
非リボソームペプチドシンターゼをコードする生合成クラスターの核酸分子およびその使用
本発明は、式(I)または(I’)の化合物の生産に関与する生合成遺伝子クラスターの1個以上の機能的フラグメントを含むポリヌクレオチドの提供に関する。本発明はまた、式(I)もしくは(I’)または式(II)〜(VII)、(XI)〜(XIV)ならびに(XVII)および(XVIII)の化合物を製造する方法も提供する。さらに、医薬組成物としてのかかる化合物の使用も本発明において提供される。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、式(I)または(I’)の化合物の生産に関与する生合成遺伝子クラスターの1個以上の機能的フラグメントを含むポリヌクレオチドの提供に関する。本発明はまた、式(I)もしくは(I’)または式(II)〜(VII)、(XI)〜(XIV)ならびに(XVII)および(XVIII)の化合物を製造する方法も提供する。さらに、医薬組成物としてのかかる化合物の使用も本発明において提供される。
【背景技術】
【0002】
微生物由来の多くの天然産物は、高等生物において見られる生物学的活性を有しており、数世紀にわたってその治療特性が探索されている。これらの天然産物のほとんどはポリケチドクラスおよび非リボソームペプチドクラスに属しており、ポリケチドシンターゼ(PKS)および非リボソームペプチドシンターゼ(NRPS)として知られるモジュラー酵素系によって合成される(Finkering and Marahiel 2004; Staunton and Weissman, 2001)。さらに、同じ経路にPKS遺伝子およびNRPS遺伝子の両方を含み、したがってこれら2つのクラスのハイブリッドである二次代謝産物を生産する経路が存在している。これらの生合成経路によって生産される天然産物は、短鎖カルボン酸およびアミノ酸のような小さな、比較的単純な構成要素から構成されている。しかし、これらの経路に由来する最終天然産物は、極めて多様であり、しばしば構造的に複雑であり、通常複数の立体中心を含んでいる。このため、これらの化合物の生産のための合成アプローチは、しばしば実行不可能であり、したがってそれらの生産には発酵が常套のアプローチで有り続けている。しかし、発酵プロセスは、代謝的に特徴付けられておらず、しばしば遺伝子の取り扱いが困難であり、そしてしばしばあまり増殖せず、目的化合物を十分なレベルでは生産しないという、微生物に対する信頼に関する固有の問題を有する。これらの問題を回避するため、これらの欠点を有さない、十分に特徴付けられた宿主生物におけるPKSまたはPRPS経路の異種発現が一つの選択肢であり得る(Wenzel and Muller, 2005に概説されている)。事実、このアプローチは、発見努力のために「静かな」または「隠れた」PKSおよびNRPS経路の発現に延長でき(Shen, 2004)、または研究室で培養できない生物からの経路の発現に使用できる。さらに、PKSおよびNRPS経路の異種宿主への移動により、親化合物の新規なアナログを作成するための二次代謝経路の効率的なバイオエンジニアリングが可能となる。
【0003】
異種発現は、一般に、PKSおよびNRPS経路がゲノム上の連続したクラスターに位置しているという事実を利用している。したがって、原理上は、これらの経路は、標準的なBACまたはコスミドベクターに比較的容易にクローニングされる。ある微生物から別の微生物へとある経路を動かすことの原理的な単純さに反して、2種の生物間の制御、コドン出現頻度または代謝における差異は、異種発現の成功に大きな課題を引き起こしている。さらに、BACライブラリー構築物のようなこの戦略を効率的に適用することができる分子ツールおよびクローニングのための組換えアプローチは、比較的最近になってからのみ利用可能となった(Wenzel and Muller, 2005)。このため、異種発現の成功例は、文献には極僅かしか存在しない。
【0004】
好適な異種宿主の選択は発現戦略を設計する際に重要な検討事項である。新たな宿主は、遺伝子操作が用意であり、研究室での取り扱いが容易であり、PKSおよびNRPS経路を用いる能力を有するべきである。例えば、新たな宿主におけるホスホパンテテイニルトランスフェラーゼの存在は、移入されたPKSおよびNRPSの活性化を促進するために必須である(Pfeifer et al. 2001)。さらに、新たな宿主が元の宿主のものと同様のコドン出現頻度を有し、移入された経路の効率的な発現が可能であることが重要である。使用される最も一般的な宿主は、大腸菌(Escherichia coli)、枯草菌(Bacillus subtilis)、シュードモナス・プチダ(Pseudomonas putida)および十分に特徴付けられたストレプトマイセス(Streptomyces)株のうちわずかに選択されたものである(Zhang and Pfeifer, 2008に概説されている)。利用されている他の宿主は、ミキソコッカス・ザンサス(Myxococcus xanthus)および糸状菌類を含む。これらの宿主株のいくつかは、変異によって主要な常在性二次代謝系がサイレンシングされて、バックグラウンド代謝プロフィールを除き、移入される生合成経路に利用可能な前駆体貯蓄の減少を防止するように、修飾されている。
【0005】
特定の経路を移動させるためには、適切な移動可能な遺伝的要素に基づく当該経路のパッケージングが必要である。まず初めにPKSまたはNRPS系の配列を、少なくともアミノ酸レベルで、より好ましくはヌクレオチドレベルで知らなければならない。典型的には、この配列を用いて、天然宿主から構築されたゲノムライブラリーから、BACまたはコスミドクローンを位置づけるプローブを設計する。これらの経路クラスターのサイズが大きいため(通常30kb以上、しばしば100kbを超える)、「ショットガン」クローニング戦略を用いるとき、それらはしばしば1個のBACまたはコスミドクローン中には捕捉されない。したがって、この経路は、しばしば、全経路を含む1個のBACまたはコスミドベクター構築物を作成するために再構築する必要がある。極めて大きな経路を発現させようとするとき、それは新たな宿主中でトランスで発現されるように2個以上の別個のベクター構築物に分割してもよい(Gu, et al. 2007)。最終的には、構築物を有する大腸菌から新たな宿主へと移動するために、ベクター構築物は、プラスミド移行性機能(例えばRK2からoriT)をも有していなければならない。構築物が新たな宿主において好適であることを確保するため、それを宿主染色体に融合させることを勧める。これを達成するため、構築物は、効率的な染色体融合のための部位を含んでいるべきである。例えば、ストレプトマイセスのファージ接着部位ΦC31が、この系における染色体挿入のためにしばしば用いられる(Binz, et al. 2008)。さらに、新たな宿主で適切に機能する新たなプロモーターを、生合成経路の前に挿入する必要がしばしばある。問題の生物2種が密接に関連しており、したがって多くの制御因子を共有している可能性がある場合、この工程を避けることができる。最後に、選択マーカー、一般には抗生物質耐性カセットが、新たな宿主における構築物(修飾BACまたはコスミド)の移行および融合が成功したかを選択するために、必要である。典型的には、これらの操作は大腸菌において行われ、しばしばRed/ET組換えの使用によってなされる(Zhang, et al. 1998)。このクローニングアプローチは、制限酵素利用操作がせいぜい挑戦的である場合に、大きなDNA構築物を含む応用に特に適用可能である。
【0006】
構築物が新たな宿主に融合されると、発酵および続く化学分析を行って、経路の発現が成功したかを決定する。ほぼ全ての場合で異種発現が成功した場合、天然宿主で観察されるものと比較してより低い力価で天然産物が生産される。この明らかな抑制にも拘わらず、異種発現の成功は、常套の株開発方法論に利用可能な多くの選択肢を有する発現プラットフォームを提供する。
【発明の概要】
【0007】
本発明は、式I
【化1】

[式中、A7のカルボキシ基とA2のヒドロキシ基の間にエステル結合があり、所望により、A5とA6の間のアミド結合の窒素原子がメチルで置換されていてもよく、
XおよびA1は各々独立して任意であり、そして
Xは任意の化学残基、特にHまたはアシル残基、特にCH3CH2CH(CH3)CO、(CH3)2CHCH2COまたは(CH3)2CHCOであり;
A1はアスパラギン酸以外の標準アミノ酸、特にグルタミンであり;
A2はスレオニンまたはセリン、特にスレオニンであり;
A3は非塩基性標準アミノ酸またはその非塩基性誘導体、特にロイシンであり;
A4はAhp、デヒドロ−AHP、プロリンまたはその誘導体、特にAhpまたはその誘導体、特にAhp誘導体3−アミノ−2ピペリドンであり;
A5はイソロイシンまたはバリン、特にイソロイシンであり;
A6はチロシンまたはその誘導体、特にチロシンであり;
A7はロイシン、イソロイシンまたはバリン、特にイソロイシンまたはバリン、特にイソロイシンである。]
のデプシペプチドの生合成に関与する生合成クラスターの同定、並びに式Iの非リボソームペプチドまたはその薬学的に許容される塩もしくは誘導体を生産するための異種発現系の開発に関する。特に、式(I’)
【化2】
[式中、A7のカルボキシ基とA2のヒドロキシ基の間にエステル結合があり、所望により、A5とA6の間のアミド結合の窒素原子がメチルで置換されていてもよく、
XはCH3CO、(CH3)2CHCO、CH3S(O)CH2CO、CH3CH2CH(CH3)COまたはC6H5COであり;
A1はグルタミンであり;
A2はスレオニンであり;
A3はロイシンであり;
A4はAhp、デヒドロ−AHP、プロリンまたは5−ヒドロキシ−プロリンであり;
A5はイソロイシンまたはバリン、特にイソロイシンであり;
A6はチロシンであり;
A7はイソロイシンまたはバリン、特にイソロイシンである。]
のデプシペプチドの生合成における生合成遺伝子クラスターの使用を見出す。
【0008】
特に本発明は、図1に示す式(II)、(III)、(IV)、(V)、(VI)、(VII)、(XI)、(XII)〜(XIV)、(XVII)および/または(XVIII)の非リボソームペプチドの生合成に関与する生合成クラスターの同定、並びに式(I)または(I’)の非リボソームペプチドまたはその薬学的に許容される塩もしくは誘導体を生産するための異種発現系の開発に関する。
【0009】
式(I)、特に式(I’)の化合物は、粘液細菌コンドロマイセス・クロカツス(Chondromyces crocatus)NPH−MB180によって生産されるデプシペプチドのファミリーに属する非リボソームポリペプチドである。これらのデプシペプチドは、極めて強力で選択的なヒトカリクレイン7(hK7)およびエラスターゼ阻害剤であることが示されている。ヒトカリクレイン7はセリンプロテアーゼ活性を有する酵素であり、アトピー性皮膚炎の処置のための潜在的標的である。この新規化合物の詳細な物理化学データならびに発酵および抽出法は、WO2009/024527として公開されているPCT出願PCT/EP08/060689に記載されている。
【0010】
本明細書において使用するとき、用語「式(I)の化合物」または「式(I)のデプシペプチド」は、上記定義の式(I)の化合物、特に図1に記載の式(II)、(III)、(IV)、(V)、(VI)、(VII)、(XI)、(XII)、(XIII)、(XIV)および/または(XVIII)の非リボソームペプチド、ならびに実質的に同じプロテアーゼ活性を保持するあらゆる誘導体を意味する。かかる誘導体の例は、WO2009/024527として公開されているPCT出願にさらに記載されている。
【0011】
本明細書において使用するとき、用語「式(I’)の化合物」または「式(I’)のデプシペプチド」は、上記定義の式(I’)の化合物、特に図1に記載の式(II)、(III)、(IV)、(V)、(VI)、(VII)、(XI)、(XII)、(XIII)、(XIV)および/または(XVIII)の非リボソームペプチド、ならびに実質的に同じプロテアーゼ活性を保持するあらゆる誘導体を意味する。
【0012】
本発明の技術的課題は、式(I)または(I’)のデプシペプチドの生合成に関与する生合成クラスターまたはその機能的部分の提供である。
この技術的課題は、特許請求の範囲に特徴付けられた態様の提供によって解決される。
本発明の別の技術的課題は、異種遺伝子発現、例えば目的の組換えタンパク質の合成に適切な抑制プロモーターの提供である。
【0013】
本発明は、第一の態様において、(1)式(I)または(I’)の化合物の生産に関与する非リボソームペプチドシンターゼ(NRPS)(以下NRPS2と称する)をコードする生合成遺伝子クラスターの1個以上の機能的フラグメントを含むポリヌクレオチドであって、以下のものを含むポリヌクレオチドの提供に関する:
(i) NRPS2ドメインをコードする配列番号1、3、5、7、9、11、13、46、48、50、52、54、56、58および60からなる群から選択される配列と、少なくとも80%、特に少なくとも85%、特に少なくとも90%、特に少なくとも95%、特に少なくとも98%の配列同一性を有するヌクレオチド配列および/またはその相補体;
(ii) NRPS2ドメインをコードする配列番号1、3、5、7、9、11、13、46、48、50、52、54、56、58または60からなる群から選択されるヌクレオチド配列の相補鎖とハイブリダイズするヌクレオチド配列および/またはその相補体;
(iii) NRPS2ドメインを示す配列番号2、4、6、8、10、12、14、47、49、51、53、55、57、59または61からなる群から選択される配列と、少なくとも60%、特に少なくとも70%、特に少なくとも80%、特に少なくとも90%、特に少なくとも95%の配列同一性を有するアミノ酸配列をコードするヌクレオチド配列および/またはその相補体;
(iv) NRPS2ドメインを示す配列番号2、4、6、8、10、12、14、47、49、51、53、55、57、59または61からなる群から選択されるアミノ酸をコードするヌクレオチド配列の相補鎖とハイブリダイズするヌクレオチド配列および/またはその相補体;
(v) 配列番号15、配列番号28からなる群から選択される配列と少なくとも80%、特に少なくとも85%、特に少なくとも90%、特に少なくとも95%、特に少なくとも98%の配列同一性を有するヌクレオチド配列および/またはその相補体;
(vi) 配列番号15、配列番号28からなる群から選択されるヌクレオチド配列の相補鎖とハイブリダイズするヌクレオチド配列および/またはその相補体;
ここで、前記(i) 〜 (vi)のヌクレオチド配列は、配列番号2、4、6、8、10、12、14、47、49、51、53、55、59および/または61の参照配列によって示される対応NRPSドメインの活性を保持する発現産物をコードする。
【0014】
第2の態様において、(2)以下の1個以上のNRPS2ドメインの活性を保持する発現産物をコードする、態様(1)に記載のポリヌクレオチドを提供する:
(i)配列番号47のチオレーションドメイン;
(ii)配列番号49のコンデンセーションドメイン;
(iii)配列番号51のプロリンのアデニレーションドメイン;
(iv)配列番号53のチオレーションドメイン;
(v)配列番号2のコンデンセーションドメイン;
(vi)配列番号4のイソロイシンのアデニレーションドメイン;
(vii)配列番号6のチオレーションドメイン;
(viii)配列番号8のコンデンセーションドメイン;
(ix)配列番号10のチロシンのアデニレーションドメイン;
(x)配列番号12のN−メチレーションドメイン;
(xi)配列番号14のチオレーションドメイン;
(xii)配列番号55のコンデンセーションドメイン;
(xiii)配列番号57のイソロイシンのアデニレーションドメイン;
(xiv)配列番号59のチオレーションドメイン;および/または
(xv)配列番号61のチオエステラーゼドメイン。
【0015】
態様(2)の特定の態様において、ポリヌクレオチドは、式(I)または(I’)の化合物を生産するNRPS2をコードし、配列番号29に示すアミノ酸配列をコードするヌクレオチド配列を含む。
【0016】
第3の態様において、(3)本発明は、式(I)または(I’)の化合物の生産に関与するNRPSであるNRPS1をコードする生合成遺伝子クラスターの1個以上の機能的フラグメントを含むポリヌクレオチドであって、以下のものを含むポリヌクレオチドに関する:
(i) NRPSドメインをコードする配列番号30、32、34、36、38、40、42および44からなる群から選択される配列と、少なくとも80%、特に少なくとも85%、特に少なくとも90%、特に少なくとも95%、特に少なくとも98%の配列同一性を有するヌクレオチド配列および/またはその相補体;
(ii) NRPSドメインをコードする配列番号30、32、34、36、38、40、42および44からなる群から選択されるヌクレオチド配列の相補鎖とハイブリダイズするヌクレオチド配列および/またはその相補体;
(iii) NRPS1ドメインを示す配列番号31、33、35、37、39、41、43、45からなる群から選択される配列と、少なくとも60%、特に少なくとも70%、特に少なくとも80%、特に少なくとも90%、特に少なくとも95%の配列同一性を有するヌクレオチド配列および/またはその相補体;
(iv) NRPS1ドメインを示す配列番号31、33、35、37、39、41、43、45からなる群から選択されるアミノ酸をコードするヌクレオチド配列の相補鎖とハイブリダイズするヌクレオチド配列および/またはその相補体;
(v) 配列番号26からなる群から選択される配列と、少なくとも80%、特に少なくとも85%、特に少なくとも90%、特に少なくとも95%、特に少なくとも98%の配列同一性を有するヌクレオチド配列および/またはその相補体;または
(vi)配列番号26からなる群から選択されるヌクレオチド配列の相補鎖とハイブリダイズするヌクレオチド配列および/またはその相補体;
(vii) ここで、前記(i) 〜 (vi)のヌクレオチド配列は、配列番号31、33、35、37、39、41、43、45の参照配列によって示される対応NRPSドメインの活性を保持する発現産物をコードする。
【0017】
第4の態様において、態様(3)に記載のポリヌクレオチドは、以下の1個以上のNRPS1ドメインの活性を保持する発現産物をコードする:
(i) 配列番号31のローディングドメイン;
(ii)配列番号33のグルタミンのアデニレーションドメイン;
(iii)配列番号35のチオレーションドメイン;
(iv)配列番号37のコンデンセーションドメイン;
(v)配列番号39のスレオニンのアデニレーションドメイン;
(vi)配列番号41のチオレーションドメイン;
(vii)配列番号43のコンデンセーションドメイン;および
(viii)配列番号45のロイシンのアデニレーションドメイン。
【0018】
態様(4)の特定の態様において、ポリヌクレオチドは、式(I)または(I’)の化合物を生産するNRPS1をコードし、配列番号27に示すアミノ酸配列をコードするヌクレオチド配列を含む。
【0019】
他の態様において、本発明は、上記1個以上のポリヌクレオチドによってコードされるポリペプチドに関する。特に、該ポリペプチドは、式(I)または(I’)の化合物の生産に適切であり、以下のものからなる群から選択されるアミノ酸配列を含む:
(i)NRPS1を示す配列番号27、第2のNRPS2を示す配列番号29、チトクロムP450を示す配列番号63;および
(ii) (i)に列挙した参照配列と60%、特に少なくとも70%、特に少なくとも80%、特に少なくとも90%、特に少なくとも95%の配列同一性を有し、実質的に同じ触媒機能を保持する、(i)に列挙したアミノ酸配列の機能的変異体。
【0020】
本発明は、さらに、上記1個以上のポリペプチドをコードするヌクレオチド配列を含むポリヌクレオチドに関する。
【0021】
さらに別の態様において、本発明は、
(i)配列番号27またはその機能的変異体をコードするヌクレオチド配列;および
(ii)配列番号29またはその機能的変異体をコードするヌクレオチド配列
を含むポリヌクレオチドを提供する。
【0022】
かかるポリヌクレオチドは、配列番号63またはその機能的変異体をコードするヌクレオチド配列をさらに含んでいてもよい。ある特定の態様において、該ポリヌクレオチドは、受託番号DSM 19329を有するコンドロマイセス・クロカツス(Chondromyces crocatus)株NPH−MB180から単離される。
【0023】
本発明は、さらに、オープンリーディングフレームが転写および翻訳配列と作動可能に連結している、上記態様のいずれかに定義のポリヌクレオチドを含む発現ベクターを提供する。
【0024】
さらなる態様において、上記態様のいずれかに定義のポリヌクレオチドまたは発現ベクターでトランスフェクトされており、それを発現する宿主細胞、特に式(I)もしくは(I’)の化合物または式(II)〜(VII)、(XI)〜(XIV)ならびに(XVII)および(XVIII)の化合物の異種生産のための宿主細胞を提供する。
【0025】
他の態様において、本発明は、式(I)もしくは(I’)または式(II)〜(VII)、(XI)〜(XIV)ならびに(XVII)および(XVIII)の化合物を製造する方法であって、上記態様に記載の宿主細胞を、当該化合物が生産される条件下で培養することを含む、製造方法に関する。
【0026】
ある態様において、本発明は、上記態様のいずれかに記載のポリペプチドまたはNRPSもしくはNRPSドメインに特異的に結合する抗体、並びに当該抗体の使用、すなわちポリペプチドまたはNRPSの精製のための使用に関する。
【0027】
ある態様において、上記態様のいずれかに定義のポリヌクレオチド、ベクター、ポリペプチド、NRPSもしくはNRPSドメインまたは抗体を含む医薬組成物を提供する。
【0028】
ある態様において、上記態様のいずれかに定義の本発明のポリヌクレオチドを含む組換え宿主細胞を好適な条件下で培養して得られるまたはそのようにして得た式(I)または(I’)のデプシペプチドを含む医薬組成物を提供する。
【0029】
ある態様において、本発明は、疾患または状態、すなわちアトピー性皮膚炎の処置および/または診断に使用するための医薬組成物の製造のための、式(I)または(I’)のデプシペプチドに関する。ある特定の態様において、式(I)または(I’)のデプシペプチドは、選択的ヒトカリクレイン(hK7)およびエラスターゼ阻害剤、特に選択的ヒトカリクレイン(hK7)の阻害剤であり、酵素活性、特にセリンプロテアーゼ活性を有する。
【0030】
本発明のさらなる態様において、式(I)または(I’)の化合物の生産に関与するNRPSをコードする生合成遺伝子クラスターであって、上記態様のいずれかに定義のポリヌクレオチドを含むものを提供する。
【0031】
本発明の他の態様において、以下の工程:
(a)コンドロマイセス・クロカツス株または関連株のゲノムDNAからなるヌクレオチドライブラリーを構築する;(b)コロニーとしてライブラリー株を培養する;(c)NRPS遺伝子クラスターを含むクローンの同定のための上記態様のいずれかに定義のポリヌクレオチドに基づくプローブ分子で、増殖したコロニーを分析する、そして(d)NRPS遺伝子クラスターを同定する;
を含む方法によって得られる、本発明の生合成遺伝子クラスターの同定のための上記態様のいずれかに定義のポリヌクレオチド配列を提供する。
【0032】
本発明の骨子は、式(I)または(I’)のデプシペプチド、特に式(II)〜(VII)、(XI)〜(XIV)ならびに(XVII)および(XVIII)のデプシペプチドの整合性に関与する生合成クラスターまたはその機能的部分の提供にある。特に利点としては、式(I)または(I’)のデプシペプチドの生合成クラスターの同定を当該デプシペプチドの異種発現に使用できることである。
【0033】
「非リボソームペプチド」は、単純アミノ酸モノマーから構成される複雑な天然産物のファミリーに属するペプチドのクラスを意味する。これは非リボソームペプチドシンテターゼ(NRPS)と呼ばれる大型多機能タンパク質によって、多くの細菌または真菌において合成される。NRPS系の特徴は、タンパク新生および非タンパク新生アミノ酸を含むペプチドを合成できることである。
【0034】
「非リボソームペプチドシンターゼ」(NRPS)は、モジュールと呼ばれる活性部位の協調的なグループに組織される、大型多機能タンパク質を意味し、ここで各モジュールは、産物長伸長および官能基の修飾の1サイクルを触媒するために必要である。モジュールの数および順序、並びに各NRPS上のモジュール内に存在するドメインのタイプは、取り込まれるアミノ酸の数、順序、選択、そして特定のタイプの伸長に関連した修飾を指示することによって、得られるペプチド産物の構造バリエーションを決定する。
【0035】
用語「ドメイン」は、触媒活性に必要なタンパク質の機能的部分を意味する。かかるドメインは、同じ触媒活性を有する異なる種由来の酵素間で保存されている。
伸長サイクルに必要な最小セットのドメインは、アデニレーション(A)、チオレーション(T)またはペプチジルキャリアタンパク質(PCP)およびコンデンセーション(C)ドメインを有するモジュールからなる。
【0036】
「アデニレーションドメイン」は、AMP誘導体中間体を介して、基質選択およびチオエステルとしてTドメインのホスホ−パンテテインアームへの共有結合性固定に関与している。
【0037】
Cドメインは、上流モジュールからのアミノアシル−またはペプチジル−S−PCPと、対応する下流モジュールのPCPに結合したアミノアシル基の間のペプチド結合の形成を触媒する。その結果は、下流モジュールのPCPドメインに固定された1個の残基によるペプチド伸長である。任意の修飾ドメインは、基質エピマー化、N−メチル化およびヘテロ環化のために存在していてもよい。このモジュールは、1または複数のポリペプチド鎖上に残っていてよい。
【0038】
多くの場合、最終産物の放出/環化に関与する最後のモジュールの最もC末端側にチオエステラーゼ(TE)ドメインが存在する。
【0039】
1.式(I)または(I’)の化合物を生産する生合成遺伝子クラスターをコードするポリヌクレオチド
下記表1は、式(I)または(I’)の化合物の生合成遺伝子クラスターのポリヌクレオチドの具体例およびそれらの対応する機能およびアミノ酸配列を記載している。
【0040】
【表1】
【表2】
【0041】
式(I)または(I’)のデプシペプチドの合成のための単離された生合成遺伝子クラスターは、NRPS1およびNRPS2とも呼ばれる非リボソームペプチドシンテターゼをコードするORF6およびORF7を含む、8個のオープンリーディングフレーム(ORF)からなる。NRPS1およびNRPS2はNRPSドメインを含み、対応する推定機能を表1に列挙する。
【0042】
用語「ポリヌクレオチド」、「ポリヌクレオチド配列」および「ポリペプチド」の意味は当該技術分野において周知であり、当該用語は、異なる定義がないかぎり、本発明の文脈に従って使用される(例えば、それぞれ配列番号1、3、5、7、9、11、13、15、16、18、20、22、24、26、28、30、32、34、36、38、40、42、44、46、48、50、52、54、56、58、60、62)。例えば、「ポリヌクレオチド配列」は、本明細書において使用するとき、天然に生じるかまたは組換えによって作成するタイプの核酸および/またはヌクレオチド配列のあらゆる形態、並びに化学的に合成した核酸/ヌクレオチド配列を意味する。この用語は、核酸アナログおよび核酸誘導体、例えばロックドDNA、PNA、チオリン酸オリゴヌクレオチドおよび置換されたリボオリゴヌクレオチド等も含む。さらに、用語「ポリヌクレオチド配列」は、ヌクレオチドまたはヌクレオチドアナログを含むあらゆる分子をも意味する。
【0043】
好ましくは、用語「ポリヌクレオチド配列」は、核酸分子、すなわちデオキシリボヌクレオチド(DNA)および/またはリボ核酸(RNA)を意味する。本発明の文脈における「ポリヌクレオチド配列」は、当業者に知られている合成的化学法または組換え技術の使用によって作成してよく、あるいは天然源から単離してもよく、あるいはそれらの組合せでもよい。DNAおよびRNAは、所望により非天然ヌクレオチドを含んでいてもよく、一本鎖でも二本鎖でもよい。「ポリヌクレオチド配列」は、センスおよびアンチセンスDNAおよびRNA、すなわちDNAおよび/またはRNAのヌクレオチドの特定の配列に相補的であるポリヌクレオチド配列も意味する。
【0044】
さらに、用語「ポリヌクレオチド配列」は、DNAもしくはRNAまたはそれらのハイブリッドまたは当該技術分野において既知のそのいずれかの修飾物を意味していてもよい(修飾物の例として例えばUS 5525711、US 4711955、US 5792608またはEP 302175参照)。ポリヌクレオチド配列は一本鎖または二本鎖、直線または環状、天然または合成であってよく、いかなるサイズによる限定もない。例えば、ポリヌクレオチド配列は、ゲノムDNA、cDNA、mRNA、アンチセンスRNA、リボザイムまたはかかるRNAもしくはキメロプラスト(chimeroplast)をコードするDNAであってよい(Gamper, Nucleic Acids Research, 2000, 28, 4332 - 4339)。当該ポリヌクレオチド配列は、プラスミドまたはウイルス性DNAもしくはRNAの形態であってもよい。「ポリヌクレオチド配列」はホスホチオエートまたはペプチド核酸(PNA)のような当該技術分野で知られている修飾を含む、オリゴヌクレオチドを意味していてもよい。
【0045】
用語「遺伝子クラスター」または「生合成遺伝子クラスター」は、式(I)または(I’)のデプシペプチドの生合成に関与する遺伝子またはその変異体の群を意味する。遺伝子クラスターまたは生合成遺伝子クラスターの遺伝子改変は、式(I)または(I’)の化合物の変異体を作成するために適用可能な核酸の突然変異誘発、不活性化または置換を含む当該技術分野で既知のあらゆる遺伝子組換え技術を意味する。遺伝子クラスターまたは生合成遺伝子クラスターの遺伝子改変は、式(I)または(I’)の化合物の遺伝的変異体を作成するために適用可能な核酸の突然変異誘発、不活性化または置換を含む当該技術分野で既知のあらゆる遺伝子組換え技術を意味する。
【0046】
DNAもしくはヌクレオチド「コーティング配列」または特定のポリペプチドもしくはタンパク質を「コードする配列」は、適切な制御配列の制御下に置かれたときにポリペプチドまたはタンパク質に転写および翻訳されるDNA配列である。
【0047】
具体的な態様において、複数の本発明のポリヌクレオチド(例えば、それぞれ配列番号1、3、5、7、9、11、13、15、16、18、20、22、24、26、28、30、32、34、36、38、40、42、44、46、48、50、52、54、56、58、60、62)を組み合わせて用いてもよい。あるいは、本発明は、配列番号1、3、5、7、9、11、13、15、16、18、20、22、24、26、28、30、32、34、36、38、40、42、44、46、48、50、52、54、56、58、60、62のフラグメントまたは機能的変異体に関する。
【0048】
ポリヌクレオチド配列の文脈において、用語「そのフラグメント」または「その機能的フラグメント」は、特に核酸分子のフラグメントまたは変異体を意味する。「ポリヌクレオチドのフラグメント」は、例えば、少なくとも1個のアミノ酸欠失を有しており、それによって野生型ポリペプチドと同じ機能(各ポリペプチドの機能は表1および図2により詳細に記載されている)を実質的に保持している本発明のポリペプチド(例えば、配列番号2、4、6、8、10、12または14に示すポリペプチド)をコードしてよい。このような短くなったポリペプチドは、本発明のポリペプチド(例えば、配列番号2、4、6、8、10、12または14、17、19、21、23、25、27、29、31、33、35、37、39、41、43、45、47、49、51、53、55、57、59、61、63に示す)の機能的フラグメントと理解されうる。
【0049】
「ポリヌクレオチドの機能的変異体」は、例えば、少なくとも1個のアミノ酸置換または付加を有しており、それによって好ましくは野生型ポリペプチドと同じ機能(各ポリペプチドの機能は表1および図2により詳細に記載されている)を保持している本発明のポリペプチド(例えば配列番号2、4、6、8、10、12または14、17、19、21、23、25、27、29、31、33、35、37、39、41、43、45、47、49、51、53、55、57、59、61、63に示すポリペプチド)をコードしうる。このような短くなったポリペプチドは、本発明のポリペプチド(例えば、配列番号2、4、6、8、10、12または14、17、19、21、23、25、27、29、31、33、35、37、39、41、43、45、47、49、51、53、55、57、59、61、63に示す)の機能的フラグメントと理解されうる。
【0050】
本発明のポリヌクレオチド/ポリペプチドの機能的変異体は、表1に記載の対応する元のポリヌクレオチド/ポリペプチド配列と、少なくとも50%、55%、60%、好ましくは少なくとも70%、より好ましくは少なくとも80%、85%、90%、95%さらに最も好ましくは少なくとも99%の配列同一性を有する。例えば、ポリペプチドは、配列番号2、4、6、8、10、12または14、17、19、21、23、25、27、29、31、33、35、37、39、41、43、45、47、49、51、53、55、57、59、61、63に示すポリペプチドと少なくとも50%、55%、60%、好ましくは少なくとも70%、より好ましくは少なくとも80%、85%、90%、95%、最も好ましくは少なくとも99%の同一性/相同性を有する。
【0051】
非リボソームペプチドシンターゼ(NRPS)または表1に記載の他のORFのヌクレオチド配列に関して、用語「フラグメント」は、本明細書において使用するとき、少なくとも7、少なくとも10、少なくとも15、少なくとも20、少なくとも30、少なくとも50、少なくとも100、少なくとも150、少なくとも200、少なくとも250、少なくとも300、少なくとも350、少なくとも400、少なくとも450、少なくとも500、少なくとも550、少なくとも600、少なくとも650または少なくとも700ヌクレオチド長を有するヌクレオチド配列を意味する。
【0052】
用語「ハイブリダイズ」は、本明細書において使用するとき、常套のハイブリダイゼーション条件下、好ましくはストリンジェントな条件下での、例えばハイブリダイゼーションSambrook and Russell (2001), Molecular Cloning: A Laboratory Manual, CSH Press, Cold Spring Harbor, NY, USAに記載のハイブリダイゼーションを意味する。さらに特定されていない限り、条件は、好ましくは非ストリンジェントである。ハイブリダイゼーション条件は、例えばSambrook (2001) loc. citに記載の、常套のプロトコルに従って確立できる。条件設定は十分に当業者の技術の範囲内であり、当該技術分野において記載のプロトコルに従って決定できる。したがって、特異的にのみハイブリダイズする配列の検出は、通常、ストリンジェントなハイブリダイゼーションおよび線条条件を必要とする。非限定的な例として、高いストリンジェントなハイブリダイゼーションは、次の条件下で生じうる:
【表3】
【0053】
ホモログまたは正確に相補的でない配列の検出のための低いストリンジェントなハイブリダイゼーション条件は、例えば6 x SSC、1% SDS、65℃に設定できる。周知のとおり、プローブの長さおよび決定する核酸の組成は、ハイブリダイゼーション条件のさらなるパラメーターを構成する。
【0054】
本明細書において提供されるポリヌクレオチド配列とハイブリダイズすることができるポリヌクレオチドも、本発明の一部であり、例えば動物のゲノムライブラリーもしくはcDNAライブラリーまたは微生物のDNAライブラリーから単離できる。好ましくは、かかるポリヌクレオチドは微生物起源のもの、特にプロテオバクテリアクラスに属する微生物、特にデルタプロテオバクテリア、特にミキソコッカス目(Myxococcales)、特にソランギウム亜目(Sorangiineae)、特にポリアンギウム科(Polyangiaceae)、とりわけコンドロマイセス属(Chondromyces)、例えばコンドロマイセス・クロカツス(Chondromyces crocatus)またはその改良株のものである。
【0055】
あるいは、かかる本発明の変異ヌクレオチド配列は、遺伝子操作または化学合成によって作成してもよい。ハイブリダイズ可能なかかるポリヌクレオチド配列は、本明細書に記載のポリヌクレオチド配列またはその一部もしくは逆相補体を用いて、例えば標準的な方法によるハイブリダイゼーション(例えばSambrook and Russell (2001), Molecular Cloning: A Laboratory Manual, CSH Press, Cold Spring Harbor, NY, USA参照)によって、同定および単離できる。列挙した配列番号またはその一部/フラグメントと同じまたは実質的に同じヌクレオチド配列を含むヌクレオチド配列は、例えばハイブリダイゼーションプローブとして使用できる。フラグメントはNRPS遺伝子クラスターの診断、配列決定またはクローニングのためのプローブまたはプライマーとしても有用でありうる。ハイブリダイゼーションプローブとして使用されるフラグメントは、通常の合成技術によって作成される合成フラグメントであってもよく、その配列は本発明のヌクレオチド配列のものと実質的に同一である。
【0056】
本明細書において使用するとき、2つの配列間の同一性パーセントは、2つの配列の最適なアラインメントのために導入する必要のあるギャップ数および各ギャップの長さを考慮し、当該配列によって共有される同一位置の数の関数である(すなわち、同一性パーセント=同一位置数/全位置数×100)である。配列の比較および2つの配列間の同一性パーセントの決定は、下記のような、数学的アルゴリズムを用いて行うことができる。
【0057】
好ましくは、同一性/相同性の程度は、列挙した配列番号に示すヌクレオチド配列とそれぞれの配列を比較して決定する。比較する配列が同じ長さを有さないとき、相同性の程度は、好ましくは、より長い配列におけるヌクレオチド残基と同一であるより短い配列のヌクレオチド残基のパーセンテージを意味する。通常は、相同性の程度は、ClustalW分析を備えたDNASTARプログラムのような既知のコンピュータープログラムを用いて決定できる。このプログラムはDNASTAR, Inc., 1228 South Park Street, Madison, WI 53715から、またはDNASTAR, Ltd., Abacus House, West Ealing, London W13 0AS UKから入手でき(support@dnastar.com)、EMBL外部ステーションのサーバーでアクセス可能である。
【0058】
特定の配列が例えば参照配列と80%同一であるかを決定するためにClustal分析法を用いる場合、好ましくは、アミノ酸配列の比較のための設定は次の通りである:マトリックス: blosum 30; オープンギャップペナルティ: 10.0; エクステンドギャップペナルティ: 0.05; ディレイダイバージェント: 40; ギャップ分離距離: 8 。ヌクレオチド配列比較については、エクステンドギャップペナルティは好ましくは5.0に設定する。
