
非ウイルス性再プログラムによる多能性幹細胞
感染性ウイルスをコードしないエピソームベクターを使って霊長類体細胞を再プログラムして多能性細胞にする方法を開示する。また、本方法で産生した多能性細胞も開示する。
【発明の詳細な説明】
【技術分野】
【0001】
(関連出願の相互参照)
本明細書は、米国仮特許出願第61/108.362(出願日2008年10月24日)を参照によりその全体を説明したものとしてここに含める。
【0002】
(連邦支援による研究または開発に関する陳述)
本発明は、NIH GM081629及びRR000167の機関を通してアメリカ合衆国政府助成を受けた。アメリカ合衆国は本発明にある一定の権利を保有する。
【背景技術】
【0003】
胚性幹細胞(ES)は、多能性、即ち、無限に増殖し、3つの胚葉全ての細胞に分化しうる能力があり、科学と医学の分野において大いに期待されている(Thomson et al., Science 282:1145−1147(1998),参照により文献全体を説明したものとして本発明に組み込む)。治療や再生医学におけるヒトES細胞の適用は、受益者の免疫系による拒絶の可能性によって複雑化する。したがって、実質的に特定受益者と遺伝的に同一のヒト多能性細胞が高く望まれる。さらに、患者特異的な治療計画を立案する際に、遺伝的同一性がES細胞を使用するためには重要である。
【0004】
出生後の霊長類個体から多能性細胞を作り出す初の試みは、体細胞性核移植(Byrne、JA et al.,Nature 450:497−502(2007)などを参照)と細胞融合(Yu,J et al.,Stem Cells 24:168−176(2006)などを参照)を採用した。しかしながら、体細胞性核移植の臨床用途は、その効率の悪さ故に非現実であり、一方、細胞融合は概4倍体細胞を生じる。2007年に、二つのグループの科学者が、出生後の霊長類個体からの体細胞を多能性幹細胞に再プログラムした(Yu et al.,Science 318:1917−1920(2007)および Takahashi et al.,Cell 131:861−872(2007)、参照により各文献全体を説明したものとして本発明に組み込む)。両グループは、能力決定導入遺伝子(potency−determining transgene)を発現するためのウイルスベクター系を使って、4個の転写因子のcDNAをヒト体細胞に搬送し、かつ、発現させた。Takahashiらの転写因子は、OCT4、SOX2、c-Myc、およびKLF4であり、Yuらの転写因子は,OCT4、SOX2、NANOG、およびLIN28であった。これらのセットの転写因子の発現は、ヒト体細胞が、形態、増殖、遺伝子発現および表面マーカー発現を含むES細胞特異的特徴を獲得するように誘導した。このような方法で再プログラムされた体細胞は、誘導多能性(iPS)幹細胞と呼ばれる。iPS細胞の存在は、胚盤胞の必要性を回避し、免疫拒絶に伴う懸念を軽減させる。
【0005】
その直後に、Lowryらが、OCT4、SOX2、c−Myc、およびKLF4(Lowry et al.,PNAS 105:2883−2888(2008))トランスジーンの異所性発現を介して患者特有のiPS細胞株を作り出した。より最近には、上皮、線維芽細胞、肝臓、胃、神経、膵臓の細胞など、ヒトとマウスのいくつかの異なる体細胞型からiPS細胞が作り出された。さらに、iPS細胞は、様々な系統の細胞に首尾良く分化した(例えば、Dimos et al.,Science321:1218−1221(2008))。
【0006】
iPS細胞を作り出す現法は、レンチウイルスに由来するようなレトロウイルス・ベクターを採用している。これらのベクターは、標的細胞のDNAに、実質的に染色体のどの座位にも安定的に組み込まれ、該DNAを永続的に変化させる。この再プログラムするベクターとゲノムの非標的相互作用は、ウイルス遺伝子の再活性化によって引き起こされた異常な細胞遺伝子発現および新生物増殖のリスクを伴う。(Okita et al.Nature 448:313−317(2007))
【0007】
それに加えて、導入遺伝子が継続して存在し、発現することが受益者の細胞生理に干渉することがある。さらに、c−Mycなどの体細胞を再プログラムするために使われた転写因子の異所性発現がプログラム細胞死(アポトーシス)を誘発することがある(Askew et al.,Oncogene 6:1915−1922(1911),Evan et al.,Cell 69:119−128(1992))。そのうえ、OCT4などの因子が継続して発現すると、その後に続くiPS細胞の分化に干渉することがある。
【0008】
再プログラムに伴う変化以上に細胞の遺伝的構造を変化させることなく体細胞を再プログラムして高い能力状態にすることが望ましい。最近、Stadtfeldらが、過渡的にOCT4、SOX2、KLF4およびc−Mycを発現した不統合のアデノウイルスを使ってマウスのiPS細胞を作り出した。今日まで、レトロウイルス・ベクターを使用せずに作り出した霊長類iPS細胞は報告されていない。
【発明の概要】
【発明が解決しようとする課題】
【0009】
本発明は、分化した霊長類体細胞を再プログラムして霊長類多能性細胞を作り出すことを広く目的とする。
【課題を解決するための手段】
【0010】
第一の態様において、本発明は、霊長類多能性細胞を作り出す方法であって、体細胞を再プログラムして多能性状態にするのに充分な1セットの導入遺伝子を霊長類体細胞に導入する工程であって、この導入遺伝子は、感染性ウイルスをコードしない少なくとも一つのエピソームのベクターで搬送される該工程と、多能性細胞を回収する工程とを含む。ここで「非ウイルス性」のベクターまたはコンストラクトとは、感染性ウイルスをコードすることの出来ないベクターまたはコンストラクトのことを示す。
【0011】
第二の態様において、本発明は、ヒト霊長類を含む霊長類の補充可能な再プログラムされた多能性細胞の濃縮された集団に関し、ここで、現存するiPS細胞と比較すると、ウイルス源またはその派生物を有する構成要素を含む前記少なくとも一つのベクターは、実質的に当該多能性細胞には存在しない。本明細書で使われるように、このことは、再プログラムされた細胞が、1細胞に付きエピソームのベクターのコピーを1個未満含むことを意味し、好ましくは細胞内にはエピソームのベクターが残留しないことを意味する。細胞分裂の際に非対称分割が起こるとベクターが希釈するので、ベクターを失った再プログラムされた細胞を容易に入手できる。本発明の別の場所で言及したように滅多にないことであるが、再プログラムするベクターが細胞のゲノムに統合されうることがある、しかし、ベクターが統合した細胞は、ベクターが存在しないように選別することによって避けることができる。さらに、現存するES細胞と比較して、本発明の霊長類多能性細胞は、胎児のまたは出生後の個体由来の体細胞と遺伝的に実質的に同一である。胎児の細胞は、例えば羊水から取得できる。濃縮された集団の細胞は、現存する霊長類ESとiPS細胞から形態学的に(即ち、円形、大きめの核小体、および、わずかな細胞質)、または成長特性(即ち、倍加時間、ES細胞は約17〜18時間の倍加時間を持つ)からは容易に区別できない。iPS細胞およびES細胞と同様に、再プログラムされた細胞は多能性細胞特異的マーカーを発現する(例えば、OCT−4、SSEA−3、SSEA−4、TRA−1−60、TRA−1−81、但し、SSEA−1を除く)。ES細胞と異なり、再プログラムされた細胞は、胚から直接得られない。本明細書で使われるように、「胚から直接得られない」とは、多能性細胞を作り出すための開始細胞型が、胎児のもしくは出生後の個体から得られた体細胞のような多分化能の細胞または最終分化した細胞のような非多能性細胞であることを意味する。iPS細胞と同様、本方法で産生された多能性細胞は、その導出の間、選択された能力決定因子(potency−determining factor)の1つもしくは複数コピーを過渡的に発現することができる。
【0012】
別途定義されない限り、本明細書で使われる全ての技術的、科学的用語はこの発明が属する技術分野の当業者に常識的に理解されている意味と同様である。本発明を実施、または、検査するために適切な材料と方法は下記するが、当業者に広く知られている、本明細書に書かれたものと同様または等価なその他の材料と方法を使っても良い。
【0013】
添付した図とともに以下の詳細をたどれば、本発明のその他の目的、利点、特徴は明らかになるであろう。
【図面の簡単な説明】
【0014】
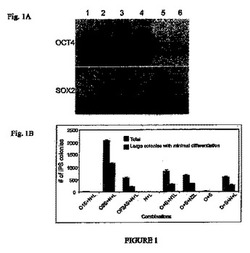
【図1A−B】本発明の再プログラムの方法の間に搬送されたベクター(単数または複数)上の導入遺伝子に連結する異なるヌクレオチド配列の再プログラム効率に関する効果を示す図である。
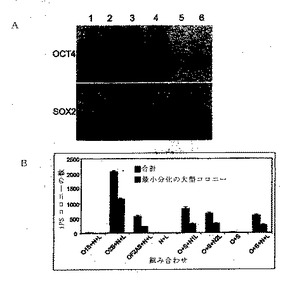
【図2A−C】ヒト新生児包皮線維芽細胞内のMyc、KLF−4、SV40大型T抗原の遺伝子発現の再プログラム効率に関する効果を示す図である。
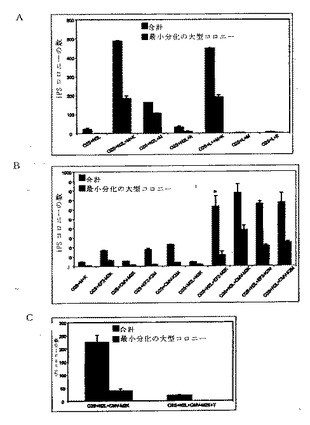
【図3A−C】本発明の方法に従って導入遺伝子を体細胞に搬送する適切なコンストラクトと、エピソームのベクター媒介導入遺伝子の時間的発現と、ヌクレオフェクション後の細胞生存に関するベクター量の効果とを示す図である。
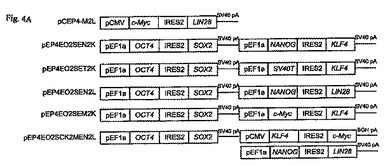
【図4A−D】エピソームのベクター媒介導入遺伝子発現を用いたヒト新生児包皮線維芽細胞の再プログラムを示す図である。
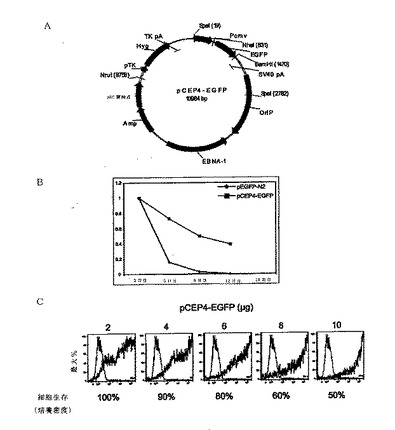
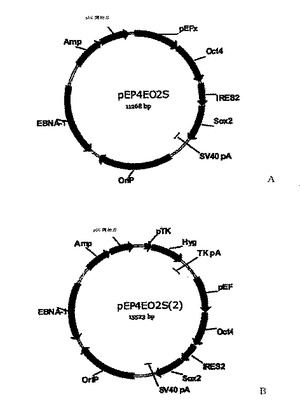
【図5A−B】本発明の再プログラム方法に有用な発現カセットを有する関連コンストラクトを示す図である。
【発明を実施するための形態】
【0015】
