
骨形成タンパク質の変異タンパク質及びその使用
【課題】骨形成タンパク質の変異タンパク質、それをコードする核酸、このような骨形成変異タンパク質の製造方法、このような変異タンパク質を含む組成物、好ましくは薬剤組成物、及びこのような変異タンパク質の、医薬の製造のための使用。
【解決手段】骨形成タンパク質の変異タンパク質に関すし、この変異タンパク質は、骨形成タンパク質の野生型と比較して、ヒトBMP−2のアミノ酸位置51に相当するアミノ酸位置にアミノ酸置換を含む。本発明の課題は、I型及びII型セリンキナーゼ受容体鎖媒介性作用の点で、骨形成タンパク質の野生型と競合しないが、依然として、ノギンタンパク質ファミリー、DANタンパク質ファミリー及びコーディンタンパク質ファミリー等のモジュレータータンパク質と相互作用能を有する。
【解決手段】骨形成タンパク質の変異タンパク質に関すし、この変異タンパク質は、骨形成タンパク質の野生型と比較して、ヒトBMP−2のアミノ酸位置51に相当するアミノ酸位置にアミノ酸置換を含む。本発明の課題は、I型及びII型セリンキナーゼ受容体鎖媒介性作用の点で、骨形成タンパク質の野生型と競合しないが、依然として、ノギンタンパク質ファミリー、DANタンパク質ファミリー及びコーディンタンパク質ファミリー等のモジュレータータンパク質と相互作用能を有する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、骨形成タンパク質の変異タンパク質、それをコードする核酸、このような骨形成変異タンパク質の製造方法、このような変異タンパク質を含む組成物、好ましくは薬剤組成物、及びこのような変異タンパク質の、医薬の製造のための使用に関する。
【背景技術】
【0002】
本発明の変異タンパク質は、骨形成タンパク質の変異タンパク質である。骨形成タンパク質は、本明細書ではBMP−2様タンパク質とも呼ばれ、TGF−βスーパーファミリーを形成する2つの主要な群の一方である。BMP−2様タンパク質とは別に、このTGF−βスーパーファミリーに属するもう一方の群のタンパク質はTGF−β−アクチン様タンパク質である。TGF−βスーパーファミリーの2つの群は、その結合機構が異なっている。一般に、ポリペプチド成長因子と構造的に関連しているTGF−βスーパーファミリーのメンバーは、細胞増殖、細胞株の運命決定、分化、移動性、接着及び細胞死をはじめとする細胞過程に関与している。こういった因子は、時間及び組織特異的パターンに従って発現され、真核生物ではほとんどすべての組織の発達、止血及び修復にとって重要である。これらの因子はまた、細胞の活性を規定する細胞内シグナルの重要な部分を占めている。
【0003】
TGF−βのシグナル伝達経路は、ごく最近解明された。TGF−βのシグナル伝達には、細胞表面の受容体セリンキナーゼ及びその基質、すなわち、SMADタンパク質が関与しており、このタンパク質は、リン酸化されると核へ移動する。リン酸化されたSMADタンパク質は、DNA結合パートナーと共同して標的遺伝子の転写を活性化する。TGF−β及びTGF−βスーパーファミリーのその他のメンバーの多機能活性は、種々の受容体とSMADタンパク質とDNA結合タンパク質との相互作用に基づいていると思われる。このシグナル伝達経路の何らかの混乱が、哺乳類、特に人間におけるいくつかの疾患の理由である。TGF−βスーパーファミリーのメンバーはいくつかの構造的特徴を共有しているが、このメンバー間の相同性はかなり限定的である場合もある。例えば、すべてのタンパク質は二量体であり、通常、ホモ二量体、すなわち、2つの同一の単量体からなるものである。また、TGF−βスーパーファミリーのメンバーは、2つの異なる種類のセリンキナーゼ受容体鎖、すなわち、I型鎖及びII型鎖からなる細胞受容体を用いる。
【0004】
I型鎖は細胞質GSボックスとセリンキナーゼとを含み、I型鎖がBRIA、BRIB又はARIである場合には、SMAD−1及びSMAD−5シグナルタンパク質を活性化する。II型鎖は、GSボックスセグメントのリン酸化によってI型受容体セリンキナーゼを活性化する。I型及びII型鎖双方の小さな受容体エクトドメインは、それぞれ約120〜150個のアミノ酸を含むが、かなり限定的な相同性しか共有していない。それにもかかわらず、TGF−βスーパーファミリーのすべての既知受容体鎖の1つの共通の特徴として、4つの保存されたジスルフィド架橋があり、さらなるジスルフィド架橋及びいくつかのアミノ酸の位置はI型又はII型受容体タンパク質のいずれかに特徴的であるようである。BMP−2様タンパク質とI型鎖との結合は、BMP−2様タンパク質のリスト(wrist)エピトープを介して起こり、II型鎖との結合は、BMP−2様タンパク質の「ナックル(knuckle)」エピトープを介して起こる。これらの骨形成タンパク質とそれぞれの受容体鎖との結合は、細胞質膜で逐次的に起こり、特定の配列は骨形成タンパク質と個々の受容体鎖との親和性に応じて変わる。例えば、溶解BMP−2は、その高親和性I型鎖、すなわち、BMPR−IA、BMPR−IBとまず結合し、おそらくはActR−Iとも結合し、続いて、低親和性II型受容体鎖と会合して活性な受容体複合体を形成する。この会合は細胞質膜で起こる。
【0005】
BMP−2のリスト又はナックルエピトープ中のいくつかのアミノ酸置換によって、不活性なBMP−2変異タンパク質となり、これは対応する受容体をもはや活性化することができない。BMP−2[A34D]などのナックルエピトープ中にアミノ酸置換を有する不活性な変異タンパク質は、それでも、野生型BMP−2と同様にI型鎖と結合できる。しかし、I型及びII型鎖の双方を含む、活性な受容体複合体を形成するための膜におけるII型鎖の会合には支障を来たす。したがって、この種の変異タンパク質は、I型鎖に対して野生型BMP−2と競合しているので、BMP−2の阻害剤として作用する。
【0006】
骨形成タンパク質及び、詳しくは、BMP−2様タンパク質は、I型鎖及びII型鎖を含む受容体が関係しているシグナル伝達経路に関与していることとは別に、モジュレータータンパク質の群と相互作用する。これらのモジュレータータンパク質は、同様にある種のシグナル伝達経路に関与している。BMP−2様タンパク質と相互作用するモジュレータータンパク質は、中でも、ノギンタンパク質ファミリー、DANタンパク質ファミリー及びコーディンタンパク質ファミリーのメンバーを含んでなる。これらのモジュレータータンパク質は、当技術分野では公知であり、例えば、Avsian-Kretchmer,O.及びHsueh, A.J. (2004)「Comparative genomicanalysis of the eight-membered ring cystine knot containingbone morphogeneticprotein antagonists.」、Mol Endocrinol、18巻、1〜12頁に記載されており、これにはBMP−モジュレータータンパク質、より詳しくは、シスチンノット含有BMPモジュレータータンパク質が記載されている。例えば、ノギンを含むノギンファミリーは、Canalis, E.、Economides, A. N.及びGazzerro, E. (2003)、「Bonemorphogeneticproteins, their antagonists, and the skeleton.」Endocr Rev巻、24、218〜235頁によって記載されており、中でも、コーディン、ベントロピン、CTGF、CYR61、クロスベインレス(Crossveinless)、プロコラーゲンIIAを含むコーディンファミリーは、例えば、Bork, P. (1993)「The modular architecture ofa new family of growth regulators related to connective tissue growth factor.」、FEBS Lett、327巻、125〜130頁、Brigstock, D. R.、Goldschmeding, R.、Katsube,K. I.、Lam, S. C.、Lau, L. F.、Lyons, K.、Naus, C.、Perbal,B.、Riser, B.、Takigawa, M.及びYeger, H. (2003)「Proposal for a unified CCNnomenclature.」Mol Pathol、56巻、127〜128頁並びにGarciaAbreu, J.、Coffinier, C.、Larrain,J.、Oelgeschlager, M.及びDeRobertis, E. M.(2002)「Chordin-like CR domains and theregulationofevolutionarily conserved extracellular signaling systems.」、Gene、287巻、39〜47頁並びに例えば、Cer1、セルベルス(Cerberus)、Dan、Dte、グレムリン(Gremlin)、PRDC、スクレロスチン(Sclerostin)を含むDANファミリーは、例えば、Pearce, J.J.、Penny, G.及びRossant, J.(1999)「Amousecerberus/Dan-related gene family.」Dev Biol、209巻、98〜110頁並びにBalemans,W.及びVan Hul, W.(2002)「Extracellular regulation of BMP signaling in vertebrates: a cocktailof modulators.」Dev Biol、250巻、231〜250頁に記載されている。
【発明の概要】
【発明が解決しようとする課題】
【0007】
したがって、本発明の基礎をなす課題は、I型及びII型セリンキナーゼ受容体鎖媒介性作用の点で、それぞれの骨形成タンパク質の野生型と競合しないが、依然として、ノギンタンパク質ファミリー、DANタンパク質ファミリー及びコーディンタンパク質ファミリーのものなどのモジュレータータンパク質と相互作用するのには適している、骨形成タンパク質、詳しくはBMP−2様タンパク質の変異タンパク質を提供することであった。
【課題を解決するための手段】
【0008】
本発明によれば、この課題は独立請求項の対象によって解決される。好ましい実施形態は従属請求項から選びとることができる。
【0009】
本発明によれば、この課題は、第1の態様で、ヒトBMP−2のアミノ酸位置51に相当するアミノ酸位置に骨形成タンパク質の野生型と比較してアミノ酸置換を含む、骨形成タンパク質の変異タンパク質によって解決される。
【図面の簡単な説明】
【0010】
【図1】(A)BMP−2及びBRIAECの複合体構造のリボン図を示す図である。(B) BMP−2及びBRIAECの接触面について、受容体BRIAがy軸の回りに180°回転している“明白な”眺望における表面表示を示す図である。(C) BMP−2及びBRIAECの接触面の表面表示を示す図である。
【図2】BMP−2−BRIA相互作用の接触面中の拡大表示を示す図である。
【図3】(A)BMP−2 L51Pの結晶構造を示す、プレヘリックスループセグメントの立体像を示す図である。
【図4】BMP−2及びBMP−2変異タンパク質に反応してATDC5(A)及びC2C12(B)細胞で測定されたアルカリホスファターゼ(ALP)活性の誘導として表されるBMP−2プロリン変異タンパク質の生物活性を示す図である。
【図5】BMP−2プロリン変異タンパク質L51Pによるノギン阻害の解除を示す図である。
【図6】ヒトBMP−2のアミノ酸位置51に相当する位置のアミノ酸残基をロイシンから好ましくはプロリンに交換し、本発明の変異タンパク質の好ましい実施形態を形成したBMP−2様タンパク質のアラインメントを示す図である。
【図7】BMPプロリン変異タンパク質と(A)BRIAEC、(B)BRIBEC又は(C)アクチビン受容体IIエクトドメイン間の相互作用、並びに(D)BMP−2とBRIAEC変異タンパク質間の相互作用のセンサーグラム(sensograms)、を示す図であって、すべてのリガンドは、120nM濃度で存在しており、センサーグラム(sensorgrams)は、アナライトであるBMP−2の濃度を飽和するために算出されたレゾナンスユニット(RU)を100%として設定することによって標準化した。
【図8】BMP−2野生型とBMP−2 L51Pの間の構造の相違を示す図であって、両構造のCα−原子がスーパーインポーズされ、中間の距離が示され、プレヘリックスループの領域は灰色で強調表示されている。
【発明を実施するための形態】
【0011】
一実施形態では、ヒトBMP−2のアミノ酸位置51に相当する位置のアミノ酸は、骨形成タンパク質の野生型ではロイシンであり、変異タンパク質では好ましくはプロリンである。
【0012】
一実施形態では、骨形成タンパク質は、hBMP−2、hBMP−4、hBMP−5、hBMP−6、hBMP−7、hBMP−8、hGDF−5、mGDF−6、mGDF−7、hBMP−10及びhGDF−2を含んでなる群から選択される。
【0013】
一実施形態では、
骨形成タンパク質は、hBMP−2であり、ヒトBMP−2のアミノ酸位置51に相当する位置はアミノ酸位置51であり、
骨形成タンパク質は、hBMP−4であり、ヒトBMP−2のアミノ酸位置51に相当する位置はアミノ酸位置53であり、
骨形成タンパク質は、hBMP−5であり、ヒトBMP−2のアミノ酸位置51に相当する位置はアミノ酸位置68であり、
骨形成タンパク質は、hBMP−6であり、ヒトBMP−2のアミノ酸位置51に相当する位置はアミノ酸位置68であり、
骨形成タンパク質は、hBMP−7であり、ヒトBMP−2のアミノ酸位置51に相当する位置はアミノ酸位置75であり、
骨形成タンパク質は、hBMP−8であり、ヒトBMP−2のアミノ酸位置51に相当する位置はアミノ酸位置75であり、
骨形成タンパク質は、hGDF−5であり、ヒトBMP−2のアミノ酸位置51に相当する位置がアミノ酸位置56であり、
骨形成タンパク質は、mGDF−6であり、ヒトBMP−2のアミノ酸位置51に相当する位置がアミノ酸位置56であり、
骨形成タンパク質は、mGDF−7であり、ヒトBMP−2のアミノ酸位置51に相当する位置はアミノ酸位置82であり、
骨形成タンパク質は、hBMP−10であり、ヒトBMP−2のアミノ酸位置51に相当する位置はアミノ酸位置44であり、
骨形成タンパク質は、hGDF−2であり、ヒトBMP−2のアミノ酸位置51に相当する位置はアミノ酸位置45である。
【0014】
一実施形態では、野生型の
hBMP−2は、配列番号1のアミノ酸配列を含み、
hBMP−4は、配列番号3のアミノ酸配列を含み、
hBMP−5は、配列番号5のアミノ酸配列を含み、
hBMP−6は、配列番号7のアミノ酸配列を含み、
hBMP−7は、配列番号9のアミノ酸配列を含み、
hBMP−8は、配列番号11のアミノ酸配列を含み、
hGDF−5は、配列番号13のアミノ酸配列を含み、
mGDF−6は、配列番号15のアミノ酸配列を含み、
mGDF−7は、配列番号17のアミノ酸配列を含み、
hBMP−10は、配列番号19のアミノ酸配列を含み、
hGDF−2は、配列番号21のアミノ酸配列を含む。
【0015】
好ましい一実施形態では、野生型の
hBMP−2は、配列番号2の核酸によってコードされており、
hBMP−4は、配列番号4の核酸によってコードされており、
hBMP−5は、配列番号6の核酸によってコードされており、
hBMP−6は、配列番号8の核酸によってコードされており、
hBMP−7は、配列番号10の核酸によってコードされており、
hBMP−8は、配列番号12の核酸によってコードされており、
hGDF−5は、配列番号14の核酸によってコードされており、
mGDF−6は、配列番号16の核酸によってコードされており、
mGDF−7は、配列番号18の核酸によってコードされており、
hBMP−10は、配列番号20の核酸によってコードされており、
hGDF−2は、配列番号22の核酸によってコードされている。
【0016】
本発明によれば、この課題は、第2の態様で、第1の骨形成タンパク質受容体と結合しておらず、かつ、ノギンタンパク質ファミリー、DANタンパク質ファミリー、コーディンタンパク質ファミリー及びシステインノット含有BMPモジュレータータンパク質を含んでなる群から選択される少なくとも1種のモジュレータータンパク質と結合している、骨形成変異タンパク質によって解決される。
【0017】
本発明によれば、この課題は、第3の態様で、骨形成変異タンパク質によって解決され、この骨形成変異タンパク質は、好ましくは第2の骨形成タンパク質受容体と相互作用するプレヘリックスループ構造を含む、本発明の第1及び第2の態様の実施形態であることが好ましい。第2の骨形成タンパク質受容体の用語は、この種の受容体と、本明細書で第1の骨形成タンパク質受容体と呼ばれる別の受容体を区別するために用いられると理解されなくてはならない。この技術用語は、本明細書で参照するためだけに取り入れられており、タンパク質及び変異タンパク質の結合をそれぞれ暗に示すものではない。その程度において、タンパク質及び変異タンパク質の結合は、それぞれ、単一の骨形成タンパク質受容体とのみ起こることが好ましい。このような単一の骨形成タンパク質受容体は、第1の骨形成タンパク質受容体又は第2の骨形成タンパク質受容体のいずれかであり得る。
【0018】
第2及び第3の態様に従う一実施形態では、第1及び/又は第2の骨形成タンパク質受容体がBRIA又はBRIBである、請求項7及び8に記載の骨形成変異タンパク質。
【0019】
第2及び第3の態様に従う一実施形態では、相互作用はアミノ酸残基、好ましくは、BRIAのGln86又はBRIBのGln67アミノ酸残基に関連している。
【0020】
本発明によれば、この課題は、第4の態様で、骨形成変異タンパク質によって解決され、この骨形成変異タンパク質は、好ましい実施形態では、第1、第2及び第3の態様に従う骨形成タンパク質であり、第2の骨形成タンパク質受容体との相互作用を有するプレヘリックスループ構造を含み、骨形成変異タンパク質のプレヘリックスループ構造と第2の骨形成タンパク質受容体との相互作用が、野生型骨形成タンパク質のプレヘリックスループ構造と第2の骨形成タンパク質受容体との相互作用とは異なる。
【0021】
いずれかの態様に従う一実施形態では、異なる相互作用又は変化は、屈折データ、好ましくは、少なくとも約2.7Åの分解能まで室温で得られているような屈折データで表される。
【0022】
いずれかの態様に従う一実施形態では、プレヘリックスループ構造が、骨形成タンパク質の野生型と比較して変異している。
【0023】
好ましい一実施形態では、野生型BMP−2の位置51のロイシンに相当するアミノ酸が変異している。
【0024】
もう1つの好ましい実施形態では、ヒトBMP−2の位置51のロイシンに相当するアミノ酸がプロリンに変異している。
【0025】
いずれかの態様に従う一実施形態では、骨形成変異タンパク質は、hBMP−2、hBMP−4、hBMP−5、hBMP−6、hBMP−7、hBMP−8、hGDF−5、mGDF−6、mGDF−7、hBMP−10及びhGDF−2を含んでなる群から選択される骨形成タンパク質の変異タンパク質である。
【0026】
好ましい一実施形態では、骨形成タンパク質はBMP−2又はプロ−BMP−2である。
【0027】
本発明によれば、この課題は、第5の態様で、配列番号23〜33のいずれかに記載のアミノ酸配列を含む骨形成タンパク質によって解決される。
【0028】
一実施形態では、骨形成変異タンパク質は、本発明のいずれかの態様に記載の骨形成変異タンパク質である。
【0029】
本発明によれば、この課題は、第6の態様で、本発明のいずれかの態様に従う、骨形成タンパク質及び骨形成変異タンパク質をそれぞれコードする核酸、並びに/又はその相補鎖によって解決される。
【0030】
本発明によれば、この課題は、第7の態様で、配列番号2、4、6、8、10、12、14、16、18、20及び22の核酸配列を含む核酸、好ましくは、本発明のいずれかの態様に従う骨形成変異タンパク質をコードする核酸、並びに/又はその相補鎖によって解決される。
【0031】
本発明によれば、この課題は、第8の態様で、本発明のいずれかの態様に従う骨形成変異タンパク質をコードする核酸であって、より好ましくは、ストリンジェントな条件下で、遺伝暗号の縮重がなければ、第6及び第7の態様に従う核酸とハイブリダイズする核酸によって解決される。
【0032】
本発明によれば、この課題は、第9の態様で、第6、第7及び第8の態様に従う核酸を含むベクターであって、好ましくは発現ベクターであるベクターによって解決される。
【0033】
本発明によれば、この課題は、第10の態様で、第6、第7及び第8の態様に従う核酸及び/又は本発明の第10の態様に従うベクターを含む細胞、好ましくは、哺乳類細胞によって解決される。
【0034】
本発明によれば、この課題は、第11の態様で、本発明の第10の態様に従う細胞を含む、宿主生物、好ましくは、哺乳類宿主生物、より好ましくは、非ヒト宿主生物によって解決される。
【0035】
本発明によれば、この課題は、第12の態様で、
a)請求項25に記載の細胞を培養ブロスで培養するステップと、
b)細胞から、及び/又は培養ブロスから骨形成変異タンパク質を調製するステップと、
を含む、本発明の第1〜第5のいずれかの態様に従う骨形成変異タンパク質の製造方法によって解決される。
【0036】
本発明によれば、この課題は、第13の態様で、本発明の第1〜第5のいずれかの態様に従う骨形成変異タンパク質と特異的に結合しているモノクローナル抗体によって解決される。
【0037】
本発明によれば、この課題は、第14の態様で、本発明の第1〜第5のいずれかの態様に従う変異タンパク質及び/又は本発明の第6〜第8のいずれかの態様に従う核酸を含む組成物によって解決される。
【0038】
本発明によれば、この課題は、第15の態様で、本発明の第1〜第5のいずれかの態様に従う変異タンパク質又は第6〜第8のいずれかの態様に従う核酸と、製薬上許容される担体とを含む薬剤組成物によって解決される。
【0039】
本発明によれば、この課題は、第16の態様で、医薬を製造するための、本発明の第1〜第5のいずれかの態様に従う骨形成変異タンパク質及び/又は第6〜第8のいずれかの態様に従う核酸の使用によって解決される。
【0040】
一実施形態では、医薬は、線維性疾患、創傷治癒、過剰血管新生、血管疾患、骨折及び骨粗しょう症を含んでなる群から選択される疾患の治療用及び/又は予防用のものである。
【0041】
好ましい一実施形態では、線維性疾患は、腎線維症、肝硬変、肺線維症及び慢性炎症、好ましくは、喘息を伴う慢性炎症を含んでなる群から選択される。
【0042】
もう1つの実施形態では、創傷治癒はケロイド、瘢痕形成及び腹膜の閉塞と関連している。
【0043】
さらなる一実施形態では、過剰血管新生は、網膜症、動脈硬化症及び/又は腫瘍と関連しているか、又は網膜症、動脈硬化症及び/又は腫瘍を伴う。
【0044】
いっそうさらなる実施形態では、骨折は難治性骨折である。
【0045】
もう1つの実施形態では、疾患は骨粗しょう症である。
【0046】
本発明によれば、この課題は、第17の態様で、形態形成タンパク質、好ましくは、本発明の第1〜第5のいずれかの態様に従う形態形成変異タンパク質の、BMP相互作用タンパク質に対する阻害剤としての使用によって解決される。
【0047】
一実施形態では、BMP相互作用タンパク質は、ノギンタンパク質ファミリー、DANタンパク質ファミリー及びコーディンタンパク質ファミリーを含んでなる群から選択される。
【0048】
本発明者らは、驚くべきことに、本発明の変異タンパク質が、ノギンファミリー、DANファミリー及びコーディンタンパク質ファミリーのものなどのモジュレータータンパク質の活性を阻害できる一方で、少なくとも1個のアミノ酸を置換することによって生じる骨形成タンパク質に対応する受容体、より詳しくは、BMP−2様タンパク質受容体、とは相互作用しないことを見出した。本発明の変異タンパク質はこれらの特徴を有するため、それらはいずれかの生物に適用することができ、この場合、前記モジュレータータンパク質が調節され、好ましくは、阻害され、ひいては、前記モジュレータータンパク質によって発揮される阻害が消滅するか減少することになり、その一方で、前記受容体鎖によって媒介される、骨形成タンパク質に関連しているその他の生物活性はもはや存在しない。骨形成タンパク質の変異タンパク質のこういった設計は、この種の骨形成タンパク質の、望ましくない受容体媒介性作用を引き起こすことなく、モジュレータータンパク質が関連しているか媒介する疾患の治療又は予防に使用できる限りにおいて、明らかに有利である。その程度において、本発明の変異タンパク質は“受容体非感受性(receptordead)”である。
【0049】
また、本発明者らは驚くべきことに、骨形成変異タンパク質のこの特有の特徴は、野生型の骨形成タンパク質中のプレヘリックスループ構造を変更することによって作出できることを見出した。本明細書においては、そうでないことが示されない場合は、骨形成タンパク質は、BMP−2様タンパク質であることが好ましい。プレヘリックスループ構造は、好ましくは骨形成変異タンパク質の構造であり、これは、二次構造表示でヘリックスの前のループであることが好ましい。ループは4〜10個、好ましくは6〜8個、より好ましくは、6個のアミノ酸残基を含むことが好ましい。ヒトBMP−2タンパク質のアミノ酸配列を例にとると、プレヘリックスループ構造はアミノ酸位置48〜アミノ酸位置53に広がっている。本明細書では、好ましい実施形態では、プレヘリックスループは、いずれかのループであって、より好ましくはヒトBMP−2のアミノ酸位置48〜53に相当するいずれかのアミノ酸配列であり、より好ましくは配列番号1のアミノ酸配列である。プレヘリックスループ構造は、好ましくは、本明細書においてI型骨形成タンパク質受容体と呼ばれる骨形成タンパク質受容体との相互作用を媒介していることが好ましい。I型骨形成タンパク質受容体はBRIAであることがいっそうより好ましい。BRIAECは、BMPR−IAの細胞外ドメインであり、以下に記載されている(Kirsch, T.、Sebald, W.、Dreyer, M.K.(2000) Nat Struct Biol、7、492〜496参照)。BRIBECは、BMPR−IBの細胞外ドメインである。ARIECは、ActR−Iの細胞外ドメインである。
【0050】
本発明の変異タンパク質は、骨形成タンパク質の野生型と比較して、構造、好ましくは二次構造及び三次構造の変化のために、I型骨形成タンパク質受容体ともはや相互作用できない。本明細書において用いられるのに好ましい変化とは、対応する骨形成タンパク質の野生型と前記I型形成タンパク質受容体との相互作用を参照することによって検出可能なものである。より詳しくは、I型骨形成タンパク質受容体がBRIAECである場合には、本発明の変異タンパク質によって最も影響を受けるアミノ酸はGln86であるようである。
