
除草剤の製造方法
【課題】市場価値のある植物の生育を阻害せずに雑草の生育を阻害する除草剤を製造する方法の提供。
【解決手段】特定な配列のアミノ酸配列からなるCDPK2酵素タンパク質等か、該酵素タンパク質を含む融合タンパク質かと、タンパク質リン酸化活性増強剤候補化合物とを用意するステップと、前記タンパク質又は融合タンパク質と、前記タンパク質リン酸化活性増強剤候補化合物とを含む反応液中で、タンパク質リン酸化活性を測定するステップとを含むタンパク質リン酸化活性増強作用の評価方法を用いる、複数の前記タンパク質リン酸化活性増強剤候補化合物のうちタンパク質リン酸化活性増強作用が強力な候補化合物をタンパク質リン酸化活性増強剤として選択するステップを含む、除草剤の製造方法。
【解決手段】特定な配列のアミノ酸配列からなるCDPK2酵素タンパク質等か、該酵素タンパク質を含む融合タンパク質かと、タンパク質リン酸化活性増強剤候補化合物とを用意するステップと、前記タンパク質又は融合タンパク質と、前記タンパク質リン酸化活性増強剤候補化合物とを含む反応液中で、タンパク質リン酸化活性を測定するステップとを含むタンパク質リン酸化活性増強作用の評価方法を用いる、複数の前記タンパク質リン酸化活性増強剤候補化合物のうちタンパク質リン酸化活性増強作用が強力な候補化合物をタンパク質リン酸化活性増強剤として選択するステップを含む、除草剤の製造方法。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、新規なタンパク質リン酸化活性増強作用の評価方法と、該タンパク質リン酸化活性増強作用を指標とする除草剤の製造方法に関する。
【背景技術】
【0002】
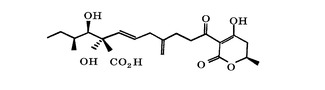
アルタナル酸(Alternaric acid)は、ジャガイモ又はトマトの胴枯れ病の原因となる真菌であるAlternaria solaniにより産生される毒素であり、図1に示す構造を有する。アルタナル酸は、宿主特異性の決定と、Alternaria solaniにより引き起こされる疾病の発病とに関わることが報告されている(非特許文献1及び2)。またジャガイモの塊茎の切片に塗布されたアルタナル酸がPhytophthora infestansの非親和性レースによる感染後の宿主植物の過敏感反応(Hypersensitive Response)の発生を遅延させることから、アルタナル酸は過敏感反応のサプレッサーであることが報告されている(非特許文献3)。アルタナル酸は除草剤としての用途があることが知られている(特許文献1)。
【特許文献1】米国特許第5,256,628号明細書
【非特許文献1】Furuichi,N.and Nishimura,S.(1984)Isolation of alternaric acid from the germination fluids of Alternaria solani. Ann.hytopathol.Soc.Jpn.50:128(Abstr.in Japanese).
【非特許文献2】Langsdorf,G.,Furuichi,N.,Doke,N.and Nishimura,S.(1990)Investigations on Alternaria solani infections:Detection of alternaric acid and a susceptibility inducing factor in the spore−germination fluid of A.solani. J.Phytopathology 128:271−282.
【非特許文献3】Furuichi,N.,Nishimura,S. and Langsdorf,G.(1992)Effect of alternaric acid,a toxin of Alternaria solani,on the hypersensitive response of potato to Phytophthora infestans. Ann.Phytopathol.Soc.Jpn.58:1−7
【0003】
過敏感反応サプレッサーで処理後のジャガイモのさまざまなタンパク質がカルシウム依存性のリン酸化により修飾されることが報告されている(非特許文献4)。また、カルシウム依存性タンパク質リン酸化酵素(以下、「CDPK」という。)は、遺伝子発現、代謝、膜輸送及び細胞構造の制御に関与する複数のアイソフォームを有する多機能酵素である(非特許文献5)。
【非特許文献4】Furuichi,N.,Anderson,A.J.,Suzuki,Y.and Takemoto,J.Y.(1994)Elicitor and suppressor of Phytophthora infestans stimulate phosphorylation of plasma membrane proteins from potato and bean tissues. In Host Specific Toxin:Biosynthesis,Receptor and Molecular Biology (Kohmoto,K.and Yoder,O.C.,eds.).pp.273−274,Tottori University Press,Tottori.
【非特許文献5】Harmon,A.C.,Gribskov,M.and Harper,J.F.(2000)CDPKs−a kinase for every Ca2+ Signal? Trends Plant Sci.5:154−159.
【0004】
最近、病原菌由来の過敏感反応サプレッサーによる過敏感反応の抑制と、前記病原菌による病気の宿主組織全体での発症とにジャガイモ由来のCDPKが関与することが発表された(特許文献2)。それによると、過敏感反応が抑制される過程は、(1)過敏感反応サプレッサーが宿主細胞壁及び形質膜を貫通する段階と、(2)前記過敏感反応サプレッサーとCDPK1の受容体部位とが結合する段階と、(3)リン酸化反応を介してCDPK2がシグナル伝達を行う段階と、(4)過敏感反応が抑制されることにより宿主組織全体が疫病に罹患する段階とを含む。
【特許文献2】公開特許公報(特開2007−68439)明細書
【0005】
本発明はアルタナル酸がCDPK2のタンパク質リン酸化活性を増強する作用を有することの発見に基づいて、新規な除草剤を開発することを意図する。
【発明の開示】
【発明が解決しようとする課題】
【0006】
従来の除草剤は市場価値のある植物の生育を阻害せずに雑草の生育を阻害することができない。特定の除草剤に対する耐性遺伝子を導入したトランスジェニック植物は該特定の除草剤によって生育が阻害されないが、市場価値のある全ての植物に前記耐性遺伝子を導入することは事実上不可能に近い。そこで、除草剤の作用機序の知見に基づいて、より強力な除草剤及び市場価値のある植物の生育を阻害せずに雑草の生育を阻害する除草剤を製造する方法を開発する必要がある。
【課題を解決するための手段】
【0007】
本発明はタンパク質リン酸化活性増強作用の評価方法を提供する。本発明のタンパク質リン酸化活性増強作用の評価方法は、
(A)(1)配列番号1のアミノ酸配列からなるCDPK2酵素タンパク質と、(2)前記アミノ酸配列において1個若しくは数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列からなり、かつ、カルシウム依存性タンパク質リン酸化活性を有するタンパク質と、(3)前記アミノ酸配列において1個若しくは数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列からなり、かつ、アルタナル酸により増強される前記タンパク質リン酸化活性を有するタンパク質とからなるグループから選択されることを特徴とするタンパク質と、(4)前記(1)ないし(3)のタンパク質からなる第1ポリペプチドと、特異的結合タグとからなる第2ポリペプチドとが連結した融合タンパク質とのうちのいずれか1つのタンパク質と、タンパク質リン酸化活性増強剤候補化合物とを用意するステップと、
(B)前記タンパク質又は融合タンパク質と、前記タンパク質リン酸化活性増強剤候補化合物とを含む反応液中で、タンパク質リン酸化活性を測定するステップとを含む。本発明の評価方法は、前記候補化合物の存在下と非存在下とにおける前記タンパク質又は融合タンパク質のタンパク質リン酸化活性を比較すること、及び、前記候補化合物存在下でのタンパク質リン酸化活性が非存在下でのタンパク質リン酸化活性よりどの程度高いかを評価することを含む場合がある。本発明のタンパク質リン酸化活性増強作用の評価方法に用いる反応液にはCa2+及びMg2+イオンを含む場合がある。
【0008】
第2ポリペプチドの特異的結合タグは、Hisタグ、mycタグ、HAタグ、インテインタグ、MBP又はGSTからなるグループから選択される場合がある。
【0009】
本発明の評価方法でタンパク質リン酸化活性増強作用が評価されるタンパク質リン酸化活性増強剤候補化合物はアルタナル酸の誘導体の場合がある。
【0010】
本発明は、本発明の評価方法を用いて複数の前記タンパク質リン酸化活性増強剤候補化合物のうちタンパク質リン酸化活性増強作用が強力な候補化合物を除草剤として選択するステップを含むことを特徴とする除草剤の製造方法を提供する。
【0011】
本発明は、本発明の評価方法によって複数の前記タンパク質リン酸化活性増強剤候補化合物のうち特定の植物種由来のCDPK2酵素に対するタンパク質リン酸化活性増強作用が他の植物種由来CDPK2酵素に対するタンパク質リン酸化活性増強作用に比べて弱い候補化合物を除草剤として選択するステップを含むことを特徴とする除草剤の製造方法を提供する。
【0012】
本発明の除草剤の製造方法において、前記特定の植物種はジャガイモの場合があり、前記他の植物種はシロイヌナズナ又はタバコの場合がある。
【0013】
本発明はタンパク質リン酸化活性増強剤候補化合物のタンパク質リン酸化活性増強作用の評価キットを提供する。本発明の評価キットは、(A)(1)配列番号1のアミノ酸配列からなるCDPK2酵素タンパク質と、(2)前記アミノ酸配列において1個若しくは数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列からなり、かつ、カルシウム依存性タンパク質リン酸化活性を有するタンパク質と、(3)前記アミノ酸配列において1個若しくは数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列からなり、かつ、アルタナル酸により増強される前記タンパク質リン酸化活性を有するタンパク質とからなるグループから選択されることを特徴とするタンパク質と、(4)前記(1)ないし(3)のタンパク質からなる第1ポリペプチドと、特異的結合タグとからなる第2ポリペプチドとが連結した融合タンパク質とのうちのいずれか1つのタンパク質とを含むことを特徴とする。
【0014】
本発明のタンパク質リン酸化活性増強作用の評価キットにおいて、前記特異的結合タグは、Hisタグ、mycタグ、HAタグ、インテインタグ、MBP又はGSTからなるグループから選択される場合がある。前記タンパク質リン酸化活性増強剤候補化合物はアルタナル酸の誘導体である場合がある。
【0015】
本明細書で用いられるところの「タンパク質」とは、2個以上のアミノ酸がペプチド結合で連結した化合物である。タンパク質は、メチル基、チオール基、リン酸、糖鎖、及び/又は、その他の共有結合による修飾を含む場合がある。本明細書で用いられるところの「ポリペプチド」とは、2個以上のアミノ酸がペプチド結合で連結した化合物である。
【0016】
本発明のタンパク質は、本発明のタンパク質をエンコードするヌクレオチド配列からなるDNAを、無生物発現系か、宿主生物及び発現ベクターを使用する発現系かで発現させることにより産生される。本発明の融合タンパク質は、第1ポリペプチドをエンコードするヌクレオチド配列からなるDNAと、第2ポリペプチドをエンコードするヌクレオチド配列からなるDNAとが連結されたDNAを、無生物発現系か、宿主生物及び発現ベクターを使用する発現系かで発現させることにより産生される。前記宿主生物は、大腸菌、枯草菌等のような原核生物と、酵母、菌類、植物、動物等のような真核生物とを含む。本発明の宿主生物及び発現ベクターを使用する発現系は、細胞や組織のような生物の一部か、生物の個体全体かの場合がある。