【0059】
配列比較によって比較される2つのヌクレオチド配列が異なる場合、同一性は、より短い配列とより短い配列にマッチするより長い配列の一部を意味する。換言すれば、比較する配列が同じ長さを有さない場合、同一性の程度は、好ましくは、より長い配列におけるヌクレオチド残基と同一であるより短い配列におけるヌクレオチド残基のパーセンテージまたはより短い配列におけるヌクレオチド残基と同一であるより長い配列におけるヌクレオチドのパーセンテージのいずれかを意味する。この文脈において、当業者は、より短い配列に「マッチ」するより長い配列の一部を、容易に決定することができる。
【0060】
一般に、どのように核酸分子を、例えば天然源から得るか、あるいは合成またはPCRのような組換え技術によって作成できるかを、当業者は知っている。これらの核酸分子は、関連文献に記載の技術を適用して得ることができる修飾または誘導体化された核酸分子を含む。
【0061】
さらに、同一性は、対応するヌクレオチド配列またはポリペプチド(例えばそれによってコードされるポリペプチド)間の機能的および/または構造的均等性が存在することを意味する。本明細書に記載の特定のヌクレオチド/アミノ酸配列と少なくとも50%、55%、60%、好ましくは少なくとも70%、より好ましくは少なくとも80%、85%、90%、95%、最も好ましくは少なくとも99%の同一性を有するヌクレオチド/アミノ酸配列は、好ましくは同じ生物学的機能を有し、これらの配列の誘導体/変異体を意味しうる。それらは天然に生じる変異体、例えば生態型、変種、種等に由来する配列または変異体であってよく、当該変異体は、天然に形成されるかあるいは計画的突然変異誘発によって作成してよい。さらに、変異体は合成的に作成された配列であってもよい。アレル変異体は天然に生じる変異体または合成によって作成した変異体または組換えDNA技術によって作成した変異体であってよい。上記ポリヌクレオチドからの逸脱は、例えば欠失、置換、付加、挿入および/または組換えによって作成できる。用語「付加」は、所定の配列の末端に少なくとも1個の核酸残基/アミノ酸を加えることを意味し、一方で「挿入」は、所定の配列中に少なくとも1個の核酸残基/アミノ酸を挿入することを意味する。
【0062】
変異体ポリペプチドおよび特に本発明のヌクレオチド配列の異なる変異体によってコードされるポリペプチドは、好ましくは、共通して有するある特徴を示す。これらは、例えば生物学的活性、分子量、免疫応答、コンホメーション等および物理的特性、例えばゲル電気泳動における泳動動態、クロマトグラフでの動態、沈降係数、溶解性、分光学的特性、安定性、至適pH、至適温度等を含む。
【0063】
ある具体的な態様において、本発明は、下記1個以上のNRPS1ドメインの活性を保持する1個以上の発現産物をコードするポリヌクレオチドを提供する:
(i)配列番号31のローディングドメイン;
(ii)配列番号33のグルタミンのアデニレーションドメイン;
(iii)配列番号35のチオレーションドメイン;
(iv)配列番号37のコンデンセーションドメイン;
(v)配列番号39のスレオニンのアデニレーションドメイン;
(vi)配列番号41のチオレーションドメイン;
(vii)配列番号43のコンデンセーションドメイン;および
(viii)配列番号45のロイシンのアデニレーションドメイン。
【0064】
特定の態様において、ポリヌクレオチドは上記全てのNRPS1ドメインの活性を保持する1個以上の発現産物をコードする。
【0065】
別の態様において、ポリヌクレオチドは、1、2または3個のアデニレーションドメインが異なるアミノ酸特異性を有する1個以上のアデニレーションドメインに置換されていることを除き、上記全てのNRPS1ドメインの活性を保持する1個以上の発現産物をコードする。
【0066】
別の具体的な態様において、本発明は、下記1個以上のNRPS2ドメインの活性を保持する1個以上の発現産物をコードするポリヌクレオチドを提供する:
(i)配列番号47のチオレーションドメイン;
(ii)配列番号49のコンデンセーションドメイン;
(iii)配列番号51のプロリンのアデニレーションドメイン;
(iv) 配列番号53のチオレーションドメイン;
(v) 配列番号2のコンデンセーションドメイン;
(vi) 配列番号4のイソロイシンのアデニレーションドメイン;
(vii) 配列番号6のチオレーションドメイン;
(viii) 配列番号8のコンデンセーションドメイン;
(ix) 配列番号10のチロシンのアデニレーションドメイン;
(x) 配列番号12のN−メチレーション ドメイン;
(xi) 配列番号14のチオレーションドメイン;
(xii) 配列番号55のコンデンセーションドメイン;
(xiii)配列番号57のイソロイシンのアデニレーションドメイン;
(xiv) 配列番号59のチオレーションドメイン;および
(xv) 配列番号61のチオエステラーゼドメイン。
【0067】
特定の態様において、ポリヌクレオチドは上記全てのNRPS2ドメインの活性を保持する1個以上の発現産物をコードする。別の態様において、ポリヌクレオチドは、1、2、3または4個のアデニレーションドメインが異なるアミノ酸特異性を有する1個以上のアデニレーションドメインに置換されていることを除き、上記全てのNRPS1ドメインの活性を保持する1個以上の発現産物をコードする。
【0068】
NRPS1をコードするORF6、NRPS2をコードするORF7およびチトクロムP450をコードするORF8は、式(I)または(I’)のデプシペプチドの生合成のためのコア酵素をコードすると推定される。したがって、さらなる局面において、本発明は、
(i) 配列番号27(NRPS1)をコードするヌクレオチド配列またはその機能的変異体;および
(ii) 配列番号29(NRPS2)をコードするヌクレオチド配列またはその機能的変異体
を含むポリヌクレオチドに関する。
【0069】
ポリヌクレオチドは、さらに、配列番号63をコードするヌクレオチド配列またはその機能的変異体を含んでいてもよい。ある具体的な態様において、これらのポリヌクレオチドは、受託番号DSM19329を有するコンドロマイセス・クロカツス(Chondromyces crocatus)株NPH−MB180から単離される。
【0070】
2.式(I)または(I’)の化合物の生産に関与するNRPSおよび他のポリペプチド
本発明はさらに、本発明のポリヌクレオチドによってコードされるポリペプチド、特に表1に記載のもの、例えばNRPS1およびNRPS2に関する。本発明はさらにそれらの機能的フラグメントおよび機能的変異体に関する。
【0071】
本発明はまた、配列番号2、4、6、8、10、12、14、17、19、21、23、25、27、29、31、33、35、37、39、41、43、45、47、49、51、53、55、57、59、61、63のポリペプチドの変異体またはその少なくとも50、75、100、150、200、300、400または500連続アミノ酸を含むフラグメントに関する。用語「変異体」は、誘導体またはこれらのポリペプチドのアナログを含む。特に、変異体は、配列番号2、4、6、8、10、12、14、17、19、21、23、25、27、29、31、33、35、37、39、41、43、45、47、49、51、53、55、57、59、61、63のポリペプチドとアミノ酸配列において、1、2、3、4、5またはそれ以上の置換、付加、欠失、融合およびトランケーション(これらは任意の組合せで存在していてもよい)によって異なっていてよい。
【0072】
変異体は天然に存在するかあるいはインビトロで作成できる。特に、かかる変異体は遺伝子操作技術、例えば部位特異的突然変異誘発、ランダム突然変異誘発、エキソヌクレアーゼIII欠失法および標準的なクローニング技術を用いて作成できる。あるいは、かかる変異体、フラグメント、アナログまたは誘導体は、化学合成または修飾法を用いて作成してもよい。
【0073】
変異体を作成する他の方法も当業者に周知である。これらには、天然単離体から得られた核酸配列を修飾して、工業的または研究的利用価値を高めた特性を有するポリペプチドをコードする核酸を作成する手法が含まれる。かかる手法において、天然単離体から得られる配列について1個以上のヌクレオチドの相違を有する多数の変異体配列が作成され、特徴付けられる。好ましくは、これらのヌクレオチドの相違は、天然単離体からの核酸によってコードされるポリペプチドに対して、アミノ酸の変化を引き起こす。
【0074】
例えば、変異体は、エラープローンPCRを用いて作成できる。エラープローンPCRにおいて、DNA増幅は、DNAポリメラーゼの忠実度が低く、したがってPCR産物の全長にわたって点突然変異が高い確率で得られるような条件下で実施される。エラープローンPCRはLeung, D.W., et al., Technique, 1:11-15 (1989)およびCaldwell, R. C. & Joyce G.F., PCR Methods Applic., 2:28-33 (1992)に記載されている。変異体は、部位特異的突然変異誘発を用いて、目的のクローン化されたDNAセグメントに部位特異的変異を作成することで作成してもよい。オリゴヌクレオチド突然変異誘発はReidhaar-Olson, J.F. & Sauer, R.T., et al., Science, 241:53-57 (1988)に記載されている。米国特許第6,361,974号及び第6,372,497号に記載のもののような定向進化戦略を用いて変異体を作成してもよい。配列番号2、4、6、8、10、12、14のポリペプチドの変異体は、配列番号2、4、6、8、10、12、14、17、19、21、23、25、27、29、31、33、35、37、39、41、43、45、47、49、51、53、55、57、59、61、63のポリペプチドのアミノ酸残基の1、2、3、4、5またはそれ以上が保存的または非保存的アミノ酸残基(好ましくは保存的アミノ酸残基)で置換されており、かかる置換アミノ酸残基は遺伝コードによってコードされるかされなくてよい、変異体であってよい。
【0075】
保存的置換は、あるポリペプチドにおける所定のアミノ酸を、同様の特性の別のアミノ酸で置換することである。典型的には、保存的置換は以下の置換として見られる:Ala、Val、LeuおよびIleのような脂肪族アミノ酸の別の脂肪族アミノ酸での置換;SerのThrでの置換またはThrのSerでの置換;AspまたはGluのような酸性残基の別の酸性残基での置換;AsnまたはGlnのようなアミド基を有する残基の別のアミド基を有する残基での置換;LysまたはArgのような塩基性残基の別の塩基性残基での交換;およびPheまたはTyrのような芳香族性残基の別の芳香族性残基での置換。
【0076】
他の変異体は、配列番号2、4、6、8、10、12、14、17、19、21、23、25、27、29、31、33、35、37、39、41、43、45、47、49、51、53、55、57、59、61、63のポリペプチドの1個以上のアミノ酸残基が置換基を含むものである。さらに別の変異体は、ポリペプチドが別の化合物、例えばポリペプチドの半減期を延長させる化合物(例えばポリエチレングリコール)と結合しているものである。さらなる変異体は、ポリペプチドにさらなるアミノ酸、例えばリーダー配列、分泌配列、前タンパク質配列またはポリペプチドの精製、富化もしくは安定化を促進する配列と融合しているものである。
【0077】
ある態様において、フラグメント、誘導体およびアナログは、配列番号2、4、6、8、10、12、14、17、19、21、23、25、27、29、31、33、35、37、39、41、43、45、47、49、51、53、55、57、59、61、63のポリペプチドと同じ生物学的機能または活性を保持している。用語「そのフラグメント」は、ポリペプチドの文脈において本明細書において使用するとき、本発明のポリヌクレオチド(例えばそれぞれ配列番号1、3、5、7、9、11、13、15、16、18、20、22、24、26、28、30、32、34、36、38、40、42、44、46、48、50、52、54、56、58、60、62)によってコードされてもよい本発明に定義のポリペプチド(例えばそれぞれ配列番号2、4、6、8、10、12または14、17、19、21、23、25、27、29、31、33、35、37、39、41、43、45、47、49、51、53、55、57、59、61、63のとおり)と実質的に同じ(生物学的)活性を有する機能的フラグメントを意味する。
【0078】
他の態様において、フラグメント、誘導体およびアナログは、少なくとも1、2、3、4、5、6または7このアデニレーションドメインが異なるアデニレーションドメインによって置換されておりそれによって異なるアミノ酸特異性が与えられていることを除き、配列番号2、4、6、8、10、12、14、17、19、21、23、25、27、29、31、33、35、37、39、41、43、45、47、49、51、53、55、57、59、61、63のポリペプチドと同じ生物学的機能または活性を保持する。
【0079】
他の態様において、フラグメント、誘導体またはアナログは、フラグメント、誘導体またはアナログから離れて全体または一部で酵素的に切断されうるポリペプチドの精製、富化、検出、安定化または分泌を促進する融合異種配列を含む。
【0080】
本発明の別の局面は、配列番号2、4、6、8、10、12、14、17、19、21、23、25、27、29、31、33、35、37、39、41、43、45、47、49、51、53、55、57、59、61、63のポリペプチドまたはその少なくとも50、75、100、150、200、300、400もしくは500連続アミノ酸を含むフラグメントの一つと、少なくとも60%、少なくとも70%、少なくとも80%、少なくとも90%または少なくとも95%の同一性を有するポリペプチドまたはそのフラグメントである。アミノ酸「同一性」は、上記のもののような保存的置換を含むと理解される。
【0081】
配列番号2、4、6、8、10、12、14、17、19、21、23、25、27、29、31、33、35、37、39、41、43、45、47、49、51、53、55、57、59、61、63のポリペプチドまたはその少なくとも少なくとも50、75、100、150、200、300、400もしくは500連続アミノ酸を含むフラグメントの一つと相同性を有するポリペプチドまたはフラグメントは、上記技術を用いてそれらをコードする核酸を単離して得ることができる。
【0082】
あるいは、生化学的富化または精製手法によって、相同なポリペプチドまたはフラグメントを得ることができる。潜在的に相同なポリペプチドまたはフラグメントの配列は、タンパク質消化、ゲル電気泳動および/またはミクロシークエンシングによって決定できる。推定される相同なポリペプチドまたはフラグメントの配列は、配列番号2、4、6、8、10、12、14、17、19、21、23、25、27、29、31、33、35、37、39、41、43、45、47、49、51、53、55、57、59、61、63のポリペプチドまたはその少なくとも50、75、100、150、200、300、400もしくは500連続アミノ酸を含むフラグメントの一つと比較できる。
【0083】
配列番号2、4、6、8、10、12、14、17、19、21、23、25、27、29、31、33、35、37、39、41、43、45、47、49、51、53、55、57、59、61、63のポリペプチドまたはその少なくとも50、75、100、150、200、300、400もしくは500連続アミノ酸を含むフラグメント、その少なくとも40、50、75、100、150、200もしくは300連続アミノ酸を含むフラグメントは、様々な用途に使用できる。例えばポリペプチドまたはそのフラグメント、誘導体もしくはアナログを用いて、本明細書の別の箇所に記載の生化学反応を触媒することができる。
【0084】
配列番号2、4、6、8、10、12、14、17、19、21、23、25、27、29、31、33、35、37、39、41、43、45、47、49、51、53、55、57、59、61、63のポリペプチドまたはその少なくとも50、75、100、150、200、300、400もしくは500連続アミノ酸を含むフラグメントを用いて、ポリペプチドまたはフラグメント、誘導体もしくはアナログに特異的に結合する抗体を作成してもよい。
【0085】
具体的な態様において、本発明のポリペプチド(例えばそれぞれ配列番号2、4、6、8、10、12または14、17、19、21、23、25、27、29、31、33、35、37、39、41、43、45、47、49、51、53、55、57、59、61、63)を組み合わせて用いてよい。
【0086】
用語「活性」または「機能性」は、本明細書において使用するとき、特に酵素活性、例えばNRPS1およびNRPS2のペプチドシンターゼ活性を誘発するポリペプチドまたはそのフラグメントの能力を意味する。当業者は本明細書に記載の機能性の(生物学的)活性はしばしば発現レベル(例えばタンパク質/mRNA)と相関すると気付くであろう。異なることが記載されていない限り、用語「発現」は、本明細書において用いるとき、本発明のポリペプチド/タンパク質(またはそのフラグメント)をコードする核酸分子の発現を意味し、一方で「活性」は、当該ポリペプチド/タンパク質の活性を意味する。本明細書に記載のポリペプチドの活性を測定する方法/アッセイは、当該技術分野において周知である。
【0087】
3.発現ベクター、組換え宿主細胞および式(I)または(I’)のデプシペプチドを製造する方法
本明細書に記載の本発明のポリヌクレオチドは、例えば式(I)または(I’)の化合物の異種発現に有用である。具体的な態様において、それらは、式(I’)の化合物の異種発現に有用である。
【0088】
したがって、さらなる局面において、本発明は、本明細書に記載の核酸分子を含むベクター、より具体的には発現ベクター、ならびに当該核酸分子および/またはベクターを含む組換え宿主細胞に関する。
【0089】
用語「ベクター」は、本明細書において使用するとき、特にプラスミド、コスミド、細菌人工染色体(BAC)、酵母人工染色体、ウイルス、バクテリオファージおよび遺伝子工学において一般に使用される他のベクターを意味する。好ましい態様において、本発明のベクターは、細胞、例えば真菌細胞、酵母もしくは細菌細胞のような微生物の細胞または動物細胞の形質転換に好適である。「発現ベクター」は、核酸を宿主細胞に導入して導入した配列の発現を引き起こすビークルを意味する。
【0090】
本明細書に記載のとおり、ポリペプチドは、コード配列が好適な宿主細胞においてコードされるポリペプチドの発現を駆動することができる配列と作動可能に連結するように、ポリペプチドをコードする核酸をベクターに挿入して、得ることができる。例えば、発現ベクターは、プロモーター、翻訳開始のためのリボソーム結合部位および転写ターミネーターを含んでいてよい。ベクターは、発現レベルを調節するための適切な配列、複製起点および選択マーカーを含んでいてもよい。細菌におけるポリペプチドまたはそのフラグメントの発現に好適なプロモーターは、E. coli lacまたはtrp プロモーター、lacl プロモーター、lacZ プロモーター、T3 プロモーター、T7 プロモーター、gpt プロモーター、ラムダ PR プロモーター、ラムダ PL プロモーター、解糖系の酵素、例えば3−ホスホグリセレートキナーゼ(PGK)をコードするオペロン由来のプロモーターおよび酸ホスファターゼプロモーターを含む。真菌プロモーターは、α因子プロモーターを含む。シュードモナス・プチダ(Pseudomonas Putida)における発現に好適なプロモーターは、ゲノムに存在する7つの16S rRNA遺伝子(PP 16SA、PP 16SB、PP 16SC、PP 16SD、PP 16SE、PP 16SF、PP 16SG)の対応転写プロモーター、抗生物質耐性決定要因の転写プロモーター、三価鉄取り込みリプレッサー(Fur)制御遺伝子の転写プロモーターを含むが、これらに限定されない。三価鉄取り込みリプレッサー(Fur)制御プロモーターのより詳細な説明は、以下に提供する。真核生物プロモーターは、CMV前初期プロモーター、HSVチミジンキナーゼプロモーター、熱ショックプロモーター、初期および後期SV40プロモーター、レトロウイルス由来のLTRおよびマウスメタロチオネイン−Iプロモーターを含む。原核生物もしくは真核生物細胞またはウイルスにおいて、遺伝子の発現を制御することが知られている他のプロモーターを用いてもよい。
【0091】
哺乳類発現ベクターは、複製起点、必要なリボソーム結合部位、ポリアデニレーション部位、スプライスドナーおよびアクセプター部位、転写終止配列および5’フランキング非転写配列を含んでいてもよい。ある態様において、SV40スプライスおよびポリアデニレーション部位由来のDNA配列を用いて、必要な非転写遺伝要素を提供することができる。
【0092】
真核生物細胞においてポリペプチドまたはそのフラグメントを発現するためのベクターは、発現レベルを上昇させるエンハンサーを含んでいてもよい。エンハンサーは、転写を増加させるプロモーターとして作用する通常約10〜約300bp長のDNAのシス−作用エレメントである。例えば、複製起点の後側の100〜270bpのSV40エンハンサー、サイトメガロウイルス初期プロモーターエンハンサー、複製起点の後側のポリオーマエンハンサーおよびアデノウイルスエンハンサーを含む。
【0093】
さらに、発現ベクターは好ましくは、ベクターを含む宿主細胞の選択が可能なように1個以上の選択マーカー遺伝子を含む。使用できる選択マーカーの例は、ジヒドロ葉酸レダクターゼをコードする遺伝子または真核細胞培養にネオマイシン耐性を与える遺伝子、大腸菌にテトラサイクリンもしくはアンピシリン耐性を与える遺伝子、ならびに出芽酵母TRP1遺伝子を含む。好適なマーカーの例は、ゲンタマイシン耐性カセットaacClである。他の選択マーカーーは、アンピシリン耐性を与えるヌクレオチドカセット(例えばbla)、クロラムフェニコール耐性を与えるヌクレオチドカセット(例えばcat)、カナマイシン耐性を与えるヌクレオチドカセット(例えばaacC2、aadBまたは他のアミノグリコシド修飾酵素)またはテトラサイクリン耐性を与えるヌクレオチドカセット(例えばtetAまたはtetB)である。
【0094】
適切なDNA配列は、様々な手段でベクターに挿入できる。一般に、DNA配列をベクターの所望の位置にライゲートし、次いで適切な制限エンドヌクレアーゼで挿入物およびベクターを消化する。あるいは、PCRにより適切な制限酵素部位をDNA配列へと設計することができる。様々なクローニング技術がAusbel et al. Current Protocols in Molecular Biology, John Wiley 503 Sons, Inc. 1997 および Sambrook et al., Molecular Cloning: A Laboratory Manual 2d Ed., Cold Spring Harbour Laboratory Press, 1989に記載されている。かかる手法および他のものは、当該技術分野の技術範囲内であると理解される。
【0095】
ベクターは例えば、プラスミド、ウイルス粒子またはファージの形態であってよい。他のベクターは、染色体の誘導体、非染色体および合成DNA配列、ウイルス、細菌プラスミド、ファージDNA、バキュロウイルス、酵母プラスミド、プラスミドおよびファージDNAの組合せに由来するベクター、ワクチンのようなウイルス性DNA,アデノウイルス、鶏痘ウイルスおよび仮性狂犬病ウイルスを含む。原核および真核宿主で使用するための多様なクローニングおよび発現ベクターは、Sambrook et al., Molecular Cloning: A Laboratory Manual, Second Edition, Cold Spring Harbor, N.Y., (1989)に記載されている。
【0096】
使用できる具体的な細菌ベクターは、周知のクローニングベクターの遺伝的要素を含む商業的に入手可能なプラスミド、例えばpBR322 (ATCC 37017)、pKK223-3 (Pharmacia Fine Chemicals, Uppsala, Sweden)、pGEM1 (Promega Biotec, Madison, WI, USA) pQE70、pQE60、pQE-9 (Qiagen)、pD10、phiX174、pBluescript(商標) II KS、pNH8A、pNH16a、pNH18A、pNH46A (Stratagene)、ptrc99a、pKK223-3、pKK233-3、pDR540、pRIT5 (Pharmacia)、pKK232-8 および pCM7を含む。具体的な真核ベクターは、pSV2CAT、pOG44、pXT1、pSG (Stratagene) pSVK3、pBPV、pMSGおよびpSVL (Pharmacia)を含む。しかし、あらゆる他のベクターが、宿主細胞において複製可能であり安定である限り、使用可能である。
【0097】
エレクトロポレーション形質転換、トランスフェクション、形質導入、ウイルス感染、遺伝子銃またはTi介在遺伝子導入を含む多様な技術のいずれかを用いて、ベクターを宿主細胞に導入してよい。適切であるとき、操作された宿主細胞は、プロモーターを活性化させ、形質転換体を選択しまたは本発明の遺伝子を増幅するために適切なように修飾された、常套の栄養培地で培養できる。好適な宿主株の形質転換および適切な細胞密度への当該宿主株の成長の後、選択されたプロモーターを適切な手段(例えば温度変化または化学的誘導)で誘導してよく、所望のポリペプチドまたはそのフラグメントを生産させるためにさらなる期間にわたって当該細胞を培養してよい。
【0098】
さらなる局面において、本発明の組換え宿主細胞は、本発明のポリヌクレオチド配列によってコードされるポリペプチドを発現できるかあるいは発現している。具体的な態様において、宿主細胞に含まれる「ポリペプチド」は、宿主細胞の起源に対して異種であってよい。本発明の宿主細胞を作成するために用いる様々な発現系の例の概略、例えば上記の具体的なものは、例えばGlorioso et al. (1999), Expression of Recombinant Genes in Eukaryotic Systems, Academic Press Inc., Burlington, USA、Paulina Balbas und Argelia Lorence (2004), Recombinant Gene Expression: Reviews and Protocols, Second Edition: Reviews and Protocols (Methods in Molecular Biology), Humana Press, USAに記載されている。
【0099】
本発明のヌクレオチド配列またはベクターによる宿主細胞の形質転換または遺伝子操作は、標準的な方法、例えばSambrook and Russell (2001), Molecular Cloning: A Laboratory Manual, CSH Press, Cold Spring Harbor, NY, USAに記載のとおりに行うことができる。さらに、本発明の宿主細胞は、特にpH値、温度、塩濃度、通気、抗生物質、ビタミン、微量元素等の観点から、使用する具体的な宿主細胞の要求を満たす栄養培地中で培養する。
【0100】
一般に、本発明の宿主細胞は、本発明のヌクレオチド配列、ベクターおよび/またはポリペプチドを含む原核または真核細胞、あるいは本発明のヌクレオチド配列、ベクターおよび/またはポリペプチドを含む細胞に由来する細胞であってよい。好ましい態様において、宿主細胞は、例えば遺伝子操作によって、そのゲノムに組み込まれた本発明のヌクレオチド配列をそれが含むように、本発明のヌクレオチド配列またはベクターを含む。かかる本発明の宿主細胞の非限定的な例(一般的な本発明の宿主細胞でもある)は、細菌、酵母、真菌、植物、動物またはヒト細胞であってよい。
【0101】
用語「宿主細胞」または「単離された宿主細胞」は、その生物が式(I)の化合物または式(I’)の化合物を生産することが知られていようといまいと、その化合物を生産するために必要な遺伝子情報を担持する微生物を意味する。該用語は、本明細書において使用するとき、天然環境に存在するままでその生物に例えば式(I)または(I’)の化合物を生産するための遺伝情報が見られる生物と、遺伝情報が組換え技術によって導入される生物に、等しく適用する。宿主細胞は原核細胞または真核細胞を含む当該技術分野において周知のあらゆる宿主細胞であってよい。適切な宿主の代表例として、次のものを挙げることができる:細菌細胞、例えば大腸菌(E. coli)、ストレプトマイセス・リビダンス(Streptomyces lividans)、ストレプトマイセス・グリセオフスカス(Streptomyces griseofuscus)、ストレプトマイセス・アムボファシエンス(Streptomyces ambofaciens)、枯草菌(Bacillus subtilis)、ネズミチフス菌(Salmonella typhimurium)、ミキソコッカス・キサンタス(Myxococcus xanthus)、ソランギウム・セルロサム(Sorangium cellulosum)、コンドロマイセス・クロカツス(Chondromyces crocatus)およびシュードモナス属、ストレプトマイセス属、バシラス属およびスタフィロコッカス属の多様な種、真菌細胞、例えば酵母、困窮細胞、例えばDrosophila S2およびSpodoptera Sf9、動物細胞、例えばCHO、COSまたはBowes黒色腫、ならびにアデノウイルス。適切な宿主の選択は当業者の能力の範囲内である。
【0102】
本明細書で意図する生物源は、プロテオバクテリアに含まれる生物、好ましくはデルタプロテオバクテリア、より好ましくはミキソコッカス目(Myxococcales)、より好ましくはソランギウム亜目(Sorangiineae)、より好ましくはポリアンギウム科(Polyangiaceae)、最も好ましくはコンドロマイセス属(Chondromyces)、例えばコンドロマイセス・クロカツス(Chondromyces crocatus)またはその改良株のものである。
【0103】
用語「組換え宿主細胞」は、本明細書において使用するとき、本発明のヌクレオチド配列で遺伝子操作されているかまたは本発明のベクターまたはポリペプチドもしくはそのフラグメントを含む宿主細胞を意味する。本発明は、異種組換え宿主細胞、すなわち天然生産株とは異なる別の株において発現される式(I)または式(I’)のデプシペプチドの生産を可能とする。例示として細菌株の使用を説明するが、本明細書に記載のとおり、あらゆる生物または発現系を用いてよい。生物の選択は当業者の必要に応じる。例えば、遺伝子操作を受け入れられる株は、デプシペプチド化合物の修飾および生産を促進するために使用できる。
【0104】
ある具体的な態様において、宿主細胞は、ミキソコッカス属またはシュードモナス属、例えばシュードモナス・プチダから選択される。あるより具体的な態様において、組換え宿主細胞、例えばシュードモナス・プチダは、NRPS1(配列番号27)およびNRPS2(配列番号29)またはその機能的変異体をコードするヌクレオチドを含む。それはさらに、配列番号63のチトクロムP450またはその機能的変異体をコードするヌクレオチド配列を含んでいてもよい。それはまた、配列番号17、配列番号18、配列番号21、配列番号23、配列番号25および配列番号27の1つ以上を含んでいてもよい。有利には、各オープンリーディングフレームは、これらのORFが組換え宿主細胞によって好適な条件下で発現されるように、機能的転写および翻訳配列の制御下に存在する。シュードモナス・プチダにおける異種発現の具体例は、下記実施例にさらに説明されている。
【0105】
上記に従って、本発明は、さらなる態様において、式(I)または(I’)の化合物を生産する方法であって、式(I)または式(I’)、例えば式(II)〜(VII)、(XI)〜(XIV)ならびに(XVII)および(XVIII)の化合物が合成される条件下で組換え宿主細胞を培養し、そして当該化合物を回収することを含む方法に関する。
【0106】
用語「条件」は、本明細書において使用するとき、式(I)の化合物または式(I’)の化合物を発現し、回収するための組換え宿主細胞の培養条件を意味する。ある具体的な態様において、組換え宿主細胞はシュードモナス・プチダである。別の具体的な態様において、組換え宿主細胞はシュードモナス・プチダであり、当該細胞は30℃未満、例えば10〜20℃、例えば約15℃で成長させる。
【0107】
別の具体的な態様において、成長培地はイソ酪酸、例えば1〜5g/lのイソ酪酸、例えば約2g/lのイソ酪酸を含む。
【0108】
例えば、本発明の組換え宿主細胞は、特に、式(I)または式(I’)のデプシペプチドの促進されたかあるいは増加した生産に好適であり得る。
【0109】
4.異種遺伝子発現における鉄制御プロモーターの使用
本発明の別の局面は、宿主細胞、例えばシュードモナス属宿主細胞、例えばシュードモナス・プチダにおける、目的の組換えタンパク質の異種遺伝子発現または合成に関する。例えば、特に組換えタンパク質発現が細菌の増殖を損ねうる場合、増殖の転換期までまたは宿主細胞が健康な集団密度もしくは最も適切な異種遺伝子発現の段階まで、異種遺伝子発現が阻害されるようにそれを制御する必要がある。本発明は、宿主細胞、例えばシュードモナス・プチダにおいて、Fur制御プロモーターによって異種遺伝子発現の制御に成功しうることを示す。かかるプロモーターの使用は、デプシペプチドの生合成遺伝子クラスターの異種発現のために、本明細書に記載されているが、本発明のFur制御プロモーターは、目的の組換えタンパク質の異種遺伝子発現または合成のために、当該技術分野においてより広い用途を有しうる。
【0110】
したがって本発明は、組換え宿主細胞、好ましくは細菌宿主細胞、例えばシュードモナス属、例えばシュードモナス・プチダにおける異種遺伝子発現を制御し、向上させる手段を提供する。
【0111】
ある態様において、本発明は、異種遺伝子発現または目的の組換えタンパク質の合成のための発現カセットに関する。かかる発現カセットは、鉄制御プロモーターと作動可能に連結した、目的の成熟組換えタンパク質をコードするオープンリーディングフレーム(以下、コード配列と称する)を少なくとも含むポリヌクレオチドである。
【0112】
「異種遺伝子発現」の文脈内で本明細書において使用するとき、用語「目的の組換えタンパク質」は、鉄制御プロモーターの制御下では天然には発現されないタンパク質を意味する。好ましい態様において、目的の組換えタンパク質は、酵素、治療用タンパク質(ホルモン、成長因子、抗凝固剤、レセプターアゴニストもしくはアンタゴニストまたはデコイレセプターを含むがこれらに限定されない)、抗体(診断用または治療用を含む)または別の標的結合足場、例えばフィブロネクチン由来のタンパク質、シングルドメイン抗体、一本鎖抗体、ナノボディー等であってよいが、これらに限定されない。
【0113】
発現カセットの文脈において本明細書において使用するとき、用語「作動可能に連結」は、プロモーターがタンパク質をコードするヌクレオチド配列の発現を制御するようにタンパク質をコードするヌクレオチド配列に連結しているプロモーターを含むポリヌクレオチド配列を意味する。
【0114】
本発明の発現カセットは、宿主細胞における目的の組換えタンパク質の好適な発現に必要な他の制御配列、例えば5’非翻訳領域、シグナルペプチド、ポリアデニレーション領域および/または他の3’非翻訳領域をさらに含んでいてよい。
【0115】
4.1 鉄制御プロモーターおよびFur制御プロモーター
ある具体的な態様において、本明細書の第4項に記載の本発明の発現カセットに使用できる鉄制御プロモーターは、培養培地中の鉄の利用能に応答して機能する鉄上流リプレッサー(Fur)またはFurリプレッサータンパク質のホモログから選択されるタンパク質によって部分的または完全に転写的に抑制される、いずれかの細菌プロモーターであり得る。それはさらに、Fur依存的および培養培地中の鉄の利用能に応答してコード配列の発現を制御するように、コード配列と作動可能に連結していてよいFurリプレッサー結合部位を含む、いずれかのプロモーターを含む。細菌Furリプレッサータンパク質の例は、当該技術分野において知られており、例えばCarpenter et al. (2009)に記載されている。
【0116】
本明細書において使用するとき、抑制条件下(すなわちリプレッサーもしくはリプレッサー刺激および/またはリプレッサー結合部位の存在下)でのプロモーター活性が、レポータージーンアッセイ、例えばlacZレポータージーンアッセイで測定したとき、非抑制条件下(すなわち、リプレッサーもしくはリプレッサー刺激および/またはリプレッサー結合部位の非存在下)でのプロモーター活性の少なくとも5倍低いとき、プロモーターは、外部の刺激もしくはシス−エレメントまたはリプレッサーに応答して抑制されている。
【0117】
Furリプレッサー結合部位は当該技術分野において既知であり、多くの細菌、例えば大腸菌、緑膿菌、ネズミチフス菌および枯草菌において発見されている(Carpenter et al. (2002))。他のFurリプレッサー結合部位は、配列番号64のFurリプレッサー結合部位コンセンサス配列とのホモロジーによって探索できる。好ましい態様において、Furリプレッサー結合部位は配列番号64〜68のいずれか1つからなる群から選択される。
【0118】
Fur制御プロモーターは当該技術分野において既知であり、多くの細菌、例えば大腸菌、緑膿菌、コレラ菌、ネズミチフス菌、枯草菌、ヘリコバクター・ピロリ(Helicobacter pylorii)、ヒト結核菌、ブラディリゾビウム・ジャポニクム(Bradyrhizobium japonicum)、リステリア菌(Listeria monocytogenes)、カンピロバクター・ジェジュニ(Campylobacter jejuni)、ストレプトマイセス・セリカラー(Streptomyces coelicolor)、ペスト菌および黄色ブドウ球菌において同定されている(Carpenter et al. (2002))。Fur制御プロモーターの例は、配列番号69〜71のいずれか1つを含むが、それらに限定されない。
【0119】
好ましい態様において、Fur制御プロモーターは、下記群から選択されるポリヌクレオチド配列である:
a) 配列番号69
b) 配列番号69と実質的に同じプロモーター活性を保持する配列番号69のフラグメント;
c) 配列番号69と少なくとも50%、60%、70%、80%、90%または95%の同一性を有する配列番号69の変異プロモーター。
【0120】
ある態様において、配列番号69のフラグメントは、配列番号65または配列番号66のFurリプレッサー結合部位の少なくとも1個および配列番号69の3’下流配列のいずれかを含むフラグメントである。
【0121】
ある態様において、かかる変異プロモーターは、配列番号65もしくは配列番号66と同一または配列番号65および配列番号66のFurリプレッサー結合部位における1、2、3、4または5個以上のヌクレオチドの変化を有する、Furリプレッサー結合部位を含む核酸であってよい。
【0122】
他の態様において、配列番号69の変異プロモーターは、配列番号69と実質的に同じ活性を保持する機能的変異体である。具体的な態様において、変異プロモーターは配列番号69と実質的に同じ活性を保持し、配列番号69と少なくとも50%同一であるが、配列番号65および配列番号66と同じかまたは配列番号65および配列番号66とアラインメントしたときそれぞれ1、2、3、4または5個以上のヌクレオチドの変化を有する2つのリプレッサー結合部位を含む機能的変異体である。
