
非対称PCR増幅法、その特別なプライマーおよび用途
本発明は、非対称PCR増幅法、その特別なプライマーおよび用途を開示し、一本鎖産物の調製のための単純で効率的なPCR増幅を提供する。本発明のPCRプライマーは、いくつかのプライマー対を含み、検出される標的配列とは無関係の核酸配列が、一方のプライマーの5’末端に付加されている。提供される非対称PCR増幅は:1)予備的に変性する工程;2)PCRの第一段階として、変性、プライマーアニーリング、伸長サイクルを繰り返す工程;3)PCR増幅の第二段階として、変性、プライマー伸長サイクルを繰り返す工程、を包含し、検出される標的配列とは無関係の核酸配列が、伸長における各対の一方のPCRプライマーの5’末端に付加されている。本発明の非対称PCR増幅によって、一本鎖産生のハイスループットを得ることができ、単一のPCR増幅または複数のPCR増幅が実施され得る。そして本方法は、核酸の検出において広範に使用され得る。
【発明の詳細な説明】
【技術分野】
【0001】
(技術分野)
本願は、PCR増幅法、PCR増幅法のためのプライマー、およびそれらの使用に関する。具体的には、本願は、核酸の検出における、非対称PCR増幅法、非対称PCR増幅のためのプライマー、およびそれらの使用に関する。
【背景技術】
【0002】
(背景)
ライフサイエンスの研究が進歩するにつれて、核酸が、遺伝情報の決定のための重要な物質であることがよく認識されるようになった。被験体のサンプルにおいて核酸配列の変化または変異を決定することによって、その被験体が、病原性微生物および/もしくはそのような微生物に対する耐性を有するか否か、その被験体が特定の疾患を有するか否か、ならびにその被験体が特定の遺伝状態にあるか否かを決定することができる。したがって、核酸分析技術には、ライフサイエンス研究の種々の領域において用途が見出され、この領域としては、病原性微生物の耐性遺伝子の試験、分類および検出、疾患の診断および予後、HLA分類、ならびにSNP検出が挙げられる。
【0003】
サンプルにおける核酸の量は通常分析には不十分であるため、検出されるためには、分析の前にその核酸を増幅することが通常必須である。増幅法としては、ポリメラーゼ連鎖反応(PCR)、逆転写ポリメラーゼ連鎖反応(RT−PCR)、鎖置換増幅(RDA)、およびローリングサイクル増幅(rolling cycle amplification)(RCA)が挙げられる(非特許文献1)。とりわけ、PCRが近年最も頻繁に使用されている。PCR産物を分析するための、多くの方法が存在する。例えば、アガロースゲル電気泳動またはPAGE電気泳動が、PCR産物の検出のために使用されている。これらの電気泳動法は、迅速かつ簡便な分析を提供する。しかしながら、これらの方法には、特異性が低いという問題があり、それゆえ遺伝子変異分析には適していない。PCR産物を分析するための別の方法は、制限酵素の使用であり、この方法は、制限された用途、、低い感受性を有し、操作が困難である。PCR産物の配列情報における正確性を保証する1つの方法は、PCR産物をクローニングすること、およびそのクローニングされた配列を配列決定することを包含する。しかしながら、この方法は、複数の工程を包含し、それゆえ高価で、実用的ではない。
【0004】
PCR産物のプローブとのハイブリダーゼーションの方法としては、以下が挙げられる:1)サザンハイブリダイゼーション、すなわち、そのPCR産物を析出させるために電気泳動を使用し、そのPCR産物をメンブレンに移し、そしてそのPCR産物を標識されたプローブとハイブリダイズさせる。この技術は、良好な特異性を生じるが、操作が複雑であり、時間を要し、そしてそれゆえ、複数の特徴についての並行分析には適していない。2)ポジティブドットハイブリダイゼーション(positive dot Hybridization)、すなわち、PCR産物を巻くまたは他の固体基板上に固定化し、その固定化されたPCR産物とプローブとをハイブリダイズさせる。この方法は、PCR産物が生成、定量および固定化されていることを必要とし、時間を要し、そして少量のサンプルにおける検出に適していない。3)リバースドットハイブリダイゼーション(reverse dot Hybridization)、すなわち、PCR産物と、既に膜または他の固体基板の表面に固定化したプローブとをハイブリダイズさせる。多数のプローブを含有する基板は、迅速に調製され得、そしてPCR反応完了後速やかにPCR産物を分析するために使用され得る。したがって、この方法は、迅速かつ簡便であり、キットおよび遺伝子チップにおける使用に適している。
【0005】
遺伝子チップ技術は、革新的である。その対称的、微小的および自動的特性に起因して、遺伝子チップ技術は、核酸分析(特に、ハイスループット核酸分析)において、重要な用途が見出されている(非特許文献2;非特許文献3;非特許文献4;および非特許文献5)。核酸チップは、特異的な条件下で、遺伝子発現プロファイルを分析するために使用されており、そして1kbまでの遺伝子領域において、一塩基多型(SNP)を決定するためにもまた使用されている。
【0006】
バイオチップ(例えば、感染性疾患の検出のために使用されるバイオチップ)を使用する伝統的な受動的核酸分析は、代表的に、3つの工程を包含する。第一の工程は、サンプル調製、すなわち、血漿、血液、唾液、尿、および糞便のようなサンプルからの核酸の調製である。そのようなサンプルから得られた核酸は、通常、直接分析されるには不十分であり、さらなる増幅(例えば、PCR増幅)が必要である。第二の工程は、核酸ハイブリダイゼーション、すなわち、その増幅された産物と、そのチップ上に固定化されたプローブとの間のハイブリダイゼーションである。第三の工程は、ハイブリダイゼーションシグナルの検出である。この工程は、代表的に、特定の標識の検出によって実施される。この標識は、増幅およびハイブリダイゼーションの間に導入され得る。検出方法は、使用される標識に依存する。例えば、蛍光検出器が蛍光標識を検出するために使用され得、一方オートラジオグラムが放射能標識を検出するのに使用され得る。ビオチンおよびストレプトアビジン標識が使用される場合、さらなる酵素的増幅が実施され得る。異なる増幅法が、実験の所望の感度に依存して使用される。例えば、チラミドシグナル増幅(Tyramide signal amplification)(TSA)および分枝DNA法(それぞれ、非特許文献6および非特許文献7に記載される)である。
【0007】
標的核酸と、バイオチップの表面上に固定化されたプローブとの間のハイブリダイゼーションは、核酸検出における中心的な工程を構成する。この標的核酸は、代表的に、PCRによって増幅され、一本鎖PCR産物に変性され、これが次いで、ストリンジェントな条件下でプローブにハイブリダイズされる。このハイブリダイズされた産物を、次いで、洗浄および検出する。ハイブリダイゼーションの間、そのPCR産物の一方の鎖のみがプローブとハイブリダイズし得る。対応する相補鎖は、PCR二本鎖産物の自己アニーリングに起因して、ハイブリダイゼーションを妨害し得る。結果として、ハイブリダイゼーションシグナルが失われ得る。オリゴヌクレオチドプローブとハイブリダイズする場合、一本鎖DNAのハイブリダイゼーション感度は、変性した二本鎖DNAよりも約5倍高いことが見出されている(非特許文献8)。したがって、遺伝子チップ上でのオリゴヌクレオチドとの高効率ハイブリダイゼーションのために、一本鎖核酸を得ることが望ましい。
【0008】
一本鎖核酸を調製するための、いくつかの方法が存在する。熱または塩基による二本鎖DNAの変性に加えて、一本鎖核酸を調製するための方法としては、以下が挙げられる。
【0009】
1.逆転写法。この方法では、T7プロモーターがPCRプライマーに加えられる。一本鎖核酸は、鋳型として精製PCR産物を使用する、T7 RNAポリメラーゼ媒介性インビトロ転写によって生成される(非特許文献9)。一本鎖核酸の収率はこの方法では非常に高いが、そのような二工程の方法は不便であり、そしてRNaseによる汚染の傾向がある。
【0010】
2.エキソヌクレアーゼ切断法(非特許文献10)。この方法において、上記PCRプライマーの一方はリン酸化されている。PCR産物がエキソヌクレアーゼによる切断に供される場合、このリン酸化されたプライマーから伸長した鎖は切断されない。次いで、このエキソヌクレアーゼは熱不活性化されなければならない。この方法は、PCR産物の精製を必要とし、エキソヌクレアーゼ活性に依存し、そしてそれゆえ不便である。
【0011】
4.変性高速液体クロマトグラフィー(DHPLC)。この方法において、上記PCRプライマーの一方はビオチンで標識されている。標識されたプライマーから伸長した鎖は、それゆえ、DHPLCにおいて他の鎖から分離され得る(非特許文献11)。所望の一本鎖は、15分以内に、二本鎖PCR産物から得ることができる。しかしながら、そのような方法は、高価な機器を必要とし、それゆえ一般的には使用され得ない。
【0012】
4.磁性ビーズ捕捉法。この方法では、ビオチンが上記PCRプライマーの一方に結合される。標識されたプライマーから伸長した鎖は、ストレプトアビジンコーティングされた磁性ビーズによって捕捉され、そしてNaOHを使用してそのビーズから分離される(非特許文献12)。この方法は、コーティングされた磁性ビーズの使用に起因して、非常に高価である。
【0013】
5.非対称PCR。上記の方法の全てがPCR反応後に余分な工程を含む一方で、非対称PCRは、PCR反応プロセスの間のDNAの調製を可能にする。本発明者らは、非対称PCRおよび磁性ビーズ捕捉法の両方が、比較的高い感度および特異性を有することを見出したが、熱変性および塩基変性法はしばしば、偽陰性の結果を生じる。これら全ての方法の中で、非対称PCRは、比較的単純かつ低価格であり、それゆえずっと実用的である(非特許文献13)。
【0014】
近年、非対称PCRのための以下のスキームが存在する。
【0015】
