
間葉幹細胞分化
本発明は、関節炎および関節損傷を処置または予防するための方法および組成物を提供する。
【発明の詳細な説明】
【技術分野】
【0001】
関連出願に対する相互参照
本出願は、2009年7月14日に出願された米国仮出願第61/225,293の35U.S.C.§1.119(e)の下での利益を請求し、その全体の内容が、参照により組み込まれる。
【背景技術】
【0002】
骨関節炎(OA)は、もっとも一般的な筋骨格障害を示す。約4000万人のアメリカ人が、近年罹患しており、この数は、高齢化や平均寿命の上昇の結果として、次の20年のうちに6000万に上昇し、身体障害の原因の第4位になることが予測されている。OAは、関節軟骨(関節の潤滑および緩衝を生む細胞およびマトリックスを含む)および関節軟骨の下部の軟骨下骨の両方を含む関節の緩やかな変性衰弱を特徴とする。近年のOA治療は、経口NSAIDsまたは選択的シクロオキシゲナーゼ2(COX−2)阻害剤での鎮痛、コルチコステロイドおよびヒアルロナンなどの剤の関節内(IA)注射、ならびに外科的アプローチを含む。
【0003】
間葉幹細胞(MSCs)は、成人関節軟骨に存在し、単離してインビトロで軟骨細胞および他の間葉細胞系列へ分化させるようにプログラムすることができる。それは、部分的に、成長因子(TGFβ、BMPs)、血清条件および細胞間接触によって制御される。
【発明の概要】
【0004】
本発明は、哺乳動物における関節炎または関節損傷を改善または予防する方法を提供する。ある実施態様において、該方法は、配列番号1または25に対して少なくとも95%の同一性を有するアミノ酸配列を含む有効量のポリペプチドを含む組成物を、哺乳動物の関節に投与し、それによって、哺乳動物における関節炎または関節損傷を改善または予防することを含む。
【0005】
ある実施態様において、該個体は、関節炎または関節損傷を有している。ある実施態様において、該個体は、関節炎または関節損傷を有していないが、関節炎または関節損傷のリスクを有する。
【0006】
ある実施態様において、該ポリペプチドは、配列番号1または25を含む。
【0007】
ある実施態様において、該アミノ酸配列は、配列番号2、3、4、26、27、または28に対して少なくとも95%の同一性を有する。ある実施態様において、該アミノ酸配列は、配列番号2、3、4、26、27、または28を含む。
【0008】
ある実施態様において、該アミノ酸配列は、配列番号5〜24のいずれかに対して少なくとも80%同一である。ある実施態様において、該アミノ酸配列は、配列番号5〜24のいずれかを含む。
【0009】
ある実施態様において、該関節炎は、骨関節炎、外傷的関節炎、および自己免疫関節炎からなる群から選択される。
【0010】
ある実施態様において、該哺乳動物はヒトである。
【0011】
ある実施態様において、該組成物は、さらにヒアルロン酸を含む。
【0012】
本発明は、軟骨基質を形成する軟骨細胞への間葉幹細胞の分化を誘導する方法も提供する。ある実施態様において、該方法は、配列番号1または25に対して少なくとも95%の同一性を有するアミノ酸配列を含む十分量のポリペプチドを間葉幹細胞に接触させ、軟骨細胞への幹細胞の分化を誘導することを含む。
【0013】
ある実施態様において、該方法は、インビトロで実行される。ある実施態様において、該方法は、インビボで実行され、該幹細胞は、哺乳動物に存在する。ある実施態様において、該哺乳動物はヒトである。
【0014】
ある実施態様において、該ポリペプチドは配列番号1または25を含む。
【0015】
ある実施態様において、該アミノ酸配列は、配列番号2、3、4、26、27、または28に対して少なくとも95%の同一性を有する。ある実施態様において、該アミノ酸配列は、配列番号2、3、4、26、27、または28を含む。
【0016】
ある実施態様において、該アミノ酸配列は、配列番号5〜24のいずれかに対して少なくとも80%同一である。ある実施態様において、該アミノ酸配列は、配列番号5〜24のいずれかを含む。
【0017】
本発明は、関節内輸送および全身性輸送のための医薬組成物も提供する。ある実施態様において、該組成物は、配列番号1または25に対して少なくとも95%の同一性を有するアミノ酸配列を含む医薬的に有効量のポリペプチドを含む。
【0018】
ある実施態様において、該組成物は、さらにヒアルロン酸を含む。
【0019】
ある実施態様において、該ポリペプチドは、配列番号1または25を含む。
【0020】
ある実施態様において、該アミノ酸配列は、配列番号2、3、4、26、27、または28に対して少なくとも95%の同一性を有する。ある実施態様において、該アミノ酸配列は、配列番号2、3、4、26、27、または28を含む。
【0021】
ある実施態様において、該アミノ酸配列は、配列番号5〜24のいずれかに対して少なくとも80%同一である。ある実施態様において、該アミノ酸配列は、配列番号5〜24のいずれかを含む。
【0022】
本発明のさらなる特徴、長所、実施態様は、次の詳細な説明および請求項の考察から説明される、または明らかであろう。さらに、先述の発明の要旨および、次の詳細な説明の両方が、例示であり、限定なしに請求項に規定された発明の範囲のさらなる説明を提供するためのものであることが、理解されるべきである。
【0023】
定義
用語「ペプチド模倣物」および「模倣物」は、天然または非天然に生じるポリペプチド(例えば、ANGPTL3)と実質的に同一の構造的および機能的な特徴を有する合成化学化合物に言及する。ペプチド類似物は、テンプレートペプチドのものと類似の特性を有する非ペプチド薬として製薬産業において広く用いられている。これらのタイプの非ペプチド化合物は、「ペプチド模倣物」または「ペプチド模倣物」と呼ばれる(Fauchere,J.Adv.Drug Res.15:29(1986);Veber and Freidinger TINS p.392(1985);およびEvans et al.J.Med.Chem.30:1229(1987)。これらは、参照によって本明細書に包含される)。ペプチド模倣物は、治療的に有用なペプチドに構造的に類似するものであり、等価のまたは強化した治療的または予防的効果を産出するために使用してもよい。一般的には、ペプチド模倣物は、目的のポリペプチドにおいて見出されるなどの模範ポリペプチド(すなわち、生物学的または薬理的活性を有するポリペプチド)に構造的に類似するが、例えば、−CH2NH−、−CH2S−、−CH2−CH2−、−CH=CH−(シスおよびトランス)、−COCH2−、−CH(OH)CH2−、および−CH2SO−からなる群から選択される結合によって任意的に交換された1または複数のペプチド結合を有する。該模倣物は、完全に合成、非天然のアミノ酸類似物からなるか、または、部分的な天然ペプチドアミノ酸および部分的な非天然アミノ酸類似物のキメラ分子であることができる。該模倣物は、模倣物の構造および/または活性を実質的に改変しない置換でもある限り、任意の量の天然アミノ酸保存的置換を包含することもできる。例えば、模倣物組成物は、目的のポリペプチドの少なくとも1つの活性を実現することができる場合には、本発明の範囲内である。
【0024】
用語「ポリペプチド」、「ペプチド」および「タンパク質」は、本明細書において、アミノ酸残基のポリマーに言及するために区別せずに用いられる。該用語は、1または複数のアミノ酸残基が天然に生じる対応のアミノ酸の人工的化学模倣物であるアミノ酸ポリマーに対して用いられ、また、天然に生じるアミノ酸ポリマーおよび非天然に生じるアミノ酸ポリマーに対して用いられる。
【0025】
用語「アミノ酸」は、天然に生じる合成アミノ酸に言及し、また、天然に生じるアミノ酸に類似する様式で機能するアミノ酸類似物、およびアミノ酸模倣物に言及する。天然に生じるアミノ酸は、遺伝的コードによってコードされるものであり、また、例えば、ヒドロキシプロリン、γ−カルボキシグルタミン酸、およびO−ホスホセリンなどの後で修飾されたアミノ酸である。アミノ酸類似物は、天然に生じるアミノ酸としての基本的化学構造、すなわち、水素、カルボキシル基、アミノ基、および、例えば、ホモセリン、ノルロイシン、メチオニンスルホキシド、メチオニンメチルスルホニウムなどのR基に結合した
α炭素、と同一の構造を有する化合物に言及する。このような類似物は、修飾されたR基(例えば、ノルロイシン)または修飾されたペプチド骨格を有するが、天然に生じるアミノ酸と同一の基本的化学構造を保持する。天然にコードされるアミノ酸は、20の共通のアミノ酸(アラニン、アルギニン、アスパラギン、アスパラギン酸、システイン、グルタミン、グルタミン酸、グリシン、ヒスチジン、イソロイシン、ロイシン、リジン、メチオニン、フェニルアラニン、プロリン、セリン、スレオニン、トリプトファン、チロシン、およびバリン)およびピロールリジンならびにセレノシステインである。
【0026】
「保存的に修飾された変異体」は、アミノ酸および核酸の両方の配列に対して用いられる。特定の核酸配列に関して、保存的に修飾された変異体は、同一である、または本質的に同一であるアミノ酸配列をコードする、またはアミノ酸配列をコードしない核酸に言及し、本質的に同一である配列に言及する。遺伝的コードの縮重のため、多数の機能的に同一である核酸が、任意の所与のタンパク質をコードする。たとえば、コドンGCA、GCC、GCGおよびGCUは、すべて、アミノ酸アラニンをコードする。したがって、アラニンがコドンによって特定される各々の位置において、該コドンは、コードされるポリペプチドを改変することなく、説明された対応コドンのいずれかに改変することができる。
このような核酸変異は、「サイレント変異」であり、保存的に修飾された変異の一種である。本明細書において、ポリペプチドをコードする各核酸配列は、核酸の各々のありうるサイレント変異も説明する。当業者は、核酸における各々のコドン(但し、通常メチオニンのための唯一のコドンであるAUG、および通常メチオニンのための唯一のコドンであるトリプトファンTGGを除く)は、機能的に同一である分子を産出するために変更することができることを認識するであろう。したがって、ポリペプチドをコードする核酸の各々のサイレント変異は、各々記述された配列において、黙示されている。
【0027】
アミノ酸配列に関して、コードされる配列における単一のアミノ酸または少数のパーセンテージのアミノ酸を改変、付加または欠失させる、核酸、ペプチド、ポリペプチド、またはタンパク質配列に対する個々の置換、欠失または付加が、改変によって化学的に類似するアミノ酸でアミノ酸を置換させる「保存的に修飾された変異体」であることを、当業者は認識するであろう。機能的に類似するアミノ酸を提供する保存的置換の一覧は、本技術分野においてよく知られている。このような保存的に修飾された変異体は、本発明の多型変異体、種間ホモログ、およびアレルに対して付加されるものであり、これらを排斥するものではない。
【0028】
次の8つの群は、互いに保存的置換であるアミノ酸をそれぞれ含む:
1)アラニン(A)、グリシン(G);
2)アスパラギン酸(D)、グルタミン酸(E);
3)アスパラギン(N)、グルタミン(Q);
4)アルギニン(R)、リジン(K);
5)イソロイシン(I)、ロイシン(L)、メチオニン(M)、バリン(V);
6)フェニルアラニン(F)、チロシン(Y)、トリプトファン(W);
7)セリン(S)、スレオニン(T);および
8)システイン(C)、メチオニン(M)(例えば、Creighton,Proteins(1984)を参照)。
【0029】
「配列同一性のパーセンテージ」は、最適にアラインされた2つの配列を、該2つの配列の最適アラインメントのために、比較窓で比較することによって測定され、ここで、比較窓におけるポリヌクレオチド配列の部分は、付加または欠失を含まない参照配列(例えば、本発明のポリペプチド)と比較して付加または欠失(すなわち、ギャップ)を含んでもよい。該パーセンテージは、同一である核酸塩基またはアミノ酸残基が、両方の配列に生じる位置の数を測定し、一致した位置の数を得て、一致した位置の数を、比較の窓における位置の総数で割り、その結果に100をかけて、配列同一性のパーセンテージを得ることによって計算される。
【0030】
2またはそれを超える核酸またはポリペプチド配列の文脈において、用語「同一である」またはパーセント「同一性」は、同一の配列である2またはそれを超える配列または部分列に言及する。2つの配列が、比較窓にわたる最大一致にわたって、または、次の配列比較アルゴリズムの1つを用いてまたは手動アラインメントおよび目視検査によって測定される指定領域にわたって、比較してアラインした場合に、特定のパーセンテージ同一である(すなわち、特定の領域にわたって、または、特定されない場合には、完全な配列にわたって、60%同一である、任意的に65%、70%、75%、80%、85%、90%、または95%同一である)アミノ酸残基またはヌクレオチドを有する場合には、2つの配列は、「実質的に同一である」。本発明は、本明細書において例示されるポリペプチド(例えば、配列番号1〜28のいずれか)にそれぞれ実質的に同一であるポリペプチド、およびその使用を提供し、その使用は、関節炎または関節損傷を処置または予防するための使用を含むが、それらに限定されない。任意的に、該同一性は、少なくとも約50ヌクレオチドの長さの領域にわたって、より好ましくは、100から500または1000またはそれを超えるヌクレオチドの長さの領域にわたって、または、参照配列の完全長にわたって、存在する。
【0031】
配列比較のためには、典型的には、1つの配列が、参照配列として作用し、試験配列は、これと比較される。配列比較アルゴリズムを用いる場合、試験および参照配列が、コンピューターに入力され、必要に応じて部分列座標が指定され、そして、配列アルゴリズムプログラムパラメーターが指定される。デフォルトプログラムパラメーターを使用することができ、また、別のパラメーター指定することができる。配列比較アルゴリズムは、続いて、プログラムパラメーターに基づき、参照配列と比較した試験配列におけるパーセント配列同一性を計算する。
【0032】
本明細書において、「比較窓」は、20〜600、普通には、約50〜約200、より普通には約100〜約150からなる群から選択される、隣接位置の数の任意の1つの断片への参照を含み、ここで、配列は、2つの配列が最適にアラインされた後、同一数の隣接位置の参照配列と比較されてもよい。比較のための配列のアラインメントの方法は、本技術分野においてよく知られている。比較のための配列の最適なアラインメントは、例えば、Smith and Waterman(1970)Adv.Appl.Math.2:482cのローカルホモロジーアルゴリズムによって、Needleman and Wunsch(1970)J.MoI.Biol.48:443のホモロジーアラインメントアルゴリズムによって、Pearson and Lipman(1988)Proc.Nat’l.Acad.Sci.USA 85:2444の類似性検索の方法によって、これらのアルゴリズムのコンピューターでの遂行によって(Wisconsin Genetics Software Package、 Genetics Computer Group、 575 Science Dr., Madison、 WIにおけるGAP、BESTFIT、FASTA、およびTFASTA)、または、手動アラインメントおよび目視検査によって(例えば、Ausubel et al., Current Protocols in Molecular Biology(1995supplement)を参照)によって行うことができる。
【0033】
パーセント配列同一性および配列類似性の測定に適したアルゴリズムの2つの例が、BLASTおよびBLAST2.0アルゴリズムであり、それぞれAltschul et al.(1977)Nuc. Acids Res.25:3389−3402およびAltschul et al.(1990)J. MoI. Biol.215:403−410に説明されている。BLAST分析を実行するためのソフトウェアは、National Center for Biotechnology Informationを通して公的に入手可能である。このアルゴリズムは、データベース配列における同一長のワードとアラインした場合にいくつかの正値閾値スコアTと一致するか、それを満たすかのいずれかである、クレリー配列におけるW長のショートワードを同定することによって、高スコア配列ペア(HSPs)をまず同定することを含む。Tは、近隣ワードスコア閾値とも呼ばれる(上記Altschul et al.,)。これらの最初の近隣ワードヒットは、それらを含むより長いHSPsを探すためのデフォルト検索の種としての役割を果たす。該ワードヒットは、累積アラインメントスコアが上昇することができる限り、各配列にそって両方向に延長される。累積スコアは、ヌクレオチド配列のためにはパラメーターM(一致残基のペアのためのリワードスコア;常に>0)およびN(非一致残基のためのペナルティスコア;常に<0)を用いて計算される。アミノ酸配列のためには、スコアリングマトリックスが、累積スコアを計算するために用いられる。それぞれの方向におけるワードヒットの延長は、累積アラインメントスコアが、その最大達成値からの数Xによって低下する場合、1または複数の負スコア残基アラインメントの蓄積のために累積スコアが、ゼロまたはそれ未満になる場合、または、いずれかの配列の末端に到達した場合に中止される。BLASTアルゴリズムパラメーターW、T、およびXは、アラインメントの感度およびスピードを決定する。(ヌクレオチド配列のための)BLASTNプログラムは、デフォルトとして、11のワード長(W)、10の期待値(E)M=5、N=−4および、両ストランドの比較を使用する。アミノ酸配列のためのBLASTPプログラムは、デフォルトとして、3のワード長、10の期待値(E)、およびBLOSUM62スコアリングマトリックス(Henikoff and Henikoff(1989)Proc.Natl.Acad.Sci.USA 89:10915を参照)、50のアラインメント(B)、10の期待値(E)M=5、N=−4、および両ストランドの比較を用いる。
【0034】
BLAST アルゴリズムは、2つの配列の間の類似性の統計的分析も実行する(例えば、Karlin and Altschul(1993)Proc.Natl.Acad.Sci.USA 90:5873−5787を参照)。BLASTアルゴリズムによって提供される類似性の1つの測定は、最小合計確率(P(N))であり、これは、2つのヌクレオチドまたはアミノ酸配列の間の一致が偶然に起こりうる確率の指標を提供する。例えば、もし、参照核酸との試験核酸の比較における最小合計確率が、約0.2未満、より好ましくは約0.01未満、および最も好ましくは約0.001未満である場合には、核酸は、参照配列と類似していると考えられる。
【0035】
本明細書において、用語「ヒアルロン酸」は、ヒアルロン酸のエステルおよびヒアルロン酸の塩を含むヒアルロン酸の誘導体を含むものとして使用され、用語ヒアルロナンも含む。該呼称は、低および高分子量型のヒアルロナンの両方、および、架橋ヒアルロナンまたはハイランも含む。このようなヒアルロナンの例は、Synvisc(商標)(Genzyme Corp.Cambridge,Mass.),ORTHO VISC(商標)(Anika therapeutics,Woburn,Mass.)、およびHYALGAN(商標)(Sanofi−Synthelabo Inc.,Malvern,Pa.)である。
【図面の簡単な説明】
【0036】
【図1】インビトロ軟骨形成の間のMSC−誘導II型コラーゲン産生の定量的評価。 hMSCs(10,000/384ウェル)をhMSC増殖培地(Millipore)において24時間プレートした。該細胞を、上記タンパク質でさらに72時間処理した。培地を無血清DMEMに交換し、追加の刺激なしにさらに14日間培養した。終わりに際して、細胞をホルマリンで固定し、洗浄し、II型コラーゲンに対する抗体で染色し、DAPIで対比染色した。II型コラーゲン染色の量は、ハイコンテントイメージング(Opera、 Perkin Elmer)によって定量化した。データは、3つの実験からそれぞれの投与量について2つプールした(n=6)。
【0037】
【図2】ANGTPL3−誘導軟骨形成の特性評価。 (A)ペレット培養(1x106細胞/ペレット)において、21日間、無血清DMEM、1×ITSおよびANGTPL3(指定の場合)中でhMSCsを培養した。培地を3日ごとに交換した。製造者の指示によるヒトTaqman特異的プローブを用いてアグレカンmRNA発現を定量化した(3つの実験から2つずつプールされたデータ(n=6))。 (B)ウシ軟骨を単離し、叩いて対称円にし、器官培養に入れた。該スライスを48時間20ng/ml TNFaおよび10ng/ml OSM(炎症性メディエーター)で処理して、ANGPTL3の存在下または非存在下で、軟骨マトリックスの分解を誘導し、マトリックス損傷の指標であるグルコサミノグルカン放出のパーセント阻害を同定した(4つのドナーからプールしたデータ、n=12)。 (C)初期正常ヒト軟骨細胞および滑膜細胞(増殖培地中、2500/384ウェル)を、プレートし、24時間37℃で培養した。24時間の期間をとおしてANGTPL3に応じた穏やかな軟骨細胞特異的細胞増殖の上昇が実証された(2つの実験からプールされたデータ、2反復/投与量(n=4))。
【0038】
【図3】mANGTPTL3。 (A)主要タンパク質構造および確認されたグリコシル化部位。 (B)mANGTPL3C末端FLD(225−455)の原子構造(1.8オングストローム)。
【0039】
【図4】ANGPTL3配列の配列アラインメント。 (A)ヒト、マウス、ウシ、およびイヌネイティブANGPTL3タンパク質の配列アラインメント; (B)ヒト、マウス、ウシ、イヌおよびウマネイティブANGPTL3タンパク質の配列アラインメント。
【0040】
【図5】外科骨関節炎モデルにおけるのインビボ効能。 C57BL/6マウス(n=12/群)からの右膝の前十字靱帯(ACL)、内側半月板脛骨靱帯(MMTL)、および内側側副靱帯(MCL)の外科的切断を行い、膝関節における不安定化を誘導し、それによってOA表現型を引き起こした。外科手術の1週間後、マウスに3〜4週間に1回ごと、関節内投与を行った。 (A)末梢血を、外科手術の28日後に収集した。循環II型コラーゲン断片(CTX−II)をELISA(Nordic biosciences)によって定量化した。mANGPTL3投与量=200ng/膝、3週間ごとの注射。 (B)脛骨プラトーの定量的評価を0〜4スケールで行った。0は正常であり、5は重度のOAである(軟骨の全層破壊)。各マウスからの2つの片が、2の観察者によって無分別に等級分けされた。mANGPTL3投与量=200ng/膝、3週間ごと注射。 (C)インキャパシタンステストによって、または、モニター装置を通して外科手術の脚と別の脚とで立ったマウスの時間のパーセンテージを測定することによって、OA痛を測定した。読み出し値は、外科手術36日後の痛応答を示す(示された濃度でANGPTL3の1週間ごと投与量の3投与量)。
【発明を実施するための形態】
【0041】
詳細な説明
I.導入
【0042】
本発明は、アンジオポイエチン様3(ANGPTL3)が間葉幹細胞における軟骨細胞分化を刺激するという発見に、部分的に基づいている。したがって、本発明は、軟骨細胞への間葉幹細胞分化を誘導する方法を提供する。また、本発明は、ANGPTL3タンパク質を関節、椎骨、椎間板、または全身に投与することによって関節炎または関節損傷を予防または改善するためのANGPTL3タンパク質の投与を提供する。
II.アンジオポイエチン様3
【0043】
