
除草剤抵抗性遺伝子
【課題】2,4-Dおよび他のフェノキシオーキシン除草剤に対してのみならず、アリールオキシフェノキシプロピオネート除草剤にもまた抵抗性である新規な植物を提供する。
【解決手段】より広くかつより強固な雑草の制御、処理の柔軟性の増加、および除草剤抵抗性管理の選択肢の改善を提供するために、本発明の1種または複数の酵素を、単独で、または別の除草剤抵抗性遺伝子、好ましくは、グリフォセート抵抗性遺伝子とともに「重ね合わせて」産生する植物を含む。
【解決手段】より広くかつより強固な雑草の制御、処理の柔軟性の増加、および除草剤抵抗性管理の選択肢の改善を提供するために、本発明の1種または複数の酵素を、単独で、または別の除草剤抵抗性遺伝子、好ましくは、グリフォセート抵抗性遺伝子とともに「重ね合わせて」産生する植物を含む。
Notice: Undefined index: DEJ in /mnt/www/gzt_disp.php on line 298
【特許請求の範囲】
【請求項1】
SEQ ID NO:9またはSEQ ID NO:9の変種のアミノ酸配列を含むAAD-1タンパク質をコードするポリヌクレオチドを含むトランスジェニック植物細胞であって、該変種がアリールオキシアルカノエートジオキシゲナーゼ活性、少なくとも1つのアミノ酸欠失または保存性置換、およびSEQ ID NO:9との少なくとも95%の配列同一性を有する、トランスジェニック植物細胞。
【請求項2】
植物細胞が双子葉植物細胞および単子葉植物細胞からなる群より選択される、請求項1に記載の細胞。
【請求項3】
植物細胞が双子葉植物細胞であり、ワタ細胞、タバコ細胞、カノーラ細胞、ダイズ細胞、およびシロイヌナズナ(Arabidopsis)細胞からなる群より選択される、請求項2に記載の細胞。
【請求項4】
植物細胞がイネ細胞およびトウモロコシ細胞からなる群より選択される単子葉植物細胞である、請求項2に記載の細胞。
【請求項5】
ポリヌクレオチドの発現がアリールオキシアルカノエート除草剤に対して細胞を耐性にする、請求項1に記載の複数の細胞を含むトランスジェニック植物。
【請求項6】
除草剤がフェノキシオーキシン除草剤である、請求項5に記載の植物。
【請求項7】
除草剤が2,4-ジクロロフェノキシ酢酸、MCPA、ジクロルプロップ、およびメコプロップからなる群より選択される、請求項5に記載の植物。
【請求項8】
除草剤がアリールオキシフェノキシプロピオネートである、請求項5に記載の植物。
【請求項9】
除草剤が、フルアジホップ、ハロキシホップ、ジクロホップ、キザロホップ、フェノキサプロップ、メタニホップ、シハロホップ、およびクロジノホップからなる群より選択される、請求項5に記載の植物。
【請求項10】
ポリヌクレオチドの発現が、フェノキシオーキシン除草剤とアリールオキシフェノキシプロピオネート除草剤の両方に対して植物を抵抗性にする、請求項5に記載の植物。
【請求項11】
第2の除草剤抵抗性遺伝子をさらに含む、請求項5に記載の植物。
【請求項12】
第2の除草剤抵抗性遺伝子が、グリフォセート、グルフォシネート、ALS阻害剤、4-ヒドロキシフェニル-ピルビン酸-ジオキシゲナーゼ(HPPD)の阻害剤、およびプロトポルフィリノーゲンオキシダーゼ(PPO)の阻害剤からなる群より選択される除草剤に対して植物を抵抗性にする、請求項11に記載の植物。
【請求項13】
フィールドが少なくとも1種の請求項5に記載の植物を含み、フェノキシオーキシン除草剤およびアリールオキシフェノキシプロピオネート除草剤からなる群より選択される第1の除草剤を、該フィールドの少なくとも一部に適用する工程を含む、フィールド中で少なくとも1種の雑草を制御する方法。
【請求項14】