【0051】
また、本発明の骨形成変異タンパク質はプレヘリックスループ構造を含み、これがBRIAECなどのI型骨形成タンパク質受容体との相互作用を媒介しており、この相互作用は骨形成タンパク質の野生型のプレヘリックスループ構造とI型骨形成タンパク質受容体との相互作用とは異なっている。これらの誘導された変化のいずれか又は変異タンパク質とそれぞれの第2の骨形成タンパク質受容体との間の異なる相互作用パターンは、骨形成タンパク質の野生型の対応する構造又は相互作用と比較して検出される。前記I型骨形成タンパク質受容体と変異タンパク質のいずれか又は双方のこのような変化は、屈折データを得ることによってモニターできることが好ましい。より好ましい実施形態では、このような屈折データは少なくとも約2.7Åの分解能まで室温で得られる。
【0052】
好ましい実施形態では、骨形成タンパク質は、BMP−2である。BMP−2は当技術分野では公知であり、本明細書に記載される配列番号1のアミノ酸配列を含むことが好ましく、配列番号2の核酸配列を含む核酸によってコードされることが好ましい。
【0053】
より詳しくは、本発明者らはまた、本発明の変異タンパク質の特有の特徴は、プレヘリックスループ構造でのアミノ酸置換、より詳しくは、骨形成タンパク質のアミノ酸位置、好ましくはヒトBMP−2のアミノ酸位置51に相当するBMP−2様タンパク質のアミノ酸位置、さらに好ましくは、配列番号1のBMP−2のアミノ酸位置でのアミノ酸置換、すなわち変異、によって付与されるということも発見した。より詳細には、変異タンパク質中で変異されているそれぞれのアミノ酸は、対応する骨形成タンパク質の野生型においてはロイシンである。より好ましくは、その交換はロイシン残基からプロリン残基である。骨形成タンパク質、特に、BMP−2様タンパク質の高い相同性、ひいては類似の二次及び三次構造のために、特定の位置が同様に活性であり、この群の骨形成タンパク質の種々のメンバー内でそれぞれ関連があり、その結果、変化、すなわち、アミノ酸置換は、前記骨形成タンパク質のいずれにおいても有効であり、したがって、それぞれの変異タンパク質は前記で概説した特徴を示す。骨形成タンパク質はhBMP−2、hBMP−4、hBMP−5、hBMP−6、hBMP−7、hBMP−8、hGDF−5、mGDF−6、mGDF−7、hBMP−10及びhGDF−2であることがより好ましく、これらは当技術分野ではそれ自体公知であり、これらのすべては同じようにBRIAECと結合する。しかし、対応する変異タンパク質、すなわち、ヒトBMP−2のアミノ酸位置51に相当するそれらの位置にアミノ酸置換を有する変異タンパク質、にそれらを変えるそれぞれの変異によって、それらを先行技術から区別している。前記の骨形成タンパク質の野生型配列は本明細書では配列番号1(hBMP−2)、配列番号3(hBMP−4)、配列番号5(hBMP−5)、配列番号7(hBMP−6)、配列番号9(hBMP−7)、配列番号11(hBMP−8)、配列番号13(hGDF−5)、配列番号15(mGDF−6)、配列番号17(mGDF−7)、配列番号19(hBMP−10)及び配列番号21(hGDF−2)として開示されており、本発明の前記骨形成タンパク質の変異タンパク質の対応するアミノ酸配列は、野生型と比較して前記の単一のアミノ酸位置で異なっており、前記アミノ酸配列は配列番号23(hBMP−2)、配列番号24(hBMP−4)、配列番号25(hBMP−5)、配列番号26(hBMP−6)、配列番号27(hBMP−7)、配列番号28(hBMP−8)、配列番号29(hGDF−5)、配列番号30(mGDF−6)、配列番号31(mGDF−7)、配列番号32(hBMP−10)及び配列番号33(hGDF−2)のものである。より詳しくは、前記のヒトBMP−2のアミノ酸位置51は、hBMP−4の場合には位置53、hBMP−5の場合には位置68、hBMP−6の場合には位置68、hBMP−7の場合には位置75、hBMP−8の場合には位置75、hGDF−5の場合には位置56、mGDF−6の場合には位置56、mGDF−7の場合には位置82、hBMP−10の場合には位置44及びhGDF−2の場合には位置45に相当する。
【0054】
本明細書において、本明細書に開示される、骨形成タンパク質及びひいては、単一のアミノ酸置換でのみ異なっていることが好ましい対応する変異タンパク質は、プレプロ型で存在する場合も、プロ型で存在する場合も、成熟タンパク質として存在する場合もある。この基本設計は、すべての骨形成タンパク質にもともと備わっているものである。例えば、BMP−2の場合には、プレプロ型は全部で396個のアミノ酸を含む。プレ配列はシグナルペプチドとして働き、新生ポリペプチド鎖の小胞体への輸送に関与している。輸送された後、タンパク質はその未変性コンホメーションに折り畳まれ、その際ジスルフィド架橋が形成される。BMP−2のプロ型は、位置20〜位置396のアミノ酸すべてを含む。したがって、プロ配列はBMP−2のヒトプレプロ型の位置20(Gly)〜アミノ酸位置282(Arg)までのアミノ酸を含む。最後に、成熟BMP−2はプレプロ型のアミノ酸283〜396を含む。成熟型は本明細書では配列番号1として表されている。
【0055】
本発明の骨形成タンパク質及びその対応する変異タンパク質は、プレプロ型、プロ型、又は成熟型で存在し、いずれかのシグナルペプチドが、プレ配列として、好ましくは、骨形成タンパク質のプレ配列として作用し、いずれの骨形成タンパク質のいずれのプロ配列であれば、特定の骨形成タンパク質のプロ配列として使用できるということは本発明の範囲内であるが、成熟骨形成変異タンパク質のN末端に結合しているプロ配列が、in vivoでそれぞれの骨形成タンパク質のN末端に結合するプロ配列に相当することがより好ましい。同じことがプレ配列にも当てはまる。
【0056】
本明細書において、用語「骨形成タンパク質」及び「骨形成変異タンパク質」とはそれぞれ、その相同分子種も含むものとして好ましく使用される。本明細書において、相同分子種とは、参照タンパク質(reference protein)として、同様の遺伝的機能及び生理学的機能を果たす別の生物由来のタンパク質である。また、より好ましい実施形態では、用語「骨形成タンパク質」及び「骨形成変異タンパク質」は、いずれかの末端切断型タンパク質及び変異タンパク質をそれぞれ含む。このような末端切断型タンパク質及び変異タンパク質は、少なくとも1個のアミノ酸残基を欠いており、N末端又はC末端のいずれかで少なくとも1個のアミノ酸残基を欠いていることが好ましい。したがって、本発明の骨形成変異タンパク質は、前記で概説された特徴、すなわち、「受容体非感受性」であり、かつ、少なくとも1種のモジュレータータンパク質、好ましくは、ノギンタンパク質ファミリー、DANタンパク質ファミリー及び/又はコーディンタンパク質ファミリーのメンバーと相互作用するのに依然として適している限りは、末端切断型変異タンパク質である。
【0057】
変異タンパク質はさらに修飾され、このような修飾はリン酸化、ペグ化及びグリコシル化を含んでなる群から選択されることが好ましいということもまた本発明の範囲内である。
【0058】
本発明の核酸は、いずれもがDNA、RNA又はそれらの誘導体、例えば、LNA若しくはPNA、として存在し得ることを理解されなくてはならない。それぞれの核酸は、一本鎖又は部分的な二本鎖若しくは完全な二本鎖のいずれかとして存在することも本発明の範囲内である。最後に、用語「核酸配列」は、例えば、配列識別子に従う配列によって、単独で又は本質的な相補的鎖であるもう一方の鎖と組み合わせて特定されるような核酸の補完物を含むことも本発明の範囲内である。
【0059】
本明細書において、用語「核酸」とは、また、本明細書に記載される核酸のいずれの断片も含み、このような断片のいずれもが、約19〜30の長さの連続するヌクレオチドを含むことが好ましく、19〜25の長さがより好ましく、21〜22の長さが最も好ましい。核酸のこの種の断片は、二本鎖構造として存在することがいっそうより好ましい。
【0060】
本発明によれば、核酸はまた、本明細書に開示される核酸の何らかのその他の形とハイブリダイズする核酸であってもよく、このようなハイブリダイゼーションは、ストリンジェントな条件で起きることが好ましい。このようなストリンジェントな条件は、中でも、Sambrook, J.、Fritsch, E.F.及びManiatis, T. (1989) Molecular Cloning. A.Laboratory Manual、第2版、9.47〜9.55頁、Cold Spring Harbour Laboratory Pressに記載されている。
【0061】
本明細書に記載される、骨形成タンパク質の野生型配列又はその変異タンパク質の配列のいずれかを発端として、骨形成タンパク質の野生型とその変異タンパク質とを区別できる核酸を設計することが可能であることは、当業者に認められよう。このような区別はハイブリダイゼーション条件のストリンジェンシーを高めることによって可能となることが好ましい。
【0062】
本明細書に開示される特定の配列だけでなく、種々の配列、より好ましくは、核酸配列も、それらが本発明の変異タンパク質をコードするのに適している限りは本発明に含まれるということも当業者に認められよう。このような配列は遺伝暗号を用いて導くことができ、事実上、遺伝暗号の冗長性によって生じるこのような配列のいずれの順列も本発明の範囲内にある。ヒトBMP−2のアミノ酸位置51にそれぞれ相当する、骨形成タンパク質及び変異タンパク質の位置が、ロイシンではなくプロリンであることが本発明の変異タンパク質の特徴となっており、この特定のアミノ酸置換については、それぞれのコドンがCCU、CCC、CCA又はCCGであると認められよう。
【0063】
本発明のベクターは、本明細書に開示されるいずれかの断片を含み、本明細書に開示されるいずれかの核酸を含む。ベクターは発現ベクターであることが好ましい。このような発現ベクターは、通常、プロモーター又はプロモーター様構造と、場合によっては、このようなプロモーター及びその他の制御エレメントによって制御されるような、ベクターにクローニングされている核酸の発現を制御するのに適しているさらなるエレメントを含む。発現ベクターは原核生物の発現ベクターか、真核生物の発現ベクターかそれぞれのシャトルベクターであることが好ましい。この種の発現ベクターの個々の設計は当業者には公知である。
【0064】
本発明の細胞は、原核細胞か真核細胞のいずれかである。原核細胞としては、原則として、大腸菌(E.coli)、枯草菌(B.subtilis)又はS.カルノサス(S.carnosus)が本発明の骨形成変異タンパク質をコードする核酸、又はその断片若しくは誘導体の発現に用いられる。
【0065】
本明細書において、真核細胞は、酵母細胞又は哺乳類細胞であることが好ましい。哺乳類細胞はマウス、ラット、モルモット、ブタ、サル又はヒト由来の細胞であることがより好ましい。
【0066】
本明細書において、宿主生物は本発明のいずれかの細胞又は多細胞生物であることが好ましい。このような多細胞生物は好ましくは、哺乳類であることが好ましく、非ヒト哺乳類であることがより好ましい。好ましい非ヒト哺乳類はマウス、ラット、モルモット、ヒツジ、ウサギ及びブタを含んでなる群から選択される。
【0067】
本発明の骨形成変異タンパク質を製造するための本発明の方法では、本発明の細胞は培養されていることが好ましい。このような培養は培養ブロスで行われることが好ましい。培養ブロスは発酵ブロス又は、培養に適し、かつ、より好ましくは培養される細胞の増殖に適しているいずれかのその他の溶液、であることが好ましい。培養ブロスは当業者には公知である。培養及び/又は細胞の増殖において、本発明の変異タンパク質は、培養ブロスから調製されることが好ましい。培養ブロスからの調製は、本明細書に記載されているように、本発明の変異タンパク質を細胞内に含ませるか又は細胞に結合させてその細胞からの調製すること、又は培養ブロスからの本発明の変異タンパク質を単離若しくは調製すること、を含む。本発明の変異タンパク質の調製又は単離方法は当業者には公知である。
【0068】
さらなる態様では、本発明は、本発明の変異タンパク質に対するモノクローナル抗体と関連している。抗体はモノクローナル抗体であることが好ましい。抗体、好ましくは、本発明の骨形成変異タンパク質と特異的に結合するモノクローナル抗体、を調製することが可能であることは、当業者に認められよう。
【0069】
用語「特異的な結合」とは、抗体が、変異タンパク質に対してと同程度に、又は以下に示すように、変異タンパク質に対して同じ結合特性で、骨形成タンパク質の野生型とは結合しておらず、対応する骨形成タンパク質の野生型と比較した変異タンパク質の結合定数が、少なくとも2倍異なっている、好ましくは少なくとも5倍、最も好ましくは少なくとも10倍異なることを意味している。
【0070】
さらなる態様では、本発明は、本発明の変異タンパク質又は本発明の核酸のいずれかを含む組成物に関連している。このような組成物は、タンパク質及び/又は核酸だけでなく、担体を含み得る。このような担体は固相、水相及び脂質相を含んでなる群から選択されることが好ましい。本明細書において、脂質相は核酸及び変異タンパク質とそれぞれ複合体を形成しているか、核酸及び変異タンパク質をそれぞれ包含している脂質又は脂質組成物であることが好ましい。脂質組成物は、その程度において当業者に公知のリポソームであることが好ましい。水相は水及び水性バッファー溶液及びヒアルロン酸を含んでなる群から選択されることが好ましい。固相は、リン酸三カルシウム、ポリラクチド、コラーゲン及び不溶性コラーゲン骨マトリックスであることが好ましい。
【0071】
さらなる態様では、本発明は薬剤組成物に関連している。このような薬剤組成物は、本発明の変異タンパク質又は本発明の核酸あるいは双方と、さらに製薬上許容される担体とを含む。製薬上許容される担体は液体又は固体担体のいずれかであることが好ましい。適した液体担体としては、中でも、水、水溶液、より好ましくは、バッファー又は脂質若しくは脂質組成物がある。好ましい固体担体は、中でも、糖、セルロース及びデンプンを含む。当業者、好ましくは、製剤の技術分野の当業者であれば、さらなる製薬上許容される担体及び担体物質をそれぞれ承知しているものと認められる。好ましい実施形態では、薬剤組成物はさらなる製薬上活性な薬剤を含む。
【0072】
さらなる態様では、本発明は、医薬の製造のための本発明の骨形成変異タンパク質及び/又は本発明の核酸の使用に関する。
【0073】
本発明者らの驚くべき発見に基づいて、すなわち、骨形成タンパク質、より詳しくは、本発明のBMP−2様タンパク質の変異タンパク質は、骨形成タンパク質のような受容体媒介性作用を示さず、すなわち「受容体非感受性」であるが、依然として、それぞれの骨形成タンパク質の野生型が、相互作用するモジュレータータンパク質と相互作用することは当業者に認められる。その程度において、本発明の変異タンパク質及びそれをコードする核酸は、骨形成タンパク質の公知の望ましくない作用が生じるという危険に出くわすことなくそれらを必要とする患者に適用でき、このことは、その阻害活性を阻害するモジュレータータンパク質と相互作用する利益よりも上回る。例えば、BMP−2の望ましくない活性の中には、腫瘍成長を促進することがある。このために、先行技術における、いずれかの変異タンパク質をも含む骨形成タンパク質の医学的使用は、骨再生などの局所投与に制限されてきた。しかし、骨形成タンパク質の活性であるそのような作用、詳しくは、その受容体との相互作用によって媒介されるもの、を欠いている本発明の変異タンパク質は、このクラスの製薬上活性を有する化合物の医学的な適用を全く初めて可能にするものである。
【0074】
本発明のさらなる態様は、本発明の医薬を使用できる、本明細書に記載された疾患のいずれかを患っているか、又はそれが進行する状態にある患者の治療方法に関している。このような方法は、変異タンパク質又はそれをコードする核酸の患者への投与を含む。患者は哺乳類であることが好ましく、ヒトであることがより好ましい。
【0075】
本発明の変異タンパク質の作用様式のために、それらは種々の疾患、すなわち、本発明の骨形成タンパク質とこれに対応する変異タンパク質との間の影響を受けない相互作用、及びいずれかのモジュレータータンパク質、を提供することによって治療又は予防できる疾患、に容易に用いることができる。モジュレータータンパク質は当技術分野では公知であり、シスチンノット含有骨形成タンパク質、より詳しくは、シスチンノット含有BMPモジュレータータンパク質、ノギンファミリー、コーディンファミリー及びDANファミリーを含んでなる群から選択されることが好ましい。シスチンノット含有BMPモジュレータータンパク質は、例えば、Avsian-Kretchmer,O.及びHsueh, A. J. (2004)Comparative genomic analysis of theeight-membered ring cystine knot-containingbone morphogenetic proteinantagonists. Mol Endocrinol、18巻、1〜12頁に記載されている。例えば、ノギンを含むノギンファミリーは、Canalis, E.、Economides, A. N.及びGazzerro,E.によって(2003) Bonemorphogeneticproteins, their antagonists, and the skeleton. Endocr Rev、24巻、218〜235頁に記載されており、中でもコーディン、ベントロプチン、CTGF、CYR61、クロスベインレス、プロコラーゲンIIAを含むコーディンファミリーは、例えば、Bork, P. (1993) The modular architecture of anew family of growthregulators related to connective tissue growth factor.FEBS Lett、327巻、125〜130頁、Brigstock, D. R.、Goldschmeding,R.、Katsube, K. I.、Lam, S. C.、Lau, L. F.、Lyons, K.、Naus, C.、Perbal, B.、Riser,B.、Takigawa, M.及びYeger, H.(2003)Proposal for a unified CCN nomenclature. Mol Pathol、56巻、127〜128頁及びGarcia Abreu, J.、Coffinier, C.、Larrain, J.、Oelgeschlager, M.及びDe Robertis, E.M.(2002) Chordin-like CR domains and theregulation ofevolutionarily conserved extracellular signaling systems. Gene、287巻、39〜47頁に記載されており、例えば、Cer1、セルベルス、Dan、Dte、グレムリン、PRDC、スクレロスチン(Sclerostin)を含むDANファミリーは、例えば、Pearce, J.J.、Penny, G.及びRossant, J. (1999) A mouse cerberus/Dan-related genefamily. Dev Biol、209巻、98〜110頁及びBalemans, W.及びVan Hul, W.(2002)Extracellular regulation of BMP singaling in vertebrates: a cocktailofmodulators. Dev Biol、250巻、231〜250頁に記載されている。これらのモジュレータータンパク質は、偏っている場合、特にそれ自身が阻害されない場合に、in vivoプロセスに対する阻害作用を有し、その結果、何らかの疾患又は病的状態をもたらす傾向がある細胞における経路及び事象に対して阻害又は刺激作用を有することがわかっている。したがって、いずれの理論にも拘束されようとは思わないが、本発明の変異タンパク質は、依然として前記モジュレータータンパク質と相互作用し、これによりモジュレータータンパク質によって刺激され、さもなければ阻害されるプロセスを許容するそれらの修飾作用をも阻害でき、モジュレータータンパク質単独の存在下で観察される程度と比較して、あらゆるレベルで、又は高いレベルで、又は低いレベルで実施され得る。
【0076】
本発明の、変異タンパク質及びそれをコードする核酸によって治療できる特に好ましい疾患としては、線維性疾患がある。本発明に従って治療できる特に好ましい疾患としては、腎線維症、肝硬変、肺線維症及び慢性炎症があり、喘息を伴う慢性炎症がより好ましく、したがって、喘息の治療も本発明に含まれる。
【0077】
再度、いずれの理論にも拘束されようとは思わないが、コーディン−ファミリーに属するCTGFは、BMP−2と、ひいては本発明の変異タンパク質と相互作用する。CTGFはTGF−βのプロ線維活性を媒介している。より詳しくは、TGF−βは、細胞外マトリックスのタンパク質の産生を刺激しており、この種のタンパク質の分解を阻害する。すなわち、このプロセスは組織の治癒に不可欠なものである。しかし、炎症性疾患、より詳しくは、慢性炎症などの慢性疾患の場合には、TGF−β活性が豊富であり、ひいては組織線維症、特に、過剰組織線維症をもたらす(Branton, M. H.及びKopp, J. B.(1999)TGF-betaand fibrosis. Microbes Infect、1巻、1349〜1365頁)。したがって、これら種々の線維性疾患は本発明の変異タンパク質の投与によって阻害できる。
【0078】
本発明に従って治療できるさらなる種類の疾患には、創傷治癒がある。この医学的状態、すなわち、創傷治癒の形はケロイド、瘢痕形成及び腹膜の閉塞であることがより好ましい。本発明に従って治療できる特定の群の患者は、好ましくは、視力に影響を及ぼす極度の瘢痕形成が同時に起きる緑内障手術を受けるか受けたことがある患者である。再度、いずれの論理的根拠にも拘束されようとは思わないが、過増殖性瘢痕形成はCTGF及びCYR61によって媒介されるようであり(Branton, M. H.及びKopp, J. B.(1999)TGF-betaand fibrosis.Microbes Infect、1巻、1349〜1365頁)、その結果、これらのモジュレータータンパク質の本発明の変異タンパク質による阻害は、瘢痕形成を避けるための、ひいては、個々の状態及び疾患をそれぞれ治療するための適した手段である。
【0079】
本発明によって治療されるもう1つの疾患又は状態は、過剰血管新生である。より詳しくは、用語「過剰血管新生」とは、過剰血管新生によって引き起こされるか、過剰血管新生と関連しているか、又は過剰血管新生を伴う、いずれの疾患も含む。この種の疾患は腫瘍、網膜症及び動脈硬化症であることが好ましい。その程度において、本発明はまた、腫瘍、網膜症の治療のための本発明の変異タンパク質の使用に関する。腫瘍は、固形癌であることが好ましい。また、血管疾患、例えば、好ましくは、動脈硬化症は、本発明によって治療できる。再度、いずれの理論にも拘束されようとは思わないが、固形癌は、1〜3mm3という体積を超えて成長する場合には十分な血管新生が必要であるようである。十分な血液供給は、腫瘍の増殖及び進行の際の律速ステップである。本発明者らの現時点での理解によれば、CYR61は腫瘍の脈管形成に関与している。より詳しくは、CYR61及びCTGFはそれぞれ、脈管形成性進行及び線維形成性進行に関与しており、それらは進行性血管閉塞に関与していると理解される(Brigstock, D. R. (2002) Regulation of angiogenesis and endothelialcell function by connective tissue growth factor(CTGF) and cysteine-rich 61(CYR61)、Angiogenesis、5巻、153〜165頁)。したがって、前記標的分子、すなわち、CYR61及びCTGFと相互作用することによって、本発明の変異タンパク質はそれぞれの疾患の予防及び治療に適している。
【0080】
それぞれ、本発明に従って治療できる、さらなる疾患及び状態としては、骨折、好ましくは、難治性骨折がある。再度、いずれの理論にも拘束されようとは思わないが、ノギン及びCTGFは、骨折の治癒に関与しているBMP調節性タンパク質であるようである。より詳しくは、ノギン及びCTGFは、骨形成を阻害し、終結させる(Yoshimura, Y.ら、(2001)、Colocalization of noggin and bone morphogenetic protein-4duringfracture healing、J Bone Miner Res、16巻、876〜884頁)。したがって、例えば、前記標的の相互作用パートナーとの競合阻害によって、本発明の変異タンパク質はこの状態の治療に影響を及ぼす。
【0081】
骨粗しょう症は、本発明に従って治療できるもう1つの疾患である。再度、いずれの理論にも拘束されようとは思わないが、この疾患に関連して、標的はスクレロスチンであるように思われる。スクレロスチンは、生物における骨形成プロセスを阻害する。スクレロスチン遺伝子(SOST)の不活性化を受けている人は、骨密度の持続的増加を示し、これは標準値の3倍にも達する場合がある。その程度において、スクレロスチンを阻害する化合物はいずれも、骨密度の低下に付随している状態、例えば、骨粗しょう症などの治療のための手段として適している(Balemans, W.及び Van Hul, W.(2002)、Extracellular regulation of BMP signaling invertebrates: a cocktailof modulators, Dev Biol、250巻、231〜250頁)。その程度において、本発明の変異タンパク質がスクレロスチンと相互作用し、ひいては、この化合物を阻害するという事実を踏まえると、それらはこの種の疾患及び状態、より好ましくは、骨粗しょう症の治療にそれぞれ適している。
【0082】
本発明を図及び実施例によってさらに説明するが、そこからはさらなる特徴、実施形態及び利点が得られる。
【0083】
実施例1:材料及び方法
(タンパク質の調製)
