
間葉系細胞の製造方法、歯の製造方法及び歯形成用間葉系細胞
【課題】特有の細胞配置を保持した歯を効率よく得ることができる歯形成用間葉系細胞を提供すること、及びこれを用いて特有の細胞配置を保持した歯を効率よく得る。
【解決手段】全能性幹細胞、例えば胚性癌腫細胞を、ジメチルスルホキシドなどの分化誘導剤の存在下で培養して、分化誘導後の細胞集団から、CD44陽性且つCD29陽性細胞又はCD44陽性且つCD106陽性細胞を歯形成用間葉系細胞として選別する。また、間葉系細胞と上皮系細胞とのいずれか一方から実質的になる第1の細胞集合体と、いずれか他方のみから実質的になる第2の細胞集合体とを、細胞の接触状態を保持可能な支持担体の内部に混合することなく密着させて配置して培養する際、上記間葉系細胞を、歯形成用間葉系細胞を含むものとすることよって、特有の細胞配置を有する歯を得る。
【解決手段】全能性幹細胞、例えば胚性癌腫細胞を、ジメチルスルホキシドなどの分化誘導剤の存在下で培養して、分化誘導後の細胞集団から、CD44陽性且つCD29陽性細胞又はCD44陽性且つCD106陽性細胞を歯形成用間葉系細胞として選別する。また、間葉系細胞と上皮系細胞とのいずれか一方から実質的になる第1の細胞集合体と、いずれか他方のみから実質的になる第2の細胞集合体とを、細胞の接触状態を保持可能な支持担体の内部に混合することなく密着させて配置して培養する際、上記間葉系細胞を、歯形成用間葉系細胞を含むものとすることよって、特有の細胞配置を有する歯を得る。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、歯を形成するために用いられる間葉系細胞の製造方法、歯の製造方法及び歯形成用間葉系細胞に関する。
【背景技術】
【0002】
歯は、最外層にエナメル質、その内層に象牙質という硬組織を有し、さらに象牙質の内側に象牙質を産生する象牙芽細胞、その更に中心部に歯髄を有する器官であって、齲蝕や歯周病等によって失われることがある器官である。一般に歯の損失については、生命に対する危惧が少ないと考えられているため、現在は主として入れ歯やインプラントにより補うことが多い。しかしながら、歯の有無は外見や食べ物の味覚に大きく影響し、また健康維持や質の高い生活を維持するという観点から歯の再生技術の開発への関心が高まって来た。
【0003】
歯は、胎児期の発生過程の誘導によって形成され、複数の細胞種によって構築された機能単位であり、器官や臓器と同じであると考えられている。そのため歯は、成体内の造血幹細胞や間葉系幹細胞のような幹細胞から細胞種が発生する幹細胞システムによって発生するのではない。このことから、現在、再生医療によって進められている幹細胞の移入のみ(幹細胞移入療法)では、歯を再生することができない。
そこで、近年、単離された歯胚細胞を用いた歯胚を再構成させて、この再構成歯胚を移植することによる歯の再生を中心とした検討が行われている。
【0004】
例えば、非特許文献1には、歯胚から単離された上皮系細胞や間葉系の歯嚢細胞などの細胞を、生体吸収性の担体と共にラットの腹腔内に移植することで歯様の組織が再生されることが開示されている。
歯胚の再生方法としては、例えば、特許文献1には、生体から単離された歯胚細胞を、線維芽細胞増殖因子等の生理活性物質の存在下で培養することが記載されている。また、特許文献2には、生体から単離された歯胚細胞及びこれらの細胞に分化可能な細胞のうち少なくとも1種類を、フィブリンを含む担体と一緒に培養することが提案されており、ここでフィブリンを含む担体は、歯胚の目的形状のものを使用して、特有の形態を有する「歯」を形成すると記載されている。
【0005】
特許文献3及び特許文献4号公報には、6ヶ月のブタの下顎骨から、象牙質を形成する歯髄由来の間葉系細胞とエナメル形成に寄与する上皮系細胞とを含む歯胚との細胞混合物を、ポリグリコール酸−ポリ酢酸共重合体からなる生分解性ポリマーを固化させた担体(Scaffold)に播種して、動物の体内へ移植し、歯を形成する方法が開示されている。ここで担体は、歯胚の目的形状のものを使用して、特有の形態を有する「歯」を形成すると記載されている。
【0006】
一方、特許文献5には、骨の欠損又は損傷を有する患者を治療するための歯の再生方法を開示している。この方法によれば、ポリグリコール酸メッシュ担体に、例えば歯胚由来の間葉系細胞を播種した後に、上皮系細胞をコラーゲンと共に重層する又は上皮細胞シートで包むことによって、骨が形成される。なお、特許文献5では、骨の形状を構築するために担体を用いている。
【0007】
また、特許文献6には、歯胚由来の上皮系細胞と間葉系細胞とをそれぞれ細胞集合体とした上で、コラーゲンゲルの内部に細胞集合体同士を密着させ、その状態を保持しながら培養することによって、歯特有の細胞配置を備えた歯を製造する技術が開示されている。
【0008】
しかしながら、上記技術では、歯又は歯胚を再生するために歯胚細胞又はこれに分化可能な細胞を生体から入手している。このように生体から採取する細胞を用いる技術では、入手可能な細胞数が充分でない場合がある。また、細胞の入手元について制限が設けられることがある。
また、組織として機能するには、組織を構成する複数種の細胞が適切な相対位置に配置(細胞配置)され、組織としての方向性を有することが必須である。
【0009】
【非特許文献1】J. Dent. Res., 2002, Vol.81(10), pp.695-700
【特許文献1】特開2004−331557号公報
【特許文献2】特開2004−357567号公報
【特許文献3】米国特許出願公開第2002/0119180号公報
【特許文献4】米国特許出願公開第2004/0219489号公報
【特許文献5】国際公開第2005/014070号パンフレット
【特許文献6】国際公開第2006/129672号パンフレット
【発明の開示】
【発明が解決しようとする課題】
【0010】
しかしながら、上記技術では、歯又は歯胚を再生するために歯胚細胞又はこれに分化可能な細胞を生体から入手している。このように生体から採取する細胞を用いる技術では、入手可能な細胞数が充分でない場合があるだけでなく、入手元について制限が設けられることがある。
また、組織として機能するには、組織を構成する複数種の細胞が適切な相対位置に配置(細胞配置)され、組織としての方向性を有することが必須である。
【0011】
従って本発明は、特有の細胞配置を保持した歯を効率よく得ることができる歯形成用間葉系細胞を提供すること、及びこれを用いて特有の細胞配置を保持した歯を効率よく得ることを目的とする。
【課題を解決するための手段】
【0012】
本発明の間葉系細胞の製造方法は、歯を形成するために用いられる間葉系細胞の製造方法であって、全能性幹細胞を分化誘導剤の存在下で培養して、CD44陽性且つCD29陽性細胞又はCD44陽性且つCD106陽性細胞を含む分化誘導処理後の細胞集団を生成すること、前記分化誘導処理後の細胞集団から、CD44陽性且つCD29陽性細胞又はCD44陽性且つCD106陽性細胞を、歯形成用間葉系細胞として選別することを含む。
本発明の歯の製造方法は、支持担体の内部に、間葉系細胞及び上皮系細胞のいずれか一方のみから実質的になる第1の細胞集合体と、いずれか他方のみから実質的になる第2の細胞集合体とを混合することなく密着させて配置すること、前記第1及び第2の細胞集合体を前記支持担体の内部で培養すること、を含み、前記間葉系細胞が上記歯形成用間葉系細胞を含む。
本発明の歯形成用間葉系細胞は、全能性幹細胞から誘導されるCD44陽性且つCD29陽性又はCD44陽性且つCD106陽性の歯形成用間葉系細胞である。
【0013】
本発明の間葉系細胞の製造方法は、全能性幹細胞に分化誘導処理を行うことによって得られたCD44陽性且つCD29陽性細胞又はCD44陽性且つCD106陽性細胞を歯形成用間葉系細胞として選別するので、この間葉系細胞を得るために生体由来の歯等を使用する必要がなく、目的とする歯を大量に作製することができる。
また、このような歯形成用間葉系細胞を用いるので、目的とする歯を効率よく製造することができる。
【発明の効果】
【0014】
本発明によれば、目的とする歯を効率よく大量に作製することができる間葉系細胞の製造方法、及びエナメル質及び象牙質による特有の細胞配置を保持した歯を、効率よく大量に製造することができる歯の製造方法を提供することができる。
【発明を実施するための最良の形態】
【0015】
[間葉系細胞の製造方法]
本発明の間葉系細胞の製造方法は、歯を形成するために用いられる間葉系細胞を製造する間葉系細胞の製造方法であって、全能性幹細胞を分化誘導剤の存在下で培養して、CD44陽性且つCD29陽性細胞又はCD44陽性且つCD106陽性細胞を含む分化誘導処理後の細胞集団を生成すること、前記分化誘導処理後の細胞集団から、CD44陽性且つCD29陽性細胞又はCD44陽性且つCD106陽性細胞を、歯形成用間葉系細胞として選別することを含むものである。
【0016】
本発明は、全能性幹細胞を分化誘導して得られたCD44陽性且つCD29陽性細胞又はCD44陽性且つCD106陽性細胞が、歯形成用間葉系細胞として使用できることを見出したものである。これにより、歯を作製するために必要な間葉系細胞を、生体由来の歯又は歯胚以外からも得ることができ、歯を形成するための間葉系細胞を得るために、生体由来の歯等を使用する必要性を排除することができる。この結果、歯胚から直接採取した細胞数が充分でない事情や、入手元における制限の有無といった事情を考慮することなく、必要な量の間葉系細胞を容易に、また場合よって大量に得ることができる。
【0017】
本発明において、「歯」とは、内側に象牙質及び外側にエナメル質の層を連続して備えた組織をいい、特に歯冠や歯根を有する方向性を備えた組織をいう。歯の方向性は、歯冠や歯根の配置によって特定することができる。歯冠や歯根は、形状や組織染色などに基づいて目視にて確認することができる。歯冠とは、エナメル質と象牙質の層構造を有する部分をいい、歯根にはエナメル質の層は存在しない。
【0018】
象牙質及びエナメル質は、当業者には、組織染色などによって形態的に容易に特定することができる。また、エナメル質は、エナメル芽細胞の存在によって特定することができ、エナメル芽細胞の存在は、アメロジェニンの有無によって確認することができる。一方、象牙質は、象牙芽細胞の存在によって特定することができ、象牙芽細胞の存在は、デンチンシアロプロテインの有無によって確認することができる。アメロジェニン及びデンチンシアロプロテインの確認はこの分野で周知の方法によって容易に実施することができ、例えば、in situ ハイブリダイゼーション、抗体染色等をあげることができる。
【0019】
また本発明における間葉系細胞は、歯を形成するために、まず歯胚を形成するために使用できる。
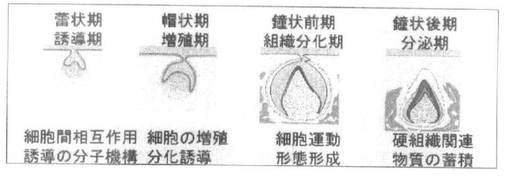
本発明において「歯胚」及び「歯芽」は、後述する発生段階に基づいて区別されたものに特に言及する場合に用いられる表現である。この場合の「歯胚」とは、将来歯になることが決定付けられた歯の初期胚であり、歯の発生ステージで一般的に用いられる蕾状期(Bud stage)から鐘状期(Bell stage)までの段階であり、特に歯の硬組織としての特徴である象牙質、エナメル質の蓄積が認められない組織である。一方、「歯芽」とは本発明で用いられる「歯胚」の段階移行の、歯の硬組織の特徴である象牙質、エナメル質の蓄積が始まった段階から歯が歯肉から萌芽して一般的に歯としての機能を発現する前の段階の組織をいう。
【0020】
歯胚から歯への発生は、図1に示されるように個体発生の過程で、蕾状期、帽状期、鐘状前期及び後期の各ステージを経て行われる。ここで、蕾状期では、上皮系細胞が間葉系細胞を包むように陥入し、鐘状前期及び鐘状後期に至ると、上皮系細胞部分が外側のエナメル質となり、間葉系細胞部分が内部に象牙質を形成するようになる。従って、上皮系細胞と間葉系細胞との細胞間相互作用によって歯胚から歯が形成される。
なお本発明において「間葉系細胞」とは、間葉組織由来の細胞を意味し、「上皮系細胞」とは上皮組織由来の細胞を意味する。
また本発明において「歯周組織」とは、歯の主として外層の形成された歯槽骨及び歯根膜をいう。歯槽骨及び歯根膜は、当業者には、組織染色などによって形態的に容易に特定することができる。
【0021】
以下に本発明にかかる歯形成用間葉系細胞について説明する。
本発明に係る歯形成用間葉系細胞を得るために用いられる全能性幹細胞は、少なくとも2以上の細胞に分化可能な多分化能を有する細胞を言い、好ましくは胚性癌腫細胞、胚性幹細胞、胚性生殖幹細胞からなる群より選択されたものを使用することができる。このうち、入手の容易性の観点から胚性癌腫細胞(以下、「EC細胞」という)であることがより好ましい。本発明に適用可能なEC細胞には、神経、睾丸、卵巣などいずれの組織に由来するものであってもよい。このようなEC細胞としては。例えばヒト由来としてはNCR−G3細胞、並びにNTERA−2細胞を挙げることができ、マウス由来としてはc−1300細胞やF9細胞、LT−2細胞、OTT6050細胞、PCC4細胞、P19細胞、METT−1細胞、STT−3細胞などの細胞株を挙げることができる。これらの細胞は、哺乳動物の霊長類(例えばヒト、サルなど)、有蹄類(例えば豚、牛、馬など)、小型哺乳類の齧歯類(例えばマウス、ラット、ウサギなど)の種々の動物に由来するものから、適宜使用目的に応じて選択することができる。例えばマウス由来の胚性癌腫細胞株としては、上記に記載した細胞株やこれらの派生クローンを挙げることができる
【0022】
全能性幹細胞の培養には、通常使用される培地を使用することができる。培養に用いられる培地としては、一般に動物細胞の培養に用いられる培地、例えばダルベッコ改変イーグル培地(DMEM)等を用いることができる。この培地には、細胞の増殖を促進するための血清を添加するか、あるいは血清に代替するものとして、例えばFGF、EGF、PDGF等の細胞増殖因子やトランスフェリン等の既知血清成分を添加してもよい。なお、血清を添加する場合の濃度は、そのときの培養状態によって適宜変更することができるが、通常10%とすることができる。細胞の培養には、通常の培養条件、例えば37℃の温度で5%CO2濃度のインキュベーター内での培養が適用される。また、適宜、ストレプトマイシン等の抗生物質を添加したものであってもよい。
【0023】
本発明では、全能性幹細胞に対して、分化誘導剤の存在下で培養することによる分化誘導処理が行われる。この分化誘導処理によって、全能性幹細胞から、CD44陽性且つCD29陽性細胞又はCD44陽性且つCD106陽性細胞を含む細胞集団が生成する。誘導処理によって得られる細胞集団には、上記のCD44陽性且つCD29陽性細胞又はCD44陽性且つCD106陽性細胞が含まれていればよく、これらの双方の細胞が含まれていてもよい。
【0024】
上記分化誘導剤としては、少なくとも全能性幹細胞からCD44発現細胞への分化誘導が可能なCD44陽性細胞誘導因子であれば使用することができる。このようなCD44陽性細胞誘導因子としては、神経堤細胞系誘導因子、神経堤細胞発生関与因子、神経堤細胞増殖促進因子を挙げることができる。神経堤細胞系誘導因子は、全能性幹細胞を神経堤細胞又はその派生細胞(平滑筋細胞、心筋細胞等)へ誘導することができる因子であり、例えば、ジメチルスルホキシド(DMSO)、Dex、cAMP、ヘキサメチレンビスアセトアミド、5’−アザシチジン、TGFβ、Noggin等を挙げることができる。神経堤細胞発生関与因子としては、レチノイン酸(RA)、BMP−4、BMP−7、Shh、Wnt、FGF2、エンドセリン−1、エントセリン−3、アクチビンβA、PDGF、VEGF等を挙げることができる。神経堤細胞増殖促進因子としては、FGF2、FGF8、FGF10、BMP−2、SCF、活性型ビタミンD3等を挙げることができる。神経堤細胞系誘導因子、神経堤細胞発生関与因子及び神経堤細胞増殖促進因子はそれぞれ明確に区別されるものではなく、また、これらを単独で又は1以上を組み合わせて使用することができる。このうち、後述する本発明の目的細胞集団を効率よく生成可能なDMSO及びRAが好ましく、これらは単独又は組み合わせて使用することができる。
【0025】
これらの分化誘導剤は、全能性幹細胞の培地に分化誘導可能な濃度、添加すればよい。ここで分化誘導可能な濃度とは、用いられる分化誘導剤の種類や全能性幹細胞の種類等によって異なる。
例えばDMSOの場合、一般に培地の容量に対して、0.5容量%〜10容量%、処理に対する細胞の生存率及び培養によって得られる目的の細胞の取得効率の観点から、好ましくは2.5容量%〜5容量%、更に好ましくは4容量%〜5容量%の濃度とすることができる。2.5容量%以上の濃度であれば、全能性幹細胞の分化を充分に誘導することができ、一方、5容量%以下の濃度であれば目的の細胞取得の効率を著しく損なうことがない。
また、RAの場合、一般に培地中で0.1μM〜10μM、処理に対する細胞の生存率及び培養によって得られる目的の細胞の取得効率の観点から、好ましくは0.5μM〜5μM、更に好ましくは0.5μM〜2μMの濃度とすることができる。0.1μM以上の濃度であれば、全能性幹細胞の分化を充分に誘導することができ、一方、10μM以下の濃度であれば目的の細胞取得の効率を著しく損なうことがない。
【0026】
全能性幹細胞に対する分化誘導処理は、上記分化誘導剤の存在下で全能性幹細胞を培養によって行われる。分化誘導処理(培養)の期間は、分化誘導剤の濃度と細胞の成育活性によって異なるが、一般に2時間〜3日、細胞の生存率、ならびに目的の細胞の入手効率の観点から好ましくは6時間〜24時間とすることができる。6時間以上とすることにより充分に分化誘導された細胞を効率よく得ることができ、24時間以下とすることによって効率を著しく損なうことがない。
【0027】
なお、分化誘導処理の期間と分化誘導剤の濃度との間には、全能性幹細胞に対する分化誘導を効果的に起こさせるために一定の関係が成立する。即ち、一般に、分化誘導剤存在下での培養期間(処理時間)は、分化誘導剤の濃度と分化誘導剤の種類によって異なるが、特定の分化誘導剤の場合には、培養日数を固定した場合、一般にその濃度には好ましい至適濃度域が存在する。また培養日数を短くした場合には、至適濃度域は高い濃度側に移行し、逆に培養日数を長くする場合には至適濃度域は低い濃度側に移行する。これらの要素を考慮すると、培養日数の調節によって幅広い濃度域での分化誘導効果が得られると共に、当業者には、誘導効果が得られる至適濃度域を容易に設定することができる。
【0028】
上記分化誘導処理によって得られた分化誘導処理後の細胞集団には、CD44陽性且つCD29陽性細胞又はCD44陽性且つCD106陽性細胞を含む細胞集団が生成する。これら双方の細胞が含まれていてもよい。この分化誘導処理後の細胞集団は、次いで、CD44陽性且つCD29陽性細胞又はCD44陽性且つCD106陽性細胞を歯形成用間葉系細胞として選別する選別処理に供される。
【0029】
細胞の選別は、上述したような本発明にかかる間葉系細胞を選別することができる手段であればいずれを用いてもよい。このような選別手段としては、遺伝子発現パターン、細胞表面抗原、形態などを基準とする選別を挙げることができ、これらの1つ又は2つ以上を用いて、全能性幹細胞を選別することができる。選別手段は、細胞選別の効率の観点から、細胞表面の発現をもとにしたセルソーターを用いることが好ましく、確実に間葉系細胞を選別するためには、複数の組み合わせ、特に、遺伝子発現パターンと細胞表現抗原の発現とを組み合わせて使用することが好ましい。
【0030】
