
類型不能HAEMOPHILUSINFLUENZAEの中耳炎単離株の遺伝子
【課題】Haemophilus influenzae(NTHi)の類型不能株の遺伝子およびその使用を提供すること。
【解決手段】Haemophilus influenzae(NTHi)の類型不能株のポリヌクレオチド配列、そのポリヌクレオチドによってコードされるポリペプチドならびにその使用。中耳または鼻咽頭のNTHi感染の間またはその感染に応じて、アップレギュレートされるNTHi遺伝子。また、NTHi関連障害を処置および予防するワクチンおよび方法を含む、これらのNTHiポリヌクレオチドおよびNTHiポリペプチドの使用。
【解決手段】Haemophilus influenzae(NTHi)の類型不能株のポリヌクレオチド配列、そのポリヌクレオチドによってコードされるポリペプチドならびにその使用。中耳または鼻咽頭のNTHi感染の間またはその感染に応じて、アップレギュレートされるNTHi遺伝子。また、NTHi関連障害を処置および予防するワクチンおよび方法を含む、これらのNTHiポリヌクレオチドおよびNTHiポリペプチドの使用。
【発明の詳細な説明】
【技術分野】
【0001】
(関連出願)
本願は、2003年3月6日出願の米国仮特許出願第60/453,134号(これは、その全体が本明細書に参考として援用される)の優先権の利益を主張する。
【0002】
(発明の分野)
本発明は、Haemophilus influenzae(NTHi)ゲノムの類型不能株のポリヌクレオチド配列、そのゲノム内に含まれるNTHi遺伝子およびそのポリヌクレオチドによってコードされるポリペプチドに関する。本発明はまた、NTHi関連障害を処置および予防するワクチンおよび方法を含む、これらのNTHiポリヌクレオチドおよびNTHiポリペプチドの使用に関する。本発明はまた、中耳または鼻咽頭のNTHi感染の間またはその感染に応じて、アップレギュレートされるNTHi遺伝子に関する。
【背景技術】
【0003】
(背景)
中耳炎(OM)は、全世界で非常に優勢な小児疾患であり、小児が救急室に運ばれる主な原因である(Infante−Rivand and Fernandez,Epidemiol.Rev.,15:444−465,1993)。最近の統計では、1990年に、OMが原因で2億4500万回の通院(physician office visit)があったことが示されている。このことは、1980年代において報告されたものより200%を超えて増大していることを示している。もはや死と関連することは稀であるが、OMと関連する罹患率は、重大である。聴覚喪失は、この疾患と関連する一般的な問題であり、しばしば、小児の行動、教育および言語能力の発達に影響を与える(Baldwin,Am.J.Otol.,14:601−604,1993;Hunteretら,Ann.Otol.Rhinol.Laryngol.Suppl.,163:59−61,1994;Teeleら,J.Infect.Dis.,162:685−694,1990)。OMのその社会経済的な影響もまた大きく、OMを診断および管理する直接的および間接的な費用は、米国単独で年間50億ドルを超えている(Kaplanら,Pediatr.Infect.Dis.J.,16:S9−11,1997)。
【0004】
抗生物質療法は、一般的かつ中耳腔換気用チューブの外科手術による置換は、滲出液を排出すること、感染を除去すること、および中耳に流体が蓄積することと関連する頭痛を緩和することという観点から好結果であったが、多剤耐性細菌の出現およびチューブの置換と関連するその侵襲的性質によって、OMの管理、より好ましくはOMの予防に対するより有効かつ容認されたアプローチが必要であることが明らかにされた。慢性OMの外科手術的管理は、小児に全身麻酔をかけている間に、鼓膜を通して中耳腔換気用チューブを挿入することを包含する。この手順は平凡であり(普及率は約13%である;Brightら,Am.J.Public Health,83(7):1026−8,1993)、蓄積した流体を中耳から排出することによって、痛みの伴う症状を軽減するという点で非常に有効であるが、これは、その手順の侵襲的性質およびその伴う危険性に起因して、数多くの批判を受けている(Bermanら,Pediatrics,93(3):353−63,1994;Brightら,前出;Cimons,ASMNews,60:527−528;Paap,Ann.Pharmacother.,30(11):1291−7,1996)。
【0005】
ワクチン開発における進歩は、7価の莢膜結合体ワクチンであるPREVNAR(登録商標)(EskolaおよびKilpi,Pedriatr.Effect.Dis.J.16:S72−78,2000)が最近認可および解放されたことによって明らかなように、Streptococcus pneumoniae(OMの主な原因となる因子)について最も促進されている(AOM)。PREVNAR(登録商標)は、侵襲性の肺炎球菌疾患に非常に有用であったが、OMについての適用範囲は、そのワクチンに含まれない血清型に起因して、数が増大したOMの症例の報告によって期待が裏切られている(6〜8%)(Blackら,Pedriatr.Infect.Dis J.,19:187−195;Eskolaら,Pedriatr.Infect.Dis J.,19:S72−78,2000;Eskolaら,N.Engl.J Med.344:403−409,2001;Snowら,Otol.Neurotol.,23:1−2,2002)。類型不可Haemopilus influenzae(NTHi)(滲出液を伴う慢性OM中で優勢なグラム陰性病原体(Klein,Pedriatr.Infect.Dis J.,16:S5−8,1997;Spinolaら,J.Infect.Dis.,154:100−109,1986))については、あまり進歩していない。NTHiに対する有効なワクチン開発の妨害は、NTHi誘導性中耳疾患の病原の現在の不完全な理解である。この遅れの一因は、その疾患が、良性鼻咽頭共生生物の宿主の免疫学的寛容性のうちの1つから進行するので、微生物が発現する毒性因子と宿主の免疫応答との間の動的な相互作用の理解が欠如していることであり、通常は無菌の中耳腔の日和見的な侵入物に対する活発な防御反応の欠如に起因している。
【0006】
現在では、小児においてNTHiがどのようにOMを引き起こすかということの理解は乏しい。OMの誘導に必要な推定毒性因子の同定は、その宿主−病原体相互作用の理解、最終的には、潜在的なワクチン候補および化学療法の標的の同定に大きく寄与する。中耳炎の管理、好ましくは、その予防により有効かつ容認されたアプローチを開発することは非常に必要とされている。ワクチン開発は、この目的を達成するための非常に有望なかつ費用効果的な方法である(Giebank,Pediatr.Infect.Dis J.,13(11):1064−8,1994:Karmaら,Int.J.Pedritr.Otorhinolaryngol.,32(Suppl.):S127−34,1995)。
【発明の概要】
【課題を解決するための手段】
【0007】
本発明は、例えば、以下の項目を提供する。
(項目1)
単離されたポリヌクレオチドであって、該ポリヌクレオチドは、配列番号577〜579、配列番号589〜614、配列番号675〜685、配列番号615、配列番号617、配列番号619、配列番号621、配列番号623、配列番号625、配列番号627、配列番号629、配列番号631、配列番号633、配列番号635、配列番号637、配列番号639、配列番号641、配列番号643、配列番号645、配列番号647、配列番号649、配列番号651、配列番号653、配列番号655、配列番号657、配列番号659、配列番号661、配列番号663、配列番号665、配列番号667、配列番号669、配列番号671、配列番号673、配列番号686、配列番号688、配列番号692、配列番号694、配列番号696、配列番号698、配列番号700、配列番号702、配列番号704、配列番号706、配列番号708、配列番号710、配列番号712、配列番号714、配列番号716、配列番号718、配列番号720、配列番号722、配列番号724、配列番号726、配列番号728、配列番号730、配列番号732、配列番号734、配列番号736、配列番号738、配列番号740、配列番号742、配列番号744、配列番号746、配列番号748、配列番号750、配列番号752、配列番号754、配列番号756、配列番号758、配列番号760、配列番号762、配列番号764、配列番号766、配列番号768または配列番号770のいずれか一つのヌクレオチド配列を含む、ポリヌクレオチド。
(項目2)
項目1に示されたヌクレオチド配列によってコードされるアミノ酸配列を含む単離されたポリペプチドまたはそのフラグメント。
(項目3)
単離されたポリペプチドであって、該ポリペプチドは、配列番号616、配列番号618、配列番号620、配列番号622、配列番号624、配列番号626、配列番号628、配列番号628、配列番号630、配列番号632、配列番号634、配列番号636、配列番号638、配列番号640、配列番号642、配列番号644、配列番号646、配列番号648、配列番号650、配列番号652、配列番号654、配列番号656、配列番号658、配列番号660、配列番号662、配列番号664、配列番号666、配列番号668、配列番号670、配列番号672、配列番号674、配列番号687、配列番号689、配列番号691、配列番号693、配列番号695、配列番号697、配列番号699、配列番号701、配列番号703、配列番号705、配列番号707、配列番号709、配列番号711、配列番号713、配列番号715、配列番号717、配列番号719、配列番号721、配列番号723、配列番号725、配列番号727、配列番号729、配列番号731、配列番号733、配列番号735、配列番号737、配列番号739、配列番号741、配列番号743、配列番号745、配列番号747、配列番号749、配列番号751、配列番号753、配列番号755、配列番号757、配列番号759、配列番号761、配列番号763、配列番号765、配列番号767、配列番号769または配列番号771のいずれか一つのアミノ酸配列を含む、ポリペプチド。
(項目4)
項目2または項目3に記載のポリペプチドおよび薬学的に受容可能なキャリアを含む組成物。
(項目5)
項目2もしくは項目3に記載のポリペプチドまたはそのフラグメントに特異的に結合する抗体。
(項目6)
項目5に記載の抗体および薬学的に受容可能なキャリアを含む組成物。
(項目7)
生物学的サンプル中のNTHi細菌を検出するための方法であって、該方法は、
(a)項目1に記載のポリヌクレオチドまたはそのフラグメントと生物学的サンプルとを接触させる工程、および
(b)該サンプル中の該ポリヌクレオチドのハイブリダイゼーションを検出する工程、を包含する、方法。
(項目8)
生物学的サンプル中のNTHi細菌を検出するための方法であって、該方法は、
(a)項目5に記載の抗体と生物学的サンプルとを接触させる工程、および
(b)該サンプル中の該抗体の結合を検出する工程、
を包含する、方法。
(項目9)
前記生物学的サンプルは、血清、唾液、耳の分泌液、血液、尿、リンパ液、および脳脊髄液からなる群より選択される、項目7または8に記載の方法。
(項目10)
NTHi細菌に対する免疫応答を誘導するための方法であって、該方法は、免疫原性的に有効量の項目2もしくは3に記載のポリペプチドまたはそのフラグメントをNTHi細菌感染の危険にある患者に投与する工程を包含する、方法。
(項目11)
項目2もしくは3に記載のポリペプチドまたはそのフラグメントおよび薬学的に適切なキャリアを含むワクチン。
(項目12)
NTHi細菌感染を防止するための方法であって、該方法は、NTHiの細胞接着をブロックする項目2または3に記載のポリペプチドをNTHi細菌感染の危険にある患者に投与する工程を包含する、方法。
(項目13)
NTHi細菌感染を防止するための方法であって、該方法は、NTHiの細胞接着をブロックする項目5に記載の抗体をNTHi細菌感染の危険にある患者に投与する工程を包含する、方法。
(項目14)
前記NTHi感染は、中耳内である、項目13に記載の方法。
(項目15)
NTHi細菌感染を処置または防止するための方法であって、該方法は、項目2もしくは3に記載のポリペプチドの発現または活性を阻害する分子を必要とする患者に投与する工程を包含する、方法。
(項目16)
項目15に記載の方法であって、前記NTHiポリペプチドは、hisB、lppB、sapA、rbsC、pure、rib、arcB,uxuA、licC、ispZ、mukF、glpR、ihfB、cspD、lav、HI1647、HI0094、HI1163、HI0665、HI1292、HI1064、HI1386、HI1462、HI1369、およびHI1598からなる群より選択されるNTHi遺伝子によってコードされるアミノ酸配列を含む、方法。
(項目17)
項目15に記載の方法であって、前記NTHiポリペプチドは、配列番号616、配列番号618、配列番号620、配列番号622、配列番号624、配列番号626、配列番号628、配列番号634、配列番号638、配列番号642、配列番号644、配列番号646、配列番号650、配列番号652、配列番号656、配列番号658、配列番号660、配列番号662、配列番号664、配列番号666、配列番号668、配列番号670、配列番号672および配列番号674からなる群より選択されるアミノ酸配列を含む、方法。
(項目18)
前記必要とする患者に投与される前記分子は、アンチセンスオリゴヌクレオチドである、
項目15に記載の方法。
(項目19)
前記必要とする患者に投与される前記分子は、抗体である、項目15に記載の方法。
(項目20)
前記必要とする患者に投与される前記分子は、低分子である、項目15に記載の方法。
(項目21)
前記NTHi感染は、中耳内である、項目15に記載の方法。
本発明は、NTHiであるH.influenzae株86−028NPのゲノム配列の同定および特徴付け、ならびにこれらによりコードされるポリペプチド配列を提供する。NTHiゲノム配列の三倍分析(3−fold analysis)を、配列番号1〜576として示される一連のコンティグ配列において設定し、その後のゲノム配列の八倍分析(8−fold analysis)を、配列番号675〜685として示される一連の11個のコンティグ配列において設定する。これらのコンティグは、稀なデータであり、当業者は、重複する配列を比較して、慣用的な方法を使用して、NTHi株86−028NPの完全ゲノムを構築することによって、これらのコンティグを組み立て得る。
【0008】
本発明はまた、本発明のNTHiポリペプチドに特異的な抗体を提供する。ヒトにおいて、またはサンプル(例えば、血清、精液、耳からの分泌液(ear fluid)、血液、尿、リンパ液および脳脊髄液)において、NTHi細菌を検出する方法が、企図される。これらの方法は、NTHiポリヌクレオチドを、特異的ポリヌクレオチドプローブで検出する工程、またはNTHiポリペプチドを特異的抗体で検出する工程を包含する。本発明はまた、NTHi細菌を検出するこれらの方法を利用する診断キットを企図する。
【0009】
本発明はまた、本発明のNTHiポリペプチドまたはそのNTHiペプチドを投与することによって、免疫応答を誘発する方法を企図する。これらの方法は、NTHiポリペプチドまたはNTHiペプチドを、NTHi感染によって引き起こされる疾患(例えば、OM)の処置および/または予防のためのワクチンとして投与する工程を包含する。以下のNTHi遺伝子は、中耳および/または鼻咽頭の感染の間またはその感染に応答してアップレギュレートされ;これらの遺伝子によってコードされるポリペプチドおよびそのペプチドは、あり得るOMワクチン候補および/または化学療法の標的として企図される:hisB、lppB、sapA、lolA、rbsC、purE、ribB、arcB、uxuA、dsbB、ureH、licC、HI1647、ispZ、radC、mukF、glpR、ihfB、argR、cspD、HI0094、HI1163、HI1063、HI0665、HI1292、HI1064。NTHi hisB遺伝子は、配列番号615のヌクレオチド配列として表され、配列番号616として表されるアミノ酸配列をコードする。NTHi sapA遺伝子は、配列番号617のヌクレオチド配列として表され、配列番号618として表されるアミノ酸配列をコードする。NTHi rbsC遺伝子は、配列番号619のヌクレオチド配列として表され、配列番号620として表されるアミノ酸配列をコードする。NTHi purE遺伝子は、配列番号621のヌクレオチド配列として表され、配列番号622として表されるアミノ酸配列をコードする。NTHi ribB遺伝子は、配列番号623のヌクレオチド配列として表され、配列番号624として表されるアミノ酸配列をコードする。NTHi arcB遺伝子は、配列番号625のヌクレオチド配列として表され、配列番号626として表されるアミノ酸配列をコードする。NTHi uxuA遺伝子は、配列番号627のヌクレオチド配列として表され、配列番号628として表されるアミノ酸配列をコードする。NTHi dsbB遺伝子は、配列番号629のヌクレオチド配列として表され、配列番号630として表されるアミノ酸配列をコードする。NTHi ureH遺伝子は、配列番号631のヌクレオチド配列として表され、配列番号632として表されるアミノ酸配列をコードする。NTHi licC遺伝子は、配列番号633のヌクレオチド配列として表され、配列番号634として表されるアミノ酸配列をコードする。NTHi HI1647遺伝子は、配列番号635のヌクレオチド配列として表され、配列番号636として表されるアミノ酸配列をコードする。NTHi ispZ遺伝子は、配列番号637のヌクレオチド配列として表され、配列番号638として表されるアミノ酸配列をコードする。NTHi radC遺伝子は、配列番号639のヌクレオチド配列として表され、配列番号640として表されるアミノ酸配列をコードする。NTHi mukF遺伝子は、配列番号641のヌクレオチド配列として表され、配列番号642として表されるアミノ酸配列をコードする。NTHi glpR遺伝子は、配列番号643のヌクレオチド配列として表され、配列番号644として表されるアミノ酸配列をコードする。NTHi ihfB遺伝子は、配列番号645のヌクレオチド配列として表され、配列番号646として表されるアミノ酸配列をコードする。NTHi argR遺伝子は、配列番号647のヌクレオチド配列として表され、配列番号648として表されるアミノ酸配列をコードする。NTHi cspD遺伝子は、配列番号649のヌクレオチド配列として表され、配列番号650として表されるアミノ酸配列をコードする。NTHi HI1163遺伝子は、配列番号651のヌクレオチド配列として表され、配列番号652として表されるアミノ酸配列をコードする。NTHi HI1063遺伝子は、配列番号653のヌクレオチド配列として表され、配列番号654として表されるアミノ酸配列をコードする。NTHi HI0665遺伝子は、配列番号655のヌクレオチド配列として表され、配列番号656として表されるアミノ酸配列をコードする。NTHi HI1292遺伝子は、配列番号657のヌクレオチド配列として表され、配列番号658として表されるアミノ酸配列をコードする。
【0010】
配列番号1〜576、配列番号675〜685として示されるポリヌクレオチド配列ならびに表4および表4Bにおいて示されるポリヌクレオチド配列中に含まれる新規なNTHi遺伝子はまた、中耳および/または鼻咽頭の感染の間にアップレギュレートされ、従って、OMワクチン候補および/または化学療法の標的をコードすると予測される。さらに、以下のNTHi遺伝子は、毒性関連遺伝子であると予測され、従って、あり得るOMワクチン候補および/または化学療法の標的をコードすることが予測される:HI1386、HI1462、HI1369、lav、HI1598。NTHi HI1386遺伝子配列は、配列番号659として表され、配列番号660として表されるアミノ酸配列をコードする。NTHi HI1462遺伝子配列は、配列番号661として表され、配列番号662として表されるアミノ酸配列をコードする。NTHi HI1369遺伝子配列は、配列番号665として表され、配列番号666として表されるアミノ酸配列をコードする。NTHi lav遺伝子配列は、配列番号663として表され、配列番号664として表されるアミノ酸配列をコードする。NTHi HI1598遺伝子配列は、配列番号669および配列番号671として表され、配列番号670および配列番号672として表されるアミノ酸配列をコードする。毒性と関連したさらなるNTHi遺伝子としては、配列番号667および配列番号673として示されるポリヌクレオチド配列が挙げられる。
【0011】
NTHi感染を処置または予防する方法として、本発明は、その感染の間にアップレギュレートまたは活性になるNTHiポリペプチドの発現または活性を阻害する分子を投与することを企図する。特に、本発明は、NTHi感染を処置または予防する方法を企図し、この方法は、NTHi感染の間にアップレギュレートされるNTHi遺伝子に特異的に結合するアンチセンスオリゴヌクレオチドを投与することによって、NTHiタンパク質発現を調節する工程を包含する。このような遺伝子としては、hisB、lppB、sapA、lolA、rbsC、purE、ribB、arcB、uxuA、dsbB、ureH、licC、HI1647、ispZ、radC、mu、glpR、ihfB、argR、cspD、HI0094、HI1163、HI1063、HI0665、HI1292、HI1064が挙げられる。本発明はまた、NTHi感染を処置または予防する方法を企図し、この方法は、これらの遺伝子によってコードされるタンパク質の活性を調節する抗体または低分子を投与する工程を包含する。配列番号1〜576、配列番号675〜685として示されるポリヌクレオチド配列ならびに表4および表4Bに示されるヌクレオチド配列に含まれる新規なNTHi遺伝子もまた、中耳および/または鼻咽頭の感染の間にアップレギュレートされ、従って、これらのポリヌクレオチド配列に特異的に結合するアンチセンスオリゴヌクレオチドもまた、企図される。
【0012】
(本発明のポリヌクレオチドおよびポリペプチド)
本発明は、NTHi株86−028NPゲノムの配列を提供する。このゲノム配列は、本明細書中で「コンティグ1〜576」として表される一連のコンティグ配列として示される。各コンティグは、その「コンティグ番号」と関連して配列識別番号を割り当てられる。従って、本発明のコンティグは、配列番号1〜576として表される。これらのコンティグポリヌクレオチド配列は、慣用的な方法を使用して、NTHi株86−028NPの完全ゲノム配列に組み立てられ得る。NTHi株82−028NPゲノムの八倍配列分析の完了の際に、そのゲノム配列は、配列番号675〜685として本明細書中で表される11のコンティグに組み立てられた。
【0013】
本発明は、NTHiポリヌクレオチド配列および配列番号1〜576、配列番号675〜685のコンティグ内に含まれるオープンリーディングフレーム、ならびに表3B、表4Bおよび表5に示されるヌクレオチド配列を提供する。本発明はまた、本発明のNTHiポリヌクレオチドによってコードされるポリペプチド配列(例えば、表3B、表4Bおよび表5に示されるアミノ酸配列)を提供する。本発明は、(a)配列番号1〜576のヌクレオチド配列の相補体;配列番号675〜685のヌクレオチド配列の相補体および本明細書中の表3B、表4Bおよび表5に示されるヌクレオチド配列の相補対;(b)上記のポリヌクレオチドの対立遺伝子改変体であるポリヌクレオチド;(c)上記のタンパク質のいずれかの種ホモログをコードするポリヌクレオチド;あるいは(d)本発明のNTHiポリペプチドの特定のドメインまたは短縮を含むポリペプチドをコードするポリヌクレオチド、にストリンジェントな条件下でハイブリダイズするポリヌクレオチドを提供する。
【0014】
本発明のNTHiポリヌクレオチドはまた、上記のポリヌクレオチドに実質的に等価なヌクレオチド配列を包含する。本発明に従うポリヌクレオチドは、上記のNTHiポリヌクレオチドに対して、例えば、少なくとも65%、少なくとも70%、少なくとも75%、少なくとも80%、81%、82%、83%、84%、85%、86%、87%、88%、または89%、より代表的には、少なくとも90%、91%、92%、93%、または94%、およびより代表的には、少なくとも95%、96%、97%、98%または99%の配列同一性を有する。
【0015】
本発明の核酸配列の範囲内に含まれるのは、配列番号1〜576、配列番号675〜685のNTHiヌクレオチド配列、ならびに本明細書中の表3B、表4Bおよび表5に示されるヌクレオチド配列、またはそれらの相補体にストリンジェントな条件下でハイブリダイズする核酸配列フラグメントであり、これらのフラグメントは、約5ヌクレオチドより大きく、好ましくは7ヌクレオチド、より好ましくは、9ヌクレオチドより大きく、最も好ましくは17ヌクレオチドより大きい。例えば、選択的な(すなわち、本発明のポリヌクレオチドのいずれか1つに特異的にハイブリダイズする)15ヌクレオチド以上、17ヌクレオチド以上、または20ヌクレオチド以上のフラグメントが、企図される。ポリヌクレオチドに特異的にハイブリダイズし得るプローブは、本発明のNTHiポリヌクレオチド配列を、遺伝子の同じファミリーにおける他のポリヌクレオチド配列から区別することができるか、またはNTHi遺伝子を他の細菌遺伝子から区別することができ、より好ましくは、独特のヌクレオチド配列に基づく。
【0016】
用語「ストリンジェント」とは、当該分野でストリンジェントとして一般に理解されている条件をいうために使用される。ハイブリダイゼーションのストリンジェンシーは、原理的に、温度、イオン強度、および変性剤(例えば、ホルムアミド)の濃度によって決定される。ハイブリダイゼーションおよび洗浄のためのストリンジェントなじょう件の例は、65〜68℃での0.015M 塩化ナトリウム、0.0015M クエン酸ナトリウムまたは42℃での0.015M 塩化ナトリウム、0.0015M クエン酸ナトリウム、および50% ホルムアミドである。Sambrookら,Molecular Cloning:A Laboratory Manual,第2版,Cold Spring Harbor Laboratory,(Cold Spring Harbor,N.Y.1989)を参照のこと。よりストリンジェントな条件(例えば、より高温、より低いイオン強度、より高いホルムアミドもしくは他の変性剤)もまた、使用され得るが、ハイブリダイゼーションの速度は影響を受ける。デオキシオリゴヌクレオチドのハイブリダイゼーションが考慮される場合において、さらなる例示的なストリンジェントなハイブリダイゼーション条件としては、37℃(14塩基オリゴについて)、48℃(17塩基オリゴについて)、55℃(20塩基オリゴについて)および60℃(23塩基オリゴについて)での6×SSC 0.05% リン酸ナトリウム中での洗浄が挙げられる。
【0017】
他の薬剤は、非特異的ハイブリダイゼーションかつ/またはバックグラウンドハイブリダイゼーションを減少させる目的で、ハイブリダイゼーション緩衝液および洗浄緩衝液中に含まれ得る。例は、0.1% ウシ血清アルブミン、0.1% ポリビニル−ピロリドン、0.1% ピロリン酸ナトリウム、0.1% ドデシル硫酸ナトリウムNaDodSO4(SDS)、フィコール、デンハルト溶液、超音波処理サケ精子DNA(または他の非相補的DNA)、および硫酸デキストランであるが、他の適切な薬剤もまた使用され得る。これらの添加物の濃度および型は、ハイブリダイゼーション条件のストリンジェンシーに実質的に影響を及ぼすことなく、変更され得る。ハイブリダイゼーション実験は、通常、pH6.8〜7.4で行われるが、代表的なイオン強度条件では、ハイブリダイゼーションの速度は、pHとはほとんど関係ない。Andersonら,Nucleic Acid Hybridisation:A Practical Approach,Ch.4,IRL Press Limited(Oxford,England)を参照のこと。ハイブリダイゼーション条件は、これらの変数を適合させ、異なる配列関連性のDNAにハイブリッドを形成させるために、当業者によって調節され得る。
【0018】
本発明の範囲内にある配列は、これらの特定の配列に限定されず、それらの対立遺伝子および種改変体もまた包含する。対立遺伝子および種改変体は、配列番号1〜576、配列番号675〜685に示される配列、および本明細書中の表3B、表4Bおよび表5に示されるヌクレオチド配列(好ましくはそれらのオープンリーディングフレーム、それらの代表的なフラグメント)、あるいは配列番号1〜576、配列番号675〜685に示される配列および本明細書中の表3B、表4Bおよび表5に示されるヌクレオチド配列と少なくとも90%同一(好ましくは95%同一)のヌクレオチド配列を、同種の別の単離体に由来する配列と比較することによって規定どおりに決定され得る。2つの配列間の同一性および類似性を決定するための好ましいコンピュータプログラムとしては、GCGプログラムパッケージ(Devereuxら、Nucl.Acid.Res.,12:387、1984;Genetics Computer Group、University of Wisconsin、Madison、WI、BLASTP、BLASTN、およびFASTA(Altschulら、J.Mol.Biol.,215:403−410、1990)が挙げられる)が挙げられるが、これらに限定されない。BLASTXプログラムは、全米バイオテクノロジー情報センター(NCBI)および他の供給元(BLAST Manual、Altschulら、NCB/NLM/NIH Bethesda、MD 20894;Altschulら、前出)から公的に入手可能である。公知のSmith Watermanアルゴリズムもまた、同一性を決定するために使用され得る。
【0019】
さらに、コドン可変性に適応するために、本発明は、同じアミノ酸配列をコードする核酸分子を包含し、本明細書中に開示される特定のオープンリーディングフレーム(ORF)もまた包含する。換言すると、ORFのコード領域において、あるコドンの、同じアミノ酸をコードする別のコドンへの置換は、明示的に包含される。
【0020】
本発明の単離されたポリペプチドとしては、以下:
配列番号1〜576、配列番号675〜685として示されるポリヌクレオチド配列に含まれるヌクレオチドならびに表3B、表4Bおよび表5に示されるヌクレオチド配列によってコードされるアミノ酸配列、あるいは対応する全長タンパク質または成熟タンパク質を含むポリペプチドが挙げられるが、これらに限定されない。本発明のポリペプチドとしては、本明細書中の表3B、表4Bおよび表5に示される配列番号616、配列番号618、配列番号620、配列番号622、配列番号624、配列番号626、配列番号628、配列番号630、配列番号632、配列番号634、配列番号636、配列番号638、配列番号640、配列番号642、配列番号644、配列番号646、配列番号648、配列番号650、配列番号652、配列番号654、配列番号656、配列番号658、配列番号660、配列番号662、配列番号664、配列番号666、配列番号668、配列番号670、配列番号672、配列番号674、配列番号687、配列番号689、配列番号691、配列番号693、配列番号695、配列番号697、配列番号699、配列番号701、配列番号703、配列番号705、配列番号707、配列番号709、配列番号711、配列番号713、配列番号715、配列番号717、配列番号719、配列番号721、配列番号723、配列番号725、配列番号727、配列番号729、配列番号731、配列番号733、配列番号735、配列番号737、配列番号739、配列番号741、配列番号743、配列番号745、配列番号747、配列番号749、配列番号751、配列番号753、配列番号755、配列番号757、配列番号759、配列番号761、配列番号763、配列番号765、配列番号767、配列番号769または配列番号771のアミノ酸配列が挙げられる。
【0021】
本発明のポリペプチドはまた、好ましくは以下:
(a)配列番号1〜576、配列番号675〜685として示されるヌクレオチド配列ならびに表3B、表4Bおよび表5に示されるヌクレオチド配列に含まれるオープンリーディングフレーム、あるいは
(b)ストリンジェントなハイブリダイゼーション条件下で(a)のポリヌクレオチドの相補体にハイブリダイズするポリヌクレオチド
によってコードされる生物学的活性または免疫学的活性を有するポリペプチドを包含する。
【0022】
本発明はまた、本発明のアミノ酸配列の生物学的また免疫学的に活性な改変体;ならびに生物学的活性および/または免疫学的活性を保持するそれらの「実質的な等価物」(例えば、少なくとも約65%、少なくとも約70%、少なくとも約75%、少なくとも約80%、少なくとも約85%、86%、87%、88%、89%、少なくとも約90%、91%、92%、93%、94%、代表的には、少なくとも約95%、96%、97%、より代表的には、少なくとも約98%、または最も代表的には、少なくとも約99%アミノ酸同一性を有するもの)を提供する。対立遺伝子改変体によってコードされるポリペプチドは、配列番号1〜576、配列番号675〜685に示されるヌクレオチド配列ならびに本明細書中で表3B、表4Bおよび表5に示されるヌクレオチド配列に含まれるポリヌクレオチドによってコードされるポリペプチド、および本明細書中で表3B、表4Bおよび表5に示されるアミノ酸配列を有するポリペプチドと比較して、類似の、増加した活性もしくは減少した活性を有し得る。
【0023】
NTHiペプチドとは、配列番号1〜576、配列番号675〜685に示されるヌクレオチド配列あるいは本明細書中で表3B、表4Bおよび表5に示されるヌクレオチド配列によってコードされるNTHiポリペプチドのフラグメント、および本明細書中で表3B、表4Bおよび表5に示されるアミノ酸配列を有するポリペプチドのフラグメントをいう。好ましいNTHiペプチドは、生物学的および/または免疫学的に活性である。
【0024】
本発明はさらに、本発明のNTHi核酸フラグメントまたは本発明の核酸フラグメントの縮重改変体によってコードされる、単離されたNTHiポリペプチドまたはNTHiペプチドを提供する。用語「変性改変体」とは、ヌクレオチド配列により本発明の核酸フラグメント(例えば、ORF)とは異なるが、遺伝暗号の縮重に依存してNTHiポリペプチド配列をコードするヌクレオチドフラグメントをいう。本発明の好ましい核酸フラグメントは、タンパク質をコードするORFである。
【0025】
本発明はまた、NTHiポリペプチドを提供し、このNTHiポリペプチドは、そのポリペプチドの生物学的活性および/または免疫学的活性に影響を及ぼさない1以上の保存的アミノ酸置換を有する。あるいは、本発明のNTHiポリペプチドは、保存的アミノ酸置換を有することを包含し、この保存的アミノ酸置換は生物学的活性を変化させても変化させなくてもよい。用語「保存的アミノ酸置換」とは、その置換位置においてアミノ酸の極性または電荷にほとんど影響を及ぼさないかもしくは全く影響を及ぼさないように、ネイティブでない残基(天然のアミノ酸および非天然のアミノ酸が挙げられる)を有する天然のアミノ酸残基の置換をいう。例えば、保存的置換は、ポリペプチド中の無極性残基の、任意の他の無極性残基との交換から生じる。さらに、ポリペプチド中の任意のネイティブな残基もまた、「アラニンスキャニング変異誘発」の方法に従って、アラニンに置換され得る。天然のアミノ酸は、以下:
塩基性:アルギニン、リジン、ヒスチジン;
酸性:グルタミン酸、アルパラギン酸;
無電荷で極性:グルタミン、アスパラギン、セリン、スレオニン、チロシン;および
無極性:フェニルアラニン、トリプトファン、システイン、グリシン、アラニン、バリン、プロリン、メチオニン、ロイシン、ノルロイシン、イソロイシン
のように、それらの側鎖に基づいて特徴付けられる。アミノ酸置換についての一般的な規定は、以下の表1に記載されている。
【0026】
【表1】

