
間葉幹細胞及びその使用法
動物間葉幹細胞を有効な量で投与することにより、動物の自己免疫疾患、アレルギー性疾患、癌あるいは炎症性疾患を治療する方法、創傷治癒を促進する方法、及び動物の器官あるいは組織における血管形成を促進する方法。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、間葉幹細胞に関する。特に本発明は、種々の組織及び器官における血管形成の促進、自己免疫疾患の治療、アレルギー反応の治療、ガンの治療、炎症性疾患の治療、及び回復の促進を含む間葉幹細胞の新規用途に関する。
【背景技術】
【0002】
間葉幹細胞(MSCs)は、骨芽細胞、筋細胞、軟骨細胞及び脂肪細胞を含む系統に容易に分化可能な多能性幹細胞である(非特許文献1、非特許文献2、非特許文献3)。MSCsの、筋肉(非特許文献4)、ニューロン様前駆体(非特許文献5、非特許文献6)、心筋細胞(非特許文献7、非特許文献8)に、また場合によっては他の細胞種に分化する能力が、In vitro研究により示されている。さらに、MSCsは、造血幹細胞及び胚性幹細胞の増殖(expansion)に有効な支持細胞層をもたらすと見られている(非特許文献9、非特許文献10)。MSCsが、損傷した骨、軟骨、半月板(meniscus)あるいは心筋の組織の修復あるいは再生に有用であり得ることが、種々の動物モデルを用いた最近の研究で示されている(非特許文献11、非特許文献12、非特許文献13、非特許文献14、非特許文献15)。複数の研究者はMSCsを用い、骨形成不全症(非特許文献16)、パーキンソン症候群(非特許文献17)、脊髄損傷(非特許文献18、非特許文献19)、及び心疾患(非特許文献20、非特許文献21)を含む動物疾患モデルへの移植に有望な結果を示した。重要なことには、骨形成不全症への臨床試験(非特許文献22、非特許文献23)及び異種骨髄移植の促進移植(enhanced engraftment)(非特許文献24、非特許文献25)においても有望な結果が報告されている。
【0003】
MSCsはその表面に主要組織適合複合体(MHC)クラスI抗原を発現するが、MHCクラスIIは制限され(非特許文献26、非特許文献27)、またB7あるいはCD40共刺激分子は発現しないが(非特許文献28)、これはこれらの細胞が低免疫原性表現型を持つことを示している(非特許文献29)。MSCsはまた、MHC非依存性方法でT細胞増殖反応を抑制する(非特許文献30、非特許文献31、非特許文献32)。これらのMSCsの免疫学的性質は、その移植を強化し、移植後に同種異系細胞を認識し拒絶する受容者の免疫システムの能力を制限することができる。MSCsは、局所刺激下で適切な細胞系に分化するための能力とともに、免疫反応を調節し、造血をサポートする因子を生成するため、細胞移植研究にとって望ましい幹細胞である(非特許文献33、非特許文献34)。
【非特許文献1】Pittenger,et al.,Science,Vol.284,pg.143(1999)
【非特許文献2】Haynesworth,et al.,Bone,Vol.13,pg.69(1992)
【非特許文献3】Prockop,Science,Vol.276,pg.71(1997)
【非特許文献4】Wakitani,et al.,Muscle Nerve,Vol.18,pg.1417(1995)
【非特許文献5】Woodbury,et al.,J.Neurosci.Res.,Vol.69,pg.908(2002)
【非特許文献6】Sanchez−Ramos,et al.,Exp.Neurol.,Vol.171,pg.109(2001)
【非特許文献7】Toma、et al.,Circulation,Vol.105,pg.93(2002)
【非特許文献8】Fakuda,Artif.Organs,Vol.25,pg.187(2001)
【非特許文献9】Eaves,et al.,Ann.N.Y.Acad.Sci.,Vol.938,pg.63(2001)
【非特許文献10】Wagers,et al.,Gene Therapy,Vol.9,pg.606(2002)
【非特許文献11】Dekok,et al.,Clin.Oral Implants Res.,Vol.14,pg.481(2003)
【非特許文献12】Wu,et al.,Transplantation,Vol.75,pg.679(2003)
【非特許文献13】Noel,et al.,Curr.Opin.Investig.Drugs,Vol.3,pg.1000(2002)
【非特許文献14】Ballas,et al.,J.Cell.Biochem.Suppl.,Vol.38,pg.20(2002)
【非特許文献15】Mackenzie,et al.,Blood Cells Mol.Dis.,Vol.27(2002)
【非特許文献16】Pereira,et al.,Proc.Nat.Acad.Sci.,Vol.95,pg.1142(1998)
【非特許文献17】Schwartz,et al.,Hum.Gene Ther.,Vol.10,pg.2539(1999)
【非特許文献18】Chopp、et al.,Neuroreport,Vol.11,pg.3001(2000)
【非特許文献19】Wu,et al.,J.Neurosci.Res.,Vol.72,pg.393(2003)
【非特許文献20】Tomita,et al.,Circulation,Vol.100,pg.247(1999)
【非特許文献21】Shake,et al.,Ann.Thorac.Surg.,Vol.73,pg.1919(2002)
【非特許文献22】Horwitz,et al.,Blood,Vol.97,pg.1227(2001)
【非特許文献23】Horowitz,et al.Proc.Nat.Acad.Sci.,Vol.99,pg.8932(2002)
【非特許文献24】Frassoni,et al.,Int.Society for Cell Therapy,SA006(abstract)(2002)
【非特許文献25】Koc,et al.,J.Clin.Oncol.,Vol.18,pg.307(2000)
【非特許文献26】Le Blanc,et al.,Exp.Hematol.,Vol.31,pg.890(2003)
【非特許文献27】Potian,et al.,J.Immunol.,Vol.171,pg.3426(2003)
【非特許文献28】Majumdar,et al.,J.Biomed.Sci.,Vol.10,pg.228(2003)
【非特許文献29】Tse,et al.,Transplantation,Vol.75,pg.389(2003)
【非特許文献30】Bartholomew,et al.,Exp.Hematol.,Vol.30,pg.42(2002)
【非特許文献31】Devine,et al.,Cancer J.,Vol.7,pg.576(2001)
【非特許文献32】DiNicola,et al.,Blood,Vol.99,pg.3838(2002)
【非特許文献33】Majumdar,et al.,Hematother.Stem Cell Res.,Vol.9,pg.841(2000)
【非特許文献34】Haynesworth,et al.,J.Cell.Physiol.,Vol.166,pg.585(1996)
【発明の開示】
【0004】
出願人は現在、樹状細胞(DC1及びDC2)、エフェクターT細胞(Th1及びTh2)、及びNK細胞を含む単離された免疫細胞集団と間葉幹細胞との相互作用を調査した。出願人は前記相互作用に基づいて、間葉幹細胞が、免疫反応プロセスのいくつかの段階を規制し得る種々の因子の産生を調節し得ることを発見した。したがって、間葉幹細胞は、免疫システムが関与する疾病、状態及び障害の治療に、あるいは、炎症又はアレルギー反応を含む疾病、状態、障害の治療に用いられ得る。前記疾病、状態及び障害は、自己免疫疾患、アレルギー、関節炎、炎症を起した創傷、円形脱毛症(はげ頭症)、歯肉炎及び歯周炎(periodontitis)を含む歯周病、及び免疫反応が関与する他の疾病、状態あるいは障害を含むがこれらに限定されない。
【0005】
さらに、間葉幹細胞は、新たな血管の形成を促進することで血管形成を促進する血管内皮増殖因子、すなわちVEGFを産生するように抹消血単核細胞(PBMCs)を刺激すると考えられる。
【0006】
さらに、間葉幹細胞は、癌抑制及びウイルス感染に対する免疫力を高めるインターフェロン−ベータ(INF−β)を産生するように樹状細胞(DCs)を刺激すると考えられる。
【0007】
本発明の一態様にしたがって、動物の自己免疫疾患を治療する方法が提供される。この方法は、動物の自己免疫疾患を治療するのに有効な量で、間葉幹細胞を動物に投与することを含む。
【0008】
本発明のこの態様の範囲は如何なる理論上の推論にも限定されるべきではないが、間葉幹細胞が自己免疫疾患を抑制する最低一つのメカニズムは、制御性T細胞(Treg細胞)及び/又は樹状細胞(DC)からインターロイキン10(IL−10)を放出することによると考えられる。
【0009】
本発明にしたがって治療され得る自己免疫疾患は、多発性硬化症、1型糖尿病、関節リウマチ、ブドウ膜炎、自己免疫性甲状腺疾患、炎症性大腸炎、自己免疫リンパ増殖症候群(ALPS)、脱髄疾患、自己免疫性脳脊髄炎、自己免疫性胃炎(AIG)、及び自己免疫性糸球体疾患(autoimmune glomerular diseases)を含むがこれらに限定されない。しかしながら、本発明の範囲がここに開示された特定の自己免疫疾患の治療に限定されないことは、理解されるべきである。
【0010】
一実施例において、間葉幹細胞が投与される動物は、哺乳類である。哺乳類は、ヒト及び非ヒト霊長類を含む霊長類であり得る。
【0011】
通常、間葉幹細胞(MSC)療法は、例えば以下の手順に基づく:MSC含有組織の収集、MSCsの単離及び増殖、及び生化学的操作あるいは遺伝子操作を伴った、あるいは伴わない、動物へのMSCsの投与。
【0012】
投与される間葉幹細胞は、均一組成物か、あるいはMSCsに富んだ混合細胞集団であり得る。均一間葉幹細胞組成物は、接着性の骨髄細胞あるいは骨膜細胞を培養することで得られ得る。また間葉幹細胞組成物は、接着性の骨髄細胞あるいは骨膜細胞を培養することで得られ得る。また間葉幹細胞は、固有のモノクローナル抗体で同定される特異的細胞表面マーカーにより同定され得る。間葉幹細胞に富む細胞集団の獲得方法は、例えば、米国特許5486359号に開示されている。間葉幹細胞の代替資源は、血、皮膚、臍帯血、筋肉、脂肪、骨、及び軟骨膜を含むがこれらに限定されない。
【0013】
間葉幹細胞は、種々の方法で投与することができる。間葉幹細胞は、静脈内、動脈内、あるいは腹腔内投与等により、全身に投与することができる。
【0014】
間葉幹細胞は、自己、同種異系、異種を含む多種多様な資源から得られ得る。
【0015】
間葉幹細胞は、自己免疫疾患を投与するのに有効な量で動物に投与される。間葉幹細胞は、約1×105cells/kgから約1×107cells/kg、好ましくは約1×106cells/kgから約5×106cells/kgの量で、投与され得る。投与される間葉幹細胞の量は、年齢、体重、患者の性別、治療される自己免疫疾患、及びその範囲と重症度を含む、種々の因子に依存する。
【0016】
間葉幹細胞は、薬学的許容担体と併せて投与され得る。例えば、間葉幹細胞は、注入用の薬学的許容液状媒体中の細胞懸濁液として投与され得る。
【0017】
本発明の他の形態にしたがって、動物の炎症反応の治療法が提供される。この方法は、動物における炎症反応を治療するのに有効な量で、間葉幹細胞を動物に投与することを含む。
【0018】
本発明のこの態様の範囲は如何なる理論上の推論にも限定されるべきではないが、間葉幹細胞がT細胞の制御性T細胞(Treg)への成熟を促進し、それにより炎症反応を制御すると考えられる。また、間葉幹細胞がTヘルパー1細胞(Th1細胞)を抑制し、それにより、例えば乾癬に関連するもの等の特定の炎症反応におけるインターフェロンγ(INF−γ)の発現を減少させるとも考えられる。
【0019】
一実施例において、治療され得る炎症反応は、乾癬に関連するものである。
【0020】
他の実施例において、間葉幹細胞は、間葉幹細胞が脳内の小膠細胞(ミクログリア)及び又は星状細胞に接触して炎症を緩和するように、動物に投与され得る。これにより、間葉幹細胞は、アルツハイマー病、パーキンソン病、脳梗塞あるいは脳細胞損傷等の疾病あるいは障害において、活性化膠細胞(活性化グリア細胞)により引き起こされる神経変性を抑制する。
【0021】
また、他の実施例において、間葉幹細胞は、間葉幹細胞が皮膚の表皮中のケラチン生成細胞及びランゲルハンス細胞に接触して乾癬、慢性皮膚炎及び接触性皮膚炎に起こり得る炎症を緩和するように、動物に投与され得る。この実施例は如何なる理論上の推論にも限定されるべきではないが、間葉幹細胞が表皮のケラチン生成細胞及びランゲルハンス細胞に接触し、T細胞レセプター及びサイトカイン分泌プロファイルを変更して、腫瘍壊死因子アルファ(TNF−α)の発現が減少し、制御性T細胞(Treg細胞)集団が増加すると考えられる。
【0022】
さらなる実施例において、間葉幹細胞は、変形性関節炎、関節リウマチ及びウェブサイトwww.arthritis.org/conditions/diseasesに掲載されている他の関節疾患を含むがこれらに限定されない関節炎及び関節炎様状態において起こる、骨中の炎症を緩和させるために用いられ得る。この実施例の範囲は如何なる理論上の推論にも限定されると意図されないが、間葉幹細胞が関節液中の記憶T細胞によるインターロイキン−17分泌を抑制し得ると考えられる。
【0023】
他の実施例において、間葉幹細胞は、炎症性大腸炎及び慢性肝炎において、それぞれ消化管及び肝臓における炎症を緩和するために用いられ得る。本発明のこの態様の範囲は如何なる理論上の推論にも限定されるとは意図されないが、間葉幹細胞がインターロイキン−10(IL−10)の分泌の増加と制御性T細胞(Treg細胞)の発生を促進すると考えられる。
【0024】
他の実施例において、間葉幹細胞は、敗血症及び、火傷、外科処置、移植を含む外傷(trauma)等の病態における、過度の好中球活性化及びマクロファージ活性化を抑制するために用いられ得る。この実施例の範囲は如何なる理論上の推論にも限定されるべきではないが、間葉幹細胞がIL−10等の抑制性サイトカインの分泌を促進し、またマクロファージ遊走阻止因子を抑制すると考えられる。
【0025】
他の実施例において、間葉幹細胞は、角膜、水晶体、色素上皮、網膜を含む目、脳、脊髄、妊娠子宮、及び胎盤、卵巣、精巣、副腎皮質、肝臓、及び毛包(hair follicles)等の免疫学的特権部位における炎症を抑えるために用いられ得る。この実施例の範囲は如何なる理論上の推論にも限定されるべきではないが、間葉幹細胞がIL−10等の抑制性サイトカインの分泌及びTreg細胞の発生を促進すると考えられる。
【0026】
また他の実施例において、間葉幹細胞は、透析及び又は糸球体腎炎の間の末期腎不全(ESRD)感染を調節するために用いられ得る。この実施例の範囲は如何なる理論上の推論にも限定されるべきではないが、間葉幹細胞が、糸球体構造形成を促進する血管内皮増殖因子、すなわちVEGFを発現するように抹消血単核細胞を誘導すると考えられる。
【0027】
さらなる実施例において、間葉幹細胞は、インフルエンザ、C型肝炎、単純ヘルペスウイルス、ワクシニアウイルス感染、及びエプスタイン・バーウイルス等のウイルス感染を抑えるために用いられ得る。この実施例の範囲は如何なる理論上の推論にも限定されるべきではないが、間葉幹細胞がインターフェロン−ベータ(IFN−β)の分泌を促進すると考えられる。