除草剤がキラルフェノキシオーキシンのR-エナンチオマーである、請求項13に記載の方法。
【請求項15】
除草剤がR-ジクロルプロップおよびR-メコプロップからなる群より選択される、請求項14に記載の方法。
【請求項16】
除草剤がアキラルフェノキシオーキシンである、請求項13に記載の方法。
【請求項17】
除草剤が2,4-DおよびMCPAからなる群より選択される、請求項16に記載の方法。
【請求項18】
第1の除草剤がアリールオキシフェノキシプロピオネートであり、植物が単子葉植物である、請求項13に記載の方法。
【請求項19】
単子葉植物が、トウモロコシ、イネ、コムギ、オオムギ、ライムギ、芝草、オートムギ、モロコシ、および牧草からなる群より選択される、請求項18に記載の方法。
【請求項20】
第1の除草剤がフェノキシオーキシンであり、植物が双子葉植物である、請求項13に記載の方法。
【請求項21】
双子葉植物が、ワタ、タバコ、カノーラ、およびダイズからなる群より選択される、請求項20に記載の方法。
【請求項22】
第2の除草剤を適用する工程を含む、請求項13に記載の方法。
【請求項23】
第1の除草剤および第2の除草剤が連続的に適用される、請求項22に記載の方法。
【請求項24】
第1の除草剤および第2の除草剤が同時に適用される、請求項22に記載の方法。
【請求項25】
第1の除草剤がフェノキシオーキシンであり、第2の除草剤がアリールオキシフェノキシプロピオネートである、請求項22に記載の方法。
【請求項26】
植物がグリフォセートに対して抵抗性である、請求項13に記載の方法。
【請求項27】
R-エナンチオマーがラセミ混合物の成分である、請求項14に記載の方法。
【請求項28】
植物が、該植物を第2の除草剤に対して抵抗性にする第2の除草剤抵抗性遺伝子をさらに含む、請求項22に記載の方法。
【請求項29】
第2の遺伝子が、改変AHAS(アセトヒドロキシ酸シンターゼ)、SurA、SurB、Csr1、Csr1-1、Csr1-2、改変EPSPS(5-エノールピルビルシキミ酸-3-ホスフェートシンターゼ)、GOX、GAT、PAT(ホスフィノスリシン-N-アセチルトランスフェラーゼ)、bar、およびジキャンバ分解酵素からなる群より選択される、請求項28に記載の方法。
【請求項30】
第2の除草剤が、グリフォセート、グルフォシネート、ジキャンバ、アセト乳酸シンターゼ阻害剤、プロトポルフィリノーゲンオキシダーゼ阻害剤、およびヒドロキシフェニル-ピルビン酸-ジオキシゲナーゼ阻害剤からなる群より選択される、請求項28に記載の方法。
【請求項31】
第2の除草剤が、イミダゾリノン、スルホニルウレアおよびトリアゾロピリミジン除草剤からなる群より選択されるアセト乳酸シンターゼ阻害剤である、請求項30に記載の方法。
【請求項32】
第2の除草剤が、イマザモックス、イマゼサピル、イマザキン、およびイマザピックからなる群より選択されるイミダゾリノンである、請求項31に記載の方法。
【請求項33】
第1の除草剤がフェノキシオーキシンであり、第2の除草剤がグリフォセートおよびグルフォシネートからなる群より選択される、請求項28に記載の方法。
【請求項34】
フェノキシオーキシンが2,4-Dであり、第2の除草剤がグリフォセートである、請求項33に記載の方法。
【請求項35】
第1の除草剤がアリールオキシフェノキシプロピオネートであり、第2の除草剤がグリフォセートである、請求項28に記載の方法。
【請求項36】
第1の除草剤が、キザロホップ、ハロキシホップ、およびシハロホップからなる群より選択される、請求項35に記載の方法。
【請求項37】
