
非ヒト科動物が正常な学習能力を有する状態であるか否かを評価する方法、及び医薬のスクリーニング方法
【課題】従来のLTPとは異なった概念を基盤とし、動物の学習能力及びその脳の状態を解析・解釈することで、非ヒト科動物が正常な学習能力を有する状態であるか否かを評価する方法の提供。
【解決手段】非ヒト科動物が正常な学習能力を有する状態であるか否かを評価する方法であって、前記動物に学習行動を行わせる学習工程と、前記動物の脳の神経細胞におけるCapZの局在状態を調べる検出工程と、前記CapZの局在状態の観察に基づき、前記脳の状態を評価する評価工程と、を含むことを特徴とする非ヒト科動物の学習能力の評価方法。
【解決手段】非ヒト科動物が正常な学習能力を有する状態であるか否かを評価する方法であって、前記動物に学習行動を行わせる学習工程と、前記動物の脳の神経細胞におけるCapZの局在状態を調べる検出工程と、前記CapZの局在状態の観察に基づき、前記脳の状態を評価する評価工程と、を含むことを特徴とする非ヒト科動物の学習能力の評価方法。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、非ヒト科動物が正常な学習能力を有する状態であるか否かを評価する方法に関する。より詳しくは、非ヒト科動物の脳の神経細胞におけるCapZの局在を調べることにより、当該動物の脳が正常に学習できる状態であるか否かを評価する方法に関する。
【背景技術】
【0002】
従来の薬物スクリーニングの場合、記憶能、学習能の判定に齧歯類の行動変化を指標とする試験法が一般的である。例えば「文脈依存的恐怖条件付け文脈学習」の場合、過去に痛みを与えられた場所に置かれた動物が、その痛みを想起して表す恐怖反応、全く動かなくなる「すくみ行動」、の時間率を学習能として評価する。
【0003】
また、神経細胞・分子レベルで記憶や学習の根本的なメカニズムを研究するために、主に生体外で切片標本を用いたLTP(Long Term Potentiation)が、研究材料として使用されている。LTPで観察されるAMPA型グルタミン酸受容体による電流の増加は、記憶の素過程であると考えられおり(非特許文献1及び2)、記憶や学習に影響を及ぼす薬物のスクリーニング等への応用研究も盛んである。ところで、記憶の素過程として現れるはずの現象(神経回路の可塑性を示す現象)は、少ない頻度でしか起こらない可能性があることに留意しなければならない。発明者らの過去の研究(非特許文献3,4)からも、記憶が形成される刺激によって変化するスパインの数は、全体の数%程度である可能性が示唆されている。
【先行技術文献】
【非特許文献】
【0004】
【非特許文献1】“Memory enhancement: the search for mechanism-based drugs.” Lynch G. Nat Neurosci. 2002 Nov;5 Suppl:1035-8.
【非特許文献2】”Learning from LTP: a comment on recent attempts to identify cellular and molecular mechanisms of memory.” Learn Mem. 1996 Sep-Oct;3(2-3):61-73.
【非特許文献3】”Experience-dependent, rapid structural changes in hippocampal pyramidal cell spines.” Kitanishi T, Ikegaya Y, Matsuki N, Yamada MK. Cereb Cortex. 2009 Nov;19(11):2572-8. Epub 2009 Feb 24.
【非特許文献4】”Contextual learning induces an increase in the number of hippocampal CA1 neurons expressing high levels of BDNF.” Chen J, Kitanishi T, Ikeda T, Matsuki N, Yamada MK. Neurobiol Learn Mem. 2007 Nov;88(4):409-15. Epub 2007 Aug 31.
【発明の概要】
【発明が解決しようとする課題】
【0005】
従来の薬物スクリーニングに用いられている、動物の学習能の判定に用いられる試験法は、健康な動物を試験対象とする場合には適当な方法であるとしても、疾患モデル動物を試験対象とする場合には、単に行動だけが変化したのか、或いは学習能が変化したのか、を弁別して解釈することは難しいという問題がある。例えば、学習能が低下していると考えられている鬱病モデル動物や統合失調症モデル動物を対象にする場合は、従来の試験法では、だるさや無気力等が原因で不動状態になっているのか、恐怖反応を示して不動状態になっているのか、を弁別することが難しい。このように、特に疾患モデル動物の場合には、学習とは異なる原因の不動も考えられるので、従来の試験法が本当に学習能を反映するのかは確実ではない。また、薬物のスクリーニングを行う場合、投与した被検薬物が副作用を与えて、体が不調になったことが原因で不動状態になっているにも関わらず、当該被検薬物して学習が達成されたために不動状態になっている、と誤って解釈される危険もある。
【0006】
したがって、従来の学習試験の不確実性を回避する方法としては、脳の記憶担当部位に起こる変化を直接解析する手法が望ましい。
【0007】
脳の記憶担当部位に起こる変化を捉えるために、従来からLTPを用いる方法が一般的に考えられている。しかし、近年の研究の進展によって、LTP形成に基づいた記憶の素過程の研究について、疑念が生じつつある。そもそも、従来のLTPの研究に用いられてきた、in vitroの神経細胞を人為的に直接発火させる刺激は、動物が活動や学習する際に神経細胞に自然に入力する刺激とは同じでなく、その人為的な刺激は生体中の自然な刺激を再現できていない可能性が高いことが明らかになりつつある。また、ここ数年間の技術進歩により、LTPを観察する手法は格段に進化しており、もし生体内でLTP形成が起これば確実に観察結果の報告があっていいはずである。それにも関わらず、LTPと同様の現象が生体内で起こったという報告は未だになされていない。
【0008】
これらのことから、今まで研究されてきたLTPと部分的に共通の分子メカニズムを持ちつつも、生体内で生じる自然な刺激によって起きる新たな現象を、記憶との相関という原点に戻って確認する必要がある。
記憶が形成されるメカニズムについて理解しようと試みた従来の実験手法は、脳全体を見る分子科学か、ランダムに選んだ一部の神経細胞を見る生理学かのどちらかであった。このような実験手法だけを用いて研究していると、記憶が形成されるメカニズムにおいて重要となりうる、少数の特定の神経細胞に起こる変化(現象)を見逃してしまう危険がある。
【0009】
本発明は、上記事情に鑑みてなされたものであり、従来のLTPとは異なった概念を基盤とし、動物の学習能力及びその脳の状態を解析・解釈することで、非ヒト科動物が正常な学習能力を有する状態であるか否かを評価する方法の提供を課題とする。
【課題を解決するための手段】
【0010】
本発明者が鋭意研究したところ、動物が学習することに伴う記憶の形成と、当該動物の脳の神経細胞におけるCapZの局在とに相関があることを見出した。この知見に基づき、以下の方法を提供する。
【0011】
本発明の請求項1に記載の非ヒト科動物の学習能力の評価方法は、非ヒト科動物が正常な学習能力を有する状態であるか否かを評価する方法であって、前記動物に学習行動を行わせる学習工程と、前記動物の脳の神経細胞におけるCapZの局在状態を調べる検出工程と、前記CapZの局在状態の観察に基づき、前記脳の状態を評価する評価工程と、を含むことを特徴とする。
本発明の請求項2に記載の評価方法は、請求項1において、前記評価工程において、CapZが一部のスパインに局在している場合、前記動物が正常な学習能力を有する状態であると評価することを特徴とする。
本発明の請求項3に記載の評価方法は、請求項1又は2において、前記非ヒト科動物が、CapZとマーカータンパク質及び/又はペプチドタグとを結合させた融合タンパク質をコードする遺伝子が導入された、トランスジェニック動物であり、前記マーカータンパク質に由来するシグナル、及び/又は前記ペプチドタグに特異的に結合する抗体に由来するシグナルを検出することにより、CapZの局在を調べることを特徴とする。
本発明の請求項4に記載の評価方法は、請求項3において、前記トランスジェニック動物における前記融合タンパク質の発現を、前記学習行動に伴って入力を受ける脳部位において、前記入力により誘導させることを特徴とする。
本発明の請求項5に記載の評価方法は、請求項1〜4の何れか一項において、前記動物が疾患モデル動物であることを特徴とする。
本発明の請求項6に記載の評価方法は、請求項1〜5の何れか一項において、CapZが、以下の(i)又は(ii)のタンパク質であることを特徴とする。
(i)配列番号1〜5の何れかで表されるアミノ酸配列を有するタンパク質。
(ii)配列番号1〜5の何れかで表されるアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列を有し、且つ神経細胞への入力依存的にスパインに局在する機能を有するタンパク質。
本発明の請求項7に記載の医薬のスクリーニング方法は、学習障害若しくは精神疾患を呈する請求項5に記載の疾患モデル動物に対して、被検物質を投与する工程と、前記動物に学習行動を行わせる学習工程と、前記動物の脳の神経細胞におけるCapZの局在を調べる検出工程と、前記CapZの局在に基づき、前記脳の状態を評価する評価工程と、
を含み、前記評価工程において、CapZが一部のスパインに局在している場合、前記被検物質を、前記学習障害若しくは精神疾患を治療又は予防する医薬として有用であると判断することを特徴とする。
【発明の効果】
【0012】
本発明の評価方法によれば、学習行動を行わせた動物の脳が、当該学習に関する記憶を蓄積(記銘)できる正常な学習能力を有するか否かを評価することができる。本発明の評価方法を用いることによって、動物の学習と記憶に関して、LTP形成とは異なる現象に基づいた新たな研究基盤を提供できる。
本発明の医薬のスクリーニング方法によれば、学習障害若しくは精神疾患を治療又は予防する医薬として有用な医薬候補となる物質を見出すことができる。
【図面の簡単な説明】
【0013】
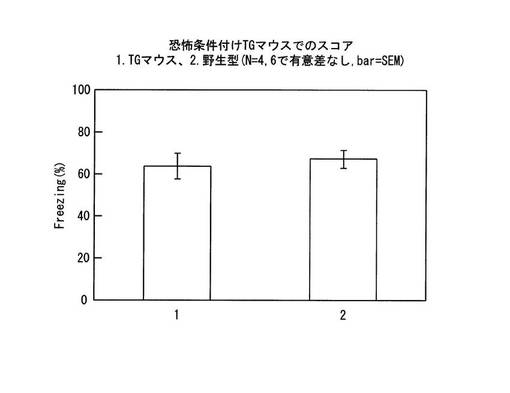
【図1】実施例1で使用したトランスジェニックマウスに導入した遺伝子の模式図である。
【図2】実施例1で使用したトランスジェニックマウスの脳切片を蛍光顕微鏡で観察して検出された、EGFP-CapZに由来する緑色蛍光シグナルを示す写真である。
【図3】図2と同じ観察視野において、Rhodamine-phalloidinの赤色蛍光で染色されたスパインを示す写真である。
【図4】左は図2と同じEGFP-CapZに由来する蛍光シグナルを示す写真であり、右は図3と同じRhodamine-phalloidin染色によって観察された蛍光シグナルを示す写真である。
【図5】図2と同じ観察視野において、MAP2が存在する樹状突起を図2のEGFP-CapZのシグナルに重ねて示した写真である。
【図6】図5と同じ観察視野において、EGFP-CapZに由来するシグナルを除き、MAP2が存在する樹状突起のみを示した写真である。
【図7】図2よりも広い視野で観察した(ズームなしで観察した)写真である。
【図8】比較例1で使用したトランスジェニックマウスの脳切片を蛍光顕微鏡で観察して検出された、EGFP-CapZに由来する緑色蛍光シグナルを示す写真である。
【図9】図8と同じ写真において、画像の全体のシグナル強度を下げた写真である。
【図10】左は図7と同一視野におけるファロイジン染色の写真であり、右は図8と同一視野におけるファロイジン染色の写真である。
【図11】実施例2で使用した野生型マウスの脳切片における、抗CapZ抗体に由来するシグナルを示す写真である。
【図12】比較例2で使用した野生型マウスの脳切片における、抗CapZ抗体に由来するシグナルを示す写真である。
【図13】比較例3で使用した疾患モデルマウスの脳切片における、抗CapZ抗体に由来するシグナルを示す写真である。
【図14】恐怖条件付けを行ったトランスジェニックマウスの学習能力を表すグラフである。
【発明を実施するための形態】
【0014】
以下、好適な実施の形態に基づき、図面を参照して本発明を説明する。
<評価方法>
本発明の非ヒト科動物の学習能力の評価方法は、非ヒト科動物が正常な学習能力を有する状態であるか否かを評価する方法であって、前記動物に学習行動を行わせる学習工程と、前記動物の脳の神経細胞におけるCapZの局在状態を調べる検出工程と、前記CapZの局在状態の観察に基づき、前記脳の状態を評価する評価工程と、を含むものである。
前記学習工程、検出工程及び評価工程以外の工程は、必要に応じて組み入れられる。
【0015】
本発明において、「動物が正常な学習能力を有する状態である(あった)」又は「動物が正常な学習能力を有する状態でない(なかった)」と評価・判断する際の根拠は、
「正常な学習能力を有することを予め別の方法で確かめている動物が、本発明の学習工程における学習行動を行うことにより記憶を蓄積すること」と「当該学習行動に伴って信号が入力される神経細胞においてCapZが一部のスパインに局在すること」が高い相関を有すること、及び
「学習障害を呈し、正常な学習能力を有さないことを別の方法で予め確かめている動物が、本発明の学習工程における学習行動を行うことにより記憶を蓄積しないこと」と「正常である場合には、当該学習行動に伴って信号が入力される神経細胞においてCapZが一部のスパインに局在しないこと」が高い相関を有すること、
を本発明者が見出したことである。
【0016】
本明細書及び特許請求の範囲において「CapZが一部のスパインに局在している」とは、学習行動を行うことに伴って信号(活動電位)が入力される脳部位において、当該脳部位に含まれる一部のスパインにおけるCapZの濃度が、そのスパインを持つ樹状突起本体におけるCapZの濃度よりも高く、且つ、当該神経回路に含まれるスパイン間で、CapZの濃度に明らかな偏りがあることを意味する。通常、CapZの濃度は、CapZに由来するシグナル強度に正の相関をもって反映される。
【0017】
本発明における非ヒト科動物としては、学習行動を行わせて、当該学習行動に関する記憶を蓄積しうる動物であれば特に制限されず、例えば、マウス、ラット、モルモット、ウサギ等のげっ歯類、サル、オランウータン等の霊長類、ブタ、ウシ、ウマ、イヌ、ネコ、イルカ等の哺乳類、インコ、ジュウシマツ等の鳥類、コイ、ゼブラフィッシュ等の魚類などが挙げられる。これらの中でも、比較的高等な学習行動を学んで記憶できる霊長類、哺乳類およびげっ歯類が好ましく、飼育が容易であり、医薬の候補化合物のスクリーニングにも用いられるげっ歯類がより好ましい。
前記動物として、内在性又は外来性のCapZを予め導入した動物を用いることができる。前記導入した動物については、後で詳述する。
なお、ヒトにおいても、本発明の評価方法を適用することは原理的には可能であるが、安全性及び倫理的な問題について、十分に検討がなされるべきである。
【0018】
[学習工程]
本発明の評価方法における学習工程は、前記動物に学習行動を行わせる工程である。