1)非対称PCRのための異なる濃度の上流プライマーおよび下流プライマーの使用。サイクルが増加するにつれて、低い濃度のプライマーが使い果たされ、一方高濃度のプライマーは、線形に増加する速度で一本鎖DNAを産生する(非特許文献14)。同様に、Zihong へらは、SNP検出のためにIGF−II遺伝子を非対照的に増幅するために、プライマーを1:10および1:20の比率において使用した(非特許文献15)。Shuangding Liらは、HLA−DRB1遺伝子を非対照的に増幅するために、プライマーを1:15の比率で使用した(非特許文献16)。そのような方法は、上流プライマーと下流プライマーとの間の比率の最適化を必要とし、非特異的増幅の可能性を増加させる。結果として、その産物は、通常、電気泳動上で拡散したバンドとして示される(非特許文献17)。
【0016】
2)非対称PCRのための異なる長さの上流プライマーおよび下流プライマーの使用。Xiaomou Pengらは、HBVのS遺伝子を非対照的に増幅するために、34ヌクレオチドの上流プライマーおよび20ヌクレオチドの下流プライマーを使用した。PCRサイクルの第二相の間に、アニーリング温度が上昇される。短いプライマーはそのような条件下ではアニーリングできず、一方長いプライマーは伸長し続け、一本鎖核酸を得る(非特許文献18)。このプライマーは遺伝子特異的であるが、長いプライマーの使用により、増幅に非特異性が導入される。したがってそのような方法は、多くのSNP部位を有する遺伝子(例えば、細菌の16S rRNA)の増幅には適していない。
【0017】
3)PCR鋳型を生成するための対称PCRの使用、ならびにPCR産物をさらに非対称増幅および標識するための1つのプライマーまたは異なる量のプライマーの使用(非特許文献19;非特許文献20;非特許文献21;非特許文献22)。これらの方法は、鋳型として精製された対称PCR産物を使用し、その後PCRサイクルを介して一本鎖を産生するために1つのプライマーを使用する。この方法は、複数工程の反応を必要とし、そしてそれゆえ、時間を要し、不便である。
【0018】
複数PCRの慣習的な方法は、複数のレベルの最適化を必要とする。これらの方法は、以下の問題点を有する。1)複数のプライマーの使用が、これらの異なるプライマーによる偽陽性の増幅を生成する。2)異なるプライマー間での競合により、標的核酸のバランスのとれていない増幅が生じる。すなわち、特定のプライマー対が効率的に増幅し、一方特定のプライマー対は非常に非効率的に増幅する。3)異なる実験を繰り返す。したがって、そのような状況下で非対称PCRをさらに実施するのは不満である。上述の方法は、それゆえ、複数の標的の同時分析には適していない。一工程の非対称PCRに対する必要性が存在する。
【非特許文献1】Andrasら、Mol.Biotechnol.,19:29−44,2001
【非特許文献2】DebouckおよびGoodfellow,Nature Genetics,1999,21(補遺):48−50
【非特許文献3】Dugganら、Nature Genetics,1999,21(補遺):10−14
【非特許文献4】Gerholdら、Trends Biochem.Sci.,1999,24:168−173
【非特許文献5】Alizadehら、Nature,2000,403:503−511
【非特許文献6】Karstenら、Nucleic Acid Res.,2002,E4
【非特許文献7】Kricka,Clin.Chem.,1999,45:453−458
【非特許文献8】Kawaiら、Anal.Biochem.1993,209:63−69
【非特許文献9】Hughesら、Nat.Biotechnol.,2001,19:342−347
【非特許文献10】HiguchiおよびOchman,Nucleic Acid Res.,1989,17:5865
【非特許文献11】DickmanおよびHornby,Anal.Biochem.,2000,284:164−167
【非特許文献12】Espelundら、Nucleic Acids Res.1990,18:6157−6158
【非特許文献13】Gaoら、Analytical Letters,2003,33:2849−2863
【非特許文献14】GyllenstenおよびErlich,Proc.Natl.Acad.Sci.U.S.A.,1988,85:7652−7656
【非特許文献15】Heら、Chinese Journal of Sports Medicine,2002,21:116−121
【非特許文献16】Liら、Chinese Journal of Experimental Hematology,2003,11:393−397
【非特許文献17】Erdoganら、Nucleic Acids Res.,2001,29:E36
【非特許文献18】Pengら、Chinese Experimental Diagnostics,2002,6:206−208
【非特許文献19】Gorelovら、Biochem.Biophys.Res.Commun.,1994,200:365−369
【非特許文献20】Scottら、Lett.Appl.未C路日おL。、1998,27:39−44
【非特許文献21】Guoら、Genome Res.,2002,12:447−457
【非特許文献22】Zhouら、Medical Animal Control,2003,19:524−527
【発明の開示】
【課題を解決するための手段】
【0019】
本発明は、一本鎖増幅産物の調製のための単純かつ効率的な非対称PCR増幅法、およびそのような方法のためのプライマーを提供する。
【0020】
本発明は、非対称PCR増幅のためのプライマーセットを提供し、このプライマーセットは複数のプライマー対を含み、各プライマー対における2つのプライマーのうちの一方は、その5’末端において、増幅される標的配列と無関係な配列を有するオリゴヌクレオチドテイルを有する。
【0021】
プライマーが適したTm値を有するために、本明細書中に記載されるオリゴヌクレオチドは、通常、8bp〜40bpであり、好ましくは15bp〜25bpである。
【0022】
異なる標的配列の間の増幅効率のバランスをとるために、非対称PCRプライマーにおいて、一般プライマーが提供される。ここで、その一般プライマーの少なくとも8個の連続するヌクレオチドが、オリゴヌクレオチドテイルにおけるヌクレオチドの同じである。好ましくは、この一般プライマーは、オリゴヌクレオチドテイルの配列と同じ配列を有する。
【0023】
一本鎖の非対称増幅および生成を促進するために、本明細書中に記載される一般プライマーの濃度は、本明細書中に記載される遺伝子特異的プライマー対の濃度よりも高い。
【0024】
したがって、本発明は、非対称PCRプライマーを使用する非対称PCR増幅法を提供する。この方法は、1)プレ変性(predenaturation);2)変性、アニーリングおよびプライマー伸長のサイクルの1つ以上を含む、第一相PCR増幅;ならびに3)変性、アニーリングおよびプライマー伸長のサイクルの1つ以上を含む第二相PCR増幅、の工程を包含し、ここで、プライマー伸長のためのプライマー対の2つのプライマーのうちの一方は、その5’末端において、標的配列とは無関係の配列を有するオリゴヌクレオチドテイルを有する。
【0025】
増幅効率を増加させるために、さらなるプライマー伸長反応が、第二相PCR増幅の後に実施される。
【0026】
上記工程2)における第一相PCR反応は、15〜25サイクルを有し、工程3)における第二相PCRのための伸長温度は、60℃〜75℃であり得る。
【0027】
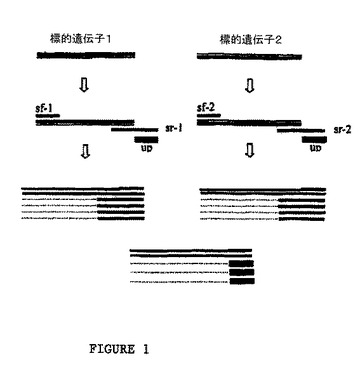
本発明の非対称PCR法は、図1にさらに図示される。図1において、sf−1/sr−1およびsf−2/sr−2が、それぞれ標的遺伝子1および標的遺伝子2に対する特異的プライマー対である。これらの中で、sr−1およびsr−2は、その5’末端において、一般配列を有する。一般プライマーに対して「Up」という場合、その配列は、sr−1およびsr−2における一般配列と同じである。非対称PCR増幅反応の間、反応混合物におけるDNAポリメラーゼ、dNTP、Mg2+、および反応プライマーは、慣習的なPCRのために使用されているものと同じであり、そして異なる反応の要求に基づいて条件は最適化され得る。
【0028】
本発明の非対称PCR増幅反応は、2つの相を有する。PCRの第一相は、慣習的なPCRと同じであり、変性、プライマーアニーリングおよびプライマー伸長の工程を包含する。このアニーリング温度は、上記遺伝子特異的プライマーのTmに基づいて調整され得る。同様に、伸長時間は、増幅されるフラグメントの長さに依存する。両方のプライマーが、慣習的な増幅の間に使用される。PCR増幅の第二相において、プライマーアニーリングのための温度は、プライマー伸長のための温度と同じである。それゆえ、この第二相は、変性およびプライマー伸長の工程のみを有する。長く、それゆえより高いTmを有する、テイルを有するプライマーのみが、アニーリングおよび伸長することができ、それによって一本鎖を産生する。
【0029】
本発明の非対称PCRにおいて使用される一般プライマーは、以下の機能を有する。PCRの第一相の間、この一般プライマーは、第二の温度サイクルの後の増幅反応に関与し得、それによって複数の増幅反応における異なる標的の増幅効率のバランスをとる。この一般プライマーの高い濃度に起因して、増幅プロセス全体を通じて異なるプライマーの濃度における差異を釣り合わせ、それによって非対称的増幅および一本鎖の作製を促進する。
【0030】
本発明の別の目的は、核酸の検出における非対称PCRプライマーの使用を提供することである。
【0031】
本発明において記載される非対称PCRプライマーは、複数のPCR増幅のために有用であり、その後、遺伝子チップまたは他のハイブリダイゼーション(例えば、メンブレンハイブリダイゼーション)が行われ、複数の標的配列を簡便に検出することを可能にする。
【0032】