本発明において選別された歯形成用間葉系細胞は、CD44陽性であることに加えて、CD29陽性又はCD106陽性の細胞性状を示すものである。CD44は、ヒアルロン酸受容体として細胞表面に発現する膜タンパク質であることが知られている。一方、CD29は、インテグリンβ1分子として細胞表面に発現する膜タンパク質であることが知られており、EC細胞では弱陽性、間葉系細胞ではmRNAレベルで発現していることがそれぞれ報告されている。またCD106は、接着分子として既知のVCAM−1分子であることが知られており、主として上皮系細胞のマーカーとして利用されることが多い。このような性状を示す細胞であれば、後述するように歯の形成において間葉系細胞として使用することができる。
本発明における歯形成用間葉系細胞は、CD44陽性に加えて、CD29及びCD106の少なくともいずれか一方の抗原が陽性となる集団であればよく、CD44、CD29、CD106の3種の抗原が共に陽性となる三重陽性細胞が含まれていてもよい。
【0031】
本発明における歯形成用間葉系細胞は、上記CD44、CD29及びCD106の細胞表現抗原に加えて、他の細胞性状に基づいて選別を行ってもよい。
他の細胞性状としては他の細胞表面抗原パターンや遺伝子発現パターンを挙げることができる。細胞表現抗原パターンとして、例えばCD14陰性、CD34陰性、CD45陰性等を挙げることができ、これらを単独又は組み合わせて使用することができる。また遺伝子発現パターンとしては、例えばSlug発現、Wnt5a発現、Lhx8発現、BMP4発現、Pax3発現、Pax9発現、Msx1発現、Oct3/4非発現、nanog非発現、Sox9非発現、Sox5非発現等を挙げることができ、これらを単独又は組み合わせて使用することができるが、これらに限定されるものではない。
【0032】
本発明の歯形成用間葉系細胞は、好ましくは、上記CD44、CD29、CD106に加えて、歯形成能の高さから、Slug遺伝子、Pax3遺伝子、Msx1遺伝子及びPax9遺伝子の群から選択された少なくとも1つの遺伝子を発現していることが好ましく、Slug遺伝子及びPax3遺伝子を共に発現していることが更に好ましく、これら4つの遺伝子をすべて発現していることが特に好ましい。
【0033】
これらの細胞表面抗原や遺伝子発現パターンの確認及びこれに基づく選別には、表面抗原を認識する各種抗体や遺伝子発現の確認が可能な核酸配列などを用いる。これらの抗体及び核酸配列は、いずれも公知であり容易に入手可能である。また、これらの表面抗原や遺伝子発現に基づく確認及び選別は、この用途に一般的に用いられる手段、例えば、各種抗体による免疫染色、フローサイトメトリー、セルソーター、RT−PCR等を用いればよい。
【0034】
また、本発明の製造方法では、上記選別手段に組み合わせて、単一の間葉系細胞とするためのクローニングを用いることができる。クローニングの方法としては、通常この目的で用いられている方法、例えば、限界希釈法、セルソーター、クローニングリングなどを用いることができる。クローニングは、上記選別手段による選別の前後のいずれで行ってもよいが、目的とする間葉系細胞を効率よく得るためには選別の後に行うことが好ましい。
【0035】
選別工程において目的とする細胞数を効率よく得るために、選別工程の前に増殖工程を設けてもよい。このような増殖工程では、分化誘導剤を含まない通常培養用の培地を用いて分化誘導後の細胞集団の培養を行う。これにより、分化誘導された目的細胞を所定の細胞数まで増殖させることができる。その結果、充分な細胞数により選別を容易に実施可能にすることができる。
このような増殖工程の期間は、分化誘導処理後の細胞数及び細胞の状態に基づいて適宜設定することができるが、目的細胞の濃縮効率の観点から一般に3〜30日、好ましくは5〜10日とすることができる。この結果、選別工程の対象となる細胞集団中に、目的とする歯形成用間葉系細胞が含まれることになり、このような細胞集団を選別工程に供することによって、本発明にかかる歯形成用間葉系細胞を効率よく且つ大量に得ることができる。
【0036】
本発明の方法で得られた間葉系細胞は、歯を形成するために用いられる。歯の形成は、本発明に係る間葉系細胞を用いるものであればいずれの方法であってもよいが、以下の本発明の歯の製造方法に用いられることが最も好ましい。
以下、本発明の歯の製造方法について説明する。
【0037】
[歯の製造方法]
本発明の歯形成用間葉系細胞を用いた好ましい歯の製造方法は、支持担体の内部に、間葉系細胞及び上皮系細胞のいずれか一方のみから実質的になる第1の細胞集合体と、いずれか他方のみから実質的になる第2の細胞集合体とを混合することなく密着させて配置すること(配置工程)、前記第1及び第2の細胞集合体を前記支持担体の内部で培養すること(培養工程)、を含み、前記間葉系細胞が上記歯形成用間葉系細胞を含むものである。
本製造方法では、間葉系細胞と上皮系細胞とを細胞集合体として支持担体の内部で混合することなく密着させた状態で生育させるので、緊密な接触状態によって細胞間相互作用を効果的に再現することができ、内側に象牙質、外側にエナメル質という歯に特有の細胞配置を有する歯を得ることができる。この際、間葉系細胞から実質的になる細胞集合体が本発明にかかる歯形成用間葉系細胞を含むので、特有の細胞配置を有する歯を効率よく大量に製造することができる。
【0038】
配置工程では、支持担体の内部に、第1の細胞集合体と第2の細胞集合体とを接触させて配置する。
ここで第1の細胞集合体及び第2の細胞集合体は、それぞれ間葉系細胞のみ、又は上皮系細胞のみから実質的に構成されているものである。ここで、間葉系細胞のみから実質的に構成されている細胞集合体は、上述した歯形成用間葉系細胞を含むものである。この歯形成用間葉系細胞を含む細胞集合体は、上述した製造方法に従った調製工程によって調製することができ、一方、上皮系細胞のみから実質的に構成されている細胞集合体は、間葉系細胞から実質的になる細胞集合体とは独立して調製することができる(第1の細胞調製工程及び第2の細胞調製工程)。
【0039】
また、「細胞集合体」とは、細胞が密集した状態をいい、組織の状態であっても、単一細胞の状態から調製された細胞集合体(細胞凝集体)であってもよい。また「実質的になる」とは、対象となる細胞以外のものをできるだけ含まないことを意味する。このため上皮系細胞からなる細胞集合体は、組織の一部又は単一細胞の集合体とすることができる。また、上皮系細胞及び間葉系細胞は共に単一細胞で構成された細胞集合体であってもよい。
第1の細胞集合体第2の細胞集合体は、いずれが上皮系細胞及び間葉系細胞であってもよく、この細胞集合体を構成する細胞の数は、動物の種類や、支持担体の種類、硬さ及び大きさによって異なるが、細胞集合体1個あたり、一般に101〜108個、好ましくは103〜108個とすることができる。
【0040】
間葉系細胞から実質的になる細胞集団は、歯形成用間葉系細胞を含んでいれば他の間葉系細胞を含んでいてもよい。
上記間葉系細胞以外の他の間葉系細胞としては、歯胚及び歯胚以外に由来する間葉系細胞を挙げることができる。歯胚以外に由来する間葉系細胞としては、生体内の他の間葉系組織に由来する細胞であり、好ましくは、血液細胞を含まない骨髄細胞や間葉系幹細胞、さらに好ましくは口腔内間葉系細胞や顎骨の内部の骨髄細胞、頭部神経堤細胞に由来する間葉系細胞、前記間葉系細胞を生み出しうる間葉系前駆細胞やその幹細胞等を挙げることができる。
【0041】
本製造方法で用いられる上皮系細胞は、生体内での細胞配置を再現して特有の構造及び方向性を有する歯を効果的に形成するために歯胚に由来するものであることが好ましく、細胞の分化段階の幼若性と均質性の観点から蕾状期から帽状期からのものであることが好ましい。
また、歯胚以外に由来する上皮系細胞であってもよく、これには、生体内の他の上皮系組織に由来する細胞を挙げることができる。好ましくは、皮膚や口腔内の粘膜や歯肉の上皮系細胞、さらに好ましくは皮膚や粘膜などの分化した、例えば角化した、あるいは錯角化した上皮系細胞を生み出しうる未熟な上皮系前駆細胞、たとえば非角化上皮系細胞やその幹細胞等を挙げることができる。
【0042】
細胞集合体を調製するために各細胞を組織から単離する場合、歯胚及び他の組織は、哺乳動物の霊長類、例えばヒト、サルなど、有蹄類、例えば豚、牛、馬など、小型哺乳類の齧歯類、例えばマウス、ラット、ウサギなどの種々の動物の顎骨等から採取することができる。歯胚及び組織の採取は、通常、組織の採取で用いられる条件をそのまま適用すればよく、無菌状態で取り出し、適当な保存液に保存すればよい。なお、ヒトの歯胚としては、第3大臼歯いわゆる親知らずの歯胚の他、胎児歯胚を挙げることができるが、自家組織の利用との観点から、親知らず歯胚を用いることが好ましい。
【0043】
組織、例えば歯胚から、上記細胞を調製する場合には、まず周囲の組織から単離された歯胚を、形状に従って歯胚間葉組織及び歯胚上皮組織に分ける。このとき、歯胚組織は顕微鏡下で構造的に見分けることが可能であるため、解剖用ハサミやピンセット等で切断、あるいは引き剥がすことによって容易に分離することができる。また、歯胚組織からの歯胚間葉組織及び歯胚上皮組織の分離は、その形状に従って注射針、タングステンニードル、ピンセット等で切断、あるいは引き剥がすことにより容易に行うことができる。
好ましくは、周囲組織から歯胚細胞を容易に分離するため及び/又は歯胚組織から上皮組織及び間葉組織を分離するために、酵素を用いてもよい。このような用途に用いられる酵素としては、ディスパーゼ、コラーゲナーゼ、トリプシン等を挙げることができる。
【0044】
細胞集合体を構成する細胞は、採取した組織から単一細胞の状態に調製してもよい。調製工程において単一の細胞に容易に分散可能とするために、酵素を用いてもよい。このような酵素としては、ディスパーゼ、コラーゲナーゼ、トリプシン等を挙げることができる。このとき、上皮組織からの上皮系細胞の分離にはコラーゲナーゼ処理後にトリプシン処理とDNase処理をすることが好ましい。他方、間葉組織からの間葉系細胞の分離には、コラーゲナーゼとトリプシンで同時に処理し、最終的にDNase処理をすることが好ましい。このときDNase処理を行うのは、酵素処理により一部の細胞がダメージをうけ、細胞膜が溶解したときに溶液中に放出されるDNAによって細胞が凝集し細胞の回収量が低下することすることを防ぐためである。
【0045】
なお、細胞集合体を構成する細胞は、それぞれ充分な細胞数を得るために、配置工程に先立って予備的な培養を経たものであってもよい。細胞の培養は、一般に動物細胞の培養に用いられる温度等の条件をそのまま用いることができる。
培養に用いられる培地としては、一般に動物細胞の培養に用いられる培地、例えばダルベッコ改変イーグル培地(DMEM)等を用いることができ、細胞の増殖を促進するための血清を添加するか、あるいは血清に代替するものとして、例えばFGF、EGF、PDGF等の細胞増殖因子やトランスフェリン等の既知血清成分を添加してもよい。なお、血清を添加する場合の濃度は、そのときの培養状態によって適宜変更することができるが、通常10容量%とすることができる。細胞の培養には、通常の培養条件、例えば37℃の温度で5%CO2濃度のインキュベーター内での培養が適用される。また、適宜、ストレプトマイシン等の抗生物質を添加したものであってもよい。
【0046】
この予備的な培養を間葉系細胞に適用する場合、上述した歯形成用間葉系細胞の増殖のための培養を兼ねたものであってもよく、選別工程後に行うもの、又はこの双方であってもよい。
また、細胞の性質を変えないという観点から、間葉系細胞の予備培養や上述した歯形成用間葉系細胞の単なる増殖のための培養では、前述した分化誘導剤を含有しない培地で行うことが好ましい。
【0047】
配置工程における細胞集合体の配置は、上記第1及び第2の細胞集合体を、細胞の接触状態を保持可能な支持担体の内部に配置する。このとき、各細胞集合体は互いに混合することがない。このように各細胞集合体を混合することなく配置するので、細胞集合体の間に境界線が形成される。このような配置形態を、本明細書中では適宜「区画化」と表現する。
【0048】
ここで用いられる支持担体としては、細胞を内部で培養可能なものであればよく、好ましくは、上記培地との混合物である。このような支持担体としては、コラーゲン、アガロースゲル、カルボキシメチルセルロース、ゼラチン、寒天、ハイドロゲル、セルマトリクス(商品名)、メビオールゲル(商品名)、マトリゲル(商品名)、エラスチン、フィブリン、ラミニン、細胞外マトリクス混合物、ポリグリコール酸(PGA)、ポリ乳酸(PLA)、乳酸/グリコール酸共重合体(PLGA)等を挙げることができる。これらの支持担体は、細胞を内部に配置したときに配置した位置をほぼ維持可能な程度な硬さを有するものであればよく、ゲル状、繊維状、固体状のものを挙げることができる。中でも細胞外マトリクス混合物等のゲルが適切な硬さや保持力を提供しやすいといった観点から、コラーゲン、アガロースゲル、カルボキシメチルセルロース、ゼラチン、寒天、ハイドロゲル、セルマトリクス、メビオールゲル、マトリゲル、細胞倍マトリクス混合物、エラスチン、フィブリン、ラミニンがより好ましい。ここで、細胞の位置を維持可能な硬さとは、通常、三次元培養として適用される硬さ、即ち、細胞の配置を保持できると共に増殖による肥大化を阻害しない硬さであればよく、容易に決定することができる。
なお、ここで支持担体は、第1及び第2の細胞集合体が担体内部で成育することができる程度の厚みを有すればよく、目的とする組織の大きさ等によって適宜設定することができる。
【0049】
また、支持担体は、細胞が分散することなく細胞の接触状態を保持可能な保持力を備えていればよい。ここでいう「接触状態」とは、各細胞集合体において、また細胞集合体間において、確実に細胞相互作用させるために緊密(高密度)の状態であることが好ましく、このような高密度の状態とは、細胞凝集体の場合には、例えば、単に触れ合うよりも強く密着した状態を維持可能な保持力で細胞を培養可能なものとすることができる。例えば、コラーゲンの場合、最終濃度2〜3mg/mlの濃度、即ちJIS-K6503-1996に準拠した方法(12.7mm径のプランジャーで4mm押し下げるのに必要な荷重として測定)によって120g〜250gのゼリー強度となる濃度での使用が適切な硬さを提供する。また、このゼリー強度は限定されず、他の種の支持担体においても、同様の評価方法によって同様の強度があれば本発明の支持担体として好ましく用いられる。また1種又は複数種の支持担体を混合することによって、目的とするゼリー強度に相当する硬さの支持担体を得てもよい。
【0050】
高密度の状態とは、組織を構成する際の密度と同等程度であることをいい、例えば、細胞集合体の場合、細胞配置時で5×107〜1×109個/ml、細胞の活性を損なわずに確実に細胞相互作用させるため好ましくは1×108〜1×109個/ml、最も好ましくは2×108〜8×108個/mlの密度をいう。このような細胞密度に細胞集合体を調製するには、細胞を遠心によって凝集させ沈殿化することが細胞の活性を損なわずに簡便に高密度化できるため好ましい。このような遠心は、細胞の生存を損ねない300〜1200×g、好ましくは500〜1000×gの遠心力に該当する回転数で3〜10分間行えばよい。300×gよりも低い遠心では、細胞の沈殿が不十分となって細胞密度が低くなる場合があり、一方、1200×gよりも高い遠心では細胞が損傷を受ける場合があるため、それぞれ好ましくない。
【0051】
遠心分離によって高密度の細胞を調製する場合には、通常、細胞遠心分離するために用いられるチューブ等の容器に単一細胞の懸濁液を調製した後に遠心分離し、沈殿物としての細胞を残して上清をできるだけ取り除けばよい。このときに使用されるチューブ等の容器は、上清を完全に除去する観点から、シリコーンコートされたものであることが好ましい。
【0052】
遠心分離による沈殿物とした場合には、沈殿物をそのまま支持担体の内部に配置すればよい。このとき、目的とする細胞以外の成分(例えば、培養液、緩衝液、支持担体等)は、細胞の容量と等量以下であることが好ましく、目的とする細胞以外の成分を含まないことを最も好ましい。このような高密度の細胞集合体では、細胞が緊密に接触しており、細胞間相互作用が効果的に発揮される。特に目的とする細胞以外の成分が極端に少ない細胞集合体を支持担体の内部に配置すると、支持担体の固化等によって更に凝集し、より一層緊密状態となる。
【0053】
組織の状態で使用する場合には、酵素処理等を行って、対象となる細胞以外の結合組織等を除去することが好ましい。目的とする細胞以外の成分が多い場合、例えば細胞の容量と等量以上になると、細胞間相互作用が充分に発揮されないため、好ましくない。
【0054】
また、第1の細胞集合体と第2の細胞集合体との接触は、第1の細胞集合体と第2の細胞集合体の接触は密接であるほど好ましく、第1の細胞集合体に対して第2の細胞集合体を押し付けて配置することが特に好ましい。また、第1の細胞集合体と第2の細胞集合体との周囲を培養液、酸素透過を阻害しない固形物で包み込むことも、細胞集合体同士の接触を密接にするのに有効である。粘度の異なる溶液に密度の高い細胞懸濁液を入れて配置させ、溶液をそのまま固化することも、細胞の接触の保持を容易に達成できるため、好ましい。このとき、第1の細胞集合体を歯胚間葉系細胞の単一細胞集合物とし、第2の細胞集合体を歯胚上皮組織とした場合には、歯胚上皮組織のエナメル結節が第1の細胞集合体に接触するように配置することが好ましいが、これに限定されない。
【0055】
支持担体がゲル状、あるいは溶液状等の場合には、配置工程の後に支持担体を固化する固化工程を設けてもよい。固化工程によって、支持担体内部に配置された細胞が支持担体内部に固定化される。支持担体の固化には、一般に用いた支持担体の固化条件をそのまま適用すればよい。例えば支持担体にコラーゲン等の固化可能な化合物を用いた場合には、通常適用される条件で、例えば培養温度下で数分〜数十分間静置させることにより、固化することができる。これにより、支持担体内部における細胞間の結合を固定化できると共に、強固なものにすることができる。
【0056】
本発明の製造方法における培養工程では、第1の細胞集合体及び第2の細胞集合体を支持担体内部で培養する。この培養工程では、互いに緊密に接触された第1の細胞集合体及び第2の細胞集合体によって細胞間相互作用が効果的に行われて、組織、即ち歯が再構成される。
培養工程は、支持担体によって第1の細胞集合体と第2の細胞集合体との接触状態が維持されて行われればよく、第1及び第2の細胞集合体を有する支持担体単独による培養であっても、他の動物細胞の存在下での培養であってもよい。
【0057】
培養期間としては、支持担体内部に配置された細胞数及び細胞集合体の状態、更には培養工程の実施条件によって異なるが、一般に、1〜300日、エナメル質を外側に有し、象牙質を内側に有する歯を形成するためには、好ましくは1〜120日、迅速に提供可能とする観点からは、好ましくは1〜60日とすることができる。更に歯周組織を備えた歯とするためには、一般に1〜300日、好ましくは1〜60日とすることができる。
【0058】
支持担体のみによる培養とした場合には、動物細胞の培養に用いられる通常の条件下での培養とすることができる。ここでの培養は、一般に動物細胞での培養条件をそのまま適用すればよく、前述した条件をそのまま適用することができる。また培養には、哺乳動物由来の血清を添加してもよく、またこれらの細胞の増殖や分化に有効であることが既知の各種細胞因子を添加してもよい。このような細胞因子としては、FGF、BMP等を挙げることができる。
【0059】
また、組織や細胞集合体のガス交換や栄養供給の観点から器官培養を用いることが好ましい。器官培養では、一般に、動物細胞の増殖に適した培地上に多孔性の膜をフロートさせ、その膜上に支持担体で包埋された細胞集合体を置いて培養を行う。ここで用いられる多孔性の膜には、0.3〜5μm程度の孔を多数有した膜であることが好ましく、具体的にはセルカルチャーインサート(商品名)、アイソポアフィルター(商品名)を挙げることができる。