本発明は、分化した霊長類体細胞を再プログラムして、再プログラムされた霊長類細胞にする新規な方法に広く関連する。再プログラムの間には存在し、しかし、再プログラムされた細胞には実質的に存在しない非ウイルス性ベクターに能力決定因子を導入することによって、再プログラムされた霊長類細胞は、その産生中に使われたベクターを実質的に有しない。本明細書で使われるように、「再プログラミング」とは、分化した体細胞を、その由来源の細胞よりも高い能力を有する脱分化した細胞に転換する遺伝的プロセスのことである。
【0016】
好都合に、本方法で産生した高い能力を有する細胞は、正倍数体多能性細胞である。ここで使われるように、「多能性細胞」は、多能性細胞特異的マーカーを発現する細胞集団を意味し、未分化細胞の細胞形態特徴(即ち、緻密なコロニー、高い核対細胞質比、および、顕著な核小体)を有し、3個全ての胚葉(例えば、内胚葉、中胚葉および外胚葉)に分化することができる。SCIDマウスなどの免疫無防備状態の動物に導入された場合、多能性細胞は、典型的に3つの全ての胚葉の細胞または組織の特徴を含む奇形腫を形成する。当業者は、当分野で通常使われている技術を利用してこれらの特徴を評価することができる。上記のThomson et al.,を参照されたい。多能性細胞は細胞培養で増殖でき、かつ、多分化能性質を示す様々な系統限定細胞集団へと分化できる。多能性細胞は、体細胞性多分化能細胞より高い能力を有する。体細胞性多分化能細胞は、比較的より分化しているが、最終分化している訳ではない。霊長類体細胞再プログラム方法の多能性産物は、ここで再プログラムされた霊長類多能性細胞、または、誘発多能性(iPS)細胞と呼ばれる。そのような細胞は、ヒトES細胞または現存するiPS細胞に関して現在予期される研究と治療上の適用における使用に適している。
【0017】
ヒトを含む霊長類の、胎性、新生児、若年性、または、成体の個々からの細胞を含む分化した体細胞は、本方法に適した開始細胞である。適した体細胞は、これらに制限されないが、骨髄細胞、上皮細胞、血管内皮細胞、線維芽細胞、造血細胞、ケラチノサイト、肝細胞、腸細胞、間葉系細胞、骨髄性前駆細胞、および、脾臓細胞を含む。別の適した体細胞は、基質に付着したCD29+、CD44+、CD166+、CD105+、CD73+およびCD31-の間葉系細胞である。代わりに、体細胞は、それら自身が増殖でき、かつ、血液幹細胞、筋/骨幹細胞、脳幹細胞および肝臓幹細胞を含む他の型の細胞に分化できる細胞であっても良い。適した体細胞は、受容性であるか、または、科学文献に一般に知られた方法を使って受容性にできて、因子をコードする遺伝物質を含む能力決定因子を取り込む。取り込み増強方法は、細胞型と発現系に依存して変えることができる。適切な伝達効率を有した受容性体細胞を調製するために使われる典型的な条件は、当業者によって良く知られている。開始体細胞は、約24時間の倍加時間を持つことができる。
【0018】
本明細書に記述したベクターは、治療上の使用の安全性を向上するために科学文献に一般的に知られたた方法を使って構成し、設計できる。この方法は、選択と強化のマーカーを含み、希望するならば、ベクター上のヌクレオチド配列の発現を最適化することができる。このベクターは、ベクターが体細胞性開始細胞内で自己複製できる構造要素を含むべきである。例えば、既知のエプシュタイン・バーウイルス oriP/Nuclear抗原(EBNA−1)組み合わせ(例、Lindner,S.E.and B.The plasmid replicon of Epstein−Barr virus:mechanistic insights intoefficient,licensed,extrachromosomal replication in human cells, Plasmid 58:1(2007)を参照、本文全体をここに説明したものとして参照により本明細書に組み込む)は、ベクターの自己複製を支持するのに充分であり、また、哺乳類、特に霊長類の細胞内で機能すると知られているその他の組み合わせを採用しても良い。本発明において使用するのに適した発現ベクターを構築する標準技術は、当業者に知られており、Sambrook Jらによる「分子クローニング:ラボラトリマニュアル」(3rd ed.Cold Spring harbor Press,Cold Spring Harbor,N.Y. 2001)などの文献に見出すことができ、参照により本文全体を説明されたものとして本明細書に取り込む。
【0019】
本発明の方法では、1セットの能力決定因子をコードする遺伝子物質が、1個または複数個の再プログラム用ベクターを介して体細胞に搬送される。適切な能力決定因子は、これらに限定されるものではないが、OCT−4、SOX2、LIN28、NANOG、c−Myc、KLF4、および、これらの組み合わせを含むことができる。体細胞内にポリヌクレオチド発現を引き起こすことができる非相同性のプロモータに機能的に結合している能力決定因子をコードするポリヌクレオチド導入遺伝子として、各能力決定因子を体細胞に導入することができる。SV40T抗原それ自体は能力決定因子ではないけれども、能力決定因子が発現されている再プログラムの間に、細胞生存を促進するのに充分な条件を持つ細胞を提供するように、都合良く体細胞に導入される。因子の発現に充分なその他の条件は、実施例に記述されている細胞培養条件を含む。
【0020】
適切な再プログラム用ベクターは、プラスミドのようなエピソームのベクターであって、感染性または有複製能ウイルスを生じるのに十分なウイルス性ゲノムの全部または一部をコードしないが、当該ベクターは一つまたは複数のウイルスから得た構造要素を含むことができる。1個または複数個の再プログラム用ベクターが、1個の体細胞に導入しうる。1個または複数個の導入遺伝子が、1個の再プログラム用ベクターに供給されうる。1個の強い、恒常的な転写性プロモータが、複数の導入遺伝子のために転写調節を提供することができ、該導入遺伝子は発現カセットして提供されうる。1個のベクター上の別々の発現カセットは、それぞれの強い恒常的なプロモータの転写調節下にあり、これらのプロモータは、同一プロモータのコピーであるか、異なるプロモータのコピーであり得る。多様な非相同性プロモータが当分野で知られており、能力決定因子の所望の発現レベルなどの要因に応じて使うことができる。下記に例示するように、標的体細胞内の異なる強度を持つ異なるプロモータを使って、別々の発現カセットを転写コントロールすることは好都合である。転写性プロモータ(単数または複数)を選択する際に別途考慮することは、プロモータが標的体細胞内で発現抑制される割合である。再プログラム法において、遺伝子(単数または複数)の産生がその役割を完了または実質的に完了した後に、1個または複数個の、導入遺伝子または導入遺伝子発現カセットの発現を減らすと好都合であることを当業者は想定する。例示的プロモータは、ヒトEF1α伸長因子プロモータ、CMVサイトメガロウイルス即時型初期プロモータ、CAGニワトリ・アルブミン・プロモータ、および、別種由来のそれぞれに対応する相同型プロモータである。ヒト体細胞内において、EF1αとCMVの両方は強いプロモータであるが、CMVプロモータはEF1αプロモータより効率的に抑制される、即ち、EF1αの制御下の導入遺伝子の発現が停止するや否や、CMVの制御下の導入遺伝子の発現が停止する。
【0021】
能力決定因子は、再プログラムの効率を調整するために変動する相対比率で、体細胞内において発現できる。例えば、OCT−4とSOX2が、単一のベクター内の単一の転写で1:1の比率でコードされる場合は、単一の細胞に吸収される比率が制御できないように2個の因子それぞれを異なるベクター上に提供する時に比べて(即ち、各因子が各単一細胞に入る比率を制御できない)、体細胞の再プログラム効率は4倍高い。好ましくは、複数の導入遺伝子が単一の転写でコードされる場合には、配列内リボソーム進入部位が、転写性プロモータから離れた導入遺伝子(単数または複数)の上流に位置する。因子の相対的比率は、搬送された因子に依存して変化させることができるが、本開示を見た当業者は、最適な比率を求めることができる。
【0022】
当業者は、複数のベクターではなく、単一のベクターで全ての因子を導入できる効率の良さを想定するだろうが、ベクター全体のサイズが増大するにつれて、ベクターを導入することがますます困難になる。当業者は、また、ベクター上の因子が時間的発現に及ぼす影響と結果として生じる再プログラム効率も想定するであろう。そのため、出願人らは、ベクターの組み合わせについて様々な因子の組み合わせを採用した。いくつかのそのような組み合わせが、本明細書に提示されて再プログラムを立証する。
【0023】
再プログラム用ベクター(単数または複数)を導入後で、かつ体細胞が再プログラム中である場合、導入された導入遺伝子が転写および翻訳される間は、標的細胞内にベクターが存続し得る。導入遺伝子発現は、再プログラムされて多能性状態になった細胞内で、好都合にも下方制御しうるか、停止しうる。再プログラム用ベクター(単数または複数)は、染色体外に残ることができる。極度に低い効率の場合、ベクター(単数または複数)は、細胞のゲノム内へと組み込める。再プログラム用ベクター(単数または複数)は、レシピエント細胞のゲノムと協調して複製し、そのため、エピソームのベクターがそのDNAを複製することが出来なくなる期間より長く、2週間ほど適度に安定している。にもかかわらず、ベクターは細胞分裂の際に均等に分配されないので、選択圧が無い場合、細胞がエピソームのベクター(単数または複数)を失い、それにより本方法ではベクター・フリーの多能性細胞を直ちに回収できる。例えば、体細胞内にoriP/EBNA−1−ベースのエピソームのプラスミドが、通常2〜3週間安定的に維持される。初期の2〜3週間に、細胞はエピソームのプラスミドを急速に失う。細胞が安定化すると、細胞は、一世代に付き〜5%の割合でエピソームのベクターを連続的に失う。
【0024】