【0028】
また他の実施例において、間葉幹細胞は、リーシュマニア(Leishmania)感染及びヘリコバクター(Helicobacter)感染等の寄生虫感染を抑えるために用いられ得る。この実施例の範囲は如何なる理論上の推論にも限定されるべきではないが、間葉幹細胞がTヘルパー2(Th2)細胞による反応を調節し、それによりβ細胞による免疫グロブリンEの産生の増加を促進すると考えられる。
【0029】
しかしながら、本発明のこの態様の範囲が、如何なる特定の炎症反応の治療にも限定されるべきではないことは理解されるべきである。
【0030】
間葉幹細胞は、前述のとおり、ヒト及び非ヒト霊長類を含む動物に投与され得る。
【0031】
また間葉幹細胞は、前述のとおり、全身に投与され得る。あるいは、変形性関節症又は関節リウマチの場合には、間葉幹細胞は関節炎部位に直接投与され得る。
【0032】
間葉幹細胞は、動物における炎症反応を治療するのに有効な量で投与される。間葉幹細胞は、約1×105cells/kgから約1×107cells/kg、好ましくは約1×106cells/kgから約5×106cells/kgの量で、投与され得る。投与されるべき間葉幹細胞の正確な投与量は、年齢、体重、患者の性別、治療される炎症反応、及びその範囲と重症度を含む、種々の因子に依存する。
【0033】
間葉幹細胞は、前述のとおり、薬学的許容担体と併せて投与され得る。
【0034】
本発明のまた別の態様にしたがって、動物の癌の治療法が提供される。前記方法は、動物の癌を治療するのに有効な量の間葉幹細胞を動物に投与することを含む。
【0035】
本発明のこの態様の範囲は如何なる理論上の推論にも限定されるべきではないが、間葉幹細胞が樹状細胞と相互作用し、IFN−β分泌をもたらし、腫瘍抑制因子として働くと考えられる。治療され得る癌は、肝細胞癌、子宮頸癌、膵臓癌、前立腺癌、線維肉腫、髄芽腫、及び星状細胞腫を含むがこれらに限定されない。しかしながら、本発明の範囲が如何なる特定の癌にも限定されるべきではないことは理解されるべきである。
【0036】
動物は、前述のとおり、ヒト及び非ヒト霊長類を含む哺乳類であり得る。
【0037】
間葉幹細胞は、動物における癌を治療するのに有効な量で動物に投与される。通常、間葉幹細胞は、約1×105cells/kgから約1×107cells/kg、好ましくは約1×106cells/kgから約5×106cells/kgの量で、投与される。投与されるべき間葉幹細胞の正確な量は、年齢、体重、患者の性別、治療される癌のタイプ、及びその範囲と重症度を含む、種々の因子に依存する。
【0038】
間葉幹細胞は、前述のとおり、薬学的許容担体と併せて投与され、また全身に投与され得る。あるいは、間葉幹細胞は治療される癌に直接投与され得る。
【0039】
本発明のさらなる別の態様にしたがって、動物のアレルギー性疾患の治療法が提供される。この方法は、動物のアレルギー性疾患を治療するのに有効な量で間葉幹細胞を動物に投与することを含む。
【0040】
本発明のこの態様の範囲は如何なる理論上の推論にも限定されるべきではないが、間葉幹細胞が、急性アレルギー反応の後に投与された場合、肥満細胞の活性化及び脱顆粒の抑制をもたらすと考えられる。また、間葉幹細胞は、好塩基球活性化を下方制御し、またTNF−α等のサイトカイン、インターロイキン−8及び単球走化性タンパク質、あるいはMCP−1等のケモカイン、ロイコトリエン等の脂質メディエータを抑制し、またヒスタミン、ヘパリン、コンドロイチン硫酸、カテプシン等の主要メディエータを抑制すると考えられる。
【0041】
治療され得るアレルギー性疾患は、ぜんそく、アレルギー性鼻炎、アトピー性皮膚炎、及び接触性皮膚炎を含むがこれらに限定されない。しかしながら、本発明の範囲が如何なる特定のアレルギー疾患にも限定されるべきでないことは、理解されるべきである。
【0042】
間葉幹細胞は、動物のアレルギー疾患を治療するのに有効な量で動物に投与される。動物は哺乳類であり得る。哺乳類は、ヒト及び非ヒト霊長類を含む霊長類であり得る。通常、間葉幹細胞は、約1×105cells/kgから約1×107cells/kg、好ましくは約1×106cells/kgから約5×106cells/kgの量で、投与される。正確な投与量は、年齢、体重、患者の性別、治療されるアレルギー疾患、及びその範囲と重症度を含む、種々の因子に依存する。
【0043】
間葉幹細胞は、前述のとおり、薬学的許容担体と併せて投与され得る。間葉幹細胞は、例えば静脈内投与あるいは動脈内投与等により、全身に投与され得る。
【0044】
本発明のさらなる態様にしたがって、動物における創傷治癒を促進する方法が提供される。この方法は、動物における創傷治癒を促進するのに有効な量の間葉幹細胞を動物に投与することを含む。
【0045】
本発明の範囲は如何なる理論上の推論にも限定されるべきではないが、前述のとおり、間葉幹細胞がTreg細胞及び樹状細胞にインターロイキン−10(IL−10)を放出させると考えられる。IL−10は、創傷における炎症を抑制あるいは調節し、それにより創傷の治癒を促進する。
【0046】
さらに、間葉幹細胞は、他の細胞タイプにより分泌因子を誘導することで、創傷治癒及び骨折治癒を促進し得る。例えば、間葉幹細胞は、末梢血単核細胞(PBMCs)による血管内皮増殖因子(VEGF)のプロスタグランジンE2(PGE2)媒介性放出、及び成長ホルモン、インスリン、インスリン様成長因子1(IGF−1)、インスリン様成長因子結合タンパク質3(IGFBP−3)、及びエンドセリン−1のPGE2−媒介性放出を誘導し得る。
【0047】
間葉幹細胞は、動物の創傷治癒を促進するのに有効な量で動物に投与される。動物は哺乳類であり得る。また哺乳類は、ヒト及び非ヒト霊長類を含む霊長類であり得る。通常、間葉幹細胞は、約1×105cells/kgから約1×107cells/kg、好ましくは約1×106cells/kgから約5×106cells/kgの量で、投与される。投与されるべき間葉幹細胞の正確な量は、年齢、体重、患者の性別、及び治療される創傷の範囲と重症度を含む、種々の因子に依存する。
【0048】
間葉幹細胞は、前述のとおり、薬学的許容担体と併せて投与され得る。間葉幹細胞は、前述のとおり、全身に投与され得る。あるいは、間葉幹細胞は、例えば間葉幹細胞を包含する包袋あるいは貯蔵容器の流体のように、直接創傷に投与され得る。
【0049】
本発明のまた別の態様にしたがって、動物の線維症を治療あるいは予防する方法が提供される。この方法は、動物の線維症を治療あるいは予防するのに有効な量の間葉幹細胞を動物に投与することを含む。
【0050】
間葉幹細胞は、肝硬変、末期腎不全に関連する腎臓の線維症、及び肺の線維症を含むがこれらに限定されない任意のタイプの動物の線維症を治療あるいは予防するために、動物に投与され得る。本発明の範囲が如何なる特定のタイプの線維症にも限定されるべきでないことは理解されるべきである。
【0051】
間葉幹細胞は、動物の線維症を治療あるいは予防するのに有効な量で動物に投与される。動物は哺乳類であり得る。また哺乳類は、ヒト及び非ヒト霊長類を含む霊長類であり得る。通常、間葉幹細胞は、約1×105cells/kgから約1×107cells/kg、好ましくは約1×106cells/kgから約5×106cells/kgの量で、投与される。投与されるべき間葉幹細胞の正確な量は、年齢、体重、患者の性別、及び治療あるいは予防される線維症の範囲と重症度を含む、種々の因子に依存する。
【0052】
間葉幹細胞は、前述のとおり、薬学的許容担体と併せて投与され得る。間葉幹細胞は、前述のとおり、全身に投与され得る。
【0053】
本発明の他の目的は、動物の組織あるいは器官における血管形成を促進することである。ここで前記組織あるいは器官は、血管形成を必要としているものである。
【0054】
したがって、本発明のさらなる態様にしたがって、動物の器官あるいは組織における血管形成を促進する方法が提供される。この方法は、動物の器官あるいは組織における血管形成を促進するのに有効な量の間葉幹細胞を動物に投与することを含む。
【0055】
血管形成は、既存の微小血管床からの新規血管の形成である。
【0056】
血管形成の誘導は、冠状動脈機能不全及び末梢動脈機能不全の治療に用いられ得る。したがって、冠状動脈疾患、虚血性心疾患、及び末梢動脈疾患の治療への非侵襲性の治癒的方法となり得る。血管形成は、心臓以外の組織及び器官における疾病及び障害の治療に、また心臓以外の器官の発達及び又は維持に関与し得る。血管形成は、内部及び外部の創傷、及び皮膚潰瘍の治療に関与し得る。また血管形成は、肺着床、及び胎盤成長、及び胎児血管系の発達に関与する。また血管形成は、軟骨吸収と骨形成の結びつけに極めて重要であり、また的確な成長板形態形成に極めて重要である。
【0057】
さらに、血管形成は、肝臓等の高代謝器官の十分な形成及び維持に必要であり、ここで密集した血管網が、十分な栄養と気体の輸送を提供するために必要である。
【0058】
間葉幹細胞は、様々な方法で血管形成を必要とする組織あるいは器官に投与することができる。間葉幹細胞は、静脈内、動脈内あるいは腹腔内投与等により、全身に投与され得る。あるいは間葉幹細胞は、例えば血管形成を必要とする組織あるいは器官に直接注入することにより、血管形成を必要とする組織あるいは器官に直接投与され得る。
【0059】
間葉幹細胞は、自己、同種異系、異種を含む多種多様な資源から得られ得る。
【0060】
本発明の範囲は如何なる理論上の推論にも限定されるべきではないが、間葉幹細胞が、動物に投与された場合、新規血管の形成を促進する血管内皮増殖因子、すなわちVEGFを産生するように末梢血単核細胞(PBMCs)を刺激すると考えられる。
【0061】
一実施例において、動物は哺乳類である。哺乳類は、ヒト及び非ヒト霊長類を含む霊長類であり得る。
【0062】
本発明にしたがって、間葉幹細胞は、血管形成を通して軽減、治療、あるいは予防することができる任意の疾病あるいは障害の治療、軽減あるいは予防に用いられ得る。したがって、例えば間葉幹細胞は、四肢、すなわち腕、脚部、手、足、及び頸部あるいは種々の器官におけるものを含む動脈閉塞を治療するために、動物に投与され得る。例えば間葉幹細胞は、脳に分布する動脈の動脈閉塞を治療し、それにより脳梗塞を治療あるいは予防するために用いられ得る。また、間葉幹細胞は、胎児期及び出生後の角膜の血管の治療に用いられ得る。また糸球体構造形成をもたらすために用いられ得る。他の実施例において、間葉幹細胞は、内部及び外部双方の創傷の治療に、また糖尿病及び鎌状赤血球貧血等の疾病により引き起こされる皮膚潰瘍を含むがこれらに限定されない、足、手、脚部あるいは腕に見られる皮膚潰瘍の治療に用いられ得る。
【0063】
さらに、血管形成は胚着床及び胎盤形成に関与するため、間葉幹細胞は、胚着床を促進するために、また流産を予防するために用いられ得る。
【0064】
さらに、間葉幹細胞は、生まれる前の動物における血管系の発達を促進するために、ヒトを含む生まれる前の動物に投与され得る。
【0065】
他の実施例において、間葉幹細胞は、軟骨吸収及び骨形成を促進するために、また的確な成長板形態形成を促進するために、生後あるいは生前の動物に投与され得る。
【0066】
間葉幹細胞は、動物の血管形成を促進するのに有効な量で投与される。間葉幹細胞は、約1×105cells/kgから約1×107cells/kg、好ましくは約1×106cells/kgから約5×106cells/kgの量で、投与され得る。投与されるべき間葉幹細胞の量は、年齢、体重、患者の性別、治療、軽減、あるいは予防される疾病あるいは障害、及びその範囲と重症度を含む、種々の因子に依存する。
【0067】
間葉幹細胞は、薬学的許容担体と併せて投与され得る。例えば、間葉幹細胞は、注入用の薬学的許容液状媒体中の細胞懸濁液として投与され得る。注入は、局所的、すなわち血管形成を必要とする組織あるいは器官に直接であっても、あるいは全身的であってもよい。
【0068】
間葉幹細胞は、治療因子をエンコードする一つあるいはそれ以上のポリヌクレオチドで遺伝子組み換えされ得る。ポリヌクレオチドは、適切な発現媒体(expression vehicles)によって間葉幹細胞に送達され得る。間葉幹細胞を遺伝子組み換えするために用いられ得る発現媒体は、レトロウイルスベクター、アデノウイルスベクター及びアデノ随伴ウイルスベクターを含むがこれらに限定されない。
【0069】
治療因子をエンコードする適切なポリヌクレオチドの選択は、治療される疾病あるいは障害、及びその範囲と重傷度を含む、種々の因子に依存する。治療因子をエンコードするポリヌクレオチド、及び適切な発現媒体は、米国特許6355239号にさらに開示されている。
【0070】
間葉幹細胞が、上述の治療に用いられた場合、増殖因子、サイトカイン、抗炎症剤等の薬剤、及び樹状細胞等の間葉幹細胞以外の細胞を含むがこれらに限定されない、この分野における通常の知識を有する者に知られた他の治療因子と併せて用いられ得ること、及び、必要に応じて、ヒアルロン酸等の細胞用可溶性担体とともに、あるいはコラーゲン、ゼラチン、あるいは他の生体適合性ポリマー等の固形物と併せて投与され得ることは理解されるべきである。
【0071】
ここに開示された方法が、多数の方法で、またこの分野においてよく知られた種々の変更及び置換を伴って実施され得ることは理解されるべきである。また、細胞タイプ間の作用機構あるいは相互作用について説明する任意の理論が本発明を任意の方法に制限すると解釈されるべきではなく、本発明の方法をより十分に理解することができるように提示されていると認識され得る。
【0072】
本発明は、図面に関して開示される。ここで、
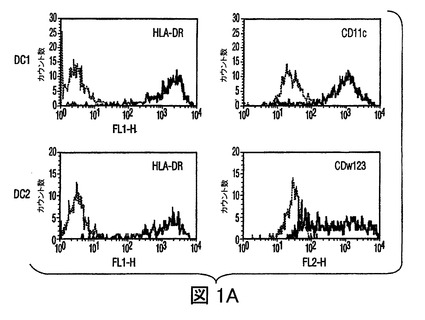
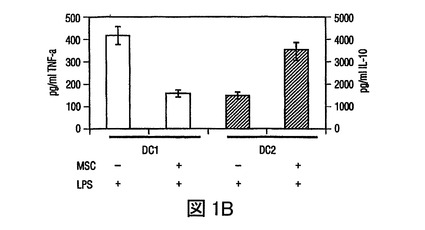
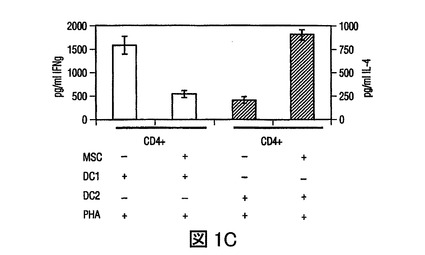
図1 MSCsの樹状細胞作用の調節
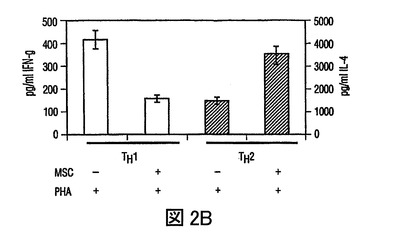
(A)HLA−DR及びCD11cに対する抗体を用いた成熟単球DC1細胞のフローサイトメトリー分析、及びHLA−DR及びCD123(IL−3レセプター)に対する抗体を用いた形質細胞様(plasmacytoid)DC2細胞のフローサイトメトリー分析。(−−−(破線)):アイソタイプ コントロール;(___(実線)):FITC/PE結合抗体。(B)MSCsは、それぞれ活性化したDC1及びDC2からのTNF−α分泌を抑制し(第一y軸)、IL−10分泌を増加させる(第二y軸)。(C)成熟DC1細胞とともに培養したMSCsは、MSCあるいはDC単独の場合と比較して、T細胞によるIFN−γ分泌(第一y軸)を抑制し、またIL−4レベルを増加させる(第二y軸)。MSCsの存在下での炎症誘発性IFN−γの産生の減少、及び抗炎症性IL−4の産生の増加は、抗炎症性表現型に関してのT細胞集団の変動を示唆した。
【0073】
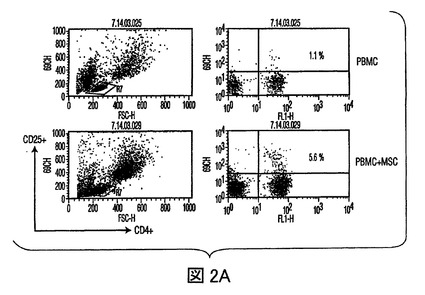
図2 MSCsの炎症誘発性エフェクターT細胞作用の抑制
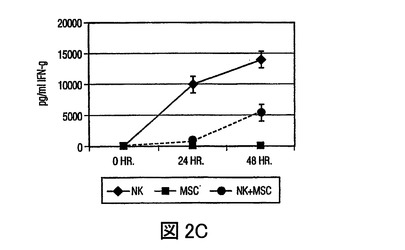