本発明の方法によって生成される非対称PCR産物は、以下の遺伝子チップ検出法において使用され得る:a)リバースハイブリダイゼーション、すなわち、非対称PCR増幅の産物とハイブリダイゼーション緩衝液とを混合し、熱変性し、そして遺伝子チップ上に固定化されたオリゴヌクレオチドプローブとハイブリダイズさせる;あるいは、b)ポジティブハイブリダイゼーション、すなわち、非対称PCR増幅の産物を精製し、その精製産物を、固体キャリアの表面上に固定化し、遺伝子チップを作製し、そしてその遺伝子チップを核酸プローブとハイブリダイズさせる。ハイブリダイゼーションの最後に、異なる標識法およびシグナル検出法が、ハイブリダイゼーションシグナルを検出し、標的配列の検出を完了するために使用され得る。
【0033】
本発明の非対称PCR法は慣習的な非対称PCR法に対して以下の利点を有する。
【0034】
1)プライマー対の一方のプライマーの5’末端において標的配列に無関係である一般配列を加えることにより、そのプライマー対における一方のプライマーが他方のプライマーよりも長くなり、それによってそのプライマー対における2つのプライマーのTmを差別化する。5’末端オリゴヌクレオチドテイルの一般配列はまた、遺伝子特異的配列の間の差異を強調しないようにし(deemphasize)、初期PCRサイクルの増幅頻度を増加させ、そしてプライマー設計における困難性を克服する。
【0035】
2)オリゴヌクレオチドテイルの配列と同じ配列を有する一般プライマーの付加により、上流プライマーと下流プライマーとの間のプライマー数におけるさらなる不均衡を作り出す。
【0036】
3)より重要なことに、本発明の非対称PCRプライマーは、複雑な最適化プロセスの必要なしで、複数のPCR増幅のために使用され得る。この利点には、2つの理由が存在する。第一に、全てのプライマー対に対する2つのプライマーのうちの1つは同じ一般配列を有するため、これらのプライマー対に対する増幅効率は似通う傾向があり、異なるプライマー間の干渉の可能性は低い。第二に、この一般プライマーは、増幅プロセスにおけるプライマーのバランスをとる。ある特定の遺伝子特異的プライマー対が低い増幅効率を有する場合、より多くの一般プライマーが、その後の増幅反応に関与する。遺伝子特異的プライマーの増幅効率が低くなるほど、その後の反応に関与する一般プライマーの数は増加し、逆もまた同様である。したがって、異なる標的分子が、ほぼ同じ効率において増幅される。この一般配列は、それゆえ、異なるPCR反応の間のバランスを生み出し、そして全ての遺伝子特異的プライマーの類似の増幅効率を保証する。
【発明を実施するための最良の形態】
【0037】
(本発明の実施例の最良の形態)
(実施例1.単一非対称PCR増幅および遺伝子チップ上でのグラム陽性細菌の決定におけるその適用)
1.PCR増幅のための遺伝子特異的プライマーおよび種々のオリゴヌクレオチドプローブ。
【0038】
プライマーおよびプローブはいずれも、Shanghai Boya Biotechnology Companyによって合成した。5’末端にTAMRA蛍光標識を有する特定のプライマーおよび5’末端に修飾アミノ基を有するプローブもまた、Shanghai Boya Biotechnology Companyから提供された。
【0039】
遺伝子特異的プライマーに対する標的配列は、細菌の16S rRNA遺伝子であった。増幅フラグメントは、約1.5kbであった。このプライマーの分類およびそのオリゴヌクレオチド配列を、表1に提供する。
【0040】
【表1】

種々のオリゴヌクレオチドプローブが、標的細菌の一本鎖増幅産物における種々の部位にハイブリダイズし得る。このプローブの配列は、細菌16S rRNA遺伝子のフラグメントと同一であった。いくつかの核酸プローブの核酸配列およびその対応する標的を、表2に提供する。
【0041】
【表2】
2.アルデヒド基を有する基板の調製
室温で一晩、酸性溶液中にガラス基板を浸す。この酸性溶液を水道水で3回洗浄し、蒸留水で3回洗浄し、脱イオン水で1回浸し洗いし、そして脱イオン水で1回洗浄する。このガラス基板を、次いで、遠心分離によって乾燥させ、110℃で15分間焼き、このガラス基板を完全に乾燥させる。このガラス基板を、次いで、95%エタノール中1% APTES(3−アミノプロピルトリエトキシシラン)に浸し、振盪床上で4時間、室温にて振盪する。このガラス基板を、次いで、1回洗浄し、95%EtOHで1回浸し洗いする。この洗浄されたガラス基板を、次いで、110℃において2時間ガスバルブを閉じて、最大減圧(−0.08Mpa〜−0.1Mpa)下で、減圧乾燥器内に置いた。このガラス基板を、次いで、室温まで冷却し、12.5%グルタルジアルデヒド(400mlの12.5%グルタルジアルデヒド溶液、100mlの50%グルタルジアルデヒド、300mlのリン酸緩衝液(1モル/L NaH2PO4 30ml、2.682g NaCl)pH7.0)に浸し、室温下で4時間振盪した。このガラス基板を、次いで、グルタルジアルデヒド溶液から取り出し、3×SSCで3回、脱イオン水で2回洗浄し、遠心分離して水を除去し、そして室温において乾燥させた。
【0042】
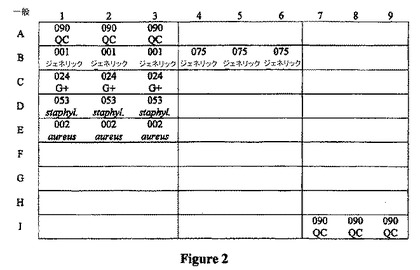
3.プローブが表面上に固定化されたガラス基板(遺伝子チップ)の調製
表2に示されるプローブを、最終濃度10μmol/Lで50% DMSOに溶解する。このプローブを、Cartesian Spotting Instrument(Cartesian Technologies,Inc.,CA,USA)(9×9、090QCはプローブPBB−0201090を示す)を使用して、図2に示すように基板上にスポットする。このスポットされたガラス基板を、室温下で一晩乾燥させ、室温において0.2%SDS中に2回、それぞれ振動条件下で浸した。このガラス基板を、次いで、脱イオン水で2回洗浄し、脱イオン水で1回浸し洗いし、そして遠心分離して水を除去した。このガラス基板を次いで、NaBH4溶液(1g NaBH4を300mlの1×PBSに溶解し、次いで100mlエタノールを加えた)に移し、5分間わずかに振盪した。このガラス基板を、次いで、脱イオン水で1回洗浄し、脱イオン水で1回浸し洗いし(それぞれ1分)、そして遠心分離して水を除去した。
【0043】
4.細菌培養および核酸抽出
本明細書中で使用されるStaphylococcus aureus 26001を、Chinese Medical Bacterial Reservation Center,Institute of Chinese Medicinal and Biological Productsから入手した。
【0044】
無菌のワークステーションにおいて、MH(Mueller Hinton寒天培地)培養スラブに細菌を播種し、シングルコロニーを分離する。このスラブを、35℃インキュベーターにおいて、24時間、上下にインキュベートした。
【0045】
10mgのG1145ガラスビーズおよび40mgのG1152ガラスビーズ(Sigma)を、無菌の1.5ml遠心チューブ内に入れ、100μlの1×TEを加える。炎によって滅菌されたピンセットを使用して、爪楊枝またはチップを掴み、その培養スラブからシングルコロニーを取り、その爪楊枝またはチップをそのチューブの底部においてガラスビーズにこすりつけ、それによって十分量の細菌をそのチューブ内に残す。その爪楊枝またはチップを、細菌殺傷溶液を含むくず入れに投げ入れ、その遠心チューブを閉じる。そのチューブを、ボルテックスミキサー(TDX−1,Beijing Tongda)において最大速度で5分間ボルテックスする。この遠心分離チューブを、次いで、95℃の水浴に5分間入れ、4℃で保管する。
【0046】
5.核酸PCR増幅
PCR系を、表3に示すように組み立てた。26001細菌懸濁液においてガラスビーズを振盪することによって調製した1μLの細菌サンプルを、系A、BおよびCの各々に加えた。総反応容量は、25μlであった。
【0047】
PCR反応を、PTC−200サイクラー(MJ Research Inc.)において実施した。系Aおよび系Bは、表4に示すような非対称PCR増幅サイクルを使用した。系Cは、表5に示されるような非対称PCRサイクルを使用した。
【0048】
【表3】
【表4】
【0049】
【表5】
6.電気泳動
アガロースゲル電気泳動を、PCR増幅産物を検出するために使用した。アガロースゲルの濃度は1.5%であった。各サンプルは2μlであり、電気泳動は、50V/cmで30分間実施した。
【0050】
7.ハイブリダイゼーション
ハイブリダイゼーション系を、表6に示す。
【0051】
【表6】
200μlの蒸留水を、HybriCassettesTM(Beijing Bo’ao Biochip,Ltd)に加え、ハイブリダイゼーション系の蒸発を防ぐ。反応物が表面上に固定化されたガラス基板およびカバースリップ(SmartCoverTM,Beijing Bo’ao Biochip,Ltd.)を、ハイブリダイゼーションカセット中に入れる。このハイブリダイゼーション系を95℃に加熱し、5分間95℃で維持し、PCR産物の変性を可能にする。この混合物を、次いで、氷水浴において速やかに冷却する。13μLのハイブリダイゼーション系を採取し、そのカバースリップ上の孔を通して、その基板とそのカバースリップとの間の空隙にそのハイブリダイゼーション系を加える。そのハイブリダイゼーションカセットを閉じ、90分間ハイブリダイゼーションさせる。次いでチップを取り出し、2×SSC、0.2%SDS溶液中に浸し、そして5分間振盪する。このチップを、次いで、脱イオン水で2回、それぞれ1分間浸し洗いし、そして遠心分離して水を除去した。2回の実験を、Staphylococcus aureus 26001サンプルを含むPCR系で実施した。
【0052】
8.チップ上でのシグナルの検出
チップ上の蛍光シグナルを、GenePix4000Bスキャナ(Axon Instruments,Inc.,CA,USA)を使用することによって検出した。波長532nm、PMT600、および出力33%。
【0053】
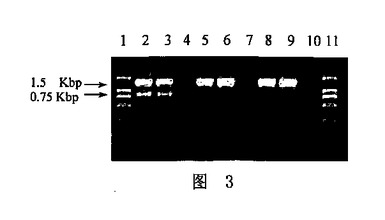
9.結果の分析