【0060】
他の動物細胞の存在下での培養の場合には、動物細胞からの各種サイトカイン等の作用を受けて、早期に特有の細胞配置を有する歯を形成することができるので、好ましい。このような他の動物細胞の存在下での培養は、単離細胞や培養細胞を用いて生体外での培養によって行ってもよい。
【0061】
また、第1及び第2の細胞集合体を有する支持担体を生体へ移植して、生体内で培養を行ってもよい。このような生体内での培養は、歯、更には歯周組織の形成を早期に行うことができるため特に好ましい。この場合、支持担体と共に第1及び第2の細胞集合体が生体内へ移植される。
【0062】
この用途に利用可能な動物は、哺乳動物、例えばヒト、豚、マウス等を好ましく挙げることができ、歯胚組織と同一の種に由来するものであることが更に好ましい。ヒト歯胚組織を移植する場合には、ヒト、又は免疫不全に改変したヒト以外の他の哺乳動物を用いることが好ましい。このような生体内成育に好適な生体部位としては、動物細胞の器官や組織をできる限り正常に発生させるためには、腎臓皮膜下、腸間膜、皮下移植、口腔内等が好ましい。
移植による成育期間としては、移植時の大きさと発生させる歯の大きさによって異なるが、一般に、3〜400日とすることができる。例えば、腎臓皮膜下への移植期間は移植する培養物の大きさと再生させる歯の大きさによっても異なるが、歯の再生と移植先で発生させる歯の大きさの観点から7〜60日間であることが好ましい。
【0063】
生体への移植を行う前に、生体外での培養(前培養)を行ってもよい。この前培養によって細胞間の結合と第1及び第2の細胞集合体同士の結合を強固にして、細胞間相互作用をより強固にすることができるため好ましい。その結果、全体の成育期間を短縮することができる。
前培養の期間は短期であっても長期であってもよい。長期間、例えば3日以上、好ましくは7日以上とした場合には、歯胚から歯芽に発生させることができるので、移植後に歯ができるまでの期間を短縮することもできるため好ましい。前培養の期間としては、例えば腎臓皮膜下へ移植を行う場合の器官培養として、好ましくは1〜7日とすることが効率よく歯を再生するために好ましい。
【0064】
本発明の製造方法によって製造された歯は、内側に象牙質、外側にエナメル質という歯としての特有の細胞配置(構造)を有するものであり、また好ましくは、更に歯の先端(歯冠)と歯根という方向性も備えているものである。少なくともこのような特有の細胞配置、好ましくは細胞配置に加えて方向性を有することによって、歯としての機能も発揮できるものである。このため、歯の代替物として広く利用することが可能である。特に、自家の歯胚に由来した間葉系細胞及び上皮系細胞を用いた場合には、拒絶反応による問題を回避しつつ使用することができる。また一般に移植抗原が適合した他人の歯胚に由来する細胞を用いる場合にも拒絶反応による問題を回避することが可能である。
【0065】
また本発明の製造方法によって製造された歯は、歯特有の細胞配置を有する歯の集合体であってもよい。
このような歯の集合体は、歯特有の細胞配置を有する複数の歯で構成されているため、個々の歯を集合体から分離して、以下に述べるように1つの歯の移植片として用いることができる。この結果、移植片としての歯を効率よく作製することができる。
【0066】
複数の歯で構成された歯の集合体を得るためには、第1及び第2の細胞集合体いずれも、複数本の歯が発生するための歯胚の再誘導を容易にするために共に単一細胞により構成されていることが好ましい。
なお、培養工程は、前記と同様に、器官培養であっても腎臓皮膜下での培養であってもよいが、得られた歯を移植片として用いる場合には、他の動物細胞との接触がなく且つ全行程in vitroで調製することができる器官培養とすることが好ましい。
【0067】
さらに本発明の製造方法では、培養期間を歯周組織が形成されるまで継続してもよい。これにより、歯そのものに加えて、歯を顎骨上で支持し、固定化する歯槽骨や歯根膜などの歯周組織も形成させることができる。この結果、移植後に実用可能な歯を提供することが可能である。
【0068】
なお、歯周組織を製造するためには、上記培養工程の後に、前記培養によって得られた歯周組織を単離する工程を設けて歯周組織のみを得てもよい。歯周組織の単離は、培養工程の過程で形成された歯周組織と歯とを分離することができる如何なる方法によって行ってもよく、ピンセット等による分離や、酵素による部分消化等を挙げることができる。
【0069】
本発明に従って得られた歯及び歯周組織は、移植片として用いられる他、歯の発生過程を解明するための研究にも好ましく利用することができ、今後の歯に関連する組織発生のために有効なリサーチツールとして利用することができる。
なお、得られた歯又は歯周組織を移植片として用いる場合には、製造方法における培養工程を、他の動物細胞との接触がなく且つ全行程in vitroで調製することができる器官培養としたものであることが好ましい。
【0070】
また、本発明には、歯の移植方法が含まれる。この移植方法では、上記歯の集合体を得る工程と、歯の複合体から個々の歯を分離する工程と、分離された歯を、移植部位での他の歯と同一の方向性となるように揃えて移植する工程とを含む。
これにより、歯の移植を、特有の細胞配置と方向性を備えた複数の歯を同時に得て、効率よく実施することができる。
【0071】
本発明による歯は、歯の欠損及び損傷を伴う各種症状、例えば、齲蝕、辺縁性歯周炎(歯槽膿漏)、歯周病による歯の欠損、事故などによる折損や脱落等の治療又は処置に適用することもできる。
即ち、本発明の治療方法は、本発明による製造方法によって得られた歯及び/又は歯周組織を、欠損及び/又は損傷部位へ移植することを含む。これにより、欠損及び/又は損傷部位の上記症状を治療及び/又は緩和することができる。
本発明の他の治療方法は、本発明における培養工程のみ、或いは、配置工程及び培養工程を、欠損及び/又は損傷部位において実施させることを含む。この場合、支持担体としては、上述したものに加えて、欠損及び/又は損傷部位の周囲組織そのものを支持担体として適用してもよい。これにより、生体内での周辺組織からのサイトカイン等によって、より迅速に欠損及び/又は損傷部位の治療等を行うことができる。
【実施例】
【0072】
以下に本発明の実施例について説明するが、これに限定されるものではない。また実施例中の%は、特に断らない限り、重量(質量)基準である。
[実施例1]
1. EC細胞の培養方法
EC細胞は、AT805細胞(129系EC細胞であるOTT6050の派生クローン、理化学研究所バイオリソースセンター細胞材料開発室より入手)を使用した。AT805細胞の培養は、10容量%牛胎児血清(FCS:JRH又はJBS、Hyclone社製)及び55μM 2−メルカプトエタノール(GIBCO社製)を添加したダルベッコ改変イーグル培地(DMEM:SIGMA社又はコージンバイオ社製)を用いて行った。EC細胞は通常、100mmディッシュあたり5〜8×105個の濃度で播種し、一日おきに全量培地交換と継代培養を繰り返した。継代培養では、細胞をHCMFバッファ(pH7.4、10mM Hepes、136.9mM NaCl、0.34mM Na2HPO3、13.9mMグルコース、5.37mM KCl)にて1回洗浄後、最終濃度0.025%のトリプシン−EDTA・2Na(GIBCO社製)を溶解した酵素溶液を5ml添加し、37℃にて1分間酵素処理を行った。その後、等量の10%FCS添加DMEMを加えて細胞を分散させた後、遠心分離によって沈殿回収した細胞を再度懸濁し、新たな培養ディッシュへ播種し、継代培養した。
【0073】
2.EC細胞の分化誘導及び細胞の回収
トリプシン処理により回収したEC細胞を、終濃度0.5〜5容量%のジメチルスルホキシド(DMSO:SIGMA社製)を添加した10容量%FCS添加DMEMで懸濁し、1.0×106個/100mmディッシュの濃度で播種した。5%CO2濃度条件下で37℃にて培養し、14時間後、HCMFバッファにて細胞を1回洗浄し、培養液を10容量%FCS添加DMEMに培地交換した。
【0074】
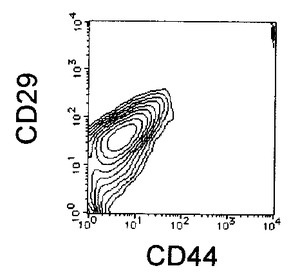
DMSOで処理したEC細胞は、3日ごとに培養液の半量を10容量%FCS添加DMEMに交換を行いながら培養を6日間(5容量%DMSOのみ19日間)継続し、適宜観察を行った。未分化細胞を含む細胞集団が増殖した時点で、HCMFバッファにて1回洗浄後、3mM EDTA、並びに0.5%BSAを含むCa2+、Mg2+不含PBS(−)(EDTA/BSA−PBS)を添加して細胞を分散し、遠心操作により細胞を沈殿として回収した。細胞を再びEDTA/BSA−PBSに懸濁し、FITC標識した抗CD44抗体(CD44−FITC:BD Pharmingen社製)及びPE標識した抗CD29抗体(CD29−PE:BD Pharmingen社製)により二重染色を行い、CD44陽性かつCD29陽性の細胞フラクションを、セルソーター、Epics ALTRA(Beckman Coulter社製)を用いて分離取得することで、未分化EC細胞が除去されたDMSO処理により分化誘導したEC細胞フラクションを得た(図2参照)。この細胞をDMSO−EC細胞と名付けた。
また、各DMSO濃度で処理した場合のDMSO−EC細胞の割合について、表1に示す。表1に示されるように、DMSO処理濃度に依存して、CD44陽性かつCD29陽性の細胞は増加することが判明した。なお、5容量%DMSOで処理したEC細胞は、ほとんどの細胞が処理直後に死滅するため、処理後の培養日数が他の処理濃度より長く必要になる一方で、培養後に得られたCD44陽性かつCD29陽性の細胞は高い割合であることが明らかになった。
以後の実験には、5容量%のDMSOを用いて得られた細胞を用いた。
【0075】
【表1】

【0076】
DMSO−EC細胞のクローン化
DMSO−EC細胞のクローン化には、クローニングリングによるコロニーの分離、あるいは限界希釈法によって行った。
クローニングリングによる細胞の取得では、DMSO−EC細胞を培養ディッシュに低濃度(約100個)で播種し、増殖によってできたコロニーの細胞数が10個以上になったところで、滅菌済みのシリコングリースをクローニングリングの一端に塗り、培地を取り除いたディッシュ底面に、細胞のコロニーを囲うようにして貼り付けた。その後、クローニングリング内でトリプシン処理を行い、細胞を回収して、増殖した細胞から順次、培養のスケールアップを行ない、クローンを取得した。
限界希釈法によるクローンの取得は、上記方法により取得したDMSO−EC細胞をHCMFバッファにて1回洗浄後、トリプシン処理により細胞を回収した。回収された細胞数を計測し、1細胞/200μl/ウエル(説明)になるように96ウエル培養プレートに播種した。3〜5日後に単一のコロニーであることを確認した。その後5日ごとに、半量の培地交換を行いながら培養を継続し、増殖した細胞から順次、培養のスケールアップを行ない、クローンを取得した。
これにより、DMSO−EC細胞由来のクローン1(DMSO−ECクローン1:Clone #1)及びクローン2(DMSO−ECクローン2:Clone #2)を得た。
【0077】
[実施例2]
DMSO−EC細胞の性状
(1)位相差顕微鏡像
上記で得られたクローン1及び2の形態と、EC細胞、DMSO−EC細胞、並びにEC細胞由来軟骨系前駆細胞であるATDC5細胞(理化学研究所バイオリソースセンター細胞材料開発室より入手)の形態をそれぞれ位相差顕微鏡で観察した。
この結果、EC細胞は、コロニー内の細胞が小さく、高い密度で増殖するほか、コロニー周辺が盛り上がっており、典型的な全能性幹細胞様の形態であることが観察された。これに対して、DMSO−ECやDMSO−ECクローン1及び2、ATDC5細胞はいずれも、コロニーを構成するひとつずつの細胞の面積が大きく、付着性細胞としての特徴である単一層を形成しており、特徴的なEC細胞の形態と明らかに異なる形態を示した。このような付着性の細胞形態は、EC細胞よりは間葉系細胞に類似している。
【0078】
(2)各種遺伝子発現
EC細胞、DMSO−EC、DMSO−ECクローン1及び2、ATDC5細胞における各種遺伝子発現をリアルタイムPCR(RT−PCR)により解析し、各種マーカー遺伝子の発現を検討した。
EC細胞、並びにDMSO−EC、DMSO−ECクローン1及び2、ATDC5細胞からの全RNAの抽出は、TRIzol Reagent(Invitrogen社製)を用いて行なった。解析する細胞の培養液を除去し、100mmの培養ディッシュに直接2mlのTRIzol Reagentを滴下し、Cell Scraper(Falcon社製)で充分混合した後、エッペンチューブに回収し、ポリトロンホモゲナイザー(ポリトロン社製)でホモジナイズを行った。その後のRNA抽出までの操作は、添付の使用説明書に従った。細胞・組織から抽出された全RNAは、DEPC処理水に溶解し、分光光度計にて濃度算定を行った後、−30℃に保存した。
【0079】
細胞から抽出した全RNAを鋳型として、リアルタイムPCR(RT−PCR)解析のためテンプレートとなるcDNAの合成を行った。cDNA合成は、ReverTra Ace(TOYOBO)を用いて行い、操作は添付された使用説明書に従って行った。
PCR及び解析は、ABI PRISM 7000(Applied Biosystems社製)を使用して行った。ポリメラーゼはSYBR Premix Ex Taq(TAKARA社製)を使用し、内在性コントロールはβ−アクチンを用いた。RT−PCR専用96ウエルプレートに1サンプルあたり、SYBR Premix Ex Taq (TAKARA社製)を13μl、20倍に希釈したcDNA反応溶液を2μl、1μMの濃度のプライマーをそれぞれ5μl、滅菌水を5μl添加し、最終容量25μlの反応系を調製し、PCR反応を常法に従って行なった。また、それぞれの遺伝子についてコントロールの反応を行って検量線を作成し、それぞれの定量化を行った。解析に使用したプライマーの配列を表2及び表3に示した。
遺伝子発現は、それぞれの細胞におけるβ−アクチンの発現量により標準化し、β−アクチン発現量を一定にした場合の相対的な発現量で比較した。結果を表4に示す。
【0080】
【表2】
【0081】
【表3】
【0082】
【表4】
【0083】
表4に示されるように、未分化幹細胞マーカー遺伝子であるOct3/4とNanogの発現は、EC細胞でのみ検出され、DMSOで処理して得られたEC細胞やそのクローン細胞、ATDC5細胞では認められなかった。神経堤細胞のマーカー遺伝子であるSlug、Pax3、Wnt1の発現は、EC細胞では認められず、SlugはEC細胞以外の細胞群の全てで認められ、Pax3とWnt1は、発現量の差異はあるものの、EC細胞とATDC5細胞以外のDMSO−EC細胞群で認められた。
【0084】
このことから、DMSO−EC細胞とそのクローン化細胞は、DMSOの添加によって未分化幹細胞から神経堤細胞へと分化誘導された細胞であることが示唆され、遺伝子発現の特性から、Oct3/4陰性、Nanog陰性、Slug陽性、Pax3陽性、Wnt1陽性の細胞であることが判明した。
一方、歯の間葉細胞のマーカー群では、EC細胞ではMsx1とBMP−7が発現しているほかはいずれの遺伝子も発現していないのに対して、DMSO−EC細胞、並びにそのクローン化細胞では、Msx1、Pax9、Wnt5a、Lhx8が特異的に高発現していた。BMP4、Runx2、Dlx1は、DMSO−EC細胞、並びにそのクローン化細胞とATDC5細胞で、発現量に多少の差異はあるものの発現が認められた。
また軟骨細胞における特異的なマーカー遺伝子であるSox9、Sox5の発現は、ATDC5細胞でのみ高発現しており、DMSO−EC細胞とそのクローン化細胞は軟骨系への分化をしていない細胞であると判断された。
従って、DMSOによって、EC細胞が神経堤細胞様ないしは間葉系細胞様の細胞へ分化誘導されたことは明らかであった。
【0085】
[実施例3]
EC細胞から分化誘導した細胞の歯形成能の評価
次に、本発明によって得られたDMSO−EC細胞、そのクローン化細胞の象牙芽細胞への分化能、並びに歯の象牙質形成能を、以下のように行って、確認した。
【0086】
(1)歯胚上皮組織の調製
C57BL/6マウス(SLCより購入)の胎齢14.5日、胚仔から下顎切歯歯胚組織を顕微鏡下で常法により摘出した。摘出した下顎切歯歯胚組織をPBS(−)で洗浄し、PBS(−)に最終濃度1.2U/mlのディスパーゼ II (Roche, Mannheim, Germany)を添加した酵素液で室温にて12.5分間処理した後、10%FCS(JRH Biosciences, Lenexa, KS)を添加したDMEM(Sigma, St. Louis, MO)で3回洗浄した。さらにDNaseI溶液 (Takara, Siga, Japan)を最終濃度70U/mlになるよう添加し歯胚組織を分散させ、25G注射針 (Terumo, Tokyo, Japan)を用いて外科的に歯胚上皮組織を分離した。
【0087】
(2)再構成歯胚の作製
再構成歯胚の作製には、上記で調製された歯胚上皮組織と、間葉系細胞として、EC細胞、DMSO−EC細胞、DMSO−ECクローン1及び2のいずれかの評価対象細胞とを用いた。
歯胚再構築に使用する評価対象細胞群を、それぞれトリプシン処理によりディッシュから回収した。シリコングリースを塗布した1.5mlマイクロチューブ(Eppendorf, Hamburg, Germany)に、10%FCS(JRH)添加DMEM(Sigma)で懸濁した評価対象細胞を入れ、遠心分離(580×g)により細胞を沈殿として回収した。遠心後の培養液の上清をできる限り除去し、再度遠心操作を行い、実体顕微鏡で観察しながら細胞の沈殿周囲に残存する培養液をGELoader Tip 0.5−20μL(エッペンドルフ社製)を用いて完全に除去し、再構成歯胚作製に用いるそれぞれの評価対象細胞を準備した。
【0088】
シリコングリースを塗布したペトリディッシュに2.4mg/mlのCellmatrix type I−A(新田ゼラチン社製)を30μl滴下してコラーゲンゲルドロップを作製した。この溶液に、上記評価対象細胞を、0.1−10μlのピペットチップ(エッペンドルフ社製)を用いて、0.2μl〜l0.3μlアプライして、高密度の細胞集合体として細胞凝集体を作製した(細胞密度:2×108個/ml)。続いて、10μlのピペットチップを用いて、歯胚上皮組織或いは歯胚間葉組織を同じゲルドロップ内にアプライし、タングステン針を用いて、分離した歯胚由来上皮組織が本来、間葉組織と接触していた面を評価対象細胞の細胞凝集体に密着させた。その後、ゲルドロップを固化させることにより、歯胚組織と評価対象細胞間の結合をより強固にすることで、高密度再構成歯胚を作製した。
【0089】
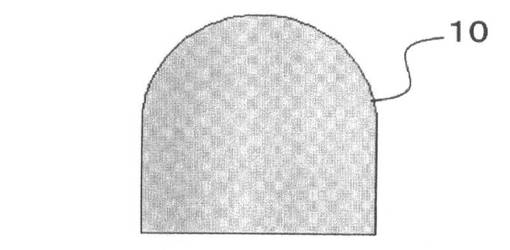
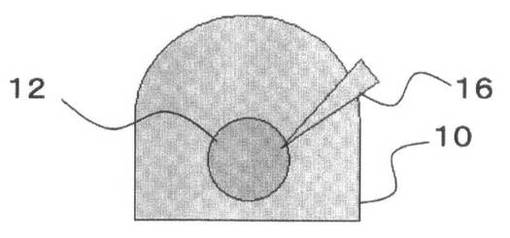
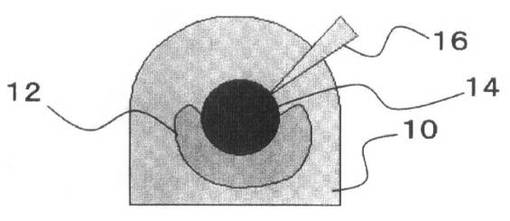
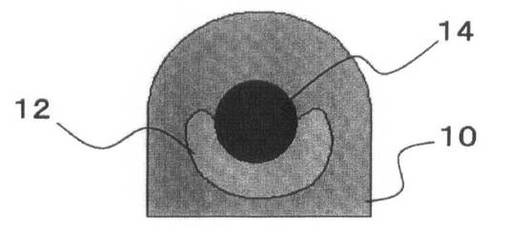
これを、図3を参照して説明する。
ピペットチップ16で先にゲルドロップ10(図3A参照)内に配置された細胞凝集体12は、ゲルドロップ10内で球体を構成する(図3B参照)。この後に他方の細胞凝集体14を押し込むことによって、球体の細胞凝集体12がつぶされて、他方の細胞凝集体14を包むようになることが多い(図3C参照)。その後にゲルドロップ10を固化させることにより、細胞間の結合が強固になる(図3D参照)。
【0090】
