
骨形成タンパク質(BMP)、BMPレセプターおよびBMP結合タンパク質ならびに緑内障の診断および処置におけるそれらの使用
【課題】 緑内障の診断および処置のための方法およびキットを提供することを、本発明の課題とする。
【解決手段】 細胞または体液から得られたサンプルにおいて、骨形態形成ファミリーメンバー遺伝子の発現の変化を検出することによる、緑内障を診断する方法を提供することによって、上記課題は解決された。この方法は、好ましくは、緑内障を有すると疑われる患者から組織または液体のサンプルを得る工程;該サンプルからDNAを抽出する工程;複数のPCRプライマーを得る工程;該プライマーを使用して、該抽出されたDNAの領域を増幅してPCR産物を得る工程;該PCR産物を分離する工程;ならびに該増幅した抽出されたDNA配列と該プライマーの配列との間の違いを同定する工程を包含する。
【解決手段】 細胞または体液から得られたサンプルにおいて、骨形態形成ファミリーメンバー遺伝子の発現の変化を検出することによる、緑内障を診断する方法を提供することによって、上記課題は解決された。この方法は、好ましくは、緑内障を有すると疑われる患者から組織または液体のサンプルを得る工程;該サンプルからDNAを抽出する工程;複数のPCRプライマーを得る工程;該プライマーを使用して、該抽出されたDNAの領域を増幅してPCR産物を得る工程;該PCR産物を分離する工程;ならびに該増幅した抽出されたDNA配列と該プライマーの配列との間の違いを同定する工程を包含する。
【発明の詳細な説明】
【技術分野】
【0001】
(1.発明の分野)
本発明は、緑内障および関連障害の診断および処置のための方法および試薬を開示する。
【背景技術】
【0002】
(2.関連分野の説明)
「緑内障」は、消耗性の眼の疾患の一群であり、米国および他の先進国において回復不能の失明の主要原因である。緑内障のもっとも一般的な形態である原発性開放隅角緑内障(「POAG」)は、虹彩と角膜との間の空間(例えば、「角」)の閉鎖無しに、房水が眼を離れる正常な能力の閉塞を起こす、小柱網の変性によって特徴づけられる(非特許文献1)。この疾患におけるこのような閉塞の特徴は、眼内圧力(「IOP」)上昇であり、もし適切にかつ適時な様式で処置されなければ、進行性の視力喪失および失明を起こす。この疾患は、40歳を越える全ての成人のうち0.4%と3.3%との間の人々に影響を及ぼすことが推測される(Leske,M.C.ら、(1986);Bengtsson,B.(1989);Strong,N.P.(1992))。さらに、この疾患の有病率は、年齢とともに増加して、75歳以上では6%を超える(Strong,N.P.、(1992))。
【0003】
IOPの上昇は、緑内障の容易に測定可能な特徴であるので、この疾患の診断は、眼内力の測定(眼圧測定)によって大まかにスクリーニングされる(Strong,N.P.(1992);Greve,M.ら、(1993))。不幸なことに、緑内障および正常の圧力範囲は重複するので、このような方法は、多数の読み取りが得られない限り、限定された値のものである(Hitchings,R.A.、(1993);Tuck,M.W.ら、(1993);Vaugham,D.ら、(1992);Vernon,S.A.、(1993))。この理由から、さらなる方法(例えば、視神経円板の直接的な検査および患者の視野損失の程度の決定)が、しばしば、診断の正確性を改善するために実施される(Greve,M.ら、(1993))。
【0004】
緑内障は、眼における3つの別個の組織に影響する。POAG関連の上昇したIOPは、小柱網(TM)(角膜と虹彩との間の角に位置する組織)での形態的変化および生物化学的な変化に起因する。栄養性房水のほとんどは、TMを通って眼の前方区画に存在する。緑内障の眼のTMにおけるTM細胞の進行性の喪失および細胞外破片の増加は、房水流出に対する耐性の増加を引き起し(Lutjen−DrecollおよびRohen 1996;Rohen 1983;Rohenら、1993;GriersonおよびCalthorpe 1988)、それによって、IOPが上昇する。IOPの上昇ならびに他の因子(例えば、虚血)は、視神経乳頭(ONH)において変性性の変化を起こし、ONHの進行性の「杯形成」(VarmaおよびMinckler 1996;HernandezおよびGong 1996;Hernandezら、1990;HernandezおよびPena 1997;Morrisonら、1990))、ならびに網膜神経節細胞(Quigleyら、2000;Quigley 1999;Quigleyら、1995;Kerriganら、1997)ならびに軸策の喪失を起こす。TM、ONHおよび網膜の神経節細胞に対する緑内障による損傷の原因となる詳細な分子機構は、未知である。
【0005】
現在の緑内障の治療は、緑内障の発病および進行の主な危険因子であるIOPを低下させることに関する。これらの治療はIOPを低下させるが、病原機構に直接的に対処せず
、そしてこの疾患は進行を続ける。緑内障を有する患者らの少なくとも半分は診断されず、患者らが緑内障を有すると診断されるまでに、患者らはすでに40%近くの網膜の神経節細胞を喪失してしまっている。したがって、緑内障のより早い検出および診断の方法が必要とされる。
この出願の発明に関連する先行技術文献情報としては、次のものがある。
【非特許文献1】Vaughan,D.ら、(1992)
【発明の開示】
【発明が解決しようとする課題】
【0006】
緑内障の重要性、および以前の診断方法の少なくとも部分的な欠点の見地において、初期段階の緑内障の診断のための改善されたより正確な方法を有することが所望される。さらに、緑内障の病原の機構に対処する新規の治療剤を有することが所望される。
【課題を解決するための手段】
【0007】
(発明の要旨)
本発明は、先行技術のこれらおよび他の欠点を、緑内障の初期診断のための方法およびキット、緑内障の処置のための方法およびキット、ならびに緑内障の処置において有用な化合物の同定のための方法およびキットを提供することによって克服する。
【0008】
ある特定の実施形態において、本発明は、細胞および体液から得られたサンプル中の骨形成タンパク質ファミリーメンバー遺伝子の発現の変化を検出することによる、緑内障の診断のための方法を提供する。この方法は、一般に以下の工程:
a)緑内障を有すると疑われる患者から組織サンプルまたは体液サンプルを得る工程;
b)このサンプルからDNAを抽出する工程;
c)複数のPCRプライマーを得る工程であって、これらプライマーの各々は、配列番号1、配列番号3、配列番号5、配列番号7、配列番号37、配列番号39、配列番号41、配列番号43、配列番号45、配列番号47または配列番号53由来の18〜1547連続するヌクレオチドからなる配列を含む、工程;
d)これらのプライマーを使用して、抽出されたDNAの領域を増幅してPCR産物を得る工程;
e)このPCR産物を分離する工程;ならびに
f)このPCR産物の配列と正常な遺伝子配列との間の違いを同定する工程、を包含し、ここで、この増幅配列と正常な遺伝子配列との間の違いは、緑内障の診断特徴である。
【0009】
一般に、本発明の方法は、個体からサンプルを得る工程およびこのサンプルからDNAを抽出する工程を包含し得る。BMP遺伝子ファミリーの特定のメンバーのための選択されたPCRプライマーは、PCR産物を得るために、抽出された遺伝子の適切な領域の増幅のために使用される。PCR産物を、評価されている特定のBMPファミリーの遺伝子(抽出されたDNA)の正常形態と変異形態との間のDNA配列の違いを効果的に同定する技術によって、分離される。同定された配列間の違いは、緑内障を示す。
【0010】
本発明の方法において使用される組織サンプルおよび液体サンプルは、血液または頬の細胞であり得る。
【0011】
代表的に、プライマー配列は、約10、15、または18ヌクレオチド〜約20ヌクレオチド、あるいは約10、15、または18ヌクレオチド〜約30ヌクレオチドの範囲の長さを有する。より長い配列(例えば、40、50、80、90、95、100、全長までさえ)が、ある実施形態についてなおさらに好ましい。少なくとも約18〜20ヌクレオチドのオリゴヌクレオチドの長さは、分子プローブとして有用であるように十分に特異的なハイブリダイゼーションを可能にするのに十分であるとして、当業者に十分受け入れられている(Lathe(1985)によって記載され、この参考文献は、この目的のために、本明細書中で参考として具体的に採用される)。好ましくは、このヌクレオチド配
列は、配列番号1、配列番号3、配列番号5、配列番号7、配列番号37、配列番号39、配列番号41、配列番号43、配列番号45、配列番号47または配列番号53の、20〜100連続するヌクレオチドからなる。プライマー配列は、その配列が公知であるBMPレセプター遺伝子の配列およびのBMP関連タンパク質の配列由来の、少なくとも10、15、または18連続するヌクレオチドの配列からなり得ることもまた企図される。
【0012】
配列番号1、配列番号3、配列番号5、配列番号7、配列番号37、配列番号39、配列番号41、配列番号43、配列番号45、配列番号47または配列番号53のいずれか1つに相補的な、10、18、20、30、50、60、65または100ヌクレオチド程度まで、および100ヌクレオチド程度を含むストレッチを有する核酸分子は、ハイブリダイゼーションプローブとしての有用性を有する。約18ヌクレオチドのヌクレオチド長を有するプライマーまたはプローブが、標的配列に対する高度に特異的なハイブリダイゼーションを提供することが、当業者に認識される。フラグメントの全サイズならびに相補的ストレッチのサイズは、特定の核酸セグメントの適用の意図した使用に最終的に依存する。より小さいフラグメントは、一般に、ハイブリダイゼーション実施形態における用途を見出し、ここで、相補的な領域の長さは変化し得る(例えば、検出したいと望む相補的配列に従って、約10、18、20もしくは30ヌクレオチドと約50、60、70、80、80もしくは100ヌクレオチドとの間、または全長まで)。
【0013】
特に好ましい実施形態において、プライマーは、配列番号1、配列番号3、配列番号5、配列番号7、配列番号37、配列番号39、配列番号41、配列番号43、配列番号45、配列番号47または配列番号53由来の連続する配列からなる。他の好ましい実施形態において、プライマーは、BMPレセプター遺伝子(ten Dijkeら、1993;Astromら、1999;Nohnoら、1995によって開示され、これらはすべて参考として採用される)または、BMP関連遺伝子(例えば、コーディン(chordin)(NCBI NM_029130)、グレムリン(gremlin)(Murphyら、1999;McMahonら、2000)、フォリスタチン(follistatin)(NCBI NM_003892)、またはバンビ(bambi)(NCBI NM_005791))由来の連続配列からなる。最も好ましくは、プライマーは、配列番号3由来の連続配列からなる。ある局面において、少なくともいくつかのプライマーは、検出可能な標識をさらに含み得る。
【0014】
他の実施形態において、本発明は、BMP2アゴニスト、BMP4アゴニスト、BMP5アゴニスト、BMP7アゴニスト、Smad1/5アゴニスト、コーディンアンタゴニスト、グレムリンアンタゴニスト、およびフォリスタチンアンタゴニストからなる群より選択される少なくとも1つの化合物からなる配列を含む組成物を、緑内障の処置を必要とする患者に投与することによる、緑内障の処置の方法を提供する。
【0015】
さらなる局面において、本発明は、緑内障の処置のための治療剤を同定する方法を提供する。治療剤は、例えば、以下によって同定され得る;
a)BMP−2A、BMP4、BMP−5またはBMP7を発現する組換え細胞の集団を含む第一の組成物を得ること;
b)候補物質を得ること;
c)この組成物およびこの候補物質をインキュベートすること;
BMP誘導性Smadシグナル伝達経路および/またはBMP調節性の遺伝子発現を開始する能力について、この化合物を試験すること;ならびにBMPのこれらの下流の効果を阻害、または刺激する候補物質を同定すること。
【0016】
本発明の別の局面は、本発明の配列および適切な試薬(例えば、タンパク質、ペプチド、または抗体自体に連結した検出可能な標識)を含む診断キットである。あるいは、検出
可能な標識は、本発明の配列に選択的にハイブリダイズする第2の配列に連結され得る。
【0017】
関連する実施形態は、本明細書中に開示される核酸配列またはペプチド配列もしくはタンパク質配列のいずれかの、薬学的に受容可能な処方物を含む治療キットを含む。このようなキットは、緑内障の診断のための、臨床的なサンプル中のBMP遺伝子およびBMPタンパク質の変化した発現の検出に有用である。
従って、本発明は、以下を提供する:
(項目1)
細胞または体液から得られたサンプルにおいて、骨形態形成ファミリーメンバー遺伝子の発現の変化を検出することによる、緑内障を診断する方法であって、該方法は、以下の工程:
g)緑内障を有すると疑われる患者から組織または液体のサンプルを得る工程;
h)該サンプルからDNAを抽出する工程;
i)複数のPCRプライマーを得る工程であって、該プライマーの各々は、配列番号1、配列番号3、配列番号5、配列番号7、配列番号37、配列番号39、配列番号41、配列番号43、配列番号45、配列番号47または配列番号53由来の、18〜1547連続するヌクレオチドからなる配列を含む、工程;
j)該プライマーを使用して、該抽出されたDNAの領域を増幅してPCR産物を得る工程;
k)該PCR産物を分離する工程;ならびに
l)該増幅した抽出されたDNA配列と該プライマーの配列との間の違いを同定する工程、を包含し、
ここで該増幅配列とプライマーとの間の違いは、緑内障の診断特徴である、方法。
(項目2)
項目1に記載の方法であって、前記組織サンプルまたは前記液体サンプルは、血液または頬の細胞である、方法。
(項目3)
項目1に記載の方法であって、前記プライマーは、配列番号1、配列番号3、配列番号5、配列番号7、配列番号37、配列番号39、配列番号41、配列番号43、配列番号45、配列番号47または配列番号53の、20〜100連続するヌクレオチドからなる配列を包む、方法。
(項目4)
項目1に記載の方法であって、前記プライマーは、配列番号3の20〜50連続するヌクレオチドからなる配列を包む、方法。
(項目5)
項目1に記載の方法であって、前記PCR産物は、SSCP、DGGE,ASOまたはRFLPによって分離される、方法。
(項目6)
緑内障を処置する方法であって、該方法は、BMP2アゴニスト、BMP4アゴニスト、BMP5アゴニスト、BMP7アゴニスト、Smad1/5アゴニスト、コーディンアンタゴニスト、グレムリンアンタゴニストおよび、フォリスタチンアンタゴニストからなる群から選択される少なくとも1つの化合物からなる配列を包む組成物を、このような処置を必要とする患者に投与する工程を包含する、方法。
(項目7)
緑内障を処置するための治療剤を同定する方法であって、該方法は以下の工程:
d)BMP−2A、BMP4、BMP−5、またはBMP7を発現する組換え細胞の集団を含む、第一の組成物を得る工程;
e)候補物質を得る工程;
f)該組成物および該候補物質をインキュベートする工程;
g)該組成物を、BMP誘導性Smadシグナル伝達経路、またはBMP調節性の遺伝子発現を開始する能力について試験する工程;ならびに
h)BMP誘導性のSmadシグナル伝達経路、または該BMP調節性の遺伝子発現を阻害または刺激する候補物質を同定する工程、
を包含する、方法。
【0018】
(好ましい実施形態の詳細な説明)
小柱網は、房水の正常な流れにおいて、重要な役割を担うことが提案されており、そして緑内障の眼における流出抵抗の主な部位であると推測されている。ヒト小柱網(HTM)細胞は、房水が眼から出る流出チャネルを裏打ちする特殊化した細胞である。細胞の変化した合成機能は、POAG、ステロイド緑内障、および他のタイプの緑内障の病因に関与し得る。
【0019】
集中的な研究の年月にもかかわらず、眼への緑内障による損傷の原因となる正確な分子機構は、知られていない。最近の研究によって、増殖因子が、緑内障に関連する眼の組織における正常な恒常性を維持することにおいて重要であり得、そして増殖因子/増殖因子レセプターの変化が、緑内障の病因において役割を担い得ることが示唆されている。増殖因子は、ポリペプチドの非常に大きなファミリーであり、細胞の増殖および分化を制御する。これらの分子は、遺伝子発現、細胞外マトリックスの組成および堆積、細胞骨格構築、ならびに細胞機能の調節に対して、種々の細胞特異的影響を有する。TMは、広範囲な様々の増殖因子、増殖因子レセプター(Tripathiら、1993a;Tripathiら、1993b;Tripathiら、1994a;Tripathiら、1994b;Wordingerら、1998;Wordingerら、1999)ならびに神経栄養ニューロトロフィン/神経栄養因子およびそれらのレセプター(Liuら、2001;Wordingerら、2000)を発現する。ONH星状細胞および篩板細胞(視神経乳頭の2つの細胞型)は、増殖因子であるニューロトロフィンおよびそのレセプターを発現する(Lambertら、2001;Penaら、1999)。房水はまた、種々の増殖因子(FGF2、EGF、TGFβ、HGFを含む)(Tripathiら、1996;Tripathiら、1991;Tripathiら,1992;HuおよびRitch、2001)ならびにニューロトロフィン(Chundruら、2000)を含む。房水のTGFβ−2およびHGFの上昇したレベルは、POAG患者において報告されている(Tripathiら、1994c;Inataniら、2001;Pichtら2001)。増殖因子は、TMおよびONHの正常な発達および/または機能を変更することによって、緑内障に関与し得る。
【0020】
本発明は、骨形成タンパク質(BMP)が、骨形成および軟骨形成を誘導するだけではなく、多数の細胞型に対する広範な影響を有する多機能的なサイトカインであり(Hogan 1996;Reddi 1997)、そしてヒト小柱網(HTM)細胞および視神経乳頭(ONH)細胞の両方によって発現される(Wordingerら、2002)という認識に部分的に由来する。BMPは、TGFβスーパーファミリーのメンバーであり、ヒトには約15〜20のBMP遺伝子、3つのBMPレセプター、そしてBMPアンタゴニストとして機能する多数のBMP関連タンパク質がある(Yamashitaら、1996)。BMPは、BMPR−IおよびBMPR−IIからなるレセプター複合体を介してシグナルを伝達する。スーパーファミリーメンバーのであるTGFβおよびTGFβR(Agarwalら、1997;Lambertら、1997)ならびにGDNFおよびGDNFR(Wordingerら、1999;Liuら、1999)は、HTM細胞およびONH細胞の両方によって発現されることが報告されている。
【0021】
BMPおよびBMPレセプターは、眼の組織で発現しているが(Obataら、199
9;Youら、1999)、以前の報告は、眼の発達に集中している。マウスにおけるBMPをコードする遺伝子の標的化された破壊が、網膜および水晶体に重篤な発達欠損を引き起こすので、BMPの機能は眼の発達に重要である(Jenaら、1997;Luoら、1995;Dudleyら、1995)。BMP−2、BMP−4およびBMP−7は、眼および網膜の発達に関与する(Jenaら、1997;FurutaおよびHogan 1998;Reddi 2000;Trousseら、2001)。BMP−6およびBMP−7もまた、低血糖または虚血損傷からニューロンを保護する役割を果たすようであり(Nonnerら、2001;Liuら、2001)、そしてBMP2は、神経節細胞のニューロトロフィン発現を増強することが示されている(Zhangら、1998)。Bmp4について不十分なハプロイド(haploinsufficient)であるヘテロ接合体ノックアウトマウスは、前方部分の発育不全、IOPの上昇、および視神経異常を含む眼の表現型を有する(Changら、2001)。ヒトの出生後の眼におけるBMPの役割に関して、非常に限られた情報が発表されている。
【0022】
Mohanおよび共同研究者(1998)は、BMP−2およびBMP−4ならびにBMPレセプターが、成体の角膜細胞において発現することを報告し、そしてBMP機能が、角膜の角膜実質細胞の増殖およびアポトーシスを含み得ることを示唆した。Youおよび共同研究者(1999)は、この研究を確認し、そしてまた、エキソビボならびに培養角膜上皮および培養基質細胞でのBMP−3、BMP−5およびBMP−7の発現を報告した。彼らは、BMPの転写レベルが、基質においてより高く、一方でレセプターのレベルが培養角膜上皮細胞においてより高いことを報告した。
【0023】
RT−PCRを使用して、本発明者らは、BMPのmRNA、BMPレセプターであるBMPR−IA、BMPR−IBおよびBMPR−IIのmRNA、ならびにBMP結合タンパク質であるグレムリン、コーディン、フォリスタチン、およびバンビのmRNAを、HTM、篩板(LC)およびONH星状細胞細胞株および組織で発見した(Wordingerら、2002)。本発明者らはさらに、HTM細胞およびONH細胞が、タンパク質BMP−2、BMP−4、BMP−5およびBMP−7を発現することを発見した。
【0024】
緑内障は、BMPシグナル伝達ファミリーのメンバーの遺伝子における遺伝的変化の特徴付けによって診断される。本明細書中で使用する場合、句「骨形成タンパク質ファミリーメンバー遺伝子」および「BMPシグナル伝達ファミリー」は、全てのBMP、BMPレセプターおよび関連するタンパク質をいう。用語「遺伝的変化」は、当業者にとって周知である。特定の遺伝子における遺伝的変化に関する障害の多数の例が存在する(例えば、Cummings 1997;Strachanら、1996;Jordeら、1999を参照のこと)。特定の遺伝子(例えば、BMP)における遺伝的変化は、当業者に周知の様々な技術を使用することによって決定され得る(例えば、SSCP、DGGE、ASO、RFLP、ヘテロニ重鎖分析、CCM、PTT、ならびにRNase切断(Birrenら、1998を参照のこと))。
【0025】
緑内障は、上昇したIOPおよび/または緑内障的な眼の神経障害を引き起こす、眼における1つ以上のBMPファミリー遺伝子の発現の変化によって生じ得る。「変化したBMP遺伝子発現」は、この遺伝子産物の発現が正常とは異なることを意味する。この用語はまた、遺伝子またはタンパク質の配列における変化もいい得る。正常なBMP遺伝子は、十分特徴づけられており(上記を参照のこと)、そしてBMPの発現は、TMおよびONHを含む様々な組織において報告されている。BMPファミリー遺伝子のコード領域の遺伝的変化は、これらのタンパク質の機能を変化し得る。コード領域外の遺伝的変化もまた、緑内障を引き起こし得る。
【0026】
特定の遺伝子のコード領域の「外の変化」が、遺伝子発現の調節において重要であるこ
とは当業者に周知である。例えば、ほとんどの遺伝子のコード領域の上流(5’)領域は、その遺伝子の発現を「促進」および調節するプロモーター領域として公知である。プロモーター領域は、遺伝子発現の活性化または抑制の原因となる、様々な転写因子およびDNA結合タンパク質によって認識される多数のヌクレオチド配列を含む。遺伝子の下流(3’)領域は、遺伝子産物のポリアデニル化を決定し得、それによってRNAプロセシングおよび遺伝子産物の翻訳を調節し得る。
【0027】
緑内障を示すBMP遺伝子の発現の変化または、遺伝子配列における変異は、当業者に周知の技術を使用して検出され得る。例えば、ほぼ全ての長さの核酸フラグメントが使用され得、その全長は、調製しやすさおよび意図するプロトコールでの使用しやすさによって好ましくは限定されることが企図される。本明細書中に開示される核酸配列はまた、核酸ハイブリダイゼーション実施形態におけるプローブまたはプライマーとしての有用性も有し得る。このように、BMP−2A(配列番号1)、BMP−4(配列番号3)、BMP−5(配列番号5)、BMP−7(配列番号7)、BMP−RIA(配列番号37)、BMP−RIB(配列番号39)、BMP−RII(配列番号41)、コーディン(配列番号43)、グレムリン(配列番号45)、フォリスタチン(配列番号47)、またはバンビ(配列番号53)の少なくとも14ヌクレオチド長連続する配列と同じ配列を有するか、またはこれらに相補的な配列からなる配列領域を含む核酸セグメントは、特定の有用性を見出すことが企図される。より長く、連続した、同一または相補的な配列(例えば、約20、30、40、50、100、200、500、1000ヌクレオチド(全ての中間の長さを含む)の配列)、約1547ヌクレオチド(BMP−2Aについて)、1946ヌクレオチド(BMP−4について)、2153ヌクレオチド(BMP−5について)および1878ヌクレオチド(BMP−7について)、2932ヌクレオチド(BMP−RIAについて)、2032ヌクレオチド(BMP−RIBについて)、3611ヌクレオチド(BMP−RIIについて)、3561ヌクレオチド(コーディンについて)、4049ヌクレオチド(グレムリンについて)、1386ヌクレオチド(フォリスタチンについて)、および1523ヌクレオチド(バンビについて)の全長配列までさえもまた、ある実施形態において使用される。