【0017】
配列番号1のアミノ酸配列からなるカルシウム依存性タンパク質リン酸化酵素CDPK2は、Rishiri品種のジャガイモから単離された。配列番号1のアミノ酸配列は、DDBJ/GenBank/EMBLのデータベースにアクセッション番号AB0551809として登録された。
【0018】
本明細書で用いられるところの「第1ポリペプチド」とは、所望の機能、すなわち、タンパク質リン酸化活性を有するポリペプチドである。本明細書で用いられるところの「第2ポリペプチド」とは、発現タンパク質の分離、精製又は検出をより容易に行うために、第1ポリペプチドとペプチド結合で連結されるポリペプチドである。第2ポリペプチドは、第1ポリペプチドのN末端又はC末端に連結される場合がある。
【0019】
本発明の第1ポリペプチドは、配列番号1のアミノ酸配列からなるRiCDPK2酵素タンパク質か、前記アミノ酸配列において1個又は2個以上のアミノ酸が欠失、置換又は付加されたアミノ酸配列からなり、かつ、カルシウム依存性タンパク質リン酸化活性を有するタンパク質かの場合がある。本発明の第1ポリペプチドのタンパク質リン酸化活性はアルタナル酸により増強される場合がある。
【0020】
本発明の酵素のタンパク質リン酸化活性は、該酵素タンパク質自体が基質になる場合の他、他のタンパク質又はポリペプチドを基質とする場合がある。本発明のカルシウム依存性タンパク質リン酸化活性とは、本発明の酵素のタンパク質リン酸化活性がCa2+イオン存在下では非存在下でよりも高い、すなわち、増強されることをいう。本発明のタンパク質リン酸化活性増強剤は、該増強剤の存在下で非存在下でよりもタンパク質リン酸化活性を増強する。本発明のタンパク質リン酸化活性増強剤候補化合物は、本発明のタンパク質リン酸化活性増強剤となる可能性がある化合物であって、本発明の評価方法によってタンパク質リン酸化活性増強作用が強力なものが本発明のタンパク質リン酸化活性増強剤として選択される。本発明のタンパク質リン酸化活性増強剤の製造方法は、本発明の評価方法を用いて本発明のタンパク質リン酸化活性増強剤候補化合物のタンパク質リン酸化活性増強作用を評価して、該作用が強力なものを本発明のタンパク質リン酸化活性増強剤として選択するステップを含む。
【0021】
第2ポリペプチドは特異的結合タグからなる。本発明で用いられるところの「特異的結合タグ」とは、所望の機能を有するポリペプチドを遺伝子組み換えによって調製する際に、前記所望の機能を有するポリペプチドとペプチド結合で連結した融合タンパク質として発現させることにより、形質転換体からの発現タンパク質の分離、精製又は検出をより容易に行うことを可能にするために、他のタンパク質、多糖類、糖脂質、核酸及びこれらの誘導体、樹脂、シリコン等と特異的に結合するポリペプチドである。特異的結合タグは、水溶液中に溶解した物質又は固体支持体と結合する場合がある。本発明の特異的結合タグは、Hisタグ、mycタグ、HAタグ、インテインタグ、MBP、GSTその他これらに類するポリペプチドが含まれるが、これらに限定されない。前記特異的結合タグは、Hisタグ、mycタグ、HAタグ、インテインタグ、MBP又はGSTからなるグループから選択される場合がある。
【0022】
本発明で用いられるところの「Hisタグ」とは、ヒスチジン残基が連続するアミノ酸配列からなるタグである。前記ヒスチジン残基の数は少なくとも5個、好ましくは6ないし10個、最も好ましくは6個である。Hisタグを連結した融合タンパク質は、Hisタグと金属イオンとキレート化合物とが錯体を形成することを利用して分離、精製される。本発明で用いられるところの「キレート化合物」とは、金属イオンと特異的に配位結合をする原子団をいう。前記キレート化合物は、ニトリロ三酢酸又はイミノ二酢酸の場合があるがこれらに限定されない。前記キレート化合物は、直接的に、又はスペーサーを介して間接的に、カラム担体等の固体支持体に固定化される。前記金属イオンは、Ni2+イオン又はCo2+イオンのような遷移金属イオンの場合があるがこれらに限定されない。Hisタグを利用するタンパク質精製用の前記金属イオンを捕捉したキレート化合物を固定化した固体支持体は、Ni−NTAアガロースとして、インビトロジェン、キアゲン社等から入手することができる。Hisタグを利用するタンパク質精製システムで商業的に入手可能なものには、Ni−NTA精製システム(インビトロジェン株式会社、カタログ番号:K95001)、Ni−NTA Fast Start Kit(株式会社キアゲン、カタログ番号:30600)等が含まれるがこれらに限定されない。
【0023】
Hisタグを連結した融合タンパク質の精製の手順は、(1)前記金属イオンを捕捉したキレート化合物を固定化した固体支持体を用意するステップと、(2)前記固体支持体と前記融合タンパク質とを接触させることにより、前記金属イオンと融合タンパク質のHisタグ部分のイミダゾール基を構成する窒素との間で配位結合を形成させ、前記融合タンパク質を前記固体支持体に捕捉するステップと、(3)前記固体支持体を洗浄して不要成分を除去するステップと、(4)遊離イミダゾール化合物を含む溶出液を添加することにより前記融合タンパク質を前記固体支持体から解離させて回収するステップとを含むのが一般的である。
【0024】
本発明で用いられるところの「mycタグ」とは、ヒトc−mycタンパク質の410番−419番の10個のアミノ酸の配列に対応する、配列番号3のアミノ酸配列からなるタグである。mycタグを連結した融合タンパク質は、mycタグと該タグに特異的に結合する抗体との抗原抗体複合体形成を利用して分離、精製される。本発明で用いられるところの「mycタグペプチド」とは、ヒトc−mycタンパク質の410番−419番の10個のアミノ酸の配列に対応する、配列番号3のアミノ酸配列からなるポリペプチドである。本発明で用いられるところの「抗体」は、ポリクローナル抗体及びモノクローナル抗体を含む。抗mycタグモノクローナル抗体は、例えば、ハイブリドーマ細胞9E10株により産生される。mycタグを利用するタンパク質精製システムで商業的に入手可能なものには、ANTI−c−Myc AGAROSE CONJUGATE(シグマアルドリッチ ジャパン株式会社、カタログ番号:A7470)、c−Myc Peptide(シグマアルドリッチ ジャパン株式会社、カタログ番号:M2435)、c−Myc Monoclonal Antibody−Agarose Beads(クロンテック、タカラバイオ株式会社、カタログ番号:3843−1)等が含まれるが、これらに限定されない。
【0025】
mycタグを連結した融合タンパク質の精製の手順は、(1)抗mycタグ抗体が固定化された固体支持体を用意するステップと、(2)前記固体支持体と前記融合タンパク質とを接触させることにより、前記融合タンパク質のmycタグ部分と前記抗体との間で複合体を形成させて、前記融合タンパク質を前記固体支持体に捕捉するステップと、(3)前記固体支持体を洗浄し不要成分を除去するステップと、(4)mycタグペプチドを含む溶出液を添加することにより前記融合タンパク質を前記固体支持体から解離させて回収するステップとを含むのが一般的である。
【0026】
本発明で用いられるところの「HAタグ」とは、ヒトインフルエンザウィルスのヘマグルチニン(HA)の98番−106番の9個のアミノ酸に対応する、配列番号4のアミノ酸配列からなるタグである。HAタグを連結した融合タンパク質は、HAタグと該タグに特異的に結合する抗体との抗原抗体複合体形成を利用して分離、精製される。HAタグに対応するモノクローナル抗体は例えば、ハイブリドーマ細胞HA−7株により産生される。HAタグを利用するタンパク質精製システムで商業的に入手可能なものには、例えばMonoclonal Anti−HA Agarose Conjugate Clone HA−7(シグマアルドリッチ ジャパン株式会社、カタログ番号:A2095)、INFLUENZA HEMAGGLUTININ(HA)PEPTIDE(シグマアルドリッチ ジャパン株式会社、カタログ番号:I2149)等が含まれるがこれらに限定されない。
【0027】
HAタグを連結した融合タンパク質は、mycタグの場合と同様の手順で精製される。
【0028】
本発明で用いられるところの「インテインタグ」とは、インテインにキチン結合ドメイン(CBD)等のリガンド特異的結合ドメインを連結したタグである。インテインタグを連結した融合タンパク質は、キチン結合ドメインとキチンとの結合形成を利用して分離、精製される。インテインの自己切断能力を利用して、ワンステップで前記タグの除去後のタンパク質として精製することができる。インテインタグを利用するタンパク質発現・精製システムで商業的に入手可能なものには、IMPACT(商標) Systems(ニュー・イングランド・バイオラボ・ジャパン株式会社、カタログ番号:E6900S)が含まれるがこれに限定されない。
【0029】
インテインタグを連結した融合タンパク質の精製の手順は、(1)インテインタグを連結した融合タンパク質の溶液を用意するステップと、(2)前記溶液とキチンが固定化された固体支持体とを接触させることによりインテインタグのキチン結合ドメインとキチンとの間で複合体を形成させ、前記融合タンパク質を前記固体支持体に捕捉させるステップと、(3)前記固体支持体を洗浄し不要成分を除去するステップと、(4)DTT又はβ−メルカプトエタノールのような還元剤の添加によりインテインの自己切断能力を活性化させて融合タンパク質からインテインタグを切断するステップと、(5)インテインタグが切断されたタンパク質を回収するステップとを含むのが一般的である。
【0030】
本発明で用いられるところの「MBP」とは、マルトース結合タンパク質である。MBPを連結した融合タンパク質は、アミロース等とMBPとの結合形成を利用して分離、精製される。MBPを利用するタンパク質発現・精製システムで商業的に入手可能なものには、例えばpMAL(商標) Protein Fusion and Purification System(ニュー・イングランド・バイオラボ・ジャパン株式会社、カタログ番号:#E8000S)が含まれるがこれに限定されない。
【0031】
本発明で用いられるところの「GST」とは、グルタチオンS転移酵素である。GSTを連結した融合タンパク質は、GSTと該酵素の基質であるグルタチオンとの親和性を利用して分離、精製される。GSTを利用するタンパク質精製システムで商業的に入手可能なものには、例えばGlutathione Sepharose 4 Fast Flow(GEヘルスケアバイオサイエンス株式会社、カタログ番号:17−5132−01)、GSTrap FF(GEヘルスケアバイオサイエンス株式会社、カタログ番号:17−5130−02)等が含まれるがこれらに限定されない。
【0032】
好ましくは第2ポリペプチドには、Hisタグ、mycタグ、HAタグ、インテインタグ、MBP又はGSTが選択される。さらに好ましくはHisタグが選択される。
【0033】
本発明の融合タンパク質は、第2ポリペプチドに加えて第3ポリペプチドが第1ポリペプチドに連結される場合がある。本明細書で用いられるところの「第3ポリペプチド」とは、第1ポリペプチドが第2ポリペプチドに連結されている場合にさらに連結されるポリペプチドであって、第1ポリペプチドを遺伝子組み換えによって調製する際に前記第3ポリペプチドと前記第1ポリペプチドとを連結した融合タンパク質として発現させることにより、形質転換体からの発現タンパク質の分離、精製又は検出をより容易に行うことを可能にする場合があるポリペプチドである。第3ポリペプチドは、形質転換体での発現タンパク質の分泌、安定化を可能にする場合がある。第3ポリペプチドは、第1ポリペプチドに対して第2ポリペプチドと同じ側か反対側かに連結される場合がある。第3ポリペプチドには、Hisタグ、mycタグ、HAタグ、インテインタグ、MBP、GSTの他、シグナルペプチド、プロテインA、Trxタグ、CBPタグ、CBDタグ、CBRタグ、luc、HSP、LacZ、IgG、アビジン、プロテインG等が含まれるがこれらに限定されない。
【0034】