(A)FITC結合CD4抗体(x軸)及びPE結合CD25抗体(y軸)を用いた、PBMCsあるいは、MSC+PBMC培養(MSC+PBMC)における非接着性画分の染色によるTReg細胞数(%)のフローサイトメトリー分析。ゲートは、バックグラウンドとしてのアイソタイプ コントロール抗体に基づいて設定された。グラフは、5つの独立した実験の代表である。(B)細胞培養上清中で、MSCsの存在下で発生したTH1細胞は減少したレベルのIFN−γを分泌し(第一y軸)、またMSCsの存在下で発生したTH2細胞は増加した量のIL−4を分泌した(第二y軸)。(C)MSCsは、24ウェルプレート中で0、24、あるいは48時間培養された精製NK細胞からのIFN−γ分泌を抑制する。示されたデータは、一つの実験における平均値±SDサイトカイン分泌であり、また3つの独立した実験の代表である。
【0074】
図3 MSCsのTreg細胞集団数の増加及びGITR発現の増加の誘導
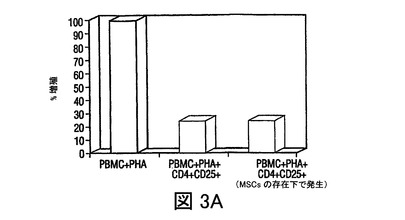
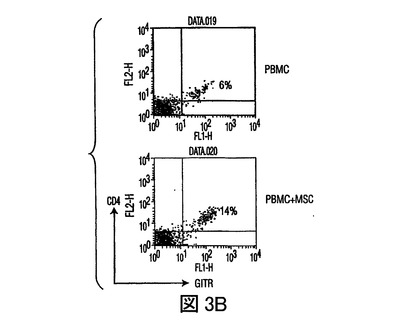
(A)PRMCあるいはMSC+PBMC(MSCのPBMCに対する比 1:10)培養(3日間さらなる刺激もなく培養された)由来のCD4+CD25+Treg細胞集団は、2ステップ磁気分離法を用いて単離された。これらの細胞は、(さらなる増殖を阻止するために)放射線照射され、混合リンパ球反応(MLR)における刺激因子として用いられた。ここで応答物質(responder)は、フィトヘムアグルチニン(PHA)の存在下(2.5mg/ml)での同種異系PBMCs(刺激因子の応答物質に対する比 1:100)であった。細胞は48時間培養され、その後3Hチミジンが添加され、24時間後に組み込まれた放射能がカウントされた。結果は、MSCsの存在下で発生したTreg集団(レーン3)が、MSCsの不在下で発生したTreg細胞(レーン2)と機能上類似していることを示した。(B)PBMCsは、MSCsの不在下(上段プロット)あるいは存在下(下段プロット)(MSCのPBMCに対する比 1:10)で、3日間培養され、その後非接着性画分が回収され、FITC標識GITR及びPE標識CD4を用いて免疫染色された。結果は、MSCsの存在下で培養された細胞のGITR発現において、2倍を超える増加を示す。
【0075】
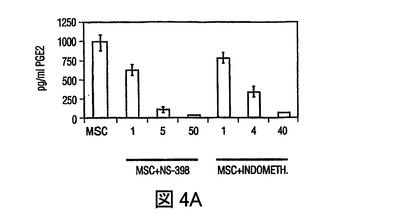
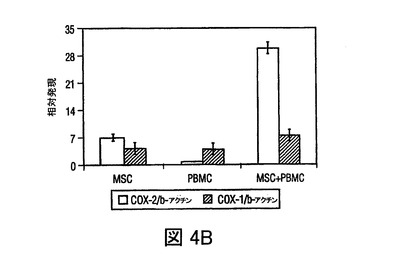
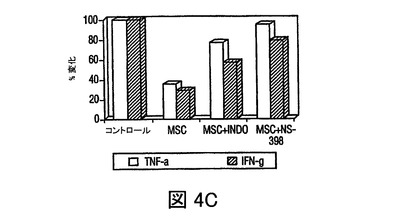
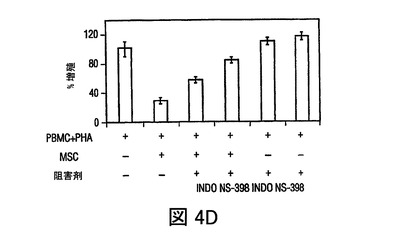
図4 MSCsのPGE2の産生、及びMSC媒介免疫調節作用を無効にするPGE2の阻害
(A)種々の濃度の、PGE2阻害剤であるNS−398あるいはインドメタシン(Indometh.)の存在下あるいは不在下で培養されたMSCsから得られた培養上清におけるPGE2分泌(平均値±SD)。阻害剤濃度はμMであり、提示されたデータは24時間培養後に得られた値である。(B)リアルタイムRT−PCRを用いたMSCs及びPBMCsにおけるCOX−1及びCOX−2発現。MSCsは、PBMCsと比較してはるかに高いレベルのCOX−2を発現し、MSCsがPBMCsの存在下で培養された場合には、MSCsのCOX−2発現の3倍以上の増加であった。3つの独立した実験のうちの1つからの代表データが示される。MSC+PBMC培養はトランスウェルチャンバープレートに設置され、ここでMSCsは下部チャンバーに播種され、PBMCsは上部チャンバーに播種された。(C)PGE2阻害剤インドメタシン(Ind.)あるいはNS−398の存在が、コントロールと比較して、活性化DCsからのTNF−α分泌と、TH1細胞からのIFN−γ分泌を増加させた。データは、MSCs及びPGE2阻害剤の不在化で発生した培養からの%変化として計算された。(D)MSC−PBMC共培養(1:10)でのPGE2阻害剤インドメタシン(Indo)及びNS−398の存在は、PHA処理したPBMCsに対するMSC媒介抗増殖促進作用を無効にする。示されたデータは、一つの実験からのものであり、また3つの独立した実験の代表である。
【0076】
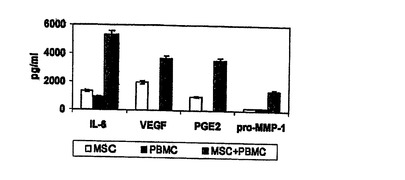
図5 同種異系PBMCsの存在下での構成的MSCサイトカイン分泌の増加
予め特性の示されたヒトMSCsを用いて、PBMCsの存在下(MSCのPBMCに対する比 1:10)(斜線バー)あるいは不在下(白抜きバー)で24時間培養したMSCsの培養上清における、サイトカインIL−6及びVEGF、脂質メディエータPGE2、及びマトリクスメタロプロテイナーゼ1(pro−MMP−1)のレベルが分析された。MSCsはIL−6、VEGF、及びPGE2を構成的に産生し、これらの因子のレベルはPBMCsとの共培養により増加した。これにより、MSCsが炎症設定における免疫機能の調節に関与し得ることを示す。
【0077】
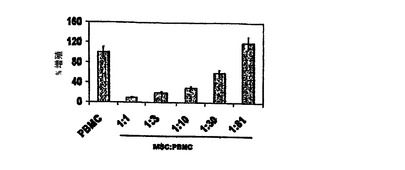
図6 MSCsの用量依存的様式でのマイトジェン誘導T細胞増殖の抑制
増加数の同種異系PBMCsが、PHAの存在下(2.5mg/ml)あるいは不在下で、96ウェルプレートに播種された定数のMSCs(2000cells/well)とともにインキュベートされ、3Hチミジン取り込みが(カウント数/分、すなわちcpmで)測定された。MSCsの存在下で、PHAで処理したPBMCsの増殖の用量依存的抑制があった。3つの独立した実験の1つからの代表的な結果が示される。類似の結果は、LeBlanc,et al.,Scand J.Immunol.,Vol.57,pg.11(2003)により報告された。
【0078】
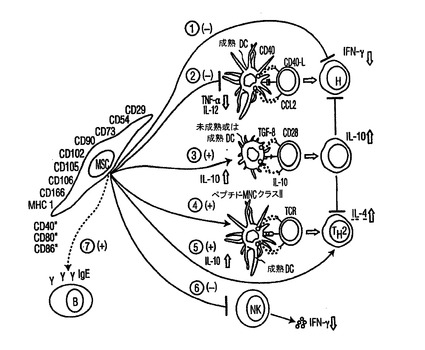
図7 MSC作用機序案の系統図
MSCsは、先天性(DCs経路 2から4;及びNK経路 6)及び適応的(T経路 1及び5、及びB経路 7)免疫システムの双方からの細胞に作用することで、その免疫調節作用を調節する。侵入病原体に対する反応で、未成熟DCsは潜在的入口部位に移動し、成熟して、(抗原特異的及び副刺激シグナルで)保護(protective)エフェクターT細胞(細胞媒介TH1免疫あるいは体液性TH2免疫)となるためのnaiveT細胞を用意(prime)する能力を獲得する。MSC−DC相互作用の間に、MSCsは、直接細胞間接触で、あるいは分泌された因子を介して、細胞媒介反応(経路2)を開始するDCsの能力を制限することによって、あるいは体液性反応(経路4)を開始する能力を促進することによって、免疫反応の結果を変更し得る。また、成熟エフェクターT細胞が存在する場合は、MSCsは、TH1(経路1)反応のバランスをTH2反応(経路5)の方へ、またおそらくIgE産生B細胞活性の増加(経路7)、GvHD及び自己免疫疾患症状の抑制について望ましい結果の方へ傾けるために、互いに相互作用し得る。MSCsは、TReg集団の発生の増加(経路3)をもたらす能力において、体制表現型をもたらし、また局所微小環境におけるバイスタンダー(bystander)炎症を弱めることで受容者ホストを救済し得る。破線(−−−)は、提案されたメカニズムを表す。
【0079】
本発明は、以下の実施例に関して開示される。しかしながら、本発明の範囲はそれらによって限定されるべきでないことは理解されるべきである。
【発明を実施するための最良の形態】
【0080】
(実施例1)
材料及び方法
ヒトMSCsの培養
Pittenger et al.,Science,Vol.284,pg.143(1999)により開示されたように、ヒトMSCsを培養した。つまり、Poietics Technologies、Div of Cambrex Biosciencesによるインフォームドコセントの後に、匿名提供者の腸骨稜から骨髄サンプルを回収した。MSCsを、1%抗生物質−抗真菌(antibiotic−antimycotic)溶液(Invitrogen,Carlsbad,California)と、10%ウシ胎仔血清(FBS,JRH BioSciences、lenexa、Kansas)とを含有する、完全ダルベッコ変法イーグル培地(complete Dulbecco‘s Modified Eagle’s Medium)−低グルコース(Life Technologies,Carlsbad,California)中で培養した。MSCsは接着性の単層として増殖し、トリプシン/EDTA(0.05%トリプシンを37℃で3分間)を用いて分離した。用いた全てのMSCsが多分化能について予め明らかにされており、また間葉系に分化(軟骨、脂質生成、及び骨形成)する能力を保持していた(Pittenger et al.,Science,Vol.284,pg.143(1999))。
【0081】
樹状細胞の単離
末梢血単核細胞(PBMCs)を、Poietics Technologies,Div of Cambrex Biosciences(Walkersville,MD)から獲得した。単球系の樹状細胞(DCs)の前駆体(CD1c+)を、Dzionek、et al.,J.Immunol.,Vol.165,pg.6037(2000)にしたがった2ステップ磁気分離法を用いて、PBMCsからポジティブセレクションした。つまり、CD1c発現B細胞が磁気ビーズを用いてCD19+細胞を磁気的に除去され、その後、ビオチン標識CD1c(BDCA1+)及び抗ビオチン抗体を用いてB細胞除去画分を標識し、それらを、メーカーの使用説明書(Miltenyi Biotech,Auburn,California)にしたがって磁気カラムを利用して非標識細胞画分から分離した。形質細胞様系統のDCsの前駆体が、陽性に標識される抗体でコートされた細胞(BDCA2+)の免疫−磁気ソーティングにより、PBMCsから単離された(Miltenyi Biotech,Auburn,California)。
【0082】
MSC−DC培養
大部分の実験において、等数のヒトMSCs及びDCsを、種々の期間培養し、細胞培養上清を回収し、さらなる実験まで−80℃で保存した。選択した実験では、MSCsを成熟DC1あるいはDC2細胞とともに(1:1 MSC:DCの比)3日間培養し、続いて混合培養(MSCsとDCs)を、増殖を阻止するために放射線照射した。続いて、抗体精製、naive、同種異系T細胞(CD4+、CD45RA+)を放射線照射MSCs/DCsに添加し、さらに6日間培養した。その後、非接着性細胞画分(精製T細胞)を培養液から回収し、2回洗浄し、さらに24時間PHAで再刺激した。その後、細胞培養上清を収集し、ELISAにより分泌されたIFN−γ及びIL−4について分析した。
【0083】
NK細胞の単離
NK細胞の精製集団を、一次試薬としてのビオチン結合モノクローナル抗体(抗CD3、CD14、CD19、CD36及び抗IgE抗体)と、二次標識試薬としてのマイクロビーズに結合した抗ビオチンモノクローナル抗体との混合物で磁気的に標識した非NK細胞を除去することで、獲得した。磁気標識された非NK細胞は、磁気領域中のMACS(Miltenyi Biotech,Auburn,California)カラムに保持され、一方NK細胞は、通過して回収された。
【0084】
Treg細胞集団の単離
Treg細胞集団を、2ステップ単離法を用いて単離した。第一に、非CD4+T細胞を、ビオチン標識抗体と抗ビオチンマイクロビーズの混合物で間接的磁気的に標識した。続いて標識した細胞を、MACSカラム(Miltenyi Biotech,Auburn,California)での分離により除去した。次に、CD4+CD25+細胞を、CD25マイクロビーズで直接標識し、濃縮前CD4+T細胞画分からポジティブセレクションにより単離した。磁気的に標識したCD4+CD25+T細胞をカラム上に保持し、磁気領域からのカラムの除去の後に溶出した。
【0085】
MSCsの存在下で発生したCD4+CD25+集団の増加が実際に抑制されたか否かを測定するために、2ステップ磁気単離法を用いて、CD4+CD25+TReg細胞集団をPBMCあるいはMSC+PBMC(MSCのPBMCに対する比 1:10)培養(3日間さらなる刺激もなく培養された)から単離した。これらの細胞を、さらなる増殖を阻止するために放射線照射し、混合リンパ球反応(MLR)における刺激因子として用いた。ここで応答物質は、PHAの存在下(2.5μg/ml)での同種異系PBMCs(刺激因子の応答物質に対する比 1:100)であった。48時間の培養を実施し、その後3Hチミジンを添加して、24時間後に組み込まれた放射能をカウントした。
【0086】
PBMCsをMSCsの不在下あるいは存在下(MSCのPBMCに対する比 1:10)で培養し、その後、非接着性画分を収集し、FITC標識グルココルチコイド誘導TNFレセプター、すなわちGITR、及びPE標識CD4を用いて免疫染色した。
【0087】
TH1/TH2細胞の発生
末梢血単核細胞(PBMCs)を、単球を除去するために、37℃で45分間2×106cells/mlで播種した。非接着性画分を、MSCsの存在下あるいは不在下で、TH1(IL−2(4ng/ml)+IL−12(5ng/ml)+抗IL−4(1μg/ml))あるいはTH2(IL−2(4ng/ml)+IL−4(4ng/ml)+抗IFN−γ(1μg/ml))条件下、プレート固定抗CD3(5μg/ml)抗体、及び抗CD28(1μg/ml)抗体の存在下で3日間インキュベートした。細胞を洗浄し、その後24時間あるいは48時間、PHA(2.5μg/ml)で再刺激した。その後、ELISA(R&D Systems,Minneapolis,Minnesota)で、培養上清中のIFN−γ及びIL−4のレベルを測定した。
【0088】
MSCsの培養上清中のVEGF、PGE2、及びpro−MMP−1のレベルの分析
予め特定されたヒトMSCsを用いて、PBMCsの存在下(MSCのPBMCに対する比 1:10)あるいは不在下で24時間培養したMSCsの培養上清中で、インターロイキン−6(IL−6)、VEGF、脂質メディエータ プロスタグランジンE2(PGE2)、及びマトリクスメタロプロテイナーゼ1(pro−MMP−1)のレベルを分析した。
【0089】
PBMCsの増殖
精製PBMCsを、Ficoll−Hypaque(Lymphoprep,Oslo,Norway)での遠心分離ロイコパック(centrifuging leukopack)(Cambrex,Walkersville,Maryland)により処理した。分離した細胞を、マイトジェンPHA(Sigma Chemicals,St.Louis,Missouri)の存在下、MSCsの存在下(定着するように、PBMC添加の3から4時間前に播種した)あるいは不在下で、48時間培養した(トリプリケイト(triplicates)で)。選択した実験においては、PBMCsを、PGE2阻害剤インドメタシン(Sigma Chemicals,St.Louis,Missouri)あるいはNS−938(Cayman Chemicals,Ann Arbor,Michigan)を含有する培地中に再懸濁した。(3H)−チミジンを添加し(200μlの培養液中に20μl)、さらなる24時間の培養の後に自動収集器を用いて細胞を収集した。MSCsあるいはPGE2阻害剤の作用効果を、PHAの存在下のコントロール反応(100%)の割合として計算した。
【0090】
量的RT−PCR