【0028】
この文脈における「中間の長さ」は、引用された範囲(例えば、14、15、16、17、18、19、20など;21、22、23など;30、31、32など;50、51、52、53など;100、101、102、103など;150、151、152、153など;200〜500;500〜1,000;1,000〜2,000の範囲の全てを含み、そして、2,001、2,002、2,050、2,051などの配列までおよびそれらを含む)の間の任意の長さを意味することが容易に理解される。
【0029】
このような核酸プローブおよび核酸プライマーがBMPコード配列に特異的にハイブリダイズする能力、ならびにプライマーがBMP配列を特異的に増幅する能力により、所与のサンプル中の相補配列の存在の検出において、それらが使用可能となる。ところが、変異種のプライマーの調製のための配列情報の使用、または他の遺伝的構築物の調製における使用のためのプライマーの使用を含む、他の使用が想定される。
【0030】
BMP−2A(配列番号1)、BMP4(配列番号3)、BMP−5(配列番号5)、BMP7(配列番号7)、BMP−RIA(配列番号37)、BMP−RIB(配列番号39)、BMP−RII(配列番号41)、コーディン(配列番号43)、グレムリン(配列番号45)、フォリスタチン(配列番号47)またはバンビ(配列番号53)と同一かもしくは相補的な、10、20、30、50、またはちょうど100〜200ヌクレオチドなどの連続するヌクレオチドストレッチからなる配列領域を有する核酸分子は、、例えばサザンブロッティングおよびノザンブロッティングに加えて、SNP評価および固相ハイブリダイゼーションアッセイにおける使用のための、ハイブリダイゼーションプロー
ブとして特に企図される。これにより、組織および細胞の両方におけるBMPの構造遺伝子または調節遺伝子の解析が可能となる。フラグメントの全サイズならびに相補的ストレッチのサイズは、特定の核酸セグメントの適用について意図する使用に最終的に依存する。より小さいフラグメントは、一般に、ハイブリダイゼーション実施形態における使用を見出す。ここで、連続する相補的な領域の長さは、変化し得る(例えば、約10ヌクレオチドと約100ヌクレオチドとの間)しかし、約1547ヌクレオチド(BMP−2Aについて)、1946ヌクレオチド(BMP−4について)、2153ヌクレオチド(BMP−5について)、ならびに1878ヌクレオチド(BMP−7について)、2932ヌクレオチド(BMP−RIAについて)、2032ヌクレオチド(BMP−RIBについて)、3611ヌクレオチド(BMP−RIIについて)、3561ヌクレオチド(コーディンについて)、4049ヌクレオチド(グレムリンについて)、1386ヌクレオチド(フォリスタチンについて)、1523ヌクレオチド(バンビについて)までの、より長い連続する相補的ストレッチが、検出したいと望む相補的配列の長さに従って使用され得る。
【0031】
約10〜14ヌクレオチド長のハイブリダイゼーションプローブの使用により、安定かつ選択的な二重鎖分子の形成が可能である。10塩基長を越えるストレッチにわたる連続する相補的配列を有する分子が一般に好ましいが、ハイブリッドの安定性および選択性を高め、それによって得られる特異的なハイブリッド分子の質および程度を改善するために、一般に、15〜20連続するヌクレオチドの遺伝子相補的なストレッチを有するか、または所望される場合、さらに長い核酸分子を設計することが好まれる。
【0032】
ハイブリダイゼーションプローブは、本明細書中に開示される任意の配列のいずれの部分からも選択され得る。配列番号1、配列番号3、配列番号5、配列番号7、配列番号37、配列番号39、配列番号41、配列番号43、配列番号45、配列番号47または配列番号53に示される配列を検討すること、ならびにプローブまたはプライマーとしての利用が望まれる、約10ヌクレオチド長から、全長配列までを含むまでの配列の任意の連続する部分を選択することが必要なだけである。プローブ配列およびプライマー配列の選択は、様々な因子によって左右され得る(例えば、例示のみとして、さらなるDNAを増幅するために、全配列の終点に向かうプライマーまたは、機能的ドメインコード配列の末端からのプライマ−の設計が望まれ得る)。
【0033】
BMP−2A(配列番号1)、BMP4(配列番号3)、BMP−5(配列番号5)、BMP7(配列番号7)、BMP−RIA(配列番号37)、BMP−RIB(配列番号39)、BMP−RII(配列番号41)、コーディン(配列番号43)、グレムリン(配列番号45)、フォリスタチン(配列番号47)またはバンビ(配列番号53)内由来の連続する配列を含む核酸セグメントを選択および調製する工程は、核酸フラグメントを調製するとして代替的に記載されている。もちろん、フラグメントはまた、他の技術によって(例えば、機械的剪断または、制限酵素消化によって)獲得され得る。小さな核酸セグメントまたはフラグメントは、例えば、自動化されたオリゴヌクレオチド合成装置の使用によって一般に実行されるような、化学的手段によるフラグメントの直接的な合成によって容易に調製され得る。また、フラグメントは、核酸再生成技術(例えば、米国特許第4,683,202号および同第4,682,195号(各々は、参照として上記に記載される)のPCRTM技術)の適用によって、組換え生成のために選択した配列を組換えベクターに導入することによって、ならびに分子生物学の分野の当業者に一般に公知の他の組換えDNA技術によって、獲得され得る。
【0034】
従って、本発明のヌクレオチド配列は、BMPの遺伝子またはcDNAの相補的なストレッチとの二重鎖分子を選択的に形成するその能力のために、使用され得る。想定される適用に依存して、標的配列に対するプローブの選択性を様々な程度にするために、様々な
程度のハイブリダイゼーションの選択性を使用することが所望される。高い選択性を必要とする適用について、ハイブリッドを形成するために、比較的ストリンジェントな条件を使用することが代表的に所望される(例えば、比較的低い塩および/または高い温度条件(例えば、50℃〜70℃の温度で0.02M〜0.15MのNaClによって提供される)を選択する)。このような選択的条件は、もしあったとしてもプローブと鋳型または標的鎖との間のミスマッチをほとんど許容せず、そして特にBMP遺伝子の解析に適切である。
【0035】
もちろん、いくつかの適用のために、例えば基礎をなす鋳型にハイブリダイズした変異プライマー鎖を使用する変異体の調製または同定を所望する場合、あるいは関連する種由来のBMPコード配列、機能的同等物などを単離することを求める場合、あまりストリンジェントでないハイブリダイゼーション条件が、ヘテロ二重鎖の形成を可能にするために代表的に必要とされる。これらの状況下で、条件(例えば、0.15M〜1.0Mの塩、20℃〜55℃の範囲にある温度)を使用することが所望され得る。それにより、交差ハイブリダイズする種は、コントロールハイブリダイゼーションに関連してポジティブにハイブリダイズするシグナルとして、容易に同定され得る。いずれの場合においても、条件は、NaCl濃度を低くすることによって、または温度の上昇と同じ様式でハイブリッド二重鎖を不安定化するように作用するホルムアミドの斬新量を添加することによって、よりストリンジェントにされ得ることが一般に理解される。このように、ハイブリダイゼーション条件は容易に操作され得、従って一般に、所望される結果に依存する選択の方法である。
【0036】
ある実施形態において、ハイブリダイゼーションを決定するための適切な手段(例えば、標識)と組み合わせて本発明の核酸配列を使用することは有益である。様々な種類の適切な指示手段は、当該分野で公知であり、これには蛍光、放射性、酵素的または他のリガンド(例えば、アビジン/ビオチン)が挙げられ、これらは、検出可能なシグナルを生じ得る。好ましい実施形態においては、放射性または他の環境的に所望されない試薬の代わりに、蛍光標識または酵素タグ(例えば、ウレアーゼ、アルカリホスファタ−ゼまたはペルオキシダーゼ)を使用することが所望される傾向にある。酵素タグの場合、ヒトの眼または吸収分光測光計で見ることができる手段を提供して、相補的核酸含有サンプルとの特異的ハイブリダイゼーションを同定するために使用され得る比色指示基質は公知である。
【0037】
一般に、本明細書に記載されたハイブリダイゼーションプローブは、溶液ハイブリダイゼーションならびに固相を使用する実施形態の両方において試薬として有用であると想定される。固相を含む実施形態において、試験DNA(またはRNA)は、吸収されるか、または他の方法で、選択されたマトリックスもしくは表面に結合する。この固定された一本鎖の核酸は、その後、所望される条件下で、選択されたプローブとの特異的なハイブリダイゼーションに供される。この選択された条件は、必要とされる特定の基準に基づく特定の環境に依存する(例えば、G+C含有量、標的核酸のタイプ、核酸の供給源、ハイブリダイゼーションプローブのサイズなどに依存する)。非特異的に結合したプローブ分子を除くためのハイブリダイズ表面の洗浄に続いて、標識によって、特異的ハイブリダイゼーションは検出されるか、またはさらに測定される。
【0038】
本発明は、BMP−2A(配列番号1)、BMP4(配列番号3)、BMP−5(配列番号5)、BMP7(配列番号7)、BMP−RIA(配列番号37)、BMP−RIB(配列番号39)、BMP−RII(配列番号41)、コーディン(配列番号43)、グレムリン(配列番号45)、フォリスタチン(配列番号47)またはバンビ(配列番号53)の特定の核酸配列コピーおよびアミノ酸に限定されないこともまた理解される。組換えベクターおよび単離されたDNAセグメントは、従って、BMPコード領域自体、遺伝子の上流領域または下流領域、選択された変化または改変を基本的なコード領域中に有す
るコード領域を様々に含み得るか、あるいは、それにもかかわらずBMPコード領域を含むより大きなペプチドをコードし得るかまたは、改変体アミノ酸配列を有する生物学的に機能的な等価なタンパク質もしくはポリペプチドをコードし得る。
【0039】
本発明のDNAセグメントは、生物学的に機能的な等価なBMPタンパク質およびポリペプチドを包含する。このような配列は、コドン縮重および機能的等価性の結果として生じ得、これらは核酸配列およびこのようにコードされるタンパク質内に天然に存在するものであることが公知である。あるいは、機能的に等価なタンパク質またはポリペプチドは、タンパク質構造における変化が、交換されるアミノ酸の特性の考慮に基づいて操作され得る組換えDNA技術の適用を通じて作製され得る。人為的に設計された変化は、部位特異的変異誘発技術の適用を通して、例えば、タンパク質の抗原性に改善を導入するため、または分子レベルでの結合活性を試験するためにBMP変異を試験するために導入され得る。
【0040】
緑内障の処置のための治療剤は、ペプチドまたはタンパク質、ペプチド模倣物、オリゴヌクレオチドまたは誘導体化オリゴヌクレオチド、あるいは薬物様の低分子であり得、その全てが、眼のBMP経路の1つ以上の局面に影響する。好ましい治療剤は以下である:(1)BMP2アゴニスト、BMP4アゴニスト、BMP5アゴニスト、またはBMP7アゴニスト;(2)コーディンアンタゴニスト、グレムリンアンタゴニスト、フォリスタチンアンタゴニスト、またはバンビアンタゴニスト;および/あるいは(3)Smad1アゴニスト、Smad5アゴニスト、および/またはSmad4アゴニスト。
【0041】
この薬剤は、当業者に周知の技術を使用することで、直接的に眼に送達され得る(例えば、局所的点眼薬または軟膏;盲嚢中、のあるいは強膜に隣接してかまたは眼内に移植された徐放デバイス;眼周囲、結膜、サブテノン(sub−Tenons)、眼房内(intracameral)または硝子体内注射)か、あるいは非経口的に(例えば:経口;静脈内、皮下または筋内注射;皮膚送達など)。以下は、本発明によって具体化される可能な処方物の例示である。
(a)局所的な眼用処方 重量%
眼のBMP−4の発現を上昇させる薬剤 0.01〜2
HPMC 0.5
塩化ナトリウム 0.8
BAC 0.01
EDTA 0.01
NaOH/HCl qs pH7.4
精製水 qs 100mL
(b)局所的な眼用処方 重量%
グレムリンアンタゴニスト 0.01〜2
HPMC 0.5
塩化ナトリウム 0.8
BAC 0.01
EDTA 0.01
NaOH/HCl qs pH7.4
精製水 qs 100mL
(c)局所的な眼用処方 重量%
Smad1/5アゴニスト 0.01〜2
HPMC 0.5
塩化ナトリウム 0.8
BAC 0.01
EDTA 0.01
NaOH/HCl qs pH7.2
精製水 qs 100mL
本発明の化合物は、眼内挿入デバイス中で処方されることがさらに企図される。
【0042】
(A.治療剤についてのアッセイ)
本発明はまた、BMPシグナル伝達経路に関与する新しい抗緑内障治療剤の発見のために有用である(図5を参照のこと)。選択的BMPリガンドは、BMP I型およびBMP II型セリン/スレオニンキナーゼレセプター(BMP−RIおよびBMP−RII)に結合し、そしてSmadタンパク質を介してシグナルを伝達する。BMPシグナルは、タンパク質−タンパク質相互作用およびタンパク質−DNA相互作用を介して、Smadによって伝搬される(AttisanoおよびTuen Lee−Hoeflich 2001)。調節的Smad1およびSmad5は、BMPレセプターへのリガンドの結合によって(リン酸化を介して)活性化される(von BubnoffおよびCho 2001)。これらの調節的Smadは、その後、Smad4と相互作用して、核に転移するヘテロマー複合体を形成する。この複合体は、どの核補助因子が存在するかに依存して、この転写複合体を認識する選択的遺伝子の転写を活性化することまたは抑制することが可能である。
【0043】
BMP/Smadシグナル伝達経路は、いくつかの機構によってネガティブに調節される。あるBMP結合性タンパク質(例えば、グレムリン、BAMBI、またはフォリスタチン)は、BMPを結合し、そしてBMPレセプターとのそれらの相互作用を阻害する。さらに、阻害性Smadタンパク質(例えば、Smad6およびSmad7)が存在し、これらはBMPレセプターを結合して不活性化させる(Kowabataら、1998;Itohら、2000;Miyazono 2000)。本発明者らは、ヒトTM細胞、ONH星状細胞および相篩板細胞が、BMPレセプター複合体についてのメッセージおよびタンパク質を発現することを発見している。従って、これらの細胞は、内因性のBMPリガンドに応答し得る。
【0044】
様々な方法が、新しい抗緑内障治療剤を発見するために使用され得、そしてこれらの技術は当業者に周知である。例えば、BMPのアゴニストまたはインヒビターとして働くペプチドまたはペプチド模倣剤は、BMP/BMPレセプター構造の分子モデリングによって発見され得る(Nickelら、2001)。BMPシグナル伝達は、Smadタンパク質の選択されたセットを含む(Kawabataら、1998;Itohら、2000;Attisenoら、2000)。選択されたBMPアゴニストおよびSmadアゴニストは、細胞ベースのアッセイを使用して発見され得る。試験細胞は、適切なBMPレセプターを発現するべきであり、適切なBMPシグナル伝達経路を所有するべきである。BMPシグナル伝達の主な影響の1つは、遺伝子発現の変化であるので、BMPアゴニストおよびSmadアゴニストは、BMP誘導性遺伝子についてのスクリーニングによって発見され得る。BMP調節性遺伝子の誘導はまた、定量的RT−PCR(Wangら、2001)、DNAマイクロアレイ、またはレポーター遺伝子構築物を使用することによって、mRNAのレベルを定量してアッセイされ得る。BMPシグナル伝達、BMP結合タンパク質(BMP関連タンパク質としても公知)(例えば、コーディン、グレムリン、およびフォリスタチン)の天然のインヒビターが存在する。タンパク質インヒビターのアンタゴニストは、リガンド結合アッセイを用いて発見され得る。例えば、試験薬剤は、組み換え精製グレムリンに付加され得、そしてグレムリンに結合するこれら薬剤は、当業者に公知の様々な技術を使用することで同定される。これらの薬剤がグレムリンアンタゴニストであるか否かを決定するために、上記で記載したアッセイと類似の細胞ベースのアッセイが使用される。
【0045】
任意の公知のインビトロおよびインビボのスクリーニングモデルは、本発明と組み合わ
せて、BMPファミリーの遺伝子に関する新しい緑内障治療を同定するために使用され得ることが企図される。このようなモデルは当業者にとって周知であり、それらの実施は決まりきった仕事になる。小さいペプチドまたはペプチド模倣物は、BMP、BMPRおよび/またはBMP結合タンパク質の遺伝子産物の構造/機能の知識に基づいて設計され得る。リガンド結合アッセイは、BMP、BMPRまたはBMP結合タンパク質に結合する低分子の検出に使用され得る。細胞ベースのアッセイにより、BMPシグナル伝達経路に対する様々な薬剤の影響を見ることができ得る。レポーター遺伝子に連結されたBMPファミリー遺伝子プロモーターを含むノック−イン細胞株は、BMPファミリーメンバーの遺伝子発現を変化させる薬剤を探索するために作製され得る。これらのアッセイは、アゴニスト分子およびアンタゴニスト分子の両方の同定に使用され得る。エキソビボアッセイ(例えば、灌流培養されたヒトの眼由来の前方区画の(Clarkら、1995a;Pangら2000))は、TM組織におけるIOPおよびBMPシグナル伝達に対する薬剤の影響を試験するために使用され得る。緑内障のげっ歯類モデルは、マウスおよびラットの、安定なBMPファミリーメンバーのトランスジェニック株、ノックアウト株、またはノックイン株を作製するための周知の技術を使用して作製され得る。これらのげっ歯類モデルは、緑内障様の表現型を変化させる薬剤についてスクリーニングするために使用され得る(例えば、IOPに対する影響を評価するための限圧測定、緑内障の目の神経学に対する影響を評価するための組織学)。
【0046】
(B.キット)
本発明は、緑内障の初期検出のための方法、化合物およびキットを提供する。このキットは、BMPポリペプチドまたはBMPタンパク質をコードする核酸セグメントを含み得る。このキットはさらに、サンプルと本発明の核酸またはペプチドとの間の相互作用を検出するための試薬を含み得る。提供された試薬は、放射性標識化、蛍光標識化、または酵素的標識化され得る。このキットは、本発明の核酸またはペプチドもしくはタンパク質と結合または相互作用し得る、公知の放射性標識された薬剤を含み得る。
【0047】
キットの試薬は、液体溶液としてか、固体支持体に結合してかまたは乾燥した粉末として提供され得る。好ましくは、試薬が液体溶液中に提供される場合、この液体溶液は水溶液である。好ましくは、提供される試薬が固体支持体に結合する場合、この固体支持体はクロマトグラフィー媒体、複数のウェルを有する試験プレート、あるいは顕微鏡スライドであり得る。提供される試薬が乾燥した粉末である場合、この粉末は、提供され得る適切な溶媒の添加によって再び構成され得る。
【0048】
なお、さらなる実施形態において、本発明は、緑内障の診断のための診断方法、および関連するキットに関する。本発明のBMP関連のペプチドおよび核酸は、患者のサンプル由来のBMP核酸における多型または変異を検出するために使用され得ることが提案される。一般に、これらの方法は、このような多型または変異を含むことが疑われるサンプルを最初に獲得する工程、本発明のペプチドまたは核酸とこのサンプルとを接触させる工程、(この場合には、複合体を形成させるのに有効な条件下であり得る)、および次いで複合体の存在を検出する工程を包含する。
【0049】
一般に、複合体形成の検出は当該分野で周知であり、多数のアプローチの適用を通して成し遂げられ得る。例えば、本発明は、ELISA、RIA、間接的蛍光技術などの適用を企図する。概して、複合体形成は、標識(例えば、放射性標識または酵素タグ(例えば、アルカリホスファターゼ、西洋ワサビペルオキシダーゼなど)の使用を通して検出される。もちろん、二次結合リガンドの使用によってさらなる利益を見出し得る。
【0050】
以下の実施例は、本発明の局面の実行において本発明者らによって使用される技術を示す。これらの技術が、本発明を実施するための好ましい実施形態の例示であることは、本
開示を鑑みて当業者に認識されるべきであり、多数の改変が、本発明の精神および意図される範囲から逸脱することなくなされ得ることが認識される。
【実施例】
【0051】
(実施例1)
細胞培養:ヒトTM細胞およびONH細胞を、ドナーの眼から記載されるように生成した(Steelyら、1992;Steelyら、2000;Wilsonら、1993;Clarkら、1994;Clarkら、1995b;Clarkら、1995c;Clarkら、1996;Clarkら、2001a;Clarkら、2001b;Dickersonら、1998;Wordingerら、1998;Wordingerら、1999;Wordingerら、2000;Wordingerら、2002;Lambertら、2001;Agarwalら、1999;Liuら、2001)。TM細胞を、年齢6日齢から90歳の範囲のドナーのTM外移植体から増殖させた。ヒト視神経乳頭星状細胞および篩板(LC)細胞を、慎重に解剖した視神経乳頭(2日齢〜90歳までの年齢のドナー)から生成し、そして以前の報告に従って特徴付けた(Lambertら、2001;Clarkら、1995a)。細胞を、以下の培地:TM細胞については10%ウシ胎仔血清(HyClone,Logan,UT)および抗生物質(Gibco BRL−Life Technologies,Grand Island,NY)を含むHam’s F10培地(JRH Biosciences、Lenexa、KS);LC細胞については10%FBSを含むダルベッコ改変イーグル培地(DMEM,HyClone);そしてONH星状細胞については5%FBSを含む星状細胞増殖培地(AGM,Clonetics,San Diego,CA)内で、コンフルエントまで増殖させた。
【0052】
RT−PCR:ヒトTM組織およびONH組織もまた、ドナーの眼から解剖した(Wordingerら、1998;Wangら、2001)。全RNAをTM細胞およびTM組織ならびにONH細胞およびONH組織から、TRIzol抽出(Gibco BRL−Life Technologies)を使用して抽出し、そしてcDNAを標準的な手順(Wordingerら、1998;Worgingerら、1999;Worgdingerら、2000;Wordingerら、2002)を使用して逆転写によって調製した。PCRプライマーを、Oligos 4.0ソフトウェアプログラムを使用して設計した(表1のプライマー対を参照のこと)。全てのプライマー対を、潜在的に混入したゲノムDNA配列の増幅が予想される産物よりも実質的に長いmRNA PCR産物を生じるように設計した。なぜなら、RNAプロセシングの間に切除されるイントロン配列が、ゲノムDNAに含まれるからである。β−アクチンPCRプライマーAGGCCAACCGCGAGAAGATGACC(上流)およびGAAGTCCAGGGCGACGTAGCAC(下流)と、55℃アニーリング温度とにより、350bpのPCR産物を得た。
【0053】
PCR反応を、Taq Start Antibody Hot Startを以下:94℃で2分間、92℃で2分間、そして(最適なアニーリング温度で30秒間、72℃で90秒間の伸長、92℃で45秒間の変性)×40サイクルのサイクル条件を使用して、記載されたように実行した(Wordingerら、1998;Wordingerら、1999;Wordingerら、2000;Lambertら、2001;Wordingerら、2002)。増幅したPCR産物を、1.5%アガロースゲルで水平な電気泳動によって試験した。RT−PCR産物の特異性を確実にするために、増幅したPCR産物内の領域にハイブリダイズする、Oligo 4.0を用いて設計したプローブを使用して、サザンブロット解析を実行した。PCR反応の特異性を証明するために、PCR産物を配列決定した。表2は、ヒトTMおよびONHで発現しているBMPファミリーのメンバーを列挙している。
【0054】
【表1】