アンジオポイエチン様3は、分泌因子のアンジオポイエチン様ファミリーのメンバーである。これは、主に肝臓に発現し、シグナルペプチド、N−末端コイルドコイルドメイン(CCD)およびC末端フィブリノゲン(FBN)様ドメインからなるアンジオポイエチンの特徴構造を有する。アンジオポイエチン様3におけるFBN様ドメインは、αV/β3インテグリンに結合し、この結合が、内皮細胞接着および遊走を誘導することが示された。
【0044】
様々なANGPTL3タンパク質が、本発明において使用することができる。本明細書において説明されるように、ネイティブANGPTL3は、一般的には、インビボで開裂され、アミノ−末端およびカルボキシル末端断片になる。本発明は、軟骨形成活性を有する様々なANGPTL3タンパク質の使用を想定する。ある実施態様において、本発明は、完全長ネイティブ(またはその変異体)ANGPTL3タンパク質アミノ酸配列の使用を提供する。ある実施態様において、本発明は、ANGPTL3配列、またはその変異体の(完全長ネイティブ配列ではない)部分を含むANGPTL3タンパク質であって、軟骨形成活性を保持するもの、すなわち、ネイティブタンパク質のアミノ酸末端側終端ではないものを提供する。ある実施態様において、本発明のANGPTL3タンパク質は、CCDドメインを持たない、および/または、有意なCCD活性を持たない。したがって、ある実施態様において、本発明のANGPTL3タンパク質は、ネイティブマウス(例えば、配列番号12)、ヒト(例えば、配列番号8)、ウシ(例えば、配列番号16)、イヌ(例えば、配列番号20)、またはウマ(例えば、配列番号24)ANGPTL3タンパク質配列、または実質的に同一である配列の少なくとも断片(例えば、少なくとも50、100、150、200、250の隣接するアミノ酸)を含むが、ネイティブANGPTL3タンパク質の少なくとも200の隣接するアミノ−末端アミノ酸を含まない。
【0045】
ある実施態様において、本発明のANGPTL3タンパク質は、フィブリノゲン−様ドメインを含む。ある実施態様において、本発明のANGPTL3タンパク質は、ネイティブマウス(例えば、配列番号12)、ヒト(例えば、配列番号8)、ウシ(例えば、配列番号16)、イヌ(例えば、配列番号20)、またはウマ(例えば、配列番号24)ANGPTL3タンパク質配列のアミノ酸207−455、207−400、207−350、225−455、225−400、225−350、241−455、241−400、241−350に対応する隣接するアミノ酸を含む、または、これらの配列と実質的同一であるが、フランキングネイティブANGPTL3タンパク質アミノ酸配列を含まない。ある実施態様において、本発明のANGPTL3タンパク質(配列番号1〜28のいずれかを含むがこれらに限定されない)は、C末端配列の少なくとも部分を欠き、例えば、C−末端からの10、20、30、40、50アミノ酸を欠く。
【0046】
上記の本発明のANGPTL3タンパク質は、上記の領域の側面に位置するネイティブANGPTL3タンパク質配列を含まなくてよいが、本発明のANGPTL3タンパク質は、非ネイティブANGPTL3タンパク質フランキング配列を含むことができる。例えば、ANGPTL3タンパク質の軟骨形成活性部分は、1または複数の異種アミノ酸に融合させて、融合タンパク質を形成させることができる。融合パートナー配列は、アミノ酸タグ非−L(例えば、D−)アミノ酸、またはインビボ半減期を伸ばすための、および/もしくはプロテアーゼ耐性を高めるための他のアミノ酸模倣物、標的化配列または他の配列を含むことができるが、それらに限定されない。
【0047】
本発明のANGPTL3タンパク質は、ネイティブANGPTL3タンパク質の変異体およびトランケーション、ならびに上記活性断片の変異体およびトランケーションを包含する。活性変異体は、本技術分野における当業者に知られているあらゆる方法によって同定することができる。ある実施態様において、活性タンパク質のアミノ酸アラインメントは、不変である位置、または保存されたアミノ酸変化を含む位置を同定するために確立されうる。配列番号1、2、3、または4は、ヒト、マウス、ウシ、およびイヌネイティブANGPTL3タンパク質のある領域(それぞれ、位置241−455、225−455および207−455、ならびにネイティブ完全長)の間に不変アミノ酸を含むコンセンサス配列を表す。配列番号25、26、27、または28は、ヒト、マウス、ウシ、イヌ、およびウマネイティブANGPTL3タンパク質のある領域(それぞれ、位置241−455、225−455および207−455、ならびにネイティブ完全長)の間において不変アミノ酸を含むコンセンサス配列を表す。したがって、ある実施態様において、本発明の軟骨形成ANGPTL3タンパク質は、配列番号1、2、3、4、25、26、27、または28を含む。ある実施態様において、本発明の軟骨形成ANGPTL3タンパク質は、配列番号5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23または24のいずれかと(配列番号の長に渡って測定して)実質的に同一であるアミノ酸配列を含む。
【0048】
本発明のANGPTL3タンパク質は、軟骨形成活性を有する。本明細書において定義されるように、軟骨形成または軟骨形成活性は、MSCsからの軟骨細胞の成長に言及する。軟骨形成活性の指標は、軟骨マトリックス産生を含むがこれらに限定されない。軟骨マトリックス産生は、様々なマーカー、例えば、Sox9、II型コラーゲン、またはグルコサミノグルカン(GAG)産生によって測定してもよい。ある実施態様において、GAG産生は、軟骨マトリックス産生のためのマーカーとして測定される。ある実施態様において、軟骨特異的タンパク質発現でのGAG産生の3倍増加は、陽性の軟骨マトリックス産生を示す。
【0049】
ある実施態様において、本発明のANGPTL3ポリペプチドは、少なくとも1つの非天然にコードされるアミノ酸を含むであろう。非天然に生じるアミノ酸を作製しタンパク質に導入する方法は、知られている。例えば、米国特許第7,083,970、および7,524,647を参照のこと。1または複数の所望の非天然アミノ酸を含むタンパク質の作製に適した直交翻訳システムの作成のための一般的な原理は、直交翻訳システムを作成するための一般的な方法として、本技術分野において知られている。例えば、“METHODS AND COMPOSITION FOR THE PRODUCTION OF ORTHOGONAL tRNA−AMINOACYL−tRNA SYNTHETASE PAIRS;”と題された国際公開公報WO2002/086075、“IN VIVO INCORPORATION OF UNNATURAL AMINO ACIDS;”と題されたWO2002/085923、“EXPANDING THE EUKARYOTIC GENETIC CODE;”と題されたWO2004/094593、“ORTHOGONAL TRANSLATION COMPONENTS FOR THE VIVO INCORPORATION OF UNNATURAL AMINO ACIDS”と題され、2004年7月7日出願のWO2005/019415、2004年7月7日出願のWO2005/007870、2004年7月7日出願のWO2005/007624、2005年10月27日出願のWO2006/110182、および“SYSTEMS FOR THE EXPRESSION OF ORTHOGONAL TRANSLATION COMPONENTS IN EUBACTERIAL HOST CELLS”と題され2007年3月7日出願のWO2007/103490を参照のこと。これらのそれぞれの内容は、参照によって全体が組み込まれる。非天然アミノ酸を組み込む直交翻訳システム、およびその産生および使用の方法の議論のためには、Wang and Schultz,(2005)“Expanding the Genetic Code.”Angewandte Chemie Int Ed 44:34−66;Xie and Schultz,(2005)“An Expanding Genetic Code.”Methods 36:227−238;Xie and Schultz,(2005)“Adding Amino Acids to the Genetic Repertoire.”Curr Opinion in Chemical Biology 9:548−554;およびWang,et al,(2006)“Expanding the Genetic Code.”Annu Rev Biophys Biomol Struct 35:225−249;Deiters,et al,(2005)“In vivo incorporation of an alkyne into proteins in Escherichia coli.”Bioorganic&Medicinal Chemistry Letters 15:1521−1524;Chin,et al.,(2002)“Addition of p−Azido−L−phenylalanine to the Genetic Code of Escherichia coli.”J Am Chem Soc 124:9026−9027;および2005年9月20日出願の国際公開公報WO2006/034332も参照のこと。これらのそれぞれの内容は、参照によって全体が組み込まれる。さらなる詳細は、米国特許第7,045,337;第7,083,970;第7,238,510;第7,129,333;第7,262,040;第7,183,082;第7,199,222;および第7,217,809に見出される。
【0050】
「非天然にコードされるアミノ酸」は、共通アミノ酸の1つではないアミノ酸またはピロールリジンもしくはセレノシステインに言及する。用語「非天然にコードされるアミノ酸」と同意語として用いられる他の用語は、「非天然アミノ酸」、「非天然アミノ酸」、「非天然に生じるアミノ酸」、および様々にハイフンで結んだ、およびハイフンで結んでいないそれらのバージョンである。用語「非天然にコードされるアミノ酸」は、天然にコードされるアミノ酸(20の共通アミノ酸、またはピロールリジンおよびセレノシステインを含むがこれらに限定されない)の修飾(例えば、翻訳後修飾)によって生じるが、それ自体増殖しているポリペプチド鎖には翻訳複合体によって天然に組み込まれないアミノ酸も含むがそれらに限定されない。このような非天然に生じるアミノ酸の例は、N−アセチルグルコサミニル−L−セリン、N−アセチルグルコサミニル−L−スレオニン、およびO−フォスフォチロシンを含むがこれらに限定されない。
【0051】
非天然にコードされるアミノ酸は、典型的には20の天然アミノ酸において用いられるものとは別の任意の置換側鎖を有する任意の構造である。本発明の非天然にコードされるアミノ酸は、典型的には、天然アミノ酸とは側鎖の構造の点でのみ異なるので、非天然にコードされるアミノ酸は、天然にまたは非天然にコードされるものを非限定的に含む他のアミノ酸と、天然に生じるポリペプチドにおいて形成されるのと同一の様式でアミド結合を形成する。しかしながら、非天然にコードされるアミノ酸は、天然アミノ酸からそれらを区別する側鎖基を有する。例えば、Rは、任意的にアルキル−、アリール−、アシル−、ケト−、アジド−、ヒドロキシル−、ヒドラジン、シアノ−、ハロ−、ヒドラジド、アルケニル、アルキニル、エーテル、チオール、セレノ−、スルホニル−、ホウ酸塩、ボロン酸、ホスホ、ホスホノ、ホスフィン、複素環、エノン、イミン、アルデヒド、エステル、チオ酸、ヒドロキシルアミン、アミノ基、もしくは同様のもの、またはその任意の組み合わせを含む。本発明における使用に適しうる他の非天然に生じる目的のアミノ酸は、光活性化可能な架橋剤を含むアミノ酸、スピン標識アミノ酸、蛍光アミノ酸、金属結合アミノ酸、金属−含有アミノ酸、放射性活性アミノ酸、新規の官能基を有するアミノ酸、他の分子と共有結合的にまたは非共有結合的に相互作用するアミノ酸、フォトケージされたおよび/または光異性化可能なアミノ酸、ビオチンまたはビオチン類似物を含むアミノ酸、糖置換セリンなどのグリコシル化アミノ酸、他の炭水化物修飾アミノ酸、ケト−含有アミノ酸、ポリエチレングリコールまたはポリエーテルを含むアミノ酸、重原子置換アミノ酸、化学的に開裂可能なおよび/または光開裂可能なアミノ酸、約5を超えるまたは約10を超える炭素を非限定的に含むポリエーテルまたは長鎖炭化水素を非限定的に含む天然アミノ酸と比較して伸張した側鎖を持つアミノ酸、炭素接続糖類含有アミノ酸、レドックス活性アミノ酸、アミノチオ酸含有アミノ酸、および1または複数の毒性部分を含むアミノ酸を含むがそれらに限定されない。
【0052】
本発明における使用に適し得て、水溶性ポリマーとの反応に有用な非天然にコードされるアミノ酸は、例えば、カルボニル、アミノオキシ、ヒドラジン、ヒドラジド、セミカルバジド、アジドおよびアルキン反応基を有するものを含むがこれらに限定されない。ある実施態様において、非天然にコードされるアミノ酸は、サッカリド部分を含む。このようなアミノ酸の例は、N−アセチル−L−グルコサミニル−L−セリン、N−アセチル−L−ガラクトサミニル−L−セリン、N−アセチル−L−グルコサミニル−L−スレオニン、N−アセチル−L−グルコサミニル−L−アスパラギンおよびO−マンノサミニル−L−セリンを含む。このようなアミノ酸の例は、天然に生じるアミノ酸とサッカリドとの間のN−またはO−結合が、天然には一般には見出されない共有結合(アルケン、オキシム、チオエーテル、アミド、および同等物を含むがこれらに限定されない)に交換されている例も含む。このようなアミノ酸の例は、2−デオキシ−グルコース、2−デオキシガラクトース、および同等物などの天然に生じるタンパク質において一般的には見出されないサッカリドも含む。
【0053】
例えば、インビボ半減期を伸張するために、本発明のANGPTL3タンパク質に(例えば、ポリペプチド鎖内において、または、N−もしくはC末端のいずれかにおいて)導入できる別の型の修飾は、PEG化または長鎖ポリエチレングリコールポリマー(PEG)の組み込みである。PEGまたはPEGの長鎖ポリマーの導入は、例えば、尿への急速ろ過を予防するために、本発明のポリペプチドの有効分子量を増大させる。ある実施態様において、ANGPTL3配列におけるリジン残基は、PEGに直接的にまたはリンカーを通して、接合される。このようなリンカーは、例えば、適切に修飾されたPEG鎖への結合のためのチオール官能基を含有するGlu残基またはアシル残基であることができる。PEG鎖を導入するための別の方法は、まずC−末端に、または溶媒露出残基に、ArgまたはLys残基の代用などのCys残基を導入する。このCys残基は、次に、例えば、マレイミド機能を含有するPEG鎖に部位特異的に付着させられる。PEGまたはPEGの長鎖ポリマーを組み込むための方法は、本技術分野においてよく知られており(例えば、Veronese,F.M.,et al.,Drug Disc.Today 10:1451−8(2005);Greenwald,R.B.,et al.,Adv.DrugDeliv.Rev.55:217−50(2003);Roberts,M.J.,et al.,Adv.DrugDeliv.Rev.,54:459−76(2002)などに説明されている)、その内容は参照によって本明細書に包含される。本技術分野において知られているポリマー接合の別の方法も、本発明において使用することができる。ある実施態様において、ポリ(2−メタクリロイルオキシエチルホスホリルコリン)(PMPC)が、本発明のANGPTL3タンパク質へのポリマー接合として導入される(例えば、WO2008/098930;Lewis,et al.,Bioconjug Chem.,19:2144−55(2008)を参照のこと)。ある実施態様において、ANGPTL3タンパク質と接合したホスホリルコリン−含有ポリマーが、本発明において使用することができる。当業者は、別の生体適合性ポリマー接合を利用することができることを容易に認識するであろう。
【0054】
(上記などの)非天然アミノ酸の組み込みを通してPEGまたはPEGポリマーを組み込むためのより最近に報告された改変ネイティブアプローチは、本発明のポリペプチドで実行することができる。このアプローチは、進化したtRNA/tRNAシンテターゼペアを利用し、アンバーサプレッサーコドンによる発現プラスミドにおいてコードされる(Deiters,A,et al.(2004).Bio−org.Med.Chem.Lett.14,5743−5)。例えば、p−アジドフェニルアラニンは、本発明のポリペプチドに組み込まれ、そして、還元剤および銅イオンの存在下で、アセチレン部分を有するPEGポリマーとの反応に供され、「ヒュスゲン[3+2]付加環化」として知られる有機反応を促進させることができる。
【0055】
ある実施態様において、本発明は、ポリペプチドのグリコシル化を改変するためのANGPTL3タンパク質の特異的な変異を想定する。このような変異は、O−連結またはN−連結グリコシル化部位を含むがそれらに限定されない1または複数のグリコシル化部位を導入または除去するように選択されることができる。ある実施態様において、本発明のANGPTL3タンパク質は、天然に生じるANGPTL3タンパク質と比較して非改変であるグリコシル化部位およびパターンを有する。ある実施態様において、ANGPTL3タンパク質の変異体は、グリコシル化部位の数および/またはタイプが、天然に生じるANGPTL3タンパク質と比較して改変されたグリコシル化変異体を含む。ある実施態様において、ポリペプチドの変異体は、ネイティブのポリペプチドと比較して数が多いまたは少ないN−連結グリコシル化部位を含む。N−連結グリコシル化部位は、配列:Asn−X−SerまたはAsn−X−Thrによって特徴付けられ、ここで、Xと指定されたアミノ酸残基は、プロリン以外の任意のアミノ酸残基であってよい。この配列を作製するためのアミノ酸残基の置換は、N−連結炭水化物鎖の付加のための潜在的な新しい部位を提供する。また、この配列を除去する置換は、存在するN−連結炭水化物鎖を除去するであろう。ある実施態様において、N−連結炭水化物鎖の再配列が、提供され、ここで、1または複数のN−連結グリコシル化部位(典型的には天然に生じるもの)が除去され、そして、1または複数の新しいN−連結部位が作製される。
【0056】
ANGPTL3タンパク質変異体の例は、天然に生じるANGPTL3タンパク質のアミノ酸配列と比較して1または複数のシステイン残基が欠失して別のアミノ酸(例えば、、セリン)に置換されたシステイン変異体を含む。ある実施態様において、不溶性封入体の単離後など、ANGPTL3タンパク質が、生物学的に活性なコンフォーメーションにリフォールドされなければならない場合には、システイン変異体は、有用でありうる。ある実施態様において、システイン変異体は、ネイティブのポリペプチドと比較して少ない数のシステイン残基を有する。ある実施態様において、システイン変異体は、非ペアシステインにからもたらされる相互作用を最小化するために、偶数のシステイン残基を有する。
【0057】
ある実施態様において、ANGPTL3タンパク質の機能的変異体または修飾されたフォームは、本発明のANGPTL3タンパク質の融合タンパク質および1または複数の融合ドメインを含む。融合ドメインのよく知られた例は、ポリヒスチジン、Glu−Glu、グルタチオンSトランスフェラーゼ(GST)、チオレドキシン、プロテインA、プロテインG、免疫グロブリン重鎖定常領域(Fc)、マルトース結合タンパク質(MBP)、またはヒト血清アルブミンを含むがこれらに限定されない。融合ドメインは、所望の特性を授与するために選択することができる。例えば、ある融合ドメインは、アフィニティクロマトグラフィによって融合タンパク質を単離するために特に有用である。親和性精製の目的のために、グルタチオン−、アミラーゼ−、およびニッケル−またはコバルト−接合樹脂などのアフィニティクロマトグラフィのための関連するマトリックスが使用される。このようなマトリックスの多くは、(HIS6)融合パートナーに有用なPharmacia GST purification system、およびQLAexpress(商標)system(Qiagen)などの「キット」の形で入手することができる。別の例として、融合ドメインは、ANGPTL3タンパク質の検出を促進するように選択されうる。このような検出ドメインの例は、様々な蛍光タンパク質(例えば、GFP)、ならびに通常は短いペプチド配列でありそれに対する特異的な抗体を入手することができる「エピトープタグ」を含む。特異的なモノクローナル抗体を容易に入手することができ、よく知られているエピトープタグは、FLAG、インフルエンザウイルス血球凝集(HA)、およびc−mycタグを含む。ある場合において、融合ドメインは、関連するプロテアーゼが融合タンパク質を部分的に消化し、それによってそれからの組換えタンパク質を遊離させるのを可能にする、Xa因子またはトロンビンなどのプロテアーゼ開裂部位を有する。そして、遊離されたタンパク質は、続いて行われるクロマトグラフィ分離によって融合ドメインから単離することができる。ある実施態様において、ANGPTL3タンパク質は、インビボでANGPTL3タンパク質を安定化するドメイン(「安定用」ドメイン)と融合している。「安定化」は、血清半減期を上昇させるあらゆることを意味し、これが破壊の減少、腎臓によるクリアランスの減少、または他の薬物動態効果によるかどうかは関係ない。免疫グロブリンのFc部分との融合は、幅広いタンパク質に所望の薬物動態特性を授与することが知られている。同様に、ヒト血清アルブミンへの融合は、所望の特性を授与することができる。選択されうる別のタイプの融合ドメインは、多量体化(例えば、二量体化、四量体化)ドメイン、および(所望の付加的な生物学的機能を授与する)機能的ドメインを含む。
III.アンジオポイエチン様3タンパク質疾患指標
【0058】
本発明のポリペプチド、組成物、および方法が、任意のタイプの関節炎または関節損傷の処置または予防のために使用されてもよいことが想定される。本発明のポリペプチド、組成物、および方法は、様々な軟骨性障害の処置または予防に使用されてもよいことがさらに想定される。ある実施態様において、本発明のタンパク質は、例えば、関節炎または関節損傷の遺伝的または家族歴がある、または以前に関節手術がある、もしくは関節手術の最中であるなどの、関節炎または関節損傷を予防するために投与される。本発明のポリペプチド、組成物、および方法で処置されるべきまたは予防されるべき病状または障害の例は、関節リウマチ、若年性慢性関節炎、骨関節炎、変性椎間板疾患、脊椎関節症、全身性硬化症(強皮症)、特発性炎症性ミオパシー(皮膚筋炎、多発性筋炎)、シェーグレン症候群、全身性血管炎、サルコイドーシス、自己免疫性溶血性貧血(免疫汎血球減少症、発作性夜間血色素尿症)、自己免疫性血小板減少症(特発性血小板減少性紫斑病、免疫介在性血小板減少症)、甲状腺炎(グレーブス病、橋本甲状腺炎、幼若リンパ球性甲状腺炎、萎縮性甲状腺炎)、糖尿病、免疫介在性腎疾患(糸球体腎炎、尿細管間質性腎炎)、多発性硬化、特発性脱髄性多発神経障害またはギラン−バール症候群、および慢性炎症性脱髄性多発神経障害などの中枢および末梢神経系の脱髄疾患、伝染性肝炎(肝炎A、B、C、D、E、および他の非肝炎ウイルス)などの肝胆道疾患、自己免疫性慢性活動性肝炎、原発性胆汁性肝硬変、肉芽腫性肝炎、および硬化性胆管炎、炎症性腸疾患(潰瘍性大腸炎:クローン病)、グルテン過敏性腸疾患、およびウィップル病、水疱性皮膚病を含む自己免疫または免疫介在性皮膚疾患、多形紅斑、および接触皮膚炎、乾癬、ぜんそく、アレルギー性鼻炎、アトピー性皮膚炎、食物過敏症、および、じんましん、などのアレルギー疾患、好酸球性肺炎、特発性肺線維症、および過敏性肺炎などの肺の免疫学的疾患、移植片拒絶および移植片対宿主病を含む移植関連疾患を含むがそれらに限定されない。本発明のある実施態様において、本発明のポリペプチド、組成物、および方法は、骨関節炎の処置のために使用されうる。