【0123】
プロモーターのプロモーター活性を測定し、配列番号69のプロモーター活性と比較するために、lacZレポータージーンアッセイのようないずれかの好適なレポータージーンアッセイを用いて、レポータージーン発現を直接、例えばmRNAレベルを測定して、あるいは間接的に、レポーター酵素活性(例えばβガラクトシダーゼ活性)を抑制および非抑制条件下で測定することがでる。試験プロモーターと配列番号66のプロモーターの間で抑制および非抑制条件下でのかかる活性に顕著な違いがない場合、該試験プロモーターは、配列番号69と実質的に同じプロモーター活性を保持すると言える。
【0124】
4.2 鉄制御プロモーターを有する発現カセットを含む発現ベクターおよび組換え宿主細胞
発現カセットを任意の好適な発現ベクターに挿入できる。Fur制御プロモーターを用いた目的の組換えタンパク質の合成の文脈において、発現ベクターは、核酸を宿主細胞に導入して目的の組換えタンパク質をコードする遺伝子の異種発現を引き起こすことのできるビークルを意味する。
【0125】
それは例えばプラスミド、バクテリオファージまたはコスミドもしくは他の人工染色体、あるいは宿主細胞における組換えタンパク質生産に一般に用いられる他のベクターに由来してよい。かかる発現ベクターは、さらに、発現カセットに加えて、宿主細胞に入るための手段および/または宿主細胞において複製するための手段、および/またはポリペプチドを細胞の表面または細胞の外側に分泌するための手段を含む。発現ベクターは、1種以上の細胞タイプ、例えば少なくとも2種の細胞タイプ、1種の原核細胞タイプおよび1種の真核細胞タイプにおいて複製されるかまたは増殖されるための手段を含んでいてもよい。
【0126】
使用できる具体的な細菌ベクターは、周知のクローニングベクターの遺伝的要素を含む商業的に入手可能なプラスミド、例えばpBR322 (ATCC 37017)、pKK223-3 (Pharmacia Fine Chemicals, Uppsala, Sweden)、pGEM1 (Promega Biotec, Madison, WI, USA) pQE70、pQE60、pQE-9 (Qiagen)、pD10、phiX174、pBluescript(商標) II KS、pNH8A、pNH16a、pNH18A、pNH46A (Stratagene)、ptrc99a、pKK223-3、pKK233-3、pDR540、pRIT5 (Pharmacia)、pKK232-8 および pCM7を含む。具体的な真核ベクターは、pSV2CAT、pOG44、pXT1、pSG (Stratagene) pSVK3、pBPV、pMSGおよびpSVL (Pharmacia)を含む。しかし、あらゆる他のベクターが、宿主細胞において複製可能であり安定である限り、使用可能である。
【0127】
エレクトロポレーション形質転換、トランスフェクション、形質導入、ウイルス感染、遺伝子銃またはTi介在遺伝子導入を含む多様な技術のいずれかを用いて、発現ベクターを宿主細胞に導入してよい。適切であるとき、操作された宿主細胞は、プロモーターを活性化させ、形質転換体を選択しまたは目的の組換えタンパク質をコードする遺伝子を増幅するために適切なように修飾された、常套の栄養培地で培養できる。
【0128】
さらなる局面において、本発明の組換え宿主細胞は、目的の組換えタンパク質を発現できるかあるいは発現している。発明の宿主細胞を作成するために用いる様々な発現系の例の概略、例えば上記の具体的なものは、例えばGlorioso et al. (1999), Expression of Recombinant Genes in Eukaryotic Systems, Academic Press Inc., Burlington, USA、Paulina Balbas und Argelia Lorence (2004), Recombinant Gene Expression: Reviews and Protocols, Second Edition: Reviews and Protocols (Methods in Molecular Biology), Humana Press, USAに記載されている。
【0129】
本発明のヌクレオチド配列またはベクターによる宿主細胞の形質転換または遺伝子操作は、標準的な方法、例えばSambrook and Russell (2001), Molecular Cloning: A Laboratory Manual, CSH Press, Cold Spring Harbor, NY, USAに記載のとおりに行うことができる。さらに、本発明の宿主細胞は、特にpH値、温度、塩濃度、通気、抗生物質、ビタミン、微量元素等の観点から、使用する具体的な宿主細胞の要求を満たす栄養培地中で培養する。
【0130】
一般に、本発明の組換え宿主細胞は、本発明の発現カセットおよび/または発現ベクターを含む原核または真核細胞、または本発明の発現カセットおよび/または本発明の発現ベクターを含む細胞由来の細胞であってよい。
【0131】
したがって本発明は、適切な成長培地条件下での異種遺伝子発現または目的の組換えタンパク質の合成のための組換え宿主細胞であって、ゲノムに融合されているかまたは自律レプリコンとして、上記本発明の発現カセットまたは発現ベクターを含む、組換え宿主細胞に関する。
【0132】
「組換え宿主細胞」は、適切な成長培地条件下での目的の組換えタンパク質の異種発現に好適なあらゆる細胞であってよい。好ましくは、かかる組換え宿主細胞は、細菌細胞である。
【0133】
好ましい態様において、組換え宿主細胞は、上段に記載の鉄制御プロモーターと作動可能に連結した目的の成熟組換えタンパク質をコードするオープンリーディングフレームを含む発現ベクターで形質転換またはトランスフェクトされている細菌宿主細胞である。より具体的な態様において、組換え宿主細胞は、本発明の発現ベクターを含むシュードモナス属、例えばシュードモナス・プチダ、最も好ましくは、シュードモナス・プチダKT2440から選択され、ここで、鉄制御プロモーターは配列番号69〜71のいずれか1つまたはそのいずれかの機能的変異プロモーターからなる群から選択される。
【0134】
本発明はさらに、例えば目的の組換えタンパク質の合成における、異種遺伝子発現のための上記発現カセット、発現ベクターおよび/または組換え宿主細胞の使用に関する。
【0135】
4.3 異種遺伝子発現方法
鉄制御プロモーターを含む本発明の組換え宿主細胞は、有利には、異種遺伝子発現、例えば目的の組換えタンパク質の合成のために使用できる。好適な宿主細胞の形質転換および当該宿主細胞の適切な細胞密度への増殖の後、Fur制御プロモーターを適切な手段(例えばFeキレート剤、Fe飢餓)で抑制でき、当該細胞をさらなる期間培養して目的のタンパク質を生産させることができる。
【0136】
したがって、ある態様において、本発明は、宿主細胞、好ましくは細菌宿主細胞、頼好ましくはシュードモナス属における異種遺伝子発現または目的の組換えタンパク質の合成のための方法であって、a) 鉄制御プロモーターを含む発現カセットを含んである宿主細胞を、抑制条件下で培養し、
b) 適切な生産段階で鉄制御プロモーターを抑制解除するために増殖条件を変化させ、
c) 抑制解除条件下で細胞を増殖させて異種遺伝子を発現させ、そして/または目的の組換えタンパク質を合成すること
を含む方法を提供する。
【0137】
具体的な態様において、抑制条件は、成長培地中で十分な濃度の鉄を提供することで得られ、抑制解除条件は、鉄不足条件を作出することで得られる。係る条件は成長期の自然な使用および鉄飢餓によって達成されうる。あるいは、かかる条件は、鉄キレート剤を培地に加えて得られる。
【0138】
あらゆる好適な鉄キレート剤を、鉄制御プロモーターの抑制解除のために使用できる。かかる鉄キレート剤の例は、エチレンジアミンテトラ酢酸(EDTA)、クエン酸塩または鉄取り込みシデロホアとして作用する既知化合物(例えばデスフェリオキサミン、エンテロバクチンまたはバシリバクチン)を含むがこれらに限定されない。好ましい態様において、かかる鉄キレート剤は、2’2’ジピリジルである。キレート剤を培地に、例えば少なくとも成長培地中の鉄濃度と等しい、好ましくはそれの少なくとも3倍の濃度で、加えてよい。
【0139】
4.4 異種遺伝子発現のための鉄制御プロモーターの使用に関する本発明の具体的な態様
態様1:宿主細胞、好ましくは細菌宿主細胞、より好ましくはシュードモナス属宿主細胞における異種遺伝子発現に好適な発現カセットであって、天然には鉄制御プロモーターを制御しない遺伝子と作動可能に連結された鉄制御プロモーターを含む、発現カセット。
【0140】
態様2:鉄制御プロモーターが、鉄取り込み制御リプレッサータンパク質(Fur)からなる群から選択されるタンパク質によって抑制される細菌プロモーター、またはFurリプレッサータンパク質によって転写的に抑制されるいずれかの相同プロモーターである、態様1に記載の発現カセット。
【0141】
態様3:Furリプレッサータンパク質によって抑制されるプロモーターが、以下のものからなる群から選択されるポリヌクレオチド配列である、態様2に記載の発現カセット:
(a) 配列番号69、
(b) 配列番号69と実質的に同じプロモーター活性を保持する配列番号69のフラグメント、
(c) 配列番号69と実質的に同じプロモーター活性を保持する、配列番号69と少なくとも50%同一性を有するポリヌクレオチド配列。
【0142】
態様4:態様1〜3のいずれかの発現カセットを含む、組換え宿主細胞。
態様5:細菌種から選択される、態様4の組換え宿主細胞。
態様6:シュードモナス属、例えばシュードモナス・プチダから選択される、態様5の組換え宿主細胞。
【0143】
態様7:宿主細胞における目的の組換えタンパク質の合成のための、鉄制御プロモーターの使用。
態様8:鉄制御プロモーターが、鉄取り込み制御リプレッサータンパク質(Fur)からなる群から選択されるタンパク質によって抑制される細菌プロモーター、またはFurリプレッサータンパク質によって転写的に抑制されるいずれかの相同プロモーターである、態様7に記載の使用。
【0144】
態様9:Furリプレッサータンパク質によって抑制されるプロモーターが、以下のものからなる群から選択されるポリヌクレオチド配列である、態様7に記載の使用:
(a) 配列番号69、
(b) 配列番号69と実質的に同じプロモーター活性を保持する配列番号69のフラグメント、
(c) 配列番号69と実質的に同じプロモーター活性を保持する、配列番号69と少なくとも50%同一性を有するポリヌクレオチド配列。
【0145】
態様10:目的の組換えタンパク質の合成が成長培地中の鉄濃度を調節することによって制御される、態様7〜9のいずれか1つに記載の使用。
態様11:目的の組換えタンパク質の合成が細菌宿主細胞、好ましくはシュードモナス属、例えばシュードモナス・プチダで行われる、態様7〜10のいずれか1つに記載の使用。
態様12:目的の組換えタンパク質の合成が鉄をキレート化して鉄制御プロモーターを制御解除するのに十分な濃度で鉄キレート剤を培地に加えることによって誘導される、態様7〜11のいずれか1つに記載の使用。
態様13:鉄キレート剤が2’2’ジピリジルである、態様12に記載の使用。
【0146】
5.異種発現によって得られるデプシペプチドおよびその使用
本発明はさらに、上記方法で得られるかまたは得た式(I)または(I’)、例えば式(II)〜(VII)、(XI)〜(XIV)ならびに(XVII)および(XVIII)の化合物に関する。
【0147】
さらなる局面において、本発明は、上記方法で得られるかまたは得た式(I)または(I’)、例えば式(II)〜(VII)、(XI)〜(XIV)ならびに(XVII)および(XVIII)の化合物を含む医薬組成物に関する。
【0148】
医薬組成物は、個々の患者の臨床状態、医薬組成物の送達部位、投与方法、投与スケジュールおよび実施者に知られている他の要因を考慮して、良好な医療実務に一致するように、製剤され、投薬される。本明細書の目的での医薬組成物の「有効量」は、したがって、かかる検討事項によって決定される。
【0149】
当業者は個体に投与される医薬組成物の有効量が、特に、化合物の性質に依存することを理解している。例えば、化合物が(ポリ)ペプチドまたはタンパク質である場合、非経腸的に投与される医薬組成物の用量あたりの総薬学的有効量は、患者体重の約1μgのタンパク質/kg/日〜10mgのタンパク質/kg/日の範囲であるが、上記の通り、これは治療上の裁量の問題である。より好ましくは、この用量は少なくとも0.01mgのタンパク質/kg/日、例えばヒトについて、約0.01〜1mgのタンパク質/kg/日である。連続的に投与する場合、医薬組成物は典型的には、1日あたり1〜4回の注射または例えばミニポンプを用いた連続皮下輸液によって、投薬速度約1μg/kg/時〜約50μg/kg/時で投与する。静脈内バッグ溶液を用いてもよい。変化が観察されるのに必要な処置の長さおよび応答が生じるのに必要な処置後の感覚は、望まれる効果に依存して変化すると考えられる。具体的な量は、当業者に周知の常套の試験によって決定されうる。
【0150】
本発明の医薬組成物は、経口、非経腸、大槽内、腹腔内、局所(粉末、軟膏、ドロップまたは経皮パッチとして)、舌下または経口もしくは経鼻スプレーとして、投与してよい。
【0151】
本発明の医薬組成物は、好ましくは、薬学的に許容される担体を含む。「薬学的に許容される担体」とは、非毒性の固体、半固体または液体増量剤、希釈剤、カプセル化材またはあらゆるタイプの添加剤を意味する。用語「非経腸」は、本明細書において使用するとき、静脈内、筋肉内、腹腔内、胸骨内、皮下および関節内注射および輸液を含む投与形態を意味する。
【0152】
医薬組成物は好適には徐放系で投与する。徐放組成物の好適な例は、造形品の形態の半透ポリマー材料、例えばフィルムまたはマイクロカプセルを含む。徐放性材料は、ポリラクチド(米国特許第3,773,919号、EP 58,481)、L−グルタミン酸とγ−エチル−L−グルタミン酸エステルのコポリマー(Sidman, U. et al., Biopolymers 22:547-556 (1983))、ポリ(2−ヒドロキシエチルメタクリレート)(R. Langer et al., J. Biomed. Mater. Res. 15:167-277 (1981)、およびR. Langer, Chem. Tech. 12:98-105 (1982))、エチレンビニルアセテート(R. Langer et al., Id.)またはポリ−D−(−)−3−ヒドロキシ酪酸(EP 133,988)を含む。徐放医薬組成物は、リポソームに封入した化合物を含んでいてもよい。リポソーム含有医薬組成物は、自体公知の方法:E 3,218,121; Epstein et al., Proc. Natl. Acad. Sci. (USA) 82:3688-3692 (1985); Hwang et al., Proc. Natl. Acad. Sci. (USA) 77:4030-4034 (1980); EP 52,322; EP 36,676; EP 88,046; EP 143,949; EP 142,641; 日本特許出願第83-118008号; 米国特許第4,485,045号および第4,544,545号;ならびにEP 102,324によって製造される。通常は、リポソームは、約30モルパーセントのコレステロール以上の脂質含有量であり、選択された割合は最適な治療のために調節される、小さな(約200〜800オングストローム)単層型のものである。
【0153】
非経腸投与用医薬組成物は、一般に、所望の純度で薬学的に許容される担体と、すなわち使用する用量および濃度で受容者に非毒性であり製剤の他の成分と適合するものと、単位投与注射形態(溶液、懸濁液またはエマルジョン)に混合して製剤する。
【0154】
一般に、医薬組成物の成分を液体担体または微粉化担体またはその両方と均一かつ密接に接触させて、製剤を製造する。次いで、所望により、製品を所望の剤形に造形する。好ましくは、担体は非経腸担体、より好ましくは受容者の血液と等張な溶液である。かかる担体ビークルの例は、水、食塩水、リンゲル液およびデキストロース溶液を含む。非水性ビークル、例えば固定油およびオレイン酸エチルならびにリポソームもここで有用である。担体は好適には、微量の添加物、例えば等張性および化学的安定性を向上させる物質を含む。かかる物質は使用する用量および濃度で受容者に非毒性であり、バッファー、例えばリン酸塩、クエン酸塩、コハク酸塩、酢酸および他の有機酸またはそれらの塩;抗酸化剤、例えばアスコルビン酸;低分子量(約10残基未満)(ポリ)ペプチド、例えばポリアルギニンまたはトリペプチド;タンパク質、例えば血清アルブミン、ゼラチンまたは免疫グロブリン;親水性ポリマー、例えばポリビニルピロリドン;アミノ酸、例えばグリシン、グルタミン酸、アスパラギン酸またはアルギニン;単糖類、二糖類および他の炭水化物、例えばセルロールまたはその誘導体、グルコース、マンノースまたはデキストリン;キレート剤、例えばEDTA;糖アルコール、例えばマンニトールまたはソルビトール;対イオン、例えばナトリウム;および/または非イオン性界面活性剤、例えばポリソルベート、ポロキサマーまたはPEGを含む。
【0155】
治療投与に使用する医薬組成物の成分は滅菌されていなければならない。滅菌は、滅菌濾過膜(例えば0.2ミクロンの膜)による濾過によって容易に達成される。医薬組成物の治療用成分は、一般に、滅菌アクセスポートを有する容器、例えば静脈内溶液バッグまたは皮下注射針により貫通可能なストッパーを有するバイアルに入れる。
【0156】
通常、医薬組成物の成分は、単位または複数投与容器、例えば密封アンプルまたはバイアルに、水溶液または再構成用凍結乾燥製剤として保存される。凍結乾燥製剤の例として、10mlバイアルを滅菌濾過した1%(w/v)水溶液5mlで満たし、得られた混合物を凍結乾燥させる。輸液溶液は、静菌注射用水を用いて、凍結乾燥化合物を再構成して製造する。
【0157】
本発明はまた、上記デプシペプチドまたはその誘導体の、医薬としての使用に関する。例えばがん、特に卵巣がんの処置または炎症性および/または過増殖性および掻痒性皮膚疾患、例えばケロイド、肥厚性瘢痕、ざ瘡、アトピー性皮膚炎、乾癬、膿疱性乾癬、酒さ、ネザートン症候群または他の掻痒性皮膚疾患、例えば結節性痒疹、老人の不特定の痒み、ならびに上皮バリアー機能不全に関連した他の疾患、例えば加齢性皮膚、炎症性腸疾患およびクローン病、ならびに膵炎またはがん、特に卵巣がん、嚢胞性線維症(CF)、慢性閉塞性肺疾患(COPD)、肺線維症、成人呼吸窮迫症候群、慢性気管支炎、遺伝性肺気腫、リウマチ性関節炎、IBD、乾癬、喘息のため。
【0158】
ある態様において、本発明は、炎症性および/または過増殖性および掻痒性皮膚疾患、例えばケロイド、肥厚性瘢痕、ざ瘡、アトピー性皮膚炎、乾癬、膿疱性乾癬、酒さ、ネザートン症候群または他の掻痒性皮膚疾患、例えば結節性痒疹、老人の不特定の痒み、ならびに上皮バリアー機能不全に関連した他の疾患、例えば加齢性皮膚、炎症性腸疾患およびクローン病、ならびに膵炎またはがん、特に卵巣がんの処置用医薬としての、上記デプシペプチドまたはその誘導体の使用に関する。
【0159】
他の態様において、本発明は、嚢胞性線維症(CF)、慢性閉塞性肺疾患(COPD)、肺線維症、成人呼吸窮迫症候群、慢性気管支炎、遺伝性肺気腫、リウマチ性関節炎、IBD、乾癬、喘息の処置用医薬としての、上記デプシペプチドまたはその誘導体の使用に関する。
【0160】
さらに別の態様において、本発明は、炎症性および/または過増殖性および掻痒性皮膚疾患、例えばケロイド、肥厚性瘢痕、ざ瘡、アトピー性皮膚炎、乾癬、膿疱性乾癬、酒さ、ネザートン症候群または他の掻痒性皮膚疾患、例えば結節性痒疹、老人の不特定の痒みの処置用医薬としての、上記デプシペプチドまたはその誘導体の使用に関する。
【0161】
6.本発明のポリペプチドに対する抗体
具体的な態様において、本発明は、本明細書に記載され、定義されている本発明のポリペプチドまたはそのフラグメントに特異的に結合する抗体およびその使用に関する。さらに、かかる抗体は、当該ポリペプチド、特に非リポソームペプチドおよび/または非リポソームペプチドシンターゼ(NRPS)の精製に使用できる。用語「抗体」は、当該技術分野において周知である。
【0162】
本発明の文脈において、用語「抗体」は、本明細書において使用するとき、特に、完全な免疫グロブリン分子および実質的に結合特異性を保持しているかかる免疫グロブリン分子の一部を意味する。さらに、この用語は、キメラおよびヒト化抗体のような修飾および/または改変抗体分子、組換えまたは合成により作成/合成した抗体およびインタクトな抗体、ならびに分離した軽鎖および重鎖、Fab、Fab/c、Fv、Fab’、F(ab’)2のようなその抗体フラグメントを意味する。用語「抗体」はまた、二重機能後退、三重機能抗体および一本鎖Fvs(scFv)または抗体融合タンパク質のような抗体構築物も含む。
【0163】
抗体を作成する技術は当該技術分野において周知であり、例えばHoward and Bethell (2000) Basic Methods in Antibody Production and Characterization, Crc. Pr. Inc.に記載されている。本発明のポリペプチドに対する抗体は、例えばポリペプチド(またはそのフラグメント)を動物に直接注射して、あるいはポリペプチド(またはそのフラグメント)を動物に投与して得ることができる。そのようにして得られた抗体は、ポリペプチド(またはそのフラグメント)それ自体に結合する。この方法において、結合が上記定義の「特異的」である限り、ポリペプチドのフラグメントを用いて全ポリペプチドに結合する抗体を作成することすらできる。
【0164】
本発明の文脈において特に好ましいものは、モノクローナル抗体である。連続細胞系培養によって作成した抗体を提供するモノクローナル抗体の作製のためのあらゆる技術を使用してよい。かかる技術の例は、ヒトモノクローナル抗体を作成するためのハイブリドーマ技術、トリオーマ技術、ヒトB細胞ハイブリドーマ技術およびEBVハイブリドーマ技術を含む(Shepherd and Dean (2000), Monoclonal Antibodies: A Practical Approach, Oxford University Press、Goding and Goding (1996), Monoclonal Antibodies: Principles and Practice - Production and Application of Monoclonal Antibodies in Cell Biology, Biochemistry and Immunology, Academic Pr Inc, USA)。
【0165】
抗体誘導体はまた、ペプチド模倣によって作成してもよい。さらに、一本鎖抗体の作製について記載されている技術(特に米国特許第4,946,778号参照)は、本発明のポリペプチドを特異的に認識する一本鎖抗体を作成するために適用できる。また、トランスジェニック動物を用いて、本発明のポリペプチドに対するヒト化抗体を発現させてもよい。
【0166】
用語「特異的に結合」は、本明細書において使用するとき、タンパク質および他の生物製剤の異種集団の存在下での非リボソームペプチドおよび/または非リボソームペプチドシンターゼ(NRPS)および抗体の存在を決定づける結合反応を意味する。したがって、指定されたアッセイ条件下では、特異的抗体とポリペプチドは互いに結合するが、サンプル中に存在する他の成分とは顕著な量で結合しない。かかる条件下での標的分析物との特異的結合は、特定の標的分析物に対する特異性について選択される結合部分を必要としうる。多様なイムノアッセイフォーマットを用いて、特定の抗原と特異的に反応する抗体を選択できる。例えば、固相ELISAイムノアッセイを通常通りに用いて、分析物と特異的に免疫応答するモノクローナル抗体を選択する。イムノアッセイフォーマットおよび特異的免疫応答を決定するために使用できる条件の説明については、Shepherd and Dean (2000), Monoclonal Antibodies: A Practical Approach, Oxford University Press および/または Howard and Bethell (2000) Basic Methods in Antibody Production and Characterization, Crc. Pr. Inc.を参照されたい。典型的には、特異的または選択的応答は、ノイズよるバックグラウンドシグナルの少なくとも2倍、より典型的にはバックグラウンドの10〜100倍以上である。
【0167】
用語「精製」は、本明細書において使用するとき、複数の混合物から1つのタイプのタンパク質を単離することを意図する一連のプロセスを意味する。タンパク質精製は目的のタンパク質の機能、構造および相互作用の特徴付けに重要である。出発物質は、非限定的な例として、生物学的組織または微生物培養であってよい。精製プロセスの多様な工程が、それを制限するマトリックスからタンパク質を開放し、タンパク質と混合物の非タンパク質部分とを分離し、そして最終的に所望のタンパク質を他の全てのタンパク質から分離することができる。分離工程は、タンパク質のサイズ、物理化学的特性および結合親和性の違いを利用する。
【0168】
以下の非限定的な図、配列および実施例を参照して、本発明をさらに説明する。
【図面の簡単な説明】
【0169】
【図1】本発明のNRPSクラスターから生合成される、コンドロマイセスNPH−MB180によって生産された確認された構造の一覧を示す。
【図2】式(I)または(I’)の化合物をコードするNRPS生合成遺伝子クラスターのドメイン構造を示し、式(II)、(III)、(VI)および(VII)〜(XVII)の化合物の生合成経路案を例示する。L、ローディングドメイン;AQ、アデニレーションドメイン(Gln);T、チオレーションドメイン;C、コンデンセーションドメイン;NM、N−メチレーションドメイン;TE、チオエステラーゼドメイン、APアデニレーションドメイン(Pro);AT、アデニレーションドメイン(Thr);AL、アデニレーションドメイン(Leu);AE、アデニレーションドメイン(Glu);AI、アデニレーションドメイン(Ile);AY、アデニレーションドメイン(Tyr)。
【図3】NRPSセグメントF 10517242における2個のアデニレーションドメインの結合ポケットとそれらの最もマッチする定義されたアデニレーションドメインを並べた、10アミノ酸残基のアラインメントを示す。
【図4】コンドロマイセスNPH−MB180に対するコンドロマイドN−メチレーションドメインのBLASTpアラインメントからの結果を示し、これは、NRPSセグメントF 10517242に位置するN−メチレーションドメインを明らかにする。N−メチレーションドメインモチーフは太字で示す。
【図5】ahp残基を形成するヒドロキシプロリンを含む化合物の推定相互変換を示す。水性条件下では、式(XVIII)で例示されるヒドロキシプロリンと、式(II)で例示されるahp含有化合物間との間で、平衡が存在する。
【図6】P.プチダ KT2440の異種発現培養抽出物のLC−MS分析による式(II)の化合物の検出。HPLCクロマトグラムはMSによって陽イオン(左パネル)および陰イオン(右パネル)検出を示す:A)式(II)参照化合物;B)6日目 LB_D培地;C)6日目 P.プチダ陰性対照。MSスペクトル:D)Aに示すHPLC実行からの式(II)参照化合物;E)6日目 LB_D培地、Bに示すHPLC実行から3.2分でピーク。
【発明を実施するための形態】
【0170】
本発明は以下のヌクレオチドおよびアミノ酸配列に関する:
配列番号1は、Val/Ileコンデンセーションドメインを示す、ドメイン1のアミノ酸配列をコードするヌクレオチド配列を示す。
配列番号2は、Val/Ileコンデンセーションドメインを示す、ドメイン1のアミノ酸配列を示す。
配列番号3は、Val/Ileアデニレーションドメインを示す、ドメイン2のアミノ酸配列をコードするヌクレオチド配列を示す。
配列番号4は、Val/Ileアデニレーションドメインを示す、ドメイン2のアミノ酸配列を示す。
配列番号5は、Val/Ileチオレーションドメインを示す、ドメイン3のアミノ酸配列をコードするヌクレオチド配列を示す。
【0171】
配列番号6は、Val/Ileチオレーションドメインを示す、ドメイン3のアミノ酸配列を示す。
配列番号7は、Tyrコンデンセーションドメインを示す、ドメイン4のアミノ酸配列をコードするヌクレオチド配列を示す。
配列番号8は、Tyrコンデンセーションドメインを示す、ドメイン4のアミノ酸配列を示す。
配列番号9は、Tyrアデニレーションドメインを示す、ドメイン5のアミノ酸配列をコードするヌクレオチド配列を示す。
配列番号10は、Tyrアデニレーションドメインを示す、ドメイン5のアミノ酸配列を示す。
【0172】
配列番号11は、Tyr 6−N−メチレーションドメインを示す、ドメイン6のアミノ酸配列をコードするヌクレオチド配列を示す。
配列番号12は、Tyr 6−N−メチレーション ドメインを示す、ドメイン6のアミノ酸配列を示す。
配列番号13は、Tyrチオレーションドメインを示す、ドメイン7のアミノ酸配列をコードするヌクレオチド配列を示す。
配列番号14は、Tyrチオレーションドメインを示す、ドメイン7のアミノ酸配列を示す。
配列番号15は、それぞれVal/IleおよびTyrのアデニレーションドメイン、コンデンセーションドメインおよびチオレーションドメインならびにTyr 6−N−メチレーションドメインを含むNRPSフラグメントをコードするヌクレオチド配列を示す。
【0173】
本明細書に記載のヌクレオチドおよびアミノ酸配列の機能および推定される役割は、上記表1および下記実施例においてさらに説明される。
【実施例】
【0174】
実施例
下記実施例は本発明を説明する:
【0175】
実施例1:NPH−MB180のゲノム配列;会合および分析。
454配列決定法(ピロホスフェートを利用した配列決定プラットフォーム)を用いてNPH−MB180の完全なゲノムを配列決定して、「ドラフト」配列を作成した。ショットガン配列決定を1回行い、次に対末端配列決定を2回行った。対末端試験は、より伝統的なショットガン法の補完的技術として用いられる。簡潔に述べると、物理的に断片化され、環化された染色体DNAフラグメントを短いDNAアダプターセクションにライゲートする配列決定法である。これにより、互いに約3kb(断片化された環化DNAの平均サイズ)離れている2つの短いリード(約150〜200bp)を与えるアダプターからの配列決定を多岐にわたって行うことができる。2つの別個のコンティグにおける2つの短いリードのオーバーラップ(ホモロジー)によって、非オーバーラップコンティグを一体に連結することができ、未定義ヌクレオチド(N)の進展によって3kb近似に基づいて見積もられたおよその長さと連結することができる。このようにして連結されたコンティグを足場と呼ぶ。全体で、1,295,834の個々のリードを実行して、配列決定された310,674,400塩基が得られた。平均リード長は、このタイプの配列決定法の標準である239塩基であった。これらのリードを集積して、リード間の配列オーバーラップに基づきコンティグを形成した。この努力により15,449,316塩基についての平均コンティグ長8,931 bpのコンティグ4,038個が得られた。対末端試験の使用は、足場を作成するためにオーバーラップし、15,029,556塩基を含む96個の足場の集積をもたらす。平均足場サイズは1,227,671塩基であり、平均足場サイズは156,557塩基であった。
【0176】
式(I)または(I’)のデプシペプチドの生合成に関与するNRPS遺伝子クラスターを同定するために、ゲノムデータを分析した。全体的なアプローチは、サーチクエリーとしてNRPSドメインを用いて96個の足場に対してBLASTサーチ(Altschul et. al. 1990; Gish, W. & States, D.J. 1993)を用いることであった。信頼されるNRPSドメインは、どのアミノ酸が非リボソームペプチドに導入されるかが特異的であり、したがって特定のNRPSクラスターを同定するための良好なマーカーであるため、アデニレーションドメインであった(Marahiel, M.A. et. al. 1997)。一般に、以下の特異性および相対順序:Gln-Thr-Val-Glu-Ile-Tyr+(N-meth.)-Ileを有するその構造アデニレーションドメイン内に含まれるNRPSクラスターを発見すると予測された。さらに、遺伝子クラスターは、イソ酪酸のようなカルボン酸で生合成を開始することができるローディングドメインによって開始すると予測され、さらにクラスターがチオエステラーゼドメインで終わると推測された。また、グルタミン酸残基の酸化を促進して3−アミノ−6−ヒドロキシ−ピペリドン残基(Ahp)を形成する他の生合成ユニットが存在している可能性があった。これらのアクセサリー遺伝子の相対位置は、このクラスター内に存在するならば、予測できなかった。さらに、この領域に存在する1個以上の転写産物を定義する転写開始および終了の位置は、この段階では予測できなかった。
【0177】
実施例2:BLAST分析によるNPH−MB180ゲノム配列における全NRPSアデニレーションドメインの同定。
NRPSクラスターを同定するために頼ったアプローチは、まずNPH−MB180ゲノムの全NRPSアデニレーションドメインを同定することであった。NRPSアデニレーションドメインは、それらが使用するアミノ酸について特異的であり、したがってこれらのドメインを分析して、式(I)または(I’)のデプシペプチドを構成するアミノ酸の含有量および相対順序に基づいて正確なNRPSクラスターを同定した。このような目的で、ゲノム配列データにおける全ての可能性のあるNRPSクラスターを同定するための一般アデニレーションドメインの例として、シクロスポリンのバリンアデニレーションドメインを用いた。これは、アミノ酸配列のホモロジーによって全てのNRPSアデニレーションドメインを同定するため、ゲノムのtBLASTn(Altschul et. al. 1990; Gish, W. & States, D.J. 1993)分析を行って達成した。このアプローチにより14個の可能性のあるNRPSクラスター(表2)を同定し、これらのクラスターを含む足場オリジナルBLASTヒット(例えばA 12171827)の開始のヌクレオチド数と共にA−N標識した。このリストから各アデニレーションドメインを同定し、ドメイン特異性を定義する保存アミノ酸残基の分析によって、各ドメインの特異性を決定した(詳細は実施例3を参照)。
【0178】
【表4】
【0179】
この第一の分析は、正確なアデニレーションドメイン組成および生合成経路をコードする予測されたクラスター(〜30kb)の全体のサイズを有するいずれかのNRPSクラスターを同定することができなかった。実際、我々の目的の経路に見られると予測した7個のアデニレーションドメインを含むNRPS経路は同定されなかった。しかし、F 10517242 がイソロイシンおよびチロシンアデニレーションドメイン(表2)を含むことを見出した。なお、この足場(足場#72)は極めて短い(〜7.4kb)が、これは目的のNRPSクラスターの一部であり、クラスターの残りの部分(配列決定ギャップ領域の残基)が配列決定されないままであると仮定した。チロシンアデニレーションドメインと部分Tドメインの間に存在するN−メチレーションドメインの発見は、この仮定にさらなるサポートを与えた(詳細は実施例4参照)。
【0180】
これらのデータに基づいて、ゲノム配列は生合成遺伝子クラスターの全体を含んでいないと結論づけた。実際、5’方向の約20kbおよび3’方向の6kbは不明であると予測できる。
【0181】
実施例3:NRPSアデニレーションドメイン特異性の予測
本明細書に記載のアデニレーションドメインの特異性を、下記一般プロトコルを用いて予測する。シクロスポリンシンターゼ(CssA)のバリンアデニレーションドメインのアミノ酸配列を目的のコンドロマイセスゲノムDNAに対してアラインメントするtBLASTn(Altschul et. al. 1990; Gish, W. & States, D.J. 1993)サーチを用いて、アデニレーションドメインを同定した。ClustalX 複数配列アラインメントソフトウェア(Higgins et. al. 1996)を用いて、Marahiel et. al. (1997)により定義された2つのコアモチーフ(A3およびA6)間のアミノ酸レベルで、翻訳されたコンドロマイセスアデニレーションドメインをGrsA(PheA)(グラミシジンSシンテターゼ)に対してアラインメントした。アデニレーションドメインの結合ポケットを定義し、したがってアミノ酸特異性を決定づける、Marahiel et al.によって報告された10個のアミノ酸を、このアラインメントで同定した。次いでRausch et. al. (2005) および Stachelhaus et. al. (1999)によって報告されたデータを用いて、この10個のアミノ酸を定義されたアデニレーションドメインアミノ酸コードと比較した。
【0182】
生合成クラスターのセグメントにおいて同定された2個のアデニレーションドメインが、イソロイシンおよびチロシンの結合ポケットを定義する10個のアミノ酸と高い相同性を示した(図3)。これらのアミノ酸特異性は絶対ではなく、同様の化学的特徴を有するアミノ酸がしばしば該ドメインを定義するアミノ酸に代えて使用される。これは、1個のNRPSオペロンに沿って合成される構造の可変性に寄与する。本件の場合、イソロイシンアデニレーションドメインはその結合ポケットにバリンも受け入れると推測され、特徴は他の「イソロイシン」アデニレーションドメインについても見られる(Rausch et. al. (2005))。実際、利用可能なNRPS予測ツール(例えば、http://www-ab.informatik.uni-tuebingen.de/software/NRPSpredictor)は一般に、アデニレーションドメインをイソロイシン特異的ともバリン特異的とも宣言することができない。
【0183】
実施例4:NRPSN−メチレーションドメインの予測
N−メチレーションドメインの存在は、下記アプローチを用いて、チロシンアデニレーションドメインと3’方向に直接隣接して位置していると予測された。コンドロマイセス・クロカツスNPH−MB180 コンドロマイドNRPSクラスターのN−メチレーションドメインのアミノ酸配列を用いて、tBLASTn(Altschul et. al. 1990; Gish, W. & States, D.J. 1993)を用いて同様のドメインのゲノムをサーチした。このアプローチを用いて、N−メチレーションドメインはNRPSセグメント内で期待値5e-43 および46%アミノ酸配列同一性を有することが同定された(図4)。さらに、このNRPSセグメントからのN−メチレーションドメインは機能的N−メチレーションドメインにおいて共通して見出される予測アミノ酸モチーフを有することを見出した(von Dohren, H. et. al. 1997; Marahiel, M.A. et. al. 1997)。このデータを確認するため、アナベナ株90アナベノペプチリド生合成クラスター由来のN−メチルトランスフェラーゼApsy−6(Rouhiainen et. al. 2000)を、上記のN−メチルトランスフェラーゼと比較した。この比較のBLASTpの結果は、これらのドメインが期待値2e-65で極めて類似していることを示し、したがって、このドメインの最初の同定が確認された。このドメインがチロシンアデニレーションドメインに直接隣接して存在していることは、NRPS遺伝子クラスターの予測された構造と一致する。さらに、N−メチレーションドメインは比較的珍しく、したがってNRPSセグメント内のこのドメインの存在は、NRPSクラスターに属するこのセグメントについて強い証拠を提供する。