アガロースゲル電気泳動の結果を、図3に示す。レーン1およびレーン11は、DL2000マーカー(Takarar)であり;レーン2、レーン3およびレーン4は、系AのPCR産物であり;レーン5、レーン6およびレーン7は、系BのPCR産物であり;レーン8、レーン9およびレーン10は、系CのPCR産物であった。これらのレーンの中で、レーン2、レーン3、レーン5、レーン6、レーン8およびレーン9は、26001細菌サンプルを含有し、一方レーン4、レーン7およびレーン10におけるPCR系は、ブランクコントロールを含有した。図3に示されるように、3つ全てのPCR系(A、BおよびC)が、約1.5kbの二本鎖標的核酸の良好な増幅をもたらしたが、一方系Aのみが明確な一本鎖PCR増幅産物を産生した。
【0054】
図4は、本明細書中に記載される実験のポジティブなハイブリダイゼーション結果を示す。図4Aは、Staphylococcus aureusの所望のハイブリダイゼーションシグナルを示し;図5は、本明細書中に記載されるハイブリダイゼーション実験の蛍光検出結果を示す。図4および図5に示されるように、系Aを使用したハイブリダイゼーション結果のみが、図4と一致するシグナルを生じ、一方他の2つの系は、部分的な結果を生じるのみであった。1つの可能な説明は、系Bおよび系Cにおいて産生された主要な産物は二本鎖PCR産物であり、これはハイブリダイゼーションの間に自己アニーリングすることができ、それによってポジティブなシグナルを弱めているか、またはポジティブなシグナルを生じることができなくなっているというものである。
【0055】
電気泳動およびチップハイブリダイゼーション結果は、本発明の非対称PCR増幅系および非対称PCR増幅温度サイクルが一本鎖増幅産物の調製において効率的であることを実証した。これは、ハイブリダイゼーション反応に特に適しており、それによってハイブリダイゼーション効率およびハイブリダイゼーションシグナルを増加させる。
【0056】
(実施例2.複数の非対称PCR増幅および遺伝子チップを使用する、細菌の薬物耐性遺伝子の検出)
プライマーおよびプローブはいずれも、Shanghai Boya Biotechnology Companyによって合成した。5’末端にTAMRA蛍光標識を有する特定のプライマーおよび5’末端に修飾アミノ基を有する特定のプローブもまた、Shanghai Boya Biotechnology Companyによって提供された。
【0057】
遺伝子特異的プライマーの分類、配列情報、標的遺伝子、および増幅されたフラグメントの長さは、表7に示す。各標的遺伝子は2つのプローブを有し、それらの配列情報は表8に示す。これらの中で、標的遺伝子tetKおよびtetMは、グラム陽性細菌由来のテトラサイクリン耐性遺伝子であった。標的遺伝子ermAおよびermCは、グラム陽性細菌由来のマクロライド−リンコサミド−ストレプトグラミンB耐性遺伝子であった。普遍的な細菌の23rRNA遺伝子の部分を、PCR増幅およびハイブリダイゼーションプロセスをモニタリングするために、内部コントロールとして使用した。
【0058】
【表7】
【0059】
【表8】
2.アルデヒド基を有するガラス基板の調製
アルデヒド基を有するガラス基板は、実施例1のように調製した。
【0060】
3.プローブが表面上に固定化されたガラス基板の調製
調製法は、実施例1と同じであった。このプローブは、表8に示す。これらのプローブを、図6に示すように(6×6、090QCは、コントロールプローブPBB−0201090を示し、残りのプローブは、プローブ番号の最後3つの数字で番号を付けた)スポットした。例えば、PBB−024654は、「654」と示される。
【0061】
4.細菌培養および核酸抽出
本実施例において使用した最近の供給元は、表9に示す。細菌の培養および核酸抽出法は、実施例2に示される。
【0062】
【表9】
5.核酸のPCR増幅
本実施例は、1回の実験における5つの異なるPCR増幅を示す。このPCR反応系を、以下に示す:1×MaterMix(Beijing Tianwei Times);0.2μmol/Lの、表7に示される5対の遺伝子特異的プライマー;1μmol/Lの一般プライマーAMB_0408047;ガラスビーズとの振動後の1μLの細菌サンプル。総反応容量は、25μLであった。
【0063】
PCRを、PTC−200(MJ Research Inc.)サーモサイクラーで実施した。この非対称PCRサーモサイクルは、表4に示した。
【0064】
6.ハイブリダイゼーションおよび検出
ハイブリダイゼーション検出系およびハイブリダイゼーション検出法、ならびにハイブリダイゼーション後の蛍光検出法は、実施例1に示した。
【0065】
7.結果の分析
図7は、8種の細菌株についての薬物耐性遺伝子検出結果を示す。図7に示されるように、異なる細菌株は、異なる種類および数の薬物耐性遺伝子を有する。Staphylococcus aureus MRSA6581およびEnterococcus faecalisは、1つの薬物耐性遺伝子のみを有する。Staphylococcus aureus B437およびStaphylococcus epidermis TR2041は、2つの薬物耐性遺伝子を有する。Staphylococcus aureus B435およびStaphylococcus aureus MRSA6460は、3つの薬物耐性遺伝子を有する。Staphylococcus aureus MRSA6437は、4つの薬物耐性遺伝子を有する。Enterococcus faeciumは、薬物耐性遺伝子を有さない。
【0066】
さらに、図7に示されるように、PCRおよびハイブリダイゼーションコントロールの23S rRNA遺伝子は、全ての細菌株に対して良好に機能し、良好な増幅および強力なハイブリダイゼーションシグナルを生じた。ポジティブおよび相同なシグナルを、1つ、2つ、3つまたは4つの薬物耐性遺伝子を有する全ての細菌から得た。このことは、本発明の非対称PCR増幅法および温度サイクルが、複数の遺伝子の並行検出に適していることを示す。
【産業上の利用可能性】
【0067】
本発明の非対称PCR増幅法は、高収率の一本鎖産物を産生し、このことはその後のハイブリダイゼーション反応を促進し、ハイブリダイゼーション効率を向上させ、そしてハイブリダイゼーションシグナルを増加させる。この方法はまた、最適化の必要性なく、類似の条件下での単一または複数のPCR増幅のために有用である。本発明のプライマーは、設計が単純である。本方法は、操作が容易であり、多因子のハイスループット遺伝子チップ法に適用可能である。本明細書中に記載される方法は、微生物判定、分類、薬物耐性遺伝子検出、疾患診断および予後、HLA分類、SNP検出ならびに遺伝子分析において用途を見出し得る。
【図面の簡単な説明】
【0068】
【図1】図1は、本発明の非対称PCRの概略図を提供する。
【図2】図2は、実施例1において使用されるプローブアレイの概略図を提供する。
【図3】図3は、実施例1のPCR産物の電気泳動写真を提供する。
【図4】図4Aは、実施例1に記載されるStaphylococcus aureus 26001サンプルにおける所望のハイブリダイゼーション結果を提供する。図4Bは、実施例1に記載されるブランクコントロールにおける所望のハイブリダイゼーション結果を提供する。
【図5】図5は、ハイブリダイゼーション後の蛍光検出の結果を提供する。
【図6】図6は、実施例2に記載されるプローブのアレイの概略図を提供する。
【図7】図7は、実施例2に記載されるハイブリダイゼーション後の蛍光検出の結果を提供する。
【技術分野】
【0001】
(技術分野)
本願は、PCR増幅法、PCR増幅法のためのプライマー、およびそれらの使用に関する。具体的には、本願は、核酸の検出における、非対称PCR増幅法、非対称PCR増幅のためのプライマー、およびそれらの使用に関する。
【背景技術】
【0002】
(背景)
ライフサイエンスの研究が進歩するにつれて、核酸が、遺伝情報の決定のための重要な物質であることがよく認識されるようになった。被験体のサンプルにおいて核酸配列の変化または変異を決定することによって、その被験体が、病原性微生物および/もしくはそのような微生物に対する耐性を有するか否か、その被験体が特定の疾患を有するか否か、ならびにその被験体が特定の遺伝状態にあるか否かを決定することができる。したがって、核酸分析技術には、ライフサイエンス研究の種々の領域において用途が見出され、この領域としては、病原性微生物の耐性遺伝子の試験、分類および検出、疾患の診断および予後、HLA分類、ならびにSNP検出が挙げられる。
【0003】
サンプルにおける核酸の量は通常分析には不十分であるため、検出されるためには、分析の前にその核酸を増幅することが通常必須である。増幅法としては、ポリメラーゼ連鎖反応(PCR)、逆転写ポリメラーゼ連鎖反応(RT−PCR)、鎖置換増幅(RDA)、およびローリングサイクル増幅(rolling cycle amplification)(RCA)が挙げられる(非特許文献1)。とりわけ、PCRが近年最も頻繁に使用されている。PCR産物を分析するための、多くの方法が存在する。例えば、アガロースゲル電気泳動またはPAGE電気泳動が、PCR産物の検出のために使用されている。これらの電気泳動法は、迅速かつ簡便な分析を提供する。しかしながら、これらの方法には、特異性が低いという問題があり、それゆえ遺伝子変異分析には適していない。PCR産物を分析するための別の方法は、制限酵素の使用であり、この方法は、制限された用途、、低い感受性を有し、操作が困難である。PCR産物の配列情報における正確性を保証する1つの方法は、PCR産物をクローニングすること、およびそのクローニングされた配列を配列決定することを包含する。しかしながら、この方法は、複数の工程を包含し、それゆえ高価で、実用的ではない。
【0004】
