
髄膜炎菌性疾患の予防および処置のための新規な免疫原性組成物
本発明は、Neisseria ORF2086タンパク質、ナイセリア菌株から単離または組み換え的に調製され得る、交差反応性免疫原性タンパク質(その免疫原性部分、その生物学的等価物を含む)、上記のものに免疫特異的に結合する抗体および上記のものを各々コードする核酸配列、ならびにNeisseria meningitidisセログループBによる感染症に対して効果的な免疫原性組成物における上記のものの使用に関する。

【発明の詳細な説明】
【技術分野】
【0001】
本発明は、Neisseria meningitidis(髄膜炎菌)(セログループA、B、C、D、W−135、X、Y、Zおよび29E)、Neisseria gonorrhoeae(淋菌)、およびNeisseria lactamicaの菌株を含むナイセリア種菌株のような菌株から単離され得るナイセリアORF2086タンパク質(サブファミリーAおよびサブファミリーB)、ならびに該タンパク質の免疫原性部分および/または生物学的等価物に関する。本発明はまた、前記タンパク質、免疫原性部分および/または生物学的等価物に免疫特異的に結合する抗体に関する。さらに、本発明は、上記タンパク質、免疫原性部分、生物学的等価物および/または抗体のいずれかをコードする核酸配列を含む単離されたポリヌクレオチドに関する。加えて、本発明は、免疫原性組成物、およびN.meningitidisによって引き起こされる髄膜炎菌感染症、特にN.meningitidisセログループBによって引き起こされる髄膜炎菌性疾患の予防、処置および/または診断におけるそれらの使用、ならびに前記組成物の調製方法に関する。本発明は、組換え型および天然供与源由来の単離型の両方、ならびに脂質化型および非脂質化型の両方に関する。
【背景技術】
【0002】
髄膜炎菌性髄膜炎は、抗生物質の有効性にもかかわらず、数時間以内に小児および若い成人を死に至らせることがある破壊的な疾患である。非特許文献1。髄膜炎は、激しい頭痛、発熱、食欲不振、光および音に対する不耐性、特に頸部における筋肉の硬直、ならびに重篤なケースでは、死に至る痙攣、嘔吐およびせん妄という結果になる髄膜の炎症として特徴付けられる。髄膜炎菌性髄膜炎の症状は突然現れ、ついにはその特徴的な出血性発疹を伴う髄膜炎菌性敗血症となる。もしなんらかの生存のチャンスがあるとすれば、迅速な診断および大量の抗生物質による速やかな処置が決定的である。2000.Bantam Medical Dictionary,Third Edition 302。
【0003】
髄膜炎菌性髄膜炎は、Neisseria meningitidis(髄膜炎菌)によって引き起こされ、この菌は、A、B、C、D、W−135、X、Y、Zおよび29Eを含むいくつかの病原性セログループに分類されているグラム陰性の莢膜形成菌である。N.meningitidisのセログループB株は、世界中の髄膜炎菌性疾患の主要な原因である。例えば、セログループBが、米国および欧州在住の幼児および小児における細菌性髄膜炎の約50%の原因であると医学文献において報告されている。N.meningitidisセログループBによって引き起こされる髄膜炎菌性疾患を予防するためのワクチンは、現在全く存在しない。
【0004】
30年以上前のGoldschneiderらの研究以来、セログループB髄膜炎菌性疾患予防のための免疫原性組成物の開発は、研究者にとっての課題である。非特許文献2;非特許文献3;非特許文献4;および非特許文献5。第二次世界大戦後、北米から事実上消滅したセログループA疾患(Achtman,M.1995,Trends in Microbiology 3(5):186−92)とは異なり、セログループBおよびC微生物により引き起こされる疾患は、経済的に発展した世界の多くにわたって依然として風土病のままである。疾患発生率は、風土病がまれな<1/100,000から、流行病の間の高リスク個体群における200/100,000までの範囲で変わる。
【0005】
多糖結合体をベースとするワクチンが、N.meningitidisセログループAおよびCに対して開発されてきており、疾患の予防に効果的であると思われる。現在、セログループA、C、Y、およびW−135由来の莢膜状多糖で作られた免疫原性組成物が利用可能である。Ambroschら、1983,Immugenicity and side−effects of a new tetravalent.Bulletin of the World Health Organization 61(2):317−23。しかしながら、この免疫原性組成物は、T細胞非依存性免疫応答を引き起こし、幼い児童においては効果的でなく、髄膜炎菌性疾患の50%以上を引き起こすセログループB菌株については全くカバーしない。
【0006】
他の研究者らも、莢膜多糖を用いて免疫原性組成物の開発を試みている。最近、セログループC莢膜材料をタンパク質に接合させることによって調製されたセログループC疾患のための免疫原性組成物が、欧州において使用が認可されている。しかしながら、セログループB莢膜はワクチン候補として適さないかもしれない。なぜならば、莢膜多糖は、発達中のヒト神経組織上の炭水化物部分との類似性を持つポリシアル酸で構成されているからである。この糖半分は、自己抗原と認識され、したがってヒトにおいて免疫原性が乏しい。
【0007】
外膜タンパク質(OMP)が、セログループB疾患のための代替ワクチン抗原として開発されてきた。PorAの2つの可変領域に結合するモノクローナル抗体により、髄膜炎菌についてのセロサブタイプ判別方式が定義される。したがって、PorAタンパク質は、髄膜炎菌株のためのセロサブタイプ判別抗原として役立ち(Abdillahら、1988,Microbial Pathogenesis 4(1):27−32)、セログループB免疫原性組成物の成分として活発に研究されている(Poolman,1996,Adv.Exp.Med.Biol.397:73−7)。なぜならば、それらは殺菌抗体を誘発することができるからである(Saukkonen,1987,Microbial Pathogenesis 3(4):261−7)。殺菌抗体は保護の指標であると考えられ、どのような新しい免疫原性組成物候補も、これらの機能的抗体を誘導するべきである。
【0008】
ヒトならびに動物における研究は、セロサブタイプ判別抗原、PorAが殺菌抗体を誘導することを示す。しかしながら、PorAに対する免疫応答は一般に、セロサブタイプ特異的である。特に、セロサブタイプ判別データは、PorAで作られた免疫原性組成物が、そのような免疫原性組成物によりカバーされる各セロサブタイプについてPorAをおそらく6〜9も要求し得ることを示す。したがって、70〜80%のセログループB菌株をカバーするために、6〜9のPorAが必要とされるであろう。かくして、このタンパク質の可変的性質は、十分な数の髄膜炎菌セロサブタイプ臨床隔離集団から保護するために、多価ワクチン組成物を必要とする。
【0009】
セログループB髄膜炎菌のための免疫原性組成物の開発は非常に困難だったので、最近いくつかのグループが、新しい免疫原性組成物候補の同定を促すため、セログループAおよびB両方を代表する菌株由来のゲノムを配列決定している。Tettelin,2000,Science,287(5459):1809−15;非特許文献1。ナイセリアのゲノムの知識があっても、新しい免疫原性組成物候補の同定は、適切な数学的アルゴリズムが現在存在しない困難なプロセスである。実際、最近の報告は、理論的な膜貫通ドメインを含む数百のオープンリーディングフレーム(「ORF」)の同定にもかかわらず、発現、精製、および表面反応性で機能的に活性な抗体の誘起の問題により、研究者は、セログループB髄膜炎菌免疫原性組成物についてわずか7つの候補にしか至っていないことを示している。上記参照。これらのうちの1つは以前から知られていた。
【先行技術文献】
【非特許文献】
【0010】
【非特許文献1】Pizzaら、Science(2000)287:1816−1820
【非特許文献2】Goldschneiderら、J.Exp.Med.(1969)129(6):1307−26
【非特許文献3】Goldschneiderら、J.Exp.Med.(1969)129(6):1327−48
【非特許文献4】Gotschlichら、J.Exp.Med.(1969)129(6):1385−95
【非特許文献5】Gotschlichら、J.Exp.Med.(1969)129(6):1367−84
【発明の概要】
【発明が解決しようとする課題】
【0011】
したがって、(1)複数のナイセリア菌株に対する殺菌抗体を誘導し、(2)複数の菌株の表面と反応し、(3)生菌攻撃に対する受動的保護を付与し、および/または(4)コロニー形成を妨げる、免疫原性組成物に対する要求が依然としてある。
【課題を解決するための手段】
【0012】
本発明の1つの実施形態は、(a)配列番号1−11の奇数番号配列のいずれかと少なくとも約95%配列同一性を有するヌクレオチド配列、または(b)配列番号2−12の偶数番号配列のいずれかのアミノ酸配列と少なくとも約95%配列同一性を有するアミノ酸配列を含むポリペプチドをコードするヌクレオチド配列を含むポリヌクレオチドを提供する。
【0013】
本発明のさらなる実施形態は、本発明のポリヌクレオチドを含むベクターを提供する。
【0014】
本発明のさらなる実施形態は、本発明のベクターを含む組換え細胞を提供する。
【0015】
本発明のさらなる実施形態は、(a)配列番号2−12の偶数番号配列のいずれかと少なくとも95%配列同一性を有するアミノ酸配列、(b)配列番号1−11の奇数番号配列のいずれかと少なくとも95%配列同一性を有するヌクレオチド配列によってコードされるアミノ酸配列、(c)(a)もしくは(b)に記載されたアミノ酸配列の少なくとも1つの免疫原性部分、あるいは(d)(a)もしくは(b)に記載されたアミノ酸配列または(c)に記載された免疫原性部分の少なくとも1つの生物学的等価物を含むポリペプチドを提供する。
【0016】
本発明のさらなる実施形態は、(a)偶数番号の配列番号2−12のいずれかのアミノ酸配列を含むポリペプチドと免疫特異的に結合するポリペプチド、または(b)(a)に記載されたポリペプチドの少なくとも1つの免疫原性部分、または(c)(a)に記載されたポリペプチドまたは(b)に記載された免疫原性フラグメントの少なくとも1つの生物学的等価物、のいずれかを含む抗体を提供する。
【0017】
本発明のさらなる実施形態は、本発明のポリヌクレオチド、ベクター、組換え細胞、ポリペプチドまたは抗体を含む組成物を提供する。
【0018】
本発明のさらなる実施形態は、(a)配列番号1−5の奇数番号配列のいずれかと少なくとも約95%配列同一性または配列番号2−6の偶数番号配列のいずれかのアミノ酸配列をコードするヌクレオチド配列と少なくとも約95%配列同一性を有するヌクレオチド配列を含む第1のポリヌクレオチドと、(b)配列番号7−11の奇数番号配列のいずれかと少なくとも約95%配列同一性または配列番号8−12の偶数番号配列のいずれかのアミノ酸配列をコードするヌクレオチド配列と少なくとも約95%配列同一性を有するヌクレオチド配列を含む第2のポリヌクレオチドと、を含む組成物を提供する。
【0019】
本発明のさらなる実施形態は、(a)配列番号2−6の偶数番号配列のいずれかと少なくとも約95%配列同一性を有するアミノ酸配列を含む第1のポリペプチドと、(b)配列番号8−12の偶数番号配列のいずれかと少なくとも約95%配列同一性を有するアミノ酸配列を含む第2のポリペプチドと、を含む組成物を提供する。
【0020】
本発明のさらなる実施形態は、(a)偶数番号の配列番号2−12のいずれかのアミノ酸配列を含むポリペプチド、(b)奇数番号の配列番号1−11のいずれかの核酸配列を含むポリヌクレオチドによりコードされたポリペプチド、(c)(a)または(b)に記載されたポリペプチドの少なくとも1つの免疫原性部分、あるいは(d)(a)もしくは(b)に記載されたポリペプチドまたは(c)に記載された免疫原性フラグメントの少なくとも1つの生物学的等価物のいずれかを、ナイセリア種から単離および精製するか、または組換えにより調製するステップを含むプロセスにより調製される組成物を提供する。
【0021】
本発明のさらなる実施形態は、哺乳動物において免疫応答を引き起こすための薬剤の調製における本発明の組成物の使用を提供する。
【0022】
本発明のさらなる実施形態は、哺乳動物における細菌性髄膜炎に対して効果的な薬剤における本発明の組成物の使用を提供する。
【0023】
本発明のさらなる実施形態は、本明細書中で記載されたポリペプチドのいずれかをコードする核酸配列を宿主細胞において発現させるステップを含む組成物を調製する方法を提供する。
【0024】
本発明のさらなる実施形態は、本明細書中で記載されたタンパク質、免疫原性部分または生物学的等価物のいずれかを含む組成物を動物に導入した後にその動物から抗体を回収するステップを含む、抗体組成物を調製する方法を提供する。
【0025】
本発明のさらなる実施形態は、有効量の本発明の組成物の1つ以上を哺乳動物に投与するステップを含む、哺乳動物において免疫応答を引き起こす方法を提供する。
【0026】
本発明のさらなる実施形態は、有効量の本発明の組成物の1つ以上を哺乳動物に投与すことを含む、哺乳動物における細菌性髄膜炎を予防または処置する方法を提供する。
【図面の簡単な説明】
【0027】
【図1A】異種菌株に対する殺菌抗体を誘導することができるナイセリア膜タンパク質抽出物を同定するための実験から得られたタンパク質画分の2つの主要なタンパク質を示すSDS−PAGEゲルを示す。
【図1B】プロテアーゼ消化および逆相N−末端配列決定によるTMAEフロースルー成分の分析による2つの主要なタンパク質の同定の実験結果を示す。
【図2】rLP2086の精製スキームおよびSDS−PAGEにより決定される等質性を示す。
【図3】LC−MS/MSおよび対応するSDS−PAGEによるTMAEフロースルー成分の分析による2つの主要なタンパク質および1つの少量タンパク質の同定の実験結果を示す。
【図4】2086タンパク質の組換え発現のSDS−PAGEゲルである。
【図5】本明細書中の実施例に記載されるプラスミドpPX7340の概略図である。
【図6】本明細書中の実施例に記載されるプラスミドpPX7328の概略図である。
【図7】本明細書中の実施例に記載されるプラスミドpPX7343の概略図である。
【図8】様々な菌株由来の2086遺伝子のN末端領域を例示する。
【図9A】ナイセリア菌株中の免疫原性成分の同定における準備段階を示すフローチャートである。
【図9B】ナイセリア菌株中の免疫原性成分の同定における最終段階を示すフローチャートである。
【図10】図10Aは、本明細書中の実施例に記載されるrP2086脂質化型を発現させるためのP4シグナル/ORF2086融合タンパク質を発現させるpBADアラビノース誘導プロモーターの概略図である。図10Bは、ORF2086非脂質化型の組換え発現のためのpET9a−T7ベクターの概略図である。
【図11】図11Aは、rLP2086タンパク質を発現するE.coli Bの全細胞ライゼートを表す写真である。図11Bは、rP2086タンパク質を発現するE.coli Bの全細胞ライゼートを示す。
【図12A】本発明の実施に従うORF2086タンパク質のサブファミリーおよびグループの構成を示す系統樹である。
【図12B】本発明の実施に従うORF2086タンパク質のサブファミリーおよびグループの構成を示す系統樹である。
【図13】rLP2086サブファミリーA抗血清についての全細胞ELISAデータのグラフである。
【図14】rLP2086サブファミリーB抗血清についての全細胞ELISAデータのグラフである。
【図15】rLP2086混合研究−WCE力価の結果のグラフである。
【図16】rLP2086/rPorA混合研究−WCE力価の結果のグラフである。
【図17】P2086サブファミリーB N.meningitidis全細胞ライゼートに対するrLP2086マウス抗血清の反応性を示すウェスタンブロットである。
【図18】P2086サブファミリーA N.meningitidisおよびN.lactamica全細胞ライゼートに対するrLP2086マウス抗血清の反応性を示すウェスタンブロットである。
【発明を実施するための形態】
【0028】
配列の概要
配列番号1 天然のリーダー配列と組み合わされた時のCDC1135菌株由来の成熟2086タンパク質についてのアミノ酸配列をコードする核酸配列。
【0029】
配列番号2 天然のリーダー配列を用いて調製されたCDC1135菌株由来の成熟2086タンパク質についてのアミノ酸配列。
【0030】
配列番号3 P4リーダー配列と組み合わされた時のCDC1135由来の成熟2086タンパク質についてのアミノ酸配列をコードするための核酸配列。
【0031】
配列番号4 P4リーダー配列を用いて調製されたCDC1135菌株由来の成熟2086タンパク質についてのアミノ酸配列。
【0032】
配列番号5 CDC1135菌株由来の成熟2086タンパク質についてのアミノ酸配列をコードする核酸配列。
【0033】
配列番号6 CDC1135菌株由来の成熟2086タンパク質についてのアミノ酸配列。
【0034】
配列番号7 天然のリーダー配列と組み合わされた時のCDC1127菌株由来の成熟2086タンパク質についてのアミノ酸配列をコードする核酸配列。
【0035】
配列番号8 天然のリーダー配列を用いて調製されたCDC1127菌株由来の成熟2086タンパク質についてのアミノ酸配列。
【0036】
配列番号9 P4リーダー配列と組み合わされた時のCDC1127由来の成熟2086タンパク質についてのアミノ酸配列をコードするための核酸配列。
【0037】
配列番号10 P4リーダー配列を用いて調製されたCDC1127菌株由来の成熟2086タンパク質についてのアミノ酸配列。
【0038】
配列番号11 CDC1127菌株由来の成熟2086タンパク質についてのアミノ酸配列をコードする核酸配列。
【0039】
配列番号12 CDC1127菌株由来の成熟2086タンパク質についてのアミノ酸配列。
【0040】
発明の詳細な説明
本発明は、2086サブファミリーAタンパク質および2086サブファミリーBタンパク質を含むナイセリアORF2086タンパク質(「2086タンパク質」)を提供する。2086タンパク質の各々は、Neisseria meningitidis(セログループA、B、C、D、W−135、X、Y、Zおよび29E)、Neisseria gonorrhoeae、およびNeisseria lactamicaの菌株を含む、天然のナイセリア菌株から単離され得るタンパク質である。2086タンパク質はまた、組換え技術を用いて調製されてもよい。
【0041】
様々な実施形態によれば、本発明は、2086タンパク質、それらの免疫原性部分、および/またはそれらの生物学的等価物、上記のもののいずれかに免疫特異的に結合する抗体、ならびに上記のもののいずれかをコードする核酸配列を含むポリヌクレオチドを提供する。本発明は、組成物、免疫原性組成物、および髄膜炎菌性感染症、特にN.meningitidisにより引き起こされる髄膜炎菌性疾患の予防、処置および/または診断におけるそれらの使用、ならびに前記組成物を調製するための方法を提供する。2086タンパク質は本明細書中で、組換え型および天然供給源由来の単離型、ならびに脂質化型および非脂質化型を含む。
【0042】
本発明は、(1)N.meningitidis、N.gonorrhoeae、および/またはN.lactamicaの菌株のような複数のナイセリア菌株に対する殺菌抗体を誘導し、(2)複数の菌株の表面と反応し、(3)生菌攻撃に対する受動的保護を付与し、および/または(4)コロニー形成を妨げる組成物、ならびに前記組成物を使用する方法および前記組成物を調製する方法を、思いがけずかつ有利に提供する。本発明の様々な実施形態が以下で説明される。
【0043】
本明細書中で記載されるように、N.meningitidisから単離されたナイセリア種ORF2086タンパク質(本明細書中で区別なく用いられる「2086タンパク質」または「ORF2086」タンパク質、あるいは非脂質化タンパク質についてはP2086およびタンパク質の脂質化バージョンについてはLP2086とも呼ばれる)をベースとした新規な免疫原性組成物候補は、抗血清の調製に伴う細胞分画、ディファレンシャル界面活性剤抽出、タンパク質精製を、複数の菌株を用いた殺菌活性アッセイと組み合わせることにより同定された。上記で引用された参照文献中で候補として開示される免疫原性組成物および診断法に代わるものとして、本発明は、タンパク質、その免疫原性部分およびその生物学的等価物のほか、前記ポリペプチド、部分、および等価物をコードする遺伝子およびそれらに免疫特異的に結合する抗体の使用による、髄膜炎菌感染の処置および/または予防のための組成物および方法に関する。
【0044】
本明細書中で用いられるように、用語「菌株非特異的(non−strain specific)」は、1つ以上のN.meningitidis菌株(例えば、異種髄膜炎菌株)に対して効果的な免疫応答を誘発する抗原の特徴を指す。用語「交差反応性(cross−reactive)」は、本明細書中で用いられるように、用語「菌株非特異的」と区別なく用いられる。用語「免疫原性菌株非特異的N.meningitidis抗原」は、本明細書中で用いられるように、N.meningitidisから単離され得る抗原を述べるが、これは、別の細菌(例えば、淋菌のような他のナイセリア菌株)から単離することも、または組換え技術を用いて調製することもできる。
【0045】
本発明の2086タンパク質は、脂質化タンパク質および非脂質化タンパク質を含む。さらに、本発明はまた、中間体化合物/組成物として各タンパク質に対応する未熟タンパク質またはプレタンパク質の使用も考慮する。
【0046】
本発明はまた、本発明の実施に従って、上記の免疫原性薬剤に免疫特異的に結合する抗体を提供する。さらに、本発明は、上記のうちのいずれかをコードする核酸配列を含む単離されたポリヌクレオチドに関する。加えて、本発明は、髄膜炎菌性髄膜炎、特にセログループB髄膜炎菌性疾患の予防、処置および/または診断における組成物および/または免疫原性組成物およびそれらの使用、ならびに前記組成物を調製するための方法を提供する。
【0047】
本発明の組成物は高度に免疫原性であり、殺菌抗体の産生を誘導できる。これらの抗体は、セログループ、セロタイプおよびセロサブタイプ異種髄膜炎菌株と交差反応性である。したがって、本発明の組成物は、異種ナイセリア菌株に対する殺菌抗体を誘導する能力を示すことにより、以前のN.meningitidisワクチンの試みの欠陥を克服する。したがって、本発明は、他にも利点はあるが、特に従来使用されていた薬剤と比較して、保護を誘導するためにより少ない成分で調合され得る免疫原性組成物を提供する。組成物またはその中野免疫原性薬剤(例えば、ポリペプチド、免疫原性部分またはフラグメント、および生物学的等価物など、ただしこれらに限定されない)は、髄膜炎菌性感染および疾患からの免疫学的保護を引き出すため、ならびに他の病原体により引き起こされた感染および/または疾患からの免疫学的保護を引き出すため、単独で、または他の抗原もしくは薬剤と組み合わせて使用され得る。これにより、複数の菌株から保護するために必要とされる抗原の数を減らすことにより、髄膜炎菌性感染に対して使用する免疫原性組成物のデザインが単純化される。実際、精製2086タンパク質は、髄膜炎菌性疾患の原因となる菌株の適切な免疫原性の有効範囲を提供するために必要とされるタンパク質の数を劇的かつ予想外に低減する。2086タンパク質は、E.coliにおいてリポタンパク質として組換えによって発現させることができ、これは、天然の髄膜炎菌におけるよりもずっと高いレベルの野生型タンパク質である。
【0048】
以下の公開国際特許出願は、参照によりその全体が本明細書に組み込まれる:PCT/US02/32369(2003年8月7日に国際公開第03/063766号として公開)およびPCT/US04/11901(2004年11月4日に国際公開第04/094596号として公開)。
【0049】
2086タンパク質は野生型菌株中には大量に存在しないが、これは殺菌抗体の標的である。これらの抗体は、PorAに応答して産生された抗体と異なり、異種セロサブタイプを発現する菌株を殺すことができる。
【0050】
2086タンパク質に対する抗体はまた、髄膜炎菌による攻撃から幼若ラットを受動的に保護する。2086タンパク質の組換え発現により、髄膜炎菌性疾患の予防のための免疫原性組成物としての2086タンパク質の使用が可能になる。臨床試験における最近の髄膜炎菌性免疫原性組成物のすべては、多くの異なるタンパク質を含む複合組成物または外膜タンパク質製剤であった。セロサブタイプ特異性をもたらすPorAタンパク質は、約70〜80%の疾患関連セロサブタイプの有効範囲を提供するために、免疫原性組成物中に6〜9の変異体を含有することを必要とする。対照的に、単一の2086タンパク質に対する抗血清単独で、西ヨーロッパおよび米国における疾病隔離集団の約65%の原因となる6つのセロサブタイプの典型を殺すことができることが本明細書中で明確に実証される。したがって、精製2086タンパク質は、髄膜炎菌性疾患の原因となるセロサブタイプの十分な免疫原性組成物の有効範囲を提供するために必要とされるタンパク質の数を低減することが見込まれる。
タンパク質、免疫原性部分および生物学等価物
本発明により提供される2086タンパク質は、単離されたタンパク質またはポリペプチドである。用語「単離された(isolated)」は、天然状態から人の手によって変更されたことを意味する。もし「単離された」組成物または物質が自然界で生ずるのであれば、それはすでにその本来の環境から変化しているかまたは取り除かれているか、あるいはその両方である。例えば、生きている動物中に天然に存在するポリペプチドまたはポリヌクレオチドは「単離され」てはいないが、同じポリペプチドまたはポリヌクレオチドがその天然の状態の共存物質から分離されていれば、本明細書中で使用される用語「単離され」と同じく「単離され」ている。したがって、本明細書中で用いられるように、用語「単離されたタンパク質(isolated protein)」は、天然供給源から単離されたタンパク質および組換え技術を用いて調製されたタンパク質、ならびに、他の抗原および/または例えば薬学的に許容される担体、バッファー、アジュバント等の添加物と組み合わされた時の上記のようなタンパク質を包含する。
【0051】
本発明の実施形態によれば、2086タンパク質は、免疫原性、非病原および非菌株特異的であることを特徴とする。2086タンパク質は、高度に可変性であり、したがって、タンパク質の免疫原性を損なうことなくアミノ酸残基の挿入、置換および/または欠失を受けてもよい。2086タンパク質は、2つのサブファミリー、すなわちサブファミリーAおよびサブファミリーBに分割され得る。
【0052】
サブファミリーA由来の2086タンパク質は、配列番号2−6の偶数番号配列のいずれかのアミノ酸配列または配列番号1−5の奇数番号配列のいずれかのヌクレオチド配列を含むポリヌクレオチドによってコードされるアミノ酸配列を含む。サブファミリーB由来の2086タンパク質は、配列番号8−12の偶数番号配列のいずれかのアミノ酸配列または配列番号7−11の奇数番号配列のいずれかのヌクレオチド配列を含むポリヌクレオチドによってコードされるアミノ酸配列を含む。
【0053】
本発明のポリペプチド配列は、基準配列(例えば、偶数番号のついた配列番号2−12)と同一、すなわち100%同一、または、基準配列と比較して同一性の%が100%未満であるようないくつかのアミノ酸変更を含み得る。そのような変更としては、少なくとも1つのアミノ酸欠失、同類置換もしくは非同類置換を含む置換、または挿入が含まれる。変更は、基準ポリペプチド配列のアミノ−またはカルボキシ−末端位置において生じてもよいし、あるいは基準アミノ酸配列中、または基準アミノ酸配列内の1つ以上の隣接するグループ中のアミノ酸間に個別に散在されて、それらの末端位置間のどこかで生じてもよい。
【0054】
したがって、本発明はまた、配列表に含まれるアミノ酸配列(すなわち、偶数番号のついた配列番号2−12)との配列同一性を有するタンパク質を提供する。本発明の様々な実施形態によれば、2086タンパク質は、配列番号2−12の偶数番号アミノ酸配列のいずれかと約95%、約96%、約97%、約98%、約99%、約99.9%またはそれ以上の同一性より大きい配列を有する。これらは、制限なく、突然変異体および対立遺伝子変異体を含んでいる。
【0055】
本発明の好ましい実施形態において、2086タンパク質または他の2086ポリペプチド(例えば、免疫学的部分および生物学的等価物)は、髄膜炎菌の相同菌株および少なくとも1つの異種菌株に対する殺菌抗体を生成する。具体的には、2086ポリペプチドに対する抗体は、髄膜炎菌による鼻腔内などでの攻撃から幼若ラットを受動的に保護する。さらなる好ましい実施形態において、2086ポリペプチドは、相同菌株および少なくとも1つの異種菌株について幼若ラットに対し上記のような保護を発揮する。ポリペプチドは、偶数番号のついた配列番号2−12中に示されるように、上記の配列概要から選択されればよく、あるいはポリペプチドは、列挙されたポリペプチドの任意の免疫学的フラグメントまたは生物学的等価物としてもよい。好ましくは、ポリペプチドは、上記の配列概要における偶数番号のついた配列番号2−12のいずれかから選ばれる。
【0056】
本発明はまた、生物学的等価物である2086ポリペプチドの対立遺伝子変異体または他の変異体に関する。適切な生物学等価物とは、(1)相同菌株ならびに少なくとも1つの異種ナイセリア菌株および/または淋菌株に対する殺菌抗体を誘導し、(2)相同菌株および少なくとも1つの異種ナイセリア菌株および/または淋菌株の表面と反応し、(3)生菌攻撃に対し受動的保護を付与し、および/または(4)コロニー形成を妨げる、能力を示すものである。
【0057】
適切な生物学的等価物は、該生物学的等価物が本発明の2086タンパク質の1つと実質的に同じ免疫抗原特性を誘導できるという前提で、本明細書中で明記された2086ポリペプチド(すなわち、偶数番号のついた配列番号2−12)の1つと少なくとも約95%、約96%、約97%、約98%、約99%または約99.9%の類似性を有する。
【0058】
あるいは、生物学的等価物は、偶数番号のついた配列番号2−12中の2086タンパク質の1つと実質的に同じ免疫抗原特性を有する。本発明の実施形態によれば、生物学的等価物は、偶数番号のついた配列番号2−12と同じ免疫抗原特性を有する。
【0059】
生物学的等価物は、本発明のタンパク質に対する変異体および修飾を生成することにより得られる。タンパク質に対するこれらの変異体および修飾は、1つ以上のアミノ酸の挿入、欠失または置換によりアミノ酸配列を改変することによって得られる。アミノ酸配列は、実質的に同じまたは改善された品質を有するポリペプチドを作り出すために、例えば置換によって修飾される。改変を導入する好ましい手段は、部位指定突然変異誘発によりポリペプチドの核酸配列のあらかじめ決められた突然変異を作るステップを含む。
【0060】
修飾および変更は、本発明のポリペプチドの構造中において行うことができ、さらにN.meningitidis免疫原性を有する分子を得ることができる。例えば、制限なく、ある一定のアミノ酸が、免疫原性をそれほど損失することなく配列中で、非保存的および保存的置換を含む他のアミノ酸で置換され得る。ポリペプチドの生物学的機能活性を定義するのはポリペプチドの相互作用的な能力および性質なので、いくつかのアミノ酸配列置換は、ポリペプチド配列(または、当然、その基礎をなすDNAコード化配列)中で行うことができ、依然として同様な特性を有するポリペプチドを得ることができる。本発明は、本明細書中のポリペプチドの構造ならびに前記ポリペプチドをコードする核酸配列へのどのような変化も考慮し、ここで、ポリペプチドは免疫原性を保持している。当業者は、本明細書中で提供される手引きに基づいて、開示されたポリペプチドおよびポリヌクレオチドをそれに応じて容易に修飾できるであろう。
【0061】
例えば、置換または欠失が許容されるある一定の可変領域が同定されている。2086共通配列は、かねて論じられたように、本発明の実施に従うタンパク質の2086ファミリーの保存および非保存領域を示す。
【0062】
そのような変更を行う際、当業者に知られているどのような技術も利用できる。例えば、アミノ酸のハイドロパシック・インデックス(hydropathic index)を、これに限定することを意図することなく、考慮することができる。ポリペプチド上に相互作用的な生物学的機能を付与する際のハイドロパシックアミノ酸インデックスの重要性は、技術分野において一般に理解されている。Kyte et al.1982.J.Mol.Bio.157:105〜132。
【0063】
同様なアミノ酸の置換はまた、特に、生物学的機能性等価ポリペプチドまたはそれによって作り出されたペプチドが免疫学的実施形態における使用を意図される場合、親水性に基づいて行われ得る。参照により本明細書に組み込まれる米国特許第4,554,101号は、ポリペプチドの最大局所平均親水性が、その隣接アミノ酸の親水性により支配されるように、その免疫原性、すなわちポリペプチドの生物学的特性と相関関係にあると述べている。
【0064】
ポリペプチドの生物学的等価物はまた、部位特異的突然変異誘発を用いて調製できる。部位特異的突然変異誘発は、第二世代ポリペプチド、すなわち基となるDNAの特定の突然変異誘発を通して、その配列から誘導された生物学的機能性等価ポリペプチドまたはペプチドの調製において有用な手法である。そのような変更は、アミノ酸置換が望ましい場合に望ましいことがあり得る。この手法はさらに、1つ以上のヌクレオチド配列変化をDNAに導入することにより、例えば、上記の問題点のうちの1つ以上を組み込んだ配列変異体の調製および試験を素早く行えるようにする。部位特異的突然変異誘発は、所望の突然変異のDNA配列をコードする特定のオリゴヌクレオチド配列、ならびに横断されている欠失接合の両側で安定な二重鎖を形成するのに十分なサイズおよび配列複雑度のプライマー配列を提供するのに十分な数の隣接ヌクレオチドの使用によって、突然変異体の産生を可能にする。典型的には、改変される配列の接合部の両側に約5〜10の残基を有する長さが約17〜25ヌクレオチドのプライマーが好ましい。
【0065】
一般に、部位特異的突然変異誘発の手法は、技術的に知られている。理解されるように、この手法は典型的には、一本鎖型および二本鎖型の両方で存在し得るファージベクターを用いる。典型的には、本明細書に従う部位指定突然変異誘発は、最初に、選ばれたN.meningitidisポリペプチド配列の全部または一部をコードするDNA配列をその配列内に含む一本鎖ベクターを得ることにより実行される。所望の変異された配列を持つオリゴヌクレオチドプライマーが(例えば、合成的に)調製される。このプライマーは次に、一本鎖ベクターにアニールされ、変異支持鎖の合成を完成するためにE.coli DNAポリメラーゼI(クレノウフラグメント)のような酵素を用いて伸長される。かくして、1つの鎖がオリジナルの非変異配列をコードし、第2の鎖が所望の突然変異を有するヘテロ二本鎖が形成される。このヘテロ二本鎖ベクターは次に、E.coliのような適切な細胞を形質転換するために用いられ、突然変異を有する組換えベクターを含むクローンが選択される。商業的に利用可能なキットには、オリゴヌクレオチドプライマーを除く必要なすべての試薬が付属している。
【0066】
2086ポリペプチドとしては、偶数番号の配列番号 2−12の1つからのアミノ酸配列を有する2086タンパク質との実質的な配列類似性および/または生物学的等価性を含む任意のタンパク質またはポリペプチドが含まれる。加えて、本発明の2086ポリペプチドは、特定の供給源に限定されない。したがって、本発明は、各種の供給源由来のポリペプチドの一般的検出および分離に備える。また、2086ポリペプチドは、当業者に知られているように、本明細書中で提供される手引きに基づいて組換え的に、または技術的に知られている任意の他の合成的方法で調製され得る。
