
非マルトース生成エキソアミラーゼ類及び澱粉の劣化を遅延することへのそれらの使用
【課題】非マルトース生成エキソアミラーゼの使用による澱粉生成物の製造方法及びベーキングされた生成物における澱粉の老化を遅延するための方法を提供する。
【解決手段】アミロペクチンの側鎖の非還元末端から4〜8個のD−グルコピラノシル単位から主として成る1又は複数の線状マルトオリゴ糖を切り取ることによって澱粉を加水分解することができるシュードモナス・サッカロフィラ由来の非マルトース生成エキソアミラーゼを澱粉基材に添加することを含む方法。
【解決手段】アミロペクチンの側鎖の非還元末端から4〜8個のD−グルコピラノシル単位から主として成る1又は複数の線状マルトオリゴ糖を切り取ることによって澱粉を加水分解することができるシュードモナス・サッカロフィラ由来の非マルトース生成エキソアミラーゼを澱粉基材に添加することを含む方法。
【発明の詳細な説明】
【技術分野】
【0001】
発明の分野
本発明は、タンパク質類、特に澱粉を分解することかできるタンパク質類に関する。
特に、本発明は、澱粉の有害な劣化を遅延することができるタンパク質の使用に関する。
有害な劣化(retrogradation)過程、たとえば老化(staling)は、典型的には、澱粉基材、特に水性澱粉懸濁液の加熱及び冷却の後に生じ、そしてゲル化された澱粉の徐々の規則状態への転位のためである。
【0002】
さらに具体的には、本発明は、アミロペクチンの有害な劣化を遅延することができるタンパク質の使用に関する。
さらに特別には、本発明は、ベーキングされたパン生成物を調製するためへのタンパク質の使用、及びそのベーキングされたパン生成物自体にも関する。
さらに特別には、本発明は、ベーキングされた澱粉質のパン生成物の劣化の遅延にも関する。
【0003】
さらに特定には、本発明は、パンドウに非マルトース生成エキソアミラーゼを添加することを含んで成る、劣化を遅延又は低下せしめた、ベーキングされた澱粉質のパン生成物の製造方法にも関する。
本発明はまた、非マルトース生成エキソアミラーゼを含んで成る、ドウ及びベーキングされた澱粉質のパン生成物のための改良された生成物にも関する。
【背景技術】
【0004】
発明の背景
澱粉はアミロペクチン及びアミロースを含んで成る。アミロペクチンは、枝分かれした及びブッシュ様構造を形成するα−(1→6)結合を通して枝分かれ点で一緒に連結される短いα−(1→4)−D−グルカン鎖を有する高度に枝分かれされた炭水化物ポリマーである。平均的に、20〜25のα−(1→4)結合されたグルコース残基ごとに1つの枝分かれ点が存在する。対照的に、アミロースは、枝だなしのα−(1→4)−D−グルカン単位から主に成る線状構造である。典型的には、澱粉は、約75%のアミロペクチン分子及び約25%のアミロース分子を含む。
【0005】
より具体的には、線状マルト−多糖類は、α−(1→4)結合により連結される2〜10単位のα−D−グルコピラノースから構成される。それらの性質、たとえば低甘味性、高い水保持機能及びスクロース結晶化の阻止[1]のために、それらの化合物は食品産業への可能性ある用途を有する。しかしながら、3以上の重合程度(すなわち、DP>3)を有するマルト−オリゴ糖の多量の調製は、短調で退屈であり、且つ費用が高い。
【0006】
バックグラウンド情報として、次のものを列挙する:DP1=グルコース、DP2=マルトース、DP3=マルトトリオース、DP4=マルトテトラオース、DP5=マルトペンタオース、DP6=マルトヘキサオーズ、DP7=マルトヘプタオース、DP8=マルトオクタオース、DP9=マルトノナオース、及びDP10=マルトデカオース。
特定の長さのマルト−オリゴ糖を生成する微生物酵素の発見は、多量のそれらのオリゴ糖の生成を可能にする[2]。
【0007】
アミラーゼは、澱粉においてα−D−(1→4)O−グリコシド結合を分解する、ヒドロラーゼとして分類される澱粉−分解酵素である。一般的に、α−アミラーゼ(E. C. 3. 2. 1. 1, α−D−(1→4)−グルカングルカノヒドロラーゼは、ランダム態様で澱粉分子内のα−D−(1→4)O−グルコシド結合を分解するエンド−作用酵素として定義される[3]。
【0008】
対照的に、エキソ−作用性アミロース分解酵素、たとえばβアミラーゼ(E. C. 3. 2. 1, α−D−(1→4)−グルカンマルトヒドロラーゼ)、及びいくつかの生成物−特異的アミラーゼは、基質の非還元末端から澱粉分子を分解する[4]。β−アミラーゼ、α−グルコシダーゼ(E. C. 3. 2. 2. 20, α−D−グルコシドグルコヒドロラーゼ)、グルコアミラーゼ(E. C. 3. 2. 1. 3, α−D−(1→4)−グルカングルコヒドロラーゼ)、及び生成物−特異的アミラーゼは、澱粉から特定の長さのマルト−オリゴ糖を生成することができる。
【0009】
特定のDPのマルト−オリゴ糖を生成するいくつかのアミラーゼ、たとえばクレビシェラ・プネウモニア(Klebsiella pneumonia)[5, 6]、バチルス・スブチリス(Bacillus subtilis)[7], B. サーキュランスG−6(B. circulans G-6)[8]、B.サーキュランスF-2 [9]、及びB.カルドベロックス(B. caldovelox)[11, 12]が、これまで同定されている。マルトペンタオース−生成アミラーゼは、B.リケニホルミス(B. licheniformis)584 [13] 及びプロイドモナスspp. (Pseudomonas spp.) [14, 15] において検出されている。さらに、マルトテトラオース−生成アミラーゼが、シュードモナス・スツゼリ(Pseudomonas stutzeri)NRRL B-3389 [16, 17]、バチルスsp. MG-4 [14] 及びシュードモナスsp. IMD353 [19] から、及びマルトトリオース−生成アミラーゼが、ストレプトミセス・グリセウス(Streptomyces griseus)NA-468 [20] 及びB. スブチリス[21] から報告されている。
【0010】
EP−B1−298,645号は、遺伝子工学技法を用いて、シュードモナス・スツゼリ又はP. サッカロフィラ(P. saccharophila)のエキソ−マルトテトラヒドロラーゼを調製するための方法を記載する。
アメリカ特許第5,204,254号は、好アルカリ性細菌(DSM5853)の天然の及び遺伝子的に修飾されたエキソ−マルトペンタヒドロラーゼを記載する。
【0011】
高いpHで活性な非常に少数の生成物−特異的アミラーゼが同定されている。同定されているそれらの例は、マルトヘキサノースを生成するバチルスsp. H−167[22, 23]、マルトペンタオースを生成する細菌単離物(163−26、DSM5853)[24]、マルトテトラオース及びより小さなマルト−オリゴ糖を生成するバチルスsp. IMD370 [25]、及び延長された加水分解期間、マルトテトラオースに転換された澱粉からマルトヘキサオースを最初に生成したバチルスsp. GM8901 [26] からのアミラーゼを包含する。
【0012】
水の存在下で加熱された澱粉粒質物は、ゲル化と呼ばれる規則不規則転移を受け、ここで液体が膨潤粒質物により取られる。ゲル化温度は、異なって澱粉で変化し、そして天然の修飾されていない澱粉に関しては、それらの生物学的な源に依存する。
冷却は、澱粉濃度に依存して、ゲル化された相を粘弾性ペースト又は弾性ゲルに転換する。この工程の間、アミロース及びアミロペクチン鎖は、より規則化された構造を形成するために再会合する。時間の経過に従って、会合がより形成され、そしてそれらはさらにより規則的に成る。アミロペクチン鎖DP−15−20の会合が熱可逆性擬似結晶構造を導くと思われる。
【0013】
有害な劣化の結果、ペースト又はゲルシステムの水保持能力が、ゲルテキスチャー及びダイエット性質に重要に関係して変更される。
ベーキングされたパン生成物の品質が貯蔵の間、徐々に劣化することは知られている。パンの中味は柔軟性及び弾性を失い、そして堅く且つ脆くなる。このいわゆる劣化は、非晶性状態から擬似結晶状態に、ベーキングの間ゲル化される澱粉の転移であることが理解されている、主に澱粉の有害な劣化のためである。パンの中味の堅さの上昇はしばしば、パンの劣化過程の測定値として使用される。
【0014】
あらたにベーキングされたパンの冷却に基づいて、アミロース画分は、時間の経過に従って、劣化し、網状構造を進行せしめる。この過程は、それが低い程度の堅さ及び改良されたスライスシング特性を有する所望のパン中心構造を創造することにおいて有益である。アミロペクチンの徐々の結晶化が、ベーキングの後、数日間、ゲル化された澱粉顆粒内で生じる。この過程において、アミロペクチンは、澱粉顆粒が埋封されているアミロース網状構造を強化すると思われる。この強化は、パン中心の高められた硬性を導く。この強化がパン劣化の主な原因の1つである。
【0015】
アミロペクチンの有害な劣化又は結晶化の割合は、アミロペクチンの側鎖の長さに依存する。これによれば、穀物アミロペクチンは、穀物アミロペクチンよりも長い平均側鎖を有する、エンドウ又はジャガイモからのアミロペクチンによりも遅い速度で劣化する。
【0016】
これは、11以下のDP、すなわち重合の程度の平均鎖長を有するアミロペクチンがまったく結晶化しない、アミロペクチンゲルシステムからの観察により支持される。さらに、DP6−9の非常に短い鎖の存在が、たぶん立体的妨害のために、周囲の長い側鎖の結晶化を阻害すると思われる。それにより、それらの短い鎖は、強い反−有害な劣化効果を有すると思われる。これによれば、アミロペクチン劣化は、DP14−24を有する側鎖のモル画分に直接的に比例し、そしてDPG6−9を有する側鎖のモル画分に反比例する。
【0017】
小麦及び他の穀類においては、アミロペクチンの外部側鎖は、DP12−19の範囲である。従って、アミロペクチン側鎖の酵素加水分解は、それらの結晶化傾向を著しく低めることができる。
グルコース生成及びマルトース生成エキソ−アミラーゼ、たとえばグルコースを開放することによって澱粉を加水分解するアミログリコシダーゼ、及び非還元鎖末端からマルトースを開放することによって澱粉を加水分解するマルトース生成エキソアミラーゼ又はβ−アミラーゼを用いることによって、パンの劣化を遅延することは、当業界において知られている。
【0018】
これに関しては、Jakubczykなど. (Zesz. Nauk. Sck. Gl. Gospod Wiejsk. Warzawie, Technol. Reino-Spozyw, 1973, 223-235)が、アミログルコシダーゼが小麦粉をベーキングすることによって得られたパンの劣化を遅延することができることを報告している。
JP−62−79745号及びJP−62−79746号は、それぞれ、バチルス・ステアロサーモフィラス(Bacillus stearothermophilus)及びバチルス・メガテリウム(Bacillus megateriumu )により生成されるβ−アミラーゼの使用がパンを包含する澱粉食品の劣化の遅延において効果的であることを言及している。
【0019】
EP−A412,607号は、ゲル化の前、不活性化されない、熱安定性エキソアミラーゼのドウへの添加により劣化性質を遅延しているパン生成物の生成方法を開示する。アミログリコシダーゼ及びβ−アミラーゼのみが、使用される適切なエキソアミラーゼとして列挙されている。エキソアミラーゼは、アミロース及びアミロペクチンの非還元末端からグルコース又はマルトースを分解することによって、ベーキングの間、アミロペクチン成分の結晶化性質を選択的に変性することができる量で存在する。EP−A−412, 607号によれば、エキソアミラーゼは、アミロースの結晶化性質に対し実質的な影響を及ぼさないで、アミロペクチンの結晶化性質を選択的に低める。
【0020】
EP−A−494,233号は、α−形状で存在し、そしてゲル化の前、不活性化されていないマルトースを開放するために遅延された劣化性質を有する、ベーキングされた生成物の生成工程へのマルトース生成エキソアミラーゼの使用を開示する。バチルス株NCIB11837からのマルトース生成α−アミラーゼのみが特別に開示される。明らかに、マルトース生成エキソアミラーゼは、段階的に、多糖鎖の非還元末端からのα−マルトース単位を除去することによって、澱粉における(1→4)−α−グルコシド結合を加水分解する。
【0021】
従って、従来技術は、一定のグルコース生成エキソアミラーゼ及びマルトース生成エキソアミラーゼが、アミロペクチン側鎖の短縮を通してアミロペクチンの有害な劣化傾向を選択的に低めることによって、非劣化効果を提供することができることを教授する。
それにもかかわらず、澱粉生成物、特にベーキングされた生成物、より特定にはパン生成物の有害な劣化を遅延するための、たとえば老化を遅延するための、異なった及び効果的な、好ましくはより効果的手段を提供する必要性がまだ存在する。
【発明の概要】
【0022】
発明の要約
本発明は、有害な劣化特性が遅延する澱粉生成物の製造方法を提供する。
本発明はまた、本発明の方法において有用である酵素も提供する。
本発明の酵素はアミラーゼ酵素である。より具体的には、本発明の酵素は非マルトース生成エキソアミラーゼ酵素である。
非マルトース生成エキソアミラーゼが、ベーキングされた生成物における老化を遅延するためにはもちろんのこと、澱粉生成物の有害な劣化を遅延するためにこれまで使用されたことはないことが注目されるべきである。
【0023】
従って、本発明の第1の観点によれば、アミロペクチンの側鎖の非還元末端から4〜8個のD−グルコピラノシル単位から主として成る1又は複数の線状マルトオリゴ糖を切り取ることによって澱粉を加水分解することができる非マルトース生成エキソアミラーゼを澱粉基材に添加することを含んで成る澱粉生成物の製造方法が提供される。
澱粉基材への非マルトース生成エキソアミラーゼの添加は、澱粉生成物の加熱の間、及び/又はその後に行うことができる。
従って、本発明の第2の観点によれば、本発明の方法により得られる、ベーキングされた生成物が提供される。
【0024】
従って、本発明の第3の観点によれば、ドウのための改良された組成物が提供され;ここで前記組成物は非マルトース生成エキソアミラーゼ、及び少なくとも1つの追加のドウ成分又はドウ添加材を含んで成る。
従って、本発明の第4の観点によれば、澱粉生成物の有害な劣化を遅延するために澱粉生成物への非マルトース生成エキソアミラーゼの使用が提供される。
従って、本発明の第5の観点によれば、新規非マルトース生成エキソアミラーゼが提供される。
本発明のそれらの及び他の観点は、請求の範囲に提供される。さらに、本発明のそれらの及び他の観点、並びにそれらの好ましい観点が下記に提供され、そして論じられる。
【図面の簡単な説明】
【0025】
【図1】図1は、45℃で2%澱粉基質において培養されたB.クラウジBT−21の液体培養物における細胞外澱粉分解活性(mU/ml)を示す。◆は可溶性澱粉、●はアミロペクチン、黒三角はコーン澱粉、黒四角は完全な玄米。バーは標準偏差を示す。
【図2】図2は、生成物−特異的アミラーゼの活性に対するpHの効果を示す。55℃でのpHの効果。
【0026】
【図3】図3は、生成物−特異的アミラーゼの活性に対する温度の効果を示す。5mMのCaCl2を伴って〔黒四角〕又はCaCl2を伴わないで〔□〕、pH9.5での温度の効果。
【図4】図4は、5mMのCaCl2を伴って、pH9.5での上昇する温度でのインキュベーション後、生成物−特異的アミラーゼの残留活性として試験される熱安定性を示す。
【0027】
【図5】図5は、55℃及びpH9.5で、1%の可溶性澱粉及び5mMのCaCl2と共に、生成物−特異的アミラーゼ(505mU/ml)をインキュベートすることによって形成される生成物(mM)を示す。○グルコース、×マルトース、□マルトトリオース、黒三角マルトテトラオース、●マルトペンタオース、黒四角マルトヘキサオース。
【0028】
【図6】図6は、PH9.5及び55℃で1%可溶性澱粉と共に生成物−特異的アミラーゼ(505mU/ml)をインキュベートすることによって得られるHPAEC−PAD形跡を示す。A)酵素を含まない可溶性澱粉、B)酵素と共に30分間インキュベーション。
【図7】図7は、既知の澱粉分解作用のアミラーゼに比較して、B.クラウジBT−21生成物−特異的アミラーゼのエンド−及びエキソ−活性の決定を示す。青色形成(最大の%)が、mMマルトースの生成に対してプロットされる。曲線の傾斜は、エンド−又はエキソ−活性の有力性を示す。
【発明を実施するための形態】
【0029】
一般的な定義
従って、本発明は、澱粉基材、特に澱粉ゲルの有害な劣化を遅延することができるタンパク質の使用に関する。
1つの好ましい観点においては、本発明は、澱粉の劣化を遅延することができるタンパク質の使用に関する。
もう1つの観点においては、本発明は、澱粉基材、たとえば澱粉ゲルの有害な劣化を遅延することができるタンパク質の使用に関する。
本発明によれば、用語“澱粉”とは、澱粉自体、又はその成分、特にアミロペクチンを意味する。
本発明によれば、用語“澱粉基材”とは、澱粉を含んで成るいずれかの適切な基材を意味する。
【0030】
用語“澱粉生成物”とは、澱粉を含むか、又は澱粉に基づくか、又は澱粉に由来するいずれかの生成物を意味する。
好ましくは、澱粉生成物は、小麦粉から得られた澱粉を含み、又はその澱粉に基づかれ、又はその澱粉に由来する。
用語“小麦粉”とは、本明細書において使用される場合、小麦粉又は他の穀物の細かく粉砕された粉と同義語である。しかしながら、好ましくは、その用語は、小麦自体から得られ、そして他の穀物からではない食料粉を意味する。従って、及び特にことわらない限り、“小麦粉”とは、本明細書において使用される場合、小麦粉自体を意味し、そして基材、たとえばドウに存在する場合も小麦粉を意味する。
【0031】
好ましい食料粉は、小麦粉又はライ麦粉、又はそれらの混合物である。しかしながら、他のタイプの食料粉、たとえば米、トウモロコシ、大麦及びアズキに由来する食料粉を含んで成るドウもまた企画される。
好ましくは、澱粉生成物は、大麦生成物である。
より好ましくは、澱粉生成物は、パン生成物である。
さらにより好ましくは、澱粉生成物は、ベーキングされた澱粉質のパン生成物である。
【0032】
用語“ベーキングされた澱粉質のパン生成物”とは、粉砕された穀物に基づく、及び食料粉、水及び発酵剤をドウ形成条件下で混合することにより得ることができるドウに基づくいずれかのベーキングされた生成物を言及することが理解される。しかしながら、追加の成分がそのドウ混合物に添加され得ることも、本発明の範囲内である。
【0033】
用語“アミラーゼ”は、その通常の意味においては、中でも、澱粉の分解を触媒することが酵素を意味する。特に、それらは澱粉においてα−D−(1→4)O−グリコシド結合を分解できるヒドロラーゼである。
用語“非マルトース生成エキソアミラーゼ酵素”とは、最初、澱粉を実質的な量のマルトースに分解しない酵素を意味する。非常に好ましい観点においては、その用語はまた、最初、澱粉を実質的な量のマルトース及びグルコースに分解しない酵素を意味する。
【0034】
本発明の前、非マルトース生成エキソアミラーゼ酵素は、澱粉基材、特に澱粉ゲルの有害な劣化を遅延することについて提案されたことはない。
本発明に従ってアミラーゼ活性を決定するための適切なアッセイは、後で提供される。便利には、このアッセイは“アミラーゼアッセイプロトコール”と呼ばれる。
従って、好ましくは、用語“非マルトース生成エキソアミラーゼ酵素”とは、本明細書に提供される“アミラーゼ活性プロトコール”に記載のような生成物測定方法に従って分析される場合、前記酵素が最初、澱粉を実質的な量のマルトースに分解しないことを意味する。
【0035】
好ましい観点においては、非マルトース生成エキソアミラーゼは、0.7単位の量の前記非マルトース生成エキソアミラーゼが、50mMの2−(N−モルホリノ)エタンスルホン酸及び2mMの塩化カルシウムを含む緩衝液1ml当たり10mgの予備煮沸されたモチ状澱粉の水溶液4mlにおいて、50℃の温度でpH6.0で15分間インキュベートされる場合、前記酵素が2〜10個のD−グルコピラノシル単位及び任意にはグルコースの1又は複数の線状マルト−オリゴ糖から成る加水分解生成物を生成し、その結果、前記加水分解生成物の少なくとも60重量%、好ましくは少なくとも70重量%、より好ましくは少なくとも80重量%そして最も好ましくは少なくとも85重量%が、3〜10個のD−グルコピラノシル単位の線状マルトオリゴ糖、好ましくは4〜8個のD−グルコピラノシル単位の線状マルトオリゴ糖から構成されることにより特徴づけることができる。
【0036】
参照の容易さ及び本発明の目的のために、50mMの2−(N−モルホリノ)エタンスルホン酸及び2mMの塩化カルシウムを含む緩衝液1mlあたり10mgの予備煮沸されたモチ状トウモロコシ澱粉の水溶液4mlにおいて、50℃の温度及びpH6.0で15分間、0.7単位の量の非マルトース生成エキソアミラーゼをインキュベートする特徴が、“モチ状トウモロコシ澱粉インキュベーション試験”として言及され得る。
【0037】
従って、好ましい非マルトース生成エキソアミラーゼは、2〜10個のD−グルコピラノシル単位及び任意にはグルコースの1又は複数の線状マルト−オリゴ糖から成る加水分解生成物を、モチ状トウモロコシ澱粉インキュベーション試験において生成する能力を有するものとして特徴づけられ;その加水分解の結果、前記加水分解生成物の少なくとも60重量%、好ましくは少なくとも70重量%、より好ましくは少なくとも80重量%及び最も好ましくは少なくとも85重量%が、3〜10個のD−グルコピラノシル単位の線状マルトオリゴ糖、好ましくは4〜8個のD−グルコピラノシル単位から成る線状マルトオリゴ糖から成る。
【0038】
モチ状トウモリコシ澱粉インキュベーション試験における加水分解生成物は、2〜10個のD−グルコピラノシル単位の1又は複数の線状マルト−オリゴ糖及び任意にはグルコースを含む。モチ状トウモロコシ澱粉インキュベーション試験における加水分解生成物はまた、他の加水分解生成物も含むことができる。それにもかかわらず、3〜10個のD−グルコピラノシル単位の線状マルトオリゴ糖の重量%の量は、2〜10個のD−グリコピラノシル単位及び任意にはグルコースの1又は複数の線状マルト−オリゴ糖から成る加水分解生成物の量に基づく。換言すれば、3〜10個のD−グルコピラノシル単位の線状マルトオリゴ糖の重量%の量は、2〜10個のD−グルコプラノシル単位の1又は複数の線状マルト−オリゴ糖及びグルコース以外の他の加水分解生成物の量に基づかない。
【0039】
加水分解生成物は、いずれかの適切な手段により分析され得る。たとえば、加水分解生成物は、パルスされた電流滴定検出及びたとえば標準として、グルコース〜マルトヘプタオースの既知の線状マルトオリゴ糖と共に、Dionex PA 100カラムを用いて、アニオン交換HPLCにより分析され得る。
参照の容易さ及び本発明の目的のために、パルスされた電流滴定検出及び標準として、グルコース〜マルトヘプタオースの既知線状マルトオリゴ糖と共にDionex PA 100 カラムを用いて、アニオン交換HPLCによる加水分解生成物の分析特徴は、“アニオン交換による分析”として言及され得る。もちろん、及びまさに示されるように、他の分析技法及び他の特定のアニオン交換技法でも十分である。
【0040】
従って、他方では、好ましい非マルトース生成エキソアミロースは、モチ状トウモロコシ澱粉インキュベーション試験において、2〜10個のD−グルコピラノシル単位及び任意にはグルコースの1又は複数の線状マルト−オリゴ糖から成る加水分解生成物を生じさせる能力を有し、その結果、前記加水分解生成物の少なくとも60重量%、好ましくは少なくとも70重量%、より好ましくは少なくとも80重量%そして最も好ましくは少なくとも85重量%が、3〜10個のD−グルコピラノシル単位の線状マルトオリゴ糖、好ましくは4〜8個のD−グルコピラノシル単位の線状マルトオリゴ糖から成ることを特徴とする。
【0041】
本明細書において使用される場合、用語“線状マルト−オリゴ糖”とは、通常、α−(1→4)結合により結合される、2〜10個のα−D−グルコプラノース単位を意味するものとして使用される。
用語“P.サッカロフィラ(Pseudomonas sacharophila)から得ることができる”とは、酵素がP.サッカロフィラから必ずしも得られる必要はないことを意味する。代わりに、酵素は組換えDNA技法の使用により調製され得る。
【0042】
P.サッカロフィラから得ることができる酵素に関しての用語“その機能的同等物”とは、その機能的同等物が他の源から得られることを意味する。機能的に同等の酵素は異なったアミノ酸配列を有するが、しかし非マルトース生成エキソアミラーゼ活性を有するであろう。その機能的に同等の酵素は異なった化学構造及び/又は化学式を有するが、しかし非マルトース生成エキソアミラーゼ活性を有するであろう。その機能的に同等の酵素は、P.サッカロフィラから得られる非マルトース生成エキソアミラーゼ酵素と正確に同じ非マルトース生成エキソアミラーゼ活性を必ずしも有する必要はない。いくつかの用途においては、好ましくは、機能的に同等の酵素は、P.サッカロフィラから得られる酵素と少なくとも同じ活性プロフィールを有する。
【0043】
用語“バチルス・クラウジ(Bacillus clausii)から得ることができる”とは、酵素がバチルス・クラウジから必ずしも得られる必要はないことを意味する。代わりに、酵素は、組換えDNA技法の使用により調製され得る。
バチルス・グラウジから得ることができる酵素に関しての用語“その機能的同等物”とは、その機能的同等物が他の源から得られることを意味する。機能的に同等の酵素は異なったアミノ酸配列を有するが、しかし非マルトース生成エキソアミラーゼ活性を有するであろう。
【0044】
その機能的に同等の酵素は異なった化学構造及び/又は化学式を有するが、しかし非マルトース生成エキソアミラーゼ活性を有するであろう。その機能的に同等の酵素は、バチルス・グラウジから得られる非マルトース生成エキソアミラーゼ酵素と正確に同じ非マルトース生成エキソアミラーゼ活性を必ずしも有する必要はない。いくつかの用途においては、好ましくは、機能的に同等の酵素は、バチルス・グラウジから得られる酵素と少なくとも同じ活性プロフィール(たとえば、図7に示される反応性プロフィール)を有する。
【0045】
一般的な解説
本発明は、非マルトース生成エキソアミアーゼが澱粉生成物、たとえばベーキングされた生成物において有害な劣化、たとえば老化(staling)を遅延し又は低めることにおいて非常に効果的である驚くべき発見に基づかれている。
本発明者は、本発明の非マルトース生成エキソアミラーゼが、グルース生成及びマルトース生成エキソアミラーゼよりも、パンにおける有害な劣化、たとえば老化を遅延することにおいてより効果的であり得ることを見出した。
有害な劣化の低下は、当業界において知られている標準の技法により測定され得る。例によれば、いくつかの技法が、セクション標題“劣化の測定のためのアッセイ”において、後で示される。
【0046】
本発明の研究において、本発明者は、十分な熱安定性を有する、十分な量の活性の非マルトース生成エキソアミラーゼ、たとえばエキソ−マルトテトラヒドロラーゼ(EC 3. 2. 1. 60)をドウ中に導入することによって、たとえば貯蔵条件下で対照のパンの有害な劣化に比較して、低められた、多くの場合、十分に低められた有害な劣化を有するベーキングされた生成物が提供されることを見出した。対照的に、非マルトース生成エキソアミラーゼの熱安定性に匹敵する熱安定性を有するマルトース生成エキソアミラーゼの同じ量の活性を組み込むことによる有害な劣化を低める効果は、有意に劣っている。
【0047】
従って、非マルトース生成エキソアミラーゼの抗劣化効果は、マルトース生成エキソアミラーゼのその効果よりもより効果的である。本発明者は、この差異が一部、アミロペクチン側鎖が短くされる程度によるかも知れないと信じている。本発明はまた、抗劣化効果が、マルトヘプタオース及び/又はマルトオクタオース及び/又はマルトへキソースを開放する本発明の非マルトース生成エキソアミラーゼを用いる場合、さらにより著しいことも信じている。
【0048】
本発明の研究において、本発明者はまた、マルトヘキサオースを生成する高いpHで活性な生成物−特異的アミラーゼを精製し、そして特徴づけた。このアミラーゼは、バチルス・クラウジBT−21のアルカリ耐性株から単離された。
さらに、本発明者は、本発明の非マルトース生成エキソアミラーゼを用いることによって得ることができる有害な劣化の遅延が非常に広範囲にわたって、用量応答性であることを見出した。これは、かなり制限され、そして強く低下する用量応答を有するマルトース生成エキソアミラーゼからの効果と対照的である。
【0049】
アミラーゼ
1つの観点において、本発明は、澱粉生成物、たとえば製パン製品を調製するためへの一定のアミラーゼの使用を提供する。この観点においては、非マルトース生成エキソアミラーゼであるアミラーゼは、澱粉生成物、特にベーキングされた澱粉質パン生成物の老化を遅延し、又は低める(すなわち、老化の速度を低める)。
好ましくは、アミラーゼは、単離された形で及び/又は実質的に純粋な形で存在する。ここで、用語“単離された”とは、酵素がその天然の環境下で存在しないことを意味する。
【0050】
上記に示されるように、本発明の非マルトース生成エキソアミラーゼ酵素は、最初に、澱粉を実質的な量のマルトースに分解しない。
本発明によれば、非マルトース生成エキソアミラーゼは、アミロペクチンの側鎖の非還元末端から、4〜8個のD−グルコピラノシル単位から主に成る線状マルトオリゴ糖を切り離すことができる。この特徴を有し、そして本発明への使用のために適切である非マルトース生成エキソアミラーゼは、アミラーゼアッセイプロトコール(前記)に示されるモデル系におけるゲル化されたモチ状トウモロコシを加水分解するそれらの能力により同定される。
【0051】
アミラーゼアッセイプロトコールにおける記載される条件下で15分間インキュベートされる場合、本発明への使用のために適切である非マルトース生成エキソアミラーゼは、2〜10個のD−グルコピラノシル単位及び任意にはグルコースの1又は複数の線状マルト−オリゴ糖から成る加水分解生成物を生成し、その結果、その加水分解生成物の生成物パターンは、少なくとも60重量%、特に少なくとも70重量%、より好ましくは少なくとも80重量%及び最も好ましくは少なくとも90重量%が、マルトース及びグルコース以外の澱粉加水分解生成物から成る。
【0052】
本発明の好ましい観点に関しては、本発明への使用のために適切である非マルトース生成エキソアミラーゼは、モチ状トウモロコシ澱粉インキュベーション試験について記載される条件下で15分間インキュベートされる場合、前記加水分解生成物をもたらし、その結果、その加水分解生成物は、少なくとも60重量%、特に少なくとも70重量%、より好ましくは少なくとも80重量%そして最も好ましくは少なくとも90重量%の、3〜10個のD−グルコピラノシル単位の線状マルトオリゴ糖、特に4〜8個のD−グルコピラノシル単位から成る線状マルトオリゴ糖の生成物パターンを有する。
【0053】
本発明のより好ましい観点においては、前記試験における前記加水分解生成物は、少なくとも60重量%、特に少なくとも70重量%、より好ましくは少なくとも80重量%そして最も好ましくは少なくとも85重量%の、4又は6個のD−グルコピラノシル単位の線状マルトオリゴ糖の生成物パターンを有する。
本発明のより好ましい観点においては、前記試験における前記加水分解生成物は、少なくとも60重量%、特に少なくとも70重量%、より好ましくは少なくとも80重量%そして最も好ましくは少なくとも85重量%の、4個のD−グルコピラノシル単位の線状マルトオリゴ糖の生成物パターンを有する。
【0054】
本発明のより好ましい観点においては、前記試験における前記加水分解生成物は、少なくとも60重量%、特に少なくとも70重量%、より好ましくは少なくとも80重量%そして最も好ましくは少なくとも85重量%の、6個のD−グルコピラノシル単位の線状マルトオリゴ糖の生成物パターンを有する。
好ましくは、非マルトース生成エキソアミラーゼは、その一次生成物をグルコース、マルトース及びマルトトリオースに転換するように該生成物を実質的に加水分解しない。その場合、前記一次生成物は、酵素に対してアミロペクチン非還元鎖末端と基質として競争し、その結果、その抗−劣化効率を低められる。
【0055】
従って、好ましくは、非マルトース生成エキソアミラーゼは、モチ状トウモロコシ澱粉インキュベーション試験に類似する条件下で(但し、15分間が300分に延長され、そして本発明の便利さのために、この変性されたモチ状トウモリコシ澱粉インキュベーション試験は“延長されたモチ状トウモロコシ澱粉インキュベーション試験”と呼ばれる)、300分間インキュベートされる場合、前記加水分解生成物をなお生成し、ここで前記加水分解生成物は、少なくとも50重量%、特に少なくとも60重量%、より好ましくは少なくとも70重量%、そして最も好ましくは少なくとも80重量%の4〜8個のD−グルコピラノシル単位の生成物パターンを有する。
【0056】
