
非小細胞肺癌関連遺伝子ANLN、およびそのRhoAとの相互作用
本発明は、癌を処置するための化合物を同定するための組成物および方法、ならびに癌の予後を予測するための方法を提供する。特に、本発明は、癌、特に非小細胞肺癌(NSCLC)などの肺癌の処置および予防に有用な、ANLNとRhoAとの間の相互作用の阻害剤を同定するための方法およびキットを提供する。本発明のスクリーニング方法によって同定された癌を処置または予防するための組成物およびそれらを癌の処置および予防に用いる方法もまた本明細書に開示する。
【発明の詳細な説明】
【技術分野】
【0001】
発明の分野
本発明は、癌、特に肺癌の処置および予防に有用な化合物を同定するための方法およびキットに関し、ならびに、それを処置および予防するための方法および組成物に関する。さらに詳しくは、本方法は、非小細胞肺癌でアップレギュレートされている癌特異的遺伝子であるANLN(全体が参照として本明細書に組み入れられるPCT公報WO2004/031413を参照されたい)が、腫瘍形成に関わる低分子量GTPaseのRasホモロジーファミリーのメンバーであるRhoAと相互作用するという発見に関する。
【背景技術】
【0002】
発明の背景
肺癌は世界中で最も一般的な癌による死因の一つであり、非小細胞肺癌(NSCLC)がこれらのケースの80%近くを占めている(Greenlee, R.T. et al. CA. Cancer J. Clin. 51:15-36 (2001)(非特許文献1))。肺癌の発症および/または進行に関わる多くの遺伝子変化が報告されているが、正確な分子メカニズムは不明のままである(Sozzi, G., Eur. J. Cancer. 37:63-73 (2001)(非特許文献2))。新たに開発された細胞毒性薬剤は進行したNSCLCを有する患者に複数の治療選択肢を与えるが、これらの新たな治療法の各々はシスプラチンをベースに使用する療法と比較して少しの生存利益を提供できるのみである(Schiller, J.H. et al. N. Engl. J. Med. 346(2):92-98 (2002)(非特許文献3); Kelly, K. et al. J. Clin. Oncol. 19(13):3210-3218 (2001)(非特許文献4))。それゆえ、分子標的薬剤および抗体の開発ならびに癌ワクチンのような新規治療戦略が熱望される。
【0003】
cDNAマイクロアレイを用いる何千もの遺伝子の発現レベルの体系的な解析は、発癌経路にかかわる分子セットを同定する有効なアプローチであり(Kikuchi, T. et al. Oncogene 22(14):2192-2205 (2003)(非特許文献5); Kakiuchi, S. et al. Mol. Cancer Res. 1:485-499 (2003)(非特許文献6); Zembutsu, H. et al. Int. J. Oncol. 23:29-39 (2003)(非特許文献7); Suzuki, C. et al. Cancer Res. 63(21):7038-7041 (2003)(非特許文献8))、それらのいくつかは新規抗癌薬および腫瘍マーカーの開発のための候補標的でありうる。
【0004】
本発明は、このような標的分子に対する当技術分野における必要性に応えるものである。特に、NSCLCの診断および処置のための新規分子標的を単離するため、本発明者らは、NSCLCのゲノム全域にわたる発現プロファイル解析を、レーザーキャプチャーマイクロダイセクションによる37個の癌組織からの腫瘍細胞の純粋精製と組み合わせて行った。これら研究の過程において、ショウジョウバエ(Drosophila)アクチン結合タンパク質(ANLN)であるアニリン(anillin)のヒトホモログをコードする遺伝子が高頻度に原発性NSCLCで過剰発現していることが観察された。ANLNは、分裂溝におけるアクチンケーブルの形成または構築に不可欠で、細胞質分裂に重要な役割を担うことが報告されている(Oegema, K. et al. J. Cell Biol. 150(3):539-551 (2000)(非特許文献9))。非小細胞肺癌に関連する差次的遺伝子発現は以前に述べられている。例えばPCT公報WO2004/031413(特許文献1)を参照されたい。
【0005】
低分子量グアノシントリホスファターゼ(GTPase)は、細胞骨格のリモデリングの調節および協調に重要な役割を担う。哺乳動物細胞におけるRasホモロジーファミリーメンバー(RHO)のうち、RhoA、Ras関連C3ボツリヌス毒素基質1(RAC1)、および細胞分裂周期42(CDC42)がその生物学的機能について広範に研究されている。RhoAは、そのエフェクターであるmDia、Rho関連コイルドコイル含有タンパク質キナーゼ1(ROCK1)、およびROCK2の動員および活性化を通じた接着斑の組み立ておよびアクチンストレスファイバーの形成に原形質膜受容体が関連しているシグナル伝達経路を調節する(Ridley, A. J. and Hall, A., Cell 70:389-399 (1992)(非特許文献10); Leung, T. et al. Mol. Cell Biol. 16:5313-5327(1996)(非特許文献11); Amano, M. et al. Science 275:1308-1311 (1997)(非特許文献12))。RHO活性は細胞の運動性のために重要であるが、効率的な遊走は、細胞環境における適切な時空間において厳密な均衡の取れたRAC1、CDC42、およびRhoAの活性化および非活性化を必要とする。RHOタンパク質はまた、遺伝子転写、細胞周期進行、または抗アポトーシス経路の制御にも関与しており(Etienne-Manneville, S. and Hall, A., Nature 420:629-635 (2002)(非特許文献13))、最近の研究によってRhoAは一部のヒト腫瘍において活性化されていることが示されている(Sahai, E. and Marshall, C., J., Nat. Rev. Cancer 2(2):133-142 (2002)(非特許文献14))が、その正確なメカニズム、特に発癌の際のRhoAシグナル伝達の上流の経路は明らかにされていない。
【0006】
【特許文献1】PCT公報WO2004/031413
【非特許文献1】Greenlee, R.T. et al. CA. Cancer J. Clin. 51:15-36 (2001)
【非特許文献2】Sozzi, G., Eur. J. Cancer. 37:63-73 (2001)
【非特許文献3】Schiller, J.H. et al. N. Engl. J. Med. 346(2):92-98 (2002)
【非特許文献4】Kelly, K. et al. J. Clin. Oncol. 19(13):3210-3218 (2001)
【非特許文献5】Kikuchi, T. et al. Oncogene 22(14):2192-2205 (2003)
【非特許文献6】Kakiuchi, S. et al. Mol. Cancer Res. 1:485-499 (2003)
【非特許文献7】Zembutsu, H. et al. Int. J. Oncol. 23:29-39 (2003)
【非特許文献8】Suzuki, C. et al. Cancer Res. 63(21):7038-7041 (2003)
【非特許文献9】Oegema, K. et al. J. Cell Biol. 150(3):539-551 (2000)
【非特許文献10】Ridley, A. J. and Hall, A., Cell 70:389-399 (1992)
【非特許文献11】Leung, T. et al. Mol. Cell Biol. 16:5313-5327(1996)
【非特許文献12】Amano, M. et al. Science 275:1308-1311 (1997)
【非特許文献13】Etienne-Manneville, S. and Hall, A., Nature 420:629-635 (2002)
【非特許文献14】Sahai, E. and Marshall, C., J., Nat. Rev. Cancer 2(2):133-142 (2002)
【発明の開示】
【0007】
発明の概要
本発明は、ANLNおよびRhoAが非小細胞肺癌細胞中で相互作用すること、さらに、ANLNの核局在が非小細胞肺癌患者における不良な予後に関係しているという知見に基づく。よって、本発明は、ANLN/RhoA相互作用を妨害することによって、またはANLNを介する細胞運動性を阻害することによって、癌、例えば非小細胞肺癌の進行を遅延または停止させる化合物を同定するための新規方法を提供する。本発明はまた、サンプル中のANLNの核局在の有無を決定することによって、癌患者について予後を判断するための方法を提供する。
【0008】
よって、癌、特に非小細胞肺癌(NSCLC)などの肺癌の処置または予防に有用な化合物をスクリーニングする方法を提供することが、本発明の目的である。一部の態様において、本方法は以下の工程を含む:
(a)試験化合物の存在下で、ANLNポリペプチドのRhoA結合ドメインを含むポリペプチドを、RhoAポリペプチドのANLN結合ドメインを含むポリペプチドと接触させる工程;
(b)ポリペプチド間の結合を検出する工程;および
(c)ポリペプチド間の結合を阻害する試験化合物を選択する工程。
【0009】
一部の態様において、RhoA結合ドメインを含むポリペプチドはANLNポリペプチドを含む。同様に、他の態様において、ANLN結合ドメインを含むポリペプチドはRhoAポリペプチドを含む。
【0010】
一部の態様において、RhoA結合ドメインを含むポリペプチドは生細胞中で発現する。
【0011】
一部の態様において、以下のものを検出する工程を含む方法によってポリペプチド間の結合が検出される:
(a)活性化RhoAの濃度;
(b)RhoAとRHOエフェクターまたはそのRhoA結合領域との間の相互作用;
(c)活性化RhoAを介する下流遺伝子発現または下流遺伝子産物活性を含む任意のシグナル複合体の活性化;
(d)DNA合成および細胞周期移行の促進;
(e)細胞遊走または任意の他の発癌表現型;
(f)アクチンストレスファイバー形成およびF-アクチン産生;ならびに
(g)細胞接着、遊走、および浸潤に重要な任意の分子との相互作用。
【0012】
本発明はまた、癌、特に非小細胞肺癌(NSCLC)などの肺癌の処置または予防に有用な化合物をスクリーニングするためのキットを提供する。一部の態様において、本キットは以下のものを含む:
(a)ANLNポリペプチドのRhoA結合ドメインを含む第一のポリペプチド;
(b)RhoAポリペプチドのANLN結合ドメインを含む第二のポリペプチド;および
(c)第一のポリペプチドと第二のポリペプチドとの間の相互作用を検出する手段(例えば試薬)。
【0013】
一部の態様において、第一のポリペプチド、すなわちRhoA結合ドメインを含むポリペプチドは、ANLNポリペプチドを含む。同様に、他の態様において、第二のポリペプチド、すなわちANLN結合ドメインを含むポリペプチドは、RhoAポリペプチドを含む。
【0014】
一部の態様において、RhoA結合ドメインを含むポリペプチドは生細胞中で発現する。
【0015】
一部の態様において、2つのポリペプチド間の相互作用を検出する手段(例えば試薬)は、以下のものを検出する:
(a)活性化RhoAの濃度;
(b)RhoAとRHOエフェクターまたはそのRhoA結合領域との間の相互作用;
(c)活性化RhoAを介する下流遺伝子を含む任意のシグナル複合体の活性化;
(d)DNA合成および細胞周期移行の促進;
(e)細胞遊走または任意の他の発癌表現型;
(f)アクチンストレスファイバー形成およびF-アクチン産生;ならびに
(g)細胞接着、遊走、および浸潤に重要な任意の分子との相互作用。
【0016】
代替的な態様において、癌、特に非小細胞肺癌(NSCLC)などの肺癌の処置または予防に有用な化合物をスクリーニングする本発明の方法は、以下の工程を含む:
(a)ANLNポリペプチドまたはその機能的同等物を発現する細胞を試験化合物と接触させる工程;
(b)細胞のANLNを介する運動性を検出する工程;および
(c)試験化合物の非存在下で検出される運動性レベルと比較して細胞の運動性を阻害する試験化合物を選択する工程。
【0017】
一部の態様において、細胞は、ANLNポリペプチドまたはその機能的同等物を発現可能にコードするポリヌクレオチドを(例えばANLNポリペプチドをコードするポリヌクレオチドに機能的に連結されたプロモーターとして)含むベクターを含む。
【0018】
同様に、癌、特に非小細胞肺癌(NSCLC)などの肺癌の処置または予防に有用な化合物をスクリーニングするための本発明のキットは、以下のものを含みうる:
(a)ANLNポリペプチドまたはその機能的同等物を発現する細胞;および
(b)細胞の運動性を検出する手段(例えば試薬)。
【0019】
本発明はさらに、対象における癌の予後、より詳しくは非小細胞肺癌(NSCLC)に罹患した患者などの肺癌患者の予後を予測する方法を提供する。一部の態様において、本方法は以下の工程を含む:
(a)NSCLCの予後を予測しようとする対象から収集された試料の核に局在するANLNを検出する工程、および
(b)核内のANLNの局在が検出された場合に予後が不良であると予測する工程。
【0020】
一部の態様において、試料の核内のANLNの局在は以下の工程によって検出される:
(a)ANLNタンパク質を認識する抗体を試料と接触させる工程;および
(b)核領域において試料に結合する抗体を検出する工程。
【0021】
本発明はまた、対象における癌の予後、特に非小細胞肺癌(NSCLC)などの肺癌の予後を予測するためのキットを提供する。一部の態様において、本キットは、(a)ANLNタンパク質を認識する抗体と、(b)核における検出用の薬剤とを含む。一部の態様において、核の検出用の薬剤はヘマトキシリン-エオシン染色染料である。
【0022】
本発明はまた、対象における癌、特に非小細胞肺癌(NSCLC)などの肺癌を処置または予防するための方法を提供する。一部の態様において、本方法は以下の工程によって選択される化合物を投与する工程を含む:
(a)試験化合物の存在下で、ANLNポリペプチドのRhoA結合ドメインを含むポリペプチドを、RhoAポリペプチドのANLN結合ドメインを含むポリペプチドと接触させる工程;
(b)ポリペプチド間の結合を検出する工程;および
(c)ポリペプチド間の結合を阻害する試験化合物を選択する工程。
【0023】
または、本方法は以下の工程を含みうる:
(a)ANLNポリペプチドまたはその機能的同等物を発現する細胞を試験化合物と接触させる工程;
(b)細胞の運動性を検出する工程;および
(c)試験化合物の非存在下で検出される運動性レベルと比較して細胞の運動性を阻害する試験化合物を選択する工程。
【0024】
本発明はまた、対象における癌、特に非小細胞肺癌(NSCLC)などの肺癌を処置または予防するための方法であって、ANLNとRhoAとの間の結合を阻害する化合物を投与する工程を含む方法を提供する。
【0025】
本発明はまた、癌、特に非小細胞肺癌(NSCLC)などの肺癌の処置または予防に有用な組成物であって、以下の工程によって選択される化合物の薬学的有効量を含む組成物を提供する:
(a)試験化合物の存在下で、ANLNポリペプチドのRhoA結合ドメインを含むポリペプチドを、RhoAポリペプチドのANLN結合ドメインを含むポリペプチドと接触させる工程;
(b)ポリペプチド間の結合を検出する工程;および
(c)ポリペプチド間の結合を阻害する試験化合物を選択する工程;
または、
(a)ANLNポリペプチドまたはその機能的同等物を発現する細胞を試験化合物と接触させる工程;
(b)細胞の運動性を検出する工程;および
(c)試験化合物の非存在下で検出される運動性レベルと比較して細胞の運動性を阻害する試験化合物を選択する工程。
【0026】
本発明はまた、癌、特に非小細胞肺癌(NSCLC)などの肺癌の処置または予防に有用な組成物であって、ANLNとRhoAとの間の結合を阻害する化合物の薬学的有効量と、薬学的に許容される担体とを含む組成物を提供する。
【0027】
本発明のこれらおよび他の目的および特徴は、添付の図面および実施例と合わせて以下の詳細な説明を読むことにより、さらに十分明らかとなるであろう。しかし、上記の発明の概要および下記の詳細な説明はいずれも好ましい態様のものであって、発明または他の代替的な発明の態様を限定するものではないことを理解されたい。
【0028】
好ましい態様の詳細な説明
I. 概説
分子標的薬は、明確な作用メカニズムによって副作用を最小限に抑えつつ、悪性細胞に高度に特異的であることが期待される。このような薬の開発のための適切な分子標的を同定するために見込みある戦略として、本発明者らは、癌細胞で過剰発現している遺伝子を選択できるゲノム全域にわたる発現解析を、RNAi技術による機能喪失効果のハイスループットスクリーニングと組み合わせた。加えて、組織マイクロアレイ方法を応用し、可能性のある標的タンパク質の検証のために何百もの保管臨床サンプルを解析した。この種の体系的なアプローチを用いて、ANLNが臨床NSCLCサンプルならびに細胞株中で高頻度に過剰発現していること、また、この遺伝子産物が肺癌細胞の成長および進行において必要不可欠な役割を担うことを本明細書において示す。
【0029】
ANLNは当初、ショウジョウバエアクチン結合タンパク質であるアニリンのヒトホモログとして特徴付けられた(Oegema, K. et al. J. Cell Biol. 150(3):539-551 (2000))。ヒトANLN cDNAは、アクチン結合ドメインおよびC末端プレクストリン相同(PH)ドメインを含む1124アミノ酸タンパク質をコードする。また、いくつかのコンセンサス核局在配列(NLS)および一つのコンセンサスSH3結合モチーフを含んでいた。ANLNは細胞質分裂の間、分裂溝に局在しており、細胞質分裂に重要な役割を担っているとされる。本明細書において、ANLNは、一定の割合の癌細胞において細胞質だけでなく核にも局在する一方で、核膜崩壊後の皮層および細胞質分裂の間の分裂溝に位置することが見出された。以前に報告されているように、ANLNは細胞周期進行に重要な役割を担っている可能性が高く、後期において、少なくとも2つの他の溝タンパク質(アクチンおよびセプチン(SEPT))との相互作用を通じて、娘細胞を分けるアクチン-ミオシン収縮環を組み立てている可能性がある(Oegema, K. et al. J. Cell Biol. 150(3):539-551 (2000))。本明細書において考察するように、ANLN-siRNAで処置したNSCLC細胞は溝退行およびより大きな細胞形態を示し、おそらく収縮環の組み立てが阻止された結果としての細胞質分裂プロセスの機能不全により、多核化したことが観察されている。本明細書において、内因性RhoA(SEQ ID NO:4(SEQ ID NO:3にコードされる))のANLNとの相互作用は細胞質中のみでなく分裂溝および中央体においても起こることが見出され、細胞成長がANLN-RhoA相互作用およびNSCLC細胞の細胞質分裂の加速を通じて促進されたことが示された。
【0030】
そこで、本発明者らは、アクチン構造の形成を制御することが知られるRHO活性化(Ridley, A. J. and Hall, A. Cell 70:389-399 (1992))に対するANLNの効果に焦点を絞り、外因性ANLNの過剰発現が哺乳動物細胞においてアクチンストレスファイバーの形成を促進したことを見出した。本明細書におけるデータは、ANLNがRhoAとの相互作用を通じてRHOシグナル伝達を活性化してアクチン細胞骨格の再構築を有意に促進し、その結果、マトリゲル浸潤アッセイおよび傷害遊走アッセイによって観察されるように、細胞遊走活性を活性化した可能性があることを示唆する。
【0031】
RASおよびRHO GTPaseは十分に研究されているシグナル伝達分子であり、RHO GTPaseは、アクチン構築、限局性複合体/接着の組み立て、細胞運動性、細胞極性、遺伝子転写、および細胞周期進行を含む多様な一連の生物学的活性の調節において重要な分子として浮上してきた。最近の研究により、RHOタンパク質がいくつかのヒト腫瘍で過剰発現していること、ならびに上皮細胞成長因子(EGF)、肝細胞成長因子(HGF)、リポホスファチジン酸(lypophosphatidic acid)(LPA)、血小板由来成長因子(PDGF)、およびトランスフォーミング成長因子β(TGFB)を含む一部の成長因子がRHOタンパク質を活性化しうること(Zondag, G. C. et al. J. Cell Biol. 149:775-781 (2000); Bhowmick, N. A., et al., Mol. Biol. Cell 12, 27-36 (2001); Liu, A. X. et al., Mol. Cell Biol. 21:6906-6912 (2001))が示されている。インテグリン、カドヘリン、および免疫グロブリンスーパーファミリーメンバーを含む細胞接着分子のいくつかのクラスもRHO活性に影響を及ぼすことが示されている(DeMali, K. A., et al., Curr. Opin. Cell Biol. 15:572-582 (2003); Braga, V.M., Curr. Opin. Cell Biol. 14, 546-556 (2002); Thompson, P. W., et al., J. Immunol. 169:1007-1013 (2002))。加えて、いくつかのグアニンヌクレオチド交換因子(GEF)がRHOタンパク質およびそれらの下流エフェクターを異常に活性化して新生物性の形質転換を起こす可能性がある(Fort, P., Prog. Mol. Subcell. Biol. 22, 159-181 (1999); Zohn, I. M., et al., Oncogene 17, 1415-1438 (1998))。本発明者らのデータにより、過剰発現したANLNによるRhoAの異常な活性化がアクチン細胞骨格の再構築を通じて哺乳動物細胞の遊走活性を促進し、癌細胞の浸潤能および転移能に寄与しうるであろうことが示唆された。
【0032】
BrdU取り込みアッセイにより、外因的に発現されたANLNによる用量依存的なNSCLC細胞のDNA合成の促進が検出され、ANLNが細胞周期進行の重要な正のレギュレーターでありうることが示唆された。RHO、RAC、およびCDC42などのRHO GTPaseは、細胞周期のG1期に様々な活性をもたらすことが知られている(Olson, M. F., et al., Science 269, 1270-1272 (1995))。インビトロ研究により、活性がサイクリンD1(CCND1)などの活性化剤ならびにCDK阻害剤(CDKI)であるp21WAF1およびp27KIP1などの阻害剤によって制御されるサイクリン依存性キナーゼ(CDK)の調節を主に介して、RHOタンパク質が細胞周期の進行を促進することができる、複数のメカニズムが明らかとなっている(Danen, E. H., et al., J. Cell Biol. 151, 1413-1422 (2000); Olson, M. F., et al., Nature 394:295-299 (1998); Adnane, J., et al., Mol. Cell. Biol. 18:6962-6970 (1998))。ANLNによるNSCLC細胞のDNA合成の促進は、これら分子経路の機能に影響を及ぼす活性化RhoAのアップレギュレーションによるものである可能性がある。
【0033】
本明細書に提示する組織マイクロアレイ実験の結果は、n-ANLN陽性腫瘍を有する肺癌患者が、陰性腫瘍を有さない患者と比べて不良な癌特異的生存を示したことを示している。核へのANLN輸送の正確な分子メカニズム、およびそれが核特異的な追加機能を有するかどうかは不明であるが、このデータは、n-ANLNが何らかの未知のシグナル伝達経路を活性化することによって肺癌細胞の非常に悪性の表現型に寄与することを示唆している。
【0034】
以上をまとめると、ANLNは、RhoAと直接相互作用してそれを活性化することが本明細書において示され、この複合体は肺癌の成長促進経路および攻撃的な特性、ならびに細胞分裂/細胞周期進行に不可欠である可能性が高い。ここで報告するデータは、このANLN-RhoA経路が新規バイオマーカーを設計するための、および肺癌に特異的な抗癌薬を開発するための、好適な分子標的となりうることを示している。治療用siRNAはこの経路を妨害する選択肢の一つであるはずである。
【0035】
II. 定義:
本明細書において用いられる冠詞(a、an、およびthe)は、特に具体的に示されていない限り「少なくとも1つ」を意味する。
【0036】
本発明の文脈において、「ANLNポリペプチド」または「ANLN」は、しばしば科学文献において「アニリン」と呼ばれるアクチン結合タンパク質を指す。例えば、全体が参照として本明細書に組み入れられるOegema et al., J. Cell Biol. 150(3):539-551 (2000)を参照されたい。ANLNポリペプチドは、ヒト(Genbankアクセッション番号AF273437)、ショウジョウバエ(Genbankアクセッション番号X89858、ショウジョウバエ遺伝子CG4530(GenBankアクセッション番号AAF47044)の産物)、線虫(C. elegans)遺伝子K10B2.5(GenBankアクセッション番号T16604)、Y43F8C.14(GenBankアクセッション番号T26874)、およびY49E10.19(GenBankアクセッション番号T27053)の産物、または他のオルソロガスなポリペプチドと実質的に同一でありうる。一部の態様において、ANLNポリペプチドは、ヒトとショウジョウバエのANLNオルソログ間で保存されているアミノ酸を含む。または、ANLNポリペプチドは、例えばOegemaらの文献の542頁に示されるような、上に列挙したタンパク質の全ての間で保存されているアミノ酸を含みうる。全長ANLNポリペプチドは、例えば、PH結合ドメイン、アクチン結合ドメイン、核局在シグナル配列、およびSH3ドメインを含みうる。
【0037】
本明細書において、「RhoAポリペプチド」または「RhoA」は、GTP結合タンパク質のRho遺伝子ファミリーを指す。RhoAの結晶構造は決定されている。例えば、全体が参照として本明細書に組み入れられるMaesaki et al., Molec. Cell 4:793-803 (1999)を参照されたい。Maesakiらは、RhoAをRhoファミリーの他のメンバーから識別するACCフィンガー構造の位置および構造を含むRhoAの一次および二次構造に関してかなりの量の情報を提供している。RhoAポリペプチドの例には、例えばSEQ ID NO:4と実質的に同一なタンパク質が含まれる。
【0038】
最近の研究により、RHOファミリータンパク質が腫瘍形成に関与することが示されている。経路はまだ不明であるが、RHOと癌との関連は本質的なものである。特にRhoAタンパク質は癌に多大に関連しているようであり、RhoAの過剰発現は、大腸、乳房、肺、精巣生殖細胞、および頭頸部扁平上皮細胞癌腫瘍に関連している。加えて、RhoAは細胞運動性および細胞極性に関与することが示唆されている。これら2つの機能に対するRhoA発現の効果は、腫瘍形成の原因である高い可能性を示唆する。
【0039】
本発明の文脈において、2つのタンパク質間の「結合の阻害」とは、タンパク質間の結合を少なくとも減少させることを指す。従って、場合によっては、サンプル中の結合対のパーセンテージは、適切な(例えば、試験化合物で処置されていない、または非癌サンプル由来の、または癌サンプル由来の)対照と比較して低下する。結合したタンパク質の量における減少は、例えば、対照サンプル中で結合した対よりも、90%、80%、70%、60%、50%、40%、25%、10%、5%、1%よりも少ない、またはそれより少なく(例えば0%)なりうる。
【0040】
用語「試験化合物」は、本明細書において詳しく述べられるような、ANLNとRhoAとの間のタンパク質-タンパク質相互作用を阻害しうる任意の(例えば化学的または組換え的に産生された)分子を指す。一部の態様において、試験化合物は1,500ダルトン未満の分子量を有し、場合によっては、1,000、800、600、500、または400ダルトン未満の分子量を有する。
【0041】
化合物の「薬学的有効量」とは、個体においてANLNを介する疾患を処置および/または改善するのに十分な量である。薬学的有効量の一例は、動物に投与したときに、ANLNとRhoAとの間の相互作用を低下させるのに必要な量でありうる。相互作用における低下とは、例えば、結合における少なくとも5%、10%、20%、30%、40%、50%、75%、80%、90%、95%、99%、または100%の変化でありうる。または、この量は、野生型ANLNについて本明細書に記載されているように、投与したときに核局在を検出可能に低下させる量を含みうる。
【0042】
「薬学的に許容される担体」という表現は、薬物の希釈剤または賦形剤として用いられる不活性な物質を指す。
【0043】
本発明の文脈において、用語「機能的に同等」とは、対象ポリペプチドが参照ポリペプチドの生物学的活性を有することを意味する。例えば、ANLNの機能的同等物は、野生型ANLNのようにインビトロでアクチンに結合して細胞運動性を誘導する能力を有するであろう。このような活性を測定するためのアッセイ法は当技術分野において周知である。
【0044】
用語「単離」および「生物学的に純粋」とは、自然の状態で見られる通常はそれに付随する成分を実質的または本質的に含まない材料を指す。しかしながら、用語「単離」は、電気泳動ゲルまたは他の分離媒体中に存在する成分を指すことを意図しない。単離された成分はそのような分離媒体を含まず、別の応用にすぐに使用可能な形態にあるか、あるいは既に新しい応用/環境に使用されている。
【0045】
「保存的に改変された変異体」という表現は、アミノ酸配列および核酸配列の両方に適用される。特定の核酸配列に関して、保存的に改変された変異体とは、同一または本質的に同一のアミノ酸配列をコードする核酸を指し、核酸がアミノ酸配列をコードしない場合は、本質的に同一の配列を指す。遺伝コードの縮重により、機能的に同一の核酸の多数が任意の所与のタンパク質をコードする。例えば、コドンGCA、GCC、GCG、およびGCUは全て、アミノ酸のアラニンをコードする。したがって、アラニンがコドンによって指定される全ての位置で、そのコドンは、コードするポリペプチドを変化させることなく任意の上記対応コドンに変えることができる。このような核酸変異は「サイレント変異」、つまり保存的に改変された変異の一種である。本明細書において、ポリペプチドをコードする全ての核酸配列は、その核酸の全ての可能なサイレント変異も表す。当業者は、核酸中の各コドン(通常メチオニンの唯一のコドンであるAUGと、通常トリプトファンの唯一のコドンであるTGGを除く)を改変して機能的に同一の分子を得ることができることを認識するであろう。よって、ポリペプチドをコードする核酸の各サイレント変異は、各開示配列に暗黙のうちに記載されている。
【0046】
アミノ酸配列に関し、コードされた配列中の1個のアミノ酸を、または少ないパーセンテージのアミノ酸を変化、付加、または欠失させる、核酸、ペプチド、ポリペプチド、またはタンパク質配列に対する置換、欠失、または付加はそれぞれ、その変化がアミノ酸を化学的に類似のアミノ酸と置換する「保存的に改変された変異体」であることを当業者は認識するであろう。機能的に類似のアミノ酸を提供する保存的置換表は当技術分野において周知である。このような保存的に改変された変異体は、本発明の多型変異体、種間ホモログ、および対立遺伝子に加えられるものであって、これらを排除するものではない。
【0047】
以下の8群はそれぞれ、互いに保存的置換であるアミノ酸を含む:
1)アラニン(A)、グリシン(G);
2)アスパラギン酸(D)、グルタミン酸(E);
3)アスパラギン(N)、グルタミン(Q);
4)アルギニン(R)、リジン(K);
5)イソロイシン(I)、ロイシン(L)、メチオニン(M)、バリン(V);
6)フェニルアラニン(F)、チロシン(Y)、トリプトファン(W);
7)セリン(S)、スレオニン(T);および
8)システイン(C)、メチオニン(M)(例えば、Creighton, Proteins (1984) を参照されたい)。
【0048】
本発明の文脈において、「配列同一性のパーセンテージ」は、2つの最適に整列された配列を比較ウィンドウにわたって比較することにより決定され、このとき比較ウィンドウ中のポリヌクレオチド配列部分は、2つの配列の最適な整列について、付加または欠失を含まない参照配列(例えば本発明のポリペプチド)と比較して付加または欠失(すなわちギャップ)を含んでもよい。このパーセンテージは、同一核酸塩基またはアミノ酸残基が両配列に現れる位置の数を決定してマッチする位置の数を出し、マッチする位置の数を比較ウィンドウ中の位置の総数で割り、その結果に100を乗じて配列同一性のパーセンテージを出すことによって計算される。
【0049】
2つもしくはそれ以上の核酸またはポリペプチド配列の文脈において、用語「同一」またはパーセント「同一性」は、同じ配列である2つまたはそれ以上の配列または部分配列を指す。2つの配列が、下記の配列比較アルゴリズムの1つを用いてまたはマニュアル整列と目視検査によって測定された比較ウィンドウまたは指定領域にわたって最大一致するように比較整列された際に、同じアミノ酸残基またはヌクレオチドを特定のパーセンテージで有する(すなわち、特定領域にわたって、あるいは特定されていない場合は全配列にわたって、60%同一性、任意で65%、70%、75%、80%、85%、90%、または95%同一性)場合、2つの配列は「実質的に同一」である。任意で、同一性は長さが少なくとも約50ヌクレオチドである領域にわたって存在し、またはより好ましくは長さが100〜500または1000ヌクレオチド以上である領域にわたって存在する。
【0050】
配列比較には、典型的には1つの配列を参照配列とし、これに対して試験配列を比較する。配列比較アルゴリズムを用いる場合、試験および参照配列をコンピュータに入力し、必要であれば部分配列座標を指定して、配列アルゴリズムプログラムパラメータを指定する。デフォルトプログラムパラメータを用いてもよく、また、代替的なパラメータを指定してもよい。そして配列比較アルゴリズムが、プログラムパラメータに基づいて、試験配列について参照配列に対するパーセント配列同一性を計算する。