【0067】
2086ポリペプチドが、さらなる構造的または機能的分析において、あるいは2086関連ポリペプチドおよび2086特異的抗体のような試薬の産生において使用するためにフラグメントに有利に切断され得ることが本発明において考慮される。これは、精製または未精製N.meningitidisポリペプチドを、エンドプロテイナーゼglu−C(Boehringer,Indianapolis,IN)のようなペプチダーゼで処理することにより達成され得る。CNBrによる処理は、ペプチドフラグメントを天然のN.meningitidis2086ポリペプチドから産生できる別の方法である。組換え手法も、2086タンパク質の特定フラグメントを産生するために用いることができる。
【0068】
本明細書中で用いられる用語「変異体(variant)」は、基準ポリヌクレオチドまたはポリペプチドとそれぞれ異なるポリヌクレオチドまたはポリペプチドであるが、本質的な特性を保持する。ポリヌクレオチドの典型的な変異体は、別の基準ポリヌクレオチドとはヌクレオチド配列が異なる。変異体のヌクレオチド配列中の変化は、基準ポリヌクレオチドによってコードされるポリペプチドのアミノ酸配列を改変することも、改変しないこともある。ヌクレオチド変化は、以下で論じられるように、基準配列によってコードされるポリペプチド中でのアミノ酸置換、付加、欠失、融合およびトランケーションという結果になり得る。ポリペプチドの典型的な変異体は、アミノ酸配列が別の基準ポリペプチドと異なる。一般に、差異は限定されており、その結果、基準ポリペプチドおよび変異体の配列は、全体的に密接に類似しており、多くの領域において同一(すなわち、生物学的に等価)である。変異体および基準ポリペプチドは、いずれかの組み合わせで1つ以上の置換、付加、欠失だけアミノ酸配列が異なり得る。置換または挿入されたアミノ酸残基は、遺伝コードによってコードされたものであっても、されてないものであってもよい。ポリヌクレオチドまたはポリペプチドの変異体は、対立遺伝子変異体のような自然に発生するものであることもあり、または自然に発生することが知られていない変異体であることもある。ポリヌクレオチドおよびポリペプチドの非自然発生変異体は、突然変異誘発手法または直接合成により作られ得る。
【0069】
当該技術において知られているように、「同一性(identity)」は、配列を比較することによって確認される、2つ以上のポリペプチド配列または2つ以上のポリヌクレオチド配列間の関係である。当該技術において、「同一性」はまた、場合によっては、そのような配列のストリング間の一致によって確認される、ポリペプチドまたはポリヌクレオチド配列間の配列関連性の程度も意味する。「同一性」および「類似性(similarity)」は、Computational Molecular Biology,Lesk,A.M.,ed.Oxford University Press,New York,1988;Biocomputing:Informatics and Genome Projects,Smith,D.W.,ed.Academic Press,New York,1993;Computer Analysis of Sequence Data,Part I,Griffin,A.M.and Griffin,H.G.,eds.,HumanaPress,New Jersey,1994;Sequence Analysis in Molecular Biology,von Heinje,G.,Academic Press,1987;およびSequence Analysis Primer,Gribskov,M.and Devreux,J.,eds.,M Stockton Press,New York,1991;およびCarillo,H.,and Lipman,D.,SIAM J.Applied Math.,48:1073(1988)に記載されるような既知の方法で容易に計算できるが、それらに限定されない。同一性を確認する好ましい方法は、試験される配列間で最も大きい一致をもたらすように設計される。同一性および類似性を確認する方法は、公的に利用可能なコンピュータプログラムにおいて体系化されている。2つの配列間の同一性および類似性を確認する好ましいコンピュータプログラム方法としては、GCGプログラムパッケージ(Devreux,J.,et al 1984)、BLASTP、BLASTN、およびFASTA(Altschul,S.F.,et al.,1990)が含まれるが、それらに限定されない。BLASTXプログラムは、NCBIおよび他の供給源(BLAST Manual,Altschul,S.,et al.,NCBI NLM NIH Bethesda,Md.20894;Altschul,S.,et al.,1990)から公的に利用可能である。よく知られているスミスウォーターマンアルゴリズムもまた、同一性を確認するために用いられ得る。
【0070】
一例として、本発明のアミノ酸配列は、基準配列、偶数番号のついた配列番号2−12と同一、すなわち100%同一であってもよく、あるいは、本発明のアミノ酸配列は、基準配列と比較して同一性の%が100%未満であるようないくつかのアミノ酸変更を含むこともできるが、それに限定されることを意図するものではない。そのような変更は、少なくとも1つのアミノ酸欠失、保存的および非保存的置換を含む置換、または挿入からなる群から選ばれ、その場合、前記変更は、基準ポリペプチド配列のアミノ−またはカルボキシ−末端位置において生じてもよいし、あるいは基準アミノ酸配列中、または基準アミノ酸配列内の1つ以上の隣接するグループ中のアミノ酸間に個別に散在されて、それらの末端位置のどこかで生じてもよい。所与の%同一性についてのアミノ酸改変数は、偶数番号の配列番号2−12中のアミノ酸の総数にそれぞれのパーセント同一性の数値パーセント(100で割る)を乗じ、次にその積を偶数番号の配列番号2−12のいずれかの中のアミノ酸の前記総数から引くことにより求められる:すなわち、
na=xa−(xa・y)
式中、naはアミノ酸改変の数であり、xaは偶数番号の配列番号2−12中のアミノ酸の総数であり、yは、例えば70%については0.70、80%については0.80、85%については0.85等であり、xaとyとのどのような非整数積も、これをxaから引く前に最も近い整数に丸められる。
【0071】
好ましい実施形態において、上記のポリペプチドは、2086タンパク質の成熟プロセス型のような、偶数番号の配列番号 2−12中に明示されるタンパク質から選ばれる。2086タンパク質または等価物等は、脂質化または非脂質化され得る。
【0072】
ORF2086は、天然のORF 2086シグナル配列を持つE.coli菌中で発現可能である。しかしながら、タンパク質の発現を改善する手段を見出すことが望ましい。本発明の実施形態によれば、リーダー配列は、タンパク質の脂質化型を産生する。例えば、以下は、発現を増強するための、莢膜非保有ヘモフィルス・インフルエンザP4タンパク質のシグナル配列の使用を説明する。
【0073】
細菌リポタンパク質のプロセシングは、シグナル配列を含む前駆体またはプロリポタンパク質の合成から始まり、それが今度は共通リポタンパク質プロセシング/修飾部位を含む。このプロリポタンパク質は当初、グラム陰性菌の内膜上またはグラム陽性菌の膜上の共通Sec系を通過する。ひとたびSec系によって膜中に置かれると、プロリポタンパク質は共通部位においてシグナルペプチダーゼIIによって開裂され、露出されたN−末端システイン残基がグリセレート化およびアシル化される。Hayashi et al.1990.Lipoproteins in bacteria.J.Bioenerg.Biomembr.Jun;22(3):451−71;Oudega et al.1993。Escherichia coli SecB,SecA,and SecY proteins are requiered for expression and mambrane insertion of the bacteriocin release protein,a small lipoprotein.J.Bacteriol.Mar;175(5):1543−7;Sankaran et al.1995.Modification of bacterialLipoproteins.Methods Enzymo.250:683−97。
【0074】
グラム陰性菌において、外膜への脂質化タンパク質の輸送は、リポタンパク質の位置2におけるソーティングシグナルに応じて、膜特異性を有する特有のABCトランスポーター系によって媒介される。Yakushi et al.2000.A new ABC transporter mediating the detachment of lipid modified proteins from membranes.Nat Cell Biolo.Apr;2(4):212−8。
【0075】
細菌リポタンパク質およびそれらのシグナル配列との融合が、細菌表面の組換えタンパク質を表示するために用いられてきた。米国特許第5,583,038号および同第6,130,085号。リポタンパク質シグナル配列を交換することにより、リポタンパク質の産生を増大することができる。De et al.2000.Purification and characterization of Streptococcus pneumoniae palmitoylated pneumococcal surface adhesin A expressed in Escherichia coli.Vaccine.Mar 6;18(17):1811−21。
【0076】
タンパク質の細菌脂質化は、タンパク質に対する免疫学的応答を増大または修正することが知られている。Erdile et al.1993.Role of attached lipid in immunogenicity of Borrelia burgdorferi OspA. Infect.Immun.Jan;61(1):81−90;Snapper et al.1995.細菌リポタンパク質は、T細胞独立タイプII抗原に対する体液性免疫応答におけるサイトカインに取って代わり得る。J.Immuno.Dec 15;155(12):5582−9。しかしながら、細菌リポタンパク質発現は、プロセシングのストリンジェンシーにより複雑化され得る。Pollitt et al.1986.Effect of amino acid substitutions at the signal peptide cleavage しte of the Escherichia coli major outer membrane lipoprotein.J.Biol.Chem.Feb 5;261(4):1835−7;Lunn et al.1987.Effect of prolipoprotein signal peptide mutations on secretion of hybrido propolipo−beta−lactamase in Escherichia coli.J.Biol.Chem.Jun 15;262(17):8318−24;Klein et al.1988.Distinctive propertiees of signal sequences from bacterial lipoproteins.Protein Eng.Apr;2(1):15−20。細菌リポタンパク質発現はまた、毒性および低発現レベルのような他の問題によって複雑化される。Gomez et al.1994.Nucleotide The Bacillus subtilis lipoprotein LplA causes cell lysis when expressed in Escherichia coli.Microbiology.Aug;140(Pt8):1839−45;Hansson et al.1995.Expression of truncated and full−length forms of the Lyme disease Borrelia outer surface protein A in Escherichia coli.Protein Expr.Purif.Feb;6(1):15−24;Yakushi et al.1997.Lethality of the covalent linkage between mislocalized major outer membrane lipoprotein and the peptidoglycan of Escherichia coli.J.Bacteriol.May;179(9):2857−62。
【0077】
型分類不能なヘモフィルス・インフルエンザ菌は、P4と呼ばれるリポタンパク質(タンパク質“e”としても知られる)を発現する。P4タンパク質の組換え型は、天然のP4シグナル配列を用いて、E.coli中で高度に発現される。米国特許第5,955,580号。天然のP4シグナル配列がE.coli中の発現ベクター中の天然のORF2086シグナル配列に置換されると、ORF2086発現レベルは増大される。
【0078】
発現を増大させるために異種P4シグナル配列を用いるというこの概念は、他の細菌リポタンパク質にまで拡張可能である。特に、細菌ゲノムの分析は、関心の対象となる可能性のある多くのORFの同定につながる。E.coliのような異種宿主細胞中でORFをその天然のシグナル配列によって発現させようとする試みにより、安定性、適合性等を含む各種のシグナル配列を用いる場合に固有の各種の問題を引き起こす。これらの問題を最小限にするため、P4シグナル配列が対象となる各ORFを発現させるために用いられる。上記で記載されるように、P4シグナル配列は、異種2086ORFの発現を改善する。発現ベクターが、対象となるORFの天然のシグナル配列を欠失させ、そしてORFにP4シグナル配列を結合することによって構成される。次に適切な宿主細胞が発現ベクターで形質変換、形質移入または感染させられ、ORFの発現は、ORFの天然のシグナル配列を用いた発現と比較して増大する。
【0079】
非脂質化型は、本来のリーダー配列を欠くタンパク質によって、または宿主細胞中の脂肪酸アシル化のための部位を特定しない配列の部分で置き換えられたリーダー配列によって産生される。
【0080】
本発明の2086タンパク質の様々な形態は、特にことわらない限り、本明細書中では「2086」タンパク質と呼ばれる。また、「2086ポリペプチド」は、特にことわらない限り、2086タンパク質ならびにその免疫原性部分または生物学的等価物を指す。
【0081】
全長単離および精製されたN.meningitidisタンパク質は、10%〜20%勾配SDSポリアクリルアミドゲル(SDS−PAGE)上で測定されると約28〜35kDaの見掛けの分子量を有する。より具体的には、このタンパク質は、質量分析法により測定されると約26,000〜30,000ダルトンの分子量を有する。
【0082】
好ましくは、2086ポリペプチドおよびそのようなポリペプチドをコードする核酸は、N.meningitidisおよび/または他の種により引き起こされる感染を予防または改善するために用いられる。
抗体
配列番号2−12のアミノ酸配列を含む本発明のタンパク質、それらのフラグメント、およびそれらの類似体、またはそれらを発現させる細胞はまた、本発明のポリペプチドについて免疫特異性の抗体を産生するための免疫原として用いられる。本発明は、免疫特異性のポリペプチドに対する抗体およびN.meningitidisの存在を検出し、受動的保護を提供あるいは細胞、組織抽出物、または生体液中のポリペプチドの量もしく濃度を測定するためのそのような抗体の使用を含む。
【0083】
本発明の抗体としては、ポリクローナル抗体、モノクローナル抗体、キメラ抗体、および抗イディオタイプ抗体が含まれる。ポリクローナル抗体は、抗原で免疫性化された動物の血清から得られた抗体分子の異種個体群である。モノクローナル抗体は、特定の抗原に対する抗体の実質的に均質な個体群である。モノクローナル抗体は、当業者に知られている方法、例えば、Kohler and Milstein,1975,Nature 256:495−497および米国特許第4,376,110号により得られ。そのような抗体は、IgG、IgM、IgE、IgA、GILDおよびそれらの任意のサブクラスを含む任意の免疫グロブリンクラスであり得る。
【0084】
キメラ抗体は、マウスモノクローナル抗体およびヒト免疫グロブリン定常領域から得られた可変領域を有するもののような、その異なる部分が異なる動物種から得られる分子である。キメラ抗体およびそれらの産生方法は、技術的に知られている
【0085】
【化1】
これらの参考文献は、参照によりその全体が本明細書に組み込まれる。
【0086】
抗イディオタイプ(抗Id)抗体は、抗体の抗原結合部位と一般に関連付けられる特有の決定基を認識する抗体である。抗Id抗体は、モノクローナル抗体の供給源と同じ種および遺伝子型の動物(例えば、マウス株)に、抗Idが調製されているモノクローナル抗体で免疫化することによって調製される。免疫化された動物は、これらのイソタイプ決定基に対する抗体(抗Id抗体)を産生することにより、免疫化する抗体のイディオタイプ決定基を認識して応答する。
【0087】
したがって、本発明のポリペプチドに対して発生したモノクローナル抗体は、適切な動物の抗Id抗体を引き起こすように使用できる。そのような免疫化されたマウス由来の脾臓細胞は、抗Idモノクローナル抗体を分泌する抗Idハイブリドーマを産生するために使用できる。さらに、抗Id抗体は、キーホール・リンペット・ヘモシアニン(KLH)のような担体に結合させて、付加的なBALB/cマウスを免疫化するために使用できる。これらのマウス由来の血清は、R−PTPaseエピトープについて特異的な最終mAbの結合特性を有する抗抗Id抗体を含んでいる。抗Id抗体は、したがって、それらのイディオタイプエピトープ、すなわち、ストレプトコッカス・ピオゲネスポリペプチドのような評価されるエピトープと構造的に類似する「イディオトープ(idiotope)」を有する。
【0088】
用語「抗体(antibody)」はまた、無傷の分子ならびにFabのようなフラグメント、一本鎖抗体および抗原に結合できる抗体の他の抗原認識フラグメントの双方を含むことを意図している。Fabフラグメントは、無傷の抗体のFcフラグメントを欠いており、循環からより迅速に消え、そして無傷の抗体よりも非特異的組織結合が少ないことがある(Wahl et al.、1983,J.Nucl.Med.24:316−325)。本発明において有用な抗体のFabおよび他のフラグメントが、無傷の抗体分子のための方法に従ってN.meningitidisポリペプチドの検出および定量に用い得ることが理解されるであろう。
【0089】
anti−iodiotypic(「抗Id」)抗体のような本発明の抗体は、上記で記載されたようなポリペプチドに特異的な抗体を免疫学的に有効量投与することを含む、哺乳動物宿主におけるナイセリア感染の処置または予防のための方法において用いられ得る。抗Id抗体はまた、さらに別の動物において、いわゆる抗抗Id抗体(anti−anti−Id antibody)を産生する免疫応答を誘発する「免疫原」として用いられ得る。抗抗Idは、抗Idを誘発した当初のmAbとエピトープ的に同一であり得る。したがって、mAbのイディオタイプ決定基に対する抗体を用いることにより、同一の特異性の抗体を発現させる他のクローンを同定することが可能である。
【0090】
抗体は、様々なやり方、例えば、タンパク質が発現されていることの確認のため、またはタンパク質がどこで発現されているかを確認するために用いられる。標識抗体(例えば、FACSのための蛍光標識化)は、無傷の細菌でインキュベートすることができ、細菌表面上の標識の存在は、例えばタンパク質の位置を確認する。
【0091】
本発明のポリペプチドに対して生成された抗体は、ポリペプチドまたエピトープ坦持フラグメント、類似体、または細胞を、常用の手順を用いて動物に投与することによって得ることができる。モノクローナル抗体の調製については、継代細胞系培養により産生された抗体を提供する任意の手法を用いることができる。
ポリヌクレオチド
本発明のタンパク質と同様に、本発明のポリヌクレオチドは、奇数番号の配列番号1−11の基準配列のうちのいずれかと同一、すなわち100%同一の核酸配列を含んでもよく、または、基準配列と比較していくつかのヌクレオチド改変を含んでもよい。そのような改変は、少なくとも1つのヌクレオチド欠失、転位および転換を含む置換、または挿入からなる群から選ばれ、前記改変は、基準ヌクレオチド配列の5’もしくは3’末端位置において、あるいは個別に基準アミノ酸配列中か基準アミノ酸配列内の1つ以上の隣接する群中のアミノ酸間に散在されて、それらの末端位置間のどこかで生じる。ヌクレオチド変更の数は、奇数番号の配列番号1−11のいずれかの中のヌクレオチド総数にそれぞれのパーセント同一性(100で割られる)を乗じ、その積を前記配列中の前記総数から引くことにより求められる。
【0092】
一例として、それに限定されることを意図することなく、奇数番号の配列番号1−11のいずれかの核酸配列、その縮退変異体またはそのフラグメントと少なくとも95%同一性を有するポリヌクレオチド配列を含む単離N.meningitidisポリヌクレオチドであり、ここでポリヌクレオチド配列は、奇数番号の配列番号1−11の核酸配列のポリヌクレオチド領域全体にわたり最高nnの核酸変更を含むことができ、nnは、改変の最大数であり、次式により計算される:
nn=xn−(xn・y)、
式中、xnは、奇数番号の配列番号1−11のいずれかの核酸の総数であり、yは、0.95の値を有し、xnとyとのどのような非整数積も、そのような積をxnから引く前に最も近い整数に丸められる。当然、yは、95%について0.95の値等を有することもできる。偶数番号配列配列番号2−12のいずれかのアミノ酸配列を含むポリペプチドをコードするポリヌクレオチド配列の改変は、このコーディング配列中にナンセンス、ミスセンスまたはフレームシフト突然変異を作り出し、それによって、そのような改変に続いてポリヌクレオチドによってコードされるポリペプチドを改変することがある。
【0093】
本発明のある一定の実施形態は、2086タンパク質に対して作られた2086タンパク質および抗体をコードするポリヌクレオチド(本明細書中では、「2086ポリヌクレオチド」または「ORF2086ポリヌクレオチド」と呼ばれる)に関する。好ましい実施形態において、本発明の単離ポリヌクレオチドは、奇数番号の配列番号1−11のうちの1つ、その縮退変異体、またはそのフラグメントから選ばれるヌクレオチド配列と少なくとも約95%の同一性を有するヌクレオチド配列を含むポリヌクレオチドである。本明細書中で定義されるように、「縮退変異体(degenarate variant)」は、遺伝コードの縮退により奇数番号の配列番号1−11(およびそのフラグメント)中に示されるヌクレオチド配列とは異なるポリヌクレオチドと定義されるが、奇数番号の配列番号1−11中に示されるヌクレオチド配列によりコードされるものと同じ2086タンパク質(例えば、偶数番号配列配列番号2−12)を依然としてコードする。
【0094】
他の実施形態において、ポリヌクレオチドは、奇数番号の配列番号1−11のうちの1つ、その縮退変異体、またはそのフラグメントから選ばれるヌクレオチド配列に対する補体である。さらに他の実施形態において、ポリヌクレオチドは、DNA、染色体DNA、cDNAおよびRNAからなる群から選ばれ、異種ヌクレオチドをさらに含み得る。
【0095】
2086ポリヌクレオチドが、天然の、合成または半合成の供給源から得られること、さらに、ヌクレオチド配列が、天然に生じる配列であり得ること、あるいは、もし常にそのような配列を含む核酸分子が上記のように2086免疫原性ポリペプチドとして発現させられ得るのであれば、ヌクレオチド配列が、そのような天然に生じる配列への一もしくは複数塩基置換、欠失、挿入および逆位を含む突然変異に関連し得ることが理解されるであろう。核酸分子は、RNA、DNA、一本鎖もしくは二本鎖、線状または共有結合環状形であり得る。ヌクレオチド配列は、それに隣接して配置された発現制御配列を有することがあり、そのような制御配列は通常、異種供給源から導かれる。一般に、本発明の核酸配列の組換え発現は、核酸配列の末端において、TAAのような停止コドン配列を用いる。
【0096】
本発明はまた、低減されたストリンジェンシー条件下、好ましくはストリンジェントな条件下、そして最も好ましくは高度にストリンジェントな条件下で、本明細書中に記載されるたポリヌクレオチドにハイブリダイズできるポリヌクレオチドも含む。ストリンジェンシー条件の例は、以下のストリンジェンシー条件表に示してあり、高度にストリンジェントな条件とは、例えば、少なくとも条件A〜Fと同じくらいストリンジェントな条件であり、ストリンジェントな条件とは、例えば、少なくとも条件G〜Lと同じくらいストリンジェント条件であり、低減されたストリンジェンシー条件とは、例えば、少なくとも条件M〜Rと同じくらいストリンジェントな条件である。
【0097】
【表1−1】
【0098】
【表1−2】
【0099】
【表1−3】
bpI:ハイブリッド長は、ハイブリダイズしているポリヌクレオチドのハイブリダイズされた領域について予想される長さである。ポリヌクレオチドを、未知の配列の標的ポリヌクレオチドにハイブリダイズする場合、ハイブリッド長は、ハイブリダイズしているポリヌクレオチドの長さであると仮定される。既知配列のポリヌクレオチドがハイブリダイズされる場合、ハイブリッド長は、ポリヌクレオチドの配列を一直線に並べ、最適配列相補性の1つ以上の領域を同定することによって確認できる。
【0100】
バッファーH:SSPE(1×SSPEは、0.15 MNaCl、10 mM NaH2PO4、および1.25 mM EDTA、pH 7.4である)は、ハイブリダイゼーションおよび洗浄バッファーにおいて、SSC(1×SSCは、0.15M NaClおよび15 mMクエン酸ナトリウムである)の代わりに使用でき、洗浄は、ハイブリダイゼーション完了後に15分間実行される。
【0101】
TB〜TR:長さが50塩基対未満であると予想されるハイブリッドのためのハイブリダイゼーション温度は、ハイブリッドの融解温度(Tm)より5〜10EC低くあるべきであり、ここで、Tmは、以下の式に従って決定される。長さが18塩基対未満のハイブリッドについては、Tm(EC)=2(A+T塩基の数)+4(G+C塩基の数)。長さが18〜49塩基対のハイブリッドについては、Tm(EC)=81.5+16.6(log10[Na+])+0.41(%G+C)−(600/N)であり、ここでNは、ハイブリッド中の塩基の数であり、[Na+]は、ハイブリダイゼーションバッファ中のナトリウムイオン濃度である(1×SSCについての[Na+]=0.165M)。
【0102】
ポリヌクレオチドハイブリダイゼーションのためのストリンジェンシー条件の付加的な例は、Sambrook,J.,E.F.Fritsch,and T.Maniatis,1989,Molecular Cloning:A Laboratory Manual,Cold Spring Harbor Laboratory Press,Cold Spring Harbor,NY,chpters 9 and 11,and Current Protocols in Molecular Biology,1995,F.M.Ausubelら、eds.,John Wiley & Sons,Inc.,sections 2.10 and 6.3−6.4に提供されており、これらは参照により本明細書に組み込まれる。
【0103】
本発明はまた、これらのポリヌクレオチドに完全に相補的なポリヌクレオチドを提供し、またアンチセンス配列も提供する。アンチセンスオリゴヌクレオチドとも呼ばれる本発明のアンチセンス配列は、本発明のポリペプチドをコードするポリヌクレオチドの発現を妨げる内部生成配列および外部投与発現の両方を含む。本発明のアンチセンス配列は、例えば、約15〜20塩基対を含む。アンチセンス配列は、例えば、上流の非翻訳配列に結合するプロモーターを妨げることにより、またはリボソームの結合を妨げることにより本発明のポリペプチドをコードする転写産物の翻訳を妨げることにより、転写を阻害するように設計することができる。
【0104】
本発明のポリヌクレオチドは、多くのやり方で(例えば、化学合成により、DNAライブラリから、生物体それ自体から)調製され、様々な形態(例えば、一本鎖、二本鎖、ベクター、プローブ、プライマー)を取ることができる。用語「ポリヌクレオチド(polynucleotide)」としては、DNAおよびRNA、ならびに修飾されたバックボーンを含むそれらの類似物が含まれる。
【0105】
本発明のさらなる実施形態によれば、本発明のポリヌクレオチドは、cDNAライブラリのようなDNAライブラリを含む。
融合タンパク質
本発明はまた、融合タンパク質に関する。「融合タンパク質(fusion protein)」は、2つの、しばしば無関係な、融合遺伝子またはそのフラグメントによりコードされたタンパク質を意味する。例えば、免疫グロブリン分子の定常領域の様々な部分を別の免疫原性タンパク質またはその部分と共に含む融合タンパク質である。多くの場合、免疫グロブリンFc領域を融合タンパク質の一部として使用することは、治療および診断での使用に有利であり、例えば、改善された薬物動態学特性という結果になる(例えば、欧州特許出願公開第0232262号参照)。他方、用途によっては、融合タンパク質が発現され、検出され、精製された後に、Fc部分を欠失させることができることが望ましいであろう。本発明の2086ポリヌクレオチドは、本発明のポリペプチドの組換え産生に用いられ、ポリヌクレオチドは、それ自体で、成熟ポリペプチドのためのコード配列、あるいは、リーダーまたは分泌配列、プレもしくはプロないしはプレプロタンパク質配列、または他の融合ペプチド部分をコードするコード配列のような、他のコード配列を持つリーディングフレーム中の成熟ポリペプチドのためのコード配列を含み得る。例えば、2086ポリペプチドまたは融合ポリペプチドの精製を容易にするマーカー配列がコードされ得る(参照によりその全体が本明細書に組み込まれる、Gentzら、1989参照)。したがって、発現産物のHisタグ精製を可能にする融合ポリペプチドをコードするポリヌクレオチドの調製が本発明の実施において考慮される。ポリヌクレオチドはまた、転写された、非翻訳配列、スプライシングおよびポリアデニル化シグナルのような、非コーディング5’および3’配列を含み得る。そのような融合ポリペプチドは、形質変換/形質移入されたまたは感染させられたもしくは以下に記載されるように組換えDNAクローニングビークルで感染させられた宿主細胞により産生されることができ、そのような融合ペプチドはその後、宿主細胞から単離され、他の宿主細胞タンパク質を実質的に含まない融合ポリペプチドを提供することができる。
免疫原性組成物
本発明の1つの態様は、少なくとも1つの2086タンパク質または前記タンパク質をコードする核酸を含む免疫原性組成物を提供する。上記のものは、(1)複数の菌株への殺菌抗体を引き出し、(2)複数の菌株の表面と反応し、(3)生菌攻撃に対する受動的保護を付与し、および/または、(4)コロニー形成を防止する能力を有する。そのような免疫原性組成物の製剤は、当業者によく知られている。ある一定の実施形態において、本発明の組成物は、薬学的に許容される担体および/または希釈剤を含む。適切な薬学的に許容される担体および/または希釈剤としては、あらゆるすべての従来の溶媒、分散媒、フィラー、固体担体、水溶液、コーティング、抗菌性および抗真菌性薬剤、等張性および吸収遅延薬剤等が含まれる。適切な薬学的に許容される担体としては、例えば、1つ以上の水、食塩水、リン酸緩衝食塩水、デキストロース、グリセリロール、エタノール等、ならびにそれらの組み合わせが含まれる。薬学的に許容される担体はさらに、湿潤または乳化剤、防腐剤またはバッファーのような抗体の保存期間または有効性を向上する少量の補助物質をさらに含み得る。薬学的に許容される担体の調製および使用は、技術的によく知られている。いずれかの従来の培地または薬剤が活性成分と不和合である限りを除き、本発明の免疫原性組成物におけるそれらの使用は考慮される。本発明のある一定の実施形態によれば、薬学的に許容される担体は、担体タンパク質である。
担体タンパク質
担体タンパク質は、好ましくは非毒性かつ非反応原性であり、さらに十分な量および純度で入手可能なタンパク質である。担体タンパク質は、標準の接合手順に適用できるべきである。本発明の特定の実施形態において、CRM197が担体タンパク質として用いられる。
【0106】
CRM197197(Wyeth,Sanford,NC)は、カザアミノ酸および酵母抽出物をベースとする培地中で生育されたコリネバクテリウム・ジフテリア菌株C7(β197)の培養物から単離されたジフテリア毒素の非毒性変異体(すなわち、トキソイド)である。CRM197は、限外濾過、硫酸アンモニウム沈降、およびイオン交換クロマトグラフィーによって精製される。CRM197を得る別の方法は、米国特許第4,925,792号に記載される。代わりに、CRM197は、米国特許第5,614,382号に従って組換え的に調製される。他のジフテリアトキソイドも、担体タンパク質としての使用に適する。
【0107】
他の実施形態において、本発明の担体タンパク質は、酵素的に不活性なストレプトコッカルC5aペプチダーゼ(SCP)(例えば、米国特許第6,270,775号、同第6,355,255号、および同第6,951,653号に記載される1つ以上のSCP変異体)である。
【0108】
他の適切な担体タンパク質としては、破傷風トキソイド、百日咳トキソイド、コレラトキソイド(例えば、国際公開第2004/083251号に記載されるCT E29H)、E.coli LT、E.coli ST、E.coli DnaKタンパク質、およびPseudomonas aeruginosa由来の外毒素Aのような不活性化された細菌毒素が含まれる。外膜複合体c(OMPC)、ポリン、トランスフェリン結合タンパク質、ニューモリシン毒素(例えば、米国特許第5,565,204号)、ニューモリシントキソイド(例えば、国際公開第2005/108580号)肺炎球菌表面タンパク質A(PspA)、肺炎球菌アドヘシンタンパク質(PsaA)、またHaemophilus influenzaeタンパク質Dのような細菌外膜タンパク質もまた用いられる。ミコバクテリアhsp−70のような細菌熱ショックタンパク質もまた用いられる。Staphylococcus epidermidisタンパク質SdrG、SitCおよびフェロクロム結合タンパク質、ならびにStaphylococcus aureusタンパク質ClfA、ClfBおよびFnbAのような他のタンパク質もまた用いられる。オボアルブミン、キーホール・リンペット・ヘモシアニン(KLH)、グルタチオンS−トランスフェラーザ(GST)、ウシ血清アルブミン(BSA)、ガラクトキナーザ(galK)、ユビキチン、β−ガラクトシダーゼ、インフルエンザNS1タンパク質、またはツベルクリンの精製タンパク質誘導体(PPD)のようなさらに他のタンパク質もまた、担体タンパク質として用いられる。例えばロタウイルスVP6またはバクテリオファージQβ由来のウイルス様粒子もまた用いられる。
アジュバント
