
院内感染に対する免疫化のための多糖ブドウ球菌表面付着因子キャリアタンパク質接合体
【課題】院内感染症に対する有効な免疫原性組成物の提供。
【解決手段】多糖抗原およびブドウ球菌表面付着因子キャリアタンパク質の両方に対する抗体反応を惹起するための、ブドウ球菌表面付着因子キャリアタンパク質に接合した院内病原菌に由来する多糖抗原を有する免疫原性多糖タンパク質接合体(またはその1以上の抗原エピトープを表すジサッカライドフラグメント)の免疫原性組成物としての使用。当該免疫原性組成物は、スタフィロコッカス・アウレウス、スタフィロコッカス・エピデルミディスまたは他の院内病原菌により引き起こされる疾患に対して免疫化するために使用される。
【解決手段】多糖抗原およびブドウ球菌表面付着因子キャリアタンパク質の両方に対する抗体反応を惹起するための、ブドウ球菌表面付着因子キャリアタンパク質に接合した院内病原菌に由来する多糖抗原を有する免疫原性多糖タンパク質接合体(またはその1以上の抗原エピトープを表すジサッカライドフラグメント)の免疫原性組成物としての使用。当該免疫原性組成物は、スタフィロコッカス・アウレウス、スタフィロコッカス・エピデルミディスまたは他の院内病原菌により引き起こされる疾患に対して免疫化するために使用される。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、院内病原菌から得られる多糖抗原(または1以上の抗原性エピトープを表すそのオリゴ糖フラグメント)およびブドウ球菌表面付着因子キャリアタンパク質を含む免疫原性多糖タンパク質接合体に関する。本発明はさらに、多糖−タンパク質接合体を含む免疫原性組成物、およびその使用に関する。
【背景技術】
【0002】
毎年、米国において病院に入院している推定40000000人のうち約2000000人が院内感染を発症している(Anonyomous 1997)。死亡率は約4.4%であるので、院内感染のため毎年88000人が死亡している。米国における院内感染の費用は、1年あたり推定4500000000ドルである(Weinstein 1998)。これらの推定値は毎年行われる31000000人の外来患者の手術(National Center for Health Statistics’ website)、1500000人の養護施設居住者、拡張看護施設、または外来治療を受けている者において発生する感染症を包含しない。
【0003】
スタフィロコッカス・アウレウス(Staphylococcus aureus)および凝集酵素陰性ブドウ球菌(CoNS)、特にエス・エピデルミディス(S.epidermidis)は、院内感染の大部分の原因であるグラム陽性日和見院内病原菌である。ブドウ球菌感染症はすべての院内感染症のほぼ25%(約500000人)の原因である(Haley,Culverら、1985)(Boyce 1997)。一部の病院では入院患者全体の1%までがエス・アウレウス感染症にかかっている(StorchおよびRajagopalan 1986)。ブドウ球菌(エス・アウレウスおよびエス・エピデルミディス)は院内血流感染症の約47%、手術部位感染症(SSI)の24%、および院内感染肺炎の17%の原因である(Anonyomous 1997)。院内エス・アウレウスおよびCoNS感染症の患者の死亡率は5%〜68%までの範囲で相当変化する(Nada,Ichiyamaら、1996);(Thylefors,Harbarthら、1998)。
【0004】
ブドウ球菌感染症は、範囲が広く、皮膚感染症、例えば、膿痂疹、できもの、創傷感染および埋め込み装置からの感染症から重度の命にかかわる感染症、例えば骨髄炎、心内膜炎および転移性合併症を伴う菌血症にまで及ぶ。このような多様性のために、ブドウ球菌に対する有効な免疫原性組成物の設計を非常に困難になる。薬剤耐性院内細菌の出現が急激に増加し、そのためにこのような設計がさらに困難になっている。メチシリン耐性エス・アウレウスは院内感染症の原因の約40%である(Boyce 1997)。最近、バンコマイシン中間耐性エス・アウレウス(VISA)が出現し、その拡大に関する不安がさらに大きくなっている。従って、院内感染症に対する有効な免疫原性組成物に対する需要が強く、そして急速に増大している。
【0005】
莢膜多糖(capsular polysaccharide)
ヘモフィルス・インフルエンゼ(Haemophilus influenzae)、ストレプトコッカス・ニューモニエ(Streptococcus pneumoniae)およびB群ストレプトコッカスをはじめとする多くの病原菌の毒性における莢膜多糖(CP)の関与は、十分に証明されている。莢膜で覆われた細菌は、白血球による食菌作用に対して耐性であり、従って血液および組織に感染し得る。莢膜多糖に対する抗体は細菌莢膜の抗食細胞特性を中和するので(Karakawa、Suttonら、1988,Thakker、Parkら、1988)、ブドウ球菌莢膜は、人間におけるブドウ球菌感染症を予防するための免疫原性組成物の開発における主要な標的である。
【0006】
エス・アウレウスの12の公知莢膜血清型のうち、血清型5(CP5)および血清型8(CP8)は全臨床分離株の約85〜90%を占める(Arbeit,Karakawaら、1984、Karakawa,Fournierら、1985、Essawi,Na’wasら、1998、Na’was、Hawwariら、1998)。ほとんどのメチシリン耐性エス・アウレウス分離株はCP5を発現する(Sompolinsky,Samraら、1985)。CP5およびCP8に対する抗体は、インビトロのヒト多形核好中球による型特異性オプソニン作用による食菌現象を誘発し、動物において保護を付与する(Karakawa,Suttonら、1988、Fattom,Sarwarら、1996)。
【0007】
ほとんどの細菌莢膜多糖は、動物および人間において不十分な免疫原である。しかしながら、精製された多糖がタンパク質キャリア分子と接合するならば、これらは免疫原性およびT細胞依存性を獲得する。いくつかの研究所は、タンパク質と共有結合したCP5およびCP8からなる免疫原性接合体を合成した。これらの接合体は、マウスおよびヒトにおいて高度に免疫原性であり、食菌作用のためにマイクロカプセル化エス・アウレウスをオプソニン化する抗体を誘発する(Fattom,Schneersonら、1993、Gilbertら、1994,Reynaud−Rondierら、1991)。シュードモナス・エルジノーサ組み換え外毒素Aに接合したCP5を含有する一価免疫原性組成物は、免疫原性であり、健康な成人および末期の腎臓病の患者において良好な耐容性を示す(Welchら、1996)。血液透析を受けている末期腎臓病の患者を含む二重盲検法において、シュードモナス・エルジノーサ組み換え外毒素Aと共有結合したCP5およびCP8からなる二価接合体ワクチンは、約40週間エス・アウレウス菌血症に対して部分的免疫を付与し、その後、防御は抗体レベルが減少するにつれて減少した(Shinefieldら、2002)。この試験の結果は、さらに完全な防御をもたらすことができる改善された免疫原性組成物が必要とされていることを示す。
【0008】
多糖付着因子(PS/A;(Tojo,Yamashitaら、1988))、ポリ−N−スクシニルβ−1−6−グルコサミン(PNSG、(McKenney,Pouliotら、1999))、ポリ−N−アセチルグルコサミン表面多糖(PNAG、(Maria−Litran,Kropecら、2002))、または多糖細胞間付着因子(PIA(Mack,Fisherら、1996))と称する別の種類の細胞外多糖は、エス・アウレウスおよびエス・エピデルミディスの両方により発現される。PIAまたはPS/Aは直鎖β−1,6−結合グルコサミングリカンである。マウスをPS/A(PNSG、PNAG)で免疫化すると腎臓の転移増殖が減少し、インビトロで若干のPS/A(PNSG、PNAG)を産生するエス・アウレウス株で攻撃した後にマウスを死から防御する(McKenney,Pouliotら、1999)。PIAは静脈内カテーテルに関連する感染症の発生機序において重要な役割を果たす(Rupp,Ulphaniら、1999;Rupp,Ulphaniら、1999;RuppおよびFey 2001;Rupp,Feyら、2001)。個々のエス・エピデルミディス細胞間の接着を促進することに加えて、PIAは赤血球と結合し、ヘマグルチニンとして作用する(Fey,Ulphaniら、1999)。
【0009】
ブドウ球菌表面付着因子
ブドウ球菌は複数の表面付着因子(接着性マトリックス分子を認識する微生物表面成分と称する)を発現し、これらには、例えば、フィブロネクチン結合タンパク質、フィブリノゲン結合タンパク質、コラーゲン結合タンパク質およびビトロネクチン結合タンパク質が含まれる。これらの付着因子は細胞外マトリックス(ECM)成分、例えば、フィブロネクチン、フィブリノゲン、コラーゲンおよびビトロネクチンを特異的に認識し、結合する。エス・アウレウスにより発現される多数の接着因子は、様々な組織に対する接着および感染の代替法を提供することにより、その病原性に寄与する。ブドウ球菌付着因子の抗体は、細菌が哺乳動物宿主組織を侵襲することを防止するか、またはオプソニン作用による食菌作用を促進することにより疾患を軽減することができる。エス・アウレウスフィブロネクチン結合タンパク質A(融合タンパク質として提供される)の一部で免疫化されたラットは、実験的心内膜炎から適度に保護された。フィブロネクチン結合タンパク質Aの抗体を惹起するように設計された同様の免疫原性組成物をエス・アウレウス乳腺炎のマウスモデルにおいて試験した。免疫化されたマウスは対照マウスよりも重度の乳腺炎の症例が少なく、免疫化されたマウスの乳腺炎から回収される細菌は対照マウスよりも少なかった。19および87kDaのフィブリノゲン結合タンパク質で免疫化されたマウスは、非免疫化対照と比較して乳腺炎の発病率が減少し、一方、コラーゲン結合タンパク質での免疫化は保護しなかった(Lee,Pier 1997)。
【発明の概要】
【発明が解決しようとする課題】
【0010】
しかしながら、多糖抗原を様々なタンパク質キャリアに接合させるためのこれらおよび他の試みにもかかわらず、院内感染を治療または予防するための有効な免疫原性組成物は現在のところない。
【課題を解決するための手段】
【0011】
本発明は、従って、院内病原菌に由来する少なくとも1種の多糖抗原、または少なくとも1種のブドウ球菌表面付着キャリアタンパク質に接合した少なくとも1種の多糖抗原(合成または天然の多糖の加水分解により調製)の1以上の抗原エピトープを表すオリゴ糖フラグメントを含む免疫原性多糖−タンパク質接合体を提供する。本発明の接合体は、院内病原菌の多糖抗原および表面付着因子キャリアタンパク質の両方に対して特異的な抗体反応を対象において惹起する際に有用な免疫原性組成物において用いられる。従って、これらの接合体は、エス・アウレウス、エス・エピデルミディスまたは他の院内病原菌により引き起こされる院内感染症に対して免疫化するために、また受動免疫化のための免疫グロブリンの生成に関しては、院内感染症の予防または重篤度の軽減のために使用することができる。
【0012】
本発明の一態様において、少なくとも1種のブドウ球菌表面付着因子キャリアタンパク質と接合した院内病原菌に由来する少なくとも1種の多糖抗原を含む免疫原性多糖タンパク質接合体が提供され、ここにおいて該接合体は、多糖抗原および表面付着因子キャリアタンパク質の両方に対する特異的抗体を生成する。
本発明のもう一つ別の態様において、少なくとも1種のブドウ球菌表面付着因子キャリアタンパク質と接合した院内病原菌から得られる少なくとも1種の多糖抗原の1以上の抗原エピトープを表すオリゴ糖フラグメントを含む免疫原性多糖タンパク質接合体が提供され、ここにおいて、該接合体は、多糖抗原および表面付着因子キャリアタンパク質の両方に対する特異的抗体を生成する。
【0013】
さらにもう一つ別の態様において、多糖抗原−表面付着因子タンパク質接合体を適当な担体または希釈剤との組み合わせにおいて含む免疫原性組成物が提供される。本発明の免疫原性組成物はさらにアジュバント、例えば、水酸化アルミニウムまたはリン酸アルミニウムを含んでもよい。
さらにもう一つ別の態様において、哺乳動物において院内感染症に対する活性免疫を誘発する方法であって、このような感染症にかかっているヒトをはじめとする哺乳動物に、免疫原性量の本発明の免疫原性組成物を投与することを含む方法が提供される。
さらにもう一つ別の態様において、院内感染症に対する免疫治療薬の調製法であって、哺乳動物を本発明の免疫原性組成物で免疫化する工程、免疫化された哺乳動物から血漿を採集する工程、および集められた血漿から抗多糖抗体および抗表面付着因子抗体を含有する高度免疫グロブリンを収穫する工程を含む方法が提供される。高度免疫グロブリンは、院内感染症に対する受動免疫を誘発するために使用できる。
本発明の接合体は、多糖抗原および表面付着因子キャリアタンパク質の両方(両方とも毒性因子である)に対する抗体を惹起し、院内病原菌により引き起こされる疾患に対する免疫を付与する明確な利点を有する。すなわち、表面付着因子タンパク質それ自体が単に多糖抗原のタンパク質キャリアとして作用するだけでなく、免疫を付与することができる。
【図面の簡単な説明】
【0014】
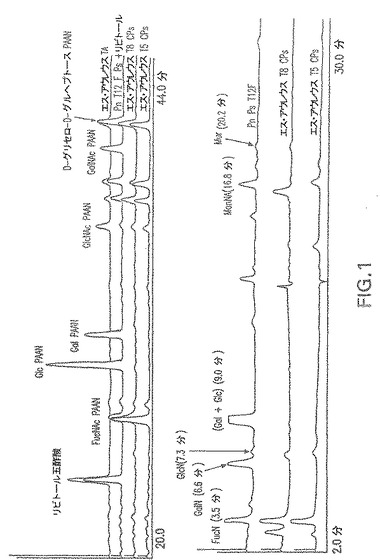
【図1】GLCおよびHPAEC−PAD分析により決定されるエス・アウレウスCP5およびCP8の組成を示す。
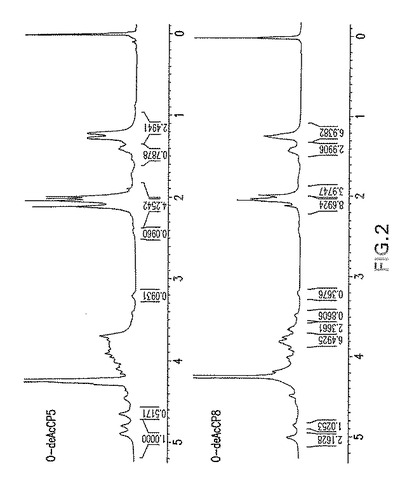
【図2】デ−O−アセチル化エス・アウレウスCP5およびCP8の1H−NMR分析を示す。
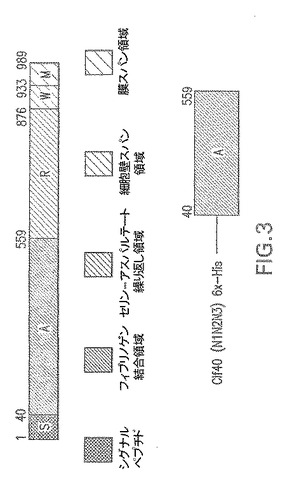
【図3】エス・アウレウス−ClfAから得られるクランピング因子の概略図である。
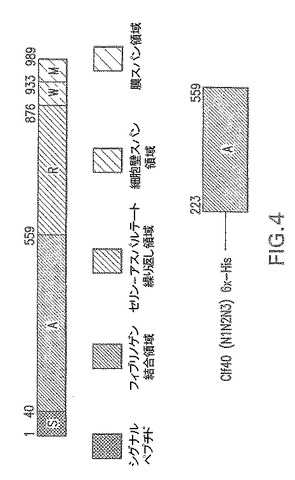
【図4】エス・アウレウスClfAに由来する組み換えタンパク質Clf40およびClf41の概略図である。
【図5】エス・エピデルミディス−SdrGから得られるクランピング因子の概略図である。
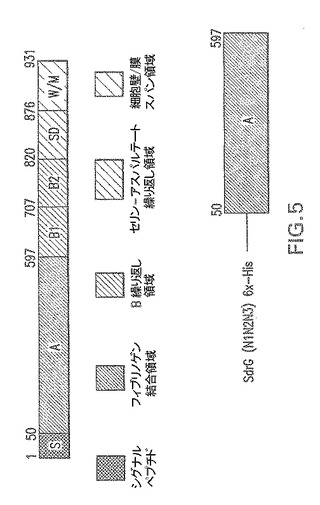
【図6】エス・エピデルミディスSdrG:SdrG(N1N2N3)およびSdrG(N2N3)に由来する組み換えタンパク質の概略図である。
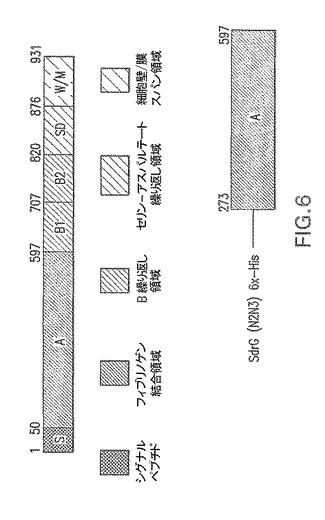
【図7】表面付着因子タンパク質のブロモアセチル化を示す。
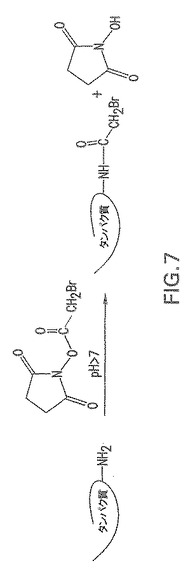
【図8】エス・アウレウスCPの3−(2−ピリジルジチオ)プロピオニルヒドラジド(PDPH)での活性化を示す。
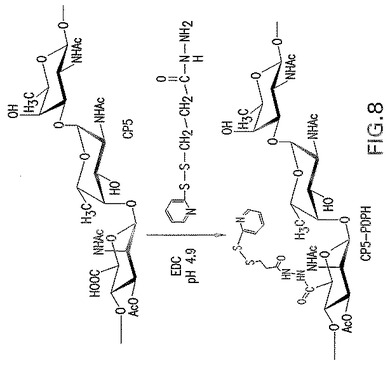
【図9】チオール化エス・アウレウスCPの表面付着因子タンパク質への接合を示す。
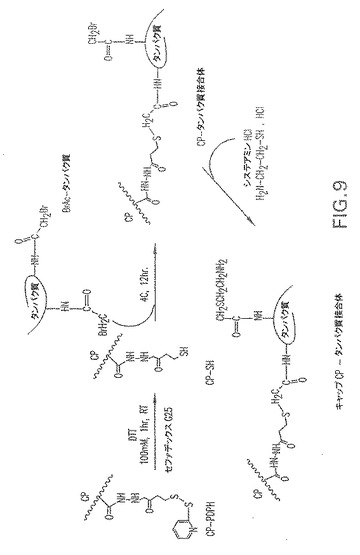
【図10】CP特異性ウサギ抗血清に関する抗原性についてのCP5−およびCP8−SdrG(N1N2N3)およびCP5−およびCP8−Clf41(N2N3)接合体の分析を示す。
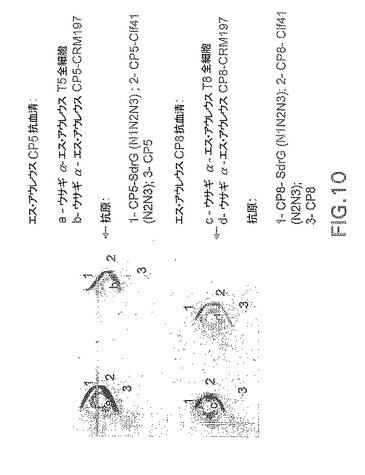
【図11】ClfA特異性ウサギ抗血清に関する抗原性についてのCP5−およびCP8−Clf41(N2N3)接合体の分析を示す。
【図12】二重免疫拡散検定による抗原性についてのCP5−およびCP8−SdrG(N2N3)6xHisおよびCP5−およびCP8−Clf40(N1N2N3)6xHis接合体の分析を示す。
【図13】Ouchterlony免疫拡散検定による抗原性についてのCP5−およびCP8−SdrG(N2N3)およびCP5−およびCP8−FnbA接合体の分析を示す。
【図14】ドットブロットによる接合体の分析を示す。
【図15A】SdrG(N2N3)に接合したエス・アウレウスCP8に対する免疫応答を示す。
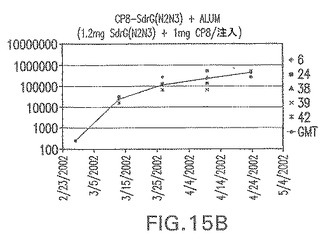
【図15B】アジュバントを含む、SdrG(N2N3)に接合したエス・アウレウスCP8に対する免疫応答を示す。
【図15C】Clf40(N1N2N3)に接合したエス・アウレウスCP8に対する免疫応答を示す。
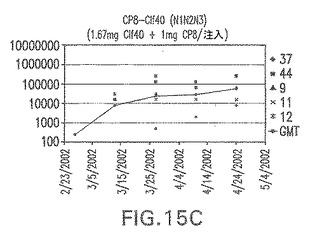
【図15D】アジュバントを含む、Clf40(N1N2N3)に接合したエス・アウレウスCP8に対する免疫応答を示す。
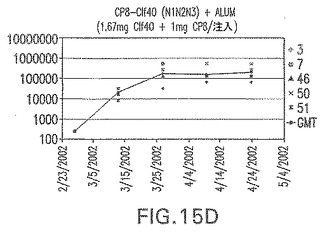
【図15E】アジュバントを含む、Clf41(N2N3)に接合したエス・アウレウスCP8に対する免疫応答を示す。
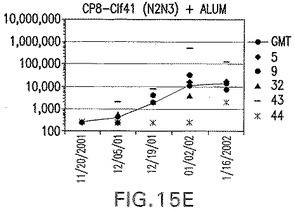
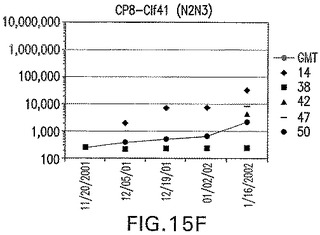
【図15F】Clf41(N2N3)に接合したエス・アウレウスCP8に対する免疫応答を示す。
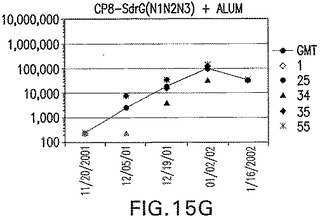
【図15G】アジュバントを含む、SdrG(N1N2N3)に接合したエス・アウレウスCP8に対する免疫応答を示す。
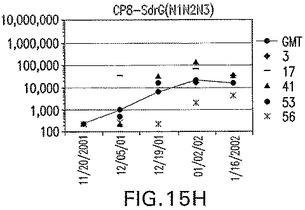
【図15H】SdrG(N1N2N3)に接合したエス・アウレウスCP8に対する免疫応答を示す。
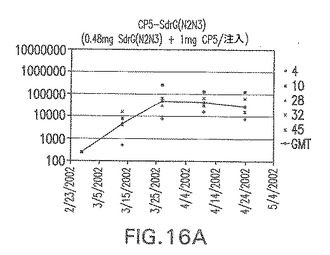
【図16A】SdrG(N2N3)に接合したエス・アウレウスCP5に対する免疫応答を示す。
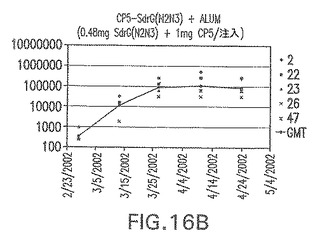
【図16B】アジュバントを含む、SdrG(N2N3)に接合したエス・アウレウスCP5に対する免疫応答を示す。
【図16C】Clf40(N1N2N3)に接合したエス・アウレウスCP5に対する免疫応答を示す。
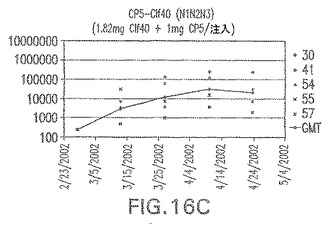
【図16D】アジュバントを含む、Clf40(N1N2N3)に接合したエス・アウレウスCP5に対する免疫応答を示す。
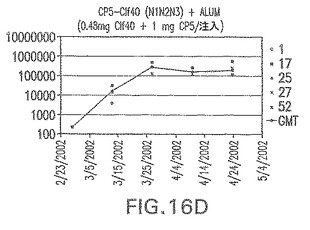
【図16E】アジュバントを含む、Clf41(N2N3)に接合したエス・アウレウスCP5に対する免疫応答を示す。
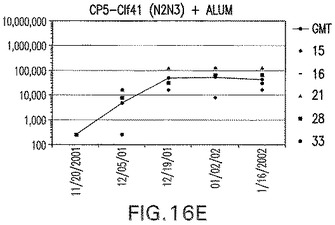
【図16F】Clf41(N2N3)に接合したエス・アウレウスCP5に対する免疫応答を示す。
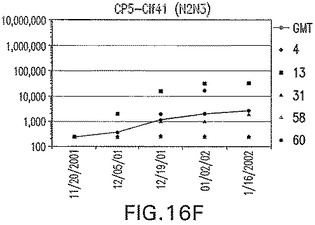
【図16G】アジュバントを含む、SdrG(N1N2N3)に接合したエス・アウレウスCP5に対する免疫応答を示す。
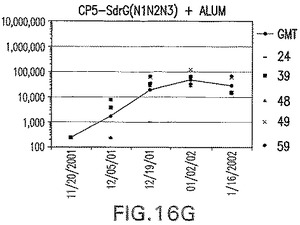
【図16H】SdrG(N1N2N3)に接合したエス・アウレウスCP5に対する免疫応答を示す。
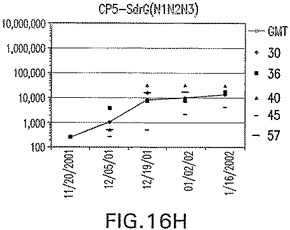
【図17A】アジュバントを含まない、未接合エス・アウレウスClf40(N1N2N3)に対する免疫応答を示す。
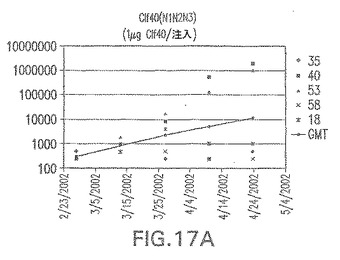
【図17B】アジュバントを含まない、CP8に接合したエス・アウレウスClf40(N1N2N3)に対する免疫応答を示す。
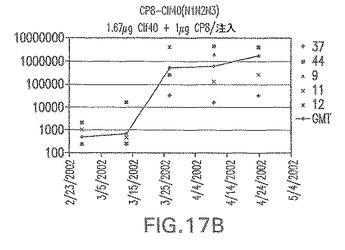
【図17C】アジュバントを含まない、CP5に接合したエス・アウレウスClf40(N1N2N3)に対する免疫応答を示す。
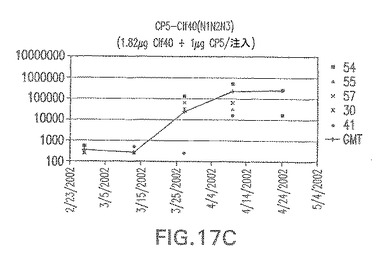
【図17D】アジュバントを含む、CP5に接合したエス・アウレウスClf40(N1N2N3)に対する免疫応答を示す。
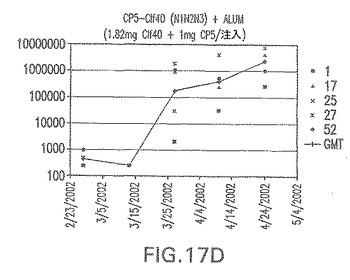
【図17E】アジュバントを含む、CP8に接合したエス・アウレウスClf40(N1N2N3)に対する免疫応答を示す。
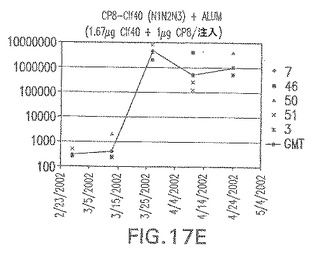
【図17F】アジュバントを含む、未接合エス・アウレウスClf40(N1N2N3)に対する免疫応答を示す。
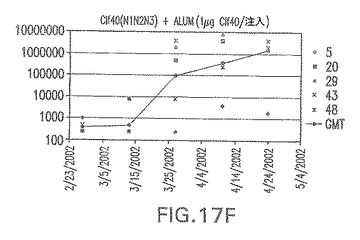
【図18A】アジュバントを含まない、CP5に接合したエス・アウレウスClf41(N2N3)に対する免疫応答を示す。