【0059】
ある実施態様において、本発明のポリペプチド、組成物、および方法は、外傷的損傷または軟骨疾患に起因して損傷した軟骨性組織における軟骨細胞増殖および軟骨産生を刺激するための方法を提供する。脊椎、肩、肘、手首、指の関節、尻、膝、足首、および足の関節などの関節表面を示す組織は、処置にとって特に重要である。処置の恩恵を受けうる疾患の例は、骨関節炎、リウマチ性関節炎、他の自己免疫疾患、または離断性骨軟骨症を含む。さらに、軟骨奇形が、ヒトにおいて小人症の形態でしばしば見られ、このことは、該ポリペプチド、組成物、および方法が、これらの患者において有用であろうことを示唆する。
【0060】
本発明のポリペプチド、組成物、および方法が、哺乳動物の処置のために有用であることが想定される。本明細書においては、「哺乳動物」は、ヒト、牛(例えば、雌牛)、馬、イヌ、羊、豚、ウサギ、ヤギ、ネコなどの家畜および家畜動物、ならびに動物園の動物スポーツ動物またはペット動物を含む、哺乳動物として分類される任意の哺乳動物を意味する。本発明のある実施態様において、哺乳動物はヒトである。
【0061】
ある実施態様において、本発明のポリペプチドは、処置されるべき哺乳動物とは異種であることができる。例えば、ウシANGPTL3タンパク質またはその断片、ウシANGPTL3タンパク質に由来するタンパク質またはペプチド(例えば、修飾されたウシANGPTL3タンパク質、ウシANGPTL3タンパク質の保存的変異体、ウシANGPTL3タンパク質に由来するペプチド模倣物)が、ヒト患者の処置において使用される。ある実施態様において、異種ANGPTL3タンパク質が、移植のための培養の軟骨細胞集団を拡張するために使用することができる。ある実施態様において、拡張した培養は、次に、処置されるべき哺乳動物にホモロジーがあるポリペプチドおよび組成物と混ぜ合わせられ、関節腔に、または、軟骨欠損に直接に、入れられる。また、本発明のポリペプチドは、同一の種に由来する。すなわち、ヒトANGPTL3タンパク質またはその断片、ヒトANGPTL3タンパク質に由来するタンパク質またはペプチド(例えば、修飾されたヒトANGPTL3タンパク質、ヒトANGPTL3タンパク質の保存的変異体、ヒトANGPTL3タンパク質に由来するペプチド模倣物)が、ヒト患者の処置において使用される。処置されつつある哺乳動物の同一種に由来するタンパク質の使用によって、不用意な免疫応答を避けてもよい。
【0062】
本発明のポリペプチドおよび組成物は、単独で、またはタンパク質の持続放出に適した担体との複合体で、関節の滑液への直接注射、全身性投与(経口または静脈内)、または軟骨欠損への直接投与によって適用することができる。本発明のポリペプチド、組成物、および方法は、自家または同種軟骨細胞移植のための培養における軟骨細胞集団を拡張するために使用することができる。該移植は、本発明のポリペプチドおよび組成物の投与からなる併用療法で任意的に投与することができる。これらの手順において、例えば、軟骨細胞は、損傷した関節の非損傷小耐力領域から関節鏡視下的に収集することができ、本発明のポリペプチドおよび組成物の存在下で、培養し、移植に先立ち、細胞の数を増加させることができる。拡張した培養は、続いて、本発明のポリペプチドおよび組成物と混ぜ合わされ、関節腔に、または欠損に直接に、入れられる。本発明のポリペプチドおよび組成物は、軟骨を形成することができる、および/または移植された軟骨細胞またはその前駆細胞を適所に保つのに役立つ、細胞を含む骨膜または軟骨膜移植片と組み合わせて使用することができる。本発明のポリペプチドおよび組成物は、関節の洗浄、骨髄の刺激、掻爬関節形成、骨軟骨病変ドリリング、または軟骨下骨の微小破壊と併用して、軟骨損傷の修復のために使用することができる。さらに、本発明のポリペプチドおよび組成物の投与に起因する軟骨の増殖の後、新たに形成された軟骨表面を適切に形成させるには、さらなる外科処置が必要であるかもしれない。
IV.医薬組成物
【0063】
上記の疾患または障害を処置するための本発明の化合物の投与量は、投与の様式、対象者の年齢、体重、および処置されるべき対象者の病状に応じて異なり、最終的には、主治医または主治獣医によって決定されるであろう。主治医または主治獣医によってこのように決定される化合物の量は、本明細書において「有効量」と呼ばれる。
【0064】
投与に適した剤形は、賦形剤を含み、水性および非水性溶液;酸化防止剤、緩衝剤、静菌剤、およびその剤形を等張にする溶質を含むことができる等張滅菌溶液;懸濁化剤、可溶化剤、増粘剤、安定剤、および防腐剤を含むことができる水性および非水性無菌懸濁液を含むがこれらに限定されない。
【0065】
本発明の文脈において、患者に投与される投与量は、時間とともに対象者における有益な応答をもたらすために十分であるべきである。投与量は、用いられる特定のタンパク質の効能、および対象者の病状、ならびに、体重または処置されるべき領域の表面領域によって決定されるであろう。投与量の多さは、特定の対象者における特定のタンパク質またはベクターの投与に伴う副作用の存在、性質、および程度によっても決定されるであろう。投与は、単一の投与量によって、または分けた投与量によって達成することができる。
V.投与の方法
【0066】
本発明のタンパク質を罹患した関節に輸送するためには、任意の方法を用いることができる。本発明の実施において、組成物は、例えば、関節内に(すなわち、関節へ)、経口的に、静脈内に投与することができる。化合物の製剤は、アンプルおよびバイアルなどのユニット投与量または多投与量密封容器において存在することができる。溶液および懸濁液は、前記の種類の滅菌粉末、顆粒、および錠剤から調製することができる。本発明のタンパク質は、所望の治療または効果に応じて、1または複数のさらなる活性剤(例えば、、ヒアルロン酸またはその塩)と組み合わせて効率的に使用することもできる。
【実施例】
【0067】
次の実施例は、請求項の発明を説明するために提供されるものであって、これを限定するためのものではない。
実施例1 軟骨形成の誘導剤のハイスループットスクリーン
【0068】
OA関節修復のための新しい非侵襲戦略を同定し明らかにするために、我々は、ヒト間葉幹細胞(MSCs)の軟骨細胞への分化を選択的に指示することができるタンパク質の非偏向ハイスループットスクリーンを開発した。このアッセイシステムは、軟骨における在住MSCをモデルとし、天然修復能力を刺激し、統合した軟骨再生を増進するメディエーターを同定する。我々は、ハイスループットによって固有の分泌タンパク質ライブラリーを試験するために、ヒトおよびマウスMSCsの細胞ベースのスクリーンを選んだ。このアプローチは、軟骨形成に影響を与える非同定およびネイティブタンパク質リガンドの迅速な同定を可能にする戦略を提供する。
【0069】
これらの分泌タンパク質を研究するために、哺乳動物産生細胞株(HEK293T)からの調整培地(CM)の産生、およびフリースタイルHEK−F細胞からの精製タンパク質の産出の2つのアプローチがとられた。これらの補完的アプローチは、組み合わさって、様々な適用において新規の標的同定を可能にする。
【0070】
MSCアッセイにおけるタンパク質のスクリーニングによって、有力候補であるアンジオポイエチン−様3、略称ANGPTL3が同定された。ANGPTL3は、CMおよび精製タンパク質の両方の2つの並行直交初期スクリーンで同定された。コンセプトCMスクリーンの証拠で、CMの移動の後、C3H10tl/2細胞を7日間インキュベートした。軟骨マトリックス産生を検出するためにアルシアンブルー染色によってアッセイし、軟骨形成は、ANGPTL3−75含有ウェルにおいて起こることが見出された。ANGPTL3−75は、ヒトMSCsの軟骨分化のための531の精製タンパク質のスクリーニングによって独立に同定された。
【0071】
初期スクリーニングアッセイの後、続いて、ANGPTL3を、6つの第2アッセイにおいて特徴付けた。これらアッセイは、
(1)マウス間葉C3H10tl/2細胞の単層培養:II型コラーゲンおよびSox9タンパク質発現(軟骨形成のマーカー)を誘導するが、オステオカルシン(骨発生のマーカー)の誘導はない;
(2)ウシ軟骨細胞におけるTNFα/オンコスタチンM(OSM)−誘導酸化窒素(NO)放出の阻害;
(3)ウシ軟骨器官培養におけるTNFα/OSM−誘導グルコサミノグルカン(GAG)放出の阻害;
(4)ヒトMSCペレット培養システムにおける軟骨マトリックス遺伝子発現(アグレカン)の誘導;
(5)初期ヒト軟骨細胞、ヒト滑膜線維芽細胞、およびヒトMSCsにおける毒性の欠如;
(6)ヒト軟骨細胞の増殖の刺激、
を含む。
【0072】
そのヒトホモログは、他の既知の軟骨形成の誘導剤と比較して、定量的イメージベースのII型コラーゲンアッセイにおいて軟骨形成のための類似の効能を保持するけれども、マウスANGPTL3タンパク質は、マウス、ヒトおよびウシ細胞タイプにおいて機能するので、さらなる特徴づけのために、潜在的に軟骨形成分化を誘導したとして(0.5−5nM)選択された(図1)。マウスおよびヒトMSCの両方が、(それぞれ)単分子層およびペレット培養で、0.5−5nM ANGPTL3で処置されて18〜21日間分化させられ、3つの軟骨特異的タンパク質:II型コラーゲン、Sox9およびアグレカンを発現する。これは、免疫組織化学染色およびTaqmanによるmRNA定量化(図2A)の両方によってアッセイされた。さらに、データは、マウスMSCsを18日間培養後、アルカリフォスファターゼの発現の減少を通し、ANGPTL3が、線維性修復応答に向けた自発傾向を阻害することを示唆する。組織損傷を予防する潜在的能力を評価するために、ウシ軟骨器官培養を、TNFαおよびオンコスタチンM(OSM)で刺激した。マトリックス変性の指標である刺激されたグルコサミノグルカン放出は、ANGPTL3での処置によって有意に阻害された(図2B)。さらに、初期ヒト軟骨細胞の処置によって、24時間内において細胞増殖において2倍の増加がもたらされたのに対し、ヒト滑膜細胞ではそうではなかったが(図2C)、このことは、軟骨に対するその作用の特異性を示唆する。そのタンパク質は、また、初期ヒト軟骨細胞、滑膜線維芽細胞およびMSCsの生存能力に対して、明白なインビトロ毒性効果を持たなかった(<100μM)(データは示さず)。
【0073】
目下の臨床試験での2つの候補間の直接的な比較において、100ng/mlのFGF18または100ng/ml BMP7の処置が、軟骨形成小節を誘導することができたが、全般的なマトリックス産生は、ANGPTL3と比較して、少なかった。FGF18は、アルシアンブルー、Sox9またはII型コラーゲン染色を増加させる能力に欠けており、このことは、本物の軟骨基質の特異性を欠くことを示す。100ng/mlのBMP7が、アルシアンブルー、Sox9およびII型コラーゲン染色を増加させたが、有意性は、類似濃度では、それほど高くなかった。
実施例2:組換え完全長ANGPTL3および変異タンパク質の発現、ならびに機能分析
【0074】
マウスANGPTL3は、51kDaタンパク質であると予測される。これは、アンジオポイエチンに類似した構造を有するが、Tie2受容体に結合する能力を欠き、区別される機能を有する、7つの同定されたアンジオポイエチン−様(ANGPTL)タンパク質のファミリーに属する。それらは、N−末端コイルドコイルドメイン(CCD)およびC末端フィブリノゲン−様ドメイン(FLD)を含む。ANGPTLタンパク質は、その微環境、および、細胞外マトリックス(ECM)との相互作用によって密接に制御されるが、その正確な相互作用部位も、パートナーも、詳細には解明されてこなかった。ANGPTL3は、肝臓によって分泌され、全身に循環する。LXR−誘導高トリグリセリド血が、ANGPTL3放出に起因することを証拠として、これは、肝臓X受容体(LXR)を通して制御される。推定ヘパリン結合モチーフを通したCCDおよびECMの間の相互作用は、ANGPTL4について報告されるのと似たように、前駆タンパク質転換酵素認識配列(R221−R224)における開裂の阻害をもたらすかもしれない。開裂は、リポタンパク質リパーゼ活性(LPL)を阻害するCCDの能力の有意な増加をもたらし、それによって、トリグリセリド血(TG)の増加をもたらす。これは、本発明に先立ち同定されたANGPTL3の主要な生物学的機能を表す。C末端FLDは、内皮細胞接着を誘導し、インビボでのラット角膜直接移植後、血管形成媒介するのに十分である。組換えANGPTL3は、精製αVβ3インテグリンに結合することができ、内皮細胞におけるFAK、MAPKおよびAKTシグナル伝達の増加を引き起こすことも実証された。これらのデータは、FLDのインテグリンとの相互作用が、血管形成を媒介することを示唆する。
【0075】
ANGPTL3の発現は、報告されてこなかったし、ヒト軟骨細胞、hMSCまたはヒト滑膜線維芽細胞におけるウェスタンブロッティングを用いる我々の研究でも観察されてこなかったし、その発現は、マウス膝関節において見出されなかった。さらに、我々のスクリーンで同定されたこの新規の軟骨形成機能に関する、関節細胞におけるいずれかの断片についての活性も報告されていない。
【0076】
我々のデータは、完全長ANGPTL3が軟骨形成および軟骨保護のための新規メディエーターであることを示す。我々は、ANGPTL3のどのドメインが、この新規軟骨形成機能に決定的かを調べた。詳細な生物物理的特性決定のために、ANGPTL3のトランケーション系および完全長タンパク質を、限定的で均一なグリコシル化を与える改変HEK−S細胞株において発現させ、精製した。質量分光学アプローチによって、すべての4つの予測されたN−連結グリコシル化(N115、N232、N296、およびN357、図3A)の場所が確認された。HEK内因的にプロセスされた断片から、CCDおよびFLDドメインの間のタンパク質分解開裂部位がR224であると測定された。完全長ANGPTL3は、質量>400kDaでのサイズ排除クロマトグラフィによって溶出し、このことは、異種グリコシル化での三量体を示す。サイズ排除および静的光散乱法によって単量体であることが示されるFLD(241−455)を結晶化した。FLDの原子構造を、1.8オングストローム解像度で測定した。FLD構造は、C−末端に向けて3つのループを向けるらせん伸長を持つコアベータシート形態を明らかにし、これは、他のFLDホモログの典型である(図3B)。ドメインのC末端部分における温度要因は、残りの構造よりも高く、このことは、この領域における高度の柔軟性を示唆する。ANGPTL2/Tie2構造(PDBコード:2GY7)との重ね合わせは、FLDのC末端部分が、タンパク質タンパク質相互作用に関与するかもしれないことを示唆する。
【0077】
産生された5つのトランケーション生成物を、複数の軟骨形成アッセイで評価した。結果は、CCD単独では活性保持がないが、FLDでは活性が維持することを示唆する。さらに、SMADlリン酸化は、完全長または変異GまたはH単独での3日間の刺激に応じて増加し、このことは、軟骨形成シグナル伝達カスケードの活性化を示唆する。
【0078】
関節内に注射されたタンパク質の全身性暴露は、滑液リンパ排出のために限られているけれども、全身性暴露を最小化することが、時には望ましい。全身性暴露がある場合には、ANGPTL3の開裂は、CCDドメインの放出を引き起こすかもしれない。CCDによるLPL活性の阻害の増加は、患者のTGレベルの改変をもたらしうる。我々の結果は、ANGPTL3の潜在的な臨床的利点を示す。たとえば、ANGPTL3における軟骨形成活性が位置する場所の切除によって、我々は、脂質代謝または血管形成特性に対する望まない全身性効果を最小化または排除するであろう。軟骨形成活性は、主としてC末端に位置し、そのため、このドメインの特異的使用は、完全長分子またはN末端のTG制御の懸念を軽減するであろう。完全長ANGPTL3は、通常、血清において存在するので、予想される免疫原性はない。C末端を使用するさらなる利点は、それは、開裂し、かつ三量体の完全長タンパク質と比較して単量体(〜29kDa)であり、そのため、循環において存在するかもしれない任意の小パーセンテージの全身性半減期を減少させ、血管形成からの任意の潜在的な効果を制限するであろうことである。
実施例3:ANGPTL3のインビボ分析
【0079】
我々は、潜在的な有害減少および関節内滞留に対処するために、完全長ANGPTL3のいくつかのインビボ評価を行った。8週齢C57BL/10マウスの左膝関節に3.6μgの完全長ANGPTL3で関節内(IA)注射した後、我々は、24時間の暴露の存在下および非存在下での発現を免疫組織化学的に評価した。滑液リンパ管へのトランス滑液流によるタンパク質および水にとっての典型的なターンオーバーが約2時間であるので、24時間まで、検出できるレベルのANGPTL3は、わずかしか滑液に留まらない、または滑液に留まらないはずである。結果は、非処置関節におけるANGPTL3の内因性発現がないが、関節軟骨および半月板において、24時間の時点でさえ細胞周囲マトリックスのタンパク質が有意に検出されることを明らかにした。さらに、細胞毒性の軟骨細胞への広がりや、インビボでの軟骨損傷は、検出されなかった。一連のラットの膝関節への3IA注射(1週間に1回を3週間)の後、臨床的には、毒性はなかった(関節膨潤も、歩き方の変化もなかった)し、ラットの関節における急性炎症反応の証拠となるものもなかった。組織学的には、滑膜炎の増加や、注射された5つのラットの関節における無制御的な増殖はなかった。マウスでのように、ANGPTL3は、軟骨マトリックスおよび周囲の軟骨細胞において検出され得た。これらの結果は、ANGPTL3は、軟骨自体に対する望ましくない効果を引き起こさず、そして、ANGPTL3は、インビボで軟骨および半月板に入り保持されることを示す。
【0080】
OAは、単一疾患実体ではなく、様々な病院的因子の帰結でありうる。ヒトにおいて、それは、しばしば、異常な生物力学的ストレスまたは遺伝的に獲得された関節軟骨または骨の異常性によって引き起こされる。したがって、「最善の」小動物OAモデルを選択することは、困難であり、複数のモデルが、任意の治療の保護特性を測定するために、探索されるべきである。我々は、慢性OAモデル(van der Kraanおよび同僚らによって説明された研究に基づきコラーゲナーゼVII誘導)およびGlassonらの仕事に基づき関節における3つの主要な靱帯(ACL、MCLおよびMMTL)の横断面を使う急性外科モデルの効能研究を完成させた。両方のモデルは、OAに共通に関係する病理的変化誘導した。プロテオグリカン染色の喪失、軟骨および骨の浸食、骨棘形成ならびに滑膜および靱帯の化生性変化が、OAの注射の4〜8週間後にはっきりと分かりうる。図5は、OAのANGPTL3外科モデルの再生能力を表す。OAについて潜在的バイオマーカーの検証を始めるため、外科モデルの間で末梢血を回収して、軟骨損傷ゆえに放出されたII型コラーゲン断片を測定した(図5A)。組織学的分析および、それに続く中間脛骨プラトーの等級付けは、200ng ANGTPL3/膝の1週間に1回3週間での処置後の軟骨マトリックスにおける再生を明らかにした(図5B)。
【0081】
8週間外科OAマウスのサブセットにおいて、3投与量のANGPTL3が、インキャパシタンス測定を通したOA−誘導痛を軽減するかどうかについて調べられた。この方法は、外科手術および非外科手術肢の間の重量配分を測定する。外科手術の後36日目に、および、PROlでの処置3週間ごとに、100ng/膝もの低さの投与が、ビヒクル処置外科手術膝と比較した有意な向上を実証した(図5C)。これらの組み合わせたデータは、ANGTPL3が、2つのOAモデル(病理的修正および痛み縮小の両方)においてインビボ効能を有することの具体的な証拠を提供し、新規のOA治療としてのその発達の進歩を裏付ける。
【0082】
本明細書に記載された実施例および実施態様は、例示のためだけのものであり、それらを考慮した様々な修飾または変化が本技術分野における当業者によって示唆されるであろうし、それらは、本出願、および添付の請求項の範囲の精神および範囲内に含まれると理解される。本明細書において引用された全ての公開、特許、および特許出願は、あらゆる目的でその全体が、参照によって本明細書に包含される。
【0083】
配列表
X = 任意のアミノ酸
丸括弧中のアミノ酸は、アミノ酸があってもなくてもよいことをを示す。
【0084】
配列番号:1
241−455 コンセンサス
XXPAXCXXXYNXGXHTSGXYXIXPXNSQXFXVYCDXXSGSXWTLIQHRXDGSQXFNETWENYXXGFGRLDGEFWLGLEKIYXIVXQSNYXLRXELXDWXD(X)KXYXEYXFXLGXHETNYTLHXXXIXGNXXXAXPEXXDLXFSTWXHXAKGXXXCPEXXSGGWWXXXXCGENNLNGKYNKXXXKXXPERXXGXXWXXQXXXLYXIKXXKMXXXPXX
【0085】
配列番号:2
225−455 コンセンサス
TTPXXXXNEXXNXXXXXXPAXCXXXYNXGXHTSGXYXIXPXNSQXFXVYCDXXSGSXWTLIQHRXDGSQXFNETWENYXXGFGRLDGEFWLGLEKIYXIVXQSNYXLRXELXDWXD(X)KXYXEYXFXLGXHETNYTLHXXXIXGNXXXAXPEXXDLXFSTWXHXAKGXXXCPEXXSGGWWXXXXCGENNLNGKYNKXXXKXXPERXXGXXWXXQXXXLYXIKXXKMXXXPXX
【0086】
配列番号:3
207−455 コンセンサス
XIXEXXEXSLSSKXRAPRTTPXXXXNEXXNXXXXXXPAXCXXXYNXGXHTSGXYXIXPXNSQXFXVYCDXXSGSXWTLIQHRXDGSQXFNETWENYXXGFGRLDGEFWLGLEKIYXIVXQSNYXLRXELXDWXD(X)KXYXEYXFXLGXHETNYTLHXXXIXGNXXXAXPEXXDLXFSTWXHXAKGXXXCPEXXSGGWWXXXXCGENNLNGKYNKXXXKXXPERXXGXXWXXQXXXLYXIKXXKMXXXPXX
【0087】
配列番号:4
完全長コンセンサス
MXTIKLXLXXXPLVIXSXXDXDXXSXDSXXXEPKSRFAMLDDVKILANGLLQLGHGLKDFVHKTKGQINDIFQKLNIFDQSFYDLSLXTXEIKEEEKELRRXTXXLQVKNEEVKNMSXELXSKXESLLEEKXXLQXKVXXLEXQLTXLIXXXXXXXEXXEVTSLKXXVEXQDNSIXXLLQXVEXQYXQLXQQXXQIKEIEXQLR(X)XXIXEXXEXSLSSKXRAPRTTPXXXXNEXXNXXXXXXPAXCXXXYNXGXHTSGXYXIXPXNSQXFXVYCDXXSGSXWTLIQHRXDGSQXFNETWENYXXGFGRLDGEFWLGLEKIYXIVXQSNYXLRXELXDWXD(X)KXYXEYXFXLGXHETNYTLHXXXIXGNXXXAXPEXXDLXFSTWXHXAKGXXXCPEXXSGGWWXXXXCGENNLNGKYNKXXXKXXPERXXGXXWXXQXXXLYXIKXXKMXXXPXX(SEXXE)
【0088】
配列番号:5
ヒトANGPTL3 241−455