細胞ペレットからの全RNAを、市販のキット(Qiagen,Valencia,California)を用いて、またメーカーの使用説明書にしたがって、処理した。混入ゲノムDNAを、DNAフリーキット(Ambion,Austin,Texas)を用いて除去した。0.5μM濃度のプライマーとともにQuantiTect SYBR Green RT−PCRキット(Qiagen,Valencia,California)を用いて、MJ Research Opticon検出システム(South San Francisco,California)で量的RT−PCRを実施した。種々の条件下で培養された細胞における発現レベルの相対的変化を、内在コントロールとしてのβアクチンを用いて、Ct値(交点)の違いにより算出した。COX−1及びCOX−2の特異的プライマーの配列は、COX−1:5’−CCG GAT GCC AGT CAG GAT GAT G−3’(フォワード)、5’−CTA GAC AGC CAG ATG CTG ACA G−3’(リバース);COX−2:5’−ATC TAC CCT CCT CAA GTC CC−3’(フォワード)、5’−TAC CAG AAG GGC AGG ATA CAG−3’(リバース)である。
【0091】
増加数の同種異系PBMCsを、PHAの存在下(2.5μg/ml)で、96ウェルプレートに播種した定数のMSCs(2000cells/well)とともに72時間インキュベートし、3Hチミジン取り込み(カウント数/分、すなわちcpmで)を測定した。
【0092】
PBMCsとMSCsは、MSC:PBMCが1:1、1:3、1:10、1:30、及び1:81の割合で培養した。
【0093】
結果
本研究において、ヒトMSCsと、樹状細胞(DC1及びDC2)、エフェクターT細胞(TH1及びTH2)、及びNK細胞を含む、単離された免疫細胞集団との相互作用を調査した。MSCsと各免疫細胞タイプとの相互作用は、MSCsが免疫反応プロセスの複数の段階を調節し得ることを示す特異的な結果であった。MSC免疫調節作用を調節し、またMSC免疫調節作用の原因となり得る分泌因子の産生が評価され、プロスタグランジン合成が関与することが示された。
【0094】
骨髄(DC1)前駆樹状細胞及び形質細胞様(DC2)前駆樹状細胞を、それぞれBDCA1+細胞及びBDCA2+細胞の免疫−磁気ソーティングにより単離し、DC1細胞についてはGM−CSF及びIL−4(それぞれ1×103IU/ml、及び1×103IU/ml)とともに、DC2細胞についてはIL−3(10ng/ml)とともにインキュベートすることで成熟させた。フローサイトメトリーを用いて、DC1細胞はHLA−DR+及びCD11c+であり、一方DC2細胞はHLA−DR+及びCD123+であった(図1A)。炎症性因子である細菌性リポ多糖(LPS,1ng/ml)の存在下で、DC1細胞は中程度のレベルのTNF−αを産生したが、MSCsが存在した場合(1:1及び1:10の比で調査された)には、TNF−α分泌の50%以上の減少があった(図1B)。他方では、DC2細胞はLPSの存在下でIL−10を産生し、そのレベルはMSC:DC2共培養(1:1)で2倍以上増加した(図1B)。したがって、MSCsは、培養中の活性化されたDCsのサイトカインプロファイルを、より寛容原生表現型に変更した。さらに、活性化したDCsは、MSCsとともに培養された場合に、IFN−γを減少させ、またnaiveCD4+T細胞により分泌されたIL−4レベルを増加させることができ(図1C)、これは前炎症性T細胞表現型から抗炎症性T細胞表現型へのMSC媒介変化を示す。
【0095】
IL−10分泌の増加が制御性細胞の発生に関与するため(Kingsley,et al.,J.Immunol.,Vol.168,pg.1080(2000))、T制御性細胞(TReg)を、PBMCs及びMSCsの共培養中で、フローサイトメトリーにより定量した。PBMCsのMSCsとの3から5日間の培養により、抗CD4抗体及び抗CD25抗体でのPBMCsの染色により測定されたTReg細胞数の増加があり(図2A)、さらにMSC誘導寛容原生反応を支持した。MSCsの存在下で発生したCD4+CD25+TReg細胞集団は、増加したレベルのグルココルチコイド誘導TNFレセプター(GITR)、TReg細胞集団上に発現する細胞表面レセプターを発現し、また同種異系T細胞増殖を抑制した(図3A、3B)ことから、実際抑制的であった。次に、T細胞分化に作用する直接の能力に関して、MSCsを調査した。抗体選別精製T細胞(CD4+Th細胞)を用いて、IFN−γ産生TH1細胞及びIL−4産生TH2細胞を、MSCsの存在下あるいは不在下で発生させた。分化の間にMSCsが存在すると、TH1細胞によるIFN−γ分泌が減少し、TH2細胞によるIL−4分泌が増加した(図2B)。Th細胞がエフェクターTH1タイプあるいはTH2タイプに分化した後(3日)に、MSCsが培養に添加された場合は、IFN−γあるいはIL−4のレベルの著しい変化は見られなかった(不図示)。これらの実験は、MSCsがエフェクターT細胞分化に直接作用し、T細胞サイトカイン分泌を体液性表現型に変化させることができることを示す。
【0096】
同様に、MSCsを精製NK細胞(CD3−、CD14−、CD19−、CD36−)と比率1:1で異なる期間(0から48時間)培養すると、培養上清中でのIFN−γ分泌の減少があった(図2C)。したがって、これはMSCsがNK細胞作用をも調節することができることを示す。
【0097】
過去の研究は、MSCsが水溶性因子によりT細胞作用を調節することを示している(LeBlanc,et al.,Exp.Hematol.,Vol.31,pg.890(2003);Tse,et al.,Transplantation,Vol.75,pg.389(2003))。MSCsが、IL−6、プロスタグランジンE2、VEGF、及びproMMP−1を含む複数の因子を構成的に分泌し、PBMCsを含む培養によりそれぞれのレベルが増加したことが観察された(図5)。DCsによる、TNF−α産生の抑制、及びIL−10産生の増加をもたらすMSC生成因子を調査するために、プロスタグランジンE2の潜在的役割を調査したが、活性化DCsによるTNF−γ産生を抑制することが認められた(Vassiliou,et al.,Cell.Immunol.,Vol.223,pg.120(2003))。MSC培養(0.5×106cells/mlの24時間培養)の条件培地は、おおよそ1000pg/mlのPGE2を含有していた(図4A)。培養上清中に、PGE2分泌の既知の誘導因子、例えばTNF−α、IFN−γあるいはIL−1βの存在は検出できなかったが(不図示)、これはMSCsによるPGE2の構成的分泌を示す。hMSCsによるPGE2分泌は、PGE2産生の既知の阻害剤、NS−398(5μM)及びインドメタシン(4μM)の存在下で、60から90%抑制された(図4A)。PGE2分泌の放出が、構成的活性シクロオキシゲナーゼ酵素1(COX−1)及び誘導性シクロオキシゲナーゼ酵素2(COX−2)の酵素活性の結果として起こるため(Harris,et al.,Trends Immunol.,Vol.23,pg.144(2002))、MSCs及びPBMCsにおけるCOX−1及びCOX−2のmRNA発現を、トランスウェル培養システムを用いて分析した。MSCsは、PBMCsと比較してはるかに高いレベルのCOX−2を発現し、MSCsとPBMCsの24時間の共培養(MSCのPBMCに対する比 1:10)の場合には、発現レベルは3倍以上増加した(図4B)。COX−1レベルのわずかな変化が見られ、これはMSC−PBMC共培養によるPGE2分泌の増加(図5)が、COX−2の上向き調節により調節されることを示す。DCs及びT細胞に対するMSCの免疫調節効果がPGE2によって調節されたか否かを調査するために、PGE2阻害剤NS−398あるいはインドメタシンの存在下で、MSCsを活性化樹状細胞(DC1)あるいはTH1細胞とともに培養した。NS−398あるいはインドメタシンの存在が、DC1sによるTNF−α分泌と、TH1細胞からのIFN−γ分泌をそれぞれ増加させたが(図4C)、これは免疫細胞タイプに対するMSCの作用が、分泌されたPGE2により調節され得ることを示す。最近の研究が、MSCsが様々な刺激で誘導されるT細胞増殖を抑制することを示した(Denicola,et al.,Blood,Vol.99,pg.3838(2002);LeBlanc,et al.,Scand.J.Immunol.,Vol.57,pg.11(2003))。MSCsがマイトジェン誘導T細胞増殖を用量依存的様式で抑制することが観察され(図6)、PGE2阻害剤NS−398(5μM)あるいはインドメタシン(4μM)が存在した場合には、MSC含有培養液中のPHA処理PBMCsによる3Hチミジン取り込みが、阻害剤を含まないコントロールと比べて、70%以上増加した(図4D)。
【0098】
要約すると、MSCの他の免疫細胞タイプとの相互作用のモデル(図7)が提案される。成熟T細胞が存在する場合には、MSCsは、それらと直接相互作用して、前炎症性IFN−γ産生を抑制し(経路1)、制御性T細胞表現型を促進し(経路3)、また抗炎症性TH2細胞を促進する(経路5)。さらに、MSCsは、PGE2を分泌し、前炎症性DC1細胞を抑制し(経路2)、また抗炎症性DC2細胞(経路4)あるいは制御性DCs(経路3)を促進することで、DCsを介したT細胞免疫反応の結果を変えることができる。TH2免疫への変化は言い換えると、B細胞活性の、IgE/IgG1サブタイプ抗体の発生の増加(経路7)への変化を示す。MSCsは、そのNK細胞からのIFN−γ分泌を抑制する能力により、NK細胞作用を調節し得る(経路6)。このMSCのモデル:免疫細胞相互作用は、複数の他の研究所で行われた実験と一致する(LeBlanc,et al.,Exp.Hematol.,Vol.31,pg.890(2003);Tse,et al.,Transplantation,Vol.75,pg.389(2003);DiNicola,et al.,Blood,Vol.99,pg.3838(2002))。提案されたメカニズムについてのさらなる調査が進行中であり、現在はMSC投与のin vivo効果を調査するために、動物研究が必要である。
【0099】
公開された特許出願、保管登録番号、及びデータベース登録番号を含む、全ての特許、文献は、各特許、文献、保管登録番号、及びデータベース登録番号が明確に、また個別に参照として組み込まれているように、同じ範囲で参照としてここに組み込まれている。
【0100】
しかしながら、本発明の範囲は、上述の特定の実施例に限定されるべきでないことは理解されるべきである。本発明は、特に開示されている以外の様式で実施され得るが、これも添付の請求の範囲の範囲内である。
【図面の簡単な説明】
【0101】
【図1A】MSCsの樹状細胞作用の調節を説明する図
【図1B】MSCsの樹状細胞作用の調節を説明する図
【図1C】MSCsの樹状細胞作用の調節を説明する図
【図2A】MSCsの炎症誘発性エフェクターT細胞作用の抑制を説明する図
【図2B】MSCsの炎症誘発性エフェクターT細胞作用の抑制を説明する図
【図2C】MSCsの炎症誘発性エフェクターT細胞作用の抑制を説明する図
【図3A】MSCsのTreg細胞集団数の増加及びGITR発現の増加の誘導を説明する図
【図3B】MSCsのTreg細胞集団数の増加及びGITR発現の増加の誘導を説明する図
【図4A】MSCsのPGE2の産生、及びMSC媒介免疫調節作用を無効にするPGE2の阻害を説明する図
【図4B】MSCsのPGE2の産生、及びMSC媒介免疫調節作用を無効にするPGE2の阻害を説明する図
【図4C】MSCsのPGE2の産生、及びMSC媒介免疫調節作用を無効にするPGE2の阻害を説明する図
【図4D】MSCsのPGE2の産生、及びMSC媒介免疫調節作用を無効にするPGE2の阻害を説明する図
【図5】同種異系PBMCsの存在下での構成的MSCサイトカイン分泌の増加を説明する図
【図6】MSCsの用量依存的様式でのマイトジェン誘導T細胞増殖の抑制を説明する図
【図7】MSC作用機序案の系統図
【技術分野】
【0001】
本発明は、間葉幹細胞に関する。特に本発明は、種々の組織及び器官における血管形成の促進、自己免疫疾患の治療、アレルギー反応の治療、ガンの治療、炎症性疾患の治療、及び回復の促進を含む間葉幹細胞の新規用途に関する。
【背景技術】
【0002】
間葉幹細胞(MSCs)は、骨芽細胞、筋細胞、軟骨細胞及び脂肪細胞を含む系統に容易に分化可能な多能性幹細胞である(非特許文献1、非特許文献2、非特許文献3)。MSCsの、筋肉(非特許文献4)、ニューロン様前駆体(非特許文献5、非特許文献6)、心筋細胞(非特許文献7、非特許文献8)に、また場合によっては他の細胞種に分化する能力が、In vitro研究により示されている。さらに、MSCsは、造血幹細胞及び胚性幹細胞の増殖(expansion)に有効な支持細胞層をもたらすと見られている(非特許文献9、非特許文献10)。MSCsが、損傷した骨、軟骨、半月板(meniscus)あるいは心筋の組織の修復あるいは再生に有用であり得ることが、種々の動物モデルを用いた最近の研究で示されている(非特許文献11、非特許文献12、非特許文献13、非特許文献14、非特許文献15)。複数の研究者はMSCsを用い、骨形成不全症(非特許文献16)、パーキンソン症候群(非特許文献17)、脊髄損傷(非特許文献18、非特許文献19)、及び心疾患(非特許文献20、非特許文献21)を含む動物疾患モデルへの移植に有望な結果を示した。重要なことには、骨形成不全症への臨床試験(非特許文献22、非特許文献23)及び異種骨髄移植の促進移植(enhanced engraftment)(非特許文献24、非特許文献25)においても有望な結果が報告されている。
【0003】
MSCsはその表面に主要組織適合複合体(MHC)クラスI抗原を発現するが、MHCクラスIIは制限され(非特許文献26、非特許文献27)、またB7あるいはCD40共刺激分子は発現しないが(非特許文献28)、これはこれらの細胞が低免疫原性表現型を持つことを示している(非特許文献29)。MSCsはまた、MHC非依存性方法でT細胞増殖反応を抑制する(非特許文献30、非特許文献31、非特許文献32)。これらのMSCsの免疫学的性質は、その移植を強化し、移植後に同種異系細胞を認識し拒絶する受容者の免疫システムの能力を制限することができる。MSCsは、局所刺激下で適切な細胞系に分化するための能力とともに、免疫反応を調節し、造血をサポートする因子を生成するため、細胞移植研究にとって望ましい幹細胞である(非特許文献33、非特許文献34)。
【非特許文献1】Pittenger,et al.,Science,Vol.284,pg.143(1999)
【非特許文献2】Haynesworth,et al.,Bone,Vol.13,pg.69(1992)
【非特許文献3】Prockop,Science,Vol.276,pg.71(1997)
【非特許文献4】Wakitani,et al.,Muscle Nerve,Vol.18,pg.1417(1995)
【非特許文献5】Woodbury,et al.,J.Neurosci.Res.,Vol.69,pg.908(2002)
【非特許文献6】Sanchez−Ramos,et al.,Exp.Neurol.,Vol.171,pg.109(2001)
【非特許文献7】Toma、et al.,Circulation,Vol.105,pg.93(2002)
【非特許文献8】Fakuda,Artif.Organs,Vol.25,pg.187(2001)
【非特許文献9】Eaves,et al.,Ann.N.Y.Acad.Sci.,Vol.938,pg.63(2001)
【非特許文献10】Wagers,et al.,Gene Therapy,Vol.9,pg.606(2002)
【非特許文献11】Dekok,et al.,Clin.Oral Implants Res.,Vol.14,pg.481(2003)
【非特許文献12】Wu,et al.,Transplantation,Vol.75,pg.679(2003)
