
骨誘導タンパク質及びペプチドの細胞内送達
本発明は、タンパク質導入ドメインと骨誘導ポリペプチドを含む融合タンパク質、ならびに、このようなポリペプチドを用いて骨形成を誘導し、そしてプロテオグリカンの合成を促進する方法を、提供する。本発明はまた、in vivoにおいて骨形成誘導能が実証されている骨誘導ポリペプチドを、提供する。
【発明の詳細な説明】
【発明の開示】
【0001】
本出願は、2003年3月24日に出願された米国仮出願第60/456,551の利益を主張する。
発明の属する分野
本発明は、概して、骨誘導タンパク質、及びこれらのタンパク質を細胞内に送達するための方法に、関する。より具体的には、本発明は、LIM石灰化タンパク質(LMP)、骨形態形成タンパク質(BMP)及びSmadタンパク質などの骨誘導タンパク質、このような骨誘導タンパク質とタンパク質導入ドメイン(PTD)との複合体、PTDと骨誘導タンパク質をコードするヌクレオチド配列を含む核酸との複合体、ならびに、これらの複合体の細胞内への導入に、関する。
【0002】
背景
組織再生は、疾患、外傷または外科手術に続く治癒過程の重要な要素である。疾患または外傷により、例えば骨欠損が生じる状況や、外科的手順(自家移植片もしくは同種移植片の挿入、骨架橋または骨融合など)が、骨欠損を補正するために行われる状況において、骨再生は、回復の主要な目標である。しかしながら、これは、常にまたは容易に達成される目標ではなく、そして多くの研究が、組織修復及び再生を促進するためのより新たな、そしてより効果的な方法に対して、供されている。
【0003】
骨架橋を作ることによって関節動作を排除することは、変性脊椎及び関節障害の治療に対する一般的な整形外科的ストラテジーである。脊椎融合の失敗は、該手順を受ける患者の45%ほどにおいて起こる可能性があり、彼らに疼痛の持続、外科手術の再施行、医療費及び全体的な治療の失敗を残す。
【0004】
細胞内及び細胞外骨誘導タンパク質は、骨成長及び修復を促進し、そして治療に用いるための潜在的な標的となる。このようなタンパク質としては、骨形態形成タンパク質及びLIM石灰化タンパク質が挙げられる。BMPは、in vivoにおいて骨成長を刺激することが示されており、そして、LMP、特にLMP−1及びLMP−3は骨誘導のより上流に影響を与え、これは、LMP−1発現の阻害が、糖質コルチコイド類またはBMP−6によって通常刺激されるであろう小結節の形成を遮断することからも明らかである。これらは、細胞表面受容体との相互作用によって作用する「細胞外」タンパク質と考えられるため、非常に高用量の骨形態形成タンパク質が、ヒトにおいて一貫した影響に達するのに必要とされる。BMPの製造費用は概して高いため、このことは、治療費用が法外に高いことを意味する可能性がある。したがって、BMPは有効性を示し、そして骨誘導に対する実行可能な療法ではあるものの、より費用効率が高く、そしておそらくさらにいっそう治療的に有効であろう代替治療法を開発することが、有益であろう。
【0005】
LMPの細胞内環境への送達は、魅力的な治療レジメンを提供する。これは、細胞に、LIM石灰化タンパク質をコードするヌクレオチド配列を含むプラスミドをトランスフェクトすることによって、遂行することが可能であり、あるいは、標的細胞に、LMPのヌクレオチド配列を保有するウイルスベクターを感染させることによって、行うことが可能である。しかしながら、これら各々の技術には限界がある。プラスミドのトランスフェクションでは、細胞をトランスフェクションのために単離し、次いでトランスフェクション後に植え込むことが一般に必要となる。ウイルスの送達では、ウイルスの細胞への侵入を容易にするために、適切な受容体を標的細胞の表面に配置することが、一般に必要となる。
【0006】
骨誘導タンパク質及びペプチドの使用、特に、細胞内メカニズムによって作用するこれらのタンパク質及びペプチドの使用に対して、多大な可能性がある。必要とされるのは、有効な細胞内骨誘導タンパク質及びペプチドを細胞内に送達する方法である。
【0007】
発明の概要
本発明は、細胞透過性の骨誘導ポリペプチドを作製する方法を提供し、該方法は、骨誘導ポリペプチドが細胞透過性ポリペプチドとの融合タンパク質の一部として発現するように配置された、細胞透過性ポリペプチドと骨誘導ポリペプチドをコードする発現コンストラクトを、適切な宿主細胞に導入することを含む。発現コンストラクトは、融合産物をコードするポリヌクレオチド配列の転写を導くように配置されたプロモーターを、一般に含有する。
【0008】
発現コンストラクトは、精製タグをさらに含んでもよい。細胞透過性ポリペプチドは、HIV−TAT、VP−22、増殖因子のシグナルペプチド配列、Pep−1及びショウジョウバエAntpペプチドからなる群より選択されてもよい。骨誘導ポリペプチドは、LMP−1、LMP−3、配列番号1、配列番号2、配列番号3、配列番号4、配列番号5、配列番号6、配列番号7、配列番号8、BMP−2、BMP−4、BMP−6、BMP−7、TGF−β1及びSmadからなる群より選択されてもよい。
【0009】
本発明は、配列番号1、配列番号2、配列番号3、配列番号4、配列番号5、配列番号6、配列番号7及び配列番号8、またはその組み合わせからなる群の中から選択される骨誘導ポリペプチドを、提供する。
【0010】
本発明はまた、哺乳動物において骨形成を誘導する方法であって、タンパク質導入ドメインと少なくとも一つの骨誘導ポリペプチドとを含む、有効量の融合ポリペプチドを投与することを含む方法を提供する。融合ポリペプチドは、移植片(インプラント)として投与されてもよく、そして、少なくとも一つの多能性前駆細胞に投与されてもよく、該細胞は、骨誘導を促進するために哺乳動物に植え込まれることが可能である。
【0011】
本発明はまた、タンパク質導入ドメインと少なくとも一つの骨誘導ポリペプチドを含む融合タンパク質をコードするポリヌクレオチドを提供し、タンパク質導入ドメインは、例えばHIV−TAT、VP−22、増殖因子のシグナルペプチド配列、Pep−1及びショウジョウバエAntpペプチドによって代表される、多様なタンパク質導入、膜移行、及びその他の同様なポリペプチドの中から選択される。骨誘導ポリペプチドは、LMP−1、LMP−3、配列番号1、配列番号2、配列番号3、配列番号4、配列番号5、配列番号6、配列番号7、配列番号8、BMP−2、BMP−4、BMP−6、BMP−7、TGF−β1及びSmadからなる群より選択されてもよい。
【0012】
哺乳動物においてプロテオグリカン合成を誘導する方法もまた、提供される。該方法は、タンパク質導入ドメインと少なくとも一つの骨誘導ポリペプチドを含む有効量の融合ポリペプチドを投与することを含む。融合ポリペプチドは、移植片として投与されてもよく、そして、少なくとも一つの多能性前駆細胞に投与されてもよい。
【0013】
骨誘導ポリペプチドと機能可能なように連結されている膜移行ペプチドを含む、単離された融合タンパク質が、本発明によって提供される。膜移行ペプチドは、HIV−TAT、VP−22、増殖因子のシグナルペプチド配列、Pep−1及びショウジョウバエAntpペプチドからなる群より選択されてもよく、そして骨誘導ポリペプチドは、LMP−1、LMP−3、配列番号1、配列番号2、配列番号3、配列番号4、配列番号5、配列番号6、配列番号7、配列番号8、BMP−2、BMP−4、BMP−6、BMP−7、TGF−β1及びSmadからなる群より選択されてもよい。
【0014】
本発明は、前駆細胞において骨芽細胞への分化を誘導する方法を提供し、該方法は、タンパク質導入ドメインと少なくとも一つの骨誘導ポリペプチドとを含む有効量の融合ポリペプチドを前駆細胞に投与することを含む。タンパク質導入ドメインは、HIV−TAT、VP−22、増殖因子のシグナルペプチド配列、Pep−1及びショウジョウバエAntpペプチドによって代表される群から選択されることが可能であり、そして骨誘導ポリペプチドは、LMP−1、LMP−3、配列番号1、配列番号2、配列番号3、配列番号4、配列番号5、配列番号6、配列番号7、配列番号8、BMP−2、BMP−4、BMP−6、BMP−7、TGF−β1及びSmadによって代表される群から選択されてもよい。
【0015】
詳細な説明
本発明者は、タンパク質導入ポリペプチドと骨誘導ポリペプチドを含む融合タンパク質を、in vivoにおいて骨の発達及び椎間板再生を促進するために効果的に用いることが可能であることを、発見した。したがって、本発明は、細胞内へ送達するための骨誘導ポリペプチド、このような骨誘導ポリペプチドとタンパク質導入配列をコードするポリヌクレオチド、ならびに、in vivoにおいて骨の発達及び椎間板再生を促進するのにこれらの融合タンパク質を利用する方法を、提供する。
【0016】
以前の研究により、LIM石灰化タンパク質のスプライスバリアント1及び3(LMP−1及びLMP−3)は骨誘導性であるが、LMP−2はこのような骨誘導の可能性を有するようにはみえないことが、実証された。ヒトLMP−1(hLMP−1)のアミノ酸配列のアミノ酸94〜133に相当する40アミノ酸配列は、LMP−1及びLMP−3の双方に共通している。したがって、発明者は、このタンパク質のこの特有な領域自体が、骨誘導の可能性を有するのではないかと推測した。この配列の重複セグメントを含むペプチドを設計し、そして、これを用いて発明者の仮説を検証した。その結果、LMP−1及びLMP−3由来のペプチドが、骨誘導の可能性を有することが、示唆される。これらのペプチドは、in vivoにおいて用いた場合に、骨形成誘導能を示した。図6は、本発明の方法において、本発明の融合タンパク質の一部として細胞内に導入された場合に、骨誘導の機能性を示したペプチドを示す。
【0017】
LMP−1を発現している細胞を胸椎及び腰椎に植え込むと、無胸腺ラットにおいて堅固な脊椎融合がもたらされることが、発明者によって示された。本明細書において開示された結果により、配列番号1、配列番号2、配列番号3、配列番号4、配列番号5、配列番号6、配列番号7及び配列番号8によって提供されるペプチドなどの、LMP−1及びLMP−3ペプチドもまた、LMP−1について実証された骨誘導の利益を提供することが、実証される。
【0018】
骨形態形成タンパク質が、ヒトにおいて一貫した効果を発揮するには、概して、非常に高用量が必要とされる。PTD融合タンパク質によるLMP−1の細胞内への送達は、本明細書に記載されるように、より費用効率的な治療の選択肢を提供する。さらに、これまでの結果により、LMP−1、及びLMP関連骨誘導ペプチドが、細胞内で作用して諸因子のカスケードを誘導し、天然の骨誘導経路を活性化することが、示唆される。
【0019】
タンパク質導入ポリペプチドは、核酸配列または治療用タンパク質の取り込み、及びそれに続く発現を、容易にする。文献において、これらは、代替的に、そしてしばしば同義的に、細胞透過性ペプチド、タンパク質導入ドメイン、膜輸送配列及び膜移行ペプチドと称されてもよい。これらは、付属するペプチド、ポリペプチドまたはタンパク質を、受容体とは独立に、細胞膜を介して細胞内部に輸送する働きをする。タンパク質導入ドメインを利用する融合タンパク質は、タンパク質導入ドメインと機能可能なように連結されている、1またはそれより多くのペプチド、ポリペプチドまたはタンパク質を含むことが可能である。本発明において、このような融合タンパク質は、タンパク質導入ドメインと少なくとも一つの骨誘導ペプチド、ポリペプチドまたはタンパク質、あるいはその組み合わせを含むことが可能である。これらのペプチドを用いて、外胚葉、間葉または造血を起源とする自己、同種異系または異種の細胞あるいは組織に形質導入し、そして、新たな組織の形成を誘導または助長するためにこれをレシピエントに注入または植え込むことが可能である。本発明の方法において、このようなポリペプチドは、例えば多能性前駆(幹)細胞などの細胞を、例えばBMP−2、BMP−4、BMP−6、BMP−7、BMP−9、BMP−12、BMP−13、アグリカン、I型コラーゲン、II型コラーゲン、バーシカン、ルミカン、フィブロモジュリン、ビグリカン及びデコリンを産生するよう誘導することが可能なタンパク質の取り込みを、容易にする。本発明のポリペプチドの有効量を、実験デザイン及び本明細書に開示した結果において示すが、しかし、本明細書にて提供される有効量の開示をもとに、当業者が決定してもよい。
【0020】
LMPタンパク質ファミリーの一つである、ヒトLIM石灰化タンパク質−1(hLMP−1)は、in vitro及びin vivoにおいて骨石灰化の有効性を増強することができる細胞内調節タンパク質である。ヒトLMP−1は、アミノ酸スペーサーによって連結された二つの特別なジンクフィンガーから構成される特徴的な構造モチーフを有するため、このように称される。LIM石灰化タンパク質のスプライスバリアント及びそれらの使用については、米国特許第6,300,127;6,444,803;及び6,521,750において、発明者によって記載されている。LMP−1、LMP−2及びLMP−3の配列もまた、これらの特許において公開されている。1997年7月22日に、指定されたベクターpCMV2/RLMP(挿入断片10−4クローン/RLMPを有するベクターpRc/CMV2である)中の、10−4/RLMP(ドブネズミLIM石灰化タンパク質のcDNA)の試料を、アメリカン・タイプ・カルチャー・コレクション(ATCC)(12301 Parklawn Drive, Rockville, Md. 20852)に寄託し、そして受入番号209153を割り当てられた。1998年3月19日に、挿入断片HLPM−1s(ホモサピエンスLIM石灰化タンパク質のcDNA)を有するベクターpHis−Aの試料を、アメリカン・タイプ・カルチャー・コレクションに寄託し、そして受入番号209698を割り当てられた。
【0021】
血清型5型のアデノウイルス(Ad5)は、末梢血及び骨髄由来細胞を含む多様な細胞及び組織にLMPを送達するために、使用されている。(Bodenら、「Adenoviral Delivery of LMP-1 Induces Consistent Spine Fusion」、第47回年会、Orthopaedic Research Society、サンフランシスコ、カリフォルニア州(2001年))。しかしながら、Ad5ウイルスは、特異的な受容体(すなわち、コクサッキーアデノウイルス受容体またはCAR)を利用するが、これは、これらの細胞において存在しないかまたは限られた量にて存在する。受容体を介したメカニズムなしにタンパク質の細胞内への送達を容易にする、細胞膜を介したタンパク質導入は、多様な種類の細胞及び組織の処理を可能にする魅力的な代替手段を、提示する。
【0022】
LMP及び他の骨誘導性タンパク質の作用は、脳、脊髄、末梢神経、骨、軟骨、椎間板、結合組織、腱及び靭帯などの多様な組織において、これらが治療の可能性を有することを、示唆する。LMPの、例えば多様な組織への送達は、例えばコラーゲン、コラーゲンとセラミックとの組み合わせ、脱石灰化骨基質、ならびに、天然または合成ポリマー類(エラスチン、フィブリン、ポリ乳酸、ポリグリコール酸、ポリカプロラクトン、フマル酸ポリプロピレン、ポリビニルアルコール、ポリエステル類、ポリエーテル類、ポリヒドロキシル類など)、ならびに構造的移植片を含む送達システムによって、遂行されることが可能である。このような基質は、注射可能で成型可能な固形移植片、構造的移植片、またはその組み合わせであってもよい。
【0023】
本発明者は、機能性骨誘導タンパク質を細胞内に送達し、そして骨形成及びプロテオグリカン合成を効率的に誘導するのに、PTDを用いることが可能であることを、発見した。このような細胞透過性ペプチド移入(CPPI)は、骨誘導タンパク質を多様な細胞種類に送達するための方法を、提供する。元来はHIV−1 TATタンパク質に由来する11アミノ酸ペプチドを用いて、骨誘導タンパク質を細胞内に送達することに成功した。TATペプチドを、pTAT−HAベクターを用いて、細菌細胞において過剰発現させることが可能である。リコンビナントヒト遺伝子を、TATペプチド配列と目的の遺伝子産物の双方を含有する融合タンパク質を産生させるように、このベクター中に挿入することが可能である。さらに、PTD/骨誘導ポリペプチドを、融合タンパク質の単離及び精製を容易にするために、polyHisタグと連結して発現させることが可能である。pTAT−HAベクター、及び、TAT融合タンパク質の精製プロトコールについては、以前にNagaharaら(Nature Medicine 第4巻,1449〜1452頁、1998年12月)によって記載されている。
【0024】
多様なPTDにおいて見つけられるペプチド配列は、コクサッキー−アデノウイルス受容体(CAR)とは独立に、細胞内への侵入を容易にすることが可能であり、それによって標的細胞への導入効率を改良し、続いて、所望の効果に達するのに必要な核酸またはタンパク質の必要量を低減させる。したがって、PTD融合タンパク質は、治療費用を低下させるのに用いられてもよい治療ツールを、提供する。
【0025】
本発明の一つの態様において、タンパク質導入ドメインと骨誘導タンパク質との融合タンパク質が、提供される。骨誘導タンパク質としては、これに限定されるものではないが、LIM石灰化タンパク質(LMP)、骨形態形成タンパク質(BMP)及びSmadタンパク質が挙げられる。本明細書では、「骨誘導タンパク質」、「骨誘導ポリペプチド」及び「骨誘導ペプチド」は、骨誘導の機能性を有する、様々な長さのペプチドまたはポリペプチド、あるいは完全長タンパク質のいずれかを指すのに、同義的に用いられてもよい。
【0026】
PTDとLIM石灰化タンパク質を含む融合タンパク質は、本発明の一つの態様として提供される。融合タンパク質は、PTDと1またはそれより多くのLIM石灰化タンパク質またはポリペプチドを含むことが可能である。有用なLIM石灰化タンパク質としては、例えば、米国特許第6,300,127;6,444,803;及び6,521,750;ならびに、係属中の米国特許出願第09/959,578(2000年4月28日出願)において開示されるようなLMPが挙げられる。好ましくは、LMPは、RLMP、HLMP−1、HLMP−1s、HLMP−2、HLMP−3またはそれに由来するペプチドである。これらのペプチドは、例えば、配列番号1、配列番号2、配列番号3、配列番号4、配列番号5、配列番号6、配列番号7、または、配列番号8にあるようなポリペプチドを含むことが可能である。
【0027】
LIM石灰化タンパク質をコードするヌクレオチド配列は、好ましくは、完全長の以下配列:
tcctcatccg ggtcttgcat gaactcggtg
に相補的な核酸分子と、標準的な条件下でハイブリダイズするか、または、完全長の以下配列:
gcccccgccc gctgacagcg ccccgcaa
に相補的な核酸分子と、高ストリンジェントな条件下でハイブリダイズするか、あるいはその双方である。
【0028】
「標準的なハイブリダイゼーション条件」は、プローブのサイズ、背景及び核酸試薬の濃度、ならびにハイブリダイゼーションの種類(in situ、サザンブロット、またはDNA−RNAハイブリッドのハイブリダイゼーション(ノーザンブロット))によって変わるであろう。「標準的なハイブリダイゼーション条件」の決定は、当該技術分野における技能水準の範囲内にある。方法としては、例えば、米国特許第5,580,775(Fremeauら)、Southern, J. Mol. Biol., 98:503(1975年)、Alwineら、Meth. Enzymol., 68:220(1979年)、及び、Sambrookら、Molecular Cloning: A Laboratory Manual,第2版、コールドスプリングハーバー・プレス、7.19-7.50(1989年)に記載されるものが挙げられる。
【0029】
1セットの標準的なハイブリダイゼーション条件は、50%ホルムアミド、5X SSPE(150nM NaCl、10mM NaH2PO4[pH7.4]、1mM EDTA[pH8.0])、5Xデンハート液(100mlの水につき、フィコール20mg、ポリビニルピロリドン20mg、及びBSA 20mg)、10%硫酸デキストラン、1%SDS、及び100μg/mlサケ精子DNA中で、42℃にて2時間、ブロットをプレハイブリダイズすることを、含む。32P−標識cDNAプローブを加え、そしてさらにハイブリダイゼーションを14時間続ける。その後、ブロットを、2X SSPE、0.1%SDS中で、22℃にて20分間、2回洗浄し、続いて、0.1X SSPE、0.1%SDS中で、65℃にて1時間洗浄する。次いで、ブロットを乾燥させ、そして増感スクリーンの存在下でX線フイルムに5日間曝露する。
【0030】
「高ストリンジェントな条件」下では、プローブはその標的配列と、これら二つの配列が実質的に同一であるならば、ハイブリダイズするであろう。実質的に同一の配列のみがハイブリダイズし、非同一配列がハイブリダイズしないであろう条件を決定するための技術は、当業者に公知である。
【0031】
本明細書では、「タンパク質」の語は、ミメティック(類似体、模倣物)を含むことを意図し、そして「アミノ酸」の語は、L型アミノ酸、D型アミノ酸及び修飾アミノ酸を含むことを意図している。これらの代替品は、分子間の既知の構造類似性を用いて、当業者が作製してもよい。また、アミノ酸配列には、記載された配列のN末端またはC末端のいずれか、あるいは両方に追加のアミノ酸を含んでもよい、任意のペプチドまたはタンパク質配列を含まれることが、意図される。「骨誘導タンパク質」の語は、完全長タンパク質のみならず、バリアントまたは生物学的に活性なポリペプチド断片も含むことが、意図される。
【0032】
一つのアミノ酸が、1より多いヌクレオチドコドンによってコードされてもよく、そしてヌクレオチド配列が、同一ペプチドをコードする代替のヌクレオチド配列を作り出すために修飾されてもよいことは、当該技術分野において周知である。したがって、本発明の代替の態様には、先に記載されたようなアミノ酸配列を含有するペプチドをコードする、代替のDNA配列が、含まれる。特許請求の範囲にあるアミノ酸配列を含有するペプチドをコードする、DNA配列は、特許請求の範囲にある配列と、特許請求の範囲にあるアミノ酸配列のN末端またはC末端に位置する他のアミノ酸との、任意の組み合わせをコードするDNA配列を、含む。当然のことながら、アミノ酸及び核酸配列は、追加の残基、特にNまたはC末端アミノ酸、あるいは5’または3’核酸配列を含んでもよく、そして、該配列は、発現したポリペプチドまたはタンパク質に骨誘導の可能性を授与する限り、本質的には、本明細書にて開示される配列に、なお記載されてもよい。
【0033】
追加の核酸塩基を、骨誘導ポリペプチドをコードする核酸配列の5’または3’のいずれかに加えてもよく、そして、プロモーター、ポリアデニル化シグナル、追加の制限酵素部位、多重クローニング部位、他のコードセグメント等などの他のDNA配列と、組み合わせてもよい。したがって、このようなポリヌクレオチドの全体的な長さは、かなり変わってもよい。
【0034】
