
間葉系幹細胞とその用途
動物に有効量の間葉系幹細胞を投与することにより、動物の自己免疫疾患、アレルギー応答、癌、炎症性疾患を治療し、動物の臓器又は組織の創傷治癒を促進し、上皮傷害を修復し、血管形成を促進する方法。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は間葉系幹細胞に関する。さらに詳しくは、本発明は、種々の組織や臓器における血管形成の促進、自己免疫疾患の治療、アレルギー応答の治療、癌の治療、炎症性疾患と障害の治療、創傷治癒の促進、炎症の治療、及び上皮傷害の修復を含む間葉系幹細胞の新規用途に関する。
【背景技術】
【0002】
本発明は、海軍省により契約第N66001−02−C−8068号で政府の援助で行われた。政府は本発明において一定の権利を有する。
【0003】
本出願は、2004年3月22日に出願された仮特許出願第60/555,118号(その内容は参照することによりその全体が本明細書に組み込まれる)に基づく優先権を請求する2005年3月15日出願の特許出願第11/080,298号の一部継続出願である。
【0004】
間葉系幹細胞(MSC)は、骨芽細胞、筋細胞、軟骨細胞、及び脂肪細胞を含む系統に容易に分化できる多能性幹細胞である(Pittenger, et al., Science, Vol. 284, pg. 143 (1999); Haynesworth, et al., Bone, Vol. 13, pg. 69 (1992); Prockop, Science, Vol. 276, pg. 71 (1997))。インビトロの試験により、MSCが筋肉(Wakitani, et al., Muscle Nerve, Vol. 18,, pg. 1417 (1995))、神経様前駆体(Woodbury, et al., J. Neurosci. Res., Vol. 69, pg. 908 (2002); Sanchez-Ramos, et al., Exp. Neurol., Vol. 171 , pg. 109 (2001))、心筋細胞(Toma, et al., Circulation, Vol. 105, pg. 93 (2002); Fakuda, Artif. Organs, Vol. 25, pg. 187 (2001))、及びおそらくは他のタイプの細胞に分化する能力が証明されている。さらにMSCは、造血幹細胞や胚幹細胞の拡張のための有効なフィーダー層となることが証明されている(Eaves, et al., Ann. N.Y. Acad. Sci., Vol. 938, pg. 63 (2001); Wagers, et al., Gene Therapy, Vol. 9, pg. 606 (2002))。種々の動物モデルを用いた最近の研究は、MSCが、傷害された骨、軟骨、半月板、又は心筋組織の修復又は再生において有用であることを証明している(DeKok, et al., Clin. Oral Implants Res., Vol. 14, pg. 481 (2003)); Wu, et al., Transplantation, Vol. 75, pg. 679 (2003); Noel, et al., Curr. Opin. Investig. Drugs, Vol. 3, pg. 1000 (2002); Ballas, et al., J. Cell. Biochem. Suppl., Vol. 38, pg. 20 (2002); Mackenzie, et al., Blood Cells Mol. Dis., Vol. 27 (2002))。何人かの研究者は、骨形成不全症(Pereira, et al., Proc. Nat. Acad. Sci., Vol. 95, pg. 1142 (1998))、パーキンソン病(Schwartz, et al., Hum. Gene Ther., Vol. 10, pg. 2539 (1999))、脊髄損傷(Chopp, et al., Neuroreport, Vol. 11 , pg. 3001 (2000); Wu, et al., J. Neurosci. Res., Vol. 72, pg. 393 (2003))、及び心臓疾患(Tomita, et al., Circulation, Vol. 100, pg. 247 (1999). Shake, et al., Ann. Thorac. Surg., Vol. 73, pg. 1919 (2002))を含む動物疾患モデルにおける移植について、MSCを使用して有望な結果を得ている。重要なことに、有望な結果はまた、骨形成不全症(Horwitz, et al., Blood, Vol. 97, pg. 1227 (2001); Horowitz, et al. Proc. Nat. Acad. Sci., Vol. 99, pg. 8932 (2002))と異種骨髄移植(Frassoni, et al., Int. Society for Cell Therapy, SA006 (要約) (2002); Koc, et al., J. Clin. Oncol., Vol. 18, pg. 307 (2000))の移植の向上の臨床治験でも報告されている。
【0005】
MSCは、その表面に主要組織適合遺伝子複合体(MHC)クラスI抗原を発現するが、MHCクラスII(Le Blanc, et al., Exp. Hematol., Vol. 31 , pg. 890 (2003); Potian, et al., J. Immunol., Vol. 171 , pg. 3426 (2003))やB7もCD40同時刺激分子(Majumdar, et al., J. Biomed. Sci., Vol. 10, pg. 228 (2003))も発現せず、これらの細胞が免疫原性表現型(Tse, et al., Transplantation, Vol. 75, pg. 389 (2003))を有することを示唆している。MSCはまたMHC非依存性にT細胞増殖応答を阻害する(Bartholomew, et al., Exp. Hematol., Vol. 30, pg. 42 (2002); Devine, et al., Cancer J., Vol. 7, pg. 576 (2001); DiNicola, et al., Blood, Vol. 99, pg. 3838 (2002))。MSCのこれらの免疫学的性質は移植を向上させ、移植後の同種異系細胞を認識し拒絶する受容者の免疫系の能力を制限しているかも知れない。局所的刺激下で適切なタイプの細胞に分化する能力とともに、免疫応答を調節し造血を支持する因子のMSCによる産生は、これらを細胞移植研究のための好ましい幹細胞にしている(Majumdar, et al., Hematother. Stem Cell Res., Vol. 9, pg. 841 (2000); Haynesworth, et al., J. Cell. Physiol., Vol. 166, pg. 585 (1996))。
【発明の開示】
【0006】
本出願人は、樹状細胞(DC1とDC2)、エフェクターT細胞(Th1とTh2)、及びNK細胞を含む、間葉系幹細胞と単離された免疫細胞集団との相互作用を調べた。かかる相互作用に基づいて本出願人は、間葉系幹細胞が、免疫応答プロセスのいくつかの工程を制御する種々の因子の産生を制御することを発見した。すなわち間葉系幹細胞は、免疫系が関与する症状や障害、炎症が関与する疾患、症状、及び障害、上皮傷害、又はアレルギー応答の治療に使用できるかも知れない。かかる疾患、症状、及び障害には、特に限定されないが、自己免疫疾患、アレルギー、関節炎、炎症を起こした創傷、円形脱毛症(禿)、歯周病(歯肉炎と歯周炎を含む)、及び免疫応答が関与する他の疾患、症状、又は障害がある。
【0007】
さらに間葉系幹細胞は、血管内皮増殖因子、又はVEGF(これは新しい血管の形成を刺激することにより血管形成を促進する)を発現し分泌する。間葉系幹細胞はまた末梢血単核細胞(PBMC)を刺激してVEGFを産生させる。
【0008】
さらに間葉系幹細胞は樹状細胞がインターフェロン−ベータ(IFN−β)を産生するのを刺激して、これが腫瘍抑制とウイルス感染に対する免疫を促進する。
【0009】
本発明のある態様において、動物の自己免疫疾患と移植片対宿主病よりなる群から選択される疾患を治療する方法が提供される。この方法は、動物の疾患を治療するために有効な量の間葉系幹細胞を動物に投与することを含む。
【0010】
本発明のこの態様の範囲はいかなる理論にも限定されないが、間葉系幹細胞が自己免疫疾患と移植片対宿主病を抑制する少なくとも1つの機構は、制御T細胞(Treg細胞)及び/又は樹状細胞(DC)からのインターロイキン−10(IL−10)の放出を引き起こすことによると考えられる。
【0011】
本発明で治療される自己免疫疾患には、特に限定されないが、多発性硬化症、1型糖尿病、慢性関節リウマチ、ブドウ膜炎、自己免疫甲状腺疾患、炎症性腸疾患、強皮症、グレーブス病、ループス、クローン病、自己免疫リンパ増殖性疾患(ALPS)、脱髄疾患、自己免疫脳脊髄炎、自己免疫胃炎(AIG)、及び自己免疫糸球体疾患がある。また上記したように、移植片対宿主病も治療される。しかし本発明の範囲は、本明細書に記載の特定の疾患の治療に限定されないことを理解されたい。
【0012】
ある実施態様において、間葉系幹細胞が投与される動物は哺乳動物である。哺乳動物は、ヒト及び非ヒト霊長類を含む霊長類である。
【0013】
一般に間葉系幹細胞(MSC)療法は、以下の順序に基づく:MSC含有組織の採取、MSCの単離と拡張、動物へのMSCの投与(生化学的又は遺伝的操作有り又は無し)。
【0014】
投与される間葉系幹細胞は均一組成物であるか、又はMSCで濃縮された混合細胞集団でもよい。均一間葉系幹細胞組成物は接着性骨髄細胞又は骨膜細胞を培養することにより得られ、間葉系幹細胞組成物は接着性骨髄細胞又は骨膜細胞を培養することにより得られ、間葉系幹細胞は、ユニークなモノクローナル抗体を用いて同定される特異的細胞表面マーカーにより同定される。間葉系幹細胞で濃縮された細胞集団を得る方法は、例えば米国特許第5,486,359号明細書に記載されている。間葉系幹細胞の別の供給源には、特に限定されないが、血液、皮膚、臍帯血、筋肉、脂肪、骨、及び軟骨膜がある。
【0015】
間葉系幹細胞は種々の方法で投与される。間葉系幹細胞は、例えば静脈内、動脈内、又は腹腔内投与により全身的に投与してもよい。
【0016】
間葉系幹細胞は、自己、同種異系、又は異種を含む範囲の供給源由来でもよい。
【0017】
間葉系幹細胞は、動物の自己免疫疾患又は移植片対宿主病を治療するために有効な量で投与される。間葉系幹細胞は約1×105細胞/kg〜約1×107細胞/kgの量で投与される。別の実施態様において間葉系幹細胞は、約1×106細胞/kg〜約5×106細胞/kgの量で投与される。投与される間葉系幹細胞の量は、患者の年齢、体重、性、治療される自己免疫疾患、及びその程度と重症度を含む種々の要因に依存する。
【0018】
間葉系幹細胞は薬学的に許容される担体とともに投与してもよい。例えば間葉系幹細胞は、注射又は局所的投与のための薬学的に許容される液体媒体又はゲル中で細胞懸濁液として投与してもよい。
【0019】
本発明の別の態様において、動物の炎症応答を治療する方法が提供される。この方法は、動物の炎症応答を治療するために有効な量の間葉系幹細胞を動物に投与することを含む。
【0020】
本発明のこの態様の範囲はいかなる理論にも限定されないが、間葉系幹細胞が、制御T細胞(Treg細胞)へのT細胞成熟を促進し、こうして炎症応答が抑制されると考えられる。また、間葉系幹細胞がTヘルパー1細胞(Th1細胞)を阻害し、それによって、乾癬に関連するもののようなある炎症反応のインターフェロン−γ(IFN−γ)の発現を低下させると考えられる。
【0021】
ある実施態様において治療される炎症応答は、乾癬に関連するものである。
【0022】
別の実施態様において間葉系幹細胞は、脳中の小神経膠細胞及び/又は星状細胞に接触して炎症を低下させるように動物に投与され、こうして間葉系幹細胞は、アルツハイマー病、パーキンソン病、卒中、又は脳細胞損傷のような疾患もしくは障害において活性化グリア細胞により引き起こされる神経変性を制限する。
【0023】
さらに別の実施態様において間葉系幹細胞は、皮膚の表皮中のケラチン細胞及びランゲルハンス細胞に間葉系幹細胞が接触して、乾癬、慢性皮膚炎、及び接触皮膚炎で起きるような炎症を低下させるように動物に投与される。本発明はいかなる理論にも限定されないが、間葉系幹細胞が表皮中のケラチン細胞及びランゲルハンス細胞に接触して、表皮中のT細胞受容体やランゲルハンス細胞の発現を変化させ、こうして腫瘍壊死因子−アルファ(TNF−α)の発現を低下させ、制御T細胞(Treg細胞)集団を上昇させると考えられる。
【0024】
さらなる実施態様において間葉系幹細胞は、特に限定されないが骨関節炎や慢性関節リウマチ、及びウェブサイトwww.arthritis.org/conditions/diseasesに記載されている他の関節炎疾患を含む関節炎や関節炎様症状で起こるような、骨の炎症を低下させるのに使用される。本発明のこの実施態様の範囲はいかなる理論にも限定されないが、間葉系幹細胞が、滑液中の記憶T細胞によるインターロイキン−17分泌を阻害すると考えられる。
【0025】
別の実施態様において間葉系幹細胞は、炎症性腸疾患や慢性肝炎におけるそれぞれ消化管及び肝臓中の炎症を抑えるために使用される。本発明のこの実施態様の範囲はいかなる理論にも限定されないが、間葉系幹細胞が、インターロイキン−10(IL−10)の分泌増加と制御T細胞(Treg細胞)の生成を促進すると考えられる。
【0026】
別の実施態様において間葉系幹細胞は、敗血症及び火傷、手術、及び移植を含む外傷のような病理症状において好中球とマクロファージの過度の活性化を阻害するために使用される。本発明のこの実施態様の範囲はいかなる理論にも限定されないが、間葉系幹細胞が、IL−10のような抑制性サイトカインの分泌を促進し、マクロファージ遊走阻止因子を阻害すると考えられる。
【0027】
別の実施態様において間葉系幹細胞は、角膜、水晶体、色素上皮細胞、及び網膜を含む眼、脳、脊髄、妊娠子宮と胎盤、卵巣、睾丸、副腎皮質、肝臓、及び毛包のような免疫学的特権部位の炎症を制御するために使用される。この実施態様の範囲はいかなる理論にも限定されないが、間葉系幹細胞が、IL−10のような抑制性サイトカインの分泌とTreg細胞の生成を促進すると考えられる。
【0028】
別の実施態様において間葉系幹細胞は、透析及び/又は糸球体腎炎の最終段階腎疾患(ESRD)に関連する組織傷害を治療するために使用される。本発明のこの実施態様の範囲はいかなる理論にも限定されないが、間葉系幹細胞が腎修復を促進すると考えられる。間葉系幹細胞はまた、血管内皮増殖因子(すなわちVEGF)を発現し分泌し、これが新しい血管形成を刺激し、傷害された腎組織の修復を助けると考えられる。
【0029】
さらなる実施態様において間葉系幹細胞は、インフルエンザ、C型肝炎、単純ヘルペスウイルス、ワクシニアウイルス感染症、エプスタインバーウイルスのようなウイルス感染症を抑制するために使用される。この実施態様の範囲はいかなる理論にも限定されないが、間葉系幹細胞がインターフェロン−ベータ(IFN−β)の分泌を促進すると考えられる。
【0030】
さらに別の実施態様において間葉系幹細胞は、リーシュマニア (Leishmania)感染症やヘリコバクター(Helicobacter)感染症のような寄生虫感染症を制御するために使用される。この実施態様の範囲はいかなる理論にも限定されないが、間葉系幹細胞がTヘルパー2(Th2)細胞による応答を仲介し、こうしてβ細胞による免疫グロブリンE(IgE)の産生増加を促進すると考えられる。
【0031】
しかし、本発明のこの態様は、特定の炎症応答の治療に限定されないことを理解されたい。
【0032】
間葉系幹細胞は、上記したようにヒト及び非ヒト霊長類を含む哺乳動物に投与される。
【0033】
間葉系幹細胞はまた、上記したように全身的に投与される。あるいは骨関節炎又は慢性関節リウマチの場合、間葉系幹細胞は関節炎関節に直接投与される。
【0034】
間葉系幹細胞は、動物の炎症応答を治療するために有効な量で投与される。間葉系幹細胞は、約1×105細胞/kg〜約1×107細胞/kgの量で投与される。別の実施態様において間葉系幹細胞は、約1×106細胞/kg〜約5×106細胞/kgの量で投与される。投与される間葉系幹細胞の正確な用量は、患者の年齢、体重、性、治療される炎症応答、及びその程度と重症度を含む種々の要因に依存する。
【0035】
上記したように、間葉系幹細胞は薬学的に許容される担体とともに投与される。
【0036】
本発明の別の態様において、動物の炎症を治療し、及び/又は上皮傷害を修復する方法が提供される。この方法は、動物の炎症及び/又は上皮傷害を治療するために有効な量の間葉系幹細胞を動物に投与することを含む。
【0037】
本発明のこの態様の範囲はいかなる理論にも限定されないが、間葉系幹細胞が、T細胞による炎症促進性サイトカインであるTNF−αとインターフェロン−γの分泌の低下と、T細胞による抗炎症性サイトカインであるインターロイキン−10(IL−10)とインターロイキン−4(IL−4)の分泌の上昇とを引き起こすと考えられている。また間葉系幹細胞は、ナチュラルキラー(NK)細胞によるインターフェロン−γ分泌の低下を引き起こすと考えられる。