PCR産物のプローブとのハイブリダーゼーションの方法としては、以下が挙げられる:1)サザンハイブリダイゼーション、すなわち、そのPCR産物を析出させるために電気泳動を使用し、そのPCR産物をメンブレンに移し、そしてそのPCR産物を標識されたプローブとハイブリダイズさせる。この技術は、良好な特異性を生じるが、操作が複雑であり、時間を要し、そしてそれゆえ、複数の特徴についての並行分析には適していない。2)ポジティブドットハイブリダイゼーション(positive dot Hybridization)、すなわち、PCR産物を巻くまたは他の固体基板上に固定化し、その固定化されたPCR産物とプローブとをハイブリダイズさせる。この方法は、PCR産物が生成、定量および固定化されていることを必要とし、時間を要し、そして少量のサンプルにおける検出に適していない。3)リバースドットハイブリダイゼーション(reverse dot Hybridization)、すなわち、PCR産物と、既に膜または他の固体基板の表面に固定化したプローブとをハイブリダイズさせる。多数のプローブを含有する基板は、迅速に調製され得、そしてPCR反応完了後速やかにPCR産物を分析するために使用され得る。したがって、この方法は、迅速かつ簡便であり、キットおよび遺伝子チップにおける使用に適している。
【0005】
遺伝子チップ技術は、革新的である。その対称的、微小的および自動的特性に起因して、遺伝子チップ技術は、核酸分析(特に、ハイスループット核酸分析)において、重要な用途が見出されている(非特許文献2;非特許文献3;非特許文献4;および非特許文献5)。核酸チップは、特異的な条件下で、遺伝子発現プロファイルを分析するために使用されており、そして1kbまでの遺伝子領域において、一塩基多型(SNP)を決定するためにもまた使用されている。
【0006】
バイオチップ(例えば、感染性疾患の検出のために使用されるバイオチップ)を使用する伝統的な受動的核酸分析は、代表的に、3つの工程を包含する。第一の工程は、サンプル調製、すなわち、血漿、血液、唾液、尿、および糞便のようなサンプルからの核酸の調製である。そのようなサンプルから得られた核酸は、通常、直接分析されるには不十分であり、さらなる増幅(例えば、PCR増幅)が必要である。第二の工程は、核酸ハイブリダイゼーション、すなわち、その増幅された産物と、そのチップ上に固定化されたプローブとの間のハイブリダイゼーションである。第三の工程は、ハイブリダイゼーションシグナルの検出である。この工程は、代表的に、特定の標識の検出によって実施される。この標識は、増幅およびハイブリダイゼーションの間に導入され得る。検出方法は、使用される標識に依存する。例えば、蛍光検出器が蛍光標識を検出するために使用され得、一方オートラジオグラムが放射能標識を検出するのに使用され得る。ビオチンおよびストレプトアビジン標識が使用される場合、さらなる酵素的増幅が実施され得る。異なる増幅法が、実験の所望の感度に依存して使用される。例えば、チラミドシグナル増幅(Tyramide signal amplification)(TSA)および分枝DNA法(それぞれ、非特許文献6および非特許文献7に記載される)である。
【0007】
標的核酸と、バイオチップの表面上に固定化されたプローブとの間のハイブリダイゼーションは、核酸検出における中心的な工程を構成する。この標的核酸は、代表的に、PCRによって増幅され、一本鎖PCR産物に変性され、これが次いで、ストリンジェントな条件下でプローブにハイブリダイズされる。このハイブリダイズされた産物を、次いで、洗浄および検出する。ハイブリダイゼーションの間、そのPCR産物の一方の鎖のみがプローブとハイブリダイズし得る。対応する相補鎖は、PCR二本鎖産物の自己アニーリングに起因して、ハイブリダイゼーションを妨害し得る。結果として、ハイブリダイゼーションシグナルが失われ得る。オリゴヌクレオチドプローブとハイブリダイズする場合、一本鎖DNAのハイブリダイゼーション感度は、変性した二本鎖DNAよりも約5倍高いことが見出されている(非特許文献8)。したがって、遺伝子チップ上でのオリゴヌクレオチドとの高効率ハイブリダイゼーションのために、一本鎖核酸を得ることが望ましい。
【0008】
一本鎖核酸を調製するための、いくつかの方法が存在する。熱または塩基による二本鎖DNAの変性に加えて、一本鎖核酸を調製するための方法としては、以下が挙げられる。
【0009】
1.逆転写法。この方法では、T7プロモーターがPCRプライマーに加えられる。一本鎖核酸は、鋳型として精製PCR産物を使用する、T7 RNAポリメラーゼ媒介性インビトロ転写によって生成される(非特許文献9)。一本鎖核酸の収率はこの方法では非常に高いが、そのような二工程の方法は不便であり、そしてRNaseによる汚染の傾向がある。
【0010】
2.エキソヌクレアーゼ切断法(非特許文献10)。この方法において、上記PCRプライマーの一方はリン酸化されている。PCR産物がエキソヌクレアーゼによる切断に供される場合、このリン酸化されたプライマーから伸長した鎖は切断されない。次いで、このエキソヌクレアーゼは熱不活性化されなければならない。この方法は、PCR産物の精製を必要とし、エキソヌクレアーゼ活性に依存し、そしてそれゆえ不便である。
【0011】
4.変性高速液体クロマトグラフィー(DHPLC)。この方法において、上記PCRプライマーの一方はビオチンで標識されている。標識されたプライマーから伸長した鎖は、それゆえ、DHPLCにおいて他の鎖から分離され得る(非特許文献11)。所望の一本鎖は、15分以内に、二本鎖PCR産物から得ることができる。しかしながら、そのような方法は、高価な機器を必要とし、それゆえ一般的には使用され得ない。
【0012】
4.磁性ビーズ捕捉法。この方法では、ビオチンが上記PCRプライマーの一方に結合される。標識されたプライマーから伸長した鎖は、ストレプトアビジンコーティングされた磁性ビーズによって捕捉され、そしてNaOHを使用してそのビーズから分離される(非特許文献12)。この方法は、コーティングされた磁性ビーズの使用に起因して、非常に高価である。
【0013】
5.非対称PCR。上記の方法の全てがPCR反応後に余分な工程を含む一方で、非対称PCRは、PCR反応プロセスの間のDNAの調製を可能にする。本発明者らは、非対称PCRおよび磁性ビーズ捕捉法の両方が、比較的高い感度および特異性を有することを見出したが、熱変性および塩基変性法はしばしば、偽陰性の結果を生じる。これら全ての方法の中で、非対称PCRは、比較的単純かつ低価格であり、それゆえずっと実用的である(非特許文献13)。
【0014】
近年、非対称PCRのための以下のスキームが存在する。
【0015】
1)非対称PCRのための異なる濃度の上流プライマーおよび下流プライマーの使用。サイクルが増加するにつれて、低い濃度のプライマーが使い果たされ、一方高濃度のプライマーは、線形に増加する速度で一本鎖DNAを産生する(非特許文献14)。同様に、Zihong へらは、SNP検出のためにIGF−II遺伝子を非対照的に増幅するために、プライマーを1:10および1:20の比率において使用した(非特許文献15)。Shuangding Liらは、HLA−DRB1遺伝子を非対照的に増幅するために、プライマーを1:15の比率で使用した(非特許文献16)。そのような方法は、上流プライマーと下流プライマーとの間の比率の最適化を必要とし、非特異的増幅の可能性を増加させる。結果として、その産物は、通常、電気泳動上で拡散したバンドとして示される(非特許文献17)。
【0016】
2)非対称PCRのための異なる長さの上流プライマーおよび下流プライマーの使用。Xiaomou Pengらは、HBVのS遺伝子を非対照的に増幅するために、34ヌクレオチドの上流プライマーおよび20ヌクレオチドの下流プライマーを使用した。PCRサイクルの第二相の間に、アニーリング温度が上昇される。短いプライマーはそのような条件下ではアニーリングできず、一方長いプライマーは伸長し続け、一本鎖核酸を得る(非特許文献18)。このプライマーは遺伝子特異的であるが、長いプライマーの使用により、増幅に非特異性が導入される。したがってそのような方法は、多くのSNP部位を有する遺伝子(例えば、細菌の16S rRNA)の増幅には適していない。
【0017】
3)PCR鋳型を生成するための対称PCRの使用、ならびにPCR産物をさらに非対称増幅および標識するための1つのプライマーまたは異なる量のプライマーの使用(非特許文献19;非特許文献20;非特許文献21;非特許文献22)。これらの方法は、鋳型として精製された対称PCR産物を使用し、その後PCRサイクルを介して一本鎖を産生するために1つのプライマーを使用する。この方法は、複数工程の反応を必要とし、そしてそれゆえ、時間を要し、不便である。
【0018】
複数PCRの慣習的な方法は、複数のレベルの最適化を必要とする。これらの方法は、以下の問題点を有する。1)複数のプライマーの使用が、これらの異なるプライマーによる偽陽性の増幅を生成する。2)異なるプライマー間での競合により、標的核酸のバランスのとれていない増幅が生じる。すなわち、特定のプライマー対が効率的に増幅し、一方特定のプライマー対は非常に非効率的に増幅する。3)異なる実験を繰り返す。したがって、そのような状況下で非対称PCRをさらに実施するのは不満である。上述の方法は、それゆえ、複数の標的の同時分析には適していない。一工程の非対称PCRに対する必要性が存在する。
【非特許文献1】Andrasら、Mol.Biotechnol.,19:29−44,2001
【非特許文献2】DebouckおよびGoodfellow,Nature Genetics,1999,21(補遺):48−50
【非特許文献3】Dugganら、Nature Genetics,1999,21(補遺):10−14
【非特許文献4】Gerholdら、Trends Biochem.Sci.,1999,24:168−173
【非特許文献5】Alizadehら、Nature,2000,403:503−511
【非特許文献6】Karstenら、Nucleic Acid Res.,2002,E4
【非特許文献7】Kricka,Clin.Chem.,1999,45:453−458
【非特許文献8】Kawaiら、Anal.Biochem.1993,209:63−69
【非特許文献9】Hughesら、Nat.Biotechnol.,2001,19:342−347
【非特許文献10】HiguchiおよびOchman,Nucleic Acid Res.,1989,17:5865
【非特許文献11】DickmanおよびHornby,Anal.Biochem.,2000,284:164−167
【非特許文献12】Espelundら、Nucleic Acids Res.1990,18:6157−6158