前記学習行動は、動物に刺激を与えることにより、当該動物がその刺激を記憶できるものであれば特に制限されない。例えば、動物を一定の条件下に置いたとき、当該動物に快適な又は不快な体性感覚を与えることによって、当該動物が前記一定の条件下に置かれた状況と前記体性感覚とを連合学習して、両者が関連すると理解できるようにする「条件付け」が挙げられる。前記「条件付け」を利用した従来方法としては、記憶・学習にかかる研究で用いられてきた「文脈依存的恐怖条件付け学習」が挙げられる。
【0019】
従来の「文脈依存的恐怖条件付け文脈学習」の代表的な実施方法としては、電気ショック(無条件刺激)と動物が居る場所から得られる視覚刺激などの中立刺激(条件刺激)を組み合わせて条件付けを成立させた動物が、再び電気ショックを与えられた場所に置かれた際に表す恐怖反応(すくみ行動)を、文脈学習として評価する方法である。具体的には,電気グリッドを設置したチャンバー内に動物をいれ、痛みを感じる電気刺激を短時間与える。条件付けの一定時間 (1 時間、24 時間など)後に、電気刺激のない条件付けチャンバー内でのすくみ行動を2〜5分間測定し、全測定時間に対するすくみ行動時間の百分率(%)を算出し、これを学習能の指標とする
【0020】
本発明の評価方法において、動物に学習行動を行わせる場合、当該学習行動が当該動物に記憶されたか否かは、「学習工程」の段階では評価・判定しない点が、従来の方法とは相違する。つまり、従来の文脈依存的恐怖条件付け学習においては、特定条件(条件刺激)と電気刺激とを組み合わせて提示した後、所定の時間後に再び特定条件を提示した場合の当該動物の反応(すくみ行動)を解析する。一方、本発明の学習工程において文脈恐怖条件付けを行う場合、前記特定条件と電気刺激とを組み合わせて提示する方法は同様に行うが、所定の時間経過後に再び特定条件を提示して当該動物の反応を解析することは行う必要はない。
すなわち、本発明の評価方法は、学習工程では、当該学習が成立して、当該動物がその学習行動に関する記憶を蓄積したか否かは評価・判定せず、後段の検出工程において、当該動物の脳の状態を直接観察し、評価工程において、当該動物がその学習行動に関する記憶を蓄積したか否かを評価・判定する。
したがって、本発明の学習工程において、動物に行わせる学習行動は、従来から用いられ、その学習行動が当該動物によって記憶されうることが予め確認されている学習行動が好ましい。そのような学習行動としては、例えば、Y字型迷路試験、ロータロッド試験、恐怖条件付け学習試験、水探索試験、新奇物質探索試験、受動回避試験、放射状迷路試験、Morris水迷路試験及び遅延見本見合わせ・非見本見合わせ試験等の従来方法における、特定条件と特定刺激の提示にかかる部分の方法が、好適なものとして例示できる。
【0021】
[検出工程]
本発明の評価方法における検出工程は、前記動物の脳の神経細胞におけるCapZの局在状態を調べる工程である。
前記CapZの局在状態を調べる方法としては、前記学習工程を経た動物の脳を所定の時間後に直接観察するか、化学的に固定して摘出し、前記学習行動に関連する脳部位を含む切片を作製して、CapZの分布を調べる方法が好適である。
【0022】
前記所定の時間は特に制限されず、学習行動に関する記憶が脳部位に蓄積されると考えられる時間とすればよい。
前記学習行動が恐怖条件付けであり、当該動物の海馬を解析する場合、前記所定の時間は4時間〜72時間が好ましく、8時間〜48時間がより好ましく、12時間〜24時間が更に好ましい。
上記範囲であることにより、当該動物の海馬に学習行動に関する記憶が蓄積されると考えられ、当該記憶と相関の高い現象としてCapZの局在を検出することができる。上記範囲の下限値未満の時間で脳を調べると、後段の評価工程における当該シグナルの解釈が困難になる場合がある。記憶は海馬で数日〜数週間は保持されると考えられるが、上記範囲の上限値を超えた時間で調べると、別の新しい記憶が上書きされることによって、当該学習依存のCapZの局在を示すシグナルを検出できなくなる恐れがある。
【0023】
前記脳を直接観察する方法、化学的に工程し摘出する方法及び脳の切片を作製する方法は公知の方法が適用できる。
脳内に多数存在する部位のうち、どの部位を調べるかは特に制限されない。学習工程で動物に行わせる学習行動と関連性の高い脳部位の状態を調べることが好ましい。例えば、空間認知に関する学習の記憶は、脳における海馬に蓄積されることが報告されている。従って、空間認知に関する学習行動を行わせた動物の脳における海馬を調べることによって、当該学習行動に関する記憶の蓄積と相関する現象を捉えることができる。
本発明の評価方法において、記憶の蓄積と相関する現象は、当該脳部位若しくは神経細胞を構成するスパインの一部に、CapZと呼ばれるタンパク質が局在する現象である。
【0024】
CapZは、F-アクチン キャッピング プロテイン(F-actin capping protein)とも呼ばれる約270〜290アミノ酸残基のタンパク質であり、当初、F-アクチンに結合する機能を有するタンパク質として同定された。その後、本発明者らによって、CapZが神経細胞のシナプス可塑性に関与している可能性が検討された。その結果、本発明者は、動物が学習行動を行うことにより記憶を蓄積する事象と、当該学習行動に伴って発火する神経細胞においてCapZの少なくとも一部が、スパインの一部に局在する事象と、が高い相関を有することを見出した。
【0025】
動物の臓器別に、異なるアイソタイプのCapZが存在する場合があり、これらのアイソタイプがホモ複合体又はヘテロ複合体を形成して機能している。例えば、マウスにはCapZ beta1、CapZ beta2及びCapZ alphaの少なくとも3種類のアイソタイプが確認されている。このうち、主にCapZ beta2及びCapZ alphaがマウスの脳内に発現している。
【0026】
本発明におけるCapZとしては、当該動物の内在性CapZであってもよいし、当該動物に遺伝子導入若しくはタンパク質導入された外来性CapZであってもよい。
前記外来性のCapZは、神経細胞において、信号の入力依存的にスパインに局在する機能及び/又はF-アクチンに結合する機能を有するものであれば特に制限されず、同種の動物に由来するCapZであってもよいし、異種の動物に由来するCapZであってもよい。また、同種若しくは異種の動物に由来するCapZのアミノ酸配列において1若しくは数残基のアミノ酸が欠失・置換・付加された変異型CapZであってもよい。さらに、CapZにマーカータンパク質及び/又はペプチドタグを結合させた融合タンパク質を、外来性CapZとして導入してもよい。
前記外来性のCapZは、由来する動物において脳内に発現しているものが好ましい。また、異種の動物に由来するCapZを導入する場合、その異種の動物は、導入する動物に近縁であるものが好ましい。
【0027】
動物からクローニングされたCapZのcDNA情報は、NCBIのデータバンクUniGeneで一括して見ることができる。CapZ alphaのID番号はUGID:905681であり、CapZ beta2のID番号はUGID:256943である。このデータバンク及びリンクされたデータバンクで検索できるCapZを前記外来性のCapZとして導入することが可能である。
【0028】
ほ乳類に属する動物間でのCapZのアミノ酸配列の相同性は大変高い。例えばヒトとマウス間で比較したアミノ酸配列の相同性は、サブユニットalphaでは98%、サブユニット betaでは100%である。このため、ほ乳類内では、異種の動物由来のCapZが遺伝子導入された動物において、導入されたCapZは機能すると考えられる。
【0029】
前記外来性又は内在性のCapZとして、マウス(Mus musculus)由来のCapZを用いる場合、当該CapZは、
配列表の配列番号1で表される272アミノ酸残基のタンパク質(CapZ beta2);
配列表の配列番号2で表される286アミノ酸残基のタンパク質(CapZ alpha);
配列番号1若しくは配列番号2のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換又は付加されたアミノ酸を有し、且つ神経細胞への入力依存的にスパインに局在する機能及び/又はF-アクチンに結合する機能を有するタンパク質;
配列番号1若しくは配列番号2のアミノ酸配列と相同性が90%以上であり且つ神経細胞への入力依存的にスパインに局在する機能及び/又はF-アクチンに結合する機能を有するタンパク質、が好ましい。
【0030】
配列番号1のアミノ酸配列(CapZ beta2)は、GenBank Accession No.NM_009798で登録されているMus musculus capping protein (actin filament) muscle Z-line, beta (Capzb), transcript variant 2, mRNAと同一の配列である。配列番号2のアミノ酸配列(CapZ alpha)は、GenBank Accession No. NM_009797.2で登録されているMus musculus capping protein (actin filament) muscle Z-line, alpha 1 (Capza1), mRNAと同一の配列である。
【0031】
前記外来性又は内在性のCapZとして、ラット(Rattus norvegicus)由来のCapZを用いる場合、当該CapZは、
配列表の配列番号3で表される272アミノ酸残基のタンパク質(CapZ beta);
配列番号3のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換又は付加されたアミノ酸を有し、且つ神経細胞への入力依存的にスパインに局在する機能及び/又はF-actinに結合する機能を有するタンパク質;
配列番号3のアミノ酸配列と相同性が90%以上であり且つ神経細胞への入力依存的にスパインに局在する機能及び/又はF-actinに結合する機能を有するタンパク質、が好ましい。
【0032】
配列番号3のアミノ酸配列(CapZ beta)は、GenBank Accession No.NM_001005903.1で登録されているRattus norvegicus capping protein (actin filament) muscle Z-line, beta (Capzb), mRNAと同一の配列である。
【0033】
前記外来性又は内在性のCapZとして、ヒト由来のCapZを用いる場合、当該CapZは
配列表の配列番号4で表される272アミノ酸残基のタンパク質(CapZ beta1);
配列表の配列番号5で表される286アミノ酸残基のタンパク質(CapZ alpha1);
配列番号4若しくは配列番号5のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換又は付加されたアミノ酸を有し、且つ神経細胞への入力依存的にスパインに局在する機能及び/又はF-アクチンに結合する機能を有するタンパク質;
配列番号4若しくは配列番号5のアミノ酸配列と相同性が90%以上であり且つ神経細胞への入力依存的にスパインに局在する機能及び/又はF-アクチンに結合する機能を有するタンパク質、が好ましい。
【0034】
配列番号4のアミノ酸配列(CapZ beta1)は、GenBank Accession No.NM_004930.3で登録されているHomo sapiens capping protein (actin filament) muscle Z-line, beta (CAPZB), transcript variant 1, mRNAと同一の配列である。配列番号5のアミノ酸配列(CapZ alpha1)は、GenBank Accession No.NM_006135.2で登録されているHomo sapiens capping protein (actin filament) muscle Z-line, alpha 1 (CAPZA1), mRNAと同一の配列である。
【0035】
作製した脳の切片標本において、どの部位にCapZが局在しているかを調べる方法は特に制限されず、従来公知の方法を適用できる。例えば、CapZに選択的(特異的)に結合する抗体を用いた免疫染色法が適用できる。当該抗体は市販のものを購入して用いても良いし、常法に従って抗CapZ抗体を作製して用いても良い。
【0036】
CapZが神経細胞のどの部位に分布しているかを調べるために、神経細胞を構成する構造体を染色することが好ましい。例えば、抗MAP2抗体を使用して当該切片を染色すると、MAP2が存在する樹状突起の位置、形状及び輪郭を観察することができる。また、ローダミン−ファロイジン(Rhodamine-phalloidine)を使用して当該切片を染色すると、F-アクチンを含むスパインの位置、形状及び輪郭を観察することができる。したがって、当該切片の観察視野において、樹状突起およびスパインの位置や大きさ等を確認し、同じ視野におけるCapZ(融合タンパク質)に由来するシグナルを検出することによって、CapZが神経細胞のどの部位に分布しているか、又はCapZが神経細胞のどの部位に局在しているか、を調べられる。
【0037】
また、より好適な別の方法として、CapZを遺伝子導入したトランスジェニック動物(以下、CapZ-TG動物ということがある。)の使用が例示できる。
前記CapZ-TG動物としては、CapZとマーカータンパク質及び/又はペプチドタグとを結合させた融合タンパク質をコードする遺伝子が導入されたものが好ましい。
前記融合タンパク質は、マーカータンパク質又はペプチドタグが結合されているので、それを利用すると、検出がより容易となるので好ましい。なお、前記CapZ-TG動物は、常法によって作製できる。
【0038】
学習工程を経た動物の脳の切片において、CapZに選択的に結合した一次抗体を検出する方法、及び前記マーカータンパク質又はペプチドタグを検出する方法は特に制限されず、公知方法が適用できる。例えば、前記一次抗体に結合する蛍光標識付きの二次抗体を使用し、蛍光顕微鏡で検出する方法や、前記マーカータンパク質が発する蛍光を顕微鏡で検出する方法が挙げられる。これらの方法で検出されるシグナルは、当該切片におけるCapZの分布を示すものである。
【0039】
前記CapZ-TG動物に導入するCapZとして、前述の外来性CapZを適用できる。当該CapZは、同種の動物からクローニングされたCapZであってもよいし、異種の動物からクローニングされたCapZであってもよい。例えば、マウスに導入するCapZは、マウス由来のCapZであってもよいし、ヒト、ラット、ウサギ等の別種のほ乳類に由来するCapZであってもよい。ここで、導入するCapZは、神経細胞において、信号の入力依存的にスパインに局在する機能及び/又はF-アクチンに結合する機能を有するものである。この機能を有する限り、導入するCapZのアミノ酸配列において1若しくは数残基のアミノ酸の欠失・置換・付加が行われていてもよい。
【0040】
前記マーカータンパク質は特に制限されず、従来公知のマーカータンパク質が適用可能であり、例えばアルカリフォスファターゼ、HRP、GFP、EGFP等が挙げられる。
前記ペプチドタグは特に制限されず、従来公知のペプチドタグが適用可能であり、例えばHAタグ、Mycタグ、Hisタグ、FLAGタグ、GSTタグ等が挙げられる。
前記マーカータンパク質に由来するシグナル、及び/又は前記ペプチドタグに特異的に結合する抗体に由来するシグナルを検出することにより、前記CapZを含む融合タンパク質の局在を調べることがより容易となるので好ましい。
【0041】
前記CapZ-TG動物を用いる場合、学習行動を行わせたことに対する反応を増幅するために、当該CapZ-TG動物における前記融合タンパク質の発現を、前記学習行動を行わせた期間に対応する、できるだけ狭い時間的な幅で誘導させることが好ましい。