【0055】
【表2】
ウェスタンイムノブロッティング:タンパク質を、溶解緩衝液を使用して培養細胞から抽出し、そしてタンパク質を、ニトロセルロースメンブレンへの電気泳動的な移動に先立って、変性ポリアクリルアミドゲル電気泳動によって分離した(Lambertら、2001)。メンブレンを、5%ミルク(BMPについて)または3%ゼラチン(BMPRについて)でブロッキングし、そして以下の一次抗原:BMP2、BMP4、BMP5、BMP7(すべてSanta Cruz,Santa Cruz,CAから)、あるいはBMP−RIA、BMP−RIB、BMP−RII(Jackson Immuno Research,West Grove,PAから)と共にインキュベートした。メンブレンを洗浄し、二次抗体(BMPについてヤギ抗マウスIgG−西洋ワサビペルオキシダーゼ、Santa Cruz;BMPレセプターについてロバ抗ヤギ−西洋ワサビペルオキシダーゼ、Jackson Immuno Research)と共にインキュベートし、そしてWesternBreeze化学発光免疫検出システム(Invitrogen,Carlsbad,CA)を使用して現像した。
【0056】
ヒトのTM細胞およびTM組織におけるBMP、BMPR mRNAの発現:ヒトのTM細胞およびTM組織においてBMP−2プライマー対、BMP−4プライマー対、BMP−5プライマー対およびBMP−7プライマー対についての予想された増幅産物は、図6に示される。特異的プローブを使用したサザンブロットによって、これらが予想されたPCR産物であることが証明された。全てのヒトTM細胞株および組織は、BMP−2、BMP−4、およびBMP−7のメッセージを発現した。ところが、BMP−5のメッセージは、ヒトTM組織サンプルにおいて検出されないくらい低いものであった(図6、レーン6およびレーン7)。cDNA無しのコントロール反応は、増幅産物を生じず、試薬およびプライマーには、DNAもRNAも混入しれていなかったことを示す(図6、レーンC)。
【0057】
図7は、ヒトのTM細胞およびTM組織においてBMP−RIAプライマー対、BMP−RIBプライマー対、およびBMP−RIIプライマー対についての予想されたサイズの増幅産物を示す。全てのヒトTM細胞およびヒトTM組織は、BMPレセプター複合体のメッセージを発現した。特異的なプローブを使用したサザンブロットによって、これらが予想されたPCR産物であることが証明された。代替的な増幅産物(350bp)は、BMP−RII PCR反応において検出された。その代替的な増幅産物は、全てのヒトTM細胞およびヒトTM組織において存在していた。この代替的なバンドは現在、選択的にスプライスされた型のレセプターであるか否かを決定するために同定中である。cDNA無しのコントロール反応は増幅産物を生じず(図7、レーンC)このことは、試薬およびプライマーには、DNAもRNAも混入していなかったことを示す。
【0058】
ヒトONH細胞およびONH組織におけるBMPおよびBMPレセプターmRNAの発現:ヒトONH星状細胞およびONH組織における、BMP−2プライマー対、BMP−4プライマー対、BMP−5プライマー対およびBMP−7プライマー対についての予想されたサイズの増幅産物は、図8に示される。全てのONH星状細胞およびONH組織は、それぞれのBMPのメッセージを発現した。ヒト脳星状細胞を、ポジティブコントロール細胞株として使用した。特異的プローブを使用したサザンブロットによって、これらが予想されたPCR産物であることが証明された。BMP−2以外の、全ての他のBMPは、ヒト脳星状細胞により発現された(図8、レーン7)。cDNA無しのコントロール反応は、増幅産物を生じず(図8、レーンC)、試薬およびプライマーには、DNAもRNAも混入していなかったことを示す。
【0059】
図9は、培養ヒトLC細胞における、BMP−2プライマー対、BMP−4プライマー対、BMP−5プライマー対およびBMP−7のプライマー対についての予想されたサイズの増幅産物を示す。全てのLC細胞株は各々のBMPのメッセージを発現した。特異的
なプローブを使用したサザンブロットによって、これらが予想されたPCR産物であることが証明された。cDNA無しのコントロール反応は増幅産物を生じず(図9、レーンC)このことは、試薬およびプライマーには、DNAもRNAも混入していなかったことを示す。
【0060】
ヒトONH星状細胞およびONH組織における、BMP−RIA、BMP−RIBおよびBMP−RIIのプライマー対についての予想されたサイズの増幅産物は、図10に示される。全てのONH星状細胞およびONH組織は、BMP−RIAおよびBMP−RIBのメッセージを発現した。特異的プローブを使用したサザンブロットによって、これらが予想されたPCR産物であることが証明された。ONH組織(図10、レーン6)以外の全てのONH星状細胞細胞株によってBMP−RIIが発現された。全てのBMPレセプターのメッセージ(図10、レーン7)が、ポジティブコントロールとして働くヒト脳星状細胞細胞株により発現された。ONH組織およびONH細胞株におけるBMP−RIIの発現には食い違いが存在するようであった。ONH組織での発現の減少は、発現の低いレベルを反映し得る。cDNA無しのコントロール反応は、増幅産物を生じず(図5、レーンC)、試薬およびプライマーにはDNAもRNAも混入していなかったことを示す。
【0061】
図11は、培養ヒトLC細胞における、BMP−RIA、BMP−RIB、およびBMP−RIIのプライマー対についての予想されたサイズの増幅産物を示す。全てのLC細胞株は、各々のBMPレセプターのメッセージを発現した。特異的なプローブを使用したサザンブロットによって、これらが予想されたPCR産物であることが証明された。cDNA無しのコントロール反応は増幅産物を生じず(図11、レーンC)このことは、試薬およびプライマーには、DNAもRNAも混入していなかったことを示す。
【0062】
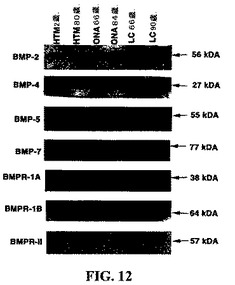
ヒトTM細胞およびTM組織ならびにONH細胞およびONH組織におけるBMPタンパク質およびBMPレセプタータンパク質の発現:図12は、ヒトTM細胞およびTM組織ならびにONH細胞およびONH組織におけるBMP−2、BMP−4、BMP−5、BMP−7、BMP−RIA、BMP−RIB、およびBMP−RIIのタンパク質の化学発光免疫ブロット検出を示す。研究した全ての細胞株は、それぞれのBMPタンパク質を発現した。BMPタンパク質は、細胞株において以下の分子量:BMP−2について54〜56kDa、BMP−4について25〜27kDa、BMP−5について55〜57kDa、およびBMP−7について77kDaで検出された。複数のバンドがBMP−2およびBMP−4について検出され、これらは、他の研究において見られるようなこれらBMPのグリコシル化された型、および部分的にグリコシル化された型を示す可能性が最も高い。ところがグリコシル化の研究は、本研究の範囲を越えているので、本研究者らはグリコシル化の研究を行っていない。BMPレセプタータンパク質は、細胞株において以下の分子量:BMP−RIAについて38kDa、BMP−RIBについて64kDa、およびBMP−RIIについて57kDaで検出された。複数のバンドがTM細胞においてBMP−RIBおよびBMP−RIIについて検出され、これらは、他の研究において見られるようなグリコシル化された型および、部分的にグリコシル化された型を示す可能性が最も高い。BMPレセプターのタンパク質の発現レベルは、ONH細胞と比較してTM細胞では低いようであった。例えば、BMP−RIIは、TM細胞において検出されず、そしてBMP−RIBは大々的に減少した。
【0063】
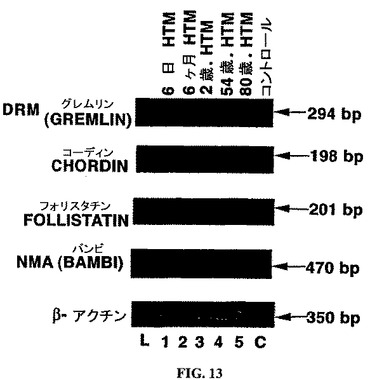
培養ヒトTM細胞および培養ヒトONH細胞におけるBMP関連タンパク質mRNAの発現:ヒトTM細胞株における、BMP関連タンパク質のプライマー対について予想されたサイズの増幅産物は、図13に示される。ヒトTM細胞株は、DRM(グレムリン)、コーディン、フォリスタチンおよびNMA(バンビ)のメッセージを発現した。特異的プローブを使用したサザンブロットによって、これらが、予想されたPCR産物であること
が証明された。細胞株間でメッセージの発現において明らかな違いは無かった。試験した全てのヒトTM細胞は、BMP関連タンパク質であるノギンおよびCer−1のmRNAを発現できなかった。cDNA無しのコントロール反応は、増幅産物を生じず、試薬およびプライマーにはDNAもRNAも混入していなかったことを示す。
【0064】
ONH星状細胞およびLC細胞株における、BMP関連タンパク質のプライマー対について予想されたサイズの増幅産物は、図14に示される。全てのONH星状細胞およびLC細胞株は、DRM(グレムリン)、フォリスタチンおよびNMA(バンビ)のメッセージを発現した。特異的プローブを使用したサザンブロットによって、これらが、予想されたPCR産物であることが証明された。大多数のLC細胞およびONH星状細胞は、コーディンのメッセージを発現した。試験した全てのヒトONH星状細胞およびLS細胞は、BMP関連タンパク質であるノギンおよびCer−1のmRNAを発現できなかった。cDNA無しのコントロール反応は、増幅産物を生じず、試薬およびプライマーには、DNAもRNAも混入していなかったことを示す。
【0065】
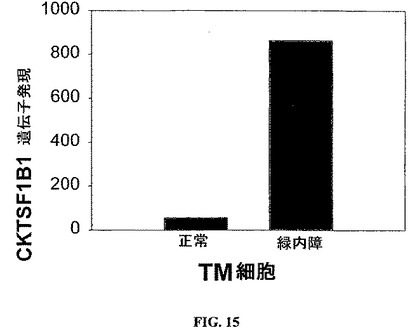
図15は、緑内障のTM細胞におけるBMPアンタゴニストのグレムリン(CKTSF1B1)の発現の上昇を示す。遺伝子発現を、Affymetrix遺伝子アレイ(Affymetrix遺伝子チップU133A)を使用して評価した。
【0066】
本明細書中で開示し、特許請求した組成物および/または方法の全ては、本開示を鑑みて過度の実験無しになされ、そして実施され得る。本発明の組成物および方法を好ましい実施形態に関して記載してきたが、本発明の概念、意図および範囲から逸脱することなく、本明細書中に記載される組成物および/または方法ならびに方法の工程または工程の順序にバリエーションが適用され得ることは、当業者にとって明らかである。より具体的には、化学的および構造的の両方で関連する特定の薬剤は、似た結果を達成するために、本明細書中に記載される薬剤の代用になり得ることが明らかである。当業者にとって明らかである全てのこのような代替物および改変物は、添付した特許請求の範囲に定義されるような、本発明の精神、範囲および概念内であると考えられる。
【0067】
(参考文献)
以下の参考文献は、例示的な手順、または本明細書中に記載される手順に対して補助的な他の詳細を提供する程度にまで、本明細書中で参考として具体的に援用される。
(書籍)
【0068】
【表3】
【図面の簡単な説明】
【0069】
図面は、本明細書の一部を成し、そして本発明のある局面をさらに実証するために含まれる。本発明は、本明細書中に示された特定の実施形態の詳細な説明と組み合わせて、1つ以上のこれらの図面の参照によって、より良く理解され得る。
【図1a】BMP2Aのヌクレオチド配列およびアミノ酸配列。
【図1b】BMP2Aのヌクレオチド配列およびアミノ酸配列。
【図1c】BMP2Aのヌクレオチド配列およびアミノ酸配列。
【図2a】BMP4のヌクレオチド配列およびアミノ酸配列。
【図2b】BMP4のヌクレオチド配列およびアミノ酸配列。
【図2c】BMP4のヌクレオチド配列およびアミノ酸配列。
【図2d】BMP4のヌクレオチド配列およびアミノ酸配列。
【図3a】BMP5のヌクレオチド配列およびアミノ酸配列。
【図3b】BMP5のヌクレオチド配列およびアミノ酸配列。
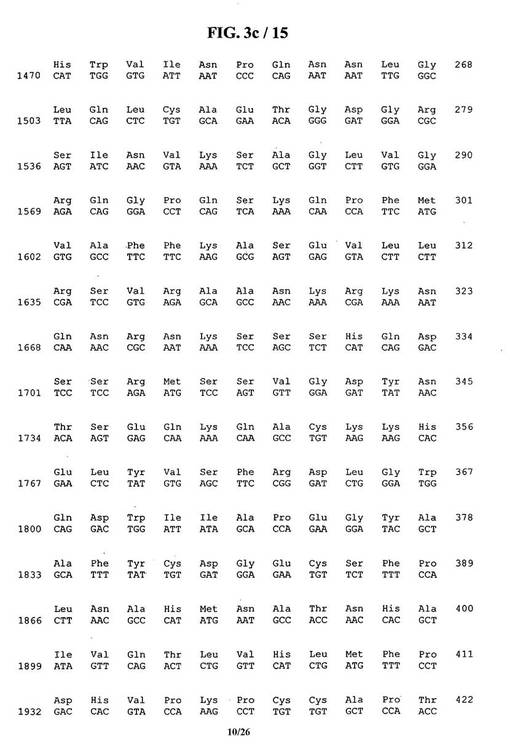
【図3c】BMP5のヌクレオチド配列およびアミノ酸配列。
【図3d】BMP5のヌクレオチド配列およびアミノ酸配列。
【図4a】BMP7のヌクレオチド配列およびアミノ酸配列。
【図4b】BMP7のヌクレオチド配列およびアミノ酸配列。
【図4c】BMP7のヌクレオチド配列およびアミノ酸配列。
【図4d】BMP7のヌクレオチド配列およびアミノ酸配列。
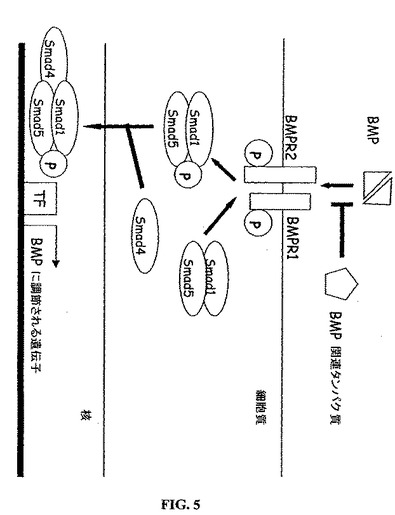
【図5】骨形成タンパク質シグナル伝達経路。骨形成タンパク質は(BMP)2量体は、セリン/スレオニンキナーゼであるBMPレセプター1および2から構成される膜複合体に結合する。調節性のSmad(Smad1/Smad5)は、リン酸化されそして補助Smad(co−Smad)(Smad4)に結合する。結果できたこのSmad複合体は、核に入り、核で転写因子(TF)に結合し、そして遺伝子の発現を調節する。BMP関連タンパク質は、BMPに結合し、そしてBMPとBMPレセプターとの相互作用を阻害することによってBMPアンタゴニストとして働く。
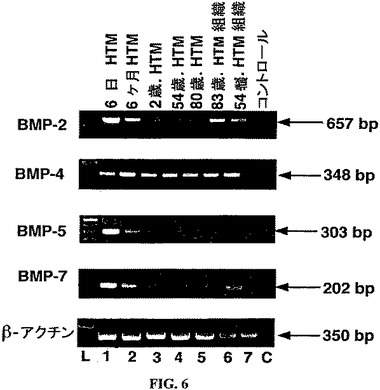
【図6】ヒトTM細胞およびヒトTM組織におけるBMP発現。ヒトTM細胞(レーン1〜5)およびヒトTM組織(レーン6〜7)におけるBMP発現のRT−PCR解析から生成されたcDNAサンプル由来のBMP PCR産物のアガロースゲルの、エチジウムブロマイド染色。L=塩基対マーカー。C=PCRのネガティブコントロールレーン。β−アクチンを、ポジティブなRT−PCR内部コントロールとして使用した。
【図7】ヒトTM細胞およびヒトTM組織のBMPレセプター発現。ヒトTM細胞(レーン1〜5)およびヒトTM組織(レーン6〜7)におけるBMPレセプター発現のRT−PCR解析から生成されたcDNAサンプル由来のPCR産物のアガロースゲルのエチジウムブロマイド染色。L=塩基対マーカー。C=PCRのネガティブコントロールレーン。β−アクチンを、ポジティブなRT−PCR内部コントロールとして使用した。
【図8】ヒトONH星状細胞、ヒトONH組織およびヒト脳星状細胞におけるBMPの発現。ヒトONH星状細胞(レーン1〜5)、ヒトONH組織(レーン6)、およびヒト脳星状細胞(レーン7)におけるBMP発現のRT−PCR解析から生成されたcDNAサンプル由来のPCR産物のアガロースゲルのエチジウムブロマイド染色。L=塩基対マーカー。C=PCRのネガティブコントロールレーン。β−アクチンを、ポジティブなRT−PCR内部コントロールとして使用した。
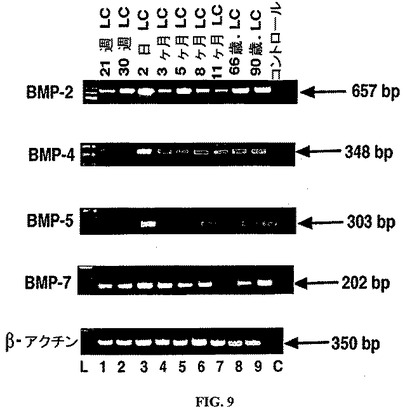
【図9】ヒト篩板細胞株におけるBMPの発現。ヒト篩板細胞(レーン1〜9)のRT−PCR解析から生成されたcDNAサンプル由来のPCR産物のアガロースゲルのエチジウムブロマイド染色。L=塩基対マーカー。C=PCRのネガティブコントロールレーン。β−アクチンを、ポジティブなRT−PCR内部コントロールとして使用した。
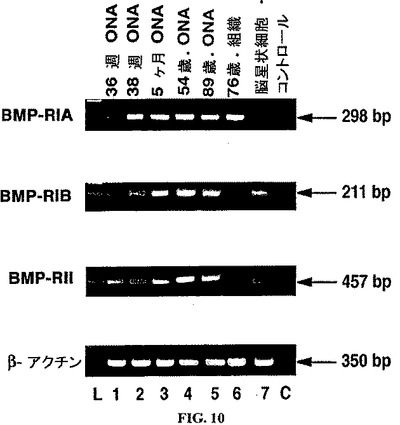
【図10】ヒトONH星状細胞、ONH組織およびヒト脳星状細胞におけるBMPレセプターの発現。ヒト視神経乳頭星状細胞(ONH)(レーン1〜5)、ヒトONH組織(レーン6)およびヒト脳星状細胞(レーン7)におけるBMPレセプター発現のRT−PCR解析から生成されたcDNAサンプル由来のPCR産物のアガロースゲルのエチジウムブロマイド染色。L=塩基対マーカー。C=PCRのネガティブコントロールレーン。β−アクチンを、ポジティブなRT−PCR内部コントロールとして使用した。
【図11】ヒト篩板細胞株におけるBMPレセプターの発現。ヒト篩板細胞(レーン1〜9)のRT−PCR解析から生成されたcDNAサンプル由来のPCR産物のアガロースゲルのエチジウムブロマイド染色。L=塩基対マーカー。C=PCRのネガティブコントロールレーン。β−アクチンを、ポジティブなRT−PCR内部コントロールとして使用した。
【図12】培養ヒトTM細胞、培養ヒト視神経乳頭星状細胞(ONA)、および培養ヒト篩板細胞におけるBMPおよびBMPレセプターの発現のウェスタンイムノブロット。ヒト小柱網細胞(レーン1〜2)、ヒトONH星状細胞(レーン3〜4)、およびヒト篩板細胞(レーン5〜6)におけるBMPタンパク質およびBMPレセプターの化学発光による検出。タンパク質サイズは、kDaで示す。
【図13】ヒトTM細胞におけるBMP関連タンパク質mRNAの発現。ヒトTM細胞(レーン1〜5)のRT−PCR解析から生成されたcDNAサンプル由来のPCR産物のアガロースゲルのエチジウムブロマイド染色。L=塩基対マーカー。C=PCRのネガティブコントロールレーン。β−アクチンを、ポジティブなRT−PCR内部コントロールとして使用した。