例によれば、本発明の方法において有用な非マルトース生成エキソアミラーゼは、それがモチ状トウモロコシ澱粉インキュベーション試験において、2〜10個のD−グルコピラノシル単位及び任意にはグルコースの1又は複数の線状マルト−オリゴ糖から成る加水分解生成物を生成する能力を有し、その結果、前記加水分解生成物の少なくとも60重量%、好ましくは少なくとも70重量%、より好ましくは少なくとも80重量%そして最も好ましくは少なくとも85重量%が、3〜10個のD−グルコピラノシル単位の線状マルトオリゴ糖、好ましくは4〜8個のD−グルコピラノシル単位の線状マルトオリゴ糖から成り、そして前記酵素がP.サッカロフィラから得ることができるものであり、又はその機能的同等物であることにより特徴づけられ得る。
【0057】
さらなる例によれば、本発明の方法において有用なもう1つの非マルトース生成エキソミラーゼは、それがモチ状トウモロコシ澱粉インキュベーション試験において、2〜10個のD−グルコピラノシル単位及び任意にはグルコースの1又は複数の線状マルト−オリゴ糖から成る加水分解生成物を生成する能力を有し、その結果、前記加水分解生成物の少なくとも60重量%、好ましくは少なくとも70重量%、より好ましくは少なくとも80重量%そして最も好ましくは少なくとも85重量%が、3〜10個のD−グルコピラノシル単位の線状マルトオリゴ糖、好ましくは4〜8個のD−グルコピラノシル単位の線状マルトオリゴ糖から成り、そして前記酵素がバチルス・クラウジから得ることができるものであり、又はその機能的同等物であり、そして前記酵素が約101,000Daの分子量を有し(ドデシル硫酸ナトリウムポリアクリルアミド電気泳動により推定される場合)、そして/又は前記酵素がpH9.5及び55℃で最適活性を有することにより特徴づけられる。
【0058】
好ましくは、本発明の使用のために適切である非マルトース生成エキソアミラーゼは、ベーキングの間、活性であり、そして約55℃の温度で開始する澱粉顆粒のゲル化の間及びその後、澱粉を加水分解する。非マルトース生成エキソアミラーゼはより熱安定性であるほど、より長い時間、活性であり、そして従って、より抗老化効果を提供するであろう。しかしながら、約85℃の温度以上でベーキングする間、非マルトース生成エキソアミラーゼは選択的に徐々に不活性化され、その結果、ベーキング工程の後、最終のパンにおいて活性は実質的に存在しない。
【0059】
従って、好ましくは、本発明への使用のために適切な非マルトース生成エキソアミラーゼは、上記のようにして調製された、50mMのMES、2mMの塩化カルシウム溶液(pH6.0)中、10mg/mlのモチ状トウモロコシ澱粉4mlを含む試験において、40, 45, 50, 55, 60, 65, 70, 75, 80, 85又は90℃で15分間インキュベートされ、そして上記のようにして加水分解生成物の開放についてアッセイされる場合、約45℃〜98℃の最適温度を有する。好ましくは、非マルトース生成エキソアミラーゼの最適温度は55℃〜95℃であり、そしてさらにより好ましくは、60℃〜90℃である。
【0060】
低い熱安定性であることが見出され得る非マルトース生成エキソアミラーゼは、より熱安定性になるようタンパク質工学を用いて改良され得、そして従って、本発明への使用のために適切である。従って、タンパク質工学により一層熱安定性になるよう変性された非マルトース生成エキソアミラーゼの使用が本発明により包含される。
【0061】
いくつかの非マルトース生成エキソアミラーゼがいくらかの程度のエンドアミラーゼ活性を有することは知られている。多くの場合、このタイプの活性は減じられるか又は排除される必要がある。なぜならば、エンドアミラーゼ活性は、枝分かれのデキストリンの蓄積のために、粘着性の又はゴム状のパンの中味(crumb)を生成することによって、最終のパン生成物の品質におそらく負の効果を与えるためである。
【0062】
従って、好ましい観点においては、本発明の非マルトオース生成エキソアミラーゼは、1単位のエキソアミラーゼ活性当たり0.5より少いエンドアミラーゼ単位(EAU)を有するであろう。
好ましくは、本発明の使用のために適切である非マルトース生成エキソアミラーゼは、1単位のエキソアミラーゼ活性当たり0.05より少いEAUを有し、そしてより好ましくは、1単位のエキソアミラーゼ活性当たり0.01より少いEAUを有する。
エンドアミラーゼ単位は、下記に示されるエンドアミラーゼアッセイプロトコールにより決定され得る。
【0063】
本発明への使用のために適切な非マルトース生成エキソアミラーゼの例は、非還元鎖末端から、それぞれマルトテトラオース、マルトペンタオース又はマルトヘキサオースの連続的な残基を除去するために、澱粉質多糖類の1, 4−α−グルコシル結合を加水分解する、エキソーマルトテトラヒドロラーゼ(E. C. 3. 2. 1. 60)、エキソ−マルトペンタヒドロラーゼ及びエキソ−マルトヘキサヒドロラーゼ(E. C. 3. 2. 1. 98)を、包含する。
【0064】
例としては、シュードモナス・サッカロフィラ及びP. スツゼリ(EP−0298645B1)のエキソ−マルトテトラヒドロラーゼ、好アルカリ性グラム−陽性細菌(アメリカ特許第5,204,254号)及びシュードモナスsp. (Shidaなど., Biosci. Biotechnol. Biochem., 1992, 56, 76-80)のエクソ−マルトペンタヒドロラーゼ、及びバチルスsp. #707(Tsukamoto など., Biochem. Biophys. Res. Commun., 1988, 151, 25-31)、B.サーキュランスF2(Taniguchi, ACS Symp., 1991, Ser. 458, 111-124)及びアエロバクター・アエロゲネス(Kainumaなど., Biochim. Biophys. Acta, 1975, 410, 333-346)のエキソ−マルトヘキサヒドロラーゼを列挙することができる。
【0065】
本発明への使用のために適切な非マルトース生成エキソアミラーゼのもう1つの例は、好アルカリ性バチルス株、GM8901からのエキソアミラーゼである[28]。これは、澱粉からマルトテトラオース、マルトペンタオース及びマルトヘキサオースを生成する非マルトース生成エキソアミラーゼである。
【0066】
さらに、本発明への使用のために適切な非マルトース生成エキソアミラーゼはまた、非還元鎖末端から、それぞれマルトヘプタオース又はマルトオクタオースの残基を除去するために、澱粉質多糖類における1, 4−α−グルコシド結合を加水分解する、エキソ−マルトヘプタヒドロラーゼ又はエキソ−マルトオクタヒドロラーゼを包含する。エクソ−マルトヘプタヒドロラーゼ及びエキソ−マルトオクタヒドロラーゼは、野生型株をスクリーニングすることによって見出され得、又はタンパク質工学により他の澱粉分解酵素から開発され得る。従って、非マルトース生成エキソアミラーゼになるよう他の澱粉分解酵素からタンパク質工学により開発された非マルトース生成エキソアミラーゼもまた、本発明への使用のために適切である。
【0067】
新規アミラーゼ
1つの観点においては、本発明はまた、本発明の澱粉生成物、たとえば製パン生成物を調製するために適切である新規アミラーゼを提供する。本発明の新規アミラーゼは、非マルトース生成エキソアミラーゼである。本発明の研究において、本発明者は、食品、特に製パン生成物の調製に使用するためのドウの調製のために適切であるこの新規アミラーゼを特性決定した。
【0068】
従って、本発明はまた、非マルトース生成エキソアミラーゼを提供し、ここで前記非マルトース生成エキソアミラーゼは、それがモチ状トウモロコシ澱粉インキュベーション試験において、2〜10個のD−グルコピラノシル単位及び任意にはグルコースの1又は複数の線状マルト−オリゴ糖から成る生成物を加水分解する能力を有し、その結果、前記加水分解生成物の少なくとも60重量%、好ましくは少なくとも70重量%、より好ましくは少なくとも80重量%そして最も好ましくは少なくとも85重量%が、3〜10個のD−グルコピラノシル単位の線状マルトオリゴ糖、好ましくは4〜8個のD−グルコピラノシル単位の線状マルトオリゴ糖から成り、そして前記酵素がバチルス・クラウジから得ることができるものであり、又はその機能的同等物であり、そして前記酵素が約101,000Daの分子量を有し(ドデシル硫酸ナトリウムポリアクリルアミド電気泳動により推定される場合)、そして/又は前記酵素がpH9.5及び55℃で活性の最適性を有することにより特徴づけられる。
【0069】
好ましくは、アミラーゼは、単離された形及び/又は実質的に純粋な形で存在する。ここで、用語“単離された”とは、酵素がその天然の環境下に存在しないことを意味する。
【0070】
抗体
本発明の酵素はまた、たとえば標準技法の使用により、抗体を生成するためにも使用され得る。従って、本発明の個々の酵素に対する抗体が生成され得る。個々の抗体は、発明の他の適切なアミラーゼ酵素をスクリーンするために使用され得る。さらに、個々の抗体は本発明の酵素を単離するためにも使用され得る。
【0071】
抗体の生成に関しては、種々の宿主、たとえばヤギ、ウサギ、ラット、マウス、等が、免疫原性質を保持する、インヒビター又はそのいずれかの部分、変異体、相同体、フラグメント又は誘導体、又はオリゴペプチドによる注射により免疫化され得る。宿主種に依存して、種々のアジュバントが免疫学的応答を高めるために使用され得る。そのようなアジュバントは次のものを包含するが、但しそれらだけには限定されない:フロイント、鉱物ゲル、たとえば水酸化アルミニウム、及び界面活性物質、たとえばリゾレシチン、プルロニックポリオール、ポリアニオン、ペプチド、油エマルジョン、キーオール・リンペット(Keyhole limpet)ヘモシアニン及びジニトロフェノール。BCG(バチルス・カルメト−グエリン)及びコリネバクテリウム・パルバム(Corynebacterium parvum)は、開発され得る可能性ある有用なヒトアジュバントである。
【0072】
酵素に対するモノクローナル抗体は、培養での連続細胞系による抗体分子の生成を提供するいずれかの技法を用いて調製され得る。それらは次の技法を包含するが、但しそれらだけには限定されない:Koehler and Milstein (1975, Nature 256: 495-497) により最初に記載されたハイブリドーマ技法、ヒトB−細胞ハイブリドーマ技法(Kosbor など(1983)Immunol Today 4: 72; Cote など(1983) Proc. Natl. Acad. Sci. 80: 2026-2030)及びEBV−ハイブリドーマ技法(Cole など(1985)Monoclonal Antibodies and Cancer Therapy, Alan R Liss Inc. pp77-96)。
【0073】
さらに、“キメラ抗体”の生成のために開発された技法、すなわち適切な抗原特異性及び生物学的活性を有する分子を得るためにヒト抗体遺伝子へのマウス抗体遺伝子のスプライシングが使用さえ得る(Morrison など(1984)Proc. Natl. Acad. Sci. 81: 6851-6855; Neuberger など(1984)Nature 312: 604-608; Takeda など. (1985) Nature 314: 452-454)。他方では、一本鎖抗体の生成について記載される技法(アメリカ特許第4946779号)が、インヒビター特異的一本鎖抗体を生成するために適合され得る。
【0074】
抗体はまた、リンパ球集団においてインビボ生成を誘発することによって、又はOrlandiなど(1989, Proc. Natl. Acad. Sci. 86: 3833-3837)及びWinter G and Milstein C (1991; Nature 349: 293-299) に開示されるようにして、高い特異的結合試薬の組換え免疫グロブリンライブラリー又はパネルをスクリーニングすることによって生成され得る。
【0075】
改良性組成物
示されるように、本発明の1つの観点は、澱粉生成物、特にドウ及び/又はドウから製造されるベーキングされた澱粉質パン生成物のための改良性組成物に関する。
改良性組成物は、本発明の非マルトース生成エキソアミラーゼ及び少なくとも1つの追加のドウ成分又はドウ添加剤を含んでなる。
本発明によれば、追加のドウ成分又はドウ添加剤は、上記に記載されるドウ成分又はドウ添加剤のいずれかであり得る。
【0076】
好都合には、その改良性組成物は、少なくとも1つの追加の成分又は添加剤と共に混合される本発明の非マルトース生成エキソアミラーゼを含んで成る乾燥微紛組成物である。しかしながら、改良性組成物はまた、本発明の非マルトース生成エキソアミラーゼ、及び水又は他の液体に溶解されているか又は分散されている少なくとも1つの追加の成分又は添加剤を含んでなる溶液調製物であり得る。改良性組成物中の酵素活性の量は、改良性組成物の一部を形成する追加の成分及び添加剤の量及びタイプに依存するであろう。
【0077】
任意には、改良性組成物は、特定のベーキングされた生成物を製造するための乾燥成分及び添加剤のすべてを含む、完全混合物、いわゆるプレ−混合物の形で存在することができる。
【0078】
澱粉生成物の調製
本発明の1つの観点によれば、その方法は、適切な非マルトース生成エキソアミラーゼ酵素、たとえば本明細書に示される新規の非マルトース生成エキソアミラーゼ酵素の1つを、澱粉基材に添加することによって、澱粉生成物を形成することを含んで成る。
澱粉基材がドウである場合、ドウは、穀物、水、本発明の非マルトース生成エキソアミラーゼ及び他の可能な成分及び添加剤を一緒に混合することによって調製される。
【0079】
追加の例によれば、澱粉生成物がベーキングされた澱粉質パン生成物(非常に好ましい態様である)である場合、その技法は、ドウ形成条件下で、いずれか適切な順序で、食料粉、水及び発酵剤を混合し、そしてさらに、適切な非マルトース生成エキソアミラーゼ酵素を添加することを含んで成る。
膨化剤(leavening agent)は、化学膨化剤、たとえば炭酸水素ナトリウム、又はサッカロミセス・セレビシアエのいずれかの株(パン酵素)であり得る。
非マルトース生成エキソアミラーゼは、いずれかのドウ成分、たとえば水又はドウ成分混合物と共に、又はいずれかの添加剤又は添加剤混合物と共に添加され得る。
【0080】
ドウは、ベーキング産業、又は食料粉ドウ基材の生成物を製造するいずれかの他の産業において共通するいずれかの従来のドウ調製物により調製され得る。
澱粉質パン生成物、たとえば精白パン、篩分けされたライ麦粉及び小麦粉から製造されたパン、ロールパン及び同様のものベーキングは、典型的には、180〜250℃の範囲のオーブン温度で、約15〜60分間、パン用ドウをベーキングすることによって達成される。ベーキング工程の間、急な温度グラジェント(200→120℃)が外部ドウ層において有力であり、ここでベーキングされた生成物の特徴的な外皮が進行する。しかしながら、蒸気発生による熱消費のために、パンの中味の温度がベーキング工程の最後で、100℃に接近する。
【0081】
非マルトース生成エキソアミラーゼは、単独の活性成分として、又は1又は複数の追加のドウ成分又はドウ添加剤と混合して酵素を含んで成る、液体調製物として、又は乾燥微紛組成物として添加され得る。
ベーキングされた生成物の性質をさらに改良し、そしてベーキングされた生成物に特徴的な性質を付与するために、追加のドウ成分及び/又はドウ添加剤がドウ中に導入され得る。典型的には、そのような追加の成分は、ドウ成分、たとえば塩、穀粒、脂肪及び油、糖、ダイエット用繊維物質、粉乳、グルテン及びドウ添加剤、たとえば乳化剤、他の酵素、親水コロイド、風味剤、酸化剤、鉱物及びビタミンを含むことができる。
【0082】
乳化剤はドウ補強剤及びパン中味の軟化剤として有用である。ドウ補強剤として、乳化剤は、残り時間に関する耐性及び加工の間、衝撃に対する耐性を提供することができる。さらに、ドウ補強剤は、発酵時間における変化に対する所定のドウの耐性を改良するであろう。ほとんどのドウ補強剤はまた、ベーキングされた製品に耐性を与えるものからの体積の上昇を意味するオーブンスプリングを改良する。最後に、ドウ補強剤は、レセピ−混合物に存在するいずれかの脂肪を乳化するであろう。
【0083】
モノグリセリドの特性を主に示すパン中味の軟化は、冷却に基づいて再結晶化せず、そして従ってパン中味の堅さに寄与しないであろうアミラーゼとの不溶性包含複合体の形成を導く澱粉のアミロース画分と乳化剤との間の相互作用に帰因する。
【0084】
追加のドウ添加剤として使用され得る適切な乳化剤は次のものを包含する:レシチン、ポリオキシエチレンステアレート、食用脂肪酸のモノ−及びジグリセリド、食用脂肪酸のモノ−及びジグリセリドの酢酸エステル、食用脂肪酸のモノ−及びジグリセリドの乳酸エステル、食用脂肪酸のモノ−及びジグリセリドのクエン酸エステル、食用脂肪酸のモノ−及びジグリセリドのジアセチル酒石酸エステル、食用脂肪酸のスクロースエステル、ナトリウムステアロイル−2−ラクチレ−ト、及びカルシウムステアロイル−2−ラクチレ−ト。
【0085】
追加のドウ添加剤として有用である他の酵素は、例として、次のものを包含する:オキシドレダクターゼ、たとえばグルコースオキシダーゼ、ヘキソースオキシダーゼ、及びアスコルベートオキシダーゼ、たとえばリパーゼ及びエステラーゼ、並びにグリコシダーゼ様α−アミラーゼ、プルラナーゼ及びキシラナーゼ。オキシドレダクターゼ、たとえばグルコースオキシダーゼ及びヘキソースオキシダーゼは、ドウ補強剤及びベーキングされた生成物の体積の調節のために使用され、そしてキシラナーゼ及び他のヘミセルラーゼは、ドウ取り扱い性質、パン中味軟化及びパン体積を改良するために添加され得る。リパーゼはドウ補強剤及びパン中身軟化剤として有用であり、そしてα−アミラーゼ及び他の澱粉分解酵素はパンの体積を調節し、そしてさらに、パン中味の堅さを低めるためにドウ中に導入され得る。
【0086】
添加される本発明の非マルトース生成エキソアミラーゼの量は通常、kg穀物当たり50〜100,000単位、好ましくはkg 穀物当たり100〜50,000単位の量で最終ドウに存在する。本発明の有用な態様においては、その量はkg穀物当たり200〜20,000単位の範囲である。
本明細書においては、1単位の非マルトース生成エキソアミラーゼは、この後に記載されるように、50mMのMES, 2mMの塩化カルシウム溶液(pH6.0)中、10mg/mlのモチ状トウモロコシ澱粉4mlを有する試験管において50℃でインキュベートされる場合、1分当たり1μモルの還元糖に等しい加水分解生成物を開放する量の酵素として定義される。
【0087】
アミラーゼにより調製される食品
本発明は、食品の製造への使用のための適切なアミラーゼを提供する。動物飼料を包含する典型的な食品は、酪農製品、食肉製品、家禽製品、魚製品及び製パン製品を包含する。
好ましくは、食品は製パン生成物、たとえば上記に記載される製パン生成物である。本発明の範囲内に組み込まれる典型的な製パン(ベーキングされた)生成物は、パン、たとえば一塊のパン、ロールパン、バン、ビザ生地、等、プレッツェル、トルティヤ、ケーキ、クッキー、ビスケット、クラッカー、等を包含する。
【0088】
アミラーゼアッセイプロトコール
次のシステムが、本発明への使用のために適切である非マルトース生成エキソアミラーゼを特徴づけるために使用される。
初期のバックグラウンド情報によれば、モチ状トウモロコシアミロペクチン(Roquette, France からWAXILYS 200 として得ることができる)は、非常に高いアミロペクチン含有率(90%以上)を有する澱粉である。
20mg/mlのモチ状トウモロコシ澱粉は、50mMのMES(2−(N−モルホリノ)エタンスルホン酸)、2mMの塩化カルシウムの緩衝液(pH6.0)において3分間煮沸され、そして続いて50℃でインキュベートされ、そして30分以内に使用される。
【0089】
1単位の非マルトース生成エキソアミラーゼは、上記に記載されるように、50mMのMES, 2mMの塩化カルシウム溶液(pH6.0)中、10mg/mlのモチ状トウモロコシ澱粉4mlを有する試験管において50℃でインキュベートされる場合、1分間当たり1μモルの還元糖に等しい加水分解生成物を開放する量の酵素として定義される。
還元糖は、標準としてマルトースを用い、及びBernfeld, Methods Enzymol., (1954), 1, 149-158のジニトロサリチル酸法又は還元糖を定量化するための当業界において知られている他の方法を用いて測定される。
【0090】
非マルトース生成エキソアミラーゼの加水分解生成物パターンは、上記のようにして調製された緩衝液中、10mg/ml のモチ状トウモロコシ澱粉4mlを有する試験管において、0.7単位の非マルトース生成エキソアミラーゼを50℃で15又は300分間、インキュベートすることによって決定される。反応は、試験管を煮沸水浴に3分間浸すことによって停止された。
【0091】
加水分解生成物は、パルスされた電流滴定検出、及び標準としてグルコース〜マルトペプタオースの既知の線状マルトオリゴ糖と共に、溶離剤として酢酸ナトリウム、水酸化ナトリウム及び水を用いて、Dionex PA100 カラムのアニオン交換HPLCにより分析され、そして定量化される。マルトオクタオース〜マルトデカオースのために使用される応答因子は、マルトヘプタオースに関して見出された応答因子である。
【0092】
エンドアミラーゼアッセイプロトコール
0.75mlの酵素溶液が、50mMのMES(2−(N−モルホリノ)エタンスルホン酸)、2mMの塩化カルシウム溶液(pH6.0)中、0.5%(w/v)のAZCL−アミロース(Megazyme, Ireland から入手できるアズリン架橋されたアミロース)6.75mlと共に50℃でインキュベートされる。5, 10, 15, 20及び25分後、それぞれ1.0mlの反応混合物が、4%(w/v)のTRIS(トリス(ヒドロキシメチル)アミノメタン)から成る停止溶液4.0mlに移される。
【0093】
停止されたサンプルは、Whatman No.1フィルターを通して濾過され、そして590nmでのその光学密度が蒸留水に対して測定される。アッセイされる酵素溶液が希釈され、その結果、得られる光学密度は時間の線状関数である。光学密度対時間についての直線の傾斜が使用され、g当たり100エンドアミラーゼ単位(EAU)を有するものとして定義される、標準GRINDAMYL(商標)A100 (Danisco Ingredients から入手できる) に対するエンドアミラーゼが計算される。
【0094】
劣化(老化を包含する)の測定のためのアッセイ
本発明の非マルトース生成エキソアミラーゼの抗老化効果の評価のために、パン中味の堅さが、Instrn 4301 Universal Food Taxture Analyzer 又は当業界において知られている類似する装置により、ベーキングの1, 3及び7日後、測定され得る。
【0095】
当業界において従来使用され、そして本発明の非マルトース生成エキソアミラーゼの澱粉劣化に対する効果を評価するために使用されるもう1つの方法は、DSC(示差走査熱量)に基づかれる。それにより、酵素と共に又は酵素なしで(対照)ベーキングされたモデルシステムドウからのパン中味における劣化されたアミロペクチンの溶融エントロピーが測定される。記載される例に適用されるDSC装置は、20〜95℃、1分当たり10℃の温度グラジエントで作動するMettler-Toledo DSC820 である。サンプルの調製のためには、10〜20mgのパン中味が計量され、そしてMetler-Toledo アルミニウムパンに移され、次に密封される。
【0096】
記載される例に使用されるモデルシステムドウは、本発明の非マルトース生成エキソアミラーゼと共に又はそれを伴わないで、標準の小麦粉及び最適量の水を含む。それらは、それぞれ6又は7分間、Brabender Farinographにより10又は50gで混合される。ドウのサンプルが、蓋付きのガラス試験管(15×0.8cm)に配置される。それらの試験管は、33℃で30分間のインキュベーションから開始して、続いて1.1℃/分のグラジエントで33℃から95℃まで、水浴においてのベーキング工程にゆだねられ、そして最終的に95℃で5分間インキュベートされる。続いて、試験管が、DSC分析の前、20℃でサーモスタットにおいて貯蔵される。
【0097】
要約
要約すれば、本発明は、アミロペクチンの非還元鎖末端から4〜8個の範囲のD−グルコピラノシル単位の線状マルトオリゴ糖を切り取ることによって澱粉を加水分解し、そして好ましくは、十分な程度の熱安定性を有する非マルトース生成エキソアミラーゼが、ベーキングされた生成物における有害な劣化を遅延し、又は低めるのに非常に効果的である驚くべき発見に基づかれている。
【0098】
寄託
次のサンプルが、1999年3月12日、公認された寄託機関DSMZ(Deutsche Sammlung von Microorganismen und Zellkulturen GmbH of Mascheroder Weg 1b, D-38124 Braunschweig)に、ブダペスト条約に従って寄託された:BT−21 DSM番号DSM12731。
本発明はまた、それらの寄託物及びそれを含んで成る具体物から誘導でき、そして/又は発現できる配列、及びその活性フラグメントも包含する。
【実施例】
【0099】
例−セクションA
例1.シュードモナス・サッカロフィラ非マルトース生成エキソアミラーゼの発酵及び生成
1.1生成:
P.サッカロフィラ株IAM No. 1544を、IAM Culture Collection, Inst. Of Mloecular and Cellular Biosciences, University of Tokyo, Japan から得た。
【0100】
前記菌株の発酵を、2Lの使用体積を有するApplikon ADIの3Lの生物反応器において、次の条件下で行った:
温度: 30℃
撹拌速度: 1000rpm
通気 1体積空気/1体積培地/分
pH: 2Mの水酸化ナトリウム及び10%(w/v)の塩酸による調節による一定のpH7.4
【0101】
培地: Bacto Trypton 20g/l
Bacto Yeast 抽出物 20g/l
澱粉 20g/l
Na2HPO4・2H2O 5.6g/l
KH2PO4 1.5g/l。
1日の発酵の後、発酵ブイヨンを遠心分離し、そして濾過し、細胞を除いた。細胞フリーブイヨンにおける非マルトース生成エキソアミラーゼの活性は、上記のようにして決定される場合、5単位/mlであった。
【0102】
1.2 P.サッカロフィラ非マルトース生成エキソアミラーゼの精製:
P.サッカロフィラの非マルトース生成エキソアミラーゼを、200mMの硫酸ナトリウム、50mMのトリエタノールアミン、2mMの塩化カルシウムを含むA−緩衝液(pH7.2)を含む、150mlのPhenyl Sepharose FF低サブカラム(Pharmacia, Sweden) を用いて、疎水性相互作用グロマトグラフィーにより、部分的に精製した。濾過された発酵ブイヨン(500ml)を、200mMの硫酸ナトリウムによりpH7.2に調節し、そしてカラム上に負荷した。非マルトース生成エキソアミラーゼを、A−緩衝液中、直線的に低下する硫酸ナトリウムグラジエントにより遊離した。エキソアミラーゼ活性を含む画分をプールした。
【0103】
プールされた画分を水により3倍に希釈し、そしてさらに、50mMのトリエタノールアミン、5mMの塩化カルシウムから成るA−緩衝液(pH7.5)により平衡化された150mlのQ−Sepharose FF (Pharmacia) カラム上でのアニオン−交換クロマトグラフィーにより精製した。非マルトース生成エキソアミラーゼを、A−緩衝液中、0〜1Mの塩化ナトリウム直線グラジエントにより溶離した。エキソアミラーゼ活性を含む画分をプールした。この部分的に精製された調製物を、下記試験のために使用した。それは、アミラーゼ活性について染色されたポリアクリルアミドゲル電気泳動システムにおいて試験される場合、14.7単位/mlの活性及びわずか1つのアミラーゼ活性の結合を有した。
【0104】
1.3 P. サッカロフィラ非マルトース生成エキソアミラーゼの特徴化:
最初に、P.サッカロフィラのエキソ−アミラーゼ(PS4と呼ばれる)をコードする遺伝子のDNA配列が、Zhouなど(Zho JH, Baba T, Takano T, Kobayash S, Arai Y (1989) FEBS Lett 1989 Sep 11; 255 (1): 37-41 “Nucleotide sequence of the maltotetraohydrolase gene from Pseudomonas sacchrophila”)により公開されている。さらに、前記DNA配列は、受託番号X16732としてGenBankから入手できる。
【0105】
本発明者は、質量分光計(MALDI−TOFF)によれば、精製されたPS4酵素のMWが57500±500D(配列からの理論的MWによれば、57741Dである)であることを決定した。
PS4の最適温度及びpHは、Zhouなど(Zhon JH, Baba T, Takano T, Kobayashi S, Arai Y (1992) Carbohydr Res 1992 Jan; 223: 255-61 “Properties of the enzyme exzyme expressed by the Pseudomonas saccharophila maltotetrahydrolase gene (mta) in Escherichia coli”)によれば、45℃及びpH6.5である。
【0106】
非マルトース生成エキソアミラーゼの加水分解パターンを、上記のようにして、10mg/mlのモチ状トウモロコシ澱粉4mlを有する試験管において、0.7単位の部分的な精製された非マルトース生成エキソアミラーゼを50℃で15又は300分間インキュベートすることによって生成される加水分解生成物を分析することによって決定した。
【0107】
15分及び300分後に検出される加水分解生成物のパターンは、表1に示されており、そしてP.サッカロフィラが本発明において定義されるような非マルトース生成エキソアミラーゼを生成し、そしてこの酵素が、それぞれ、15分及び30分の加水分解の後、85.8重量%及び93.0重量%を占める有力な生成物としてマルトテトラオースを開放することを示す。
【0108】
【表1】

【0109】
例2.P. サッカロフィラ非マルトース生成エキソアミラーゼのベーキング試験
ベーキング試験を、P.サッカロフィラ非マルトース生成エキソアミラーゼの抗固化効果を試験するために設定した。Danish Toast Bread のためのレセピーを使用した。それは、食料粉(2000g)、乾燥酵母(30g)、糖(30g)、塩(30g)及び水(400g Brabender Units (BU)のドウコンシステンシーに対応する約1200g+使用される乾燥酵母を補足するための追加の水60g)を含み、そしてそれらを、遅い速度で2分間及び早い速度で12分間、Hobartミキサー(モデルA−200)において混合する。
【0110】
ドウ温度は、混合の最後で26℃である。ドウを30℃で10分間、放置し、この後、ドウを750gのドウ部分に分割する。そのドウ部分を、33℃の温度及び85%の相対湿度で試験キャビネットにおいて5分間、放置する。次に、ドウ部分を、次の設定でGlimek成形機(タイプLR−67)上で成形し:1:4、2:2、3:14及び4:12、この後、成形されたドウ部分を、ベーキング用缶に移し、そして33℃の温度及び85%の相対湿度で50分間、試験キャビネットにおいて試験する。最終的に、試験されたドウ部分を、Wachtel オーブン(モデルAE416/38COM)において、220℃の温度で40分間、10秒間の蒸気を伴ってベーキングする。
【0111】
P.サッカロフィラ非マルトースエキソアミラーゼの部分的に精製された調製物を、食料粉1kg当たり1470単位で、導入されるドウに添加した。ベーキングした後、非マルトース生成エキソアミラーゼを有するか又は有さないパンを20℃に冷却し、そしてその後、プラスチックバックにおいて20℃で貯蔵した。堅さを、ベーキングの後3日目、1つのパンからの10枚のスライスの平均として及び7日目、2個のパンからのそれぞれ10枚のスライスの平均として、それぞれ3日目、Instron 4301 Universal Food Texture Analyzer により決定した。表2は、酵素を有するパンにおいて低い堅さが、両日で観察されたことを示す。
【0112】
【表2】
【0113】
表3は、7日目、P.サッカロフィラ非マルトース生成エキソアミラーゼの抗固化効果が95%の信頼レベルで統計学的に有意であることを示す。
【表3】
【0114】
The StatAdvisor:
ANOVA表は、次の2種の成分に7日目での堅さの変動を分ける:グループ間成分及びグループ内成分。この場合、72.0に等しいF−比率は、グループ内推定値に対するグルループ間推定値の比率である。F−試験のP−値は0.05以下であるので、95.0%の信頼レベルで、1つの酵素レベルからもう1つの酵素レベルでの平均堅さ間に統計学的に有意な差異が存在する。平均が他の平均と有意に異なるかどうかを決定するためには、Tabalar Options の列挙からMaltiple Range Testを選択すること。
【0115】
例3.