【非特許文献13】Gaoら、Analytical Letters,2003,33:2849−2863
【非特許文献14】GyllenstenおよびErlich,Proc.Natl.Acad.Sci.U.S.A.,1988,85:7652−7656
【非特許文献15】Heら、Chinese Journal of Sports Medicine,2002,21:116−121
【非特許文献16】Liら、Chinese Journal of Experimental Hematology,2003,11:393−397
【非特許文献17】Erdoganら、Nucleic Acids Res.,2001,29:E36
【非特許文献18】Pengら、Chinese Experimental Diagnostics,2002,6:206−208
【非特許文献19】Gorelovら、Biochem.Biophys.Res.Commun.,1994,200:365−369
【非特許文献20】Scottら、Lett.Appl.未C路日おL。、1998,27:39−44
【非特許文献21】Guoら、Genome Res.,2002,12:447−457
【非特許文献22】Zhouら、Medical Animal Control,2003,19:524−527
【発明の開示】
【課題を解決するための手段】
【0019】
本発明は、一本鎖増幅産物の調製のための単純かつ効率的な非対称PCR増幅法、およびそのような方法のためのプライマーを提供する。
【0020】
本発明は、非対称PCR増幅のためのプライマーセットを提供し、このプライマーセットは複数のプライマー対を含み、各プライマー対における2つのプライマーのうちの一方は、その5’末端において、増幅される標的配列と無関係な配列を有するオリゴヌクレオチドテイルを有する。
【0021】
プライマーが適したTm値を有するために、本明細書中に記載されるオリゴヌクレオチドは、通常、8bp〜40bpであり、好ましくは15bp〜25bpである。
【0022】
異なる標的配列の間の増幅効率のバランスをとるために、非対称PCRプライマーにおいて、一般プライマーが提供される。ここで、その一般プライマーの少なくとも8個の連続するヌクレオチドが、オリゴヌクレオチドテイルにおけるヌクレオチドの同じである。好ましくは、この一般プライマーは、オリゴヌクレオチドテイルの配列と同じ配列を有する。
【0023】
一本鎖の非対称増幅および生成を促進するために、本明細書中に記載される一般プライマーの濃度は、本明細書中に記載される遺伝子特異的プライマー対の濃度よりも高い。
【0024】
したがって、本発明は、非対称PCRプライマーを使用する非対称PCR増幅法を提供する。この方法は、1)プレ変性(predenaturation);2)変性、アニーリングおよびプライマー伸長のサイクルの1つ以上を含む、第一相PCR増幅;ならびに3)変性、アニーリングおよびプライマー伸長のサイクルの1つ以上を含む第二相PCR増幅、の工程を包含し、ここで、プライマー伸長のためのプライマー対の2つのプライマーのうちの一方は、その5’末端において、標的配列とは無関係の配列を有するオリゴヌクレオチドテイルを有する。
【0025】
増幅効率を増加させるために、さらなるプライマー伸長反応が、第二相PCR増幅の後に実施される。
【0026】
上記工程2)における第一相PCR反応は、15〜25サイクルを有し、工程3)における第二相PCRのための伸長温度は、60℃〜75℃であり得る。
【0027】
本発明の非対称PCR法は、図1にさらに図示される。図1において、sf−1/sr−1およびsf−2/sr−2が、それぞれ標的遺伝子1および標的遺伝子2に対する特異的プライマー対である。これらの中で、sr−1およびsr−2は、その5’末端において、一般配列を有する。一般プライマーに対して「Up」という場合、その配列は、sr−1およびsr−2における一般配列と同じである。非対称PCR増幅反応の間、反応混合物におけるDNAポリメラーゼ、dNTP、Mg2+、および反応プライマーは、慣習的なPCRのために使用されているものと同じであり、そして異なる反応の要求に基づいて条件は最適化され得る。
【0028】
本発明の非対称PCR増幅反応は、2つの相を有する。PCRの第一相は、慣習的なPCRと同じであり、変性、プライマーアニーリングおよびプライマー伸長の工程を包含する。このアニーリング温度は、上記遺伝子特異的プライマーのTmに基づいて調整され得る。同様に、伸長時間は、増幅されるフラグメントの長さに依存する。両方のプライマーが、慣習的な増幅の間に使用される。PCR増幅の第二相において、プライマーアニーリングのための温度は、プライマー伸長のための温度と同じである。それゆえ、この第二相は、変性およびプライマー伸長の工程のみを有する。長く、それゆえより高いTmを有する、テイルを有するプライマーのみが、アニーリングおよび伸長することができ、それによって一本鎖を産生する。
【0029】
本発明の非対称PCRにおいて使用される一般プライマーは、以下の機能を有する。PCRの第一相の間、この一般プライマーは、第二の温度サイクルの後の増幅反応に関与し得、それによって複数の増幅反応における異なる標的の増幅効率のバランスをとる。この一般プライマーの高い濃度に起因して、増幅プロセス全体を通じて異なるプライマーの濃度における差異を釣り合わせ、それによって非対称的増幅および一本鎖の作製を促進する。
【0030】
本発明の別の目的は、核酸の検出における非対称PCRプライマーの使用を提供することである。
【0031】
本発明において記載される非対称PCRプライマーは、複数のPCR増幅のために有用であり、その後、遺伝子チップまたは他のハイブリダイゼーション(例えば、メンブレンハイブリダイゼーション)が行われ、複数の標的配列を簡便に検出することを可能にする。
【0032】
本発明の方法によって生成される非対称PCR産物は、以下の遺伝子チップ検出法において使用され得る:a)リバースハイブリダイゼーション、すなわち、非対称PCR増幅の産物とハイブリダイゼーション緩衝液とを混合し、熱変性し、そして遺伝子チップ上に固定化されたオリゴヌクレオチドプローブとハイブリダイズさせる;あるいは、b)ポジティブハイブリダイゼーション、すなわち、非対称PCR増幅の産物を精製し、その精製産物を、固体キャリアの表面上に固定化し、遺伝子チップを作製し、そしてその遺伝子チップを核酸プローブとハイブリダイズさせる。ハイブリダイゼーションの最後に、異なる標識法およびシグナル検出法が、ハイブリダイゼーションシグナルを検出し、標的配列の検出を完了するために使用され得る。
【0033】
本発明の非対称PCR法は慣習的な非対称PCR法に対して以下の利点を有する。
【0034】
1)プライマー対の一方のプライマーの5’末端において標的配列に無関係である一般配列を加えることにより、そのプライマー対における一方のプライマーが他方のプライマーよりも長くなり、それによってそのプライマー対における2つのプライマーのTmを差別化する。5’末端オリゴヌクレオチドテイルの一般配列はまた、遺伝子特異的配列の間の差異を強調しないようにし(deemphasize)、初期PCRサイクルの増幅頻度を増加させ、そしてプライマー設計における困難性を克服する。
【0035】
2)オリゴヌクレオチドテイルの配列と同じ配列を有する一般プライマーの付加により、上流プライマーと下流プライマーとの間のプライマー数におけるさらなる不均衡を作り出す。
【0036】
3)より重要なことに、本発明の非対称PCRプライマーは、複雑な最適化プロセスの必要なしで、複数のPCR増幅のために使用され得る。この利点には、2つの理由が存在する。第一に、全てのプライマー対に対する2つのプライマーのうちの1つは同じ一般配列を有するため、これらのプライマー対に対する増幅効率は似通う傾向があり、異なるプライマー間の干渉の可能性は低い。第二に、この一般プライマーは、増幅プロセスにおけるプライマーのバランスをとる。ある特定の遺伝子特異的プライマー対が低い増幅効率を有する場合、より多くの一般プライマーが、その後の増幅反応に関与する。遺伝子特異的プライマーの増幅効率が低くなるほど、その後の反応に関与する一般プライマーの数は増加し、逆もまた同様である。したがって、異なる標的分子が、ほぼ同じ効率において増幅される。この一般配列は、それゆえ、異なるPCR反応の間のバランスを生み出し、そして全ての遺伝子特異的プライマーの類似の増幅効率を保証する。
【発明を実施するための最良の形態】
【0037】
(本発明の実施例の最良の形態)
(実施例1.単一非対称PCR増幅および遺伝子チップ上でのグラム陽性細菌の決定におけるその適用)
1.PCR増幅のための遺伝子特異的プライマーおよび種々のオリゴヌクレオチドプローブ。
【0038】
プライマーおよびプローブはいずれも、Shanghai Boya Biotechnology Companyによって合成した。5’末端にTAMRA蛍光標識を有する特定のプライマーおよび5’末端に修飾アミノ基を有するプローブもまた、Shanghai Boya Biotechnology Companyから提供された。
【0039】
遺伝子特異的プライマーに対する標的配列は、細菌の16S rRNA遺伝子であった。増幅フラグメントは、約1.5kbであった。このプライマーの分類およびそのオリゴヌクレオチド配列を、表1に提供する。
【0040】
【表1】
種々のオリゴヌクレオチドプローブが、標的細菌の一本鎖増幅産物における種々の部位にハイブリダイズし得る。このプローブの配列は、細菌16S rRNA遺伝子のフラグメントと同一であった。いくつかの核酸プローブの核酸配列およびその対応する標的を、表2に提供する。
【0041】
【表2】
2.アルデヒド基を有する基板の調製