【0184】
実施例5:全生合成NRPS遺伝子クラスターの同定
式(I’)のデプシペプチドの生産に関与する完全な非リボソームペプチド生合成遺伝子を同定し、特徴付けた。足場D 942267に挿入した足場F 10517242からなる足場に、生合成遺伝子を構築した(表1)。足場F 10517242に直接隣接するヌクレオチドの配列分析によって、元のゲノムアセンブリへのこの挿入調節が保証されることが示された後に、これらの足場の組合せを行った。この構築物をPCRで確認し、次いで足場連結領域を介してDNA配列決定した。この足場は式(I’)のデプシペプチドの生合成、修飾および細胞外輸送に関与している可能性がある8個の近接オープンリーディングフレームを含む。さらに、可能性のある分泌型プロテアーゼは、最終的にはデプシペプチドの天然の細胞標的であり得るこれらのオープンリーディングフレーム内に位置し、プロテアーゼ阻害剤であることが示された。これらのORFおよび対応するNRPSドメインの配置は、図2に示す。
【0185】
核非リボソームペプチドオープンリーディングフレーム(ORF6およびORF7)の直前に5個のORFが存在する。ORF1およびORF2は、ソランギウム・セルロサム(Sorangium cellulosum)から報告された2個の異なる特徴付けられていないタンパク質とそれぞれ類似する。これらのタンパク質は仮説機能を有さないが、重要なことに、それらはポリアンギウム科(Polyangiaceae)においてのみ見られるようである。さらに、ORF2と類似するソランギウム(Sorangium)タンパク質は、S. cellulosumゲノムにおいて少なくとも5回発見されている。これらのタンパク質は、ほぼ完全なヌクレオチド配列連結性に基づいて、ORF3と共に転写されると考えられる。ORF3はセリンプロテアーゼ、特にスブチリシン群に属するものと高い配列ホモロジーを有する。我々は、デプシペプチドが高度に特異的なセリンプロテアーゼ阻害剤であることを生化学的に決定し、したがって、デプシペプチドがORF3セリンプロテアーゼの阻害剤であることは真実味がある。逆に、ORF3は、コンドロマイセス株へのデプシペプチド耐性付与に関連しているかもしれない。ORF4およびORF5は、ABCトランスレポータータイプのシデロフォアパーミアーゼおよび一般的な環状ペプチドパーミアーゼと類似する。このパーミアーゼ系は細胞膜を通過するデプシペプチドの排出に関与している可能性がある。実際、これらのORFの5つ全てが細胞膜転位プロセスに関与しており、実際のプロテアーゼとしてそれはプロテアーゼ阻害剤に結合するため、「セリンプロテアーゼ様」ORF3はセリンプロテアーゼファミリーとだけ類似性を有する可能性がある。
【0186】
コアデプシペプチド生合成クラスターはORF6に属し、ORF7へと続く。これらの2個のORFは合わせて15kb長以上である。全NRPS生合成クラスター同様、それらはコンデンセーションドメイン、次いでアデニレーションドメイン、次いでチオレーションドメインからなる一般トポロジーを有する機能的ドメインへと崩壊しうる(Marahiel et. al. 1997)。この3個のドメインモジュールは、通常、ペプチドに取り込まれる各アミノ酸につき、NRPSクラスターで複数回反復される。デプシペプチド生合成クラスターは、ペプチドコアに含まれる7個のアミノ酸からなる7個のかかるモジュラー反復を有するこのパターンに従う。アデニレーションドメインは成長ペプチドにアミノ酸特異性を与え、それらが受け入れ、次いで取り込むアミノ酸を同定するために分析できる。
【0187】
ORF6および7に存在する7個のアデニレーションドメインの予測されるアミノ酸特異性は、1つの例外を有するが、デプシペプチドの最終構造と一般に一致する。第4アデニレーションドメイン(ドメイン 7.3)は、プロリンを成長ペプチドにこの位置で受け入れ、取り込むが、最終ペプチドは非標準アミノ酸、3−アミノ−6−ヒドロキシピペリドン(ahp)をこの位置で含むと予測される。ahpはアナベナ株90によって生産される関連アナベナペプトリドを含む様々なデプシペプチドに存在する(Rouhiainen et. al. 2000)。ahp形成は、アナベナペプトリドにおいて、鎖の4位にグルタミンが取り込まれた後、前のアミノ酸のアミンに戻る反応を起こしてahpを形成することで生じると主張されている(Rouhiainen et. al. 2000)。しかし、ahp特異的アデニレーションドメインは文献にも記載されている(Rausch et al. 2005)。式(II)〜(VII)、(XI)〜(XIII)および(XVII)のMB180株から単離したahp含有デプシペプチドにおいて、我々は、最初にプロリンが成長ペプチドに4位において取り込まれ、次いでオキシドレダクターゼの作用によりahpが形成されるというahp形成の新たなプロセスを仮定する。実際、驚くべきことに、チトクロムP450遺伝子(ORF8)がデプシペプチド生合成クラスターにおいて見出されており、それはNRPS生合成クラスターの直後に位置しており、プロリン残基をヒドロキシル化する変換を触媒している可能性がある。
【0188】
重要なことに、この位置でプロリンを含むデプシペプチドアナログがMB180株から単離されている(式(XIV))。また、数日間水性環境中でインキュベーションすると、ahp含有デプシペプチド(例えば式(II))から自然に5−ヒドロキシプロリンを有するアナログ(式(XVIII))が形成されることも示された。この5−ヒドロキシプロリン形態とahp形態の相互変換が可逆性であることも我々は示した。他のデプシペプチドもこの戦略に従うかは不明であるが、これが我々のMB180株によって使用されるahp形成戦略である可能性がある。
【0189】
デプシペプチド生合成クラスターは、スターターユニットで生合成を開始するローディングドメインを有するORF6で開始する。式(I’)のデプシペプチドにおけるX残基の構造バリエーションに基づき、スターターユニットとして、カルボン酸、例えばCH3CH2CH(CH3)COOH、(CH3)2CHCOOH、C6H5COOH、CH3S(O)CH2COOHまたはCH3COOHが想定できる。
【0190】
非リボソープペプチドにとって小さな酸性残基で開始することは一般的であるが、残基の選択はペプチド毎に大きく異なる。しかし、複雑なカルボン酸スターターユニットは、非リボソープペプチドの間でも比較的稀である。デプシペプチド生合成を開始するために用いるローディングドメインは、構造においても使用するスターターユニットにおいてもアナベナペプトリドローディングドメインと異なっている。実際、デプシペプチドローディングは、標準コンデンセーションドメインと極めて密接に関連しているが、アナベナペプトリドのホルミル基ローディングドメインは前述のホルミルトランスフェラーゼと極めて類似している(Rouhiainen et. al. 2000)。カルボン酸スターターユニットがドメイン6.2によって特定されるグルタミンアミノ酸のアルファアミノ基に濃縮された後、鎖が1回に1アミノ酸成長し続け、次いでNRPS生合成装置を通る(図2)。
【0191】
デプシペプチド生合成装置は、ペプチド結合の2級アミンをメチル化する比較的稀なメチルトランスフェラーゼドメイン(ドメイン7.10)に遭遇するまで、1回に1アミノ酸、狂いなく、単純なNRPSペプチドから、ペプチドを合成する。この場合、これはチロシン由来アミノ基に3級アミンをもたらす。おそらく、このメチル化は、チロシンが成長ペプチドに加えられた後であるが、次のおよび最終アミノ酸が加えられる前に生じる。このことは、N−メチラーゼドメインがチロシン特異的アデニレーションドメインの直後に位置することによって強く示唆されている。
【0192】
最後に、ペプチドが最終チオレーションドメインから除去され、スレオニンのアルコールと末端イソロイシンのアルファケト基の間でエステル結合が形成されて環化する。これは、ORF7に位置する最終ドメインである標準チオエステラーゼドメイン(ドメイン7.15)によって行われる。ahp形成がこのチオエステラーゼ段階の前後いずれに生じるのかは不明である。しかし、この生合成クラスターに含まれる遺伝子は、式(I’)のデプシペプチドの全構造を明らかにするのに十分である。
【0193】
実施例6:シュードモナス・プチダKT2440におけるデプシペプチドの異種発現
ここで、我々は、シュードモナス・プチダKT2440における式(I)または(I’)のデプシペプチドの異種発現を行うためのアプローチの一例を説明する。この宿主は天然生産株C. crocatusと比較して様々な利点、例えば速やかで予測可能な増殖、遺伝子ツールの利用可能性および大規模発酵におけるバリデートされた使用を有する。さらに、この宿主は、C. crocatusと同様のゲノムGC%を有し、天然NRPS系を有する;これら2つの特徴は、異種発現戦略を設計する際に重要な検討事項である。
【0194】
大きな挿入を受け入れることができるコスミドpWEB-TNC(Epicenter Biotechnologies, Madison WI, USA)に生合成遺伝子クラスターをクローン化した;本質は生合成遺伝子クラスターが30kb長を超えることである。生合成遺伝子クラスターのクローニングは、まず、約30〜40kbの直線DNAフラグメントを作成するために生合成遺伝子クラスターの境界の外側を切断する、適切な制限酵素を同定することによって行った。ゲノム配列データの分析により、この目的のために酵素XmnIが適切であり、完全ゲノムDNAフラグメント消化を行うとこのサイズ範囲の異なる15個のDNAフラグメントを作成することが示された。これらの15個のDNAフラグメントのうち、1個の39kbのフラグメントが生合成クラスターを含むと予測された。アガロースゲル電気泳動により、これらの15個のDNAフラグメントを他の染色体消化フラグメントから分離した。ガイドとして適切なサイズのDNA標準を用いて所望のサイズ範囲の15個のDNAフラグメントをゲルから切り出し、製造業者の指示書に従ってコスミドpWEB-TNCにクローン化した。完全な生合成クラスターを含むコスミドクローンは、コロニーPCRで同定し、DNA配列決定で確認した。コスミドまたはBACベクターを用いて完全ゲノムのランダムショットガンライブラリーを作成し、次いで放射線標識化プローブを用いたクローンライブラリーへのコロニーハイブリダイゼーションにより、目的の生合成クラスターを含むクローンライブラリーのメンバーを同定するという別のアプローチも存在する。
【0195】
クローン化した生合成経路を得た後、コスミドクローンに挿入して異種発現を成功させるためにはいくつかの遺伝子成分が必要であった。これらの成分は、i) 異種宿主への移入が成功したことを同定することができる選択マーカー、ii) 異種宿主において機能するプロモーター、iii) 異種宿主への染色体組込みのための部位および iv) pRK2013 oriT配列によって与えられるプラスミド接合伝達性機能(RK2トランスファー機能で使用するため)を含む。本実施例のためにシュードモナス・プチダKT2440に使用するため選択した選択マーカーは、ゲンタマイシン耐性カセットaacCI(Blondelet-Rouault et al. 1997)であった。他の選択マーカーは以下の薬剤耐性を与えるヌクレオチドカセットを含む:アンピシリン(例えばbla)、クロラムフェニコール(例えばcat)、カナマイシン(例えばaacC2、aadBまたは他のアミノグリコシド修飾酵素)またはテトラサイクリン(例えばtetAおよびtetB)。シュードモナス・プチダKT2440における異種発現を駆動するためのプロモーターとして、ここで、フマラーゼC−1(PP 0944)遺伝子プロモーターの使用を記載する(実施例8も参照)。転写プロモーターの選択は、上記抗生物質耐性決定因子のいずれかの転写プロモーターまたはシュードモナス・プチダKT2440において機能的であるいずれかの転写プロモーター、例えばシュードモナス・プチダKT2440ゲノムに存在する7個の16S rRNA遺伝子の転写プロモーター(PP 16SA、PP 16SB、PP 16SC、PP 16SD、PP 16SE、PP 16SF、PP 16SG)、シュードモナス・プチダKT2440鉄取り込みリプレッサー(Fur)制御遺伝子のいずれかの転写プロモーター(fagA (PP 0943)または他のfumC ホモログ、fumC-2 [PP 1755]のプロモーターを含む)、シデロフォアまたはシデロフォア様化合物の生合成および輸送に関与するプロモーター(pvdE [PP 4216]、fpvA [PP 4217]を含む)または遺伝子PP 4243もしくはPP 0946の転写プロモーターを含んでいてよいが、それらに限定されない。ここに記載のフマラーゼC−1プロモーターの使用を含むP. putida由来のプロモーターは、我々の戦略において、RecA介在染色体組込み事象を介してP. putida宿主に染色体組込みの部位を提供することによって、第二の目的に供される。効果的な染色体組込みを促進するために、1046 bpのプロモーター領域がコスミド構築物内に含まれた。下流生合成クラスター遺伝子に転写の促進を可能とするため、プロモーター要素は意図した挿入の3’末端に位置した。プラスミド接合はpSET152からのori Tヌクレオチド配列の取り込みによって促進された。oriT配列は、RK2転写機能がトランスで提供されるとき、コスミドの接合伝達を成功させるために必要かつ十分である。これら3つの遺伝子成分は、バックボーンとしてpUC19を用いて、順次クローン化した(5’−ゲンタマイシン耐性−oriT−fumC1プロモーター−3’)。この異種発現カセットは標準的な分子生物学の実技を用いて作成した。
【0196】
完了後、pUC19から生合成遺伝子クラスターを含むコスミドクローンに異種発現カセットを移した。この挿入は、プロモーター要素を含む挿入物の3’末端が生合成遺伝子クラスターの第一オープンリーディングフレームの翻訳開始コドンから20塩基対離れて位置するように行い、それによってプロモーター要素と生合成遺伝子クラスターの転写融合体を作成した。プロモーターは、遺伝子クラスターの転写を駆動することおよび生合成タンパク質の翻訳を開始するため生合成遺伝子クラスター内に位置する天然リボソーム結合部位に依拠することを意図した。この挿入は、Chaveroche et al. 2000に従って、ラムダREDリコンビナーゼ機能によって介在される異種組換えの使用によって行った。簡潔には、生合成遺伝子クラスターの意図した挿入部位とホモロジーを有する100 ntの隣接領域(PCRプライマーに設計)を有する上記構築物からなるPCR産物を作成した。これらの100 ntの隣接領域は、長フランキングホモロジーPCR(Moore et al. 2005)によって、存在する100 ntの隣接領域に600 nt長のPCR作成隣接領域を加えることにより、さらに延長した。プラスミドpKOBEGhyg(HindIII制限部位にクローン化されたpKOBEGプラスミドのハイグロマイシンカセット含有構築物)からのラムダREDタンパク質を予め発現していた大腸菌EPI100エレクトロコンピテント細胞に、600 ntのホモロジー隣接領域を有する異種発現カセットをエレクトロポレーションした。15μg/mlのゲンタマイシンを補ったLauria培養液寒天で、コスミドへの組込みに成功したトランスコンジュゲートを選択した。このようにして作成した異種発現構築物をPCRおよびDNA配列決定で確認した。効率が低くなるが、コスミドクローンへの異種発現カセットの挿入は、伝統的な制限酵素を利用したクローニング戦略によって行ってもよい。
【0197】
大腸菌ヘルパー株HB101 (pRK2013)に依拠してRK2トランスファー機能を提供する確立された方法(Stanisich and Holloway, 1969)を用いた三親接合により、シュードモナス・プチダKT2440に異種発現構築物を接合により移した。P. putidaトランスコンジュゲートを選択するため75μg/mlのゲンタマイシンならびに大腸菌ドナーおよびヘルパー株の増殖を防止するため25μg/mlのイルガサンを補ったLauria培養液寒天でP. putidaトランスコンジュゲートを選択した。Transconjugates that had successfully integrated into the P. putida chromosome at the fumC-1上流プロモーター領域でのP. putida染色体への組込みに成功したトランスコンジュゲートを、PCR、サザンハイブリダイゼーションおよびDNA配列分析により確認した。
【0198】
2g/Lのイソ酪酸および100μMの2,2,ジピリジル(培地pHは7.0に調節)を含むLauria培養液中、15℃、200rpmの一定回転撹拌で増殖させて、式(II)の化合物の生産を確認した。化学抽出は、6日目に粗発酵液5mLについて1:1の酢酸エチルで行い、次いで30℃で乾燥濃縮し、メタノールで20X最終濃度に再構成した。オンラインDAD、MSおよびMS/MS検出を組み合わせたC−18カラムを用いたHPLC分離により分析を行った。式(II)の化合物は、MSおよびMS/MS検出を用いてはっきりと確認された(図6)。
【0199】
実施例7:5−ヒドロキシプロリンから3−アミノ−6−ヒドロキシ−2−ピペリドン(ahp)への転位のメカニズム
式(I’)のデプシペプチドの重要な生合成経路は、プロリンがデプシペプチド鎖の4位アミノ酸で取り込まれることを示唆している。これはプロリンに代えてahpまたはデヒドロahpを含む式(XIV)の化合物と一致している。我々は、プロリンをヒドロキシル化して、式(XVIII)で例示される5−ヒドロキシプロリンを有する化合物を製造していると仮定するチトクロムP450酵素(orf 8)を同定した。式(XVIII)の化合物は、ahp含有デプシペプチド(例えば式(II))を水性環境中に数日インキュベーションすると自然に形成された(図5)。この5−ヒドロキシプロリン形態とahp形態の相互変換が可逆性であること、そして50℃の水中で10日後にモル比約9:1(ahp:5−ヒドロキシプロリン)で平衡に達することも我々は示した。
【0200】
実施例8:デプシペプチド生合成用遺伝子クラスターの異種遺伝子発現のための、シュードモナス・プチダKT2440由来Fur制御fumC-1プロモーターの使用
宿主シュードモナス・プチダKT2440におけるデプシペプチドの生合成遺伝子の異種発現を成功させるために、異種宿主の遺伝子クラスターの前に置く好適なプロモーターを見つける必要があった。異種宿主、シュードモナス・プチダKT2440からのfur制御プロモーターを選択した(配列番号69)。ほとんどではないが多くの細菌において、標準の複合成長培地(たとえばLB)を用いた場合、成長の転換期は、成長培地中の鉄制限の発生と一致する。我々は、この成長段階で一般にほとんどの二次代謝産物が作成されることが知られているため、健康な集団密度に細胞が達することができるように成長の転換期まで異種宿主におけるデプシペプチドの生合成遺伝子クラスターの転写を遅延させることが有利であると考えた。鉄制限に応答して活性化される遺伝子は、しばしば鉄取り込みリプレッサー(Fur)によって制御される。このメタロレギュレーターは、鉄が十分な条件下で制御遺伝子のプロモーター領域と直接結合して物理的にRNAポリメラーゼ結合を防止することによって、遺伝子セットを抑制するFeセンサーとして作用する(Barton et. al. 1996)。鉄が不充分な条件下では、Furはプロモーター領域から遊離して、遺伝子の転写が生じる。したがって、Fur制御プロモーターを用いて、転換期まで異種遺伝子の発現を抑制することができる。
【0201】
我々は、十分な鉄レベルに対して低い鉄レベルに応答して発現される遺伝子の公表されているプロテオーム(Heim et al. 2003)から、シュードモナス・プチダKT2440における潜在的なFur制御遺伝子を同定し、緑膿菌Furリプレッサーコンセンサス部位「gataatgataatcattatc」(配列番号64)(Barton et al. 1996)を用いてそれらの遺伝子の前のプロモーター領域を探索した。Barton et alからの鉄制御プロテオームの試験によって測定したとき、シュードモナス・プチダKT2440において最も高度にアップレギュレートされた遺伝子産物の一つが、2つのP. putidaフマラーゼ酵素の一方をコードするfumC-1の遺伝子産物であった。さらなる調査により、この遺伝子はFur制御であることが既に示されていることが分かった(Hassett et. al. 1997)。したがって我々は、公表されたデータに基づいて、このプロモーター領域が強く、鉄依存的に作用し、鉄レベルが細胞中で低いときにオンになることを期待した。これらの特徴は、fumC-1プロモーター領域をシュードモナス・プチダKT2440における異種遺伝子発現の目的に使用する理想的な候補たらしめた。実施例6および図6に示すように、全生合成遺伝子クラスターの異種遺伝子発現の成功によってかかる仮説が確認された。
【0202】
鉄が不充分な条件は、発酵成長培地中の鉄濃度と同等乃至3倍以上のモル濃度で鉄キレート剤2’2’ジピリジルを発酵培養に加えて得られる。これによりFur制御遺伝子を、2’2’ジピリジルの添加により制御可能な方法で、アップレギュレートできる。例えば、我々は、成長培地LBを用いる異種発現発酵培養において300μMの2’2’ジピリジルを用いた。他の鉄キレート剤、例えばエチレンジアミンテトラ酢酸(EDTA)、クエン酸塩または鉄取り込みシデロフォアとして作用することが知られている化合物(例えばデスフェリオキサミン、エンテロバクチンまたはバシリバクチン)も、発酵培地に鉄が不充分な条件を作り出すための同様の方法において使用できた。あるいは、所定の発酵培地を用いて、鉄レベルを注意深く制御することができた。
【0203】
ここに記載したfumC-1プロモーターの使用の成功と同じように、他のFur制御プロモーターを使用できた。同様にFurリプレッサー結合部位を含むため、例えばFpvA および OmpR-1 の発現を制御するプロモーターを使用できた。かかるプロモーターは、下記実施例9に更に詳細に記載する。Feが不充分な条件下でアップレギュレートされるあらゆる遺伝子の前の他のFur結合部位は、ここに記載のバイオインフォマティクスアプローチを用いるか、あるいはBaichoo et al. (2002)に記載のように、プロモーター領域のDNAに対する精製Furタンパク質の電気泳動移動度シフトアッセイを用いて、同定された。Furファミリーは細菌ドメインに広く拡がっており、プロモーター領域およびそれらのそれぞれのFur結合部位は、一般に、属特異的であり、しばしば種特異的である。したがって、シュードモナス・プチダKT2440のFur制御プロモーター領域が他のシュードモナス種でも機能することは明らかである。
【0204】
実施例9:Fur制御プロモーター
シュードモナス・プチダKT2440からのFur制御プロモーター。Furリプレッサー結合部位は下線で示し、緑膿菌Furリプレッサーコンセンサス部位gataatgataatcattatc (配列番号64)(Barton et al. 1996)に対するコンセンサスヌクレオチド類似性サーチにより同定した。
【0205】
fumC-1 Fur制御プロモーター領域(Furリプレッサー部位には下線を付す)
atcaggccgcgctgattcgccgtatggggcgcgggctgctggtgaccgaactgatggggcatggcttgaacatggtgacgggggactattcccgtggtgcggcggggttctgggtcgagaatggcgagattcagcatgccgtacaggaagtcaccatcgccggaaacatgaaggacatgttccagcagattgtcgcgatcggtagcgatcttgaaacccgtagcaatattcatacgggctcggtgttgatcgagcggatgaccgttgctggtagctgatctttagcctgcgccggccctttcgcgggtaaacccgctcctacacggtggtggacgtacatcggggttggacacaggccgttgtaggagcgggttcacccgcgaagaggccggaacagcactacacctttccctgcaaatccgaagacccggccctcgcgccgggtttttatttcatcacctttttcttgaagtgattctatttatcacttaataatgaatatcattatccagtaacccggcgatgatgttcatgaaatccgtcctccgcgaactgccctacctggaaaactggcgctggctcagccggcgcattcgctgtgcgctcgaccccgacgagccgcgcctgatcgagcattacctggccgaaggccgctatctggtgtgctgcaccgaaacctcgccatggacggtggcgctgacagcgtttcgcctgctgctggataccgcctgcgatcgcatgctcccctggcattggcgttgtctgtgcctggaccaggcgtggcgccctctgctggacctgcgcaacctcgaccgccaggaacagaaccaacgctggcaaccctacgccttgcagttggccaattgccgtctgctgccttcgatttctcccgatgaactgatgcaaggatttgatgatgagtgatacccgtatcgagcg (配列番号69)
FpvA Fur制御プロモーター領域(Furリプレッサー部位には下線を付す)
tccggcgaattttctacacagagctgctgccggacctcaagcgcctgggcaagaccatca
tcgtgataagccacgacgaccgctacttcgacgtcgccgaccagctcatccacatggcgg
caggcaaggtccaacaggagaaccgcgtcgcagattgcatttaatttttccggttttggc
cgatgagtgcgtcccaatcaataacaagaattaatactattaacatctgacactcaaggg
ctttgaaaaa (配列番号70)
OmpR-1 Fur制御プロモーター領域(Furリプレッサー部位には下線を付す)
caggtagcgcaggcgctcttccaggtggcgcaactgagtgtcgtcaaggctaccggtcac
ttccttgcgatagcgggcgatgaagggcacggtcgagccttcgtccaacaggctcacggc
cgcctcgacctgctgcgggcgtacgcccagttcctcggcgatacggctgttgatgctgtc
catgtaaaccacctgacatttgtgaatacgggggtcgcctgtgggctttttgcccggcgg
cgctggatgaaagccgcgcattatacccatcgcaaacggcttgcggtgatggcgcccggc
cagccggaactggcgccgggggaaaaatctgctaacaatgctcacgcaacgtgcagcaat
ggctacgccataatgcgcggcgatatcagaggagttattc (配列番号71)
fumC-1 プロモーターのFurリプレッサー結合部位
aaacatgaaggacatgttc (配列番号65)
aataatgaatatcattatc (配列番号66)
fpvA プロモーターFurリプレッサー結合部位
aataacaagaattaatact (配列番号67)
ompR-1 プロモーターのFurリプレッサー結合部位
cataatgcgcggcgatatc (配列番号68)
【0206】
Fur制御プロモーターおよびそれらのFurリプレッサー部位は、多くの非シュードモナス種から記載され、特徴付けられており、そしてCarpenter et al. (2009)に列挙され、レビューされている。Fur結合は異なる属間で大きく異なりうる。例えば、大腸菌のコンセンサスFur結合部位はGATAATGATAATCATTATC(de Lorenzo et al. 1987)であるが、枯草菌のコンセンサスFur結合部位はTGATAATTATTATCA(Baichoo and Helmann, 2002)である。
【0207】
参考文献
Altschul, S.F., Gish, W., Miller, W., Myers, E.W. & Lipman, D.J. (1990) "Basic local alignment search tool." J. Mol. Biol. 215:403-410.
Baichoo N, Helmann JD. (2002) Recognition of DNA by Fur: a reinterpretation of the Fur box consensus sequence. J Bacteriol. 184(21):5826-32.
Baichoo N, Wang T, Ye R, Helmann JD. (2002) Global analysis of the Bacillus subtilis Fur regulon and the iron starvation stimulon. Mol Microbiol. 45(6):1613-29.
Barton HA, Johnson Z, Cox CD, Vasil AI, Vasil ML. (1996) Ferric uptake regulator mutants of Pseudomonas aeruginosa with distinct alterations in the iron-dependent repression of exotoxin A and siderophores in aerobic and microaerobic environments. Mol Microbiol. 21(5):1001-17.
Binz, T.M., Wenzel, S.C., Schbell, H., Bechthold, A., Mueller, R. (2008) Heterologous expression and genetic engineering of the phenalinolactone biosynthetic gene cluster by using Red/ET recombineering. ChemBioChem. 9: 447-454.
Carpenter BM, Whitmire JM, Merrell DS. (2009) This is not your mother's repressor: the complex role of fur in pathogenesis. Infect Immun. 77(7):2590-601.
de Lorenzo V, Wee S, Herrero M, Neilands JB. (1987) Operator sequences of the aerobactin operon of plasmid ColV-K30 binding the ferric uptake regulation (fur) repressor. J Bacteriol. 169(6):2624-30.
Garrity, P. A., Ligation-Mediated PCR, in PCR 2 A Practical Appraoch, McPherson, M. J. et. al. (Eds.) pp. 309-322 Oxford University Press, New York (1995).
Gish, W. & States, D.J. (1993) "Identification of protein coding regions by database similarity search." Nature Genet. 3:266-272.
Gu, J.Q., Nguyen, K.T., Gandhi, C., Rajgarhia, V., Baltz, R.H., Brian, P., Chu, M. (2007) Structural characterization of daptomycin analogues A21978C1-3(d-Asn11) produced by a recombinant Streptomyces roseosporus strain. J. Nat. Prod. 70: 233-240.
Hassett DJ, Howell ML, Ochsner UA, Vasil ML, Johnson Z, Dean GE. (1997) An operon containing fumC and sodA encoding fumarase C and manganese superoxide dismutase is controlled by the ferric uptake regulator in Pseudomonas aeruginosa: fur mutants produce elevated alginate levels. J Bacteriol. 179(5):1452-9.
Heim S, Ferrer M, Heuer H, Regenhardt D, Nimtz M, Timmis KN. (2003) Proteome reference map of Pseudomonas putida strain KT2440 for genome expression profiling: distinct responses of KT2440 and Pseudomonas aeruginosa strain PAO1 to iron deprivation and a new form of superoxide dismutase. Environ Microbiol. 5(12):1257-69.
Higgins D.G., Thompson J.D., Gibson T.J. (1996). Using CLUSTAL for multiple sequence alignments. Methods Enzymol., 266, 383-402.
Finking, R., Marahiel, MA., (2004) Biosynthesis of nonribosomal polypeptides. Annu Rev. Microbiol. 58: 453-488.
Marahiel, M. A., Stachelhaus, T., Mootz, H.D. (1997) Modular peptide synthetases involved in nonribosomal peptide synthesis. Cem. Rev. 97:2651-2673.
Pfeifer, B.A., Admiraal, S.J., Gramajo, H., Cane, D.E., Khosla, C. (2001) Biosynthesis of complex polyketides in a metabolically engineered strain of E. coli. Science. 291: 1790-1792.
Rausch, C., Weber, T., Kohlbacher, O. Wohlleben, W., Huson, D.H. (2005) Specificity prediction of adenylation domains in nonribosomal peptide synthetases (NRPS) using transductive support vector machines (TSVMs). Nuc. Acids Res. 33: 5799-5808.
Rouhiainen, L., Paulin, L., Suomalainen, S., Hyytiainen, H., Buikema, W., Haselkorn, R., Sivonen, K. (2000) Genes encoding synthetases of cyclic depsipeptides, anabaenopeptilides, in Anabaena strain 90. Mol. Microbiol. 37: 156-167.
Sambrook, J., Maniatis, T. (1989) Molecular Cloning: A Laboratory Manual. Cold Spring Harbor Laboratory Press. Cold Spring Harbor NY.
Shaying Zhao (Ed.), Marvin Stodolsky, Marvin Stodolsky (Ed.) (2003) Bacterial Artificial Chromosomes (Methods in Molecular Biology Series v255-256): Library Construction, Physical Mapping, and Sequencing, Vol. 1 Springer-Verlag New York, LLC.
Shen, B. (2004) Accessing natural products by combinatorial biosynthesis. Sci STKE. Pe14.
Stachelhaus, T. Mootz, H.D., Marahiel, M.A. (1999) The specificity-conferring code of adenylation domains in nonribosomal peptide synthetases. Chem. Biol. 6: 493-505.
Staunton, J., Weissman, K.J. (2001) Polyketide biosynthesis: a millennium review. Nat. Prod. Rep. 18:380-416.
Wenzel, S.C., Mueller, R. (2005) Recent developments towards the heterologous expression of complex bacterial natural product biosynthetic pathways. Curr. Op. Biotechnol. 16: 594-606.
Von Dohren, H., Keller, U., Vater, J., and Zocher, R. (1997) Multifunctional peptide synthetases. Chem. Rev. 97:2675-2705.
Zhang, Y., Muyrers, J. Testa, G., Stewart, F. (1998) A new logic for DNA engineering using recombination in Escherichia coli. Nat. Genet. 20: 123-128.
Zhuang, H., Yong, W., Pfeifer, B.A. (2008) Bacterial hosts for natural product production. Molecular Pharmaceuticals. 5: 212-225.