(3)再構成した歯胚の器官培養
ゲル中で作製した高密度再構成歯胚は、CO2インキュベーターに10分間静置してCellmatrix type I-A (Nitta Gelatin)を固形化した。10容量%FCS(JRH社製)、0.1mg/mlのL−アスコルビン酸(Sigma社製)、2mMのL−グルタミン(GIBCO社製)添加したDMEM(Sigma社製)に、セルカルチャーインサート(ポアサイズが0.4ミクロンのPETメンブレン;BD社製)が接するように培養容器を準備した。培養容器のセルカルチャーインサートの膜上に、再構成歯胚を、支持担体である周囲のゲルと共に移して、器官培養した。
【0091】
器官培養により歯の発生を解析する場合には、通常、14日間の培養を行なった。器官培養した場合には、移植後10〜30日目に再構成歯胚を摘出し、4%パラホルムアルデヒド−リン酸緩衝液で6時間固定した後、4.5%EDTA溶液(pH7.4)を用いて24時間の中性脱灰を行った。その後、常法によりパラフィン包埋して、10μmの切片を作製した。組織学的解析のためには常法に従い、ヘマトキシリン−エオジン染色(HE染色)を行った。
【0092】
器官培養による評価
14.5日齢、下顎切歯歯胚上皮組織とEC細胞の高密度再構成歯胚では、EC細胞の異常増殖により再構成歯胚は肥大化し、HE染色においても歯の間葉細胞に由来する象牙芽細胞や象牙質は観察されなかった。
これに対し、DMSO−EC細胞、DMSO−ECクローン1及び2では、上皮組織との相互作用が誘導され、位相差顕微鏡像においても歯胚の器官培養と同様に組織誘導が観察された。またさらに、DMSO−EC細胞、DMSO−ECクローン1及び2ではいずれも、外側にエナメル質、内側に象牙質を有する再構成歯胚を形成できることがHE染色像から確認でき、歯に特有の組織構造が形成可能であることが示された。
これらのことから、DMSO−EC細胞、DMSO−ECクローン1及び2は、歯胚上皮細胞との相互作用によって、歯の間葉組織である象牙芽細胞へ分化し、歯に特有の象牙質を産生できることは明らかであった。また高密度再構成歯胚法によって、器官培養においても、外側にエナメル質、内側に象牙質、内部に歯髄細胞、歯の先端と歯根を有する歯に特有の組織構造を有する歯を形成できることが明らかであった。
【0093】
[実施例4]
腎皮膜下移植方法
次に、DMSO−EC細胞、DMSO−ECクローン1及び2の細胞と14.5日齢胎児下顎切歯歯胚上皮細胞とから、高密度再構成歯胚を作製し、得られた再構成歯胚をC57BL6マウス(日本クレア社から入手)腎皮膜下に移植して、歯形成能を評価した。
実施例3と同様にして作製した再構成歯胚を、48時間から96時間、器官培養を行った後、周囲のゲルごと8週齢のNOD−SCIDマウス(チャールズリバー社から入手)の腎皮膜下に移植して異所的な歯の発生を進行させ、解析を行った。
腎皮膜下に移植した場合には、移植後10〜30日目に周囲の腎組織ごと再構成歯胚を摘出した。摘出組織を、4%パラホルムアルデヒド−リン酸緩衝液で6時間固定した後、4.5%EDTA溶液(pH7.4)を用いて24時間の中性脱灰を行った。その後、常法によりパラフィン包埋して、10μmの切片を作製した。組織学的解析のためには常法に従い、HE染色を行った。
【0094】
(1)腎皮膜下移植による評価
腎皮膜下移植では、14.5日齢胎児切歯歯胚を移植した場合には、内側に象牙質及び外側にエナメル質を有する特徴的な構造の歯を16日の期間で腎皮膜下移植により発生させることができた(図4参照)。14.5日齢胎児切歯歯胚に由来する歯胚上皮組織と歯胚間葉細胞(図4A参照)、歯胚上皮組織とDMSO−EC細胞(図4B参照)、歯胚上皮組織とDMSO−ECクローン(クローン#1、図4C参照)による再構成歯胚では、腎皮膜下移植によって移植後14日目には、正常歯胚をそのまま腎皮膜下へ移植した場合と同様に、外側のエナメル芽細胞、エナメル質、その内側に象牙質、象牙芽細胞が容易に認められた。できた歯は、先端と歯根を有しており、正常発生の歯と同様の構造を有していた。
歯形成の頻度は、EC細胞及びATDC細胞が0%であるのに対して、DMSO−EC細胞では51.3±8.8%、DMSO−ECクローン1では23.3%±16.7%、クローン2では33.3%±47.1%であった。
【0095】
これらのことから、DMSO−EC細胞、DMSO−ECクローン1及び2は、歯の間葉由来組織である象牙芽細胞へと分化し、歯の硬組織である象牙質を産生することが可能であることが明らかになった。さらに高密度再構成歯胚法によって、腎皮膜下移植においても、DMSO−EC細胞、DMSO−ECクローン1及び2によって特有の組織構造を有する歯を形成できることが明らかになった。
【0096】
(2)In situハイブリダイゼーション
14.5日齢胎児切歯歯胚に由来する歯胚上皮組織とDMSO−EC細胞を腎皮膜下へ移植し、14日後に摘出した組織について、In situハイブリダイゼーションにより、エナメル質の構成分子であるアメロジェニンと、象牙質の構成要素であるデンチンシアロホスホプロテイン(DSPP)のmRNAの発現、並びに歯根膜特異的な遺伝子であるペチロスタチンmRNAの発現を解析した。
【0097】
腎皮膜下移植後14日目の再構成歯胚を摘出し、4%パラホルムアルデヒド−リン酸緩衝液で6時間固定した後、4.5%EDTA溶液(pH7.4)を用いて24時間の中性脱灰を行った。その後、12.5%スクロース溶液、25%スクロース溶液へと順次浸漬した後、OCT compound(SAKURA Finetechnica社製)に包埋した。包埋後はクリオスタット(Leica社製)で10μmの切片を作製した。
【0098】
上記切片を、最終濃度2μg/mlのプロテアーゼK(Nacalai tesque, kyoto, Japan)を含むPBS(−)で10分間処理し、最終濃度4%パラホルムアルデヒド(Nacalai tesque)を含むPBS(−)で10分間固定した。それぞれ最終濃度で1.325%トリエタノールアミン、0.0175NのHCl(Wako)、0.25%無水酢酸を含むジエチルピロカーボネート処理(DEPC)水で10分間処理した後、PBS(−)で5分間、3回洗浄した。1.5% (v/v) トリエタノールアミン(Nacalai tesque)、0.33NのHCl(Wako)、0.25%(v/v)の無水酢酸(Nacalai tesque) in DEPC水で10分間処理した後、2×SSCで10分間、2回洗浄した。歯根膜特異的遺伝子であるペリオスチン (Genbank accession no. NM_015784) プローブは、センスプライマー (-7; ggctgaagatggttcctctc:配列番号35)とアンチセンスプライマー (573; gtacattgaaggaataacca:配列番号36)を用いてPCRにより取得したcDNA断片を、DIG標識して用いた。
【0099】
定法に従ってin situ ハイブリダイゼーションを行い、抗DIG−アルカリホルファターゼ(AP)Fabフラグメント(Roche)とNBT/BCIPStock Solution(Roche)で発色させ、Axio Imager A.1 (Zeiss)とAxioCam MRc5 (Zeiss)で解析を行なった。
その結果から、HE染色像におけるエナメル芽細胞ではアメロジェニンmRNAの発現が、象牙芽細胞ではDSPP mRNAの発現がそれぞれ認められた。またペリオスタチンmRNAの発現が認められた。このことから硬組織形成に関与する分子のmRNAは、それぞれが適切にその産生細胞において明らかに発現していた。また歯根膜で発現するペリオスチンが検出されたことから歯周組織が形成されていることが示唆された。
【0100】
[実施例5]
レチノイン酸による誘導
(1)CD44陽性かつCD29陽性の細胞の取得のためのEC細胞の分化誘導の方法
実施例1と同様にして、トリプシン処理にて回収されたAT805細胞を1.0×106個/100mmディッシュの濃度で播種し、翌日、終濃度0〜10.0μMのレチノイン酸(RA)(SIGMA社製)を添加した10容量%FCS DMEMに置換して刺激を加えた。培養72時間後、HCMFバッファにて2回洗浄を行い、全量10容量%FCS DMEMで培地交換を行った。
【0101】
(2)RA処理後のCD44陽性かつCD29陽性の細胞の取得
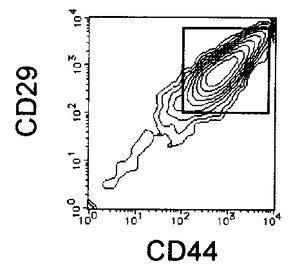
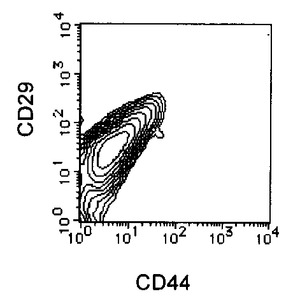
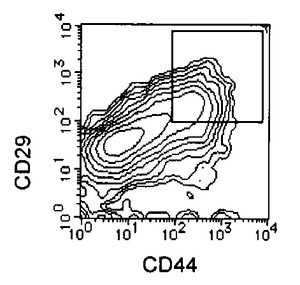
3日毎に10容量%FCS DMEMを用いて半量培地交換を行いながら培養を7日間継続し、適宜観察を行った。RA処理後増殖してきた、未分化細胞を含む細胞集団をHCMFにて1回洗浄後、3mMのEDTA−0.5% BSA−PBS(−)を添加し、37℃でインキュベートして回収した。回収後の細胞はCD44 FITC(BD Pharmingen)及びCD29(BD Pharmingen)PEにより二重染色を行い、CD44陽性かつCD29陽性の細胞をEpics ALTRA(Beckman Coulter)を用いて分離取得することで、未分化細胞を除いた分化した細胞集団のみを得た。この細胞をRA−EC細胞と名付けた。結果を図5に示す。
また、各RA濃度で処理した場合のRA−EC細胞の割合について表5に示す。表5に示されるようにRA濃度に依存して、CD44陽性且つCD29陽性細胞が変化することが判明した。
またRA処理後のCD44陽性且つCD29陽性細胞について実施例2(2)と同様に遺伝子の発現について確認したところ、DMSO−EC細胞と同様に、Oct3/4陰性、Nanog陰性、Slug陽性、Pax3陽性、Wnt1陽性の細胞であることが判明した。
【0102】
【表5】
【0103】
(3)RA処理後のCD44陽性かつCD29陽性の細胞による歯形成能の評価
上記のようにして得られたCD44陽性且つCD29陽性細胞(RA濃度2μM)を用いて、実施例3と同様にして歯胚再構築及び器官培養を行い、評価した。器官培養14日後には、歯胚上皮細胞との相互作用によって、歯の間葉組織である象牙芽細胞へ分化した。また高密度再構成歯胚法によって、外側にエナメル質、内側に象牙質、内部に歯髄細胞を有する歯に特有の組織構造を有する歯が形成された(図6参照)。
またRA処理後のCD44陽性且つCD29陽性細胞について実施例4(2)と同様にしてアメロジェニン及びDSPP、ペルオスタチンの発現を確認したところ、DMSO−ECと同様に各mRNAの発現が認められた。このことから、硬組織、歯根膜を含む歯周組織が形成されていることが示唆された。
【0104】
[実施例6]
(1)CD44陽性かつCD106陽性の細胞の取得のためのEC細胞の分化誘導の方法
実施例1と同様にしてトリプシン処理により回収したAT805細胞を、1.0×106個/100mmディッシュの濃度で播種し、翌日、終濃度2μMのRA(SIGMA社製)を添加した10容量%FCS DMEMに置換して刺激を加えた。培養72時間後、HCMFバッファにて2回洗浄を行い、全量10容量%FCS DMEMで培地交換を行った。翌日、HCMFバッファにて2回洗浄を行い、死細胞を除去した後、さらに10容量%FCS DMEMで培養を継続した。以降1日おきに培地交換を行い、7日間培養した。
【0105】
(2)CD44陽性かつCD106陽性の細胞の取得
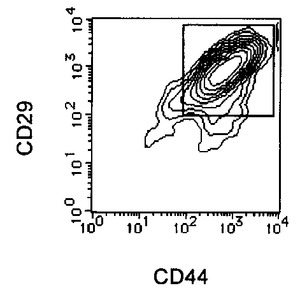
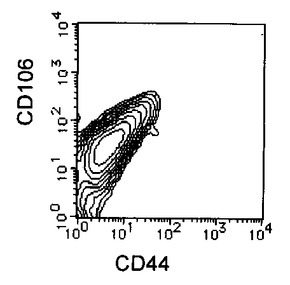
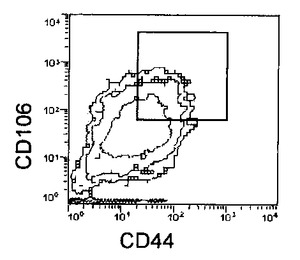
上記処理により取得された未分化細胞を含む細胞集団はHCMFにて1回洗浄後、3mM EDTA−0.5% BSA−PBS(−)を添加し、37℃でインキュベートして回収した。回収後の細胞はCD44 FITC(BD Pharmingen)及び抗CD106抗体(CD106(BD Pharmingen)PE)により二重染色を行い、CD44陽性且つCD106陽性フラクションをEpics ALTRA(Beckman Coulter)を用いて分離取得することで、未分化細胞を除いた分化した細胞集団のみを得た。結果を図7に示す。
【0106】
(3)RA処理後のCD44陽性かつCD106陽性の細胞による歯形成能の評価
上記のようにして得られたCD44陽性且つCD106陽性細胞を用いて、実施例3と同様にして歯胚再構築及び器官培養を行い、評価した。器官培養14日後には、歯胚上皮細胞との相互作用によって、歯の間葉組織である象牙芽細胞へ分化した。また高密度再構成歯胚法によって、外側にエナメル質、内側に象牙質内部に歯髄細胞を有する歯に特有の組織構造を有する歯が形成された。
またこれらのCD44陽性且つCD106陽性のDMSO−EC細胞について実施例2(2)と同様に遺伝子の発現について確認した。この結果、CD44陽性且つCD29陽性DMSO−EC細胞と同様に、Oct3/4陰性、Nanog陰性、Slug陽性、Pax3陽性、Wnt1陽性の細胞であることが示された。
更に、実施例4(2)と同様にしてアメロジェニン及びDSPP、ペルオスタチンの発現を確認したところ、DMSO−ECと同様に各mRNAの発現が認められた。このことから、硬組織、歯根膜を含む歯周組織が形成されていることが示唆された。
【0107】
[実施例7]
実施例6(2)及び(3)と同様にして、DMSO−EC細胞についてCD44 FITC及びCD106 PEを用いて二重染色したところ、CD44陽性且つCD106陽性細胞の存在が示された。これらのCD44陽性且つCD106陽性のDMSO−EC細胞について実施例2と同様に遺伝子の発現について確認したところ、CD44陽性且つCD29陽性DMSO−EC細胞と同様に、Oct3/4陰性、Nanog陰性、Slug陽性、Pax3陽性、Wnt1陽性の細胞であることが判明した。
ここで得られたCD44陽性且つCD106陽性細胞の細胞集団を用いて、実施例3及び実施例4(2)と同様に歯胚再構築、器官培養及びmRNAの発現を行い評価した。その結果、歯胚上皮細胞との相互作用による特有の組織構造を有する歯が形成された。また実施例4(2)と同様にしてアメロジェニン及びDSPP、ペルオスタチンの発現が認められた。このことから、硬組織、歯根膜を含む歯周組織の形成が可能であることが示唆された。
【0108】
このように全能性幹細胞からCD44陽性且つCD29陽性細胞又はCD44陽性且つCD106陽性細胞を分化誘導することができ、これらの細胞を上皮系細胞と共に区画化して培養することにより、歯胚を再構成して、エナメル質及び象牙質を有する特有の構造を示す歯を提供することができた。
従って、本発明によれば、全能性幹細胞から歯形成用間葉系細胞を得ることができ、効率よく大量に歯を作製することができる。
【図面の簡単な説明】
【0109】
【図1】歯胚の形成を模式的に表した概念図である。
【図2A】本発明の実施例1にかかるEC細胞のCD44及びCD29の二重染色の様子を示す図である。
【図2B】本発明の実施例1にかかるDMSO処理後6日目の細胞のCD44及びCD29の二重染色の様子を示す図である。
【図2C】本発明の実施例1にかかる、選別されたCD44陽性且つCD29陽性細胞のCD44及びCD29の二重染色の様子を示す図である。
【図3A】本発明の実施例にかかる、歯胚由来間葉系細胞と上皮系細胞を用いた歯胚再構築の手順を概念的に示した図であり、細胞配置前のゲルパックの様子を示した図である。
【図3B】本発明の実施例にかかる、歯胚由来間葉系細胞と上皮系細胞を用いた歯胚再構築の手順を概念的に示した図であり、第一の細胞集合体をゲルパック内に配置した様子を示した図である。
【図3C】本発明の実施例にかかる、歯胚由来間葉系細胞と上皮系細胞を用いた歯胚再構築の手順を概念的に示した図であり、第二の細胞集合体をゲルパック内に配置した様子を示した図である。
【図3D】本発明の実施例にかかる、歯胚由来間葉系細胞と上皮系細胞を用いた歯胚再構築の手順を概念的に示した図であり、第一の細胞集合体及び第二の細胞集合体が配置されたゲルパックを固化した様子を示した図である。
【図4A】本発明の実施例4にかかる歯胚上皮組織と歯胚間葉細胞から作製した歯胚のHE染色像である(倍率:100倍)。
【図4B】本発明の実施例4にかかる歯胚上皮組織とDMSO−EC細胞から作製した歯胚のHE染色像である(倍率:100倍)。
【図4C】本発明の実施例4にかかる歯胚上皮組織とDMSO−ECクローン化細胞から作製した歯胚のHE染色像である(倍率:100倍)。
【図5A】本発明の実施例5にかかるEC細胞のCD44及びCD29の二重染色の様子を示す図である。
【図5B】本発明の実施例5にかかるRA処理後7日目の細胞のCD44及びCD29の二重染色の様子を示す図である。
【図5C】本発明の実施例5にかかる、選別されたCD44陽性且つCD29陽性細胞のCD44及びCD29の二重染色の様子を示す図である。
【図6】本発明の実施例5においてRA処理により得られたCD44陽性且つCD106陽性細胞と歯胚上皮組織とから作製した歯胚のHE染色像である(倍率:200倍、バーは50μm)。
【図7A】本発明の実施例6にかかるEC細胞のCD44及びCD106の二重染色の様子を示す図である。
【図7B】本発明の実施例6にかかるRA処理後7日目の細胞のCD44及びCD106の二重染色の様子を示す図である。
【図7C】本発明の実施例6にかかる選別されたCD44陽性且つCD106陽性細胞のCD44及びCD106の二重染色の様子を示す図である。
【符号の説明】
【0110】
10 ゲルパック(支持担体)
12 細胞凝集塊(第1の細胞集合体)
14 細胞凝集塊(第2の細胞集合体)
16 ピペットチップ
【技術分野】
【0001】
本発明は、歯を形成するために用いられる間葉系細胞の製造方法、歯の製造方法及び歯形成用間葉系細胞に関する。
【背景技術】
【0002】
歯は、最外層にエナメル質、その内層に象牙質という硬組織を有し、さらに象牙質の内側に象牙質を産生する象牙芽細胞、その更に中心部に歯髄を有する器官であって、齲蝕や歯周病等によって失われることがある器官である。一般に歯の損失については、生命に対する危惧が少ないと考えられているため、現在は主として入れ歯やインプラントにより補うことが多い。しかしながら、歯の有無は外見や食べ物の味覚に大きく影響し、また健康維持や質の高い生活を維持するという観点から歯の再生技術の開発への関心が高まって来た。
【0003】
歯は、胎児期の発生過程の誘導によって形成され、複数の細胞種によって構築された機能単位であり、器官や臓器と同じであると考えられている。そのため歯は、成体内の造血幹細胞や間葉系幹細胞のような幹細胞から細胞種が発生する幹細胞システムによって発生するのではない。このことから、現在、再生医療によって進められている幹細胞の移入のみ(幹細胞移入療法)では、歯を再生することができない。
そこで、近年、単離された歯胚細胞を用いた歯胚を再構成させて、この再構成歯胚を移植することによる歯の再生を中心とした検討が行われている。
【0004】
例えば、非特許文献1には、歯胚から単離された上皮系細胞や間葉系の歯嚢細胞などの細胞を、生体吸収性の担体と共にラットの腹腔内に移植することで歯様の組織が再生されることが開示されている。
歯胚の再生方法としては、例えば、特許文献1には、生体から単離された歯胚細胞を、線維芽細胞増殖因子等の生理活性物質の存在下で培養することが記載されている。また、特許文献2には、生体から単離された歯胚細胞及びこれらの細胞に分化可能な細胞のうち少なくとも1種類を、フィブリンを含む担体と一緒に培養することが提案されており、ここでフィブリンを含む担体は、歯胚の目的形状のものを使用して、特有の形態を有する「歯」を形成すると記載されている。
【0005】