BMP−2変異タンパク質及びBMP受容体エクトドメインタンパク質の調製を、先に記載された通りに実施した(Kirschら、2000b;Ruppertら、1996)。BMP−2又はBRIAECの変異体は、組換えPCRによって得た(Kirschら、2000a)。結晶化に用いたBMP−2野生型及び変異タンパク質タンパク質は、EMD SO3−カラム(Merck)を用いる陽イオン交換クロマトグラフィーによって精製した。画分をSDS−PAGEによって分析し、プールした画分を水に対して透析し、凍結乾燥した。ESI FT−CR質量分析及び分析用逆相HPLCによってタンパク質の均質性を調べた。リフォールディングしたBMP−2変異タンパク質の結合活性を、BIAコア方法を用いるBMPI型及びII型受容体の結合親和性の測定によって評価した。構造解析のためのBMP−2BRIAEC複合体の調製は公開されている通りに実施した(Kirschら、2000b)。
【0084】
(タンパク質の結晶化)
結晶化のために、BMP−2 D53P及びBMP−2 L51Pを、それぞれ5mg/ml及び8mg/mlの濃度で水に溶解した。BMP−2 L51Pは、室温で、15%t−ブタノール、0.1M硫酸リチウム及び0.1Mクエン酸ナトリウムpH5.0から水滴蒸気拡散法によって結晶化した。BMP−2−BRIAECの複合体の結晶化は、基本的には公開された様に実施した(Kirschら、2000b)が、100Kで測定するための結晶は、グルコースを抗凍結剤として用いる共結晶化によって得た。データ収集のための高品質結晶は、11mg/mlのタンパク質濃度において、10℃で、0.75M酢酸ナトリウム、0.1MイミダゾールpH7.0及び30%グルコースから成長させた。
【0085】
(X線データ収集)
BMP−2−BRIAECという複合体のデータは、Swiss Light Source(Paul Scherrer Institute、スイス)のビームラインX06SA PXを用いて、100Kでの単結晶から得た。結晶を90°回転させて(フレームあたり1°)2つのデータセットを測定し、1°に対して10秒曝露した高分解能セットからは1.9Åという最高分解能を得て、1°あたり1秒の曝露時間で記録することによって低分解能データを得た。データはソフトウェアMOSFLMバージョン6.2.1を用いて加工し、完全なものにし(Leslie、1992)、2つのデータセットをマージし(オーバーラップ:低分解能については20〜5.0Å、高分解能については20〜1.9Å)、ソフトウェアSCALA CCP4バージョン4.2.1(Collaborative Computational Project、1994;Evans、1993)を用いてスケーリングし、加工統計データの要約を表A付録に示す。BMP−2変異タンパク質L51Pの回析データは、Rigaku RU−300(50 kV、100mA、0.3mmフィラメント)、Osmic ConfocalBlue optics及びRigaku R−AXIS IV++イメージプレートシステムからなるhome−sourceで室温で得た。曝露時間は、0.5°回転あたり5分に設定し、結晶は2.5Åまで回析した。放射線損傷を最小化ために獲得戦略をソフトウェアCrystalClear(Rigaku)を用いて決定したが、相応な完全性を実現するために、2種の結晶からデータを集め、ソフトウェアMOSFLM及びSCALAを用いてマージし、スケーリングした(Evans、1993;Leslie、1992)。
【0086】
(構造決定)
BMP−2−BRIAECという複合体の構造を、低分解能構造(PDBエントリー1ES7)を出発モデルとして用いてリファインメントした。BMP−2−BRIAEC複合体を、空間群P65で結晶化し、これは非対称単位にBMP−2二量体と2つのBRIAEC分子を保持していた。容器定数のわずかな変化のために、回転及び翻訳検索とそれに続く剛体リファインメントからなる分子置換ステップを、ソフトウェアCNS1.1を用いて実施した(Brungerら、1998)。リファインメントのためにプログラムREFMAC5(Murshudovら、1997)を続いて用い、それに続いてソフトウェアQUANTA2000(Accelrys)を用いて座標を手動で再建した。分解能を20〜1.9Åという最終分解能範囲まで徐々に高めた。データの異方性を明らかにするために、各鎖、すなわち、BMP−2単量体及び各BRIAEC分子に対して1つのTLSグループを定義した(Winnら、2001)。リファインメントの進行は、5%の反射を含む試験データセットを用いる相互検証によってモニターした(Brunger、1992)。複合体構造は、基本的には対称であるので、本発明者らは非結晶学的対称拘束を用いることを試みたが、続くリファインメントステップでは、RfreeをNCS拘束を破らずに低下させることができなかった。したがって、NCS拘束はリファインメントを通じて用いなかった。最終ラウンドのリファインメントでは、fobs−fcalc相違電子密度マップを用いて185の水分子を同定した。最終最小化サイクルによって、R−因子は20.7、Rfreeに対しては22.9となった。BMP−2 L51Pの構造分析はBMP−2−BRIAEC複合体のプロトコールに従ったが、焼き鈍し法及びエネルギー最小化のためにCNS 1.1を用いた。BMP−2野生型(PDBエントリー3BMP)の構造を出発モデルとして用いた。モデルの手動での再建は、σA加重した2fobs−fcalc及びfobs−fcalc電子密度マップを基にし、最終ラウンドのリファインメントでは、13の水及び2つのMPD分子を加えた。BMP−2 L51Pの最終構造のR−因子は、21.5であり、Rfreeに対しては23.5である。
【0087】
(相互作用分析)
タンパク質をビオチン化し、ストレプトアビジンコートしたセンサーチップCM5に固定化した(Kirschら、2000a)。BRIA、BRIB及びARIIの細胞外ドメインを記載された通りに調製した(Kirschら、2000a)。ノギン/Fc、コーディン及びグレムリンをR&Dシステム(Wiesbaden−Nordenstadt)から得た。分析物との相互作用をBIAコア2000システムで測定した(Kirschら、2000a)。センサーグラム(Biaevaluation software 2.0)の評価から複合体形成(kon)及び解離(koff)の速度定数が得られた。6〜12回の測定から平均値を得、平均標準偏差はkoffについては35%及びkonについては12%であった。見掛けの解離定数KDはKD=koff/konとして算出するか、平衡結合の線量依存性を評価することによるかのいずれかとした。特別の条件を適用して、BMP−2結合後に遊離ノギンを再生しなくてはならなかった。残存する複合体は、1M塩化ナトリウム及び6M尿素の存在下で酸性pH3でのみ解離することができた。幸にも、タンパク質は、センサーに固定されたタンパク質Aにノギン−Fc融合タンパク質を固定するのに使用する独立した単一サイクルの実験によって確立される厳しい処理の後にも天然の結合特性を保持していた。
【0088】
(細胞株における生物活性)
アルカリホスファターゼ(ALP)活性は、血清を欠乏させたATDC5(Riken Bioresource Center、細胞番号RCB0565)及びC2C12(ATCC番号CRL−1772)細胞で測定した(Kirschら、2000a)。
【0089】
実施例2:BMP−2−BRIAEC複合体における水素結合パターン
BMP−2−BRIAEC複合体の高分解能構造は、非対称単位に2つのBRIAエクトドメインと結合している1つの完全なBMP−2二量体を含んでいる(図1A)。厳しい非結晶学的対称性を適用することに失敗したということは、リガンド−受容体複合体の対称性は完全ではないということを示すものである。しかし、2つのセグメント(BMP−2二量体と1つのBRIAEC分子)間の相違は小さく、すべての原子についてr.m.s.偏差は0.9Åであり、すべてのCα−原子については0.4Åである。詳細な分析により、2つのBMP−2/BRIAEC接触面の構造は同一であることが示される。非結晶学的対称性に関連する分子間のすべての相違は、結晶格子における充填環境の相違によるものである。これらのセグメントにおいて高温因子は、すべての相違は骨格柔軟性によるものであり得るということを示唆する。したがって、対称性に関連しているタンパク質セグメントにおける構造変動は、個々の分子間の実際の相違には反映されないだけでなく、あり得る高次構造並びにBMP−2及びBRIAポリペプチド鎖間の動力学を反映しない。
【0090】
BMP−2/BRIA接触面は成長因子/受容体接触面の新規パラダイムを表す。BMP−2の結合エピトープは、高度に疎水性であり、2310Å2という全部の埋もれた表面のほぼ60%が疎水性残基で作られている(図1b)。ヒト成長ホルモン(de Vosら、1992)、インターロイキン−4(Hageら、1999)、エリスロポエチン(Syedら、1998)又はインターロイキン−6(Boulangerら、2003)のようなその他の成長因子の結合エピトープと比較すると、このレベルの疎水性はかなり高い。アクチビン受容体IIエクトドメインに結合しているBMP−7の複合体構造(Greenwaldら、2003)、TGF−β受容体IIとの複合体中のTGF−β3(Hartら、2002)及びアクチビン受容体IIBと結合しているアクチビンA(Thompsonら、2003)に示されるように、極端に疎水性接触面は、TGF−βスーパーファミリーのメンバーに特徴的であるようである。対照的に、BRIAECの結合エピトープは、結合接触面に広がる極性及び電荷を有する残基の広がりのためにあまり疎水性ではない(30%)(図1B)。
【0091】
BMP−2−BRIAEC複合体のリファインメントされた構造の1.9Åというより高い分解能によって、低分解能でこれまでの構造に可能であったようなより詳細な分析が可能となる。各BMP−2/受容体接触面において、プログラムHBPLUS(McDonald及びThornton、1994)によって10の分子間水素結合(H−結合)が同定される(表1)。BMP−2は5個の、その受容体は4個の主鎖原子を水素結合に提供している。H−結合に携わっている原子を、表1に表されるように2つの構造エレメントにまとめる:リガンドBMP−2では、残基Phe49〜Asn59からなるプレヘリックスループ、受容体BRIAでは、短いα−ヘリックス及びフランキング配列(Asp84〜Arg97)。2つの水素結合、BMP−2 Asp53(N)−BRIAECCys77(O)及びBMP−2 Ser69(O)−BRIAEC Gln94(N)が、主鎖原子の間でのみ形成され、2つの水素結合は、側鎖間でのみ生じ、4つの混合水素結合は側鎖と主鎖原子とを含む(表1)。10の水素結合のうち8は、正真正銘の水素結合に対して予測されるような(Xuら、1997)ドナー−アクセプター距離(2.8±0.14Å)、φ結合角(137±11°)を有する。しかし、結合パラメータの幾何学的評価に純粋に基づいた個々の水素結合の強さの分類はこの複合体には可能ではないようである。
【0092】
【表1】

【0093】
カッコ内の数字は、ドナー−アクセプター原子の距離及び非結晶学的対称性で関連している二次接触面の角度を表す。
a)斜線をつけた行中のH−結合は、前記の幾何統計データから排除した。
b)N−O−Cはドナー−アクセプター原子であり;統計データ(Xuら、1997)から、この角度は、MC−MC水素結合については149°±15°並びにSC−MC及びSC−SC H−結合については129°±18°である。
c)MC(主鎖)及びSC(側鎖)ドナー/アクセプター原子
d)BMP−2(イタリック体)及びBRIA(太字)における分析したH結合を乱す置換。
【0094】
接触面構造から機能的に重要なH−結合を予測できるかどうかを調査するために、残基部分の接近可能性を算出した(図1C)。複合体におけるこの残基の接近可能性は、コアに向かった、又は接触面の表面に向かった残基の位置と関連がある(Chakrabarti及びJanin、2002)。10個の界面のH−結合のうち8個は、表面に位置する残基の間に生じる。受容体Gln86とBMP−2主鎖のLeu51アミド及びカルボニルをつなぐ2個のH−結合だけが、複合体形成の際に完全に埋もれる(図1C)。したがって、この相互作用は、結合のホットスポットを表す可能性があるが(Bogan及びThorn、1998)、その他のH−結合及び疎水性相互作用は、結合親和性には寄与していないか、ほんのわずかしか寄与していない可能性がある。
【0095】
接触面における水分子の調査によって、結合エピトープの脱溶媒和がその他のタンパク質−タンパク質接触面と比較してかなり効果的であるということが明らかになっている。複合体形成の際、各リガンド−受容体接触面には4個の水分子しか完全に埋もれない。これらの埋もれた水分子の低温因子(すなわち、これらの埋もれた水分子の値は、40〜50Å2の範囲であり、近接するタンパク質原子が32Å2というB因子を有する)は、外部の水との交換が遅いものであり得ることを示唆する。驚くべきことに、これらの水分子は、ホットスポットBRIAEC Gln86に近接している。3個の水分子は、側鎖アミド窒素原子に近いBRIAEC Gln86の「膜遠位側」に位置している。これらの水素結合している水分子は、BRIAEC Lys97との接触面の大きな割れ目を埋めており、この窪みへの入口を塞いでいる。単一で単離された水分子が、BRIAEC Gln86の側鎖カルボニル酸素原子の隣の窪んだ空間に位置している。この窪みはBMP−2の残基Phe49及びPro50によって外側から塞がれている。BMP−2結合の際の受容体Gln86の不完全な脱溶媒和は、疎水性環境における強力な水素結合の形成を支持している可能性がある。
【0096】
実施例3:水素結合の突然変異解析
BMP−2及び2種のアミノ酸置換を含むBRIAEC変異タンパク質を用いる相互作用分析によって、結合親和性にとっての水素結合の重要性を解析した。第1に、アラニン置換を導入して、側鎖結合を乱した(実施例2の表1参照)。BMP−2 W28A変異タンパク質は単離できなかったので、Trp28を、その側鎖も水素結合能を欠いているフェニルアラニンで置換した。(1個の電荷を有する残基の置換をBMP−2 S69R変異タンパク質で行った)。第2に、プロリン置換を導入して、主鎖アミド基による水素結合を消滅させた。
【0097】
表2に表されるように、固定化した受容体BRIAECを用いるBMP−2変異タンパク質の相互作用解析によって、置換W28F、D53A及びY103Aは結合親和性に対して小さな作用しか有していないことが示された。S69R変異タンパク質に電荷を有する残基を挿入した後に結合親和性のわずかに大きな低下(17倍)が観察された。際立ったことに、L51P及びD53P置換は著しく低い親和性をもたらす。BRIAECとL51Pの間の見掛けの解離定数KDは、BMP−2のものより7100倍よりも高く、D53Pについては53倍高かった(表2及び図7)。両変異タンパク質とも、BRIB受容体に対する親和性の同様の低下を有し、このことはBMP−2の同様の決定基が2つのI型受容体の結合に用いられていることを示す。このことは、2つのプロリン変異タンパク質において乱された主鎖水素結合は主要な結合決定基であるが、W28、D53、S69及びY103という側鎖結合はBRIA受容体との結合にはあまり重要なものでないということを示している。
【0098】
II型受容体ARIIに対するプロリン変異タンパク質の親和性は、野生型と比較して変わらない(表2)。II型受容体アクチビン受容体IIBとBMP受容体IIとの相互作用についても同じことが観察されている(データは示していない)。このことにより、置換されたプロリンによる局所構造の変化は、II型受容体結合の並列するエピトープ(ナックルエピトープ)には伝わっていないので、変異タンパク質の全体構造は変わっていないということが確認される。
【0099】
【表2】
【0100】
BRIA受容体の結合決定基を同定するために、3種のエクトドメイン変異タンパク質を調製した。受容体Q94P変異タンパク質では、野生型ではBMP−2 Ser69骨格カルボニルに水素結合を供与する骨格窒素アミドが乱されている。受容体Q86A変異タンパク質は、野生型ではBMP−2 L51骨格アミド及びカルボニルとH結合を形成する側鎖カルボキシアミド基を欠いている(図2)。S90A置換によって水素結合をBMP2 Val26骨格カルボニルに供与するセリンヒドロキシル基が消滅する。すべての受容体変異タンパク質は野生型エクトドメインと同様に大腸菌で発現された後に高度に精製され得る。バイオセンサーに固定化した場合、受容体Q94P変異タンパク質は、野生型エクトドメインと比較して親和性が3倍しか低下しないでBMP−2と結合したが(表2)、このことは、変異タンパク質中で失われた主鎖/主鎖結合は結合にはわずかにしか寄与していないということを示す。S90A変異タンパク質において親和性の同様の小さな変化が観察されている。
【0101】
しかし、受容体Q86A置換は、BMP−2相互作用の際に86倍増加したKDをもたらした。しかし、BMP−2 L51P変異タンパク質は、影響を受けるH−結合が主要な結合決定基に相当するということをすでに実証していたので(前記参照)、親和性のこのような大きな低下は予測されたものであった。最も重要なことは、関与しているドナー及びアクセプター群の残基部分の接近可能性を分析することによって推測されるホットスポットと突然変異解析によって同定されたホットスポットとが一致することである(図1C)。受容体Q86A変異タンパク質における親和性の喪失は、BMP−2 L51P変異タンパク質におけるものよりは深刻なものではない(表2)。受容体中の隣接側鎖、例えば、Lys79又はGlu81又はAsp89が、失われたGln86のカルボキサミド基を部分的に補っている可能性がある(図1b)。他方、BMP−2 L51Pに導入されたプロリンは、立体障害を引き起こすか、例えば、BMP−2 Asp53骨格アミドと受容体Cys77カルボニルの間の局所的な構造転位によって隣接する結合を阻害し、それによって置換の作用をさらに悪化させる(図2)。
【0102】
実施例4:プロリンBMP−2変異タンパク質L51Pの構造
BMP−2 L51Pの結合親和性の大きな低下を、BRIAEC Q86Aに対して観察されたものと比較することによって、結合親和性の喪失は、単一の水素結合の喪失だけに起因するわけではない可能性があることが示唆される。さらに、結合エピトープにおける大きな構造変化がタンパク質相互作用を乱す可能性がある。したがって、BMP−2 L51P及びまたD53P変異タンパク質を結晶化試験に付した。BMP−2 L51Pは野生型BMP−2と同一条件下で結晶化でき、同一の結晶パラメータ、例えば、野生型BMP−2の結晶について観察されたものと同一の空間群、R32、及びほぼ同一の容器定数を有していた。明らかに、プロリン残基はLeu51の代わりに受け入れられ、その疎水性側鎖がBMP−2のタンパク質コア中に向かっている。変異タンパク質D53Pは、そのような条件下では結晶化せず、D53Pの結晶は、エタノール又は2−メチル−2,4−ペンタジオールのいずれかから成長し、極めて低い分解能までしか回析せず、さらには分析しなかった。
【0103】
BMP−2 L51Pの回析データは室温で2.7Åという分解能まで得た。突然変異部位は別として、BMP−2野生型とL51Pの構造は基本的に同一であり、表3に表される構造決定法の誤差限度内にあり、すべてのCα原子位置のr.m.s.偏差は0.6Åである。残基Pro48〜His54を含み、位置51の突然変異部位も保持するプレヘリックスループに明確な変化が観察される。残基Pro48、Phe49、Pro50、Leu/Pro51、Ala52及びHis54について、野生型と変異タンパク質のCα位置間の距離は1Åを越え、最大の相違はAla52のCαについての1.8Åである(表3)。両構造、BMP−2野生型とL51Pの全体的な位置の誤差は、σAプロットから算出されるように約0.4であるので、プレヘリックスループの高次構造の変化は小さいが有意なものと考えられる。
【0104】
【表3】
【0105】
この変化は野生型のロイシンのものと比較したプロリン残基の骨格の形状の変化によるものであり得る。しかし、結合していない高次構造及び結合している高次構造におけるBMP−2の構造を比較すると、Cα位置間の相違はBMP−2野生型と変異タンパク質L51P間で観察されるものと同じ大きさであり、このことは、プレヘリックスループの高次構造は前もって形成されているものでも、固定されているものでもなく、複合体形成の際に受容体表面に適応するということを示す。
【0106】
Pro51の側鎖は野生型BMP−2中のLeu51と同様の方向を向いており、このことは疎水性コアのパッキングは突然変異によって影響を受けないということを示す。さらに、Leu51からプロリンへの交換は、残基51の骨格カルボニルの方向を変えず、それゆえに、BRIA Gln86と残基51との1個の水素結合が依然として複合体に形成され得る(図2、3A)。突然変異によって、結合していないタンパク質に導入された、かなり小さい構造の相違を考慮すると、結合親和性の大きな変化は不可解なものに思われる。
【0107】
L51PとBMP−2−BRIAECという複合体の構造の重ね合わせにより、プロリン残基の環が、BMP−2 Leu51アミドとBRIAEC Gln86の側鎖カルボニルの間の水素結合に用いられていた容積を元は占めていることが示される(図3A)。プロリン環の大きな空間要求が、BMP−2Pro51の側鎖とBRIAEC Gln86の間に立体障害を引き起こす(図3A)。Pro51を受け入れるには、その側鎖はさらに押し戻される必要がある。基本的に環の方向にアミド窒素を固定する堅い環構造であるために、もし環構造が押し戻されると、これは、Pro50の骨格カルボニルがBRIAEC Gln86の側鎖の方向に移動するφ骨格ねじれ角の回りの回転に相当する。結果として、1つの立体障害を除去することによって、別の1つが引き起こされることとなり、これによって、おそらくは、突然変異L51Pによる結合エネルギーの大きな喪失が説明される。
【0108】
実施例5:プロリン変異タンパク質及び生物活性
BMP−2プロリン変異タンパク質の生物活性は、C2C12細胞及びわずかにより感受性の高いATDC5細胞株を用いて、アルカリホスファターゼ(ALP)活性の用量依存的誘導によって決定した。最大応答の50%(ED50)をもたらす野生型BMP−2の用量は、C2C12細胞では25nMであった。ATDC5細胞を用いた場合、ED50は8nMであり、このことはこれらの細胞の高い反応性を示す。対照的に、BMP−2 L51Pには、C2C12(>1600nM)又はATDC5細胞(>1500nM)で測定可能な生物活性はない。BMP−2 D53P変異タンパク質は、C2C12及びATDC5細胞に対して、330nM及び140nMというED50値をそれぞれ示す。これらの値は、野生型BMP−2のED50値と比較して約13〜17倍高い。したがって、2種のBMP−2プロリン変異タンパク質におけるBRIA受容体親和性の異なる低下は生物活性の喪失と平行している(図4)。
【0109】
BMP−2 Leu51のプロリン置換は、I型受容体との相互作用にのみ影響を及ぼしており、他方でII型受容体の親和性は変わっていないので、L51P変異タンパク質は、主としてドミナントネガティブ阻害剤として機能している可能性がある。しかし、これは、250nMまでの濃度の変異タンパク質を、20nM BMP−2で刺激した細胞に適用した場合には観察されなかった(データは示していない)。相対的に、ATDC5細胞における定常ALP活性は、アンタゴニストBMP−2 A34D変異タンパク質によって阻害される(Kirschら、2000a)が、L51Pによっては阻害されない(図4A)。
【0110】
実施例6:結合タンパク質のアンタゴニストとしてのプロリン変異タンパク質
BMP−2 L51PがBRIAECと結合しないことは、その他の結合タンパク質との相互作用もプロリン置換によって影響を受け得るかどうかという疑問を提起した。特に注目されるものは、BMPと高親和性で結合し、BMP−7との複合体中のその構造が最近決定された(Groppeら、2002)ノギンである。もう1つの重要な結合タンパク質はコーディンであり、これはBMP−2と2つのシステインリッチドメインによって相互作用する。さらにもう1つの種類の結合タンパク質は、いわゆるDNAドメインによってBMP−2と相互作用するグレムリンに相当する。3種のタンパク質の各々を、バイオセンサーの表面に固定し、表4に表されるようにBMP−2変異タンパク質相互作用について分析した。
【0111】
【表4】
【0112】
ノギンとの複合体についての2種のBMP−2プロリン変異タンパク質の解離定数KDはBMP−2野生型のものと同様であり、0.5〜2.5nMの範囲であった。したがって、ノギンのN末端アミノ酸がBMP−2 Leu51(BMP−7 Leu75)及びAsp53(BMP−7 Ser77、プレヘリックスループ)に対応するBMP−7領域と接触していることが示され、ノギンはBMP−2とのBRIA結合を妨げるという事実があるにもかかわらず、BMP−2 Leu51及びAsp53はノギンに対する主要な結合決定基ではない。
【0113】
コーディンドメインCR1及びCR3並びにグレムリンに対するBMP−2の親和性は、ノギンに対してよりも10〜30倍低い(表4)。BMP−2野生型又は2種のプロリン変異タンパク質に対しても再度、同様の解離定数KDが認められ、このことは、これらのタンパク質の結合がプロリン置換によって影響を受けないことを示す。したがって、両結合タンパク質に対するBMP−2エピトープはBRIA受容体に対するものとは異なる。
【0114】
実施例7:プロリン変異タンパク質L51Pは、C2C12細胞においてノギン阻害を解除する
BMP−2は飢餓前骨髄芽球性細胞株C2C12において、アルカリホスファターゼの発現を誘導する。このBMP応答は、等モル濃度のノギン−Fc融合タンパク質によってバックグラウンドレベルまで阻害される(図5)。ノギンによるBMP−2活性の阻害は、L51P変異タンパク質によって用量依存的に解除される。ほぼ等モル濃度の変異タンパク質L51Pとノギンで、ノギンの阻害作用は、80%よりも多くまで中和される。このことは、受容体非感受性L51P変異タンパク質はノギンタンパク質の効率的な阻害剤に相当するということを示す。この結果は、BMP−2 L51P置換後のI型受容体結合の混乱とノギン結合の保持を示す物理的結合研究と一致する。
【0115】
参照文献
不要な参照を避けるために、本明細書に記載した参照文献の包括的な一覧を以下に示す。これらの参照文献のいずれの開示内容も参照により本明細書に組み込まれると理解されなくてはならない。
【0116】
Balemans, W. and Van Hul, W. (2002)Extracellularregulation of BMP signaling in vertebrates: a cocktail ofmodulators. Dev Biol,250, 231-250.