GIPAECTTIYNRGEHTSGMYAIRPSNSQVFHVYCDVISGSPWTLIQHRIDGSQNFNETWENYKYGFGRLDGEFWLGLEKIYSIVKQSNYVLRIELEDWKDNKHYIEYSFYLGNHETNYTLHLVAITGNVPNAIPENKDLVFSTWDHKAKGHFNCPEGYSGGWWWHDECGENNLNGKYNKPRAKSKPERRRGLSWKSQNGRLYSIKSTKMLIHPTD
【0089】
配列番号:6
ヒトANGPTL3 225−455
TTPFLQLNEIRNVKHDGIPAECTTIYNRGEHTSGMYAIRPSNSQVFHVYCDVISGSPWTLIQHRIDGSQNFNETWENYKYGFGRLDGEFWLGLEKIYSIVKQSNYVLRIELEDWKDNKHYIEYSFYLGNHETNYTLHLVAITGNVPNAIPENKDLVFSTWDHKAKGHFNCPEGYSGGWWWHDECGENNLNGKYNKPRAKSKPERRRGLSWKSQNGRLYSIKSTKMLIHPTD
【0090】
配列番号:7
ヒトANGPTL3 207−455
IQEPTEISLSSKPRAPRTTPFLQLNEIRNVKHDGIPAECTTIYNRGEHTSGMYAIRPSNSQVFHVYCDVISGSPWTLIQHRIDGSQNFNETWENYKYGFGRLDGEFWLGLEKIYSIVKQSNYVLRIELEDWKDNKHYIEYSFYLGNHETNYTLHLVAITGNVPNAIPENKDLVFSTWDHKAKGHFNCPEGYSGGWWWHDECGENNLNGKYNKPRAKSKPERRRGLSWKSQNGRLYSIKSTKMLIHPTD
【0091】
配列番号:8
完全長ヒトANGPTL3 gi|7656888|ref|NP_055310.1|[Homo sapiens]
MFTIKLLLFIVPLVISSRIDQDNSSFDSLSPEPKSRFAMLDDVKILANGLLQLGHGLKDFVHKTKGQINDIFQKLNIFDQSFYDLSLQTSEIKEEEKELRRTTYKLQVKNEEVKNMSLELNSKLESLLEEKILLQQKVKYLEEQLTNLIQNQPETPEHPEVTSLKTFVEKQDNSIKDLLQTVEDQYKQLNQQHSQIKEIENQLRRTSIQEPTEISLSSKPRAPRTTPFLQLNEIRNVKHDGIPAECTTIYNRGEHTSGMYAIRPSNSQVFHVYCDVISGSPWTLIQHRIDGSQNFNETWENYKYGFGRLDGEFWLGLEKIYSIVKQSNYVLRIELEDWKDNKHYIEYSFYLGNHETNYTLHLVAITGNVPNAIPENKDLVFSTWDHKAKGHFNCPEGYSGGWWWHDECGENNLNGKYNKPRAKSKPERRRGLSWKSQNGRLYSIKSTKMLIHPTDSESFE
【0092】
配列番号:9
マウスANGPTL3 241−455
DLPADCSAVYNRGEHTSGVYTIKPRNSQGFNVYCDTQSGSPWTLIQHRKDGSQDFNETWENYEKGFGRLDGEFWLGLEKIYAIVQQSNYILRLELQDWKDSKHYVEYSFHLGSHETNYTLHVAEIAGNIPGALPEHTDLMFSTWNHRAKGQLYCPESYSGGWWWNDICGENNLNGKYNKPRTKSRPERRRGIYWRPQSRKLYAIKSSKMMLQPTT
【0093】
配列番号:10
マウスANGPTL3 225−455
TTPPLQLNETENTEQDDLPADCSAVYNRGEHTSGVYTIKPRNSQGFNVYCDTQSGSPWTLIQHRKDGSQDFNETWENYEKGFGRLDGEFWLGLEKIYAIVQQSNYILRLELQDWKDSKHYVEYSFHLGSHETNYTLHVAEIAGNIPGALPEHTDLMFSTWNHRAKGQLYCPESYSGGWWWNDICGENNLNGKYNKPRTKSRPERRRGIYWRPQSRKLYAIKSSKMMLQPTT
【0094】
配列番号:11
マウスANGPTL3 207−455
IQEPSENSLSSKSRAPRTTPPLQLNETENTEQDDLPADCSAVYNRGEHTSGVYTIKPRNSQGFNVYCDTQSGSPWTLIQHRKDGSQDFNETWENYEKGFGRLDGEFWLGLEKIYAIVQQSNYILRLELQDWKDSKHYVEYSFHLGSHETNYTLHVAEIAGNIPGALPEHTDLMFSTWNHRAKGQLYCPESYSGGWWWNDICGENNLNGKYNKPRTKSRPERRRGIYWRPQSRKLYAIKSSKMMLQPTT
【0095】
配列番号:12
完全長マウスANGPTL3 gi|33469117|ref|NP_038941.1|[Mus musculus]
MHTIKLFLFWPLVIASRVDPDLSSFDSAPSEPKSRFAMLDDVKILANGLLQLGHGLKDFVHKTKGQINDIFQKLNIFDQSFYDLSLRTNEIKEEEKELRRTTSTLQVKNEEVKNMSVELNSKLESLLEEKTALQHKVRALEEQLTNLILSPAGAQEHPEVTSLKSFVEQQDNSIRELLQSVEEQYKQLSQQHMQIKEIEKQLRKTGIQEPSENSLSSKSRAPRTTPPLQLNETENTEQDDLPADCSAVYNRGEHTSGVYTIKPRNSQGFNVYCDTQSGSPWTLIQHRKDGSQDFNETWENYEKGFGRLDGEFWLGLEKIYAIVQQSNYILRLELQDWKDSKHYVEYSFHLGSHETNYTLHVAEIAGNIPGALPEHTDLMFSTWNHRAKGQLYCPESYSGGWWWNDICGENNLNGKYNKPRTKSRPERRRGIYWRPQSRKLYAIKSSKMMLQPTT
【0096】
配列番号:13
ウシANGPTL3 241−454
DIPADCTIIYNQGKHTSGIYSIRPSNSQVFNVYCDVKSGSSWTLIQHRIDGSQNFNETWENYKYGFGRLDGEFWLGLEKIYSIVMQSNYILRIELEDWKDKYYTEYSFHLGDHETNYTLHLAEISGNGPKAFPEHKDLMFSTWDHKAKGHFNCPESNSGGWWYHDVCGENNLNGKYNKPKAKAKPERKEGICWKSQDGRLYSIKATKMLIHPSD
【0097】
配列番号:14
ウシANGPTL3 225−454
TTPSFHSNETKNVEHDDIPADCTIIYNQGKHTSGIYSIRPSNSQVFNVYCDVKSGSSWTLIQHRIDGSQNFNETWENYKYGFGRLDGEFWLGLEKIYSIVMQSNYILRIELEDWKDKYYTEYSFHLGDHETNYTLHLAEISGNGPKAFPEHKDLMFSTWDHKAKGHFNCPESNSGGWWYHDVCGENNLNGKYNKPKAKAKPERKEGICWKSQDGRLYSIKATKMLIHPSD
【0098】
配列番号:15
ウシANGPTL3 207−454
IKESTEISLSSKPRAPRTTPSFHSNETKNVEHDDIPADCTIIYNQGKHTSGIYSIRPSNSQVFNVYCDVKSGSSWTLIQHRIDGSQNFNETWENYKYGFGRLDGEFWLGLEKIYSIVMQSNYILRIELEDWKDKYYTEYSFHLGDHETNYTLHLAEISGNGPKAFPEHKDLMFSTWDHKAKGHFNCPESNSGGWWYHDVCGENNLNGKYNKPKAKAKPERKEGICWKSQDGRLYSIKATKMLIHPSD
【0099】
配列番号:16
完全長ウシANGPTL3 gi|122692391|ref|NP_001073814.1|[Bos taurus]
MYTIKLFLIIAPLVISSRTDQDYTSLDSISPEPKSRFAMLDDVKILANGLLQLGHGLKDFVHKTKGQINDIFQKLNIFDQSFYDLSLQTNEIKEEEKELRRATSKLQVKNEEVKNMSLELDSKLESLLEEKILLQQKVRYLEDQLTDLIKNQPQIQEYLEVTSLKTLVEQQDNSIKDLLQIVEEQYRQLNQQQSQIKEIENQLRRTGIKESTEISLSSKPRAPRTTPSFHSNETKNVEHDDIPADCTIIYNQGKHTSGIYSIRPSNSQVFNVYCDVKSGSSWTLIQHRIDGSQNFNETWENYKYGFGRLDGEFWLGLEKIYSIVMQSNYILRIELEDWKDKYYTEYSFHLGDHETNYTLHLAEISGNGPKAFPEHKDLMFSTWDHKAKGHFNCPESNSGGWWYHDVCGENNLNGKYNKPKAKAKPERKEGICWKSQDGRLYSIKATKMLIHPSDSENSE
【0100】
配列番号:17
イヌANGPTL3 240−454
DIPANCTTIYNRGEHTSGIYSIRPSNSQVFNVYCDVKSGSSWTLIQHRIDGSQNFNETWENYRYGFGRLDGEFWLGLEKIYSIVKQSNYILRIELEDWNDNKHYIEYFFHLGNHETNYTLHLVEITGNILNALPEHKDLVFSTWDHKAKGHVNCPESYSGGWWWHNVCGENNLNGKYNKQRAKTKPERRRGLYWKSQNGRLYSIKSTKMLIHPID
【0101】
配列番号:18
イヌANGPTL3 224−454
TTPFLHLNETKNVEHNDIPANCTTIYNRGEHTSGIYSIRPSNSQVFNVYCDVKSGSSWTLIQHRIDGSQNFNETWENYRYGFGRLDGEFWLGLEKIYSIVKQSNYILRIELEDWNDNKHYIEYFFHLGNHETNYTLHLVEITGNILNALPEHKDLVFSTWDHKAKGHVNCPESYSGGWWWHNVCGENNLNGKYNKQRAKTKPERRRGLYWKSQNGRLYSIKSTKMLIHPID
【0102】
配列番号:19
イヌANGPTL3 206−454
IQESTENSLSSKPRAPRTTPFLHLNETKNVEHNDIPANCTTIYNRGEHTSGIYSIRPSNSQVFNVYCDVKSGSSWTLIQHRIDGSQNFNETWENYRYGFGRLDGEFWLGLEKIYSIVKQSNYILRIELEDWNDNKHYIEYFFHLGNHETNYTLHLVEITGNILNALPEHKDLVFSTWDHKAKGHVNCPESYSGGWWWHNVCGENNLNGKYNKQRAKTKPERRRGLYWKSQNGRLYSIKSTKMLIHPID
【0103】
配列番号:20
完全長イヌANGPTL3 gi|57086505|ref|XP_536686.1|[Canis familiaris]
MYTIKLFLFIIPLVISSKIDRDYSSYDSVSPEPKSRFAMLDDVKILANGLLQLGHGLKDFVHKTKGQINDIFQKLNIFDQSFYDLSLQTNEIKEEEKELRRTTSKLQVKNEEVKNMSLELNSKVESLLEEKILLQQKVRYLEKQLTSLIKNQPEIQEHPEVTSLKTFVEQQDNSIKDLLQTVEEQYRQLNQQHSQIKEIENQLRNVIQESTENSLSSKPRAPRTTPFLHLNETKNVEHNDIPANCTTIYNRGEHTSGIYSIRPSNSQVFNVYCDVKSGSSWTLIQHRIDGSQNFNETWENYRYGFGRLDGEFWLGLEKIYSIVKQSNYILRIELEDWNDNKHYIEYFFHLGNHETNYTLHLVEITGNILNALPEHKDLVFSTWDHKAKGHVNCPESYSGGWWWHNVCGENNLNGKYNKQRAKTKPERRRGLYWKSQNGRLYSIKSTKMLIHPIDSESSE
【0104】
配列番号:21
ウマANGPTL3 241−455
DFPADCTTIYNRGEHTSGIYSIKPSNSQVFNVYCDVISGSSWILIQRRIDGSQNFNETWQNYKYGFGRLDFEFWLGLEKIYSIVKRSNYILRIELEDWKDNKHTIEYSFHLGNHETNYTLHLVEITGNVPNALPEHKDLVFSTWDHKAKGQLNCLESYSGGWWWHDVCGGDNPNGKYNKPRSKTKPERRRGICWKSQNGRLYTIKSTKMLIHPID
【0105】
配列番号:22
ウマANGPTL3 225−455
TTPSFHLNETKDVEHDDFPADCTTIYNRGEHTSGIYSIKPSNSQVFNVYCDVISGSSWILIQRRIDGSQNFNETWQNYKYGFGRLDFEFWLGLEKIYSIVKRSNYILRIELEDWKDNKHTIEYSFHLGNHETNYTLHLVEITGNVPNALPEHKDLVFSTWDHKAKGQLNCLESYSGGWWWHDVCGGDNPNGKYNKPRSKTKPERRRGICWKSQNGRLYTIKSTKMLIHPID
【0106】
配列番号:23
ウマANGPTL3 207−455
IQESTENSLSSKPRAPRTTPSFHLNETKDVEHDDFPADCTTIYNRGEHTSGIYSIKPSNSQVFNVYCDVISGSSWILIQRRIDGSQNFNETWQNYKYGFGRLDFEFWLGLEKIYSIVKRSNYILRIELEDWKDNKHTIEYSFHLGNHETNYTLHLVEITGNVPNALPEHKDLVFSTWDHKAKGQLNCLESYSGGWWWHDVCGGDNPNGKYNKPRSKTKPERRRGICWKSQNGRLYTIKSTKMLIHPID
【0107】
配列番号:24
完全長ウマANGPTL3[equus caballus]
MYTIKLFLVIAPLVISSRIDQDYSSLDSIPPEPKSRFAMLDDVKILANGLLQLGHGLKDFVHKTKGQINDIFQKLNIFDQSFYALSLQTNEIKEEEKELRRTTSKLQVKNEEVKNMSLELNSKLESLLEEKSLLQQKVKYLEEQLTKLIKNQPEIQEHPEVTSLKTFVEQQDNSIKDLLQTMEEQYRQLNQQHSQIKEIENQLRRTGIQESTENSLSSKPRAPRTTPSFHLNETKDVEHDDFPADCTTIYNRGEHTSGIYSIKPSNSQVFNVYCDVISGSSWILIQRRIDGSQNFNETWQNYKYGFGRLDFEFWLGLEKIYSIVKRSNYILRIELEDWKDNKHTIEYSFHLGNHETNYTLHLVEITGNVPNALPEHKDLVFSTWDHKAKGQLNCLESYSGGWWWHDVCGGDNPNGKYNKPRSKTKPERRRGICWKSQNGRLYTIKSTKMLIHPIDSESFELRQIKKPMN
【0108】
配列番号:25
241−455 コンセンサス
XXPAXCXXXYNXGXHTSGXYXIXPXNSQXFXVYCDXXSGSXWXLIQXRXDGSQXFNETWXNYXXGFGRLDXEFWLGLEKIYXIVXXSNYXLEXELXDWXD(X)KXXXEYXFXLGXHETNYTLHXXXIXGNXXXAXPEXXDLXFSTWXHXAKGXXXCXEXXSGGWWXXXXCGXXNXNGKYNKXXXKXXPERXXGXXWXXQXXXLYXIKXXKMXXXPXX
【0109】
配列番号:26
225−455 コンセンサス
TTPXXXXNEXXXXXXXXXPAXCXXXYNXGXHTSGXYXIXPXNSQXFXVYCDXXSGSXWXLIQXRXDGSQXFNETWXNYXXGFGRLDXEFWLGLEKIYXIVXXSNYXLEXELXDWXD(X)KXXXEYXFXLGXHETNYTLHXXXIXGNXXXAXPEXXDLXFSTWXHXAKGXXXCXEXXSGGWWXXXXCGXXNXNGKYNKXXXKXXPERXXGXXWXXQXXXLYXIKXXKMXXXPXX
【0110】
配列番号:27
207−455 コンセンサス
XIXEXXEXSLSSKXRAPRTTPXXXXNEXXXXXXXXXPAXCXXXYNXGXHTSGXYXIXPXNSQXFXVYCDXXSGSXWXLIQXRXDGSQXFNETWXNYXXGFGRLDXEFWLGLEKIYXIVXXSNYXLEXELXDWXD(X)KXXXEYXFXLGXHETNYTLHXXXIXGNXXXAXPEXXDLXFSTWXHXAKGXXXCXEXXSGGWWXXXXCGXXNXNGKYNKXXXKXXPERXXGXXWXXQXXXLYXIKXXKMXXXPXX
【0111】
配列番号:28
完全長コンセンサス
MXTIKLXLXXXPLVIXSXXDXDXXSXDSXXXEPKSRFAMLDDVKILANGLLQLGHGLKDFVHKTKGQINDIFQKLNIFDQSFYXLSLXTXEIKEEEKELRRXTXXLQVKNEEVKNMSXELXSKXESLLEEKXXLQXKVXXLEXQLTXLIXXXXXXXEXXEVTSLKXXVEXQDNSIXXLLQXXEXQYXQLXQQXXQIKEIEXQLR(X)XXIXEXXEXSLSSKXRAPRTTPXXXXNEXXXXXXXXXPAXCXXXYNXGXHTSGXYXIXPXNSQXFXVYCDXXSGSXWXLIQXRXDGSQXFNETWXNYXXGFGRLDXEFWLGLEKIYXIVXXSNYXLEXELXDWXD(X)KXXXEYXFXLGXHETNYTLHXXXIXGNXXXAXPEXXDLXFSTWXHXAKGXXXCXEXXSGGWWXXXXCGXXNXNGKYNKXXXKXXPERXXGXXWXXQXXXLYXIKXXKMXXXPXX(SEXXEXXXXXXXXX)
【技術分野】
【0001】
関連出願に対する相互参照
本出願は、2009年7月14日に出願された米国仮出願第61/225,293の35U.S.C.§1.119(e)の下での利益を請求し、その全体の内容が、参照により組み込まれる。
【背景技術】
【0002】
骨関節炎(OA)は、もっとも一般的な筋骨格障害を示す。約4000万人のアメリカ人が、近年罹患しており、この数は、高齢化や平均寿命の上昇の結果として、次の20年のうちに6000万に上昇し、身体障害の原因の第4位になることが予測されている。OAは、関節軟骨(関節の潤滑および緩衝を生む細胞およびマトリックスを含む)および関節軟骨の下部の軟骨下骨の両方を含む関節の緩やかな変性衰弱を特徴とする。近年のOA治療は、経口NSAIDsまたは選択的シクロオキシゲナーゼ2(COX−2)阻害剤での鎮痛、コルチコステロイドおよびヒアルロナンなどの剤の関節内(IA)注射、ならびに外科的アプローチを含む。
【0003】
間葉幹細胞(MSCs)は、成人関節軟骨に存在し、単離してインビトロで軟骨細胞および他の間葉細胞系列へ分化させるようにプログラムすることができる。それは、部分的に、成長因子(TGFβ、BMPs)、血清条件および細胞間接触によって制御される。
【発明の概要】
【0004】
本発明は、哺乳動物における関節炎または関節損傷を改善または予防する方法を提供する。ある実施態様において、該方法は、配列番号1または25に対して少なくとも95%の同一性を有するアミノ酸配列を含む有効量のポリペプチドを含む組成物を、哺乳動物の関節に投与し、それによって、哺乳動物における関節炎または関節損傷を改善または予防することを含む。
【0005】
ある実施態様において、該個体は、関節炎または関節損傷を有している。ある実施態様において、該個体は、関節炎または関節損傷を有していないが、関節炎または関節損傷のリスクを有する。
【0006】
ある実施態様において、該ポリペプチドは、配列番号1または25を含む。
【0007】
ある実施態様において、該アミノ酸配列は、配列番号2、3、4、26、27、または28に対して少なくとも95%の同一性を有する。ある実施態様において、該アミノ酸配列は、配列番号2、3、4、26、27、または28を含む。
【0008】
ある実施態様において、該アミノ酸配列は、配列番号5〜24のいずれかに対して少なくとも80%同一である。ある実施態様において、該アミノ酸配列は、配列番号5〜24のいずれかを含む。
【0009】
ある実施態様において、該関節炎は、骨関節炎、外傷的関節炎、および自己免疫関節炎からなる群から選択される。
【0010】
ある実施態様において、該哺乳動物はヒトである。
【0011】
ある実施態様において、該組成物は、さらにヒアルロン酸を含む。
【0012】
本発明は、軟骨基質を形成する軟骨細胞への間葉幹細胞の分化を誘導する方法も提供する。ある実施態様において、該方法は、配列番号1または25に対して少なくとも95%の同一性を有するアミノ酸配列を含む十分量のポリペプチドを間葉幹細胞に接触させ、軟骨細胞への幹細胞の分化を誘導することを含む。
【0013】
ある実施態様において、該方法は、インビトロで実行される。ある実施態様において、該方法は、インビボで実行され、該幹細胞は、哺乳動物に存在する。ある実施態様において、該哺乳動物はヒトである。
【0014】
ある実施態様において、該ポリペプチドは配列番号1または25を含む。
【0015】
ある実施態様において、該アミノ酸配列は、配列番号2、3、4、26、27、または28に対して少なくとも95%の同一性を有する。ある実施態様において、該アミノ酸配列は、配列番号2、3、4、26、27、または28を含む。
【0016】
ある実施態様において、該アミノ酸配列は、配列番号5〜24のいずれかに対して少なくとも80%同一である。ある実施態様において、該アミノ酸配列は、配列番号5〜24のいずれかを含む。
【0017】
本発明は、関節内輸送および全身性輸送のための医薬組成物も提供する。ある実施態様において、該組成物は、配列番号1または25に対して少なくとも95%の同一性を有するアミノ酸配列を含む医薬的に有効量のポリペプチドを含む。
【0018】
ある実施態様において、該組成物は、さらにヒアルロン酸を含む。
【0019】
ある実施態様において、該ポリペプチドは、配列番号1または25を含む。
【0020】
ある実施態様において、該アミノ酸配列は、配列番号2、3、4、26、27、または28に対して少なくとも95%の同一性を有する。ある実施態様において、該アミノ酸配列は、配列番号2、3、4、26、27、または28を含む。
【0021】
ある実施態様において、該アミノ酸配列は、配列番号5〜24のいずれかに対して少なくとも80%同一である。ある実施態様において、該アミノ酸配列は、配列番号5〜24のいずれかを含む。
【0022】
本発明のさらなる特徴、長所、実施態様は、次の詳細な説明および請求項の考察から説明される、または明らかであろう。さらに、先述の発明の要旨および、次の詳細な説明の両方が、例示であり、限定なしに請求項に規定された発明の範囲のさらなる説明を提供するためのものであることが、理解されるべきである。
【0023】
定義
用語「ペプチド模倣物」および「模倣物」は、天然または非天然に生じるポリペプチド(例えば、ANGPTL3)と実質的に同一の構造的および機能的な特徴を有する合成化学化合物に言及する。ペプチド類似物は、テンプレートペプチドのものと類似の特性を有する非ペプチド薬として製薬産業において広く用いられている。これらのタイプの非ペプチド化合物は、「ペプチド模倣物」または「ペプチド模倣物」と呼ばれる(Fauchere,J.Adv.Drug Res.15:29(1986);Veber and Freidinger TINS p.392(1985);およびEvans et al.J.Med.Chem.30:1229(1987)。これらは、参照によって本明細書に包含される)。ペプチド模倣物は、治療的に有用なペプチドに構造的に類似するものであり、等価のまたは強化した治療的または予防的効果を産出するために使用してもよい。一般的には、ペプチド模倣物は、目的のポリペプチドにおいて見出されるなどの模範ポリペプチド(すなわち、生物学的または薬理的活性を有するポリペプチド)に構造的に類似するが、例えば、−CH2NH−、−CH2S−、−CH2−CH2−、−CH=CH−(シスおよびトランス)、−COCH2−、−CH(OH)CH2−、および−CH2SO−からなる群から選択される結合によって任意的に交換された1または複数のペプチド結合を有する。該模倣物は、完全に合成、非天然のアミノ酸類似物からなるか、または、部分的な天然ペプチドアミノ酸および部分的な非天然アミノ酸類似物のキメラ分子であることができる。該模倣物は、模倣物の構造および/または活性を実質的に改変しない置換でもある限り、任意の量の天然アミノ酸保存的置換を包含することもできる。例えば、模倣物組成物は、目的のポリペプチドの少なくとも1つの活性を実現することができる場合には、本発明の範囲内である。