特許文献3及び特許文献4号公報には、6ヶ月のブタの下顎骨から、象牙質を形成する歯髄由来の間葉系細胞とエナメル形成に寄与する上皮系細胞とを含む歯胚との細胞混合物を、ポリグリコール酸−ポリ酢酸共重合体からなる生分解性ポリマーを固化させた担体(Scaffold)に播種して、動物の体内へ移植し、歯を形成する方法が開示されている。ここで担体は、歯胚の目的形状のものを使用して、特有の形態を有する「歯」を形成すると記載されている。
【0006】
一方、特許文献5には、骨の欠損又は損傷を有する患者を治療するための歯の再生方法を開示している。この方法によれば、ポリグリコール酸メッシュ担体に、例えば歯胚由来の間葉系細胞を播種した後に、上皮系細胞をコラーゲンと共に重層する又は上皮細胞シートで包むことによって、骨が形成される。なお、特許文献5では、骨の形状を構築するために担体を用いている。
【0007】
また、特許文献6には、歯胚由来の上皮系細胞と間葉系細胞とをそれぞれ細胞集合体とした上で、コラーゲンゲルの内部に細胞集合体同士を密着させ、その状態を保持しながら培養することによって、歯特有の細胞配置を備えた歯を製造する技術が開示されている。
【0008】
しかしながら、上記技術では、歯又は歯胚を再生するために歯胚細胞又はこれに分化可能な細胞を生体から入手している。このように生体から採取する細胞を用いる技術では、入手可能な細胞数が充分でない場合がある。また、細胞の入手元について制限が設けられることがある。
また、組織として機能するには、組織を構成する複数種の細胞が適切な相対位置に配置(細胞配置)され、組織としての方向性を有することが必須である。
【0009】
【非特許文献1】J. Dent. Res., 2002, Vol.81(10), pp.695-700
【特許文献1】特開2004−331557号公報
【特許文献2】特開2004−357567号公報
【特許文献3】米国特許出願公開第2002/0119180号公報
【特許文献4】米国特許出願公開第2004/0219489号公報
【特許文献5】国際公開第2005/014070号パンフレット
【特許文献6】国際公開第2006/129672号パンフレット
【発明の開示】
【発明が解決しようとする課題】
【0010】
しかしながら、上記技術では、歯又は歯胚を再生するために歯胚細胞又はこれに分化可能な細胞を生体から入手している。このように生体から採取する細胞を用いる技術では、入手可能な細胞数が充分でない場合があるだけでなく、入手元について制限が設けられることがある。
また、組織として機能するには、組織を構成する複数種の細胞が適切な相対位置に配置(細胞配置)され、組織としての方向性を有することが必須である。
【0011】
従って本発明は、特有の細胞配置を保持した歯を効率よく得ることができる歯形成用間葉系細胞を提供すること、及びこれを用いて特有の細胞配置を保持した歯を効率よく得ることを目的とする。
【課題を解決するための手段】
【0012】
本発明の間葉系細胞の製造方法は、歯を形成するために用いられる間葉系細胞の製造方法であって、全能性幹細胞を分化誘導剤の存在下で培養して、CD44陽性且つCD29陽性細胞又はCD44陽性且つCD106陽性細胞を含む分化誘導処理後の細胞集団を生成すること、前記分化誘導処理後の細胞集団から、CD44陽性且つCD29陽性細胞又はCD44陽性且つCD106陽性細胞を、歯形成用間葉系細胞として選別することを含む。
本発明の歯の製造方法は、支持担体の内部に、間葉系細胞及び上皮系細胞のいずれか一方のみから実質的になる第1の細胞集合体と、いずれか他方のみから実質的になる第2の細胞集合体とを混合することなく密着させて配置すること、前記第1及び第2の細胞集合体を前記支持担体の内部で培養すること、を含み、前記間葉系細胞が上記歯形成用間葉系細胞を含む。
本発明の歯形成用間葉系細胞は、全能性幹細胞から誘導されるCD44陽性且つCD29陽性又はCD44陽性且つCD106陽性の歯形成用間葉系細胞である。
【0013】
本発明の間葉系細胞の製造方法は、全能性幹細胞に分化誘導処理を行うことによって得られたCD44陽性且つCD29陽性細胞又はCD44陽性且つCD106陽性細胞を歯形成用間葉系細胞として選別するので、この間葉系細胞を得るために生体由来の歯等を使用する必要がなく、目的とする歯を大量に作製することができる。
また、このような歯形成用間葉系細胞を用いるので、目的とする歯を効率よく製造することができる。
【発明の効果】
【0014】
本発明によれば、目的とする歯を効率よく大量に作製することができる間葉系細胞の製造方法、及びエナメル質及び象牙質による特有の細胞配置を保持した歯を、効率よく大量に製造することができる歯の製造方法を提供することができる。
【発明を実施するための最良の形態】
【0015】
[間葉系細胞の製造方法]
本発明の間葉系細胞の製造方法は、歯を形成するために用いられる間葉系細胞を製造する間葉系細胞の製造方法であって、全能性幹細胞を分化誘導剤の存在下で培養して、CD44陽性且つCD29陽性細胞又はCD44陽性且つCD106陽性細胞を含む分化誘導処理後の細胞集団を生成すること、前記分化誘導処理後の細胞集団から、CD44陽性且つCD29陽性細胞又はCD44陽性且つCD106陽性細胞を、歯形成用間葉系細胞として選別することを含むものである。
【0016】
本発明は、全能性幹細胞を分化誘導して得られたCD44陽性且つCD29陽性細胞又はCD44陽性且つCD106陽性細胞が、歯形成用間葉系細胞として使用できることを見出したものである。これにより、歯を作製するために必要な間葉系細胞を、生体由来の歯又は歯胚以外からも得ることができ、歯を形成するための間葉系細胞を得るために、生体由来の歯等を使用する必要性を排除することができる。この結果、歯胚から直接採取した細胞数が充分でない事情や、入手元における制限の有無といった事情を考慮することなく、必要な量の間葉系細胞を容易に、また場合よって大量に得ることができる。
【0017】
本発明において、「歯」とは、内側に象牙質及び外側にエナメル質の層を連続して備えた組織をいい、特に歯冠や歯根を有する方向性を備えた組織をいう。歯の方向性は、歯冠や歯根の配置によって特定することができる。歯冠や歯根は、形状や組織染色などに基づいて目視にて確認することができる。歯冠とは、エナメル質と象牙質の層構造を有する部分をいい、歯根にはエナメル質の層は存在しない。
【0018】
象牙質及びエナメル質は、当業者には、組織染色などによって形態的に容易に特定することができる。また、エナメル質は、エナメル芽細胞の存在によって特定することができ、エナメル芽細胞の存在は、アメロジェニンの有無によって確認することができる。一方、象牙質は、象牙芽細胞の存在によって特定することができ、象牙芽細胞の存在は、デンチンシアロプロテインの有無によって確認することができる。アメロジェニン及びデンチンシアロプロテインの確認はこの分野で周知の方法によって容易に実施することができ、例えば、in situ ハイブリダイゼーション、抗体染色等をあげることができる。
【0019】
また本発明における間葉系細胞は、歯を形成するために、まず歯胚を形成するために使用できる。
本発明において「歯胚」及び「歯芽」は、後述する発生段階に基づいて区別されたものに特に言及する場合に用いられる表現である。この場合の「歯胚」とは、将来歯になることが決定付けられた歯の初期胚であり、歯の発生ステージで一般的に用いられる蕾状期(Bud stage)から鐘状期(Bell stage)までの段階であり、特に歯の硬組織としての特徴である象牙質、エナメル質の蓄積が認められない組織である。一方、「歯芽」とは本発明で用いられる「歯胚」の段階移行の、歯の硬組織の特徴である象牙質、エナメル質の蓄積が始まった段階から歯が歯肉から萌芽して一般的に歯としての機能を発現する前の段階の組織をいう。
【0020】
歯胚から歯への発生は、図1に示されるように個体発生の過程で、蕾状期、帽状期、鐘状前期及び後期の各ステージを経て行われる。ここで、蕾状期では、上皮系細胞が間葉系細胞を包むように陥入し、鐘状前期及び鐘状後期に至ると、上皮系細胞部分が外側のエナメル質となり、間葉系細胞部分が内部に象牙質を形成するようになる。従って、上皮系細胞と間葉系細胞との細胞間相互作用によって歯胚から歯が形成される。
なお本発明において「間葉系細胞」とは、間葉組織由来の細胞を意味し、「上皮系細胞」とは上皮組織由来の細胞を意味する。
また本発明において「歯周組織」とは、歯の主として外層の形成された歯槽骨及び歯根膜をいう。歯槽骨及び歯根膜は、当業者には、組織染色などによって形態的に容易に特定することができる。
【0021】
以下に本発明にかかる歯形成用間葉系細胞について説明する。
本発明に係る歯形成用間葉系細胞を得るために用いられる全能性幹細胞は、少なくとも2以上の細胞に分化可能な多分化能を有する細胞を言い、好ましくは胚性癌腫細胞、胚性幹細胞、胚性生殖幹細胞からなる群より選択されたものを使用することができる。このうち、入手の容易性の観点から胚性癌腫細胞(以下、「EC細胞」という)であることがより好ましい。本発明に適用可能なEC細胞には、神経、睾丸、卵巣などいずれの組織に由来するものであってもよい。このようなEC細胞としては。例えばヒト由来としてはNCR−G3細胞、並びにNTERA−2細胞を挙げることができ、マウス由来としてはc−1300細胞やF9細胞、LT−2細胞、OTT6050細胞、PCC4細胞、P19細胞、METT−1細胞、STT−3細胞などの細胞株を挙げることができる。これらの細胞は、哺乳動物の霊長類(例えばヒト、サルなど)、有蹄類(例えば豚、牛、馬など)、小型哺乳類の齧歯類(例えばマウス、ラット、ウサギなど)の種々の動物に由来するものから、適宜使用目的に応じて選択することができる。例えばマウス由来の胚性癌腫細胞株としては、上記に記載した細胞株やこれらの派生クローンを挙げることができる
【0022】
全能性幹細胞の培養には、通常使用される培地を使用することができる。培養に用いられる培地としては、一般に動物細胞の培養に用いられる培地、例えばダルベッコ改変イーグル培地(DMEM)等を用いることができる。この培地には、細胞の増殖を促進するための血清を添加するか、あるいは血清に代替するものとして、例えばFGF、EGF、PDGF等の細胞増殖因子やトランスフェリン等の既知血清成分を添加してもよい。なお、血清を添加する場合の濃度は、そのときの培養状態によって適宜変更することができるが、通常10%とすることができる。細胞の培養には、通常の培養条件、例えば37℃の温度で5%CO2濃度のインキュベーター内での培養が適用される。また、適宜、ストレプトマイシン等の抗生物質を添加したものであってもよい。
【0023】
本発明では、全能性幹細胞に対して、分化誘導剤の存在下で培養することによる分化誘導処理が行われる。この分化誘導処理によって、全能性幹細胞から、CD44陽性且つCD29陽性細胞又はCD44陽性且つCD106陽性細胞を含む細胞集団が生成する。誘導処理によって得られる細胞集団には、上記のCD44陽性且つCD29陽性細胞又はCD44陽性且つCD106陽性細胞が含まれていればよく、これらの双方の細胞が含まれていてもよい。
【0024】
上記分化誘導剤としては、少なくとも全能性幹細胞からCD44発現細胞への分化誘導が可能なCD44陽性細胞誘導因子であれば使用することができる。このようなCD44陽性細胞誘導因子としては、神経堤細胞系誘導因子、神経堤細胞発生関与因子、神経堤細胞増殖促進因子を挙げることができる。神経堤細胞系誘導因子は、全能性幹細胞を神経堤細胞又はその派生細胞(平滑筋細胞、心筋細胞等)へ誘導することができる因子であり、例えば、ジメチルスルホキシド(DMSO)、Dex、cAMP、ヘキサメチレンビスアセトアミド、5’−アザシチジン、TGFβ、Noggin等を挙げることができる。神経堤細胞発生関与因子としては、レチノイン酸(RA)、BMP−4、BMP−7、Shh、Wnt、FGF2、エンドセリン−1、エントセリン−3、アクチビンβA、PDGF、VEGF等を挙げることができる。神経堤細胞増殖促進因子としては、FGF2、FGF8、FGF10、BMP−2、SCF、活性型ビタミンD3等を挙げることができる。神経堤細胞系誘導因子、神経堤細胞発生関与因子及び神経堤細胞増殖促進因子はそれぞれ明確に区別されるものではなく、また、これらを単独で又は1以上を組み合わせて使用することができる。このうち、後述する本発明の目的細胞集団を効率よく生成可能なDMSO及びRAが好ましく、これらは単独又は組み合わせて使用することができる。
【0025】
これらの分化誘導剤は、全能性幹細胞の培地に分化誘導可能な濃度、添加すればよい。ここで分化誘導可能な濃度とは、用いられる分化誘導剤の種類や全能性幹細胞の種類等によって異なる。
例えばDMSOの場合、一般に培地の容量に対して、0.5容量%〜10容量%、処理に対する細胞の生存率及び培養によって得られる目的の細胞の取得効率の観点から、好ましくは2.5容量%〜5容量%、更に好ましくは4容量%〜5容量%の濃度とすることができる。2.5容量%以上の濃度であれば、全能性幹細胞の分化を充分に誘導することができ、一方、5容量%以下の濃度であれば目的の細胞取得の効率を著しく損なうことがない。
また、RAの場合、一般に培地中で0.1μM〜10μM、処理に対する細胞の生存率及び培養によって得られる目的の細胞の取得効率の観点から、好ましくは0.5μM〜5μM、更に好ましくは0.5μM〜2μMの濃度とすることができる。0.1μM以上の濃度であれば、全能性幹細胞の分化を充分に誘導することができ、一方、10μM以下の濃度であれば目的の細胞取得の効率を著しく損なうことがない。
【0026】
全能性幹細胞に対する分化誘導処理は、上記分化誘導剤の存在下で全能性幹細胞を培養によって行われる。分化誘導処理(培養)の期間は、分化誘導剤の濃度と細胞の成育活性によって異なるが、一般に2時間〜3日、細胞の生存率、ならびに目的の細胞の入手効率の観点から好ましくは6時間〜24時間とすることができる。6時間以上とすることにより充分に分化誘導された細胞を効率よく得ることができ、24時間以下とすることによって効率を著しく損なうことがない。
【0027】
なお、分化誘導処理の期間と分化誘導剤の濃度との間には、全能性幹細胞に対する分化誘導を効果的に起こさせるために一定の関係が成立する。即ち、一般に、分化誘導剤存在下での培養期間(処理時間)は、分化誘導剤の濃度と分化誘導剤の種類によって異なるが、特定の分化誘導剤の場合には、培養日数を固定した場合、一般にその濃度には好ましい至適濃度域が存在する。また培養日数を短くした場合には、至適濃度域は高い濃度側に移行し、逆に培養日数を長くする場合には至適濃度域は低い濃度側に移行する。これらの要素を考慮すると、培養日数の調節によって幅広い濃度域での分化誘導効果が得られると共に、当業者には、誘導効果が得られる至適濃度域を容易に設定することができる。
【0028】
上記分化誘導処理によって得られた分化誘導処理後の細胞集団には、CD44陽性且つCD29陽性細胞又はCD44陽性且つCD106陽性細胞を含む細胞集団が生成する。これら双方の細胞が含まれていてもよい。この分化誘導処理後の細胞集団は、次いで、CD44陽性且つCD29陽性細胞又はCD44陽性且つCD106陽性細胞を歯形成用間葉系細胞として選別する選別処理に供される。
【0029】
細胞の選別は、上述したような本発明にかかる間葉系細胞を選別することができる手段であればいずれを用いてもよい。このような選別手段としては、遺伝子発現パターン、細胞表面抗原、形態などを基準とする選別を挙げることができ、これらの1つ又は2つ以上を用いて、全能性幹細胞を選別することができる。選別手段は、細胞選別の効率の観点から、細胞表面の発現をもとにしたセルソーターを用いることが好ましく、確実に間葉系細胞を選別するためには、複数の組み合わせ、特に、遺伝子発現パターンと細胞表現抗原の発現とを組み合わせて使用することが好ましい。
【0030】
本発明において選別された歯形成用間葉系細胞は、CD44陽性であることに加えて、CD29陽性又はCD106陽性の細胞性状を示すものである。CD44は、ヒアルロン酸受容体として細胞表面に発現する膜タンパク質であることが知られている。一方、CD29は、インテグリンβ1分子として細胞表面に発現する膜タンパク質であることが知られており、EC細胞では弱陽性、間葉系細胞ではmRNAレベルで発現していることがそれぞれ報告されている。またCD106は、接着分子として既知のVCAM−1分子であることが知られており、主として上皮系細胞のマーカーとして利用されることが多い。このような性状を示す細胞であれば、後述するように歯の形成において間葉系細胞として使用することができる。
本発明における歯形成用間葉系細胞は、CD44陽性に加えて、CD29及びCD106の少なくともいずれか一方の抗原が陽性となる集団であればよく、CD44、CD29、CD106の3種の抗原が共に陽性となる三重陽性細胞が含まれていてもよい。
【0031】
本発明における歯形成用間葉系細胞は、上記CD44、CD29及びCD106の細胞表現抗原に加えて、他の細胞性状に基づいて選別を行ってもよい。
他の細胞性状としては他の細胞表面抗原パターンや遺伝子発現パターンを挙げることができる。細胞表現抗原パターンとして、例えばCD14陰性、CD34陰性、CD45陰性等を挙げることができ、これらを単独又は組み合わせて使用することができる。また遺伝子発現パターンとしては、例えばSlug発現、Wnt5a発現、Lhx8発現、BMP4発現、Pax3発現、Pax9発現、Msx1発現、Oct3/4非発現、nanog非発現、Sox9非発現、Sox5非発現等を挙げることができ、これらを単独又は組み合わせて使用することができるが、これらに限定されるものではない。
【0032】
本発明の歯形成用間葉系細胞は、好ましくは、上記CD44、CD29、CD106に加えて、歯形成能の高さから、Slug遺伝子、Pax3遺伝子、Msx1遺伝子及びPax9遺伝子の群から選択された少なくとも1つの遺伝子を発現していることが好ましく、Slug遺伝子及びPax3遺伝子を共に発現していることが更に好ましく、これら4つの遺伝子をすべて発現していることが特に好ましい。
【0033】
これらの細胞表面抗原や遺伝子発現パターンの確認及びこれに基づく選別には、表面抗原を認識する各種抗体や遺伝子発現の確認が可能な核酸配列などを用いる。これらの抗体及び核酸配列は、いずれも公知であり容易に入手可能である。また、これらの表面抗原や遺伝子発現に基づく確認及び選別は、この用途に一般的に用いられる手段、例えば、各種抗体による免疫染色、フローサイトメトリー、セルソーター、RT−PCR等を用いればよい。
【0034】
また、本発明の製造方法では、上記選別手段に組み合わせて、単一の間葉系細胞とするためのクローニングを用いることができる。クローニングの方法としては、通常この目的で用いられている方法、例えば、限界希釈法、セルソーター、クローニングリングなどを用いることができる。クローニングは、上記選別手段による選別の前後のいずれで行ってもよいが、目的とする間葉系細胞を効率よく得るためには選別の後に行うことが好ましい。
【0035】
選別工程において目的とする細胞数を効率よく得るために、選別工程の前に増殖工程を設けてもよい。このような増殖工程では、分化誘導剤を含まない通常培養用の培地を用いて分化誘導後の細胞集団の培養を行う。これにより、分化誘導された目的細胞を所定の細胞数まで増殖させることができる。その結果、充分な細胞数により選別を容易に実施可能にすることができる。
このような増殖工程の期間は、分化誘導処理後の細胞数及び細胞の状態に基づいて適宜設定することができるが、目的細胞の濃縮効率の観点から一般に3〜30日、好ましくは5〜10日とすることができる。この結果、選別工程の対象となる細胞集団中に、目的とする歯形成用間葉系細胞が含まれることになり、このような細胞集団を選別工程に供することによって、本発明にかかる歯形成用間葉系細胞を効率よく且つ大量に得ることができる。