Bogan, A.A. and Thorn, K.S. (1998)Anatomy of hot spots inprotein interfaces. J Mol Biol, 280, 1-9.
Boulanger, M.J., Chow, D.C., Brevnova,E.E. and Garcia,K.C. (2003) Hexameric structure and assembly of theinterleukin-6/IL-6alpha-receptor/gp130 complex. Science, 300, 2101-2104.
Brunger, A.T. (1992) Free R value: anovel statisticalquantity for assessing the accuracy of crystal structures.Nature, 355,472-475.
Brunger, A.T., Adams, P.D., Clore, G.M.,DeLano, W.L.,Gros, P., Grosse-Kunstleve, R.W., Jiang, J.S., Kuszewski, J.,Nilges, M., Pannu, N.S., Read, R.J., Rice, L.M., Simonson, T. and Warren, G.L.(1998) Crystallography & NMR system: A new software suite formacromolecular structure determination. Acta Crystallogr DBiol Crystallogr, 54(Pt 5), 905-921.
Chakrabarti, P. and Janin, J. (2002)Dissectingprotein-protein recognition sites. Proteins, 47, 334-343.
Collaborative Computational Project, N.(1994) The CCP4Suite: Programs for Protein Crystallography. Acta Crystallogsect D, 50.
de Vos, A.M., Ultsch, M. and Kossiakoff,A.A. (1992) Humangrowth hormone and extracellular domain of its receptor:crystal structure ofthe complex. Science, 255, 306-312.
Evans, P.R. (1993) Data reduction.Proceedings of CCP4Study Weekend, pp. 114-122.
Greenwald, J., Groppe, J., Gray, P.,Wiater, E.,Kwiatkowski, W., Vale, W. and Choe, S. (2003) The BMP7/ActRII extracellulardomaincomplex provides new insights into the cooperative nature of receptorassembly.Mol Cell, 11, 605-617.
Groppe, J., Greenwald, J., Wiater, E.,Rodriguez-Leon, J.,Economides, A.N., Kwiatkowski, W., Affolter, M., Vale, W.W.,Belmonte, J.C. andChoe, S. (2002) Structural basis of BMP signalling inhibitionby the cystineknot protein Noggin. Nature, 420, 636-642.
Hage, T., Sebald, W. and Reinemer, P.(1999) Crystal structure of the interleukin-4/receptor alpha chaincomplexreveals a mosaic binding interface. Cell, 97, 271-281.
Hart, P.J., Deep, S., Taylor, A.B.,Shu,Z., Hinck, C.S. and Hinck, A.P. (2002) Crystal structure of the human TbetaR2ectodomain--TGF-beta3 complex. Nat Struct Biol,9, 203-208.
Kirsch, T., Nickel, J. and Sebald, W.(2000a) BMP-2 antagonistsemerge from alterations in the low-affinity bindingepitope for receptor BMPR-II. Embo J, 19, 3314-3324.
Kirsch, T., Nickel, J. and Sebald, W.(2000b) Isolation ofrecombinant BMP receptor IA ectodomain and its 2:1 complexwith BMP-2. FEBSLett, 468, 215-219.
Leslie, A.G.W. (1992) Recent changes tothe MOSFLM packagefor processing film and image plate data. Joint CCP4 +ESF-EAMCB Newsletter onProtein Crystallography, 26.
McDonald, I.K. and Thornton,J.M. (1994)Satisfying hydrogen bonding potential in proteins. J Mol Biol, 238,777-793.
Murshudov, G.N., Vagin, A.A. and Dodson,E.J. (1997)Refinement of Macromolecular Structures by the Maximum-LikelihoodMethod. ActaCrystallog sect D, D53, 240-25
5.
Ruppert, R., Hoffmann, E. and Sebald, W.(1996) Human bonemorphogenetic protein2 contains a heparin-binding site whichmodifies itsbiological activity. Eur J Biochem, 237, 295-302.
Syed, R.S., Reid, S.W., Li, C.,Cheetham, J.C., Aoki, K.H.,Liu, B., Zhan, H., Osslund, T.D., Chirino, A.J.,Zhang, J., Finer-Moore, J.,Elliott, S., Sitney, K.,Katz, B.A., Matthews, D.J.,
Thompson, T.B., Woodruff, T.K. andJardetzky, T.S. (2003)Structures of an ActRIIB:activin A complex reveal a novelbinding mode forTGF-beta ligand:receptor interactions. Embo J, 22, 1555-1566.
Winn, M.D., Isupov, M.N. and Murshudov,G.N. (2001) Use ofTLS parameters to model anisotropic displacements inmacromolecular refinement.Acta Crystallogr D Biol Crystallogr, 57, 122-133.
Xu, D., Tsai, C.J. and Nussinov, R.(1997) Hydrogen bondsand salt bridges across protein-protein interfaces.Protein Eng, 10, 999-1012.
【0117】
本明細書に開示した本発明の特徴、特許請求の範囲及び/又は図面は、個別に、及びいずれかの組合せでの双方で、種々の形の本発明を理解する材料であり得る。
【技術分野】
【0001】
本発明は、骨形成タンパク質の変異タンパク質、それをコードする核酸、このような骨形成変異タンパク質の製造方法、このような変異タンパク質を含む組成物、好ましくは薬剤組成物、及びこのような変異タンパク質の、医薬の製造のための使用に関する。
【背景技術】
【0002】
本発明の変異タンパク質は、骨形成タンパク質の変異タンパク質である。骨形成タンパク質は、本明細書ではBMP−2様タンパク質とも呼ばれ、TGF−βスーパーファミリーを形成する2つの主要な群の一方である。BMP−2様タンパク質とは別に、このTGF−βスーパーファミリーに属するもう一方の群のタンパク質はTGF−β−アクチン様タンパク質である。TGF−βスーパーファミリーの2つの群は、その結合機構が異なっている。一般に、ポリペプチド成長因子と構造的に関連しているTGF−βスーパーファミリーのメンバーは、細胞増殖、細胞株の運命決定、分化、移動性、接着及び細胞死をはじめとする細胞過程に関与している。こういった因子は、時間及び組織特異的パターンに従って発現され、真核生物ではほとんどすべての組織の発達、止血及び修復にとって重要である。これらの因子はまた、細胞の活性を規定する細胞内シグナルの重要な部分を占めている。
【0003】
TGF−βのシグナル伝達経路は、ごく最近解明された。TGF−βのシグナル伝達には、細胞表面の受容体セリンキナーゼ及びその基質、すなわち、SMADタンパク質が関与しており、このタンパク質は、リン酸化されると核へ移動する。リン酸化されたSMADタンパク質は、DNA結合パートナーと共同して標的遺伝子の転写を活性化する。TGF−β及びTGF−βスーパーファミリーのその他のメンバーの多機能活性は、種々の受容体とSMADタンパク質とDNA結合タンパク質との相互作用に基づいていると思われる。このシグナル伝達経路の何らかの混乱が、哺乳類、特に人間におけるいくつかの疾患の理由である。TGF−βスーパーファミリーのメンバーはいくつかの構造的特徴を共有しているが、このメンバー間の相同性はかなり限定的である場合もある。例えば、すべてのタンパク質は二量体であり、通常、ホモ二量体、すなわち、2つの同一の単量体からなるものである。また、TGF−βスーパーファミリーのメンバーは、2つの異なる種類のセリンキナーゼ受容体鎖、すなわち、I型鎖及びII型鎖からなる細胞受容体を用いる。
【0004】
I型鎖は細胞質GSボックスとセリンキナーゼとを含み、I型鎖がBRIA、BRIB又はARIである場合には、SMAD−1及びSMAD−5シグナルタンパク質を活性化する。II型鎖は、GSボックスセグメントのリン酸化によってI型受容体セリンキナーゼを活性化する。I型及びII型鎖双方の小さな受容体エクトドメインは、それぞれ約120〜150個のアミノ酸を含むが、かなり限定的な相同性しか共有していない。それにもかかわらず、TGF−βスーパーファミリーのすべての既知受容体鎖の1つの共通の特徴として、4つの保存されたジスルフィド架橋があり、さらなるジスルフィド架橋及びいくつかのアミノ酸の位置はI型又はII型受容体タンパク質のいずれかに特徴的であるようである。BMP−2様タンパク質とI型鎖との結合は、BMP−2様タンパク質のリスト(wrist)エピトープを介して起こり、II型鎖との結合は、BMP−2様タンパク質の「ナックル(knuckle)」エピトープを介して起こる。これらの骨形成タンパク質とそれぞれの受容体鎖との結合は、細胞質膜で逐次的に起こり、特定の配列は骨形成タンパク質と個々の受容体鎖との親和性に応じて変わる。例えば、溶解BMP−2は、その高親和性I型鎖、すなわち、BMPR−IA、BMPR−IBとまず結合し、おそらくはActR−Iとも結合し、続いて、低親和性II型受容体鎖と会合して活性な受容体複合体を形成する。この会合は細胞質膜で起こる。
【0005】
BMP−2のリスト又はナックルエピトープ中のいくつかのアミノ酸置換によって、不活性なBMP−2変異タンパク質となり、これは対応する受容体をもはや活性化することができない。BMP−2[A34D]などのナックルエピトープ中にアミノ酸置換を有する不活性な変異タンパク質は、それでも、野生型BMP−2と同様にI型鎖と結合できる。しかし、I型及びII型鎖の双方を含む、活性な受容体複合体を形成するための膜におけるII型鎖の会合には支障を来たす。したがって、この種の変異タンパク質は、I型鎖に対して野生型BMP−2と競合しているので、BMP−2の阻害剤として作用する。
【0006】
骨形成タンパク質及び、詳しくは、BMP−2様タンパク質は、I型鎖及びII型鎖を含む受容体が関係しているシグナル伝達経路に関与していることとは別に、モジュレータータンパク質の群と相互作用する。これらのモジュレータータンパク質は、同様にある種のシグナル伝達経路に関与している。BMP−2様タンパク質と相互作用するモジュレータータンパク質は、中でも、ノギンタンパク質ファミリー、DANタンパク質ファミリー及びコーディンタンパク質ファミリーのメンバーを含んでなる。これらのモジュレータータンパク質は、当技術分野では公知であり、例えば、Avsian-Kretchmer,O.及びHsueh, A.J. (2004)「Comparative genomicanalysis of the eight-membered ring cystine knot containingbone morphogeneticprotein antagonists.」、Mol Endocrinol、18巻、1〜12頁に記載されており、これにはBMP−モジュレータータンパク質、より詳しくは、シスチンノット含有BMPモジュレータータンパク質が記載されている。例えば、ノギンを含むノギンファミリーは、Canalis, E.、Economides, A. N.及びGazzerro, E. (2003)、「Bonemorphogeneticproteins, their antagonists, and the skeleton.」Endocr Rev巻、24、218〜235頁によって記載されており、中でも、コーディン、ベントロピン、CTGF、CYR61、クロスベインレス(Crossveinless)、プロコラーゲンIIAを含むコーディンファミリーは、例えば、Bork, P. (1993)「The modular architecture ofa new family of growth regulators related to connective tissue growth factor.」、FEBS Lett、327巻、125〜130頁、Brigstock, D. R.、Goldschmeding, R.、Katsube,K. I.、Lam, S. C.、Lau, L. F.、Lyons, K.、Naus, C.、Perbal,B.、Riser, B.、Takigawa, M.及びYeger, H. (2003)「Proposal for a unified CCNnomenclature.」Mol Pathol、56巻、127〜128頁並びにGarciaAbreu, J.、Coffinier, C.、Larrain,J.、Oelgeschlager, M.及びDeRobertis, E. M.(2002)「Chordin-like CR domains and theregulationofevolutionarily conserved extracellular signaling systems.」、Gene、287巻、39〜47頁並びに例えば、Cer1、セルベルス(Cerberus)、Dan、Dte、グレムリン(Gremlin)、PRDC、スクレロスチン(Sclerostin)を含むDANファミリーは、例えば、Pearce, J.J.、Penny, G.及びRossant, J.(1999)「Amousecerberus/Dan-related gene family.」Dev Biol、209巻、98〜110頁並びにBalemans,W.及びVan Hul, W.(2002)「Extracellular regulation of BMP signaling in vertebrates: a cocktailof modulators.」Dev Biol、250巻、231〜250頁に記載されている。
【発明の概要】
【発明が解決しようとする課題】
【0007】
したがって、本発明の基礎をなす課題は、I型及びII型セリンキナーゼ受容体鎖媒介性作用の点で、それぞれの骨形成タンパク質の野生型と競合しないが、依然として、ノギンタンパク質ファミリー、DANタンパク質ファミリー及びコーディンタンパク質ファミリーのものなどのモジュレータータンパク質と相互作用するのには適している、骨形成タンパク質、詳しくはBMP−2様タンパク質の変異タンパク質を提供することであった。
【課題を解決するための手段】
【0008】
本発明によれば、この課題は独立請求項の対象によって解決される。好ましい実施形態は従属請求項から選びとることができる。
【0009】
本発明によれば、この課題は、第1の態様で、ヒトBMP−2のアミノ酸位置51に相当するアミノ酸位置に骨形成タンパク質の野生型と比較してアミノ酸置換を含む、骨形成タンパク質の変異タンパク質によって解決される。
【図面の簡単な説明】
【0010】
【図1】(A)BMP−2及びBRIAECの複合体構造のリボン図を示す図である。(B) BMP−2及びBRIAECの接触面について、受容体BRIAがy軸の回りに180°回転している“明白な”眺望における表面表示を示す図である。(C) BMP−2及びBRIAECの接触面の表面表示を示す図である。
【図2】BMP−2−BRIA相互作用の接触面中の拡大表示を示す図である。
【図3】(A)BMP−2 L51Pの結晶構造を示す、プレヘリックスループセグメントの立体像を示す図である。
【図4】BMP−2及びBMP−2変異タンパク質に反応してATDC5(A)及びC2C12(B)細胞で測定されたアルカリホスファターゼ(ALP)活性の誘導として表されるBMP−2プロリン変異タンパク質の生物活性を示す図である。
【図5】BMP−2プロリン変異タンパク質L51Pによるノギン阻害の解除を示す図である。
【図6】ヒトBMP−2のアミノ酸位置51に相当する位置のアミノ酸残基をロイシンから好ましくはプロリンに交換し、本発明の変異タンパク質の好ましい実施形態を形成したBMP−2様タンパク質のアラインメントを示す図である。
【図7】BMPプロリン変異タンパク質と(A)BRIAEC、(B)BRIBEC又は(C)アクチビン受容体IIエクトドメイン間の相互作用、並びに(D)BMP−2とBRIAEC変異タンパク質間の相互作用のセンサーグラム(sensograms)、を示す図であって、すべてのリガンドは、120nM濃度で存在しており、センサーグラム(sensorgrams)は、アナライトであるBMP−2の濃度を飽和するために算出されたレゾナンスユニット(RU)を100%として設定することによって標準化した。
【図8】BMP−2野生型とBMP−2 L51Pの間の構造の相違を示す図であって、両構造のCα−原子がスーパーインポーズされ、中間の距離が示され、プレヘリックスループの領域は灰色で強調表示されている。
【発明を実施するための形態】
【0011】
一実施形態では、ヒトBMP−2のアミノ酸位置51に相当する位置のアミノ酸は、骨形成タンパク質の野生型ではロイシンであり、変異タンパク質では好ましくはプロリンである。
【0012】
一実施形態では、骨形成タンパク質は、hBMP−2、hBMP−4、hBMP−5、hBMP−6、hBMP−7、hBMP−8、hGDF−5、mGDF−6、mGDF−7、hBMP−10及びhGDF−2を含んでなる群から選択される。
【0013】
一実施形態では、
骨形成タンパク質は、hBMP−2であり、ヒトBMP−2のアミノ酸位置51に相当する位置はアミノ酸位置51であり、
骨形成タンパク質は、hBMP−4であり、ヒトBMP−2のアミノ酸位置51に相当する位置はアミノ酸位置53であり、
骨形成タンパク質は、hBMP−5であり、ヒトBMP−2のアミノ酸位置51に相当する位置はアミノ酸位置68であり、
骨形成タンパク質は、hBMP−6であり、ヒトBMP−2のアミノ酸位置51に相当する位置はアミノ酸位置68であり、
骨形成タンパク質は、hBMP−7であり、ヒトBMP−2のアミノ酸位置51に相当する位置はアミノ酸位置75であり、
骨形成タンパク質は、hBMP−8であり、ヒトBMP−2のアミノ酸位置51に相当する位置はアミノ酸位置75であり、
骨形成タンパク質は、hGDF−5であり、ヒトBMP−2のアミノ酸位置51に相当する位置がアミノ酸位置56であり、
骨形成タンパク質は、mGDF−6であり、ヒトBMP−2のアミノ酸位置51に相当する位置がアミノ酸位置56であり、
骨形成タンパク質は、mGDF−7であり、ヒトBMP−2のアミノ酸位置51に相当する位置はアミノ酸位置82であり、
骨形成タンパク質は、hBMP−10であり、ヒトBMP−2のアミノ酸位置51に相当する位置はアミノ酸位置44であり、
骨形成タンパク質は、hGDF−2であり、ヒトBMP−2のアミノ酸位置51に相当する位置はアミノ酸位置45である。
【0014】
一実施形態では、野生型の
hBMP−2は、配列番号1のアミノ酸配列を含み、
hBMP−4は、配列番号3のアミノ酸配列を含み、
hBMP−5は、配列番号5のアミノ酸配列を含み、
hBMP−6は、配列番号7のアミノ酸配列を含み、
hBMP−7は、配列番号9のアミノ酸配列を含み、
hBMP−8は、配列番号11のアミノ酸配列を含み、
hGDF−5は、配列番号13のアミノ酸配列を含み、
mGDF−6は、配列番号15のアミノ酸配列を含み、
mGDF−7は、配列番号17のアミノ酸配列を含み、
hBMP−10は、配列番号19のアミノ酸配列を含み、
hGDF−2は、配列番号21のアミノ酸配列を含む。
【0015】
好ましい一実施形態では、野生型の
hBMP−2は、配列番号2の核酸によってコードされており、
hBMP−4は、配列番号4の核酸によってコードされており、
hBMP−5は、配列番号6の核酸によってコードされており、
hBMP−6は、配列番号8の核酸によってコードされており、
hBMP−7は、配列番号10の核酸によってコードされており、
hBMP−8は、配列番号12の核酸によってコードされており、
hGDF−5は、配列番号14の核酸によってコードされており、
mGDF−6は、配列番号16の核酸によってコードされており、
mGDF−7は、配列番号18の核酸によってコードされており、
hBMP−10は、配列番号20の核酸によってコードされており、
hGDF−2は、配列番号22の核酸によってコードされている。
【0016】
本発明によれば、この課題は、第2の態様で、第1の骨形成タンパク質受容体と結合しておらず、かつ、ノギンタンパク質ファミリー、DANタンパク質ファミリー、コーディンタンパク質ファミリー及びシステインノット含有BMPモジュレータータンパク質を含んでなる群から選択される少なくとも1種のモジュレータータンパク質と結合している、骨形成変異タンパク質によって解決される。
【0017】