【図1A】
【図1B】
【図2A】
【図2B】
【図2C】
【図2D】
【図2E】
【図2F】
【図2G】
【技術分野】
【0001】
本発明は、式(I)または(I’)の化合物の生産に関与する生合成遺伝子クラスターの1個以上の機能的フラグメントを含むポリヌクレオチドの提供に関する。本発明はまた、式(I)もしくは(I’)または式(II)〜(VII)、(XI)〜(XIV)ならびに(XVII)および(XVIII)の化合物を製造する方法も提供する。さらに、医薬組成物としてのかかる化合物の使用も本発明において提供される。
【背景技術】
【0002】
微生物由来の多くの天然産物は、高等生物において見られる生物学的活性を有しており、数世紀にわたってその治療特性が探索されている。これらの天然産物のほとんどはポリケチドクラスおよび非リボソームペプチドクラスに属しており、ポリケチドシンターゼ(PKS)および非リボソームペプチドシンターゼ(NRPS)として知られるモジュラー酵素系によって合成される(Finkering and Marahiel 2004; Staunton and Weissman, 2001)。さらに、同じ経路にPKS遺伝子およびNRPS遺伝子の両方を含み、したがってこれら2つのクラスのハイブリッドである二次代謝産物を生産する経路が存在している。これらの生合成経路によって生産される天然産物は、短鎖カルボン酸およびアミノ酸のような小さな、比較的単純な構成要素から構成されている。しかし、これらの経路に由来する最終天然産物は、極めて多様であり、しばしば構造的に複雑であり、通常複数の立体中心を含んでいる。このため、これらの化合物の生産のための合成アプローチは、しばしば実行不可能であり、したがってそれらの生産には発酵が常套のアプローチで有り続けている。しかし、発酵プロセスは、代謝的に特徴付けられておらず、しばしば遺伝子の取り扱いが困難であり、そしてしばしばあまり増殖せず、目的化合物を十分なレベルでは生産しないという、微生物に対する信頼に関する固有の問題を有する。これらの問題を回避するため、これらの欠点を有さない、十分に特徴付けられた宿主生物におけるPKSまたはPRPS経路の異種発現が一つの選択肢であり得る(Wenzel and Muller, 2005に概説されている)。事実、このアプローチは、発見努力のために「静かな」または「隠れた」PKSおよびNRPS経路の発現に延長でき(Shen, 2004)、または研究室で培養できない生物からの経路の発現に使用できる。さらに、PKSおよびNRPS経路の異種宿主への移動により、親化合物の新規なアナログを作成するための二次代謝経路の効率的なバイオエンジニアリングが可能となる。
【0003】
異種発現は、一般に、PKSおよびNRPS経路がゲノム上の連続したクラスターに位置しているという事実を利用している。したがって、原理上は、これらの経路は、標準的なBACまたはコスミドベクターに比較的容易にクローニングされる。ある微生物から別の微生物へとある経路を動かすことの原理的な単純さに反して、2種の生物間の制御、コドン出現頻度または代謝における差異は、異種発現の成功に大きな課題を引き起こしている。さらに、BACライブラリー構築物のようなこの戦略を効率的に適用することができる分子ツールおよびクローニングのための組換えアプローチは、比較的最近になってからのみ利用可能となった(Wenzel and Muller, 2005)。このため、異種発現の成功例は、文献には極僅かしか存在しない。
【0004】
好適な異種宿主の選択は発現戦略を設計する際に重要な検討事項である。新たな宿主は、遺伝子操作が用意であり、研究室での取り扱いが容易であり、PKSおよびNRPS経路を用いる能力を有するべきである。例えば、新たな宿主におけるホスホパンテテイニルトランスフェラーゼの存在は、移入されたPKSおよびNRPSの活性化を促進するために必須である(Pfeifer et al. 2001)。さらに、新たな宿主が元の宿主のものと同様のコドン出現頻度を有し、移入された経路の効率的な発現が可能であることが重要である。使用される最も一般的な宿主は、大腸菌(Escherichia coli)、枯草菌(Bacillus subtilis)、シュードモナス・プチダ(Pseudomonas putida)および十分に特徴付けられたストレプトマイセス(Streptomyces)株のうちわずかに選択されたものである(Zhang and Pfeifer, 2008に概説されている)。利用されている他の宿主は、ミキソコッカス・ザンサス(Myxococcus xanthus)および糸状菌類を含む。これらの宿主株のいくつかは、変異によって主要な常在性二次代謝系がサイレンシングされて、バックグラウンド代謝プロフィールを除き、移入される生合成経路に利用可能な前駆体貯蓄の減少を防止するように、修飾されている。
【0005】
特定の経路を移動させるためには、適切な移動可能な遺伝的要素に基づく当該経路のパッケージングが必要である。まず初めにPKSまたはNRPS系の配列を、少なくともアミノ酸レベルで、より好ましくはヌクレオチドレベルで知らなければならない。典型的には、この配列を用いて、天然宿主から構築されたゲノムライブラリーから、BACまたはコスミドクローンを位置づけるプローブを設計する。これらの経路クラスターのサイズが大きいため(通常30kb以上、しばしば100kbを超える)、「ショットガン」クローニング戦略を用いるとき、それらはしばしば1個のBACまたはコスミドクローン中には捕捉されない。したがって、この経路は、しばしば、全経路を含む1個のBACまたはコスミドベクター構築物を作成するために再構築する必要がある。極めて大きな経路を発現させようとするとき、それは新たな宿主中でトランスで発現されるように2個以上の別個のベクター構築物に分割してもよい(Gu, et al. 2007)。最終的には、構築物を有する大腸菌から新たな宿主へと移動するために、ベクター構築物は、プラスミド移行性機能(例えばRK2からoriT)をも有していなければならない。構築物が新たな宿主において好適であることを確保するため、それを宿主染色体に融合させることを勧める。これを達成するため、構築物は、効率的な染色体融合のための部位を含んでいるべきである。例えば、ストレプトマイセスのファージ接着部位ΦC31が、この系における染色体挿入のためにしばしば用いられる(Binz, et al. 2008)。さらに、新たな宿主で適切に機能する新たなプロモーターを、生合成経路の前に挿入する必要がしばしばある。問題の生物2種が密接に関連しており、したがって多くの制御因子を共有している可能性がある場合、この工程を避けることができる。最後に、選択マーカー、一般には抗生物質耐性カセットが、新たな宿主における構築物(修飾BACまたはコスミド)の移行および融合が成功したかを選択するために、必要である。典型的には、これらの操作は大腸菌において行われ、しばしばRed/ET組換えの使用によってなされる(Zhang, et al. 1998)。このクローニングアプローチは、制限酵素利用操作がせいぜい挑戦的である場合に、大きなDNA構築物を含む応用に特に適用可能である。
【0006】
構築物が新たな宿主に融合されると、発酵および続く化学分析を行って、経路の発現が成功したかを決定する。ほぼ全ての場合で異種発現が成功した場合、天然宿主で観察されるものと比較してより低い力価で天然産物が生産される。この明らかな抑制にも拘わらず、異種発現の成功は、常套の株開発方法論に利用可能な多くの選択肢を有する発現プラットフォームを提供する。
【発明の概要】
【0007】
本発明は、式I
【化1】
[式中、A7のカルボキシ基とA2のヒドロキシ基の間にエステル結合があり、所望により、A5とA6の間のアミド結合の窒素原子がメチルで置換されていてもよく、
XおよびA1は各々独立して任意であり、そして
Xは任意の化学残基、特にHまたはアシル残基、特にCH3CH2CH(CH3)CO、(CH3)2CHCH2COまたは(CH3)2CHCOであり;
A1はアスパラギン酸以外の標準アミノ酸、特にグルタミンであり;
A2はスレオニンまたはセリン、特にスレオニンであり;
A3は非塩基性標準アミノ酸またはその非塩基性誘導体、特にロイシンであり;
A4はAhp、デヒドロ−AHP、プロリンまたはその誘導体、特にAhpまたはその誘導体、特にAhp誘導体3−アミノ−2ピペリドンであり;
A5はイソロイシンまたはバリン、特にイソロイシンであり;
A6はチロシンまたはその誘導体、特にチロシンであり;
A7はロイシン、イソロイシンまたはバリン、特にイソロイシンまたはバリン、特にイソロイシンである。]
のデプシペプチドの生合成に関与する生合成クラスターの同定、並びに式Iの非リボソームペプチドまたはその薬学的に許容される塩もしくは誘導体を生産するための異種発現系の開発に関する。特に、式(I’)
【化2】
[式中、A7のカルボキシ基とA2のヒドロキシ基の間にエステル結合があり、所望により、A5とA6の間のアミド結合の窒素原子がメチルで置換されていてもよく、
XはCH3CO、(CH3)2CHCO、CH3S(O)CH2CO、CH3CH2CH(CH3)COまたはC6H5COであり;
A1はグルタミンであり;
A2はスレオニンであり;
A3はロイシンであり;
A4はAhp、デヒドロ−AHP、プロリンまたは5−ヒドロキシ−プロリンであり;
A5はイソロイシンまたはバリン、特にイソロイシンであり;
A6はチロシンであり;
A7はイソロイシンまたはバリン、特にイソロイシンである。]
のデプシペプチドの生合成における生合成遺伝子クラスターの使用を見出す。
【0008】
特に本発明は、図1に示す式(II)、(III)、(IV)、(V)、(VI)、(VII)、(XI)、(XII)〜(XIV)、(XVII)および/または(XVIII)の非リボソームペプチドの生合成に関与する生合成クラスターの同定、並びに式(I)または(I’)の非リボソームペプチドまたはその薬学的に許容される塩もしくは誘導体を生産するための異種発現系の開発に関する。
【0009】
式(I)、特に式(I’)の化合物は、粘液細菌コンドロマイセス・クロカツス(Chondromyces crocatus)NPH−MB180によって生産されるデプシペプチドのファミリーに属する非リボソームポリペプチドである。これらのデプシペプチドは、極めて強力で選択的なヒトカリクレイン7(hK7)およびエラスターゼ阻害剤であることが示されている。ヒトカリクレイン7はセリンプロテアーゼ活性を有する酵素であり、アトピー性皮膚炎の処置のための潜在的標的である。この新規化合物の詳細な物理化学データならびに発酵および抽出法は、WO2009/024527として公開されているPCT出願PCT/EP08/060689に記載されている。
【0010】
本明細書において使用するとき、用語「式(I)の化合物」または「式(I)のデプシペプチド」は、上記定義の式(I)の化合物、特に図1に記載の式(II)、(III)、(IV)、(V)、(VI)、(VII)、(XI)、(XII)、(XIII)、(XIV)および/または(XVIII)の非リボソームペプチド、ならびに実質的に同じプロテアーゼ活性を保持するあらゆる誘導体を意味する。かかる誘導体の例は、WO2009/024527として公開されているPCT出願にさらに記載されている。
【0011】
本明細書において使用するとき、用語「式(I’)の化合物」または「式(I’)のデプシペプチド」は、上記定義の式(I’)の化合物、特に図1に記載の式(II)、(III)、(IV)、(V)、(VI)、(VII)、(XI)、(XII)、(XIII)、(XIV)および/または(XVIII)の非リボソームペプチド、ならびに実質的に同じプロテアーゼ活性を保持するあらゆる誘導体を意味する。
【0012】
本発明の技術的課題は、式(I)または(I’)のデプシペプチドの生合成に関与する生合成クラスターまたはその機能的部分の提供である。
この技術的課題は、特許請求の範囲に特徴付けられた態様の提供によって解決される。
本発明の別の技術的課題は、異種遺伝子発現、例えば目的の組換えタンパク質の合成に適切な抑制プロモーターの提供である。
【0013】
本発明は、第一の態様において、(1)式(I)または(I’)の化合物の生産に関与する非リボソームペプチドシンターゼ(NRPS)(以下NRPS2と称する)をコードする生合成遺伝子クラスターの1個以上の機能的フラグメントを含むポリヌクレオチドであって、以下のものを含むポリヌクレオチドの提供に関する:
(i) NRPS2ドメインをコードする配列番号1、3、5、7、9、11、13、46、48、50、52、54、56、58および60からなる群から選択される配列と、少なくとも80%、特に少なくとも85%、特に少なくとも90%、特に少なくとも95%、特に少なくとも98%の配列同一性を有するヌクレオチド配列および/またはその相補体;
(ii) NRPS2ドメインをコードする配列番号1、3、5、7、9、11、13、46、48、50、52、54、56、58または60からなる群から選択されるヌクレオチド配列の相補鎖とハイブリダイズするヌクレオチド配列および/またはその相補体;
(iii) NRPS2ドメインを示す配列番号2、4、6、8、10、12、14、47、49、51、53、55、57、59または61からなる群から選択される配列と、少なくとも60%、特に少なくとも70%、特に少なくとも80%、特に少なくとも90%、特に少なくとも95%の配列同一性を有するアミノ酸配列をコードするヌクレオチド配列および/またはその相補体;
(iv) NRPS2ドメインを示す配列番号2、4、6、8、10、12、14、47、49、51、53、55、57、59または61からなる群から選択されるアミノ酸をコードするヌクレオチド配列の相補鎖とハイブリダイズするヌクレオチド配列および/またはその相補体;
(v) 配列番号15、配列番号28からなる群から選択される配列と少なくとも80%、特に少なくとも85%、特に少なくとも90%、特に少なくとも95%、特に少なくとも98%の配列同一性を有するヌクレオチド配列および/またはその相補体;
(vi) 配列番号15、配列番号28からなる群から選択されるヌクレオチド配列の相補鎖とハイブリダイズするヌクレオチド配列および/またはその相補体;
ここで、前記(i) 〜 (vi)のヌクレオチド配列は、配列番号2、4、6、8、10、12、14、47、49、51、53、55、59および/または61の参照配列によって示される対応NRPSドメインの活性を保持する発現産物をコードする。
【0014】
第2の態様において、(2)以下の1個以上のNRPS2ドメインの活性を保持する発現産物をコードする、態様(1)に記載のポリヌクレオチドを提供する:
(i)配列番号47のチオレーションドメイン;
(ii)配列番号49のコンデンセーションドメイン;
(iii)配列番号51のプロリンのアデニレーションドメイン;
(iv)配列番号53のチオレーションドメイン;
(v)配列番号2のコンデンセーションドメイン;
(vi)配列番号4のイソロイシンのアデニレーションドメイン;
(vii)配列番号6のチオレーションドメイン;
(viii)配列番号8のコンデンセーションドメイン;
(ix)配列番号10のチロシンのアデニレーションドメイン;
(x)配列番号12のN−メチレーションドメイン;
(xi)配列番号14のチオレーションドメイン;
(xii)配列番号55のコンデンセーションドメイン;
(xiii)配列番号57のイソロイシンのアデニレーションドメイン;
(xiv)配列番号59のチオレーションドメイン;および/または
(xv)配列番号61のチオエステラーゼドメイン。
【0015】
態様(2)の特定の態様において、ポリヌクレオチドは、式(I)または(I’)の化合物を生産するNRPS2をコードし、配列番号29に示すアミノ酸配列をコードするヌクレオチド配列を含む。
【0016】
第3の態様において、(3)本発明は、式(I)または(I’)の化合物の生産に関与するNRPSであるNRPS1をコードする生合成遺伝子クラスターの1個以上の機能的フラグメントを含むポリヌクレオチドであって、以下のものを含むポリヌクレオチドに関する:
(i) NRPSドメインをコードする配列番号30、32、34、36、38、40、42および44からなる群から選択される配列と、少なくとも80%、特に少なくとも85%、特に少なくとも90%、特に少なくとも95%、特に少なくとも98%の配列同一性を有するヌクレオチド配列および/またはその相補体;
(ii) NRPSドメインをコードする配列番号30、32、34、36、38、40、42および44からなる群から選択されるヌクレオチド配列の相補鎖とハイブリダイズするヌクレオチド配列および/またはその相補体;
(iii) NRPS1ドメインを示す配列番号31、33、35、37、39、41、43、45からなる群から選択される配列と、少なくとも60%、特に少なくとも70%、特に少なくとも80%、特に少なくとも90%、特に少なくとも95%の配列同一性を有するヌクレオチド配列および/またはその相補体;
(iv) NRPS1ドメインを示す配列番号31、33、35、37、39、41、43、45からなる群から選択されるアミノ酸をコードするヌクレオチド配列の相補鎖とハイブリダイズするヌクレオチド配列および/またはその相補体;
(v) 配列番号26からなる群から選択される配列と、少なくとも80%、特に少なくとも85%、特に少なくとも90%、特に少なくとも95%、特に少なくとも98%の配列同一性を有するヌクレオチド配列および/またはその相補体;または
(vi)配列番号26からなる群から選択されるヌクレオチド配列の相補鎖とハイブリダイズするヌクレオチド配列および/またはその相補体;
(vii) ここで、前記(i) 〜 (vi)のヌクレオチド配列は、配列番号31、33、35、37、39、41、43、45の参照配列によって示される対応NRPSドメインの活性を保持する発現産物をコードする。
【0017】
第4の態様において、態様(3)に記載のポリヌクレオチドは、以下の1個以上のNRPS1ドメインの活性を保持する発現産物をコードする:
(i) 配列番号31のローディングドメイン;
(ii)配列番号33のグルタミンのアデニレーションドメイン;
(iii)配列番号35のチオレーションドメイン;
(iv)配列番号37のコンデンセーションドメイン;
(v)配列番号39のスレオニンのアデニレーションドメイン;
(vi)配列番号41のチオレーションドメイン;
(vii)配列番号43のコンデンセーションドメイン;および
(viii)配列番号45のロイシンのアデニレーションドメイン。
【0018】
態様(4)の特定の態様において、ポリヌクレオチドは、式(I)または(I’)の化合物を生産するNRPS1をコードし、配列番号27に示すアミノ酸配列をコードするヌクレオチド配列を含む。
【0019】
他の態様において、本発明は、上記1個以上のポリヌクレオチドによってコードされるポリペプチドに関する。特に、該ポリペプチドは、式(I)または(I’)の化合物の生産に適切であり、以下のものからなる群から選択されるアミノ酸配列を含む:
(i)NRPS1を示す配列番号27、第2のNRPS2を示す配列番号29、チトクロムP450を示す配列番号63;および
(ii) (i)に列挙した参照配列と60%、特に少なくとも70%、特に少なくとも80%、特に少なくとも90%、特に少なくとも95%の配列同一性を有し、実質的に同じ触媒機能を保持する、(i)に列挙したアミノ酸配列の機能的変異体。
【0020】
本発明は、さらに、上記1個以上のポリペプチドをコードするヌクレオチド配列を含むポリヌクレオチドに関する。
【0021】
さらに別の態様において、本発明は、
(i)配列番号27またはその機能的変異体をコードするヌクレオチド配列;および
(ii)配列番号29またはその機能的変異体をコードするヌクレオチド配列
を含むポリヌクレオチドを提供する。
【0022】
かかるポリヌクレオチドは、配列番号63またはその機能的変異体をコードするヌクレオチド配列をさらに含んでいてもよい。ある特定の態様において、該ポリヌクレオチドは、受託番号DSM 19329を有するコンドロマイセス・クロカツス(Chondromyces crocatus)株NPH−MB180から単離される。
【0023】
本発明は、さらに、オープンリーディングフレームが転写および翻訳配列と作動可能に連結している、上記態様のいずれかに定義のポリヌクレオチドを含む発現ベクターを提供する。
【0024】
さらなる態様において、上記態様のいずれかに定義のポリヌクレオチドまたは発現ベクターでトランスフェクトされており、それを発現する宿主細胞、特に式(I)もしくは(I’)の化合物または式(II)〜(VII)、(XI)〜(XIV)ならびに(XVII)および(XVIII)の化合物の異種生産のための宿主細胞を提供する。
【0025】
他の態様において、本発明は、式(I)もしくは(I’)または式(II)〜(VII)、(XI)〜(XIV)ならびに(XVII)および(XVIII)の化合物を製造する方法であって、上記態様に記載の宿主細胞を、当該化合物が生産される条件下で培養することを含む、製造方法に関する。
【0026】
ある態様において、本発明は、上記態様のいずれかに記載のポリペプチドまたはNRPSもしくはNRPSドメインに特異的に結合する抗体、並びに当該抗体の使用、すなわちポリペプチドまたはNRPSの精製のための使用に関する。
【0027】
ある態様において、上記態様のいずれかに定義のポリヌクレオチド、ベクター、ポリペプチド、NRPSもしくはNRPSドメインまたは抗体を含む医薬組成物を提供する。
【0028】
ある態様において、上記態様のいずれかに定義の本発明のポリヌクレオチドを含む組換え宿主細胞を好適な条件下で培養して得られるまたはそのようにして得た式(I)または(I’)のデプシペプチドを含む医薬組成物を提供する。
【0029】
ある態様において、本発明は、疾患または状態、すなわちアトピー性皮膚炎の処置および/または診断に使用するための医薬組成物の製造のための、式(I)または(I’)のデプシペプチドに関する。ある特定の態様において、式(I)または(I’)のデプシペプチドは、選択的ヒトカリクレイン(hK7)およびエラスターゼ阻害剤、特に選択的ヒトカリクレイン(hK7)の阻害剤であり、酵素活性、特にセリンプロテアーゼ活性を有する。
【0030】
本発明のさらなる態様において、式(I)または(I’)の化合物の生産に関与するNRPSをコードする生合成遺伝子クラスターであって、上記態様のいずれかに定義のポリヌクレオチドを含むものを提供する。
【0031】
本発明の他の態様において、以下の工程:
(a)コンドロマイセス・クロカツス株または関連株のゲノムDNAからなるヌクレオチドライブラリーを構築する;(b)コロニーとしてライブラリー株を培養する;(c)NRPS遺伝子クラスターを含むクローンの同定のための上記態様のいずれかに定義のポリヌクレオチドに基づくプローブ分子で、増殖したコロニーを分析する、そして(d)NRPS遺伝子クラスターを同定する;
を含む方法によって得られる、本発明の生合成遺伝子クラスターの同定のための上記態様のいずれかに定義のポリヌクレオチド配列を提供する。
【0032】
本発明の骨子は、式(I)または(I’)のデプシペプチド、特に式(II)〜(VII)、(XI)〜(XIV)ならびに(XVII)および(XVIII)のデプシペプチドの整合性に関与する生合成クラスターまたはその機能的部分の提供にある。特に利点としては、式(I)または(I’)のデプシペプチドの生合成クラスターの同定を当該デプシペプチドの異種発現に使用できることである。
【0033】
「非リボソームペプチド」は、単純アミノ酸モノマーから構成される複雑な天然産物のファミリーに属するペプチドのクラスを意味する。これは非リボソームペプチドシンテターゼ(NRPS)と呼ばれる大型多機能タンパク質によって、多くの細菌または真菌において合成される。NRPS系の特徴は、タンパク新生および非タンパク新生アミノ酸を含むペプチドを合成できることである。
【0034】
「非リボソームペプチドシンターゼ」(NRPS)は、モジュールと呼ばれる活性部位の協調的なグループに組織される、大型多機能タンパク質を意味し、ここで各モジュールは、産物長伸長および官能基の修飾の1サイクルを触媒するために必要である。モジュールの数および順序、並びに各NRPS上のモジュール内に存在するドメインのタイプは、取り込まれるアミノ酸の数、順序、選択、そして特定のタイプの伸長に関連した修飾を指示することによって、得られるペプチド産物の構造バリエーションを決定する。
【0035】
用語「ドメイン」は、触媒活性に必要なタンパク質の機能的部分を意味する。かかるドメインは、同じ触媒活性を有する異なる種由来の酵素間で保存されている。
伸長サイクルに必要な最小セットのドメインは、アデニレーション(A)、チオレーション(T)またはペプチジルキャリアタンパク質(PCP)およびコンデンセーション(C)ドメインを有するモジュールからなる。
【0036】
「アデニレーションドメイン」は、AMP誘導体中間体を介して、基質選択およびチオエステルとしてTドメインのホスホ−パンテテインアームへの共有結合性固定に関与している。
【0037】
Cドメインは、上流モジュールからのアミノアシル−またはペプチジル−S−PCPと、対応する下流モジュールのPCPに結合したアミノアシル基の間のペプチド結合の形成を触媒する。その結果は、下流モジュールのPCPドメインに固定された1個の残基によるペプチド伸長である。任意の修飾ドメインは、基質エピマー化、N−メチル化およびヘテロ環化のために存在していてもよい。このモジュールは、1または複数のポリペプチド鎖上に残っていてよい。
【0038】
多くの場合、最終産物の放出/環化に関与する最後のモジュールの最もC末端側にチオエステラーゼ(TE)ドメインが存在する。
【0039】
1.式(I)または(I’)の化合物を生産する生合成遺伝子クラスターをコードするポリヌクレオチド
下記表1は、式(I)または(I’)の化合物の生合成遺伝子クラスターのポリヌクレオチドの具体例およびそれらの対応する機能およびアミノ酸配列を記載している。
【0040】
【表1】
【表2】
【0041】
式(I)または(I’)のデプシペプチドの合成のための単離された生合成遺伝子クラスターは、NRPS1およびNRPS2とも呼ばれる非リボソームペプチドシンテターゼをコードするORF6およびORF7を含む、8個のオープンリーディングフレーム(ORF)からなる。NRPS1およびNRPS2はNRPSドメインを含み、対応する推定機能を表1に列挙する。
【0042】
用語「ポリヌクレオチド」、「ポリヌクレオチド配列」および「ポリペプチド」の意味は当該技術分野において周知であり、当該用語は、異なる定義がないかぎり、本発明の文脈に従って使用される(例えば、それぞれ配列番号1、3、5、7、9、11、13、15、16、18、20、22、24、26、28、30、32、34、36、38、40、42、44、46、48、50、52、54、56、58、60、62)。例えば、「ポリヌクレオチド配列」は、本明細書において使用するとき、天然に生じるかまたは組換えによって作成するタイプの核酸および/またはヌクレオチド配列のあらゆる形態、並びに化学的に合成した核酸/ヌクレオチド配列を意味する。この用語は、核酸アナログおよび核酸誘導体、例えばロックドDNA、PNA、チオリン酸オリゴヌクレオチドおよび置換されたリボオリゴヌクレオチド等も含む。さらに、用語「ポリヌクレオチド配列」は、ヌクレオチドまたはヌクレオチドアナログを含むあらゆる分子をも意味する。
【0043】
好ましくは、用語「ポリヌクレオチド配列」は、核酸分子、すなわちデオキシリボヌクレオチド(DNA)および/またはリボ核酸(RNA)を意味する。本発明の文脈における「ポリヌクレオチド配列」は、当業者に知られている合成的化学法または組換え技術の使用によって作成してよく、あるいは天然源から単離してもよく、あるいはそれらの組合せでもよい。DNAおよびRNAは、所望により非天然ヌクレオチドを含んでいてもよく、一本鎖でも二本鎖でもよい。「ポリヌクレオチド配列」は、センスおよびアンチセンスDNAおよびRNA、すなわちDNAおよび/またはRNAのヌクレオチドの特定の配列に相補的であるポリヌクレオチド配列も意味する。
【0044】
さらに、用語「ポリヌクレオチド配列」は、DNAもしくはRNAまたはそれらのハイブリッドまたは当該技術分野において既知のそのいずれかの修飾物を意味していてもよい(修飾物の例として例えばUS 5525711、US 4711955、US 5792608またはEP 302175参照)。ポリヌクレオチド配列は一本鎖または二本鎖、直線または環状、天然または合成であってよく、いかなるサイズによる限定もない。例えば、ポリヌクレオチド配列は、ゲノムDNA、cDNA、mRNA、アンチセンスRNA、リボザイムまたはかかるRNAもしくはキメロプラスト(chimeroplast)をコードするDNAであってよい(Gamper, Nucleic Acids Research, 2000, 28, 4332 - 4339)。当該ポリヌクレオチド配列は、プラスミドまたはウイルス性DNAもしくはRNAの形態であってもよい。「ポリヌクレオチド配列」はホスホチオエートまたはペプチド核酸(PNA)のような当該技術分野で知られている修飾を含む、オリゴヌクレオチドを意味していてもよい。
【0045】
用語「遺伝子クラスター」または「生合成遺伝子クラスター」は、式(I)または(I’)のデプシペプチドの生合成に関与する遺伝子またはその変異体の群を意味する。遺伝子クラスターまたは生合成遺伝子クラスターの遺伝子改変は、式(I)または(I’)の化合物の変異体を作成するために適用可能な核酸の突然変異誘発、不活性化または置換を含む当該技術分野で既知のあらゆる遺伝子組換え技術を意味する。遺伝子クラスターまたは生合成遺伝子クラスターの遺伝子改変は、式(I)または(I’)の化合物の遺伝的変異体を作成するために適用可能な核酸の突然変異誘発、不活性化または置換を含む当該技術分野で既知のあらゆる遺伝子組換え技術を意味する。
【0046】
DNAもしくはヌクレオチド「コーティング配列」または特定のポリペプチドもしくはタンパク質を「コードする配列」は、適切な制御配列の制御下に置かれたときにポリペプチドまたはタンパク質に転写および翻訳されるDNA配列である。
【0047】
具体的な態様において、複数の本発明のポリヌクレオチド(例えば、それぞれ配列番号1、3、5、7、9、11、13、15、16、18、20、22、24、26、28、30、32、34、36、38、40、42、44、46、48、50、52、54、56、58、60、62)を組み合わせて用いてもよい。あるいは、本発明は、配列番号1、3、5、7、9、11、13、15、16、18、20、22、24、26、28、30、32、34、36、38、40、42、44、46、48、50、52、54、56、58、60、62のフラグメントまたは機能的変異体に関する。
【0048】
ポリヌクレオチド配列の文脈において、用語「そのフラグメント」または「その機能的フラグメント」は、特に核酸分子のフラグメントまたは変異体を意味する。「ポリヌクレオチドのフラグメント」は、例えば、少なくとも1個のアミノ酸欠失を有しており、それによって野生型ポリペプチドと同じ機能(各ポリペプチドの機能は表1および図2により詳細に記載されている)を実質的に保持している本発明のポリペプチド(例えば、配列番号2、4、6、8、10、12または14に示すポリペプチド)をコードしてよい。このような短くなったポリペプチドは、本発明のポリペプチド(例えば、配列番号2、4、6、8、10、12または14、17、19、21、23、25、27、29、31、33、35、37、39、41、43、45、47、49、51、53、55、57、59、61、63に示す)の機能的フラグメントと理解されうる。
【0049】
「ポリヌクレオチドの機能的変異体」は、例えば、少なくとも1個のアミノ酸置換または付加を有しており、それによって好ましくは野生型ポリペプチドと同じ機能(各ポリペプチドの機能は表1および図2により詳細に記載されている)を保持している本発明のポリペプチド(例えば配列番号2、4、6、8、10、12または14、17、19、21、23、25、27、29、31、33、35、37、39、41、43、45、47、49、51、53、55、57、59、61、63に示すポリペプチド)をコードしうる。このような短くなったポリペプチドは、本発明のポリペプチド(例えば、配列番号2、4、6、8、10、12または14、17、19、21、23、25、27、29、31、33、35、37、39、41、43、45、47、49、51、53、55、57、59、61、63に示す)の機能的フラグメントと理解されうる。
【0050】
本発明のポリヌクレオチド/ポリペプチドの機能的変異体は、表1に記載の対応する元のポリヌクレオチド/ポリペプチド配列と、少なくとも50%、55%、60%、好ましくは少なくとも70%、より好ましくは少なくとも80%、85%、90%、95%さらに最も好ましくは少なくとも99%の配列同一性を有する。例えば、ポリペプチドは、配列番号2、4、6、8、10、12または14、17、19、21、23、25、27、29、31、33、35、37、39、41、43、45、47、49、51、53、55、57、59、61、63に示すポリペプチドと少なくとも50%、55%、60%、好ましくは少なくとも70%、より好ましくは少なくとも80%、85%、90%、95%、最も好ましくは少なくとも99%の同一性/相同性を有する。
【0051】
非リボソームペプチドシンターゼ(NRPS)または表1に記載の他のORFのヌクレオチド配列に関して、用語「フラグメント」は、本明細書において使用するとき、少なくとも7、少なくとも10、少なくとも15、少なくとも20、少なくとも30、少なくとも50、少なくとも100、少なくとも150、少なくとも200、少なくとも250、少なくとも300、少なくとも350、少なくとも400、少なくとも450、少なくとも500、少なくとも550、少なくとも600、少なくとも650または少なくとも700ヌクレオチド長を有するヌクレオチド配列を意味する。
【0052】
用語「ハイブリダイズ」は、本明細書において使用するとき、常套のハイブリダイゼーション条件下、好ましくはストリンジェントな条件下での、例えばハイブリダイゼーションSambrook and Russell (2001), Molecular Cloning: A Laboratory Manual, CSH Press, Cold Spring Harbor, NY, USAに記載のハイブリダイゼーションを意味する。さらに特定されていない限り、条件は、好ましくは非ストリンジェントである。ハイブリダイゼーション条件は、例えばSambrook (2001) loc. citに記載の、常套のプロトコルに従って確立できる。条件設定は十分に当業者の技術の範囲内であり、当該技術分野において記載のプロトコルに従って決定できる。したがって、特異的にのみハイブリダイズする配列の検出は、通常、ストリンジェントなハイブリダイゼーションおよび線条条件を必要とする。非限定的な例として、高いストリンジェントなハイブリダイゼーションは、次の条件下で生じうる:
【表3】
【0053】
ホモログまたは正確に相補的でない配列の検出のための低いストリンジェントなハイブリダイゼーション条件は、例えば6 x SSC、1% SDS、65℃に設定できる。周知のとおり、プローブの長さおよび決定する核酸の組成は、ハイブリダイゼーション条件のさらなるパラメーターを構成する。
【0054】
本明細書において提供されるポリヌクレオチド配列とハイブリダイズすることができるポリヌクレオチドも、本発明の一部であり、例えば動物のゲノムライブラリーもしくはcDNAライブラリーまたは微生物のDNAライブラリーから単離できる。好ましくは、かかるポリヌクレオチドは微生物起源のもの、特にプロテオバクテリアクラスに属する微生物、特にデルタプロテオバクテリア、特にミキソコッカス目(Myxococcales)、特にソランギウム亜目(Sorangiineae)、特にポリアンギウム科(Polyangiaceae)、とりわけコンドロマイセス属(Chondromyces)、例えばコンドロマイセス・クロカツス(Chondromyces crocatus)またはその改良株のものである。
【0055】
あるいは、かかる本発明の変異ヌクレオチド配列は、遺伝子操作または化学合成によって作成してもよい。ハイブリダイズ可能なかかるポリヌクレオチド配列は、本明細書に記載のポリヌクレオチド配列またはその一部もしくは逆相補体を用いて、例えば標準的な方法によるハイブリダイゼーション(例えばSambrook and Russell (2001), Molecular Cloning: A Laboratory Manual, CSH Press, Cold Spring Harbor, NY, USA参照)によって、同定および単離できる。列挙した配列番号またはその一部/フラグメントと同じまたは実質的に同じヌクレオチド配列を含むヌクレオチド配列は、例えばハイブリダイゼーションプローブとして使用できる。フラグメントはNRPS遺伝子クラスターの診断、配列決定またはクローニングのためのプローブまたはプライマーとしても有用でありうる。ハイブリダイゼーションプローブとして使用されるフラグメントは、通常の合成技術によって作成される合成フラグメントであってもよく、その配列は本発明のヌクレオチド配列のものと実質的に同一である。
【0056】
本明細書において使用するとき、2つの配列間の同一性パーセントは、2つの配列の最適なアラインメントのために導入する必要のあるギャップ数および各ギャップの長さを考慮し、当該配列によって共有される同一位置の数の関数である(すなわち、同一性パーセント=同一位置数/全位置数×100)である。配列の比較および2つの配列間の同一性パーセントの決定は、下記のような、数学的アルゴリズムを用いて行うことができる。
【0057】
好ましくは、同一性/相同性の程度は、列挙した配列番号に示すヌクレオチド配列とそれぞれの配列を比較して決定する。比較する配列が同じ長さを有さないとき、相同性の程度は、好ましくは、より長い配列におけるヌクレオチド残基と同一であるより短い配列のヌクレオチド残基のパーセンテージを意味する。通常は、相同性の程度は、ClustalW分析を備えたDNASTARプログラムのような既知のコンピュータープログラムを用いて決定できる。このプログラムはDNASTAR, Inc., 1228 South Park Street, Madison, WI 53715から、またはDNASTAR, Ltd., Abacus House, West Ealing, London W13 0AS UKから入手でき(support@dnastar.com)、EMBL外部ステーションのサーバーでアクセス可能である。
【0058】
特定の配列が例えば参照配列と80%同一であるかを決定するためにClustal分析法を用いる場合、好ましくは、アミノ酸配列の比較のための設定は次の通りである:マトリックス: blosum 30; オープンギャップペナルティ: 10.0; エクステンドギャップペナルティ: 0.05; ディレイダイバージェント: 40; ギャップ分離距離: 8 。ヌクレオチド配列比較については、エクステンドギャップペナルティは好ましくは5.0に設定する。
【0059】
配列比較によって比較される2つのヌクレオチド配列が異なる場合、同一性は、より短い配列とより短い配列にマッチするより長い配列の一部を意味する。換言すれば、比較する配列が同じ長さを有さない場合、同一性の程度は、好ましくは、より長い配列におけるヌクレオチド残基と同一であるより短い配列におけるヌクレオチド残基のパーセンテージまたはより短い配列におけるヌクレオチド残基と同一であるより長い配列におけるヌクレオチドのパーセンテージのいずれかを意味する。この文脈において、当業者は、より短い配列に「マッチ」するより長い配列の一部を、容易に決定することができる。
【0060】
一般に、どのように核酸分子を、例えば天然源から得るか、あるいは合成またはPCRのような組換え技術によって作成できるかを、当業者は知っている。これらの核酸分子は、関連文献に記載の技術を適用して得ることができる修飾または誘導体化された核酸分子を含む。
【0061】
さらに、同一性は、対応するヌクレオチド配列またはポリペプチド(例えばそれによってコードされるポリペプチド)間の機能的および/または構造的均等性が存在することを意味する。本明細書に記載の特定のヌクレオチド/アミノ酸配列と少なくとも50%、55%、60%、好ましくは少なくとも70%、より好ましくは少なくとも80%、85%、90%、95%、最も好ましくは少なくとも99%の同一性を有するヌクレオチド/アミノ酸配列は、好ましくは同じ生物学的機能を有し、これらの配列の誘導体/変異体を意味しうる。それらは天然に生じる変異体、例えば生態型、変種、種等に由来する配列または変異体であってよく、当該変異体は、天然に形成されるかあるいは計画的突然変異誘発によって作成してよい。さらに、変異体は合成的に作成された配列であってもよい。アレル変異体は天然に生じる変異体または合成によって作成した変異体または組換えDNA技術によって作成した変異体であってよい。上記ポリヌクレオチドからの逸脱は、例えば欠失、置換、付加、挿入および/または組換えによって作成できる。用語「付加」は、所定の配列の末端に少なくとも1個の核酸残基/アミノ酸を加えることを意味し、一方で「挿入」は、所定の配列中に少なくとも1個の核酸残基/アミノ酸を挿入することを意味する。
【0062】
変異体ポリペプチドおよび特に本発明のヌクレオチド配列の異なる変異体によってコードされるポリペプチドは、好ましくは、共通して有するある特徴を示す。これらは、例えば生物学的活性、分子量、免疫応答、コンホメーション等および物理的特性、例えばゲル電気泳動における泳動動態、クロマトグラフでの動態、沈降係数、溶解性、分光学的特性、安定性、至適pH、至適温度等を含む。
【0063】
ある具体的な態様において、本発明は、下記1個以上のNRPS1ドメインの活性を保持する1個以上の発現産物をコードするポリヌクレオチドを提供する:
(i)配列番号31のローディングドメイン;
(ii)配列番号33のグルタミンのアデニレーションドメイン;
(iii)配列番号35のチオレーションドメイン;
(iv)配列番号37のコンデンセーションドメイン;
(v)配列番号39のスレオニンのアデニレーションドメイン;
(vi)配列番号41のチオレーションドメイン;
(vii)配列番号43のコンデンセーションドメイン;および
(viii)配列番号45のロイシンのアデニレーションドメイン。
【0064】
特定の態様において、ポリヌクレオチドは上記全てのNRPS1ドメインの活性を保持する1個以上の発現産物をコードする。
【0065】
別の態様において、ポリヌクレオチドは、1、2または3個のアデニレーションドメインが異なるアミノ酸特異性を有する1個以上のアデニレーションドメインに置換されていることを除き、上記全てのNRPS1ドメインの活性を保持する1個以上の発現産物をコードする。
【0066】
別の具体的な態様において、本発明は、下記1個以上のNRPS2ドメインの活性を保持する1個以上の発現産物をコードするポリヌクレオチドを提供する:
(i)配列番号47のチオレーションドメイン;
(ii)配列番号49のコンデンセーションドメイン;
(iii)配列番号51のプロリンのアデニレーションドメイン;
(iv) 配列番号53のチオレーションドメイン;
(v) 配列番号2のコンデンセーションドメイン;
(vi) 配列番号4のイソロイシンのアデニレーションドメイン;
(vii) 配列番号6のチオレーションドメイン;
(viii) 配列番号8のコンデンセーションドメイン;
(ix) 配列番号10のチロシンのアデニレーションドメイン;
(x) 配列番号12のN−メチレーション ドメイン;
(xi) 配列番号14のチオレーションドメイン;
(xii) 配列番号55のコンデンセーションドメイン;
(xiii)配列番号57のイソロイシンのアデニレーションドメイン;
(xiv) 配列番号59のチオレーションドメイン;および
(xv) 配列番号61のチオエステラーゼドメイン。
【0067】
特定の態様において、ポリヌクレオチドは上記全てのNRPS2ドメインの活性を保持する1個以上の発現産物をコードする。別の態様において、ポリヌクレオチドは、1、2、3または4個のアデニレーションドメインが異なるアミノ酸特異性を有する1個以上のアデニレーションドメインに置換されていることを除き、上記全てのNRPS1ドメインの活性を保持する1個以上の発現産物をコードする。
【0068】
NRPS1をコードするORF6、NRPS2をコードするORF7およびチトクロムP450をコードするORF8は、式(I)または(I’)のデプシペプチドの生合成のためのコア酵素をコードすると推定される。したがって、さらなる局面において、本発明は、
(i) 配列番号27(NRPS1)をコードするヌクレオチド配列またはその機能的変異体;および
(ii) 配列番号29(NRPS2)をコードするヌクレオチド配列またはその機能的変異体
を含むポリヌクレオチドに関する。
【0069】
ポリヌクレオチドは、さらに、配列番号63をコードするヌクレオチド配列またはその機能的変異体を含んでいてもよい。ある具体的な態様において、これらのポリヌクレオチドは、受託番号DSM19329を有するコンドロマイセス・クロカツス(Chondromyces crocatus)株NPH−MB180から単離される。
【0070】
2.式(I)または(I’)の化合物の生産に関与するNRPSおよび他のポリペプチド
本発明はさらに、本発明のポリヌクレオチドによってコードされるポリペプチド、特に表1に記載のもの、例えばNRPS1およびNRPS2に関する。本発明はさらにそれらの機能的フラグメントおよび機能的変異体に関する。
【0071】
本発明はまた、配列番号2、4、6、8、10、12、14、17、19、21、23、25、27、29、31、33、35、37、39、41、43、45、47、49、51、53、55、57、59、61、63のポリペプチドの変異体またはその少なくとも50、75、100、150、200、300、400または500連続アミノ酸を含むフラグメントに関する。用語「変異体」は、誘導体またはこれらのポリペプチドのアナログを含む。特に、変異体は、配列番号2、4、6、8、10、12、14、17、19、21、23、25、27、29、31、33、35、37、39、41、43、45、47、49、51、53、55、57、59、61、63のポリペプチドとアミノ酸配列において、1、2、3、4、5またはそれ以上の置換、付加、欠失、融合およびトランケーション(これらは任意の組合せで存在していてもよい)によって異なっていてよい。
【0072】
変異体は天然に存在するかあるいはインビトロで作成できる。特に、かかる変異体は遺伝子操作技術、例えば部位特異的突然変異誘発、ランダム突然変異誘発、エキソヌクレアーゼIII欠失法および標準的なクローニング技術を用いて作成できる。あるいは、かかる変異体、フラグメント、アナログまたは誘導体は、化学合成または修飾法を用いて作成してもよい。
【0073】
変異体を作成する他の方法も当業者に周知である。これらには、天然単離体から得られた核酸配列を修飾して、工業的または研究的利用価値を高めた特性を有するポリペプチドをコードする核酸を作成する手法が含まれる。かかる手法において、天然単離体から得られる配列について1個以上のヌクレオチドの相違を有する多数の変異体配列が作成され、特徴付けられる。好ましくは、これらのヌクレオチドの相違は、天然単離体からの核酸によってコードされるポリペプチドに対して、アミノ酸の変化を引き起こす。
【0074】
例えば、変異体は、エラープローンPCRを用いて作成できる。エラープローンPCRにおいて、DNA増幅は、DNAポリメラーゼの忠実度が低く、したがってPCR産物の全長にわたって点突然変異が高い確率で得られるような条件下で実施される。エラープローンPCRはLeung, D.W., et al., Technique, 1:11-15 (1989)およびCaldwell, R. C. & Joyce G.F., PCR Methods Applic., 2:28-33 (1992)に記載されている。変異体は、部位特異的突然変異誘発を用いて、目的のクローン化されたDNAセグメントに部位特異的変異を作成することで作成してもよい。オリゴヌクレオチド突然変異誘発はReidhaar-Olson, J.F. & Sauer, R.T., et al., Science, 241:53-57 (1988)に記載されている。米国特許第6,361,974号及び第6,372,497号に記載のもののような定向進化戦略を用いて変異体を作成してもよい。配列番号2、4、6、8、10、12、14のポリペプチドの変異体は、配列番号2、4、6、8、10、12、14、17、19、21、23、25、27、29、31、33、35、37、39、41、43、45、47、49、51、53、55、57、59、61、63のポリペプチドのアミノ酸残基の1、2、3、4、5またはそれ以上が保存的または非保存的アミノ酸残基(好ましくは保存的アミノ酸残基)で置換されており、かかる置換アミノ酸残基は遺伝コードによってコードされるかされなくてよい、変異体であってよい。