具体的には、前記融合タンパク質の発現を特定のプロモーターによって制御することが好ましい。前記プロモーターとして、神経活動入力依存的に発現誘導されるArcプロモーター、c-Fos プロモーター等が好適なものとして例示できる。また、所望のタイミングで発現を制御できる、テトラサイクリン誘導プロモーター、cre依存性プロモーター構造なども好適なものとして例示できる。
【0042】
例示した中でも、Arcプロモーターがより好ましい。Arcプロモーターを用いることによって、例えばCapZを遺伝子導入したトランスジェニックマウス(以下、CapZ-TGマウスと呼ぶことがある。)に空間認識に関する学習行動を行わせた場合、当該CapZ-TGマウスの脳における海馬のCA1野の放線層に属する神経細胞に信号が入力され、当該入力に伴って、前記融合タンパク質を当該神経細胞において発現させることができる。すなわち、学習行動に関する記憶を蓄積すると考えられる神経細胞において、融合タンパク質の発現が当該学習行動を行わせることを引き金としてなされる。このメカニズムを利用して、当該学習行動に関する記憶の蓄積の際、新たに発現した融合タンパク質の分布を、もともと存在していた内在性のCapZと区別することができる。つまり、当該学習行動を行わせる以前に蓄積されていた記憶に関するCapZと、当該学習行動を行わせたことにより発現したEGFP-CapZ(融合タンパク質)とを区別することが可能となる。この結果、新たに発現した融合タンパク質の分布が、当該学習行動に関する記憶の蓄積におけるCapZの、学習マーカーとして果たす役割をよりよく反映していると考えられる。
【0043】
[評価工程]
本発明の評価方法における評価工程は、前記CapZの局在状態の観察に基づき、前記脳の状態を評価する工程である。
評価工程において、検出工程で検出したCapZに由来するシグナルの分布を解析して、前記CapZが一部のスパインに局在している場合、前記動物が正常な学習能力を有する状態である、又は前記動物が少なくとも学習工程において正常な学習能力を有する状態であった、と評価・判断する。
また、評価工程において、検出工程で検出したCapZに由来するシグナルの分布を解析して、前記CapZが一部のスパインに局在していない場合、前記動物が正常な学習能力を有する状態ではない(異常な状態である)、又は前記動物が少なくとも学習工程において正常な学習能力を有する状態ではなかった(異常な状態であった)、と評価・判断する。
【0044】
本明細書では、前記学習行動を行うことに伴って信号が入力される脳部位に含まれるスパインのうち、CapZのシグナルが強く検出されたスパインの割合を「CapZのスパイン局在率」又は単に「局在率」と呼ぶ。局在率は、脳切片を観察した画像全体から、ファロイジン染色により定義される各スパイン内の、CapZ強度を取得して、その強度分布(ヒストグラム)が最尤値付近にまとまらず、突出して高い値を有する強度が分布に占める割合で規定できる。
【0045】
観察した神経細胞及び脳部位において、スパインは多数存在することが普通である。ここで、「CapZの一部がスパインに局在する」とは、複数のスパインの全てにおいてCapZの濃度が高まるのではなく、一部のスパインにおいてのみCapZの濃度が高まることを意味する。
最近の研究では、記憶に関する神経細胞のシナプス可塑性の変化は、一部の神経細胞にのみ起きることが示唆されている。これと符号するように、本発明者の検討によれば、正常な学習能力を有する野生型のマウスに学習行動を行わせて24時間経過した脳の海馬においては、特定の神経回路をなす神経細胞群が有する全スパインのうち、1〜5%のスパインにのみCapZのシグナルが強く検出されている。
【0046】
本発明において、当該動物が正常な学習能力を有するか否かを局在率に基づいて判定することができる。具体的には、動物に学習行動を行わせた後、所定時間経過時の脳の切片において、学習行動に関する記憶が形成されると考えられる脳部位の神経細胞群が有するスパインのうち、所定割合のスパインにのみCapZのシグナルが強く検出された場合、すなわち局在率が高い場合に、当該動物は正常な学習能力を有する、と判定できる。
前記所定時間としては、4時間〜72時間が好ましく、8時間〜48時間がより好ましく、12時間〜24時間がさらに好ましい。この範囲であると、学習行動に関する記憶が神経細胞に蓄積(形成)されたことをより確実に判定できる。
前記局在率としては、0.01〜10%が好ましく、0.1〜10%がより好ましく、0.1〜5%がさらに好ましい。この範囲であると、学習行動に関する記憶が蓄積された場合(正常な学習能力を有する場合)と蓄積されていない場合(正常な学習能力を有さない場合)とをより確実に区別して判定できる。
【0047】
本発明の評価方法において、前記動物として、疾患の状態を呈する疾患モデル動物を使用できる。前記疾患としては、ダウン症、統合失調症、認知症、公知の学習障害を呈する疾患又は公知の精神疾患が挙げられる。このような疾患モデル動物は、公知のものを適用できる。
また、前記疾患モデル動物として、前記融合タンパク質をコードする遺伝子が導入されたCapZ-TG動物を疾患モデル動物となるように加工して用いてもよい。この疾患モデル化CapZ-TG動物を用いることにより、当該動物中のCapZを検出することがより容易となるので好ましい。以下では、「疾患モデル動物」は、特に明記しない限り「疾患モデル化CapZ-TG動物」を含むものとする。
【0048】
前記疾患モデル動物の作製方法は特に制限されず、例えば、受精卵に当該疾患関連遺伝子を導入する方法、当該疾患関連遺伝子を欠損または置換したノックアウトマウスを常法により作製する方法、疾患関連遺伝子をウイルスベクターや電気穿孔法などで神経細胞に導入する方法、脳に作用し疾患様症状を呈する薬物を投与する方法、脳に部分的に損傷を与える方法、疾患様症状を誘導する強度のストレスを与える方法などが挙げられる。
【0049】
前記疾患関連遺伝子としては、例えばダウン症、統合失調症、認知症、公知の学習障害を呈する疾患又は公知の精神疾患に関連する遺伝子が挙げられる。
前記疾患モデル動物は、当該疾患の症状又は当該疾患に近い症状を呈する。また、前記疾患モデル化CapZ-TG動物は、当該疾患の症状又は当該疾患に近い症状を呈し、且つ前記融合タンパク質が導入された動物である。
例えばダウン症に関連する遺伝子を含む染色体が導入された疾患モデルマウスは、学習障害を呈することが知られている。
疾患モデル動物と健康な動物とを用いることによって、当該疾患に関連する学習障害若しくは精神疾患を呈する脳の状態と、正常な学習能力を有する脳の状態とを、CapZ(融合タンパク質)の分布の違いに基づいて比較検討することができる。
【0050】
<医薬のスクリーニング方法>
本発明の医薬のスクリーニング方法は、学習障害若しくは精神疾患を呈する前記疾患モデル動物に対して、被検物質を投与する工程と、前記動物に学習行動を行わせる学習工程と、前記動物の脳の神経細胞におけるCapZの局在を調べる検出工程と、前記CapZの局在に基づき、前記脳の状態を評価する評価工程と、を含み、前記評価工程において、前記CapZが一部のスパインに局在している場合、前記被検物質を、前記学習障害若しくは精神疾患を治療又は予防する医薬として有用であると判断する方法である。
【0051】
本発明の医薬のスクリーニング方法における学習工程は、予め動物に被検物質を投与することを除いて、前述の本発明の評価方法における学習工程と同様に行えばよい。
前記動物は、学習障害若しくは精神疾患を呈する前述の疾患モデル動物が好ましい。
【0052】
医薬のスクリーニング方法に用いる疾患モデル動物としては、前記学習行動を行わせても当該学習行動に関連する記憶が正常に蓄積されないこと又は学習記憶が正常に成立しないこと、が予め別の方法で示唆又は確認されているものが好ましい。さらに、前記疾患モデル動物としては、前記学習行動を行わせても、学習記憶に関連する脳部位においてCapZが正常に局在しないことが、予め確認されているものが好ましい。
また、前記疾患モデル動物としては、前記学習行動を行わせても、当該学習行動に関連する記憶が正常に蓄積されないこと又は学習記憶が正常に成立しないこと、が予め別の方法で示唆又は確認されていて、且つ同じ学習行動を行わせた場合、その学習記憶に関連する脳部位においてCapZが正常に局在しないことが、予め確認されているものがより好ましい。ただし、本方法においては、従来の学習試験での効果の判定とは違う結果となる可能性がある。例えば、恐怖条件付けのテストの際、被験物質での腹痛のため動けない動物は記憶していなくてもすくみ反応をしたと誤解されて解釈される場合や、恐怖を記憶しているにも関わらず、錯乱して動いてしまい(すくみ反応を示さず)、記憶していないと判断される場合など、従来の記憶試験では判断が誤ってしまうケースにおいても、本発明の医薬のスクリーニング方法によれば、正確に判断できる可能性があることに十分留意する。記憶など認知の障害が、体のだるさ(倦怠感)等の他の症状にマスクされがちな統合失調症や鬱病のモデルの場合には特に留意する。
【0053】
前記被検物質としては、疾患モデル動物に対して毒性が低いものが好ましく、さらに当該疾患を改善すると見込まれるものがより好ましい。
前記被検物質の投与量および投与方法は特に制限されず、薬理学的に許容される投与量で公知の方法で投与すればよい。
【0054】
本発明の医薬のスクリーニング方法における検出工程は、前述の本発明の評価方法における学習工程と同じ様に行えばよい。前記疾患モデル化CapZ-TG動物を用いると、CapZに結合されたマーカータンパク質又はペプチドタグを利用することにより、CapZの検出がより容易となるので好ましい。
【0055】
本発明の医薬のスクリーニング方法における評価工程は、前記CapZの局在に基づき、前記被検物質が投与された疾患モデル動物の脳の状態を評価する工程である。
【0056】
前記評価は、前述の本発明にかかる評価方法と同様に行うことができる。CapZ-TG動物を含む健康な動物が示す、当該CapZの局在状態(一部のスパインに局在する)を正常の指標とし、被検物質を投与する前の疾患モデル動物が示す当該CapZの局在状態(一部のスパインに局在しない)を異常の指標とし、被検物質を投与した疾患モデル動物が示す当該CapZの局在状態が異常から正常へ近づいた場合、又は被検物質を投与した疾患モデル動物が示す当該CapZの前記局在率が上昇した場合、当該疾患モデル動物の異常であった学習能力が正常に回復した若しくは正常に近づいたと評価できる。
この評価に基づき、当該被検物質を、当該疾患モデル動物が呈する学習障害若しくは精神疾患を治療又は予防する医薬として有用であると判断できる。
【0057】
医薬のスクリーニング方法の評価工程において、前段の検出工程で検出した前記CapZが一部のスパインに局在している場合、当該疾患モデル動物の異常であった学習能力が正常に回復した若しくは正常に近づいたと評価し、前記被検物質を、当該疾患モデル動物が呈する学習障害若しくは精神疾患を治療又は予防する医薬として有用であると判断する。
また、当該評価工程において、前段の検出工程で検出した前記CapZの前記局在率が上昇した場合、当該疾患モデル動物の異常であった学習能力が正常に回復した若しくは正常に近づいたと評価し、前記被検物質が有する、当該疾患モデル動物が呈する学習障害若しくは精神疾患を治療又は予防する作用効果の程度を判断することもできる。
【0058】
<本発明と、従来の記憶に関する研究との関係>
記憶などの脳の高次機能を発揮するためには、各神経細胞は個性を持ち、役割を分担して働く必要がある。もし、役割分担をせずに大多数の神経細胞が同期して活動した場合は、てんかん発作のような痙攣を起こすはずである。しかし、記憶に関する従来の研究では、どの神経細胞のどの部分が、どのように変化して働いているのかを解析する手法はなかった。
【0059】
脳の一部に人工的に長期増強を起こす刺激をした後に長期増強部位にCapZの濃度(免疫組織染色強度)が上昇すること、全てのスパインに均一に存在するのではなく染色強度が高いスパインが一部にあること、はすでに発明者らが下記の論文(6)で報告している。
Genes Cells. 2010 Jun;15(7):737-47. Epub 2010 Jun 10. “Activity-dependent localization in spines of the F-actin capping protein CapZ screened in a rat model of dementia.” Kitanishi T, Sakai J, Kojima S, Saitoh Y, Inokuchi K, Fukaya M, Watanabe M, Matsuki N, Yamada MK.
しかし、この論文の実験では二つの問題点があった。1.刺激が人工的であるため、学習に関連した入力と同じである保証がない。2.免疫組織染色の場合、抗体の浸透性の問題があり、とりわけシナプス後部では存在しても染色されない分子があることが知られている。そこで、本発明者は、免疫染色とは異なる方法として、CapZとEGFPを融合タンパク質として発現するトランスジェニックマウス(CapZ-TGマウス)を作製し、脳の自然な活動に伴ってCapZの局在を検出しうることを見出した。具体的には、そのCapZ-TGマウスに学習行動を行わせ(文脈依存恐怖条件づけを行い)、記憶が固定されているはずの24時間後に脳切片を作成して、文脈と恐怖の連合記憶に必要な背側海馬を観察すると、学習行動を行わせていない動物の脳に比べて特徴的なパターンが現れる。それは、背側海馬CA1野の一部のスパインに、他と比べて正常な分散範囲を超えた(目立って高い)EGFP強度をもつものがあるパターンである。また、そうしたスパインを多数持つ樹状突起があることも分かった。この、EGFP-CapZ融合タンパク質発現トランスジェニックマウスでの結果により、免疫組織染色での問題点であった、抗体の浸透性からみかけだけスパイン間の存在量に差が出た可能性は否定された。
【0060】
そもそも、神経細胞間での情報の伝達は、少数の神経細胞によって担われていると考えられる。その根拠として、神経細胞の活動電位の発生について、どのような学習行動を行った場合でも最大10%の神経細胞しか活動しないと考えられる実験事実がある(Henze et al. JNP)。そのような少数の神経活動による情報処理を前提とすると、機能的な神経回路では情報入力が、川をながれる水のように、ある部分だけを伝播する状態が形成される必要がある。そして、学習に関する記憶が蓄積されるためには、神経のつながりにおいて一部が周りと異なった状態となって、新しく入力を受けやすい部分、水の流れやすくなる部分が生じる必要がある。その変化は「少数」でないと拡散してしまう。
発明者らの過去の実験から、人工的に長期増強を起こす刺激をした実験において長期増強部位(神経伝達効率が上がった部位)にCapZの濃度が上昇したことがわかっている。そこから類推すると、EGFP-CapZ強度の高いスパインには長期増強が起きてcoding(記憶情報の流れやすい部位の形成)をしている可能性が高い。また、そのスパインが集積している樹状突起があれば神経細胞はcoding neuronとして働きうる。本発明者が見出したEGFP-CapZ強度の高いスパインが出現する頻度は少数であり、特定の記憶に関連して活動する神経細胞が少数であることから考えて妥当である。
(参考文献;J Neurophysiol. 2000 Jul;84(1):390-400. “Intracellular features predicted by extracellular recordings in the hippocampus in vivo.” Henze DA, Borhegyi Z, Csicsvari J, Mamiya A, Harris KD, Buzsa’ki G.)