本発明によれば、この課題は、第3の態様で、骨形成変異タンパク質によって解決され、この骨形成変異タンパク質は、好ましくは第2の骨形成タンパク質受容体と相互作用するプレヘリックスループ構造を含む、本発明の第1及び第2の態様の実施形態であることが好ましい。第2の骨形成タンパク質受容体の用語は、この種の受容体と、本明細書で第1の骨形成タンパク質受容体と呼ばれる別の受容体を区別するために用いられると理解されなくてはならない。この技術用語は、本明細書で参照するためだけに取り入れられており、タンパク質及び変異タンパク質の結合をそれぞれ暗に示すものではない。その程度において、タンパク質及び変異タンパク質の結合は、それぞれ、単一の骨形成タンパク質受容体とのみ起こることが好ましい。このような単一の骨形成タンパク質受容体は、第1の骨形成タンパク質受容体又は第2の骨形成タンパク質受容体のいずれかであり得る。
【0018】
第2及び第3の態様に従う一実施形態では、第1及び/又は第2の骨形成タンパク質受容体がBRIA又はBRIBである、請求項7及び8に記載の骨形成変異タンパク質。
【0019】
第2及び第3の態様に従う一実施形態では、相互作用はアミノ酸残基、好ましくは、BRIAのGln86又はBRIBのGln67アミノ酸残基に関連している。
【0020】
本発明によれば、この課題は、第4の態様で、骨形成変異タンパク質によって解決され、この骨形成変異タンパク質は、好ましい実施形態では、第1、第2及び第3の態様に従う骨形成タンパク質であり、第2の骨形成タンパク質受容体との相互作用を有するプレヘリックスループ構造を含み、骨形成変異タンパク質のプレヘリックスループ構造と第2の骨形成タンパク質受容体との相互作用が、野生型骨形成タンパク質のプレヘリックスループ構造と第2の骨形成タンパク質受容体との相互作用とは異なる。
【0021】
いずれかの態様に従う一実施形態では、異なる相互作用又は変化は、屈折データ、好ましくは、少なくとも約2.7Åの分解能まで室温で得られているような屈折データで表される。
【0022】
いずれかの態様に従う一実施形態では、プレヘリックスループ構造が、骨形成タンパク質の野生型と比較して変異している。
【0023】
好ましい一実施形態では、野生型BMP−2の位置51のロイシンに相当するアミノ酸が変異している。
【0024】
もう1つの好ましい実施形態では、ヒトBMP−2の位置51のロイシンに相当するアミノ酸がプロリンに変異している。
【0025】
いずれかの態様に従う一実施形態では、骨形成変異タンパク質は、hBMP−2、hBMP−4、hBMP−5、hBMP−6、hBMP−7、hBMP−8、hGDF−5、mGDF−6、mGDF−7、hBMP−10及びhGDF−2を含んでなる群から選択される骨形成タンパク質の変異タンパク質である。
【0026】
好ましい一実施形態では、骨形成タンパク質はBMP−2又はプロ−BMP−2である。
【0027】
本発明によれば、この課題は、第5の態様で、配列番号23〜33のいずれかに記載のアミノ酸配列を含む骨形成タンパク質によって解決される。
【0028】
一実施形態では、骨形成変異タンパク質は、本発明のいずれかの態様に記載の骨形成変異タンパク質である。
【0029】
本発明によれば、この課題は、第6の態様で、本発明のいずれかの態様に従う、骨形成タンパク質及び骨形成変異タンパク質をそれぞれコードする核酸、並びに/又はその相補鎖によって解決される。
【0030】
本発明によれば、この課題は、第7の態様で、配列番号2、4、6、8、10、12、14、16、18、20及び22の核酸配列を含む核酸、好ましくは、本発明のいずれかの態様に従う骨形成変異タンパク質をコードする核酸、並びに/又はその相補鎖によって解決される。
【0031】
本発明によれば、この課題は、第8の態様で、本発明のいずれかの態様に従う骨形成変異タンパク質をコードする核酸であって、より好ましくは、ストリンジェントな条件下で、遺伝暗号の縮重がなければ、第6及び第7の態様に従う核酸とハイブリダイズする核酸によって解決される。
【0032】
本発明によれば、この課題は、第9の態様で、第6、第7及び第8の態様に従う核酸を含むベクターであって、好ましくは発現ベクターであるベクターによって解決される。
【0033】
本発明によれば、この課題は、第10の態様で、第6、第7及び第8の態様に従う核酸及び/又は本発明の第10の態様に従うベクターを含む細胞、好ましくは、哺乳類細胞によって解決される。
【0034】
本発明によれば、この課題は、第11の態様で、本発明の第10の態様に従う細胞を含む、宿主生物、好ましくは、哺乳類宿主生物、より好ましくは、非ヒト宿主生物によって解決される。
【0035】
本発明によれば、この課題は、第12の態様で、
a)請求項25に記載の細胞を培養ブロスで培養するステップと、
b)細胞から、及び/又は培養ブロスから骨形成変異タンパク質を調製するステップと、
を含む、本発明の第1〜第5のいずれかの態様に従う骨形成変異タンパク質の製造方法によって解決される。
【0036】
本発明によれば、この課題は、第13の態様で、本発明の第1〜第5のいずれかの態様に従う骨形成変異タンパク質と特異的に結合しているモノクローナル抗体によって解決される。
【0037】
本発明によれば、この課題は、第14の態様で、本発明の第1〜第5のいずれかの態様に従う変異タンパク質及び/又は本発明の第6〜第8のいずれかの態様に従う核酸を含む組成物によって解決される。
【0038】
本発明によれば、この課題は、第15の態様で、本発明の第1〜第5のいずれかの態様に従う変異タンパク質又は第6〜第8のいずれかの態様に従う核酸と、製薬上許容される担体とを含む薬剤組成物によって解決される。
【0039】
本発明によれば、この課題は、第16の態様で、医薬を製造するための、本発明の第1〜第5のいずれかの態様に従う骨形成変異タンパク質及び/又は第6〜第8のいずれかの態様に従う核酸の使用によって解決される。
【0040】
一実施形態では、医薬は、線維性疾患、創傷治癒、過剰血管新生、血管疾患、骨折及び骨粗しょう症を含んでなる群から選択される疾患の治療用及び/又は予防用のものである。
【0041】
好ましい一実施形態では、線維性疾患は、腎線維症、肝硬変、肺線維症及び慢性炎症、好ましくは、喘息を伴う慢性炎症を含んでなる群から選択される。
【0042】
もう1つの実施形態では、創傷治癒はケロイド、瘢痕形成及び腹膜の閉塞と関連している。
【0043】
さらなる一実施形態では、過剰血管新生は、網膜症、動脈硬化症及び/又は腫瘍と関連しているか、又は網膜症、動脈硬化症及び/又は腫瘍を伴う。
【0044】
いっそうさらなる実施形態では、骨折は難治性骨折である。
【0045】
もう1つの実施形態では、疾患は骨粗しょう症である。
【0046】
本発明によれば、この課題は、第17の態様で、形態形成タンパク質、好ましくは、本発明の第1〜第5のいずれかの態様に従う形態形成変異タンパク質の、BMP相互作用タンパク質に対する阻害剤としての使用によって解決される。
【0047】
一実施形態では、BMP相互作用タンパク質は、ノギンタンパク質ファミリー、DANタンパク質ファミリー及びコーディンタンパク質ファミリーを含んでなる群から選択される。
【0048】
本発明者らは、驚くべきことに、本発明の変異タンパク質が、ノギンファミリー、DANファミリー及びコーディンタンパク質ファミリーのものなどのモジュレータータンパク質の活性を阻害できる一方で、少なくとも1個のアミノ酸を置換することによって生じる骨形成タンパク質に対応する受容体、より詳しくは、BMP−2様タンパク質受容体、とは相互作用しないことを見出した。本発明の変異タンパク質はこれらの特徴を有するため、それらはいずれかの生物に適用することができ、この場合、前記モジュレータータンパク質が調節され、好ましくは、阻害され、ひいては、前記モジュレータータンパク質によって発揮される阻害が消滅するか減少することになり、その一方で、前記受容体鎖によって媒介される、骨形成タンパク質に関連しているその他の生物活性はもはや存在しない。骨形成タンパク質の変異タンパク質のこういった設計は、この種の骨形成タンパク質の、望ましくない受容体媒介性作用を引き起こすことなく、モジュレータータンパク質が関連しているか媒介する疾患の治療又は予防に使用できる限りにおいて、明らかに有利である。その程度において、本発明の変異タンパク質は“受容体非感受性(receptordead)”である。
【0049】
また、本発明者らは驚くべきことに、骨形成変異タンパク質のこの特有の特徴は、野生型の骨形成タンパク質中のプレヘリックスループ構造を変更することによって作出できることを見出した。本明細書においては、そうでないことが示されない場合は、骨形成タンパク質は、BMP−2様タンパク質であることが好ましい。プレヘリックスループ構造は、好ましくは骨形成変異タンパク質の構造であり、これは、二次構造表示でヘリックスの前のループであることが好ましい。ループは4〜10個、好ましくは6〜8個、より好ましくは、6個のアミノ酸残基を含むことが好ましい。ヒトBMP−2タンパク質のアミノ酸配列を例にとると、プレヘリックスループ構造はアミノ酸位置48〜アミノ酸位置53に広がっている。本明細書では、好ましい実施形態では、プレヘリックスループは、いずれかのループであって、より好ましくはヒトBMP−2のアミノ酸位置48〜53に相当するいずれかのアミノ酸配列であり、より好ましくは配列番号1のアミノ酸配列である。プレヘリックスループ構造は、好ましくは、本明細書においてI型骨形成タンパク質受容体と呼ばれる骨形成タンパク質受容体との相互作用を媒介していることが好ましい。I型骨形成タンパク質受容体はBRIAであることがいっそうより好ましい。BRIAECは、BMPR−IAの細胞外ドメインであり、以下に記載されている(Kirsch, T.、Sebald, W.、Dreyer, M.K.(2000) Nat Struct Biol、7、492〜496参照)。BRIBECは、BMPR−IBの細胞外ドメインである。ARIECは、ActR−Iの細胞外ドメインである。
【0050】
本発明の変異タンパク質は、骨形成タンパク質の野生型と比較して、構造、好ましくは二次構造及び三次構造の変化のために、I型骨形成タンパク質受容体ともはや相互作用できない。本明細書において用いられるのに好ましい変化とは、対応する骨形成タンパク質の野生型と前記I型形成タンパク質受容体との相互作用を参照することによって検出可能なものである。より詳しくは、I型骨形成タンパク質受容体がBRIAECである場合には、本発明の変異タンパク質によって最も影響を受けるアミノ酸はGln86であるようである。
【0051】
また、本発明の骨形成変異タンパク質はプレヘリックスループ構造を含み、これがBRIAECなどのI型骨形成タンパク質受容体との相互作用を媒介しており、この相互作用は骨形成タンパク質の野生型のプレヘリックスループ構造とI型骨形成タンパク質受容体との相互作用とは異なっている。これらの誘導された変化のいずれか又は変異タンパク質とそれぞれの第2の骨形成タンパク質受容体との間の異なる相互作用パターンは、骨形成タンパク質の野生型の対応する構造又は相互作用と比較して検出される。前記I型骨形成タンパク質受容体と変異タンパク質のいずれか又は双方のこのような変化は、屈折データを得ることによってモニターできることが好ましい。より好ましい実施形態では、このような屈折データは少なくとも約2.7Åの分解能まで室温で得られる。
【0052】
好ましい実施形態では、骨形成タンパク質は、BMP−2である。BMP−2は当技術分野では公知であり、本明細書に記載される配列番号1のアミノ酸配列を含むことが好ましく、配列番号2の核酸配列を含む核酸によってコードされることが好ましい。
【0053】
より詳しくは、本発明者らはまた、本発明の変異タンパク質の特有の特徴は、プレヘリックスループ構造でのアミノ酸置換、より詳しくは、骨形成タンパク質のアミノ酸位置、好ましくはヒトBMP−2のアミノ酸位置51に相当するBMP−2様タンパク質のアミノ酸位置、さらに好ましくは、配列番号1のBMP−2のアミノ酸位置でのアミノ酸置換、すなわち変異、によって付与されるということも発見した。より詳細には、変異タンパク質中で変異されているそれぞれのアミノ酸は、対応する骨形成タンパク質の野生型においてはロイシンである。より好ましくは、その交換はロイシン残基からプロリン残基である。骨形成タンパク質、特に、BMP−2様タンパク質の高い相同性、ひいては類似の二次及び三次構造のために、特定の位置が同様に活性であり、この群の骨形成タンパク質の種々のメンバー内でそれぞれ関連があり、その結果、変化、すなわち、アミノ酸置換は、前記骨形成タンパク質のいずれにおいても有効であり、したがって、それぞれの変異タンパク質は前記で概説した特徴を示す。骨形成タンパク質はhBMP−2、hBMP−4、hBMP−5、hBMP−6、hBMP−7、hBMP−8、hGDF−5、mGDF−6、mGDF−7、hBMP−10及びhGDF−2であることがより好ましく、これらは当技術分野ではそれ自体公知であり、これらのすべては同じようにBRIAECと結合する。しかし、対応する変異タンパク質、すなわち、ヒトBMP−2のアミノ酸位置51に相当するそれらの位置にアミノ酸置換を有する変異タンパク質、にそれらを変えるそれぞれの変異によって、それらを先行技術から区別している。前記の骨形成タンパク質の野生型配列は本明細書では配列番号1(hBMP−2)、配列番号3(hBMP−4)、配列番号5(hBMP−5)、配列番号7(hBMP−6)、配列番号9(hBMP−7)、配列番号11(hBMP−8)、配列番号13(hGDF−5)、配列番号15(mGDF−6)、配列番号17(mGDF−7)、配列番号19(hBMP−10)及び配列番号21(hGDF−2)として開示されており、本発明の前記骨形成タンパク質の変異タンパク質の対応するアミノ酸配列は、野生型と比較して前記の単一のアミノ酸位置で異なっており、前記アミノ酸配列は配列番号23(hBMP−2)、配列番号24(hBMP−4)、配列番号25(hBMP−5)、配列番号26(hBMP−6)、配列番号27(hBMP−7)、配列番号28(hBMP−8)、配列番号29(hGDF−5)、配列番号30(mGDF−6)、配列番号31(mGDF−7)、配列番号32(hBMP−10)及び配列番号33(hGDF−2)のものである。より詳しくは、前記のヒトBMP−2のアミノ酸位置51は、hBMP−4の場合には位置53、hBMP−5の場合には位置68、hBMP−6の場合には位置68、hBMP−7の場合には位置75、hBMP−8の場合には位置75、hGDF−5の場合には位置56、mGDF−6の場合には位置56、mGDF−7の場合には位置82、hBMP−10の場合には位置44及びhGDF−2の場合には位置45に相当する。
【0054】
本明細書において、本明細書に開示される、骨形成タンパク質及びひいては、単一のアミノ酸置換でのみ異なっていることが好ましい対応する変異タンパク質は、プレプロ型で存在する場合も、プロ型で存在する場合も、成熟タンパク質として存在する場合もある。この基本設計は、すべての骨形成タンパク質にもともと備わっているものである。例えば、BMP−2の場合には、プレプロ型は全部で396個のアミノ酸を含む。プレ配列はシグナルペプチドとして働き、新生ポリペプチド鎖の小胞体への輸送に関与している。輸送された後、タンパク質はその未変性コンホメーションに折り畳まれ、その際ジスルフィド架橋が形成される。BMP−2のプロ型は、位置20〜位置396のアミノ酸すべてを含む。したがって、プロ配列はBMP−2のヒトプレプロ型の位置20(Gly)〜アミノ酸位置282(Arg)までのアミノ酸を含む。最後に、成熟BMP−2はプレプロ型のアミノ酸283〜396を含む。成熟型は本明細書では配列番号1として表されている。
【0055】
本発明の骨形成タンパク質及びその対応する変異タンパク質は、プレプロ型、プロ型、又は成熟型で存在し、いずれかのシグナルペプチドが、プレ配列として、好ましくは、骨形成タンパク質のプレ配列として作用し、いずれの骨形成タンパク質のいずれのプロ配列であれば、特定の骨形成タンパク質のプロ配列として使用できるということは本発明の範囲内であるが、成熟骨形成変異タンパク質のN末端に結合しているプロ配列が、in vivoでそれぞれの骨形成タンパク質のN末端に結合するプロ配列に相当することがより好ましい。同じことがプレ配列にも当てはまる。
【0056】
本明細書において、用語「骨形成タンパク質」及び「骨形成変異タンパク質」とはそれぞれ、その相同分子種も含むものとして好ましく使用される。本明細書において、相同分子種とは、参照タンパク質(reference protein)として、同様の遺伝的機能及び生理学的機能を果たす別の生物由来のタンパク質である。また、より好ましい実施形態では、用語「骨形成タンパク質」及び「骨形成変異タンパク質」は、いずれかの末端切断型タンパク質及び変異タンパク質をそれぞれ含む。このような末端切断型タンパク質及び変異タンパク質は、少なくとも1個のアミノ酸残基を欠いており、N末端又はC末端のいずれかで少なくとも1個のアミノ酸残基を欠いていることが好ましい。したがって、本発明の骨形成変異タンパク質は、前記で概説された特徴、すなわち、「受容体非感受性」であり、かつ、少なくとも1種のモジュレータータンパク質、好ましくは、ノギンタンパク質ファミリー、DANタンパク質ファミリー及び/又はコーディンタンパク質ファミリーのメンバーと相互作用するのに依然として適している限りは、末端切断型変異タンパク質である。
【0057】
変異タンパク質はさらに修飾され、このような修飾はリン酸化、ペグ化及びグリコシル化を含んでなる群から選択されることが好ましいということもまた本発明の範囲内である。
【0058】
本発明の核酸は、いずれもがDNA、RNA又はそれらの誘導体、例えば、LNA若しくはPNA、として存在し得ることを理解されなくてはならない。それぞれの核酸は、一本鎖又は部分的な二本鎖若しくは完全な二本鎖のいずれかとして存在することも本発明の範囲内である。最後に、用語「核酸配列」は、例えば、配列識別子に従う配列によって、単独で又は本質的な相補的鎖であるもう一方の鎖と組み合わせて特定されるような核酸の補完物を含むことも本発明の範囲内である。
【0059】
本明細書において、用語「核酸」とは、また、本明細書に記載される核酸のいずれの断片も含み、このような断片のいずれもが、約19〜30の長さの連続するヌクレオチドを含むことが好ましく、19〜25の長さがより好ましく、21〜22の長さが最も好ましい。核酸のこの種の断片は、二本鎖構造として存在することがいっそうより好ましい。
【0060】
本発明によれば、核酸はまた、本明細書に開示される核酸の何らかのその他の形とハイブリダイズする核酸であってもよく、このようなハイブリダイゼーションは、ストリンジェントな条件で起きることが好ましい。このようなストリンジェントな条件は、中でも、Sambrook, J.、Fritsch, E.F.及びManiatis, T. (1989) Molecular Cloning. A.Laboratory Manual、第2版、9.47〜9.55頁、Cold Spring Harbour Laboratory Pressに記載されている。
【0061】
本明細書に記載される、骨形成タンパク質の野生型配列又はその変異タンパク質の配列のいずれかを発端として、骨形成タンパク質の野生型とその変異タンパク質とを区別できる核酸を設計することが可能であることは、当業者に認められよう。このような区別はハイブリダイゼーション条件のストリンジェンシーを高めることによって可能となることが好ましい。
【0062】
本明細書に開示される特定の配列だけでなく、種々の配列、より好ましくは、核酸配列も、それらが本発明の変異タンパク質をコードするのに適している限りは本発明に含まれるということも当業者に認められよう。このような配列は遺伝暗号を用いて導くことができ、事実上、遺伝暗号の冗長性によって生じるこのような配列のいずれの順列も本発明の範囲内にある。ヒトBMP−2のアミノ酸位置51にそれぞれ相当する、骨形成タンパク質及び変異タンパク質の位置が、ロイシンではなくプロリンであることが本発明の変異タンパク質の特徴となっており、この特定のアミノ酸置換については、それぞれのコドンがCCU、CCC、CCA又はCCGであると認められよう。
【0063】
本発明のベクターは、本明細書に開示されるいずれかの断片を含み、本明細書に開示されるいずれかの核酸を含む。ベクターは発現ベクターであることが好ましい。このような発現ベクターは、通常、プロモーター又はプロモーター様構造と、場合によっては、このようなプロモーター及びその他の制御エレメントによって制御されるような、ベクターにクローニングされている核酸の発現を制御するのに適しているさらなるエレメントを含む。発現ベクターは原核生物の発現ベクターか、真核生物の発現ベクターかそれぞれのシャトルベクターであることが好ましい。この種の発現ベクターの個々の設計は当業者には公知である。
【0064】
本発明の細胞は、原核細胞か真核細胞のいずれかである。原核細胞としては、原則として、大腸菌(E.coli)、枯草菌(B.subtilis)又はS.カルノサス(S.carnosus)が本発明の骨形成変異タンパク質をコードする核酸、又はその断片若しくは誘導体の発現に用いられる。
【0065】
本明細書において、真核細胞は、酵母細胞又は哺乳類細胞であることが好ましい。哺乳類細胞はマウス、ラット、モルモット、ブタ、サル又はヒト由来の細胞であることがより好ましい。
【0066】
本明細書において、宿主生物は本発明のいずれかの細胞又は多細胞生物であることが好ましい。このような多細胞生物は好ましくは、哺乳類であることが好ましく、非ヒト哺乳類であることがより好ましい。好ましい非ヒト哺乳類はマウス、ラット、モルモット、ヒツジ、ウサギ及びブタを含んでなる群から選択される。
【0067】
本発明の骨形成変異タンパク質を製造するための本発明の方法では、本発明の細胞は培養されていることが好ましい。このような培養は培養ブロスで行われることが好ましい。培養ブロスは発酵ブロス又は、培養に適し、かつ、より好ましくは培養される細胞の増殖に適しているいずれかのその他の溶液、であることが好ましい。培養ブロスは当業者には公知である。培養及び/又は細胞の増殖において、本発明の変異タンパク質は、培養ブロスから調製されることが好ましい。培養ブロスからの調製は、本明細書に記載されているように、本発明の変異タンパク質を細胞内に含ませるか又は細胞に結合させてその細胞からの調製すること、又は培養ブロスからの本発明の変異タンパク質を単離若しくは調製すること、を含む。本発明の変異タンパク質の調製又は単離方法は当業者には公知である。
【0068】
さらなる態様では、本発明は、本発明の変異タンパク質に対するモノクローナル抗体と関連している。抗体はモノクローナル抗体であることが好ましい。抗体、好ましくは、本発明の骨形成変異タンパク質と特異的に結合するモノクローナル抗体、を調製することが可能であることは、当業者に認められよう。
【0069】
用語「特異的な結合」とは、抗体が、変異タンパク質に対してと同程度に、又は以下に示すように、変異タンパク質に対して同じ結合特性で、骨形成タンパク質の野生型とは結合しておらず、対応する骨形成タンパク質の野生型と比較した変異タンパク質の結合定数が、少なくとも2倍異なっている、好ましくは少なくとも5倍、最も好ましくは少なくとも10倍異なることを意味している。
【0070】
さらなる態様では、本発明は、本発明の変異タンパク質又は本発明の核酸のいずれかを含む組成物に関連している。このような組成物は、タンパク質及び/又は核酸だけでなく、担体を含み得る。このような担体は固相、水相及び脂質相を含んでなる群から選択されることが好ましい。本明細書において、脂質相は核酸及び変異タンパク質とそれぞれ複合体を形成しているか、核酸及び変異タンパク質をそれぞれ包含している脂質又は脂質組成物であることが好ましい。脂質組成物は、その程度において当業者に公知のリポソームであることが好ましい。水相は水及び水性バッファー溶液及びヒアルロン酸を含んでなる群から選択されることが好ましい。固相は、リン酸三カルシウム、ポリラクチド、コラーゲン及び不溶性コラーゲン骨マトリックスであることが好ましい。
【0071】