本明細書中で記載されるような免疫原性組成物はまた、ある一定の実施形態において、1つ以上のアジュバントを含む。アジュバントは、免疫原または抗原と共に投与された時に、免疫応答を増大させる物質である。いくつかのサイトカインおよびリンホカインは、免疫調整活性を有することが示されており、したがってアジュバントとして有用であり、インターロイキン1−α、1−β、2、4、5、6、7、8、10、12(例えば、米国特許第5,723,127号参照)、13、14、15、16、17および 18(ならびにその突然変異体);インターフェロン−α、βおよびγ;顆粒状マクロファージコロニー刺激因子(GM−CSF)(例えば、米国特許第5,078,996号およびATCC受入番号39900参照)、マクロファージコロニー刺激因子(M−CSF);顆粒状コロニー刺激因子(G−CSF)、ならびに腫瘍壊死因子αおよびβが含まれるが、これらに限定されない。本明細書中で記載される免疫原性組成物と共に有用な他のアジュバントとしては、MCP−1、MIP−1α、MIP−1βおよびRANETSを制限無く含むケモカイン;セレクチン、例えば、L−セレクチン、P−セレクチンおよびE−セレクチンのような接着分子;ムチン様分子、例えば、CD34、GlyCAM−1およびMadCAM−1;LFA−1、VLA−1、Mac−1およびp150.95のようなインテグリンファミリーの一つ;PECAM、ICAMs、例えば、ICAM−1、ICAM−2およびICAM−3、CD2およびLFA−3のような免疫グロブリンスーパーファミリーの一つ;CD40およびCD40Lのような共刺激分子;血管成長因子、神経成長因子、神経芽細胞成長因子、表皮成長因子、B7.2、PDGF、BL−1、ならびに血管内皮成長因子を含む成長因子;Fas、TNF受容体、Flt、Apo−1、p55、WSL−1、DR3、TRAMP、Apo−3、AIR、LARD、NGRF、DR4、DR5、KILLER、TRAIL−R2、TRICK2、およびDR6を含む受容体分子;ならびにカスパーゼ(ICE)が含まれる。
【0109】
免疫応答を強化するために用いられる適切なアジュバントは、制限なく、MPL(登録商標)(3−O−デアシル化モノホスホリル脂質A,Corixa,Hamilton,MT)をさらに含み、これは米国特許第4,912,094号に記載される。合成脂質A類似体またはアミノアルキルグルコサミンホスフェート化合物(AGP)あるいはその誘導体または類似物も、アジュバントとしての使用に有用であり、これらは、Corixa(Hamilton,MT)から利用可能であり、米国特許第6,113,918号に記載される。1つのそのようなAGPは、2−[(R)−3−テトラデカノイルオキシテトラデカノイルアミノ]エチル2−デオキシ−4−O−ホスホノ−3−O−[(R)−3−テトラデカノイルオキシテトラデカノイル]−2−[(R)−3−テトラデカノイルオキシテトラデカノイル−アミノ]−b−D−グルコピラノシドであり、これも、529として知られている(以前は、RC529として知られていた)。この529アジュバントは、水性形態(AF)または安定エマルション(SE)として配合される。
【0110】
さらに他のアジュバントとしては、N−アセチル−ムラニル−L−トレオニル−D−イソグルタミン(thr−MDP)、N−アセチル−ノルムラニル−L−アラニン−2−(1’−2’ジパルミトイル−sn−グリセロ−3−ヒドロキシホスホリルオキシ)−エチルアミン(MTP−PE)のようなムラミルペプチド;MF59(国際公開第90/14837号)(5%スクワレン、0.5%Tween80、および0.5%Span85を含み(種々の量のMTP−PEを任意に含み))、モデル110Yマイクロフルイダイザー(Microfluidics,Newton,MA)のようなマイクロフルイダイザーを用いてサブミクロン粒子状に処方される)、およびSAF(10%スクワレン、0.4%Tween80、5%プルロニックブロックポリマーL121、およびthr−MDPを含み、サブミクロンエマルションに微小液滴化されるか、より大きい粒径のエマルションを生成するために渦撹拌される)のような水中油型エマルション;不完全フロイントアジュバント(IFA);水酸化アルミニウム、リン酸アルミニウム、硫酸アルミニウムのようなアルミニウム塩(みょうばん);アンフィゲン(Amphigen)、アブリジン、L121/スクワレン、D−ラクチド−ポリアクチド/グリコシド;プルロニックポリオール;死菌Bordetella;米国特許第5,057,540号に記載のStimulon(商標)QS−21(Antigenics,Framingham,MA.)、米国特許第5,254,339号に記載のISCOMATRIX(CSL Limited,Parkville,Australia)、および免疫刺激複合体(ISCOMS)のようなサポニン;Mycobacterium tuberculosis;細菌リポ多糖;CpGモチーフを含むオリゴヌクレオチドのような合成ポリヌクレオチド(例えば、米国特許第6,207,646号);欧州特許第1,296,713号および同第1,326,634号に記載のIC−31(Intercell AG,Vienna,Austria);百日咳毒素(PT)またはその突然変異体、コレラ毒素またはその突然変異体(例えば、国際出願第00/18434号、同第02/098368号および同第02/098369号);あるいはE.coli熱不安定毒素(LT)、特にLT−K63、LT−R72、PT−K9/G129;例えば、国際出願第93/13302号および同第92/19265号参照、が含まれる。
投与様式
そのような免疫原性組成物は、非経口的に、例えば、皮下注射か筋肉内注射により、ならびに経口的または鼻腔内的に投与され得る。筋肉内免疫化の方法は、WolffらおよびSedegahらにより記載されている。他の投与様式は、例えば、経口製剤、肺製剤、座薬、および経皮適用を用いるが、それらに限定されない。経口製剤としては、例えば、製薬グレードのマンニトール、乳糖、デンプン、ステアリン酸マグネシウム、サッカリンナトリウム、セルロース、炭酸マグネシウム等の通常用いられる賦形剤が含まれるが、それらに限定されない。
【0111】
本発明の免疫原性組成物は、ISCOMS(免疫刺激性複合体)、ISCOMS含有CTB、リポソームの形態で送達されるか、あるいは吸着に適したサイズの微小球を形成するためにアクリレートまたはポリ(DL−ラクチド−グリコシド)のような化合物中にカプセル化され得る。本発明のタンパク質はまた、油性エマルションに組み入れられ得る。
多重抗原
本発明のタンパク質、ポリヌクレオチドおよび等価物を含む免疫原性薬剤は、免疫原性組成物中の唯一の能動免疫原として投与されてもよく、代わりに、組成物は、他のNeisseria sp.免疫原性ポリペプチド、または1つ以上の他の微生物病原体(例えば、ウイルス、プリオン、細菌、真菌、ただしこれらに限定されない)の免疫学的活性なタンパク質もしくは莢膜多糖を含む他の能動免疫原を含んでもよい。組成物は、選ばれた徴候について要求されるように、1つ以上の所望のタンパク質、フラグメントまたは製薬化合物を含み得る。同様に、免疫原性組成物中に1つ以上の核酸を使用する本発明の組成物は、上記のように、同じ多様なタンパク質群をコードする核酸も含み得る。
【0112】
任意の多重抗原または多価免疫原性組成物が本発明により考慮される。例えば、本発明の組成物は、粘膜送達に適した形態で、2つ以上の2086タンパク質の組み合わせ、2086タンパク質と1つ以上のPorAタンパク質との組み合わせ、2086タンパク質と髄膜炎菌セログループA、C、YおよびW135多糖および/または多糖結合体との組み合わせ、2086タンパク質と髄膜炎菌および肺炎球菌組み合わせとの組み合わせ、または上記のいずれかの組み合わせを含み得る。当業者は、そのような多重抗原または多価免疫学的組成物を容易に配合できるであろう。
【0113】
本発明はまた、病原体に対し有用な任意の組成物を本発明の組成物の中でまたは本発明の組成物と共に組み合わせることができる、多重免疫化処方計画も考慮する。例えば、制限無く、患者は、多重免疫化処方計画の一環として、本発明の免疫原性組成物およびS.Pneumoniaeに対して免疫化するための別の免疫学的組成物を投与され得る。当業者は、多重免疫化処方計画を開発および実施するために、本発明の免疫原性組成物と併用するための免疫原性組成物を容易に選択することができるであろう。
【0114】
本発明の特定の実施形態は、S.Pneumoniae感染の予防または改善のための組成物中でまたは処置処方計画の一環として、1つ以上の本発明のポリペプチドまたはそれをコードする核酸の使用に関する。2086ポリペプチドまたは2086ポリヌクレオチドを、S.Pneumoniae感染に対して使用するための任意の免疫原性組成物と組み合わせることができる。また、2086ポリペプチドまたは2086ポリヌクレオチドを、任意の他のタンパク質または多糖ベースの髄膜炎菌ワクチンと組み合わせることができる。
【0115】
2086ポリペプチド、フラグメントおよび等価物は、接合免疫原性組成物の一部として用いることができ、その場合、1つ以上のタンパク質またはポリペプチドは、いくつかのセロタイプに対しておよび/またはいくつかの疾患に対して免疫原性特性を有する組成物を生成するために、担体タンパク質に接合する。代わりに、2086ポリペプチドの1つが、他の免疫原性ポリペプチドのための担体タンパク質として用いられ得る。
【0116】
本発明はまた、哺乳動物における免疫応答を引き起こす方法に関し、この方法は本発明の免疫原性組成物を前記哺乳動物に提供するステップを含む。この免疫原性組成物は、そのような組成物中に含まれる免疫学的有効量のポリペプチドがN.meningitidis感染に対して所望の免疫応答を引き起こすように、処置される動物またはヒトにおいて抗原性の組成物である。好ましい実施形態は、ヒトにおけるN.meningitidis感染の改善または予防を含む処置のための方法に関し、この方法は、免疫学的有効量の組成物をヒトに投与するステップを含む。
【0117】
本明細書中で用いられるように、文言「免疫学的有効量」は、処置された個体の免疫系に細菌感染の臨床的衝撃を低減する応答を少なくとも引き起こさせるのに十分な単一用量か、一連の用量の一環としての、哺乳動物宿主(好ましくはヒト)への投与を意味する。これは、細菌負荷の最小限の減少から細菌感染の予防にまで及び得る。理想的には、処置された個体は、細菌感染のより深刻な臨床的徴候を示さない。投薬量は、個体の特定の条件に応じて変わり得る。この量は、常用の試行により、さもなければ当業者に既知の手段により決定できる。
【0118】
本発明の別の特定の態様は、本発明のタンパク質、またはその免疫原性部分を発現させるベクターまたはプラスミドを免疫原性組成物として用いることに関する。したがって、さらなる態様として、本発明は、哺乳動物において免疫応答を引き起こす方法を提供し、この方法は、少なくとも1つの単離2086ポリペプチドを発現させるベクターまたはプラスミドを哺乳動物に提供するステップを含む。本発明のタンパク質は、生きたベクターを用いて、特に、異物ポリペプチドとしてポリペプチドまたは免疫原性部分の発現に必要な遺伝物質を含んでいる、生きた組換え細菌、ウイルスまたは他の生きた薬剤を用いて哺乳動物に送達され得る。
【0119】
本発明のさらなる実施によれば、哺乳動物において細菌性髄膜炎を診断するための方法が提供され、この方法は、哺乳動物または該哺乳動物の組織試料中の免疫複合体の存在を検出するステップを含み、前記哺乳動物または組織試料は、偶数番号配列配列番号2−12のいずれかのアミノ酸配列を含む少なくとも1つのポリペプチドと免疫特異的に結合する抗体を含む抗体組成物と接触させられ、哺乳動物または組織サンプルは、免疫複合体の形成に適した条件下で抗体組成物と接触させられる。
ウイルスベクターおよび非ウイルスベクター
特にin vitoroおよびin vivo細胞アッセイのための好ましいベクターは、レンチウイルス、レトロウイルス、ヘルペスウイルス、アデノウイルス、アデノ随伴ウイルス、ワクシニアウイルス、バキュロウイルス、および望ましい細胞指向性を持つ他の組換えウイルスのようなウイルスベクターである。したがって、2086タンパク質またはその免疫原性フラグメントをコードする核酸は、ウイルスベクターを用いてまたはDNAの直接導入により、in vivo、ex vivo、またはin vitroで導入され得る。標的組織中での発現は、ウイルスベクターまたは受容体リガンドにより、または組織特異性プロモーターを用いて、あるいはその両方により、トランスジェニックベクターを特定の細胞特定に向けることにより行われ得る。標的遺伝子送達は、国際公開第95/28494号に記載され、これは、参照によりその全体が本明細書に組み入れられる。
【0120】
in vivoまたはex vivoターゲティングおよび療法手順において一般的に用いられるウイルスベクターは、DNAベースのベクターおよびレトロウイルスベクターである。ウイルスベクターを調製し、使用する方法は、技術的に知られている(例えば、Miller and Rosman,BioTechniques,1992,7:980−990)。好ましくは、ウイルスベクターは複製欠陥性であり、すなわち、それらのウイルスベクターは、標的細胞中で自律的に複製できない。好ましくは、複製欠陥性ウイルスは最小のウイルスであり、すなわち、そのウイルスは、ウイルス粒子を産生するゲノムを被包するのに必要なそのゲノム配列のみを保持する。
【0121】
DNAウイルスベクターとしては、単純ヘルペスウイルス(HSV)、乳頭腫ウイルス、エプスタイン−バールウイルス(EBV)、アデノウイルス、アデノ随伴ウイルス(AAV)等のような、ただしこれらに限定されない弱毒性または欠陥DNAウイルスが含まれる。ウイルス遺伝子を完全にまたはほぼ完全に欠く欠陥ウイルスが好ましい。欠陥ウイルスは、細胞内への導入後に感染性はない。欠陥ウイルスベクターの使用は、ベクターが他細胞に感染し得るという不安なしに、特定の局部的領域における細胞への投与ができる。したがって、特定の組織を特異的に標的とし得る。具体的なベクターの例としては、各々が参照によりその全体が本明細書に組み入れられる、欠陥ヘルペスウイルス1(HSV1)ベクター(Kaplittら、Molec.Cell.Neurosci.,1991,2:320−330)、糖タンパク質L遺伝子を欠く欠陥ヘルペスウイルスベクター、または他の欠陥ヘルペスウイルスベクター(国際公開第94/21807号および同92/05263号);Stratford−Perricaudet et al.により記載されたベクターのような、弱毒アデノウイルスベクター(J.Clin.Invest.,1992,90:626−630;La Salleら、Science,1993,259:988−990も参照);および欠陥アデノ随伴ウイルスベクター(Samulskiら、J.Viol.,1987,61:3096−3101;Samulskiら、J.Viol.,1989,63:3822−3828;Lebkowskiら、Mol.Cell.Biol.,1988,8:3988−3996)が含まれるが、それらに限定されない。
【0122】
Avigen,Inc.(Alameda,CA;AAVベクター)、Cell Genesys(Foster City,CA;レトロウイルス、アデノウイルス、AAVベクター、およびレンチウイルスベクター)、Clontech(レトロウイルスおよびバキュロウイスルベクター)、Genovo,Inc.(Sharon Hill,PA;アデノウイルスおよびAAVベクター)、Genvec(アデノウイルスベクター)、IntroGene(Leiden,Netherland;アデノウイルスベクター)、Molecular Medicine(レトロウイルス、アデノウイルス、AAV、およびヘルペスウイルスベクター)、Norgen(アデノウイルスベクター)、Oxford BioMedica(Oxford,United Kingdom;レンチウイルスベクター)、およびTransgene(Strasbourg,France;アデノウイルス、ワクシニア、レトロウイルス、およびレンチウイルスベクター)を含む様々な企業がウイルスベクターを商業的に製造しているが、これらに限定されるものではない。
【0123】
アデノウイルスベクター。アデノウイルスは、本発明の核酸を各種の細胞型に効率的に送達するように修飾され得る真核DNAウイルスである。アデノウイルスの様々なセロタイプが存在する。これらのセロタイプのうち、本発明の範囲内で、2型または5型ヒトアデノウイルス(Ad2またはAd5)あるいは動物起源アデノウイルス(国際公開第94/26914号参照)の使用が優先される。本発明の範囲内で用いられ得るそれらの動物起源アデノウイルスとしては、イヌ、ウシ、マウス(例:Mav1,Beardら、Viroloy,1990,75−81)、ヒツジ、ブタ、トリ、およびサル(例:SAV)起源が含まれる。好ましくは、動物起源アデノウイルスは、イヌアデノウイルス、より好ましくはCAV2アデノウイルス(例えば、マンハッタンまたはA26/61株、ATCC VR−800)である。様々な複製欠陥アデノウイルスおよび最小アデノウイルスベクターが記載されている(国際公開第94/26914号、同第95/02697号、同第94/28938号、同第94/28152号、同第94/12649号、同第95/02697号、同第96/22378号)。本発明による複製欠陥組換えアデノウイルスは、当業者に知られている任意の手法で調製され得る(Levreroら、Gene,1991,101:195;欧州特許出願公開第185573号;Graham,EMBO J.,1984,3:2917;Grahamら、J.Gen.Virol.,1977,36:59)。組換えアデノウイルスは、当業者に周知の標準的分子生物学的手法を用いて回収および精製される。
【0124】
アデノ随伴ウイルス。アデノ随伴ウイルス(AAV)は、それらが感染する細胞のゲノムに安定的かつ部位特異的なやり方で同化できる比較的小サイズのDNAウイルスである。これらのウイルスは、細胞の成長、形態、または分化へのどのような効果も引き起こすことなく広範囲の細胞に感染することができ、ヒトの病理学に関与しないと思われる。AAVゲノムは、クローン化され、配列決定され、特性決定されている。遺伝子をin vivoおよびin vitroで転移するためのAAV由来ベクターの使用が記載されている(国際公開第91/18088号および同第93/09239号;米国特許第4,797,368号および同第5,139,941号;欧州特許出願公開第488528号参照)。本発明による複製欠陥語組換えAAVは、2つのAAV逆方向末端反復(ITR)領域により隣接された対象となる核酸配列を含むプラスミドを、AAVキャプシド形成遺伝子(repおよびcap遺伝子)を持つプラスミドと、ヒトヘルパーウイルス(例えば、アデノウイルス)で感染させた細胞系に同時形質移入することにより調製できる。産生されたAAV組換え体は次に、標準的手法によって精製される。
【0125】
レトロウイルスベクター。本発明の別の実施において、例えば、各々が参照によりその全体が組み込まれる、米国特許第5,399,346号;Mannら、Cell,1983,33:153;米国特許第4,650,764および同第4,980,289号、Markowitzら、J.Viol.,1988,62:1120;米国特許第5,124,263号;欧州特許出願公開第453 242号および同第178 220号;Bersteinら、Genet.Eng.,1985,7:235;McCormick,BioTechnology,1985,3:689;国際公開第95/07358号;およびKuoら、Blood,1993,82:845に記載されるように、核酸をレトロウイルスベクターに導入できる。レトロウイルスは、分裂細胞に感染する組込みウイルスである。レトロウイルスゲノムは、2つのLTR、キャプシド形成配列および3つのコード化領域(gag、polおよびenv)を含む。組換えレトロウイルスベクターにおいて、gag、polおよびenv遺伝子は一般に、全部または部分的に欠失され、対象となる異種核酸配列で置き換えられる。これらのベクターは、HIV、MoMuLV(「マウスモロニー白血病ウイルス」)、MSV(「マウスモロニー肉腫ウイルス」)、HaSV(「ハーベイ肉腫ウイルス」);SNV(「脾臓壊死ウイルス」);RSV(「ラウス肉腫ウイルス」)およびフレンドウイルスのような、異なる型のレトロウイルスから構築できる。適切なパッケージング細胞系が先行技術において記載されており、特に細胞系PA317(米国特許第4,861,719号);PsiCRIP細胞系(国際公開第90/02806号)およびGP+envAm−12細胞系(国際公開第89/07150号)である。加えて、組換えレトロウイルスベクターは、転写活性を抑制するためのLTR内の修飾、ならびにgag遺伝子の一部を含み得る広いキャプシド形成配列も含み得る(Benderら、J.Viol.,1987,61:1639)。組換えレトロウイルスベクターは、当業者に知られている標準的手法により精製される。
【0126】
レトロウイルスベクターは、感染性粒子として機能するように、または単一ラウンドの形質移入を受けるように構成され得る。前者の場合、ウイルスは、発がん性形質転換の原因となるものを除くその遺伝子すべてを保持し、異種遺伝子を発現させるように修飾される。非感染性ウイルスベクターは、ウイルスパッケージングシグナルを抹消するが、異種遺伝子およびパッケージングシグナルを含むように設計された同時導入ウイルスをパッケージするために必要な構造遺伝子を保持するように操作される。したがって、産生されるウイルス粒子は、付加的なウイルスを産生することができない。
【0127】
レトロウイルスベクターはまた、DNAウイルスにより導入されてもよく、それにより、1サイクルのレトロウイルス複製が可能になり、形質移入効率が増幅される(国際公開第95/22617号、同第95/26411号、同第96/39036号および同第97/19182号参照)。
【0128】
レンチウイルスベクター。本発明の別の実施において、レンチウイルスベクターは、脳、網膜、筋肉、肝臓および血液を含むいくつかの組織型における導入遺伝子の直接送達および持続的発現のための薬剤として用いられ得る。ベクターは、これらの組織中の分裂および非分裂細胞を効率的に形質導入し、対象となる遺伝子の長期発現をもたらし得る。概要については、Naldini,Curr.Opin.Biotechnol.,1998,9:457−63を参照、また、Zufferey,et al.,J.Viol.,1998,72:9873−80も参照されたい。レンチウイルスパッケージング細胞系は、利用可能であり、当該技術分野において一般に知られている。レンチウイルスパッケージング細胞系は、遺伝子治療のための高力価レンチウイルスベクターの産生を容易にする。一つの例が、少なくとも3ないし4日間、106IU/mLより大きい力価のウイルス粒子を生成できるテトラサイクリン誘導VSV−Gプソイド型レンチウイルスパッケージング細胞系である(Kafri,et al.,J.Viol.,1999,73:576−584)。誘導細胞系により産生されたベクターは、in vitroおよびin vivoで非分裂細胞を効率的に形質導入するために必要に応じて濃縮され得る。
【0129】
非ウイルスベクター。本発明の別の実施において、ベクターは、in vivoでリポフェクションにより裸のDNAとして、または他の形質移入促進薬剤(ペプチド、ポリマー等)を用いて導入され得る。合成カチオン性脂質は、マーカーをコードする遺伝子のin vivo形質移入のためのリポソームを調製するために用いられ得る(Felgner,et al.,Proc.Natl.Acad.Sci.U.S.A.,1987,84:7413−7417;Felgner and Ringold,Science,1989,337:387−388;Mackey,et al.,Proc.Natl.Acad.Sci.U.S.A.,1988,85:8027−8031;Ulmerら、Science,1993,259:1745−1748)。核酸の転移に有用な脂質化合物および組成物は、国際出願第95/18863号、同第96/17823号、および米国特許第5,459,127号に記載される。ターゲティングの目的で、脂質を他の分子に化学的に結合させることができる(上記、Mackey,et al.参照)。標的ペプチド、例えば、ホルモンまたは神経伝達物質、および抗体のようなタンパク質、または非ペプチド分子を、リポソームに化学的に結合させることができるであろう。
【0130】
カチオン性オリゴペプチド(例えば、国際公開第95/21931号)、DNA結合タンパク質から誘導されたペプチド(例えば、国際公開第96/25508号)、またはカチオン性ポリマー(例えば、国際公開第95/21931号)のような他の分子もまた、in vivoで核酸の形質移入を促進するのに有用である。
【0131】
ベクターを裸のDNAプラスミドとしてin vivoで導入することも可能である。ワクチン目的または遺伝子治療のための裸のDNAベクターは、技術的に知られている方法、例えば、電気穿孔法、顕微注射、細胞融合、DEAEデキストラン、リン酸カルシウム沈降、遺伝子銃の使用、またはDNAベクター輸送体の使用により所望の宿主細胞に導入され得る(例えば、Wuら、J.Biol.Chem.,1992,267:963−967;Wu and wu,J.Biol.Chem.,1988,263:14621−14624;カナダ特許出願公開第2,012,311号;Williamsら、Proc.Natl.Acad.Sci.USA,1991,88:2726−2730)。受容体媒介DNA送達方法もまた用いられ得る(Curielら、Hum.Gene Ther.,1992,3:147−154;Wu and wu,J.Biol.Chem.,1987,262:4429−4432)。米国特許第5,580,859号および同第5,589,466号は、哺乳動物における、形質移入助長薬剤の無い、外因性DNA配列の送達を開示している。最近、エレクトロトランスファー(electrotranfer)と呼ばれる比較的低電圧、高効率のin vivo DNA転移手法が記載されている(Mirら、C.P.Acad.Sci.,1988,321:893;国際公開第99/01157号;同第99/01158号;同第99/01175号)。したがって、本発明の付加的な実施形態は、ヒトにおける免疫応答を引き起こす方法に関し、この方法は、本発明の2086ポリペプチドをコードするある量のDNA分子を、任意に形質導入促進薬剤と共に、前記ヒトに投与するステップを含み、前記ポリペプチドは、発現された場合に免疫原性を保持し、免疫原性組成物中に組み込まれてヒトに投与された場合に、N.meningitidisのようなNeisseria sp.病原体にその後ヒトが感染した際に、増大した疾患を引き起こすことなく保護を提供する。形質移入促進薬剤は、技術的に知られており、参照により本明細書に組み込まれるブピビカイン、ならびに他の局部麻酔薬(例えば、米国特許第5,739,118号参照)およびカチオン性ポリアミン(国際公開第96/10038号に公開される)を含む。
【0132】
本発明はまた、抗体に関し、この抗体は、上記で記載されるように2086ポリペプチドに特異的な、モノクローナル抗体かポリクローナル抗体であり得る。そのような抗体は、当業者によく知られた方法により産生され得る。
細菌発現系およびプラスミド
本発明はまた、ベクターまたはプラスミドのような組換えDNA分子を提供し、これは、プロモーター配列およびイニシエーター配列ならびに本発明のポリペプチドをコードするヌクレオチド配列を有する発現制御配列を含み、ヌクレオチド配列は、プロモーターおよびイニシエーター配列に対して3’に位置する。さらに別の態様において、本発明は、プロモーター配列およびイニシエーター配列を有する発現制御配列、ならびに2086ポリペプチドをコードするヌクレオチド配列を含有する、2086ポリペプチドを発現することができる組換えDNAクローニングビヒクルを提供し、ヌクレオチド配列は、プロモーターおよびイニシエーター配列に対し3’に位置する。さらなる態様において、上記で記載されるような組換えDNAクローニングビークルおよび/または組換えDNA分子を含む宿主細胞が提供される。適切な発現制御配列および宿主細胞/クローニングビークルの組み合わせは、技術的によく知られており、Sambrookら(1989)に例証として記載されている。
【0133】
ひとたび本発明の所望のポリペプチドを発現する組換えDNAクローニングビークルおよび/または宿主細胞が、対応する2086ポリヌクレオチドを含有するプラスミドにより形質転換、形質移入または感染させることにより構築されたら、クローニングビークルまたは宿主細胞は、ポリペプチドが発現されるような条件下で培養される。ポリペプチドは次に、当業者によく知られた手法により、汚染宿主細胞成分を実質的に含まずに単離される。
【0134】
以下の実施例は、本発明の好ましい実施形態を実証するために含まれる。以下の実施例中で開示される手法が、本発明の実施においてうまく機能することが本発明者らに発見された手法を代表するものであり、したがって本発明の実施のための好ましい態様を構成することが当業者により理解されるべきである。しかしながら、当業者は、本開示を考慮して、開示される特定の実施形態において多くの変更がなされ、それでもなお本発明の趣旨および範囲を逸脱することなく同様なまたは類似の結果を得ることができると理解するべきである。
【実施例】
【0135】
(実施例1)
異種菌株に対する殺菌抗体を引き出すことが可能なナイセリア膜タンパク質抽出物の同定:
以下の表IIを参照すると、LOS(脂質オリゴ糖)除去外膜タンパク質調製物は、殺菌抗体を引き出すことが示されている。これらの抗体は、それぞれの菌株のPorAに向けられることが多くある。セログループB髄膜炎菌株8529(B:15:P1.7b,3)由来のLOS除去外膜調製物は、このように普通ではない。なぜならば、それらの調製物は、いくつかの異種菌株に対する殺菌抗体を引き出すからである。
【0136】
【表2】
異種殺菌抗体を引き出す原因となる抗原の単離および特性決定を容易にするため、本発明者らは、どの界面活性剤が抗原を最適に抽出したかを同定しようと努めた。
菌株および培養条件
凍結したガラスびんからのN.meningitidis菌株8529を、GCプレート上に筋状に塗った。(髄膜炎菌株8529は、The RIVM,Bilthoven,The Netherlandsから受領した)。プレートを、36C/5%CO2で7.5時間インキュベートした。いくつかのコロニーを用い、50mLの修正フランツ培地+GCサプリメントを含むフラスコに接種した。フラスコを36℃にてエアシェーカー中でインキュベートし、200RPMで4.5時間撹拌した。5mLを用いて、450mLの修正フランツ培地+GCサプリメントを含むフェルンバッハフラスコに接種した。フラスコを36℃にてエアシェーカー中でインキュベートし、100RPMで11時間撹拌した。450mL全部を用いて、10L発酵槽中の8.5Lの修正フランツ培地+GCサプリメントに接種した。
修正フランツ培地の組成:
グルタミン酸 1.3g/L
システイン 0.02
リン酸水素二ナトリウム・7水和物 10
塩化カリウム 0.09
塩化ナトリウム 6
塩化アンモニウム 1.25
透析酵母抽出物(YE) 40ml
(5容量のdH2Oに対して一晩透析し、次いでオートクレーブした25%YE溶液)
GCサプリメント100X、フィルタ殺菌
デキストロース 400g/L
グルタミン酸 10
コカルボキシラーゼ 0.02
硝酸第二鉄 0.5
以下のパラメータを発酵中制御した:温度=36℃;pH=7.4;溶存酸素=20%。数滴のP−2000消泡剤を添加して発泡を制御した。培養物を定常期まで生育させた。細胞を、OD650=5.25において遠心分離により収集した。一般に、全部で100〜300グラムの湿潤細胞ペーストが、〜8.5Lの培養物から収集される。
異種殺菌抗体を引き出す髄膜炎菌由来の外膜タンパク質画分の部分的精製:
湿重量100グラムの細胞を、10mM HEPES−NaOH、pH7.4、1mN Na2EDTAで湿重量の5倍の体積に縣濁させ、そしてチャンバを備える110Yマイクロフルイダイザーを約18,000psiで通過させて溶解させた。細胞ライゼートを清澄にし、300,000×gで1時間の遠心分離により細胞エンベロープを単離した。細胞エンベロープを、同じバッファーでホモジナイザーを用いて縣濁し、続いて上記のように遠心分離して2回洗浄した。次に細胞エンベロープを、10mM HEPES−NaOH、pH7.4、1mN MgCl2中の1%(w/v)Triton X−100 320mLで抽出した。以下の表IIIを参照すると、Triton X−100およびZwittergent3−14を用いた逐次ディファレンシャル界面活性剤抽出、続いてのマウスの免疫化の結果により、Triton抽出物が、対象となる候補を最適に抽出したと確認することができた。次に、表IIIに列記された5つの菌株のうち4つに対して殺菌抗体応答を引き出すこのTriton X−100抽出物を、BioRad Rotophorユニット中での調製用等電点分離(IEF)により分画した。両性電解質濃度は、1%pH4〜6と混合された1%pH3〜10であった。表IIIに示されるように、いくつかの画分は、異種殺菌性応答を引き出すことが見出された。IEFから得られた画分は、5.5〜7.8のpH範囲で集束したものであり、殺菌アッセイにより確認されるように大部分の菌株に対する異種応答を引き出した。プールしたIEF画分を濃縮し、両性電解質をエタノール沈降により除去した。約5.5〜7.8のpH範囲で得られたタンパク質のいくらかを陰イオン交換カラムに吸着させ、吸着タンパク質および非吸着タンパク質でマウスを免疫化した後に得られる殺菌活性を比較することにより、さらなる精製が達成された。表IIを再度参照すると、多くのタンパク質が陰イオン交換樹脂に吸着された一方、カラムにより吸着されなかったタンパク質はより多くの異種殺菌抗体を引き出した。
【0137】
【表3】
NT:試験せず
*臨床分離株539は、同じ発生から単離された8529の相同菌株である。
【0138】
図1Aに示されるように、SDS−PAGEにより確認されるように、非吸着画分中に2つの主要タンパク質が存在した。これらのタンパク質を同定するため、2種類の分析を実行した。一方の分析は、限定タンパク質分解(図1Aおよび図1B参照)を実行し、続いてペプチドの単離および直接的タンパク質配列決定を行うものであった。他方の分析は、SDS−PAGEを実行し、続いてゲル切り出し、タンパク質分解消化、およびLS−MS/MS(液体クロマトグラフィータンデム質量分析)を行い、(図3参照)対象となる調製物の成分についての質量スペクトル情報を得ることであった。(後に本節中で説明されるペプチドマッピングおよび配列決定方法を参照)
N.meningitidis A Sangerゲノム配列は、ZaugurskyおよびRussel,2001,BioTechniques,31:636−659に記載される方法およびアルゴリズムを用いて分析した。このマイニング分析により、12,000以上の考え得るオープンリーディングフレーム(ORF)がもたらされた。上述の直接配列データおよび質量スペクトルデータは、非吸着画分の主要成分が、Sangerデータベースの分析中に存在するいくつかのORFの生成物であることを示した。この方法論により同定された3つの主なタンパク質は、ORF4431、5163および2086に対応する(図1Bおよび3参照)。
【0139】
ORF4431は、画分中で同定された最も支配的なタンパク質であったが、組換え脂質化4431に対するマウス抗体は殺菌性ではなく、動物モデルにおいて防御応答をもたらさなかった。ORF5163の付加的な分析は進行中である。
【0140】
本明細書中で記載される調製物の2番目に支配的な成分は、ORF2086の生成物に対応する。
免疫原性方法:
抗血清の調製:
言及される場合を除き、タンパク質組成物/ワクチンは、25μgの総タンパク質で製剤し、20μgQS−21をアジュバントとした。0.2mLの用量を皮下(臀部)注射により、6〜8週齢雌スイスウェブスターマウスに0週目および4週目に投与した。0週目および4週目に血液を採取し、最終瀉血を6週目に実行した。
殺菌アッセイ:
殺菌アッセイは、本質的に記載されたように実行した(Mountzouros and Howell,2000,J.Clin.Microbiol.38(8):2878−2884参照)。SBAについての補体媒介抗体依存性の殺菌力価は、アッセイに導入された標的分子の≧50%を致死させる試験血清の最高希釈の逆数として表した(BC50力価)。
2086タンパク質の同定に用いた方法:
臭化シアン開裂およびフラグメントの直接的配列決定:
陰イオン交換非吸着画分(AEUF)の臭化シアン開裂。AEUFを90%冷エタノールにより沈降させ、70%蟻酸中の10mg/mL臭化シアンを用いて1mg/mLのタンパク質濃度に可溶化した。反応は、暗所において室温で夜通し実行した。開裂生成物を高速真空によって乾燥させ、ペレットを、HE/0.1%還元TX−100で可溶化した。SDS−PAGE、続いてN末端アミノ酸配列決定を用いて、この画分の成分を同定した。