本発明の評価方法においては、正常な分散範囲を超えて、CapZが一部のスパインに局在する局在率が高い場合に、学習ができた脳と定義して、脳機能の評価に用いる。
【実施例】
【0061】
<実施例1>
[非ヒト科動物の準備]
非ヒト科動物として、トランスジェニックマウス(以下、CapZ-TGマウスと呼ぶ。)を準備して用いた。CapZ-TGマウスの作製は、以下の方法でEGFPを融合した CapZを発現させるために行った。導入遺伝子は、遺伝子組み換えにより、以下の(1)〜(8)のDNA配列又はアミノ酸配列をコードするDNA配列を順に連結して作製した(図1)。
(1)mouse Arc promoter (SmaI-NaeI断片;7.1kb)
(2)Tetracyclin-responsive transcriptional activator (Clontech社;pTet-off由来)
(3)P2A配列(TSATNFSLLKQAGDVEENPGPVAT;人工合成)
(4)EGFP (Clontech社;pEGFP-N1由来)
(5)Spacer(SGRTQISSS;人工合成)
(6)HAtag(YPYDVPDYA;人工合成)
(7)mouse CapZ beta 2 完全長272a.a.(Met-Cys)(RT-PCRにより単離;Genbank Accession No. NM_009798;Mus musculus capping protein (actin filament) muscle Z-line, beta (Capzb), transcript variant 2, mRNA)
(8)SV40-polyA(Clontech社;pEGFP-N1由来)
【0062】
(1)のマウスArcプロモーターは、マウスゲノムの該当部分(Ch15、 74.50M-74.51Mの間)を含むBACクローン(B6Ng01-232C07など)をBACライブラリ(http://dna.brc.riken.jp/ja/MSMbac.htmlなど)から入手し、SmaI-NaeI消化物の7.1kb付近からクローニングできる。
(3)のP2A配列は、異なる二つのたんぱく質(この場合は(2)と(4)-(7)融合タンパク質)を一連のcDNAから等量合成するためのシグナルである(Donnelly et al. J Gen Virol 2001, 82:1013-1025.)。なお、(2)は、本CapZ-TGマウスで将来別の遺伝子発現をする場合に備えた転写因子である。
(7)CapZ はalphaとbetaの二量体でbetaには1と2のSplicing Variantがある。脳ではbeta2が主要であるため、これを用いている。
【0063】
上記の作製した導入遺伝子を、制限酵素を用いてベクター部分を除きつつ直線化したのち、フェノールクロロフォルム法及びエタノール沈殿により精製した。さらに、ゲル電気泳動を行い、ベクター部分を含まないよう該当長のバンドを切り出して、キアゲン社のgel extraction kitを用いた精製を二回繰り返して高度に精製した。
精製した導入遺伝子を受精卵の核に注入し、正常に発生した胚を借り腹に移植した。産仔42匹のうち、CapZ-TGマウスとして7lineが同定された。各lineの導入コピー数は(1,3,10,10,10,10,100)であったため、実験には100コピー導入されたlineを用いた。
【0064】
作製したCapZ-TGマウスでは、Arcプロモーターが有るので、最近活動した神経細胞に選択的に、EGFP-HA-CapZ融合タンパク質を発現させることができる。
【0065】
[学習行動を行わせる工程]
作製したCapZ-TGマウスに、以下の方法で、学習行動として文脈恐怖条件付けを行った。
小原医科産業の文脈恐怖条件付け装置(ショックジェネレーター型番:SGA-2010、実験チャンバー:型番CL-3002)及び防音箱(型番CL-4210C)を使用した。まず、防音された装置のチャンバー内にマウスを5分間置いた。この5分間のうち、1分経過時に1回、4分経過時に1回、合計2回の電気ショック(0.2mA、2秒間)を、マウスの足に与えた。電気ショックの様子をカメラを通して観察し、確認した。5分の条件付けセッションの後、マウスを通常飼育しているケージに戻した。
この学習行動後において、電気ショックを受けたマウスが、「特定のチャンバーに入れられること」と「電気ショックを受けること」とを結び付ける文脈依存性恐怖条件を学習・記憶することが一般に知られているが、該当マウスでは記憶のテストはしていない。
この工程とは別に、24時間後にテストを行うことによって、当該CapZ-TGマウスが上記文脈恐怖条件を学習・記憶できることは、従来方法により別途確認している。すなわち、この確認において、CapZ-TGマウスと野生型マウスとで、正常な学習能力が発揮されることに有意差はなかった(図14参照)。テストでは常法に従い、ショックを与えたチャンバーに5分間マウスを置き、恐怖反応であるFreezingを起こしていた時間的割合(%)を記録した。記録はチャンバー上部のカメラから0.5秒おきに撮影した画像のマウス部分(黒)に一定のずれが生じないことが2秒続いた場合にFreezingとカウントした。上記小原医科産業の装置により画像を取得しPCに転送後、ソフトウェアTimeFZ1により自動測定した。
【0066】
[CapZ-TGマウスの脳の神経細胞におけるCapZの局在状態を調べる検出工程]
文脈恐怖条件付け後24時間経過したCapZ-TGマウスを麻酔し、開胸後30秒以内(通常10秒以内)に左心室に針を刺し、PBSを2ml、次いで2%パラホルムアルデヒドを含む0.1Mリン酸緩衝液(ともに氷冷しておいたもの)を20〜30ml(マウスの最高血圧を超えない約100mmHgで)全身灌流して固定し、脳を取出して正中で半割し、さらに1時間、4℃室で十分に振とうしながら後固定した。固定したマウスの脳をビブラトームで冠状断切片とした。以下の観察には空間学習に関わるとされる背側海馬(座標Bregma1.4mm-2.2mmの範囲)のCA1放線層、内側(lateral, 1.5mm以内)を用いた。以下ではCA1放線層の当該部位についての結果を示すが、CA1内の他の部位(網状分子層、多形層)、CA3においても同様の傾向が見られた。なお、固定処理によりタンパク質は分解・移動などの変化をしない状態になる。また蛍光タンパク質EGFPの蛍光はこの固定法では消失しない。
【0067】
得られた切片に、0.1%TritonX100で15分間の膜透過処理を行い、5%山羊血清で30分間の非特異的吸着を防ぐ処置をしたのち、1%山羊血清入りPBSで希釈した抗MAP2抗体の一次抗体(1:3000、chicken IgY、 Phosphosolutions社製;型番1100)を4℃で一晩振とうしながら反応させた。反応後、PBSで3回洗浄した。次に、二次抗体としてfluorescence(Alexa647)-labeled anti-chicken IgY(1:1000、 Invitrogen社製;型番A21463)を1%山羊血清入りPBSに希釈して、切片に反応させると同時にRhodamine-phalloidin (0.7 unit / ml, Invitrogen社製;型番R415)を加えて、室温で1時間または4℃で3時間反応させることにより、ファロイジンと抗MAP2抗体による二重染色を行った。
なお、抗体液は反応前に遠心(15,000×g、5分)し、上清部分を用いることで変性して凝集した抗体などによる擬染色を起こさぬよう注意を払った。
染色した切片をレーザー共焦点顕微鏡:Olympus社FV1000(倒立顕微鏡:IX81、対物レンズ:UPLSAPO 100×O NA1.40) を用いて、最も遅いスキャンスピードで観察し、出力を660V以内に抑えた光電子増倍管で画像を取得した。画像取得時は画像化できる最大強度を超えないよう、最も明るいサンプルにあわせてレーザー強度を調節した。その結果を図2〜10に示す。
【0068】
取得した画像の処理方法は次の通りである。
<画像処理法>:画像を処理する際は実質的に切り取った値がでないよう、比較する画像の中でもっとも幅の広い値にあわせて画像内ピクセル強度の最大値と最小値に基づいて強度を拡大表示した。例えば「0-255階調の8bit画像で 10-210、8-150の二枚の画像があるときは双方8-210を0-255に引き延ばす。」と、処理できる。なお、表示した図については原則的には原図を掲載し、4倍ズームで図を取得した図2〜6,11〜13については見易くするためにメディアンフィルターをかけた。
メディアンフィルターは測定器に由来するちらつきノイズを消すために一般的に用いられており、周囲の点の中間値を中央値に採用して突出した値をならす原理による。ここでは半径1ピクセルに設定したので3×3の点を用いていることになる。
【0069】
図2は、EGFP‐CapZのEGFP由来の緑色蛍光シグナルを示す。図2のように、学習行動を行わせた後のCapZ-TGマウスの海馬CA1野の放線層におけるEGFP‐CapZのシグナルは、一部に局在している。この図中のバーは2マイクロメーターであり、シグナルが検出された部分は、直径が約1マイクロメーター程度までで、大きさのばらつきがあることがわかる。
【0070】
図3は、図2と同じ視野においてRhodamine-phalloidinの赤色蛍光で染色されたスパインを示す。スパインは繊維状アクチンが多いため、Rhodamine-phalloidinで染色されると考えられている。脳の興奮性神経細胞の樹状突起上には、このスパイン(棘突起)と呼ばれる、頭部の直径が1マイクロメーター程度で、大きさのばらつきがある突起状の構造が密に存在することが知られる。スパインは、興奮性シグナルを受け取るシナプス後部がある部分である。神経細胞が記憶すべき情報を、あるスパインで受け取った場合、次なる想起刺激に対応すべく、そのスパインは他のスパインに比べて、より情報を伝え易くするように変化するはずである。すなわち、記憶形成に伴って、何らかの変化が起こる部分が、一部のスパインに存在しなければならない。
【0071】
図4の左は図2と同じEGFP‐CapZのシグナルを示し、図4の右は図3と同じRhodamine-phalloidin染色を示す。両者は、同一視野から別の蛍光シグナルとして、同時に取得した。矢印は同じ位置にあるので、EGFP‐CapZのシグナル(左)がRhodamine-phalloidin染色されたスパイン(右)のうちの一部だけに強く局在することが明確にわかる。
【0072】
図5は、MAP2が存在する樹状突起が、図2のEGFP-CapZのシグナルに重ねて示されている。粒状のEGFP-CapZのシグナルが、筋状の樹状突起に沿って、樹状突起から突出する棘のように存在している。この図は、EGFP‐CapZのシグナルがスパインにあることをさらに補強している。なお、樹状突起とスパインとの間に隙間があるように見えるものもあるが、これは、スパインのネック部分にはCapZが存在しないためであると考えられる。スパインにはマッシュルーム型のものがあり、ネックとよばれる細い首状の構造を持つ。ネックは、長いものでは2マイクロメーター(図5に示したバーの長さ)を超えるものもある。
【0073】
図6は、図5を解釈する際の参考とするために、図5と同じ視野において、EGFP-CapZのシグナルを除き、MAP2が存在する樹状突起のみを示している。丸く見える部分はスパインではなく、画面手前に向かう細い樹状突起の切断面と思われる。
【0074】
以上の観察から、文脈恐怖条件付けを行ったマウス脳の前記学習に関する脳部位の神経細胞では、EGFP-CapZの濃度が局所的に高い箇所があることが明らかである。その局所的に高い濃度でEGFP-CapZが主に存在する箇所は、Rhodamine-phalloidinで染色されているためスパインであると考えられる。さらに、当該脳部位に観察されるスパインの全てにおいてEGFP-CapZのシグナル強度が高くなるのではなく、一部のスパインにおいてのみEGFP-CapZのシグナル強度が高くなっている。つまり、学習行動を行わせた(文脈恐怖条件付けを行った)マウス脳では、CapZが一部のスパインに多く局在することが理解される。
前記脳部位に観察されるスパインのうち、EGFP-CapZのシグナル強度が顕著に高くなったスパインの割合(局在率)は、1〜5%であると見積もられた。
【0075】
図7は、図2〜5の脳と同じ条件で作製された標本である。すなわち、CapZ-TGマウスに学習行動を行わせた(文脈恐怖条件付けを行った)24時間後に固定した脳での海馬CA1野の放線層からのEGFP-CapZシグナルを示す。スケールバーは10マイクロメーターであり、広い範囲のシグナルをとっても、EGFP-CapZが存在する部分は疎であることがわかる。
【0076】
<比較例1>
上記CapZ-TGマウスを用いて、文脈恐怖条件付けを行わずに、実施例1と同じ方法でマウスを処理した。得られた切片を同様に観察した。その結果を図8〜9にと示す。
【0077】
図8は、学習行動を行わせなかったマウス脳の海馬CA1野の放線層におけるEGFP-CapZの分布パターンを示す。図9は、図8と同じ画像の全体の強度を下げた写真である。図8の強度を低くなるよう撮影しても、実施例の図7の写真のようにはならないことを示している。つまり、実施例1の図7と比較例1の図8〜9を比較することによって、図7では、図8〜9と異なり、「CapZが“一部の”スパインに局在」していることが明らかである。ここで「CapZが“一部の”スパインに局在する」とは、スパインに検出されるCapZに由来するシグナルの強度が樹状突起の幹の部分(スパイン以外)に比べて強いことを意味するだけでなく、同時に、同じ領域(特定の脳部位、ここでは空間記憶に関わる背側海馬)でのスパイン間で、当該シグナルの強度に偏りがあることを意味し、これが学習後の特徴と考えられる。なお、EGFPのシグナル強度は例えばpH(主に固定液の)や観察深度によっても変化するので、絶対強度の比較での議論は現時点では難しい。
【0078】
図10の左は、図7と同一視野でのファロイジン染色(スパイン染色)の写真であり、図10の右は図8と同一視野でのファロイジン染色(スパイン染色)の写真であり、両者間でスパイン密度には変化がないことがわかる。つまり、図7で検出されたCapZに由来するシグナルの偏りは、スパイン自体の消滅によるものではなく、各スパインごとのCapZの存在量が調節されたことによりCapZの濃度が異なっているためと考えられる。
【0079】
以上の観察により、学習行動を行わせなかった(文脈恐怖条件付けを行わなかった)マウス脳の神経細胞では、EGFP-CapZは全体のスパインに同様の濃度で存在して見え、濃度が他のスパインと比べて偏って高くなった一部のスパインは見出されなかった。
【0080】
学習行動を行わせた場合のEGFP-CapZのシグナルの分布が、上記のように一部のスパインを残して無くなって見える状態になる理由としては、一部のスパイン以外の領域に存在したCapZが分解されているためであると解釈できる。これを支持する知見として、CapZ分子に分解シグナルがあり、結合タンパクBAG3とHsc70が共存していないと、ユビキチン非依存にプロテアソームで分解されることが知られている。
(参考文献:Circ Res. 2010 Nov 12;107(10):1220-31. BAG3 and Hsc70 interact with actin capping protein CapZ to maintain myofibrillar integrity under mechanical stress.Hishiya A, Kitazawa T, Takayama S.)
【0081】
実施例1及び比較例1の結果から、学習行動を行わせたことによってマウス脳内に蓄積した記憶と、当該学習タスクによってマウス脳内に新たに発現したEGFP-CapZが一部のスパインに局在して検出されたこと(一部のスパインに偏って強く検出されたこと)は、高い相関を示している。すなわち、学習行動に伴ってEGFP-CapZが、その学習行動に関する神経細胞の一部のスパインに選択的に局在したことは、学習行動に関する記憶が当該スパインに記銘されたことを示していると考えられる。
【0082】
<実施例2>
CapZ-TGマウスの代わりに野生型のマウスを用いたこと、及びEGFPの蛍光の代わりに抗CapZ beta2抗体(1:300、mouse monoclonal IgG、The Developmental Studies Hybridoma Bank製;型番3F2.3-s, betaのうちValiant2にのみ存在するC末端配列部分に対する抗体)を抗MAP2抗体と同時に反応させ、その二次抗体としてfluorescence(Alexa488)-labeled anti-chicken IgG(1: 1000、Invitrogen社製;型番A11029)を検出に用いたこと以外は、実施例1と同じ方法で行った。その結果を図11に示す。なお用いた抗CapZ beta2抗体はimmunoblotting実験で脳のタンパク質内で非常に高い特異性を示すことは確認している(非特許文献4のsupplementary figure1で単一バンド)。
【0083】
図11において、白く示した領域が、抗CapZ抗体に由来するシグナルを示す。バーは1マイクロメーターであり、図2で見られたものとほぼ同様のサイズのシグナルが得られていることがわかる。なお、このシグナルの部位は図3〜6で示したのと同様に、一部のスパインに局在するものであることは確認している。この観察により、実施例1の図7と同じ結論が導かれた。すなわち、野生型で免疫染色を行う方法によっても、一部のスパインに強いCapZシグナルが見られることがわかった。
【0084】
<比較例2−1>
文脈恐怖条件付けを行わずに、実施例2と同じ方法でマウスを処理した。得られた切片を同様に観察した。その結果を図12に示す。
図12において、白く示した領域が、抗CapZ抗体に由来するシグナルを示す。この観察により、学習行動を行わせなかった(文脈恐怖条件付けを行わなかった)マウス脳では、実施例1の図8と同様に比較的均一なシグナルが観察される。つまりCapZが一部のスパインに局在している現象は見出されなかった。
【0085】
<比較例2−2>
ダウン症モデルマウス(ダウン症で3本になっているヒトの21番染色体に相同な、マウス16番染色体の一部を付加して遺伝的に部分的Trisomyにしている)、Ts1Cje mouseを用いて、実施例2と同様の文脈恐怖条件付けを行い、実施例2と同様に解析した。その結果を図13に示す。
使用したマウスは、作成者であるDr. C.J.Epsteinに許可を得た後、理化学研究所の山川和弘先生より分与いただいた。このマウスは、学習障害を呈することが以下の参考文献に報告されている。
(参考文献;Proc Natl Acad Sci U S A. 1998 May 26;95(11):6256-61. “Ts1Cje, a partial trisomy 16 mouse model for Down syndrome, exhibits learning and behavioral abnormalities.” Sago H, Carlson EJ, Smith DJ, Kilbridge J, Rubin EM, Mobley WC, Epstein CJ, Huang TT.)
【0086】
図13において、白く示した領域が、抗CapZ beta2抗体に由来するシグナルを示す。
この観察により、学習行動を行わせたにも関わらず、学習障害を持つダウン症モデルマウス脳では、CapZの濃度が全体に高めに測定されたが、各スパインで検出されるシグナルに偏りは少なく、一部のスパインにだけシグナルが強く検出されることはなかった。
【産業上の利用可能性】
【0087】
本発明の非ヒト科動物の学習能力の評価方法は、当該動物が学習能力を有するか否かの客観的な指標を提供できる。この評価方法を用いて、当該動物に投与した薬物が、学習能力に及ぼす影響を評価することができる。従って、学習障害や精神疾患の治療又は予防に有用な薬物のスクリーニングを行う際に、本発明の評価方法を利用できる。
【技術分野】
【0001】
本発明は、非ヒト科動物が正常な学習能力を有する状態であるか否かを評価する方法に関する。より詳しくは、非ヒト科動物の脳の神経細胞におけるCapZの局在を調べることにより、当該動物の脳が正常に学習できる状態であるか否かを評価する方法に関する。
【背景技術】
【0002】
従来の薬物スクリーニングの場合、記憶能、学習能の判定に齧歯類の行動変化を指標とする試験法が一般的である。例えば「文脈依存的恐怖条件付け文脈学習」の場合、過去に痛みを与えられた場所に置かれた動物が、その痛みを想起して表す恐怖反応、全く動かなくなる「すくみ行動」、の時間率を学習能として評価する。
【0003】
また、神経細胞・分子レベルで記憶や学習の根本的なメカニズムを研究するために、主に生体外で切片標本を用いたLTP(Long Term Potentiation)が、研究材料として使用されている。LTPで観察されるAMPA型グルタミン酸受容体による電流の増加は、記憶の素過程であると考えられおり(非特許文献1及び2)、記憶や学習に影響を及ぼす薬物のスクリーニング等への応用研究も盛んである。ところで、記憶の素過程として現れるはずの現象(神経回路の可塑性を示す現象)は、少ない頻度でしか起こらない可能性があることに留意しなければならない。発明者らの過去の研究(非特許文献3,4)からも、記憶が形成される刺激によって変化するスパインの数は、全体の数%程度である可能性が示唆されている。
【先行技術文献】
【非特許文献】
【0004】
【非特許文献1】“Memory enhancement: the search for mechanism-based drugs.” Lynch G. Nat Neurosci. 2002 Nov;5 Suppl:1035-8.
【非特許文献2】”Learning from LTP: a comment on recent attempts to identify cellular and molecular mechanisms of memory.” Learn Mem. 1996 Sep-Oct;3(2-3):61-73.
【非特許文献3】”Experience-dependent, rapid structural changes in hippocampal pyramidal cell spines.” Kitanishi T, Ikegaya Y, Matsuki N, Yamada MK. Cereb Cortex. 2009 Nov;19(11):2572-8. Epub 2009 Feb 24.