次のことは、E.コリMC1061におけるシュードモナス・サッカロフィラからの非マルトース生成エキソアミラーゼをコードするmta遺伝子の本発明のクローニング及び発現を記載する。
この観点においては、P.サッカロフィラIAM1520を、2mlのLB培地において増殖し、そして細胞を、20,000xgで10分間の遠心分離により収穫した。全DNAを、わずかに変更されたminiprep プロトコールを用いて単離した。細胞を、300μlの再懸濁緩衝液(50mMのトリス−HCl、pH8.0:10mMのEDTA;100μg/mlのRNアーゼA)に再懸濁し、この後、細胞を、Fastprep FP120 (BIO101; California)を用いて粉砕した。
【0116】
粉砕に続いて、300μlの溶解緩衝液(200mMのNaOH;1%のSDS)及び300μlの中和緩衝液(3.0Mの酢酸カリウム、pH5.5)を添加した。20.000xgで、4℃で15分間の遠心分離の後、上清液を集め、そして0.6体積のイソプロパノールを添加した。DNAを、4℃で、20,000xgでの30分間の遠心分離により沈殿せしめ、70%エタノールにより洗浄し、そして100μlのTE(10mMのトリス−HCl, pH8.0, 1mMのEDTA)に再溶解した。
【0117】
PCR増幅のために、次の4種の異なったPCRプライマーを企画した:
#1 ATG ACG AGG TCC TTG TTT TTC 位置213−233
#2 GCT CCT GAT ACG ACA GCG 位置2403−2386
#3 GCC ATG GAT CAG GCC GGC AAG AGC CCG 位置663−683
#4 TGG ATC CTC AGA ACG AGC CGC TGG T 位置2258−2238
【0118】
前記位置は、GenBank 受託番号X16732号に見出されるmtaについての配列に関する。上方の番号のプライマーはまず、アンチセンスプライマーであり、そしてその配列は相補的配列である。鋳型DNAに対して相補的でないヌクレオチドは太字で示され、そして導入された制限部位は下線で示される。プライマー#3は、ユニークNco1部位を導入し、そしてプライマー#4はBamHI部位を導入し、この部位は、発現ベクターpBAD/gIII (Invitrogen) における次のクローニングのために使用される。
【0119】
第1のPCR増幅を、次のプライマーの組み合わせ:反応1:2190bpのフラグメントを付与する#1+#2を用いて、製造業者の説明書及び次の増幅プロトコールに従って、Expand DNA ポリメタ−ゼ(Boehringer Mannheim; Germany)、及び鋳型としての50〜150ngのゲノムIAM 1520 DNAを用いて行った:35サイクルの間94℃で2分(94℃で1分、58℃で2分、72℃で2分)、及び最終的に、72℃で5分。
【0120】
2190bpのフラグメントを、“gene Clean Kit (BIO 101; California)を用いて、ゲルから単離した。そのフラグメントを、上記と同じ増幅プロトコールを用いて、第2のPCRにおいて、次のプライマーの組み合わせを用いて、鋳型DNAとして使用した:
反応2:1605bpのグラグメントを付与する#3+#4.
【0121】
1605bpのフラグメントを精製し、そして製造業者の説明書に従って、PCR−BLUNTベクター(Invitrogen)中にクローン化した。そのクローン化されたフラグメントの配列を、普遍及び逆プライマー及び4種のラベルされた内部プライマーを用いて、単一色素配列決定技法及びALF配列決定機(Pharmacia;Sweden)により配列決定することによって確かめた:
【0122】
CAT CGT AGA GCA CCT CCA 999-982
GAT CAT CAA GGA CTG GTC C 1382-1400
CTT GAG AGC GAA GTC GAA C 1439-1421
GAC TTC ATC CGC CAG CTG AT 1689-1780
前記位置は、GenBan受託番号X16732号に見出されるmtaについての配列を示す。上方の番号のプライマーはまず、アンチセンスプライマーであり、そしてその配列は相補的配列である。
【0123】
配列を確かめた後、mta遺伝子を発現ベクターpBAD/gIII(Invitroge)中にクローン化した。Mta 遺伝子は、BamHIによる消化、続いてクレノウフラグメントによるブラント化及びNcoIによる消化によりpCR−BLUNTから開放し、そして1620bpのフラグメントを精製した。発現ベクターpBAD/gIIIをNcoI及びPmeIにより消化し、そして精製した。連結の後、得られる発現構造体を用いて、E.コリMC1061細胞を形質転換し、そしてタンパク質を、pBAD/gIIIマニュアル(Invitrogen)に従って発現した。
【0124】
例4.澱粉劣化に対する非マルトース生成及びマルトース生成エキソアミラーゼの効果の比較
サツマイモβ−アミラーゼ(EC 3. 2. 1. 2; 製品番号A7005号としてSigmaから得ることができる)は、澱粉の非還元末端からマルトースを開放するマルトース生成エキソアミラーゼである。
このマルトース生成エキソアミラーゼの熱安定性は、50mMのクエン酸ナトリウム、5mMの塩化カルシウム溶液(pH6.5)において45〜75℃の温度で15分間インキュベートした後、残留活性により示されるように、P.サッカロフィラ非マルトース生成エキソアミラーゼの熱安定性に類似する(表4)。
【表4】
澱粉劣化に対する両酵素の効果を、“Assys for measurement of retrogradation and staling”に記載されるように、モデルシステムドウからのベーキングされ、そして貯蔵された生成物のDSC分析により試験した。この試験に関しては、“アミラーゼアッセイプロトコール”に従ってアッセイされた、485単位のP.サッカロフィラ非マルトース生成エキソアミラーゼ及び735単位のサツマイモβ−アミラーゼをドウに使用した。ドウを、50mMのクエン酸ナトリウム、5mMの塩化カルシウム溶液(pH6.5)30.7ml又は添加される酸素、及び50gの標準デンマーク小麦粉(Danisco 98022)から調製した。
【0125】
7日間の貯蔵の後、劣化されたアミロペクチンの量を、その溶融エントロピーを測定することによって定量化した。統計学的分析によれば、両酵素は澱粉劣化を有意に低め(表5);すなわちP.サッカロフィラ非マルトース生成エキソアミラーゼは、7日目、劣化されたアミロペクチンの量を86%(1.77 J/g)に低め、ところがサツマイモβ−アミラーゼは、それを96%(1.96 J/g)に低めることが見出された。結論的には、P.サッカロフィラ非マルトース生成エキソアミラーゼは、比較できる熱安定性のマルトース生成アミラーゼよりも、劣化及び老化を低めるために、明らかにより効果的である。
【0126】
【表5】
【0127】
この表は、平均が他の平均と有意に異なるかどうかを決定するために、複数の比較方法を利用する。アウトプットの低部半分は、個々の対の平均間の推定される差異を示す。星印は3個の対に続いて配置され、これは、それらの対が95.0%の信頼レベルで統計学的に有意な差異を示すことを示唆する。平均間を区別するために現在使用される方法は、Fisher’s最少有意差異(LSD)方法である。
【0128】
例−セクションB
材料及び方法:
材料:コーンからのアミロペクチン及びアミロース、コーン澱粉、カルボキシメチルセルロース(CMC)、ウシ血清アルブミン(BSA)、デキストラン、プルラン、マルトース、マルトトリオース、及びマルトテトラオース〜マルトデカオースの混合物を、Sigma Chemical Co., St. Louis, USAから得た。可溶性澱粉を、Merck, Darmstadt, Germany から得た。酵素抽出物及びトリプトンは、Difco Laboratories, Detroit, USAから得られた。
【0129】
Neue Allgemeine Reisgesellschaft mbH, Hamburg, Germany からの完全な玄米が使用された。医薬品種のα−、β−及びγ−シクロデキストリンは、Wacker Chemie Danmark Aps, Glostrup, Denmark から得られた。マルトテトラオースは、前記のようにして調製された[32]。すべての化学物質は、特にことわらない限り、分析用銘柄のものであった。
B.クラウジBT−21の単離:前記株は、DSMZ(Deutsche Sammlung von Mikroorganismen and Zellkulturen GmbH, Braunshweig, Germany)により同定された、Assens, Denmark により集められた土壌サンプルから単離される。
【0130】
酵素の生成:B.クラウジBT−21を、ジャガイモからの2.0%可溶性澱粉トウモロコシからのアミロペクチン、コーン澱粉又は完全な玄米、0.5%酵母抽出物、0.5%トリプトン、0.1%KH2PO4, 0.1%Na2HPO4, 0.02%MgSO4・7H2O, 0.02% CaCl2・2H2O 及び0.1%(NH4)2 SO4から成る最適化された液体培地において増殖させた。オートクレーブの後、無菌Na2CO3溶液を添加し、1%の最終濃度にした(約pH10)。グリセロール中の胞子懸濁液(−80℃で貯蔵される)を用いて、100mlの実際の培地を接種し、そして200rpmでの振盪インキュベーターにおいて45℃で18時間インキュベートした(New Brunswick Scientific, Edison, N.J., USA)。
【0131】
この培養物2mlを用いて、200mlの培地を有する振盪フラスコを接種し、そして振盪インキュベーターにおいて45℃でインキュベートした。アリコートを規則的な間隔で採取し、そして600nmでのODを測定し、培地における菌類の増殖を決定した。サンプル(4ml)を、9600rpmで4℃で10分間、遠心分離し、そしてpH及びアミラーゼ活性を決定した。すべての増殖実験は三重反復して行われた。平均値(X=(Σni=1Xi)/m)及び標準偏差値(std.=√(Σni=1(Xi-X)/(n-1))を決定した。
【0132】
生成物−特異的アミラーゼの精製:完全な玄米上でのB.クラウジBT−21の52時間の増殖の後、細胞及び完全な米穀粒を、9600rpm及び4℃での15分間の遠心分離により細胞外流体(1000ml)から除いた。生成物−特異的アミラーゼを、エポキシ−活性化されたセファロース6Bマトリックス(Pharmacia Biotech, Uppsala, Sweden)にβ−シクロデキストリンを共有結合することによって調製された親和性ゲルを用いて精製した[33]。細胞外細胞−フリー上清液を、12gのゲルと共に、4℃で1時間、振盪しながらインキュベートした。
【0133】
次に、上清液を、9600rpm及び4℃で10分間の遠心分離により除去した。結合されなかったタンパク質を、50mMのリン酸緩衝液(pH8.0)75mlによりゲルを洗浄し、続いて遠心分離することによって除去した。洗浄段階を7度反復した。結合されたタンパク質を、10mMのα―シクロデキストリンを含む50mMの燐酸緩衝液(pH8.0)45mlにより溶出し、続いて遠心分離した。溶出段階は4回反復された。α−シクロデキストリンを、β−及びγ−シクロデキストリンはBradford (1976)のタンパク質決定方法を妨害するので、酵素の溶出のために使用した[34]。
【0134】
次に、α−シクロデキストリンを、4℃で撹拌しながら、10mMのトリエタノールアミン(pH7.5)5Lに対する透析(6〜8kDaのSpectra/Por透析膜、The Spedtrum Companies, Gardena, CA, USA)により除去した。緩衝液を2時間後に交換し、続いてさらに12時間、透析した。透析バッグをCMCに配置し、サンプルを濃縮した。10mlを、FPLC−システム(Pharmacia, Uppsala, Sweden)を用いて、HiTrap Qカラム(Pharmacia Biotech, Uppsala, Sweden)に適用した。
【0135】
タンパク質を、10mMのトリエタノールアミン(pH7.5)25mlにより1.0ml/分の速度で、続いて、10mMのトリエタノールアミン(pH7.5)中、20mMのNaClのグラジエントにより溶出した。酵素を0.5MのNaClにより溶出した。タンパク質の含有率を、BIO−RAD Protein Assay (Bio-Rad Laboratories, Hercules, CA, USA)を用いてBradford, (1976)の方法[34]により評価した。BSAを標準として利用した。
【0136】
ゲル電気泳動:15μlのサンプルを、[35]により記載されるようにして、天然のトリス−グリシンゲル(10%)により分析した。次に、ゲルを、pH6.5での50mMのリン酸緩衝液に配置し、そして30分間、振盪した。1%(w/v)の可溶性澱粉溶液を、ゲルと共に45分間、振盪しながらインキュベートした。緩衝溶液により洗浄した後、ゲルをヨウ素溶液(4mMのI2, 160mM のKI)と共にインキュベートし、そして緩衝液により脱色した。脱色されバンドは、澱粉加水分解活性を示した。
【0137】
SDS−PAGE(10%)を、[36]に従って行い、続いて銀染色[37]した。SDS−PAGE広範囲分子量標準(Bio-Rad Laboratories, hercules, CA, USA)を用いた。
酵素アッセイ:0.1Mの硼酸緩衝液(pH10.0)中、2mlの可溶性澱粉溶液(1.25%)を、0.5mlの酵素溶液と共に45℃で2時間インキュベートした。反応を、その混合物を10分間、煮沸することによって停止した。還元糖の形成を、CuSO4/ビシンコネート アッセイ[38]により決定し、そして形成されるmMマルトース同等物として計算した。1単位の活性は、pH10.0及び45℃で1分当たり1μモルのマルトース同等物を生成する酵素の量に対応する。
【0138】
酵素特徴化:温度最適値の決定のために、精製された酵素を、0.1Mの硼酸緩衝液(pH10.0)(0.5mMのCaCl2を含むか又は含まない)中、1%の最終濃度の可溶性澱粉において、30℃〜90℃の温度で15分間インキュベートした。温度安定性の決定を、5mMのCaCl2を含む50mMのグリシン−NaOH緩衝液(pH9.5)において、精製された酵素を30分間、30, 40, 50, 60, 70, 80及び90℃でインキュベートすることによって行った。
【0139】
残留活性を、50mMのグリシン−NaOH緩衝液(pH9.5)中、1%の最終濃度の可溶性澱粉において、加熱処理された酵素を、55℃で15分間インキュベートすることによって決定した。PH最適値を、異なった緩衝液中、1%の最終濃度の可溶性澱粉において、精製された酵素を55℃で15分間インキュベートすることによって決定した。使用される緩衝液は、50mMのクエン酸塩(pH4.0〜6.0)、50mMのトリス−マレエート(pH6.5〜8.5)及び50mMのグリシン−NaOH(pH9.0〜11.0)であった。
【0140】
I2−KI溶液(0.02%のI2及び0.2%のKI)を、Fuwa、1954[39]に従って調製した。澱粉−ヨウ素青色形成を、次の変法により二重反復して測定した。サンプル500μlを、異なった時間間隔で、可溶性澱粉の酵素加水分解から回収した。次に、250μlのHCl及び250μlのI2−KI溶液を添加し、そして混合した。脱イオン水(4.0ml)を添加し、そして混合を反復した。青色の形成を、600nmで分光光学的に測定した。
【0141】
精製された酵素による異なった基質の加水分解を、ジャガイモからの可溶性澱粉、トウモロコシからのアミロペクチン、デキストラン、プルラン(1%)、アミロース(0.1%)、及び10mMのα−、β−及びγ−シクロデキストリンにより試験した。基質を、5mMのCaCl2を含む、50mMのグリシン−NaOH緩衝液(pH9.5)に溶解し、そして精製された酵素を添加した(505mU/ml)。種々の基質を55℃でインキュベートし、そしてサンプルを下記のようにして分析した。
【0142】
精製された酵素によるマルト−オリゴ糖の加水分解を、2mMの最終濃度でマルトース、マルトトリオース及びマルトテトラオースにより、及びマルトテトラオース〜マルトドテカオースの混合物(5mM)により試験した。マルト−オリゴ糖を、5mMのCaCl2を含む50mMの硼酸緩衝液(pH9.5)に溶解し、そして精製された酵素を添加した(147mU/ml)。基質を55℃でインキュベートし、そしてサンプルを異なった時間間隔で採取した。反応を10分間の煮沸により停止し、そしてサンプルを下記のようにして分析した。
【0143】
加水分解生成物の分析:加水分解生成物を、パルスされた電流滴定検出を有する高性能アニオン交換クロマトグラフィー(HPAEC−PAD)を用いて検出した。CarboPac PA-1カラム(Dionex Corporation, Sunnyvale, CA, USA)を、100mMのNaOH中、30分間にわたっての0〜60%の1.0Mの酢酸ナトリウムグラジエント及びDionex DX-300又はDX-500システム上での1.0ml/分の流速と共に使用した。澱粉加水分生成物を、グルコース、マルトース、マルトトリオース、マルトテトラオース、マルトペンタオース及びマルトヘキサオースとそれらの保持時間とを比較することによって同定した。相同の線状マルト−オリゴ糖の保持時間は、重合の程度と共に上昇するので、中間DPの線状マルト−オリゴ糖は容易に同定され得る[40, 41]。
【0144】
結果及び論議:
B.グラウジBT−21の同定:その脂肪酸組成によれば、前記菌株は、バチルス属に対する類似性を示した。16SrDNAの部分配列決定は、B.クラウジに対して99.4%の類似性を示した。アルカリ耐性株の生理学的性質がこの同定を確認した。
B.グラウジBT−21によるアミラーゼ活性の生成:ジャガイモからの可溶性澱粉、コーン澱粉、トウモロコシ及び玄米からのアミロペクチンは、基材において異なったレベルの細胞外アミラーゼ活性をもたらした。
【0145】
コーン澱粉は、ジャガイモ澱粉に比較してより脂質を含み、そしてリンを含まないが、トウモロコシからのアミロペクチンは、α−D−(1→6)O−グリコシド結合を含む高い枝分かれ鎖構造を有する。それらの3種のタイプの澱粉は、加熱ゲル化の後、酵素のために入手でき、ところが完全な玄米は米穀粒に封入される低い入手可能性の澱粉を含む。異なった澱粉基質を含む液体培養物の細胞外流体におけるアミラーゼ活性が図1に示される。最高の澱粉分解活性が基質として完全な玄米により得られた。これは、低い入手可能性澱粉基質の存在がB.クラウジBT−21による細胞外澱粉分解活性の高められた生成をもたらした。
【0146】
類似する結果が、酵素の精製の前、細胞外流体から除去することが困難である小麦ふすまにより得られた。炭素源、たとえばガラクトース、グリコーゲン及びイヌリンはこれまで、B.リケニホルミスによるアミラーゼ生成のために適切であるものとして報告されており[27]、そして可溶性澱粉がB.ステアロサーモフィラスによるアミラーゼの生成のために最良の基質として見出された[28]。しかしながら、それらの研究のどれも、低い入手可能性の澱粉基質を包含していない。
生成物−特異的アミラーゼの精製:酵素を、β−CD Sepharose 6Bによる親和性クロマトグラフィー、続くアニオン交換クロマトグラフィーにより精製した(表6)。
【0147】
【表6】
【0148】
活性染色された天然のPAGEは、細胞外流体における3種の澱粉分解活性の存在を示した。生成物−特異的酵素を、β−CD親和性クロマトグラフィー、続くアニオン−交換クロマトグラフィー処理の後、他の澱粉分解活性から完全に分離した。精製された酵素調製物のSDS−PAGEは、生成物−特異的アミラーゼが相同性に精製され、そして約101kDaの見掛け分子量を有することを示した。シクロデキストリン セファロース6B親和性グロマトグラフィーによる他のアミラーゼの除去の後、最終精製段階としてα−アミラーゼの精製のためにこれまで使用して来た[29]。8.7%の酵素回収率、及び生成物−特異的アミラーゼのために得られた18.5の精製因子は、それらの著者により報告される値に類似した。
【0149】
生成物−特異的アミラーゼの特徴化:精製された酵素は、pH9.5での活性の最適値を示し(図2)、そしてその活性のための最適温度は、5mMのCaCl2の存在下で、又は不在下で55℃であった(図3)。5mMのCaCl2の存在下でpH9.5での酵素の熱安定性が図4に示される。55℃以上の温度で、酵素は、30分のインキュベーション期間の間、その最大活性の75%を失った。マルトヘキサオース[23]、マルトペンタオース[24]及びマルトテトラオース[25]を形成する5種の他の生成物−特異的アミラーゼはまた、アルカリpH最適性を示す。
【0150】
それらの酵素の分子量は、59, 73及び80kDa[23], 180kDa[24]及び97kDa[26]であることが推定され、そしてB.グラウジBT−21からの生成物−特異的アミラーゼは101kDaの推定分子量を示した。生成物−特異的アミラーゼ及びα−アミラーゼは、50〜65kDaの範囲の低い分子量を示す[3, 16, 17, 19及び21]。約55℃の温度適度は、上記生成物−特異的アミラーゼについて報告される最適度に類似した[23, 24及び26]。
【0151】
精製された生成物−特異的アミラーゼは、低いDPの主な初期生成物として、1時間のインキュベートの後、可溶性澱粉を、主にマルトヘキサオース及びマルトペンタオース(合計の加水分解された生成物の52%及び19%)に加水分解した(図5)。2時間のインキュベーションの後、マルトヘキサオースの量、及び4時間後、マルトペンタオースの量は低下し、そしてマルトテトラオース、マルトトリオース、マルトース及びグルコースの量は低下した。それらの生成物は延長された加水分解の後、蓄積し、このことは、それらがさらに加水分解されなかったことを示す。
【0152】
24時間の澱粉加水分解の後、マルト−オリゴ糖の量は3%であり、マルトヘキサオースの量は(3%)マルトヘキサオース、(4%)マルトペンタオース、(41%)マルトテトラオース、(13%)マルトトリオース、(16%マルトース及び(4%)グルコースであった。30分の澱粉加水分解の後に得られる、パルスされた電気滴定検出を有する高性能アニオン−交換クロマトグラフィー(HPAEC−PAD)形跡は(図6)、マルトヘキサオース(DP6)よりも大きな澱粉加水分解生成物が不在であることを示す。マルトヘキサオース−形成生成物−特異的アミラーゼによる可溶性澱粉の加水分解の時間経過研究はまた、マルトヘキサオースが加水分解の初期段階において、選択的に生成されたことも示す[23]。
【0153】
Kimなど(1995)[26]は、マルトテトラオース−形成生成物−特異的アミラーゼによる澱粉の1時間の加水分解の後の初期加水分解生成物が主にマルトヘキサオース(54%)であり、続いてマルトテトラオース及びマルトースの量の徐々の上昇が存在し、そしてマルトヘキサオースの量が低下したことも見出した。20時間後、マルト−オリゴ糖の組成が、0.6%のマルトヘキサオース、1.3%のマルトペンタオース、53.2%のマルトテトラオース、8.3%のマルトトリオース、27.6%マルトース及び9%のグルコースに変化した。
澱粉加水分解の間、酵素の作用のモードをさらに試験するために、異なった基質を、精製された酵素と共にインキュベートした(表7)。
【0154】
【表7】
【0155】
澱粉は、アミロース(20〜30%)及びアミロペクチン(80〜70%)から構成される。アミラーゼはα−D−(1→4)O−グルコシド結合された線状グルカンであり、そしてアミロペクチンは分子におけるα−D−(1→6)O−グリコシド結合の存在のために枝分かれ鎖のグルカンである。生成物−特異的アミラーゼは容易に、アミロペクチンを加水分解し、それは、可溶性澱粉(7%及び17.8%)及びアミロース(6.3%及び20%)に比較して、マルトペンタオース(9.2%)及びマルトヘキサオース(24.8%)の形成により示された。
【0156】
酵素は、プルラン、すなわちマルトトリオースを鎖から構成されるα−(1→6)O−グリコシド結合グルカン、又はデキストラン、すなわち0〜3個の主鎖単位に結合される技を有するα−(1→6)O−グリコシド結合グルカンを加水分解しなかった。α−、β−又はγ−シクロデキストリン、すなわち6.7及び8個のグルコース単位から構成される環状マルト−オリゴ糖はまた、24時間のインキュベーションの後でさえ、加水分解されなかった。
【0157】