【0075】
保存的置換は、あるポリペプチドにおける所定のアミノ酸を、同様の特性の別のアミノ酸で置換することである。典型的には、保存的置換は以下の置換として見られる:Ala、Val、LeuおよびIleのような脂肪族アミノ酸の別の脂肪族アミノ酸での置換;SerのThrでの置換またはThrのSerでの置換;AspまたはGluのような酸性残基の別の酸性残基での置換;AsnまたはGlnのようなアミド基を有する残基の別のアミド基を有する残基での置換;LysまたはArgのような塩基性残基の別の塩基性残基での交換;およびPheまたはTyrのような芳香族性残基の別の芳香族性残基での置換。
【0076】
他の変異体は、配列番号2、4、6、8、10、12、14、17、19、21、23、25、27、29、31、33、35、37、39、41、43、45、47、49、51、53、55、57、59、61、63のポリペプチドの1個以上のアミノ酸残基が置換基を含むものである。さらに別の変異体は、ポリペプチドが別の化合物、例えばポリペプチドの半減期を延長させる化合物(例えばポリエチレングリコール)と結合しているものである。さらなる変異体は、ポリペプチドにさらなるアミノ酸、例えばリーダー配列、分泌配列、前タンパク質配列またはポリペプチドの精製、富化もしくは安定化を促進する配列と融合しているものである。
【0077】
ある態様において、フラグメント、誘導体およびアナログは、配列番号2、4、6、8、10、12、14、17、19、21、23、25、27、29、31、33、35、37、39、41、43、45、47、49、51、53、55、57、59、61、63のポリペプチドと同じ生物学的機能または活性を保持している。用語「そのフラグメント」は、ポリペプチドの文脈において本明細書において使用するとき、本発明のポリヌクレオチド(例えばそれぞれ配列番号1、3、5、7、9、11、13、15、16、18、20、22、24、26、28、30、32、34、36、38、40、42、44、46、48、50、52、54、56、58、60、62)によってコードされてもよい本発明に定義のポリペプチド(例えばそれぞれ配列番号2、4、6、8、10、12または14、17、19、21、23、25、27、29、31、33、35、37、39、41、43、45、47、49、51、53、55、57、59、61、63のとおり)と実質的に同じ(生物学的)活性を有する機能的フラグメントを意味する。
【0078】
他の態様において、フラグメント、誘導体およびアナログは、少なくとも1、2、3、4、5、6または7このアデニレーションドメインが異なるアデニレーションドメインによって置換されておりそれによって異なるアミノ酸特異性が与えられていることを除き、配列番号2、4、6、8、10、12、14、17、19、21、23、25、27、29、31、33、35、37、39、41、43、45、47、49、51、53、55、57、59、61、63のポリペプチドと同じ生物学的機能または活性を保持する。
【0079】
他の態様において、フラグメント、誘導体またはアナログは、フラグメント、誘導体またはアナログから離れて全体または一部で酵素的に切断されうるポリペプチドの精製、富化、検出、安定化または分泌を促進する融合異種配列を含む。
【0080】
本発明の別の局面は、配列番号2、4、6、8、10、12、14、17、19、21、23、25、27、29、31、33、35、37、39、41、43、45、47、49、51、53、55、57、59、61、63のポリペプチドまたはその少なくとも50、75、100、150、200、300、400もしくは500連続アミノ酸を含むフラグメントの一つと、少なくとも60%、少なくとも70%、少なくとも80%、少なくとも90%または少なくとも95%の同一性を有するポリペプチドまたはそのフラグメントである。アミノ酸「同一性」は、上記のもののような保存的置換を含むと理解される。
【0081】
配列番号2、4、6、8、10、12、14、17、19、21、23、25、27、29、31、33、35、37、39、41、43、45、47、49、51、53、55、57、59、61、63のポリペプチドまたはその少なくとも少なくとも50、75、100、150、200、300、400もしくは500連続アミノ酸を含むフラグメントの一つと相同性を有するポリペプチドまたはフラグメントは、上記技術を用いてそれらをコードする核酸を単離して得ることができる。
【0082】
あるいは、生化学的富化または精製手法によって、相同なポリペプチドまたはフラグメントを得ることができる。潜在的に相同なポリペプチドまたはフラグメントの配列は、タンパク質消化、ゲル電気泳動および/またはミクロシークエンシングによって決定できる。推定される相同なポリペプチドまたはフラグメントの配列は、配列番号2、4、6、8、10、12、14、17、19、21、23、25、27、29、31、33、35、37、39、41、43、45、47、49、51、53、55、57、59、61、63のポリペプチドまたはその少なくとも50、75、100、150、200、300、400もしくは500連続アミノ酸を含むフラグメントの一つと比較できる。
【0083】
配列番号2、4、6、8、10、12、14、17、19、21、23、25、27、29、31、33、35、37、39、41、43、45、47、49、51、53、55、57、59、61、63のポリペプチドまたはその少なくとも50、75、100、150、200、300、400もしくは500連続アミノ酸を含むフラグメント、その少なくとも40、50、75、100、150、200もしくは300連続アミノ酸を含むフラグメントは、様々な用途に使用できる。例えばポリペプチドまたはそのフラグメント、誘導体もしくはアナログを用いて、本明細書の別の箇所に記載の生化学反応を触媒することができる。
【0084】
配列番号2、4、6、8、10、12、14、17、19、21、23、25、27、29、31、33、35、37、39、41、43、45、47、49、51、53、55、57、59、61、63のポリペプチドまたはその少なくとも50、75、100、150、200、300、400もしくは500連続アミノ酸を含むフラグメントを用いて、ポリペプチドまたはフラグメント、誘導体もしくはアナログに特異的に結合する抗体を作成してもよい。
【0085】
具体的な態様において、本発明のポリペプチド(例えばそれぞれ配列番号2、4、6、8、10、12または14、17、19、21、23、25、27、29、31、33、35、37、39、41、43、45、47、49、51、53、55、57、59、61、63)を組み合わせて用いてよい。
【0086】
用語「活性」または「機能性」は、本明細書において使用するとき、特に酵素活性、例えばNRPS1およびNRPS2のペプチドシンターゼ活性を誘発するポリペプチドまたはそのフラグメントの能力を意味する。当業者は本明細書に記載の機能性の(生物学的)活性はしばしば発現レベル(例えばタンパク質/mRNA)と相関すると気付くであろう。異なることが記載されていない限り、用語「発現」は、本明細書において用いるとき、本発明のポリペプチド/タンパク質(またはそのフラグメント)をコードする核酸分子の発現を意味し、一方で「活性」は、当該ポリペプチド/タンパク質の活性を意味する。本明細書に記載のポリペプチドの活性を測定する方法/アッセイは、当該技術分野において周知である。
【0087】
3.発現ベクター、組換え宿主細胞および式(I)または(I’)のデプシペプチドを製造する方法
本明細書に記載の本発明のポリヌクレオチドは、例えば式(I)または(I’)の化合物の異種発現に有用である。具体的な態様において、それらは、式(I’)の化合物の異種発現に有用である。
【0088】
したがって、さらなる局面において、本発明は、本明細書に記載の核酸分子を含むベクター、より具体的には発現ベクター、ならびに当該核酸分子および/またはベクターを含む組換え宿主細胞に関する。
【0089】
用語「ベクター」は、本明細書において使用するとき、特にプラスミド、コスミド、細菌人工染色体(BAC)、酵母人工染色体、ウイルス、バクテリオファージおよび遺伝子工学において一般に使用される他のベクターを意味する。好ましい態様において、本発明のベクターは、細胞、例えば真菌細胞、酵母もしくは細菌細胞のような微生物の細胞または動物細胞の形質転換に好適である。「発現ベクター」は、核酸を宿主細胞に導入して導入した配列の発現を引き起こすビークルを意味する。
【0090】
本明細書に記載のとおり、ポリペプチドは、コード配列が好適な宿主細胞においてコードされるポリペプチドの発現を駆動することができる配列と作動可能に連結するように、ポリペプチドをコードする核酸をベクターに挿入して、得ることができる。例えば、発現ベクターは、プロモーター、翻訳開始のためのリボソーム結合部位および転写ターミネーターを含んでいてよい。ベクターは、発現レベルを調節するための適切な配列、複製起点および選択マーカーを含んでいてもよい。細菌におけるポリペプチドまたはそのフラグメントの発現に好適なプロモーターは、E. coli lacまたはtrp プロモーター、lacl プロモーター、lacZ プロモーター、T3 プロモーター、T7 プロモーター、gpt プロモーター、ラムダ PR プロモーター、ラムダ PL プロモーター、解糖系の酵素、例えば3−ホスホグリセレートキナーゼ(PGK)をコードするオペロン由来のプロモーターおよび酸ホスファターゼプロモーターを含む。真菌プロモーターは、α因子プロモーターを含む。シュードモナス・プチダ(Pseudomonas Putida)における発現に好適なプロモーターは、ゲノムに存在する7つの16S rRNA遺伝子(PP 16SA、PP 16SB、PP 16SC、PP 16SD、PP 16SE、PP 16SF、PP 16SG)の対応転写プロモーター、抗生物質耐性決定要因の転写プロモーター、三価鉄取り込みリプレッサー(Fur)制御遺伝子の転写プロモーターを含むが、これらに限定されない。三価鉄取り込みリプレッサー(Fur)制御プロモーターのより詳細な説明は、以下に提供する。真核生物プロモーターは、CMV前初期プロモーター、HSVチミジンキナーゼプロモーター、熱ショックプロモーター、初期および後期SV40プロモーター、レトロウイルス由来のLTRおよびマウスメタロチオネイン−Iプロモーターを含む。原核生物もしくは真核生物細胞またはウイルスにおいて、遺伝子の発現を制御することが知られている他のプロモーターを用いてもよい。
【0091】
哺乳類発現ベクターは、複製起点、必要なリボソーム結合部位、ポリアデニレーション部位、スプライスドナーおよびアクセプター部位、転写終止配列および5’フランキング非転写配列を含んでいてもよい。ある態様において、SV40スプライスおよびポリアデニレーション部位由来のDNA配列を用いて、必要な非転写遺伝要素を提供することができる。
【0092】
真核生物細胞においてポリペプチドまたはそのフラグメントを発現するためのベクターは、発現レベルを上昇させるエンハンサーを含んでいてもよい。エンハンサーは、転写を増加させるプロモーターとして作用する通常約10〜約300bp長のDNAのシス−作用エレメントである。例えば、複製起点の後側の100〜270bpのSV40エンハンサー、サイトメガロウイルス初期プロモーターエンハンサー、複製起点の後側のポリオーマエンハンサーおよびアデノウイルスエンハンサーを含む。
【0093】
さらに、発現ベクターは好ましくは、ベクターを含む宿主細胞の選択が可能なように1個以上の選択マーカー遺伝子を含む。使用できる選択マーカーの例は、ジヒドロ葉酸レダクターゼをコードする遺伝子または真核細胞培養にネオマイシン耐性を与える遺伝子、大腸菌にテトラサイクリンもしくはアンピシリン耐性を与える遺伝子、ならびに出芽酵母TRP1遺伝子を含む。好適なマーカーの例は、ゲンタマイシン耐性カセットaacClである。他の選択マーカーーは、アンピシリン耐性を与えるヌクレオチドカセット(例えばbla)、クロラムフェニコール耐性を与えるヌクレオチドカセット(例えばcat)、カナマイシン耐性を与えるヌクレオチドカセット(例えばaacC2、aadBまたは他のアミノグリコシド修飾酵素)またはテトラサイクリン耐性を与えるヌクレオチドカセット(例えばtetAまたはtetB)である。
【0094】
適切なDNA配列は、様々な手段でベクターに挿入できる。一般に、DNA配列をベクターの所望の位置にライゲートし、次いで適切な制限エンドヌクレアーゼで挿入物およびベクターを消化する。あるいは、PCRにより適切な制限酵素部位をDNA配列へと設計することができる。様々なクローニング技術がAusbel et al. Current Protocols in Molecular Biology, John Wiley 503 Sons, Inc. 1997 および Sambrook et al., Molecular Cloning: A Laboratory Manual 2d Ed., Cold Spring Harbour Laboratory Press, 1989に記載されている。かかる手法および他のものは、当該技術分野の技術範囲内であると理解される。
【0095】
ベクターは例えば、プラスミド、ウイルス粒子またはファージの形態であってよい。他のベクターは、染色体の誘導体、非染色体および合成DNA配列、ウイルス、細菌プラスミド、ファージDNA、バキュロウイルス、酵母プラスミド、プラスミドおよびファージDNAの組合せに由来するベクター、ワクチンのようなウイルス性DNA,アデノウイルス、鶏痘ウイルスおよび仮性狂犬病ウイルスを含む。原核および真核宿主で使用するための多様なクローニングおよび発現ベクターは、Sambrook et al., Molecular Cloning: A Laboratory Manual, Second Edition, Cold Spring Harbor, N.Y., (1989)に記載されている。
【0096】
使用できる具体的な細菌ベクターは、周知のクローニングベクターの遺伝的要素を含む商業的に入手可能なプラスミド、例えばpBR322 (ATCC 37017)、pKK223-3 (Pharmacia Fine Chemicals, Uppsala, Sweden)、pGEM1 (Promega Biotec, Madison, WI, USA) pQE70、pQE60、pQE-9 (Qiagen)、pD10、phiX174、pBluescript(商標) II KS、pNH8A、pNH16a、pNH18A、pNH46A (Stratagene)、ptrc99a、pKK223-3、pKK233-3、pDR540、pRIT5 (Pharmacia)、pKK232-8 および pCM7を含む。具体的な真核ベクターは、pSV2CAT、pOG44、pXT1、pSG (Stratagene) pSVK3、pBPV、pMSGおよびpSVL (Pharmacia)を含む。しかし、あらゆる他のベクターが、宿主細胞において複製可能であり安定である限り、使用可能である。
【0097】
エレクトロポレーション形質転換、トランスフェクション、形質導入、ウイルス感染、遺伝子銃またはTi介在遺伝子導入を含む多様な技術のいずれかを用いて、ベクターを宿主細胞に導入してよい。適切であるとき、操作された宿主細胞は、プロモーターを活性化させ、形質転換体を選択しまたは本発明の遺伝子を増幅するために適切なように修飾された、常套の栄養培地で培養できる。好適な宿主株の形質転換および適切な細胞密度への当該宿主株の成長の後、選択されたプロモーターを適切な手段(例えば温度変化または化学的誘導)で誘導してよく、所望のポリペプチドまたはそのフラグメントを生産させるためにさらなる期間にわたって当該細胞を培養してよい。
【0098】
さらなる局面において、本発明の組換え宿主細胞は、本発明のポリヌクレオチド配列によってコードされるポリペプチドを発現できるかあるいは発現している。具体的な態様において、宿主細胞に含まれる「ポリペプチド」は、宿主細胞の起源に対して異種であってよい。本発明の宿主細胞を作成するために用いる様々な発現系の例の概略、例えば上記の具体的なものは、例えばGlorioso et al. (1999), Expression of Recombinant Genes in Eukaryotic Systems, Academic Press Inc., Burlington, USA、Paulina Balbas und Argelia Lorence (2004), Recombinant Gene Expression: Reviews and Protocols, Second Edition: Reviews and Protocols (Methods in Molecular Biology), Humana Press, USAに記載されている。
【0099】
本発明のヌクレオチド配列またはベクターによる宿主細胞の形質転換または遺伝子操作は、標準的な方法、例えばSambrook and Russell (2001), Molecular Cloning: A Laboratory Manual, CSH Press, Cold Spring Harbor, NY, USAに記載のとおりに行うことができる。さらに、本発明の宿主細胞は、特にpH値、温度、塩濃度、通気、抗生物質、ビタミン、微量元素等の観点から、使用する具体的な宿主細胞の要求を満たす栄養培地中で培養する。
【0100】
一般に、本発明の宿主細胞は、本発明のヌクレオチド配列、ベクターおよび/またはポリペプチドを含む原核または真核細胞、あるいは本発明のヌクレオチド配列、ベクターおよび/またはポリペプチドを含む細胞に由来する細胞であってよい。好ましい態様において、宿主細胞は、例えば遺伝子操作によって、そのゲノムに組み込まれた本発明のヌクレオチド配列をそれが含むように、本発明のヌクレオチド配列またはベクターを含む。かかる本発明の宿主細胞の非限定的な例(一般的な本発明の宿主細胞でもある)は、細菌、酵母、真菌、植物、動物またはヒト細胞であってよい。
【0101】
用語「宿主細胞」または「単離された宿主細胞」は、その生物が式(I)の化合物または式(I’)の化合物を生産することが知られていようといまいと、その化合物を生産するために必要な遺伝子情報を担持する微生物を意味する。該用語は、本明細書において使用するとき、天然環境に存在するままでその生物に例えば式(I)または(I’)の化合物を生産するための遺伝情報が見られる生物と、遺伝情報が組換え技術によって導入される生物に、等しく適用する。宿主細胞は原核細胞または真核細胞を含む当該技術分野において周知のあらゆる宿主細胞であってよい。適切な宿主の代表例として、次のものを挙げることができる:細菌細胞、例えば大腸菌(E. coli)、ストレプトマイセス・リビダンス(Streptomyces lividans)、ストレプトマイセス・グリセオフスカス(Streptomyces griseofuscus)、ストレプトマイセス・アムボファシエンス(Streptomyces ambofaciens)、枯草菌(Bacillus subtilis)、ネズミチフス菌(Salmonella typhimurium)、ミキソコッカス・キサンタス(Myxococcus xanthus)、ソランギウム・セルロサム(Sorangium cellulosum)、コンドロマイセス・クロカツス(Chondromyces crocatus)およびシュードモナス属、ストレプトマイセス属、バシラス属およびスタフィロコッカス属の多様な種、真菌細胞、例えば酵母、困窮細胞、例えばDrosophila S2およびSpodoptera Sf9、動物細胞、例えばCHO、COSまたはBowes黒色腫、ならびにアデノウイルス。適切な宿主の選択は当業者の能力の範囲内である。
【0102】
本明細書で意図する生物源は、プロテオバクテリアに含まれる生物、好ましくはデルタプロテオバクテリア、より好ましくはミキソコッカス目(Myxococcales)、より好ましくはソランギウム亜目(Sorangiineae)、より好ましくはポリアンギウム科(Polyangiaceae)、最も好ましくはコンドロマイセス属(Chondromyces)、例えばコンドロマイセス・クロカツス(Chondromyces crocatus)またはその改良株のものである。
【0103】
用語「組換え宿主細胞」は、本明細書において使用するとき、本発明のヌクレオチド配列で遺伝子操作されているかまたは本発明のベクターまたはポリペプチドもしくはそのフラグメントを含む宿主細胞を意味する。本発明は、異種組換え宿主細胞、すなわち天然生産株とは異なる別の株において発現される式(I)または式(I’)のデプシペプチドの生産を可能とする。例示として細菌株の使用を説明するが、本明細書に記載のとおり、あらゆる生物または発現系を用いてよい。生物の選択は当業者の必要に応じる。例えば、遺伝子操作を受け入れられる株は、デプシペプチド化合物の修飾および生産を促進するために使用できる。
【0104】
ある具体的な態様において、宿主細胞は、ミキソコッカス属またはシュードモナス属、例えばシュードモナス・プチダから選択される。あるより具体的な態様において、組換え宿主細胞、例えばシュードモナス・プチダは、NRPS1(配列番号27)およびNRPS2(配列番号29)またはその機能的変異体をコードするヌクレオチドを含む。それはさらに、配列番号63のチトクロムP450またはその機能的変異体をコードするヌクレオチド配列を含んでいてもよい。それはまた、配列番号17、配列番号18、配列番号21、配列番号23、配列番号25および配列番号27の1つ以上を含んでいてもよい。有利には、各オープンリーディングフレームは、これらのORFが組換え宿主細胞によって好適な条件下で発現されるように、機能的転写および翻訳配列の制御下に存在する。シュードモナス・プチダにおける異種発現の具体例は、下記実施例にさらに説明されている。
【0105】
上記に従って、本発明は、さらなる態様において、式(I)または(I’)の化合物を生産する方法であって、式(I)または式(I’)、例えば式(II)〜(VII)、(XI)〜(XIV)ならびに(XVII)および(XVIII)の化合物が合成される条件下で組換え宿主細胞を培養し、そして当該化合物を回収することを含む方法に関する。
【0106】
用語「条件」は、本明細書において使用するとき、式(I)の化合物または式(I’)の化合物を発現し、回収するための組換え宿主細胞の培養条件を意味する。ある具体的な態様において、組換え宿主細胞はシュードモナス・プチダである。別の具体的な態様において、組換え宿主細胞はシュードモナス・プチダであり、当該細胞は30℃未満、例えば10〜20℃、例えば約15℃で成長させる。
【0107】
別の具体的な態様において、成長培地はイソ酪酸、例えば1〜5g/lのイソ酪酸、例えば約2g/lのイソ酪酸を含む。
【0108】
例えば、本発明の組換え宿主細胞は、特に、式(I)または式(I’)のデプシペプチドの促進されたかあるいは増加した生産に好適であり得る。
【0109】
4.異種遺伝子発現における鉄制御プロモーターの使用
本発明の別の局面は、宿主細胞、例えばシュードモナス属宿主細胞、例えばシュードモナス・プチダにおける、目的の組換えタンパク質の異種遺伝子発現または合成に関する。例えば、特に組換えタンパク質発現が細菌の増殖を損ねうる場合、増殖の転換期までまたは宿主細胞が健康な集団密度もしくは最も適切な異種遺伝子発現の段階まで、異種遺伝子発現が阻害されるようにそれを制御する必要がある。本発明は、宿主細胞、例えばシュードモナス・プチダにおいて、Fur制御プロモーターによって異種遺伝子発現の制御に成功しうることを示す。かかるプロモーターの使用は、デプシペプチドの生合成遺伝子クラスターの異種発現のために、本明細書に記載されているが、本発明のFur制御プロモーターは、目的の組換えタンパク質の異種遺伝子発現または合成のために、当該技術分野においてより広い用途を有しうる。
【0110】
したがって本発明は、組換え宿主細胞、好ましくは細菌宿主細胞、例えばシュードモナス属、例えばシュードモナス・プチダにおける異種遺伝子発現を制御し、向上させる手段を提供する。
【0111】
ある態様において、本発明は、異種遺伝子発現または目的の組換えタンパク質の合成のための発現カセットに関する。かかる発現カセットは、鉄制御プロモーターと作動可能に連結した、目的の成熟組換えタンパク質をコードするオープンリーディングフレーム(以下、コード配列と称する)を少なくとも含むポリヌクレオチドである。
【0112】
「異種遺伝子発現」の文脈内で本明細書において使用するとき、用語「目的の組換えタンパク質」は、鉄制御プロモーターの制御下では天然には発現されないタンパク質を意味する。好ましい態様において、目的の組換えタンパク質は、酵素、治療用タンパク質(ホルモン、成長因子、抗凝固剤、レセプターアゴニストもしくはアンタゴニストまたはデコイレセプターを含むがこれらに限定されない)、抗体(診断用または治療用を含む)または別の標的結合足場、例えばフィブロネクチン由来のタンパク質、シングルドメイン抗体、一本鎖抗体、ナノボディー等であってよいが、これらに限定されない。
【0113】
発現カセットの文脈において本明細書において使用するとき、用語「作動可能に連結」は、プロモーターがタンパク質をコードするヌクレオチド配列の発現を制御するようにタンパク質をコードするヌクレオチド配列に連結しているプロモーターを含むポリヌクレオチド配列を意味する。
【0114】
本発明の発現カセットは、宿主細胞における目的の組換えタンパク質の好適な発現に必要な他の制御配列、例えば5’非翻訳領域、シグナルペプチド、ポリアデニレーション領域および/または他の3’非翻訳領域をさらに含んでいてよい。
【0115】
4.1 鉄制御プロモーターおよびFur制御プロモーター
ある具体的な態様において、本明細書の第4項に記載の本発明の発現カセットに使用できる鉄制御プロモーターは、培養培地中の鉄の利用能に応答して機能する鉄上流リプレッサー(Fur)またはFurリプレッサータンパク質のホモログから選択されるタンパク質によって部分的または完全に転写的に抑制される、いずれかの細菌プロモーターであり得る。それはさらに、Fur依存的および培養培地中の鉄の利用能に応答してコード配列の発現を制御するように、コード配列と作動可能に連結していてよいFurリプレッサー結合部位を含む、いずれかのプロモーターを含む。細菌Furリプレッサータンパク質の例は、当該技術分野において知られており、例えばCarpenter et al. (2009)に記載されている。
【0116】
本明細書において使用するとき、抑制条件下(すなわちリプレッサーもしくはリプレッサー刺激および/またはリプレッサー結合部位の存在下)でのプロモーター活性が、レポータージーンアッセイ、例えばlacZレポータージーンアッセイで測定したとき、非抑制条件下(すなわち、リプレッサーもしくはリプレッサー刺激および/またはリプレッサー結合部位の非存在下)でのプロモーター活性の少なくとも5倍低いとき、プロモーターは、外部の刺激もしくはシス−エレメントまたはリプレッサーに応答して抑制されている。
【0117】
Furリプレッサー結合部位は当該技術分野において既知であり、多くの細菌、例えば大腸菌、緑膿菌、ネズミチフス菌および枯草菌において発見されている(Carpenter et al. (2002))。他のFurリプレッサー結合部位は、配列番号64のFurリプレッサー結合部位コンセンサス配列とのホモロジーによって探索できる。好ましい態様において、Furリプレッサー結合部位は配列番号64〜68のいずれか1つからなる群から選択される。
【0118】
Fur制御プロモーターは当該技術分野において既知であり、多くの細菌、例えば大腸菌、緑膿菌、コレラ菌、ネズミチフス菌、枯草菌、ヘリコバクター・ピロリ(Helicobacter pylorii)、ヒト結核菌、ブラディリゾビウム・ジャポニクム(Bradyrhizobium japonicum)、リステリア菌(Listeria monocytogenes)、カンピロバクター・ジェジュニ(Campylobacter jejuni)、ストレプトマイセス・セリカラー(Streptomyces coelicolor)、ペスト菌および黄色ブドウ球菌において同定されている(Carpenter et al. (2002))。Fur制御プロモーターの例は、配列番号69〜71のいずれか1つを含むが、それらに限定されない。
【0119】
好ましい態様において、Fur制御プロモーターは、下記群から選択されるポリヌクレオチド配列である:
a) 配列番号69
b) 配列番号69と実質的に同じプロモーター活性を保持する配列番号69のフラグメント;
c) 配列番号69と少なくとも50%、60%、70%、80%、90%または95%の同一性を有する配列番号69の変異プロモーター。
【0120】
ある態様において、配列番号69のフラグメントは、配列番号65または配列番号66のFurリプレッサー結合部位の少なくとも1個および配列番号69の3’下流配列のいずれかを含むフラグメントである。
【0121】
ある態様において、かかる変異プロモーターは、配列番号65もしくは配列番号66と同一または配列番号65および配列番号66のFurリプレッサー結合部位における1、2、3、4または5個以上のヌクレオチドの変化を有する、Furリプレッサー結合部位を含む核酸であってよい。
【0122】
他の態様において、配列番号69の変異プロモーターは、配列番号69と実質的に同じ活性を保持する機能的変異体である。具体的な態様において、変異プロモーターは配列番号69と実質的に同じ活性を保持し、配列番号69と少なくとも50%同一であるが、配列番号65および配列番号66と同じかまたは配列番号65および配列番号66とアラインメントしたときそれぞれ1、2、3、4または5個以上のヌクレオチドの変化を有する2つのリプレッサー結合部位を含む機能的変異体である。
【0123】
プロモーターのプロモーター活性を測定し、配列番号69のプロモーター活性と比較するために、lacZレポータージーンアッセイのようないずれかの好適なレポータージーンアッセイを用いて、レポータージーン発現を直接、例えばmRNAレベルを測定して、あるいは間接的に、レポーター酵素活性(例えばβガラクトシダーゼ活性)を抑制および非抑制条件下で測定することがでる。試験プロモーターと配列番号66のプロモーターの間で抑制および非抑制条件下でのかかる活性に顕著な違いがない場合、該試験プロモーターは、配列番号69と実質的に同じプロモーター活性を保持すると言える。
【0124】
4.2 鉄制御プロモーターを有する発現カセットを含む発現ベクターおよび組換え宿主細胞
発現カセットを任意の好適な発現ベクターに挿入できる。Fur制御プロモーターを用いた目的の組換えタンパク質の合成の文脈において、発現ベクターは、核酸を宿主細胞に導入して目的の組換えタンパク質をコードする遺伝子の異種発現を引き起こすことのできるビークルを意味する。
【0125】
それは例えばプラスミド、バクテリオファージまたはコスミドもしくは他の人工染色体、あるいは宿主細胞における組換えタンパク質生産に一般に用いられる他のベクターに由来してよい。かかる発現ベクターは、さらに、発現カセットに加えて、宿主細胞に入るための手段および/または宿主細胞において複製するための手段、および/またはポリペプチドを細胞の表面または細胞の外側に分泌するための手段を含む。発現ベクターは、1種以上の細胞タイプ、例えば少なくとも2種の細胞タイプ、1種の原核細胞タイプおよび1種の真核細胞タイプにおいて複製されるかまたは増殖されるための手段を含んでいてもよい。
【0126】
使用できる具体的な細菌ベクターは、周知のクローニングベクターの遺伝的要素を含む商業的に入手可能なプラスミド、例えばpBR322 (ATCC 37017)、pKK223-3 (Pharmacia Fine Chemicals, Uppsala, Sweden)、pGEM1 (Promega Biotec, Madison, WI, USA) pQE70、pQE60、pQE-9 (Qiagen)、pD10、phiX174、pBluescript(商標) II KS、pNH8A、pNH16a、pNH18A、pNH46A (Stratagene)、ptrc99a、pKK223-3、pKK233-3、pDR540、pRIT5 (Pharmacia)、pKK232-8 および pCM7を含む。具体的な真核ベクターは、pSV2CAT、pOG44、pXT1、pSG (Stratagene) pSVK3、pBPV、pMSGおよびpSVL (Pharmacia)を含む。しかし、あらゆる他のベクターが、宿主細胞において複製可能であり安定である限り、使用可能である。
【0127】
エレクトロポレーション形質転換、トランスフェクション、形質導入、ウイルス感染、遺伝子銃またはTi介在遺伝子導入を含む多様な技術のいずれかを用いて、発現ベクターを宿主細胞に導入してよい。適切であるとき、操作された宿主細胞は、プロモーターを活性化させ、形質転換体を選択しまたは目的の組換えタンパク質をコードする遺伝子を増幅するために適切なように修飾された、常套の栄養培地で培養できる。
【0128】
さらなる局面において、本発明の組換え宿主細胞は、目的の組換えタンパク質を発現できるかあるいは発現している。発明の宿主細胞を作成するために用いる様々な発現系の例の概略、例えば上記の具体的なものは、例えばGlorioso et al. (1999), Expression of Recombinant Genes in Eukaryotic Systems, Academic Press Inc., Burlington, USA、Paulina Balbas und Argelia Lorence (2004), Recombinant Gene Expression: Reviews and Protocols, Second Edition: Reviews and Protocols (Methods in Molecular Biology), Humana Press, USAに記載されている。
【0129】
本発明のヌクレオチド配列またはベクターによる宿主細胞の形質転換または遺伝子操作は、標準的な方法、例えばSambrook and Russell (2001), Molecular Cloning: A Laboratory Manual, CSH Press, Cold Spring Harbor, NY, USAに記載のとおりに行うことができる。さらに、本発明の宿主細胞は、特にpH値、温度、塩濃度、通気、抗生物質、ビタミン、微量元素等の観点から、使用する具体的な宿主細胞の要求を満たす栄養培地中で培養する。
【0130】
一般に、本発明の組換え宿主細胞は、本発明の発現カセットおよび/または発現ベクターを含む原核または真核細胞、または本発明の発現カセットおよび/または本発明の発現ベクターを含む細胞由来の細胞であってよい。
【0131】
したがって本発明は、適切な成長培地条件下での異種遺伝子発現または目的の組換えタンパク質の合成のための組換え宿主細胞であって、ゲノムに融合されているかまたは自律レプリコンとして、上記本発明の発現カセットまたは発現ベクターを含む、組換え宿主細胞に関する。
【0132】
「組換え宿主細胞」は、適切な成長培地条件下での目的の組換えタンパク質の異種発現に好適なあらゆる細胞であってよい。好ましくは、かかる組換え宿主細胞は、細菌細胞である。
【0133】
好ましい態様において、組換え宿主細胞は、上段に記載の鉄制御プロモーターと作動可能に連結した目的の成熟組換えタンパク質をコードするオープンリーディングフレームを含む発現ベクターで形質転換またはトランスフェクトされている細菌宿主細胞である。より具体的な態様において、組換え宿主細胞は、本発明の発現ベクターを含むシュードモナス属、例えばシュードモナス・プチダ、最も好ましくは、シュードモナス・プチダKT2440から選択され、ここで、鉄制御プロモーターは配列番号69〜71のいずれか1つまたはそのいずれかの機能的変異プロモーターからなる群から選択される。
【0134】
本発明はさらに、例えば目的の組換えタンパク質の合成における、異種遺伝子発現のための上記発現カセット、発現ベクターおよび/または組換え宿主細胞の使用に関する。
【0135】
4.3 異種遺伝子発現方法
鉄制御プロモーターを含む本発明の組換え宿主細胞は、有利には、異種遺伝子発現、例えば目的の組換えタンパク質の合成のために使用できる。好適な宿主細胞の形質転換および当該宿主細胞の適切な細胞密度への増殖の後、Fur制御プロモーターを適切な手段(例えばFeキレート剤、Fe飢餓)で抑制でき、当該細胞をさらなる期間培養して目的のタンパク質を生産させることができる。
【0136】
したがって、ある態様において、本発明は、宿主細胞、好ましくは細菌宿主細胞、頼好ましくはシュードモナス属における異種遺伝子発現または目的の組換えタンパク質の合成のための方法であって、a) 鉄制御プロモーターを含む発現カセットを含んである宿主細胞を、抑制条件下で培養し、
b) 適切な生産段階で鉄制御プロモーターを抑制解除するために増殖条件を変化させ、
c) 抑制解除条件下で細胞を増殖させて異種遺伝子を発現させ、そして/または目的の組換えタンパク質を合成すること
を含む方法を提供する。
【0137】
具体的な態様において、抑制条件は、成長培地中で十分な濃度の鉄を提供することで得られ、抑制解除条件は、鉄不足条件を作出することで得られる。係る条件は成長期の自然な使用および鉄飢餓によって達成されうる。あるいは、かかる条件は、鉄キレート剤を培地に加えて得られる。
【0138】
あらゆる好適な鉄キレート剤を、鉄制御プロモーターの抑制解除のために使用できる。かかる鉄キレート剤の例は、エチレンジアミンテトラ酢酸(EDTA)、クエン酸塩または鉄取り込みシデロホアとして作用する既知化合物(例えばデスフェリオキサミン、エンテロバクチンまたはバシリバクチン)を含むがこれらに限定されない。好ましい態様において、かかる鉄キレート剤は、2’2’ジピリジルである。キレート剤を培地に、例えば少なくとも成長培地中の鉄濃度と等しい、好ましくはそれの少なくとも3倍の濃度で、加えてよい。
【0139】
4.4 異種遺伝子発現のための鉄制御プロモーターの使用に関する本発明の具体的な態様
態様1:宿主細胞、好ましくは細菌宿主細胞、より好ましくはシュードモナス属宿主細胞における異種遺伝子発現に好適な発現カセットであって、天然には鉄制御プロモーターを制御しない遺伝子と作動可能に連結された鉄制御プロモーターを含む、発現カセット。
【0140】
態様2:鉄制御プロモーターが、鉄取り込み制御リプレッサータンパク質(Fur)からなる群から選択されるタンパク質によって抑制される細菌プロモーター、またはFurリプレッサータンパク質によって転写的に抑制されるいずれかの相同プロモーターである、態様1に記載の発現カセット。
【0141】
態様3:Furリプレッサータンパク質によって抑制されるプロモーターが、以下のものからなる群から選択されるポリヌクレオチド配列である、態様2に記載の発現カセット:
(a) 配列番号69、
(b) 配列番号69と実質的に同じプロモーター活性を保持する配列番号69のフラグメント、
(c) 配列番号69と実質的に同じプロモーター活性を保持する、配列番号69と少なくとも50%同一性を有するポリヌクレオチド配列。
【0142】
態様4:態様1〜3のいずれかの発現カセットを含む、組換え宿主細胞。
態様5:細菌種から選択される、態様4の組換え宿主細胞。
態様6:シュードモナス属、例えばシュードモナス・プチダから選択される、態様5の組換え宿主細胞。
【0143】
態様7:宿主細胞における目的の組換えタンパク質の合成のための、鉄制御プロモーターの使用。
態様8:鉄制御プロモーターが、鉄取り込み制御リプレッサータンパク質(Fur)からなる群から選択されるタンパク質によって抑制される細菌プロモーター、またはFurリプレッサータンパク質によって転写的に抑制されるいずれかの相同プロモーターである、態様7に記載の使用。
【0144】
態様9:Furリプレッサータンパク質によって抑制されるプロモーターが、以下のものからなる群から選択されるポリヌクレオチド配列である、態様7に記載の使用:
(a) 配列番号69、
(b) 配列番号69と実質的に同じプロモーター活性を保持する配列番号69のフラグメント、
(c) 配列番号69と実質的に同じプロモーター活性を保持する、配列番号69と少なくとも50%同一性を有するポリヌクレオチド配列。
【0145】
態様10:目的の組換えタンパク質の合成が成長培地中の鉄濃度を調節することによって制御される、態様7〜9のいずれか1つに記載の使用。
態様11:目的の組換えタンパク質の合成が細菌宿主細胞、好ましくはシュードモナス属、例えばシュードモナス・プチダで行われる、態様7〜10のいずれか1つに記載の使用。
態様12:目的の組換えタンパク質の合成が鉄をキレート化して鉄制御プロモーターを制御解除するのに十分な濃度で鉄キレート剤を培地に加えることによって誘導される、態様7〜11のいずれか1つに記載の使用。
態様13:鉄キレート剤が2’2’ジピリジルである、態様12に記載の使用。
【0146】
5.異種発現によって得られるデプシペプチドおよびその使用
本発明はさらに、上記方法で得られるかまたは得た式(I)または(I’)、例えば式(II)〜(VII)、(XI)〜(XIV)ならびに(XVII)および(XVIII)の化合物に関する。
【0147】
さらなる局面において、本発明は、上記方法で得られるかまたは得た式(I)または(I’)、例えば式(II)〜(VII)、(XI)〜(XIV)ならびに(XVII)および(XVIII)の化合物を含む医薬組成物に関する。
【0148】
医薬組成物は、個々の患者の臨床状態、医薬組成物の送達部位、投与方法、投与スケジュールおよび実施者に知られている他の要因を考慮して、良好な医療実務に一致するように、製剤され、投薬される。本明細書の目的での医薬組成物の「有効量」は、したがって、かかる検討事項によって決定される。
【0149】
当業者は個体に投与される医薬組成物の有効量が、特に、化合物の性質に依存することを理解している。例えば、化合物が(ポリ)ペプチドまたはタンパク質である場合、非経腸的に投与される医薬組成物の用量あたりの総薬学的有効量は、患者体重の約1μgのタンパク質/kg/日〜10mgのタンパク質/kg/日の範囲であるが、上記の通り、これは治療上の裁量の問題である。より好ましくは、この用量は少なくとも0.01mgのタンパク質/kg/日、例えばヒトについて、約0.01〜1mgのタンパク質/kg/日である。連続的に投与する場合、医薬組成物は典型的には、1日あたり1〜4回の注射または例えばミニポンプを用いた連続皮下輸液によって、投薬速度約1μg/kg/時〜約50μg/kg/時で投与する。静脈内バッグ溶液を用いてもよい。変化が観察されるのに必要な処置の長さおよび応答が生じるのに必要な処置後の感覚は、望まれる効果に依存して変化すると考えられる。具体的な量は、当業者に周知の常套の試験によって決定されうる。
【0150】
本発明の医薬組成物は、経口、非経腸、大槽内、腹腔内、局所(粉末、軟膏、ドロップまたは経皮パッチとして)、舌下または経口もしくは経鼻スプレーとして、投与してよい。
【0151】
本発明の医薬組成物は、好ましくは、薬学的に許容される担体を含む。「薬学的に許容される担体」とは、非毒性の固体、半固体または液体増量剤、希釈剤、カプセル化材またはあらゆるタイプの添加剤を意味する。用語「非経腸」は、本明細書において使用するとき、静脈内、筋肉内、腹腔内、胸骨内、皮下および関節内注射および輸液を含む投与形態を意味する。
【0152】
医薬組成物は好適には徐放系で投与する。徐放組成物の好適な例は、造形品の形態の半透ポリマー材料、例えばフィルムまたはマイクロカプセルを含む。徐放性材料は、ポリラクチド(米国特許第3,773,919号、EP 58,481)、L−グルタミン酸とγ−エチル−L−グルタミン酸エステルのコポリマー(Sidman, U. et al., Biopolymers 22:547-556 (1983))、ポリ(2−ヒドロキシエチルメタクリレート)(R. Langer et al., J. Biomed. Mater. Res. 15:167-277 (1981)、およびR. Langer, Chem. Tech. 12:98-105 (1982))、エチレンビニルアセテート(R. Langer et al., Id.)またはポリ−D−(−)−3−ヒドロキシ酪酸(EP 133,988)を含む。徐放医薬組成物は、リポソームに封入した化合物を含んでいてもよい。リポソーム含有医薬組成物は、自体公知の方法:E 3,218,121; Epstein et al., Proc. Natl. Acad. Sci. (USA) 82:3688-3692 (1985); Hwang et al., Proc. Natl. Acad. Sci. (USA) 77:4030-4034 (1980); EP 52,322; EP 36,676; EP 88,046; EP 143,949; EP 142,641; 日本特許出願第83-118008号; 米国特許第4,485,045号および第4,544,545号;ならびにEP 102,324によって製造される。通常は、リポソームは、約30モルパーセントのコレステロール以上の脂質含有量であり、選択された割合は最適な治療のために調節される、小さな(約200〜800オングストローム)単層型のものである。
【0153】
非経腸投与用医薬組成物は、一般に、所望の純度で薬学的に許容される担体と、すなわち使用する用量および濃度で受容者に非毒性であり製剤の他の成分と適合するものと、単位投与注射形態(溶液、懸濁液またはエマルジョン)に混合して製剤する。
【0154】
一般に、医薬組成物の成分を液体担体または微粉化担体またはその両方と均一かつ密接に接触させて、製剤を製造する。次いで、所望により、製品を所望の剤形に造形する。好ましくは、担体は非経腸担体、より好ましくは受容者の血液と等張な溶液である。かかる担体ビークルの例は、水、食塩水、リンゲル液およびデキストロース溶液を含む。非水性ビークル、例えば固定油およびオレイン酸エチルならびにリポソームもここで有用である。担体は好適には、微量の添加物、例えば等張性および化学的安定性を向上させる物質を含む。かかる物質は使用する用量および濃度で受容者に非毒性であり、バッファー、例えばリン酸塩、クエン酸塩、コハク酸塩、酢酸および他の有機酸またはそれらの塩;抗酸化剤、例えばアスコルビン酸;低分子量(約10残基未満)(ポリ)ペプチド、例えばポリアルギニンまたはトリペプチド;タンパク質、例えば血清アルブミン、ゼラチンまたは免疫グロブリン;親水性ポリマー、例えばポリビニルピロリドン;アミノ酸、例えばグリシン、グルタミン酸、アスパラギン酸またはアルギニン;単糖類、二糖類および他の炭水化物、例えばセルロールまたはその誘導体、グルコース、マンノースまたはデキストリン;キレート剤、例えばEDTA;糖アルコール、例えばマンニトールまたはソルビトール;対イオン、例えばナトリウム;および/または非イオン性界面活性剤、例えばポリソルベート、ポロキサマーまたはPEGを含む。
【0155】
治療投与に使用する医薬組成物の成分は滅菌されていなければならない。滅菌は、滅菌濾過膜(例えば0.2ミクロンの膜)による濾過によって容易に達成される。医薬組成物の治療用成分は、一般に、滅菌アクセスポートを有する容器、例えば静脈内溶液バッグまたは皮下注射針により貫通可能なストッパーを有するバイアルに入れる。
【0156】
通常、医薬組成物の成分は、単位または複数投与容器、例えば密封アンプルまたはバイアルに、水溶液または再構成用凍結乾燥製剤として保存される。凍結乾燥製剤の例として、10mlバイアルを滅菌濾過した1%(w/v)水溶液5mlで満たし、得られた混合物を凍結乾燥させる。輸液溶液は、静菌注射用水を用いて、凍結乾燥化合物を再構成して製造する。
【0157】
本発明はまた、上記デプシペプチドまたはその誘導体の、医薬としての使用に関する。例えばがん、特に卵巣がんの処置または炎症性および/または過増殖性および掻痒性皮膚疾患、例えばケロイド、肥厚性瘢痕、ざ瘡、アトピー性皮膚炎、乾癬、膿疱性乾癬、酒さ、ネザートン症候群または他の掻痒性皮膚疾患、例えば結節性痒疹、老人の不特定の痒み、ならびに上皮バリアー機能不全に関連した他の疾患、例えば加齢性皮膚、炎症性腸疾患およびクローン病、ならびに膵炎またはがん、特に卵巣がん、嚢胞性線維症(CF)、慢性閉塞性肺疾患(COPD)、肺線維症、成人呼吸窮迫症候群、慢性気管支炎、遺伝性肺気腫、リウマチ性関節炎、IBD、乾癬、喘息のため。
【0158】
ある態様において、本発明は、炎症性および/または過増殖性および掻痒性皮膚疾患、例えばケロイド、肥厚性瘢痕、ざ瘡、アトピー性皮膚炎、乾癬、膿疱性乾癬、酒さ、ネザートン症候群または他の掻痒性皮膚疾患、例えば結節性痒疹、老人の不特定の痒み、ならびに上皮バリアー機能不全に関連した他の疾患、例えば加齢性皮膚、炎症性腸疾患およびクローン病、ならびに膵炎またはがん、特に卵巣がんの処置用医薬としての、上記デプシペプチドまたはその誘導体の使用に関する。
【0159】
他の態様において、本発明は、嚢胞性線維症(CF)、慢性閉塞性肺疾患(COPD)、肺線維症、成人呼吸窮迫症候群、慢性気管支炎、遺伝性肺気腫、リウマチ性関節炎、IBD、乾癬、喘息の処置用医薬としての、上記デプシペプチドまたはその誘導体の使用に関する。