【非特許文献4】”Contextual learning induces an increase in the number of hippocampal CA1 neurons expressing high levels of BDNF.” Chen J, Kitanishi T, Ikeda T, Matsuki N, Yamada MK. Neurobiol Learn Mem. 2007 Nov;88(4):409-15. Epub 2007 Aug 31.
【発明の概要】
【発明が解決しようとする課題】
【0005】
従来の薬物スクリーニングに用いられている、動物の学習能の判定に用いられる試験法は、健康な動物を試験対象とする場合には適当な方法であるとしても、疾患モデル動物を試験対象とする場合には、単に行動だけが変化したのか、或いは学習能が変化したのか、を弁別して解釈することは難しいという問題がある。例えば、学習能が低下していると考えられている鬱病モデル動物や統合失調症モデル動物を対象にする場合は、従来の試験法では、だるさや無気力等が原因で不動状態になっているのか、恐怖反応を示して不動状態になっているのか、を弁別することが難しい。このように、特に疾患モデル動物の場合には、学習とは異なる原因の不動も考えられるので、従来の試験法が本当に学習能を反映するのかは確実ではない。また、薬物のスクリーニングを行う場合、投与した被検薬物が副作用を与えて、体が不調になったことが原因で不動状態になっているにも関わらず、当該被検薬物して学習が達成されたために不動状態になっている、と誤って解釈される危険もある。
【0006】
したがって、従来の学習試験の不確実性を回避する方法としては、脳の記憶担当部位に起こる変化を直接解析する手法が望ましい。
【0007】
脳の記憶担当部位に起こる変化を捉えるために、従来からLTPを用いる方法が一般的に考えられている。しかし、近年の研究の進展によって、LTP形成に基づいた記憶の素過程の研究について、疑念が生じつつある。そもそも、従来のLTPの研究に用いられてきた、in vitroの神経細胞を人為的に直接発火させる刺激は、動物が活動や学習する際に神経細胞に自然に入力する刺激とは同じでなく、その人為的な刺激は生体中の自然な刺激を再現できていない可能性が高いことが明らかになりつつある。また、ここ数年間の技術進歩により、LTPを観察する手法は格段に進化しており、もし生体内でLTP形成が起これば確実に観察結果の報告があっていいはずである。それにも関わらず、LTPと同様の現象が生体内で起こったという報告は未だになされていない。
【0008】
これらのことから、今まで研究されてきたLTPと部分的に共通の分子メカニズムを持ちつつも、生体内で生じる自然な刺激によって起きる新たな現象を、記憶との相関という原点に戻って確認する必要がある。
記憶が形成されるメカニズムについて理解しようと試みた従来の実験手法は、脳全体を見る分子科学か、ランダムに選んだ一部の神経細胞を見る生理学かのどちらかであった。このような実験手法だけを用いて研究していると、記憶が形成されるメカニズムにおいて重要となりうる、少数の特定の神経細胞に起こる変化(現象)を見逃してしまう危険がある。
【0009】
本発明は、上記事情に鑑みてなされたものであり、従来のLTPとは異なった概念を基盤とし、動物の学習能力及びその脳の状態を解析・解釈することで、非ヒト科動物が正常な学習能力を有する状態であるか否かを評価する方法の提供を課題とする。
【課題を解決するための手段】
【0010】
本発明者が鋭意研究したところ、動物が学習することに伴う記憶の形成と、当該動物の脳の神経細胞におけるCapZの局在とに相関があることを見出した。この知見に基づき、以下の方法を提供する。
【0011】
本発明の請求項1に記載の非ヒト科動物の学習能力の評価方法は、非ヒト科動物が正常な学習能力を有する状態であるか否かを評価する方法であって、前記動物に学習行動を行わせる学習工程と、前記動物の脳の神経細胞におけるCapZの局在状態を調べる検出工程と、前記CapZの局在状態の観察に基づき、前記脳の状態を評価する評価工程と、を含むことを特徴とする。
本発明の請求項2に記載の評価方法は、請求項1において、前記評価工程において、CapZが一部のスパインに局在している場合、前記動物が正常な学習能力を有する状態であると評価することを特徴とする。
本発明の請求項3に記載の評価方法は、請求項1又は2において、前記非ヒト科動物が、CapZとマーカータンパク質及び/又はペプチドタグとを結合させた融合タンパク質をコードする遺伝子が導入された、トランスジェニック動物であり、前記マーカータンパク質に由来するシグナル、及び/又は前記ペプチドタグに特異的に結合する抗体に由来するシグナルを検出することにより、CapZの局在を調べることを特徴とする。
本発明の請求項4に記載の評価方法は、請求項3において、前記トランスジェニック動物における前記融合タンパク質の発現を、前記学習行動に伴って入力を受ける脳部位において、前記入力により誘導させることを特徴とする。
本発明の請求項5に記載の評価方法は、請求項1〜4の何れか一項において、前記動物が疾患モデル動物であることを特徴とする。
本発明の請求項6に記載の評価方法は、請求項1〜5の何れか一項において、CapZが、以下の(i)又は(ii)のタンパク質であることを特徴とする。
(i)配列番号1〜5の何れかで表されるアミノ酸配列を有するタンパク質。
(ii)配列番号1〜5の何れかで表されるアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列を有し、且つ神経細胞への入力依存的にスパインに局在する機能を有するタンパク質。
本発明の請求項7に記載の医薬のスクリーニング方法は、学習障害若しくは精神疾患を呈する請求項5に記載の疾患モデル動物に対して、被検物質を投与する工程と、前記動物に学習行動を行わせる学習工程と、前記動物の脳の神経細胞におけるCapZの局在を調べる検出工程と、前記CapZの局在に基づき、前記脳の状態を評価する評価工程と、
を含み、前記評価工程において、CapZが一部のスパインに局在している場合、前記被検物質を、前記学習障害若しくは精神疾患を治療又は予防する医薬として有用であると判断することを特徴とする。
【発明の効果】
【0012】
本発明の評価方法によれば、学習行動を行わせた動物の脳が、当該学習に関する記憶を蓄積(記銘)できる正常な学習能力を有するか否かを評価することができる。本発明の評価方法を用いることによって、動物の学習と記憶に関して、LTP形成とは異なる現象に基づいた新たな研究基盤を提供できる。
本発明の医薬のスクリーニング方法によれば、学習障害若しくは精神疾患を治療又は予防する医薬として有用な医薬候補となる物質を見出すことができる。
【図面の簡単な説明】
【0013】
【図1】実施例1で使用したトランスジェニックマウスに導入した遺伝子の模式図である。
【図2】実施例1で使用したトランスジェニックマウスの脳切片を蛍光顕微鏡で観察して検出された、EGFP-CapZに由来する緑色蛍光シグナルを示す写真である。
【図3】図2と同じ観察視野において、Rhodamine-phalloidinの赤色蛍光で染色されたスパインを示す写真である。
【図4】左は図2と同じEGFP-CapZに由来する蛍光シグナルを示す写真であり、右は図3と同じRhodamine-phalloidin染色によって観察された蛍光シグナルを示す写真である。
【図5】図2と同じ観察視野において、MAP2が存在する樹状突起を図2のEGFP-CapZのシグナルに重ねて示した写真である。
【図6】図5と同じ観察視野において、EGFP-CapZに由来するシグナルを除き、MAP2が存在する樹状突起のみを示した写真である。
【図7】図2よりも広い視野で観察した(ズームなしで観察した)写真である。
【図8】比較例1で使用したトランスジェニックマウスの脳切片を蛍光顕微鏡で観察して検出された、EGFP-CapZに由来する緑色蛍光シグナルを示す写真である。
【図9】図8と同じ写真において、画像の全体のシグナル強度を下げた写真である。
【図10】左は図7と同一視野におけるファロイジン染色の写真であり、右は図8と同一視野におけるファロイジン染色の写真である。
【図11】実施例2で使用した野生型マウスの脳切片における、抗CapZ抗体に由来するシグナルを示す写真である。
【図12】比較例2で使用した野生型マウスの脳切片における、抗CapZ抗体に由来するシグナルを示す写真である。
【図13】比較例3で使用した疾患モデルマウスの脳切片における、抗CapZ抗体に由来するシグナルを示す写真である。
【図14】恐怖条件付けを行ったトランスジェニックマウスの学習能力を表すグラフである。
【発明を実施するための形態】
【0014】
以下、好適な実施の形態に基づき、図面を参照して本発明を説明する。
<評価方法>
本発明の非ヒト科動物の学習能力の評価方法は、非ヒト科動物が正常な学習能力を有する状態であるか否かを評価する方法であって、前記動物に学習行動を行わせる学習工程と、前記動物の脳の神経細胞におけるCapZの局在状態を調べる検出工程と、前記CapZの局在状態の観察に基づき、前記脳の状態を評価する評価工程と、を含むものである。
前記学習工程、検出工程及び評価工程以外の工程は、必要に応じて組み入れられる。
【0015】
本発明において、「動物が正常な学習能力を有する状態である(あった)」又は「動物が正常な学習能力を有する状態でない(なかった)」と評価・判断する際の根拠は、
「正常な学習能力を有することを予め別の方法で確かめている動物が、本発明の学習工程における学習行動を行うことにより記憶を蓄積すること」と「当該学習行動に伴って信号が入力される神経細胞においてCapZが一部のスパインに局在すること」が高い相関を有すること、及び
「学習障害を呈し、正常な学習能力を有さないことを別の方法で予め確かめている動物が、本発明の学習工程における学習行動を行うことにより記憶を蓄積しないこと」と「正常である場合には、当該学習行動に伴って信号が入力される神経細胞においてCapZが一部のスパインに局在しないこと」が高い相関を有すること、
を本発明者が見出したことである。
【0016】
本明細書及び特許請求の範囲において「CapZが一部のスパインに局在している」とは、学習行動を行うことに伴って信号(活動電位)が入力される脳部位において、当該脳部位に含まれる一部のスパインにおけるCapZの濃度が、そのスパインを持つ樹状突起本体におけるCapZの濃度よりも高く、且つ、当該神経回路に含まれるスパイン間で、CapZの濃度に明らかな偏りがあることを意味する。通常、CapZの濃度は、CapZに由来するシグナル強度に正の相関をもって反映される。
【0017】
本発明における非ヒト科動物としては、学習行動を行わせて、当該学習行動に関する記憶を蓄積しうる動物であれば特に制限されず、例えば、マウス、ラット、モルモット、ウサギ等のげっ歯類、サル、オランウータン等の霊長類、ブタ、ウシ、ウマ、イヌ、ネコ、イルカ等の哺乳類、インコ、ジュウシマツ等の鳥類、コイ、ゼブラフィッシュ等の魚類などが挙げられる。これらの中でも、比較的高等な学習行動を学んで記憶できる霊長類、哺乳類およびげっ歯類が好ましく、飼育が容易であり、医薬の候補化合物のスクリーニングにも用いられるげっ歯類がより好ましい。
前記動物として、内在性又は外来性のCapZを予め導入した動物を用いることができる。前記導入した動物については、後で詳述する。
なお、ヒトにおいても、本発明の評価方法を適用することは原理的には可能であるが、安全性及び倫理的な問題について、十分に検討がなされるべきである。
【0018】
[学習工程]
本発明の評価方法における学習工程は、前記動物に学習行動を行わせる工程である。
前記学習行動は、動物に刺激を与えることにより、当該動物がその刺激を記憶できるものであれば特に制限されない。例えば、動物を一定の条件下に置いたとき、当該動物に快適な又は不快な体性感覚を与えることによって、当該動物が前記一定の条件下に置かれた状況と前記体性感覚とを連合学習して、両者が関連すると理解できるようにする「条件付け」が挙げられる。前記「条件付け」を利用した従来方法としては、記憶・学習にかかる研究で用いられてきた「文脈依存的恐怖条件付け学習」が挙げられる。
【0019】
従来の「文脈依存的恐怖条件付け文脈学習」の代表的な実施方法としては、電気ショック(無条件刺激)と動物が居る場所から得られる視覚刺激などの中立刺激(条件刺激)を組み合わせて条件付けを成立させた動物が、再び電気ショックを与えられた場所に置かれた際に表す恐怖反応(すくみ行動)を、文脈学習として評価する方法である。具体的には,電気グリッドを設置したチャンバー内に動物をいれ、痛みを感じる電気刺激を短時間与える。条件付けの一定時間 (1 時間、24 時間など)後に、電気刺激のない条件付けチャンバー内でのすくみ行動を2〜5分間測定し、全測定時間に対するすくみ行動時間の百分率(%)を算出し、これを学習能の指標とする
【0020】
本発明の評価方法において、動物に学習行動を行わせる場合、当該学習行動が当該動物に記憶されたか否かは、「学習工程」の段階では評価・判定しない点が、従来の方法とは相違する。つまり、従来の文脈依存的恐怖条件付け学習においては、特定条件(条件刺激)と電気刺激とを組み合わせて提示した後、所定の時間後に再び特定条件を提示した場合の当該動物の反応(すくみ行動)を解析する。一方、本発明の学習工程において文脈恐怖条件付けを行う場合、前記特定条件と電気刺激とを組み合わせて提示する方法は同様に行うが、所定の時間経過後に再び特定条件を提示して当該動物の反応を解析することは行う必要はない。
すなわち、本発明の評価方法は、学習工程では、当該学習が成立して、当該動物がその学習行動に関する記憶を蓄積したか否かは評価・判定せず、後段の検出工程において、当該動物の脳の状態を直接観察し、評価工程において、当該動物がその学習行動に関する記憶を蓄積したか否かを評価・判定する。
したがって、本発明の学習工程において、動物に行わせる学習行動は、従来から用いられ、その学習行動が当該動物によって記憶されうることが予め確認されている学習行動が好ましい。そのような学習行動としては、例えば、Y字型迷路試験、ロータロッド試験、恐怖条件付け学習試験、水探索試験、新奇物質探索試験、受動回避試験、放射状迷路試験、Morris水迷路試験及び遅延見本見合わせ・非見本見合わせ試験等の従来方法における、特定条件と特定刺激の提示にかかる部分の方法が、好適なものとして例示できる。
【0021】
[検出工程]
本発明の評価方法における検出工程は、前記動物の脳の神経細胞におけるCapZの局在状態を調べる工程である。
前記CapZの局在状態を調べる方法としては、前記学習工程を経た動物の脳を所定の時間後に直接観察するか、化学的に固定して摘出し、前記学習行動に関連する脳部位を含む切片を作製して、CapZの分布を調べる方法が好適である。
【0022】
前記所定の時間は特に制限されず、学習行動に関する記憶が脳部位に蓄積されると考えられる時間とすればよい。
前記学習行動が恐怖条件付けであり、当該動物の海馬を解析する場合、前記所定の時間は4時間〜72時間が好ましく、8時間〜48時間がより好ましく、12時間〜24時間が更に好ましい。
上記範囲であることにより、当該動物の海馬に学習行動に関する記憶が蓄積されると考えられ、当該記憶と相関の高い現象としてCapZの局在を検出することができる。上記範囲の下限値未満の時間で脳を調べると、後段の評価工程における当該シグナルの解釈が困難になる場合がある。記憶は海馬で数日〜数週間は保持されると考えられるが、上記範囲の上限値を超えた時間で調べると、別の新しい記憶が上書きされることによって、当該学習依存のCapZの局在を示すシグナルを検出できなくなる恐れがある。
【0023】
前記脳を直接観察する方法、化学的に工程し摘出する方法及び脳の切片を作製する方法は公知の方法が適用できる。
脳内に多数存在する部位のうち、どの部位を調べるかは特に制限されない。学習工程で動物に行わせる学習行動と関連性の高い脳部位の状態を調べることが好ましい。例えば、空間認知に関する学習の記憶は、脳における海馬に蓄積されることが報告されている。従って、空間認知に関する学習行動を行わせた動物の脳における海馬を調べることによって、当該学習行動に関する記憶の蓄積と相関する現象を捉えることができる。
本発明の評価方法において、記憶の蓄積と相関する現象は、当該脳部位若しくは神経細胞を構成するスパインの一部に、CapZと呼ばれるタンパク質が局在する現象である。
【0024】
CapZは、F-アクチン キャッピング プロテイン(F-actin capping protein)とも呼ばれる約270〜290アミノ酸残基のタンパク質であり、当初、F-アクチンに結合する機能を有するタンパク質として同定された。その後、本発明者らによって、CapZが神経細胞のシナプス可塑性に関与している可能性が検討された。その結果、本発明者は、動物が学習行動を行うことにより記憶を蓄積する事象と、当該学習行動に伴って発火する神経細胞においてCapZの少なくとも一部が、スパインの一部に局在する事象と、が高い相関を有することを見出した。
【0025】
動物の臓器別に、異なるアイソタイプのCapZが存在する場合があり、これらのアイソタイプがホモ複合体又はヘテロ複合体を形成して機能している。例えば、マウスにはCapZ beta1、CapZ beta2及びCapZ alphaの少なくとも3種類のアイソタイプが確認されている。このうち、主にCapZ beta2及びCapZ alphaがマウスの脳内に発現している。