本方法で産生した多能性細胞は、多能性細胞増殖をサポートする任意の培地で培養することができる。培地として、ここに書かれたものに限定されるものではないが、TeSR(商標)(StemCell Technologies,Inc.;Vancouver,Canada),mTeSR(商標)(StemCell Technologies, Inc.)and StemLine(登録商標)serum−free medium(Sigma;St.Louis.,Mo.)、または、マウス胚性線維芽細胞(MEF)馴化培地などの馴化培地などがある。本明細書で使われるように、「限定培地」とは、既知の原料由来の既知の化学組成物または成分からなる生化学的に定義した成分のみを含む、生化学的に定義した処方を意味する。本明細書で使われるように、「馴化培地」とは、培地で培養された細胞由来の可溶性因子をさらに補充した増殖培地のことを意味する。別の方法として、細胞は培地内のMEF上で維持しても良い。
【0025】
本発明は、以下のこれらに限定されることのない実施例を熟慮することにより十分に理解できる。
【0026】
実施例1
発現カセットの設計と作製
プライマーとして、かつ表1に収載の配列内リボソーム進入部位から得られるコード領域の最初と最後の20〜22の塩基を使用して、して、導入遺伝子の一部または全てから得られるオープン・リーディング・フレーム(ORF)をポリメラーゼ連鎖反応(PCR)法で直接増幅するという従来の方法を使って、適した発現カセットを作製した。SV40T抗原とヒトテロメラーゼ逆転写酵素、プラスミドpBABE−puro SV40 LT、および、pBABE−hygro−hTERT(ヒトテロメラーゼ逆転写酵素)の供給源は、プラスミド1390および1773としてそれぞれAddgene,Inc,Cambridge、MAから市販されている。 IRES1とIRES2、プラスミドpIRESpuro3とプラスミドpIRES2EGFPの供給源は、Clontech Laboratories,Inc.,Mountain View,CA.から市販されている。
口蹄疫ウイルスセグメント2を化学的に合成した。インフレームの発現カセットは、下記表1記載のコードを使って記述する。例えば、「E−O2S」とは、OCT4とSOX2コード領域(両者の間にIRES2)の上流に位置するEF1αプロモータを有する発現カセットを意味する。同様に、C−M2Kとは、c−MycとKlf4コード領域(両者の間にIRES2)の上流に位置するCMVプロモータを有する発現カセットを意味する。
いくつかのコンストラクトでは、上記のカセットは、一連の再プログラム中では使わなかった。O2Sとは異なるO2S変異体発現カセット(「O2S(2)」)を採用した。この変異体は、TKプロモータ−Hyg −TK polyAカセット(図5Aおよび図5Bを比較せよ)を含んでいた。指示した構造を有するカセットは、様々な因子の発現レベルを実験的に測定することによる再プログラムの方法で、後に使用するために選択した。
EF2(配列番号:12)として設計されたプロモータは、既知のEF1αプロモータ(配列番号:11)とはわずかに異なる変異体であったが、活性化中のEF1αとは異ならず、また後に以下のエピソームのベクターの再プログラムの試行には使用しなかった。F2Aは、単一のトランスクリプトから発現した異なるコード領域を同時翻訳するのを促進するペプチド・リンカーである。F2Aを試用したが、後のエピソームのベクターを使った再プログラム試行には使わなかった。IRES1を試用したが、後のエピソームのベクターを使った再プログラム試行には使わなかった。
【0027】
ORF上流または下流の発現に関して、様々なプロモータ、IRES配列、および、導入遺伝子の配置の相対的効果を、以前に記述(Yu et al.,上記参照)したように、形質移入後ヒト体細胞を再プログラムできるかをテストするために、様々な導入遺伝子発現カセットをpSin4、修飾したレンチウイルス・ベースのベクターへと別々にクローニングして評価した。293FT細胞に、下記に示すようにSuperFect(Qiagen,Valencia,CA)を使いIRES1またはIRES2で連結されたOCT4とSOX2を発現するレンチウイルスのプラスミドベクターで形質移入した。細胞は、形質移入2日後に収集した。図1Aは、293FT細胞内のOCT−4とSOX2のウエスタンブロット法による解析を示す。レーン1、pSIN4−EF2−OCT4−IRES1−SOX2、レーン2,pSIN4−EF2−OCT4−IRES2−SOX2、レーン3,pSIN4−EF2−OCT4−F2A−SOX2、レーン4、pSIN4−EF2−OCT4−IRES1−PURO、レーン5、pSIN4−EF2−SOX2−IRES1−PURO、レーン6、プラスミド無し(コントロール)。マウス抗ヒトOCT4モノクローナル抗体(1:500,Santa Cruz ,Biotechnology Inc.,Santa Cruz,CA,sc−5279)とヤギ抗ヒトSOX2ポリクローナル抗体(1:500,R&D Systems,Minneapolis,MN AF2018)を使用して、OCT4とSOX2それぞれの相対的発現を検出した。
【0028】
図1Bは、OCT4ノックイン・ヒト胚性幹細胞(米国特許出願第2006/0128018号、および、Zwaka and Thomson,Nature Biotechnology 21:319−321(2003)、参照により全体を説明したものとして各文献を本発明に取り込む)由来の(Yu et al.,上記参照)0.2×106の間葉系細胞内で連結させた能力決定因子を使った再プログラムを示す。
この系統は、ネオマイシン選択(ジェネテシン:100μg/ml,Invitrogen Corp.)下で維持した。形質導入16日後にヒトiPS細胞コロニーを計数した。遺伝子組み合わせは、pSIN4−EF2−OCT4−IRES1−SOX2(O1S)、pSIN4−EF2−OCT4−IRES2−SOX2(O2S)、pSIN4−EF2−OCT4−F2A−SOX2(OF2AS)、pSIN4−EF2−NANOG−IRES1−LIN28(N1L)pSIN4−EF2−NANOG−IRES2−LIN28(N2L)、pSIN4−EF2−OCT4−IRES1−PURO(O)、pSIN4−EF2−SOX2−IRES1−PURO(S)、pSIN4−EF2−NANOG−IRES1−PURO(N)、pSIN4−EF2−LIN28−IRES1−PURO(L)であった。レンチウイルスのプラスミドベクターの各々の略語は、ベクターの名称の後に、括弧で囲んでいる。
【0029】
実施例2
レンチウイルス・コンストラクトを使用したヒト新生児包皮線維芽細胞の再プログラム化
予備的再プログラムの実験は、レンチウイルスベクターをヒト新生児の包皮線維芽細胞に導入することによって行った。図2Aは、OCT4、SOX2、LIN28、および、さらにc−MYCを導入する場合、NANOGが再プログラム効率にかなりのプラス効果をもたらすことを示し、 また、OCT4、SOX2、および、LIN28との組み合わせでは、c−MYCまたはKLF4が無い場合でもNANOGが再プログラムを補助できることを示す。使用したレンチウイルスのコンストラクトは、pSIN4−EF2−OCT4−IRES2−SOX2(O2S)、pSIN4−EF2−NANOG−IRES2−LIN28 (N2L)、pSIN4−EF2−LIN28−IRES1−PURO(L)、pSIN4−CMV−c−Myc−IRES1−PURO(M)、pSIN4−EF2−KLF4−IRES1−PURO(K)であった。形質移入20日後、アルカリ・ホスファターゼ-陽性ヒトiPS細胞コロニーを計数した。iPS細胞コロニーの数は、2.5×104個のヒト新生児包皮線維芽細胞(継代数9)の入力由来であった。薄灰色の棒は、再プログラムされて形成した、代表的なヒトES細胞形態を有するコロニーの合計数を表し、濃い灰色棒は、分化が最も少なかった大型コロニーの数を表す。
【0030】
図2Bは、連結した能力決定因子を用いた再プログラムを証明する。使用したレンチウイルスのコンストラクトは、pSIN4−EF2−c−Myc−IRES2−KLF4(EF2−M2K)、pSIN4−CMV−c−Myc−IRES2−KLF4(CMV−M2K)、pSIN4−EF2−KLF4−IRES2−c−Myc(EF2−K2M)、pSIN4−CMV−KLF4−IRES2−c−Myc(CMV−K2M)、pSIN4−CMV−c−Myc−IRES2−LIN28(M2L)、pSIN4−EF2−NANOG−IRES2−KLF4(N2K)であった。
【0031】
形質導入14日後、アルカリ・ホスファターゼ-陽性ヒトiPS細胞コロニーを計数した。iPS細胞コロニーの数は、約7.0×104個のヒト新生児包皮線維芽細胞(継代数12)の入力由来であった。アステリスクは、多くのアルカリ・ホスファターゼ−陽性コロニーが形態学的に緩やかに現れたことを示す。
【0032】
図2Cは、SV40大型T抗原遺伝子発現の再プログラムへの効果を示す。SV40大型T抗原は、マウスの線維芽細胞内にc−Mycが誘発するのを妨ぎ(Hermeking et al.,PNAS 91:10412−10416(1994))、再プログラム効率を高める(Hanna et al.,Cell 133:250−264(2008);Mali et al.,Stem Cells doi:10.1634/stemcells.2008−0346(2008))。遺伝子組み合わせ略名は、図2B内のものと同じであり、さらにSV40大型T抗原(T)を追加する。c−Mycも細胞増殖を促進する。形質導入12日後、アルカリ・ホスファターゼ-陽性ヒトiPS細胞コロニーを計数した。iPS細胞コロニーの数は、約〜3.5×104個のヒト新生児包皮線維芽細胞(継代数17)の入力由来であった。図2Cは、T抗原がレンチウイルス・ベースの再プログラムの間に達成したレベルで存在するならば、iPS細胞導出の最終段階を抑制することを実証する。対照的に、下記を参照されたいが、T抗原は、一時的発現時間に存在し、かつ/または、エピソームのベクターを使って再プログラム中に達成したレベルで存在するならば、この効果をもたない。さらに加えて、T抗原はc−Myc−誘導アポトーシスを妨げないがc−Myc−誘導細胞増殖に逆の影響を与える。
【0033】
実施例3.