【0036】
本発明の方法で得られた間葉系細胞は、歯を形成するために用いられる。歯の形成は、本発明に係る間葉系細胞を用いるものであればいずれの方法であってもよいが、以下の本発明の歯の製造方法に用いられることが最も好ましい。
以下、本発明の歯の製造方法について説明する。
【0037】
[歯の製造方法]
本発明の歯形成用間葉系細胞を用いた好ましい歯の製造方法は、支持担体の内部に、間葉系細胞及び上皮系細胞のいずれか一方のみから実質的になる第1の細胞集合体と、いずれか他方のみから実質的になる第2の細胞集合体とを混合することなく密着させて配置すること(配置工程)、前記第1及び第2の細胞集合体を前記支持担体の内部で培養すること(培養工程)、を含み、前記間葉系細胞が上記歯形成用間葉系細胞を含むものである。
本製造方法では、間葉系細胞と上皮系細胞とを細胞集合体として支持担体の内部で混合することなく密着させた状態で生育させるので、緊密な接触状態によって細胞間相互作用を効果的に再現することができ、内側に象牙質、外側にエナメル質という歯に特有の細胞配置を有する歯を得ることができる。この際、間葉系細胞から実質的になる細胞集合体が本発明にかかる歯形成用間葉系細胞を含むので、特有の細胞配置を有する歯を効率よく大量に製造することができる。
【0038】
配置工程では、支持担体の内部に、第1の細胞集合体と第2の細胞集合体とを接触させて配置する。
ここで第1の細胞集合体及び第2の細胞集合体は、それぞれ間葉系細胞のみ、又は上皮系細胞のみから実質的に構成されているものである。ここで、間葉系細胞のみから実質的に構成されている細胞集合体は、上述した歯形成用間葉系細胞を含むものである。この歯形成用間葉系細胞を含む細胞集合体は、上述した製造方法に従った調製工程によって調製することができ、一方、上皮系細胞のみから実質的に構成されている細胞集合体は、間葉系細胞から実質的になる細胞集合体とは独立して調製することができる(第1の細胞調製工程及び第2の細胞調製工程)。
【0039】
また、「細胞集合体」とは、細胞が密集した状態をいい、組織の状態であっても、単一細胞の状態から調製された細胞集合体(細胞凝集体)であってもよい。また「実質的になる」とは、対象となる細胞以外のものをできるだけ含まないことを意味する。このため上皮系細胞からなる細胞集合体は、組織の一部又は単一細胞の集合体とすることができる。また、上皮系細胞及び間葉系細胞は共に単一細胞で構成された細胞集合体であってもよい。
第1の細胞集合体第2の細胞集合体は、いずれが上皮系細胞及び間葉系細胞であってもよく、この細胞集合体を構成する細胞の数は、動物の種類や、支持担体の種類、硬さ及び大きさによって異なるが、細胞集合体1個あたり、一般に101〜108個、好ましくは103〜108個とすることができる。
【0040】
間葉系細胞から実質的になる細胞集団は、歯形成用間葉系細胞を含んでいれば他の間葉系細胞を含んでいてもよい。
上記間葉系細胞以外の他の間葉系細胞としては、歯胚及び歯胚以外に由来する間葉系細胞を挙げることができる。歯胚以外に由来する間葉系細胞としては、生体内の他の間葉系組織に由来する細胞であり、好ましくは、血液細胞を含まない骨髄細胞や間葉系幹細胞、さらに好ましくは口腔内間葉系細胞や顎骨の内部の骨髄細胞、頭部神経堤細胞に由来する間葉系細胞、前記間葉系細胞を生み出しうる間葉系前駆細胞やその幹細胞等を挙げることができる。
【0041】
本製造方法で用いられる上皮系細胞は、生体内での細胞配置を再現して特有の構造及び方向性を有する歯を効果的に形成するために歯胚に由来するものであることが好ましく、細胞の分化段階の幼若性と均質性の観点から蕾状期から帽状期からのものであることが好ましい。
また、歯胚以外に由来する上皮系細胞であってもよく、これには、生体内の他の上皮系組織に由来する細胞を挙げることができる。好ましくは、皮膚や口腔内の粘膜や歯肉の上皮系細胞、さらに好ましくは皮膚や粘膜などの分化した、例えば角化した、あるいは錯角化した上皮系細胞を生み出しうる未熟な上皮系前駆細胞、たとえば非角化上皮系細胞やその幹細胞等を挙げることができる。
【0042】
細胞集合体を調製するために各細胞を組織から単離する場合、歯胚及び他の組織は、哺乳動物の霊長類、例えばヒト、サルなど、有蹄類、例えば豚、牛、馬など、小型哺乳類の齧歯類、例えばマウス、ラット、ウサギなどの種々の動物の顎骨等から採取することができる。歯胚及び組織の採取は、通常、組織の採取で用いられる条件をそのまま適用すればよく、無菌状態で取り出し、適当な保存液に保存すればよい。なお、ヒトの歯胚としては、第3大臼歯いわゆる親知らずの歯胚の他、胎児歯胚を挙げることができるが、自家組織の利用との観点から、親知らず歯胚を用いることが好ましい。
【0043】
組織、例えば歯胚から、上記細胞を調製する場合には、まず周囲の組織から単離された歯胚を、形状に従って歯胚間葉組織及び歯胚上皮組織に分ける。このとき、歯胚組織は顕微鏡下で構造的に見分けることが可能であるため、解剖用ハサミやピンセット等で切断、あるいは引き剥がすことによって容易に分離することができる。また、歯胚組織からの歯胚間葉組織及び歯胚上皮組織の分離は、その形状に従って注射針、タングステンニードル、ピンセット等で切断、あるいは引き剥がすことにより容易に行うことができる。
好ましくは、周囲組織から歯胚細胞を容易に分離するため及び/又は歯胚組織から上皮組織及び間葉組織を分離するために、酵素を用いてもよい。このような用途に用いられる酵素としては、ディスパーゼ、コラーゲナーゼ、トリプシン等を挙げることができる。
【0044】
細胞集合体を構成する細胞は、採取した組織から単一細胞の状態に調製してもよい。調製工程において単一の細胞に容易に分散可能とするために、酵素を用いてもよい。このような酵素としては、ディスパーゼ、コラーゲナーゼ、トリプシン等を挙げることができる。このとき、上皮組織からの上皮系細胞の分離にはコラーゲナーゼ処理後にトリプシン処理とDNase処理をすることが好ましい。他方、間葉組織からの間葉系細胞の分離には、コラーゲナーゼとトリプシンで同時に処理し、最終的にDNase処理をすることが好ましい。このときDNase処理を行うのは、酵素処理により一部の細胞がダメージをうけ、細胞膜が溶解したときに溶液中に放出されるDNAによって細胞が凝集し細胞の回収量が低下することすることを防ぐためである。
【0045】
なお、細胞集合体を構成する細胞は、それぞれ充分な細胞数を得るために、配置工程に先立って予備的な培養を経たものであってもよい。細胞の培養は、一般に動物細胞の培養に用いられる温度等の条件をそのまま用いることができる。
培養に用いられる培地としては、一般に動物細胞の培養に用いられる培地、例えばダルベッコ改変イーグル培地(DMEM)等を用いることができ、細胞の増殖を促進するための血清を添加するか、あるいは血清に代替するものとして、例えばFGF、EGF、PDGF等の細胞増殖因子やトランスフェリン等の既知血清成分を添加してもよい。なお、血清を添加する場合の濃度は、そのときの培養状態によって適宜変更することができるが、通常10容量%とすることができる。細胞の培養には、通常の培養条件、例えば37℃の温度で5%CO2濃度のインキュベーター内での培養が適用される。また、適宜、ストレプトマイシン等の抗生物質を添加したものであってもよい。
【0046】
この予備的な培養を間葉系細胞に適用する場合、上述した歯形成用間葉系細胞の増殖のための培養を兼ねたものであってもよく、選別工程後に行うもの、又はこの双方であってもよい。
また、細胞の性質を変えないという観点から、間葉系細胞の予備培養や上述した歯形成用間葉系細胞の単なる増殖のための培養では、前述した分化誘導剤を含有しない培地で行うことが好ましい。
【0047】
配置工程における細胞集合体の配置は、上記第1及び第2の細胞集合体を、細胞の接触状態を保持可能な支持担体の内部に配置する。このとき、各細胞集合体は互いに混合することがない。このように各細胞集合体を混合することなく配置するので、細胞集合体の間に境界線が形成される。このような配置形態を、本明細書中では適宜「区画化」と表現する。
【0048】
ここで用いられる支持担体としては、細胞を内部で培養可能なものであればよく、好ましくは、上記培地との混合物である。このような支持担体としては、コラーゲン、アガロースゲル、カルボキシメチルセルロース、ゼラチン、寒天、ハイドロゲル、セルマトリクス(商品名)、メビオールゲル(商品名)、マトリゲル(商品名)、エラスチン、フィブリン、ラミニン、細胞外マトリクス混合物、ポリグリコール酸(PGA)、ポリ乳酸(PLA)、乳酸/グリコール酸共重合体(PLGA)等を挙げることができる。これらの支持担体は、細胞を内部に配置したときに配置した位置をほぼ維持可能な程度な硬さを有するものであればよく、ゲル状、繊維状、固体状のものを挙げることができる。中でも細胞外マトリクス混合物等のゲルが適切な硬さや保持力を提供しやすいといった観点から、コラーゲン、アガロースゲル、カルボキシメチルセルロース、ゼラチン、寒天、ハイドロゲル、セルマトリクス、メビオールゲル、マトリゲル、細胞倍マトリクス混合物、エラスチン、フィブリン、ラミニンがより好ましい。ここで、細胞の位置を維持可能な硬さとは、通常、三次元培養として適用される硬さ、即ち、細胞の配置を保持できると共に増殖による肥大化を阻害しない硬さであればよく、容易に決定することができる。
なお、ここで支持担体は、第1及び第2の細胞集合体が担体内部で成育することができる程度の厚みを有すればよく、目的とする組織の大きさ等によって適宜設定することができる。
【0049】
また、支持担体は、細胞が分散することなく細胞の接触状態を保持可能な保持力を備えていればよい。ここでいう「接触状態」とは、各細胞集合体において、また細胞集合体間において、確実に細胞相互作用させるために緊密(高密度)の状態であることが好ましく、このような高密度の状態とは、細胞凝集体の場合には、例えば、単に触れ合うよりも強く密着した状態を維持可能な保持力で細胞を培養可能なものとすることができる。例えば、コラーゲンの場合、最終濃度2〜3mg/mlの濃度、即ちJIS-K6503-1996に準拠した方法(12.7mm径のプランジャーで4mm押し下げるのに必要な荷重として測定)によって120g〜250gのゼリー強度となる濃度での使用が適切な硬さを提供する。また、このゼリー強度は限定されず、他の種の支持担体においても、同様の評価方法によって同様の強度があれば本発明の支持担体として好ましく用いられる。また1種又は複数種の支持担体を混合することによって、目的とするゼリー強度に相当する硬さの支持担体を得てもよい。
【0050】
高密度の状態とは、組織を構成する際の密度と同等程度であることをいい、例えば、細胞集合体の場合、細胞配置時で5×107〜1×109個/ml、細胞の活性を損なわずに確実に細胞相互作用させるため好ましくは1×108〜1×109個/ml、最も好ましくは2×108〜8×108個/mlの密度をいう。このような細胞密度に細胞集合体を調製するには、細胞を遠心によって凝集させ沈殿化することが細胞の活性を損なわずに簡便に高密度化できるため好ましい。このような遠心は、細胞の生存を損ねない300〜1200×g、好ましくは500〜1000×gの遠心力に該当する回転数で3〜10分間行えばよい。300×gよりも低い遠心では、細胞の沈殿が不十分となって細胞密度が低くなる場合があり、一方、1200×gよりも高い遠心では細胞が損傷を受ける場合があるため、それぞれ好ましくない。
【0051】
遠心分離によって高密度の細胞を調製する場合には、通常、細胞遠心分離するために用いられるチューブ等の容器に単一細胞の懸濁液を調製した後に遠心分離し、沈殿物としての細胞を残して上清をできるだけ取り除けばよい。このときに使用されるチューブ等の容器は、上清を完全に除去する観点から、シリコーンコートされたものであることが好ましい。
【0052】
遠心分離による沈殿物とした場合には、沈殿物をそのまま支持担体の内部に配置すればよい。このとき、目的とする細胞以外の成分(例えば、培養液、緩衝液、支持担体等)は、細胞の容量と等量以下であることが好ましく、目的とする細胞以外の成分を含まないことを最も好ましい。このような高密度の細胞集合体では、細胞が緊密に接触しており、細胞間相互作用が効果的に発揮される。特に目的とする細胞以外の成分が極端に少ない細胞集合体を支持担体の内部に配置すると、支持担体の固化等によって更に凝集し、より一層緊密状態となる。
【0053】
組織の状態で使用する場合には、酵素処理等を行って、対象となる細胞以外の結合組織等を除去することが好ましい。目的とする細胞以外の成分が多い場合、例えば細胞の容量と等量以上になると、細胞間相互作用が充分に発揮されないため、好ましくない。
【0054】
また、第1の細胞集合体と第2の細胞集合体との接触は、第1の細胞集合体と第2の細胞集合体の接触は密接であるほど好ましく、第1の細胞集合体に対して第2の細胞集合体を押し付けて配置することが特に好ましい。また、第1の細胞集合体と第2の細胞集合体との周囲を培養液、酸素透過を阻害しない固形物で包み込むことも、細胞集合体同士の接触を密接にするのに有効である。粘度の異なる溶液に密度の高い細胞懸濁液を入れて配置させ、溶液をそのまま固化することも、細胞の接触の保持を容易に達成できるため、好ましい。このとき、第1の細胞集合体を歯胚間葉系細胞の単一細胞集合物とし、第2の細胞集合体を歯胚上皮組織とした場合には、歯胚上皮組織のエナメル結節が第1の細胞集合体に接触するように配置することが好ましいが、これに限定されない。
【0055】
支持担体がゲル状、あるいは溶液状等の場合には、配置工程の後に支持担体を固化する固化工程を設けてもよい。固化工程によって、支持担体内部に配置された細胞が支持担体内部に固定化される。支持担体の固化には、一般に用いた支持担体の固化条件をそのまま適用すればよい。例えば支持担体にコラーゲン等の固化可能な化合物を用いた場合には、通常適用される条件で、例えば培養温度下で数分〜数十分間静置させることにより、固化することができる。これにより、支持担体内部における細胞間の結合を固定化できると共に、強固なものにすることができる。
【0056】
本発明の製造方法における培養工程では、第1の細胞集合体及び第2の細胞集合体を支持担体内部で培養する。この培養工程では、互いに緊密に接触された第1の細胞集合体及び第2の細胞集合体によって細胞間相互作用が効果的に行われて、組織、即ち歯が再構成される。
培養工程は、支持担体によって第1の細胞集合体と第2の細胞集合体との接触状態が維持されて行われればよく、第1及び第2の細胞集合体を有する支持担体単独による培養であっても、他の動物細胞の存在下での培養であってもよい。
【0057】
培養期間としては、支持担体内部に配置された細胞数及び細胞集合体の状態、更には培養工程の実施条件によって異なるが、一般に、1〜300日、エナメル質を外側に有し、象牙質を内側に有する歯を形成するためには、好ましくは1〜120日、迅速に提供可能とする観点からは、好ましくは1〜60日とすることができる。更に歯周組織を備えた歯とするためには、一般に1〜300日、好ましくは1〜60日とすることができる。
【0058】
支持担体のみによる培養とした場合には、動物細胞の培養に用いられる通常の条件下での培養とすることができる。ここでの培養は、一般に動物細胞での培養条件をそのまま適用すればよく、前述した条件をそのまま適用することができる。また培養には、哺乳動物由来の血清を添加してもよく、またこれらの細胞の増殖や分化に有効であることが既知の各種細胞因子を添加してもよい。このような細胞因子としては、FGF、BMP等を挙げることができる。
【0059】
また、組織や細胞集合体のガス交換や栄養供給の観点から器官培養を用いることが好ましい。器官培養では、一般に、動物細胞の増殖に適した培地上に多孔性の膜をフロートさせ、その膜上に支持担体で包埋された細胞集合体を置いて培養を行う。ここで用いられる多孔性の膜には、0.3〜5μm程度の孔を多数有した膜であることが好ましく、具体的にはセルカルチャーインサート(商品名)、アイソポアフィルター(商品名)を挙げることができる。
【0060】
他の動物細胞の存在下での培養の場合には、動物細胞からの各種サイトカイン等の作用を受けて、早期に特有の細胞配置を有する歯を形成することができるので、好ましい。このような他の動物細胞の存在下での培養は、単離細胞や培養細胞を用いて生体外での培養によって行ってもよい。
【0061】
また、第1及び第2の細胞集合体を有する支持担体を生体へ移植して、生体内で培養を行ってもよい。このような生体内での培養は、歯、更には歯周組織の形成を早期に行うことができるため特に好ましい。この場合、支持担体と共に第1及び第2の細胞集合体が生体内へ移植される。
【0062】
この用途に利用可能な動物は、哺乳動物、例えばヒト、豚、マウス等を好ましく挙げることができ、歯胚組織と同一の種に由来するものであることが更に好ましい。ヒト歯胚組織を移植する場合には、ヒト、又は免疫不全に改変したヒト以外の他の哺乳動物を用いることが好ましい。このような生体内成育に好適な生体部位としては、動物細胞の器官や組織をできる限り正常に発生させるためには、腎臓皮膜下、腸間膜、皮下移植、口腔内等が好ましい。
移植による成育期間としては、移植時の大きさと発生させる歯の大きさによって異なるが、一般に、3〜400日とすることができる。例えば、腎臓皮膜下への移植期間は移植する培養物の大きさと再生させる歯の大きさによっても異なるが、歯の再生と移植先で発生させる歯の大きさの観点から7〜60日間であることが好ましい。
【0063】
生体への移植を行う前に、生体外での培養(前培養)を行ってもよい。この前培養によって細胞間の結合と第1及び第2の細胞集合体同士の結合を強固にして、細胞間相互作用をより強固にすることができるため好ましい。その結果、全体の成育期間を短縮することができる。
前培養の期間は短期であっても長期であってもよい。長期間、例えば3日以上、好ましくは7日以上とした場合には、歯胚から歯芽に発生させることができるので、移植後に歯ができるまでの期間を短縮することもできるため好ましい。前培養の期間としては、例えば腎臓皮膜下へ移植を行う場合の器官培養として、好ましくは1〜7日とすることが効率よく歯を再生するために好ましい。
【0064】
本発明の製造方法によって製造された歯は、内側に象牙質、外側にエナメル質という歯としての特有の細胞配置(構造)を有するものであり、また好ましくは、更に歯の先端(歯冠)と歯根という方向性も備えているものである。少なくともこのような特有の細胞配置、好ましくは細胞配置に加えて方向性を有することによって、歯としての機能も発揮できるものである。このため、歯の代替物として広く利用することが可能である。特に、自家の歯胚に由来した間葉系細胞及び上皮系細胞を用いた場合には、拒絶反応による問題を回避しつつ使用することができる。また一般に移植抗原が適合した他人の歯胚に由来する細胞を用いる場合にも拒絶反応による問題を回避することが可能である。
【0065】
また本発明の製造方法によって製造された歯は、歯特有の細胞配置を有する歯の集合体であってもよい。
このような歯の集合体は、歯特有の細胞配置を有する複数の歯で構成されているため、個々の歯を集合体から分離して、以下に述べるように1つの歯の移植片として用いることができる。この結果、移植片としての歯を効率よく作製することができる。
【0066】
複数の歯で構成された歯の集合体を得るためには、第1及び第2の細胞集合体いずれも、複数本の歯が発生するための歯胚の再誘導を容易にするために共に単一細胞により構成されていることが好ましい。