【0026】
本発明におけるCapZとしては、当該動物の内在性CapZであってもよいし、当該動物に遺伝子導入若しくはタンパク質導入された外来性CapZであってもよい。
前記外来性のCapZは、神経細胞において、信号の入力依存的にスパインに局在する機能及び/又はF-アクチンに結合する機能を有するものであれば特に制限されず、同種の動物に由来するCapZであってもよいし、異種の動物に由来するCapZであってもよい。また、同種若しくは異種の動物に由来するCapZのアミノ酸配列において1若しくは数残基のアミノ酸が欠失・置換・付加された変異型CapZであってもよい。さらに、CapZにマーカータンパク質及び/又はペプチドタグを結合させた融合タンパク質を、外来性CapZとして導入してもよい。
前記外来性のCapZは、由来する動物において脳内に発現しているものが好ましい。また、異種の動物に由来するCapZを導入する場合、その異種の動物は、導入する動物に近縁であるものが好ましい。
【0027】
動物からクローニングされたCapZのcDNA情報は、NCBIのデータバンクUniGeneで一括して見ることができる。CapZ alphaのID番号はUGID:905681であり、CapZ beta2のID番号はUGID:256943である。このデータバンク及びリンクされたデータバンクで検索できるCapZを前記外来性のCapZとして導入することが可能である。
【0028】
ほ乳類に属する動物間でのCapZのアミノ酸配列の相同性は大変高い。例えばヒトとマウス間で比較したアミノ酸配列の相同性は、サブユニットalphaでは98%、サブユニット betaでは100%である。このため、ほ乳類内では、異種の動物由来のCapZが遺伝子導入された動物において、導入されたCapZは機能すると考えられる。
【0029】
前記外来性又は内在性のCapZとして、マウス(Mus musculus)由来のCapZを用いる場合、当該CapZは、
配列表の配列番号1で表される272アミノ酸残基のタンパク質(CapZ beta2);
配列表の配列番号2で表される286アミノ酸残基のタンパク質(CapZ alpha);
配列番号1若しくは配列番号2のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換又は付加されたアミノ酸を有し、且つ神経細胞への入力依存的にスパインに局在する機能及び/又はF-アクチンに結合する機能を有するタンパク質;
配列番号1若しくは配列番号2のアミノ酸配列と相同性が90%以上であり且つ神経細胞への入力依存的にスパインに局在する機能及び/又はF-アクチンに結合する機能を有するタンパク質、が好ましい。
【0030】
配列番号1のアミノ酸配列(CapZ beta2)は、GenBank Accession No.NM_009798で登録されているMus musculus capping protein (actin filament) muscle Z-line, beta (Capzb), transcript variant 2, mRNAと同一の配列である。配列番号2のアミノ酸配列(CapZ alpha)は、GenBank Accession No. NM_009797.2で登録されているMus musculus capping protein (actin filament) muscle Z-line, alpha 1 (Capza1), mRNAと同一の配列である。
【0031】
前記外来性又は内在性のCapZとして、ラット(Rattus norvegicus)由来のCapZを用いる場合、当該CapZは、
配列表の配列番号3で表される272アミノ酸残基のタンパク質(CapZ beta);
配列番号3のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換又は付加されたアミノ酸を有し、且つ神経細胞への入力依存的にスパインに局在する機能及び/又はF-actinに結合する機能を有するタンパク質;
配列番号3のアミノ酸配列と相同性が90%以上であり且つ神経細胞への入力依存的にスパインに局在する機能及び/又はF-actinに結合する機能を有するタンパク質、が好ましい。
【0032】
配列番号3のアミノ酸配列(CapZ beta)は、GenBank Accession No.NM_001005903.1で登録されているRattus norvegicus capping protein (actin filament) muscle Z-line, beta (Capzb), mRNAと同一の配列である。
【0033】
前記外来性又は内在性のCapZとして、ヒト由来のCapZを用いる場合、当該CapZは
配列表の配列番号4で表される272アミノ酸残基のタンパク質(CapZ beta1);
配列表の配列番号5で表される286アミノ酸残基のタンパク質(CapZ alpha1);
配列番号4若しくは配列番号5のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換又は付加されたアミノ酸を有し、且つ神経細胞への入力依存的にスパインに局在する機能及び/又はF-アクチンに結合する機能を有するタンパク質;
配列番号4若しくは配列番号5のアミノ酸配列と相同性が90%以上であり且つ神経細胞への入力依存的にスパインに局在する機能及び/又はF-アクチンに結合する機能を有するタンパク質、が好ましい。
【0034】
配列番号4のアミノ酸配列(CapZ beta1)は、GenBank Accession No.NM_004930.3で登録されているHomo sapiens capping protein (actin filament) muscle Z-line, beta (CAPZB), transcript variant 1, mRNAと同一の配列である。配列番号5のアミノ酸配列(CapZ alpha1)は、GenBank Accession No.NM_006135.2で登録されているHomo sapiens capping protein (actin filament) muscle Z-line, alpha 1 (CAPZA1), mRNAと同一の配列である。
【0035】
作製した脳の切片標本において、どの部位にCapZが局在しているかを調べる方法は特に制限されず、従来公知の方法を適用できる。例えば、CapZに選択的(特異的)に結合する抗体を用いた免疫染色法が適用できる。当該抗体は市販のものを購入して用いても良いし、常法に従って抗CapZ抗体を作製して用いても良い。
【0036】
CapZが神経細胞のどの部位に分布しているかを調べるために、神経細胞を構成する構造体を染色することが好ましい。例えば、抗MAP2抗体を使用して当該切片を染色すると、MAP2が存在する樹状突起の位置、形状及び輪郭を観察することができる。また、ローダミン−ファロイジン(Rhodamine-phalloidine)を使用して当該切片を染色すると、F-アクチンを含むスパインの位置、形状及び輪郭を観察することができる。したがって、当該切片の観察視野において、樹状突起およびスパインの位置や大きさ等を確認し、同じ視野におけるCapZ(融合タンパク質)に由来するシグナルを検出することによって、CapZが神経細胞のどの部位に分布しているか、又はCapZが神経細胞のどの部位に局在しているか、を調べられる。
【0037】
また、より好適な別の方法として、CapZを遺伝子導入したトランスジェニック動物(以下、CapZ-TG動物ということがある。)の使用が例示できる。
前記CapZ-TG動物としては、CapZとマーカータンパク質及び/又はペプチドタグとを結合させた融合タンパク質をコードする遺伝子が導入されたものが好ましい。
前記融合タンパク質は、マーカータンパク質又はペプチドタグが結合されているので、それを利用すると、検出がより容易となるので好ましい。なお、前記CapZ-TG動物は、常法によって作製できる。
【0038】
学習工程を経た動物の脳の切片において、CapZに選択的に結合した一次抗体を検出する方法、及び前記マーカータンパク質又はペプチドタグを検出する方法は特に制限されず、公知方法が適用できる。例えば、前記一次抗体に結合する蛍光標識付きの二次抗体を使用し、蛍光顕微鏡で検出する方法や、前記マーカータンパク質が発する蛍光を顕微鏡で検出する方法が挙げられる。これらの方法で検出されるシグナルは、当該切片におけるCapZの分布を示すものである。
【0039】
前記CapZ-TG動物に導入するCapZとして、前述の外来性CapZを適用できる。当該CapZは、同種の動物からクローニングされたCapZであってもよいし、異種の動物からクローニングされたCapZであってもよい。例えば、マウスに導入するCapZは、マウス由来のCapZであってもよいし、ヒト、ラット、ウサギ等の別種のほ乳類に由来するCapZであってもよい。ここで、導入するCapZは、神経細胞において、信号の入力依存的にスパインに局在する機能及び/又はF-アクチンに結合する機能を有するものである。この機能を有する限り、導入するCapZのアミノ酸配列において1若しくは数残基のアミノ酸の欠失・置換・付加が行われていてもよい。
【0040】
前記マーカータンパク質は特に制限されず、従来公知のマーカータンパク質が適用可能であり、例えばアルカリフォスファターゼ、HRP、GFP、EGFP等が挙げられる。
前記ペプチドタグは特に制限されず、従来公知のペプチドタグが適用可能であり、例えばHAタグ、Mycタグ、Hisタグ、FLAGタグ、GSTタグ等が挙げられる。
前記マーカータンパク質に由来するシグナル、及び/又は前記ペプチドタグに特異的に結合する抗体に由来するシグナルを検出することにより、前記CapZを含む融合タンパク質の局在を調べることがより容易となるので好ましい。
【0041】
前記CapZ-TG動物を用いる場合、学習行動を行わせたことに対する反応を増幅するために、当該CapZ-TG動物における前記融合タンパク質の発現を、前記学習行動を行わせた期間に対応する、できるだけ狭い時間的な幅で誘導させることが好ましい。
具体的には、前記融合タンパク質の発現を特定のプロモーターによって制御することが好ましい。前記プロモーターとして、神経活動入力依存的に発現誘導されるArcプロモーター、c-Fos プロモーター等が好適なものとして例示できる。また、所望のタイミングで発現を制御できる、テトラサイクリン誘導プロモーター、cre依存性プロモーター構造なども好適なものとして例示できる。
【0042】
例示した中でも、Arcプロモーターがより好ましい。Arcプロモーターを用いることによって、例えばCapZを遺伝子導入したトランスジェニックマウス(以下、CapZ-TGマウスと呼ぶことがある。)に空間認識に関する学習行動を行わせた場合、当該CapZ-TGマウスの脳における海馬のCA1野の放線層に属する神経細胞に信号が入力され、当該入力に伴って、前記融合タンパク質を当該神経細胞において発現させることができる。すなわち、学習行動に関する記憶を蓄積すると考えられる神経細胞において、融合タンパク質の発現が当該学習行動を行わせることを引き金としてなされる。このメカニズムを利用して、当該学習行動に関する記憶の蓄積の際、新たに発現した融合タンパク質の分布を、もともと存在していた内在性のCapZと区別することができる。つまり、当該学習行動を行わせる以前に蓄積されていた記憶に関するCapZと、当該学習行動を行わせたことにより発現したEGFP-CapZ(融合タンパク質)とを区別することが可能となる。この結果、新たに発現した融合タンパク質の分布が、当該学習行動に関する記憶の蓄積におけるCapZの、学習マーカーとして果たす役割をよりよく反映していると考えられる。
【0043】
[評価工程]
本発明の評価方法における評価工程は、前記CapZの局在状態の観察に基づき、前記脳の状態を評価する工程である。
評価工程において、検出工程で検出したCapZに由来するシグナルの分布を解析して、前記CapZが一部のスパインに局在している場合、前記動物が正常な学習能力を有する状態である、又は前記動物が少なくとも学習工程において正常な学習能力を有する状態であった、と評価・判断する。
また、評価工程において、検出工程で検出したCapZに由来するシグナルの分布を解析して、前記CapZが一部のスパインに局在していない場合、前記動物が正常な学習能力を有する状態ではない(異常な状態である)、又は前記動物が少なくとも学習工程において正常な学習能力を有する状態ではなかった(異常な状態であった)、と評価・判断する。
【0044】
本明細書では、前記学習行動を行うことに伴って信号が入力される脳部位に含まれるスパインのうち、CapZのシグナルが強く検出されたスパインの割合を「CapZのスパイン局在率」又は単に「局在率」と呼ぶ。局在率は、脳切片を観察した画像全体から、ファロイジン染色により定義される各スパイン内の、CapZ強度を取得して、その強度分布(ヒストグラム)が最尤値付近にまとまらず、突出して高い値を有する強度が分布に占める割合で規定できる。
【0045】
観察した神経細胞及び脳部位において、スパインは多数存在することが普通である。ここで、「CapZの一部がスパインに局在する」とは、複数のスパインの全てにおいてCapZの濃度が高まるのではなく、一部のスパインにおいてのみCapZの濃度が高まることを意味する。
最近の研究では、記憶に関する神経細胞のシナプス可塑性の変化は、一部の神経細胞にのみ起きることが示唆されている。これと符号するように、本発明者の検討によれば、正常な学習能力を有する野生型のマウスに学習行動を行わせて24時間経過した脳の海馬においては、特定の神経回路をなす神経細胞群が有する全スパインのうち、1〜5%のスパインにのみCapZのシグナルが強く検出されている。
【0046】
本発明において、当該動物が正常な学習能力を有するか否かを局在率に基づいて判定することができる。具体的には、動物に学習行動を行わせた後、所定時間経過時の脳の切片において、学習行動に関する記憶が形成されると考えられる脳部位の神経細胞群が有するスパインのうち、所定割合のスパインにのみCapZのシグナルが強く検出された場合、すなわち局在率が高い場合に、当該動物は正常な学習能力を有する、と判定できる。
前記所定時間としては、4時間〜72時間が好ましく、8時間〜48時間がより好ましく、12時間〜24時間がさらに好ましい。この範囲であると、学習行動に関する記憶が神経細胞に蓄積(形成)されたことをより確実に判定できる。
前記局在率としては、0.01〜10%が好ましく、0.1〜10%がより好ましく、0.1〜5%がさらに好ましい。この範囲であると、学習行動に関する記憶が蓄積された場合(正常な学習能力を有する場合)と蓄積されていない場合(正常な学習能力を有さない場合)とをより確実に区別して判定できる。
【0047】
本発明の評価方法において、前記動物として、疾患の状態を呈する疾患モデル動物を使用できる。前記疾患としては、ダウン症、統合失調症、認知症、公知の学習障害を呈する疾患又は公知の精神疾患が挙げられる。このような疾患モデル動物は、公知のものを適用できる。
また、前記疾患モデル動物として、前記融合タンパク質をコードする遺伝子が導入されたCapZ-TG動物を疾患モデル動物となるように加工して用いてもよい。この疾患モデル化CapZ-TG動物を用いることにより、当該動物中のCapZを検出することがより容易となるので好ましい。以下では、「疾患モデル動物」は、特に明記しない限り「疾患モデル化CapZ-TG動物」を含むものとする。
【0048】
前記疾患モデル動物の作製方法は特に制限されず、例えば、受精卵に当該疾患関連遺伝子を導入する方法、当該疾患関連遺伝子を欠損または置換したノックアウトマウスを常法により作製する方法、疾患関連遺伝子をウイルスベクターや電気穿孔法などで神経細胞に導入する方法、脳に作用し疾患様症状を呈する薬物を投与する方法、脳に部分的に損傷を与える方法、疾患様症状を誘導する強度のストレスを与える方法などが挙げられる。
【0049】
前記疾患関連遺伝子としては、例えばダウン症、統合失調症、認知症、公知の学習障害を呈する疾患又は公知の精神疾患に関連する遺伝子が挙げられる。
前記疾患モデル動物は、当該疾患の症状又は当該疾患に近い症状を呈する。また、前記疾患モデル化CapZ-TG動物は、当該疾患の症状又は当該疾患に近い症状を呈し、且つ前記融合タンパク質が導入された動物である。
例えばダウン症に関連する遺伝子を含む染色体が導入された疾患モデルマウスは、学習障害を呈することが知られている。
疾患モデル動物と健康な動物とを用いることによって、当該疾患に関連する学習障害若しくは精神疾患を呈する脳の状態と、正常な学習能力を有する脳の状態とを、CapZ(融合タンパク質)の分布の違いに基づいて比較検討することができる。
【0050】
<医薬のスクリーニング方法>
本発明の医薬のスクリーニング方法は、学習障害若しくは精神疾患を呈する前記疾患モデル動物に対して、被検物質を投与する工程と、前記動物に学習行動を行わせる学習工程と、前記動物の脳の神経細胞におけるCapZの局在を調べる検出工程と、前記CapZの局在に基づき、前記脳の状態を評価する評価工程と、を含み、前記評価工程において、前記CapZが一部のスパインに局在している場合、前記被検物質を、前記学習障害若しくは精神疾患を治療又は予防する医薬として有用であると判断する方法である。