【0024】
用語「ポリペプチド」、「ペプチド」および「タンパク質」は、本明細書において、アミノ酸残基のポリマーに言及するために区別せずに用いられる。該用語は、1または複数のアミノ酸残基が天然に生じる対応のアミノ酸の人工的化学模倣物であるアミノ酸ポリマーに対して用いられ、また、天然に生じるアミノ酸ポリマーおよび非天然に生じるアミノ酸ポリマーに対して用いられる。
【0025】
用語「アミノ酸」は、天然に生じる合成アミノ酸に言及し、また、天然に生じるアミノ酸に類似する様式で機能するアミノ酸類似物、およびアミノ酸模倣物に言及する。天然に生じるアミノ酸は、遺伝的コードによってコードされるものであり、また、例えば、ヒドロキシプロリン、γ−カルボキシグルタミン酸、およびO−ホスホセリンなどの後で修飾されたアミノ酸である。アミノ酸類似物は、天然に生じるアミノ酸としての基本的化学構造、すなわち、水素、カルボキシル基、アミノ基、および、例えば、ホモセリン、ノルロイシン、メチオニンスルホキシド、メチオニンメチルスルホニウムなどのR基に結合した
α炭素、と同一の構造を有する化合物に言及する。このような類似物は、修飾されたR基(例えば、ノルロイシン)または修飾されたペプチド骨格を有するが、天然に生じるアミノ酸と同一の基本的化学構造を保持する。天然にコードされるアミノ酸は、20の共通のアミノ酸(アラニン、アルギニン、アスパラギン、アスパラギン酸、システイン、グルタミン、グルタミン酸、グリシン、ヒスチジン、イソロイシン、ロイシン、リジン、メチオニン、フェニルアラニン、プロリン、セリン、スレオニン、トリプトファン、チロシン、およびバリン)およびピロールリジンならびにセレノシステインである。
【0026】
「保存的に修飾された変異体」は、アミノ酸および核酸の両方の配列に対して用いられる。特定の核酸配列に関して、保存的に修飾された変異体は、同一である、または本質的に同一であるアミノ酸配列をコードする、またはアミノ酸配列をコードしない核酸に言及し、本質的に同一である配列に言及する。遺伝的コードの縮重のため、多数の機能的に同一である核酸が、任意の所与のタンパク質をコードする。たとえば、コドンGCA、GCC、GCGおよびGCUは、すべて、アミノ酸アラニンをコードする。したがって、アラニンがコドンによって特定される各々の位置において、該コドンは、コードされるポリペプチドを改変することなく、説明された対応コドンのいずれかに改変することができる。
このような核酸変異は、「サイレント変異」であり、保存的に修飾された変異の一種である。本明細書において、ポリペプチドをコードする各核酸配列は、核酸の各々のありうるサイレント変異も説明する。当業者は、核酸における各々のコドン(但し、通常メチオニンのための唯一のコドンであるAUG、および通常メチオニンのための唯一のコドンであるトリプトファンTGGを除く)は、機能的に同一である分子を産出するために変更することができることを認識するであろう。したがって、ポリペプチドをコードする核酸の各々のサイレント変異は、各々記述された配列において、黙示されている。
【0027】
アミノ酸配列に関して、コードされる配列における単一のアミノ酸または少数のパーセンテージのアミノ酸を改変、付加または欠失させる、核酸、ペプチド、ポリペプチド、またはタンパク質配列に対する個々の置換、欠失または付加が、改変によって化学的に類似するアミノ酸でアミノ酸を置換させる「保存的に修飾された変異体」であることを、当業者は認識するであろう。機能的に類似するアミノ酸を提供する保存的置換の一覧は、本技術分野においてよく知られている。このような保存的に修飾された変異体は、本発明の多型変異体、種間ホモログ、およびアレルに対して付加されるものであり、これらを排斥するものではない。
【0028】
次の8つの群は、互いに保存的置換であるアミノ酸をそれぞれ含む:
1)アラニン(A)、グリシン(G);
2)アスパラギン酸(D)、グルタミン酸(E);
3)アスパラギン(N)、グルタミン(Q);
4)アルギニン(R)、リジン(K);
5)イソロイシン(I)、ロイシン(L)、メチオニン(M)、バリン(V);
6)フェニルアラニン(F)、チロシン(Y)、トリプトファン(W);
7)セリン(S)、スレオニン(T);および
8)システイン(C)、メチオニン(M)(例えば、Creighton,Proteins(1984)を参照)。
【0029】
「配列同一性のパーセンテージ」は、最適にアラインされた2つの配列を、該2つの配列の最適アラインメントのために、比較窓で比較することによって測定され、ここで、比較窓におけるポリヌクレオチド配列の部分は、付加または欠失を含まない参照配列(例えば、本発明のポリペプチド)と比較して付加または欠失(すなわち、ギャップ)を含んでもよい。該パーセンテージは、同一である核酸塩基またはアミノ酸残基が、両方の配列に生じる位置の数を測定し、一致した位置の数を得て、一致した位置の数を、比較の窓における位置の総数で割り、その結果に100をかけて、配列同一性のパーセンテージを得ることによって計算される。
【0030】
2またはそれを超える核酸またはポリペプチド配列の文脈において、用語「同一である」またはパーセント「同一性」は、同一の配列である2またはそれを超える配列または部分列に言及する。2つの配列が、比較窓にわたる最大一致にわたって、または、次の配列比較アルゴリズムの1つを用いてまたは手動アラインメントおよび目視検査によって測定される指定領域にわたって、比較してアラインした場合に、特定のパーセンテージ同一である(すなわち、特定の領域にわたって、または、特定されない場合には、完全な配列にわたって、60%同一である、任意的に65%、70%、75%、80%、85%、90%、または95%同一である)アミノ酸残基またはヌクレオチドを有する場合には、2つの配列は、「実質的に同一である」。本発明は、本明細書において例示されるポリペプチド(例えば、配列番号1〜28のいずれか)にそれぞれ実質的に同一であるポリペプチド、およびその使用を提供し、その使用は、関節炎または関節損傷を処置または予防するための使用を含むが、それらに限定されない。任意的に、該同一性は、少なくとも約50ヌクレオチドの長さの領域にわたって、より好ましくは、100から500または1000またはそれを超えるヌクレオチドの長さの領域にわたって、または、参照配列の完全長にわたって、存在する。
【0031】
配列比較のためには、典型的には、1つの配列が、参照配列として作用し、試験配列は、これと比較される。配列比較アルゴリズムを用いる場合、試験および参照配列が、コンピューターに入力され、必要に応じて部分列座標が指定され、そして、配列アルゴリズムプログラムパラメーターが指定される。デフォルトプログラムパラメーターを使用することができ、また、別のパラメーター指定することができる。配列比較アルゴリズムは、続いて、プログラムパラメーターに基づき、参照配列と比較した試験配列におけるパーセント配列同一性を計算する。
【0032】
本明細書において、「比較窓」は、20〜600、普通には、約50〜約200、より普通には約100〜約150からなる群から選択される、隣接位置の数の任意の1つの断片への参照を含み、ここで、配列は、2つの配列が最適にアラインされた後、同一数の隣接位置の参照配列と比較されてもよい。比較のための配列のアラインメントの方法は、本技術分野においてよく知られている。比較のための配列の最適なアラインメントは、例えば、Smith and Waterman(1970)Adv.Appl.Math.2:482cのローカルホモロジーアルゴリズムによって、Needleman and Wunsch(1970)J.MoI.Biol.48:443のホモロジーアラインメントアルゴリズムによって、Pearson and Lipman(1988)Proc.Nat’l.Acad.Sci.USA 85:2444の類似性検索の方法によって、これらのアルゴリズムのコンピューターでの遂行によって(Wisconsin Genetics Software Package、 Genetics Computer Group、 575 Science Dr., Madison、 WIにおけるGAP、BESTFIT、FASTA、およびTFASTA)、または、手動アラインメントおよび目視検査によって(例えば、Ausubel et al., Current Protocols in Molecular Biology(1995supplement)を参照)によって行うことができる。
【0033】
パーセント配列同一性および配列類似性の測定に適したアルゴリズムの2つの例が、BLASTおよびBLAST2.0アルゴリズムであり、それぞれAltschul et al.(1977)Nuc. Acids Res.25:3389−3402およびAltschul et al.(1990)J. MoI. Biol.215:403−410に説明されている。BLAST分析を実行するためのソフトウェアは、National Center for Biotechnology Informationを通して公的に入手可能である。このアルゴリズムは、データベース配列における同一長のワードとアラインした場合にいくつかの正値閾値スコアTと一致するか、それを満たすかのいずれかである、クレリー配列におけるW長のショートワードを同定することによって、高スコア配列ペア(HSPs)をまず同定することを含む。Tは、近隣ワードスコア閾値とも呼ばれる(上記Altschul et al.,)。これらの最初の近隣ワードヒットは、それらを含むより長いHSPsを探すためのデフォルト検索の種としての役割を果たす。該ワードヒットは、累積アラインメントスコアが上昇することができる限り、各配列にそって両方向に延長される。累積スコアは、ヌクレオチド配列のためにはパラメーターM(一致残基のペアのためのリワードスコア;常に>0)およびN(非一致残基のためのペナルティスコア;常に<0)を用いて計算される。アミノ酸配列のためには、スコアリングマトリックスが、累積スコアを計算するために用いられる。それぞれの方向におけるワードヒットの延長は、累積アラインメントスコアが、その最大達成値からの数Xによって低下する場合、1または複数の負スコア残基アラインメントの蓄積のために累積スコアが、ゼロまたはそれ未満になる場合、または、いずれかの配列の末端に到達した場合に中止される。BLASTアルゴリズムパラメーターW、T、およびXは、アラインメントの感度およびスピードを決定する。(ヌクレオチド配列のための)BLASTNプログラムは、デフォルトとして、11のワード長(W)、10の期待値(E)M=5、N=−4および、両ストランドの比較を使用する。アミノ酸配列のためのBLASTPプログラムは、デフォルトとして、3のワード長、10の期待値(E)、およびBLOSUM62スコアリングマトリックス(Henikoff and Henikoff(1989)Proc.Natl.Acad.Sci.USA 89:10915を参照)、50のアラインメント(B)、10の期待値(E)M=5、N=−4、および両ストランドの比較を用いる。
【0034】
BLAST アルゴリズムは、2つの配列の間の類似性の統計的分析も実行する(例えば、Karlin and Altschul(1993)Proc.Natl.Acad.Sci.USA 90:5873−5787を参照)。BLASTアルゴリズムによって提供される類似性の1つの測定は、最小合計確率(P(N))であり、これは、2つのヌクレオチドまたはアミノ酸配列の間の一致が偶然に起こりうる確率の指標を提供する。例えば、もし、参照核酸との試験核酸の比較における最小合計確率が、約0.2未満、より好ましくは約0.01未満、および最も好ましくは約0.001未満である場合には、核酸は、参照配列と類似していると考えられる。
【0035】
本明細書において、用語「ヒアルロン酸」は、ヒアルロン酸のエステルおよびヒアルロン酸の塩を含むヒアルロン酸の誘導体を含むものとして使用され、用語ヒアルロナンも含む。該呼称は、低および高分子量型のヒアルロナンの両方、および、架橋ヒアルロナンまたはハイランも含む。このようなヒアルロナンの例は、Synvisc(商標)(Genzyme Corp.Cambridge,Mass.),ORTHO VISC(商標)(Anika therapeutics,Woburn,Mass.)、およびHYALGAN(商標)(Sanofi−Synthelabo Inc.,Malvern,Pa.)である。
【図面の簡単な説明】
【0036】
【図1】インビトロ軟骨形成の間のMSC−誘導II型コラーゲン産生の定量的評価。 hMSCs(10,000/384ウェル)をhMSC増殖培地(Millipore)において24時間プレートした。該細胞を、上記タンパク質でさらに72時間処理した。培地を無血清DMEMに交換し、追加の刺激なしにさらに14日間培養した。終わりに際して、細胞をホルマリンで固定し、洗浄し、II型コラーゲンに対する抗体で染色し、DAPIで対比染色した。II型コラーゲン染色の量は、ハイコンテントイメージング(Opera、 Perkin Elmer)によって定量化した。データは、3つの実験からそれぞれの投与量について2つプールした(n=6)。
【0037】
【図2】ANGTPL3−誘導軟骨形成の特性評価。 (A)ペレット培養(1x106細胞/ペレット)において、21日間、無血清DMEM、1×ITSおよびANGTPL3(指定の場合)中でhMSCsを培養した。培地を3日ごとに交換した。製造者の指示によるヒトTaqman特異的プローブを用いてアグレカンmRNA発現を定量化した(3つの実験から2つずつプールされたデータ(n=6))。 (B)ウシ軟骨を単離し、叩いて対称円にし、器官培養に入れた。該スライスを48時間20ng/ml TNFaおよび10ng/ml OSM(炎症性メディエーター)で処理して、ANGPTL3の存在下または非存在下で、軟骨マトリックスの分解を誘導し、マトリックス損傷の指標であるグルコサミノグルカン放出のパーセント阻害を同定した(4つのドナーからプールしたデータ、n=12)。 (C)初期正常ヒト軟骨細胞および滑膜細胞(増殖培地中、2500/384ウェル)を、プレートし、24時間37℃で培養した。24時間の期間をとおしてANGTPL3に応じた穏やかな軟骨細胞特異的細胞増殖の上昇が実証された(2つの実験からプールされたデータ、2反復/投与量(n=4))。
【0038】
【図3】mANGTPTL3。 (A)主要タンパク質構造および確認されたグリコシル化部位。 (B)mANGTPL3C末端FLD(225−455)の原子構造(1.8オングストローム)。
【0039】
【図4】ANGPTL3配列の配列アラインメント。 (A)ヒト、マウス、ウシ、およびイヌネイティブANGPTL3タンパク質の配列アラインメント; (B)ヒト、マウス、ウシ、イヌおよびウマネイティブANGPTL3タンパク質の配列アラインメント。
【0040】
【図5】外科骨関節炎モデルにおけるのインビボ効能。 C57BL/6マウス(n=12/群)からの右膝の前十字靱帯(ACL)、内側半月板脛骨靱帯(MMTL)、および内側側副靱帯(MCL)の外科的切断を行い、膝関節における不安定化を誘導し、それによってOA表現型を引き起こした。外科手術の1週間後、マウスに3〜4週間に1回ごと、関節内投与を行った。 (A)末梢血を、外科手術の28日後に収集した。循環II型コラーゲン断片(CTX−II)をELISA(Nordic biosciences)によって定量化した。mANGPTL3投与量=200ng/膝、3週間ごとの注射。 (B)脛骨プラトーの定量的評価を0〜4スケールで行った。0は正常であり、5は重度のOAである(軟骨の全層破壊)。各マウスからの2つの片が、2の観察者によって無分別に等級分けされた。mANGPTL3投与量=200ng/膝、3週間ごと注射。 (C)インキャパシタンステストによって、または、モニター装置を通して外科手術の脚と別の脚とで立ったマウスの時間のパーセンテージを測定することによって、OA痛を測定した。読み出し値は、外科手術36日後の痛応答を示す(示された濃度でANGPTL3の1週間ごと投与量の3投与量)。
【発明を実施するための形態】
【0041】
詳細な説明
I.導入
【0042】
本発明は、アンジオポイエチン様3(ANGPTL3)が間葉幹細胞における軟骨細胞分化を刺激するという発見に、部分的に基づいている。したがって、本発明は、軟骨細胞への間葉幹細胞分化を誘導する方法を提供する。また、本発明は、ANGPTL3タンパク質を関節、椎骨、椎間板、または全身に投与することによって関節炎または関節損傷を予防または改善するためのANGPTL3タンパク質の投与を提供する。
II.アンジオポイエチン様3
【0043】
アンジオポイエチン様3は、分泌因子のアンジオポイエチン様ファミリーのメンバーである。これは、主に肝臓に発現し、シグナルペプチド、N−末端コイルドコイルドメイン(CCD)およびC末端フィブリノゲン(FBN)様ドメインからなるアンジオポイエチンの特徴構造を有する。アンジオポイエチン様3におけるFBN様ドメインは、αV/β3インテグリンに結合し、この結合が、内皮細胞接着および遊走を誘導することが示された。
【0044】
様々なANGPTL3タンパク質が、本発明において使用することができる。本明細書において説明されるように、ネイティブANGPTL3は、一般的には、インビボで開裂され、アミノ−末端およびカルボキシル末端断片になる。本発明は、軟骨形成活性を有する様々なANGPTL3タンパク質の使用を想定する。ある実施態様において、本発明は、完全長ネイティブ(またはその変異体)ANGPTL3タンパク質アミノ酸配列の使用を提供する。ある実施態様において、本発明は、ANGPTL3配列、またはその変異体の(完全長ネイティブ配列ではない)部分を含むANGPTL3タンパク質であって、軟骨形成活性を保持するもの、すなわち、ネイティブタンパク質のアミノ酸末端側終端ではないものを提供する。ある実施態様において、本発明のANGPTL3タンパク質は、CCDドメインを持たない、および/または、有意なCCD活性を持たない。したがって、ある実施態様において、本発明のANGPTL3タンパク質は、ネイティブマウス(例えば、配列番号12)、ヒト(例えば、配列番号8)、ウシ(例えば、配列番号16)、イヌ(例えば、配列番号20)、またはウマ(例えば、配列番号24)ANGPTL3タンパク質配列、または実質的に同一である配列の少なくとも断片(例えば、少なくとも50、100、150、200、250の隣接するアミノ酸)を含むが、ネイティブANGPTL3タンパク質の少なくとも200の隣接するアミノ−末端アミノ酸を含まない。
【0045】
ある実施態様において、本発明のANGPTL3タンパク質は、フィブリノゲン−様ドメインを含む。ある実施態様において、本発明のANGPTL3タンパク質は、ネイティブマウス(例えば、配列番号12)、ヒト(例えば、配列番号8)、ウシ(例えば、配列番号16)、イヌ(例えば、配列番号20)、またはウマ(例えば、配列番号24)ANGPTL3タンパク質配列のアミノ酸207−455、207−400、207−350、225−455、225−400、225−350、241−455、241−400、241−350に対応する隣接するアミノ酸を含む、または、これらの配列と実質的同一であるが、フランキングネイティブANGPTL3タンパク質アミノ酸配列を含まない。ある実施態様において、本発明のANGPTL3タンパク質(配列番号1〜28のいずれかを含むがこれらに限定されない)は、C末端配列の少なくとも部分を欠き、例えば、C−末端からの10、20、30、40、50アミノ酸を欠く。
【0046】
上記の本発明のANGPTL3タンパク質は、上記の領域の側面に位置するネイティブANGPTL3タンパク質配列を含まなくてよいが、本発明のANGPTL3タンパク質は、非ネイティブANGPTL3タンパク質フランキング配列を含むことができる。例えば、ANGPTL3タンパク質の軟骨形成活性部分は、1または複数の異種アミノ酸に融合させて、融合タンパク質を形成させることができる。融合パートナー配列は、アミノ酸タグ非−L(例えば、D−)アミノ酸、またはインビボ半減期を伸ばすための、および/もしくはプロテアーゼ耐性を高めるための他のアミノ酸模倣物、標的化配列または他の配列を含むことができるが、それらに限定されない。
【0047】
本発明のANGPTL3タンパク質は、ネイティブANGPTL3タンパク質の変異体およびトランケーション、ならびに上記活性断片の変異体およびトランケーションを包含する。活性変異体は、本技術分野における当業者に知られているあらゆる方法によって同定することができる。ある実施態様において、活性タンパク質のアミノ酸アラインメントは、不変である位置、または保存されたアミノ酸変化を含む位置を同定するために確立されうる。配列番号1、2、3、または4は、ヒト、マウス、ウシ、およびイヌネイティブANGPTL3タンパク質のある領域(それぞれ、位置241−455、225−455および207−455、ならびにネイティブ完全長)の間に不変アミノ酸を含むコンセンサス配列を表す。配列番号25、26、27、または28は、ヒト、マウス、ウシ、イヌ、およびウマネイティブANGPTL3タンパク質のある領域(それぞれ、位置241−455、225−455および207−455、ならびにネイティブ完全長)の間において不変アミノ酸を含むコンセンサス配列を表す。したがって、ある実施態様において、本発明の軟骨形成ANGPTL3タンパク質は、配列番号1、2、3、4、25、26、27、または28を含む。ある実施態様において、本発明の軟骨形成ANGPTL3タンパク質は、配列番号5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23または24のいずれかと(配列番号の長に渡って測定して)実質的に同一であるアミノ酸配列を含む。
【0048】
本発明のANGPTL3タンパク質は、軟骨形成活性を有する。本明細書において定義されるように、軟骨形成または軟骨形成活性は、MSCsからの軟骨細胞の成長に言及する。軟骨形成活性の指標は、軟骨マトリックス産生を含むがこれらに限定されない。軟骨マトリックス産生は、様々なマーカー、例えば、Sox9、II型コラーゲン、またはグルコサミノグルカン(GAG)産生によって測定してもよい。ある実施態様において、GAG産生は、軟骨マトリックス産生のためのマーカーとして測定される。ある実施態様において、軟骨特異的タンパク質発現でのGAG産生の3倍増加は、陽性の軟骨マトリックス産生を示す。
【0049】