デキストランに基づいて得られた結果は、α−(1→6)O−グリコシド結合が生成物−特異的アミラーゼにより分解されなかったことを示した。プルランに対する活性の欠失は、生成物−特異的アミラーゼが3個のグルコース単位に続くα−(1→6)O−グルコシド結合を回避することができず、又は3個のグルコース単位のいずれかを結合し得ないことを示した。α−、β−又はγ−シクロデキストリンに対する加水分解の欠失は、生成物−特異的アミラーゼがエキソタイプの分解機構により澱粉を加水分解したことを示した[30]。HPAEC−PAD形跡(図6)はまた、DP6よりも長い澱粉加水分解生成物が不在であるので、エキソータイプの分解機構を示した。
【0158】
可溶性澱粉に対する酵素分解作用をさらに試験するために、澱粉−ヨウ素青色形成を、還元糖の生成に対してプロットした(図7)。曲線の傾斜は、澱粉分解澱粉加水分解の有力な型の分解機構を示す[31]。エンド−作用性酵素は、エキソ−作用性酵素に比較される小さな値を有する傾斜を生成するであろう。傾斜の小さな値は、A.オリザエからのα−アミラーゼにより示される、ランダム澱粉分解活性による澱粉−ヨウ素青色複合体の早い還元の結果である(傾斜は−61である)。P.スツゼリからの細胞外酵素調製物は、大きな傾斜値により示される有力なエキソ−作用分解機構のための証拠を示した(傾斜は−13である)。精製された生成物−特異的アミラーゼは、エキソ−活性を示す、−6に傾斜値を示した。
【0159】
生成物−特異的アミラーゼによる、低いDPを有する基質の加水分解の作用のモードを、そのような基質と共にインキュベートすることによって試験した。マルトース、マルトトリオース及びマルトテトラオースは、精製されたB.クラウジBT−21アミラーゼにより加水分解されなかった。これは、それらの生成物が蓄積し、そして従って、加水分解の最終生成物として見なされる、可溶性澱粉に対して得られる結果を確認する。
【0160】
DP4〜DP10のマルト−オリゴ糖の混合物に対する生成物−特異的アミラーゼ活性を、時間経過実験により研究した。HPAEC−PADにより得られるピーク領域の変化は、マルト−オリゴ糖の形成又は加水分解に対応する。マルトヘキサオース(DP6)の形成、及びDP7, DP8, DP9及びDP10の量の同時低下が、酵素のマルトヘキサオース形成能力を確認した。しかしながら、定常状態条件が達成され、そして澱粉加水分解により見出されるようなDP6のさらなる分解が加水分解の7日後でさえ、検出されなかった。DP6の濃度は、澱粉加水分解で得られる濃度よりも低く、そしてこのことは、一定量のマルトヘキサオースが生成するマルトテトラオース及びマルトースの形成のために必要とされた。
【0161】
B.クラウジBT−21生成物特異的アミラーゼによる澱粉加水分解は、2段階工程に類似することが見出された。この工程は、主にマルトヘキサオース及び少量のマルトペンタオースへの澱粉の初期加水分解を包含し、それらは、集中的な加水分解の後、蓄積する主要マルトテトラオース及びマルトースにさらに加水分解された。マルトテトラオース及びマルトースへの第2加水分解段階は、濃度依存性が進行する第2段階のための調節体と思われるので、大きな基質のマルトヘキサオースへの予備加水分解により制限されると思われた。
【0162】
ベーキング実験:
ベーキング実験を、生成物−特異的アミラーゼにより行った。ドウを、10gの標準デンマークの小麦粉(Danisco 98078)、及び40単位の酵素(セクションBの材料及び方法に記載されるように、45℃及びpH10でアッセイされる)を有さない(対照)又は有する、0.2MのNaOH−グリシン緩衝液(pH10)6.2mlにより調製し、ベーキングし、そして“劣化及び老化の測定のためのアッセイ”に従って、貯蔵の後、DSCにより分析した。表8に示されるように、酵素は、それが有意な抗老化効果有することを示すベーキングの7日後に見出される劣化されるアミロペクチンの量を有意に低める。
【0163】
【表8】
【0164】
ベーキングの後、凍結されたベーキングされた生成物のパン中味サンプルを、蒸留水により抽出し(1gのベーキングされた生成物/10gの水を、1時間撹拌し、そして遠心分離した)、そして上記のようにしてHPAEC−PADにより分析し、ドウのベーキングの間、酵素により形成される澱粉加水分解生成物を検出した。対照に対する、マルトテトラオース、マルトペンタオース、マルトヘキサオース及びマルトヘプタオースの蓄積が、この酵素の活性の結果として見出された。
【0165】
発明の要約
本発明は製パン製品を製造するための方法、及びそのような方法への使用のために適切なアミラーゼを開示する。
本発明の好ましい態様は、次の番号付けされた文章により示される。
1.遅延された老化性質を有するベーキングされた澱粉質パン生成物を製造する方法が提供され、ここで前記方法は、ベーキング工程の間、アミロペクチンの側鎖の非還元末端から、4〜8個のD−グルコピラノシル単位から主に成る線状マルトオリゴ糖を分解することによって、澱粉を加水分解することができる有効量の非マルトース生成エキソアミラーゼのドウ成分、ドウ添加物又はドウへの添加を含んで成る。
【0166】
2.前記食料粉が小麦粉又はライ麦粉、又は小麦粉及びライ麦粉の混合物である前記1の方法。
3.前記非マルトース生成エキソアミラーゼが、kg食料粉当たり50〜100,000単位、好ましくはkg食料粉当たり100〜50,000単位の範囲で存在する量で添加される前記1又は2の方法。
4.前記非マルトース生成エキソアミラーゼがkg食料粉当たり200〜20,000単位の範囲で存在する量で添加される前記3の方法。
【0167】
5.前記非マルトース生成エキソアミラーゼが単位エキソアミラーゼ活性当たり0.5以下のエンドアミラーゼ単位(EAU)のエンドアミラーゼ活性を有する前記1〜4のいずれかに記載の方法。
6.前記非マルトース生成エキソアミラーゼが単位エキソアミラーゼ活性当たり0.05以下のエンドアミラーゼ単位(EAU)のエンドアミラーゼ活性を有する前記1〜4のいずれかに記載の方法。
7.前記非マルトース生成エキソアミラーゼが単位エキソアミラーゼ活性当たり0.01以下のエンドアミラーゼ単位(EAU)のエンドアミラーゼ活性を有する前記1〜4のいずれかに記載の方法。
【0168】
8.前記非マルトース生成エキソアミラーゼが、0.7単位の量の前記非マルトース生成エキソアミラーゼが、50mMの2−(N−モルホリノ)エタンスルホン酸及び2mMの塩化カルシウムを含む緩衝溶液1ml当たり10mgの予備煮沸されたトウモロコシ澱粉の水溶液4mlにおいて、50℃の温度で、pH6.0で15分間インキュベートされる場合、それが、少なくとも60重量%、好ましくは少なくとも70重量%、より好ましくは少なくとも80重量%及び最も好ましくは少なくとも90重量%の、グルコース、マルトース、及び3〜10個のD−グルコピラノシル単位の線状マルトオリゴ糖、好ましくは4〜8個のD−グルコピラノシル単位の線状マルトオリゴ糖から成る加水分解生成物を生成することを特徴として、ここで前記加水分解生成物は、パルスされた電流滴定検出、及び標準としてグルコース〜マルトヘプタオースの既知の線状マルトオリゴ糖と共にDionex PA100カラムを用いてアニオン交換HPLCにより分析される前記1〜4のいずれかに記載の方法。
【0169】
9.前記非マルトース生成エキソアミラーゼが、前記8記載の条件下でインキュベートされ、そして前記加水分解生成物が請求項8記載のようにして分析される場合、それが少なくとも60重量%、好ましくは少なくとも70重量%、より好ましくは少なくとも80重量%及び最も好ましくは少なくとも90重量%のマルトテトラオースを生成する前記8の方法。
10.前記非マルトース生成エキソアミラーゼが、前記8記載の条件下でインキュベートされ、そして前記加水分解生成物が請求項8記載のようにして分析される場合、それが少なくとも60重量%、好ましくは少なくとも70重量%、より好ましくは少なくとも80重量%及び最も好ましくは少なくとも90重量%のマルトペンタオースを生成する前記8の方法。
【0170】
11.前記非マルトース生成エキソアミラーゼが、前記8記載の条件下でインキュベートされ、そして前記加水分解生成物が請求項8記載のようにして分析される場合、それが少なくとも60重量%、好ましくは少なくとも70重量%、より好ましくは少なくとも80重量%及び最も好ましくは少なくとも90重量%のマルトヘキサオースを生成する前記8の方法。
12.前記非マルトース生成エキソアミラーゼが、前記8記載の条件下でインキュベートされ、そして前記加水分解生成物が請求項8記載のようにして分析される場合、それが少なくとも60重量%、好ましくは少なくとも70重量%、より好ましくは少なくとも80重量%及び最も好ましくは少なくとも90重量%のマルヘプタオースを生成する前記8の方法。
【0171】
13.前記非マルトース生成エキソアミラーゼが、請求項8記載の条件下でインキュベートされ、そして前記加水分解生成物が請求項8記載のようにして分析される場合、それが少なくとも60重量%、好ましくは少なくとも70重量%、より好ましくは少なくとも80重量%及び最も好ましくは少なくとも90重量%のマルトオクタオースを生成する前記8の方法。
14.少なくとも1つの乳化剤がドウ成分、ドウ添加剤又はドウに添加される請求項1〜4のいずれか1項記載の方法。
【0172】
15.前記乳化剤が、レシチン、ポリオキシエチレンステアレート、食用脂肪酸のモノ−及びジグリセリド、食用脂肪酸のモノ−及びジグリセリドの酢酸エステル、食用脂肪酸のモノ−及びジグリセリドの乳酸エステル、食用脂肪酸のモノ−及びジグリセリドのクエン酸エステル、食用脂肪酸のモノ−及びジグリセリドのジアセチル酒石酸エステル、食用脂肪酸のスクロースエステル、ナトリウムステアロイル−2−ラクチレ−ト及びカルシウムステアロイル−2−ラクチレ−トから成る群から選択される前記14の方法。
【0173】
16.少なくとも1つの追加の酵素が、ドウ成分、ドウ添加剤又はドウに添加される前記1〜4のいずれかに記載の方法。
17.前記追加の酵素が、セルラーゼ、ヘミセルラーゼ、キシラナーゼ、オキシドレダクターゼ及びプロテアーゼから成る群から選択される前記16の方法。
18.ベーキング工程の間、アミロペクチンの側鎖の非還元末端から、4〜8個のD−グルコピラノシル単位から主として成る線状マルトオリゴ糖を分解することによって、澱粉を加水分解できる非マルトース生成エキソアミラーゼ、少なくとも1つの追加のドウ成分又はドウ添加剤から成るドウから製造されるドウ及びベーキングされた澱粉質パン生成物のための改良生成物。
【0174】
19.前記非マルトース生成エキソアミラーゼがエキソアミラーゼ活性1単位当たり0.5以下のエンドアミラーゼ単位(EAU)のエンドアミラーゼ活性を有する前記18の改良性組成物。
20.前記非マルトース生成エキソアミラーゼがエキソアミラーゼ活性1単位当たり0.05以下のエンドアミラーゼ単位(EAU)のエンドアミラーゼ活性を有する前記18の改良性組成物。
21.前記非マルトース生成エキソアミラーゼがエキソアミラーゼ活性1単位当たり0.01以下のエンドアミラーゼ単位(EAU)のエンドアミラーゼ活性を有する前記18の改良性組成物。
【0175】
22.0.7単位の量の前記非マルトース生成エキソアミラーゼが、50mMの2−(N−モルホリノ)エタンスルホン酸及び2mMの塩化カルシウムを含む緩衝溶液1ml当たり10mgの予備煮沸されたモチ状トウモロコシ澱粉の水溶液4mlにおいて、50℃の温度で、pH6.0で15分間インキュベートされる場合、それが、少なくとも60重量%、好ましくは少なくとも70重量%、より好ましくは少なくとも80重量%及び最も好ましくは少なくとも90重量%の、グルコース、マルトース、及び3〜10個のD−グルコピラノシル単位の線状マルトオリゴ糖、好ましくは4〜8個のD−グルコピラノシル単位の線状マルトオリゴ糖から成る加水分解生成物を生成することを特徴として、ここで前記加水分解生成物は、パルスされた電流滴定検出、及び標準としてグルコース〜マルトヘプタオースの既知の線状マルトオリゴ糖と共にDionex PA100カラムを用いてアニオン交換HPLCにより分析される前記18の改良性組成物。
【0176】
23.前記非マルトース生成エキソアミラーゼが、請求項8記載の条件下でインキュベートされ、そして加水分解生成物が請求項8記載のようにして分析される場合、少なくとも60重量%、好ましくは少なくとも70重量%、より好ましくは少なくとも80重量%及び最も好ましくは少なくとも90重量%のマルトテトラオースを生成する前記22の改良性組成物。
24.前記非マルトース生成エキソアミラーゼが、請求項8記載の条件下でインキュベートされ、そして加水分解生成物が請求項8記載のようにして分析される場合、少なくとも60重量%、好ましくは少なくとも70重量%、より好ましくは少なくとも80重量%及び最も好ましくは少なくとも90重量%のマルトペンタオースを生成する前記22の改良性組成物。
【0177】
25.前記非マルトース生成エキソアミラーゼが、請求項8記載の条件下でインキュベートされ、そして加水分解生成物が請求項8記載のようにして分析される場合、少なくとも60重量%、好ましくは少なくとも70重量%、より好ましくは少なくとも80重量%及び最も好ましくは少なくとも90重量%のマルトヘキサオースを生成する前記22の改良性組成物。
26.前記非マルトース生成エキソアミラーゼが、請求項8記載の条件下でインキュベートされ、そして加水分解生成物が請求項8記載のようにして分析される場合、少なくとも60重量%、好ましくは少なくとも70重量%、より好ましくは少なくとも80重量%及び最も好ましくは少なくとも90重量%のマルトヘプタオースを生成する前記22の改良性組成物。
【0178】
27.前記非マルトース生成エキソアミラーゼが、請求項8記載の条件下でインキュベートされ、そして加水分解生成物が請求項8記載のようにして分析される場合、少なくとも60重量%、好ましくは少なくとも70重量%、より好ましくは少なくとも80重量%及び最も好ましくは少なくとも90重量%のマルトオクタオースを生成する前記22の改良性組成物。
28.乳化剤及びヒドロコロイドから成る群から選択された少なくとも1つの添加剤を含んで成る前記19〜27のいずれかに記載の改良性組成物。
29.少なくとも1つの乳化剤がドウ成分、ドウ添加剤又はドウに添加される前記28の改良性組成物。
【0179】
30.前記ヒドロコロイドが、アルギネート、カラゲニン、ペクチン及び植物ガムから成る群から選択される前記28の改良性組成物。
31.セルラーゼ、ヘミセルラーゼ、キシラナーゼ、オキシドレダクターゼ及びプロテアーゼから成る群から選択された少なくとも1つの追加の酵素を含んで成る前記19〜30のいずれかに記載の改良性組成物。
32.請求項1〜17のいずれか1項記載の方法により得ることができるベーキングされた澱粉質パン生成物。
【0180】
本明細書に言及されるすべての出版物は、引用により組み込まれる。本発明の記載される方法及びシステムの種々の修飾及び変更は、当業者に明らかであり、そして本発明の範囲内で行われるであろう。本発明は特定の好ましい態様で記載されて来たが、本発明はそのような特定の態様に限定されるものではない。実際、生化学又は関連する分野の当業者に明らかである、本発明を実施するための記載される態様の種々の修飾は、本発明の範囲内で行われる。
【0181】
参考文献
[1]. K.H. Park, Food Sci. Ind. 25 (1992) 73-82.
[2]. M. Okada and T. Nakakuki, Oligosaccharides: production, properties and application, in F.W. Schenck and R.E. Hebeda (Eds.), Starch hydrolysis products worldwide technology, production and application, VCH Publishers, New York, 1992, pp 335-366.
[3]. W.M.Fogarty, Microbial amylases, in W.M. Fogarty (Ed.), Microbial enzymes and biotechnology, Applied Science, London, 1983, pp. 1-92.
【0182】
[4]. W.M. Fogarty and C.T. Kelly, Starch-degrading enzymes of microbial origin, in M.J. Bull (Ed.), Progress in industrial microbiology, Vol. 15, Elsevier Scientific 1978, pp. 87-150.
[5]. K. Kainuma, S. Kobayashi, T. Ito, and S. Suzuki, FEBS Letters, 26 (1972) 281-285.
[6]. N. Monma, T. Nakakuki, and K. Kainuma, Agric. Biol. Chem., 47 (1983) 1769-1774.
[7]. J.F. Kennedy and C.A. White, Starch/Starke 31 (1979) 93-99.
[8]. Y. Takasaki, Agric. Biol. Chem. 46 (1982) 1539-1547.
【0183】
[9]. H. Taniguchi, C.M. Jae, N. Yoshigi, and Y. Maruyama, Agric. Biol. Chem. 47 (1983) 511-519.
[10]. H. Taniguchi, Maltohexaose-producing amylase of Bacillus circulans F-2 in R.B. Friendman (Ed.) Biotechnology of amylodextrin oligosaccharides. ACS Symp. Ser. 458. American Chemical Society, Washington DC, 1991, pp 111-124.
[11]. F. Bealin-Kelly, C.T. Kelly, and W.M. Fogarty, Biochem. Soc. Trans., 18 (1990) 310-311.
[12]. W.M. Fogarty, F. Bealin-Kelly, C.T.Kelly, and E.M. Appl. Microbiol. Biotechnol., 36 (1991) 184-489.
【0184】
[13]. N. Saito, Archives. Biochem. Biophys., 155 (1973) 290-298.
[14]. H. Okemoto, S. Kobayashi, M. Monma, H. Hashimoto, K. Hara, and K. Kainuma, Appl. Microbiol. Biotechnol., 25 (1986) 137-142.
[15]. O. Shida, T. Takano, H. Takagi, K. Kadowaki, and S. Kobayashi, Biosci., Biotechnol., Biochem. 56 (1992) 76-80.
[16]. (There is no ref. [16])
[17]. Y. Sakano, Y. Kashiwagi, and T. Kobayashi, Agric. Biol. Chem., 46 (1982) 639-646.
[18]. Y. Takasaki, H. Shinohara, M. Tsuruhisa, S. Hayashi, K. Imada, Agric. Biol. Chem. 55 (1991) 1715-1720.
【0185】
[19]. W.M. Fogarty, C.T. Kelly, A.C. Bourke, and E.M. Doyle, Biotechnol. Lett. 16 (1994) 473-478.
[20]. K. Wako, S. Hashimoo, S. Kubomura, A. Yokota, K. Aikawa, and J. Kamaeda, J. Jap. Soc. Starch. Sci 26 (1979) 175-181.
[21]. Y. Takasaki, Agric. Biol. Chem. 49 (1985) 1091-1097.
[22]. (There is no ref. [22])
[23]. T. Hayashi, T. Akiba, and K. Horikoshi, Appl. Microbiol. Biotechnol. 28 (1988b) 281-285.
[24]. G. Schmid, A. Candussio, and A. Bock, U.S. Patent 5 304 723 (1994).
【0186】
[25]. M.A. Mc Tigue, C.T. Kelly, E.M. Doyle, and W.M. Fogarty, Enzyme Microb. Technol., 17 (1995) 570-573.
[26]. T.U. Kim, B.G. Gu, J.Y. Jeong, S.M. Byun, and Y.C. Shin, appl. Environm. Microbiol. 61 (1995) 3105-3112.
[27]. A.K. Chandra, S. Medda, and A.K. Bhadra, J. Ferment. Technol., 58, (1980), 1-10.
[28]. R.A.K. Srivastava and J.N. Baruah, Appl. Environ. Mocrobiol. 52 (1986) 179-184.
[29]. V. Planchot an P. Colonna, Carbohydr. Res., 272 (1995) 97-109.
【0187】
[30]. J.F. Robyt and W.J. Whelan, The α−amylases in J.A. Radley (Ed.) Starch and its derivatives, 4.ed. Chapman and Hall, London, 1968, pp. 430-476.
[31]. M. Ohnishi and K. Hiromi, General considerations for conditions and methods of amylase assay in The Amylase Research Society of Japan (Ed.) Handbook of amylases and related enzymes. Their Sources, isolation methods, properties and applications. Pergamon, Oxford, 1988, pp. 10-14.
[32]. L. Duedahl-Olesen, W. Zimmemamm, and J.A. Delcour, Cereal Chemistry (1999) in press.
【0188】
[33]. K.L. Larsen, L. Duedahl-Olesen, H.J.S. Christensen, F. Mathiesen, L.H. Pedersen, and W. Zimmermann, Carbohydr. Res. 310 (1998) 211-219.
[34]. M.M. Bradford, Anal. Chem., 72 (1976) 248-254.
[35]. Novex, Precast Gel Instructions. NOVEX, San Diego, CA, USA.
[36]. Mini Protean II Electrophoresis Cell Instructions manual. Bio-Rad laboratories, Hercules, CA, U.S.A.
[37]. H. Blum, H. Beier, and H.J. Gross, Electrophoresis (1987) 93-99.
[38]. L.H. Pedersen, H.J.S. Christensen, F. Mathiesen, K.L. Larsen, and W. Zimmermamm, Starch, 49 (1997) 250-253.
【0189】
[39]. H.J. Fuwa, J. Biochem., 41 (1954) 583-603.
[40]. Y.C. Lee, J. Chormatogr. A, 720 (1996) 137-149.