非ウイルス性エピソームのベクターを使ったヒト新生児包皮線維芽細胞の再プログラム
ヒト新生児包皮線維芽細胞(Cat# CRL−2097(商標),ATCC)は、10%熱失活したウシ胎仔血清(FBS,HyClone Laboratories,Logan,UT)、2mMのグルタマックス、0.1mMの非必須アミノ酸、および、0.1mMのβ−メルカプトエタノールを添加した包皮線維芽細胞培地(DMEM(Cat# 11965,Invitrogen))で維持した。
【0034】
実施例1および以下の表3で詳細に示されるように構成された導入遺伝子発現カセットとして提供された能力決定因子の様々な組み合わせは、エピソームのコンストラクトpCEP4−EGFP(図3Aに示すように)を使って体細胞に導入されて、様々な効率を持つ再プログラムをもたらした。pCEP4−EGFPは、市販の哺乳類エピソームの発現ベクターpCEP4(Invitrogen Corp., Carlsbad, CA)から、EGFPコード領域をpCEP4 BamHI部位とNheI部位との間に挿入することによって創出した。表2のエピソームのベクターは、指定した発現ベクターをpCEP4−EGFPもしくは関連するバックボーンのないPcmv(指定されたpEP4)に挿入することによって創出した。発現カセットが挿入されたクローニング部位については図3Aと表2の脚注を参照せよ。
【0035】
製造元の指示に従い、ヒト真皮線維芽細胞キット(Normal Human Dermal Fibroblasts,Amaxa,Inc.Cat.No.VPD−1001)を使用して、単一のヌクレオフェクション・イベント経由で線維芽細胞にベクターを導入した。ヌクレオフェクション後、照射を受けたマウス胚性線維芽細胞(MEF)が播種された3個の10cmの皿の上に形質移入した線維芽細胞(それぞれ〜0.8から1.0×106細胞)を直ちにプレートした。包皮線維芽細胞培地は一日おきに交換した。4日後、包皮線維芽細胞培地をヒト胚性幹細胞培地(DMEM/F12培地に以下を補充、20%ノックアウト血清リプレーサー、0.1mMの非必須アミノ酸(全てInvitrogen Corp.製)、1mMのグルタマックス、0.1mMのβメルカプトエタノール、および、前記した100ng/mlのゼブラフィッシュタンパク・塩基性線維芽細胞増殖因子(zbFGF)(Amit et al., Developmental Biology 227:271−278 (2006); Ludwig et al., Nature Methods 3:637−646(2006)、ここで、それぞれは、参照により全体を説明したものとしてここに組み入れる)と交換した。プレーティング8〜10日後、播種されたMEFが、再プログラム培地を維持出来なくなったとき、照射されたMEFで馴化したヒト胚性幹細胞培地を代わりに使用した。適切な時期(形質移入2〜3週間後)に、ヒトiPSコロニー発生の徴候としてアルカリ・ホスファターゼにより培養物を着色した。
【0036】
導入遺伝子コンストラクトを導入するための適切なパラメータを決定するために、pEGFP−N2(対照)とpCEP4−EGFPのエピソームのベクター由来のEGFPを293FT細胞に導入して評価した後、EGFPレベルを経時的に測定することによって、時間的発現について初めに評価した(図3B)。
【0037】
ヒト新生児包皮線維芽細胞生存物について、導入された導入遺伝子コンストラクトの量の効果も、予備的実験で評価した。図3Cは、ヌクレオフェクション後翌日の細胞コンフルエンスから推定した、ヌクレオフェクションの効率性とヒト新生児包皮線維芽細胞の生存物について、使用されたpCEP4−EGFPのエピソームのベクターの量の効果を示す。ヌクレオフェクションされた約1×106個の包皮線維芽細胞を6ウェル・プレートの各ウェルにプレートした。灰色線は形質移入されなかった対照の線維芽細胞を表し、黒色線は形質移入された線維芽細胞を表す。
【0038】
図4Aは、様々な発現カセットを含む表3からの模式的な導入遺伝子発現コンストラクトを示し、ある組み合わせをヒト新生児包皮線維芽細胞に導入すると、線維芽細胞から多能性細胞への再プログラムが生じる。導入されるエピソームの再プログラム用ベクターの3つの組み合わせが、次の再プログラムされた多能性細胞を生じさせた。(1)pEP4−E−O2S−E−T2K、pEP4−E−O2S−E−N2K、およびpCEP4−C−M2L、(2)pEP4−E−O2S−C−K2M−E−N2L、およびpEP4−E−O2S−E−T2K、ならびに、(3)pEP4−E−O2S−E−N2L、pEP4−E−O2S−E−T2K、およびpEP4−E−O2S−E−M2K。表3は、成功した組み合わせで使われた各ベクターの量を示す。各成功した再プログラムの組み合わせ内の1個のベクターは、EF1αプロモータの制御の元、T抗原をコードした。
【0039】
図4Bは、エピソームのベクターの形質移入18日後に観察した形態変化をもつ典型的なコロニーの明視野顕微鏡のイメージを示す。図4Cは、エピソームのベクターの形質移入18日後のアルカリホスファターゼ−陽性コロニーの明視野顕微鏡イメージを示す。
【0040】
形質移入25〜30日後、ヒトiPS細胞コロニーに形態的に類似した多くの非iPS 細胞コロニーがある故に、再プログラム培養物を一度新鮮な10cmのMEF皿(1:3比)に移した。そして、その後の解析のためにコロニーを選択した。図4Dは、形質移入再プログラム培養後28日目の第一継代6日後のヒトiPS細胞コロニーの明視野顕微鏡イメージを示す。スケールバーは、0.1mmを表す。再プログラムされた細胞は、後の解析のために、前述した馴化培地(Xu et al.,Nat.Biotechnol.19:971(2001)、参照により全体を説明したものとして本明細書に組み込む)を用い、マトリゲル(BD Biosciences,Bedford,MA)のフィーダ・フリー培養で維持した。
【0041】
好都合にも、再プログラムされた新生児包皮線維芽細胞の内1%を大きく上回る再プログラム効率が達成でき、4個の遺伝子の組み合わせを使用して達成した場合に比べ、有意に短い再プログラム時間であった。
【0042】
本開示に記述した本発明のいくつかの改作は当業者においては通常の最適化事項であり、本発明の精神もしくは添付の請求項の範囲を逸脱することなく実施できることを理解されたい。明細書で上述した出版物と特許は、参照により本明細書に組み込まれる。本発明の記述した方法とシステムの様々な改変と変形は、本発明の範囲と精神を逸脱することなく、当業者には明らかである。しかし、上記した本発明の実施例と実施形態は、例証的であり、本発明を限定するわけではない。本発明は、下記の請求項の範囲に付随する実施例と実施形態の全ての改変体を含む。
【技術分野】
【0001】
(関連出願の相互参照)
本明細書は、米国仮特許出願第61/108.362(出願日2008年10月24日)を参照によりその全体を説明したものとしてここに含める。
【0002】
(連邦支援による研究または開発に関する陳述)
本発明は、NIH GM081629及びRR000167の機関を通してアメリカ合衆国政府助成を受けた。アメリカ合衆国は本発明にある一定の権利を保有する。
【背景技術】
【0003】
胚性幹細胞(ES)は、多能性、即ち、無限に増殖し、3つの胚葉全ての細胞に分化しうる能力があり、科学と医学の分野において大いに期待されている(Thomson et al., Science 282:1145−1147(1998),参照により文献全体を説明したものとして本発明に組み込む)。治療や再生医学におけるヒトES細胞の適用は、受益者の免疫系による拒絶の可能性によって複雑化する。したがって、実質的に特定受益者と遺伝的に同一のヒト多能性細胞が高く望まれる。さらに、患者特異的な治療計画を立案する際に、遺伝的同一性がES細胞を使用するためには重要である。
【0004】
出生後の霊長類個体から多能性細胞を作り出す初の試みは、体細胞性核移植(Byrne、JA et al.,Nature 450:497−502(2007)などを参照)と細胞融合(Yu,J et al.,Stem Cells 24:168−176(2006)などを参照)を採用した。しかしながら、体細胞性核移植の臨床用途は、その効率の悪さ故に非現実であり、一方、細胞融合は概4倍体細胞を生じる。2007年に、二つのグループの科学者が、出生後の霊長類個体からの体細胞を多能性幹細胞に再プログラムした(Yu et al.,Science 318:1917−1920(2007)および Takahashi et al.,Cell 131:861−872(2007)、参照により各文献全体を説明したものとして本発明に組み込む)。両グループは、能力決定導入遺伝子(potency−determining transgene)を発現するためのウイルスベクター系を使って、4個の転写因子のcDNAをヒト体細胞に搬送し、かつ、発現させた。Takahashiらの転写因子は、OCT4、SOX2、c-Myc、およびKLF4であり、Yuらの転写因子は,OCT4、SOX2、NANOG、およびLIN28であった。これらのセットの転写因子の発現は、ヒト体細胞が、形態、増殖、遺伝子発現および表面マーカー発現を含むES細胞特異的特徴を獲得するように誘導した。このような方法で再プログラムされた体細胞は、誘導多能性(iPS)幹細胞と呼ばれる。iPS細胞の存在は、胚盤胞の必要性を回避し、免疫拒絶に伴う懸念を軽減させる。
【0005】
その直後に、Lowryらが、OCT4、SOX2、c−Myc、およびKLF4(Lowry et al.,PNAS 105:2883−2888(2008))トランスジーンの異所性発現を介して患者特有のiPS細胞株を作り出した。より最近には、上皮、線維芽細胞、肝臓、胃、神経、膵臓の細胞など、ヒトとマウスのいくつかの異なる体細胞型からiPS細胞が作り出された。さらに、iPS細胞は、様々な系統の細胞に首尾良く分化した(例えば、Dimos et al.,Science321:1218−1221(2008))。
【0006】
iPS細胞を作り出す現法は、レンチウイルスに由来するようなレトロウイルス・ベクターを採用している。これらのベクターは、標的細胞のDNAに、実質的に染色体のどの座位にも安定的に組み込まれ、該DNAを永続的に変化させる。この再プログラムするベクターとゲノムの非標的相互作用は、ウイルス遺伝子の再活性化によって引き起こされた異常な細胞遺伝子発現および新生物増殖のリスクを伴う。(Okita et al.Nature 448:313−317(2007))
【0007】
それに加えて、導入遺伝子が継続して存在し、発現することが受益者の細胞生理に干渉することがある。さらに、c−Mycなどの体細胞を再プログラムするために使われた転写因子の異所性発現がプログラム細胞死(アポトーシス)を誘発することがある(Askew et al.,Oncogene 6:1915−1922(1911),Evan et al.,Cell 69:119−128(1992))。そのうえ、OCT4などの因子が継続して発現すると、その後に続くiPS細胞の分化に干渉することがある。
【0008】
再プログラムに伴う変化以上に細胞の遺伝的構造を変化させることなく体細胞を再プログラムして高い能力状態にすることが望ましい。最近、Stadtfeldらが、過渡的にOCT4、SOX2、KLF4およびc−Mycを発現した不統合のアデノウイルスを使ってマウスのiPS細胞を作り出した。今日まで、レトロウイルス・ベクターを使用せずに作り出した霊長類iPS細胞は報告されていない。
【発明の概要】
【発明が解決しようとする課題】
【0009】
本発明は、分化した霊長類体細胞を再プログラムして霊長類多能性細胞を作り出すことを広く目的とする。
【課題を解決するための手段】
【0010】
第一の態様において、本発明は、霊長類多能性細胞を作り出す方法であって、体細胞を再プログラムして多能性状態にするのに充分な1セットの導入遺伝子を霊長類体細胞に導入する工程であって、この導入遺伝子は、感染性ウイルスをコードしない少なくとも一つのエピソームのベクターで搬送される該工程と、多能性細胞を回収する工程とを含む。ここで「非ウイルス性」のベクターまたはコンストラクトとは、感染性ウイルスをコードすることの出来ないベクターまたはコンストラクトのことを示す。