室温で一晩、酸性溶液中にガラス基板を浸す。この酸性溶液を水道水で3回洗浄し、蒸留水で3回洗浄し、脱イオン水で1回浸し洗いし、そして脱イオン水で1回洗浄する。このガラス基板を、次いで、遠心分離によって乾燥させ、110℃で15分間焼き、このガラス基板を完全に乾燥させる。このガラス基板を、次いで、95%エタノール中1% APTES(3−アミノプロピルトリエトキシシラン)に浸し、振盪床上で4時間、室温にて振盪する。このガラス基板を、次いで、1回洗浄し、95%EtOHで1回浸し洗いする。この洗浄されたガラス基板を、次いで、110℃において2時間ガスバルブを閉じて、最大減圧(−0.08Mpa〜−0.1Mpa)下で、減圧乾燥器内に置いた。このガラス基板を、次いで、室温まで冷却し、12.5%グルタルジアルデヒド(400mlの12.5%グルタルジアルデヒド溶液、100mlの50%グルタルジアルデヒド、300mlのリン酸緩衝液(1モル/L NaH2PO4 30ml、2.682g NaCl)pH7.0)に浸し、室温下で4時間振盪した。このガラス基板を、次いで、グルタルジアルデヒド溶液から取り出し、3×SSCで3回、脱イオン水で2回洗浄し、遠心分離して水を除去し、そして室温において乾燥させた。
【0042】
3.プローブが表面上に固定化されたガラス基板(遺伝子チップ)の調製
表2に示されるプローブを、最終濃度10μmol/Lで50% DMSOに溶解する。このプローブを、Cartesian Spotting Instrument(Cartesian Technologies,Inc.,CA,USA)(9×9、090QCはプローブPBB−0201090を示す)を使用して、図2に示すように基板上にスポットする。このスポットされたガラス基板を、室温下で一晩乾燥させ、室温において0.2%SDS中に2回、それぞれ振動条件下で浸した。このガラス基板を、次いで、脱イオン水で2回洗浄し、脱イオン水で1回浸し洗いし、そして遠心分離して水を除去した。このガラス基板を次いで、NaBH4溶液(1g NaBH4を300mlの1×PBSに溶解し、次いで100mlエタノールを加えた)に移し、5分間わずかに振盪した。このガラス基板を、次いで、脱イオン水で1回洗浄し、脱イオン水で1回浸し洗いし(それぞれ1分)、そして遠心分離して水を除去した。
【0043】
4.細菌培養および核酸抽出
本明細書中で使用されるStaphylococcus aureus 26001を、Chinese Medical Bacterial Reservation Center,Institute of Chinese Medicinal and Biological Productsから入手した。
【0044】
無菌のワークステーションにおいて、MH(Mueller Hinton寒天培地)培養スラブに細菌を播種し、シングルコロニーを分離する。このスラブを、35℃インキュベーターにおいて、24時間、上下にインキュベートした。
【0045】
10mgのG1145ガラスビーズおよび40mgのG1152ガラスビーズ(Sigma)を、無菌の1.5ml遠心チューブ内に入れ、100μlの1×TEを加える。炎によって滅菌されたピンセットを使用して、爪楊枝またはチップを掴み、その培養スラブからシングルコロニーを取り、その爪楊枝またはチップをそのチューブの底部においてガラスビーズにこすりつけ、それによって十分量の細菌をそのチューブ内に残す。その爪楊枝またはチップを、細菌殺傷溶液を含むくず入れに投げ入れ、その遠心チューブを閉じる。そのチューブを、ボルテックスミキサー(TDX−1,Beijing Tongda)において最大速度で5分間ボルテックスする。この遠心分離チューブを、次いで、95℃の水浴に5分間入れ、4℃で保管する。
【0046】
5.核酸PCR増幅
PCR系を、表3に示すように組み立てた。26001細菌懸濁液においてガラスビーズを振盪することによって調製した1μLの細菌サンプルを、系A、BおよびCの各々に加えた。総反応容量は、25μlであった。
【0047】
PCR反応を、PTC−200サイクラー(MJ Research Inc.)において実施した。系Aおよび系Bは、表4に示すような非対称PCR増幅サイクルを使用した。系Cは、表5に示されるような非対称PCRサイクルを使用した。
【0048】
【表3】
【表4】
【0049】
【表5】
6.電気泳動
アガロースゲル電気泳動を、PCR増幅産物を検出するために使用した。アガロースゲルの濃度は1.5%であった。各サンプルは2μlであり、電気泳動は、50V/cmで30分間実施した。
【0050】
7.ハイブリダイゼーション
ハイブリダイゼーション系を、表6に示す。
【0051】
【表6】
200μlの蒸留水を、HybriCassettesTM(Beijing Bo’ao Biochip,Ltd)に加え、ハイブリダイゼーション系の蒸発を防ぐ。反応物が表面上に固定化されたガラス基板およびカバースリップ(SmartCoverTM,Beijing Bo’ao Biochip,Ltd.)を、ハイブリダイゼーションカセット中に入れる。このハイブリダイゼーション系を95℃に加熱し、5分間95℃で維持し、PCR産物の変性を可能にする。この混合物を、次いで、氷水浴において速やかに冷却する。13μLのハイブリダイゼーション系を採取し、そのカバースリップ上の孔を通して、その基板とそのカバースリップとの間の空隙にそのハイブリダイゼーション系を加える。そのハイブリダイゼーションカセットを閉じ、90分間ハイブリダイゼーションさせる。次いでチップを取り出し、2×SSC、0.2%SDS溶液中に浸し、そして5分間振盪する。このチップを、次いで、脱イオン水で2回、それぞれ1分間浸し洗いし、そして遠心分離して水を除去した。2回の実験を、Staphylococcus aureus 26001サンプルを含むPCR系で実施した。
【0052】
8.チップ上でのシグナルの検出
チップ上の蛍光シグナルを、GenePix4000Bスキャナ(Axon Instruments,Inc.,CA,USA)を使用することによって検出した。波長532nm、PMT600、および出力33%。
【0053】
9.結果の分析
アガロースゲル電気泳動の結果を、図3に示す。レーン1およびレーン11は、DL2000マーカー(Takarar)であり;レーン2、レーン3およびレーン4は、系AのPCR産物であり;レーン5、レーン6およびレーン7は、系BのPCR産物であり;レーン8、レーン9およびレーン10は、系CのPCR産物であった。これらのレーンの中で、レーン2、レーン3、レーン5、レーン6、レーン8およびレーン9は、26001細菌サンプルを含有し、一方レーン4、レーン7およびレーン10におけるPCR系は、ブランクコントロールを含有した。図3に示されるように、3つ全てのPCR系(A、BおよびC)が、約1.5kbの二本鎖標的核酸の良好な増幅をもたらしたが、一方系Aのみが明確な一本鎖PCR増幅産物を産生した。
【0054】
図4は、本明細書中に記載される実験のポジティブなハイブリダイゼーション結果を示す。図4Aは、Staphylococcus aureusの所望のハイブリダイゼーションシグナルを示し;図5は、本明細書中に記載されるハイブリダイゼーション実験の蛍光検出結果を示す。図4および図5に示されるように、系Aを使用したハイブリダイゼーション結果のみが、図4と一致するシグナルを生じ、一方他の2つの系は、部分的な結果を生じるのみであった。1つの可能な説明は、系Bおよび系Cにおいて産生された主要な産物は二本鎖PCR産物であり、これはハイブリダイゼーションの間に自己アニーリングすることができ、それによってポジティブなシグナルを弱めているか、またはポジティブなシグナルを生じることができなくなっているというものである。
【0055】
電気泳動およびチップハイブリダイゼーション結果は、本発明の非対称PCR増幅系および非対称PCR増幅温度サイクルが一本鎖増幅産物の調製において効率的であることを実証した。これは、ハイブリダイゼーション反応に特に適しており、それによってハイブリダイゼーション効率およびハイブリダイゼーションシグナルを増加させる。
【0056】
(実施例2.複数の非対称PCR増幅および遺伝子チップを使用する、細菌の薬物耐性遺伝子の検出)
プライマーおよびプローブはいずれも、Shanghai Boya Biotechnology Companyによって合成した。5’末端にTAMRA蛍光標識を有する特定のプライマーおよび5’末端に修飾アミノ基を有する特定のプローブもまた、Shanghai Boya Biotechnology Companyによって提供された。
【0057】
遺伝子特異的プライマーの分類、配列情報、標的遺伝子、および増幅されたフラグメントの長さは、表7に示す。各標的遺伝子は2つのプローブを有し、それらの配列情報は表8に示す。これらの中で、標的遺伝子tetKおよびtetMは、グラム陽性細菌由来のテトラサイクリン耐性遺伝子であった。標的遺伝子ermAおよびermCは、グラム陽性細菌由来のマクロライド−リンコサミド−ストレプトグラミンB耐性遺伝子であった。普遍的な細菌の23rRNA遺伝子の部分を、PCR増幅およびハイブリダイゼーションプロセスをモニタリングするために、内部コントロールとして使用した。
【0058】
【表7】
【0059】
【表8】
2.アルデヒド基を有するガラス基板の調製
アルデヒド基を有するガラス基板は、実施例1のように調製した。
【0060】
3.プローブが表面上に固定化されたガラス基板の調製
調製法は、実施例1と同じであった。このプローブは、表8に示す。これらのプローブを、図6に示すように(6×6、090QCは、コントロールプローブPBB−0201090を示し、残りのプローブは、プローブ番号の最後3つの数字で番号を付けた)スポットした。例えば、PBB−024654は、「654」と示される。
【0061】
4.細菌培養および核酸抽出
本実施例において使用した最近の供給元は、表9に示す。細菌の培養および核酸抽出法は、実施例2に示される。
【0062】
【表9】
5.核酸のPCR増幅
本実施例は、1回の実験における5つの異なるPCR増幅を示す。このPCR反応系を、以下に示す:1×MaterMix(Beijing Tianwei Times);0.2μmol/Lの、表7に示される5対の遺伝子特異的プライマー;1μmol/Lの一般プライマーAMB_0408047;ガラスビーズとの振動後の1μLの細菌サンプル。総反応容量は、25μLであった。
【0063】
PCRを、PTC−200(MJ Research Inc.)サーモサイクラーで実施した。この非対称PCRサーモサイクルは、表4に示した。
