
除去されたエフェクター機能を伴う抗体Fc突然変異
Fc領域に変異を有する抗体及び他のFc含有分子は、Fcγ受容体との結合と、得られる活性とを低減し、種々の疾患及び傷害の治療に使用され得る。

【発明の詳細な説明】
【技術分野】
【0001】
本発明は、Fcγ受容体に特異的に結合する能力か、又は表面標的抗原のFc受容体媒介性架橋結合によって免疫細胞による分裂応答を活性化させる能力を実質的に失うように突然変異される、ヒト抗体IgG2定常領域(Fc領域)に関する。本発明は、突然変異IgG2定常領域が組み込まれ得る、新規の抗体も提供する。
【背景技術】
【0002】
細胞表面抗原を標的とする抗体は、不必要な免疫活性化、及び免疫細胞とのFc受容体(FcR)結合と補体の活性化とに関連するエフェクター機能を引き起こす。標的配位子機能を標的にし、かつ活性化又は中和するが、必要な局所的細胞又は組織を傷つける又は破壊することのないように意図された治療用抗体及びFc融合構築物として、縮小された(除去された)エフェクター機能を伴うFc突然変異が模索されている。
【0003】
ヒトIgGアイソタイプ(成熟γグロブリンクラスG抗体のサブクラス、IgG1、IgG2、IgG3、及びIgG4)は、例えば、抗体依存性細胞傷害(ADCC、例えば、IgG1及びIgG3)、抗体依存性細胞貪食作用(ADCP、例えば、IgG1、IgG2、IgG3、及びIgG4)、及び補体依存性細胞傷害(CDC、例えば、IgG1、IgG3)といった免疫機能を回復させる、差異的な能力を示す。そのような免疫機能のアイソタイプ特異的結合は、別個の免疫細胞上のFc受容体に対する選択性と、C1qに結合し膜攻撃複合体(MAC)のアセンブリを活性化する能力とに基づく。種々のアイソタイプの中で、Fcγ受容体(例えば、FcγRI、FcγRIIa/b/c、FcγRIIIa/b)に対する相対的親和性は、IgG1及びIgG3に対して高いが、しかしながら、IgG2に対しては親和性が最小であり(FcγRIIa 131H多型に限定される)、またIgG4はFcγRIに対してある程度の親和性しか有さない。比較配列分析及び共結晶構造を用いて、受容体結合に対して主に接触する残基は、より低いヒンジ及びCH2領域にまたがるアミノ酸残基にマッピングされている。標準的タンパク質工学技術を用いて、Fc受容体及びC1q補体成分に対する抗体調製物の親和性を強化又は低減することにおいて、いくらかの成功が収められている。
【0004】
アイソタイプの中で、IgG2はFc受容体のファミリーに結合する能力が最少である。IgG2を起点として、減少されたエフェクター機能を伴いつつ、しかしFcRn結合、長期安定性、及び低免疫原性を保持する突然変異誘発物質を見出す試みがなされている。この類の改善された突然変異は、安全性を保持しつつ、改善された抗体治療薬を提供することができる。
【発明の概要】
【課題を解決するための手段】
【0005】
本発明は、例えば、FC領域を含有し細胞表面配位子を標的とするような、抗体又は抗体様治療薬の技術作業において有用な、修飾されたグリコシル化免疫グロブリン定常ドメインを提供する。本発明の組成物は、減少されたFcγR結合能力を示すが保存されたFcRn結合を有する、IgG 2Fc突然変異である。これらのIgG Fc突然変異は、免疫エフェクター機能のFc関連結合及び補体媒介性細胞傷害を最小にしつつ、可溶性又は細胞表面抗原を治療標的化することを可能にする。1つの態様において、IgG 2Fc突然変異は、EU番号付け方式に従うV234A、G237A、P238Sを含む。別の態様において、IgG2 Fc突然変異は、EU番号付け方式に従うV234A、G237A、H268Q、又はH268A、V309L、A330S、P331Sを含む。特定の態様において、IgG2 Fc突然変異は、EU番号付け方式に従うV234A、G237A、P238S、H268A、V309L、A330S、P331S、及び任意でP233Sを含む。
【0006】
1つの実施形態において、IgG2 Fc突然変異組成物は、i)抗体依存性細胞傷害(ADCC)、ii)補体依存性細胞傷害(CDC)、iii)抗体依存性細胞貪食作用(ADCP)、iv)FcR媒介性細胞活性化(例えば、FcR架橋結合を介するサイトカイン放出)、及びv)FcR媒介性血小板活性化/枯渇といった、免疫及びエフェクター機能に関連するFcγRの活性化に由来する潜在毒性が、最小化又は排除されながら、治療用抗体(又はFc融合体)半減期の保持がFcRnとの相互作用を通して保存される効能において、使用される。1つの態様において、IgG2 Fc突然変異は、例えば基底細胞神経節といった神経疾患に関与する細胞上の配位子;B細胞又はT細胞活性化に、あるいは線維芽細胞又は幹細胞といった組織修復又は治癒に関与する細胞に関係する免疫系疾患;を標的とする、多価結合剤等の結合剤の治療用抗体又はFc融合体に組み込まれる。
【0007】
別の実施形態において、IgG2 Fc突然変異は医薬組成物を含む。別の実施形態において、IgG2 Fc突然変異は薬剤活性分子の一部を含む。IgG2 Fc突然変異又は活性IgG2 Fc突然変異含有分子を含む医薬組成物は、マクロファージ又は単核細胞の遊走及び集結によって特徴付けられる疾患の治療に有用である。1つの態様において、IgG2 Fc突然変異含有分子は、神経組織、内分泌組織、血管組織、心臓組織、滑膜組織、皮膚組織、又は粘膜組織内の標的に結合するのに有用である。本発明のIgG2 Fc突然変異の多くの使用のうちの1つは、移植片対宿主疾患;宿主対移植片疾患;臓器移植拒絶反応;骨髄移植拒絶反応;血管炎、自己免疫性溶血性貧血、自己免疫性血小板減少症、及び関節炎等の自己免疫;胎児/新生児同種免疫性血小板減少症等の同種免疫;ぜんそく及びアレルギー;クローン病又は強皮症等の慢性若しくは急性炎症性疾病;アルツハイマー病、冠状動脈閉塞症、の治療におけるものである。
【図面の簡単な説明】
【0008】
【図1】それぞれの残基に対応するEU番号付けを示す、野生型ヒトIgG2(配列番号:1)、IgG4(配列番号:2)、及びIgG1(配列番号:3)のアミノ酸配列の整列を示し、IgG2のヒンジ領域はEU残基218で始まる。
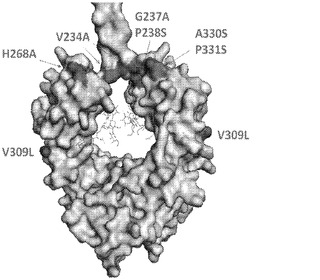
【図2】修飾された残基の表面位置(EU番号付け)を示す、Fc断片の構造を示す。
【図3A】それぞれのアイソタイプと、AlphaScreenビーズアッセイプラットフォームを使用して、ヒトIgG1野生型アイソタイプの抗体との抗Her2/neu結合抗体として構築した突然変異との、競合を示すグラフである。FcgRI(図示せず)、FcgRIIa(A)、FcgRIIb(B)、FcgRIIIa(図示せず)、及びFcRn(C)。
【図3B】それぞれのアイソタイプと、AlphaScreenビーズアッセイプラットフォームを使用して、ヒトIgG1野生型アイソタイプの抗体との抗Her2/neu結合抗体として構築した突然変異との、競合を示すグラフである。FcgRI(図示せず)、FcgRIIa(A)、FcgRIIb(B)、FcgRIIIa(図示せず)、及びFcRn(C)。
【図3C】それぞれのアイソタイプと、AlphaScreenビーズアッセイプラットフォームを使用して、ヒトIgG1野生型アイソタイプの抗体との抗Her2/neu結合抗体として構築した突然変異との、競合を示すグラフである。FcgRI(図示せず)、FcgRIIa(A)、FcgRIIb(B)、FcgRIIIa(図示せず)、及びFcRn(C)。
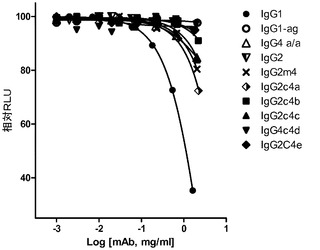
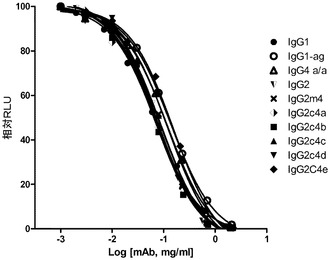
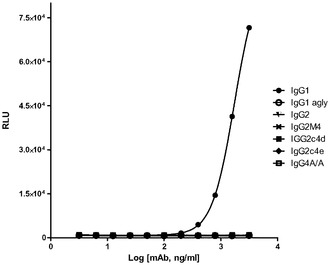
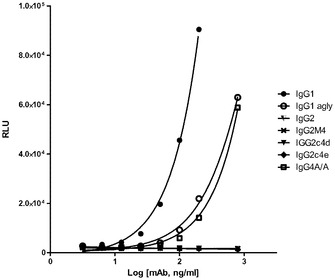
【図4A】それぞれのアイソタイプと、AlphaScreenビーズアッセイを使用して、抗Her2/neu結合抗体として構築した突然変異との、直接結合を示すグラフである。FcgRIIIa(A)、FcgRI(B)、及びFcgRIIa(C)。
【図4B】それぞれのアイソタイプと、AlphaScreenビーズアッセイを使用して、抗Her2/neu結合抗体として構築した突然変異との、直接結合を示すグラフである。FcgRIIIa(A)、FcgRI(B)、及びFcgRIIa(C)。
【図4C】それぞれのアイソタイプと、AlphaScreenビーズアッセイを使用して、抗Her2/neu結合抗体として構築した突然変異との、直接結合を示すグラフである。FcgRIIIa(A)、FcgRI(B)、及びFcgRIIa(C)。
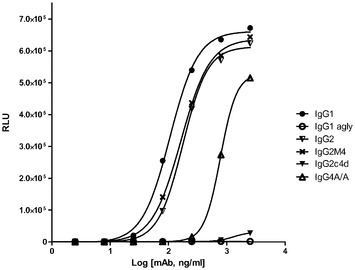
【図5】ADCCアッセイの結果、及び選択されたアイソタイプと、抗Her2/neu結合抗体として構築し、またヒトPBMC(25倍過剰)とSK−Br3乳癌細胞とを標的として用いた突然変異と、を示す。
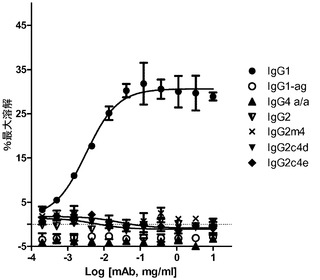
【図6】ヒト補体とWIL2−Sリンパ腫細胞とを標的として用いた、選択された抗CD20構築物の、CDCアッセイの結果を示す。
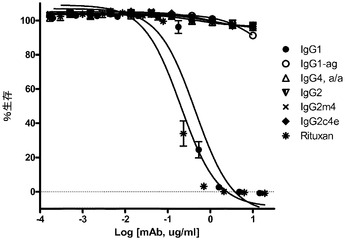
【図7】Sk−Br3標的細胞とGM−CSF分化マクロファージとを用いた、選択された抗Her2/neu構築物の、フローサイトメトリー分析からのADCPアッセイの結果を示す。
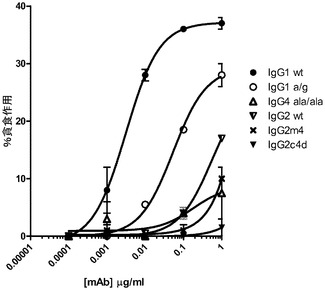
【図8】IgG2野生型及び6つの突然変異を含む、種々の構築物に対するIgGアイソタイプとのビーズ結合を伴うPBMCの刺激後24時間からの、サイトカイン(TNFα)放出を示すグラフ。
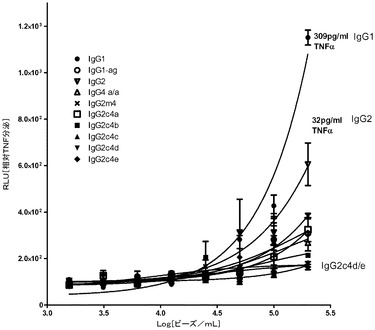
【図9】種々のFc領域を伴う、又はFc領域(Fab’)2が欠損する抗CD20結合ドメイン抗体構築物を注射されたカニクイザル群(Cynomologous monkeys)における、時間経過に伴う平均循環B細胞数のグラフ。
【0009】
【表1】
【発明を実施するための形態】
【0010】
略語
ADCC=抗体依存性細胞傷害、ADCP=抗体依存性細胞貪食作用、CDC=補体依存性細胞傷害、IgG=免疫グロブリンG、ITAM=免疫受容体チロシン活性化モチーフ、ITIM=免疫受容体チロシン阻害モチーフ、Mab=モノクローナル抗体、FDCR=Fc依存性サイトカイン放出、FcγR、FcgR、又はFcγR=Fcγ受容体。
【0011】
用語の定義&説明
「抗体依存性細胞媒介性細胞傷害」又は「ADCC」とは、細胞傷害性細胞(例えばナチュラルキラー(NK)細胞、好中球及びマクロファージ)上に存在するFc受容体(FcR)上に結合した、分泌されたIgが、これらの細胞傷害性エフェクター細胞が抗原を有する標的細胞に特異的に結合してその後標的細胞を細胞毒で殺傷することを可能にするような、細胞傷害の形態を意味する。標的細胞の表面を対象とする配位子特異的高親和性IgG抗体は、細胞傷害性細胞を刺激し、またそのような死滅に絶対に必要である。標的細胞の溶解は細胞外であり、直接の細胞間接触を必要とし、また補体に関与しない。
【0012】
任意の特定の抗体の、ADCCによる標的細胞の溶解を媒介する能力が測定され得る。ADCC活性を評価するために、目的の抗体は、標的細胞の細胞溶解をもたらす抗原抗体複合体によって活性化され得る免疫エフェクター細胞と共同して、標的配位子を示す、標的細胞に添加される。細胞溶解は、通常、溶解した細胞からの標識(例えば、放射性基質、蛍光染料、又は天然細胞内タンパク質)の放出によって検出される。そのようなアッセイに有用なエフェクター細胞としては、末梢血単核球(PBMC)及びナチュラルキラー(NK)細胞が挙げられる。生体外ADCCアッセイの具体的な例は、Wisecarverら、1985,19:211、Bruggemannら、1987,J Exp Med 166:1351、Wilkinsonら、2001,J Immunol Methods 258:183、Patelら、1995,J Immunol Methods 184:29に説明される(これらのそれぞれは、参照によって組み込まれる)。別の方法として、又は追加として、目的の抗体のADCC活性は、生体内で、例えば、その内容が参照によってその全体において組み込まれるClynesら、1998,PNAS USA 95:652に開示されるような動物モデルにおいて、評価されてもよい。
【0013】
「補体指向性細胞傷害」又はCDCは、補体カスケードが、抗体Fcに結合する補体成分C1qによって活性化される、細胞傷害の形態を指す。
【0014】
本明細書で使用するとき、「Fc」、「Fc含有タンパク質」又は「Fc含有分子」という用語は、少なくとも免疫グロブリンCH2及びCH3ドメインを有する単量体、二量体、又はヘテロ二量体タンパク質を指す。CH2及びCH3ドメインは、タンパク質/分子(例えば、抗体)の二量体領域の少なくとも一部を形成することができる。
【0015】
本明細書で使用される場合の「モノクローナル抗体」という用語は、動物抗体の少なくとも1つの種の重鎖又は軽鎖抗体可変ドメインのうちの少なくとも1つに対して実質的に相同性を保持する、少なくとも1つの配位子結合ドメインを含む、Fc含有タンパク質の特定の形態である。
【0016】
「野生型ヒトIgG2 Fc領域」とは、カバットのEU番号付けに従う、ヒトIgG重鎖の残基K218から残基K447までである、配列番号:1のアミノ酸配列又はその断片を有する、ヒトIgG Fc領域を指す。定常領域内のアミノ酸は、ヒトIgG1抗体、EUIgG1(配列番号:3)との整列によって番号付けされる(Cunninghamら、1970、J.Biol. Chem.,9:3161〜70を参照)。すなわち、抗体の重鎖及び軽鎖は、アミノ酸配列同一性を最大化するようにEUの重鎖及び軽鎖と整列され、抗体内のそれぞれのアミノ酸は、EUにおける対応するアミノ酸と同じ番号を割り当てられる。EU番号付け方式は、当該技術分野において従来使用されている(概略的には、Kabatら、Sequences of Protein of Immunological Interest,NIH Publication No.91−3242,US Department of Health and Human Services(1991)を参照)。この慣習に従って、説明される「野生型IgG2」定常領域は、236位でアミノ酸を欠損する(図1、配列番号:1)。
【0017】
概論
本発明は、サイトカイン放出を引き起す能力がない、又は標的配位子を示し標的とされる細胞を囲む細胞及び組織を損傷若しくは死滅させる能力がないという観点から改善された安全性を有する、治療用抗体、Fc融合体、及び類似のバイオ医薬品生物薬剤の製造に使用するためのFcドメインを同定することへの関心によって動機付けられている。