【0038】
本発明のこの態様で治療される炎症及び/又は上皮傷害は、自己免疫疾患、移植臓器の拒絶、火傷、切傷、裂傷、皮膚潰瘍形成と糖尿病性潰瘍形成を含む潰瘍形成を含むがこれらに限定されない、種々の疾患と障害により引き起こされる炎症及び/又は上皮傷害がある。
【0039】
ある実施態様において間葉系幹細胞は、慢性関節リウマチ、クローン病、1型糖尿病、多発性硬化症、強皮症、グレーブス病、ループス、炎症性腸疾患、自己免疫胃炎(AIG)、及び自己免疫糸球体疾患を含むがこれらに限定されない、自己免疫疾患により生じる上皮傷害を修復するために、動物に投与される。間葉系幹細胞はまた、移植片対宿主病(GVHD)により生じる上皮傷害を修復する。
【0040】
本発明のこの態様は、移植片対宿主病により生じる上皮傷害の修復、及びさらに詳しくは、皮膚及び/又は消化管系に影響を与えるグレードIIIとIVの移植片対宿主病を含む重症の移植片対宿主病により生じる上皮傷害の修復に適用される。本出願人は特に、重症の移植片対宿主病、特にグレードIIIとIVの消化管移植片対宿主病に罹った患者に間葉系幹細胞が投与されると、間葉系幹細胞の投与により患者の皮膚及び/又は潰瘍形成した小腸上皮組織を修復させることを発見した。
【0041】
別の実施態様において間葉系幹細胞は、移植臓器又は組織の拒絶により引き起こされる、腎臓、心臓、及び肺を含むがこれらに限定されない移植臓器又は組織に対する上皮傷害を修復するために動物に投与される。
【0042】
さらに別の実施態様において間葉系幹細胞は、火傷、切傷、裂傷、皮膚潰瘍形成と糖尿病性潰瘍形成を含む潰瘍形成により引き起こされる上皮傷害を修復するために動物に投与される。
【0043】
間葉系幹細胞は、上記したようにヒト及び非ヒト霊長類を含む哺乳動物に投与される。
【0044】
間葉系幹細胞はまた、上記したように全身的に投与される。
【0045】
間葉系幹細胞は、動物の上皮傷害を修復するために有効な量で投与される。間葉系幹細胞は、約1×105細胞/kg〜約1×107細胞/kgの量で投与される。別の実施態様において間葉系幹細胞は、約1×106細胞/kg〜約5×106細胞/kgの量で投与される。投与される間葉系幹細胞の正確な用量は、患者の年齢、体重、性、修復される上皮傷害のタイプ、及びその程度と重症度を含む種々の要因に依存する。
【0046】
本発明の別の態様において、動物の癌を治療する方法が提供される。この方法は、動物の癌を治療するために有効な量の間葉系幹細胞を動物に投与することを含む。
【0047】
本発明のこの態様の範囲はいかなる理論にも限定されないが、間葉系幹細胞が樹状細胞と相互作用してIFN−βを分泌させ、これが次に腫瘍サプレッサーとして作用すると考えられる。治療される癌には、特に限定されないが、肝細胞癌、子宮頚癌、膵臓癌、前立腺癌、線維肉腫、髄芽細胞腫、及び星状細胞腫がある。しかし本発明の範囲は特定の種類の癌に限定されないことを理解されたい。
【0048】
動物は上記したようにヒト及び非ヒト霊長類を含む哺乳動物である。
【0049】
間葉系幹細胞は、動物の癌を治療するために有効な量で投与される。一般に間葉系幹細胞は、約1×105細胞/kg〜約1×107細胞/kgの量で投与される。別の実施態様において間葉系幹細胞は、約1×106細胞/kg〜約5×106細胞/kgの量で投与される。投与される間葉系幹細胞の正確な用量は、患者の年齢、体重、性、治療される癌のタイプ、及びその程度と重症度を含む種々の要因に依存する。
【0050】
上記したように、間葉系幹細胞は薬学的に許容される担体とともに投与され、全身的に投与される。あるいは間葉系幹細胞は、治療される癌に直接投与される。
【0051】
本発明の別の態様において、動物のアレルギー疾患又は障害を治療する方法が提供される。この方法は、動物のアレルギー疾患又は障害を治療するために有効な量の間葉系幹細胞を動物に投与することを含む。
【0052】
本発明のこの態様の範囲はいかなる理論にも限定されないが、間葉系幹細胞が急性アレルギー応答後に投与されると、肥満細胞活性化と脱顆粒の阻害を提供すると考えられる。また間葉系幹細胞は好塩基球活性化をダウンレギュレートし、サイトカイン、例えばTNF−α、ケモカイン、例えばインターロイキン−8、単球化学誘引性タンパク質(すなわちMCP−1)、脂質メディエーター、例えばロイコトリエンを阻害し、主要なメディエーター、例えばヒスタミン、ヘパリン、コンドロイチン硫酸、及びカテプシンを阻害すると考えられる。
【0053】
治療されるアレルギー疾患又は障害には、特に限定されないが、喘息、アレルギー性鼻炎、アトピー性皮膚炎、及び接触皮膚炎がある。しかし、本発明の範囲は、特定のアレルギー疾患又は障害に限定されないことを理解されたい。
【0054】
間葉系幹細胞は、動物のアレルギー疾患又は障害を治療するために有効な量で投与される。動物は哺乳動物でもよい。哺乳動物は、ヒト及び非ヒト霊長類を含む霊長類でもよい。一般に間葉系幹細胞は、約1×105細胞/kg〜約1×107細胞/kgの量で投与される。別の実施態様において間葉系幹細胞は、約1×106細胞/kg〜約5×106細胞/kgの量で投与される。投与される間葉系幹細胞の正確な用量は、患者の年齢、体重、性、治療されるアレルギー疾患又は障害、及びその程度と重症度を含む種々の要因に依存する。
【0055】
上記したように、間葉系幹細胞は薬学的に許容される担体とともに投与される。間葉系幹細胞は、例えば静脈内又は動脈内投与のように全身的に投与される。
【0056】
本発明のさらなる態様において、動物の創傷治癒を促進する方法が提供される。この方法は、動物の創傷治癒を促進するために有効な量の間葉系幹細胞を動物に投与することを含む。
【0057】
本発明の範囲はいかなる理論にも限定されないが、上記したように間葉系幹細胞は、Treg細胞と樹状細胞にインターロイキン−1−10(IL−10)を放出させる。IL−10は、創傷の炎症を制限又は抑制し、こうして創傷治癒を促進する。
【0058】
さらに間葉系幹細胞は、他のタイプの細胞により分泌因子を誘導することにより、創傷治癒と骨折治癒とを促進する。例えば間葉系幹細胞は、末梢血単核細胞(PBMC)による血管内皮増殖因子(VEGF)のプロスタグランジンE2(PGE2)介在放出、ならびに成長ホルモン、インスリン、インスリン様増殖因子1(IGF−1)インスリン様増殖因子結合タンパク質−3(IGFBP−3)、及びエンドテリン−1のPGE2介在放出を誘導する。
【0059】
治癒される創傷には、特に限定されないが、切傷、裂傷、火傷、及び皮膚潰瘍形成から生じるものがある。
【0060】
間葉系幹細胞は、動物の創傷治癒を促進するために有効な量で動物に投与される。動物は哺乳動物でもよく、哺乳動物はヒト及び非ヒト霊長類を含む霊長類でもよい。一般に間葉系幹細胞は、約1×105細胞/kg〜約1×107細胞/kgの量で投与される。別の実施態様において間葉系幹細胞は、約1×106細胞/kg〜約5×106細胞/kgの量で投与される。投与される間葉系幹細胞の正確な用量は、患者の年齢、体重、性、治療される創傷、及びその程度と重症度を含む種々の要因に依存する。
【0061】
上記したように、間葉系幹細胞は薬学的に許容される担体とともに投与される。間葉系幹細胞は、上記したように全身的に投与される。あるいは間葉系幹細胞は、間葉系幹細胞を含有する包帯又はリザーバー上の液体で、創傷に直接投与される。
【0062】
本発明のさらに別の態様において、動物の繊維症を治療または予防する方法が提供される。この方法は、動物の繊維症を治療または予防するために有効な量の間葉系幹細胞を動物に投与することを含む。
【0063】
間葉系幹細胞は、肝硬変、最終段階腎疾患に関連する腎臓の繊維症、および肺の繊維症を含むがこれらに限定されない動物の任意の種類の繊維症、成人型呼吸窮迫症候群(ARDS)および慢性閉塞性肺疾患(COPD)を含むがこれらに限定されない肺の繊維症を治療または予防するために動物に投与される。本発明の範囲は特定の種類の繊維症に限定されないことを理解されたい。
【0064】
間葉系幹細胞は、動物の繊維症を治療または予防するために有効な量で動物に投与される。動物は哺乳動物でもよく、哺乳動物はヒト及び非ヒト霊長類を含む霊長類でもよい。一般に間葉系幹細胞は、約1×105細胞/kg〜約1×107細胞/kgの量で投与される。別の実施態様において間葉系幹細胞は、約1×106細胞/kg〜約5×106細胞/kgの量で投与される。投与される間葉系幹細胞の正確な用量は、患者の年齢、体重、性、治療または予防される繊維症の程度と重症度を含む種々の要因に依存する。
【0065】
間葉系幹細胞は、上記したように薬学的に許容される担体とともに投与される。間葉系幹細胞は上記したように全身的に投与される。
【0066】
本発明の別の目的は、動物の組織又は臓器の血管形成であって、該組織又は臓器は血管形成が必要である、血管形成を促進することである。
【0067】
本発明のさらなる態様において、動物の臓器又は組織の血管形成を促進する方法が提供される。この方法は、動物の臓器又は組織の血管形成を促進するために有効な量の間葉系幹細胞を動物に投与することを含む。
【0068】
血管形成は、既存の微小血管床からの新しい血管の形成である。
【0069】
血管形成の誘導は、冠動脈及び末梢動脈不全症を治療するために使用され、冠動脈疾患、虚血性心疾患、及び末梢血動脈疾患の治療のための非侵襲的及び治療的アプローチでもよい。血管形成は心臓以外の組織や臓器の疾患と障害の治療、ならびに心臓以外の臓器の成長及び/又は維持において役割を果たす。血管形成は、内部及び外部創傷ならびに皮膚潰瘍の治療において役割を果たす。血管形成はまた、胚の着床、胎盤成長、ならびに胚血管の成長において役割を果たす。血管形成はまた、軟骨再吸収と骨形成との連結に必須であり、正しい成長板の形態形成に必須である。
【0070】
さらに血管形成は、充分な栄養物質と気体輸送を提供するために必要な高密度の血管ネットワークが必要な高代謝臓器、例えば肝臓の構築と維持の成功に必要である。
【0071】
間葉系幹細胞は、種々の方法により血管形成の必要な組織又は臓器に投与することができる。間葉系幹細胞は、例えば静脈内、動脈内、又は腹腔内投与により全身的に投与されるか、又は間葉系幹細胞は、例えば血管形成の必要な組織又は臓器への直接投与により、血管形成の必要な組織又は臓器に直接投与される。
【0072】
間葉系幹細胞は、自己、同種異系、又は異種を含む範囲の供給源由来でもよい。
【0073】
本発明の範囲はいかなる理論にも限定されないが、間葉系幹細胞は動物に投与されると、末梢血単核細胞(PBMC)を刺激して血管内皮増殖因子(すなわちVEGF)を産生させ、これが新しい血管の形成を刺激すると考えられる。
【0074】
ある実施態様において動物は哺乳動物である。哺乳動物は、ヒト及び非ヒト霊長類を含む霊長類でもよい。
【0075】
本発明の間葉系幹細胞は、血管形成を介して緩和、治療、又は予防できる疾患もしくは障害の治療、緩和、又は予防に使用される。例えば間葉系幹細胞は、四肢(すなわち腕、脚、手、及び足)、ならびに首又は種々の臓器中の閉塞動脈を治療するために動物に投与される。例えば間葉系幹細胞は、脳の需要を満たす閉塞動脈を治療、従って卒中を治療又は予防するために使用される。また間葉系幹細胞は、胚及び生後角膜の血管を治療するために使用され、かつ糸球体構造形成を提供するために使用される。別の実施態様において間葉系幹細胞は、内部及び外部の創傷の治療、ならびに糖尿病や鎌状赤血球貧血のような疾患により引き起こされる皮膚潰瘍を含むがこれらに限定されない足、手、脚、又は腕の皮膚潰瘍の治療に使用される。
【0076】
さらに血管形成は胚の着床と胎盤形成に関与するため、間葉系幹細胞は胚の着床を促進し流産を防ぐために使用される。
【0077】
さらに間葉系幹細胞は、胎内の動物の血管の成長を促進するために、ヒトを含む胎内の動物に投与される。
【0078】
別の実施態様において間葉系幹細胞は、軟骨再吸収と骨形成を促進し、正しい成長板の形態形成を促進するために、生後又は生前の動物に投与される。
【0079】
間葉系幹細胞は、動物の血管形成を促進するために有効な量で投与される。間葉系幹細胞は、約1×105細胞/kg〜約1×107細胞/kgの量で投与される。別の実施態様において間葉系幹細胞は、約1×106細胞/kg〜約5×106細胞/kgの量で投与される。投与される間葉系幹細胞の量は、患者の年齢、体重、性、治療、緩和、又は予防される疾患もしくは障害、及びその程度と重症度を含む種々の要因に依存する。
【0080】
間葉系幹細胞は薬学的に許容される担体とともに投与してもよい。例えば間葉系幹細胞は、注射用のための薬学的に許容される液体媒体中で細胞懸濁液として投与してもよい。注射は、局所的、すなわち血管形成の必要な組織又は臓器に直接、又は全身性でもよい。
【0081】
間葉系幹細胞は、治療薬をコードする1つ又はそれ以上のポリヌクレオチドを用いて遺伝子操作してもよい。ポリヌクレオチドは、適切な発現ビヒクルを介して間葉系幹細胞に提供される。間葉系幹細胞を遺伝子操作するために使用される発現ビヒクルには、特に限定されないが、レトロウイルスベクター、アデノウイルスベクター、及びアデノ関連ウイルスベクターがある。
【0082】
治療薬をコードする適切なポリヌクレオチドの選択は、治療される疾患もしくは障害、及びその程度と重症度を含む種々の要因に依存する。治療薬をコードするポリヌクレオチド及び適切な発現ビヒクルは、さらに米国特許第6,355,239号明細書に記載されている。
【0083】
間葉系幹細胞は上記治療で使用される時には、増殖因子、サイトカイン、抗炎症剤のような薬剤を含むがこれらに限定されない当業者に公知の他の治療薬、と間葉系幹細胞以外の細胞、例えば樹状細胞とを組合せて使用されるか、あるいは細胞のための可溶性担体、例えばヒアルロン酸、又は適宜固体マトリックス、例えば、コラーゲン、ゼラチン、又は他の生体適合性ポリマーと組合せて投与される。
【0084】
本明細書に記載の方法は、多くの方法でかつ当該分野で公知の種々の修飾及び置換を用いて行われる。細胞タイプ間の作用又は相互作用のモードについて提唱されている理論は、決して本発明を限定するものではなく、本発明の方法をよりよく理解できるように提供されていることを理解されたい。
【0085】
本発明を図面により説明する。
【0086】
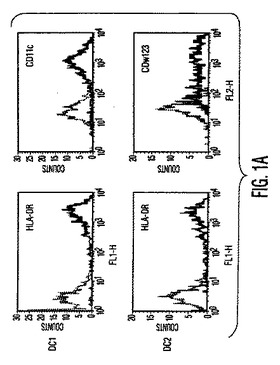
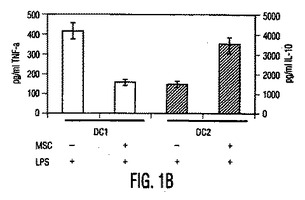
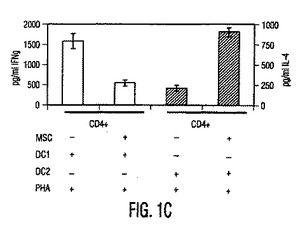
図1。MSCは樹状細胞機能を調節する。(A)HLA−DRとCD11cに対する抗体を使用する成熟単球DC1細胞のフローサイトメトリー解析と、HLA−DRとCD123(IL−3受容体)に対する抗体を使用する形質球様DC2細胞のフローサイトメトリー解析。(破線):アイソタイプ対照;(実線):FITC/PE結合抗体。(B)MSCは、活性化DC1とDC2からの、それぞれTNF−α分泌(第1y軸)を阻害し、IL−10分泌(第2y軸)を上昇させる。(C)成熟DC1細胞で培養したMSCは、MSCもしくはDC単独と比較して、T細胞によるIFN−γ分泌(第1y軸)を阻害しIL−4レベル(第2y軸)を上昇させる。MSCの存在下での炎症促進性IFN−γの産生低下と抗炎症性IL−4の産生増加は、T細胞集団の抗炎症性表現型への移動を示した。
【0087】
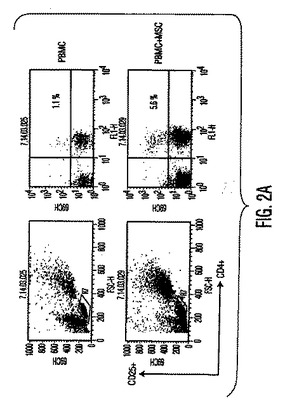
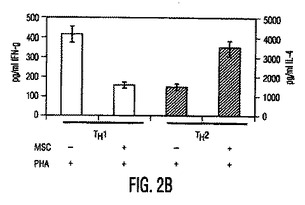
図2。MSCは炎症促進性エフェクターT細胞機能を阻害する。(A)FITC結合CD4(x軸)抗体とPE結合CD25(y軸)抗体を用いる、MSC+PBMC培養物(MSC+PBMC)中のPBMC又は非接着性画分を染色することによるTReg細胞数(%)のフローサイトメトリー解析。ゲートはバックグランドとしてのアイソタイプ対照抗体に基づく。グラフは5つの独立した実験の典型である。(B)細胞培養上清中で、MSCの存在下で生成したTH1細胞は低レベルのIFN−γ(第1y軸)を分泌し、MSCの存在下で生成したTH2細胞は増加量のIL−4(第2y軸)を分泌した。(C)MSCは、24ウェルプレート中で0、24、又は48時間培養した精製NK細胞からのIFN−γ分泌を阻害する。示したデータは1つの実験の平均±SDサイトカイン分泌であり、3つの独立した実験の典型である。
【0088】
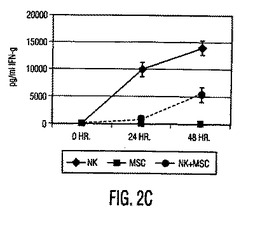
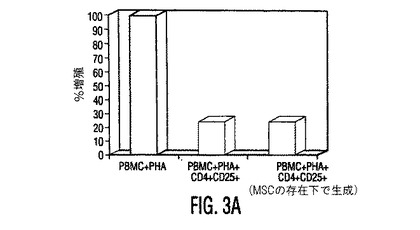
図3。MSCはTreg細胞集団の数とGITR発現を上昇させる。(A)CD4+CD25+Treg細胞集団を、PBMC又はMSC+PBMC(MSC対PBMC比は1:10)培養物(さらなる刺激無しで3日間培養した)から2工程磁性単離法を使用して単離した。これらの細胞に放射線照射し(さらなる増殖を阻止するため)、混合リンパ球反応(MLR)において刺激細胞として使用し、ここで応答細胞は植物性血球凝集素(PHA)(2.5mg/ml)の存在下の同種異系PBMC(刺激細胞対応答細胞比は1:100)であった。細胞を48時間培養し、次に3Hチミジンを加え、24時間後、取り込まれた放射活性を計測した。結果は、MSCの存在下で生成したTreg集団(レーン3)は、MSCの非存在下で生成したTreg細胞(レーン2)と機能的に同等であることを示した。(B)PBMCをMSCの非存在下(上のプロット)又は存在下(下のプロット)で3日間培養(MSC対PBMC比は1:10)し、次に非接着画分を採取し、FITC標識GITRとPE標識CD4で免疫染色した。結果は、MSCの存在下で培養した細胞でGITR発現の2倍以上の上昇を示す。