本発明のタンパク質リン酸化活性増強作用の評価方法のステップ(A)で用意されるタンパク質リン酸化活性増強剤候補化合物は、ステップ(B)のリン酸化活性の測定に供試できるものであればよく、特に限定されない。前記タンパク質リン酸化活性増強剤候補化合物は、アルタナル酸の誘導体の場合がある。
【0035】
本発明のタンパク質リン酸化活性増強作用の評価方法のステップ(B)では、該ステップのリン酸化活性測定において前記リン酸化活性増強剤誘導体候補化合物によるリン酸化活性を増強させ、該候補化合物間のリン酸化活性の差異をより明確に把握するために、Ca2+及びMg2+イオンを添加する場合がある。前記Ca2+及びMg2+イオンの濃度は、リン酸化活性測定に問題が生じない濃度であれば特に限定されない。Ca2+イオンの濃度は100μMの場合がある。Mg2+イオンの濃度は900μMの場合がある。
【0036】
本発明のタンパク質リン酸化活性増強作用の評価方法のステップ(B)のタンパク質リン酸化活性は、CDPK2酵素によるタンパク質リン酸基転移反応を適切に測定できるいずれかの方法で測定される。例えば、CDPK2酵素とATPとを混合してリン酸化反応を行わせた後、リン酸基の脱離により生成したADPをホスホクレアチンとクレアチンキナーゼとを使用して定量することによりタンパク質リン酸化活性を測定することができる。また、CDPK2酵素とγ位のリン原子が放射性同位体であるATP(例えば[γ−32P]ATP)とを混合して反応させた後、タンパク質に結合した放射能を定量することによりタンパク質リン酸化活性を定量することができる。リン酸化活性測定システムで商業的に入手可能なものには、Protein Kinase C Enzyme Assay(GEヘルスケアバイオサイエンス株式会社、カタログ番号:RPN77)、SignaTECT(商標)CaM KII Assay System(プロメガ株式会社、カタログ番号:V8161)等が含まれるが、これらに限定されない。
【0037】
本発明のタンパク質リン酸化活性増強剤の製造方法では、前記リン酸化活性増強剤の評価方法のステップ(B)に供試された複数のリン酸化活性増強剤候補化合物の中から、測定されたリン酸化活性が高い候補化合物をタンパク質リン酸化活性増強剤として選択する場合がある。
【0038】
本発明のタンパク質リン酸化活性増強剤候補化合物はアルタナル酸の誘導体の場合がある。アルタナル酸は除草剤であるから、本発明のタンパク質リン酸化活性増強剤の製造方法により除草剤が製造される場合がある。したがって、本発明は除草剤の製造方法を提供する。アルタナル酸の誘導体は、アルタナル酸の各原子にさまざまな修飾を加えることによって合成される。
【0039】
本発明の除草剤の製造方法における前記リン酸化活性増強剤候補化合物を選択するステップでは、前記リン酸化活性増強剤の評価方法のステップ(2)に供試された複数の前記タンパク質リン酸化活性増強剤候補化合物のうち特定の植物種由来のCDPK2酵素に対するタンパク質リン酸化活性増強作用が他の植物種由来CDPK2酵素に対するタンパク質リン酸化活性増強作用に比べて弱い候補化合物を選択するステップを含む場合がある。このようにして選択されたリン酸化活性増強剤は前記特定の植物種の生育を阻害しない除草剤の場合がある。前記特定の植物種がジャガイモの場合には、前記他の植物種はジャガイモ畑に繁茂しうるジャガイモ以外のいずれかの植物であり、本発明の除草剤の製造方法はジャガイモの生育を阻害せずに雑草に有効な除草剤を製造することができる。
【0040】
本発明において雑草とは、作物、花卉園芸等として市場価値がある植物以外の植物をいい、ジャガイモについて雑草とはジャガイモ以外の植物をいう。本発明の雑草には、ウキクサ(duckweeds)、アカザ(redroot pigweed)、イヌホオズキ(black nightshade)、白花ようしゅちょうせんあさがお(jimsonweed)、タデ科雑草(northen jointvetch)、マメ科雑草、ウマゴヤシ、クローバー等が含まれるが、これらに限定されない。前記雑草のモデル植物としてシロイヌナズナ又はタバコを用いる場合がある。シロイヌナズナ及びタバコ由来のCDPK2タンパク質のアミノ酸配列をそれぞれ配列番号3及び4に示す。本発明の除草剤の製造方法では、シロイヌナズナ又はタバコ由来のCDPK2タンパク質を用いる本発明の評価方法によってタンパク質リン酸化活性増強作用が強力な候補化合物を選択し、該候補化合物のうち、ジャガイモ由来のRiCDPK2タンパク質を用いる本発明の評価方法によってタンパク質リン酸化活性増強作用が弱い候補化合物をさらに選択することにより、ジャガイモの生育を阻害しないでジャガイモ以外の植物の生育を阻害する除草剤を製造することができる。
【発明を実施するための最良の形態】
【0041】
以下に示す実施例によって本発明をより詳細に説明するが、これらの実施例は本発明の詳細な説明のために一例として示すものであり、これらの実施例によって本発明の範囲が制限されると理解するべきではない。
【実施例1】
【0042】
1.トマト葉に及ぼすアルタナル酸処理の影響の検討
1.1 方法
(生物学的材料の調製)
トマト植物として、品種「Fukuju II」を供試した。ジャガイモ植物として、品種「Rishiri」の塊茎を4°Cで保存し供試した。Alternaria solaniを、ジャガイモ−ブドウ糖培地にて保存し供試した。
【0043】
(アルタナル酸の精製)
Langsdorf,G.,Furuichi,N.and Nishimura,S.(1989)Evaluation of Alternaric acid in pathogenesis of Alternaria solani.In Host−specific Toxins: Recognition and Specificity Factors in Plant Disease(Kohmoto,K.and Durbin,R.D.eds.).pp.45−58,Tottori University Press,Tottori.及びFuruichi,N.,Nishimura,S.and Langsdorf,G.(1992)Effect of alternaric acid,a toxin of Alternaria solani,on the hypersensitive response of potato to Phytophthora infestans.Ann.Phytopathol.Soc.Jpn.58:1−7に記載の方法に従って、アルタナル酸(Alternaric acid)をA.solaniの培養液から精製した。具体的には、A.solaniを、ジャガイモブドウ糖培地で25°C、25日間増殖させ、A.solani培養液を得た。該培養液は和光ゲルC200シリカゲルを用いてシリカゲルカラムクロマトグラフィー法で分画し、クロロホルム:エタノール(体積比19:1)の混合液で溶出した。アルタナル酸は0.5%アセトンを含む水溶液に溶解して保存した。
【0044】
(トマト葉のアルタナル酸処理)
完全に成長したFukujuII品種のトマトの複葉を0.1%次亜塩素酸ナトリウムで消毒し、蒸留水で複数回リンスした後、植物1本あたり2枚の葉の表面を15mm2未満の面積だけ均一に針で穿孔した。その後、前記葉をアルタナル酸を0、0.1、0.25、2.5及び25μMそれぞれ添加した0.5%アセトンを含む水溶液30μLで処理し、昼14時間、23°C、無菌条件下でインキュベーションした。葉のネクローシスの有無はアルタナル酸処理の24時間後に評価した。
【0045】
1.2 結果
図2Aはそれぞれの濃度のアルタナル酸で処理された葉表面の写真である。図2A−a、図2A−b、図2A−c、図2A−dはそれぞれ、25μM、2.5μM、0.25μM、0.1μMのアルタナル酸溶液で処理したトマトの葉である。図2A−eは対照実験のアルタナル酸を含まない0.5%アセトンを含む水溶液で処理したトマトの葉である。アルタナル酸での処理による特徴的な症状として、トマトの葉の葉脈につづく葉脈間のネクローシスと、広範な白化とが観察された。トマト葉でのネクローシスの重篤度は、処理液中のアルタナル酸の濃度に依存し(図2A)、0.25μMより高い濃度のアルタナル酸溶液で処理した場合には処理後24時間以内に静脈のネクローシスが肉眼で観察され(図2A−a、図2A−b、図2A−c)、25μMのアルタナル酸溶液で処理したときに最も重度なネクローシスが観察された(図2A−a)。しかし、0.1μMのアルタナル酸溶液処理でも葉の処理部位の黄変がみられた(図2A−d)。これは、A.solaniをジャガイモの葉に感染させたときの徴候に類似していた。なお、本条件下での処理の24時間後に顕微鏡で観察したところ、アルタナル酸で処理されたトマト細胞の過敏感細胞死の発生は、観察されなかった。これにより、アルタナル酸は宿主植物の過敏感反応を抑制することが確認された。本実施例により、A.solaniは宿主植物組織の過敏感反応を抑制することによりネクローシスを進行させ感染を成立させることと、A.solaniが生成するアルタナル酸が感染成立に関与する成分であることとが確認された。
【実施例2】
【0046】
2.トマト葉に及ぼすA.solaniの接種の影響の検討
2.1 方法
アルタナル酸処理の実験と同じように消毒、リンス及び穿孔されたトマトの葉をA.solani生菌で処理して感染させた。
【0047】
2.2 結果
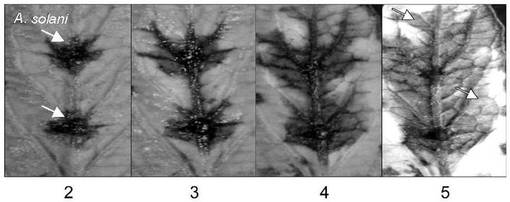
図2B−2、2B−3、2B−4及び2B−5は、それぞれA.solani感染後2、3,4及び5日後のトマト葉表面の写真である。図2Bから、A.solaniは接種2日後にトマト葉で症状を引き起こし(図2B−2)、感染後5日間にネクローシスが広がった(図2B−5)。これによりA.solani生菌がネクローシスを誘発し感染を成立させることが確認された。
【実施例3】
【0048】
3.RiCDPK2cDNAの単離及びRiCDPK2融合タンパク質の発現
3.1 方法
(cDNAライブラリーの調製)
全RNAをRishiri品種のジャガイモの葉及び根茎から単離し、オリゴdTセルロースを用いるクロマトグラフィによってポリA+−RNAを精製した。該ポリA+−RNAからオリゴdTプライマー及びcDNA synthesis System Plus(アマシャムバイオサイエンス株式会社)を使って2本鎖cDNAを合成した。前記2本鎖cDNAとラムダgt11ファージをベクターとからcDNAライブラリを構築した。得られたcDNAライブラリからDNAを抽出して、これを鋳型として、配列番号3及び4に列挙されたヌクレオチド配列を有するプライマーを用いてRishiri品種のジャガイモCDPK2(以下、「RiCDPK2」という。)をエンコードするcDNAを増幅した。
【0049】
(RiCDPK2融合タンパク質発現ベクターの構築)
配列番号1のアミノ酸配列を有するRiCDPK2タンパク質のC末端に6個のヒスチジン残基からなるHisタグが連結した融合タンパク質(以下、「His−RiCDPK2」という。)が発現できるようにRiCDPK2のcDNAをpCRT7CT TOPOベクターにクローン化した。Sambrook,J.及びRussell,D.W.(2001、Molecular Cloning:A Laboratory Manual,Vol.1,Cold Spring Harbor Laboratory Press,New York.)に記載の方法に従って、His−RiCDPK2を発現するpCTT7CT TOPOコンストラクトで大腸菌(BL21、pLysS)細胞を形質転換し、His−RiCDPK2を発現させた。
【0050】
(His−RiCDPK2の調製)