なお、培養工程は、前記と同様に、器官培養であっても腎臓皮膜下での培養であってもよいが、得られた歯を移植片として用いる場合には、他の動物細胞との接触がなく且つ全行程in vitroで調製することができる器官培養とすることが好ましい。
【0067】
さらに本発明の製造方法では、培養期間を歯周組織が形成されるまで継続してもよい。これにより、歯そのものに加えて、歯を顎骨上で支持し、固定化する歯槽骨や歯根膜などの歯周組織も形成させることができる。この結果、移植後に実用可能な歯を提供することが可能である。
【0068】
なお、歯周組織を製造するためには、上記培養工程の後に、前記培養によって得られた歯周組織を単離する工程を設けて歯周組織のみを得てもよい。歯周組織の単離は、培養工程の過程で形成された歯周組織と歯とを分離することができる如何なる方法によって行ってもよく、ピンセット等による分離や、酵素による部分消化等を挙げることができる。
【0069】
本発明に従って得られた歯及び歯周組織は、移植片として用いられる他、歯の発生過程を解明するための研究にも好ましく利用することができ、今後の歯に関連する組織発生のために有効なリサーチツールとして利用することができる。
なお、得られた歯又は歯周組織を移植片として用いる場合には、製造方法における培養工程を、他の動物細胞との接触がなく且つ全行程in vitroで調製することができる器官培養としたものであることが好ましい。
【0070】
また、本発明には、歯の移植方法が含まれる。この移植方法では、上記歯の集合体を得る工程と、歯の複合体から個々の歯を分離する工程と、分離された歯を、移植部位での他の歯と同一の方向性となるように揃えて移植する工程とを含む。
これにより、歯の移植を、特有の細胞配置と方向性を備えた複数の歯を同時に得て、効率よく実施することができる。
【0071】
本発明による歯は、歯の欠損及び損傷を伴う各種症状、例えば、齲蝕、辺縁性歯周炎(歯槽膿漏)、歯周病による歯の欠損、事故などによる折損や脱落等の治療又は処置に適用することもできる。
即ち、本発明の治療方法は、本発明による製造方法によって得られた歯及び/又は歯周組織を、欠損及び/又は損傷部位へ移植することを含む。これにより、欠損及び/又は損傷部位の上記症状を治療及び/又は緩和することができる。
本発明の他の治療方法は、本発明における培養工程のみ、或いは、配置工程及び培養工程を、欠損及び/又は損傷部位において実施させることを含む。この場合、支持担体としては、上述したものに加えて、欠損及び/又は損傷部位の周囲組織そのものを支持担体として適用してもよい。これにより、生体内での周辺組織からのサイトカイン等によって、より迅速に欠損及び/又は損傷部位の治療等を行うことができる。
【実施例】
【0072】
以下に本発明の実施例について説明するが、これに限定されるものではない。また実施例中の%は、特に断らない限り、重量(質量)基準である。
[実施例1]
1. EC細胞の培養方法
EC細胞は、AT805細胞(129系EC細胞であるOTT6050の派生クローン、理化学研究所バイオリソースセンター細胞材料開発室より入手)を使用した。AT805細胞の培養は、10容量%牛胎児血清(FCS:JRH又はJBS、Hyclone社製)及び55μM 2−メルカプトエタノール(GIBCO社製)を添加したダルベッコ改変イーグル培地(DMEM:SIGMA社又はコージンバイオ社製)を用いて行った。EC細胞は通常、100mmディッシュあたり5〜8×105個の濃度で播種し、一日おきに全量培地交換と継代培養を繰り返した。継代培養では、細胞をHCMFバッファ(pH7.4、10mM Hepes、136.9mM NaCl、0.34mM Na2HPO3、13.9mMグルコース、5.37mM KCl)にて1回洗浄後、最終濃度0.025%のトリプシン−EDTA・2Na(GIBCO社製)を溶解した酵素溶液を5ml添加し、37℃にて1分間酵素処理を行った。その後、等量の10%FCS添加DMEMを加えて細胞を分散させた後、遠心分離によって沈殿回収した細胞を再度懸濁し、新たな培養ディッシュへ播種し、継代培養した。
【0073】
2.EC細胞の分化誘導及び細胞の回収
トリプシン処理により回収したEC細胞を、終濃度0.5〜5容量%のジメチルスルホキシド(DMSO:SIGMA社製)を添加した10容量%FCS添加DMEMで懸濁し、1.0×106個/100mmディッシュの濃度で播種した。5%CO2濃度条件下で37℃にて培養し、14時間後、HCMFバッファにて細胞を1回洗浄し、培養液を10容量%FCS添加DMEMに培地交換した。
【0074】
DMSOで処理したEC細胞は、3日ごとに培養液の半量を10容量%FCS添加DMEMに交換を行いながら培養を6日間(5容量%DMSOのみ19日間)継続し、適宜観察を行った。未分化細胞を含む細胞集団が増殖した時点で、HCMFバッファにて1回洗浄後、3mM EDTA、並びに0.5%BSAを含むCa2+、Mg2+不含PBS(−)(EDTA/BSA−PBS)を添加して細胞を分散し、遠心操作により細胞を沈殿として回収した。細胞を再びEDTA/BSA−PBSに懸濁し、FITC標識した抗CD44抗体(CD44−FITC:BD Pharmingen社製)及びPE標識した抗CD29抗体(CD29−PE:BD Pharmingen社製)により二重染色を行い、CD44陽性かつCD29陽性の細胞フラクションを、セルソーター、Epics ALTRA(Beckman Coulter社製)を用いて分離取得することで、未分化EC細胞が除去されたDMSO処理により分化誘導したEC細胞フラクションを得た(図2参照)。この細胞をDMSO−EC細胞と名付けた。
また、各DMSO濃度で処理した場合のDMSO−EC細胞の割合について、表1に示す。表1に示されるように、DMSO処理濃度に依存して、CD44陽性かつCD29陽性の細胞は増加することが判明した。なお、5容量%DMSOで処理したEC細胞は、ほとんどの細胞が処理直後に死滅するため、処理後の培養日数が他の処理濃度より長く必要になる一方で、培養後に得られたCD44陽性かつCD29陽性の細胞は高い割合であることが明らかになった。
以後の実験には、5容量%のDMSOを用いて得られた細胞を用いた。
【0075】
【表1】
【0076】
DMSO−EC細胞のクローン化
DMSO−EC細胞のクローン化には、クローニングリングによるコロニーの分離、あるいは限界希釈法によって行った。
クローニングリングによる細胞の取得では、DMSO−EC細胞を培養ディッシュに低濃度(約100個)で播種し、増殖によってできたコロニーの細胞数が10個以上になったところで、滅菌済みのシリコングリースをクローニングリングの一端に塗り、培地を取り除いたディッシュ底面に、細胞のコロニーを囲うようにして貼り付けた。その後、クローニングリング内でトリプシン処理を行い、細胞を回収して、増殖した細胞から順次、培養のスケールアップを行ない、クローンを取得した。
限界希釈法によるクローンの取得は、上記方法により取得したDMSO−EC細胞をHCMFバッファにて1回洗浄後、トリプシン処理により細胞を回収した。回収された細胞数を計測し、1細胞/200μl/ウエル(説明)になるように96ウエル培養プレートに播種した。3〜5日後に単一のコロニーであることを確認した。その後5日ごとに、半量の培地交換を行いながら培養を継続し、増殖した細胞から順次、培養のスケールアップを行ない、クローンを取得した。
これにより、DMSO−EC細胞由来のクローン1(DMSO−ECクローン1:Clone #1)及びクローン2(DMSO−ECクローン2:Clone #2)を得た。
【0077】
[実施例2]
DMSO−EC細胞の性状
(1)位相差顕微鏡像
上記で得られたクローン1及び2の形態と、EC細胞、DMSO−EC細胞、並びにEC細胞由来軟骨系前駆細胞であるATDC5細胞(理化学研究所バイオリソースセンター細胞材料開発室より入手)の形態をそれぞれ位相差顕微鏡で観察した。
この結果、EC細胞は、コロニー内の細胞が小さく、高い密度で増殖するほか、コロニー周辺が盛り上がっており、典型的な全能性幹細胞様の形態であることが観察された。これに対して、DMSO−ECやDMSO−ECクローン1及び2、ATDC5細胞はいずれも、コロニーを構成するひとつずつの細胞の面積が大きく、付着性細胞としての特徴である単一層を形成しており、特徴的なEC細胞の形態と明らかに異なる形態を示した。このような付着性の細胞形態は、EC細胞よりは間葉系細胞に類似している。
【0078】
(2)各種遺伝子発現
EC細胞、DMSO−EC、DMSO−ECクローン1及び2、ATDC5細胞における各種遺伝子発現をリアルタイムPCR(RT−PCR)により解析し、各種マーカー遺伝子の発現を検討した。
EC細胞、並びにDMSO−EC、DMSO−ECクローン1及び2、ATDC5細胞からの全RNAの抽出は、TRIzol Reagent(Invitrogen社製)を用いて行なった。解析する細胞の培養液を除去し、100mmの培養ディッシュに直接2mlのTRIzol Reagentを滴下し、Cell Scraper(Falcon社製)で充分混合した後、エッペンチューブに回収し、ポリトロンホモゲナイザー(ポリトロン社製)でホモジナイズを行った。その後のRNA抽出までの操作は、添付の使用説明書に従った。細胞・組織から抽出された全RNAは、DEPC処理水に溶解し、分光光度計にて濃度算定を行った後、−30℃に保存した。
【0079】
細胞から抽出した全RNAを鋳型として、リアルタイムPCR(RT−PCR)解析のためテンプレートとなるcDNAの合成を行った。cDNA合成は、ReverTra Ace(TOYOBO)を用いて行い、操作は添付された使用説明書に従って行った。
PCR及び解析は、ABI PRISM 7000(Applied Biosystems社製)を使用して行った。ポリメラーゼはSYBR Premix Ex Taq(TAKARA社製)を使用し、内在性コントロールはβ−アクチンを用いた。RT−PCR専用96ウエルプレートに1サンプルあたり、SYBR Premix Ex Taq (TAKARA社製)を13μl、20倍に希釈したcDNA反応溶液を2μl、1μMの濃度のプライマーをそれぞれ5μl、滅菌水を5μl添加し、最終容量25μlの反応系を調製し、PCR反応を常法に従って行なった。また、それぞれの遺伝子についてコントロールの反応を行って検量線を作成し、それぞれの定量化を行った。解析に使用したプライマーの配列を表2及び表3に示した。
遺伝子発現は、それぞれの細胞におけるβ−アクチンの発現量により標準化し、β−アクチン発現量を一定にした場合の相対的な発現量で比較した。結果を表4に示す。
【0080】
【表2】
【0081】
【表3】
【0082】
【表4】
【0083】
表4に示されるように、未分化幹細胞マーカー遺伝子であるOct3/4とNanogの発現は、EC細胞でのみ検出され、DMSOで処理して得られたEC細胞やそのクローン細胞、ATDC5細胞では認められなかった。神経堤細胞のマーカー遺伝子であるSlug、Pax3、Wnt1の発現は、EC細胞では認められず、SlugはEC細胞以外の細胞群の全てで認められ、Pax3とWnt1は、発現量の差異はあるものの、EC細胞とATDC5細胞以外のDMSO−EC細胞群で認められた。
【0084】
このことから、DMSO−EC細胞とそのクローン化細胞は、DMSOの添加によって未分化幹細胞から神経堤細胞へと分化誘導された細胞であることが示唆され、遺伝子発現の特性から、Oct3/4陰性、Nanog陰性、Slug陽性、Pax3陽性、Wnt1陽性の細胞であることが判明した。
一方、歯の間葉細胞のマーカー群では、EC細胞ではMsx1とBMP−7が発現しているほかはいずれの遺伝子も発現していないのに対して、DMSO−EC細胞、並びにそのクローン化細胞では、Msx1、Pax9、Wnt5a、Lhx8が特異的に高発現していた。BMP4、Runx2、Dlx1は、DMSO−EC細胞、並びにそのクローン化細胞とATDC5細胞で、発現量に多少の差異はあるものの発現が認められた。
また軟骨細胞における特異的なマーカー遺伝子であるSox9、Sox5の発現は、ATDC5細胞でのみ高発現しており、DMSO−EC細胞とそのクローン化細胞は軟骨系への分化をしていない細胞であると判断された。
従って、DMSOによって、EC細胞が神経堤細胞様ないしは間葉系細胞様の細胞へ分化誘導されたことは明らかであった。
【0085】
[実施例3]
EC細胞から分化誘導した細胞の歯形成能の評価
次に、本発明によって得られたDMSO−EC細胞、そのクローン化細胞の象牙芽細胞への分化能、並びに歯の象牙質形成能を、以下のように行って、確認した。
【0086】
(1)歯胚上皮組織の調製
C57BL/6マウス(SLCより購入)の胎齢14.5日、胚仔から下顎切歯歯胚組織を顕微鏡下で常法により摘出した。摘出した下顎切歯歯胚組織をPBS(−)で洗浄し、PBS(−)に最終濃度1.2U/mlのディスパーゼ II (Roche, Mannheim, Germany)を添加した酵素液で室温にて12.5分間処理した後、10%FCS(JRH Biosciences, Lenexa, KS)を添加したDMEM(Sigma, St. Louis, MO)で3回洗浄した。さらにDNaseI溶液 (Takara, Siga, Japan)を最終濃度70U/mlになるよう添加し歯胚組織を分散させ、25G注射針 (Terumo, Tokyo, Japan)を用いて外科的に歯胚上皮組織を分離した。
【0087】
(2)再構成歯胚の作製
再構成歯胚の作製には、上記で調製された歯胚上皮組織と、間葉系細胞として、EC細胞、DMSO−EC細胞、DMSO−ECクローン1及び2のいずれかの評価対象細胞とを用いた。
歯胚再構築に使用する評価対象細胞群を、それぞれトリプシン処理によりディッシュから回収した。シリコングリースを塗布した1.5mlマイクロチューブ(Eppendorf, Hamburg, Germany)に、10%FCS(JRH)添加DMEM(Sigma)で懸濁した評価対象細胞を入れ、遠心分離(580×g)により細胞を沈殿として回収した。遠心後の培養液の上清をできる限り除去し、再度遠心操作を行い、実体顕微鏡で観察しながら細胞の沈殿周囲に残存する培養液をGELoader Tip 0.5−20μL(エッペンドルフ社製)を用いて完全に除去し、再構成歯胚作製に用いるそれぞれの評価対象細胞を準備した。
【0088】
シリコングリースを塗布したペトリディッシュに2.4mg/mlのCellmatrix type I−A(新田ゼラチン社製)を30μl滴下してコラーゲンゲルドロップを作製した。この溶液に、上記評価対象細胞を、0.1−10μlのピペットチップ(エッペンドルフ社製)を用いて、0.2μl〜l0.3μlアプライして、高密度の細胞集合体として細胞凝集体を作製した(細胞密度:2×108個/ml)。続いて、10μlのピペットチップを用いて、歯胚上皮組織或いは歯胚間葉組織を同じゲルドロップ内にアプライし、タングステン針を用いて、分離した歯胚由来上皮組織が本来、間葉組織と接触していた面を評価対象細胞の細胞凝集体に密着させた。その後、ゲルドロップを固化させることにより、歯胚組織と評価対象細胞間の結合をより強固にすることで、高密度再構成歯胚を作製した。
【0089】
これを、図3を参照して説明する。
ピペットチップ16で先にゲルドロップ10(図3A参照)内に配置された細胞凝集体12は、ゲルドロップ10内で球体を構成する(図3B参照)。この後に他方の細胞凝集体14を押し込むことによって、球体の細胞凝集体12がつぶされて、他方の細胞凝集体14を包むようになることが多い(図3C参照)。その後にゲルドロップ10を固化させることにより、細胞間の結合が強固になる(図3D参照)。
【0090】
(3)再構成した歯胚の器官培養
ゲル中で作製した高密度再構成歯胚は、CO2インキュベーターに10分間静置してCellmatrix type I-A (Nitta Gelatin)を固形化した。10容量%FCS(JRH社製)、0.1mg/mlのL−アスコルビン酸(Sigma社製)、2mMのL−グルタミン(GIBCO社製)添加したDMEM(Sigma社製)に、セルカルチャーインサート(ポアサイズが0.4ミクロンのPETメンブレン;BD社製)が接するように培養容器を準備した。培養容器のセルカルチャーインサートの膜上に、再構成歯胚を、支持担体である周囲のゲルと共に移して、器官培養した。
【0091】
器官培養により歯の発生を解析する場合には、通常、14日間の培養を行なった。器官培養した場合には、移植後10〜30日目に再構成歯胚を摘出し、4%パラホルムアルデヒド−リン酸緩衝液で6時間固定した後、4.5%EDTA溶液(pH7.4)を用いて24時間の中性脱灰を行った。その後、常法によりパラフィン包埋して、10μmの切片を作製した。組織学的解析のためには常法に従い、ヘマトキシリン−エオジン染色(HE染色)を行った。
【0092】
器官培養による評価
14.5日齢、下顎切歯歯胚上皮組織とEC細胞の高密度再構成歯胚では、EC細胞の異常増殖により再構成歯胚は肥大化し、HE染色においても歯の間葉細胞に由来する象牙芽細胞や象牙質は観察されなかった。
これに対し、DMSO−EC細胞、DMSO−ECクローン1及び2では、上皮組織との相互作用が誘導され、位相差顕微鏡像においても歯胚の器官培養と同様に組織誘導が観察された。またさらに、DMSO−EC細胞、DMSO−ECクローン1及び2ではいずれも、外側にエナメル質、内側に象牙質を有する再構成歯胚を形成できることがHE染色像から確認でき、歯に特有の組織構造が形成可能であることが示された。
これらのことから、DMSO−EC細胞、DMSO−ECクローン1及び2は、歯胚上皮細胞との相互作用によって、歯の間葉組織である象牙芽細胞へ分化し、歯に特有の象牙質を産生できることは明らかであった。また高密度再構成歯胚法によって、器官培養においても、外側にエナメル質、内側に象牙質、内部に歯髄細胞、歯の先端と歯根を有する歯に特有の組織構造を有する歯を形成できることが明らかであった。
【0093】
[実施例4]
腎皮膜下移植方法
次に、DMSO−EC細胞、DMSO−ECクローン1及び2の細胞と14.5日齢胎児下顎切歯歯胚上皮細胞とから、高密度再構成歯胚を作製し、得られた再構成歯胚をC57BL6マウス(日本クレア社から入手)腎皮膜下に移植して、歯形成能を評価した。
実施例3と同様にして作製した再構成歯胚を、48時間から96時間、器官培養を行った後、周囲のゲルごと8週齢のNOD−SCIDマウス(チャールズリバー社から入手)の腎皮膜下に移植して異所的な歯の発生を進行させ、解析を行った。
腎皮膜下に移植した場合には、移植後10〜30日目に周囲の腎組織ごと再構成歯胚を摘出した。摘出組織を、4%パラホルムアルデヒド−リン酸緩衝液で6時間固定した後、4.5%EDTA溶液(pH7.4)を用いて24時間の中性脱灰を行った。その後、常法によりパラフィン包埋して、10μmの切片を作製した。組織学的解析のためには常法に従い、HE染色を行った。
【0094】
(1)腎皮膜下移植による評価
腎皮膜下移植では、14.5日齢胎児切歯歯胚を移植した場合には、内側に象牙質及び外側にエナメル質を有する特徴的な構造の歯を16日の期間で腎皮膜下移植により発生させることができた(図4参照)。14.5日齢胎児切歯歯胚に由来する歯胚上皮組織と歯胚間葉細胞(図4A参照)、歯胚上皮組織とDMSO−EC細胞(図4B参照)、歯胚上皮組織とDMSO−ECクローン(クローン#1、図4C参照)による再構成歯胚では、腎皮膜下移植によって移植後14日目には、正常歯胚をそのまま腎皮膜下へ移植した場合と同様に、外側のエナメル芽細胞、エナメル質、その内側に象牙質、象牙芽細胞が容易に認められた。できた歯は、先端と歯根を有しており、正常発生の歯と同様の構造を有していた。
歯形成の頻度は、EC細胞及びATDC細胞が0%であるのに対して、DMSO−EC細胞では51.3±8.8%、DMSO−ECクローン1では23.3%±16.7%、クローン2では33.3%±47.1%であった。