ある実施態様において、本発明のANGPTL3ポリペプチドは、少なくとも1つの非天然にコードされるアミノ酸を含むであろう。非天然に生じるアミノ酸を作製しタンパク質に導入する方法は、知られている。例えば、米国特許第7,083,970、および7,524,647を参照のこと。1または複数の所望の非天然アミノ酸を含むタンパク質の作製に適した直交翻訳システムの作成のための一般的な原理は、直交翻訳システムを作成するための一般的な方法として、本技術分野において知られている。例えば、“METHODS AND COMPOSITION FOR THE PRODUCTION OF ORTHOGONAL tRNA−AMINOACYL−tRNA SYNTHETASE PAIRS;”と題された国際公開公報WO2002/086075、“IN VIVO INCORPORATION OF UNNATURAL AMINO ACIDS;”と題されたWO2002/085923、“EXPANDING THE EUKARYOTIC GENETIC CODE;”と題されたWO2004/094593、“ORTHOGONAL TRANSLATION COMPONENTS FOR THE VIVO INCORPORATION OF UNNATURAL AMINO ACIDS”と題され、2004年7月7日出願のWO2005/019415、2004年7月7日出願のWO2005/007870、2004年7月7日出願のWO2005/007624、2005年10月27日出願のWO2006/110182、および“SYSTEMS FOR THE EXPRESSION OF ORTHOGONAL TRANSLATION COMPONENTS IN EUBACTERIAL HOST CELLS”と題され2007年3月7日出願のWO2007/103490を参照のこと。これらのそれぞれの内容は、参照によって全体が組み込まれる。非天然アミノ酸を組み込む直交翻訳システム、およびその産生および使用の方法の議論のためには、Wang and Schultz,(2005)“Expanding the Genetic Code.”Angewandte Chemie Int Ed 44:34−66;Xie and Schultz,(2005)“An Expanding Genetic Code.”Methods 36:227−238;Xie and Schultz,(2005)“Adding Amino Acids to the Genetic Repertoire.”Curr Opinion in Chemical Biology 9:548−554;およびWang,et al,(2006)“Expanding the Genetic Code.”Annu Rev Biophys Biomol Struct 35:225−249;Deiters,et al,(2005)“In vivo incorporation of an alkyne into proteins in Escherichia coli.”Bioorganic&Medicinal Chemistry Letters 15:1521−1524;Chin,et al.,(2002)“Addition of p−Azido−L−phenylalanine to the Genetic Code of Escherichia coli.”J Am Chem Soc 124:9026−9027;および2005年9月20日出願の国際公開公報WO2006/034332も参照のこと。これらのそれぞれの内容は、参照によって全体が組み込まれる。さらなる詳細は、米国特許第7,045,337;第7,083,970;第7,238,510;第7,129,333;第7,262,040;第7,183,082;第7,199,222;および第7,217,809に見出される。
【0050】
「非天然にコードされるアミノ酸」は、共通アミノ酸の1つではないアミノ酸またはピロールリジンもしくはセレノシステインに言及する。用語「非天然にコードされるアミノ酸」と同意語として用いられる他の用語は、「非天然アミノ酸」、「非天然アミノ酸」、「非天然に生じるアミノ酸」、および様々にハイフンで結んだ、およびハイフンで結んでいないそれらのバージョンである。用語「非天然にコードされるアミノ酸」は、天然にコードされるアミノ酸(20の共通アミノ酸、またはピロールリジンおよびセレノシステインを含むがこれらに限定されない)の修飾(例えば、翻訳後修飾)によって生じるが、それ自体増殖しているポリペプチド鎖には翻訳複合体によって天然に組み込まれないアミノ酸も含むがそれらに限定されない。このような非天然に生じるアミノ酸の例は、N−アセチルグルコサミニル−L−セリン、N−アセチルグルコサミニル−L−スレオニン、およびO−フォスフォチロシンを含むがこれらに限定されない。
【0051】
非天然にコードされるアミノ酸は、典型的には20の天然アミノ酸において用いられるものとは別の任意の置換側鎖を有する任意の構造である。本発明の非天然にコードされるアミノ酸は、典型的には、天然アミノ酸とは側鎖の構造の点でのみ異なるので、非天然にコードされるアミノ酸は、天然にまたは非天然にコードされるものを非限定的に含む他のアミノ酸と、天然に生じるポリペプチドにおいて形成されるのと同一の様式でアミド結合を形成する。しかしながら、非天然にコードされるアミノ酸は、天然アミノ酸からそれらを区別する側鎖基を有する。例えば、Rは、任意的にアルキル−、アリール−、アシル−、ケト−、アジド−、ヒドロキシル−、ヒドラジン、シアノ−、ハロ−、ヒドラジド、アルケニル、アルキニル、エーテル、チオール、セレノ−、スルホニル−、ホウ酸塩、ボロン酸、ホスホ、ホスホノ、ホスフィン、複素環、エノン、イミン、アルデヒド、エステル、チオ酸、ヒドロキシルアミン、アミノ基、もしくは同様のもの、またはその任意の組み合わせを含む。本発明における使用に適しうる他の非天然に生じる目的のアミノ酸は、光活性化可能な架橋剤を含むアミノ酸、スピン標識アミノ酸、蛍光アミノ酸、金属結合アミノ酸、金属−含有アミノ酸、放射性活性アミノ酸、新規の官能基を有するアミノ酸、他の分子と共有結合的にまたは非共有結合的に相互作用するアミノ酸、フォトケージされたおよび/または光異性化可能なアミノ酸、ビオチンまたはビオチン類似物を含むアミノ酸、糖置換セリンなどのグリコシル化アミノ酸、他の炭水化物修飾アミノ酸、ケト−含有アミノ酸、ポリエチレングリコールまたはポリエーテルを含むアミノ酸、重原子置換アミノ酸、化学的に開裂可能なおよび/または光開裂可能なアミノ酸、約5を超えるまたは約10を超える炭素を非限定的に含むポリエーテルまたは長鎖炭化水素を非限定的に含む天然アミノ酸と比較して伸張した側鎖を持つアミノ酸、炭素接続糖類含有アミノ酸、レドックス活性アミノ酸、アミノチオ酸含有アミノ酸、および1または複数の毒性部分を含むアミノ酸を含むがそれらに限定されない。
【0052】
本発明における使用に適し得て、水溶性ポリマーとの反応に有用な非天然にコードされるアミノ酸は、例えば、カルボニル、アミノオキシ、ヒドラジン、ヒドラジド、セミカルバジド、アジドおよびアルキン反応基を有するものを含むがこれらに限定されない。ある実施態様において、非天然にコードされるアミノ酸は、サッカリド部分を含む。このようなアミノ酸の例は、N−アセチル−L−グルコサミニル−L−セリン、N−アセチル−L−ガラクトサミニル−L−セリン、N−アセチル−L−グルコサミニル−L−スレオニン、N−アセチル−L−グルコサミニル−L−アスパラギンおよびO−マンノサミニル−L−セリンを含む。このようなアミノ酸の例は、天然に生じるアミノ酸とサッカリドとの間のN−またはO−結合が、天然には一般には見出されない共有結合(アルケン、オキシム、チオエーテル、アミド、および同等物を含むがこれらに限定されない)に交換されている例も含む。このようなアミノ酸の例は、2−デオキシ−グルコース、2−デオキシガラクトース、および同等物などの天然に生じるタンパク質において一般的には見出されないサッカリドも含む。
【0053】
例えば、インビボ半減期を伸張するために、本発明のANGPTL3タンパク質に(例えば、ポリペプチド鎖内において、または、N−もしくはC末端のいずれかにおいて)導入できる別の型の修飾は、PEG化または長鎖ポリエチレングリコールポリマー(PEG)の組み込みである。PEGまたはPEGの長鎖ポリマーの導入は、例えば、尿への急速ろ過を予防するために、本発明のポリペプチドの有効分子量を増大させる。ある実施態様において、ANGPTL3配列におけるリジン残基は、PEGに直接的にまたはリンカーを通して、接合される。このようなリンカーは、例えば、適切に修飾されたPEG鎖への結合のためのチオール官能基を含有するGlu残基またはアシル残基であることができる。PEG鎖を導入するための別の方法は、まずC−末端に、または溶媒露出残基に、ArgまたはLys残基の代用などのCys残基を導入する。このCys残基は、次に、例えば、マレイミド機能を含有するPEG鎖に部位特異的に付着させられる。PEGまたはPEGの長鎖ポリマーを組み込むための方法は、本技術分野においてよく知られており(例えば、Veronese,F.M.,et al.,Drug Disc.Today 10:1451−8(2005);Greenwald,R.B.,et al.,Adv.DrugDeliv.Rev.55:217−50(2003);Roberts,M.J.,et al.,Adv.DrugDeliv.Rev.,54:459−76(2002)などに説明されている)、その内容は参照によって本明細書に包含される。本技術分野において知られているポリマー接合の別の方法も、本発明において使用することができる。ある実施態様において、ポリ(2−メタクリロイルオキシエチルホスホリルコリン)(PMPC)が、本発明のANGPTL3タンパク質へのポリマー接合として導入される(例えば、WO2008/098930;Lewis,et al.,Bioconjug Chem.,19:2144−55(2008)を参照のこと)。ある実施態様において、ANGPTL3タンパク質と接合したホスホリルコリン−含有ポリマーが、本発明において使用することができる。当業者は、別の生体適合性ポリマー接合を利用することができることを容易に認識するであろう。
【0054】
(上記などの)非天然アミノ酸の組み込みを通してPEGまたはPEGポリマーを組み込むためのより最近に報告された改変ネイティブアプローチは、本発明のポリペプチドで実行することができる。このアプローチは、進化したtRNA/tRNAシンテターゼペアを利用し、アンバーサプレッサーコドンによる発現プラスミドにおいてコードされる(Deiters,A,et al.(2004).Bio−org.Med.Chem.Lett.14,5743−5)。例えば、p−アジドフェニルアラニンは、本発明のポリペプチドに組み込まれ、そして、還元剤および銅イオンの存在下で、アセチレン部分を有するPEGポリマーとの反応に供され、「ヒュスゲン[3+2]付加環化」として知られる有機反応を促進させることができる。
【0055】
ある実施態様において、本発明は、ポリペプチドのグリコシル化を改変するためのANGPTL3タンパク質の特異的な変異を想定する。このような変異は、O−連結またはN−連結グリコシル化部位を含むがそれらに限定されない1または複数のグリコシル化部位を導入または除去するように選択されることができる。ある実施態様において、本発明のANGPTL3タンパク質は、天然に生じるANGPTL3タンパク質と比較して非改変であるグリコシル化部位およびパターンを有する。ある実施態様において、ANGPTL3タンパク質の変異体は、グリコシル化部位の数および/またはタイプが、天然に生じるANGPTL3タンパク質と比較して改変されたグリコシル化変異体を含む。ある実施態様において、ポリペプチドの変異体は、ネイティブのポリペプチドと比較して数が多いまたは少ないN−連結グリコシル化部位を含む。N−連結グリコシル化部位は、配列:Asn−X−SerまたはAsn−X−Thrによって特徴付けられ、ここで、Xと指定されたアミノ酸残基は、プロリン以外の任意のアミノ酸残基であってよい。この配列を作製するためのアミノ酸残基の置換は、N−連結炭水化物鎖の付加のための潜在的な新しい部位を提供する。また、この配列を除去する置換は、存在するN−連結炭水化物鎖を除去するであろう。ある実施態様において、N−連結炭水化物鎖の再配列が、提供され、ここで、1または複数のN−連結グリコシル化部位(典型的には天然に生じるもの)が除去され、そして、1または複数の新しいN−連結部位が作製される。
【0056】
ANGPTL3タンパク質変異体の例は、天然に生じるANGPTL3タンパク質のアミノ酸配列と比較して1または複数のシステイン残基が欠失して別のアミノ酸(例えば、、セリン)に置換されたシステイン変異体を含む。ある実施態様において、不溶性封入体の単離後など、ANGPTL3タンパク質が、生物学的に活性なコンフォーメーションにリフォールドされなければならない場合には、システイン変異体は、有用でありうる。ある実施態様において、システイン変異体は、ネイティブのポリペプチドと比較して少ない数のシステイン残基を有する。ある実施態様において、システイン変異体は、非ペアシステインにからもたらされる相互作用を最小化するために、偶数のシステイン残基を有する。
【0057】
ある実施態様において、ANGPTL3タンパク質の機能的変異体または修飾されたフォームは、本発明のANGPTL3タンパク質の融合タンパク質および1または複数の融合ドメインを含む。融合ドメインのよく知られた例は、ポリヒスチジン、Glu−Glu、グルタチオンSトランスフェラーゼ(GST)、チオレドキシン、プロテインA、プロテインG、免疫グロブリン重鎖定常領域(Fc)、マルトース結合タンパク質(MBP)、またはヒト血清アルブミンを含むがこれらに限定されない。融合ドメインは、所望の特性を授与するために選択することができる。例えば、ある融合ドメインは、アフィニティクロマトグラフィによって融合タンパク質を単離するために特に有用である。親和性精製の目的のために、グルタチオン−、アミラーゼ−、およびニッケル−またはコバルト−接合樹脂などのアフィニティクロマトグラフィのための関連するマトリックスが使用される。このようなマトリックスの多くは、(HIS6)融合パートナーに有用なPharmacia GST purification system、およびQLAexpress(商標)system(Qiagen)などの「キット」の形で入手することができる。別の例として、融合ドメインは、ANGPTL3タンパク質の検出を促進するように選択されうる。このような検出ドメインの例は、様々な蛍光タンパク質(例えば、GFP)、ならびに通常は短いペプチド配列でありそれに対する特異的な抗体を入手することができる「エピトープタグ」を含む。特異的なモノクローナル抗体を容易に入手することができ、よく知られているエピトープタグは、FLAG、インフルエンザウイルス血球凝集(HA)、およびc−mycタグを含む。ある場合において、融合ドメインは、関連するプロテアーゼが融合タンパク質を部分的に消化し、それによってそれからの組換えタンパク質を遊離させるのを可能にする、Xa因子またはトロンビンなどのプロテアーゼ開裂部位を有する。そして、遊離されたタンパク質は、続いて行われるクロマトグラフィ分離によって融合ドメインから単離することができる。ある実施態様において、ANGPTL3タンパク質は、インビボでANGPTL3タンパク質を安定化するドメイン(「安定用」ドメイン)と融合している。「安定化」は、血清半減期を上昇させるあらゆることを意味し、これが破壊の減少、腎臓によるクリアランスの減少、または他の薬物動態効果によるかどうかは関係ない。免疫グロブリンのFc部分との融合は、幅広いタンパク質に所望の薬物動態特性を授与することが知られている。同様に、ヒト血清アルブミンへの融合は、所望の特性を授与することができる。選択されうる別のタイプの融合ドメインは、多量体化(例えば、二量体化、四量体化)ドメイン、および(所望の付加的な生物学的機能を授与する)機能的ドメインを含む。
III.アンジオポイエチン様3タンパク質疾患指標
【0058】
本発明のポリペプチド、組成物、および方法が、任意のタイプの関節炎または関節損傷の処置または予防のために使用されてもよいことが想定される。本発明のポリペプチド、組成物、および方法は、様々な軟骨性障害の処置または予防に使用されてもよいことがさらに想定される。ある実施態様において、本発明のタンパク質は、例えば、関節炎または関節損傷の遺伝的または家族歴がある、または以前に関節手術がある、もしくは関節手術の最中であるなどの、関節炎または関節損傷を予防するために投与される。本発明のポリペプチド、組成物、および方法で処置されるべきまたは予防されるべき病状または障害の例は、関節リウマチ、若年性慢性関節炎、骨関節炎、変性椎間板疾患、脊椎関節症、全身性硬化症(強皮症)、特発性炎症性ミオパシー(皮膚筋炎、多発性筋炎)、シェーグレン症候群、全身性血管炎、サルコイドーシス、自己免疫性溶血性貧血(免疫汎血球減少症、発作性夜間血色素尿症)、自己免疫性血小板減少症(特発性血小板減少性紫斑病、免疫介在性血小板減少症)、甲状腺炎(グレーブス病、橋本甲状腺炎、幼若リンパ球性甲状腺炎、萎縮性甲状腺炎)、糖尿病、免疫介在性腎疾患(糸球体腎炎、尿細管間質性腎炎)、多発性硬化、特発性脱髄性多発神経障害またはギラン−バール症候群、および慢性炎症性脱髄性多発神経障害などの中枢および末梢神経系の脱髄疾患、伝染性肝炎(肝炎A、B、C、D、E、および他の非肝炎ウイルス)などの肝胆道疾患、自己免疫性慢性活動性肝炎、原発性胆汁性肝硬変、肉芽腫性肝炎、および硬化性胆管炎、炎症性腸疾患(潰瘍性大腸炎:クローン病)、グルテン過敏性腸疾患、およびウィップル病、水疱性皮膚病を含む自己免疫または免疫介在性皮膚疾患、多形紅斑、および接触皮膚炎、乾癬、ぜんそく、アレルギー性鼻炎、アトピー性皮膚炎、食物過敏症、および、じんましん、などのアレルギー疾患、好酸球性肺炎、特発性肺線維症、および過敏性肺炎などの肺の免疫学的疾患、移植片拒絶および移植片対宿主病を含む移植関連疾患を含むがそれらに限定されない。本発明のある実施態様において、本発明のポリペプチド、組成物、および方法は、骨関節炎の処置のために使用されうる。
【0059】
ある実施態様において、本発明のポリペプチド、組成物、および方法は、外傷的損傷または軟骨疾患に起因して損傷した軟骨性組織における軟骨細胞増殖および軟骨産生を刺激するための方法を提供する。脊椎、肩、肘、手首、指の関節、尻、膝、足首、および足の関節などの関節表面を示す組織は、処置にとって特に重要である。処置の恩恵を受けうる疾患の例は、骨関節炎、リウマチ性関節炎、他の自己免疫疾患、または離断性骨軟骨症を含む。さらに、軟骨奇形が、ヒトにおいて小人症の形態でしばしば見られ、このことは、該ポリペプチド、組成物、および方法が、これらの患者において有用であろうことを示唆する。
【0060】
本発明のポリペプチド、組成物、および方法が、哺乳動物の処置のために有用であることが想定される。本明細書においては、「哺乳動物」は、ヒト、牛(例えば、雌牛)、馬、イヌ、羊、豚、ウサギ、ヤギ、ネコなどの家畜および家畜動物、ならびに動物園の動物スポーツ動物またはペット動物を含む、哺乳動物として分類される任意の哺乳動物を意味する。本発明のある実施態様において、哺乳動物はヒトである。
【0061】
ある実施態様において、本発明のポリペプチドは、処置されるべき哺乳動物とは異種であることができる。例えば、ウシANGPTL3タンパク質またはその断片、ウシANGPTL3タンパク質に由来するタンパク質またはペプチド(例えば、修飾されたウシANGPTL3タンパク質、ウシANGPTL3タンパク質の保存的変異体、ウシANGPTL3タンパク質に由来するペプチド模倣物)が、ヒト患者の処置において使用される。ある実施態様において、異種ANGPTL3タンパク質が、移植のための培養の軟骨細胞集団を拡張するために使用することができる。ある実施態様において、拡張した培養は、次に、処置されるべき哺乳動物にホモロジーがあるポリペプチドおよび組成物と混ぜ合わせられ、関節腔に、または、軟骨欠損に直接に、入れられる。また、本発明のポリペプチドは、同一の種に由来する。すなわち、ヒトANGPTL3タンパク質またはその断片、ヒトANGPTL3タンパク質に由来するタンパク質またはペプチド(例えば、修飾されたヒトANGPTL3タンパク質、ヒトANGPTL3タンパク質の保存的変異体、ヒトANGPTL3タンパク質に由来するペプチド模倣物)が、ヒト患者の処置において使用される。処置されつつある哺乳動物の同一種に由来するタンパク質の使用によって、不用意な免疫応答を避けてもよい。
【0062】
本発明のポリペプチドおよび組成物は、単独で、またはタンパク質の持続放出に適した担体との複合体で、関節の滑液への直接注射、全身性投与(経口または静脈内)、または軟骨欠損への直接投与によって適用することができる。本発明のポリペプチド、組成物、および方法は、自家または同種軟骨細胞移植のための培養における軟骨細胞集団を拡張するために使用することができる。該移植は、本発明のポリペプチドおよび組成物の投与からなる併用療法で任意的に投与することができる。これらの手順において、例えば、軟骨細胞は、損傷した関節の非損傷小耐力領域から関節鏡視下的に収集することができ、本発明のポリペプチドおよび組成物の存在下で、培養し、移植に先立ち、細胞の数を増加させることができる。拡張した培養は、続いて、本発明のポリペプチドおよび組成物と混ぜ合わされ、関節腔に、または欠損に直接に、入れられる。本発明のポリペプチドおよび組成物は、軟骨を形成することができる、および/または移植された軟骨細胞またはその前駆細胞を適所に保つのに役立つ、細胞を含む骨膜または軟骨膜移植片と組み合わせて使用することができる。本発明のポリペプチドおよび組成物は、関節の洗浄、骨髄の刺激、掻爬関節形成、骨軟骨病変ドリリング、または軟骨下骨の微小破壊と併用して、軟骨損傷の修復のために使用することができる。さらに、本発明のポリペプチドおよび組成物の投与に起因する軟骨の増殖の後、新たに形成された軟骨表面を適切に形成させるには、さらなる外科処置が必要であるかもしれない。
IV.医薬組成物
【0063】
上記の疾患または障害を処置するための本発明の化合物の投与量は、投与の様式、対象者の年齢、体重、および処置されるべき対象者の病状に応じて異なり、最終的には、主治医または主治獣医によって決定されるであろう。主治医または主治獣医によってこのように決定される化合物の量は、本明細書において「有効量」と呼ばれる。
【0064】
投与に適した剤形は、賦形剤を含み、水性および非水性溶液;酸化防止剤、緩衝剤、静菌剤、およびその剤形を等張にする溶質を含むことができる等張滅菌溶液;懸濁化剤、可溶化剤、増粘剤、安定剤、および防腐剤を含むことができる水性および非水性無菌懸濁液を含むがこれらに限定されない。
【0065】
本発明の文脈において、患者に投与される投与量は、時間とともに対象者における有益な応答をもたらすために十分であるべきである。投与量は、用いられる特定のタンパク質の効能、および対象者の病状、ならびに、体重または処置されるべき領域の表面領域によって決定されるであろう。投与量の多さは、特定の対象者における特定のタンパク質またはベクターの投与に伴う副作用の存在、性質、および程度によっても決定されるであろう。投与は、単一の投与量によって、または分けた投与量によって達成することができる。
V.投与の方法
【0066】
本発明のタンパク質を罹患した関節に輸送するためには、任意の方法を用いることができる。本発明の実施において、組成物は、例えば、関節内に(すなわち、関節へ)、経口的に、静脈内に投与することができる。化合物の製剤は、アンプルおよびバイアルなどのユニット投与量または多投与量密封容器において存在することができる。溶液および懸濁液は、前記の種類の滅菌粉末、顆粒、および錠剤から調製することができる。本発明のタンパク質は、所望の治療または効果に応じて、1または複数のさらなる活性剤(例えば、、ヒアルロン酸またはその塩)と組み合わせて効率的に使用することもできる。
【実施例】
【0067】
次の実施例は、請求項の発明を説明するために提供されるものであって、これを限定するためのものではない。