発現したHis−RiCDPK2の精製は、Xpress(商標)Systemタンパク質精製プロトコール(インビトロジェン株式会社)に従って行なった。アンピシリン70μg/mlと形質転換された大腸菌細胞とを含むLuria−Bertani(LB)培地に0.6mMのIPTGを添加することによりタンパク質発現を誘導した。前記大腸菌細胞を24時間、25°Cで振とう培養し、RPR−20ローター(株式会社日立ハイテクノロジーズ)で4°C、10分間、4000rpm遠心分離して回収した。回収した大腸菌をグアニジウムを含む溶菌バッファーで可溶化し、RPR−20ローターで4°C、15分間、6500rpmの遠心分離により上清を回収した。該上清を前記タンパク質精製プロトコールに従ってヒスチジン結合樹脂カラムに通し、His−RiCDPK2を溶出した。溶出されたHis−RiCDPK2を10mMのTris−HCl(pH8.0)及び0.1%のTriton X−100を含む水溶液に対して透析した。透析後のHis−RiCDPK2溶液中のタンパク質濃度は、Bradfordの方法(Bradford,M.M.(1976)A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein−dye binding.Analytical Biochem.72:248−254.)に従って、ウシ血清アルブミン(BSA)を標準としてBio−Rad assay kitを使用することにより決定した。
【0051】
(SDS−PAGE及びウエスタンイムノブロット分析)
His−RiCDPK2の純度の確認のために、Leammli,U.K.et al.,Nature Vol.227,pp.680−681(1970)に記載の方法に従って、ドデシル硫酸ナトリウム−ポリアクリルアミドゲル電気泳動をminiPAGE System(アトー株式会社)を用いて分離し、クマシーブリリアントブルー(CBB)G−250により染色し検出した。ウエスタンイムノブロット分析のため、電気泳動で分離されたタンパク質を、ポリビニリデンジフルオリド(PVDF)膜(Immoblin(商標)P、孔径0.45μm、日本ミリポア株式会社)に写しとった。前記膜を、ブロッキング用緩衝液(3%(w/v)BSA、TBS(20mM TrisHCl)でブロッキングし、希釈用緩衝液(1%のBSA及び0.1%のTriton X−100を添加したTBS)で1/5000に希釈した抗His(C末端)−マウスモノクローナルIgG2b抗体とともに終夜インキュベーションを行なった。前記膜をTBST(0.1%のTriton X−100を添加したTBS)で2回洗浄した後、250倍に希釈したアルカリホスフェターゼ結合ラビット抗マウス抗体(DAKO D0314、ダコ ジャパン株式会社)とともに室温にて1時間インキュベーションを行なった。前記膜をTBSTで洗浄した後、アルカリホスファターゼ緩衝液(0.1M Tris−HCl、0.1M NaCl、5mM MgCl2、pH9.5)で洗浄し、AP結合基質キット(バイオラッドラボラトリーズ株式会社)を用いて、抗原−抗体複合体を検出した。
【0052】
3.2 結果
RiCDPK2のヌクレオチド配列を配列番号2に示す。RiCDPK2がエンコードするタンパク質のアミノ酸配列を配列番号1に示す。His−RiCDPK2は配列番号1のRiCDPK2タンパク質のN末端に6個のヒスチジン残基からなるHisタグが連結した融合タンパク質である。IPTG誘導によりHis−RiCDPK2を過剰発現させた大腸菌の培養200mLから、約100μgのHis−RiCDPK2が精製された。His−RiCDPK2融合タンパク質は主に不溶性タンパク質画分で回収された。精製後の純度はSDS−PAGEの泳動バンドから判断して約90%であった。
【実施例4】
【0053】
4.Ca2+イオン存在下及び非存在下でのHis−RiCDPK2の酵素活性に対するアルタナル酸の影響の検討
4.1 方法
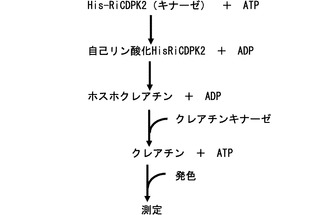
His−RiCDPK2の酵素活性は、Furuichi,N.、Anderson,A.J.、Suzuki,Y.及びTakemoto,J.Y.(1994、Elicitor and suppressor of Phytophthora infestans stimulate phosphorylation of plasma membrane proteins from potato and bean tissues.In Host Specific Toxin:Biosynthesis,Receptor and Molecular Biology(Kohmoto,K.and Yoder,O.C.,eds.).pp.273−274、Tottori University Press、Tottori)に記載の方法に従って実施された。図3に酵素活性のアッセイ系の概略を示す。簡潔には、96ウェルマイクロタイタープレートを使用して、1処理当たり1個のウェルに155μLの反応混合液(8.5mM Tris−HCl(pH7.1)、5mM ホスホクレアチン、0.4単位 クレアチンホスホキナーゼ(シグマ アルドリッチ ジャパン株式会社)、1.5μg His−RiCDPK2)を添加した。アルタナル酸及びCa2+イオンの効果を調べる実験にはそれぞれ25μMのアルタナル酸及び0.1mM CaCl2を前記反応混合液に添加した。酵素反応は0.9mM ATPを添加することによって開始され、30°C、10分間継続された。酵素反応は、0.2% 1−ナフトール(和光純薬工業株式会社)及び0.06% 2,3−ブタンジオン(和光純薬工業株式会社)を含むストックアルカリ溶液(1.5M NaOH、0.7M NaHCO3)を各ウェルに添加して発色させた。酵素活性の定量は、マイクロプレートリーダー(バイオラッド3500、バイオラッドラボラトリーズ株式会社)を用いて595nmの吸光度を10分ごとには50分後まで測定することによって実施された。吸光度の測定と次の測定との間、酵素反応に用いるマイクロタイタープレートは30°Cに保温された。
【0054】
4.2 結果
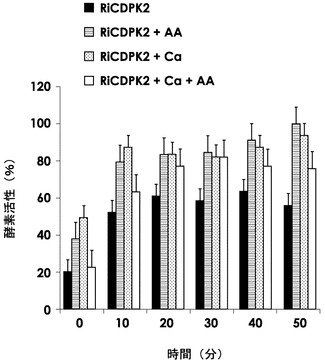
図4は、アルタナル酸及びCa2+イオンの存在下におけるHis−RiCDPK2のキナーゼ活性を、キナーゼ反応で生じるADP量に対応する吸光度の相対値で表した棒グラフである。アルタナル酸存在下で50分間反応させた場合の吸光度を100%として、アルタナル酸及びCa2+イオンとも非存在下(黒ベタ塗り)か、あるいは、アルタナル酸単独存在下(横線ハッチング)か、Ca2+イオン単独存在下(網掛けハッチング)か、アルタナル酸及びCa2+イオンの両方の存在下(白)かのそれぞれの2回の異なる実験の結果の0分から50分までの10分おきの平均と標準偏差の上限とを示す。図4のグラフから、His−RiCDPK2のキナーゼ活性がCa2+イオン存在下で増強されることから、カルシウム依存性タンパク質リン酸化活性があること、及び、アルタナル酸がHis−RiCDPK2のキナーゼ活性を増強することが示された。
【実施例5】
【0055】
5.Ca2+及びMg2+イオン存在下及び非存在下でのHis−RiCDPK2の酵素活性に対するアルタナル酸の影響の検討
5.1 方法
アルタナル酸がMg2+イオン存在下でもHis−RiCDPK2のタンパク質リン酸化活性に影響を与えるかどうかを調べるために、25μMのアルタナル酸、100μMのCaCl2及び900μMのMgCl2を前記反応混合液に添加することを除いて、実施例4に記載のアッセイ系と同じ条件で実験を行なった。
【0056】
5.2 結果
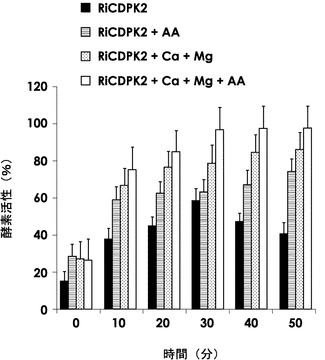
図5は、アルタナル酸、Ca2+及びMg2+イオンの存在下におけるHis−RiCDPK2のキナーゼ活性を、キナーゼ反応で生じるADP量に対応する吸光度の相対値で表した棒グラフである。アルタナル酸、Ca2+及びMg2+イオン存在下で50分間反応させた場合の吸光度を100%として、アルタナル酸及びCa2+イオンとも非存在下(黒ベタ塗り)か、あるいは、アルタナル酸単独存在下(横線ハッチング)か、Ca2+及びMg2+イオン存在下(網掛けハッチング)か、アルタナル酸とCa2+及びMg2+イオンとの存在下(白)かのそれぞれの2回の異なる実験の結果の0分から50分までの10分おきの平均と標準偏差の上限とを示す。図5のグラフから、His−RiCDPK2のキナーゼ活性がCa2+及びMg2+イオン存在下で増強されること、及び、Ca2+及びMg2+イオンの有無にかかわらずアルタナル酸がHis−RiCDPK2のキナーゼ活性を増強することが示された。
【図面の簡単な説明】
【0057】
【図1】アルタナル酸の化学構造式。
【図2A】異なる濃度のアルタナル酸で処理された葉表面の写真。
【図2B】A.solani感染後2ないし5日後のトマト葉表面の写真。
【図3】酵素活性のアッセイ系の模式図。
【図4】アルタナル酸及びCa2+イオンの存在下におけるHis−RiCDPK2のキナーゼ活性を、キナーゼ反応で生じるADP量に対応する吸光度の相対値で表した棒グラフ。
【図5】アルタナル酸、Ca2+及びMg2+イオンの存在下におけるHis−RiCDPK2のキナーゼ活性を、キナーゼ反応で生じるADP量に対応する吸光度の相対値で表した棒グラフ。
【技術分野】
【0001】
本発明は、新規なタンパク質リン酸化活性増強作用の評価方法と、該タンパク質リン酸化活性増強作用を指標とする除草剤の製造方法に関する。
【背景技術】
【0002】
アルタナル酸(Alternaric acid)は、ジャガイモ又はトマトの胴枯れ病の原因となる真菌であるAlternaria solaniにより産生される毒素であり、図1に示す構造を有する。アルタナル酸は、宿主特異性の決定と、Alternaria solaniにより引き起こされる疾病の発病とに関わることが報告されている(非特許文献1及び2)。またジャガイモの塊茎の切片に塗布されたアルタナル酸がPhytophthora infestansの非親和性レースによる感染後の宿主植物の過敏感反応(Hypersensitive Response)の発生を遅延させることから、アルタナル酸は過敏感反応のサプレッサーであることが報告されている(非特許文献3)。アルタナル酸は除草剤としての用途があることが知られている(特許文献1)。
【特許文献1】米国特許第5,256,628号明細書
【非特許文献1】Furuichi,N.and Nishimura,S.(1984)Isolation of alternaric acid from the germination fluids of Alternaria solani. Ann.hytopathol.Soc.Jpn.50:128(Abstr.in Japanese).
【非特許文献2】Langsdorf,G.,Furuichi,N.,Doke,N.and Nishimura,S.(1990)Investigations on Alternaria solani infections:Detection of alternaric acid and a susceptibility inducing factor in the spore−germination fluid of A.solani. J.Phytopathology 128:271−282.
【非特許文献3】Furuichi,N.,Nishimura,S. and Langsdorf,G.(1992)Effect of alternaric acid,a toxin of Alternaria solani,on the hypersensitive response of potato to Phytophthora infestans. Ann.Phytopathol.Soc.Jpn.58:1−7
【0003】
過敏感反応サプレッサーで処理後のジャガイモのさまざまなタンパク質がカルシウム依存性のリン酸化により修飾されることが報告されている(非特許文献4)。また、カルシウム依存性タンパク質リン酸化酵素(以下、「CDPK」という。)は、遺伝子発現、代謝、膜輸送及び細胞構造の制御に関与する複数のアイソフォームを有する多機能酵素である(非特許文献5)。
【非特許文献4】Furuichi,N.,Anderson,A.J.,Suzuki,Y.and Takemoto,J.Y.(1994)Elicitor and suppressor of Phytophthora infestans stimulate phosphorylation of plasma membrane proteins from potato and bean tissues. In Host Specific Toxin:Biosynthesis,Receptor and Molecular Biology (Kohmoto,K.and Yoder,O.C.,eds.).pp.273−274,Tottori University Press,Tottori.