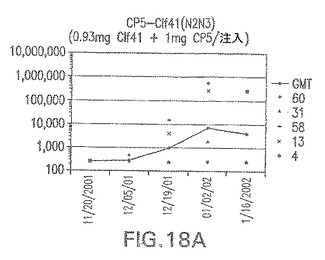
【図18B】アジュバントを含まない、CP8に接合したエス・アウレウスClf41(N2N3)に対する免疫応答を示す。
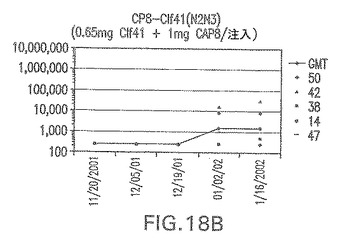
【図18C】アジュバントを含まない、未接合エス・アウレウスClf41(N2N3)に対する免疫応答を示す。
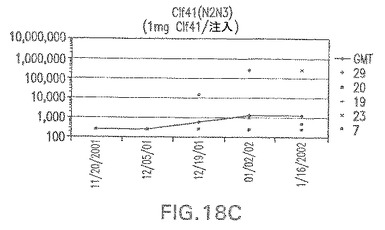
【図18D】アジュバントを含む、CP5に接合したエス・アウレウスClf41(N2N3)に対する免疫応答を示す。
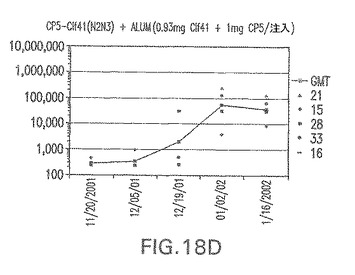
【図18E】アジュバントを含む、CP8に接合したエス・アウレウスClf41(N2N3)に対する免疫応答を示す。
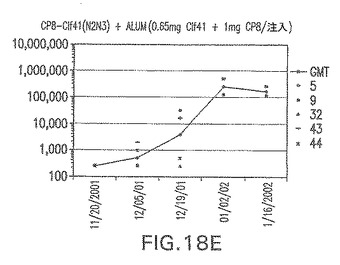
【図18F】アジュバントを含む、未接合エス・アウレウスClf41(N2N3)に対する免疫応答を示す。
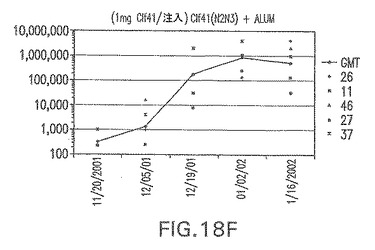
【図19A】アジュバントを含まない、CP5に接合したエス・エピデルミディスSdrG(N1N2N3)に対する免疫応答を示す。
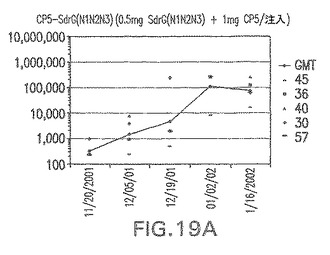
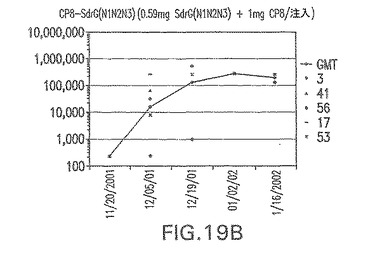
【図19B】アジュバントを含まない、CP8に接合したエス・エピデルミディスSdrG(N1N2N3)に対する免疫応答を示す。
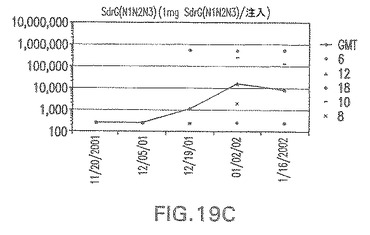
【図19C】アジュバントを含まない、未接合エス・エピデルミディスSdrG(N1N2N3)に対する免疫応答を示す。
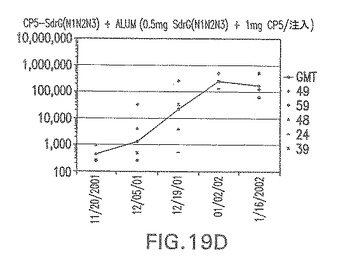
【図19D】アジュバントを含む、CP5に接合したエス・エピデルミディスSdrG(N1N2N3)に対する免疫応答を示す。
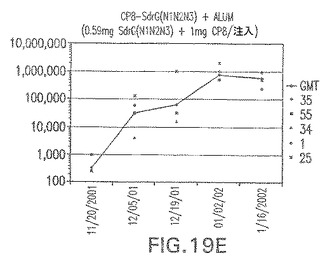
【図19E】アジュバントを含む、CP8に接合したエス・エピデルミディスSdrG(N1N2N3)に対する免疫応答を示す。
【図19F】アジュバントを含む、未接合エス・エピデルミディスSdrG(N1N2N3)に対する免疫応答を示す。
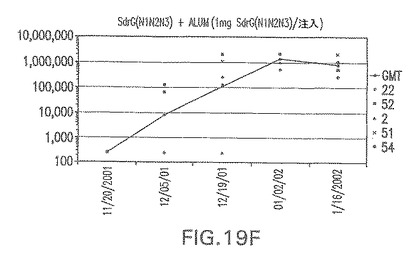
【図20A】アジュバントを含まない、CP5に接合したエス・エピデルミディスSdrG(N2N3)に対する免疫応答を示す。
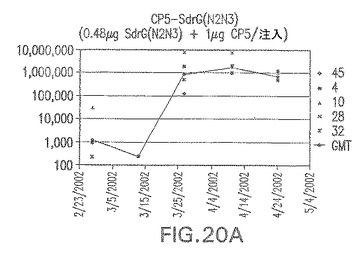
【図20B】アジュバントを含まない、CP8に接合したエス・エピデルミディスSdrG(N2N3)に対する免疫応答を示す。
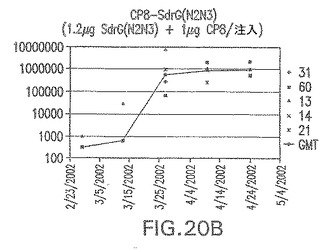
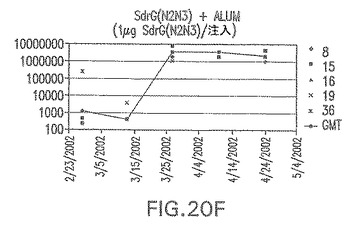
【図20C】アジュバントを含まない、未接合エス・エピデルミディスSdrG(N2N3)に対する免疫応答を示す。
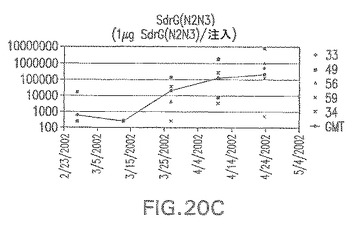
【図20D】アジュバントを含む、CP5に接合したエス・エピデルミディスSdrG(N2N3)に対する免疫応答を示す。
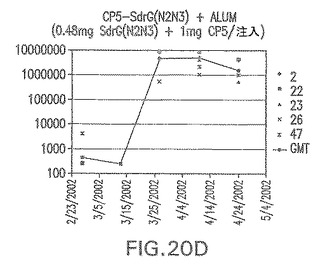
【図20E】アジュバントを含む、CP8に接合したエス・エピデルミディスSdrG(N2N3)に対する免疫応答を示す。
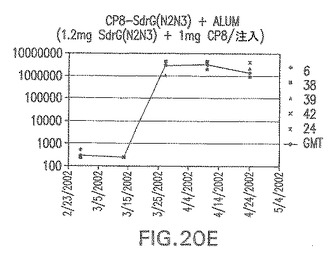
【図20F】アジュバントを含む、未接合エス・エピデルミディスSdrG(N2N3)に対する免疫応答を示す。
【発明を実施するための形態】
【0015】
院内感染症は複数の毒性因子を含む。従って、一つの毒性決定因子のみを含有する免疫原性組成物と比較して、免疫原性組成物中に成分として含まれる毒性決定因子の組み合わせが防御を増大させる可能性が非常に高い。本発明の多糖抗原は、これらに限定されないが、スタフィロコッカス・アウレウス(Staphylococcus aureus)、スタフィロコッカス・エピデルミディス(Staphylococcus epidermidis)および他の凝固酵素陰性ブドウ球菌(CoNS)、エンテロコッカス種(Enterococcus spp.)、カンジダ・アルビカンス(Candida albicans)、エンテロバクター種(Enterobacter spp.)、ヘモフィルス・インフルエンゼ(Haemophilus influenzae)、クレブシエラ・ニューモニエ(Klebsiella pneumoniae)、エシェリキア・コリ(Escherichia coli)、およびシュードモナス・エルジノーサ(Pseudomonas aeruginosa)をはじめとする様々な院内病原微生物に由来する。これらの抗原は全身性感染症における毒性因子であり、不十分な免疫源である。その免疫原性は、キャリアタンパク質への接合により向上させることができる。本発明の目的に関して、表面付着因子タンパク質は接着性マトリックス分子を認識する微生物表面成分である。これらはInhibitex Inc.(Alpharetta,GA、USA)からMSCRAMMの登録商標で入手可能である。以下に記載するように、ブドウ球菌表面付着因子キャリアタンパク質を多糖抗原のキャリアタンパク質として利用することにより、多糖がT細胞依存性抗原に変換され、かくして抗多糖IgG応答が誘発される。さらに、接合体は感染症に対して防御し、哺乳動物宿主組織への細菌の付着の防止を助ける抗表面付着因子キャリアタンパク質抗体を誘発する。タンパク質−サッカライド接合法の化学反応はキャリアタンパク質抗体の免疫原性エピトープに対して有害な影響を及ぼし得ることが知られているが、驚くべきことに、本発明において、このような影響は見られず、タンパク質は保護エピトープに対して反応を惹起できる状態のままである。
【0016】
細菌細胞表面上の表面付着因子タンパク質および宿主組織内のリガンドは厳重に相互作用し、その結果、細菌が宿主に付着する。付着はしばしば細菌の生存に必要であり、細菌が宿主防御メカニズムおよび抗生物質の攻撃をくぐりぬける助けをする。細菌がうまく宿主組織に付着し、コロニー化すると、その生理機能は劇的に変化し、毒素および酵素などの損傷成分が分泌される。さらに、付着性細菌はしばしばバイオフィルムを生成し、ほとんどの抗生物質の殺菌効果に対して素速く耐性になる。
表面付着因子タンパク質の代表例としては、フィブロネクチン結合タンパク質、フィブリノゲン結合タンパク質、コラーゲン結合タンパク質およびビトロネクチン結合タンパク質が挙げられる。これらの付着因子は細胞外マトリックス成分フィブリノゲン、フィブロネクチン、コラーゲンおよびビトロネクチンを特異的に認識し、結合する。
【0017】
フィブロネクチン結合タンパク質
フィブロネクチン(Fn)は、動物のECMおよび体液において見いだされる440kDa糖タンパクである。フィブロネクチンの第一の生物学的機能は、適当なインテグリンを発現する細胞の付着に関して基質として作用するその能力に関連するようである。いくつかの細菌種がフィブロネクチンと特異的に結合し、フィブロネクチン含有低質に付着することが証明されている。ほとんどのエス・アウレウス分離株はFnと結合するが、様々な程度で結合し、このことは細菌表面上に発現される表面付着因子分子の数の多様性を反映する。Fnとエス・アウレウス間の相互作用は非常に特異性である(Kuusela 1978)。Fn結合は、110kDaの分子量を有する、FnBP−AおよびFnBP−Bと称する2つの表面が露出したタンパク質により媒介される。第一Fn結合部位は、3〜5回繰り返す35〜40アミノ酸のモチーフからなる。これらの遺伝子はクローンされ、配列決定されている(Jonsson 1991)。
【0018】
WO−A−85/05553は、フィブロネクチン−、フィブリノゲン、コラーゲン、および/またはラミニン結合能力を有する細菌細胞表面タンパク質を開示している。
米国特許第5320951号および第5571514号(Hookら)は、フィブロネクチン結合タンパク質A(fngA)遺伝子配列、およびこの配列に基づく生成物および方法を開示している。
【0019】
米国特許第5175096号(Hookら)は、fnbBの遺伝子配列、ハイブリッドDNA分子(fnbB)およびこの配列に基づく生物学的産物および方法を開示している。
米国特許第5652217号は、定義された配列を有するエス・アウレウスから得られるハイブリッドDNA分子によりエンコードされる結合活性を有する単離および精製タンパク質を開示している。
米国特許第5440014号は、ブドウ球菌感染症により引き起こされる乳腺炎に対する反芻動物の免疫化、傷の治療、タンパク質レセプターのブロッキング、他の動物の免疫化、または診断検定における使用のために用いることができるエス・アウレウスのフィブロネクチン結合タンパク質のD3相同性単位内のフィブロネクチン結合ペプチドを開示している。
【0020】
米国特許第5189015号は、哺乳動物におけるフィブロネクチンと結合する能力を有するエス・アウレウス細菌株のコロニー化の予防的治療のための方法であって、フィブロネクチンと結合する能力を有するエス・アウレウス細菌株により引き起こされる感染症の発生を予防するために、治療を必要とする哺乳動物に、予防的に有効な量のフィブロネクチン結合特性を有するタンパク質を投与することを含む方法を開示し、ここにおいて、タンパク質は87kDa〜165kDaの分子量を有する。
米国特許第5416021号は、ストレプトコッカス・ディスガラクティエ(Streptococcus dysgalactiae)から得られるフィブロネクチン結合タンパク質をエンコードするDNAとイー・コリに含まれるエス・ディスガラクティエから得られるフィブロネクチン結合タンパク質をエンコードするDNAを含むプラスミド、エス・ディスガラクティエからのフィブロネクチン結合タンパク質をエンコードするDNAおよびエス・ディスガラクティエからのフィブロネクチンをエンコードするDNAにより形質転換されたイー・コリ微生物を開示している。
【0021】
コラーゲン結合タンパク質
コラーゲンは軟骨の主な構成成分である。コラーゲン(Cn)結合タンパク質は通常、ブドウ球菌株により発現される。エス・アウレウスのコラーゲン結合表面付着因子タンパク質は、ブドウ球菌感染症における発病のメカニズムの重要な部分を構成するプロセスにおいて軟骨に付着する(Switalski 1993)。エス・アウレウスによるコラーゲン結合は、これだけではないが、少なくとも関節炎および敗血症に関与することが判明している。133、110および87kDaの分子量を有するコラーゲン付着因子(CNA)(Patti,J.ら、1992)が同定されている。異なる分子量を有するCNAを発現する株は、そのコラーゲン結合能力に差はない(Switalski 1993)。
敗血症性関節炎または骨髄炎と診断された患者の関節から回収されるブドウ球菌株は、ほぼ必ずと言っていいほどコラーゲン結合タンパク質を発現するが、創傷感染から得られる分離株でこの付着因子を発現するものは著しく少ない(Switalskiら 1993)。同様に、骨髄炎の患者から単離されたエス・アウレウス株はしばしば骨特異性タンパク質、骨シアロタンパク質(BSP)を認識する表面付着因子タンパク質を有する(Rydenら、1987)。関節空間内の関節軟骨のエス・アウレウスコロニー化は、敗血症性関節炎の発症に寄与する重要な因子であるらしい。
【0022】
WO92/07002は、コラーゲン結合活性を有するタンパク質またはポリペプチドをコードするエス・アウレウスからのヌクレオチド配列および該ヌクレオチド配列を含むプラスミドまたはファージを含むハイブリッドDNA分子を開示している。
コラーゲン結合タンパク質を発現するイー・コリ株、組み換えDNAにより形質転換された微生物、コラーゲン結合タンパク質またはポリペプチドを産生する方法、およびコラーゲン結合タンパク質またはポリペプチドのタンパク質配列も開示されている。
エス・アウレウスコラーゲン結合タンパク質をエンコードする遺伝子cnaのクローニング、配列決定、および発現が報告されている(Patti,Jら、1992)。
cna遺伝子は、グラム陽性菌から単離される表面タンパク質に特徴的な構造特性を含む133kDa付着因子をエンコードする。
【0023】
最近、リガンド結合部位がコラーゲン結合タンパク質のN−末端側の半分内に特定された(Patti,Jら、1993)。表面付着因子タンパク質の異なるセグメントに対応する組み換えタンパク質のコラーゲン結合活性を分析することにより、相当のコラーゲン結合活性を有する168アミノ酸長のタンパク質フラグメント(アミン酸残基151〜318に対応)が同定された。NまたはC末端においてこのタンパク質を切断すると、リガンド結合活性が失われるが、円偏光二色性分光分析により示されるようにタンパク質において構造変化が起きた。
Pattiら(1995)は、cna遺伝子によりエンコードされるエス・アウレウス付着因子におけるコラーゲン結合エピトープを開示している。その研究において、著者らは前記タンパク質の配列に由来するペプチドを合成し、抗体を産生するためにこれらを使用した。これらの抗体の内のいくつかはタンパク質のコラーゲンに対する結合を阻害する。
【0024】
WO97/43314は、コラーゲン結合タンパク質のある同定されたエピトープ(M55、M33、およびM17)は保護抗体を生成させるために使用できることを開示している。
該出願は、エス・アウレウスコラーゲン結合タンパク質に対するコラーゲンの結合と相互作用するか、または完全にブロックする組成物を同定するために必要な重要な情報を提供するコラーゲン結合タンパク質の結晶構造も開示している。エス・アウレウスコラーゲン結合タンパク質および25−アミノ酸ペプチドにおけるリガンド結合部位は、125I標識II型コラーゲンに対するエス・アウレウスの結合を直接阻害することを特徴とする。
【0025】
フィブリノゲン結合タンパク質
フィブリンは血餅の主成分であり、フィブリノゲン/フィブリンは埋め込まれた生体適合物質上に沈着する主な血漿タンパク質の一つである。細菌のフィブリノゲン/フィブリンへの付着が装置に関連する感染症の開始に重要であるということを示唆する証拠がかなり存在する。例えば、Vaudauxら(1989)により示されるように、エス・アウレウスはインビトロでフィブリノゲンでコートされたプラスチックに用量に依存した方法で付着する。加えて、血餅または心臓弁に対する損傷を模倣したモデルにおいて、Hermannら(1993)は、エス・アウレウスはフィブリノゲンブリッジを介して表面に付着する血小板に強力に結合することを示した。エス・アウレウスはインビトロで形成された血餅中のフィブリノゲンに直接付着することができ、ブリッジとして作用する血漿から沈着したフィブリノゲンにより培養された内皮細胞に付着することができる(Moreillonら、1995、Cheungら、1991)。VaudauxらおよびMoreillonらにより示されるように、フィブリノゲン結合タンパク質クランピング因子(ClfA)が欠損した突然変異体は、心内膜炎のラットモデルにおいて、インビトロでのフィブリノゲン、外植カテーテル、血餅、および損傷を受けた心臓弁に対する付着が減少した(Vaudauxら、1995、Moreillonら、1995)。
【0026】
フィブリノゲンの付着因子(「クランピング因子」と称することが多い)はエス・アウレウス細胞の表面上に位置する。溶液中での細菌とフィブリノゲン間の相互作用の結果、細菌細胞が即座に凝集する。フィブリノゲン上の結合部位はダイマーフィブリノゲン糖タンパク質のγ鎖のC末端に位置する。親和力は非常に高く、凝集は低濃度のフィブリノゲンで起こる。科学者らは最近、クランピング因子が固相フィブリノゲン、血餅、および損傷を受けた心臓弁への付着も促進することを示している(McDevittら、1994;Vaudauxら、1995;Moreillonら、1995)。
エス・アウレウスにおける2つの遺伝子が2つのフィブリノゲン結合タンパク質ClfAおよびClfBをコードすることが見いだされた。遺伝子clfAはクローンされ、配列決定され、92kDaのポリペプチドをコードすることが見いだされた。ClfAはフィブリノゲンのγ鎖と結合し、ClfBはαおよびβ鎖と結合する(Eidhinら、1998)。ClfBは推定分子量88kDa、見かけの分子量124kDaを有する、可溶性および固定化フィブリノゲンの両方に結合し、クランピング因子として作用する細胞壁に結合したタンパク質である。
【0027】
クランピング因子タンパク質の遺伝子(ClfAと表示)はクローンされ、配列決定され、分子レベルで詳細に分析された(McDevittら、1994、McDevittら、1995)。予想されるタンパク質は、933アミノ酸から構成される。39残基のシグナル配列はN末端にあり、その後に520残基の領域(領域A)が続き、これはフィブリノゲン結合ドメインを含有する。ジペプチドセリン−アスパルテートの154の繰り返しからなる308残基領域(領域R)がその後に続く。R領域配列は、18塩基対繰り返しGAY TCN GAY TCN GAY AGY(Yはピリミジンであり、Nは任意の塩基である)によりエンコードされる。ClfAのC末端は、タンパク質を細胞壁、膜アンカー、およびC末端の正に荷電した残基に固定するLPDTGモチーフなどのグラム陽性菌の多くの表面タンパク質中に存在する特徴を有する。
【0028】
血小板インテグリンαIIbβ3はフィブリノゲンのγ鎖のC末端を認識する。これは凝集中の血液凝固の開始における重大な事象である。ClfAは血小板凝集をブロックでき、γ鎖のC末端に対応するペプチド(198−411)はインテグリンおよびフィブリノゲンと相互作用するClfAの両方をブロックできるので、ClfAおよびαIIbβ3はフィブリノゲンγ鎖上の正確に同じ部位を認識するようである(McDevittら、1997)。αIIbβ3のフィブリノゲン結合部位は、「EFハンド」と称するCa2+結合決定因子の近くにあるか、または重複する。ClfA領域AはいくつかのEFハンド様モチーフを有する。3〜5mMの範囲のCa2+の濃度は、これらのClfA−フィブリノゲン相互作用をブロックし、ClfAタンパク質の二次構造を変化させる。ClfA EFハンドに影響を及ぼす突然変異はフィブリノゲンとの相互作用を軽減または防止する。Ca2+およびフィブリノゲンγ鎖はClfA領域Aの同じ部位に結合するか、または重複するようである。
【0029】
白血球インテグリンのα鎖、αMβ2は、200アミノ酸(AまたはIドメイン)の挿入を有し、これはリガンド結合活性の原因である。Iドメインにおける新規金属イオン依存性付着部位(MIDAS)モチーフはリガンド結合に必要とされる。認識されるリガンドにはフィブリノゲンが含まれる。フィブリノゲン上の結合部位はγ鎖にある(残基190〜202)。カンジダ・アルビカンスは、白血球インテグリンとよく似た性質を有する表面タンパク質、αIntlpを有することが近年報告された。表面タンパク質は、MIDASモチーフを包含するMβ2のIドメインと相同性であるアミノ酸配列を有する。さらに、Intlpはフィブリノゲンと結合する。
【0030】
ClfA領域Aはさらに、αIntlpと相同性である配列もある程度示す。ClfA領域A配列の調査により有効なMIDASモチーフが明らかになった。ClfAにおけるMIDASモチーフのDxSxS部分における推定カチオン配位残基における突然変異の結果、フィブリノゲン結合が著しく減少する。αMβ2のγ鎖結合部位に対応するペプチド(190−202)は、O’ConnellらによりClfA−フィブリノゲン相互作用を阻害することが示されている(O’Connell、1998)。したがって、ClfAは2つの別個の部位でフィブリノゲンのγ鎖と結合できるようである。ClfA上のリガンド結合部位は真核細胞インテグリンにより採用されるものと類似し、二価カチオン結合EFハンドおよびMIDASモチーフを含む。
【0031】
推定分子量約88kDaおよび見かけの分子量約124kDaを有するフィブリノゲン結合タンパク質ClfBも公知である。ClfBは細胞壁結合タンパク質であり、可溶性および固定化フィブリノゲンの両方と結合する。加えて、ClfBはフィブリノゲンのαおよびβ鎖の両方と結合し、クランピング因子として作用する。