【0089】
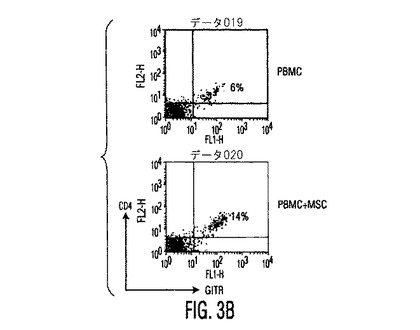
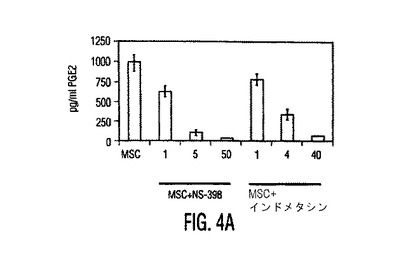
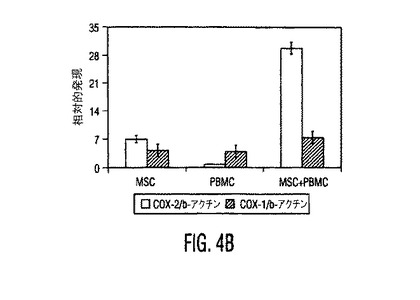
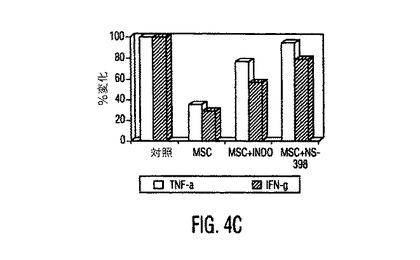
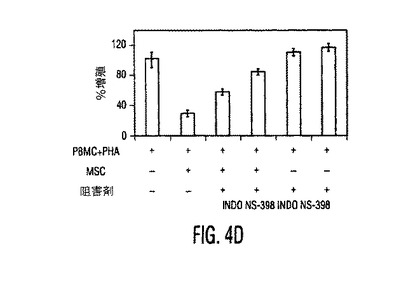
図4。MSCはPGE2を産生し、PGE2を阻止するとMSC性免疫調節作用が逆転する。(A)種々の濃度のPGE2ブロッカーであるNS−398又はインドメタシン(Indometh.)の存在下又は非存在下で培養したMSCから得られた培養上清中のPGE2分泌(平均±SD)。インヒビター濃度はμMであり、示したデータは24時間培養後に得られた値である。(B)リアルタイムRT−PCRを使用したMSCとPBMC中のCOX−1とCOX−2発現。MSCはPBMCと比較して有意に高レベルのCOX−2を発現し、MSCをPBMCの存在下で培養すると、MSC中のCOX−2発現の3倍を超える上昇があった。1〜3つの独立した実験からの典型的データを示す。MSC+PBMC培養物をトランスウェルチャンバープレート中で準備し、ここでMSCは下のチャンバーに蒔き、PBMCは上のチャンバーに蒔いた。(C)PGE2ブロッカーであるインドメタシン(Ind.)又はNS−398の存在は、対照と比較して、活性化DC(白バー)からのTNF−α分泌と、TH1細胞(斜線バー)からのIFN−γ分泌を上昇させる。データは、MSCとPGE2インヒビターの非存在下で生成した培養物からの%変化として計算した。(D)MSC−PBMC同時培養物(1:10)中のPGE2ブロッカーであるインドメタシン(Indo)又はNS−398の存在は、PHA処理PBMCに対するMSC性抗増殖作用を逆転させる。示したデータは1つの実験からであり、3つの独立した実験の典型である。
【0090】
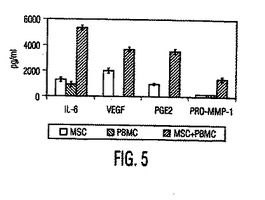
図5。構成性MSCサイトカイン分泌は同種異系PBMCの存在下で上昇する。すでに性状解析したヒトMSCを使用して、PBMCの存在下(斜線)又は非存在下(白い棒)(MSC対PBMC比は1:10)で24時間培養したMSCの培養上清中のサイトカインIL−6とVEGF、脂質メディエーターPGE2、及びマトリックス金属プロテアーゼ1(pro−MMP−1)のレベルを分析した。MSCはIL−6、VEGF、及びPGE2を構成性に産生し、これらの因子のレベルはPBMCとの同時培養で上昇し、従ってMSCが炎症性状況で免疫機能の調節において役割を果たすことを示唆する。
【0091】
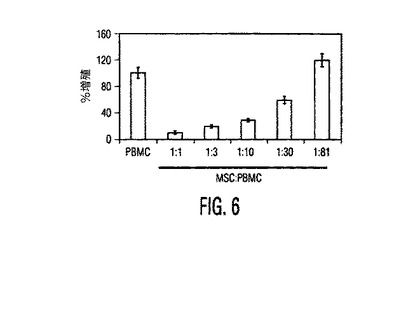
図6。MSCは分裂促進剤誘導性のT細胞増殖を用量依存的に阻害する。増加する数の同種異系PBMCを、PHA(2.5mg/ml)の存在下又は非存在下で96ウェルプレートに蒔いた一定数のMSC(2,000細胞/ウェル)とインキュベートし、3Hチミジン取り込みを測定した(1分当たりのカウント、cpm)。MSCの存在下でPHA処理PBMCの増殖の用量依存性阻害があった。3つの独立した実験の1つからの典型的結果を示す。同様の結果が、LeBlanc, et al., Scand J. Immunol., Vol. 57, pg. 11 (2003)により報告された。
【0092】
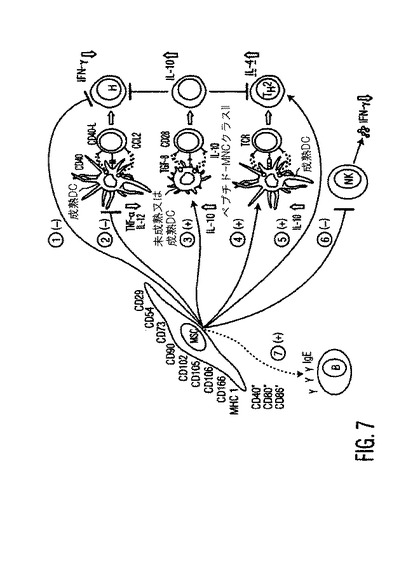
図7。提唱されているMSC作用機構の模式図
MSCは、先天的(DC経路2〜4;及びNK経路6)免疫系と適応(T経路1と5、及びB経路7)免疫系の両方からの細胞に影響を与えることにより免疫調節作用を仲介する。浸入病原体に応答して未成熟DCは、侵入可能部位に移動し、成熟し、未処理のT細胞を刺激して(抗原特異的及び同時刺激シグナルにより)、防御性エフェクターT細胞(細胞性TH1又は体液性TH2免疫)になる能力を獲得する。MSC−DC相互作用中に、MSCは直接の細胞−細胞接触により又は分泌された因子により、細胞性応答(経路2)を開始するDCの能力を制限することにより、又は体液性応答(経路4)を開始する能力を促進することにより、免疫応答の結果を変化させる。また成熟エフェクターT細胞が存在する時、MSCはこれらと相互作用してTH1(経路1)応答のバランスをTH2応答(経路5)に向けて、おそらくIgE産生B細胞活性(経路7)の上昇(これは、GvHDと自己免疫疾患症状の抑制に好ましい結果である)に向ける。MSCはTReg集団(経路3)の生成を増加させる能力により、寛容表現型となり、局所的微小環境におけるバイスタンダー(bystander)炎症を緩和することにより受容体宿主を助ける。点線(----)は提唱された機構である。
【0093】
本発明を以下の実施例で説明する;しかし本発明の範囲は決してこれらにより限定されるものではない。
【0094】
実施例1
【0095】
材料と方法
【0096】
ヒトMSCの培養
ヒトMSCを、Pittenger et al., Science, Vol. 284, pg. 143 (1999)に記載されたように培養した。すなわち、Poietics Technologies, Div of Cambrex Biosciencesによるインフォームドコンセントの後、匿名のドナーの腸骨稜から骨髄試料を採取した。1%抗生物質−抗真菌剤溶液(Invitrogen, Carlsbad, California)と10%牛胎児血清(FBS, JRH BioSciences, Lenexa, Kansas)を含有する完全ダルベッコー改変イーグル培地−低グルコース(Life Technologies, Carlsbad, California)中でMSCを培養した。MSCは接着性単層として増殖し、トリプシン/EDTAを用いて剥がした(0.05%トリプシンで37℃で3分)。使用したすべてのMSCは多分化性についてすでに性状解析され、間葉性系統(軟骨細胞性、脂肪細胞生成性、及び骨形成性)に分化する能力を保持した(Pittenger, et al., Science, Vol. 284, pg. 143 (1999))。
【0097】
樹状細胞の単離
末梢血単核細胞(PBMC)は、Poietics Technologies, Div of Cambrex Biosciences (Walkersville, MD)から得た。単球系統(CD1c+)の樹状細胞(DC)の前駆体を、Dzionek, et. al., J. Immunol., Vol. 165, pg. 6037 (2000)に従う2工程磁性分離法を使用して、PBMCから陽性選択した。すなわち、CD1c発現B細胞を磁性ビーズを使用してCD19+細胞を磁性的に枯渇させ、次にB細胞枯渇画分をビオチン標識CD1c(BDCA1+)と抗ビオチン抗体とで標識し、製造業者(Miltenyi Biotech, Auburn, California)の説明書に従って磁性カラムを使用して、非標識細胞画分からこれらを分離した。陽性標識した抗体コーティング細胞(BDCA2+)(Miltenyi Biotech, Auburn, California)の免疫−磁性分類により、PBMCから形質細胞様系統のDCの前駆体を単離した。
【0098】
MSC−DC培養
ほとんどの実験でヒトMSCとDCを同じ数で種々の時間培養し、細胞培養物上清を採取し、さらに評価するまで−80℃で保存した。選択された実験で、MSCを成熟DC1もしくはDC2細胞(1:1 MSC:DC比)と3日間培養し、次に一緒にした培養物(MSCとDC)に放射線照射してさらなる増殖を止めた。次に抗体で精製した未処理の同種異系T細胞(CD4+、CD45RA+)を放射線照射MSC/DCに加え、さらに6日間培養した。次に培養物から非接着性細胞画分(精製T細胞)を採取し、2回洗浄し、PHAでさらに24時間再刺激し、次に細胞培養上清を採取し、分離されたIFN−γとIL−4についてELISAにより分析した。
【0099】
NK細胞の単離
ビオチン結合モノクローナル抗体のカクテル(抗−CD3、−CD14、−CD19、−CD36、及び抗IgE抗体)を1次試薬とし、マイクロビーズに結合した抗ビオチンモノクローナル抗体を2次標識試薬として、磁性標識した非NK細胞を枯渇させることにより、NK細胞の精製集団が得られた。磁性標識した非NK細胞は、磁界中でMACS(Miltenyi Biotech, Auburn, California)カラム中に保持され、NK細胞は通過し、採取された。
【0100】
TReg細胞集団の単離
2工程単離法を使用してTReg細胞集団を単離した。まずビオチン標識抗体のカクテルと抗ビオチンマイクロビーズを用いて、非CD4+ T細胞を間接に磁性標識した。次に標識した細胞をMACSカラム(Miltenyi Biotech, Auburn, California)で分離して枯渇させた。次にCD4+CD25+細胞をCD25マイクロビーズで直接標識し、あらかじめ濃縮したCD4+ T細胞画分から陽性選択により単離した。磁性標識したCD4+CD25+細胞はカラムに保持され、磁界からカラムを取り出した後溶出した。
【0101】
MSCの存在下で生成されたCD4+CD25+集団の増加が自然界で抑制性であるかどうかを調べるために、CD4+CD25+ Treg細胞集団を、2工程磁性単離法を使用してPBMC又はMSC+PBMC(MSC対PBMC比は1:10)培養物(さらなる刺激無しで3日間培養した)から単離した。これらの細胞に放射線照射してさらなる増殖を阻止し、混合リンパ球反応(MLR)で刺激細胞として使用し、ここで応答細胞はPHA(2.5μg/ml)の存在下で同種異系PBMC(刺激細胞対応答細胞の比は1:100)であった。48時間培養を行い。次に3Hチミジンを加えた。24時間後に取り込まれた放射活性を計測した。
【0102】
MSCの非存在下又は存在下でPBMCを培養(MSC対PBMC比は1:10)し、次に非接着性画分を採取し、FITC標識グルココルチコイド誘導性TNF受容体(すなわちGITR)、及びPE標識CD4で免疫染色した。
【0103】
TH1/TH2細胞の作製
2×106細胞/mlで37℃で45分、末梢血単核細胞(PBMC)をプレートに蒔き、単球を除去した。非接着性画分をプレート結合抗CD3(5μg/ml)と抗CD28(1μg/ml)抗体の存在下で、TH1(IL−2(4ng/ml)+IL−12(5ng/ml)+抗IL−4(1μg/ml))条件又はTH2(IL−2(4ng/ml)+IL−4(4ng/ml)+抗IFN−γ(1μg/ml))条件で3日間、MSCの存在下又は非存在下でインキュベートした。細胞を洗浄し、次にPHA(2.5μg/ml)でさらに24もしくは48時間再刺激し、次に培養上清中のIFN−γとIL−4のレベルをELISA(R&D Systems, Minneapolis, Minnesota)により測定した。
【0104】
MSCの培養上清中のVEGF、PGE2、及びpro−MMP−1のレベルの分析
すでに性状解析されたヒトMSCを使用して、インターロイキン−6(IL−6)、VEGF、脂質メディエータープロスタグランジンE2(PGE2)、及びマトリックス金属プロテアーゼ1(pro−MMP−1)のレベルを、PBMCの存在下又は非存在下(MSC対PBMC比は1:10)で24時間培養したMSCの培養上清で分析した。
【0105】
PBMCの増殖
フィコール−ハイパーク(Lymphoprep, Oslo, Norway)上でleukopack(Cambrex, Walkersville, Maryland)を遠心分離して、精製PBMCを調製した。分離した細胞をMSC(PBMC添加の3〜4時間前にプレートに蒔いて沈降させた)の存在下又は非存在下で、分裂促進剤PHA(Sigma Chemicals, St. Louis, Missouri)の存在下で48時間培養した(三重測定)。選択された実験で、PBMCをPGE2インヒビターであるインドメタシン(Sigma Chemicals, St. Louis, Missouri)又はNS−938(Cayman Chemicals, Ann Arbor, Michigan)を含有する培地に再懸濁した。(3H)チミジンを加え(200μlの培養物に20μl)、さらに24時間培養後自動ハーベスターを使用して細胞を採取した。MSC又はPGE2ブロッカーの作用を、PHAの存在下での対照応答(100%)のパーセントとして計算した。
【0106】
定量的RT−PCR
市販のキット(Qiagen, Valencia, California)を使用して製造業者の説明書に従って、細胞ペレットから総RNAを調製した。混入しているゲノムDNAをDNAフリーキット(Ambion, Austin, Texas)を使用して除去した。QuantiTect SYBR Green RT-PCR キット(Qiagen, Valencia, California)を使用して、0.5μM濃度のプライマーを用いて、MJ Research Opticon検出系(South San Francisco, California)で定量的RT−PCRを行った。異なる条件で培養した細胞中の発現レベルの相対的変化を、β−アクチンを内部対照として使用してCt値(交差点)の差により計算した。COX−1及びCOX−2特異的プライマーの配列は以下の通りであった:COX−1:5'-CCG GAT GCC AGT CAG GAT GAT G-3' (フォワード)、 5'-CTA GAC AGC CAG ATG CTG ACA G-3' (リバース);COX-2:5'-ATC TAC CCT CCT CAA GTC CC-3'(フォワード)、 5'-TAC CAG AAG GGC AGG ATA CAG-3' (リバース)。
【0107】
増加する数の同種異系PBMCを、96ウェルプレート上に蒔いた一定数のMSC(2,000細胞/ウェル)と、PHA(2.5μg/ml)の存在下で72時間インキュベートし、3Hチミジンの取り込み(1分当たりのカウント、cpm)を測定した。PBMCとMSCは、MSC:PBMCが1:1、1:3、1:10、1:30、及び1:81の比で培養した。
【0108】
結果
本試験では、ヒトMSCと、樹状細胞(DC1とDC2)、エフェクターT細胞(TH1とTH2)及びNK細胞を含む単離した免疫細胞集団との相互作用を調べた。MSCと各免疫細胞タイプとの相互作用は特定の結果を有し、MSCが免疫応答プロセスのいくつかの工程を調節することを示唆した。MSC免疫調節作用を調節し、これの原因かも知れない分泌因子の産生を評価し、プロスタグランジン合成が示唆された。
【0109】
それぞれBDCA1+とBDCA2+細胞の免疫磁性分類により、骨髄性(DC1)と形質細胞性(DC2)前駆樹状細胞を単離し、DC1細胞についてはGM−CSFとIL−4(それぞれ1×103IU/mlと1×103IU/ml)と、DC2細胞についてはIL−3(10ng/ml)とインキュベートして成熟させた。フローサイトメトリーを使用すると、DC1細胞はHLA−DR+であり、一方DC2細胞はHLA−DR+とCD123+であった(図1A)。炎症性物質である細菌リポ多糖(LPS、1ng/ml)の存在下で、DC1細胞は中レベルのTNF−αを産生したが、MSCが存在する時(調べた比率1:1と1:10)は、TNF−α分泌の>50%低下があった(図1B)。一方DC2細胞はLPSの存在下でIL−10を産生し、そのレベルは、MSC:DC2同時培養(1:1)で2倍以上上昇した(図1B)。従ってMSCは培養物中の活性化DCのサイトカインプロフィールをより寛容原性表現型に修飾した。さらに活性化DCはMSCと培養するとIFN−γを低下させ、かつ未処理のCD4+T細胞により分泌されるIL−4レベルを上昇(図1C)させることができ、炎症促進性から抗炎症性T細胞表現型へのMSC性シフトを示唆した。
【0110】
上昇したIL−10分泌は制御細胞の生成において役割を果たす(Kingsley, et al., J. Immunol., Vol. 168, pg. 1080 (2002))ため、T制御細胞(TReg)は、PBMCとMSCの同時培養でフローサイトメトリーにより定量した。PBMCとMSCを3〜5日間培養すると、抗CD4と抗CD25抗体を用いてPBMCを染色して測定したTReg細胞数が増加し(図2A)、これはMSC誘導性の寛容原性応答をさらに支持した。MSCの存在下で生成するCD4+CD25+ TReg細胞集団は、グルココルチコイド誘導性のTNF受容体(GITR)(TReg細胞集団上で発現される細胞表面受容体)のレベル上昇を示し、自然界では同種異系T細胞増殖を抑制したため抑制性であった(図3A、3 B)。次にT細胞分化に影響を与えるその直接能力についてMSCを調べた。抗体で選択した精製T細胞(CD4+ Th細胞)を使用して、IFN−γ産生TH1細胞とIL−4産生TH2細胞を、MSCの存在下又は非存在下で作製した。分化中にMSCが存在する時に、TH1細胞によるIFN−γ分泌が低下し、TH2細胞によるIL−4分泌が上昇した(図2B)。T細胞がエフェクターTH1又はTH2型(データは示していない)に分化(3日目)した後にMSCを培養物に加えると、IFN−γとIL−4レベルに有意な変化はなかった。これらの実験は、MSCがエフェクターT細胞分化に直接影響を与え、T細胞サイトカイン分泌を体液性表現型に変化させることを示唆する。