【非特許文献5】Harmon,A.C.,Gribskov,M.and Harper,J.F.(2000)CDPKs−a kinase for every Ca2+ Signal? Trends Plant Sci.5:154−159.
【0004】
最近、病原菌由来の過敏感反応サプレッサーによる過敏感反応の抑制と、前記病原菌による病気の宿主組織全体での発症とにジャガイモ由来のCDPKが関与することが発表された(特許文献2)。それによると、過敏感反応が抑制される過程は、(1)過敏感反応サプレッサーが宿主細胞壁及び形質膜を貫通する段階と、(2)前記過敏感反応サプレッサーとCDPK1の受容体部位とが結合する段階と、(3)リン酸化反応を介してCDPK2がシグナル伝達を行う段階と、(4)過敏感反応が抑制されることにより宿主組織全体が疫病に罹患する段階とを含む。
【特許文献2】公開特許公報(特開2007−68439)明細書
【0005】
本発明はアルタナル酸がCDPK2のタンパク質リン酸化活性を増強する作用を有することの発見に基づいて、新規な除草剤を開発することを意図する。
【発明の開示】
【発明が解決しようとする課題】
【0006】
従来の除草剤は市場価値のある植物の生育を阻害せずに雑草の生育を阻害することができない。特定の除草剤に対する耐性遺伝子を導入したトランスジェニック植物は該特定の除草剤によって生育が阻害されないが、市場価値のある全ての植物に前記耐性遺伝子を導入することは事実上不可能に近い。そこで、除草剤の作用機序の知見に基づいて、より強力な除草剤及び市場価値のある植物の生育を阻害せずに雑草の生育を阻害する除草剤を製造する方法を開発する必要がある。
【課題を解決するための手段】
【0007】
本発明はタンパク質リン酸化活性増強作用の評価方法を提供する。本発明のタンパク質リン酸化活性増強作用の評価方法は、
(A)(1)配列番号1のアミノ酸配列からなるCDPK2酵素タンパク質と、(2)前記アミノ酸配列において1個若しくは数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列からなり、かつ、カルシウム依存性タンパク質リン酸化活性を有するタンパク質と、(3)前記アミノ酸配列において1個若しくは数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列からなり、かつ、アルタナル酸により増強される前記タンパク質リン酸化活性を有するタンパク質とからなるグループから選択されることを特徴とするタンパク質と、(4)前記(1)ないし(3)のタンパク質からなる第1ポリペプチドと、特異的結合タグとからなる第2ポリペプチドとが連結した融合タンパク質とのうちのいずれか1つのタンパク質と、タンパク質リン酸化活性増強剤候補化合物とを用意するステップと、
(B)前記タンパク質又は融合タンパク質と、前記タンパク質リン酸化活性増強剤候補化合物とを含む反応液中で、タンパク質リン酸化活性を測定するステップとを含む。本発明の評価方法は、前記候補化合物の存在下と非存在下とにおける前記タンパク質又は融合タンパク質のタンパク質リン酸化活性を比較すること、及び、前記候補化合物存在下でのタンパク質リン酸化活性が非存在下でのタンパク質リン酸化活性よりどの程度高いかを評価することを含む場合がある。本発明のタンパク質リン酸化活性増強作用の評価方法に用いる反応液にはCa2+及びMg2+イオンを含む場合がある。
【0008】
第2ポリペプチドの特異的結合タグは、Hisタグ、mycタグ、HAタグ、インテインタグ、MBP又はGSTからなるグループから選択される場合がある。
【0009】
本発明の評価方法でタンパク質リン酸化活性増強作用が評価されるタンパク質リン酸化活性増強剤候補化合物はアルタナル酸の誘導体の場合がある。
【0010】
本発明は、本発明の評価方法を用いて複数の前記タンパク質リン酸化活性増強剤候補化合物のうちタンパク質リン酸化活性増強作用が強力な候補化合物を除草剤として選択するステップを含むことを特徴とする除草剤の製造方法を提供する。
【0011】
本発明は、本発明の評価方法によって複数の前記タンパク質リン酸化活性増強剤候補化合物のうち特定の植物種由来のCDPK2酵素に対するタンパク質リン酸化活性増強作用が他の植物種由来CDPK2酵素に対するタンパク質リン酸化活性増強作用に比べて弱い候補化合物を除草剤として選択するステップを含むことを特徴とする除草剤の製造方法を提供する。
【0012】
本発明の除草剤の製造方法において、前記特定の植物種はジャガイモの場合があり、前記他の植物種はシロイヌナズナ又はタバコの場合がある。
【0013】
本発明はタンパク質リン酸化活性増強剤候補化合物のタンパク質リン酸化活性増強作用の評価キットを提供する。本発明の評価キットは、(A)(1)配列番号1のアミノ酸配列からなるCDPK2酵素タンパク質と、(2)前記アミノ酸配列において1個若しくは数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列からなり、かつ、カルシウム依存性タンパク質リン酸化活性を有するタンパク質と、(3)前記アミノ酸配列において1個若しくは数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列からなり、かつ、アルタナル酸により増強される前記タンパク質リン酸化活性を有するタンパク質とからなるグループから選択されることを特徴とするタンパク質と、(4)前記(1)ないし(3)のタンパク質からなる第1ポリペプチドと、特異的結合タグとからなる第2ポリペプチドとが連結した融合タンパク質とのうちのいずれか1つのタンパク質とを含むことを特徴とする。
【0014】
本発明のタンパク質リン酸化活性増強作用の評価キットにおいて、前記特異的結合タグは、Hisタグ、mycタグ、HAタグ、インテインタグ、MBP又はGSTからなるグループから選択される場合がある。前記タンパク質リン酸化活性増強剤候補化合物はアルタナル酸の誘導体である場合がある。
【0015】
本明細書で用いられるところの「タンパク質」とは、2個以上のアミノ酸がペプチド結合で連結した化合物である。タンパク質は、メチル基、チオール基、リン酸、糖鎖、及び/又は、その他の共有結合による修飾を含む場合がある。本明細書で用いられるところの「ポリペプチド」とは、2個以上のアミノ酸がペプチド結合で連結した化合物である。
【0016】
本発明のタンパク質は、本発明のタンパク質をエンコードするヌクレオチド配列からなるDNAを、無生物発現系か、宿主生物及び発現ベクターを使用する発現系かで発現させることにより産生される。本発明の融合タンパク質は、第1ポリペプチドをエンコードするヌクレオチド配列からなるDNAと、第2ポリペプチドをエンコードするヌクレオチド配列からなるDNAとが連結されたDNAを、無生物発現系か、宿主生物及び発現ベクターを使用する発現系かで発現させることにより産生される。前記宿主生物は、大腸菌、枯草菌等のような原核生物と、酵母、菌類、植物、動物等のような真核生物とを含む。本発明の宿主生物及び発現ベクターを使用する発現系は、細胞や組織のような生物の一部か、生物の個体全体かの場合がある。
【0017】
配列番号1のアミノ酸配列からなるカルシウム依存性タンパク質リン酸化酵素CDPK2は、Rishiri品種のジャガイモから単離された。配列番号1のアミノ酸配列は、DDBJ/GenBank/EMBLのデータベースにアクセッション番号AB0551809として登録された。
【0018】
本明細書で用いられるところの「第1ポリペプチド」とは、所望の機能、すなわち、タンパク質リン酸化活性を有するポリペプチドである。本明細書で用いられるところの「第2ポリペプチド」とは、発現タンパク質の分離、精製又は検出をより容易に行うために、第1ポリペプチドとペプチド結合で連結されるポリペプチドである。第2ポリペプチドは、第1ポリペプチドのN末端又はC末端に連結される場合がある。
【0019】
本発明の第1ポリペプチドは、配列番号1のアミノ酸配列からなるRiCDPK2酵素タンパク質か、前記アミノ酸配列において1個又は2個以上のアミノ酸が欠失、置換又は付加されたアミノ酸配列からなり、かつ、カルシウム依存性タンパク質リン酸化活性を有するタンパク質かの場合がある。本発明の第1ポリペプチドのタンパク質リン酸化活性はアルタナル酸により増強される場合がある。
【0020】
本発明の酵素のタンパク質リン酸化活性は、該酵素タンパク質自体が基質になる場合の他、他のタンパク質又はポリペプチドを基質とする場合がある。本発明のカルシウム依存性タンパク質リン酸化活性とは、本発明の酵素のタンパク質リン酸化活性がCa2+イオン存在下では非存在下でよりも高い、すなわち、増強されることをいう。本発明のタンパク質リン酸化活性増強剤は、該増強剤の存在下で非存在下でよりもタンパク質リン酸化活性を増強する。本発明のタンパク質リン酸化活性増強剤候補化合物は、本発明のタンパク質リン酸化活性増強剤となる可能性がある化合物であって、本発明の評価方法によってタンパク質リン酸化活性増強作用が強力なものが本発明のタンパク質リン酸化活性増強剤として選択される。本発明のタンパク質リン酸化活性増強剤の製造方法は、本発明の評価方法を用いて本発明のタンパク質リン酸化活性増強剤候補化合物のタンパク質リン酸化活性増強作用を評価して、該作用が強力なものを本発明のタンパク質リン酸化活性増強剤として選択するステップを含む。
【0021】
第2ポリペプチドは特異的結合タグからなる。本発明で用いられるところの「特異的結合タグ」とは、所望の機能を有するポリペプチドを遺伝子組み換えによって調製する際に、前記所望の機能を有するポリペプチドとペプチド結合で連結した融合タンパク質として発現させることにより、形質転換体からの発現タンパク質の分離、精製又は検出をより容易に行うことを可能にするために、他のタンパク質、多糖類、糖脂質、核酸及びこれらの誘導体、樹脂、シリコン等と特異的に結合するポリペプチドである。特異的結合タグは、水溶液中に溶解した物質又は固体支持体と結合する場合がある。本発明の特異的結合タグは、Hisタグ、mycタグ、HAタグ、インテインタグ、MBP、GSTその他これらに類するポリペプチドが含まれるが、これらに限定されない。前記特異的結合タグは、Hisタグ、mycタグ、HAタグ、インテインタグ、MBP又はGSTからなるグループから選択される場合がある。
【0022】
本発明で用いられるところの「Hisタグ」とは、ヒスチジン残基が連続するアミノ酸配列からなるタグである。前記ヒスチジン残基の数は少なくとも5個、好ましくは6ないし10個、最も好ましくは6個である。Hisタグを連結した融合タンパク質は、Hisタグと金属イオンとキレート化合物とが錯体を形成することを利用して分離、精製される。本発明で用いられるところの「キレート化合物」とは、金属イオンと特異的に配位結合をする原子団をいう。前記キレート化合物は、ニトリロ三酢酸又はイミノ二酢酸の場合があるがこれらに限定されない。前記キレート化合物は、直接的に、又はスペーサーを介して間接的に、カラム担体等の固体支持体に固定化される。前記金属イオンは、Ni2+イオン又はCo2+イオンのような遷移金属イオンの場合があるがこれらに限定されない。Hisタグを利用するタンパク質精製用の前記金属イオンを捕捉したキレート化合物を固定化した固体支持体は、Ni−NTAアガロースとして、インビトロジェン、キアゲン社等から入手することができる。Hisタグを利用するタンパク質精製システムで商業的に入手可能なものには、Ni−NTA精製システム(インビトロジェン株式会社、カタログ番号:K95001)、Ni−NTA Fast Start Kit(株式会社キアゲン、カタログ番号:30600)等が含まれるがこれらに限定されない。
【0023】
Hisタグを連結した融合タンパク質の精製の手順は、(1)前記金属イオンを捕捉したキレート化合物を固定化した固体支持体を用意するステップと、(2)前記固体支持体と前記融合タンパク質とを接触させることにより、前記金属イオンと融合タンパク質のHisタグ部分のイミダゾール基を構成する窒素との間で配位結合を形成させ、前記融合タンパク質を前記固体支持体に捕捉するステップと、(3)前記固体支持体を洗浄して不要成分を除去するステップと、(4)遊離イミダゾール化合物を含む溶出液を添加することにより前記融合タンパク質を前記固体支持体から解離させて回収するステップとを含むのが一般的である。