【0018】
ヒトIgG4アイソタイプ抗体及びFc融合タンパク質は、顕著なADCC(NKにより、FcgRIIIaだけを発現する)を引き起こさないが、しかし、マクロファージ(FcγRI、IIa、及びIIIaを発現する)による貪食作用(ADCP)を誘発し、かつ可能であれば、免疫複合体において特定の免疫細胞上の活性化FcγRの分布に起因する場合に、単核細胞を活性化する能力を、保持する。残留活性を最小化する試みにより、野生型IgG4 Fc(配列番号:2)にV234A、L235A(ala/ala)を有する突然変異の、開発及び使用がもたらされた。
【0019】
Armourらは、IgG2において多重点突然変異を生成し、FcgRI(Armourら、1999)、並びにFcgRIIa及びIIIa(Armourら、2003)との結合を最小化した。Muellerらの特許出願(PCT国際公開特許WO97/11971号)に開示される追加の突然変異は、IgG2 CH2及びIgG4 CH3ドメインからなる、IgG2/IgG4のハイブリッドに由来し、特に、残基330及び331は、本発明のIgG2突然変異内のIgG4に由来する。免疫活性化作用を減少させるための、弱化又は発現抑制の試みはまた、IgG2又はIgG4又は2つのサブクラス間を交換するドメインを使用し、IgG1(配列番号:3)内に最小のエフェクター機能を伴うmAbを生成させた(Taoら、1991)。Strohlの特許出願(米国特許出願US2007/0148167号)は、EU位置268、309、330、及び331位残基でのIgG2における4つの突然変異を開示している。Shieldsら、(2001)は、追加の置換物、H268Aも開示している。
【0020】
本発明は、カバット/EU番号付け方式に従うIgG4残基H268Q又はA、V309L、A330S、及びP331Sを伴う、IgG2定常領域(Fc)の多数の位置における置換の初めての実証である。多数の残基置換の指向選択は、測定可能なエフェクター機能を欠く治療用実体の提供の可能性に加え、予想外に抗体工学で用いるための機能的Fcドメインを提供し、融合ポリペプチドとして使用した。
【0021】
多置換IgG2突然変異は、Alpha Screen競合アッセイ及びSPR/Biacore分析によって評価された、ヒトFcR(FcγRI、FcγRIIa、FcγRIIb、FcγRIIIa、及びFcRn)に対するそれらの相対親和性に基づき選択された。これらの突然変異は、更に、CDC、ADCC、及びADCPを誘発し、またPBMCによるTNF−α分泌を引き起こす能力について、適切な細胞システムにおいて試験され順位付けられた。本明細書で提供される一連の実験データにおいて、IgG2突然変異が、既知の調整物、又は野生型IgG1及びIgG2に加えてIgG1 ag及びIgG4 Ala/Alaを含む突然変異と、比較された。いくつかの生体外生物検定におけるこれらの突然変異の更なる分析は、最小の検出不可能な活性レベルと、大いに除去されたFcRに対する結合親和性とを実証した。これらの試験に基づき、IgG2 Fc突然変異は、IgG2c4dとして名付けられ、FcRに対する検出可能な親和性又は結合活性(単量体対二重〜多重配位子結合)を有しておらず、かつ前述の種々のエフェクター/免疫活性化生物検定において活性を有していないことが確認されている。IgG2c4d Fcは、アラニン、セリン、及びロイシンの置換物:V234A、G237A、P238S、H28A、V309L、A330S、P331S(EU番号付け)を含む。7つの残基置換されたIgG2であるIgG2c4dは、FcRに結合する能力、エフェクター機能を媒介する能力、又はFc媒介性サイトカイン放出に関与する能力がないという点において、第1の真に「発現抑制された」Fcであると考えられ得る。
【0022】
本発見に基づき、IgG2c4dの7つの突然変異の一部が使用されるか、又は他のアミノ酸突然変異と組み合わされることができ、又は本突然変異は、本明細書で教示されるようなエフェクター機能の類似又は選択的発現抑制を達成するために別のIgGアイソタイプにおいて使用され、また当該技術分野において既知であるものと組み合わされることができる。
【0023】
本明細書で具体的に例示されるように、三つ組のV234A、G237A、P238S置換物は、分子のFcRn結合親和性を維持しつつ、FcγRIIa結合親和性及びFcγRIIa結合活性を低減し、かつADCP及びFc依存性サイトカイン放出が除去される。
【0024】
【表2】
【0025】
【表3】
【0026】
【表4】
【0027】
変性Fc含有分子を作製する方法
置換のための部位は、自然抗体Fcの構造的特性、保持されたFcRn結合、良好な安定性を有し、並びに補体カスケード、細胞溶解、細胞貪食作用、又はサイトカイン放出などを刺激する能力が減少された組成物を製造したいという要望に基づき、選択された。
【0028】
IgGクラス抗体は二価であるため、機能的に関連する重可変ドメイン及び軽可変ドメインからなる2つの完全なFvドメインを有する。二価性は、結合活性作用と、同一又は別個の細胞上の標的抗原又はFc受容体を架橋結合し、それによって、抗体の生物活性に由来する非標的特異的な受容体結合のスペクトルを誘発する能力とを提供する。この理由により、本発明のFc突然変異は「結合活性関連」内で試験され、すなわち、IgGのFc突然変異は表面上で多量体形成され、かつ特定のFc受容体の多量体との相互作用に対して試験された。
【0029】
突然変異の生物学的特徴
Fc含有タンパク質は、いくつかの周知の生体外アッセイによって、機能性を比較され得る。具体的には、Fcγ受容体のFcγRI、FcγRII、及びFcγRIIIファミリーのメンバーに対する親和性が対象となる。これらの測定値は、受容体の組換え可溶型又は受容体の細胞結合型を使用して作製され得る。加えて、IgGの長期の循環半減期に関与する受容体、FcRnに対する親和性は、例えば、組換え可溶性FcRnを使用する「ALPHASCREEN」の、配位子結合型ビーズフォーマットを使用して、測定することができる。高処理量スクリーニングで使用される、ALPHASCREENは、結合等の分子現象の検出を可能にする均質アッセイ技術である。コーティングされた「供与体」及び「受容体」ビーズが、アッセイ技術の基礎である。ビーズに基づくアッセイとして、ALPHASCREENは、大いに増幅された信号を産生するように働く化学反応のカスケードをもたらす、近接近したビーズの相互作用を通して機能する。例えば、競合結合測定といった、直接又は非直接的測定は、タンパク質間の相対親和性及び結合活性を評価するために、適用され得る。
【0030】
ヒトIgGアイソタイプ(例えば、IgG1、IgG2、IgG3、及びIgG4)の自然進化は、それぞれに、抗体依存性細胞傷害(ADCC、例えば、IgG1及びIgG3)、抗体依存性細胞貪食作用(ADCP、例えば、IgG1、IgG2、IgG3、及びIgG4)、及び補体依存性細胞傷害(CDC、例えば、IgG1、IgG3)等の、免疫機能を回復させる能力の異なるスペクトルを示させることを可能にしている。これらの機能のアイソタイプ特異的結合は、別個の免疫細胞上に存在するFc受容体に対する異なる選択性と、C1qに結合し、かつエフェクターマクロファージ上の特定受容体結合補体成分を通じて、CDC及びCDP(補体依存性貪食作用)をもたらす膜攻撃複合体(MAC)のアセンブリを活性化する能力と、に基づく。ヒトアイソタイプの、補体カスケードの初期成分、C1qに結合する能力の優先順位は、IgG1>IgG3>IgG2>IgG4であり、しかし微生物感染においてはIgG2及びIgG4による補体活性化が十分に立証されている。
【0031】
ADCCアッセイ及びCDCアッセイ等の細胞に基づいた機能性アッセイは、特定の変異構造の可能な機能性の結果に関する見識を提供する。抗体依存性細胞媒介性細胞傷害(ADCC)は細胞媒介性反応であり、Fc受容体(FcR)を発現する非特異性の細胞傷害性細胞(例えば、ナチュラルキラー(NK)細胞、好中球、及びマクロファージ)は、標的細胞上に結合した抗体を認識し、その後標的細胞の溶解を引き起こす。1つの実施形態において、ADCCアッセイは、主要エフェクター細胞としてNK細胞を有し、これらの細胞によって発現されることが知られている唯一のFcγ型受容体である、FcγRIIIA上の機能的作用を反映するように構成される。
【0032】
過酸化物又は炎症媒介放出等の細胞応答を測定できるため、貪食作用アッセイを使用して、異なる突然変異の免疫エフェクター機能を比較することもできる。生体内モデルは、Fc変異型の研究において有用であることが証明されている。例えば、抗CD3抗体の突然変異を使用し、マウスにおいて、Fcドメインが、Fcγ受容体等の特異的配位子に関与することに依存する活性であるT細胞活性化を測定することで、実証されている。マクロファージの抗体指向性活性化は、抗体依存性細胞貪食作用(ADCP)を媒介し、オプソニン化された標的細胞がマクロファージによって飲み込まれ消化されるようにする。高レベルのFcRを発現する生体外で分化されたマクロファージは、IFNγ又はGM−CSFを使用して単核細胞に対する全FcR(FcγRI,FcγRIIa,FcγRIIIa)の上昇したレベルを発現する(to expressed)ことによって、M1表現型へと分化され得る。そのようなアッセイは、当該抗体工学分野における技術者にとって既知である。
【0033】
抗体を作製する方法
所定の組換え処理を使用して、Fc突然変異の生成及び試験の出発点として使用されるヒトIgG2定常ドメインに対する、配列内の定方向突然変異が作成された。当該技術分野の技術者は、コード配列内に変化をもたらす種々の技術が、回収及び試験のための種々の宿主細胞における、所望のアミノ酸配列の発現に好適なベクターを作成するために、使用され得るということを理解するであろう。
【0034】
宿主細胞選択又は宿主細胞工学
本明細書で説明されるように、組換えFc含有タンパク質又はモノクローナル抗体の発現のために選択される宿主細胞は、免疫グロブリンCH2ドメイン内でタンパク質を修飾するオリゴ糖部分の組成物における変化を含むが、しかしそれに限定されず、最終的な組成物にとって重要な寄与因子である。したがって、本発明の1つの態様は、所望の治療用タンパク質を発現する産生細胞の使用及び/又は開発のために適切な、宿主細胞の選択を含む。
【0035】
更に、宿主細胞は、哺乳類起源であるか、又はCOS−1、COS−7、HEK293、BHK21、CHO、BSC−1、Hep G2,653、SP2/0、293、HeLa、骨髄腫、リンパ腫、酵母、昆虫又は植物細胞、又はそれらの任意の誘導体、不死化細胞、若しくは形質転換細胞から選択され得る。
【0036】
別の方法としては、宿主細胞は、例えば、天然又は人口の大腸菌種、クレブシェラ菌種、又はシュードモナス菌種等の、原核細胞又は有機体といった、ポリペプチドをグリコシル化する能力がない種又は有機体から選択されてもよい。
【0037】
抗体
本出願で説明される抗体は、非限定的に、ヒト、マウス、ウサギ、ラット、げっ歯類、霊長類、ヤギ、又はそれらの任意の組み合わせ等を含むか、又はそれらに由来することができ、また、単離したヒト、霊長類、げっ歯類、哺乳類、キメラ、ヒト化、及び/又はCDR移植等の抗体、免疫グロブリン、開裂産物及び他の指定部分、並びにそれらの変異を含む。
【0038】
本明細書で説明される抗体、Fc含有タンパク質、又はFc断片は、当該技術分野において周知であるいくつかの方法で、得ることができる。1つの態様において、抗体は、マウス又は他の動物を、標的ペプチド、細胞、又は組織抽出物によって免疫化して調製されるハイブリドーマから、都合よく得られる。抗体はしたがって、当該技術分野において周知である任意のハイブリドーマ技術を使用して得ることができ、例えば参照によって本明細書に全体が組み込まれる、Harlow及びLane、antibodies,a Laboratory Manual,Cold Spring Harbor,NY(1989)を参照されたい。
【0039】
抗体若しくはFc融合タンパク質、又はそれらの成分及びドメインは、例えば、ファージライブラリー等の、そのようなドメイン又は成分のライブラリーから選択し、得ることもできる。ファージライブラリーは、免疫化された動物又はヒトのB細胞からのような、ランダムオリゴヌクレオチドのライブラリー又は目的の配列を含有するポリヌクレオチドのライブラリーを挿入することによって、作成できる(Smith,G.P.1985.Science 228:1315〜1317)。抗体ファージライブラリーは、1つのファージ内に重鎖(H)及び軽鎖(L)の可変領域対を含有し、単一鎖Fv断片又はFab断片の発現を可能にする(Hoogenboomら、2000,Immunol.Today 21(8)371〜8)。ファージミドライブラリの多様性は、ライブラリーのモノクローナル抗体の免疫特異性を増大及び/又は変化させ、追加の望ましいヒトモノクローナル抗体を産生しその後同定するように、操作され得る。例えば、遺伝子を符号化する重鎖(H)及び軽鎖(L)の免疫グロブリン分子は、組み立てられた免疫グロブリン分子における新規のHL対を作成するように、無作為に混合(入れ替え)され得る。更に、遺伝子をコードするH鎖及びL鎖の一方又は双方は、免疫グロブリンポリペプチドの可変領域の相補性決定領域(CDR)において変異誘発され、かつその後に望ましい親和性及び中和能力について篩分けされ得る。抗体ライブラリーはまた、1つ以上のヒトフレームワーク配列を選択し、かつヒト抗体レパートリーから得られるか又は設計変更を通じて得られるCDRカセットの収集を導入することによって、合成的に作成され得る(Kretzschmar及びvon Ruden 2000,Current Opinion in Biotechnology,13:598〜602)。多様性の位置はCDRに限定されないが、可変領域のフレームワークセグメントを含むこともでき、又はペプチド等の抗体可変領域以外を含んでもよい。
【0040】
抗体可変領域以外を含み得る標的結合成分の他のライブラリーは、リボソームディスプレイ、酵母ディスプレイ、及び細菌ディスプレイである。リボソームディスプレイは、タンパク質のRNAとの付着を保ちつつ、mRNAをそれらの同属タンパク質へと翻訳する方法である。核酸コード配列は、RT−PCRによって回収される(Mattheakis,L.C.ら、1994.Proc.Natl.Acad.Sci.USA 91,9022)。酵母ディスプレイは、膜結合αアグルチニン酵母接着受容体の融合タンパク質の構築物aga1及びaga2に基づき、接合型システムの一部である(Broderら、1997.Nature Biotechnology,15:553〜7)。細菌ディスプレイは、細胞膜又は細胞壁に関係している、標的と排出された細菌タンパク質との融合に基づく(Chen及びGeorgiou 2002.Biotechnol Bioeng,79:496〜503)。