【0051】
本発明の医薬のスクリーニング方法における学習工程は、予め動物に被検物質を投与することを除いて、前述の本発明の評価方法における学習工程と同様に行えばよい。
前記動物は、学習障害若しくは精神疾患を呈する前述の疾患モデル動物が好ましい。
【0052】
医薬のスクリーニング方法に用いる疾患モデル動物としては、前記学習行動を行わせても当該学習行動に関連する記憶が正常に蓄積されないこと又は学習記憶が正常に成立しないこと、が予め別の方法で示唆又は確認されているものが好ましい。さらに、前記疾患モデル動物としては、前記学習行動を行わせても、学習記憶に関連する脳部位においてCapZが正常に局在しないことが、予め確認されているものが好ましい。
また、前記疾患モデル動物としては、前記学習行動を行わせても、当該学習行動に関連する記憶が正常に蓄積されないこと又は学習記憶が正常に成立しないこと、が予め別の方法で示唆又は確認されていて、且つ同じ学習行動を行わせた場合、その学習記憶に関連する脳部位においてCapZが正常に局在しないことが、予め確認されているものがより好ましい。ただし、本方法においては、従来の学習試験での効果の判定とは違う結果となる可能性がある。例えば、恐怖条件付けのテストの際、被験物質での腹痛のため動けない動物は記憶していなくてもすくみ反応をしたと誤解されて解釈される場合や、恐怖を記憶しているにも関わらず、錯乱して動いてしまい(すくみ反応を示さず)、記憶していないと判断される場合など、従来の記憶試験では判断が誤ってしまうケースにおいても、本発明の医薬のスクリーニング方法によれば、正確に判断できる可能性があることに十分留意する。記憶など認知の障害が、体のだるさ(倦怠感)等の他の症状にマスクされがちな統合失調症や鬱病のモデルの場合には特に留意する。
【0053】
前記被検物質としては、疾患モデル動物に対して毒性が低いものが好ましく、さらに当該疾患を改善すると見込まれるものがより好ましい。
前記被検物質の投与量および投与方法は特に制限されず、薬理学的に許容される投与量で公知の方法で投与すればよい。
【0054】
本発明の医薬のスクリーニング方法における検出工程は、前述の本発明の評価方法における学習工程と同じ様に行えばよい。前記疾患モデル化CapZ-TG動物を用いると、CapZに結合されたマーカータンパク質又はペプチドタグを利用することにより、CapZの検出がより容易となるので好ましい。
【0055】
本発明の医薬のスクリーニング方法における評価工程は、前記CapZの局在に基づき、前記被検物質が投与された疾患モデル動物の脳の状態を評価する工程である。
【0056】
前記評価は、前述の本発明にかかる評価方法と同様に行うことができる。CapZ-TG動物を含む健康な動物が示す、当該CapZの局在状態(一部のスパインに局在する)を正常の指標とし、被検物質を投与する前の疾患モデル動物が示す当該CapZの局在状態(一部のスパインに局在しない)を異常の指標とし、被検物質を投与した疾患モデル動物が示す当該CapZの局在状態が異常から正常へ近づいた場合、又は被検物質を投与した疾患モデル動物が示す当該CapZの前記局在率が上昇した場合、当該疾患モデル動物の異常であった学習能力が正常に回復した若しくは正常に近づいたと評価できる。
この評価に基づき、当該被検物質を、当該疾患モデル動物が呈する学習障害若しくは精神疾患を治療又は予防する医薬として有用であると判断できる。
【0057】
医薬のスクリーニング方法の評価工程において、前段の検出工程で検出した前記CapZが一部のスパインに局在している場合、当該疾患モデル動物の異常であった学習能力が正常に回復した若しくは正常に近づいたと評価し、前記被検物質を、当該疾患モデル動物が呈する学習障害若しくは精神疾患を治療又は予防する医薬として有用であると判断する。
また、当該評価工程において、前段の検出工程で検出した前記CapZの前記局在率が上昇した場合、当該疾患モデル動物の異常であった学習能力が正常に回復した若しくは正常に近づいたと評価し、前記被検物質が有する、当該疾患モデル動物が呈する学習障害若しくは精神疾患を治療又は予防する作用効果の程度を判断することもできる。
【0058】
<本発明と、従来の記憶に関する研究との関係>
記憶などの脳の高次機能を発揮するためには、各神経細胞は個性を持ち、役割を分担して働く必要がある。もし、役割分担をせずに大多数の神経細胞が同期して活動した場合は、てんかん発作のような痙攣を起こすはずである。しかし、記憶に関する従来の研究では、どの神経細胞のどの部分が、どのように変化して働いているのかを解析する手法はなかった。
【0059】
脳の一部に人工的に長期増強を起こす刺激をした後に長期増強部位にCapZの濃度(免疫組織染色強度)が上昇すること、全てのスパインに均一に存在するのではなく染色強度が高いスパインが一部にあること、はすでに発明者らが下記の論文(6)で報告している。
Genes Cells. 2010 Jun;15(7):737-47. Epub 2010 Jun 10. “Activity-dependent localization in spines of the F-actin capping protein CapZ screened in a rat model of dementia.” Kitanishi T, Sakai J, Kojima S, Saitoh Y, Inokuchi K, Fukaya M, Watanabe M, Matsuki N, Yamada MK.
しかし、この論文の実験では二つの問題点があった。1.刺激が人工的であるため、学習に関連した入力と同じである保証がない。2.免疫組織染色の場合、抗体の浸透性の問題があり、とりわけシナプス後部では存在しても染色されない分子があることが知られている。そこで、本発明者は、免疫染色とは異なる方法として、CapZとEGFPを融合タンパク質として発現するトランスジェニックマウス(CapZ-TGマウス)を作製し、脳の自然な活動に伴ってCapZの局在を検出しうることを見出した。具体的には、そのCapZ-TGマウスに学習行動を行わせ(文脈依存恐怖条件づけを行い)、記憶が固定されているはずの24時間後に脳切片を作成して、文脈と恐怖の連合記憶に必要な背側海馬を観察すると、学習行動を行わせていない動物の脳に比べて特徴的なパターンが現れる。それは、背側海馬CA1野の一部のスパインに、他と比べて正常な分散範囲を超えた(目立って高い)EGFP強度をもつものがあるパターンである。また、そうしたスパインを多数持つ樹状突起があることも分かった。この、EGFP-CapZ融合タンパク質発現トランスジェニックマウスでの結果により、免疫組織染色での問題点であった、抗体の浸透性からみかけだけスパイン間の存在量に差が出た可能性は否定された。
【0060】
そもそも、神経細胞間での情報の伝達は、少数の神経細胞によって担われていると考えられる。その根拠として、神経細胞の活動電位の発生について、どのような学習行動を行った場合でも最大10%の神経細胞しか活動しないと考えられる実験事実がある(Henze et al. JNP)。そのような少数の神経活動による情報処理を前提とすると、機能的な神経回路では情報入力が、川をながれる水のように、ある部分だけを伝播する状態が形成される必要がある。そして、学習に関する記憶が蓄積されるためには、神経のつながりにおいて一部が周りと異なった状態となって、新しく入力を受けやすい部分、水の流れやすくなる部分が生じる必要がある。その変化は「少数」でないと拡散してしまう。
発明者らの過去の実験から、人工的に長期増強を起こす刺激をした実験において長期増強部位(神経伝達効率が上がった部位)にCapZの濃度が上昇したことがわかっている。そこから類推すると、EGFP-CapZ強度の高いスパインには長期増強が起きてcoding(記憶情報の流れやすい部位の形成)をしている可能性が高い。また、そのスパインが集積している樹状突起があれば神経細胞はcoding neuronとして働きうる。本発明者が見出したEGFP-CapZ強度の高いスパインが出現する頻度は少数であり、特定の記憶に関連して活動する神経細胞が少数であることから考えて妥当である。
(参考文献;J Neurophysiol. 2000 Jul;84(1):390-400. “Intracellular features predicted by extracellular recordings in the hippocampus in vivo.” Henze DA, Borhegyi Z, Csicsvari J, Mamiya A, Harris KD, Buzsa’ki G.)
本発明の評価方法においては、正常な分散範囲を超えて、CapZが一部のスパインに局在する局在率が高い場合に、学習ができた脳と定義して、脳機能の評価に用いる。
【実施例】
【0061】
<実施例1>
[非ヒト科動物の準備]
非ヒト科動物として、トランスジェニックマウス(以下、CapZ-TGマウスと呼ぶ。)を準備して用いた。CapZ-TGマウスの作製は、以下の方法でEGFPを融合した CapZを発現させるために行った。導入遺伝子は、遺伝子組み換えにより、以下の(1)〜(8)のDNA配列又はアミノ酸配列をコードするDNA配列を順に連結して作製した(図1)。
(1)mouse Arc promoter (SmaI-NaeI断片;7.1kb)
(2)Tetracyclin-responsive transcriptional activator (Clontech社;pTet-off由来)
(3)P2A配列(TSATNFSLLKQAGDVEENPGPVAT;人工合成)
(4)EGFP (Clontech社;pEGFP-N1由来)
(5)Spacer(SGRTQISSS;人工合成)
(6)HAtag(YPYDVPDYA;人工合成)
(7)mouse CapZ beta 2 完全長272a.a.(Met-Cys)(RT-PCRにより単離;Genbank Accession No. NM_009798;Mus musculus capping protein (actin filament) muscle Z-line, beta (Capzb), transcript variant 2, mRNA)
(8)SV40-polyA(Clontech社;pEGFP-N1由来)
【0062】
(1)のマウスArcプロモーターは、マウスゲノムの該当部分(Ch15、 74.50M-74.51Mの間)を含むBACクローン(B6Ng01-232C07など)をBACライブラリ(http://dna.brc.riken.jp/ja/MSMbac.htmlなど)から入手し、SmaI-NaeI消化物の7.1kb付近からクローニングできる。
(3)のP2A配列は、異なる二つのたんぱく質(この場合は(2)と(4)-(7)融合タンパク質)を一連のcDNAから等量合成するためのシグナルである(Donnelly et al. J Gen Virol 2001, 82:1013-1025.)。なお、(2)は、本CapZ-TGマウスで将来別の遺伝子発現をする場合に備えた転写因子である。
(7)CapZ はalphaとbetaの二量体でbetaには1と2のSplicing Variantがある。脳ではbeta2が主要であるため、これを用いている。
【0063】
上記の作製した導入遺伝子を、制限酵素を用いてベクター部分を除きつつ直線化したのち、フェノールクロロフォルム法及びエタノール沈殿により精製した。さらに、ゲル電気泳動を行い、ベクター部分を含まないよう該当長のバンドを切り出して、キアゲン社のgel extraction kitを用いた精製を二回繰り返して高度に精製した。
精製した導入遺伝子を受精卵の核に注入し、正常に発生した胚を借り腹に移植した。産仔42匹のうち、CapZ-TGマウスとして7lineが同定された。各lineの導入コピー数は(1,3,10,10,10,10,100)であったため、実験には100コピー導入されたlineを用いた。
【0064】
作製したCapZ-TGマウスでは、Arcプロモーターが有るので、最近活動した神経細胞に選択的に、EGFP-HA-CapZ融合タンパク質を発現させることができる。
【0065】
[学習行動を行わせる工程]
作製したCapZ-TGマウスに、以下の方法で、学習行動として文脈恐怖条件付けを行った。
小原医科産業の文脈恐怖条件付け装置(ショックジェネレーター型番:SGA-2010、実験チャンバー:型番CL-3002)及び防音箱(型番CL-4210C)を使用した。まず、防音された装置のチャンバー内にマウスを5分間置いた。この5分間のうち、1分経過時に1回、4分経過時に1回、合計2回の電気ショック(0.2mA、2秒間)を、マウスの足に与えた。電気ショックの様子をカメラを通して観察し、確認した。5分の条件付けセッションの後、マウスを通常飼育しているケージに戻した。
この学習行動後において、電気ショックを受けたマウスが、「特定のチャンバーに入れられること」と「電気ショックを受けること」とを結び付ける文脈依存性恐怖条件を学習・記憶することが一般に知られているが、該当マウスでは記憶のテストはしていない。
この工程とは別に、24時間後にテストを行うことによって、当該CapZ-TGマウスが上記文脈恐怖条件を学習・記憶できることは、従来方法により別途確認している。すなわち、この確認において、CapZ-TGマウスと野生型マウスとで、正常な学習能力が発揮されることに有意差はなかった(図14参照)。テストでは常法に従い、ショックを与えたチャンバーに5分間マウスを置き、恐怖反応であるFreezingを起こしていた時間的割合(%)を記録した。記録はチャンバー上部のカメラから0.5秒おきに撮影した画像のマウス部分(黒)に一定のずれが生じないことが2秒続いた場合にFreezingとカウントした。上記小原医科産業の装置により画像を取得しPCに転送後、ソフトウェアTimeFZ1により自動測定した。
【0066】
[CapZ-TGマウスの脳の神経細胞におけるCapZの局在状態を調べる検出工程]
文脈恐怖条件付け後24時間経過したCapZ-TGマウスを麻酔し、開胸後30秒以内(通常10秒以内)に左心室に針を刺し、PBSを2ml、次いで2%パラホルムアルデヒドを含む0.1Mリン酸緩衝液(ともに氷冷しておいたもの)を20〜30ml(マウスの最高血圧を超えない約100mmHgで)全身灌流して固定し、脳を取出して正中で半割し、さらに1時間、4℃室で十分に振とうしながら後固定した。固定したマウスの脳をビブラトームで冠状断切片とした。以下の観察には空間学習に関わるとされる背側海馬(座標Bregma1.4mm-2.2mmの範囲)のCA1放線層、内側(lateral, 1.5mm以内)を用いた。以下ではCA1放線層の当該部位についての結果を示すが、CA1内の他の部位(網状分子層、多形層)、CA3においても同様の傾向が見られた。なお、固定処理によりタンパク質は分解・移動などの変化をしない状態になる。また蛍光タンパク質EGFPの蛍光はこの固定法では消失しない。
【0067】
得られた切片に、0.1%TritonX100で15分間の膜透過処理を行い、5%山羊血清で30分間の非特異的吸着を防ぐ処置をしたのち、1%山羊血清入りPBSで希釈した抗MAP2抗体の一次抗体(1:3000、chicken IgY、 Phosphosolutions社製;型番1100)を4℃で一晩振とうしながら反応させた。反応後、PBSで3回洗浄した。次に、二次抗体としてfluorescence(Alexa647)-labeled anti-chicken IgY(1:1000、 Invitrogen社製;型番A21463)を1%山羊血清入りPBSに希釈して、切片に反応させると同時にRhodamine-phalloidin (0.7 unit / ml, Invitrogen社製;型番R415)を加えて、室温で1時間または4℃で3時間反応させることにより、ファロイジンと抗MAP2抗体による二重染色を行った。
なお、抗体液は反応前に遠心(15,000×g、5分)し、上清部分を用いることで変性して凝集した抗体などによる擬染色を起こさぬよう注意を払った。
染色した切片をレーザー共焦点顕微鏡:Olympus社FV1000(倒立顕微鏡:IX81、対物レンズ:UPLSAPO 100×O NA1.40) を用いて、最も遅いスキャンスピードで観察し、出力を660V以内に抑えた光電子増倍管で画像を取得した。画像取得時は画像化できる最大強度を超えないよう、最も明るいサンプルにあわせてレーザー強度を調節した。その結果を図2〜10に示す。
【0068】
取得した画像の処理方法は次の通りである。
<画像処理法>:画像を処理する際は実質的に切り取った値がでないよう、比較する画像の中でもっとも幅の広い値にあわせて画像内ピクセル強度の最大値と最小値に基づいて強度を拡大表示した。例えば「0-255階調の8bit画像で 10-210、8-150の二枚の画像があるときは双方8-210を0-255に引き延ばす。」と、処理できる。なお、表示した図については原則的には原図を掲載し、4倍ズームで図を取得した図2〜6,11〜13については見易くするためにメディアンフィルターをかけた。
メディアンフィルターは測定器に由来するちらつきノイズを消すために一般的に用いられており、周囲の点の中間値を中央値に採用して突出した値をならす原理による。ここでは半径1ピクセルに設定したので3×3の点を用いていることになる。
【0069】