【0024】
本発明で用いられるところの「mycタグ」とは、ヒトc−mycタンパク質の410番−419番の10個のアミノ酸の配列に対応する、配列番号3のアミノ酸配列からなるタグである。mycタグを連結した融合タンパク質は、mycタグと該タグに特異的に結合する抗体との抗原抗体複合体形成を利用して分離、精製される。本発明で用いられるところの「mycタグペプチド」とは、ヒトc−mycタンパク質の410番−419番の10個のアミノ酸の配列に対応する、配列番号3のアミノ酸配列からなるポリペプチドである。本発明で用いられるところの「抗体」は、ポリクローナル抗体及びモノクローナル抗体を含む。抗mycタグモノクローナル抗体は、例えば、ハイブリドーマ細胞9E10株により産生される。mycタグを利用するタンパク質精製システムで商業的に入手可能なものには、ANTI−c−Myc AGAROSE CONJUGATE(シグマアルドリッチ ジャパン株式会社、カタログ番号:A7470)、c−Myc Peptide(シグマアルドリッチ ジャパン株式会社、カタログ番号:M2435)、c−Myc Monoclonal Antibody−Agarose Beads(クロンテック、タカラバイオ株式会社、カタログ番号:3843−1)等が含まれるが、これらに限定されない。
【0025】
mycタグを連結した融合タンパク質の精製の手順は、(1)抗mycタグ抗体が固定化された固体支持体を用意するステップと、(2)前記固体支持体と前記融合タンパク質とを接触させることにより、前記融合タンパク質のmycタグ部分と前記抗体との間で複合体を形成させて、前記融合タンパク質を前記固体支持体に捕捉するステップと、(3)前記固体支持体を洗浄し不要成分を除去するステップと、(4)mycタグペプチドを含む溶出液を添加することにより前記融合タンパク質を前記固体支持体から解離させて回収するステップとを含むのが一般的である。
【0026】
本発明で用いられるところの「HAタグ」とは、ヒトインフルエンザウィルスのヘマグルチニン(HA)の98番−106番の9個のアミノ酸に対応する、配列番号4のアミノ酸配列からなるタグである。HAタグを連結した融合タンパク質は、HAタグと該タグに特異的に結合する抗体との抗原抗体複合体形成を利用して分離、精製される。HAタグに対応するモノクローナル抗体は例えば、ハイブリドーマ細胞HA−7株により産生される。HAタグを利用するタンパク質精製システムで商業的に入手可能なものには、例えばMonoclonal Anti−HA Agarose Conjugate Clone HA−7(シグマアルドリッチ ジャパン株式会社、カタログ番号:A2095)、INFLUENZA HEMAGGLUTININ(HA)PEPTIDE(シグマアルドリッチ ジャパン株式会社、カタログ番号:I2149)等が含まれるがこれらに限定されない。
【0027】
HAタグを連結した融合タンパク質は、mycタグの場合と同様の手順で精製される。
【0028】
本発明で用いられるところの「インテインタグ」とは、インテインにキチン結合ドメイン(CBD)等のリガンド特異的結合ドメインを連結したタグである。インテインタグを連結した融合タンパク質は、キチン結合ドメインとキチンとの結合形成を利用して分離、精製される。インテインの自己切断能力を利用して、ワンステップで前記タグの除去後のタンパク質として精製することができる。インテインタグを利用するタンパク質発現・精製システムで商業的に入手可能なものには、IMPACT(商標) Systems(ニュー・イングランド・バイオラボ・ジャパン株式会社、カタログ番号:E6900S)が含まれるがこれに限定されない。
【0029】
インテインタグを連結した融合タンパク質の精製の手順は、(1)インテインタグを連結した融合タンパク質の溶液を用意するステップと、(2)前記溶液とキチンが固定化された固体支持体とを接触させることによりインテインタグのキチン結合ドメインとキチンとの間で複合体を形成させ、前記融合タンパク質を前記固体支持体に捕捉させるステップと、(3)前記固体支持体を洗浄し不要成分を除去するステップと、(4)DTT又はβ−メルカプトエタノールのような還元剤の添加によりインテインの自己切断能力を活性化させて融合タンパク質からインテインタグを切断するステップと、(5)インテインタグが切断されたタンパク質を回収するステップとを含むのが一般的である。
【0030】
本発明で用いられるところの「MBP」とは、マルトース結合タンパク質である。MBPを連結した融合タンパク質は、アミロース等とMBPとの結合形成を利用して分離、精製される。MBPを利用するタンパク質発現・精製システムで商業的に入手可能なものには、例えばpMAL(商標) Protein Fusion and Purification System(ニュー・イングランド・バイオラボ・ジャパン株式会社、カタログ番号:#E8000S)が含まれるがこれに限定されない。
【0031】
本発明で用いられるところの「GST」とは、グルタチオンS転移酵素である。GSTを連結した融合タンパク質は、GSTと該酵素の基質であるグルタチオンとの親和性を利用して分離、精製される。GSTを利用するタンパク質精製システムで商業的に入手可能なものには、例えばGlutathione Sepharose 4 Fast Flow(GEヘルスケアバイオサイエンス株式会社、カタログ番号:17−5132−01)、GSTrap FF(GEヘルスケアバイオサイエンス株式会社、カタログ番号:17−5130−02)等が含まれるがこれらに限定されない。
【0032】
好ましくは第2ポリペプチドには、Hisタグ、mycタグ、HAタグ、インテインタグ、MBP又はGSTが選択される。さらに好ましくはHisタグが選択される。
【0033】
本発明の融合タンパク質は、第2ポリペプチドに加えて第3ポリペプチドが第1ポリペプチドに連結される場合がある。本明細書で用いられるところの「第3ポリペプチド」とは、第1ポリペプチドが第2ポリペプチドに連結されている場合にさらに連結されるポリペプチドであって、第1ポリペプチドを遺伝子組み換えによって調製する際に前記第3ポリペプチドと前記第1ポリペプチドとを連結した融合タンパク質として発現させることにより、形質転換体からの発現タンパク質の分離、精製又は検出をより容易に行うことを可能にする場合があるポリペプチドである。第3ポリペプチドは、形質転換体での発現タンパク質の分泌、安定化を可能にする場合がある。第3ポリペプチドは、第1ポリペプチドに対して第2ポリペプチドと同じ側か反対側かに連結される場合がある。第3ポリペプチドには、Hisタグ、mycタグ、HAタグ、インテインタグ、MBP、GSTの他、シグナルペプチド、プロテインA、Trxタグ、CBPタグ、CBDタグ、CBRタグ、luc、HSP、LacZ、IgG、アビジン、プロテインG等が含まれるがこれらに限定されない。
【0034】
本発明のタンパク質リン酸化活性増強作用の評価方法のステップ(A)で用意されるタンパク質リン酸化活性増強剤候補化合物は、ステップ(B)のリン酸化活性の測定に供試できるものであればよく、特に限定されない。前記タンパク質リン酸化活性増強剤候補化合物は、アルタナル酸の誘導体の場合がある。
【0035】
本発明のタンパク質リン酸化活性増強作用の評価方法のステップ(B)では、該ステップのリン酸化活性測定において前記リン酸化活性増強剤誘導体候補化合物によるリン酸化活性を増強させ、該候補化合物間のリン酸化活性の差異をより明確に把握するために、Ca2+及びMg2+イオンを添加する場合がある。前記Ca2+及びMg2+イオンの濃度は、リン酸化活性測定に問題が生じない濃度であれば特に限定されない。Ca2+イオンの濃度は100μMの場合がある。Mg2+イオンの濃度は900μMの場合がある。
【0036】
本発明のタンパク質リン酸化活性増強作用の評価方法のステップ(B)のタンパク質リン酸化活性は、CDPK2酵素によるタンパク質リン酸基転移反応を適切に測定できるいずれかの方法で測定される。例えば、CDPK2酵素とATPとを混合してリン酸化反応を行わせた後、リン酸基の脱離により生成したADPをホスホクレアチンとクレアチンキナーゼとを使用して定量することによりタンパク質リン酸化活性を測定することができる。また、CDPK2酵素とγ位のリン原子が放射性同位体であるATP(例えば[γ−32P]ATP)とを混合して反応させた後、タンパク質に結合した放射能を定量することによりタンパク質リン酸化活性を定量することができる。リン酸化活性測定システムで商業的に入手可能なものには、Protein Kinase C Enzyme Assay(GEヘルスケアバイオサイエンス株式会社、カタログ番号:RPN77)、SignaTECT(商標)CaM KII Assay System(プロメガ株式会社、カタログ番号:V8161)等が含まれるが、これらに限定されない。
【0037】
本発明のタンパク質リン酸化活性増強剤の製造方法では、前記リン酸化活性増強剤の評価方法のステップ(B)に供試された複数のリン酸化活性増強剤候補化合物の中から、測定されたリン酸化活性が高い候補化合物をタンパク質リン酸化活性増強剤として選択する場合がある。
【0038】
本発明のタンパク質リン酸化活性増強剤候補化合物はアルタナル酸の誘導体の場合がある。アルタナル酸は除草剤であるから、本発明のタンパク質リン酸化活性増強剤の製造方法により除草剤が製造される場合がある。したがって、本発明は除草剤の製造方法を提供する。アルタナル酸の誘導体は、アルタナル酸の各原子にさまざまな修飾を加えることによって合成される。
【0039】
本発明の除草剤の製造方法における前記リン酸化活性増強剤候補化合物を選択するステップでは、前記リン酸化活性増強剤の評価方法のステップ(2)に供試された複数の前記タンパク質リン酸化活性増強剤候補化合物のうち特定の植物種由来のCDPK2酵素に対するタンパク質リン酸化活性増強作用が他の植物種由来CDPK2酵素に対するタンパク質リン酸化活性増強作用に比べて弱い候補化合物を選択するステップを含む場合がある。このようにして選択されたリン酸化活性増強剤は前記特定の植物種の生育を阻害しない除草剤の場合がある。前記特定の植物種がジャガイモの場合には、前記他の植物種はジャガイモ畑に繁茂しうるジャガイモ以外のいずれかの植物であり、本発明の除草剤の製造方法はジャガイモの生育を阻害せずに雑草に有効な除草剤を製造することができる。
【0040】
本発明において雑草とは、作物、花卉園芸等として市場価値がある植物以外の植物をいい、ジャガイモについて雑草とはジャガイモ以外の植物をいう。本発明の雑草には、ウキクサ(duckweeds)、アカザ(redroot pigweed)、イヌホオズキ(black nightshade)、白花ようしゅちょうせんあさがお(jimsonweed)、タデ科雑草(northen jointvetch)、マメ科雑草、ウマゴヤシ、クローバー等が含まれるが、これらに限定されない。前記雑草のモデル植物としてシロイヌナズナ又はタバコを用いる場合がある。シロイヌナズナ及びタバコ由来のCDPK2タンパク質のアミノ酸配列をそれぞれ配列番号3及び4に示す。本発明の除草剤の製造方法では、シロイヌナズナ又はタバコ由来のCDPK2タンパク質を用いる本発明の評価方法によってタンパク質リン酸化活性増強作用が強力な候補化合物を選択し、該候補化合物のうち、ジャガイモ由来のRiCDPK2タンパク質を用いる本発明の評価方法によってタンパク質リン酸化活性増強作用が弱い候補化合物をさらに選択することにより、ジャガイモの生育を阻害しないでジャガイモ以外の植物の生育を阻害する除草剤を製造することができる。
【発明を実施するための最良の形態】
【0041】
以下に示す実施例によって本発明をより詳細に説明するが、これらの実施例は本発明の詳細な説明のために一例として示すものであり、これらの実施例によって本発明の範囲が制限されると理解するべきではない。
【実施例1】
【0042】
1.トマト葉に及ぼすアルタナル酸処理の影響の検討
1.1 方法
(生物学的材料の調製)
トマト植物として、品種「Fukuju II」を供試した。ジャガイモ植物として、品種「Rishiri」の塊茎を4°Cで保存し供試した。Alternaria solaniを、ジャガイモ−ブドウ糖培地にて保存し供試した。