NTHiポリペプチドをコードするポリヌクレオチドに対して相補的なアンチセンスポリヌクレオチドもまた、提供される。
【0027】
本発明は、本発明のポリヌクレオチドが、増幅または発現のためにベクターに挿入され得ることを包含する。発現のために、ポリヌクレオチドは、必要に応じて、適切な発現制御配列(例えば、プロモーター配列およびポリアデニル化シグナル配列)に連結される。本発明のポリヌクレオチドを含む細胞が、さらに提供される。例示的な真菌宿主としては、E.Coli、Bacillus、Streptomyces、Pseudomonas、SalmonellaおよびSerratiaのような細菌が挙げられる。
【0028】
用語「単離された」とは、それが天然に存在する環境から取り出され、かつそれが天然に存在する環境の他の成分を実質的に含まない実体をいう。例えば、ポリペプチドは、他の細胞タンパク質から分離されるか、またはDNAは、それが天然に生じるゲノムにおいてそれに隣接する他のDNAから分離される。
【0029】
(抗体および免疫応答を誘発するための方法)
本発明は、NTHiポリペプチドに特有の(すなわち、NTHiポリペプチドに特異的な)抗原性エピトープに結合する抗体を提供する。また、複数のH.influenzaeサブタイプ間で共通だが、任意の他の抗原性エピトープに関して特有である抗原性エピトープに結合する抗体が提供される。その抗体は、ポリクローナル抗体、モノクローナル抗体、それらの特有のエピトープに結合する能力を保持する抗体フラグメント(例えば、Fvフラグメント、FabフラグメントおよびF(ab)2フラグメント)、一本鎖抗体およびヒト抗体もしくはヒト化抗体であり得る。抗体は、当該分野で標準的な技術によって生成され得る。
【0030】
当該分野において、H.influenzaeの莢膜多糖類に対する抗体が、インビトロアッセイで細菌を殺す能力を示すことが公知である。これらの抗体はまた、動物モデル系において、H.influenzaeを用いるチャレンジから保護することが公知である。これらの研究は、莢膜多糖類に対する抗体がヒトにおいて防御免疫反応を誘導する可能性を有することを示す。本発明は、本発明のNTHiポリペプチドおよびそれらのフラグメントに特異的な抗体を提供し、それは、H.influenzae細菌を殺す活性およびヒトをNTHi感染から守る活性の両方を示す。本発明はまた、本発明のNTHiポリペプチドに特異的な抗体を提供し、それは、毒性を減少し、付着を阻害し、細胞分裂を阻害し、そして/あるいはH.influenzae細菌の上皮への侵入を阻害するかまたはH.influenzae細菌の食作用を促進する。
【0031】
インビトロ補体媒介性殺菌アッセイシステム(Musherら、Infect.Immun.39:297−304、1983;Andersonら、J.Clin.Invest.51:31−38、1972)が、抗NTHi抗体の殺菌活性を測定するために使用され得る。NTHiポリペプチドおよびNTHiペプチドの防御抗体反応を誘導する活性におけるさらなるデータは、本明細書中に記載されるチンチラモデル系のような感染の動物モデルを使用することによって、生成され得る。
【0032】
NTHiのエピトープに対して予備形成された抗体(例えば、NTHi、OMP、LOSまたは非被膜タンパク質に対する抗体)の投与を介する受動免疫治療によって宿主に短期間の防御を与えることもまた、可能である。従って、意図されたワクチン処方物が、受動免疫治療で使用するための抗体を産生するために使用され得る。ヒト免疫グロブリンがヒト医薬では好ましい。なぜなら、異種免疫グロブリンは、その外来の免疫原成分に対して免疫応答を誘発し得るからである。そのような受動免疫付与は、特別な危険性に曝された非免疫性の個体の即時的保護のための緊急性を根拠に使用され得る。あるいは、これらの抗体は、抗イディオタイプ抗体の産生に使用され得、それは次にNTHiエピトープに対する免疫応答を刺激するための抗体として使用され得る。
【0033】
本発明は、個体においてNTHiに対する免疫応答を誘導する方法を包含する。これらの方法は、NYHi細菌を殺す免疫応答、および細胞へのH.influenzae付着をブロックする免疫応答を包含する。1つの実施形態において、その方法は、本発明のNTHiタンパク質またはNTHiペプチドを含む組成物の免疫原性用量を投与する工程を包含する。別の実施形態において、その方法は、本発明のNTHiタンパク質またはNTHiペプチドを発現する細胞を含む組成物の免疫原性用量を投与する工程を包含する。なお別の実施形態において、その方法は、本発明のNTHiタンパク質またはNTHiペプチドをコードするポリヌクレオチドを含む組成物の免疫原性用量を投与する工程を包含する。そのポリヌクレオチドは、任意の他の核酸とは無関係の裸の(naked)ポリヌクレオチドであっても、プラスミドまたはウイルスベクター(例えば、アデノ随伴ウイルスベクターまたはアデノウイルスベクター)のようなベクター中に存在していてもよい。組成物の投与は、当該分野で標準的な経路(例えば、非経口、静脈内、経口、口腔、鼻、肺、直腸、または膣)によってであり得る。その方法は、単一の個体において組み合わせて使用され得る。その方法は、個体のNTHi感染前または感染後に使用され得る。
【0034】
「免疫学的用量」は、上記個体をNTHi感染(特に、中耳および/または鼻咽頭もしくは下気道のNTHi感染)から守るために抗体免疫応答および/またはT細胞免疫応答を発生させるのに十分な用量である。そのような免疫学的応答が細菌複製を遅延させる方法もまた、提供される。本発明のさらなる局面は、免疫学的組成物に関し、それは、個体に取り込まれた場合に免疫学的応答を誘導し得るかまたは誘導する。その免疫学的応答は、治療的または予防的に使用され得、そしてCTLまたはCD4+ T細胞から生じるような抗体免疫または細胞免疫の形態を取り得る。NTHiタンパク質またはその抗原性ペプチドは、補タンパク質と融合され得、その補タンパク質は、それ自体抗体を産生し得ないが、第1のタンパク質を安定化し得、そして免疫原性特性および保護特性を有する融合タンパク質を生成し得る。従って、融合組換えタンパク質は、好ましくは、免疫原性の補タンパク質(例えば、グルタチオン−S−トランスフェラーゼ(GST)またはβ−ガラクトシダーゼ、タンパク質を可溶化させその生成および精製を促進する比較的大きな補タンパク質)をさらに含む。さらに、補タンパク質は、免疫系の一般的な刺激を提供するという意味でアジュバントとして作用し得る。補タンパク質は、第1のタンパク質のアミノ末端またはカルボキシ末端のいずれかに結合し得る。本発明のポリヌクレオチドまたはその免疫原性ペプチドによってコードされるNTHiポリペプチドを含む組成物(特にワクチン組成物)、および方法が、本発明によって提供される。
【0035】
本発明は、それに対応して、NTHi感染に対する免疫応答を誘導するのに適切な組成物を提供し、ここで、誘導される抗体は、宿主細胞へのNTHi細菌の結合をブロックする。その組成物は、本発明のNTHiタンパク質もしくはNTHiペプチド、NTHiポリペプチドを発現する細胞、またはそのポリペプチドをコードするポリヌクレオチドを含む。その組成物はまた、キャリアおよびアジュバントのような他の成分を含み得る。
【0036】
中耳および/または鼻咽頭のNTHi感染においてアップレギュレートされる遺伝子、ならびにNTHi毒性に関連する遺伝子が、本明細書中に記載される。これらのNTHi遺伝子によってコードされるそれらのポリペプチドおよびペプチドは、OMのようなNTHi感染に関連する障害を処置または予防するために免疫応答を誘導するのに有用であることが意図される。これらの遺伝子によってコードされるポリペプチドのいくつかとしては、以下が挙げられる:ヒスチジン生合成タンパク質、リポタンパク質B、ペプチドABCトランスポーター、ペリプラスムSapA前駆体、外膜リポタンパク質キャリアタンパク質前駆体、リボース輸送系パーミアーゼタンパク質、ホスホリボシルアミノイミダゾールカルボキシラーゼ触媒性サブユニット、PurE、ホスホリボシルアミノイミダゾールカルボキシラーゼ触媒性サブユニット、オルニチンカルバモイルトランスフェラーゼ、マンノネートデヒドラターゼ、ジスルフィドオキシドレダクターゼ、ウレアーゼ補タンパク質、ホスホコリンシチジリルトランスフェラーゼ、推定ピリドキシン生合成タンパク質、一重項酸素耐性タンパク質、細胞内隔壁タンパク質(intracellular septation protein)、DNA修復タンパク質、MukFタンパク質、グリセロール−3−ホスフェートレギュロンリプレッサー、融合宿主因子βサブユニット、アルギニンリプレッサー、低温ショック様タンパク質、ストレス反応タンパク質、LicA、MukF、RadAおよびHI0094、HI1163、HI0665、HI1292、HI1064、HI186、HI0352遺伝子によってコードされるそれらの仮想タンパク質。NTHi OMP、LOSおよび非被膜タンパク質もまた、NTHi感染に関連する障害の予防および処置のための免疫応答を誘導すると意図される。
【0037】
本発明の組成物の「免疫原性用量」は、投与前に検出可能な免疫応答と比較してか、または投与前の標準的な免疫応答と比較して、投与後に、検出可能な体液性免疫応答および/または細胞性免疫応答を生じる用量である。本発明は、方法から生じる免疫応答が、予防的および/または治療的であり得ることを意図する。
【0038】
本発明は、個体において宿主細胞に対するNTHi細菌の結合をブロックする方法を包含する。その方法は、NTHi細胞の付着の結合をブロックする本発明の抗体またはポリペプチドを投与する工程を包含する。あるいは、NTHi細胞付着の結合をブロックする1以上の低分子の投与が意図される。インビトロアッセイは、本発明の抗体、ポリペプチドまたは低分子がNTHi細胞付着をブロックする能力を示すために使用され得る。
【0039】
NTHi細胞の付着をブロックする本発明の抗体、本発明のポリペプチドおよび/または本発明の低分子を含む薬学的組成物が提供される。その薬学的組成物は、上記の活性成分のうちの1つ単独からなっても、上記の活性成分の組み合わせを含んでも、細菌感染を処置するために使用されるさらなる活性成分を含んでもよい。その薬学的組成物は、薬学的に有効なキャリアのような1以上のさらなる成分を含み得る。薬学的組成物の投与の投薬量および頻度は、標準的な技術によって決定され、例えば、個体の体重および年齢、投与経路、ならびに症状の重篤度に依存する。薬学的組成物の投与は、当該分野で標準的な経路(例えば、非経口、静脈内、経口、口腔、鼻、肺、直腸、または膣)によってであり得る。
【0040】
個体におけるNTHi感染を検出するための方法もまた、提供される。1つの実施形態において、これらの方法は、サンプル中の本発明のNTHiポリヌクレオチドに特異的に結合するプライマーまたはプローブを使用して、そのポリヌクレオチドを検出する工程を包含する。そのポリヌクレオチドの検出は、例えば、ハイブリダイゼーションおよびPCRに関連する当該分野での多くの技術的慣用法によって達成され得る。
【0041】
本発明の抗体はまた、H.influenzaeの感染が疑われる個体の種々の体液中でNTHi抗原(NTHiポリペプチドおよびそのペプチド)を検出するための診断アッセイにおける用途のための試薬を提供するのに使用され得る。別の実施形態において、本発明のNTHiタンパク質およびNTHiペプチドは、種々の患者の組織および体液中でのNTHiの検出のためのイムノアッセイにおける抗原として使用され得る。これらの組織および流体としては、血液、血漿、耳滲出液(ear fluid)、脊髄液、唾、尿、リンパ液、および脳髄液が挙げられるが、これらの限定されない。本発明の抗原は、当該分野で公知のイムノアッセイのいずれかにおいて使用され得る。これらのイムノアッセイとしては、ラジオイムノアッセイ、ELISAアッセイ、サンドウィッチアッセイ、沈降素反応、ゲル拡散沈降素反応、免疫拡散アッセイ、凝集アッセイ、発光イムノアッセイ、プロテインAイムノアッセイおよび免疫電気泳動アッセイが挙げられるが、これらの限定されない。
【0042】
(ワクチン化学療法標的)
本発明の1つの局面は、個体(特に哺乳類)における免疫応答を誘導するための方法に関連し、この方法は、NTHi抗原タンパク質またはそれらの抗原性ペプチドをその個体に接種する工程を包含する。
【0043】
本発明はまた、本発明の免疫原性組換えNTHiタンパク質またはNTHiペプチドを適切なキャリアとともに含むワクチン処方物を提供する。ワクチン候補および/または化学療法の標的として企図される、NTHiポリペプチドおよびそれらのペプチドとしては、以下が挙げられるが、これらに限定されない:ヒスチジン生合成タンパク質、リポタンパク質B、ペプチドABCトランスポータ、ペリプラズムSapA前駆体、外膜リポタンパク質キャリアタンパク質前駆体、リボース輸送系透過酵素タンパク質、ホスホリボシルアミノイミダゾールカルボキシレース触媒サブユニット、PurE、3,4−ジヒドロキシ(hydroxt)−2−ブトン4−ホスフェートシンターゼ、オミニチンカルバモリルトランスフェラーゼ、マンノエートデヒドラターゼ、ジスルフィドオキヒドレダクターゼ、ウレーゼ、オミチンカルバモリルトランスフェラーゼ(omithine carbamolytransferase)、マンノネートデヒドラターゼ(mannonate dehydratase)、ジスフィドオキシドレダクターゼ(disulfide oxidoreductase)、ウレアーゼ修飾タンパク質(urease accessory protein)、ホスホコリンシチジリトランスフェラーゼ(phospshocholine cytidylytransferase)、推定ピリドキシン生合成タンパク質、一重項酸素耐性タンパク質(singlet oxygen resistance protein)、細胞内隔膜タンパク質(intracellular septation protein)、DNA修復タンパク質、MUKFタンパク質、グリセロール−3−ホスフェートレギュロンリプレッサー、組込み宿主因子βサブユニット、アルギニンリプレッサー、冷ショック様タンパク質、ストレス応答性タンパク質、LicA、RadA、ならびにHI0094遺伝子、HI1163遺伝子、HI0665遺伝子、HI1292遺伝子、HI1064遺伝子、HI1386遺伝子、HI0352遺伝子によってコードされるそれらの推定タンパク質、NTHiOMP、NTHi LOSおよびNTHi非カプセル状タンパク質および配列番号1〜576、配列番号675〜685に示されるヌクレオチド配列、ならびに本明細書中の表3B、表4および表5に示されるヌクレオチド配列において存在するその新規NTHiポリヌクレオチド配列によってコードされるポリペプチド、ならびに本明細書中の表3B、表4B、および表5に示されるアミノ酸配列を有するポリペプチド。
【0044】
このタンパク質は、胃の中で分解され得るので、好ましくは、このタンパク質は、非経口的に投与され、この投与としては、例えば、皮下、筋肉内、静脈内、皮内である投与が挙げられる。非経口投与にとって適切な処方物としては、抗酸化剤、緩衝液、静菌剤、およびその個体の体液、好ましくは血液と等張である処方物を与える溶質を含み得る、水溶性および非水溶性の滅菌注入溶液;ならびに懸濁剤または濃化剤を含み得る水性および非水性の滅菌懸濁物が挙げられる。これらの処方物は、単位用量の容器または複数回用量の容器中に提供され得る。これらの容器としては、例えば、密閉したアンプルおよびバイアルである。そして、これらの処方物は、使用直前に、滅菌液体キャリアの添加のみが必要とされる凍結乾燥条件で貯蔵され得る。このワクチン処方物はまた、その処方物の免疫原性を増強するためのアジュバント系(例えば、水中油系および当該分野で公知の系)を包含し得る。この投与は、そのワクチンの比活性に依存し、そして、慣用的な実験によって容易に決定されされ得る。
【0045】
(A.ペプチドワクチン)
ペプチド治療剤(例えば、ペプチドワクチン)は、当該分野において周知であり、薬学分野において使用が増加している。このようなペプチド化合物の非経口投与の一貫した欠点は、破壊または変性が速いことである。注入ポンプあんらびびワックスまたはオイル移植物は、ペプチド様治療剤の存在を長引かせることおよびこのような薬剤の完全性を保存することの両方のために、治療剤の長期投与のために使用されている。さらに、ペプチド様薬剤は、(ペプチド様薬剤の各エピトープに対する特定の参照とともに)長期間、天然の状態の構成を理想的に維持し、さらに、チャレンジされた動物または免疫されたヒトにおいて免疫原性応答を誘発するために適した様式で提示されるべきである。
【0046】
本発明のNTHi抗原性ペプチドは、多くの従来の方法で調製され得る。短いペプチド配列は、標準的な手段を使用する化学合成によって調製され得る。固相技術(例えば、Eriksonら、The Proteins(1976)v.2,Academic Press,New York,p255を参照のこと)が特に簡便である。自動固相合成機は、市販される。さらに、配列の改変は、適切な残基の置換、付加または省略によって容易になされる。例えば、システイン残基は、カルボキシ末端に付加されて、キャリアタンパク質への簡便な連結のためのスルフヒドリル基を提供し得るか、またはスペーサー要素(例えば、追加のグリシン残基)が、C末端の連結アミノ酸とペプチドの残りとの間の配列内に組み込まれ得る。短いNTHiペプチドはまた、組換え技術によって産生され得る。この長さのペプチドについてのコード配列は、化学技術(例えば、Matteucciら、J Am Chem Soc.,103:3185(1981)に記載されるホスホトリエステル法)によって容易に合成され得る。
【0047】
本明細書中で企図されるNTHiペプチド配列のうちのいくつかは、免疫原性であるには小さすぎると考えられ得、これらは、それらにこの特質を与えるためにキャリア物質に連結され得る。当該分野で公知のこのような連結を作製するための任意の方法が使用され得る。連結は、一方の官能基末端にジスルフィド連結を生成し、そして他方のペプチド連結を生じるヘテロ二官能性剤(例えば、ジスルフィドアミド形成剤(例えば、N−スクシジミジル−3−(2−ピリジルジチオ)プロプリオネート(SPDP)(例えば、Jansenら、,Immun.Rev.62:185,1982を参照のこと)))およびジスルフィド連結以外のチオエーテルを形成する二官能性連結剤(例えば、6−マレイミドカプロン酸、2−ブロモ酢酸、2−ヨード酢酸、4−(N−マレイミド−メチル)シクロヘキサン−1−カルボン酸など)、ならびにカルボキシル基をスクシンイミドまたは1−ヒドロキシ−2−ニトロ−4−スルホン酸(スクシンイミジル4−(N−マレイミド−メチル)シクロヘキサン−1−カルボキシレート(SMCC)のようなナトリウム塩に対して)と組み合わせることによってカルボキシル基を活性化するカップリング剤を用いて形成され得る。
【0048】
(B.ワクチン組成物および投与)
プライミング用量の免疫原(これは、免疫原への一回以上のブースター曝露が続く)は、有効ワクチンであることが必要であり得る(Krampら、Infect.Immun.,25:771−773,1979;Davisら、Immunology Letters,14:341−8 1986 1987)。同時投与される場合、免疫応答を有利に増強し得るタンパク質またはポリペプチドの例としては、サイトカイン(例えば、IL−2、Il−12、GM−CSF)、サイトカイン誘導分子(例えば、Leaf)または同時刺激分子が挙げられる。ヘルパー(HTL)エピトープは、細胞内標的化シグナルに連結され得、CTLエピトープから別々に発現され得る。これにより、CTLエピトープとは異なる細胞区画へのHTLエピトープの指向が可能になる。必要な場合、これは、HTLエピトープのMHCクラスII経路へのより効率的な進入を容易にし得、それによって、VTL誘導を改善する。CTL誘導と対照的に、免疫抑制分子(例えば、TGF−β)の同時発現による免疫応答を特異的に減少することは、特定の疾患において有利であり得る。
【0049】
理想的には、免疫原は、2つ特性を示す;対応する抗体の形成を刺激する能力、およびこれらの抗体と特異的に反応する傾向。免疫原は、抗体の部位を組み合わせることによって認識可能な免疫原の最も小さな部分である1つ以上のエピトープを有する。特定の場合において、免疫原、免疫原の画分または免疫原が提示される条件は、促進するには不適切であり、不十分な免疫を生じる。これは、しばしば、免疫原として使用されるペプチドまたは他の低分子の場合である。他の物質(例えば、免疫調節剤(例えば、インターロイキンのようなサイトカイン)は、同様にワクチンにおいて組み合わされ得る。
【0050】
上記のワクチンの分野は、免疫原と組み合わせて使用した場合に免疫応答を増強する特定の物質(アジュバントと呼ばれる)の使用を認識する。アジュバントは、そのアジュバントを使用することなく惹起されるよりも速いかまたは大きい免疫応答を惹起するために、さらに使用される。さらに、アジュバントは、アジュバントを含まずに必要とされるよりも少ない免疫原を使用して免疫学的応答を生成するため、免疫学的防御を与える特定の抗体物質の生成を増加するため、またはその免疫応答(例えば、体液性免疫応答、細胞性免疫応答)の成分を増強するために、使用され得る。公知のアジュバントとしては、エマルジョン(例えば、フロイントアジュバントおよび他のエマルジョン)、Bordetella pertussis、MF59、Quillaja saponaria由来の精製サポニン(QS21)、アルミニウム塩(例えば、水酸化アルミニウム、リン酸アルミニウム、およびミョウバン)、リン酸カルシウム(および他の金属塩)、ゲル(例えば、水酸化アルミニウム塩)、マイコバクテリア生成物(ムラミルジペプチドを含む)、固体物質、粒子(例えば、リポソームおよびビロソーム)が挙げられる。アジュバントとして使用されることが公知である天然生成物および細菌生成物の例としては、モノホスホリルリピドA(MPL)、RC−59(合成MPL様アシル化単糖)、OM−174(これは、E.coli由来のリピドA誘導体である)、ホロ毒素(例えば、コレラ毒素(CT)もしくはその誘導体のうちの1つ、破傷風毒素(PT)、およびE.coliの易熱性毒素(LT)、もしくはその誘導体のうちの1つ)、ならびにCpGオリゴヌクレオチドが挙げられる。アジュバント活性は、多数の要因(例えば、キャリア効果、デポー(depot)形成、変化したリンパ球動員、Tリンパ球の刺激、Bリンパ球の直接刺激、およびマクロファージの刺激)によって影響を受け得る。
【0051】
ワクチンは、代表的には、注射可能な物質として、液体溶液または液体懸濁液のいずれかとして、調製される。注射前に液体中の溶液または懸濁液のために適切な固体形態もまた、調製され得る。その調製物はまた、乳濁され得る。その活性な免疫原性成分は、しばしば、賦形剤(これは、薬学的に受容可能であり、そしてその活性成分と適合性である)と混合される。適切な賦形剤は、例えば、水、生理食塩水、デキストロース、グリセロール、エタノールなど、およびそれらの組み合わせである。さらに、望ましい場合、そのワクチンは、少量の補助物質(例えば、湿潤剤または乳化剤、pH緩衝剤、またはアジュバント(これは、そのワクチンの有効性を増強する)を含み得る。このワクチンは、注射(例えば、皮下注射または筋肉内注射のいずれか)によって、従来のように非経口投与される。他の投与様式のために適切なさらなる処方物としては、坐剤が挙げられ、時には、経口処方物が挙げられる。坐剤について、従来の結合剤およびキャリアとしては、例えば、ポリアルカレングリコールまたはトリグリセリドが挙げられ得る。そのような坐剤は、0.5%〜10%、好ましくは1〜2%の範囲で上記活性成分を含む混合物から形成され得る。経口処方物は、通常使用される賦形剤(例えば、薬剤グレードのマンニトール、ラクトース、デンプン、ステアリン酸マグネシウム、サッカリンナトリウム、セルロース、炭酸マグネシウムなど)を含む。これらの組成物は、溶液、懸濁物、錠剤、ピル剤、カプセル剤、徐放処方物または散剤の形態を採り得、10%〜95%(好ましくは25〜70%)の活性成分を含む。
【0052】
ワクチンはまた、ジェット注射器、微小針、エレクトロポレーション、ソノポレーション、微小カプセル化、ポリマーまたはリポソームを使用して経皮経路を介して、噴霧器、エアロゾル、および鼻スプレーを使用して経粘膜経路および鼻腔内経路を介して、投与され得る。天然ポリマーまたは合成ポリマー(例えば、デンプン、アルギネート、およびキトサン、D−ポリL−ラクテート(PLA)、D−ポリDL−乳酸−co−グリコール酸ミクロスフェア、ポリカプロラクトン、ポリオルトエステル、ポリ無水物、およびポリホスファゼン、ポリホスファタザン)を使用するマイクロカプセル化が、経皮投与および経粘膜投与の両方のために有用である。合成ポリ−オルニテート、ポリ−リジン、およびポリ−アルギニンまたは両親媒性ペプチドを含む、ポリマー性複合体は、経皮送達系のために有用である。さらに、それらの両親媒性の性質に起因して、リポソームは、経皮ワクチン送達系、経粘膜ワクチン送達系、および鼻腔内ワクチン送達系のために企図される。ワクチン送達のために使用される一般的脂質としては、N−(1)2,3−(ジオレイル−ジヒドロキシプロピル)−N,N,N−トリメチルアンモニウム−メチルスルフェート(DOTAP)、ジオレイルオキシ−プロピル−トリメチルアンモニウムクロリド(DOTMA)、ジミリスチルオキシプロピル−3−ジメチル−ヒドロキシエチルアンモニウム(DMRIE)、ジメチルジオクタデシルアンモニウムブロミド(DDAB)、および9N(N’,N’−ジメチルアミノエタン)カルバモイルコレステロール(DC−Chol)が、挙げられる。ヘルパー脂質とリポソームとの組み合わせは、皮膚を通してのそのリポソームの取り込みを増強する。これらのヘルパー脂質としては、ジオレイルホスファチジルエタノールアミン(DOPE)、ジラウロイルホスファチジルエタノールアミン(DLPE)、ジミリストイルホスファチジルエタノールアミン(DMPE)、ジパルミトイルホスファチジルエタノールアミン(DPPE)が挙げられる。さらに、トリテルペノイドグリコシドまたはチリセッケンボク(Quillaja saponaria)由来のサポニンは、鼻腔内ワクチン送達および経粘膜ワクチン送達のための有用なアジュバントとして企図されている。
【0053】
タンパク質は、中性形態または塩形態としてワクチンに処方され得る。薬学的に受容可能な塩としては、酸付加塩(ペプチドの遊離アミノ基と共に形成される)が挙げられ、これは、例えば、塩酸もしくはリン酸のような無機酸、または酢酸、シュウ酸、酒石酸、マンデル酸のような有機酸と共に形成される。遊離のカルボキシ基と共に形成される塩はまた、例えば、ナトリウム、カリウム、アンモニウム、カルシウム、または水酸化第2鉄のような無機塩、およびイソプロピルアミン、トリメチルアミン、2−エチルアミノエタノール、ヒスチジン、およびプロカインのような有機塩に由来し得る。
【0054】
ワクチンは、投薬処方物に適合する様式、そして治療的に有効かつ免疫原性な量で投与される。投与される量は、処置される被験体、被験体の免疫系が抗体を合成する能力、および所望される防御の程度に依存する。投与するのに必要な活性成分の正確な量は、開業医の判断に依存し、各個体で特有である。しかし、適切な投薬量範囲は、1個体当たり数百マイクログラムの活性成分のオーダーである。初回投与およびブースター投与のための適切なレジメンもまた可変性であるが、初回投与後1〜3ヶ月の間隔に次いでその後の注射もしくは他の投与に代表される。
【0055】
本明細書中に記載されるようにワクチン組成物で免疫化する際に、宿主の免疫系は、所望の抗原に特異的な大量のCTLを産生することによってワクチンに応答し、そしてその宿主は、少なくとも部分的に免疫化されて感染を遅らせるかまたは発生している慢性的感染に耐性を示す。本発明のNTHiポリペプチドまたはNTHiペプチドを含むワクチン組成物は、細菌感染に感受性であるかもしくはそれ以外では細菌感染の危険性を有する患者に投与されて、その抗原に対する免疫応答を誘発する。従って、患者自身の免疫応答能が増強される。そのような量は、「免疫原性的に有効な用量」として規定される。この使用において、正確な量は、この場合もまた、患者の健康状態および年齢、投与形式、処方物の性質などに依存するが、一般的には、70kgの患者1人当たり約1.0μg〜約5000、より一般には70kgの体重当たり約10〜500mgである。治療目的または免疫目的のために、本発明のNTHiポリペプチドまたはNTHiペプチドはまた、弱毒化したウイルス宿主(例えば、ワクチンまたは鶏痘)によって発現され得る。このアプローチは、本発明のペプチドをコードするヌクレオチド配列を発現するためのベクターとして、ワクチンウイルスの使用を含む。急性的にかもしくは慢性的に感染した宿主への導入、または非感染宿主への導入の際、組換えワクチンウイルスは、免疫原性ペプチドを発現し、それによって、宿主CTL応答を促進する。
【0056】
体液性免疫応答は、多くの周知の方法(例えば、単一放射状免疫拡散アッセイ(SRID)、酵素免疫アッセイ(EIA)、および赤血球凝集阻害アッセイ(HAI)によって測定され得る。詳細には、SRIDは、試験される免疫原を含むゲル(例えば、アガロース)の層を利用する。ウェルが、そのゲル中に切除され、試験される血清が、そのウェル中に配置される。そのゲルの外から中への抗体の拡散は、沈殿環の形成をもたらす。その沈殿環の領域は、試験される血清中の抗体濃度に比例する。EIA(ELISA(酵素結合イムノアッセイとしても公知)は、サンプル中の全抗体を決定するために使用される。その免疫原は、マイクロタイタープレートの表面に吸着される。その試験血清は、そのプレートに暴露され、その後、酵素結合した免疫グロブリン(例えば、IgG)に暴露される。そのプレートに付着する酵素活性は、従来の任意の手段(例えば、分光光度法)によって定量され、その活性は、その試験サンプル中に存在する免疫原に対する抗体の濃度に比例する。HAIは、免疫原(例えば、ウイルスタンパク質)がニワトリ赤血球(など)を凝集する能力を利用する。このアッセイは、中和抗体(すなわち、赤血球凝集を阻害可能な抗体)を検出する。その試験血清の希釈物が、標準濃度の免疫原とともにインキュベートされ、その後、上記赤血球が添加される。中和抗体の存在は、その免疫原による赤血球の凝集を阻害する。細胞性免疫応答を測定するための試験としては、遅延型過敏症の決定または標的免疫原に対するリンパ球の増殖応答の測定が挙げられる。
【0057】
(分類不能型インフルエンザ菌(Haemophilus influenzae)(NTHi))
H.influenzaeは、小さい非運動性のグラム陰性細菌である。他のH.influenzae株とは異なり、分類不能型H.influenzae(NTHi)下部は、多糖莢膜を欠き、時には、「非カプセル型」と示される」NTHi株は、カプセル型株とは遺伝的に別個であり、b型H.influenzae単離株よりも異種である。NTHiは、ヒト宿主に対して複雑した抗原群を提示する。防御を惹起し得る可能な抗原としては、OMP、リポ多糖、リポタンパク質、接着タンパク質、および非莢膜タンパク質が挙げられる。
【0058】
ヒトは、H.influenzaeのための唯一の宿主である。NTHi株は、呼吸管上部(鼻咽頭および口腔咽頭部後部を含む)、呼吸管下部および女性生殖管に一般的に存在する。NTHiは、ヒトにおいて広範なスペクトルの疾患を引き起こす。その疾患としては、中耳炎、肺炎、静脈洞炎、敗血症、心内膜炎、喉頭蓋炎、化膿性関節炎、髄膜炎、分娩後感染症および新生児感染症、分娩後敗血症および新生児敗血症、急性卵管炎および慢性卵管炎、喉頭蓋炎、心膜炎、蜂巣炎、骨髄炎、心内膜炎、胆嚢炎、腹内感染、泌尿器管感染、乳様突起炎、大動脈移植片感染、結膜炎、ブラジル紫斑熱、潜在性菌血症、ならびに基礎となる肺疾患(例えば、慢性肺炎、気管支炎、および嚢胞性線維症)の憎悪が挙げられるが、これらに限定されない。
【0059】
NTHiの疫学的研究は、この株が、外膜タンパク質プロフィール(Barenkampら、Infect.Immun.36:535〜40,1982)、酵素アロタイプ(Musserら、Infect.Immun.52:183〜191,1986)および他の一般的に使用される疫学的ツールに関して、不均質であることを示した。NTHiをサブタイプに分けるいくつかの試みが存在したが、それらの方法論はいずれも、完全には満足のいくものではなかった。NTHiの外膜タンパク質の組成は、約20種のタンパク質からなる。すべてのNTHi株は、2種の共通するOMPを含み、これらは、分子量が30,000ダルトンおよび16,600ダルトンである。NTHi株は、32,000〜42,000ダルトンの範囲内にある2種のOMPに基づいてサブタイプ分けされ得る。そのNTHiリポ多糖プロフィールは、腸内グラム陰性細菌とは基本的に異なり、20,000ダルトン未満の範囲の1〜4つの別個のバンドへと分かれる。
【0060】
プロトタイプNTHi単離株は、慢性中耳炎に罹患していた子供から回収された、低継代単離株86−028NPである。この株は、インビトロで(Bakaletzら、Infect.Immun.53:331〜5,1988;Holmesら、Microb.Pathog.23:157〜66,1997)ならびにチンチラOMモデル(本明細書中に記載される)(Bakaletzら、Vaccine 15:955〜61,1997;Suzukiら、Infect.Immun.62:1710〜8,1994;DeMariaら、Infect.Immun.64:5187〜92,1996)で、充分に特徴付けられている。この86−028NP株は、本明細書中に記載されるように、チンチラ中耳炎モデルにおいて発現がアップレギュレートされる遺伝子を同定するため、およびそのチンチラの中耳におけるNTHiの生存のために必要な遺伝子を同定するために、使用された。
【0061】
(DFI戦略)
差次的蛍光誘導(DFI)戦略が、本明細書中において、チンチラ動物モデルにおいてOMの間に誘導されるNTHi遺伝子を同定するために使用された。いくつかの方法が、感染の間の生物の毒性に寄与する細菌遺伝子を同定するために開発されている。そのような方法としては、細菌プロモーターが、宿主における生存のために必要な必須栄養素の合成のために必要とされる遺伝子の発現を調節する、インビボ発現方法(IVET);変異した場合に微生物のビルレンス特性を変化させる遺伝子のタグ特異的同定を可能にする、サイン(signature)タグ化変異誘発(STM);転写的に活性ない電子について全体的にスクリーニングするためのDNAマイクロアレイ技術、および転写的に活性なプロモーターについて選択するためにFACS分析を使用するDFI(Chiangら、Ann.Rev.Microbiol.53:129〜154,1999)。DFIは、基礎発現レベルに関わらず差次的に調節される遺伝子の同定を可能にし、かつインビトロでの増殖のために必須の遺伝子は排除しない、高スループット方法である。
【0062】
DFIは、多くの微生物において首尾良く利用されている。例えば、GFPレポーター系およびフローサイトメトリーが、マクロファージと相互作用する際の微生物遺伝子発現を研究するために使用された(Dhandayuthapaniら、Mol.Microbiol.17:901〜912,1995)。インビボ増殖を刺激する環境にSalmonellaeが供された場合、および培養マクロファージ様細胞によりインターナライズされた場合に、転写が増加される遺伝子を同定するために、プロモータートラップ系が、使用された(ValdivaおよびFalkow,Mol.Microbiol.22:367〜378,1996;ValdiviaおよびFalkow,Science,277:2007〜2011,1997;ValdiviaおよびFalkow,Curr.Opin.Microbiol.1:359〜363,1998)。さらに、DFIは、感染を模倣する変化したインビトロ状態下で増殖した場合にS.pneumoniaeおよびS.aureusにおいて発現されるプロモーターを同定するために、使用された。さらに、DFIは、Bacillus cereusにおいて、環境刺激に応答した遺伝子調節を研究するために(DunnおよびHandelsman、Gene,226:297〜305,1999)、S.pneumoniaeにおいてコンピテンス刺激ペプチドに応答した遺伝子調節を研究するために(Bartilsonら、Mol.Microbiol.39:126〜135,2001)、Bartonella henselae(LeeおよびFalkow,Infect.Immun.66:3964〜3967,1998)、Listeria monocytogenes(Wilsonら、Infect.Immun.69:5016〜5024,2001)、Brucella abortus(Eskraら、Infect.Immun.69:7736〜7742,2001)、およびEscherichia coli(Badgerら、Mol.Microbiol.36:174〜182,2000)における宿主細胞との相互作用および宿主細胞の侵襲の際の遺伝子調節を研究するために、使用された。
【0063】