当然のことながら、ポリペプチドの「バリアント」は、天然のタンパク質とは完全に同一ではない。骨誘導ポリペプチドまたはタンパク質のバリアントは、例えば、1またはそれより多くのアミノ酸を挿入、欠失または置換することによりアミノ酸配列を変えることによって、得ることが可能である。ポリペプチドまたはタンパク質のアミノ酸配列を、例えば、置換によって修飾して、天然ポリペプチドと比べて同一または改良された質を実質的に有するポリペプチドを作り出すことが、可能である。置換は、保存的置換であってもよい。「保存的置換」とは、アミノ酸を、極性/非極性の性質、電荷またはサイズにおいて類似した側鎖を有する別のアミノ酸に置換することである。20の必須アノミ酸は、非極性側鎖を有するもの(アラニン、バリン、ロイシン、イソロイシン、プロリン、フェニルアラニン及びトリプトファン)、非荷電性極性側鎖を有するもの(メチオニン、グリシン、セリン、トレオニン、シスチン、チロシン、アスパラギン及びグルタミン)、酸性側鎖を有するもの(アスパラギン酸及びグルタミン酸)、及び塩基性側鎖を有するもの(リジン、アルギニン及びヒスチジン)として分類されることが可能である。保存性置換には、例えば、AspをGlu、AsnまたはGlnに;HisをLys、ArgまたはPheに;AsnをGln、AspまたはGluに;及び、SerをCys、ThrまたはGlyに置換することが含まれてもよい。例えば、アラニンは、保存的置換を行うのにしばしば用いられる。
【0035】
当業者は、バリアントポリペプチドを、生物学的活性に影響を及ぼすために、ポリペプチド構造中の1またはそれより多くの位置において第一のアミノ酸を第二のアミノ酸に置換することによって、得ることが可能である。アミノ酸置換は、例えば、生物学的活性の増加をもたらすポリペプチドの立体構造変化を誘導する可能性がある。
【0036】
当業者はまた、アミノ酸の親水性指数または疎水・親水性指数に基づいて、アミノ酸配列における置換を行ってもよい。したがって、本発明のバリアントアミノ酸分子は、LMP−1、LMP−2、LMP−3、配列番号1、配列番号2、配列番号3、配列番号4、配列番号5、配列番号6、配列番号7、あるいは、配列番号8にあるようなポリペプチドのアミノ酸配列を含むポリペプチドのアミノ酸配列と、100%よりは少ないが、しかし少なくとも約50%、そして好ましくは少なくとも約80%〜約90%の、アミノ酸配列相同性または同一性を有する。したがって、バリアント骨誘導ポリペプチドまたはタンパク質のアミノ酸配列は、本質的には、天然の骨誘導ポリペプチドまたはタンパク質のアミノ酸配列に相当する。本明細書では、「本質的には〜に相当する」の語は、LMP−1、LMP−2、LMP−3、配列番号1、配列番号2、配列番号3、配列番号4、配列番号5、配列番号6、配列番号7、あるいは、配列番号8にあるようなポリペプチドを含む骨誘導ポリペプチドまたはタンパク質によって生じる活性に類似した、生物学的活性及び酵素活性を誘発するであろうポリペプチド配列に向けたものであり、このような活性は、天然の骨誘導タンパク質の少なくとも約70%であり、そしてより好ましくは、天然の骨誘導タンパク質活性の100%を超える。
【0037】
骨誘導タンパク質のバリアントには、LMP−1、LMP−2、LMP−3、配列番号1、配列番号2、配列番号3、配列番号4、配列番号5、配列番号6、配列番号7または配列番号8を含む骨誘導タンパク質を含む、相当する骨誘導タンパク質中には存在しないアミノ酸残基が含まれてもよく、あるいは、LMP−1、LMP−2、LMP−3、配列番号1、配列番号2、配列番号3、配列番号4、配列番号5、配列番号6、配列番号7または配列番号8を含む骨誘導タンパク質と比べて、欠失が含まれてもよい。バリアントはまた、LMP−1、LMP−2、LMP−3、配列番号1、配列番号2、配列番号3、配列番号4、配列番号5、配列番号6、配列番号7または配列番号8を含む、相当するタンパク質と比べて、切断された「断片」であってもよく、該断片は、完全長のタンパク質またはポリペプチドのほんの一部である。
【0038】
骨形態形成タンパク質(BMP)は、タンパク質のTGF−βスーパーファミリーのメンバーである。BMPは、異所性骨または軟骨形成を誘導することが示されている。本発明にしたがって、PTDと骨形態形成タンパク質との融合タンパク質もまた、提供される。BMPとしては、例えば、BMP−2、BMP−3、BMP−3b、BMP−4、BMP−5、BMP−6、BMP−7、BMP−8、BMP−9、BMP−10、BMP−11、BMP−12、BMP−13、BMP−14、BMP−15、GDF−1、GDF−3、GDF−8及びGDF−9が挙げられる。骨形態形成タンパク質であるBMP−2、BMP−4、BMP−5、BMP−6、BMP−7、BMP−8またはBMP−9は、本発明の方法において、特に有用である可能性がある。
【0039】
Smadタンパク質は、細胞外TGF−β関連因子の受容体からのシグナル伝達を媒介する、細胞内タンパク質である(Heldinら、”TGF-β Signalling from Cell Membrane to Nucleus through SMAD Proteins”、Nature、第390巻(1997年))。Smadタンパク質は、BMPがその受容体に結合することによって、活性化される(すなわち、リン酸化される)ことが可能である。活性化されると、Smadタンパク質は、それが遺伝子発現を調節する場所である核に移行する。PTDとSmadタンパク質との融合タンパク質もまた、本発明において提供される。Smad−1、Smad−2、Smad−3、Smad−4、Smad−5、Smad−6、Smad−7またはSmad−8は、本発明におけるようなタンパク質導入ドメインとの融合タンパク質として送達される場合、骨誘導を促進するのに特に有用である可能性がある。
【0040】
本発明にしたがったタンパク質導入ドメインは、付属もしくは付随のタンパク質、ペプチドまたは核酸を細胞内部に送達するために、細胞の原形質膜を越えることが可能な、任意のペプチド、ミメティック、またはペプチドの核酸(PNA)配列であることが可能である。発明者は、骨誘導タンパク質を、骨誘導及びプロテオグリカン合成を促進する能力を損なうことなく、細胞内に(例えば、融合タンパク質の一部分として)送達可能であることを、実証した。PTDとしては、例えば、ショウジョウバエのホメオティック転写因子であるアンテナペディア(Antp)由来のポリペプチド、単純ヘルペスウイルス(HSV)タンパク質VP22、カポジ線維芽細胞増殖因子(K−FGF)などの増殖因子由来シグナルペプチド配列(Linら、J. Biol. Chem.、第270巻、14255〜14258頁、1995年)、K−FGFシグナルペプチド配列由来の膜移行配列(Rojasら、Nat. Biotech.、第16巻、370〜375頁、1998年)、及びヒト免疫不全ウイルス(HIV)−1転写活性化因子TAT(Fawellら、Proc. Natl. Acad. Sci. USA、第91巻、664〜668頁、1994年)が挙げられる。PTDについては、米国特許第5,652,122、及びSchwarzeら、「Protein Transduction: Unrestricted Delivery into all Cells」、Trends in Cell Biology、第10巻(2000年)において、開示されている。発明者は、HIV−TATのPTDが本発明において特に有用であることを、見いだした。
【0041】
プロモーターと機能可能なように連結されている、融合タンパク質(タンパク質導入ドメイン(PTD)と骨誘導タンパク質を含む)をコードするヌクレオチド配列を含む核酸もまた、提供される。該核酸は、ベクター(例えば、プラスミドなどの発現ベクター)の一部であることが可能である。骨誘導タンパク質は、例えば、LIM石灰化タンパク質、骨形態形成タンパク質、Smadタンパク質及び骨誘導ペプチド、ならびにそれに由来するポリペプチドを含むことが可能である。骨誘導ペプチド及びポリペプチドの例としては、配列番号1、配列番号2、配列番号3、配列番号4、配列番号5、配列番号6、配列番号7及び配列番号8が挙げられる。
【0042】
骨誘導タンパク質を細胞内に送達する方法もまた、本発明によって提供される。本発明の方法において、少なくとも一つの骨誘導タンパク質を、導入によって細胞内に送達することが可能であり、ここで、タンパク質導入ドメイン(PTD)と骨誘導タンパク質を含む融合タンパク質は、融合タンパク質が細胞内に送達されるように細胞と接触し、タンパク質導入ドメインまたは細胞透過性ペプチドによって、その送達が容易となる。
【0043】
骨誘導タンパク質を送達可能な細胞としては、例えば、骨(すなわち、骨形成)及び非骨細胞が挙げられる。このような細胞は、例えば、軟膜細胞、幹細胞(例えば、間葉幹細胞、多能(multipotent)性及び多分化性(pluripotent)幹細胞)、椎間板細胞(disc cell)(例えば、線維輪細胞(nucleus cell)及び髄核細胞(annulus cell))、間葉細胞、造血性細胞、内皮細胞及び筋細胞を含んでもよい。幹細胞は、自己または同種異系の組織由来であることが可能である。
【0044】
タンパク質導入ドメイン(PTD)と骨誘導タンパク質との融合タンパク質を導入されているかまたはこれを発現する細胞もまた、提供される。このような細胞は、これに限定されないが、軟膜細胞、幹細胞(例えば、間葉幹細胞及び多分化性(pluripotential)幹細胞)、椎間板細胞(例えば、線維輪細胞及び髄核細胞)、間葉細胞、造血性細胞、内皮細胞及び筋細胞を含んでもよい。本明細書において記載されるような、PTDと骨誘導タンパク質との融合タンパク質を含有する細胞は、哺乳動物体内に植え込まれて、骨形成を誘導することが可能である。LMPを骨誘導タンパク質として用いた骨形成誘導法については、例えば、米国特許第6,300,127に記載されている。例えば、2002年11月13日に出願され、係属中の米国特許第10/292,951に記載のように、PTDと骨誘導タンパク質との融合タンパク質を含む細胞を、椎間板内に植え込んで、プロテオグリカン及び/またはコラーゲン合成を刺激してもよい。
【0045】
PTDと、骨誘導タンパク質をコードするヌクレオチド配列を含む核酸との複合体もまた、提供される。PTD/核酸複合体を、例えば、骨形成または椎間板再生を促進するために、骨誘導タンパク質の過剰発現を導くのに用いることが可能である。ヌクレオチド配列によってコードされる骨誘導タンパク質は、これに限定されないが、LMP、BMP及びSmadタンパク質を含むことが可能である。ペプチドを核酸と化学的に結合させるための方法は、当該技術分野において公知である。一つのこのような方法は、米国特許第5,652,122に記載されている。核酸は、プロモーターと機能可能に連結されている骨誘導タンパク質をコードするヌクレオチド配列を含む、発現ベクターの形であることが可能である。
【0046】
本発明の方法を、1またはそれより多くの骨形態形成タンパク質の発現、または、2003年3月7日に出願された同時係属中の米国特許出願第10/382,844に記載されるように、増殖因子−βタンパク質の細胞内での形質転換を誘導するのに、用いることが可能である。例えば、BMP−2、BMP−4、BMP−6、BMP−7、TGF−β1及びその組み合わせからなる群より選択される、1またはそれより多くのタンパク質の発現は、細胞を、PTDと骨誘導タンパク質を含む融合タンパク質と本発明にしたがって接触させることにより、誘導可能である。加えて、BMP−2、BMP−4、BMP−6、BMP−7、TGF−β1及びその組み合わせからなる群より選択される、1またはそれより多くのタンパク質を過剰発現する細胞もまた、本発明にしたがって提供される。細胞は、非限定的に、幹細胞、軟膜細胞、骨髄細胞、末梢血細胞または脂肪細胞を含む、任意の体細胞であることが可能である。細胞は、自己または同種異系組織に由来する幹細胞であることが可能である。
【0047】
腎臓においては、例えば、細胞を、本発明において記載されるようなPTD/LMP融合ポリペプチドの投与によって刺激して、BMP−7を産生させてもよく、あるいは、幹細胞を、PTD/LMP融合ポリペプチドで処理して、これにBMP−7の発現を誘導させてもよい。ひとたび腎臓に移植されるならば、そのBMP−7産生は、外因性BMP−7の投与を介してこれまで利用可能でなかった治療効用を提供することが可能である。BMP−7は、慢性腎障害の回復に関連した、重度に損傷した腎尿細管上皮細胞の修復を、誘導する(Zeisbergら、Nat. Med. 9:964-968、2000年)。マウスにおいて、BMP−7を投与することによって、慢性腎疾患の一合併症である無形成骨障害の治療に成功したことが、報告されている(Lundら、J Am Soc Nephrol 15(2):359-369、2004年)。慢性腎障害及び線維症のマウス遺伝モデルにおいて、外因性ヒトリコンビナントBMP−7の投与は、腎機能及び生存性を向上させた(Zeisbergら、Am J Physiol Renal Physiol 285(6):F1060-7、2003年)。BMP−7が、糖尿病誘導性の腎肥大を部分的に回復させることもまた、示されている(Wangら、Kidney Int 63(6):2037-2049、2003年)。Daviesらは、BMP−7の欠損が、慢性腎不全における病態生理学的因子であると、判断した(J Am Soc Nephrol 14(6):1559-1567、2003年)。これらの試験は、LMP−1が細胞によるBMP−7産生を誘導し、そしてLMPペプチド(配列番号1、配列番号2、配列番号3、配列番号4、配列番号5、配列番号6、配列番号7及び配列番号8)がLMP−1タンパク質と同じように機能するという、発明者による発見と併せて、タンパク質導入ドメインとLMP−1もしくはLMP−3タンパク質または関連ペプチドを含む融合ポリペプチドが、腎疾患の治療において有効である可能性があることを、実証する。本発明の融合ポリペプチドを用いてBMP−7の発現を誘導された幹細胞は、この目的に対して特に有用である可能性がある。
【0048】
BMP−4及びBMP−7もまた、ヒト骨髄腫細胞において、増殖を阻害し、そしてアポトーシスを誘導することが示されている。BMPの治療への使用は、骨髄腫の骨疾患及び骨髄腫細胞の増殖に対して、提案されている。本発明の方法は、LMP−1、LMP−3、または、配列番号1、配列番号2、配列番号3、配列番号4、配列番号5、配列番号6、配列番号7及び配列番号8を含むペプチドを、例えば注射、移植片、またはPTD/LMP融合ポリペプチドで処理された幹細胞の植え込みによって投与することにより、in vivoにおいてBMP−4とBMP−7双方の産生を刺激する方法を、提供する。幹細胞移植片を、植え込み型薬物送達装置などのPTD/LMP源とともに組み合わせると、特に有効である可能性がある。
【0049】
幹細胞または多能性前駆細胞は、骨芽細胞を作り出す細胞源を提供することが可能である。これらの細胞を、種々の分化段階において単離し、そして特定系統の経路にて分化を誘導してもよい。細胞を、骨粗鬆症または骨形成不全症などの骨疾患、ならびに非治癒骨折を治療するのに用いてもよい。核結合因子α1(Cbfa1)は、骨形成に必要であることが実証されている。BMP−2、BMP−4及びBMP−7は、骨芽細胞の分化を誘導することが知られており、Cbfa1の発現を亢進させる。BMP−8及びSmad−3は、骨芽細胞の分化中に亢進している。TGF−β/BMP−Smadのシグナル伝達の活性化は、Cbfa1の発現及び骨芽細胞の分化を促進することが、示されている。本発明は、例えば、ヒト骨髄由来中胚葉前駆細胞などの細胞において骨芽細胞の分化を促進するために、機能性BMP、LMP、Smadタンパク質またはその組み合わせを含む融合タンパク質を提供する。適切な細胞には、例えば、Jiangら(Nature、第418巻、41〜49頁、2002年)によって記載された細胞のような、多能性細胞が含まれてもよい。適切な骨誘導タンパク質またはポリペプチド、あるいはその組み合わせの投与は、細胞を植え込む前にex vivoで、あるいは、植え込みまたは注射後にin vivoで、行うことが可能である。
【0050】
In vivo投与に関して、本発明の骨誘導タンパク質を、例えばPTDまたは細胞透過性ペプチドによって近隣の細胞の内部に送達可能なように、標的部位に注射することが可能である。あるいは、PTD/骨誘導ポリペプチドと組み合わせたキャリアーを含む移植片を、用いてもよい。移植片は、周囲組織に放出するためにPTD/骨誘導ポリペプチドを入れる貯蔵所を含有してもよく、あるいは、1またはそれより多くのPTD/骨誘導ポリペプチドコンストラクトを含有する溶液中に浸された多孔性の組成物を含んでもよい。ヒドロゲル、持続放出カプセルもしくは球(spheres)、リポソーム、ミクロスフェア、ナノスフェア、生分解性ポリマー、または他のこのような薬物送達システムもまた、本発明のペプチド及びタンパク質を標的細胞及び組織に送達するのに用いられてよい。例えば、米国特許第6,475,516(DiCosmoら)は、抗生物質などのリポソーム治療剤が詰め込まれたヒドロゲルを提供し、該ヒドロゲルは、移植片などの留置された医療装置の表面と共有結合している。
【0051】
椎間板変性の特徴は、椎間板においてプロテオグリカン類、特に硫酸化グリコサミノグリカン類(sGAG)及びアグリカンの産生が減少することである。椎間板の主要なプロテオグリカンであるアグリカンの産生速度の減少は、椎間板変性における重要な要素である。椎間板の機能においてプロテオグリカン類は中心的な役割を果たすため、椎間板の正常なプロテオグリカン産生を回復させることは、椎間板変性のいかなる生物学的治療においても、非常に重要である可能性がある。
【0052】
発明者は、LMP−1の過剰発現または細胞内投与が、in vitro及びin vivoにおいて椎間板細胞のプロテオグリカン産生を増加させることを実証する実験を、行った。LMP−1の過剰発現は、in vitro及びin vivoにおいて、BMP−2及びBMP−7のmRNAの亢進を誘導する。これらのBMP−2及びBMP−7を特異的に阻害するノギンは、AdLMP−1によるプロテオグリカンの亢進を阻害し、このことは、LMP−1によって誘導されるプロテオグリカンの亢進が、BMPの亢進によって媒介されることを示唆する。したがって、遺伝子治療またはタンパク質治療によるLMP−1の投与(例えば、PTD複合体による送達)を、椎間板におけるプロテオグリカンの産生を刺激し、そして椎間板再生において治療的な役割を果たすのに用いることが可能である。
【0053】
TGF−β1、IGF−1及びEGFなどのサイトカイン類は、椎間板細胞の有糸分裂、及びある程度プロテオグリカン産生を刺激することが示されている。BMP−2及びBMP−7などの他のサイトカイン類もまた、プロテオグリカン産生を刺激するのに有効であることが示されている。しかしながら、サイトカイン類は小さな水溶性分子であるため、これらは椎間板から急速に拡散して出て行くか、または他の調節因子によって不活化される。
LIM石灰化タンパク質−1(LMP−1)は、白血球及び骨芽細胞から複数の異なるBMPの分泌を誘導することが知られている、細胞内調節分子である。LMP−1、LMP−2、LMP−3、あるいはLMP−1またはLMP−3由来骨誘導ペプチド、あるいはその組み合わせを、特にPTD/核酸複合体によって細胞内に送達することによって、BMP産生を細胞内部から刺激することが可能である。適切な骨誘導ペプチドは、例えば、配列番号1、配列番号2、配列番号3、配列番号4、配列番号5、配列番号6、配列番号7、または、配列番号8にあるようなポリペプチドを含む。
【0054】
本発明は、以下の限定されない実施例を参照することによって、より十分に理解されるだろう。
(実施例)
HIV−Tatのタンパク質導入ドメインとLMP−1タンパク質を含む、(His)6TAT−LMPタンパク質複合体の合成及び使用について、以下に記載する。pTAT−HA−ベクターを、ワシントン大学(セントルイス、ミズーリ州)から、物質譲渡契約(material transfer agreement)に基づいて得た。
【0055】
pCDNA3.1/hLMP−1を以下のプライマー:
正:5’−CCATGGAflCCflCAAAGTAGTGC−3’
逆:5’−CAGGGCGGGCGGCTGGTAG−3’
を用いたPCRの鋳型として利用することによって、NcoI制限酵素部位を、hLMP−1の5’末端に付加した。反応を、95℃にて2分間[95℃、30秒間;66℃、30秒間;72℃、1分間]x25、及び72℃にて10分間、行った。PCR産物を、PCRII−TOPOベクター(インビトロジェン社)中にクローニングし、そして適切なクローンを、配列決定によって同定した。
【0056】
(His)6TAT−LMPベクターの構築を、NcoI及びClaIを用いてプラスミドクローンを制限エンドヌクレアーゼ消化し、そしてその結果として得られた産物をアガロースゲル電気泳動及び電気溶出によって精製することにより、行った。hLMP−1の完全長配列を、ClaI及びEcoRIを用いてpCDNA3.1/hLMP−1ベクターを制限酵素消化し、そしてその結果として得られた産物をアガロースゲル電気泳動及び電気溶出によって精製することにより、単離した。pTAT−HA−ベクターもまた、NcoI及びEcoRIで制限酵素消化を受け、そしてその結果として得られた線状化ベクターを、アガロースゲル電気泳動及び電気溶出によって精製した。次いで、産物を、標準的な手順によって、16℃にて一晩かけて連結させた。正しく連結された産物(5’hLMP−1+3’hLMP−1+線状化pTAT−HA−ベクター=(His)6TAT−LMPベクター)を、続いて行ったアガロースゲル電気泳動及び分子量決定によって、決定した。
【0057】
連結された産物を、BL21(DE3)コンピテント細胞にトランスフェクトし、そして適切なクローンを制限酵素解析によって同定した。
(His)6TAT−LMPタンパク質の合成及び回収
(His)6TAT−LMP融合コンストラクトの陽性クローンを含有する、適切なBL21(DE3)大腸菌(Eschericia coli)コロニーを、0.8OD(600nm)まで増殖させ、そしてタンパク質産生を、100μMのIPTGで37℃にて4時間誘導した。誘導された細胞を標準的な方法によって回収し、そして超音波処理(4x20秒、それぞれ2分間の休止期間を伴う、4℃)によって溶解した(20mM PO4バッファー、pH7.2、8M尿素、100mM NaCl、20mMイミダゾール)。
【0058】
溶解物を、遠心分離(10000xg)によって澄明にし、そしてその結果として得られた上清を、Ni2+セファロースアフィニティカラム(インビトロジェン社)に、重力流の条件下で載せた。カラムを洗浄し(20mM PO4バッファー、pH6.0、8M尿素、250mM NaCl、20mMイミダゾール)、次いで、結合しているタンパク質を溶出させた(20mM PO4バッファー、pH4.0、8M尿素、500mM NaCl)。
【0059】