【0043】
(アルタナル酸の精製)
Langsdorf,G.,Furuichi,N.and Nishimura,S.(1989)Evaluation of Alternaric acid in pathogenesis of Alternaria solani.In Host−specific Toxins: Recognition and Specificity Factors in Plant Disease(Kohmoto,K.and Durbin,R.D.eds.).pp.45−58,Tottori University Press,Tottori.及びFuruichi,N.,Nishimura,S.and Langsdorf,G.(1992)Effect of alternaric acid,a toxin of Alternaria solani,on the hypersensitive response of potato to Phytophthora infestans.Ann.Phytopathol.Soc.Jpn.58:1−7に記載の方法に従って、アルタナル酸(Alternaric acid)をA.solaniの培養液から精製した。具体的には、A.solaniを、ジャガイモブドウ糖培地で25°C、25日間増殖させ、A.solani培養液を得た。該培養液は和光ゲルC200シリカゲルを用いてシリカゲルカラムクロマトグラフィー法で分画し、クロロホルム:エタノール(体積比19:1)の混合液で溶出した。アルタナル酸は0.5%アセトンを含む水溶液に溶解して保存した。
【0044】
(トマト葉のアルタナル酸処理)
完全に成長したFukujuII品種のトマトの複葉を0.1%次亜塩素酸ナトリウムで消毒し、蒸留水で複数回リンスした後、植物1本あたり2枚の葉の表面を15mm2未満の面積だけ均一に針で穿孔した。その後、前記葉をアルタナル酸を0、0.1、0.25、2.5及び25μMそれぞれ添加した0.5%アセトンを含む水溶液30μLで処理し、昼14時間、23°C、無菌条件下でインキュベーションした。葉のネクローシスの有無はアルタナル酸処理の24時間後に評価した。
【0045】
1.2 結果
図2Aはそれぞれの濃度のアルタナル酸で処理された葉表面の写真である。図2A−a、図2A−b、図2A−c、図2A−dはそれぞれ、25μM、2.5μM、0.25μM、0.1μMのアルタナル酸溶液で処理したトマトの葉である。図2A−eは対照実験のアルタナル酸を含まない0.5%アセトンを含む水溶液で処理したトマトの葉である。アルタナル酸での処理による特徴的な症状として、トマトの葉の葉脈につづく葉脈間のネクローシスと、広範な白化とが観察された。トマト葉でのネクローシスの重篤度は、処理液中のアルタナル酸の濃度に依存し(図2A)、0.25μMより高い濃度のアルタナル酸溶液で処理した場合には処理後24時間以内に静脈のネクローシスが肉眼で観察され(図2A−a、図2A−b、図2A−c)、25μMのアルタナル酸溶液で処理したときに最も重度なネクローシスが観察された(図2A−a)。しかし、0.1μMのアルタナル酸溶液処理でも葉の処理部位の黄変がみられた(図2A−d)。これは、A.solaniをジャガイモの葉に感染させたときの徴候に類似していた。なお、本条件下での処理の24時間後に顕微鏡で観察したところ、アルタナル酸で処理されたトマト細胞の過敏感細胞死の発生は、観察されなかった。これにより、アルタナル酸は宿主植物の過敏感反応を抑制することが確認された。本実施例により、A.solaniは宿主植物組織の過敏感反応を抑制することによりネクローシスを進行させ感染を成立させることと、A.solaniが生成するアルタナル酸が感染成立に関与する成分であることとが確認された。
【実施例2】
【0046】
2.トマト葉に及ぼすA.solaniの接種の影響の検討
2.1 方法
アルタナル酸処理の実験と同じように消毒、リンス及び穿孔されたトマトの葉をA.solani生菌で処理して感染させた。
【0047】
2.2 結果
図2B−2、2B−3、2B−4及び2B−5は、それぞれA.solani感染後2、3,4及び5日後のトマト葉表面の写真である。図2Bから、A.solaniは接種2日後にトマト葉で症状を引き起こし(図2B−2)、感染後5日間にネクローシスが広がった(図2B−5)。これによりA.solani生菌がネクローシスを誘発し感染を成立させることが確認された。
【実施例3】
【0048】
3.RiCDPK2cDNAの単離及びRiCDPK2融合タンパク質の発現
3.1 方法
(cDNAライブラリーの調製)
全RNAをRishiri品種のジャガイモの葉及び根茎から単離し、オリゴdTセルロースを用いるクロマトグラフィによってポリA+−RNAを精製した。該ポリA+−RNAからオリゴdTプライマー及びcDNA synthesis System Plus(アマシャムバイオサイエンス株式会社)を使って2本鎖cDNAを合成した。前記2本鎖cDNAとラムダgt11ファージをベクターとからcDNAライブラリを構築した。得られたcDNAライブラリからDNAを抽出して、これを鋳型として、配列番号3及び4に列挙されたヌクレオチド配列を有するプライマーを用いてRishiri品種のジャガイモCDPK2(以下、「RiCDPK2」という。)をエンコードするcDNAを増幅した。
【0049】
(RiCDPK2融合タンパク質発現ベクターの構築)
配列番号1のアミノ酸配列を有するRiCDPK2タンパク質のC末端に6個のヒスチジン残基からなるHisタグが連結した融合タンパク質(以下、「His−RiCDPK2」という。)が発現できるようにRiCDPK2のcDNAをpCRT7CT TOPOベクターにクローン化した。Sambrook,J.及びRussell,D.W.(2001、Molecular Cloning:A Laboratory Manual,Vol.1,Cold Spring Harbor Laboratory Press,New York.)に記載の方法に従って、His−RiCDPK2を発現するpCTT7CT TOPOコンストラクトで大腸菌(BL21、pLysS)細胞を形質転換し、His−RiCDPK2を発現させた。
【0050】
(His−RiCDPK2の調製)
発現したHis−RiCDPK2の精製は、Xpress(商標)Systemタンパク質精製プロトコール(インビトロジェン株式会社)に従って行なった。アンピシリン70μg/mlと形質転換された大腸菌細胞とを含むLuria−Bertani(LB)培地に0.6mMのIPTGを添加することによりタンパク質発現を誘導した。前記大腸菌細胞を24時間、25°Cで振とう培養し、RPR−20ローター(株式会社日立ハイテクノロジーズ)で4°C、10分間、4000rpm遠心分離して回収した。回収した大腸菌をグアニジウムを含む溶菌バッファーで可溶化し、RPR−20ローターで4°C、15分間、6500rpmの遠心分離により上清を回収した。該上清を前記タンパク質精製プロトコールに従ってヒスチジン結合樹脂カラムに通し、His−RiCDPK2を溶出した。溶出されたHis−RiCDPK2を10mMのTris−HCl(pH8.0)及び0.1%のTriton X−100を含む水溶液に対して透析した。透析後のHis−RiCDPK2溶液中のタンパク質濃度は、Bradfordの方法(Bradford,M.M.(1976)A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein−dye binding.Analytical Biochem.72:248−254.)に従って、ウシ血清アルブミン(BSA)を標準としてBio−Rad assay kitを使用することにより決定した。
【0051】
(SDS−PAGE及びウエスタンイムノブロット分析)
His−RiCDPK2の純度の確認のために、Leammli,U.K.et al.,Nature Vol.227,pp.680−681(1970)に記載の方法に従って、ドデシル硫酸ナトリウム−ポリアクリルアミドゲル電気泳動をminiPAGE System(アトー株式会社)を用いて分離し、クマシーブリリアントブルー(CBB)G−250により染色し検出した。ウエスタンイムノブロット分析のため、電気泳動で分離されたタンパク質を、ポリビニリデンジフルオリド(PVDF)膜(Immoblin(商標)P、孔径0.45μm、日本ミリポア株式会社)に写しとった。前記膜を、ブロッキング用緩衝液(3%(w/v)BSA、TBS(20mM TrisHCl)でブロッキングし、希釈用緩衝液(1%のBSA及び0.1%のTriton X−100を添加したTBS)で1/5000に希釈した抗His(C末端)−マウスモノクローナルIgG2b抗体とともに終夜インキュベーションを行なった。前記膜をTBST(0.1%のTriton X−100を添加したTBS)で2回洗浄した後、250倍に希釈したアルカリホスフェターゼ結合ラビット抗マウス抗体(DAKO D0314、ダコ ジャパン株式会社)とともに室温にて1時間インキュベーションを行なった。前記膜をTBSTで洗浄した後、アルカリホスファターゼ緩衝液(0.1M Tris−HCl、0.1M NaCl、5mM MgCl2、pH9.5)で洗浄し、AP結合基質キット(バイオラッドラボラトリーズ株式会社)を用いて、抗原−抗体複合体を検出した。
【0052】
3.2 結果
RiCDPK2のヌクレオチド配列を配列番号2に示す。RiCDPK2がエンコードするタンパク質のアミノ酸配列を配列番号1に示す。His−RiCDPK2は配列番号1のRiCDPK2タンパク質のN末端に6個のヒスチジン残基からなるHisタグが連結した融合タンパク質である。IPTG誘導によりHis−RiCDPK2を過剰発現させた大腸菌の培養200mLから、約100μgのHis−RiCDPK2が精製された。His−RiCDPK2融合タンパク質は主に不溶性タンパク質画分で回収された。精製後の純度はSDS−PAGEの泳動バンドから判断して約90%であった。
【実施例4】
【0053】
4.Ca2+イオン存在下及び非存在下でのHis−RiCDPK2の酵素活性に対するアルタナル酸の影響の検討
4.1 方法
His−RiCDPK2の酵素活性は、Furuichi,N.、Anderson,A.J.、Suzuki,Y.及びTakemoto,J.Y.(1994、Elicitor and suppressor of Phytophthora infestans stimulate phosphorylation of plasma membrane proteins from potato and bean tissues.In Host Specific Toxin:Biosynthesis,Receptor and Molecular Biology(Kohmoto,K.and Yoder,O.C.,eds.).pp.273−274、Tottori University Press、Tottori)に記載の方法に従って実施された。図3に酵素活性のアッセイ系の概略を示す。簡潔には、96ウェルマイクロタイタープレートを使用して、1処理当たり1個のウェルに155μLの反応混合液(8.5mM Tris−HCl(pH7.1)、5mM ホスホクレアチン、0.4単位 クレアチンホスホキナーゼ(シグマ アルドリッチ ジャパン株式会社)、1.5μg His−RiCDPK2)を添加した。アルタナル酸及びCa2+イオンの効果を調べる実験にはそれぞれ25μMのアルタナル酸及び0.1mM CaCl2を前記反応混合液に添加した。酵素反応は0.9mM ATPを添加することによって開始され、30°C、10分間継続された。酵素反応は、0.2% 1−ナフトール(和光純薬工業株式会社)及び0.06% 2,3−ブタンジオン(和光純薬工業株式会社)を含むストックアルカリ溶液(1.5M NaOH、0.7M NaHCO3)を各ウェルに添加して発色させた。酵素活性の定量は、マイクロプレートリーダー(バイオラッド3500、バイオラッドラボラトリーズ株式会社)を用いて595nmの吸光度を10分ごとには50分後まで測定することによって実施された。吸光度の測定と次の測定との間、酵素反応に用いるマイクロタイタープレートは30°Cに保温された。