細胞外マトリックスと結合するフィブリノゲン結合ClfAおよびClfBに関連するタンパク質が見いだされている。SdrC、SdrDおよびSdrEタンパク質は一次配列および構造機構においてClfAおよびClfBタンパク質に関連し、また細胞表面上に局在化する。細胞表面上に局在化するこれらのタンパク質のA領域に関して、タンパク質は血漿中のタンパク質、細胞外マトリックスまたは宿主細胞の表面上の分子と相互作用する。SdrCは細胞外マトリックスタンパク質、例えばビトロネクチンと結合することができる。SdrEも細胞外マトリックスと結合することができ;例えばSdrEは骨シアロタンパク質(BSP)と結合する。
【0032】
SdrC、SdrD、SdrE、ClfAおよびClfBのA領域において、コンセンサスTYTFTDYVDモチーフを誘導するために使用できる高度に保存されたアミノ酸配列が存在することが見いだされた。このモチーフは細菌感染症に対して広範囲の免疫を付与する多成分ワクチンにおいて使用でき、また広範囲の受動免疫を付与するモノクローナルまたはポリクローナル抗体を産生するために使用できる。別の具体例において、免疫を付与するために、または保護抗体を誘発するために、SdrおよびClfタンパク質ファミリーに由来する様々な配列モチーフの任意の組み合わせ(T/I)(Y/F)(T/V)(F)(T)(D/N)(Y)(V)(D/N)を使用できる。
【0033】
MHC−II類似タンパク質
フィブリノゲン、フィブロネクチンおよびコラーゲンに加えて、エス・アウレウス株は、その多くが付着性マトリックスタンパク質のファミリーに属するビトロネクチンなどの他の付着性真核細胞タンパク質に関連する(Chatwalら、1987)。米国特許第5648240号は、約70kDaの分子量を有するエス・アウレウスの広範囲の付着因子をエンコードする遺伝子を含むDNAセグメントを開示している。この付着因子は、フィブロネクチンまたはビトロネクチンと結合することができ、約30アミノ酸のMHC II様単位を含む。このタンパク質の結合特異性をさらに分析すると、これは合成ペプチドと結合する点でMHC II抗原と機能的に類似していることが明らかになる。従って、細胞外マトリックスタンパク質に対する細菌付着を媒介することに加えて、宿主の免疫系を抑制することにより、ブドウ球菌感染症に関与し得る。
【0034】
スタフィロコッカス・エピデルミディスからのSdrタンパク質
スタフィロコッカス・エピデルミディスは凝固酵素陰性菌であり、人間の皮膚の一般的な常在菌であり、しばしば異物感染症を引き起こす。病原性は、人工弁、整形外科用装置、ならびに静脈内および腹膜透析カテーテルなどの留置医療装置と生物体がまず付着し、その後、その上にバイオフィルムを形成する生物体の能力により促進される。装置に関連する感染症は、医療の成功を危うくし、患者の死亡率を著しく増大させる。従って、エス・エピデルミディス感染症の発生を制御または防止できるワクチンを開発できることは、同時に凝固酵素陽性および凝固酵素陰性菌の両方を包含する広範囲の細菌からの感染症を予防または治療できる複合ワクチンの開発と同様に非常に重要である。
エス・エピデルミディスにより発現される3つのSdr(セリン−アスパルテート(SD)繰り返し領域)タンパク質は、SdrF、SdrGおよびSdrHと表示され、これらのタンパク質のアミノ酸配列およびその核酸配列はWO00/12131(出典明示により本発明の一部として参照される)に示されている。
【0035】
本発明に従って、前記の表面付着因子タンパク質の少なくとも1つと接合した少なくとも1種の多糖抗原を含む免疫原性組成物として有用な接合体が提供される。加えて、多糖抗原および表面付着因子タンパク質に対する抗体は通常の手段を用いて産生される。従って、SdrGなどの表面付着因子タンパク質を含む免疫原性組成物は、凝固酵素陽性菌および凝固酵素陰性菌の両方に由来するものを含む広範囲の細菌感染症を治療するために使用される。
本発明の接合体の他の成分は、院内病原菌に由来する少なくとも1種の多糖抗原を含む。このような院内病原菌としては、これらに限定されないが、スタフィロコッカス・アウレウス(Staphylococcus aureus)、スタフィロコッカス・エピデルミディス(Staphylococcus epidermidis)および他の凝固酵素陰性ブドウ球菌(CoNS)、エンテロコッカス種(Enterococcus spp.、カンジダ・アルビカンス(Candida albicans)、エンテロバクター種(Enterobacter spp.)、ヘモフィルス・インフルエンゼ(Haemophilus influenzae)、クレブシエラ・ニューモニエ(Klebsiella pneumoniae)、エシェリキア・コリ(Escherichia coli)およびシュードモナス・エルジノーサ(Pseudomonas aeruginosa)が挙げられる。
【0036】
本発明の一具体例において、多糖抗原は、エス・アウレウスCP5およびCP8の少なくとも1つを含む。
本発明のもう一つ別の具体例において、多糖抗原は、エス・アウレウスおよび/またはエス・エピデルミディスにより発現されるようなPS/A、PNSG、PNAGおよびPIAのうちの少なくとも1つを含む。
【0037】
免疫原性組成物の調製および使用
免疫原性組成物は、本明細書に開示されるような多糖抗原−表面付着因子タンパク質接合体から調製される。免疫原性組成物は、多糖抗原および表面付着因子キャリアタンパク質の両方に対する抗体を産生する免疫応答を惹起する。
免疫原性組成物は、本明細書において開示されるようなオリゴ糖抗原−表面付着因子タンパク質接合体からも調製される。免疫原性組成物は、オリゴ糖抗原および表面付着因子キャリアタンパク質の両方に対する抗体を産生する免疫応答を惹起する。
【0038】
免疫原性組成物としての使用に好適な、本発明において提供される接合体は、これらに限定されないが、以下のものを包含する:
(i)クランピング因子A(ClfA)などのエス・アウレウスのフィブリノゲン結合タンパク質またはペプチドと接合するCP5、またはその有用なフラグメント、またはこれと十分高度に相同性であるタンパク質またはフラグメント;または
(ii)クランピング因子A(ClfA)などのエス・アウレウスのフィブリノゲン結合タンパク質またはペプチドと接合するCP8、またはその有用なフラグメント、またはこれと十分高度に相同性であるタンパク質またはフラグメント;または
(iii)クランピング因子A(ClfA)などのエス・アウレウスのフィブリノゲン結合タンパク質またはペプチドと接合するPIA、またはその有用なフラグメント、またはこれと十分高度に相同性であるタンパク質またはフラグメント;または
(iv)SdrGなどのエス・エピデルミディスのフィブリノゲン結合タンパク質またはペプチドと接合するCP5、またはその有用なフラグメント、またはこれと十分高度に相同性であるタンパク質またはフラグメント;または
(v)SdrGなどのエス・エピデルミディスのフィブリノゲン結合タンパク質またはペプチドと接合するCP8、またはその有用なフラグメント、またはこれと十分高度に相同性であるタンパク質またはフラグメント;または
(vi)SdrGなどのエス・エピデルミディスのフィブリノゲン結合タンパク質またはペプチドと接合するPIA、またはその有用なフラグメント、またはこれと十分高度に相同性であるタンパク質またはフラグメント。
【0039】
各例において、任意の接合体(i)〜(vi)から調製された免疫原性組成物は、エス・アウレウスなどの凝固酵素陽性菌、ならびにエス・エピデルミディスなどの凝固酵素陰性菌からの感染症に対して患者を免疫化するために有用である。
表面付着因子キャリアタンパク質がフィブリノゲン結合タンパク質である前記の接合体(i)〜(vi)に加えて、本発明はさらに、表面付着因子キャリアタンパク質が、例えばフィブロネクチン結合タンパク質、コラーゲン結合タンパク質およびビトロネクチン結合タンパク質などの任意のブドウ球菌表面付着因子タンパク質である接合体を包含する。本発明はさらに、多糖抗原がPS/A、PNAGまたはPNSG、またはエス・アウレウス、エス・エピデルミディスおよび他のCoNs、エンテロコッカス種、カンジダ・アルビカンス、エンテロバクター種、ヘモフィルス・インフルエンゼ、クレブシエラ・ニューモニエ、エシェリキア・コリ、およびシュードモナス・エルジノーサなどの院内病原微生物からの他の多糖抗原であり得ることも意図する。
【0040】
多糖をタンパク質に接合するための多くの方法が当該分野において公知であり、本発明における使用に適している。一般に、多糖を活性化させるか、または他の方法で接合しやすくする、すなわち、少なくとも1つの部分をタンパク質または他の分子と共有結合できるようにしなければならない。多くのこのような方法は当該分野において公知である。例えば、米国特許第4356170号(Jennings)は、多糖上にアルデヒド基を生成させるために過ヨウ素酸を使用し、次いでシアノボロヒドリドを用いて還元的アミノ化を行うことを記載している。米国特許第4663160号は(Tsayら)もアルデヒド基を生成させるために過ヨウ素酸を使用したが、次にシアノボロヒドリドなどの還元剤の存在下でのシッフ塩基反応で4〜12の炭素部分(縮合剤の存在下で調製)で誘導化されたタンパク質に多糖を結合させた。米国特許第4619828号(Gordon)は、多糖を活性化するために臭化シアンを使用し、次いで4〜8個の炭素原子のスペーサーブリッジによりタンパク質と接合させた。さらに他の接合法も当該分野において公知である。
【0041】
本発明の一具体例において、CPはリンカー3−(2−ピリジルジチオ)−プロピオニルヒドラジド(PDPH)で活性化され、これによりCPにおけるN−アセチルマンノースアミノウロン酸のカルボジイミド活性化カルボキシレート基がPDPHのヒドラジド基とカップリングする(図8)。MSCRMAMMキャリアタンパク質はブロモ酢酸のN−ヒドロキシスクシイミドエステルでのリシン残基のブロモアセチル化により活性化される(図7)。PDPH−チオール化CPを次いで、チオールでブロモアセチル化タンパク質における臭素を置換することにより活性化表面付着因子タンパク質に接合させると、結果として安定なチオエーテル結合が得られる(図9)。
CP−CONHNHCOCH2CH2SCH2CONH−表面付着因子タンパク質
【0042】
本発明のCP−表面抗原タンパク質接合体を含む免疫原性組成物をマウスにおいて試験し、不十分な免疫原性未接合CPと比較して、向上された免疫原性を有することが示された(図15〜20)。加えて、莢膜多糖特異性抗体ならびにCP−表面付着因子接合免疫原性組成物により誘発されたClfAおよびSdrG特異性抗体はどちらも対応する抗原を発現する生株と結合することが示された(表5および6)。これらの結果を考慮すると、本発明の免疫原性組成物はエス・アウレウスまたはエス・エピデルミディスなどの病原体により引き起こされる院内感染症に対して有用であると考えられる。また、CP−表面付着因子接合体により誘発される抗体が免疫原性組成物として傷に投与されるか、またはインビトロまたはインビボで医療装置またはポリマー生体適合物質をコートするために使用される場合、組成物は創傷部位または生体適合物質へのブドウ球菌菌の結合を防止または抑制するであろう。本発明に従って処理された接合体は、患者を院内感染に対して保護するための免疫原性組成物の調製において使用される。本明細書において用いられる「対象」とは、温血哺乳動物であり、例えば、ヒト、霊長類、ウマ、ウシ、イヌおよびネコを包含する。
【0043】
注射可能な液体溶液または懸濁液を調製するために、接合体を免疫学的に許容される希釈剤または担体に通常の方法で添加することができる。
本発明の免疫原性組成物は、典型的には、任意の適当な医薬的に許容される担体、例えば生理食塩水または他の注射可能な液体中に接合体を分散させることにより形成される。本明細書において用いられる場合、「医薬的に許容される担体」なる語句は、医薬投与に適合するありとあらゆる溶媒、分散媒体、コーティング、抗菌および抗真菌剤、等張および吸収遅延剤などを包含することを意図される。医薬的に活性な物質に関するかかる媒体および薬剤の使用は当該分野において周知である。通常の媒体または薬剤が活性化合物と不適合である場合を除いて、かかる媒体を本発明の組成物において用いることができる。例えば、接合体調製物をリン酸ナトリウム緩衝塩溶液(PBS)(pH7.0〜8.0)中に多糖1mlあたり1〜100μgの濃度で懸濁させる。本発明の免疫原性組成物の投与は、これらに限定されないが、非経口(例えば、皮下、腹腔内、筋肉内、静脈内、皮内)、経口および鼻内を包含する任意の周知の方法により行うことができる。免疫原性組成物の好ましい投与方法は非経口投与である。非経口投与に用いられる溶液または懸濁液は、次の成分を含む:無菌希釈剤、例えば、注射用水、塩溶液、不揮発性油、ポリエチレングリコール、グリセリン、ポリプロピレングリコールまたは他の合成溶媒;抗菌剤、例えばベンジルアルコールまたはメチルパラベン;酸化防止剤、例えばアスコルビン酸または重亜硫酸ナトリウム;キレート化剤、例えばエチレンジアミン四酢酸;緩衝剤、例えば酢酸塩、クエン酸塩またはリン酸塩および等張性を調節するための薬剤、例えば塩化ナトリウムまたはデキストロース。pHは酸または塩基、例えば、塩酸または水酸化ナトリウムで調節することができる。非経口製剤はアンプル、使い捨て注射器あるいはガラスまたはプラスチック製の複数回投与用バイアル中に封入することができる。
【0044】
注射可能な用途に適した免疫原性組成物は、無菌水性溶液(水溶性の場合)または分散液および無菌注射可能な溶液または分散液の即時調製用無菌粉末を含む。あらゆる場合において、組成物は無菌でなければならず、容易に注射可能である程度に流動性でなければならない。製造および貯蔵条件下で安定でなければならず、細菌および真菌などの微生物の汚染作用に対して保護されなければならない。担体は、例えば、水、エタノール、ポリオール(例えば、グリセロール、プロピレングリコール、および液体ポリエチレングリコールなど)、およびその適当な混合物を含有する溶媒または分散媒体である。適当な流動性は、例えば、レシチンなどのコーティングの使用、分散液の場合は必要とされる粒子サイズの維持、および界面活性剤の使用により維持される。微生物の作用の防止は、様々な抗菌剤および抗真菌剤、例えば、パラベン、クロロブタノール、フェノール、アスコルビン酸、チメロサールなどにより達成される。多くの場合において、等張剤、例えば、糖、ポリアルコール、例えばマンニトール、ソルビトール、および塩化ナトリウムを組成物中に含めることが好ましい。組成物中に吸収を遅らせる薬剤、例えばアルミニウムモノステアレートおよびゼラチンを含めることにより注射可能な組成物の吸収が延長される。
【0045】
無菌注射可能な溶液は、本発明の接合体を必要とされる量、適当な溶媒中に、単独または必要に応じて前記の成分と組み合わせて配合し、続いて濾過滅菌することにより調製される。一般に、分散液は、活性化合物を、塩基性分散媒体および前記のもののうち必要とされる他の成分を含有する無菌ビヒクル中に配合することにより調製される。無菌注射可能な溶液を調製するための無菌粉末の場合、好ましい調製法は真空乾燥および凍結乾燥であり、これにより活性成分の粉末と、あらかじめ無菌濾過されたその溶液からの任意のさらなる所望の成分が得られる。
ある具体例において、免疫原性組成物は1以上のアジュバントを含む。本明細書において定義されるように、「アジュバント」は本発明の免疫原性組成物の免疫原性を向上させる働きをする物質である。従って、アジュバントは免疫反応の追加免疫(boost)のために投与され、当業者には周知である。
【0046】
組成物の有効性を向上させるための好ましいアジュバントとしては、これらに限定されないが、次のものが挙げられる:
(1)アルミニウム塩(alum)、例えば水酸化アルミニウム、リン酸アルミニウム、硫酸アルミニウムなど;
(2)水中油エマルジョン処方(他の特異的免疫刺激剤、例えばムラミルペプチド(以下参照)または細菌細胞壁成分の存在下または非存在下)、例えば
(a)ミクロフルイダイザー、例えば110型ミクロフルイダイザー(Microfluidics,Newton,MA)を用いて1ミクロン以下の粒子に処方された5%スクアレン、0.5%Tween80、および0.5%Span85(任意に様々な量のMTP−PE(必要ではないが、以下参照)を含有してもよい)を含有するMF59(PCT公開番号WO90/14837)、
(b)10%スクアレン、0.4%Tween80、5%プルロンブロックされたポリマーL121、およびthr−MDP(以下参照)(ミクロ流動化されて1ミクロン以下のエマルジョンにされているか、または撹拌されて大きな粒子サイズのエマルジョンが生じるかのいずれか)を含有するSAF、
(c)2%スクアレン、0.2%Tween80、および米国特許第4912094号(Corixa)に記載されている3−デアシル化モノホスホリピッドA(MPL)、トレハロースジミコレート(TDM)、および細胞壁骨格(CWS)、好ましくはMPL+CWS(Detox)からなる群からの1以上の細菌細胞壁成分を含有するRibiアジュバント系(RAS)(Corixa,Hamilton,MT);
(3)サポニンアジュバント、例えばQuil AまたはSTIMULON QS−21(Antigenics,Framingham,MA)(米国特許第5,057,540号)またはこれから生成する粒子、例えばISCOM(免疫刺激複合体);
(4)細菌リポ多糖、合成リピッドA類似体、例えばアミノアルキルグルコサミンホスフェート化合物(AGP)、またはその誘導体または類似体(Corixaから入手可能で、米国特許第6,113,918号に記載されている);例えばAGPは2−[(R)−3−テトラデカノイルオキシテトラデカノイルアミノ]エチル2−デオキシ−4−O−ホスホノ−3−O−[(R)−3−テトラデカノイルオキシテトラデカノイル]−2−[(R)−3−テトラデカノイルオキシテトラデカノイルアミノ]−b−D−グルコピラノシド(529(以前はRC529と呼んだ)をも呼ばれ、水性形態または安定なエマルジョンとして処方される)、合成ポリヌクレオチド、例えばCpGモチーフを含有するオリゴヌクレオチド(米国特許第6,207,646号);
(5)サイトカイン、例えばインターロイキン(例えば、IL−1、IL−2、IL−4、IL−5、IL−6、IL−7、IL−12、IL−15、IL−18など)、インターフェロン(例えば、ガンマインターフェロン)、顆粒球マクロファージコロニー刺激因子((GM−CSF)、マクロファージコロニー刺激因子(M−CSF)、腫瘍壊死因子(TNF)など;
(6)コレラ毒素(CT)(野生型または突然変異形態、例えば公開国際特許出願番号WO00/18434に従ってアミノ酸29位でグルタミン酸が別のアミノ酸、好ましくはヒスチジンにより置換されている場合(WO02/098368およびWO02/098369も参照))などの細菌ADP−リボシル化トキシンの無毒化突然変異体、狂犬病毒素(PT)、またはイー・コリ易熱性毒素(LT)、特にLT−K63、LT−R72、CT−S109、PT−K9/G129(例えば、WO93/13302およびWO92/19265参照);および
(7)組成物の有効性を向上させるために免疫刺激剤として作用する他の物質。
【0047】
前記のように、ムラミルペプチドとしては、これらに限定されないが、N−アセチル−ムラミル−L−スレオニル−D−イソグルタミン(thr−MDP)、N−アセチル−ノルムラミル−L−アラニン−2−(1’−2’−ジパルミトイル−sn−グリセロ−3−ヒドロキシホスホリルオキシ)−エチルアミン(MTP−PE)などが挙げられる。
本発明の免疫原性組成物は、免疫原性反応を誘発するために十分な量で投与される。用量は、該免疫原性組成物を受容する個体の大きさ、体重または年齢に基づいて調節することができる。個体における抗体応答は、抗体価または殺菌活性について分析することによりモニターすることができ、必要に応じて追加免疫して反応を向上させることができる。
【0048】
本発明の免疫原性組成物は、体液性免疫応答を誘発するために対象に投与される。対象は次に免疫原組成物に応答して生成されたイムノグロブリンの供給源(高度免疫イムノグロブリン)として作用する。免疫化された対象は血漿を供与し、これから通常の血漿分画技術により高イムノグロブリンが得られ、院内感染に対する耐性を付与するため、または院内感染を治療するために別の対象に投与される。
【0049】
(実施例)
前記開示は一般に本発明を説明する。以下の実施例を参照することによりさらに完全に理解することができる。これらの実施例は、説明の目的のみで記載したものであり、本発明の範囲を制限することを意図するものではない。
【0050】
実施例1
エス・アウレウスCP5およびCP8多糖の精製