【0051】
本明細書において用いられる「比較ウィンドウ」は、20〜600、一般的には約50〜約200、より一般的には約100〜約150からなる群より選択される連続位置の数のいずれか1つのセグメントに対する参照を含み、この内部において、ある配列が同じ数の連続位置の参照配列と最適に整列された後、これら2つの配列が比較されうる。比較するために配列を整列する方法は当技術分野において周知である。例えば、Smith and Waterman (1981) Adv. Appl. Math. 2:482-489の局所相同性アルゴリズム、Needleman and Wunsch (1970) J. Mol. Biol. 48:443の相同性整列アルゴリズム、Pearson and Lipman (1988) Proc. Nat'l. Acad. Sci. USA 85:2444の類似性検索法、これらアルゴリズムのコンピュータによる実施(GAP、BESTFIT、FASTA、およびTFASTA、Wisconsin Genetics Software Package, Genetics Computer Group, 575 Science Dr., Madison, WI)、またはマニュアル整列と目視検査(例えば、Ausubel et al., Current Protocols in Molecular Biology (1995 supplement) を参照されたい)によって比較用に配列を最適に整列できる。
【0052】
パーセント配列同一性および配列類似性を決定するために適したアルゴリズムの2つの例がBLASTアルゴリズムおよびBLAST 2.0アルゴリズムであり、これらはそれぞれAltschul et al. (1997) Nuc. Acids Res. 25:3389-3402、およびAltschul et al. (1990) J. Mol. Biol. 215:403-410に記載されている。BLAST解析を行うためのソフトウェアは、National Center for Biotechnology Informationを通じて公的に利用可能である。このアルゴリズムは、データベース配列中の同じ長さのワード(word)と整列させた場合にいくつかの正の値の閾値スコアTにマッチするかまたはそれを満たす、クエリー(query)配列中の長さの短いワードWを同定することによって、高スコア配列対(HSP)をまず同定することを含む。Tは近傍ワードスコア閾値(neighborhood word score threshold)(Altschul et al.、上記)と呼ばれる。これら初期近傍ワードヒットが、これらを含むさらに長いHSPを見つけるための検索を開始するためのシード(seed)となる。ワードヒットは、累積整列スコアを増加できる限り、各配列に沿って両方向に延長される。累積スコアは、ヌクレオチド配列に関しては、パラメータM(マッチする残基対に対するリワード(reward)スコア;常に>0)およびパラメータN(ミスマッチ残基に対するペナルティ(penalty)スコア;常に<0)を用いて計算される。アミノ酸配列に関しては、スコアリングマトリックスを用いて累積スコアを計算する。各方向におけるワードヒットの延長は以下の場合に停止される:累積整列スコアが最大達成値から量X分減少する場合;累積スコアが、1つまたは複数の負のスコアの残基整列の蓄積によってゼロ以下になる場合;あるいは、いずれかの配列の終わりに達する場合。BLASTアルゴリズムパラメータW、T、およびXは、整列の感度と速度を決定する。BLASTNプログラム(ヌクレオチド配列用)はデフォルトとして、word length(ワード長)(W)11、expectation(期待値)(E)10、M=5、N=-4、およびcomparison of both strands(両鎖の比較)を用いる。アミノ酸配列については、BLASTPプログラムはデフォルトとして、word length 3、expectation(E)10、BLOSUM62 scoring matrix(スコアリング マトリックス)(Henikoff and Henikoff (1989) Proc. Natl. Acad. Sci. USA 89:10915を参照されたい)alignment(アライメント)(B)50、expectation(E)10、M=5、N=-4、およびcomparison of both strandsを用いる。
【0053】
BLASTアルゴリズムはまた、2つの配列間の類似性の統計学的解析を行う(例えば、Karlin and Altschul (1993) Proc. Natl. Acad. Sci. USA 90:5873-7を参照されたい)。BLASTアルゴリズムによって提供される類似性の1つの尺度が最小合計確率(smallest sum probability, P(N))であり、2つのヌクレオチドまたはアミノ酸配列間のマッチが偶然に起こる確率の指標を提供する。例えば、試験核酸の参照核酸との比較における最小合計確率が約0.2未満、より好ましくは約0.01未満、最も好ましくは約0.001未満である場合、この核酸は参照配列に類似すると見なされる。
【0054】
用語「有機低分子」は、薬剤に一般的に用いられる有機分子に相当するサイズの分子を指す。この用語は生物学的高分子(例えば、タンパク質、核酸など)は除外する。好ましい有機低分子はサイズが約5000Daまで、例えば、2000Daまで、あるいは約1000Daまでの範囲である。
【0055】
用語「標識」および「検出可能な標識」は、分光学的、光化学的、生化学的、免疫化学的、電気的、光学的、または化学的手段によって検出可能な任意の組成物を指すために本明細書において用いられる。このような標識としては、標識されたストレプトアビジン結合体での染色のためのビオチン、磁気ビーズ(例えばDYNABEADS(商標))、蛍光色素(例えば、フルオレセイン、テキサスレッド、ローダミン、緑色蛍光タンパク質など)、放射標識(例えば、3H、125I、.35S、14C、または32P)、酵素(例えば、ホースラディッシュペルオキシダーゼ、アルカリホスファターゼ、およびELISAで一般的に用いられる他のもの)、および金コロイドまたは着色ガラスもしくはプラスチック(例えば、ポリスチレン、ポリプロピレン、ラテックスなどの)ビーズなどの熱量測定標識が挙げられる。このような標識の使用を教示する特許としては、米国特許第3,817,837号;第3,850,752号;第3,939,350号;第3,996,345号;第4,277,437号;第4,275,149号;および第4,366,241号が挙げられる。このような標識を検出する手段は当業者に周知である。したがって、例えば、放射標識は感光フィルムまたはシンチレーションカウンターを用いて検出することができ、蛍光マーカーは放出された光を検出する光検出器を用いて検出しうる。酵素標識は典型的には、酵素に基質を提供して酵素の基質に対する作用によって産生された反応産物を検出することによって検出され、熱量測定標識は着色された標識を単純に視覚化することによって検出される。
【0056】
本明細書において用いられる用語「抗体」は、天然抗体および非天然抗体を包含し、例えば、単鎖抗体、キメラ、二機能性、およびヒト化抗体、ならびにそれらの抗原結合断片(例えば、Fab'、F(ab')2、Fab、Fv、およびrIgG)を含む。Pierce Catalog and Handbook, 1994-1995 (Pierce Chemical Co., Rockford, IL) も参照されたい。例えば、Kuby, J., Immunology, 3rd Ed., W.H. Freeman & Co., New York (1998) も参照されたい。このような非天然抗体は固相ペプチド合成を用いて構築することができ、組換え的に産生することも、あるいは例えば、参照として本明細書に組み入れられる、Huse et al., Science 246:1275-1281 (1989) に記載されているように、可変重鎖と可変軽鎖とからなるコンビナトリアルライブラリーをスクリーニングすることによって得ることもできる。例えば、キメラ、ヒト化、CDR移植、単鎖、および二機能性抗体を作成するこれらおよび他の方法は当業者に周知である(Winter and Harris, Immunol. Today 14:243-246 (1993); Ward et al., Nature 341:544-546 (1989); Harlow and Lane, Antibodies: A Laboratory Manual, 1988; Hilyard et al., Protein Engineering: A practical approach (IRL Press 1992); Borrebeck, Antibody Engineering, 2d ed. (Oxford University Press 1995);これらは各々参照として本明細書に組み入れられる)。
【0057】
用語「抗体」は、ポリクローナル抗体およびモノクローナル抗体の両方を含む。この用語はまた、キメラ抗体(例えばヒト化マウス抗体)およびヘテロ結合抗体(heteroconjugate antibody)(例えば二重特異性抗体)などの遺伝的に操作された形態を含む。この用語はまた、組換え単鎖Fv断片(scFv)も指す。抗体という用語はまた、二価または二重特異性分子、二重特異性抗体(diabody)、三重特異性抗体(triabody)、および四重特異性抗体(tetrabody)を含む。二価および二重特異性分子は、例えば、Kostelny et al. (1992) J Immunol 148:1547, Pack and Pluckthun (1992) Biochemistry 31:1579, Holliger et al. (1993) Proc Natl Acad Sci U S A. 90:6444, Gruber et al. (1994) J Immunol 152:5368, Zhu et al. (1997) Protein Sci 6:781, Hu et al. (1996) Cancer Res. 56:3055, Adams et al. (1993) Cancer Res. 53:4026、および McCartney, et al. (1995) Protein Eng. 8:301に記載されている。
【0058】
典型的には、抗体は重鎖および軽鎖を有する。重鎖および軽鎖はそれぞれ定常領域および可変領域(これら領域は「ドメイン」としても知られる)を含む。軽鎖および重鎖可変領域は、「相補性決定領域」または「CDR」とも呼ばれる、3つの超可変領域によって中断される4つの「フレームワーク」領域を含む。フレームワーク領域およびCDRの範囲は規定されている。異なる軽鎖または重鎖のフレームワーク領域の配列は種内で比較的保存されている。抗体のフレームワーク領域、すなわち構成軽鎖と重鎖のフレームワーク領域を合わせたものは、CDRを3次元空間に位置付け、整列させる。
【0059】
CDRは主として抗原エピトープへの結合を担っている。各鎖のCDRは、典型的にはN末端から順に番号がつけられてCDR1、CDR2、およびCDR3と呼ばれ、また典型的には、その特定のCDRが位置する鎖によって同定される。したがって、VH CDR3はそれが見出される抗体の重鎖の可変ドメインに位置し、VL CDR1はそれが見出される抗体の軽鎖の可変ドメインのCDR1である。「VH」に関しては抗体の免疫グロブリン重鎖の可変領域を指し、Fv、scFv、またはFabの重鎖を含む。「VL」に関しては免疫グロブリン軽鎖の可変領域を指し、Fv、scFv、dsFv、またはFabの軽鎖を含む。
【0060】
「単鎖Fv」または「scFv」という表現は、通常の2鎖抗体の重鎖および軽鎖の可変ドメインが連結して1つの鎖を形成した抗体を指す。典型的にはリンカーペプチドが2つの鎖の間に挿入されており、適正なフォールディングと活性な結合部位の形成とを可能にする。
【0061】
「キメラ抗体」は、(a)異なるまたは変えられた、クラス、エフェクター機能、および/または種の定常領域に、あるいはキメラ抗体に新しい特性を付与する全く異なる分子、例えば、酵素、毒素、ホルモン、成長因子、薬物などに、抗原結合部位(可変領域)が結合するように、定常領域またはその一部分が変えられた、置き換えられた、または交換された;あるいは(b)可変領域またはその一部分が、異なるまたは変えられた抗原特異性を有する可変領域に変えられた、置き換えられた、または交換された、免疫グロブリン分子である。
【0062】
「ヒト化抗体」は、非ヒト免疫グロブリンに由来する最小配列を含む免疫グロブリン分子である。ヒト化抗体には、所望の特異性、親和性、および能力を有するマウス、ラット、またはウサギなどの非ヒト種(ドナー抗体)の相補性決定領域(CDR)に由来する残基でヒト免疫グロブリン(レシピエント抗体)のCDR由来の残基が置き換えられているヒト免疫グロブリンが含まれる。一部の例において、ヒト免疫グロブリンのFvフレームワーク残基は、対応する非ヒト残基で置き換えられる。ヒト化抗体はまた、レシピエント抗体にも、導入されたCDRもしくはフレームワーク配列にも見られない残基を含みうる。一般に、ヒト化抗体は少なくとも1つ、典型的には2つの可変ドメインの実質的に全てを含み、これらCDR領域の全てまたは実質的に全ては非ヒト免疫グロブリンのものに対応し、フレームワーク(FR)領域の全てまたは実質的に全てはヒト免疫グロブリンコンセンサス配列のものである。ヒト化抗体は最適にはまた、免疫グロブリン定常領域(Fc)、典型的にはヒト免疫グロブリンのものの少なくとも一部分を含む(Jones et al., Nature 321:522-525 (1986); Riechmann et al., Nature 332:323-327 (1988)、 および Presta, Curr. Op. Struct. Biol. 2:593-596 (1992))。ヒト化は本質的には、Winterと共同研究者(Jones et al., Nature 321:522-525 (1986); Riechmann et al., Nature 332:323-327 (1988); Verhoeyen et al., Science 239:1534-1536 (1988))の方法に従って、齧歯類のCDRまたはCDR配列でヒト抗体の対応配列を置換することによって行うことができる。従って、このようなヒト化抗体はキメラ抗体(米国特許第4,816,567号)であり、実質的に完全でないヒト可変ドメインが非ヒト種由来の対応配列によって置換されている。
【0063】
用語「エピトープ」および「抗原決定基」は、抗体が結合する抗原上の部位を指す。エピトープは連続アミノ酸から、またはタンパク質の三次フォールディングによって近接して並べられた非連続アミノ酸から形成されうる。連続アミノ酸から形成されたエピトープは典型的には変性溶媒にさらされても保持され、三次フォールディングによって形成されたエピトープは典型的には変性溶媒処置で失われる。エピトープは典型的には、独自の空間的高次構造中に少なくとも3、より一般的には少なくとも5または8〜10個のアミノ酸を含む。エピトープの空間的高次構造を決定する方法には、例えばx線結晶解析および2次元核磁気共鳴が含まれる。例えば、Epitope Mapping Protocols in Methods in Molecular Biology, Vol. 66, Glenn E. Morris, Ed (1996) を参照されたい。
【0064】
用語「ポリペプチド」、「ペプチド」、および「タンパク質」は、アミノ酸残基のポリマーを指して本明細書において相互に交換可能に用いられる。これら用語は、天然アミノ酸ポリマー、修飾残基を含むもの、および非天然アミノ酸ポリマーに適用されるのに加え、1つまたは複数のアミノ酸残基が対応する天然アミノ酸の人工的な化学的模倣体であるアミノ酸ポリマーにも適用される。
【0065】
用語「アミノ酸」は、天然および合成アミノ酸、ならびに天然アミノ酸と同様に機能するアミノ酸アナログおよびアミノ酸模倣体を指す。天然アミノ酸とは、遺伝コードによってコードされるもの、ならびに例えば、ヒドロキシプロリン、γ-カルボキシグルタメート、およびO-ホスホセリンで後に修飾されるアミノ酸である。アミノ酸アナログとは、天然アミノ酸と同じ基本化学構造、例えば、水素、カルボキシル基、アミノ基、およびR基に結合されたα炭素、例えば、ホモセリン、ノルロイシン、メチオニンスルホキシド、メチオニンメチルスルホニウム、を有する化合物を指す。このようなアナログは修飾されたR基(例えばノルロイシン)または修飾されたペプチド骨格を有しうるが、天然アミノ酸と同じ基本化学構造を保持する。アミノ酸模倣体とは、アミノ酸の一般的な化学構造とは異なる構造を有するが天然アミノ酸と同様に機能する化学的な化合物を指す。
【0066】
アミノ酸は本明細書において、一般的に知られる3文字表記で表されることもあり、またIUPAC-IUB Biochemical Nomenclature Commission推奨の1文字表記で表されることもある。ヌクレオチドは同様に、一般的に認められた1文字コードで表されうる。
【0067】
用語「組換え」は、例えば細胞、あるいは核酸、タンパク質、またはベクターなどへの言及と共に用いられる場合、細胞、核酸、タンパク質、またはベクターが、異種核酸またはタンパク質の導入、あるいは元来の核酸またはタンパク質の変化によって修飾されていること、あるいは細胞がそのように修飾された細胞に由来することを示す。したがって例えば、組換え細胞は元来の(非組換え)形態の細胞では見られない遺伝子を発現するか、または元来の遺伝子を異常に発現する、より少なく発現する、または全く発現しない。本明細書における用語「組換え核酸」とは、一般に核酸の操作によって、例えばポリメラーゼおよびエンドヌクレアーゼを用いて、最初からインビトロで形成された、天然に普通では見られない形態にある核酸を意味する。このようにして異なる配列の機能的な連結が達成される。このように、通常は繋がっていないDNA分子を連結してインビトロで形成した線状形態の単離核酸または発現ベクターは両方とも、本発明の目的に関して組換え型と見なされる。組換え核酸を作製して宿主細胞または生物体に再び導入すると、これは非組換え的に、すなわちインビトロ操作ではなく宿主細胞のインビボ細胞機構を用いて、複製する;しかし、このような核酸は組換え的に産生されると続いて非組換え的に複製されるが、それでも本発明の目的に関して組換え型と見なされることが理解される。同様に、「組換えタンパク質」は、組換え技術を用いて、すなわち上記のような組換え核酸の発現を通じて作製されたタンパク質である。
【0068】
特に定義しない限り、本明細書において用いられる技術的および科学的用語は全て本発明の属する技術分野の当業者によって一般的に理解されるものと同じ意味を有する。矛盾する場合は、本明細書が定義を含めて統御する。
【0069】
III. ANLNを介する疾患を処置する化合物の産生および同定
ANLNがRhoAと相互作用し、ANLN発現が癌患者における不良な予後に関連するという本明細書に提供される事項を考慮し、本発明の一局面は、ANLNとRhoAとの間の結合を減少または予防する試験化合物を同定することに関する。さらに、ANLNの発現が細胞運動性の増加と関連するという本明細書において提供される事項を考慮し、本発明は、ANLNを介する運動性を阻害する試験化合物を同定する方法を提供する。
【0070】
ANLN/RhoA結合を決定するための方法には、2つのタンパク質の相互作用を決定するための任意の方法が含まれる。このようなアッセイとしては、従来のアプローチ、例えば、架橋、共免疫沈降、および勾配またはクロマトグラフィーカラムを通した同時精製が挙げられるが、これらに限定されない。加えて、タンパク質-タンパク質相互作用は、Fieldsと共同研究者によって述べられている酵母ベースの遺伝学的システム(Fields and Song, Nature 340:245-246 (1989); Chien et al., Proc. Natl. Acad. Sci. USA 88, 9578-9582 (1991))を用い、Chevray and Nathans (Proc. Natl. Acad. Sci. USA 89:5789-5793 (1992))に開示されているようにモニターすることができる。酵母GALAなどの多くの転写アクチベーターは2個の物理的に別個のモジュラードメインからなり、片方がDNA結合ドメインとして働き、もう片方が転写活性化ドメインとして機能する。上記刊行物に記載の酵母発現システム(一般に「ツーハイブリッドシステム」と呼ばれる)はこの特性を利用して2つのハイブリッドタンパク質を使用しており、片方は標的タンパク質がGAL4のDNA結合ドメインに融合しており、もう片方は候補活性化タンパク質が活性化ドメインに融合している。GALA活性化プロモーターの制御下にあるGAL1-lacZレポーター遺伝子の発現は、タンパク質-タンパク質相互作用を介するGAL4活性の再構成に依存する。相互作用するポリペプチドを含むコロニーは、β-ガラクトシダーゼに対する色素産生性基質で検出される。ツーハイブリッド技術を用いて2つの特異的なタンパク質間のタンパク質-タンパク質相互作用を同定するための完全なキット(MATCHMAKER(商標))がClontechから市販されている。このシステムを拡張して、特異的タンパク質相互作用に関わるタンパク質ドメインをマッピングすることもできるし、これら相互作用に非常に重要なアミノ酸残基を正確に特定することもできる。
【0071】
本出願では「ANLN」および「RhoA」と言及していても、両者の相互作用が解析または操作される場合、これらタンパク質の片方または両方の結合部分を、これらタンパク質の全長コピーのかわりに用いることができることが理解される。RhoAに結合するANLNの断片は、ANLNの標準的な欠失解析および/または突然変異誘発を用いてRhoAに結合する断片を同定することによって容易に同定しうる。類似の解析を用いてRhoAのANLN結合断片を同定しうる。
【0072】
ANLNを介する細胞運動性を阻害する試験化合物を同定する方法は、ANLNを発現する細胞を化合物と接触させ、細胞運動性を観察することによって行いうる。正常または癌組織由来の任意の哺乳動物細胞を用いることができ、例えば、NIH3T3、COS-7、HEK293、SAEC、BEAS-2B細胞などが挙げられる。細胞運動性は、マトリゲル浸潤アッセイなどの標準的なアッセイ法を用いて測定することができる。
【0073】
本明細書に開示されるように、例えば、タンパク質(抗体を含む)、ムテイン(mutein)、ポリヌクレオチド、核酸アプタマー、ならびにペプチドおよび非ペプチド有機低分子を含む任意の試験化合物が、本発明の試験化合物となりうる。試験化合物は、天然の供給源から単離されてもよく、合成的もしくは組換え的に、またはこれらの任意の組み合わせで調製されてもよい。
【0074】
例えばペプチドは、"Solid Phase Peptide Synthesis" by G. Barany and R. B. Merrifield in Peptides, Vol. 2, edited by E. Gross and J. Meienhoffer, Academic Press, New York, N.Y., pp. 100-118 (1980) に記載されているような固相技術を用いて合成的に産生しうる。同様に核酸も、Beaucage, S.L., & Iyer, R.P. (1992) Tetrahedron, 48, 2223-2311; および Matthes et al., EMBO J., 3:801-805 (1984) に記載されているように固相技術を用いて合成することができる。
【0075】
阻害ペプチドが同定されれば、インビボでのペプチドの安定性を増加させるために、様々なアミノ酸模倣体または非天然アミノ酸で本発明のペプチドを修飾することが特に有用である。安定性は数々の方法でアッセイできる。例えば、ペプチダーゼおよび様々な生物学的媒体、例えばヒト血漿および血清などを用いて安定性が試験されている。例えば、Verhoef et al., Eur. J. Drug Metab Pharmacokin. 11:291-302 (1986) を参照されたい。当技術分野で公知の他の有用なペプチド修飾にはグリコシル化およびアセチル化が含まれる。
【0076】
組換えおよび化学的合成技術は両方とも、本発明の試験化合物を産生するために用いうる。例えば、核酸の試験化合物は適切なベクターへ挿入して産生することができ、これをコンピテント細胞にトランスフェクトして増やすことができる。または、核酸はPCR技術または適した宿主中での発現を用いて増幅しうる(Sambrook et al., Molecular Cloning: A Laboratory Manual, 1989, Cold Spring Harbor Laboratory, New York, USA参照)。
【0077】
ペプチドおよびタンパク質も、当技術分野において周知の組換え技術を用いて、例えば、Morrison, J. Bact., 132:349-351 (1977)および Clark-Curtiss & Curtiss, Methods in Enzymology, 101:347-362 (1983) に記載されているように適した宿主細胞を組換えDNAコンストラクトで形質転換することによって、発現しうる。
【0078】
抗ANLNおよび抗RhoA抗体
本発明の一部の局面において、試験化合物は抗ANLNまたは抗RhoA抗体である。一部の態様において、これら抗体は、ヒト化抗体を含むがこれに限定されないキメラである。場合によっては、本発明の抗体の態様は、これらタンパク質の片方がもう一方に会合する境界面でANLNまたはRhoAのいずれかに結合する。一部の態様において、これら抗体は、生理的条件下で少なくとも約105 mol-1、106 mol-1以上、107 mol-1以上、108 mol-1以上、または109 mol-1以上のKaでANLNまたはRhoAに結合する。このような抗体は、例えばChemicon, Inc. (Temecula CA) などの商業的供給元から購入することができ、あるいは実質的に精製されたANLNまたはRhoAタンパク質、例えばヒトタンパク質またはその抗原性断片などを、免疫原として用いて作製することができる。提供された免疫原からモノクローナルおよびポリクローナル抗体の両方を調製する方法は、当技術分野において周知である。精製技術、および特定の免疫原に対する抗体を同定するための方法については、例えば、その全体が参照として本明細書に組み入れられるPCT/US02/07144 (WO/03/077838) を参照されたい。例えば親和性カラムを形成するために抗体親和性マトリックスを用いる抗体精製法も当技術分野において周知で、市販されている(AntibodyShop, Copenhagen, Denmark)。ANLN/RhoAの会合を阻害しうる抗体の同定は、試験化合物について全般的に以下に詳述するものと同じ試験アッセイを用いて行われる。
【0079】
変換酵素
変換酵素は本発明の試験化合物となりうる。本発明の文脈において、変換酵素とは、ANLN、RhoA、または両方に対して共有結合性翻訳後修飾を行う分子触媒である。本発明の変換酵素は、ANLNおよび/またはRhoAの1つまたは複数のアミノ酸残基を、修飾タンパク質の構造中のアロステリック変化を引き起こすように、あるいはANLNとRhoAとの間の結合を妨害するようにANLN/RhoA分子結合部位の化学的性質もしくは修飾タンパク質の構造を変えて、共有結合的に修飾する。本明細書において、これら2分子間の結合の妨害とは、結合のKaにおいて、30℃、イオン強度0.1、界面活性剤の非存在下で測定したこれらタンパク質間の結合のKaと比べて、少なくとも25%、30%、40%、50%、60%、70%またはそれ以上低下させることを指す。本発明の例示的な変換酵素には、キナーゼ、ホスファターゼ、アミダーゼ、アセチラーゼ、グリコシダーゼなどが含まれる。
【0080】
試験化合物ライブラリーの構築
試験化合物ライブラリーの構築は当技術分野において周知であるが、本セクションでは、試験化合物の同定と、ANLN/RhoA相互作用および/またはANLNを介する細胞運動性の有効な阻害剤のスクリーニングのためのこのような化合物のライブラリーの構築とにおけるさらなるガイダンスを提供する。
【0081】
分子モデリング
試験化合物ライブラリーの構築は、求められる特性を有することが知られている化合物の分子構造、ならびに/または阻害する標的分子、すなわちANLNおよびRhoAの分子構造の知識によって促進される。さらなる評価に適した試験化合物の予備スクリーニングに対する1つのアプローチが、試験化合物とその標的との間の相互作用のコンピュータモデリングである。本発明において、ANLNおよび/またはRhoAの間の相互作用をモデリングすることにより、相互作用自体の詳細への洞察が提供され、また、可能性のある、相互作用の分子阻害剤を含む、相互作用を阻害するために可能な方法が示唆される。
【0082】
コンピュータモデリングテクノロジーにより、選択した分子の3次元原子構造の視覚化と、その分子と相互作用するであろう新しい化合物の合理的設計とが可能である。3次元構築物は典型的には、選択した分子のx線結晶構造解析またはNMRイメージングのデータに依存する。分子動態は力場データを必要とする。コンピュータグラフィックシステムは、新しい化合物がどのように標的分子と関連するかを予測することができ、結合特異性を完全にするように化合物および標的分子の構造を実験操作することが可能である。片方または両方に小さな変化を起こしたときに分子-化合物間相互作用がどうなるかの予測には、通常ユーザーが使いやすい、分子設計プログラムとユーザーとの間のメニュー駆動型インターフェースと連係した、分子力学ソフトウェアおよび計算集約型コンピュータが必要である。
【0083】
一般的に述べた上記の分子モデリングシステムの一例は、CHARMmおよびQUANTAプログラム(Polygen Corporation, Waltham, Mass)からなる。CHARMmはエネルギー最小化および分子動態機能を行う。QUANTAは分子構造の構築、グラフィックモデリング、および解析を行う。QUANTAによって、分子の互いの挙動の相互的な構築、修飾、視覚化、および解析が可能である。
【0084】
特定のタンパク質と相互作用する薬物のコンピュータモデリングが多くの論文に概説されている。例えば、Rotivinen, et al. Acta Pharmaceutica Fennica 97, 159-166 (1988); Ripka, New Scientist 54-57 (Jun. 16, 1988); McKinlay and Rossmann, Annu. Rev. Pharmacol. Toxiciol. 29, 111-122 (1989); Perry and Davies, Prog Clin Biol Res.291:189-93(1989); Lewis and Dean, Proc. R. Soc. Lond. 236, 125-140 and 141-162 (1989);および、核酸成分に対するモデル受容体に関し、Askew, et al., J. Am. Chem. Soc. 111, 1082-1090 (1989)。
【0085】
化学物質をスクリーニングし、画像として描写する他のコンピュータプログラムが、BioDesign, Inc., Pasadena, Calif.、Allelix, Inc, Mississauga, Ontario, Canada、およびHypercube, Inc., Cambridge, Ontarioなどの会社から利用可能である。例えば、DesJarlais et al. (1988) J. Med. Chem. 31:722; Meng et al. (1992) J. Computer Chem. 13:505; Meng et al. (1993) Proteins 17:266; Shoichet et al. (1993) Science 259:1445を参照されたい。
【0086】
ANLN/RhoA相互作用の推定阻害剤が同定されれば、以下に詳述するように、同定された推定阻害剤の化学的構造に基づき、コンビナトリアルケミカル技術を使用して任意の数の変異体を構築することができる。こうして得られる推定阻害剤または「試験化合物」のライブラリーを本発明の方法を用いてスクリーニングし、ANLN/RhoAの会合を阻害するライブラリーの試験化合物を同定することができる。
【0087】
コンビナトリアルケミカル合成
試験化合物のコンビナトリアルライブラリーは、ANLN/RhoA相互作用の公知の阻害剤に存在するコア構造の知識を含む合理的薬物設計プログラムの一部として産生しうる。このアプローチは、ライブラリーを妥当なサイズに維持することが可能であり、ハイスループットスクリーニングを促進する。または、単純な、特に短いポリマー分子ライブラリーは、ライブラリーを構成する分子ファミリーの全ての並べ換えを単純に合成することによって構築しうる。この後者のアプローチの一例は、すべてのペプチドが6アミノ酸長のライブラリーであろう。このようなペプチドライブラリーは、全ての6アミノ酸配列の並べ換えを含みうる。このタイプのライブラリーは直線的コンビナトリアルケミカルライブラリーと呼ばれる。
【0088】