【0011】
第二の態様において、本発明は、ヒト霊長類を含む霊長類の補充可能な再プログラムされた多能性細胞の濃縮された集団に関し、ここで、現存するiPS細胞と比較すると、ウイルス源またはその派生物を有する構成要素を含む前記少なくとも一つのベクターは、実質的に当該多能性細胞には存在しない。本明細書で使われるように、このことは、再プログラムされた細胞が、1細胞に付きエピソームのベクターのコピーを1個未満含むことを意味し、好ましくは細胞内にはエピソームのベクターが残留しないことを意味する。細胞分裂の際に非対称分割が起こるとベクターが希釈するので、ベクターを失った再プログラムされた細胞を容易に入手できる。本発明の別の場所で言及したように滅多にないことであるが、再プログラムするベクターが細胞のゲノムに統合されうることがある、しかし、ベクターが統合した細胞は、ベクターが存在しないように選別することによって避けることができる。さらに、現存するES細胞と比較して、本発明の霊長類多能性細胞は、胎児のまたは出生後の個体由来の体細胞と遺伝的に実質的に同一である。胎児の細胞は、例えば羊水から取得できる。濃縮された集団の細胞は、現存する霊長類ESとiPS細胞から形態学的に(即ち、円形、大きめの核小体、および、わずかな細胞質)、または成長特性(即ち、倍加時間、ES細胞は約17〜18時間の倍加時間を持つ)からは容易に区別できない。iPS細胞およびES細胞と同様に、再プログラムされた細胞は多能性細胞特異的マーカーを発現する(例えば、OCT−4、SSEA−3、SSEA−4、TRA−1−60、TRA−1−81、但し、SSEA−1を除く)。ES細胞と異なり、再プログラムされた細胞は、胚から直接得られない。本明細書で使われるように、「胚から直接得られない」とは、多能性細胞を作り出すための開始細胞型が、胎児のもしくは出生後の個体から得られた体細胞のような多分化能の細胞または最終分化した細胞のような非多能性細胞であることを意味する。iPS細胞と同様、本方法で産生された多能性細胞は、その導出の間、選択された能力決定因子(potency−determining factor)の1つもしくは複数コピーを過渡的に発現することができる。
【0012】
別途定義されない限り、本明細書で使われる全ての技術的、科学的用語はこの発明が属する技術分野の当業者に常識的に理解されている意味と同様である。本発明を実施、または、検査するために適切な材料と方法は下記するが、当業者に広く知られている、本明細書に書かれたものと同様または等価なその他の材料と方法を使っても良い。
【0013】
添付した図とともに以下の詳細をたどれば、本発明のその他の目的、利点、特徴は明らかになるであろう。
【図面の簡単な説明】
【0014】
【図1A−B】本発明の再プログラムの方法の間に搬送されたベクター(単数または複数)上の導入遺伝子に連結する異なるヌクレオチド配列の再プログラム効率に関する効果を示す図である。
【図2A−C】ヒト新生児包皮線維芽細胞内のMyc、KLF−4、SV40大型T抗原の遺伝子発現の再プログラム効率に関する効果を示す図である。
【図3A−C】本発明の方法に従って導入遺伝子を体細胞に搬送する適切なコンストラクトと、エピソームのベクター媒介導入遺伝子の時間的発現と、ヌクレオフェクション後の細胞生存に関するベクター量の効果とを示す図である。
【図4A−D】エピソームのベクター媒介導入遺伝子発現を用いたヒト新生児包皮線維芽細胞の再プログラムを示す図である。
【図5A−B】本発明の再プログラム方法に有用な発現カセットを有する関連コンストラクトを示す図である。
【発明を実施するための形態】
【0015】
本発明は、分化した霊長類体細胞を再プログラムして、再プログラムされた霊長類細胞にする新規な方法に広く関連する。再プログラムの間には存在し、しかし、再プログラムされた細胞には実質的に存在しない非ウイルス性ベクターに能力決定因子を導入することによって、再プログラムされた霊長類細胞は、その産生中に使われたベクターを実質的に有しない。本明細書で使われるように、「再プログラミング」とは、分化した体細胞を、その由来源の細胞よりも高い能力を有する脱分化した細胞に転換する遺伝的プロセスのことである。
【0016】
好都合に、本方法で産生した高い能力を有する細胞は、正倍数体多能性細胞である。ここで使われるように、「多能性細胞」は、多能性細胞特異的マーカーを発現する細胞集団を意味し、未分化細胞の細胞形態特徴(即ち、緻密なコロニー、高い核対細胞質比、および、顕著な核小体)を有し、3個全ての胚葉(例えば、内胚葉、中胚葉および外胚葉)に分化することができる。SCIDマウスなどの免疫無防備状態の動物に導入された場合、多能性細胞は、典型的に3つの全ての胚葉の細胞または組織の特徴を含む奇形腫を形成する。当業者は、当分野で通常使われている技術を利用してこれらの特徴を評価することができる。上記のThomson et al.,を参照されたい。多能性細胞は細胞培養で増殖でき、かつ、多分化能性質を示す様々な系統限定細胞集団へと分化できる。多能性細胞は、体細胞性多分化能細胞より高い能力を有する。体細胞性多分化能細胞は、比較的より分化しているが、最終分化している訳ではない。霊長類体細胞再プログラム方法の多能性産物は、ここで再プログラムされた霊長類多能性細胞、または、誘発多能性(iPS)細胞と呼ばれる。そのような細胞は、ヒトES細胞または現存するiPS細胞に関して現在予期される研究と治療上の適用における使用に適している。
【0017】
ヒトを含む霊長類の、胎性、新生児、若年性、または、成体の個々からの細胞を含む分化した体細胞は、本方法に適した開始細胞である。適した体細胞は、これらに制限されないが、骨髄細胞、上皮細胞、血管内皮細胞、線維芽細胞、造血細胞、ケラチノサイト、肝細胞、腸細胞、間葉系細胞、骨髄性前駆細胞、および、脾臓細胞を含む。別の適した体細胞は、基質に付着したCD29+、CD44+、CD166+、CD105+、CD73+およびCD31-の間葉系細胞である。代わりに、体細胞は、それら自身が増殖でき、かつ、血液幹細胞、筋/骨幹細胞、脳幹細胞および肝臓幹細胞を含む他の型の細胞に分化できる細胞であっても良い。適した体細胞は、受容性であるか、または、科学文献に一般に知られた方法を使って受容性にできて、因子をコードする遺伝物質を含む能力決定因子を取り込む。取り込み増強方法は、細胞型と発現系に依存して変えることができる。適切な伝達効率を有した受容性体細胞を調製するために使われる典型的な条件は、当業者によって良く知られている。開始体細胞は、約24時間の倍加時間を持つことができる。
【0018】
本明細書に記述したベクターは、治療上の使用の安全性を向上するために科学文献に一般的に知られたた方法を使って構成し、設計できる。この方法は、選択と強化のマーカーを含み、希望するならば、ベクター上のヌクレオチド配列の発現を最適化することができる。このベクターは、ベクターが体細胞性開始細胞内で自己複製できる構造要素を含むべきである。例えば、既知のエプシュタイン・バーウイルス oriP/Nuclear抗原(EBNA−1)組み合わせ(例、Lindner,S.E.and B.The plasmid replicon of Epstein−Barr virus:mechanistic insights intoefficient,licensed,extrachromosomal replication in human cells, Plasmid 58:1(2007)を参照、本文全体をここに説明したものとして参照により本明細書に組み込む)は、ベクターの自己複製を支持するのに充分であり、また、哺乳類、特に霊長類の細胞内で機能すると知られているその他の組み合わせを採用しても良い。本発明において使用するのに適した発現ベクターを構築する標準技術は、当業者に知られており、Sambrook Jらによる「分子クローニング:ラボラトリマニュアル」(3rd ed.Cold Spring harbor Press,Cold Spring Harbor,N.Y. 2001)などの文献に見出すことができ、参照により本文全体を説明されたものとして本明細書に取り込む。
【0019】
本発明の方法では、1セットの能力決定因子をコードする遺伝子物質が、1個または複数個の再プログラム用ベクターを介して体細胞に搬送される。適切な能力決定因子は、これらに限定されるものではないが、OCT−4、SOX2、LIN28、NANOG、c−Myc、KLF4、および、これらの組み合わせを含むことができる。体細胞内にポリヌクレオチド発現を引き起こすことができる非相同性のプロモータに機能的に結合している能力決定因子をコードするポリヌクレオチド導入遺伝子として、各能力決定因子を体細胞に導入することができる。SV40T抗原それ自体は能力決定因子ではないけれども、能力決定因子が発現されている再プログラムの間に、細胞生存を促進するのに充分な条件を持つ細胞を提供するように、都合良く体細胞に導入される。因子の発現に充分なその他の条件は、実施例に記述されている細胞培養条件を含む。
【0020】
適切な再プログラム用ベクターは、プラスミドのようなエピソームのベクターであって、感染性または有複製能ウイルスを生じるのに十分なウイルス性ゲノムの全部または一部をコードしないが、当該ベクターは一つまたは複数のウイルスから得た構造要素を含むことができる。1個または複数個の再プログラム用ベクターが、1個の体細胞に導入しうる。1個または複数個の導入遺伝子が、1個の再プログラム用ベクターに供給されうる。1個の強い、恒常的な転写性プロモータが、複数の導入遺伝子のために転写調節を提供することができ、該導入遺伝子は発現カセットして提供されうる。1個のベクター上の別々の発現カセットは、それぞれの強い恒常的なプロモータの転写調節下にあり、これらのプロモータは、同一プロモータのコピーであるか、異なるプロモータのコピーであり得る。多様な非相同性プロモータが当分野で知られており、能力決定因子の所望の発現レベルなどの要因に応じて使うことができる。下記に例示するように、標的体細胞内の異なる強度を持つ異なるプロモータを使って、別々の発現カセットを転写コントロールすることは好都合である。転写性プロモータ(単数または複数)を選択する際に別途考慮することは、プロモータが標的体細胞内で発現抑制される割合である。再プログラム法において、遺伝子(単数または複数)の産生がその役割を完了または実質的に完了した後に、1個または複数個の、導入遺伝子または導入遺伝子発現カセットの発現を減らすと好都合であることを当業者は想定する。例示的プロモータは、ヒトEF1α伸長因子プロモータ、CMVサイトメガロウイルス即時型初期プロモータ、CAGニワトリ・アルブミン・プロモータ、および、別種由来のそれぞれに対応する相同型プロモータである。ヒト体細胞内において、EF1αとCMVの両方は強いプロモータであるが、CMVプロモータはEF1αプロモータより効率的に抑制される、即ち、EF1αの制御下の導入遺伝子の発現が停止するや否や、CMVの制御下の導入遺伝子の発現が停止する。
【0021】
能力決定因子は、再プログラムの効率を調整するために変動する相対比率で、体細胞内において発現できる。例えば、OCT−4とSOX2が、単一のベクター内の単一の転写で1:1の比率でコードされる場合は、単一の細胞に吸収される比率が制御できないように2個の因子それぞれを異なるベクター上に提供する時に比べて(即ち、各因子が各単一細胞に入る比率を制御できない)、体細胞の再プログラム効率は4倍高い。好ましくは、複数の導入遺伝子が単一の転写でコードされる場合には、配列内リボソーム進入部位が、転写性プロモータから離れた導入遺伝子(単数または複数)の上流に位置する。因子の相対的比率は、搬送された因子に依存して変化させることができるが、本開示を見た当業者は、最適な比率を求めることができる。
【0022】