【0041】
ハイブリドーマ技術との比較において、ファージ及び他の抗体ディスプレイ方法は、生体外の抗原標的に対する選択を操作する機会を与え、かつ宿主の抗原への作用の可能性を制限せず逆もまた同様である。
【0042】
本発明はまた、単離したポリヌクレオチドとして、又は原核細胞、真核性、又は繊維状のファージ発現、組成物の分泌及び/又はそれらの定方向突然変異誘発物質又は組成物のディスプレイと適合するベクターを含む発現ベクターの部分として、本発明の組成物をコードするする核酸を提供する。
【0043】
Fc含有分子の使用
上述された方法のうちのいずれかによって生成される組成物(抗体、Fc融合体、Fc断片)は、ヒトの疾患、又は細胞、組織、器官、体液、若しくは概して宿主における特定の病理を、診断、治療、検出、又は調節するために使用され得る。本明細書で教示されるように、Fcγ受容体結合及び指定のエフェクター機能を低減又は除去するために、抗体、Fc融合タンパク質、又はFc断片の、Fc部分を修正することは、抗体が元の標的指向化性質を保持するが、優れた活性スペクトル、生物物理学的性質、安定性、及び宿主の身体内に持続させる能力を有する抗体及びFc構築物を提供する。
【0044】
本発明によって提供される組成物を用いた治療に使用できる疾患又は病理としては、例えばアルツハイマー病であるが、それに限定されない神経性疼痛を含む、神経疾患;皮膚病;代謝病;変形性関節炎;及び火傷又は怪我から生じる症状;心筋梗塞、うっ血性心不全、脳卒中、虚血性脳卒中、及び出血を含むがそれに限定されない、心臓血管疾患;並びにリウマチ性疾患、乾癬、及び強皮症を含む一般的な免疫介在性疾患が挙げられるが、それらに限定されない。
【0045】
本発明は一般論として記述されてきているが、本発明の実施形態は、特許請求の範囲を限定するように解釈されるべきではない以下の実施例で更に開示される。
【実施例】
【0046】
実施例1.Fc突然変異の構築及び試験
表4に示されるヒトIgG2抗体に由来する突然変異を伴う一連の構築物は、標準的組換え方法を使用して構築された。完全な可変ドメインを伴う抗体に対して、抗HER2及び抗CD20抗体の既知のCDR配列が使用され、示されたようなアイソタイプ及びFc突然変異が構築された。抗体突然変異は、標準的クローン化及び発現手順を使用し、293T細胞内に一時的に発現された。MAbは、次の実験的分析の前に、タンパク質Aカラムを使用して、95%を超える均質性まで浄化された。
【0047】
【表5】
【0048】
親和性のビアコア(Biacore)研究
Biacore 3000光学バイオセンサー(Biacore AB,Uppsala,Sweden、現在は、GE Healthcareの一部である)を使用して、表面プラズモン共鳴実験が実行された。実験は、3 mMのEDTAと0.01%の界面活性剤P20とを含有するD−PBS緩衝剤中、摂氏25度で実行された。受容体とFc突然変異との相互作用を分析するために、マウスの抗His IgG(R&D systems cat#MAB050)をCM−5センサーチップへ共有結合させることによって、捕捉表面が生成された。抗His Abは、10mMの酢酸ナトリウム緩衝剤pH 4.5(Biacore AB)の中に希釈され、アミン連結化学剤製造者の説明書を使用し、CM−5チップ(〜3000 RU)のカルボキシメチル化デキストラン表面と連結された。表面上に残留する反応基は、エタノールアミンHCLを使用して非活性化された。動力学的実験を実行するために、FcγRI、FcγRII、及びFcγRIIIの、165、351、及び208反応単位(RU)がそれぞれ、この表面上で捕捉された。受容体捕捉の後に、野生型又はFc突然変異の一連の希釈物が(4倍希釈工程で4000nMから3.9nMまで)、30uL/分で注入された。会合相を3分間観察した。その後に、20分間の緩衝剤流入が続き、結合解離を観察した。捕捉表面は、100mMのリン酸の9秒パルスを100uL/分で使用し、その後の移動緩衝剤の注入によって再生された。
【0049】
参照のために、Scrubberソフトウェアバージョン1.1g(BioLogic Software)を使用して、データの二重参照減算(double reference subtraction)を実行し、信号及び機器ノイズ(Myszka 1999)への緩衝剤寄与を修正した。この初期データ処理の後、単純1:1結合モデルを仮定とし、BIA評価ソフトウェア、バージョン4.0.1(Biacore、AB)を使用して、データの動力学的分析を実行した。AlphaScreen及び結合研究。
【0050】
種々のFcγRとのIgGの競合結合及び直接結合の双方が、均質ビーズ系結合アッセイ、AlphaScreen(商標)(PerkinElmer,Waltham,MA)を使用して評価された。手短に言うと、わずかな修正を伴って、以前に説明されたように実験が実行された(Lazarら、2006 Proc Natl Acad Sci USA 103(11):4005〜10)。FcγRI、IIaは、R&Dシステムから取得された。FcγRIIIa及びFcRnは、クローン作製、発現、及び浄化された。IgGFc突然変異が、NLSビオチン、Pierce、2:1比率を使用してビオチン化された、CNT06234(非特異的制御ヒトIgG1サブクラス抗体)、又は抗Her2/neu(ヒトIgG2抗体)に対する競合結合について、試験された。
【0051】
競合結合研究において、ビオチン化抗体が、200ng/mLの最終アッセイ濃度に添加され、次にそれぞれの実験図に規定されている指定された最終濃度に、競合試験抗体が添加された。FcγRが、200ng/mL最終濃度で96穴プレートへ添加され、次に、ストレプトアビジン供与体及びNiキレート受容体ビーズが連続的に添加された。プレートを密封し室温で振盪した後、プレートはEnvision plate readerを使用して読み取られ、GraphPad Prismにおいてデータがグラフ化されプロットされた。結合活性研究は、試験IgG分子が2:1比率でビオチニル化され、かつ直接FcR結合が競合の不在のもとに対照抗体に対して評価されたということ以外は、競合研究と同様に実行された。
【0052】
結果
SPR/Biacore分析によって評価され、センサグラムから得られたヒトFcR(FcγRI、FcγRIIa、FcγRIIb、及びFcγRIIIa)に対するIgG変異型の相対親和性を、表5に示す。
【0053】
【表6】
【0054】
これらの数字は、1つの実験のグローバル・フィットのために生成されたパラメーターに対応する。
【0055】
これらの4つのデータセットに対して、単純1:1結合モデル・キネティック・フィットを使用してフィッティングを実行することによって、親和性が獲得された。他の全てに対して、単純1:1結合定常状態親和性解析を使用してフィッティングを実行することによって、親和性が獲得された。
【0056】
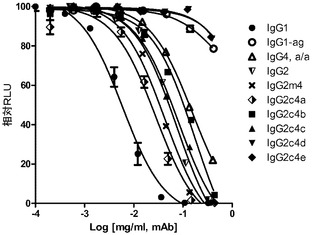
ヒトIgG1(CNTO6234)を用いたAlpha Screen(商標)競合アッセイを使用して測定された、結合活性関連におけるヒトFcR(FcγRIIa、FcγRIIb、及びFcRn)に対する、すなわち高密度の標的分野に結合する二価抗体に対するIgG突然変異の相対親和性を、図3A〜Cにそれぞれ示す。
【0057】
実験データは、IgG1 ag及びIgG4 Ala/Alaと同様に、IgG2突然変異が、FcRn(生体内半減期をもたらす新生児Fc受容体)と結合するそれらの能力を保持しつつ、IgG1と比較して大幅に減少したFcγ受容体との結合親和性を示すということを実証した。具体的には、IgG2に対する競合結合における、また高親和性から最低親和性まで順位付けされたFcgRIIaとの結合におけるその順序は、以下の通りである。IgG1>IgG2c4a>IgG2m4>IgG2=IgGc4c>IgG2c4b>IgG4ala/ala>IgG4agly>IgG2c4d。この順序は、FcgRIIbとの結合におけるIgG1(CNTO6234)に対する競合において、一致している。pH 6.4でのFcRnとの結合における、IgG1(CNTO6234)に対する更なる競合結合分析は、全てのアイソタイプ及び突然変異がFcRnと比較的均等に結合することを示した。重要なことには、IgG2c4d及びIgG1 aglyは、仮にあったとしても、FcgRIIa及びFcgRIIbとで最小の検出可能な結合を示す。
【0058】
実施例2:ADCC及びCDC
CDCは、3つの経路カテゴリーにおいて始まる。抗体依存性(古典経路)、多糖類依存性(レクチン依存性)、及び外来性表面構造(代替経路)であり、その全ては、タンパク質分解工程のカスケードを産生し、ついには標的細胞又は微細物の溶解に至る膜攻撃複合体のアセンブリをもたらす(W.E.Paul Immunologyを参照)。実験1で説明されたアイソタイプ及びFc突然変異の部分集合が、抗CD20抗体の可変ドメインを使用して調製され、ヒト血清存在下におけるWIL2−S B細胞リンパ腫(lymophoma)標的細胞を溶解させる能力が評価された。WIL2−S細胞のCDCを媒介することで知られる治療用抗CD20である、市販のRituxan(登録商標)が、溶解に対する陽性対照として使用された。CDC分析のために、Wil2−S標的細胞が96穴プレート内に播種され、ヒト血清補体(1/6希釈)とともに培養され、AlamarBlueを使用して相対的細胞生存率が評価された。
【0059】
ADCCアッセイは、実験1に説明されたように、またSkBr3乳癌細胞を標的として、異なるIgGアイソタイプ又は変異型の種々のFcドメインと組み合わされた、抗HER2/neu可変ドメインを使用して実行された。アッセイは、以前に説明されたように、細胞溶解検出のEuTDA法(PerkinElmer,Waltham,MA)を使用して実行された。TDAを負荷したSkBr3乳癌標的細胞が、U−bottom 96穴プレートに播種され、指定の抗体濃度でオプソニン化され、また37℃でロイコパックから単離された25倍過剰のPBMCとともに共培養された。3時間後、プレートが遠心分離され、製造者の説明書に従って上清がTDA放出について分析された。生データが正規化され、GraphPad Prismを使用してプロットされた。CDC分析のために、Wil2−S標的細胞が、96穴プレート内に一穴あたり50,000個で播種され、ヒト血清補体(1/6希釈)とともに培養され、AlamarBlueを使用して相対的細胞生存率が評価された。
【0060】
結果
NK細胞上のFcgRIIIaに主として関わるADCCは、IgG2、IgG2m4、IgG2c4d及びe、IgG1 agly、並びにIgG4 ala/ala(図5)を含む、試験された一連の構築物に関して非検出可能であり、AlphaScreen(商標)アッセイ(図4A)における結合活性の欠損と同様に、より高い親和性FcgRIIIa(V)受容体に見られる減少した結合特性と一致した。乳癌細胞上のHER2/neu等の高密度標的の使用は、FcgRIIIa(40uMより低い)に対するFc親和性が標的細胞溶解を誘発するには不十分にしか出現しないということを実証した。同様に、IgG1を除いて、抗体又はFc突然変異のいずれも、試験された抗CD20構築物内のWIL2−S標的細胞に対する、有意なCDCレベルを示さなかった(図6)。試験されたIgGサブクラス及び突然変異のうち、IgG1のみが、検出可能なCDCレベルを示し、これは、C1q結合とは無関係に、残留する突然変異のいずれもがCDCを引き起こすことはできないということを示唆している(図6)。先行の試みにより、IgG2は、C1q結合及び古典経路の活性化を通じて最小のCDCを有すると指摘されているが、我々は、先行文献の観察(Idusogie,Prestaら、2000 J Immunol 164(8):4178〜84)と一致する抗CD20関連において、有意な活性レベルを観察しなかった。更に、抗CD20及びヒト血清を用いて、残留補体活性を有するIgG1 aglyの先行文献の指摘も検出されなかった(Dorai,Muellerら、1991 Hybridoma 10(2):211〜7)。この矛盾に対する説明は、低いレベルの補体活性化がIgG2によって媒介され得るが、活性化は、オプソニン化された細胞を溶解させるのに十分な膜攻撃複合体(MAC)のアセンブリを始動させるには不十分であるかもしれない、ということである。
【0061】
これらのデータは、補体活性化のレベルにかかわらず、ついには標的細胞溶解に至る膜攻撃複合体のアセンブリが、結合型IgG1以外のアイソタイプの抗体に対しては、欠乏しているか又は不十分であるということを示唆する。
【0062】
実施例3:抗体依存性細胞貪食作用
抗HER2/neu結合Fc突然変異が、オプソニン化された標的乳癌細胞であるSk−Br3及びマクロファージを使用して、抗体依存性細胞貪食作用(ADCP)を媒介する能力について評価された。
【0063】
抗体依存性細胞貪食作用(ADCP)
末梢血単核球が、標準Ficoll−Paque(GE Healthcare)密度勾配調製によってロイコパック(Biological Specialty Corporation)から単離され、細胞が等分され、かつ窒素内で貯蔵された。PBMCは解凍され、製造者の説明書に従って、CD14陽性細胞が、CD16枯渇を伴わないCD14 Isolation kit(Stem Cell Technologies)を使用する陰性除去(negative depletion)によって単離された。細胞は、0.1×106細胞/cm2で、20ng/mLのGM−CSF(R&D Systems)の存在下で、RPMI/5%の加熱不活性化したFBS/50μg/mLゲンタマイシン内で、7日間平板培養され、単核細胞由来マクロファージが生成された。SK−BR−3腫瘍細胞は、製造者の説明書に従って、PKH67(Sigma)で標識付けされた。標的細胞は洗浄され、かつ4エフェクター細胞に対して1標的細胞の比率で単核細胞由来マクロファージを用いて、抗体の存在下で、37℃で4時間5% CO2培養器内で培養された。培養後、細胞はAccutase(Millipore)によって分離され、またマクロファージは、AlexaFluor−647(Invitrogen)と複合化された抗CD11b抗体(BD Biosciences)で標識付けされた。フローサイトメトリーによって細胞が分析され、腫瘍細胞単独(PKH67pos、CD11bneg)、マクロファージ単独(PKH67neg、CD11bpos)、及び貪食された腫瘍細胞(PKH67pos、CD11bpos)が測定された。パーセント貪食作用が、次の等式によって決定された。(貪食された腫瘍細胞)/(貪食された腫瘍細胞+腫瘍細胞単独)×100%。細胞は、FACS Calibur(Becton Dickinson)上に取得され、その結果はFloJo Software(Tree Star)によって分析された。