コンビナトリアルケミカルライブラリーの調製は当業者に周知であり、化学的または生物学的合成によって生成されうる。コンビナトリアルケミカルライブラリーには、ペプチドライブラリー(例えば、米国特許第5,010,175号, Furka, Int. J. Pept. Prot. Res. 37:487-493 (1991) および Houghten et al., Nature 354:84-86 (1991) を参照されたい)が挙げられるが、これに限定されない。化学的多様性ライブラリーを生成するための他の化学も用いることができる。このような化学としては以下のものが挙げられるが、これらに限定されない:ペプチド(例えばPCT公報WO91/19735)、コードされるペプチド(例えばPCT公報WO93/20242)、ランダムバイオオリゴマー(例えばPCT公報WO92/00091)、ベンゾジアゼピン(例えば米国特許第5,288,514号)、ヒダントイン、ベンゾジアゼピン、およびジペプチドなどのディバーソマー(diversomer)(DeWitt et al., Proc. Natl. Acad. Sci. USA 90:6909-6913 (1993))、ビニル性(vinylogous)ポリペプチド(Hagihara et al., J. Amer. Chem. Soc. 114:6568 (1992))、グルコース骨格を有する非ペプチド性ペプチド模倣体(Hirschmann et al., J. Amer. Chem. Soc. 114:9217-9218 (1992))、低分子化合物ライブラリーの類似の有機合成(Chen et al., J. Amer. Chem. Soc. 116:2661 (1994))、オリゴカルバメート(Cho et al., Science 261:1303 (1993))、および/またはペプチジルホスホネート(Campbell et al., J. Org. Chem. 59:658 (1994))、核酸ライブラリー(Ausubel、およびSambrook(全て上記)を参照されたい)、ペプチド核酸ライブラリー(例えば米国特許第5,539,083号を参照されたい)、抗体ライブラリー(例えばVaughan et al., Nature Biotechnology, 14(3):309-314 (1996) およびPCT/US96/10287を参照されたい)、炭水化物ライブラリー(例えばLiang et al., Science, 274:1520-1522 (1996) および米国特許第5,593,853号を参照されたい)、有機低分子ライブラリー(例えば、ベンゾジアゼピン、Baum C&EN, Jan 18, page 33 (1993); イソプレノイド、米国特許第5,569,588号; チアゾリジノン(thiazolidinone)およびメタチアザノン(metathiazanone)、米国特許第5,549,974号; ピロリジン、米国特許第5,525,735号および第5,519,134号; モルホリノ化合物、米国特許第5,506,337号; ベンゾジアゼピン、米国特許第5,288,514号などを参照されたい)。
【0089】
ファージディスプレイ
別のアプローチは、組換えバクテリオファージを用いてライブラリーを産生する。この「ファージ法」(Scott and Smith, Science 249:386-390, 1990; Cwirla, et al, Proc. Natl. Acad. Sci., 87:6378-6382, 1990; Devlin et al., Science, 249:404-406, 1990)を用いて、非常に大きなライブラリーを構築することができる(例えば106〜108化学単位)。第二のアプローチは主に化学的方法を用いるものであり、Geysen法(Geysen et al., Molecular Immunology 23:709-715, 1986; Geysen et al. J. Immunologic Method 102:259-274, 1987)およびFodorらの方法(Science 251:767-773, 1991)がその例である。Furka et al. (14th International Congress of Biochemistry, Volume #5, Abstract FR:013, 1988; Furka, Int. J. Peptide Protein Res. 37:487-493, 1991)、(米国特許第4,631,211号) 、および (米国特許第5,010,175号)が、アゴニストまたはアンタゴニストとして試験されうるペプチドの混合物を産生するための方法を記載している。
【0090】
コンビナトリアルライブラリー調製用の装置が市販されている(例えば、357 MPS, 390 MPS, Advanced ChemTech, Louisville KY, Symphony, Rainin, Woburn, MA, 433A Applied Biosystems, Foster City, CA, 9050 Plus, Millipore, Bedford, MAを参照されたい)。また、多数のコンビナトリアルライブラリー自体が市販されている(例えばComGenex, Princeton, N.J., Tripos, Inc., St. Louis, MO, 3D Pharmaceuticals, Exton, PA, Martek Biosciences, Columbia, MDなどを参照されたい)。
【0091】
試験化合物ライブラリーのスクリーニング
本発明のスクリーニング方法は、ANLN/RhoAの会合またはANLNを介する細胞運動性を妨害する確率が高い試験化合物の効率的で迅速な同定を提供する。一般に、ANLN/RhoAの会合またはANLNを介する細胞運動性を妨害する試験化合物の能力を決定する方法はいずれも本発明と共に使用するのに適する。例えば、ELISA形式の競合および非競合阻害アッセイを利用しうる。対照実験を行ってシステムの最大結合能を決定すべきである(例えば、下記の例においては、結合させたANLNをRhoAと接触させ、ANLNに結合するRhoAの量を決定する)。
【0092】
競合アッセイ形式
本発明の試験化合物をスクリーニングするために競合アッセイを用いうる。例として、競合ELISA形式は、固体支持体に結合されたANLN(またはRhoA)を含みうる。この結合されたANLN(またはRhoA)は、RhoA(またはANLN)および試験化合物とインキュベートされる。試験化合物および/またはRhoA(もしくはANLN)をANLN(またはRhoA)に結合させるのに十分な時間の後、基質を洗浄して非結合物質を除く。そしてANLN(またはRhoA)に結合したRhoA(またはANLN)の量を決定する。これは、当技術分野で公知の様々な方法のいずれか、例えば、検出可能な標識でタグ付されたRhoA(またはANLN)種を用いて、あるいは洗浄した基質を標識化抗RhoA(またはANLN)抗体と接触させて、遂行しうる。ANLN(またはRhoA)に結合したRhoA(またはANLN)の量は、RhoA/ANLNの会合を妨害する試験化合物の能力に反比例するであろう。抗体を含むがそれに限定されないタンパク質の標識は、Harlow & Lane, Antibodies, A Laboratory Manual (1988) に記載されている。
【0093】
あるバリエーションでは、ANLN(またはRhoA)は親和性タグで標識される。標識されたANLN(またはRhoA)を次に、試験化合物およびRhoA(またはANLN)とインキュベートし、免疫沈降する。そして免疫沈降物を抗RhoA(またはANLN)抗体を用いたウェスタンブロッティングに供する。前記の競合アッセイ形式のように、ANLN(またはRhoA)と会合することが見出されたRhoA(またはANLN)の量は、ANLN/RhoAの会合を妨害する試験化合物の能力と反比例する。
【0094】
非競合アッセイ形式
非競合結合アッセイも、本明細書に記載されるもののような競合アッセイを用いたスクリーニングに容易に適用できない形式で構築された試験化合物ライブラリーの初期スクリーニングとして有用でありうる。このようなライブラリーの一例は、ファージディスプレイライブラリーである(例えば、Barrett, et al. (1992) Anal. Biochem 204, 357-364を参照されたい)。
【0095】
ファージライブラリーは、数多くの異なる組換えペプチドの作業量(working quantity)を素速く産生しうる点において有用である。ファージライブラリーはそれ自体は本発明の競合アッセイに向いてはいないが、非競合形式で効率的にスクリーニングされ、どの組換えペプチド試験化合物がANLNまたはRhoAに結合するかを決定することができる。次いで、結合すると同定された試験化合物を産生し、競合アッセイ形式を用いてスクリーニングすることができる。ファージおよび細胞ディスプレイライブラリーの産生およびスクリーニングは当技術分野において周知であり、例えば、Ladner et al., WO 88/06630; Fuchs et al. (1991) Biotechnology 9:1369-1372; Goward et al. (1993) TIBS 18:136-140; Charbit et al. (1986) EMBO J 5, 3029-3037. Cull et al. (1992) PNAS USA 89:1865-1869; Cwirla, et al. (1990) Proc. Natl. Acad. Sci. U.S.A. 87, 6378-6382に考察されている。
【0096】
例示的な非競合アッセイは、成分の1つ(ANLNまたはRhoA)の添加なしで、競合アッセイに関して記載したものと類似の手順に従う。しかしながら、非競合形式はANLNまたはRhoAに対する試験化合物の結合を決定するので、ANLNおよびRhoAの両方に対する試験化合物の能力を各候補について決定する必要がある。したがって、例として、固定化されたANLNへの試験化合物の結合は、結合していない試験化合物を洗い流し、結合した試験化合物を支持体から溶出させた後に、例えば、質量分析、タンパク質測定(BradfordもしくはLowryアッセイ法、または280nmの吸光度測定)によって溶出物を解析することによって、決定しうる。または、溶出工程を省いて、支持体表面での有機層の分光学的特性における変化をモニターすることによって試験化合物の結合を決定してもよい。表面の分光学的特性をモニターするための方法としては以下のものが含まれるが、これらに限定されない:吸光度、反射率、透過率、複屈折、屈折率、回折、表面プラズモン共鳴、偏光解析法、共鳴ミラー法、格子共役導波管(grating coupled waveguide)技術、および多極共鳴分光法(これらは全て当業者に公知である)。溶出工程の必要を省くために、標識された試験化合物もアッセイに用いうる。この場合、非結合物質を洗い流した後に支持体に会合している標識の量が試験化合物の結合に直接比例する。
【0097】
多くの周知のロボットシステムが液相化学用に開発されてきた。これらシステムとしては、Takeda Chemical Industries, LTD.(Osaka, Japan)によって開発された自動合成器のような自動ワークステーションが挙げられ、多くのロボットシステムがロボットアーム(Zymate II, Zymark Corporation, Hopkinton, Mass.; Orca, Hewlett Packard, Palo Alto, Calif.)を利用しており、化学者によって行われるマニュアル合成操作を模倣する。上記装置はいずれも、本発明と共に使用するのに適している。本明細書において述べられているように操作可能になるようなこれら装置に対する修飾(もしあれば)の性質および実施は、関連技術分野の当業者には明らかであろう。加えて、多数のコンビナトリアルライブラリーはそれ自体が市販されている(例えば、ComGenex, Princeton, N.J., Asinex, Moscow, Ru, Tripos, Inc., St. Louis, MO, ChemStar, Ltd, Moscow, RU, 3D Pharmaceuticals, Exton, PA, Martek Biosciences, Columbia, MDなどを参照されたい)。
【0098】
変換酵素のスクリーニング
変換酵素である試験化合物は、アッセイする変換酵素に特異的なコファクターおよび補助基質を用いて非競合形式でアッセイしうる。このようなコファクターおよび補助基質は、調べる変換酵素のタイプが与えられれば、当業者に公知である。
【0099】
変換酵素の1つの例示的なスクリーニング手順は、まず、変換酵素に特徴的なタンパク質の共有結合性修飾を行うのに必要なコファクターおよび補助基質の存在下、好ましくは生理的条件下で、ANLNおよび/またはRhoAを変換酵素と接触させることを含む。修飾されたタンパク質を次に、結合パートナーに結合する能力(すなわちANLNのRhoAに対する結合)について試験する。次に、修飾されたタンパク質の結合パートナーに対する結合を、未修飾の対照対の結合と比較し、上記のKaにおいて要求される変化が達成されたかどうかを決定する。
【0100】
アッセイを行う上でタンパク質の検出を促進するために、当業者に周知の技術を用いて、1つまたは複数のタンパク質を上記のような検出可能な標識で標識してもよい。
【0101】
スクリーニング方法
上記のスクリーニングの態様は、さらなる調査に適する試験化合物のハイスループットな決定に適する。本発明のスクリーニング方法は、以下のものの1つまたは複数の検出を含みうる:
(a)活性化RhoAの濃度;
(b)RhoAと、RHOエフェクター(例えばROCK(Maekawa, M. et.al. Science vol 285. 895-898(1999))、Rhophilin-2(Jeremy W. Peck et.al. Journal of Biological Chemistry 277, 43924-32(2002))、またはそのRhoA結合領域との間の相互作用;
(c)活性化RhoAを介する下流遺伝子発現(例えばROCK(Stephan A.K. Harvey et.al. Investigative Ophthalmology & Visual Science 45, 2168-76(2004))、c-JUN(Maria Julia Marinissen et.al. Molecular Cell 14, 29-41(2004))、または下流遺伝子産物活性(例えばERK(Laboureau J, et.al. Experimental Dermatology 13, 70-7(2004))、GATA-4(Yanazume T, et.al. The Journal of Biological Chemistry 277, 8618-25(2002))を含む任意のシグナル複合体の活性化;
(d)DNA合成および細胞周期移行の促進;
(e)細胞遊走または任意の他の発癌表現型細胞接着、細胞浸潤(Shibata, T et.al. American Journal of Pathology 164, 2269-78(2004), Selma Cetin et.al. The Journal of Biological Chemistry 279, 24592-600(2004));
(f)アクチンストレスファイバー形成およびF-アクチン産生;ならびに
(g)細胞接着、遊走、および浸潤に重要な任意の分子との相互作用(Spred(Miyoshi, K. et.al. Oncogene 23, 5567-5576(2004))、Smurf1(Hong-Rui Wang et.al. Science 302, 1775-9(2003)))。
【0102】
または、調査中の試験化合物を増殖中の細胞に加えて、試験化合物を与えていない対照集団の増殖を基準として処置細胞の増殖をモニターしてもよい。試験化合物のスクリーニングに適した細胞株は、本明細書に提供される教示から当業者には明らかであろう。
【0103】
インビボ試験に関しては、認められた動物モデルに試験化合物を投与すればよい。
【0104】
IV. 同定された試験化合物からの薬剤の製剤化
よって、本発明は、癌、特に非小細胞肺癌などの肺癌、ならびにANLNおよび/もしくはRhoAの発現レベルおよび活性の上昇を示す細胞、ならびに/またはANLNの核局在によって特徴付けられる他の癌の予防または処置に有用な薬剤および方法を含む。これら薬剤および方法は、ANLN/RhoA相互作用を阻害するとして同定された本発明の少なくとも1つの試験化合物を、異常な細胞増殖の減衰または停止を達成するのに有効な量で含む。より具体的には、治療的に有効な量とは、処置対象の発症を防ぐのに、あるいはその既存の症状を緩和するのに、有効な量を意味する。
【0105】
本発明の方法で処置される個体は、例えばマーカータンパク質ANLNの発現の上昇またはANLNの核局在を示すことによって特徴付けられる非小細胞肺癌を含む癌に罹患した任意の個体でありうる。このような個体は、例えば、ヒト、イヌ、ネコ、ウマ、ウシ、もしくはヤギを含む哺乳類などの脊椎動物、あるいは任意の他の動物、特に商業的に重要な動物または家畜化された動物でありうる。本発明の目的のためには、マーカータンパク質の発現の上昇とは、一方または両方のマーカータンパク質について、マーカータンパク質の正常細胞での平均濃度よりも、少なくとも10%、好ましくは15%、20%、25%、30%、35%、40%、45%、50%、55%、またはそれよりも多い、平均細胞マーカータンパク質濃度を指す。
【0106】
治療用量範囲の決定
本発明の薬剤のための有効な用量範囲の決定は、特に本明細書に提供される詳述された開示を鑑みれば、十分に当業者の能力の範囲内である。試験化合物の治療的に有効な用量は、まず細胞培養アッセイおよび/または動物モデルから推定できる。例えば、動物モデルにおいて用量を定め、細胞培養で決定されるIC50(50%の細胞が所望の効果を示す用量)を含む循環濃度範囲を達成することができる。また、試験化合物の毒性および治療有効性は、例えば、LD50(集団の50%に致死的な用量)およびED50(集団の50%に治療有効量)を決定するための、細胞培養または実験動物における標準的な薬学的手順によって決定することができる。毒性効果と治療効果との間の用量比が、治療指数(すなわちLD50とED50の比)である。高い治療指数を示す化合物を用いることができる。これら細胞培養アッセイおよび動物試験から得られたデータを、ヒトに用いる用量範囲を定める際に用いうる。このような化合物の用量は、毒性がほとんどないかまたは全くないED50を含む循環濃度の範囲内に収まるであろう。用量は、使用される用量形態および利用される投与経路に従い、この範囲内で変わりうる。厳密な処方、投与経路、および用量は、患者の状態を考慮して個別の医師によって選択されうる。例えばFingl et al., (1975), in "The Pharmacological Basis of Therapeutics", Ch. 1 p1を参照されたい。用量および間隔は、所望の効果を維持するのに十分な活性試験化合物の血漿レベルを提供するように個別に調整しうる。
【0107】
薬学的に許容される添加物
哺乳類(例えばヒト)に投与する薬剤は、薬学的に許容される添加物、または担体を含みうる。適した添加物およびそれらの製剤は、Remington's Pharmaceutical Sciences, 16th ed., (1980), Mack Publishing Co., edited by Oslo et al.に記載されている。水溶性の調製物には、製剤を等張にするために、適切な量の薬学的に許容される塩が製剤中に典型的に用いられる。薬学的に許容される等張添加物の例としては、生理的食塩水、リンゲル液、ハンクス液、およびデキストロース溶液などの液体が含まれる。等張添加物は、注射用製剤に特に重要である。
【0108】
経粘膜投与には、透過させる障壁に適切な浸透剤を製剤に用いる。このような浸透剤は当技術分野において一般的に知られている。
【0109】
添加物は製剤の適正なpHを維持するために用いうる。最適な有効期間のためには、試験化合物を含む溶液のpHは好ましくは約5〜約8、より好ましくは約7〜約7.5の範囲にわたる。製剤はまた、凍結乾燥粉末、または、マトリックスが成形品、例えば、フィルム、リポソーム、もしくは微粒子の形態にある、固体疎水性ポリマーの半透性マトリックスなどの徐放性調製物を含む本発明に適した他の選択的な添加物を含みうる。例えば、投与経路、投与される試験化合物の濃度、または、処置がタンパク質、試験化合物をコードする核酸、もしくは試験化合物を分泌することができる細胞を有効成分として含む薬剤を用いるかどうかによって、特定の添加物がより好ましい場合があることが当業者には明らかであろう。
【0110】
本発明の薬学的組成物は、それ自体公知の様式で、例えば、通常の混合、溶解、顆粒化、糖衣丸形成、研和、乳化、カプセル化、閉じ込め(entrapping)、または凍結乾燥プロセスによって、製造しうる。適正な製剤は、選択された投与経路に依存する。
【0111】
経口投与には、担体によって、本発明の化合物の処置患者による経口摂取のために、錠剤、丸剤、糖衣丸、カプセル、液体、ゲル、シロップ、スラリー、懸濁液などとして製剤化することが可能である。経口用の薬学的調製物は、試験化合物を固体の拡散性添加物と共に製剤化し、得られた混合物を任意ですりつぶして、所望であれば適した補助剤を加えた後に顆粒の混合物を加工して錠剤または糖衣丸コアを得ることによって得ることができる。適した添加物は特に、ラクトース、スクロース、マンニトール、またはソルビトールを含む糖類などの増量剤;例えば、トウモロコシデンプン、コムギデンプン、イネデンプン、ジャガイモデンプン、ゼラチン、トラガカントゴム、メチルセルロース、ヒドロキシプロピルメチルセルロース、カルボキシメチルセルロースナトリウム、および/またはポリビニルピロリドン(PVP)などのセルロース調製物である。所望であれば、架橋ポリビニルピロリドン、寒天、またはアルギン酸またはアルギン酸ナトリウムなどのそれらの塩などの崩壊剤を加えてもよい。
【0112】
本発明の化合物の多くは薬学的に適合性の対イオンとの塩として提供してもよい。薬学的に適合性の塩は、用途によって、塩酸、硫酸、酢酸、乳酸、酒石酸、リンゴ酸、コハク酸などを含むがこれらに限定されない多くの酸と共に形成しうる。塩は、水性または対応する遊離塩基形態である他のプロトン性溶媒に、より可溶性である傾向がある。
【0113】
許容可能な添加物に加えて、本発明の製剤は、同定された試験化合物以外の治療剤を含みうる。例えば、製剤は、抗炎症剤、鎮痛剤、化学療法剤、粘液溶解薬(例えばn-アセチル-システイン)などを含みうる。薬剤自体に他の治療剤を含むことに加え、本発明の薬剤はまた、1つまたは複数の他の薬理的物質に連続的にまたは同時に投与しうる。薬剤および薬理的物質の量は、例えば、どのタイプの薬理的物質を用いるか、また、処置される疾患、ならびにスケジュールおよび投与経路に依存する。
【0114】
本発明の薬剤の投与に続いて、熟練した施術者に周知の様々な方法で哺乳類の生理的状態をモニターすることができる。
【0115】
遺伝子療法
ANLN/RhoAの会合の阻害物質(disruptor)として同定されたタンパク質およびペプチド試験化合物は、癌、例えば非小細胞肺癌に苦しむ患者に遺伝子療法を用いて治療的に送達しうる。遺伝子療法技術に適用できる例示的試験化合物としては、変換酵素や、立体またはアロステリック阻害によってANLN/RhoAの会合を直接変化させるペプチドが含まれるが、これらに限定されない。一部の局面において、遺伝子療法の態様は、本発明の適した同定された試験化合物をコードする核酸配列を含む。一部の態様において、核酸配列は、標的細胞における試験化合物の発現に必要なそれらの調節エレメントを含む。核酸は、標的細胞のゲノムに安定に挿入されるように準備されうる(例えば、相同組換えカセットベクターの記載について、Thomas, K. R. and Capecchi, M. R. (1987) Cell 51:503を参照されたい)。
【0116】
患者への核酸の送達は直接的でもよいし(この場合、患者は核酸または核酸を含むベクターに直接さらされる)、間接的でもよい(この場合、細胞がまずインビトロで核酸によって形質転換されてから患者に移植される)。これら2つのアプローチはそれぞれインビボまたはエクスビボ遺伝子療法として知られている。
【0117】
遺伝子療法の方法の全般的概説については、Goldspiel et al., 1993 Clinical Pharmacy 12:488-505; Wu and Wu, 1991 Biotherapy 3:87-95; Tolstoshev, 1993 Ann. Rev. Pharmacol. Toxicol. 33:573-596; Mulligan, 1993 Science 260:926-932; および Morgan and Anderson, 1993 Ann. Rev. Biochem. 62:191-217; 1993 TIBTECH 11(5):155-215を参照されたい。用いうる組換えDNA技術の分野で一般的に知られる方法は、Ausubel et al. (eds.), 1993, Current Protocols in Molecular Biology, John Wiley & Sons, NY; および Kriegler, 1990, Gene Transfer and Expression, A Laboratory Manual, Stockton Press, NYに記載されている。
【0118】
V. スクリーニング、予後、および処置キット
本発明はまた、癌、特に非小細胞肺癌(NSCLC)などの肺癌の処置または予防に有用な化合物をスクリーニングするための材料を含む製品またはキットも提供する。このような製品は、本明細書に記載される材料の、1つまたは複数のラベルされた容器を、使用説明と共に含みうる。適した容器には、例えば、ボトル、バイアル、および試験管が含まれる。容器は、ガラスまたはプラスチックなどの様々な材料から形成されうる。
【0119】
一つの態様において、スクリーニングキットは以下のものを含む:(a)ANLNポリペプチドのRhoA結合ドメインを含む第一のポリペプチド;(b)RhoAポリペプチドのANLN結合ドメインを含む第二のポリペプチド、および(c)第一のポリペプチドと第二のポリペプチドとの間の相互作用を検出する手段(例えば試薬)。
【0120】
一部の態様において、第一のポリペプチド、すなわちRhoA結合ドメインを含むポリペプチドは、ANLNポリペプチドを含む。同様に、他の態様において、第二のポリペプチド、すなわちANLN結合ドメインを含むポリペプチドは、RhoAポリペプチドを含む。
【0121】
一部の態様において、RhoA結合ドメインを含むポリペプチドは生細胞中で発現される。
【0122】
一部の態様において、2つのポリペプチド間の相互作用を検出する手段(例えば試薬)は、以下のものを検出することができる:
(1)活性化RhoAの濃度;
(2)RhoAとRHOエフェクターまたはそのRhoA結合領域との間の相互作用;
(3)活性化RhoAを介する下流遺伝子を含む任意のシグナル複合体の活性化;
(4)DNA合成および細胞周期移行の促進;
(5)細胞遊走または任意の他の発癌表現型;
(6)アクチンストレスファイバー形成およびF-アクチン産生;ならびに
(7)細胞接着、遊走、および浸潤に重要な任意の分子との相互作用。
【0123】
別の態様において、スクリーニングキットは以下のものを含みうる:(a)ANLNポリペプチドまたはその機能的同等物を発現する細胞;および(b)細胞の運動性を検出する手段(例えば試薬)。
【0124】
本発明はまた、癌対象、例えば非小細胞肺癌(NSCLC)などの肺癌に罹患した対象の予後を予測するためのキットを提供する。一部の態様において、このような予後キットは以下のものを含む:(a)ANLNタンパク質を認識する抗体、および(b)核における検出用の薬剤。一部の態様において、核の検出用の薬剤はヘマトキシリン-エオシン染色染料である。使用説明は、NSCLCの予後を予測しようとする対象から収集した試料の核に局在するANLNが検出されることが、不良な予後の指標であることを示すであろう。
【0125】
本発明はさらに、本明細書に記載の病理学的状態を処置するのに有用な物質を含む製品およびキットを提供する。このような製品は、本明細書に記載されるような薬剤の容器をラベルと共に含みうる。上記のように、適した容器には、例えば、ボトル、バイアル、および試験管が含まれる。容器は、ガラスまたはプラスチックなどの様々な材料から形成されうる。本発明の文脈において、容器とは、細胞増殖性疾患、例えば非小細胞肺癌を処置するために有効な活性物質を有する組成物を保持するものである。組成物中の活性物質は、インビボでANLN/RhoAの会合を阻害しうる同定された試験化合物(例えば、抗体、低分子など)でるかあもしれない。容器上のラベルは、異常な細胞増殖によって特徴付けられる1つまたは複数の状態を処置するために組成物が用いられることが示されうる。ラベルはまた、本明細書に記載するもののような投与およびモニタリング技術に関する指示を示しうる。
【0126】
上記の容器に加え、本発明の処置キットは、任意で薬学的に許容される希釈剤を収容する第二の容器を含んでもよい。さらに、使用説明書と共に、他のバッファー、希釈剤、フィルター、針、シリンジ、およびパッケージ挿入物を含む、商業的およびユーザーの観点から望ましい他の材料を含みうる。
【0127】
所望であれば、組成物は、有効成分を含む1つまたは複数の単位用量形態を含みうるパックまたはディスペンサー装置で提供されうる。パックは、例えば、ブリスターパック(blister pack)などの金属またはプラスチックのフォイルを含みうる。パックまたはディスペンサー装置は投与のための説明書を伴いうる。また、適合性の薬学的担体中に製剤化された本発明の化合物を含む組成物は、指示された状態の処置のために調製し、適切な容器に入れてラベルされうる。
【0128】
VI. 癌の診断および予後
本方法は、数々のタイプの癌の診断、予後、分類、および処置に用いることができる。原発性、転移性、および再発癌などの任意の進行段階の癌を検出することができる。診断、予後、分類のための例示的な癌には、例えば、非小細胞肺癌(NSCLC)などの肺癌が含まれる。
【0129】
本発明は、癌を有する哺乳類の予後を判断するための方法を提供する。このような方法は、予後が不良な非小細胞肺癌患者においてANLNの核局在が最も高頻度に起こるという発見に基づく。従って、ある個体由来のサンプル(例えば生検)が、細胞質よりはむしろ核にANLNが局在する細胞を含むかどうかを測定することによって、その個体の予後を予測することが可能である。
【0130】
本明細書に記載のように、ANLNの局在は完全またはほぼ完全に細胞質中であることもあり、また細胞の核および細胞質の両方に局在することもある。このように、「核局在」は、ANLNが細胞質および核の両方において検出できる状況を含む。
【0131】
核局在は、当技術分野において公知の任意の方法によって測定しうる。一部の態様において、免疫組織化学解析を用いて、痰検査または任意の生検などの、どの病院でも利用可能な手術または最小限の浸潤的技術によって得られる組織または細胞学的サンプルにおいて細胞中のANLNを検出する。不良な予後の判断は、さらなる処置を決定するため、例えば、生存の質を落とすようなさらなる処置を中止したり、以前に用いられたものとは異なる方法で癌を処置したり、より積極的に癌を処置したりするために用いられうる。すなわち、ANLNによる予後の予測は最終的に、組織サンプリングのルーチン的手順のみを用いて、従来の疾患の臨床段階の情報がなくてさえも、各癌患者にとって最も適切な処置を臨床医が前もって選ぶことを可能にするはずである。
【0132】
さらに、本方法を用いて処置経過の有効性を評価しうる。例えば、ANLNが核に局在することが見出された生物学的サンプルが得られた癌を有する哺乳類において、時間を追ってANLNの核局在をモニターすることによって抗癌処置の有効性を評価することができる。例えば、処置前または処置の初期に哺乳類から採取されたサンプルにおけるレベルと比べて、処置後の哺乳類から採取された生物学的サンプルにおけるANLN局在が減少していれば、処置が有効であることが示される。
【0133】
上記のように、本発明はまた、細胞のANLNの位置を検出するためのキットも提供する。このようなキットの成分の例には、免疫組織化学試薬、例えば、ANLNに結合する抗体、および細胞内で抗体を検出するための検出可能な標識が含まれる。
【0134】
以下に、実施例を参照して本発明をさらに詳細に説明する。しかしながら、以下の材料、方法、および例は、本発明の局面を例示するのみであって、本発明の範囲を限定する意図は全くない。このように、本明細書に記載のものと類似または同等の方法および材料を本発明の実施または試験に用いることができる。
【0135】
実施例
上記に提供した開示から理解できるように、本発明は広範な応用を有する。よって、以下の実施例は例示目的のために提供するものであって、いずれにしても本発明を限定するものと見なす意図はない。当業者は、本質的に類似の結果を得るために変化または修飾しうる様々な重要でないパラメータを容易に認識するであろう。
【0136】
実施例1 - 材料および方法
(a)細胞株および臨床サンプル
本明細書の実施例において用いられた23個のヒト肺癌細胞株は以下の通りである:肺腺癌(ADC);A549、LC319、PC-3、PC-9、PC-14、A427、NCI-H1373、細気管支肺胞癌(BAC);NCI-H1666、NCI-H1781、肺扁平上皮細胞癌(SCC);RERF-LC-AI、SK-MES-1、EBC-1、LU61、NCI-H520、NCI-H1703、NCI-H2170、肺腺扁平上皮癌(ASC);NCI-H226、NCI-H647、肺大細胞癌(LCC);LX1、小細胞肺癌(SCLC);DMS114、DMS273、SBC-3、SBC-5。細胞は全て、10%ウシ胎児血清(FCS)を加えた適切な培地中で、単層で生育させ、37℃、5%CO2の加湿空気雰囲気下で維持した。ヒト小気道上皮細胞(SAEC)を正常対照として用い、Cambrex Bio Science Inc.(Walkersville, MD)から購入した最適培地(SAGM)で生育させた。37人の患者からのインフォームドコンセントが得られた原発性NSCLCサンプル(このうち22個がADC、14個がSCC、1個がASCに分類された)を前もって得た(Kikuchi, T., et al., Oncogene. 22:2192-205 (2003))。
【0137】
手術を受けた患者からインフォームドコンセントを得て、組織マイクロアレイでの免疫染色に用いる合計285個のホルマリンで固定した原発性NSCLC(I〜IIIA期)および隣接正常肺組織サンプルを得た。
【0138】
(b)候補遺伝子の選択および半定量的RT-PCRによる解析
遺伝子発現プロファイル解析に基づき、50%を超える腫瘍中で正常肺の5倍以上の発現レベルを示す遺伝子を検査するために選択した。ANLN転写物がそれら遺伝子のリストに含まれ、続いて半定量的RT-PCRによって過剰発現していることが確認された。βアクチン(ACTB)の発現レベルを定量対照として用いて、臨床肺癌細胞のmRNAから調製された各一本鎖cDNAの適切な希釈物を調製した。増幅用のプライマーセットは以下の通りである:
ACTBには、ACTB-F (5’-GAGGTGATAGCATTGCTTTCG-3’(SEQ ID No.5))および
ACTB-R (5’-CAAGTCAGTGTACAGGTAAGC-3’(SEQ ID No.6))、ならびに
ANLNには、ANLN-F1 (5’-GCTGCGTAGCTTACAGACTTAGC-3’(SEQ ID No.7))および