DFIは、感染の細胞培養モデルまたはインビボ環境を刺激するように設計されたインビトロ条件において活性なプロモーターを同定するために首尾良く使用されているが、動物全体において特定の生物学的ニッチにおいて調節されるプロモーターを同定するためには、DFIはほとんど適用されていない。このことは、おそらく、インビボ環境から選別することに関連する多数の難問に起因する。宿主の免疫応答、感染部位からの細菌細胞の蔓延おおび/または除去、ならびに上皮細胞に対する細菌の付着(おそらく、バイオフィルム形成を介する)は、細菌を、生存動物から回収するために接近不能にし得る。これらの要因は、とりわけ、細菌がこれらの難問を感知して応答する場合に、微小環境の複雑性および遺伝子発現の不均一性に寄与する。最近、DFIは、最近が、呼吸器管感染のマウスモデルおよびOMのアレチネズミ感染モデルにおいてスクリーニングされた場合に、S.pneumoniaeにおいて発現されるプロモーターを同定するために使用されている(Marraら、Infect.Immun.70:1422〜33,2002;Marraら、Microbiol.148:1483〜91,2002)。
【0064】
(動物モデル)
上記チンチラモデルは、OMについての広範に受容されている実験モデルである。特に、NTHi誘導性OMのチンチラモデルは、十分に特徴付けられており(Bakaletzら、J.Infect.Dis.168:865〜872,1993;BakaletzおよびHolmes,Clin.Diagn.Lab.Immunol.4:223〜225,1997;SuzukiおよびBakaletz,Infect.Immun.62:1710〜1718,1994)、OMに対する痘苗原としての、いくつかのNTHi外膜タンパク質、外膜タンパク質の組み合わせ、キメラ合成ペプチドワクチン成分、およびアジュバント処方物の防御効力を決定するために使用されている(Bakaletzら、Vaccine 15:955〜961,1997;Bakaletzら、Infect.Immun.67:2746〜2762,1999;Kennedyら、Infect.Immun.68:2756〜2765,2000)。
【0065】
特に、アデノウイルスが、チンチラをH.influenzae誘導性中耳炎に対して罹りやすくする独特のインビボモデルが存在し、これにより、NTHiの生物学的評価についての関連する細胞、組織、および器官の培養系の確立が可能になった(Bakaletzら、J.Infect.Dis.168:865〜72,1993;Suzukiら、Infect.Immunity 62:1710〜8,1994)。アデノウイルス感染単独が、鼓室中への誘導された血清抗体の漏出について評価するために使用され(Bakaletzら、Clin.Diagnostic Lab Immunol.4(2):223〜5,1997)、NTHiとともに共病原体として使用されて、中耳炎に対する痘苗原として種々のNTHi外膜タンパク質、OMPの組み合わせ、キメラ合成ペプチドワクチン成分、およびアジュバント処方物を標的とするいくつかの能動的免疫化レジメンおよび受動的免疫化レジメンの防御効力が決定された(Bakaletzら、Infect Immunity,67(6):2746〜62,1999;Kennedyら、Infect Immun.68(5):2756〜65、2000;Novotnyら、Infect Immunity 68(4):2119〜28,2000;Poolmanら、Vaccine 19(補遺1):S108〜15,2000。
【0066】
(中耳炎のNTHi感染に応答してインビボでアップレギュレートされる遺伝子)
中耳炎のNTHi感染に応答して差次的に調節されるプロモーターを同定するために、プロモータートラップライブラリーが、構築され、そして選別パラメーターが、規定された。上記プロモータートラップライブラリーの一部が、上記チンチラ中耳に直接接種され、OM発症が、ビデオ耳鏡検査法およびティンパノメトリによって24時間目および48時間目にモニターされた。さらに、中耳液が、感染の24時間後および48時間後に回収された。二色FACS分析が、浸出に関連する他の細胞および破片から、GFPを発現している細菌を単離するために使用された。単離後、gfpmut3遺伝子の5’側にあるHaemophilus挿入物のDNA配列が、決定され、分析された。この様式で、本発明者らは、NTHiがAOMの間にチンチラ中耳の環境を感知して応答する場合にアップレギュレートされる遺伝子を同定した。以下の遺伝子が、同定され、NTHi感染の間にそれらの遺伝子がアップレギュレートされることに起因して、それらの遺伝子は、NTHi感染および毒性において一定の役割を果たし得る。
【0067】
下記実施例7に記載されるように、上記のDFI手順およびその後のgfp発現クローンのFACS分析の後、可能性のあるインビボで調節されるプロモーターを含む52候補クローンが、単離された。これらのクローンが制御する遺伝子は、一般的内容およびその細胞中での機能に基づいて分類された。それらとしては、一般的代謝プロセス、環境情報処理および膜輸送、膜タンパク質、および仮想タンパク質が挙げられる。これらの52個のクローンのうち8個が、NTHi株86−028NPについて独特である配列を含む。重要なことには、3つのクローンが、1つよりも多い動物において独立したスクリーニングから単離され、それによって、単離方法が確認された。
【0068】
上記のFACSデータを独立して確認するために、本発明者らは、定量的RT−PCRによる候補遺伝子の相対的発現を決定した。親株86−028NPを、これらの研究のために使用した。従って、遺伝子調節に対するプラスミドコピー数の影響を伴わない野生型遺伝子発現が、分析され、これにより、FACSによる擬陽性クローン同定を同定することが可能になった。H.influenzae株Rdにおいて同定された配列に類似する配列を含む44個の候補クローンのうち、インビトロおよびインビボでの遺伝子発現の定量的比較により、チンチラ中耳中に存在する環境的合図にNTHiが応答する場合に、26個の遺伝子についてのアップレギュレートされた遺伝子発現(60%)が確認された。この分析により、膜輸送、環境情報処理、細胞代謝、遺伝子調節、ならびに未知の機能を有する仮想タンパク質に関与する遺伝子の発現を駆動する、インビボで調節されるプロモーターが同定された。(実施例6における表4参照)。
【0069】
定量的RT−PCRは、lolA発現の2倍の増加を示し、これは、内膜から外膜へのリポタンパク質輸送を可能にした。細菌は、中耳環境ちゅうで迅速に増殖して、48時間以内に、中耳液1ml当たり5.0×108 GFU NTHiに達する。その細菌は、その環境を感知して応答し、増殖および生存のために必要な栄養素を獲得または合成する。リボース糖輸送における膜成分をコードする遺伝子であるrbsC(配列番号619)は、インビトロで増殖する細胞と比較して、インビボでの発現の5倍増加を示した。さらに、代謝プロセスに関与する多くの遺伝子は、インビトロで増殖する細胞と比較して、インビボでの遺伝子発現の劇的な増加を示す。これらとしては、リボフラビン合成遺伝子ribB(配列番号623)、プリンヌクレオチド生合成遺伝子purE(配列番号621)、オルニチンカルバモイルトランスフェラーゼarcB(配列番号625)(これは、尿素サイクルを介するアルギニン分解に関与する)、およびuxuA(配列番号627)(これは、D−グルクロネートの取り込みおよびグリセルアルデヒド3リン酸への変換に必要な、マンノエートヒドロラーゼをコードする)が、挙げられる。さらに、より弱い程度ではあるが、ヒスチジン生合成についての遺伝子(hisB;配列番号615)、DNA修復についての遺伝子(radC;配列番号639)および推定細胞内中隔形成膜貫通タンパク質についての遺伝子(ispZ;配列番号637)が、アップレギュレートされた。
【0070】
ジスルフィド結合形成は、細菌における多くの分泌タンパク質のフォールディングおよびアセンブリのために重要である。原核生物において、DsbAおよびDsbBは、ジスルフィドの形成を担う酸化経路を構成する。DsbBは、DNAを再酸化する。これは、フォールディングしていないポリペプチドへとジスルフィド結合を直接提供する。DsbBは、酸化キノンからジスルフィドを新規に生成することが示されている(ColletおよびBardwell、Mol.Microbiol.44:1〜8,2002)。H.influenzae株Rdにおいて、DsbAは、形質転換のためのコンピテンスのために必要である(Tomb,Proc.Natl.Acad.Sci.U.S.A.,89,10252〜10256,1992)。本明細書中で、dsbB遺伝子(配列番号629)の転写の約3倍の増加が、示された。これは、中耳環境において増殖するNTHiについてのジスルフィド交換のための重要な役割に光を当てる。
【0071】
中耳の細菌コロニー形成(通常の滅菌環境)は、宿主炎症応答を生じ、引き続いて、好中球浸潤を生じる。細菌は、この宿主応答と戦うために、多くの戦略を引き起こす。NTHiは、ureH(配列番号631)(酸耐性に関与することが示されている(Youngら、J.Bacterol.,178:6487−6495,1996)、Helicobacter中の活性ウレアーゼの発現に必要な遺伝子のホモログ)の遺伝子発現を増加する(4倍)。最近、ウレアーゼ活性が、感染で生じるpHの低下を中和することによって、慢性Actinobacillus pleuropneumoniae完成においてある役割を果たし得ることが報告されている(Baltesら、Infect.Immun.,69:472−478,2000;Baltersら、Infect.Immun.,69:472−478,2001;BosseおよびMacInnes,Can.J.Vet。Res.,64:145−150)。中耳浸出液からのNTHi単離物に対する生物型分析は、87%がウレアーゼ陽性であることを実証した(DeMariaら、J.Clin.Microbiol.,20:1102−1104,1984)。しかし、NTHi毒性におけるウレアーゼの役割は未知である。同様に、産物が、S.pneumoniaeにおけるピリドキシン生合成タンパク質に対して88%の配列同一性を示し、抗酸化剤として機能し得る推定一重項酸素耐性タンパク質に60%の相同生を示す遺伝子の発現が増加した。ホスホリルコリン(ChoP)は、NTHiの病因に関わっている(Weiserら、Infect.Immun.65:943−950,1997)。NTHiは、相変動によってChoP発現を調節し、細胞表面上にLOSを装飾する。ChoPは、抗微生物ペプチドに対する減少した感受性を介して気道におけるNTHi残存に寄与し得(Lyseckoら、Infect.Immun.,68:1664−1671,2000)、そしてC反応性タンパク質(CRP)により媒介される血清殺傷に対する感受性を変化させる(Weiserら、J.Exp.Med.187:631−640,1998)。鼻咽腔および中耳腔の微環境は、ChoP+菌株がチンチラ鼻咽腔のより多くのコロニー形成を示すので、ChoP+表現型を選択し得る(Tongら、Infect.Immun.,68:4593−4597,2000)。licC遺伝子(配列番号633)の発現もまた増加した。licC遺伝子は、ホスホリルコリン誘導体化LOSの生合成におけるある役割を果たすホスホリルコリンシチジリルトランスフェラーゼをコードする(Rockら、J.Bacterol.,183:4927−4931,2001)。
【0072】
産物が引き続いて遺伝子発現またはDNA複製を調節するセットがまた、インビボ誘導遺伝子に含まれる。これらの遺伝子は、glpリプレッサー、glpR(配列番号643)、アルギニンリプレッサー遺伝子、argR(配列番号647)、および組込み宿主因子(IHF)βサブユニット、ihfB(配列番号645)によるグリセロール代謝の転写調節を含む。IHFは、特定の配列においてDNAに結合するヒストン様タンパク質(複製、部位特異的組換えおよび転写に関与し、多数のオペロンの活性を変化させる補助因子)である(Goosenおよびvan de Putte,Mol.Microbiol.161−7,1995)。さらに、CspDは、E.coliにおける静止期誘導ストレス応答の間のDNA複製を阻害し(Yamanakaら、Mol.Microbiol.,39:1572−1584,2001)、そしてmukF(配列番号641)遺伝子タンパク質ホモログは、細胞分離の前により緻密な形態へのヌクレイド構造のリモデリングに寄与する(SawitzkeおよびAustin,Proc.Natl.Acad.Sci.U.S.A.,62:1710−1718,2000)。本明細書中に記載されるDFI戦略は、未知の機能についてインビボで誘導されるプロモーターを同定した。仮想タンパク質HI0094は、初期OMの間、遺伝子発現の8倍の増加を示したが、その役割は未知のままである。HI1163(配列番号651)は、E.coliの推定オキシダーゼである仮想YdiJタンパク質と58%のアミノ酸同一性を示した。
【0073】
高密度トランスポゾン突然変異誘発戦略は、高栄養培地上で増殖するために必須のH.influenzae遺伝子を同定するために使用された(Akerleyら,Proc.Natl.Acad.Sci.U.S.A.,99:966−971,2002)。本明細書において記載されるスクリーニングにおいて、Akerleyら(上述)に記載された必須の遺伝子セットに含まれる6つの遺伝子が同定された(hisB、lppB、lolX、ispZ、mukFおよび未知のHI0665)。最近、2種類のヒト気道由来上皮細胞株との相互作用によって発現される、分類不可能なH.influenzaeの遺伝子が同定された。これらの遺伝子は、代謝プロセス、ストレス応答、遺伝子発現、細胞エンベロープ生合成、DNA関連プロセス、細胞分裂、および機能未知のORFがコードするタンパク質に関与する遺伝子を含んだ(Ulsenら,Mol.Microbiol.,45:485−500,2002)。同様に、ストレス応答遺伝子、cspD(配列番号649)、プリンおよびリボフラビン生合成に関与する遺伝子、ならびに機能未知のタンパク質、vapAが、本明細書において記載されるスクリーニングで同定された。vapAの発現は、インビトロで検出され、さらにvapA遺伝子発現は、インビボで2倍に増加した。これらの独自のアプローチは、NTHi誘導性OMにおいてアップレギュレートされ、したがってNTHiの感染およびビルレンスに役割を果たす可能性があり;かつワクチン治療およびアンチセンス治療、ならびにNTHi関連障害の処置の、他の治療法の潜在的な候補であり得る、既知の遺伝子を同定した。
DFI戦略は、その上、機能未知の遺伝子のための、インビボで誘導されるプロモーターの同定という結果を生じた。仮説的タンパク質、HI0094は、初期OMの間、遺伝子発現における8倍の増加を示したが、その機能は未知のままである。HI1163(配列番号651)は、E.Coliの仮説的YdiJタンパク質(推定上のオキシダーゼ)と58%のアミノ酸同一性を示した。したがって、これらの仮説的遺伝子は、NTHi感染によって誘導されるOMに役割を果たす可能性がある。
【図面の簡単な説明】
【0074】
【図1】図1は、Haemophilus単離物のパネルにおけるLKP遺伝子領域を示す。86−028NP株の配列は、この領域において、NTHiR3001株における配列と同一である。これらのNTHiの両方は、血球凝集性線毛をコードするhif遺伝子クラスターを欠損している。
【図2】図2は、Haemophilus単離物のパネルにおけるrfaD領域を示す。86−028NP株のゲノムのrfaD領域における遺伝子の配置は、Rd株のゲノムの配置と類似するが、検査したほとんどのNTHiのゲノムにおいて見られるこれらの遺伝子の配置とは異なる。
【図3A】図3A〜図3Mは、表4に示されるヌクレオチド配列(配列番号589〜614)を提示する。これらは、OM感染の間にアップレギュレートされたと同定された(実施例6を参照のこと)。配列番号1〜576として提示される、既知の遺伝子に対応するヌクレオチド(nt.)およびコンティグ配列に対応するヌクレオチド(nt.)もまた、示される。
【図3B】図3A〜図3Mは、表4に示されるヌクレオチド配列(配列番号589〜614)を提示する。これらは、OM感染の間にアップレギュレートされたと同定された(実施例6を参照のこと)。配列番号1〜576として提示される、既知の遺伝子に対応するヌクレオチド(nt.)およびコンティグ配列に対応するヌクレオチド(nt.)もまた、示される。
【図3C】図3A〜図3Mは、表4に示されるヌクレオチド配列(配列番号589〜614)を提示する。これらは、OM感染の間にアップレギュレートされたと同定された(実施例6を参照のこと)。配列番号1〜576として提示される、既知の遺伝子に対応するヌクレオチド(nt.)およびコンティグ配列に対応するヌクレオチド(nt.)もまた、示される。
【図3D】図3A〜図3Mは、表4に示されるヌクレオチド配列(配列番号589〜614)を提示する。これらは、OM感染の間にアップレギュレートされたと同定された(実施例6を参照のこと)。配列番号1〜576として提示される、既知の遺伝子に対応するヌクレオチド(nt.)およびコンティグ配列に対応するヌクレオチド(nt.)もまた、示される。
【図3E】図3A〜図3Mは、表4に示されるヌクレオチド配列(配列番号589〜614)を提示する。これらは、OM感染の間にアップレギュレートされたと同定された(実施例6を参照のこと)。配列番号1〜576として提示される、既知の遺伝子に対応するヌクレオチド(nt.)およびコンティグ配列に対応するヌクレオチド(nt.)もまた、示される。
【図3F】図3A〜図3Mは、表4に示されるヌクレオチド配列(配列番号589〜614)を提示する。これらは、OM感染の間にアップレギュレートされたと同定された(実施例6を参照のこと)。配列番号1〜576として提示される、既知の遺伝子に対応するヌクレオチド(nt.)およびコンティグ配列に対応するヌクレオチド(nt.)もまた、示される。
【図3G】図3A〜図3Mは、表4に示されるヌクレオチド配列(配列番号589〜614)を提示する。これらは、OM感染の間にアップレギュレートされたと同定された(実施例6を参照のこと)。配列番号1〜576として提示される、既知の遺伝子に対応するヌクレオチド(nt.)およびコンティグ配列に対応するヌクレオチド(nt.)もまた、示される。
【図3H】図3A〜図3Mは、表4に示されるヌクレオチド配列(配列番号589〜614)を提示する。これらは、OM感染の間にアップレギュレートされたと同定された(実施例6を参照のこと)。配列番号1〜576として提示される、既知の遺伝子に対応するヌクレオチド(nt.)およびコンティグ配列に対応するヌクレオチド(nt.)もまた、示される。
【図3I】図3A〜図3Mは、表4に示されるヌクレオチド配列(配列番号589〜614)を提示する。これらは、OM感染の間にアップレギュレートされたと同定された(実施例6を参照のこと)。配列番号1〜576として提示される、既知の遺伝子に対応するヌクレオチド(nt.)およびコンティグ配列に対応するヌクレオチド(nt.)もまた、示される。
【図3J】図3A〜図3Mは、表4に示されるヌクレオチド配列(配列番号589〜614)を提示する。これらは、OM感染の間にアップレギュレートされたと同定された(実施例6を参照のこと)。配列番号1〜576として提示される、既知の遺伝子に対応するヌクレオチド(nt.)およびコンティグ配列に対応するヌクレオチド(nt.)もまた、示される。
【図3K】図3A〜図3Mは、表4に示されるヌクレオチド配列(配列番号589〜614)を提示する。これらは、OM感染の間にアップレギュレートされたと同定された(実施例6を参照のこと)。配列番号1〜576として提示される、既知の遺伝子に対応するヌクレオチド(nt.)およびコンティグ配列に対応するヌクレオチド(nt.)もまた、示される。
【図3L】図3A〜図3Mは、表4に示されるヌクレオチド配列(配列番号589〜614)を提示する。これらは、OM感染の間にアップレギュレートされたと同定された(実施例6を参照のこと)。配列番号1〜576として提示される、既知の遺伝子に対応するヌクレオチド(nt.)およびコンティグ配列に対応するヌクレオチド(nt.)もまた、示される。
【図3M】図3A〜図3Mは、表4に示されるヌクレオチド配列(配列番号589〜614)を提示する。これらは、OM感染の間にアップレギュレートされたと同定された(実施例6を参照のこと)。配列番号1〜576として提示される、既知の遺伝子に対応するヌクレオチド(nt.)およびコンティグ配列に対応するヌクレオチド(nt.)もまた、示される。
【発明を実施するための形態】
【0075】
(詳細な説明)
以下の実施例は、本発明を説明する。ここで、実施例1は、NTHiゲノムの配列を記載し、実施例2は、同定されたコンティグおよび最初の遺伝子発見を記載し、実施例3は、NTHiプロモータートラップライブラリーの構築を記載し、実施例4は、GFPを発現する86−028NP誘導体の分析を記載し、実施例5は、中耳の流体由来の細菌の直接標識を示し、実施例6は、急性中耳炎においてインビボで誘導されたプロモーターの同定を記載し、実施例7は、ビルレンス関連遺伝子の同定を記載し、そして実施例8は、固有のNTHi遺伝子配列の同定を記載する。
【実施例】
【0076】
(実施例1)
(分類不可能なHaemophilus influenzaeゲノムの配列)
NTHi86−028NP株は、コロンバスのChildren’s Hospitalにおいて鼓膜切開術および慢性OMのための管挿入を受けた小児科患者から得られた、最小限に継代された臨床単離物である(Bakaletzら,Infection and Immunity,56(2):331−335,1988)。86−028NP株は、2002年10月16日にAmerican Type Tissue Collection(Manassas,VA 20108 USA)に寄託され、そして受託番号PTA−4764を割当てられた。
【0077】
NTHiにおけるビルレンス決定因子の同定により広範に近づくための試みにおいて、NTHi 86−028NP株のゲノムを3倍の適用範囲(3−fold coverage)でシークエンシングした。染色体DNA、86−028NP株からPuregeneプロトコールを用いて調製し、そしてHydroshear装置(Gene Machines)によって2kb〜4kbのサイズにせん断した。Chissoeら(Methods:a Companion to Methods of Enzymology 3:55−65,1991)およびSambrookら(Molecular Cloning:a Laboratory Manual,第2版 Cold Springs Harbor,NY,1989)に記載されたように、せん断したDNAを、エタノール沈殿し、クレノウ酵素の混合物を用いて末端修復し、そしてアガロースゲル電気泳動によってサイズ選別して、2kb〜4kbのフラグメントを得た。これらのフラグメントを、SmaI制限酵素認識部位(ホスファターゼ処理されている)を用いてpUC18ベクター中にクローニングし、そしてE.coli XL−1 Blueに形質転換し、アンピシリン耐性について選択する。X−galを含有するLB−Ampプレート上での青/白スクリーニングによって、インサートを含むコロニーを同定し、そして1.5mlのTB−Amp(TB=Ternfic Broth)ブロスを含む96ディープウェルプレートに移す。ディープウェルプレート培養物を、37℃で一晩増殖させる。テンプレート調製、シークエンシングおよびコンティグアセンブリを行った。
【0078】
Chissoeら(上述)に記載されるように、Beckman Biomek2000自動ロボットワークステーション上で、自動テンプレート調製を行った。簡単に言うと、上記で調製されたクローンを含む各96ディープウェルプレートを遠心し、細胞をペレット化した。上清をデカントし、細胞を(必要な場合)−20℃で凍結した。Chissoeら(上述)に記載されるように、4つの96ディープウェルブロックをBiomekテーブル上に配置し、液体ハンドリングロボットを使用して、代表的なSDS−NaOH溶解プロトコールの自動バージョンを用いてテンプレートを調製した。最終的なエタノール沈殿テンプレートを、50μlのddH2Oに各々溶解し、そしてDNAシークエンシングのために使用した。
【0079】
Robbins Hydra 96ロボットを用いて、上記テンプレートを(96ウェルプレートから)384ウェルプレートへと再整列化することによって、シークエンシング反応を実行した。PE Big−DyeTMターミネーターおよびユニバーサルプライマー(M13フォワードおよびリバース)を用いてサイクルシークエンシング反応を実行し、Sephadex G50カラムを通して浄化し、そしてPE Biosystems 3700キャピラリー電気泳動DNAシークエンサー上で、製造者の指示に従って分析した。シークエンシングの読み(8219)を、576のコンティグ(本明細書における配列番号1〜576)に組み立てた。3倍(3−fold)シークエンシングについての統計を、表2Aに示す。アセンブリ17の全固有配列は、1.74Mbである。
【0080】
(表2A)
【0081】
【表2A】
続いて、NTHiゲノムの8倍(8−fold)シークエンシング分析を行った。8倍シークエンシングは、NTHiゲノムを11のコンティグに組み立てた。コンティグ5、8、9、10、12〜18を、本明細書において配列番号675〜685として示す。8倍シークエンシングについての統計を、表2Bに示す。
【0082】
(表2B)
【0083】
【表2B】
(実施例2)
(コンティグ説明および最初の遺伝子発見)
3倍配列分析によって同定された、長さ5000bp以上を有する88個のコンティグ中75個は、BLASTNを介して、H.influenzae Rd株に対する有意な類似性を示す。H.influenzae 86−028NP株における遺伝子順とH.influenzae Rd株における遺伝子順との間の潜在的な関係を可視化するため、86−028NP 3倍コンティグセットおよびRd遺伝子セットを、BLASTNを用いて双方向性に比較した。結果を、ヒットしたRd遺伝子の遺伝子座標に基づいてコンティグをソーティングし、Rayら(Bioinformatics 17:1105−12,2001)に記載されるように、見出された最小の座標で各コンティグを固定することによって、遺伝子順 対 コンティグ空間にプロットした。この様式で比較し、完全に既知のゲノムに対して同一の遺伝子順を有するゲノムの不完全なアセンブリを、単調増加する階段状の形態で表示した。
【0084】
BLASTXを使用して、Rd株ゲノムにおける遺伝子に対して相同性を有する配列、およびH.influefzzae Rd株に見出されない遺伝子に対して相同性を有する配列に対するヒットを同定した。
【0085】
Rd株配列へのヒットをデータセットから除去し、そして他のセットを表3Aにまとめた。データは以下のように提示される:コンティグ番号(=配列番号)、カラム1;各ヒットについてのEスコア、カラム2;引用されたコンティグのアミノ酸翻訳部分に対する相同性を有したタンパク質の名前、カラム3;上記ホモログを産生する生物体、カラム4;そして、カラム3に引用されたタンパク質の各々についてのGenbankタンパク質識別子、カラム5;コンティグ内の対応するヌクレオチド(配列番号によって参照される)。ほとんどの例においていくつかのホモログが同定されたが、明確化のため、最も相同性の高いタンパク質を表3Aに引用する。
【0086】
表3Aに列挙した遺伝子のいくつかについての配列を、NTHiゲノムの8倍シークエンシング内で同定した。表3Bは、11コンティグ内のこれらの遺伝子の配置、全長オープンリーディングフレーム配列(配列番号によって同定される)、上記オープンリーディングフレームによってコードされる生成されたアミノ酸配列、およびBLASTXによって同定された高相同性を有する遺伝子(表3Aで列挙されたようなもの)を列挙する。
【0087】
NTHiおよびRd株中における比較的短い遺伝子の配置を検査するため、詳細に記載されている2つの遺伝子クラスター中の遺伝子順を比較した。第一に、血液凝集性線毛(LKP)遺伝子領域中に存在する遺伝子を検査した(Mhlanga−Mutangaduraら,J Bacteriol.180(17):4693−703,1998)。線毛遺伝子クラスターは、purE遺伝子とpepN遺伝子との間に位置する。これらのフラグメントのみを、図1に記載する。血清型b株、Eaganは、hifABCDE遺伝子クラスターを含み、そして血液凝集性線毛を生成する。Rd株は、hicAB遺伝子およびhifABCDE遺伝子クラスターを欠損している。一般的に、既に検査した分類不可能な株は、hicAB遺伝子を含むが、血液凝集性線毛をコードするhif遺伝子を含まない。86−028NP株配列(本明細書において記載される)は、この領域において、NTHi R3001株の配列と同一である(図1)。rfaD遺伝子は、エンドトキシンの生合成に関与する酵素をコードする。さらに、NTHi 2019株由来のrfaD遺伝子は、Nicholsら(Infect Immunity 65(4):1377−86,1997)によって特徴付けられている。2019株において、rfaD遺伝子は、エンドトキシン生合成に関与する別の酵素をコードするrfaF遺伝子のすぐ上流にある。Rd株における遺伝子配置は、様々である;rfaD遺伝子およびrfaF遺伝子は、およそ11kbの配列によって分離されている。検査された最も分類不可能な株は、2019株において見られる遺伝子配置を含んでいた。対照的に、86−028NP株は、Rd株に見られる遺伝子配置と同一の遺伝子配置を有する(図2)。
【0088】
現行のアセンブリの全体的な分析は、遺伝子の内容および順序が、Rd株における内容および順序と類似することを示す。より詳細な分析は、Pasteurellaceaにおいて以前見られなかったかなりの数のNTHi遺伝子が存在すること、そしてNTHi遺伝子の内容および順序がRd株において見られる内容および順序と異なるいくつかの領域が存在することを、明らかにした。したがって、現行のデータは、86−028NP株ゲノムが、Rd様性質と非Rd様性質との複雑なモザイクを含むことを示唆する。
【0089】
DFI戦略はまた、遺伝子発現を増加させる新規のNTHi配列を同定した。他の生物体(主にグラム陰性細菌)のORFに対して相同性を有する遺伝子または遺伝子フラグメントを含む、これらの新規コンティグ配列のリストを、表3Aに示す。例えば、コンティグ442(配列番号442)のヌクレオチド配列、ヌクレオチド1498〜1845は、H.influenzae Rd株リポタンパク質B(LppB)のアミノ酸1〜116をコードする配列に対して、高度に相同性である。この遺伝子は、静止期生存遺伝子、surEと、ビルレンスに関連している、最近同定された細菌リポタンパク質、LppB/NlpDに対して高度に相同性である、43kDの抗原外膜リポタンパク質をコードする遺伝子との間に位置する(Padmalayamら,Infect.Immun.,68:4972−4979,2000)。最近、Zhangおよび共同研究者らは、Thermotoga maritimaにおいて、nlpD遺伝子およびsurE遺伝子の発現が、細胞増殖の静止期の間に誘導されることを示した(Zhangら,Structure(Camb),9:1095−1106,2001)。したがって、中耳でのストレス誘導条件下において、このNTHiリポタンパク質が発現される。
【0090】
【表3A−1】
【表3A−2】
【表3A−3】
【表3A−4】
【表3A−5】
【表3A−6】
【表3A−7】
【表3A−8】
【表3A−9】
【表3A−10】
【表3A−11】
【表3A−12】
【表3A−13】
【表3A−14】
【表3A−15】
【表3A−16】
【表3A−17】
【表3A−18】
【表3B−1】
【表3B−2】
【表3B−3】
(実施例3)
(NTHiプロモータートラップライブラリーの構築)
NTHiの潜在的なビルレンス決定因子を同定するために、細菌遺伝子発現を、NTHi誘導中耳炎(OM)のチンチラモデルの1つの特定の解剖学的くぼみ(niche)における初期疾患の進行の間の微分蛍光誘導(DFI)によって、モニターした。NTHi菌株86−028NP由来のゲノムDNAフラグメントを、プロモータートラップライブラリーを使用して、プロモーターレスgfpmut3遺伝子の上流にクローニングした。プラスミドpGZRS39A(Actinobacillus pleuropneumoniaeから単離したpGZES−1の誘導体)は、A.pleuropneumoniae−Escherichia coliシャトルベクターである。このプラスミドは、A.pleuropneumoniae由来の複製起点、pUC19由来のlacZα遺伝子およびTn903由来のカナマイシン耐性遺伝子を含む(Westら、Genes,160:81−86,1995)。
【0091】
プロモータートラップベクターを、GTP変異gfpmut3遺伝子を、EcoRIフラグメントに対するBamHIとして、pGZRS−39A内にクローニングしてpRSM2167を形成することによって構築した。この変異GTP遺伝子は、2つのアミノ酸変化(S65GおよびS72A)を含み、これは、488nmで励起される場合、蛍光発光を増強する。この変異体はまた、高い溶解性および発色団形成の速い動力学を有する(Cormackら、Gene,173:33−38,1996)。このプラスミドは、NTHi菌株86−028NP内へのエレクトロポレーションによって形質転換され、親のプラスミド菌株86−028NP/pRSM2167を生成する。
【0092】
ランダムゲノムDNAフラグメント(実施例1に記載される)を、puroもタープローブベクター内への連結のために調製した。ゲノムDNAを、製造業者のプロトコルに従って、PuregeneDNA単離キット(Gentra Systems,Minneapolis,MN)を使用して、菌株86−028NPから単離した。制限障壁に起因して、プラスミドDNAを単離し、これをライブラリー作製のために使用することが必要であった。この単離されたDNAを37℃で1時間、Sau3AI(NEB,Beverly,MA;0.25単位/μg DNA)を用いて部分的に消化し、ゲル電気泳動によって分離し、そしてDNAフラグメント(0.5〜1.5kbのサイズ)を、Qiagenゲル抽出キットを使用して、回収した。ベクター調製のために、pRSM2167を、製造業者のプロトコルに従って、Wizard Plus Maxiprep DNA精製システム(Promega,Madison WI)を使用して、一晩の培養物から単離した。
【0093】
プラスミドDNAを、BamHI消化によって線形化し、そして5’リン酸基を、ウシ腸アルカリホスファターゼ(CIAP;GibcoBRL Life Technologies)を用いる処置によって除去した。ゲノムDNAフラグメントを、線形化されホスファターゼで処理されたベクターを用いて連結し、そして改変プロトコルに従ってエレクトロポレーションのための調製したコンピテントNTHi菌株86−028NP内エレクトロポレートした(Mitchellら、Nucleic Acids Res.,19:3625−3628,1991)。プラスミドDNAをNTHi菌株86−028NP内にエレクトロポレーションで戻した場合、形質転換効率は、千倍改善した。簡単に述べると、細胞を、sBHI(脳心臓注入物)ブロスにおいて37℃、220rpmで、OD600=0.3まで増殖させた。細胞を、30分間、氷において冷やし、引き続いて、4℃で、等量の0.5×SG(1×SG:15%グリセロール、272mMスクロース)を用いて洗浄した。洗浄を合計3回繰り返した。引き続いて、細胞を、1×SG中で、100×濃縮用量に希釈した。細胞を、200オーム、2.5kVおよび25μFで、BioRad Gene Pulser IIセットを使用して、エレクトロポレーションし、次いで、1mlの予め温めたsBHI中に希釈し、37℃、5%CO2で2時間インキュベートし、そして形質転換体の一晩の増殖のためにチョコレート寒天にプレートした。
【0094】
形質転換体を選択し、20%グリセロール(容積/容積)を含む脱脂乳中、1000クローンのプール中に凍結させた。68,000メンバーのgfpプロモータープローブライブラリーを作製した。ClarkeおよびCarbon(Cell,9:91−99,1976)の確率計算を使用して、菌株86−028NP DNAの300bpフラグメントのライブラリー(1.8×106bp/ゲノム)に表される所定のDNA配列の99%確率を達成するために、27,629クローンのライブラリーを必要とした。従って、本発明のライブラリーは、86−028NPゲノムの2.5倍の適用範囲を表す。
【0095】
ライブラリーの質を評価するために、50クローンをランダムに選択し、チョコレート寒天で一晩増殖させ、そしてプラスミドを単離し、そして挿入DNAを配列決定した。選択されたクローンの大部分(64%)は、200bpと500bpとの間の範囲挿入サイズを有したが、32%は、500bpを超えた。挿入物の大部分は、独特のH.influenzae菌株Rdオープンリーディングフレーム(ORF)と相同性を示し、そして15クローンが、菌株86−028NP DNAに対して独特の配列を有した。菌株Rdに対して相同性を有するこれらのクローンのうち、60%が、正しい方向であり、その36%が、ORFの上流に配列を含んだ。クローンの大部分が500bp未満の挿入サイズを有したが、小さな挿入サイズと増加したGFP発現との間に相関は見出されなかった。実際、4つのクローンは、インビトロにおいて、わずかから中程度の蛍光を示し、これらの3つが、200塩基対〜500塩基対の間の挿入サイズを有し、1つが、700塩基対よりも長い挿入物を有した。