【0111】
同様に、MSCを精製NK細胞(CD3−、CD14−、CD19−、CD36−)と1:1の比率で異なる時間(0〜48時間)培養すると、培養上清中でIFN−γ分泌が低下(図2C)し、従ってMSCがNK細胞機能も調節できることを示唆する。
【0112】
以前の研究は、MSCが可溶性因子によりT細胞機能を修飾することを示している(LeBlanc, et al., Exp. Hematol., Vol. 31 , pg. 890 (2003); Tse, et al., Transplantation, Vol. 75, pg. 389 (2003))。MSCは、IL−6、プロスタグランジンE2、VEGF、及びpro−MMP−1を含むいくつかの因子を構成的に分泌し、それぞれのレベルがPBMCとの培養により上昇することが観察された(図5)。DCによるTNF−αの阻害とIL−10産生の上昇を引き起こすMSC由来因子を調べるために、プロスタグランジンE2の役割の可能性を調べた(これは活性化DCによるTNF−α産生を阻害することが証明されているため)(Vassiliou, et al., Cell. Immunol., Vol. 223, pg. 120 (2003))。MSC培養物(0.5×106細胞/mlの24時間培養物)からの調整培地は約1000pg/mlのPGE2を含有した(図4A)。培養上清中にPGE2分泌の公知のインデューサー、例えば、TNF−α、IFN−γ、又はIL−1βの検出可能な存在は無く(データは示していない)、MSCによるPGE2の構成性分泌を示している。hMSCによるPGE2分泌は、PGE2産生の公知のインヒビター[NS−398(5μM)及びインドメタシン(4μM)]の存在下で60〜90%阻害された(図4A)。構成的活性なシクロオキシゲナーゼ酵素1(COX−1)と誘導性シクロオキシゲナーゼ酵素2(COX−2)の酵素活性の結果としてPGE2分泌の放出が起きるため(Harris, et al., Trends Immunol., Vol. 23, pg. 144 (2002))、トランスウェル培養系を使用してMSCとPBMC中のCOX−1とCOX−2のmRNA発現を分析した。MSCは、PBMCと比較して有意に高レベルのCOX−2を発現し、その発現レベル上昇は、MSCとPBMC(MSCとPBMCの比は1:10)と24時間同時培養すると>3倍であった(図4B)。COX−1レベルにわずかな変化が見られ、MSC−PBMC同時培養でのPGE2分泌の上昇(図5)はCOX−2のアップレギュレーションにより仲介されることを示唆している。DCとT細胞に対するMSCの免疫調節作用がPGE2により仲介されるかどうかを調べるために、PGE2インヒビターであるNS−398又はインドメタシンの存在下でMSCを活性化樹状細胞(DC1)又はTH1細胞と培養した。NS−398又はインドメタシンの存在は、それぞれDC1によるTNF−α分泌と、TH1細胞によるIFN−γ分泌を上昇させ(図4C)、免疫細胞タイプへのMSCの作用は分泌されたPGE2により仲介されることを示唆した。最近の研究は、種々の刺激により誘導されるT細胞増殖をMSCが阻害することを証明している(DeNicola, et al., Blood, Vol. 99, pg. 3838 (2002); LeBlanc, et al., Scand. J. Immunol., Vol. 57, pg. 11 (2003))。MSCは分裂促進剤誘導性のT細胞増殖を用量依存性に阻害(図6)し、PGE2インヒビターであるNS−398(5μM)又はインドメタシン(4μM)が存在すると、インヒビターの無い対照と比較して、培養物を含有するMSC中のPHA処理PBMCにより(3H)チミジン取り込みが>70%上昇することが観察された(図4D)。
【0113】
要約すると、MSCと他の免疫細胞種との相互作用のモデル(図7)が提唱される。成熟T細胞が存在すると、MSCが直接これらと相互作用し、炎症促進IFN−γ産生(経路1)を阻害し、制御性T細胞表現型(経路3)を増加させ、抗炎症性TH2細胞(経路5)を増加する。さらにMSCは、PGE2を分泌することにより、炎症促進DC1細胞を阻害(経路2)することにより、及び抗炎症性DC2細胞(経路4)又は制御性DC(経路3)を増加させることにより、DCを介するT細胞免疫応答の結果を変化させることができる。TH2免疫へのシフトは、IgE/IgG1サブタイプ抗体の生成上昇(経路7)へのB細胞活性の変化を示唆する。MSCはNK細胞からのIFN−γ分泌を阻害する能力により、NK細胞機能を修飾するようである(経路6)。このMSC:免疫細胞相互作用のモデルは、いくつかの他の実験室で行われた実験と一致する(LeBlanc, et al., Exp. Hematol., Vol. 31 , pg. 890 (2003); Tse, et al., Transplantation, Vol. 75, pg. 389 (2003); DiNicola, et al., Blood, Vol. 99, pg. 3838 (2002))。提唱された機構のさらなる研究が進行中であり、MSC投与のインビボ作用を調べるために動物試験が必要である。
【0114】
実施例2
【0115】
重症の段階IVの消化管移植片対宿主病(GVHD)に罹っている33才の女性患者に間葉系幹細胞を投与した。この患者は他のすべてのGVHD治療に抵抗性であった。患者の結腸の内視鏡観察は、治療前に潰瘍形成と炎症の部位を示した。患者の結腸の組織検査は、治療前にGVHDが患者の腸陰窩の大部分を破壊していることを証明した。
【0116】
患者は、体重1kg当たり3×106細胞の量でPlasma Lyte Aの50ml中で同種異系間葉系幹細胞の静脈内注入を受けた。
【0117】
注入の2週間後患者を評価した。注入後2週間で、患者の結腸の内視鏡観察は、治療前に見えていた炎症部位と潰瘍形成が消失したことを示した。さらに患者の結腸の生検は、腸陰窩の有意な再生を示した。すなわち患者への間葉系幹細胞の投与により、消化管移植片対宿主病の炎症性成分の有意な低下と、新しい機能的小腸組織の再生が起きた。
【0118】
すべての特許、刊行物(公開された特許出願、寄託受け入れ番号、及びデータベース受け入れ番号を含む)は、すべての特許、刊行物(公開された特許出願、寄託受け入れ番号、及びデータベース受け入れ番号を含む)が具体的かつ個々に取り込まれているのと同じ程度に、参照することにより本明細書に組み込まれる。
【0119】
しかし本発明の範囲は本明細書に記載の具体例に限定されるものではないことを理解されたい。本発明は、本明細書の具体的な記載とは別に実施することができ、それでも添付の特許請求の範囲内にある。
【図面の簡単な説明】
【0120】
【図1A】図1Aは、MSCは樹状細胞機能を調節することを示す。HLA−DRとCD11cに対する抗体を使用する成熟単球DC1細胞のフローサイトメトリー解析と、HLA−DRとCD123(IL−3受容体)に対する抗体を使用する形質球様DC2細胞のフローサイトメトリー解析。(破線):アイソタイプ対照;(実線):FITC/PE結合抗体。
【図1B】図1Bは、MSCが、活性化DC1とDC2からの、それぞれTNF−α分泌(第1y軸)を阻害し、IL−10分泌(第2y軸)を上昇させる、ことを示す。
【図1C】図1Cは、成熟DC1細胞で培養したMSCが、MSCもしくはDC単独と比較して、T細胞によるIFN−γ分泌(第1y軸)を阻害しIL−4レベル(第2y軸)を上昇させる、ことを示す。MSCの存在下での炎症促進性IFN−γの産生低下と抗炎症性IL−4の産生増加は、T細胞集団の抗炎症性表現型への移動を示した。
【図2A】図2Aは、MSCは炎症促進性エフェクターT細胞機能を阻害することを示す。FITC結合CD4(x軸)抗体とPE結合CD25(y軸)抗体を用いる、MSC+PBMC培養物(MSC+PBMC)中のPBMC又は非接着性画分を染色することによるTReg細胞数(%)のフローサイトメトリー解析。ゲートはバックグランドとしてのアイソタイプ対照抗体に基づく。グラフは5つの独立した実験の典型である。
【図2B】図2Bは、細胞培養上清中で、MSCの存在下で生成したTH1細胞は低レベルのIFN−γ(第1y軸)を分泌し、MSCの存在下で生成したTH2細胞は増加量のIL−4(第2y軸)を分泌した、ことを示す。
【図2C】図2Cは、MSCが、24ウェルプレート中で0、24、又は48時間培養した精製NK細胞からのIFN−γ分泌を阻害する、ことを示す。示したデータは1つの実験の平均±SDサイトカイン分泌であり、3つの独立した実験の典型である。
【図3A】図3Aは、MSCはTreg細胞集団の数とGITR発現を上昇させることを示す。CD4+CD25+Treg細胞集団を、PBMC又はMSC+PBMC(MSC対PBMC比は1:10)培養物(さらなる刺激無しで3日間培養した)から2工程磁性単離法を使用して単離した。これらの細胞に放射線照射し(さらなる増殖を阻止するため)、混合リンパ球反応(MLR)において刺激細胞として使用し、ここで応答細胞は植物性血球凝集素(PHA)(2.5mg/ml)の存在下の同種異系PBMC(刺激細胞対応答細胞比は1:100)であった。細胞を48時間培養し、次に3Hチミジンを加え、24時間後、取り込まれた放射活性を計測した。結果は、MSCの存在下で生成したTreg集団(レーン3)は、MSCの非存在下で生成したTreg細胞(レーン2)と機能的に同等であることを示した。
【図3B】図3Bは、PBMCをMSCの非存在下(上のプロット)又は存在下(下のプロット)で3日間培養(MSC対PBMC比は1:10)し、次に非接着画分を採取し、FITC標識GITRとPE標識CD4で免疫染色した結果を示す。結果は、MSCの存在下で培養した細胞でGITR発現の2倍以上の上昇を示す。
【図4A】図4Aは、MSCはPGE2を産生し、PGE2を阻止するとMSC性免疫調節作用が逆転することを示す。種々の濃度のPGE2ブロッカーであるNS−398又はインドメタシン(Indometh.)の存在下又は非存在下で培養したMSCから得られた培養上清中のPGE2分泌(平均±SD)。インヒビター濃度はμMであり、示したデータは24時間培養後に得られた値である。
【図4B】図4Bは、リアルタイムRT−PCRを使用したMSCとPBMC中のCOX−1とCOX−2発現を示す。MSCはPBMCと比較して有意に高レベルのCOX−2を発現し、MSCをPBMCの存在下で培養すると、MSC中のCOX−2発現の3倍を超える上昇があった。1〜3つの独立した実験からの典型的データを示す。MSC+PBMC培養物をトランスウェルチャンバープレート中で準備し、ここでMSCは下のチャンバーに蒔き、PBMCは上のチャンバーに蒔いた。
【図4C】図4Cは、(C)PGE2ブロッカーであるインドメタシン(Ind.)又はNS−398の存在は、対照と比較して、活性化DC(白バー)からのTNF−α分泌と、TH1細胞(斜線バー)からのIFN−γ分泌を上昇させることを示す。データは、MSCとPGE2インヒビターの非存在下で生成した培養物からの%変化として計算した。
【図4D】図4Dは、MSC−PBMC同時培養物(1:10)中のPGE2ブロッカーであるインドメタシン(Indo)又はNS−398の存在は、PHA処理PBMCに対するMSC性抗増殖作用を逆転させることを示す。示したデータは1つの実験からであり、3つの独立した実験の典型である。
【図5】図5は、構成性MSCサイトカイン分泌は同種異系PBMCの存在下で上昇することを示す。すでに性状解析したヒトMSCを使用して、PBMCの存在下(斜線バー)又は非存在下(白バー)(MSC対PBMC比は1:10)で24時間培養したMSCの培養上清中のサイトカインIL−6とVEGF、脂質メディエーターPGE2、及びマトリックス金属プロテアーゼ1(pro−MMP−1)のレベルを分析した。MSCはIL−6、VEGF、及びPGE2を構成性に産生し、これらの因子のレベルはPBMCとの同時培養で上昇し、従ってMSCが炎症性状況で免疫機能の調節において役割を果たすことを示唆する。
【図6】図6は、MSCは分裂促進剤誘導性のT細胞増殖を用量依存的に阻害することを示す。増加する数の同種異系PBMCを、PHA(2.5mg/ml)の存在下又は非存在下で96ウェルプレートに蒔いた一定数のMSC(2,000細胞/ウェル)とインキュベートし、3Hチミジン取り込みを測定した(1分当たりのカウント、cpm)。MSCの存在下でPHA処理PBMCの増殖の用量依存性阻害があった。3つの独立した実験の1つからの典型的結果を示す。同様の結果が、LeBlanc, et al., Scand J. Immunol., Vol. 57, pg. 11 (2003)により報告された。
【図7】図7は、提唱されているMSC作用機構の模式図を示す。MSCは、先天的(DC経路2〜4;及びNK経路6)免疫系と適応(T経路1と5、及びB経路7)免疫系の両方からの細胞に影響を与えることにより免疫調節作用を仲介する。浸入病原体に応答して未成熟DCは、侵入可能部位に移動し、成熟し、未処理のT細胞を刺激して(抗原特異的及び同時刺激シグナルにより)、防御性エフェクターT細胞(細胞性TH1又は体液性TH2免疫)になる能力を獲得する。MSC−DC相互作用中に、MSCは直接の細胞−細胞接触により又は分泌された因子により、細胞性応答(経路2)を開始するDCの能力を制限することにより、又は体液性応答(経路4)を開始する能力を促進することにより、免疫応答の結果を変化させる。また成熟エフェクターT細胞が存在する時、MSCはこれらと相互作用してTH1(経路1)応答のバランスをTH2応答(経路5)に向けて、おそらくIgE産生B細胞活性(経路7)の上昇(これは、GvHDと自己免疫疾患症状の抑制に好ましい結果である)に向ける。MSCはTReg集団(経路3)の生成を増加させる能力により、寛容表現型となり、局所的微小環境におけるバイスタンダー(bystander)炎症を緩和することにより受容体宿主を助ける。点線は提唱された機構である。
【技術分野】
【0001】
本発明は間葉系幹細胞に関する。さらに詳しくは、本発明は、種々の組織や臓器における血管形成の促進、自己免疫疾患の治療、アレルギー応答の治療、癌の治療、炎症性疾患と障害の治療、創傷治癒の促進、炎症の治療、及び上皮傷害の修復を含む間葉系幹細胞の新規用途に関する。
【背景技術】
【0002】
本発明は、海軍省により契約第N66001−02−C−8068号で政府の援助で行われた。政府は本発明において一定の権利を有する。
【0003】
本出願は、2004年3月22日に出願された仮特許出願第60/555,118号(その内容は参照することによりその全体が本明細書に組み込まれる)に基づく優先権を請求する2005年3月15日出願の特許出願第11/080,298号の一部継続出願である。
【0004】
間葉系幹細胞(MSC)は、骨芽細胞、筋細胞、軟骨細胞、及び脂肪細胞を含む系統に容易に分化できる多能性幹細胞である(Pittenger, et al., Science, Vol. 284, pg. 143 (1999); Haynesworth, et al., Bone, Vol. 13, pg. 69 (1992); Prockop, Science, Vol. 276, pg. 71 (1997))。インビトロの試験により、MSCが筋肉(Wakitani, et al., Muscle Nerve, Vol. 18,, pg. 1417 (1995))、神経様前駆体(Woodbury, et al., J. Neurosci. Res., Vol. 69, pg. 908 (2002); Sanchez-Ramos, et al., Exp. Neurol., Vol. 171 , pg. 109 (2001))、心筋細胞(Toma, et al., Circulation, Vol. 105, pg. 93 (2002); Fakuda, Artif. Organs, Vol. 25, pg. 187 (2001))、及びおそらくは他のタイプの細胞に分化する能力が証明されている。さらにMSCは、造血幹細胞や胚幹細胞の拡張のための有効なフィーダー層となることが証明されている(Eaves, et al., Ann. N.Y. Acad. Sci., Vol. 938, pg. 63 (2001); Wagers, et al., Gene Therapy, Vol. 9, pg. 606 (2002))。種々の動物モデルを用いた最近の研究は、MSCが、傷害された骨、軟骨、半月板、又は心筋組織の修復又は再生において有用であることを証明している(DeKok, et al., Clin. Oral Implants Res., Vol. 14, pg. 481 (2003)); Wu, et al., Transplantation, Vol. 75, pg. 679 (2003); Noel, et al., Curr. Opin. Investig. Drugs, Vol. 3, pg. 1000 (2002); Ballas, et al., J. Cell. Biochem. Suppl., Vol. 38, pg. 20 (2002); Mackenzie, et al., Blood Cells Mol. Dis., Vol. 27 (2002))。何人かの研究者は、骨形成不全症(Pereira, et al., Proc. Nat. Acad. Sci., Vol. 95, pg. 1142 (1998))、パーキンソン病(Schwartz, et al., Hum. Gene Ther., Vol. 10, pg. 2539 (1999))、脊髄損傷(Chopp, et al., Neuroreport, Vol. 11 , pg. 3001 (2000); Wu, et al., J. Neurosci. Res., Vol. 72, pg. 393 (2003))、及び心臓疾患(Tomita, et al., Circulation, Vol. 100, pg. 247 (1999). Shake, et al., Ann. Thorac. Surg., Vol. 73, pg. 1919 (2002))を含む動物疾患モデルにおける移植について、MSCを使用して有望な結果を得ている。重要なことに、有望な結果はまた、骨形成不全症(Horwitz, et al., Blood, Vol. 97, pg. 1227 (2001); Horowitz, et al. Proc. Nat. Acad. Sci., Vol. 99, pg. 8932 (2002))と異種骨髄移植(Frassoni, et al., Int. Society for Cell Therapy, SA006 (要約) (2002); Koc, et al., J. Clin. Oncol., Vol. 18, pg. 307 (2000))の移植の向上の臨床治験でも報告されている。