[41]. R.N. Ammerall, G.A. Delgado, F.L. Tenbarge, and R.B. Freidmann, Carbohydr. Res. 215 (1991) 179-192。
【技術分野】
【0001】
発明の分野
本発明は、タンパク質類、特に澱粉を分解することかできるタンパク質類に関する。
特に、本発明は、澱粉の有害な劣化を遅延することができるタンパク質の使用に関する。
有害な劣化(retrogradation)過程、たとえば老化(staling)は、典型的には、澱粉基材、特に水性澱粉懸濁液の加熱及び冷却の後に生じ、そしてゲル化された澱粉の徐々の規則状態への転位のためである。
【0002】
さらに具体的には、本発明は、アミロペクチンの有害な劣化を遅延することができるタンパク質の使用に関する。
さらに特別には、本発明は、ベーキングされたパン生成物を調製するためへのタンパク質の使用、及びそのベーキングされたパン生成物自体にも関する。
さらに特別には、本発明は、ベーキングされた澱粉質のパン生成物の劣化の遅延にも関する。
【0003】
さらに特定には、本発明は、パンドウに非マルトース生成エキソアミラーゼを添加することを含んで成る、劣化を遅延又は低下せしめた、ベーキングされた澱粉質のパン生成物の製造方法にも関する。
本発明はまた、非マルトース生成エキソアミラーゼを含んで成る、ドウ及びベーキングされた澱粉質のパン生成物のための改良された生成物にも関する。
【背景技術】
【0004】
発明の背景
澱粉はアミロペクチン及びアミロースを含んで成る。アミロペクチンは、枝分かれした及びブッシュ様構造を形成するα−(1→6)結合を通して枝分かれ点で一緒に連結される短いα−(1→4)−D−グルカン鎖を有する高度に枝分かれされた炭水化物ポリマーである。平均的に、20〜25のα−(1→4)結合されたグルコース残基ごとに1つの枝分かれ点が存在する。対照的に、アミロースは、枝だなしのα−(1→4)−D−グルカン単位から主に成る線状構造である。典型的には、澱粉は、約75%のアミロペクチン分子及び約25%のアミロース分子を含む。
【0005】
より具体的には、線状マルト−多糖類は、α−(1→4)結合により連結される2〜10単位のα−D−グルコピラノースから構成される。それらの性質、たとえば低甘味性、高い水保持機能及びスクロース結晶化の阻止[1]のために、それらの化合物は食品産業への可能性ある用途を有する。しかしながら、3以上の重合程度(すなわち、DP>3)を有するマルト−オリゴ糖の多量の調製は、短調で退屈であり、且つ費用が高い。
【0006】
バックグラウンド情報として、次のものを列挙する:DP1=グルコース、DP2=マルトース、DP3=マルトトリオース、DP4=マルトテトラオース、DP5=マルトペンタオース、DP6=マルトヘキサオーズ、DP7=マルトヘプタオース、DP8=マルトオクタオース、DP9=マルトノナオース、及びDP10=マルトデカオース。
特定の長さのマルト−オリゴ糖を生成する微生物酵素の発見は、多量のそれらのオリゴ糖の生成を可能にする[2]。
【0007】
アミラーゼは、澱粉においてα−D−(1→4)O−グリコシド結合を分解する、ヒドロラーゼとして分類される澱粉−分解酵素である。一般的に、α−アミラーゼ(E. C. 3. 2. 1. 1, α−D−(1→4)−グルカングルカノヒドロラーゼは、ランダム態様で澱粉分子内のα−D−(1→4)O−グルコシド結合を分解するエンド−作用酵素として定義される[3]。
【0008】
対照的に、エキソ−作用性アミロース分解酵素、たとえばβアミラーゼ(E. C. 3. 2. 1, α−D−(1→4)−グルカンマルトヒドロラーゼ)、及びいくつかの生成物−特異的アミラーゼは、基質の非還元末端から澱粉分子を分解する[4]。β−アミラーゼ、α−グルコシダーゼ(E. C. 3. 2. 2. 20, α−D−グルコシドグルコヒドロラーゼ)、グルコアミラーゼ(E. C. 3. 2. 1. 3, α−D−(1→4)−グルカングルコヒドロラーゼ)、及び生成物−特異的アミラーゼは、澱粉から特定の長さのマルト−オリゴ糖を生成することができる。
【0009】
特定のDPのマルト−オリゴ糖を生成するいくつかのアミラーゼ、たとえばクレビシェラ・プネウモニア(Klebsiella pneumonia)[5, 6]、バチルス・スブチリス(Bacillus subtilis)[7], B. サーキュランスG−6(B. circulans G-6)[8]、B.サーキュランスF-2 [9]、及びB.カルドベロックス(B. caldovelox)[11, 12]が、これまで同定されている。マルトペンタオース−生成アミラーゼは、B.リケニホルミス(B. licheniformis)584 [13] 及びプロイドモナスspp. (Pseudomonas spp.) [14, 15] において検出されている。さらに、マルトテトラオース−生成アミラーゼが、シュードモナス・スツゼリ(Pseudomonas stutzeri)NRRL B-3389 [16, 17]、バチルスsp. MG-4 [14] 及びシュードモナスsp. IMD353 [19] から、及びマルトトリオース−生成アミラーゼが、ストレプトミセス・グリセウス(Streptomyces griseus)NA-468 [20] 及びB. スブチリス[21] から報告されている。
【0010】
EP−B1−298,645号は、遺伝子工学技法を用いて、シュードモナス・スツゼリ又はP. サッカロフィラ(P. saccharophila)のエキソ−マルトテトラヒドロラーゼを調製するための方法を記載する。
アメリカ特許第5,204,254号は、好アルカリ性細菌(DSM5853)の天然の及び遺伝子的に修飾されたエキソ−マルトペンタヒドロラーゼを記載する。
【0011】
高いpHで活性な非常に少数の生成物−特異的アミラーゼが同定されている。同定されているそれらの例は、マルトヘキサノースを生成するバチルスsp. H−167[22, 23]、マルトペンタオースを生成する細菌単離物(163−26、DSM5853)[24]、マルトテトラオース及びより小さなマルト−オリゴ糖を生成するバチルスsp. IMD370 [25]、及び延長された加水分解期間、マルトテトラオースに転換された澱粉からマルトヘキサオースを最初に生成したバチルスsp. GM8901 [26] からのアミラーゼを包含する。
【0012】
水の存在下で加熱された澱粉粒質物は、ゲル化と呼ばれる規則不規則転移を受け、ここで液体が膨潤粒質物により取られる。ゲル化温度は、異なって澱粉で変化し、そして天然の修飾されていない澱粉に関しては、それらの生物学的な源に依存する。
冷却は、澱粉濃度に依存して、ゲル化された相を粘弾性ペースト又は弾性ゲルに転換する。この工程の間、アミロース及びアミロペクチン鎖は、より規則化された構造を形成するために再会合する。時間の経過に従って、会合がより形成され、そしてそれらはさらにより規則的に成る。アミロペクチン鎖DP−15−20の会合が熱可逆性擬似結晶構造を導くと思われる。
【0013】
有害な劣化の結果、ペースト又はゲルシステムの水保持能力が、ゲルテキスチャー及びダイエット性質に重要に関係して変更される。
ベーキングされたパン生成物の品質が貯蔵の間、徐々に劣化することは知られている。パンの中味は柔軟性及び弾性を失い、そして堅く且つ脆くなる。このいわゆる劣化は、非晶性状態から擬似結晶状態に、ベーキングの間ゲル化される澱粉の転移であることが理解されている、主に澱粉の有害な劣化のためである。パンの中味の堅さの上昇はしばしば、パンの劣化過程の測定値として使用される。
【0014】
あらたにベーキングされたパンの冷却に基づいて、アミロース画分は、時間の経過に従って、劣化し、網状構造を進行せしめる。この過程は、それが低い程度の堅さ及び改良されたスライスシング特性を有する所望のパン中心構造を創造することにおいて有益である。アミロペクチンの徐々の結晶化が、ベーキングの後、数日間、ゲル化された澱粉顆粒内で生じる。この過程において、アミロペクチンは、澱粉顆粒が埋封されているアミロース網状構造を強化すると思われる。この強化は、パン中心の高められた硬性を導く。この強化がパン劣化の主な原因の1つである。
【0015】
アミロペクチンの有害な劣化又は結晶化の割合は、アミロペクチンの側鎖の長さに依存する。これによれば、穀物アミロペクチンは、穀物アミロペクチンよりも長い平均側鎖を有する、エンドウ又はジャガイモからのアミロペクチンによりも遅い速度で劣化する。
【0016】
これは、11以下のDP、すなわち重合の程度の平均鎖長を有するアミロペクチンがまったく結晶化しない、アミロペクチンゲルシステムからの観察により支持される。さらに、DP6−9の非常に短い鎖の存在が、たぶん立体的妨害のために、周囲の長い側鎖の結晶化を阻害すると思われる。それにより、それらの短い鎖は、強い反−有害な劣化効果を有すると思われる。これによれば、アミロペクチン劣化は、DP14−24を有する側鎖のモル画分に直接的に比例し、そしてDPG6−9を有する側鎖のモル画分に反比例する。
【0017】
小麦及び他の穀類においては、アミロペクチンの外部側鎖は、DP12−19の範囲である。従って、アミロペクチン側鎖の酵素加水分解は、それらの結晶化傾向を著しく低めることができる。
グルコース生成及びマルトース生成エキソ−アミラーゼ、たとえばグルコースを開放することによって澱粉を加水分解するアミログリコシダーゼ、及び非還元鎖末端からマルトースを開放することによって澱粉を加水分解するマルトース生成エキソアミラーゼ又はβ−アミラーゼを用いることによって、パンの劣化を遅延することは、当業界において知られている。
【0018】
これに関しては、Jakubczykなど. (Zesz. Nauk. Sck. Gl. Gospod Wiejsk. Warzawie, Technol. Reino-Spozyw, 1973, 223-235)が、アミログルコシダーゼが小麦粉をベーキングすることによって得られたパンの劣化を遅延することができることを報告している。
JP−62−79745号及びJP−62−79746号は、それぞれ、バチルス・ステアロサーモフィラス(Bacillus stearothermophilus)及びバチルス・メガテリウム(Bacillus megateriumu )により生成されるβ−アミラーゼの使用がパンを包含する澱粉食品の劣化の遅延において効果的であることを言及している。
【0019】
EP−A412,607号は、ゲル化の前、不活性化されない、熱安定性エキソアミラーゼのドウへの添加により劣化性質を遅延しているパン生成物の生成方法を開示する。アミログリコシダーゼ及びβ−アミラーゼのみが、使用される適切なエキソアミラーゼとして列挙されている。エキソアミラーゼは、アミロース及びアミロペクチンの非還元末端からグルコース又はマルトースを分解することによって、ベーキングの間、アミロペクチン成分の結晶化性質を選択的に変性することができる量で存在する。EP−A−412, 607号によれば、エキソアミラーゼは、アミロースの結晶化性質に対し実質的な影響を及ぼさないで、アミロペクチンの結晶化性質を選択的に低める。
【0020】
EP−A−494,233号は、α−形状で存在し、そしてゲル化の前、不活性化されていないマルトースを開放するために遅延された劣化性質を有する、ベーキングされた生成物の生成工程へのマルトース生成エキソアミラーゼの使用を開示する。バチルス株NCIB11837からのマルトース生成α−アミラーゼのみが特別に開示される。明らかに、マルトース生成エキソアミラーゼは、段階的に、多糖鎖の非還元末端からのα−マルトース単位を除去することによって、澱粉における(1→4)−α−グルコシド結合を加水分解する。
【0021】
従って、従来技術は、一定のグルコース生成エキソアミラーゼ及びマルトース生成エキソアミラーゼが、アミロペクチン側鎖の短縮を通してアミロペクチンの有害な劣化傾向を選択的に低めることによって、非劣化効果を提供することができることを教授する。
それにもかかわらず、澱粉生成物、特にベーキングされた生成物、より特定にはパン生成物の有害な劣化を遅延するための、たとえば老化を遅延するための、異なった及び効果的な、好ましくはより効果的手段を提供する必要性がまだ存在する。
【発明の概要】
【0022】
発明の要約
本発明は、有害な劣化特性が遅延する澱粉生成物の製造方法を提供する。
本発明はまた、本発明の方法において有用である酵素も提供する。
本発明の酵素はアミラーゼ酵素である。より具体的には、本発明の酵素は非マルトース生成エキソアミラーゼ酵素である。
非マルトース生成エキソアミラーゼが、ベーキングされた生成物における老化を遅延するためにはもちろんのこと、澱粉生成物の有害な劣化を遅延するためにこれまで使用されたことはないことが注目されるべきである。
【0023】
従って、本発明の第1の観点によれば、アミロペクチンの側鎖の非還元末端から4〜8個のD−グルコピラノシル単位から主として成る1又は複数の線状マルトオリゴ糖を切り取ることによって澱粉を加水分解することができる非マルトース生成エキソアミラーゼを澱粉基材に添加することを含んで成る澱粉生成物の製造方法が提供される。
澱粉基材への非マルトース生成エキソアミラーゼの添加は、澱粉生成物の加熱の間、及び/又はその後に行うことができる。
従って、本発明の第2の観点によれば、本発明の方法により得られる、ベーキングされた生成物が提供される。
【0024】
従って、本発明の第3の観点によれば、ドウのための改良された組成物が提供され;ここで前記組成物は非マルトース生成エキソアミラーゼ、及び少なくとも1つの追加のドウ成分又はドウ添加材を含んで成る。
従って、本発明の第4の観点によれば、澱粉生成物の有害な劣化を遅延するために澱粉生成物への非マルトース生成エキソアミラーゼの使用が提供される。
従って、本発明の第5の観点によれば、新規非マルトース生成エキソアミラーゼが提供される。
本発明のそれらの及び他の観点は、請求の範囲に提供される。さらに、本発明のそれらの及び他の観点、並びにそれらの好ましい観点が下記に提供され、そして論じられる。
【図面の簡単な説明】
【0025】
【図1】図1は、45℃で2%澱粉基質において培養されたB.クラウジBT−21の液体培養物における細胞外澱粉分解活性(mU/ml)を示す。◆は可溶性澱粉、●はアミロペクチン、黒三角はコーン澱粉、黒四角は完全な玄米。バーは標準偏差を示す。
【図2】図2は、生成物−特異的アミラーゼの活性に対するpHの効果を示す。55℃でのpHの効果。
【0026】
【図3】図3は、生成物−特異的アミラーゼの活性に対する温度の効果を示す。5mMのCaCl2を伴って〔黒四角〕又はCaCl2を伴わないで〔□〕、pH9.5での温度の効果。
【図4】図4は、5mMのCaCl2を伴って、pH9.5での上昇する温度でのインキュベーション後、生成物−特異的アミラーゼの残留活性として試験される熱安定性を示す。
【0027】
【図5】図5は、55℃及びpH9.5で、1%の可溶性澱粉及び5mMのCaCl2と共に、生成物−特異的アミラーゼ(505mU/ml)をインキュベートすることによって形成される生成物(mM)を示す。○グルコース、×マルトース、□マルトトリオース、黒三角マルトテトラオース、●マルトペンタオース、黒四角マルトヘキサオース。
【0028】
【図6】図6は、PH9.5及び55℃で1%可溶性澱粉と共に生成物−特異的アミラーゼ(505mU/ml)をインキュベートすることによって得られるHPAEC−PAD形跡を示す。A)酵素を含まない可溶性澱粉、B)酵素と共に30分間インキュベーション。
【図7】図7は、既知の澱粉分解作用のアミラーゼに比較して、B.クラウジBT−21生成物−特異的アミラーゼのエンド−及びエキソ−活性の決定を示す。青色形成(最大の%)が、mMマルトースの生成に対してプロットされる。曲線の傾斜は、エンド−又はエキソ−活性の有力性を示す。
【発明を実施するための形態】
【0029】
一般的な定義
従って、本発明は、澱粉基材、特に澱粉ゲルの有害な劣化を遅延することができるタンパク質の使用に関する。
1つの好ましい観点においては、本発明は、澱粉の劣化を遅延することができるタンパク質の使用に関する。
もう1つの観点においては、本発明は、澱粉基材、たとえば澱粉ゲルの有害な劣化を遅延することができるタンパク質の使用に関する。
本発明によれば、用語“澱粉”とは、澱粉自体、又はその成分、特にアミロペクチンを意味する。
本発明によれば、用語“澱粉基材”とは、澱粉を含んで成るいずれかの適切な基材を意味する。
【0030】
用語“澱粉生成物”とは、澱粉を含むか、又は澱粉に基づくか、又は澱粉に由来するいずれかの生成物を意味する。
好ましくは、澱粉生成物は、小麦粉から得られた澱粉を含み、又はその澱粉に基づかれ、又はその澱粉に由来する。
用語“小麦粉”とは、本明細書において使用される場合、小麦粉又は他の穀物の細かく粉砕された粉と同義語である。しかしながら、好ましくは、その用語は、小麦自体から得られ、そして他の穀物からではない食料粉を意味する。従って、及び特にことわらない限り、“小麦粉”とは、本明細書において使用される場合、小麦粉自体を意味し、そして基材、たとえばドウに存在する場合も小麦粉を意味する。
【0031】
好ましい食料粉は、小麦粉又はライ麦粉、又はそれらの混合物である。しかしながら、他のタイプの食料粉、たとえば米、トウモロコシ、大麦及びアズキに由来する食料粉を含んで成るドウもまた企画される。
好ましくは、澱粉生成物は、大麦生成物である。
より好ましくは、澱粉生成物は、パン生成物である。
さらにより好ましくは、澱粉生成物は、ベーキングされた澱粉質のパン生成物である。
【0032】
用語“ベーキングされた澱粉質のパン生成物”とは、粉砕された穀物に基づく、及び食料粉、水及び発酵剤をドウ形成条件下で混合することにより得ることができるドウに基づくいずれかのベーキングされた生成物を言及することが理解される。しかしながら、追加の成分がそのドウ混合物に添加され得ることも、本発明の範囲内である。
【0033】
用語“アミラーゼ”は、その通常の意味においては、中でも、澱粉の分解を触媒することが酵素を意味する。特に、それらは澱粉においてα−D−(1→4)O−グリコシド結合を分解できるヒドロラーゼである。
用語“非マルトース生成エキソアミラーゼ酵素”とは、最初、澱粉を実質的な量のマルトースに分解しない酵素を意味する。非常に好ましい観点においては、その用語はまた、最初、澱粉を実質的な量のマルトース及びグルコースに分解しない酵素を意味する。
【0034】
本発明の前、非マルトース生成エキソアミラーゼ酵素は、澱粉基材、特に澱粉ゲルの有害な劣化を遅延することについて提案されたことはない。
本発明に従ってアミラーゼ活性を決定するための適切なアッセイは、後で提供される。便利には、このアッセイは“アミラーゼアッセイプロトコール”と呼ばれる。
従って、好ましくは、用語“非マルトース生成エキソアミラーゼ酵素”とは、本明細書に提供される“アミラーゼ活性プロトコール”に記載のような生成物測定方法に従って分析される場合、前記酵素が最初、澱粉を実質的な量のマルトースに分解しないことを意味する。
【0035】
好ましい観点においては、非マルトース生成エキソアミラーゼは、0.7単位の量の前記非マルトース生成エキソアミラーゼが、50mMの2−(N−モルホリノ)エタンスルホン酸及び2mMの塩化カルシウムを含む緩衝液1ml当たり10mgの予備煮沸されたモチ状澱粉の水溶液4mlにおいて、50℃の温度でpH6.0で15分間インキュベートされる場合、前記酵素が2〜10個のD−グルコピラノシル単位及び任意にはグルコースの1又は複数の線状マルト−オリゴ糖から成る加水分解生成物を生成し、その結果、前記加水分解生成物の少なくとも60重量%、好ましくは少なくとも70重量%、より好ましくは少なくとも80重量%そして最も好ましくは少なくとも85重量%が、3〜10個のD−グルコピラノシル単位の線状マルトオリゴ糖、好ましくは4〜8個のD−グルコピラノシル単位の線状マルトオリゴ糖から構成されることにより特徴づけることができる。
【0036】
参照の容易さ及び本発明の目的のために、50mMの2−(N−モルホリノ)エタンスルホン酸及び2mMの塩化カルシウムを含む緩衝液1mlあたり10mgの予備煮沸されたモチ状トウモロコシ澱粉の水溶液4mlにおいて、50℃の温度及びpH6.0で15分間、0.7単位の量の非マルトース生成エキソアミラーゼをインキュベートする特徴が、“モチ状トウモロコシ澱粉インキュベーション試験”として言及され得る。
【0037】
従って、好ましい非マルトース生成エキソアミラーゼは、2〜10個のD−グルコピラノシル単位及び任意にはグルコースの1又は複数の線状マルト−オリゴ糖から成る加水分解生成物を、モチ状トウモロコシ澱粉インキュベーション試験において生成する能力を有するものとして特徴づけられ;その加水分解の結果、前記加水分解生成物の少なくとも60重量%、好ましくは少なくとも70重量%、より好ましくは少なくとも80重量%及び最も好ましくは少なくとも85重量%が、3〜10個のD−グルコピラノシル単位の線状マルトオリゴ糖、好ましくは4〜8個のD−グルコピラノシル単位から成る線状マルトオリゴ糖から成る。
【0038】
モチ状トウモリコシ澱粉インキュベーション試験における加水分解生成物は、2〜10個のD−グルコピラノシル単位の1又は複数の線状マルト−オリゴ糖及び任意にはグルコースを含む。モチ状トウモロコシ澱粉インキュベーション試験における加水分解生成物はまた、他の加水分解生成物も含むことができる。それにもかかわらず、3〜10個のD−グルコピラノシル単位の線状マルトオリゴ糖の重量%の量は、2〜10個のD−グリコピラノシル単位及び任意にはグルコースの1又は複数の線状マルト−オリゴ糖から成る加水分解生成物の量に基づく。換言すれば、3〜10個のD−グルコピラノシル単位の線状マルトオリゴ糖の重量%の量は、2〜10個のD−グルコプラノシル単位の1又は複数の線状マルト−オリゴ糖及びグルコース以外の他の加水分解生成物の量に基づかない。
【0039】
加水分解生成物は、いずれかの適切な手段により分析され得る。たとえば、加水分解生成物は、パルスされた電流滴定検出及びたとえば標準として、グルコース〜マルトヘプタオースの既知の線状マルトオリゴ糖と共に、Dionex PA 100カラムを用いて、アニオン交換HPLCにより分析され得る。
参照の容易さ及び本発明の目的のために、パルスされた電流滴定検出及び標準として、グルコース〜マルトヘプタオースの既知線状マルトオリゴ糖と共にDionex PA 100 カラムを用いて、アニオン交換HPLCによる加水分解生成物の分析特徴は、“アニオン交換による分析”として言及され得る。もちろん、及びまさに示されるように、他の分析技法及び他の特定のアニオン交換技法でも十分である。
【0040】
従って、他方では、好ましい非マルトース生成エキソアミロースは、モチ状トウモロコシ澱粉インキュベーション試験において、2〜10個のD−グルコピラノシル単位及び任意にはグルコースの1又は複数の線状マルト−オリゴ糖から成る加水分解生成物を生じさせる能力を有し、その結果、前記加水分解生成物の少なくとも60重量%、好ましくは少なくとも70重量%、より好ましくは少なくとも80重量%そして最も好ましくは少なくとも85重量%が、3〜10個のD−グルコピラノシル単位の線状マルトオリゴ糖、好ましくは4〜8個のD−グルコピラノシル単位の線状マルトオリゴ糖から成ることを特徴とする。
【0041】
本明細書において使用される場合、用語“線状マルト−オリゴ糖”とは、通常、α−(1→4)結合により結合される、2〜10個のα−D−グルコプラノース単位を意味するものとして使用される。
用語“P.サッカロフィラ(Pseudomonas sacharophila)から得ることができる”とは、酵素がP.サッカロフィラから必ずしも得られる必要はないことを意味する。代わりに、酵素は組換えDNA技法の使用により調製され得る。
【0042】
P.サッカロフィラから得ることができる酵素に関しての用語“その機能的同等物”とは、その機能的同等物が他の源から得られることを意味する。機能的に同等の酵素は異なったアミノ酸配列を有するが、しかし非マルトース生成エキソアミラーゼ活性を有するであろう。その機能的に同等の酵素は異なった化学構造及び/又は化学式を有するが、しかし非マルトース生成エキソアミラーゼ活性を有するであろう。その機能的に同等の酵素は、P.サッカロフィラから得られる非マルトース生成エキソアミラーゼ酵素と正確に同じ非マルトース生成エキソアミラーゼ活性を必ずしも有する必要はない。いくつかの用途においては、好ましくは、機能的に同等の酵素は、P.サッカロフィラから得られる酵素と少なくとも同じ活性プロフィールを有する。
【0043】
用語“バチルス・クラウジ(Bacillus clausii)から得ることができる”とは、酵素がバチルス・クラウジから必ずしも得られる必要はないことを意味する。代わりに、酵素は、組換えDNA技法の使用により調製され得る。
バチルス・グラウジから得ることができる酵素に関しての用語“その機能的同等物”とは、その機能的同等物が他の源から得られることを意味する。機能的に同等の酵素は異なったアミノ酸配列を有するが、しかし非マルトース生成エキソアミラーゼ活性を有するであろう。
【0044】
その機能的に同等の酵素は異なった化学構造及び/又は化学式を有するが、しかし非マルトース生成エキソアミラーゼ活性を有するであろう。その機能的に同等の酵素は、バチルス・グラウジから得られる非マルトース生成エキソアミラーゼ酵素と正確に同じ非マルトース生成エキソアミラーゼ活性を必ずしも有する必要はない。いくつかの用途においては、好ましくは、機能的に同等の酵素は、バチルス・グラウジから得られる酵素と少なくとも同じ活性プロフィール(たとえば、図7に示される反応性プロフィール)を有する。
【0045】
一般的な解説
本発明は、非マルトース生成エキソアミアーゼが澱粉生成物、たとえばベーキングされた生成物において有害な劣化、たとえば老化(staling)を遅延し又は低めることにおいて非常に効果的である驚くべき発見に基づかれている。
本発明者は、本発明の非マルトース生成エキソアミラーゼが、グルース生成及びマルトース生成エキソアミラーゼよりも、パンにおける有害な劣化、たとえば老化を遅延することにおいてより効果的であり得ることを見出した。
有害な劣化の低下は、当業界において知られている標準の技法により測定され得る。例によれば、いくつかの技法が、セクション標題“劣化の測定のためのアッセイ”において、後で示される。
【0046】
本発明の研究において、本発明者は、十分な熱安定性を有する、十分な量の活性の非マルトース生成エキソアミラーゼ、たとえばエキソ−マルトテトラヒドロラーゼ(EC 3. 2. 1. 60)をドウ中に導入することによって、たとえば貯蔵条件下で対照のパンの有害な劣化に比較して、低められた、多くの場合、十分に低められた有害な劣化を有するベーキングされた生成物が提供されることを見出した。対照的に、非マルトース生成エキソアミラーゼの熱安定性に匹敵する熱安定性を有するマルトース生成エキソアミラーゼの同じ量の活性を組み込むことによる有害な劣化を低める効果は、有意に劣っている。
【0047】
従って、非マルトース生成エキソアミラーゼの抗劣化効果は、マルトース生成エキソアミラーゼのその効果よりもより効果的である。本発明者は、この差異が一部、アミロペクチン側鎖が短くされる程度によるかも知れないと信じている。本発明はまた、抗劣化効果が、マルトヘプタオース及び/又はマルトオクタオース及び/又はマルトへキソースを開放する本発明の非マルトース生成エキソアミラーゼを用いる場合、さらにより著しいことも信じている。
【0048】
本発明の研究において、本発明者はまた、マルトヘキサオースを生成する高いpHで活性な生成物−特異的アミラーゼを精製し、そして特徴づけた。このアミラーゼは、バチルス・クラウジBT−21のアルカリ耐性株から単離された。
さらに、本発明者は、本発明の非マルトース生成エキソアミラーゼを用いることによって得ることができる有害な劣化の遅延が非常に広範囲にわたって、用量応答性であることを見出した。これは、かなり制限され、そして強く低下する用量応答を有するマルトース生成エキソアミラーゼからの効果と対照的である。
【0049】
アミラーゼ
1つの観点において、本発明は、澱粉生成物、たとえば製パン製品を調製するためへの一定のアミラーゼの使用を提供する。この観点においては、非マルトース生成エキソアミラーゼであるアミラーゼは、澱粉生成物、特にベーキングされた澱粉質パン生成物の老化を遅延し、又は低める(すなわち、老化の速度を低める)。
好ましくは、アミラーゼは、単離された形で及び/又は実質的に純粋な形で存在する。ここで、用語“単離された”とは、酵素がその天然の環境下で存在しないことを意味する。
【0050】
上記に示されるように、本発明の非マルトース生成エキソアミラーゼ酵素は、最初に、澱粉を実質的な量のマルトースに分解しない。
本発明によれば、非マルトース生成エキソアミラーゼは、アミロペクチンの側鎖の非還元末端から、4〜8個のD−グルコピラノシル単位から主に成る線状マルトオリゴ糖を切り離すことができる。この特徴を有し、そして本発明への使用のために適切である非マルトース生成エキソアミラーゼは、アミラーゼアッセイプロトコール(前記)に示されるモデル系におけるゲル化されたモチ状トウモロコシを加水分解するそれらの能力により同定される。
【0051】
アミラーゼアッセイプロトコールにおける記載される条件下で15分間インキュベートされる場合、本発明への使用のために適切である非マルトース生成エキソアミラーゼは、2〜10個のD−グルコピラノシル単位及び任意にはグルコースの1又は複数の線状マルト−オリゴ糖から成る加水分解生成物を生成し、その結果、その加水分解生成物の生成物パターンは、少なくとも60重量%、特に少なくとも70重量%、より好ましくは少なくとも80重量%及び最も好ましくは少なくとも90重量%が、マルトース及びグルコース以外の澱粉加水分解生成物から成る。
【0052】
本発明の好ましい観点に関しては、本発明への使用のために適切である非マルトース生成エキソアミラーゼは、モチ状トウモロコシ澱粉インキュベーション試験について記載される条件下で15分間インキュベートされる場合、前記加水分解生成物をもたらし、その結果、その加水分解生成物は、少なくとも60重量%、特に少なくとも70重量%、より好ましくは少なくとも80重量%そして最も好ましくは少なくとも90重量%の、3〜10個のD−グルコピラノシル単位の線状マルトオリゴ糖、特に4〜8個のD−グルコピラノシル単位から成る線状マルトオリゴ糖の生成物パターンを有する。
【0053】
本発明のより好ましい観点においては、前記試験における前記加水分解生成物は、少なくとも60重量%、特に少なくとも70重量%、より好ましくは少なくとも80重量%そして最も好ましくは少なくとも85重量%の、4又は6個のD−グルコピラノシル単位の線状マルトオリゴ糖の生成物パターンを有する。
本発明のより好ましい観点においては、前記試験における前記加水分解生成物は、少なくとも60重量%、特に少なくとも70重量%、より好ましくは少なくとも80重量%そして最も好ましくは少なくとも85重量%の、4個のD−グルコピラノシル単位の線状マルトオリゴ糖の生成物パターンを有する。
【0054】
本発明のより好ましい観点においては、前記試験における前記加水分解生成物は、少なくとも60重量%、特に少なくとも70重量%、より好ましくは少なくとも80重量%そして最も好ましくは少なくとも85重量%の、6個のD−グルコピラノシル単位の線状マルトオリゴ糖の生成物パターンを有する。
好ましくは、非マルトース生成エキソアミラーゼは、その一次生成物をグルコース、マルトース及びマルトトリオースに転換するように該生成物を実質的に加水分解しない。その場合、前記一次生成物は、酵素に対してアミロペクチン非還元鎖末端と基質として競争し、その結果、その抗−劣化効率を低められる。
【0055】
従って、好ましくは、非マルトース生成エキソアミラーゼは、モチ状トウモロコシ澱粉インキュベーション試験に類似する条件下で(但し、15分間が300分に延長され、そして本発明の便利さのために、この変性されたモチ状トウモリコシ澱粉インキュベーション試験は“延長されたモチ状トウモロコシ澱粉インキュベーション試験”と呼ばれる)、300分間インキュベートされる場合、前記加水分解生成物をなお生成し、ここで前記加水分解生成物は、少なくとも50重量%、特に少なくとも60重量%、より好ましくは少なくとも70重量%、そして最も好ましくは少なくとも80重量%の4〜8個のD−グルコピラノシル単位の生成物パターンを有する。
【0056】
例によれば、本発明の方法において有用な非マルトース生成エキソアミラーゼは、それがモチ状トウモロコシ澱粉インキュベーション試験において、2〜10個のD−グルコピラノシル単位及び任意にはグルコースの1又は複数の線状マルト−オリゴ糖から成る加水分解生成物を生成する能力を有し、その結果、前記加水分解生成物の少なくとも60重量%、好ましくは少なくとも70重量%、より好ましくは少なくとも80重量%そして最も好ましくは少なくとも85重量%が、3〜10個のD−グルコピラノシル単位の線状マルトオリゴ糖、好ましくは4〜8個のD−グルコピラノシル単位の線状マルトオリゴ糖から成り、そして前記酵素がP.サッカロフィラから得ることができるものであり、又はその機能的同等物であることにより特徴づけられ得る。
【0057】
さらなる例によれば、本発明の方法において有用なもう1つの非マルトース生成エキソミラーゼは、それがモチ状トウモロコシ澱粉インキュベーション試験において、2〜10個のD−グルコピラノシル単位及び任意にはグルコースの1又は複数の線状マルト−オリゴ糖から成る加水分解生成物を生成する能力を有し、その結果、前記加水分解生成物の少なくとも60重量%、好ましくは少なくとも70重量%、より好ましくは少なくとも80重量%そして最も好ましくは少なくとも85重量%が、3〜10個のD−グルコピラノシル単位の線状マルトオリゴ糖、好ましくは4〜8個のD−グルコピラノシル単位の線状マルトオリゴ糖から成り、そして前記酵素がバチルス・クラウジから得ることができるものであり、又はその機能的同等物であり、そして前記酵素が約101,000Daの分子量を有し(ドデシル硫酸ナトリウムポリアクリルアミド電気泳動により推定される場合)、そして/又は前記酵素がpH9.5及び55℃で最適活性を有することにより特徴づけられる。
【0058】
好ましくは、本発明の使用のために適切である非マルトース生成エキソアミラーゼは、ベーキングの間、活性であり、そして約55℃の温度で開始する澱粉顆粒のゲル化の間及びその後、澱粉を加水分解する。非マルトース生成エキソアミラーゼはより熱安定性であるほど、より長い時間、活性であり、そして従って、より抗老化効果を提供するであろう。しかしながら、約85℃の温度以上でベーキングする間、非マルトース生成エキソアミラーゼは選択的に徐々に不活性化され、その結果、ベーキング工程の後、最終のパンにおいて活性は実質的に存在しない。