溶出物を、0%バッファーBから100%バッファーBまでの直線勾配(バッファーA:20mM炭酸ナトリウム、pH11、8M尿素;バッファーB:20mM炭酸ナトリウム、pH11、8M尿素、2M塩化ナトリウム)を用いた陰イオン交換クロマトグラフィ(Hitrap Q HP、5mL、ファルマシア社)にかけた(5mL/分にて40分間)。溶出画分(5mL)を、SDS−PAGE及びウエスタンブロット解析によって、hLMP−1の存在について解析した。
【0060】
hLMP−1陽性画分を、疎水性相互作用クロマトグラフィ(Hitrap(商標登録)フェニル−セファロース、5mL、ファルマシア社)にかけた。溶出(バッファーA:20mM炭酸ナトリウム、pH10.5、1.5M硫酸アンモニウム;バッファーB:20mM炭酸ナトリウム、pH10.5)を、0%〜100%のバッファーBの直線勾配で、5mL/分にて20分間行った。溶出画分(5mL)を、SDS−PAGE及びウエスタンブロット解析によって、hLMP−1の存在について解析した。(His)6TAT−LMP融合タンパク質に陽性の画分を、使用時まで凍結乾燥させておいた。
【0061】
新規の骨形成のための(His)6TAT−LMP融合タンパク質の使用
凍結乾燥させた画分(55pg)を、40mM KOH中に再懸濁した(ストック溶液;1.0μM)。ヒト軟膜細胞を、Viggeswarapuら、J.Bone Joint Surg.、第83(3)巻、364頁(2001年)に記載されるように調製した。これらの細胞を、アルファMEM(ギブコ社)に溶かした(His)6TAT−LMP融合タンパク質5μLと混合し、そして37℃にて30分間インキュベートした。
【0062】
次いで、適切な容量の、(His)6TAT−LMP融合タンパク質を含有するヒト軟膜細胞を、植え込み用に、多孔性コラーゲン基質に載せた。In vivoにおける新規の骨形成を実証するため、ラットへの植え込み用に、100μLの細胞懸濁液を、無菌ピペットによって無菌の5x5mmI型ヒトコラーゲンディスクに載せた。脊椎の融合を促進するための移植用に、同様な量の細胞懸濁液を、10x25mmシートに載せた。ディスクを、4〜5週齢の無胸腺ラット(rnu−/rnu−)の胸/腹部に、外科的に皮下に植え込んだ。動物を4週間後に屠殺し、この時点でディスクを切除し、70%エタノール中で固定し、そしてX線撮影及び脱灰していない組織の検査(5μmの切片を作製し、そしてゴールドナー・トリクロムで染色)によって解析した。
【0063】
ウサギにおいて、後外側腰椎の関節固定を行い、そして、脊椎の各側が、4x106個の導入された軟膜細胞を受け入れるように、キャリアー基質を植え込んだ(すなわち、15%ヒドロキシアパタイト/85%リン酸三カルシウムのコラーゲンスポンジ)。4週間後に、ウサギを屠殺し、そしてその腰椎を切除した。脊椎融合の状態を、残存する運動(融合の失敗を示唆)を検出するための、盲検化された手による運動分節の触診、X線撮影、CTスキャン及び脱灰していない組織の検査によって、評価した。
【0064】
ラットとウサギの双方において、X線撮影により、LMP−1をトランスフェクトされたヒト軟膜細胞を含有する元々のコラーゲンディスクまたはシートの形に一致した、高レベルの石灰化骨形成が示された。石灰化骨形成は、対照において認められず、そして元々のコラーゲンディスクまたはシートは、吸収を受けているようにみえた。組織像は、LMP−1を導入された移植片において、骨芽細胞の並んだ新たな骨梁を示したが、対照においては、骨はみられず、そしてキャリアーは部分的に再吸収された。
【0065】
In vivo骨形成を誘導するための骨誘導ペプチドの使用
プロトコールは、基本的には上記のとおりである。ペプチドを、4個の別々の移植片(3個はウサギ用)に、250μl/ディスク、6M/mlにて載せた。移植片は、コラーゲンディスクを含んだ。移植片を、Harlan無胸腺ラットまたはニュージーランドホワイトウサギの胸部内に置いた。試験された用量範囲は5nM、10nM、12.5nM、15nM、17.5nM、20nM、22.5nM及び25nMであった。結果を、図6に示す。
【0066】
sGAG合成のLMP−1刺激
Sprague−Dawleyラットから得た腰椎間板細胞を用いたin vitro実験を、種々の用量(感染多重度0、5、10、25、50)のLMP−1ヌクレオチド配列の挿入断片を含有するアデノウイルス(AdLMP−1)で細胞を処理し、そしてこれを単層にて6日間培養することによって行い、in vitroにおけるLMP−1過剰発現の影響を判定した。DMMB法を用いて、培地中の硫酸化グリコサミノグリカン(sGAG)を定量した。リアルタイムPCRを用いて、アグリカン、過剰発現LMP−1、BMP−2、4、6及び7のmRNAレベルを定量した。直接ELISA法を用いて、培地中のBMP−2、4、6及び7のレベルを定量した。LMP−1による椎間板細胞のプロテオグリカン産生亢進にBMPが関与することを実証するため、BMP−2、4、6及び7の活性を特異的に遮断する分子(ノギン)を、異なる濃度で、AdLMP−1処理細胞に加えた。
【0067】
In vivo遺伝子治療実験を、ニュージーランドホワイトウサギにおいて行った。腰椎間板に、AdLMP−1(実験用)、またはマーカー遺伝子を保有するアデノウイルス(AdLacZ−対照)のいずれかを、異なる3用量(106、107及び108pfu/椎間板)にて注射した。3週間後に、注射された椎間板を回収し、そして総LMP−1(内因性)、過剰発現LMP−1、アグリカン、BMP−2及びBMP−7のmRNAレベルを測定した。
【0068】
感染多重度(MOI)25のAdLMP−1は、最大レベルのsGAG亢進を誘導するのに十分であった。6日間の単層培養期間が、AdLMP−1処理(MOI 25)後に最大レベルのsGAG亢進に達するのに必要とされた。アグリカンのmRNAは、対照と比べて2倍増加した。細胞をアルギン酸中で培養した場合、sGAG産生に対するAdLMP−1処理の効果は、3週間持続した。AdLMP−1処理後6日に、BMP−2及びBMP−7のmRNAレベルは、それぞれ対照の3.0±0.2及び2.8±0.3倍(p<0.01)有意に増加した。BMP−4及びBMP−6のmRNAレベルは、変わらなかった。培地中のBMP−2及びBMP7のタンパク質レベルは、対照と比べて有意に増加した(p<0.01)。対照的に、BMP−4及びBMP−6のタンパク質レベルは、上昇しなかった。3200ng/mlのノギンは、AdLMP−1によるプロテオグリカンの亢進を完全に遮断した。内因性レベルのLMP−1 mRNAが、腰髄核中に検出された。107pfu/椎間板のAdLMP−1を注射されたin vivo椎間板では、対照と比べてLMP−1、BMP−2及びBMP−7のmRNAレベルが有意に上昇した。
【0069】
In vitro実験を、Sprague−Dawley(SD)ラットの腰椎間板細胞を用いて行った。両側スチューデントt検定を用いて、実験群を対照群と比較した。図中の誤差バーは、1SEM(標準誤差)を示す。
【0070】
ヒトLMP−1遺伝子のcDNAを保有する複製欠損性5型アデノウイルスを用いた(AdLMP−1)。SD線維輪細胞を用いた単層培養実験を行い、ウイルス用量と過剰発現LMP−1 mRNA発現との関連性を、ウイルスによって送達されるLMP−1 cDNAのみに特異的なプライマーを用いたリアルタイムPCR法を用いて、判定した。SD線維輪細胞を用いた単層培養実験を行い、ウイルス用量とsGAG産生との関連を判定した。sGAGレベルを、DMMB法を用いて測定した。AdLMP−1の最適用量を、sGAGの産生をプラトーレベルに導く最低用量として定めた。最適用量(25MOI)を用いて、経時的な実験(9日間)を行い、sGAGの産生がプラトーレベルに達するのに必要な最短期間(6日間)を決定した。先に決定された最適ウイルス量及び時間(25MOIのAdLMP−1、及び処理後6日)を用いて、AdLMP−1処理された線維輪細胞のアグリカンのmRNAレベルを、対照と比較した。髄核細胞が、線維輪細胞と同じように反応することを確かめるため、sGAG産生及び細胞数に対するAdLMP−1の影響を、処理後6日に、25MOIにて、線維輪細胞及び髄核細胞の双方について判定した。AdLMP−1処理のより長期の影響を検討するため、アルギン酸中で培養されたSD線維輪細胞のアルギン酸中のsGAGの蓄積を、1、2及び3週間後に判定した。すべての実験を、少なくとも2回繰り返した。
【0071】
SD線維輪細胞を用いた単層培養実験を、MOI 25のAdLMP−1を用いて行った。簡潔に記すと、LMP−1、BMP−2、BMP−4、BMP−6及びBMP−7のmRNAレベルを、AdLMP−1処理後第0.5、1、3及び6日に、リアルタイムPCR解析を用いて測定した。BMP−2、BMP−4、BMP−6及びBMP−7のレベルを、直接ELISA法を用いて、処理後6日に培地中にて測定した。BMP活性を遮断する影響を判定するために、ノギンを、AdLMP−1処理開始時に培地に加えた。AdLMP−1で処理後の、種々の濃度のノギン存在下または非存在下でのsGAGレベルの変化を、6日間の実験において測定した。
【0072】
アグリカン、BMP−2及びBMP−7に対するLMP−1投与(AdLMP−1による)のin vivo影響を判定するために、4例のニュージーランドホワイト(NZW)ウサギ(3〜4kg)を用いた。前部腰椎間板L2/3、L3/4、L4/5及びL5/6を、左後腹膜からのアプローチを介して露出させた。107プラーク形成単位(plaque-forming units、pfu)の実験用ウイルス(AdLMP−1)または対照ウイルス(AdGFP−緑色蛍光タンパク質のcDNAを挿入断片として有する、5型アデノウイルス)のいずれかを、露出させた椎間板髄核のそれぞれに、交互に注射した(すなわち、各ウサギにおいて、二つの椎間板にはAdLMP−1を注射し、そして二つの椎間板にはAdGFPを注射)。ウイルスを、30Gハミルトンシリンジを介して、10μlのリン酸緩衝生理食塩液中にて送達することによって、投与した。3週間後に、注射された腰椎間板から得た髄核組織を回収した。2例のウサギから得た髄核組織を、対照または実験椎間板群のいずれかにプールして、さらなる解析に十分なmRNAを得た。逆転写及びリアルタイムPCRを用いて、総LMP−1、BMP−7及びアグリカンのmRNAレベルを定量した。総LMP−1のプライマーは、内因性及び過剰発現LMP−1の双方を同定するように、設計された。
【0073】
第二のin vivo実証においては、種々の用量のAdLMP−1ウイルスを用いて、用量反応の関係性を確立した。異なる3用量(106、107、108pfu)のAdLMP−1及び単一用量(107pfu)のAdGFPを、投与した。この実験では、それぞれの動物のすべての椎間板に、先の実験におけるような交互のウイルス種ではなく、単一用量のウイルスを注射した。8例のNZWウサギ(3〜4kg)を、4つのウイルス群に2例ずつのウサギで、用いた。ウサギを3週間後に屠殺し、そして髄核を回収した。各処理群内から回収した組織をプールし、mRNAを単離し、そしてこれを用いて対応するcDNAを作製した。リアルタイムPCRを用いて、総LMP−1、過剰発現LMP−1、BMP−2、BMP−7及びアグリカンのmRNAレベルを定量した。
【0074】
様々な用量(MOI 0、10、25及び50)でウイルス処理した12時間後における、過剰発現LMP−1のmRNAの相対量を、過剰発現LMP−1のmRNAの最低検出可能レベル(AdLMP−1 MOI 5)を基準に正規化した。AdLMP−1の用量を増加させると、過剰発現LMP−1のmRNAに、MOI 5と比べて統計学的に有意な増加が生じた。非処理対照群またはAdLacZ対照群において、検出可能なレベルの過剰発現LMP−1のmRNAはみられず、このことは、AdLMP−1が、線維輪細胞において用量相関的にLMP−1の過剰発現を誘導することを示唆した。
【0075】
線維輪細胞の培地中sGAG濃度を、様々な用量のAdLMP−1による処理後6日に、測定した。第6日における培地中sGAG濃度は、培地交換からsGAG測定までの3日間の期間中に細胞によって産生された総sGAGの指標であり、したがって、この期間中のsGAG産生速度の指標である。データを、処理と非処理対照間の割合として表す。LMP−1の投与(MOI 25のAdLMP−1による)は、最も高いsGAGレベルを提供し、これは、対照の3.1±0.2倍(p<0.01)であった。MOI 25またはMOI 50で処理された細胞間では、培地中sGAGレベルに有意な差はなかった。このMOI 25のAdLMP−1の用量は、最大のsGAG応答に達することが可能な最低用量であったため、発明者は、実験の残りに用いる用量としてMOI 25を選択した。
【0076】
時間経過実験を行って、MOI 25のAdLMP−1で処理後の実験の長さを変えた影響を、判定した。線維輪細胞をLMP−1(MOI 25のAdLMP−1)で処理し、そして3日間の増殖期間中のsGAG産生を測定した。結果を、各時点におけるDNA含量によって正規化し、そして同時点の非処理対照に対する割合として表した。sGAGレベルは、第3日に1.6±0.2(p<0.01)に、第6日に2.9±0.1(p<0.01)に、第9日には2.8±0.1(p<0.01)に増加した。第6日から第9日までの間に、sGAGレベルの増加はみられず、このことは、第6日までにプラトーレベルに達したことを示唆した。
【0077】
LMP−1によるアグリカン合成の誘導
アグリカンは椎間板の主なプロテオグリカンであるため、アグリカンのコアタンパク質のmRNAレベルを、AdLMP−1で処理後6日に測定した。定量性リアルタイムPCR法を用い、そしてデータを非処理対照に対する割合として表した。AdLMP−1処理(MOI 25)後に、アグリカンのmRNAレベルは、対照の2.1±0.1(p<0.01)倍であった。AdLacZ処理(MOI 25)後に、アグリカンのmRNAレベルは変わらず、対照の1.0±0.16倍であった。これらの結果は、sGAGの実験と併せて、LMP−1が椎間板細胞のプロテオグリカン産生を刺激することを、実証した。
【0078】
最適なAdLMP−1活性に必要な用量及びタイミングを培養系において確立したことから、次いで、発明者は、AdLMP−1によって投与されるLMP−1が線維輪細胞及び髄核細胞に及ぼす影響を、比較した。ラット線維輪細胞及び髄核細胞を、MOI 25のAdLMP−1で処理し、そして各細胞種のsGAG濃度を、第6日に測定した。sGAG濃度を、細胞数の軽微な変化を明らかにするために、DNA含量によって正規化した。線維輪細胞及び髄核細胞種の非処理対照を比較するため、結果を、非処理対照を基準に正規化しなかった。非処理の線維輪細胞及び髄核細胞の結果は、それぞれ0.6±0.04 sGAG/DNA、及び1.0±0.12 sGAG/DNAであった。この差は統計学的に有意であり(p<0.05)、そして、非処理髄核細胞が、線維輪細胞よりも一細胞につきより多くのsGAGを産生することを、示唆した。AdLMP−1処理後、線維輪細胞及び髄核細胞の結果は、それぞれ1.5±0.08 sGAG/DNA(p<0.01)、及び2.4±0.1 sGAG/DNA(p<0.01)であった。これらの値は、そのそれぞれの非処理対照と比べて、統計学的に有意な増加を示した。非処理細胞からAdLMP−1処理細胞への相対的な増加は、線維輪細胞と髄核細胞との間で同様であった。第6日におけるAdLMP−1処理及び非処理の細胞数を、DNA含量を測定することによって決定した。線維輪細胞のDNA含量は、非処理対照と比べて1.2倍増加したが(p<0.01)、しかし、髄核細胞のDNA含量は、AdLMP−1による影響を受けなかった。このことは、LMP−1が、軽度であるがしかし有意な線維輪細胞数の増加を誘導するが、髄核細胞では誘導しないことを、示唆した。
【0079】
LMP−1の投与(AdLMP−1による)がin vitroにおけるsGAG産生に及ぼす影響を、数週間の期間、試験するために、発明者は、アルギン酸培養系を用いた。アルギン酸は、in vitro培養液中で長期間、軟骨細胞の表現型を維持するのに重要な細胞の三次元マトリックスを、提供する。単層で増殖させた線維輪細胞を、MOI 25のAdLMP−1で処理し、次いで24時間後にアルギン酸培養液に移した。細胞を、1、2及び3週間の期間、培養した。1週間後に、AdLMP−1群におけるsGAGレベルは、非処理対照の1.5±0.06(p<0.01)倍であった。2週間後に、AdLMP−1処理群におけるsGAGレベルは、非処理対照の2.9±0.3(p<0.01)倍に増加した。この差は、3週間後において維持され;AdLMP−1処理群におけるsGAGレベルは、対照の2.9±0.1(p<0.01)倍であった。このことは、AdLMP−1が、少なくとも3週間、培養液中で、アルギン酸中のsGAG増加を維持するのに効果的であることを、示唆した。
【0080】
プロテオグリカン産生に対するLMP−1過剰発現の効果を実証したことから、我々は、LMP−1の過剰発現がこの効果を誘導するメカニズムを検討した。過剰発現LMP−1及びBMP(BMP−2、4、6及び7)のmRNAの変化の時間経過を、測定した。これらのBMPは、白血球及び骨芽細胞において、LMP−1の過剰発現によって刺激されることが、これまでに示されていた。
【0081】
線維輪細胞をMOI 25のAdLMP−1で処理後の、過剰発現LMP−1の時間依存的な変化を、測定した。データを、非処理対照に対する割合としてでなく、過剰発現LMP−1の最大mRNAレベル(第6日)の百分率として表したが、これは、対照において過剰発現LMP−1のmRNAが検出可能でなかったためである。LMP−1のmRNAは、処理後12時間に検出可能であり、そして調査した最終時点(第6日)まで増加し続けた。このことは、過剰発現LMP−1が、LMP−1投与後早ければ12時間後には下流の遺伝子に影響を与え得ることを、示唆した。
【0082】
MOI 25のAdLMP−1で処理後の、BMPのmRNAレベルの時間経過も、リアルタイムPCRによって測定し、そして各時点において非処理対照に対する割合として算出した。BMP−2のmRNAは早期に亢進し、LMP−1処理後12時間に、統計学的に有意な(p<0.01)増加に達した。BMP−2のmRNA増加は、第3日までにプラトーレベルに達した。BMP−7のmRNAはBMP−2よりも遅れて亢進し、第3日(p<0.01)及び第6日(p<0.01)に、統計学的に有意な増加に達した。BMP−4及びBMP−6のmRNAレベルには、いずれも非処理対照のものと有意な差はなかった。
【0083】
LMP−1のmRNAの過剰発現が、線維輪細胞においてBMP−2及びBMP−7の亢進をもたらすことを確立したことから、発明者は、このmRNA増加が、BMPタンパク質の培養液への分泌増加と相関するかどうかを検討した。ELISAアッセイを、調製された培地中のBMP−2、4、6及び7タンパク質レベルを定量するために行った。単層にて培養された線維輪細胞を、MOI 25のAdLMP−1で処理した。培地を第3日に一度交換し、そして培地を第6日に解析した。したがって、被験試料は、培養最終3日間(第4〜6日)に分泌されたBMPを含有した。培地中のBMP−2及びBMP−7のタンパク質レベルは、それぞれ、非処理対照の3.5±0.4(p<0.01)倍及び2.5±0.3(p<0.05)倍であった。BMP−4及びBMP−6のタンパク質レベルには、非処理対照のものと有意な差はなく、このことは、mRNAについて得られた結果と一致した。
【0084】
BMP−2及びBMP−7のmRNA及びタンパク質が、LMP−1によって亢進したことから、発明者は、これらのBMPの遮断が、LMP−1によって誘導されるsGAG産生増加を妨げるかどうかを、検討した。単層においてMOI 25のAdLMP−1とともに培養された線維輪細胞を、BMP阻害剤であるノギンで、培地中にて処理し、そしてsGAGレベルを第6日に測定した。AdLMP−1のみで処理された細胞では、sGAGレベルが増加したが(非処理対照の2.7±0.3倍)、3200ng/mlのノギン及びMOI 25のAdLMP−1で処理された細胞では、sGAGレベルは変わらず(非処理対照の1.1±0.1倍)、このことは、ノギンがAdLMP−1の影響を完全に遮断することを示唆した。ノギン(3200ng/ml)のみで処理された細胞では、sGAGレベルはほとんど変わらないままであり(非処理対照の0.8±0.1倍)、このことは、ノギンによる毒性的影響がないことを示唆した。AdLMP−1によって誘導されるsGAG産生に及ぼすノギンの阻害的影響は、濃度依存性であった。
【0085】
対照の椎間板(AdGFPを注射された椎間板)におけるアグリカン、BMP−7及びLMP−1のmRNAの内因性レベルを、髄核において検出した。内因性mRNAレベルを用いて、AdLMP−1を注射された椎間板におけるアグリカン、BMP−7及びLMP−1のmRNAの増加を、算出した。
【0086】
AdLMP−1を注射された椎間板は、AdGFPを注射された椎間板よりも、総LMP−1のmRNAを、830%高いレベルで発現した。AdLMP−1によるLMP−1の投与は、対照群よりも1100%、BMP−7のmRNAレベルの増加(p<0.05)をもたらした。アグリカンのmRNAレベルは、対照群よりも66%増加した(p<0.05)。
【0087】
BMP−2、BMP−7、LMP−1及びアグリカンのmRNAの内因性レベルも、検出した。AdLMP−1の用量増加と総LMP−1のmRNAとの相関がみられた。AdLMP−1によるLMP−1の投与は、107pfu/椎間板の用量で最大限に、BMP−2及びBMP−7のmRNAレベルを有意に増加させた(p<0.05)。107pfu/椎間板の用量のAdLMP−1は、対照群より50%と(p<0.05)、アグリカンのmRNAを最も増加させた。
【0088】
LMP−1の椎間板細胞への投与は、sGAGの産生増加及びアグリカンのmRNAレベルの増加をもたらした。LMP−1の投与は、in vitroにおいてBMP−2及びBMP7のmRNAならびにタンパク質レベルの増加をもたらし、そしてLMP−1の投与がプロテオグリカン産生亢進に及ぼす影響を、BMP阻害剤であるノギンの投与によって遮断することが可能であった。
【0089】
11ヶ月齢のSprague−Dawleyラットを屠殺し、そして腰椎及び尾から得た椎間板組織を、無菌条件下で回収した。線維輪及び髄核を、別々に、解剖しそして小さい角切りにした。椎間板組織を、100単位/mlペニシリン及び100mg/mlストレプトマイシンを含有する、ダルベッコ変法イーグル培地及びハムF12培地(DMEM/F−12;ギブコBRL社、グランドアイランド、ニューヨーク州、米国)中に入れた。椎間板組織を、培地中で、0.2%プロナーゼ(シグマケミカル社、セントルイス、ミズーリ州、米国)を用いて37℃にて1時間処理し、次いで、0.025%コラゲナーゼ(シグマケミカル社、セントルイス、ミズーリ州、米国)を用いて37℃にて6時間処理した。単離された細胞を洗浄し、そして70mmメッシュ(ファルコン社、フランクリンレイクス、ニュージャージー州、米国)を介してろ過し、10%胎児ウシ血清(FBS)、100単位/mlペニシリン、100mg/mlストレプトマイシン、2mM L−グルタミン及び50mg/mlアスコルビン酸塩を含有するDMEM/F12培地12mlを入れた75cm2フラスコに、加えた。細胞を、加湿された5%CO2中で37℃にて、増殖させた。培養液を、およそ8日間、2日毎に交換した。