【0005】
MSCは、その表面に主要組織適合遺伝子複合体(MHC)クラスI抗原を発現するが、MHCクラスII(Le Blanc, et al., Exp. Hematol., Vol. 31 , pg. 890 (2003); Potian, et al., J. Immunol., Vol. 171 , pg. 3426 (2003))やB7もCD40同時刺激分子(Majumdar, et al., J. Biomed. Sci., Vol. 10, pg. 228 (2003))も発現せず、これらの細胞が免疫原性表現型(Tse, et al., Transplantation, Vol. 75, pg. 389 (2003))を有することを示唆している。MSCはまたMHC非依存性にT細胞増殖応答を阻害する(Bartholomew, et al., Exp. Hematol., Vol. 30, pg. 42 (2002); Devine, et al., Cancer J., Vol. 7, pg. 576 (2001); DiNicola, et al., Blood, Vol. 99, pg. 3838 (2002))。MSCのこれらの免疫学的性質は移植を向上させ、移植後の同種異系細胞を認識し拒絶する受容者の免疫系の能力を制限しているかも知れない。局所的刺激下で適切なタイプの細胞に分化する能力とともに、免疫応答を調節し造血を支持する因子のMSCによる産生は、これらを細胞移植研究のための好ましい幹細胞にしている(Majumdar, et al., Hematother. Stem Cell Res., Vol. 9, pg. 841 (2000); Haynesworth, et al., J. Cell. Physiol., Vol. 166, pg. 585 (1996))。
【発明の開示】
【0006】
本出願人は、樹状細胞(DC1とDC2)、エフェクターT細胞(Th1とTh2)、及びNK細胞を含む、間葉系幹細胞と単離された免疫細胞集団との相互作用を調べた。かかる相互作用に基づいて本出願人は、間葉系幹細胞が、免疫応答プロセスのいくつかの工程を制御する種々の因子の産生を制御することを発見した。すなわち間葉系幹細胞は、免疫系が関与する症状や障害、炎症が関与する疾患、症状、及び障害、上皮傷害、又はアレルギー応答の治療に使用できるかも知れない。かかる疾患、症状、及び障害には、特に限定されないが、自己免疫疾患、アレルギー、関節炎、炎症を起こした創傷、円形脱毛症(禿)、歯周病(歯肉炎と歯周炎を含む)、及び免疫応答が関与する他の疾患、症状、又は障害がある。
【0007】
さらに間葉系幹細胞は、血管内皮増殖因子、又はVEGF(これは新しい血管の形成を刺激することにより血管形成を促進する)を発現し分泌する。間葉系幹細胞はまた末梢血単核細胞(PBMC)を刺激してVEGFを産生させる。
【0008】
さらに間葉系幹細胞は樹状細胞がインターフェロン−ベータ(IFN−β)を産生するのを刺激して、これが腫瘍抑制とウイルス感染に対する免疫を促進する。
【0009】
本発明のある態様において、動物の自己免疫疾患と移植片対宿主病よりなる群から選択される疾患を治療する方法が提供される。この方法は、動物の疾患を治療するために有効な量の間葉系幹細胞を動物に投与することを含む。
【0010】
本発明のこの態様の範囲はいかなる理論にも限定されないが、間葉系幹細胞が自己免疫疾患と移植片対宿主病を抑制する少なくとも1つの機構は、制御T細胞(Treg細胞)及び/又は樹状細胞(DC)からのインターロイキン−10(IL−10)の放出を引き起こすことによると考えられる。
【0011】
本発明で治療される自己免疫疾患には、特に限定されないが、多発性硬化症、1型糖尿病、慢性関節リウマチ、ブドウ膜炎、自己免疫甲状腺疾患、炎症性腸疾患、強皮症、グレーブス病、ループス、クローン病、自己免疫リンパ増殖性疾患(ALPS)、脱髄疾患、自己免疫脳脊髄炎、自己免疫胃炎(AIG)、及び自己免疫糸球体疾患がある。また上記したように、移植片対宿主病も治療される。しかし本発明の範囲は、本明細書に記載の特定の疾患の治療に限定されないことを理解されたい。
【0012】
ある実施態様において、間葉系幹細胞が投与される動物は哺乳動物である。哺乳動物は、ヒト及び非ヒト霊長類を含む霊長類である。
【0013】
一般に間葉系幹細胞(MSC)療法は、以下の順序に基づく:MSC含有組織の採取、MSCの単離と拡張、動物へのMSCの投与(生化学的又は遺伝的操作有り又は無し)。
【0014】
投与される間葉系幹細胞は均一組成物であるか、又はMSCで濃縮された混合細胞集団でもよい。均一間葉系幹細胞組成物は接着性骨髄細胞又は骨膜細胞を培養することにより得られ、間葉系幹細胞組成物は接着性骨髄細胞又は骨膜細胞を培養することにより得られ、間葉系幹細胞は、ユニークなモノクローナル抗体を用いて同定される特異的細胞表面マーカーにより同定される。間葉系幹細胞で濃縮された細胞集団を得る方法は、例えば米国特許第5,486,359号明細書に記載されている。間葉系幹細胞の別の供給源には、特に限定されないが、血液、皮膚、臍帯血、筋肉、脂肪、骨、及び軟骨膜がある。
【0015】
間葉系幹細胞は種々の方法で投与される。間葉系幹細胞は、例えば静脈内、動脈内、又は腹腔内投与により全身的に投与してもよい。
【0016】
間葉系幹細胞は、自己、同種異系、又は異種を含む範囲の供給源由来でもよい。
【0017】
間葉系幹細胞は、動物の自己免疫疾患又は移植片対宿主病を治療するために有効な量で投与される。間葉系幹細胞は約1×105細胞/kg〜約1×107細胞/kgの量で投与される。別の実施態様において間葉系幹細胞は、約1×106細胞/kg〜約5×106細胞/kgの量で投与される。投与される間葉系幹細胞の量は、患者の年齢、体重、性、治療される自己免疫疾患、及びその程度と重症度を含む種々の要因に依存する。
【0018】
間葉系幹細胞は薬学的に許容される担体とともに投与してもよい。例えば間葉系幹細胞は、注射又は局所的投与のための薬学的に許容される液体媒体又はゲル中で細胞懸濁液として投与してもよい。
【0019】
本発明の別の態様において、動物の炎症応答を治療する方法が提供される。この方法は、動物の炎症応答を治療するために有効な量の間葉系幹細胞を動物に投与することを含む。
【0020】
本発明のこの態様の範囲はいかなる理論にも限定されないが、間葉系幹細胞が、制御T細胞(Treg細胞)へのT細胞成熟を促進し、こうして炎症応答が抑制されると考えられる。また、間葉系幹細胞がTヘルパー1細胞(Th1細胞)を阻害し、それによって、乾癬に関連するもののようなある炎症反応のインターフェロン−γ(IFN−γ)の発現を低下させると考えられる。
【0021】
ある実施態様において治療される炎症応答は、乾癬に関連するものである。
【0022】
別の実施態様において間葉系幹細胞は、脳中の小神経膠細胞及び/又は星状細胞に接触して炎症を低下させるように動物に投与され、こうして間葉系幹細胞は、アルツハイマー病、パーキンソン病、卒中、又は脳細胞損傷のような疾患もしくは障害において活性化グリア細胞により引き起こされる神経変性を制限する。
【0023】
さらに別の実施態様において間葉系幹細胞は、皮膚の表皮中のケラチン細胞及びランゲルハンス細胞に間葉系幹細胞が接触して、乾癬、慢性皮膚炎、及び接触皮膚炎で起きるような炎症を低下させるように動物に投与される。本発明はいかなる理論にも限定されないが、間葉系幹細胞が表皮中のケラチン細胞及びランゲルハンス細胞に接触して、表皮中のT細胞受容体やランゲルハンス細胞の発現を変化させ、こうして腫瘍壊死因子−アルファ(TNF−α)の発現を低下させ、制御T細胞(Treg細胞)集団を上昇させると考えられる。
【0024】
さらなる実施態様において間葉系幹細胞は、特に限定されないが骨関節炎や慢性関節リウマチ、及びウェブサイトwww.arthritis.org/conditions/diseasesに記載されている他の関節炎疾患を含む関節炎や関節炎様症状で起こるような、骨の炎症を低下させるのに使用される。本発明のこの実施態様の範囲はいかなる理論にも限定されないが、間葉系幹細胞が、滑液中の記憶T細胞によるインターロイキン−17分泌を阻害すると考えられる。
【0025】
別の実施態様において間葉系幹細胞は、炎症性腸疾患や慢性肝炎におけるそれぞれ消化管及び肝臓中の炎症を抑えるために使用される。本発明のこの実施態様の範囲はいかなる理論にも限定されないが、間葉系幹細胞が、インターロイキン−10(IL−10)の分泌増加と制御T細胞(Treg細胞)の生成を促進すると考えられる。
【0026】
別の実施態様において間葉系幹細胞は、敗血症及び火傷、手術、及び移植を含む外傷のような病理症状において好中球とマクロファージの過度の活性化を阻害するために使用される。本発明のこの実施態様の範囲はいかなる理論にも限定されないが、間葉系幹細胞が、IL−10のような抑制性サイトカインの分泌を促進し、マクロファージ遊走阻止因子を阻害すると考えられる。
【0027】
別の実施態様において間葉系幹細胞は、角膜、水晶体、色素上皮細胞、及び網膜を含む眼、脳、脊髄、妊娠子宮と胎盤、卵巣、睾丸、副腎皮質、肝臓、及び毛包のような免疫学的特権部位の炎症を制御するために使用される。この実施態様の範囲はいかなる理論にも限定されないが、間葉系幹細胞が、IL−10のような抑制性サイトカインの分泌とTreg細胞の生成を促進すると考えられる。
【0028】
別の実施態様において間葉系幹細胞は、透析及び/又は糸球体腎炎の最終段階腎疾患(ESRD)に関連する組織傷害を治療するために使用される。本発明のこの実施態様の範囲はいかなる理論にも限定されないが、間葉系幹細胞が腎修復を促進すると考えられる。間葉系幹細胞はまた、血管内皮増殖因子(すなわちVEGF)を発現し分泌し、これが新しい血管形成を刺激し、傷害された腎組織の修復を助けると考えられる。
【0029】
さらなる実施態様において間葉系幹細胞は、インフルエンザ、C型肝炎、単純ヘルペスウイルス、ワクシニアウイルス感染症、エプスタインバーウイルスのようなウイルス感染症を抑制するために使用される。この実施態様の範囲はいかなる理論にも限定されないが、間葉系幹細胞がインターフェロン−ベータ(IFN−β)の分泌を促進すると考えられる。
【0030】
さらに別の実施態様において間葉系幹細胞は、リーシュマニア (Leishmania)感染症やヘリコバクター(Helicobacter)感染症のような寄生虫感染症を制御するために使用される。この実施態様の範囲はいかなる理論にも限定されないが、間葉系幹細胞がTヘルパー2(Th2)細胞による応答を仲介し、こうしてβ細胞による免疫グロブリンE(IgE)の産生増加を促進すると考えられる。
【0031】
しかし、本発明のこの態様は、特定の炎症応答の治療に限定されないことを理解されたい。
【0032】
間葉系幹細胞は、上記したようにヒト及び非ヒト霊長類を含む哺乳動物に投与される。
【0033】
間葉系幹細胞はまた、上記したように全身的に投与される。あるいは骨関節炎又は慢性関節リウマチの場合、間葉系幹細胞は関節炎関節に直接投与される。
【0034】
間葉系幹細胞は、動物の炎症応答を治療するために有効な量で投与される。間葉系幹細胞は、約1×105細胞/kg〜約1×107細胞/kgの量で投与される。別の実施態様において間葉系幹細胞は、約1×106細胞/kg〜約5×106細胞/kgの量で投与される。投与される間葉系幹細胞の正確な用量は、患者の年齢、体重、性、治療される炎症応答、及びその程度と重症度を含む種々の要因に依存する。
【0035】
上記したように、間葉系幹細胞は薬学的に許容される担体とともに投与される。
【0036】
本発明の別の態様において、動物の炎症を治療し、及び/又は上皮傷害を修復する方法が提供される。この方法は、動物の炎症及び/又は上皮傷害を治療するために有効な量の間葉系幹細胞を動物に投与することを含む。
【0037】
本発明のこの態様の範囲はいかなる理論にも限定されないが、間葉系幹細胞が、T細胞による炎症促進性サイトカインであるTNF−αとインターフェロン−γの分泌の低下と、T細胞による抗炎症性サイトカインであるインターロイキン−10(IL−10)とインターロイキン−4(IL−4)の分泌の上昇とを引き起こすと考えられている。また間葉系幹細胞は、ナチュラルキラー(NK)細胞によるインターフェロン−γ分泌の低下を引き起こすと考えられる。
【0038】
本発明のこの態様で治療される炎症及び/又は上皮傷害は、自己免疫疾患、移植臓器の拒絶、火傷、切傷、裂傷、皮膚潰瘍形成と糖尿病性潰瘍形成を含む潰瘍形成を含むがこれらに限定されない、種々の疾患と障害により引き起こされる炎症及び/又は上皮傷害がある。
【0039】
ある実施態様において間葉系幹細胞は、慢性関節リウマチ、クローン病、1型糖尿病、多発性硬化症、強皮症、グレーブス病、ループス、炎症性腸疾患、自己免疫胃炎(AIG)、及び自己免疫糸球体疾患を含むがこれらに限定されない、自己免疫疾患により生じる上皮傷害を修復するために、動物に投与される。間葉系幹細胞はまた、移植片対宿主病(GVHD)により生じる上皮傷害を修復する。
【0040】
本発明のこの態様は、移植片対宿主病により生じる上皮傷害の修復、及びさらに詳しくは、皮膚及び/又は消化管系に影響を与えるグレードIIIとIVの移植片対宿主病を含む重症の移植片対宿主病により生じる上皮傷害の修復に適用される。本出願人は特に、重症の移植片対宿主病、特にグレードIIIとIVの消化管移植片対宿主病に罹った患者に間葉系幹細胞が投与されると、間葉系幹細胞の投与により患者の皮膚及び/又は潰瘍形成した小腸上皮組織を修復させることを発見した。
【0041】
別の実施態様において間葉系幹細胞は、移植臓器又は組織の拒絶により引き起こされる、腎臓、心臓、及び肺を含むがこれらに限定されない移植臓器又は組織に対する上皮傷害を修復するために動物に投与される。
【0042】
さらに別の実施態様において間葉系幹細胞は、火傷、切傷、裂傷、皮膚潰瘍形成と糖尿病性潰瘍形成を含む潰瘍形成により引き起こされる上皮傷害を修復するために動物に投与される。
【0043】
間葉系幹細胞は、上記したようにヒト及び非ヒト霊長類を含む哺乳動物に投与される。
【0044】
間葉系幹細胞はまた、上記したように全身的に投与される。
【0045】
間葉系幹細胞は、動物の上皮傷害を修復するために有効な量で投与される。間葉系幹細胞は、約1×105細胞/kg〜約1×107細胞/kgの量で投与される。別の実施態様において間葉系幹細胞は、約1×106細胞/kg〜約5×106細胞/kgの量で投与される。投与される間葉系幹細胞の正確な用量は、患者の年齢、体重、性、修復される上皮傷害のタイプ、及びその程度と重症度を含む種々の要因に依存する。
【0046】
本発明の別の態様において、動物の癌を治療する方法が提供される。この方法は、動物の癌を治療するために有効な量の間葉系幹細胞を動物に投与することを含む。
【0047】
本発明のこの態様の範囲はいかなる理論にも限定されないが、間葉系幹細胞が樹状細胞と相互作用してIFN−βを分泌させ、これが次に腫瘍サプレッサーとして作用すると考えられる。治療される癌には、特に限定されないが、肝細胞癌、子宮頚癌、膵臓癌、前立腺癌、線維肉腫、髄芽細胞腫、及び星状細胞腫がある。しかし本発明の範囲は特定の種類の癌に限定されないことを理解されたい。
【0048】
動物は上記したようにヒト及び非ヒト霊長類を含む哺乳動物である。
【0049】
間葉系幹細胞は、動物の癌を治療するために有効な量で投与される。一般に間葉系幹細胞は、約1×105細胞/kg〜約1×107細胞/kgの量で投与される。別の実施態様において間葉系幹細胞は、約1×106細胞/kg〜約5×106細胞/kgの量で投与される。投与される間葉系幹細胞の正確な用量は、患者の年齢、体重、性、治療される癌のタイプ、及びその程度と重症度を含む種々の要因に依存する。
【0050】