第2の除草剤が、アミドスルフロン、ベンスルフロン、クロリムロン、クロルスルフロン、シノスルフロン、フルピルスルフロン、フォラムスルフロン、ハロスルフロン、ニコスルフロン、プリミスルフロン、プロスルフロン、リムスルフロン、スルホメツロン、スルホスルフロン、チフェンスルフロン、トリアスルフロン、トリフロキシスルフロン、およびトリフルスルフロンからなる群より選択されるスルホニルウレアである、請求項31に記載の方法。
【請求項38】
第3の除草剤を適用する工程をさらに含む、請求項28に記載の方法。
【請求項39】
除草剤が2,4-D;キザロホップ;およびグルフォシネートである、請求項38に記載の方法。
【請求項40】
第1の除草剤をフィールドに適用する工程、および該第1の除草剤の適用の14日以内に該フィールドに種子を播種する工程を含み、該種子が請求項1の細胞を含み、該第1の除草剤が、フェノキシオーキシンおよびアリールオキシフェノキシプロピオネートからなる群より選択される、フィールドにおいて雑草を制御する方法。
【請求項41】
第1の除草剤が、酸、無機塩、有機塩、エステル、R-エナンチオ特異的異性体、またはラセミ混合物の成分である、請求項40に記載の方法。
【請求項42】
種子が、第2の除草剤に対して植物を抵抗性にする第二の種子を含み、播種の前にフィールドに第2の除草剤を適用する工程をさらに含む、請求項40に記載の方法。
【請求項43】
第2の除草剤が、グリフォセート、グラモゾン、およびグルフォシネートからなる群より選択される、請求項42に記載の方法。
【請求項44】
アリールオキシアルカノエートジオキシゲナーゼ活性を有するタンパク質をコードし、該タンパク質をコードする核酸分子が、SEQ ID NO:3、SEQ ID NO:4、およびSEQ ID NO:5からなる群より選択される配列の全長相補体と、ストリンジェントな条件下でハイブリダイズする、植物中での発現のために最適化されたポリヌクレオチド。
【請求項45】
双子葉植物または単子葉植物中での発現のために最適化されている、請求項44に記載のポリヌクレオチド。
【請求項46】
SEQ ID NO:5を含む、請求項44に記載のポリヌクレオチド。
【請求項47】
フェノキシオーキシンおよびアリールオキシフェノキシプロピオネートからなる群より選択される除草剤を酵素により分解するタンパク質をコードする単離されたポリヌクレオチドであって、該タンパク質をコードする核酸分子が、SEQ ID NO:3、SEQ ID NO:4、およびSEQ ID NO:5からなる群より選択される配列の全長相補体とストリンジェントな条件下でハイブリダイズし、該ポリヌクレオチドが植物細胞中で機能的であるプロモーターに作動可能に連結されている、単離されたポリヌクレオチド。
【請求項48】
プロモーターが植物プロモーターである、請求項47に記載のポリヌクレオチド。
【請求項49】
プロモーターが、キャッサバ葉脈モザイクウイルスプロモーターである、請求項47に記載のポリヌクレオチド。
【請求項50】
複数の植物細胞を、請求項44または47に記載のポリヌクレオチドを用いる形質転換に供する工程、および非形質転換細胞を殺傷またはその成長を阻害しながら、該ポリヌクレオチドを発現する形質転換細胞を成長させることを可能にする除草剤の濃度で、該細胞を成長させる工程を含み、該除草剤が、フェノキシオーキシンおよびアリールオキシフェノキシアルカノエートからなる群より選択される、形質転換植物細胞を選択するための方法。
【請求項51】
細胞が植物の細胞であり、形質転換植物を選択するために使用される、請求項50に記載の方法。
【請求項52】
請求項50に記載の方法において選択マーカーとして使用される、請求項47または50に記載のポリヌクレオチド。
【請求項53】