【0096】
ライブラリーの画分(約1000クローン)を、チョコレート寒天で増殖させ、PBS中で収集し、GFP蛍光についてフローサイトメトリーによって分析した。挿入DNAの無いプロモータートラップベクターを含む菌株86−028NP/pRSM2169と比較して、ライブラリークローンのプールは、増加した蛍光強度を示す。従って、ライブラリーは、種々のレベルの活性でプロモーターを有するクローンを含む。
【0097】
(実施例4)
(GFPを発現する86−028NP誘導体の分析)
gfp発現細菌を同定および分類するために必要なFACSプロモーターを確立するために、種々のレベルのgfp発現を示すパネル単離物を利用した。バックグラウンド蛍光を、菌株86−028NP/pRSM2169(ネガティブコントロール)を使用して評価し、従って、任意の観察された蛍光は、lacZプロモーター駆動gfp発現に起因している。しかし、この菌株は、検出可能なレベルのGFPを生成せず、実際、親の菌株86−028NPと比較した場合、増加した蛍光を示さない。高レベルのgfp発現単離物を、外膜タンパク質P2発現に対する強力なプロモーターを含む500bpフラグメントを、Sai−BamHI消化pRSM2167にクローニングすることによって作製した。このプラスミドを、エレクトロポレーションによって86−028NP内に形質転換し、高レベルのgfp発現菌株86−028NP/pRSM2211(高い蛍光コントロール)を生成する。この菌株は、菌株86−028NP/pRSM2169と比較して、GFP蛍光において約100倍増加を示した。中程度の蛍光誘導体クローン、86−028NP/pKMM4B5(中程度の蛍光コントロール)を、FACS分析によって単離し、そして予備的実験および細胞選別のためのコントロールとして使用した。インビトロでgfp発現を駆動するプロモーターを含むDNAフラグメントは、他の生物のDNAに対して公知の相同を有さない、菌株86−028NPに対して独特である。このクローンは、菌株86−028NP/pRSM2169と比較して、蛍光において約10倍の増加を示す。
【0098】
コントロール菌株をチョコレート寒天上での増殖物から再懸濁させ、4℃で30分間、交差反応性Phycoprobe R−PE抗ヒトIgG(H+L)抗体(100μlPBS中、10μg/ml;Biomeda Corp)を用いて標識した。未結合の抗体を除去するための3回の連続洗浄に続いて、細菌をFACS分析のために、300μlDPBS中で再懸濁させた。これらのコントロール調製物を使用して、488nmを発するアルゴンレーザーを備えるCoulter Epics Eliteフローサイトメーター(Coulter Corp.)を使用して、適切なサイズおよび蛍光ゲートを設定した。細菌を、ログフォワードアングル(log forward angle)および側方散乱検出に基づくサイズ、ならびに細菌のFITC/PE標識による選別のためにゲート(gate)した。選別された細胞を、冷却したsBHI中に収集し、そしてチョコレート寒天上にプレートした。一晩の増殖後、細胞を、第2回の感染のために収集するか、または個々に選択し、一晩増殖させ、インビトロで増殖する場合、蛍光について個々のクローンによってスクリーニングし、そしてプラスミド単離の前に、20%(容積/容積)グリセロールを含む脱脂乳中で凍結させ、そして挿入物DNAを配列決定した。コントロール菌株の選別効率を、Coulter EPICSフローサイトメーター(Coulter Corp.)を使用して確認した。
【0099】
多くのプラスミドを、抗生物質選択の非存在下でインビトロで迅速に分離した。従って、本明細書中で使用されるプロモータートラップベクターがこの事象の傾向があるか否かを評価するために、菌株86−028NP/pRSM2211(高い蛍光コントロール)の単一のコロニーを、チョコレート寒天上で単離し、抗生物質選択の非存在下で20回継代させた。抗生物質の存在下で増殖された細菌と比較した場合、蛍光強度の有意な減少は、観察されなかった。さらに、プラスミドをインビボでの抗生物質選択の非存在下で維持する。チンチラから収集された細菌含有中耳分泌液を、カナマイシン無しまたは有りでチョコレート寒天上にプレートした場合、類似の細菌カウントが観察された。これらのデータは、プロモータートラップベクターが抗生物質選択の非存在下で安定に維持されたことを実証する。
【0100】
プラスミドの安定性に問題に加えて、宿主−病原体相互作用を研究するためのレポーターとしてのGFPの使用についての初期の研究は、GFPが、細菌細胞−表面動力学に対して最小の効果を有する低い毒性で、細胞質タンパク質として連続的に合成され得ることを実証した(Chalfieら、Science,263:802−805,1994)。高いレベルのgfp発現誘導体の構築は、NTHiに対するGFP毒性の評価を可能にした。野生型菌株(86−028NP)および高いGFP産生菌株86−028NP/pRSM2211の両方の増殖曲線を、類似の条件下で増殖した場合と比較した。増殖速度は類似し、GFP発現が細胞に対して毒性でないことを示す。
【0101】
86−028NP gfp発現誘導体を使用して、効率的な細胞選別についてのパラメーターを規定した。菌株86−028NP/pRSM2169を、中程度gfp発現誘導体、菌株86−028NP/pKMM4B5と、100:1の比で混合し、細菌集団の合計に対して小さな割合のgfp発現クローンを含むことが予期されるインビボ環境をシミュレートした。この混合物をFACS分析に供し、1.8%最大蛍光集団および52%最小蛍光集団を収集する。選別された集団のフローサイトメトリー分析は、細菌集団の65%まで、菌株86−028NP/pKMM4B5の富化を示し、これは、ネガティブ集団での選別の場合に観察されない減少である。引き続く回の選別は、この中程度蛍光集団をさらに富化させることをが期待される。ネガティブ選別において蛍光細菌の量を減少させることができないことは、ネガティブ選別についてのゲート設定のサイズに起因した。GFP−ネガティブ細胞を、10%の最小蛍光集団でゲーティングすることによって富化した。
【0102】
(実施例5)
(中耳分泌液からの細菌の直接的標識)
類似の戦略(実施例5に記載されるとおり)を、AOMの間にチンチラの中耳から得られた浸出液から蛍光クローンを選別するために適用した。インビボの微分蛍光誘導(DFI)を使用する本発明者らの能力は、非蛍光細菌、蛍光および非経口細胞細片、ならびに真核生物細胞からgfp発現細菌を選別する本発明者らの能力に依存した。
【0103】
耳鏡検査法またはティンパノメトリのいずれによっても中耳感染の証拠のない健常な成体チンチラ(Chinchilla lanigera)を使用して、インビボのプロモーター活性についてライブラリーをスクリーニングした。NTHi/pRSM2169ライブラリーの2つのプール(それぞれ1000クローン)を、カナマイシンを含むチョコレート寒天上で一晩増殖させた。このライブラリーを組合せ、冷却10mM滅菌PBS中に3.3×106CFU/mlまで希釈し、300μl(1.0×106CFU;500CFU/クローン)を使用して、左および右のチンチラトランスブラーキャビティー(transbullar cavity)を播種した(2000クローン/耳)。OM発生(development)を、24時間および48時間で、ビデオ耳鏡検査法およびティンパノメトリーによってモニターした。細菌を中耳腔において増殖させ、予期されるように、48時間で、接種材料用量の500倍の濃度に達した(Bakaletzら、Infect.Immunity 67:2746−62,1999)。宿主環境に対するこの細菌の適応は、炎症応答を生じ、これは、紅斑、脈管拡大および鼓膜の隆起、多形核球細胞(PMS)の浸潤、ならびに回収される浸出液の耳鏡検査および顕微鏡検査によって観察されるような中耳腔の分泌液の蓄積によって示される。24時間後および48時間後、中耳分泌液を上鼓室タップによって回収し、そしてFACSのために調製した。
【0104】
この分析が中耳分泌液中で回収可能な細菌に制限されたことに注意することが重要である。いくつかの場合において、FACS分析のために細菌を収集するために、中耳腔を洗浄することが重要であった。従って、この分析は、NTHiが粘膜にゆるく付着する場合、アップレギュレートされる遺伝子を含む。NTHiは、OMのチンチラモデルにおいて中耳腔内に生物膜を形成することが観察された(Erhlichら、JAMA,287;1710−5,2002)。本明細書中に記載されるプロトコルがプランクトンの集団から回収されるクローンを選択するので、細菌が粘膜生体膜と関連する場合、遺伝子がアップレギュレートされるクローンを回収することは予期されない。しかし、中耳粘膜の均質化および引き続く細菌細胞の単離によって、本発明者らは、これらのクローンを回収し得る。いくつかのGFP発現クローンがこの浸出液において回収されたが、剥離した細胞として浸出液中にまたは凝集物中に存在する真核生物細胞に付着したこともあり得る。これらの細菌は、選別効率を損なうことなく浸出液から回収することは困難である。従って、中耳分泌液を粘液溶解剤を用いて処理し、次いで、遠心分離して、標識化する前に、大きな凝集物および真核生物細胞を除去した。
【0105】
必要な場合に、チンチラの中耳の分泌液を、滅菌した生理食塩水を用いて250μlまで希釈した。等量のDPBS中のN−アセチル−L−システイン(0.5%;w/v)(pH 7.4)を、粘液溶解剤として室温で5分間添加した(MiyamotoおよびBakaletz,Microb.Pathog.,21:343〜356 1996)。分泌液を、細胞の残屑、赤血球、および炎症性細胞を除去するために遠心分離し(300×g、5分間)、そして細菌を含有する上清を、新しいチューブへ移した。細菌を、NTHi菌株86−028NPに由来する、全OMP調製物に対して指向されるチンチラ抗血清(1:50希釈)と共に、4℃で45分間インキュベートし、遠心分離(2000×g、5分間)によってペレット化し、そして0.05%ウシ血清アルブミンを含有する冷DPBSを用いて2回洗浄した。続いて、細菌を、交差反応性フィコプローブ(phycoprobe)R−PE抗ヒトIgG(H+L)抗体(100μlPBS中10μg/ml;Biomeda Corp)を用いて、4℃で30分間標識した。非結合の抗体を除去するために連続して3回洗浄し、続いて、細胞を、FACS分析のために300μl DPBSに再懸濁した。
【0106】
(実施例6)
(急性中耳炎におけるインビボで誘導したプロモーターの同定)
プロモータートラップライブラリーを用いて形質転換したH.influenzae 86−028NPを、チョコレート寒天で一晩増殖させた。インビトロでgfpを発現したプロモーターを含有するこれらクローンを選択するために、上記ライブラリーを、1回のFACS分析に供し(実施例6に記載するように)、低レベル量のGFPを発現しているこれらのクローンのみを回収した。これらのクローンを、プールして、チンチラの中耳の骨胞を経由して(transbullarly)接種するために用いた。注射の24時間および48時間の後、細菌含有滲出液を、鼓室または鼓膜の上部の穿刺によって除去した。細菌をR−PE標識化抗体を用いて間接的に標識し、蛍光タグ化された細菌のゲーティングによるが、発現もしている細菌について選別することによってFACS分析に供した。これらのクローンを、さらに富化するために動物を再感染させるために用いた。最終回の選別の後、単一のコロニー単離体を、蛍光の欠如についてインビトロでスクリーニングした。
【0107】
インビトロで蛍光を発していなかった、FACS分析によって単離したこれらのクローン(インビボでGFP蛍光について陽性)を、プラスミド単離および挿入したDNA配列の同定のために調製した。これらのクローンを、カナマイシンを含有するチョコレート寒天プレートで一晩増殖させ、製造者のプロトコルに従ってQiaprep Miniprep Kit(Qiagen)を用いてプラスミド単離のために調製した。プラスミド挿入DNAを、gfpmut3遺伝子に相補的であり、かつ挿入DNAの下流であるプライマー5’−TGCCCATTAACATCACCATCTA−3’(配列番号588)を用いて配列決定した。配列決定反応を、製造者のプロトコルに従って、ABI prism BigDye(登録商標)ターミネーター・サイクル・シークエンシング・レディー・リアクション・キット(Applied Biosystems)を使用して、GeneAmp PCR System 9700 (Applied Biosystems)を用いて実施した。この配列を次いで、96ウェル・マルチスクリーンHVプレート(Millipore)中のセファデックスG−50を通過させることによって精製し、続いてABI Prism3100 DNAアナライザー(Applied Biosystems)で分析した。
【0108】
挿入配列を、H.influenzae菌株Rdの注釈をつけた全配列と比較した。菌株Rdとヌクレオチド相同性を有さないこれらの挿入物を続いて、BLASTNおよびBLASTXアルゴリズムを用いて分析した。さらに配列分析を、DNASTAR(Madison,Wisc)を用いて実施した。正しい向きでかつ推定ORFの5’側配列を含む挿入物は、NTHi細菌が、チンチラの中耳に存在する場合、優先的に活性化した推定プロモーターを含有した。
【0109】
インビボで調節される推定プロモーターを有する52個のクローンを、単離した。H.influenzae菌株Rdに同定された配列と類似する配列を含む44個の候補クローンの、インビトロおよびインビボでの遺伝子発現の定量的な比較によって、NTHiが、チンチラの中耳内に存在する周囲のきっかけに応答する場合、26個の遺伝子(60%)がアップレギュレートした遺伝子発現があることを確認した。これらの遺伝子を以下の表4Aに要約した。遺伝子発現を駆動するインビボ調節プロモーターは、膜輸送、周囲の情報のプロセシング、細胞の代謝、遺伝子調節、および未知の機能を有する仮想タンパク質に関係していると予測される。
【0110】
インビボで推定プロモーター候補の誘導を確認するために、NTHi菌株86−028NPが、インビトロで中間対数期まで、またはインビボで48時間増殖させた場合に、メッセンジャーRNA発現の相対量を、比較した。このRNAを、製造者のプロトコルに従って、TRIzol LS試薬(Gibco Life Technologies)を用いて単離した。DNAを、製造者のプロトコルに従って、DNAフリー・キット(Ambion)を用いてRNA調製物から除去した。DNase I処理したRNAサンプルを、Qiagen RNeasyカラムを通過させることによって精製した。RNAの純度および完全性を、260/280nm分光光度計読み取りおよびAgilent 2100 Bioanalyzer(Agilent Technologies)によってそれぞれ、調べた。
【0111】
FACSデータを独立に確認するために、本発明者らは、定量的RT−PCRによって候補遺伝子の相対的な発現を決定した。親株86−028NPを、これらの研究に用いた。製造者の説明書に従って、ワン・ステップQuantiTectSYBR Green RT−PCRキット(Qiagen)を用いるリアル・タイム定量的RT−PCRによって、転写レベルを調べた。簡単に言えば、FACS分析によって同定した推定上のインビボで誘導されるプロモーターのオープン・リーディング・フレームの下流に作製したプライマーを用いて、遺伝子特異的mRNAを、ABI Prism 7700シークエンス・ディテクション・システム(Applied Biosystems)でのRT−PCRによって、逆転写し、増幅した。産物の量を、ジャイレース(gyr)遺伝子のフラグメントを増幅することによって、既知量の細菌のゲノムDNA(102〜107個のゲノムコピーDNA)について作製した検量線を用いて計算した。RNA調製物(−RTコントロール)中にDNAが存在しないことおよび鋳型RNAのないコントロールサンプル中にプライマーダイマーが無いことを確認するために、コントロールを、同時に分析した。さらに、RT−PCR産物を、ゲル電気泳動によって分析した。全ての場合において、単一の産物を、適切な塩基対サイズで観察した。サンプル間の細菌RNAの量を、インビトロで本発明者らが試験した種々の増殖条件の下、構成的に発現することを示す、gyr発現に対して標準化した。既知量の細菌ゲノムDNA(102〜107個のゲノムコピーDNA)を用いて、ジャイレース(gyr)遺伝子のフラグメントを増幅することによるRT−PCR定量のための標準曲線を作製した。ジャイレースは、種々の増殖条件の下インビトロで構成的に発現し、そのためサンプル間の全細菌RNAレベルの標準化のために用いた。インビボでの相対的な遺伝子発現を、インビトロでの遺伝子発現と比較し、倍率増加(fold−increase)データを、表4に要約する。
【0112】
NTHiゲノムの8倍の配列決定は、表4Aに列挙した大部分の遺伝子についての全長オープン・リーディング・フレームを同定した。表4Bは、NTHiゲノムおよび対応するアミノ酸配列内の全長ヌクレオチド配列を提供する。遺伝子の誘導倍率は、チンチラの中耳内に存在する周囲のきっかけに起因し、遺伝子の産物または機能を、便宜のために表4Bに繰り返す。
【0113】
【表4A】
【0114】
【表4B−1】
【0115】
【表4B−2】
(実施例7)
(ビルレンス関連遺伝子の同定)
多くの細菌種において、ビルレンス関連遺伝子のサブセットは、短いリピートの複製における誤りによって調節される。これらのリピートは、遺伝子の5’またはコード配列内であり得、これらの存在は、ビルレンスと関連することを示す遺伝子の制御発現の指標である。リピートの付加または欠失は、特定のビルレンス決定因子の発現または発現を欠く結果を生じる。
【0116】
NTHi H.influenzae菌株86−028NPコンティグセットは、短いオリゴヌクレオチド・リピートについて問い合わせた。このリピート周辺の領域を、このリピートに関連する遺伝子を同定するために分析した。表5に、同定したリピートおよび各リピートに関連するORF(BLASTによって同定した)を列挙する。
【0117】
さらに配列分析によって、ビルレンス関連遺伝子の全長ヌクレオチド配列およびORFによってコードされる対応するアミノ酸配列を同定した。誘導したアミノ酸配列は、列挙したGenbank配列と高い相同性である
【0118】
【表5−1】
【0119】
【表5−2】
(実施例8)
(独特のNTHi遺伝子配列の同定)
NTHi毒性に関連する遺伝子をまた、NTHi細菌属が、組織に感染している場合の遺伝子発現レベルとNTHiを、人工的な検査室培地で増殖させた場合の同一遺伝子発現レベルとを比較することによって同定した。これらの新規遺伝子を、実施例4〜6の上記プロモーター・トラップ技術を用いて同定し、続いて公知のRdゲノムとの比較によってこれらの遺伝子がNTHi菌株86−028NPに特有であることを示した。
【0120】
このスクリーニング方法を用いて同定したDNA配列を、配列番号577〜580として示す。これらの配列は、H.influenzae Rd.ゲノム配列と相同性を有する遺伝子または遺伝子フラグメントを含まなかった。これらは、完全に新規配列であるが、チンチラの中耳におけるNTHi感染の間の遺伝子の発現レベルに起因して、これら遺伝子の発現は、NTHi毒性に関係する可能性がある。
【技術分野】
【0001】
(関連出願)
本願は、2003年3月6日出願の米国仮特許出願第60/453,134号(これは、その全体が本明細書に参考として援用される)の優先権の利益を主張する。
【0002】
(発明の分野)
本発明は、Haemophilus influenzae(NTHi)ゲノムの類型不能株のポリヌクレオチド配列、そのゲノム内に含まれるNTHi遺伝子およびそのポリヌクレオチドによってコードされるポリペプチドに関する。本発明はまた、NTHi関連障害を処置および予防するワクチンおよび方法を含む、これらのNTHiポリヌクレオチドおよびNTHiポリペプチドの使用に関する。本発明はまた、中耳または鼻咽頭のNTHi感染の間またはその感染に応じて、アップレギュレートされるNTHi遺伝子に関する。
【背景技術】
【0003】
(背景)
中耳炎(OM)は、全世界で非常に優勢な小児疾患であり、小児が救急室に運ばれる主な原因である(Infante−Rivand and Fernandez,Epidemiol.Rev.,15:444−465,1993)。最近の統計では、1990年に、OMが原因で2億4500万回の通院(physician office visit)があったことが示されている。このことは、1980年代において報告されたものより200%を超えて増大していることを示している。もはや死と関連することは稀であるが、OMと関連する罹患率は、重大である。聴覚喪失は、この疾患と関連する一般的な問題であり、しばしば、小児の行動、教育および言語能力の発達に影響を与える(Baldwin,Am.J.Otol.,14:601−604,1993;Hunteretら,Ann.Otol.Rhinol.Laryngol.Suppl.,163:59−61,1994;Teeleら,J.Infect.Dis.,162:685−694,1990)。OMのその社会経済的な影響もまた大きく、OMを診断および管理する直接的および間接的な費用は、米国単独で年間50億ドルを超えている(Kaplanら,Pediatr.Infect.Dis.J.,16:S9−11,1997)。
【0004】
抗生物質療法は、一般的かつ中耳腔換気用チューブの外科手術による置換は、滲出液を排出すること、感染を除去すること、および中耳に流体が蓄積することと関連する頭痛を緩和することという観点から好結果であったが、多剤耐性細菌の出現およびチューブの置換と関連するその侵襲的性質によって、OMの管理、より好ましくはOMの予防に対するより有効かつ容認されたアプローチが必要であることが明らかにされた。慢性OMの外科手術的管理は、小児に全身麻酔をかけている間に、鼓膜を通して中耳腔換気用チューブを挿入することを包含する。この手順は平凡であり(普及率は約13%である;Brightら,Am.J.Public Health,83(7):1026−8,1993)、蓄積した流体を中耳から排出することによって、痛みの伴う症状を軽減するという点で非常に有効であるが、これは、その手順の侵襲的性質およびその伴う危険性に起因して、数多くの批判を受けている(Bermanら,Pediatrics,93(3):353−63,1994;Brightら,前出;Cimons,ASMNews,60:527−528;Paap,Ann.Pharmacother.,30(11):1291−7,1996)。
【0005】
ワクチン開発における進歩は、7価の莢膜結合体ワクチンであるPREVNAR(登録商標)(EskolaおよびKilpi,Pedriatr.Effect.Dis.J.16:S72−78,2000)が最近認可および解放されたことによって明らかなように、Streptococcus pneumoniae(OMの主な原因となる因子)について最も促進されている(AOM)。PREVNAR(登録商標)は、侵襲性の肺炎球菌疾患に非常に有用であったが、OMについての適用範囲は、そのワクチンに含まれない血清型に起因して、数が増大したOMの症例の報告によって期待が裏切られている(6〜8%)(Blackら,Pedriatr.Infect.Dis J.,19:187−195;Eskolaら,Pedriatr.Infect.Dis J.,19:S72−78,2000;Eskolaら,N.Engl.J Med.344:403−409,2001;Snowら,Otol.Neurotol.,23:1−2,2002)。類型不可Haemopilus influenzae(NTHi)(滲出液を伴う慢性OM中で優勢なグラム陰性病原体(Klein,Pedriatr.Infect.Dis J.,16:S5−8,1997;Spinolaら,J.Infect.Dis.,154:100−109,1986))については、あまり進歩していない。NTHiに対する有効なワクチン開発の妨害は、NTHi誘導性中耳疾患の病原の現在の不完全な理解である。この遅れの一因は、その疾患が、良性鼻咽頭共生生物の宿主の免疫学的寛容性のうちの1つから進行するので、微生物が発現する毒性因子と宿主の免疫応答との間の動的な相互作用の理解が欠如していることであり、通常は無菌の中耳腔の日和見的な侵入物に対する活発な防御反応の欠如に起因している。
【0006】
現在では、小児においてNTHiがどのようにOMを引き起こすかということの理解は乏しい。OMの誘導に必要な推定毒性因子の同定は、その宿主−病原体相互作用の理解、最終的には、潜在的なワクチン候補および化学療法の標的の同定に大きく寄与する。中耳炎の管理、好ましくは、その予防により有効かつ容認されたアプローチを開発することは非常に必要とされている。ワクチン開発は、この目的を達成するための非常に有望なかつ費用効果的な方法である(Giebank,Pediatr.Infect.Dis J.,13(11):1064−8,1994:Karmaら,Int.J.Pedritr.Otorhinolaryngol.,32(Suppl.):S127−34,1995)。
【発明の概要】
【課題を解決するための手段】
【0007】
本発明は、例えば、以下の項目を提供する。
(項目1)
単離されたポリヌクレオチドであって、該ポリヌクレオチドは、配列番号577〜579、配列番号589〜614、配列番号675〜685、配列番号615、配列番号617、配列番号619、配列番号621、配列番号623、配列番号625、配列番号627、配列番号629、配列番号631、配列番号633、配列番号635、配列番号637、配列番号639、配列番号641、配列番号643、配列番号645、配列番号647、配列番号649、配列番号651、配列番号653、配列番号655、配列番号657、配列番号659、配列番号661、配列番号663、配列番号665、配列番号667、配列番号669、配列番号671、配列番号673、配列番号686、配列番号688、配列番号692、配列番号694、配列番号696、配列番号698、配列番号700、配列番号702、配列番号704、配列番号706、配列番号708、配列番号710、配列番号712、配列番号714、配列番号716、配列番号718、配列番号720、配列番号722、配列番号724、配列番号726、配列番号728、配列番号730、配列番号732、配列番号734、配列番号736、配列番号738、配列番号740、配列番号742、配列番号744、配列番号746、配列番号748、配列番号750、配列番号752、配列番号754、配列番号756、配列番号758、配列番号760、配列番号762、配列番号764、配列番号766、配列番号768または配列番号770のいずれか一つのヌクレオチド配列を含む、ポリヌクレオチド。
(項目2)
項目1に示されたヌクレオチド配列によってコードされるアミノ酸配列を含む単離されたポリペプチドまたはそのフラグメント。
(項目3)
単離されたポリペプチドであって、該ポリペプチドは、配列番号616、配列番号618、配列番号620、配列番号622、配列番号624、配列番号626、配列番号628、配列番号628、配列番号630、配列番号632、配列番号634、配列番号636、配列番号638、配列番号640、配列番号642、配列番号644、配列番号646、配列番号648、配列番号650、配列番号652、配列番号654、配列番号656、配列番号658、配列番号660、配列番号662、配列番号664、配列番号666、配列番号668、配列番号670、配列番号672、配列番号674、配列番号687、配列番号689、配列番号691、配列番号693、配列番号695、配列番号697、配列番号699、配列番号701、配列番号703、配列番号705、配列番号707、配列番号709、配列番号711、配列番号713、配列番号715、配列番号717、配列番号719、配列番号721、配列番号723、配列番号725、配列番号727、配列番号729、配列番号731、配列番号733、配列番号735、配列番号737、配列番号739、配列番号741、配列番号743、配列番号745、配列番号747、配列番号749、配列番号751、配列番号753、配列番号755、配列番号757、配列番号759、配列番号761、配列番号763、配列番号765、配列番号767、配列番号769または配列番号771のいずれか一つのアミノ酸配列を含む、ポリペプチド。
(項目4)
項目2または項目3に記載のポリペプチドおよび薬学的に受容可能なキャリアを含む組成物。
(項目5)
項目2もしくは項目3に記載のポリペプチドまたはそのフラグメントに特異的に結合する抗体。
(項目6)
項目5に記載の抗体および薬学的に受容可能なキャリアを含む組成物。
(項目7)
生物学的サンプル中のNTHi細菌を検出するための方法であって、該方法は、
(a)項目1に記載のポリヌクレオチドまたはそのフラグメントと生物学的サンプルとを接触させる工程、および
(b)該サンプル中の該ポリヌクレオチドのハイブリダイゼーションを検出する工程、を包含する、方法。
(項目8)
生物学的サンプル中のNTHi細菌を検出するための方法であって、該方法は、
(a)項目5に記載の抗体と生物学的サンプルとを接触させる工程、および
(b)該サンプル中の該抗体の結合を検出する工程、
を包含する、方法。
(項目9)
前記生物学的サンプルは、血清、唾液、耳の分泌液、血液、尿、リンパ液、および脳脊髄液からなる群より選択される、項目7または8に記載の方法。
(項目10)
NTHi細菌に対する免疫応答を誘導するための方法であって、該方法は、免疫原性的に有効量の項目2もしくは3に記載のポリペプチドまたはそのフラグメントをNTHi細菌感染の危険にある患者に投与する工程を包含する、方法。
(項目11)
項目2もしくは3に記載のポリペプチドまたはそのフラグメントおよび薬学的に適切なキャリアを含むワクチン。
(項目12)
NTHi細菌感染を防止するための方法であって、該方法は、NTHiの細胞接着をブロックする項目2または3に記載のポリペプチドをNTHi細菌感染の危険にある患者に投与する工程を包含する、方法。
(項目13)
NTHi細菌感染を防止するための方法であって、該方法は、NTHiの細胞接着をブロックする項目5に記載の抗体をNTHi細菌感染の危険にある患者に投与する工程を包含する、方法。
(項目14)
前記NTHi感染は、中耳内である、項目13に記載の方法。
(項目15)
NTHi細菌感染を処置または防止するための方法であって、該方法は、項目2もしくは3に記載のポリペプチドの発現または活性を阻害する分子を必要とする患者に投与する工程を包含する、方法。
(項目16)
項目15に記載の方法であって、前記NTHiポリペプチドは、hisB、lppB、sapA、rbsC、pure、rib、arcB,uxuA、licC、ispZ、mukF、glpR、ihfB、cspD、lav、HI1647、HI0094、HI1163、HI0665、HI1292、HI1064、HI1386、HI1462、HI1369、およびHI1598からなる群より選択されるNTHi遺伝子によってコードされるアミノ酸配列を含む、方法。
(項目17)
項目15に記載の方法であって、前記NTHiポリペプチドは、配列番号616、配列番号618、配列番号620、配列番号622、配列番号624、配列番号626、配列番号628、配列番号634、配列番号638、配列番号642、配列番号644、配列番号646、配列番号650、配列番号652、配列番号656、配列番号658、配列番号660、配列番号662、配列番号664、配列番号666、配列番号668、配列番号670、配列番号672および配列番号674からなる群より選択されるアミノ酸配列を含む、方法。
(項目18)
前記必要とする患者に投与される前記分子は、アンチセンスオリゴヌクレオチドである、
項目15に記載の方法。
(項目19)
前記必要とする患者に投与される前記分子は、抗体である、項目15に記載の方法。
(項目20)
前記必要とする患者に投与される前記分子は、低分子である、項目15に記載の方法。
(項目21)
前記NTHi感染は、中耳内である、項目15に記載の方法。
本発明は、NTHiであるH.influenzae株86−028NPのゲノム配列の同定および特徴付け、ならびにこれらによりコードされるポリペプチド配列を提供する。NTHiゲノム配列の三倍分析(3−fold analysis)を、配列番号1〜576として示される一連のコンティグ配列において設定し、その後のゲノム配列の八倍分析(8−fold analysis)を、配列番号675〜685として示される一連の11個のコンティグ配列において設定する。これらのコンティグは、稀なデータであり、当業者は、重複する配列を比較して、慣用的な方法を使用して、NTHi株86−028NPの完全ゲノムを構築することによって、これらのコンティグを組み立て得る。
【0008】
本発明はまた、本発明のNTHiポリペプチドに特異的な抗体を提供する。ヒトにおいて、またはサンプル(例えば、血清、精液、耳からの分泌液(ear fluid)、血液、尿、リンパ液および脳脊髄液)において、NTHi細菌を検出する方法が、企図される。これらの方法は、NTHiポリヌクレオチドを、特異的ポリヌクレオチドプローブで検出する工程、またはNTHiポリペプチドを特異的抗体で検出する工程を包含する。本発明はまた、NTHi細菌を検出するこれらの方法を利用する診断キットを企図する。
【0009】
本発明はまた、本発明のNTHiポリペプチドまたはそのNTHiペプチドを投与することによって、免疫応答を誘発する方法を企図する。これらの方法は、NTHiポリペプチドまたはNTHiペプチドを、NTHi感染によって引き起こされる疾患(例えば、OM)の処置および/または予防のためのワクチンとして投与する工程を包含する。以下のNTHi遺伝子は、中耳および/または鼻咽頭の感染の間またはその感染に応答してアップレギュレートされ;これらの遺伝子によってコードされるポリペプチドおよびそのペプチドは、あり得るOMワクチン候補および/または化学療法の標的として企図される:hisB、lppB、sapA、lolA、rbsC、purE、ribB、arcB、uxuA、dsbB、ureH、licC、HI1647、ispZ、radC、mukF、glpR、ihfB、argR、cspD、HI0094、HI1163、HI1063、HI0665、HI1292、HI1064。NTHi hisB遺伝子は、配列番号615のヌクレオチド配列として表され、配列番号616として表されるアミノ酸配列をコードする。NTHi sapA遺伝子は、配列番号617のヌクレオチド配列として表され、配列番号618として表されるアミノ酸配列をコードする。NTHi rbsC遺伝子は、配列番号619のヌクレオチド配列として表され、配列番号620として表されるアミノ酸配列をコードする。NTHi purE遺伝子は、配列番号621のヌクレオチド配列として表され、配列番号622として表されるアミノ酸配列をコードする。NTHi ribB遺伝子は、配列番号623のヌクレオチド配列として表され、配列番号624として表されるアミノ酸配列をコードする。NTHi arcB遺伝子は、配列番号625のヌクレオチド配列として表され、配列番号626として表されるアミノ酸配列をコードする。NTHi uxuA遺伝子は、配列番号627のヌクレオチド配列として表され、配列番号628として表されるアミノ酸配列をコードする。NTHi dsbB遺伝子は、配列番号629のヌクレオチド配列として表され、配列番号630として表されるアミノ酸配列をコードする。NTHi ureH遺伝子は、配列番号631のヌクレオチド配列として表され、配列番号632として表されるアミノ酸配列をコードする。NTHi licC遺伝子は、配列番号633のヌクレオチド配列として表され、配列番号634として表されるアミノ酸配列をコードする。NTHi HI1647遺伝子は、配列番号635のヌクレオチド配列として表され、配列番号636として表されるアミノ酸配列をコードする。NTHi ispZ遺伝子は、配列番号637のヌクレオチド配列として表され、配列番号638として表されるアミノ酸配列をコードする。NTHi radC遺伝子は、配列番号639のヌクレオチド配列として表され、配列番号640として表されるアミノ酸配列をコードする。NTHi mukF遺伝子は、配列番号641のヌクレオチド配列として表され、配列番号642として表されるアミノ酸配列をコードする。NTHi glpR遺伝子は、配列番号643のヌクレオチド配列として表され、配列番号644として表されるアミノ酸配列をコードする。NTHi ihfB遺伝子は、配列番号645のヌクレオチド配列として表され、配列番号646として表されるアミノ酸配列をコードする。NTHi argR遺伝子は、配列番号647のヌクレオチド配列として表され、配列番号648として表されるアミノ酸配列をコードする。NTHi cspD遺伝子は、配列番号649のヌクレオチド配列として表され、配列番号650として表されるアミノ酸配列をコードする。NTHi HI1163遺伝子は、配列番号651のヌクレオチド配列として表され、配列番号652として表されるアミノ酸配列をコードする。NTHi HI1063遺伝子は、配列番号653のヌクレオチド配列として表され、配列番号654として表されるアミノ酸配列をコードする。NTHi HI0665遺伝子は、配列番号655のヌクレオチド配列として表され、配列番号656として表されるアミノ酸配列をコードする。NTHi HI1292遺伝子は、配列番号657のヌクレオチド配列として表され、配列番号658として表されるアミノ酸配列をコードする。