上記したように、間葉系幹細胞は薬学的に許容される担体とともに投与され、全身的に投与される。あるいは間葉系幹細胞は、治療される癌に直接投与される。
【0051】
本発明の別の態様において、動物のアレルギー疾患又は障害を治療する方法が提供される。この方法は、動物のアレルギー疾患又は障害を治療するために有効な量の間葉系幹細胞を動物に投与することを含む。
【0052】
本発明のこの態様の範囲はいかなる理論にも限定されないが、間葉系幹細胞が急性アレルギー応答後に投与されると、肥満細胞活性化と脱顆粒の阻害を提供すると考えられる。また間葉系幹細胞は好塩基球活性化をダウンレギュレートし、サイトカイン、例えばTNF−α、ケモカイン、例えばインターロイキン−8、単球化学誘引性タンパク質(すなわちMCP−1)、脂質メディエーター、例えばロイコトリエンを阻害し、主要なメディエーター、例えばヒスタミン、ヘパリン、コンドロイチン硫酸、及びカテプシンを阻害すると考えられる。
【0053】
治療されるアレルギー疾患又は障害には、特に限定されないが、喘息、アレルギー性鼻炎、アトピー性皮膚炎、及び接触皮膚炎がある。しかし、本発明の範囲は、特定のアレルギー疾患又は障害に限定されないことを理解されたい。
【0054】
間葉系幹細胞は、動物のアレルギー疾患又は障害を治療するために有効な量で投与される。動物は哺乳動物でもよい。哺乳動物は、ヒト及び非ヒト霊長類を含む霊長類でもよい。一般に間葉系幹細胞は、約1×105細胞/kg〜約1×107細胞/kgの量で投与される。別の実施態様において間葉系幹細胞は、約1×106細胞/kg〜約5×106細胞/kgの量で投与される。投与される間葉系幹細胞の正確な用量は、患者の年齢、体重、性、治療されるアレルギー疾患又は障害、及びその程度と重症度を含む種々の要因に依存する。
【0055】
上記したように、間葉系幹細胞は薬学的に許容される担体とともに投与される。間葉系幹細胞は、例えば静脈内又は動脈内投与のように全身的に投与される。
【0056】
本発明のさらなる態様において、動物の創傷治癒を促進する方法が提供される。この方法は、動物の創傷治癒を促進するために有効な量の間葉系幹細胞を動物に投与することを含む。
【0057】
本発明の範囲はいかなる理論にも限定されないが、上記したように間葉系幹細胞は、Treg細胞と樹状細胞にインターロイキン−1−10(IL−10)を放出させる。IL−10は、創傷の炎症を制限又は抑制し、こうして創傷治癒を促進する。
【0058】
さらに間葉系幹細胞は、他のタイプの細胞により分泌因子を誘導することにより、創傷治癒と骨折治癒とを促進する。例えば間葉系幹細胞は、末梢血単核細胞(PBMC)による血管内皮増殖因子(VEGF)のプロスタグランジンE2(PGE2)介在放出、ならびに成長ホルモン、インスリン、インスリン様増殖因子1(IGF−1)インスリン様増殖因子結合タンパク質−3(IGFBP−3)、及びエンドテリン−1のPGE2介在放出を誘導する。
【0059】
治癒される創傷には、特に限定されないが、切傷、裂傷、火傷、及び皮膚潰瘍形成から生じるものがある。
【0060】
間葉系幹細胞は、動物の創傷治癒を促進するために有効な量で動物に投与される。動物は哺乳動物でもよく、哺乳動物はヒト及び非ヒト霊長類を含む霊長類でもよい。一般に間葉系幹細胞は、約1×105細胞/kg〜約1×107細胞/kgの量で投与される。別の実施態様において間葉系幹細胞は、約1×106細胞/kg〜約5×106細胞/kgの量で投与される。投与される間葉系幹細胞の正確な用量は、患者の年齢、体重、性、治療される創傷、及びその程度と重症度を含む種々の要因に依存する。
【0061】
上記したように、間葉系幹細胞は薬学的に許容される担体とともに投与される。間葉系幹細胞は、上記したように全身的に投与される。あるいは間葉系幹細胞は、間葉系幹細胞を含有する包帯又はリザーバー上の液体で、創傷に直接投与される。
【0062】
本発明のさらに別の態様において、動物の繊維症を治療または予防する方法が提供される。この方法は、動物の繊維症を治療または予防するために有効な量の間葉系幹細胞を動物に投与することを含む。
【0063】
間葉系幹細胞は、肝硬変、最終段階腎疾患に関連する腎臓の繊維症、および肺の繊維症を含むがこれらに限定されない動物の任意の種類の繊維症、成人型呼吸窮迫症候群(ARDS)および慢性閉塞性肺疾患(COPD)を含むがこれらに限定されない肺の繊維症を治療または予防するために動物に投与される。本発明の範囲は特定の種類の繊維症に限定されないことを理解されたい。
【0064】
間葉系幹細胞は、動物の繊維症を治療または予防するために有効な量で動物に投与される。動物は哺乳動物でもよく、哺乳動物はヒト及び非ヒト霊長類を含む霊長類でもよい。一般に間葉系幹細胞は、約1×105細胞/kg〜約1×107細胞/kgの量で投与される。別の実施態様において間葉系幹細胞は、約1×106細胞/kg〜約5×106細胞/kgの量で投与される。投与される間葉系幹細胞の正確な用量は、患者の年齢、体重、性、治療または予防される繊維症の程度と重症度を含む種々の要因に依存する。
【0065】
間葉系幹細胞は、上記したように薬学的に許容される担体とともに投与される。間葉系幹細胞は上記したように全身的に投与される。
【0066】
本発明の別の目的は、動物の組織又は臓器の血管形成であって、該組織又は臓器は血管形成が必要である、血管形成を促進することである。
【0067】
本発明のさらなる態様において、動物の臓器又は組織の血管形成を促進する方法が提供される。この方法は、動物の臓器又は組織の血管形成を促進するために有効な量の間葉系幹細胞を動物に投与することを含む。
【0068】
血管形成は、既存の微小血管床からの新しい血管の形成である。
【0069】
血管形成の誘導は、冠動脈及び末梢動脈不全症を治療するために使用され、冠動脈疾患、虚血性心疾患、及び末梢血動脈疾患の治療のための非侵襲的及び治療的アプローチでもよい。血管形成は心臓以外の組織や臓器の疾患と障害の治療、ならびに心臓以外の臓器の成長及び/又は維持において役割を果たす。血管形成は、内部及び外部創傷ならびに皮膚潰瘍の治療において役割を果たす。血管形成はまた、胚の着床、胎盤成長、ならびに胚血管の成長において役割を果たす。血管形成はまた、軟骨再吸収と骨形成との連結に必須であり、正しい成長板の形態形成に必須である。
【0070】
さらに血管形成は、充分な栄養物質と気体輸送を提供するために必要な高密度の血管ネットワークが必要な高代謝臓器、例えば肝臓の構築と維持の成功に必要である。
【0071】
間葉系幹細胞は、種々の方法により血管形成の必要な組織又は臓器に投与することができる。間葉系幹細胞は、例えば静脈内、動脈内、又は腹腔内投与により全身的に投与されるか、又は間葉系幹細胞は、例えば血管形成の必要な組織又は臓器への直接投与により、血管形成の必要な組織又は臓器に直接投与される。
【0072】
間葉系幹細胞は、自己、同種異系、又は異種を含む範囲の供給源由来でもよい。
【0073】
本発明の範囲はいかなる理論にも限定されないが、間葉系幹細胞は動物に投与されると、末梢血単核細胞(PBMC)を刺激して血管内皮増殖因子(すなわちVEGF)を産生させ、これが新しい血管の形成を刺激すると考えられる。
【0074】
ある実施態様において動物は哺乳動物である。哺乳動物は、ヒト及び非ヒト霊長類を含む霊長類でもよい。
【0075】
本発明の間葉系幹細胞は、血管形成を介して緩和、治療、又は予防できる疾患もしくは障害の治療、緩和、又は予防に使用される。例えば間葉系幹細胞は、四肢(すなわち腕、脚、手、及び足)、ならびに首又は種々の臓器中の閉塞動脈を治療するために動物に投与される。例えば間葉系幹細胞は、脳の需要を満たす閉塞動脈を治療、従って卒中を治療又は予防するために使用される。また間葉系幹細胞は、胚及び生後角膜の血管を治療するために使用され、かつ糸球体構造形成を提供するために使用される。別の実施態様において間葉系幹細胞は、内部及び外部の創傷の治療、ならびに糖尿病や鎌状赤血球貧血のような疾患により引き起こされる皮膚潰瘍を含むがこれらに限定されない足、手、脚、又は腕の皮膚潰瘍の治療に使用される。
【0076】
さらに血管形成は胚の着床と胎盤形成に関与するため、間葉系幹細胞は胚の着床を促進し流産を防ぐために使用される。
【0077】
さらに間葉系幹細胞は、胎内の動物の血管の成長を促進するために、ヒトを含む胎内の動物に投与される。
【0078】
別の実施態様において間葉系幹細胞は、軟骨再吸収と骨形成を促進し、正しい成長板の形態形成を促進するために、生後又は生前の動物に投与される。
【0079】
間葉系幹細胞は、動物の血管形成を促進するために有効な量で投与される。間葉系幹細胞は、約1×105細胞/kg〜約1×107細胞/kgの量で投与される。別の実施態様において間葉系幹細胞は、約1×106細胞/kg〜約5×106細胞/kgの量で投与される。投与される間葉系幹細胞の量は、患者の年齢、体重、性、治療、緩和、又は予防される疾患もしくは障害、及びその程度と重症度を含む種々の要因に依存する。
【0080】
間葉系幹細胞は薬学的に許容される担体とともに投与してもよい。例えば間葉系幹細胞は、注射用のための薬学的に許容される液体媒体中で細胞懸濁液として投与してもよい。注射は、局所的、すなわち血管形成の必要な組織又は臓器に直接、又は全身性でもよい。
【0081】
間葉系幹細胞は、治療薬をコードする1つ又はそれ以上のポリヌクレオチドを用いて遺伝子操作してもよい。ポリヌクレオチドは、適切な発現ビヒクルを介して間葉系幹細胞に提供される。間葉系幹細胞を遺伝子操作するために使用される発現ビヒクルには、特に限定されないが、レトロウイルスベクター、アデノウイルスベクター、及びアデノ関連ウイルスベクターがある。
【0082】
治療薬をコードする適切なポリヌクレオチドの選択は、治療される疾患もしくは障害、及びその程度と重症度を含む種々の要因に依存する。治療薬をコードするポリヌクレオチド及び適切な発現ビヒクルは、さらに米国特許第6,355,239号明細書に記載されている。
【0083】
間葉系幹細胞は上記治療で使用される時には、増殖因子、サイトカイン、抗炎症剤のような薬剤を含むがこれらに限定されない当業者に公知の他の治療薬、と間葉系幹細胞以外の細胞、例えば樹状細胞とを組合せて使用されるか、あるいは細胞のための可溶性担体、例えばヒアルロン酸、又は適宜固体マトリックス、例えば、コラーゲン、ゼラチン、又は他の生体適合性ポリマーと組合せて投与される。
【0084】
本明細書に記載の方法は、多くの方法でかつ当該分野で公知の種々の修飾及び置換を用いて行われる。細胞タイプ間の作用又は相互作用のモードについて提唱されている理論は、決して本発明を限定するものではなく、本発明の方法をよりよく理解できるように提供されていることを理解されたい。
【0085】
本発明を図面により説明する。
【0086】
図1。MSCは樹状細胞機能を調節する。(A)HLA−DRとCD11cに対する抗体を使用する成熟単球DC1細胞のフローサイトメトリー解析と、HLA−DRとCD123(IL−3受容体)に対する抗体を使用する形質球様DC2細胞のフローサイトメトリー解析。(破線):アイソタイプ対照;(実線):FITC/PE結合抗体。(B)MSCは、活性化DC1とDC2からの、それぞれTNF−α分泌(第1y軸)を阻害し、IL−10分泌(第2y軸)を上昇させる。(C)成熟DC1細胞で培養したMSCは、MSCもしくはDC単独と比較して、T細胞によるIFN−γ分泌(第1y軸)を阻害しIL−4レベル(第2y軸)を上昇させる。MSCの存在下での炎症促進性IFN−γの産生低下と抗炎症性IL−4の産生増加は、T細胞集団の抗炎症性表現型への移動を示した。
【0087】
図2。MSCは炎症促進性エフェクターT細胞機能を阻害する。(A)FITC結合CD4(x軸)抗体とPE結合CD25(y軸)抗体を用いる、MSC+PBMC培養物(MSC+PBMC)中のPBMC又は非接着性画分を染色することによるTReg細胞数(%)のフローサイトメトリー解析。ゲートはバックグランドとしてのアイソタイプ対照抗体に基づく。グラフは5つの独立した実験の典型である。(B)細胞培養上清中で、MSCの存在下で生成したTH1細胞は低レベルのIFN−γ(第1y軸)を分泌し、MSCの存在下で生成したTH2細胞は増加量のIL−4(第2y軸)を分泌した。(C)MSCは、24ウェルプレート中で0、24、又は48時間培養した精製NK細胞からのIFN−γ分泌を阻害する。示したデータは1つの実験の平均±SDサイトカイン分泌であり、3つの独立した実験の典型である。
【0088】
図3。MSCはTreg細胞集団の数とGITR発現を上昇させる。(A)CD4+CD25+Treg細胞集団を、PBMC又はMSC+PBMC(MSC対PBMC比は1:10)培養物(さらなる刺激無しで3日間培養した)から2工程磁性単離法を使用して単離した。これらの細胞に放射線照射し(さらなる増殖を阻止するため)、混合リンパ球反応(MLR)において刺激細胞として使用し、ここで応答細胞は植物性血球凝集素(PHA)(2.5mg/ml)の存在下の同種異系PBMC(刺激細胞対応答細胞比は1:100)であった。細胞を48時間培養し、次に3Hチミジンを加え、24時間後、取り込まれた放射活性を計測した。結果は、MSCの存在下で生成したTreg集団(レーン3)は、MSCの非存在下で生成したTreg細胞(レーン2)と機能的に同等であることを示した。(B)PBMCをMSCの非存在下(上のプロット)又は存在下(下のプロット)で3日間培養(MSC対PBMC比は1:10)し、次に非接着画分を採取し、FITC標識GITRとPE標識CD4で免疫染色した。結果は、MSCの存在下で培養した細胞でGITR発現の2倍以上の上昇を示す。
【0089】
図4。MSCはPGE2を産生し、PGE2を阻止するとMSC性免疫調節作用が逆転する。(A)種々の濃度のPGE2ブロッカーであるNS−398又はインドメタシン(Indometh.)の存在下又は非存在下で培養したMSCから得られた培養上清中のPGE2分泌(平均±SD)。インヒビター濃度はμMであり、示したデータは24時間培養後に得られた値である。(B)リアルタイムRT−PCRを使用したMSCとPBMC中のCOX−1とCOX−2発現。MSCはPBMCと比較して有意に高レベルのCOX−2を発現し、MSCをPBMCの存在下で培養すると、MSC中のCOX−2発現の3倍を超える上昇があった。1〜3つの独立した実験からの典型的データを示す。MSC+PBMC培養物をトランスウェルチャンバープレート中で準備し、ここでMSCは下のチャンバーに蒔き、PBMCは上のチャンバーに蒔いた。(C)PGE2ブロッカーであるインドメタシン(Ind.)又はNS−398の存在は、対照と比較して、活性化DC(白バー)からのTNF−α分泌と、TH1細胞(斜線バー)からのIFN−γ分泌を上昇させる。データは、MSCとPGE2インヒビターの非存在下で生成した培養物からの%変化として計算した。(D)MSC−PBMC同時培養物(1:10)中のPGE2ブロッカーであるインドメタシン(Indo)又はNS−398の存在は、PHA処理PBMCに対するMSC性抗増殖作用を逆転させる。示したデータは1つの実験からであり、3つの独立した実験の典型である。
【0090】
図5。構成性MSCサイトカイン分泌は同種異系PBMCの存在下で上昇する。すでに性状解析したヒトMSCを使用して、PBMCの存在下(斜線)又は非存在下(白い棒)(MSC対PBMC比は1:10)で24時間培養したMSCの培養上清中のサイトカインIL−6とVEGF、脂質メディエーターPGE2、及びマトリックス金属プロテアーゼ1(pro−MMP−1)のレベルを分析した。MSCはIL−6、VEGF、及びPGE2を構成性に産生し、これらの因子のレベルはPBMCとの同時培養で上昇し、従ってMSCが炎症性状況で免疫機能の調節において役割を果たすことを示唆する。
【0091】
図6。MSCは分裂促進剤誘導性のT細胞増殖を用量依存的に阻害する。増加する数の同種異系PBMCを、PHA(2.5mg/ml)の存在下又は非存在下で96ウェルプレートに蒔いた一定数のMSC(2,000細胞/ウェル)とインキュベートし、3Hチミジン取り込みを測定した(1分当たりのカウント、cpm)。MSCの存在下でPHA処理PBMCの増殖の用量依存性阻害があった。3つの独立した実験の1つからの典型的結果を示す。同様の結果が、LeBlanc, et al., Scand J. Immunol., Vol. 57, pg. 11 (2003)により報告された。
【0092】
図7。提唱されているMSC作用機構の模式図
MSCは、先天的(DC経路2〜4;及びNK経路6)免疫系と適応(T経路1と5、及びB経路7)免疫系の両方からの細胞に影響を与えることにより免疫調節作用を仲介する。浸入病原体に応答して未成熟DCは、侵入可能部位に移動し、成熟し、未処理のT細胞を刺激して(抗原特異的及び同時刺激シグナルにより)、防御性エフェクターT細胞(細胞性TH1又は体液性TH2免疫)になる能力を獲得する。MSC−DC相互作用中に、MSCは直接の細胞−細胞接触により又は分泌された因子により、細胞性応答(経路2)を開始するDCの能力を制限することにより、又は体液性応答(経路4)を開始する能力を促進することにより、免疫応答の結果を変化させる。また成熟エフェクターT細胞が存在する時、MSCはこれらと相互作用してTH1(経路1)応答のバランスをTH2応答(経路5)に向けて、おそらくIgE産生B細胞活性(経路7)の上昇(これは、GvHDと自己免疫疾患症状の抑制に好ましい結果である)に向ける。MSCはTReg集団(経路3)の生成を増加させる能力により、寛容表現型となり、局所的微小環境におけるバイスタンダー(bystander)炎症を緩和することにより受容体宿主を助ける。点線(----)は提唱された機構である。