【0090】
二つの異なるウイルスを用いた。CMVプロモーターによって駆動される、ヒトLMP−1 cDNAを含有する複製欠損性5型アデノウイルス(AdLMP−1)を、実験用ウイルスとして用いた。対照ウイルスは、lacZのcDNAを含有する同様な複製欠損性5型アデノウイルスからなった。
【0091】
椎間板細胞の初代培養物がコンフルエントになったとき、細胞を、400,000細胞/ウェルにて6ウェルプレートに継代培養した。3日後に、細胞を、ヒトLMP−1遺伝子またはLacZ遺伝子のいずれかのcDNAを含有するアデノウイルス(それぞれ、AdLMP−1とAdLacZ)で、処理した。細胞数を、血球計数器を用いて対照ウェルをカウントすることによって、第0日に測定した。ウイルス用量を、感染多重度(MOI)、すなわちプラーク形成単位(pfu)/細胞の数として、表した。これは、本質的には、一つの椎間板細胞が曝露されたリコンビナントアデノウイルスのプラーク形成単位の数である。培養された細胞を、各実験において指定されたように、異なるMOI(0、10、25、50)のAdLMP−1またはAdLacZを用いて、300mlの0%FBS添加DMEM/F−12中で37℃にて30分間処理した。次いで、培養容量を、1%FBS、100単位/mlペニシリン、100mg/mlストレプトマイシン、2mM L−グルタミン及び50mg/mlビタミンCを含有するDMEM/F−12培地を用いて、2.0mlに上昇させた。培地を、実験期間中、3日毎に交換した。
【0092】
培養液の硫酸化グリコサミノグリカン(sGAG)含量を、1,9−ジメチルメチレンブルー(DMMB)法を用いてアッセイした。培養液2mlを、セントリコンYM−50遠心フィルター(ミリポア・Co.、ベッドフォード、マサチューセッツ州、米国)を用いて、遠心分離し(5000xGで30分間)、sGAGを濃縮した。試料溶液(20ml)を、96ウェルのマイクロタイタープレート中で、200mlのDMMB色素溶液と穏やかに混合し、そして光学濃度(OD)を、520nm波長フィルターにて直ちに検査した。標準曲線を、硫酸コンドロイチン(シグマケミカル社、セントルイス、ミズーリ州、米国)の連続希釈系列を用いて、作図した。培地中の総sGAGを、DNA含量によって正規化し、そして非処理対照に対する割合として表した。
【0093】
細胞数を、各ウェルのDNA含量によって決定し、そして、先に記載したように、DNA含量を、ヘキスト色素33258(Polysciences社、Warrington、ペンシルバニア州、米国)法を用いて測定した。培養された細胞を、パパイン(10単位/ml)に曝露することによって、プレートから除いた。次いで、培養された細胞をペレット状にし、そして60℃にて3時間インキュベートした。20μlのパパイン消化物のアリコットを、200mlのヘキスト色素33258溶液と、96ウェルのフルオロプレート中で混合した。発光及び励起スペクトルを、ルミネセンス分光器LS 50B(パーキン−エルマー社、Wellesly、マサチューセッツ州、米国)において、それぞれ456nm及び365nmにて測定した。標準曲線を、子ウシ胸腺DNA(シグマケミカル社、セントルイス、ミズーリ州、米国)の既知の濃度を用いて、各測定時点について作製した。
【0094】
アルギン酸ビーズ培養は、長期培養において軟骨細胞の表現型を維持するのに有用である。この方法は、3週間の培養においてAdLMP−1の影響を判定するためのものである。細胞を、上記のように、単層培養において処理した。1日後に、細胞を、トリプシン処理によって剥離させ、そして培地で2回洗浄した。単離された細胞を、0.6%低粘度無菌アルギン酸塩(シグマケミカル社、セントルイス、ミズーリ州、米国)溶液中に、600,000細胞/mlにて再懸濁した。アルギン酸ビーズを形成させるため、細胞を、10mlプラスチックシリンジに取り付けた21ゲージ針を介して、1滴ずつ、0.102M CaCl+0.15M NaCl溶液中に加えた。10分後に、新たに形成されたビーズ(およそ12,000細胞/ビーズを含有)を、無菌0.9%NaCl溶液で3回、続いてDMEM/F−12で2回、洗浄した。線維輪細胞を含有するビーズを、1%FBS、100単位/mlペニシリン、100mg/mlストレプトマイシン、2mM L−グルタミン及び50mg/mlビタミンCを含有するDMEM/F−12培地で、6ウェルプレートにおいて別々に培養した。培地を、異なる期間中(1、2及び3週間)、2日毎に交換した。アルギン酸ビーズを、350mlのクエン酸ナトリウムバッファー(55mmol/L Na−クエン酸、50mmol/L EDTA、150mmol/L NaCl、pH7.4)に溶解した。細胞を、遠心分離によってペレット状にし、そして溶解液中のsGAG含量を、上記のDMMB法で測定した。細胞ペレットに残存するsGAG含量は、懸濁液中のものと比べて、無視できるほどであった。結果を、溶解液のsGAGを用いて、非処理対照群の倍数増加として記載した。
mRNAレベルの定量
リアルタイムPCRを用いて、BMP−2、BMP−4、BMP−6、BMP−7及び過剰発現LMP−1のmRNAレベルを、定量的に測定した。遺伝子のすべてのプライマーを、アガロースゲル上で産物のサイズを決定することによって、そしてアンプリコンをDNA配列決定することによって、確認した。18Sのレベルを各試料において測定し、内在性対照として用いた。
【0095】
各試料の総RNAを、グラニジウム・チオシアネート−フェノール−クロロホルム技術を用いた単一工程の方法によって、抽出した。単離RNAの濃度を、分光光度計(DU−500;ベックマン社、フラトン、カリフォルニア州、米国)を用いて、260nm波長にて測定した。RNAを、DNA分解酵素1(アンビオン・Inc.、テキサス州、米国)で処理して、試料のDNA混入を除去した。逆転写を、総RNA2mg;トリ骨髄芽球症ウイルス逆転写酵素(プロメガ社、マディソン、ウィスコンシン州、米国)30U;デオキシアデノシン三リン酸(dATP)、デオキシシチジン三リン酸(dCTP)、デオキシグアニジン三リン酸(dGTP)、デオキシチミジン三リン酸(dTTP)各1mM;及び、オリゴ(dT)15プライマー1mgを用いて、42℃にて45分間、40ml容量中で行った。PCRを、Amplitaq(商標登録)DNAポリメラーゼを用いて、30サイクル(95℃、30秒間;62℃、30秒間;72℃、45秒間)行った。DNAの混入がないことを確認するため、逆転写酵素非存在下で処理したRNA試料も、PCRにかけ:PCR産物がないことで、DNA混入していないと確認した。
【0096】
リアルタイムPCRは、迅速で、信頼性があり、そして再現可能な、特定のmRNAの定量性検出法であると、報告されている。SYBR(商標登録)グリーン・リアルタイムPCRキット(Applied Biosystems社、フォスターシティ、カリフォルニア州、米国)を用いたリアルタイムPCR法を用いて、BMP−2、BMP−4、BMP−6、BMP−7及びアグリカンの定量性mRNA解析を行った。25マイクロリットル(25ml)の反応容量は、cDNA 5ml、各プライマー(BMP−2、−4、−6、−7、及び18S)3.75ピコモル、及びSYBR(商標登録)グリーン・マスターミックス(2x、バイオラッド社、ヘラクレス、カリフォルニア州、米国)を含んだ。過剰発現LMP−1及び18SのmRNAレベルを定量するために、TaqMan(商標登録)リアルタイムPCRキット(Applied Biosystems社、フォスターシティ、カリフォルニア州、米国)を用いたリアルタイムPCR法も、行った。25マイクロリットル(25ml)の反応容量には、cDNA 5ml、各プライマー3.75ピコモル、及びTaqMan(商標登録)PCRマスターミックス(2x、バイオラッド社、ヘラクレス、カリフォルニア州、米国)が含まれた。
【0097】
リアルタイムPCRを、GeneAmp(商標登録);5700配列検出システム(Applied Biosystems社、フォスターシティ、カリフォルニア州、米国)を用いて、以下の3工程のプロトコールで行った;工程1:50℃にて2分間、工程2:95℃にて10分間、及び、工程3:(95℃にて15秒間、60℃にて1分間)x45サイクル。増幅の特異性を確認するため、PCR産物を、転移曲線解析にかけた。先に記載のように、各反応のthreshold cycle(Ct)を、競合的_ΔΔCt法を用いて、18Sを基準として標準化した。
【0098】
BMP2、4、6及び7のELISAアッセイ
BMPの標準曲線を、0.05mol/L重炭酸バッファーに溶解した、ヒトBMP2、4、6及び7(Genetics Institute、ケンブリッジ、マサチューセッツ州)の漸増させた濃度(0.1ng/100μL/ウェル〜1000ng/100μL/ウェル)を用いて、作図した。100マイクロリットルの試料を、トリプリケートで各ウェルに加えた。4℃にて一晩インキュベートした後、プレートを、0.5%Tween 20(PBST)添加0.01Mリン酸緩衝生理食塩液で3回洗浄し、そして非反応部位を、1%ウシアルブミン(シグマ社、セントルイス、ミズーリ州)を用いて、室温にて1時間ブロッキングした。プレートをPBSTで洗浄後に、一次抗体(1:1000)を100マイクロリットルにて各ウェルに分注し、そして室温にて2時間インキュベートした。BMP2、4及び6に対するヤギポリクローナル抗体(サンタクルーズ・Inc、サンタクルーズ、カリフォルニア州)、ならびBMP7に対するウサギ抗体(シグマ社、セントルイス、ミズーリ州)が、用いられた。プレートをPBSTで洗浄し、次いで、アルカリホスファターゼ結合抗ヤギIgG及び抗ウサギIgG(シグマ社、セントルイス、ミズーリ州)を用いて、室温にて1時間、それぞれインキュベートした。基質であるp−ニトロフェニルリン酸塩(シグマ社、セントルイス、ミズーリ州)を用いて発色させ、20分後に3N NaOHで反応を止めた。呈色を、Elx 800−マイクロプレートリーダー(バイオ−テック・インスツルメント社、Winooska、バーモント州)を用いて405nmにて吸光度差を測定することによって、定量した。
【0099】
結果を定量化するため、線形回帰プロットを、各標準品について作製した。すべての場合において、試料濃度を、標準品と同じ吸光度の対応値に準じて、標準品の線形回帰プロットから外挿した。
【0100】
ノギン糖タンパク質によるBMPの阻害
ノギンは、BMP−2、4、6及び7と高特異的に結合し、そしてこれらのBMPがその同族受容体を活性化させることを妨げる、糖タンパク質である。マウスノギン(ノギン−FC・シグマケミカル社、セントルイス、ミズーリ州、米国)の形態を実験において用い、AdLMP−1処理後にBMPを特異的に遮断する影響を、判定した。異なるノギン濃度(100、200、400、800、1600及び3200ng/ml)を、AdLMP−1(MOI 25)処理後第0日及び第3日に、細胞に載せた。第6日に、上記のDMMB法を用いて、条件培地を、sGAGの産生を検討するためにアッセイした。結果は、LMP−1の影響が、ノギンの投与によって遮断され得ることを示唆し、このことは、LMP−1活性が、これによるBMPの誘導に部分的に関連することを、実証した。
【図面の簡単な説明】
【0101】
【図1】図1は、線維輪細胞をLMP−1(MOI 25のAdLMP−1)で処理後6日の、培地中BMPタンパク質レベル(量)のグラフである。BMP−2及びBMP−7のタンパク質レベルは有意に増加したが、しかしBMP−4及びBMP−6のタンパク質レベルは、非処理対照細胞の培地と有意な差がなかった。各結果を、非処理細胞の値に比例した割合として表す。7試料の平均値及び標準誤差を、報告する(*p<0.05 **p<0.01 vs.NT)
【図2】図2は、線維輪細胞をLMP−1(MOI 25のAdLMP−1)で処理後の、経時的なBMPのmRNAレベルを示す。BMP−2のmRNAレベルは、AdLMP−1で処理後早ければ12時間に有意に亢進し、そしてAdLMP−1処理後3日にプラトーに達した。BMP−7のmRNAレベルは、AdLMP−1で処理後3日に、有意に増加した。各結果を、同時点における非処理細胞の値に対する割合として表す。6試料の平均値及び標準誤差を、報告する(対応する時点について、**p<0.01 vs.NT)
【図3】図3は、リアルタイムPCRによって測定された、LMP−1(AdLMP−1)で処理後の線維輪細胞中のアグリカンmRNAレベルを、グラフに描いたものである。アグリカンのmRNAレベルは、MOI 25のAdLMP−1で処理後6日に、非処理細胞と比べて有意に増加した。アグリカンのmRNAレベルは、AdLacZで処理された細胞において、非処理細胞と比べて変わらなかった。各結果を、非処理細胞の値に対する割合として、表す。9試料の平均値及び標準誤差を、報告する。AdLMP−1:MOI 25、AdLacZ:MOI 25(**p<0.01 vs.NT)。
【図4】図4は、Harlan無胸腺ラットにおける、PTD/LMP−1融合タンパク質によるLMP−1投与のパラメータ及び結果を要約した表である。
【図5】図5は、ニュージーランドホワイトウサギにおける、PTD/LMP−1融合タンパク質によるLMP−1投与のパラメータ及び結果を要約した表である。
【図6】図6は、Harlan無胸腺ラット、及び示された箇所ではニュージーランドホワイトウサギにおける、PTD/ペプチド融合タンパク質による、示された骨誘導ペプチド投与の結果を要約した表である。骨成長は、X線によって検出され、そして、示された箇所では触診も可能であった。
【発明の開示】
【0001】
本出願は、2003年3月24日に出願された米国仮出願第60/456,551の利益を主張する。
発明の属する分野
本発明は、概して、骨誘導タンパク質、及びこれらのタンパク質を細胞内に送達するための方法に、関する。より具体的には、本発明は、LIM石灰化タンパク質(LMP)、骨形態形成タンパク質(BMP)及びSmadタンパク質などの骨誘導タンパク質、このような骨誘導タンパク質とタンパク質導入ドメイン(PTD)との複合体、PTDと骨誘導タンパク質をコードするヌクレオチド配列を含む核酸との複合体、ならびに、これらの複合体の細胞内への導入に、関する。
【0002】
背景
組織再生は、疾患、外傷または外科手術に続く治癒過程の重要な要素である。疾患または外傷により、例えば骨欠損が生じる状況や、外科的手順(自家移植片もしくは同種移植片の挿入、骨架橋または骨融合など)が、骨欠損を補正するために行われる状況において、骨再生は、回復の主要な目標である。しかしながら、これは、常にまたは容易に達成される目標ではなく、そして多くの研究が、組織修復及び再生を促進するためのより新たな、そしてより効果的な方法に対して、供されている。
【0003】
骨架橋を作ることによって関節動作を排除することは、変性脊椎及び関節障害の治療に対する一般的な整形外科的ストラテジーである。脊椎融合の失敗は、該手順を受ける患者の45%ほどにおいて起こる可能性があり、彼らに疼痛の持続、外科手術の再施行、医療費及び全体的な治療の失敗を残す。
【0004】
細胞内及び細胞外骨誘導タンパク質は、骨成長及び修復を促進し、そして治療に用いるための潜在的な標的となる。このようなタンパク質としては、骨形態形成タンパク質及びLIM石灰化タンパク質が挙げられる。BMPは、in vivoにおいて骨成長を刺激することが示されており、そして、LMP、特にLMP−1及びLMP−3は骨誘導のより上流に影響を与え、これは、LMP−1発現の阻害が、糖質コルチコイド類またはBMP−6によって通常刺激されるであろう小結節の形成を遮断することからも明らかである。これらは、細胞表面受容体との相互作用によって作用する「細胞外」タンパク質と考えられるため、非常に高用量の骨形態形成タンパク質が、ヒトにおいて一貫した影響に達するのに必要とされる。BMPの製造費用は概して高いため、このことは、治療費用が法外に高いことを意味する可能性がある。したがって、BMPは有効性を示し、そして骨誘導に対する実行可能な療法ではあるものの、より費用効率が高く、そしておそらくさらにいっそう治療的に有効であろう代替治療法を開発することが、有益であろう。
【0005】
LMPの細胞内環境への送達は、魅力的な治療レジメンを提供する。これは、細胞に、LIM石灰化タンパク質をコードするヌクレオチド配列を含むプラスミドをトランスフェクトすることによって、遂行することが可能であり、あるいは、標的細胞に、LMPのヌクレオチド配列を保有するウイルスベクターを感染させることによって、行うことが可能である。しかしながら、これら各々の技術には限界がある。プラスミドのトランスフェクションでは、細胞をトランスフェクションのために単離し、次いでトランスフェクション後に植え込むことが一般に必要となる。ウイルスの送達では、ウイルスの細胞への侵入を容易にするために、適切な受容体を標的細胞の表面に配置することが、一般に必要となる。
【0006】
骨誘導タンパク質及びペプチドの使用、特に、細胞内メカニズムによって作用するこれらのタンパク質及びペプチドの使用に対して、多大な可能性がある。必要とされるのは、有効な細胞内骨誘導タンパク質及びペプチドを細胞内に送達する方法である。
【0007】
発明の概要
本発明は、細胞透過性の骨誘導ポリペプチドを作製する方法を提供し、該方法は、骨誘導ポリペプチドが細胞透過性ポリペプチドとの融合タンパク質の一部として発現するように配置された、細胞透過性ポリペプチドと骨誘導ポリペプチドをコードする発現コンストラクトを、適切な宿主細胞に導入することを含む。発現コンストラクトは、融合産物をコードするポリヌクレオチド配列の転写を導くように配置されたプロモーターを、一般に含有する。
【0008】
発現コンストラクトは、精製タグをさらに含んでもよい。細胞透過性ポリペプチドは、HIV−TAT、VP−22、増殖因子のシグナルペプチド配列、Pep−1及びショウジョウバエAntpペプチドからなる群より選択されてもよい。骨誘導ポリペプチドは、LMP−1、LMP−3、配列番号1、配列番号2、配列番号3、配列番号4、配列番号5、配列番号6、配列番号7、配列番号8、BMP−2、BMP−4、BMP−6、BMP−7、TGF−β1及びSmadからなる群より選択されてもよい。
【0009】
本発明は、配列番号1、配列番号2、配列番号3、配列番号4、配列番号5、配列番号6、配列番号7及び配列番号8、またはその組み合わせからなる群の中から選択される骨誘導ポリペプチドを、提供する。
【0010】
本発明はまた、哺乳動物において骨形成を誘導する方法であって、タンパク質導入ドメインと少なくとも一つの骨誘導ポリペプチドとを含む、有効量の融合ポリペプチドを投与することを含む方法を提供する。融合ポリペプチドは、移植片(インプラント)として投与されてもよく、そして、少なくとも一つの多能性前駆細胞に投与されてもよく、該細胞は、骨誘導を促進するために哺乳動物に植え込まれることが可能である。
【0011】
本発明はまた、タンパク質導入ドメインと少なくとも一つの骨誘導ポリペプチドを含む融合タンパク質をコードするポリヌクレオチドを提供し、タンパク質導入ドメインは、例えばHIV−TAT、VP−22、増殖因子のシグナルペプチド配列、Pep−1及びショウジョウバエAntpペプチドによって代表される、多様なタンパク質導入、膜移行、及びその他の同様なポリペプチドの中から選択される。骨誘導ポリペプチドは、LMP−1、LMP−3、配列番号1、配列番号2、配列番号3、配列番号4、配列番号5、配列番号6、配列番号7、配列番号8、BMP−2、BMP−4、BMP−6、BMP−7、TGF−β1及びSmadからなる群より選択されてもよい。
【0012】
哺乳動物においてプロテオグリカン合成を誘導する方法もまた、提供される。該方法は、タンパク質導入ドメインと少なくとも一つの骨誘導ポリペプチドを含む有効量の融合ポリペプチドを投与することを含む。融合ポリペプチドは、移植片として投与されてもよく、そして、少なくとも一つの多能性前駆細胞に投与されてもよい。
【0013】
骨誘導ポリペプチドと機能可能なように連結されている膜移行ペプチドを含む、単離された融合タンパク質が、本発明によって提供される。膜移行ペプチドは、HIV−TAT、VP−22、増殖因子のシグナルペプチド配列、Pep−1及びショウジョウバエAntpペプチドからなる群より選択されてもよく、そして骨誘導ポリペプチドは、LMP−1、LMP−3、配列番号1、配列番号2、配列番号3、配列番号4、配列番号5、配列番号6、配列番号7、配列番号8、BMP−2、BMP−4、BMP−6、BMP−7、TGF−β1及びSmadからなる群より選択されてもよい。
【0014】
本発明は、前駆細胞において骨芽細胞への分化を誘導する方法を提供し、該方法は、タンパク質導入ドメインと少なくとも一つの骨誘導ポリペプチドとを含む有効量の融合ポリペプチドを前駆細胞に投与することを含む。タンパク質導入ドメインは、HIV−TAT、VP−22、増殖因子のシグナルペプチド配列、Pep−1及びショウジョウバエAntpペプチドによって代表される群から選択されることが可能であり、そして骨誘導ポリペプチドは、LMP−1、LMP−3、配列番号1、配列番号2、配列番号3、配列番号4、配列番号5、配列番号6、配列番号7、配列番号8、BMP−2、BMP−4、BMP−6、BMP−7、TGF−β1及びSmadによって代表される群から選択されてもよい。
【0015】
詳細な説明
本発明者は、タンパク質導入ポリペプチドと骨誘導ポリペプチドを含む融合タンパク質を、in vivoにおいて骨の発達及び椎間板再生を促進するために効果的に用いることが可能であることを、発見した。したがって、本発明は、細胞内へ送達するための骨誘導ポリペプチド、このような骨誘導ポリペプチドとタンパク質導入配列をコードするポリヌクレオチド、ならびに、in vivoにおいて骨の発達及び椎間板再生を促進するのにこれらの融合タンパク質を利用する方法を、提供する。
【0016】
以前の研究により、LIM石灰化タンパク質のスプライスバリアント1及び3(LMP−1及びLMP−3)は骨誘導性であるが、LMP−2はこのような骨誘導の可能性を有するようにはみえないことが、実証された。ヒトLMP−1(hLMP−1)のアミノ酸配列のアミノ酸94〜133に相当する40アミノ酸配列は、LMP−1及びLMP−3の双方に共通している。したがって、発明者は、このタンパク質のこの特有な領域自体が、骨誘導の可能性を有するのではないかと推測した。この配列の重複セグメントを含むペプチドを設計し、そして、これを用いて発明者の仮説を検証した。その結果、LMP−1及びLMP−3由来のペプチドが、骨誘導の可能性を有することが、示唆される。これらのペプチドは、in vivoにおいて用いた場合に、骨形成誘導能を示した。図6は、本発明の方法において、本発明の融合タンパク質の一部として細胞内に導入された場合に、骨誘導の機能性を示したペプチドを示す。