【図14】ヒト篩板細胞およびONH星状細胞におけるBMP関連タンパク質mRMAの発現。篩板(LC)細胞(レーン1〜7)およびONH星状細胞(ONA)(レーン8〜11)のRT−PCR解析から生成されたcDNAサンプル由来のPCR産物のアガロースゲルのエチジウムブロマイド染色。L=塩基対マーカー。C=PCRのネガティブコントロールレーン。β−アクチンは、ポジティブなRT−PCRインターナルコントロールとして使用された。
【図15】緑内障TM細胞における、BMPアンタゴニストのグレムリン(CKTSF1B1)発現の上昇の例示。遺伝子発現を、Affymetrix遺伝子アレイ(Affymetrix遺伝子チップU133A)を使用して評価した。(配列表)
【化1】
【化2】
【化3】
【化4】
【化5】
【化6】
【化7】
【化8】
【化9】
【化10】
【化11】
【化12】
【化13】
【化14】
【化15】
【化16】
【化17】
【化18】
【化19】
【化20】
【化21】
【化22】
【化23】
【化24】
【化25】
【化26】
【化27】
【化28】
【化29】
【化30】
【化31】
【化32】
【化33】
【化34】
【化35】
【化36】
【化37】
【化38】
【化39】
【技術分野】
【0001】
(1.発明の分野)
本発明は、緑内障および関連障害の診断および処置のための方法および試薬を開示する。
【背景技術】
【0002】
(2.関連分野の説明)
「緑内障」は、消耗性の眼の疾患の一群であり、米国および他の先進国において回復不能の失明の主要原因である。緑内障のもっとも一般的な形態である原発性開放隅角緑内障(「POAG」)は、虹彩と角膜との間の空間(例えば、「角」)の閉鎖無しに、房水が眼を離れる正常な能力の閉塞を起こす、小柱網の変性によって特徴づけられる(非特許文献1)。この疾患におけるこのような閉塞の特徴は、眼内圧力(「IOP」)上昇であり、もし適切にかつ適時な様式で処置されなければ、進行性の視力喪失および失明を起こす。この疾患は、40歳を越える全ての成人のうち0.4%と3.3%との間の人々に影響を及ぼすことが推測される(Leske,M.C.ら、(1986);Bengtsson,B.(1989);Strong,N.P.(1992))。さらに、この疾患の有病率は、年齢とともに増加して、75歳以上では6%を超える(Strong,N.P.、(1992))。
【0003】
IOPの上昇は、緑内障の容易に測定可能な特徴であるので、この疾患の診断は、眼内力の測定(眼圧測定)によって大まかにスクリーニングされる(Strong,N.P.(1992);Greve,M.ら、(1993))。不幸なことに、緑内障および正常の圧力範囲は重複するので、このような方法は、多数の読み取りが得られない限り、限定された値のものである(Hitchings,R.A.、(1993);Tuck,M.W.ら、(1993);Vaugham,D.ら、(1992);Vernon,S.A.、(1993))。この理由から、さらなる方法(例えば、視神経円板の直接的な検査および患者の視野損失の程度の決定)が、しばしば、診断の正確性を改善するために実施される(Greve,M.ら、(1993))。
【0004】
緑内障は、眼における3つの別個の組織に影響する。POAG関連の上昇したIOPは、小柱網(TM)(角膜と虹彩との間の角に位置する組織)での形態的変化および生物化学的な変化に起因する。栄養性房水のほとんどは、TMを通って眼の前方区画に存在する。緑内障の眼のTMにおけるTM細胞の進行性の喪失および細胞外破片の増加は、房水流出に対する耐性の増加を引き起し(Lutjen−DrecollおよびRohen 1996;Rohen 1983;Rohenら、1993;GriersonおよびCalthorpe 1988)、それによって、IOPが上昇する。IOPの上昇ならびに他の因子(例えば、虚血)は、視神経乳頭(ONH)において変性性の変化を起こし、ONHの進行性の「杯形成」(VarmaおよびMinckler 1996;HernandezおよびGong 1996;Hernandezら、1990;HernandezおよびPena 1997;Morrisonら、1990))、ならびに網膜神経節細胞(Quigleyら、2000;Quigley 1999;Quigleyら、1995;Kerriganら、1997)ならびに軸策の喪失を起こす。TM、ONHおよび網膜の神経節細胞に対する緑内障による損傷の原因となる詳細な分子機構は、未知である。
【0005】
現在の緑内障の治療は、緑内障の発病および進行の主な危険因子であるIOPを低下させることに関する。これらの治療はIOPを低下させるが、病原機構に直接的に対処せず
、そしてこの疾患は進行を続ける。緑内障を有する患者らの少なくとも半分は診断されず、患者らが緑内障を有すると診断されるまでに、患者らはすでに40%近くの網膜の神経節細胞を喪失してしまっている。したがって、緑内障のより早い検出および診断の方法が必要とされる。
この出願の発明に関連する先行技術文献情報としては、次のものがある。
【非特許文献1】Vaughan,D.ら、(1992)
【発明の開示】
【発明が解決しようとする課題】
【0006】
緑内障の重要性、および以前の診断方法の少なくとも部分的な欠点の見地において、初期段階の緑内障の診断のための改善されたより正確な方法を有することが所望される。さらに、緑内障の病原の機構に対処する新規の治療剤を有することが所望される。
【課題を解決するための手段】
【0007】
(発明の要旨)
本発明は、先行技術のこれらおよび他の欠点を、緑内障の初期診断のための方法およびキット、緑内障の処置のための方法およびキット、ならびに緑内障の処置において有用な化合物の同定のための方法およびキットを提供することによって克服する。
【0008】
ある特定の実施形態において、本発明は、細胞および体液から得られたサンプル中の骨形成タンパク質ファミリーメンバー遺伝子の発現の変化を検出することによる、緑内障の診断のための方法を提供する。この方法は、一般に以下の工程:
a)緑内障を有すると疑われる患者から組織サンプルまたは体液サンプルを得る工程;
b)このサンプルからDNAを抽出する工程;
c)複数のPCRプライマーを得る工程であって、これらプライマーの各々は、配列番号1、配列番号3、配列番号5、配列番号7、配列番号37、配列番号39、配列番号41、配列番号43、配列番号45、配列番号47または配列番号53由来の18〜1547連続するヌクレオチドからなる配列を含む、工程;
d)これらのプライマーを使用して、抽出されたDNAの領域を増幅してPCR産物を得る工程;
e)このPCR産物を分離する工程;ならびに
f)このPCR産物の配列と正常な遺伝子配列との間の違いを同定する工程、を包含し、ここで、この増幅配列と正常な遺伝子配列との間の違いは、緑内障の診断特徴である。
【0009】
一般に、本発明の方法は、個体からサンプルを得る工程およびこのサンプルからDNAを抽出する工程を包含し得る。BMP遺伝子ファミリーの特定のメンバーのための選択されたPCRプライマーは、PCR産物を得るために、抽出された遺伝子の適切な領域の増幅のために使用される。PCR産物を、評価されている特定のBMPファミリーの遺伝子(抽出されたDNA)の正常形態と変異形態との間のDNA配列の違いを効果的に同定する技術によって、分離される。同定された配列間の違いは、緑内障を示す。
【0010】
本発明の方法において使用される組織サンプルおよび液体サンプルは、血液または頬の細胞であり得る。
【0011】
代表的に、プライマー配列は、約10、15、または18ヌクレオチド〜約20ヌクレオチド、あるいは約10、15、または18ヌクレオチド〜約30ヌクレオチドの範囲の長さを有する。より長い配列(例えば、40、50、80、90、95、100、全長までさえ)が、ある実施形態についてなおさらに好ましい。少なくとも約18〜20ヌクレオチドのオリゴヌクレオチドの長さは、分子プローブとして有用であるように十分に特異的なハイブリダイゼーションを可能にするのに十分であるとして、当業者に十分受け入れられている(Lathe(1985)によって記載され、この参考文献は、この目的のために、本明細書中で参考として具体的に採用される)。好ましくは、このヌクレオチド配
列は、配列番号1、配列番号3、配列番号5、配列番号7、配列番号37、配列番号39、配列番号41、配列番号43、配列番号45、配列番号47または配列番号53の、20〜100連続するヌクレオチドからなる。プライマー配列は、その配列が公知であるBMPレセプター遺伝子の配列およびのBMP関連タンパク質の配列由来の、少なくとも10、15、または18連続するヌクレオチドの配列からなり得ることもまた企図される。
【0012】
配列番号1、配列番号3、配列番号5、配列番号7、配列番号37、配列番号39、配列番号41、配列番号43、配列番号45、配列番号47または配列番号53のいずれか1つに相補的な、10、18、20、30、50、60、65または100ヌクレオチド程度まで、および100ヌクレオチド程度を含むストレッチを有する核酸分子は、ハイブリダイゼーションプローブとしての有用性を有する。約18ヌクレオチドのヌクレオチド長を有するプライマーまたはプローブが、標的配列に対する高度に特異的なハイブリダイゼーションを提供することが、当業者に認識される。フラグメントの全サイズならびに相補的ストレッチのサイズは、特定の核酸セグメントの適用の意図した使用に最終的に依存する。より小さいフラグメントは、一般に、ハイブリダイゼーション実施形態における用途を見出し、ここで、相補的な領域の長さは変化し得る(例えば、検出したいと望む相補的配列に従って、約10、18、20もしくは30ヌクレオチドと約50、60、70、80、80もしくは100ヌクレオチドとの間、または全長まで)。
【0013】
特に好ましい実施形態において、プライマーは、配列番号1、配列番号3、配列番号5、配列番号7、配列番号37、配列番号39、配列番号41、配列番号43、配列番号45、配列番号47または配列番号53由来の連続する配列からなる。他の好ましい実施形態において、プライマーは、BMPレセプター遺伝子(ten Dijkeら、1993;Astromら、1999;Nohnoら、1995によって開示され、これらはすべて参考として採用される)または、BMP関連遺伝子(例えば、コーディン(chordin)(NCBI NM_029130)、グレムリン(gremlin)(Murphyら、1999;McMahonら、2000)、フォリスタチン(follistatin)(NCBI NM_003892)、またはバンビ(bambi)(NCBI NM_005791))由来の連続配列からなる。最も好ましくは、プライマーは、配列番号3由来の連続配列からなる。ある局面において、少なくともいくつかのプライマーは、検出可能な標識をさらに含み得る。
【0014】
他の実施形態において、本発明は、BMP2アゴニスト、BMP4アゴニスト、BMP5アゴニスト、BMP7アゴニスト、Smad1/5アゴニスト、コーディンアンタゴニスト、グレムリンアンタゴニスト、およびフォリスタチンアンタゴニストからなる群より選択される少なくとも1つの化合物からなる配列を含む組成物を、緑内障の処置を必要とする患者に投与することによる、緑内障の処置の方法を提供する。
【0015】
さらなる局面において、本発明は、緑内障の処置のための治療剤を同定する方法を提供する。治療剤は、例えば、以下によって同定され得る;
a)BMP−2A、BMP4、BMP−5またはBMP7を発現する組換え細胞の集団を含む第一の組成物を得ること;
b)候補物質を得ること;
c)この組成物およびこの候補物質をインキュベートすること;
BMP誘導性Smadシグナル伝達経路および/またはBMP調節性の遺伝子発現を開始する能力について、この化合物を試験すること;ならびにBMPのこれらの下流の効果を阻害、または刺激する候補物質を同定すること。
【0016】
本発明の別の局面は、本発明の配列および適切な試薬(例えば、タンパク質、ペプチド、または抗体自体に連結した検出可能な標識)を含む診断キットである。あるいは、検出
可能な標識は、本発明の配列に選択的にハイブリダイズする第2の配列に連結され得る。
【0017】
関連する実施形態は、本明細書中に開示される核酸配列またはペプチド配列もしくはタンパク質配列のいずれかの、薬学的に受容可能な処方物を含む治療キットを含む。このようなキットは、緑内障の診断のための、臨床的なサンプル中のBMP遺伝子およびBMPタンパク質の変化した発現の検出に有用である。
従って、本発明は、以下を提供する:
(項目1)
細胞または体液から得られたサンプルにおいて、骨形態形成ファミリーメンバー遺伝子の発現の変化を検出することによる、緑内障を診断する方法であって、該方法は、以下の工程:
g)緑内障を有すると疑われる患者から組織または液体のサンプルを得る工程;
h)該サンプルからDNAを抽出する工程;
i)複数のPCRプライマーを得る工程であって、該プライマーの各々は、配列番号1、配列番号3、配列番号5、配列番号7、配列番号37、配列番号39、配列番号41、配列番号43、配列番号45、配列番号47または配列番号53由来の、18〜1547連続するヌクレオチドからなる配列を含む、工程;
j)該プライマーを使用して、該抽出されたDNAの領域を増幅してPCR産物を得る工程;
k)該PCR産物を分離する工程;ならびに
l)該増幅した抽出されたDNA配列と該プライマーの配列との間の違いを同定する工程、を包含し、
ここで該増幅配列とプライマーとの間の違いは、緑内障の診断特徴である、方法。
(項目2)
項目1に記載の方法であって、前記組織サンプルまたは前記液体サンプルは、血液または頬の細胞である、方法。
(項目3)
項目1に記載の方法であって、前記プライマーは、配列番号1、配列番号3、配列番号5、配列番号7、配列番号37、配列番号39、配列番号41、配列番号43、配列番号45、配列番号47または配列番号53の、20〜100連続するヌクレオチドからなる配列を包む、方法。
(項目4)
項目1に記載の方法であって、前記プライマーは、配列番号3の20〜50連続するヌクレオチドからなる配列を包む、方法。
(項目5)
項目1に記載の方法であって、前記PCR産物は、SSCP、DGGE,ASOまたはRFLPによって分離される、方法。
(項目6)
緑内障を処置する方法であって、該方法は、BMP2アゴニスト、BMP4アゴニスト、BMP5アゴニスト、BMP7アゴニスト、Smad1/5アゴニスト、コーディンアンタゴニスト、グレムリンアンタゴニストおよび、フォリスタチンアンタゴニストからなる群から選択される少なくとも1つの化合物からなる配列を包む組成物を、このような処置を必要とする患者に投与する工程を包含する、方法。
(項目7)
緑内障を処置するための治療剤を同定する方法であって、該方法は以下の工程:
d)BMP−2A、BMP4、BMP−5、またはBMP7を発現する組換え細胞の集団を含む、第一の組成物を得る工程;
e)候補物質を得る工程;
f)該組成物および該候補物質をインキュベートする工程;
g)該組成物を、BMP誘導性Smadシグナル伝達経路、またはBMP調節性の遺伝子発現を開始する能力について試験する工程;ならびに
h)BMP誘導性のSmadシグナル伝達経路、または該BMP調節性の遺伝子発現を阻害または刺激する候補物質を同定する工程、
を包含する、方法。
【0018】
(好ましい実施形態の詳細な説明)
小柱網は、房水の正常な流れにおいて、重要な役割を担うことが提案されており、そして緑内障の眼における流出抵抗の主な部位であると推測されている。ヒト小柱網(HTM)細胞は、房水が眼から出る流出チャネルを裏打ちする特殊化した細胞である。細胞の変化した合成機能は、POAG、ステロイド緑内障、および他のタイプの緑内障の病因に関与し得る。
【0019】
集中的な研究の年月にもかかわらず、眼への緑内障による損傷の原因となる正確な分子機構は、知られていない。最近の研究によって、増殖因子が、緑内障に関連する眼の組織における正常な恒常性を維持することにおいて重要であり得、そして増殖因子/増殖因子レセプターの変化が、緑内障の病因において役割を担い得ることが示唆されている。増殖因子は、ポリペプチドの非常に大きなファミリーであり、細胞の増殖および分化を制御する。これらの分子は、遺伝子発現、細胞外マトリックスの組成および堆積、細胞骨格構築、ならびに細胞機能の調節に対して、種々の細胞特異的影響を有する。TMは、広範囲な様々の増殖因子、増殖因子レセプター(Tripathiら、1993a;Tripathiら、1993b;Tripathiら、1994a;Tripathiら、1994b;Wordingerら、1998;Wordingerら、1999)ならびに神経栄養ニューロトロフィン/神経栄養因子およびそれらのレセプター(Liuら、2001;Wordingerら、2000)を発現する。ONH星状細胞および篩板細胞(視神経乳頭の2つの細胞型)は、増殖因子であるニューロトロフィンおよびそのレセプターを発現する(Lambertら、2001;Penaら、1999)。房水はまた、種々の増殖因子(FGF2、EGF、TGFβ、HGFを含む)(Tripathiら、1996;Tripathiら、1991;Tripathiら,1992;HuおよびRitch、2001)ならびにニューロトロフィン(Chundruら、2000)を含む。房水のTGFβ−2およびHGFの上昇したレベルは、POAG患者において報告されている(Tripathiら、1994c;Inataniら、2001;Pichtら2001)。増殖因子は、TMおよびONHの正常な発達および/または機能を変更することによって、緑内障に関与し得る。