【0064】
単離した単核細胞が、GM−CSFを使用して生体外で分化され、更に、フローサイトメトリック分析によってFcRの発現レベルについて特徴付けられた。他者の先行研究で述べられているように、GM−CSF活性化マクロファージは、親単核細胞に対して、全FcR(FcγRI、FcγRIIa、FcγRIIIa)の上昇したレベルを発現した(データは図示せず)。抗HER2/neu IgG Fc突然変異構築物は、その後、M1マクロファージを使用して貪食作用アッセイで試験された。
【0065】
結果
それぞれのmAbの存在下での、SkBr3細胞を伴う4時間の共培養後、かなりのレベルのADCPがIgG1に対して出現したが、しかしながら、非グリコシル化IgG1、IgG2、IgG2m4、及びIgG4 S>P ala/alaに対しては、より高濃度の抗体において最小レベルのADCPも観察された(図7)。対照的に、IgG2c4dは、検出可能なレベルのADCPを示さなかった。この観察結果は、以前に実証されたFcRに対する種々のIgGの結合プロファイルと一致している。例えば、IgG1、IgG agly、及びIgG4 ala/alaは、多数の配位子ディスプレイビーズ(例えば、AlphaScreen(登録商標)システムを使用して)の使用においてFcγRIとの結合を実証したが、これら3つの全てはADCPも実証した。IgG2及びIgG2M4及びIgG4 ala/alaは、より高濃度で最小のADCPを示した。IgG2及びIgG2m4によるFcγRIIaの結合は、FcγRIIaの寄与を示唆する結合活性研究によって示されるように、それ自体としては、有意なレベルのADCPを生じさせるには不十分であるかもしれない。BiaCore及びAlphaScreen(商標)の結果(表5、並びに図4B及び4C)は更に、IgG aglyがFcγRIとの結合を保持し、IgG4 ala/alaはFcγRI及びFcγRIIaの双方との結合活性を示すが、しかしADCPは、IgG1 aglyに対してIgG4 ala/alaよりも幾分強固であるということを示した。IgG1 aglyは、FcγRIIaに対し、また更には高度に類似した阻害物質FcγRIIb(細胞外ドメイン(doman)内での、配列同一性>95%に基づく)に拡大して、仮にあったとしても最小の結合性を有するため、FcγRI信号伝達を通してのITAMの活性化は、FcγRIIb活性化に関連するITIMを通しての信号伝達によって相殺されない。対照的に、IgG4 S>P ala/alaは抑制された貪食作用を示し、FcγRIIbの活性化に起因する可能性が高い。最後に、単量体又は結合活性に基づく、種々のFcRとのIgG2c4dの検出可能な結合の完全な欠損は更に、このFc主鎖の固有の無効にされた貪食能力を立証する。
【0066】
実施例4:抗体媒介性サイトカイン放出
免疫細胞上のFcRのFc結合は、架橋結合されるとサイトカイン放出を促進する。免疫細胞上のFcRの結合活性系結合を模倣するために、mAbがポリスチレンビーズへ結合された。
【0067】
抗HER2/neu IgG突然変異を使用したサイトカイン放出が、IgGをラテックスビーズへ直接結合した後、一晩培養してから、実行された。洗浄されたビーズが、1mLあたり約1500個〜250,000個の指定の種々の濃度で単離したヒトPBMCへ添加され、一晩培養した後、共培養上清を除去し、PerkinElmer(Waltham,MA)からの標準AlphaELISAキットを使用して、分泌されたTNFαが測定された。
【0068】
試験されたIgGアイソタイプ及びFc突然変異は分化能力を有し、PBMCからのFc受容体媒介性TNFα分泌を通じてサイトカイン放出を刺激する(図8)。従って、種々のアイソタイプ及びそれらのFc突然変異による、高レベルから低レベルまでのTNF分泌のレベルは、以下の通りである。IgG1>IgG2>IgG2m4>IgG2c4a>IgG1agly>IgG4ala/ala>IgG2c4b>IgG2c4c>IgG4c4d及びe留意すべき点として、IgG4d及びeFc突然変異の双方は、仮にあったとしても、検出可能なカイトカイン(TNF)放出を誘発する最小の能力を持つ。
【0069】
実施例5:生体外B細胞枯渇
生体内での発現抑制レベルに関するアイソタイプ及び突然変異の能力をより良く理解するために、ヘパリン化ヒト全血液の存在下で、WIL2−S B細胞の生体外枯渇が測定された。抗CD20 IgG1は、全てのエフェクター機能(ADCC、CDC、ADCP)に関与することが知られているため、PMN(好中球、好塩基球等)、ヒト補体、及び過剰IgGの存在を含む全血液システムは、それぞれの変異型によって与えられる「発現抑制」のレベルを表していると見なされた。
【0070】
手短に言うと、ヒト全血液は、ADCCアッセイのための標的細胞の機能を果たすエフェクター細胞及びWIL2−S細胞を提供した。標的細胞は、BATDA(PerkinElmer)を用いて摂氏37度で30分間事前標識付けされ、2回洗浄されDMEM/5%の加熱不活性化したFBS内で再懸濁され、その後50μLの標的(1穴あたり2×104細胞)が、96穴U−bottomプレートの穴へ添加された。種々の濃度の抗体を伴って又は伴わないで追加の50μLが添加され、細胞は室温で20分間培養された。次に、ヒト全血液(100μL)が穴へ添加された。全ての試料は3組で実行された。プレートは、200gで3分間遠心分離され、5% CO2培養器において摂氏37度で3時間培養され、次に、200gで3分間再び遠心分離された。1穴あたり合計20μLの上清が除去され、細胞溶解が、200μLのDELPHIA Europium系試薬(PerkinElmer)の添加によって測定された。Envision 2101 Multilabel Reader(PerkinElmer)を使用して、蛍光性が測定された。データは、Triton X−100(Sigma Aldrich)で細胞を処理することによって得られた最大の細胞傷害に対して正規化され、また標的細胞単独からのBATDAの自然放出によって最小対照が決定された。データは、GraphPad Prism v5.01を使用して、S字状用量−応答モデルにフィッティングされた。
【0071】
ヒト血液の存在下でのWIL2−Sの共培養は、IgG1を使用して、またある程度はIgG2及びIgG2M4によって、標識付けされたWIL2−Sのエフェクター媒介による重度の枯渇を明らかにした。留意すべき点として、(Fab’)2及びFab抗CD20断片の双方は、エフェクター機能を回復する能力がある血清内の開裂IgG自己抗体(例えば、Breskiら、2008 J Immunol 181:3183〜3192を参照)の存在を示し得る、あるレベルのWIL2−S枯渇を誘発した。先行のADCC、CDC、及びADCPデータと一致して、IgG2c4eに伴って、大幅な又は検出可能なレベルのサイトカイン放出は、観察されなかった。
【0072】
実施例6:CD20標的化を伴う生体内B細胞生存
IgG2c4eに関連する生体内エフェクター機能が、抗CD20の確立されたカニクイザルB細胞(Cynomologous B-cell)枯渇モデル(Reffら、1994,Blood 83:435〜445)を使用して評価された。
【0073】
カニクイザル(Cynomologous monkeys)(n=3/群)は、0.2mg/Kg又は2mg/Kgでの、IgG1か又はIgG2Σかの単回ボーラス静脈内投与の7日前に、生理食塩水を注入された。注入後の所定の日に、全血液試料からのB細胞レベルが、抗CD20及び抗CD3をそれぞれマーカーとして使用するB及びT細胞のフローサイトメトリー分析によって測定された。それぞれの群に対する平均B細胞レベル(CD20+/CD3−)が、注入後3週間の間プロットされた(図9)。IgG1の低投与量(0.2mg/kg)は、注入後1日で全B細胞の完全に近い枯渇(99%)を誘発したのに対し、抗CD20 IgG2c4eによっては有意な枯渇は誘発されなかった(群内平均15%)。B細胞レベルは、抗CD20 IgG2c4eで処理された動物に対しては、後続の数日に渡って相対的に正常なままであり、またIgG1で処理された動物に対しては、後続の数週間の間で、徐々に回復へと向かうB細胞レベルの傾向が観察された。留意すべき点として、IgG2c4e及びIgG1の双方とも、2mg/Kgのより高い投与量では、完全に近いB細胞枯渇を誘発した。
【0074】
抗CD20媒介性B細胞枯渇は、ADCC、CDC、及びアポトーシスを含むいくつかのメカニズムによって媒介されると考えられる。IgG2c4eによる、より高い投与量(2mg/Kg)でのB細胞の枯渇を示すサルのB細胞枯渇データを考慮し、単離したB細胞内の抗体によって誘発されるアポトーシスのレベルを測定することによって、B細胞枯渇の基本メカニズムが更に評価された。
【0075】
単離したB細胞は、0、0.26、2.6、及び26μg/mL濃度のIgG1、IgG2c4e、(Fab’)2、及びIgG2と、非結合性の対照mAb(BM21)とで、4時間処理され、Annexin V陽性及び7AAD陰性細胞が、フローサイトメトリーによって定量化された。0.2及び2mg/Kgのボーラス注入後の、概算の最大生体内血清IgG濃度を反映するように、2.6及び26μg/mLの特定の最終濃度が選択された。
【0076】
これらの3つの全ての結合抗体に対して、アポトーシスの投与量依存性誘発が、(Fab’)2を含む全てのIgGに対して観察され、抗CD20媒介性架橋結合は、より高い投与量での細胞死滅の誘発に十分であるが、しかし、IgG2c4eに対するより低い投与量では十分ではないことが示された。留意すべき点として、(Fab’)2はまた、Fcの不在下でも有意なアポトーシスを示し、抗IgG媒介性アポトーシスは、以前に観察されたように、Fc媒介性架橋結合とは無関係に誘発され得るという考えを裏付けた。
【0077】
したがって、細胞上の標的抗原の架橋結合に関連する正常な機能は、修飾されたFc変異型によって除去されない。
【技術分野】
【0001】
本発明は、Fcγ受容体に特異的に結合する能力か、又は表面標的抗原のFc受容体媒介性架橋結合によって免疫細胞による分裂応答を活性化させる能力を実質的に失うように突然変異される、ヒト抗体IgG2定常領域(Fc領域)に関する。本発明は、突然変異IgG2定常領域が組み込まれ得る、新規の抗体も提供する。
【背景技術】
【0002】
細胞表面抗原を標的とする抗体は、不必要な免疫活性化、及び免疫細胞とのFc受容体(FcR)結合と補体の活性化とに関連するエフェクター機能を引き起こす。標的配位子機能を標的にし、かつ活性化又は中和するが、必要な局所的細胞又は組織を傷つける又は破壊することのないように意図された治療用抗体及びFc融合構築物として、縮小された(除去された)エフェクター機能を伴うFc突然変異が模索されている。
【0003】
ヒトIgGアイソタイプ(成熟γグロブリンクラスG抗体のサブクラス、IgG1、IgG2、IgG3、及びIgG4)は、例えば、抗体依存性細胞傷害(ADCC、例えば、IgG1及びIgG3)、抗体依存性細胞貪食作用(ADCP、例えば、IgG1、IgG2、IgG3、及びIgG4)、及び補体依存性細胞傷害(CDC、例えば、IgG1、IgG3)といった免疫機能を回復させる、差異的な能力を示す。そのような免疫機能のアイソタイプ特異的結合は、別個の免疫細胞上のFc受容体に対する選択性と、C1qに結合し膜攻撃複合体(MAC)のアセンブリを活性化する能力とに基づく。種々のアイソタイプの中で、Fcγ受容体(例えば、FcγRI、FcγRIIa/b/c、FcγRIIIa/b)に対する相対的親和性は、IgG1及びIgG3に対して高いが、しかしながら、IgG2に対しては親和性が最小であり(FcγRIIa 131H多型に限定される)、またIgG4はFcγRIに対してある程度の親和性しか有さない。比較配列分析及び共結晶構造を用いて、受容体結合に対して主に接触する残基は、より低いヒンジ及びCH2領域にまたがるアミノ酸残基にマッピングされている。標準的タンパク質工学技術を用いて、Fc受容体及びC1q補体成分に対する抗体調製物の親和性を強化又は低減することにおいて、いくらかの成功が収められている。
【0004】
アイソタイプの中で、IgG2はFc受容体のファミリーに結合する能力が最少である。IgG2を起点として、減少されたエフェクター機能を伴いつつ、しかしFcRn結合、長期安定性、及び低免疫原性を保持する突然変異誘発物質を見出す試みがなされている。この類の改善された突然変異は、安全性を保持しつつ、改善された抗体治療薬を提供することができる。
【発明の概要】
【課題を解決するための手段】
【0005】
本発明は、例えば、FC領域を含有し細胞表面配位子を標的とするような、抗体又は抗体様治療薬の技術作業において有用な、修飾されたグリコシル化免疫グロブリン定常ドメインを提供する。本発明の組成物は、減少されたFcγR結合能力を示すが保存されたFcRn結合を有する、IgG 2Fc突然変異である。これらのIgG Fc突然変異は、免疫エフェクター機能のFc関連結合及び補体媒介性細胞傷害を最小にしつつ、可溶性又は細胞表面抗原を治療標的化することを可能にする。1つの態様において、IgG 2Fc突然変異は、EU番号付け方式に従うV234A、G237A、P238Sを含む。別の態様において、IgG2 Fc突然変異は、EU番号付け方式に従うV234A、G237A、H268Q、又はH268A、V309L、A330S、P331Sを含む。特定の態様において、IgG2 Fc突然変異は、EU番号付け方式に従うV234A、G237A、P238S、H268A、V309L、A330S、P331S、及び任意でP233Sを含む。
【0006】
1つの実施形態において、IgG2 Fc突然変異組成物は、i)抗体依存性細胞傷害(ADCC)、ii)補体依存性細胞傷害(CDC)、iii)抗体依存性細胞貪食作用(ADCP)、iv)FcR媒介性細胞活性化(例えば、FcR架橋結合を介するサイトカイン放出)、及びv)FcR媒介性血小板活性化/枯渇といった、免疫及びエフェクター機能に関連するFcγRの活性化に由来する潜在毒性が、最小化又は排除されながら、治療用抗体(又はFc融合体)半減期の保持がFcRnとの相互作用を通して保存される効能において、使用される。1つの態様において、IgG2 Fc突然変異は、例えば基底細胞神経節といった神経疾患に関与する細胞上の配位子;B細胞又はT細胞活性化に、あるいは線維芽細胞又は幹細胞といった組織修復又は治癒に関与する細胞に関係する免疫系疾患;を標的とする、多価結合剤等の結合剤の治療用抗体又はFc融合体に組み込まれる。