植物から試料を収集する工程、およびポリヌクレオチドの存在について該試料をアッセイする工程を含む、植物が請求項47または50に記載のポリヌクレオチドを含むか否かを検出する方法。
【請求項54】
ポリヌクレオチドによってコードされるタンパク質の存在について、試料をアッセイする工程を含む、請求項53に記載の方法。
【請求項55】
ポリヌクレオチドの存在について検出するためのPCRプライマーまたはプローブを使用する工程を含む、請求項53に記載の方法。
【請求項56】
タンパク質の存在について検出するための抗体を使用する工程を含む、請求項54に記載の方法。
【請求項57】
請求項1に記載の植物細胞を含む種子。
【請求項58】
請求項57に記載の種子から生長した植物。
【請求項59】
請求項5に記載の植物の一部分、子孫、または無性繁殖物。
【請求項60】
除草剤抵抗性雑草を処理または予防するために使用される、請求項13に記載の方法。
【請求項61】
バチルス チューリンジエンシス(Bacillus thuringiensis)、フォトラブダス(Photorhabdus)、およびジェノラブダズ(Xenorhabdus)からなる群より選択される生物に由来する昆虫抵抗性遺伝子をさらに含む、請求項5に記載の植物。
【請求項62】
真菌抵抗性、ストレス耐性、収量の増加、オイルプロファイルの改善、繊維品質の改善、ウイルス抵抗性、成熟の遅延、低温耐性、および塩耐性からなる群より選択される農学的な形質についての遺伝子をさらに含む、請求項5に記載の植物。
【請求項63】
2,4-D除草剤に対してR-特異的アリールオキシアルカノエートジオキシゲナーゼ活性を有する、核酸によってコードされる酵素を、作物の少なくとも1つの植物細胞に導入する工程を含む、穀物に対して2,4-D除草剤抵抗性を付与する方法。
【請求項64】
グリフォセート耐性作物植物が請求項47に記載のポリヌクレオチドを含み、フィールドの少なくとも一部に、アリールオキシアルカノエート除草剤を適用する工程を含む、該植物のフィールドにおいてグリフォセート抵抗性雑草を制御する方法。
【請求項65】
除草剤がフェノキシオーキシンである、請求項64に記載の方法。
【請求項66】
除草剤がアリールオキシフェノキシプロピオネートである、請求項64に記載の方法。
【請求項67】
植物が双子葉植物である、請求項64に記載の方法。
【請求項68】
植物が単子葉植物である、請求項64に記載の方法。
【請求項1】
SEQ ID NO:9またはSEQ ID NO:9の変種のアミノ酸配列を含むAAD-1タンパク質をコードするポリヌクレオチドを含むトランスジェニック植物細胞であって、該変種がアリールオキシアルカノエートジオキシゲナーゼ活性、少なくとも1つのアミノ酸欠失または保存性置換、およびSEQ ID NO:9との少なくとも95%の配列同一性を有する、トランスジェニック植物細胞。
【請求項2】
植物細胞が双子葉植物細胞および単子葉植物細胞からなる群より選択される、請求項1に記載の細胞。
【請求項3】
植物細胞が双子葉植物細胞であり、ワタ細胞、タバコ細胞、カノーラ細胞、ダイズ細胞、およびシロイヌナズナ(Arabidopsis)細胞からなる群より選択される、請求項2に記載の細胞。
【請求項4】
植物細胞がイネ細胞およびトウモロコシ細胞からなる群より選択される単子葉植物細胞である、請求項2に記載の細胞。
【請求項5】
ポリヌクレオチドの発現がアリールオキシアルカノエート除草剤に対して細胞を耐性にする、請求項1に記載の複数の細胞を含むトランスジェニック植物。
【請求項6】
除草剤がフェノキシオーキシン除草剤である、請求項5に記載の植物。
【請求項7】
除草剤が2,4-ジクロロフェノキシ酢酸、MCPA、ジクロルプロップ、およびメコプロップからなる群より選択される、請求項5に記載の植物。