【0054】
4.2 結果
図4は、アルタナル酸及びCa2+イオンの存在下におけるHis−RiCDPK2のキナーゼ活性を、キナーゼ反応で生じるADP量に対応する吸光度の相対値で表した棒グラフである。アルタナル酸存在下で50分間反応させた場合の吸光度を100%として、アルタナル酸及びCa2+イオンとも非存在下(黒ベタ塗り)か、あるいは、アルタナル酸単独存在下(横線ハッチング)か、Ca2+イオン単独存在下(網掛けハッチング)か、アルタナル酸及びCa2+イオンの両方の存在下(白)かのそれぞれの2回の異なる実験の結果の0分から50分までの10分おきの平均と標準偏差の上限とを示す。図4のグラフから、His−RiCDPK2のキナーゼ活性がCa2+イオン存在下で増強されることから、カルシウム依存性タンパク質リン酸化活性があること、及び、アルタナル酸がHis−RiCDPK2のキナーゼ活性を増強することが示された。
【実施例5】
【0055】
5.Ca2+及びMg2+イオン存在下及び非存在下でのHis−RiCDPK2の酵素活性に対するアルタナル酸の影響の検討
5.1 方法
アルタナル酸がMg2+イオン存在下でもHis−RiCDPK2のタンパク質リン酸化活性に影響を与えるかどうかを調べるために、25μMのアルタナル酸、100μMのCaCl2及び900μMのMgCl2を前記反応混合液に添加することを除いて、実施例4に記載のアッセイ系と同じ条件で実験を行なった。
【0056】
5.2 結果
図5は、アルタナル酸、Ca2+及びMg2+イオンの存在下におけるHis−RiCDPK2のキナーゼ活性を、キナーゼ反応で生じるADP量に対応する吸光度の相対値で表した棒グラフである。アルタナル酸、Ca2+及びMg2+イオン存在下で50分間反応させた場合の吸光度を100%として、アルタナル酸及びCa2+イオンとも非存在下(黒ベタ塗り)か、あるいは、アルタナル酸単独存在下(横線ハッチング)か、Ca2+及びMg2+イオン存在下(網掛けハッチング)か、アルタナル酸とCa2+及びMg2+イオンとの存在下(白)かのそれぞれの2回の異なる実験の結果の0分から50分までの10分おきの平均と標準偏差の上限とを示す。図5のグラフから、His−RiCDPK2のキナーゼ活性がCa2+及びMg2+イオン存在下で増強されること、及び、Ca2+及びMg2+イオンの有無にかかわらずアルタナル酸がHis−RiCDPK2のキナーゼ活性を増強することが示された。
【図面の簡単な説明】
【0057】
【図1】アルタナル酸の化学構造式。
【図2A】異なる濃度のアルタナル酸で処理された葉表面の写真。
【図2B】A.solani感染後2ないし5日後のトマト葉表面の写真。
【図3】酵素活性のアッセイ系の模式図。
【図4】アルタナル酸及びCa2+イオンの存在下におけるHis−RiCDPK2のキナーゼ活性を、キナーゼ反応で生じるADP量に対応する吸光度の相対値で表した棒グラフ。
【図5】アルタナル酸、Ca2+及びMg2+イオンの存在下におけるHis−RiCDPK2のキナーゼ活性を、キナーゼ反応で生じるADP量に対応する吸光度の相対値で表した棒グラフ。
【特許請求の範囲】
【請求項1】
(A)(1)配列番号1のアミノ酸配列からなるCDPK2酵素タンパク質と、
(2)前記アミノ酸配列において1個若しくは数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列からなり、かつ、カルシウム依存性タンパク質リン酸化活性を有するタンパク質と、
(3)前記アミノ酸配列において1個若しくは数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列からなり、かつ、アルタナル酸により増強される前記タンパク質リン酸化活性を有するタンパク質とからなるグループから選択されることを特徴とするタンパク質と、
(4)前記(1)ないし(3)のタンパク質からなる第1ポリペプチドと、特異的結合タグとからなる第2ポリペプチドとが連結した融合タンパク質とのうちのいずれか1つのタンパク質と、タンパク質リン酸化活性増強剤候補化合物とを用意するステップと、
(B)前記タンパク質又は融合タンパク質と、前記タンパク質リン酸化活性増強剤候補化合物とを含む反応液中で、タンパク質リン酸化活性を測定するステップとを含むことを特徴とするタンパク質リン酸化活性増強作用の評価方法。
【請求項2】
前記特異的結合タグは、Hisタグ、mycタグ、HAタグ、インテインタグ、MBP又はGSTからなるグループから選択されることを特徴とする請求項1に記載のタンパク質リン酸化活性増強作用の評価方法。
【請求項3】
前記反応液はCa2+及びMg2+イオンを含むことを特徴とする請求項1又は2に記載の評価方法。
【請求項4】
前記タンパク質リン酸化活性増強剤候補化合物はアルタナル酸の誘導体であることを特徴とする請求項1ないし3のいずれかに記載の評価方法。
【請求項5】
請求項1ないし4のいずれかに記載の評価方法を用いて、複数の前記タンパク質リン酸化活性増強剤候補化合物のうちタンパク質リン酸化活性増強作用が強力な候補化合物をタンパク質リン酸化活性増強剤として選択するステップを含むことを特徴とするタンパク質リン酸化活性増強剤の製造方法。
【請求項6】
請求項1ないし4のいずれかに記載の評価方法によって、複数の前記タンパク質リン酸化活性増強剤候補化合物のうち特定の植物種由来のCDPK2酵素に対するタンパク質リン酸化活性増強作用が他の植物種由来CDPK2酵素に対するタンパク質リン酸化活性増強作用に比べて弱い候補化合物を該特定の植物の生育を阻害しない除草剤として選択するステップを含むことを特徴とする除草剤の製造方法。
【請求項7】
前記特定の植物種はジャガイモであることを特徴とする請求項6に記載の除草剤の製造方法。
【請求項8】
前記他の植物種はシロイヌナズナ又はタバコであることを特徴とする請求項6又は7に記載の除草剤の製造方法。
【請求項9】
(A)(1)配列番号1のアミノ酸配列からなるCDPK2酵素タンパク質と、
(2)前記アミノ酸配列において1個若しくは数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列からなり、かつ、カルシウム依存性タンパク質リン酸化活性を有するタンパク質と、
(3)前記アミノ酸配列において1個若しくは数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列からなり、かつ、アルタナル酸により増強される前記タンパク質リン酸化活性を有するタンパク質とからなるグループから選択されることを特徴とするタンパク質と、
(4)前記(1)ないし(3)のタンパク質からなる第1ポリペプチドと、特異的結合タグとからなる第2ポリペプチドとが連結した融合タンパク質とのうちのいずれか1つのタンパク質とを含むことを特徴とするタンパク質リン酸化活性増強剤候補化合物のタンパク質リン酸化活性増強作用の評価キット。
【請求項10】
前記特異的結合タグは、Hisタグ、mycタグ、HAタグ、インテインタグ、MBP又はGSTからなるグループから選択されることを特徴とする請求項9に記載の評価キット。
【請求項11】
前記タンパク質リン酸化活性増強剤候補化合物はアルタナル酸の誘導体であることを特徴とする請求項9又は10に記載の評価キット。
【請求項1】
(A)(1)配列番号1のアミノ酸配列からなるCDPK2酵素タンパク質と、
(2)前記アミノ酸配列において1個若しくは数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列からなり、かつ、カルシウム依存性タンパク質リン酸化活性を有するタンパク質と、
(3)前記アミノ酸配列において1個若しくは数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列からなり、かつ、アルタナル酸により増強される前記タンパク質リン酸化活性を有するタンパク質とからなるグループから選択されることを特徴とするタンパク質と、
(4)前記(1)ないし(3)のタンパク質からなる第1ポリペプチドと、特異的結合タグとからなる第2ポリペプチドとが連結した融合タンパク質とのうちのいずれか1つのタンパク質と、タンパク質リン酸化活性増強剤候補化合物とを用意するステップと、
(B)前記タンパク質又は融合タンパク質と、前記タンパク質リン酸化活性増強剤候補化合物とを含む反応液中で、タンパク質リン酸化活性を測定するステップとを含むことを特徴とするタンパク質リン酸化活性増強作用の評価方法。
【請求項2】
前記特異的結合タグは、Hisタグ、mycタグ、HAタグ、インテインタグ、MBP又はGSTからなるグループから選択されることを特徴とする請求項1に記載のタンパク質リン酸化活性増強作用の評価方法。
【請求項3】
前記反応液はCa2+及びMg2+イオンを含むことを特徴とする請求項1又は2に記載の評価方法。
【請求項4】
前記タンパク質リン酸化活性増強剤候補化合物はアルタナル酸の誘導体であることを特徴とする請求項1ないし3のいずれかに記載の評価方法。
【請求項5】
請求項1ないし4のいずれかに記載の評価方法を用いて、複数の前記タンパク質リン酸化活性増強剤候補化合物のうちタンパク質リン酸化活性増強作用が強力な候補化合物をタンパク質リン酸化活性増強剤として選択するステップを含むことを特徴とするタンパク質リン酸化活性増強剤の製造方法。
【請求項6】
請求項1ないし4のいずれかに記載の評価方法によって、複数の前記タンパク質リン酸化活性増強剤候補化合物のうち特定の植物種由来のCDPK2酵素に対するタンパク質リン酸化活性増強作用が他の植物種由来CDPK2酵素に対するタンパク質リン酸化活性増強作用に比べて弱い候補化合物を該特定の植物の生育を阻害しない除草剤として選択するステップを含むことを特徴とする除草剤の製造方法。
【請求項7】
前記特定の植物種はジャガイモであることを特徴とする請求項6に記載の除草剤の製造方法。
【請求項8】
前記他の植物種はシロイヌナズナ又はタバコであることを特徴とする請求項6又は7に記載の除草剤の製造方法。
【請求項9】
(A)(1)配列番号1のアミノ酸配列からなるCDPK2酵素タンパク質と、
(2)前記アミノ酸配列において1個若しくは数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列からなり、かつ、カルシウム依存性タンパク質リン酸化活性を有するタンパク質と、
(3)前記アミノ酸配列において1個若しくは数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列からなり、かつ、アルタナル酸により増強される前記タンパク質リン酸化活性を有するタンパク質とからなるグループから選択されることを特徴とするタンパク質と、
(4)前記(1)ないし(3)のタンパク質からなる第1ポリペプチドと、特異的結合タグとからなる第2ポリペプチドとが連結した融合タンパク質とのうちのいずれか1つのタンパク質とを含むことを特徴とするタンパク質リン酸化活性増強剤候補化合物のタンパク質リン酸化活性増強作用の評価キット。
【請求項10】
前記特異的結合タグは、Hisタグ、mycタグ、HAタグ、インテインタグ、MBP又はGSTからなるグループから選択されることを特徴とする請求項9に記載の評価キット。
【請求項11】
前記タンパク質リン酸化活性増強剤候補化合物はアルタナル酸の誘導体であることを特徴とする請求項9又は10に記載の評価キット。
【図1】


【図3】
【図4】
【図5】
【図2A】

【図2B】
【図3】
【図4】
【図5】
【図2A】
【図2B】
【公開番号】特開2009−50223(P2009−50223A)
【公開日】平成21年3月12日(2009.3.12)
【国際特許分類】
【出願番号】特願2007−221693(P2007−221693)
【出願日】平成19年8月28日(2007.8.28)
【出願人】(304027279)国立大学法人 新潟大学 (310)
【Fターム(参考)】
【公開日】平成21年3月12日(2009.3.12)
【国際特許分類】
【出願日】平成19年8月28日(2007.8.28)
【出願人】(304027279)国立大学法人 新潟大学 (310)
【Fターム(参考)】
[ Back to top ]