ANLN-R1 (5’-AAGGCGTTTAAAGGTG ATAGGTG-3’(SEQ ID No.8))。
【0139】
反応は全て、GeneAmp PCRシステム9700(Applied Biosystems, Foster City, CA)での、94℃で2分間の初期変性に続き、95℃で30秒間、58〜62℃で30秒間、および72℃で45秒間の21サイクル(ACTBについて)または30サイクル(ANLNについて)を含んだ。
【0140】
(c)ノーザンブロット解析
ヒト複数組織ブロット(Human multiple-tissue blot)(BD Biosciences Clontech, Palo Alto, CA)をANLNの32P標識PCR産物とハイブリダイズさせた。ANLNの全長cDNAを以下のプライマーを用いてRT-PCRで調製した:
ANLN-F2 (5’- CCCAAGCTTGGGGCCACCATGGATCCGTTTACGGAGAAAC -3’(SEQ ID No.9))および
ANLN-R2 (5’- TGCTCTAGAGCAAGGCTTTCCAATAGGTTTGTAG -3’(SEQ ID No.10))。
プレハイブリダイゼーション、ハイブリダイゼーション、および洗浄は供給業者の推奨に従って行った。増感スクリーンを用いて室温で96時間、ブロットをオートラジオグラフィーにかけた。
【0141】
(d)ウェスタンブロット解析
コドン位置428〜718のGST融合ヒトANLNタンパク質でウサギを免疫化して、ヒトANLNタンパク質に特異的なウサギ抗体を作成した。抗体は標準的な手順で精製した。ECL Western Blotting System(Amersham Biosciences, Uppsala, Sweden)を用いた。細胞は、プラスミドトランスフェクション後24時間、無血清培地中で維持し、適切な量の溶解バッファー(150mM NaCl、50mM Tris-HCl, pH8.0、1%Nonidet P-40、0.1%SDS、0.5%デオキシコール酸Na、に加えてプロテアーゼ阻害剤)に溶解させた。SDS-PAGEを行い、PAGEで分離されたタンパク質をニトロセルロース膜(Amersham Biosciences)上にエレクトロブロットし、抗体と共にインキュベートした。ヒツジ抗マウスIgG-HRP抗体(Amersham Biosciences)およびヤギ抗ウサギIgG-HRP抗体(Amersham Biosciences)をこれら実験の二次抗体として供した。
【0142】
(e)免疫細胞化学解析
培養細胞をPBS(-)で2回洗浄し、4%パラホルムアルデヒド溶液中、60分間室温で固定し、0.1%Triton X-100を含むPBS(-)で1.5分間透過化した。一次抗体反応の前に、細胞をブロッキング溶液(PBS(-)中3%BSA)で60分間覆って非特異的な抗体結合をブロックした。そして、細胞をヒトANLNに対する抗体と共にインキュベートした。内因性ANLNを明らかにするために、FITC(Cappel, Durham, NC, USA)またはローダミン(Cappel)に結合したヤギ抗ウサギ二次抗体で染色し、レーザー共焦点顕微鏡(TSC SP2 AOBS: Leica Microsystems, Wetzlar, Germany)で観察した。アクチンフィラメントを可視化するため、Alexa59結合ファロイジン(Molecular Probes, Eugene, OR, USA)も、二次抗体とのインキュベーション後に加えた。
【0143】
(f)アンチセンスS-オリゴヌクレオチド
6ウェルディッシュにプレーティングした2 X 105個のNSCLC細胞株A549を、リポフェクタミン試薬(40nM)(Invitrogen, Carlsbad, CA, USA)を用いて、ANLN遺伝子に対応する合成S-オリゴヌクレオチド(0.2μM)でトランスフェクトし、10%FCSを含む培地中で2日間維持した。細胞生存率を、それぞれ三回MTTアッセイによって評価した。S-オリゴヌクレオチドの配列は以下の通り:
アンチセンス1(AS1)5’-CTCCGTAAACGGATCCAT-3’(SEQ ID No.11);
リバース1(R1)5’-TACCTAGGCAAATGCCTC-3’(SEQ ID No.12)、
アンチセンス2(AS2)5’-CGGATCCATCGCCCCAGG-3’(SEQ ID No.13);
リバース2(R2)5’-GGACCCCGCTACCTAGGC-3’(SEQ ID No.14)。
MTTアッセイは、別に記載されているように(Suzuki, C., et al., Cancer Res. 63:7038-41 (2003))行った。
【0144】
(g)RNA干渉アッセイ
別に報告されているように(Shimokawa T, et al., Cancer Res. 2003;63:6116-20.)、ベクターベースのRNA干渉(RNAi)システムpsiH1BX3.0は哺乳動物細胞におけるsiRNAの合成を誘導するために確立された。30μlのリポフェクタミン 2000(Invitrogen)を用いて、10μgのsiRNA発現ベクターをNSCLC細胞株LC319およびA549にトランスフェクトした。このアッセイでは、90%を超えるトランスフェクト細胞がこの合成siRNAを発現し、内因性のANLN発現が有効に抑制された。トランスフェクト細胞を、適切な濃度のジェネテシン(G418)存在下で5日間培養した。細胞数および生存率を三回Giemsa染色およびMTTアッセイによって測定した。RNAiの合成オリゴヌクレオチドの標的配列は以下の通り:
対照(LUC(ルシフェラーゼ:フォチナス ピラリス(Photinus pyralis)ルシフェラーゼ遺伝子)5’-CGTACGCGGAATACTTCGA-3’(SEQ ID No.15);
SCR(スクランブル:5Sおよび16S rRNAをコードするミドリムシ葉緑体遺伝子)5’-GCGCGCTTTGTAGGATTCG-3’(SEQ ID No.16));
siRNA-ANLN-1(si-1)5’-CCAGTTGAGTCGACATCTG-3’(SEQ ID No.17);
siRNA-ANLN-2(si-2)5’-GCAGCAGATACCATCAGTG-3’(SEQ ID No.18)。
本発明者らのRNAiシステムを実証するため、個々の対照siRNAを半定量的RT-PCRによって試験し、COS-7細胞に一過的にトランスフェクトした対応標的遺伝子の発現の低下を確認した。また、このアッセイに用いた細胞株において、対照ではなく機能的siRNAによるANLN発現のダウンレギュレーションが確認された。
【0145】
(h)フローサイトメトリー
細胞を5 X 105細胞/100mmディッシュの密度でプレーティングし、siRNA発現ベクターでトランスフェクトして、適切な濃度のジェネテシンの存在下で培養した。トランスフェクション5日後、細胞をトリプシン処理し、PBS中に収集して、70%冷エタノール中で30分間固定した。100μg/ml RNase(Sigma-Aldrich Co.)で処置した後、細胞をPBS中50μg/mlヨウ化プロピジウム(Sigma-Aldrich Co.)で染色した。フローサイトメトリーをBecton Dickinson FACScanで行い、ModFitソフトウェア(Verity Software House, Inc., Topsham, ME, USA)によって解析した。少なくとも20,000個の非ゲート化(ungated)細胞から決定した細胞をDNA含量について解析した。
【0146】
(i)BrdU取り込みアッセイ
ANLNおよびモック(mock)プラスミドを発現するように設計されたプラスミドでトランスフェクトした肺癌細胞(LC319およびA549細胞)を、無血清培地中で4時間培養した。次に培地を10%FCSと10μM BrdUを含むRPMI1640と交換した。これらの細胞を20時間インキュベートし、さらに固定して、取り込まれたBrdUを市販のキット(Cell Proliferation ELISA, BrdU; Roche Diagnostics, Basel, Switzerland)を用いて測定するために用いた。
【0147】
(j)マトリゲル浸潤アッセイ
ANLNまたはモックプラスミドを発現するように設計されたプラスミドでトランスフェクトしたNIH3T3およびCOS-7細胞を、10%FCSを含むDMEM中で融合性の段階まで生育させた。細胞をトリプシン処理によって回収し、続いて血清またはプロテイナーゼ阻害剤を加えないDMEM中で洗浄した。細胞をDMEM中で1 x 105/mlで懸濁した。細胞懸濁液を調製する前に、マトリゲル基質(Becton Dickinson Labware, Bedford, MA, USA)の乾燥層をDMEMで2時間、室温で再水和した。10%FCSを含むDMEM(0.75ml)を24ウェルマトリゲル浸潤チャンバーの各下部チャンバーに加え、0.5ml(5 x 104細胞)の細胞懸濁液を上部チャンバーの各インサートに加えた。インサートのプレートを22時間、37℃でインキュベートした。インキュベーション後チャンバーを処理し、供給業者(Becton Dickinson Labware)の指示通りに、マトリゲルで被覆されたインサートを通って浸潤している細胞を固定してGiemsaによって染色した。
【0148】
(k)傷害遊走アッセイ
ANLNまたはモックプラスミドを発現するように設計されたプラスミドでトランスフェクトされたNIH3T3細胞を無血清DMEM中に懸濁し、10μg/mlのフィブロネクチンで被覆された2ウェルチャンバー(Becton Dickinson Labware)の個々のウェルにプレーティングした。4時間インキュベーション後、各チャンバーの底からP200ピペットチップで接着細胞を線状に掻き取って傷をつけ、培地を10%FCSを含むDMEMで交換した。細胞を増殖させ、傷害区域に48時間遊走させた。そして、傷害区域内の細胞数を顕微鏡(DP50, OLYMPUS, Tokyo, Japan)を用いて数えた。
【0149】
(l)RHO活性化の検出
ANLNによるRHOの活性化を、EZ-Detect(商標)Rho Activation Kit(PIERCE, Rockford, IL, USA)を用いて検出した。簡単に述べると、ANLN発現プラスミドまたはモックプラスミドでトランスフェクトしたLC319細胞を24時間培養した。次に細胞を洗浄し、溶解バッファーで溶解した。16,000xgで遠心分離した後、溶解物を、活性化RHOをアフィニティー沈降させるためにGST-rhotekin(RTKN)-RBDおよび「SwellGel Immobilized Glutathione Disc」と混合した。活性化RHOを含むGSTプルダウン沈殿剤を洗浄し、サンプルバッファーで煮沸して、抗RHO(-A、-B、および-C)および抗ANLN抗体を用いるウェスタンブロット解析に供した。
【0150】
(m)免疫組織化学および組織マイクロアレイ
ホルマリンで固定した肺癌を用いた腫瘍組織マイクロアレイを、以前に公開されているように(Kononen, J., et al., Nat. Med. 4: 844-847 (1998); Chin, S. F., et al., Mol. Pathol., 56:275-79 (2003); Callagy, G., et al., Diagn. Mol. Pathol., 12:27-34 (2003))構築した。サンプリング用の組織区域は、スライド上の対応するHE染色されたセクションとの視覚的配列に基づいて選択した。ドナー腫瘍ブロックから採取された3、4、または5個の組織コア(直径0.6mm;高さ3〜4mm)を、組織マイクロアレイヤー(Beecher Instruments, Hummingbird Court Sun Prairie, WI, USA)を用いてレシピエントパラフィンブロック中に配置した。また、各ケースから正常組織のコアも切り抜いた。得られたマイクロアレイブロックの5μmセクションを免疫組織化学解析に用いた。ANLNの染色パターンを、なし(absent)または陽性として半定量的に評価し、また、臨床経過観察データの予備知識のない3人の独立した研究者が、核または細胞質ANLN(n-ANLN、c-ANLN)に応じて定性的に評価した。n-またはc-ANLN染色腫瘍細胞が10%未満のケースはそれぞれのタイプのANLN陰性と判定した。評価者が独立して陽性と規定したケースのみを陽性として認めた。
【0151】
組織マイクロアレイ上の臨床サンプル中のANLNタンパク質の存在を調査するため、ENVISION+ Kit/ホースラディッシュペルオキシダーゼ(HRP)(DakoCytomation, Glostrup, Denmark)を用いてセクションを染色した。簡単に述べると、内因性ペルオキシダーゼおよびタンパク質をブロッキングした後に、抗ヒトANLN抗体を添加し、HRP標識抗ウサギIgGを二次抗体としてセクションをインキュベートした。基質-色素体を加え、試料をヘマトキシリンで対比染色した。
【0152】
(n)統計的解析
組織マイクロアレイ解析によって決定されるANLNタンパク質の発現レベルとの相関について、年齢、性別、および病理学的TNM段階などの臨床病理学的変数を調べた。手術日からNSCLCに関連した死亡時または最終経過観察時までの腫瘍特異的生存曲線を計算した。各関連変数およびANLN発現についてそれぞれKaplan-Meier曲線を計算した。Log-rank検定を用いて患者サブグループ間の生存期間の差を解析した。Cox比例ハザード回帰モデルで一変量および多変量解析を行い、臨床病理学的変数と癌関連死亡率との関連を決定した。まず、年齢、性別、pT分類、およびpN分類を含む可能な予後因子と死との関連を、1回につき1つの因子を考慮して解析した。次に、多変量Cox解析を、P値<0.05のエントリーレベルを満たす任意のおよび全ての変数と協調して、常にANLN発現をモデルに強制する逆方向(段階的)手順で行った。モデルが因子を加え続けていくと、独立因子はP値<0.05のエグジット(exit)レベルを超えなかった。
【0153】
実施例2 - NSCLC組織および細胞株ならびに正常組織におけるANLNの過剰発現
cDNAマイクロアレイによる解析で37個のNSCLCのうちの50%以上において5倍以上の発現を示す遺伝子は、以前にスクリーニングされている(Kikuchi, T. et al. Oncogene 22(14):2192-2205 (2003))。スクリーニングされた23,040個の遺伝子のうち、ANLN転写物は、NSCLCにおいて頻繁に過剰発現しているとして同定され、半定量的RT-PCR実験によって12個の代表的なNSCLC症例において過剰発現が確認された(図1a)。加えて、高レベルのANLN(Genbankアクセッション番号NM_018685)発現が23個の肺癌細胞株全てにおいて観察された一方、正常小気道上皮由来細胞(SAEC)においてはPCR産物は検出されなかった(図1b)。ANLN cDNAをブローブとして用いたノーザンブロッティング解析によって、約4.0kbの転写物が、調べた24個の正常ヒト組織のうち精巣および脊髄においてのみ弱いバンドとして同定された(図1c)。NSCLC細胞株A549、LC319、およびNCI-H522におけるANLNタンパク質の発現は、抗ANLN抗体を用いたウェスタンブロッティング解析によって内因性ANLNタンパク質の発現を調べることにより確認された。
【0154】
実施例3 - ANLNの細胞内局在およびアクチンストレスファイバー形成
肺癌細胞におけるANLNの細胞内局在を確認するため、免疫細胞化学解析を行った。肺癌細胞株LC319およびA549で内因的に発現されたANLNは、様々な細胞内局在パターンを示した(図2a)。ANLNタンパク質は、核および/または細胞質(n-ANLNおよびc-ANLN)、ならびに核膜崩壊後の皮層、細胞質分裂中の分裂溝、および分裂終期の後期の中央体において観察された。細胞質における線維様染色もかなりの数の細胞において観察された。これら細胞におけるストレスファイバー上の内因性ANLNおよびF-アクチンの共存が、抗ANLN抗体およびファロイジンを用いた免疫染色によって確認された(図2b)。アクチン細胞骨格は哺乳動物細胞の細胞質分裂および形態に重要な役割を担うことが知られていることから、次に、アクチンストレスファイバー形成に対するANLNの効果を、ANLN発現プラスミドをLC319およびNIH3T3細胞にトランスフェクションすることによって調べた。ファロイジンで検出された線維様染色はANLNのトランスフェクション後に有意に増加し、外因性ANLNの過剰発現がこれら細胞において多くのストレスファイバーを誘導することが示唆された(図2c)。
【0155】
実施例4 - NSCLC細胞の成長に対するANLNの効果
ANLNが肺癌細胞の成長または生存に不可欠であるかどうかを評価するため、ANLN配列(Genbankアクセッション番号NM_018685、SEQ ID NO:1(SEQ ID NO:2をコードする)に対応する2対のリバース(対照)およびアンチセンスS-オリゴヌクレオチドを合成して、高レベルのANLN発現を示したA549細胞にそれぞれトランスフェクトした。2つの異なるアンチセンスS-オリゴヌクレオチド(AS1およびAS2)のそれぞれの導入は、対応対照ヌクレオチド(R1およびR2)と比較して細胞生存度を低下させ、ANLNが癌細胞の成長および/または生存に不可欠であることが示唆された(データ示さず)。アンチセンスS-オリゴヌクレオチドによる成長抑制効果がANLN特異的であることをさらに確認するため、ANLNに対するsiRNA(siRNA-ANLN-1および-2)および2個の対照プラスミド(ルシフェラーゼ(LUC)またはスクランブル(SCR)に対するsiRNA)を発現するようにプラスミドを設計、構築し、それぞれをLC319およびA549細胞にトランスフェクトした。siRNA-ANLN-1または-2でトランスフェクトした細胞におけるANLN転写物の量は、2個の対照siRNAのいずれかでトランスフェクトしたものと比べて有意に低下した(図3a);siRNA-ANLN-1または-2のトランスフェクションによってもまた、コロニー形成およびMTTアッセイによって測定したところコロニー数および細胞生存度が有意に低下した(図3b、c)。さらに、siRNA-ANLN-1で処置した細胞は、複数の核を有する、より大きな細胞形態を示した(図3d)。この表現型の分子メカニズムをさらに明らかにするため、siRNA-ANLN-1でトランスフェクトしたLC319細胞を用いてフローサイトメトリーを行い、siRNA-ANLN-1でトランスフェクトした細胞において4N〜16NのDNA含量を有する細胞の比率が、対照siRNA(LUC)でトランスフェクトした細胞におけるものよりも有意に高いことを見出した(図3e)。
【0156】
細胞周期進行の調節に対するANLNの効果をさらに調べるため、ANLN発現プラスミドで一過的にトランスフェクトしたLC319およびA549細胞を用いて、BrdU取り込みアッセイを行った。DNA合成は、ANLN発現の誘導によって両細胞株において用量依存的に増強されているようであった(図4a)。
【0157】
実施例5 - 細胞運動性に対するANLNの効果
ストレスファイバー上のANLNタンパク質およびF-アクチンが共存し、外因性ANLN発現の誘導が哺乳動物細胞においてアクチンストレスファイバーの形成を促進することが免疫細胞化学解析により示されたので、続いてマトリゲル浸潤アッセイを行い、ANLNが細胞運動性に役割を担っている可能性があるかどうかを判定した。ANLN発現ベクターでトランスフェクトしたNIH3T3およびCOS-7細胞のマトリゲルを通じた浸潤は、対照細胞と比較して有意に促進された(図4b、4c)。ANLNまたはモックプラスミドを発現するように設計されたプラスミドでトランスフェクトしたNIH3T3細胞を用いた傷害遊走アッセイもまた、ANLN発現細胞の遊走が有意に活性化されたことを示した。
【0158】
ANLNによる細胞運動性の活性化のメカニズムを明らかにするため、以下のアッセイを実施した。低分子量GTPase RhoAはアクチン構造物の形成を制御することが知られているので、まずRhoAとANLNとの間の可能な相互作用を調べた。図5aに示すとおり、肺癌細胞株LC319細胞の細胞質および分裂溝における内因性RhoAおよびANLNの共存が、免疫細胞化学解析によって検出された。内因性RhoAの外因性発現ANLNとの直接的関連が免疫沈降アッセイによって確認された(図5b)。
【0159】
他のGTPaseと同様に、RHOはGTPに結合しているときは活性で、GDPに結合しているときは不活性である。GTPと結合すると、RHOはrhotekin(RTKN)などの下流エフェクターと相互作用する。これら事実に基づいて、ANLNの活性型RHOとの相互作用を調べた。具体的には、GST融合RTKNを用いてGSTプルダウンアッセイを行い、GTP-RHO(活性型)およびANLNを含む複合体をアフィニティー沈降させた;ANLNおよびRHOを含む免疫複合体が、いずれかの抗体を用いたウェスタンブロッティング解析によって検出された。ANLNを発現するように設計されたプラスミドでLC319細胞をトランスフェクトすると、RHO活性化の誘導、ならびにANLNと活性型RHOとの間の直接的相互作用が観察された(図5c、上部および下部パネル)。
【0160】
実施例6 - 核ANLN発現は不良な予後に関連する
外科的に切除された285個のNSCLC組織からなる組織マイクロアレイを用い、抗ANLNポリクローナル抗体で免疫組織化学解析を行った。この研究により、285ケースのうち267(93.6%)個がc-ANLN(細胞質性)のみで陽性に染色され、128(44.9%)個がc-ANLNおよびn-ANLN(核性)の両方で陽性であり、n-ANLNのみで陽性であるケースはなかったことが示された(図6a)。そうすると問題は、ANLN発現がNSCLCの臨床結果と関連するのかどうかであった。統計解析では、c-またはn-ANLN発現のpT-またはpN-因子との有意な相関は認められなかった。しかし、Kaplan-Meier法を用い、n-ANLN発現と腫瘍特異的5年生存率との間に顕著な関連が見出された(Log-rank検定でP<0.0001)(図6b)。一変量解析を用いると、pT、pN、性別、およびn-ANLN発現が、NSCLC患者の不良な腫瘍特異的生存と有意に関連していた。さらに、Cox比例ハザードモデルを用いた多変量解析によって、n-ANLN染色は独立した予後因子であることが決定された(P=0.001)。
【0161】
上記の例は本発明を例示するために提供されているが、その範囲を限定することを意図するものではない。本発明の他の変形は当業者には容易に明らかであり、添付の請求の範囲に包含される。
【0162】
産業上の利用可能性
本明細書で示したように、ANLNのRhoAとの相互作用、およびこの相互作用の阻害により癌細胞の細胞増殖が阻害された。このように、ANLNとRhoAの結合を阻害し、その活性を妨げる薬剤は、抗癌剤、特にNSCLCのような肺癌の治療のための抗癌剤として、治療的に有用であると考えられる。
【0163】
本明細書に引用される全ての刊行物、データベース、Genbank配列、特許、および特許出願は、これによって参照として本明細書に組み入れられる。
【0164】
本発明は詳細にその具体的な態様を参照して記載されているが、本発明の要旨および範囲から逸脱することなく様々な変化および修飾をその内に施すことができることは当業者には明らかであり、その境界および範囲は添付の請求の範囲によって定められる。
【図面の簡単な説明】
【0165】
【図1】原発性NSCLCおよび肺癌細胞株におけるANLN発現を示す。パート(a)は、半定量的RT-PCRによって調べた、NSCLCの臨床サンプルおよび正常肺組織におけるANLNの発現を表す。パート(b)は、肺癌細胞株におけるANLNの発現を表す。パート(c)は、ノーザンブロット解析によって検出した正常組織におけるANLNの発現を表す。
【図2】ANLNの細胞内局在およびアクチンストレスファイバー形成を示す。パート(a)は、NSCLC細胞株LC319における内因性ANLNの細胞内局在を表す。核(n-ANLN)、細胞質(c-ANLN)、および分裂溝におけるANLN発現を、ローダミン結合二次抗体を用いた免疫細胞化学染色によって検出した。パート(b)は、FITC結合二次抗体およびAlexa594結合ファロイジンでそれぞれ検出されたLC319細胞における内因性c-ANLNおよびF-アクチンの共存を表す。パート(c)においては、LC319細胞を解析の24時間前にANLN発現プラスミドで一過的にトランスフェクトし、ANLNについてはFITC免疫染色によって、およびAlexa594-ファロイジン染色によってANLNおよびF-アクチン分布を評価した。外因性ANLN発現およびそれらの共存によるストレスファイバー形成の誘導がLC319細胞において観察された。
【図3】ANLNに対するsiRNAによるNSCLC細胞の成長阻害を示す。パート(a)は、半定量的RT-PCRによって解析した、LC319細胞におけるルシフェラーゼ(LUC)またはスクランブル(SCR)に対するsiRNA-ANLN-1(si-1)、-2(si-2)、または対照siRNAに応答したANLNの発現を表す。パート(b)は、ANLN(si-1および-2)または対照プラスミド(si-LUCおよび-SCR)の特異的siRNAでトランスフェクトしたLC319細胞のコロニー形成アッセイを表す。パート(c)は、si-1、-2、-LUC、または-SCRに応答した、MTTアッセイによって評価したLC319細胞の生存度を表す。パート(d)は、si-1または-LUCでトランスフェクトしたLC319細胞の顕微鏡観察を表す。矢印は、多核で、より大きな細胞形態を示すsiRNA-ANLN-1(si-1)で処置した細胞を示す。パート(e)は、si-1または-LUCでトランスフェクトしたLC319細胞のフローサイトメトリー解析の結果を表す。si-1でトランスフェクトした細胞において4N〜16NのDNA含量を有する細胞の比率は、対照siRNA(si-LUC)でトランスフェクトした細胞におけるものよりも有意に高かった。アッセイは3回、三つ組ウェルにおいて行った。
【図4】ANLNによるDNA合成の促進および細胞運動性の活性化を示す。パート(a)は、BrdUと20時間インキュベーションした後の、ANLNで一過的にトランスフェクトしたLC319またはA549細胞におけるBrdU取り込みを表す。ANLN発現プラスミドで一過的にトランスフェクトしたLC319およびA549細胞の両方において、用量依存的にDNA合成が促進されたようであった。パート(b)および(c)は、ヒトANLN発現プラスミドをトランスフェクトした際の、マトリゲル基質中でのNIH3T3およびCOS-7細胞の浸潤性の促進を示すマトリゲル浸潤アッセイの結果を表す。具体的には、パート(b)はGiemsa染色(X100)を示し、パート(c)はマトリゲル被覆フィルターを通過して遊走した細胞の数を表す。アッセイは3回、三つ組ウェルにおいて行った。
【図5】ANLNのRhoAとの相互作用およびそのRHO活性化の調節を示す。パート(a)は、抗ANLN抗体(FITC)および抗RhoA抗体(ローダミン)を用いた免疫細胞化学染色によって検出した内因性ANLNおよびRhoAの共存を表す。パート(b)は、肺癌細胞株LC319抽出物からの外因性ANLNおよび内因性RhoAの免疫沈降(IP)を表す。LC319細胞はモックまたはANLNでトランスフェクトし、抗ANLN抗体を用いたANLN-IP、続いて抗RhoA抗体を用いた免疫ブロッティング(IB)に供した(上部)。細胞溶解物のアリコートを免疫ブロッティングに直接供して、下3つのパネルに示すように、各タンパク質の発現を確認した。パート(c)は、ANLNの直接結合によって誘導されたRHO活性化を表す。(上3つのパネル)LC319細胞をモックまたはANLNでトランスフェクトし、免疫ブロッティングに直接供して各タンパク質の発現を確認した。(下2つのパネル)細胞溶解物のアリコートをGST-RTKN-RBDと共にインキュベートしてGSTプルダウンアッセイに供し、次に抗RHOおよび抗ANLN抗体を用いた免疫ブロッティングに供した。最初のパネルは外因性ANLN発現によって活性化されたRHOのレベルを示し、二番目のパネルは活性化RHOとのANLNの直接的相互作用を示す。
【図6】n-ANLNの過剰発現がNSCLCにおける不良な結果に関連することを示す。パート(a)は、組織マイクロアレイに対して抗ANLNポリクローナル抗体を用いた、外科的に切除されたNSCLC組織由来の代表的サンプルの免疫組織化学評価の結果を表す(ADC、SCC、X100)。c-ANLNは細胞質に局在するANLNを示し、n-ANLNは核のものを示す。矢印はn-ANLNを発現する細胞の例を示す。パート(b)は、n-ANLN発現によるNSCLC患者における腫瘍特異的生存の組織マイクロアレイおよびKaplan-Meier解析の結果を表す(P<0.0001;Log-rank検定)。
【技術分野】
【0001】
発明の分野
本発明は、癌、特に肺癌の処置および予防に有用な化合物を同定するための方法およびキットに関し、ならびに、それを処置および予防するための方法および組成物に関する。さらに詳しくは、本方法は、非小細胞肺癌でアップレギュレートされている癌特異的遺伝子であるANLN(全体が参照として本明細書に組み入れられるPCT公報WO2004/031413を参照されたい)が、腫瘍形成に関わる低分子量GTPaseのRasホモロジーファミリーのメンバーであるRhoAと相互作用するという発見に関する。
【背景技術】
【0002】
発明の背景
肺癌は世界中で最も一般的な癌による死因の一つであり、非小細胞肺癌(NSCLC)がこれらのケースの80%近くを占めている(Greenlee, R.T. et al. CA. Cancer J. Clin. 51:15-36 (2001)(非特許文献1))。肺癌の発症および/または進行に関わる多くの遺伝子変化が報告されているが、正確な分子メカニズムは不明のままである(Sozzi, G., Eur. J. Cancer. 37:63-73 (2001)(非特許文献2))。新たに開発された細胞毒性薬剤は進行したNSCLCを有する患者に複数の治療選択肢を与えるが、これらの新たな治療法の各々はシスプラチンをベースに使用する療法と比較して少しの生存利益を提供できるのみである(Schiller, J.H. et al. N. Engl. J. Med. 346(2):92-98 (2002)(非特許文献3); Kelly, K. et al. J. Clin. Oncol. 19(13):3210-3218 (2001)(非特許文献4))。それゆえ、分子標的薬剤および抗体の開発ならびに癌ワクチンのような新規治療戦略が熱望される。
【0003】
cDNAマイクロアレイを用いる何千もの遺伝子の発現レベルの体系的な解析は、発癌経路にかかわる分子セットを同定する有効なアプローチであり(Kikuchi, T. et al. Oncogene 22(14):2192-2205 (2003)(非特許文献5); Kakiuchi, S. et al. Mol. Cancer Res. 1:485-499 (2003)(非特許文献6); Zembutsu, H. et al. Int. J. Oncol. 23:29-39 (2003)(非特許文献7); Suzuki, C. et al. Cancer Res. 63(21):7038-7041 (2003)(非特許文献8))、それらのいくつかは新規抗癌薬および腫瘍マーカーの開発のための候補標的でありうる。
【0004】
本発明は、このような標的分子に対する当技術分野における必要性に応えるものである。特に、NSCLCの診断および処置のための新規分子標的を単離するため、本発明者らは、NSCLCのゲノム全域にわたる発現プロファイル解析を、レーザーキャプチャーマイクロダイセクションによる37個の癌組織からの腫瘍細胞の純粋精製と組み合わせて行った。これら研究の過程において、ショウジョウバエ(Drosophila)アクチン結合タンパク質(ANLN)であるアニリン(anillin)のヒトホモログをコードする遺伝子が高頻度に原発性NSCLCで過剰発現していることが観察された。ANLNは、分裂溝におけるアクチンケーブルの形成または構築に不可欠で、細胞質分裂に重要な役割を担うことが報告されている(Oegema, K. et al. J. Cell Biol. 150(3):539-551 (2000)(非特許文献9))。非小細胞肺癌に関連する差次的遺伝子発現は以前に述べられている。例えばPCT公報WO2004/031413(特許文献1)を参照されたい。
【0005】
低分子量グアノシントリホスファターゼ(GTPase)は、細胞骨格のリモデリングの調節および協調に重要な役割を担う。哺乳動物細胞におけるRasホモロジーファミリーメンバー(RHO)のうち、RhoA、Ras関連C3ボツリヌス毒素基質1(RAC1)、および細胞分裂周期42(CDC42)がその生物学的機能について広範に研究されている。RhoAは、そのエフェクターであるmDia、Rho関連コイルドコイル含有タンパク質キナーゼ1(ROCK1)、およびROCK2の動員および活性化を通じた接着斑の組み立ておよびアクチンストレスファイバーの形成に原形質膜受容体が関連しているシグナル伝達経路を調節する(Ridley, A. J. and Hall, A., Cell 70:389-399 (1992)(非特許文献10); Leung, T. et al. Mol. Cell Biol. 16:5313-5327(1996)(非特許文献11); Amano, M. et al. Science 275:1308-1311 (1997)(非特許文献12))。RHO活性は細胞の運動性のために重要であるが、効率的な遊走は、細胞環境における適切な時空間において厳密な均衡の取れたRAC1、CDC42、およびRhoAの活性化および非活性化を必要とする。RHOタンパク質はまた、遺伝子転写、細胞周期進行、または抗アポトーシス経路の制御にも関与しており(Etienne-Manneville, S. and Hall, A., Nature 420:629-635 (2002)(非特許文献13))、最近の研究によってRhoAは一部のヒト腫瘍において活性化されていることが示されている(Sahai, E. and Marshall, C., J., Nat. Rev. Cancer 2(2):133-142 (2002)(非特許文献14))が、その正確なメカニズム、特に発癌の際のRhoAシグナル伝達の上流の経路は明らかにされていない。
【0006】
【特許文献1】PCT公報WO2004/031413
【非特許文献1】Greenlee, R.T. et al. CA. Cancer J. Clin. 51:15-36 (2001)