成分を同定するためのプロテアーゼ消化/逆相/N末端配列決定:
AEUFを、GluC(V8)、LysCまたはArgCで消化した。タンパク質と酵素の比は、30μgタンパク質と1μg酵素であった。消化は夜通し37℃にて実行した。消化されたタンパク質混合物(30μg)を、7ミクロンAquapore RF−300カラムに通し、0.1%トリフルオロ酢酸中の10〜95%アセトニトリルの勾配で溶出し、ピークを手作業で収集した。無タンパク質ブランクも走らせ、これからのピークを試料クロマトグラムから減じた。試料ランにおいてのみ生じるピークを質量分析器により分析し、明確な質量を与える試料をN末端アミノ酸配列決定について分析した。
N末端アミノ酸配列決定:
ブロットから切り出されたバンドの場合、タンパク質試料を、SDSゲルからPVDF膜に移し、アミドブラック(脱イオン水中の10%酢酸、0.1%アミドブラック)で染色し、10%酢酸中で脱色した。次に所望のタンパク質バンドを、メタノールで洗浄した円刃刀またはミニ−Exactoナイフを用いて全10レーンから切り出し、Applied Biosystems477Aタンパク質シークエンサーの反応カートリッジに入れた。溶液中の試料の直接配列決定の場合、Prosorbカートリッジを集め、PVDFを60μLのメタノールで湿らせた。PVDFを50μLの脱イオン水で洗浄し、試料(50μL)をPVDFに置いた。50μLの脱イオン水を使用して試料を洗浄した後、Prosorb PVDFを押し抜き、乾燥し、Applied Biosystems477Aタンパク質シークエンサーの反応カートリッジに入れた。両方の方法について、Applied Biosystems N末端シークエンサーを、最適ブロット条件下で12サイクル以上(1サイクル ブランク、1サイクル 標準、および所望の残基同定のための10サイクル以上)走らせ、PTH−アミノ酸検出をApplied Biosystems 120A PTHアナライザーで行った。サイクルは、アナログチャートレコーダーおよび計測器ソフトウェアを介してデジタル式の両方で収集した。アミノ酸の帰属は、アナログおよびデジタルデータの両方を使用して、PTH−アミノ酸の標準セットとアナライザー上でのそれら個々の保持時間の比較により行った(システイン残基は変換中に破壊され、検出されなかった)。複数の配列情報を単一の残基から得ることが可能であり、一次対二次帰属はシグナル強度に基づいて行われた。
LC−MS/MS
IEFにより精製されたタンパク質試料を、SDSポリアクリルアミドゲル電気泳動によりさらに分析した。タンパク質は、クーマシーブルー染色により視覚化し、対象となるバンドを手動で切り出し、その後還元し、アルキル化し、そして自動in−gelトリプシン消化ロボット(1)を用いてin situでトリプシンにより消化(Promega,Madison,WI)した。消化後、ペプチド抽出物を、Savant Speed Vac Concentrator(ThermoQuest,Holdbrook,NY)を使用して、最終容積10〜20μLに濃縮した。
【0141】
ペプチド抽出物を、自動マイクロエレクトロスプレイ逆相HPLCで分析した。簡単に言えば、マイクロエレクトロスプレイインターフェースは、10umC18逆相ビーズ(YMC,Wilmington,NC)で長さ10cmに充填した、Picofrit石英ガラススプレイニードル、長さ50cm、ID75um、8umオリフィス直径(New Objective,Cambridge MA)で構成された。Picofritニードルは、質量分析器の前に位置する手製基部に保持された光ファイバーホルダー(Melles Griot,Irvine,CA)に取り付けた。カラムの後部を、チタンユニオンを介して配管し、エレクトロスプレイインターフェースに電気的接続を供給した。ユニオンを、ある長さの石英ガラスキャピラリー(FSC)管と共に、HPLC溶媒ポンプ(ABI 140C,Perkin−Elmer,Norwalk,CT)に連結されたFAMOSオートサンプラー(LC−Packings,San Francisco,CA)に連結した。HPLC溶媒ポンプは、50μL/分の流量を送達し、これをPEEKマイクロタイトスプリッティングティー(Upchurch Scientific,Oak Harbor,WA)を使用して250nL/分に低下させ、FSC移送ラインを用いてオートサンプラーに送った。LCポンプおよびオートサンプラーは各々、それらの内部ユーザープログラムを使用して制御された。試料をプラスチック製オートサンプラーバイアルに入れ、密封し、5μL試料ループを使用して注入した。
マイクロキャピラリーHPLC−質量分析:
in−gel消化物由来の抽出ペプチドを、マイクロエレクトロスプレイHPLCシステムにより0〜50%溶媒Bの50分間勾配(A:0.1M HOAc、B:90% MeCN/0.1M HOAc)を用いて分離した。ペプチド分析は、スプレイ電位1.5kVで作動し、加熱キャピラリー温度150℃を使用したFinnigan LCQイオントラップ質量分析器(ThermoQuest,San Jose,CA)で行った。データは、機器と共に提供されたデータ取得ソフトウェアを使用して、自動MS/MSモードで得た。取得方法は、1MSスキャン(375〜1200m/z)、続いてMSスキャン中の最も豊富な3種のイオンのMS/MSスキャンを包含した。動的排除関数および同位体排除関数を使用して分析されるペプチドイオンの数を増加させた(設定:3amu=排除幅、3分=排除期間、30秒=排除前期間、3amu=同位体排除幅)。MS/MSデータの自動分析は、N.meningitidisの完全ゲノム由来のタンパク質データベース(Sangerより)を使用して、Finnigan Bioworksデータ解析パッケージに含まれたSEQUESTコンピュータアルゴリズム(ThermoQuest,San Jose,CA)を使用して実行した。研究の結果を図3に示す。
(実施例2)
組換え脂質化P2086(RLP2086)のクローニング:
A.)天然のリーダー配列:
供給源物質:
ORF2086遺伝子を、8529と呼ばれるセログループB Neisseria meningitidis株の臨床分離菌からPCRにより増幅させた。この菌株のセログループ、セロタイプおよびセロサブタイプは、括弧内に示される;8529(B:15、P1:7b、3)。この髄膜炎菌株は、The RIVM,Bilthoven,The Netherlandsから得た。
PCR増幅およびクローニング戦略:
ORF2086の目視検査は、この遺伝子が潜在的なリポタンパク質シグナル配列を有することを示した。特許権によって保護されている隠れマルコフモデルリポタンパク質アルゴリズム(Hidden Markov Model Lipoprotein algorithm)を使用した付加的な解析は、ORF2086がリポタンパク質シグナル配列を含むことを確認した。より自然に近い立体配座でP2086を組換え発現させるために、オリゴヌクレオチドプライマーは、リポタンパク質シグナル配列を持つ全長遺伝子を増幅するように設計し、そしてN.meningitidis A ORF2086のSanger配列の分析に基づいた。2086遺伝子を、ポリメラーゼ連鎖反応(PCR)[ABI2400サーマルサイクラー,Applied Biosystems,Foster City,CA]によりN.meningitidis菌株8529から増幅した。正しいサイズの増幅産物を結合し、pCR2.1−TOPO(Invitrogen)にクローニングした。プラスミドDNAを、NdeIおよびBamHIで制限消化し、ゲル精製し、pET−27b(+)ベクター(Novagen)に結合した。
【0142】
本明細書に記載のオリゴヌクレオチドプライマーを、β−シアノエチルホスホルアミダイト・ケミストリー(Applied Biosystems,Foster City CA)を使用して、PerSeptive Biosystemsオリゴヌクレオチドシンセサイザー(Applied Biosystems,Foster City CA)で合成した。
天然のリーダー配列を使用したrLP2086タンパク質発現:
図5を参照すると、プラスミドpPX7340をBLR(DE3)pLysS宿主細胞(Life Sciences)に形質転換/形質移入、または感染させた。1つの形質転換体を選択し、2%グルコース、カナマイシン(30μg/mL)、クロラムフェニコール(30μg/mL)、およびテトラサイクリン(12μg/mL)を含有するテリフィックブロス(Terrific Broth)50mLに接種した。一晩培養した培養物のOD600は6.0であった。一晩培養した培養物を、1%グリセロールおよび同じ抗生物質を含有するテリフィックブロス1リットルで希釈した。出発OD600は、0.4であった。2時間後、OD600は1.6であり、誘導前試料を採取した。OD600=1に等価の細胞は遠心分離し、上澄みを除去した。全細胞ペレットを150μLのTris−EDTAバッファーおよび150μLの2xSDS−PAGE試料バッファーに再懸濁した。IPTGを添加し、最終濃度1mMとした。3.5時間後、記載されるように誘導後試料を採取し、SDS−PAGEで分析した(図4参照)。
rLP2086の精製:
rLP2086を、ディファレンシャル界面活性剤抽出後にE.coliから可溶化した。天然環境にあるP2086とは異なり、rLP2086は、Triton X−100またはZwittergent 3−12によって著しく可溶化されなかった。rLP2086のバルクをサルコシルで可溶化し、それはN.meningitidisにおける相互作用とは異なってE.coliの外膜成分と相互作用することを示唆した。ひとたび可溶化されると、rLP2086は天然タンパク質と同様に精製され、その中に混在する多くのE.coliタンパク質はアニオン交換樹脂にpH=8で吸着させることにより除去した。その理論的pIより2分の1pH単位上であるにもかかわらず、rLP2086はpH8で吸着されないままであった。それ以上の精製は、pH4.5におけるカチオン交換樹脂へのrLP2086の吸着により達成された。
【0143】
rLP2086の均一性をSDS−PAGE後の図2に示す。rLP2086の質量はMALDI−TOF質量スペクトル分析により27,836であると確認された。この質量は理論的質量の27,100と736異なり、これは細菌リポタンパク質に共通なN末端脂質修飾の質量に近似する。天然およびrLP2086は共に外膜リポタンパク質であると考えられる。N末端配列決定の試みは妨げられ、このことは末端修飾と一致する。
精製方法:
P2086を発現するBLR DE3 pLysS細胞の凍結ペレットを、20mL/g細胞湿重量で10mM HEPES−NaOH/1mM EDTA/1μg/mL Pefabloc SCプロテアーゼ阻害剤(Roche)pH7.4(HEP)に再懸濁し、マイクロフルイダイザー(Microfluidics Corporation Model 110Y)により溶解した。細胞ライゼートは150,000×gで1時間遠心分離した。ペレットはHEPで2回洗浄し、2回遠心分離し、得られた膜ペレットは一晩凍結した。該ペレットは10mM HEPES−NaOH/1mM MgCl2/1%TX−100 pH7.4で30分間可溶化し、続いて150,000xgで30分間遠心分離した。これを3回繰り返した。膜ペレットは上記のように50mM Tris−HCl/5mM EDTA/1%Zwittergent3−12 pH8で2回洗浄し、続いて50mM Tris−HCl/5mM EDTA/1%Zwittergent3−14 pH8および50mM Tris−HCl/5mM EDTA/1%Zwittergent3−14/0.5M NaCl pH8のそれぞれで2回洗浄した。
【0144】
次にrLP2086を、50mM Tris−HCl/5mM EDTA/1%サルコシル pH8で可溶化した。このサルコシル抽出物を1%Zwittergent3−14(Z3−14)に調整し、30倍過剰の50mM Tris−HCl/5mM EDTA/1%Z3−14に対して2回透析した。透析したrLP2086抽出物は90%エタノールで沈殿させ、残存するサルコシルを除去し、50mM Tris−HCl/5mM EDTA/1%Z3−14 pH8(TEZ)で可溶化した。不溶な物質は遠心分離により除去し、上澄みは陰イオン交換クロマトグラフィーカラムを通し、rLP2086を非結合画分に集めた。次に非結合画分を30倍過剰の25mM NaAc/1%Z3−14pH4.5に対して2回透析し、カチオン交換クロマトグラフィーカラムを通した。rLP2086は0〜0.3M NaCl勾配で溶出し、SDS−PAGE(クーマシー染色)により分析した。rLP2086プールはレーザーデンシトメトリーにより84%純粋であることが確認された。
rLP2086サブファミリーBに対する抗血清の表面反応性および殺菌活性:
表VIIを参照すると、サブファミリーB菌株8526由来の精製rLP2086に対する抗血清は、全細胞ELISAにより試験した10種の2086サブファミリーB菌株すべてに対して表面反応性を示した。殺菌活性は異種セロサブタイプ抗原、PorAを発現する10種の2086サブファミリーB菌株のうち9種に対して検出された。これらの菌株は西ヨーロッパ、アメリカ、オーストラリア、およびニュージーランド全体にわたってセログループB髄膜炎菌性疾患を引き起こす代表的な菌株である。殺菌アッセイで致死されなかった唯一の菌株、870227は、全細胞ELISAにより抗rLP2086(サブファミリーB)血清と強く反応し、この菌株がP2086に共通のエピトープを持つタンパク質を発現することを示した。
【0145】
表VIIに挙げた2086サブファミリーA菌株も同様に、全細胞ELISAにより表面反応性を試験した。これらの菌株3種の中の2種は非常に低レベルの反応性を有することが分かり、いくつかの2086サブファミリーA菌株はrLP2086サブファミリーBに対して産生された抗体と交差反応性ではない可能性があることを示した。菌株8529から2086サブファミリーB遺伝子を同定するために使用されるPCR増幅手順もまた、菌株870446、NMBおよび6557に対して実行した。2086サブファミリーBのPCR増幅産物は全く検出されなかった。
免疫原性方法:
抗血清の調製:
ワクチンを、前に実施例1に記載されるように製剤した。しかしながら、10μg投与量を使用した。
全細胞酵素結合免疫吸着アッセイ(ELISA):
N.meningitidis全細胞懸濁液を、無菌0.01M ホスフェート、0.137M NaCl、0.002M KCl(PBS)中、620nmにおける吸光度0.1に希釈した。この懸濁液から、0.1mLをNunc Bac T96ウェルプレート(Cat# 2−69620)の各々のウェルに添加した。細胞は室温で3日間、プレート上で乾燥し、その後覆い、反転させ、4℃で保存した。プレートは洗浄バッファー(0.01M Tris−HCl、0.139M NaCl/KCl、0.1%ドデシルポリ(オキシエチレレングリコールエーテル)n n=23(Brij−35(登録商標)、ICI Americas,Inc.,Wilmington,Delawareから入手可能)、pH7.0〜7.4)で3回洗浄した。抗血清の希釈物を、PBS、0.05%Tween−20/アジドで調製し、そして0.1mLを被覆したプレートに移した。プレートを2時間37℃でインキュベートした。プレートは洗浄バッファーで3回洗浄した。ヤギ−抗マウスIgG AP(Southern Biotech)をPBS/0.05%Tween−20で1:1500に希釈し、0.1mLを各々のウェルに添加し、プレートを37℃で2時間インキュベートした。プレートは(上記のように)洗浄した。基質溶液は、1M ジエタノールアミン/0.5mM MgCl2中にp−ニトロフェニルホスフェート(Sigma)を1mg/mLに溶解することにより調製した。基質はウェルあたり0.1mLでプレートに添加し、室温で1時間インキュベートした。反応は50μL/ウェルの3N NaOHで停止させ、プレートは、690nmを基準として、405nmで読み取った。
B.)P4リーダー配列:
PCR増幅およびクローニング法:
rLP2086発現を最適化するために、2086遺伝子を型分類不能なHaemophilus influenzaeのP4シグナル配列の後にクローニングした(Greenら、1991)。リポタンパク質クローニングに使用されたプライマーが表IVに列挙してあり、化合物番号:5658、5660、6473、6543および6385によって識別される。ORF2086は、以下の化合物番号5658および5660を持つプライマーを使用してN.meningitidisセログループB菌株8529から増幅された。ORF2086は、以下の化合物番号6385および5660を持つプライマーを使用してN.meningitidisセログループB菌株CDC1573から増幅された。ORF2086は、以下の化合物番号6473および6543を持つプライマーを使用してN.meningitidisセログループB菌株2996から増幅された。N末端(5’)プライマーは、2086遺伝子の成熟領域(システインのすぐ下流のアミノ酸位置3のセリン残基において開始)に相同であるように設計した。制限部位BamHI(GGATTC)はそれぞれのN−末端プライマーの5’末端に組み込まれ、アミノ酸位置2において成熟タンパク質にグリシン残基が挿入された。C末端(3’)プライマーは2086遺伝子のC−末端に相同であるように設計され、終止コドンおよびクローニングのためにSphI部位を含有した。各N.meningitidis B菌株から増幅されたフラグメントを、中間ベクターにクローニングし、配列分析によってスクリーニングした。
【0146】
正しいクローン由来のプラスミドDNAを、BamHIおよびSphI制限酵素(New England Biolabs,(NEB))で消化した。pLP339という名称のベクター(出願者の譲受人により供給された)を発現ベクターとして選択した。このベクターは、pBAD18−Cmバックボーン(Beckwithら、1995)を使用し、P4リポタンパク質シグナル配列および型分類不能なヘモフィルス インフルエンザのP4遺伝子(Greenら、1991)を含有する。pLP339ベクターは、制限酵素BamHIにより部分的に消化され、その後SphIで消化された。増幅された2086フラグメント(BamHI/SphI)は各々別個にpLP339ベクター(部分的BamHI/SphI)に結合された。このクローニング戦略は、成熟2086遺伝子をP4リポタンパク質シグナル配列の後に配置する。BamHI部位はP4シグナル配列と2086遺伝子間のクローニング接合部に残る(図7に示したプラスミド構築物を参照されたい)。以下のものはBamHIクローニング接合部における配列の例である:
[P4シグナル配列]−TGT GGA TCC−[残存2086成熟核酸配列]
[P4シグナル配列]−Cys Gly Ser−[残存2086成熟アミノ酸配列]
図7を参照すると、各増幅されたフラグメントはP4リーダー配列を含有する改変pBAD18−Cmベクターにクローニングされた。発酵はrP4LP2086(組換えP4脂質化2086)を発現する組換えE.coli BLR pPX7343で行い、付加的グルコースの添加により細胞密度増大を試みた。発酵装置は、1%グルコースを補足したSambrookの完全M9最小培地10Lで満たした。
【0147】
発酵装置中のグルコースの初期濃度は45g/Lであった。初期ODが約0.25になるように発酵装置に菌を接種した。約OD25で、付加的な20g/Lグルコースを添加した。培養物はOD63.4、グルコース枯渇において1%アラビノースで誘導した。発酵は、誘導後3時間まで継続した。試料は誘導後t=0、1、2、3に保存し、タンパク質はBSAを用いて定量した。t=3において、タンパク質収量は約0.35g/L、そして7%総細胞タンパク質であった。約10Lの培養物から全部で895グラムの湿細胞ペーストが採取された。
【0148】
rP4LP2086の精製は、上記の実施例2、節Aに記載のものと同じ方法を使用して行った。
【0149】
本明細書に記載のオリゴヌクレオチドプライマーを、−シアノエチルホスホルアミダイト・ケミストリー(Applied Biosystems,Foster City CA)を使用して、PerSeptive Biosystemsオリゴヌクレオチドシンセサイザー(Applied Biosystems,Foster City CA)で合成した。ORF2086遺伝子ファミリーのPCR増幅に使用されるプライマーは表IVに挙げられ、それは本発明のプライマーの非限定的な例を示す。
【0150】
【表4】
(実施例3)
非脂質化成熟2086タンパク質の発生遺伝学:
2086タンパク質の免疫原性をさらに評価するため、P2086の非脂質化型のクローニングおよび発現を行った。
ORF2086のPCR遺伝子増幅:
様々な染色由来の2086遺伝子が、参照により本明細書に組み込まれる国際出願PCT/US02/32369号(2003年8月7日、国際公開第03/063766号として公開)、および国際出願PCT/US04/11901号(2004年11月4日、国際公開第04/094596号として公開)に記載されるように、プライマーで増幅され得る。
【0151】
これらのプライマーの特徴としては、各プライマーにおける合成BglII制限部位、化合物番号6406および6474における合成NdeI制限部位が含まれ、そして3種すべてのリーディングフレームの停止コドンは化合物番号5135および6605に存在する。プライマー番号6406および6474は、第2のアミノ末端コドン(ACG)に融合したATG(Met)を持つ2086遺伝子を増幅し、成熟2086ポリペプチドの単一アミノ酸置換(TGC Cysを置換)を表す。
【0152】
PCRクローニングベクターは、TOPO−PCR2.1、Invitrogen,Valencia,CAであった。
【0153】
非脂質化2086タンパク質を発現するために使用されたベクターはNovagen,Madison,WIのpET9aであった。
【0154】
E.coliクローニング菌株は、Top10,Invitrogen,Carlsbad,CAであった。
【0155】
E.coli発現菌株は、BLR(DE3)pLysS,Novagen,Madison,WIであった。
【0156】
クローニング目的のための培地は、グリセロールの代わりの1%無菌グルコース、および適切な抗生物質(アンピシリンまたはカナマイシン)を含む、Sambrookら、によるテリフィックブロス液または寒天培地であった。
【0157】
プラスミド精製は、Qiagen Spin Miniprepキット(Valencia,CA)により行った。
非脂質化2086発現のための産生菌株または細胞系統の調製:
2086遺伝子は、髄膜炎菌株8529に由来する染色体DNAからポリメラーゼ連鎖反応(PCR)[AmpliTaq and ABI2400サーマルサイクラー,Applied Biosystems,Foster City,CA]により増幅した。2086遺伝子のPCR増幅は、それぞれの反応において化合物番号6474および5135によって同定された2種のオリゴヌレオチドプライマーを使用した。増幅された2086PCR産物はTOPO−PCR2.1クローニングベクターに直接クローニングし、100μg/mlアンピシリンおよび20μg/ml X−Galを補ったテリフィックブロス寒天培地で選択した。白色コロニーを選択し、培養した。プラスミドDNAは、Qiagen miniprepキットを使用して調製し、該プラスミドをPCRフラグメントインサートに関してスクリーニングした。PCRインサートプラスミドを、DNA配列決定にかけた(ABI377シークエンサーでのBig Dye化学,Applied Biosystems,Foster City,CA)。
【0158】
適切なDNA配列を示すプラスミドを、BglII制限酵素で消化し、BglIIフラグメントを、GeneClean II精製キット(Bio101、Carlsbad,CA)を使用してゲル精製した。精製BglIIフラグメントを、発現ベクターpET9aのBamHI部位にクローニングした。pET9a/2086クローンは30μg/mlのカナマイシンを補ったテリフィックブロスプレート上で選択した。カナマイシン耐性クローンを培養し、miniprepプラスミドDNAを調製した。プラスミドはBamHI部位における2086遺伝子の適切な配向に関してスクリーニングした。適切に配置されたプラスミドは、2086遺伝子のアミノ末端へのT7−抗原の融合物(rP2086T7)を表す。これらのrP2086T7遺伝子融合物を、BLR(DE3)pLysSに形質転換し、テリフィックブロス/Kanプレート上で選択し、テリフィックブロスで培養し、rP2086T7融合タンパク質を発現するように1mM IPTG(イソプロピルβ−D−チオガラクトピラノシド)により誘導した。rP2086T7融合タンパク質は高レベルで発現された。
【0159】
次に、これらの融合プラスミドをNdeI制限消化にかけ、それによりT7抗原を欠失し、ベクターにより提供されたATGスタートに成熟2086遺伝子を直接連結する。これらのNdeI欠失プラスミドを、Top10細胞に形質転換し、テリフィックブロス/Kanプレート上で選択した。候補クローンを培養し、ミニプレッププラスミドDNAを調製した。プラスミドDNAをDNA配列決定にかけ、欠失および2086遺伝子配列の完全性を確認した。これらのプラスミドはpPX7328と呼ぶプラスミドマップによって表される(図6)。適切なDNA配列を表すプラスミドを、BLR(DE3)pLysSに形質転換し、テリフィックブロス/Kanプレート上で選択し、テリフィックブロスで培養し、IPTGにより2086タンパク質を発現するように誘導した。pET9aベクターは、T7タグが除去されると、菌株BLR(DE3)pLysSに成熟2086タンパク質を発現できなかった。
非脂質化2086タンパク質の産生:
精製プラスミドDNAを用い、発現菌株BLR(DE3)pLysSを形質転換した。プラスミドを持つBLR(DE3)pLysS細胞は、カナマイシン耐性で、1mM IPTGの添加により高レベルのPorAタンパク質を発現するように誘導することができる。rP2086T7融合タンパク質は、E.coli細胞系統BLR(DE3)pLysS中に不溶性封入体として、全タンパク質の約40%において発現させることができる。この精製融合タンパク質は、マウスを免疫するために用いられ、異種髄膜炎菌株に対する有意レベルの殺菌抗体を生成した(表V参照)。
2086非脂質化遺伝子変異誘発:
PCRプライマー変異誘発は2086遺伝子の5’末端で行われた。成熟rP2086T7の高発現レベルを示しつつT7−タグを除去できるかどうかを確かめるための発現研究が進行中である。
非脂質化rP2086T7の精製:
非脂質化rP2086T7を発現するE.coliBLR(DE3)pLysS細胞を、10mM HEPES−NaOH/5mM EDTA/1mM Pefabloc SC pH7.4中で、マイクロフルイダイザーにより溶解した。細胞ライゼートを18,000xgで30分間遠心分離した。封入体ペレットを50mM Tris−HCl/5mM EDTA/1%Triton X−100 pH8で3回洗浄し、都度24,000xgで30分間遠心分離した。次に封入体ペレットを50mM Tris−HCl/5mM EDTA/1%Zwittergent3−14 pH8で2回洗浄し、都度24,000xgで15分間遠心分離した。続いて封入体ペレットを50mM Tris−HCl/5mM EDTA/4M 尿素 pH8で2時間可溶化し、その後不溶性物質を遠心分離により除去した。上澄み(可溶化rP2086T7)を4つの等しい試料に分けた。保存溶液を使用して、1つは50mM Tris−HCl/5mM EDTA/250mM NaCl/2M 尿素 pH8(界面活性剤を含まない)に調整し、1つは50mM Tris−HCl/5mM EDTA/250mM NaCl/2M 尿素/1%水素化Triton X−100 pH8(TX−100)に調整し、1つは50mM Tris−HCl/5mM EDTA/250mM NaCl/2M 尿素/1%Zwittergent3−12 pH8(Z3−12)に調整し、そして1つは50mM Tris−HCl/5mM EDTA/250mM NaCl/2M 尿素/1%Zwittergent3−14 pH8(Z3−14)に調整した。尿素を除去するために、尿素を含まないそれぞれのバッファーに対して試料を透析した。次に尿素を含まずに60mM NaClを含むそれぞれのバッファーに対して試料を透析し、NaCl濃度を減少させた。不溶性物質は2,000xgで15分間遠心分離することにより除去し、得られた上澄み(再生rP2086T7)をその後の実験に使用した。rP2086T7の均一性は、クーマシー染色したSDS−PAGEおよびレーザーデンシトメトリーを使用して測定し、91〜95%であることが見出された。
【0160】
免疫原性手順−実施例2に記載の通り
この精製融合タンパク質を使用してマウスを免疫化し、異種髄膜炎菌株に対する有意レベルの殺菌抗体を生成した。(以下の表V参照):
【0161】
【表5】
(* rLP2086によるマウスの免疫化によって生成された陽性対照血清)
(実施例4)
ORF2086のキメラクローンの開発
菌株CDC−1573由来の2086タンパク質N末端領域は、菌株8529および2996由来の2086遺伝子には存在しない反復部分を含有する(図8参照)。この反復部分は、E.coliベースの2つの発現系(pETおよびpBAD)由来の高レベルの組換え2086タンパク質発現に関与すると考えられる。pETおよびpBAD発現系におけるCDC−1573 2086遺伝子由来の組換えタンパク質発現レベルは、同じ系を使用した菌株8529および2996による2086遺伝子由来の組換え体発現レベルに比較して有意に良好であった。3種すべての菌株に由来する2086遺伝子のN末端領域は、この反復部分以外は比較的相同である。したがって、pETおよびpBAD系を使用した場合、菌株8529および2996由来の2086遺伝子にCDC−1573N末端を融合することにより、これらの遺伝子から発現される組換え2086タンパク質レベルが増大すると考えることは妥当である。
材料および方法:
菌株8529および2996由来の染色体DNAを精製し、キメラ2086遺伝子のPCR増幅のための鋳型として使用した。化合物番号6721および5135を持つPCRプライマーを使用して菌株8529由来のキメラ2086遺伝子を増幅し、化合物番号6721および6605を持つPCRプライマーを使用して、菌株2996由来のキメラ2086遺伝子を増幅した。PCR産物は、InvitrogenのPCR2.1 TOPOベクターに直接クローニングし、次にDNA配列分析によりスクリーニングして、無傷のキメラ2086遺伝子を同定した。次にその遺伝子をBglIIによりPCR2.1ベクターから開裂し、BglIIフラグメントをpET9aプラスミドのBamHI部位に挿入した。プラスミドインサートが適切な配向であることをスクリーニングした後、NdeI消化にかけた。線状NdeIフラグメントは自己連結し、pET9aベクターによって与えられたT7−タグ配列を含有する小NdeIフラグメントの欠失が達成された。この欠失は、キメラ2086遺伝子の5’末端にT7プロモーターを直接結合する。NdeI欠失プラスミドをE.coli菌株BL21(DE3)に形質転換し、カナマイシン耐性コロニーをIPTG誘導によるキメラ2086タンパク質発現に関してスクリーニングした。
【0162】
初期の研究は、pET9a系で発現される場合、菌株2996由来のキメラ2086遺伝子は、天然の2996/2086遺伝子に比較して2倍の組換えタンパク質を発現することを示す。pBAD系は現在のところ試験されていない。
【0163】
ただ1回の実験が行われただけであるが、データは、キメラ2086遺伝子に由来する、増大した用途があることを示唆する。菌株8529および2996由来2086遺伝子に対するCDC−1573N−末端融合物の生成により、増大した組換え2086タンパク質発現がもたらされる。
(実施例5)
N.meningitidis菌株の2086PCRスクリーニング:
臨床分離菌間の2086遺伝子の保存を確かめるために、88のN.meningitidis菌株にPCR増幅を行った。
【0164】
ORF2086の当初のPCR同定は、化合物番号:4623、4624および4625により同定された表IV(先の実施例2参照)に列挙されるプライマーを使用した。これらのプライマーは、SangerのN.meningitidisセログループA配列に基づいて設計された。多数の菌株のスクリーニングを容易にするため、内部プライマーが2086遺伝子に関して設計された。全部で88のN.meningitidis菌株を、化合物番号5005および5007により同定された、新規に設計された内部2086プライマーを用いて、PCRによりスクリーニングした。これらのプライマーを用い、本出願人らは、88のN.meningitidis菌株の中の63(約70%)を同定することができた(表VI−A参照)。
【0165】
SangerのN.meningitidisセログループA配列およびTIGRのN.meningitidisセログループB配列中の2086遺伝子を囲む伸長領域を調べ、並べた。プライマーは、2086遺伝子の上流および下流領域に対応するように設計した。目的は、配列比較のためにこれらのプライマーを使用して、多様なN.meningitidis菌株から全長2086遺伝子より大きいものを増幅することであった。化合物No.6470および6472を使用した1種の菌株(6577)のPCR増幅は、低収率の産物という結果になった。菌株6557増幅産物をクローニングし、プラスミドDNAを配列分析にかけた。結果は、以前見られたよりも配列可変性の大きい新型の2086遺伝子を示した。菌株6557由来の2086遺伝子は、配列決定された他の菌株に対してアミノ酸レベルで約75%同一であった。興味深いことに、菌株6557は、上述の2086PCRスクリーニングにより以前陰性とされた菌株30%の中の1種であった。
【0166】
菌株6557内のC末端可変領域に特異的な内部プライマーが設計された。これらのプライマーを使用して、2086PCRスクリーニングによる試験で以前陰性とされた約30%の菌株中のより可変性の2086遺伝子をスクリーニングした。これらの新規に同定された内部2086プライマー(化合物番号6495および6496により識別)を使用して、入手可能なすべてのN.meningitidis菌株(n=88)をPCRによりスクリーニングした。2086に関してPCRにより以前陰性とされた約30%のN.meningitidis菌株だけがこのスクリーニングでPCR陽性であった。以前のPCR陰性(約30%)菌株から増幅された遺伝子の組は、新型2086遺伝子または2086遺伝子の第2ファミリーを表すべきであり、本明細書では2086サブファミリーAと呼ぶ。8529由来のプライマーにより約70%の菌株から増幅された2086遺伝子の組は、本明細書ではサブファミリーBと呼ぶ。
【0167】
PCR増幅研究に使用されるN.meningitidis菌株は、以下の表、表VI Aおよび表VI Bから選択された。表に列挙された菌株は、前に本明細書中で開示されたものに加えて、N.meningitidis菌株の例として提供されるが、それらに限定されない。表VI Aに列挙された菌株は、2086タンパク質サブファミリーAに分類され、表VI Bに列挙された菌株は、2086タンパク質サブファミリーBに分類される。各表中に列挙された菌株は、セロサブタイプにより分類される。菌株は表中に示すような以下の4つの供与源から入手可能である:MPHL−Manchester Public Health Laboratory,Manchester,UK;RIVM,Bilthoven,The Netherlands;University of Iowa,College of Medicine,Department of Microbiology,Iowa City,IA;およびWalter Reed Army Institute of Research,Washington,D.C.