【0020】
本発明は、骨形成タンパク質(BMP)が、骨形成および軟骨形成を誘導するだけではなく、多数の細胞型に対する広範な影響を有する多機能的なサイトカインであり(Hogan 1996;Reddi 1997)、そしてヒト小柱網(HTM)細胞および視神経乳頭(ONH)細胞の両方によって発現される(Wordingerら、2002)という認識に部分的に由来する。BMPは、TGFβスーパーファミリーのメンバーであり、ヒトには約15〜20のBMP遺伝子、3つのBMPレセプター、そしてBMPアンタゴニストとして機能する多数のBMP関連タンパク質がある(Yamashitaら、1996)。BMPは、BMPR−IおよびBMPR−IIからなるレセプター複合体を介してシグナルを伝達する。スーパーファミリーメンバーのであるTGFβおよびTGFβR(Agarwalら、1997;Lambertら、1997)ならびにGDNFおよびGDNFR(Wordingerら、1999;Liuら、1999)は、HTM細胞およびONH細胞の両方によって発現されることが報告されている。
【0021】
BMPおよびBMPレセプターは、眼の組織で発現しているが(Obataら、199
9;Youら、1999)、以前の報告は、眼の発達に集中している。マウスにおけるBMPをコードする遺伝子の標的化された破壊が、網膜および水晶体に重篤な発達欠損を引き起こすので、BMPの機能は眼の発達に重要である(Jenaら、1997;Luoら、1995;Dudleyら、1995)。BMP−2、BMP−4およびBMP−7は、眼および網膜の発達に関与する(Jenaら、1997;FurutaおよびHogan 1998;Reddi 2000;Trousseら、2001)。BMP−6およびBMP−7もまた、低血糖または虚血損傷からニューロンを保護する役割を果たすようであり(Nonnerら、2001;Liuら、2001)、そしてBMP2は、神経節細胞のニューロトロフィン発現を増強することが示されている(Zhangら、1998)。Bmp4について不十分なハプロイド(haploinsufficient)であるヘテロ接合体ノックアウトマウスは、前方部分の発育不全、IOPの上昇、および視神経異常を含む眼の表現型を有する(Changら、2001)。ヒトの出生後の眼におけるBMPの役割に関して、非常に限られた情報が発表されている。
【0022】
Mohanおよび共同研究者(1998)は、BMP−2およびBMP−4ならびにBMPレセプターが、成体の角膜細胞において発現することを報告し、そしてBMP機能が、角膜の角膜実質細胞の増殖およびアポトーシスを含み得ることを示唆した。Youおよび共同研究者(1999)は、この研究を確認し、そしてまた、エキソビボならびに培養角膜上皮および培養基質細胞でのBMP−3、BMP−5およびBMP−7の発現を報告した。彼らは、BMPの転写レベルが、基質においてより高く、一方でレセプターのレベルが培養角膜上皮細胞においてより高いことを報告した。
【0023】
RT−PCRを使用して、本発明者らは、BMPのmRNA、BMPレセプターであるBMPR−IA、BMPR−IBおよびBMPR−IIのmRNA、ならびにBMP結合タンパク質であるグレムリン、コーディン、フォリスタチン、およびバンビのmRNAを、HTM、篩板(LC)およびONH星状細胞細胞株および組織で発見した(Wordingerら、2002)。本発明者らはさらに、HTM細胞およびONH細胞が、タンパク質BMP−2、BMP−4、BMP−5およびBMP−7を発現することを発見した。
【0024】
緑内障は、BMPシグナル伝達ファミリーのメンバーの遺伝子における遺伝的変化の特徴付けによって診断される。本明細書中で使用する場合、句「骨形成タンパク質ファミリーメンバー遺伝子」および「BMPシグナル伝達ファミリー」は、全てのBMP、BMPレセプターおよび関連するタンパク質をいう。用語「遺伝的変化」は、当業者にとって周知である。特定の遺伝子における遺伝的変化に関する障害の多数の例が存在する(例えば、Cummings 1997;Strachanら、1996;Jordeら、1999を参照のこと)。特定の遺伝子(例えば、BMP)における遺伝的変化は、当業者に周知の様々な技術を使用することによって決定され得る(例えば、SSCP、DGGE、ASO、RFLP、ヘテロニ重鎖分析、CCM、PTT、ならびにRNase切断(Birrenら、1998を参照のこと))。
【0025】
緑内障は、上昇したIOPおよび/または緑内障的な眼の神経障害を引き起こす、眼における1つ以上のBMPファミリー遺伝子の発現の変化によって生じ得る。「変化したBMP遺伝子発現」は、この遺伝子産物の発現が正常とは異なることを意味する。この用語はまた、遺伝子またはタンパク質の配列における変化もいい得る。正常なBMP遺伝子は、十分特徴づけられており(上記を参照のこと)、そしてBMPの発現は、TMおよびONHを含む様々な組織において報告されている。BMPファミリー遺伝子のコード領域の遺伝的変化は、これらのタンパク質の機能を変化し得る。コード領域外の遺伝的変化もまた、緑内障を引き起こし得る。
【0026】
特定の遺伝子のコード領域の「外の変化」が、遺伝子発現の調節において重要であるこ
とは当業者に周知である。例えば、ほとんどの遺伝子のコード領域の上流(5’)領域は、その遺伝子の発現を「促進」および調節するプロモーター領域として公知である。プロモーター領域は、遺伝子発現の活性化または抑制の原因となる、様々な転写因子およびDNA結合タンパク質によって認識される多数のヌクレオチド配列を含む。遺伝子の下流(3’)領域は、遺伝子産物のポリアデニル化を決定し得、それによってRNAプロセシングおよび遺伝子産物の翻訳を調節し得る。
【0027】
緑内障を示すBMP遺伝子の発現の変化または、遺伝子配列における変異は、当業者に周知の技術を使用して検出され得る。例えば、ほぼ全ての長さの核酸フラグメントが使用され得、その全長は、調製しやすさおよび意図するプロトコールでの使用しやすさによって好ましくは限定されることが企図される。本明細書中に開示される核酸配列はまた、核酸ハイブリダイゼーション実施形態におけるプローブまたはプライマーとしての有用性も有し得る。このように、BMP−2A(配列番号1)、BMP−4(配列番号3)、BMP−5(配列番号5)、BMP−7(配列番号7)、BMP−RIA(配列番号37)、BMP−RIB(配列番号39)、BMP−RII(配列番号41)、コーディン(配列番号43)、グレムリン(配列番号45)、フォリスタチン(配列番号47)、またはバンビ(配列番号53)の少なくとも14ヌクレオチド長連続する配列と同じ配列を有するか、またはこれらに相補的な配列からなる配列領域を含む核酸セグメントは、特定の有用性を見出すことが企図される。より長く、連続した、同一または相補的な配列(例えば、約20、30、40、50、100、200、500、1000ヌクレオチド(全ての中間の長さを含む)の配列)、約1547ヌクレオチド(BMP−2Aについて)、1946ヌクレオチド(BMP−4について)、2153ヌクレオチド(BMP−5について)および1878ヌクレオチド(BMP−7について)、2932ヌクレオチド(BMP−RIAについて)、2032ヌクレオチド(BMP−RIBについて)、3611ヌクレオチド(BMP−RIIについて)、3561ヌクレオチド(コーディンについて)、4049ヌクレオチド(グレムリンについて)、1386ヌクレオチド(フォリスタチンについて)、および1523ヌクレオチド(バンビについて)の全長配列までさえもまた、ある実施形態において使用される。
【0028】
この文脈における「中間の長さ」は、引用された範囲(例えば、14、15、16、17、18、19、20など;21、22、23など;30、31、32など;50、51、52、53など;100、101、102、103など;150、151、152、153など;200〜500;500〜1,000;1,000〜2,000の範囲の全てを含み、そして、2,001、2,002、2,050、2,051などの配列までおよびそれらを含む)の間の任意の長さを意味することが容易に理解される。
【0029】
このような核酸プローブおよび核酸プライマーがBMPコード配列に特異的にハイブリダイズする能力、ならびにプライマーがBMP配列を特異的に増幅する能力により、所与のサンプル中の相補配列の存在の検出において、それらが使用可能となる。ところが、変異種のプライマーの調製のための配列情報の使用、または他の遺伝的構築物の調製における使用のためのプライマーの使用を含む、他の使用が想定される。
【0030】
BMP−2A(配列番号1)、BMP4(配列番号3)、BMP−5(配列番号5)、BMP7(配列番号7)、BMP−RIA(配列番号37)、BMP−RIB(配列番号39)、BMP−RII(配列番号41)、コーディン(配列番号43)、グレムリン(配列番号45)、フォリスタチン(配列番号47)またはバンビ(配列番号53)と同一かもしくは相補的な、10、20、30、50、またはちょうど100〜200ヌクレオチドなどの連続するヌクレオチドストレッチからなる配列領域を有する核酸分子は、、例えばサザンブロッティングおよびノザンブロッティングに加えて、SNP評価および固相ハイブリダイゼーションアッセイにおける使用のための、ハイブリダイゼーションプロー
ブとして特に企図される。これにより、組織および細胞の両方におけるBMPの構造遺伝子または調節遺伝子の解析が可能となる。フラグメントの全サイズならびに相補的ストレッチのサイズは、特定の核酸セグメントの適用について意図する使用に最終的に依存する。より小さいフラグメントは、一般に、ハイブリダイゼーション実施形態における使用を見出す。ここで、連続する相補的な領域の長さは、変化し得る(例えば、約10ヌクレオチドと約100ヌクレオチドとの間)しかし、約1547ヌクレオチド(BMP−2Aについて)、1946ヌクレオチド(BMP−4について)、2153ヌクレオチド(BMP−5について)、ならびに1878ヌクレオチド(BMP−7について)、2932ヌクレオチド(BMP−RIAについて)、2032ヌクレオチド(BMP−RIBについて)、3611ヌクレオチド(BMP−RIIについて)、3561ヌクレオチド(コーディンについて)、4049ヌクレオチド(グレムリンについて)、1386ヌクレオチド(フォリスタチンについて)、1523ヌクレオチド(バンビについて)までの、より長い連続する相補的ストレッチが、検出したいと望む相補的配列の長さに従って使用され得る。
【0031】
約10〜14ヌクレオチド長のハイブリダイゼーションプローブの使用により、安定かつ選択的な二重鎖分子の形成が可能である。10塩基長を越えるストレッチにわたる連続する相補的配列を有する分子が一般に好ましいが、ハイブリッドの安定性および選択性を高め、それによって得られる特異的なハイブリッド分子の質および程度を改善するために、一般に、15〜20連続するヌクレオチドの遺伝子相補的なストレッチを有するか、または所望される場合、さらに長い核酸分子を設計することが好まれる。
【0032】
ハイブリダイゼーションプローブは、本明細書中に開示される任意の配列のいずれの部分からも選択され得る。配列番号1、配列番号3、配列番号5、配列番号7、配列番号37、配列番号39、配列番号41、配列番号43、配列番号45、配列番号47または配列番号53に示される配列を検討すること、ならびにプローブまたはプライマーとしての利用が望まれる、約10ヌクレオチド長から、全長配列までを含むまでの配列の任意の連続する部分を選択することが必要なだけである。プローブ配列およびプライマー配列の選択は、様々な因子によって左右され得る(例えば、例示のみとして、さらなるDNAを増幅するために、全配列の終点に向かうプライマーまたは、機能的ドメインコード配列の末端からのプライマ−の設計が望まれ得る)。
【0033】
BMP−2A(配列番号1)、BMP4(配列番号3)、BMP−5(配列番号5)、BMP7(配列番号7)、BMP−RIA(配列番号37)、BMP−RIB(配列番号39)、BMP−RII(配列番号41)、コーディン(配列番号43)、グレムリン(配列番号45)、フォリスタチン(配列番号47)またはバンビ(配列番号53)内由来の連続する配列を含む核酸セグメントを選択および調製する工程は、核酸フラグメントを調製するとして代替的に記載されている。もちろん、フラグメントはまた、他の技術によって(例えば、機械的剪断または、制限酵素消化によって)獲得され得る。小さな核酸セグメントまたはフラグメントは、例えば、自動化されたオリゴヌクレオチド合成装置の使用によって一般に実行されるような、化学的手段によるフラグメントの直接的な合成によって容易に調製され得る。また、フラグメントは、核酸再生成技術(例えば、米国特許第4,683,202号および同第4,682,195号(各々は、参照として上記に記載される)のPCRTM技術)の適用によって、組換え生成のために選択した配列を組換えベクターに導入することによって、ならびに分子生物学の分野の当業者に一般に公知の他の組換えDNA技術によって、獲得され得る。
【0034】
従って、本発明のヌクレオチド配列は、BMPの遺伝子またはcDNAの相補的なストレッチとの二重鎖分子を選択的に形成するその能力のために、使用され得る。想定される適用に依存して、標的配列に対するプローブの選択性を様々な程度にするために、様々な
程度のハイブリダイゼーションの選択性を使用することが所望される。高い選択性を必要とする適用について、ハイブリッドを形成するために、比較的ストリンジェントな条件を使用することが代表的に所望される(例えば、比較的低い塩および/または高い温度条件(例えば、50℃〜70℃の温度で0.02M〜0.15MのNaClによって提供される)を選択する)。このような選択的条件は、もしあったとしてもプローブと鋳型または標的鎖との間のミスマッチをほとんど許容せず、そして特にBMP遺伝子の解析に適切である。
【0035】
もちろん、いくつかの適用のために、例えば基礎をなす鋳型にハイブリダイズした変異プライマー鎖を使用する変異体の調製または同定を所望する場合、あるいは関連する種由来のBMPコード配列、機能的同等物などを単離することを求める場合、あまりストリンジェントでないハイブリダイゼーション条件が、ヘテロ二重鎖の形成を可能にするために代表的に必要とされる。これらの状況下で、条件(例えば、0.15M〜1.0Mの塩、20℃〜55℃の範囲にある温度)を使用することが所望され得る。それにより、交差ハイブリダイズする種は、コントロールハイブリダイゼーションに関連してポジティブにハイブリダイズするシグナルとして、容易に同定され得る。いずれの場合においても、条件は、NaCl濃度を低くすることによって、または温度の上昇と同じ様式でハイブリッド二重鎖を不安定化するように作用するホルムアミドの斬新量を添加することによって、よりストリンジェントにされ得ることが一般に理解される。このように、ハイブリダイゼーション条件は容易に操作され得、従って一般に、所望される結果に依存する選択の方法である。
【0036】
ある実施形態において、ハイブリダイゼーションを決定するための適切な手段(例えば、標識)と組み合わせて本発明の核酸配列を使用することは有益である。様々な種類の適切な指示手段は、当該分野で公知であり、これには蛍光、放射性、酵素的または他のリガンド(例えば、アビジン/ビオチン)が挙げられ、これらは、検出可能なシグナルを生じ得る。好ましい実施形態においては、放射性または他の環境的に所望されない試薬の代わりに、蛍光標識または酵素タグ(例えば、ウレアーゼ、アルカリホスファタ−ゼまたはペルオキシダーゼ)を使用することが所望される傾向にある。酵素タグの場合、ヒトの眼または吸収分光測光計で見ることができる手段を提供して、相補的核酸含有サンプルとの特異的ハイブリダイゼーションを同定するために使用され得る比色指示基質は公知である。
【0037】