【非特許文献13】Noel,et al.,Curr.Opin.Investig.Drugs,Vol.3,pg.1000(2002)
【非特許文献14】Ballas,et al.,J.Cell.Biochem.Suppl.,Vol.38,pg.20(2002)
【非特許文献15】Mackenzie,et al.,Blood Cells Mol.Dis.,Vol.27(2002)
【非特許文献16】Pereira,et al.,Proc.Nat.Acad.Sci.,Vol.95,pg.1142(1998)
【非特許文献17】Schwartz,et al.,Hum.Gene Ther.,Vol.10,pg.2539(1999)
【非特許文献18】Chopp、et al.,Neuroreport,Vol.11,pg.3001(2000)
【非特許文献19】Wu,et al.,J.Neurosci.Res.,Vol.72,pg.393(2003)
【非特許文献20】Tomita,et al.,Circulation,Vol.100,pg.247(1999)
【非特許文献21】Shake,et al.,Ann.Thorac.Surg.,Vol.73,pg.1919(2002)
【非特許文献22】Horwitz,et al.,Blood,Vol.97,pg.1227(2001)
【非特許文献23】Horowitz,et al.Proc.Nat.Acad.Sci.,Vol.99,pg.8932(2002)
【非特許文献24】Frassoni,et al.,Int.Society for Cell Therapy,SA006(abstract)(2002)
【非特許文献25】Koc,et al.,J.Clin.Oncol.,Vol.18,pg.307(2000)
【非特許文献26】Le Blanc,et al.,Exp.Hematol.,Vol.31,pg.890(2003)
【非特許文献27】Potian,et al.,J.Immunol.,Vol.171,pg.3426(2003)
【非特許文献28】Majumdar,et al.,J.Biomed.Sci.,Vol.10,pg.228(2003)
【非特許文献29】Tse,et al.,Transplantation,Vol.75,pg.389(2003)
【非特許文献30】Bartholomew,et al.,Exp.Hematol.,Vol.30,pg.42(2002)
【非特許文献31】Devine,et al.,Cancer J.,Vol.7,pg.576(2001)
【非特許文献32】DiNicola,et al.,Blood,Vol.99,pg.3838(2002)
【非特許文献33】Majumdar,et al.,Hematother.Stem Cell Res.,Vol.9,pg.841(2000)
【非特許文献34】Haynesworth,et al.,J.Cell.Physiol.,Vol.166,pg.585(1996)
【発明の開示】
【0004】
出願人は現在、樹状細胞(DC1及びDC2)、エフェクターT細胞(Th1及びTh2)、及びNK細胞を含む単離された免疫細胞集団と間葉幹細胞との相互作用を調査した。出願人は前記相互作用に基づいて、間葉幹細胞が、免疫反応プロセスのいくつかの段階を規制し得る種々の因子の産生を調節し得ることを発見した。したがって、間葉幹細胞は、免疫システムが関与する疾病、状態及び障害の治療に、あるいは、炎症又はアレルギー反応を含む疾病、状態、障害の治療に用いられ得る。前記疾病、状態及び障害は、自己免疫疾患、アレルギー、関節炎、炎症を起した創傷、円形脱毛症(はげ頭症)、歯肉炎及び歯周炎(periodontitis)を含む歯周病、及び免疫反応が関与する他の疾病、状態あるいは障害を含むがこれらに限定されない。
【0005】
さらに、間葉幹細胞は、新たな血管の形成を促進することで血管形成を促進する血管内皮増殖因子、すなわちVEGFを産生するように抹消血単核細胞(PBMCs)を刺激すると考えられる。
【0006】
さらに、間葉幹細胞は、癌抑制及びウイルス感染に対する免疫力を高めるインターフェロン−ベータ(INF−β)を産生するように樹状細胞(DCs)を刺激すると考えられる。
【0007】
本発明の一態様にしたがって、動物の自己免疫疾患を治療する方法が提供される。この方法は、動物の自己免疫疾患を治療するのに有効な量で、間葉幹細胞を動物に投与することを含む。
【0008】
本発明のこの態様の範囲は如何なる理論上の推論にも限定されるべきではないが、間葉幹細胞が自己免疫疾患を抑制する最低一つのメカニズムは、制御性T細胞(Treg細胞)及び/又は樹状細胞(DC)からインターロイキン10(IL−10)を放出することによると考えられる。
【0009】
本発明にしたがって治療され得る自己免疫疾患は、多発性硬化症、1型糖尿病、関節リウマチ、ブドウ膜炎、自己免疫性甲状腺疾患、炎症性大腸炎、自己免疫リンパ増殖症候群(ALPS)、脱髄疾患、自己免疫性脳脊髄炎、自己免疫性胃炎(AIG)、及び自己免疫性糸球体疾患(autoimmune glomerular diseases)を含むがこれらに限定されない。しかしながら、本発明の範囲がここに開示された特定の自己免疫疾患の治療に限定されないことは、理解されるべきである。
【0010】
一実施例において、間葉幹細胞が投与される動物は、哺乳類である。哺乳類は、ヒト及び非ヒト霊長類を含む霊長類であり得る。
【0011】
通常、間葉幹細胞(MSC)療法は、例えば以下の手順に基づく:MSC含有組織の収集、MSCsの単離及び増殖、及び生化学的操作あるいは遺伝子操作を伴った、あるいは伴わない、動物へのMSCsの投与。
【0012】
投与される間葉幹細胞は、均一組成物か、あるいはMSCsに富んだ混合細胞集団であり得る。均一間葉幹細胞組成物は、接着性の骨髄細胞あるいは骨膜細胞を培養することで得られ得る。また間葉幹細胞組成物は、接着性の骨髄細胞あるいは骨膜細胞を培養することで得られ得る。また間葉幹細胞は、固有のモノクローナル抗体で同定される特異的細胞表面マーカーにより同定され得る。間葉幹細胞に富む細胞集団の獲得方法は、例えば、米国特許5486359号に開示されている。間葉幹細胞の代替資源は、血、皮膚、臍帯血、筋肉、脂肪、骨、及び軟骨膜を含むがこれらに限定されない。
【0013】
間葉幹細胞は、種々の方法で投与することができる。間葉幹細胞は、静脈内、動脈内、あるいは腹腔内投与等により、全身に投与することができる。
【0014】
間葉幹細胞は、自己、同種異系、異種を含む多種多様な資源から得られ得る。
【0015】
間葉幹細胞は、自己免疫疾患を投与するのに有効な量で動物に投与される。間葉幹細胞は、約1×105cells/kgから約1×107cells/kg、好ましくは約1×106cells/kgから約5×106cells/kgの量で、投与され得る。投与される間葉幹細胞の量は、年齢、体重、患者の性別、治療される自己免疫疾患、及びその範囲と重症度を含む、種々の因子に依存する。
【0016】
間葉幹細胞は、薬学的許容担体と併せて投与され得る。例えば、間葉幹細胞は、注入用の薬学的許容液状媒体中の細胞懸濁液として投与され得る。
【0017】
本発明の他の形態にしたがって、動物の炎症反応の治療法が提供される。この方法は、動物における炎症反応を治療するのに有効な量で、間葉幹細胞を動物に投与することを含む。
【0018】
本発明のこの態様の範囲は如何なる理論上の推論にも限定されるべきではないが、間葉幹細胞がT細胞の制御性T細胞(Treg)への成熟を促進し、それにより炎症反応を制御すると考えられる。また、間葉幹細胞がTヘルパー1細胞(Th1細胞)を抑制し、それにより、例えば乾癬に関連するもの等の特定の炎症反応におけるインターフェロンγ(INF−γ)の発現を減少させるとも考えられる。
【0019】
一実施例において、治療され得る炎症反応は、乾癬に関連するものである。
【0020】
他の実施例において、間葉幹細胞は、間葉幹細胞が脳内の小膠細胞(ミクログリア)及び又は星状細胞に接触して炎症を緩和するように、動物に投与され得る。これにより、間葉幹細胞は、アルツハイマー病、パーキンソン病、脳梗塞あるいは脳細胞損傷等の疾病あるいは障害において、活性化膠細胞(活性化グリア細胞)により引き起こされる神経変性を抑制する。
【0021】
また、他の実施例において、間葉幹細胞は、間葉幹細胞が皮膚の表皮中のケラチン生成細胞及びランゲルハンス細胞に接触して乾癬、慢性皮膚炎及び接触性皮膚炎に起こり得る炎症を緩和するように、動物に投与され得る。この実施例は如何なる理論上の推論にも限定されるべきではないが、間葉幹細胞が表皮のケラチン生成細胞及びランゲルハンス細胞に接触し、T細胞レセプター及びサイトカイン分泌プロファイルを変更して、腫瘍壊死因子アルファ(TNF−α)の発現が減少し、制御性T細胞(Treg細胞)集団が増加すると考えられる。
【0022】
さらなる実施例において、間葉幹細胞は、変形性関節炎、関節リウマチ及びウェブサイトwww.arthritis.org/conditions/diseasesに掲載されている他の関節疾患を含むがこれらに限定されない関節炎及び関節炎様状態において起こる、骨中の炎症を緩和させるために用いられ得る。この実施例の範囲は如何なる理論上の推論にも限定されると意図されないが、間葉幹細胞が関節液中の記憶T細胞によるインターロイキン−17分泌を抑制し得ると考えられる。
【0023】
他の実施例において、間葉幹細胞は、炎症性大腸炎及び慢性肝炎において、それぞれ消化管及び肝臓における炎症を緩和するために用いられ得る。本発明のこの態様の範囲は如何なる理論上の推論にも限定されるとは意図されないが、間葉幹細胞がインターロイキン−10(IL−10)の分泌の増加と制御性T細胞(Treg細胞)の発生を促進すると考えられる。
【0024】
他の実施例において、間葉幹細胞は、敗血症及び、火傷、外科処置、移植を含む外傷(trauma)等の病態における、過度の好中球活性化及びマクロファージ活性化を抑制するために用いられ得る。この実施例の範囲は如何なる理論上の推論にも限定されるべきではないが、間葉幹細胞がIL−10等の抑制性サイトカインの分泌を促進し、またマクロファージ遊走阻止因子を抑制すると考えられる。
【0025】
他の実施例において、間葉幹細胞は、角膜、水晶体、色素上皮、網膜を含む目、脳、脊髄、妊娠子宮、及び胎盤、卵巣、精巣、副腎皮質、肝臓、及び毛包(hair follicles)等の免疫学的特権部位における炎症を抑えるために用いられ得る。この実施例の範囲は如何なる理論上の推論にも限定されるべきではないが、間葉幹細胞がIL−10等の抑制性サイトカインの分泌及びTreg細胞の発生を促進すると考えられる。
【0026】
また他の実施例において、間葉幹細胞は、透析及び又は糸球体腎炎の間の末期腎不全(ESRD)感染を調節するために用いられ得る。この実施例の範囲は如何なる理論上の推論にも限定されるべきではないが、間葉幹細胞が、糸球体構造形成を促進する血管内皮増殖因子、すなわちVEGFを発現するように抹消血単核細胞を誘導すると考えられる。
【0027】
さらなる実施例において、間葉幹細胞は、インフルエンザ、C型肝炎、単純ヘルペスウイルス、ワクシニアウイルス感染、及びエプスタイン・バーウイルス等のウイルス感染を抑えるために用いられ得る。この実施例の範囲は如何なる理論上の推論にも限定されるべきではないが、間葉幹細胞がインターフェロン−ベータ(IFN−β)の分泌を促進すると考えられる。
【0028】
また他の実施例において、間葉幹細胞は、リーシュマニア(Leishmania)感染及びヘリコバクター(Helicobacter)感染等の寄生虫感染を抑えるために用いられ得る。この実施例の範囲は如何なる理論上の推論にも限定されるべきではないが、間葉幹細胞がTヘルパー2(Th2)細胞による反応を調節し、それによりβ細胞による免疫グロブリンEの産生の増加を促進すると考えられる。
【0029】
しかしながら、本発明のこの態様の範囲が、如何なる特定の炎症反応の治療にも限定されるべきではないことは理解されるべきである。
【0030】
間葉幹細胞は、前述のとおり、ヒト及び非ヒト霊長類を含む動物に投与され得る。
【0031】
また間葉幹細胞は、前述のとおり、全身に投与され得る。あるいは、変形性関節症又は関節リウマチの場合には、間葉幹細胞は関節炎部位に直接投与され得る。
【0032】
間葉幹細胞は、動物における炎症反応を治療するのに有効な量で投与される。間葉幹細胞は、約1×105cells/kgから約1×107cells/kg、好ましくは約1×106cells/kgから約5×106cells/kgの量で、投与され得る。投与されるべき間葉幹細胞の正確な投与量は、年齢、体重、患者の性別、治療される炎症反応、及びその範囲と重症度を含む、種々の因子に依存する。
【0033】
間葉幹細胞は、前述のとおり、薬学的許容担体と併せて投与され得る。
【0034】
本発明のまた別の態様にしたがって、動物の癌の治療法が提供される。前記方法は、動物の癌を治療するのに有効な量の間葉幹細胞を動物に投与することを含む。
【0035】
本発明のこの態様の範囲は如何なる理論上の推論にも限定されるべきではないが、間葉幹細胞が樹状細胞と相互作用し、IFN−β分泌をもたらし、腫瘍抑制因子として働くと考えられる。治療され得る癌は、肝細胞癌、子宮頸癌、膵臓癌、前立腺癌、線維肉腫、髄芽腫、及び星状細胞腫を含むがこれらに限定されない。しかしながら、本発明の範囲が如何なる特定の癌にも限定されるべきではないことは理解されるべきである。
【0036】
動物は、前述のとおり、ヒト及び非ヒト霊長類を含む哺乳類であり得る。
【0037】
間葉幹細胞は、動物における癌を治療するのに有効な量で動物に投与される。通常、間葉幹細胞は、約1×105cells/kgから約1×107cells/kg、好ましくは約1×106cells/kgから約5×106cells/kgの量で、投与される。投与されるべき間葉幹細胞の正確な量は、年齢、体重、患者の性別、治療される癌のタイプ、及びその範囲と重症度を含む、種々の因子に依存する。
【0038】
間葉幹細胞は、前述のとおり、薬学的許容担体と併せて投与され、また全身に投与され得る。あるいは、間葉幹細胞は治療される癌に直接投与され得る。
【0039】
本発明のさらなる別の態様にしたがって、動物のアレルギー性疾患の治療法が提供される。この方法は、動物のアレルギー性疾患を治療するのに有効な量で間葉幹細胞を動物に投与することを含む。
【0040】
本発明のこの態様の範囲は如何なる理論上の推論にも限定されるべきではないが、間葉幹細胞が、急性アレルギー反応の後に投与された場合、肥満細胞の活性化及び脱顆粒の抑制をもたらすと考えられる。また、間葉幹細胞は、好塩基球活性化を下方制御し、またTNF−α等のサイトカイン、インターロイキン−8及び単球走化性タンパク質、あるいはMCP−1等のケモカイン、ロイコトリエン等の脂質メディエータを抑制し、またヒスタミン、ヘパリン、コンドロイチン硫酸、カテプシン等の主要メディエータを抑制すると考えられる。
【0041】
治療され得るアレルギー性疾患は、ぜんそく、アレルギー性鼻炎、アトピー性皮膚炎、及び接触性皮膚炎を含むがこれらに限定されない。しかしながら、本発明の範囲が如何なる特定のアレルギー疾患にも限定されるべきでないことは、理解されるべきである。
【0042】
間葉幹細胞は、動物のアレルギー疾患を治療するのに有効な量で動物に投与される。動物は哺乳類であり得る。哺乳類は、ヒト及び非ヒト霊長類を含む霊長類であり得る。通常、間葉幹細胞は、約1×105cells/kgから約1×107cells/kg、好ましくは約1×106cells/kgから約5×106cells/kgの量で、投与される。正確な投与量は、年齢、体重、患者の性別、治療されるアレルギー疾患、及びその範囲と重症度を含む、種々の因子に依存する。