【0007】
別の実施形態において、IgG2 Fc突然変異は医薬組成物を含む。別の実施形態において、IgG2 Fc突然変異は薬剤活性分子の一部を含む。IgG2 Fc突然変異又は活性IgG2 Fc突然変異含有分子を含む医薬組成物は、マクロファージ又は単核細胞の遊走及び集結によって特徴付けられる疾患の治療に有用である。1つの態様において、IgG2 Fc突然変異含有分子は、神経組織、内分泌組織、血管組織、心臓組織、滑膜組織、皮膚組織、又は粘膜組織内の標的に結合するのに有用である。本発明のIgG2 Fc突然変異の多くの使用のうちの1つは、移植片対宿主疾患;宿主対移植片疾患;臓器移植拒絶反応;骨髄移植拒絶反応;血管炎、自己免疫性溶血性貧血、自己免疫性血小板減少症、及び関節炎等の自己免疫;胎児/新生児同種免疫性血小板減少症等の同種免疫;ぜんそく及びアレルギー;クローン病又は強皮症等の慢性若しくは急性炎症性疾病;アルツハイマー病、冠状動脈閉塞症、の治療におけるものである。
【図面の簡単な説明】
【0008】
【図1】それぞれの残基に対応するEU番号付けを示す、野生型ヒトIgG2(配列番号:1)、IgG4(配列番号:2)、及びIgG1(配列番号:3)のアミノ酸配列の整列を示し、IgG2のヒンジ領域はEU残基218で始まる。
【図2】修飾された残基の表面位置(EU番号付け)を示す、Fc断片の構造を示す。
【図3A】それぞれのアイソタイプと、AlphaScreenビーズアッセイプラットフォームを使用して、ヒトIgG1野生型アイソタイプの抗体との抗Her2/neu結合抗体として構築した突然変異との、競合を示すグラフである。FcgRI(図示せず)、FcgRIIa(A)、FcgRIIb(B)、FcgRIIIa(図示せず)、及びFcRn(C)。
【図3B】それぞれのアイソタイプと、AlphaScreenビーズアッセイプラットフォームを使用して、ヒトIgG1野生型アイソタイプの抗体との抗Her2/neu結合抗体として構築した突然変異との、競合を示すグラフである。FcgRI(図示せず)、FcgRIIa(A)、FcgRIIb(B)、FcgRIIIa(図示せず)、及びFcRn(C)。
【図3C】それぞれのアイソタイプと、AlphaScreenビーズアッセイプラットフォームを使用して、ヒトIgG1野生型アイソタイプの抗体との抗Her2/neu結合抗体として構築した突然変異との、競合を示すグラフである。FcgRI(図示せず)、FcgRIIa(A)、FcgRIIb(B)、FcgRIIIa(図示せず)、及びFcRn(C)。
【図4A】それぞれのアイソタイプと、AlphaScreenビーズアッセイを使用して、抗Her2/neu結合抗体として構築した突然変異との、直接結合を示すグラフである。FcgRIIIa(A)、FcgRI(B)、及びFcgRIIa(C)。
【図4B】それぞれのアイソタイプと、AlphaScreenビーズアッセイを使用して、抗Her2/neu結合抗体として構築した突然変異との、直接結合を示すグラフである。FcgRIIIa(A)、FcgRI(B)、及びFcgRIIa(C)。
【図4C】それぞれのアイソタイプと、AlphaScreenビーズアッセイを使用して、抗Her2/neu結合抗体として構築した突然変異との、直接結合を示すグラフである。FcgRIIIa(A)、FcgRI(B)、及びFcgRIIa(C)。
【図5】ADCCアッセイの結果、及び選択されたアイソタイプと、抗Her2/neu結合抗体として構築し、またヒトPBMC(25倍過剰)とSK−Br3乳癌細胞とを標的として用いた突然変異と、を示す。
【図6】ヒト補体とWIL2−Sリンパ腫細胞とを標的として用いた、選択された抗CD20構築物の、CDCアッセイの結果を示す。
【図7】Sk−Br3標的細胞とGM−CSF分化マクロファージとを用いた、選択された抗Her2/neu構築物の、フローサイトメトリー分析からのADCPアッセイの結果を示す。
【図8】IgG2野生型及び6つの突然変異を含む、種々の構築物に対するIgGアイソタイプとのビーズ結合を伴うPBMCの刺激後24時間からの、サイトカイン(TNFα)放出を示すグラフ。
【図9】種々のFc領域を伴う、又はFc領域(Fab’)2が欠損する抗CD20結合ドメイン抗体構築物を注射されたカニクイザル群(Cynomologous monkeys)における、時間経過に伴う平均循環B細胞数のグラフ。
【0009】
【表1】
【発明を実施するための形態】
【0010】
略語
ADCC=抗体依存性細胞傷害、ADCP=抗体依存性細胞貪食作用、CDC=補体依存性細胞傷害、IgG=免疫グロブリンG、ITAM=免疫受容体チロシン活性化モチーフ、ITIM=免疫受容体チロシン阻害モチーフ、Mab=モノクローナル抗体、FDCR=Fc依存性サイトカイン放出、FcγR、FcgR、又はFcγR=Fcγ受容体。
【0011】
用語の定義&説明
「抗体依存性細胞媒介性細胞傷害」又は「ADCC」とは、細胞傷害性細胞(例えばナチュラルキラー(NK)細胞、好中球及びマクロファージ)上に存在するFc受容体(FcR)上に結合した、分泌されたIgが、これらの細胞傷害性エフェクター細胞が抗原を有する標的細胞に特異的に結合してその後標的細胞を細胞毒で殺傷することを可能にするような、細胞傷害の形態を意味する。標的細胞の表面を対象とする配位子特異的高親和性IgG抗体は、細胞傷害性細胞を刺激し、またそのような死滅に絶対に必要である。標的細胞の溶解は細胞外であり、直接の細胞間接触を必要とし、また補体に関与しない。
【0012】
任意の特定の抗体の、ADCCによる標的細胞の溶解を媒介する能力が測定され得る。ADCC活性を評価するために、目的の抗体は、標的細胞の細胞溶解をもたらす抗原抗体複合体によって活性化され得る免疫エフェクター細胞と共同して、標的配位子を示す、標的細胞に添加される。細胞溶解は、通常、溶解した細胞からの標識(例えば、放射性基質、蛍光染料、又は天然細胞内タンパク質)の放出によって検出される。そのようなアッセイに有用なエフェクター細胞としては、末梢血単核球(PBMC)及びナチュラルキラー(NK)細胞が挙げられる。生体外ADCCアッセイの具体的な例は、Wisecarverら、1985,19:211、Bruggemannら、1987,J Exp Med 166:1351、Wilkinsonら、2001,J Immunol Methods 258:183、Patelら、1995,J Immunol Methods 184:29に説明される(これらのそれぞれは、参照によって組み込まれる)。別の方法として、又は追加として、目的の抗体のADCC活性は、生体内で、例えば、その内容が参照によってその全体において組み込まれるClynesら、1998,PNAS USA 95:652に開示されるような動物モデルにおいて、評価されてもよい。
【0013】
「補体指向性細胞傷害」又はCDCは、補体カスケードが、抗体Fcに結合する補体成分C1qによって活性化される、細胞傷害の形態を指す。
【0014】
本明細書で使用するとき、「Fc」、「Fc含有タンパク質」又は「Fc含有分子」という用語は、少なくとも免疫グロブリンCH2及びCH3ドメインを有する単量体、二量体、又はヘテロ二量体タンパク質を指す。CH2及びCH3ドメインは、タンパク質/分子(例えば、抗体)の二量体領域の少なくとも一部を形成することができる。
【0015】
本明細書で使用される場合の「モノクローナル抗体」という用語は、動物抗体の少なくとも1つの種の重鎖又は軽鎖抗体可変ドメインのうちの少なくとも1つに対して実質的に相同性を保持する、少なくとも1つの配位子結合ドメインを含む、Fc含有タンパク質の特定の形態である。
【0016】
「野生型ヒトIgG2 Fc領域」とは、カバットのEU番号付けに従う、ヒトIgG重鎖の残基K218から残基K447までである、配列番号:1のアミノ酸配列又はその断片を有する、ヒトIgG Fc領域を指す。定常領域内のアミノ酸は、ヒトIgG1抗体、EUIgG1(配列番号:3)との整列によって番号付けされる(Cunninghamら、1970、J.Biol. Chem.,9:3161〜70を参照)。すなわち、抗体の重鎖及び軽鎖は、アミノ酸配列同一性を最大化するようにEUの重鎖及び軽鎖と整列され、抗体内のそれぞれのアミノ酸は、EUにおける対応するアミノ酸と同じ番号を割り当てられる。EU番号付け方式は、当該技術分野において従来使用されている(概略的には、Kabatら、Sequences of Protein of Immunological Interest,NIH Publication No.91−3242,US Department of Health and Human Services(1991)を参照)。この慣習に従って、説明される「野生型IgG2」定常領域は、236位でアミノ酸を欠損する(図1、配列番号:1)。
【0017】
概論
本発明は、サイトカイン放出を引き起す能力がない、又は標的配位子を示し標的とされる細胞を囲む細胞及び組織を損傷若しくは死滅させる能力がないという観点から改善された安全性を有する、治療用抗体、Fc融合体、及び類似のバイオ医薬品生物薬剤の製造に使用するためのFcドメインを同定することへの関心によって動機付けられている。
【0018】
ヒトIgG4アイソタイプ抗体及びFc融合タンパク質は、顕著なADCC(NKにより、FcgRIIIaだけを発現する)を引き起こさないが、しかし、マクロファージ(FcγRI、IIa、及びIIIaを発現する)による貪食作用(ADCP)を誘発し、かつ可能であれば、免疫複合体において特定の免疫細胞上の活性化FcγRの分布に起因する場合に、単核細胞を活性化する能力を、保持する。残留活性を最小化する試みにより、野生型IgG4 Fc(配列番号:2)にV234A、L235A(ala/ala)を有する突然変異の、開発及び使用がもたらされた。
【0019】
Armourらは、IgG2において多重点突然変異を生成し、FcgRI(Armourら、1999)、並びにFcgRIIa及びIIIa(Armourら、2003)との結合を最小化した。Muellerらの特許出願(PCT国際公開特許WO97/11971号)に開示される追加の突然変異は、IgG2 CH2及びIgG4 CH3ドメインからなる、IgG2/IgG4のハイブリッドに由来し、特に、残基330及び331は、本発明のIgG2突然変異内のIgG4に由来する。免疫活性化作用を減少させるための、弱化又は発現抑制の試みはまた、IgG2又はIgG4又は2つのサブクラス間を交換するドメインを使用し、IgG1(配列番号:3)内に最小のエフェクター機能を伴うmAbを生成させた(Taoら、1991)。Strohlの特許出願(米国特許出願US2007/0148167号)は、EU位置268、309、330、及び331位残基でのIgG2における4つの突然変異を開示している。Shieldsら、(2001)は、追加の置換物、H268Aも開示している。
【0020】
本発明は、カバット/EU番号付け方式に従うIgG4残基H268Q又はA、V309L、A330S、及びP331Sを伴う、IgG2定常領域(Fc)の多数の位置における置換の初めての実証である。多数の残基置換の指向選択は、測定可能なエフェクター機能を欠く治療用実体の提供の可能性に加え、予想外に抗体工学で用いるための機能的Fcドメインを提供し、融合ポリペプチドとして使用した。
【0021】
多置換IgG2突然変異は、Alpha Screen競合アッセイ及びSPR/Biacore分析によって評価された、ヒトFcR(FcγRI、FcγRIIa、FcγRIIb、FcγRIIIa、及びFcRn)に対するそれらの相対親和性に基づき選択された。これらの突然変異は、更に、CDC、ADCC、及びADCPを誘発し、またPBMCによるTNF−α分泌を引き起こす能力について、適切な細胞システムにおいて試験され順位付けられた。本明細書で提供される一連の実験データにおいて、IgG2突然変異が、既知の調整物、又は野生型IgG1及びIgG2に加えてIgG1 ag及びIgG4 Ala/Alaを含む突然変異と、比較された。いくつかの生体外生物検定におけるこれらの突然変異の更なる分析は、最小の検出不可能な活性レベルと、大いに除去されたFcRに対する結合親和性とを実証した。これらの試験に基づき、IgG2 Fc突然変異は、IgG2c4dとして名付けられ、FcRに対する検出可能な親和性又は結合活性(単量体対二重〜多重配位子結合)を有しておらず、かつ前述の種々のエフェクター/免疫活性化生物検定において活性を有していないことが確認されている。IgG2c4d Fcは、アラニン、セリン、及びロイシンの置換物:V234A、G237A、P238S、H28A、V309L、A330S、P331S(EU番号付け)を含む。7つの残基置換されたIgG2であるIgG2c4dは、FcRに結合する能力、エフェクター機能を媒介する能力、又はFc媒介性サイトカイン放出に関与する能力がないという点において、第1の真に「発現抑制された」Fcであると考えられ得る。
【0022】
本発見に基づき、IgG2c4dの7つの突然変異の一部が使用されるか、又は他のアミノ酸突然変異と組み合わされることができ、又は本突然変異は、本明細書で教示されるようなエフェクター機能の類似又は選択的発現抑制を達成するために別のIgGアイソタイプにおいて使用され、また当該技術分野において既知であるものと組み合わされることができる。
【0023】
本明細書で具体的に例示されるように、三つ組のV234A、G237A、P238S置換物は、分子のFcRn結合親和性を維持しつつ、FcγRIIa結合親和性及びFcγRIIa結合活性を低減し、かつADCP及びFc依存性サイトカイン放出が除去される。
【0024】
【表2】
【0025】
【表3】
【0026】
【表4】
【0027】
変性Fc含有分子を作製する方法
置換のための部位は、自然抗体Fcの構造的特性、保持されたFcRn結合、良好な安定性を有し、並びに補体カスケード、細胞溶解、細胞貪食作用、又はサイトカイン放出などを刺激する能力が減少された組成物を製造したいという要望に基づき、選択された。
【0028】
IgGクラス抗体は二価であるため、機能的に関連する重可変ドメイン及び軽可変ドメインからなる2つの完全なFvドメインを有する。二価性は、結合活性作用と、同一又は別個の細胞上の標的抗原又はFc受容体を架橋結合し、それによって、抗体の生物活性に由来する非標的特異的な受容体結合のスペクトルを誘発する能力とを提供する。この理由により、本発明のFc突然変異は「結合活性関連」内で試験され、すなわち、IgGのFc突然変異は表面上で多量体形成され、かつ特定のFc受容体の多量体との相互作用に対して試験された。
【0029】
突然変異の生物学的特徴