【請求項8】
除草剤がアリールオキシフェノキシプロピオネートである、請求項5に記載の植物。
【請求項9】
除草剤が、フルアジホップ、ハロキシホップ、ジクロホップ、キザロホップ、フェノキサプロップ、メタニホップ、シハロホップ、およびクロジノホップからなる群より選択される、請求項5に記載の植物。
【請求項10】
ポリヌクレオチドの発現が、フェノキシオーキシン除草剤とアリールオキシフェノキシプロピオネート除草剤の両方に対して植物を抵抗性にする、請求項5に記載の植物。
【請求項11】
第2の除草剤抵抗性遺伝子をさらに含む、請求項5に記載の植物。
【請求項12】
第2の除草剤抵抗性遺伝子が、グリフォセート、グルフォシネート、ALS阻害剤、4-ヒドロキシフェニル-ピルビン酸-ジオキシゲナーゼ(HPPD)の阻害剤、およびプロトポルフィリノーゲンオキシダーゼ(PPO)の阻害剤からなる群より選択される除草剤に対して植物を抵抗性にする、請求項11に記載の植物。
【請求項13】
フィールドが少なくとも1種の請求項5に記載の植物を含み、フェノキシオーキシン除草剤およびアリールオキシフェノキシプロピオネート除草剤からなる群より選択される第1の除草剤を、該フィールドの少なくとも一部に適用する工程を含む、フィールド中で少なくとも1種の雑草を制御する方法。
【請求項14】
除草剤がキラルフェノキシオーキシンのR-エナンチオマーである、請求項13に記載の方法。
【請求項15】
除草剤がR-ジクロルプロップおよびR-メコプロップからなる群より選択される、請求項14に記載の方法。
【請求項16】
除草剤がアキラルフェノキシオーキシンである、請求項13に記載の方法。
【請求項17】
除草剤が2,4-DおよびMCPAからなる群より選択される、請求項16に記載の方法。
【請求項18】
第1の除草剤がアリールオキシフェノキシプロピオネートであり、植物が単子葉植物である、請求項13に記載の方法。
【請求項19】
単子葉植物が、トウモロコシ、イネ、コムギ、オオムギ、ライムギ、芝草、オートムギ、モロコシ、および牧草からなる群より選択される、請求項18に記載の方法。
【請求項20】
第1の除草剤がフェノキシオーキシンであり、植物が双子葉植物である、請求項13に記載の方法。
【請求項21】
双子葉植物が、ワタ、タバコ、カノーラ、およびダイズからなる群より選択される、請求項20に記載の方法。
【請求項22】
第2の除草剤を適用する工程を含む、請求項13に記載の方法。
【請求項23】
第1の除草剤および第2の除草剤が連続的に適用される、請求項22に記載の方法。
【請求項24】
第1の除草剤および第2の除草剤が同時に適用される、請求項22に記載の方法。
【請求項25】
第1の除草剤がフェノキシオーキシンであり、第2の除草剤がアリールオキシフェノキシプロピオネートである、請求項22に記載の方法。
【請求項26】
植物がグリフォセートに対して抵抗性である、請求項13に記載の方法。
【請求項27】
R-エナンチオマーがラセミ混合物の成分である、請求項14に記載の方法。
【請求項28】
植物が、該植物を第2の除草剤に対して抵抗性にする第2の除草剤抵抗性遺伝子をさらに含む、請求項22に記載の方法。
【請求項29】
第2の遺伝子が、改変AHAS(アセトヒドロキシ酸シンターゼ)、SurA、SurB、Csr1、Csr1-1、Csr1-2、改変EPSPS(5-エノールピルビルシキミ酸-3-ホスフェートシンターゼ)、GOX、GAT、PAT(ホスフィノスリシン-N-アセチルトランスフェラーゼ)、bar、およびジキャンバ分解酵素からなる群より選択される、請求項28に記載の方法。
【請求項30】