【0064】
6.ハイブリダイゼーションおよび検出
ハイブリダイゼーション検出系およびハイブリダイゼーション検出法、ならびにハイブリダイゼーション後の蛍光検出法は、実施例1に示した。
【0065】
7.結果の分析
図7は、8種の細菌株についての薬物耐性遺伝子検出結果を示す。図7に示されるように、異なる細菌株は、異なる種類および数の薬物耐性遺伝子を有する。Staphylococcus aureus MRSA6581およびEnterococcus faecalisは、1つの薬物耐性遺伝子のみを有する。Staphylococcus aureus B437およびStaphylococcus epidermis TR2041は、2つの薬物耐性遺伝子を有する。Staphylococcus aureus B435およびStaphylococcus aureus MRSA6460は、3つの薬物耐性遺伝子を有する。Staphylococcus aureus MRSA6437は、4つの薬物耐性遺伝子を有する。Enterococcus faeciumは、薬物耐性遺伝子を有さない。
【0066】
さらに、図7に示されるように、PCRおよびハイブリダイゼーションコントロールの23S rRNA遺伝子は、全ての細菌株に対して良好に機能し、良好な増幅および強力なハイブリダイゼーションシグナルを生じた。ポジティブおよび相同なシグナルを、1つ、2つ、3つまたは4つの薬物耐性遺伝子を有する全ての細菌から得た。このことは、本発明の非対称PCR増幅法および温度サイクルが、複数の遺伝子の並行検出に適していることを示す。
【産業上の利用可能性】
【0067】
本発明の非対称PCR増幅法は、高収率の一本鎖産物を産生し、このことはその後のハイブリダイゼーション反応を促進し、ハイブリダイゼーション効率を向上させ、そしてハイブリダイゼーションシグナルを増加させる。この方法はまた、最適化の必要性なく、類似の条件下での単一または複数のPCR増幅のために有用である。本発明のプライマーは、設計が単純である。本方法は、操作が容易であり、多因子のハイスループット遺伝子チップ法に適用可能である。本明細書中に記載される方法は、微生物判定、分類、薬物耐性遺伝子検出、疾患診断および予後、HLA分類、SNP検出ならびに遺伝子分析において用途を見出し得る。
【図面の簡単な説明】
【0068】
【図1】図1は、本発明の非対称PCRの概略図を提供する。
【図2】図2は、実施例1において使用されるプローブアレイの概略図を提供する。
【図3】図3は、実施例1のPCR産物の電気泳動写真を提供する。
【図4】図4Aは、実施例1に記載されるStaphylococcus aureus 26001サンプルにおける所望のハイブリダイゼーション結果を提供する。図4Bは、実施例1に記載されるブランクコントロールにおける所望のハイブリダイゼーション結果を提供する。
【図5】図5は、ハイブリダイゼーション後の蛍光検出の結果を提供する。
【図6】図6は、実施例2に記載されるプローブのアレイの概略図を提供する。
【図7】図7は、実施例2に記載されるハイブリダイゼーション後の蛍光検出の結果を提供する。
【特許請求の範囲】
【請求項1】
複数のPCRプライマー対を含む非対称PCR増幅のためのプライマーセットであって、各プライマー対における一方のプライマーは、5’末端において、増幅される標的配列に無関係な配列を有するオリゴヌクレオチドテイルを有する、プライマーセット。
【請求項2】
前記オリゴヌクレオチドテイルは8オリゴヌクレオチド長〜40オリゴヌクレオチド長である、請求項1に記載のプライマーセット。
【請求項3】
前記オリゴヌクレオチドテイルは15ヌクレオチド長〜25ヌクレオチド長である、請求項2に記載のプライマーセット。
【請求項4】
一般プライマーをさらに含む、請求項1、2または3のいずれか1項に記載のプライマーセットであって、該一般プライマーは、前記オリゴヌクレオチドテイルのヌクレオチドと同じ、少なくとも8個の連続するヌクレオチドを有する、プライマーセット。
【請求項5】
前記一般プライマーの配列は、前記オリゴヌクレオチドテイルの配列と同じである、請求項4に記載のプライマーセット。
【請求項6】
前記一般プライマーの濃度は、前記プライマー対の濃度よりも高い、請求項4に記載のプライマーセット。
【請求項7】
非対称PCR増幅法であって、該方法は、
1)プレ変性;
2)変性、アニーリングおよびプライマー伸長のサイクルを1つ以上含む、第一相のPCR増幅;ならびに
3)変性およびプライマー伸長のサイクルの1つ以上を1つ以上含む、第二相のPCR増幅、
を包含し、
伸長のための各プライマー対における一方のプライマーは、その5’末端において、増幅される標的配列に無関係な配列を有するオリゴヌクレオチドを有する、方法。
【請求項8】
前記第二相のPCRの後にさらなる伸長工程をさらに包含する、請求項7に記載の方法。
【請求項9】
前記第一相のPCR増幅のための前記PCR増幅サイクルは、8回〜25回である、請求項7または8に記載の方法。
【請求項10】
前記オリゴヌクレオチドテイルは8ヌクレオチド長〜40ヌクレオチド長である、請求項7または8に記載の方法。
【請求項11】
前記PCRのためのプライマーはさらに一般プライマーを含み、該一般プライマーは、前記オリゴヌクレオチドテイルのヌクレオチドと同じ、少なくとも8個の連続したヌクレオチドを有する、請求項7または8に記載の方法。
【請求項12】
前記一般プライマーの配列は、前記オリゴヌクレオチドテイルのプライマーと同一である、請求項11に記載の方法。
【請求項13】
前記一般プライマーの濃度は、前記プライマー対の濃度よりも高い、請求項11に記載の方法。
【請求項14】
前記第二相のPCRのための伸長温度は、60〜75℃である、請求項7または8に記載の方法。
【請求項15】
核酸の検出のための、請求項1〜6のいずれか1項に記載の非対称PCR増幅プライマーセットの使用。
【請求項16】
前記核酸検出は、遺伝子チップまたはメンブレンハイブリダイゼーションを使用する、請求項15に記載の使用。
【請求項1】
複数のPCRプライマー対を含む非対称PCR増幅のためのプライマーセットであって、各プライマー対における一方のプライマーは、5’末端において、増幅される標的配列に無関係な配列を有するオリゴヌクレオチドテイルを有する、プライマーセット。
【請求項2】
前記オリゴヌクレオチドテイルは8オリゴヌクレオチド長〜40オリゴヌクレオチド長である、請求項1に記載のプライマーセット。
【請求項3】
前記オリゴヌクレオチドテイルは15ヌクレオチド長〜25ヌクレオチド長である、請求項2に記載のプライマーセット。
【請求項4】
一般プライマーをさらに含む、請求項1、2または3のいずれか1項に記載のプライマーセットであって、該一般プライマーは、前記オリゴヌクレオチドテイルのヌクレオチドと同じ、少なくとも8個の連続するヌクレオチドを有する、プライマーセット。
【請求項5】
前記一般プライマーの配列は、前記オリゴヌクレオチドテイルの配列と同じである、請求項4に記載のプライマーセット。
【請求項6】
前記一般プライマーの濃度は、前記プライマー対の濃度よりも高い、請求項4に記載のプライマーセット。
【請求項7】
非対称PCR増幅法であって、該方法は、
1)プレ変性;
2)変性、アニーリングおよびプライマー伸長のサイクルを1つ以上含む、第一相のPCR増幅;ならびに
3)変性およびプライマー伸長のサイクルの1つ以上を1つ以上含む、第二相のPCR増幅、
を包含し、
伸長のための各プライマー対における一方のプライマーは、その5’末端において、増幅される標的配列に無関係な配列を有するオリゴヌクレオチドを有する、方法。
【請求項8】
前記第二相のPCRの後にさらなる伸長工程をさらに包含する、請求項7に記載の方法。
【請求項9】
前記第一相のPCR増幅のための前記PCR増幅サイクルは、8回〜25回である、請求項7または8に記載の方法。
【請求項10】
前記オリゴヌクレオチドテイルは8ヌクレオチド長〜40ヌクレオチド長である、請求項7または8に記載の方法。
【請求項11】
前記PCRのためのプライマーはさらに一般プライマーを含み、該一般プライマーは、前記オリゴヌクレオチドテイルのヌクレオチドと同じ、少なくとも8個の連続したヌクレオチドを有する、請求項7または8に記載の方法。
【請求項12】
前記一般プライマーの配列は、前記オリゴヌクレオチドテイルのプライマーと同一である、請求項11に記載の方法。
【請求項13】
前記一般プライマーの濃度は、前記プライマー対の濃度よりも高い、請求項11に記載の方法。
【請求項14】
前記第二相のPCRのための伸長温度は、60〜75℃である、請求項7または8に記載の方法。
【請求項15】
核酸の検出のための、請求項1〜6のいずれか1項に記載の非対称PCR増幅プライマーセットの使用。
【請求項16】
前記核酸検出は、遺伝子チップまたはメンブレンハイブリダイゼーションを使用する、請求項15に記載の使用。
【図1】

【図2】
【図3】
【図4A】

【図4B】

【図5】

【図6】

【図7】

【図2】
【図3】
【図4A】
【図4B】
【図5】
【図6】
【図7】
【公表番号】特表2008−510471(P2008−510471A)
【公表日】平成20年4月10日(2008.4.10)
【国際特許分類】
【出願番号】特願2007−528557(P2007−528557)
【出願日】平成16年11月22日(2004.11.22)
【国際出願番号】PCT/CN2004/001330
【国際公開番号】WO2006/021131
【国際公開日】平成18年3月2日(2006.3.2)
【出願人】(507060099)
【出願人】(507061421)清▲華▼大学 (3)
【Fターム(参考)】
【公表日】平成20年4月10日(2008.4.10)
【国際特許分類】
【出願日】平成16年11月22日(2004.11.22)
【国際出願番号】PCT/CN2004/001330
【国際公開番号】WO2006/021131
【国際公開日】平成18年3月2日(2006.3.2)
【出願人】(507060099)
【出願人】(507061421)清▲華▼大学 (3)
【Fターム(参考)】
[ Back to top ]