【0093】
本発明を以下の実施例で説明する;しかし本発明の範囲は決してこれらにより限定されるものではない。
【0094】
実施例1
【0095】
材料と方法
【0096】
ヒトMSCの培養
ヒトMSCを、Pittenger et al., Science, Vol. 284, pg. 143 (1999)に記載されたように培養した。すなわち、Poietics Technologies, Div of Cambrex Biosciencesによるインフォームドコンセントの後、匿名のドナーの腸骨稜から骨髄試料を採取した。1%抗生物質−抗真菌剤溶液(Invitrogen, Carlsbad, California)と10%牛胎児血清(FBS, JRH BioSciences, Lenexa, Kansas)を含有する完全ダルベッコー改変イーグル培地−低グルコース(Life Technologies, Carlsbad, California)中でMSCを培養した。MSCは接着性単層として増殖し、トリプシン/EDTAを用いて剥がした(0.05%トリプシンで37℃で3分)。使用したすべてのMSCは多分化性についてすでに性状解析され、間葉性系統(軟骨細胞性、脂肪細胞生成性、及び骨形成性)に分化する能力を保持した(Pittenger, et al., Science, Vol. 284, pg. 143 (1999))。
【0097】
樹状細胞の単離
末梢血単核細胞(PBMC)は、Poietics Technologies, Div of Cambrex Biosciences (Walkersville, MD)から得た。単球系統(CD1c+)の樹状細胞(DC)の前駆体を、Dzionek, et. al., J. Immunol., Vol. 165, pg. 6037 (2000)に従う2工程磁性分離法を使用して、PBMCから陽性選択した。すなわち、CD1c発現B細胞を磁性ビーズを使用してCD19+細胞を磁性的に枯渇させ、次にB細胞枯渇画分をビオチン標識CD1c(BDCA1+)と抗ビオチン抗体とで標識し、製造業者(Miltenyi Biotech, Auburn, California)の説明書に従って磁性カラムを使用して、非標識細胞画分からこれらを分離した。陽性標識した抗体コーティング細胞(BDCA2+)(Miltenyi Biotech, Auburn, California)の免疫−磁性分類により、PBMCから形質細胞様系統のDCの前駆体を単離した。
【0098】
MSC−DC培養
ほとんどの実験でヒトMSCとDCを同じ数で種々の時間培養し、細胞培養物上清を採取し、さらに評価するまで−80℃で保存した。選択された実験で、MSCを成熟DC1もしくはDC2細胞(1:1 MSC:DC比)と3日間培養し、次に一緒にした培養物(MSCとDC)に放射線照射してさらなる増殖を止めた。次に抗体で精製した未処理の同種異系T細胞(CD4+、CD45RA+)を放射線照射MSC/DCに加え、さらに6日間培養した。次に培養物から非接着性細胞画分(精製T細胞)を採取し、2回洗浄し、PHAでさらに24時間再刺激し、次に細胞培養上清を採取し、分離されたIFN−γとIL−4についてELISAにより分析した。
【0099】
NK細胞の単離
ビオチン結合モノクローナル抗体のカクテル(抗−CD3、−CD14、−CD19、−CD36、及び抗IgE抗体)を1次試薬とし、マイクロビーズに結合した抗ビオチンモノクローナル抗体を2次標識試薬として、磁性標識した非NK細胞を枯渇させることにより、NK細胞の精製集団が得られた。磁性標識した非NK細胞は、磁界中でMACS(Miltenyi Biotech, Auburn, California)カラム中に保持され、NK細胞は通過し、採取された。
【0100】
TReg細胞集団の単離
2工程単離法を使用してTReg細胞集団を単離した。まずビオチン標識抗体のカクテルと抗ビオチンマイクロビーズを用いて、非CD4+ T細胞を間接に磁性標識した。次に標識した細胞をMACSカラム(Miltenyi Biotech, Auburn, California)で分離して枯渇させた。次にCD4+CD25+細胞をCD25マイクロビーズで直接標識し、あらかじめ濃縮したCD4+ T細胞画分から陽性選択により単離した。磁性標識したCD4+CD25+細胞はカラムに保持され、磁界からカラムを取り出した後溶出した。
【0101】
MSCの存在下で生成されたCD4+CD25+集団の増加が自然界で抑制性であるかどうかを調べるために、CD4+CD25+ Treg細胞集団を、2工程磁性単離法を使用してPBMC又はMSC+PBMC(MSC対PBMC比は1:10)培養物(さらなる刺激無しで3日間培養した)から単離した。これらの細胞に放射線照射してさらなる増殖を阻止し、混合リンパ球反応(MLR)で刺激細胞として使用し、ここで応答細胞はPHA(2.5μg/ml)の存在下で同種異系PBMC(刺激細胞対応答細胞の比は1:100)であった。48時間培養を行い。次に3Hチミジンを加えた。24時間後に取り込まれた放射活性を計測した。
【0102】
MSCの非存在下又は存在下でPBMCを培養(MSC対PBMC比は1:10)し、次に非接着性画分を採取し、FITC標識グルココルチコイド誘導性TNF受容体(すなわちGITR)、及びPE標識CD4で免疫染色した。
【0103】
TH1/TH2細胞の作製
2×106細胞/mlで37℃で45分、末梢血単核細胞(PBMC)をプレートに蒔き、単球を除去した。非接着性画分をプレート結合抗CD3(5μg/ml)と抗CD28(1μg/ml)抗体の存在下で、TH1(IL−2(4ng/ml)+IL−12(5ng/ml)+抗IL−4(1μg/ml))条件又はTH2(IL−2(4ng/ml)+IL−4(4ng/ml)+抗IFN−γ(1μg/ml))条件で3日間、MSCの存在下又は非存在下でインキュベートした。細胞を洗浄し、次にPHA(2.5μg/ml)でさらに24もしくは48時間再刺激し、次に培養上清中のIFN−γとIL−4のレベルをELISA(R&D Systems, Minneapolis, Minnesota)により測定した。
【0104】
MSCの培養上清中のVEGF、PGE2、及びpro−MMP−1のレベルの分析
すでに性状解析されたヒトMSCを使用して、インターロイキン−6(IL−6)、VEGF、脂質メディエータープロスタグランジンE2(PGE2)、及びマトリックス金属プロテアーゼ1(pro−MMP−1)のレベルを、PBMCの存在下又は非存在下(MSC対PBMC比は1:10)で24時間培養したMSCの培養上清で分析した。
【0105】
PBMCの増殖
フィコール−ハイパーク(Lymphoprep, Oslo, Norway)上でleukopack(Cambrex, Walkersville, Maryland)を遠心分離して、精製PBMCを調製した。分離した細胞をMSC(PBMC添加の3〜4時間前にプレートに蒔いて沈降させた)の存在下又は非存在下で、分裂促進剤PHA(Sigma Chemicals, St. Louis, Missouri)の存在下で48時間培養した(三重測定)。選択された実験で、PBMCをPGE2インヒビターであるインドメタシン(Sigma Chemicals, St. Louis, Missouri)又はNS−938(Cayman Chemicals, Ann Arbor, Michigan)を含有する培地に再懸濁した。(3H)チミジンを加え(200μlの培養物に20μl)、さらに24時間培養後自動ハーベスターを使用して細胞を採取した。MSC又はPGE2ブロッカーの作用を、PHAの存在下での対照応答(100%)のパーセントとして計算した。
【0106】
定量的RT−PCR
市販のキット(Qiagen, Valencia, California)を使用して製造業者の説明書に従って、細胞ペレットから総RNAを調製した。混入しているゲノムDNAをDNAフリーキット(Ambion, Austin, Texas)を使用して除去した。QuantiTect SYBR Green RT-PCR キット(Qiagen, Valencia, California)を使用して、0.5μM濃度のプライマーを用いて、MJ Research Opticon検出系(South San Francisco, California)で定量的RT−PCRを行った。異なる条件で培養した細胞中の発現レベルの相対的変化を、β−アクチンを内部対照として使用してCt値(交差点)の差により計算した。COX−1及びCOX−2特異的プライマーの配列は以下の通りであった:COX−1:5'-CCG GAT GCC AGT CAG GAT GAT G-3' (フォワード)、 5'-CTA GAC AGC CAG ATG CTG ACA G-3' (リバース);COX-2:5'-ATC TAC CCT CCT CAA GTC CC-3'(フォワード)、 5'-TAC CAG AAG GGC AGG ATA CAG-3' (リバース)。
【0107】
増加する数の同種異系PBMCを、96ウェルプレート上に蒔いた一定数のMSC(2,000細胞/ウェル)と、PHA(2.5μg/ml)の存在下で72時間インキュベートし、3Hチミジンの取り込み(1分当たりのカウント、cpm)を測定した。PBMCとMSCは、MSC:PBMCが1:1、1:3、1:10、1:30、及び1:81の比で培養した。
【0108】
結果
本試験では、ヒトMSCと、樹状細胞(DC1とDC2)、エフェクターT細胞(TH1とTH2)及びNK細胞を含む単離した免疫細胞集団との相互作用を調べた。MSCと各免疫細胞タイプとの相互作用は特定の結果を有し、MSCが免疫応答プロセスのいくつかの工程を調節することを示唆した。MSC免疫調節作用を調節し、これの原因かも知れない分泌因子の産生を評価し、プロスタグランジン合成が示唆された。
【0109】
それぞれBDCA1+とBDCA2+細胞の免疫磁性分類により、骨髄性(DC1)と形質細胞性(DC2)前駆樹状細胞を単離し、DC1細胞についてはGM−CSFとIL−4(それぞれ1×103IU/mlと1×103IU/ml)と、DC2細胞についてはIL−3(10ng/ml)とインキュベートして成熟させた。フローサイトメトリーを使用すると、DC1細胞はHLA−DR+であり、一方DC2細胞はHLA−DR+とCD123+であった(図1A)。炎症性物質である細菌リポ多糖(LPS、1ng/ml)の存在下で、DC1細胞は中レベルのTNF−αを産生したが、MSCが存在する時(調べた比率1:1と1:10)は、TNF−α分泌の>50%低下があった(図1B)。一方DC2細胞はLPSの存在下でIL−10を産生し、そのレベルは、MSC:DC2同時培養(1:1)で2倍以上上昇した(図1B)。従ってMSCは培養物中の活性化DCのサイトカインプロフィールをより寛容原性表現型に修飾した。さらに活性化DCはMSCと培養するとIFN−γを低下させ、かつ未処理のCD4+T細胞により分泌されるIL−4レベルを上昇(図1C)させることができ、炎症促進性から抗炎症性T細胞表現型へのMSC性シフトを示唆した。
【0110】
上昇したIL−10分泌は制御細胞の生成において役割を果たす(Kingsley, et al., J. Immunol., Vol. 168, pg. 1080 (2002))ため、T制御細胞(TReg)は、PBMCとMSCの同時培養でフローサイトメトリーにより定量した。PBMCとMSCを3〜5日間培養すると、抗CD4と抗CD25抗体を用いてPBMCを染色して測定したTReg細胞数が増加し(図2A)、これはMSC誘導性の寛容原性応答をさらに支持した。MSCの存在下で生成するCD4+CD25+ TReg細胞集団は、グルココルチコイド誘導性のTNF受容体(GITR)(TReg細胞集団上で発現される細胞表面受容体)のレベル上昇を示し、自然界では同種異系T細胞増殖を抑制したため抑制性であった(図3A、3 B)。次にT細胞分化に影響を与えるその直接能力についてMSCを調べた。抗体で選択した精製T細胞(CD4+ Th細胞)を使用して、IFN−γ産生TH1細胞とIL−4産生TH2細胞を、MSCの存在下又は非存在下で作製した。分化中にMSCが存在する時に、TH1細胞によるIFN−γ分泌が低下し、TH2細胞によるIL−4分泌が上昇した(図2B)。T細胞がエフェクターTH1又はTH2型(データは示していない)に分化(3日目)した後にMSCを培養物に加えると、IFN−γとIL−4レベルに有意な変化はなかった。これらの実験は、MSCがエフェクターT細胞分化に直接影響を与え、T細胞サイトカイン分泌を体液性表現型に変化させることを示唆する。
【0111】
同様に、MSCを精製NK細胞(CD3−、CD14−、CD19−、CD36−)と1:1の比率で異なる時間(0〜48時間)培養すると、培養上清中でIFN−γ分泌が低下(図2C)し、従ってMSCがNK細胞機能も調節できることを示唆する。
【0112】
以前の研究は、MSCが可溶性因子によりT細胞機能を修飾することを示している(LeBlanc, et al., Exp. Hematol., Vol. 31 , pg. 890 (2003); Tse, et al., Transplantation, Vol. 75, pg. 389 (2003))。MSCは、IL−6、プロスタグランジンE2、VEGF、及びpro−MMP−1を含むいくつかの因子を構成的に分泌し、それぞれのレベルがPBMCとの培養により上昇することが観察された(図5)。DCによるTNF−αの阻害とIL−10産生の上昇を引き起こすMSC由来因子を調べるために、プロスタグランジンE2の役割の可能性を調べた(これは活性化DCによるTNF−α産生を阻害することが証明されているため)(Vassiliou, et al., Cell. Immunol., Vol. 223, pg. 120 (2003))。MSC培養物(0.5×106細胞/mlの24時間培養物)からの調整培地は約1000pg/mlのPGE2を含有した(図4A)。培養上清中にPGE2分泌の公知のインデューサー、例えば、TNF−α、IFN−γ、又はIL−1βの検出可能な存在は無く(データは示していない)、MSCによるPGE2の構成性分泌を示している。hMSCによるPGE2分泌は、PGE2産生の公知のインヒビター[NS−398(5μM)及びインドメタシン(4μM)]の存在下で60〜90%阻害された(図4A)。構成的活性なシクロオキシゲナーゼ酵素1(COX−1)と誘導性シクロオキシゲナーゼ酵素2(COX−2)の酵素活性の結果としてPGE2分泌の放出が起きるため(Harris, et al., Trends Immunol., Vol. 23, pg. 144 (2002))、トランスウェル培養系を使用してMSCとPBMC中のCOX−1とCOX−2のmRNA発現を分析した。MSCは、PBMCと比較して有意に高レベルのCOX−2を発現し、その発現レベル上昇は、MSCとPBMC(MSCとPBMCの比は1:10)と24時間同時培養すると>3倍であった(図4B)。COX−1レベルにわずかな変化が見られ、MSC−PBMC同時培養でのPGE2分泌の上昇(図5)はCOX−2のアップレギュレーションにより仲介されることを示唆している。DCとT細胞に対するMSCの免疫調節作用がPGE2により仲介されるかどうかを調べるために、PGE2インヒビターであるNS−398又はインドメタシンの存在下でMSCを活性化樹状細胞(DC1)又はTH1細胞と培養した。NS−398又はインドメタシンの存在は、それぞれDC1によるTNF−α分泌と、TH1細胞によるIFN−γ分泌を上昇させ(図4C)、免疫細胞タイプへのMSCの作用は分泌されたPGE2により仲介されることを示唆した。最近の研究は、種々の刺激により誘導されるT細胞増殖をMSCが阻害することを証明している(DeNicola, et al., Blood, Vol. 99, pg. 3838 (2002); LeBlanc, et al., Scand. J. Immunol., Vol. 57, pg. 11 (2003))。MSCは分裂促進剤誘導性のT細胞増殖を用量依存性に阻害(図6)し、PGE2インヒビターであるNS−398(5μM)又はインドメタシン(4μM)が存在すると、インヒビターの無い対照と比較して、培養物を含有するMSC中のPHA処理PBMCにより(3H)チミジン取り込みが>70%上昇することが観察された(図4D)。
【0113】
要約すると、MSCと他の免疫細胞種との相互作用のモデル(図7)が提唱される。成熟T細胞が存在すると、MSCが直接これらと相互作用し、炎症促進IFN−γ産生(経路1)を阻害し、制御性T細胞表現型(経路3)を増加させ、抗炎症性TH2細胞(経路5)を増加する。さらにMSCは、PGE2を分泌することにより、炎症促進DC1細胞を阻害(経路2)することにより、及び抗炎症性DC2細胞(経路4)又は制御性DC(経路3)を増加させることにより、DCを介するT細胞免疫応答の結果を変化させることができる。TH2免疫へのシフトは、IgE/IgG1サブタイプ抗体の生成上昇(経路7)へのB細胞活性の変化を示唆する。MSCはNK細胞からのIFN−γ分泌を阻害する能力により、NK細胞機能を修飾するようである(経路6)。このMSC:免疫細胞相互作用のモデルは、いくつかの他の実験室で行われた実験と一致する(LeBlanc, et al., Exp. Hematol., Vol. 31 , pg. 890 (2003); Tse, et al., Transplantation, Vol. 75, pg. 389 (2003); DiNicola, et al., Blood, Vol. 99, pg. 3838 (2002))。提唱された機構のさらなる研究が進行中であり、MSC投与のインビボ作用を調べるために動物試験が必要である。