Fc含有タンパク質は、いくつかの周知の生体外アッセイによって、機能性を比較され得る。具体的には、Fcγ受容体のFcγRI、FcγRII、及びFcγRIIIファミリーのメンバーに対する親和性が対象となる。これらの測定値は、受容体の組換え可溶型又は受容体の細胞結合型を使用して作製され得る。加えて、IgGの長期の循環半減期に関与する受容体、FcRnに対する親和性は、例えば、組換え可溶性FcRnを使用する「ALPHASCREEN」の、配位子結合型ビーズフォーマットを使用して、測定することができる。高処理量スクリーニングで使用される、ALPHASCREENは、結合等の分子現象の検出を可能にする均質アッセイ技術である。コーティングされた「供与体」及び「受容体」ビーズが、アッセイ技術の基礎である。ビーズに基づくアッセイとして、ALPHASCREENは、大いに増幅された信号を産生するように働く化学反応のカスケードをもたらす、近接近したビーズの相互作用を通して機能する。例えば、競合結合測定といった、直接又は非直接的測定は、タンパク質間の相対親和性及び結合活性を評価するために、適用され得る。
【0030】
ヒトIgGアイソタイプ(例えば、IgG1、IgG2、IgG3、及びIgG4)の自然進化は、それぞれに、抗体依存性細胞傷害(ADCC、例えば、IgG1及びIgG3)、抗体依存性細胞貪食作用(ADCP、例えば、IgG1、IgG2、IgG3、及びIgG4)、及び補体依存性細胞傷害(CDC、例えば、IgG1、IgG3)等の、免疫機能を回復させる能力の異なるスペクトルを示させることを可能にしている。これらの機能のアイソタイプ特異的結合は、別個の免疫細胞上に存在するFc受容体に対する異なる選択性と、C1qに結合し、かつエフェクターマクロファージ上の特定受容体結合補体成分を通じて、CDC及びCDP(補体依存性貪食作用)をもたらす膜攻撃複合体(MAC)のアセンブリを活性化する能力と、に基づく。ヒトアイソタイプの、補体カスケードの初期成分、C1qに結合する能力の優先順位は、IgG1>IgG3>IgG2>IgG4であり、しかし微生物感染においてはIgG2及びIgG4による補体活性化が十分に立証されている。
【0031】
ADCCアッセイ及びCDCアッセイ等の細胞に基づいた機能性アッセイは、特定の変異構造の可能な機能性の結果に関する見識を提供する。抗体依存性細胞媒介性細胞傷害(ADCC)は細胞媒介性反応であり、Fc受容体(FcR)を発現する非特異性の細胞傷害性細胞(例えば、ナチュラルキラー(NK)細胞、好中球、及びマクロファージ)は、標的細胞上に結合した抗体を認識し、その後標的細胞の溶解を引き起こす。1つの実施形態において、ADCCアッセイは、主要エフェクター細胞としてNK細胞を有し、これらの細胞によって発現されることが知られている唯一のFcγ型受容体である、FcγRIIIA上の機能的作用を反映するように構成される。
【0032】
過酸化物又は炎症媒介放出等の細胞応答を測定できるため、貪食作用アッセイを使用して、異なる突然変異の免疫エフェクター機能を比較することもできる。生体内モデルは、Fc変異型の研究において有用であることが証明されている。例えば、抗CD3抗体の突然変異を使用し、マウスにおいて、Fcドメインが、Fcγ受容体等の特異的配位子に関与することに依存する活性であるT細胞活性化を測定することで、実証されている。マクロファージの抗体指向性活性化は、抗体依存性細胞貪食作用(ADCP)を媒介し、オプソニン化された標的細胞がマクロファージによって飲み込まれ消化されるようにする。高レベルのFcRを発現する生体外で分化されたマクロファージは、IFNγ又はGM−CSFを使用して単核細胞に対する全FcR(FcγRI,FcγRIIa,FcγRIIIa)の上昇したレベルを発現する(to expressed)ことによって、M1表現型へと分化され得る。そのようなアッセイは、当該抗体工学分野における技術者にとって既知である。
【0033】
抗体を作製する方法
所定の組換え処理を使用して、Fc突然変異の生成及び試験の出発点として使用されるヒトIgG2定常ドメインに対する、配列内の定方向突然変異が作成された。当該技術分野の技術者は、コード配列内に変化をもたらす種々の技術が、回収及び試験のための種々の宿主細胞における、所望のアミノ酸配列の発現に好適なベクターを作成するために、使用され得るということを理解するであろう。
【0034】
宿主細胞選択又は宿主細胞工学
本明細書で説明されるように、組換えFc含有タンパク質又はモノクローナル抗体の発現のために選択される宿主細胞は、免疫グロブリンCH2ドメイン内でタンパク質を修飾するオリゴ糖部分の組成物における変化を含むが、しかしそれに限定されず、最終的な組成物にとって重要な寄与因子である。したがって、本発明の1つの態様は、所望の治療用タンパク質を発現する産生細胞の使用及び/又は開発のために適切な、宿主細胞の選択を含む。
【0035】
更に、宿主細胞は、哺乳類起源であるか、又はCOS−1、COS−7、HEK293、BHK21、CHO、BSC−1、Hep G2,653、SP2/0、293、HeLa、骨髄腫、リンパ腫、酵母、昆虫又は植物細胞、又はそれらの任意の誘導体、不死化細胞、若しくは形質転換細胞から選択され得る。
【0036】
別の方法としては、宿主細胞は、例えば、天然又は人口の大腸菌種、クレブシェラ菌種、又はシュードモナス菌種等の、原核細胞又は有機体といった、ポリペプチドをグリコシル化する能力がない種又は有機体から選択されてもよい。
【0037】
抗体
本出願で説明される抗体は、非限定的に、ヒト、マウス、ウサギ、ラット、げっ歯類、霊長類、ヤギ、又はそれらの任意の組み合わせ等を含むか、又はそれらに由来することができ、また、単離したヒト、霊長類、げっ歯類、哺乳類、キメラ、ヒト化、及び/又はCDR移植等の抗体、免疫グロブリン、開裂産物及び他の指定部分、並びにそれらの変異を含む。
【0038】
本明細書で説明される抗体、Fc含有タンパク質、又はFc断片は、当該技術分野において周知であるいくつかの方法で、得ることができる。1つの態様において、抗体は、マウス又は他の動物を、標的ペプチド、細胞、又は組織抽出物によって免疫化して調製されるハイブリドーマから、都合よく得られる。抗体はしたがって、当該技術分野において周知である任意のハイブリドーマ技術を使用して得ることができ、例えば参照によって本明細書に全体が組み込まれる、Harlow及びLane、antibodies,a Laboratory Manual,Cold Spring Harbor,NY(1989)を参照されたい。
【0039】
抗体若しくはFc融合タンパク質、又はそれらの成分及びドメインは、例えば、ファージライブラリー等の、そのようなドメイン又は成分のライブラリーから選択し、得ることもできる。ファージライブラリーは、免疫化された動物又はヒトのB細胞からのような、ランダムオリゴヌクレオチドのライブラリー又は目的の配列を含有するポリヌクレオチドのライブラリーを挿入することによって、作成できる(Smith,G.P.1985.Science 228:1315〜1317)。抗体ファージライブラリーは、1つのファージ内に重鎖(H)及び軽鎖(L)の可変領域対を含有し、単一鎖Fv断片又はFab断片の発現を可能にする(Hoogenboomら、2000,Immunol.Today 21(8)371〜8)。ファージミドライブラリの多様性は、ライブラリーのモノクローナル抗体の免疫特異性を増大及び/又は変化させ、追加の望ましいヒトモノクローナル抗体を産生しその後同定するように、操作され得る。例えば、遺伝子を符号化する重鎖(H)及び軽鎖(L)の免疫グロブリン分子は、組み立てられた免疫グロブリン分子における新規のHL対を作成するように、無作為に混合(入れ替え)され得る。更に、遺伝子をコードするH鎖及びL鎖の一方又は双方は、免疫グロブリンポリペプチドの可変領域の相補性決定領域(CDR)において変異誘発され、かつその後に望ましい親和性及び中和能力について篩分けされ得る。抗体ライブラリーはまた、1つ以上のヒトフレームワーク配列を選択し、かつヒト抗体レパートリーから得られるか又は設計変更を通じて得られるCDRカセットの収集を導入することによって、合成的に作成され得る(Kretzschmar及びvon Ruden 2000,Current Opinion in Biotechnology,13:598〜602)。多様性の位置はCDRに限定されないが、可変領域のフレームワークセグメントを含むこともでき、又はペプチド等の抗体可変領域以外を含んでもよい。
【0040】
抗体可変領域以外を含み得る標的結合成分の他のライブラリーは、リボソームディスプレイ、酵母ディスプレイ、及び細菌ディスプレイである。リボソームディスプレイは、タンパク質のRNAとの付着を保ちつつ、mRNAをそれらの同属タンパク質へと翻訳する方法である。核酸コード配列は、RT−PCRによって回収される(Mattheakis,L.C.ら、1994.Proc.Natl.Acad.Sci.USA 91,9022)。酵母ディスプレイは、膜結合αアグルチニン酵母接着受容体の融合タンパク質の構築物aga1及びaga2に基づき、接合型システムの一部である(Broderら、1997.Nature Biotechnology,15:553〜7)。細菌ディスプレイは、細胞膜又は細胞壁に関係している、標的と排出された細菌タンパク質との融合に基づく(Chen及びGeorgiou 2002.Biotechnol Bioeng,79:496〜503)。
【0041】
ハイブリドーマ技術との比較において、ファージ及び他の抗体ディスプレイ方法は、生体外の抗原標的に対する選択を操作する機会を与え、かつ宿主の抗原への作用の可能性を制限せず逆もまた同様である。
【0042】
本発明はまた、単離したポリヌクレオチドとして、又は原核細胞、真核性、又は繊維状のファージ発現、組成物の分泌及び/又はそれらの定方向突然変異誘発物質又は組成物のディスプレイと適合するベクターを含む発現ベクターの部分として、本発明の組成物をコードするする核酸を提供する。
【0043】
Fc含有分子の使用
上述された方法のうちのいずれかによって生成される組成物(抗体、Fc融合体、Fc断片)は、ヒトの疾患、又は細胞、組織、器官、体液、若しくは概して宿主における特定の病理を、診断、治療、検出、又は調節するために使用され得る。本明細書で教示されるように、Fcγ受容体結合及び指定のエフェクター機能を低減又は除去するために、抗体、Fc融合タンパク質、又はFc断片の、Fc部分を修正することは、抗体が元の標的指向化性質を保持するが、優れた活性スペクトル、生物物理学的性質、安定性、及び宿主の身体内に持続させる能力を有する抗体及びFc構築物を提供する。
【0044】
本発明によって提供される組成物を用いた治療に使用できる疾患又は病理としては、例えばアルツハイマー病であるが、それに限定されない神経性疼痛を含む、神経疾患;皮膚病;代謝病;変形性関節炎;及び火傷又は怪我から生じる症状;心筋梗塞、うっ血性心不全、脳卒中、虚血性脳卒中、及び出血を含むがそれに限定されない、心臓血管疾患;並びにリウマチ性疾患、乾癬、及び強皮症を含む一般的な免疫介在性疾患が挙げられるが、それらに限定されない。
【0045】
本発明は一般論として記述されてきているが、本発明の実施形態は、特許請求の範囲を限定するように解釈されるべきではない以下の実施例で更に開示される。
【実施例】
【0046】
実施例1.Fc突然変異の構築及び試験
表4に示されるヒトIgG2抗体に由来する突然変異を伴う一連の構築物は、標準的組換え方法を使用して構築された。完全な可変ドメインを伴う抗体に対して、抗HER2及び抗CD20抗体の既知のCDR配列が使用され、示されたようなアイソタイプ及びFc突然変異が構築された。抗体突然変異は、標準的クローン化及び発現手順を使用し、293T細胞内に一時的に発現された。MAbは、次の実験的分析の前に、タンパク質Aカラムを使用して、95%を超える均質性まで浄化された。
【0047】
【表5】
【0048】
親和性のビアコア(Biacore)研究
Biacore 3000光学バイオセンサー(Biacore AB,Uppsala,Sweden、現在は、GE Healthcareの一部である)を使用して、表面プラズモン共鳴実験が実行された。実験は、3 mMのEDTAと0.01%の界面活性剤P20とを含有するD−PBS緩衝剤中、摂氏25度で実行された。受容体とFc突然変異との相互作用を分析するために、マウスの抗His IgG(R&D systems cat#MAB050)をCM−5センサーチップへ共有結合させることによって、捕捉表面が生成された。抗His Abは、10mMの酢酸ナトリウム緩衝剤pH 4.5(Biacore AB)の中に希釈され、アミン連結化学剤製造者の説明書を使用し、CM−5チップ(〜3000 RU)のカルボキシメチル化デキストラン表面と連結された。表面上に残留する反応基は、エタノールアミンHCLを使用して非活性化された。動力学的実験を実行するために、FcγRI、FcγRII、及びFcγRIIIの、165、351、及び208反応単位(RU)がそれぞれ、この表面上で捕捉された。受容体捕捉の後に、野生型又はFc突然変異の一連の希釈物が(4倍希釈工程で4000nMから3.9nMまで)、30uL/分で注入された。会合相を3分間観察した。その後に、20分間の緩衝剤流入が続き、結合解離を観察した。捕捉表面は、100mMのリン酸の9秒パルスを100uL/分で使用し、その後の移動緩衝剤の注入によって再生された。
【0049】
参照のために、Scrubberソフトウェアバージョン1.1g(BioLogic Software)を使用して、データの二重参照減算(double reference subtraction)を実行し、信号及び機器ノイズ(Myszka 1999)への緩衝剤寄与を修正した。この初期データ処理の後、単純1:1結合モデルを仮定とし、BIA評価ソフトウェア、バージョン4.0.1(Biacore、AB)を使用して、データの動力学的分析を実行した。AlphaScreen及び結合研究。
【0050】
種々のFcγRとのIgGの競合結合及び直接結合の双方が、均質ビーズ系結合アッセイ、AlphaScreen(商標)(PerkinElmer,Waltham,MA)を使用して評価された。手短に言うと、わずかな修正を伴って、以前に説明されたように実験が実行された(Lazarら、2006 Proc Natl Acad Sci USA 103(11):4005〜10)。FcγRI、IIaは、R&Dシステムから取得された。FcγRIIIa及びFcRnは、クローン作製、発現、及び浄化された。IgGFc突然変異が、NLSビオチン、Pierce、2:1比率を使用してビオチン化された、CNT06234(非特異的制御ヒトIgG1サブクラス抗体)、又は抗Her2/neu(ヒトIgG2抗体)に対する競合結合について、試験された。