第2の除草剤が、グリフォセート、グルフォシネート、ジキャンバ、アセト乳酸シンターゼ阻害剤、プロトポルフィリノーゲンオキシダーゼ阻害剤、およびヒドロキシフェニル-ピルビン酸-ジオキシゲナーゼ阻害剤からなる群より選択される、請求項28に記載の方法。
【請求項31】
第2の除草剤が、イミダゾリノン、スルホニルウレアおよびトリアゾロピリミジン除草剤からなる群より選択されるアセト乳酸シンターゼ阻害剤である、請求項30に記載の方法。
【請求項32】
第2の除草剤が、イマザモックス、イマゼサピル、イマザキン、およびイマザピックからなる群より選択されるイミダゾリノンである、請求項31に記載の方法。
【請求項33】
第1の除草剤がフェノキシオーキシンであり、第2の除草剤がグリフォセートおよびグルフォシネートからなる群より選択される、請求項28に記載の方法。
【請求項34】
フェノキシオーキシンが2,4-Dであり、第2の除草剤がグリフォセートである、請求項33に記載の方法。
【請求項35】
第1の除草剤がアリールオキシフェノキシプロピオネートであり、第2の除草剤がグリフォセートである、請求項28に記載の方法。
【請求項36】
第1の除草剤が、キザロホップ、ハロキシホップ、およびシハロホップからなる群より選択される、請求項35に記載の方法。
【請求項37】
第2の除草剤が、アミドスルフロン、ベンスルフロン、クロリムロン、クロルスルフロン、シノスルフロン、フルピルスルフロン、フォラムスルフロン、ハロスルフロン、ニコスルフロン、プリミスルフロン、プロスルフロン、リムスルフロン、スルホメツロン、スルホスルフロン、チフェンスルフロン、トリアスルフロン、トリフロキシスルフロン、およびトリフルスルフロンからなる群より選択されるスルホニルウレアである、請求項31に記載の方法。
【請求項38】
第3の除草剤を適用する工程をさらに含む、請求項28に記載の方法。
【請求項39】
除草剤が2,4-D;キザロホップ;およびグルフォシネートである、請求項38に記載の方法。
【請求項40】
第1の除草剤をフィールドに適用する工程、および該第1の除草剤の適用の14日以内に該フィールドに種子を播種する工程を含み、該種子が請求項1の細胞を含み、該第1の除草剤が、フェノキシオーキシンおよびアリールオキシフェノキシプロピオネートからなる群より選択される、フィールドにおいて雑草を制御する方法。
【請求項41】
第1の除草剤が、酸、無機塩、有機塩、エステル、R-エナンチオ特異的異性体、またはラセミ混合物の成分である、請求項40に記載の方法。
【請求項42】
種子が、第2の除草剤に対して植物を抵抗性にする第二の種子を含み、播種の前にフィールドに第2の除草剤を適用する工程をさらに含む、請求項40に記載の方法。
【請求項43】
第2の除草剤が、グリフォセート、グラモゾン、およびグルフォシネートからなる群より選択される、請求項42に記載の方法。
【請求項44】
アリールオキシアルカノエートジオキシゲナーゼ活性を有するタンパク質をコードし、該タンパク質をコードする核酸分子が、SEQ ID NO:3、SEQ ID NO:4、およびSEQ ID NO:5からなる群より選択される配列の全長相補体と、ストリンジェントな条件下でハイブリダイズする、植物中での発現のために最適化されたポリヌクレオチド。
【請求項45】
双子葉植物または単子葉植物中での発現のために最適化されている、請求項44に記載のポリヌクレオチド。
【請求項46】
SEQ ID NO:5を含む、請求項44に記載のポリヌクレオチド。
【請求項47】