【0160】
さらに別の態様において、本発明は、炎症性および/または過増殖性および掻痒性皮膚疾患、例えばケロイド、肥厚性瘢痕、ざ瘡、アトピー性皮膚炎、乾癬、膿疱性乾癬、酒さ、ネザートン症候群または他の掻痒性皮膚疾患、例えば結節性痒疹、老人の不特定の痒みの処置用医薬としての、上記デプシペプチドまたはその誘導体の使用に関する。
【0161】
6.本発明のポリペプチドに対する抗体
具体的な態様において、本発明は、本明細書に記載され、定義されている本発明のポリペプチドまたはそのフラグメントに特異的に結合する抗体およびその使用に関する。さらに、かかる抗体は、当該ポリペプチド、特に非リポソームペプチドおよび/または非リポソームペプチドシンターゼ(NRPS)の精製に使用できる。用語「抗体」は、当該技術分野において周知である。
【0162】
本発明の文脈において、用語「抗体」は、本明細書において使用するとき、特に、完全な免疫グロブリン分子および実質的に結合特異性を保持しているかかる免疫グロブリン分子の一部を意味する。さらに、この用語は、キメラおよびヒト化抗体のような修飾および/または改変抗体分子、組換えまたは合成により作成/合成した抗体およびインタクトな抗体、ならびに分離した軽鎖および重鎖、Fab、Fab/c、Fv、Fab’、F(ab’)2のようなその抗体フラグメントを意味する。用語「抗体」はまた、二重機能後退、三重機能抗体および一本鎖Fvs(scFv)または抗体融合タンパク質のような抗体構築物も含む。
【0163】
抗体を作成する技術は当該技術分野において周知であり、例えばHoward and Bethell (2000) Basic Methods in Antibody Production and Characterization, Crc. Pr. Inc.に記載されている。本発明のポリペプチドに対する抗体は、例えばポリペプチド(またはそのフラグメント)を動物に直接注射して、あるいはポリペプチド(またはそのフラグメント)を動物に投与して得ることができる。そのようにして得られた抗体は、ポリペプチド(またはそのフラグメント)それ自体に結合する。この方法において、結合が上記定義の「特異的」である限り、ポリペプチドのフラグメントを用いて全ポリペプチドに結合する抗体を作成することすらできる。
【0164】
本発明の文脈において特に好ましいものは、モノクローナル抗体である。連続細胞系培養によって作成した抗体を提供するモノクローナル抗体の作製のためのあらゆる技術を使用してよい。かかる技術の例は、ヒトモノクローナル抗体を作成するためのハイブリドーマ技術、トリオーマ技術、ヒトB細胞ハイブリドーマ技術およびEBVハイブリドーマ技術を含む(Shepherd and Dean (2000), Monoclonal Antibodies: A Practical Approach, Oxford University Press、Goding and Goding (1996), Monoclonal Antibodies: Principles and Practice - Production and Application of Monoclonal Antibodies in Cell Biology, Biochemistry and Immunology, Academic Pr Inc, USA)。
【0165】
抗体誘導体はまた、ペプチド模倣によって作成してもよい。さらに、一本鎖抗体の作製について記載されている技術(特に米国特許第4,946,778号参照)は、本発明のポリペプチドを特異的に認識する一本鎖抗体を作成するために適用できる。また、トランスジェニック動物を用いて、本発明のポリペプチドに対するヒト化抗体を発現させてもよい。
【0166】
用語「特異的に結合」は、本明細書において使用するとき、タンパク質および他の生物製剤の異種集団の存在下での非リボソームペプチドおよび/または非リボソームペプチドシンターゼ(NRPS)および抗体の存在を決定づける結合反応を意味する。したがって、指定されたアッセイ条件下では、特異的抗体とポリペプチドは互いに結合するが、サンプル中に存在する他の成分とは顕著な量で結合しない。かかる条件下での標的分析物との特異的結合は、特定の標的分析物に対する特異性について選択される結合部分を必要としうる。多様なイムノアッセイフォーマットを用いて、特定の抗原と特異的に反応する抗体を選択できる。例えば、固相ELISAイムノアッセイを通常通りに用いて、分析物と特異的に免疫応答するモノクローナル抗体を選択する。イムノアッセイフォーマットおよび特異的免疫応答を決定するために使用できる条件の説明については、Shepherd and Dean (2000), Monoclonal Antibodies: A Practical Approach, Oxford University Press および/または Howard and Bethell (2000) Basic Methods in Antibody Production and Characterization, Crc. Pr. Inc.を参照されたい。典型的には、特異的または選択的応答は、ノイズよるバックグラウンドシグナルの少なくとも2倍、より典型的にはバックグラウンドの10〜100倍以上である。
【0167】
用語「精製」は、本明細書において使用するとき、複数の混合物から1つのタイプのタンパク質を単離することを意図する一連のプロセスを意味する。タンパク質精製は目的のタンパク質の機能、構造および相互作用の特徴付けに重要である。出発物質は、非限定的な例として、生物学的組織または微生物培養であってよい。精製プロセスの多様な工程が、それを制限するマトリックスからタンパク質を開放し、タンパク質と混合物の非タンパク質部分とを分離し、そして最終的に所望のタンパク質を他の全てのタンパク質から分離することができる。分離工程は、タンパク質のサイズ、物理化学的特性および結合親和性の違いを利用する。
【0168】
以下の非限定的な図、配列および実施例を参照して、本発明をさらに説明する。
【図面の簡単な説明】
【0169】
【図1】本発明のNRPSクラスターから生合成される、コンドロマイセスNPH−MB180によって生産された確認された構造の一覧を示す。
【図2】式(I)または(I’)の化合物をコードするNRPS生合成遺伝子クラスターのドメイン構造を示し、式(II)、(III)、(VI)および(VII)〜(XVII)の化合物の生合成経路案を例示する。L、ローディングドメイン;AQ、アデニレーションドメイン(Gln);T、チオレーションドメイン;C、コンデンセーションドメイン;NM、N−メチレーションドメイン;TE、チオエステラーゼドメイン、APアデニレーションドメイン(Pro);AT、アデニレーションドメイン(Thr);AL、アデニレーションドメイン(Leu);AE、アデニレーションドメイン(Glu);AI、アデニレーションドメイン(Ile);AY、アデニレーションドメイン(Tyr)。
【図3】NRPSセグメントF 10517242における2個のアデニレーションドメインの結合ポケットとそれらの最もマッチする定義されたアデニレーションドメインを並べた、10アミノ酸残基のアラインメントを示す。
【図4】コンドロマイセスNPH−MB180に対するコンドロマイドN−メチレーションドメインのBLASTpアラインメントからの結果を示し、これは、NRPSセグメントF 10517242に位置するN−メチレーションドメインを明らかにする。N−メチレーションドメインモチーフは太字で示す。
【図5】ahp残基を形成するヒドロキシプロリンを含む化合物の推定相互変換を示す。水性条件下では、式(XVIII)で例示されるヒドロキシプロリンと、式(II)で例示されるahp含有化合物間との間で、平衡が存在する。
【図6】P.プチダ KT2440の異種発現培養抽出物のLC−MS分析による式(II)の化合物の検出。HPLCクロマトグラムはMSによって陽イオン(左パネル)および陰イオン(右パネル)検出を示す:A)式(II)参照化合物;B)6日目 LB_D培地;C)6日目 P.プチダ陰性対照。MSスペクトル:D)Aに示すHPLC実行からの式(II)参照化合物;E)6日目 LB_D培地、Bに示すHPLC実行から3.2分でピーク。
【発明を実施するための形態】
【0170】
本発明は以下のヌクレオチドおよびアミノ酸配列に関する:
配列番号1は、Val/Ileコンデンセーションドメインを示す、ドメイン1のアミノ酸配列をコードするヌクレオチド配列を示す。
配列番号2は、Val/Ileコンデンセーションドメインを示す、ドメイン1のアミノ酸配列を示す。
配列番号3は、Val/Ileアデニレーションドメインを示す、ドメイン2のアミノ酸配列をコードするヌクレオチド配列を示す。
配列番号4は、Val/Ileアデニレーションドメインを示す、ドメイン2のアミノ酸配列を示す。
配列番号5は、Val/Ileチオレーションドメインを示す、ドメイン3のアミノ酸配列をコードするヌクレオチド配列を示す。
【0171】
配列番号6は、Val/Ileチオレーションドメインを示す、ドメイン3のアミノ酸配列を示す。
配列番号7は、Tyrコンデンセーションドメインを示す、ドメイン4のアミノ酸配列をコードするヌクレオチド配列を示す。
配列番号8は、Tyrコンデンセーションドメインを示す、ドメイン4のアミノ酸配列を示す。
配列番号9は、Tyrアデニレーションドメインを示す、ドメイン5のアミノ酸配列をコードするヌクレオチド配列を示す。
配列番号10は、Tyrアデニレーションドメインを示す、ドメイン5のアミノ酸配列を示す。
【0172】
配列番号11は、Tyr 6−N−メチレーションドメインを示す、ドメイン6のアミノ酸配列をコードするヌクレオチド配列を示す。
配列番号12は、Tyr 6−N−メチレーション ドメインを示す、ドメイン6のアミノ酸配列を示す。
配列番号13は、Tyrチオレーションドメインを示す、ドメイン7のアミノ酸配列をコードするヌクレオチド配列を示す。
配列番号14は、Tyrチオレーションドメインを示す、ドメイン7のアミノ酸配列を示す。
配列番号15は、それぞれVal/IleおよびTyrのアデニレーションドメイン、コンデンセーションドメインおよびチオレーションドメインならびにTyr 6−N−メチレーションドメインを含むNRPSフラグメントをコードするヌクレオチド配列を示す。
【0173】
本明細書に記載のヌクレオチドおよびアミノ酸配列の機能および推定される役割は、上記表1および下記実施例においてさらに説明される。
【実施例】
【0174】
実施例
下記実施例は本発明を説明する:
【0175】
実施例1:NPH−MB180のゲノム配列;会合および分析。
454配列決定法(ピロホスフェートを利用した配列決定プラットフォーム)を用いてNPH−MB180の完全なゲノムを配列決定して、「ドラフト」配列を作成した。ショットガン配列決定を1回行い、次に対末端配列決定を2回行った。対末端試験は、より伝統的なショットガン法の補完的技術として用いられる。簡潔に述べると、物理的に断片化され、環化された染色体DNAフラグメントを短いDNAアダプターセクションにライゲートする配列決定法である。これにより、互いに約3kb(断片化された環化DNAの平均サイズ)離れている2つの短いリード(約150〜200bp)を与えるアダプターからの配列決定を多岐にわたって行うことができる。2つの別個のコンティグにおける2つの短いリードのオーバーラップ(ホモロジー)によって、非オーバーラップコンティグを一体に連結することができ、未定義ヌクレオチド(N)の進展によって3kb近似に基づいて見積もられたおよその長さと連結することができる。このようにして連結されたコンティグを足場と呼ぶ。全体で、1,295,834の個々のリードを実行して、配列決定された310,674,400塩基が得られた。平均リード長は、このタイプの配列決定法の標準である239塩基であった。これらのリードを集積して、リード間の配列オーバーラップに基づきコンティグを形成した。この努力により15,449,316塩基についての平均コンティグ長8,931 bpのコンティグ4,038個が得られた。対末端試験の使用は、足場を作成するためにオーバーラップし、15,029,556塩基を含む96個の足場の集積をもたらす。平均足場サイズは1,227,671塩基であり、平均足場サイズは156,557塩基であった。
【0176】
式(I)または(I’)のデプシペプチドの生合成に関与するNRPS遺伝子クラスターを同定するために、ゲノムデータを分析した。全体的なアプローチは、サーチクエリーとしてNRPSドメインを用いて96個の足場に対してBLASTサーチ(Altschul et. al. 1990; Gish, W. & States, D.J. 1993)を用いることであった。信頼されるNRPSドメインは、どのアミノ酸が非リボソームペプチドに導入されるかが特異的であり、したがって特定のNRPSクラスターを同定するための良好なマーカーであるため、アデニレーションドメインであった(Marahiel, M.A. et. al. 1997)。一般に、以下の特異性および相対順序:Gln-Thr-Val-Glu-Ile-Tyr+(N-meth.)-Ileを有するその構造アデニレーションドメイン内に含まれるNRPSクラスターを発見すると予測された。さらに、遺伝子クラスターは、イソ酪酸のようなカルボン酸で生合成を開始することができるローディングドメインによって開始すると予測され、さらにクラスターがチオエステラーゼドメインで終わると推測された。また、グルタミン酸残基の酸化を促進して3−アミノ−6−ヒドロキシ−ピペリドン残基(Ahp)を形成する他の生合成ユニットが存在している可能性があった。これらのアクセサリー遺伝子の相対位置は、このクラスター内に存在するならば、予測できなかった。さらに、この領域に存在する1個以上の転写産物を定義する転写開始および終了の位置は、この段階では予測できなかった。
【0177】
実施例2:BLAST分析によるNPH−MB180ゲノム配列における全NRPSアデニレーションドメインの同定。
NRPSクラスターを同定するために頼ったアプローチは、まずNPH−MB180ゲノムの全NRPSアデニレーションドメインを同定することであった。NRPSアデニレーションドメインは、それらが使用するアミノ酸について特異的であり、したがってこれらのドメインを分析して、式(I)または(I’)のデプシペプチドを構成するアミノ酸の含有量および相対順序に基づいて正確なNRPSクラスターを同定した。このような目的で、ゲノム配列データにおける全ての可能性のあるNRPSクラスターを同定するための一般アデニレーションドメインの例として、シクロスポリンのバリンアデニレーションドメインを用いた。これは、アミノ酸配列のホモロジーによって全てのNRPSアデニレーションドメインを同定するため、ゲノムのtBLASTn(Altschul et. al. 1990; Gish, W. & States, D.J. 1993)分析を行って達成した。このアプローチにより14個の可能性のあるNRPSクラスター(表2)を同定し、これらのクラスターを含む足場オリジナルBLASTヒット(例えばA 12171827)の開始のヌクレオチド数と共にA−N標識した。このリストから各アデニレーションドメインを同定し、ドメイン特異性を定義する保存アミノ酸残基の分析によって、各ドメインの特異性を決定した(詳細は実施例3を参照)。
【0178】
【表4】
【0179】
この第一の分析は、正確なアデニレーションドメイン組成および生合成経路をコードする予測されたクラスター(〜30kb)の全体のサイズを有するいずれかのNRPSクラスターを同定することができなかった。実際、我々の目的の経路に見られると予測した7個のアデニレーションドメインを含むNRPS経路は同定されなかった。しかし、F 10517242 がイソロイシンおよびチロシンアデニレーションドメイン(表2)を含むことを見出した。なお、この足場(足場#72)は極めて短い(〜7.4kb)が、これは目的のNRPSクラスターの一部であり、クラスターの残りの部分(配列決定ギャップ領域の残基)が配列決定されないままであると仮定した。チロシンアデニレーションドメインと部分Tドメインの間に存在するN−メチレーションドメインの発見は、この仮定にさらなるサポートを与えた(詳細は実施例4参照)。
【0180】
これらのデータに基づいて、ゲノム配列は生合成遺伝子クラスターの全体を含んでいないと結論づけた。実際、5’方向の約20kbおよび3’方向の6kbは不明であると予測できる。
【0181】
実施例3:NRPSアデニレーションドメイン特異性の予測
本明細書に記載のアデニレーションドメインの特異性を、下記一般プロトコルを用いて予測する。シクロスポリンシンターゼ(CssA)のバリンアデニレーションドメインのアミノ酸配列を目的のコンドロマイセスゲノムDNAに対してアラインメントするtBLASTn(Altschul et. al. 1990; Gish, W. & States, D.J. 1993)サーチを用いて、アデニレーションドメインを同定した。ClustalX 複数配列アラインメントソフトウェア(Higgins et. al. 1996)を用いて、Marahiel et. al. (1997)により定義された2つのコアモチーフ(A3およびA6)間のアミノ酸レベルで、翻訳されたコンドロマイセスアデニレーションドメインをGrsA(PheA)(グラミシジンSシンテターゼ)に対してアラインメントした。アデニレーションドメインの結合ポケットを定義し、したがってアミノ酸特異性を決定づける、Marahiel et al.によって報告された10個のアミノ酸を、このアラインメントで同定した。次いでRausch et. al. (2005) および Stachelhaus et. al. (1999)によって報告されたデータを用いて、この10個のアミノ酸を定義されたアデニレーションドメインアミノ酸コードと比較した。
【0182】
生合成クラスターのセグメントにおいて同定された2個のアデニレーションドメインが、イソロイシンおよびチロシンの結合ポケットを定義する10個のアミノ酸と高い相同性を示した(図3)。これらのアミノ酸特異性は絶対ではなく、同様の化学的特徴を有するアミノ酸がしばしば該ドメインを定義するアミノ酸に代えて使用される。これは、1個のNRPSオペロンに沿って合成される構造の可変性に寄与する。本件の場合、イソロイシンアデニレーションドメインはその結合ポケットにバリンも受け入れると推測され、特徴は他の「イソロイシン」アデニレーションドメインについても見られる(Rausch et. al. (2005))。実際、利用可能なNRPS予測ツール(例えば、http://www-ab.informatik.uni-tuebingen.de/software/NRPSpredictor)は一般に、アデニレーションドメインをイソロイシン特異的ともバリン特異的とも宣言することができない。
【0183】
実施例4:NRPSN−メチレーションドメインの予測
N−メチレーションドメインの存在は、下記アプローチを用いて、チロシンアデニレーションドメインと3’方向に直接隣接して位置していると予測された。コンドロマイセス・クロカツスNPH−MB180 コンドロマイドNRPSクラスターのN−メチレーションドメインのアミノ酸配列を用いて、tBLASTn(Altschul et. al. 1990; Gish, W. & States, D.J. 1993)を用いて同様のドメインのゲノムをサーチした。このアプローチを用いて、N−メチレーションドメインはNRPSセグメント内で期待値5e-43 および46%アミノ酸配列同一性を有することが同定された(図4)。さらに、このNRPSセグメントからのN−メチレーションドメインは機能的N−メチレーションドメインにおいて共通して見出される予測アミノ酸モチーフを有することを見出した(von Dohren, H. et. al. 1997; Marahiel, M.A. et. al. 1997)。このデータを確認するため、アナベナ株90アナベノペプチリド生合成クラスター由来のN−メチルトランスフェラーゼApsy−6(Rouhiainen et. al. 2000)を、上記のN−メチルトランスフェラーゼと比較した。この比較のBLASTpの結果は、これらのドメインが期待値2e-65で極めて類似していることを示し、したがって、このドメインの最初の同定が確認された。このドメインがチロシンアデニレーションドメインに直接隣接して存在していることは、NRPS遺伝子クラスターの予測された構造と一致する。さらに、N−メチレーションドメインは比較的珍しく、したがってNRPSセグメント内のこのドメインの存在は、NRPSクラスターに属するこのセグメントについて強い証拠を提供する。
【0184】
実施例5:全生合成NRPS遺伝子クラスターの同定
式(I’)のデプシペプチドの生産に関与する完全な非リボソームペプチド生合成遺伝子を同定し、特徴付けた。足場D 942267に挿入した足場F 10517242からなる足場に、生合成遺伝子を構築した(表1)。足場F 10517242に直接隣接するヌクレオチドの配列分析によって、元のゲノムアセンブリへのこの挿入調節が保証されることが示された後に、これらの足場の組合せを行った。この構築物をPCRで確認し、次いで足場連結領域を介してDNA配列決定した。この足場は式(I’)のデプシペプチドの生合成、修飾および細胞外輸送に関与している可能性がある8個の近接オープンリーディングフレームを含む。さらに、可能性のある分泌型プロテアーゼは、最終的にはデプシペプチドの天然の細胞標的であり得るこれらのオープンリーディングフレーム内に位置し、プロテアーゼ阻害剤であることが示された。これらのORFおよび対応するNRPSドメインの配置は、図2に示す。
【0185】
核非リボソームペプチドオープンリーディングフレーム(ORF6およびORF7)の直前に5個のORFが存在する。ORF1およびORF2は、ソランギウム・セルロサム(Sorangium cellulosum)から報告された2個の異なる特徴付けられていないタンパク質とそれぞれ類似する。これらのタンパク質は仮説機能を有さないが、重要なことに、それらはポリアンギウム科(Polyangiaceae)においてのみ見られるようである。さらに、ORF2と類似するソランギウム(Sorangium)タンパク質は、S. cellulosumゲノムにおいて少なくとも5回発見されている。これらのタンパク質は、ほぼ完全なヌクレオチド配列連結性に基づいて、ORF3と共に転写されると考えられる。ORF3はセリンプロテアーゼ、特にスブチリシン群に属するものと高い配列ホモロジーを有する。我々は、デプシペプチドが高度に特異的なセリンプロテアーゼ阻害剤であることを生化学的に決定し、したがって、デプシペプチドがORF3セリンプロテアーゼの阻害剤であることは真実味がある。逆に、ORF3は、コンドロマイセス株へのデプシペプチド耐性付与に関連しているかもしれない。ORF4およびORF5は、ABCトランスレポータータイプのシデロフォアパーミアーゼおよび一般的な環状ペプチドパーミアーゼと類似する。このパーミアーゼ系は細胞膜を通過するデプシペプチドの排出に関与している可能性がある。実際、これらのORFの5つ全てが細胞膜転位プロセスに関与しており、実際のプロテアーゼとしてそれはプロテアーゼ阻害剤に結合するため、「セリンプロテアーゼ様」ORF3はセリンプロテアーゼファミリーとだけ類似性を有する可能性がある。
【0186】
コアデプシペプチド生合成クラスターはORF6に属し、ORF7へと続く。これらの2個のORFは合わせて15kb長以上である。全NRPS生合成クラスター同様、それらはコンデンセーションドメイン、次いでアデニレーションドメイン、次いでチオレーションドメインからなる一般トポロジーを有する機能的ドメインへと崩壊しうる(Marahiel et. al. 1997)。この3個のドメインモジュールは、通常、ペプチドに取り込まれる各アミノ酸につき、NRPSクラスターで複数回反復される。デプシペプチド生合成クラスターは、ペプチドコアに含まれる7個のアミノ酸からなる7個のかかるモジュラー反復を有するこのパターンに従う。アデニレーションドメインは成長ペプチドにアミノ酸特異性を与え、それらが受け入れ、次いで取り込むアミノ酸を同定するために分析できる。
【0187】
ORF6および7に存在する7個のアデニレーションドメインの予測されるアミノ酸特異性は、1つの例外を有するが、デプシペプチドの最終構造と一般に一致する。第4アデニレーションドメイン(ドメイン 7.3)は、プロリンを成長ペプチドにこの位置で受け入れ、取り込むが、最終ペプチドは非標準アミノ酸、3−アミノ−6−ヒドロキシピペリドン(ahp)をこの位置で含むと予測される。ahpはアナベナ株90によって生産される関連アナベナペプトリドを含む様々なデプシペプチドに存在する(Rouhiainen et. al. 2000)。ahp形成は、アナベナペプトリドにおいて、鎖の4位にグルタミンが取り込まれた後、前のアミノ酸のアミンに戻る反応を起こしてahpを形成することで生じると主張されている(Rouhiainen et. al. 2000)。しかし、ahp特異的アデニレーションドメインは文献にも記載されている(Rausch et al. 2005)。式(II)〜(VII)、(XI)〜(XIII)および(XVII)のMB180株から単離したahp含有デプシペプチドにおいて、我々は、最初にプロリンが成長ペプチドに4位において取り込まれ、次いでオキシドレダクターゼの作用によりahpが形成されるというahp形成の新たなプロセスを仮定する。実際、驚くべきことに、チトクロムP450遺伝子(ORF8)がデプシペプチド生合成クラスターにおいて見出されており、それはNRPS生合成クラスターの直後に位置しており、プロリン残基をヒドロキシル化する変換を触媒している可能性がある。
【0188】
重要なことに、この位置でプロリンを含むデプシペプチドアナログがMB180株から単離されている(式(XIV))。また、数日間水性環境中でインキュベーションすると、ahp含有デプシペプチド(例えば式(II))から自然に5−ヒドロキシプロリンを有するアナログ(式(XVIII))が形成されることも示された。この5−ヒドロキシプロリン形態とahp形態の相互変換が可逆性であることも我々は示した。他のデプシペプチドもこの戦略に従うかは不明であるが、これが我々のMB180株によって使用されるahp形成戦略である可能性がある。
【0189】
デプシペプチド生合成クラスターは、スターターユニットで生合成を開始するローディングドメインを有するORF6で開始する。式(I’)のデプシペプチドにおけるX残基の構造バリエーションに基づき、スターターユニットとして、カルボン酸、例えばCH3CH2CH(CH3)COOH、(CH3)2CHCOOH、C6H5COOH、CH3S(O)CH2COOHまたはCH3COOHが想定できる。
【0190】
非リボソープペプチドにとって小さな酸性残基で開始することは一般的であるが、残基の選択はペプチド毎に大きく異なる。しかし、複雑なカルボン酸スターターユニットは、非リボソープペプチドの間でも比較的稀である。デプシペプチド生合成を開始するために用いるローディングドメインは、構造においても使用するスターターユニットにおいてもアナベナペプトリドローディングドメインと異なっている。実際、デプシペプチドローディングは、標準コンデンセーションドメインと極めて密接に関連しているが、アナベナペプトリドのホルミル基ローディングドメインは前述のホルミルトランスフェラーゼと極めて類似している(Rouhiainen et. al. 2000)。カルボン酸スターターユニットがドメイン6.2によって特定されるグルタミンアミノ酸のアルファアミノ基に濃縮された後、鎖が1回に1アミノ酸成長し続け、次いでNRPS生合成装置を通る(図2)。
【0191】
デプシペプチド生合成装置は、ペプチド結合の2級アミンをメチル化する比較的稀なメチルトランスフェラーゼドメイン(ドメイン7.10)に遭遇するまで、1回に1アミノ酸、狂いなく、単純なNRPSペプチドから、ペプチドを合成する。この場合、これはチロシン由来アミノ基に3級アミンをもたらす。おそらく、このメチル化は、チロシンが成長ペプチドに加えられた後であるが、次のおよび最終アミノ酸が加えられる前に生じる。このことは、N−メチラーゼドメインがチロシン特異的アデニレーションドメインの直後に位置することによって強く示唆されている。
【0192】
最後に、ペプチドが最終チオレーションドメインから除去され、スレオニンのアルコールと末端イソロイシンのアルファケト基の間でエステル結合が形成されて環化する。これは、ORF7に位置する最終ドメインである標準チオエステラーゼドメイン(ドメイン7.15)によって行われる。ahp形成がこのチオエステラーゼ段階の前後いずれに生じるのかは不明である。しかし、この生合成クラスターに含まれる遺伝子は、式(I’)のデプシペプチドの全構造を明らかにするのに十分である。
【0193】
実施例6:シュードモナス・プチダKT2440におけるデプシペプチドの異種発現
ここで、我々は、シュードモナス・プチダKT2440における式(I)または(I’)のデプシペプチドの異種発現を行うためのアプローチの一例を説明する。この宿主は天然生産株C. crocatusと比較して様々な利点、例えば速やかで予測可能な増殖、遺伝子ツールの利用可能性および大規模発酵におけるバリデートされた使用を有する。さらに、この宿主は、C. crocatusと同様のゲノムGC%を有し、天然NRPS系を有する;これら2つの特徴は、異種発現戦略を設計する際に重要な検討事項である。
【0194】
大きな挿入を受け入れることができるコスミドpWEB-TNC(Epicenter Biotechnologies, Madison WI, USA)に生合成遺伝子クラスターをクローン化した;本質は生合成遺伝子クラスターが30kb長を超えることである。生合成遺伝子クラスターのクローニングは、まず、約30〜40kbの直線DNAフラグメントを作成するために生合成遺伝子クラスターの境界の外側を切断する、適切な制限酵素を同定することによって行った。ゲノム配列データの分析により、この目的のために酵素XmnIが適切であり、完全ゲノムDNAフラグメント消化を行うとこのサイズ範囲の異なる15個のDNAフラグメントを作成することが示された。これらの15個のDNAフラグメントのうち、1個の39kbのフラグメントが生合成クラスターを含むと予測された。アガロースゲル電気泳動により、これらの15個のDNAフラグメントを他の染色体消化フラグメントから分離した。ガイドとして適切なサイズのDNA標準を用いて所望のサイズ範囲の15個のDNAフラグメントをゲルから切り出し、製造業者の指示書に従ってコスミドpWEB-TNCにクローン化した。完全な生合成クラスターを含むコスミドクローンは、コロニーPCRで同定し、DNA配列決定で確認した。コスミドまたはBACベクターを用いて完全ゲノムのランダムショットガンライブラリーを作成し、次いで放射線標識化プローブを用いたクローンライブラリーへのコロニーハイブリダイゼーションにより、目的の生合成クラスターを含むクローンライブラリーのメンバーを同定するという別のアプローチも存在する。
【0195】
クローン化した生合成経路を得た後、コスミドクローンに挿入して異種発現を成功させるためにはいくつかの遺伝子成分が必要であった。これらの成分は、i) 異種宿主への移入が成功したことを同定することができる選択マーカー、ii) 異種宿主において機能するプロモーター、iii) 異種宿主への染色体組込みのための部位および iv) pRK2013 oriT配列によって与えられるプラスミド接合伝達性機能(RK2トランスファー機能で使用するため)を含む。本実施例のためにシュードモナス・プチダKT2440に使用するため選択した選択マーカーは、ゲンタマイシン耐性カセットaacCI(Blondelet-Rouault et al. 1997)であった。他の選択マーカーは以下の薬剤耐性を与えるヌクレオチドカセットを含む:アンピシリン(例えばbla)、クロラムフェニコール(例えばcat)、カナマイシン(例えばaacC2、aadBまたは他のアミノグリコシド修飾酵素)またはテトラサイクリン(例えばtetAおよびtetB)。シュードモナス・プチダKT2440における異種発現を駆動するためのプロモーターとして、ここで、フマラーゼC−1(PP 0944)遺伝子プロモーターの使用を記載する(実施例8も参照)。転写プロモーターの選択は、上記抗生物質耐性決定因子のいずれかの転写プロモーターまたはシュードモナス・プチダKT2440において機能的であるいずれかの転写プロモーター、例えばシュードモナス・プチダKT2440ゲノムに存在する7個の16S rRNA遺伝子の転写プロモーター(PP 16SA、PP 16SB、PP 16SC、PP 16SD、PP 16SE、PP 16SF、PP 16SG)、シュードモナス・プチダKT2440鉄取り込みリプレッサー(Fur)制御遺伝子のいずれかの転写プロモーター(fagA (PP 0943)または他のfumC ホモログ、fumC-2 [PP 1755]のプロモーターを含む)、シデロフォアまたはシデロフォア様化合物の生合成および輸送に関与するプロモーター(pvdE [PP 4216]、fpvA [PP 4217]を含む)または遺伝子PP 4243もしくはPP 0946の転写プロモーターを含んでいてよいが、それらに限定されない。ここに記載のフマラーゼC−1プロモーターの使用を含むP. putida由来のプロモーターは、我々の戦略において、RecA介在染色体組込み事象を介してP. putida宿主に染色体組込みの部位を提供することによって、第二の目的に供される。効果的な染色体組込みを促進するために、1046 bpのプロモーター領域がコスミド構築物内に含まれた。下流生合成クラスター遺伝子に転写の促進を可能とするため、プロモーター要素は意図した挿入の3’末端に位置した。プラスミド接合はpSET152からのori Tヌクレオチド配列の取り込みによって促進された。oriT配列は、RK2転写機能がトランスで提供されるとき、コスミドの接合伝達を成功させるために必要かつ十分である。これら3つの遺伝子成分は、バックボーンとしてpUC19を用いて、順次クローン化した(5’−ゲンタマイシン耐性−oriT−fumC1プロモーター−3’)。この異種発現カセットは標準的な分子生物学の実技を用いて作成した。
【0196】
完了後、pUC19から生合成遺伝子クラスターを含むコスミドクローンに異種発現カセットを移した。この挿入は、プロモーター要素を含む挿入物の3’末端が生合成遺伝子クラスターの第一オープンリーディングフレームの翻訳開始コドンから20塩基対離れて位置するように行い、それによってプロモーター要素と生合成遺伝子クラスターの転写融合体を作成した。プロモーターは、遺伝子クラスターの転写を駆動することおよび生合成タンパク質の翻訳を開始するため生合成遺伝子クラスター内に位置する天然リボソーム結合部位に依拠することを意図した。この挿入は、Chaveroche et al. 2000に従って、ラムダREDリコンビナーゼ機能によって介在される異種組換えの使用によって行った。簡潔には、生合成遺伝子クラスターの意図した挿入部位とホモロジーを有する100 ntの隣接領域(PCRプライマーに設計)を有する上記構築物からなるPCR産物を作成した。これらの100 ntの隣接領域は、長フランキングホモロジーPCR(Moore et al. 2005)によって、存在する100 ntの隣接領域に600 nt長のPCR作成隣接領域を加えることにより、さらに延長した。プラスミドpKOBEGhyg(HindIII制限部位にクローン化されたpKOBEGプラスミドのハイグロマイシンカセット含有構築物)からのラムダREDタンパク質を予め発現していた大腸菌EPI100エレクトロコンピテント細胞に、600 ntのホモロジー隣接領域を有する異種発現カセットをエレクトロポレーションした。15μg/mlのゲンタマイシンを補ったLauria培養液寒天で、コスミドへの組込みに成功したトランスコンジュゲートを選択した。このようにして作成した異種発現構築物をPCRおよびDNA配列決定で確認した。効率が低くなるが、コスミドクローンへの異種発現カセットの挿入は、伝統的な制限酵素を利用したクローニング戦略によって行ってもよい。
【0197】
大腸菌ヘルパー株HB101 (pRK2013)に依拠してRK2トランスファー機能を提供する確立された方法(Stanisich and Holloway, 1969)を用いた三親接合により、シュードモナス・プチダKT2440に異種発現構築物を接合により移した。P. putidaトランスコンジュゲートを選択するため75μg/mlのゲンタマイシンならびに大腸菌ドナーおよびヘルパー株の増殖を防止するため25μg/mlのイルガサンを補ったLauria培養液寒天でP. putidaトランスコンジュゲートを選択した。Transconjugates that had successfully integrated into the P. putida chromosome at the fumC-1上流プロモーター領域でのP. putida染色体への組込みに成功したトランスコンジュゲートを、PCR、サザンハイブリダイゼーションおよびDNA配列分析により確認した。
【0198】
2g/Lのイソ酪酸および100μMの2,2,ジピリジル(培地pHは7.0に調節)を含むLauria培養液中、15℃、200rpmの一定回転撹拌で増殖させて、式(II)の化合物の生産を確認した。化学抽出は、6日目に粗発酵液5mLについて1:1の酢酸エチルで行い、次いで30℃で乾燥濃縮し、メタノールで20X最終濃度に再構成した。オンラインDAD、MSおよびMS/MS検出を組み合わせたC−18カラムを用いたHPLC分離により分析を行った。式(II)の化合物は、MSおよびMS/MS検出を用いてはっきりと確認された(図6)。
【0199】
実施例7:5−ヒドロキシプロリンから3−アミノ−6−ヒドロキシ−2−ピペリドン(ahp)への転位のメカニズム
式(I’)のデプシペプチドの重要な生合成経路は、プロリンがデプシペプチド鎖の4位アミノ酸で取り込まれることを示唆している。これはプロリンに代えてahpまたはデヒドロahpを含む式(XIV)の化合物と一致している。我々は、プロリンをヒドロキシル化して、式(XVIII)で例示される5−ヒドロキシプロリンを有する化合物を製造していると仮定するチトクロムP450酵素(orf 8)を同定した。式(XVIII)の化合物は、ahp含有デプシペプチド(例えば式(II))を水性環境中に数日インキュベーションすると自然に形成された(図5)。この5−ヒドロキシプロリン形態とahp形態の相互変換が可逆性であること、そして50℃の水中で10日後にモル比約9:1(ahp:5−ヒドロキシプロリン)で平衡に達することも我々は示した。
【0200】
実施例8:デプシペプチド生合成用遺伝子クラスターの異種遺伝子発現のための、シュードモナス・プチダKT2440由来Fur制御fumC-1プロモーターの使用
宿主シュードモナス・プチダKT2440におけるデプシペプチドの生合成遺伝子の異種発現を成功させるために、異種宿主の遺伝子クラスターの前に置く好適なプロモーターを見つける必要があった。異種宿主、シュードモナス・プチダKT2440からのfur制御プロモーターを選択した(配列番号69)。ほとんどではないが多くの細菌において、標準の複合成長培地(たとえばLB)を用いた場合、成長の転換期は、成長培地中の鉄制限の発生と一致する。我々は、この成長段階で一般にほとんどの二次代謝産物が作成されることが知られているため、健康な集団密度に細胞が達することができるように成長の転換期まで異種宿主におけるデプシペプチドの生合成遺伝子クラスターの転写を遅延させることが有利であると考えた。鉄制限に応答して活性化される遺伝子は、しばしば鉄取り込みリプレッサー(Fur)によって制御される。このメタロレギュレーターは、鉄が十分な条件下で制御遺伝子のプロモーター領域と直接結合して物理的にRNAポリメラーゼ結合を防止することによって、遺伝子セットを抑制するFeセンサーとして作用する(Barton et. al. 1996)。鉄が不充分な条件下では、Furはプロモーター領域から遊離して、遺伝子の転写が生じる。したがって、Fur制御プロモーターを用いて、転換期まで異種遺伝子の発現を抑制することができる。
【0201】
我々は、十分な鉄レベルに対して低い鉄レベルに応答して発現される遺伝子の公表されているプロテオーム(Heim et al. 2003)から、シュードモナス・プチダKT2440における潜在的なFur制御遺伝子を同定し、緑膿菌Furリプレッサーコンセンサス部位「gataatgataatcattatc」(配列番号64)(Barton et al. 1996)を用いてそれらの遺伝子の前のプロモーター領域を探索した。Barton et alからの鉄制御プロテオームの試験によって測定したとき、シュードモナス・プチダKT2440において最も高度にアップレギュレートされた遺伝子産物の一つが、2つのP. putidaフマラーゼ酵素の一方をコードするfumC-1の遺伝子産物であった。さらなる調査により、この遺伝子はFur制御であることが既に示されていることが分かった(Hassett et. al. 1997)。したがって我々は、公表されたデータに基づいて、このプロモーター領域が強く、鉄依存的に作用し、鉄レベルが細胞中で低いときにオンになることを期待した。これらの特徴は、fumC-1プロモーター領域をシュードモナス・プチダKT2440における異種遺伝子発現の目的に使用する理想的な候補たらしめた。実施例6および図6に示すように、全生合成遺伝子クラスターの異種遺伝子発現の成功によってかかる仮説が確認された。
【0202】
鉄が不充分な条件は、発酵成長培地中の鉄濃度と同等乃至3倍以上のモル濃度で鉄キレート剤2’2’ジピリジルを発酵培養に加えて得られる。これによりFur制御遺伝子を、2’2’ジピリジルの添加により制御可能な方法で、アップレギュレートできる。例えば、我々は、成長培地LBを用いる異種発現発酵培養において300μMの2’2’ジピリジルを用いた。他の鉄キレート剤、例えばエチレンジアミンテトラ酢酸(EDTA)、クエン酸塩または鉄取り込みシデロフォアとして作用することが知られている化合物(例えばデスフェリオキサミン、エンテロバクチンまたはバシリバクチン)も、発酵培地に鉄が不充分な条件を作り出すための同様の方法において使用できた。あるいは、所定の発酵培地を用いて、鉄レベルを注意深く制御することができた。
【0203】
ここに記載したfumC-1プロモーターの使用の成功と同じように、他のFur制御プロモーターを使用できた。同様にFurリプレッサー結合部位を含むため、例えばFpvA および OmpR-1 の発現を制御するプロモーターを使用できた。かかるプロモーターは、下記実施例9に更に詳細に記載する。Feが不充分な条件下でアップレギュレートされるあらゆる遺伝子の前の他のFur結合部位は、ここに記載のバイオインフォマティクスアプローチを用いるか、あるいはBaichoo et al. (2002)に記載のように、プロモーター領域のDNAに対する精製Furタンパク質の電気泳動移動度シフトアッセイを用いて、同定された。Furファミリーは細菌ドメインに広く拡がっており、プロモーター領域およびそれらのそれぞれのFur結合部位は、一般に、属特異的であり、しばしば種特異的である。したがって、シュードモナス・プチダKT2440のFur制御プロモーター領域が他のシュードモナス種でも機能することは明らかである。
【0204】
実施例9:Fur制御プロモーター
シュードモナス・プチダKT2440からのFur制御プロモーター。Furリプレッサー結合部位は下線で示し、緑膿菌Furリプレッサーコンセンサス部位gataatgataatcattatc (配列番号64)(Barton et al. 1996)に対するコンセンサスヌクレオチド類似性サーチにより同定した。
【0205】
fumC-1 Fur制御プロモーター領域(Furリプレッサー部位には下線を付す)
atcaggccgcgctgattcgccgtatggggcgcgggctgctggtgaccgaactgatggggcatggcttgaacatggtgacgggggactattcccgtggtgcggcggggttctgggtcgagaatggcgagattcagcatgccgtacaggaagtcaccatcgccggaaacatgaaggacatgttccagcagattgtcgcgatcggtagcgatcttgaaacccgtagcaatattcatacgggctcggtgttgatcgagcggatgaccgttgctggtagctgatctttagcctgcgccggccctttcgcgggtaaacccgctcctacacggtggtggacgtacatcggggttggacacaggccgttgtaggagcgggttcacccgcgaagaggccggaacagcactacacctttccctgcaaatccgaagacccggccctcgcgccgggtttttatttcatcacctttttcttgaagtgattctatttatcacttaataatgaatatcattatccagtaacccggcgatgatgttcatgaaatccgtcctccgcgaactgccctacctggaaaactggcgctggctcagccggcgcattcgctgtgcgctcgaccccgacgagccgcgcctgatcgagcattacctggccgaaggccgctatctggtgtgctgcaccgaaacctcgccatggacggtggcgctgacagcgtttcgcctgctgctggataccgcctgcgatcgcatgctcccctggcattggcgttgtctgtgcctggaccaggcgtggcgccctctgctggacctgcgcaacctcgaccgccaggaacagaaccaacgctggcaaccctacgccttgcagttggccaattgccgtctgctgccttcgatttctcccgatgaactgatgcaaggatttgatgatgagtgatacccgtatcgagcg (配列番号69)
FpvA Fur制御プロモーター領域(Furリプレッサー部位には下線を付す)
tccggcgaattttctacacagagctgctgccggacctcaagcgcctgggcaagaccatca
tcgtgataagccacgacgaccgctacttcgacgtcgccgaccagctcatccacatggcgg
caggcaaggtccaacaggagaaccgcgtcgcagattgcatttaatttttccggttttggc
cgatgagtgcgtcccaatcaataacaagaattaatactattaacatctgacactcaaggg
ctttgaaaaa (配列番号70)
OmpR-1 Fur制御プロモーター領域(Furリプレッサー部位には下線を付す)
caggtagcgcaggcgctcttccaggtggcgcaactgagtgtcgtcaaggctaccggtcac
ttccttgcgatagcgggcgatgaagggcacggtcgagccttcgtccaacaggctcacggc
cgcctcgacctgctgcgggcgtacgcccagttcctcggcgatacggctgttgatgctgtc
catgtaaaccacctgacatttgtgaatacgggggtcgcctgtgggctttttgcccggcgg
cgctggatgaaagccgcgcattatacccatcgcaaacggcttgcggtgatggcgcccggc
cagccggaactggcgccgggggaaaaatctgctaacaatgctcacgcaacgtgcagcaat
ggctacgccataatgcgcggcgatatcagaggagttattc (配列番号71)
fumC-1 プロモーターのFurリプレッサー結合部位
aaacatgaaggacatgttc (配列番号65)
aataatgaatatcattatc (配列番号66)
fpvA プロモーターFurリプレッサー結合部位
aataacaagaattaatact (配列番号67)
ompR-1 プロモーターのFurリプレッサー結合部位
cataatgcgcggcgatatc (配列番号68)
【0206】
Fur制御プロモーターおよびそれらのFurリプレッサー部位は、多くの非シュードモナス種から記載され、特徴付けられており、そしてCarpenter et al. (2009)に列挙され、レビューされている。Fur結合は異なる属間で大きく異なりうる。例えば、大腸菌のコンセンサスFur結合部位はGATAATGATAATCATTATC(de Lorenzo et al. 1987)であるが、枯草菌のコンセンサスFur結合部位はTGATAATTATTATCA(Baichoo and Helmann, 2002)である。
【0207】
参考文献
Altschul, S.F., Gish, W., Miller, W., Myers, E.W. & Lipman, D.J. (1990) "Basic local alignment search tool." J. Mol. Biol. 215:403-410.