一般に、本明細書に記載されたハイブリダイゼーションプローブは、溶液ハイブリダイゼーションならびに固相を使用する実施形態の両方において試薬として有用であると想定される。固相を含む実施形態において、試験DNA(またはRNA)は、吸収されるか、または他の方法で、選択されたマトリックスもしくは表面に結合する。この固定された一本鎖の核酸は、その後、所望される条件下で、選択されたプローブとの特異的なハイブリダイゼーションに供される。この選択された条件は、必要とされる特定の基準に基づく特定の環境に依存する(例えば、G+C含有量、標的核酸のタイプ、核酸の供給源、ハイブリダイゼーションプローブのサイズなどに依存する)。非特異的に結合したプローブ分子を除くためのハイブリダイズ表面の洗浄に続いて、標識によって、特異的ハイブリダイゼーションは検出されるか、またはさらに測定される。
【0038】
本発明は、BMP−2A(配列番号1)、BMP4(配列番号3)、BMP−5(配列番号5)、BMP7(配列番号7)、BMP−RIA(配列番号37)、BMP−RIB(配列番号39)、BMP−RII(配列番号41)、コーディン(配列番号43)、グレムリン(配列番号45)、フォリスタチン(配列番号47)またはバンビ(配列番号53)の特定の核酸配列コピーおよびアミノ酸に限定されないこともまた理解される。組換えベクターおよび単離されたDNAセグメントは、従って、BMPコード領域自体、遺伝子の上流領域または下流領域、選択された変化または改変を基本的なコード領域中に有す
るコード領域を様々に含み得るか、あるいは、それにもかかわらずBMPコード領域を含むより大きなペプチドをコードし得るかまたは、改変体アミノ酸配列を有する生物学的に機能的な等価なタンパク質もしくはポリペプチドをコードし得る。
【0039】
本発明のDNAセグメントは、生物学的に機能的な等価なBMPタンパク質およびポリペプチドを包含する。このような配列は、コドン縮重および機能的等価性の結果として生じ得、これらは核酸配列およびこのようにコードされるタンパク質内に天然に存在するものであることが公知である。あるいは、機能的に等価なタンパク質またはポリペプチドは、タンパク質構造における変化が、交換されるアミノ酸の特性の考慮に基づいて操作され得る組換えDNA技術の適用を通じて作製され得る。人為的に設計された変化は、部位特異的変異誘発技術の適用を通して、例えば、タンパク質の抗原性に改善を導入するため、または分子レベルでの結合活性を試験するためにBMP変異を試験するために導入され得る。
【0040】
緑内障の処置のための治療剤は、ペプチドまたはタンパク質、ペプチド模倣物、オリゴヌクレオチドまたは誘導体化オリゴヌクレオチド、あるいは薬物様の低分子であり得、その全てが、眼のBMP経路の1つ以上の局面に影響する。好ましい治療剤は以下である:(1)BMP2アゴニスト、BMP4アゴニスト、BMP5アゴニスト、またはBMP7アゴニスト;(2)コーディンアンタゴニスト、グレムリンアンタゴニスト、フォリスタチンアンタゴニスト、またはバンビアンタゴニスト;および/あるいは(3)Smad1アゴニスト、Smad5アゴニスト、および/またはSmad4アゴニスト。
【0041】
この薬剤は、当業者に周知の技術を使用することで、直接的に眼に送達され得る(例えば、局所的点眼薬または軟膏;盲嚢中、のあるいは強膜に隣接してかまたは眼内に移植された徐放デバイス;眼周囲、結膜、サブテノン(sub−Tenons)、眼房内(intracameral)または硝子体内注射)か、あるいは非経口的に(例えば:経口;静脈内、皮下または筋内注射;皮膚送達など)。以下は、本発明によって具体化される可能な処方物の例示である。
(a)局所的な眼用処方 重量%
眼のBMP−4の発現を上昇させる薬剤 0.01〜2
HPMC 0.5
塩化ナトリウム 0.8
BAC 0.01
EDTA 0.01
NaOH/HCl qs pH7.4
精製水 qs 100mL
(b)局所的な眼用処方 重量%
グレムリンアンタゴニスト 0.01〜2
HPMC 0.5
塩化ナトリウム 0.8
BAC 0.01
EDTA 0.01
NaOH/HCl qs pH7.4
精製水 qs 100mL
(c)局所的な眼用処方 重量%
Smad1/5アゴニスト 0.01〜2
HPMC 0.5
塩化ナトリウム 0.8
BAC 0.01
EDTA 0.01
NaOH/HCl qs pH7.2
精製水 qs 100mL
本発明の化合物は、眼内挿入デバイス中で処方されることがさらに企図される。
【0042】
(A.治療剤についてのアッセイ)
本発明はまた、BMPシグナル伝達経路に関与する新しい抗緑内障治療剤の発見のために有用である(図5を参照のこと)。選択的BMPリガンドは、BMP I型およびBMP II型セリン/スレオニンキナーゼレセプター(BMP−RIおよびBMP−RII)に結合し、そしてSmadタンパク質を介してシグナルを伝達する。BMPシグナルは、タンパク質−タンパク質相互作用およびタンパク質−DNA相互作用を介して、Smadによって伝搬される(AttisanoおよびTuen Lee−Hoeflich 2001)。調節的Smad1およびSmad5は、BMPレセプターへのリガンドの結合によって(リン酸化を介して)活性化される(von BubnoffおよびCho 2001)。これらの調節的Smadは、その後、Smad4と相互作用して、核に転移するヘテロマー複合体を形成する。この複合体は、どの核補助因子が存在するかに依存して、この転写複合体を認識する選択的遺伝子の転写を活性化することまたは抑制することが可能である。
【0043】
BMP/Smadシグナル伝達経路は、いくつかの機構によってネガティブに調節される。あるBMP結合性タンパク質(例えば、グレムリン、BAMBI、またはフォリスタチン)は、BMPを結合し、そしてBMPレセプターとのそれらの相互作用を阻害する。さらに、阻害性Smadタンパク質(例えば、Smad6およびSmad7)が存在し、これらはBMPレセプターを結合して不活性化させる(Kowabataら、1998;Itohら、2000;Miyazono 2000)。本発明者らは、ヒトTM細胞、ONH星状細胞および相篩板細胞が、BMPレセプター複合体についてのメッセージおよびタンパク質を発現することを発見している。従って、これらの細胞は、内因性のBMPリガンドに応答し得る。
【0044】
様々な方法が、新しい抗緑内障治療剤を発見するために使用され得、そしてこれらの技術は当業者に周知である。例えば、BMPのアゴニストまたはインヒビターとして働くペプチドまたはペプチド模倣剤は、BMP/BMPレセプター構造の分子モデリングによって発見され得る(Nickelら、2001)。BMPシグナル伝達は、Smadタンパク質の選択されたセットを含む(Kawabataら、1998;Itohら、2000;Attisenoら、2000)。選択されたBMPアゴニストおよびSmadアゴニストは、細胞ベースのアッセイを使用して発見され得る。試験細胞は、適切なBMPレセプターを発現するべきであり、適切なBMPシグナル伝達経路を所有するべきである。BMPシグナル伝達の主な影響の1つは、遺伝子発現の変化であるので、BMPアゴニストおよびSmadアゴニストは、BMP誘導性遺伝子についてのスクリーニングによって発見され得る。BMP調節性遺伝子の誘導はまた、定量的RT−PCR(Wangら、2001)、DNAマイクロアレイ、またはレポーター遺伝子構築物を使用することによって、mRNAのレベルを定量してアッセイされ得る。BMPシグナル伝達、BMP結合タンパク質(BMP関連タンパク質としても公知)(例えば、コーディン、グレムリン、およびフォリスタチン)の天然のインヒビターが存在する。タンパク質インヒビターのアンタゴニストは、リガンド結合アッセイを用いて発見され得る。例えば、試験薬剤は、組み換え精製グレムリンに付加され得、そしてグレムリンに結合するこれら薬剤は、当業者に公知の様々な技術を使用することで同定される。これらの薬剤がグレムリンアンタゴニストであるか否かを決定するために、上記で記載したアッセイと類似の細胞ベースのアッセイが使用される。
【0045】
任意の公知のインビトロおよびインビボのスクリーニングモデルは、本発明と組み合わ
せて、BMPファミリーの遺伝子に関する新しい緑内障治療を同定するために使用され得ることが企図される。このようなモデルは当業者にとって周知であり、それらの実施は決まりきった仕事になる。小さいペプチドまたはペプチド模倣物は、BMP、BMPRおよび/またはBMP結合タンパク質の遺伝子産物の構造/機能の知識に基づいて設計され得る。リガンド結合アッセイは、BMP、BMPRまたはBMP結合タンパク質に結合する低分子の検出に使用され得る。細胞ベースのアッセイにより、BMPシグナル伝達経路に対する様々な薬剤の影響を見ることができ得る。レポーター遺伝子に連結されたBMPファミリー遺伝子プロモーターを含むノック−イン細胞株は、BMPファミリーメンバーの遺伝子発現を変化させる薬剤を探索するために作製され得る。これらのアッセイは、アゴニスト分子およびアンタゴニスト分子の両方の同定に使用され得る。エキソビボアッセイ(例えば、灌流培養されたヒトの眼由来の前方区画の(Clarkら、1995a;Pangら2000))は、TM組織におけるIOPおよびBMPシグナル伝達に対する薬剤の影響を試験するために使用され得る。緑内障のげっ歯類モデルは、マウスおよびラットの、安定なBMPファミリーメンバーのトランスジェニック株、ノックアウト株、またはノックイン株を作製するための周知の技術を使用して作製され得る。これらのげっ歯類モデルは、緑内障様の表現型を変化させる薬剤についてスクリーニングするために使用され得る(例えば、IOPに対する影響を評価するための限圧測定、緑内障の目の神経学に対する影響を評価するための組織学)。
【0046】
(B.キット)
本発明は、緑内障の初期検出のための方法、化合物およびキットを提供する。このキットは、BMPポリペプチドまたはBMPタンパク質をコードする核酸セグメントを含み得る。このキットはさらに、サンプルと本発明の核酸またはペプチドとの間の相互作用を検出するための試薬を含み得る。提供された試薬は、放射性標識化、蛍光標識化、または酵素的標識化され得る。このキットは、本発明の核酸またはペプチドもしくはタンパク質と結合または相互作用し得る、公知の放射性標識された薬剤を含み得る。
【0047】
キットの試薬は、液体溶液としてか、固体支持体に結合してかまたは乾燥した粉末として提供され得る。好ましくは、試薬が液体溶液中に提供される場合、この液体溶液は水溶液である。好ましくは、提供される試薬が固体支持体に結合する場合、この固体支持体はクロマトグラフィー媒体、複数のウェルを有する試験プレート、あるいは顕微鏡スライドであり得る。提供される試薬が乾燥した粉末である場合、この粉末は、提供され得る適切な溶媒の添加によって再び構成され得る。
【0048】
なお、さらなる実施形態において、本発明は、緑内障の診断のための診断方法、および関連するキットに関する。本発明のBMP関連のペプチドおよび核酸は、患者のサンプル由来のBMP核酸における多型または変異を検出するために使用され得ることが提案される。一般に、これらの方法は、このような多型または変異を含むことが疑われるサンプルを最初に獲得する工程、本発明のペプチドまたは核酸とこのサンプルとを接触させる工程、(この場合には、複合体を形成させるのに有効な条件下であり得る)、および次いで複合体の存在を検出する工程を包含する。
【0049】
一般に、複合体形成の検出は当該分野で周知であり、多数のアプローチの適用を通して成し遂げられ得る。例えば、本発明は、ELISA、RIA、間接的蛍光技術などの適用を企図する。概して、複合体形成は、標識(例えば、放射性標識または酵素タグ(例えば、アルカリホスファターゼ、西洋ワサビペルオキシダーゼなど)の使用を通して検出される。もちろん、二次結合リガンドの使用によってさらなる利益を見出し得る。
【0050】
以下の実施例は、本発明の局面の実行において本発明者らによって使用される技術を示す。これらの技術が、本発明を実施するための好ましい実施形態の例示であることは、本
開示を鑑みて当業者に認識されるべきであり、多数の改変が、本発明の精神および意図される範囲から逸脱することなくなされ得ることが認識される。
【実施例】
【0051】
(実施例1)
細胞培養:ヒトTM細胞およびONH細胞を、ドナーの眼から記載されるように生成した(Steelyら、1992;Steelyら、2000;Wilsonら、1993;Clarkら、1994;Clarkら、1995b;Clarkら、1995c;Clarkら、1996;Clarkら、2001a;Clarkら、2001b;Dickersonら、1998;Wordingerら、1998;Wordingerら、1999;Wordingerら、2000;Wordingerら、2002;Lambertら、2001;Agarwalら、1999;Liuら、2001)。TM細胞を、年齢6日齢から90歳の範囲のドナーのTM外移植体から増殖させた。ヒト視神経乳頭星状細胞および篩板(LC)細胞を、慎重に解剖した視神経乳頭(2日齢〜90歳までの年齢のドナー)から生成し、そして以前の報告に従って特徴付けた(Lambertら、2001;Clarkら、1995a)。細胞を、以下の培地:TM細胞については10%ウシ胎仔血清(HyClone,Logan,UT)および抗生物質(Gibco BRL−Life Technologies,Grand Island,NY)を含むHam’s F10培地(JRH Biosciences、Lenexa、KS);LC細胞については10%FBSを含むダルベッコ改変イーグル培地(DMEM,HyClone);そしてONH星状細胞については5%FBSを含む星状細胞増殖培地(AGM,Clonetics,San Diego,CA)内で、コンフルエントまで増殖させた。
【0052】
RT−PCR:ヒトTM組織およびONH組織もまた、ドナーの眼から解剖した(Wordingerら、1998;Wangら、2001)。全RNAをTM細胞およびTM組織ならびにONH細胞およびONH組織から、TRIzol抽出(Gibco BRL−Life Technologies)を使用して抽出し、そしてcDNAを標準的な手順(Wordingerら、1998;Worgingerら、1999;Worgdingerら、2000;Wordingerら、2002)を使用して逆転写によって調製した。PCRプライマーを、Oligos 4.0ソフトウェアプログラムを使用して設計した(表1のプライマー対を参照のこと)。全てのプライマー対を、潜在的に混入したゲノムDNA配列の増幅が予想される産物よりも実質的に長いmRNA PCR産物を生じるように設計した。なぜなら、RNAプロセシングの間に切除されるイントロン配列が、ゲノムDNAに含まれるからである。β−アクチンPCRプライマーAGGCCAACCGCGAGAAGATGACC(上流)およびGAAGTCCAGGGCGACGTAGCAC(下流)と、55℃アニーリング温度とにより、350bpのPCR産物を得た。
【0053】
PCR反応を、Taq Start Antibody Hot Startを以下:94℃で2分間、92℃で2分間、そして(最適なアニーリング温度で30秒間、72℃で90秒間の伸長、92℃で45秒間の変性)×40サイクルのサイクル条件を使用して、記載されたように実行した(Wordingerら、1998;Wordingerら、1999;Wordingerら、2000;Lambertら、2001;Wordingerら、2002)。増幅したPCR産物を、1.5%アガロースゲルで水平な電気泳動によって試験した。RT−PCR産物の特異性を確実にするために、増幅したPCR産物内の領域にハイブリダイズする、Oligo 4.0を用いて設計したプローブを使用して、サザンブロット解析を実行した。PCR反応の特異性を証明するために、PCR産物を配列決定した。表2は、ヒトTMおよびONHで発現しているBMPファミリーのメンバーを列挙している。
【0054】
【表1】
【0055】
【表2】
ウェスタンイムノブロッティング:タンパク質を、溶解緩衝液を使用して培養細胞から抽出し、そしてタンパク質を、ニトロセルロースメンブレンへの電気泳動的な移動に先立って、変性ポリアクリルアミドゲル電気泳動によって分離した(Lambertら、2001)。メンブレンを、5%ミルク(BMPについて)または3%ゼラチン(BMPRについて)でブロッキングし、そして以下の一次抗原:BMP2、BMP4、BMP5、BMP7(すべてSanta Cruz,Santa Cruz,CAから)、あるいはBMP−RIA、BMP−RIB、BMP−RII(Jackson Immuno Research,West Grove,PAから)と共にインキュベートした。メンブレンを洗浄し、二次抗体(BMPについてヤギ抗マウスIgG−西洋ワサビペルオキシダーゼ、Santa Cruz;BMPレセプターについてロバ抗ヤギ−西洋ワサビペルオキシダーゼ、Jackson Immuno Research)と共にインキュベートし、そしてWesternBreeze化学発光免疫検出システム(Invitrogen,Carlsbad,CA)を使用して現像した。