フェノキシオーキシンおよびアリールオキシフェノキシプロピオネートからなる群より選択される除草剤を酵素により分解するタンパク質をコードする単離されたポリヌクレオチドであって、該タンパク質をコードする核酸分子が、SEQ ID NO:3、SEQ ID NO:4、およびSEQ ID NO:5からなる群より選択される配列の全長相補体とストリンジェントな条件下でハイブリダイズし、該ポリヌクレオチドが植物細胞中で機能的であるプロモーターに作動可能に連結されている、単離されたポリヌクレオチド。
【請求項48】
プロモーターが植物プロモーターである、請求項47に記載のポリヌクレオチド。
【請求項49】
プロモーターが、キャッサバ葉脈モザイクウイルスプロモーターである、請求項47に記載のポリヌクレオチド。
【請求項50】
複数の植物細胞を、請求項44または47に記載のポリヌクレオチドを用いる形質転換に供する工程、および非形質転換細胞を殺傷またはその成長を阻害しながら、該ポリヌクレオチドを発現する形質転換細胞を成長させることを可能にする除草剤の濃度で、該細胞を成長させる工程を含み、該除草剤が、フェノキシオーキシンおよびアリールオキシフェノキシアルカノエートからなる群より選択される、形質転換植物細胞を選択するための方法。
【請求項51】
細胞が植物の細胞であり、形質転換植物を選択するために使用される、請求項50に記載の方法。
【請求項52】
請求項50に記載の方法において選択マーカーとして使用される、請求項47または50に記載のポリヌクレオチド。
【請求項53】
植物から試料を収集する工程、およびポリヌクレオチドの存在について該試料をアッセイする工程を含む、植物が請求項47または50に記載のポリヌクレオチドを含むか否かを検出する方法。
【請求項54】
ポリヌクレオチドによってコードされるタンパク質の存在について、試料をアッセイする工程を含む、請求項53に記載の方法。
【請求項55】
ポリヌクレオチドの存在について検出するためのPCRプライマーまたはプローブを使用する工程を含む、請求項53に記載の方法。
【請求項56】
タンパク質の存在について検出するための抗体を使用する工程を含む、請求項54に記載の方法。
【請求項57】
請求項1に記載の植物細胞を含む種子。
【請求項58】
請求項57に記載の種子から生長した植物。
【請求項59】
請求項5に記載の植物の一部分、子孫、または無性繁殖物。
【請求項60】
除草剤抵抗性雑草を処理または予防するために使用される、請求項13に記載の方法。
【請求項61】
バチルス チューリンジエンシス(Bacillus thuringiensis)、フォトラブダス(Photorhabdus)、およびジェノラブダズ(Xenorhabdus)からなる群より選択される生物に由来する昆虫抵抗性遺伝子をさらに含む、請求項5に記載の植物。
【請求項62】
真菌抵抗性、ストレス耐性、収量の増加、オイルプロファイルの改善、繊維品質の改善、ウイルス抵抗性、成熟の遅延、低温耐性、および塩耐性からなる群より選択される農学的な形質についての遺伝子をさらに含む、請求項5に記載の植物。
【請求項63】
2,4-D除草剤に対してR-特異的アリールオキシアルカノエートジオキシゲナーゼ活性を有する、核酸によってコードされる酵素を、作物の少なくとも1つの植物細胞に導入する工程を含む、穀物に対して2,4-D除草剤抵抗性を付与する方法。
【請求項64】
グリフォセート耐性作物植物が請求項47に記載のポリヌクレオチドを含み、フィールドの少なくとも一部に、アリールオキシアルカノエート除草剤を適用する工程を含む、該植物のフィールドにおいてグリフォセート抵抗性雑草を制御する方法。
【請求項65】
除草剤がフェノキシオーキシンである、請求項64に記載の方法。
【請求項66】
除草剤がアリールオキシフェノキシプロピオネートである、請求項64に記載の方法。
【請求項67】
植物が双子葉植物である、請求項64に記載の方法。
【請求項68】
植物が単子葉植物である、請求項64に記載の方法。