【0168】
【表6A】
【0169】
【表6B−1】
【0170】
【表6B−2】
その他の菌株は、感染した個体から分離菌として容易に入手可能である。
(実施例6)
髄膜炎菌株に対するrLP2086抗血清の反応性:
以下の表、表VIIは、上述のrLP2086の交差反応性および交差保護能力を示す。表に示すように、rLP2086は、全細胞ELISA(WCE)力価、殺菌アッセイ(BCA)および幼若ラット(IR)アッセイを含む、多様な手法を使用して処理および分析して、2086タンパク質に対して引き起こされたポリクローナル抗体の細菌表面反応性を確認した。
【0171】
【表7】
+ 菌血病において10分の1減少より大
− 菌血病において10分の1減少より小
(実施例7)
ORF2086タンパク質を発現させるための種々の構築物を調製した。以下の表、表VIIIは、例を示し、本発明の実施を例示するために提供されるr2086構築物表であるが、それらに限定されない。
【0172】
【表8】
(実施例8)
LOS除去外膜タンパク質を用いたさらなる研究から、異種セロサブタイプを発現する菌株に殺菌抗体を誘導することができる、PorA以外の外膜タンパク質(単数または複数)を産生する付加的な菌株を同定した。以下に、本発明の一実施態様による付加的なタンパク質、具体的には髄膜炎菌免疫原性組成物に必要なタンパク質の数を減少させることができる外膜リポタンパク質を同定するためのさらなる研究を記載する。これらのさらなる研究は、以前の実施例に記載された研究を補足する。
【0173】
複数菌株に対する免疫および殺菌アッセイと共に、細胞下分画、ディファレンシャル界面活性剤抽出、等電点分離法、およびイオン交換クロマトグラフィーを使用して、対象となる小グループのタンパク質を同定した。主成分の直接配列決定は、N末端が遮断されていることを示した。内部タンパク質配列は、化学的およびタンパク質分解消化に由来するポリペプチドの直接配列決定により得た。グループA髄膜炎菌株のゲノム配列は、Sanger Centerからダウンロードし、既存および本出願者らが所有するアルゴリズムを使用して本出願者らのバイオインフォマティックグループが分析し、検索データベースを調製した。ペプチド配列データは、ORF2086が関心のあるものであることを示した。このorfに基づいたプライマーを使用して、菌株8529からP2086遺伝子をPCRした。遺伝子配列の分析、N末端が遮断されているという事実、およびその細胞下局在は、P2086が脂質化外膜タンパク質(LP2086)であることを示した。rLP2086−8529および別の髄膜炎菌株由来の変異体は、H.インフルエンザP4シグナル配列を使用してE.coli中でリポタンパク質として組換え発現された。これらの組換えタンパク質を、ディファレンシャル界面活性剤抽出によりE.coli膜から単離し、イオン交換クロマトグラフィーを使用して精製し、マウスを免疫するために使用した。マウス抗LP2086血清は、N.meningitidisのいくつかの異なるセロサブタイプ株に対する殺菌活性の促進が可能であった。多くのN.meningitidis菌株由来のP2086遺伝子のさらなる分析は、これらの配列がサブファミリーAおよびサブファミリーBと呼ばれる2種のグループに分類されることを示した。(図12参照)。サブファミリーBタンパク質に対して誘導された抗血清はサブファミリーBタンパク質を発現する9種の菌株、およびサブファミリーAタンパク質を発現する1種の菌株に対して殺菌性であった。サブファミリーA抗血清はサブファミリーA系統に対して殺菌性であった。1種のrPorAと1種のrLP2086の混合物は、いずれかのタンパク質だけによって誘導されたもの以上の広いワクチン適用範囲を持つ相補的抗体を誘導した。
【0174】
これらの観察は以下の結論に至る。rLP2086抗原は、異種PorAおよび異種P2086タンパク質を発現する髄膜炎菌株に対して殺菌抗体を誘導することができる。抗原のP2086ファミリーは、単独または別の髄膜炎菌抗原との組み合わせのいずれかで、有用なワクチンであるか、または免疫原性であり得る。
【0175】
以下、先の研究について詳細に説明する。可溶性外膜タンパク質(sOMPs)の複合体混合物は、異種PorAタンパク質を発現する菌株に対してPorA非依存的殺菌抗体を誘導することが見出された。ディファレンシャル界面活性剤抽出、等電点分離法およびイオン交換クロマトグラフィー、続くマウス免疫化の工程を使用して免疫学的に活性な成分を追跡した。
【0176】
各ステップで、髄膜炎菌性疾患の世界的疫学を代表するセロサブタイプ抗原を含有するいくつかの菌株に対して、血清は表面反応性および殺菌活性についてアッセイされた。
【0177】
この分離および免疫化の工程を使用して、グループB N.meningitidisに関する新規な交差反応性免疫原性候補を同定した。
【0178】
PorA欠乏菌株の調製−porA染色体座位を、菌株2996由来のプラスミドpPX7016にクローニングした。プラスミド内では、porAプロモーター、S/Dボックスおよび初めの38のN末端コドンが欠失し、自己含有KanR発現カセットにより置換されている。該プラスミドは制限酵素で線状にされ、セロサブタイプ菌株PI:5,2;PI:9;PI:7,16;PI:15;PI:4;PI:3&PI:10に自然に形質転換された。カナマイシン耐性形質転換体を選択し、ELISAにおいてセロサブタイプ特異的モノクローナル抗体によりPorAの損失をスクリーニングした。
【0179】
殺菌アッセイ:Mountzouros,K.T.and Howell,A.P.Detection of Complement−Medicated Antibody−Dependent Bactericidal Activity in a Fluorescence−Based Serum Bactericidal Assay for Group b Neisseria meningitidis.J Clin Microbiol.2000;38:2878−2884を参照されたい。
【0180】
全細胞酵素結合免疫吸着アッセイ(ELISA):N.meningitidis全細胞懸濁液を、滅菌0.01Mホスフェート、0.137M NaCl、0.002M KCl(PBS)中で、620nmにおける光学密度度0.1に希釈した。この懸濁液から、Nunc Bac T96ウェルプレート(Cat#2−69620)の各ウェルに0.1mL添加した。細胞を37℃で一晩、プレート上で乾燥させ、その後被覆し、反転させ、4℃で保存した。プレートは、洗浄バッファー(0.01M Tris−HCl、0.139M NaCl/KCl、0.1% Brij−35、pH7.0〜7.4)で3回洗浄した。抗血清の希釈物は、PBS、0.05%Tween−20/アジドで調製し、0.1mLを被覆プレートに移し、37℃で2時間インキュベートした。プレートは洗浄バッファー中で3回洗浄した。ヤギ抗マウスIgG AP(Southern Biotech)をPBS/0.05%Tween−20で1:1500に希釈し、0.1mLを各ウェルに添加し、プレートを37℃で2時間インキュベートした。プレートを(上記の通り)洗浄した。基質溶液は、ジエタノールアミンで、p−ニトロフェニルホスフェート(Sigma)を1mg/mlに希釈することにより調製した。ウェルあたり0.1mLの基質をプレートに添加し、室温で1時間インキュベートした。反応は50ul/ウェルの3N NaOHで停止し、690nmを参照として405nmでプレートを読んだ。
【0181】
組換えPorA誘導:BLR(DE3)/pET9a菌株を、Kan−30および2%グルコースを補ったハイソイ(HySoy)ブロス(Sheffield Products)中で、37℃で一晩培養した。翌朝、O/N培養物をハイソイブロス、Kan−30および1%グリセロールで希釈し、37℃で1時間培養した。これらの培養物にIPTGを最終濃度1mMで添加することにより誘導した。培養物はさらに2〜3時間培養し、その後採取した。
【0182】
組換えPorA精製:rPorAを、E.coli封入体から8M尿素で可溶化し、尿素を全く含まないバッファーに対して透析して再生した。再生されたrPorAを透析ろ過により濃縮し、バッファーはG25カラムでNaPO4 pH6に交換した。次に、透析したrPorAをカチオン交換カラム(S Fractogel)に流し、1M NaClで溶出した。
【0183】
菌株8529(P1.7−2,3)のsOMPsは、異種セロサブタイプを発現する菌株に対し、マウスにおいてPorA非依存性殺菌活性を誘導する。以下の表、表IXは、調べた菌株における殺菌活性を示す。
【0184】
【表9−1】
【0185】
【表9−2】
sOMPsの調製:N.meningitidis膜を、TX−100、Zwittergent3−14,およびZwittergent3−14+0.5N NaClで抽出した。上記のsOMPを、Zwittergent3−14/0.5M NaCl抽出物中で可溶化した。抽出は当業者に周知の技術、例えば、参照により本明細書に組み込まれる米国特許第6,355,253号を使用して実行する。
【0186】
免疫原性:雌スイスウェブスターマウスを、0週目および4週目に20ug QS−21をアジュバントとして添加した25ug総タンパク質量で免疫化した。瀉血およびデータ分析を6週目に行った。
【0187】
1 殺菌性(BC50)力価(生細胞数を50%減少させる抗血清の希釈度の逆数として表される)。0週目の正常マウス血清のBC50力価は<25であった。
【0188】
2 NST=セロタイプ分類不能
以下の表、表Xはおよび表XIは、サブファミリーAおよびサブファミリーB両方の組換え脂質化P2086(rLP2086)の精製および特徴の概要を示す。
サブファミリーA rLP2086精製
【0189】
【表10】
サブファミリーB rLP2086精製
【0190】
【表11】
精製法:すべての変異体は、TX−100によりE.coli膜から可溶化された(サルコシルまたは尿素で可溶化されたrLP2086−8529を除く)。さらなる精製は、Tris−HClまたはNaPO4バッファー中でアニオン交換(TMAE)、サイズ排除および/またはカチオン交換(S Fractogel)クロマトグラフィーの組み合わせにより行った。
【0191】
1 菌株8529由来のP2086と比較したアミノ酸相同性
2 SDS−PAGEおよびコロイド状クーマシー染色バンド(Simply Blue染色)のレーザーデンシトメトリーにより確認した純度
同種および異種菌株に対して試験された、サブファミリーBメンバー、rLP2086−8529の免疫原性。
【0192】
以下の表XIIは、同種および異種菌株に対して試験された、サブファミリーBメンバー、rLP2086−8529の免疫原性を示す。
【0193】
【表12】
ワクチン接種手順:6〜8週齢雌スイスウェブスターマウスを、0週目および4週目に10ug rLP2086−8529+20ug QS−21で免疫化した。データ分析は6週目の瀉血により行った。
【0194】
a rLP2086−8529と比較したP2086のアミノ酸相同性
b 吸光度=0.1における希釈度の逆数として表したエンドポイント力価
c 生細胞数を50%減少させる抗血清の希釈度の逆数として表したBC50力価。0週目の正常マウス血清のBC50力価は<10であった。
【0195】
表XIIIは、同種および異種菌株に対して試験した、サブファミリーBメンバー、rLP2086−2996の免疫原性を示す。
【0196】
【表13】
ワクチン接種手順:6〜8週齢雌スイスウェブスターマウスを、0週目および4週目に10ug rLP2086−2996+20ug QS−21で免疫化した。データ分析は6週目の瀉血で行った。
【0197】
a rLP2086−2996と比較したP2086のアミノ酸相同性
b 吸光度=0.1における希釈度の逆数として表したエンドポイント力価
c 生細胞数を50%減少させる抗血清の希釈度の逆数として表した殺菌性(BC50)力価。0週目の正常マウス血清のBC50力価は<10であった。
【0198】
以下の表XIVは、rLP2086およびrPorAに対する抗血清は、混合され殺菌活性をアッセイした場合に相補的であることを示す。
【0199】
【表14】
ワクチン接種手順:6〜8週齢雌スイスウェブスターマウスを、0週目および4週目に10ug rLP2086−8529/20ug QS−21か、15ug rPorA/100ug MPLのいずれかで免疫化した。データ分析は、6週目の瀉血に関して実行した。
【0200】
a 生細胞数を50%減少させる抗血清の希釈度の逆数として表した殺菌性(BC50)力価。0週目の正常マウス血清のBC50力価は<10であった。
【0201】
以下の表XVは、rLP2086サブファミリーと2種のrPorAとの混合物がマウスに殺菌抗体を誘導することを示す。
【0202】
【表15】
ワクチン接種手順:6〜8週齢雌スイスウェブスターマウスを、0週目および4週目に各タンパク質10ug+20ug QS−21で免疫化した。データ分析は6週目の瀉血で行った。
【0203】
a 生細胞数を50%減少させる抗血清の希釈度の逆数として表した殺菌性(BC50)力価。0週目の正常マウス血清のBC50力価は<10であった。
【0204】
b SfA−サブファミリーA、SfB−サブファミリーB
c 適切な1価対照:rLP2086−8529、rLP2086−2996、rP1.5−1,2−2またはrP1.22−1,14−1抗血清
以下に上記の研究の結果を概説する。抗rLP2086抗血清は、13/16の試験菌株に対して殺菌性である。異なるセロサブタイプを発現する11種の菌株は抗P2086血清により致死する。抗rLP2086血清の殺菌活性は抗rPorA血清に相補的である。P2086とPorAとの混合物は、マウスに相補的殺菌抗体を誘導する。ディファレンシャル界面活性剤抽出、精製および免疫、ならびに多くの菌株に対する機能的抗体アッセイを使用して、新規ワクチン候補を同定することができる。P2086は、P2086およびrPorAの両方において異種の菌株に対し殺菌抗体を誘導するワクチン候補として同定されている。したがって、タンパク質の2086ファミリーは、単独で、または別の髄膜炎菌抗原と組み合わせて有用なワクチンであり得る。
(実施例9)
変化するセログループの髄膜炎菌株を、ORF2086遺伝子の存在についてPCRによりスクリーニングした。結局、100種以上の髄膜炎菌株をスクリーニングした。
【0205】
C末端可変領域に特異的な2組の内部PCRプライマーを利用して、サブファミリーAおよびB遺伝子配列を識別した。約350bpのPCR増幅産物の存在は、2086遺伝子配列が染色体上に存在することを示した。すべての菌株は、予想されたサイズの単一のPCR産物を生じた。全長ORF2086遺伝子のヌクレオチド配列を決定し、整列させ(DNAStar MegAlign)、系統樹を調製するために使用した。
【0206】
2086遺伝子は、pBADアラビノース誘導性プロモーター系においてrLP2086リポタンパク質として組換え発現され、またはIPTG誘導性pET系においてrLP2086非脂質化タンパク質として組換え発現された。これらの組換えタンパク質は、E.coli Bにおいて発現させた。精製組換えタンパク質を使用してマウスを免疫化し、マウス抗血清を、その血清IgG力価および多様な髄膜炎菌株に対する殺菌活性についてアッセイした。
【0207】
ORF2086を、以下のものうちの1つからPCRにより増幅した:全髄膜炎菌細胞、精製染色体DNAまたはプラスミドDNA鋳型。
【0208】
ORF2086遺伝子をベクターpLP339にクローニングし、これはヘモフィルスP4リーダー配列をORF2086遺伝子の5’末端に融合する。E.coli菌株BLRは、pBAD/ORF2086クローン由来のrP2086の脂質化型の組換え発現のための宿主菌株として使用した。(図10A参照)。pBADアラビノース誘導性プロモーターは、P4シグナル/ORF2086融合タンパク質の発現を操作して、rP2086の脂質化型を発現させる。シグナル配列を欠くP2086遺伝子を、高活性T7ファージプロモーターの後にpET9aベクターにクローニングされた。E.coli菌株BL21(DE3)は、pET9a/ORF2086クローン由来のORF2086の非脂質化型の組換え発現のための宿主菌株として使用した。E.coli菌株BL21におけるDE3溶原菌は、lacUV5プロモーターの制御下で、IPTGの添加によりに誘導されてT7RNAポリメラーゼを発現することができる。WCE;FEMS Micro.Lett.,48(1987)367−371およびBCA;J.Clin.Microbiol.,38(2000)2878−2884を参照されたい。
【0209】
遺伝子、ORF2086は、異なるN.meningitidis菌株からクローニングおよび配列決定された。ヌクレオチド配列を整列させ(DNAStar MegAlign)、系統樹調製に使用した。この樹は、ORF2086遺伝子ヌクレオチド配列の2種の異なるサブファミリーを明らかにする。2つの遺伝子サブファミリーは、それらの5’末端において類似するが、それらの3’末端近くではかなりの変化を含む。かなりの変動性があると考えられるが、遺伝子のある種の重要な領域は、異なる菌株間で高度に相同性である。理論により拘束されることを意図することなく、これらの保存された領域は、タンパク質に対して機能的連続性を提供することができ、そしてワクチン標的として利用されることになる交差防御エピトープを表し得る。
【0210】
2086遺伝子は、いくつかのセログループB髄膜炎菌株からクローニングされ、脂質化シグナル配列を伴って、またはそれを伴わずに発現された。T7−タグに融合された非脂質化型は、最高レベルで発現した。T7−タグ配列は、mRNAに安定性を提供し、翻訳されたポリペプチドのレベルを有意に向上し得る。この融合タンパク質は、封入体内に沈着すると考えられ、公知のプロトコルにより容易に精製および再生できる。P2086の脂質化型および非脂質化型は、全細胞タンパク質の約5〜8%で発現され、全タンパク質の約50%としてrP2086を発現するT7−タグ融合物を例外とする。タンパク質の非脂質化型は、可溶性で細胞質中に局在すると考えられる。タンパク質の脂質化型は、膜画分に結合すると考えられ、界面活性剤により可溶化される。天然の脂質化型のタンパク質は、抗原提示に関して優れた三次構造を有することができ、および/または付着した脂質は、より強い免疫原性反応を刺激するアジュバントとして作用し得る。
【0211】
試験したすべてのN.meningitidisB菌株は、1つ2086様遺伝子を有すると考えられる。2086遺伝子の少なくとも2種のファミリーが提示される。2086ホモログは、PCRスクリーニングにより以下のものに同定されている:
N.meningitidisA、B、C,W135、Y
N.lactamica
N.gonorrhoeaeFA1090
いくつかのORF2086遺伝子は、クローニングされ、組換え発現されている
P2086の脂質化バージョンは、種々の髄膜炎菌株から発現された。
【0212】
これらの組換えタンパク質は精製され、マウスに予防接種するために使用されている。
【0213】
結果として生じる抗血清は殺菌性である。
【0214】
P2086の非脂質化バージョンは、様々な菌株から発現された。
【0215】
rLP2086は、rP2086より強い免疫反応を一貫して誘発する。
【0216】
rLP2086はまた、同種および異種髄膜炎菌株の両方に対して高められた殺菌活性を示す。
(実施例11)
【0217】
以下は、P2086が髄膜炎菌株において発現され、そしていくつかの菌株においてP2086発現の付加的な具体例を提供することをさらに示す。
【0218】
細胞ライゼートを、SDS試料バッファーに再懸濁されたプレート培養物由来の細胞で調製し、98℃で4分間加熱し。試料を、ウェルあたり約30〜50ug総タンパク質で、10〜20%プレキャストゲル(ICN)に負荷し、175Vで流した。ゲルをニトロセルロース膜に移し、次にそれを30分間、Tris緩衝生理食塩水(Blotto)中の5%粉末ミルクでブロッキングした。使用した一次抗体は、マウスにおいて個々のrLP2086変異体に対して誘導されたポリクローナル抗血清のプールであった。
【0219】
図17および18を参照すると、ウェスタンブロットは、P2086サブファミリーAおよびB全細胞ライゼートに対するrLP2086マウス抗血清の反応性を示す。サブファミリーA細胞ライゼートブロットの場合、使用される抗血清は、rLP2086−2996、−870446および−250771に対して誘導され、rLP2086−250771はBlottoで1/500に希釈され、他はBlottoで1/1000に希釈された。サブファミリーB細胞ライゼートブロットの場合、使用される抗血清は、rLP2086−8529(Blottoで1/1000に希釈)、−CDC1573、−M982および−880049(これら3種はBlottoで1/500に希釈)に対して誘導された。一次抗血清およびブロットを、4℃で一晩インキュベートした。ブロットを洗浄し、ヤギ抗マウスAP二次をBlottoで1/500に希釈して添加し、ブロットを30分間室温でインキュベートした。洗浄後、ブロットをBCIP/NBT膜ホスファターゼ基質系(KPL)を使用して展開した。
参考文献一覧
本明細書中で参照した参考文献が以下に記載され、それらは、参照によりその全体が本明細書に組み込まれる:
【0220】
【化2】
【0221】
【化3】
【0222】
【化4】
【0223】
【化5】
【0224】
【化6】
【0225】
【化7】
【0226】
【化8】
【0227】
【化9】
【0228】
【化10】
【0229】
【化11】
【0230】
【化12】
【0231】
【化13】
本発明は今や十分に説明されているが、本明細書中で示された本発明の趣旨または範囲を逸脱することなく、本発明に対し多くの変更および修正を行い得ることが当業者には明らかであろう。上記は、本発明の好ましい実施態様をいくつかの考え得る代替案と共に説明している。しかしながら、これらの実施態様は、単に例のためであり、本発明はそれらに限定されない。
【技術分野】
【0001】
本発明は、Neisseria meningitidis(髄膜炎菌)(セログループA、B、C、D、W−135、X、Y、Zおよび29E)、Neisseria gonorrhoeae(淋菌)、およびNeisseria lactamicaの菌株を含むナイセリア種菌株のような菌株から単離され得るナイセリアORF2086タンパク質(サブファミリーAおよびサブファミリーB)、ならびに該タンパク質の免疫原性部分および/または生物学的等価物に関する。本発明はまた、前記タンパク質、免疫原性部分および/または生物学的等価物に免疫特異的に結合する抗体に関する。さらに、本発明は、上記タンパク質、免疫原性部分、生物学的等価物および/または抗体のいずれかをコードする核酸配列を含む単離されたポリヌクレオチドに関する。加えて、本発明は、免疫原性組成物、およびN.meningitidisによって引き起こされる髄膜炎菌感染症、特にN.meningitidisセログループBによって引き起こされる髄膜炎菌性疾患の予防、処置および/または診断におけるそれらの使用、ならびに前記組成物の調製方法に関する。本発明は、組換え型および天然供与源由来の単離型の両方、ならびに脂質化型および非脂質化型の両方に関する。
【背景技術】
【0002】
髄膜炎菌性髄膜炎は、抗生物質の有効性にもかかわらず、数時間以内に小児および若い成人を死に至らせることがある破壊的な疾患である。非特許文献1。髄膜炎は、激しい頭痛、発熱、食欲不振、光および音に対する不耐性、特に頸部における筋肉の硬直、ならびに重篤なケースでは、死に至る痙攣、嘔吐およびせん妄という結果になる髄膜の炎症として特徴付けられる。髄膜炎菌性髄膜炎の症状は突然現れ、ついにはその特徴的な出血性発疹を伴う髄膜炎菌性敗血症となる。もしなんらかの生存のチャンスがあるとすれば、迅速な診断および大量の抗生物質による速やかな処置が決定的である。2000.Bantam Medical Dictionary,Third Edition 302。
【0003】
髄膜炎菌性髄膜炎は、Neisseria meningitidis(髄膜炎菌)によって引き起こされ、この菌は、A、B、C、D、W−135、X、Y、Zおよび29Eを含むいくつかの病原性セログループに分類されているグラム陰性の莢膜形成菌である。N.meningitidisのセログループB株は、世界中の髄膜炎菌性疾患の主要な原因である。例えば、セログループBが、米国および欧州在住の幼児および小児における細菌性髄膜炎の約50%の原因であると医学文献において報告されている。N.meningitidisセログループBによって引き起こされる髄膜炎菌性疾患を予防するためのワクチンは、現在全く存在しない。
【0004】
30年以上前のGoldschneiderらの研究以来、セログループB髄膜炎菌性疾患予防のための免疫原性組成物の開発は、研究者にとっての課題である。非特許文献2;非特許文献3;非特許文献4;および非特許文献5。第二次世界大戦後、北米から事実上消滅したセログループA疾患(Achtman,M.1995,Trends in Microbiology 3(5):186−92)とは異なり、セログループBおよびC微生物により引き起こされる疾患は、経済的に発展した世界の多くにわたって依然として風土病のままである。疾患発生率は、風土病がまれな<1/100,000から、流行病の間の高リスク個体群における200/100,000までの範囲で変わる。
【0005】
多糖結合体をベースとするワクチンが、N.meningitidisセログループAおよびCに対して開発されてきており、疾患の予防に効果的であると思われる。現在、セログループA、C、Y、およびW−135由来の莢膜状多糖で作られた免疫原性組成物が利用可能である。Ambroschら、1983,Immugenicity and side−effects of a new tetravalent.Bulletin of the World Health Organization 61(2):317−23。しかしながら、この免疫原性組成物は、T細胞非依存性免疫応答を引き起こし、幼い児童においては効果的でなく、髄膜炎菌性疾患の50%以上を引き起こすセログループB菌株については全くカバーしない。
【0006】
他の研究者らも、莢膜多糖を用いて免疫原性組成物の開発を試みている。最近、セログループC莢膜材料をタンパク質に接合させることによって調製されたセログループC疾患のための免疫原性組成物が、欧州において使用が認可されている。しかしながら、セログループB莢膜はワクチン候補として適さないかもしれない。なぜならば、莢膜多糖は、発達中のヒト神経組織上の炭水化物部分との類似性を持つポリシアル酸で構成されているからである。この糖半分は、自己抗原と認識され、したがってヒトにおいて免疫原性が乏しい。
【0007】
外膜タンパク質(OMP)が、セログループB疾患のための代替ワクチン抗原として開発されてきた。PorAの2つの可変領域に結合するモノクローナル抗体により、髄膜炎菌についてのセロサブタイプ判別方式が定義される。したがって、PorAタンパク質は、髄膜炎菌株のためのセロサブタイプ判別抗原として役立ち(Abdillahら、1988,Microbial Pathogenesis 4(1):27−32)、セログループB免疫原性組成物の成分として活発に研究されている(Poolman,1996,Adv.Exp.Med.Biol.397:73−7)。なぜならば、それらは殺菌抗体を誘発することができるからである(Saukkonen,1987,Microbial Pathogenesis 3(4):261−7)。殺菌抗体は保護の指標であると考えられ、どのような新しい免疫原性組成物候補も、これらの機能的抗体を誘導するべきである。
【0008】
ヒトならびに動物における研究は、セロサブタイプ判別抗原、PorAが殺菌抗体を誘導することを示す。しかしながら、PorAに対する免疫応答は一般に、セロサブタイプ特異的である。特に、セロサブタイプ判別データは、PorAで作られた免疫原性組成物が、そのような免疫原性組成物によりカバーされる各セロサブタイプについてPorAをおそらく6〜9も要求し得ることを示す。したがって、70〜80%のセログループB菌株をカバーするために、6〜9のPorAが必要とされるであろう。かくして、このタンパク質の可変的性質は、十分な数の髄膜炎菌セロサブタイプ臨床隔離集団から保護するために、多価ワクチン組成物を必要とする。
【0009】
セログループB髄膜炎菌のための免疫原性組成物の開発は非常に困難だったので、最近いくつかのグループが、新しい免疫原性組成物候補の同定を促すため、セログループAおよびB両方を代表する菌株由来のゲノムを配列決定している。Tettelin,2000,Science,287(5459):1809−15;非特許文献1。ナイセリアのゲノムの知識があっても、新しい免疫原性組成物候補の同定は、適切な数学的アルゴリズムが現在存在しない困難なプロセスである。実際、最近の報告は、理論的な膜貫通ドメインを含む数百のオープンリーディングフレーム(「ORF」)の同定にもかかわらず、発現、精製、および表面反応性で機能的に活性な抗体の誘起の問題により、研究者は、セログループB髄膜炎菌免疫原性組成物についてわずか7つの候補にしか至っていないことを示している。上記参照。これらのうちの1つは以前から知られていた。
【先行技術文献】
【非特許文献】
【0010】
【非特許文献1】Pizzaら、Science(2000)287:1816−1820
【非特許文献2】Goldschneiderら、J.Exp.Med.(1969)129(6):1307−26
【非特許文献3】Goldschneiderら、J.Exp.Med.(1969)129(6):1327−48
【非特許文献4】Gotschlichら、J.Exp.Med.(1969)129(6):1385−95
【非特許文献5】Gotschlichら、J.Exp.Med.(1969)129(6):1367−84
【発明の概要】
【発明が解決しようとする課題】
【0011】
したがって、(1)複数のナイセリア菌株に対する殺菌抗体を誘導し、(2)複数の菌株の表面と反応し、(3)生菌攻撃に対する受動的保護を付与し、および/または(4)コロニー形成を妨げる、免疫原性組成物に対する要求が依然としてある。
【課題を解決するための手段】
【0012】
本発明の1つの実施形態は、(a)配列番号1−11の奇数番号配列のいずれかと少なくとも約95%配列同一性を有するヌクレオチド配列、または(b)配列番号2−12の偶数番号配列のいずれかのアミノ酸配列と少なくとも約95%配列同一性を有するアミノ酸配列を含むポリペプチドをコードするヌクレオチド配列を含むポリヌクレオチドを提供する。
【0013】
本発明のさらなる実施形態は、本発明のポリヌクレオチドを含むベクターを提供する。
【0014】
本発明のさらなる実施形態は、本発明のベクターを含む組換え細胞を提供する。
【0015】
本発明のさらなる実施形態は、(a)配列番号2−12の偶数番号配列のいずれかと少なくとも95%配列同一性を有するアミノ酸配列、(b)配列番号1−11の奇数番号配列のいずれかと少なくとも95%配列同一性を有するヌクレオチド配列によってコードされるアミノ酸配列、(c)(a)もしくは(b)に記載されたアミノ酸配列の少なくとも1つの免疫原性部分、あるいは(d)(a)もしくは(b)に記載されたアミノ酸配列または(c)に記載された免疫原性部分の少なくとも1つの生物学的等価物を含むポリペプチドを提供する。
【0016】