実施例1 軟骨形成の誘導剤のハイスループットスクリーン
【0068】
OA関節修復のための新しい非侵襲戦略を同定し明らかにするために、我々は、ヒト間葉幹細胞(MSCs)の軟骨細胞への分化を選択的に指示することができるタンパク質の非偏向ハイスループットスクリーンを開発した。このアッセイシステムは、軟骨における在住MSCをモデルとし、天然修復能力を刺激し、統合した軟骨再生を増進するメディエーターを同定する。我々は、ハイスループットによって固有の分泌タンパク質ライブラリーを試験するために、ヒトおよびマウスMSCsの細胞ベースのスクリーンを選んだ。このアプローチは、軟骨形成に影響を与える非同定およびネイティブタンパク質リガンドの迅速な同定を可能にする戦略を提供する。
【0069】
これらの分泌タンパク質を研究するために、哺乳動物産生細胞株(HEK293T)からの調整培地(CM)の産生、およびフリースタイルHEK−F細胞からの精製タンパク質の産出の2つのアプローチがとられた。これらの補完的アプローチは、組み合わさって、様々な適用において新規の標的同定を可能にする。
【0070】
MSCアッセイにおけるタンパク質のスクリーニングによって、有力候補であるアンジオポイエチン−様3、略称ANGPTL3が同定された。ANGPTL3は、CMおよび精製タンパク質の両方の2つの並行直交初期スクリーンで同定された。コンセプトCMスクリーンの証拠で、CMの移動の後、C3H10tl/2細胞を7日間インキュベートした。軟骨マトリックス産生を検出するためにアルシアンブルー染色によってアッセイし、軟骨形成は、ANGPTL3−75含有ウェルにおいて起こることが見出された。ANGPTL3−75は、ヒトMSCsの軟骨分化のための531の精製タンパク質のスクリーニングによって独立に同定された。
【0071】
初期スクリーニングアッセイの後、続いて、ANGPTL3を、6つの第2アッセイにおいて特徴付けた。これらアッセイは、
(1)マウス間葉C3H10tl/2細胞の単層培養:II型コラーゲンおよびSox9タンパク質発現(軟骨形成のマーカー)を誘導するが、オステオカルシン(骨発生のマーカー)の誘導はない;
(2)ウシ軟骨細胞におけるTNFα/オンコスタチンM(OSM)−誘導酸化窒素(NO)放出の阻害;
(3)ウシ軟骨器官培養におけるTNFα/OSM−誘導グルコサミノグルカン(GAG)放出の阻害;
(4)ヒトMSCペレット培養システムにおける軟骨マトリックス遺伝子発現(アグレカン)の誘導;
(5)初期ヒト軟骨細胞、ヒト滑膜線維芽細胞、およびヒトMSCsにおける毒性の欠如;
(6)ヒト軟骨細胞の増殖の刺激、
を含む。
【0072】
そのヒトホモログは、他の既知の軟骨形成の誘導剤と比較して、定量的イメージベースのII型コラーゲンアッセイにおいて軟骨形成のための類似の効能を保持するけれども、マウスANGPTL3タンパク質は、マウス、ヒトおよびウシ細胞タイプにおいて機能するので、さらなる特徴づけのために、潜在的に軟骨形成分化を誘導したとして(0.5−5nM)選択された(図1)。マウスおよびヒトMSCの両方が、(それぞれ)単分子層およびペレット培養で、0.5−5nM ANGPTL3で処置されて18〜21日間分化させられ、3つの軟骨特異的タンパク質:II型コラーゲン、Sox9およびアグレカンを発現する。これは、免疫組織化学染色およびTaqmanによるmRNA定量化(図2A)の両方によってアッセイされた。さらに、データは、マウスMSCsを18日間培養後、アルカリフォスファターゼの発現の減少を通し、ANGPTL3が、線維性修復応答に向けた自発傾向を阻害することを示唆する。組織損傷を予防する潜在的能力を評価するために、ウシ軟骨器官培養を、TNFαおよびオンコスタチンM(OSM)で刺激した。マトリックス変性の指標である刺激されたグルコサミノグルカン放出は、ANGPTL3での処置によって有意に阻害された(図2B)。さらに、初期ヒト軟骨細胞の処置によって、24時間内において細胞増殖において2倍の増加がもたらされたのに対し、ヒト滑膜細胞ではそうではなかったが(図2C)、このことは、軟骨に対するその作用の特異性を示唆する。そのタンパク質は、また、初期ヒト軟骨細胞、滑膜線維芽細胞およびMSCsの生存能力に対して、明白なインビトロ毒性効果を持たなかった(<100μM)(データは示さず)。
【0073】
目下の臨床試験での2つの候補間の直接的な比較において、100ng/mlのFGF18または100ng/ml BMP7の処置が、軟骨形成小節を誘導することができたが、全般的なマトリックス産生は、ANGPTL3と比較して、少なかった。FGF18は、アルシアンブルー、Sox9またはII型コラーゲン染色を増加させる能力に欠けており、このことは、本物の軟骨基質の特異性を欠くことを示す。100ng/mlのBMP7が、アルシアンブルー、Sox9およびII型コラーゲン染色を増加させたが、有意性は、類似濃度では、それほど高くなかった。
実施例2:組換え完全長ANGPTL3および変異タンパク質の発現、ならびに機能分析
【0074】
マウスANGPTL3は、51kDaタンパク質であると予測される。これは、アンジオポイエチンに類似した構造を有するが、Tie2受容体に結合する能力を欠き、区別される機能を有する、7つの同定されたアンジオポイエチン−様(ANGPTL)タンパク質のファミリーに属する。それらは、N−末端コイルドコイルドメイン(CCD)およびC末端フィブリノゲン−様ドメイン(FLD)を含む。ANGPTLタンパク質は、その微環境、および、細胞外マトリックス(ECM)との相互作用によって密接に制御されるが、その正確な相互作用部位も、パートナーも、詳細には解明されてこなかった。ANGPTL3は、肝臓によって分泌され、全身に循環する。LXR−誘導高トリグリセリド血が、ANGPTL3放出に起因することを証拠として、これは、肝臓X受容体(LXR)を通して制御される。推定ヘパリン結合モチーフを通したCCDおよびECMの間の相互作用は、ANGPTL4について報告されるのと似たように、前駆タンパク質転換酵素認識配列(R221−R224)における開裂の阻害をもたらすかもしれない。開裂は、リポタンパク質リパーゼ活性(LPL)を阻害するCCDの能力の有意な増加をもたらし、それによって、トリグリセリド血(TG)の増加をもたらす。これは、本発明に先立ち同定されたANGPTL3の主要な生物学的機能を表す。C末端FLDは、内皮細胞接着を誘導し、インビボでのラット角膜直接移植後、血管形成媒介するのに十分である。組換えANGPTL3は、精製αVβ3インテグリンに結合することができ、内皮細胞におけるFAK、MAPKおよびAKTシグナル伝達の増加を引き起こすことも実証された。これらのデータは、FLDのインテグリンとの相互作用が、血管形成を媒介することを示唆する。
【0075】
ANGPTL3の発現は、報告されてこなかったし、ヒト軟骨細胞、hMSCまたはヒト滑膜線維芽細胞におけるウェスタンブロッティングを用いる我々の研究でも観察されてこなかったし、その発現は、マウス膝関節において見出されなかった。さらに、我々のスクリーンで同定されたこの新規の軟骨形成機能に関する、関節細胞におけるいずれかの断片についての活性も報告されていない。
【0076】
我々のデータは、完全長ANGPTL3が軟骨形成および軟骨保護のための新規メディエーターであることを示す。我々は、ANGPTL3のどのドメインが、この新規軟骨形成機能に決定的かを調べた。詳細な生物物理的特性決定のために、ANGPTL3のトランケーション系および完全長タンパク質を、限定的で均一なグリコシル化を与える改変HEK−S細胞株において発現させ、精製した。質量分光学アプローチによって、すべての4つの予測されたN−連結グリコシル化(N115、N232、N296、およびN357、図3A)の場所が確認された。HEK内因的にプロセスされた断片から、CCDおよびFLDドメインの間のタンパク質分解開裂部位がR224であると測定された。完全長ANGPTL3は、質量>400kDaでのサイズ排除クロマトグラフィによって溶出し、このことは、異種グリコシル化での三量体を示す。サイズ排除および静的光散乱法によって単量体であることが示されるFLD(241−455)を結晶化した。FLDの原子構造を、1.8オングストローム解像度で測定した。FLD構造は、C−末端に向けて3つのループを向けるらせん伸長を持つコアベータシート形態を明らかにし、これは、他のFLDホモログの典型である(図3B)。ドメインのC末端部分における温度要因は、残りの構造よりも高く、このことは、この領域における高度の柔軟性を示唆する。ANGPTL2/Tie2構造(PDBコード:2GY7)との重ね合わせは、FLDのC末端部分が、タンパク質タンパク質相互作用に関与するかもしれないことを示唆する。
【0077】
産生された5つのトランケーション生成物を、複数の軟骨形成アッセイで評価した。結果は、CCD単独では活性保持がないが、FLDでは活性が維持することを示唆する。さらに、SMADlリン酸化は、完全長または変異GまたはH単独での3日間の刺激に応じて増加し、このことは、軟骨形成シグナル伝達カスケードの活性化を示唆する。
【0078】
関節内に注射されたタンパク質の全身性暴露は、滑液リンパ排出のために限られているけれども、全身性暴露を最小化することが、時には望ましい。全身性暴露がある場合には、ANGPTL3の開裂は、CCDドメインの放出を引き起こすかもしれない。CCDによるLPL活性の阻害の増加は、患者のTGレベルの改変をもたらしうる。我々の結果は、ANGPTL3の潜在的な臨床的利点を示す。たとえば、ANGPTL3における軟骨形成活性が位置する場所の切除によって、我々は、脂質代謝または血管形成特性に対する望まない全身性効果を最小化または排除するであろう。軟骨形成活性は、主としてC末端に位置し、そのため、このドメインの特異的使用は、完全長分子またはN末端のTG制御の懸念を軽減するであろう。完全長ANGPTL3は、通常、血清において存在するので、予想される免疫原性はない。C末端を使用するさらなる利点は、それは、開裂し、かつ三量体の完全長タンパク質と比較して単量体(〜29kDa)であり、そのため、循環において存在するかもしれない任意の小パーセンテージの全身性半減期を減少させ、血管形成からの任意の潜在的な効果を制限するであろうことである。
実施例3:ANGPTL3のインビボ分析
【0079】
我々は、潜在的な有害減少および関節内滞留に対処するために、完全長ANGPTL3のいくつかのインビボ評価を行った。8週齢C57BL/10マウスの左膝関節に3.6μgの完全長ANGPTL3で関節内(IA)注射した後、我々は、24時間の暴露の存在下および非存在下での発現を免疫組織化学的に評価した。滑液リンパ管へのトランス滑液流によるタンパク質および水にとっての典型的なターンオーバーが約2時間であるので、24時間まで、検出できるレベルのANGPTL3は、わずかしか滑液に留まらない、または滑液に留まらないはずである。結果は、非処置関節におけるANGPTL3の内因性発現がないが、関節軟骨および半月板において、24時間の時点でさえ細胞周囲マトリックスのタンパク質が有意に検出されることを明らかにした。さらに、細胞毒性の軟骨細胞への広がりや、インビボでの軟骨損傷は、検出されなかった。一連のラットの膝関節への3IA注射(1週間に1回を3週間)の後、臨床的には、毒性はなかった(関節膨潤も、歩き方の変化もなかった)し、ラットの関節における急性炎症反応の証拠となるものもなかった。組織学的には、滑膜炎の増加や、注射された5つのラットの関節における無制御的な増殖はなかった。マウスでのように、ANGPTL3は、軟骨マトリックスおよび周囲の軟骨細胞において検出され得た。これらの結果は、ANGPTL3は、軟骨自体に対する望ましくない効果を引き起こさず、そして、ANGPTL3は、インビボで軟骨および半月板に入り保持されることを示す。
【0080】
OAは、単一疾患実体ではなく、様々な病院的因子の帰結でありうる。ヒトにおいて、それは、しばしば、異常な生物力学的ストレスまたは遺伝的に獲得された関節軟骨または骨の異常性によって引き起こされる。したがって、「最善の」小動物OAモデルを選択することは、困難であり、複数のモデルが、任意の治療の保護特性を測定するために、探索されるべきである。我々は、慢性OAモデル(van der Kraanおよび同僚らによって説明された研究に基づきコラーゲナーゼVII誘導)およびGlassonらの仕事に基づき関節における3つの主要な靱帯(ACL、MCLおよびMMTL)の横断面を使う急性外科モデルの効能研究を完成させた。両方のモデルは、OAに共通に関係する病理的変化誘導した。プロテオグリカン染色の喪失、軟骨および骨の浸食、骨棘形成ならびに滑膜および靱帯の化生性変化が、OAの注射の4〜8週間後にはっきりと分かりうる。図5は、OAのANGPTL3外科モデルの再生能力を表す。OAについて潜在的バイオマーカーの検証を始めるため、外科モデルの間で末梢血を回収して、軟骨損傷ゆえに放出されたII型コラーゲン断片を測定した(図5A)。組織学的分析および、それに続く中間脛骨プラトーの等級付けは、200ng ANGTPL3/膝の1週間に1回3週間での処置後の軟骨マトリックスにおける再生を明らかにした(図5B)。
【0081】
8週間外科OAマウスのサブセットにおいて、3投与量のANGPTL3が、インキャパシタンス測定を通したOA−誘導痛を軽減するかどうかについて調べられた。この方法は、外科手術および非外科手術肢の間の重量配分を測定する。外科手術の後36日目に、および、PROlでの処置3週間ごとに、100ng/膝もの低さの投与が、ビヒクル処置外科手術膝と比較した有意な向上を実証した(図5C)。これらの組み合わせたデータは、ANGTPL3が、2つのOAモデル(病理的修正および痛み縮小の両方)においてインビボ効能を有することの具体的な証拠を提供し、新規のOA治療としてのその発達の進歩を裏付ける。
【0082】
本明細書に記載された実施例および実施態様は、例示のためだけのものであり、それらを考慮した様々な修飾または変化が本技術分野における当業者によって示唆されるであろうし、それらは、本出願、および添付の請求項の範囲の精神および範囲内に含まれると理解される。本明細書において引用された全ての公開、特許、および特許出願は、あらゆる目的でその全体が、参照によって本明細書に包含される。
【0083】
配列表
X = 任意のアミノ酸
丸括弧中のアミノ酸は、アミノ酸があってもなくてもよいことをを示す。
【0084】
配列番号:1
241−455 コンセンサス
XXPAXCXXXYNXGXHTSGXYXIXPXNSQXFXVYCDXXSGSXWTLIQHRXDGSQXFNETWENYXXGFGRLDGEFWLGLEKIYXIVXQSNYXLRXELXDWXD(X)KXYXEYXFXLGXHETNYTLHXXXIXGNXXXAXPEXXDLXFSTWXHXAKGXXXCPEXXSGGWWXXXXCGENNLNGKYNKXXXKXXPERXXGXXWXXQXXXLYXIKXXKMXXXPXX
【0085】
配列番号:2
225−455 コンセンサス
TTPXXXXNEXXNXXXXXXPAXCXXXYNXGXHTSGXYXIXPXNSQXFXVYCDXXSGSXWTLIQHRXDGSQXFNETWENYXXGFGRLDGEFWLGLEKIYXIVXQSNYXLRXELXDWXD(X)KXYXEYXFXLGXHETNYTLHXXXIXGNXXXAXPEXXDLXFSTWXHXAKGXXXCPEXXSGGWWXXXXCGENNLNGKYNKXXXKXXPERXXGXXWXXQXXXLYXIKXXKMXXXPXX
【0086】
配列番号:3
207−455 コンセンサス
XIXEXXEXSLSSKXRAPRTTPXXXXNEXXNXXXXXXPAXCXXXYNXGXHTSGXYXIXPXNSQXFXVYCDXXSGSXWTLIQHRXDGSQXFNETWENYXXGFGRLDGEFWLGLEKIYXIVXQSNYXLRXELXDWXD(X)KXYXEYXFXLGXHETNYTLHXXXIXGNXXXAXPEXXDLXFSTWXHXAKGXXXCPEXXSGGWWXXXXCGENNLNGKYNKXXXKXXPERXXGXXWXXQXXXLYXIKXXKMXXXPXX
【0087】
配列番号:4
完全長コンセンサス
MXTIKLXLXXXPLVIXSXXDXDXXSXDSXXXEPKSRFAMLDDVKILANGLLQLGHGLKDFVHKTKGQINDIFQKLNIFDQSFYDLSLXTXEIKEEEKELRRXTXXLQVKNEEVKNMSXELXSKXESLLEEKXXLQXKVXXLEXQLTXLIXXXXXXXEXXEVTSLKXXVEXQDNSIXXLLQXVEXQYXQLXQQXXQIKEIEXQLR(X)XXIXEXXEXSLSSKXRAPRTTPXXXXNEXXNXXXXXXPAXCXXXYNXGXHTSGXYXIXPXNSQXFXVYCDXXSGSXWTLIQHRXDGSQXFNETWENYXXGFGRLDGEFWLGLEKIYXIVXQSNYXLRXELXDWXD(X)KXYXEYXFXLGXHETNYTLHXXXIXGNXXXAXPEXXDLXFSTWXHXAKGXXXCPEXXSGGWWXXXXCGENNLNGKYNKXXXKXXPERXXGXXWXXQXXXLYXIKXXKMXXXPXX(SEXXE)
【0088】
配列番号:5
ヒトANGPTL3 241−455
GIPAECTTIYNRGEHTSGMYAIRPSNSQVFHVYCDVISGSPWTLIQHRIDGSQNFNETWENYKYGFGRLDGEFWLGLEKIYSIVKQSNYVLRIELEDWKDNKHYIEYSFYLGNHETNYTLHLVAITGNVPNAIPENKDLVFSTWDHKAKGHFNCPEGYSGGWWWHDECGENNLNGKYNKPRAKSKPERRRGLSWKSQNGRLYSIKSTKMLIHPTD
【0089】
配列番号:6
ヒトANGPTL3 225−455
TTPFLQLNEIRNVKHDGIPAECTTIYNRGEHTSGMYAIRPSNSQVFHVYCDVISGSPWTLIQHRIDGSQNFNETWENYKYGFGRLDGEFWLGLEKIYSIVKQSNYVLRIELEDWKDNKHYIEYSFYLGNHETNYTLHLVAITGNVPNAIPENKDLVFSTWDHKAKGHFNCPEGYSGGWWWHDECGENNLNGKYNKPRAKSKPERRRGLSWKSQNGRLYSIKSTKMLIHPTD
【0090】
配列番号:7
ヒトANGPTL3 207−455
IQEPTEISLSSKPRAPRTTPFLQLNEIRNVKHDGIPAECTTIYNRGEHTSGMYAIRPSNSQVFHVYCDVISGSPWTLIQHRIDGSQNFNETWENYKYGFGRLDGEFWLGLEKIYSIVKQSNYVLRIELEDWKDNKHYIEYSFYLGNHETNYTLHLVAITGNVPNAIPENKDLVFSTWDHKAKGHFNCPEGYSGGWWWHDECGENNLNGKYNKPRAKSKPERRRGLSWKSQNGRLYSIKSTKMLIHPTD
【0091】
配列番号:8
完全長ヒトANGPTL3 gi|7656888|ref|NP_055310.1|[Homo sapiens]
MFTIKLLLFIVPLVISSRIDQDNSSFDSLSPEPKSRFAMLDDVKILANGLLQLGHGLKDFVHKTKGQINDIFQKLNIFDQSFYDLSLQTSEIKEEEKELRRTTYKLQVKNEEVKNMSLELNSKLESLLEEKILLQQKVKYLEEQLTNLIQNQPETPEHPEVTSLKTFVEKQDNSIKDLLQTVEDQYKQLNQQHSQIKEIENQLRRTSIQEPTEISLSSKPRAPRTTPFLQLNEIRNVKHDGIPAECTTIYNRGEHTSGMYAIRPSNSQVFHVYCDVISGSPWTLIQHRIDGSQNFNETWENYKYGFGRLDGEFWLGLEKIYSIVKQSNYVLRIELEDWKDNKHYIEYSFYLGNHETNYTLHLVAITGNVPNAIPENKDLVFSTWDHKAKGHFNCPEGYSGGWWWHDECGENNLNGKYNKPRAKSKPERRRGLSWKSQNGRLYSIKSTKMLIHPTDSESFE
【0092】
配列番号:9
マウスANGPTL3 241−455
DLPADCSAVYNRGEHTSGVYTIKPRNSQGFNVYCDTQSGSPWTLIQHRKDGSQDFNETWENYEKGFGRLDGEFWLGLEKIYAIVQQSNYILRLELQDWKDSKHYVEYSFHLGSHETNYTLHVAEIAGNIPGALPEHTDLMFSTWNHRAKGQLYCPESYSGGWWWNDICGENNLNGKYNKPRTKSRPERRRGIYWRPQSRKLYAIKSSKMMLQPTT
【0093】
配列番号:10
マウスANGPTL3 225−455
TTPPLQLNETENTEQDDLPADCSAVYNRGEHTSGVYTIKPRNSQGFNVYCDTQSGSPWTLIQHRKDGSQDFNETWENYEKGFGRLDGEFWLGLEKIYAIVQQSNYILRLELQDWKDSKHYVEYSFHLGSHETNYTLHVAEIAGNIPGALPEHTDLMFSTWNHRAKGQLYCPESYSGGWWWNDICGENNLNGKYNKPRTKSRPERRRGIYWRPQSRKLYAIKSSKMMLQPTT
【0094】
配列番号:11
マウスANGPTL3 207−455
IQEPSENSLSSKSRAPRTTPPLQLNETENTEQDDLPADCSAVYNRGEHTSGVYTIKPRNSQGFNVYCDTQSGSPWTLIQHRKDGSQDFNETWENYEKGFGRLDGEFWLGLEKIYAIVQQSNYILRLELQDWKDSKHYVEYSFHLGSHETNYTLHVAEIAGNIPGALPEHTDLMFSTWNHRAKGQLYCPESYSGGWWWNDICGENNLNGKYNKPRTKSRPERRRGIYWRPQSRKLYAIKSSKMMLQPTT
【0095】
配列番号:12
完全長マウスANGPTL3 gi|33469117|ref|NP_038941.1|[Mus musculus]
MHTIKLFLFWPLVIASRVDPDLSSFDSAPSEPKSRFAMLDDVKILANGLLQLGHGLKDFVHKTKGQINDIFQKLNIFDQSFYDLSLRTNEIKEEEKELRRTTSTLQVKNEEVKNMSVELNSKLESLLEEKTALQHKVRALEEQLTNLILSPAGAQEHPEVTSLKSFVEQQDNSIRELLQSVEEQYKQLSQQHMQIKEIEKQLRKTGIQEPSENSLSSKSRAPRTTPPLQLNETENTEQDDLPADCSAVYNRGEHTSGVYTIKPRNSQGFNVYCDTQSGSPWTLIQHRKDGSQDFNETWENYEKGFGRLDGEFWLGLEKIYAIVQQSNYILRLELQDWKDSKHYVEYSFHLGSHETNYTLHVAEIAGNIPGALPEHTDLMFSTWNHRAKGQLYCPESYSGGWWWNDICGENNLNGKYNKPRTKSRPERRRGIYWRPQSRKLYAIKSSKMMLQPTT
【0096】
配列番号:13
ウシANGPTL3 241−454
DIPADCTIIYNQGKHTSGIYSIRPSNSQVFNVYCDVKSGSSWTLIQHRIDGSQNFNETWENYKYGFGRLDGEFWLGLEKIYSIVMQSNYILRIELEDWKDKYYTEYSFHLGDHETNYTLHLAEISGNGPKAFPEHKDLMFSTWDHKAKGHFNCPESNSGGWWYHDVCGENNLNGKYNKPKAKAKPERKEGICWKSQDGRLYSIKATKMLIHPSD