さらなる態様では、本発明は薬剤組成物に関連している。このような薬剤組成物は、本発明の変異タンパク質又は本発明の核酸あるいは双方と、さらに製薬上許容される担体とを含む。製薬上許容される担体は液体又は固体担体のいずれかであることが好ましい。適した液体担体としては、中でも、水、水溶液、より好ましくは、バッファー又は脂質若しくは脂質組成物がある。好ましい固体担体は、中でも、糖、セルロース及びデンプンを含む。当業者、好ましくは、製剤の技術分野の当業者であれば、さらなる製薬上許容される担体及び担体物質をそれぞれ承知しているものと認められる。好ましい実施形態では、薬剤組成物はさらなる製薬上活性な薬剤を含む。
【0072】
さらなる態様では、本発明は、医薬の製造のための本発明の骨形成変異タンパク質及び/又は本発明の核酸の使用に関する。
【0073】
本発明者らの驚くべき発見に基づいて、すなわち、骨形成タンパク質、より詳しくは、本発明のBMP−2様タンパク質の変異タンパク質は、骨形成タンパク質のような受容体媒介性作用を示さず、すなわち「受容体非感受性」であるが、依然として、それぞれの骨形成タンパク質の野生型が、相互作用するモジュレータータンパク質と相互作用することは当業者に認められる。その程度において、本発明の変異タンパク質及びそれをコードする核酸は、骨形成タンパク質の公知の望ましくない作用が生じるという危険に出くわすことなくそれらを必要とする患者に適用でき、このことは、その阻害活性を阻害するモジュレータータンパク質と相互作用する利益よりも上回る。例えば、BMP−2の望ましくない活性の中には、腫瘍成長を促進することがある。このために、先行技術における、いずれかの変異タンパク質をも含む骨形成タンパク質の医学的使用は、骨再生などの局所投与に制限されてきた。しかし、骨形成タンパク質の活性であるそのような作用、詳しくは、その受容体との相互作用によって媒介されるもの、を欠いている本発明の変異タンパク質は、このクラスの製薬上活性を有する化合物の医学的な適用を全く初めて可能にするものである。
【0074】
本発明のさらなる態様は、本発明の医薬を使用できる、本明細書に記載された疾患のいずれかを患っているか、又はそれが進行する状態にある患者の治療方法に関している。このような方法は、変異タンパク質又はそれをコードする核酸の患者への投与を含む。患者は哺乳類であることが好ましく、ヒトであることがより好ましい。
【0075】
本発明の変異タンパク質の作用様式のために、それらは種々の疾患、すなわち、本発明の骨形成タンパク質とこれに対応する変異タンパク質との間の影響を受けない相互作用、及びいずれかのモジュレータータンパク質、を提供することによって治療又は予防できる疾患、に容易に用いることができる。モジュレータータンパク質は当技術分野では公知であり、シスチンノット含有骨形成タンパク質、より詳しくは、シスチンノット含有BMPモジュレータータンパク質、ノギンファミリー、コーディンファミリー及びDANファミリーを含んでなる群から選択されることが好ましい。シスチンノット含有BMPモジュレータータンパク質は、例えば、Avsian-Kretchmer,O.及びHsueh, A. J. (2004)Comparative genomic analysis of theeight-membered ring cystine knot-containingbone morphogenetic proteinantagonists. Mol Endocrinol、18巻、1〜12頁に記載されている。例えば、ノギンを含むノギンファミリーは、Canalis, E.、Economides, A. N.及びGazzerro,E.によって(2003) Bonemorphogeneticproteins, their antagonists, and the skeleton. Endocr Rev、24巻、218〜235頁に記載されており、中でもコーディン、ベントロプチン、CTGF、CYR61、クロスベインレス、プロコラーゲンIIAを含むコーディンファミリーは、例えば、Bork, P. (1993) The modular architecture of anew family of growthregulators related to connective tissue growth factor.FEBS Lett、327巻、125〜130頁、Brigstock, D. R.、Goldschmeding,R.、Katsube, K. I.、Lam, S. C.、Lau, L. F.、Lyons, K.、Naus, C.、Perbal, B.、Riser,B.、Takigawa, M.及びYeger, H.(2003)Proposal for a unified CCN nomenclature. Mol Pathol、56巻、127〜128頁及びGarcia Abreu, J.、Coffinier, C.、Larrain, J.、Oelgeschlager, M.及びDe Robertis, E.M.(2002) Chordin-like CR domains and theregulation ofevolutionarily conserved extracellular signaling systems. Gene、287巻、39〜47頁に記載されており、例えば、Cer1、セルベルス、Dan、Dte、グレムリン、PRDC、スクレロスチン(Sclerostin)を含むDANファミリーは、例えば、Pearce, J.J.、Penny, G.及びRossant, J. (1999) A mouse cerberus/Dan-related genefamily. Dev Biol、209巻、98〜110頁及びBalemans, W.及びVan Hul, W.(2002)Extracellular regulation of BMP singaling in vertebrates: a cocktailofmodulators. Dev Biol、250巻、231〜250頁に記載されている。これらのモジュレータータンパク質は、偏っている場合、特にそれ自身が阻害されない場合に、in vivoプロセスに対する阻害作用を有し、その結果、何らかの疾患又は病的状態をもたらす傾向がある細胞における経路及び事象に対して阻害又は刺激作用を有することがわかっている。したがって、いずれの理論にも拘束されようとは思わないが、本発明の変異タンパク質は、依然として前記モジュレータータンパク質と相互作用し、これによりモジュレータータンパク質によって刺激され、さもなければ阻害されるプロセスを許容するそれらの修飾作用をも阻害でき、モジュレータータンパク質単独の存在下で観察される程度と比較して、あらゆるレベルで、又は高いレベルで、又は低いレベルで実施され得る。
【0076】
本発明の、変異タンパク質及びそれをコードする核酸によって治療できる特に好ましい疾患としては、線維性疾患がある。本発明に従って治療できる特に好ましい疾患としては、腎線維症、肝硬変、肺線維症及び慢性炎症があり、喘息を伴う慢性炎症がより好ましく、したがって、喘息の治療も本発明に含まれる。
【0077】
再度、いずれの理論にも拘束されようとは思わないが、コーディン−ファミリーに属するCTGFは、BMP−2と、ひいては本発明の変異タンパク質と相互作用する。CTGFはTGF−βのプロ線維活性を媒介している。より詳しくは、TGF−βは、細胞外マトリックスのタンパク質の産生を刺激しており、この種のタンパク質の分解を阻害する。すなわち、このプロセスは組織の治癒に不可欠なものである。しかし、炎症性疾患、より詳しくは、慢性炎症などの慢性疾患の場合には、TGF−β活性が豊富であり、ひいては組織線維症、特に、過剰組織線維症をもたらす(Branton, M. H.及びKopp, J. B.(1999)TGF-betaand fibrosis. Microbes Infect、1巻、1349〜1365頁)。したがって、これら種々の線維性疾患は本発明の変異タンパク質の投与によって阻害できる。
【0078】
本発明に従って治療できるさらなる種類の疾患には、創傷治癒がある。この医学的状態、すなわち、創傷治癒の形はケロイド、瘢痕形成及び腹膜の閉塞であることがより好ましい。本発明に従って治療できる特定の群の患者は、好ましくは、視力に影響を及ぼす極度の瘢痕形成が同時に起きる緑内障手術を受けるか受けたことがある患者である。再度、いずれの論理的根拠にも拘束されようとは思わないが、過増殖性瘢痕形成はCTGF及びCYR61によって媒介されるようであり(Branton, M. H.及びKopp, J. B.(1999)TGF-betaand fibrosis.Microbes Infect、1巻、1349〜1365頁)、その結果、これらのモジュレータータンパク質の本発明の変異タンパク質による阻害は、瘢痕形成を避けるための、ひいては、個々の状態及び疾患をそれぞれ治療するための適した手段である。
【0079】
本発明によって治療されるもう1つの疾患又は状態は、過剰血管新生である。より詳しくは、用語「過剰血管新生」とは、過剰血管新生によって引き起こされるか、過剰血管新生と関連しているか、又は過剰血管新生を伴う、いずれの疾患も含む。この種の疾患は腫瘍、網膜症及び動脈硬化症であることが好ましい。その程度において、本発明はまた、腫瘍、網膜症の治療のための本発明の変異タンパク質の使用に関する。腫瘍は、固形癌であることが好ましい。また、血管疾患、例えば、好ましくは、動脈硬化症は、本発明によって治療できる。再度、いずれの理論にも拘束されようとは思わないが、固形癌は、1〜3mm3という体積を超えて成長する場合には十分な血管新生が必要であるようである。十分な血液供給は、腫瘍の増殖及び進行の際の律速ステップである。本発明者らの現時点での理解によれば、CYR61は腫瘍の脈管形成に関与している。より詳しくは、CYR61及びCTGFはそれぞれ、脈管形成性進行及び線維形成性進行に関与しており、それらは進行性血管閉塞に関与していると理解される(Brigstock, D. R. (2002) Regulation of angiogenesis and endothelialcell function by connective tissue growth factor(CTGF) and cysteine-rich 61(CYR61)、Angiogenesis、5巻、153〜165頁)。したがって、前記標的分子、すなわち、CYR61及びCTGFと相互作用することによって、本発明の変異タンパク質はそれぞれの疾患の予防及び治療に適している。
【0080】
それぞれ、本発明に従って治療できる、さらなる疾患及び状態としては、骨折、好ましくは、難治性骨折がある。再度、いずれの理論にも拘束されようとは思わないが、ノギン及びCTGFは、骨折の治癒に関与しているBMP調節性タンパク質であるようである。より詳しくは、ノギン及びCTGFは、骨形成を阻害し、終結させる(Yoshimura, Y.ら、(2001)、Colocalization of noggin and bone morphogenetic protein-4duringfracture healing、J Bone Miner Res、16巻、876〜884頁)。したがって、例えば、前記標的の相互作用パートナーとの競合阻害によって、本発明の変異タンパク質はこの状態の治療に影響を及ぼす。
【0081】
骨粗しょう症は、本発明に従って治療できるもう1つの疾患である。再度、いずれの理論にも拘束されようとは思わないが、この疾患に関連して、標的はスクレロスチンであるように思われる。スクレロスチンは、生物における骨形成プロセスを阻害する。スクレロスチン遺伝子(SOST)の不活性化を受けている人は、骨密度の持続的増加を示し、これは標準値の3倍にも達する場合がある。その程度において、スクレロスチンを阻害する化合物はいずれも、骨密度の低下に付随している状態、例えば、骨粗しょう症などの治療のための手段として適している(Balemans, W.及び Van Hul, W.(2002)、Extracellular regulation of BMP signaling invertebrates: a cocktailof modulators, Dev Biol、250巻、231〜250頁)。その程度において、本発明の変異タンパク質がスクレロスチンと相互作用し、ひいては、この化合物を阻害するという事実を踏まえると、それらはこの種の疾患及び状態、より好ましくは、骨粗しょう症の治療にそれぞれ適している。
【0082】
本発明を図及び実施例によってさらに説明するが、そこからはさらなる特徴、実施形態及び利点が得られる。
【0083】
実施例1:材料及び方法
(タンパク質の調製)
BMP−2変異タンパク質及びBMP受容体エクトドメインタンパク質の調製を、先に記載された通りに実施した(Kirschら、2000b;Ruppertら、1996)。BMP−2又はBRIAECの変異体は、組換えPCRによって得た(Kirschら、2000a)。結晶化に用いたBMP−2野生型及び変異タンパク質タンパク質は、EMD SO3−カラム(Merck)を用いる陽イオン交換クロマトグラフィーによって精製した。画分をSDS−PAGEによって分析し、プールした画分を水に対して透析し、凍結乾燥した。ESI FT−CR質量分析及び分析用逆相HPLCによってタンパク質の均質性を調べた。リフォールディングしたBMP−2変異タンパク質の結合活性を、BIAコア方法を用いるBMPI型及びII型受容体の結合親和性の測定によって評価した。構造解析のためのBMP−2BRIAEC複合体の調製は公開されている通りに実施した(Kirschら、2000b)。
【0084】
(タンパク質の結晶化)
結晶化のために、BMP−2 D53P及びBMP−2 L51Pを、それぞれ5mg/ml及び8mg/mlの濃度で水に溶解した。BMP−2 L51Pは、室温で、15%t−ブタノール、0.1M硫酸リチウム及び0.1Mクエン酸ナトリウムpH5.0から水滴蒸気拡散法によって結晶化した。BMP−2−BRIAECの複合体の結晶化は、基本的には公開された様に実施した(Kirschら、2000b)が、100Kで測定するための結晶は、グルコースを抗凍結剤として用いる共結晶化によって得た。データ収集のための高品質結晶は、11mg/mlのタンパク質濃度において、10℃で、0.75M酢酸ナトリウム、0.1MイミダゾールpH7.0及び30%グルコースから成長させた。
【0085】
(X線データ収集)
BMP−2−BRIAECという複合体のデータは、Swiss Light Source(Paul Scherrer Institute、スイス)のビームラインX06SA PXを用いて、100Kでの単結晶から得た。結晶を90°回転させて(フレームあたり1°)2つのデータセットを測定し、1°に対して10秒曝露した高分解能セットからは1.9Åという最高分解能を得て、1°あたり1秒の曝露時間で記録することによって低分解能データを得た。データはソフトウェアMOSFLMバージョン6.2.1を用いて加工し、完全なものにし(Leslie、1992)、2つのデータセットをマージし(オーバーラップ:低分解能については20〜5.0Å、高分解能については20〜1.9Å)、ソフトウェアSCALA CCP4バージョン4.2.1(Collaborative Computational Project、1994;Evans、1993)を用いてスケーリングし、加工統計データの要約を表A付録に示す。BMP−2変異タンパク質L51Pの回析データは、Rigaku RU−300(50 kV、100mA、0.3mmフィラメント)、Osmic ConfocalBlue optics及びRigaku R−AXIS IV++イメージプレートシステムからなるhome−sourceで室温で得た。曝露時間は、0.5°回転あたり5分に設定し、結晶は2.5Åまで回析した。放射線損傷を最小化ために獲得戦略をソフトウェアCrystalClear(Rigaku)を用いて決定したが、相応な完全性を実現するために、2種の結晶からデータを集め、ソフトウェアMOSFLM及びSCALAを用いてマージし、スケーリングした(Evans、1993;Leslie、1992)。
【0086】
(構造決定)
BMP−2−BRIAECという複合体の構造を、低分解能構造(PDBエントリー1ES7)を出発モデルとして用いてリファインメントした。BMP−2−BRIAEC複合体を、空間群P65で結晶化し、これは非対称単位にBMP−2二量体と2つのBRIAEC分子を保持していた。容器定数のわずかな変化のために、回転及び翻訳検索とそれに続く剛体リファインメントからなる分子置換ステップを、ソフトウェアCNS1.1を用いて実施した(Brungerら、1998)。リファインメントのためにプログラムREFMAC5(Murshudovら、1997)を続いて用い、それに続いてソフトウェアQUANTA2000(Accelrys)を用いて座標を手動で再建した。分解能を20〜1.9Åという最終分解能範囲まで徐々に高めた。データの異方性を明らかにするために、各鎖、すなわち、BMP−2単量体及び各BRIAEC分子に対して1つのTLSグループを定義した(Winnら、2001)。リファインメントの進行は、5%の反射を含む試験データセットを用いる相互検証によってモニターした(Brunger、1992)。複合体構造は、基本的には対称であるので、本発明者らは非結晶学的対称拘束を用いることを試みたが、続くリファインメントステップでは、RfreeをNCS拘束を破らずに低下させることができなかった。したがって、NCS拘束はリファインメントを通じて用いなかった。最終ラウンドのリファインメントでは、fobs−fcalc相違電子密度マップを用いて185の水分子を同定した。最終最小化サイクルによって、R−因子は20.7、Rfreeに対しては22.9となった。BMP−2 L51Pの構造分析はBMP−2−BRIAEC複合体のプロトコールに従ったが、焼き鈍し法及びエネルギー最小化のためにCNS 1.1を用いた。BMP−2野生型(PDBエントリー3BMP)の構造を出発モデルとして用いた。モデルの手動での再建は、σA加重した2fobs−fcalc及びfobs−fcalc電子密度マップを基にし、最終ラウンドのリファインメントでは、13の水及び2つのMPD分子を加えた。BMP−2 L51Pの最終構造のR−因子は、21.5であり、Rfreeに対しては23.5である。
【0087】
(相互作用分析)
タンパク質をビオチン化し、ストレプトアビジンコートしたセンサーチップCM5に固定化した(Kirschら、2000a)。BRIA、BRIB及びARIIの細胞外ドメインを記載された通りに調製した(Kirschら、2000a)。ノギン/Fc、コーディン及びグレムリンをR&Dシステム(Wiesbaden−Nordenstadt)から得た。分析物との相互作用をBIAコア2000システムで測定した(Kirschら、2000a)。センサーグラム(Biaevaluation software 2.0)の評価から複合体形成(kon)及び解離(koff)の速度定数が得られた。6〜12回の測定から平均値を得、平均標準偏差はkoffについては35%及びkonについては12%であった。見掛けの解離定数KDはKD=koff/konとして算出するか、平衡結合の線量依存性を評価することによるかのいずれかとした。特別の条件を適用して、BMP−2結合後に遊離ノギンを再生しなくてはならなかった。残存する複合体は、1M塩化ナトリウム及び6M尿素の存在下で酸性pH3でのみ解離することができた。幸にも、タンパク質は、センサーに固定されたタンパク質Aにノギン−Fc融合タンパク質を固定するのに使用する独立した単一サイクルの実験によって確立される厳しい処理の後にも天然の結合特性を保持していた。
【0088】
(細胞株における生物活性)
アルカリホスファターゼ(ALP)活性は、血清を欠乏させたATDC5(Riken Bioresource Center、細胞番号RCB0565)及びC2C12(ATCC番号CRL−1772)細胞で測定した(Kirschら、2000a)。
【0089】
実施例2:BMP−2−BRIAEC複合体における水素結合パターン
BMP−2−BRIAEC複合体の高分解能構造は、非対称単位に2つのBRIAエクトドメインと結合している1つの完全なBMP−2二量体を含んでいる(図1A)。厳しい非結晶学的対称性を適用することに失敗したということは、リガンド−受容体複合体の対称性は完全ではないということを示すものである。しかし、2つのセグメント(BMP−2二量体と1つのBRIAEC分子)間の相違は小さく、すべての原子についてr.m.s.偏差は0.9Åであり、すべてのCα−原子については0.4Åである。詳細な分析により、2つのBMP−2/BRIAEC接触面の構造は同一であることが示される。非結晶学的対称性に関連する分子間のすべての相違は、結晶格子における充填環境の相違によるものである。これらのセグメントにおいて高温因子は、すべての相違は骨格柔軟性によるものであり得るということを示唆する。したがって、対称性に関連しているタンパク質セグメントにおける構造変動は、個々の分子間の実際の相違には反映されないだけでなく、あり得る高次構造並びにBMP−2及びBRIAポリペプチド鎖間の動力学を反映しない。
【0090】
BMP−2/BRIA接触面は成長因子/受容体接触面の新規パラダイムを表す。BMP−2の結合エピトープは、高度に疎水性であり、2310Å2という全部の埋もれた表面のほぼ60%が疎水性残基で作られている(図1b)。ヒト成長ホルモン(de Vosら、1992)、インターロイキン−4(Hageら、1999)、エリスロポエチン(Syedら、1998)又はインターロイキン−6(Boulangerら、2003)のようなその他の成長因子の結合エピトープと比較すると、このレベルの疎水性はかなり高い。アクチビン受容体IIエクトドメインに結合しているBMP−7の複合体構造(Greenwaldら、2003)、TGF−β受容体IIとの複合体中のTGF−β3(Hartら、2002)及びアクチビン受容体IIBと結合しているアクチビンA(Thompsonら、2003)に示されるように、極端に疎水性接触面は、TGF−βスーパーファミリーのメンバーに特徴的であるようである。対照的に、BRIAECの結合エピトープは、結合接触面に広がる極性及び電荷を有する残基の広がりのためにあまり疎水性ではない(30%)(図1B)。
【0091】
BMP−2−BRIAEC複合体のリファインメントされた構造の1.9Åというより高い分解能によって、低分解能でこれまでの構造に可能であったようなより詳細な分析が可能となる。各BMP−2/受容体接触面において、プログラムHBPLUS(McDonald及びThornton、1994)によって10の分子間水素結合(H−結合)が同定される(表1)。BMP−2は5個の、その受容体は4個の主鎖原子を水素結合に提供している。H−結合に携わっている原子を、表1に表されるように2つの構造エレメントにまとめる:リガンドBMP−2では、残基Phe49〜Asn59からなるプレヘリックスループ、受容体BRIAでは、短いα−ヘリックス及びフランキング配列(Asp84〜Arg97)。2つの水素結合、BMP−2 Asp53(N)−BRIAECCys77(O)及びBMP−2 Ser69(O)−BRIAEC Gln94(N)が、主鎖原子の間でのみ形成され、2つの水素結合は、側鎖間でのみ生じ、4つの混合水素結合は側鎖と主鎖原子とを含む(表1)。10の水素結合のうち8は、正真正銘の水素結合に対して予測されるような(Xuら、1997)ドナー−アクセプター距離(2.8±0.14Å)、φ結合角(137±11°)を有する。しかし、結合パラメータの幾何学的評価に純粋に基づいた個々の水素結合の強さの分類はこの複合体には可能ではないようである。
【0092】
【表1】
【0093】
カッコ内の数字は、ドナー−アクセプター原子の距離及び非結晶学的対称性で関連している二次接触面の角度を表す。
a)斜線をつけた行中のH−結合は、前記の幾何統計データから排除した。
b)N−O−Cはドナー−アクセプター原子であり;統計データ(Xuら、1997)から、この角度は、MC−MC水素結合については149°±15°並びにSC−MC及びSC−SC H−結合については129°±18°である。