【0043】
間葉幹細胞は、前述のとおり、薬学的許容担体と併せて投与され得る。間葉幹細胞は、例えば静脈内投与あるいは動脈内投与等により、全身に投与され得る。
【0044】
本発明のさらなる態様にしたがって、動物における創傷治癒を促進する方法が提供される。この方法は、動物における創傷治癒を促進するのに有効な量の間葉幹細胞を動物に投与することを含む。
【0045】
本発明の範囲は如何なる理論上の推論にも限定されるべきではないが、前述のとおり、間葉幹細胞がTreg細胞及び樹状細胞にインターロイキン−10(IL−10)を放出させると考えられる。IL−10は、創傷における炎症を抑制あるいは調節し、それにより創傷の治癒を促進する。
【0046】
さらに、間葉幹細胞は、他の細胞タイプにより分泌因子を誘導することで、創傷治癒及び骨折治癒を促進し得る。例えば、間葉幹細胞は、末梢血単核細胞(PBMCs)による血管内皮増殖因子(VEGF)のプロスタグランジンE2(PGE2)媒介性放出、及び成長ホルモン、インスリン、インスリン様成長因子1(IGF−1)、インスリン様成長因子結合タンパク質3(IGFBP−3)、及びエンドセリン−1のPGE2−媒介性放出を誘導し得る。
【0047】
間葉幹細胞は、動物の創傷治癒を促進するのに有効な量で動物に投与される。動物は哺乳類であり得る。また哺乳類は、ヒト及び非ヒト霊長類を含む霊長類であり得る。通常、間葉幹細胞は、約1×105cells/kgから約1×107cells/kg、好ましくは約1×106cells/kgから約5×106cells/kgの量で、投与される。投与されるべき間葉幹細胞の正確な量は、年齢、体重、患者の性別、及び治療される創傷の範囲と重症度を含む、種々の因子に依存する。
【0048】
間葉幹細胞は、前述のとおり、薬学的許容担体と併せて投与され得る。間葉幹細胞は、前述のとおり、全身に投与され得る。あるいは、間葉幹細胞は、例えば間葉幹細胞を包含する包袋あるいは貯蔵容器の流体のように、直接創傷に投与され得る。
【0049】
本発明のまた別の態様にしたがって、動物の線維症を治療あるいは予防する方法が提供される。この方法は、動物の線維症を治療あるいは予防するのに有効な量の間葉幹細胞を動物に投与することを含む。
【0050】
間葉幹細胞は、肝硬変、末期腎不全に関連する腎臓の線維症、及び肺の線維症を含むがこれらに限定されない任意のタイプの動物の線維症を治療あるいは予防するために、動物に投与され得る。本発明の範囲が如何なる特定のタイプの線維症にも限定されるべきでないことは理解されるべきである。
【0051】
間葉幹細胞は、動物の線維症を治療あるいは予防するのに有効な量で動物に投与される。動物は哺乳類であり得る。また哺乳類は、ヒト及び非ヒト霊長類を含む霊長類であり得る。通常、間葉幹細胞は、約1×105cells/kgから約1×107cells/kg、好ましくは約1×106cells/kgから約5×106cells/kgの量で、投与される。投与されるべき間葉幹細胞の正確な量は、年齢、体重、患者の性別、及び治療あるいは予防される線維症の範囲と重症度を含む、種々の因子に依存する。
【0052】
間葉幹細胞は、前述のとおり、薬学的許容担体と併せて投与され得る。間葉幹細胞は、前述のとおり、全身に投与され得る。
【0053】
本発明の他の目的は、動物の組織あるいは器官における血管形成を促進することである。ここで前記組織あるいは器官は、血管形成を必要としているものである。
【0054】
したがって、本発明のさらなる態様にしたがって、動物の器官あるいは組織における血管形成を促進する方法が提供される。この方法は、動物の器官あるいは組織における血管形成を促進するのに有効な量の間葉幹細胞を動物に投与することを含む。
【0055】
血管形成は、既存の微小血管床からの新規血管の形成である。
【0056】
血管形成の誘導は、冠状動脈機能不全及び末梢動脈機能不全の治療に用いられ得る。したがって、冠状動脈疾患、虚血性心疾患、及び末梢動脈疾患の治療への非侵襲性の治癒的方法となり得る。血管形成は、心臓以外の組織及び器官における疾病及び障害の治療に、また心臓以外の器官の発達及び又は維持に関与し得る。血管形成は、内部及び外部の創傷、及び皮膚潰瘍の治療に関与し得る。また血管形成は、肺着床、及び胎盤成長、及び胎児血管系の発達に関与する。また血管形成は、軟骨吸収と骨形成の結びつけに極めて重要であり、また的確な成長板形態形成に極めて重要である。
【0057】
さらに、血管形成は、肝臓等の高代謝器官の十分な形成及び維持に必要であり、ここで密集した血管網が、十分な栄養と気体の輸送を提供するために必要である。
【0058】
間葉幹細胞は、様々な方法で血管形成を必要とする組織あるいは器官に投与することができる。間葉幹細胞は、静脈内、動脈内あるいは腹腔内投与等により、全身に投与され得る。あるいは間葉幹細胞は、例えば血管形成を必要とする組織あるいは器官に直接注入することにより、血管形成を必要とする組織あるいは器官に直接投与され得る。
【0059】
間葉幹細胞は、自己、同種異系、異種を含む多種多様な資源から得られ得る。
【0060】
本発明の範囲は如何なる理論上の推論にも限定されるべきではないが、間葉幹細胞が、動物に投与された場合、新規血管の形成を促進する血管内皮増殖因子、すなわちVEGFを産生するように末梢血単核細胞(PBMCs)を刺激すると考えられる。
【0061】
一実施例において、動物は哺乳類である。哺乳類は、ヒト及び非ヒト霊長類を含む霊長類であり得る。
【0062】
本発明にしたがって、間葉幹細胞は、血管形成を通して軽減、治療、あるいは予防することができる任意の疾病あるいは障害の治療、軽減あるいは予防に用いられ得る。したがって、例えば間葉幹細胞は、四肢、すなわち腕、脚部、手、足、及び頸部あるいは種々の器官におけるものを含む動脈閉塞を治療するために、動物に投与され得る。例えば間葉幹細胞は、脳に分布する動脈の動脈閉塞を治療し、それにより脳梗塞を治療あるいは予防するために用いられ得る。また、間葉幹細胞は、胎児期及び出生後の角膜の血管の治療に用いられ得る。また糸球体構造形成をもたらすために用いられ得る。他の実施例において、間葉幹細胞は、内部及び外部双方の創傷の治療に、また糖尿病及び鎌状赤血球貧血等の疾病により引き起こされる皮膚潰瘍を含むがこれらに限定されない、足、手、脚部あるいは腕に見られる皮膚潰瘍の治療に用いられ得る。
【0063】
さらに、血管形成は胚着床及び胎盤形成に関与するため、間葉幹細胞は、胚着床を促進するために、また流産を予防するために用いられ得る。
【0064】
さらに、間葉幹細胞は、生まれる前の動物における血管系の発達を促進するために、ヒトを含む生まれる前の動物に投与され得る。
【0065】
他の実施例において、間葉幹細胞は、軟骨吸収及び骨形成を促進するために、また的確な成長板形態形成を促進するために、生後あるいは生前の動物に投与され得る。
【0066】
間葉幹細胞は、動物の血管形成を促進するのに有効な量で投与される。間葉幹細胞は、約1×105cells/kgから約1×107cells/kg、好ましくは約1×106cells/kgから約5×106cells/kgの量で、投与され得る。投与されるべき間葉幹細胞の量は、年齢、体重、患者の性別、治療、軽減、あるいは予防される疾病あるいは障害、及びその範囲と重症度を含む、種々の因子に依存する。
【0067】
間葉幹細胞は、薬学的許容担体と併せて投与され得る。例えば、間葉幹細胞は、注入用の薬学的許容液状媒体中の細胞懸濁液として投与され得る。注入は、局所的、すなわち血管形成を必要とする組織あるいは器官に直接であっても、あるいは全身的であってもよい。
【0068】
間葉幹細胞は、治療因子をエンコードする一つあるいはそれ以上のポリヌクレオチドで遺伝子組み換えされ得る。ポリヌクレオチドは、適切な発現媒体(expression vehicles)によって間葉幹細胞に送達され得る。間葉幹細胞を遺伝子組み換えするために用いられ得る発現媒体は、レトロウイルスベクター、アデノウイルスベクター及びアデノ随伴ウイルスベクターを含むがこれらに限定されない。
【0069】
治療因子をエンコードする適切なポリヌクレオチドの選択は、治療される疾病あるいは障害、及びその範囲と重傷度を含む、種々の因子に依存する。治療因子をエンコードするポリヌクレオチド、及び適切な発現媒体は、米国特許6355239号にさらに開示されている。
【0070】
間葉幹細胞が、上述の治療に用いられた場合、増殖因子、サイトカイン、抗炎症剤等の薬剤、及び樹状細胞等の間葉幹細胞以外の細胞を含むがこれらに限定されない、この分野における通常の知識を有する者に知られた他の治療因子と併せて用いられ得ること、及び、必要に応じて、ヒアルロン酸等の細胞用可溶性担体とともに、あるいはコラーゲン、ゼラチン、あるいは他の生体適合性ポリマー等の固形物と併せて投与され得ることは理解されるべきである。
【0071】
ここに開示された方法が、多数の方法で、またこの分野においてよく知られた種々の変更及び置換を伴って実施され得ることは理解されるべきである。また、細胞タイプ間の作用機構あるいは相互作用について説明する任意の理論が本発明を任意の方法に制限すると解釈されるべきではなく、本発明の方法をより十分に理解することができるように提示されていると認識され得る。
【0072】
本発明は、図面に関して開示される。ここで、
図1 MSCsの樹状細胞作用の調節
(A)HLA−DR及びCD11cに対する抗体を用いた成熟単球DC1細胞のフローサイトメトリー分析、及びHLA−DR及びCD123(IL−3レセプター)に対する抗体を用いた形質細胞様(plasmacytoid)DC2細胞のフローサイトメトリー分析。(−−−(破線)):アイソタイプ コントロール;(___(実線)):FITC/PE結合抗体。(B)MSCsは、それぞれ活性化したDC1及びDC2からのTNF−α分泌を抑制し(第一y軸)、IL−10分泌を増加させる(第二y軸)。(C)成熟DC1細胞とともに培養したMSCsは、MSCあるいはDC単独の場合と比較して、T細胞によるIFN−γ分泌(第一y軸)を抑制し、またIL−4レベルを増加させる(第二y軸)。MSCsの存在下での炎症誘発性IFN−γの産生の減少、及び抗炎症性IL−4の産生の増加は、抗炎症性表現型に関してのT細胞集団の変動を示唆した。
【0073】
図2 MSCsの炎症誘発性エフェクターT細胞作用の抑制
(A)FITC結合CD4抗体(x軸)及びPE結合CD25抗体(y軸)を用いた、PBMCsあるいは、MSC+PBMC培養(MSC+PBMC)における非接着性画分の染色によるTReg細胞数(%)のフローサイトメトリー分析。ゲートは、バックグラウンドとしてのアイソタイプ コントロール抗体に基づいて設定された。グラフは、5つの独立した実験の代表である。(B)細胞培養上清中で、MSCsの存在下で発生したTH1細胞は減少したレベルのIFN−γを分泌し(第一y軸)、またMSCsの存在下で発生したTH2細胞は増加した量のIL−4を分泌した(第二y軸)。(C)MSCsは、24ウェルプレート中で0、24、あるいは48時間培養された精製NK細胞からのIFN−γ分泌を抑制する。示されたデータは、一つの実験における平均値±SDサイトカイン分泌であり、また3つの独立した実験の代表である。
【0074】
図3 MSCsのTreg細胞集団数の増加及びGITR発現の増加の誘導
(A)PRMCあるいはMSC+PBMC(MSCのPBMCに対する比 1:10)培養(3日間さらなる刺激もなく培養された)由来のCD4+CD25+Treg細胞集団は、2ステップ磁気分離法を用いて単離された。これらの細胞は、(さらなる増殖を阻止するために)放射線照射され、混合リンパ球反応(MLR)における刺激因子として用いられた。ここで応答物質(responder)は、フィトヘムアグルチニン(PHA)の存在下(2.5mg/ml)での同種異系PBMCs(刺激因子の応答物質に対する比 1:100)であった。細胞は48時間培養され、その後3Hチミジンが添加され、24時間後に組み込まれた放射能がカウントされた。結果は、MSCsの存在下で発生したTreg集団(レーン3)が、MSCsの不在下で発生したTreg細胞(レーン2)と機能上類似していることを示した。(B)PBMCsは、MSCsの不在下(上段プロット)あるいは存在下(下段プロット)(MSCのPBMCに対する比 1:10)で、3日間培養され、その後非接着性画分が回収され、FITC標識GITR及びPE標識CD4を用いて免疫染色された。結果は、MSCsの存在下で培養された細胞のGITR発現において、2倍を超える増加を示す。
【0075】
図4 MSCsのPGE2の産生、及びMSC媒介免疫調節作用を無効にするPGE2の阻害
(A)種々の濃度の、PGE2阻害剤であるNS−398あるいはインドメタシン(Indometh.)の存在下あるいは不在下で培養されたMSCsから得られた培養上清におけるPGE2分泌(平均値±SD)。阻害剤濃度はμMであり、提示されたデータは24時間培養後に得られた値である。(B)リアルタイムRT−PCRを用いたMSCs及びPBMCsにおけるCOX−1及びCOX−2発現。MSCsは、PBMCsと比較してはるかに高いレベルのCOX−2を発現し、MSCsがPBMCsの存在下で培養された場合には、MSCsのCOX−2発現の3倍以上の増加であった。3つの独立した実験のうちの1つからの代表データが示される。MSC+PBMC培養はトランスウェルチャンバープレートに設置され、ここでMSCsは下部チャンバーに播種され、PBMCsは上部チャンバーに播種された。(C)PGE2阻害剤インドメタシン(Ind.)あるいはNS−398の存在が、コントロールと比較して、活性化DCsからのTNF−α分泌と、TH1細胞からのIFN−γ分泌を増加させた。データは、MSCs及びPGE2阻害剤の不在化で発生した培養からの%変化として計算された。(D)MSC−PBMC共培養(1:10)でのPGE2阻害剤インドメタシン(Indo)及びNS−398の存在は、PHA処理したPBMCsに対するMSC媒介抗増殖促進作用を無効にする。示されたデータは、一つの実験からのものであり、また3つの独立した実験の代表である。
【0076】
図5 同種異系PBMCsの存在下での構成的MSCサイトカイン分泌の増加
予め特性の示されたヒトMSCsを用いて、PBMCsの存在下(MSCのPBMCに対する比 1:10)(斜線バー)あるいは不在下(白抜きバー)で24時間培養したMSCsの培養上清における、サイトカインIL−6及びVEGF、脂質メディエータPGE2、及びマトリクスメタロプロテイナーゼ1(pro−MMP−1)のレベルが分析された。MSCsはIL−6、VEGF、及びPGE2を構成的に産生し、これらの因子のレベルはPBMCsとの共培養により増加した。これにより、MSCsが炎症設定における免疫機能の調節に関与し得ることを示す。
【0077】
図6 MSCsの用量依存的様式でのマイトジェン誘導T細胞増殖の抑制
増加数の同種異系PBMCsが、PHAの存在下(2.5mg/ml)あるいは不在下で、96ウェルプレートに播種された定数のMSCs(2000cells/well)とともにインキュベートされ、3Hチミジン取り込みが(カウント数/分、すなわちcpmで)測定された。MSCsの存在下で、PHAで処理したPBMCsの増殖の用量依存的抑制があった。3つの独立した実験の1つからの代表的な結果が示される。類似の結果は、LeBlanc,et al.,Scand J.Immunol.,Vol.57,pg.11(2003)により報告された。
【0078】
図7 MSC作用機序案の系統図