当業者は、複数のベクターではなく、単一のベクターで全ての因子を導入できる効率の良さを想定するだろうが、ベクター全体のサイズが増大するにつれて、ベクターを導入することがますます困難になる。当業者は、また、ベクター上の因子が時間的発現に及ぼす影響と結果として生じる再プログラム効率も想定するであろう。そのため、出願人らは、ベクターの組み合わせについて様々な因子の組み合わせを採用した。いくつかのそのような組み合わせが、本明細書に提示されて再プログラムを立証する。
【0023】
再プログラム用ベクター(単数または複数)を導入後で、かつ体細胞が再プログラム中である場合、導入された導入遺伝子が転写および翻訳される間は、標的細胞内にベクターが存続し得る。導入遺伝子発現は、再プログラムされて多能性状態になった細胞内で、好都合にも下方制御しうるか、停止しうる。再プログラム用ベクター(単数または複数)は、染色体外に残ることができる。極度に低い効率の場合、ベクター(単数または複数)は、細胞のゲノム内へと組み込める。再プログラム用ベクター(単数または複数)は、レシピエント細胞のゲノムと協調して複製し、そのため、エピソームのベクターがそのDNAを複製することが出来なくなる期間より長く、2週間ほど適度に安定している。にもかかわらず、ベクターは細胞分裂の際に均等に分配されないので、選択圧が無い場合、細胞がエピソームのベクター(単数または複数)を失い、それにより本方法ではベクター・フリーの多能性細胞を直ちに回収できる。例えば、体細胞内にoriP/EBNA−1−ベースのエピソームのプラスミドが、通常2〜3週間安定的に維持される。初期の2〜3週間に、細胞はエピソームのプラスミドを急速に失う。細胞が安定化すると、細胞は、一世代に付き〜5%の割合でエピソームのベクターを連続的に失う。
【0024】
本方法で産生した多能性細胞は、多能性細胞増殖をサポートする任意の培地で培養することができる。培地として、ここに書かれたものに限定されるものではないが、TeSR(商標)(StemCell Technologies,Inc.;Vancouver,Canada),mTeSR(商標)(StemCell Technologies, Inc.)and StemLine(登録商標)serum−free medium(Sigma;St.Louis.,Mo.)、または、マウス胚性線維芽細胞(MEF)馴化培地などの馴化培地などがある。本明細書で使われるように、「限定培地」とは、既知の原料由来の既知の化学組成物または成分からなる生化学的に定義した成分のみを含む、生化学的に定義した処方を意味する。本明細書で使われるように、「馴化培地」とは、培地で培養された細胞由来の可溶性因子をさらに補充した増殖培地のことを意味する。別の方法として、細胞は培地内のMEF上で維持しても良い。
【0025】
本発明は、以下のこれらに限定されることのない実施例を熟慮することにより十分に理解できる。
【0026】
実施例1
発現カセットの設計と作製
プライマーとして、かつ表1に収載の配列内リボソーム進入部位から得られるコード領域の最初と最後の20〜22の塩基を使用して、して、導入遺伝子の一部または全てから得られるオープン・リーディング・フレーム(ORF)をポリメラーゼ連鎖反応(PCR)法で直接増幅するという従来の方法を使って、適した発現カセットを作製した。SV40T抗原とヒトテロメラーゼ逆転写酵素、プラスミドpBABE−puro SV40 LT、および、pBABE−hygro−hTERT(ヒトテロメラーゼ逆転写酵素)の供給源は、プラスミド1390および1773としてそれぞれAddgene,Inc,Cambridge、MAから市販されている。 IRES1とIRES2、プラスミドpIRESpuro3とプラスミドpIRES2EGFPの供給源は、Clontech Laboratories,Inc.,Mountain View,CA.から市販されている。
口蹄疫ウイルスセグメント2を化学的に合成した。インフレームの発現カセットは、下記表1記載のコードを使って記述する。例えば、「E−O2S」とは、OCT4とSOX2コード領域(両者の間にIRES2)の上流に位置するEF1αプロモータを有する発現カセットを意味する。同様に、C−M2Kとは、c−MycとKlf4コード領域(両者の間にIRES2)の上流に位置するCMVプロモータを有する発現カセットを意味する。
いくつかのコンストラクトでは、上記のカセットは、一連の再プログラム中では使わなかった。O2Sとは異なるO2S変異体発現カセット(「O2S(2)」)を採用した。この変異体は、TKプロモータ−Hyg −TK polyAカセット(図5Aおよび図5Bを比較せよ)を含んでいた。指示した構造を有するカセットは、様々な因子の発現レベルを実験的に測定することによる再プログラムの方法で、後に使用するために選択した。
EF2(配列番号:12)として設計されたプロモータは、既知のEF1αプロモータ(配列番号:11)とはわずかに異なる変異体であったが、活性化中のEF1αとは異ならず、また後に以下のエピソームのベクターの再プログラムの試行には使用しなかった。F2Aは、単一のトランスクリプトから発現した異なるコード領域を同時翻訳するのを促進するペプチド・リンカーである。F2Aを試用したが、後のエピソームのベクターを使った再プログラム試行には使わなかった。IRES1を試用したが、後のエピソームのベクターを使った再プログラム試行には使わなかった。
【0027】
ORF上流または下流の発現に関して、様々なプロモータ、IRES配列、および、導入遺伝子の配置の相対的効果を、以前に記述(Yu et al.,上記参照)したように、形質移入後ヒト体細胞を再プログラムできるかをテストするために、様々な導入遺伝子発現カセットをpSin4、修飾したレンチウイルス・ベースのベクターへと別々にクローニングして評価した。293FT細胞に、下記に示すようにSuperFect(Qiagen,Valencia,CA)を使いIRES1またはIRES2で連結されたOCT4とSOX2を発現するレンチウイルスのプラスミドベクターで形質移入した。細胞は、形質移入2日後に収集した。図1Aは、293FT細胞内のOCT−4とSOX2のウエスタンブロット法による解析を示す。レーン1、pSIN4−EF2−OCT4−IRES1−SOX2、レーン2,pSIN4−EF2−OCT4−IRES2−SOX2、レーン3,pSIN4−EF2−OCT4−F2A−SOX2、レーン4、pSIN4−EF2−OCT4−IRES1−PURO、レーン5、pSIN4−EF2−SOX2−IRES1−PURO、レーン6、プラスミド無し(コントロール)。マウス抗ヒトOCT4モノクローナル抗体(1:500,Santa Cruz ,Biotechnology Inc.,Santa Cruz,CA,sc−5279)とヤギ抗ヒトSOX2ポリクローナル抗体(1:500,R&D Systems,Minneapolis,MN AF2018)を使用して、OCT4とSOX2それぞれの相対的発現を検出した。
【0028】
図1Bは、OCT4ノックイン・ヒト胚性幹細胞(米国特許出願第2006/0128018号、および、Zwaka and Thomson,Nature Biotechnology 21:319−321(2003)、参照により全体を説明したものとして各文献を本発明に取り込む)由来の(Yu et al.,上記参照)0.2×106の間葉系細胞内で連結させた能力決定因子を使った再プログラムを示す。
この系統は、ネオマイシン選択(ジェネテシン:100μg/ml,Invitrogen Corp.)下で維持した。形質導入16日後にヒトiPS細胞コロニーを計数した。遺伝子組み合わせは、pSIN4−EF2−OCT4−IRES1−SOX2(O1S)、pSIN4−EF2−OCT4−IRES2−SOX2(O2S)、pSIN4−EF2−OCT4−F2A−SOX2(OF2AS)、pSIN4−EF2−NANOG−IRES1−LIN28(N1L)pSIN4−EF2−NANOG−IRES2−LIN28(N2L)、pSIN4−EF2−OCT4−IRES1−PURO(O)、pSIN4−EF2−SOX2−IRES1−PURO(S)、pSIN4−EF2−NANOG−IRES1−PURO(N)、pSIN4−EF2−LIN28−IRES1−PURO(L)であった。レンチウイルスのプラスミドベクターの各々の略語は、ベクターの名称の後に、括弧で囲んでいる。
【0029】
実施例2
レンチウイルス・コンストラクトを使用したヒト新生児包皮線維芽細胞の再プログラム化
予備的再プログラムの実験は、レンチウイルスベクターをヒト新生児の包皮線維芽細胞に導入することによって行った。図2Aは、OCT4、SOX2、LIN28、および、さらにc−MYCを導入する場合、NANOGが再プログラム効率にかなりのプラス効果をもたらすことを示し、 また、OCT4、SOX2、および、LIN28との組み合わせでは、c−MYCまたはKLF4が無い場合でもNANOGが再プログラムを補助できることを示す。使用したレンチウイルスのコンストラクトは、pSIN4−EF2−OCT4−IRES2−SOX2(O2S)、pSIN4−EF2−NANOG−IRES2−LIN28 (N2L)、pSIN4−EF2−LIN28−IRES1−PURO(L)、pSIN4−CMV−c−Myc−IRES1−PURO(M)、pSIN4−EF2−KLF4−IRES1−PURO(K)であった。形質移入20日後、アルカリ・ホスファターゼ-陽性ヒトiPS細胞コロニーを計数した。iPS細胞コロニーの数は、2.5×104個のヒト新生児包皮線維芽細胞(継代数9)の入力由来であった。薄灰色の棒は、再プログラムされて形成した、代表的なヒトES細胞形態を有するコロニーの合計数を表し、濃い灰色棒は、分化が最も少なかった大型コロニーの数を表す。
【0030】
図2Bは、連結した能力決定因子を用いた再プログラムを証明する。使用したレンチウイルスのコンストラクトは、pSIN4−EF2−c−Myc−IRES2−KLF4(EF2−M2K)、pSIN4−CMV−c−Myc−IRES2−KLF4(CMV−M2K)、pSIN4−EF2−KLF4−IRES2−c−Myc(EF2−K2M)、pSIN4−CMV−KLF4−IRES2−c−Myc(CMV−K2M)、pSIN4−CMV−c−Myc−IRES2−LIN28(M2L)、pSIN4−EF2−NANOG−IRES2−KLF4(N2K)であった。
【0031】
形質導入14日後、アルカリ・ホスファターゼ-陽性ヒトiPS細胞コロニーを計数した。iPS細胞コロニーの数は、約7.0×104個のヒト新生児包皮線維芽細胞(継代数12)の入力由来であった。アステリスクは、多くのアルカリ・ホスファターゼ−陽性コロニーが形態学的に緩やかに現れたことを示す。
【0032】
図2Cは、SV40大型T抗原遺伝子発現の再プログラムへの効果を示す。SV40大型T抗原は、マウスの線維芽細胞内にc−Mycが誘発するのを妨ぎ(Hermeking et al.,PNAS 91:10412−10416(1994))、再プログラム効率を高める(Hanna et al.,Cell 133:250−264(2008);Mali et al.,Stem Cells doi:10.1634/stemcells.2008−0346(2008))。遺伝子組み合わせ略名は、図2B内のものと同じであり、さらにSV40大型T抗原(T)を追加する。c−Mycも細胞増殖を促進する。形質導入12日後、アルカリ・ホスファターゼ-陽性ヒトiPS細胞コロニーを計数した。iPS細胞コロニーの数は、約〜3.5×104個のヒト新生児包皮線維芽細胞(継代数17)の入力由来であった。図2Cは、T抗原がレンチウイルス・ベースの再プログラムの間に達成したレベルで存在するならば、iPS細胞導出の最終段階を抑制することを実証する。対照的に、下記を参照されたいが、T抗原は、一時的発現時間に存在し、かつ/または、エピソームのベクターを使って再プログラム中に達成したレベルで存在するならば、この効果をもたない。さらに加えて、T抗原はc−Myc−誘導アポトーシスを妨げないがc−Myc−誘導細胞増殖に逆の影響を与える。
【0033】
実施例3.