【0051】
競合結合研究において、ビオチン化抗体が、200ng/mLの最終アッセイ濃度に添加され、次にそれぞれの実験図に規定されている指定された最終濃度に、競合試験抗体が添加された。FcγRが、200ng/mL最終濃度で96穴プレートへ添加され、次に、ストレプトアビジン供与体及びNiキレート受容体ビーズが連続的に添加された。プレートを密封し室温で振盪した後、プレートはEnvision plate readerを使用して読み取られ、GraphPad Prismにおいてデータがグラフ化されプロットされた。結合活性研究は、試験IgG分子が2:1比率でビオチニル化され、かつ直接FcR結合が競合の不在のもとに対照抗体に対して評価されたということ以外は、競合研究と同様に実行された。
【0052】
結果
SPR/Biacore分析によって評価され、センサグラムから得られたヒトFcR(FcγRI、FcγRIIa、FcγRIIb、及びFcγRIIIa)に対するIgG変異型の相対親和性を、表5に示す。
【0053】
【表6】
【0054】
これらの数字は、1つの実験のグローバル・フィットのために生成されたパラメーターに対応する。
【0055】
これらの4つのデータセットに対して、単純1:1結合モデル・キネティック・フィットを使用してフィッティングを実行することによって、親和性が獲得された。他の全てに対して、単純1:1結合定常状態親和性解析を使用してフィッティングを実行することによって、親和性が獲得された。
【0056】
ヒトIgG1(CNTO6234)を用いたAlpha Screen(商標)競合アッセイを使用して測定された、結合活性関連におけるヒトFcR(FcγRIIa、FcγRIIb、及びFcRn)に対する、すなわち高密度の標的分野に結合する二価抗体に対するIgG突然変異の相対親和性を、図3A〜Cにそれぞれ示す。
【0057】
実験データは、IgG1 ag及びIgG4 Ala/Alaと同様に、IgG2突然変異が、FcRn(生体内半減期をもたらす新生児Fc受容体)と結合するそれらの能力を保持しつつ、IgG1と比較して大幅に減少したFcγ受容体との結合親和性を示すということを実証した。具体的には、IgG2に対する競合結合における、また高親和性から最低親和性まで順位付けされたFcgRIIaとの結合におけるその順序は、以下の通りである。IgG1>IgG2c4a>IgG2m4>IgG2=IgGc4c>IgG2c4b>IgG4ala/ala>IgG4agly>IgG2c4d。この順序は、FcgRIIbとの結合におけるIgG1(CNTO6234)に対する競合において、一致している。pH 6.4でのFcRnとの結合における、IgG1(CNTO6234)に対する更なる競合結合分析は、全てのアイソタイプ及び突然変異がFcRnと比較的均等に結合することを示した。重要なことには、IgG2c4d及びIgG1 aglyは、仮にあったとしても、FcgRIIa及びFcgRIIbとで最小の検出可能な結合を示す。
【0058】
実施例2:ADCC及びCDC
CDCは、3つの経路カテゴリーにおいて始まる。抗体依存性(古典経路)、多糖類依存性(レクチン依存性)、及び外来性表面構造(代替経路)であり、その全ては、タンパク質分解工程のカスケードを産生し、ついには標的細胞又は微細物の溶解に至る膜攻撃複合体のアセンブリをもたらす(W.E.Paul Immunologyを参照)。実験1で説明されたアイソタイプ及びFc突然変異の部分集合が、抗CD20抗体の可変ドメインを使用して調製され、ヒト血清存在下におけるWIL2−S B細胞リンパ腫(lymophoma)標的細胞を溶解させる能力が評価された。WIL2−S細胞のCDCを媒介することで知られる治療用抗CD20である、市販のRituxan(登録商標)が、溶解に対する陽性対照として使用された。CDC分析のために、Wil2−S標的細胞が96穴プレート内に播種され、ヒト血清補体(1/6希釈)とともに培養され、AlamarBlueを使用して相対的細胞生存率が評価された。
【0059】
ADCCアッセイは、実験1に説明されたように、またSkBr3乳癌細胞を標的として、異なるIgGアイソタイプ又は変異型の種々のFcドメインと組み合わされた、抗HER2/neu可変ドメインを使用して実行された。アッセイは、以前に説明されたように、細胞溶解検出のEuTDA法(PerkinElmer,Waltham,MA)を使用して実行された。TDAを負荷したSkBr3乳癌標的細胞が、U−bottom 96穴プレートに播種され、指定の抗体濃度でオプソニン化され、また37℃でロイコパックから単離された25倍過剰のPBMCとともに共培養された。3時間後、プレートが遠心分離され、製造者の説明書に従って上清がTDA放出について分析された。生データが正規化され、GraphPad Prismを使用してプロットされた。CDC分析のために、Wil2−S標的細胞が、96穴プレート内に一穴あたり50,000個で播種され、ヒト血清補体(1/6希釈)とともに培養され、AlamarBlueを使用して相対的細胞生存率が評価された。
【0060】
結果
NK細胞上のFcgRIIIaに主として関わるADCCは、IgG2、IgG2m4、IgG2c4d及びe、IgG1 agly、並びにIgG4 ala/ala(図5)を含む、試験された一連の構築物に関して非検出可能であり、AlphaScreen(商標)アッセイ(図4A)における結合活性の欠損と同様に、より高い親和性FcgRIIIa(V)受容体に見られる減少した結合特性と一致した。乳癌細胞上のHER2/neu等の高密度標的の使用は、FcgRIIIa(40uMより低い)に対するFc親和性が標的細胞溶解を誘発するには不十分にしか出現しないということを実証した。同様に、IgG1を除いて、抗体又はFc突然変異のいずれも、試験された抗CD20構築物内のWIL2−S標的細胞に対する、有意なCDCレベルを示さなかった(図6)。試験されたIgGサブクラス及び突然変異のうち、IgG1のみが、検出可能なCDCレベルを示し、これは、C1q結合とは無関係に、残留する突然変異のいずれもがCDCを引き起こすことはできないということを示唆している(図6)。先行の試みにより、IgG2は、C1q結合及び古典経路の活性化を通じて最小のCDCを有すると指摘されているが、我々は、先行文献の観察(Idusogie,Prestaら、2000 J Immunol 164(8):4178〜84)と一致する抗CD20関連において、有意な活性レベルを観察しなかった。更に、抗CD20及びヒト血清を用いて、残留補体活性を有するIgG1 aglyの先行文献の指摘も検出されなかった(Dorai,Muellerら、1991 Hybridoma 10(2):211〜7)。この矛盾に対する説明は、低いレベルの補体活性化がIgG2によって媒介され得るが、活性化は、オプソニン化された細胞を溶解させるのに十分な膜攻撃複合体(MAC)のアセンブリを始動させるには不十分であるかもしれない、ということである。
【0061】
これらのデータは、補体活性化のレベルにかかわらず、ついには標的細胞溶解に至る膜攻撃複合体のアセンブリが、結合型IgG1以外のアイソタイプの抗体に対しては、欠乏しているか又は不十分であるということを示唆する。
【0062】
実施例3:抗体依存性細胞貪食作用
抗HER2/neu結合Fc突然変異が、オプソニン化された標的乳癌細胞であるSk−Br3及びマクロファージを使用して、抗体依存性細胞貪食作用(ADCP)を媒介する能力について評価された。
【0063】
抗体依存性細胞貪食作用(ADCP)
末梢血単核球が、標準Ficoll−Paque(GE Healthcare)密度勾配調製によってロイコパック(Biological Specialty Corporation)から単離され、細胞が等分され、かつ窒素内で貯蔵された。PBMCは解凍され、製造者の説明書に従って、CD14陽性細胞が、CD16枯渇を伴わないCD14 Isolation kit(Stem Cell Technologies)を使用する陰性除去(negative depletion)によって単離された。細胞は、0.1×106細胞/cm2で、20ng/mLのGM−CSF(R&D Systems)の存在下で、RPMI/5%の加熱不活性化したFBS/50μg/mLゲンタマイシン内で、7日間平板培養され、単核細胞由来マクロファージが生成された。SK−BR−3腫瘍細胞は、製造者の説明書に従って、PKH67(Sigma)で標識付けされた。標的細胞は洗浄され、かつ4エフェクター細胞に対して1標的細胞の比率で単核細胞由来マクロファージを用いて、抗体の存在下で、37℃で4時間5% CO2培養器内で培養された。培養後、細胞はAccutase(Millipore)によって分離され、またマクロファージは、AlexaFluor−647(Invitrogen)と複合化された抗CD11b抗体(BD Biosciences)で標識付けされた。フローサイトメトリーによって細胞が分析され、腫瘍細胞単独(PKH67pos、CD11bneg)、マクロファージ単独(PKH67neg、CD11bpos)、及び貪食された腫瘍細胞(PKH67pos、CD11bpos)が測定された。パーセント貪食作用が、次の等式によって決定された。(貪食された腫瘍細胞)/(貪食された腫瘍細胞+腫瘍細胞単独)×100%。細胞は、FACS Calibur(Becton Dickinson)上に取得され、その結果はFloJo Software(Tree Star)によって分析された。
【0064】
単離した単核細胞が、GM−CSFを使用して生体外で分化され、更に、フローサイトメトリック分析によってFcRの発現レベルについて特徴付けられた。他者の先行研究で述べられているように、GM−CSF活性化マクロファージは、親単核細胞に対して、全FcR(FcγRI、FcγRIIa、FcγRIIIa)の上昇したレベルを発現した(データは図示せず)。抗HER2/neu IgG Fc突然変異構築物は、その後、M1マクロファージを使用して貪食作用アッセイで試験された。
【0065】
結果
それぞれのmAbの存在下での、SkBr3細胞を伴う4時間の共培養後、かなりのレベルのADCPがIgG1に対して出現したが、しかしながら、非グリコシル化IgG1、IgG2、IgG2m4、及びIgG4 S>P ala/alaに対しては、より高濃度の抗体において最小レベルのADCPも観察された(図7)。対照的に、IgG2c4dは、検出可能なレベルのADCPを示さなかった。この観察結果は、以前に実証されたFcRに対する種々のIgGの結合プロファイルと一致している。例えば、IgG1、IgG agly、及びIgG4 ala/alaは、多数の配位子ディスプレイビーズ(例えば、AlphaScreen(登録商標)システムを使用して)の使用においてFcγRIとの結合を実証したが、これら3つの全てはADCPも実証した。IgG2及びIgG2M4及びIgG4 ala/alaは、より高濃度で最小のADCPを示した。IgG2及びIgG2m4によるFcγRIIaの結合は、FcγRIIaの寄与を示唆する結合活性研究によって示されるように、それ自体としては、有意なレベルのADCPを生じさせるには不十分であるかもしれない。BiaCore及びAlphaScreen(商標)の結果(表5、並びに図4B及び4C)は更に、IgG aglyがFcγRIとの結合を保持し、IgG4 ala/alaはFcγRI及びFcγRIIaの双方との結合活性を示すが、しかしADCPは、IgG1 aglyに対してIgG4 ala/alaよりも幾分強固であるということを示した。IgG1 aglyは、FcγRIIaに対し、また更には高度に類似した阻害物質FcγRIIb(細胞外ドメイン(doman)内での、配列同一性>95%に基づく)に拡大して、仮にあったとしても最小の結合性を有するため、FcγRI信号伝達を通してのITAMの活性化は、FcγRIIb活性化に関連するITIMを通しての信号伝達によって相殺されない。対照的に、IgG4 S>P ala/alaは抑制された貪食作用を示し、FcγRIIbの活性化に起因する可能性が高い。最後に、単量体又は結合活性に基づく、種々のFcRとのIgG2c4dの検出可能な結合の完全な欠損は更に、このFc主鎖の固有の無効にされた貪食能力を立証する。
【0066】
実施例4:抗体媒介性サイトカイン放出
免疫細胞上のFcRのFc結合は、架橋結合されるとサイトカイン放出を促進する。免疫細胞上のFcRの結合活性系結合を模倣するために、mAbがポリスチレンビーズへ結合された。
【0067】
抗HER2/neu IgG突然変異を使用したサイトカイン放出が、IgGをラテックスビーズへ直接結合した後、一晩培養してから、実行された。洗浄されたビーズが、1mLあたり約1500個〜250,000個の指定の種々の濃度で単離したヒトPBMCへ添加され、一晩培養した後、共培養上清を除去し、PerkinElmer(Waltham,MA)からの標準AlphaELISAキットを使用して、分泌されたTNFαが測定された。
【0068】
試験されたIgGアイソタイプ及びFc突然変異は分化能力を有し、PBMCからのFc受容体媒介性TNFα分泌を通じてサイトカイン放出を刺激する(図8)。従って、種々のアイソタイプ及びそれらのFc突然変異による、高レベルから低レベルまでのTNF分泌のレベルは、以下の通りである。IgG1>IgG2>IgG2m4>IgG2c4a>IgG1agly>IgG4ala/ala>IgG2c4b>IgG2c4c>IgG4c4d及びe留意すべき点として、IgG4d及びeFc突然変異の双方は、仮にあったとしても、検出可能なカイトカイン(TNF)放出を誘発する最小の能力を持つ。
【0069】
実施例5:生体外B細胞枯渇
生体内での発現抑制レベルに関するアイソタイプ及び突然変異の能力をより良く理解するために、ヘパリン化ヒト全血液の存在下で、WIL2−S B細胞の生体外枯渇が測定された。抗CD20 IgG1は、全てのエフェクター機能(ADCC、CDC、ADCP)に関与することが知られているため、PMN(好中球、好塩基球等)、ヒト補体、及び過剰IgGの存在を含む全血液システムは、それぞれの変異型によって与えられる「発現抑制」のレベルを表していると見なされた。
【0070】