c)MC(主鎖)及びSC(側鎖)ドナー/アクセプター原子
d)BMP−2(イタリック体)及びBRIA(太字)における分析したH結合を乱す置換。
【0094】
接触面構造から機能的に重要なH−結合を予測できるかどうかを調査するために、残基部分の接近可能性を算出した(図1C)。複合体におけるこの残基の接近可能性は、コアに向かった、又は接触面の表面に向かった残基の位置と関連がある(Chakrabarti及びJanin、2002)。10個の界面のH−結合のうち8個は、表面に位置する残基の間に生じる。受容体Gln86とBMP−2主鎖のLeu51アミド及びカルボニルをつなぐ2個のH−結合だけが、複合体形成の際に完全に埋もれる(図1C)。したがって、この相互作用は、結合のホットスポットを表す可能性があるが(Bogan及びThorn、1998)、その他のH−結合及び疎水性相互作用は、結合親和性には寄与していないか、ほんのわずかしか寄与していない可能性がある。
【0095】
接触面における水分子の調査によって、結合エピトープの脱溶媒和がその他のタンパク質−タンパク質接触面と比較してかなり効果的であるということが明らかになっている。複合体形成の際、各リガンド−受容体接触面には4個の水分子しか完全に埋もれない。これらの埋もれた水分子の低温因子(すなわち、これらの埋もれた水分子の値は、40〜50Å2の範囲であり、近接するタンパク質原子が32Å2というB因子を有する)は、外部の水との交換が遅いものであり得ることを示唆する。驚くべきことに、これらの水分子は、ホットスポットBRIAEC Gln86に近接している。3個の水分子は、側鎖アミド窒素原子に近いBRIAEC Gln86の「膜遠位側」に位置している。これらの水素結合している水分子は、BRIAEC Lys97との接触面の大きな割れ目を埋めており、この窪みへの入口を塞いでいる。単一で単離された水分子が、BRIAEC Gln86の側鎖カルボニル酸素原子の隣の窪んだ空間に位置している。この窪みはBMP−2の残基Phe49及びPro50によって外側から塞がれている。BMP−2結合の際の受容体Gln86の不完全な脱溶媒和は、疎水性環境における強力な水素結合の形成を支持している可能性がある。
【0096】
実施例3:水素結合の突然変異解析
BMP−2及び2種のアミノ酸置換を含むBRIAEC変異タンパク質を用いる相互作用分析によって、結合親和性にとっての水素結合の重要性を解析した。第1に、アラニン置換を導入して、側鎖結合を乱した(実施例2の表1参照)。BMP−2 W28A変異タンパク質は単離できなかったので、Trp28を、その側鎖も水素結合能を欠いているフェニルアラニンで置換した。(1個の電荷を有する残基の置換をBMP−2 S69R変異タンパク質で行った)。第2に、プロリン置換を導入して、主鎖アミド基による水素結合を消滅させた。
【0097】
表2に表されるように、固定化した受容体BRIAECを用いるBMP−2変異タンパク質の相互作用解析によって、置換W28F、D53A及びY103Aは結合親和性に対して小さな作用しか有していないことが示された。S69R変異タンパク質に電荷を有する残基を挿入した後に結合親和性のわずかに大きな低下(17倍)が観察された。際立ったことに、L51P及びD53P置換は著しく低い親和性をもたらす。BRIAECとL51Pの間の見掛けの解離定数KDは、BMP−2のものより7100倍よりも高く、D53Pについては53倍高かった(表2及び図7)。両変異タンパク質とも、BRIB受容体に対する親和性の同様の低下を有し、このことはBMP−2の同様の決定基が2つのI型受容体の結合に用いられていることを示す。このことは、2つのプロリン変異タンパク質において乱された主鎖水素結合は主要な結合決定基であるが、W28、D53、S69及びY103という側鎖結合はBRIA受容体との結合にはあまり重要なものでないということを示している。
【0098】
II型受容体ARIIに対するプロリン変異タンパク質の親和性は、野生型と比較して変わらない(表2)。II型受容体アクチビン受容体IIBとBMP受容体IIとの相互作用についても同じことが観察されている(データは示していない)。このことにより、置換されたプロリンによる局所構造の変化は、II型受容体結合の並列するエピトープ(ナックルエピトープ)には伝わっていないので、変異タンパク質の全体構造は変わっていないということが確認される。
【0099】
【表2】
【0100】
BRIA受容体の結合決定基を同定するために、3種のエクトドメイン変異タンパク質を調製した。受容体Q94P変異タンパク質では、野生型ではBMP−2 Ser69骨格カルボニルに水素結合を供与する骨格窒素アミドが乱されている。受容体Q86A変異タンパク質は、野生型ではBMP−2 L51骨格アミド及びカルボニルとH結合を形成する側鎖カルボキシアミド基を欠いている(図2)。S90A置換によって水素結合をBMP2 Val26骨格カルボニルに供与するセリンヒドロキシル基が消滅する。すべての受容体変異タンパク質は野生型エクトドメインと同様に大腸菌で発現された後に高度に精製され得る。バイオセンサーに固定化した場合、受容体Q94P変異タンパク質は、野生型エクトドメインと比較して親和性が3倍しか低下しないでBMP−2と結合したが(表2)、このことは、変異タンパク質中で失われた主鎖/主鎖結合は結合にはわずかにしか寄与していないということを示す。S90A変異タンパク質において親和性の同様の小さな変化が観察されている。
【0101】
しかし、受容体Q86A置換は、BMP−2相互作用の際に86倍増加したKDをもたらした。しかし、BMP−2 L51P変異タンパク質は、影響を受けるH−結合が主要な結合決定基に相当するということをすでに実証していたので(前記参照)、親和性のこのような大きな低下は予測されたものであった。最も重要なことは、関与しているドナー及びアクセプター群の残基部分の接近可能性を分析することによって推測されるホットスポットと突然変異解析によって同定されたホットスポットとが一致することである(図1C)。受容体Q86A変異タンパク質における親和性の喪失は、BMP−2 L51P変異タンパク質におけるものよりは深刻なものではない(表2)。受容体中の隣接側鎖、例えば、Lys79又はGlu81又はAsp89が、失われたGln86のカルボキサミド基を部分的に補っている可能性がある(図1b)。他方、BMP−2 L51Pに導入されたプロリンは、立体障害を引き起こすか、例えば、BMP−2 Asp53骨格アミドと受容体Cys77カルボニルの間の局所的な構造転位によって隣接する結合を阻害し、それによって置換の作用をさらに悪化させる(図2)。
【0102】
実施例4:プロリンBMP−2変異タンパク質L51Pの構造
BMP−2 L51Pの結合親和性の大きな低下を、BRIAEC Q86Aに対して観察されたものと比較することによって、結合親和性の喪失は、単一の水素結合の喪失だけに起因するわけではない可能性があることが示唆される。さらに、結合エピトープにおける大きな構造変化がタンパク質相互作用を乱す可能性がある。したがって、BMP−2 L51P及びまたD53P変異タンパク質を結晶化試験に付した。BMP−2 L51Pは野生型BMP−2と同一条件下で結晶化でき、同一の結晶パラメータ、例えば、野生型BMP−2の結晶について観察されたものと同一の空間群、R32、及びほぼ同一の容器定数を有していた。明らかに、プロリン残基はLeu51の代わりに受け入れられ、その疎水性側鎖がBMP−2のタンパク質コア中に向かっている。変異タンパク質D53Pは、そのような条件下では結晶化せず、D53Pの結晶は、エタノール又は2−メチル−2,4−ペンタジオールのいずれかから成長し、極めて低い分解能までしか回析せず、さらには分析しなかった。
【0103】
BMP−2 L51Pの回析データは室温で2.7Åという分解能まで得た。突然変異部位は別として、BMP−2野生型とL51Pの構造は基本的に同一であり、表3に表される構造決定法の誤差限度内にあり、すべてのCα原子位置のr.m.s.偏差は0.6Åである。残基Pro48〜His54を含み、位置51の突然変異部位も保持するプレヘリックスループに明確な変化が観察される。残基Pro48、Phe49、Pro50、Leu/Pro51、Ala52及びHis54について、野生型と変異タンパク質のCα位置間の距離は1Åを越え、最大の相違はAla52のCαについての1.8Åである(表3)。両構造、BMP−2野生型とL51Pの全体的な位置の誤差は、σAプロットから算出されるように約0.4であるので、プレヘリックスループの高次構造の変化は小さいが有意なものと考えられる。
【0104】
【表3】
【0105】
この変化は野生型のロイシンのものと比較したプロリン残基の骨格の形状の変化によるものであり得る。しかし、結合していない高次構造及び結合している高次構造におけるBMP−2の構造を比較すると、Cα位置間の相違はBMP−2野生型と変異タンパク質L51P間で観察されるものと同じ大きさであり、このことは、プレヘリックスループの高次構造は前もって形成されているものでも、固定されているものでもなく、複合体形成の際に受容体表面に適応するということを示す。
【0106】
Pro51の側鎖は野生型BMP−2中のLeu51と同様の方向を向いており、このことは疎水性コアのパッキングは突然変異によって影響を受けないということを示す。さらに、Leu51からプロリンへの交換は、残基51の骨格カルボニルの方向を変えず、それゆえに、BRIA Gln86と残基51との1個の水素結合が依然として複合体に形成され得る(図2、3A)。突然変異によって、結合していないタンパク質に導入された、かなり小さい構造の相違を考慮すると、結合親和性の大きな変化は不可解なものに思われる。
【0107】
L51PとBMP−2−BRIAECという複合体の構造の重ね合わせにより、プロリン残基の環が、BMP−2 Leu51アミドとBRIAEC Gln86の側鎖カルボニルの間の水素結合に用いられていた容積を元は占めていることが示される(図3A)。プロリン環の大きな空間要求が、BMP−2Pro51の側鎖とBRIAEC Gln86の間に立体障害を引き起こす(図3A)。Pro51を受け入れるには、その側鎖はさらに押し戻される必要がある。基本的に環の方向にアミド窒素を固定する堅い環構造であるために、もし環構造が押し戻されると、これは、Pro50の骨格カルボニルがBRIAEC Gln86の側鎖の方向に移動するφ骨格ねじれ角の回りの回転に相当する。結果として、1つの立体障害を除去することによって、別の1つが引き起こされることとなり、これによって、おそらくは、突然変異L51Pによる結合エネルギーの大きな喪失が説明される。
【0108】
実施例5:プロリン変異タンパク質及び生物活性
BMP−2プロリン変異タンパク質の生物活性は、C2C12細胞及びわずかにより感受性の高いATDC5細胞株を用いて、アルカリホスファターゼ(ALP)活性の用量依存的誘導によって決定した。最大応答の50%(ED50)をもたらす野生型BMP−2の用量は、C2C12細胞では25nMであった。ATDC5細胞を用いた場合、ED50は8nMであり、このことはこれらの細胞の高い反応性を示す。対照的に、BMP−2 L51Pには、C2C12(>1600nM)又はATDC5細胞(>1500nM)で測定可能な生物活性はない。BMP−2 D53P変異タンパク質は、C2C12及びATDC5細胞に対して、330nM及び140nMというED50値をそれぞれ示す。これらの値は、野生型BMP−2のED50値と比較して約13〜17倍高い。したがって、2種のBMP−2プロリン変異タンパク質におけるBRIA受容体親和性の異なる低下は生物活性の喪失と平行している(図4)。
【0109】
BMP−2 Leu51のプロリン置換は、I型受容体との相互作用にのみ影響を及ぼしており、他方でII型受容体の親和性は変わっていないので、L51P変異タンパク質は、主としてドミナントネガティブ阻害剤として機能している可能性がある。しかし、これは、250nMまでの濃度の変異タンパク質を、20nM BMP−2で刺激した細胞に適用した場合には観察されなかった(データは示していない)。相対的に、ATDC5細胞における定常ALP活性は、アンタゴニストBMP−2 A34D変異タンパク質によって阻害される(Kirschら、2000a)が、L51Pによっては阻害されない(図4A)。
【0110】
実施例6:結合タンパク質のアンタゴニストとしてのプロリン変異タンパク質
BMP−2 L51PがBRIAECと結合しないことは、その他の結合タンパク質との相互作用もプロリン置換によって影響を受け得るかどうかという疑問を提起した。特に注目されるものは、BMPと高親和性で結合し、BMP−7との複合体中のその構造が最近決定された(Groppeら、2002)ノギンである。もう1つの重要な結合タンパク質はコーディンであり、これはBMP−2と2つのシステインリッチドメインによって相互作用する。さらにもう1つの種類の結合タンパク質は、いわゆるDNAドメインによってBMP−2と相互作用するグレムリンに相当する。3種のタンパク質の各々を、バイオセンサーの表面に固定し、表4に表されるようにBMP−2変異タンパク質相互作用について分析した。
【0111】
【表4】
【0112】
ノギンとの複合体についての2種のBMP−2プロリン変異タンパク質の解離定数KDはBMP−2野生型のものと同様であり、0.5〜2.5nMの範囲であった。したがって、ノギンのN末端アミノ酸がBMP−2 Leu51(BMP−7 Leu75)及びAsp53(BMP−7 Ser77、プレヘリックスループ)に対応するBMP−7領域と接触していることが示され、ノギンはBMP−2とのBRIA結合を妨げるという事実があるにもかかわらず、BMP−2 Leu51及びAsp53はノギンに対する主要な結合決定基ではない。
【0113】
コーディンドメインCR1及びCR3並びにグレムリンに対するBMP−2の親和性は、ノギンに対してよりも10〜30倍低い(表4)。BMP−2野生型又は2種のプロリン変異タンパク質に対しても再度、同様の解離定数KDが認められ、このことは、これらのタンパク質の結合がプロリン置換によって影響を受けないことを示す。したがって、両結合タンパク質に対するBMP−2エピトープはBRIA受容体に対するものとは異なる。
【0114】
実施例7:プロリン変異タンパク質L51Pは、C2C12細胞においてノギン阻害を解除する
BMP−2は飢餓前骨髄芽球性細胞株C2C12において、アルカリホスファターゼの発現を誘導する。このBMP応答は、等モル濃度のノギン−Fc融合タンパク質によってバックグラウンドレベルまで阻害される(図5)。ノギンによるBMP−2活性の阻害は、L51P変異タンパク質によって用量依存的に解除される。ほぼ等モル濃度の変異タンパク質L51Pとノギンで、ノギンの阻害作用は、80%よりも多くまで中和される。このことは、受容体非感受性L51P変異タンパク質はノギンタンパク質の効率的な阻害剤に相当するということを示す。この結果は、BMP−2 L51P置換後のI型受容体結合の混乱とノギン結合の保持を示す物理的結合研究と一致する。
【0115】
参照文献
不要な参照を避けるために、本明細書に記載した参照文献の包括的な一覧を以下に示す。これらの参照文献のいずれの開示内容も参照により本明細書に組み込まれると理解されなくてはならない。
【0116】
Balemans, W. and Van Hul, W. (2002)Extracellularregulation of BMP signaling in vertebrates: a cocktail ofmodulators. Dev Biol,250, 231-250.
Bogan, A.A. and Thorn, K.S. (1998)Anatomy of hot spots inprotein interfaces. J Mol Biol, 280, 1-9.
Boulanger, M.J., Chow, D.C., Brevnova,E.E. and Garcia,K.C. (2003) Hexameric structure and assembly of theinterleukin-6/IL-6alpha-receptor/gp130 complex. Science, 300, 2101-2104.
Brunger, A.T. (1992) Free R value: anovel statisticalquantity for assessing the accuracy of crystal structures.Nature, 355,472-475.
Brunger, A.T., Adams, P.D., Clore, G.M.,DeLano, W.L.,Gros, P., Grosse-Kunstleve, R.W., Jiang, J.S., Kuszewski, J.,Nilges, M., Pannu, N.S., Read, R.J., Rice, L.M., Simonson, T. and Warren, G.L.(1998) Crystallography & NMR system: A new software suite formacromolecular structure determination. Acta Crystallogr DBiol Crystallogr, 54(Pt 5), 905-921.
Chakrabarti, P. and Janin, J. (2002)Dissectingprotein-protein recognition sites. Proteins, 47, 334-343.
Collaborative Computational Project, N.(1994) The CCP4Suite: Programs for Protein Crystallography. Acta Crystallogsect D, 50.
de Vos, A.M., Ultsch, M. and Kossiakoff,A.A. (1992) Humangrowth hormone and extracellular domain of its receptor:crystal structure ofthe complex. Science, 255, 306-312.
Evans, P.R. (1993) Data reduction.Proceedings of CCP4Study Weekend, pp. 114-122.
Greenwald, J., Groppe, J., Gray, P.,Wiater, E.,Kwiatkowski, W., Vale, W. and Choe, S. (2003) The BMP7/ActRII extracellulardomaincomplex provides new insights into the cooperative nature of receptorassembly.Mol Cell, 11, 605-617.
Groppe, J., Greenwald, J., Wiater, E.,Rodriguez-Leon, J.,Economides, A.N., Kwiatkowski, W., Affolter, M., Vale, W.W.,Belmonte, J.C. andChoe, S. (2002) Structural basis of BMP signalling inhibitionby the cystineknot protein Noggin. Nature, 420, 636-642.
Hage, T., Sebald, W. and Reinemer, P.(1999) Crystal structure of the interleukin-4/receptor alpha chaincomplexreveals a mosaic binding interface. Cell, 97, 271-281.
Hart, P.J., Deep, S., Taylor, A.B.,Shu,Z., Hinck, C.S. and Hinck, A.P. (2002) Crystal structure of the human TbetaR2ectodomain--TGF-beta3 complex. Nat Struct Biol,9, 203-208.
Kirsch, T., Nickel, J. and Sebald, W.(2000a) BMP-2 antagonistsemerge from alterations in the low-affinity bindingepitope for receptor BMPR-II. Embo J, 19, 3314-3324.
Kirsch, T., Nickel, J. and Sebald, W.(2000b) Isolation ofrecombinant BMP receptor IA ectodomain and its 2:1 complexwith BMP-2. FEBSLett, 468, 215-219.
Leslie, A.G.W. (1992) Recent changes tothe MOSFLM packagefor processing film and image plate data. Joint CCP4 +ESF-EAMCB Newsletter onProtein Crystallography, 26.
McDonald, I.K. and Thornton,J.M. (1994)Satisfying hydrogen bonding potential in proteins. J Mol Biol, 238,777-793.
Murshudov, G.N., Vagin, A.A. and Dodson,E.J. (1997)Refinement of Macromolecular Structures by the Maximum-LikelihoodMethod. ActaCrystallog sect D, D53, 240-25
5.
Ruppert, R., Hoffmann, E. and Sebald, W.(1996) Human bonemorphogenetic protein2 contains a heparin-binding site whichmodifies itsbiological activity. Eur J Biochem, 237, 295-302.
Syed, R.S., Reid, S.W., Li, C.,Cheetham, J.C., Aoki, K.H.,Liu, B., Zhan, H., Osslund, T.D., Chirino, A.J.,Zhang, J., Finer-Moore, J.,Elliott, S., Sitney, K.,Katz, B.A., Matthews, D.J.,
Thompson, T.B., Woodruff, T.K. andJardetzky, T.S. (2003)Structures of an ActRIIB:activin A complex reveal a novelbinding mode forTGF-beta ligand:receptor interactions. Embo J, 22, 1555-1566.
Winn, M.D., Isupov, M.N. and Murshudov,G.N. (2001) Use ofTLS parameters to model anisotropic displacements inmacromolecular refinement.Acta Crystallogr D Biol Crystallogr, 57, 122-133.
Xu, D., Tsai, C.J. and Nussinov, R.(1997) Hydrogen bondsand salt bridges across protein-protein interfaces.Protein Eng, 10, 999-1012.