【0059】
従って、好ましくは、本発明への使用のために適切な非マルトース生成エキソアミラーゼは、上記のようにして調製された、50mMのMES、2mMの塩化カルシウム溶液(pH6.0)中、10mg/mlのモチ状トウモロコシ澱粉4mlを含む試験において、40, 45, 50, 55, 60, 65, 70, 75, 80, 85又は90℃で15分間インキュベートされ、そして上記のようにして加水分解生成物の開放についてアッセイされる場合、約45℃〜98℃の最適温度を有する。好ましくは、非マルトース生成エキソアミラーゼの最適温度は55℃〜95℃であり、そしてさらにより好ましくは、60℃〜90℃である。
【0060】
低い熱安定性であることが見出され得る非マルトース生成エキソアミラーゼは、より熱安定性になるようタンパク質工学を用いて改良され得、そして従って、本発明への使用のために適切である。従って、タンパク質工学により一層熱安定性になるよう変性された非マルトース生成エキソアミラーゼの使用が本発明により包含される。
【0061】
いくつかの非マルトース生成エキソアミラーゼがいくらかの程度のエンドアミラーゼ活性を有することは知られている。多くの場合、このタイプの活性は減じられるか又は排除される必要がある。なぜならば、エンドアミラーゼ活性は、枝分かれのデキストリンの蓄積のために、粘着性の又はゴム状のパンの中味(crumb)を生成することによって、最終のパン生成物の品質におそらく負の効果を与えるためである。
【0062】
従って、好ましい観点においては、本発明の非マルトオース生成エキソアミラーゼは、1単位のエキソアミラーゼ活性当たり0.5より少いエンドアミラーゼ単位(EAU)を有するであろう。
好ましくは、本発明の使用のために適切である非マルトース生成エキソアミラーゼは、1単位のエキソアミラーゼ活性当たり0.05より少いEAUを有し、そしてより好ましくは、1単位のエキソアミラーゼ活性当たり0.01より少いEAUを有する。
エンドアミラーゼ単位は、下記に示されるエンドアミラーゼアッセイプロトコールにより決定され得る。
【0063】
本発明への使用のために適切な非マルトース生成エキソアミラーゼの例は、非還元鎖末端から、それぞれマルトテトラオース、マルトペンタオース又はマルトヘキサオースの連続的な残基を除去するために、澱粉質多糖類の1, 4−α−グルコシル結合を加水分解する、エキソーマルトテトラヒドロラーゼ(E. C. 3. 2. 1. 60)、エキソ−マルトペンタヒドロラーゼ及びエキソ−マルトヘキサヒドロラーゼ(E. C. 3. 2. 1. 98)を、包含する。
【0064】
例としては、シュードモナス・サッカロフィラ及びP. スツゼリ(EP−0298645B1)のエキソ−マルトテトラヒドロラーゼ、好アルカリ性グラム−陽性細菌(アメリカ特許第5,204,254号)及びシュードモナスsp. (Shidaなど., Biosci. Biotechnol. Biochem., 1992, 56, 76-80)のエクソ−マルトペンタヒドロラーゼ、及びバチルスsp. #707(Tsukamoto など., Biochem. Biophys. Res. Commun., 1988, 151, 25-31)、B.サーキュランスF2(Taniguchi, ACS Symp., 1991, Ser. 458, 111-124)及びアエロバクター・アエロゲネス(Kainumaなど., Biochim. Biophys. Acta, 1975, 410, 333-346)のエキソ−マルトヘキサヒドロラーゼを列挙することができる。
【0065】
本発明への使用のために適切な非マルトース生成エキソアミラーゼのもう1つの例は、好アルカリ性バチルス株、GM8901からのエキソアミラーゼである[28]。これは、澱粉からマルトテトラオース、マルトペンタオース及びマルトヘキサオースを生成する非マルトース生成エキソアミラーゼである。
【0066】
さらに、本発明への使用のために適切な非マルトース生成エキソアミラーゼはまた、非還元鎖末端から、それぞれマルトヘプタオース又はマルトオクタオースの残基を除去するために、澱粉質多糖類における1, 4−α−グルコシド結合を加水分解する、エキソ−マルトヘプタヒドロラーゼ又はエキソ−マルトオクタヒドロラーゼを包含する。エクソ−マルトヘプタヒドロラーゼ及びエキソ−マルトオクタヒドロラーゼは、野生型株をスクリーニングすることによって見出され得、又はタンパク質工学により他の澱粉分解酵素から開発され得る。従って、非マルトース生成エキソアミラーゼになるよう他の澱粉分解酵素からタンパク質工学により開発された非マルトース生成エキソアミラーゼもまた、本発明への使用のために適切である。
【0067】
新規アミラーゼ
1つの観点においては、本発明はまた、本発明の澱粉生成物、たとえば製パン生成物を調製するために適切である新規アミラーゼを提供する。本発明の新規アミラーゼは、非マルトース生成エキソアミラーゼである。本発明の研究において、本発明者は、食品、特に製パン生成物の調製に使用するためのドウの調製のために適切であるこの新規アミラーゼを特性決定した。
【0068】
従って、本発明はまた、非マルトース生成エキソアミラーゼを提供し、ここで前記非マルトース生成エキソアミラーゼは、それがモチ状トウモロコシ澱粉インキュベーション試験において、2〜10個のD−グルコピラノシル単位及び任意にはグルコースの1又は複数の線状マルト−オリゴ糖から成る生成物を加水分解する能力を有し、その結果、前記加水分解生成物の少なくとも60重量%、好ましくは少なくとも70重量%、より好ましくは少なくとも80重量%そして最も好ましくは少なくとも85重量%が、3〜10個のD−グルコピラノシル単位の線状マルトオリゴ糖、好ましくは4〜8個のD−グルコピラノシル単位の線状マルトオリゴ糖から成り、そして前記酵素がバチルス・クラウジから得ることができるものであり、又はその機能的同等物であり、そして前記酵素が約101,000Daの分子量を有し(ドデシル硫酸ナトリウムポリアクリルアミド電気泳動により推定される場合)、そして/又は前記酵素がpH9.5及び55℃で活性の最適性を有することにより特徴づけられる。
【0069】
好ましくは、アミラーゼは、単離された形及び/又は実質的に純粋な形で存在する。ここで、用語“単離された”とは、酵素がその天然の環境下に存在しないことを意味する。
【0070】
抗体
本発明の酵素はまた、たとえば標準技法の使用により、抗体を生成するためにも使用され得る。従って、本発明の個々の酵素に対する抗体が生成され得る。個々の抗体は、発明の他の適切なアミラーゼ酵素をスクリーンするために使用され得る。さらに、個々の抗体は本発明の酵素を単離するためにも使用され得る。
【0071】
抗体の生成に関しては、種々の宿主、たとえばヤギ、ウサギ、ラット、マウス、等が、免疫原性質を保持する、インヒビター又はそのいずれかの部分、変異体、相同体、フラグメント又は誘導体、又はオリゴペプチドによる注射により免疫化され得る。宿主種に依存して、種々のアジュバントが免疫学的応答を高めるために使用され得る。そのようなアジュバントは次のものを包含するが、但しそれらだけには限定されない:フロイント、鉱物ゲル、たとえば水酸化アルミニウム、及び界面活性物質、たとえばリゾレシチン、プルロニックポリオール、ポリアニオン、ペプチド、油エマルジョン、キーオール・リンペット(Keyhole limpet)ヘモシアニン及びジニトロフェノール。BCG(バチルス・カルメト−グエリン)及びコリネバクテリウム・パルバム(Corynebacterium parvum)は、開発され得る可能性ある有用なヒトアジュバントである。
【0072】
酵素に対するモノクローナル抗体は、培養での連続細胞系による抗体分子の生成を提供するいずれかの技法を用いて調製され得る。それらは次の技法を包含するが、但しそれらだけには限定されない:Koehler and Milstein (1975, Nature 256: 495-497) により最初に記載されたハイブリドーマ技法、ヒトB−細胞ハイブリドーマ技法(Kosbor など(1983)Immunol Today 4: 72; Cote など(1983) Proc. Natl. Acad. Sci. 80: 2026-2030)及びEBV−ハイブリドーマ技法(Cole など(1985)Monoclonal Antibodies and Cancer Therapy, Alan R Liss Inc. pp77-96)。
【0073】
さらに、“キメラ抗体”の生成のために開発された技法、すなわち適切な抗原特異性及び生物学的活性を有する分子を得るためにヒト抗体遺伝子へのマウス抗体遺伝子のスプライシングが使用さえ得る(Morrison など(1984)Proc. Natl. Acad. Sci. 81: 6851-6855; Neuberger など(1984)Nature 312: 604-608; Takeda など. (1985) Nature 314: 452-454)。他方では、一本鎖抗体の生成について記載される技法(アメリカ特許第4946779号)が、インヒビター特異的一本鎖抗体を生成するために適合され得る。
【0074】
抗体はまた、リンパ球集団においてインビボ生成を誘発することによって、又はOrlandiなど(1989, Proc. Natl. Acad. Sci. 86: 3833-3837)及びWinter G and Milstein C (1991; Nature 349: 293-299) に開示されるようにして、高い特異的結合試薬の組換え免疫グロブリンライブラリー又はパネルをスクリーニングすることによって生成され得る。
【0075】
改良性組成物
示されるように、本発明の1つの観点は、澱粉生成物、特にドウ及び/又はドウから製造されるベーキングされた澱粉質パン生成物のための改良性組成物に関する。
改良性組成物は、本発明の非マルトース生成エキソアミラーゼ及び少なくとも1つの追加のドウ成分又はドウ添加剤を含んでなる。
本発明によれば、追加のドウ成分又はドウ添加剤は、上記に記載されるドウ成分又はドウ添加剤のいずれかであり得る。
【0076】
好都合には、その改良性組成物は、少なくとも1つの追加の成分又は添加剤と共に混合される本発明の非マルトース生成エキソアミラーゼを含んで成る乾燥微紛組成物である。しかしながら、改良性組成物はまた、本発明の非マルトース生成エキソアミラーゼ、及び水又は他の液体に溶解されているか又は分散されている少なくとも1つの追加の成分又は添加剤を含んでなる溶液調製物であり得る。改良性組成物中の酵素活性の量は、改良性組成物の一部を形成する追加の成分及び添加剤の量及びタイプに依存するであろう。
【0077】
任意には、改良性組成物は、特定のベーキングされた生成物を製造するための乾燥成分及び添加剤のすべてを含む、完全混合物、いわゆるプレ−混合物の形で存在することができる。
【0078】
澱粉生成物の調製
本発明の1つの観点によれば、その方法は、適切な非マルトース生成エキソアミラーゼ酵素、たとえば本明細書に示される新規の非マルトース生成エキソアミラーゼ酵素の1つを、澱粉基材に添加することによって、澱粉生成物を形成することを含んで成る。
澱粉基材がドウである場合、ドウは、穀物、水、本発明の非マルトース生成エキソアミラーゼ及び他の可能な成分及び添加剤を一緒に混合することによって調製される。
【0079】
追加の例によれば、澱粉生成物がベーキングされた澱粉質パン生成物(非常に好ましい態様である)である場合、その技法は、ドウ形成条件下で、いずれか適切な順序で、食料粉、水及び発酵剤を混合し、そしてさらに、適切な非マルトース生成エキソアミラーゼ酵素を添加することを含んで成る。
膨化剤(leavening agent)は、化学膨化剤、たとえば炭酸水素ナトリウム、又はサッカロミセス・セレビシアエのいずれかの株(パン酵素)であり得る。
非マルトース生成エキソアミラーゼは、いずれかのドウ成分、たとえば水又はドウ成分混合物と共に、又はいずれかの添加剤又は添加剤混合物と共に添加され得る。
【0080】
ドウは、ベーキング産業、又は食料粉ドウ基材の生成物を製造するいずれかの他の産業において共通するいずれかの従来のドウ調製物により調製され得る。
澱粉質パン生成物、たとえば精白パン、篩分けされたライ麦粉及び小麦粉から製造されたパン、ロールパン及び同様のものベーキングは、典型的には、180〜250℃の範囲のオーブン温度で、約15〜60分間、パン用ドウをベーキングすることによって達成される。ベーキング工程の間、急な温度グラジェント(200→120℃)が外部ドウ層において有力であり、ここでベーキングされた生成物の特徴的な外皮が進行する。しかしながら、蒸気発生による熱消費のために、パンの中味の温度がベーキング工程の最後で、100℃に接近する。
【0081】
非マルトース生成エキソアミラーゼは、単独の活性成分として、又は1又は複数の追加のドウ成分又はドウ添加剤と混合して酵素を含んで成る、液体調製物として、又は乾燥微紛組成物として添加され得る。
ベーキングされた生成物の性質をさらに改良し、そしてベーキングされた生成物に特徴的な性質を付与するために、追加のドウ成分及び/又はドウ添加剤がドウ中に導入され得る。典型的には、そのような追加の成分は、ドウ成分、たとえば塩、穀粒、脂肪及び油、糖、ダイエット用繊維物質、粉乳、グルテン及びドウ添加剤、たとえば乳化剤、他の酵素、親水コロイド、風味剤、酸化剤、鉱物及びビタミンを含むことができる。
【0082】
乳化剤はドウ補強剤及びパン中味の軟化剤として有用である。ドウ補強剤として、乳化剤は、残り時間に関する耐性及び加工の間、衝撃に対する耐性を提供することができる。さらに、ドウ補強剤は、発酵時間における変化に対する所定のドウの耐性を改良するであろう。ほとんどのドウ補強剤はまた、ベーキングされた製品に耐性を与えるものからの体積の上昇を意味するオーブンスプリングを改良する。最後に、ドウ補強剤は、レセピ−混合物に存在するいずれかの脂肪を乳化するであろう。
【0083】
モノグリセリドの特性を主に示すパン中味の軟化は、冷却に基づいて再結晶化せず、そして従ってパン中味の堅さに寄与しないであろうアミラーゼとの不溶性包含複合体の形成を導く澱粉のアミロース画分と乳化剤との間の相互作用に帰因する。
【0084】
追加のドウ添加剤として使用され得る適切な乳化剤は次のものを包含する:レシチン、ポリオキシエチレンステアレート、食用脂肪酸のモノ−及びジグリセリド、食用脂肪酸のモノ−及びジグリセリドの酢酸エステル、食用脂肪酸のモノ−及びジグリセリドの乳酸エステル、食用脂肪酸のモノ−及びジグリセリドのクエン酸エステル、食用脂肪酸のモノ−及びジグリセリドのジアセチル酒石酸エステル、食用脂肪酸のスクロースエステル、ナトリウムステアロイル−2−ラクチレ−ト、及びカルシウムステアロイル−2−ラクチレ−ト。
【0085】
追加のドウ添加剤として有用である他の酵素は、例として、次のものを包含する:オキシドレダクターゼ、たとえばグルコースオキシダーゼ、ヘキソースオキシダーゼ、及びアスコルベートオキシダーゼ、たとえばリパーゼ及びエステラーゼ、並びにグリコシダーゼ様α−アミラーゼ、プルラナーゼ及びキシラナーゼ。オキシドレダクターゼ、たとえばグルコースオキシダーゼ及びヘキソースオキシダーゼは、ドウ補強剤及びベーキングされた生成物の体積の調節のために使用され、そしてキシラナーゼ及び他のヘミセルラーゼは、ドウ取り扱い性質、パン中味軟化及びパン体積を改良するために添加され得る。リパーゼはドウ補強剤及びパン中身軟化剤として有用であり、そしてα−アミラーゼ及び他の澱粉分解酵素はパンの体積を調節し、そしてさらに、パン中味の堅さを低めるためにドウ中に導入され得る。
【0086】
添加される本発明の非マルトース生成エキソアミラーゼの量は通常、kg穀物当たり50〜100,000単位、好ましくはkg 穀物当たり100〜50,000単位の量で最終ドウに存在する。本発明の有用な態様においては、その量はkg穀物当たり200〜20,000単位の範囲である。
本明細書においては、1単位の非マルトース生成エキソアミラーゼは、この後に記載されるように、50mMのMES, 2mMの塩化カルシウム溶液(pH6.0)中、10mg/mlのモチ状トウモロコシ澱粉4mlを有する試験管において50℃でインキュベートされる場合、1分当たり1μモルの還元糖に等しい加水分解生成物を開放する量の酵素として定義される。
【0087】
アミラーゼにより調製される食品
本発明は、食品の製造への使用のための適切なアミラーゼを提供する。動物飼料を包含する典型的な食品は、酪農製品、食肉製品、家禽製品、魚製品及び製パン製品を包含する。
好ましくは、食品は製パン生成物、たとえば上記に記載される製パン生成物である。本発明の範囲内に組み込まれる典型的な製パン(ベーキングされた)生成物は、パン、たとえば一塊のパン、ロールパン、バン、ビザ生地、等、プレッツェル、トルティヤ、ケーキ、クッキー、ビスケット、クラッカー、等を包含する。
【0088】
アミラーゼアッセイプロトコール
次のシステムが、本発明への使用のために適切である非マルトース生成エキソアミラーゼを特徴づけるために使用される。
初期のバックグラウンド情報によれば、モチ状トウモロコシアミロペクチン(Roquette, France からWAXILYS 200 として得ることができる)は、非常に高いアミロペクチン含有率(90%以上)を有する澱粉である。
20mg/mlのモチ状トウモロコシ澱粉は、50mMのMES(2−(N−モルホリノ)エタンスルホン酸)、2mMの塩化カルシウムの緩衝液(pH6.0)において3分間煮沸され、そして続いて50℃でインキュベートされ、そして30分以内に使用される。
【0089】
1単位の非マルトース生成エキソアミラーゼは、上記に記載されるように、50mMのMES, 2mMの塩化カルシウム溶液(pH6.0)中、10mg/mlのモチ状トウモロコシ澱粉4mlを有する試験管において50℃でインキュベートされる場合、1分間当たり1μモルの還元糖に等しい加水分解生成物を開放する量の酵素として定義される。
還元糖は、標準としてマルトースを用い、及びBernfeld, Methods Enzymol., (1954), 1, 149-158のジニトロサリチル酸法又は還元糖を定量化するための当業界において知られている他の方法を用いて測定される。
【0090】
非マルトース生成エキソアミラーゼの加水分解生成物パターンは、上記のようにして調製された緩衝液中、10mg/ml のモチ状トウモロコシ澱粉4mlを有する試験管において、0.7単位の非マルトース生成エキソアミラーゼを50℃で15又は300分間、インキュベートすることによって決定される。反応は、試験管を煮沸水浴に3分間浸すことによって停止された。
【0091】
加水分解生成物は、パルスされた電流滴定検出、及び標準としてグルコース〜マルトペプタオースの既知の線状マルトオリゴ糖と共に、溶離剤として酢酸ナトリウム、水酸化ナトリウム及び水を用いて、Dionex PA100 カラムのアニオン交換HPLCにより分析され、そして定量化される。マルトオクタオース〜マルトデカオースのために使用される応答因子は、マルトヘプタオースに関して見出された応答因子である。
【0092】
エンドアミラーゼアッセイプロトコール
0.75mlの酵素溶液が、50mMのMES(2−(N−モルホリノ)エタンスルホン酸)、2mMの塩化カルシウム溶液(pH6.0)中、0.5%(w/v)のAZCL−アミロース(Megazyme, Ireland から入手できるアズリン架橋されたアミロース)6.75mlと共に50℃でインキュベートされる。5, 10, 15, 20及び25分後、それぞれ1.0mlの反応混合物が、4%(w/v)のTRIS(トリス(ヒドロキシメチル)アミノメタン)から成る停止溶液4.0mlに移される。
【0093】
停止されたサンプルは、Whatman No.1フィルターを通して濾過され、そして590nmでのその光学密度が蒸留水に対して測定される。アッセイされる酵素溶液が希釈され、その結果、得られる光学密度は時間の線状関数である。光学密度対時間についての直線の傾斜が使用され、g当たり100エンドアミラーゼ単位(EAU)を有するものとして定義される、標準GRINDAMYL(商標)A100 (Danisco Ingredients から入手できる) に対するエンドアミラーゼが計算される。
【0094】
劣化(老化を包含する)の測定のためのアッセイ
本発明の非マルトース生成エキソアミラーゼの抗老化効果の評価のために、パン中味の堅さが、Instrn 4301 Universal Food Taxture Analyzer 又は当業界において知られている類似する装置により、ベーキングの1, 3及び7日後、測定され得る。
【0095】
当業界において従来使用され、そして本発明の非マルトース生成エキソアミラーゼの澱粉劣化に対する効果を評価するために使用されるもう1つの方法は、DSC(示差走査熱量)に基づかれる。それにより、酵素と共に又は酵素なしで(対照)ベーキングされたモデルシステムドウからのパン中味における劣化されたアミロペクチンの溶融エントロピーが測定される。記載される例に適用されるDSC装置は、20〜95℃、1分当たり10℃の温度グラジエントで作動するMettler-Toledo DSC820 である。サンプルの調製のためには、10〜20mgのパン中味が計量され、そしてMetler-Toledo アルミニウムパンに移され、次に密封される。
【0096】
記載される例に使用されるモデルシステムドウは、本発明の非マルトース生成エキソアミラーゼと共に又はそれを伴わないで、標準の小麦粉及び最適量の水を含む。それらは、それぞれ6又は7分間、Brabender Farinographにより10又は50gで混合される。ドウのサンプルが、蓋付きのガラス試験管(15×0.8cm)に配置される。それらの試験管は、33℃で30分間のインキュベーションから開始して、続いて1.1℃/分のグラジエントで33℃から95℃まで、水浴においてのベーキング工程にゆだねられ、そして最終的に95℃で5分間インキュベートされる。続いて、試験管が、DSC分析の前、20℃でサーモスタットにおいて貯蔵される。
【0097】
要約
要約すれば、本発明は、アミロペクチンの非還元鎖末端から4〜8個の範囲のD−グルコピラノシル単位の線状マルトオリゴ糖を切り取ることによって澱粉を加水分解し、そして好ましくは、十分な程度の熱安定性を有する非マルトース生成エキソアミラーゼが、ベーキングされた生成物における有害な劣化を遅延し、又は低めるのに非常に効果的である驚くべき発見に基づかれている。
【0098】
寄託
次のサンプルが、1999年3月12日、公認された寄託機関DSMZ(Deutsche Sammlung von Microorganismen und Zellkulturen GmbH of Mascheroder Weg 1b, D-38124 Braunschweig)に、ブダペスト条約に従って寄託された:BT−21 DSM番号DSM12731。
本発明はまた、それらの寄託物及びそれを含んで成る具体物から誘導でき、そして/又は発現できる配列、及びその活性フラグメントも包含する。
【実施例】
【0099】
例−セクションA
例1.シュードモナス・サッカロフィラ非マルトース生成エキソアミラーゼの発酵及び生成
1.1生成:
P.サッカロフィラ株IAM No. 1544を、IAM Culture Collection, Inst. Of Mloecular and Cellular Biosciences, University of Tokyo, Japan から得た。
【0100】
前記菌株の発酵を、2Lの使用体積を有するApplikon ADIの3Lの生物反応器において、次の条件下で行った:
温度: 30℃
撹拌速度: 1000rpm
通気 1体積空気/1体積培地/分
pH: 2Mの水酸化ナトリウム及び10%(w/v)の塩酸による調節による一定のpH7.4
【0101】
培地: Bacto Trypton 20g/l
Bacto Yeast 抽出物 20g/l
澱粉 20g/l
Na2HPO4・2H2O 5.6g/l
KH2PO4 1.5g/l。
1日の発酵の後、発酵ブイヨンを遠心分離し、そして濾過し、細胞を除いた。細胞フリーブイヨンにおける非マルトース生成エキソアミラーゼの活性は、上記のようにして決定される場合、5単位/mlであった。
【0102】
1.2 P.サッカロフィラ非マルトース生成エキソアミラーゼの精製:
P.サッカロフィラの非マルトース生成エキソアミラーゼを、200mMの硫酸ナトリウム、50mMのトリエタノールアミン、2mMの塩化カルシウムを含むA−緩衝液(pH7.2)を含む、150mlのPhenyl Sepharose FF低サブカラム(Pharmacia, Sweden) を用いて、疎水性相互作用グロマトグラフィーにより、部分的に精製した。濾過された発酵ブイヨン(500ml)を、200mMの硫酸ナトリウムによりpH7.2に調節し、そしてカラム上に負荷した。非マルトース生成エキソアミラーゼを、A−緩衝液中、直線的に低下する硫酸ナトリウムグラジエントにより遊離した。エキソアミラーゼ活性を含む画分をプールした。
【0103】
プールされた画分を水により3倍に希釈し、そしてさらに、50mMのトリエタノールアミン、5mMの塩化カルシウムから成るA−緩衝液(pH7.5)により平衡化された150mlのQ−Sepharose FF (Pharmacia) カラム上でのアニオン−交換クロマトグラフィーにより精製した。非マルトース生成エキソアミラーゼを、A−緩衝液中、0〜1Mの塩化ナトリウム直線グラジエントにより溶離した。エキソアミラーゼ活性を含む画分をプールした。この部分的に精製された調製物を、下記試験のために使用した。それは、アミラーゼ活性について染色されたポリアクリルアミドゲル電気泳動システムにおいて試験される場合、14.7単位/mlの活性及びわずか1つのアミラーゼ活性の結合を有した。
【0104】
1.3 P. サッカロフィラ非マルトース生成エキソアミラーゼの特徴化:
最初に、P.サッカロフィラのエキソ−アミラーゼ(PS4と呼ばれる)をコードする遺伝子のDNA配列が、Zhouなど(Zho JH, Baba T, Takano T, Kobayash S, Arai Y (1989) FEBS Lett 1989 Sep 11; 255 (1): 37-41 “Nucleotide sequence of the maltotetraohydrolase gene from Pseudomonas sacchrophila”)により公開されている。さらに、前記DNA配列は、受託番号X16732としてGenBankから入手できる。
【0105】
本発明者は、質量分光計(MALDI−TOFF)によれば、精製されたPS4酵素のMWが57500±500D(配列からの理論的MWによれば、57741Dである)であることを決定した。
PS4の最適温度及びpHは、Zhouなど(Zhon JH, Baba T, Takano T, Kobayashi S, Arai Y (1992) Carbohydr Res 1992 Jan; 223: 255-61 “Properties of the enzyme exzyme expressed by the Pseudomonas saccharophila maltotetrahydrolase gene (mta) in Escherichia coli”)によれば、45℃及びpH6.5である。
【0106】
非マルトース生成エキソアミラーゼの加水分解パターンを、上記のようにして、10mg/mlのモチ状トウモロコシ澱粉4mlを有する試験管において、0.7単位の部分的な精製された非マルトース生成エキソアミラーゼを50℃で15又は300分間インキュベートすることによって生成される加水分解生成物を分析することによって決定した。
【0107】
15分及び300分後に検出される加水分解生成物のパターンは、表1に示されており、そしてP.サッカロフィラが本発明において定義されるような非マルトース生成エキソアミラーゼを生成し、そしてこの酵素が、それぞれ、15分及び30分の加水分解の後、85.8重量%及び93.0重量%を占める有力な生成物としてマルトテトラオースを開放することを示す。
【0108】
【表1】
【0109】
例2.P. サッカロフィラ非マルトース生成エキソアミラーゼのベーキング試験
ベーキング試験を、P.サッカロフィラ非マルトース生成エキソアミラーゼの抗固化効果を試験するために設定した。Danish Toast Bread のためのレセピーを使用した。それは、食料粉(2000g)、乾燥酵母(30g)、糖(30g)、塩(30g)及び水(400g Brabender Units (BU)のドウコンシステンシーに対応する約1200g+使用される乾燥酵母を補足するための追加の水60g)を含み、そしてそれらを、遅い速度で2分間及び早い速度で12分間、Hobartミキサー(モデルA−200)において混合する。
【0110】
ドウ温度は、混合の最後で26℃である。ドウを30℃で10分間、放置し、この後、ドウを750gのドウ部分に分割する。そのドウ部分を、33℃の温度及び85%の相対湿度で試験キャビネットにおいて5分間、放置する。次に、ドウ部分を、次の設定でGlimek成形機(タイプLR−67)上で成形し:1:4、2:2、3:14及び4:12、この後、成形されたドウ部分を、ベーキング用缶に移し、そして33℃の温度及び85%の相対湿度で50分間、試験キャビネットにおいて試験する。最終的に、試験されたドウ部分を、Wachtel オーブン(モデルAE416/38COM)において、220℃の温度で40分間、10秒間の蒸気を伴ってベーキングする。
【0111】
P.サッカロフィラ非マルトースエキソアミラーゼの部分的に精製された調製物を、食料粉1kg当たり1470単位で、導入されるドウに添加した。ベーキングした後、非マルトース生成エキソアミラーゼを有するか又は有さないパンを20℃に冷却し、そしてその後、プラスチックバックにおいて20℃で貯蔵した。堅さを、ベーキングの後3日目、1つのパンからの10枚のスライスの平均として及び7日目、2個のパンからのそれぞれ10枚のスライスの平均として、それぞれ3日目、Instron 4301 Universal Food Texture Analyzer により決定した。表2は、酵素を有するパンにおいて低い堅さが、両日で観察されたことを示す。
【0112】
【表2】
【0113】
表3は、7日目、P.サッカロフィラ非マルトース生成エキソアミラーゼの抗固化効果が95%の信頼レベルで統計学的に有意であることを示す。
【表3】
【0114】
The StatAdvisor:
ANOVA表は、次の2種の成分に7日目での堅さの変動を分ける:グループ間成分及びグループ内成分。この場合、72.0に等しいF−比率は、グループ内推定値に対するグルループ間推定値の比率である。F−試験のP−値は0.05以下であるので、95.0%の信頼レベルで、1つの酵素レベルからもう1つの酵素レベルでの平均堅さ間に統計学的に有意な差異が存在する。平均が他の平均と有意に異なるかどうかを決定するためには、Tabalar Options の列挙からMaltiple Range Testを選択すること。
【0115】
例3.
次のことは、E.コリMC1061におけるシュードモナス・サッカロフィラからの非マルトース生成エキソアミラーゼをコードするmta遺伝子の本発明のクローニング及び発現を記載する。
この観点においては、P.サッカロフィラIAM1520を、2mlのLB培地において増殖し、そして細胞を、20,000xgで10分間の遠心分離により収穫した。全DNAを、わずかに変更されたminiprep プロトコールを用いて単離した。細胞を、300μlの再懸濁緩衝液(50mMのトリス−HCl、pH8.0:10mMのEDTA;100μg/mlのRNアーゼA)に再懸濁し、この後、細胞を、Fastprep FP120 (BIO101; California)を用いて粉砕した。
【0116】
粉砕に続いて、300μlの溶解緩衝液(200mMのNaOH;1%のSDS)及び300μlの中和緩衝液(3.0Mの酢酸カリウム、pH5.5)を添加した。20.000xgで、4℃で15分間の遠心分離の後、上清液を集め、そして0.6体積のイソプロパノールを添加した。DNAを、4℃で、20,000xgでの30分間の遠心分離により沈殿せしめ、70%エタノールにより洗浄し、そして100μlのTE(10mMのトリス−HCl, pH8.0, 1mMのEDTA)に再溶解した。
【0117】
PCR増幅のために、次の4種の異なったPCRプライマーを企画した:
#1 ATG ACG AGG TCC TTG TTT TTC 位置213−233
#2 GCT CCT GAT ACG ACA GCG 位置2403−2386
#3 GCC ATG GAT CAG GCC GGC AAG AGC CCG 位置663−683
#4 TGG ATC CTC AGA ACG AGC CGC TGG T 位置2258−2238
【0118】
前記位置は、GenBank 受託番号X16732号に見出されるmtaについての配列に関する。上方の番号のプライマーはまず、アンチセンスプライマーであり、そしてその配列は相補的配列である。鋳型DNAに対して相補的でないヌクレオチドは太字で示され、そして導入された制限部位は下線で示される。プライマー#3は、ユニークNco1部位を導入し、そしてプライマー#4はBamHI部位を導入し、この部位は、発現ベクターpBAD/gIII (Invitrogen) における次のクローニングのために使用される。
【0119】
第1のPCR増幅を、次のプライマーの組み合わせ:反応1:2190bpのフラグメントを付与する#1+#2を用いて、製造業者の説明書及び次の増幅プロトコールに従って、Expand DNA ポリメタ−ゼ(Boehringer Mannheim; Germany)、及び鋳型としての50〜150ngのゲノムIAM 1520 DNAを用いて行った:35サイクルの間94℃で2分(94℃で1分、58℃で2分、72℃で2分)、及び最終的に、72℃で5分。
【0120】
2190bpのフラグメントを、“gene Clean Kit (BIO 101; California)を用いて、ゲルから単離した。そのフラグメントを、上記と同じ増幅プロトコールを用いて、第2のPCRにおいて、次のプライマーの組み合わせを用いて、鋳型DNAとして使用した:
反応2:1605bpのグラグメントを付与する#3+#4.