MSCsは、先天性(DCs経路 2から4;及びNK経路 6)及び適応的(T経路 1及び5、及びB経路 7)免疫システムの双方からの細胞に作用することで、その免疫調節作用を調節する。侵入病原体に対する反応で、未成熟DCsは潜在的入口部位に移動し、成熟して、(抗原特異的及び副刺激シグナルで)保護(protective)エフェクターT細胞(細胞媒介TH1免疫あるいは体液性TH2免疫)となるためのnaiveT細胞を用意(prime)する能力を獲得する。MSC−DC相互作用の間に、MSCsは、直接細胞間接触で、あるいは分泌された因子を介して、細胞媒介反応(経路2)を開始するDCsの能力を制限することによって、あるいは体液性反応(経路4)を開始する能力を促進することによって、免疫反応の結果を変更し得る。また、成熟エフェクターT細胞が存在する場合は、MSCsは、TH1(経路1)反応のバランスをTH2反応(経路5)の方へ、またおそらくIgE産生B細胞活性の増加(経路7)、GvHD及び自己免疫疾患症状の抑制について望ましい結果の方へ傾けるために、互いに相互作用し得る。MSCsは、TReg集団の発生の増加(経路3)をもたらす能力において、体制表現型をもたらし、また局所微小環境におけるバイスタンダー(bystander)炎症を弱めることで受容者ホストを救済し得る。破線(−−−)は、提案されたメカニズムを表す。
【0079】
本発明は、以下の実施例に関して開示される。しかしながら、本発明の範囲はそれらによって限定されるべきでないことは理解されるべきである。
【発明を実施するための最良の形態】
【0080】
(実施例1)
材料及び方法
ヒトMSCsの培養
Pittenger et al.,Science,Vol.284,pg.143(1999)により開示されたように、ヒトMSCsを培養した。つまり、Poietics Technologies、Div of Cambrex Biosciencesによるインフォームドコセントの後に、匿名提供者の腸骨稜から骨髄サンプルを回収した。MSCsを、1%抗生物質−抗真菌(antibiotic−antimycotic)溶液(Invitrogen,Carlsbad,California)と、10%ウシ胎仔血清(FBS,JRH BioSciences、lenexa、Kansas)とを含有する、完全ダルベッコ変法イーグル培地(complete Dulbecco‘s Modified Eagle’s Medium)−低グルコース(Life Technologies,Carlsbad,California)中で培養した。MSCsは接着性の単層として増殖し、トリプシン/EDTA(0.05%トリプシンを37℃で3分間)を用いて分離した。用いた全てのMSCsが多分化能について予め明らかにされており、また間葉系に分化(軟骨、脂質生成、及び骨形成)する能力を保持していた(Pittenger et al.,Science,Vol.284,pg.143(1999))。
【0081】
樹状細胞の単離
末梢血単核細胞(PBMCs)を、Poietics Technologies,Div of Cambrex Biosciences(Walkersville,MD)から獲得した。単球系の樹状細胞(DCs)の前駆体(CD1c+)を、Dzionek、et al.,J.Immunol.,Vol.165,pg.6037(2000)にしたがった2ステップ磁気分離法を用いて、PBMCsからポジティブセレクションした。つまり、CD1c発現B細胞が磁気ビーズを用いてCD19+細胞を磁気的に除去され、その後、ビオチン標識CD1c(BDCA1+)及び抗ビオチン抗体を用いてB細胞除去画分を標識し、それらを、メーカーの使用説明書(Miltenyi Biotech,Auburn,California)にしたがって磁気カラムを利用して非標識細胞画分から分離した。形質細胞様系統のDCsの前駆体が、陽性に標識される抗体でコートされた細胞(BDCA2+)の免疫−磁気ソーティングにより、PBMCsから単離された(Miltenyi Biotech,Auburn,California)。
【0082】
MSC−DC培養
大部分の実験において、等数のヒトMSCs及びDCsを、種々の期間培養し、細胞培養上清を回収し、さらなる実験まで−80℃で保存した。選択した実験では、MSCsを成熟DC1あるいはDC2細胞とともに(1:1 MSC:DCの比)3日間培養し、続いて混合培養(MSCsとDCs)を、増殖を阻止するために放射線照射した。続いて、抗体精製、naive、同種異系T細胞(CD4+、CD45RA+)を放射線照射MSCs/DCsに添加し、さらに6日間培養した。その後、非接着性細胞画分(精製T細胞)を培養液から回収し、2回洗浄し、さらに24時間PHAで再刺激した。その後、細胞培養上清を収集し、ELISAにより分泌されたIFN−γ及びIL−4について分析した。
【0083】
NK細胞の単離
NK細胞の精製集団を、一次試薬としてのビオチン結合モノクローナル抗体(抗CD3、CD14、CD19、CD36及び抗IgE抗体)と、二次標識試薬としてのマイクロビーズに結合した抗ビオチンモノクローナル抗体との混合物で磁気的に標識した非NK細胞を除去することで、獲得した。磁気標識された非NK細胞は、磁気領域中のMACS(Miltenyi Biotech,Auburn,California)カラムに保持され、一方NK細胞は、通過して回収された。
【0084】
Treg細胞集団の単離
Treg細胞集団を、2ステップ単離法を用いて単離した。第一に、非CD4+T細胞を、ビオチン標識抗体と抗ビオチンマイクロビーズの混合物で間接的磁気的に標識した。続いて標識した細胞を、MACSカラム(Miltenyi Biotech,Auburn,California)での分離により除去した。次に、CD4+CD25+細胞を、CD25マイクロビーズで直接標識し、濃縮前CD4+T細胞画分からポジティブセレクションにより単離した。磁気的に標識したCD4+CD25+T細胞をカラム上に保持し、磁気領域からのカラムの除去の後に溶出した。
【0085】
MSCsの存在下で発生したCD4+CD25+集団の増加が実際に抑制されたか否かを測定するために、2ステップ磁気単離法を用いて、CD4+CD25+TReg細胞集団をPBMCあるいはMSC+PBMC(MSCのPBMCに対する比 1:10)培養(3日間さらなる刺激もなく培養された)から単離した。これらの細胞を、さらなる増殖を阻止するために放射線照射し、混合リンパ球反応(MLR)における刺激因子として用いた。ここで応答物質は、PHAの存在下(2.5μg/ml)での同種異系PBMCs(刺激因子の応答物質に対する比 1:100)であった。48時間の培養を実施し、その後3Hチミジンを添加して、24時間後に組み込まれた放射能をカウントした。
【0086】
PBMCsをMSCsの不在下あるいは存在下(MSCのPBMCに対する比 1:10)で培養し、その後、非接着性画分を収集し、FITC標識グルココルチコイド誘導TNFレセプター、すなわちGITR、及びPE標識CD4を用いて免疫染色した。
【0087】
TH1/TH2細胞の発生
末梢血単核細胞(PBMCs)を、単球を除去するために、37℃で45分間2×106cells/mlで播種した。非接着性画分を、MSCsの存在下あるいは不在下で、TH1(IL−2(4ng/ml)+IL−12(5ng/ml)+抗IL−4(1μg/ml))あるいはTH2(IL−2(4ng/ml)+IL−4(4ng/ml)+抗IFN−γ(1μg/ml))条件下、プレート固定抗CD3(5μg/ml)抗体、及び抗CD28(1μg/ml)抗体の存在下で3日間インキュベートした。細胞を洗浄し、その後24時間あるいは48時間、PHA(2.5μg/ml)で再刺激した。その後、ELISA(R&D Systems,Minneapolis,Minnesota)で、培養上清中のIFN−γ及びIL−4のレベルを測定した。
【0088】
MSCsの培養上清中のVEGF、PGE2、及びpro−MMP−1のレベルの分析
予め特定されたヒトMSCsを用いて、PBMCsの存在下(MSCのPBMCに対する比 1:10)あるいは不在下で24時間培養したMSCsの培養上清中で、インターロイキン−6(IL−6)、VEGF、脂質メディエータ プロスタグランジンE2(PGE2)、及びマトリクスメタロプロテイナーゼ1(pro−MMP−1)のレベルを分析した。
【0089】
PBMCsの増殖
精製PBMCsを、Ficoll−Hypaque(Lymphoprep,Oslo,Norway)での遠心分離ロイコパック(centrifuging leukopack)(Cambrex,Walkersville,Maryland)により処理した。分離した細胞を、マイトジェンPHA(Sigma Chemicals,St.Louis,Missouri)の存在下、MSCsの存在下(定着するように、PBMC添加の3から4時間前に播種した)あるいは不在下で、48時間培養した(トリプリケイト(triplicates)で)。選択した実験においては、PBMCsを、PGE2阻害剤インドメタシン(Sigma Chemicals,St.Louis,Missouri)あるいはNS−938(Cayman Chemicals,Ann Arbor,Michigan)を含有する培地中に再懸濁した。(3H)−チミジンを添加し(200μlの培養液中に20μl)、さらなる24時間の培養の後に自動収集器を用いて細胞を収集した。MSCsあるいはPGE2阻害剤の作用効果を、PHAの存在下のコントロール反応(100%)の割合として計算した。
【0090】
量的RT−PCR
細胞ペレットからの全RNAを、市販のキット(Qiagen,Valencia,California)を用いて、またメーカーの使用説明書にしたがって、処理した。混入ゲノムDNAを、DNAフリーキット(Ambion,Austin,Texas)を用いて除去した。0.5μM濃度のプライマーとともにQuantiTect SYBR Green RT−PCRキット(Qiagen,Valencia,California)を用いて、MJ Research Opticon検出システム(South San Francisco,California)で量的RT−PCRを実施した。種々の条件下で培養された細胞における発現レベルの相対的変化を、内在コントロールとしてのβアクチンを用いて、Ct値(交点)の違いにより算出した。COX−1及びCOX−2の特異的プライマーの配列は、COX−1:5’−CCG GAT GCC AGT CAG GAT GAT G−3’(フォワード)、5’−CTA GAC AGC CAG ATG CTG ACA G−3’(リバース);COX−2:5’−ATC TAC CCT CCT CAA GTC CC−3’(フォワード)、5’−TAC CAG AAG GGC AGG ATA CAG−3’(リバース)である。
【0091】
増加数の同種異系PBMCsを、PHAの存在下(2.5μg/ml)で、96ウェルプレートに播種した定数のMSCs(2000cells/well)とともに72時間インキュベートし、3Hチミジン取り込み(カウント数/分、すなわちcpmで)を測定した。
【0092】
PBMCsとMSCsは、MSC:PBMCが1:1、1:3、1:10、1:30、及び1:81の割合で培養した。
【0093】
結果
本研究において、ヒトMSCsと、樹状細胞(DC1及びDC2)、エフェクターT細胞(TH1及びTH2)、及びNK細胞を含む、単離された免疫細胞集団との相互作用を調査した。MSCsと各免疫細胞タイプとの相互作用は、MSCsが免疫反応プロセスの複数の段階を調節し得ることを示す特異的な結果であった。MSC免疫調節作用を調節し、またMSC免疫調節作用の原因となり得る分泌因子の産生が評価され、プロスタグランジン合成が関与することが示された。
【0094】
骨髄(DC1)前駆樹状細胞及び形質細胞様(DC2)前駆樹状細胞を、それぞれBDCA1+細胞及びBDCA2+細胞の免疫−磁気ソーティングにより単離し、DC1細胞についてはGM−CSF及びIL−4(それぞれ1×103IU/ml、及び1×103IU/ml)とともに、DC2細胞についてはIL−3(10ng/ml)とともにインキュベートすることで成熟させた。フローサイトメトリーを用いて、DC1細胞はHLA−DR+及びCD11c+であり、一方DC2細胞はHLA−DR+及びCD123+であった(図1A)。炎症性因子である細菌性リポ多糖(LPS,1ng/ml)の存在下で、DC1細胞は中程度のレベルのTNF−αを産生したが、MSCsが存在した場合(1:1及び1:10の比で調査された)には、TNF−α分泌の50%以上の減少があった(図1B)。他方では、DC2細胞はLPSの存在下でIL−10を産生し、そのレベルはMSC:DC2共培養(1:1)で2倍以上増加した(図1B)。したがって、MSCsは、培養中の活性化されたDCsのサイトカインプロファイルを、より寛容原生表現型に変更した。さらに、活性化したDCsは、MSCsとともに培養された場合に、IFN−γを減少させ、またnaiveCD4+T細胞により分泌されたIL−4レベルを増加させることができ(図1C)、これは前炎症性T細胞表現型から抗炎症性T細胞表現型へのMSC媒介変化を示す。
【0095】
IL−10分泌の増加が制御性細胞の発生に関与するため(Kingsley,et al.,J.Immunol.,Vol.168,pg.1080(2000))、T制御性細胞(TReg)を、PBMCs及びMSCsの共培養中で、フローサイトメトリーにより定量した。PBMCsのMSCsとの3から5日間の培養により、抗CD4抗体及び抗CD25抗体でのPBMCsの染色により測定されたTReg細胞数の増加があり(図2A)、さらにMSC誘導寛容原生反応を支持した。MSCsの存在下で発生したCD4+CD25+TReg細胞集団は、増加したレベルのグルココルチコイド誘導TNFレセプター(GITR)、TReg細胞集団上に発現する細胞表面レセプターを発現し、また同種異系T細胞増殖を抑制した(図3A、3B)ことから、実際抑制的であった。次に、T細胞分化に作用する直接の能力に関して、MSCsを調査した。抗体選別精製T細胞(CD4+Th細胞)を用いて、IFN−γ産生TH1細胞及びIL−4産生TH2細胞を、MSCsの存在下あるいは不在下で発生させた。分化の間にMSCsが存在すると、TH1細胞によるIFN−γ分泌が減少し、TH2細胞によるIL−4分泌が増加した(図2B)。Th細胞がエフェクターTH1タイプあるいはTH2タイプに分化した後(3日)に、MSCsが培養に添加された場合は、IFN−γあるいはIL−4のレベルの著しい変化は見られなかった(不図示)。これらの実験は、MSCsがエフェクターT細胞分化に直接作用し、T細胞サイトカイン分泌を体液性表現型に変化させることができることを示す。
【0096】
同様に、MSCsを精製NK細胞(CD3−、CD14−、CD19−、CD36−)と比率1:1で異なる期間(0から48時間)培養すると、培養上清中でのIFN−γ分泌の減少があった(図2C)。したがって、これはMSCsがNK細胞作用をも調節することができることを示す。
【0097】