手短に言うと、ヒト全血液は、ADCCアッセイのための標的細胞の機能を果たすエフェクター細胞及びWIL2−S細胞を提供した。標的細胞は、BATDA(PerkinElmer)を用いて摂氏37度で30分間事前標識付けされ、2回洗浄されDMEM/5%の加熱不活性化したFBS内で再懸濁され、その後50μLの標的(1穴あたり2×104細胞)が、96穴U−bottomプレートの穴へ添加された。種々の濃度の抗体を伴って又は伴わないで追加の50μLが添加され、細胞は室温で20分間培養された。次に、ヒト全血液(100μL)が穴へ添加された。全ての試料は3組で実行された。プレートは、200gで3分間遠心分離され、5% CO2培養器において摂氏37度で3時間培養され、次に、200gで3分間再び遠心分離された。1穴あたり合計20μLの上清が除去され、細胞溶解が、200μLのDELPHIA Europium系試薬(PerkinElmer)の添加によって測定された。Envision 2101 Multilabel Reader(PerkinElmer)を使用して、蛍光性が測定された。データは、Triton X−100(Sigma Aldrich)で細胞を処理することによって得られた最大の細胞傷害に対して正規化され、また標的細胞単独からのBATDAの自然放出によって最小対照が決定された。データは、GraphPad Prism v5.01を使用して、S字状用量−応答モデルにフィッティングされた。
【0071】
ヒト血液の存在下でのWIL2−Sの共培養は、IgG1を使用して、またある程度はIgG2及びIgG2M4によって、標識付けされたWIL2−Sのエフェクター媒介による重度の枯渇を明らかにした。留意すべき点として、(Fab’)2及びFab抗CD20断片の双方は、エフェクター機能を回復する能力がある血清内の開裂IgG自己抗体(例えば、Breskiら、2008 J Immunol 181:3183〜3192を参照)の存在を示し得る、あるレベルのWIL2−S枯渇を誘発した。先行のADCC、CDC、及びADCPデータと一致して、IgG2c4eに伴って、大幅な又は検出可能なレベルのサイトカイン放出は、観察されなかった。
【0072】
実施例6:CD20標的化を伴う生体内B細胞生存
IgG2c4eに関連する生体内エフェクター機能が、抗CD20の確立されたカニクイザルB細胞(Cynomologous B-cell)枯渇モデル(Reffら、1994,Blood 83:435〜445)を使用して評価された。
【0073】
カニクイザル(Cynomologous monkeys)(n=3/群)は、0.2mg/Kg又は2mg/Kgでの、IgG1か又はIgG2Σかの単回ボーラス静脈内投与の7日前に、生理食塩水を注入された。注入後の所定の日に、全血液試料からのB細胞レベルが、抗CD20及び抗CD3をそれぞれマーカーとして使用するB及びT細胞のフローサイトメトリー分析によって測定された。それぞれの群に対する平均B細胞レベル(CD20+/CD3−)が、注入後3週間の間プロットされた(図9)。IgG1の低投与量(0.2mg/kg)は、注入後1日で全B細胞の完全に近い枯渇(99%)を誘発したのに対し、抗CD20 IgG2c4eによっては有意な枯渇は誘発されなかった(群内平均15%)。B細胞レベルは、抗CD20 IgG2c4eで処理された動物に対しては、後続の数日に渡って相対的に正常なままであり、またIgG1で処理された動物に対しては、後続の数週間の間で、徐々に回復へと向かうB細胞レベルの傾向が観察された。留意すべき点として、IgG2c4e及びIgG1の双方とも、2mg/Kgのより高い投与量では、完全に近いB細胞枯渇を誘発した。
【0074】
抗CD20媒介性B細胞枯渇は、ADCC、CDC、及びアポトーシスを含むいくつかのメカニズムによって媒介されると考えられる。IgG2c4eによる、より高い投与量(2mg/Kg)でのB細胞の枯渇を示すサルのB細胞枯渇データを考慮し、単離したB細胞内の抗体によって誘発されるアポトーシスのレベルを測定することによって、B細胞枯渇の基本メカニズムが更に評価された。
【0075】
単離したB細胞は、0、0.26、2.6、及び26μg/mL濃度のIgG1、IgG2c4e、(Fab’)2、及びIgG2と、非結合性の対照mAb(BM21)とで、4時間処理され、Annexin V陽性及び7AAD陰性細胞が、フローサイトメトリーによって定量化された。0.2及び2mg/Kgのボーラス注入後の、概算の最大生体内血清IgG濃度を反映するように、2.6及び26μg/mLの特定の最終濃度が選択された。
【0076】
これらの3つの全ての結合抗体に対して、アポトーシスの投与量依存性誘発が、(Fab’)2を含む全てのIgGに対して観察され、抗CD20媒介性架橋結合は、より高い投与量での細胞死滅の誘発に十分であるが、しかし、IgG2c4eに対するより低い投与量では十分ではないことが示された。留意すべき点として、(Fab’)2はまた、Fcの不在下でも有意なアポトーシスを示し、抗IgG媒介性アポトーシスは、以前に観察されたように、Fc媒介性架橋結合とは無関係に誘発され得るという考えを裏付けた。
【0077】
したがって、細胞上の標的抗原の架橋結合に関連する正常な機能は、修飾されたFc変異型によって除去されない。
【特許請求の範囲】
【請求項1】
野生型Fcと比較して少なくとも1つのFcγ受容体に対する低下した親和性を有するFc含有分子であって、突然変異したIgG定常領域を伴う抗体Fcドメインを含み、EU番号付け方式によって定義されるアミノ酸残基233、234、235、237、及び238が、PAAAP(配列番号:4)、PAAAS(配列番号:5)、及びSAAAS(配列番号:6)から選択される配列を含む、Fc含有分子。
【請求項2】
前記Fcドメインが更に、EU番号付け方式によって定義される突然変異H268A又はH268Q、V309L、A330S、及びP331Sを含む、請求項1に記載のFc含有分子。
【請求項3】
前記ドメインが、FcRnに特異的に結合することができる、請求項1に記載のFc含有分子。
【請求項4】
前記Fcドメイン配列が、ヒトIgG2重鎖CH2ドメインと少なくとも90%同一である、請求項1に記載のFc含有分子。
【請求項5】
前記Fc含有分子が、抗体又はFc融合タンパク質である、請求項1に記載のFc含有分子。
【請求項6】
残基228がSからPへ突然変異される、請求項1に記載のFc含有分子。
【請求項7】
組換えポリペプチド系結合分子であって、(i)標的分子に結合することができる結合ドメインと、(ii)ヒト免疫グロブリン重鎖のCH2及びCH3定常ドメインの全て又は一部に実質的に相同であるアミノ酸配列を有するFcドメインであって、EU番号付け方式によって定義される残基233、234、235、237、及び238が、PAAAP、PAAAS、及びSAAASから選択されるアミノ酸配列を含む、Fcドメインと、を含み、前記結合分子が、著しい補体依存性溶解又は前記標的の細胞媒介性破壊を引き起こすことなしに、前記標的分子に結合することができる、結合分子。
【請求項8】
前記Fcドメインが、FcRnに特異的に結合することができる、請求項7に記載の結合分子。
【請求項9】
前記結合ドメインが、抗体、酵素、ホルモン、受容体、サイトカイン、免疫細胞表面抗原、配位子、及び接着分子の結合部位から選択される、請求項8に記載の結合分子。
【請求項10】
前記分子が結合活性を示す、請求項9に記載の結合分子。
【請求項11】
前記結合ドメインが、神経組織、内分泌組織、血管組織、心臓組織、滑膜組織、皮膚組織、又は粘膜組織内で標的と特異的に結合する、請求項7〜10のいずれか一項に記載の結合分子。
【請求項12】
マクロファージの遊走及び集結によって特徴付けられる疾患を治療するための方法であって、請求項1〜7のいずれかに記載のFc含有タンパク質調製物を、対象又は患者に投与する工程を含む、方法。
【請求項13】
移植片対宿主疾患;宿主対移植片疾患;臓器移植拒絶反応;骨髄移植拒絶反応;自己免疫、血管炎、自己免疫性溶血性貧血、自己免疫性血小板減少症及び関節炎;胎児/新生児同種免疫性血小板減少症等の同種免疫;ぜんそく及びアレルギー;慢性若しくは急性炎症性疾病、クローン病又は強皮症;アルツハイマー病、又は冠状動脈閉塞を治療する方法であって、請求項1〜7のいずれかに記載のFc含有分子を対象又は患者に投与する工程を含む、方法。
【請求項14】
請求項1〜7のいずれかに記載のFc含有分子を対象又は患者に投与する工程を含む、症状を治療するための方法であって、前記結合分子が、患者に投与されるか、又は任意で前記患者が胎児である場合に前記患者の母親に投与される、方法。
【請求項15】
野生型Fcと比較してFcγ受容体に対する低下した親和性を有するFc含有分子であって、IgG2定常領域に基づく抗体Fcドメインを含み、EU番号付け方式によって定義されるアミノ酸残基233、234、235、237、及び238が、PAAAP、PAAAS、及びSAAASから選択される配列を含み、かつEU番号付け方式によって定義される突然変異H268A又はH268Q、V309L、A330S、及びP331Sを更に含む、Fc含有分子。
【請求項16】
IgG2系Fc含有分子のFcγ受容体への結合を、野生型IgG2系Fcと比較して変化させる方法であって、EU番号付け方式によって定義される残基233、234、235、237、及び238でのIgG2定常領域に基づくFcドメインの配列を、PAAAP、PAAAS、及びSAAASから選択される配列を含み、かつ突然変異H268A又はH268Q、V309L、A330S、及びP331Sを含むように、変化させる工程を含む、方法。
【請求項17】
本明細書に記載されたいずれかの発明。
【請求項1】
野生型Fcと比較して少なくとも1つのFcγ受容体に対する低下した親和性を有するFc含有分子であって、突然変異したIgG定常領域を伴う抗体Fcドメインを含み、EU番号付け方式によって定義されるアミノ酸残基233、234、235、237、及び238が、PAAAP(配列番号:4)、PAAAS(配列番号:5)、及びSAAAS(配列番号:6)から選択される配列を含む、Fc含有分子。
【請求項2】
前記Fcドメインが更に、EU番号付け方式によって定義される突然変異H268A又はH268Q、V309L、A330S、及びP331Sを含む、請求項1に記載のFc含有分子。
【請求項3】
前記ドメインが、FcRnに特異的に結合することができる、請求項1に記載のFc含有分子。
【請求項4】
前記Fcドメイン配列が、ヒトIgG2重鎖CH2ドメインと少なくとも90%同一である、請求項1に記載のFc含有分子。
【請求項5】
前記Fc含有分子が、抗体又はFc融合タンパク質である、請求項1に記載のFc含有分子。
【請求項6】
残基228がSからPへ突然変異される、請求項1に記載のFc含有分子。
【請求項7】
組換えポリペプチド系結合分子であって、(i)標的分子に結合することができる結合ドメインと、(ii)ヒト免疫グロブリン重鎖のCH2及びCH3定常ドメインの全て又は一部に実質的に相同であるアミノ酸配列を有するFcドメインであって、EU番号付け方式によって定義される残基233、234、235、237、及び238が、PAAAP、PAAAS、及びSAAASから選択されるアミノ酸配列を含む、Fcドメインと、を含み、前記結合分子が、著しい補体依存性溶解又は前記標的の細胞媒介性破壊を引き起こすことなしに、前記標的分子に結合することができる、結合分子。
【請求項8】
前記Fcドメインが、FcRnに特異的に結合することができる、請求項7に記載の結合分子。
【請求項9】
前記結合ドメインが、抗体、酵素、ホルモン、受容体、サイトカイン、免疫細胞表面抗原、配位子、及び接着分子の結合部位から選択される、請求項8に記載の結合分子。
【請求項10】
前記分子が結合活性を示す、請求項9に記載の結合分子。
【請求項11】
前記結合ドメインが、神経組織、内分泌組織、血管組織、心臓組織、滑膜組織、皮膚組織、又は粘膜組織内で標的と特異的に結合する、請求項7〜10のいずれか一項に記載の結合分子。
【請求項12】
マクロファージの遊走及び集結によって特徴付けられる疾患を治療するための方法であって、請求項1〜7のいずれかに記載のFc含有タンパク質調製物を、対象又は患者に投与する工程を含む、方法。
【請求項13】
移植片対宿主疾患;宿主対移植片疾患;臓器移植拒絶反応;骨髄移植拒絶反応;自己免疫、血管炎、自己免疫性溶血性貧血、自己免疫性血小板減少症及び関節炎;胎児/新生児同種免疫性血小板減少症等の同種免疫;ぜんそく及びアレルギー;慢性若しくは急性炎症性疾病、クローン病又は強皮症;アルツハイマー病、又は冠状動脈閉塞を治療する方法であって、請求項1〜7のいずれかに記載のFc含有分子を対象又は患者に投与する工程を含む、方法。
【請求項14】
請求項1〜7のいずれかに記載のFc含有分子を対象又は患者に投与する工程を含む、症状を治療するための方法であって、前記結合分子が、患者に投与されるか、又は任意で前記患者が胎児である場合に前記患者の母親に投与される、方法。
【請求項15】
野生型Fcと比較してFcγ受容体に対する低下した親和性を有するFc含有分子であって、IgG2定常領域に基づく抗体Fcドメインを含み、EU番号付け方式によって定義されるアミノ酸残基233、234、235、237、及び238が、PAAAP、PAAAS、及びSAAASから選択される配列を含み、かつEU番号付け方式によって定義される突然変異H268A又はH268Q、V309L、A330S、及びP331Sを更に含む、Fc含有分子。
【請求項16】
IgG2系Fc含有分子のFcγ受容体への結合を、野生型IgG2系Fcと比較して変化させる方法であって、EU番号付け方式によって定義される残基233、234、235、237、及び238でのIgG2定常領域に基づくFcドメインの配列を、PAAAP、PAAAS、及びSAAASから選択される配列を含み、かつ突然変異H268A又はH268Q、V309L、A330S、及びP331Sを含むように、変化させる工程を含む、方法。
【請求項17】
本明細書に記載されたいずれかの発明。
【図1】


【図2】
【図3A】
【図3B】
【図3C】
【図4A】
【図4B】
【図4C】
【図5】
【図6】
【図7】
【図8】
【図9】
【図2】
【図3A】
【図3B】
【図3C】
【図4A】
【図4B】
【図4C】
【図5】
【図6】
【図7】
【図8】
【図9】
【公表番号】特表2013−512258(P2013−512258A)
【公表日】平成25年4月11日(2013.4.11)
【国際特許分類】
【出願番号】特願2012−541220(P2012−541220)
【出願日】平成22年11月29日(2010.11.29)
【国際出願番号】PCT/US2010/058188
【国際公開番号】WO2011/066501
【国際公開日】平成23年6月3日(2011.6.3)
【出願人】(509087759)ヤンセン バイオテツク,インコーポレーテツド (77)
【Fターム(参考)】
【公表日】平成25年4月11日(2013.4.11)
【国際特許分類】
【出願日】平成22年11月29日(2010.11.29)
【国際出願番号】PCT/US2010/058188
【国際公開番号】WO2011/066501
【国際公開日】平成23年6月3日(2011.6.3)
【出願人】(509087759)ヤンセン バイオテツク,インコーポレーテツド (77)
【Fターム(参考)】
[ Back to top ]