【0121】
1605bpのフラグメントを精製し、そして製造業者の説明書に従って、PCR−BLUNTベクター(Invitrogen)中にクローン化した。そのクローン化されたフラグメントの配列を、普遍及び逆プライマー及び4種のラベルされた内部プライマーを用いて、単一色素配列決定技法及びALF配列決定機(Pharmacia;Sweden)により配列決定することによって確かめた:
【0122】
CAT CGT AGA GCA CCT CCA 999-982
GAT CAT CAA GGA CTG GTC C 1382-1400
CTT GAG AGC GAA GTC GAA C 1439-1421
GAC TTC ATC CGC CAG CTG AT 1689-1780
前記位置は、GenBan受託番号X16732号に見出されるmtaについての配列を示す。上方の番号のプライマーはまず、アンチセンスプライマーであり、そしてその配列は相補的配列である。
【0123】
配列を確かめた後、mta遺伝子を発現ベクターpBAD/gIII(Invitroge)中にクローン化した。Mta 遺伝子は、BamHIによる消化、続いてクレノウフラグメントによるブラント化及びNcoIによる消化によりpCR−BLUNTから開放し、そして1620bpのフラグメントを精製した。発現ベクターpBAD/gIIIをNcoI及びPmeIにより消化し、そして精製した。連結の後、得られる発現構造体を用いて、E.コリMC1061細胞を形質転換し、そしてタンパク質を、pBAD/gIIIマニュアル(Invitrogen)に従って発現した。
【0124】
例4.澱粉劣化に対する非マルトース生成及びマルトース生成エキソアミラーゼの効果の比較
サツマイモβ−アミラーゼ(EC 3. 2. 1. 2; 製品番号A7005号としてSigmaから得ることができる)は、澱粉の非還元末端からマルトースを開放するマルトース生成エキソアミラーゼである。
このマルトース生成エキソアミラーゼの熱安定性は、50mMのクエン酸ナトリウム、5mMの塩化カルシウム溶液(pH6.5)において45〜75℃の温度で15分間インキュベートした後、残留活性により示されるように、P.サッカロフィラ非マルトース生成エキソアミラーゼの熱安定性に類似する(表4)。
【表4】
澱粉劣化に対する両酵素の効果を、“Assys for measurement of retrogradation and staling”に記載されるように、モデルシステムドウからのベーキングされ、そして貯蔵された生成物のDSC分析により試験した。この試験に関しては、“アミラーゼアッセイプロトコール”に従ってアッセイされた、485単位のP.サッカロフィラ非マルトース生成エキソアミラーゼ及び735単位のサツマイモβ−アミラーゼをドウに使用した。ドウを、50mMのクエン酸ナトリウム、5mMの塩化カルシウム溶液(pH6.5)30.7ml又は添加される酸素、及び50gの標準デンマーク小麦粉(Danisco 98022)から調製した。
【0125】
7日間の貯蔵の後、劣化されたアミロペクチンの量を、その溶融エントロピーを測定することによって定量化した。統計学的分析によれば、両酵素は澱粉劣化を有意に低め(表5);すなわちP.サッカロフィラ非マルトース生成エキソアミラーゼは、7日目、劣化されたアミロペクチンの量を86%(1.77 J/g)に低め、ところがサツマイモβ−アミラーゼは、それを96%(1.96 J/g)に低めることが見出された。結論的には、P.サッカロフィラ非マルトース生成エキソアミラーゼは、比較できる熱安定性のマルトース生成アミラーゼよりも、劣化及び老化を低めるために、明らかにより効果的である。
【0126】
【表5】
【0127】
この表は、平均が他の平均と有意に異なるかどうかを決定するために、複数の比較方法を利用する。アウトプットの低部半分は、個々の対の平均間の推定される差異を示す。星印は3個の対に続いて配置され、これは、それらの対が95.0%の信頼レベルで統計学的に有意な差異を示すことを示唆する。平均間を区別するために現在使用される方法は、Fisher’s最少有意差異(LSD)方法である。
【0128】
例−セクションB
材料及び方法:
材料:コーンからのアミロペクチン及びアミロース、コーン澱粉、カルボキシメチルセルロース(CMC)、ウシ血清アルブミン(BSA)、デキストラン、プルラン、マルトース、マルトトリオース、及びマルトテトラオース〜マルトデカオースの混合物を、Sigma Chemical Co., St. Louis, USAから得た。可溶性澱粉を、Merck, Darmstadt, Germany から得た。酵素抽出物及びトリプトンは、Difco Laboratories, Detroit, USAから得られた。
【0129】
Neue Allgemeine Reisgesellschaft mbH, Hamburg, Germany からの完全な玄米が使用された。医薬品種のα−、β−及びγ−シクロデキストリンは、Wacker Chemie Danmark Aps, Glostrup, Denmark から得られた。マルトテトラオースは、前記のようにして調製された[32]。すべての化学物質は、特にことわらない限り、分析用銘柄のものであった。
B.クラウジBT−21の単離:前記株は、DSMZ(Deutsche Sammlung von Mikroorganismen and Zellkulturen GmbH, Braunshweig, Germany)により同定された、Assens, Denmark により集められた土壌サンプルから単離される。
【0130】
酵素の生成:B.クラウジBT−21を、ジャガイモからの2.0%可溶性澱粉トウモロコシからのアミロペクチン、コーン澱粉又は完全な玄米、0.5%酵母抽出物、0.5%トリプトン、0.1%KH2PO4, 0.1%Na2HPO4, 0.02%MgSO4・7H2O, 0.02% CaCl2・2H2O 及び0.1%(NH4)2 SO4から成る最適化された液体培地において増殖させた。オートクレーブの後、無菌Na2CO3溶液を添加し、1%の最終濃度にした(約pH10)。グリセロール中の胞子懸濁液(−80℃で貯蔵される)を用いて、100mlの実際の培地を接種し、そして200rpmでの振盪インキュベーターにおいて45℃で18時間インキュベートした(New Brunswick Scientific, Edison, N.J., USA)。
【0131】
この培養物2mlを用いて、200mlの培地を有する振盪フラスコを接種し、そして振盪インキュベーターにおいて45℃でインキュベートした。アリコートを規則的な間隔で採取し、そして600nmでのODを測定し、培地における菌類の増殖を決定した。サンプル(4ml)を、9600rpmで4℃で10分間、遠心分離し、そしてpH及びアミラーゼ活性を決定した。すべての増殖実験は三重反復して行われた。平均値(X=(Σni=1Xi)/m)及び標準偏差値(std.=√(Σni=1(Xi-X)/(n-1))を決定した。
【0132】
生成物−特異的アミラーゼの精製:完全な玄米上でのB.クラウジBT−21の52時間の増殖の後、細胞及び完全な米穀粒を、9600rpm及び4℃での15分間の遠心分離により細胞外流体(1000ml)から除いた。生成物−特異的アミラーゼを、エポキシ−活性化されたセファロース6Bマトリックス(Pharmacia Biotech, Uppsala, Sweden)にβ−シクロデキストリンを共有結合することによって調製された親和性ゲルを用いて精製した[33]。細胞外細胞−フリー上清液を、12gのゲルと共に、4℃で1時間、振盪しながらインキュベートした。
【0133】
次に、上清液を、9600rpm及び4℃で10分間の遠心分離により除去した。結合されなかったタンパク質を、50mMのリン酸緩衝液(pH8.0)75mlによりゲルを洗浄し、続いて遠心分離することによって除去した。洗浄段階を7度反復した。結合されたタンパク質を、10mMのα―シクロデキストリンを含む50mMの燐酸緩衝液(pH8.0)45mlにより溶出し、続いて遠心分離した。溶出段階は4回反復された。α−シクロデキストリンを、β−及びγ−シクロデキストリンはBradford (1976)のタンパク質決定方法を妨害するので、酵素の溶出のために使用した[34]。
【0134】
次に、α−シクロデキストリンを、4℃で撹拌しながら、10mMのトリエタノールアミン(pH7.5)5Lに対する透析(6〜8kDaのSpectra/Por透析膜、The Spedtrum Companies, Gardena, CA, USA)により除去した。緩衝液を2時間後に交換し、続いてさらに12時間、透析した。透析バッグをCMCに配置し、サンプルを濃縮した。10mlを、FPLC−システム(Pharmacia, Uppsala, Sweden)を用いて、HiTrap Qカラム(Pharmacia Biotech, Uppsala, Sweden)に適用した。
【0135】
タンパク質を、10mMのトリエタノールアミン(pH7.5)25mlにより1.0ml/分の速度で、続いて、10mMのトリエタノールアミン(pH7.5)中、20mMのNaClのグラジエントにより溶出した。酵素を0.5MのNaClにより溶出した。タンパク質の含有率を、BIO−RAD Protein Assay (Bio-Rad Laboratories, Hercules, CA, USA)を用いてBradford, (1976)の方法[34]により評価した。BSAを標準として利用した。
【0136】
ゲル電気泳動:15μlのサンプルを、[35]により記載されるようにして、天然のトリス−グリシンゲル(10%)により分析した。次に、ゲルを、pH6.5での50mMのリン酸緩衝液に配置し、そして30分間、振盪した。1%(w/v)の可溶性澱粉溶液を、ゲルと共に45分間、振盪しながらインキュベートした。緩衝溶液により洗浄した後、ゲルをヨウ素溶液(4mMのI2, 160mM のKI)と共にインキュベートし、そして緩衝液により脱色した。脱色されバンドは、澱粉加水分解活性を示した。
【0137】
SDS−PAGE(10%)を、[36]に従って行い、続いて銀染色[37]した。SDS−PAGE広範囲分子量標準(Bio-Rad Laboratories, hercules, CA, USA)を用いた。
酵素アッセイ:0.1Mの硼酸緩衝液(pH10.0)中、2mlの可溶性澱粉溶液(1.25%)を、0.5mlの酵素溶液と共に45℃で2時間インキュベートした。反応を、その混合物を10分間、煮沸することによって停止した。還元糖の形成を、CuSO4/ビシンコネート アッセイ[38]により決定し、そして形成されるmMマルトース同等物として計算した。1単位の活性は、pH10.0及び45℃で1分当たり1μモルのマルトース同等物を生成する酵素の量に対応する。
【0138】
酵素特徴化:温度最適値の決定のために、精製された酵素を、0.1Mの硼酸緩衝液(pH10.0)(0.5mMのCaCl2を含むか又は含まない)中、1%の最終濃度の可溶性澱粉において、30℃〜90℃の温度で15分間インキュベートした。温度安定性の決定を、5mMのCaCl2を含む50mMのグリシン−NaOH緩衝液(pH9.5)において、精製された酵素を30分間、30, 40, 50, 60, 70, 80及び90℃でインキュベートすることによって行った。
【0139】
残留活性を、50mMのグリシン−NaOH緩衝液(pH9.5)中、1%の最終濃度の可溶性澱粉において、加熱処理された酵素を、55℃で15分間インキュベートすることによって決定した。PH最適値を、異なった緩衝液中、1%の最終濃度の可溶性澱粉において、精製された酵素を55℃で15分間インキュベートすることによって決定した。使用される緩衝液は、50mMのクエン酸塩(pH4.0〜6.0)、50mMのトリス−マレエート(pH6.5〜8.5)及び50mMのグリシン−NaOH(pH9.0〜11.0)であった。
【0140】
I2−KI溶液(0.02%のI2及び0.2%のKI)を、Fuwa、1954[39]に従って調製した。澱粉−ヨウ素青色形成を、次の変法により二重反復して測定した。サンプル500μlを、異なった時間間隔で、可溶性澱粉の酵素加水分解から回収した。次に、250μlのHCl及び250μlのI2−KI溶液を添加し、そして混合した。脱イオン水(4.0ml)を添加し、そして混合を反復した。青色の形成を、600nmで分光光学的に測定した。
【0141】
精製された酵素による異なった基質の加水分解を、ジャガイモからの可溶性澱粉、トウモロコシからのアミロペクチン、デキストラン、プルラン(1%)、アミロース(0.1%)、及び10mMのα−、β−及びγ−シクロデキストリンにより試験した。基質を、5mMのCaCl2を含む、50mMのグリシン−NaOH緩衝液(pH9.5)に溶解し、そして精製された酵素を添加した(505mU/ml)。種々の基質を55℃でインキュベートし、そしてサンプルを下記のようにして分析した。
【0142】
精製された酵素によるマルト−オリゴ糖の加水分解を、2mMの最終濃度でマルトース、マルトトリオース及びマルトテトラオースにより、及びマルトテトラオース〜マルトドテカオースの混合物(5mM)により試験した。マルト−オリゴ糖を、5mMのCaCl2を含む50mMの硼酸緩衝液(pH9.5)に溶解し、そして精製された酵素を添加した(147mU/ml)。基質を55℃でインキュベートし、そしてサンプルを異なった時間間隔で採取した。反応を10分間の煮沸により停止し、そしてサンプルを下記のようにして分析した。
【0143】
加水分解生成物の分析:加水分解生成物を、パルスされた電流滴定検出を有する高性能アニオン交換クロマトグラフィー(HPAEC−PAD)を用いて検出した。CarboPac PA-1カラム(Dionex Corporation, Sunnyvale, CA, USA)を、100mMのNaOH中、30分間にわたっての0〜60%の1.0Mの酢酸ナトリウムグラジエント及びDionex DX-300又はDX-500システム上での1.0ml/分の流速と共に使用した。澱粉加水分生成物を、グルコース、マルトース、マルトトリオース、マルトテトラオース、マルトペンタオース及びマルトヘキサオースとそれらの保持時間とを比較することによって同定した。相同の線状マルト−オリゴ糖の保持時間は、重合の程度と共に上昇するので、中間DPの線状マルト−オリゴ糖は容易に同定され得る[40, 41]。
【0144】
結果及び論議:
B.グラウジBT−21の同定:その脂肪酸組成によれば、前記菌株は、バチルス属に対する類似性を示した。16SrDNAの部分配列決定は、B.クラウジに対して99.4%の類似性を示した。アルカリ耐性株の生理学的性質がこの同定を確認した。
B.グラウジBT−21によるアミラーゼ活性の生成:ジャガイモからの可溶性澱粉、コーン澱粉、トウモロコシ及び玄米からのアミロペクチンは、基材において異なったレベルの細胞外アミラーゼ活性をもたらした。
【0145】
コーン澱粉は、ジャガイモ澱粉に比較してより脂質を含み、そしてリンを含まないが、トウモロコシからのアミロペクチンは、α−D−(1→6)O−グリコシド結合を含む高い枝分かれ鎖構造を有する。それらの3種のタイプの澱粉は、加熱ゲル化の後、酵素のために入手でき、ところが完全な玄米は米穀粒に封入される低い入手可能性の澱粉を含む。異なった澱粉基質を含む液体培養物の細胞外流体におけるアミラーゼ活性が図1に示される。最高の澱粉分解活性が基質として完全な玄米により得られた。これは、低い入手可能性澱粉基質の存在がB.クラウジBT−21による細胞外澱粉分解活性の高められた生成をもたらした。
【0146】
類似する結果が、酵素の精製の前、細胞外流体から除去することが困難である小麦ふすまにより得られた。炭素源、たとえばガラクトース、グリコーゲン及びイヌリンはこれまで、B.リケニホルミスによるアミラーゼ生成のために適切であるものとして報告されており[27]、そして可溶性澱粉がB.ステアロサーモフィラスによるアミラーゼの生成のために最良の基質として見出された[28]。しかしながら、それらの研究のどれも、低い入手可能性の澱粉基質を包含していない。
生成物−特異的アミラーゼの精製:酵素を、β−CD Sepharose 6Bによる親和性クロマトグラフィー、続くアニオン交換クロマトグラフィーにより精製した(表6)。
【0147】
【表6】
【0148】
活性染色された天然のPAGEは、細胞外流体における3種の澱粉分解活性の存在を示した。生成物−特異的酵素を、β−CD親和性クロマトグラフィー、続くアニオン−交換クロマトグラフィー処理の後、他の澱粉分解活性から完全に分離した。精製された酵素調製物のSDS−PAGEは、生成物−特異的アミラーゼが相同性に精製され、そして約101kDaの見掛け分子量を有することを示した。シクロデキストリン セファロース6B親和性グロマトグラフィーによる他のアミラーゼの除去の後、最終精製段階としてα−アミラーゼの精製のためにこれまで使用して来た[29]。8.7%の酵素回収率、及び生成物−特異的アミラーゼのために得られた18.5の精製因子は、それらの著者により報告される値に類似した。
【0149】
生成物−特異的アミラーゼの特徴化:精製された酵素は、pH9.5での活性の最適値を示し(図2)、そしてその活性のための最適温度は、5mMのCaCl2の存在下で、又は不在下で55℃であった(図3)。5mMのCaCl2の存在下でpH9.5での酵素の熱安定性が図4に示される。55℃以上の温度で、酵素は、30分のインキュベーション期間の間、その最大活性の75%を失った。マルトヘキサオース[23]、マルトペンタオース[24]及びマルトテトラオース[25]を形成する5種の他の生成物−特異的アミラーゼはまた、アルカリpH最適性を示す。
【0150】
それらの酵素の分子量は、59, 73及び80kDa[23], 180kDa[24]及び97kDa[26]であることが推定され、そしてB.グラウジBT−21からの生成物−特異的アミラーゼは101kDaの推定分子量を示した。生成物−特異的アミラーゼ及びα−アミラーゼは、50〜65kDaの範囲の低い分子量を示す[3, 16, 17, 19及び21]。約55℃の温度適度は、上記生成物−特異的アミラーゼについて報告される最適度に類似した[23, 24及び26]。
【0151】
精製された生成物−特異的アミラーゼは、低いDPの主な初期生成物として、1時間のインキュベートの後、可溶性澱粉を、主にマルトヘキサオース及びマルトペンタオース(合計の加水分解された生成物の52%及び19%)に加水分解した(図5)。2時間のインキュベーションの後、マルトヘキサオースの量、及び4時間後、マルトペンタオースの量は低下し、そしてマルトテトラオース、マルトトリオース、マルトース及びグルコースの量は低下した。それらの生成物は延長された加水分解の後、蓄積し、このことは、それらがさらに加水分解されなかったことを示す。
【0152】
24時間の澱粉加水分解の後、マルト−オリゴ糖の量は3%であり、マルトヘキサオースの量は(3%)マルトヘキサオース、(4%)マルトペンタオース、(41%)マルトテトラオース、(13%)マルトトリオース、(16%マルトース及び(4%)グルコースであった。30分の澱粉加水分解の後に得られる、パルスされた電気滴定検出を有する高性能アニオン−交換クロマトグラフィー(HPAEC−PAD)形跡は(図6)、マルトヘキサオース(DP6)よりも大きな澱粉加水分解生成物が不在であることを示す。マルトヘキサオース−形成生成物−特異的アミラーゼによる可溶性澱粉の加水分解の時間経過研究はまた、マルトヘキサオースが加水分解の初期段階において、選択的に生成されたことも示す[23]。
【0153】
Kimなど(1995)[26]は、マルトテトラオース−形成生成物−特異的アミラーゼによる澱粉の1時間の加水分解の後の初期加水分解生成物が主にマルトヘキサオース(54%)であり、続いてマルトテトラオース及びマルトースの量の徐々の上昇が存在し、そしてマルトヘキサオースの量が低下したことも見出した。20時間後、マルト−オリゴ糖の組成が、0.6%のマルトヘキサオース、1.3%のマルトペンタオース、53.2%のマルトテトラオース、8.3%のマルトトリオース、27.6%マルトース及び9%のグルコースに変化した。
澱粉加水分解の間、酵素の作用のモードをさらに試験するために、異なった基質を、精製された酵素と共にインキュベートした(表7)。
【0154】
【表7】
【0155】
澱粉は、アミロース(20〜30%)及びアミロペクチン(80〜70%)から構成される。アミラーゼはα−D−(1→4)O−グルコシド結合された線状グルカンであり、そしてアミロペクチンは分子におけるα−D−(1→6)O−グリコシド結合の存在のために枝分かれ鎖のグルカンである。生成物−特異的アミラーゼは容易に、アミロペクチンを加水分解し、それは、可溶性澱粉(7%及び17.8%)及びアミロース(6.3%及び20%)に比較して、マルトペンタオース(9.2%)及びマルトヘキサオース(24.8%)の形成により示された。
【0156】
酵素は、プルラン、すなわちマルトトリオースを鎖から構成されるα−(1→6)O−グリコシド結合グルカン、又はデキストラン、すなわち0〜3個の主鎖単位に結合される技を有するα−(1→6)O−グリコシド結合グルカンを加水分解しなかった。α−、β−又はγ−シクロデキストリン、すなわち6.7及び8個のグルコース単位から構成される環状マルト−オリゴ糖はまた、24時間のインキュベーションの後でさえ、加水分解されなかった。
【0157】
デキストランに基づいて得られた結果は、α−(1→6)O−グリコシド結合が生成物−特異的アミラーゼにより分解されなかったことを示した。プルランに対する活性の欠失は、生成物−特異的アミラーゼが3個のグルコース単位に続くα−(1→6)O−グルコシド結合を回避することができず、又は3個のグルコース単位のいずれかを結合し得ないことを示した。α−、β−又はγ−シクロデキストリンに対する加水分解の欠失は、生成物−特異的アミラーゼがエキソタイプの分解機構により澱粉を加水分解したことを示した[30]。HPAEC−PAD形跡(図6)はまた、DP6よりも長い澱粉加水分解生成物が不在であるので、エキソータイプの分解機構を示した。
【0158】
可溶性澱粉に対する酵素分解作用をさらに試験するために、澱粉−ヨウ素青色形成を、還元糖の生成に対してプロットした(図7)。曲線の傾斜は、澱粉分解澱粉加水分解の有力な型の分解機構を示す[31]。エンド−作用性酵素は、エキソ−作用性酵素に比較される小さな値を有する傾斜を生成するであろう。傾斜の小さな値は、A.オリザエからのα−アミラーゼにより示される、ランダム澱粉分解活性による澱粉−ヨウ素青色複合体の早い還元の結果である(傾斜は−61である)。P.スツゼリからの細胞外酵素調製物は、大きな傾斜値により示される有力なエキソ−作用分解機構のための証拠を示した(傾斜は−13である)。精製された生成物−特異的アミラーゼは、エキソ−活性を示す、−6に傾斜値を示した。
【0159】
生成物−特異的アミラーゼによる、低いDPを有する基質の加水分解の作用のモードを、そのような基質と共にインキュベートすることによって試験した。マルトース、マルトトリオース及びマルトテトラオースは、精製されたB.クラウジBT−21アミラーゼにより加水分解されなかった。これは、それらの生成物が蓄積し、そして従って、加水分解の最終生成物として見なされる、可溶性澱粉に対して得られる結果を確認する。
【0160】
DP4〜DP10のマルト−オリゴ糖の混合物に対する生成物−特異的アミラーゼ活性を、時間経過実験により研究した。HPAEC−PADにより得られるピーク領域の変化は、マルト−オリゴ糖の形成又は加水分解に対応する。マルトヘキサオース(DP6)の形成、及びDP7, DP8, DP9及びDP10の量の同時低下が、酵素のマルトヘキサオース形成能力を確認した。しかしながら、定常状態条件が達成され、そして澱粉加水分解により見出されるようなDP6のさらなる分解が加水分解の7日後でさえ、検出されなかった。DP6の濃度は、澱粉加水分解で得られる濃度よりも低く、そしてこのことは、一定量のマルトヘキサオースが生成するマルトテトラオース及びマルトースの形成のために必要とされた。
【0161】
B.クラウジBT−21生成物特異的アミラーゼによる澱粉加水分解は、2段階工程に類似することが見出された。この工程は、主にマルトヘキサオース及び少量のマルトペンタオースへの澱粉の初期加水分解を包含し、それらは、集中的な加水分解の後、蓄積する主要マルトテトラオース及びマルトースにさらに加水分解された。マルトテトラオース及びマルトースへの第2加水分解段階は、濃度依存性が進行する第2段階のための調節体と思われるので、大きな基質のマルトヘキサオースへの予備加水分解により制限されると思われた。
【0162】
ベーキング実験:
ベーキング実験を、生成物−特異的アミラーゼにより行った。ドウを、10gの標準デンマークの小麦粉(Danisco 98078)、及び40単位の酵素(セクションBの材料及び方法に記載されるように、45℃及びpH10でアッセイされる)を有さない(対照)又は有する、0.2MのNaOH−グリシン緩衝液(pH10)6.2mlにより調製し、ベーキングし、そして“劣化及び老化の測定のためのアッセイ”に従って、貯蔵の後、DSCにより分析した。表8に示されるように、酵素は、それが有意な抗老化効果有することを示すベーキングの7日後に見出される劣化されるアミロペクチンの量を有意に低める。
【0163】
【表8】
【0164】
ベーキングの後、凍結されたベーキングされた生成物のパン中味サンプルを、蒸留水により抽出し(1gのベーキングされた生成物/10gの水を、1時間撹拌し、そして遠心分離した)、そして上記のようにしてHPAEC−PADにより分析し、ドウのベーキングの間、酵素により形成される澱粉加水分解生成物を検出した。対照に対する、マルトテトラオース、マルトペンタオース、マルトヘキサオース及びマルトヘプタオースの蓄積が、この酵素の活性の結果として見出された。
【0165】
発明の要約
本発明は製パン製品を製造するための方法、及びそのような方法への使用のために適切なアミラーゼを開示する。
本発明の好ましい態様は、次の番号付けされた文章により示される。
1.遅延された老化性質を有するベーキングされた澱粉質パン生成物を製造する方法が提供され、ここで前記方法は、ベーキング工程の間、アミロペクチンの側鎖の非還元末端から、4〜8個のD−グルコピラノシル単位から主に成る線状マルトオリゴ糖を分解することによって、澱粉を加水分解することができる有効量の非マルトース生成エキソアミラーゼのドウ成分、ドウ添加物又はドウへの添加を含んで成る。
【0166】
2.前記食料粉が小麦粉又はライ麦粉、又は小麦粉及びライ麦粉の混合物である前記1の方法。
3.前記非マルトース生成エキソアミラーゼが、kg食料粉当たり50〜100,000単位、好ましくはkg食料粉当たり100〜50,000単位の範囲で存在する量で添加される前記1又は2の方法。
4.前記非マルトース生成エキソアミラーゼがkg食料粉当たり200〜20,000単位の範囲で存在する量で添加される前記3の方法。
【0167】
5.前記非マルトース生成エキソアミラーゼが単位エキソアミラーゼ活性当たり0.5以下のエンドアミラーゼ単位(EAU)のエンドアミラーゼ活性を有する前記1〜4のいずれかに記載の方法。
6.前記非マルトース生成エキソアミラーゼが単位エキソアミラーゼ活性当たり0.05以下のエンドアミラーゼ単位(EAU)のエンドアミラーゼ活性を有する前記1〜4のいずれかに記載の方法。
7.前記非マルトース生成エキソアミラーゼが単位エキソアミラーゼ活性当たり0.01以下のエンドアミラーゼ単位(EAU)のエンドアミラーゼ活性を有する前記1〜4のいずれかに記載の方法。
【0168】
8.前記非マルトース生成エキソアミラーゼが、0.7単位の量の前記非マルトース生成エキソアミラーゼが、50mMの2−(N−モルホリノ)エタンスルホン酸及び2mMの塩化カルシウムを含む緩衝溶液1ml当たり10mgの予備煮沸されたトウモロコシ澱粉の水溶液4mlにおいて、50℃の温度で、pH6.0で15分間インキュベートされる場合、それが、少なくとも60重量%、好ましくは少なくとも70重量%、より好ましくは少なくとも80重量%及び最も好ましくは少なくとも90重量%の、グルコース、マルトース、及び3〜10個のD−グルコピラノシル単位の線状マルトオリゴ糖、好ましくは4〜8個のD−グルコピラノシル単位の線状マルトオリゴ糖から成る加水分解生成物を生成することを特徴として、ここで前記加水分解生成物は、パルスされた電流滴定検出、及び標準としてグルコース〜マルトヘプタオースの既知の線状マルトオリゴ糖と共にDionex PA100カラムを用いてアニオン交換HPLCにより分析される前記1〜4のいずれかに記載の方法。
【0169】
9.前記非マルトース生成エキソアミラーゼが、前記8記載の条件下でインキュベートされ、そして前記加水分解生成物が請求項8記載のようにして分析される場合、それが少なくとも60重量%、好ましくは少なくとも70重量%、より好ましくは少なくとも80重量%及び最も好ましくは少なくとも90重量%のマルトテトラオースを生成する前記8の方法。
10.前記非マルトース生成エキソアミラーゼが、前記8記載の条件下でインキュベートされ、そして前記加水分解生成物が請求項8記載のようにして分析される場合、それが少なくとも60重量%、好ましくは少なくとも70重量%、より好ましくは少なくとも80重量%及び最も好ましくは少なくとも90重量%のマルトペンタオースを生成する前記8の方法。
【0170】
11.前記非マルトース生成エキソアミラーゼが、前記8記載の条件下でインキュベートされ、そして前記加水分解生成物が請求項8記載のようにして分析される場合、それが少なくとも60重量%、好ましくは少なくとも70重量%、より好ましくは少なくとも80重量%及び最も好ましくは少なくとも90重量%のマルトヘキサオースを生成する前記8の方法。
12.前記非マルトース生成エキソアミラーゼが、前記8記載の条件下でインキュベートされ、そして前記加水分解生成物が請求項8記載のようにして分析される場合、それが少なくとも60重量%、好ましくは少なくとも70重量%、より好ましくは少なくとも80重量%及び最も好ましくは少なくとも90重量%のマルヘプタオースを生成する前記8の方法。
【0171】
13.前記非マルトース生成エキソアミラーゼが、請求項8記載の条件下でインキュベートされ、そして前記加水分解生成物が請求項8記載のようにして分析される場合、それが少なくとも60重量%、好ましくは少なくとも70重量%、より好ましくは少なくとも80重量%及び最も好ましくは少なくとも90重量%のマルトオクタオースを生成する前記8の方法。
14.少なくとも1つの乳化剤がドウ成分、ドウ添加剤又はドウに添加される請求項1〜4のいずれか1項記載の方法。
【0172】
15.前記乳化剤が、レシチン、ポリオキシエチレンステアレート、食用脂肪酸のモノ−及びジグリセリド、食用脂肪酸のモノ−及びジグリセリドの酢酸エステル、食用脂肪酸のモノ−及びジグリセリドの乳酸エステル、食用脂肪酸のモノ−及びジグリセリドのクエン酸エステル、食用脂肪酸のモノ−及びジグリセリドのジアセチル酒石酸エステル、食用脂肪酸のスクロースエステル、ナトリウムステアロイル−2−ラクチレ−ト及びカルシウムステアロイル−2−ラクチレ−トから成る群から選択される前記14の方法。
【0173】
16.少なくとも1つの追加の酵素が、ドウ成分、ドウ添加剤又はドウに添加される前記1〜4のいずれかに記載の方法。
17.前記追加の酵素が、セルラーゼ、ヘミセルラーゼ、キシラナーゼ、オキシドレダクターゼ及びプロテアーゼから成る群から選択される前記16の方法。
18.ベーキング工程の間、アミロペクチンの側鎖の非還元末端から、4〜8個のD−グルコピラノシル単位から主として成る線状マルトオリゴ糖を分解することによって、澱粉を加水分解できる非マルトース生成エキソアミラーゼ、少なくとも1つの追加のドウ成分又はドウ添加剤から成るドウから製造されるドウ及びベーキングされた澱粉質パン生成物のための改良生成物。
【0174】
19.前記非マルトース生成エキソアミラーゼがエキソアミラーゼ活性1単位当たり0.5以下のエンドアミラーゼ単位(EAU)のエンドアミラーゼ活性を有する前記18の改良性組成物。
20.前記非マルトース生成エキソアミラーゼがエキソアミラーゼ活性1単位当たり0.05以下のエンドアミラーゼ単位(EAU)のエンドアミラーゼ活性を有する前記18の改良性組成物。
21.前記非マルトース生成エキソアミラーゼがエキソアミラーゼ活性1単位当たり0.01以下のエンドアミラーゼ単位(EAU)のエンドアミラーゼ活性を有する前記18の改良性組成物。
【0175】
22.0.7単位の量の前記非マルトース生成エキソアミラーゼが、50mMの2−(N−モルホリノ)エタンスルホン酸及び2mMの塩化カルシウムを含む緩衝溶液1ml当たり10mgの予備煮沸されたモチ状トウモロコシ澱粉の水溶液4mlにおいて、50℃の温度で、pH6.0で15分間インキュベートされる場合、それが、少なくとも60重量%、好ましくは少なくとも70重量%、より好ましくは少なくとも80重量%及び最も好ましくは少なくとも90重量%の、グルコース、マルトース、及び3〜10個のD−グルコピラノシル単位の線状マルトオリゴ糖、好ましくは4〜8個のD−グルコピラノシル単位の線状マルトオリゴ糖から成る加水分解生成物を生成することを特徴として、ここで前記加水分解生成物は、パルスされた電流滴定検出、及び標準としてグルコース〜マルトヘプタオースの既知の線状マルトオリゴ糖と共にDionex PA100カラムを用いてアニオン交換HPLCにより分析される前記18の改良性組成物。
【0176】
23.前記非マルトース生成エキソアミラーゼが、請求項8記載の条件下でインキュベートされ、そして加水分解生成物が請求項8記載のようにして分析される場合、少なくとも60重量%、好ましくは少なくとも70重量%、より好ましくは少なくとも80重量%及び最も好ましくは少なくとも90重量%のマルトテトラオースを生成する前記22の改良性組成物。
24.前記非マルトース生成エキソアミラーゼが、請求項8記載の条件下でインキュベートされ、そして加水分解生成物が請求項8記載のようにして分析される場合、少なくとも60重量%、好ましくは少なくとも70重量%、より好ましくは少なくとも80重量%及び最も好ましくは少なくとも90重量%のマルトペンタオースを生成する前記22の改良性組成物。
【0177】
25.前記非マルトース生成エキソアミラーゼが、請求項8記載の条件下でインキュベートされ、そして加水分解生成物が請求項8記載のようにして分析される場合、少なくとも60重量%、好ましくは少なくとも70重量%、より好ましくは少なくとも80重量%及び最も好ましくは少なくとも90重量%のマルトヘキサオースを生成する前記22の改良性組成物。
26.前記非マルトース生成エキソアミラーゼが、請求項8記載の条件下でインキュベートされ、そして加水分解生成物が請求項8記載のようにして分析される場合、少なくとも60重量%、好ましくは少なくとも70重量%、より好ましくは少なくとも80重量%及び最も好ましくは少なくとも90重量%のマルトヘプタオースを生成する前記22の改良性組成物。
【0178】
27.前記非マルトース生成エキソアミラーゼが、請求項8記載の条件下でインキュベートされ、そして加水分解生成物が請求項8記載のようにして分析される場合、少なくとも60重量%、好ましくは少なくとも70重量%、より好ましくは少なくとも80重量%及び最も好ましくは少なくとも90重量%のマルトオクタオースを生成する前記22の改良性組成物。
28.乳化剤及びヒドロコロイドから成る群から選択された少なくとも1つの添加剤を含んで成る前記19〜27のいずれかに記載の改良性組成物。
29.少なくとも1つの乳化剤がドウ成分、ドウ添加剤又はドウに添加される前記28の改良性組成物。
【0179】
30.前記ヒドロコロイドが、アルギネート、カラゲニン、ペクチン及び植物ガムから成る群から選択される前記28の改良性組成物。
31.セルラーゼ、ヘミセルラーゼ、キシラナーゼ、オキシドレダクターゼ及びプロテアーゼから成る群から選択された少なくとも1つの追加の酵素を含んで成る前記19〜30のいずれかに記載の改良性組成物。
32.請求項1〜17のいずれか1項記載の方法により得ることができるベーキングされた澱粉質パン生成物。
【0180】
本明細書に言及されるすべての出版物は、引用により組み込まれる。本発明の記載される方法及びシステムの種々の修飾及び変更は、当業者に明らかであり、そして本発明の範囲内で行われるであろう。本発明は特定の好ましい態様で記載されて来たが、本発明はそのような特定の態様に限定されるものではない。実際、生化学又は関連する分野の当業者に明らかである、本発明を実施するための記載される態様の種々の修飾は、本発明の範囲内で行われる。
【0181】
参考文献
[1]. K.H. Park, Food Sci. Ind. 25 (1992) 73-82.