【0095】
これらのことから、DMSO−EC細胞、DMSO−ECクローン1及び2は、歯の間葉由来組織である象牙芽細胞へと分化し、歯の硬組織である象牙質を産生することが可能であることが明らかになった。さらに高密度再構成歯胚法によって、腎皮膜下移植においても、DMSO−EC細胞、DMSO−ECクローン1及び2によって特有の組織構造を有する歯を形成できることが明らかになった。
【0096】
(2)In situハイブリダイゼーション
14.5日齢胎児切歯歯胚に由来する歯胚上皮組織とDMSO−EC細胞を腎皮膜下へ移植し、14日後に摘出した組織について、In situハイブリダイゼーションにより、エナメル質の構成分子であるアメロジェニンと、象牙質の構成要素であるデンチンシアロホスホプロテイン(DSPP)のmRNAの発現、並びに歯根膜特異的な遺伝子であるペチロスタチンmRNAの発現を解析した。
【0097】
腎皮膜下移植後14日目の再構成歯胚を摘出し、4%パラホルムアルデヒド−リン酸緩衝液で6時間固定した後、4.5%EDTA溶液(pH7.4)を用いて24時間の中性脱灰を行った。その後、12.5%スクロース溶液、25%スクロース溶液へと順次浸漬した後、OCT compound(SAKURA Finetechnica社製)に包埋した。包埋後はクリオスタット(Leica社製)で10μmの切片を作製した。
【0098】
上記切片を、最終濃度2μg/mlのプロテアーゼK(Nacalai tesque, kyoto, Japan)を含むPBS(−)で10分間処理し、最終濃度4%パラホルムアルデヒド(Nacalai tesque)を含むPBS(−)で10分間固定した。それぞれ最終濃度で1.325%トリエタノールアミン、0.0175NのHCl(Wako)、0.25%無水酢酸を含むジエチルピロカーボネート処理(DEPC)水で10分間処理した後、PBS(−)で5分間、3回洗浄した。1.5% (v/v) トリエタノールアミン(Nacalai tesque)、0.33NのHCl(Wako)、0.25%(v/v)の無水酢酸(Nacalai tesque) in DEPC水で10分間処理した後、2×SSCで10分間、2回洗浄した。歯根膜特異的遺伝子であるペリオスチン (Genbank accession no. NM_015784) プローブは、センスプライマー (-7; ggctgaagatggttcctctc:配列番号35)とアンチセンスプライマー (573; gtacattgaaggaataacca:配列番号36)を用いてPCRにより取得したcDNA断片を、DIG標識して用いた。
【0099】
定法に従ってin situ ハイブリダイゼーションを行い、抗DIG−アルカリホルファターゼ(AP)Fabフラグメント(Roche)とNBT/BCIPStock Solution(Roche)で発色させ、Axio Imager A.1 (Zeiss)とAxioCam MRc5 (Zeiss)で解析を行なった。
その結果から、HE染色像におけるエナメル芽細胞ではアメロジェニンmRNAの発現が、象牙芽細胞ではDSPP mRNAの発現がそれぞれ認められた。またペリオスタチンmRNAの発現が認められた。このことから硬組織形成に関与する分子のmRNAは、それぞれが適切にその産生細胞において明らかに発現していた。また歯根膜で発現するペリオスチンが検出されたことから歯周組織が形成されていることが示唆された。
【0100】
[実施例5]
レチノイン酸による誘導
(1)CD44陽性かつCD29陽性の細胞の取得のためのEC細胞の分化誘導の方法
実施例1と同様にして、トリプシン処理にて回収されたAT805細胞を1.0×106個/100mmディッシュの濃度で播種し、翌日、終濃度0〜10.0μMのレチノイン酸(RA)(SIGMA社製)を添加した10容量%FCS DMEMに置換して刺激を加えた。培養72時間後、HCMFバッファにて2回洗浄を行い、全量10容量%FCS DMEMで培地交換を行った。
【0101】
(2)RA処理後のCD44陽性かつCD29陽性の細胞の取得
3日毎に10容量%FCS DMEMを用いて半量培地交換を行いながら培養を7日間継続し、適宜観察を行った。RA処理後増殖してきた、未分化細胞を含む細胞集団をHCMFにて1回洗浄後、3mMのEDTA−0.5% BSA−PBS(−)を添加し、37℃でインキュベートして回収した。回収後の細胞はCD44 FITC(BD Pharmingen)及びCD29(BD Pharmingen)PEにより二重染色を行い、CD44陽性かつCD29陽性の細胞をEpics ALTRA(Beckman Coulter)を用いて分離取得することで、未分化細胞を除いた分化した細胞集団のみを得た。この細胞をRA−EC細胞と名付けた。結果を図5に示す。
また、各RA濃度で処理した場合のRA−EC細胞の割合について表5に示す。表5に示されるようにRA濃度に依存して、CD44陽性且つCD29陽性細胞が変化することが判明した。
またRA処理後のCD44陽性且つCD29陽性細胞について実施例2(2)と同様に遺伝子の発現について確認したところ、DMSO−EC細胞と同様に、Oct3/4陰性、Nanog陰性、Slug陽性、Pax3陽性、Wnt1陽性の細胞であることが判明した。
【0102】
【表5】
【0103】
(3)RA処理後のCD44陽性かつCD29陽性の細胞による歯形成能の評価
上記のようにして得られたCD44陽性且つCD29陽性細胞(RA濃度2μM)を用いて、実施例3と同様にして歯胚再構築及び器官培養を行い、評価した。器官培養14日後には、歯胚上皮細胞との相互作用によって、歯の間葉組織である象牙芽細胞へ分化した。また高密度再構成歯胚法によって、外側にエナメル質、内側に象牙質、内部に歯髄細胞を有する歯に特有の組織構造を有する歯が形成された(図6参照)。
またRA処理後のCD44陽性且つCD29陽性細胞について実施例4(2)と同様にしてアメロジェニン及びDSPP、ペルオスタチンの発現を確認したところ、DMSO−ECと同様に各mRNAの発現が認められた。このことから、硬組織、歯根膜を含む歯周組織が形成されていることが示唆された。
【0104】
[実施例6]
(1)CD44陽性かつCD106陽性の細胞の取得のためのEC細胞の分化誘導の方法
実施例1と同様にしてトリプシン処理により回収したAT805細胞を、1.0×106個/100mmディッシュの濃度で播種し、翌日、終濃度2μMのRA(SIGMA社製)を添加した10容量%FCS DMEMに置換して刺激を加えた。培養72時間後、HCMFバッファにて2回洗浄を行い、全量10容量%FCS DMEMで培地交換を行った。翌日、HCMFバッファにて2回洗浄を行い、死細胞を除去した後、さらに10容量%FCS DMEMで培養を継続した。以降1日おきに培地交換を行い、7日間培養した。
【0105】
(2)CD44陽性かつCD106陽性の細胞の取得
上記処理により取得された未分化細胞を含む細胞集団はHCMFにて1回洗浄後、3mM EDTA−0.5% BSA−PBS(−)を添加し、37℃でインキュベートして回収した。回収後の細胞はCD44 FITC(BD Pharmingen)及び抗CD106抗体(CD106(BD Pharmingen)PE)により二重染色を行い、CD44陽性且つCD106陽性フラクションをEpics ALTRA(Beckman Coulter)を用いて分離取得することで、未分化細胞を除いた分化した細胞集団のみを得た。結果を図7に示す。
【0106】
(3)RA処理後のCD44陽性かつCD106陽性の細胞による歯形成能の評価
上記のようにして得られたCD44陽性且つCD106陽性細胞を用いて、実施例3と同様にして歯胚再構築及び器官培養を行い、評価した。器官培養14日後には、歯胚上皮細胞との相互作用によって、歯の間葉組織である象牙芽細胞へ分化した。また高密度再構成歯胚法によって、外側にエナメル質、内側に象牙質内部に歯髄細胞を有する歯に特有の組織構造を有する歯が形成された。
またこれらのCD44陽性且つCD106陽性のDMSO−EC細胞について実施例2(2)と同様に遺伝子の発現について確認した。この結果、CD44陽性且つCD29陽性DMSO−EC細胞と同様に、Oct3/4陰性、Nanog陰性、Slug陽性、Pax3陽性、Wnt1陽性の細胞であることが示された。
更に、実施例4(2)と同様にしてアメロジェニン及びDSPP、ペルオスタチンの発現を確認したところ、DMSO−ECと同様に各mRNAの発現が認められた。このことから、硬組織、歯根膜を含む歯周組織が形成されていることが示唆された。
【0107】
[実施例7]
実施例6(2)及び(3)と同様にして、DMSO−EC細胞についてCD44 FITC及びCD106 PEを用いて二重染色したところ、CD44陽性且つCD106陽性細胞の存在が示された。これらのCD44陽性且つCD106陽性のDMSO−EC細胞について実施例2と同様に遺伝子の発現について確認したところ、CD44陽性且つCD29陽性DMSO−EC細胞と同様に、Oct3/4陰性、Nanog陰性、Slug陽性、Pax3陽性、Wnt1陽性の細胞であることが判明した。
ここで得られたCD44陽性且つCD106陽性細胞の細胞集団を用いて、実施例3及び実施例4(2)と同様に歯胚再構築、器官培養及びmRNAの発現を行い評価した。その結果、歯胚上皮細胞との相互作用による特有の組織構造を有する歯が形成された。また実施例4(2)と同様にしてアメロジェニン及びDSPP、ペルオスタチンの発現が認められた。このことから、硬組織、歯根膜を含む歯周組織の形成が可能であることが示唆された。
【0108】
このように全能性幹細胞からCD44陽性且つCD29陽性細胞又はCD44陽性且つCD106陽性細胞を分化誘導することができ、これらの細胞を上皮系細胞と共に区画化して培養することにより、歯胚を再構成して、エナメル質及び象牙質を有する特有の構造を示す歯を提供することができた。
従って、本発明によれば、全能性幹細胞から歯形成用間葉系細胞を得ることができ、効率よく大量に歯を作製することができる。
【図面の簡単な説明】
【0109】
【図1】歯胚の形成を模式的に表した概念図である。
【図2A】本発明の実施例1にかかるEC細胞のCD44及びCD29の二重染色の様子を示す図である。
【図2B】本発明の実施例1にかかるDMSO処理後6日目の細胞のCD44及びCD29の二重染色の様子を示す図である。
【図2C】本発明の実施例1にかかる、選別されたCD44陽性且つCD29陽性細胞のCD44及びCD29の二重染色の様子を示す図である。
【図3A】本発明の実施例にかかる、歯胚由来間葉系細胞と上皮系細胞を用いた歯胚再構築の手順を概念的に示した図であり、細胞配置前のゲルパックの様子を示した図である。
【図3B】本発明の実施例にかかる、歯胚由来間葉系細胞と上皮系細胞を用いた歯胚再構築の手順を概念的に示した図であり、第一の細胞集合体をゲルパック内に配置した様子を示した図である。
【図3C】本発明の実施例にかかる、歯胚由来間葉系細胞と上皮系細胞を用いた歯胚再構築の手順を概念的に示した図であり、第二の細胞集合体をゲルパック内に配置した様子を示した図である。
【図3D】本発明の実施例にかかる、歯胚由来間葉系細胞と上皮系細胞を用いた歯胚再構築の手順を概念的に示した図であり、第一の細胞集合体及び第二の細胞集合体が配置されたゲルパックを固化した様子を示した図である。
【図4A】本発明の実施例4にかかる歯胚上皮組織と歯胚間葉細胞から作製した歯胚のHE染色像である(倍率:100倍)。
【図4B】本発明の実施例4にかかる歯胚上皮組織とDMSO−EC細胞から作製した歯胚のHE染色像である(倍率:100倍)。
【図4C】本発明の実施例4にかかる歯胚上皮組織とDMSO−ECクローン化細胞から作製した歯胚のHE染色像である(倍率:100倍)。
【図5A】本発明の実施例5にかかるEC細胞のCD44及びCD29の二重染色の様子を示す図である。
【図5B】本発明の実施例5にかかるRA処理後7日目の細胞のCD44及びCD29の二重染色の様子を示す図である。
【図5C】本発明の実施例5にかかる、選別されたCD44陽性且つCD29陽性細胞のCD44及びCD29の二重染色の様子を示す図である。
【図6】本発明の実施例5においてRA処理により得られたCD44陽性且つCD106陽性細胞と歯胚上皮組織とから作製した歯胚のHE染色像である(倍率:200倍、バーは50μm)。
【図7A】本発明の実施例6にかかるEC細胞のCD44及びCD106の二重染色の様子を示す図である。
【図7B】本発明の実施例6にかかるRA処理後7日目の細胞のCD44及びCD106の二重染色の様子を示す図である。
【図7C】本発明の実施例6にかかる選別されたCD44陽性且つCD106陽性細胞のCD44及びCD106の二重染色の様子を示す図である。
【符号の説明】
【0110】
10 ゲルパック(支持担体)
12 細胞凝集塊(第1の細胞集合体)
14 細胞凝集塊(第2の細胞集合体)
16 ピペットチップ
【特許請求の範囲】
【請求項1】
歯を形成するために用いられる間葉系細胞の製造方法であって、
全能性幹細胞を分化誘導剤の存在下で培養して、CD44陽性且つCD29陽性細胞又はCD44陽性且つCD106陽性細胞を含む分化誘導処理後の細胞集団を生成すること、
前記分化誘導処理後の細胞集団から、CD44陽性且つCD29陽性細胞又はCD44陽性且つCD106陽性細胞を、歯形成用間葉系細胞として選別すること
を含む製造方法。
【請求項2】
前記歯形成用間葉系細胞が更に、Slug遺伝子、Pax3遺伝子、Msx1遺伝子及びPax9遺伝子からなる群より選択された少なくとも1つを発現しているものである請求項1記載の製造方法。
【請求項3】
前記歯形成用間葉系細胞が更に、Slug遺伝子及びPax3遺伝子を共に発現しているものである請求項1記載の製造方法。
【請求項4】
前記全能性幹細胞が、胚性幹細胞、胚性癌腫細胞、胚性生殖幹細胞からなる群より選択された少なくとも1つである請求項1〜請求項3のいずれか1項記載の製造方法。
【請求項5】
前記全能性幹細胞が、胚性癌腫細胞であることを特徴とする請求項4記載の製造方法。
【請求項6】
前記分化誘導剤が、ジメチルスルホキシド及びレチノイン酸から選択された少なくとも一方である請求項1〜請求項5のいずれか1項記載の製造方法。
【請求項7】
支持担体の内部に、間葉系細胞及び上皮系細胞のいずれか一方のみから実質的になる第1の細胞集合体と、いずれか他方のみから実質的になる第2の細胞集合体とを混合することなく密着させて配置すること、
前記第1及び第2の細胞集合体を前記支持担体の内部で培養すること、
を含み、前記間葉系細胞が請求項1〜請求項6のいずれか1項記載の上記歯形成用間葉系細胞を含むものである歯の製造方法。
【請求項8】
前記培養が、歯周組織が形成するまで継続するものである請求項7記載の歯の製造方法。
【請求項9】
前記上皮系細胞が歯胚由来のものである請求項7又は8記載の歯の製造方法。
【請求項10】
全能性幹細胞から誘導されるCD44陽性且つCD29陽性又はCD44陽性且つCD106陽性の歯形成用間葉系細胞。
【請求項11】
前記歯形成用間葉系細胞が更に、Slug遺伝子、Pax3遺伝子、Msx1遺伝子及びPax9遺伝子からなる群より選択された少なくとも1つを発現しているものである請求項10記載の歯形成用間葉系細胞。
【請求項12】
前記歯形成用間葉系細胞が更に、Slug遺伝子及びPax3遺伝子を共に発現しているものである請求項10記載の歯形成用間葉系細胞。
【請求項13】
前記全能性幹細胞が、胚性幹細胞、胚性癌腫細胞、胚性生殖幹細胞からなる群より選択された少なくとも1つである請求項10〜請求項12のいずれか1項記載の歯形成用間葉系細胞。
【請求項1】
歯を形成するために用いられる間葉系細胞の製造方法であって、
全能性幹細胞を分化誘導剤の存在下で培養して、CD44陽性且つCD29陽性細胞又はCD44陽性且つCD106陽性細胞を含む分化誘導処理後の細胞集団を生成すること、
前記分化誘導処理後の細胞集団から、CD44陽性且つCD29陽性細胞又はCD44陽性且つCD106陽性細胞を、歯形成用間葉系細胞として選別すること
を含む製造方法。
【請求項2】
前記歯形成用間葉系細胞が更に、Slug遺伝子、Pax3遺伝子、Msx1遺伝子及びPax9遺伝子からなる群より選択された少なくとも1つを発現しているものである請求項1記載の製造方法。
【請求項3】
前記歯形成用間葉系細胞が更に、Slug遺伝子及びPax3遺伝子を共に発現しているものである請求項1記載の製造方法。
【請求項4】
前記全能性幹細胞が、胚性幹細胞、胚性癌腫細胞、胚性生殖幹細胞からなる群より選択された少なくとも1つである請求項1〜請求項3のいずれか1項記載の製造方法。
【請求項5】
前記全能性幹細胞が、胚性癌腫細胞であることを特徴とする請求項4記載の製造方法。
【請求項6】
前記分化誘導剤が、ジメチルスルホキシド及びレチノイン酸から選択された少なくとも一方である請求項1〜請求項5のいずれか1項記載の製造方法。
【請求項7】
支持担体の内部に、間葉系細胞及び上皮系細胞のいずれか一方のみから実質的になる第1の細胞集合体と、いずれか他方のみから実質的になる第2の細胞集合体とを混合することなく密着させて配置すること、
前記第1及び第2の細胞集合体を前記支持担体の内部で培養すること、
を含み、前記間葉系細胞が請求項1〜請求項6のいずれか1項記載の上記歯形成用間葉系細胞を含むものである歯の製造方法。
【請求項8】
前記培養が、歯周組織が形成するまで継続するものである請求項7記載の歯の製造方法。
【請求項9】
前記上皮系細胞が歯胚由来のものである請求項7又は8記載の歯の製造方法。
【請求項10】
全能性幹細胞から誘導されるCD44陽性且つCD29陽性又はCD44陽性且つCD106陽性の歯形成用間葉系細胞。
【請求項11】
前記歯形成用間葉系細胞が更に、Slug遺伝子、Pax3遺伝子、Msx1遺伝子及びPax9遺伝子からなる群より選択された少なくとも1つを発現しているものである請求項10記載の歯形成用間葉系細胞。
【請求項12】
前記歯形成用間葉系細胞が更に、Slug遺伝子及びPax3遺伝子を共に発現しているものである請求項10記載の歯形成用間葉系細胞。
【請求項13】
前記全能性幹細胞が、胚性幹細胞、胚性癌腫細胞、胚性生殖幹細胞からなる群より選択された少なくとも1つである請求項10〜請求項12のいずれか1項記載の歯形成用間葉系細胞。
【図2A】

【図2B】

【図2C】
【図5A】
【図5B】
【図5C】
【図7A】
【図7B】
【図7C】

【図1】
【図3A】
【図3B】
【図3C】
【図3D】
【図4A】
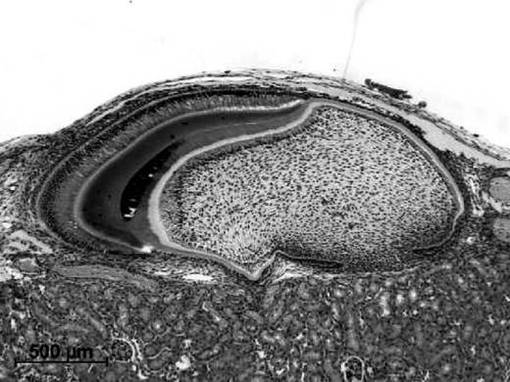
【図4B】
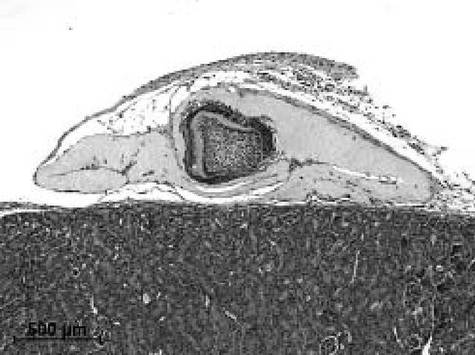
【図4C】
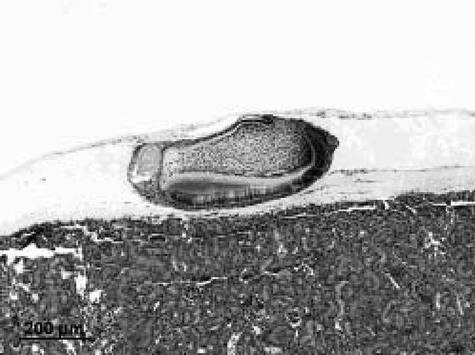
【図6】
【図2B】
【図2C】
【図5A】
【図5B】
【図5C】
【図7A】
【図7B】
【図7C】
【図1】
【図3A】
【図3B】
【図3C】
【図3D】
【図4A】
【図4B】
【図4C】
【図6】
【公開番号】特開2008−200033(P2008−200033A)
【公開日】平成20年9月4日(2008.9.4)
【国際特許分類】
【出願番号】特願2008−8772(P2008−8772)
【出願日】平成20年1月18日(2008.1.18)
【出願人】(803000115)学校法人東京理科大学 (545)
【Fターム(参考)】
【公開日】平成20年9月4日(2008.9.4)
【国際特許分類】
【出願日】平成20年1月18日(2008.1.18)
【出願人】(803000115)学校法人東京理科大学 (545)
【Fターム(参考)】
[ Back to top ]