非ウイルス性エピソームのベクターを使ったヒト新生児包皮線維芽細胞の再プログラム
ヒト新生児包皮線維芽細胞(Cat# CRL−2097(商標),ATCC)は、10%熱失活したウシ胎仔血清(FBS,HyClone Laboratories,Logan,UT)、2mMのグルタマックス、0.1mMの非必須アミノ酸、および、0.1mMのβ−メルカプトエタノールを添加した包皮線維芽細胞培地(DMEM(Cat# 11965,Invitrogen))で維持した。
【0034】
実施例1および以下の表3で詳細に示されるように構成された導入遺伝子発現カセットとして提供された能力決定因子の様々な組み合わせは、エピソームのコンストラクトpCEP4−EGFP(図3Aに示すように)を使って体細胞に導入されて、様々な効率を持つ再プログラムをもたらした。pCEP4−EGFPは、市販の哺乳類エピソームの発現ベクターpCEP4(Invitrogen Corp., Carlsbad, CA)から、EGFPコード領域をpCEP4 BamHI部位とNheI部位との間に挿入することによって創出した。表2のエピソームのベクターは、指定した発現ベクターをpCEP4−EGFPもしくは関連するバックボーンのないPcmv(指定されたpEP4)に挿入することによって創出した。発現カセットが挿入されたクローニング部位については図3Aと表2の脚注を参照せよ。
【0035】
製造元の指示に従い、ヒト真皮線維芽細胞キット(Normal Human Dermal Fibroblasts,Amaxa,Inc.Cat.No.VPD−1001)を使用して、単一のヌクレオフェクション・イベント経由で線維芽細胞にベクターを導入した。ヌクレオフェクション後、照射を受けたマウス胚性線維芽細胞(MEF)が播種された3個の10cmの皿の上に形質移入した線維芽細胞(それぞれ〜0.8から1.0×106細胞)を直ちにプレートした。包皮線維芽細胞培地は一日おきに交換した。4日後、包皮線維芽細胞培地をヒト胚性幹細胞培地(DMEM/F12培地に以下を補充、20%ノックアウト血清リプレーサー、0.1mMの非必須アミノ酸(全てInvitrogen Corp.製)、1mMのグルタマックス、0.1mMのβメルカプトエタノール、および、前記した100ng/mlのゼブラフィッシュタンパク・塩基性線維芽細胞増殖因子(zbFGF)(Amit et al., Developmental Biology 227:271−278 (2006); Ludwig et al., Nature Methods 3:637−646(2006)、ここで、それぞれは、参照により全体を説明したものとしてここに組み入れる)と交換した。プレーティング8〜10日後、播種されたMEFが、再プログラム培地を維持出来なくなったとき、照射されたMEFで馴化したヒト胚性幹細胞培地を代わりに使用した。適切な時期(形質移入2〜3週間後)に、ヒトiPSコロニー発生の徴候としてアルカリ・ホスファターゼにより培養物を着色した。
【0036】
導入遺伝子コンストラクトを導入するための適切なパラメータを決定するために、pEGFP−N2(対照)とpCEP4−EGFPのエピソームのベクター由来のEGFPを293FT細胞に導入して評価した後、EGFPレベルを経時的に測定することによって、時間的発現について初めに評価した(図3B)。
【0037】
ヒト新生児包皮線維芽細胞生存物について、導入された導入遺伝子コンストラクトの量の効果も、予備的実験で評価した。図3Cは、ヌクレオフェクション後翌日の細胞コンフルエンスから推定した、ヌクレオフェクションの効率性とヒト新生児包皮線維芽細胞の生存物について、使用されたpCEP4−EGFPのエピソームのベクターの量の効果を示す。ヌクレオフェクションされた約1×106個の包皮線維芽細胞を6ウェル・プレートの各ウェルにプレートした。灰色線は形質移入されなかった対照の線維芽細胞を表し、黒色線は形質移入された線維芽細胞を表す。
【0038】
図4Aは、様々な発現カセットを含む表3からの模式的な導入遺伝子発現コンストラクトを示し、ある組み合わせをヒト新生児包皮線維芽細胞に導入すると、線維芽細胞から多能性細胞への再プログラムが生じる。導入されるエピソームの再プログラム用ベクターの3つの組み合わせが、次の再プログラムされた多能性細胞を生じさせた。(1)pEP4−E−O2S−E−T2K、pEP4−E−O2S−E−N2K、およびpCEP4−C−M2L、(2)pEP4−E−O2S−C−K2M−E−N2L、およびpEP4−E−O2S−E−T2K、ならびに、(3)pEP4−E−O2S−E−N2L、pEP4−E−O2S−E−T2K、およびpEP4−E−O2S−E−M2K。表3は、成功した組み合わせで使われた各ベクターの量を示す。各成功した再プログラムの組み合わせ内の1個のベクターは、EF1αプロモータの制御の元、T抗原をコードした。
【0039】
図4Bは、エピソームのベクターの形質移入18日後に観察した形態変化をもつ典型的なコロニーの明視野顕微鏡のイメージを示す。図4Cは、エピソームのベクターの形質移入18日後のアルカリホスファターゼ−陽性コロニーの明視野顕微鏡イメージを示す。
【0040】
形質移入25〜30日後、ヒトiPS細胞コロニーに形態的に類似した多くの非iPS 細胞コロニーがある故に、再プログラム培養物を一度新鮮な10cmのMEF皿(1:3比)に移した。そして、その後の解析のためにコロニーを選択した。図4Dは、形質移入再プログラム培養後28日目の第一継代6日後のヒトiPS細胞コロニーの明視野顕微鏡イメージを示す。スケールバーは、0.1mmを表す。再プログラムされた細胞は、後の解析のために、前述した馴化培地(Xu et al.,Nat.Biotechnol.19:971(2001)、参照により全体を説明したものとして本明細書に組み込む)を用い、マトリゲル(BD Biosciences,Bedford,MA)のフィーダ・フリー培養で維持した。
【0041】
好都合にも、再プログラムされた新生児包皮線維芽細胞の内1%を大きく上回る再プログラム効率が達成でき、4個の遺伝子の組み合わせを使用して達成した場合に比べ、有意に短い再プログラム時間であった。
【0042】
本開示に記述した本発明のいくつかの改作は当業者においては通常の最適化事項であり、本発明の精神もしくは添付の請求項の範囲を逸脱することなく実施できることを理解されたい。明細書で上述した出版物と特許は、参照により本明細書に組み込まれる。本発明の記述した方法とシステムの様々な改変と変形は、本発明の範囲と精神を逸脱することなく、当業者には明らかである。しかし、上記した本発明の実施例と実施形態は、例証的であり、本発明を限定するわけではない。本発明は、下記の請求項の範囲に付随する実施例と実施形態の全ての改変体を含む。
【特許請求の範囲】
【請求項1】
霊長類の体細胞を再プログラムする方法であって、
前記細胞を再プログラムするのに充分な条件の下で前記霊長類体細胞を複数の能力決定因子に曝露する工程と、
前記曝露された細胞を培養して前記霊長類体細胞より高い能力レベルを有する再プログラムされた細胞を得る工程と、
を含み、
前記再プログラムされた細胞は、前記体細胞に曝露する前記能力決定因子に関連するどのような構成要素も実質的に含まない再プログラム方法。
【請求項2】
前記霊長類体細胞が胎児または出生後の個体から得られたものである、請求項1に記載の方法。
【請求項3】
前記霊長類が、ヒトである、請求項1に記載の方法。
【請求項4】
前記再プログラムされた細胞が、胎児または出生後の個体と遺伝的に同一である、請求項1に記載の方法。
【請求項5】
前記曝露する工程が、1つまたは複数個の能力決定因子をコードする少なくとも1個のベクターを前記霊長類の体細胞に導入する工程を含む、請求項1に記載の方法。
【請求項6】
前記ベクターが非ウイルス性エピソームのベクターである、請求項5に記載の方法。
【請求項7】
前記1つまたは複数個の能力決定因子が、ヒトOCT−4、ヒトSOX2、ヒトLIN28、ヒトNANOG、ヒトc−Myc、およびヒトKLF4を含み、前記ベクターがc−Mycを含む場合、前記方法はさらに前記霊長類体細胞をSV40T抗原に曝露する工程を含む、請求項5に記載の方法。
【請求項8】
前記少なくとも1つのベクターが単一ベクターである、請求項5に記載の方法。
【請求項9】
前記単一ベクターが、少なくとも1つの配列内リボソーム進入部位を含む、請求項に記載の方法。
【請求項10】
前記単一ベクターが、順番に、第一のプロモータ、OCT4、IRES2、SOX2、第二のプロモータ、SV40T抗原、IRES2、およびKLF4を含む、請求項8に記載の方法。
【請求項11】
前記ベクターが、順番に、第一のプロモータ、OCT4、IRES2、SOX2、第二のプロモータ、c−Myc、IRES2、KLF4、第三のプロモータ、NANOG、IRES2、およびLN28を含む、請求項8に記載の方法。
【請求項12】
第一のベクターが、順番に、第一のプロモータ、OCT4、IRES2、SOX2、第二のプロモータ、SV40T抗原、IRES2、およびKLF4を含み、第二のベクターが、順番に、第三のプロモータ、OCT4、IRES2、SOX2、第四のプロモータ、NANOG、IRES2、KLF4を含み、第三のベクターが、順番に、第五のプロモータ、c−Myc、IRES2、LN28を含み、前記プロモータは同一である必要がない、請求項5に記載の方法。
【請求項13】
前記第一の、第二の、第三の、および第四の各プロモータが伸長因子1α(EF1α)遺伝子プロモータである、請求項12に記載の方法。
【請求項14】