Baichoo N, Helmann JD. (2002) Recognition of DNA by Fur: a reinterpretation of the Fur box consensus sequence. J Bacteriol. 184(21):5826-32.
Baichoo N, Wang T, Ye R, Helmann JD. (2002) Global analysis of the Bacillus subtilis Fur regulon and the iron starvation stimulon. Mol Microbiol. 45(6):1613-29.
Barton HA, Johnson Z, Cox CD, Vasil AI, Vasil ML. (1996) Ferric uptake regulator mutants of Pseudomonas aeruginosa with distinct alterations in the iron-dependent repression of exotoxin A and siderophores in aerobic and microaerobic environments. Mol Microbiol. 21(5):1001-17.
Binz, T.M., Wenzel, S.C., Schbell, H., Bechthold, A., Mueller, R. (2008) Heterologous expression and genetic engineering of the phenalinolactone biosynthetic gene cluster by using Red/ET recombineering. ChemBioChem. 9: 447-454.
Carpenter BM, Whitmire JM, Merrell DS. (2009) This is not your mother's repressor: the complex role of fur in pathogenesis. Infect Immun. 77(7):2590-601.
de Lorenzo V, Wee S, Herrero M, Neilands JB. (1987) Operator sequences of the aerobactin operon of plasmid ColV-K30 binding the ferric uptake regulation (fur) repressor. J Bacteriol. 169(6):2624-30.
Garrity, P. A., Ligation-Mediated PCR, in PCR 2 A Practical Appraoch, McPherson, M. J. et. al. (Eds.) pp. 309-322 Oxford University Press, New York (1995).
Gish, W. & States, D.J. (1993) "Identification of protein coding regions by database similarity search." Nature Genet. 3:266-272.
Gu, J.Q., Nguyen, K.T., Gandhi, C., Rajgarhia, V., Baltz, R.H., Brian, P., Chu, M. (2007) Structural characterization of daptomycin analogues A21978C1-3(d-Asn11) produced by a recombinant Streptomyces roseosporus strain. J. Nat. Prod. 70: 233-240.
Hassett DJ, Howell ML, Ochsner UA, Vasil ML, Johnson Z, Dean GE. (1997) An operon containing fumC and sodA encoding fumarase C and manganese superoxide dismutase is controlled by the ferric uptake regulator in Pseudomonas aeruginosa: fur mutants produce elevated alginate levels. J Bacteriol. 179(5):1452-9.
Heim S, Ferrer M, Heuer H, Regenhardt D, Nimtz M, Timmis KN. (2003) Proteome reference map of Pseudomonas putida strain KT2440 for genome expression profiling: distinct responses of KT2440 and Pseudomonas aeruginosa strain PAO1 to iron deprivation and a new form of superoxide dismutase. Environ Microbiol. 5(12):1257-69.
Higgins D.G., Thompson J.D., Gibson T.J. (1996). Using CLUSTAL for multiple sequence alignments. Methods Enzymol., 266, 383-402.
Finking, R., Marahiel, MA., (2004) Biosynthesis of nonribosomal polypeptides. Annu Rev. Microbiol. 58: 453-488.
Marahiel, M. A., Stachelhaus, T., Mootz, H.D. (1997) Modular peptide synthetases involved in nonribosomal peptide synthesis. Cem. Rev. 97:2651-2673.
Pfeifer, B.A., Admiraal, S.J., Gramajo, H., Cane, D.E., Khosla, C. (2001) Biosynthesis of complex polyketides in a metabolically engineered strain of E. coli. Science. 291: 1790-1792.
Rausch, C., Weber, T., Kohlbacher, O. Wohlleben, W., Huson, D.H. (2005) Specificity prediction of adenylation domains in nonribosomal peptide synthetases (NRPS) using transductive support vector machines (TSVMs). Nuc. Acids Res. 33: 5799-5808.
Rouhiainen, L., Paulin, L., Suomalainen, S., Hyytiainen, H., Buikema, W., Haselkorn, R., Sivonen, K. (2000) Genes encoding synthetases of cyclic depsipeptides, anabaenopeptilides, in Anabaena strain 90. Mol. Microbiol. 37: 156-167.
Sambrook, J., Maniatis, T. (1989) Molecular Cloning: A Laboratory Manual. Cold Spring Harbor Laboratory Press. Cold Spring Harbor NY.
Shaying Zhao (Ed.), Marvin Stodolsky, Marvin Stodolsky (Ed.) (2003) Bacterial Artificial Chromosomes (Methods in Molecular Biology Series v255-256): Library Construction, Physical Mapping, and Sequencing, Vol. 1 Springer-Verlag New York, LLC.
Shen, B. (2004) Accessing natural products by combinatorial biosynthesis. Sci STKE. Pe14.
Stachelhaus, T. Mootz, H.D., Marahiel, M.A. (1999) The specificity-conferring code of adenylation domains in nonribosomal peptide synthetases. Chem. Biol. 6: 493-505.
Staunton, J., Weissman, K.J. (2001) Polyketide biosynthesis: a millennium review. Nat. Prod. Rep. 18:380-416.
Wenzel, S.C., Mueller, R. (2005) Recent developments towards the heterologous expression of complex bacterial natural product biosynthetic pathways. Curr. Op. Biotechnol. 16: 594-606.
Von Dohren, H., Keller, U., Vater, J., and Zocher, R. (1997) Multifunctional peptide synthetases. Chem. Rev. 97:2675-2705.
Zhang, Y., Muyrers, J. Testa, G., Stewart, F. (1998) A new logic for DNA engineering using recombination in Escherichia coli. Nat. Genet. 20: 123-128.
Zhuang, H., Yong, W., Pfeifer, B.A. (2008) Bacterial hosts for natural product production. Molecular Pharmaceuticals. 5: 212-225.
【図1A】
【図1B】
【図2A】
【図2B】
【図2C】
【図2D】
【図2E】
【図2F】
【図2G】
【特許請求の範囲】
【請求項1】
式(I)または(I’)の化合物の生産に関与する非リボソームペプチドシンターゼであるNRPS2をコードする生合成遺伝子クラスターの1個以上の機能的フラグメントを含むポリヌクレオチドであって、以下のものを含むポリヌクレオチド:
(i) NRPS2ドメインをコードする配列番号1、3、5、7、9、11、13、46、48、50、52、54、56、58および60からなる群から選択される配列と、少なくとも80%、特に少なくとも85%、特に少なくとも90%、特に少なくとも95%、特に少なくとも98%の配列同一性を有するヌクレオチド配列および/またはその相補体;
(ii) NRPS2ドメインをコードする配列番号1、3、5、7、9、11、13、46、48、50、52、54、56、58または60からなる群から選択されるヌクレオチド配列の相補鎖とハイブリダイズするヌクレオチド配列および/またはその相補体;
(iii) NRPS2ドメインを示す配列番号2、4、6、8、10、12、14、47、49、51、53、55、57、59または61からなる群から選択される配列と、少なくとも60%、特に少なくとも70%、特に少なくとも80%、特に少なくとも90%、特に少なくとも95%の配列同一性を有するアミノ酸配列をコードするヌクレオチド配列および/またはその相補体;
(iv) NRPS2ドメインを示す配列番号2、4、6、8、10、12、14、47、49、51、53、55、57、59または61からなる群から選択されるアミノ酸をコードするヌクレオチド配列の相補鎖とハイブリダイズするヌクレオチド配列および/またはその相補体;
(v) 配列番号15、配列番号28からなる群から選択される配列と少なくとも80%、特に少なくとも85%、特に少なくとも90%、特に少なくとも95%、特に少なくとも98%の配列同一性を有するヌクレオチド配列および/またはその相補体;
(vi) 配列番号15、配列番号28からなる群から選択されるヌクレオチド配列の相補鎖とハイブリダイズするヌクレオチド配列および/またはその相補体;
ここで、前記(i) 〜 (vi)のヌクレオチド配列は、配列番号2、4、6、8、10、12、14、47、49、51、53、55、59および/または61の参照配列によって示される対応NRPSドメインの活性を保持する発現産物をコードする。
【請求項2】
以下の1個以上のNRPS2ドメインの活性を保持する発現産物をコードする、請求項1に記載のポリヌクレオチド:
(i)配列番号47のチオレーションドメイン;
(ii)配列番号49のコンデンセーションドメイン;
(iii)配列番号51のプロリンのアデニレーションドメイン;
(iv)配列番号53のチオレーションドメイン;
(v)配列番号2のコンデンセーションドメイン;
(vi)配列番号4のイソロイシンのアデニレーションドメイン;
(vii)配列番号6のチオレーションドメイン;
(viii)配列番号8のコンデンセーションドメイン;
(ix)配列番号10のチロシンのアデニレーションドメイン;
(x)配列番号12のN−メチレーションドメイン;
(xi)配列番号14のチオレーションドメイン;
(xii)配列番号55のコンデンセーションドメイン;
(xiii)配列番号57のイソロイシンのアデニレーションドメイン;
(xiv)配列番号59のチオレーションドメイン;および
(xv)配列番号61のチオエステラーゼドメイン。
【請求項3】
式(I)または(I’)の化合物の生産に関与する非リボソームペプチドシンターゼであるNRPS1をコードする生合成遺伝子クラスターの1個以上の機能的フラグメントを含むポリヌクレオチドであって、以下のものを含むポリヌクレオチド:
(i) NRPS1ドメインをコードする配列番号30、32、34、36、38、40、42および44からなる群から選択される配列と、少なくとも80%、特に少なくとも85%、特に少なくとも90%、特に少なくとも95%、特に少なくとも98%の配列同一性を有するヌクレオチド配列および/またはその相補体;
(ii) NRPS1ドメインをコードする配列番号30、32、34、36、38、40、42および44からなる群から選択されるヌクレオチド配列の相補鎖とハイブリダイズするヌクレオチド配列および/またはその相補体;
(iii) NRPS1ドメインを示す配列番号31、33、35、37、39、41、43、45からなる群から選択される配列と、少なくとも60%、特に少なくとも70%、特に少なくとも80%、特に少なくとも90%、特に少なくとも95%の配列同一性を有するヌクレオチド配列および/またはその相補体;
(iv) NRPS1ドメインを示す配列番号31、33、35、37、39、41、43、45からなる群から選択されるアミノ酸をコードするヌクレオチド配列の相補鎖とハイブリダイズするヌクレオチド配列および/またはその相補体;
(v) 配列番号26からなる群から選択される配列と、少なくとも80%、特に少なくとも85%、特に少なくとも90%、特に少なくとも95%、特に少なくとも98%の配列同一性を有するヌクレオチド配列および/またはその相補体;または
(vi)配列番号26からなる群から選択されるヌクレオチド配列の相補鎖とハイブリダイズするヌクレオチド配列および/またはその相補体;
(vii) ここで、前記(i) 〜 (vi)のヌクレオチド配列は、配列番号31、33、35、37、39、41、43、45の参照配列によって示される対応NRPSドメインの活性を保持する発現産物をコードする。
【請求項4】
以下の1個以上のNRPS1ドメインの活性を保持する発現産物をコードする、請求項3に記載のポリヌクレオチド:
(i) 配列番号31のローディングドメイン;
(ii)配列番号33のグルタミンのアデニレーションドメイン;
(iii)配列番号35のチオレーションドメイン;
(iv)配列番号37のコンデンセーションドメイン;
(v)配列番号39のスレオニンのアデニレーションドメイン;
(vi)配列番号41のチオレーションドメイン;
(vii)配列番号43のコンデンセーションドメイン;および
(viii)配列番号45のロイシンのアデニレーションドメイン。
【請求項5】
式(I)または(I’)の化合物を生産するNRPS2をコードする請求項1または2に記載のポリヌクレオチドであって、配列番号29に示すアミノ酸配列をコードするヌクレオチド配列を含む、ポリヌクレオチド。
【請求項6】
式(I)または(I’)の化合物を生産するNRPS1をコードする請求項3または4に記載のポリヌクレオチドであって、配列番号27に示すアミノ酸配列をコードするヌクレオチド配列を含む、ポリヌクレオチド。
【請求項7】
請求項1〜6のいずれかの1個以上のポリヌクレオチドによってコードされるポリペプチド。
【請求項8】
式(I)または(I’)の化合物を生産するポリペプチドであって、以下のものからなる群から選択されるアミノ酸配列を含むポリペプチド:
(i)NRPS1を示す配列番号27、第2のNRPS2を示す配列番号29、チトクロムP450を示す配列番号63;および
(ii) (i)に列挙した参照配列と60%、特に少なくとも70%、特に少なくとも80%、特に少なくとも90%、特に少なくとも95%の配列同一性を有し、実質的に同じ触媒機能を保持する、(i)に列挙したアミノ酸配列の機能的変異体。
【請求項9】
請求項8の1個以上のポリペプチドをコードするヌクレオチド配列を含むポリヌクレオチド。
【請求項10】
(i)配列番号27またはその機能的変異体をコードするヌクレオチド配列;および
(ii)配列番号29またはその機能的変異体をコードするヌクレオチド配列
を含む、請求項9に記載のポリヌクレオチド。
【請求項11】
さらに配列番号63またはその機能的変異体をコードするヌクレオチド配列を含む、請求項10に記載のポリヌクレオチド。
【請求項12】
受託番号DSM 19329を有するコンドロマイセス・クロカツス(Chondromyces crocatus)株NPH−MB180から単離された、請求項9〜11のいずれか1項に記載のポリヌクレオチド。
【請求項13】
オープンリーディングフレームが転写および翻訳配列と作動可能に連結している、請求項9〜12のいずれか1項のポリヌクレオチドを含む発現ベクター。
【請求項14】
請求項1〜6および9〜12のいずれか1項の組換えポリヌクレオチドの1個以上または請求項13の発現ベクターを含む宿主細胞であって、組換えポリヌクレオチドが宿主細胞のゲノム中で天然には見出されない、宿主細胞。
【請求項15】
配列番号16、配列番号18、配列番号20、配列番号22および配列番号24からなる群から選択される式(I)または(I’)の化合物の生産に必要な1個以上のオープンリーディングフレームまたはそれらの機能的変異体をさらに含む、請求項14に記載の宿主細胞。
【請求項16】
式(I)もしくは(I’)または式(II)〜(VII)、(XI)〜(XIV)ならびに(XVII)および(XVIII)の化合物を生産するための、請求項14または15に記載の宿主細胞。
【請求項17】
組換えポリペプチドが遺伝子発現に最適化するために修飾されている、請求項14〜16のいずれか1項に記載の宿主細胞。
【請求項18】
ポリヌクレオチドのコドン出現頻度が宿主細胞の豊富なタンパク質のコドン出現頻度に調節されている、請求項14〜17のいずれか1項に記載の宿主細胞。
【請求項19】
宿主細胞がミキソコッカス目(Myxococcales)またはシュードモナス属(Pseudomonas)もしくはストレプトマイセス属(Streptomyces)から選択される、請求項14〜18のいずれか1項に記載の宿主細胞。
【請求項20】
宿主細胞がシュードモナス・プチダ(Pseudomonas putida)から選択される、請求項19に記載の宿主細胞。
【請求項21】
式(I)または(I’)の化合物の生産に必要な遺伝子を最早発現しない、変異微生物。
【請求項22】
配列番号63に示すチトクロムP450をコードする遺伝子を最早発現しない、請求項21に記載の変異微生物。
【請求項23】
式(I)もしくは(I’)または式(II)〜(VII)、(XI)〜(XIV)もしくは(XVII)〜(XVIII)の化合物を製造する方法であって、請求項16〜20のいずれか1項の宿主細胞を、当該化合物が生産される条件下で培養することを含む、製造方法。
【請求項24】
請求項23の方法によって得られる、式(I)もしくは(I’)または式(II)〜(VII)、(XI)〜(XIV)もしくは(XVII)〜(XVIII)の化合物。
【請求項25】
薬物として使用するための、請求項24に記載の化合物。
【請求項26】
アトピー性皮膚炎の処置に使用するための、請求項25に記載の化合物。
【請求項1】
式(I)または(I’)の化合物の生産に関与する非リボソームペプチドシンターゼであるNRPS2をコードする生合成遺伝子クラスターの1個以上の機能的フラグメントを含むポリヌクレオチドであって、以下のものを含むポリヌクレオチド:
(i) NRPS2ドメインをコードする配列番号1、3、5、7、9、11、13、46、48、50、52、54、56、58および60からなる群から選択される配列と、少なくとも80%、特に少なくとも85%、特に少なくとも90%、特に少なくとも95%、特に少なくとも98%の配列同一性を有するヌクレオチド配列および/またはその相補体;
(ii) NRPS2ドメインをコードする配列番号1、3、5、7、9、11、13、46、48、50、52、54、56、58または60からなる群から選択されるヌクレオチド配列の相補鎖とハイブリダイズするヌクレオチド配列および/またはその相補体;
(iii) NRPS2ドメインを示す配列番号2、4、6、8、10、12、14、47、49、51、53、55、57、59または61からなる群から選択される配列と、少なくとも60%、特に少なくとも70%、特に少なくとも80%、特に少なくとも90%、特に少なくとも95%の配列同一性を有するアミノ酸配列をコードするヌクレオチド配列および/またはその相補体;
(iv) NRPS2ドメインを示す配列番号2、4、6、8、10、12、14、47、49、51、53、55、57、59または61からなる群から選択されるアミノ酸をコードするヌクレオチド配列の相補鎖とハイブリダイズするヌクレオチド配列および/またはその相補体;
(v) 配列番号15、配列番号28からなる群から選択される配列と少なくとも80%、特に少なくとも85%、特に少なくとも90%、特に少なくとも95%、特に少なくとも98%の配列同一性を有するヌクレオチド配列および/またはその相補体;
(vi) 配列番号15、配列番号28からなる群から選択されるヌクレオチド配列の相補鎖とハイブリダイズするヌクレオチド配列および/またはその相補体;
ここで、前記(i) 〜 (vi)のヌクレオチド配列は、配列番号2、4、6、8、10、12、14、47、49、51、53、55、59および/または61の参照配列によって示される対応NRPSドメインの活性を保持する発現産物をコードする。
【請求項2】
以下の1個以上のNRPS2ドメインの活性を保持する発現産物をコードする、請求項1に記載のポリヌクレオチド:
(i)配列番号47のチオレーションドメイン;
(ii)配列番号49のコンデンセーションドメイン;
(iii)配列番号51のプロリンのアデニレーションドメイン;
(iv)配列番号53のチオレーションドメイン;
(v)配列番号2のコンデンセーションドメイン;
(vi)配列番号4のイソロイシンのアデニレーションドメイン;
(vii)配列番号6のチオレーションドメイン;
(viii)配列番号8のコンデンセーションドメイン;
(ix)配列番号10のチロシンのアデニレーションドメイン;
(x)配列番号12のN−メチレーションドメイン;
(xi)配列番号14のチオレーションドメイン;
(xii)配列番号55のコンデンセーションドメイン;
(xiii)配列番号57のイソロイシンのアデニレーションドメイン;
(xiv)配列番号59のチオレーションドメイン;および
(xv)配列番号61のチオエステラーゼドメイン。
【請求項3】
式(I)または(I’)の化合物の生産に関与する非リボソームペプチドシンターゼであるNRPS1をコードする生合成遺伝子クラスターの1個以上の機能的フラグメントを含むポリヌクレオチドであって、以下のものを含むポリヌクレオチド:
(i) NRPS1ドメインをコードする配列番号30、32、34、36、38、40、42および44からなる群から選択される配列と、少なくとも80%、特に少なくとも85%、特に少なくとも90%、特に少なくとも95%、特に少なくとも98%の配列同一性を有するヌクレオチド配列および/またはその相補体;
(ii) NRPS1ドメインをコードする配列番号30、32、34、36、38、40、42および44からなる群から選択されるヌクレオチド配列の相補鎖とハイブリダイズするヌクレオチド配列および/またはその相補体;
(iii) NRPS1ドメインを示す配列番号31、33、35、37、39、41、43、45からなる群から選択される配列と、少なくとも60%、特に少なくとも70%、特に少なくとも80%、特に少なくとも90%、特に少なくとも95%の配列同一性を有するヌクレオチド配列および/またはその相補体;
(iv) NRPS1ドメインを示す配列番号31、33、35、37、39、41、43、45からなる群から選択されるアミノ酸をコードするヌクレオチド配列の相補鎖とハイブリダイズするヌクレオチド配列および/またはその相補体;
(v) 配列番号26からなる群から選択される配列と、少なくとも80%、特に少なくとも85%、特に少なくとも90%、特に少なくとも95%、特に少なくとも98%の配列同一性を有するヌクレオチド配列および/またはその相補体;または
(vi)配列番号26からなる群から選択されるヌクレオチド配列の相補鎖とハイブリダイズするヌクレオチド配列および/またはその相補体;
(vii) ここで、前記(i) 〜 (vi)のヌクレオチド配列は、配列番号31、33、35、37、39、41、43、45の参照配列によって示される対応NRPSドメインの活性を保持する発現産物をコードする。
【請求項4】
以下の1個以上のNRPS1ドメインの活性を保持する発現産物をコードする、請求項3に記載のポリヌクレオチド:
(i) 配列番号31のローディングドメイン;
(ii)配列番号33のグルタミンのアデニレーションドメイン;
(iii)配列番号35のチオレーションドメイン;
(iv)配列番号37のコンデンセーションドメイン;
(v)配列番号39のスレオニンのアデニレーションドメイン;
(vi)配列番号41のチオレーションドメイン;
(vii)配列番号43のコンデンセーションドメイン;および
(viii)配列番号45のロイシンのアデニレーションドメイン。
【請求項5】
式(I)または(I’)の化合物を生産するNRPS2をコードする請求項1または2に記載のポリヌクレオチドであって、配列番号29に示すアミノ酸配列をコードするヌクレオチド配列を含む、ポリヌクレオチド。
【請求項6】
式(I)または(I’)の化合物を生産するNRPS1をコードする請求項3または4に記載のポリヌクレオチドであって、配列番号27に示すアミノ酸配列をコードするヌクレオチド配列を含む、ポリヌクレオチド。
【請求項7】
請求項1〜6のいずれかの1個以上のポリヌクレオチドによってコードされるポリペプチド。
【請求項8】
式(I)または(I’)の化合物を生産するポリペプチドであって、以下のものからなる群から選択されるアミノ酸配列を含むポリペプチド:
(i)NRPS1を示す配列番号27、第2のNRPS2を示す配列番号29、チトクロムP450を示す配列番号63;および
(ii) (i)に列挙した参照配列と60%、特に少なくとも70%、特に少なくとも80%、特に少なくとも90%、特に少なくとも95%の配列同一性を有し、実質的に同じ触媒機能を保持する、(i)に列挙したアミノ酸配列の機能的変異体。
【請求項9】
請求項8の1個以上のポリペプチドをコードするヌクレオチド配列を含むポリヌクレオチド。
【請求項10】
(i)配列番号27またはその機能的変異体をコードするヌクレオチド配列;および
(ii)配列番号29またはその機能的変異体をコードするヌクレオチド配列
を含む、請求項9に記載のポリヌクレオチド。
【請求項11】
さらに配列番号63またはその機能的変異体をコードするヌクレオチド配列を含む、請求項10に記載のポリヌクレオチド。
【請求項12】
受託番号DSM 19329を有するコンドロマイセス・クロカツス(Chondromyces crocatus)株NPH−MB180から単離された、請求項9〜11のいずれか1項に記載のポリヌクレオチド。
【請求項13】
オープンリーディングフレームが転写および翻訳配列と作動可能に連結している、請求項9〜12のいずれか1項のポリヌクレオチドを含む発現ベクター。
【請求項14】
請求項1〜6および9〜12のいずれか1項の組換えポリヌクレオチドの1個以上または請求項13の発現ベクターを含む宿主細胞であって、組換えポリヌクレオチドが宿主細胞のゲノム中で天然には見出されない、宿主細胞。
【請求項15】
配列番号16、配列番号18、配列番号20、配列番号22および配列番号24からなる群から選択される式(I)または(I’)の化合物の生産に必要な1個以上のオープンリーディングフレームまたはそれらの機能的変異体をさらに含む、請求項14に記載の宿主細胞。
【請求項16】
式(I)もしくは(I’)または式(II)〜(VII)、(XI)〜(XIV)ならびに(XVII)および(XVIII)の化合物を生産するための、請求項14または15に記載の宿主細胞。
【請求項17】
組換えポリペプチドが遺伝子発現に最適化するために修飾されている、請求項14〜16のいずれか1項に記載の宿主細胞。
【請求項18】
ポリヌクレオチドのコドン出現頻度が宿主細胞の豊富なタンパク質のコドン出現頻度に調節されている、請求項14〜17のいずれか1項に記載の宿主細胞。
【請求項19】
宿主細胞がミキソコッカス目(Myxococcales)またはシュードモナス属(Pseudomonas)もしくはストレプトマイセス属(Streptomyces)から選択される、請求項14〜18のいずれか1項に記載の宿主細胞。
【請求項20】
宿主細胞がシュードモナス・プチダ(Pseudomonas putida)から選択される、請求項19に記載の宿主細胞。
【請求項21】
式(I)または(I’)の化合物の生産に必要な遺伝子を最早発現しない、変異微生物。
【請求項22】
配列番号63に示すチトクロムP450をコードする遺伝子を最早発現しない、請求項21に記載の変異微生物。
【請求項23】
式(I)もしくは(I’)または式(II)〜(VII)、(XI)〜(XIV)もしくは(XVII)〜(XVIII)の化合物を製造する方法であって、請求項16〜20のいずれか1項の宿主細胞を、当該化合物が生産される条件下で培養することを含む、製造方法。
【請求項24】
請求項23の方法によって得られる、式(I)もしくは(I’)または式(II)〜(VII)、(XI)〜(XIV)もしくは(XVII)〜(XVIII)の化合物。
【請求項25】
薬物として使用するための、請求項24に記載の化合物。
【請求項26】
アトピー性皮膚炎の処置に使用するための、請求項25に記載の化合物。
【図3】


【図4】

【図5】
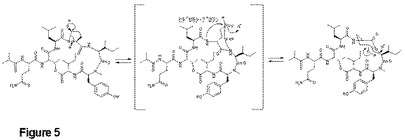
【図6】
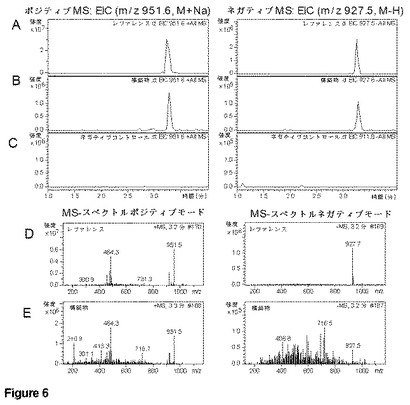
【図4】
【図5】
【図6】
【公表番号】特表2012−517801(P2012−517801A)
【公表日】平成24年8月9日(2012.8.9)
【国際特許分類】
【出願番号】特願2011−549554(P2011−549554)
【出願日】平成22年2月11日(2010.2.11)
【国際出願番号】PCT/EP2010/051696
【国際公開番号】WO2010/092109
【国際公開日】平成22年8月19日(2010.8.19)
【出願人】(504389991)ノバルティス アーゲー (806)
【Fターム(参考)】
【公表日】平成24年8月9日(2012.8.9)
【国際特許分類】
【出願日】平成22年2月11日(2010.2.11)
【国際出願番号】PCT/EP2010/051696
【国際公開番号】WO2010/092109
【国際公開日】平成22年8月19日(2010.8.19)
【出願人】(504389991)ノバルティス アーゲー (806)
【Fターム(参考)】
[ Back to top ]