【0117】
本明細書に開示した本発明の特徴、特許請求の範囲及び/又は図面は、個別に、及びいずれかの組合せでの双方で、種々の形の本発明を理解する材料であり得る。
【特許請求の範囲】
【請求項1】
骨形成タンパク質の変異タンパク質であって、前記変異タンパク質は、骨形成タンパク質の野生型と比較して、ヒトBMP−2のアミノ酸位置51に相当するアミノ酸位置にアミノ酸置換を含む、変異タンパク質。
【請求項2】
前記ヒトBMP−2のアミノ酸位置51に相当する位置のアミノ酸は、前記骨形成タンパク質の野生型ではロイシンであり、前記変異タンパク質ではプロリンであることが好ましい、請求項1記載の変異タンパク質。
【請求項3】
前記骨形成タンパク質は、hBMP−2、hBMP−4、hBMP−5、hBMP−6、hBMP−7、hBMP−8、hGDF−5、mGDF−6、mGDF−7、hBMP−10及びhGDF−2を含んでなる群から選択される、請求項1及び2記載の変異タンパク質。
【請求項4】
前記骨形成タンパク質は、hBMP−2であり、ヒトBMP−2のアミノ酸位置51に相当する位置は、アミノ酸位置51であり、
前記骨形成タンパク質は、hBMP−4であり、ヒトBMP−2のアミノ酸位置51に相当する位置は、アミノ酸位置53であり、
骨形成タンパク質は、hBMP−5であり、ヒトBMP−2のアミノ酸位置51に相当する位置は、アミノ酸位置68であり、
骨形成タンパク質は、hBMP−6であり、ヒトBMP−2のアミノ酸位置51に相当する位置は、アミノ酸位置68であり、
骨形成タンパク質は、hBMP−7であり、ヒトBMP−2のアミノ酸位置51に相当する位置は、アミノ酸位置75であり、
骨形成タンパク質は、hBMP−8であり、ヒトBMP−2のアミノ酸位置51に相当する位置は、アミノ酸位置75であり、
骨形成タンパク質は、hGDF−5であり、ヒトBMP−2のアミノ酸位置51に相当する位置は、アミノ酸位置56であり、
骨形成タンパク質は、mGDF−6であり、ヒトBMP−2のアミノ酸位置51に相当する位置は、アミノ酸位置56であり、
骨形成タンパク質は、mGDF−7であり、ヒトBMP−2のアミノ酸位置51に相当する位置は、アミノ酸位置82であり、
骨形成タンパク質は、hBMP−10であり、ヒトBMP−2のアミノ酸位置51に相当する位置は、アミノ酸位置44であり、
骨形成タンパク質は、hGDF−2であり、ヒトBMP−2のアミノ酸位置51に相当する位置は、アミノ酸位置45である、
請求項1〜3のいずれか一項記載の変異タンパク質。
【請求項5】
hBMP−2の野生型は、配列番号1のアミノ酸配列を含み、
hBMP−4の野生型は、配列番号3のアミノ酸配列を含み、
hBMP−5の野生型は、配列番号5のアミノ酸配列を含み、
hBMP−6の野生型は、配列番号7のアミノ酸配列を含み、
hBMP−7の野生型は、配列番号9のアミノ酸配列を含み、
hBMP−8の野生型は、配列番号11のアミノ酸配列を含み、
hGDF−5の野生型は、配列番号13のアミノ酸配列を含み、
mGDF−6の野生型は、配列番号15のアミノ酸配列を含み、
mGDF−7の野生型は、配列番号17のアミノ酸配列を含み、
hBMP−10の野生型は、配列番号19のアミノ酸配列を含み、
hGDF−2の野生型は、配列番号21のアミノ酸配列を含む、
請求項1〜4のいずれか一項記載の変異タンパク質。
【請求項6】
hBMP−2の野生型は、配列番号2の核酸によってコードされており、
hBMP−4の野生型は、配列番号4の核酸によってコードされており、
hBMP−5の野生型は、配列番号6の核酸によってコードされており、
hBMP−6の野生型は、配列番号8の核酸によってコードされており、
hBMP−7の野生型は、配列番号10の核酸によってコードされており、
hBMP−8の野生型は、配列番号12の核酸によってコードされており、
hGDF−5の野生型は、配列番号14の核酸によってコードされており、
mGDF−6の野生型は、配列番号16の核酸によってコードされており、
mGDF−7の野生型は、配列番号18の核酸によってコードされており、
hBMP−10の野生型は、配列番号20の核酸によってコードされており、
hGDF−2の野生型は、配列番号22の核酸によってコードされている、
請求項5記載の変異タンパク質。
【請求項7】
前記変異タンパク質は、第1の骨形成タンパク質受容体に結合していないが、少なくとも1種のモジュレータータンパク質と結合しており、
前記モジュレータータンパク質は、ノギンタンパク質ファミリー、DANタンパク質ファミリー、コーディンタンパク質ファミリー及びシステインノット含有BMPモジュレータータンパク質、を含んでなる群から選択される、
骨形成変異タンパク質。
【請求項8】
第2の骨形成タンパク質受容体と相互作用するプレヘリックスループ構造を含む、好ましくは請求項1記載の、骨形成変異タンパク質。
【請求項9】
前記第1及び/又は第2の骨形成タンパク質受容体は、BRIA又はBRIBである、請求項7及び8記載の骨形成変異タンパク質。
【請求項10】
前記相互作用は、アミノ酸残基、好ましくは、BRIAのGln86又はBRIBのGln67アミノ酸残基に関連している、請求項8及び9記載の骨形成変異タンパク質。
【請求項11】
第2の骨形成タンパク質受容体と相互作用するプレヘリックスループ構造を含み、前記骨形成変異タンパク質のプレヘリックスループ構造と、前記第2の骨形成タンパク質受容体との相互作用は、野生型骨形成タンパク質のプレヘリックスループ構造と、前記第2の骨形成タンパク質受容体との相互作用とは異なる、好ましくは請求項6〜10のいずれか一項記載の骨形成変異タンパク質。
【請求項12】
異なる相互作用又は変化は、屈折データ、好ましくは、少なくとも約2.7Åの分解能まで室温で得られている屈折データ、で表される、請求項8〜11のいずれか一項記載の骨形成変異タンパク質。
【請求項13】
前記プレヘリックスループ構造は、骨形成タンパク質の野生型と比較して変異している、請求項7〜12のいずれか一項記載の骨形成変異タンパク質。
【請求項14】
前記野生型BMP−2の位置51のロイシンに相当するアミノ酸は、変異している、請求項13記載の骨形成変異タンパク質。
【請求項15】
ヒトBMP−2の位置51のロイシンに相当するアミノ酸は、プロリンに変異している、請求項13又は14記載の骨形成変異タンパク質。
【請求項16】
hBMP−2、hBMP−4、hBMP−5、hBMP−6、hBMP−7、hBMP−8、hGDF−5、mGDF−6、mGDF−7、hBMP−10及びhGDF−2を含んでなる群から選択される骨形成タンパク質の変異タンパク質である、請求項7〜15のいずれか一項記載の骨形成変異タンパク質。
【請求項17】
BMP−2又はプロ−BMP−2である、請求項16記載の骨形成変異タンパク質。
【請求項18】
配列番号23〜33のいずれかのアミノ酸配列を含む、骨形成タンパク質。
【請求項19】
請求項7〜18のいずれか一項記載の骨形成変異タンパク質である、請求項18記載の骨形成変異タンパク質。
【請求項20】
請求項1〜19のいずれか一項記載の骨形成変異タンパク質をコードする核酸及び/又はその相補鎖。
【請求項21】
配列番号2、4、6、8、10、12、14、16、18、20及び22の核酸配列を含む核酸であって、好ましくは、請求項1〜19のいずれか一項記載の骨形成変異タンパク質をコードする核酸、及び/又はその相補鎖。
【請求項22】
前記核酸は、遺伝暗号の縮重がなければ、請求項20又は21記載の核酸とハイブリダイズし、より好ましくは、ストリンジェントな条件下で請求項20又は21記載の核酸とハイブリダイズする、請求項1〜19のいずれか一項記載の骨形成変異タンパク質をコードする核酸。
【請求項23】
ベクターは、発現ベクターであることが好ましい、請求項20〜22のいずれか一項記載の核酸を含むベクター。
【請求項24】
請求項20〜22のいずれか一項記載の核酸、及び/又は請求項23記載のベクター、を含む細胞、好ましくは、哺乳類細胞。
【請求項25】
請求項24記載の細胞を含む宿主生物、好ましくは哺乳類宿主生物、より好ましくは非ヒト宿主生物。
【請求項26】
請求項1〜19のいずれか一項記載の骨形成変異タンパク質の製造方法であって、
a)培養ブロスで請求項25記載の細胞を培養するステップと、
b)前記細胞から、及び/又は前記培養ブロスから、前記骨形成変異タンパク質を調製するステップと、
を含む方法。
【請求項27】
請求項1〜19のいずれか一項記載の骨形成変異タンパク質と特異的に結合するモノクローナル抗体。
【請求項28】
請求項1〜19のいずれか一項記載の変異タンパク質及び/又は請求項20〜22のいずれか一項記載の核酸、を含む組成物。
【請求項29】
請求項1〜19のいずれか一項記載の変異タンパク質又は請求項20〜22のいずれか一項記載の核酸と、製薬上許容される担体とを含む薬剤組成物。
【請求項30】
請求項1〜19のいずれか一項記載の骨形成変異タンパク質及び/又は請求項20〜22のいずれか一項記載の核酸、の医薬の製造のための使用。
【請求項31】
前記医薬は、線維性疾患、創傷治癒、過剰血管新生、血管疾患、骨折及び骨粗しょう症を含んでなる群から選択される疾患の治療用及び/又は予防用である、請求項30記載の使用。
【請求項32】
前記線維性疾患は、腎線維症、肝硬変、肺線維症及び慢性炎症、好ましくは、喘息を伴う慢性炎症を含んでなる群から選択される、請求項31記載の使用。
【請求項33】
前記創傷治癒は、ケロイド、瘢痕形成及び腹膜の閉塞と関連している、請求項31記載の使用。
【請求項34】
前記過剰血管新生は、網膜症、動脈硬化症及び/又は腫瘍と関連しているか、又は網膜症、動脈硬化症及び/又は腫瘍を伴う、請求項31に記載使用。
【請求項35】
前記骨折は、難治性骨折である、請求項31記載の使用。
【請求項36】
前記疾患は、骨粗しょう症である、請求項31記載の使用。
【請求項37】
BMP相互作用タンパク質に対する阻害剤としての、請求項1〜19のいずれか一項に記載の形態形成変異タンパク質の使用。
【請求項38】
前記BMP相互作用タンパク質は、ノギンタンパク質ファミリー、DANタンパク質ファミリー及びコーディンタンパク質ファミリーを含んでなる群から選択される、請求項37記載の使用。
【請求項1】
骨形成タンパク質の変異タンパク質であって、前記変異タンパク質は、骨形成タンパク質の野生型と比較して、ヒトBMP−2のアミノ酸位置51に相当するアミノ酸位置にアミノ酸置換を含む、変異タンパク質。
【請求項2】
前記ヒトBMP−2のアミノ酸位置51に相当する位置のアミノ酸は、前記骨形成タンパク質の野生型ではロイシンであり、前記変異タンパク質ではプロリンであることが好ましい、請求項1記載の変異タンパク質。
【請求項3】
前記骨形成タンパク質は、hBMP−2、hBMP−4、hBMP−5、hBMP−6、hBMP−7、hBMP−8、hGDF−5、mGDF−6、mGDF−7、hBMP−10及びhGDF−2を含んでなる群から選択される、請求項1及び2記載の変異タンパク質。
【請求項4】
前記骨形成タンパク質は、hBMP−2であり、ヒトBMP−2のアミノ酸位置51に相当する位置は、アミノ酸位置51であり、
前記骨形成タンパク質は、hBMP−4であり、ヒトBMP−2のアミノ酸位置51に相当する位置は、アミノ酸位置53であり、
骨形成タンパク質は、hBMP−5であり、ヒトBMP−2のアミノ酸位置51に相当する位置は、アミノ酸位置68であり、
骨形成タンパク質は、hBMP−6であり、ヒトBMP−2のアミノ酸位置51に相当する位置は、アミノ酸位置68であり、
骨形成タンパク質は、hBMP−7であり、ヒトBMP−2のアミノ酸位置51に相当する位置は、アミノ酸位置75であり、
骨形成タンパク質は、hBMP−8であり、ヒトBMP−2のアミノ酸位置51に相当する位置は、アミノ酸位置75であり、
骨形成タンパク質は、hGDF−5であり、ヒトBMP−2のアミノ酸位置51に相当する位置は、アミノ酸位置56であり、
骨形成タンパク質は、mGDF−6であり、ヒトBMP−2のアミノ酸位置51に相当する位置は、アミノ酸位置56であり、
骨形成タンパク質は、mGDF−7であり、ヒトBMP−2のアミノ酸位置51に相当する位置は、アミノ酸位置82であり、
骨形成タンパク質は、hBMP−10であり、ヒトBMP−2のアミノ酸位置51に相当する位置は、アミノ酸位置44であり、
骨形成タンパク質は、hGDF−2であり、ヒトBMP−2のアミノ酸位置51に相当する位置は、アミノ酸位置45である、
請求項1〜3のいずれか一項記載の変異タンパク質。
【請求項5】
hBMP−2の野生型は、配列番号1のアミノ酸配列を含み、
hBMP−4の野生型は、配列番号3のアミノ酸配列を含み、
hBMP−5の野生型は、配列番号5のアミノ酸配列を含み、
hBMP−6の野生型は、配列番号7のアミノ酸配列を含み、
hBMP−7の野生型は、配列番号9のアミノ酸配列を含み、
hBMP−8の野生型は、配列番号11のアミノ酸配列を含み、
hGDF−5の野生型は、配列番号13のアミノ酸配列を含み、
mGDF−6の野生型は、配列番号15のアミノ酸配列を含み、
mGDF−7の野生型は、配列番号17のアミノ酸配列を含み、
hBMP−10の野生型は、配列番号19のアミノ酸配列を含み、
hGDF−2の野生型は、配列番号21のアミノ酸配列を含む、
請求項1〜4のいずれか一項記載の変異タンパク質。
【請求項6】
hBMP−2の野生型は、配列番号2の核酸によってコードされており、
hBMP−4の野生型は、配列番号4の核酸によってコードされており、
hBMP−5の野生型は、配列番号6の核酸によってコードされており、
hBMP−6の野生型は、配列番号8の核酸によってコードされており、
hBMP−7の野生型は、配列番号10の核酸によってコードされており、
hBMP−8の野生型は、配列番号12の核酸によってコードされており、
hGDF−5の野生型は、配列番号14の核酸によってコードされており、
mGDF−6の野生型は、配列番号16の核酸によってコードされており、
mGDF−7の野生型は、配列番号18の核酸によってコードされており、
hBMP−10の野生型は、配列番号20の核酸によってコードされており、
hGDF−2の野生型は、配列番号22の核酸によってコードされている、
請求項5記載の変異タンパク質。
【請求項7】
前記変異タンパク質は、第1の骨形成タンパク質受容体に結合していないが、少なくとも1種のモジュレータータンパク質と結合しており、
前記モジュレータータンパク質は、ノギンタンパク質ファミリー、DANタンパク質ファミリー、コーディンタンパク質ファミリー及びシステインノット含有BMPモジュレータータンパク質、を含んでなる群から選択される、
骨形成変異タンパク質。
【請求項8】
第2の骨形成タンパク質受容体と相互作用するプレヘリックスループ構造を含む、好ましくは請求項1記載の、骨形成変異タンパク質。
【請求項9】
前記第1及び/又は第2の骨形成タンパク質受容体は、BRIA又はBRIBである、請求項7及び8記載の骨形成変異タンパク質。
【請求項10】
前記相互作用は、アミノ酸残基、好ましくは、BRIAのGln86又はBRIBのGln67アミノ酸残基に関連している、請求項8及び9記載の骨形成変異タンパク質。
【請求項11】
第2の骨形成タンパク質受容体と相互作用するプレヘリックスループ構造を含み、前記骨形成変異タンパク質のプレヘリックスループ構造と、前記第2の骨形成タンパク質受容体との相互作用は、野生型骨形成タンパク質のプレヘリックスループ構造と、前記第2の骨形成タンパク質受容体との相互作用とは異なる、好ましくは請求項6〜10のいずれか一項記載の骨形成変異タンパク質。
【請求項12】
異なる相互作用又は変化は、屈折データ、好ましくは、少なくとも約2.7Åの分解能まで室温で得られている屈折データ、で表される、請求項8〜11のいずれか一項記載の骨形成変異タンパク質。
【請求項13】
前記プレヘリックスループ構造は、骨形成タンパク質の野生型と比較して変異している、請求項7〜12のいずれか一項記載の骨形成変異タンパク質。
【請求項14】
前記野生型BMP−2の位置51のロイシンに相当するアミノ酸は、変異している、請求項13記載の骨形成変異タンパク質。
【請求項15】
ヒトBMP−2の位置51のロイシンに相当するアミノ酸は、プロリンに変異している、請求項13又は14記載の骨形成変異タンパク質。
【請求項16】
hBMP−2、hBMP−4、hBMP−5、hBMP−6、hBMP−7、hBMP−8、hGDF−5、mGDF−6、mGDF−7、hBMP−10及びhGDF−2を含んでなる群から選択される骨形成タンパク質の変異タンパク質である、請求項7〜15のいずれか一項記載の骨形成変異タンパク質。
【請求項17】
BMP−2又はプロ−BMP−2である、請求項16記載の骨形成変異タンパク質。
【請求項18】
配列番号23〜33のいずれかのアミノ酸配列を含む、骨形成タンパク質。
【請求項19】
請求項7〜18のいずれか一項記載の骨形成変異タンパク質である、請求項18記載の骨形成変異タンパク質。
【請求項20】
請求項1〜19のいずれか一項記載の骨形成変異タンパク質をコードする核酸及び/又はその相補鎖。
【請求項21】
配列番号2、4、6、8、10、12、14、16、18、20及び22の核酸配列を含む核酸であって、好ましくは、請求項1〜19のいずれか一項記載の骨形成変異タンパク質をコードする核酸、及び/又はその相補鎖。
【請求項22】
前記核酸は、遺伝暗号の縮重がなければ、請求項20又は21記載の核酸とハイブリダイズし、より好ましくは、ストリンジェントな条件下で請求項20又は21記載の核酸とハイブリダイズする、請求項1〜19のいずれか一項記載の骨形成変異タンパク質をコードする核酸。
【請求項23】
ベクターは、発現ベクターであることが好ましい、請求項20〜22のいずれか一項記載の核酸を含むベクター。
【請求項24】
請求項20〜22のいずれか一項記載の核酸、及び/又は請求項23記載のベクター、を含む細胞、好ましくは、哺乳類細胞。
【請求項25】
請求項24記載の細胞を含む宿主生物、好ましくは哺乳類宿主生物、より好ましくは非ヒト宿主生物。
【請求項26】
請求項1〜19のいずれか一項記載の骨形成変異タンパク質の製造方法であって、
a)培養ブロスで請求項25記載の細胞を培養するステップと、
b)前記細胞から、及び/又は前記培養ブロスから、前記骨形成変異タンパク質を調製するステップと、
を含む方法。
【請求項27】
請求項1〜19のいずれか一項記載の骨形成変異タンパク質と特異的に結合するモノクローナル抗体。
【請求項28】
請求項1〜19のいずれか一項記載の変異タンパク質及び/又は請求項20〜22のいずれか一項記載の核酸、を含む組成物。
【請求項29】
請求項1〜19のいずれか一項記載の変異タンパク質又は請求項20〜22のいずれか一項記載の核酸と、製薬上許容される担体とを含む薬剤組成物。
【請求項30】
請求項1〜19のいずれか一項記載の骨形成変異タンパク質及び/又は請求項20〜22のいずれか一項記載の核酸、の医薬の製造のための使用。
【請求項31】
前記医薬は、線維性疾患、創傷治癒、過剰血管新生、血管疾患、骨折及び骨粗しょう症を含んでなる群から選択される疾患の治療用及び/又は予防用である、請求項30記載の使用。
【請求項32】
前記線維性疾患は、腎線維症、肝硬変、肺線維症及び慢性炎症、好ましくは、喘息を伴う慢性炎症を含んでなる群から選択される、請求項31記載の使用。
【請求項33】
前記創傷治癒は、ケロイド、瘢痕形成及び腹膜の閉塞と関連している、請求項31記載の使用。
【請求項34】
前記過剰血管新生は、網膜症、動脈硬化症及び/又は腫瘍と関連しているか、又は網膜症、動脈硬化症及び/又は腫瘍を伴う、請求項31に記載使用。
【請求項35】
前記骨折は、難治性骨折である、請求項31記載の使用。
【請求項36】
前記疾患は、骨粗しょう症である、請求項31記載の使用。
【請求項37】
BMP相互作用タンパク質に対する阻害剤としての、請求項1〜19のいずれか一項に記載の形態形成変異タンパク質の使用。
【請求項38】
前記BMP相互作用タンパク質は、ノギンタンパク質ファミリー、DANタンパク質ファミリー及びコーディンタンパク質ファミリーを含んでなる群から選択される、請求項37記載の使用。
【図3】

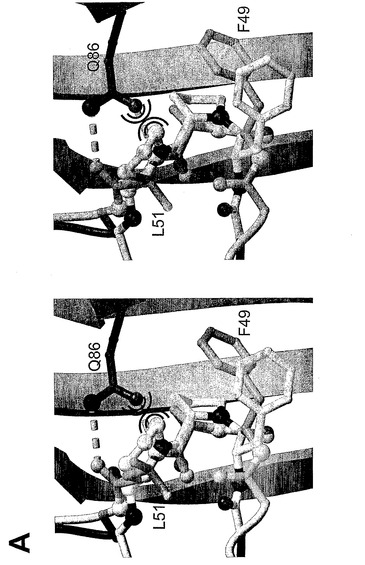
【図4】
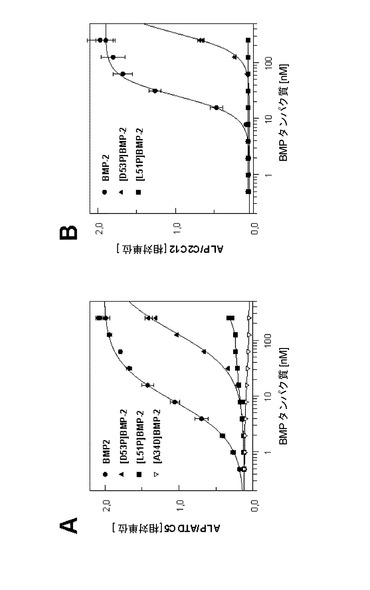
【図5】
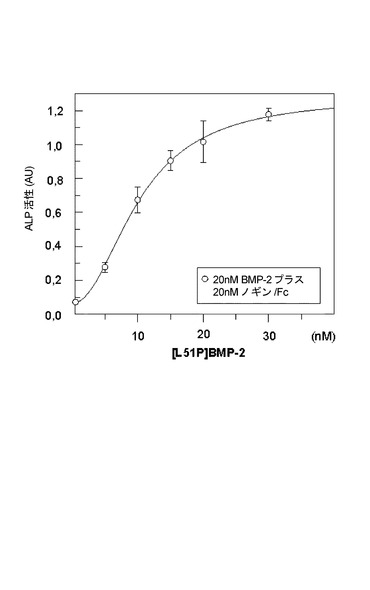
【図6】

【図7】
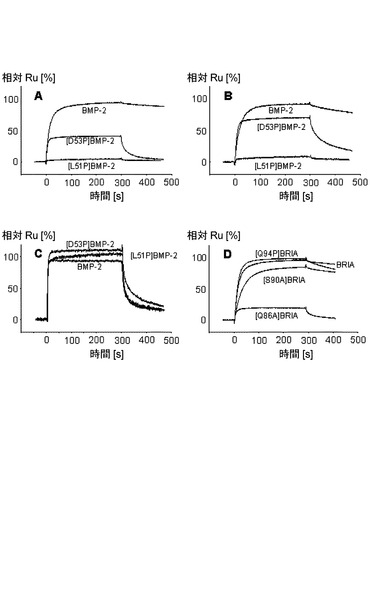
【図8】
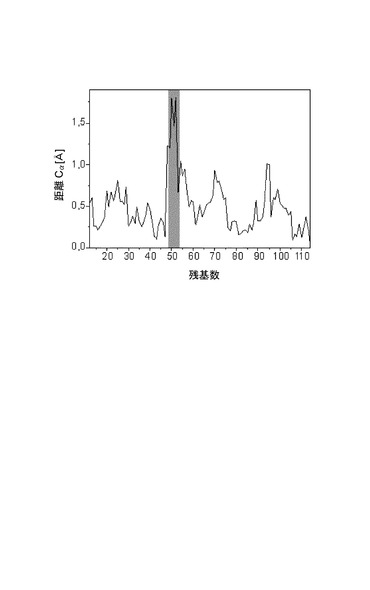
【図1】
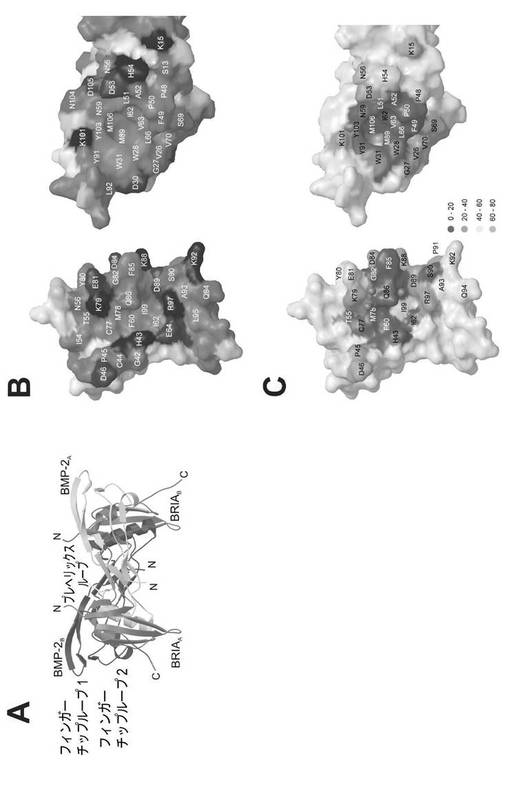
【図2】
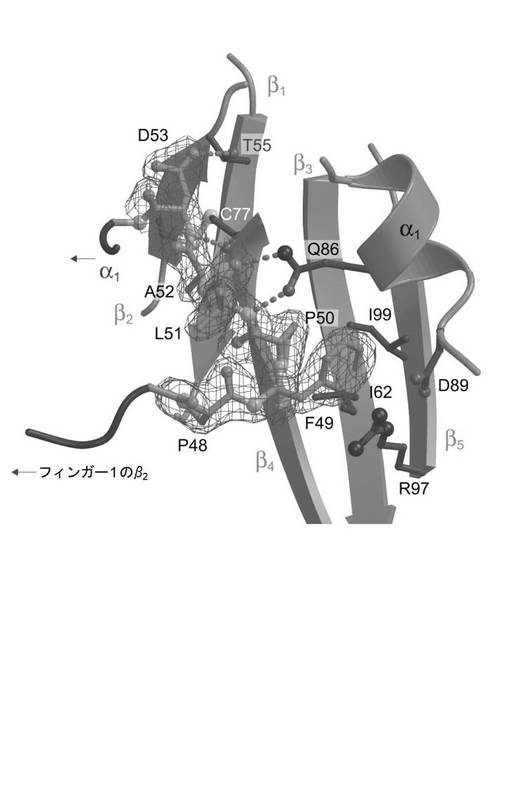
【図4】
【図5】
【図6】
【図7】
【図8】
【図1】
【図2】
【公開番号】特開2012−235784(P2012−235784A)
【公開日】平成24年12月6日(2012.12.6)
【国際特許分類】
【外国語出願】
【出願番号】特願2012−152866(P2012−152866)
【出願日】平成24年7月6日(2012.7.6)
【分割の表示】特願2007−501236(P2007−501236)の分割
【原出願日】平成17年3月4日(2005.3.4)
【出願人】(506299995)
【Fターム(参考)】
【公開日】平成24年12月6日(2012.12.6)
【国際特許分類】
【出願番号】特願2012−152866(P2012−152866)
【出願日】平成24年7月6日(2012.7.6)
【分割の表示】特願2007−501236(P2007−501236)の分割
【原出願日】平成17年3月4日(2005.3.4)
【出願人】(506299995)
【Fターム(参考)】
[ Back to top ]