【非特許文献2】Sozzi, G., Eur. J. Cancer. 37:63-73 (2001)
【非特許文献3】Schiller, J.H. et al. N. Engl. J. Med. 346(2):92-98 (2002)
【非特許文献4】Kelly, K. et al. J. Clin. Oncol. 19(13):3210-3218 (2001)
【非特許文献5】Kikuchi, T. et al. Oncogene 22(14):2192-2205 (2003)
【非特許文献6】Kakiuchi, S. et al. Mol. Cancer Res. 1:485-499 (2003)
【非特許文献7】Zembutsu, H. et al. Int. J. Oncol. 23:29-39 (2003)
【非特許文献8】Suzuki, C. et al. Cancer Res. 63(21):7038-7041 (2003)
【非特許文献9】Oegema, K. et al. J. Cell Biol. 150(3):539-551 (2000)
【非特許文献10】Ridley, A. J. and Hall, A., Cell 70:389-399 (1992)
【非特許文献11】Leung, T. et al. Mol. Cell Biol. 16:5313-5327(1996)
【非特許文献12】Amano, M. et al. Science 275:1308-1311 (1997)
【非特許文献13】Etienne-Manneville, S. and Hall, A., Nature 420:629-635 (2002)
【非特許文献14】Sahai, E. and Marshall, C., J., Nat. Rev. Cancer 2(2):133-142 (2002)
【発明の開示】
【0007】
発明の概要
本発明は、ANLNおよびRhoAが非小細胞肺癌細胞中で相互作用すること、さらに、ANLNの核局在が非小細胞肺癌患者における不良な予後に関係しているという知見に基づく。よって、本発明は、ANLN/RhoA相互作用を妨害することによって、またはANLNを介する細胞運動性を阻害することによって、癌、例えば非小細胞肺癌の進行を遅延または停止させる化合物を同定するための新規方法を提供する。本発明はまた、サンプル中のANLNの核局在の有無を決定することによって、癌患者について予後を判断するための方法を提供する。
【0008】
よって、癌、特に非小細胞肺癌(NSCLC)などの肺癌の処置または予防に有用な化合物をスクリーニングする方法を提供することが、本発明の目的である。一部の態様において、本方法は以下の工程を含む:
(a)試験化合物の存在下で、ANLNポリペプチドのRhoA結合ドメインを含むポリペプチドを、RhoAポリペプチドのANLN結合ドメインを含むポリペプチドと接触させる工程;
(b)ポリペプチド間の結合を検出する工程;および
(c)ポリペプチド間の結合を阻害する試験化合物を選択する工程。
【0009】
一部の態様において、RhoA結合ドメインを含むポリペプチドはANLNポリペプチドを含む。同様に、他の態様において、ANLN結合ドメインを含むポリペプチドはRhoAポリペプチドを含む。
【0010】
一部の態様において、RhoA結合ドメインを含むポリペプチドは生細胞中で発現する。
【0011】
一部の態様において、以下のものを検出する工程を含む方法によってポリペプチド間の結合が検出される:
(a)活性化RhoAの濃度;
(b)RhoAとRHOエフェクターまたはそのRhoA結合領域との間の相互作用;
(c)活性化RhoAを介する下流遺伝子発現または下流遺伝子産物活性を含む任意のシグナル複合体の活性化;
(d)DNA合成および細胞周期移行の促進;
(e)細胞遊走または任意の他の発癌表現型;
(f)アクチンストレスファイバー形成およびF-アクチン産生;ならびに
(g)細胞接着、遊走、および浸潤に重要な任意の分子との相互作用。
【0012】
本発明はまた、癌、特に非小細胞肺癌(NSCLC)などの肺癌の処置または予防に有用な化合物をスクリーニングするためのキットを提供する。一部の態様において、本キットは以下のものを含む:
(a)ANLNポリペプチドのRhoA結合ドメインを含む第一のポリペプチド;
(b)RhoAポリペプチドのANLN結合ドメインを含む第二のポリペプチド;および
(c)第一のポリペプチドと第二のポリペプチドとの間の相互作用を検出する手段(例えば試薬)。
【0013】
一部の態様において、第一のポリペプチド、すなわちRhoA結合ドメインを含むポリペプチドは、ANLNポリペプチドを含む。同様に、他の態様において、第二のポリペプチド、すなわちANLN結合ドメインを含むポリペプチドは、RhoAポリペプチドを含む。
【0014】
一部の態様において、RhoA結合ドメインを含むポリペプチドは生細胞中で発現する。
【0015】
一部の態様において、2つのポリペプチド間の相互作用を検出する手段(例えば試薬)は、以下のものを検出する:
(a)活性化RhoAの濃度;
(b)RhoAとRHOエフェクターまたはそのRhoA結合領域との間の相互作用;
(c)活性化RhoAを介する下流遺伝子を含む任意のシグナル複合体の活性化;
(d)DNA合成および細胞周期移行の促進;
(e)細胞遊走または任意の他の発癌表現型;
(f)アクチンストレスファイバー形成およびF-アクチン産生;ならびに
(g)細胞接着、遊走、および浸潤に重要な任意の分子との相互作用。
【0016】
代替的な態様において、癌、特に非小細胞肺癌(NSCLC)などの肺癌の処置または予防に有用な化合物をスクリーニングする本発明の方法は、以下の工程を含む:
(a)ANLNポリペプチドまたはその機能的同等物を発現する細胞を試験化合物と接触させる工程;
(b)細胞のANLNを介する運動性を検出する工程;および
(c)試験化合物の非存在下で検出される運動性レベルと比較して細胞の運動性を阻害する試験化合物を選択する工程。
【0017】
一部の態様において、細胞は、ANLNポリペプチドまたはその機能的同等物を発現可能にコードするポリヌクレオチドを(例えばANLNポリペプチドをコードするポリヌクレオチドに機能的に連結されたプロモーターとして)含むベクターを含む。
【0018】
同様に、癌、特に非小細胞肺癌(NSCLC)などの肺癌の処置または予防に有用な化合物をスクリーニングするための本発明のキットは、以下のものを含みうる:
(a)ANLNポリペプチドまたはその機能的同等物を発現する細胞;および
(b)細胞の運動性を検出する手段(例えば試薬)。
【0019】
本発明はさらに、対象における癌の予後、より詳しくは非小細胞肺癌(NSCLC)に罹患した患者などの肺癌患者の予後を予測する方法を提供する。一部の態様において、本方法は以下の工程を含む:
(a)NSCLCの予後を予測しようとする対象から収集された試料の核に局在するANLNを検出する工程、および
(b)核内のANLNの局在が検出された場合に予後が不良であると予測する工程。
【0020】
一部の態様において、試料の核内のANLNの局在は以下の工程によって検出される:
(a)ANLNタンパク質を認識する抗体を試料と接触させる工程;および
(b)核領域において試料に結合する抗体を検出する工程。
【0021】
本発明はまた、対象における癌の予後、特に非小細胞肺癌(NSCLC)などの肺癌の予後を予測するためのキットを提供する。一部の態様において、本キットは、(a)ANLNタンパク質を認識する抗体と、(b)核における検出用の薬剤とを含む。一部の態様において、核の検出用の薬剤はヘマトキシリン-エオシン染色染料である。
【0022】
本発明はまた、対象における癌、特に非小細胞肺癌(NSCLC)などの肺癌を処置または予防するための方法を提供する。一部の態様において、本方法は以下の工程によって選択される化合物を投与する工程を含む:
(a)試験化合物の存在下で、ANLNポリペプチドのRhoA結合ドメインを含むポリペプチドを、RhoAポリペプチドのANLN結合ドメインを含むポリペプチドと接触させる工程;
(b)ポリペプチド間の結合を検出する工程;および
(c)ポリペプチド間の結合を阻害する試験化合物を選択する工程。
【0023】
または、本方法は以下の工程を含みうる:
(a)ANLNポリペプチドまたはその機能的同等物を発現する細胞を試験化合物と接触させる工程;
(b)細胞の運動性を検出する工程;および
(c)試験化合物の非存在下で検出される運動性レベルと比較して細胞の運動性を阻害する試験化合物を選択する工程。
【0024】
本発明はまた、対象における癌、特に非小細胞肺癌(NSCLC)などの肺癌を処置または予防するための方法であって、ANLNとRhoAとの間の結合を阻害する化合物を投与する工程を含む方法を提供する。
【0025】
本発明はまた、癌、特に非小細胞肺癌(NSCLC)などの肺癌の処置または予防に有用な組成物であって、以下の工程によって選択される化合物の薬学的有効量を含む組成物を提供する:
(a)試験化合物の存在下で、ANLNポリペプチドのRhoA結合ドメインを含むポリペプチドを、RhoAポリペプチドのANLN結合ドメインを含むポリペプチドと接触させる工程;
(b)ポリペプチド間の結合を検出する工程;および
(c)ポリペプチド間の結合を阻害する試験化合物を選択する工程;
または、
(a)ANLNポリペプチドまたはその機能的同等物を発現する細胞を試験化合物と接触させる工程;
(b)細胞の運動性を検出する工程;および
(c)試験化合物の非存在下で検出される運動性レベルと比較して細胞の運動性を阻害する試験化合物を選択する工程。
【0026】
本発明はまた、癌、特に非小細胞肺癌(NSCLC)などの肺癌の処置または予防に有用な組成物であって、ANLNとRhoAとの間の結合を阻害する化合物の薬学的有効量と、薬学的に許容される担体とを含む組成物を提供する。
【0027】
本発明のこれらおよび他の目的および特徴は、添付の図面および実施例と合わせて以下の詳細な説明を読むことにより、さらに十分明らかとなるであろう。しかし、上記の発明の概要および下記の詳細な説明はいずれも好ましい態様のものであって、発明または他の代替的な発明の態様を限定するものではないことを理解されたい。
【0028】
好ましい態様の詳細な説明
I. 概説
分子標的薬は、明確な作用メカニズムによって副作用を最小限に抑えつつ、悪性細胞に高度に特異的であることが期待される。このような薬の開発のための適切な分子標的を同定するために見込みある戦略として、本発明者らは、癌細胞で過剰発現している遺伝子を選択できるゲノム全域にわたる発現解析を、RNAi技術による機能喪失効果のハイスループットスクリーニングと組み合わせた。加えて、組織マイクロアレイ方法を応用し、可能性のある標的タンパク質の検証のために何百もの保管臨床サンプルを解析した。この種の体系的なアプローチを用いて、ANLNが臨床NSCLCサンプルならびに細胞株中で高頻度に過剰発現していること、また、この遺伝子産物が肺癌細胞の成長および進行において必要不可欠な役割を担うことを本明細書において示す。
【0029】
ANLNは当初、ショウジョウバエアクチン結合タンパク質であるアニリンのヒトホモログとして特徴付けられた(Oegema, K. et al. J. Cell Biol. 150(3):539-551 (2000))。ヒトANLN cDNAは、アクチン結合ドメインおよびC末端プレクストリン相同(PH)ドメインを含む1124アミノ酸タンパク質をコードする。また、いくつかのコンセンサス核局在配列(NLS)および一つのコンセンサスSH3結合モチーフを含んでいた。ANLNは細胞質分裂の間、分裂溝に局在しており、細胞質分裂に重要な役割を担っているとされる。本明細書において、ANLNは、一定の割合の癌細胞において細胞質だけでなく核にも局在する一方で、核膜崩壊後の皮層および細胞質分裂の間の分裂溝に位置することが見出された。以前に報告されているように、ANLNは細胞周期進行に重要な役割を担っている可能性が高く、後期において、少なくとも2つの他の溝タンパク質(アクチンおよびセプチン(SEPT))との相互作用を通じて、娘細胞を分けるアクチン-ミオシン収縮環を組み立てている可能性がある(Oegema, K. et al. J. Cell Biol. 150(3):539-551 (2000))。本明細書において考察するように、ANLN-siRNAで処置したNSCLC細胞は溝退行およびより大きな細胞形態を示し、おそらく収縮環の組み立てが阻止された結果としての細胞質分裂プロセスの機能不全により、多核化したことが観察されている。本明細書において、内因性RhoA(SEQ ID NO:4(SEQ ID NO:3にコードされる))のANLNとの相互作用は細胞質中のみでなく分裂溝および中央体においても起こることが見出され、細胞成長がANLN-RhoA相互作用およびNSCLC細胞の細胞質分裂の加速を通じて促進されたことが示された。
【0030】
そこで、本発明者らは、アクチン構造の形成を制御することが知られるRHO活性化(Ridley, A. J. and Hall, A. Cell 70:389-399 (1992))に対するANLNの効果に焦点を絞り、外因性ANLNの過剰発現が哺乳動物細胞においてアクチンストレスファイバーの形成を促進したことを見出した。本明細書におけるデータは、ANLNがRhoAとの相互作用を通じてRHOシグナル伝達を活性化してアクチン細胞骨格の再構築を有意に促進し、その結果、マトリゲル浸潤アッセイおよび傷害遊走アッセイによって観察されるように、細胞遊走活性を活性化した可能性があることを示唆する。
【0031】
RASおよびRHO GTPaseは十分に研究されているシグナル伝達分子であり、RHO GTPaseは、アクチン構築、限局性複合体/接着の組み立て、細胞運動性、細胞極性、遺伝子転写、および細胞周期進行を含む多様な一連の生物学的活性の調節において重要な分子として浮上してきた。最近の研究により、RHOタンパク質がいくつかのヒト腫瘍で過剰発現していること、ならびに上皮細胞成長因子(EGF)、肝細胞成長因子(HGF)、リポホスファチジン酸(lypophosphatidic acid)(LPA)、血小板由来成長因子(PDGF)、およびトランスフォーミング成長因子β(TGFB)を含む一部の成長因子がRHOタンパク質を活性化しうること(Zondag, G. C. et al. J. Cell Biol. 149:775-781 (2000); Bhowmick, N. A., et al., Mol. Biol. Cell 12, 27-36 (2001); Liu, A. X. et al., Mol. Cell Biol. 21:6906-6912 (2001))が示されている。インテグリン、カドヘリン、および免疫グロブリンスーパーファミリーメンバーを含む細胞接着分子のいくつかのクラスもRHO活性に影響を及ぼすことが示されている(DeMali, K. A., et al., Curr. Opin. Cell Biol. 15:572-582 (2003); Braga, V.M., Curr. Opin. Cell Biol. 14, 546-556 (2002); Thompson, P. W., et al., J. Immunol. 169:1007-1013 (2002))。加えて、いくつかのグアニンヌクレオチド交換因子(GEF)がRHOタンパク質およびそれらの下流エフェクターを異常に活性化して新生物性の形質転換を起こす可能性がある(Fort, P., Prog. Mol. Subcell. Biol. 22, 159-181 (1999); Zohn, I. M., et al., Oncogene 17, 1415-1438 (1998))。本発明者らのデータにより、過剰発現したANLNによるRhoAの異常な活性化がアクチン細胞骨格の再構築を通じて哺乳動物細胞の遊走活性を促進し、癌細胞の浸潤能および転移能に寄与しうるであろうことが示唆された。
【0032】
BrdU取り込みアッセイにより、外因的に発現されたANLNによる用量依存的なNSCLC細胞のDNA合成の促進が検出され、ANLNが細胞周期進行の重要な正のレギュレーターでありうることが示唆された。RHO、RAC、およびCDC42などのRHO GTPaseは、細胞周期のG1期に様々な活性をもたらすことが知られている(Olson, M. F., et al., Science 269, 1270-1272 (1995))。インビトロ研究により、活性がサイクリンD1(CCND1)などの活性化剤ならびにCDK阻害剤(CDKI)であるp21WAF1およびp27KIP1などの阻害剤によって制御されるサイクリン依存性キナーゼ(CDK)の調節を主に介して、RHOタンパク質が細胞周期の進行を促進することができる、複数のメカニズムが明らかとなっている(Danen, E. H., et al., J. Cell Biol. 151, 1413-1422 (2000); Olson, M. F., et al., Nature 394:295-299 (1998); Adnane, J., et al., Mol. Cell. Biol. 18:6962-6970 (1998))。ANLNによるNSCLC細胞のDNA合成の促進は、これら分子経路の機能に影響を及ぼす活性化RhoAのアップレギュレーションによるものである可能性がある。
【0033】
本明細書に提示する組織マイクロアレイ実験の結果は、n-ANLN陽性腫瘍を有する肺癌患者が、陰性腫瘍を有さない患者と比べて不良な癌特異的生存を示したことを示している。核へのANLN輸送の正確な分子メカニズム、およびそれが核特異的な追加機能を有するかどうかは不明であるが、このデータは、n-ANLNが何らかの未知のシグナル伝達経路を活性化することによって肺癌細胞の非常に悪性の表現型に寄与することを示唆している。
【0034】
以上をまとめると、ANLNは、RhoAと直接相互作用してそれを活性化することが本明細書において示され、この複合体は肺癌の成長促進経路および攻撃的な特性、ならびに細胞分裂/細胞周期進行に不可欠である可能性が高い。ここで報告するデータは、このANLN-RhoA経路が新規バイオマーカーを設計するための、および肺癌に特異的な抗癌薬を開発するための、好適な分子標的となりうることを示している。治療用siRNAはこの経路を妨害する選択肢の一つであるはずである。
【0035】
II. 定義:
本明細書において用いられる冠詞(a、an、およびthe)は、特に具体的に示されていない限り「少なくとも1つ」を意味する。
【0036】
本発明の文脈において、「ANLNポリペプチド」または「ANLN」は、しばしば科学文献において「アニリン」と呼ばれるアクチン結合タンパク質を指す。例えば、全体が参照として本明細書に組み入れられるOegema et al., J. Cell Biol. 150(3):539-551 (2000)を参照されたい。ANLNポリペプチドは、ヒト(Genbankアクセッション番号AF273437)、ショウジョウバエ(Genbankアクセッション番号X89858、ショウジョウバエ遺伝子CG4530(GenBankアクセッション番号AAF47044)の産物)、線虫(C. elegans)遺伝子K10B2.5(GenBankアクセッション番号T16604)、Y43F8C.14(GenBankアクセッション番号T26874)、およびY49E10.19(GenBankアクセッション番号T27053)の産物、または他のオルソロガスなポリペプチドと実質的に同一でありうる。一部の態様において、ANLNポリペプチドは、ヒトとショウジョウバエのANLNオルソログ間で保存されているアミノ酸を含む。または、ANLNポリペプチドは、例えばOegemaらの文献の542頁に示されるような、上に列挙したタンパク質の全ての間で保存されているアミノ酸を含みうる。全長ANLNポリペプチドは、例えば、PH結合ドメイン、アクチン結合ドメイン、核局在シグナル配列、およびSH3ドメインを含みうる。
【0037】
本明細書において、「RhoAポリペプチド」または「RhoA」は、GTP結合タンパク質のRho遺伝子ファミリーを指す。RhoAの結晶構造は決定されている。例えば、全体が参照として本明細書に組み入れられるMaesaki et al., Molec. Cell 4:793-803 (1999)を参照されたい。Maesakiらは、RhoAをRhoファミリーの他のメンバーから識別するACCフィンガー構造の位置および構造を含むRhoAの一次および二次構造に関してかなりの量の情報を提供している。RhoAポリペプチドの例には、例えばSEQ ID NO:4と実質的に同一なタンパク質が含まれる。
【0038】
最近の研究により、RHOファミリータンパク質が腫瘍形成に関与することが示されている。経路はまだ不明であるが、RHOと癌との関連は本質的なものである。特にRhoAタンパク質は癌に多大に関連しているようであり、RhoAの過剰発現は、大腸、乳房、肺、精巣生殖細胞、および頭頸部扁平上皮細胞癌腫瘍に関連している。加えて、RhoAは細胞運動性および細胞極性に関与することが示唆されている。これら2つの機能に対するRhoA発現の効果は、腫瘍形成の原因である高い可能性を示唆する。
【0039】
本発明の文脈において、2つのタンパク質間の「結合の阻害」とは、タンパク質間の結合を少なくとも減少させることを指す。従って、場合によっては、サンプル中の結合対のパーセンテージは、適切な(例えば、試験化合物で処置されていない、または非癌サンプル由来の、または癌サンプル由来の)対照と比較して低下する。結合したタンパク質の量における減少は、例えば、対照サンプル中で結合した対よりも、90%、80%、70%、60%、50%、40%、25%、10%、5%、1%よりも少ない、またはそれより少なく(例えば0%)なりうる。
【0040】
用語「試験化合物」は、本明細書において詳しく述べられるような、ANLNとRhoAとの間のタンパク質-タンパク質相互作用を阻害しうる任意の(例えば化学的または組換え的に産生された)分子を指す。一部の態様において、試験化合物は1,500ダルトン未満の分子量を有し、場合によっては、1,000、800、600、500、または400ダルトン未満の分子量を有する。
【0041】
化合物の「薬学的有効量」とは、個体においてANLNを介する疾患を処置および/または改善するのに十分な量である。薬学的有効量の一例は、動物に投与したときに、ANLNとRhoAとの間の相互作用を低下させるのに必要な量でありうる。相互作用における低下とは、例えば、結合における少なくとも5%、10%、20%、30%、40%、50%、75%、80%、90%、95%、99%、または100%の変化でありうる。または、この量は、野生型ANLNについて本明細書に記載されているように、投与したときに核局在を検出可能に低下させる量を含みうる。
【0042】
「薬学的に許容される担体」という表現は、薬物の希釈剤または賦形剤として用いられる不活性な物質を指す。
【0043】
本発明の文脈において、用語「機能的に同等」とは、対象ポリペプチドが参照ポリペプチドの生物学的活性を有することを意味する。例えば、ANLNの機能的同等物は、野生型ANLNのようにインビトロでアクチンに結合して細胞運動性を誘導する能力を有するであろう。このような活性を測定するためのアッセイ法は当技術分野において周知である。
【0044】
用語「単離」および「生物学的に純粋」とは、自然の状態で見られる通常はそれに付随する成分を実質的または本質的に含まない材料を指す。しかしながら、用語「単離」は、電気泳動ゲルまたは他の分離媒体中に存在する成分を指すことを意図しない。単離された成分はそのような分離媒体を含まず、別の応用にすぐに使用可能な形態にあるか、あるいは既に新しい応用/環境に使用されている。
【0045】
「保存的に改変された変異体」という表現は、アミノ酸配列および核酸配列の両方に適用される。特定の核酸配列に関して、保存的に改変された変異体とは、同一または本質的に同一のアミノ酸配列をコードする核酸を指し、核酸がアミノ酸配列をコードしない場合は、本質的に同一の配列を指す。遺伝コードの縮重により、機能的に同一の核酸の多数が任意の所与のタンパク質をコードする。例えば、コドンGCA、GCC、GCG、およびGCUは全て、アミノ酸のアラニンをコードする。したがって、アラニンがコドンによって指定される全ての位置で、そのコドンは、コードするポリペプチドを変化させることなく任意の上記対応コドンに変えることができる。このような核酸変異は「サイレント変異」、つまり保存的に改変された変異の一種である。本明細書において、ポリペプチドをコードする全ての核酸配列は、その核酸の全ての可能なサイレント変異も表す。当業者は、核酸中の各コドン(通常メチオニンの唯一のコドンであるAUGと、通常トリプトファンの唯一のコドンであるTGGを除く)を改変して機能的に同一の分子を得ることができることを認識するであろう。よって、ポリペプチドをコードする核酸の各サイレント変異は、各開示配列に暗黙のうちに記載されている。
【0046】
アミノ酸配列に関し、コードされた配列中の1個のアミノ酸を、または少ないパーセンテージのアミノ酸を変化、付加、または欠失させる、核酸、ペプチド、ポリペプチド、またはタンパク質配列に対する置換、欠失、または付加はそれぞれ、その変化がアミノ酸を化学的に類似のアミノ酸と置換する「保存的に改変された変異体」であることを当業者は認識するであろう。機能的に類似のアミノ酸を提供する保存的置換表は当技術分野において周知である。このような保存的に改変された変異体は、本発明の多型変異体、種間ホモログ、および対立遺伝子に加えられるものであって、これらを排除するものではない。
【0047】
以下の8群はそれぞれ、互いに保存的置換であるアミノ酸を含む:
1)アラニン(A)、グリシン(G);
2)アスパラギン酸(D)、グルタミン酸(E);
3)アスパラギン(N)、グルタミン(Q);
4)アルギニン(R)、リジン(K);
5)イソロイシン(I)、ロイシン(L)、メチオニン(M)、バリン(V);
6)フェニルアラニン(F)、チロシン(Y)、トリプトファン(W);
7)セリン(S)、スレオニン(T);および
8)システイン(C)、メチオニン(M)(例えば、Creighton, Proteins (1984) を参照されたい)。
【0048】
本発明の文脈において、「配列同一性のパーセンテージ」は、2つの最適に整列された配列を比較ウィンドウにわたって比較することにより決定され、このとき比較ウィンドウ中のポリヌクレオチド配列部分は、2つの配列の最適な整列について、付加または欠失を含まない参照配列(例えば本発明のポリペプチド)と比較して付加または欠失(すなわちギャップ)を含んでもよい。このパーセンテージは、同一核酸塩基またはアミノ酸残基が両配列に現れる位置の数を決定してマッチする位置の数を出し、マッチする位置の数を比較ウィンドウ中の位置の総数で割り、その結果に100を乗じて配列同一性のパーセンテージを出すことによって計算される。
【0049】
2つもしくはそれ以上の核酸またはポリペプチド配列の文脈において、用語「同一」またはパーセント「同一性」は、同じ配列である2つまたはそれ以上の配列または部分配列を指す。2つの配列が、下記の配列比較アルゴリズムの1つを用いてまたはマニュアル整列と目視検査によって測定された比較ウィンドウまたは指定領域にわたって最大一致するように比較整列された際に、同じアミノ酸残基またはヌクレオチドを特定のパーセンテージで有する(すなわち、特定領域にわたって、あるいは特定されていない場合は全配列にわたって、60%同一性、任意で65%、70%、75%、80%、85%、90%、または95%同一性)場合、2つの配列は「実質的に同一」である。任意で、同一性は長さが少なくとも約50ヌクレオチドである領域にわたって存在し、またはより好ましくは長さが100〜500または1000ヌクレオチド以上である領域にわたって存在する。
【0050】
配列比較には、典型的には1つの配列を参照配列とし、これに対して試験配列を比較する。配列比較アルゴリズムを用いる場合、試験および参照配列をコンピュータに入力し、必要であれば部分配列座標を指定して、配列アルゴリズムプログラムパラメータを指定する。デフォルトプログラムパラメータを用いてもよく、また、代替的なパラメータを指定してもよい。そして配列比較アルゴリズムが、プログラムパラメータに基づいて、試験配列について参照配列に対するパーセント配列同一性を計算する。
【0051】
本明細書において用いられる「比較ウィンドウ」は、20〜600、一般的には約50〜約200、より一般的には約100〜約150からなる群より選択される連続位置の数のいずれか1つのセグメントに対する参照を含み、この内部において、ある配列が同じ数の連続位置の参照配列と最適に整列された後、これら2つの配列が比較されうる。比較するために配列を整列する方法は当技術分野において周知である。例えば、Smith and Waterman (1981) Adv. Appl. Math. 2:482-489の局所相同性アルゴリズム、Needleman and Wunsch (1970) J. Mol. Biol. 48:443の相同性整列アルゴリズム、Pearson and Lipman (1988) Proc. Nat'l. Acad. Sci. USA 85:2444の類似性検索法、これらアルゴリズムのコンピュータによる実施(GAP、BESTFIT、FASTA、およびTFASTA、Wisconsin Genetics Software Package, Genetics Computer Group, 575 Science Dr., Madison, WI)、またはマニュアル整列と目視検査(例えば、Ausubel et al., Current Protocols in Molecular Biology (1995 supplement) を参照されたい)によって比較用に配列を最適に整列できる。
【0052】
パーセント配列同一性および配列類似性を決定するために適したアルゴリズムの2つの例がBLASTアルゴリズムおよびBLAST 2.0アルゴリズムであり、これらはそれぞれAltschul et al. (1997) Nuc. Acids Res. 25:3389-3402、およびAltschul et al. (1990) J. Mol. Biol. 215:403-410に記載されている。BLAST解析を行うためのソフトウェアは、National Center for Biotechnology Informationを通じて公的に利用可能である。このアルゴリズムは、データベース配列中の同じ長さのワード(word)と整列させた場合にいくつかの正の値の閾値スコアTにマッチするかまたはそれを満たす、クエリー(query)配列中の長さの短いワードWを同定することによって、高スコア配列対(HSP)をまず同定することを含む。Tは近傍ワードスコア閾値(neighborhood word score threshold)(Altschul et al.、上記)と呼ばれる。これら初期近傍ワードヒットが、これらを含むさらに長いHSPを見つけるための検索を開始するためのシード(seed)となる。ワードヒットは、累積整列スコアを増加できる限り、各配列に沿って両方向に延長される。累積スコアは、ヌクレオチド配列に関しては、パラメータM(マッチする残基対に対するリワード(reward)スコア;常に>0)およびパラメータN(ミスマッチ残基に対するペナルティ(penalty)スコア;常に<0)を用いて計算される。アミノ酸配列に関しては、スコアリングマトリックスを用いて累積スコアを計算する。各方向におけるワードヒットの延長は以下の場合に停止される:累積整列スコアが最大達成値から量X分減少する場合;累積スコアが、1つまたは複数の負のスコアの残基整列の蓄積によってゼロ以下になる場合;あるいは、いずれかの配列の終わりに達する場合。BLASTアルゴリズムパラメータW、T、およびXは、整列の感度と速度を決定する。BLASTNプログラム(ヌクレオチド配列用)はデフォルトとして、word length(ワード長)(W)11、expectation(期待値)(E)10、M=5、N=-4、およびcomparison of both strands(両鎖の比較)を用いる。アミノ酸配列については、BLASTPプログラムはデフォルトとして、word length 3、expectation(E)10、BLOSUM62 scoring matrix(スコアリング マトリックス)(Henikoff and Henikoff (1989) Proc. Natl. Acad. Sci. USA 89:10915を参照されたい)alignment(アライメント)(B)50、expectation(E)10、M=5、N=-4、およびcomparison of both strandsを用いる。
【0053】
BLASTアルゴリズムはまた、2つの配列間の類似性の統計学的解析を行う(例えば、Karlin and Altschul (1993) Proc. Natl. Acad. Sci. USA 90:5873-7を参照されたい)。BLASTアルゴリズムによって提供される類似性の1つの尺度が最小合計確率(smallest sum probability, P(N))であり、2つのヌクレオチドまたはアミノ酸配列間のマッチが偶然に起こる確率の指標を提供する。例えば、試験核酸の参照核酸との比較における最小合計確率が約0.2未満、より好ましくは約0.01未満、最も好ましくは約0.001未満である場合、この核酸は参照配列に類似すると見なされる。
【0054】
用語「有機低分子」は、薬剤に一般的に用いられる有機分子に相当するサイズの分子を指す。この用語は生物学的高分子(例えば、タンパク質、核酸など)は除外する。好ましい有機低分子はサイズが約5000Daまで、例えば、2000Daまで、あるいは約1000Daまでの範囲である。
【0055】
用語「標識」および「検出可能な標識」は、分光学的、光化学的、生化学的、免疫化学的、電気的、光学的、または化学的手段によって検出可能な任意の組成物を指すために本明細書において用いられる。このような標識としては、標識されたストレプトアビジン結合体での染色のためのビオチン、磁気ビーズ(例えばDYNABEADS(商標))、蛍光色素(例えば、フルオレセイン、テキサスレッド、ローダミン、緑色蛍光タンパク質など)、放射標識(例えば、3H、125I、.35S、14C、または32P)、酵素(例えば、ホースラディッシュペルオキシダーゼ、アルカリホスファターゼ、およびELISAで一般的に用いられる他のもの)、および金コロイドまたは着色ガラスもしくはプラスチック(例えば、ポリスチレン、ポリプロピレン、ラテックスなどの)ビーズなどの熱量測定標識が挙げられる。このような標識の使用を教示する特許としては、米国特許第3,817,837号;第3,850,752号;第3,939,350号;第3,996,345号;第4,277,437号;第4,275,149号;および第4,366,241号が挙げられる。このような標識を検出する手段は当業者に周知である。したがって、例えば、放射標識は感光フィルムまたはシンチレーションカウンターを用いて検出することができ、蛍光マーカーは放出された光を検出する光検出器を用いて検出しうる。酵素標識は典型的には、酵素に基質を提供して酵素の基質に対する作用によって産生された反応産物を検出することによって検出され、熱量測定標識は着色された標識を単純に視覚化することによって検出される。