本発明のさらなる実施形態は、(a)偶数番号の配列番号2−12のいずれかのアミノ酸配列を含むポリペプチドと免疫特異的に結合するポリペプチド、または(b)(a)に記載されたポリペプチドの少なくとも1つの免疫原性部分、または(c)(a)に記載されたポリペプチドまたは(b)に記載された免疫原性フラグメントの少なくとも1つの生物学的等価物、のいずれかを含む抗体を提供する。
【0017】
本発明のさらなる実施形態は、本発明のポリヌクレオチド、ベクター、組換え細胞、ポリペプチドまたは抗体を含む組成物を提供する。
【0018】
本発明のさらなる実施形態は、(a)配列番号1−5の奇数番号配列のいずれかと少なくとも約95%配列同一性または配列番号2−6の偶数番号配列のいずれかのアミノ酸配列をコードするヌクレオチド配列と少なくとも約95%配列同一性を有するヌクレオチド配列を含む第1のポリヌクレオチドと、(b)配列番号7−11の奇数番号配列のいずれかと少なくとも約95%配列同一性または配列番号8−12の偶数番号配列のいずれかのアミノ酸配列をコードするヌクレオチド配列と少なくとも約95%配列同一性を有するヌクレオチド配列を含む第2のポリヌクレオチドと、を含む組成物を提供する。
【0019】
本発明のさらなる実施形態は、(a)配列番号2−6の偶数番号配列のいずれかと少なくとも約95%配列同一性を有するアミノ酸配列を含む第1のポリペプチドと、(b)配列番号8−12の偶数番号配列のいずれかと少なくとも約95%配列同一性を有するアミノ酸配列を含む第2のポリペプチドと、を含む組成物を提供する。
【0020】
本発明のさらなる実施形態は、(a)偶数番号の配列番号2−12のいずれかのアミノ酸配列を含むポリペプチド、(b)奇数番号の配列番号1−11のいずれかの核酸配列を含むポリヌクレオチドによりコードされたポリペプチド、(c)(a)または(b)に記載されたポリペプチドの少なくとも1つの免疫原性部分、あるいは(d)(a)もしくは(b)に記載されたポリペプチドまたは(c)に記載された免疫原性フラグメントの少なくとも1つの生物学的等価物のいずれかを、ナイセリア種から単離および精製するか、または組換えにより調製するステップを含むプロセスにより調製される組成物を提供する。
【0021】
本発明のさらなる実施形態は、哺乳動物において免疫応答を引き起こすための薬剤の調製における本発明の組成物の使用を提供する。
【0022】
本発明のさらなる実施形態は、哺乳動物における細菌性髄膜炎に対して効果的な薬剤における本発明の組成物の使用を提供する。
【0023】
本発明のさらなる実施形態は、本明細書中で記載されたポリペプチドのいずれかをコードする核酸配列を宿主細胞において発現させるステップを含む組成物を調製する方法を提供する。
【0024】
本発明のさらなる実施形態は、本明細書中で記載されたタンパク質、免疫原性部分または生物学的等価物のいずれかを含む組成物を動物に導入した後にその動物から抗体を回収するステップを含む、抗体組成物を調製する方法を提供する。
【0025】
本発明のさらなる実施形態は、有効量の本発明の組成物の1つ以上を哺乳動物に投与するステップを含む、哺乳動物において免疫応答を引き起こす方法を提供する。
【0026】
本発明のさらなる実施形態は、有効量の本発明の組成物の1つ以上を哺乳動物に投与すことを含む、哺乳動物における細菌性髄膜炎を予防または処置する方法を提供する。
【図面の簡単な説明】
【0027】
【図1A】異種菌株に対する殺菌抗体を誘導することができるナイセリア膜タンパク質抽出物を同定するための実験から得られたタンパク質画分の2つの主要なタンパク質を示すSDS−PAGEゲルを示す。
【図1B】プロテアーゼ消化および逆相N−末端配列決定によるTMAEフロースルー成分の分析による2つの主要なタンパク質の同定の実験結果を示す。
【図2】rLP2086の精製スキームおよびSDS−PAGEにより決定される等質性を示す。
【図3】LC−MS/MSおよび対応するSDS−PAGEによるTMAEフロースルー成分の分析による2つの主要なタンパク質および1つの少量タンパク質の同定の実験結果を示す。
【図4】2086タンパク質の組換え発現のSDS−PAGEゲルである。
【図5】本明細書中の実施例に記載されるプラスミドpPX7340の概略図である。
【図6】本明細書中の実施例に記載されるプラスミドpPX7328の概略図である。
【図7】本明細書中の実施例に記載されるプラスミドpPX7343の概略図である。
【図8】様々な菌株由来の2086遺伝子のN末端領域を例示する。
【図9A】ナイセリア菌株中の免疫原性成分の同定における準備段階を示すフローチャートである。
【図9B】ナイセリア菌株中の免疫原性成分の同定における最終段階を示すフローチャートである。
【図10】図10Aは、本明細書中の実施例に記載されるrP2086脂質化型を発現させるためのP4シグナル/ORF2086融合タンパク質を発現させるpBADアラビノース誘導プロモーターの概略図である。図10Bは、ORF2086非脂質化型の組換え発現のためのpET9a−T7ベクターの概略図である。
【図11】図11Aは、rLP2086タンパク質を発現するE.coli Bの全細胞ライゼートを表す写真である。図11Bは、rP2086タンパク質を発現するE.coli Bの全細胞ライゼートを示す。
【図12A】本発明の実施に従うORF2086タンパク質のサブファミリーおよびグループの構成を示す系統樹である。
【図12B】本発明の実施に従うORF2086タンパク質のサブファミリーおよびグループの構成を示す系統樹である。
【図13】rLP2086サブファミリーA抗血清についての全細胞ELISAデータのグラフである。
【図14】rLP2086サブファミリーB抗血清についての全細胞ELISAデータのグラフである。
【図15】rLP2086混合研究−WCE力価の結果のグラフである。
【図16】rLP2086/rPorA混合研究−WCE力価の結果のグラフである。
【図17】P2086サブファミリーB N.meningitidis全細胞ライゼートに対するrLP2086マウス抗血清の反応性を示すウェスタンブロットである。
【図18】P2086サブファミリーA N.meningitidisおよびN.lactamica全細胞ライゼートに対するrLP2086マウス抗血清の反応性を示すウェスタンブロットである。
【発明を実施するための形態】
【0028】
配列の概要
配列番号1 天然のリーダー配列と組み合わされた時のCDC1135菌株由来の成熟2086タンパク質についてのアミノ酸配列をコードする核酸配列。
【0029】
配列番号2 天然のリーダー配列を用いて調製されたCDC1135菌株由来の成熟2086タンパク質についてのアミノ酸配列。
【0030】
配列番号3 P4リーダー配列と組み合わされた時のCDC1135由来の成熟2086タンパク質についてのアミノ酸配列をコードするための核酸配列。
【0031】
配列番号4 P4リーダー配列を用いて調製されたCDC1135菌株由来の成熟2086タンパク質についてのアミノ酸配列。
【0032】
配列番号5 CDC1135菌株由来の成熟2086タンパク質についてのアミノ酸配列をコードする核酸配列。
【0033】
配列番号6 CDC1135菌株由来の成熟2086タンパク質についてのアミノ酸配列。
【0034】
配列番号7 天然のリーダー配列と組み合わされた時のCDC1127菌株由来の成熟2086タンパク質についてのアミノ酸配列をコードする核酸配列。
【0035】
配列番号8 天然のリーダー配列を用いて調製されたCDC1127菌株由来の成熟2086タンパク質についてのアミノ酸配列。
【0036】
配列番号9 P4リーダー配列と組み合わされた時のCDC1127由来の成熟2086タンパク質についてのアミノ酸配列をコードするための核酸配列。
【0037】
配列番号10 P4リーダー配列を用いて調製されたCDC1127菌株由来の成熟2086タンパク質についてのアミノ酸配列。
【0038】
配列番号11 CDC1127菌株由来の成熟2086タンパク質についてのアミノ酸配列をコードする核酸配列。
【0039】
配列番号12 CDC1127菌株由来の成熟2086タンパク質についてのアミノ酸配列。
【0040】
発明の詳細な説明
本発明は、2086サブファミリーAタンパク質および2086サブファミリーBタンパク質を含むナイセリアORF2086タンパク質(「2086タンパク質」)を提供する。2086タンパク質の各々は、Neisseria meningitidis(セログループA、B、C、D、W−135、X、Y、Zおよび29E)、Neisseria gonorrhoeae、およびNeisseria lactamicaの菌株を含む、天然のナイセリア菌株から単離され得るタンパク質である。2086タンパク質はまた、組換え技術を用いて調製されてもよい。
【0041】
様々な実施形態によれば、本発明は、2086タンパク質、それらの免疫原性部分、および/またはそれらの生物学的等価物、上記のもののいずれかに免疫特異的に結合する抗体、ならびに上記のもののいずれかをコードする核酸配列を含むポリヌクレオチドを提供する。本発明は、組成物、免疫原性組成物、および髄膜炎菌性感染症、特にN.meningitidisにより引き起こされる髄膜炎菌性疾患の予防、処置および/または診断におけるそれらの使用、ならびに前記組成物を調製するための方法を提供する。2086タンパク質は本明細書中で、組換え型および天然供給源由来の単離型、ならびに脂質化型および非脂質化型を含む。
【0042】
本発明は、(1)N.meningitidis、N.gonorrhoeae、および/またはN.lactamicaの菌株のような複数のナイセリア菌株に対する殺菌抗体を誘導し、(2)複数の菌株の表面と反応し、(3)生菌攻撃に対する受動的保護を付与し、および/または(4)コロニー形成を妨げる組成物、ならびに前記組成物を使用する方法および前記組成物を調製する方法を、思いがけずかつ有利に提供する。本発明の様々な実施形態が以下で説明される。
【0043】
本明細書中で記載されるように、N.meningitidisから単離されたナイセリア種ORF2086タンパク質(本明細書中で区別なく用いられる「2086タンパク質」または「ORF2086」タンパク質、あるいは非脂質化タンパク質についてはP2086およびタンパク質の脂質化バージョンについてはLP2086とも呼ばれる)をベースとした新規な免疫原性組成物候補は、抗血清の調製に伴う細胞分画、ディファレンシャル界面活性剤抽出、タンパク質精製を、複数の菌株を用いた殺菌活性アッセイと組み合わせることにより同定された。上記で引用された参照文献中で候補として開示される免疫原性組成物および診断法に代わるものとして、本発明は、タンパク質、その免疫原性部分およびその生物学的等価物のほか、前記ポリペプチド、部分、および等価物をコードする遺伝子およびそれらに免疫特異的に結合する抗体の使用による、髄膜炎菌感染の処置および/または予防のための組成物および方法に関する。
【0044】
本明細書中で用いられるように、用語「菌株非特異的(non−strain specific)」は、1つ以上のN.meningitidis菌株(例えば、異種髄膜炎菌株)に対して効果的な免疫応答を誘発する抗原の特徴を指す。用語「交差反応性(cross−reactive)」は、本明細書中で用いられるように、用語「菌株非特異的」と区別なく用いられる。用語「免疫原性菌株非特異的N.meningitidis抗原」は、本明細書中で用いられるように、N.meningitidisから単離され得る抗原を述べるが、これは、別の細菌(例えば、淋菌のような他のナイセリア菌株)から単離することも、または組換え技術を用いて調製することもできる。
【0045】
本発明の2086タンパク質は、脂質化タンパク質および非脂質化タンパク質を含む。さらに、本発明はまた、中間体化合物/組成物として各タンパク質に対応する未熟タンパク質またはプレタンパク質の使用も考慮する。
【0046】
本発明はまた、本発明の実施に従って、上記の免疫原性薬剤に免疫特異的に結合する抗体を提供する。さらに、本発明は、上記のうちのいずれかをコードする核酸配列を含む単離されたポリヌクレオチドに関する。加えて、本発明は、髄膜炎菌性髄膜炎、特にセログループB髄膜炎菌性疾患の予防、処置および/または診断における組成物および/または免疫原性組成物およびそれらの使用、ならびに前記組成物を調製するための方法を提供する。
【0047】
本発明の組成物は高度に免疫原性であり、殺菌抗体の産生を誘導できる。これらの抗体は、セログループ、セロタイプおよびセロサブタイプ異種髄膜炎菌株と交差反応性である。したがって、本発明の組成物は、異種ナイセリア菌株に対する殺菌抗体を誘導する能力を示すことにより、以前のN.meningitidisワクチンの試みの欠陥を克服する。したがって、本発明は、他にも利点はあるが、特に従来使用されていた薬剤と比較して、保護を誘導するためにより少ない成分で調合され得る免疫原性組成物を提供する。組成物またはその中野免疫原性薬剤(例えば、ポリペプチド、免疫原性部分またはフラグメント、および生物学的等価物など、ただしこれらに限定されない)は、髄膜炎菌性感染および疾患からの免疫学的保護を引き出すため、ならびに他の病原体により引き起こされた感染および/または疾患からの免疫学的保護を引き出すため、単独で、または他の抗原もしくは薬剤と組み合わせて使用され得る。これにより、複数の菌株から保護するために必要とされる抗原の数を減らすことにより、髄膜炎菌性感染に対して使用する免疫原性組成物のデザインが単純化される。実際、精製2086タンパク質は、髄膜炎菌性疾患の原因となる菌株の適切な免疫原性の有効範囲を提供するために必要とされるタンパク質の数を劇的かつ予想外に低減する。2086タンパク質は、E.coliにおいてリポタンパク質として組換えによって発現させることができ、これは、天然の髄膜炎菌におけるよりもずっと高いレベルの野生型タンパク質である。
【0048】
以下の公開国際特許出願は、参照によりその全体が本明細書に組み込まれる:PCT/US02/32369(2003年8月7日に国際公開第03/063766号として公開)およびPCT/US04/11901(2004年11月4日に国際公開第04/094596号として公開)。
【0049】
2086タンパク質は野生型菌株中には大量に存在しないが、これは殺菌抗体の標的である。これらの抗体は、PorAに応答して産生された抗体と異なり、異種セロサブタイプを発現する菌株を殺すことができる。
【0050】
2086タンパク質に対する抗体はまた、髄膜炎菌による攻撃から幼若ラットを受動的に保護する。2086タンパク質の組換え発現により、髄膜炎菌性疾患の予防のための免疫原性組成物としての2086タンパク質の使用が可能になる。臨床試験における最近の髄膜炎菌性免疫原性組成物のすべては、多くの異なるタンパク質を含む複合組成物または外膜タンパク質製剤であった。セロサブタイプ特異性をもたらすPorAタンパク質は、約70〜80%の疾患関連セロサブタイプの有効範囲を提供するために、免疫原性組成物中に6〜9の変異体を含有することを必要とする。対照的に、単一の2086タンパク質に対する抗血清単独で、西ヨーロッパおよび米国における疾病隔離集団の約65%の原因となる6つのセロサブタイプの典型を殺すことができることが本明細書中で明確に実証される。したがって、精製2086タンパク質は、髄膜炎菌性疾患の原因となるセロサブタイプの十分な免疫原性組成物の有効範囲を提供するために必要とされるタンパク質の数を低減することが見込まれる。
タンパク質、免疫原性部分および生物学等価物
本発明により提供される2086タンパク質は、単離されたタンパク質またはポリペプチドである。用語「単離された(isolated)」は、天然状態から人の手によって変更されたことを意味する。もし「単離された」組成物または物質が自然界で生ずるのであれば、それはすでにその本来の環境から変化しているかまたは取り除かれているか、あるいはその両方である。例えば、生きている動物中に天然に存在するポリペプチドまたはポリヌクレオチドは「単離され」てはいないが、同じポリペプチドまたはポリヌクレオチドがその天然の状態の共存物質から分離されていれば、本明細書中で使用される用語「単離され」と同じく「単離され」ている。したがって、本明細書中で用いられるように、用語「単離されたタンパク質(isolated protein)」は、天然供給源から単離されたタンパク質および組換え技術を用いて調製されたタンパク質、ならびに、他の抗原および/または例えば薬学的に許容される担体、バッファー、アジュバント等の添加物と組み合わされた時の上記のようなタンパク質を包含する。
【0051】
本発明の実施形態によれば、2086タンパク質は、免疫原性、非病原および非菌株特異的であることを特徴とする。2086タンパク質は、高度に可変性であり、したがって、タンパク質の免疫原性を損なうことなくアミノ酸残基の挿入、置換および/または欠失を受けてもよい。2086タンパク質は、2つのサブファミリー、すなわちサブファミリーAおよびサブファミリーBに分割され得る。
【0052】
サブファミリーA由来の2086タンパク質は、配列番号2−6の偶数番号配列のいずれかのアミノ酸配列または配列番号1−5の奇数番号配列のいずれかのヌクレオチド配列を含むポリヌクレオチドによってコードされるアミノ酸配列を含む。サブファミリーB由来の2086タンパク質は、配列番号8−12の偶数番号配列のいずれかのアミノ酸配列または配列番号7−11の奇数番号配列のいずれかのヌクレオチド配列を含むポリヌクレオチドによってコードされるアミノ酸配列を含む。
【0053】
本発明のポリペプチド配列は、基準配列(例えば、偶数番号のついた配列番号2−12)と同一、すなわち100%同一、または、基準配列と比較して同一性の%が100%未満であるようないくつかのアミノ酸変更を含み得る。そのような変更としては、少なくとも1つのアミノ酸欠失、同類置換もしくは非同類置換を含む置換、または挿入が含まれる。変更は、基準ポリペプチド配列のアミノ−またはカルボキシ−末端位置において生じてもよいし、あるいは基準アミノ酸配列中、または基準アミノ酸配列内の1つ以上の隣接するグループ中のアミノ酸間に個別に散在されて、それらの末端位置間のどこかで生じてもよい。
【0054】
したがって、本発明はまた、配列表に含まれるアミノ酸配列(すなわち、偶数番号のついた配列番号2−12)との配列同一性を有するタンパク質を提供する。本発明の様々な実施形態によれば、2086タンパク質は、配列番号2−12の偶数番号アミノ酸配列のいずれかと約95%、約96%、約97%、約98%、約99%、約99.9%またはそれ以上の同一性より大きい配列を有する。これらは、制限なく、突然変異体および対立遺伝子変異体を含んでいる。
【0055】
本発明の好ましい実施形態において、2086タンパク質または他の2086ポリペプチド(例えば、免疫学的部分および生物学的等価物)は、髄膜炎菌の相同菌株および少なくとも1つの異種菌株に対する殺菌抗体を生成する。具体的には、2086ポリペプチドに対する抗体は、髄膜炎菌による鼻腔内などでの攻撃から幼若ラットを受動的に保護する。さらなる好ましい実施形態において、2086ポリペプチドは、相同菌株および少なくとも1つの異種菌株について幼若ラットに対し上記のような保護を発揮する。ポリペプチドは、偶数番号のついた配列番号2−12中に示されるように、上記の配列概要から選択されればよく、あるいはポリペプチドは、列挙されたポリペプチドの任意の免疫学的フラグメントまたは生物学的等価物としてもよい。好ましくは、ポリペプチドは、上記の配列概要における偶数番号のついた配列番号2−12のいずれかから選ばれる。
【0056】
本発明はまた、生物学的等価物である2086ポリペプチドの対立遺伝子変異体または他の変異体に関する。適切な生物学等価物とは、(1)相同菌株ならびに少なくとも1つの異種ナイセリア菌株および/または淋菌株に対する殺菌抗体を誘導し、(2)相同菌株および少なくとも1つの異種ナイセリア菌株および/または淋菌株の表面と反応し、(3)生菌攻撃に対し受動的保護を付与し、および/または(4)コロニー形成を妨げる、能力を示すものである。
【0057】
適切な生物学的等価物は、該生物学的等価物が本発明の2086タンパク質の1つと実質的に同じ免疫抗原特性を誘導できるという前提で、本明細書中で明記された2086ポリペプチド(すなわち、偶数番号のついた配列番号2−12)の1つと少なくとも約95%、約96%、約97%、約98%、約99%または約99.9%の類似性を有する。
【0058】
あるいは、生物学的等価物は、偶数番号のついた配列番号2−12中の2086タンパク質の1つと実質的に同じ免疫抗原特性を有する。本発明の実施形態によれば、生物学的等価物は、偶数番号のついた配列番号2−12と同じ免疫抗原特性を有する。
【0059】
生物学的等価物は、本発明のタンパク質に対する変異体および修飾を生成することにより得られる。タンパク質に対するこれらの変異体および修飾は、1つ以上のアミノ酸の挿入、欠失または置換によりアミノ酸配列を改変することによって得られる。アミノ酸配列は、実質的に同じまたは改善された品質を有するポリペプチドを作り出すために、例えば置換によって修飾される。改変を導入する好ましい手段は、部位指定突然変異誘発によりポリペプチドの核酸配列のあらかじめ決められた突然変異を作るステップを含む。
【0060】
修飾および変更は、本発明のポリペプチドの構造中において行うことができ、さらにN.meningitidis免疫原性を有する分子を得ることができる。例えば、制限なく、ある一定のアミノ酸が、免疫原性をそれほど損失することなく配列中で、非保存的および保存的置換を含む他のアミノ酸で置換され得る。ポリペプチドの生物学的機能活性を定義するのはポリペプチドの相互作用的な能力および性質なので、いくつかのアミノ酸配列置換は、ポリペプチド配列(または、当然、その基礎をなすDNAコード化配列)中で行うことができ、依然として同様な特性を有するポリペプチドを得ることができる。本発明は、本明細書中のポリペプチドの構造ならびに前記ポリペプチドをコードする核酸配列へのどのような変化も考慮し、ここで、ポリペプチドは免疫原性を保持している。当業者は、本明細書中で提供される手引きに基づいて、開示されたポリペプチドおよびポリヌクレオチドをそれに応じて容易に修飾できるであろう。
【0061】
例えば、置換または欠失が許容されるある一定の可変領域が同定されている。2086共通配列は、かねて論じられたように、本発明の実施に従うタンパク質の2086ファミリーの保存および非保存領域を示す。
【0062】
そのような変更を行う際、当業者に知られているどのような技術も利用できる。例えば、アミノ酸のハイドロパシック・インデックス(hydropathic index)を、これに限定することを意図することなく、考慮することができる。ポリペプチド上に相互作用的な生物学的機能を付与する際のハイドロパシックアミノ酸インデックスの重要性は、技術分野において一般に理解されている。Kyte et al.1982.J.Mol.Bio.157:105〜132。
【0063】
同様なアミノ酸の置換はまた、特に、生物学的機能性等価ポリペプチドまたはそれによって作り出されたペプチドが免疫学的実施形態における使用を意図される場合、親水性に基づいて行われ得る。参照により本明細書に組み込まれる米国特許第4,554,101号は、ポリペプチドの最大局所平均親水性が、その隣接アミノ酸の親水性により支配されるように、その免疫原性、すなわちポリペプチドの生物学的特性と相関関係にあると述べている。
【0064】
ポリペプチドの生物学的等価物はまた、部位特異的突然変異誘発を用いて調製できる。部位特異的突然変異誘発は、第二世代ポリペプチド、すなわち基となるDNAの特定の突然変異誘発を通して、その配列から誘導された生物学的機能性等価ポリペプチドまたはペプチドの調製において有用な手法である。そのような変更は、アミノ酸置換が望ましい場合に望ましいことがあり得る。この手法はさらに、1つ以上のヌクレオチド配列変化をDNAに導入することにより、例えば、上記の問題点のうちの1つ以上を組み込んだ配列変異体の調製および試験を素早く行えるようにする。部位特異的突然変異誘発は、所望の突然変異のDNA配列をコードする特定のオリゴヌクレオチド配列、ならびに横断されている欠失接合の両側で安定な二重鎖を形成するのに十分なサイズおよび配列複雑度のプライマー配列を提供するのに十分な数の隣接ヌクレオチドの使用によって、突然変異体の産生を可能にする。典型的には、改変される配列の接合部の両側に約5〜10の残基を有する長さが約17〜25ヌクレオチドのプライマーが好ましい。
【0065】
一般に、部位特異的突然変異誘発の手法は、技術的に知られている。理解されるように、この手法は典型的には、一本鎖型および二本鎖型の両方で存在し得るファージベクターを用いる。典型的には、本明細書に従う部位指定突然変異誘発は、最初に、選ばれたN.meningitidisポリペプチド配列の全部または一部をコードするDNA配列をその配列内に含む一本鎖ベクターを得ることにより実行される。所望の変異された配列を持つオリゴヌクレオチドプライマーが(例えば、合成的に)調製される。このプライマーは次に、一本鎖ベクターにアニールされ、変異支持鎖の合成を完成するためにE.coli DNAポリメラーゼI(クレノウフラグメント)のような酵素を用いて伸長される。かくして、1つの鎖がオリジナルの非変異配列をコードし、第2の鎖が所望の突然変異を有するヘテロ二本鎖が形成される。このヘテロ二本鎖ベクターは次に、E.coliのような適切な細胞を形質転換するために用いられ、突然変異を有する組換えベクターを含むクローンが選択される。商業的に利用可能なキットには、オリゴヌクレオチドプライマーを除く必要なすべての試薬が付属している。
【0066】
2086ポリペプチドとしては、偶数番号の配列番号 2−12の1つからのアミノ酸配列を有する2086タンパク質との実質的な配列類似性および/または生物学的等価性を含む任意のタンパク質またはポリペプチドが含まれる。加えて、本発明の2086ポリペプチドは、特定の供給源に限定されない。したがって、本発明は、各種の供給源由来のポリペプチドの一般的検出および分離に備える。また、2086ポリペプチドは、当業者に知られているように、本明細書中で提供される手引きに基づいて組換え的に、または技術的に知られている任意の他の合成的方法で調製され得る。
【0067】
2086ポリペプチドが、さらなる構造的または機能的分析において、あるいは2086関連ポリペプチドおよび2086特異的抗体のような試薬の産生において使用するためにフラグメントに有利に切断され得ることが本発明において考慮される。これは、精製または未精製N.meningitidisポリペプチドを、エンドプロテイナーゼglu−C(Boehringer,Indianapolis,IN)のようなペプチダーゼで処理することにより達成され得る。CNBrによる処理は、ペプチドフラグメントを天然のN.meningitidis2086ポリペプチドから産生できる別の方法である。組換え手法も、2086タンパク質の特定フラグメントを産生するために用いることができる。
【0068】
本明細書中で用いられる用語「変異体(variant)」は、基準ポリヌクレオチドまたはポリペプチドとそれぞれ異なるポリヌクレオチドまたはポリペプチドであるが、本質的な特性を保持する。ポリヌクレオチドの典型的な変異体は、別の基準ポリヌクレオチドとはヌクレオチド配列が異なる。変異体のヌクレオチド配列中の変化は、基準ポリヌクレオチドによってコードされるポリペプチドのアミノ酸配列を改変することも、改変しないこともある。ヌクレオチド変化は、以下で論じられるように、基準配列によってコードされるポリペプチド中でのアミノ酸置換、付加、欠失、融合およびトランケーションという結果になり得る。ポリペプチドの典型的な変異体は、アミノ酸配列が別の基準ポリペプチドと異なる。一般に、差異は限定されており、その結果、基準ポリペプチドおよび変異体の配列は、全体的に密接に類似しており、多くの領域において同一(すなわち、生物学的に等価)である。変異体および基準ポリペプチドは、いずれかの組み合わせで1つ以上の置換、付加、欠失だけアミノ酸配列が異なり得る。置換または挿入されたアミノ酸残基は、遺伝コードによってコードされたものであっても、されてないものであってもよい。ポリヌクレオチドまたはポリペプチドの変異体は、対立遺伝子変異体のような自然に発生するものであることもあり、または自然に発生することが知られていない変異体であることもある。ポリヌクレオチドおよびポリペプチドの非自然発生変異体は、突然変異誘発手法または直接合成により作られ得る。
【0069】
当該技術において知られているように、「同一性(identity)」は、配列を比較することによって確認される、2つ以上のポリペプチド配列または2つ以上のポリヌクレオチド配列間の関係である。当該技術において、「同一性」はまた、場合によっては、そのような配列のストリング間の一致によって確認される、ポリペプチドまたはポリヌクレオチド配列間の配列関連性の程度も意味する。「同一性」および「類似性(similarity)」は、Computational Molecular Biology,Lesk,A.M.,ed.Oxford University Press,New York,1988;Biocomputing:Informatics and Genome Projects,Smith,D.W.,ed.Academic Press,New York,1993;Computer Analysis of Sequence Data,Part I,Griffin,A.M.and Griffin,H.G.,eds.,HumanaPress,New Jersey,1994;Sequence Analysis in Molecular Biology,von Heinje,G.,Academic Press,1987;およびSequence Analysis Primer,Gribskov,M.and Devreux,J.,eds.,M Stockton Press,New York,1991;およびCarillo,H.,and Lipman,D.,SIAM J.Applied Math.,48:1073(1988)に記載されるような既知の方法で容易に計算できるが、それらに限定されない。同一性を確認する好ましい方法は、試験される配列間で最も大きい一致をもたらすように設計される。同一性および類似性を確認する方法は、公的に利用可能なコンピュータプログラムにおいて体系化されている。2つの配列間の同一性および類似性を確認する好ましいコンピュータプログラム方法としては、GCGプログラムパッケージ(Devreux,J.,et al 1984)、BLASTP、BLASTN、およびFASTA(Altschul,S.F.,et al.,1990)が含まれるが、それらに限定されない。BLASTXプログラムは、NCBIおよび他の供給源(BLAST Manual,Altschul,S.,et al.,NCBI NLM NIH Bethesda,Md.20894;Altschul,S.,et al.,1990)から公的に利用可能である。よく知られているスミスウォーターマンアルゴリズムもまた、同一性を確認するために用いられ得る。
【0070】
一例として、本発明のアミノ酸配列は、基準配列、偶数番号のついた配列番号2−12と同一、すなわち100%同一であってもよく、あるいは、本発明のアミノ酸配列は、基準配列と比較して同一性の%が100%未満であるようないくつかのアミノ酸変更を含むこともできるが、それに限定されることを意図するものではない。そのような変更は、少なくとも1つのアミノ酸欠失、保存的および非保存的置換を含む置換、または挿入からなる群から選ばれ、その場合、前記変更は、基準ポリペプチド配列のアミノ−またはカルボキシ−末端位置において生じてもよいし、あるいは基準アミノ酸配列中、または基準アミノ酸配列内の1つ以上の隣接するグループ中のアミノ酸間に個別に散在されて、それらの末端位置のどこかで生じてもよい。所与の%同一性についてのアミノ酸改変数は、偶数番号の配列番号2−12中のアミノ酸の総数にそれぞれのパーセント同一性の数値パーセント(100で割る)を乗じ、次にその積を偶数番号の配列番号2−12のいずれかの中のアミノ酸の前記総数から引くことにより求められる:すなわち、