過去の研究は、MSCsが水溶性因子によりT細胞作用を調節することを示している(LeBlanc,et al.,Exp.Hematol.,Vol.31,pg.890(2003);Tse,et al.,Transplantation,Vol.75,pg.389(2003))。MSCsが、IL−6、プロスタグランジンE2、VEGF、及びproMMP−1を含む複数の因子を構成的に分泌し、PBMCsを含む培養によりそれぞれのレベルが増加したことが観察された(図5)。DCsによる、TNF−α産生の抑制、及びIL−10産生の増加をもたらすMSC生成因子を調査するために、プロスタグランジンE2の潜在的役割を調査したが、活性化DCsによるTNF−γ産生を抑制することが認められた(Vassiliou,et al.,Cell.Immunol.,Vol.223,pg.120(2003))。MSC培養(0.5×106cells/mlの24時間培養)の条件培地は、おおよそ1000pg/mlのPGE2を含有していた(図4A)。培養上清中に、PGE2分泌の既知の誘導因子、例えばTNF−α、IFN−γあるいはIL−1βの存在は検出できなかったが(不図示)、これはMSCsによるPGE2の構成的分泌を示す。hMSCsによるPGE2分泌は、PGE2産生の既知の阻害剤、NS−398(5μM)及びインドメタシン(4μM)の存在下で、60から90%抑制された(図4A)。PGE2分泌の放出が、構成的活性シクロオキシゲナーゼ酵素1(COX−1)及び誘導性シクロオキシゲナーゼ酵素2(COX−2)の酵素活性の結果として起こるため(Harris,et al.,Trends Immunol.,Vol.23,pg.144(2002))、MSCs及びPBMCsにおけるCOX−1及びCOX−2のmRNA発現を、トランスウェル培養システムを用いて分析した。MSCsは、PBMCsと比較してはるかに高いレベルのCOX−2を発現し、MSCsとPBMCsの24時間の共培養(MSCのPBMCに対する比 1:10)の場合には、発現レベルは3倍以上増加した(図4B)。COX−1レベルのわずかな変化が見られ、これはMSC−PBMC共培養によるPGE2分泌の増加(図5)が、COX−2の上向き調節により調節されることを示す。DCs及びT細胞に対するMSCの免疫調節効果がPGE2によって調節されたか否かを調査するために、PGE2阻害剤NS−398あるいはインドメタシンの存在下で、MSCsを活性化樹状細胞(DC1)あるいはTH1細胞とともに培養した。NS−398あるいはインドメタシンの存在が、DC1sによるTNF−α分泌と、TH1細胞からのIFN−γ分泌をそれぞれ増加させたが(図4C)、これは免疫細胞タイプに対するMSCの作用が、分泌されたPGE2により調節され得ることを示す。最近の研究が、MSCsが様々な刺激で誘導されるT細胞増殖を抑制することを示した(Denicola,et al.,Blood,Vol.99,pg.3838(2002);LeBlanc,et al.,Scand.J.Immunol.,Vol.57,pg.11(2003))。MSCsがマイトジェン誘導T細胞増殖を用量依存的様式で抑制することが観察され(図6)、PGE2阻害剤NS−398(5μM)あるいはインドメタシン(4μM)が存在した場合には、MSC含有培養液中のPHA処理PBMCsによる3Hチミジン取り込みが、阻害剤を含まないコントロールと比べて、70%以上増加した(図4D)。
【0098】
要約すると、MSCの他の免疫細胞タイプとの相互作用のモデル(図7)が提案される。成熟T細胞が存在する場合には、MSCsは、それらと直接相互作用して、前炎症性IFN−γ産生を抑制し(経路1)、制御性T細胞表現型を促進し(経路3)、また抗炎症性TH2細胞を促進する(経路5)。さらに、MSCsは、PGE2を分泌し、前炎症性DC1細胞を抑制し(経路2)、また抗炎症性DC2細胞(経路4)あるいは制御性DCs(経路3)を促進することで、DCsを介したT細胞免疫反応の結果を変えることができる。TH2免疫への変化は言い換えると、B細胞活性の、IgE/IgG1サブタイプ抗体の発生の増加(経路7)への変化を示す。MSCsは、そのNK細胞からのIFN−γ分泌を抑制する能力により、NK細胞作用を調節し得る(経路6)。このMSCのモデル:免疫細胞相互作用は、複数の他の研究所で行われた実験と一致する(LeBlanc,et al.,Exp.Hematol.,Vol.31,pg.890(2003);Tse,et al.,Transplantation,Vol.75,pg.389(2003);DiNicola,et al.,Blood,Vol.99,pg.3838(2002))。提案されたメカニズムについてのさらなる調査が進行中であり、現在はMSC投与のin vivo効果を調査するために、動物研究が必要である。
【0099】
公開された特許出願、保管登録番号、及びデータベース登録番号を含む、全ての特許、文献は、各特許、文献、保管登録番号、及びデータベース登録番号が明確に、また個別に参照として組み込まれているように、同じ範囲で参照としてここに組み込まれている。
【0100】
しかしながら、本発明の範囲は、上述の特定の実施例に限定されるべきでないことは理解されるべきである。本発明は、特に開示されている以外の様式で実施され得るが、これも添付の請求の範囲の範囲内である。
【図面の簡単な説明】
【0101】
【図1A】MSCsの樹状細胞作用の調節を説明する図
【図1B】MSCsの樹状細胞作用の調節を説明する図
【図1C】MSCsの樹状細胞作用の調節を説明する図
【図2A】MSCsの炎症誘発性エフェクターT細胞作用の抑制を説明する図
【図2B】MSCsの炎症誘発性エフェクターT細胞作用の抑制を説明する図
【図2C】MSCsの炎症誘発性エフェクターT細胞作用の抑制を説明する図
【図3A】MSCsのTreg細胞集団数の増加及びGITR発現の増加の誘導を説明する図
【図3B】MSCsのTreg細胞集団数の増加及びGITR発現の増加の誘導を説明する図
【図4A】MSCsのPGE2の産生、及びMSC媒介免疫調節作用を無効にするPGE2の阻害を説明する図
【図4B】MSCsのPGE2の産生、及びMSC媒介免疫調節作用を無効にするPGE2の阻害を説明する図
【図4C】MSCsのPGE2の産生、及びMSC媒介免疫調節作用を無効にするPGE2の阻害を説明する図
【図4D】MSCsのPGE2の産生、及びMSC媒介免疫調節作用を無効にするPGE2の阻害を説明する図
【図5】同種異系PBMCsの存在下での構成的MSCサイトカイン分泌の増加を説明する図
【図6】MSCsの用量依存的様式でのマイトジェン誘導T細胞増殖の抑制を説明する図
【図7】MSC作用機序案の系統図
【特許請求の範囲】
【請求項1】
動物の心臓以外の器官あるいは組織における血管形成の促進方法であって、
前記動物に、前記動物の心臓以外の器官あるいは組織における血管形成を促進するのに有効な量の間葉幹細胞を投与することを含むことを特徴とする促進方法。
【請求項2】
請求項1に記載の促進方法において、
前記動物は、哺乳類であることを特徴とする促進方法。
【請求項3】
請求項2に記載の促進方法において、
前記哺乳類は、霊長類であることを特徴とする促進方法。
【請求項4】
請求項3に記載の促進方法において、
前記霊長類は、ヒトであることを特徴とする促進方法。
【請求項5】
請求項1に記載の促進方法において、
前記間葉幹細胞は、約1×105cells/kgから約1×107cells/kgの量で投与されることを特徴とする促進方法。
【請求項6】
請求項5に記載の促進方法において、
前記間葉幹細胞は、約1×106cells/kgから約5×106cells/kgの量で投与されることを特徴とする促進方法。
【請求項7】
請求項1に記載の促進方法において、
前記間葉幹細胞は、全身に投与されることを特徴とする促進方法。
【請求項8】
請求項1に記載の促進方法において、
前記間葉幹細胞は、静脈内に投与されることを特徴とする促進方法。
【請求項9】
請求項1に記載の促進方法において、
前記間葉幹細胞は、前記動物の心臓以外の器官あるいは組織に直接注入により投与されることを特徴とする促進方法。
【請求項10】
動物の自己免疫疾患の治療方法であって、
前記動物に、前記動物の前記自己免疫疾患を治療するのに有効な量の間葉幹細胞を投与することを含むことを特徴とする治療方法。
【請求項11】
請求項10に記載の治療方法において、
前記動物は、哺乳類であることを特徴とする治療方法。
【請求項12】
請求項11に記載の治療方法において、
前記哺乳類は、ヒトであることを特徴とする治療方法。
【請求項13】
請求項10に記載の治療方法において、
前記自己免疫疾患は、多発性硬化症であることを特徴とする治療方法。
【請求項14】
動物の炎症反応の治療方法であって、
前記動物に、前記動物の前記炎症反応を治療するのに有効な量の間葉幹細胞を投与することを含むことを特徴とする治療方法。
【請求項15】
請求項14に記載の治療方法において、
前記動物は、哺乳類であることを特徴とする治療方法。
【請求項16】
請求項15に記載の治療方法において、
前記哺乳類は、ヒトであることを特徴とする治療方法。
【請求項17】
請求項14に記載の治療方法において、
前記炎症反応は、乾癬に関連するものであることを特徴とする治療方法。
【請求項18】
動物の癌の治療方法であって、
前記動物に、前記動物の癌を治療するのに有効な量の間葉幹細胞を投与することを含むことを特徴とする治療方法。
【請求項19】
請求項18に記載の治療方法において、
前記動物は、哺乳類であることを特徴とする治療方法。
【請求項20】
請求項19に記載の治療方法において、
前記哺乳類は、ヒトであることを特徴とする治療方法。
【請求項21】
動物のアレルギー性疾患の治療方法であって、
前記動物に、前記動物の前記アレルギー性疾患を治療するのに有効な量の間葉幹細胞を投与することを含むことを特徴とする治療方法。
【請求項22】
請求項21に記載の治療方法において、
前記動物は、哺乳類であることを特徴とする治療方法。
【請求項23】
請求項22に記載の治療方法において、
前記哺乳類は、ヒトであることを特徴とする治療方法。
【請求項24】
請求項21に記載の治療方法において、
前記アレルギー性疾患は、関節炎であることを特徴とする治療方法。
【請求項25】
動物の創傷治癒の促進方法であって、
前記動物に、前記動物の創傷治癒を促進するのに有効な量の間葉幹細胞を投与することを含むことを特徴とする促進方法。
【請求項26】
請求項25に記載の促進方法において、
前記動物は、哺乳類であることを特徴とする促進方法。
【請求項27】
請求項26に記載の促進方法において、
前記哺乳類は、ヒトであることを特徴とする促進方法。
【請求項28】
動物の線維症の予防方法であって、
前記動物に、前記動物の線維症を予防するのに有効な量の間葉幹細胞を投与することを含むことを特徴とする予防方法。
【請求項1】
動物の心臓以外の器官あるいは組織における血管形成の促進方法であって、
前記動物に、前記動物の心臓以外の器官あるいは組織における血管形成を促進するのに有効な量の間葉幹細胞を投与することを含むことを特徴とする促進方法。
【請求項2】
請求項1に記載の促進方法において、
前記動物は、哺乳類であることを特徴とする促進方法。
【請求項3】
請求項2に記載の促進方法において、
前記哺乳類は、霊長類であることを特徴とする促進方法。
【請求項4】
請求項3に記載の促進方法において、
前記霊長類は、ヒトであることを特徴とする促進方法。
【請求項5】
請求項1に記載の促進方法において、
前記間葉幹細胞は、約1×105cells/kgから約1×107cells/kgの量で投与されることを特徴とする促進方法。
【請求項6】
請求項5に記載の促進方法において、
前記間葉幹細胞は、約1×106cells/kgから約5×106cells/kgの量で投与されることを特徴とする促進方法。
【請求項7】
請求項1に記載の促進方法において、
前記間葉幹細胞は、全身に投与されることを特徴とする促進方法。
【請求項8】
請求項1に記載の促進方法において、
前記間葉幹細胞は、静脈内に投与されることを特徴とする促進方法。
【請求項9】
請求項1に記載の促進方法において、
前記間葉幹細胞は、前記動物の心臓以外の器官あるいは組織に直接注入により投与されることを特徴とする促進方法。
【請求項10】
動物の自己免疫疾患の治療方法であって、
前記動物に、前記動物の前記自己免疫疾患を治療するのに有効な量の間葉幹細胞を投与することを含むことを特徴とする治療方法。
【請求項11】
請求項10に記載の治療方法において、
前記動物は、哺乳類であることを特徴とする治療方法。
【請求項12】
請求項11に記載の治療方法において、
前記哺乳類は、ヒトであることを特徴とする治療方法。
【請求項13】
請求項10に記載の治療方法において、
前記自己免疫疾患は、多発性硬化症であることを特徴とする治療方法。
【請求項14】
動物の炎症反応の治療方法であって、
前記動物に、前記動物の前記炎症反応を治療するのに有効な量の間葉幹細胞を投与することを含むことを特徴とする治療方法。
【請求項15】
請求項14に記載の治療方法において、
前記動物は、哺乳類であることを特徴とする治療方法。
【請求項16】
請求項15に記載の治療方法において、
前記哺乳類は、ヒトであることを特徴とする治療方法。
【請求項17】
請求項14に記載の治療方法において、
前記炎症反応は、乾癬に関連するものであることを特徴とする治療方法。
【請求項18】
動物の癌の治療方法であって、
前記動物に、前記動物の癌を治療するのに有効な量の間葉幹細胞を投与することを含むことを特徴とする治療方法。
【請求項19】
請求項18に記載の治療方法において、
前記動物は、哺乳類であることを特徴とする治療方法。
【請求項20】
請求項19に記載の治療方法において、
前記哺乳類は、ヒトであることを特徴とする治療方法。
【請求項21】
動物のアレルギー性疾患の治療方法であって、
前記動物に、前記動物の前記アレルギー性疾患を治療するのに有効な量の間葉幹細胞を投与することを含むことを特徴とする治療方法。
【請求項22】
請求項21に記載の治療方法において、
前記動物は、哺乳類であることを特徴とする治療方法。
【請求項23】
請求項22に記載の治療方法において、
前記哺乳類は、ヒトであることを特徴とする治療方法。
【請求項24】
請求項21に記載の治療方法において、
前記アレルギー性疾患は、関節炎であることを特徴とする治療方法。
【請求項25】
動物の創傷治癒の促進方法であって、
前記動物に、前記動物の創傷治癒を促進するのに有効な量の間葉幹細胞を投与することを含むことを特徴とする促進方法。
【請求項26】
請求項25に記載の促進方法において、
前記動物は、哺乳類であることを特徴とする促進方法。
【請求項27】
請求項26に記載の促進方法において、
前記哺乳類は、ヒトであることを特徴とする促進方法。
【請求項28】
動物の線維症の予防方法であって、
前記動物に、前記動物の線維症を予防するのに有効な量の間葉幹細胞を投与することを含むことを特徴とする予防方法。
【図1A】


【図1B】
【図1C】
【図2A】
【図2B】
【図2C】
【図3A】
【図3B】
【図4A】
【図4B】
【図4C】
【図4D】
【図5】
【図6】
【図7】
【図1B】
【図1C】
【図2A】
【図2B】
【図2C】
【図3A】
【図3B】
【図4A】
【図4B】
【図4C】
【図4D】
【図5】
【図6】
【図7】
【公表番号】特表2007−530543(P2007−530543A)
【公表日】平成19年11月1日(2007.11.1)
【国際特許分類】
【出願番号】特願2007−505005(P2007−505005)
【出願日】平成17年3月15日(2005.3.15)
【国際出願番号】PCT/US2005/008506
【国際公開番号】WO2005/093044
【国際公開日】平成17年10月6日(2005.10.6)
【出願人】(500430486)オシリス セラピューティクス,インコーポレイテッド (10)
【Fターム(参考)】
【公表日】平成19年11月1日(2007.11.1)
【国際特許分類】
【出願日】平成17年3月15日(2005.3.15)
【国際出願番号】PCT/US2005/008506
【国際公開番号】WO2005/093044
【国際公開日】平成17年10月6日(2005.10.6)
【出願人】(500430486)オシリス セラピューティクス,インコーポレイテッド (10)
【Fターム(参考)】
[ Back to top ]