【0017】
LMP−1を発現している細胞を胸椎及び腰椎に植え込むと、無胸腺ラットにおいて堅固な脊椎融合がもたらされることが、発明者によって示された。本明細書において開示された結果により、配列番号1、配列番号2、配列番号3、配列番号4、配列番号5、配列番号6、配列番号7及び配列番号8によって提供されるペプチドなどの、LMP−1及びLMP−3ペプチドもまた、LMP−1について実証された骨誘導の利益を提供することが、実証される。
【0018】
骨形態形成タンパク質が、ヒトにおいて一貫した効果を発揮するには、概して、非常に高用量が必要とされる。PTD融合タンパク質によるLMP−1の細胞内への送達は、本明細書に記載されるように、より費用効率的な治療の選択肢を提供する。さらに、これまでの結果により、LMP−1、及びLMP関連骨誘導ペプチドが、細胞内で作用して諸因子のカスケードを誘導し、天然の骨誘導経路を活性化することが、示唆される。
【0019】
タンパク質導入ポリペプチドは、核酸配列または治療用タンパク質の取り込み、及びそれに続く発現を、容易にする。文献において、これらは、代替的に、そしてしばしば同義的に、細胞透過性ペプチド、タンパク質導入ドメイン、膜輸送配列及び膜移行ペプチドと称されてもよい。これらは、付属するペプチド、ポリペプチドまたはタンパク質を、受容体とは独立に、細胞膜を介して細胞内部に輸送する働きをする。タンパク質導入ドメインを利用する融合タンパク質は、タンパク質導入ドメインと機能可能なように連結されている、1またはそれより多くのペプチド、ポリペプチドまたはタンパク質を含むことが可能である。本発明において、このような融合タンパク質は、タンパク質導入ドメインと少なくとも一つの骨誘導ペプチド、ポリペプチドまたはタンパク質、あるいはその組み合わせを含むことが可能である。これらのペプチドを用いて、外胚葉、間葉または造血を起源とする自己、同種異系または異種の細胞あるいは組織に形質導入し、そして、新たな組織の形成を誘導または助長するためにこれをレシピエントに注入または植え込むことが可能である。本発明の方法において、このようなポリペプチドは、例えば多能性前駆(幹)細胞などの細胞を、例えばBMP−2、BMP−4、BMP−6、BMP−7、BMP−9、BMP−12、BMP−13、アグリカン、I型コラーゲン、II型コラーゲン、バーシカン、ルミカン、フィブロモジュリン、ビグリカン及びデコリンを産生するよう誘導することが可能なタンパク質の取り込みを、容易にする。本発明のポリペプチドの有効量を、実験デザイン及び本明細書に開示した結果において示すが、しかし、本明細書にて提供される有効量の開示をもとに、当業者が決定してもよい。
【0020】
LMPタンパク質ファミリーの一つである、ヒトLIM石灰化タンパク質−1(hLMP−1)は、in vitro及びin vivoにおいて骨石灰化の有効性を増強することができる細胞内調節タンパク質である。ヒトLMP−1は、アミノ酸スペーサーによって連結された二つの特別なジンクフィンガーから構成される特徴的な構造モチーフを有するため、このように称される。LIM石灰化タンパク質のスプライスバリアント及びそれらの使用については、米国特許第6,300,127;6,444,803;及び6,521,750において、発明者によって記載されている。LMP−1、LMP−2及びLMP−3の配列もまた、これらの特許において公開されている。1997年7月22日に、指定されたベクターpCMV2/RLMP(挿入断片10−4クローン/RLMPを有するベクターpRc/CMV2である)中の、10−4/RLMP(ドブネズミLIM石灰化タンパク質のcDNA)の試料を、アメリカン・タイプ・カルチャー・コレクション(ATCC)(12301 Parklawn Drive, Rockville, Md. 20852)に寄託し、そして受入番号209153を割り当てられた。1998年3月19日に、挿入断片HLPM−1s(ホモサピエンスLIM石灰化タンパク質のcDNA)を有するベクターpHis−Aの試料を、アメリカン・タイプ・カルチャー・コレクションに寄託し、そして受入番号209698を割り当てられた。
【0021】
血清型5型のアデノウイルス(Ad5)は、末梢血及び骨髄由来細胞を含む多様な細胞及び組織にLMPを送達するために、使用されている。(Bodenら、「Adenoviral Delivery of LMP-1 Induces Consistent Spine Fusion」、第47回年会、Orthopaedic Research Society、サンフランシスコ、カリフォルニア州(2001年))。しかしながら、Ad5ウイルスは、特異的な受容体(すなわち、コクサッキーアデノウイルス受容体またはCAR)を利用するが、これは、これらの細胞において存在しないかまたは限られた量にて存在する。受容体を介したメカニズムなしにタンパク質の細胞内への送達を容易にする、細胞膜を介したタンパク質導入は、多様な種類の細胞及び組織の処理を可能にする魅力的な代替手段を、提示する。
【0022】
LMP及び他の骨誘導性タンパク質の作用は、脳、脊髄、末梢神経、骨、軟骨、椎間板、結合組織、腱及び靭帯などの多様な組織において、これらが治療の可能性を有することを、示唆する。LMPの、例えば多様な組織への送達は、例えばコラーゲン、コラーゲンとセラミックとの組み合わせ、脱石灰化骨基質、ならびに、天然または合成ポリマー類(エラスチン、フィブリン、ポリ乳酸、ポリグリコール酸、ポリカプロラクトン、フマル酸ポリプロピレン、ポリビニルアルコール、ポリエステル類、ポリエーテル類、ポリヒドロキシル類など)、ならびに構造的移植片を含む送達システムによって、遂行されることが可能である。このような基質は、注射可能で成型可能な固形移植片、構造的移植片、またはその組み合わせであってもよい。
【0023】
本発明者は、機能性骨誘導タンパク質を細胞内に送達し、そして骨形成及びプロテオグリカン合成を効率的に誘導するのに、PTDを用いることが可能であることを、発見した。このような細胞透過性ペプチド移入(CPPI)は、骨誘導タンパク質を多様な細胞種類に送達するための方法を、提供する。元来はHIV−1 TATタンパク質に由来する11アミノ酸ペプチドを用いて、骨誘導タンパク質を細胞内に送達することに成功した。TATペプチドを、pTAT−HAベクターを用いて、細菌細胞において過剰発現させることが可能である。リコンビナントヒト遺伝子を、TATペプチド配列と目的の遺伝子産物の双方を含有する融合タンパク質を産生させるように、このベクター中に挿入することが可能である。さらに、PTD/骨誘導ポリペプチドを、融合タンパク質の単離及び精製を容易にするために、polyHisタグと連結して発現させることが可能である。pTAT−HAベクター、及び、TAT融合タンパク質の精製プロトコールについては、以前にNagaharaら(Nature Medicine 第4巻,1449〜1452頁、1998年12月)によって記載されている。
【0024】
多様なPTDにおいて見つけられるペプチド配列は、コクサッキー−アデノウイルス受容体(CAR)とは独立に、細胞内への侵入を容易にすることが可能であり、それによって標的細胞への導入効率を改良し、続いて、所望の効果に達するのに必要な核酸またはタンパク質の必要量を低減させる。したがって、PTD融合タンパク質は、治療費用を低下させるのに用いられてもよい治療ツールを、提供する。
【0025】
本発明の一つの態様において、タンパク質導入ドメインと骨誘導タンパク質との融合タンパク質が、提供される。骨誘導タンパク質としては、これに限定されるものではないが、LIM石灰化タンパク質(LMP)、骨形態形成タンパク質(BMP)及びSmadタンパク質が挙げられる。本明細書では、「骨誘導タンパク質」、「骨誘導ポリペプチド」及び「骨誘導ペプチド」は、骨誘導の機能性を有する、様々な長さのペプチドまたはポリペプチド、あるいは完全長タンパク質のいずれかを指すのに、同義的に用いられてもよい。
【0026】
PTDとLIM石灰化タンパク質を含む融合タンパク質は、本発明の一つの態様として提供される。融合タンパク質は、PTDと1またはそれより多くのLIM石灰化タンパク質またはポリペプチドを含むことが可能である。有用なLIM石灰化タンパク質としては、例えば、米国特許第6,300,127;6,444,803;及び6,521,750;ならびに、係属中の米国特許出願第09/959,578(2000年4月28日出願)において開示されるようなLMPが挙げられる。好ましくは、LMPは、RLMP、HLMP−1、HLMP−1s、HLMP−2、HLMP−3またはそれに由来するペプチドである。これらのペプチドは、例えば、配列番号1、配列番号2、配列番号3、配列番号4、配列番号5、配列番号6、配列番号7、または、配列番号8にあるようなポリペプチドを含むことが可能である。
【0027】
LIM石灰化タンパク質をコードするヌクレオチド配列は、好ましくは、完全長の以下配列:
tcctcatccg ggtcttgcat gaactcggtg
に相補的な核酸分子と、標準的な条件下でハイブリダイズするか、または、完全長の以下配列:
gcccccgccc gctgacagcg ccccgcaa
に相補的な核酸分子と、高ストリンジェントな条件下でハイブリダイズするか、あるいはその双方である。
【0028】
「標準的なハイブリダイゼーション条件」は、プローブのサイズ、背景及び核酸試薬の濃度、ならびにハイブリダイゼーションの種類(in situ、サザンブロット、またはDNA−RNAハイブリッドのハイブリダイゼーション(ノーザンブロット))によって変わるであろう。「標準的なハイブリダイゼーション条件」の決定は、当該技術分野における技能水準の範囲内にある。方法としては、例えば、米国特許第5,580,775(Fremeauら)、Southern, J. Mol. Biol., 98:503(1975年)、Alwineら、Meth. Enzymol., 68:220(1979年)、及び、Sambrookら、Molecular Cloning: A Laboratory Manual,第2版、コールドスプリングハーバー・プレス、7.19-7.50(1989年)に記載されるものが挙げられる。
【0029】
1セットの標準的なハイブリダイゼーション条件は、50%ホルムアミド、5X SSPE(150nM NaCl、10mM NaH2PO4[pH7.4]、1mM EDTA[pH8.0])、5Xデンハート液(100mlの水につき、フィコール20mg、ポリビニルピロリドン20mg、及びBSA 20mg)、10%硫酸デキストラン、1%SDS、及び100μg/mlサケ精子DNA中で、42℃にて2時間、ブロットをプレハイブリダイズすることを、含む。32P−標識cDNAプローブを加え、そしてさらにハイブリダイゼーションを14時間続ける。その後、ブロットを、2X SSPE、0.1%SDS中で、22℃にて20分間、2回洗浄し、続いて、0.1X SSPE、0.1%SDS中で、65℃にて1時間洗浄する。次いで、ブロットを乾燥させ、そして増感スクリーンの存在下でX線フイルムに5日間曝露する。
【0030】
「高ストリンジェントな条件」下では、プローブはその標的配列と、これら二つの配列が実質的に同一であるならば、ハイブリダイズするであろう。実質的に同一の配列のみがハイブリダイズし、非同一配列がハイブリダイズしないであろう条件を決定するための技術は、当業者に公知である。
【0031】
本明細書では、「タンパク質」の語は、ミメティック(類似体、模倣物)を含むことを意図し、そして「アミノ酸」の語は、L型アミノ酸、D型アミノ酸及び修飾アミノ酸を含むことを意図している。これらの代替品は、分子間の既知の構造類似性を用いて、当業者が作製してもよい。また、アミノ酸配列には、記載された配列のN末端またはC末端のいずれか、あるいは両方に追加のアミノ酸を含んでもよい、任意のペプチドまたはタンパク質配列を含まれることが、意図される。「骨誘導タンパク質」の語は、完全長タンパク質のみならず、バリアントまたは生物学的に活性なポリペプチド断片も含むことが、意図される。
【0032】
一つのアミノ酸が、1より多いヌクレオチドコドンによってコードされてもよく、そしてヌクレオチド配列が、同一ペプチドをコードする代替のヌクレオチド配列を作り出すために修飾されてもよいことは、当該技術分野において周知である。したがって、本発明の代替の態様には、先に記載されたようなアミノ酸配列を含有するペプチドをコードする、代替のDNA配列が、含まれる。特許請求の範囲にあるアミノ酸配列を含有するペプチドをコードする、DNA配列は、特許請求の範囲にある配列と、特許請求の範囲にあるアミノ酸配列のN末端またはC末端に位置する他のアミノ酸との、任意の組み合わせをコードするDNA配列を、含む。当然のことながら、アミノ酸及び核酸配列は、追加の残基、特にNまたはC末端アミノ酸、あるいは5’または3’核酸配列を含んでもよく、そして、該配列は、発現したポリペプチドまたはタンパク質に骨誘導の可能性を授与する限り、本質的には、本明細書にて開示される配列に、なお記載されてもよい。
【0033】
追加の核酸塩基を、骨誘導ポリペプチドをコードする核酸配列の5’または3’のいずれかに加えてもよく、そして、プロモーター、ポリアデニル化シグナル、追加の制限酵素部位、多重クローニング部位、他のコードセグメント等などの他のDNA配列と、組み合わせてもよい。したがって、このようなポリヌクレオチドの全体的な長さは、かなり変わってもよい。
【0034】
当然のことながら、ポリペプチドの「バリアント」は、天然のタンパク質とは完全に同一ではない。骨誘導ポリペプチドまたはタンパク質のバリアントは、例えば、1またはそれより多くのアミノ酸を挿入、欠失または置換することによりアミノ酸配列を変えることによって、得ることが可能である。ポリペプチドまたはタンパク質のアミノ酸配列を、例えば、置換によって修飾して、天然ポリペプチドと比べて同一または改良された質を実質的に有するポリペプチドを作り出すことが、可能である。置換は、保存的置換であってもよい。「保存的置換」とは、アミノ酸を、極性/非極性の性質、電荷またはサイズにおいて類似した側鎖を有する別のアミノ酸に置換することである。20の必須アノミ酸は、非極性側鎖を有するもの(アラニン、バリン、ロイシン、イソロイシン、プロリン、フェニルアラニン及びトリプトファン)、非荷電性極性側鎖を有するもの(メチオニン、グリシン、セリン、トレオニン、シスチン、チロシン、アスパラギン及びグルタミン)、酸性側鎖を有するもの(アスパラギン酸及びグルタミン酸)、及び塩基性側鎖を有するもの(リジン、アルギニン及びヒスチジン)として分類されることが可能である。保存性置換には、例えば、AspをGlu、AsnまたはGlnに;HisをLys、ArgまたはPheに;AsnをGln、AspまたはGluに;及び、SerをCys、ThrまたはGlyに置換することが含まれてもよい。例えば、アラニンは、保存的置換を行うのにしばしば用いられる。
【0035】
当業者は、バリアントポリペプチドを、生物学的活性に影響を及ぼすために、ポリペプチド構造中の1またはそれより多くの位置において第一のアミノ酸を第二のアミノ酸に置換することによって、得ることが可能である。アミノ酸置換は、例えば、生物学的活性の増加をもたらすポリペプチドの立体構造変化を誘導する可能性がある。
【0036】
当業者はまた、アミノ酸の親水性指数または疎水・親水性指数に基づいて、アミノ酸配列における置換を行ってもよい。したがって、本発明のバリアントアミノ酸分子は、LMP−1、LMP−2、LMP−3、配列番号1、配列番号2、配列番号3、配列番号4、配列番号5、配列番号6、配列番号7、あるいは、配列番号8にあるようなポリペプチドのアミノ酸配列を含むポリペプチドのアミノ酸配列と、100%よりは少ないが、しかし少なくとも約50%、そして好ましくは少なくとも約80%〜約90%の、アミノ酸配列相同性または同一性を有する。したがって、バリアント骨誘導ポリペプチドまたはタンパク質のアミノ酸配列は、本質的には、天然の骨誘導ポリペプチドまたはタンパク質のアミノ酸配列に相当する。本明細書では、「本質的には〜に相当する」の語は、LMP−1、LMP−2、LMP−3、配列番号1、配列番号2、配列番号3、配列番号4、配列番号5、配列番号6、配列番号7、あるいは、配列番号8にあるようなポリペプチドを含む骨誘導ポリペプチドまたはタンパク質によって生じる活性に類似した、生物学的活性及び酵素活性を誘発するであろうポリペプチド配列に向けたものであり、このような活性は、天然の骨誘導タンパク質の少なくとも約70%であり、そしてより好ましくは、天然の骨誘導タンパク質活性の100%を超える。
【0037】
骨誘導タンパク質のバリアントには、LMP−1、LMP−2、LMP−3、配列番号1、配列番号2、配列番号3、配列番号4、配列番号5、配列番号6、配列番号7または配列番号8を含む骨誘導タンパク質を含む、相当する骨誘導タンパク質中には存在しないアミノ酸残基が含まれてもよく、あるいは、LMP−1、LMP−2、LMP−3、配列番号1、配列番号2、配列番号3、配列番号4、配列番号5、配列番号6、配列番号7または配列番号8を含む骨誘導タンパク質と比べて、欠失が含まれてもよい。バリアントはまた、LMP−1、LMP−2、LMP−3、配列番号1、配列番号2、配列番号3、配列番号4、配列番号5、配列番号6、配列番号7または配列番号8を含む、相当するタンパク質と比べて、切断された「断片」であってもよく、該断片は、完全長のタンパク質またはポリペプチドのほんの一部である。
【0038】
骨形態形成タンパク質(BMP)は、タンパク質のTGF−βスーパーファミリーのメンバーである。BMPは、異所性骨または軟骨形成を誘導することが示されている。本発明にしたがって、PTDと骨形態形成タンパク質との融合タンパク質もまた、提供される。BMPとしては、例えば、BMP−2、BMP−3、BMP−3b、BMP−4、BMP−5、BMP−6、BMP−7、BMP−8、BMP−9、BMP−10、BMP−11、BMP−12、BMP−13、BMP−14、BMP−15、GDF−1、GDF−3、GDF−8及びGDF−9が挙げられる。骨形態形成タンパク質であるBMP−2、BMP−4、BMP−5、BMP−6、BMP−7、BMP−8またはBMP−9は、本発明の方法において、特に有用である可能性がある。
【0039】
Smadタンパク質は、細胞外TGF−β関連因子の受容体からのシグナル伝達を媒介する、細胞内タンパク質である(Heldinら、”TGF-β Signalling from Cell Membrane to Nucleus through SMAD Proteins”、Nature、第390巻(1997年))。Smadタンパク質は、BMPがその受容体に結合することによって、活性化される(すなわち、リン酸化される)ことが可能である。活性化されると、Smadタンパク質は、それが遺伝子発現を調節する場所である核に移行する。PTDとSmadタンパク質との融合タンパク質もまた、本発明において提供される。Smad−1、Smad−2、Smad−3、Smad−4、Smad−5、Smad−6、Smad−7またはSmad−8は、本発明におけるようなタンパク質導入ドメインとの融合タンパク質として送達される場合、骨誘導を促進するのに特に有用である可能性がある。
【0040】
本発明にしたがったタンパク質導入ドメインは、付属もしくは付随のタンパク質、ペプチドまたは核酸を細胞内部に送達するために、細胞の原形質膜を越えることが可能な、任意のペプチド、ミメティック、またはペプチドの核酸(PNA)配列であることが可能である。発明者は、骨誘導タンパク質を、骨誘導及びプロテオグリカン合成を促進する能力を損なうことなく、細胞内に(例えば、融合タンパク質の一部分として)送達可能であることを、実証した。PTDとしては、例えば、ショウジョウバエのホメオティック転写因子であるアンテナペディア(Antp)由来のポリペプチド、単純ヘルペスウイルス(HSV)タンパク質VP22、カポジ線維芽細胞増殖因子(K−FGF)などの増殖因子由来シグナルペプチド配列(Linら、J. Biol. Chem.、第270巻、14255〜14258頁、1995年)、K−FGFシグナルペプチド配列由来の膜移行配列(Rojasら、Nat. Biotech.、第16巻、370〜375頁、1998年)、及びヒト免疫不全ウイルス(HIV)−1転写活性化因子TAT(Fawellら、Proc. Natl. Acad. Sci. USA、第91巻、664〜668頁、1994年)が挙げられる。PTDについては、米国特許第5,652,122、及びSchwarzeら、「Protein Transduction: Unrestricted Delivery into all Cells」、Trends in Cell Biology、第10巻(2000年)において、開示されている。発明者は、HIV−TATのPTDが本発明において特に有用であることを、見いだした。
【0041】
プロモーターと機能可能なように連結されている、融合タンパク質(タンパク質導入ドメイン(PTD)と骨誘導タンパク質を含む)をコードするヌクレオチド配列を含む核酸もまた、提供される。該核酸は、ベクター(例えば、プラスミドなどの発現ベクター)の一部であることが可能である。骨誘導タンパク質は、例えば、LIM石灰化タンパク質、骨形態形成タンパク質、Smadタンパク質及び骨誘導ペプチド、ならびにそれに由来するポリペプチドを含むことが可能である。骨誘導ペプチド及びポリペプチドの例としては、配列番号1、配列番号2、配列番号3、配列番号4、配列番号5、配列番号6、配列番号7及び配列番号8が挙げられる。
【0042】
骨誘導タンパク質を細胞内に送達する方法もまた、本発明によって提供される。本発明の方法において、少なくとも一つの骨誘導タンパク質を、導入によって細胞内に送達することが可能であり、ここで、タンパク質導入ドメイン(PTD)と骨誘導タンパク質を含む融合タンパク質は、融合タンパク質が細胞内に送達されるように細胞と接触し、タンパク質導入ドメインまたは細胞透過性ペプチドによって、その送達が容易となる。
【0043】
骨誘導タンパク質を送達可能な細胞としては、例えば、骨(すなわち、骨形成)及び非骨細胞が挙げられる。このような細胞は、例えば、軟膜細胞、幹細胞(例えば、間葉幹細胞、多能(multipotent)性及び多分化性(pluripotent)幹細胞)、椎間板細胞(disc cell)(例えば、線維輪細胞(nucleus cell)及び髄核細胞(annulus cell))、間葉細胞、造血性細胞、内皮細胞及び筋細胞を含んでもよい。幹細胞は、自己または同種異系の組織由来であることが可能である。
【0044】
タンパク質導入ドメイン(PTD)と骨誘導タンパク質との融合タンパク質を導入されているかまたはこれを発現する細胞もまた、提供される。このような細胞は、これに限定されないが、軟膜細胞、幹細胞(例えば、間葉幹細胞及び多分化性(pluripotential)幹細胞)、椎間板細胞(例えば、線維輪細胞及び髄核細胞)、間葉細胞、造血性細胞、内皮細胞及び筋細胞を含んでもよい。本明細書において記載されるような、PTDと骨誘導タンパク質との融合タンパク質を含有する細胞は、哺乳動物体内に植え込まれて、骨形成を誘導することが可能である。LMPを骨誘導タンパク質として用いた骨形成誘導法については、例えば、米国特許第6,300,127に記載されている。例えば、2002年11月13日に出願され、係属中の米国特許第10/292,951に記載のように、PTDと骨誘導タンパク質との融合タンパク質を含む細胞を、椎間板内に植え込んで、プロテオグリカン及び/またはコラーゲン合成を刺激してもよい。