na=xa−(xa・y)
式中、naはアミノ酸改変の数であり、xaは偶数番号の配列番号2−12中のアミノ酸の総数であり、yは、例えば70%については0.70、80%については0.80、85%については0.85等であり、xaとyとのどのような非整数積も、これをxaから引く前に最も近い整数に丸められる。
【0071】
好ましい実施形態において、上記のポリペプチドは、2086タンパク質の成熟プロセス型のような、偶数番号の配列番号 2−12中に明示されるタンパク質から選ばれる。2086タンパク質または等価物等は、脂質化または非脂質化され得る。
【0072】
ORF2086は、天然のORF 2086シグナル配列を持つE.coli菌中で発現可能である。しかしながら、タンパク質の発現を改善する手段を見出すことが望ましい。本発明の実施形態によれば、リーダー配列は、タンパク質の脂質化型を産生する。例えば、以下は、発現を増強するための、莢膜非保有ヘモフィルス・インフルエンザP4タンパク質のシグナル配列の使用を説明する。
【0073】
細菌リポタンパク質のプロセシングは、シグナル配列を含む前駆体またはプロリポタンパク質の合成から始まり、それが今度は共通リポタンパク質プロセシング/修飾部位を含む。このプロリポタンパク質は当初、グラム陰性菌の内膜上またはグラム陽性菌の膜上の共通Sec系を通過する。ひとたびSec系によって膜中に置かれると、プロリポタンパク質は共通部位においてシグナルペプチダーゼIIによって開裂され、露出されたN−末端システイン残基がグリセレート化およびアシル化される。Hayashi et al.1990.Lipoproteins in bacteria.J.Bioenerg.Biomembr.Jun;22(3):451−71;Oudega et al.1993。Escherichia coli SecB,SecA,and SecY proteins are requiered for expression and mambrane insertion of the bacteriocin release protein,a small lipoprotein.J.Bacteriol.Mar;175(5):1543−7;Sankaran et al.1995.Modification of bacterialLipoproteins.Methods Enzymo.250:683−97。
【0074】
グラム陰性菌において、外膜への脂質化タンパク質の輸送は、リポタンパク質の位置2におけるソーティングシグナルに応じて、膜特異性を有する特有のABCトランスポーター系によって媒介される。Yakushi et al.2000.A new ABC transporter mediating the detachment of lipid modified proteins from membranes.Nat Cell Biolo.Apr;2(4):212−8。
【0075】
細菌リポタンパク質およびそれらのシグナル配列との融合が、細菌表面の組換えタンパク質を表示するために用いられてきた。米国特許第5,583,038号および同第6,130,085号。リポタンパク質シグナル配列を交換することにより、リポタンパク質の産生を増大することができる。De et al.2000.Purification and characterization of Streptococcus pneumoniae palmitoylated pneumococcal surface adhesin A expressed in Escherichia coli.Vaccine.Mar 6;18(17):1811−21。
【0076】
タンパク質の細菌脂質化は、タンパク質に対する免疫学的応答を増大または修正することが知られている。Erdile et al.1993.Role of attached lipid in immunogenicity of Borrelia burgdorferi OspA. Infect.Immun.Jan;61(1):81−90;Snapper et al.1995.細菌リポタンパク質は、T細胞独立タイプII抗原に対する体液性免疫応答におけるサイトカインに取って代わり得る。J.Immuno.Dec 15;155(12):5582−9。しかしながら、細菌リポタンパク質発現は、プロセシングのストリンジェンシーにより複雑化され得る。Pollitt et al.1986.Effect of amino acid substitutions at the signal peptide cleavage しte of the Escherichia coli major outer membrane lipoprotein.J.Biol.Chem.Feb 5;261(4):1835−7;Lunn et al.1987.Effect of prolipoprotein signal peptide mutations on secretion of hybrido propolipo−beta−lactamase in Escherichia coli.J.Biol.Chem.Jun 15;262(17):8318−24;Klein et al.1988.Distinctive propertiees of signal sequences from bacterial lipoproteins.Protein Eng.Apr;2(1):15−20。細菌リポタンパク質発現はまた、毒性および低発現レベルのような他の問題によって複雑化される。Gomez et al.1994.Nucleotide The Bacillus subtilis lipoprotein LplA causes cell lysis when expressed in Escherichia coli.Microbiology.Aug;140(Pt8):1839−45;Hansson et al.1995.Expression of truncated and full−length forms of the Lyme disease Borrelia outer surface protein A in Escherichia coli.Protein Expr.Purif.Feb;6(1):15−24;Yakushi et al.1997.Lethality of the covalent linkage between mislocalized major outer membrane lipoprotein and the peptidoglycan of Escherichia coli.J.Bacteriol.May;179(9):2857−62。
【0077】
型分類不能なヘモフィルス・インフルエンザ菌は、P4と呼ばれるリポタンパク質(タンパク質“e”としても知られる)を発現する。P4タンパク質の組換え型は、天然のP4シグナル配列を用いて、E.coli中で高度に発現される。米国特許第5,955,580号。天然のP4シグナル配列がE.coli中の発現ベクター中の天然のORF2086シグナル配列に置換されると、ORF2086発現レベルは増大される。
【0078】
発現を増大させるために異種P4シグナル配列を用いるというこの概念は、他の細菌リポタンパク質にまで拡張可能である。特に、細菌ゲノムの分析は、関心の対象となる可能性のある多くのORFの同定につながる。E.coliのような異種宿主細胞中でORFをその天然のシグナル配列によって発現させようとする試みにより、安定性、適合性等を含む各種のシグナル配列を用いる場合に固有の各種の問題を引き起こす。これらの問題を最小限にするため、P4シグナル配列が対象となる各ORFを発現させるために用いられる。上記で記載されるように、P4シグナル配列は、異種2086ORFの発現を改善する。発現ベクターが、対象となるORFの天然のシグナル配列を欠失させ、そしてORFにP4シグナル配列を結合することによって構成される。次に適切な宿主細胞が発現ベクターで形質変換、形質移入または感染させられ、ORFの発現は、ORFの天然のシグナル配列を用いた発現と比較して増大する。
【0079】
非脂質化型は、本来のリーダー配列を欠くタンパク質によって、または宿主細胞中の脂肪酸アシル化のための部位を特定しない配列の部分で置き換えられたリーダー配列によって産生される。
【0080】
本発明の2086タンパク質の様々な形態は、特にことわらない限り、本明細書中では「2086」タンパク質と呼ばれる。また、「2086ポリペプチド」は、特にことわらない限り、2086タンパク質ならびにその免疫原性部分または生物学的等価物を指す。
【0081】
全長単離および精製されたN.meningitidisタンパク質は、10%〜20%勾配SDSポリアクリルアミドゲル(SDS−PAGE)上で測定されると約28〜35kDaの見掛けの分子量を有する。より具体的には、このタンパク質は、質量分析法により測定されると約26,000〜30,000ダルトンの分子量を有する。
【0082】
好ましくは、2086ポリペプチドおよびそのようなポリペプチドをコードする核酸は、N.meningitidisおよび/または他の種により引き起こされる感染を予防または改善するために用いられる。
抗体
配列番号2−12のアミノ酸配列を含む本発明のタンパク質、それらのフラグメント、およびそれらの類似体、またはそれらを発現させる細胞はまた、本発明のポリペプチドについて免疫特異性の抗体を産生するための免疫原として用いられる。本発明は、免疫特異性のポリペプチドに対する抗体およびN.meningitidisの存在を検出し、受動的保護を提供あるいは細胞、組織抽出物、または生体液中のポリペプチドの量もしく濃度を測定するためのそのような抗体の使用を含む。
【0083】
本発明の抗体としては、ポリクローナル抗体、モノクローナル抗体、キメラ抗体、および抗イディオタイプ抗体が含まれる。ポリクローナル抗体は、抗原で免疫性化された動物の血清から得られた抗体分子の異種個体群である。モノクローナル抗体は、特定の抗原に対する抗体の実質的に均質な個体群である。モノクローナル抗体は、当業者に知られている方法、例えば、Kohler and Milstein,1975,Nature 256:495−497および米国特許第4,376,110号により得られ。そのような抗体は、IgG、IgM、IgE、IgA、GILDおよびそれらの任意のサブクラスを含む任意の免疫グロブリンクラスであり得る。
【0084】
キメラ抗体は、マウスモノクローナル抗体およびヒト免疫グロブリン定常領域から得られた可変領域を有するもののような、その異なる部分が異なる動物種から得られる分子である。キメラ抗体およびそれらの産生方法は、技術的に知られている
【0085】
【化1】
これらの参考文献は、参照によりその全体が本明細書に組み込まれる。
【0086】
抗イディオタイプ(抗Id)抗体は、抗体の抗原結合部位と一般に関連付けられる特有の決定基を認識する抗体である。抗Id抗体は、モノクローナル抗体の供給源と同じ種および遺伝子型の動物(例えば、マウス株)に、抗Idが調製されているモノクローナル抗体で免疫化することによって調製される。免疫化された動物は、これらのイソタイプ決定基に対する抗体(抗Id抗体)を産生することにより、免疫化する抗体のイディオタイプ決定基を認識して応答する。
【0087】
したがって、本発明のポリペプチドに対して発生したモノクローナル抗体は、適切な動物の抗Id抗体を引き起こすように使用できる。そのような免疫化されたマウス由来の脾臓細胞は、抗Idモノクローナル抗体を分泌する抗Idハイブリドーマを産生するために使用できる。さらに、抗Id抗体は、キーホール・リンペット・ヘモシアニン(KLH)のような担体に結合させて、付加的なBALB/cマウスを免疫化するために使用できる。これらのマウス由来の血清は、R−PTPaseエピトープについて特異的な最終mAbの結合特性を有する抗抗Id抗体を含んでいる。抗Id抗体は、したがって、それらのイディオタイプエピトープ、すなわち、ストレプトコッカス・ピオゲネスポリペプチドのような評価されるエピトープと構造的に類似する「イディオトープ(idiotope)」を有する。
【0088】
用語「抗体(antibody)」はまた、無傷の分子ならびにFabのようなフラグメント、一本鎖抗体および抗原に結合できる抗体の他の抗原認識フラグメントの双方を含むことを意図している。Fabフラグメントは、無傷の抗体のFcフラグメントを欠いており、循環からより迅速に消え、そして無傷の抗体よりも非特異的組織結合が少ないことがある(Wahl et al.、1983,J.Nucl.Med.24:316−325)。本発明において有用な抗体のFabおよび他のフラグメントが、無傷の抗体分子のための方法に従ってN.meningitidisポリペプチドの検出および定量に用い得ることが理解されるであろう。
【0089】
anti−iodiotypic(「抗Id」)抗体のような本発明の抗体は、上記で記載されたようなポリペプチドに特異的な抗体を免疫学的に有効量投与することを含む、哺乳動物宿主におけるナイセリア感染の処置または予防のための方法において用いられ得る。抗Id抗体はまた、さらに別の動物において、いわゆる抗抗Id抗体(anti−anti−Id antibody)を産生する免疫応答を誘発する「免疫原」として用いられ得る。抗抗Idは、抗Idを誘発した当初のmAbとエピトープ的に同一であり得る。したがって、mAbのイディオタイプ決定基に対する抗体を用いることにより、同一の特異性の抗体を発現させる他のクローンを同定することが可能である。
【0090】
抗体は、様々なやり方、例えば、タンパク質が発現されていることの確認のため、またはタンパク質がどこで発現されているかを確認するために用いられる。標識抗体(例えば、FACSのための蛍光標識化)は、無傷の細菌でインキュベートすることができ、細菌表面上の標識の存在は、例えばタンパク質の位置を確認する。
【0091】
本発明のポリペプチドに対して生成された抗体は、ポリペプチドまたエピトープ坦持フラグメント、類似体、または細胞を、常用の手順を用いて動物に投与することによって得ることができる。モノクローナル抗体の調製については、継代細胞系培養により産生された抗体を提供する任意の手法を用いることができる。
ポリヌクレオチド
本発明のタンパク質と同様に、本発明のポリヌクレオチドは、奇数番号の配列番号1−11の基準配列のうちのいずれかと同一、すなわち100%同一の核酸配列を含んでもよく、または、基準配列と比較していくつかのヌクレオチド改変を含んでもよい。そのような改変は、少なくとも1つのヌクレオチド欠失、転位および転換を含む置換、または挿入からなる群から選ばれ、前記改変は、基準ヌクレオチド配列の5’もしくは3’末端位置において、あるいは個別に基準アミノ酸配列中か基準アミノ酸配列内の1つ以上の隣接する群中のアミノ酸間に散在されて、それらの末端位置間のどこかで生じる。ヌクレオチド変更の数は、奇数番号の配列番号1−11のいずれかの中のヌクレオチド総数にそれぞれのパーセント同一性(100で割られる)を乗じ、その積を前記配列中の前記総数から引くことにより求められる。
【0092】
一例として、それに限定されることを意図することなく、奇数番号の配列番号1−11のいずれかの核酸配列、その縮退変異体またはそのフラグメントと少なくとも95%同一性を有するポリヌクレオチド配列を含む単離N.meningitidisポリヌクレオチドであり、ここでポリヌクレオチド配列は、奇数番号の配列番号1−11の核酸配列のポリヌクレオチド領域全体にわたり最高nnの核酸変更を含むことができ、nnは、改変の最大数であり、次式により計算される:
nn=xn−(xn・y)、
式中、xnは、奇数番号の配列番号1−11のいずれかの核酸の総数であり、yは、0.95の値を有し、xnとyとのどのような非整数積も、そのような積をxnから引く前に最も近い整数に丸められる。当然、yは、95%について0.95の値等を有することもできる。偶数番号配列配列番号2−12のいずれかのアミノ酸配列を含むポリペプチドをコードするポリヌクレオチド配列の改変は、このコーディング配列中にナンセンス、ミスセンスまたはフレームシフト突然変異を作り出し、それによって、そのような改変に続いてポリヌクレオチドによってコードされるポリペプチドを改変することがある。
【0093】
本発明のある一定の実施形態は、2086タンパク質に対して作られた2086タンパク質および抗体をコードするポリヌクレオチド(本明細書中では、「2086ポリヌクレオチド」または「ORF2086ポリヌクレオチド」と呼ばれる)に関する。好ましい実施形態において、本発明の単離ポリヌクレオチドは、奇数番号の配列番号1−11のうちの1つ、その縮退変異体、またはそのフラグメントから選ばれるヌクレオチド配列と少なくとも約95%の同一性を有するヌクレオチド配列を含むポリヌクレオチドである。本明細書中で定義されるように、「縮退変異体(degenarate variant)」は、遺伝コードの縮退により奇数番号の配列番号1−11(およびそのフラグメント)中に示されるヌクレオチド配列とは異なるポリヌクレオチドと定義されるが、奇数番号の配列番号1−11中に示されるヌクレオチド配列によりコードされるものと同じ2086タンパク質(例えば、偶数番号配列配列番号2−12)を依然としてコードする。
【0094】
他の実施形態において、ポリヌクレオチドは、奇数番号の配列番号1−11のうちの1つ、その縮退変異体、またはそのフラグメントから選ばれるヌクレオチド配列に対する補体である。さらに他の実施形態において、ポリヌクレオチドは、DNA、染色体DNA、cDNAおよびRNAからなる群から選ばれ、異種ヌクレオチドをさらに含み得る。
【0095】
2086ポリヌクレオチドが、天然の、合成または半合成の供給源から得られること、さらに、ヌクレオチド配列が、天然に生じる配列であり得ること、あるいは、もし常にそのような配列を含む核酸分子が上記のように2086免疫原性ポリペプチドとして発現させられ得るのであれば、ヌクレオチド配列が、そのような天然に生じる配列への一もしくは複数塩基置換、欠失、挿入および逆位を含む突然変異に関連し得ることが理解されるであろう。核酸分子は、RNA、DNA、一本鎖もしくは二本鎖、線状または共有結合環状形であり得る。ヌクレオチド配列は、それに隣接して配置された発現制御配列を有することがあり、そのような制御配列は通常、異種供給源から導かれる。一般に、本発明の核酸配列の組換え発現は、核酸配列の末端において、TAAのような停止コドン配列を用いる。
【0096】
本発明はまた、低減されたストリンジェンシー条件下、好ましくはストリンジェントな条件下、そして最も好ましくは高度にストリンジェントな条件下で、本明細書中に記載されるたポリヌクレオチドにハイブリダイズできるポリヌクレオチドも含む。ストリンジェンシー条件の例は、以下のストリンジェンシー条件表に示してあり、高度にストリンジェントな条件とは、例えば、少なくとも条件A〜Fと同じくらいストリンジェントな条件であり、ストリンジェントな条件とは、例えば、少なくとも条件G〜Lと同じくらいストリンジェント条件であり、低減されたストリンジェンシー条件とは、例えば、少なくとも条件M〜Rと同じくらいストリンジェントな条件である。
【0097】
【表1−1】
【0098】
【表1−2】
【0099】
【表1−3】
bpI:ハイブリッド長は、ハイブリダイズしているポリヌクレオチドのハイブリダイズされた領域について予想される長さである。ポリヌクレオチドを、未知の配列の標的ポリヌクレオチドにハイブリダイズする場合、ハイブリッド長は、ハイブリダイズしているポリヌクレオチドの長さであると仮定される。既知配列のポリヌクレオチドがハイブリダイズされる場合、ハイブリッド長は、ポリヌクレオチドの配列を一直線に並べ、最適配列相補性の1つ以上の領域を同定することによって確認できる。
【0100】
バッファーH:SSPE(1×SSPEは、0.15 MNaCl、10 mM NaH2PO4、および1.25 mM EDTA、pH 7.4である)は、ハイブリダイゼーションおよび洗浄バッファーにおいて、SSC(1×SSCは、0.15M NaClおよび15 mMクエン酸ナトリウムである)の代わりに使用でき、洗浄は、ハイブリダイゼーション完了後に15分間実行される。
【0101】
TB〜TR:長さが50塩基対未満であると予想されるハイブリッドのためのハイブリダイゼーション温度は、ハイブリッドの融解温度(Tm)より5〜10EC低くあるべきであり、ここで、Tmは、以下の式に従って決定される。長さが18塩基対未満のハイブリッドについては、Tm(EC)=2(A+T塩基の数)+4(G+C塩基の数)。長さが18〜49塩基対のハイブリッドについては、Tm(EC)=81.5+16.6(log10[Na+])+0.41(%G+C)−(600/N)であり、ここでNは、ハイブリッド中の塩基の数であり、[Na+]は、ハイブリダイゼーションバッファ中のナトリウムイオン濃度である(1×SSCについての[Na+]=0.165M)。
【0102】
ポリヌクレオチドハイブリダイゼーションのためのストリンジェンシー条件の付加的な例は、Sambrook,J.,E.F.Fritsch,and T.Maniatis,1989,Molecular Cloning:A Laboratory Manual,Cold Spring Harbor Laboratory Press,Cold Spring Harbor,NY,chpters 9 and 11,and Current Protocols in Molecular Biology,1995,F.M.Ausubelら、eds.,John Wiley & Sons,Inc.,sections 2.10 and 6.3−6.4に提供されており、これらは参照により本明細書に組み込まれる。
【0103】
本発明はまた、これらのポリヌクレオチドに完全に相補的なポリヌクレオチドを提供し、またアンチセンス配列も提供する。アンチセンスオリゴヌクレオチドとも呼ばれる本発明のアンチセンス配列は、本発明のポリペプチドをコードするポリヌクレオチドの発現を妨げる内部生成配列および外部投与発現の両方を含む。本発明のアンチセンス配列は、例えば、約15〜20塩基対を含む。アンチセンス配列は、例えば、上流の非翻訳配列に結合するプロモーターを妨げることにより、またはリボソームの結合を妨げることにより本発明のポリペプチドをコードする転写産物の翻訳を妨げることにより、転写を阻害するように設計することができる。
【0104】
本発明のポリヌクレオチドは、多くのやり方で(例えば、化学合成により、DNAライブラリから、生物体それ自体から)調製され、様々な形態(例えば、一本鎖、二本鎖、ベクター、プローブ、プライマー)を取ることができる。用語「ポリヌクレオチド(polynucleotide)」としては、DNAおよびRNA、ならびに修飾されたバックボーンを含むそれらの類似物が含まれる。
【0105】
本発明のさらなる実施形態によれば、本発明のポリヌクレオチドは、cDNAライブラリのようなDNAライブラリを含む。
融合タンパク質
本発明はまた、融合タンパク質に関する。「融合タンパク質(fusion protein)」は、2つの、しばしば無関係な、融合遺伝子またはそのフラグメントによりコードされたタンパク質を意味する。例えば、免疫グロブリン分子の定常領域の様々な部分を別の免疫原性タンパク質またはその部分と共に含む融合タンパク質である。多くの場合、免疫グロブリンFc領域を融合タンパク質の一部として使用することは、治療および診断での使用に有利であり、例えば、改善された薬物動態学特性という結果になる(例えば、欧州特許出願公開第0232262号参照)。他方、用途によっては、融合タンパク質が発現され、検出され、精製された後に、Fc部分を欠失させることができることが望ましいであろう。本発明の2086ポリヌクレオチドは、本発明のポリペプチドの組換え産生に用いられ、ポリヌクレオチドは、それ自体で、成熟ポリペプチドのためのコード配列、あるいは、リーダーまたは分泌配列、プレもしくはプロないしはプレプロタンパク質配列、または他の融合ペプチド部分をコードするコード配列のような、他のコード配列を持つリーディングフレーム中の成熟ポリペプチドのためのコード配列を含み得る。例えば、2086ポリペプチドまたは融合ポリペプチドの精製を容易にするマーカー配列がコードされ得る(参照によりその全体が本明細書に組み込まれる、Gentzら、1989参照)。したがって、発現産物のHisタグ精製を可能にする融合ポリペプチドをコードするポリヌクレオチドの調製が本発明の実施において考慮される。ポリヌクレオチドはまた、転写された、非翻訳配列、スプライシングおよびポリアデニル化シグナルのような、非コーディング5’および3’配列を含み得る。そのような融合ポリペプチドは、形質変換/形質移入されたまたは感染させられたもしくは以下に記載されるように組換えDNAクローニングビークルで感染させられた宿主細胞により産生されることができ、そのような融合ペプチドはその後、宿主細胞から単離され、他の宿主細胞タンパク質を実質的に含まない融合ポリペプチドを提供することができる。
免疫原性組成物
本発明の1つの態様は、少なくとも1つの2086タンパク質または前記タンパク質をコードする核酸を含む免疫原性組成物を提供する。上記のものは、(1)複数の菌株への殺菌抗体を引き出し、(2)複数の菌株の表面と反応し、(3)生菌攻撃に対する受動的保護を付与し、および/または、(4)コロニー形成を防止する能力を有する。そのような免疫原性組成物の製剤は、当業者によく知られている。ある一定の実施形態において、本発明の組成物は、薬学的に許容される担体および/または希釈剤を含む。適切な薬学的に許容される担体および/または希釈剤としては、あらゆるすべての従来の溶媒、分散媒、フィラー、固体担体、水溶液、コーティング、抗菌性および抗真菌性薬剤、等張性および吸収遅延薬剤等が含まれる。適切な薬学的に許容される担体としては、例えば、1つ以上の水、食塩水、リン酸緩衝食塩水、デキストロース、グリセリロール、エタノール等、ならびにそれらの組み合わせが含まれる。薬学的に許容される担体はさらに、湿潤または乳化剤、防腐剤またはバッファーのような抗体の保存期間または有効性を向上する少量の補助物質をさらに含み得る。薬学的に許容される担体の調製および使用は、技術的によく知られている。いずれかの従来の培地または薬剤が活性成分と不和合である限りを除き、本発明の免疫原性組成物におけるそれらの使用は考慮される。本発明のある一定の実施形態によれば、薬学的に許容される担体は、担体タンパク質である。
担体タンパク質
担体タンパク質は、好ましくは非毒性かつ非反応原性であり、さらに十分な量および純度で入手可能なタンパク質である。担体タンパク質は、標準の接合手順に適用できるべきである。本発明の特定の実施形態において、CRM197が担体タンパク質として用いられる。
【0106】
CRM197197(Wyeth,Sanford,NC)は、カザアミノ酸および酵母抽出物をベースとする培地中で生育されたコリネバクテリウム・ジフテリア菌株C7(β197)の培養物から単離されたジフテリア毒素の非毒性変異体(すなわち、トキソイド)である。CRM197は、限外濾過、硫酸アンモニウム沈降、およびイオン交換クロマトグラフィーによって精製される。CRM197を得る別の方法は、米国特許第4,925,792号に記載される。代わりに、CRM197は、米国特許第5,614,382号に従って組換え的に調製される。他のジフテリアトキソイドも、担体タンパク質としての使用に適する。
【0107】
他の実施形態において、本発明の担体タンパク質は、酵素的に不活性なストレプトコッカルC5aペプチダーゼ(SCP)(例えば、米国特許第6,270,775号、同第6,355,255号、および同第6,951,653号に記載される1つ以上のSCP変異体)である。
【0108】
他の適切な担体タンパク質としては、破傷風トキソイド、百日咳トキソイド、コレラトキソイド(例えば、国際公開第2004/083251号に記載されるCT E29H)、E.coli LT、E.coli ST、E.coli DnaKタンパク質、およびPseudomonas aeruginosa由来の外毒素Aのような不活性化された細菌毒素が含まれる。外膜複合体c(OMPC)、ポリン、トランスフェリン結合タンパク質、ニューモリシン毒素(例えば、米国特許第5,565,204号)、ニューモリシントキソイド(例えば、国際公開第2005/108580号)肺炎球菌表面タンパク質A(PspA)、肺炎球菌アドヘシンタンパク質(PsaA)、またHaemophilus influenzaeタンパク質Dのような細菌外膜タンパク質もまた用いられる。ミコバクテリアhsp−70のような細菌熱ショックタンパク質もまた用いられる。Staphylococcus epidermidisタンパク質SdrG、SitCおよびフェロクロム結合タンパク質、ならびにStaphylococcus aureusタンパク質ClfA、ClfBおよびFnbAのような他のタンパク質もまた用いられる。オボアルブミン、キーホール・リンペット・ヘモシアニン(KLH)、グルタチオンS−トランスフェラーザ(GST)、ウシ血清アルブミン(BSA)、ガラクトキナーザ(galK)、ユビキチン、β−ガラクトシダーゼ、インフルエンザNS1タンパク質、またはツベルクリンの精製タンパク質誘導体(PPD)のようなさらに他のタンパク質もまた、担体タンパク質として用いられる。例えばロタウイルスVP6またはバクテリオファージQβ由来のウイルス様粒子もまた用いられる。
アジュバント
本明細書中で記載されるような免疫原性組成物はまた、ある一定の実施形態において、1つ以上のアジュバントを含む。アジュバントは、免疫原または抗原と共に投与された時に、免疫応答を増大させる物質である。いくつかのサイトカインおよびリンホカインは、免疫調整活性を有することが示されており、したがってアジュバントとして有用であり、インターロイキン1−α、1−β、2、4、5、6、7、8、10、12(例えば、米国特許第5,723,127号参照)、13、14、15、16、17および 18(ならびにその突然変異体);インターフェロン−α、βおよびγ;顆粒状マクロファージコロニー刺激因子(GM−CSF)(例えば、米国特許第5,078,996号およびATCC受入番号39900参照)、マクロファージコロニー刺激因子(M−CSF);顆粒状コロニー刺激因子(G−CSF)、ならびに腫瘍壊死因子αおよびβが含まれるが、これらに限定されない。本明細書中で記載される免疫原性組成物と共に有用な他のアジュバントとしては、MCP−1、MIP−1α、MIP−1βおよびRANETSを制限無く含むケモカイン;セレクチン、例えば、L−セレクチン、P−セレクチンおよびE−セレクチンのような接着分子;ムチン様分子、例えば、CD34、GlyCAM−1およびMadCAM−1;LFA−1、VLA−1、Mac−1およびp150.95のようなインテグリンファミリーの一つ;PECAM、ICAMs、例えば、ICAM−1、ICAM−2およびICAM−3、CD2およびLFA−3のような免疫グロブリンスーパーファミリーの一つ;CD40およびCD40Lのような共刺激分子;血管成長因子、神経成長因子、神経芽細胞成長因子、表皮成長因子、B7.2、PDGF、BL−1、ならびに血管内皮成長因子を含む成長因子;Fas、TNF受容体、Flt、Apo−1、p55、WSL−1、DR3、TRAMP、Apo−3、AIR、LARD、NGRF、DR4、DR5、KILLER、TRAIL−R2、TRICK2、およびDR6を含む受容体分子;ならびにカスパーゼ(ICE)が含まれる。