CP5およびCP8それぞれの精製のためにエス・アウレウス株(ATCC#49521)およびWright(ATCC#49521)を使用した。既に公開されている方法(Fournier,Vannら、1984;Fournier,Hannonら、1987)を変更した方法により細胞から多糖を精製した。2%NaClで補足されたColumiaブロス中で成長させた細胞を3時間37℃で、リソスタフィン(175U/gの細胞)で、RNAseおよびDNAse(それぞれ0.1mg/g)で4時間、37℃で消化し、続いてプロナーゼ(1mg/gの細胞)で3時間、37℃で消化した。10mM CaCl2の存在下、25%および75%エタノールで連続して沈降させることにより酵素消化物から粗CPを調製した。0.05〜0.5M NaClの直線的勾配を用い、Q−セファロースカラム上でアニオン交換クロマトグラフィーによりCPを次いでペレットから精製した。残留するテイコ酸を0.05M NalO4で酸化した。透析後、CPを次いでSephacrylS300(Amersham Pharmacia Biotech,Piscataway,NJ)カラム上で寸法排除クロマトグラフィーによりさらに精製した。フラクション中のCPの存在をエス・アウレウスCP5およびCP8特異性抗血清との反応性により決定した。
【0051】
PIAを、Mackら(Mack,Fischerら、1996)により記載されているように、熱抽出した静止期のエス・エピデルミディス細胞から精製し、PIA含有培養上清と組み合わせた。抽出された物質および培養上清を、10K膜を用いて濃縮し、処理して、核酸および残存するタンパク質を除去した。ゲル濾過または透析濾過を用いて粗PIAを分画した。アニオン交換クロマトグラフィーによりPIA抗原陽性物質をさらに分画し、PIAフラクション含有エステル結合サクシネートを精製した。非スクシニル化および部分的非N−アセチル化PIAを含有するフロースルーフラクションを、カチオン交換クロマトグラフィーにより精製した。PS/A(PNSG、PNAG)を(Maria−Litran,Kropecら、2002)またはMcKenney,Pouliotら、1999により記載されているようにして精製した。
【0052】
実施例2
エス・アウレウスCP5およびCP8の分析
精製されたCP5およびCP8の化学的特徴化により、両多糖は事実上核酸および残存するタンパク質が無いことが示された(表1)。
HPAECクロマトグラフィーにより決定された糖組成により、CP5およびCP8におけるFucρNAcおよびManρNacAの存在が明らかになった(図1)。O−デアシル化多糖の1H NMRスペクトル(図2)は、既に公開されているスペクトル(Vann,Moreauら、1987;Moreau,Richardsら、1990)と類似しており、3種の単糖:2−アセトアミド−2−デオキシ−D−マンヌロン酸、2−アセトアミド−2−デオキシ−L−フコースおよび2−アセトアミド−2−デオキシ−D−フコースの構造および存在が確認された。
精製されたCP5、CP8およびTAは、対応する全細胞抗血清と反応した場合に二重免疫拡散検定における1つの沈降素のバンドにより確かめられるように免疫学的に異なっていた(データは省略)。
【0053】
実施例3
表面付着因子タンパク質の精製
評価した表面付着因子タンパク質は:
エス・アウレウスClf40(N1N2N3)−クランピング因子Aの完全長Aドメイン(アミノ酸(AA)40−559)−図3。
エス・アウレウスClf41(N2N3)−Clf40のポストプロテアーゼ部位フラグメント(AA223−559)−図4。
エス・エピデルミディスSdrG(N1N2N3)−SdrGの完全長Aドメイン(AA50−597)−図5。
エス・エピデルミディスSdrG(N2N3)−SdrGのポストプロテアーゼ部位フラグメント(AA273−597)−図6。
コレラの表面付着因子タンパク質は、Inhibitex,Inc.,Alpharetta,GA,USAから入手した。
【0054】
表面付着因子タンパク質のHistagマイナスバージョンをイー・コリプラスミド宿主株から精製した。イー・コリpLP1134BL21(DE3)をエス・アウレウスClf41(N2,N3)について使用し、pLP1135B21(DE3)をエス・エピデルミディスSdrG(N2,N3)精製に使用した。両タンパク質を、硫酸アンモニウム沈殿およびその後のセファクリルQ−セファロースカラム(Amersham Pharmacia Biotech,Piscataway,NJ)上でのイオン交換クロマトグラフィーにより細胞溶解物の可溶性フラクションから単離した。最終物質の純度はSDS−PAGEにより測定すると90%より高かった。
【0055】
過剰発現されたエス・アウレウスClf40(N1,N2,N3)またはClf41(N2,N3)、エス・エピデルミディスSdrG(N1,N2,N3)またはSdrG(N2,N3)を含有するイー・コリ細胞を、約13000psiで、Microfuidics M110−Yミクロフルイダイザーを1回通過させて可溶化させた。細胞片を17000rpmで30分間4℃で遠心分離することにより精製した。過剰発現されたタンパク質を、AKTAexplorer(XKカラムキレート化セファロースファーストフローおよびQセファロースHP樹脂(Amersham Pharmacia Biotech,Piscataway,NJ))を用いて上清から精製した。粗His標識されたタンパク質を、0.1M NiCl2を装填したキレート化セファロースファーストフローを用いたアフィニティ工程により上清から精製した。粗溶解物を、25mM Tris、pH8.0、0.5M NaCl、5mMイミダゾールで平衡化させたカラム上にロードし、カラムを5カラム体積の緩衝剤で洗浄することにより未結合タンパク質をカラムから溶出させた。未結合タンパク質を次いで250mM Tris、pH8.0、0.5M NaCl、500mMイミダゾール緩衝液で溶出させ、バルクで集めた。タンパク質を次いでQセファロースHPカラム上イオン交換クロマトグラフィーにより残存する不純物から精製した。
【0056】
実施例4
エス・アウレウスCP5−およびCP−8表面付着因子キャリアタンパク質接合体免疫原性組成物の合成
エス・アウレウスCP5およびCP8多糖を、リンカーを含有するチオール基を多糖に導入し、ハロアセチル基をタンパク質キャリアに導入した後、本発明により提供された表面付着因子キャリアタンパク質とチオエーテル結合を介して別々に結合させた。アミン基をブロモ酢酸のN−ヒドロキシスクシンイミドエステルと反応させることにより、ブロモアセチル基を表面付着因子タンパク質中に導入した(図7)。チオール化CPを生成させるために、莢膜多糖におけるN−アセチルマンノースアミノウロン酸のカルボジイミド活性化カルボキシレート基をスルフヒドリル反応性ヒドラジドヘテロ二官能性リンカー3−(2−ピリジルジチオ)−プロピオニルヒドラジド(PDPH、図8)のヒドラジド基とカップリングさせた。ジチオトレイトール(DTT)での還元により生成し、Sdphadex G25カラム上SECにより精製されたPDPH−チオール化CPのチオールを、活性化タンパク質のブロモアセチル基と反応させると、CPとタンパク質間の臭素置換により形成された共有チオエーテル結合が生じた(図9)。未反応ブロモアセチル基をシステアミン塩酸塩(2−アミノエタンチオール塩酸塩)で「キャップ」した。反応混合物を次いでAmicon XM100膜上で濃縮した。
【0057】
実施例5
CP−およびCP−8表面付着因子キャリアタンパク質接合体免疫原性組成物の特徴化
4Nトリフルオロ酢酸(TFA)での加水分解後、CarboPac−PA1カラム上HPAEC−PADクロマトグラフィーによるCPの定量化により、接合免疫原性組成物をCPおよび表面付着因子キャリアタンパク質含量について分析した。タンパク質含量をLowry比色検定により決定した。接合免疫原性組成物の分子量を寸法排除クロマトグラフィーおよび多角度レーザー光散乱(MALLS)の組み合わせにより測定した。結果を表2および3に記載する。接合CPおよび表面付着因子タンパク質の抗原性を二重免疫拡散法(図10〜13)およびドットブロット分析(図14)により決定した。結果は、CPの表面付着因子タンパク質への接合は、CPまたはタンパク質のいずれかの抗原性を変更しないことを示す。CPのタンパク質への接合は、接合体がニトロセルロース膜に結合する能力により確認された。未接合CPはニトロセルロース膜に結合しなかった。
【0058】
実施例6
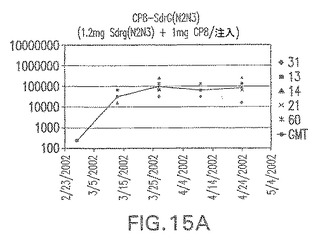
マウスにおけるCP−表面付着因子キャリアタンパク質接合体免疫原性組成物の免疫原性
接合免疫原性組成物を、CP5およびCP8および表面付着因子タンパク質キャリアに対するIgG応答を誘発する能力について試験した。Swiss−Websterマウスを1マイクログラム用量(CP基準)で2週間間隔で3回、皮下的に(SC)免疫化した。接合免疫原性組成物の免疫原性を、アジュバントとしての100マイクログラムのリン酸アルミニウムの存在下または不在下で試験した。各タンパク質免疫原性組成物候補を同様のプロトコルを用いて同様に評価した。エス・アウレウスCPおよび表面付着因子タンパク質に対する免疫応答を標準的抗原ELISAによる各注射の1週間後に分析した(以下の実施例7および8参照)。
【0059】
実施例7
エス・アウレウスCP5およびCP8−表面付着因子キャリアタンパク質接合体免疫原性組成物で免疫化されたマウスにおけるCPの抗体反応
結果(図15および16)は、CPの表面付着因子タンパク質への共有結合の結果、莢膜多糖(CP)特異性IgG反応が誘発されることを示す。このことは、CP−T細胞非依存性免疫反応はCPの表面付着因子キャリアタンパク質へのカップリング後にT細胞依存性免疫反応に変換されたことを示す。接合免疫原性組成物のリン酸アルミニウムへの吸着は、タンパク質キャリアとしてSdrG(N2N3)を投与されたマウスを除いて約10倍に増大した。CP5−およびCP8−SdrG(N2N3)接合体のアジュバントへの吸着の結果、CPに対する免疫応答は増大しなかったが、CPの抗体反応はこの実験においてアジュバントと混合された(吸着されていないが)他の表面付着因子タンパク質接合体と同様に良好であった。ClfAおよびSdrGのN1ドメインの欠失は、これらのタンパク質のキャリア特性に対して影響を及ぼさなかった。
【0060】
実施例8
エス・アウレウスCP5およびCP8−表面付着因子キャリアタンパク質接合体でワクチン接種されたマウスにおける表面付着因子タンパク質抗体反応
接合表面付着因子タンパク質は、未接合のものと比較して類似した力価の表面付着因子タンパク質特異性抗体を誘発した(図17〜20)。このことにより、表面付着因子タンパク質のCPに対する接合により抗原性エピトープが修飾されないことが確認される。未接合ClfAまたはCP−ClfA接合体のリン酸アルミニウムへの吸着の結果、アジュバントを含まない同じ免疫原性組成物で免疫化されたマウスと比較して、マウスにおけるClfA抗体価が増大した。未接合SdrGで免疫化されたマウスは、CP−SdrG接合免疫原性組成物で免疫化されたマウスと比較して低いSdrG抗体価で反応した。未接合SdrGのリン酸アルミニウムへの吸着の結果、alumなしで投与されたCP−SdrG接合体により誘発されたレベルと比較してSdrG抗体価が増大した。CP−SdrG接合体のalumへの吸着はSdrG抗体価を増大させなかった。
【0061】
実施例9
CP表面付着因子キャリアタンパク質接合体により誘発された抗体による生菌上に発現されたCPおよび表面付着因子キャリアタンパク質の認識
マウスにおけるCP−表面付着因子タンパク質接合体により誘発された抗体の生菌に対する結合をフローサイトメトリー分析により試験した。この検定において使用したエス・アウレウス株を表4に示す。SdrG接合体に対して誘発された抗体の分析について、SdrGを発現するエル・ラクチス(L.lactis)を使用した。結果は(表5および6)、莢膜多糖特異性抗体ならびに対応するCP5−およびCP8−表面付着因子タンパク質接合体により誘発されるClfAまたはSdrG特異性抗体はどちらも対応する抗原を発現する生株に結合することを示す。これにより、CPの表面付着因子タンパク質に対する接合は、CP上に存在する自然に発現されたエピトープおよび表面付着因子タンパク質抗原に対する免疫反応を変更しないことが示される。
【0062】
実施例10
フローサイトメトリー分析法
使用したエス・アウレウス株は次の通りであった:Newman、NewmanのClfAノックアウト突然変異体(Newman ClfA::emr)およびWright(ATCC49525)。ClfA発現を最大にするために、エス・アウレウス菌をトリプシンソイブロス中で静止期まで成長させた。莢膜発現を最大にするために、エス・アウレウス菌をColumbia2%NaCl寒天(BD Microbiology,Sparks,MD)上で一夜成長させた。Newman ClfA::emr株を、5μg/mlのエリスロマイシンの存在下で成長させて、ノックアウト突然変異を維持した。SdrG抗原認識を評価するためにSdrGを発現する組み換えラクトコッカス・ラクチス(Lactococcus lactis(L.Lactis))株を使用した。5μg/mlのエリスロマイシンの存在下、M17ブロス中、エル・ラクチス株を後期対数期まで成長させた。
【0063】
細菌培養物を収穫し、10mlの冷1×PBS(Invitrogen Corp, Rockville,MD)中で2回洗浄し、分析前に氷上で保存した。細菌濃度を、UV−可視記録分光光度計(Ultrospec3000、Pharmacia Biotech,Cambridge,England)を用いて1×PBSでD600nm=2.0に調節した。非特異性およびプロテインAによるマウスIgGの細胞表面に対する結合を排除するために、細菌調製物を全部、氷上で、1×PBS(Invitrogen Corp.,Rockville,MD)中10mlの1:50希釈度(2.32mg IgG)のウサギIgG(Sigma、St.Louis、Missouri)中で30分間インキュベートした。ClfA結合の不在下での8型莢膜認識を評価するために、さらに30分間高力価ClfA特異性ウサギ抗血清(Inhibitex,Alpharetta,GA)(1:100希釈)でインキュベーションすることにより、ClfAエピトープをエス・アウレウス株Wright上でブロックした。インキュベーションのブロッキング後、細菌を、3000rpmで10分間遠心分離することにより、10mlの冷1×PBS中で2回洗浄した。細菌ペレットを1×PBS中2.5%BSA(Invitrogen Corp.,Rockville,MD)(PBSA)中に再懸濁させ、氷上で保存した。
【0064】
検定をtitertube(BioRad Labs,Hercules,CA)中で行った。試験動物から得られるプレブリード(prebleed)および高力価抗血清をPBSA中に希釈し、0.5mlの各血清希釈物を、20μlの細菌懸濁液を含有する適当な試験管に添加した。全試験管を撹拌し、氷上で30分間インキュベートした。インキュベーション後、各試験管を撹拌し、次いで3000RPMで10分間遠心分離した。細菌ペレットを2回0.5mlの冷PBSA中で洗浄した。各ペレットを抗マウスIgG(H&L)(Rockland Labs,Gilbertsville,Pa)のPE接合F(ab’)2フラグメントの0.5mlの1:200希釈物中に再懸濁させた。細菌を再懸濁させ、撹拌することにより混合した。試験管を氷上で30分間インキュベートし、15分間隔で2回撹拌した。このインキュベーション後、細菌をPBSA中最終再懸濁液で洗浄した。FACS分析まで試験管を氷上で保存した。
【0065】
各titertubeを12×75mmポリスチレン試験管に移し、B−D FACSCalibur(BD Biosciences,Mansfield,MA)フローサイトメーターを用いて分析した。結果を、所定の抗血清の蛍光強度が同じ希釈度でのプレブリードで得られるシグナルより大きいならばプラスと採点した。結果を表7に示す。
前記考察および実施例は単にある具体例の詳細な説明を提示するためのものであると理解すべきである。従って、当業者には本発明の精神および範囲から逸脱することなく様々な修正および等価物を調製することが可能であることは明らかである。
本出願中に記載した全ての文献、他の刊行物、特許および特許出願は全体として本発明の一部として参照される。
【0066】
【表1】

【0067】
【表2】
【0068】
【表3】
【0069】
【表4】
【0070】
【表5−1】
【表5−2】
【0071】
【表6−1】
【表6−2】
【0072】
【表7】
*ClfA=ClfA AドメインのN1,N2,N3またはN2,N3領域
SdrG=SdrG AドメインのN1,N2,N3領域
【0073】
参考文献:
【0074】
【0075】
【0076】
【0077】
【0078】
【技術分野】
【0001】
本発明は、院内病原菌から得られる多糖抗原(または1以上の抗原性エピトープを表すそのオリゴ糖フラグメント)およびブドウ球菌表面付着因子キャリアタンパク質を含む免疫原性多糖タンパク質接合体に関する。本発明はさらに、多糖−タンパク質接合体を含む免疫原性組成物、およびその使用に関する。
【背景技術】
【0002】
毎年、米国において病院に入院している推定40000000人のうち約2000000人が院内感染を発症している(Anonyomous 1997)。死亡率は約4.4%であるので、院内感染のため毎年88000人が死亡している。米国における院内感染の費用は、1年あたり推定4500000000ドルである(Weinstein 1998)。これらの推定値は毎年行われる31000000人の外来患者の手術(National Center for Health Statistics’ website)、1500000人の養護施設居住者、拡張看護施設、または外来治療を受けている者において発生する感染症を包含しない。
【0003】
スタフィロコッカス・アウレウス(Staphylococcus aureus)および凝集酵素陰性ブドウ球菌(CoNS)、特にエス・エピデルミディス(S.epidermidis)は、院内感染の大部分の原因であるグラム陽性日和見院内病原菌である。ブドウ球菌感染症はすべての院内感染症のほぼ25%(約500000人)の原因である(Haley,Culverら、1985)(Boyce 1997)。一部の病院では入院患者全体の1%までがエス・アウレウス感染症にかかっている(StorchおよびRajagopalan 1986)。ブドウ球菌(エス・アウレウスおよびエス・エピデルミディス)は院内血流感染症の約47%、手術部位感染症(SSI)の24%、および院内感染肺炎の17%の原因である(Anonyomous 1997)。院内エス・アウレウスおよびCoNS感染症の患者の死亡率は5%〜68%までの範囲で相当変化する(Nada,Ichiyamaら、1996);(Thylefors,Harbarthら、1998)。
【0004】
ブドウ球菌感染症は、範囲が広く、皮膚感染症、例えば、膿痂疹、できもの、創傷感染および埋め込み装置からの感染症から重度の命にかかわる感染症、例えば骨髄炎、心内膜炎および転移性合併症を伴う菌血症にまで及ぶ。このような多様性のために、ブドウ球菌に対する有効な免疫原性組成物の設計を非常に困難になる。薬剤耐性院内細菌の出現が急激に増加し、そのためにこのような設計がさらに困難になっている。メチシリン耐性エス・アウレウスは院内感染症の原因の約40%である(Boyce 1997)。最近、バンコマイシン中間耐性エス・アウレウス(VISA)が出現し、その拡大に関する不安がさらに大きくなっている。従って、院内感染症に対する有効な免疫原性組成物に対する需要が強く、そして急速に増大している。
【0005】
莢膜多糖(capsular polysaccharide)
ヘモフィルス・インフルエンゼ(Haemophilus influenzae)、ストレプトコッカス・ニューモニエ(Streptococcus pneumoniae)およびB群ストレプトコッカスをはじめとする多くの病原菌の毒性における莢膜多糖(CP)の関与は、十分に証明されている。莢膜で覆われた細菌は、白血球による食菌作用に対して耐性であり、従って血液および組織に感染し得る。莢膜多糖に対する抗体は細菌莢膜の抗食細胞特性を中和するので(Karakawa、Suttonら、1988,Thakker、Parkら、1988)、ブドウ球菌莢膜は、人間におけるブドウ球菌感染症を予防するための免疫原性組成物の開発における主要な標的である。
【0006】