【0056】
ヒトのTM細胞およびTM組織におけるBMP、BMPR mRNAの発現:ヒトのTM細胞およびTM組織においてBMP−2プライマー対、BMP−4プライマー対、BMP−5プライマー対およびBMP−7プライマー対についての予想された増幅産物は、図6に示される。特異的プローブを使用したサザンブロットによって、これらが予想されたPCR産物であることが証明された。全てのヒトTM細胞株および組織は、BMP−2、BMP−4、およびBMP−7のメッセージを発現した。ところが、BMP−5のメッセージは、ヒトTM組織サンプルにおいて検出されないくらい低いものであった(図6、レーン6およびレーン7)。cDNA無しのコントロール反応は、増幅産物を生じず、試薬およびプライマーには、DNAもRNAも混入しれていなかったことを示す(図6、レーンC)。
【0057】
図7は、ヒトのTM細胞およびTM組織においてBMP−RIAプライマー対、BMP−RIBプライマー対、およびBMP−RIIプライマー対についての予想されたサイズの増幅産物を示す。全てのヒトTM細胞およびヒトTM組織は、BMPレセプター複合体のメッセージを発現した。特異的なプローブを使用したサザンブロットによって、これらが予想されたPCR産物であることが証明された。代替的な増幅産物(350bp)は、BMP−RII PCR反応において検出された。その代替的な増幅産物は、全てのヒトTM細胞およびヒトTM組織において存在していた。この代替的なバンドは現在、選択的にスプライスされた型のレセプターであるか否かを決定するために同定中である。cDNA無しのコントロール反応は増幅産物を生じず(図7、レーンC)このことは、試薬およびプライマーには、DNAもRNAも混入していなかったことを示す。
【0058】
ヒトONH細胞およびONH組織におけるBMPおよびBMPレセプターmRNAの発現:ヒトONH星状細胞およびONH組織における、BMP−2プライマー対、BMP−4プライマー対、BMP−5プライマー対およびBMP−7プライマー対についての予想されたサイズの増幅産物は、図8に示される。全てのONH星状細胞およびONH組織は、それぞれのBMPのメッセージを発現した。ヒト脳星状細胞を、ポジティブコントロール細胞株として使用した。特異的プローブを使用したサザンブロットによって、これらが予想されたPCR産物であることが証明された。BMP−2以外の、全ての他のBMPは、ヒト脳星状細胞により発現された(図8、レーン7)。cDNA無しのコントロール反応は、増幅産物を生じず(図8、レーンC)、試薬およびプライマーには、DNAもRNAも混入していなかったことを示す。
【0059】
図9は、培養ヒトLC細胞における、BMP−2プライマー対、BMP−4プライマー対、BMP−5プライマー対およびBMP−7のプライマー対についての予想されたサイズの増幅産物を示す。全てのLC細胞株は各々のBMPのメッセージを発現した。特異的
なプローブを使用したサザンブロットによって、これらが予想されたPCR産物であることが証明された。cDNA無しのコントロール反応は増幅産物を生じず(図9、レーンC)このことは、試薬およびプライマーには、DNAもRNAも混入していなかったことを示す。
【0060】
ヒトONH星状細胞およびONH組織における、BMP−RIA、BMP−RIBおよびBMP−RIIのプライマー対についての予想されたサイズの増幅産物は、図10に示される。全てのONH星状細胞およびONH組織は、BMP−RIAおよびBMP−RIBのメッセージを発現した。特異的プローブを使用したサザンブロットによって、これらが予想されたPCR産物であることが証明された。ONH組織(図10、レーン6)以外の全てのONH星状細胞細胞株によってBMP−RIIが発現された。全てのBMPレセプターのメッセージ(図10、レーン7)が、ポジティブコントロールとして働くヒト脳星状細胞細胞株により発現された。ONH組織およびONH細胞株におけるBMP−RIIの発現には食い違いが存在するようであった。ONH組織での発現の減少は、発現の低いレベルを反映し得る。cDNA無しのコントロール反応は、増幅産物を生じず(図5、レーンC)、試薬およびプライマーにはDNAもRNAも混入していなかったことを示す。
【0061】
図11は、培養ヒトLC細胞における、BMP−RIA、BMP−RIB、およびBMP−RIIのプライマー対についての予想されたサイズの増幅産物を示す。全てのLC細胞株は、各々のBMPレセプターのメッセージを発現した。特異的なプローブを使用したサザンブロットによって、これらが予想されたPCR産物であることが証明された。cDNA無しのコントロール反応は増幅産物を生じず(図11、レーンC)このことは、試薬およびプライマーには、DNAもRNAも混入していなかったことを示す。
【0062】
ヒトTM細胞およびTM組織ならびにONH細胞およびONH組織におけるBMPタンパク質およびBMPレセプタータンパク質の発現:図12は、ヒトTM細胞およびTM組織ならびにONH細胞およびONH組織におけるBMP−2、BMP−4、BMP−5、BMP−7、BMP−RIA、BMP−RIB、およびBMP−RIIのタンパク質の化学発光免疫ブロット検出を示す。研究した全ての細胞株は、それぞれのBMPタンパク質を発現した。BMPタンパク質は、細胞株において以下の分子量:BMP−2について54〜56kDa、BMP−4について25〜27kDa、BMP−5について55〜57kDa、およびBMP−7について77kDaで検出された。複数のバンドがBMP−2およびBMP−4について検出され、これらは、他の研究において見られるようなこれらBMPのグリコシル化された型、および部分的にグリコシル化された型を示す可能性が最も高い。ところがグリコシル化の研究は、本研究の範囲を越えているので、本研究者らはグリコシル化の研究を行っていない。BMPレセプタータンパク質は、細胞株において以下の分子量:BMP−RIAについて38kDa、BMP−RIBについて64kDa、およびBMP−RIIについて57kDaで検出された。複数のバンドがTM細胞においてBMP−RIBおよびBMP−RIIについて検出され、これらは、他の研究において見られるようなグリコシル化された型および、部分的にグリコシル化された型を示す可能性が最も高い。BMPレセプターのタンパク質の発現レベルは、ONH細胞と比較してTM細胞では低いようであった。例えば、BMP−RIIは、TM細胞において検出されず、そしてBMP−RIBは大々的に減少した。
【0063】
培養ヒトTM細胞および培養ヒトONH細胞におけるBMP関連タンパク質mRNAの発現:ヒトTM細胞株における、BMP関連タンパク質のプライマー対について予想されたサイズの増幅産物は、図13に示される。ヒトTM細胞株は、DRM(グレムリン)、コーディン、フォリスタチンおよびNMA(バンビ)のメッセージを発現した。特異的プローブを使用したサザンブロットによって、これらが、予想されたPCR産物であること
が証明された。細胞株間でメッセージの発現において明らかな違いは無かった。試験した全てのヒトTM細胞は、BMP関連タンパク質であるノギンおよびCer−1のmRNAを発現できなかった。cDNA無しのコントロール反応は、増幅産物を生じず、試薬およびプライマーにはDNAもRNAも混入していなかったことを示す。
【0064】
ONH星状細胞およびLC細胞株における、BMP関連タンパク質のプライマー対について予想されたサイズの増幅産物は、図14に示される。全てのONH星状細胞およびLC細胞株は、DRM(グレムリン)、フォリスタチンおよびNMA(バンビ)のメッセージを発現した。特異的プローブを使用したサザンブロットによって、これらが、予想されたPCR産物であることが証明された。大多数のLC細胞およびONH星状細胞は、コーディンのメッセージを発現した。試験した全てのヒトONH星状細胞およびLS細胞は、BMP関連タンパク質であるノギンおよびCer−1のmRNAを発現できなかった。cDNA無しのコントロール反応は、増幅産物を生じず、試薬およびプライマーには、DNAもRNAも混入していなかったことを示す。
【0065】
図15は、緑内障のTM細胞におけるBMPアンタゴニストのグレムリン(CKTSF1B1)の発現の上昇を示す。遺伝子発現を、Affymetrix遺伝子アレイ(Affymetrix遺伝子チップU133A)を使用して評価した。
【0066】
本明細書中で開示し、特許請求した組成物および/または方法の全ては、本開示を鑑みて過度の実験無しになされ、そして実施され得る。本発明の組成物および方法を好ましい実施形態に関して記載してきたが、本発明の概念、意図および範囲から逸脱することなく、本明細書中に記載される組成物および/または方法ならびに方法の工程または工程の順序にバリエーションが適用され得ることは、当業者にとって明らかである。より具体的には、化学的および構造的の両方で関連する特定の薬剤は、似た結果を達成するために、本明細書中に記載される薬剤の代用になり得ることが明らかである。当業者にとって明らかである全てのこのような代替物および改変物は、添付した特許請求の範囲に定義されるような、本発明の精神、範囲および概念内であると考えられる。
【0067】
(参考文献)
以下の参考文献は、例示的な手順、または本明細書中に記載される手順に対して補助的な他の詳細を提供する程度にまで、本明細書中で参考として具体的に援用される。
(書籍)
【0068】
【表3】
【図面の簡単な説明】
【0069】
図面は、本明細書の一部を成し、そして本発明のある局面をさらに実証するために含まれる。本発明は、本明細書中に示された特定の実施形態の詳細な説明と組み合わせて、1つ以上のこれらの図面の参照によって、より良く理解され得る。
【図1a】BMP2Aのヌクレオチド配列およびアミノ酸配列。
【図1b】BMP2Aのヌクレオチド配列およびアミノ酸配列。
【図1c】BMP2Aのヌクレオチド配列およびアミノ酸配列。
【図2a】BMP4のヌクレオチド配列およびアミノ酸配列。
【図2b】BMP4のヌクレオチド配列およびアミノ酸配列。
【図2c】BMP4のヌクレオチド配列およびアミノ酸配列。
【図2d】BMP4のヌクレオチド配列およびアミノ酸配列。
【図3a】BMP5のヌクレオチド配列およびアミノ酸配列。
【図3b】BMP5のヌクレオチド配列およびアミノ酸配列。
【図3c】BMP5のヌクレオチド配列およびアミノ酸配列。
【図3d】BMP5のヌクレオチド配列およびアミノ酸配列。
【図4a】BMP7のヌクレオチド配列およびアミノ酸配列。
【図4b】BMP7のヌクレオチド配列およびアミノ酸配列。
【図4c】BMP7のヌクレオチド配列およびアミノ酸配列。
【図4d】BMP7のヌクレオチド配列およびアミノ酸配列。
【図5】骨形成タンパク質シグナル伝達経路。骨形成タンパク質は(BMP)2量体は、セリン/スレオニンキナーゼであるBMPレセプター1および2から構成される膜複合体に結合する。調節性のSmad(Smad1/Smad5)は、リン酸化されそして補助Smad(co−Smad)(Smad4)に結合する。結果できたこのSmad複合体は、核に入り、核で転写因子(TF)に結合し、そして遺伝子の発現を調節する。BMP関連タンパク質は、BMPに結合し、そしてBMPとBMPレセプターとの相互作用を阻害することによってBMPアンタゴニストとして働く。
【図6】ヒトTM細胞およびヒトTM組織におけるBMP発現。ヒトTM細胞(レーン1〜5)およびヒトTM組織(レーン6〜7)におけるBMP発現のRT−PCR解析から生成されたcDNAサンプル由来のBMP PCR産物のアガロースゲルの、エチジウムブロマイド染色。L=塩基対マーカー。C=PCRのネガティブコントロールレーン。β−アクチンを、ポジティブなRT−PCR内部コントロールとして使用した。
【図7】ヒトTM細胞およびヒトTM組織のBMPレセプター発現。ヒトTM細胞(レーン1〜5)およびヒトTM組織(レーン6〜7)におけるBMPレセプター発現のRT−PCR解析から生成されたcDNAサンプル由来のPCR産物のアガロースゲルのエチジウムブロマイド染色。L=塩基対マーカー。C=PCRのネガティブコントロールレーン。β−アクチンを、ポジティブなRT−PCR内部コントロールとして使用した。
【図8】ヒトONH星状細胞、ヒトONH組織およびヒト脳星状細胞におけるBMPの発現。ヒトONH星状細胞(レーン1〜5)、ヒトONH組織(レーン6)、およびヒト脳星状細胞(レーン7)におけるBMP発現のRT−PCR解析から生成されたcDNAサンプル由来のPCR産物のアガロースゲルのエチジウムブロマイド染色。L=塩基対マーカー。C=PCRのネガティブコントロールレーン。β−アクチンを、ポジティブなRT−PCR内部コントロールとして使用した。
【図9】ヒト篩板細胞株におけるBMPの発現。ヒト篩板細胞(レーン1〜9)のRT−PCR解析から生成されたcDNAサンプル由来のPCR産物のアガロースゲルのエチジウムブロマイド染色。L=塩基対マーカー。C=PCRのネガティブコントロールレーン。β−アクチンを、ポジティブなRT−PCR内部コントロールとして使用した。
【図10】ヒトONH星状細胞、ONH組織およびヒト脳星状細胞におけるBMPレセプターの発現。ヒト視神経乳頭星状細胞(ONH)(レーン1〜5)、ヒトONH組織(レーン6)およびヒト脳星状細胞(レーン7)におけるBMPレセプター発現のRT−PCR解析から生成されたcDNAサンプル由来のPCR産物のアガロースゲルのエチジウムブロマイド染色。L=塩基対マーカー。C=PCRのネガティブコントロールレーン。β−アクチンを、ポジティブなRT−PCR内部コントロールとして使用した。
【図11】ヒト篩板細胞株におけるBMPレセプターの発現。ヒト篩板細胞(レーン1〜9)のRT−PCR解析から生成されたcDNAサンプル由来のPCR産物のアガロースゲルのエチジウムブロマイド染色。L=塩基対マーカー。C=PCRのネガティブコントロールレーン。β−アクチンを、ポジティブなRT−PCR内部コントロールとして使用した。
【図12】培養ヒトTM細胞、培養ヒト視神経乳頭星状細胞(ONA)、および培養ヒト篩板細胞におけるBMPおよびBMPレセプターの発現のウェスタンイムノブロット。ヒト小柱網細胞(レーン1〜2)、ヒトONH星状細胞(レーン3〜4)、およびヒト篩板細胞(レーン5〜6)におけるBMPタンパク質およびBMPレセプターの化学発光による検出。タンパク質サイズは、kDaで示す。
【図13】ヒトTM細胞におけるBMP関連タンパク質mRNAの発現。ヒトTM細胞(レーン1〜5)のRT−PCR解析から生成されたcDNAサンプル由来のPCR産物のアガロースゲルのエチジウムブロマイド染色。L=塩基対マーカー。C=PCRのネガティブコントロールレーン。β−アクチンを、ポジティブなRT−PCR内部コントロールとして使用した。
【図14】ヒト篩板細胞およびONH星状細胞におけるBMP関連タンパク質mRMAの発現。篩板(LC)細胞(レーン1〜7)およびONH星状細胞(ONA)(レーン8〜11)のRT−PCR解析から生成されたcDNAサンプル由来のPCR産物のアガロースゲルのエチジウムブロマイド染色。L=塩基対マーカー。C=PCRのネガティブコントロールレーン。β−アクチンは、ポジティブなRT−PCRインターナルコントロールとして使用された。
【図15】緑内障TM細胞における、BMPアンタゴニストのグレムリン(CKTSF1B1)発現の上昇の例示。遺伝子発現を、Affymetrix遺伝子アレイ(Affymetrix遺伝子チップU133A)を使用して評価した。(配列表)
【化1】
【化2】
【化3】
【化4】
【化5】
【化6】
【化7】
【化8】
【化9】
【化10】
【化11】
【化12】
【化13】
【化14】
【化15】
【化16】
【化17】
【化18】
【化19】
【化20】
【化21】
【化22】
【化23】
【化24】
【化25】
【化26】
【化27】
【化28】
【化29】
【化30】
【化31】
【化32】
【化33】
【化34】
【化35】
【化36】
【化37】
【化38】
【化39】
【特許請求の範囲】
【請求項1】
細胞または体液から得られたサンプルにおいて、骨形態形成ファミリーメンバー遺伝子の発現の変化を検出することによる、緑内障を診断する方法。
【請求項1】
細胞または体液から得られたサンプルにおいて、骨形態形成ファミリーメンバー遺伝子の発現の変化を検出することによる、緑内障を診断する方法。
【図5】