【0114】
実施例2
【0115】
重症の段階IVの消化管移植片対宿主病(GVHD)に罹っている33才の女性患者に間葉系幹細胞を投与した。この患者は他のすべてのGVHD治療に抵抗性であった。患者の結腸の内視鏡観察は、治療前に潰瘍形成と炎症の部位を示した。患者の結腸の組織検査は、治療前にGVHDが患者の腸陰窩の大部分を破壊していることを証明した。
【0116】
患者は、体重1kg当たり3×106細胞の量でPlasma Lyte Aの50ml中で同種異系間葉系幹細胞の静脈内注入を受けた。
【0117】
注入の2週間後患者を評価した。注入後2週間で、患者の結腸の内視鏡観察は、治療前に見えていた炎症部位と潰瘍形成が消失したことを示した。さらに患者の結腸の生検は、腸陰窩の有意な再生を示した。すなわち患者への間葉系幹細胞の投与により、消化管移植片対宿主病の炎症性成分の有意な低下と、新しい機能的小腸組織の再生が起きた。
【0118】
すべての特許、刊行物(公開された特許出願、寄託受け入れ番号、及びデータベース受け入れ番号を含む)は、すべての特許、刊行物(公開された特許出願、寄託受け入れ番号、及びデータベース受け入れ番号を含む)が具体的かつ個々に取り込まれているのと同じ程度に、参照することにより本明細書に組み込まれる。
【0119】
しかし本発明の範囲は本明細書に記載の具体例に限定されるものではないことを理解されたい。本発明は、本明細書の具体的な記載とは別に実施することができ、それでも添付の特許請求の範囲内にある。
【図面の簡単な説明】
【0120】
【図1A】図1Aは、MSCは樹状細胞機能を調節することを示す。HLA−DRとCD11cに対する抗体を使用する成熟単球DC1細胞のフローサイトメトリー解析と、HLA−DRとCD123(IL−3受容体)に対する抗体を使用する形質球様DC2細胞のフローサイトメトリー解析。(破線):アイソタイプ対照;(実線):FITC/PE結合抗体。
【図1B】図1Bは、MSCが、活性化DC1とDC2からの、それぞれTNF−α分泌(第1y軸)を阻害し、IL−10分泌(第2y軸)を上昇させる、ことを示す。
【図1C】図1Cは、成熟DC1細胞で培養したMSCが、MSCもしくはDC単独と比較して、T細胞によるIFN−γ分泌(第1y軸)を阻害しIL−4レベル(第2y軸)を上昇させる、ことを示す。MSCの存在下での炎症促進性IFN−γの産生低下と抗炎症性IL−4の産生増加は、T細胞集団の抗炎症性表現型への移動を示した。
【図2A】図2Aは、MSCは炎症促進性エフェクターT細胞機能を阻害することを示す。FITC結合CD4(x軸)抗体とPE結合CD25(y軸)抗体を用いる、MSC+PBMC培養物(MSC+PBMC)中のPBMC又は非接着性画分を染色することによるTReg細胞数(%)のフローサイトメトリー解析。ゲートはバックグランドとしてのアイソタイプ対照抗体に基づく。グラフは5つの独立した実験の典型である。
【図2B】図2Bは、細胞培養上清中で、MSCの存在下で生成したTH1細胞は低レベルのIFN−γ(第1y軸)を分泌し、MSCの存在下で生成したTH2細胞は増加量のIL−4(第2y軸)を分泌した、ことを示す。
【図2C】図2Cは、MSCが、24ウェルプレート中で0、24、又は48時間培養した精製NK細胞からのIFN−γ分泌を阻害する、ことを示す。示したデータは1つの実験の平均±SDサイトカイン分泌であり、3つの独立した実験の典型である。
【図3A】図3Aは、MSCはTreg細胞集団の数とGITR発現を上昇させることを示す。CD4+CD25+Treg細胞集団を、PBMC又はMSC+PBMC(MSC対PBMC比は1:10)培養物(さらなる刺激無しで3日間培養した)から2工程磁性単離法を使用して単離した。これらの細胞に放射線照射し(さらなる増殖を阻止するため)、混合リンパ球反応(MLR)において刺激細胞として使用し、ここで応答細胞は植物性血球凝集素(PHA)(2.5mg/ml)の存在下の同種異系PBMC(刺激細胞対応答細胞比は1:100)であった。細胞を48時間培養し、次に3Hチミジンを加え、24時間後、取り込まれた放射活性を計測した。結果は、MSCの存在下で生成したTreg集団(レーン3)は、MSCの非存在下で生成したTreg細胞(レーン2)と機能的に同等であることを示した。
【図3B】図3Bは、PBMCをMSCの非存在下(上のプロット)又は存在下(下のプロット)で3日間培養(MSC対PBMC比は1:10)し、次に非接着画分を採取し、FITC標識GITRとPE標識CD4で免疫染色した結果を示す。結果は、MSCの存在下で培養した細胞でGITR発現の2倍以上の上昇を示す。
【図4A】図4Aは、MSCはPGE2を産生し、PGE2を阻止するとMSC性免疫調節作用が逆転することを示す。種々の濃度のPGE2ブロッカーであるNS−398又はインドメタシン(Indometh.)の存在下又は非存在下で培養したMSCから得られた培養上清中のPGE2分泌(平均±SD)。インヒビター濃度はμMであり、示したデータは24時間培養後に得られた値である。
【図4B】図4Bは、リアルタイムRT−PCRを使用したMSCとPBMC中のCOX−1とCOX−2発現を示す。MSCはPBMCと比較して有意に高レベルのCOX−2を発現し、MSCをPBMCの存在下で培養すると、MSC中のCOX−2発現の3倍を超える上昇があった。1〜3つの独立した実験からの典型的データを示す。MSC+PBMC培養物をトランスウェルチャンバープレート中で準備し、ここでMSCは下のチャンバーに蒔き、PBMCは上のチャンバーに蒔いた。
【図4C】図4Cは、(C)PGE2ブロッカーであるインドメタシン(Ind.)又はNS−398の存在は、対照と比較して、活性化DC(白バー)からのTNF−α分泌と、TH1細胞(斜線バー)からのIFN−γ分泌を上昇させることを示す。データは、MSCとPGE2インヒビターの非存在下で生成した培養物からの%変化として計算した。
【図4D】図4Dは、MSC−PBMC同時培養物(1:10)中のPGE2ブロッカーであるインドメタシン(Indo)又はNS−398の存在は、PHA処理PBMCに対するMSC性抗増殖作用を逆転させることを示す。示したデータは1つの実験からであり、3つの独立した実験の典型である。
【図5】図5は、構成性MSCサイトカイン分泌は同種異系PBMCの存在下で上昇することを示す。すでに性状解析したヒトMSCを使用して、PBMCの存在下(斜線バー)又は非存在下(白バー)(MSC対PBMC比は1:10)で24時間培養したMSCの培養上清中のサイトカインIL−6とVEGF、脂質メディエーターPGE2、及びマトリックス金属プロテアーゼ1(pro−MMP−1)のレベルを分析した。MSCはIL−6、VEGF、及びPGE2を構成性に産生し、これらの因子のレベルはPBMCとの同時培養で上昇し、従ってMSCが炎症性状況で免疫機能の調節において役割を果たすことを示唆する。
【図6】図6は、MSCは分裂促進剤誘導性のT細胞増殖を用量依存的に阻害することを示す。増加する数の同種異系PBMCを、PHA(2.5mg/ml)の存在下又は非存在下で96ウェルプレートに蒔いた一定数のMSC(2,000細胞/ウェル)とインキュベートし、3Hチミジン取り込みを測定した(1分当たりのカウント、cpm)。MSCの存在下でPHA処理PBMCの増殖の用量依存性阻害があった。3つの独立した実験の1つからの典型的結果を示す。同様の結果が、LeBlanc, et al., Scand J. Immunol., Vol. 57, pg. 11 (2003)により報告された。
【図7】図7は、提唱されているMSC作用機構の模式図を示す。MSCは、先天的(DC経路2〜4;及びNK経路6)免疫系と適応(T経路1と5、及びB経路7)免疫系の両方からの細胞に影響を与えることにより免疫調節作用を仲介する。浸入病原体に応答して未成熟DCは、侵入可能部位に移動し、成熟し、未処理のT細胞を刺激して(抗原特異的及び同時刺激シグナルにより)、防御性エフェクターT細胞(細胞性TH1又は体液性TH2免疫)になる能力を獲得する。MSC−DC相互作用中に、MSCは直接の細胞−細胞接触により又は分泌された因子により、細胞性応答(経路2)を開始するDCの能力を制限することにより、又は体液性応答(経路4)を開始する能力を促進することにより、免疫応答の結果を変化させる。また成熟エフェクターT細胞が存在する時、MSCはこれらと相互作用してTH1(経路1)応答のバランスをTH2応答(経路5)に向けて、おそらくIgE産生B細胞活性(経路7)の上昇(これは、GvHDと自己免疫疾患症状の抑制に好ましい結果である)に向ける。MSCはTReg集団(経路3)の生成を増加させる能力により、寛容表現型となり、局所的微小環境におけるバイスタンダー(bystander)炎症を緩和することにより受容体宿主を助ける。点線は提唱された機構である。
【特許請求の範囲】
【請求項1】
心臓以外の動物の臓器又は組織中の血管形成を促進する方法であって、
該動物の心臓以外の臓器又は組織中の血管形成を促進するために有効な量の間葉系幹細胞を該動物に投与することを含む、前記方法。
【請求項2】
前記動物が哺乳動物である、請求項1記載の方法。
【請求項3】
前記動物が霊長類である、請求項2記載の方法。
【請求項4】
前記霊長類がヒトである、請求項3記載の方法。
【請求項5】
前記間葉系幹細胞が、約1×105細胞/kg〜約1×107細胞/kgの量で投与される、請求項1記載の方法。
【請求項6】
前記間葉系幹細胞が、約1×106細胞/kg〜約5×106細胞/kgの量で投与される、請求項5記載の方法。
【請求項7】
前記間葉系幹細胞が全身的に投与される、請求項1記載の方法。
【請求項8】
前記間葉系幹細胞が静脈内に投与される、請求項1記載の方法。
【請求項9】
前記間葉系幹細胞が、前記動物の心臓以外の臓器又は組織への直接注入により投与される、請求項1記載の方法。
【請求項10】
動物の自己免疫疾患及び移植片対宿主反応からなる群より選ばれる疾患を治療する方法であって、以下のステップ:
該動物の疾患を治療するために有効な量の間葉系幹細胞を該動物に投与すること、
を含む、前記方法。
【請求項11】
前記動物が哺乳動物である、請求項10記載の方法。
【請求項12】
前記哺乳動物がヒトである、請求項11記載の方法。
【請求項13】
前記疾患が多発性硬化症である、請求項10記載の方法。
【請求項14】
動物の炎症応答を治療するために有効な量の間葉系幹細胞を該動物に投与することを含む、動物の炎症応答の治療方法。
【請求項15】
前記動物が哺乳動物である、請求項14記載の方法。
【請求項16】
前記哺乳動物がヒトである、請求項15記載の方法。
【請求項17】
前記炎症応答が乾癬と関連している、請求項14記載の方法。
【請求項18】
動物の癌を治療するために有効な量の間葉系幹細胞を該動物に投与することを含む、動物の癌の治療方法。
【請求項19】
前記動物が哺乳動物である、請求項18記載の方法。
【請求項20】
前記哺乳動物がヒトである、請求項19記載の方法。
【請求項21】
動物のアレルギー疾患又は障害を治療するために有効な量の間葉系幹細胞を該動物に投与することを含む、動物のアレルギー疾患又は障害の治療方法。
【請求項22】
前記動物が哺乳動物である、請求項21記載の方法。
【請求項23】
前記哺乳動物がヒトである、請求項22記載の方法。
【請求項24】
前記のアレルギー疾患又は障害が関節炎である、請求項14記載の方法。
【請求項25】
動物の創傷治癒を促進するために有効な量の間葉系幹細胞を該動物に投与することを含む、動物の創傷治癒の促進方法。
【請求項26】
前記動物が哺乳動物である、請求項25記載の方法。
【請求項27】
前記哺乳動物がヒトである、請求項26記載の方法。
【請求項28】
動物の繊維症を予防するために有効な量の間葉系幹細胞を該動物に投与することを含む、動物の繊維症の予防方法。
【請求項29】
動物の上皮傷害を修復するために有効な量の間葉系幹細胞を該動物に投与することを含む、動物の上皮傷害の修復方法。
【請求項30】
前記動物が哺乳動物である、請求項29記載の方法。
【請求項31】
前記哺乳動物がヒトである、請求項30記載の方法。
【請求項32】
前記上皮傷害が、移植片対宿主病の結果である、請求項29記載の方法。
【請求項1】
心臓以外の動物の臓器又は組織中の血管形成を促進する方法であって、
該動物の心臓以外の臓器又は組織中の血管形成を促進するために有効な量の間葉系幹細胞を該動物に投与することを含む、前記方法。
【請求項2】
前記動物が哺乳動物である、請求項1記載の方法。
【請求項3】
前記動物が霊長類である、請求項2記載の方法。
【請求項4】
前記霊長類がヒトである、請求項3記載の方法。
【請求項5】
前記間葉系幹細胞が、約1×105細胞/kg〜約1×107細胞/kgの量で投与される、請求項1記載の方法。
【請求項6】
前記間葉系幹細胞が、約1×106細胞/kg〜約5×106細胞/kgの量で投与される、請求項5記載の方法。
【請求項7】
前記間葉系幹細胞が全身的に投与される、請求項1記載の方法。
【請求項8】
前記間葉系幹細胞が静脈内に投与される、請求項1記載の方法。
【請求項9】
前記間葉系幹細胞が、前記動物の心臓以外の臓器又は組織への直接注入により投与される、請求項1記載の方法。
【請求項10】
動物の自己免疫疾患及び移植片対宿主反応からなる群より選ばれる疾患を治療する方法であって、以下のステップ:
該動物の疾患を治療するために有効な量の間葉系幹細胞を該動物に投与すること、
を含む、前記方法。
【請求項11】
前記動物が哺乳動物である、請求項10記載の方法。
【請求項12】
前記哺乳動物がヒトである、請求項11記載の方法。
【請求項13】
前記疾患が多発性硬化症である、請求項10記載の方法。
【請求項14】
動物の炎症応答を治療するために有効な量の間葉系幹細胞を該動物に投与することを含む、動物の炎症応答の治療方法。
【請求項15】
前記動物が哺乳動物である、請求項14記載の方法。
【請求項16】
前記哺乳動物がヒトである、請求項15記載の方法。
【請求項17】
前記炎症応答が乾癬と関連している、請求項14記載の方法。
【請求項18】
動物の癌を治療するために有効な量の間葉系幹細胞を該動物に投与することを含む、動物の癌の治療方法。
【請求項19】
前記動物が哺乳動物である、請求項18記載の方法。
【請求項20】
前記哺乳動物がヒトである、請求項19記載の方法。
【請求項21】
動物のアレルギー疾患又は障害を治療するために有効な量の間葉系幹細胞を該動物に投与することを含む、動物のアレルギー疾患又は障害の治療方法。
【請求項22】
前記動物が哺乳動物である、請求項21記載の方法。
【請求項23】
前記哺乳動物がヒトである、請求項22記載の方法。
【請求項24】
前記のアレルギー疾患又は障害が関節炎である、請求項14記載の方法。
【請求項25】
動物の創傷治癒を促進するために有効な量の間葉系幹細胞を該動物に投与することを含む、動物の創傷治癒の促進方法。
【請求項26】
前記動物が哺乳動物である、請求項25記載の方法。
【請求項27】
前記哺乳動物がヒトである、請求項26記載の方法。
【請求項28】
動物の繊維症を予防するために有効な量の間葉系幹細胞を該動物に投与することを含む、動物の繊維症の予防方法。
【請求項29】
動物の上皮傷害を修復するために有効な量の間葉系幹細胞を該動物に投与することを含む、動物の上皮傷害の修復方法。
【請求項30】
前記動物が哺乳動物である、請求項29記載の方法。
【請求項31】
前記哺乳動物がヒトである、請求項30記載の方法。
【請求項32】
前記上皮傷害が、移植片対宿主病の結果である、請求項29記載の方法。
【図1A】


【図1B】
【図1C】
【図2A】
【図2B】
【図2C】
【図3A】
【図3B】
【図4A】
【図4B】
【図4C】
【図4D】
【図5】
【図6】
【図7】
【図1B】
【図1C】
【図2A】
【図2B】
【図2C】
【図3A】
【図3B】
【図4A】
【図4B】
【図4C】
【図4D】
【図5】
【図6】
【図7】
【公表番号】特表2010−505764(P2010−505764A)
【公表日】平成22年2月25日(2010.2.25)
【国際特許分類】
【出願番号】特願2009−530402(P2009−530402)
【出願日】平成19年9月26日(2007.9.26)
【国際出願番号】PCT/US2007/020724
【国際公開番号】WO2008/042174
【国際公開日】平成20年4月10日(2008.4.10)
【出願人】(508241152)オシリス セラピューティクス,インコーポレイティド (2)
【Fターム(参考)】
【公表日】平成22年2月25日(2010.2.25)
【国際特許分類】
【出願日】平成19年9月26日(2007.9.26)
【国際出願番号】PCT/US2007/020724
【国際公開番号】WO2008/042174
【国際公開日】平成20年4月10日(2008.4.10)
【出願人】(508241152)オシリス セラピューティクス,インコーポレイティド (2)
【Fターム(参考)】
[ Back to top ]