図2は、EGFP‐CapZのEGFP由来の緑色蛍光シグナルを示す。図2のように、学習行動を行わせた後のCapZ-TGマウスの海馬CA1野の放線層におけるEGFP‐CapZのシグナルは、一部に局在している。この図中のバーは2マイクロメーターであり、シグナルが検出された部分は、直径が約1マイクロメーター程度までで、大きさのばらつきがあることがわかる。
【0070】
図3は、図2と同じ視野においてRhodamine-phalloidinの赤色蛍光で染色されたスパインを示す。スパインは繊維状アクチンが多いため、Rhodamine-phalloidinで染色されると考えられている。脳の興奮性神経細胞の樹状突起上には、このスパイン(棘突起)と呼ばれる、頭部の直径が1マイクロメーター程度で、大きさのばらつきがある突起状の構造が密に存在することが知られる。スパインは、興奮性シグナルを受け取るシナプス後部がある部分である。神経細胞が記憶すべき情報を、あるスパインで受け取った場合、次なる想起刺激に対応すべく、そのスパインは他のスパインに比べて、より情報を伝え易くするように変化するはずである。すなわち、記憶形成に伴って、何らかの変化が起こる部分が、一部のスパインに存在しなければならない。
【0071】
図4の左は図2と同じEGFP‐CapZのシグナルを示し、図4の右は図3と同じRhodamine-phalloidin染色を示す。両者は、同一視野から別の蛍光シグナルとして、同時に取得した。矢印は同じ位置にあるので、EGFP‐CapZのシグナル(左)がRhodamine-phalloidin染色されたスパイン(右)のうちの一部だけに強く局在することが明確にわかる。
【0072】
図5は、MAP2が存在する樹状突起が、図2のEGFP-CapZのシグナルに重ねて示されている。粒状のEGFP-CapZのシグナルが、筋状の樹状突起に沿って、樹状突起から突出する棘のように存在している。この図は、EGFP‐CapZのシグナルがスパインにあることをさらに補強している。なお、樹状突起とスパインとの間に隙間があるように見えるものもあるが、これは、スパインのネック部分にはCapZが存在しないためであると考えられる。スパインにはマッシュルーム型のものがあり、ネックとよばれる細い首状の構造を持つ。ネックは、長いものでは2マイクロメーター(図5に示したバーの長さ)を超えるものもある。
【0073】
図6は、図5を解釈する際の参考とするために、図5と同じ視野において、EGFP-CapZのシグナルを除き、MAP2が存在する樹状突起のみを示している。丸く見える部分はスパインではなく、画面手前に向かう細い樹状突起の切断面と思われる。
【0074】
以上の観察から、文脈恐怖条件付けを行ったマウス脳の前記学習に関する脳部位の神経細胞では、EGFP-CapZの濃度が局所的に高い箇所があることが明らかである。その局所的に高い濃度でEGFP-CapZが主に存在する箇所は、Rhodamine-phalloidinで染色されているためスパインであると考えられる。さらに、当該脳部位に観察されるスパインの全てにおいてEGFP-CapZのシグナル強度が高くなるのではなく、一部のスパインにおいてのみEGFP-CapZのシグナル強度が高くなっている。つまり、学習行動を行わせた(文脈恐怖条件付けを行った)マウス脳では、CapZが一部のスパインに多く局在することが理解される。
前記脳部位に観察されるスパインのうち、EGFP-CapZのシグナル強度が顕著に高くなったスパインの割合(局在率)は、1〜5%であると見積もられた。
【0075】
図7は、図2〜5の脳と同じ条件で作製された標本である。すなわち、CapZ-TGマウスに学習行動を行わせた(文脈恐怖条件付けを行った)24時間後に固定した脳での海馬CA1野の放線層からのEGFP-CapZシグナルを示す。スケールバーは10マイクロメーターであり、広い範囲のシグナルをとっても、EGFP-CapZが存在する部分は疎であることがわかる。
【0076】
<比較例1>
上記CapZ-TGマウスを用いて、文脈恐怖条件付けを行わずに、実施例1と同じ方法でマウスを処理した。得られた切片を同様に観察した。その結果を図8〜9にと示す。
【0077】
図8は、学習行動を行わせなかったマウス脳の海馬CA1野の放線層におけるEGFP-CapZの分布パターンを示す。図9は、図8と同じ画像の全体の強度を下げた写真である。図8の強度を低くなるよう撮影しても、実施例の図7の写真のようにはならないことを示している。つまり、実施例1の図7と比較例1の図8〜9を比較することによって、図7では、図8〜9と異なり、「CapZが“一部の”スパインに局在」していることが明らかである。ここで「CapZが“一部の”スパインに局在する」とは、スパインに検出されるCapZに由来するシグナルの強度が樹状突起の幹の部分(スパイン以外)に比べて強いことを意味するだけでなく、同時に、同じ領域(特定の脳部位、ここでは空間記憶に関わる背側海馬)でのスパイン間で、当該シグナルの強度に偏りがあることを意味し、これが学習後の特徴と考えられる。なお、EGFPのシグナル強度は例えばpH(主に固定液の)や観察深度によっても変化するので、絶対強度の比較での議論は現時点では難しい。
【0078】
図10の左は、図7と同一視野でのファロイジン染色(スパイン染色)の写真であり、図10の右は図8と同一視野でのファロイジン染色(スパイン染色)の写真であり、両者間でスパイン密度には変化がないことがわかる。つまり、図7で検出されたCapZに由来するシグナルの偏りは、スパイン自体の消滅によるものではなく、各スパインごとのCapZの存在量が調節されたことによりCapZの濃度が異なっているためと考えられる。
【0079】
以上の観察により、学習行動を行わせなかった(文脈恐怖条件付けを行わなかった)マウス脳の神経細胞では、EGFP-CapZは全体のスパインに同様の濃度で存在して見え、濃度が他のスパインと比べて偏って高くなった一部のスパインは見出されなかった。
【0080】
学習行動を行わせた場合のEGFP-CapZのシグナルの分布が、上記のように一部のスパインを残して無くなって見える状態になる理由としては、一部のスパイン以外の領域に存在したCapZが分解されているためであると解釈できる。これを支持する知見として、CapZ分子に分解シグナルがあり、結合タンパクBAG3とHsc70が共存していないと、ユビキチン非依存にプロテアソームで分解されることが知られている。
(参考文献:Circ Res. 2010 Nov 12;107(10):1220-31. BAG3 and Hsc70 interact with actin capping protein CapZ to maintain myofibrillar integrity under mechanical stress.Hishiya A, Kitazawa T, Takayama S.)
【0081】
実施例1及び比較例1の結果から、学習行動を行わせたことによってマウス脳内に蓄積した記憶と、当該学習タスクによってマウス脳内に新たに発現したEGFP-CapZが一部のスパインに局在して検出されたこと(一部のスパインに偏って強く検出されたこと)は、高い相関を示している。すなわち、学習行動に伴ってEGFP-CapZが、その学習行動に関する神経細胞の一部のスパインに選択的に局在したことは、学習行動に関する記憶が当該スパインに記銘されたことを示していると考えられる。
【0082】
<実施例2>
CapZ-TGマウスの代わりに野生型のマウスを用いたこと、及びEGFPの蛍光の代わりに抗CapZ beta2抗体(1:300、mouse monoclonal IgG、The Developmental Studies Hybridoma Bank製;型番3F2.3-s, betaのうちValiant2にのみ存在するC末端配列部分に対する抗体)を抗MAP2抗体と同時に反応させ、その二次抗体としてfluorescence(Alexa488)-labeled anti-chicken IgG(1: 1000、Invitrogen社製;型番A11029)を検出に用いたこと以外は、実施例1と同じ方法で行った。その結果を図11に示す。なお用いた抗CapZ beta2抗体はimmunoblotting実験で脳のタンパク質内で非常に高い特異性を示すことは確認している(非特許文献4のsupplementary figure1で単一バンド)。
【0083】
図11において、白く示した領域が、抗CapZ抗体に由来するシグナルを示す。バーは1マイクロメーターであり、図2で見られたものとほぼ同様のサイズのシグナルが得られていることがわかる。なお、このシグナルの部位は図3〜6で示したのと同様に、一部のスパインに局在するものであることは確認している。この観察により、実施例1の図7と同じ結論が導かれた。すなわち、野生型で免疫染色を行う方法によっても、一部のスパインに強いCapZシグナルが見られることがわかった。
【0084】
<比較例2−1>
文脈恐怖条件付けを行わずに、実施例2と同じ方法でマウスを処理した。得られた切片を同様に観察した。その結果を図12に示す。
図12において、白く示した領域が、抗CapZ抗体に由来するシグナルを示す。この観察により、学習行動を行わせなかった(文脈恐怖条件付けを行わなかった)マウス脳では、実施例1の図8と同様に比較的均一なシグナルが観察される。つまりCapZが一部のスパインに局在している現象は見出されなかった。
【0085】
<比較例2−2>
ダウン症モデルマウス(ダウン症で3本になっているヒトの21番染色体に相同な、マウス16番染色体の一部を付加して遺伝的に部分的Trisomyにしている)、Ts1Cje mouseを用いて、実施例2と同様の文脈恐怖条件付けを行い、実施例2と同様に解析した。その結果を図13に示す。
使用したマウスは、作成者であるDr. C.J.Epsteinに許可を得た後、理化学研究所の山川和弘先生より分与いただいた。このマウスは、学習障害を呈することが以下の参考文献に報告されている。
(参考文献;Proc Natl Acad Sci U S A. 1998 May 26;95(11):6256-61. “Ts1Cje, a partial trisomy 16 mouse model for Down syndrome, exhibits learning and behavioral abnormalities.” Sago H, Carlson EJ, Smith DJ, Kilbridge J, Rubin EM, Mobley WC, Epstein CJ, Huang TT.)
【0086】
図13において、白く示した領域が、抗CapZ beta2抗体に由来するシグナルを示す。
この観察により、学習行動を行わせたにも関わらず、学習障害を持つダウン症モデルマウス脳では、CapZの濃度が全体に高めに測定されたが、各スパインで検出されるシグナルに偏りは少なく、一部のスパインにだけシグナルが強く検出されることはなかった。
【産業上の利用可能性】
【0087】
本発明の非ヒト科動物の学習能力の評価方法は、当該動物が学習能力を有するか否かの客観的な指標を提供できる。この評価方法を用いて、当該動物に投与した薬物が、学習能力に及ぼす影響を評価することができる。従って、学習障害や精神疾患の治療又は予防に有用な薬物のスクリーニングを行う際に、本発明の評価方法を利用できる。
【特許請求の範囲】
【請求項1】
非ヒト科動物が正常な学習能力を有する状態であるか否かを評価する方法であって、
前記動物に学習行動を行わせる学習工程と、
前記動物の脳の神経細胞におけるCapZの局在状態を調べる検出工程と、
前記CapZの局在状態の観察に基づき、前記脳の状態を評価する評価工程と、
を含むことを特徴とする非ヒト科動物の学習能力の評価方法。
【請求項2】
前記評価工程において、
CapZが一部のスパインに局在している場合、前記動物が正常な学習能力を有する状態であると評価することを特徴とする請求項1に記載の評価方法。
【請求項3】
前記非ヒト科動物が、CapZとマーカータンパク質及び/又はペプチドタグとを結合させた融合タンパク質をコードする遺伝子が導入された、トランスジェニック動物であり、
前記マーカータンパク質に由来するシグナル、及び/又は前記ペプチドタグに特異的に結合する抗体に由来するシグナルを検出することにより、CapZの局在を調べることを特徴とする請求項1又は2に記載の評価方法。
【請求項4】
前記トランスジェニック動物における前記融合タンパク質の発現を、
前記学習行動に伴って入力を受ける脳部位において、前記入力により誘導させることを特徴とする請求項3に記載の評価方法。
【請求項5】
前記動物が疾患モデル動物であることを特徴とする請求項1〜4の何れか一項に記載の評価方法。
【請求項6】
CapZが、以下の(i)又は(ii)のタンパク質であることを特徴とする請求項1〜5の何れか一項に記載の評価方法。
(i)配列番号1〜5の何れかで表されるアミノ酸配列を有するタンパク質。
(ii)配列番号1〜5の何れかで表されるアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列を有し、且つ神経細胞への入力依存的にスパインに局在する機能を有するタンパク質。
【請求項7】
学習障害若しくは精神疾患を呈する請求項5に記載の疾患モデル動物に対して、被検物質を投与する工程と、
前記動物に学習行動を行わせる学習工程と、
前記動物の脳の神経細胞におけるCapZの局在を調べる検出工程と、
前記CapZの局在に基づき、前記脳の状態を評価する評価工程と、
を含み、
前記評価工程において、
CapZが一部のスパインに局在している場合、前記被検物質を、前記学習障害若しくは精神疾患を治療又は予防する医薬として有用であると判断することを特徴とする医薬のスクリーニング方法。
【請求項1】
非ヒト科動物が正常な学習能力を有する状態であるか否かを評価する方法であって、
前記動物に学習行動を行わせる学習工程と、
前記動物の脳の神経細胞におけるCapZの局在状態を調べる検出工程と、
前記CapZの局在状態の観察に基づき、前記脳の状態を評価する評価工程と、
を含むことを特徴とする非ヒト科動物の学習能力の評価方法。
【請求項2】
前記評価工程において、
CapZが一部のスパインに局在している場合、前記動物が正常な学習能力を有する状態であると評価することを特徴とする請求項1に記載の評価方法。
【請求項3】
前記非ヒト科動物が、CapZとマーカータンパク質及び/又はペプチドタグとを結合させた融合タンパク質をコードする遺伝子が導入された、トランスジェニック動物であり、
前記マーカータンパク質に由来するシグナル、及び/又は前記ペプチドタグに特異的に結合する抗体に由来するシグナルを検出することにより、CapZの局在を調べることを特徴とする請求項1又は2に記載の評価方法。
【請求項4】
前記トランスジェニック動物における前記融合タンパク質の発現を、
前記学習行動に伴って入力を受ける脳部位において、前記入力により誘導させることを特徴とする請求項3に記載の評価方法。
【請求項5】
前記動物が疾患モデル動物であることを特徴とする請求項1〜4の何れか一項に記載の評価方法。
【請求項6】
CapZが、以下の(i)又は(ii)のタンパク質であることを特徴とする請求項1〜5の何れか一項に記載の評価方法。
(i)配列番号1〜5の何れかで表されるアミノ酸配列を有するタンパク質。
(ii)配列番号1〜5の何れかで表されるアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列を有し、且つ神経細胞への入力依存的にスパインに局在する機能を有するタンパク質。
【請求項7】
学習障害若しくは精神疾患を呈する請求項5に記載の疾患モデル動物に対して、被検物質を投与する工程と、
前記動物に学習行動を行わせる学習工程と、
前記動物の脳の神経細胞におけるCapZの局在を調べる検出工程と、
前記CapZの局在に基づき、前記脳の状態を評価する評価工程と、
を含み、
前記評価工程において、
CapZが一部のスパインに局在している場合、前記被検物質を、前記学習障害若しくは精神疾患を治療又は予防する医薬として有用であると判断することを特徴とする医薬のスクリーニング方法。
【図1】



【図2】

【図3】

【図4】

【図5】

【図6】

【図7】

【図8】

【図9】

【図10】

【図11】

【図12】

【図13】

【図14】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【公開番号】特開2013−40894(P2013−40894A)
【公開日】平成25年2月28日(2013.2.28)
【国際特許分類】
【出願番号】特願2011−179605(P2011−179605)
【出願日】平成23年8月19日(2011.8.19)
【国等の委託研究の成果に係る記載事項】(出願人による申告)平成21年度、独立行政法人科学技術振興機構さきがけ研究「脳情報の解読と制御」委託研究、産業技術力強化法第19条の適用を受ける特許出願
【出願人】(512185176)
【Fターム(参考)】
【公開日】平成25年2月28日(2013.2.28)
【国際特許分類】
【出願日】平成23年8月19日(2011.8.19)
【国等の委託研究の成果に係る記載事項】(出願人による申告)平成21年度、独立行政法人科学技術振興機構さきがけ研究「脳情報の解読と制御」委託研究、産業技術力強化法第19条の適用を受ける特許出願
【出願人】(512185176)
【Fターム(参考)】
[ Back to top ]