【0045】
PTDと、骨誘導タンパク質をコードするヌクレオチド配列を含む核酸との複合体もまた、提供される。PTD/核酸複合体を、例えば、骨形成または椎間板再生を促進するために、骨誘導タンパク質の過剰発現を導くのに用いることが可能である。ヌクレオチド配列によってコードされる骨誘導タンパク質は、これに限定されないが、LMP、BMP及びSmadタンパク質を含むことが可能である。ペプチドを核酸と化学的に結合させるための方法は、当該技術分野において公知である。一つのこのような方法は、米国特許第5,652,122に記載されている。核酸は、プロモーターと機能可能に連結されている骨誘導タンパク質をコードするヌクレオチド配列を含む、発現ベクターの形であることが可能である。
【0046】
本発明の方法を、1またはそれより多くの骨形態形成タンパク質の発現、または、2003年3月7日に出願された同時係属中の米国特許出願第10/382,844に記載されるように、増殖因子−βタンパク質の細胞内での形質転換を誘導するのに、用いることが可能である。例えば、BMP−2、BMP−4、BMP−6、BMP−7、TGF−β1及びその組み合わせからなる群より選択される、1またはそれより多くのタンパク質の発現は、細胞を、PTDと骨誘導タンパク質を含む融合タンパク質と本発明にしたがって接触させることにより、誘導可能である。加えて、BMP−2、BMP−4、BMP−6、BMP−7、TGF−β1及びその組み合わせからなる群より選択される、1またはそれより多くのタンパク質を過剰発現する細胞もまた、本発明にしたがって提供される。細胞は、非限定的に、幹細胞、軟膜細胞、骨髄細胞、末梢血細胞または脂肪細胞を含む、任意の体細胞であることが可能である。細胞は、自己または同種異系組織に由来する幹細胞であることが可能である。
【0047】
腎臓においては、例えば、細胞を、本発明において記載されるようなPTD/LMP融合ポリペプチドの投与によって刺激して、BMP−7を産生させてもよく、あるいは、幹細胞を、PTD/LMP融合ポリペプチドで処理して、これにBMP−7の発現を誘導させてもよい。ひとたび腎臓に移植されるならば、そのBMP−7産生は、外因性BMP−7の投与を介してこれまで利用可能でなかった治療効用を提供することが可能である。BMP−7は、慢性腎障害の回復に関連した、重度に損傷した腎尿細管上皮細胞の修復を、誘導する(Zeisbergら、Nat. Med. 9:964-968、2000年)。マウスにおいて、BMP−7を投与することによって、慢性腎疾患の一合併症である無形成骨障害の治療に成功したことが、報告されている(Lundら、J Am Soc Nephrol 15(2):359-369、2004年)。慢性腎障害及び線維症のマウス遺伝モデルにおいて、外因性ヒトリコンビナントBMP−7の投与は、腎機能及び生存性を向上させた(Zeisbergら、Am J Physiol Renal Physiol 285(6):F1060-7、2003年)。BMP−7が、糖尿病誘導性の腎肥大を部分的に回復させることもまた、示されている(Wangら、Kidney Int 63(6):2037-2049、2003年)。Daviesらは、BMP−7の欠損が、慢性腎不全における病態生理学的因子であると、判断した(J Am Soc Nephrol 14(6):1559-1567、2003年)。これらの試験は、LMP−1が細胞によるBMP−7産生を誘導し、そしてLMPペプチド(配列番号1、配列番号2、配列番号3、配列番号4、配列番号5、配列番号6、配列番号7及び配列番号8)がLMP−1タンパク質と同じように機能するという、発明者による発見と併せて、タンパク質導入ドメインとLMP−1もしくはLMP−3タンパク質または関連ペプチドを含む融合ポリペプチドが、腎疾患の治療において有効である可能性があることを、実証する。本発明の融合ポリペプチドを用いてBMP−7の発現を誘導された幹細胞は、この目的に対して特に有用である可能性がある。
【0048】
BMP−4及びBMP−7もまた、ヒト骨髄腫細胞において、増殖を阻害し、そしてアポトーシスを誘導することが示されている。BMPの治療への使用は、骨髄腫の骨疾患及び骨髄腫細胞の増殖に対して、提案されている。本発明の方法は、LMP−1、LMP−3、または、配列番号1、配列番号2、配列番号3、配列番号4、配列番号5、配列番号6、配列番号7及び配列番号8を含むペプチドを、例えば注射、移植片、またはPTD/LMP融合ポリペプチドで処理された幹細胞の植え込みによって投与することにより、in vivoにおいてBMP−4とBMP−7双方の産生を刺激する方法を、提供する。幹細胞移植片を、植え込み型薬物送達装置などのPTD/LMP源とともに組み合わせると、特に有効である可能性がある。
【0049】
幹細胞または多能性前駆細胞は、骨芽細胞を作り出す細胞源を提供することが可能である。これらの細胞を、種々の分化段階において単離し、そして特定系統の経路にて分化を誘導してもよい。細胞を、骨粗鬆症または骨形成不全症などの骨疾患、ならびに非治癒骨折を治療するのに用いてもよい。核結合因子α1(Cbfa1)は、骨形成に必要であることが実証されている。BMP−2、BMP−4及びBMP−7は、骨芽細胞の分化を誘導することが知られており、Cbfa1の発現を亢進させる。BMP−8及びSmad−3は、骨芽細胞の分化中に亢進している。TGF−β/BMP−Smadのシグナル伝達の活性化は、Cbfa1の発現及び骨芽細胞の分化を促進することが、示されている。本発明は、例えば、ヒト骨髄由来中胚葉前駆細胞などの細胞において骨芽細胞の分化を促進するために、機能性BMP、LMP、Smadタンパク質またはその組み合わせを含む融合タンパク質を提供する。適切な細胞には、例えば、Jiangら(Nature、第418巻、41〜49頁、2002年)によって記載された細胞のような、多能性細胞が含まれてもよい。適切な骨誘導タンパク質またはポリペプチド、あるいはその組み合わせの投与は、細胞を植え込む前にex vivoで、あるいは、植え込みまたは注射後にin vivoで、行うことが可能である。
【0050】
In vivo投与に関して、本発明の骨誘導タンパク質を、例えばPTDまたは細胞透過性ペプチドによって近隣の細胞の内部に送達可能なように、標的部位に注射することが可能である。あるいは、PTD/骨誘導ポリペプチドと組み合わせたキャリアーを含む移植片を、用いてもよい。移植片は、周囲組織に放出するためにPTD/骨誘導ポリペプチドを入れる貯蔵所を含有してもよく、あるいは、1またはそれより多くのPTD/骨誘導ポリペプチドコンストラクトを含有する溶液中に浸された多孔性の組成物を含んでもよい。ヒドロゲル、持続放出カプセルもしくは球(spheres)、リポソーム、ミクロスフェア、ナノスフェア、生分解性ポリマー、または他のこのような薬物送達システムもまた、本発明のペプチド及びタンパク質を標的細胞及び組織に送達するのに用いられてよい。例えば、米国特許第6,475,516(DiCosmoら)は、抗生物質などのリポソーム治療剤が詰め込まれたヒドロゲルを提供し、該ヒドロゲルは、移植片などの留置された医療装置の表面と共有結合している。
【0051】
椎間板変性の特徴は、椎間板においてプロテオグリカン類、特に硫酸化グリコサミノグリカン類(sGAG)及びアグリカンの産生が減少することである。椎間板の主要なプロテオグリカンであるアグリカンの産生速度の減少は、椎間板変性における重要な要素である。椎間板の機能においてプロテオグリカン類は中心的な役割を果たすため、椎間板の正常なプロテオグリカン産生を回復させることは、椎間板変性のいかなる生物学的治療においても、非常に重要である可能性がある。
【0052】
発明者は、LMP−1の過剰発現または細胞内投与が、in vitro及びin vivoにおいて椎間板細胞のプロテオグリカン産生を増加させることを実証する実験を、行った。LMP−1の過剰発現は、in vitro及びin vivoにおいて、BMP−2及びBMP−7のmRNAの亢進を誘導する。これらのBMP−2及びBMP−7を特異的に阻害するノギンは、AdLMP−1によるプロテオグリカンの亢進を阻害し、このことは、LMP−1によって誘導されるプロテオグリカンの亢進が、BMPの亢進によって媒介されることを示唆する。したがって、遺伝子治療またはタンパク質治療によるLMP−1の投与(例えば、PTD複合体による送達)を、椎間板におけるプロテオグリカンの産生を刺激し、そして椎間板再生において治療的な役割を果たすのに用いることが可能である。
【0053】
TGF−β1、IGF−1及びEGFなどのサイトカイン類は、椎間板細胞の有糸分裂、及びある程度プロテオグリカン産生を刺激することが示されている。BMP−2及びBMP−7などの他のサイトカイン類もまた、プロテオグリカン産生を刺激するのに有効であることが示されている。しかしながら、サイトカイン類は小さな水溶性分子であるため、これらは椎間板から急速に拡散して出て行くか、または他の調節因子によって不活化される。
LIM石灰化タンパク質−1(LMP−1)は、白血球及び骨芽細胞から複数の異なるBMPの分泌を誘導することが知られている、細胞内調節分子である。LMP−1、LMP−2、LMP−3、あるいはLMP−1またはLMP−3由来骨誘導ペプチド、あるいはその組み合わせを、特にPTD/核酸複合体によって細胞内に送達することによって、BMP産生を細胞内部から刺激することが可能である。適切な骨誘導ペプチドは、例えば、配列番号1、配列番号2、配列番号3、配列番号4、配列番号5、配列番号6、配列番号7、または、配列番号8にあるようなポリペプチドを含む。
【0054】
本発明は、以下の限定されない実施例を参照することによって、より十分に理解されるだろう。
(実施例)
HIV−Tatのタンパク質導入ドメインとLMP−1タンパク質を含む、(His)6TAT−LMPタンパク質複合体の合成及び使用について、以下に記載する。pTAT−HA−ベクターを、ワシントン大学(セントルイス、ミズーリ州)から、物質譲渡契約(material transfer agreement)に基づいて得た。
【0055】
pCDNA3.1/hLMP−1を以下のプライマー:
正:5’−CCATGGAflCCflCAAAGTAGTGC−3’
逆:5’−CAGGGCGGGCGGCTGGTAG−3’
を用いたPCRの鋳型として利用することによって、NcoI制限酵素部位を、hLMP−1の5’末端に付加した。反応を、95℃にて2分間[95℃、30秒間;66℃、30秒間;72℃、1分間]x25、及び72℃にて10分間、行った。PCR産物を、PCRII−TOPOベクター(インビトロジェン社)中にクローニングし、そして適切なクローンを、配列決定によって同定した。
【0056】
(His)6TAT−LMPベクターの構築を、NcoI及びClaIを用いてプラスミドクローンを制限エンドヌクレアーゼ消化し、そしてその結果として得られた産物をアガロースゲル電気泳動及び電気溶出によって精製することにより、行った。hLMP−1の完全長配列を、ClaI及びEcoRIを用いてpCDNA3.1/hLMP−1ベクターを制限酵素消化し、そしてその結果として得られた産物をアガロースゲル電気泳動及び電気溶出によって精製することにより、単離した。pTAT−HA−ベクターもまた、NcoI及びEcoRIで制限酵素消化を受け、そしてその結果として得られた線状化ベクターを、アガロースゲル電気泳動及び電気溶出によって精製した。次いで、産物を、標準的な手順によって、16℃にて一晩かけて連結させた。正しく連結された産物(5’hLMP−1+3’hLMP−1+線状化pTAT−HA−ベクター=(His)6TAT−LMPベクター)を、続いて行ったアガロースゲル電気泳動及び分子量決定によって、決定した。
【0057】
連結された産物を、BL21(DE3)コンピテント細胞にトランスフェクトし、そして適切なクローンを制限酵素解析によって同定した。
(His)6TAT−LMPタンパク質の合成及び回収
(His)6TAT−LMP融合コンストラクトの陽性クローンを含有する、適切なBL21(DE3)大腸菌(Eschericia coli)コロニーを、0.8OD(600nm)まで増殖させ、そしてタンパク質産生を、100μMのIPTGで37℃にて4時間誘導した。誘導された細胞を標準的な方法によって回収し、そして超音波処理(4x20秒、それぞれ2分間の休止期間を伴う、4℃)によって溶解した(20mM PO4バッファー、pH7.2、8M尿素、100mM NaCl、20mMイミダゾール)。
【0058】
溶解物を、遠心分離(10000xg)によって澄明にし、そしてその結果として得られた上清を、Ni2+セファロースアフィニティカラム(インビトロジェン社)に、重力流の条件下で載せた。カラムを洗浄し(20mM PO4バッファー、pH6.0、8M尿素、250mM NaCl、20mMイミダゾール)、次いで、結合しているタンパク質を溶出させた(20mM PO4バッファー、pH4.0、8M尿素、500mM NaCl)。
【0059】
溶出物を、0%バッファーBから100%バッファーBまでの直線勾配(バッファーA:20mM炭酸ナトリウム、pH11、8M尿素;バッファーB:20mM炭酸ナトリウム、pH11、8M尿素、2M塩化ナトリウム)を用いた陰イオン交換クロマトグラフィ(Hitrap Q HP、5mL、ファルマシア社)にかけた(5mL/分にて40分間)。溶出画分(5mL)を、SDS−PAGE及びウエスタンブロット解析によって、hLMP−1の存在について解析した。
【0060】
hLMP−1陽性画分を、疎水性相互作用クロマトグラフィ(Hitrap(商標登録)フェニル−セファロース、5mL、ファルマシア社)にかけた。溶出(バッファーA:20mM炭酸ナトリウム、pH10.5、1.5M硫酸アンモニウム;バッファーB:20mM炭酸ナトリウム、pH10.5)を、0%〜100%のバッファーBの直線勾配で、5mL/分にて20分間行った。溶出画分(5mL)を、SDS−PAGE及びウエスタンブロット解析によって、hLMP−1の存在について解析した。(His)6TAT−LMP融合タンパク質に陽性の画分を、使用時まで凍結乾燥させておいた。
【0061】
新規の骨形成のための(His)6TAT−LMP融合タンパク質の使用
凍結乾燥させた画分(55pg)を、40mM KOH中に再懸濁した(ストック溶液;1.0μM)。ヒト軟膜細胞を、Viggeswarapuら、J.Bone Joint Surg.、第83(3)巻、364頁(2001年)に記載されるように調製した。これらの細胞を、アルファMEM(ギブコ社)に溶かした(His)6TAT−LMP融合タンパク質5μLと混合し、そして37℃にて30分間インキュベートした。
【0062】
次いで、適切な容量の、(His)6TAT−LMP融合タンパク質を含有するヒト軟膜細胞を、植え込み用に、多孔性コラーゲン基質に載せた。In vivoにおける新規の骨形成を実証するため、ラットへの植え込み用に、100μLの細胞懸濁液を、無菌ピペットによって無菌の5x5mmI型ヒトコラーゲンディスクに載せた。脊椎の融合を促進するための移植用に、同様な量の細胞懸濁液を、10x25mmシートに載せた。ディスクを、4〜5週齢の無胸腺ラット(rnu−/rnu−)の胸/腹部に、外科的に皮下に植え込んだ。動物を4週間後に屠殺し、この時点でディスクを切除し、70%エタノール中で固定し、そしてX線撮影及び脱灰していない組織の検査(5μmの切片を作製し、そしてゴールドナー・トリクロムで染色)によって解析した。
【0063】
ウサギにおいて、後外側腰椎の関節固定を行い、そして、脊椎の各側が、4x106個の導入された軟膜細胞を受け入れるように、キャリアー基質を植え込んだ(すなわち、15%ヒドロキシアパタイト/85%リン酸三カルシウムのコラーゲンスポンジ)。4週間後に、ウサギを屠殺し、そしてその腰椎を切除した。脊椎融合の状態を、残存する運動(融合の失敗を示唆)を検出するための、盲検化された手による運動分節の触診、X線撮影、CTスキャン及び脱灰していない組織の検査によって、評価した。
【0064】
ラットとウサギの双方において、X線撮影により、LMP−1をトランスフェクトされたヒト軟膜細胞を含有する元々のコラーゲンディスクまたはシートの形に一致した、高レベルの石灰化骨形成が示された。石灰化骨形成は、対照において認められず、そして元々のコラーゲンディスクまたはシートは、吸収を受けているようにみえた。組織像は、LMP−1を導入された移植片において、骨芽細胞の並んだ新たな骨梁を示したが、対照においては、骨はみられず、そしてキャリアーは部分的に再吸収された。
【0065】
In vivo骨形成を誘導するための骨誘導ペプチドの使用
プロトコールは、基本的には上記のとおりである。ペプチドを、4個の別々の移植片(3個はウサギ用)に、250μl/ディスク、6M/mlにて載せた。移植片は、コラーゲンディスクを含んだ。移植片を、Harlan無胸腺ラットまたはニュージーランドホワイトウサギの胸部内に置いた。試験された用量範囲は5nM、10nM、12.5nM、15nM、17.5nM、20nM、22.5nM及び25nMであった。結果を、図6に示す。
【0066】
sGAG合成のLMP−1刺激
Sprague−Dawleyラットから得た腰椎間板細胞を用いたin vitro実験を、種々の用量(感染多重度0、5、10、25、50)のLMP−1ヌクレオチド配列の挿入断片を含有するアデノウイルス(AdLMP−1)で細胞を処理し、そしてこれを単層にて6日間培養することによって行い、in vitroにおけるLMP−1過剰発現の影響を判定した。DMMB法を用いて、培地中の硫酸化グリコサミノグリカン(sGAG)を定量した。リアルタイムPCRを用いて、アグリカン、過剰発現LMP−1、BMP−2、4、6及び7のmRNAレベルを定量した。直接ELISA法を用いて、培地中のBMP−2、4、6及び7のレベルを定量した。LMP−1による椎間板細胞のプロテオグリカン産生亢進にBMPが関与することを実証するため、BMP−2、4、6及び7の活性を特異的に遮断する分子(ノギン)を、異なる濃度で、AdLMP−1処理細胞に加えた。
【0067】
In vivo遺伝子治療実験を、ニュージーランドホワイトウサギにおいて行った。腰椎間板に、AdLMP−1(実験用)、またはマーカー遺伝子を保有するアデノウイルス(AdLacZ−対照)のいずれかを、異なる3用量(106、107及び108pfu/椎間板)にて注射した。3週間後に、注射された椎間板を回収し、そして総LMP−1(内因性)、過剰発現LMP−1、アグリカン、BMP−2及びBMP−7のmRNAレベルを測定した。
【0068】
感染多重度(MOI)25のAdLMP−1は、最大レベルのsGAG亢進を誘導するのに十分であった。6日間の単層培養期間が、AdLMP−1処理(MOI 25)後に最大レベルのsGAG亢進に達するのに必要とされた。アグリカンのmRNAは、対照と比べて2倍増加した。細胞をアルギン酸中で培養した場合、sGAG産生に対するAdLMP−1処理の効果は、3週間持続した。AdLMP−1処理後6日に、BMP−2及びBMP−7のmRNAレベルは、それぞれ対照の3.0±0.2及び2.8±0.3倍(p<0.01)有意に増加した。BMP−4及びBMP−6のmRNAレベルは、変わらなかった。培地中のBMP−2及びBMP7のタンパク質レベルは、対照と比べて有意に増加した(p<0.01)。対照的に、BMP−4及びBMP−6のタンパク質レベルは、上昇しなかった。3200ng/mlのノギンは、AdLMP−1によるプロテオグリカンの亢進を完全に遮断した。内因性レベルのLMP−1 mRNAが、腰髄核中に検出された。107pfu/椎間板のAdLMP−1を注射されたin vivo椎間板では、対照と比べてLMP−1、BMP−2及びBMP−7のmRNAレベルが有意に上昇した。
【0069】
In vitro実験を、Sprague−Dawley(SD)ラットの腰椎間板細胞を用いて行った。両側スチューデントt検定を用いて、実験群を対照群と比較した。図中の誤差バーは、1SEM(標準誤差)を示す。
【0070】
ヒトLMP−1遺伝子のcDNAを保有する複製欠損性5型アデノウイルスを用いた(AdLMP−1)。SD線維輪細胞を用いた単層培養実験を行い、ウイルス用量と過剰発現LMP−1 mRNA発現との関連性を、ウイルスによって送達されるLMP−1 cDNAのみに特異的なプライマーを用いたリアルタイムPCR法を用いて、判定した。SD線維輪細胞を用いた単層培養実験を行い、ウイルス用量とsGAG産生との関連を判定した。sGAGレベルを、DMMB法を用いて測定した。AdLMP−1の最適用量を、sGAGの産生をプラトーレベルに導く最低用量として定めた。最適用量(25MOI)を用いて、経時的な実験(9日間)を行い、sGAGの産生がプラトーレベルに達するのに必要な最短期間(6日間)を決定した。先に決定された最適ウイルス量及び時間(25MOIのAdLMP−1、及び処理後6日)を用いて、AdLMP−1処理された線維輪細胞のアグリカンのmRNAレベルを、対照と比較した。髄核細胞が、線維輪細胞と同じように反応することを確かめるため、sGAG産生及び細胞数に対するAdLMP−1の影響を、処理後6日に、25MOIにて、線維輪細胞及び髄核細胞の双方について判定した。AdLMP−1処理のより長期の影響を検討するため、アルギン酸中で培養されたSD線維輪細胞のアルギン酸中のsGAGの蓄積を、1、2及び3週間後に判定した。すべての実験を、少なくとも2回繰り返した。
【0071】
SD線維輪細胞を用いた単層培養実験を、MOI 25のAdLMP−1を用いて行った。簡潔に記すと、LMP−1、BMP−2、BMP−4、BMP−6及びBMP−7のmRNAレベルを、AdLMP−1処理後第0.5、1、3及び6日に、リアルタイムPCR解析を用いて測定した。BMP−2、BMP−4、BMP−6及びBMP−7のレベルを、直接ELISA法を用いて、処理後6日に培地中にて測定した。BMP活性を遮断する影響を判定するために、ノギンを、AdLMP−1処理開始時に培地に加えた。AdLMP−1で処理後の、種々の濃度のノギン存在下または非存在下でのsGAGレベルの変化を、6日間の実験において測定した。
【0072】