【0010】
配列番号1〜576、配列番号675〜685として示されるポリヌクレオチド配列ならびに表4および表4Bにおいて示されるポリヌクレオチド配列中に含まれる新規なNTHi遺伝子はまた、中耳および/または鼻咽頭の感染の間にアップレギュレートされ、従って、OMワクチン候補および/または化学療法の標的をコードすると予測される。さらに、以下のNTHi遺伝子は、毒性関連遺伝子であると予測され、従って、あり得るOMワクチン候補および/または化学療法の標的をコードすることが予測される:HI1386、HI1462、HI1369、lav、HI1598。NTHi HI1386遺伝子配列は、配列番号659として表され、配列番号660として表されるアミノ酸配列をコードする。NTHi HI1462遺伝子配列は、配列番号661として表され、配列番号662として表されるアミノ酸配列をコードする。NTHi HI1369遺伝子配列は、配列番号665として表され、配列番号666として表されるアミノ酸配列をコードする。NTHi lav遺伝子配列は、配列番号663として表され、配列番号664として表されるアミノ酸配列をコードする。NTHi HI1598遺伝子配列は、配列番号669および配列番号671として表され、配列番号670および配列番号672として表されるアミノ酸配列をコードする。毒性と関連したさらなるNTHi遺伝子としては、配列番号667および配列番号673として示されるポリヌクレオチド配列が挙げられる。
【0011】
NTHi感染を処置または予防する方法として、本発明は、その感染の間にアップレギュレートまたは活性になるNTHiポリペプチドの発現または活性を阻害する分子を投与することを企図する。特に、本発明は、NTHi感染を処置または予防する方法を企図し、この方法は、NTHi感染の間にアップレギュレートされるNTHi遺伝子に特異的に結合するアンチセンスオリゴヌクレオチドを投与することによって、NTHiタンパク質発現を調節する工程を包含する。このような遺伝子としては、hisB、lppB、sapA、lolA、rbsC、purE、ribB、arcB、uxuA、dsbB、ureH、licC、HI1647、ispZ、radC、mu、glpR、ihfB、argR、cspD、HI0094、HI1163、HI1063、HI0665、HI1292、HI1064が挙げられる。本発明はまた、NTHi感染を処置または予防する方法を企図し、この方法は、これらの遺伝子によってコードされるタンパク質の活性を調節する抗体または低分子を投与する工程を包含する。配列番号1〜576、配列番号675〜685として示されるポリヌクレオチド配列ならびに表4および表4Bに示されるヌクレオチド配列に含まれる新規なNTHi遺伝子もまた、中耳および/または鼻咽頭の感染の間にアップレギュレートされ、従って、これらのポリヌクレオチド配列に特異的に結合するアンチセンスオリゴヌクレオチドもまた、企図される。
【0012】
(本発明のポリヌクレオチドおよびポリペプチド)
本発明は、NTHi株86−028NPゲノムの配列を提供する。このゲノム配列は、本明細書中で「コンティグ1〜576」として表される一連のコンティグ配列として示される。各コンティグは、その「コンティグ番号」と関連して配列識別番号を割り当てられる。従って、本発明のコンティグは、配列番号1〜576として表される。これらのコンティグポリヌクレオチド配列は、慣用的な方法を使用して、NTHi株86−028NPの完全ゲノム配列に組み立てられ得る。NTHi株82−028NPゲノムの八倍配列分析の完了の際に、そのゲノム配列は、配列番号675〜685として本明細書中で表される11のコンティグに組み立てられた。
【0013】
本発明は、NTHiポリヌクレオチド配列および配列番号1〜576、配列番号675〜685のコンティグ内に含まれるオープンリーディングフレーム、ならびに表3B、表4Bおよび表5に示されるヌクレオチド配列を提供する。本発明はまた、本発明のNTHiポリヌクレオチドによってコードされるポリペプチド配列(例えば、表3B、表4Bおよび表5に示されるアミノ酸配列)を提供する。本発明は、(a)配列番号1〜576のヌクレオチド配列の相補体;配列番号675〜685のヌクレオチド配列の相補体および本明細書中の表3B、表4Bおよび表5に示されるヌクレオチド配列の相補対;(b)上記のポリヌクレオチドの対立遺伝子改変体であるポリヌクレオチド;(c)上記のタンパク質のいずれかの種ホモログをコードするポリヌクレオチド;あるいは(d)本発明のNTHiポリペプチドの特定のドメインまたは短縮を含むポリペプチドをコードするポリヌクレオチド、にストリンジェントな条件下でハイブリダイズするポリヌクレオチドを提供する。
【0014】
本発明のNTHiポリヌクレオチドはまた、上記のポリヌクレオチドに実質的に等価なヌクレオチド配列を包含する。本発明に従うポリヌクレオチドは、上記のNTHiポリヌクレオチドに対して、例えば、少なくとも65%、少なくとも70%、少なくとも75%、少なくとも80%、81%、82%、83%、84%、85%、86%、87%、88%、または89%、より代表的には、少なくとも90%、91%、92%、93%、または94%、およびより代表的には、少なくとも95%、96%、97%、98%または99%の配列同一性を有する。
【0015】
本発明の核酸配列の範囲内に含まれるのは、配列番号1〜576、配列番号675〜685のNTHiヌクレオチド配列、ならびに本明細書中の表3B、表4Bおよび表5に示されるヌクレオチド配列、またはそれらの相補体にストリンジェントな条件下でハイブリダイズする核酸配列フラグメントであり、これらのフラグメントは、約5ヌクレオチドより大きく、好ましくは7ヌクレオチド、より好ましくは、9ヌクレオチドより大きく、最も好ましくは17ヌクレオチドより大きい。例えば、選択的な(すなわち、本発明のポリヌクレオチドのいずれか1つに特異的にハイブリダイズする)15ヌクレオチド以上、17ヌクレオチド以上、または20ヌクレオチド以上のフラグメントが、企図される。ポリヌクレオチドに特異的にハイブリダイズし得るプローブは、本発明のNTHiポリヌクレオチド配列を、遺伝子の同じファミリーにおける他のポリヌクレオチド配列から区別することができるか、またはNTHi遺伝子を他の細菌遺伝子から区別することができ、より好ましくは、独特のヌクレオチド配列に基づく。
【0016】
用語「ストリンジェント」とは、当該分野でストリンジェントとして一般に理解されている条件をいうために使用される。ハイブリダイゼーションのストリンジェンシーは、原理的に、温度、イオン強度、および変性剤(例えば、ホルムアミド)の濃度によって決定される。ハイブリダイゼーションおよび洗浄のためのストリンジェントなじょう件の例は、65〜68℃での0.015M 塩化ナトリウム、0.0015M クエン酸ナトリウムまたは42℃での0.015M 塩化ナトリウム、0.0015M クエン酸ナトリウム、および50% ホルムアミドである。Sambrookら,Molecular Cloning:A Laboratory Manual,第2版,Cold Spring Harbor Laboratory,(Cold Spring Harbor,N.Y.1989)を参照のこと。よりストリンジェントな条件(例えば、より高温、より低いイオン強度、より高いホルムアミドもしくは他の変性剤)もまた、使用され得るが、ハイブリダイゼーションの速度は影響を受ける。デオキシオリゴヌクレオチドのハイブリダイゼーションが考慮される場合において、さらなる例示的なストリンジェントなハイブリダイゼーション条件としては、37℃(14塩基オリゴについて)、48℃(17塩基オリゴについて)、55℃(20塩基オリゴについて)および60℃(23塩基オリゴについて)での6×SSC 0.05% リン酸ナトリウム中での洗浄が挙げられる。
【0017】
他の薬剤は、非特異的ハイブリダイゼーションかつ/またはバックグラウンドハイブリダイゼーションを減少させる目的で、ハイブリダイゼーション緩衝液および洗浄緩衝液中に含まれ得る。例は、0.1% ウシ血清アルブミン、0.1% ポリビニル−ピロリドン、0.1% ピロリン酸ナトリウム、0.1% ドデシル硫酸ナトリウムNaDodSO4(SDS)、フィコール、デンハルト溶液、超音波処理サケ精子DNA(または他の非相補的DNA)、および硫酸デキストランであるが、他の適切な薬剤もまた使用され得る。これらの添加物の濃度および型は、ハイブリダイゼーション条件のストリンジェンシーに実質的に影響を及ぼすことなく、変更され得る。ハイブリダイゼーション実験は、通常、pH6.8〜7.4で行われるが、代表的なイオン強度条件では、ハイブリダイゼーションの速度は、pHとはほとんど関係ない。Andersonら,Nucleic Acid Hybridisation:A Practical Approach,Ch.4,IRL Press Limited(Oxford,England)を参照のこと。ハイブリダイゼーション条件は、これらの変数を適合させ、異なる配列関連性のDNAにハイブリッドを形成させるために、当業者によって調節され得る。
【0018】
本発明の範囲内にある配列は、これらの特定の配列に限定されず、それらの対立遺伝子および種改変体もまた包含する。対立遺伝子および種改変体は、配列番号1〜576、配列番号675〜685に示される配列、および本明細書中の表3B、表4Bおよび表5に示されるヌクレオチド配列(好ましくはそれらのオープンリーディングフレーム、それらの代表的なフラグメント)、あるいは配列番号1〜576、配列番号675〜685に示される配列および本明細書中の表3B、表4Bおよび表5に示されるヌクレオチド配列と少なくとも90%同一(好ましくは95%同一)のヌクレオチド配列を、同種の別の単離体に由来する配列と比較することによって規定どおりに決定され得る。2つの配列間の同一性および類似性を決定するための好ましいコンピュータプログラムとしては、GCGプログラムパッケージ(Devereuxら、Nucl.Acid.Res.,12:387、1984;Genetics Computer Group、University of Wisconsin、Madison、WI、BLASTP、BLASTN、およびFASTA(Altschulら、J.Mol.Biol.,215:403−410、1990)が挙げられる)が挙げられるが、これらに限定されない。BLASTXプログラムは、全米バイオテクノロジー情報センター(NCBI)および他の供給元(BLAST Manual、Altschulら、NCB/NLM/NIH Bethesda、MD 20894;Altschulら、前出)から公的に入手可能である。公知のSmith Watermanアルゴリズムもまた、同一性を決定するために使用され得る。
【0019】
さらに、コドン可変性に適応するために、本発明は、同じアミノ酸配列をコードする核酸分子を包含し、本明細書中に開示される特定のオープンリーディングフレーム(ORF)もまた包含する。換言すると、ORFのコード領域において、あるコドンの、同じアミノ酸をコードする別のコドンへの置換は、明示的に包含される。
【0020】
本発明の単離されたポリペプチドとしては、以下:
配列番号1〜576、配列番号675〜685として示されるポリヌクレオチド配列に含まれるヌクレオチドならびに表3B、表4Bおよび表5に示されるヌクレオチド配列によってコードされるアミノ酸配列、あるいは対応する全長タンパク質または成熟タンパク質を含むポリペプチドが挙げられるが、これらに限定されない。本発明のポリペプチドとしては、本明細書中の表3B、表4Bおよび表5に示される配列番号616、配列番号618、配列番号620、配列番号622、配列番号624、配列番号626、配列番号628、配列番号630、配列番号632、配列番号634、配列番号636、配列番号638、配列番号640、配列番号642、配列番号644、配列番号646、配列番号648、配列番号650、配列番号652、配列番号654、配列番号656、配列番号658、配列番号660、配列番号662、配列番号664、配列番号666、配列番号668、配列番号670、配列番号672、配列番号674、配列番号687、配列番号689、配列番号691、配列番号693、配列番号695、配列番号697、配列番号699、配列番号701、配列番号703、配列番号705、配列番号707、配列番号709、配列番号711、配列番号713、配列番号715、配列番号717、配列番号719、配列番号721、配列番号723、配列番号725、配列番号727、配列番号729、配列番号731、配列番号733、配列番号735、配列番号737、配列番号739、配列番号741、配列番号743、配列番号745、配列番号747、配列番号749、配列番号751、配列番号753、配列番号755、配列番号757、配列番号759、配列番号761、配列番号763、配列番号765、配列番号767、配列番号769または配列番号771のアミノ酸配列が挙げられる。
【0021】
本発明のポリペプチドはまた、好ましくは以下:
(a)配列番号1〜576、配列番号675〜685として示されるヌクレオチド配列ならびに表3B、表4Bおよび表5に示されるヌクレオチド配列に含まれるオープンリーディングフレーム、あるいは
(b)ストリンジェントなハイブリダイゼーション条件下で(a)のポリヌクレオチドの相補体にハイブリダイズするポリヌクレオチド
によってコードされる生物学的活性または免疫学的活性を有するポリペプチドを包含する。
【0022】
本発明はまた、本発明のアミノ酸配列の生物学的また免疫学的に活性な改変体;ならびに生物学的活性および/または免疫学的活性を保持するそれらの「実質的な等価物」(例えば、少なくとも約65%、少なくとも約70%、少なくとも約75%、少なくとも約80%、少なくとも約85%、86%、87%、88%、89%、少なくとも約90%、91%、92%、93%、94%、代表的には、少なくとも約95%、96%、97%、より代表的には、少なくとも約98%、または最も代表的には、少なくとも約99%アミノ酸同一性を有するもの)を提供する。対立遺伝子改変体によってコードされるポリペプチドは、配列番号1〜576、配列番号675〜685に示されるヌクレオチド配列ならびに本明細書中で表3B、表4Bおよび表5に示されるヌクレオチド配列に含まれるポリヌクレオチドによってコードされるポリペプチド、および本明細書中で表3B、表4Bおよび表5に示されるアミノ酸配列を有するポリペプチドと比較して、類似の、増加した活性もしくは減少した活性を有し得る。
【0023】
NTHiペプチドとは、配列番号1〜576、配列番号675〜685に示されるヌクレオチド配列あるいは本明細書中で表3B、表4Bおよび表5に示されるヌクレオチド配列によってコードされるNTHiポリペプチドのフラグメント、および本明細書中で表3B、表4Bおよび表5に示されるアミノ酸配列を有するポリペプチドのフラグメントをいう。好ましいNTHiペプチドは、生物学的および/または免疫学的に活性である。
【0024】
本発明はさらに、本発明のNTHi核酸フラグメントまたは本発明の核酸フラグメントの縮重改変体によってコードされる、単離されたNTHiポリペプチドまたはNTHiペプチドを提供する。用語「変性改変体」とは、ヌクレオチド配列により本発明の核酸フラグメント(例えば、ORF)とは異なるが、遺伝暗号の縮重に依存してNTHiポリペプチド配列をコードするヌクレオチドフラグメントをいう。本発明の好ましい核酸フラグメントは、タンパク質をコードするORFである。
【0025】
本発明はまた、NTHiポリペプチドを提供し、このNTHiポリペプチドは、そのポリペプチドの生物学的活性および/または免疫学的活性に影響を及ぼさない1以上の保存的アミノ酸置換を有する。あるいは、本発明のNTHiポリペプチドは、保存的アミノ酸置換を有することを包含し、この保存的アミノ酸置換は生物学的活性を変化させても変化させなくてもよい。用語「保存的アミノ酸置換」とは、その置換位置においてアミノ酸の極性または電荷にほとんど影響を及ぼさないかもしくは全く影響を及ぼさないように、ネイティブでない残基(天然のアミノ酸および非天然のアミノ酸が挙げられる)を有する天然のアミノ酸残基の置換をいう。例えば、保存的置換は、ポリペプチド中の無極性残基の、任意の他の無極性残基との交換から生じる。さらに、ポリペプチド中の任意のネイティブな残基もまた、「アラニンスキャニング変異誘発」の方法に従って、アラニンに置換され得る。天然のアミノ酸は、以下:
塩基性:アルギニン、リジン、ヒスチジン;
酸性:グルタミン酸、アルパラギン酸;
無電荷で極性:グルタミン、アスパラギン、セリン、スレオニン、チロシン;および
無極性:フェニルアラニン、トリプトファン、システイン、グリシン、アラニン、バリン、プロリン、メチオニン、ロイシン、ノルロイシン、イソロイシン
のように、それらの側鎖に基づいて特徴付けられる。アミノ酸置換についての一般的な規定は、以下の表1に記載されている。
【0026】
【表1】
NTHiポリペプチドをコードするポリヌクレオチドに対して相補的なアンチセンスポリヌクレオチドもまた、提供される。
【0027】
本発明は、本発明のポリヌクレオチドが、増幅または発現のためにベクターに挿入され得ることを包含する。発現のために、ポリヌクレオチドは、必要に応じて、適切な発現制御配列(例えば、プロモーター配列およびポリアデニル化シグナル配列)に連結される。本発明のポリヌクレオチドを含む細胞が、さらに提供される。例示的な真菌宿主としては、E.Coli、Bacillus、Streptomyces、Pseudomonas、SalmonellaおよびSerratiaのような細菌が挙げられる。
【0028】
用語「単離された」とは、それが天然に存在する環境から取り出され、かつそれが天然に存在する環境の他の成分を実質的に含まない実体をいう。例えば、ポリペプチドは、他の細胞タンパク質から分離されるか、またはDNAは、それが天然に生じるゲノムにおいてそれに隣接する他のDNAから分離される。
【0029】
(抗体および免疫応答を誘発するための方法)
本発明は、NTHiポリペプチドに特有の(すなわち、NTHiポリペプチドに特異的な)抗原性エピトープに結合する抗体を提供する。また、複数のH.influenzaeサブタイプ間で共通だが、任意の他の抗原性エピトープに関して特有である抗原性エピトープに結合する抗体が提供される。その抗体は、ポリクローナル抗体、モノクローナル抗体、それらの特有のエピトープに結合する能力を保持する抗体フラグメント(例えば、Fvフラグメント、FabフラグメントおよびF(ab)2フラグメント)、一本鎖抗体およびヒト抗体もしくはヒト化抗体であり得る。抗体は、当該分野で標準的な技術によって生成され得る。
【0030】
当該分野において、H.influenzaeの莢膜多糖類に対する抗体が、インビトロアッセイで細菌を殺す能力を示すことが公知である。これらの抗体はまた、動物モデル系において、H.influenzaeを用いるチャレンジから保護することが公知である。これらの研究は、莢膜多糖類に対する抗体がヒトにおいて防御免疫反応を誘導する可能性を有することを示す。本発明は、本発明のNTHiポリペプチドおよびそれらのフラグメントに特異的な抗体を提供し、それは、H.influenzae細菌を殺す活性およびヒトをNTHi感染から守る活性の両方を示す。本発明はまた、本発明のNTHiポリペプチドに特異的な抗体を提供し、それは、毒性を減少し、付着を阻害し、細胞分裂を阻害し、そして/あるいはH.influenzae細菌の上皮への侵入を阻害するかまたはH.influenzae細菌の食作用を促進する。
【0031】
インビトロ補体媒介性殺菌アッセイシステム(Musherら、Infect.Immun.39:297−304、1983;Andersonら、J.Clin.Invest.51:31−38、1972)が、抗NTHi抗体の殺菌活性を測定するために使用され得る。NTHiポリペプチドおよびNTHiペプチドの防御抗体反応を誘導する活性におけるさらなるデータは、本明細書中に記載されるチンチラモデル系のような感染の動物モデルを使用することによって、生成され得る。
【0032】
NTHiのエピトープに対して予備形成された抗体(例えば、NTHi、OMP、LOSまたは非被膜タンパク質に対する抗体)の投与を介する受動免疫治療によって宿主に短期間の防御を与えることもまた、可能である。従って、意図されたワクチン処方物が、受動免疫治療で使用するための抗体を産生するために使用され得る。ヒト免疫グロブリンがヒト医薬では好ましい。なぜなら、異種免疫グロブリンは、その外来の免疫原成分に対して免疫応答を誘発し得るからである。そのような受動免疫付与は、特別な危険性に曝された非免疫性の個体の即時的保護のための緊急性を根拠に使用され得る。あるいは、これらの抗体は、抗イディオタイプ抗体の産生に使用され得、それは次にNTHiエピトープに対する免疫応答を刺激するための抗体として使用され得る。
【0033】
本発明は、個体においてNTHiに対する免疫応答を誘導する方法を包含する。これらの方法は、NYHi細菌を殺す免疫応答、および細胞へのH.influenzae付着をブロックする免疫応答を包含する。1つの実施形態において、その方法は、本発明のNTHiタンパク質またはNTHiペプチドを含む組成物の免疫原性用量を投与する工程を包含する。別の実施形態において、その方法は、本発明のNTHiタンパク質またはNTHiペプチドを発現する細胞を含む組成物の免疫原性用量を投与する工程を包含する。なお別の実施形態において、その方法は、本発明のNTHiタンパク質またはNTHiペプチドをコードするポリヌクレオチドを含む組成物の免疫原性用量を投与する工程を包含する。そのポリヌクレオチドは、任意の他の核酸とは無関係の裸の(naked)ポリヌクレオチドであっても、プラスミドまたはウイルスベクター(例えば、アデノ随伴ウイルスベクターまたはアデノウイルスベクター)のようなベクター中に存在していてもよい。組成物の投与は、当該分野で標準的な経路(例えば、非経口、静脈内、経口、口腔、鼻、肺、直腸、または膣)によってであり得る。その方法は、単一の個体において組み合わせて使用され得る。その方法は、個体のNTHi感染前または感染後に使用され得る。
【0034】
「免疫学的用量」は、上記個体をNTHi感染(特に、中耳および/または鼻咽頭もしくは下気道のNTHi感染)から守るために抗体免疫応答および/またはT細胞免疫応答を発生させるのに十分な用量である。そのような免疫学的応答が細菌複製を遅延させる方法もまた、提供される。本発明のさらなる局面は、免疫学的組成物に関し、それは、個体に取り込まれた場合に免疫学的応答を誘導し得るかまたは誘導する。その免疫学的応答は、治療的または予防的に使用され得、そしてCTLまたはCD4+ T細胞から生じるような抗体免疫または細胞免疫の形態を取り得る。NTHiタンパク質またはその抗原性ペプチドは、補タンパク質と融合され得、その補タンパク質は、それ自体抗体を産生し得ないが、第1のタンパク質を安定化し得、そして免疫原性特性および保護特性を有する融合タンパク質を生成し得る。従って、融合組換えタンパク質は、好ましくは、免疫原性の補タンパク質(例えば、グルタチオン−S−トランスフェラーゼ(GST)またはβ−ガラクトシダーゼ、タンパク質を可溶化させその生成および精製を促進する比較的大きな補タンパク質)をさらに含む。さらに、補タンパク質は、免疫系の一般的な刺激を提供するという意味でアジュバントとして作用し得る。補タンパク質は、第1のタンパク質のアミノ末端またはカルボキシ末端のいずれかに結合し得る。本発明のポリヌクレオチドまたはその免疫原性ペプチドによってコードされるNTHiポリペプチドを含む組成物(特にワクチン組成物)、および方法が、本発明によって提供される。
【0035】
本発明は、それに対応して、NTHi感染に対する免疫応答を誘導するのに適切な組成物を提供し、ここで、誘導される抗体は、宿主細胞へのNTHi細菌の結合をブロックする。その組成物は、本発明のNTHiタンパク質もしくはNTHiペプチド、NTHiポリペプチドを発現する細胞、またはそのポリペプチドをコードするポリヌクレオチドを含む。その組成物はまた、キャリアおよびアジュバントのような他の成分を含み得る。
【0036】
中耳および/または鼻咽頭のNTHi感染においてアップレギュレートされる遺伝子、ならびにNTHi毒性に関連する遺伝子が、本明細書中に記載される。これらのNTHi遺伝子によってコードされるそれらのポリペプチドおよびペプチドは、OMのようなNTHi感染に関連する障害を処置または予防するために免疫応答を誘導するのに有用であることが意図される。これらの遺伝子によってコードされるポリペプチドのいくつかとしては、以下が挙げられる:ヒスチジン生合成タンパク質、リポタンパク質B、ペプチドABCトランスポーター、ペリプラスムSapA前駆体、外膜リポタンパク質キャリアタンパク質前駆体、リボース輸送系パーミアーゼタンパク質、ホスホリボシルアミノイミダゾールカルボキシラーゼ触媒性サブユニット、PurE、ホスホリボシルアミノイミダゾールカルボキシラーゼ触媒性サブユニット、オルニチンカルバモイルトランスフェラーゼ、マンノネートデヒドラターゼ、ジスルフィドオキシドレダクターゼ、ウレアーゼ補タンパク質、ホスホコリンシチジリルトランスフェラーゼ、推定ピリドキシン生合成タンパク質、一重項酸素耐性タンパク質、細胞内隔壁タンパク質(intracellular septation protein)、DNA修復タンパク質、MukFタンパク質、グリセロール−3−ホスフェートレギュロンリプレッサー、融合宿主因子βサブユニット、アルギニンリプレッサー、低温ショック様タンパク質、ストレス反応タンパク質、LicA、MukF、RadAおよびHI0094、HI1163、HI0665、HI1292、HI1064、HI186、HI0352遺伝子によってコードされるそれらの仮想タンパク質。NTHi OMP、LOSおよび非被膜タンパク質もまた、NTHi感染に関連する障害の予防および処置のための免疫応答を誘導すると意図される。
【0037】
本発明の組成物の「免疫原性用量」は、投与前に検出可能な免疫応答と比較してか、または投与前の標準的な免疫応答と比較して、投与後に、検出可能な体液性免疫応答および/または細胞性免疫応答を生じる用量である。本発明は、方法から生じる免疫応答が、予防的および/または治療的であり得ることを意図する。
【0038】
本発明は、個体において宿主細胞に対するNTHi細菌の結合をブロックする方法を包含する。その方法は、NTHi細胞の付着の結合をブロックする本発明の抗体またはポリペプチドを投与する工程を包含する。あるいは、NTHi細胞付着の結合をブロックする1以上の低分子の投与が意図される。インビトロアッセイは、本発明の抗体、ポリペプチドまたは低分子がNTHi細胞付着をブロックする能力を示すために使用され得る。
【0039】
NTHi細胞の付着をブロックする本発明の抗体、本発明のポリペプチドおよび/または本発明の低分子を含む薬学的組成物が提供される。その薬学的組成物は、上記の活性成分のうちの1つ単独からなっても、上記の活性成分の組み合わせを含んでも、細菌感染を処置するために使用されるさらなる活性成分を含んでもよい。その薬学的組成物は、薬学的に有効なキャリアのような1以上のさらなる成分を含み得る。薬学的組成物の投与の投薬量および頻度は、標準的な技術によって決定され、例えば、個体の体重および年齢、投与経路、ならびに症状の重篤度に依存する。薬学的組成物の投与は、当該分野で標準的な経路(例えば、非経口、静脈内、経口、口腔、鼻、肺、直腸、または膣)によってであり得る。
【0040】
個体におけるNTHi感染を検出するための方法もまた、提供される。1つの実施形態において、これらの方法は、サンプル中の本発明のNTHiポリヌクレオチドに特異的に結合するプライマーまたはプローブを使用して、そのポリヌクレオチドを検出する工程を包含する。そのポリヌクレオチドの検出は、例えば、ハイブリダイゼーションおよびPCRに関連する当該分野での多くの技術的慣用法によって達成され得る。
【0041】
本発明の抗体はまた、H.influenzaeの感染が疑われる個体の種々の体液中でNTHi抗原(NTHiポリペプチドおよびそのペプチド)を検出するための診断アッセイにおける用途のための試薬を提供するのに使用され得る。別の実施形態において、本発明のNTHiタンパク質およびNTHiペプチドは、種々の患者の組織および体液中でのNTHiの検出のためのイムノアッセイにおける抗原として使用され得る。これらの組織および流体としては、血液、血漿、耳滲出液(ear fluid)、脊髄液、唾、尿、リンパ液、および脳髄液が挙げられるが、これらの限定されない。本発明の抗原は、当該分野で公知のイムノアッセイのいずれかにおいて使用され得る。これらのイムノアッセイとしては、ラジオイムノアッセイ、ELISAアッセイ、サンドウィッチアッセイ、沈降素反応、ゲル拡散沈降素反応、免疫拡散アッセイ、凝集アッセイ、発光イムノアッセイ、プロテインAイムノアッセイおよび免疫電気泳動アッセイが挙げられるが、これらの限定されない。
【0042】
(ワクチン化学療法標的)
本発明の1つの局面は、個体(特に哺乳類)における免疫応答を誘導するための方法に関連し、この方法は、NTHi抗原タンパク質またはそれらの抗原性ペプチドをその個体に接種する工程を包含する。
【0043】
本発明はまた、本発明の免疫原性組換えNTHiタンパク質またはNTHiペプチドを適切なキャリアとともに含むワクチン処方物を提供する。ワクチン候補および/または化学療法の標的として企図される、NTHiポリペプチドおよびそれらのペプチドとしては、以下が挙げられるが、これらに限定されない:ヒスチジン生合成タンパク質、リポタンパク質B、ペプチドABCトランスポータ、ペリプラズムSapA前駆体、外膜リポタンパク質キャリアタンパク質前駆体、リボース輸送系透過酵素タンパク質、ホスホリボシルアミノイミダゾールカルボキシレース触媒サブユニット、PurE、3,4−ジヒドロキシ(hydroxt)−2−ブトン4−ホスフェートシンターゼ、オミニチンカルバモリルトランスフェラーゼ、マンノエートデヒドラターゼ、ジスルフィドオキヒドレダクターゼ、ウレーゼ、オミチンカルバモリルトランスフェラーゼ(omithine carbamolytransferase)、マンノネートデヒドラターゼ(mannonate dehydratase)、ジスフィドオキシドレダクターゼ(disulfide oxidoreductase)、ウレアーゼ修飾タンパク質(urease accessory protein)、ホスホコリンシチジリトランスフェラーゼ(phospshocholine cytidylytransferase)、推定ピリドキシン生合成タンパク質、一重項酸素耐性タンパク質(singlet oxygen resistance protein)、細胞内隔膜タンパク質(intracellular septation protein)、DNA修復タンパク質、MUKFタンパク質、グリセロール−3−ホスフェートレギュロンリプレッサー、組込み宿主因子βサブユニット、アルギニンリプレッサー、冷ショック様タンパク質、ストレス応答性タンパク質、LicA、RadA、ならびにHI0094遺伝子、HI1163遺伝子、HI0665遺伝子、HI1292遺伝子、HI1064遺伝子、HI1386遺伝子、HI0352遺伝子によってコードされるそれらの推定タンパク質、NTHiOMP、NTHi LOSおよびNTHi非カプセル状タンパク質および配列番号1〜576、配列番号675〜685に示されるヌクレオチド配列、ならびに本明細書中の表3B、表4および表5に示されるヌクレオチド配列において存在するその新規NTHiポリヌクレオチド配列によってコードされるポリペプチド、ならびに本明細書中の表3B、表4B、および表5に示されるアミノ酸配列を有するポリペプチド。
【0044】
このタンパク質は、胃の中で分解され得るので、好ましくは、このタンパク質は、非経口的に投与され、この投与としては、例えば、皮下、筋肉内、静脈内、皮内である投与が挙げられる。非経口投与にとって適切な処方物としては、抗酸化剤、緩衝液、静菌剤、およびその個体の体液、好ましくは血液と等張である処方物を与える溶質を含み得る、水溶性および非水溶性の滅菌注入溶液;ならびに懸濁剤または濃化剤を含み得る水性および非水性の滅菌懸濁物が挙げられる。これらの処方物は、単位用量の容器または複数回用量の容器中に提供され得る。これらの容器としては、例えば、密閉したアンプルおよびバイアルである。そして、これらの処方物は、使用直前に、滅菌液体キャリアの添加のみが必要とされる凍結乾燥条件で貯蔵され得る。このワクチン処方物はまた、その処方物の免疫原性を増強するためのアジュバント系(例えば、水中油系および当該分野で公知の系)を包含し得る。この投与は、そのワクチンの比活性に依存し、そして、慣用的な実験によって容易に決定されされ得る。
【0045】
(A.ペプチドワクチン)
ペプチド治療剤(例えば、ペプチドワクチン)は、当該分野において周知であり、薬学分野において使用が増加している。このようなペプチド化合物の非経口投与の一貫した欠点は、破壊または変性が速いことである。注入ポンプあんらびびワックスまたはオイル移植物は、ペプチド様治療剤の存在を長引かせることおよびこのような薬剤の完全性を保存することの両方のために、治療剤の長期投与のために使用されている。さらに、ペプチド様薬剤は、(ペプチド様薬剤の各エピトープに対する特定の参照とともに)長期間、天然の状態の構成を理想的に維持し、さらに、チャレンジされた動物または免疫されたヒトにおいて免疫原性応答を誘発するために適した様式で提示されるべきである。
【0046】
本発明のNTHi抗原性ペプチドは、多くの従来の方法で調製され得る。短いペプチド配列は、標準的な手段を使用する化学合成によって調製され得る。固相技術(例えば、Eriksonら、The Proteins(1976)v.2,Academic Press,New York,p255を参照のこと)が特に簡便である。自動固相合成機は、市販される。さらに、配列の改変は、適切な残基の置換、付加または省略によって容易になされる。例えば、システイン残基は、カルボキシ末端に付加されて、キャリアタンパク質への簡便な連結のためのスルフヒドリル基を提供し得るか、またはスペーサー要素(例えば、追加のグリシン残基)が、C末端の連結アミノ酸とペプチドの残りとの間の配列内に組み込まれ得る。短いNTHiペプチドはまた、組換え技術によって産生され得る。この長さのペプチドについてのコード配列は、化学技術(例えば、Matteucciら、J Am Chem Soc.,103:3185(1981)に記載されるホスホトリエステル法)によって容易に合成され得る。
【0047】
本明細書中で企図されるNTHiペプチド配列のうちのいくつかは、免疫原性であるには小さすぎると考えられ得、これらは、それらにこの特質を与えるためにキャリア物質に連結され得る。当該分野で公知のこのような連結を作製するための任意の方法が使用され得る。連結は、一方の官能基末端にジスルフィド連結を生成し、そして他方のペプチド連結を生じるヘテロ二官能性剤(例えば、ジスルフィドアミド形成剤(例えば、N−スクシジミジル−3−(2−ピリジルジチオ)プロプリオネート(SPDP)(例えば、Jansenら、,Immun.Rev.62:185,1982を参照のこと)))およびジスルフィド連結以外のチオエーテルを形成する二官能性連結剤(例えば、6−マレイミドカプロン酸、2−ブロモ酢酸、2−ヨード酢酸、4−(N−マレイミド−メチル)シクロヘキサン−1−カルボン酸など)、ならびにカルボキシル基をスクシンイミドまたは1−ヒドロキシ−2−ニトロ−4−スルホン酸(スクシンイミジル4−(N−マレイミド−メチル)シクロヘキサン−1−カルボキシレート(SMCC)のようなナトリウム塩に対して)と組み合わせることによってカルボキシル基を活性化するカップリング剤を用いて形成され得る。
【0048】
(B.ワクチン組成物および投与)
プライミング用量の免疫原(これは、免疫原への一回以上のブースター曝露が続く)は、有効ワクチンであることが必要であり得る(Krampら、Infect.Immun.,25:771−773,1979;Davisら、Immunology Letters,14:341−8 1986 1987)。同時投与される場合、免疫応答を有利に増強し得るタンパク質またはポリペプチドの例としては、サイトカイン(例えば、IL−2、Il−12、GM−CSF)、サイトカイン誘導分子(例えば、Leaf)または同時刺激分子が挙げられる。ヘルパー(HTL)エピトープは、細胞内標的化シグナルに連結され得、CTLエピトープから別々に発現され得る。これにより、CTLエピトープとは異なる細胞区画へのHTLエピトープの指向が可能になる。必要な場合、これは、HTLエピトープのMHCクラスII経路へのより効率的な進入を容易にし得、それによって、VTL誘導を改善する。CTL誘導と対照的に、免疫抑制分子(例えば、TGF−β)の同時発現による免疫応答を特異的に減少することは、特定の疾患において有利であり得る。