エス・アウレウスの12の公知莢膜血清型のうち、血清型5(CP5)および血清型8(CP8)は全臨床分離株の約85〜90%を占める(Arbeit,Karakawaら、1984、Karakawa,Fournierら、1985、Essawi,Na’wasら、1998、Na’was、Hawwariら、1998)。ほとんどのメチシリン耐性エス・アウレウス分離株はCP5を発現する(Sompolinsky,Samraら、1985)。CP5およびCP8に対する抗体は、インビトロのヒト多形核好中球による型特異性オプソニン作用による食菌現象を誘発し、動物において保護を付与する(Karakawa,Suttonら、1988、Fattom,Sarwarら、1996)。
【0007】
ほとんどの細菌莢膜多糖は、動物および人間において不十分な免疫原である。しかしながら、精製された多糖がタンパク質キャリア分子と接合するならば、これらは免疫原性およびT細胞依存性を獲得する。いくつかの研究所は、タンパク質と共有結合したCP5およびCP8からなる免疫原性接合体を合成した。これらの接合体は、マウスおよびヒトにおいて高度に免疫原性であり、食菌作用のためにマイクロカプセル化エス・アウレウスをオプソニン化する抗体を誘発する(Fattom,Schneersonら、1993、Gilbertら、1994,Reynaud−Rondierら、1991)。シュードモナス・エルジノーサ組み換え外毒素Aに接合したCP5を含有する一価免疫原性組成物は、免疫原性であり、健康な成人および末期の腎臓病の患者において良好な耐容性を示す(Welchら、1996)。血液透析を受けている末期腎臓病の患者を含む二重盲検法において、シュードモナス・エルジノーサ組み換え外毒素Aと共有結合したCP5およびCP8からなる二価接合体ワクチンは、約40週間エス・アウレウス菌血症に対して部分的免疫を付与し、その後、防御は抗体レベルが減少するにつれて減少した(Shinefieldら、2002)。この試験の結果は、さらに完全な防御をもたらすことができる改善された免疫原性組成物が必要とされていることを示す。
【0008】
多糖付着因子(PS/A;(Tojo,Yamashitaら、1988))、ポリ−N−スクシニルβ−1−6−グルコサミン(PNSG、(McKenney,Pouliotら、1999))、ポリ−N−アセチルグルコサミン表面多糖(PNAG、(Maria−Litran,Kropecら、2002))、または多糖細胞間付着因子(PIA(Mack,Fisherら、1996))と称する別の種類の細胞外多糖は、エス・アウレウスおよびエス・エピデルミディスの両方により発現される。PIAまたはPS/Aは直鎖β−1,6−結合グルコサミングリカンである。マウスをPS/A(PNSG、PNAG)で免疫化すると腎臓の転移増殖が減少し、インビトロで若干のPS/A(PNSG、PNAG)を産生するエス・アウレウス株で攻撃した後にマウスを死から防御する(McKenney,Pouliotら、1999)。PIAは静脈内カテーテルに関連する感染症の発生機序において重要な役割を果たす(Rupp,Ulphaniら、1999;Rupp,Ulphaniら、1999;RuppおよびFey 2001;Rupp,Feyら、2001)。個々のエス・エピデルミディス細胞間の接着を促進することに加えて、PIAは赤血球と結合し、ヘマグルチニンとして作用する(Fey,Ulphaniら、1999)。
【0009】
ブドウ球菌表面付着因子
ブドウ球菌は複数の表面付着因子(接着性マトリックス分子を認識する微生物表面成分と称する)を発現し、これらには、例えば、フィブロネクチン結合タンパク質、フィブリノゲン結合タンパク質、コラーゲン結合タンパク質およびビトロネクチン結合タンパク質が含まれる。これらの付着因子は細胞外マトリックス(ECM)成分、例えば、フィブロネクチン、フィブリノゲン、コラーゲンおよびビトロネクチンを特異的に認識し、結合する。エス・アウレウスにより発現される多数の接着因子は、様々な組織に対する接着および感染の代替法を提供することにより、その病原性に寄与する。ブドウ球菌付着因子の抗体は、細菌が哺乳動物宿主組織を侵襲することを防止するか、またはオプソニン作用による食菌作用を促進することにより疾患を軽減することができる。エス・アウレウスフィブロネクチン結合タンパク質A(融合タンパク質として提供される)の一部で免疫化されたラットは、実験的心内膜炎から適度に保護された。フィブロネクチン結合タンパク質Aの抗体を惹起するように設計された同様の免疫原性組成物をエス・アウレウス乳腺炎のマウスモデルにおいて試験した。免疫化されたマウスは対照マウスよりも重度の乳腺炎の症例が少なく、免疫化されたマウスの乳腺炎から回収される細菌は対照マウスよりも少なかった。19および87kDaのフィブリノゲン結合タンパク質で免疫化されたマウスは、非免疫化対照と比較して乳腺炎の発病率が減少し、一方、コラーゲン結合タンパク質での免疫化は保護しなかった(Lee,Pier 1997)。
【発明の概要】
【発明が解決しようとする課題】
【0010】
しかしながら、多糖抗原を様々なタンパク質キャリアに接合させるためのこれらおよび他の試みにもかかわらず、院内感染を治療または予防するための有効な免疫原性組成物は現在のところない。
【課題を解決するための手段】
【0011】
本発明は、従って、院内病原菌に由来する少なくとも1種の多糖抗原、または少なくとも1種のブドウ球菌表面付着キャリアタンパク質に接合した少なくとも1種の多糖抗原(合成または天然の多糖の加水分解により調製)の1以上の抗原エピトープを表すオリゴ糖フラグメントを含む免疫原性多糖−タンパク質接合体を提供する。本発明の接合体は、院内病原菌の多糖抗原および表面付着因子キャリアタンパク質の両方に対して特異的な抗体反応を対象において惹起する際に有用な免疫原性組成物において用いられる。従って、これらの接合体は、エス・アウレウス、エス・エピデルミディスまたは他の院内病原菌により引き起こされる院内感染症に対して免疫化するために、また受動免疫化のための免疫グロブリンの生成に関しては、院内感染症の予防または重篤度の軽減のために使用することができる。
【0012】
本発明の一態様において、少なくとも1種のブドウ球菌表面付着因子キャリアタンパク質と接合した院内病原菌に由来する少なくとも1種の多糖抗原を含む免疫原性多糖タンパク質接合体が提供され、ここにおいて該接合体は、多糖抗原および表面付着因子キャリアタンパク質の両方に対する特異的抗体を生成する。
本発明のもう一つ別の態様において、少なくとも1種のブドウ球菌表面付着因子キャリアタンパク質と接合した院内病原菌から得られる少なくとも1種の多糖抗原の1以上の抗原エピトープを表すオリゴ糖フラグメントを含む免疫原性多糖タンパク質接合体が提供され、ここにおいて、該接合体は、多糖抗原および表面付着因子キャリアタンパク質の両方に対する特異的抗体を生成する。
【0013】
さらにもう一つ別の態様において、多糖抗原−表面付着因子タンパク質接合体を適当な担体または希釈剤との組み合わせにおいて含む免疫原性組成物が提供される。本発明の免疫原性組成物はさらにアジュバント、例えば、水酸化アルミニウムまたはリン酸アルミニウムを含んでもよい。
さらにもう一つ別の態様において、哺乳動物において院内感染症に対する活性免疫を誘発する方法であって、このような感染症にかかっているヒトをはじめとする哺乳動物に、免疫原性量の本発明の免疫原性組成物を投与することを含む方法が提供される。
さらにもう一つ別の態様において、院内感染症に対する免疫治療薬の調製法であって、哺乳動物を本発明の免疫原性組成物で免疫化する工程、免疫化された哺乳動物から血漿を採集する工程、および集められた血漿から抗多糖抗体および抗表面付着因子抗体を含有する高度免疫グロブリンを収穫する工程を含む方法が提供される。高度免疫グロブリンは、院内感染症に対する受動免疫を誘発するために使用できる。
本発明の接合体は、多糖抗原および表面付着因子キャリアタンパク質の両方(両方とも毒性因子である)に対する抗体を惹起し、院内病原菌により引き起こされる疾患に対する免疫を付与する明確な利点を有する。すなわち、表面付着因子タンパク質それ自体が単に多糖抗原のタンパク質キャリアとして作用するだけでなく、免疫を付与することができる。
【図面の簡単な説明】
【0014】
【図1】GLCおよびHPAEC−PAD分析により決定されるエス・アウレウスCP5およびCP8の組成を示す。
【図2】デ−O−アセチル化エス・アウレウスCP5およびCP8の1H−NMR分析を示す。
【図3】エス・アウレウス−ClfAから得られるクランピング因子の概略図である。
【図4】エス・アウレウスClfAに由来する組み換えタンパク質Clf40およびClf41の概略図である。
【図5】エス・エピデルミディス−SdrGから得られるクランピング因子の概略図である。
【図6】エス・エピデルミディスSdrG:SdrG(N1N2N3)およびSdrG(N2N3)に由来する組み換えタンパク質の概略図である。
【図7】表面付着因子タンパク質のブロモアセチル化を示す。
【図8】エス・アウレウスCPの3−(2−ピリジルジチオ)プロピオニルヒドラジド(PDPH)での活性化を示す。
【図9】チオール化エス・アウレウスCPの表面付着因子タンパク質への接合を示す。
【図10】CP特異性ウサギ抗血清に関する抗原性についてのCP5−およびCP8−SdrG(N1N2N3)およびCP5−およびCP8−Clf41(N2N3)接合体の分析を示す。
【図11】ClfA特異性ウサギ抗血清に関する抗原性についてのCP5−およびCP8−Clf41(N2N3)接合体の分析を示す。
【図12】二重免疫拡散検定による抗原性についてのCP5−およびCP8−SdrG(N2N3)6xHisおよびCP5−およびCP8−Clf40(N1N2N3)6xHis接合体の分析を示す。
【図13】Ouchterlony免疫拡散検定による抗原性についてのCP5−およびCP8−SdrG(N2N3)およびCP5−およびCP8−FnbA接合体の分析を示す。
【図14】ドットブロットによる接合体の分析を示す。
【図15A】SdrG(N2N3)に接合したエス・アウレウスCP8に対する免疫応答を示す。
【図15B】アジュバントを含む、SdrG(N2N3)に接合したエス・アウレウスCP8に対する免疫応答を示す。
【図15C】Clf40(N1N2N3)に接合したエス・アウレウスCP8に対する免疫応答を示す。
【図15D】アジュバントを含む、Clf40(N1N2N3)に接合したエス・アウレウスCP8に対する免疫応答を示す。
【図15E】アジュバントを含む、Clf41(N2N3)に接合したエス・アウレウスCP8に対する免疫応答を示す。
【図15F】Clf41(N2N3)に接合したエス・アウレウスCP8に対する免疫応答を示す。
【図15G】アジュバントを含む、SdrG(N1N2N3)に接合したエス・アウレウスCP8に対する免疫応答を示す。
【図15H】SdrG(N1N2N3)に接合したエス・アウレウスCP8に対する免疫応答を示す。
【図16A】SdrG(N2N3)に接合したエス・アウレウスCP5に対する免疫応答を示す。
【図16B】アジュバントを含む、SdrG(N2N3)に接合したエス・アウレウスCP5に対する免疫応答を示す。
【図16C】Clf40(N1N2N3)に接合したエス・アウレウスCP5に対する免疫応答を示す。
【図16D】アジュバントを含む、Clf40(N1N2N3)に接合したエス・アウレウスCP5に対する免疫応答を示す。
【図16E】アジュバントを含む、Clf41(N2N3)に接合したエス・アウレウスCP5に対する免疫応答を示す。
【図16F】Clf41(N2N3)に接合したエス・アウレウスCP5に対する免疫応答を示す。
【図16G】アジュバントを含む、SdrG(N1N2N3)に接合したエス・アウレウスCP5に対する免疫応答を示す。
【図16H】SdrG(N1N2N3)に接合したエス・アウレウスCP5に対する免疫応答を示す。
【図17A】アジュバントを含まない、未接合エス・アウレウスClf40(N1N2N3)に対する免疫応答を示す。
【図17B】アジュバントを含まない、CP8に接合したエス・アウレウスClf40(N1N2N3)に対する免疫応答を示す。
【図17C】アジュバントを含まない、CP5に接合したエス・アウレウスClf40(N1N2N3)に対する免疫応答を示す。
【図17D】アジュバントを含む、CP5に接合したエス・アウレウスClf40(N1N2N3)に対する免疫応答を示す。
【図17E】アジュバントを含む、CP8に接合したエス・アウレウスClf40(N1N2N3)に対する免疫応答を示す。
【図17F】アジュバントを含む、未接合エス・アウレウスClf40(N1N2N3)に対する免疫応答を示す。
【図18A】アジュバントを含まない、CP5に接合したエス・アウレウスClf41(N2N3)に対する免疫応答を示す。
【図18B】アジュバントを含まない、CP8に接合したエス・アウレウスClf41(N2N3)に対する免疫応答を示す。
【図18C】アジュバントを含まない、未接合エス・アウレウスClf41(N2N3)に対する免疫応答を示す。
【図18D】アジュバントを含む、CP5に接合したエス・アウレウスClf41(N2N3)に対する免疫応答を示す。
【図18E】アジュバントを含む、CP8に接合したエス・アウレウスClf41(N2N3)に対する免疫応答を示す。
【図18F】アジュバントを含む、未接合エス・アウレウスClf41(N2N3)に対する免疫応答を示す。
【図19A】アジュバントを含まない、CP5に接合したエス・エピデルミディスSdrG(N1N2N3)に対する免疫応答を示す。
【図19B】アジュバントを含まない、CP8に接合したエス・エピデルミディスSdrG(N1N2N3)に対する免疫応答を示す。
【図19C】アジュバントを含まない、未接合エス・エピデルミディスSdrG(N1N2N3)に対する免疫応答を示す。
【図19D】アジュバントを含む、CP5に接合したエス・エピデルミディスSdrG(N1N2N3)に対する免疫応答を示す。
【図19E】アジュバントを含む、CP8に接合したエス・エピデルミディスSdrG(N1N2N3)に対する免疫応答を示す。
【図19F】アジュバントを含む、未接合エス・エピデルミディスSdrG(N1N2N3)に対する免疫応答を示す。
【図20A】アジュバントを含まない、CP5に接合したエス・エピデルミディスSdrG(N2N3)に対する免疫応答を示す。
【図20B】アジュバントを含まない、CP8に接合したエス・エピデルミディスSdrG(N2N3)に対する免疫応答を示す。
【図20C】アジュバントを含まない、未接合エス・エピデルミディスSdrG(N2N3)に対する免疫応答を示す。
【図20D】アジュバントを含む、CP5に接合したエス・エピデルミディスSdrG(N2N3)に対する免疫応答を示す。
【図20E】アジュバントを含む、CP8に接合したエス・エピデルミディスSdrG(N2N3)に対する免疫応答を示す。
【図20F】アジュバントを含む、未接合エス・エピデルミディスSdrG(N2N3)に対する免疫応答を示す。
【発明を実施するための形態】
【0015】
院内感染症は複数の毒性因子を含む。従って、一つの毒性決定因子のみを含有する免疫原性組成物と比較して、免疫原性組成物中に成分として含まれる毒性決定因子の組み合わせが防御を増大させる可能性が非常に高い。本発明の多糖抗原は、これらに限定されないが、スタフィロコッカス・アウレウス(Staphylococcus aureus)、スタフィロコッカス・エピデルミディス(Staphylococcus epidermidis)および他の凝固酵素陰性ブドウ球菌(CoNS)、エンテロコッカス種(Enterococcus spp.)、カンジダ・アルビカンス(Candida albicans)、エンテロバクター種(Enterobacter spp.)、ヘモフィルス・インフルエンゼ(Haemophilus influenzae)、クレブシエラ・ニューモニエ(Klebsiella pneumoniae)、エシェリキア・コリ(Escherichia coli)、およびシュードモナス・エルジノーサ(Pseudomonas aeruginosa)をはじめとする様々な院内病原微生物に由来する。これらの抗原は全身性感染症における毒性因子であり、不十分な免疫源である。その免疫原性は、キャリアタンパク質への接合により向上させることができる。本発明の目的に関して、表面付着因子タンパク質は接着性マトリックス分子を認識する微生物表面成分である。これらはInhibitex Inc.(Alpharetta,GA、USA)からMSCRAMMの登録商標で入手可能である。以下に記載するように、ブドウ球菌表面付着因子キャリアタンパク質を多糖抗原のキャリアタンパク質として利用することにより、多糖がT細胞依存性抗原に変換され、かくして抗多糖IgG応答が誘発される。さらに、接合体は感染症に対して防御し、哺乳動物宿主組織への細菌の付着の防止を助ける抗表面付着因子キャリアタンパク質抗体を誘発する。タンパク質−サッカライド接合法の化学反応はキャリアタンパク質抗体の免疫原性エピトープに対して有害な影響を及ぼし得ることが知られているが、驚くべきことに、本発明において、このような影響は見られず、タンパク質は保護エピトープに対して反応を惹起できる状態のままである。
【0016】
細菌細胞表面上の表面付着因子タンパク質および宿主組織内のリガンドは厳重に相互作用し、その結果、細菌が宿主に付着する。付着はしばしば細菌の生存に必要であり、細菌が宿主防御メカニズムおよび抗生物質の攻撃をくぐりぬける助けをする。細菌がうまく宿主組織に付着し、コロニー化すると、その生理機能は劇的に変化し、毒素および酵素などの損傷成分が分泌される。さらに、付着性細菌はしばしばバイオフィルムを生成し、ほとんどの抗生物質の殺菌効果に対して素速く耐性になる。
表面付着因子タンパク質の代表例としては、フィブロネクチン結合タンパク質、フィブリノゲン結合タンパク質、コラーゲン結合タンパク質およびビトロネクチン結合タンパク質が挙げられる。これらの付着因子は細胞外マトリックス成分フィブリノゲン、フィブロネクチン、コラーゲンおよびビトロネクチンを特異的に認識し、結合する。
【0017】
フィブロネクチン結合タンパク質
フィブロネクチン(Fn)は、動物のECMおよび体液において見いだされる440kDa糖タンパクである。フィブロネクチンの第一の生物学的機能は、適当なインテグリンを発現する細胞の付着に関して基質として作用するその能力に関連するようである。いくつかの細菌種がフィブロネクチンと特異的に結合し、フィブロネクチン含有低質に付着することが証明されている。ほとんどのエス・アウレウス分離株はFnと結合するが、様々な程度で結合し、このことは細菌表面上に発現される表面付着因子分子の数の多様性を反映する。Fnとエス・アウレウス間の相互作用は非常に特異性である(Kuusela 1978)。Fn結合は、110kDaの分子量を有する、FnBP−AおよびFnBP−Bと称する2つの表面が露出したタンパク質により媒介される。第一Fn結合部位は、3〜5回繰り返す35〜40アミノ酸のモチーフからなる。これらの遺伝子はクローンされ、配列決定されている(Jonsson 1991)。
【0018】
WO−A−85/05553は、フィブロネクチン−、フィブリノゲン、コラーゲン、および/またはラミニン結合能力を有する細菌細胞表面タンパク質を開示している。
米国特許第5320951号および第5571514号(Hookら)は、フィブロネクチン結合タンパク質A(fngA)遺伝子配列、およびこの配列に基づく生成物および方法を開示している。
【0019】
米国特許第5175096号(Hookら)は、fnbBの遺伝子配列、ハイブリッドDNA分子(fnbB)およびこの配列に基づく生物学的産物および方法を開示している。
米国特許第5652217号は、定義された配列を有するエス・アウレウスから得られるハイブリッドDNA分子によりエンコードされる結合活性を有する単離および精製タンパク質を開示している。
米国特許第5440014号は、ブドウ球菌感染症により引き起こされる乳腺炎に対する反芻動物の免疫化、傷の治療、タンパク質レセプターのブロッキング、他の動物の免疫化、または診断検定における使用のために用いることができるエス・アウレウスのフィブロネクチン結合タンパク質のD3相同性単位内のフィブロネクチン結合ペプチドを開示している。
【0020】
米国特許第5189015号は、哺乳動物におけるフィブロネクチンと結合する能力を有するエス・アウレウス細菌株のコロニー化の予防的治療のための方法であって、フィブロネクチンと結合する能力を有するエス・アウレウス細菌株により引き起こされる感染症の発生を予防するために、治療を必要とする哺乳動物に、予防的に有効な量のフィブロネクチン結合特性を有するタンパク質を投与することを含む方法を開示し、ここにおいて、タンパク質は87kDa〜165kDaの分子量を有する。