アグリカン、BMP−2及びBMP−7に対するLMP−1投与(AdLMP−1による)のin vivo影響を判定するために、4例のニュージーランドホワイト(NZW)ウサギ(3〜4kg)を用いた。前部腰椎間板L2/3、L3/4、L4/5及びL5/6を、左後腹膜からのアプローチを介して露出させた。107プラーク形成単位(plaque-forming units、pfu)の実験用ウイルス(AdLMP−1)または対照ウイルス(AdGFP−緑色蛍光タンパク質のcDNAを挿入断片として有する、5型アデノウイルス)のいずれかを、露出させた椎間板髄核のそれぞれに、交互に注射した(すなわち、各ウサギにおいて、二つの椎間板にはAdLMP−1を注射し、そして二つの椎間板にはAdGFPを注射)。ウイルスを、30Gハミルトンシリンジを介して、10μlのリン酸緩衝生理食塩液中にて送達することによって、投与した。3週間後に、注射された腰椎間板から得た髄核組織を回収した。2例のウサギから得た髄核組織を、対照または実験椎間板群のいずれかにプールして、さらなる解析に十分なmRNAを得た。逆転写及びリアルタイムPCRを用いて、総LMP−1、BMP−7及びアグリカンのmRNAレベルを定量した。総LMP−1のプライマーは、内因性及び過剰発現LMP−1の双方を同定するように、設計された。
【0073】
第二のin vivo実証においては、種々の用量のAdLMP−1ウイルスを用いて、用量反応の関係性を確立した。異なる3用量(106、107、108pfu)のAdLMP−1及び単一用量(107pfu)のAdGFPを、投与した。この実験では、それぞれの動物のすべての椎間板に、先の実験におけるような交互のウイルス種ではなく、単一用量のウイルスを注射した。8例のNZWウサギ(3〜4kg)を、4つのウイルス群に2例ずつのウサギで、用いた。ウサギを3週間後に屠殺し、そして髄核を回収した。各処理群内から回収した組織をプールし、mRNAを単離し、そしてこれを用いて対応するcDNAを作製した。リアルタイムPCRを用いて、総LMP−1、過剰発現LMP−1、BMP−2、BMP−7及びアグリカンのmRNAレベルを定量した。
【0074】
様々な用量(MOI 0、10、25及び50)でウイルス処理した12時間後における、過剰発現LMP−1のmRNAの相対量を、過剰発現LMP−1のmRNAの最低検出可能レベル(AdLMP−1 MOI 5)を基準に正規化した。AdLMP−1の用量を増加させると、過剰発現LMP−1のmRNAに、MOI 5と比べて統計学的に有意な増加が生じた。非処理対照群またはAdLacZ対照群において、検出可能なレベルの過剰発現LMP−1のmRNAはみられず、このことは、AdLMP−1が、線維輪細胞において用量相関的にLMP−1の過剰発現を誘導することを示唆した。
【0075】
線維輪細胞の培地中sGAG濃度を、様々な用量のAdLMP−1による処理後6日に、測定した。第6日における培地中sGAG濃度は、培地交換からsGAG測定までの3日間の期間中に細胞によって産生された総sGAGの指標であり、したがって、この期間中のsGAG産生速度の指標である。データを、処理と非処理対照間の割合として表す。LMP−1の投与(MOI 25のAdLMP−1による)は、最も高いsGAGレベルを提供し、これは、対照の3.1±0.2倍(p<0.01)であった。MOI 25またはMOI 50で処理された細胞間では、培地中sGAGレベルに有意な差はなかった。このMOI 25のAdLMP−1の用量は、最大のsGAG応答に達することが可能な最低用量であったため、発明者は、実験の残りに用いる用量としてMOI 25を選択した。
【0076】
時間経過実験を行って、MOI 25のAdLMP−1で処理後の実験の長さを変えた影響を、判定した。線維輪細胞をLMP−1(MOI 25のAdLMP−1)で処理し、そして3日間の増殖期間中のsGAG産生を測定した。結果を、各時点におけるDNA含量によって正規化し、そして同時点の非処理対照に対する割合として表した。sGAGレベルは、第3日に1.6±0.2(p<0.01)に、第6日に2.9±0.1(p<0.01)に、第9日には2.8±0.1(p<0.01)に増加した。第6日から第9日までの間に、sGAGレベルの増加はみられず、このことは、第6日までにプラトーレベルに達したことを示唆した。
【0077】
LMP−1によるアグリカン合成の誘導
アグリカンは椎間板の主なプロテオグリカンであるため、アグリカンのコアタンパク質のmRNAレベルを、AdLMP−1で処理後6日に測定した。定量性リアルタイムPCR法を用い、そしてデータを非処理対照に対する割合として表した。AdLMP−1処理(MOI 25)後に、アグリカンのmRNAレベルは、対照の2.1±0.1(p<0.01)倍であった。AdLacZ処理(MOI 25)後に、アグリカンのmRNAレベルは変わらず、対照の1.0±0.16倍であった。これらの結果は、sGAGの実験と併せて、LMP−1が椎間板細胞のプロテオグリカン産生を刺激することを、実証した。
【0078】
最適なAdLMP−1活性に必要な用量及びタイミングを培養系において確立したことから、次いで、発明者は、AdLMP−1によって投与されるLMP−1が線維輪細胞及び髄核細胞に及ぼす影響を、比較した。ラット線維輪細胞及び髄核細胞を、MOI 25のAdLMP−1で処理し、そして各細胞種のsGAG濃度を、第6日に測定した。sGAG濃度を、細胞数の軽微な変化を明らかにするために、DNA含量によって正規化した。線維輪細胞及び髄核細胞種の非処理対照を比較するため、結果を、非処理対照を基準に正規化しなかった。非処理の線維輪細胞及び髄核細胞の結果は、それぞれ0.6±0.04 sGAG/DNA、及び1.0±0.12 sGAG/DNAであった。この差は統計学的に有意であり(p<0.05)、そして、非処理髄核細胞が、線維輪細胞よりも一細胞につきより多くのsGAGを産生することを、示唆した。AdLMP−1処理後、線維輪細胞及び髄核細胞の結果は、それぞれ1.5±0.08 sGAG/DNA(p<0.01)、及び2.4±0.1 sGAG/DNA(p<0.01)であった。これらの値は、そのそれぞれの非処理対照と比べて、統計学的に有意な増加を示した。非処理細胞からAdLMP−1処理細胞への相対的な増加は、線維輪細胞と髄核細胞との間で同様であった。第6日におけるAdLMP−1処理及び非処理の細胞数を、DNA含量を測定することによって決定した。線維輪細胞のDNA含量は、非処理対照と比べて1.2倍増加したが(p<0.01)、しかし、髄核細胞のDNA含量は、AdLMP−1による影響を受けなかった。このことは、LMP−1が、軽度であるがしかし有意な線維輪細胞数の増加を誘導するが、髄核細胞では誘導しないことを、示唆した。
【0079】
LMP−1の投与(AdLMP−1による)がin vitroにおけるsGAG産生に及ぼす影響を、数週間の期間、試験するために、発明者は、アルギン酸培養系を用いた。アルギン酸は、in vitro培養液中で長期間、軟骨細胞の表現型を維持するのに重要な細胞の三次元マトリックスを、提供する。単層で増殖させた線維輪細胞を、MOI 25のAdLMP−1で処理し、次いで24時間後にアルギン酸培養液に移した。細胞を、1、2及び3週間の期間、培養した。1週間後に、AdLMP−1群におけるsGAGレベルは、非処理対照の1.5±0.06(p<0.01)倍であった。2週間後に、AdLMP−1処理群におけるsGAGレベルは、非処理対照の2.9±0.3(p<0.01)倍に増加した。この差は、3週間後において維持され;AdLMP−1処理群におけるsGAGレベルは、対照の2.9±0.1(p<0.01)倍であった。このことは、AdLMP−1が、少なくとも3週間、培養液中で、アルギン酸中のsGAG増加を維持するのに効果的であることを、示唆した。
【0080】
プロテオグリカン産生に対するLMP−1過剰発現の効果を実証したことから、我々は、LMP−1の過剰発現がこの効果を誘導するメカニズムを検討した。過剰発現LMP−1及びBMP(BMP−2、4、6及び7)のmRNAの変化の時間経過を、測定した。これらのBMPは、白血球及び骨芽細胞において、LMP−1の過剰発現によって刺激されることが、これまでに示されていた。
【0081】
線維輪細胞をMOI 25のAdLMP−1で処理後の、過剰発現LMP−1の時間依存的な変化を、測定した。データを、非処理対照に対する割合としてでなく、過剰発現LMP−1の最大mRNAレベル(第6日)の百分率として表したが、これは、対照において過剰発現LMP−1のmRNAが検出可能でなかったためである。LMP−1のmRNAは、処理後12時間に検出可能であり、そして調査した最終時点(第6日)まで増加し続けた。このことは、過剰発現LMP−1が、LMP−1投与後早ければ12時間後には下流の遺伝子に影響を与え得ることを、示唆した。
【0082】
MOI 25のAdLMP−1で処理後の、BMPのmRNAレベルの時間経過も、リアルタイムPCRによって測定し、そして各時点において非処理対照に対する割合として算出した。BMP−2のmRNAは早期に亢進し、LMP−1処理後12時間に、統計学的に有意な(p<0.01)増加に達した。BMP−2のmRNA増加は、第3日までにプラトーレベルに達した。BMP−7のmRNAはBMP−2よりも遅れて亢進し、第3日(p<0.01)及び第6日(p<0.01)に、統計学的に有意な増加に達した。BMP−4及びBMP−6のmRNAレベルには、いずれも非処理対照のものと有意な差はなかった。
【0083】
LMP−1のmRNAの過剰発現が、線維輪細胞においてBMP−2及びBMP−7の亢進をもたらすことを確立したことから、発明者は、このmRNA増加が、BMPタンパク質の培養液への分泌増加と相関するかどうかを検討した。ELISAアッセイを、調製された培地中のBMP−2、4、6及び7タンパク質レベルを定量するために行った。単層にて培養された線維輪細胞を、MOI 25のAdLMP−1で処理した。培地を第3日に一度交換し、そして培地を第6日に解析した。したがって、被験試料は、培養最終3日間(第4〜6日)に分泌されたBMPを含有した。培地中のBMP−2及びBMP−7のタンパク質レベルは、それぞれ、非処理対照の3.5±0.4(p<0.01)倍及び2.5±0.3(p<0.05)倍であった。BMP−4及びBMP−6のタンパク質レベルには、非処理対照のものと有意な差はなく、このことは、mRNAについて得られた結果と一致した。
【0084】
BMP−2及びBMP−7のmRNA及びタンパク質が、LMP−1によって亢進したことから、発明者は、これらのBMPの遮断が、LMP−1によって誘導されるsGAG産生増加を妨げるかどうかを、検討した。単層においてMOI 25のAdLMP−1とともに培養された線維輪細胞を、BMP阻害剤であるノギンで、培地中にて処理し、そしてsGAGレベルを第6日に測定した。AdLMP−1のみで処理された細胞では、sGAGレベルが増加したが(非処理対照の2.7±0.3倍)、3200ng/mlのノギン及びMOI 25のAdLMP−1で処理された細胞では、sGAGレベルは変わらず(非処理対照の1.1±0.1倍)、このことは、ノギンがAdLMP−1の影響を完全に遮断することを示唆した。ノギン(3200ng/ml)のみで処理された細胞では、sGAGレベルはほとんど変わらないままであり(非処理対照の0.8±0.1倍)、このことは、ノギンによる毒性的影響がないことを示唆した。AdLMP−1によって誘導されるsGAG産生に及ぼすノギンの阻害的影響は、濃度依存性であった。
【0085】
対照の椎間板(AdGFPを注射された椎間板)におけるアグリカン、BMP−7及びLMP−1のmRNAの内因性レベルを、髄核において検出した。内因性mRNAレベルを用いて、AdLMP−1を注射された椎間板におけるアグリカン、BMP−7及びLMP−1のmRNAの増加を、算出した。
【0086】
AdLMP−1を注射された椎間板は、AdGFPを注射された椎間板よりも、総LMP−1のmRNAを、830%高いレベルで発現した。AdLMP−1によるLMP−1の投与は、対照群よりも1100%、BMP−7のmRNAレベルの増加(p<0.05)をもたらした。アグリカンのmRNAレベルは、対照群よりも66%増加した(p<0.05)。
【0087】
BMP−2、BMP−7、LMP−1及びアグリカンのmRNAの内因性レベルも、検出した。AdLMP−1の用量増加と総LMP−1のmRNAとの相関がみられた。AdLMP−1によるLMP−1の投与は、107pfu/椎間板の用量で最大限に、BMP−2及びBMP−7のmRNAレベルを有意に増加させた(p<0.05)。107pfu/椎間板の用量のAdLMP−1は、対照群より50%と(p<0.05)、アグリカンのmRNAを最も増加させた。
【0088】
LMP−1の椎間板細胞への投与は、sGAGの産生増加及びアグリカンのmRNAレベルの増加をもたらした。LMP−1の投与は、in vitroにおいてBMP−2及びBMP7のmRNAならびにタンパク質レベルの増加をもたらし、そしてLMP−1の投与がプロテオグリカン産生亢進に及ぼす影響を、BMP阻害剤であるノギンの投与によって遮断することが可能であった。
【0089】
11ヶ月齢のSprague−Dawleyラットを屠殺し、そして腰椎及び尾から得た椎間板組織を、無菌条件下で回収した。線維輪及び髄核を、別々に、解剖しそして小さい角切りにした。椎間板組織を、100単位/mlペニシリン及び100mg/mlストレプトマイシンを含有する、ダルベッコ変法イーグル培地及びハムF12培地(DMEM/F−12;ギブコBRL社、グランドアイランド、ニューヨーク州、米国)中に入れた。椎間板組織を、培地中で、0.2%プロナーゼ(シグマケミカル社、セントルイス、ミズーリ州、米国)を用いて37℃にて1時間処理し、次いで、0.025%コラゲナーゼ(シグマケミカル社、セントルイス、ミズーリ州、米国)を用いて37℃にて6時間処理した。単離された細胞を洗浄し、そして70mmメッシュ(ファルコン社、フランクリンレイクス、ニュージャージー州、米国)を介してろ過し、10%胎児ウシ血清(FBS)、100単位/mlペニシリン、100mg/mlストレプトマイシン、2mM L−グルタミン及び50mg/mlアスコルビン酸塩を含有するDMEM/F12培地12mlを入れた75cm2フラスコに、加えた。細胞を、加湿された5%CO2中で37℃にて、増殖させた。培養液を、およそ8日間、2日毎に交換した。
【0090】
二つの異なるウイルスを用いた。CMVプロモーターによって駆動される、ヒトLMP−1 cDNAを含有する複製欠損性5型アデノウイルス(AdLMP−1)を、実験用ウイルスとして用いた。対照ウイルスは、lacZのcDNAを含有する同様な複製欠損性5型アデノウイルスからなった。
【0091】
椎間板細胞の初代培養物がコンフルエントになったとき、細胞を、400,000細胞/ウェルにて6ウェルプレートに継代培養した。3日後に、細胞を、ヒトLMP−1遺伝子またはLacZ遺伝子のいずれかのcDNAを含有するアデノウイルス(それぞれ、AdLMP−1とAdLacZ)で、処理した。細胞数を、血球計数器を用いて対照ウェルをカウントすることによって、第0日に測定した。ウイルス用量を、感染多重度(MOI)、すなわちプラーク形成単位(pfu)/細胞の数として、表した。これは、本質的には、一つの椎間板細胞が曝露されたリコンビナントアデノウイルスのプラーク形成単位の数である。培養された細胞を、各実験において指定されたように、異なるMOI(0、10、25、50)のAdLMP−1またはAdLacZを用いて、300mlの0%FBS添加DMEM/F−12中で37℃にて30分間処理した。次いで、培養容量を、1%FBS、100単位/mlペニシリン、100mg/mlストレプトマイシン、2mM L−グルタミン及び50mg/mlビタミンCを含有するDMEM/F−12培地を用いて、2.0mlに上昇させた。培地を、実験期間中、3日毎に交換した。
【0092】
培養液の硫酸化グリコサミノグリカン(sGAG)含量を、1,9−ジメチルメチレンブルー(DMMB)法を用いてアッセイした。培養液2mlを、セントリコンYM−50遠心フィルター(ミリポア・Co.、ベッドフォード、マサチューセッツ州、米国)を用いて、遠心分離し(5000xGで30分間)、sGAGを濃縮した。試料溶液(20ml)を、96ウェルのマイクロタイタープレート中で、200mlのDMMB色素溶液と穏やかに混合し、そして光学濃度(OD)を、520nm波長フィルターにて直ちに検査した。標準曲線を、硫酸コンドロイチン(シグマケミカル社、セントルイス、ミズーリ州、米国)の連続希釈系列を用いて、作図した。培地中の総sGAGを、DNA含量によって正規化し、そして非処理対照に対する割合として表した。
【0093】
細胞数を、各ウェルのDNA含量によって決定し、そして、先に記載したように、DNA含量を、ヘキスト色素33258(Polysciences社、Warrington、ペンシルバニア州、米国)法を用いて測定した。培養された細胞を、パパイン(10単位/ml)に曝露することによって、プレートから除いた。次いで、培養された細胞をペレット状にし、そして60℃にて3時間インキュベートした。20μlのパパイン消化物のアリコットを、200mlのヘキスト色素33258溶液と、96ウェルのフルオロプレート中で混合した。発光及び励起スペクトルを、ルミネセンス分光器LS 50B(パーキン−エルマー社、Wellesly、マサチューセッツ州、米国)において、それぞれ456nm及び365nmにて測定した。標準曲線を、子ウシ胸腺DNA(シグマケミカル社、セントルイス、ミズーリ州、米国)の既知の濃度を用いて、各測定時点について作製した。
【0094】
アルギン酸ビーズ培養は、長期培養において軟骨細胞の表現型を維持するのに有用である。この方法は、3週間の培養においてAdLMP−1の影響を判定するためのものである。細胞を、上記のように、単層培養において処理した。1日後に、細胞を、トリプシン処理によって剥離させ、そして培地で2回洗浄した。単離された細胞を、0.6%低粘度無菌アルギン酸塩(シグマケミカル社、セントルイス、ミズーリ州、米国)溶液中に、600,000細胞/mlにて再懸濁した。アルギン酸ビーズを形成させるため、細胞を、10mlプラスチックシリンジに取り付けた21ゲージ針を介して、1滴ずつ、0.102M CaCl+0.15M NaCl溶液中に加えた。10分後に、新たに形成されたビーズ(およそ12,000細胞/ビーズを含有)を、無菌0.9%NaCl溶液で3回、続いてDMEM/F−12で2回、洗浄した。線維輪細胞を含有するビーズを、1%FBS、100単位/mlペニシリン、100mg/mlストレプトマイシン、2mM L−グルタミン及び50mg/mlビタミンCを含有するDMEM/F−12培地で、6ウェルプレートにおいて別々に培養した。培地を、異なる期間中(1、2及び3週間)、2日毎に交換した。アルギン酸ビーズを、350mlのクエン酸ナトリウムバッファー(55mmol/L Na−クエン酸、50mmol/L EDTA、150mmol/L NaCl、pH7.4)に溶解した。細胞を、遠心分離によってペレット状にし、そして溶解液中のsGAG含量を、上記のDMMB法で測定した。細胞ペレットに残存するsGAG含量は、懸濁液中のものと比べて、無視できるほどであった。結果を、溶解液のsGAGを用いて、非処理対照群の倍数増加として記載した。
mRNAレベルの定量
リアルタイムPCRを用いて、BMP−2、BMP−4、BMP−6、BMP−7及び過剰発現LMP−1のmRNAレベルを、定量的に測定した。遺伝子のすべてのプライマーを、アガロースゲル上で産物のサイズを決定することによって、そしてアンプリコンをDNA配列決定することによって、確認した。18Sのレベルを各試料において測定し、内在性対照として用いた。
【0095】
各試料の総RNAを、グラニジウム・チオシアネート−フェノール−クロロホルム技術を用いた単一工程の方法によって、抽出した。単離RNAの濃度を、分光光度計(DU−500;ベックマン社、フラトン、カリフォルニア州、米国)を用いて、260nm波長にて測定した。RNAを、DNA分解酵素1(アンビオン・Inc.、テキサス州、米国)で処理して、試料のDNA混入を除去した。逆転写を、総RNA2mg;トリ骨髄芽球症ウイルス逆転写酵素(プロメガ社、マディソン、ウィスコンシン州、米国)30U;デオキシアデノシン三リン酸(dATP)、デオキシシチジン三リン酸(dCTP)、デオキシグアニジン三リン酸(dGTP)、デオキシチミジン三リン酸(dTTP)各1mM;及び、オリゴ(dT)15プライマー1mgを用いて、42℃にて45分間、40ml容量中で行った。PCRを、Amplitaq(商標登録)DNAポリメラーゼを用いて、30サイクル(95℃、30秒間;62℃、30秒間;72℃、45秒間)行った。DNAの混入がないことを確認するため、逆転写酵素非存在下で処理したRNA試料も、PCRにかけ:PCR産物がないことで、DNA混入していないと確認した。
【0096】
リアルタイムPCRは、迅速で、信頼性があり、そして再現可能な、特定のmRNAの定量性検出法であると、報告されている。SYBR(商標登録)グリーン・リアルタイムPCRキット(Applied Biosystems社、フォスターシティ、カリフォルニア州、米国)を用いたリアルタイムPCR法を用いて、BMP−2、BMP−4、BMP−6、BMP−7及びアグリカンの定量性mRNA解析を行った。25マイクロリットル(25ml)の反応容量は、cDNA 5ml、各プライマー(BMP−2、−4、−6、−7、及び18S)3.75ピコモル、及びSYBR(商標登録)グリーン・マスターミックス(2x、バイオラッド社、ヘラクレス、カリフォルニア州、米国)を含んだ。過剰発現LMP−1及び18SのmRNAレベルを定量するために、TaqMan(商標登録)リアルタイムPCRキット(Applied Biosystems社、フォスターシティ、カリフォルニア州、米国)を用いたリアルタイムPCR法も、行った。25マイクロリットル(25ml)の反応容量には、cDNA 5ml、各プライマー3.75ピコモル、及びTaqMan(商標登録)PCRマスターミックス(2x、バイオラッド社、ヘラクレス、カリフォルニア州、米国)が含まれた。
【0097】
リアルタイムPCRを、GeneAmp(商標登録);5700配列検出システム(Applied Biosystems社、フォスターシティ、カリフォルニア州、米国)を用いて、以下の3工程のプロトコールで行った;工程1:50℃にて2分間、工程2:95℃にて10分間、及び、工程3:(95℃にて15秒間、60℃にて1分間)x45サイクル。増幅の特異性を確認するため、PCR産物を、転移曲線解析にかけた。先に記載のように、各反応のthreshold cycle(Ct)を、競合的_ΔΔCt法を用いて、18Sを基準として標準化した。
【0098】
BMP2、4、6及び7のELISAアッセイ
BMPの標準曲線を、0.05mol/L重炭酸バッファーに溶解した、ヒトBMP2、4、6及び7(Genetics Institute、ケンブリッジ、マサチューセッツ州)の漸増させた濃度(0.1ng/100μL/ウェル〜1000ng/100μL/ウェル)を用いて、作図した。100マイクロリットルの試料を、トリプリケートで各ウェルに加えた。4℃にて一晩インキュベートした後、プレートを、0.5%Tween 20(PBST)添加0.01Mリン酸緩衝生理食塩液で3回洗浄し、そして非反応部位を、1%ウシアルブミン(シグマ社、セントルイス、ミズーリ州)を用いて、室温にて1時間ブロッキングした。プレートをPBSTで洗浄後に、一次抗体(1:1000)を100マイクロリットルにて各ウェルに分注し、そして室温にて2時間インキュベートした。BMP2、4及び6に対するヤギポリクローナル抗体(サンタクルーズ・Inc、サンタクルーズ、カリフォルニア州)、ならびBMP7に対するウサギ抗体(シグマ社、セントルイス、ミズーリ州)が、用いられた。プレートをPBSTで洗浄し、次いで、アルカリホスファターゼ結合抗ヤギIgG及び抗ウサギIgG(シグマ社、セントルイス、ミズーリ州)を用いて、室温にて1時間、それぞれインキュベートした。基質であるp−ニトロフェニルリン酸塩(シグマ社、セントルイス、ミズーリ州)を用いて発色させ、20分後に3N NaOHで反応を止めた。呈色を、Elx 800−マイクロプレートリーダー(バイオ−テック・インスツルメント社、Winooska、バーモント州)を用いて405nmにて吸光度差を測定することによって、定量した。