【0049】
理想的には、免疫原は、2つ特性を示す;対応する抗体の形成を刺激する能力、およびこれらの抗体と特異的に反応する傾向。免疫原は、抗体の部位を組み合わせることによって認識可能な免疫原の最も小さな部分である1つ以上のエピトープを有する。特定の場合において、免疫原、免疫原の画分または免疫原が提示される条件は、促進するには不適切であり、不十分な免疫を生じる。これは、しばしば、免疫原として使用されるペプチドまたは他の低分子の場合である。他の物質(例えば、免疫調節剤(例えば、インターロイキンのようなサイトカイン)は、同様にワクチンにおいて組み合わされ得る。
【0050】
上記のワクチンの分野は、免疫原と組み合わせて使用した場合に免疫応答を増強する特定の物質(アジュバントと呼ばれる)の使用を認識する。アジュバントは、そのアジュバントを使用することなく惹起されるよりも速いかまたは大きい免疫応答を惹起するために、さらに使用される。さらに、アジュバントは、アジュバントを含まずに必要とされるよりも少ない免疫原を使用して免疫学的応答を生成するため、免疫学的防御を与える特定の抗体物質の生成を増加するため、またはその免疫応答(例えば、体液性免疫応答、細胞性免疫応答)の成分を増強するために、使用され得る。公知のアジュバントとしては、エマルジョン(例えば、フロイントアジュバントおよび他のエマルジョン)、Bordetella pertussis、MF59、Quillaja saponaria由来の精製サポニン(QS21)、アルミニウム塩(例えば、水酸化アルミニウム、リン酸アルミニウム、およびミョウバン)、リン酸カルシウム(および他の金属塩)、ゲル(例えば、水酸化アルミニウム塩)、マイコバクテリア生成物(ムラミルジペプチドを含む)、固体物質、粒子(例えば、リポソームおよびビロソーム)が挙げられる。アジュバントとして使用されることが公知である天然生成物および細菌生成物の例としては、モノホスホリルリピドA(MPL)、RC−59(合成MPL様アシル化単糖)、OM−174(これは、E.coli由来のリピドA誘導体である)、ホロ毒素(例えば、コレラ毒素(CT)もしくはその誘導体のうちの1つ、破傷風毒素(PT)、およびE.coliの易熱性毒素(LT)、もしくはその誘導体のうちの1つ)、ならびにCpGオリゴヌクレオチドが挙げられる。アジュバント活性は、多数の要因(例えば、キャリア効果、デポー(depot)形成、変化したリンパ球動員、Tリンパ球の刺激、Bリンパ球の直接刺激、およびマクロファージの刺激)によって影響を受け得る。
【0051】
ワクチンは、代表的には、注射可能な物質として、液体溶液または液体懸濁液のいずれかとして、調製される。注射前に液体中の溶液または懸濁液のために適切な固体形態もまた、調製され得る。その調製物はまた、乳濁され得る。その活性な免疫原性成分は、しばしば、賦形剤(これは、薬学的に受容可能であり、そしてその活性成分と適合性である)と混合される。適切な賦形剤は、例えば、水、生理食塩水、デキストロース、グリセロール、エタノールなど、およびそれらの組み合わせである。さらに、望ましい場合、そのワクチンは、少量の補助物質(例えば、湿潤剤または乳化剤、pH緩衝剤、またはアジュバント(これは、そのワクチンの有効性を増強する)を含み得る。このワクチンは、注射(例えば、皮下注射または筋肉内注射のいずれか)によって、従来のように非経口投与される。他の投与様式のために適切なさらなる処方物としては、坐剤が挙げられ、時には、経口処方物が挙げられる。坐剤について、従来の結合剤およびキャリアとしては、例えば、ポリアルカレングリコールまたはトリグリセリドが挙げられ得る。そのような坐剤は、0.5%〜10%、好ましくは1〜2%の範囲で上記活性成分を含む混合物から形成され得る。経口処方物は、通常使用される賦形剤(例えば、薬剤グレードのマンニトール、ラクトース、デンプン、ステアリン酸マグネシウム、サッカリンナトリウム、セルロース、炭酸マグネシウムなど)を含む。これらの組成物は、溶液、懸濁物、錠剤、ピル剤、カプセル剤、徐放処方物または散剤の形態を採り得、10%〜95%(好ましくは25〜70%)の活性成分を含む。
【0052】
ワクチンはまた、ジェット注射器、微小針、エレクトロポレーション、ソノポレーション、微小カプセル化、ポリマーまたはリポソームを使用して経皮経路を介して、噴霧器、エアロゾル、および鼻スプレーを使用して経粘膜経路および鼻腔内経路を介して、投与され得る。天然ポリマーまたは合成ポリマー(例えば、デンプン、アルギネート、およびキトサン、D−ポリL−ラクテート(PLA)、D−ポリDL−乳酸−co−グリコール酸ミクロスフェア、ポリカプロラクトン、ポリオルトエステル、ポリ無水物、およびポリホスファゼン、ポリホスファタザン)を使用するマイクロカプセル化が、経皮投与および経粘膜投与の両方のために有用である。合成ポリ−オルニテート、ポリ−リジン、およびポリ−アルギニンまたは両親媒性ペプチドを含む、ポリマー性複合体は、経皮送達系のために有用である。さらに、それらの両親媒性の性質に起因して、リポソームは、経皮ワクチン送達系、経粘膜ワクチン送達系、および鼻腔内ワクチン送達系のために企図される。ワクチン送達のために使用される一般的脂質としては、N−(1)2,3−(ジオレイル−ジヒドロキシプロピル)−N,N,N−トリメチルアンモニウム−メチルスルフェート(DOTAP)、ジオレイルオキシ−プロピル−トリメチルアンモニウムクロリド(DOTMA)、ジミリスチルオキシプロピル−3−ジメチル−ヒドロキシエチルアンモニウム(DMRIE)、ジメチルジオクタデシルアンモニウムブロミド(DDAB)、および9N(N’,N’−ジメチルアミノエタン)カルバモイルコレステロール(DC−Chol)が、挙げられる。ヘルパー脂質とリポソームとの組み合わせは、皮膚を通してのそのリポソームの取り込みを増強する。これらのヘルパー脂質としては、ジオレイルホスファチジルエタノールアミン(DOPE)、ジラウロイルホスファチジルエタノールアミン(DLPE)、ジミリストイルホスファチジルエタノールアミン(DMPE)、ジパルミトイルホスファチジルエタノールアミン(DPPE)が挙げられる。さらに、トリテルペノイドグリコシドまたはチリセッケンボク(Quillaja saponaria)由来のサポニンは、鼻腔内ワクチン送達および経粘膜ワクチン送達のための有用なアジュバントとして企図されている。
【0053】
タンパク質は、中性形態または塩形態としてワクチンに処方され得る。薬学的に受容可能な塩としては、酸付加塩(ペプチドの遊離アミノ基と共に形成される)が挙げられ、これは、例えば、塩酸もしくはリン酸のような無機酸、または酢酸、シュウ酸、酒石酸、マンデル酸のような有機酸と共に形成される。遊離のカルボキシ基と共に形成される塩はまた、例えば、ナトリウム、カリウム、アンモニウム、カルシウム、または水酸化第2鉄のような無機塩、およびイソプロピルアミン、トリメチルアミン、2−エチルアミノエタノール、ヒスチジン、およびプロカインのような有機塩に由来し得る。
【0054】
ワクチンは、投薬処方物に適合する様式、そして治療的に有効かつ免疫原性な量で投与される。投与される量は、処置される被験体、被験体の免疫系が抗体を合成する能力、および所望される防御の程度に依存する。投与するのに必要な活性成分の正確な量は、開業医の判断に依存し、各個体で特有である。しかし、適切な投薬量範囲は、1個体当たり数百マイクログラムの活性成分のオーダーである。初回投与およびブースター投与のための適切なレジメンもまた可変性であるが、初回投与後1〜3ヶ月の間隔に次いでその後の注射もしくは他の投与に代表される。
【0055】
本明細書中に記載されるようにワクチン組成物で免疫化する際に、宿主の免疫系は、所望の抗原に特異的な大量のCTLを産生することによってワクチンに応答し、そしてその宿主は、少なくとも部分的に免疫化されて感染を遅らせるかまたは発生している慢性的感染に耐性を示す。本発明のNTHiポリペプチドまたはNTHiペプチドを含むワクチン組成物は、細菌感染に感受性であるかもしくはそれ以外では細菌感染の危険性を有する患者に投与されて、その抗原に対する免疫応答を誘発する。従って、患者自身の免疫応答能が増強される。そのような量は、「免疫原性的に有効な用量」として規定される。この使用において、正確な量は、この場合もまた、患者の健康状態および年齢、投与形式、処方物の性質などに依存するが、一般的には、70kgの患者1人当たり約1.0μg〜約5000、より一般には70kgの体重当たり約10〜500mgである。治療目的または免疫目的のために、本発明のNTHiポリペプチドまたはNTHiペプチドはまた、弱毒化したウイルス宿主(例えば、ワクチンまたは鶏痘)によって発現され得る。このアプローチは、本発明のペプチドをコードするヌクレオチド配列を発現するためのベクターとして、ワクチンウイルスの使用を含む。急性的にかもしくは慢性的に感染した宿主への導入、または非感染宿主への導入の際、組換えワクチンウイルスは、免疫原性ペプチドを発現し、それによって、宿主CTL応答を促進する。
【0056】
体液性免疫応答は、多くの周知の方法(例えば、単一放射状免疫拡散アッセイ(SRID)、酵素免疫アッセイ(EIA)、および赤血球凝集阻害アッセイ(HAI)によって測定され得る。詳細には、SRIDは、試験される免疫原を含むゲル(例えば、アガロース)の層を利用する。ウェルが、そのゲル中に切除され、試験される血清が、そのウェル中に配置される。そのゲルの外から中への抗体の拡散は、沈殿環の形成をもたらす。その沈殿環の領域は、試験される血清中の抗体濃度に比例する。EIA(ELISA(酵素結合イムノアッセイとしても公知)は、サンプル中の全抗体を決定するために使用される。その免疫原は、マイクロタイタープレートの表面に吸着される。その試験血清は、そのプレートに暴露され、その後、酵素結合した免疫グロブリン(例えば、IgG)に暴露される。そのプレートに付着する酵素活性は、従来の任意の手段(例えば、分光光度法)によって定量され、その活性は、その試験サンプル中に存在する免疫原に対する抗体の濃度に比例する。HAIは、免疫原(例えば、ウイルスタンパク質)がニワトリ赤血球(など)を凝集する能力を利用する。このアッセイは、中和抗体(すなわち、赤血球凝集を阻害可能な抗体)を検出する。その試験血清の希釈物が、標準濃度の免疫原とともにインキュベートされ、その後、上記赤血球が添加される。中和抗体の存在は、その免疫原による赤血球の凝集を阻害する。細胞性免疫応答を測定するための試験としては、遅延型過敏症の決定または標的免疫原に対するリンパ球の増殖応答の測定が挙げられる。
【0057】
(分類不能型インフルエンザ菌(Haemophilus influenzae)(NTHi))
H.influenzaeは、小さい非運動性のグラム陰性細菌である。他のH.influenzae株とは異なり、分類不能型H.influenzae(NTHi)下部は、多糖莢膜を欠き、時には、「非カプセル型」と示される」NTHi株は、カプセル型株とは遺伝的に別個であり、b型H.influenzae単離株よりも異種である。NTHiは、ヒト宿主に対して複雑した抗原群を提示する。防御を惹起し得る可能な抗原としては、OMP、リポ多糖、リポタンパク質、接着タンパク質、および非莢膜タンパク質が挙げられる。
【0058】
ヒトは、H.influenzaeのための唯一の宿主である。NTHi株は、呼吸管上部(鼻咽頭および口腔咽頭部後部を含む)、呼吸管下部および女性生殖管に一般的に存在する。NTHiは、ヒトにおいて広範なスペクトルの疾患を引き起こす。その疾患としては、中耳炎、肺炎、静脈洞炎、敗血症、心内膜炎、喉頭蓋炎、化膿性関節炎、髄膜炎、分娩後感染症および新生児感染症、分娩後敗血症および新生児敗血症、急性卵管炎および慢性卵管炎、喉頭蓋炎、心膜炎、蜂巣炎、骨髄炎、心内膜炎、胆嚢炎、腹内感染、泌尿器管感染、乳様突起炎、大動脈移植片感染、結膜炎、ブラジル紫斑熱、潜在性菌血症、ならびに基礎となる肺疾患(例えば、慢性肺炎、気管支炎、および嚢胞性線維症)の憎悪が挙げられるが、これらに限定されない。
【0059】
NTHiの疫学的研究は、この株が、外膜タンパク質プロフィール(Barenkampら、Infect.Immun.36:535〜40,1982)、酵素アロタイプ(Musserら、Infect.Immun.52:183〜191,1986)および他の一般的に使用される疫学的ツールに関して、不均質であることを示した。NTHiをサブタイプに分けるいくつかの試みが存在したが、それらの方法論はいずれも、完全には満足のいくものではなかった。NTHiの外膜タンパク質の組成は、約20種のタンパク質からなる。すべてのNTHi株は、2種の共通するOMPを含み、これらは、分子量が30,000ダルトンおよび16,600ダルトンである。NTHi株は、32,000〜42,000ダルトンの範囲内にある2種のOMPに基づいてサブタイプ分けされ得る。そのNTHiリポ多糖プロフィールは、腸内グラム陰性細菌とは基本的に異なり、20,000ダルトン未満の範囲の1〜4つの別個のバンドへと分かれる。
【0060】
プロトタイプNTHi単離株は、慢性中耳炎に罹患していた子供から回収された、低継代単離株86−028NPである。この株は、インビトロで(Bakaletzら、Infect.Immun.53:331〜5,1988;Holmesら、Microb.Pathog.23:157〜66,1997)ならびにチンチラOMモデル(本明細書中に記載される)(Bakaletzら、Vaccine 15:955〜61,1997;Suzukiら、Infect.Immun.62:1710〜8,1994;DeMariaら、Infect.Immun.64:5187〜92,1996)で、充分に特徴付けられている。この86−028NP株は、本明細書中に記載されるように、チンチラ中耳炎モデルにおいて発現がアップレギュレートされる遺伝子を同定するため、およびそのチンチラの中耳におけるNTHiの生存のために必要な遺伝子を同定するために、使用された。
【0061】
(DFI戦略)
差次的蛍光誘導(DFI)戦略が、本明細書中において、チンチラ動物モデルにおいてOMの間に誘導されるNTHi遺伝子を同定するために使用された。いくつかの方法が、感染の間の生物の毒性に寄与する細菌遺伝子を同定するために開発されている。そのような方法としては、細菌プロモーターが、宿主における生存のために必要な必須栄養素の合成のために必要とされる遺伝子の発現を調節する、インビボ発現方法(IVET);変異した場合に微生物のビルレンス特性を変化させる遺伝子のタグ特異的同定を可能にする、サイン(signature)タグ化変異誘発(STM);転写的に活性ない電子について全体的にスクリーニングするためのDNAマイクロアレイ技術、および転写的に活性なプロモーターについて選択するためにFACS分析を使用するDFI(Chiangら、Ann.Rev.Microbiol.53:129〜154,1999)。DFIは、基礎発現レベルに関わらず差次的に調節される遺伝子の同定を可能にし、かつインビトロでの増殖のために必須の遺伝子は排除しない、高スループット方法である。
【0062】
DFIは、多くの微生物において首尾良く利用されている。例えば、GFPレポーター系およびフローサイトメトリーが、マクロファージと相互作用する際の微生物遺伝子発現を研究するために使用された(Dhandayuthapaniら、Mol.Microbiol.17:901〜912,1995)。インビボ増殖を刺激する環境にSalmonellaeが供された場合、および培養マクロファージ様細胞によりインターナライズされた場合に、転写が増加される遺伝子を同定するために、プロモータートラップ系が、使用された(ValdivaおよびFalkow,Mol.Microbiol.22:367〜378,1996;ValdiviaおよびFalkow,Science,277:2007〜2011,1997;ValdiviaおよびFalkow,Curr.Opin.Microbiol.1:359〜363,1998)。さらに、DFIは、感染を模倣する変化したインビトロ状態下で増殖した場合にS.pneumoniaeおよびS.aureusにおいて発現されるプロモーターを同定するために、使用された。さらに、DFIは、Bacillus cereusにおいて、環境刺激に応答した遺伝子調節を研究するために(DunnおよびHandelsman、Gene,226:297〜305,1999)、S.pneumoniaeにおいてコンピテンス刺激ペプチドに応答した遺伝子調節を研究するために(Bartilsonら、Mol.Microbiol.39:126〜135,2001)、Bartonella henselae(LeeおよびFalkow,Infect.Immun.66:3964〜3967,1998)、Listeria monocytogenes(Wilsonら、Infect.Immun.69:5016〜5024,2001)、Brucella abortus(Eskraら、Infect.Immun.69:7736〜7742,2001)、およびEscherichia coli(Badgerら、Mol.Microbiol.36:174〜182,2000)における宿主細胞との相互作用および宿主細胞の侵襲の際の遺伝子調節を研究するために、使用された。
【0063】
DFIは、感染の細胞培養モデルまたはインビボ環境を刺激するように設計されたインビトロ条件において活性なプロモーターを同定するために首尾良く使用されているが、動物全体において特定の生物学的ニッチにおいて調節されるプロモーターを同定するためには、DFIはほとんど適用されていない。このことは、おそらく、インビボ環境から選別することに関連する多数の難問に起因する。宿主の免疫応答、感染部位からの細菌細胞の蔓延おおび/または除去、ならびに上皮細胞に対する細菌の付着(おそらく、バイオフィルム形成を介する)は、細菌を、生存動物から回収するために接近不能にし得る。これらの要因は、とりわけ、細菌がこれらの難問を感知して応答する場合に、微小環境の複雑性および遺伝子発現の不均一性に寄与する。最近、DFIは、最近が、呼吸器管感染のマウスモデルおよびOMのアレチネズミ感染モデルにおいてスクリーニングされた場合に、S.pneumoniaeにおいて発現されるプロモーターを同定するために使用されている(Marraら、Infect.Immun.70:1422〜33,2002;Marraら、Microbiol.148:1483〜91,2002)。
【0064】
(動物モデル)
上記チンチラモデルは、OMについての広範に受容されている実験モデルである。特に、NTHi誘導性OMのチンチラモデルは、十分に特徴付けられており(Bakaletzら、J.Infect.Dis.168:865〜872,1993;BakaletzおよびHolmes,Clin.Diagn.Lab.Immunol.4:223〜225,1997;SuzukiおよびBakaletz,Infect.Immun.62:1710〜1718,1994)、OMに対する痘苗原としての、いくつかのNTHi外膜タンパク質、外膜タンパク質の組み合わせ、キメラ合成ペプチドワクチン成分、およびアジュバント処方物の防御効力を決定するために使用されている(Bakaletzら、Vaccine 15:955〜961,1997;Bakaletzら、Infect.Immun.67:2746〜2762,1999;Kennedyら、Infect.Immun.68:2756〜2765,2000)。
【0065】
特に、アデノウイルスが、チンチラをH.influenzae誘導性中耳炎に対して罹りやすくする独特のインビボモデルが存在し、これにより、NTHiの生物学的評価についての関連する細胞、組織、および器官の培養系の確立が可能になった(Bakaletzら、J.Infect.Dis.168:865〜72,1993;Suzukiら、Infect.Immunity 62:1710〜8,1994)。アデノウイルス感染単独が、鼓室中への誘導された血清抗体の漏出について評価するために使用され(Bakaletzら、Clin.Diagnostic Lab Immunol.4(2):223〜5,1997)、NTHiとともに共病原体として使用されて、中耳炎に対する痘苗原として種々のNTHi外膜タンパク質、OMPの組み合わせ、キメラ合成ペプチドワクチン成分、およびアジュバント処方物を標的とするいくつかの能動的免疫化レジメンおよび受動的免疫化レジメンの防御効力が決定された(Bakaletzら、Infect Immunity,67(6):2746〜62,1999;Kennedyら、Infect Immun.68(5):2756〜65、2000;Novotnyら、Infect Immunity 68(4):2119〜28,2000;Poolmanら、Vaccine 19(補遺1):S108〜15,2000。
【0066】
(中耳炎のNTHi感染に応答してインビボでアップレギュレートされる遺伝子)
中耳炎のNTHi感染に応答して差次的に調節されるプロモーターを同定するために、プロモータートラップライブラリーが、構築され、そして選別パラメーターが、規定された。上記プロモータートラップライブラリーの一部が、上記チンチラ中耳に直接接種され、OM発症が、ビデオ耳鏡検査法およびティンパノメトリによって24時間目および48時間目にモニターされた。さらに、中耳液が、感染の24時間後および48時間後に回収された。二色FACS分析が、浸出に関連する他の細胞および破片から、GFPを発現している細菌を単離するために使用された。単離後、gfpmut3遺伝子の5’側にあるHaemophilus挿入物のDNA配列が、決定され、分析された。この様式で、本発明者らは、NTHiがAOMの間にチンチラ中耳の環境を感知して応答する場合にアップレギュレートされる遺伝子を同定した。以下の遺伝子が、同定され、NTHi感染の間にそれらの遺伝子がアップレギュレートされることに起因して、それらの遺伝子は、NTHi感染および毒性において一定の役割を果たし得る。
【0067】
下記実施例7に記載されるように、上記のDFI手順およびその後のgfp発現クローンのFACS分析の後、可能性のあるインビボで調節されるプロモーターを含む52候補クローンが、単離された。これらのクローンが制御する遺伝子は、一般的内容およびその細胞中での機能に基づいて分類された。それらとしては、一般的代謝プロセス、環境情報処理および膜輸送、膜タンパク質、および仮想タンパク質が挙げられる。これらの52個のクローンのうち8個が、NTHi株86−028NPについて独特である配列を含む。重要なことには、3つのクローンが、1つよりも多い動物において独立したスクリーニングから単離され、それによって、単離方法が確認された。
【0068】
上記のFACSデータを独立して確認するために、本発明者らは、定量的RT−PCRによる候補遺伝子の相対的発現を決定した。親株86−028NPを、これらの研究のために使用した。従って、遺伝子調節に対するプラスミドコピー数の影響を伴わない野生型遺伝子発現が、分析され、これにより、FACSによる擬陽性クローン同定を同定することが可能になった。H.influenzae株Rdにおいて同定された配列に類似する配列を含む44個の候補クローンのうち、インビトロおよびインビボでの遺伝子発現の定量的比較により、チンチラ中耳中に存在する環境的合図にNTHiが応答する場合に、26個の遺伝子についてのアップレギュレートされた遺伝子発現(60%)が確認された。この分析により、膜輸送、環境情報処理、細胞代謝、遺伝子調節、ならびに未知の機能を有する仮想タンパク質に関与する遺伝子の発現を駆動する、インビボで調節されるプロモーターが同定された。(実施例6における表4参照)。
【0069】
定量的RT−PCRは、lolA発現の2倍の増加を示し、これは、内膜から外膜へのリポタンパク質輸送を可能にした。細菌は、中耳環境ちゅうで迅速に増殖して、48時間以内に、中耳液1ml当たり5.0×108 GFU NTHiに達する。その細菌は、その環境を感知して応答し、増殖および生存のために必要な栄養素を獲得または合成する。リボース糖輸送における膜成分をコードする遺伝子であるrbsC(配列番号619)は、インビトロで増殖する細胞と比較して、インビボでの発現の5倍増加を示した。さらに、代謝プロセスに関与する多くの遺伝子は、インビトロで増殖する細胞と比較して、インビボでの遺伝子発現の劇的な増加を示す。これらとしては、リボフラビン合成遺伝子ribB(配列番号623)、プリンヌクレオチド生合成遺伝子purE(配列番号621)、オルニチンカルバモイルトランスフェラーゼarcB(配列番号625)(これは、尿素サイクルを介するアルギニン分解に関与する)、およびuxuA(配列番号627)(これは、D−グルクロネートの取り込みおよびグリセルアルデヒド3リン酸への変換に必要な、マンノエートヒドロラーゼをコードする)が、挙げられる。さらに、より弱い程度ではあるが、ヒスチジン生合成についての遺伝子(hisB;配列番号615)、DNA修復についての遺伝子(radC;配列番号639)および推定細胞内中隔形成膜貫通タンパク質についての遺伝子(ispZ;配列番号637)が、アップレギュレートされた。
【0070】
ジスルフィド結合形成は、細菌における多くの分泌タンパク質のフォールディングおよびアセンブリのために重要である。原核生物において、DsbAおよびDsbBは、ジスルフィドの形成を担う酸化経路を構成する。DsbBは、DNAを再酸化する。これは、フォールディングしていないポリペプチドへとジスルフィド結合を直接提供する。DsbBは、酸化キノンからジスルフィドを新規に生成することが示されている(ColletおよびBardwell、Mol.Microbiol.44:1〜8,2002)。H.influenzae株Rdにおいて、DsbAは、形質転換のためのコンピテンスのために必要である(Tomb,Proc.Natl.Acad.Sci.U.S.A.,89,10252〜10256,1992)。本明細書中で、dsbB遺伝子(配列番号629)の転写の約3倍の増加が、示された。これは、中耳環境において増殖するNTHiについてのジスルフィド交換のための重要な役割に光を当てる。
【0071】
中耳の細菌コロニー形成(通常の滅菌環境)は、宿主炎症応答を生じ、引き続いて、好中球浸潤を生じる。細菌は、この宿主応答と戦うために、多くの戦略を引き起こす。NTHiは、ureH(配列番号631)(酸耐性に関与することが示されている(Youngら、J.Bacterol.,178:6487−6495,1996)、Helicobacter中の活性ウレアーゼの発現に必要な遺伝子のホモログ)の遺伝子発現を増加する(4倍)。最近、ウレアーゼ活性が、感染で生じるpHの低下を中和することによって、慢性Actinobacillus pleuropneumoniae完成においてある役割を果たし得ることが報告されている(Baltesら、Infect.Immun.,69:472−478,2000;Baltersら、Infect.Immun.,69:472−478,2001;BosseおよびMacInnes,Can.J.Vet。Res.,64:145−150)。中耳浸出液からのNTHi単離物に対する生物型分析は、87%がウレアーゼ陽性であることを実証した(DeMariaら、J.Clin.Microbiol.,20:1102−1104,1984)。しかし、NTHi毒性におけるウレアーゼの役割は未知である。同様に、産物が、S.pneumoniaeにおけるピリドキシン生合成タンパク質に対して88%の配列同一性を示し、抗酸化剤として機能し得る推定一重項酸素耐性タンパク質に60%の相同生を示す遺伝子の発現が増加した。ホスホリルコリン(ChoP)は、NTHiの病因に関わっている(Weiserら、Infect.Immun.65:943−950,1997)。NTHiは、相変動によってChoP発現を調節し、細胞表面上にLOSを装飾する。ChoPは、抗微生物ペプチドに対する減少した感受性を介して気道におけるNTHi残存に寄与し得(Lyseckoら、Infect.Immun.,68:1664−1671,2000)、そしてC反応性タンパク質(CRP)により媒介される血清殺傷に対する感受性を変化させる(Weiserら、J.Exp.Med.187:631−640,1998)。鼻咽腔および中耳腔の微環境は、ChoP+菌株がチンチラ鼻咽腔のより多くのコロニー形成を示すので、ChoP+表現型を選択し得る(Tongら、Infect.Immun.,68:4593−4597,2000)。licC遺伝子(配列番号633)の発現もまた増加した。licC遺伝子は、ホスホリルコリン誘導体化LOSの生合成におけるある役割を果たすホスホリルコリンシチジリルトランスフェラーゼをコードする(Rockら、J.Bacterol.,183:4927−4931,2001)。
【0072】
産物が引き続いて遺伝子発現またはDNA複製を調節するセットがまた、インビボ誘導遺伝子に含まれる。これらの遺伝子は、glpリプレッサー、glpR(配列番号643)、アルギニンリプレッサー遺伝子、argR(配列番号647)、および組込み宿主因子(IHF)βサブユニット、ihfB(配列番号645)によるグリセロール代謝の転写調節を含む。IHFは、特定の配列においてDNAに結合するヒストン様タンパク質(複製、部位特異的組換えおよび転写に関与し、多数のオペロンの活性を変化させる補助因子)である(Goosenおよびvan de Putte,Mol.Microbiol.161−7,1995)。さらに、CspDは、E.coliにおける静止期誘導ストレス応答の間のDNA複製を阻害し(Yamanakaら、Mol.Microbiol.,39:1572−1584,2001)、そしてmukF(配列番号641)遺伝子タンパク質ホモログは、細胞分離の前により緻密な形態へのヌクレイド構造のリモデリングに寄与する(SawitzkeおよびAustin,Proc.Natl.Acad.Sci.U.S.A.,62:1710−1718,2000)。本明細書中に記載されるDFI戦略は、未知の機能についてインビボで誘導されるプロモーターを同定した。仮想タンパク質HI0094は、初期OMの間、遺伝子発現の8倍の増加を示したが、その役割は未知のままである。HI1163(配列番号651)は、E.coliの推定オキシダーゼである仮想YdiJタンパク質と58%のアミノ酸同一性を示した。
【0073】
高密度トランスポゾン突然変異誘発戦略は、高栄養培地上で増殖するために必須のH.influenzae遺伝子を同定するために使用された(Akerleyら,Proc.Natl.Acad.Sci.U.S.A.,99:966−971,2002)。本明細書において記載されるスクリーニングにおいて、Akerleyら(上述)に記載された必須の遺伝子セットに含まれる6つの遺伝子が同定された(hisB、lppB、lolX、ispZ、mukFおよび未知のHI0665)。最近、2種類のヒト気道由来上皮細胞株との相互作用によって発現される、分類不可能なH.influenzaeの遺伝子が同定された。これらの遺伝子は、代謝プロセス、ストレス応答、遺伝子発現、細胞エンベロープ生合成、DNA関連プロセス、細胞分裂、および機能未知のORFがコードするタンパク質に関与する遺伝子を含んだ(Ulsenら,Mol.Microbiol.,45:485−500,2002)。同様に、ストレス応答遺伝子、cspD(配列番号649)、プリンおよびリボフラビン生合成に関与する遺伝子、ならびに機能未知のタンパク質、vapAが、本明細書において記載されるスクリーニングで同定された。vapAの発現は、インビトロで検出され、さらにvapA遺伝子発現は、インビボで2倍に増加した。これらの独自のアプローチは、NTHi誘導性OMにおいてアップレギュレートされ、したがってNTHiの感染およびビルレンスに役割を果たす可能性があり;かつワクチン治療およびアンチセンス治療、ならびにNTHi関連障害の処置の、他の治療法の潜在的な候補であり得る、既知の遺伝子を同定した。
DFI戦略は、その上、機能未知の遺伝子のための、インビボで誘導されるプロモーターの同定という結果を生じた。仮説的タンパク質、HI0094は、初期OMの間、遺伝子発現における8倍の増加を示したが、その機能は未知のままである。HI1163(配列番号651)は、E.Coliの仮説的YdiJタンパク質(推定上のオキシダーゼ)と58%のアミノ酸同一性を示した。したがって、これらの仮説的遺伝子は、NTHi感染によって誘導されるOMに役割を果たす可能性がある。
【図面の簡単な説明】
【0074】
【図1】図1は、Haemophilus単離物のパネルにおけるLKP遺伝子領域を示す。86−028NP株の配列は、この領域において、NTHiR3001株における配列と同一である。これらのNTHiの両方は、血球凝集性線毛をコードするhif遺伝子クラスターを欠損している。
【図2】図2は、Haemophilus単離物のパネルにおけるrfaD領域を示す。86−028NP株のゲノムのrfaD領域における遺伝子の配置は、Rd株のゲノムの配置と類似するが、検査したほとんどのNTHiのゲノムにおいて見られるこれらの遺伝子の配置とは異なる。
【図3A】図3A〜図3Mは、表4に示されるヌクレオチド配列(配列番号589〜614)を提示する。これらは、OM感染の間にアップレギュレートされたと同定された(実施例6を参照のこと)。配列番号1〜576として提示される、既知の遺伝子に対応するヌクレオチド(nt.)およびコンティグ配列に対応するヌクレオチド(nt.)もまた、示される。
【図3B】図3A〜図3Mは、表4に示されるヌクレオチド配列(配列番号589〜614)を提示する。これらは、OM感染の間にアップレギュレートされたと同定された(実施例6を参照のこと)。配列番号1〜576として提示される、既知の遺伝子に対応するヌクレオチド(nt.)およびコンティグ配列に対応するヌクレオチド(nt.)もまた、示される。
【図3C】図3A〜図3Mは、表4に示されるヌクレオチド配列(配列番号589〜614)を提示する。これらは、OM感染の間にアップレギュレートされたと同定された(実施例6を参照のこと)。配列番号1〜576として提示される、既知の遺伝子に対応するヌクレオチド(nt.)およびコンティグ配列に対応するヌクレオチド(nt.)もまた、示される。
【図3D】図3A〜図3Mは、表4に示されるヌクレオチド配列(配列番号589〜614)を提示する。これらは、OM感染の間にアップレギュレートされたと同定された(実施例6を参照のこと)。配列番号1〜576として提示される、既知の遺伝子に対応するヌクレオチド(nt.)およびコンティグ配列に対応するヌクレオチド(nt.)もまた、示される。
【図3E】図3A〜図3Mは、表4に示されるヌクレオチド配列(配列番号589〜614)を提示する。これらは、OM感染の間にアップレギュレートされたと同定された(実施例6を参照のこと)。配列番号1〜576として提示される、既知の遺伝子に対応するヌクレオチド(nt.)およびコンティグ配列に対応するヌクレオチド(nt.)もまた、示される。
【図3F】図3A〜図3Mは、表4に示されるヌクレオチド配列(配列番号589〜614)を提示する。これらは、OM感染の間にアップレギュレートされたと同定された(実施例6を参照のこと)。配列番号1〜576として提示される、既知の遺伝子に対応するヌクレオチド(nt.)およびコンティグ配列に対応するヌクレオチド(nt.)もまた、示される。
【図3G】図3A〜図3Mは、表4に示されるヌクレオチド配列(配列番号589〜614)を提示する。これらは、OM感染の間にアップレギュレートされたと同定された(実施例6を参照のこと)。配列番号1〜576として提示される、既知の遺伝子に対応するヌクレオチド(nt.)およびコンティグ配列に対応するヌクレオチド(nt.)もまた、示される。
【図3H】図3A〜図3Mは、表4に示されるヌクレオチド配列(配列番号589〜614)を提示する。これらは、OM感染の間にアップレギュレートされたと同定された(実施例6を参照のこと)。配列番号1〜576として提示される、既知の遺伝子に対応するヌクレオチド(nt.)およびコンティグ配列に対応するヌクレオチド(nt.)もまた、示される。
【図3I】図3A〜図3Mは、表4に示されるヌクレオチド配列(配列番号589〜614)を提示する。これらは、OM感染の間にアップレギュレートされたと同定された(実施例6を参照のこと)。配列番号1〜576として提示される、既知の遺伝子に対応するヌクレオチド(nt.)およびコンティグ配列に対応するヌクレオチド(nt.)もまた、示される。
【図3J】図3A〜図3Mは、表4に示されるヌクレオチド配列(配列番号589〜614)を提示する。これらは、OM感染の間にアップレギュレートされたと同定された(実施例6を参照のこと)。配列番号1〜576として提示される、既知の遺伝子に対応するヌクレオチド(nt.)およびコンティグ配列に対応するヌクレオチド(nt.)もまた、示される。
【図3K】図3A〜図3Mは、表4に示されるヌクレオチド配列(配列番号589〜614)を提示する。これらは、OM感染の間にアップレギュレートされたと同定された(実施例6を参照のこと)。配列番号1〜576として提示される、既知の遺伝子に対応するヌクレオチド(nt.)およびコンティグ配列に対応するヌクレオチド(nt.)もまた、示される。