米国特許第5416021号は、ストレプトコッカス・ディスガラクティエ(Streptococcus dysgalactiae)から得られるフィブロネクチン結合タンパク質をエンコードするDNAとイー・コリに含まれるエス・ディスガラクティエから得られるフィブロネクチン結合タンパク質をエンコードするDNAを含むプラスミド、エス・ディスガラクティエからのフィブロネクチン結合タンパク質をエンコードするDNAおよびエス・ディスガラクティエからのフィブロネクチンをエンコードするDNAにより形質転換されたイー・コリ微生物を開示している。
【0021】
コラーゲン結合タンパク質
コラーゲンは軟骨の主な構成成分である。コラーゲン(Cn)結合タンパク質は通常、ブドウ球菌株により発現される。エス・アウレウスのコラーゲン結合表面付着因子タンパク質は、ブドウ球菌感染症における発病のメカニズムの重要な部分を構成するプロセスにおいて軟骨に付着する(Switalski 1993)。エス・アウレウスによるコラーゲン結合は、これだけではないが、少なくとも関節炎および敗血症に関与することが判明している。133、110および87kDaの分子量を有するコラーゲン付着因子(CNA)(Patti,J.ら、1992)が同定されている。異なる分子量を有するCNAを発現する株は、そのコラーゲン結合能力に差はない(Switalski 1993)。
敗血症性関節炎または骨髄炎と診断された患者の関節から回収されるブドウ球菌株は、ほぼ必ずと言っていいほどコラーゲン結合タンパク質を発現するが、創傷感染から得られる分離株でこの付着因子を発現するものは著しく少ない(Switalskiら 1993)。同様に、骨髄炎の患者から単離されたエス・アウレウス株はしばしば骨特異性タンパク質、骨シアロタンパク質(BSP)を認識する表面付着因子タンパク質を有する(Rydenら、1987)。関節空間内の関節軟骨のエス・アウレウスコロニー化は、敗血症性関節炎の発症に寄与する重要な因子であるらしい。
【0022】
WO92/07002は、コラーゲン結合活性を有するタンパク質またはポリペプチドをコードするエス・アウレウスからのヌクレオチド配列および該ヌクレオチド配列を含むプラスミドまたはファージを含むハイブリッドDNA分子を開示している。
コラーゲン結合タンパク質を発現するイー・コリ株、組み換えDNAにより形質転換された微生物、コラーゲン結合タンパク質またはポリペプチドを産生する方法、およびコラーゲン結合タンパク質またはポリペプチドのタンパク質配列も開示されている。
エス・アウレウスコラーゲン結合タンパク質をエンコードする遺伝子cnaのクローニング、配列決定、および発現が報告されている(Patti,Jら、1992)。
cna遺伝子は、グラム陽性菌から単離される表面タンパク質に特徴的な構造特性を含む133kDa付着因子をエンコードする。
【0023】
最近、リガンド結合部位がコラーゲン結合タンパク質のN−末端側の半分内に特定された(Patti,Jら、1993)。表面付着因子タンパク質の異なるセグメントに対応する組み換えタンパク質のコラーゲン結合活性を分析することにより、相当のコラーゲン結合活性を有する168アミノ酸長のタンパク質フラグメント(アミン酸残基151〜318に対応)が同定された。NまたはC末端においてこのタンパク質を切断すると、リガンド結合活性が失われるが、円偏光二色性分光分析により示されるようにタンパク質において構造変化が起きた。
Pattiら(1995)は、cna遺伝子によりエンコードされるエス・アウレウス付着因子におけるコラーゲン結合エピトープを開示している。その研究において、著者らは前記タンパク質の配列に由来するペプチドを合成し、抗体を産生するためにこれらを使用した。これらの抗体の内のいくつかはタンパク質のコラーゲンに対する結合を阻害する。
【0024】
WO97/43314は、コラーゲン結合タンパク質のある同定されたエピトープ(M55、M33、およびM17)は保護抗体を生成させるために使用できることを開示している。
該出願は、エス・アウレウスコラーゲン結合タンパク質に対するコラーゲンの結合と相互作用するか、または完全にブロックする組成物を同定するために必要な重要な情報を提供するコラーゲン結合タンパク質の結晶構造も開示している。エス・アウレウスコラーゲン結合タンパク質および25−アミノ酸ペプチドにおけるリガンド結合部位は、125I標識II型コラーゲンに対するエス・アウレウスの結合を直接阻害することを特徴とする。
【0025】
フィブリノゲン結合タンパク質
フィブリンは血餅の主成分であり、フィブリノゲン/フィブリンは埋め込まれた生体適合物質上に沈着する主な血漿タンパク質の一つである。細菌のフィブリノゲン/フィブリンへの付着が装置に関連する感染症の開始に重要であるということを示唆する証拠がかなり存在する。例えば、Vaudauxら(1989)により示されるように、エス・アウレウスはインビトロでフィブリノゲンでコートされたプラスチックに用量に依存した方法で付着する。加えて、血餅または心臓弁に対する損傷を模倣したモデルにおいて、Hermannら(1993)は、エス・アウレウスはフィブリノゲンブリッジを介して表面に付着する血小板に強力に結合することを示した。エス・アウレウスはインビトロで形成された血餅中のフィブリノゲンに直接付着することができ、ブリッジとして作用する血漿から沈着したフィブリノゲンにより培養された内皮細胞に付着することができる(Moreillonら、1995、Cheungら、1991)。VaudauxらおよびMoreillonらにより示されるように、フィブリノゲン結合タンパク質クランピング因子(ClfA)が欠損した突然変異体は、心内膜炎のラットモデルにおいて、インビトロでのフィブリノゲン、外植カテーテル、血餅、および損傷を受けた心臓弁に対する付着が減少した(Vaudauxら、1995、Moreillonら、1995)。
【0026】
フィブリノゲンの付着因子(「クランピング因子」と称することが多い)はエス・アウレウス細胞の表面上に位置する。溶液中での細菌とフィブリノゲン間の相互作用の結果、細菌細胞が即座に凝集する。フィブリノゲン上の結合部位はダイマーフィブリノゲン糖タンパク質のγ鎖のC末端に位置する。親和力は非常に高く、凝集は低濃度のフィブリノゲンで起こる。科学者らは最近、クランピング因子が固相フィブリノゲン、血餅、および損傷を受けた心臓弁への付着も促進することを示している(McDevittら、1994;Vaudauxら、1995;Moreillonら、1995)。
エス・アウレウスにおける2つの遺伝子が2つのフィブリノゲン結合タンパク質ClfAおよびClfBをコードすることが見いだされた。遺伝子clfAはクローンされ、配列決定され、92kDaのポリペプチドをコードすることが見いだされた。ClfAはフィブリノゲンのγ鎖と結合し、ClfBはαおよびβ鎖と結合する(Eidhinら、1998)。ClfBは推定分子量88kDa、見かけの分子量124kDaを有する、可溶性および固定化フィブリノゲンの両方に結合し、クランピング因子として作用する細胞壁に結合したタンパク質である。
【0027】
クランピング因子タンパク質の遺伝子(ClfAと表示)はクローンされ、配列決定され、分子レベルで詳細に分析された(McDevittら、1994、McDevittら、1995)。予想されるタンパク質は、933アミノ酸から構成される。39残基のシグナル配列はN末端にあり、その後に520残基の領域(領域A)が続き、これはフィブリノゲン結合ドメインを含有する。ジペプチドセリン−アスパルテートの154の繰り返しからなる308残基領域(領域R)がその後に続く。R領域配列は、18塩基対繰り返しGAY TCN GAY TCN GAY AGY(Yはピリミジンであり、Nは任意の塩基である)によりエンコードされる。ClfAのC末端は、タンパク質を細胞壁、膜アンカー、およびC末端の正に荷電した残基に固定するLPDTGモチーフなどのグラム陽性菌の多くの表面タンパク質中に存在する特徴を有する。
【0028】
血小板インテグリンαIIbβ3はフィブリノゲンのγ鎖のC末端を認識する。これは凝集中の血液凝固の開始における重大な事象である。ClfAは血小板凝集をブロックでき、γ鎖のC末端に対応するペプチド(198−411)はインテグリンおよびフィブリノゲンと相互作用するClfAの両方をブロックできるので、ClfAおよびαIIbβ3はフィブリノゲンγ鎖上の正確に同じ部位を認識するようである(McDevittら、1997)。αIIbβ3のフィブリノゲン結合部位は、「EFハンド」と称するCa2+結合決定因子の近くにあるか、または重複する。ClfA領域AはいくつかのEFハンド様モチーフを有する。3〜5mMの範囲のCa2+の濃度は、これらのClfA−フィブリノゲン相互作用をブロックし、ClfAタンパク質の二次構造を変化させる。ClfA EFハンドに影響を及ぼす突然変異はフィブリノゲンとの相互作用を軽減または防止する。Ca2+およびフィブリノゲンγ鎖はClfA領域Aの同じ部位に結合するか、または重複するようである。
【0029】
白血球インテグリンのα鎖、αMβ2は、200アミノ酸(AまたはIドメイン)の挿入を有し、これはリガンド結合活性の原因である。Iドメインにおける新規金属イオン依存性付着部位(MIDAS)モチーフはリガンド結合に必要とされる。認識されるリガンドにはフィブリノゲンが含まれる。フィブリノゲン上の結合部位はγ鎖にある(残基190〜202)。カンジダ・アルビカンスは、白血球インテグリンとよく似た性質を有する表面タンパク質、αIntlpを有することが近年報告された。表面タンパク質は、MIDASモチーフを包含するMβ2のIドメインと相同性であるアミノ酸配列を有する。さらに、Intlpはフィブリノゲンと結合する。
【0030】
ClfA領域Aはさらに、αIntlpと相同性である配列もある程度示す。ClfA領域A配列の調査により有効なMIDASモチーフが明らかになった。ClfAにおけるMIDASモチーフのDxSxS部分における推定カチオン配位残基における突然変異の結果、フィブリノゲン結合が著しく減少する。αMβ2のγ鎖結合部位に対応するペプチド(190−202)は、O’ConnellらによりClfA−フィブリノゲン相互作用を阻害することが示されている(O’Connell、1998)。したがって、ClfAは2つの別個の部位でフィブリノゲンのγ鎖と結合できるようである。ClfA上のリガンド結合部位は真核細胞インテグリンにより採用されるものと類似し、二価カチオン結合EFハンドおよびMIDASモチーフを含む。
【0031】
推定分子量約88kDaおよび見かけの分子量約124kDaを有するフィブリノゲン結合タンパク質ClfBも公知である。ClfBは細胞壁結合タンパク質であり、可溶性および固定化フィブリノゲンの両方と結合する。加えて、ClfBはフィブリノゲンのαおよびβ鎖の両方と結合し、クランピング因子として作用する。
細胞外マトリックスと結合するフィブリノゲン結合ClfAおよびClfBに関連するタンパク質が見いだされている。SdrC、SdrDおよびSdrEタンパク質は一次配列および構造機構においてClfAおよびClfBタンパク質に関連し、また細胞表面上に局在化する。細胞表面上に局在化するこれらのタンパク質のA領域に関して、タンパク質は血漿中のタンパク質、細胞外マトリックスまたは宿主細胞の表面上の分子と相互作用する。SdrCは細胞外マトリックスタンパク質、例えばビトロネクチンと結合することができる。SdrEも細胞外マトリックスと結合することができ;例えばSdrEは骨シアロタンパク質(BSP)と結合する。
【0032】
SdrC、SdrD、SdrE、ClfAおよびClfBのA領域において、コンセンサスTYTFTDYVDモチーフを誘導するために使用できる高度に保存されたアミノ酸配列が存在することが見いだされた。このモチーフは細菌感染症に対して広範囲の免疫を付与する多成分ワクチンにおいて使用でき、また広範囲の受動免疫を付与するモノクローナルまたはポリクローナル抗体を産生するために使用できる。別の具体例において、免疫を付与するために、または保護抗体を誘発するために、SdrおよびClfタンパク質ファミリーに由来する様々な配列モチーフの任意の組み合わせ(T/I)(Y/F)(T/V)(F)(T)(D/N)(Y)(V)(D/N)を使用できる。
【0033】
MHC−II類似タンパク質
フィブリノゲン、フィブロネクチンおよびコラーゲンに加えて、エス・アウレウス株は、その多くが付着性マトリックスタンパク質のファミリーに属するビトロネクチンなどの他の付着性真核細胞タンパク質に関連する(Chatwalら、1987)。米国特許第5648240号は、約70kDaの分子量を有するエス・アウレウスの広範囲の付着因子をエンコードする遺伝子を含むDNAセグメントを開示している。この付着因子は、フィブロネクチンまたはビトロネクチンと結合することができ、約30アミノ酸のMHC II様単位を含む。このタンパク質の結合特異性をさらに分析すると、これは合成ペプチドと結合する点でMHC II抗原と機能的に類似していることが明らかになる。従って、細胞外マトリックスタンパク質に対する細菌付着を媒介することに加えて、宿主の免疫系を抑制することにより、ブドウ球菌感染症に関与し得る。
【0034】
スタフィロコッカス・エピデルミディスからのSdrタンパク質
スタフィロコッカス・エピデルミディスは凝固酵素陰性菌であり、人間の皮膚の一般的な常在菌であり、しばしば異物感染症を引き起こす。病原性は、人工弁、整形外科用装置、ならびに静脈内および腹膜透析カテーテルなどの留置医療装置と生物体がまず付着し、その後、その上にバイオフィルムを形成する生物体の能力により促進される。装置に関連する感染症は、医療の成功を危うくし、患者の死亡率を著しく増大させる。従って、エス・エピデルミディス感染症の発生を制御または防止できるワクチンを開発できることは、同時に凝固酵素陽性および凝固酵素陰性菌の両方を包含する広範囲の細菌からの感染症を予防または治療できる複合ワクチンの開発と同様に非常に重要である。
エス・エピデルミディスにより発現される3つのSdr(セリン−アスパルテート(SD)繰り返し領域)タンパク質は、SdrF、SdrGおよびSdrHと表示され、これらのタンパク質のアミノ酸配列およびその核酸配列はWO00/12131(出典明示により本発明の一部として参照される)に示されている。
【0035】
本発明に従って、前記の表面付着因子タンパク質の少なくとも1つと接合した少なくとも1種の多糖抗原を含む免疫原性組成物として有用な接合体が提供される。加えて、多糖抗原および表面付着因子タンパク質に対する抗体は通常の手段を用いて産生される。従って、SdrGなどの表面付着因子タンパク質を含む免疫原性組成物は、凝固酵素陽性菌および凝固酵素陰性菌の両方に由来するものを含む広範囲の細菌感染症を治療するために使用される。
本発明の接合体の他の成分は、院内病原菌に由来する少なくとも1種の多糖抗原を含む。このような院内病原菌としては、これらに限定されないが、スタフィロコッカス・アウレウス(Staphylococcus aureus)、スタフィロコッカス・エピデルミディス(Staphylococcus epidermidis)および他の凝固酵素陰性ブドウ球菌(CoNS)、エンテロコッカス種(Enterococcus spp.、カンジダ・アルビカンス(Candida albicans)、エンテロバクター種(Enterobacter spp.)、ヘモフィルス・インフルエンゼ(Haemophilus influenzae)、クレブシエラ・ニューモニエ(Klebsiella pneumoniae)、エシェリキア・コリ(Escherichia coli)およびシュードモナス・エルジノーサ(Pseudomonas aeruginosa)が挙げられる。
【0036】
本発明の一具体例において、多糖抗原は、エス・アウレウスCP5およびCP8の少なくとも1つを含む。
本発明のもう一つ別の具体例において、多糖抗原は、エス・アウレウスおよび/またはエス・エピデルミディスにより発現されるようなPS/A、PNSG、PNAGおよびPIAのうちの少なくとも1つを含む。
【0037】
免疫原性組成物の調製および使用
免疫原性組成物は、本明細書に開示されるような多糖抗原−表面付着因子タンパク質接合体から調製される。免疫原性組成物は、多糖抗原および表面付着因子キャリアタンパク質の両方に対する抗体を産生する免疫応答を惹起する。
免疫原性組成物は、本明細書において開示されるようなオリゴ糖抗原−表面付着因子タンパク質接合体からも調製される。免疫原性組成物は、オリゴ糖抗原および表面付着因子キャリアタンパク質の両方に対する抗体を産生する免疫応答を惹起する。
【0038】
免疫原性組成物としての使用に好適な、本発明において提供される接合体は、これらに限定されないが、以下のものを包含する:
(i)クランピング因子A(ClfA)などのエス・アウレウスのフィブリノゲン結合タンパク質またはペプチドと接合するCP5、またはその有用なフラグメント、またはこれと十分高度に相同性であるタンパク質またはフラグメント;または
(ii)クランピング因子A(ClfA)などのエス・アウレウスのフィブリノゲン結合タンパク質またはペプチドと接合するCP8、またはその有用なフラグメント、またはこれと十分高度に相同性であるタンパク質またはフラグメント;または
(iii)クランピング因子A(ClfA)などのエス・アウレウスのフィブリノゲン結合タンパク質またはペプチドと接合するPIA、またはその有用なフラグメント、またはこれと十分高度に相同性であるタンパク質またはフラグメント;または
(iv)SdrGなどのエス・エピデルミディスのフィブリノゲン結合タンパク質またはペプチドと接合するCP5、またはその有用なフラグメント、またはこれと十分高度に相同性であるタンパク質またはフラグメント;または
(v)SdrGなどのエス・エピデルミディスのフィブリノゲン結合タンパク質またはペプチドと接合するCP8、またはその有用なフラグメント、またはこれと十分高度に相同性であるタンパク質またはフラグメント;または
(vi)SdrGなどのエス・エピデルミディスのフィブリノゲン結合タンパク質またはペプチドと接合するPIA、またはその有用なフラグメント、またはこれと十分高度に相同性であるタンパク質またはフラグメント。
【0039】
各例において、任意の接合体(i)〜(vi)から調製された免疫原性組成物は、エス・アウレウスなどの凝固酵素陽性菌、ならびにエス・エピデルミディスなどの凝固酵素陰性菌からの感染症に対して患者を免疫化するために有用である。
表面付着因子キャリアタンパク質がフィブリノゲン結合タンパク質である前記の接合体(i)〜(vi)に加えて、本発明はさらに、表面付着因子キャリアタンパク質が、例えばフィブロネクチン結合タンパク質、コラーゲン結合タンパク質およびビトロネクチン結合タンパク質などの任意のブドウ球菌表面付着因子タンパク質である接合体を包含する。本発明はさらに、多糖抗原がPS/A、PNAGまたはPNSG、またはエス・アウレウス、エス・エピデルミディスおよび他のCoNs、エンテロコッカス種、カンジダ・アルビカンス、エンテロバクター種、ヘモフィルス・インフルエンゼ、クレブシエラ・ニューモニエ、エシェリキア・コリ、およびシュードモナス・エルジノーサなどの院内病原微生物からの他の多糖抗原であり得ることも意図する。
【0040】
多糖をタンパク質に接合するための多くの方法が当該分野において公知であり、本発明における使用に適している。一般に、多糖を活性化させるか、または他の方法で接合しやすくする、すなわち、少なくとも1つの部分をタンパク質または他の分子と共有結合できるようにしなければならない。多くのこのような方法は当該分野において公知である。例えば、米国特許第4356170号(Jennings)は、多糖上にアルデヒド基を生成させるために過ヨウ素酸を使用し、次いでシアノボロヒドリドを用いて還元的アミノ化を行うことを記載している。米国特許第4663160号は(Tsayら)もアルデヒド基を生成させるために過ヨウ素酸を使用したが、次にシアノボロヒドリドなどの還元剤の存在下でのシッフ塩基反応で4〜12の炭素部分(縮合剤の存在下で調製)で誘導化されたタンパク質に多糖を結合させた。米国特許第4619828号(Gordon)は、多糖を活性化するために臭化シアンを使用し、次いで4〜8個の炭素原子のスペーサーブリッジによりタンパク質と接合させた。さらに他の接合法も当該分野において公知である。
【0041】
本発明の一具体例において、CPはリンカー3−(2−ピリジルジチオ)−プロピオニルヒドラジド(PDPH)で活性化され、これによりCPにおけるN−アセチルマンノースアミノウロン酸のカルボジイミド活性化カルボキシレート基がPDPHのヒドラジド基とカップリングする(図8)。MSCRMAMMキャリアタンパク質はブロモ酢酸のN−ヒドロキシスクシイミドエステルでのリシン残基のブロモアセチル化により活性化される(図7)。PDPH−チオール化CPを次いで、チオールでブロモアセチル化タンパク質における臭素を置換することにより活性化表面付着因子タンパク質に接合させると、結果として安定なチオエーテル結合が得られる(図9)。