【0056】
本明細書において用いられる用語「抗体」は、天然抗体および非天然抗体を包含し、例えば、単鎖抗体、キメラ、二機能性、およびヒト化抗体、ならびにそれらの抗原結合断片(例えば、Fab'、F(ab')2、Fab、Fv、およびrIgG)を含む。Pierce Catalog and Handbook, 1994-1995 (Pierce Chemical Co., Rockford, IL) も参照されたい。例えば、Kuby, J., Immunology, 3rd Ed., W.H. Freeman & Co., New York (1998) も参照されたい。このような非天然抗体は固相ペプチド合成を用いて構築することができ、組換え的に産生することも、あるいは例えば、参照として本明細書に組み入れられる、Huse et al., Science 246:1275-1281 (1989) に記載されているように、可変重鎖と可変軽鎖とからなるコンビナトリアルライブラリーをスクリーニングすることによって得ることもできる。例えば、キメラ、ヒト化、CDR移植、単鎖、および二機能性抗体を作成するこれらおよび他の方法は当業者に周知である(Winter and Harris, Immunol. Today 14:243-246 (1993); Ward et al., Nature 341:544-546 (1989); Harlow and Lane, Antibodies: A Laboratory Manual, 1988; Hilyard et al., Protein Engineering: A practical approach (IRL Press 1992); Borrebeck, Antibody Engineering, 2d ed. (Oxford University Press 1995);これらは各々参照として本明細書に組み入れられる)。
【0057】
用語「抗体」は、ポリクローナル抗体およびモノクローナル抗体の両方を含む。この用語はまた、キメラ抗体(例えばヒト化マウス抗体)およびヘテロ結合抗体(heteroconjugate antibody)(例えば二重特異性抗体)などの遺伝的に操作された形態を含む。この用語はまた、組換え単鎖Fv断片(scFv)も指す。抗体という用語はまた、二価または二重特異性分子、二重特異性抗体(diabody)、三重特異性抗体(triabody)、および四重特異性抗体(tetrabody)を含む。二価および二重特異性分子は、例えば、Kostelny et al. (1992) J Immunol 148:1547, Pack and Pluckthun (1992) Biochemistry 31:1579, Holliger et al. (1993) Proc Natl Acad Sci U S A. 90:6444, Gruber et al. (1994) J Immunol 152:5368, Zhu et al. (1997) Protein Sci 6:781, Hu et al. (1996) Cancer Res. 56:3055, Adams et al. (1993) Cancer Res. 53:4026、および McCartney, et al. (1995) Protein Eng. 8:301に記載されている。
【0058】
典型的には、抗体は重鎖および軽鎖を有する。重鎖および軽鎖はそれぞれ定常領域および可変領域(これら領域は「ドメイン」としても知られる)を含む。軽鎖および重鎖可変領域は、「相補性決定領域」または「CDR」とも呼ばれる、3つの超可変領域によって中断される4つの「フレームワーク」領域を含む。フレームワーク領域およびCDRの範囲は規定されている。異なる軽鎖または重鎖のフレームワーク領域の配列は種内で比較的保存されている。抗体のフレームワーク領域、すなわち構成軽鎖と重鎖のフレームワーク領域を合わせたものは、CDRを3次元空間に位置付け、整列させる。
【0059】
CDRは主として抗原エピトープへの結合を担っている。各鎖のCDRは、典型的にはN末端から順に番号がつけられてCDR1、CDR2、およびCDR3と呼ばれ、また典型的には、その特定のCDRが位置する鎖によって同定される。したがって、VH CDR3はそれが見出される抗体の重鎖の可変ドメインに位置し、VL CDR1はそれが見出される抗体の軽鎖の可変ドメインのCDR1である。「VH」に関しては抗体の免疫グロブリン重鎖の可変領域を指し、Fv、scFv、またはFabの重鎖を含む。「VL」に関しては免疫グロブリン軽鎖の可変領域を指し、Fv、scFv、dsFv、またはFabの軽鎖を含む。
【0060】
「単鎖Fv」または「scFv」という表現は、通常の2鎖抗体の重鎖および軽鎖の可変ドメインが連結して1つの鎖を形成した抗体を指す。典型的にはリンカーペプチドが2つの鎖の間に挿入されており、適正なフォールディングと活性な結合部位の形成とを可能にする。
【0061】
「キメラ抗体」は、(a)異なるまたは変えられた、クラス、エフェクター機能、および/または種の定常領域に、あるいはキメラ抗体に新しい特性を付与する全く異なる分子、例えば、酵素、毒素、ホルモン、成長因子、薬物などに、抗原結合部位(可変領域)が結合するように、定常領域またはその一部分が変えられた、置き換えられた、または交換された;あるいは(b)可変領域またはその一部分が、異なるまたは変えられた抗原特異性を有する可変領域に変えられた、置き換えられた、または交換された、免疫グロブリン分子である。
【0062】
「ヒト化抗体」は、非ヒト免疫グロブリンに由来する最小配列を含む免疫グロブリン分子である。ヒト化抗体には、所望の特異性、親和性、および能力を有するマウス、ラット、またはウサギなどの非ヒト種(ドナー抗体)の相補性決定領域(CDR)に由来する残基でヒト免疫グロブリン(レシピエント抗体)のCDR由来の残基が置き換えられているヒト免疫グロブリンが含まれる。一部の例において、ヒト免疫グロブリンのFvフレームワーク残基は、対応する非ヒト残基で置き換えられる。ヒト化抗体はまた、レシピエント抗体にも、導入されたCDRもしくはフレームワーク配列にも見られない残基を含みうる。一般に、ヒト化抗体は少なくとも1つ、典型的には2つの可変ドメインの実質的に全てを含み、これらCDR領域の全てまたは実質的に全ては非ヒト免疫グロブリンのものに対応し、フレームワーク(FR)領域の全てまたは実質的に全てはヒト免疫グロブリンコンセンサス配列のものである。ヒト化抗体は最適にはまた、免疫グロブリン定常領域(Fc)、典型的にはヒト免疫グロブリンのものの少なくとも一部分を含む(Jones et al., Nature 321:522-525 (1986); Riechmann et al., Nature 332:323-327 (1988)、 および Presta, Curr. Op. Struct. Biol. 2:593-596 (1992))。ヒト化は本質的には、Winterと共同研究者(Jones et al., Nature 321:522-525 (1986); Riechmann et al., Nature 332:323-327 (1988); Verhoeyen et al., Science 239:1534-1536 (1988))の方法に従って、齧歯類のCDRまたはCDR配列でヒト抗体の対応配列を置換することによって行うことができる。従って、このようなヒト化抗体はキメラ抗体(米国特許第4,816,567号)であり、実質的に完全でないヒト可変ドメインが非ヒト種由来の対応配列によって置換されている。
【0063】
用語「エピトープ」および「抗原決定基」は、抗体が結合する抗原上の部位を指す。エピトープは連続アミノ酸から、またはタンパク質の三次フォールディングによって近接して並べられた非連続アミノ酸から形成されうる。連続アミノ酸から形成されたエピトープは典型的には変性溶媒にさらされても保持され、三次フォールディングによって形成されたエピトープは典型的には変性溶媒処置で失われる。エピトープは典型的には、独自の空間的高次構造中に少なくとも3、より一般的には少なくとも5または8〜10個のアミノ酸を含む。エピトープの空間的高次構造を決定する方法には、例えばx線結晶解析および2次元核磁気共鳴が含まれる。例えば、Epitope Mapping Protocols in Methods in Molecular Biology, Vol. 66, Glenn E. Morris, Ed (1996) を参照されたい。
【0064】
用語「ポリペプチド」、「ペプチド」、および「タンパク質」は、アミノ酸残基のポリマーを指して本明細書において相互に交換可能に用いられる。これら用語は、天然アミノ酸ポリマー、修飾残基を含むもの、および非天然アミノ酸ポリマーに適用されるのに加え、1つまたは複数のアミノ酸残基が対応する天然アミノ酸の人工的な化学的模倣体であるアミノ酸ポリマーにも適用される。
【0065】
用語「アミノ酸」は、天然および合成アミノ酸、ならびに天然アミノ酸と同様に機能するアミノ酸アナログおよびアミノ酸模倣体を指す。天然アミノ酸とは、遺伝コードによってコードされるもの、ならびに例えば、ヒドロキシプロリン、γ-カルボキシグルタメート、およびO-ホスホセリンで後に修飾されるアミノ酸である。アミノ酸アナログとは、天然アミノ酸と同じ基本化学構造、例えば、水素、カルボキシル基、アミノ基、およびR基に結合されたα炭素、例えば、ホモセリン、ノルロイシン、メチオニンスルホキシド、メチオニンメチルスルホニウム、を有する化合物を指す。このようなアナログは修飾されたR基(例えばノルロイシン)または修飾されたペプチド骨格を有しうるが、天然アミノ酸と同じ基本化学構造を保持する。アミノ酸模倣体とは、アミノ酸の一般的な化学構造とは異なる構造を有するが天然アミノ酸と同様に機能する化学的な化合物を指す。
【0066】
アミノ酸は本明細書において、一般的に知られる3文字表記で表されることもあり、またIUPAC-IUB Biochemical Nomenclature Commission推奨の1文字表記で表されることもある。ヌクレオチドは同様に、一般的に認められた1文字コードで表されうる。
【0067】
用語「組換え」は、例えば細胞、あるいは核酸、タンパク質、またはベクターなどへの言及と共に用いられる場合、細胞、核酸、タンパク質、またはベクターが、異種核酸またはタンパク質の導入、あるいは元来の核酸またはタンパク質の変化によって修飾されていること、あるいは細胞がそのように修飾された細胞に由来することを示す。したがって例えば、組換え細胞は元来の(非組換え)形態の細胞では見られない遺伝子を発現するか、または元来の遺伝子を異常に発現する、より少なく発現する、または全く発現しない。本明細書における用語「組換え核酸」とは、一般に核酸の操作によって、例えばポリメラーゼおよびエンドヌクレアーゼを用いて、最初からインビトロで形成された、天然に普通では見られない形態にある核酸を意味する。このようにして異なる配列の機能的な連結が達成される。このように、通常は繋がっていないDNA分子を連結してインビトロで形成した線状形態の単離核酸または発現ベクターは両方とも、本発明の目的に関して組換え型と見なされる。組換え核酸を作製して宿主細胞または生物体に再び導入すると、これは非組換え的に、すなわちインビトロ操作ではなく宿主細胞のインビボ細胞機構を用いて、複製する;しかし、このような核酸は組換え的に産生されると続いて非組換え的に複製されるが、それでも本発明の目的に関して組換え型と見なされることが理解される。同様に、「組換えタンパク質」は、組換え技術を用いて、すなわち上記のような組換え核酸の発現を通じて作製されたタンパク質である。
【0068】
特に定義しない限り、本明細書において用いられる技術的および科学的用語は全て本発明の属する技術分野の当業者によって一般的に理解されるものと同じ意味を有する。矛盾する場合は、本明細書が定義を含めて統御する。
【0069】
III. ANLNを介する疾患を処置する化合物の産生および同定
ANLNがRhoAと相互作用し、ANLN発現が癌患者における不良な予後に関連するという本明細書に提供される事項を考慮し、本発明の一局面は、ANLNとRhoAとの間の結合を減少または予防する試験化合物を同定することに関する。さらに、ANLNの発現が細胞運動性の増加と関連するという本明細書において提供される事項を考慮し、本発明は、ANLNを介する運動性を阻害する試験化合物を同定する方法を提供する。
【0070】
ANLN/RhoA結合を決定するための方法には、2つのタンパク質の相互作用を決定するための任意の方法が含まれる。このようなアッセイとしては、従来のアプローチ、例えば、架橋、共免疫沈降、および勾配またはクロマトグラフィーカラムを通した同時精製が挙げられるが、これらに限定されない。加えて、タンパク質-タンパク質相互作用は、Fieldsと共同研究者によって述べられている酵母ベースの遺伝学的システム(Fields and Song, Nature 340:245-246 (1989); Chien et al., Proc. Natl. Acad. Sci. USA 88, 9578-9582 (1991))を用い、Chevray and Nathans (Proc. Natl. Acad. Sci. USA 89:5789-5793 (1992))に開示されているようにモニターすることができる。酵母GALAなどの多くの転写アクチベーターは2個の物理的に別個のモジュラードメインからなり、片方がDNA結合ドメインとして働き、もう片方が転写活性化ドメインとして機能する。上記刊行物に記載の酵母発現システム(一般に「ツーハイブリッドシステム」と呼ばれる)はこの特性を利用して2つのハイブリッドタンパク質を使用しており、片方は標的タンパク質がGAL4のDNA結合ドメインに融合しており、もう片方は候補活性化タンパク質が活性化ドメインに融合している。GALA活性化プロモーターの制御下にあるGAL1-lacZレポーター遺伝子の発現は、タンパク質-タンパク質相互作用を介するGAL4活性の再構成に依存する。相互作用するポリペプチドを含むコロニーは、β-ガラクトシダーゼに対する色素産生性基質で検出される。ツーハイブリッド技術を用いて2つの特異的なタンパク質間のタンパク質-タンパク質相互作用を同定するための完全なキット(MATCHMAKER(商標))がClontechから市販されている。このシステムを拡張して、特異的タンパク質相互作用に関わるタンパク質ドメインをマッピングすることもできるし、これら相互作用に非常に重要なアミノ酸残基を正確に特定することもできる。
【0071】
本出願では「ANLN」および「RhoA」と言及していても、両者の相互作用が解析または操作される場合、これらタンパク質の片方または両方の結合部分を、これらタンパク質の全長コピーのかわりに用いることができることが理解される。RhoAに結合するANLNの断片は、ANLNの標準的な欠失解析および/または突然変異誘発を用いてRhoAに結合する断片を同定することによって容易に同定しうる。類似の解析を用いてRhoAのANLN結合断片を同定しうる。
【0072】
ANLNを介する細胞運動性を阻害する試験化合物を同定する方法は、ANLNを発現する細胞を化合物と接触させ、細胞運動性を観察することによって行いうる。正常または癌組織由来の任意の哺乳動物細胞を用いることができ、例えば、NIH3T3、COS-7、HEK293、SAEC、BEAS-2B細胞などが挙げられる。細胞運動性は、マトリゲル浸潤アッセイなどの標準的なアッセイ法を用いて測定することができる。
【0073】
本明細書に開示されるように、例えば、タンパク質(抗体を含む)、ムテイン(mutein)、ポリヌクレオチド、核酸アプタマー、ならびにペプチドおよび非ペプチド有機低分子を含む任意の試験化合物が、本発明の試験化合物となりうる。試験化合物は、天然の供給源から単離されてもよく、合成的もしくは組換え的に、またはこれらの任意の組み合わせで調製されてもよい。
【0074】
例えばペプチドは、"Solid Phase Peptide Synthesis" by G. Barany and R. B. Merrifield in Peptides, Vol. 2, edited by E. Gross and J. Meienhoffer, Academic Press, New York, N.Y., pp. 100-118 (1980) に記載されているような固相技術を用いて合成的に産生しうる。同様に核酸も、Beaucage, S.L., & Iyer, R.P. (1992) Tetrahedron, 48, 2223-2311; および Matthes et al., EMBO J., 3:801-805 (1984) に記載されているように固相技術を用いて合成することができる。
【0075】
阻害ペプチドが同定されれば、インビボでのペプチドの安定性を増加させるために、様々なアミノ酸模倣体または非天然アミノ酸で本発明のペプチドを修飾することが特に有用である。安定性は数々の方法でアッセイできる。例えば、ペプチダーゼおよび様々な生物学的媒体、例えばヒト血漿および血清などを用いて安定性が試験されている。例えば、Verhoef et al., Eur. J. Drug Metab Pharmacokin. 11:291-302 (1986) を参照されたい。当技術分野で公知の他の有用なペプチド修飾にはグリコシル化およびアセチル化が含まれる。
【0076】
組換えおよび化学的合成技術は両方とも、本発明の試験化合物を産生するために用いうる。例えば、核酸の試験化合物は適切なベクターへ挿入して産生することができ、これをコンピテント細胞にトランスフェクトして増やすことができる。または、核酸はPCR技術または適した宿主中での発現を用いて増幅しうる(Sambrook et al., Molecular Cloning: A Laboratory Manual, 1989, Cold Spring Harbor Laboratory, New York, USA参照)。
【0077】
ペプチドおよびタンパク質も、当技術分野において周知の組換え技術を用いて、例えば、Morrison, J. Bact., 132:349-351 (1977)および Clark-Curtiss & Curtiss, Methods in Enzymology, 101:347-362 (1983) に記載されているように適した宿主細胞を組換えDNAコンストラクトで形質転換することによって、発現しうる。
【0078】
抗ANLNおよび抗RhoA抗体
本発明の一部の局面において、試験化合物は抗ANLNまたは抗RhoA抗体である。一部の態様において、これら抗体は、ヒト化抗体を含むがこれに限定されないキメラである。場合によっては、本発明の抗体の態様は、これらタンパク質の片方がもう一方に会合する境界面でANLNまたはRhoAのいずれかに結合する。一部の態様において、これら抗体は、生理的条件下で少なくとも約105 mol-1、106 mol-1以上、107 mol-1以上、108 mol-1以上、または109 mol-1以上のKaでANLNまたはRhoAに結合する。このような抗体は、例えばChemicon, Inc. (Temecula CA) などの商業的供給元から購入することができ、あるいは実質的に精製されたANLNまたはRhoAタンパク質、例えばヒトタンパク質またはその抗原性断片などを、免疫原として用いて作製することができる。提供された免疫原からモノクローナルおよびポリクローナル抗体の両方を調製する方法は、当技術分野において周知である。精製技術、および特定の免疫原に対する抗体を同定するための方法については、例えば、その全体が参照として本明細書に組み入れられるPCT/US02/07144 (WO/03/077838) を参照されたい。例えば親和性カラムを形成するために抗体親和性マトリックスを用いる抗体精製法も当技術分野において周知で、市販されている(AntibodyShop, Copenhagen, Denmark)。ANLN/RhoAの会合を阻害しうる抗体の同定は、試験化合物について全般的に以下に詳述するものと同じ試験アッセイを用いて行われる。
【0079】
変換酵素
変換酵素は本発明の試験化合物となりうる。本発明の文脈において、変換酵素とは、ANLN、RhoA、または両方に対して共有結合性翻訳後修飾を行う分子触媒である。本発明の変換酵素は、ANLNおよび/またはRhoAの1つまたは複数のアミノ酸残基を、修飾タンパク質の構造中のアロステリック変化を引き起こすように、あるいはANLNとRhoAとの間の結合を妨害するようにANLN/RhoA分子結合部位の化学的性質もしくは修飾タンパク質の構造を変えて、共有結合的に修飾する。本明細書において、これら2分子間の結合の妨害とは、結合のKaにおいて、30℃、イオン強度0.1、界面活性剤の非存在下で測定したこれらタンパク質間の結合のKaと比べて、少なくとも25%、30%、40%、50%、60%、70%またはそれ以上低下させることを指す。本発明の例示的な変換酵素には、キナーゼ、ホスファターゼ、アミダーゼ、アセチラーゼ、グリコシダーゼなどが含まれる。
【0080】
試験化合物ライブラリーの構築
試験化合物ライブラリーの構築は当技術分野において周知であるが、本セクションでは、試験化合物の同定と、ANLN/RhoA相互作用および/またはANLNを介する細胞運動性の有効な阻害剤のスクリーニングのためのこのような化合物のライブラリーの構築とにおけるさらなるガイダンスを提供する。
【0081】
分子モデリング
試験化合物ライブラリーの構築は、求められる特性を有することが知られている化合物の分子構造、ならびに/または阻害する標的分子、すなわちANLNおよびRhoAの分子構造の知識によって促進される。さらなる評価に適した試験化合物の予備スクリーニングに対する1つのアプローチが、試験化合物とその標的との間の相互作用のコンピュータモデリングである。本発明において、ANLNおよび/またはRhoAの間の相互作用をモデリングすることにより、相互作用自体の詳細への洞察が提供され、また、可能性のある、相互作用の分子阻害剤を含む、相互作用を阻害するために可能な方法が示唆される。
【0082】
コンピュータモデリングテクノロジーにより、選択した分子の3次元原子構造の視覚化と、その分子と相互作用するであろう新しい化合物の合理的設計とが可能である。3次元構築物は典型的には、選択した分子のx線結晶構造解析またはNMRイメージングのデータに依存する。分子動態は力場データを必要とする。コンピュータグラフィックシステムは、新しい化合物がどのように標的分子と関連するかを予測することができ、結合特異性を完全にするように化合物および標的分子の構造を実験操作することが可能である。片方または両方に小さな変化を起こしたときに分子-化合物間相互作用がどうなるかの予測には、通常ユーザーが使いやすい、分子設計プログラムとユーザーとの間のメニュー駆動型インターフェースと連係した、分子力学ソフトウェアおよび計算集約型コンピュータが必要である。
【0083】
一般的に述べた上記の分子モデリングシステムの一例は、CHARMmおよびQUANTAプログラム(Polygen Corporation, Waltham, Mass)からなる。CHARMmはエネルギー最小化および分子動態機能を行う。QUANTAは分子構造の構築、グラフィックモデリング、および解析を行う。QUANTAによって、分子の互いの挙動の相互的な構築、修飾、視覚化、および解析が可能である。
【0084】
特定のタンパク質と相互作用する薬物のコンピュータモデリングが多くの論文に概説されている。例えば、Rotivinen, et al. Acta Pharmaceutica Fennica 97, 159-166 (1988); Ripka, New Scientist 54-57 (Jun. 16, 1988); McKinlay and Rossmann, Annu. Rev. Pharmacol. Toxiciol. 29, 111-122 (1989); Perry and Davies, Prog Clin Biol Res.291:189-93(1989); Lewis and Dean, Proc. R. Soc. Lond. 236, 125-140 and 141-162 (1989);および、核酸成分に対するモデル受容体に関し、Askew, et al., J. Am. Chem. Soc. 111, 1082-1090 (1989)。
【0085】
化学物質をスクリーニングし、画像として描写する他のコンピュータプログラムが、BioDesign, Inc., Pasadena, Calif.、Allelix, Inc, Mississauga, Ontario, Canada、およびHypercube, Inc., Cambridge, Ontarioなどの会社から利用可能である。例えば、DesJarlais et al. (1988) J. Med. Chem. 31:722; Meng et al. (1992) J. Computer Chem. 13:505; Meng et al. (1993) Proteins 17:266; Shoichet et al. (1993) Science 259:1445を参照されたい。
【0086】
ANLN/RhoA相互作用の推定阻害剤が同定されれば、以下に詳述するように、同定された推定阻害剤の化学的構造に基づき、コンビナトリアルケミカル技術を使用して任意の数の変異体を構築することができる。こうして得られる推定阻害剤または「試験化合物」のライブラリーを本発明の方法を用いてスクリーニングし、ANLN/RhoAの会合を阻害するライブラリーの試験化合物を同定することができる。
【0087】
コンビナトリアルケミカル合成
試験化合物のコンビナトリアルライブラリーは、ANLN/RhoA相互作用の公知の阻害剤に存在するコア構造の知識を含む合理的薬物設計プログラムの一部として産生しうる。このアプローチは、ライブラリーを妥当なサイズに維持することが可能であり、ハイスループットスクリーニングを促進する。または、単純な、特に短いポリマー分子ライブラリーは、ライブラリーを構成する分子ファミリーの全ての並べ換えを単純に合成することによって構築しうる。この後者のアプローチの一例は、すべてのペプチドが6アミノ酸長のライブラリーであろう。このようなペプチドライブラリーは、全ての6アミノ酸配列の並べ換えを含みうる。このタイプのライブラリーは直線的コンビナトリアルケミカルライブラリーと呼ばれる。
【0088】
コンビナトリアルケミカルライブラリーの調製は当業者に周知であり、化学的または生物学的合成によって生成されうる。コンビナトリアルケミカルライブラリーには、ペプチドライブラリー(例えば、米国特許第5,010,175号, Furka, Int. J. Pept. Prot. Res. 37:487-493 (1991) および Houghten et al., Nature 354:84-86 (1991) を参照されたい)が挙げられるが、これに限定されない。化学的多様性ライブラリーを生成するための他の化学も用いることができる。このような化学としては以下のものが挙げられるが、これらに限定されない:ペプチド(例えばPCT公報WO91/19735)、コードされるペプチド(例えばPCT公報WO93/20242)、ランダムバイオオリゴマー(例えばPCT公報WO92/00091)、ベンゾジアゼピン(例えば米国特許第5,288,514号)、ヒダントイン、ベンゾジアゼピン、およびジペプチドなどのディバーソマー(diversomer)(DeWitt et al., Proc. Natl. Acad. Sci. USA 90:6909-6913 (1993))、ビニル性(vinylogous)ポリペプチド(Hagihara et al., J. Amer. Chem. Soc. 114:6568 (1992))、グルコース骨格を有する非ペプチド性ペプチド模倣体(Hirschmann et al., J. Amer. Chem. Soc. 114:9217-9218 (1992))、低分子化合物ライブラリーの類似の有機合成(Chen et al., J. Amer. Chem. Soc. 116:2661 (1994))、オリゴカルバメート(Cho et al., Science 261:1303 (1993))、および/またはペプチジルホスホネート(Campbell et al., J. Org. Chem. 59:658 (1994))、核酸ライブラリー(Ausubel、およびSambrook(全て上記)を参照されたい)、ペプチド核酸ライブラリー(例えば米国特許第5,539,083号を参照されたい)、抗体ライブラリー(例えばVaughan et al., Nature Biotechnology, 14(3):309-314 (1996) およびPCT/US96/10287を参照されたい)、炭水化物ライブラリー(例えばLiang et al., Science, 274:1520-1522 (1996) および米国特許第5,593,853号を参照されたい)、有機低分子ライブラリー(例えば、ベンゾジアゼピン、Baum C&EN, Jan 18, page 33 (1993); イソプレノイド、米国特許第5,569,588号; チアゾリジノン(thiazolidinone)およびメタチアザノン(metathiazanone)、米国特許第5,549,974号; ピロリジン、米国特許第5,525,735号および第5,519,134号; モルホリノ化合物、米国特許第5,506,337号; ベンゾジアゼピン、米国特許第5,288,514号などを参照されたい)。
【0089】
ファージディスプレイ
別のアプローチは、組換えバクテリオファージを用いてライブラリーを産生する。この「ファージ法」(Scott and Smith, Science 249:386-390, 1990; Cwirla, et al, Proc. Natl. Acad. Sci., 87:6378-6382, 1990; Devlin et al., Science, 249:404-406, 1990)を用いて、非常に大きなライブラリーを構築することができる(例えば106〜108化学単位)。第二のアプローチは主に化学的方法を用いるものであり、Geysen法(Geysen et al., Molecular Immunology 23:709-715, 1986; Geysen et al. J. Immunologic Method 102:259-274, 1987)およびFodorらの方法(Science 251:767-773, 1991)がその例である。Furka et al. (14th International Congress of Biochemistry, Volume #5, Abstract FR:013, 1988; Furka, Int. J. Peptide Protein Res. 37:487-493, 1991)、(米国特許第4,631,211号) 、および (米国特許第5,010,175号)が、アゴニストまたはアンタゴニストとして試験されうるペプチドの混合物を産生するための方法を記載している。
【0090】
コンビナトリアルライブラリー調製用の装置が市販されている(例えば、357 MPS, 390 MPS, Advanced ChemTech, Louisville KY, Symphony, Rainin, Woburn, MA, 433A Applied Biosystems, Foster City, CA, 9050 Plus, Millipore, Bedford, MAを参照されたい)。また、多数のコンビナトリアルライブラリー自体が市販されている(例えばComGenex, Princeton, N.J., Tripos, Inc., St. Louis, MO, 3D Pharmaceuticals, Exton, PA, Martek Biosciences, Columbia, MDなどを参照されたい)。
【0091】
試験化合物ライブラリーのスクリーニング
本発明のスクリーニング方法は、ANLN/RhoAの会合またはANLNを介する細胞運動性を妨害する確率が高い試験化合物の効率的で迅速な同定を提供する。一般に、ANLN/RhoAの会合またはANLNを介する細胞運動性を妨害する試験化合物の能力を決定する方法はいずれも本発明と共に使用するのに適する。例えば、ELISA形式の競合および非競合阻害アッセイを利用しうる。対照実験を行ってシステムの最大結合能を決定すべきである(例えば、下記の例においては、結合させたANLNをRhoAと接触させ、ANLNに結合するRhoAの量を決定する)。
【0092】
競合アッセイ形式
本発明の試験化合物をスクリーニングするために競合アッセイを用いうる。例として、競合ELISA形式は、固体支持体に結合されたANLN(またはRhoA)を含みうる。この結合されたANLN(またはRhoA)は、RhoA(またはANLN)および試験化合物とインキュベートされる。試験化合物および/またはRhoA(もしくはANLN)をANLN(またはRhoA)に結合させるのに十分な時間の後、基質を洗浄して非結合物質を除く。そしてANLN(またはRhoA)に結合したRhoA(またはANLN)の量を決定する。これは、当技術分野で公知の様々な方法のいずれか、例えば、検出可能な標識でタグ付されたRhoA(またはANLN)種を用いて、あるいは洗浄した基質を標識化抗RhoA(またはANLN)抗体と接触させて、遂行しうる。ANLN(またはRhoA)に結合したRhoA(またはANLN)の量は、RhoA/ANLNの会合を妨害する試験化合物の能力に反比例するであろう。抗体を含むがそれに限定されないタンパク質の標識は、Harlow & Lane, Antibodies, A Laboratory Manual (1988) に記載されている。
【0093】
あるバリエーションでは、ANLN(またはRhoA)は親和性タグで標識される。標識されたANLN(またはRhoA)を次に、試験化合物およびRhoA(またはANLN)とインキュベートし、免疫沈降する。そして免疫沈降物を抗RhoA(またはANLN)抗体を用いたウェスタンブロッティングに供する。前記の競合アッセイ形式のように、ANLN(またはRhoA)と会合することが見出されたRhoA(またはANLN)の量は、ANLN/RhoAの会合を妨害する試験化合物の能力と反比例する。