【図3L】図3A〜図3Mは、表4に示されるヌクレオチド配列(配列番号589〜614)を提示する。これらは、OM感染の間にアップレギュレートされたと同定された(実施例6を参照のこと)。配列番号1〜576として提示される、既知の遺伝子に対応するヌクレオチド(nt.)およびコンティグ配列に対応するヌクレオチド(nt.)もまた、示される。
【図3M】図3A〜図3Mは、表4に示されるヌクレオチド配列(配列番号589〜614)を提示する。これらは、OM感染の間にアップレギュレートされたと同定された(実施例6を参照のこと)。配列番号1〜576として提示される、既知の遺伝子に対応するヌクレオチド(nt.)およびコンティグ配列に対応するヌクレオチド(nt.)もまた、示される。
【発明を実施するための形態】
【0075】
(詳細な説明)
以下の実施例は、本発明を説明する。ここで、実施例1は、NTHiゲノムの配列を記載し、実施例2は、同定されたコンティグおよび最初の遺伝子発見を記載し、実施例3は、NTHiプロモータートラップライブラリーの構築を記載し、実施例4は、GFPを発現する86−028NP誘導体の分析を記載し、実施例5は、中耳の流体由来の細菌の直接標識を示し、実施例6は、急性中耳炎においてインビボで誘導されたプロモーターの同定を記載し、実施例7は、ビルレンス関連遺伝子の同定を記載し、そして実施例8は、固有のNTHi遺伝子配列の同定を記載する。
【実施例】
【0076】
(実施例1)
(分類不可能なHaemophilus influenzaeゲノムの配列)
NTHi86−028NP株は、コロンバスのChildren’s Hospitalにおいて鼓膜切開術および慢性OMのための管挿入を受けた小児科患者から得られた、最小限に継代された臨床単離物である(Bakaletzら,Infection and Immunity,56(2):331−335,1988)。86−028NP株は、2002年10月16日にAmerican Type Tissue Collection(Manassas,VA 20108 USA)に寄託され、そして受託番号PTA−4764を割当てられた。
【0077】
NTHiにおけるビルレンス決定因子の同定により広範に近づくための試みにおいて、NTHi 86−028NP株のゲノムを3倍の適用範囲(3−fold coverage)でシークエンシングした。染色体DNA、86−028NP株からPuregeneプロトコールを用いて調製し、そしてHydroshear装置(Gene Machines)によって2kb〜4kbのサイズにせん断した。Chissoeら(Methods:a Companion to Methods of Enzymology 3:55−65,1991)およびSambrookら(Molecular Cloning:a Laboratory Manual,第2版 Cold Springs Harbor,NY,1989)に記載されたように、せん断したDNAを、エタノール沈殿し、クレノウ酵素の混合物を用いて末端修復し、そしてアガロースゲル電気泳動によってサイズ選別して、2kb〜4kbのフラグメントを得た。これらのフラグメントを、SmaI制限酵素認識部位(ホスファターゼ処理されている)を用いてpUC18ベクター中にクローニングし、そしてE.coli XL−1 Blueに形質転換し、アンピシリン耐性について選択する。X−galを含有するLB−Ampプレート上での青/白スクリーニングによって、インサートを含むコロニーを同定し、そして1.5mlのTB−Amp(TB=Ternfic Broth)ブロスを含む96ディープウェルプレートに移す。ディープウェルプレート培養物を、37℃で一晩増殖させる。テンプレート調製、シークエンシングおよびコンティグアセンブリを行った。
【0078】
Chissoeら(上述)に記載されるように、Beckman Biomek2000自動ロボットワークステーション上で、自動テンプレート調製を行った。簡単に言うと、上記で調製されたクローンを含む各96ディープウェルプレートを遠心し、細胞をペレット化した。上清をデカントし、細胞を(必要な場合)−20℃で凍結した。Chissoeら(上述)に記載されるように、4つの96ディープウェルブロックをBiomekテーブル上に配置し、液体ハンドリングロボットを使用して、代表的なSDS−NaOH溶解プロトコールの自動バージョンを用いてテンプレートを調製した。最終的なエタノール沈殿テンプレートを、50μlのddH2Oに各々溶解し、そしてDNAシークエンシングのために使用した。
【0079】
Robbins Hydra 96ロボットを用いて、上記テンプレートを(96ウェルプレートから)384ウェルプレートへと再整列化することによって、シークエンシング反応を実行した。PE Big−DyeTMターミネーターおよびユニバーサルプライマー(M13フォワードおよびリバース)を用いてサイクルシークエンシング反応を実行し、Sephadex G50カラムを通して浄化し、そしてPE Biosystems 3700キャピラリー電気泳動DNAシークエンサー上で、製造者の指示に従って分析した。シークエンシングの読み(8219)を、576のコンティグ(本明細書における配列番号1〜576)に組み立てた。3倍(3−fold)シークエンシングについての統計を、表2Aに示す。アセンブリ17の全固有配列は、1.74Mbである。
【0080】
(表2A)
【0081】
【表2A】
続いて、NTHiゲノムの8倍(8−fold)シークエンシング分析を行った。8倍シークエンシングは、NTHiゲノムを11のコンティグに組み立てた。コンティグ5、8、9、10、12〜18を、本明細書において配列番号675〜685として示す。8倍シークエンシングについての統計を、表2Bに示す。
【0082】
(表2B)
【0083】
【表2B】
(実施例2)
(コンティグ説明および最初の遺伝子発見)
3倍配列分析によって同定された、長さ5000bp以上を有する88個のコンティグ中75個は、BLASTNを介して、H.influenzae Rd株に対する有意な類似性を示す。H.influenzae 86−028NP株における遺伝子順とH.influenzae Rd株における遺伝子順との間の潜在的な関係を可視化するため、86−028NP 3倍コンティグセットおよびRd遺伝子セットを、BLASTNを用いて双方向性に比較した。結果を、ヒットしたRd遺伝子の遺伝子座標に基づいてコンティグをソーティングし、Rayら(Bioinformatics 17:1105−12,2001)に記載されるように、見出された最小の座標で各コンティグを固定することによって、遺伝子順 対 コンティグ空間にプロットした。この様式で比較し、完全に既知のゲノムに対して同一の遺伝子順を有するゲノムの不完全なアセンブリを、単調増加する階段状の形態で表示した。
【0084】
BLASTXを使用して、Rd株ゲノムにおける遺伝子に対して相同性を有する配列、およびH.influefzzae Rd株に見出されない遺伝子に対して相同性を有する配列に対するヒットを同定した。
【0085】
Rd株配列へのヒットをデータセットから除去し、そして他のセットを表3Aにまとめた。データは以下のように提示される:コンティグ番号(=配列番号)、カラム1;各ヒットについてのEスコア、カラム2;引用されたコンティグのアミノ酸翻訳部分に対する相同性を有したタンパク質の名前、カラム3;上記ホモログを産生する生物体、カラム4;そして、カラム3に引用されたタンパク質の各々についてのGenbankタンパク質識別子、カラム5;コンティグ内の対応するヌクレオチド(配列番号によって参照される)。ほとんどの例においていくつかのホモログが同定されたが、明確化のため、最も相同性の高いタンパク質を表3Aに引用する。
【0086】
表3Aに列挙した遺伝子のいくつかについての配列を、NTHiゲノムの8倍シークエンシング内で同定した。表3Bは、11コンティグ内のこれらの遺伝子の配置、全長オープンリーディングフレーム配列(配列番号によって同定される)、上記オープンリーディングフレームによってコードされる生成されたアミノ酸配列、およびBLASTXによって同定された高相同性を有する遺伝子(表3Aで列挙されたようなもの)を列挙する。
【0087】
NTHiおよびRd株中における比較的短い遺伝子の配置を検査するため、詳細に記載されている2つの遺伝子クラスター中の遺伝子順を比較した。第一に、血液凝集性線毛(LKP)遺伝子領域中に存在する遺伝子を検査した(Mhlanga−Mutangaduraら,J Bacteriol.180(17):4693−703,1998)。線毛遺伝子クラスターは、purE遺伝子とpepN遺伝子との間に位置する。これらのフラグメントのみを、図1に記載する。血清型b株、Eaganは、hifABCDE遺伝子クラスターを含み、そして血液凝集性線毛を生成する。Rd株は、hicAB遺伝子およびhifABCDE遺伝子クラスターを欠損している。一般的に、既に検査した分類不可能な株は、hicAB遺伝子を含むが、血液凝集性線毛をコードするhif遺伝子を含まない。86−028NP株配列(本明細書において記載される)は、この領域において、NTHi R3001株の配列と同一である(図1)。rfaD遺伝子は、エンドトキシンの生合成に関与する酵素をコードする。さらに、NTHi 2019株由来のrfaD遺伝子は、Nicholsら(Infect Immunity 65(4):1377−86,1997)によって特徴付けられている。2019株において、rfaD遺伝子は、エンドトキシン生合成に関与する別の酵素をコードするrfaF遺伝子のすぐ上流にある。Rd株における遺伝子配置は、様々である;rfaD遺伝子およびrfaF遺伝子は、およそ11kbの配列によって分離されている。検査された最も分類不可能な株は、2019株において見られる遺伝子配置を含んでいた。対照的に、86−028NP株は、Rd株に見られる遺伝子配置と同一の遺伝子配置を有する(図2)。
【0088】
現行のアセンブリの全体的な分析は、遺伝子の内容および順序が、Rd株における内容および順序と類似することを示す。より詳細な分析は、Pasteurellaceaにおいて以前見られなかったかなりの数のNTHi遺伝子が存在すること、そしてNTHi遺伝子の内容および順序がRd株において見られる内容および順序と異なるいくつかの領域が存在することを、明らかにした。したがって、現行のデータは、86−028NP株ゲノムが、Rd様性質と非Rd様性質との複雑なモザイクを含むことを示唆する。
【0089】
DFI戦略はまた、遺伝子発現を増加させる新規のNTHi配列を同定した。他の生物体(主にグラム陰性細菌)のORFに対して相同性を有する遺伝子または遺伝子フラグメントを含む、これらの新規コンティグ配列のリストを、表3Aに示す。例えば、コンティグ442(配列番号442)のヌクレオチド配列、ヌクレオチド1498〜1845は、H.influenzae Rd株リポタンパク質B(LppB)のアミノ酸1〜116をコードする配列に対して、高度に相同性である。この遺伝子は、静止期生存遺伝子、surEと、ビルレンスに関連している、最近同定された細菌リポタンパク質、LppB/NlpDに対して高度に相同性である、43kDの抗原外膜リポタンパク質をコードする遺伝子との間に位置する(Padmalayamら,Infect.Immun.,68:4972−4979,2000)。最近、Zhangおよび共同研究者らは、Thermotoga maritimaにおいて、nlpD遺伝子およびsurE遺伝子の発現が、細胞増殖の静止期の間に誘導されることを示した(Zhangら,Structure(Camb),9:1095−1106,2001)。したがって、中耳でのストレス誘導条件下において、このNTHiリポタンパク質が発現される。
【0090】
【表3A−1】
【表3A−2】
【表3A−3】
【表3A−4】
【表3A−5】
【表3A−6】
【表3A−7】
【表3A−8】
【表3A−9】
【表3A−10】
【表3A−11】
【表3A−12】
【表3A−13】
【表3A−14】
【表3A−15】
【表3A−16】
【表3A−17】
【表3A−18】
【表3B−1】
【表3B−2】
【表3B−3】
(実施例3)
(NTHiプロモータートラップライブラリーの構築)
NTHiの潜在的なビルレンス決定因子を同定するために、細菌遺伝子発現を、NTHi誘導中耳炎(OM)のチンチラモデルの1つの特定の解剖学的くぼみ(niche)における初期疾患の進行の間の微分蛍光誘導(DFI)によって、モニターした。NTHi菌株86−028NP由来のゲノムDNAフラグメントを、プロモータートラップライブラリーを使用して、プロモーターレスgfpmut3遺伝子の上流にクローニングした。プラスミドpGZRS39A(Actinobacillus pleuropneumoniaeから単離したpGZES−1の誘導体)は、A.pleuropneumoniae−Escherichia coliシャトルベクターである。このプラスミドは、A.pleuropneumoniae由来の複製起点、pUC19由来のlacZα遺伝子およびTn903由来のカナマイシン耐性遺伝子を含む(Westら、Genes,160:81−86,1995)。
【0091】
プロモータートラップベクターを、GTP変異gfpmut3遺伝子を、EcoRIフラグメントに対するBamHIとして、pGZRS−39A内にクローニングしてpRSM2167を形成することによって構築した。この変異GTP遺伝子は、2つのアミノ酸変化(S65GおよびS72A)を含み、これは、488nmで励起される場合、蛍光発光を増強する。この変異体はまた、高い溶解性および発色団形成の速い動力学を有する(Cormackら、Gene,173:33−38,1996)。このプラスミドは、NTHi菌株86−028NP内へのエレクトロポレーションによって形質転換され、親のプラスミド菌株86−028NP/pRSM2167を生成する。
【0092】
ランダムゲノムDNAフラグメント(実施例1に記載される)を、puroもタープローブベクター内への連結のために調製した。ゲノムDNAを、製造業者のプロトコルに従って、PuregeneDNA単離キット(Gentra Systems,Minneapolis,MN)を使用して、菌株86−028NPから単離した。制限障壁に起因して、プラスミドDNAを単離し、これをライブラリー作製のために使用することが必要であった。この単離されたDNAを37℃で1時間、Sau3AI(NEB,Beverly,MA;0.25単位/μg DNA)を用いて部分的に消化し、ゲル電気泳動によって分離し、そしてDNAフラグメント(0.5〜1.5kbのサイズ)を、Qiagenゲル抽出キットを使用して、回収した。ベクター調製のために、pRSM2167を、製造業者のプロトコルに従って、Wizard Plus Maxiprep DNA精製システム(Promega,Madison WI)を使用して、一晩の培養物から単離した。
【0093】
プラスミドDNAを、BamHI消化によって線形化し、そして5’リン酸基を、ウシ腸アルカリホスファターゼ(CIAP;GibcoBRL Life Technologies)を用いる処置によって除去した。ゲノムDNAフラグメントを、線形化されホスファターゼで処理されたベクターを用いて連結し、そして改変プロトコルに従ってエレクトロポレーションのための調製したコンピテントNTHi菌株86−028NP内エレクトロポレートした(Mitchellら、Nucleic Acids Res.,19:3625−3628,1991)。プラスミドDNAをNTHi菌株86−028NP内にエレクトロポレーションで戻した場合、形質転換効率は、千倍改善した。簡単に述べると、細胞を、sBHI(脳心臓注入物)ブロスにおいて37℃、220rpmで、OD600=0.3まで増殖させた。細胞を、30分間、氷において冷やし、引き続いて、4℃で、等量の0.5×SG(1×SG:15%グリセロール、272mMスクロース)を用いて洗浄した。洗浄を合計3回繰り返した。引き続いて、細胞を、1×SG中で、100×濃縮用量に希釈した。細胞を、200オーム、2.5kVおよび25μFで、BioRad Gene Pulser IIセットを使用して、エレクトロポレーションし、次いで、1mlの予め温めたsBHI中に希釈し、37℃、5%CO2で2時間インキュベートし、そして形質転換体の一晩の増殖のためにチョコレート寒天にプレートした。
【0094】
形質転換体を選択し、20%グリセロール(容積/容積)を含む脱脂乳中、1000クローンのプール中に凍結させた。68,000メンバーのgfpプロモータープローブライブラリーを作製した。ClarkeおよびCarbon(Cell,9:91−99,1976)の確率計算を使用して、菌株86−028NP DNAの300bpフラグメントのライブラリー(1.8×106bp/ゲノム)に表される所定のDNA配列の99%確率を達成するために、27,629クローンのライブラリーを必要とした。従って、本発明のライブラリーは、86−028NPゲノムの2.5倍の適用範囲を表す。
【0095】
ライブラリーの質を評価するために、50クローンをランダムに選択し、チョコレート寒天で一晩増殖させ、そしてプラスミドを単離し、そして挿入DNAを配列決定した。選択されたクローンの大部分(64%)は、200bpと500bpとの間の範囲挿入サイズを有したが、32%は、500bpを超えた。挿入物の大部分は、独特のH.influenzae菌株Rdオープンリーディングフレーム(ORF)と相同性を示し、そして15クローンが、菌株86−028NP DNAに対して独特の配列を有した。菌株Rdに対して相同性を有するこれらのクローンのうち、60%が、正しい方向であり、その36%が、ORFの上流に配列を含んだ。クローンの大部分が500bp未満の挿入サイズを有したが、小さな挿入サイズと増加したGFP発現との間に相関は見出されなかった。実際、4つのクローンは、インビトロにおいて、わずかから中程度の蛍光を示し、これらの3つが、200塩基対〜500塩基対の間の挿入サイズを有し、1つが、700塩基対よりも長い挿入物を有した。
【0096】
ライブラリーの画分(約1000クローン)を、チョコレート寒天で増殖させ、PBS中で収集し、GFP蛍光についてフローサイトメトリーによって分析した。挿入DNAの無いプロモータートラップベクターを含む菌株86−028NP/pRSM2169と比較して、ライブラリークローンのプールは、増加した蛍光強度を示す。従って、ライブラリーは、種々のレベルの活性でプロモーターを有するクローンを含む。
【0097】
(実施例4)
(GFPを発現する86−028NP誘導体の分析)
gfp発現細菌を同定および分類するために必要なFACSプロモーターを確立するために、種々のレベルのgfp発現を示すパネル単離物を利用した。バックグラウンド蛍光を、菌株86−028NP/pRSM2169(ネガティブコントロール)を使用して評価し、従って、任意の観察された蛍光は、lacZプロモーター駆動gfp発現に起因している。しかし、この菌株は、検出可能なレベルのGFPを生成せず、実際、親の菌株86−028NPと比較した場合、増加した蛍光を示さない。高レベルのgfp発現単離物を、外膜タンパク質P2発現に対する強力なプロモーターを含む500bpフラグメントを、Sai−BamHI消化pRSM2167にクローニングすることによって作製した。このプラスミドを、エレクトロポレーションによって86−028NP内に形質転換し、高レベルのgfp発現菌株86−028NP/pRSM2211(高い蛍光コントロール)を生成する。この菌株は、菌株86−028NP/pRSM2169と比較して、GFP蛍光において約100倍増加を示した。中程度の蛍光誘導体クローン、86−028NP/pKMM4B5(中程度の蛍光コントロール)を、FACS分析によって単離し、そして予備的実験および細胞選別のためのコントロールとして使用した。インビトロでgfp発現を駆動するプロモーターを含むDNAフラグメントは、他の生物のDNAに対して公知の相同を有さない、菌株86−028NPに対して独特である。このクローンは、菌株86−028NP/pRSM2169と比較して、蛍光において約10倍の増加を示す。
【0098】
コントロール菌株をチョコレート寒天上での増殖物から再懸濁させ、4℃で30分間、交差反応性Phycoprobe R−PE抗ヒトIgG(H+L)抗体(100μlPBS中、10μg/ml;Biomeda Corp)を用いて標識した。未結合の抗体を除去するための3回の連続洗浄に続いて、細菌をFACS分析のために、300μlDPBS中で再懸濁させた。これらのコントロール調製物を使用して、488nmを発するアルゴンレーザーを備えるCoulter Epics Eliteフローサイトメーター(Coulter Corp.)を使用して、適切なサイズおよび蛍光ゲートを設定した。細菌を、ログフォワードアングル(log forward angle)および側方散乱検出に基づくサイズ、ならびに細菌のFITC/PE標識による選別のためにゲート(gate)した。選別された細胞を、冷却したsBHI中に収集し、そしてチョコレート寒天上にプレートした。一晩の増殖後、細胞を、第2回の感染のために収集するか、または個々に選択し、一晩増殖させ、インビトロで増殖する場合、蛍光について個々のクローンによってスクリーニングし、そしてプラスミド単離の前に、20%(容積/容積)グリセロールを含む脱脂乳中で凍結させ、そして挿入物DNAを配列決定した。コントロール菌株の選別効率を、Coulter EPICSフローサイトメーター(Coulter Corp.)を使用して確認した。
【0099】
多くのプラスミドを、抗生物質選択の非存在下でインビトロで迅速に分離した。従って、本明細書中で使用されるプロモータートラップベクターがこの事象の傾向があるか否かを評価するために、菌株86−028NP/pRSM2211(高い蛍光コントロール)の単一のコロニーを、チョコレート寒天上で単離し、抗生物質選択の非存在下で20回継代させた。抗生物質の存在下で増殖された細菌と比較した場合、蛍光強度の有意な減少は、観察されなかった。さらに、プラスミドをインビボでの抗生物質選択の非存在下で維持する。チンチラから収集された細菌含有中耳分泌液を、カナマイシン無しまたは有りでチョコレート寒天上にプレートした場合、類似の細菌カウントが観察された。これらのデータは、プロモータートラップベクターが抗生物質選択の非存在下で安定に維持されたことを実証する。
【0100】
プラスミドの安定性に問題に加えて、宿主−病原体相互作用を研究するためのレポーターとしてのGFPの使用についての初期の研究は、GFPが、細菌細胞−表面動力学に対して最小の効果を有する低い毒性で、細胞質タンパク質として連続的に合成され得ることを実証した(Chalfieら、Science,263:802−805,1994)。高いレベルのgfp発現誘導体の構築は、NTHiに対するGFP毒性の評価を可能にした。野生型菌株(86−028NP)および高いGFP産生菌株86−028NP/pRSM2211の両方の増殖曲線を、類似の条件下で増殖した場合と比較した。増殖速度は類似し、GFP発現が細胞に対して毒性でないことを示す。
【0101】
86−028NP gfp発現誘導体を使用して、効率的な細胞選別についてのパラメーターを規定した。菌株86−028NP/pRSM2169を、中程度gfp発現誘導体、菌株86−028NP/pKMM4B5と、100:1の比で混合し、細菌集団の合計に対して小さな割合のgfp発現クローンを含むことが予期されるインビボ環境をシミュレートした。この混合物をFACS分析に供し、1.8%最大蛍光集団および52%最小蛍光集団を収集する。選別された集団のフローサイトメトリー分析は、細菌集団の65%まで、菌株86−028NP/pKMM4B5の富化を示し、これは、ネガティブ集団での選別の場合に観察されない減少である。引き続く回の選別は、この中程度蛍光集団をさらに富化させることをが期待される。ネガティブ選別において蛍光細菌の量を減少させることができないことは、ネガティブ選別についてのゲート設定のサイズに起因した。GFP−ネガティブ細胞を、10%の最小蛍光集団でゲーティングすることによって富化した。
【0102】
(実施例5)
(中耳分泌液からの細菌の直接的標識)
類似の戦略(実施例5に記載されるとおり)を、AOMの間にチンチラの中耳から得られた浸出液から蛍光クローンを選別するために適用した。インビボの微分蛍光誘導(DFI)を使用する本発明者らの能力は、非蛍光細菌、蛍光および非経口細胞細片、ならびに真核生物細胞からgfp発現細菌を選別する本発明者らの能力に依存した。
【0103】
耳鏡検査法またはティンパノメトリのいずれによっても中耳感染の証拠のない健常な成体チンチラ(Chinchilla lanigera)を使用して、インビボのプロモーター活性についてライブラリーをスクリーニングした。NTHi/pRSM2169ライブラリーの2つのプール(それぞれ1000クローン)を、カナマイシンを含むチョコレート寒天上で一晩増殖させた。このライブラリーを組合せ、冷却10mM滅菌PBS中に3.3×106CFU/mlまで希釈し、300μl(1.0×106CFU;500CFU/クローン)を使用して、左および右のチンチラトランスブラーキャビティー(transbullar cavity)を播種した(2000クローン/耳)。OM発生(development)を、24時間および48時間で、ビデオ耳鏡検査法およびティンパノメトリーによってモニターした。細菌を中耳腔において増殖させ、予期されるように、48時間で、接種材料用量の500倍の濃度に達した(Bakaletzら、Infect.Immunity 67:2746−62,1999)。宿主環境に対するこの細菌の適応は、炎症応答を生じ、これは、紅斑、脈管拡大および鼓膜の隆起、多形核球細胞(PMS)の浸潤、ならびに回収される浸出液の耳鏡検査および顕微鏡検査によって観察されるような中耳腔の分泌液の蓄積によって示される。24時間後および48時間後、中耳分泌液を上鼓室タップによって回収し、そしてFACSのために調製した。
【0104】
この分析が中耳分泌液中で回収可能な細菌に制限されたことに注意することが重要である。いくつかの場合において、FACS分析のために細菌を収集するために、中耳腔を洗浄することが重要であった。従って、この分析は、NTHiが粘膜にゆるく付着する場合、アップレギュレートされる遺伝子を含む。NTHiは、OMのチンチラモデルにおいて中耳腔内に生物膜を形成することが観察された(Erhlichら、JAMA,287;1710−5,2002)。本明細書中に記載されるプロトコルがプランクトンの集団から回収されるクローンを選択するので、細菌が粘膜生体膜と関連する場合、遺伝子がアップレギュレートされるクローンを回収することは予期されない。しかし、中耳粘膜の均質化および引き続く細菌細胞の単離によって、本発明者らは、これらのクローンを回収し得る。いくつかのGFP発現クローンがこの浸出液において回収されたが、剥離した細胞として浸出液中にまたは凝集物中に存在する真核生物細胞に付着したこともあり得る。これらの細菌は、選別効率を損なうことなく浸出液から回収することは困難である。従って、中耳分泌液を粘液溶解剤を用いて処理し、次いで、遠心分離して、標識化する前に、大きな凝集物および真核生物細胞を除去した。
【0105】
必要な場合に、チンチラの中耳の分泌液を、滅菌した生理食塩水を用いて250μlまで希釈した。等量のDPBS中のN−アセチル−L−システイン(0.5%;w/v)(pH 7.4)を、粘液溶解剤として室温で5分間添加した(MiyamotoおよびBakaletz,Microb.Pathog.,21:343〜356 1996)。分泌液を、細胞の残屑、赤血球、および炎症性細胞を除去するために遠心分離し(300×g、5分間)、そして細菌を含有する上清を、新しいチューブへ移した。細菌を、NTHi菌株86−028NPに由来する、全OMP調製物に対して指向されるチンチラ抗血清(1:50希釈)と共に、4℃で45分間インキュベートし、遠心分離(2000×g、5分間)によってペレット化し、そして0.05%ウシ血清アルブミンを含有する冷DPBSを用いて2回洗浄した。続いて、細菌を、交差反応性フィコプローブ(phycoprobe)R−PE抗ヒトIgG(H+L)抗体(100μlPBS中10μg/ml;Biomeda Corp)を用いて、4℃で30分間標識した。非結合の抗体を除去するために連続して3回洗浄し、続いて、細胞を、FACS分析のために300μl DPBSに再懸濁した。
【0106】
(実施例6)
(急性中耳炎におけるインビボで誘導したプロモーターの同定)
プロモータートラップライブラリーを用いて形質転換したH.influenzae 86−028NPを、チョコレート寒天で一晩増殖させた。インビトロでgfpを発現したプロモーターを含有するこれらクローンを選択するために、上記ライブラリーを、1回のFACS分析に供し(実施例6に記載するように)、低レベル量のGFPを発現しているこれらのクローンのみを回収した。これらのクローンを、プールして、チンチラの中耳の骨胞を経由して(transbullarly)接種するために用いた。注射の24時間および48時間の後、細菌含有滲出液を、鼓室または鼓膜の上部の穿刺によって除去した。細菌をR−PE標識化抗体を用いて間接的に標識し、蛍光タグ化された細菌のゲーティングによるが、発現もしている細菌について選別することによってFACS分析に供した。これらのクローンを、さらに富化するために動物を再感染させるために用いた。最終回の選別の後、単一のコロニー単離体を、蛍光の欠如についてインビトロでスクリーニングした。
【0107】
インビトロで蛍光を発していなかった、FACS分析によって単離したこれらのクローン(インビボでGFP蛍光について陽性)を、プラスミド単離および挿入したDNA配列の同定のために調製した。これらのクローンを、カナマイシンを含有するチョコレート寒天プレートで一晩増殖させ、製造者のプロトコルに従ってQiaprep Miniprep Kit(Qiagen)を用いてプラスミド単離のために調製した。プラスミド挿入DNAを、gfpmut3遺伝子に相補的であり、かつ挿入DNAの下流であるプライマー5’−TGCCCATTAACATCACCATCTA−3’(配列番号588)を用いて配列決定した。配列決定反応を、製造者のプロトコルに従って、ABI prism BigDye(登録商標)ターミネーター・サイクル・シークエンシング・レディー・リアクション・キット(Applied Biosystems)を使用して、GeneAmp PCR System 9700 (Applied Biosystems)を用いて実施した。この配列を次いで、96ウェル・マルチスクリーンHVプレート(Millipore)中のセファデックスG−50を通過させることによって精製し、続いてABI Prism3100 DNAアナライザー(Applied Biosystems)で分析した。
【0108】
挿入配列を、H.influenzae菌株Rdの注釈をつけた全配列と比較した。菌株Rdとヌクレオチド相同性を有さないこれらの挿入物を続いて、BLASTNおよびBLASTXアルゴリズムを用いて分析した。さらに配列分析を、DNASTAR(Madison,Wisc)を用いて実施した。正しい向きでかつ推定ORFの5’側配列を含む挿入物は、NTHi細菌が、チンチラの中耳に存在する場合、優先的に活性化した推定プロモーターを含有した。
【0109】
インビボで調節される推定プロモーターを有する52個のクローンを、単離した。H.influenzae菌株Rdに同定された配列と類似する配列を含む44個の候補クローンの、インビトロおよびインビボでの遺伝子発現の定量的な比較によって、NTHiが、チンチラの中耳内に存在する周囲のきっかけに応答する場合、26個の遺伝子(60%)がアップレギュレートした遺伝子発現があることを確認した。これらの遺伝子を以下の表4Aに要約した。遺伝子発現を駆動するインビボ調節プロモーターは、膜輸送、周囲の情報のプロセシング、細胞の代謝、遺伝子調節、および未知の機能を有する仮想タンパク質に関係していると予測される。
【0110】
インビボで推定プロモーター候補の誘導を確認するために、NTHi菌株86−028NPが、インビトロで中間対数期まで、またはインビボで48時間増殖させた場合に、メッセンジャーRNA発現の相対量を、比較した。このRNAを、製造者のプロトコルに従って、TRIzol LS試薬(Gibco Life Technologies)を用いて単離した。DNAを、製造者のプロトコルに従って、DNAフリー・キット(Ambion)を用いてRNA調製物から除去した。DNase I処理したRNAサンプルを、Qiagen RNeasyカラムを通過させることによって精製した。RNAの純度および完全性を、260/280nm分光光度計読み取りおよびAgilent 2100 Bioanalyzer(Agilent Technologies)によってそれぞれ、調べた。
【0111】
FACSデータを独立に確認するために、本発明者らは、定量的RT−PCRによって候補遺伝子の相対的な発現を決定した。親株86−028NPを、これらの研究に用いた。製造者の説明書に従って、ワン・ステップQuantiTectSYBR Green RT−PCRキット(Qiagen)を用いるリアル・タイム定量的RT−PCRによって、転写レベルを調べた。簡単に言えば、FACS分析によって同定した推定上のインビボで誘導されるプロモーターのオープン・リーディング・フレームの下流に作製したプライマーを用いて、遺伝子特異的mRNAを、ABI Prism 7700シークエンス・ディテクション・システム(Applied Biosystems)でのRT−PCRによって、逆転写し、増幅した。産物の量を、ジャイレース(gyr)遺伝子のフラグメントを増幅することによって、既知量の細菌のゲノムDNA(102〜107個のゲノムコピーDNA)について作製した検量線を用いて計算した。RNA調製物(−RTコントロール)中にDNAが存在しないことおよび鋳型RNAのないコントロールサンプル中にプライマーダイマーが無いことを確認するために、コントロールを、同時に分析した。さらに、RT−PCR産物を、ゲル電気泳動によって分析した。全ての場合において、単一の産物を、適切な塩基対サイズで観察した。サンプル間の細菌RNAの量を、インビトロで本発明者らが試験した種々の増殖条件の下、構成的に発現することを示す、gyr発現に対して標準化した。既知量の細菌ゲノムDNA(102〜107個のゲノムコピーDNA)を用いて、ジャイレース(gyr)遺伝子のフラグメントを増幅することによるRT−PCR定量のための標準曲線を作製した。ジャイレースは、種々の増殖条件の下インビトロで構成的に発現し、そのためサンプル間の全細菌RNAレベルの標準化のために用いた。インビボでの相対的な遺伝子発現を、インビトロでの遺伝子発現と比較し、倍率増加(fold−increase)データを、表4に要約する。
【0112】
NTHiゲノムの8倍の配列決定は、表4Aに列挙した大部分の遺伝子についての全長オープン・リーディング・フレームを同定した。表4Bは、NTHiゲノムおよび対応するアミノ酸配列内の全長ヌクレオチド配列を提供する。遺伝子の誘導倍率は、チンチラの中耳内に存在する周囲のきっかけに起因し、遺伝子の産物または機能を、便宜のために表4Bに繰り返す。
【0113】
【表4A】
【0114】
【表4B−1】
【0115】
【表4B−2】
(実施例7)
(ビルレンス関連遺伝子の同定)
多くの細菌種において、ビルレンス関連遺伝子のサブセットは、短いリピートの複製における誤りによって調節される。これらのリピートは、遺伝子の5’またはコード配列内であり得、これらの存在は、ビルレンスと関連することを示す遺伝子の制御発現の指標である。リピートの付加または欠失は、特定のビルレンス決定因子の発現または発現を欠く結果を生じる。
【0116】
NTHi H.influenzae菌株86−028NPコンティグセットは、短いオリゴヌクレオチド・リピートについて問い合わせた。このリピート周辺の領域を、このリピートに関連する遺伝子を同定するために分析した。表5に、同定したリピートおよび各リピートに関連するORF(BLASTによって同定した)を列挙する。
【0117】
さらに配列分析によって、ビルレンス関連遺伝子の全長ヌクレオチド配列およびORFによってコードされる対応するアミノ酸配列を同定した。誘導したアミノ酸配列は、列挙したGenbank配列と高い相同性である
【0118】
【表5−1】
【0119】
【表5−2】
(実施例8)
(独特のNTHi遺伝子配列の同定)
NTHi毒性に関連する遺伝子をまた、NTHi細菌属が、組織に感染している場合の遺伝子発現レベルとNTHiを、人工的な検査室培地で増殖させた場合の同一遺伝子発現レベルとを比較することによって同定した。これらの新規遺伝子を、実施例4〜6の上記プロモーター・トラップ技術を用いて同定し、続いて公知のRdゲノムとの比較によってこれらの遺伝子がNTHi菌株86−028NPに特有であることを示した。
【0120】
このスクリーニング方法を用いて同定したDNA配列を、配列番号577〜580として示す。これらの配列は、H.influenzae Rd.ゲノム配列と相同性を有する遺伝子または遺伝子フラグメントを含まなかった。これらは、完全に新規配列であるが、チンチラの中耳におけるNTHi感染の間の遺伝子の発現レベルに起因して、これら遺伝子の発現は、NTHi毒性に関係する可能性がある。
【特許請求の範囲】
【請求項1】
明細書中に記載の発明。
【請求項1】
明細書中に記載の発明。
【図1】