CP−CONHNHCOCH2CH2SCH2CONH−表面付着因子タンパク質
【0042】
本発明のCP−表面抗原タンパク質接合体を含む免疫原性組成物をマウスにおいて試験し、不十分な免疫原性未接合CPと比較して、向上された免疫原性を有することが示された(図15〜20)。加えて、莢膜多糖特異性抗体ならびにCP−表面付着因子接合免疫原性組成物により誘発されたClfAおよびSdrG特異性抗体はどちらも対応する抗原を発現する生株と結合することが示された(表5および6)。これらの結果を考慮すると、本発明の免疫原性組成物はエス・アウレウスまたはエス・エピデルミディスなどの病原体により引き起こされる院内感染症に対して有用であると考えられる。また、CP−表面付着因子接合体により誘発される抗体が免疫原性組成物として傷に投与されるか、またはインビトロまたはインビボで医療装置またはポリマー生体適合物質をコートするために使用される場合、組成物は創傷部位または生体適合物質へのブドウ球菌菌の結合を防止または抑制するであろう。本発明に従って処理された接合体は、患者を院内感染に対して保護するための免疫原性組成物の調製において使用される。本明細書において用いられる「対象」とは、温血哺乳動物であり、例えば、ヒト、霊長類、ウマ、ウシ、イヌおよびネコを包含する。
【0043】
注射可能な液体溶液または懸濁液を調製するために、接合体を免疫学的に許容される希釈剤または担体に通常の方法で添加することができる。
本発明の免疫原性組成物は、典型的には、任意の適当な医薬的に許容される担体、例えば生理食塩水または他の注射可能な液体中に接合体を分散させることにより形成される。本明細書において用いられる場合、「医薬的に許容される担体」なる語句は、医薬投与に適合するありとあらゆる溶媒、分散媒体、コーティング、抗菌および抗真菌剤、等張および吸収遅延剤などを包含することを意図される。医薬的に活性な物質に関するかかる媒体および薬剤の使用は当該分野において周知である。通常の媒体または薬剤が活性化合物と不適合である場合を除いて、かかる媒体を本発明の組成物において用いることができる。例えば、接合体調製物をリン酸ナトリウム緩衝塩溶液(PBS)(pH7.0〜8.0)中に多糖1mlあたり1〜100μgの濃度で懸濁させる。本発明の免疫原性組成物の投与は、これらに限定されないが、非経口(例えば、皮下、腹腔内、筋肉内、静脈内、皮内)、経口および鼻内を包含する任意の周知の方法により行うことができる。免疫原性組成物の好ましい投与方法は非経口投与である。非経口投与に用いられる溶液または懸濁液は、次の成分を含む:無菌希釈剤、例えば、注射用水、塩溶液、不揮発性油、ポリエチレングリコール、グリセリン、ポリプロピレングリコールまたは他の合成溶媒;抗菌剤、例えばベンジルアルコールまたはメチルパラベン;酸化防止剤、例えばアスコルビン酸または重亜硫酸ナトリウム;キレート化剤、例えばエチレンジアミン四酢酸;緩衝剤、例えば酢酸塩、クエン酸塩またはリン酸塩および等張性を調節するための薬剤、例えば塩化ナトリウムまたはデキストロース。pHは酸または塩基、例えば、塩酸または水酸化ナトリウムで調節することができる。非経口製剤はアンプル、使い捨て注射器あるいはガラスまたはプラスチック製の複数回投与用バイアル中に封入することができる。
【0044】
注射可能な用途に適した免疫原性組成物は、無菌水性溶液(水溶性の場合)または分散液および無菌注射可能な溶液または分散液の即時調製用無菌粉末を含む。あらゆる場合において、組成物は無菌でなければならず、容易に注射可能である程度に流動性でなければならない。製造および貯蔵条件下で安定でなければならず、細菌および真菌などの微生物の汚染作用に対して保護されなければならない。担体は、例えば、水、エタノール、ポリオール(例えば、グリセロール、プロピレングリコール、および液体ポリエチレングリコールなど)、およびその適当な混合物を含有する溶媒または分散媒体である。適当な流動性は、例えば、レシチンなどのコーティングの使用、分散液の場合は必要とされる粒子サイズの維持、および界面活性剤の使用により維持される。微生物の作用の防止は、様々な抗菌剤および抗真菌剤、例えば、パラベン、クロロブタノール、フェノール、アスコルビン酸、チメロサールなどにより達成される。多くの場合において、等張剤、例えば、糖、ポリアルコール、例えばマンニトール、ソルビトール、および塩化ナトリウムを組成物中に含めることが好ましい。組成物中に吸収を遅らせる薬剤、例えばアルミニウムモノステアレートおよびゼラチンを含めることにより注射可能な組成物の吸収が延長される。
【0045】
無菌注射可能な溶液は、本発明の接合体を必要とされる量、適当な溶媒中に、単独または必要に応じて前記の成分と組み合わせて配合し、続いて濾過滅菌することにより調製される。一般に、分散液は、活性化合物を、塩基性分散媒体および前記のもののうち必要とされる他の成分を含有する無菌ビヒクル中に配合することにより調製される。無菌注射可能な溶液を調製するための無菌粉末の場合、好ましい調製法は真空乾燥および凍結乾燥であり、これにより活性成分の粉末と、あらかじめ無菌濾過されたその溶液からの任意のさらなる所望の成分が得られる。
ある具体例において、免疫原性組成物は1以上のアジュバントを含む。本明細書において定義されるように、「アジュバント」は本発明の免疫原性組成物の免疫原性を向上させる働きをする物質である。従って、アジュバントは免疫反応の追加免疫(boost)のために投与され、当業者には周知である。
【0046】
組成物の有効性を向上させるための好ましいアジュバントとしては、これらに限定されないが、次のものが挙げられる:
(1)アルミニウム塩(alum)、例えば水酸化アルミニウム、リン酸アルミニウム、硫酸アルミニウムなど;
(2)水中油エマルジョン処方(他の特異的免疫刺激剤、例えばムラミルペプチド(以下参照)または細菌細胞壁成分の存在下または非存在下)、例えば
(a)ミクロフルイダイザー、例えば110型ミクロフルイダイザー(Microfluidics,Newton,MA)を用いて1ミクロン以下の粒子に処方された5%スクアレン、0.5%Tween80、および0.5%Span85(任意に様々な量のMTP−PE(必要ではないが、以下参照)を含有してもよい)を含有するMF59(PCT公開番号WO90/14837)、
(b)10%スクアレン、0.4%Tween80、5%プルロンブロックされたポリマーL121、およびthr−MDP(以下参照)(ミクロ流動化されて1ミクロン以下のエマルジョンにされているか、または撹拌されて大きな粒子サイズのエマルジョンが生じるかのいずれか)を含有するSAF、
(c)2%スクアレン、0.2%Tween80、および米国特許第4912094号(Corixa)に記載されている3−デアシル化モノホスホリピッドA(MPL)、トレハロースジミコレート(TDM)、および細胞壁骨格(CWS)、好ましくはMPL+CWS(Detox)からなる群からの1以上の細菌細胞壁成分を含有するRibiアジュバント系(RAS)(Corixa,Hamilton,MT);
(3)サポニンアジュバント、例えばQuil AまたはSTIMULON QS−21(Antigenics,Framingham,MA)(米国特許第5,057,540号)またはこれから生成する粒子、例えばISCOM(免疫刺激複合体);
(4)細菌リポ多糖、合成リピッドA類似体、例えばアミノアルキルグルコサミンホスフェート化合物(AGP)、またはその誘導体または類似体(Corixaから入手可能で、米国特許第6,113,918号に記載されている);例えばAGPは2−[(R)−3−テトラデカノイルオキシテトラデカノイルアミノ]エチル2−デオキシ−4−O−ホスホノ−3−O−[(R)−3−テトラデカノイルオキシテトラデカノイル]−2−[(R)−3−テトラデカノイルオキシテトラデカノイルアミノ]−b−D−グルコピラノシド(529(以前はRC529と呼んだ)をも呼ばれ、水性形態または安定なエマルジョンとして処方される)、合成ポリヌクレオチド、例えばCpGモチーフを含有するオリゴヌクレオチド(米国特許第6,207,646号);
(5)サイトカイン、例えばインターロイキン(例えば、IL−1、IL−2、IL−4、IL−5、IL−6、IL−7、IL−12、IL−15、IL−18など)、インターフェロン(例えば、ガンマインターフェロン)、顆粒球マクロファージコロニー刺激因子((GM−CSF)、マクロファージコロニー刺激因子(M−CSF)、腫瘍壊死因子(TNF)など;
(6)コレラ毒素(CT)(野生型または突然変異形態、例えば公開国際特許出願番号WO00/18434に従ってアミノ酸29位でグルタミン酸が別のアミノ酸、好ましくはヒスチジンにより置換されている場合(WO02/098368およびWO02/098369も参照))などの細菌ADP−リボシル化トキシンの無毒化突然変異体、狂犬病毒素(PT)、またはイー・コリ易熱性毒素(LT)、特にLT−K63、LT−R72、CT−S109、PT−K9/G129(例えば、WO93/13302およびWO92/19265参照);および
(7)組成物の有効性を向上させるために免疫刺激剤として作用する他の物質。
【0047】
前記のように、ムラミルペプチドとしては、これらに限定されないが、N−アセチル−ムラミル−L−スレオニル−D−イソグルタミン(thr−MDP)、N−アセチル−ノルムラミル−L−アラニン−2−(1’−2’−ジパルミトイル−sn−グリセロ−3−ヒドロキシホスホリルオキシ)−エチルアミン(MTP−PE)などが挙げられる。
本発明の免疫原性組成物は、免疫原性反応を誘発するために十分な量で投与される。用量は、該免疫原性組成物を受容する個体の大きさ、体重または年齢に基づいて調節することができる。個体における抗体応答は、抗体価または殺菌活性について分析することによりモニターすることができ、必要に応じて追加免疫して反応を向上させることができる。
【0048】
本発明の免疫原性組成物は、体液性免疫応答を誘発するために対象に投与される。対象は次に免疫原組成物に応答して生成されたイムノグロブリンの供給源(高度免疫イムノグロブリン)として作用する。免疫化された対象は血漿を供与し、これから通常の血漿分画技術により高イムノグロブリンが得られ、院内感染に対する耐性を付与するため、または院内感染を治療するために別の対象に投与される。
【0049】
(実施例)
前記開示は一般に本発明を説明する。以下の実施例を参照することによりさらに完全に理解することができる。これらの実施例は、説明の目的のみで記載したものであり、本発明の範囲を制限することを意図するものではない。
【0050】
実施例1
エス・アウレウスCP5およびCP8多糖の精製
CP5およびCP8それぞれの精製のためにエス・アウレウス株(ATCC#49521)およびWright(ATCC#49521)を使用した。既に公開されている方法(Fournier,Vannら、1984;Fournier,Hannonら、1987)を変更した方法により細胞から多糖を精製した。2%NaClで補足されたColumiaブロス中で成長させた細胞を3時間37℃で、リソスタフィン(175U/gの細胞)で、RNAseおよびDNAse(それぞれ0.1mg/g)で4時間、37℃で消化し、続いてプロナーゼ(1mg/gの細胞)で3時間、37℃で消化した。10mM CaCl2の存在下、25%および75%エタノールで連続して沈降させることにより酵素消化物から粗CPを調製した。0.05〜0.5M NaClの直線的勾配を用い、Q−セファロースカラム上でアニオン交換クロマトグラフィーによりCPを次いでペレットから精製した。残留するテイコ酸を0.05M NalO4で酸化した。透析後、CPを次いでSephacrylS300(Amersham Pharmacia Biotech,Piscataway,NJ)カラム上で寸法排除クロマトグラフィーによりさらに精製した。フラクション中のCPの存在をエス・アウレウスCP5およびCP8特異性抗血清との反応性により決定した。
【0051】
PIAを、Mackら(Mack,Fischerら、1996)により記載されているように、熱抽出した静止期のエス・エピデルミディス細胞から精製し、PIA含有培養上清と組み合わせた。抽出された物質および培養上清を、10K膜を用いて濃縮し、処理して、核酸および残存するタンパク質を除去した。ゲル濾過または透析濾過を用いて粗PIAを分画した。アニオン交換クロマトグラフィーによりPIA抗原陽性物質をさらに分画し、PIAフラクション含有エステル結合サクシネートを精製した。非スクシニル化および部分的非N−アセチル化PIAを含有するフロースルーフラクションを、カチオン交換クロマトグラフィーにより精製した。PS/A(PNSG、PNAG)を(Maria−Litran,Kropecら、2002)またはMcKenney,Pouliotら、1999により記載されているようにして精製した。
【0052】
実施例2
エス・アウレウスCP5およびCP8の分析
精製されたCP5およびCP8の化学的特徴化により、両多糖は事実上核酸および残存するタンパク質が無いことが示された(表1)。
HPAECクロマトグラフィーにより決定された糖組成により、CP5およびCP8におけるFucρNAcおよびManρNacAの存在が明らかになった(図1)。O−デアシル化多糖の1H NMRスペクトル(図2)は、既に公開されているスペクトル(Vann,Moreauら、1987;Moreau,Richardsら、1990)と類似しており、3種の単糖:2−アセトアミド−2−デオキシ−D−マンヌロン酸、2−アセトアミド−2−デオキシ−L−フコースおよび2−アセトアミド−2−デオキシ−D−フコースの構造および存在が確認された。
精製されたCP5、CP8およびTAは、対応する全細胞抗血清と反応した場合に二重免疫拡散検定における1つの沈降素のバンドにより確かめられるように免疫学的に異なっていた(データは省略)。
【0053】
実施例3
表面付着因子タンパク質の精製
評価した表面付着因子タンパク質は:
エス・アウレウスClf40(N1N2N3)−クランピング因子Aの完全長Aドメイン(アミノ酸(AA)40−559)−図3。
エス・アウレウスClf41(N2N3)−Clf40のポストプロテアーゼ部位フラグメント(AA223−559)−図4。
エス・エピデルミディスSdrG(N1N2N3)−SdrGの完全長Aドメイン(AA50−597)−図5。
エス・エピデルミディスSdrG(N2N3)−SdrGのポストプロテアーゼ部位フラグメント(AA273−597)−図6。
コレラの表面付着因子タンパク質は、Inhibitex,Inc.,Alpharetta,GA,USAから入手した。
【0054】
表面付着因子タンパク質のHistagマイナスバージョンをイー・コリプラスミド宿主株から精製した。イー・コリpLP1134BL21(DE3)をエス・アウレウスClf41(N2,N3)について使用し、pLP1135B21(DE3)をエス・エピデルミディスSdrG(N2,N3)精製に使用した。両タンパク質を、硫酸アンモニウム沈殿およびその後のセファクリルQ−セファロースカラム(Amersham Pharmacia Biotech,Piscataway,NJ)上でのイオン交換クロマトグラフィーにより細胞溶解物の可溶性フラクションから単離した。最終物質の純度はSDS−PAGEにより測定すると90%より高かった。
【0055】
過剰発現されたエス・アウレウスClf40(N1,N2,N3)またはClf41(N2,N3)、エス・エピデルミディスSdrG(N1,N2,N3)またはSdrG(N2,N3)を含有するイー・コリ細胞を、約13000psiで、Microfuidics M110−Yミクロフルイダイザーを1回通過させて可溶化させた。細胞片を17000rpmで30分間4℃で遠心分離することにより精製した。過剰発現されたタンパク質を、AKTAexplorer(XKカラムキレート化セファロースファーストフローおよびQセファロースHP樹脂(Amersham Pharmacia Biotech,Piscataway,NJ))を用いて上清から精製した。粗His標識されたタンパク質を、0.1M NiCl2を装填したキレート化セファロースファーストフローを用いたアフィニティ工程により上清から精製した。粗溶解物を、25mM Tris、pH8.0、0.5M NaCl、5mMイミダゾールで平衡化させたカラム上にロードし、カラムを5カラム体積の緩衝剤で洗浄することにより未結合タンパク質をカラムから溶出させた。未結合タンパク質を次いで250mM Tris、pH8.0、0.5M NaCl、500mMイミダゾール緩衝液で溶出させ、バルクで集めた。タンパク質を次いでQセファロースHPカラム上イオン交換クロマトグラフィーにより残存する不純物から精製した。
【0056】
実施例4
エス・アウレウスCP5−およびCP−8表面付着因子キャリアタンパク質接合体免疫原性組成物の合成
エス・アウレウスCP5およびCP8多糖を、リンカーを含有するチオール基を多糖に導入し、ハロアセチル基をタンパク質キャリアに導入した後、本発明により提供された表面付着因子キャリアタンパク質とチオエーテル結合を介して別々に結合させた。アミン基をブロモ酢酸のN−ヒドロキシスクシンイミドエステルと反応させることにより、ブロモアセチル基を表面付着因子タンパク質中に導入した(図7)。チオール化CPを生成させるために、莢膜多糖におけるN−アセチルマンノースアミノウロン酸のカルボジイミド活性化カルボキシレート基をスルフヒドリル反応性ヒドラジドヘテロ二官能性リンカー3−(2−ピリジルジチオ)−プロピオニルヒドラジド(PDPH、図8)のヒドラジド基とカップリングさせた。ジチオトレイトール(DTT)での還元により生成し、Sdphadex G25カラム上SECにより精製されたPDPH−チオール化CPのチオールを、活性化タンパク質のブロモアセチル基と反応させると、CPとタンパク質間の臭素置換により形成された共有チオエーテル結合が生じた(図9)。未反応ブロモアセチル基をシステアミン塩酸塩(2−アミノエタンチオール塩酸塩)で「キャップ」した。反応混合物を次いでAmicon XM100膜上で濃縮した。
【0057】
実施例5
CP−およびCP−8表面付着因子キャリアタンパク質接合体免疫原性組成物の特徴化
4Nトリフルオロ酢酸(TFA)での加水分解後、CarboPac−PA1カラム上HPAEC−PADクロマトグラフィーによるCPの定量化により、接合免疫原性組成物をCPおよび表面付着因子キャリアタンパク質含量について分析した。タンパク質含量をLowry比色検定により決定した。接合免疫原性組成物の分子量を寸法排除クロマトグラフィーおよび多角度レーザー光散乱(MALLS)の組み合わせにより測定した。結果を表2および3に記載する。接合CPおよび表面付着因子タンパク質の抗原性を二重免疫拡散法(図10〜13)およびドットブロット分析(図14)により決定した。結果は、CPの表面付着因子タンパク質への接合は、CPまたはタンパク質のいずれかの抗原性を変更しないことを示す。CPのタンパク質への接合は、接合体がニトロセルロース膜に結合する能力により確認された。未接合CPはニトロセルロース膜に結合しなかった。
【0058】
実施例6
マウスにおけるCP−表面付着因子キャリアタンパク質接合体免疫原性組成物の免疫原性
接合免疫原性組成物を、CP5およびCP8および表面付着因子タンパク質キャリアに対するIgG応答を誘発する能力について試験した。Swiss−Websterマウスを1マイクログラム用量(CP基準)で2週間間隔で3回、皮下的に(SC)免疫化した。接合免疫原性組成物の免疫原性を、アジュバントとしての100マイクログラムのリン酸アルミニウムの存在下または不在下で試験した。各タンパク質免疫原性組成物候補を同様のプロトコルを用いて同様に評価した。エス・アウレウスCPおよび表面付着因子タンパク質に対する免疫応答を標準的抗原ELISAによる各注射の1週間後に分析した(以下の実施例7および8参照)。
【0059】
実施例7
エス・アウレウスCP5およびCP8−表面付着因子キャリアタンパク質接合体免疫原性組成物で免疫化されたマウスにおけるCPの抗体反応
結果(図15および16)は、CPの表面付着因子タンパク質への共有結合の結果、莢膜多糖(CP)特異性IgG反応が誘発されることを示す。このことは、CP−T細胞非依存性免疫反応はCPの表面付着因子キャリアタンパク質へのカップリング後にT細胞依存性免疫反応に変換されたことを示す。接合免疫原性組成物のリン酸アルミニウムへの吸着は、タンパク質キャリアとしてSdrG(N2N3)を投与されたマウスを除いて約10倍に増大した。CP5−およびCP8−SdrG(N2N3)接合体のアジュバントへの吸着の結果、CPに対する免疫応答は増大しなかったが、CPの抗体反応はこの実験においてアジュバントと混合された(吸着されていないが)他の表面付着因子タンパク質接合体と同様に良好であった。ClfAおよびSdrGのN1ドメインの欠失は、これらのタンパク質のキャリア特性に対して影響を及ぼさなかった。