【0094】
非競合アッセイ形式
非競合結合アッセイも、本明細書に記載されるもののような競合アッセイを用いたスクリーニングに容易に適用できない形式で構築された試験化合物ライブラリーの初期スクリーニングとして有用でありうる。このようなライブラリーの一例は、ファージディスプレイライブラリーである(例えば、Barrett, et al. (1992) Anal. Biochem 204, 357-364を参照されたい)。
【0095】
ファージライブラリーは、数多くの異なる組換えペプチドの作業量(working quantity)を素速く産生しうる点において有用である。ファージライブラリーはそれ自体は本発明の競合アッセイに向いてはいないが、非競合形式で効率的にスクリーニングされ、どの組換えペプチド試験化合物がANLNまたはRhoAに結合するかを決定することができる。次いで、結合すると同定された試験化合物を産生し、競合アッセイ形式を用いてスクリーニングすることができる。ファージおよび細胞ディスプレイライブラリーの産生およびスクリーニングは当技術分野において周知であり、例えば、Ladner et al., WO 88/06630; Fuchs et al. (1991) Biotechnology 9:1369-1372; Goward et al. (1993) TIBS 18:136-140; Charbit et al. (1986) EMBO J 5, 3029-3037. Cull et al. (1992) PNAS USA 89:1865-1869; Cwirla, et al. (1990) Proc. Natl. Acad. Sci. U.S.A. 87, 6378-6382に考察されている。
【0096】
例示的な非競合アッセイは、成分の1つ(ANLNまたはRhoA)の添加なしで、競合アッセイに関して記載したものと類似の手順に従う。しかしながら、非競合形式はANLNまたはRhoAに対する試験化合物の結合を決定するので、ANLNおよびRhoAの両方に対する試験化合物の能力を各候補について決定する必要がある。したがって、例として、固定化されたANLNへの試験化合物の結合は、結合していない試験化合物を洗い流し、結合した試験化合物を支持体から溶出させた後に、例えば、質量分析、タンパク質測定(BradfordもしくはLowryアッセイ法、または280nmの吸光度測定)によって溶出物を解析することによって、決定しうる。または、溶出工程を省いて、支持体表面での有機層の分光学的特性における変化をモニターすることによって試験化合物の結合を決定してもよい。表面の分光学的特性をモニターするための方法としては以下のものが含まれるが、これらに限定されない:吸光度、反射率、透過率、複屈折、屈折率、回折、表面プラズモン共鳴、偏光解析法、共鳴ミラー法、格子共役導波管(grating coupled waveguide)技術、および多極共鳴分光法(これらは全て当業者に公知である)。溶出工程の必要を省くために、標識された試験化合物もアッセイに用いうる。この場合、非結合物質を洗い流した後に支持体に会合している標識の量が試験化合物の結合に直接比例する。
【0097】
多くの周知のロボットシステムが液相化学用に開発されてきた。これらシステムとしては、Takeda Chemical Industries, LTD.(Osaka, Japan)によって開発された自動合成器のような自動ワークステーションが挙げられ、多くのロボットシステムがロボットアーム(Zymate II, Zymark Corporation, Hopkinton, Mass.; Orca, Hewlett Packard, Palo Alto, Calif.)を利用しており、化学者によって行われるマニュアル合成操作を模倣する。上記装置はいずれも、本発明と共に使用するのに適している。本明細書において述べられているように操作可能になるようなこれら装置に対する修飾(もしあれば)の性質および実施は、関連技術分野の当業者には明らかであろう。加えて、多数のコンビナトリアルライブラリーはそれ自体が市販されている(例えば、ComGenex, Princeton, N.J., Asinex, Moscow, Ru, Tripos, Inc., St. Louis, MO, ChemStar, Ltd, Moscow, RU, 3D Pharmaceuticals, Exton, PA, Martek Biosciences, Columbia, MDなどを参照されたい)。
【0098】
変換酵素のスクリーニング
変換酵素である試験化合物は、アッセイする変換酵素に特異的なコファクターおよび補助基質を用いて非競合形式でアッセイしうる。このようなコファクターおよび補助基質は、調べる変換酵素のタイプが与えられれば、当業者に公知である。
【0099】
変換酵素の1つの例示的なスクリーニング手順は、まず、変換酵素に特徴的なタンパク質の共有結合性修飾を行うのに必要なコファクターおよび補助基質の存在下、好ましくは生理的条件下で、ANLNおよび/またはRhoAを変換酵素と接触させることを含む。修飾されたタンパク質を次に、結合パートナーに結合する能力(すなわちANLNのRhoAに対する結合)について試験する。次に、修飾されたタンパク質の結合パートナーに対する結合を、未修飾の対照対の結合と比較し、上記のKaにおいて要求される変化が達成されたかどうかを決定する。
【0100】
アッセイを行う上でタンパク質の検出を促進するために、当業者に周知の技術を用いて、1つまたは複数のタンパク質を上記のような検出可能な標識で標識してもよい。
【0101】
スクリーニング方法
上記のスクリーニングの態様は、さらなる調査に適する試験化合物のハイスループットな決定に適する。本発明のスクリーニング方法は、以下のものの1つまたは複数の検出を含みうる:
(a)活性化RhoAの濃度;
(b)RhoAと、RHOエフェクター(例えばROCK(Maekawa, M. et.al. Science vol 285. 895-898(1999))、Rhophilin-2(Jeremy W. Peck et.al. Journal of Biological Chemistry 277, 43924-32(2002))、またはそのRhoA結合領域との間の相互作用;
(c)活性化RhoAを介する下流遺伝子発現(例えばROCK(Stephan A.K. Harvey et.al. Investigative Ophthalmology & Visual Science 45, 2168-76(2004))、c-JUN(Maria Julia Marinissen et.al. Molecular Cell 14, 29-41(2004))、または下流遺伝子産物活性(例えばERK(Laboureau J, et.al. Experimental Dermatology 13, 70-7(2004))、GATA-4(Yanazume T, et.al. The Journal of Biological Chemistry 277, 8618-25(2002))を含む任意のシグナル複合体の活性化;
(d)DNA合成および細胞周期移行の促進;
(e)細胞遊走または任意の他の発癌表現型細胞接着、細胞浸潤(Shibata, T et.al. American Journal of Pathology 164, 2269-78(2004), Selma Cetin et.al. The Journal of Biological Chemistry 279, 24592-600(2004));
(f)アクチンストレスファイバー形成およびF-アクチン産生;ならびに
(g)細胞接着、遊走、および浸潤に重要な任意の分子との相互作用(Spred(Miyoshi, K. et.al. Oncogene 23, 5567-5576(2004))、Smurf1(Hong-Rui Wang et.al. Science 302, 1775-9(2003)))。
【0102】
または、調査中の試験化合物を増殖中の細胞に加えて、試験化合物を与えていない対照集団の増殖を基準として処置細胞の増殖をモニターしてもよい。試験化合物のスクリーニングに適した細胞株は、本明細書に提供される教示から当業者には明らかであろう。
【0103】
インビボ試験に関しては、認められた動物モデルに試験化合物を投与すればよい。
【0104】
IV. 同定された試験化合物からの薬剤の製剤化
よって、本発明は、癌、特に非小細胞肺癌などの肺癌、ならびにANLNおよび/もしくはRhoAの発現レベルおよび活性の上昇を示す細胞、ならびに/またはANLNの核局在によって特徴付けられる他の癌の予防または処置に有用な薬剤および方法を含む。これら薬剤および方法は、ANLN/RhoA相互作用を阻害するとして同定された本発明の少なくとも1つの試験化合物を、異常な細胞増殖の減衰または停止を達成するのに有効な量で含む。より具体的には、治療的に有効な量とは、処置対象の発症を防ぐのに、あるいはその既存の症状を緩和するのに、有効な量を意味する。
【0105】
本発明の方法で処置される個体は、例えばマーカータンパク質ANLNの発現の上昇またはANLNの核局在を示すことによって特徴付けられる非小細胞肺癌を含む癌に罹患した任意の個体でありうる。このような個体は、例えば、ヒト、イヌ、ネコ、ウマ、ウシ、もしくはヤギを含む哺乳類などの脊椎動物、あるいは任意の他の動物、特に商業的に重要な動物または家畜化された動物でありうる。本発明の目的のためには、マーカータンパク質の発現の上昇とは、一方または両方のマーカータンパク質について、マーカータンパク質の正常細胞での平均濃度よりも、少なくとも10%、好ましくは15%、20%、25%、30%、35%、40%、45%、50%、55%、またはそれよりも多い、平均細胞マーカータンパク質濃度を指す。
【0106】
治療用量範囲の決定
本発明の薬剤のための有効な用量範囲の決定は、特に本明細書に提供される詳述された開示を鑑みれば、十分に当業者の能力の範囲内である。試験化合物の治療的に有効な用量は、まず細胞培養アッセイおよび/または動物モデルから推定できる。例えば、動物モデルにおいて用量を定め、細胞培養で決定されるIC50(50%の細胞が所望の効果を示す用量)を含む循環濃度範囲を達成することができる。また、試験化合物の毒性および治療有効性は、例えば、LD50(集団の50%に致死的な用量)およびED50(集団の50%に治療有効量)を決定するための、細胞培養または実験動物における標準的な薬学的手順によって決定することができる。毒性効果と治療効果との間の用量比が、治療指数(すなわちLD50とED50の比)である。高い治療指数を示す化合物を用いることができる。これら細胞培養アッセイおよび動物試験から得られたデータを、ヒトに用いる用量範囲を定める際に用いうる。このような化合物の用量は、毒性がほとんどないかまたは全くないED50を含む循環濃度の範囲内に収まるであろう。用量は、使用される用量形態および利用される投与経路に従い、この範囲内で変わりうる。厳密な処方、投与経路、および用量は、患者の状態を考慮して個別の医師によって選択されうる。例えばFingl et al., (1975), in "The Pharmacological Basis of Therapeutics", Ch. 1 p1を参照されたい。用量および間隔は、所望の効果を維持するのに十分な活性試験化合物の血漿レベルを提供するように個別に調整しうる。
【0107】
薬学的に許容される添加物
哺乳類(例えばヒト)に投与する薬剤は、薬学的に許容される添加物、または担体を含みうる。適した添加物およびそれらの製剤は、Remington's Pharmaceutical Sciences, 16th ed., (1980), Mack Publishing Co., edited by Oslo et al.に記載されている。水溶性の調製物には、製剤を等張にするために、適切な量の薬学的に許容される塩が製剤中に典型的に用いられる。薬学的に許容される等張添加物の例としては、生理的食塩水、リンゲル液、ハンクス液、およびデキストロース溶液などの液体が含まれる。等張添加物は、注射用製剤に特に重要である。
【0108】
経粘膜投与には、透過させる障壁に適切な浸透剤を製剤に用いる。このような浸透剤は当技術分野において一般的に知られている。
【0109】
添加物は製剤の適正なpHを維持するために用いうる。最適な有効期間のためには、試験化合物を含む溶液のpHは好ましくは約5〜約8、より好ましくは約7〜約7.5の範囲にわたる。製剤はまた、凍結乾燥粉末、または、マトリックスが成形品、例えば、フィルム、リポソーム、もしくは微粒子の形態にある、固体疎水性ポリマーの半透性マトリックスなどの徐放性調製物を含む本発明に適した他の選択的な添加物を含みうる。例えば、投与経路、投与される試験化合物の濃度、または、処置がタンパク質、試験化合物をコードする核酸、もしくは試験化合物を分泌することができる細胞を有効成分として含む薬剤を用いるかどうかによって、特定の添加物がより好ましい場合があることが当業者には明らかであろう。
【0110】
本発明の薬学的組成物は、それ自体公知の様式で、例えば、通常の混合、溶解、顆粒化、糖衣丸形成、研和、乳化、カプセル化、閉じ込め(entrapping)、または凍結乾燥プロセスによって、製造しうる。適正な製剤は、選択された投与経路に依存する。
【0111】
経口投与には、担体によって、本発明の化合物の処置患者による経口摂取のために、錠剤、丸剤、糖衣丸、カプセル、液体、ゲル、シロップ、スラリー、懸濁液などとして製剤化することが可能である。経口用の薬学的調製物は、試験化合物を固体の拡散性添加物と共に製剤化し、得られた混合物を任意ですりつぶして、所望であれば適した補助剤を加えた後に顆粒の混合物を加工して錠剤または糖衣丸コアを得ることによって得ることができる。適した添加物は特に、ラクトース、スクロース、マンニトール、またはソルビトールを含む糖類などの増量剤;例えば、トウモロコシデンプン、コムギデンプン、イネデンプン、ジャガイモデンプン、ゼラチン、トラガカントゴム、メチルセルロース、ヒドロキシプロピルメチルセルロース、カルボキシメチルセルロースナトリウム、および/またはポリビニルピロリドン(PVP)などのセルロース調製物である。所望であれば、架橋ポリビニルピロリドン、寒天、またはアルギン酸またはアルギン酸ナトリウムなどのそれらの塩などの崩壊剤を加えてもよい。
【0112】
本発明の化合物の多くは薬学的に適合性の対イオンとの塩として提供してもよい。薬学的に適合性の塩は、用途によって、塩酸、硫酸、酢酸、乳酸、酒石酸、リンゴ酸、コハク酸などを含むがこれらに限定されない多くの酸と共に形成しうる。塩は、水性または対応する遊離塩基形態である他のプロトン性溶媒に、より可溶性である傾向がある。
【0113】
許容可能な添加物に加えて、本発明の製剤は、同定された試験化合物以外の治療剤を含みうる。例えば、製剤は、抗炎症剤、鎮痛剤、化学療法剤、粘液溶解薬(例えばn-アセチル-システイン)などを含みうる。薬剤自体に他の治療剤を含むことに加え、本発明の薬剤はまた、1つまたは複数の他の薬理的物質に連続的にまたは同時に投与しうる。薬剤および薬理的物質の量は、例えば、どのタイプの薬理的物質を用いるか、また、処置される疾患、ならびにスケジュールおよび投与経路に依存する。
【0114】
本発明の薬剤の投与に続いて、熟練した施術者に周知の様々な方法で哺乳類の生理的状態をモニターすることができる。
【0115】
遺伝子療法
ANLN/RhoAの会合の阻害物質(disruptor)として同定されたタンパク質およびペプチド試験化合物は、癌、例えば非小細胞肺癌に苦しむ患者に遺伝子療法を用いて治療的に送達しうる。遺伝子療法技術に適用できる例示的試験化合物としては、変換酵素や、立体またはアロステリック阻害によってANLN/RhoAの会合を直接変化させるペプチドが含まれるが、これらに限定されない。一部の局面において、遺伝子療法の態様は、本発明の適した同定された試験化合物をコードする核酸配列を含む。一部の態様において、核酸配列は、標的細胞における試験化合物の発現に必要なそれらの調節エレメントを含む。核酸は、標的細胞のゲノムに安定に挿入されるように準備されうる(例えば、相同組換えカセットベクターの記載について、Thomas, K. R. and Capecchi, M. R. (1987) Cell 51:503を参照されたい)。
【0116】
患者への核酸の送達は直接的でもよいし(この場合、患者は核酸または核酸を含むベクターに直接さらされる)、間接的でもよい(この場合、細胞がまずインビトロで核酸によって形質転換されてから患者に移植される)。これら2つのアプローチはそれぞれインビボまたはエクスビボ遺伝子療法として知られている。
【0117】
遺伝子療法の方法の全般的概説については、Goldspiel et al., 1993 Clinical Pharmacy 12:488-505; Wu and Wu, 1991 Biotherapy 3:87-95; Tolstoshev, 1993 Ann. Rev. Pharmacol. Toxicol. 33:573-596; Mulligan, 1993 Science 260:926-932; および Morgan and Anderson, 1993 Ann. Rev. Biochem. 62:191-217; 1993 TIBTECH 11(5):155-215を参照されたい。用いうる組換えDNA技術の分野で一般的に知られる方法は、Ausubel et al. (eds.), 1993, Current Protocols in Molecular Biology, John Wiley & Sons, NY; および Kriegler, 1990, Gene Transfer and Expression, A Laboratory Manual, Stockton Press, NYに記載されている。
【0118】
V. スクリーニング、予後、および処置キット
本発明はまた、癌、特に非小細胞肺癌(NSCLC)などの肺癌の処置または予防に有用な化合物をスクリーニングするための材料を含む製品またはキットも提供する。このような製品は、本明細書に記載される材料の、1つまたは複数のラベルされた容器を、使用説明と共に含みうる。適した容器には、例えば、ボトル、バイアル、および試験管が含まれる。容器は、ガラスまたはプラスチックなどの様々な材料から形成されうる。
【0119】
一つの態様において、スクリーニングキットは以下のものを含む:(a)ANLNポリペプチドのRhoA結合ドメインを含む第一のポリペプチド;(b)RhoAポリペプチドのANLN結合ドメインを含む第二のポリペプチド、および(c)第一のポリペプチドと第二のポリペプチドとの間の相互作用を検出する手段(例えば試薬)。
【0120】
一部の態様において、第一のポリペプチド、すなわちRhoA結合ドメインを含むポリペプチドは、ANLNポリペプチドを含む。同様に、他の態様において、第二のポリペプチド、すなわちANLN結合ドメインを含むポリペプチドは、RhoAポリペプチドを含む。
【0121】
一部の態様において、RhoA結合ドメインを含むポリペプチドは生細胞中で発現される。
【0122】
一部の態様において、2つのポリペプチド間の相互作用を検出する手段(例えば試薬)は、以下のものを検出することができる:
(1)活性化RhoAの濃度;
(2)RhoAとRHOエフェクターまたはそのRhoA結合領域との間の相互作用;
(3)活性化RhoAを介する下流遺伝子を含む任意のシグナル複合体の活性化;
(4)DNA合成および細胞周期移行の促進;
(5)細胞遊走または任意の他の発癌表現型;
(6)アクチンストレスファイバー形成およびF-アクチン産生;ならびに
(7)細胞接着、遊走、および浸潤に重要な任意の分子との相互作用。
【0123】
別の態様において、スクリーニングキットは以下のものを含みうる:(a)ANLNポリペプチドまたはその機能的同等物を発現する細胞;および(b)細胞の運動性を検出する手段(例えば試薬)。
【0124】
本発明はまた、癌対象、例えば非小細胞肺癌(NSCLC)などの肺癌に罹患した対象の予後を予測するためのキットを提供する。一部の態様において、このような予後キットは以下のものを含む:(a)ANLNタンパク質を認識する抗体、および(b)核における検出用の薬剤。一部の態様において、核の検出用の薬剤はヘマトキシリン-エオシン染色染料である。使用説明は、NSCLCの予後を予測しようとする対象から収集した試料の核に局在するANLNが検出されることが、不良な予後の指標であることを示すであろう。
【0125】
本発明はさらに、本明細書に記載の病理学的状態を処置するのに有用な物質を含む製品およびキットを提供する。このような製品は、本明細書に記載されるような薬剤の容器をラベルと共に含みうる。上記のように、適した容器には、例えば、ボトル、バイアル、および試験管が含まれる。容器は、ガラスまたはプラスチックなどの様々な材料から形成されうる。本発明の文脈において、容器とは、細胞増殖性疾患、例えば非小細胞肺癌を処置するために有効な活性物質を有する組成物を保持するものである。組成物中の活性物質は、インビボでANLN/RhoAの会合を阻害しうる同定された試験化合物(例えば、抗体、低分子など)でるかあもしれない。容器上のラベルは、異常な細胞増殖によって特徴付けられる1つまたは複数の状態を処置するために組成物が用いられることが示されうる。ラベルはまた、本明細書に記載するもののような投与およびモニタリング技術に関する指示を示しうる。
【0126】
上記の容器に加え、本発明の処置キットは、任意で薬学的に許容される希釈剤を収容する第二の容器を含んでもよい。さらに、使用説明書と共に、他のバッファー、希釈剤、フィルター、針、シリンジ、およびパッケージ挿入物を含む、商業的およびユーザーの観点から望ましい他の材料を含みうる。
【0127】
所望であれば、組成物は、有効成分を含む1つまたは複数の単位用量形態を含みうるパックまたはディスペンサー装置で提供されうる。パックは、例えば、ブリスターパック(blister pack)などの金属またはプラスチックのフォイルを含みうる。パックまたはディスペンサー装置は投与のための説明書を伴いうる。また、適合性の薬学的担体中に製剤化された本発明の化合物を含む組成物は、指示された状態の処置のために調製し、適切な容器に入れてラベルされうる。
【0128】
VI. 癌の診断および予後
本方法は、数々のタイプの癌の診断、予後、分類、および処置に用いることができる。原発性、転移性、および再発癌などの任意の進行段階の癌を検出することができる。診断、予後、分類のための例示的な癌には、例えば、非小細胞肺癌(NSCLC)などの肺癌が含まれる。
【0129】
本発明は、癌を有する哺乳類の予後を判断するための方法を提供する。このような方法は、予後が不良な非小細胞肺癌患者においてANLNの核局在が最も高頻度に起こるという発見に基づく。従って、ある個体由来のサンプル(例えば生検)が、細胞質よりはむしろ核にANLNが局在する細胞を含むかどうかを測定することによって、その個体の予後を予測することが可能である。
【0130】
本明細書に記載のように、ANLNの局在は完全またはほぼ完全に細胞質中であることもあり、また細胞の核および細胞質の両方に局在することもある。このように、「核局在」は、ANLNが細胞質および核の両方において検出できる状況を含む。
【0131】
核局在は、当技術分野において公知の任意の方法によって測定しうる。一部の態様において、免疫組織化学解析を用いて、痰検査または任意の生検などの、どの病院でも利用可能な手術または最小限の浸潤的技術によって得られる組織または細胞学的サンプルにおいて細胞中のANLNを検出する。不良な予後の判断は、さらなる処置を決定するため、例えば、生存の質を落とすようなさらなる処置を中止したり、以前に用いられたものとは異なる方法で癌を処置したり、より積極的に癌を処置したりするために用いられうる。すなわち、ANLNによる予後の予測は最終的に、組織サンプリングのルーチン的手順のみを用いて、従来の疾患の臨床段階の情報がなくてさえも、各癌患者にとって最も適切な処置を臨床医が前もって選ぶことを可能にするはずである。
【0132】
さらに、本方法を用いて処置経過の有効性を評価しうる。例えば、ANLNが核に局在することが見出された生物学的サンプルが得られた癌を有する哺乳類において、時間を追ってANLNの核局在をモニターすることによって抗癌処置の有効性を評価することができる。例えば、処置前または処置の初期に哺乳類から採取されたサンプルにおけるレベルと比べて、処置後の哺乳類から採取された生物学的サンプルにおけるANLN局在が減少していれば、処置が有効であることが示される。
【0133】
上記のように、本発明はまた、細胞のANLNの位置を検出するためのキットも提供する。このようなキットの成分の例には、免疫組織化学試薬、例えば、ANLNに結合する抗体、および細胞内で抗体を検出するための検出可能な標識が含まれる。
【0134】
以下に、実施例を参照して本発明をさらに詳細に説明する。しかしながら、以下の材料、方法、および例は、本発明の局面を例示するのみであって、本発明の範囲を限定する意図は全くない。このように、本明細書に記載のものと類似または同等の方法および材料を本発明の実施または試験に用いることができる。
【0135】
実施例
上記に提供した開示から理解できるように、本発明は広範な応用を有する。よって、以下の実施例は例示目的のために提供するものであって、いずれにしても本発明を限定するものと見なす意図はない。当業者は、本質的に類似の結果を得るために変化または修飾しうる様々な重要でないパラメータを容易に認識するであろう。
【0136】
実施例1 - 材料および方法
(a)細胞株および臨床サンプル
本明細書の実施例において用いられた23個のヒト肺癌細胞株は以下の通りである:肺腺癌(ADC);A549、LC319、PC-3、PC-9、PC-14、A427、NCI-H1373、細気管支肺胞癌(BAC);NCI-H1666、NCI-H1781、肺扁平上皮細胞癌(SCC);RERF-LC-AI、SK-MES-1、EBC-1、LU61、NCI-H520、NCI-H1703、NCI-H2170、肺腺扁平上皮癌(ASC);NCI-H226、NCI-H647、肺大細胞癌(LCC);LX1、小細胞肺癌(SCLC);DMS114、DMS273、SBC-3、SBC-5。細胞は全て、10%ウシ胎児血清(FCS)を加えた適切な培地中で、単層で生育させ、37℃、5%CO2の加湿空気雰囲気下で維持した。ヒト小気道上皮細胞(SAEC)を正常対照として用い、Cambrex Bio Science Inc.(Walkersville, MD)から購入した最適培地(SAGM)で生育させた。37人の患者からのインフォームドコンセントが得られた原発性NSCLCサンプル(このうち22個がADC、14個がSCC、1個がASCに分類された)を前もって得た(Kikuchi, T., et al., Oncogene. 22:2192-205 (2003))。
【0137】
手術を受けた患者からインフォームドコンセントを得て、組織マイクロアレイでの免疫染色に用いる合計285個のホルマリンで固定した原発性NSCLC(I〜IIIA期)および隣接正常肺組織サンプルを得た。
【0138】
(b)候補遺伝子の選択および半定量的RT-PCRによる解析
遺伝子発現プロファイル解析に基づき、50%を超える腫瘍中で正常肺の5倍以上の発現レベルを示す遺伝子を検査するために選択した。ANLN転写物がそれら遺伝子のリストに含まれ、続いて半定量的RT-PCRによって過剰発現していることが確認された。βアクチン(ACTB)の発現レベルを定量対照として用いて、臨床肺癌細胞のmRNAから調製された各一本鎖cDNAの適切な希釈物を調製した。増幅用のプライマーセットは以下の通りである:
ACTBには、ACTB-F (5’-GAGGTGATAGCATTGCTTTCG-3’(SEQ ID No.5))および
ACTB-R (5’-CAAGTCAGTGTACAGGTAAGC-3’(SEQ ID No.6))、ならびに
ANLNには、ANLN-F1 (5’-GCTGCGTAGCTTACAGACTTAGC-3’(SEQ ID No.7))および
ANLN-R1 (5’-AAGGCGTTTAAAGGTG ATAGGTG-3’(SEQ ID No.8))。
【0139】
反応は全て、GeneAmp PCRシステム9700(Applied Biosystems, Foster City, CA)での、94℃で2分間の初期変性に続き、95℃で30秒間、58〜62℃で30秒間、および72℃で45秒間の21サイクル(ACTBについて)または30サイクル(ANLNについて)を含んだ。
【0140】
(c)ノーザンブロット解析
ヒト複数組織ブロット(Human multiple-tissue blot)(BD Biosciences Clontech, Palo Alto, CA)をANLNの32P標識PCR産物とハイブリダイズさせた。ANLNの全長cDNAを以下のプライマーを用いてRT-PCRで調製した:
ANLN-F2 (5’- CCCAAGCTTGGGGCCACCATGGATCCGTTTACGGAGAAAC -3’(SEQ ID No.9))および
ANLN-R2 (5’- TGCTCTAGAGCAAGGCTTTCCAATAGGTTTGTAG -3’(SEQ ID No.10))。
プレハイブリダイゼーション、ハイブリダイゼーション、および洗浄は供給業者の推奨に従って行った。増感スクリーンを用いて室温で96時間、ブロットをオートラジオグラフィーにかけた。
【0141】
(d)ウェスタンブロット解析
コドン位置428〜718のGST融合ヒトANLNタンパク質でウサギを免疫化して、ヒトANLNタンパク質に特異的なウサギ抗体を作成した。抗体は標準的な手順で精製した。ECL Western Blotting System(Amersham Biosciences, Uppsala, Sweden)を用いた。細胞は、プラスミドトランスフェクション後24時間、無血清培地中で維持し、適切な量の溶解バッファー(150mM NaCl、50mM Tris-HCl, pH8.0、1%Nonidet P-40、0.1%SDS、0.5%デオキシコール酸Na、に加えてプロテアーゼ阻害剤)に溶解させた。SDS-PAGEを行い、PAGEで分離されたタンパク質をニトロセルロース膜(Amersham Biosciences)上にエレクトロブロットし、抗体と共にインキュベートした。ヒツジ抗マウスIgG-HRP抗体(Amersham Biosciences)およびヤギ抗ウサギIgG-HRP抗体(Amersham Biosciences)をこれら実験の二次抗体として供した。
【0142】
(e)免疫細胞化学解析
培養細胞をPBS(-)で2回洗浄し、4%パラホルムアルデヒド溶液中、60分間室温で固定し、0.1%Triton X-100を含むPBS(-)で1.5分間透過化した。一次抗体反応の前に、細胞をブロッキング溶液(PBS(-)中3%BSA)で60分間覆って非特異的な抗体結合をブロックした。そして、細胞をヒトANLNに対する抗体と共にインキュベートした。内因性ANLNを明らかにするために、FITC(Cappel, Durham, NC, USA)またはローダミン(Cappel)に結合したヤギ抗ウサギ二次抗体で染色し、レーザー共焦点顕微鏡(TSC SP2 AOBS: Leica Microsystems, Wetzlar, Germany)で観察した。アクチンフィラメントを可視化するため、Alexa59結合ファロイジン(Molecular Probes, Eugene, OR, USA)も、二次抗体とのインキュベーション後に加えた。
【0143】
(f)アンチセンスS-オリゴヌクレオチド
6ウェルディッシュにプレーティングした2 X 105個のNSCLC細胞株A549を、リポフェクタミン試薬(40nM)(Invitrogen, Carlsbad, CA, USA)を用いて、ANLN遺伝子に対応する合成S-オリゴヌクレオチド(0.2μM)でトランスフェクトし、10%FCSを含む培地中で2日間維持した。細胞生存率を、それぞれ三回MTTアッセイによって評価した。S-オリゴヌクレオチドの配列は以下の通り:
アンチセンス1(AS1)5’-CTCCGTAAACGGATCCAT-3’(SEQ ID No.11);
リバース1(R1)5’-TACCTAGGCAAATGCCTC-3’(SEQ ID No.12)、
アンチセンス2(AS2)5’-CGGATCCATCGCCCCAGG-3’(SEQ ID No.13);
リバース2(R2)5’-GGACCCCGCTACCTAGGC-3’(SEQ ID No.14)。
MTTアッセイは、別に記載されているように(Suzuki, C., et al., Cancer Res. 63:7038-41 (2003))行った。
【0144】
(g)RNA干渉アッセイ
別に報告されているように(Shimokawa T, et al., Cancer Res. 2003;63:6116-20.)、ベクターベースのRNA干渉(RNAi)システムpsiH1BX3.0は哺乳動物細胞におけるsiRNAの合成を誘導するために確立された。30μlのリポフェクタミン 2000(Invitrogen)を用いて、10μgのsiRNA発現ベクターをNSCLC細胞株LC319およびA549にトランスフェクトした。このアッセイでは、90%を超えるトランスフェクト細胞がこの合成siRNAを発現し、内因性のANLN発現が有効に抑制された。トランスフェクト細胞を、適切な濃度のジェネテシン(G418)存在下で5日間培養した。細胞数および生存率を三回Giemsa染色およびMTTアッセイによって測定した。RNAiの合成オリゴヌクレオチドの標的配列は以下の通り:
対照(LUC(ルシフェラーゼ:フォチナス ピラリス(Photinus pyralis)ルシフェラーゼ遺伝子)5’-CGTACGCGGAATACTTCGA-3’(SEQ ID No.15);
SCR(スクランブル:5Sおよび16S rRNAをコードするミドリムシ葉緑体遺伝子)5’-GCGCGCTTTGTAGGATTCG-3’(SEQ ID No.16));
siRNA-ANLN-1(si-1)5’-CCAGTTGAGTCGACATCTG-3’(SEQ ID No.17);
siRNA-ANLN-2(si-2)5’-GCAGCAGATACCATCAGTG-3’(SEQ ID No.18)。
本発明者らのRNAiシステムを実証するため、個々の対照siRNAを半定量的RT-PCRによって試験し、COS-7細胞に一過的にトランスフェクトした対応標的遺伝子の発現の低下を確認した。また、このアッセイに用いた細胞株において、対照ではなく機能的siRNAによるANLN発現のダウンレギュレーションが確認された。