【0099】
結果を定量化するため、線形回帰プロットを、各標準品について作製した。すべての場合において、試料濃度を、標準品と同じ吸光度の対応値に準じて、標準品の線形回帰プロットから外挿した。
【0100】
ノギン糖タンパク質によるBMPの阻害
ノギンは、BMP−2、4、6及び7と高特異的に結合し、そしてこれらのBMPがその同族受容体を活性化させることを妨げる、糖タンパク質である。マウスノギン(ノギン−FC・シグマケミカル社、セントルイス、ミズーリ州、米国)の形態を実験において用い、AdLMP−1処理後にBMPを特異的に遮断する影響を、判定した。異なるノギン濃度(100、200、400、800、1600及び3200ng/ml)を、AdLMP−1(MOI 25)処理後第0日及び第3日に、細胞に載せた。第6日に、上記のDMMB法を用いて、条件培地を、sGAGの産生を検討するためにアッセイした。結果は、LMP−1の影響が、ノギンの投与によって遮断され得ることを示唆し、このことは、LMP−1活性が、これによるBMPの誘導に部分的に関連することを、実証した。
【図面の簡単な説明】
【0101】
【図1】図1は、線維輪細胞をLMP−1(MOI 25のAdLMP−1)で処理後6日の、培地中BMPタンパク質レベル(量)のグラフである。BMP−2及びBMP−7のタンパク質レベルは有意に増加したが、しかしBMP−4及びBMP−6のタンパク質レベルは、非処理対照細胞の培地と有意な差がなかった。各結果を、非処理細胞の値に比例した割合として表す。7試料の平均値及び標準誤差を、報告する(*p<0.05 **p<0.01 vs.NT)
【図2】図2は、線維輪細胞をLMP−1(MOI 25のAdLMP−1)で処理後の、経時的なBMPのmRNAレベルを示す。BMP−2のmRNAレベルは、AdLMP−1で処理後早ければ12時間に有意に亢進し、そしてAdLMP−1処理後3日にプラトーに達した。BMP−7のmRNAレベルは、AdLMP−1で処理後3日に、有意に増加した。各結果を、同時点における非処理細胞の値に対する割合として表す。6試料の平均値及び標準誤差を、報告する(対応する時点について、**p<0.01 vs.NT)
【図3】図3は、リアルタイムPCRによって測定された、LMP−1(AdLMP−1)で処理後の線維輪細胞中のアグリカンmRNAレベルを、グラフに描いたものである。アグリカンのmRNAレベルは、MOI 25のAdLMP−1で処理後6日に、非処理細胞と比べて有意に増加した。アグリカンのmRNAレベルは、AdLacZで処理された細胞において、非処理細胞と比べて変わらなかった。各結果を、非処理細胞の値に対する割合として、表す。9試料の平均値及び標準誤差を、報告する。AdLMP−1:MOI 25、AdLacZ:MOI 25(**p<0.01 vs.NT)。
【図4】図4は、Harlan無胸腺ラットにおける、PTD/LMP−1融合タンパク質によるLMP−1投与のパラメータ及び結果を要約した表である。
【図5】図5は、ニュージーランドホワイトウサギにおける、PTD/LMP−1融合タンパク質によるLMP−1投与のパラメータ及び結果を要約した表である。
【図6】図6は、Harlan無胸腺ラット、及び示された箇所ではニュージーランドホワイトウサギにおける、PTD/ペプチド融合タンパク質による、示された骨誘導ペプチド投与の結果を要約した表である。骨成長は、X線によって検出され、そして、示された箇所では触診も可能であった。
【特許請求の範囲】
【請求項1】
細胞透過性骨誘導ポリペプチドを作製する方法であって、
a) 細胞透過性ポリペプチドをコードするポリヌクレオチドと;
b) 該細胞透過性ポリペプチドと機能可能に連結されている骨誘導ポリペプチドをコードするポリヌクレオチドであって、該骨誘導ポリペプチドが、該細胞透過性ポリペプチドとの融合タンパク質の一部として発現するように配置されたポリヌクレオチドと;
c) 該ポリヌクレオチドの転写を導くように配置されたプロモーターと;
を含む発現コンストラクトを、適切な宿主細胞中に導入することを含む、上記方法。
【請求項2】
発現コンストラクトがさらに精製タグを含む、請求項1に記載の方法。
【請求項3】
細胞透過性ポリペプチドが、HIV−TAT、VP−22、増殖因子のシグナルペプチド配列、Pep−1及びショウジョウバエAntpペプチドからなる群より選択される、請求項1に記載の方法。
【請求項4】
細胞透過性ポリペプチドが、HIV−TATタンパク質導入ドメインである、請求項1に記載の方法。
【請求項5】
骨誘導ポリペプチドが、LMP−1、LMP−3、配列番号1、配列番号2、配列番号3、配列番号4、配列番号5、配列番号6、配列番号7、配列番号8、BMP−2、BMP−4、BMP−6、BMP−7、TGF−β1及びSmadからなる群より選択される、請求項1に記載の方法。
【請求項6】
骨誘導ポリペプチドが、LMP−1、LMP−3、配列番号1、配列番号2、配列番号3、配列番号4、配列番号5、配列番号6、配列番号7及び配列番号8からなる群より選択される、請求項1に記載の方法。
【請求項7】
哺乳動物において骨形成を誘導する方法であって、タンパク質導入ドメインと少なくとも一つの骨誘導ポリペプチドとを含む、有効量の融合ポリペプチドを投与することを含む、上記方法。
【請求項8】
タンパク質導入ドメインが、HIV−TAT、VP−22、増殖因子のシグナルペプチド配列、Pep−1及びショウジョウバエAntpペプチドからなる群より選択される、請求項7に記載の方法。
【請求項9】
タンパク質導入ドメインが、HIV−TATタンパク質導入ドメインである、請求項7に記載の方法。
【請求項10】
骨誘導ポリペプチドが、LMP−1、LMP−3、配列番号1、配列番号2、配列番号3、配列番号4、配列番号5、配列番号6、配列番号7、配列番号8、BMP−2、BMP−4、BMP−6、BMP−7、TGF−β1及びSmadからなる群より選択される、請求項7に記載の方法。
【請求項11】
骨誘導ポリペプチドが、LMP−1、LMP−3、配列番号1、配列番号2、配列番号3、配列番号4、配列番号5、配列番号6、配列番号7及び配列番号8からなる群より選択される、請求項7に記載の方法。
【請求項12】
融合ポリペプチドが、移植片として投与される、請求項7に記載の方法。
【請求項13】
融合ポリペプチドが、ヒドロゲルによって投与される、請求項7に記載の方法。
【請求項14】
融合ポリペプチドが、少なくとも一つの多能性前駆細胞に投与される、請求項7に記載の方法。
【請求項15】
少なくとも一つの多能性前駆細胞が、哺乳動物内に植え込まれる、請求項14に記載の方法。
【請求項16】
タンパク質導入ドメインと少なくとも一つの骨誘導ポリペプチドとを含む融合タンパク質をコードする、ポリヌクレオチド。
【請求項17】
タンパク質導入ドメインが、HIV−TAT、VP−22、増殖因子のシグナルペプチド配列、Pep−1及びショウジョウバエAntpペプチドからなる群より選択される、請求項16に記載のポリヌクレオチド。
【請求項18】
タンパク質導入ドメインが、HIV−TATタンパク質導入ドメインである、請求項16に記載のポリヌクレオチド。
【請求項19】
骨誘導ポリペプチドが、LMP−1、LMP−3、配列番号1、配列番号2、配列番号3、配列番号4、配列番号5、配列番号6、配列番号7、配列番号8、BMP−2、BMP−4、BMP−6、BMP−7、TGF−β1及びSmadからなる群より選択される、請求項16に記載のポリヌクレオチド。
【請求項20】
骨誘導ポリペプチドが、LMP−1、LMP−3、配列番号1、配列番号2、配列番号3、配列番号4、配列番号5、配列番号6、配列番号7及び配列番号8からなる群より選択される、請求項16に記載のポリヌクレオチド。
【請求項21】
哺乳動物においてプロテオグリカンの合成を誘導する方法であって、タンパク質導入ドメインと少なくとも一つの骨誘導ポリペプチドとを含む有効量の融合ポリペプチドを投与することを含む、方法。
【請求項22】
タンパク質導入ドメインが、HIV−TAT、VP−22、増殖因子のシグナルペプチド配列、Pep−1及びショウジョウバエAntpペプチドからなる群より選択される、請求項21に記載の方法。
【請求項23】
タンパク質導入ドメインが、HIV−TATタンパク質導入ドメインである、請求項21に記載の方法。
【請求項24】
骨誘導ポリペプチドが、LMP−1、LMP−3、配列番号1、配列番号2、配列番号3、配列番号4、配列番号5、配列番号6、配列番号7、配列番号8、BMP−2、及びBMP−7からなる群より選択される、請求項21に記載の方法。
【請求項25】
骨誘導ポリペプチドが、LMP−1、LMP−3、配列番号1、配列番号2、配列番号3、配列番号4、配列番号5、配列番号6、配列番号7及び配列番号8からなる群より選択される、請求項21に記載のポリヌクレオチド。
【請求項26】
融合ポリペプチドが、移植片として投与される、請求項21に記載の方法。
【請求項27】
融合ポリペプチドが、ヒドロゲルによって投与される、請求項21に記載の方法。
【請求項28】
融合ポリペプチドが、少なくとも一つの多能性前駆細胞に投与される、請求項21に記載の方法。
【請求項29】
少なくとも一つの多能性前駆細胞が、哺乳動物内に植え込まれる、請求項21に記載の方法。
【請求項30】
プロテオグリカンがアグリカンである、請求項21に記載の方法。
【請求項31】
骨誘導ポリペプチドと機能可能に連結されている膜移行ペプチドを含む、単離された融合ポリペプチド。
【請求項32】
膜移行ペプチドが、HIV−TAT、VP−22、増殖因子のシグナルペプチド配列、Pep−1及びショウジョウバエAntpペプチドからなる群より選択される、請求項31に記載の方法。
【請求項33】
膜移行ペプチドが、HIV−TATタンパク質導入ドメインである、請求項31に記載の方法。
【請求項34】
骨誘導ポリペプチドが、LMP−1、LMP−3、配列番号1、配列番号2、配列番号3、配列番号4、配列番号5、配列番号6、配列番号7、配列番号8、BMP−2、BMP−4、BMP−6、BMP−7、TGF−β1及びSmadからなる群より選択される、請求項31に記載のポリヌクレオチド。
【請求項35】
骨誘導ポリペプチドが、LMP−1、LMP−3、配列番号1、配列番号2、配列番号3、配列番号4、配列番号5、配列番号6、配列番号7及び配列番号8からなる群より選択される、請求項31に記載のポリヌクレオチド。
【請求項36】
前駆細胞において骨芽細胞の分化を誘導する方法であって、タンパク質導入ドメインと少なくとも一つの骨誘導ポリペプチドとを含む有効量の融合ポリペプチドを前駆細胞に投与することを含む、上記方法。
【請求項37】
タンパク質導入ドメインが、HIV−TAT、VP−22、増殖因子のシグナルペプチド配列、Pep−1及びショウジョウバエAntpペプチドからなる群より選択される、請求項36に記載の方法。
【請求項38】
タンパク質導入ドメインが、HIV−TATタンパク質導入ドメインである、請求項36に記載の方法。
【請求項39】
骨誘導ポリペプチドが、LMP−1、LMP−3、配列番号1、配列番号2、配列番号3、配列番号4、配列番号5、配列番号6、配列番号7、配列番号8、BMP−2、BMP−4、BMP−6、BMP−7、TGF−β1及びSmadからなる群より選択される、請求項36に記載の方法。
【請求項40】
骨誘導ポリペプチドが、LMP−1、LMP−3、配列番号1、配列番号2、配列番号3、配列番号4、配列番号5、配列番号6、配列番号7及び配列番号8からなる群より選択される、請求項36に記載の方法。
【請求項41】
配列番号1、配列番号2、配列番号3、配列番号4、配列番号5、配列番号6、配列番号7及び配列番号8、またはその組み合わせからなる群の中から選択される、骨誘導ポリペプチド。
【請求項42】
標準的な条件下で、配列:
tcctcatccg ggtcttgcat gaactcggtg
に相補的な核酸分子とハイブリダイズする、骨誘導ポリペプチド。
【請求項43】
高ストリンジェントな条件下で、配列:
gcccccgccc gctgacagcg ccccgcaa
に相補的な核酸分子とハイブリダイズする、骨誘導ポリペプチド。
【請求項1】
細胞透過性骨誘導ポリペプチドを作製する方法であって、
a) 細胞透過性ポリペプチドをコードするポリヌクレオチドと;
b) 該細胞透過性ポリペプチドと機能可能に連結されている骨誘導ポリペプチドをコードするポリヌクレオチドであって、該骨誘導ポリペプチドが、該細胞透過性ポリペプチドとの融合タンパク質の一部として発現するように配置されたポリヌクレオチドと;
c) 該ポリヌクレオチドの転写を導くように配置されたプロモーターと;
を含む発現コンストラクトを、適切な宿主細胞中に導入することを含む、上記方法。
【請求項2】
発現コンストラクトがさらに精製タグを含む、請求項1に記載の方法。
【請求項3】
細胞透過性ポリペプチドが、HIV−TAT、VP−22、増殖因子のシグナルペプチド配列、Pep−1及びショウジョウバエAntpペプチドからなる群より選択される、請求項1に記載の方法。
【請求項4】
細胞透過性ポリペプチドが、HIV−TATタンパク質導入ドメインである、請求項1に記載の方法。
【請求項5】
骨誘導ポリペプチドが、LMP−1、LMP−3、配列番号1、配列番号2、配列番号3、配列番号4、配列番号5、配列番号6、配列番号7、配列番号8、BMP−2、BMP−4、BMP−6、BMP−7、TGF−β1及びSmadからなる群より選択される、請求項1に記載の方法。
【請求項6】
骨誘導ポリペプチドが、LMP−1、LMP−3、配列番号1、配列番号2、配列番号3、配列番号4、配列番号5、配列番号6、配列番号7及び配列番号8からなる群より選択される、請求項1に記載の方法。
【請求項7】
哺乳動物において骨形成を誘導する方法であって、タンパク質導入ドメインと少なくとも一つの骨誘導ポリペプチドとを含む、有効量の融合ポリペプチドを投与することを含む、上記方法。
【請求項8】
タンパク質導入ドメインが、HIV−TAT、VP−22、増殖因子のシグナルペプチド配列、Pep−1及びショウジョウバエAntpペプチドからなる群より選択される、請求項7に記載の方法。
【請求項9】
タンパク質導入ドメインが、HIV−TATタンパク質導入ドメインである、請求項7に記載の方法。
【請求項10】
骨誘導ポリペプチドが、LMP−1、LMP−3、配列番号1、配列番号2、配列番号3、配列番号4、配列番号5、配列番号6、配列番号7、配列番号8、BMP−2、BMP−4、BMP−6、BMP−7、TGF−β1及びSmadからなる群より選択される、請求項7に記載の方法。
【請求項11】
骨誘導ポリペプチドが、LMP−1、LMP−3、配列番号1、配列番号2、配列番号3、配列番号4、配列番号5、配列番号6、配列番号7及び配列番号8からなる群より選択される、請求項7に記載の方法。
【請求項12】
融合ポリペプチドが、移植片として投与される、請求項7に記載の方法。
【請求項13】
融合ポリペプチドが、ヒドロゲルによって投与される、請求項7に記載の方法。
【請求項14】
融合ポリペプチドが、少なくとも一つの多能性前駆細胞に投与される、請求項7に記載の方法。
【請求項15】
少なくとも一つの多能性前駆細胞が、哺乳動物内に植え込まれる、請求項14に記載の方法。
【請求項16】
タンパク質導入ドメインと少なくとも一つの骨誘導ポリペプチドとを含む融合タンパク質をコードする、ポリヌクレオチド。
【請求項17】
タンパク質導入ドメインが、HIV−TAT、VP−22、増殖因子のシグナルペプチド配列、Pep−1及びショウジョウバエAntpペプチドからなる群より選択される、請求項16に記載のポリヌクレオチド。
【請求項18】
タンパク質導入ドメインが、HIV−TATタンパク質導入ドメインである、請求項16に記載のポリヌクレオチド。
【請求項19】
骨誘導ポリペプチドが、LMP−1、LMP−3、配列番号1、配列番号2、配列番号3、配列番号4、配列番号5、配列番号6、配列番号7、配列番号8、BMP−2、BMP−4、BMP−6、BMP−7、TGF−β1及びSmadからなる群より選択される、請求項16に記載のポリヌクレオチド。
【請求項20】
骨誘導ポリペプチドが、LMP−1、LMP−3、配列番号1、配列番号2、配列番号3、配列番号4、配列番号5、配列番号6、配列番号7及び配列番号8からなる群より選択される、請求項16に記載のポリヌクレオチド。
【請求項21】
哺乳動物においてプロテオグリカンの合成を誘導する方法であって、タンパク質導入ドメインと少なくとも一つの骨誘導ポリペプチドとを含む有効量の融合ポリペプチドを投与することを含む、方法。
【請求項22】
タンパク質導入ドメインが、HIV−TAT、VP−22、増殖因子のシグナルペプチド配列、Pep−1及びショウジョウバエAntpペプチドからなる群より選択される、請求項21に記載の方法。
【請求項23】
タンパク質導入ドメインが、HIV−TATタンパク質導入ドメインである、請求項21に記載の方法。
【請求項24】
骨誘導ポリペプチドが、LMP−1、LMP−3、配列番号1、配列番号2、配列番号3、配列番号4、配列番号5、配列番号6、配列番号7、配列番号8、BMP−2、及びBMP−7からなる群より選択される、請求項21に記載の方法。
【請求項25】
骨誘導ポリペプチドが、LMP−1、LMP−3、配列番号1、配列番号2、配列番号3、配列番号4、配列番号5、配列番号6、配列番号7及び配列番号8からなる群より選択される、請求項21に記載のポリヌクレオチド。
【請求項26】
融合ポリペプチドが、移植片として投与される、請求項21に記載の方法。
【請求項27】
融合ポリペプチドが、ヒドロゲルによって投与される、請求項21に記載の方法。
【請求項28】
融合ポリペプチドが、少なくとも一つの多能性前駆細胞に投与される、請求項21に記載の方法。
【請求項29】
少なくとも一つの多能性前駆細胞が、哺乳動物内に植え込まれる、請求項21に記載の方法。
【請求項30】
プロテオグリカンがアグリカンである、請求項21に記載の方法。
【請求項31】
骨誘導ポリペプチドと機能可能に連結されている膜移行ペプチドを含む、単離された融合ポリペプチド。
【請求項32】
膜移行ペプチドが、HIV−TAT、VP−22、増殖因子のシグナルペプチド配列、Pep−1及びショウジョウバエAntpペプチドからなる群より選択される、請求項31に記載の方法。
【請求項33】
膜移行ペプチドが、HIV−TATタンパク質導入ドメインである、請求項31に記載の方法。
【請求項34】
骨誘導ポリペプチドが、LMP−1、LMP−3、配列番号1、配列番号2、配列番号3、配列番号4、配列番号5、配列番号6、配列番号7、配列番号8、BMP−2、BMP−4、BMP−6、BMP−7、TGF−β1及びSmadからなる群より選択される、請求項31に記載のポリヌクレオチド。
【請求項35】
骨誘導ポリペプチドが、LMP−1、LMP−3、配列番号1、配列番号2、配列番号3、配列番号4、配列番号5、配列番号6、配列番号7及び配列番号8からなる群より選択される、請求項31に記載のポリヌクレオチド。
【請求項36】
前駆細胞において骨芽細胞の分化を誘導する方法であって、タンパク質導入ドメインと少なくとも一つの骨誘導ポリペプチドとを含む有効量の融合ポリペプチドを前駆細胞に投与することを含む、上記方法。
【請求項37】
タンパク質導入ドメインが、HIV−TAT、VP−22、増殖因子のシグナルペプチド配列、Pep−1及びショウジョウバエAntpペプチドからなる群より選択される、請求項36に記載の方法。
【請求項38】
タンパク質導入ドメインが、HIV−TATタンパク質導入ドメインである、請求項36に記載の方法。
【請求項39】
骨誘導ポリペプチドが、LMP−1、LMP−3、配列番号1、配列番号2、配列番号3、配列番号4、配列番号5、配列番号6、配列番号7、配列番号8、BMP−2、BMP−4、BMP−6、BMP−7、TGF−β1及びSmadからなる群より選択される、請求項36に記載の方法。
【請求項40】
骨誘導ポリペプチドが、LMP−1、LMP−3、配列番号1、配列番号2、配列番号3、配列番号4、配列番号5、配列番号6、配列番号7及び配列番号8からなる群より選択される、請求項36に記載の方法。
【請求項41】
配列番号1、配列番号2、配列番号3、配列番号4、配列番号5、配列番号6、配列番号7及び配列番号8、またはその組み合わせからなる群の中から選択される、骨誘導ポリペプチド。
【請求項42】
標準的な条件下で、配列:
tcctcatccg ggtcttgcat gaactcggtg
に相補的な核酸分子とハイブリダイズする、骨誘導ポリペプチド。
【請求項43】
高ストリンジェントな条件下で、配列:
gcccccgccc gctgacagcg ccccgcaa
に相補的な核酸分子とハイブリダイズする、骨誘導ポリペプチド。
【図1】


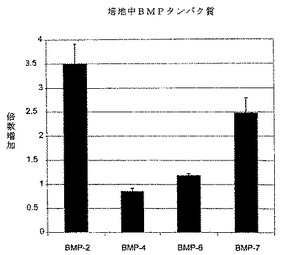
【図2】
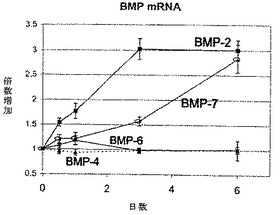
【図3】
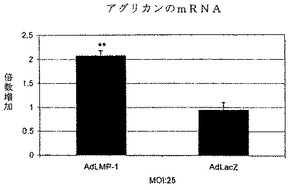
【図4】
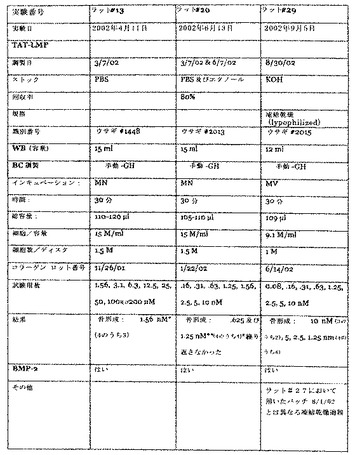
【図5】
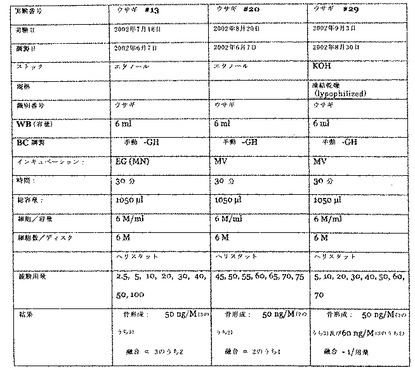
【図6】
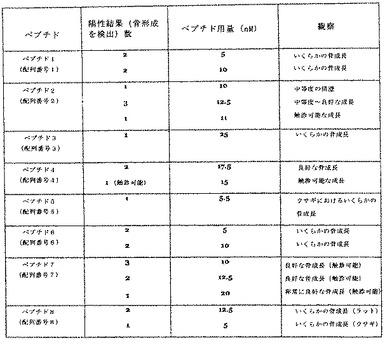
【図2】
【図3】
【図4】
【図5】
【図6】
【公表番号】特表2007−505621(P2007−505621A)
【公表日】平成19年3月15日(2007.3.15)
【国際特許分類】
【出願番号】特願2006−526862(P2006−526862)
【出願日】平成16年4月13日(2004.4.13)
【国際出願番号】PCT/US2004/009127
【国際公開番号】WO2005/111058
【国際公開日】平成17年11月24日(2005.11.24)
【出願人】(504228047)
【Fターム(参考)】
【公表日】平成19年3月15日(2007.3.15)
【国際特許分類】
【出願日】平成16年4月13日(2004.4.13)
【国際出願番号】PCT/US2004/009127
【国際公開番号】WO2005/111058
【国際公開日】平成17年11月24日(2005.11.24)
【出願人】(504228047)
【Fターム(参考)】
[ Back to top ]