【0097】
配列番号:14
ウシANGPTL3 225−454
TTPSFHSNETKNVEHDDIPADCTIIYNQGKHTSGIYSIRPSNSQVFNVYCDVKSGSSWTLIQHRIDGSQNFNETWENYKYGFGRLDGEFWLGLEKIYSIVMQSNYILRIELEDWKDKYYTEYSFHLGDHETNYTLHLAEISGNGPKAFPEHKDLMFSTWDHKAKGHFNCPESNSGGWWYHDVCGENNLNGKYNKPKAKAKPERKEGICWKSQDGRLYSIKATKMLIHPSD
【0098】
配列番号:15
ウシANGPTL3 207−454
IKESTEISLSSKPRAPRTTPSFHSNETKNVEHDDIPADCTIIYNQGKHTSGIYSIRPSNSQVFNVYCDVKSGSSWTLIQHRIDGSQNFNETWENYKYGFGRLDGEFWLGLEKIYSIVMQSNYILRIELEDWKDKYYTEYSFHLGDHETNYTLHLAEISGNGPKAFPEHKDLMFSTWDHKAKGHFNCPESNSGGWWYHDVCGENNLNGKYNKPKAKAKPERKEGICWKSQDGRLYSIKATKMLIHPSD
【0099】
配列番号:16
完全長ウシANGPTL3 gi|122692391|ref|NP_001073814.1|[Bos taurus]
MYTIKLFLIIAPLVISSRTDQDYTSLDSISPEPKSRFAMLDDVKILANGLLQLGHGLKDFVHKTKGQINDIFQKLNIFDQSFYDLSLQTNEIKEEEKELRRATSKLQVKNEEVKNMSLELDSKLESLLEEKILLQQKVRYLEDQLTDLIKNQPQIQEYLEVTSLKTLVEQQDNSIKDLLQIVEEQYRQLNQQQSQIKEIENQLRRTGIKESTEISLSSKPRAPRTTPSFHSNETKNVEHDDIPADCTIIYNQGKHTSGIYSIRPSNSQVFNVYCDVKSGSSWTLIQHRIDGSQNFNETWENYKYGFGRLDGEFWLGLEKIYSIVMQSNYILRIELEDWKDKYYTEYSFHLGDHETNYTLHLAEISGNGPKAFPEHKDLMFSTWDHKAKGHFNCPESNSGGWWYHDVCGENNLNGKYNKPKAKAKPERKEGICWKSQDGRLYSIKATKMLIHPSDSENSE
【0100】
配列番号:17
イヌANGPTL3 240−454
DIPANCTTIYNRGEHTSGIYSIRPSNSQVFNVYCDVKSGSSWTLIQHRIDGSQNFNETWENYRYGFGRLDGEFWLGLEKIYSIVKQSNYILRIELEDWNDNKHYIEYFFHLGNHETNYTLHLVEITGNILNALPEHKDLVFSTWDHKAKGHVNCPESYSGGWWWHNVCGENNLNGKYNKQRAKTKPERRRGLYWKSQNGRLYSIKSTKMLIHPID
【0101】
配列番号:18
イヌANGPTL3 224−454
TTPFLHLNETKNVEHNDIPANCTTIYNRGEHTSGIYSIRPSNSQVFNVYCDVKSGSSWTLIQHRIDGSQNFNETWENYRYGFGRLDGEFWLGLEKIYSIVKQSNYILRIELEDWNDNKHYIEYFFHLGNHETNYTLHLVEITGNILNALPEHKDLVFSTWDHKAKGHVNCPESYSGGWWWHNVCGENNLNGKYNKQRAKTKPERRRGLYWKSQNGRLYSIKSTKMLIHPID
【0102】
配列番号:19
イヌANGPTL3 206−454
IQESTENSLSSKPRAPRTTPFLHLNETKNVEHNDIPANCTTIYNRGEHTSGIYSIRPSNSQVFNVYCDVKSGSSWTLIQHRIDGSQNFNETWENYRYGFGRLDGEFWLGLEKIYSIVKQSNYILRIELEDWNDNKHYIEYFFHLGNHETNYTLHLVEITGNILNALPEHKDLVFSTWDHKAKGHVNCPESYSGGWWWHNVCGENNLNGKYNKQRAKTKPERRRGLYWKSQNGRLYSIKSTKMLIHPID
【0103】
配列番号:20
完全長イヌANGPTL3 gi|57086505|ref|XP_536686.1|[Canis familiaris]
MYTIKLFLFIIPLVISSKIDRDYSSYDSVSPEPKSRFAMLDDVKILANGLLQLGHGLKDFVHKTKGQINDIFQKLNIFDQSFYDLSLQTNEIKEEEKELRRTTSKLQVKNEEVKNMSLELNSKVESLLEEKILLQQKVRYLEKQLTSLIKNQPEIQEHPEVTSLKTFVEQQDNSIKDLLQTVEEQYRQLNQQHSQIKEIENQLRNVIQESTENSLSSKPRAPRTTPFLHLNETKNVEHNDIPANCTTIYNRGEHTSGIYSIRPSNSQVFNVYCDVKSGSSWTLIQHRIDGSQNFNETWENYRYGFGRLDGEFWLGLEKIYSIVKQSNYILRIELEDWNDNKHYIEYFFHLGNHETNYTLHLVEITGNILNALPEHKDLVFSTWDHKAKGHVNCPESYSGGWWWHNVCGENNLNGKYNKQRAKTKPERRRGLYWKSQNGRLYSIKSTKMLIHPIDSESSE
【0104】
配列番号:21
ウマANGPTL3 241−455
DFPADCTTIYNRGEHTSGIYSIKPSNSQVFNVYCDVISGSSWILIQRRIDGSQNFNETWQNYKYGFGRLDFEFWLGLEKIYSIVKRSNYILRIELEDWKDNKHTIEYSFHLGNHETNYTLHLVEITGNVPNALPEHKDLVFSTWDHKAKGQLNCLESYSGGWWWHDVCGGDNPNGKYNKPRSKTKPERRRGICWKSQNGRLYTIKSTKMLIHPID
【0105】
配列番号:22
ウマANGPTL3 225−455
TTPSFHLNETKDVEHDDFPADCTTIYNRGEHTSGIYSIKPSNSQVFNVYCDVISGSSWILIQRRIDGSQNFNETWQNYKYGFGRLDFEFWLGLEKIYSIVKRSNYILRIELEDWKDNKHTIEYSFHLGNHETNYTLHLVEITGNVPNALPEHKDLVFSTWDHKAKGQLNCLESYSGGWWWHDVCGGDNPNGKYNKPRSKTKPERRRGICWKSQNGRLYTIKSTKMLIHPID
【0106】
配列番号:23
ウマANGPTL3 207−455
IQESTENSLSSKPRAPRTTPSFHLNETKDVEHDDFPADCTTIYNRGEHTSGIYSIKPSNSQVFNVYCDVISGSSWILIQRRIDGSQNFNETWQNYKYGFGRLDFEFWLGLEKIYSIVKRSNYILRIELEDWKDNKHTIEYSFHLGNHETNYTLHLVEITGNVPNALPEHKDLVFSTWDHKAKGQLNCLESYSGGWWWHDVCGGDNPNGKYNKPRSKTKPERRRGICWKSQNGRLYTIKSTKMLIHPID
【0107】
配列番号:24
完全長ウマANGPTL3[equus caballus]
MYTIKLFLVIAPLVISSRIDQDYSSLDSIPPEPKSRFAMLDDVKILANGLLQLGHGLKDFVHKTKGQINDIFQKLNIFDQSFYALSLQTNEIKEEEKELRRTTSKLQVKNEEVKNMSLELNSKLESLLEEKSLLQQKVKYLEEQLTKLIKNQPEIQEHPEVTSLKTFVEQQDNSIKDLLQTMEEQYRQLNQQHSQIKEIENQLRRTGIQESTENSLSSKPRAPRTTPSFHLNETKDVEHDDFPADCTTIYNRGEHTSGIYSIKPSNSQVFNVYCDVISGSSWILIQRRIDGSQNFNETWQNYKYGFGRLDFEFWLGLEKIYSIVKRSNYILRIELEDWKDNKHTIEYSFHLGNHETNYTLHLVEITGNVPNALPEHKDLVFSTWDHKAKGQLNCLESYSGGWWWHDVCGGDNPNGKYNKPRSKTKPERRRGICWKSQNGRLYTIKSTKMLIHPIDSESFELRQIKKPMN
【0108】
配列番号:25
241−455 コンセンサス
XXPAXCXXXYNXGXHTSGXYXIXPXNSQXFXVYCDXXSGSXWXLIQXRXDGSQXFNETWXNYXXGFGRLDXEFWLGLEKIYXIVXXSNYXLEXELXDWXD(X)KXXXEYXFXLGXHETNYTLHXXXIXGNXXXAXPEXXDLXFSTWXHXAKGXXXCXEXXSGGWWXXXXCGXXNXNGKYNKXXXKXXPERXXGXXWXXQXXXLYXIKXXKMXXXPXX
【0109】
配列番号:26
225−455 コンセンサス
TTPXXXXNEXXXXXXXXXPAXCXXXYNXGXHTSGXYXIXPXNSQXFXVYCDXXSGSXWXLIQXRXDGSQXFNETWXNYXXGFGRLDXEFWLGLEKIYXIVXXSNYXLEXELXDWXD(X)KXXXEYXFXLGXHETNYTLHXXXIXGNXXXAXPEXXDLXFSTWXHXAKGXXXCXEXXSGGWWXXXXCGXXNXNGKYNKXXXKXXPERXXGXXWXXQXXXLYXIKXXKMXXXPXX
【0110】
配列番号:27
207−455 コンセンサス
XIXEXXEXSLSSKXRAPRTTPXXXXNEXXXXXXXXXPAXCXXXYNXGXHTSGXYXIXPXNSQXFXVYCDXXSGSXWXLIQXRXDGSQXFNETWXNYXXGFGRLDXEFWLGLEKIYXIVXXSNYXLEXELXDWXD(X)KXXXEYXFXLGXHETNYTLHXXXIXGNXXXAXPEXXDLXFSTWXHXAKGXXXCXEXXSGGWWXXXXCGXXNXNGKYNKXXXKXXPERXXGXXWXXQXXXLYXIKXXKMXXXPXX
【0111】
配列番号:28
完全長コンセンサス
MXTIKLXLXXXPLVIXSXXDXDXXSXDSXXXEPKSRFAMLDDVKILANGLLQLGHGLKDFVHKTKGQINDIFQKLNIFDQSFYXLSLXTXEIKEEEKELRRXTXXLQVKNEEVKNMSXELXSKXESLLEEKXXLQXKVXXLEXQLTXLIXXXXXXXEXXEVTSLKXXVEXQDNSIXXLLQXXEXQYXQLXQQXXQIKEIEXQLR(X)XXIXEXXEXSLSSKXRAPRTTPXXXXNEXXXXXXXXXPAXCXXXYNXGXHTSGXYXIXPXNSQXFXVYCDXXSGSXWXLIQXRXDGSQXFNETWXNYXXGFGRLDXEFWLGLEKIYXIVXXSNYXLEXELXDWXD(X)KXXXEYXFXLGXHETNYTLHXXXIXGNXXXAXPEXXDLXFSTWXHXAKGXXXCXEXXSGGWWXXXXCGXXNXNGKYNKXXXKXXPERXXGXXWXXQXXXLYXIKXXKMXXXPXX(SEXXEXXXXXXXXX)
【特許請求の範囲】
【請求項1】
哺乳動物における関節炎または関節損傷を改善または予防する方法であって、哺乳動物の関節に配列番号1または25に対して少なくとも95%の同一性を有するアミノ酸配列を含む有効量のポリペプチドを含む組成物を投与し、それにより、哺乳動物における関節炎または関節損傷を改善または予防することを含む方法。
【請求項2】
該ポリペプチドが、PEG化されている、請求項1の方法。
【請求項3】
該ポリペプチドが、ヒト血清アルブミン(HSA)、免疫グロブリン重鎖定常領域(Fc)、ポリヒスチジン、グルタチオンSトランスフェラーゼ(GST)、チオレドキシン、プロテインA、プロテインG、またはマルトース結合タンパク質(MBP)に融合している、請求項1の方法。
【請求項4】
該ポリペプチドが、1、2、3、4、またはそれを超える非天然アミノ酸を含む、請求項1の方法。
【請求項5】
個体が、関節炎または関節損傷を有している、請求項1の方法。
【請求項6】
個体が、関節炎または関節損傷を有していないが、関節炎または関節損傷のリスクを有する、請求項1の方法。
【請求項7】
該ポリペプチドが配列番号1または25を含む、請求項1の方法。
【請求項8】
該アミノ酸配列が、配列番号2、3、4、26、27、または28に対して少なくとも95%の同一性を有する、請求項1の方法。
【請求項9】
該アミノ酸配列が、配列番号2、3、4、26、27、または28を含む、請求項1の方法。
【請求項10】
該アミノ酸配列が、配列番号5、9、13、17、または21に対して少なくとも80%同一である、請求項1の方法。
【請求項11】
該アミノ酸配列が、配列番号5、9、13、17、または21を含む、請求項1の方法。
【請求項12】
該関節炎が、骨関節炎、外傷関節炎、および自己免疫関節炎からなる群から選択される、請求項1の方法。
【請求項13】
該哺乳動物が、ヒト、イヌ、またはネコである、請求項1の方法。
【請求項14】
該組成物が、さらにヒアルロン酸を含む、請求項1の方法。
【請求項15】
間葉幹細胞の軟骨細胞への分化を誘導する方法であって、配列番号1または25に対して少なくとも95%の同一性を有するアミノ酸配列を含む十分量のポリペプチドを間葉幹細胞に接触させて、軟骨細胞への幹細胞の分化を誘導すること、を含む方法。
【請求項16】
該ポリペプチドがPEG化されている、請求項15の方法。
【請求項17】
該ポリペプチドが、ヒト血清アルブミン(HSA)、免疫グロブリン重鎖定常領域(Fc)、ポリヒスチジン、グルタチオンSトランスフェラーゼ(GST)、チオレドキシン、プロテインA、プロテインG、またはマルトース結合タンパク質(MBP)に融合している、請求項15の方法。
【請求項18】
該ポリペプチドが、1、2、3、4、またはそれを超える非天然アミノ酸を含む、請求項15の方法。
【請求項19】
該方法がインビトロで実行される、請求項15の方法。
【請求項20】
該方法がインビボで実行され、該幹細胞が哺乳動物において存在する、請求項15の方法。
【請求項21】
該哺乳動物が、ヒト、イヌ、またはネコである、請求項15の方法。
【請求項22】
該ポリペプチドが、配列番号1または25を含む、請求項15の方法。
【請求項23】
該アミノ酸配列が、配列番号2、3、4、26、27、または28に対して少なくとも95%の同一性を有する、請求項15の方法。
【請求項24】
該アミノ酸配列が、配列番号2、3、4、26、27、または28を含む、請求項15の方法。
【請求項25】
該アミノ酸配列が、配列番号5、9、13、17、または21に対して少なくとも80%同一である、請求項15の方法。
【請求項26】
該アミノ酸配列が、配列番号5、9、13、17、または21を含む、請求項15の方法。
【請求項27】
関節内輸送のための医薬組成物であって、配列番号1または25に対して少なくとも95%の同一性を有するアミノ酸配列を含む医薬的に有効量のポリペプチドを含む、組成物。
【請求項28】
該ポリペプチドがPEG化されている、請求項27の組成物。
【請求項29】
該ポリペプチドが、ヒト血清アルブミン(HSA)、免疫グロブリン重鎖定常領域(Fc)、ポリヒスチジン、グルタチオンSトランスフェラーゼ(GST)、チオレドキシン、プロテインA、プロテインG、またはマルトース結合タンパク質(MBP)に融合している、請求項27の組成物。
【請求項30】
該ポリペプチドが、1、2、3、4、またはそれを超える非天然アミノ酸を含む、請求項27の組成物。
【請求項31】
さらにヒアルロン酸を含む、請求項27の組成物。
【請求項32】
該ポリペプチドが、配列番号1または25を含む、請求項27の組成物。
【請求項33】
該アミノ酸配列が、配列番号2、3、4、26、27、または28に対して少なくとも95%の同一性を有する、請求項27の組成物。
【請求項34】
該アミノ酸配列が、配列番号2、3、4、26、27、または28を含む、請求項27の組成物。
【請求項35】
該アミノ酸配列が、配列番号5、9、13、17、または21に対して少なくとも80%同一である、請求項27の組成物。
【請求項36】
該アミノ酸配列が、配列番号5、9、13、17、または21を含む、請求項27の組成物。
【請求項1】
哺乳動物における関節炎または関節損傷を改善または予防する方法であって、哺乳動物の関節に配列番号1または25に対して少なくとも95%の同一性を有するアミノ酸配列を含む有効量のポリペプチドを含む組成物を投与し、それにより、哺乳動物における関節炎または関節損傷を改善または予防することを含む方法。
【請求項2】
該ポリペプチドが、PEG化されている、請求項1の方法。
【請求項3】
該ポリペプチドが、ヒト血清アルブミン(HSA)、免疫グロブリン重鎖定常領域(Fc)、ポリヒスチジン、グルタチオンSトランスフェラーゼ(GST)、チオレドキシン、プロテインA、プロテインG、またはマルトース結合タンパク質(MBP)に融合している、請求項1の方法。
【請求項4】
該ポリペプチドが、1、2、3、4、またはそれを超える非天然アミノ酸を含む、請求項1の方法。
【請求項5】
個体が、関節炎または関節損傷を有している、請求項1の方法。
【請求項6】
個体が、関節炎または関節損傷を有していないが、関節炎または関節損傷のリスクを有する、請求項1の方法。
【請求項7】
該ポリペプチドが配列番号1または25を含む、請求項1の方法。
【請求項8】
該アミノ酸配列が、配列番号2、3、4、26、27、または28に対して少なくとも95%の同一性を有する、請求項1の方法。
【請求項9】
該アミノ酸配列が、配列番号2、3、4、26、27、または28を含む、請求項1の方法。
【請求項10】
該アミノ酸配列が、配列番号5、9、13、17、または21に対して少なくとも80%同一である、請求項1の方法。
【請求項11】
該アミノ酸配列が、配列番号5、9、13、17、または21を含む、請求項1の方法。
【請求項12】
該関節炎が、骨関節炎、外傷関節炎、および自己免疫関節炎からなる群から選択される、請求項1の方法。
【請求項13】
該哺乳動物が、ヒト、イヌ、またはネコである、請求項1の方法。
【請求項14】
該組成物が、さらにヒアルロン酸を含む、請求項1の方法。
【請求項15】
間葉幹細胞の軟骨細胞への分化を誘導する方法であって、配列番号1または25に対して少なくとも95%の同一性を有するアミノ酸配列を含む十分量のポリペプチドを間葉幹細胞に接触させて、軟骨細胞への幹細胞の分化を誘導すること、を含む方法。
【請求項16】
該ポリペプチドがPEG化されている、請求項15の方法。
【請求項17】
該ポリペプチドが、ヒト血清アルブミン(HSA)、免疫グロブリン重鎖定常領域(Fc)、ポリヒスチジン、グルタチオンSトランスフェラーゼ(GST)、チオレドキシン、プロテインA、プロテインG、またはマルトース結合タンパク質(MBP)に融合している、請求項15の方法。
【請求項18】
該ポリペプチドが、1、2、3、4、またはそれを超える非天然アミノ酸を含む、請求項15の方法。
【請求項19】
該方法がインビトロで実行される、請求項15の方法。
【請求項20】
該方法がインビボで実行され、該幹細胞が哺乳動物において存在する、請求項15の方法。
【請求項21】
該哺乳動物が、ヒト、イヌ、またはネコである、請求項15の方法。
【請求項22】
該ポリペプチドが、配列番号1または25を含む、請求項15の方法。
【請求項23】
該アミノ酸配列が、配列番号2、3、4、26、27、または28に対して少なくとも95%の同一性を有する、請求項15の方法。
【請求項24】
該アミノ酸配列が、配列番号2、3、4、26、27、または28を含む、請求項15の方法。
【請求項25】
該アミノ酸配列が、配列番号5、9、13、17、または21に対して少なくとも80%同一である、請求項15の方法。
【請求項26】
該アミノ酸配列が、配列番号5、9、13、17、または21を含む、請求項15の方法。
【請求項27】
関節内輸送のための医薬組成物であって、配列番号1または25に対して少なくとも95%の同一性を有するアミノ酸配列を含む医薬的に有効量のポリペプチドを含む、組成物。
【請求項28】
該ポリペプチドがPEG化されている、請求項27の組成物。
【請求項29】
該ポリペプチドが、ヒト血清アルブミン(HSA)、免疫グロブリン重鎖定常領域(Fc)、ポリヒスチジン、グルタチオンSトランスフェラーゼ(GST)、チオレドキシン、プロテインA、プロテインG、またはマルトース結合タンパク質(MBP)に融合している、請求項27の組成物。
【請求項30】
該ポリペプチドが、1、2、3、4、またはそれを超える非天然アミノ酸を含む、請求項27の組成物。
【請求項31】
さらにヒアルロン酸を含む、請求項27の組成物。
【請求項32】
該ポリペプチドが、配列番号1または25を含む、請求項27の組成物。
【請求項33】
該アミノ酸配列が、配列番号2、3、4、26、27、または28に対して少なくとも95%の同一性を有する、請求項27の組成物。
【請求項34】
該アミノ酸配列が、配列番号2、3、4、26、27、または28を含む、請求項27の組成物。
【請求項35】
該アミノ酸配列が、配列番号5、9、13、17、または21に対して少なくとも80%同一である、請求項27の組成物。
【請求項36】
該アミノ酸配列が、配列番号5、9、13、17、または21を含む、請求項27の組成物。
【図1】


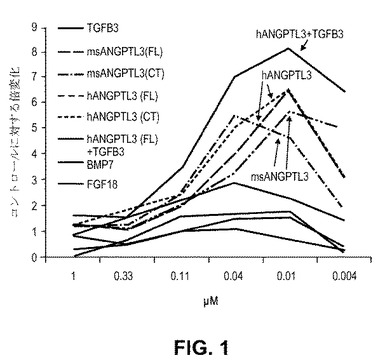
【図2】
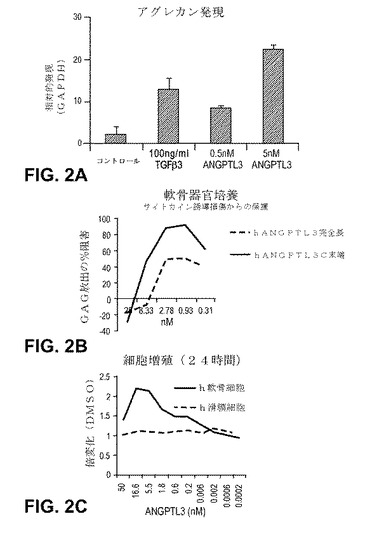
【図3A】
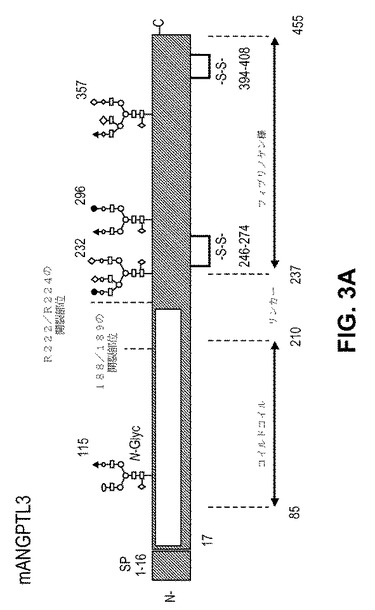
【図3B】
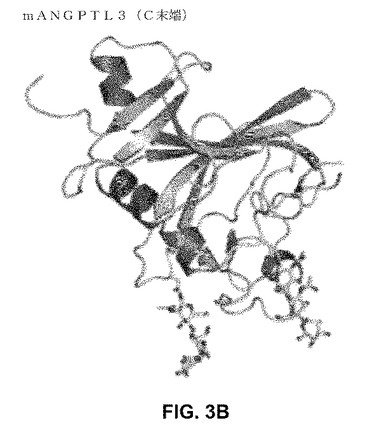
【図4A−1】

【図4A−2】

【図4B−1】

【図4B−2】

【図5】

【図2】
【図3A】
【図3B】
【図4A−1】
【図4A−2】
【図4B−1】
【図4B−2】
【図5】
【公表番号】特表2012−533549(P2012−533549A)
【公表日】平成24年12月27日(2012.12.27)
【国際特許分類】
【出願番号】特願2012−520725(P2012−520725)
【出願日】平成22年7月13日(2010.7.13)
【国際出願番号】PCT/US2010/041850
【国際公開番号】WO2011/008773
【国際公開日】平成23年1月20日(2011.1.20)
【出願人】(399038620)ザ スクリプス リサーチ インスティチュート (51)
【出願人】(503261524)アイアールエム・リミテッド・ライアビリティ・カンパニー (158)
【氏名又は名称原語表記】IRM,LLC
【Fターム(参考)】
【公表日】平成24年12月27日(2012.12.27)
【国際特許分類】
【出願日】平成22年7月13日(2010.7.13)
【国際出願番号】PCT/US2010/041850
【国際公開番号】WO2011/008773
【国際公開日】平成23年1月20日(2011.1.20)
【出願人】(399038620)ザ スクリプス リサーチ インスティチュート (51)
【出願人】(503261524)アイアールエム・リミテッド・ライアビリティ・カンパニー (158)
【氏名又は名称原語表記】IRM,LLC
【Fターム(参考)】
[ Back to top ]