【0145】
(h)フローサイトメトリー
細胞を5 X 105細胞/100mmディッシュの密度でプレーティングし、siRNA発現ベクターでトランスフェクトして、適切な濃度のジェネテシンの存在下で培養した。トランスフェクション5日後、細胞をトリプシン処理し、PBS中に収集して、70%冷エタノール中で30分間固定した。100μg/ml RNase(Sigma-Aldrich Co.)で処置した後、細胞をPBS中50μg/mlヨウ化プロピジウム(Sigma-Aldrich Co.)で染色した。フローサイトメトリーをBecton Dickinson FACScanで行い、ModFitソフトウェア(Verity Software House, Inc., Topsham, ME, USA)によって解析した。少なくとも20,000個の非ゲート化(ungated)細胞から決定した細胞をDNA含量について解析した。
【0146】
(i)BrdU取り込みアッセイ
ANLNおよびモック(mock)プラスミドを発現するように設計されたプラスミドでトランスフェクトした肺癌細胞(LC319およびA549細胞)を、無血清培地中で4時間培養した。次に培地を10%FCSと10μM BrdUを含むRPMI1640と交換した。これらの細胞を20時間インキュベートし、さらに固定して、取り込まれたBrdUを市販のキット(Cell Proliferation ELISA, BrdU; Roche Diagnostics, Basel, Switzerland)を用いて測定するために用いた。
【0147】
(j)マトリゲル浸潤アッセイ
ANLNまたはモックプラスミドを発現するように設計されたプラスミドでトランスフェクトしたNIH3T3およびCOS-7細胞を、10%FCSを含むDMEM中で融合性の段階まで生育させた。細胞をトリプシン処理によって回収し、続いて血清またはプロテイナーゼ阻害剤を加えないDMEM中で洗浄した。細胞をDMEM中で1 x 105/mlで懸濁した。細胞懸濁液を調製する前に、マトリゲル基質(Becton Dickinson Labware, Bedford, MA, USA)の乾燥層をDMEMで2時間、室温で再水和した。10%FCSを含むDMEM(0.75ml)を24ウェルマトリゲル浸潤チャンバーの各下部チャンバーに加え、0.5ml(5 x 104細胞)の細胞懸濁液を上部チャンバーの各インサートに加えた。インサートのプレートを22時間、37℃でインキュベートした。インキュベーション後チャンバーを処理し、供給業者(Becton Dickinson Labware)の指示通りに、マトリゲルで被覆されたインサートを通って浸潤している細胞を固定してGiemsaによって染色した。
【0148】
(k)傷害遊走アッセイ
ANLNまたはモックプラスミドを発現するように設計されたプラスミドでトランスフェクトされたNIH3T3細胞を無血清DMEM中に懸濁し、10μg/mlのフィブロネクチンで被覆された2ウェルチャンバー(Becton Dickinson Labware)の個々のウェルにプレーティングした。4時間インキュベーション後、各チャンバーの底からP200ピペットチップで接着細胞を線状に掻き取って傷をつけ、培地を10%FCSを含むDMEMで交換した。細胞を増殖させ、傷害区域に48時間遊走させた。そして、傷害区域内の細胞数を顕微鏡(DP50, OLYMPUS, Tokyo, Japan)を用いて数えた。
【0149】
(l)RHO活性化の検出
ANLNによるRHOの活性化を、EZ-Detect(商標)Rho Activation Kit(PIERCE, Rockford, IL, USA)を用いて検出した。簡単に述べると、ANLN発現プラスミドまたはモックプラスミドでトランスフェクトしたLC319細胞を24時間培養した。次に細胞を洗浄し、溶解バッファーで溶解した。16,000xgで遠心分離した後、溶解物を、活性化RHOをアフィニティー沈降させるためにGST-rhotekin(RTKN)-RBDおよび「SwellGel Immobilized Glutathione Disc」と混合した。活性化RHOを含むGSTプルダウン沈殿剤を洗浄し、サンプルバッファーで煮沸して、抗RHO(-A、-B、および-C)および抗ANLN抗体を用いるウェスタンブロット解析に供した。
【0150】
(m)免疫組織化学および組織マイクロアレイ
ホルマリンで固定した肺癌を用いた腫瘍組織マイクロアレイを、以前に公開されているように(Kononen, J., et al., Nat. Med. 4: 844-847 (1998); Chin, S. F., et al., Mol. Pathol., 56:275-79 (2003); Callagy, G., et al., Diagn. Mol. Pathol., 12:27-34 (2003))構築した。サンプリング用の組織区域は、スライド上の対応するHE染色されたセクションとの視覚的配列に基づいて選択した。ドナー腫瘍ブロックから採取された3、4、または5個の組織コア(直径0.6mm;高さ3〜4mm)を、組織マイクロアレイヤー(Beecher Instruments, Hummingbird Court Sun Prairie, WI, USA)を用いてレシピエントパラフィンブロック中に配置した。また、各ケースから正常組織のコアも切り抜いた。得られたマイクロアレイブロックの5μmセクションを免疫組織化学解析に用いた。ANLNの染色パターンを、なし(absent)または陽性として半定量的に評価し、また、臨床経過観察データの予備知識のない3人の独立した研究者が、核または細胞質ANLN(n-ANLN、c-ANLN)に応じて定性的に評価した。n-またはc-ANLN染色腫瘍細胞が10%未満のケースはそれぞれのタイプのANLN陰性と判定した。評価者が独立して陽性と規定したケースのみを陽性として認めた。
【0151】
組織マイクロアレイ上の臨床サンプル中のANLNタンパク質の存在を調査するため、ENVISION+ Kit/ホースラディッシュペルオキシダーゼ(HRP)(DakoCytomation, Glostrup, Denmark)を用いてセクションを染色した。簡単に述べると、内因性ペルオキシダーゼおよびタンパク質をブロッキングした後に、抗ヒトANLN抗体を添加し、HRP標識抗ウサギIgGを二次抗体としてセクションをインキュベートした。基質-色素体を加え、試料をヘマトキシリンで対比染色した。
【0152】
(n)統計的解析
組織マイクロアレイ解析によって決定されるANLNタンパク質の発現レベルとの相関について、年齢、性別、および病理学的TNM段階などの臨床病理学的変数を調べた。手術日からNSCLCに関連した死亡時または最終経過観察時までの腫瘍特異的生存曲線を計算した。各関連変数およびANLN発現についてそれぞれKaplan-Meier曲線を計算した。Log-rank検定を用いて患者サブグループ間の生存期間の差を解析した。Cox比例ハザード回帰モデルで一変量および多変量解析を行い、臨床病理学的変数と癌関連死亡率との関連を決定した。まず、年齢、性別、pT分類、およびpN分類を含む可能な予後因子と死との関連を、1回につき1つの因子を考慮して解析した。次に、多変量Cox解析を、P値<0.05のエントリーレベルを満たす任意のおよび全ての変数と協調して、常にANLN発現をモデルに強制する逆方向(段階的)手順で行った。モデルが因子を加え続けていくと、独立因子はP値<0.05のエグジット(exit)レベルを超えなかった。
【0153】
実施例2 - NSCLC組織および細胞株ならびに正常組織におけるANLNの過剰発現
cDNAマイクロアレイによる解析で37個のNSCLCのうちの50%以上において5倍以上の発現を示す遺伝子は、以前にスクリーニングされている(Kikuchi, T. et al. Oncogene 22(14):2192-2205 (2003))。スクリーニングされた23,040個の遺伝子のうち、ANLN転写物は、NSCLCにおいて頻繁に過剰発現しているとして同定され、半定量的RT-PCR実験によって12個の代表的なNSCLC症例において過剰発現が確認された(図1a)。加えて、高レベルのANLN(Genbankアクセッション番号NM_018685)発現が23個の肺癌細胞株全てにおいて観察された一方、正常小気道上皮由来細胞(SAEC)においてはPCR産物は検出されなかった(図1b)。ANLN cDNAをブローブとして用いたノーザンブロッティング解析によって、約4.0kbの転写物が、調べた24個の正常ヒト組織のうち精巣および脊髄においてのみ弱いバンドとして同定された(図1c)。NSCLC細胞株A549、LC319、およびNCI-H522におけるANLNタンパク質の発現は、抗ANLN抗体を用いたウェスタンブロッティング解析によって内因性ANLNタンパク質の発現を調べることにより確認された。
【0154】
実施例3 - ANLNの細胞内局在およびアクチンストレスファイバー形成
肺癌細胞におけるANLNの細胞内局在を確認するため、免疫細胞化学解析を行った。肺癌細胞株LC319およびA549で内因的に発現されたANLNは、様々な細胞内局在パターンを示した(図2a)。ANLNタンパク質は、核および/または細胞質(n-ANLNおよびc-ANLN)、ならびに核膜崩壊後の皮層、細胞質分裂中の分裂溝、および分裂終期の後期の中央体において観察された。細胞質における線維様染色もかなりの数の細胞において観察された。これら細胞におけるストレスファイバー上の内因性ANLNおよびF-アクチンの共存が、抗ANLN抗体およびファロイジンを用いた免疫染色によって確認された(図2b)。アクチン細胞骨格は哺乳動物細胞の細胞質分裂および形態に重要な役割を担うことが知られていることから、次に、アクチンストレスファイバー形成に対するANLNの効果を、ANLN発現プラスミドをLC319およびNIH3T3細胞にトランスフェクションすることによって調べた。ファロイジンで検出された線維様染色はANLNのトランスフェクション後に有意に増加し、外因性ANLNの過剰発現がこれら細胞において多くのストレスファイバーを誘導することが示唆された(図2c)。
【0155】
実施例4 - NSCLC細胞の成長に対するANLNの効果
ANLNが肺癌細胞の成長または生存に不可欠であるかどうかを評価するため、ANLN配列(Genbankアクセッション番号NM_018685、SEQ ID NO:1(SEQ ID NO:2をコードする)に対応する2対のリバース(対照)およびアンチセンスS-オリゴヌクレオチドを合成して、高レベルのANLN発現を示したA549細胞にそれぞれトランスフェクトした。2つの異なるアンチセンスS-オリゴヌクレオチド(AS1およびAS2)のそれぞれの導入は、対応対照ヌクレオチド(R1およびR2)と比較して細胞生存度を低下させ、ANLNが癌細胞の成長および/または生存に不可欠であることが示唆された(データ示さず)。アンチセンスS-オリゴヌクレオチドによる成長抑制効果がANLN特異的であることをさらに確認するため、ANLNに対するsiRNA(siRNA-ANLN-1および-2)および2個の対照プラスミド(ルシフェラーゼ(LUC)またはスクランブル(SCR)に対するsiRNA)を発現するようにプラスミドを設計、構築し、それぞれをLC319およびA549細胞にトランスフェクトした。siRNA-ANLN-1または-2でトランスフェクトした細胞におけるANLN転写物の量は、2個の対照siRNAのいずれかでトランスフェクトしたものと比べて有意に低下した(図3a);siRNA-ANLN-1または-2のトランスフェクションによってもまた、コロニー形成およびMTTアッセイによって測定したところコロニー数および細胞生存度が有意に低下した(図3b、c)。さらに、siRNA-ANLN-1で処置した細胞は、複数の核を有する、より大きな細胞形態を示した(図3d)。この表現型の分子メカニズムをさらに明らかにするため、siRNA-ANLN-1でトランスフェクトしたLC319細胞を用いてフローサイトメトリーを行い、siRNA-ANLN-1でトランスフェクトした細胞において4N〜16NのDNA含量を有する細胞の比率が、対照siRNA(LUC)でトランスフェクトした細胞におけるものよりも有意に高いことを見出した(図3e)。
【0156】
細胞周期進行の調節に対するANLNの効果をさらに調べるため、ANLN発現プラスミドで一過的にトランスフェクトしたLC319およびA549細胞を用いて、BrdU取り込みアッセイを行った。DNA合成は、ANLN発現の誘導によって両細胞株において用量依存的に増強されているようであった(図4a)。
【0157】
実施例5 - 細胞運動性に対するANLNの効果
ストレスファイバー上のANLNタンパク質およびF-アクチンが共存し、外因性ANLN発現の誘導が哺乳動物細胞においてアクチンストレスファイバーの形成を促進することが免疫細胞化学解析により示されたので、続いてマトリゲル浸潤アッセイを行い、ANLNが細胞運動性に役割を担っている可能性があるかどうかを判定した。ANLN発現ベクターでトランスフェクトしたNIH3T3およびCOS-7細胞のマトリゲルを通じた浸潤は、対照細胞と比較して有意に促進された(図4b、4c)。ANLNまたはモックプラスミドを発現するように設計されたプラスミドでトランスフェクトしたNIH3T3細胞を用いた傷害遊走アッセイもまた、ANLN発現細胞の遊走が有意に活性化されたことを示した。
【0158】
ANLNによる細胞運動性の活性化のメカニズムを明らかにするため、以下のアッセイを実施した。低分子量GTPase RhoAはアクチン構造物の形成を制御することが知られているので、まずRhoAとANLNとの間の可能な相互作用を調べた。図5aに示すとおり、肺癌細胞株LC319細胞の細胞質および分裂溝における内因性RhoAおよびANLNの共存が、免疫細胞化学解析によって検出された。内因性RhoAの外因性発現ANLNとの直接的関連が免疫沈降アッセイによって確認された(図5b)。
【0159】
他のGTPaseと同様に、RHOはGTPに結合しているときは活性で、GDPに結合しているときは不活性である。GTPと結合すると、RHOはrhotekin(RTKN)などの下流エフェクターと相互作用する。これら事実に基づいて、ANLNの活性型RHOとの相互作用を調べた。具体的には、GST融合RTKNを用いてGSTプルダウンアッセイを行い、GTP-RHO(活性型)およびANLNを含む複合体をアフィニティー沈降させた;ANLNおよびRHOを含む免疫複合体が、いずれかの抗体を用いたウェスタンブロッティング解析によって検出された。ANLNを発現するように設計されたプラスミドでLC319細胞をトランスフェクトすると、RHO活性化の誘導、ならびにANLNと活性型RHOとの間の直接的相互作用が観察された(図5c、上部および下部パネル)。
【0160】
実施例6 - 核ANLN発現は不良な予後に関連する
外科的に切除された285個のNSCLC組織からなる組織マイクロアレイを用い、抗ANLNポリクローナル抗体で免疫組織化学解析を行った。この研究により、285ケースのうち267(93.6%)個がc-ANLN(細胞質性)のみで陽性に染色され、128(44.9%)個がc-ANLNおよびn-ANLN(核性)の両方で陽性であり、n-ANLNのみで陽性であるケースはなかったことが示された(図6a)。そうすると問題は、ANLN発現がNSCLCの臨床結果と関連するのかどうかであった。統計解析では、c-またはn-ANLN発現のpT-またはpN-因子との有意な相関は認められなかった。しかし、Kaplan-Meier法を用い、n-ANLN発現と腫瘍特異的5年生存率との間に顕著な関連が見出された(Log-rank検定でP<0.0001)(図6b)。一変量解析を用いると、pT、pN、性別、およびn-ANLN発現が、NSCLC患者の不良な腫瘍特異的生存と有意に関連していた。さらに、Cox比例ハザードモデルを用いた多変量解析によって、n-ANLN染色は独立した予後因子であることが決定された(P=0.001)。
【0161】
上記の例は本発明を例示するために提供されているが、その範囲を限定することを意図するものではない。本発明の他の変形は当業者には容易に明らかであり、添付の請求の範囲に包含される。
【0162】
産業上の利用可能性
本明細書で示したように、ANLNのRhoAとの相互作用、およびこの相互作用の阻害により癌細胞の細胞増殖が阻害された。このように、ANLNとRhoAの結合を阻害し、その活性を妨げる薬剤は、抗癌剤、特にNSCLCのような肺癌の治療のための抗癌剤として、治療的に有用であると考えられる。
【0163】
本明細書に引用される全ての刊行物、データベース、Genbank配列、特許、および特許出願は、これによって参照として本明細書に組み入れられる。
【0164】
本発明は詳細にその具体的な態様を参照して記載されているが、本発明の要旨および範囲から逸脱することなく様々な変化および修飾をその内に施すことができることは当業者には明らかであり、その境界および範囲は添付の請求の範囲によって定められる。
【図面の簡単な説明】
【0165】
【図1】原発性NSCLCおよび肺癌細胞株におけるANLN発現を示す。パート(a)は、半定量的RT-PCRによって調べた、NSCLCの臨床サンプルおよび正常肺組織におけるANLNの発現を表す。パート(b)は、肺癌細胞株におけるANLNの発現を表す。パート(c)は、ノーザンブロット解析によって検出した正常組織におけるANLNの発現を表す。
【図2】ANLNの細胞内局在およびアクチンストレスファイバー形成を示す。パート(a)は、NSCLC細胞株LC319における内因性ANLNの細胞内局在を表す。核(n-ANLN)、細胞質(c-ANLN)、および分裂溝におけるANLN発現を、ローダミン結合二次抗体を用いた免疫細胞化学染色によって検出した。パート(b)は、FITC結合二次抗体およびAlexa594結合ファロイジンでそれぞれ検出されたLC319細胞における内因性c-ANLNおよびF-アクチンの共存を表す。パート(c)においては、LC319細胞を解析の24時間前にANLN発現プラスミドで一過的にトランスフェクトし、ANLNについてはFITC免疫染色によって、およびAlexa594-ファロイジン染色によってANLNおよびF-アクチン分布を評価した。外因性ANLN発現およびそれらの共存によるストレスファイバー形成の誘導がLC319細胞において観察された。
【図3】ANLNに対するsiRNAによるNSCLC細胞の成長阻害を示す。パート(a)は、半定量的RT-PCRによって解析した、LC319細胞におけるルシフェラーゼ(LUC)またはスクランブル(SCR)に対するsiRNA-ANLN-1(si-1)、-2(si-2)、または対照siRNAに応答したANLNの発現を表す。パート(b)は、ANLN(si-1および-2)または対照プラスミド(si-LUCおよび-SCR)の特異的siRNAでトランスフェクトしたLC319細胞のコロニー形成アッセイを表す。パート(c)は、si-1、-2、-LUC、または-SCRに応答した、MTTアッセイによって評価したLC319細胞の生存度を表す。パート(d)は、si-1または-LUCでトランスフェクトしたLC319細胞の顕微鏡観察を表す。矢印は、多核で、より大きな細胞形態を示すsiRNA-ANLN-1(si-1)で処置した細胞を示す。パート(e)は、si-1または-LUCでトランスフェクトしたLC319細胞のフローサイトメトリー解析の結果を表す。si-1でトランスフェクトした細胞において4N〜16NのDNA含量を有する細胞の比率は、対照siRNA(si-LUC)でトランスフェクトした細胞におけるものよりも有意に高かった。アッセイは3回、三つ組ウェルにおいて行った。
【図4】ANLNによるDNA合成の促進および細胞運動性の活性化を示す。パート(a)は、BrdUと20時間インキュベーションした後の、ANLNで一過的にトランスフェクトしたLC319またはA549細胞におけるBrdU取り込みを表す。ANLN発現プラスミドで一過的にトランスフェクトしたLC319およびA549細胞の両方において、用量依存的にDNA合成が促進されたようであった。パート(b)および(c)は、ヒトANLN発現プラスミドをトランスフェクトした際の、マトリゲル基質中でのNIH3T3およびCOS-7細胞の浸潤性の促進を示すマトリゲル浸潤アッセイの結果を表す。具体的には、パート(b)はGiemsa染色(X100)を示し、パート(c)はマトリゲル被覆フィルターを通過して遊走した細胞の数を表す。アッセイは3回、三つ組ウェルにおいて行った。
【図5】ANLNのRhoAとの相互作用およびそのRHO活性化の調節を示す。パート(a)は、抗ANLN抗体(FITC)および抗RhoA抗体(ローダミン)を用いた免疫細胞化学染色によって検出した内因性ANLNおよびRhoAの共存を表す。パート(b)は、肺癌細胞株LC319抽出物からの外因性ANLNおよび内因性RhoAの免疫沈降(IP)を表す。LC319細胞はモックまたはANLNでトランスフェクトし、抗ANLN抗体を用いたANLN-IP、続いて抗RhoA抗体を用いた免疫ブロッティング(IB)に供した(上部)。細胞溶解物のアリコートを免疫ブロッティングに直接供して、下3つのパネルに示すように、各タンパク質の発現を確認した。パート(c)は、ANLNの直接結合によって誘導されたRHO活性化を表す。(上3つのパネル)LC319細胞をモックまたはANLNでトランスフェクトし、免疫ブロッティングに直接供して各タンパク質の発現を確認した。(下2つのパネル)細胞溶解物のアリコートをGST-RTKN-RBDと共にインキュベートしてGSTプルダウンアッセイに供し、次に抗RHOおよび抗ANLN抗体を用いた免疫ブロッティングに供した。最初のパネルは外因性ANLN発現によって活性化されたRHOのレベルを示し、二番目のパネルは活性化RHOとのANLNの直接的相互作用を示す。
【図6】n-ANLNの過剰発現がNSCLCにおける不良な結果に関連することを示す。パート(a)は、組織マイクロアレイに対して抗ANLNポリクローナル抗体を用いた、外科的に切除されたNSCLC組織由来の代表的サンプルの免疫組織化学評価の結果を表す(ADC、SCC、X100)。c-ANLNは細胞質に局在するANLNを示し、n-ANLNは核のものを示す。矢印はn-ANLNを発現する細胞の例を示す。パート(b)は、n-ANLN発現によるNSCLC患者における腫瘍特異的生存の組織マイクロアレイおよびKaplan-Meier解析の結果を表す(P<0.0001;Log-rank検定)。
【特許請求の範囲】
【請求項1】
以下の工程を含む、癌の処置または予防に有用な化合物をスクリーニングする方法:
(a)試験化合物の存在下で、ANLNポリペプチドのRhoA結合ドメインを含むポリペプチドを、RhoAポリペプチドのANLN結合ドメインを含むポリペプチドと接触させる工程;
(b)ポリペプチド間の結合を検出する工程;および
(c)ポリペプチド間の結合を阻害する試験化合物を選択する工程。
【請求項2】
RhoA結合ドメインを含むポリペプチドがANLNポリペプチドを含む、請求項1記載の方法。
【請求項3】
ANLN結合ドメインを含むポリペプチドがRhoAポリペプチドを含む、請求項1記載の方法。
【請求項4】
RhoA結合ドメインを含むポリペプチドが生細胞中で発現される、請求項1記載の方法。
【請求項5】
以下の工程からなる群より選択される工程を含む方法によってポリペプチド間の結合が検出される、請求項1記載の方法:
(a)活性化RhoAの濃度を検出する工程;
(b)RhoAとRHOエフェクターまたはそのRhoA結合領域との間の相互作用を検出する工程;
(c)活性化RhoAを介する下流遺伝子発現または下流遺伝子産物活性を含む任意のシグナル複合体の活性化を検出する工程;
(d)DNA合成および細胞周期移行の促進を検出する工程;
(e)細胞遊走または任意の他の発癌表現型を検出する工程;
(f)アクチンストレスファイバー形成およびF-アクチン産生を検出する工程;ならびに
(g)細胞接着、遊走、および浸潤に重要な任意の分子との相互作用を検出する工程。
【請求項6】
以下のものを含む、癌の処置または予防に有用な化合物をスクリーニングするためのキット:
(a)ANLNポリペプチドのRhoA結合ドメインを含むポリペプチド;
(b)RhoAポリペプチドのANLN結合ドメインを含むポリペプチド;および
(c)ポリペプチド間の相互作用を検出する手段。
【請求項7】
RhoA結合ドメインを含むポリペプチドがANLNポリペプチドを含む、請求項6記載のキット。
【請求項8】
ANLN結合ドメインを含むポリペプチドがRhoAポリペプチドを含む、請求項6記載のキット。
【請求項9】
RhoA結合ドメインを含むポリペプチドが生細胞中で発現される、請求項6記載のキット。
【請求項10】
要素a)とb)との間の相互作用を検出する手段が以下のものを検出する、請求項6記載のキット:
(a)活性化RhoAの濃度;
(b)RhoAとRHOエフェクターまたはそのRhoA結合領域との間の相互作用;
(c)活性化RhoAを介する下流遺伝子を含む任意のシグナル複合体の活性化;
(d)DNA合成および細胞周期移行の促進;
(e)細胞遊走または任意の他の発癌表現型;
(f)アクチンストレスファイバー形成およびF-アクチン産生;ならびに
(g)細胞接着、遊走、および浸潤に重要な任意の分子との相互作用。
【請求項11】
以下の工程を含む、癌の処置または予防に有用な化合物をスクリーニングする方法:
(a)ANLNポリペプチドまたはその機能的同等物を発現する細胞を試験化合物と接触させる工程;
(b)細胞のANLNを介する運動性を検出する工程;および
(c)試験化合物の非存在下で検出される運動性レベルと比較して細胞の運動性を阻害する試験化合物を選択する工程。
【請求項12】
細胞が、ANLNポリペプチドまたはその機能的同等物を発現可能にコードするポリヌクレオチドを含むベクターを含む、請求項11記載の方法。
【請求項13】
以下のものを含む、癌の処置または予防に有用な化合物をスクリーニングするためのキット:
(a)ANLNポリペプチドまたはその機能的同等物を発現する細胞;および
(b)細胞の運動性を検出する手段。
【請求項14】
以下の工程を含む、対象における非小細胞肺癌(NSCLC)の予後を予測する方法:
(a)NSCLC予後を予測しようとする対象から収集された試料の核に局在するANLNタンパク質を検出する工程;および
(b)核内のANLNタンパク質の局在が検出された場合に予後が不良であると予測する工程。
【請求項15】
以下の工程によって試料の核内のANLNの局在が検出される、請求項14記載の方法:
(a)ANLNタンパク質を認識する抗体を試料と接触させる工程;および
(b)核領域において試料に結合する抗体を検出する工程。
【請求項16】
ANLNタンパク質を認識する抗体と、核における検出用の薬剤とを含む、非小細胞肺癌(NSCLC)に罹患した対象の予後を予測するためのキット。
【請求項17】
核における検出用の薬剤がヘマトキシリン-エオシン染色染料である、請求項16記載のキット。
【請求項18】
請求項1または12記載の方法によって選択される化合物を投与する工程を含む、対象における癌を処置または予防するための方法。
【請求項19】
ANLNとRhoAとの間の結合を阻害する化合物を投与する工程を含む、対象における癌を処置または予防するための方法。
【請求項20】
請求項1または12記載の方法によって選択される化合物の薬学的有効量と、薬学的に許容される担体とを含む、癌を処置または予防するための組成物。
【請求項21】
ANLNとRhoAとの間の結合を阻害する化合物の薬学的有効量と、薬学的に許容される担体とを含む、癌を処置または予防するための組成物。
【請求項1】
以下の工程を含む、癌の処置または予防に有用な化合物をスクリーニングする方法:
(a)試験化合物の存在下で、ANLNポリペプチドのRhoA結合ドメインを含むポリペプチドを、RhoAポリペプチドのANLN結合ドメインを含むポリペプチドと接触させる工程;
(b)ポリペプチド間の結合を検出する工程;および
(c)ポリペプチド間の結合を阻害する試験化合物を選択する工程。
【請求項2】
RhoA結合ドメインを含むポリペプチドがANLNポリペプチドを含む、請求項1記載の方法。
【請求項3】
ANLN結合ドメインを含むポリペプチドがRhoAポリペプチドを含む、請求項1記載の方法。
【請求項4】
RhoA結合ドメインを含むポリペプチドが生細胞中で発現される、請求項1記載の方法。
【請求項5】
以下の工程からなる群より選択される工程を含む方法によってポリペプチド間の結合が検出される、請求項1記載の方法:
(a)活性化RhoAの濃度を検出する工程;
(b)RhoAとRHOエフェクターまたはそのRhoA結合領域との間の相互作用を検出する工程;
(c)活性化RhoAを介する下流遺伝子発現または下流遺伝子産物活性を含む任意のシグナル複合体の活性化を検出する工程;
(d)DNA合成および細胞周期移行の促進を検出する工程;
(e)細胞遊走または任意の他の発癌表現型を検出する工程;
(f)アクチンストレスファイバー形成およびF-アクチン産生を検出する工程;ならびに
(g)細胞接着、遊走、および浸潤に重要な任意の分子との相互作用を検出する工程。
【請求項6】
以下のものを含む、癌の処置または予防に有用な化合物をスクリーニングするためのキット:
(a)ANLNポリペプチドのRhoA結合ドメインを含むポリペプチド;
(b)RhoAポリペプチドのANLN結合ドメインを含むポリペプチド;および
(c)ポリペプチド間の相互作用を検出する手段。
【請求項7】
RhoA結合ドメインを含むポリペプチドがANLNポリペプチドを含む、請求項6記載のキット。
【請求項8】
ANLN結合ドメインを含むポリペプチドがRhoAポリペプチドを含む、請求項6記載のキット。
【請求項9】
RhoA結合ドメインを含むポリペプチドが生細胞中で発現される、請求項6記載のキット。
【請求項10】
要素a)とb)との間の相互作用を検出する手段が以下のものを検出する、請求項6記載のキット:
(a)活性化RhoAの濃度;
(b)RhoAとRHOエフェクターまたはそのRhoA結合領域との間の相互作用;
(c)活性化RhoAを介する下流遺伝子を含む任意のシグナル複合体の活性化;
(d)DNA合成および細胞周期移行の促進;
(e)細胞遊走または任意の他の発癌表現型;
(f)アクチンストレスファイバー形成およびF-アクチン産生;ならびに
(g)細胞接着、遊走、および浸潤に重要な任意の分子との相互作用。
【請求項11】
以下の工程を含む、癌の処置または予防に有用な化合物をスクリーニングする方法:
(a)ANLNポリペプチドまたはその機能的同等物を発現する細胞を試験化合物と接触させる工程;
(b)細胞のANLNを介する運動性を検出する工程;および
(c)試験化合物の非存在下で検出される運動性レベルと比較して細胞の運動性を阻害する試験化合物を選択する工程。
【請求項12】
細胞が、ANLNポリペプチドまたはその機能的同等物を発現可能にコードするポリヌクレオチドを含むベクターを含む、請求項11記載の方法。
【請求項13】
以下のものを含む、癌の処置または予防に有用な化合物をスクリーニングするためのキット:
(a)ANLNポリペプチドまたはその機能的同等物を発現する細胞;および
(b)細胞の運動性を検出する手段。
【請求項14】
以下の工程を含む、対象における非小細胞肺癌(NSCLC)の予後を予測する方法:
(a)NSCLC予後を予測しようとする対象から収集された試料の核に局在するANLNタンパク質を検出する工程;および
(b)核内のANLNタンパク質の局在が検出された場合に予後が不良であると予測する工程。
【請求項15】
以下の工程によって試料の核内のANLNの局在が検出される、請求項14記載の方法:
(a)ANLNタンパク質を認識する抗体を試料と接触させる工程;および
(b)核領域において試料に結合する抗体を検出する工程。
【請求項16】
ANLNタンパク質を認識する抗体と、核における検出用の薬剤とを含む、非小細胞肺癌(NSCLC)に罹患した対象の予後を予測するためのキット。
【請求項17】
核における検出用の薬剤がヘマトキシリン-エオシン染色染料である、請求項16記載のキット。
【請求項18】
請求項1または12記載の方法によって選択される化合物を投与する工程を含む、対象における癌を処置または予防するための方法。
【請求項19】
ANLNとRhoAとの間の結合を阻害する化合物を投与する工程を含む、対象における癌を処置または予防するための方法。
【請求項20】
請求項1または12記載の方法によって選択される化合物の薬学的有効量と、薬学的に許容される担体とを含む、癌を処置または予防するための組成物。
【請求項21】
ANLNとRhoAとの間の結合を阻害する化合物の薬学的有効量と、薬学的に許容される担体とを含む、癌を処置または予防するための組成物。
【図1】



【図2】
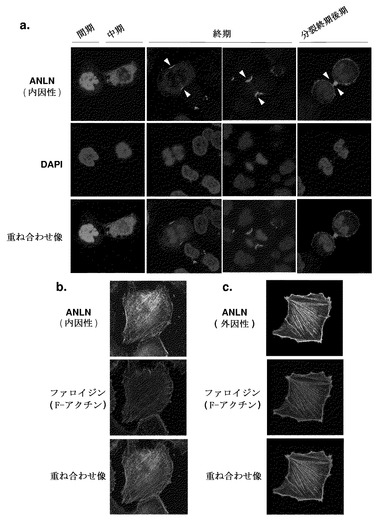
【図3】
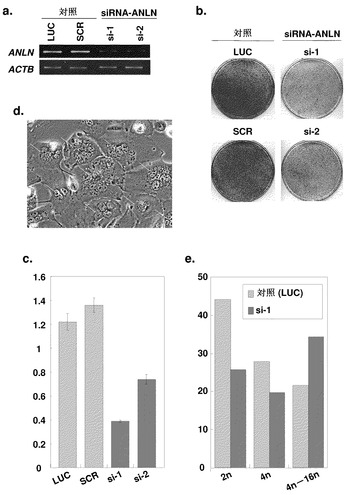
【図4】
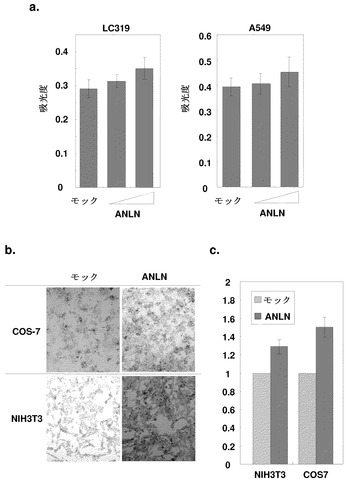
【図5】
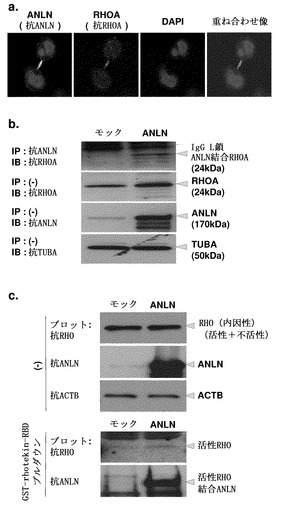
【図6A】
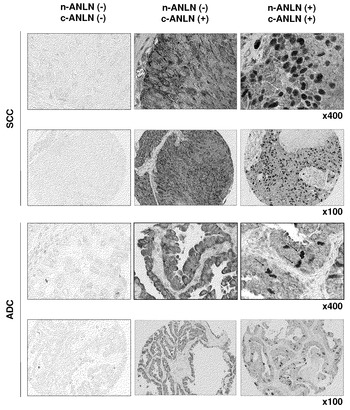
【図6B】
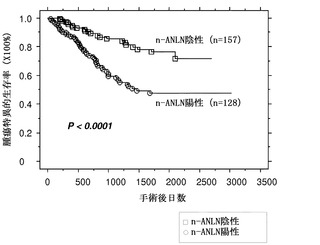
【図2】
【図3】
【図4】
【図5】
【図6A】
【図6B】
【公表番号】特表2008−510126(P2008−510126A)
【公表日】平成20年4月3日(2008.4.3)
【国際特許分類】
【出願番号】特願2007−506605(P2007−506605)
【出願日】平成17年8月9日(2005.8.9)
【国際出願番号】PCT/JP2005/014887
【国際公開番号】WO2006/016697
【国際公開日】平成18年2月16日(2006.2.16)
【出願人】(502240113)オンコセラピー・サイエンス株式会社 (142)
【出願人】(504137912)国立大学法人 東京大学 (1,942)
【Fターム(参考)】
【公表日】平成20年4月3日(2008.4.3)
【国際特許分類】
【出願日】平成17年8月9日(2005.8.9)
【国際出願番号】PCT/JP2005/014887
【国際公開番号】WO2006/016697
【国際公開日】平成18年2月16日(2006.2.16)
【出願人】(502240113)オンコセラピー・サイエンス株式会社 (142)
【出願人】(504137912)国立大学法人 東京大学 (1,942)
【Fターム(参考)】
[ Back to top ]