第一のベクターが、順番に、第一のプロモータ、OCT4、IRES2、SOX2、第二のプロモータ、KLF4、IRES2、c−Myc、第三のプロモータ、NANOG、IRES2、およびLN28を含み、第二のベクターが、順番に、第四のプロモータ、OCT4、IRES2、SOX2、第五のプロモータ、SV40T抗原、IRES2、およびKLF4を含み、前記プロモータは同一である必要がない、請求項5に記載の方法。
【請求項15】
前記第一の、第三の、第四の、および第五の、各プロモータが、EF1α遺伝子プロモータであり、また第二のプロモータが、サイトメガロウイルス・前初期遺伝子(CMV)プロモータである、請求項14に記載の方法。
【請求項16】
第一のベクターが、順番に、第一のプロモータ、OCT4、IRES2、SOX2、第二のプロモータ、NANOG、IRES2、およびLN28を含み、第二のベクターが、順番に、第三のプロモータ、OCT4、IRES2、SOX2、第四のプロモータ、SV40T抗原、IRES2、およびKLF4を含み、ならびに、第三のベクターが、順番に、第五のプロモータ、OCT4、IRES2、SOX2、第六のプロモータ、c−Myc、IRES2、およびKLF4を含み、前記プロモータは同一である必要がない、請求項5に記載の方法。
【請求項17】
霊長類多能性細胞の濃縮された集団であって、
能力決定因子を発現するのに充分な条件の下で霊長類体細胞を能力決定因子に曝露することにより、前記体細胞を再プログラムして前記霊長類体細胞を産生する工程を含む方法により産生され、
前記細胞が、前記体細胞に曝露する前記能力決定因子に関連するどのような構成要素も実質的に含まない霊長類多能性細胞の濃縮された集団。
【請求項18】
前記霊長類体細胞が、胎児または出生後の個体から得られたものである、請求項17に記載の前記集団。
【請求項19】
霊長類が、ヒトである、請求項17に記載の前記集団。
【請求項20】
前記多能性細胞が、実質的に出生後の個体と遺伝的に同一である、請求項17に記載の集団。
【請求項21】
前記曝露する工程が、1つまたは複数個の能力決定因子をコードする少なくとも1個のベクターを前記霊長類体細胞に導入する工程を含む、請求項17に記載の集団。
【請求項22】
前記ベクターが、非ウイルス性のエピソームのベクターである、請求項21に記載の集団。
【請求項23】
1つまたは複数個の能力決定因子が、ヒトOCT−4、ヒトSOX2、ヒトLIN28、ヒトNANOG、ヒトc−Myc、およびヒトKLF4を含み、前記ベクターが、c−Mycを含む場合、前記方法は、さらに前記霊長類体細胞をSV40T抗原に曝露する工程を含む、請求項21に記載の集団。
【請求項24】
多能性の誘導に関連するどのような構成要素も実質的に含まない霊長類誘導多能性細胞。
【請求項1】
霊長類の体細胞を再プログラムする方法であって、
前記細胞を再プログラムするのに充分な条件の下で前記霊長類体細胞を複数の能力決定因子に曝露する工程と、
前記曝露された細胞を培養して前記霊長類体細胞より高い能力レベルを有する再プログラムされた細胞を得る工程と、
を含み、
前記再プログラムされた細胞は、前記体細胞に曝露する前記能力決定因子に関連するどのような構成要素も実質的に含まない再プログラム方法。
【請求項2】
前記霊長類体細胞が胎児または出生後の個体から得られたものである、請求項1に記載の方法。
【請求項3】
前記霊長類が、ヒトである、請求項1に記載の方法。
【請求項4】
前記再プログラムされた細胞が、胎児または出生後の個体と遺伝的に同一である、請求項1に記載の方法。
【請求項5】
前記曝露する工程が、1つまたは複数個の能力決定因子をコードする少なくとも1個のベクターを前記霊長類の体細胞に導入する工程を含む、請求項1に記載の方法。
【請求項6】
前記ベクターが非ウイルス性エピソームのベクターである、請求項5に記載の方法。
【請求項7】
前記1つまたは複数個の能力決定因子が、ヒトOCT−4、ヒトSOX2、ヒトLIN28、ヒトNANOG、ヒトc−Myc、およびヒトKLF4を含み、前記ベクターがc−Mycを含む場合、前記方法はさらに前記霊長類体細胞をSV40T抗原に曝露する工程を含む、請求項5に記載の方法。
【請求項8】
前記少なくとも1つのベクターが単一ベクターである、請求項5に記載の方法。
【請求項9】
前記単一ベクターが、少なくとも1つの配列内リボソーム進入部位を含む、請求項に記載の方法。
【請求項10】
前記単一ベクターが、順番に、第一のプロモータ、OCT4、IRES2、SOX2、第二のプロモータ、SV40T抗原、IRES2、およびKLF4を含む、請求項8に記載の方法。
【請求項11】
前記ベクターが、順番に、第一のプロモータ、OCT4、IRES2、SOX2、第二のプロモータ、c−Myc、IRES2、KLF4、第三のプロモータ、NANOG、IRES2、およびLN28を含む、請求項8に記載の方法。
【請求項12】
第一のベクターが、順番に、第一のプロモータ、OCT4、IRES2、SOX2、第二のプロモータ、SV40T抗原、IRES2、およびKLF4を含み、第二のベクターが、順番に、第三のプロモータ、OCT4、IRES2、SOX2、第四のプロモータ、NANOG、IRES2、KLF4を含み、第三のベクターが、順番に、第五のプロモータ、c−Myc、IRES2、LN28を含み、前記プロモータは同一である必要がない、請求項5に記載の方法。
【請求項13】
前記第一の、第二の、第三の、および第四の各プロモータが伸長因子1α(EF1α)遺伝子プロモータである、請求項12に記載の方法。
【請求項14】
第一のベクターが、順番に、第一のプロモータ、OCT4、IRES2、SOX2、第二のプロモータ、KLF4、IRES2、c−Myc、第三のプロモータ、NANOG、IRES2、およびLN28を含み、第二のベクターが、順番に、第四のプロモータ、OCT4、IRES2、SOX2、第五のプロモータ、SV40T抗原、IRES2、およびKLF4を含み、前記プロモータは同一である必要がない、請求項5に記載の方法。
【請求項15】
前記第一の、第三の、第四の、および第五の、各プロモータが、EF1α遺伝子プロモータであり、また第二のプロモータが、サイトメガロウイルス・前初期遺伝子(CMV)プロモータである、請求項14に記載の方法。
【請求項16】
第一のベクターが、順番に、第一のプロモータ、OCT4、IRES2、SOX2、第二のプロモータ、NANOG、IRES2、およびLN28を含み、第二のベクターが、順番に、第三のプロモータ、OCT4、IRES2、SOX2、第四のプロモータ、SV40T抗原、IRES2、およびKLF4を含み、ならびに、第三のベクターが、順番に、第五のプロモータ、OCT4、IRES2、SOX2、第六のプロモータ、c−Myc、IRES2、およびKLF4を含み、前記プロモータは同一である必要がない、請求項5に記載の方法。
【請求項17】
霊長類多能性細胞の濃縮された集団であって、
能力決定因子を発現するのに充分な条件の下で霊長類体細胞を能力決定因子に曝露することにより、前記体細胞を再プログラムして前記霊長類体細胞を産生する工程を含む方法により産生され、
前記細胞が、前記体細胞に曝露する前記能力決定因子に関連するどのような構成要素も実質的に含まない霊長類多能性細胞の濃縮された集団。
【請求項18】
前記霊長類体細胞が、胎児または出生後の個体から得られたものである、請求項17に記載の前記集団。
【請求項19】
霊長類が、ヒトである、請求項17に記載の前記集団。
【請求項20】
前記多能性細胞が、実質的に出生後の個体と遺伝的に同一である、請求項17に記載の集団。
【請求項21】
前記曝露する工程が、1つまたは複数個の能力決定因子をコードする少なくとも1個のベクターを前記霊長類体細胞に導入する工程を含む、請求項17に記載の集団。
【請求項22】
前記ベクターが、非ウイルス性のエピソームのベクターである、請求項21に記載の集団。
【請求項23】
1つまたは複数個の能力決定因子が、ヒトOCT−4、ヒトSOX2、ヒトLIN28、ヒトNANOG、ヒトc−Myc、およびヒトKLF4を含み、前記ベクターが、c−Mycを含む場合、前記方法は、さらに前記霊長類体細胞をSV40T抗原に曝露する工程を含む、請求項21に記載の集団。
【請求項24】
多能性の誘導に関連するどのような構成要素も実質的に含まない霊長類誘導多能性細胞。
【図1A−B】


【図2A−C】
【図3A−C】
【図4A】
【図4B】

【図4C】

【図4D】

【図5A−B】
【図2A−C】
【図3A−C】
【図4A】
【図4B】
【図4C】
【図4D】
【図5A−B】
【公表番号】特表2012−506702(P2012−506702A)
【公表日】平成24年3月22日(2012.3.22)
【国際特許分類】
【出願番号】特願2011−533384(P2011−533384)
【出願日】平成21年10月23日(2009.10.23)
【国際出願番号】PCT/US2009/061935
【国際公開番号】WO2010/048567
【国際公開日】平成22年4月29日(2010.4.29)
【出願人】(500517248)ウイスコンシン アラムニ リサーチ ファンデーション (18)
【Fターム(参考)】
【公表日】平成24年3月22日(2012.3.22)
【国際特許分類】
【出願日】平成21年10月23日(2009.10.23)
【国際出願番号】PCT/US2009/061935
【国際公開番号】WO2010/048567
【国際公開日】平成22年4月29日(2010.4.29)
【出願人】(500517248)ウイスコンシン アラムニ リサーチ ファンデーション (18)
【Fターム(参考)】
[ Back to top ]