【図1】


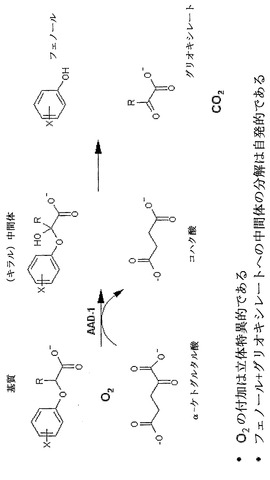
【図2】
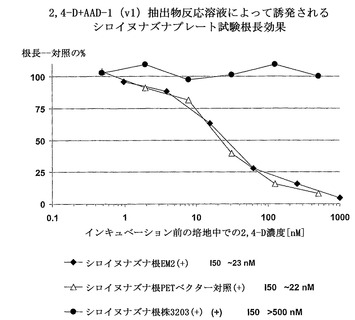
【図3】
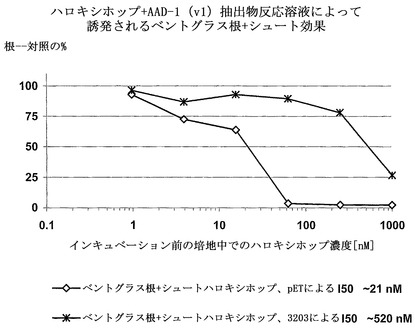
【図4】
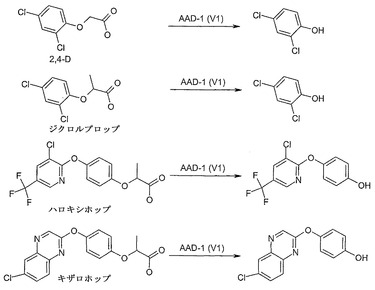
【図5】
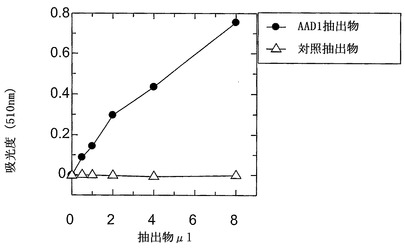
【図6】
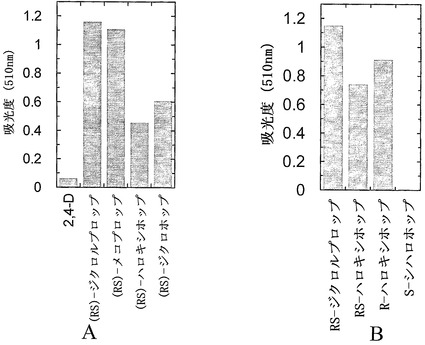
【図7】
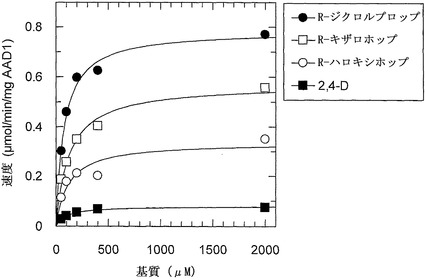
【図8】
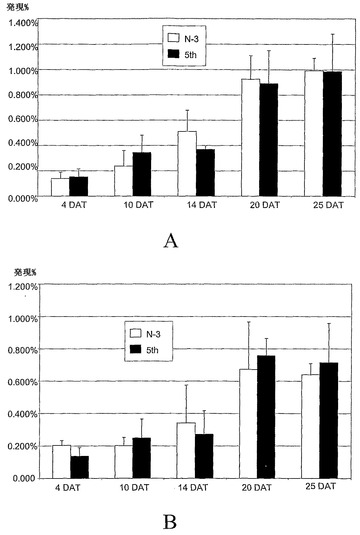
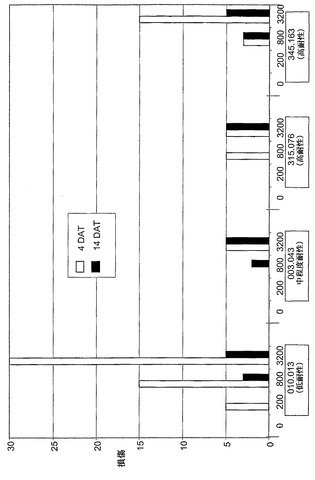
【図9A】
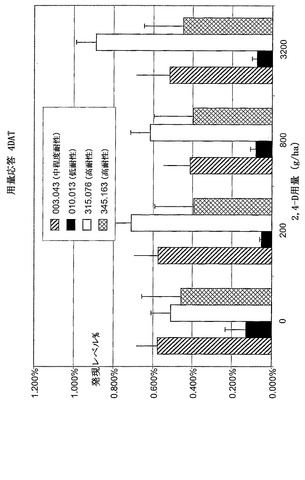
【図9B】
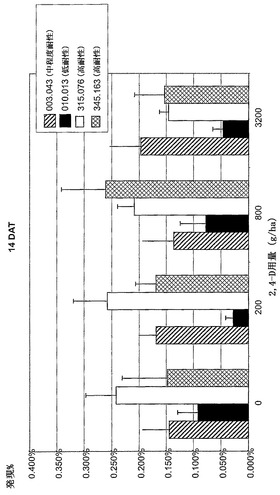
【図9C】
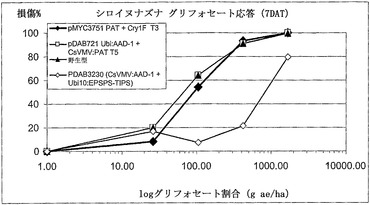
【図10】
【図11】
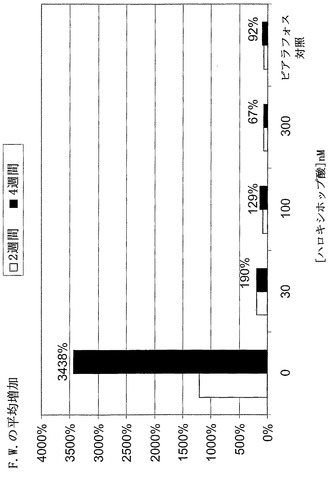
【図12】
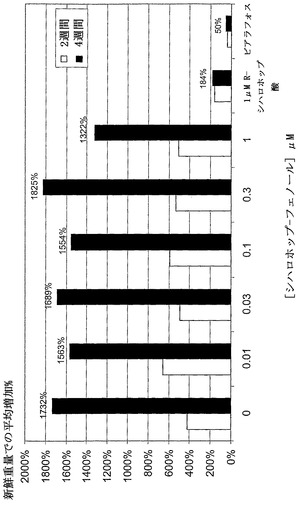
【図13】
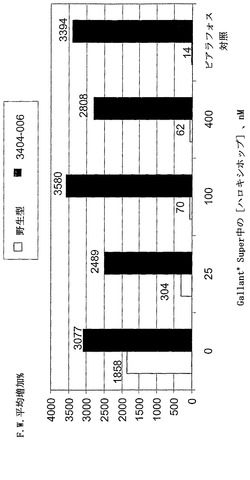
【図14】
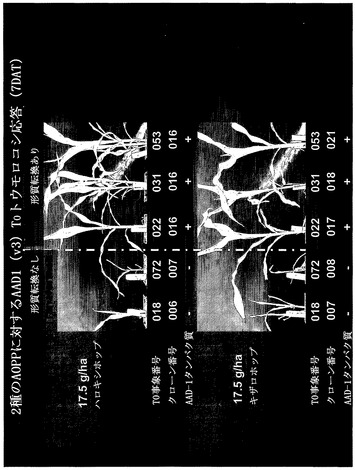
【図15】
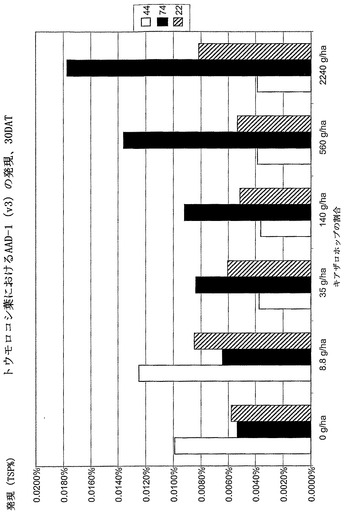
【図16】

【図17】

【図18】
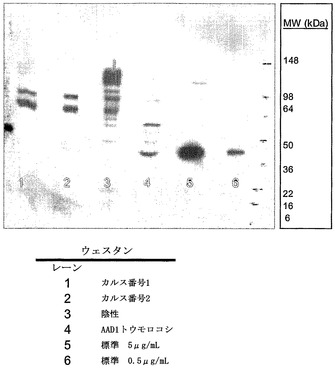
【図19】
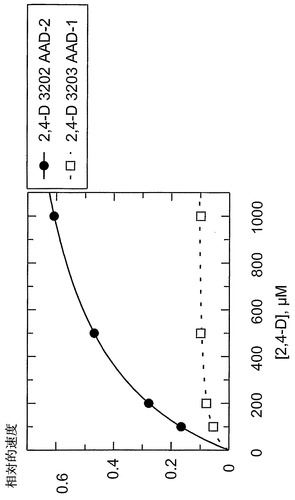
【図20】

【図21】
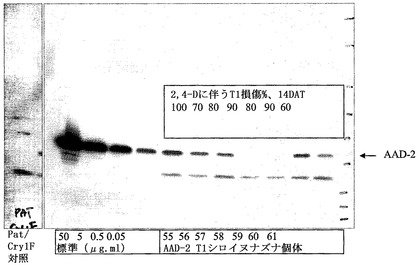
【図22】
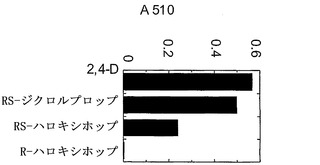
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9A】
【図9B】
【図9C】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【図19】
【図20】
【図21】
【図22】
【公開番号】特開2011−200240(P2011−200240A)
【公開日】平成23年10月13日(2011.10.13)
【国際特許分類】
【出願番号】特願2011−109120(P2011−109120)
【出願日】平成23年5月16日(2011.5.16)
【分割の表示】特願2007−510994(P2007−510994)の分割
【原出願日】平成17年5月2日(2005.5.2)
【出願人】(505412443)ダウ アグロサイエンシズ リミテッド ライアビリティー カンパニー (10)
【Fターム(参考)】
【公開日】平成23年10月13日(2011.10.13)
【国際特許分類】
【出願日】平成23年5月16日(2011.5.16)
【分割の表示】特願2007−510994(P2007−510994)の分割
【原出願日】平成17年5月2日(2005.5.2)
【出願人】(505412443)ダウ アグロサイエンシズ リミテッド ライアビリティー カンパニー (10)
【Fターム(参考)】
[ Back to top ]