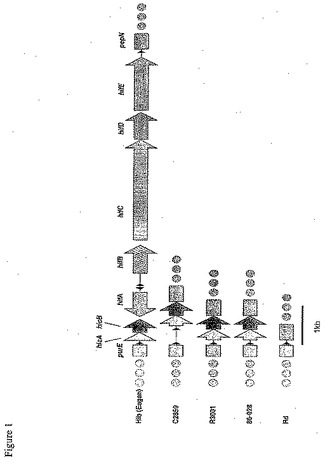
【図2】
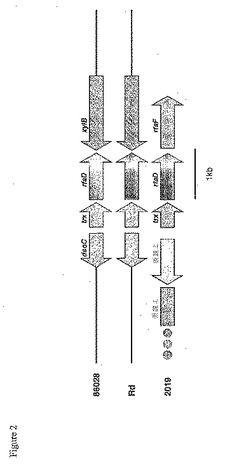
【図3A】

【図3B】

【図3C】

【図3D】

【図3E】

【図3F】

【図3G】

【図3H】

【図3I】

【図3J】

【図3K】

【図3L】

【図3M】

【図2】
【図3A】
【図3B】
【図3C】
【図3D】
【図3E】
【図3F】
【図3G】
【図3H】
【図3I】
【図3J】
【図3K】
【図3L】
【図3M】
【公開番号】特開2010−279359(P2010−279359A)
【公開日】平成22年12月16日(2010.12.16)
【国際特許分類】
【出願番号】特願2010−139968(P2010−139968)
【出願日】平成22年6月18日(2010.6.18)
【分割の表示】特願2006−506942(P2006−506942)の分割
【原出願日】平成16年3月5日(2004.3.5)
【出願人】(502200830)ネイションワイド チルドレンズ ホスピタル, インコーポレイテッド (11)
【Fターム(参考)】
【公開日】平成22年12月16日(2010.12.16)
【国際特許分類】
【出願日】平成22年6月18日(2010.6.18)
【分割の表示】特願2006−506942(P2006−506942)の分割
【原出願日】平成16年3月5日(2004.3.5)
【出願人】(502200830)ネイションワイド チルドレンズ ホスピタル, インコーポレイテッド (11)
【Fターム(参考)】
[ Back to top ]