[2]. M. Okada and T. Nakakuki, Oligosaccharides: production, properties and application, in F.W. Schenck and R.E. Hebeda (Eds.), Starch hydrolysis products worldwide technology, production and application, VCH Publishers, New York, 1992, pp 335-366.
[3]. W.M.Fogarty, Microbial amylases, in W.M. Fogarty (Ed.), Microbial enzymes and biotechnology, Applied Science, London, 1983, pp. 1-92.
【0182】
[4]. W.M. Fogarty and C.T. Kelly, Starch-degrading enzymes of microbial origin, in M.J. Bull (Ed.), Progress in industrial microbiology, Vol. 15, Elsevier Scientific 1978, pp. 87-150.
[5]. K. Kainuma, S. Kobayashi, T. Ito, and S. Suzuki, FEBS Letters, 26 (1972) 281-285.
[6]. N. Monma, T. Nakakuki, and K. Kainuma, Agric. Biol. Chem., 47 (1983) 1769-1774.
[7]. J.F. Kennedy and C.A. White, Starch/Starke 31 (1979) 93-99.
[8]. Y. Takasaki, Agric. Biol. Chem. 46 (1982) 1539-1547.
【0183】
[9]. H. Taniguchi, C.M. Jae, N. Yoshigi, and Y. Maruyama, Agric. Biol. Chem. 47 (1983) 511-519.
[10]. H. Taniguchi, Maltohexaose-producing amylase of Bacillus circulans F-2 in R.B. Friendman (Ed.) Biotechnology of amylodextrin oligosaccharides. ACS Symp. Ser. 458. American Chemical Society, Washington DC, 1991, pp 111-124.
[11]. F. Bealin-Kelly, C.T. Kelly, and W.M. Fogarty, Biochem. Soc. Trans., 18 (1990) 310-311.
[12]. W.M. Fogarty, F. Bealin-Kelly, C.T.Kelly, and E.M. Appl. Microbiol. Biotechnol., 36 (1991) 184-489.
【0184】
[13]. N. Saito, Archives. Biochem. Biophys., 155 (1973) 290-298.
[14]. H. Okemoto, S. Kobayashi, M. Monma, H. Hashimoto, K. Hara, and K. Kainuma, Appl. Microbiol. Biotechnol., 25 (1986) 137-142.
[15]. O. Shida, T. Takano, H. Takagi, K. Kadowaki, and S. Kobayashi, Biosci., Biotechnol., Biochem. 56 (1992) 76-80.
[16]. (There is no ref. [16])
[17]. Y. Sakano, Y. Kashiwagi, and T. Kobayashi, Agric. Biol. Chem., 46 (1982) 639-646.
[18]. Y. Takasaki, H. Shinohara, M. Tsuruhisa, S. Hayashi, K. Imada, Agric. Biol. Chem. 55 (1991) 1715-1720.
【0185】
[19]. W.M. Fogarty, C.T. Kelly, A.C. Bourke, and E.M. Doyle, Biotechnol. Lett. 16 (1994) 473-478.
[20]. K. Wako, S. Hashimoo, S. Kubomura, A. Yokota, K. Aikawa, and J. Kamaeda, J. Jap. Soc. Starch. Sci 26 (1979) 175-181.
[21]. Y. Takasaki, Agric. Biol. Chem. 49 (1985) 1091-1097.
[22]. (There is no ref. [22])
[23]. T. Hayashi, T. Akiba, and K. Horikoshi, Appl. Microbiol. Biotechnol. 28 (1988b) 281-285.
[24]. G. Schmid, A. Candussio, and A. Bock, U.S. Patent 5 304 723 (1994).
【0186】
[25]. M.A. Mc Tigue, C.T. Kelly, E.M. Doyle, and W.M. Fogarty, Enzyme Microb. Technol., 17 (1995) 570-573.
[26]. T.U. Kim, B.G. Gu, J.Y. Jeong, S.M. Byun, and Y.C. Shin, appl. Environm. Microbiol. 61 (1995) 3105-3112.
[27]. A.K. Chandra, S. Medda, and A.K. Bhadra, J. Ferment. Technol., 58, (1980), 1-10.
[28]. R.A.K. Srivastava and J.N. Baruah, Appl. Environ. Mocrobiol. 52 (1986) 179-184.
[29]. V. Planchot an P. Colonna, Carbohydr. Res., 272 (1995) 97-109.
【0187】
[30]. J.F. Robyt and W.J. Whelan, The α−amylases in J.A. Radley (Ed.) Starch and its derivatives, 4.ed. Chapman and Hall, London, 1968, pp. 430-476.
[31]. M. Ohnishi and K. Hiromi, General considerations for conditions and methods of amylase assay in The Amylase Research Society of Japan (Ed.) Handbook of amylases and related enzymes. Their Sources, isolation methods, properties and applications. Pergamon, Oxford, 1988, pp. 10-14.
[32]. L. Duedahl-Olesen, W. Zimmemamm, and J.A. Delcour, Cereal Chemistry (1999) in press.
【0188】
[33]. K.L. Larsen, L. Duedahl-Olesen, H.J.S. Christensen, F. Mathiesen, L.H. Pedersen, and W. Zimmermann, Carbohydr. Res. 310 (1998) 211-219.
[34]. M.M. Bradford, Anal. Chem., 72 (1976) 248-254.
[35]. Novex, Precast Gel Instructions. NOVEX, San Diego, CA, USA.
[36]. Mini Protean II Electrophoresis Cell Instructions manual. Bio-Rad laboratories, Hercules, CA, U.S.A.
[37]. H. Blum, H. Beier, and H.J. Gross, Electrophoresis (1987) 93-99.
[38]. L.H. Pedersen, H.J.S. Christensen, F. Mathiesen, K.L. Larsen, and W. Zimmermamm, Starch, 49 (1997) 250-253.
【0189】
[39]. H.J. Fuwa, J. Biochem., 41 (1954) 583-603.
[40]. Y.C. Lee, J. Chormatogr. A, 720 (1996) 137-149.
[41]. R.N. Ammerall, G.A. Delgado, F.L. Tenbarge, and R.B. Freidmann, Carbohydr. Res. 215 (1991) 179-192。
【特許請求の範囲】
【請求項1】
澱粉生成物の製造方法であって、アミロペクチンの側鎖の非還元末端から4〜8個のD−グルコピラノシル単位から主として成る1又は複数の線状マルトオリゴ糖を切り取ることによって澱粉を加水分解することができる非マルトース生成エキソアミラーゼを澱粉基材に添加することを含んで成る方法。
【請求項2】
前記澱粉基材が食料粉を含んで成り、前記食料粉が小麦粉もしくはライ麦粉、又はそれらの混合物である請求項1記載の方法。
【請求項3】
前記非マルトース生成エキソアミラーゼが、50〜100,000単位/kg食料粉、好ましくは100〜50,000単位/kg食料粉、より好ましくは200〜20,000単位/kg食料粉の範囲の量で添加される請求項2記載の方法。
【請求項4】
前記非マルトース生成エキソアミラーゼが単位エキソアミラーゼ活性当たり0.5より少いエンドアミラーゼ単位(EAU)のエンドアミラーゼ活性を有し、好ましくは単位エキソアミラーゼ活性当たり0.05より少いエンドアミラーゼ単位(EAU)のエンドアミラーゼ活性を有し、より好ましくは単位エキソアミラーゼ活性当たり0.01より少いエンドアミラーゼ単位(EAU)のエンドアミラーゼ活性を有する請求項1〜3のいずれか1項記載の方法。
【請求項5】
前記非マルトース生成エキソアミラーゼが、モチ状トウモロコシ澱粉インキュベーション試験において、2〜10個のD−グルコピラノシル単位及び任意にはグルコースの1又は複数の線状マルト−オリゴ糖から成る加水分解生成物を生成する能力を有し、その結果、少なくとも60重量%、好ましくは少なくとも70重量%、より好ましくは少なくとも80重量%そして最も好ましくは少なくとも85重量%の前記加水分解生成物が、3〜10個のD−グルコピラノシル単位の線状マルトオリゴ糖、好ましくは4〜8個のD−グルコピラノシル単位の線状マルトオリゴ糖から成ることをさらに特徴とする請求項1〜4のいずれか1項記載の方法。
【請求項6】
少なくとも60%、好ましくは少なくとも70%、より好ましくは少なくとも80%そして最も好ましくは少なくとも85%の前記加水分解生成物が、マルトテトラオース、マルトペンタオース、マルトヘキサオール、マルトヘプタオース又はマルトオクタオースである請求項5記載の方法。
【請求項7】
少なくとも60%、好ましくは少なくとも70%、より好ましくは少なくとも80%、及び最も好ましくは少なくとも80%の前記加水分解生成物がマルトテトラオースである請求項6記載の方法。
【請求項8】
前記酵素がシュードモナス・サッカロフィラ(Pseudomonas sacharophila)から得られるもの又はその機能的同等物である請求項7記載の方法。
【請求項9】
前記加水分解生成物の少なくとも60%、好ましくは少なくとも70%、より好ましくは少なくとも80%そして最も好ましくは少なくとも85%がマルトへキサオースである請求項6記載の方法。
【請求項10】
前記酵素が、バチルス・グラウジ(Bacillus clausii)から得られるもの又は機能的同等物である請求項9記載の方法。
【請求項11】
前記酵素が、約101,000Da(ドデシル硫酸ナトリウムポリアクリルアミド電気泳動により推定される場合)の分子量を有する請求項10記載の方法。
【請求項12】
前記酵素が、pH9.5及び55℃で最適活性を有する請求項10又は11記載の方法。
【請求項13】
前記澱粉生成物がドウである請求項1〜12のいずれか1項記載の方法。
【請求項14】
前記澱粉生成物がベーキングされたドウである請求項1〜13のいずれか1項記載の方法。
【請求項15】
前記澱粉生成物がベーキングされた澱粉質のパン生成物の調製のためのものである請求項1〜14のいずれか1項記載の方法。
【請求項16】
前記澱粉生成物がベーキングされる請求項1〜15のいずれか1項記載の方法。
【請求項17】
請求項16記載の方法により得られるベーキングされた生成物。
【請求項18】
請求項1〜17のいずれか1項記載の非マルトース生成エキソアミラーゼ、及び少なくとも1つの追加のドウ成分又はドウ添加剤を含んで成る、ドウのための改良性組成物。
【請求項19】
約101,000Da(ドデシル硫酸ナトリウムポリアクリルアミド電気泳動により推定される場合)の分子量、及び/又はpH9.5及び55℃で最適活性を有する、バチルス・クラウジから得られる非マルトース生成エキソアミラーゼ又はその機能的同等物。
【請求項20】
前記非マルトース生成エキソアミラーゼが、モチ状トウモロコシ澱粉インキュベーション試験において、2〜10個のD−グルコピラノシル単位及び任意にはグルコースの1又は複数の線状マルト−オリゴ糖から成る加水分解生成物を生成する能力を有し、その結果、前記加水分解生成物の少なくとも60重量%、好ましくは少なくとも70重量%、より好ましくは少なくとも80重量%そして最も好ましくは少なくとも85重量%が、3〜10個のD−グルコピラノシル単位の線状マルトオリゴ糖、好ましくは4〜8個のD−グルコピラノシル単位の線状マルトオリゴ糖から成ることをさらに特徴とする請求項19記載の非マルトース生成エキソアミラーゼ。
【請求項21】
澱粉生成物の劣化を遅延するために前記澱粉生成物への非マルトース生成エキソアミラーゼの使用。
【請求項22】
本明細書に及び請求項1に実質的に記載されるような方法、改良性組成物、酵素又は使用。
【請求項1】
澱粉生成物の製造方法であって、アミロペクチンの側鎖の非還元末端から4〜8個のD−グルコピラノシル単位から主として成る1又は複数の線状マルトオリゴ糖を切り取ることによって澱粉を加水分解することができる非マルトース生成エキソアミラーゼを澱粉基材に添加することを含んで成る方法。
【請求項2】
前記澱粉基材が食料粉を含んで成り、前記食料粉が小麦粉もしくはライ麦粉、又はそれらの混合物である請求項1記載の方法。
【請求項3】
前記非マルトース生成エキソアミラーゼが、50〜100,000単位/kg食料粉、好ましくは100〜50,000単位/kg食料粉、より好ましくは200〜20,000単位/kg食料粉の範囲の量で添加される請求項2記載の方法。
【請求項4】
前記非マルトース生成エキソアミラーゼが単位エキソアミラーゼ活性当たり0.5より少いエンドアミラーゼ単位(EAU)のエンドアミラーゼ活性を有し、好ましくは単位エキソアミラーゼ活性当たり0.05より少いエンドアミラーゼ単位(EAU)のエンドアミラーゼ活性を有し、より好ましくは単位エキソアミラーゼ活性当たり0.01より少いエンドアミラーゼ単位(EAU)のエンドアミラーゼ活性を有する請求項1〜3のいずれか1項記載の方法。
【請求項5】
前記非マルトース生成エキソアミラーゼが、モチ状トウモロコシ澱粉インキュベーション試験において、2〜10個のD−グルコピラノシル単位及び任意にはグルコースの1又は複数の線状マルト−オリゴ糖から成る加水分解生成物を生成する能力を有し、その結果、少なくとも60重量%、好ましくは少なくとも70重量%、より好ましくは少なくとも80重量%そして最も好ましくは少なくとも85重量%の前記加水分解生成物が、3〜10個のD−グルコピラノシル単位の線状マルトオリゴ糖、好ましくは4〜8個のD−グルコピラノシル単位の線状マルトオリゴ糖から成ることをさらに特徴とする請求項1〜4のいずれか1項記載の方法。
【請求項6】
少なくとも60%、好ましくは少なくとも70%、より好ましくは少なくとも80%そして最も好ましくは少なくとも85%の前記加水分解生成物が、マルトテトラオース、マルトペンタオース、マルトヘキサオール、マルトヘプタオース又はマルトオクタオースである請求項5記載の方法。
【請求項7】
少なくとも60%、好ましくは少なくとも70%、より好ましくは少なくとも80%、及び最も好ましくは少なくとも80%の前記加水分解生成物がマルトテトラオースである請求項6記載の方法。
【請求項8】
前記酵素がシュードモナス・サッカロフィラ(Pseudomonas sacharophila)から得られるもの又はその機能的同等物である請求項7記載の方法。
【請求項9】
前記加水分解生成物の少なくとも60%、好ましくは少なくとも70%、より好ましくは少なくとも80%そして最も好ましくは少なくとも85%がマルトへキサオースである請求項6記載の方法。
【請求項10】
前記酵素が、バチルス・グラウジ(Bacillus clausii)から得られるもの又は機能的同等物である請求項9記載の方法。
【請求項11】
前記酵素が、約101,000Da(ドデシル硫酸ナトリウムポリアクリルアミド電気泳動により推定される場合)の分子量を有する請求項10記載の方法。
【請求項12】
前記酵素が、pH9.5及び55℃で最適活性を有する請求項10又は11記載の方法。
【請求項13】
前記澱粉生成物がドウである請求項1〜12のいずれか1項記載の方法。
【請求項14】
前記澱粉生成物がベーキングされたドウである請求項1〜13のいずれか1項記載の方法。
【請求項15】
前記澱粉生成物がベーキングされた澱粉質のパン生成物の調製のためのものである請求項1〜14のいずれか1項記載の方法。
【請求項16】
前記澱粉生成物がベーキングされる請求項1〜15のいずれか1項記載の方法。
【請求項17】
請求項16記載の方法により得られるベーキングされた生成物。
【請求項18】
請求項1〜17のいずれか1項記載の非マルトース生成エキソアミラーゼ、及び少なくとも1つの追加のドウ成分又はドウ添加剤を含んで成る、ドウのための改良性組成物。
【請求項19】
約101,000Da(ドデシル硫酸ナトリウムポリアクリルアミド電気泳動により推定される場合)の分子量、及び/又はpH9.5及び55℃で最適活性を有する、バチルス・クラウジから得られる非マルトース生成エキソアミラーゼ又はその機能的同等物。
【請求項20】
前記非マルトース生成エキソアミラーゼが、モチ状トウモロコシ澱粉インキュベーション試験において、2〜10個のD−グルコピラノシル単位及び任意にはグルコースの1又は複数の線状マルト−オリゴ糖から成る加水分解生成物を生成する能力を有し、その結果、前記加水分解生成物の少なくとも60重量%、好ましくは少なくとも70重量%、より好ましくは少なくとも80重量%そして最も好ましくは少なくとも85重量%が、3〜10個のD−グルコピラノシル単位の線状マルトオリゴ糖、好ましくは4〜8個のD−グルコピラノシル単位の線状マルトオリゴ糖から成ることをさらに特徴とする請求項19記載の非マルトース生成エキソアミラーゼ。
【請求項21】
澱粉生成物の劣化を遅延するために前記澱粉生成物への非マルトース生成エキソアミラーゼの使用。
【請求項22】
本明細書に及び請求項1に実質的に記載されるような方法、改良性組成物、酵素又は使用。
【図1】

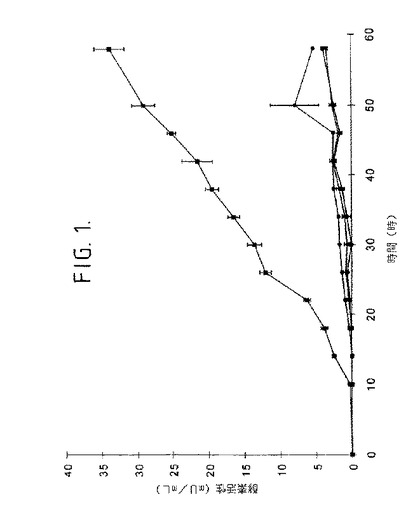
【図2】
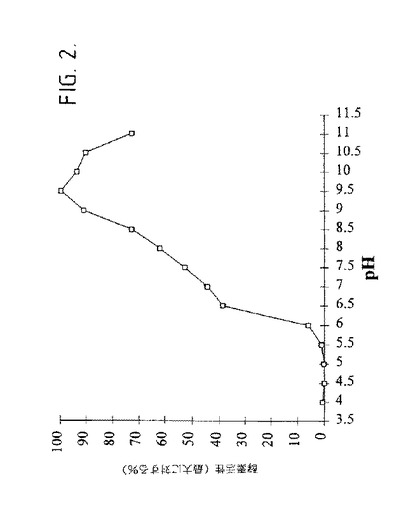
【図3】
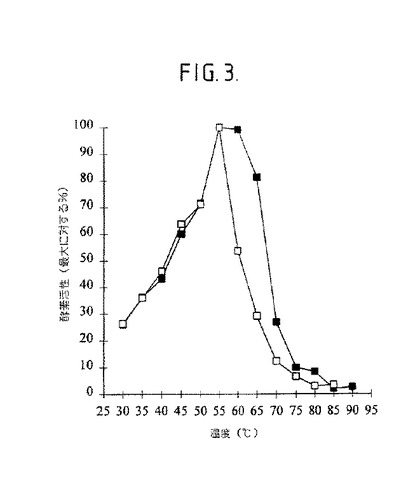
【図4】
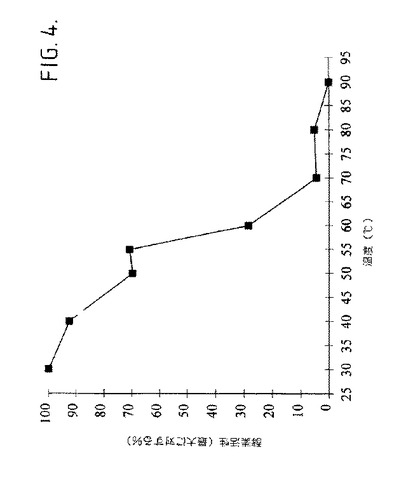
【図5】
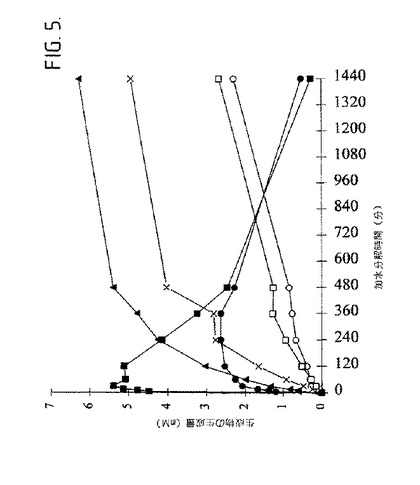
【図6】
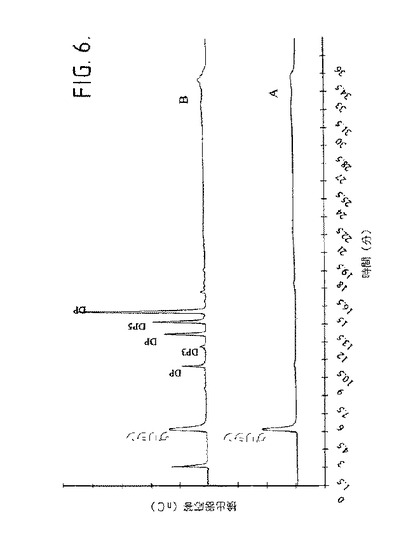
【図7】
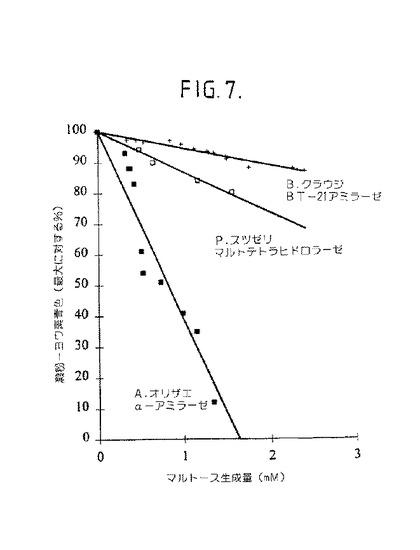
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【公開番号】特開2011−15688(P2011−15688A)
【公開日】平成23年1月27日(2011.1.27)
【国際特許分類】
【外国語出願】
【出願番号】特願2010−185039(P2010−185039)
【出願日】平成22年8月20日(2010.8.20)
【分割の表示】特願2000−541287(P2000−541287)の分割
【原出願日】平成11年3月30日(1999.3.30)
【出願人】(500457852)ダニスコ アクティーゼルスカブ (4)
【Fターム(参考)】
【公開日】平成23年1月27日(2011.1.27)
【国際特許分類】
【出願番号】特願2010−185039(P2010−185039)
【出願日】平成22年8月20日(2010.8.20)
【分割の表示】特願2000−541287(P2000−541287)の分割
【原出願日】平成11年3月30日(1999.3.30)
【出願人】(500457852)ダニスコ アクティーゼルスカブ (4)
【Fターム(参考)】
[ Back to top ]