【0060】
実施例8
エス・アウレウスCP5およびCP8−表面付着因子キャリアタンパク質接合体でワクチン接種されたマウスにおける表面付着因子タンパク質抗体反応
接合表面付着因子タンパク質は、未接合のものと比較して類似した力価の表面付着因子タンパク質特異性抗体を誘発した(図17〜20)。このことにより、表面付着因子タンパク質のCPに対する接合により抗原性エピトープが修飾されないことが確認される。未接合ClfAまたはCP−ClfA接合体のリン酸アルミニウムへの吸着の結果、アジュバントを含まない同じ免疫原性組成物で免疫化されたマウスと比較して、マウスにおけるClfA抗体価が増大した。未接合SdrGで免疫化されたマウスは、CP−SdrG接合免疫原性組成物で免疫化されたマウスと比較して低いSdrG抗体価で反応した。未接合SdrGのリン酸アルミニウムへの吸着の結果、alumなしで投与されたCP−SdrG接合体により誘発されたレベルと比較してSdrG抗体価が増大した。CP−SdrG接合体のalumへの吸着はSdrG抗体価を増大させなかった。
【0061】
実施例9
CP表面付着因子キャリアタンパク質接合体により誘発された抗体による生菌上に発現されたCPおよび表面付着因子キャリアタンパク質の認識
マウスにおけるCP−表面付着因子タンパク質接合体により誘発された抗体の生菌に対する結合をフローサイトメトリー分析により試験した。この検定において使用したエス・アウレウス株を表4に示す。SdrG接合体に対して誘発された抗体の分析について、SdrGを発現するエル・ラクチス(L.lactis)を使用した。結果は(表5および6)、莢膜多糖特異性抗体ならびに対応するCP5−およびCP8−表面付着因子タンパク質接合体により誘発されるClfAまたはSdrG特異性抗体はどちらも対応する抗原を発現する生株に結合することを示す。これにより、CPの表面付着因子タンパク質に対する接合は、CP上に存在する自然に発現されたエピトープおよび表面付着因子タンパク質抗原に対する免疫反応を変更しないことが示される。
【0062】
実施例10
フローサイトメトリー分析法
使用したエス・アウレウス株は次の通りであった:Newman、NewmanのClfAノックアウト突然変異体(Newman ClfA::emr)およびWright(ATCC49525)。ClfA発現を最大にするために、エス・アウレウス菌をトリプシンソイブロス中で静止期まで成長させた。莢膜発現を最大にするために、エス・アウレウス菌をColumbia2%NaCl寒天(BD Microbiology,Sparks,MD)上で一夜成長させた。Newman ClfA::emr株を、5μg/mlのエリスロマイシンの存在下で成長させて、ノックアウト突然変異を維持した。SdrG抗原認識を評価するためにSdrGを発現する組み換えラクトコッカス・ラクチス(Lactococcus lactis(L.Lactis))株を使用した。5μg/mlのエリスロマイシンの存在下、M17ブロス中、エル・ラクチス株を後期対数期まで成長させた。
【0063】
細菌培養物を収穫し、10mlの冷1×PBS(Invitrogen Corp, Rockville,MD)中で2回洗浄し、分析前に氷上で保存した。細菌濃度を、UV−可視記録分光光度計(Ultrospec3000、Pharmacia Biotech,Cambridge,England)を用いて1×PBSでD600nm=2.0に調節した。非特異性およびプロテインAによるマウスIgGの細胞表面に対する結合を排除するために、細菌調製物を全部、氷上で、1×PBS(Invitrogen Corp.,Rockville,MD)中10mlの1:50希釈度(2.32mg IgG)のウサギIgG(Sigma、St.Louis、Missouri)中で30分間インキュベートした。ClfA結合の不在下での8型莢膜認識を評価するために、さらに30分間高力価ClfA特異性ウサギ抗血清(Inhibitex,Alpharetta,GA)(1:100希釈)でインキュベーションすることにより、ClfAエピトープをエス・アウレウス株Wright上でブロックした。インキュベーションのブロッキング後、細菌を、3000rpmで10分間遠心分離することにより、10mlの冷1×PBS中で2回洗浄した。細菌ペレットを1×PBS中2.5%BSA(Invitrogen Corp.,Rockville,MD)(PBSA)中に再懸濁させ、氷上で保存した。
【0064】
検定をtitertube(BioRad Labs,Hercules,CA)中で行った。試験動物から得られるプレブリード(prebleed)および高力価抗血清をPBSA中に希釈し、0.5mlの各血清希釈物を、20μlの細菌懸濁液を含有する適当な試験管に添加した。全試験管を撹拌し、氷上で30分間インキュベートした。インキュベーション後、各試験管を撹拌し、次いで3000RPMで10分間遠心分離した。細菌ペレットを2回0.5mlの冷PBSA中で洗浄した。各ペレットを抗マウスIgG(H&L)(Rockland Labs,Gilbertsville,Pa)のPE接合F(ab’)2フラグメントの0.5mlの1:200希釈物中に再懸濁させた。細菌を再懸濁させ、撹拌することにより混合した。試験管を氷上で30分間インキュベートし、15分間隔で2回撹拌した。このインキュベーション後、細菌をPBSA中最終再懸濁液で洗浄した。FACS分析まで試験管を氷上で保存した。
【0065】
各titertubeを12×75mmポリスチレン試験管に移し、B−D FACSCalibur(BD Biosciences,Mansfield,MA)フローサイトメーターを用いて分析した。結果を、所定の抗血清の蛍光強度が同じ希釈度でのプレブリードで得られるシグナルより大きいならばプラスと採点した。結果を表7に示す。
前記考察および実施例は単にある具体例の詳細な説明を提示するためのものであると理解すべきである。従って、当業者には本発明の精神および範囲から逸脱することなく様々な修正および等価物を調製することが可能であることは明らかである。
本出願中に記載した全ての文献、他の刊行物、特許および特許出願は全体として本発明の一部として参照される。
【0066】
【表1】
【0067】
【表2】
【0068】
【表3】
【0069】
【表4】
【0070】
【表5−1】
【表5−2】
【0071】
【表6−1】
【表6−2】
【0072】
【表7】
*ClfA=ClfA AドメインのN1,N2,N3またはN2,N3領域
SdrG=SdrG AドメインのN1,N2,N3領域
【0073】
参考文献:
【0074】
【0075】
【0076】
【0077】
【0078】
【特許請求の範囲】
【請求項1】
院内病原菌から由来する少なくとも1種の多糖抗原および少なくとも1種のブドウ球菌表面付着因子キャリアタンパク質を含む免疫原性多糖タンパク質接合体であって、多糖抗原およびブドウ球菌表面付着因子キャリアタンパク質の両方に対する特異性抗体を産生する接合体。
【請求項2】
院内病原菌から由来する少なくとも1種の多糖抗原の1またはそれ以上の抗原エピトープを提示するオリゴ糖フラグメントおよび少なくとも1種のブドウ球菌表面付着因子キャリアタンパク質を含む免疫原性多糖タンパク質接合体であって、多糖抗原およびブドウ球菌表面付着因子キャリアタンパク質の両方に対する特異性抗体を産生する接合体。
【請求項3】
多糖抗原が、スタフィロコッカス・アウレウス(Staphylococcus aureus)、凝固酵素陰性ブドウ球菌(CoNS)、エンテロコッカス種(Enterococcus spp.)、カンジダ・アルビカンス(Candida albicans)、エンテロバクター種(Enterobacter spp.)、ヘモフィルス・インフルエンゼ(Haemophilus influenzae)、クレブシエラ・ニューモニエ(Klebsiella pneumoniae)、エシェリキア・コリ(Escherichia coli)、およびシュードモナス・エルジノーサ(Pseudomonas aeruginosa)からなる群から選択される院内病原菌から由来する請求項1または2記載の接合体。
【請求項4】
多糖抗原が、スタフィロコッカス・アウレウスまたはCoNSから由来する請求項3記載の接合体。
【請求項5】
CoNSがスタフィロコッカス・エピデルミディス(Staphylococcus epidermidis)である請求項4記載の接合体。
【請求項6】
多糖抗原が、スタフィロコッカス・アウレウス5型(CP5)または8型(CP8)から由来する請求項4記載の接合体。
【請求項7】
多糖抗原が、スタフィロコッカス・アウレウスまたはスタフィロコッカス・エピデルミディスにより発現される多糖細胞間付着因子(PIA)、多糖付着因子(PS/A)、ポリ−N−スクシニルβ−1−6−グルコサミン(PNSG)、またはポリ−N−アセチル−β−1−6−グルコサミン(PNAG)である請求項3記載の接合体。
【請求項8】
ブドウ球菌表面付着因子キャリアタンパク質が、フィブリノゲン結合タンパク質、フィブロネクチン結合タンパク質、コラーゲン結合タンパク質およびビトロネクチン結合タンパク質からなる群から選択される請求項1〜7のいずれか一つに記載の接合体。
【請求項9】
ブドウ球菌表面付着因子キャリアタンパク質が、スタフィロコッカス・アウレウスのフィブリノゲン結合タンパク質(クランピング因子A[ClfA])である請求項8記載の接合体。
【請求項10】
ブドウ球菌表面付着因子キャリアタンパク質がスタフィロコッカス・エピデルミディスのフィブリノゲン結合タンパク質(SdrG)である請求項8記載の接合体。
【請求項11】
ブドウ球菌表面付着因子キャリアタンパク質がスタフィロコッカス・アウレウスのフィブロネクチン結合タンパク質である請求項8記載の接合体。
【請求項12】
ブドウ球菌表面付着因子キャリアタンパク質がスタフィロコッカス・アウレウスのコラーゲン結合タンパク質である請求項8記載の接合体。
【請求項13】
ブドウ球菌表面付着因子キャリアタンパク質がスタフィロコッカス・アウレウスのビトロネクチン結合タンパク質である請求項8記載の接合体。
【請求項14】
多糖抗原がリンカーを介してブドウ球菌表面付着因子キャリアタンパク質と結合している請求項1〜13のいずれか1つに記載の接合体。
【請求項15】
ブドウ球菌表面付着因子キャリアタンパク質がInhibitex Inc、Alpharetta、GAから入手したMSCRAMM(登録商標)である請求項1〜14のいずれか1つに記載の接合体。
【請求項16】
リンカーが3−(2−ピリジルジチオ)−プロピオニルヒドラジド(PDPH)である請求項14記載の接合体。
【請求項17】
免疫学的に許容される担体または希釈剤中に請求項1〜16のいずれか1つに記載の接合体を含む免疫原性組成物。
【請求項18】
さらにアジュバントを含む請求項17記載の免疫原性組成物。
【請求項19】
接合体がClfAに接合したCP5を含み、その接合体がCP5およびClfAの両方に対する特異性反応性抗体を産生する請求項17または18のいずれかに記載の免疫原性組成物。
【請求項20】
接合体がClfAに接合したCP8を含み、その接合体がCP8およびClfAの両方に対する特異性反応性抗体を産生する請求項17に記載の免疫原性組成物。
【請求項21】
接合体がClfAに接合したPIA、PS/A、PNAGまたはPNSGの任意の多糖を含み、その接合体が多糖およびClfAに対する特異性反応性抗体を産生する請求項17記載の免疫原性組成物。
【請求項22】
接合体がSdrGに接合したCP5を含み、その接合体がCP5およびSdrGの両方に対する特異性反応性抗体を生成する請求項17記載の免疫原性組成物。
【請求項23】
接合体がSdrGと接合したCP8を含み、その接合体がCP8およびSdrGの両方に対する特異性反応性抗体を産生する請求項17記載の免疫原性組成物。
【請求項24】
接合体がSdrGに接合したPIA、PS/A、PNAGまたはPNSGの任意の多糖を含み、その接合体が多糖およびSdrGに対する特異性反応性抗体を産生する請求項17記載の免疫原性組成物。
【請求項1】
院内病原菌から由来する少なくとも1種の多糖抗原および少なくとも1種のブドウ球菌表面付着因子キャリアタンパク質を含む免疫原性多糖タンパク質接合体であって、多糖抗原およびブドウ球菌表面付着因子キャリアタンパク質の両方に対する特異性抗体を産生する接合体。
【請求項2】
院内病原菌から由来する少なくとも1種の多糖抗原の1またはそれ以上の抗原エピトープを提示するオリゴ糖フラグメントおよび少なくとも1種のブドウ球菌表面付着因子キャリアタンパク質を含む免疫原性多糖タンパク質接合体であって、多糖抗原およびブドウ球菌表面付着因子キャリアタンパク質の両方に対する特異性抗体を産生する接合体。
【請求項3】
多糖抗原が、スタフィロコッカス・アウレウス(Staphylococcus aureus)、凝固酵素陰性ブドウ球菌(CoNS)、エンテロコッカス種(Enterococcus spp.)、カンジダ・アルビカンス(Candida albicans)、エンテロバクター種(Enterobacter spp.)、ヘモフィルス・インフルエンゼ(Haemophilus influenzae)、クレブシエラ・ニューモニエ(Klebsiella pneumoniae)、エシェリキア・コリ(Escherichia coli)、およびシュードモナス・エルジノーサ(Pseudomonas aeruginosa)からなる群から選択される院内病原菌から由来する請求項1または2記載の接合体。
【請求項4】
多糖抗原が、スタフィロコッカス・アウレウスまたはCoNSから由来する請求項3記載の接合体。
【請求項5】
CoNSがスタフィロコッカス・エピデルミディス(Staphylococcus epidermidis)である請求項4記載の接合体。
【請求項6】
多糖抗原が、スタフィロコッカス・アウレウス5型(CP5)または8型(CP8)から由来する請求項4記載の接合体。
【請求項7】
多糖抗原が、スタフィロコッカス・アウレウスまたはスタフィロコッカス・エピデルミディスにより発現される多糖細胞間付着因子(PIA)、多糖付着因子(PS/A)、ポリ−N−スクシニルβ−1−6−グルコサミン(PNSG)、またはポリ−N−アセチル−β−1−6−グルコサミン(PNAG)である請求項3記載の接合体。
【請求項8】
ブドウ球菌表面付着因子キャリアタンパク質が、フィブリノゲン結合タンパク質、フィブロネクチン結合タンパク質、コラーゲン結合タンパク質およびビトロネクチン結合タンパク質からなる群から選択される請求項1〜7のいずれか一つに記載の接合体。
【請求項9】
ブドウ球菌表面付着因子キャリアタンパク質が、スタフィロコッカス・アウレウスのフィブリノゲン結合タンパク質(クランピング因子A[ClfA])である請求項8記載の接合体。
【請求項10】
ブドウ球菌表面付着因子キャリアタンパク質がスタフィロコッカス・エピデルミディスのフィブリノゲン結合タンパク質(SdrG)である請求項8記載の接合体。
【請求項11】
ブドウ球菌表面付着因子キャリアタンパク質がスタフィロコッカス・アウレウスのフィブロネクチン結合タンパク質である請求項8記載の接合体。
【請求項12】
ブドウ球菌表面付着因子キャリアタンパク質がスタフィロコッカス・アウレウスのコラーゲン結合タンパク質である請求項8記載の接合体。
【請求項13】
ブドウ球菌表面付着因子キャリアタンパク質がスタフィロコッカス・アウレウスのビトロネクチン結合タンパク質である請求項8記載の接合体。
【請求項14】
多糖抗原がリンカーを介してブドウ球菌表面付着因子キャリアタンパク質と結合している請求項1〜13のいずれか1つに記載の接合体。
【請求項15】
ブドウ球菌表面付着因子キャリアタンパク質がInhibitex Inc、Alpharetta、GAから入手したMSCRAMM(登録商標)である請求項1〜14のいずれか1つに記載の接合体。
【請求項16】
リンカーが3−(2−ピリジルジチオ)−プロピオニルヒドラジド(PDPH)である請求項14記載の接合体。
【請求項17】
免疫学的に許容される担体または希釈剤中に請求項1〜16のいずれか1つに記載の接合体を含む免疫原性組成物。
【請求項18】
さらにアジュバントを含む請求項17記載の免疫原性組成物。
【請求項19】
接合体がClfAに接合したCP5を含み、その接合体がCP5およびClfAの両方に対する特異性反応性抗体を産生する請求項17または18のいずれかに記載の免疫原性組成物。
【請求項20】
接合体がClfAに接合したCP8を含み、その接合体がCP8およびClfAの両方に対する特異性反応性抗体を産生する請求項17に記載の免疫原性組成物。
【請求項21】
接合体がClfAに接合したPIA、PS/A、PNAGまたはPNSGの任意の多糖を含み、その接合体が多糖およびClfAに対する特異性反応性抗体を産生する請求項17記載の免疫原性組成物。
【請求項22】
接合体がSdrGに接合したCP5を含み、その接合体がCP5およびSdrGの両方に対する特異性反応性抗体を生成する請求項17記載の免疫原性組成物。
【請求項23】
接合体がSdrGと接合したCP8を含み、その接合体がCP8およびSdrGの両方に対する特異性反応性抗体を産生する請求項17記載の免疫原性組成物。
【請求項24】
接合体がSdrGに接合したPIA、PS/A、PNAGまたはPNSGの任意の多糖を含み、その接合体が多糖およびSdrGに対する特異性反応性抗体を産生する請求項17記載の免疫原性組成物。
【図1】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】

【図13】

【図14】

【図15A】
【図15B】
【図15C】
【図15D】
【図15E】
【図15F】
【図15G】
【図15H】
【図16A】
【図16B】
【図16C】
【図16D】
【図16E】
【図16F】
【図16G】
【図16H】
【図17A】
【図17B】
【図17C】
【図17D】
【図17E】
【図17F】
【図18A】
【図18B】
【図18C】
【図18D】
【図18E】
【図18F】
【図19A】
【図19B】
【図19C】
【図19D】
【図19E】
【図19F】
【図20A】
【図20B】
【図20C】
【図20D】
【図20E】
【図20F】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15A】
【図15B】
【図15C】
【図15D】
【図15E】
【図15F】
【図15G】
【図15H】
【図16A】
【図16B】
【図16C】
【図16D】
【図16E】
【図16F】
【図16G】
【図16H】
【図17A】
【図17B】
【図17C】
【図17D】
【図17E】
【図17F】
【図18A】
【図18B】
【図18C】
【図18D】
【図18E】
【図18F】
【図19A】
【図19B】
【図19C】
【図19D】
【図19E】
【図19F】
【図20A】
【図20B】
【図20C】
【図20D】
【図20E】
【図20F】
【公開番号】特開2012−51922(P2012−51922A)
【公開日】平成24年3月15日(2012.3.15)
【国際特許分類】
【外国語出願】
【出願番号】特願2011−228995(P2011−228995)
【出願日】平成23年10月18日(2011.10.18)
【分割の表示】特願2006−509140(P2006−509140)の分割
【原出願日】平成16年3月4日(2004.3.4)
【出願人】(591000791)ワイス・ホールディングズ・コーポレイション (43)
【出願人】(505338501)インヒビテックス・インコーポレイテッド (3)
【氏名又は名称原語表記】INHIBITEX, INC.
【Fターム(参考)】
【公開日】平成24年3月15日(2012.3.15)
【国際特許分類】
【出願番号】特願2011−228995(P2011−228995)
【出願日】平成23年10月18日(2011.10.18)
【分割の表示】特願2006−509140(P2006−509140)の分割
【原出願日】平成16年3月4日(2004.3.4)
【出願人】(591000791)ワイス・ホールディングズ・コーポレイション (43)
【出願人】(505338501)インヒビテックス・インコーポレイテッド (3)
【氏名又は名称原語表記】INHIBITEX, INC.
【Fターム(参考)】
[ Back to top ]