【図6】
【図7】

【図8】

【図9】
【図10】
【図11】

【図12】
【図13】
【図14】

【図15】
【図1a】

【図1b】
【図1c】

【図2a】

【図2b】

【図2c】

【図2d】

【図3a】

【図3b】

【図3c】
【図3d】

【図4a】

【図4b】

【図4c】

【図4d】

【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図1a】
【図1b】
【図1c】
【図2a】
【図2b】
【図2c】
【図2d】
【図3a】
【図3b】
【図3c】
【図3d】
【図4a】
【図4b】
【図4c】
【図4d】
【公開番号】特開2008−301834(P2008−301834A)
【公開日】平成20年12月18日(2008.12.18)
【国際特許分類】
【出願番号】特願2008−230352(P2008−230352)
【出願日】平成20年9月8日(2008.9.8)
【分割の表示】特願2003−556021(P2003−556021)の分割
【原出願日】平成14年10月31日(2002.10.31)
【出願人】(399054697)アルコン,インコーポレイテッド (102)
【出願人】(504168673)ユニバーシティー オブ ノース テキサス ヘルス サイエンス センター (1)
【Fターム(参考)】
【公開日】平成20年12月18日(2008.12.18)
【国際特許分類】
【出願日】平成20年9月8日(2008.9.8)
【分割の表示】特願2003−556021(P2003−556021)の分割
【原出願日】平成14年10月31日(2002.10.31)
【出願人】(399054697)アルコン,インコーポレイテッド (102)
【出願人】(504168673)ユニバーシティー オブ ノース テキサス ヘルス サイエンス センター (1)
【Fターム(参考)】
[ Back to top ]