【0109】
免疫応答を強化するために用いられる適切なアジュバントは、制限なく、MPL(登録商標)(3−O−デアシル化モノホスホリル脂質A,Corixa,Hamilton,MT)をさらに含み、これは米国特許第4,912,094号に記載される。合成脂質A類似体またはアミノアルキルグルコサミンホスフェート化合物(AGP)あるいはその誘導体または類似物も、アジュバントとしての使用に有用であり、これらは、Corixa(Hamilton,MT)から利用可能であり、米国特許第6,113,918号に記載される。1つのそのようなAGPは、2−[(R)−3−テトラデカノイルオキシテトラデカノイルアミノ]エチル2−デオキシ−4−O−ホスホノ−3−O−[(R)−3−テトラデカノイルオキシテトラデカノイル]−2−[(R)−3−テトラデカノイルオキシテトラデカノイル−アミノ]−b−D−グルコピラノシドであり、これも、529として知られている(以前は、RC529として知られていた)。この529アジュバントは、水性形態(AF)または安定エマルション(SE)として配合される。
【0110】
さらに他のアジュバントとしては、N−アセチル−ムラニル−L−トレオニル−D−イソグルタミン(thr−MDP)、N−アセチル−ノルムラニル−L−アラニン−2−(1’−2’ジパルミトイル−sn−グリセロ−3−ヒドロキシホスホリルオキシ)−エチルアミン(MTP−PE)のようなムラミルペプチド;MF59(国際公開第90/14837号)(5%スクワレン、0.5%Tween80、および0.5%Span85を含み(種々の量のMTP−PEを任意に含み))、モデル110Yマイクロフルイダイザー(Microfluidics,Newton,MA)のようなマイクロフルイダイザーを用いてサブミクロン粒子状に処方される)、およびSAF(10%スクワレン、0.4%Tween80、5%プルロニックブロックポリマーL121、およびthr−MDPを含み、サブミクロンエマルションに微小液滴化されるか、より大きい粒径のエマルションを生成するために渦撹拌される)のような水中油型エマルション;不完全フロイントアジュバント(IFA);水酸化アルミニウム、リン酸アルミニウム、硫酸アルミニウムのようなアルミニウム塩(みょうばん);アンフィゲン(Amphigen)、アブリジン、L121/スクワレン、D−ラクチド−ポリアクチド/グリコシド;プルロニックポリオール;死菌Bordetella;米国特許第5,057,540号に記載のStimulon(商標)QS−21(Antigenics,Framingham,MA.)、米国特許第5,254,339号に記載のISCOMATRIX(CSL Limited,Parkville,Australia)、および免疫刺激複合体(ISCOMS)のようなサポニン;Mycobacterium tuberculosis;細菌リポ多糖;CpGモチーフを含むオリゴヌクレオチドのような合成ポリヌクレオチド(例えば、米国特許第6,207,646号);欧州特許第1,296,713号および同第1,326,634号に記載のIC−31(Intercell AG,Vienna,Austria);百日咳毒素(PT)またはその突然変異体、コレラ毒素またはその突然変異体(例えば、国際出願第00/18434号、同第02/098368号および同第02/098369号);あるいはE.coli熱不安定毒素(LT)、特にLT−K63、LT−R72、PT−K9/G129;例えば、国際出願第93/13302号および同第92/19265号参照、が含まれる。
投与様式
そのような免疫原性組成物は、非経口的に、例えば、皮下注射か筋肉内注射により、ならびに経口的または鼻腔内的に投与され得る。筋肉内免疫化の方法は、WolffらおよびSedegahらにより記載されている。他の投与様式は、例えば、経口製剤、肺製剤、座薬、および経皮適用を用いるが、それらに限定されない。経口製剤としては、例えば、製薬グレードのマンニトール、乳糖、デンプン、ステアリン酸マグネシウム、サッカリンナトリウム、セルロース、炭酸マグネシウム等の通常用いられる賦形剤が含まれるが、それらに限定されない。
【0111】
本発明の免疫原性組成物は、ISCOMS(免疫刺激性複合体)、ISCOMS含有CTB、リポソームの形態で送達されるか、あるいは吸着に適したサイズの微小球を形成するためにアクリレートまたはポリ(DL−ラクチド−グリコシド)のような化合物中にカプセル化され得る。本発明のタンパク質はまた、油性エマルションに組み入れられ得る。
多重抗原
本発明のタンパク質、ポリヌクレオチドおよび等価物を含む免疫原性薬剤は、免疫原性組成物中の唯一の能動免疫原として投与されてもよく、代わりに、組成物は、他のNeisseria sp.免疫原性ポリペプチド、または1つ以上の他の微生物病原体(例えば、ウイルス、プリオン、細菌、真菌、ただしこれらに限定されない)の免疫学的活性なタンパク質もしくは莢膜多糖を含む他の能動免疫原を含んでもよい。組成物は、選ばれた徴候について要求されるように、1つ以上の所望のタンパク質、フラグメントまたは製薬化合物を含み得る。同様に、免疫原性組成物中に1つ以上の核酸を使用する本発明の組成物は、上記のように、同じ多様なタンパク質群をコードする核酸も含み得る。
【0112】
任意の多重抗原または多価免疫原性組成物が本発明により考慮される。例えば、本発明の組成物は、粘膜送達に適した形態で、2つ以上の2086タンパク質の組み合わせ、2086タンパク質と1つ以上のPorAタンパク質との組み合わせ、2086タンパク質と髄膜炎菌セログループA、C、YおよびW135多糖および/または多糖結合体との組み合わせ、2086タンパク質と髄膜炎菌および肺炎球菌組み合わせとの組み合わせ、または上記のいずれかの組み合わせを含み得る。当業者は、そのような多重抗原または多価免疫学的組成物を容易に配合できるであろう。
【0113】
本発明はまた、病原体に対し有用な任意の組成物を本発明の組成物の中でまたは本発明の組成物と共に組み合わせることができる、多重免疫化処方計画も考慮する。例えば、制限無く、患者は、多重免疫化処方計画の一環として、本発明の免疫原性組成物およびS.Pneumoniaeに対して免疫化するための別の免疫学的組成物を投与され得る。当業者は、多重免疫化処方計画を開発および実施するために、本発明の免疫原性組成物と併用するための免疫原性組成物を容易に選択することができるであろう。
【0114】
本発明の特定の実施形態は、S.Pneumoniae感染の予防または改善のための組成物中でまたは処置処方計画の一環として、1つ以上の本発明のポリペプチドまたはそれをコードする核酸の使用に関する。2086ポリペプチドまたは2086ポリヌクレオチドを、S.Pneumoniae感染に対して使用するための任意の免疫原性組成物と組み合わせることができる。また、2086ポリペプチドまたは2086ポリヌクレオチドを、任意の他のタンパク質または多糖ベースの髄膜炎菌ワクチンと組み合わせることができる。
【0115】
2086ポリペプチド、フラグメントおよび等価物は、接合免疫原性組成物の一部として用いることができ、その場合、1つ以上のタンパク質またはポリペプチドは、いくつかのセロタイプに対しておよび/またはいくつかの疾患に対して免疫原性特性を有する組成物を生成するために、担体タンパク質に接合する。代わりに、2086ポリペプチドの1つが、他の免疫原性ポリペプチドのための担体タンパク質として用いられ得る。
【0116】
本発明はまた、哺乳動物における免疫応答を引き起こす方法に関し、この方法は本発明の免疫原性組成物を前記哺乳動物に提供するステップを含む。この免疫原性組成物は、そのような組成物中に含まれる免疫学的有効量のポリペプチドがN.meningitidis感染に対して所望の免疫応答を引き起こすように、処置される動物またはヒトにおいて抗原性の組成物である。好ましい実施形態は、ヒトにおけるN.meningitidis感染の改善または予防を含む処置のための方法に関し、この方法は、免疫学的有効量の組成物をヒトに投与するステップを含む。
【0117】
本明細書中で用いられるように、文言「免疫学的有効量」は、処置された個体の免疫系に細菌感染の臨床的衝撃を低減する応答を少なくとも引き起こさせるのに十分な単一用量か、一連の用量の一環としての、哺乳動物宿主(好ましくはヒト)への投与を意味する。これは、細菌負荷の最小限の減少から細菌感染の予防にまで及び得る。理想的には、処置された個体は、細菌感染のより深刻な臨床的徴候を示さない。投薬量は、個体の特定の条件に応じて変わり得る。この量は、常用の試行により、さもなければ当業者に既知の手段により決定できる。
【0118】
本発明の別の特定の態様は、本発明のタンパク質、またはその免疫原性部分を発現させるベクターまたはプラスミドを免疫原性組成物として用いることに関する。したがって、さらなる態様として、本発明は、哺乳動物において免疫応答を引き起こす方法を提供し、この方法は、少なくとも1つの単離2086ポリペプチドを発現させるベクターまたはプラスミドを哺乳動物に提供するステップを含む。本発明のタンパク質は、生きたベクターを用いて、特に、異物ポリペプチドとしてポリペプチドまたは免疫原性部分の発現に必要な遺伝物質を含んでいる、生きた組換え細菌、ウイルスまたは他の生きた薬剤を用いて哺乳動物に送達され得る。
【0119】
本発明のさらなる実施によれば、哺乳動物において細菌性髄膜炎を診断するための方法が提供され、この方法は、哺乳動物または該哺乳動物の組織試料中の免疫複合体の存在を検出するステップを含み、前記哺乳動物または組織試料は、偶数番号配列配列番号2−12のいずれかのアミノ酸配列を含む少なくとも1つのポリペプチドと免疫特異的に結合する抗体を含む抗体組成物と接触させられ、哺乳動物または組織サンプルは、免疫複合体の形成に適した条件下で抗体組成物と接触させられる。
ウイルスベクターおよび非ウイルスベクター
特にin vitoroおよびin vivo細胞アッセイのための好ましいベクターは、レンチウイルス、レトロウイルス、ヘルペスウイルス、アデノウイルス、アデノ随伴ウイルス、ワクシニアウイルス、バキュロウイルス、および望ましい細胞指向性を持つ他の組換えウイルスのようなウイルスベクターである。したがって、2086タンパク質またはその免疫原性フラグメントをコードする核酸は、ウイルスベクターを用いてまたはDNAの直接導入により、in vivo、ex vivo、またはin vitroで導入され得る。標的組織中での発現は、ウイルスベクターまたは受容体リガンドにより、または組織特異性プロモーターを用いて、あるいはその両方により、トランスジェニックベクターを特定の細胞特定に向けることにより行われ得る。標的遺伝子送達は、国際公開第95/28494号に記載され、これは、参照によりその全体が本明細書に組み入れられる。
【0120】
in vivoまたはex vivoターゲティングおよび療法手順において一般的に用いられるウイルスベクターは、DNAベースのベクターおよびレトロウイルスベクターである。ウイルスベクターを調製し、使用する方法は、技術的に知られている(例えば、Miller and Rosman,BioTechniques,1992,7:980−990)。好ましくは、ウイルスベクターは複製欠陥性であり、すなわち、それらのウイルスベクターは、標的細胞中で自律的に複製できない。好ましくは、複製欠陥性ウイルスは最小のウイルスであり、すなわち、そのウイルスは、ウイルス粒子を産生するゲノムを被包するのに必要なそのゲノム配列のみを保持する。
【0121】
DNAウイルスベクターとしては、単純ヘルペスウイルス(HSV)、乳頭腫ウイルス、エプスタイン−バールウイルス(EBV)、アデノウイルス、アデノ随伴ウイルス(AAV)等のような、ただしこれらに限定されない弱毒性または欠陥DNAウイルスが含まれる。ウイルス遺伝子を完全にまたはほぼ完全に欠く欠陥ウイルスが好ましい。欠陥ウイルスは、細胞内への導入後に感染性はない。欠陥ウイルスベクターの使用は、ベクターが他細胞に感染し得るという不安なしに、特定の局部的領域における細胞への投与ができる。したがって、特定の組織を特異的に標的とし得る。具体的なベクターの例としては、各々が参照によりその全体が本明細書に組み入れられる、欠陥ヘルペスウイルス1(HSV1)ベクター(Kaplittら、Molec.Cell.Neurosci.,1991,2:320−330)、糖タンパク質L遺伝子を欠く欠陥ヘルペスウイルスベクター、または他の欠陥ヘルペスウイルスベクター(国際公開第94/21807号および同92/05263号);Stratford−Perricaudet et al.により記載されたベクターのような、弱毒アデノウイルスベクター(J.Clin.Invest.,1992,90:626−630;La Salleら、Science,1993,259:988−990も参照);および欠陥アデノ随伴ウイルスベクター(Samulskiら、J.Viol.,1987,61:3096−3101;Samulskiら、J.Viol.,1989,63:3822−3828;Lebkowskiら、Mol.Cell.Biol.,1988,8:3988−3996)が含まれるが、それらに限定されない。
【0122】
Avigen,Inc.(Alameda,CA;AAVベクター)、Cell Genesys(Foster City,CA;レトロウイルス、アデノウイルス、AAVベクター、およびレンチウイルスベクター)、Clontech(レトロウイルスおよびバキュロウイスルベクター)、Genovo,Inc.(Sharon Hill,PA;アデノウイルスおよびAAVベクター)、Genvec(アデノウイルスベクター)、IntroGene(Leiden,Netherland;アデノウイルスベクター)、Molecular Medicine(レトロウイルス、アデノウイルス、AAV、およびヘルペスウイルスベクター)、Norgen(アデノウイルスベクター)、Oxford BioMedica(Oxford,United Kingdom;レンチウイルスベクター)、およびTransgene(Strasbourg,France;アデノウイルス、ワクシニア、レトロウイルス、およびレンチウイルスベクター)を含む様々な企業がウイルスベクターを商業的に製造しているが、これらに限定されるものではない。
【0123】
アデノウイルスベクター。アデノウイルスは、本発明の核酸を各種の細胞型に効率的に送達するように修飾され得る真核DNAウイルスである。アデノウイルスの様々なセロタイプが存在する。これらのセロタイプのうち、本発明の範囲内で、2型または5型ヒトアデノウイルス(Ad2またはAd5)あるいは動物起源アデノウイルス(国際公開第94/26914号参照)の使用が優先される。本発明の範囲内で用いられ得るそれらの動物起源アデノウイルスとしては、イヌ、ウシ、マウス(例:Mav1,Beardら、Viroloy,1990,75−81)、ヒツジ、ブタ、トリ、およびサル(例:SAV)起源が含まれる。好ましくは、動物起源アデノウイルスは、イヌアデノウイルス、より好ましくはCAV2アデノウイルス(例えば、マンハッタンまたはA26/61株、ATCC VR−800)である。様々な複製欠陥アデノウイルスおよび最小アデノウイルスベクターが記載されている(国際公開第94/26914号、同第95/02697号、同第94/28938号、同第94/28152号、同第94/12649号、同第95/02697号、同第96/22378号)。本発明による複製欠陥組換えアデノウイルスは、当業者に知られている任意の手法で調製され得る(Levreroら、Gene,1991,101:195;欧州特許出願公開第185573号;Graham,EMBO J.,1984,3:2917;Grahamら、J.Gen.Virol.,1977,36:59)。組換えアデノウイルスは、当業者に周知の標準的分子生物学的手法を用いて回収および精製される。
【0124】
アデノ随伴ウイルス。アデノ随伴ウイルス(AAV)は、それらが感染する細胞のゲノムに安定的かつ部位特異的なやり方で同化できる比較的小サイズのDNAウイルスである。これらのウイルスは、細胞の成長、形態、または分化へのどのような効果も引き起こすことなく広範囲の細胞に感染することができ、ヒトの病理学に関与しないと思われる。AAVゲノムは、クローン化され、配列決定され、特性決定されている。遺伝子をin vivoおよびin vitroで転移するためのAAV由来ベクターの使用が記載されている(国際公開第91/18088号および同第93/09239号;米国特許第4,797,368号および同第5,139,941号;欧州特許出願公開第488528号参照)。本発明による複製欠陥語組換えAAVは、2つのAAV逆方向末端反復(ITR)領域により隣接された対象となる核酸配列を含むプラスミドを、AAVキャプシド形成遺伝子(repおよびcap遺伝子)を持つプラスミドと、ヒトヘルパーウイルス(例えば、アデノウイルス)で感染させた細胞系に同時形質移入することにより調製できる。産生されたAAV組換え体は次に、標準的手法によって精製される。
【0125】
レトロウイルスベクター。本発明の別の実施において、例えば、各々が参照によりその全体が組み込まれる、米国特許第5,399,346号;Mannら、Cell,1983,33:153;米国特許第4,650,764および同第4,980,289号、Markowitzら、J.Viol.,1988,62:1120;米国特許第5,124,263号;欧州特許出願公開第453 242号および同第178 220号;Bersteinら、Genet.Eng.,1985,7:235;McCormick,BioTechnology,1985,3:689;国際公開第95/07358号;およびKuoら、Blood,1993,82:845に記載されるように、核酸をレトロウイルスベクターに導入できる。レトロウイルスは、分裂細胞に感染する組込みウイルスである。レトロウイルスゲノムは、2つのLTR、キャプシド形成配列および3つのコード化領域(gag、polおよびenv)を含む。組換えレトロウイルスベクターにおいて、gag、polおよびenv遺伝子は一般に、全部または部分的に欠失され、対象となる異種核酸配列で置き換えられる。これらのベクターは、HIV、MoMuLV(「マウスモロニー白血病ウイルス」)、MSV(「マウスモロニー肉腫ウイルス」)、HaSV(「ハーベイ肉腫ウイルス」);SNV(「脾臓壊死ウイルス」);RSV(「ラウス肉腫ウイルス」)およびフレンドウイルスのような、異なる型のレトロウイルスから構築できる。適切なパッケージング細胞系が先行技術において記載されており、特に細胞系PA317(米国特許第4,861,719号);PsiCRIP細胞系(国際公開第90/02806号)およびGP+envAm−12細胞系(国際公開第89/07150号)である。加えて、組換えレトロウイルスベクターは、転写活性を抑制するためのLTR内の修飾、ならびにgag遺伝子の一部を含み得る広いキャプシド形成配列も含み得る(Benderら、J.Viol.,1987,61:1639)。組換えレトロウイルスベクターは、当業者に知られている標準的手法により精製される。
【0126】
レトロウイルスベクターは、感染性粒子として機能するように、または単一ラウンドの形質移入を受けるように構成され得る。前者の場合、ウイルスは、発がん性形質転換の原因となるものを除くその遺伝子すべてを保持し、異種遺伝子を発現させるように修飾される。非感染性ウイルスベクターは、ウイルスパッケージングシグナルを抹消するが、異種遺伝子およびパッケージングシグナルを含むように設計された同時導入ウイルスをパッケージするために必要な構造遺伝子を保持するように操作される。したがって、産生されるウイルス粒子は、付加的なウイルスを産生することができない。
【0127】
レトロウイルスベクターはまた、DNAウイルスにより導入されてもよく、それにより、1サイクルのレトロウイルス複製が可能になり、形質移入効率が増幅される(国際公開第95/22617号、同第95/26411号、同第96/39036号および同第97/19182号参照)。
【0128】
レンチウイルスベクター。本発明の別の実施において、レンチウイルスベクターは、脳、網膜、筋肉、肝臓および血液を含むいくつかの組織型における導入遺伝子の直接送達および持続的発現のための薬剤として用いられ得る。ベクターは、これらの組織中の分裂および非分裂細胞を効率的に形質導入し、対象となる遺伝子の長期発現をもたらし得る。概要については、Naldini,Curr.Opin.Biotechnol.,1998,9:457−63を参照、また、Zufferey,et al.,J.Viol.,1998,72:9873−80も参照されたい。レンチウイルスパッケージング細胞系は、利用可能であり、当該技術分野において一般に知られている。レンチウイルスパッケージング細胞系は、遺伝子治療のための高力価レンチウイルスベクターの産生を容易にする。一つの例が、少なくとも3ないし4日間、106IU/mLより大きい力価のウイルス粒子を生成できるテトラサイクリン誘導VSV−Gプソイド型レンチウイルスパッケージング細胞系である(Kafri,et al.,J.Viol.,1999,73:576−584)。誘導細胞系により産生されたベクターは、in vitroおよびin vivoで非分裂細胞を効率的に形質導入するために必要に応じて濃縮され得る。
【0129】
非ウイルスベクター。本発明の別の実施において、ベクターは、in vivoでリポフェクションにより裸のDNAとして、または他の形質移入促進薬剤(ペプチド、ポリマー等)を用いて導入され得る。合成カチオン性脂質は、マーカーをコードする遺伝子のin vivo形質移入のためのリポソームを調製するために用いられ得る(Felgner,et al.,Proc.Natl.Acad.Sci.U.S.A.,1987,84:7413−7417;Felgner and Ringold,Science,1989,337:387−388;Mackey,et al.,Proc.Natl.Acad.Sci.U.S.A.,1988,85:8027−8031;Ulmerら、Science,1993,259:1745−1748)。核酸の転移に有用な脂質化合物および組成物は、国際出願第95/18863号、同第96/17823号、および米国特許第5,459,127号に記載される。ターゲティングの目的で、脂質を他の分子に化学的に結合させることができる(上記、Mackey,et al.参照)。標的ペプチド、例えば、ホルモンまたは神経伝達物質、および抗体のようなタンパク質、または非ペプチド分子を、リポソームに化学的に結合させることができるであろう。
【0130】
カチオン性オリゴペプチド(例えば、国際公開第95/21931号)、DNA結合タンパク質から誘導されたペプチド(例えば、国際公開第96/25508号)、またはカチオン性ポリマー(例えば、国際公開第95/21931号)のような他の分子もまた、in vivoで核酸の形質移入を促進するのに有用である。
【0131】
ベクターを裸のDNAプラスミドとしてin vivoで導入することも可能である。ワクチン目的または遺伝子治療のための裸のDNAベクターは、技術的に知られている方法、例えば、電気穿孔法、顕微注射、細胞融合、DEAEデキストラン、リン酸カルシウム沈降、遺伝子銃の使用、またはDNAベクター輸送体の使用により所望の宿主細胞に導入され得る(例えば、Wuら、J.Biol.Chem.,1992,267:963−967;Wu and wu,J.Biol.Chem.,1988,263:14621−14624;カナダ特許出願公開第2,012,311号;Williamsら、Proc.Natl.Acad.Sci.USA,1991,88:2726−2730)。受容体媒介DNA送達方法もまた用いられ得る(Curielら、Hum.Gene Ther.,1992,3:147−154;Wu and wu,J.Biol.Chem.,1987,262:4429−4432)。米国特許第5,580,859号および同第5,589,466号は、哺乳動物における、形質移入助長薬剤の無い、外因性DNA配列の送達を開示している。最近、エレクトロトランスファー(electrotranfer)と呼ばれる比較的低電圧、高効率のin vivo DNA転移手法が記載されている(Mirら、C.P.Acad.Sci.,1988,321:893;国際公開第99/01157号;同第99/01158号;同第99/01175号)。したがって、本発明の付加的な実施形態は、ヒトにおける免疫応答を引き起こす方法に関し、この方法は、本発明の2086ポリペプチドをコードするある量のDNA分子を、任意に形質導入促進薬剤と共に、前記ヒトに投与するステップを含み、前記ポリペプチドは、発現された場合に免疫原性を保持し、免疫原性組成物中に組み込まれてヒトに投与された場合に、N.meningitidisのようなNeisseria sp.病原体にその後ヒトが感染した際に、増大した疾患を引き起こすことなく保護を提供する。形質移入促進薬剤は、技術的に知られており、参照により本明細書に組み込まれるブピビカイン、ならびに他の局部麻酔薬(例えば、米国特許第5,739,118号参照)およびカチオン性ポリアミン(国際公開第96/10038号に公開される)を含む。
【0132】
本発明はまた、抗体に関し、この抗体は、上記で記載されるように2086ポリペプチドに特異的な、モノクローナル抗体かポリクローナル抗体であり得る。そのような抗体は、当業者によく知られた方法により産生され得る。
細菌発現系およびプラスミド
本発明はまた、ベクターまたはプラスミドのような組換えDNA分子を提供し、これは、プロモーター配列およびイニシエーター配列ならびに本発明のポリペプチドをコードするヌクレオチド配列を有する発現制御配列を含み、ヌクレオチド配列は、プロモーターおよびイニシエーター配列に対して3’に位置する。さらに別の態様において、本発明は、プロモーター配列およびイニシエーター配列を有する発現制御配列、ならびに2086ポリペプチドをコードするヌクレオチド配列を含有する、2086ポリペプチドを発現することができる組換えDNAクローニングビヒクルを提供し、ヌクレオチド配列は、プロモーターおよびイニシエーター配列に対し3’に位置する。さらなる態様において、上記で記載されるような組換えDNAクローニングビークルおよび/または組換えDNA分子を含む宿主細胞が提供される。適切な発現制御配列および宿主細胞/クローニングビークルの組み合わせは、技術的によく知られており、Sambrookら(1989)に例証として記載されている。
【0133】
ひとたび本発明の所望のポリペプチドを発現する組換えDNAクローニングビークルおよび/または宿主細胞が、対応する2086ポリヌクレオチドを含有するプラスミドにより形質転換、形質移入または感染させることにより構築されたら、クローニングビークルまたは宿主細胞は、ポリペプチドが発現されるような条件下で培養される。ポリペプチドは次に、当業者によく知られた手法により、汚染宿主細胞成分を実質的に含まずに単離される。
【0134】
以下の実施例は、本発明の好ましい実施形態を実証するために含まれる。以下の実施例中で開示される手法が、本発明の実施においてうまく機能することが本発明者らに発見された手法を代表するものであり、したがって本発明の実施のための好ましい態様を構成することが当業者により理解されるべきである。しかしながら、当業者は、本開示を考慮して、開示される特定の実施形態において多くの変更がなされ、それでもなお本発明の趣旨および範囲を逸脱することなく同様なまたは類似の結果を得ることができると理解するべきである。
【実施例】
【0135】
(実施例1)
異種菌株に対する殺菌抗体を引き出すことが可能なナイセリア膜タンパク質抽出物の同定:
以下の表IIを参照すると、LOS(脂質オリゴ糖)除去外膜タンパク質調製物は、殺菌抗体を引き出すことが示されている。これらの抗体は、それぞれの菌株のPorAに向けられることが多くある。セログループB髄膜炎菌株8529(B:15:P1.7b,3)由来のLOS除去外膜調製物は、このように普通ではない。なぜならば、それらの調製物は、いくつかの異種菌株に対する殺菌抗体を引き出すからである。
【0136】
【表2】
異種殺菌抗体を引き出す原因となる抗原の単離および特性決定を容易にするため、本発明者らは、どの界面活性剤が抗原を最適に抽出したかを同定しようと努めた。
菌株および培養条件
凍結したガラスびんからのN.meningitidis菌株8529を、GCプレート上に筋状に塗った。(髄膜炎菌株8529は、The RIVM,Bilthoven,The Netherlandsから受領した)。プレートを、36C/5%CO2で7.5時間インキュベートした。いくつかのコロニーを用い、50mLの修正フランツ培地+GCサプリメントを含むフラスコに接種した。フラスコを36℃にてエアシェーカー中でインキュベートし、200RPMで4.5時間撹拌した。5mLを用いて、450mLの修正フランツ培地+GCサプリメントを含むフェルンバッハフラスコに接種した。フラスコを36℃にてエアシェーカー中でインキュベートし、100RPMで11時間撹拌した。450mL全部を用いて、10L発酵槽中の8.5Lの修正フランツ培地+GCサプリメントに接種した。
修正フランツ培地の組成:
グルタミン酸 1.3g/L
システイン 0.02
リン酸水素二ナトリウム・7水和物 10
塩化カリウム 0.09
塩化ナトリウム 6
塩化アンモニウム 1.25
透析酵母抽出物(YE) 40ml
(5容量のdH2Oに対して一晩透析し、次いでオートクレーブした25%YE溶液)
GCサプリメント100X、フィルタ殺菌
デキストロース 400g/L
グルタミン酸 10
コカルボキシラーゼ 0.02
硝酸第二鉄 0.5
以下のパラメータを発酵中制御した:温度=36℃;pH=7.4;溶存酸素=20%。数滴のP−2000消泡剤を添加して発泡を制御した。培養物を定常期まで生育させた。細胞を、OD650=5.25において遠心分離により収集した。一般に、全部で100〜300グラムの湿潤細胞ペーストが、〜8.5Lの培養物から収集される。
異種殺菌抗体を引き出す髄膜炎菌由来の外膜タンパク質画分の部分的精製:
湿重量100グラムの細胞を、10mM HEPES−NaOH、pH7.4、1mN Na2EDTAで湿重量の5倍の体積に縣濁させ、そしてチャンバを備える110Yマイクロフルイダイザーを約18,000psiで通過させて溶解させた。細胞ライゼートを清澄にし、300,000×gで1時間の遠心分離により細胞エンベロープを単離した。細胞エンベロープを、同じバッファーでホモジナイザーを用いて縣濁し、続いて上記のように遠心分離して2回洗浄した。次に細胞エンベロープを、10mM HEPES−NaOH、pH7.4、1mN MgCl2中の1%(w/v)Triton X−100 320mLで抽出した。以下の表IIIを参照すると、Triton X−100およびZwittergent3−14を用いた逐次ディファレンシャル界面活性剤抽出、続いてのマウスの免疫化の結果により、Triton抽出物が、対象となる候補を最適に抽出したと確認することができた。次に、表IIIに列記された5つの菌株のうち4つに対して殺菌抗体応答を引き出すこのTriton X−100抽出物を、BioRad Rotophorユニット中での調製用等電点分離(IEF)により分画した。両性電解質濃度は、1%pH4〜6と混合された1%pH3〜10であった。表IIIに示されるように、いくつかの画分は、異種殺菌性応答を引き出すことが見出された。IEFから得られた画分は、5.5〜7.8のpH範囲で集束したものであり、殺菌アッセイにより確認されるように大部分の菌株に対する異種応答を引き出した。プールしたIEF画分を濃縮し、両性電解質をエタノール沈降により除去した。約5.5〜7.8のpH範囲で得られたタンパク質のいくらかを陰イオン交換カラムに吸着させ、吸着タンパク質および非吸着タンパク質でマウスを免疫化した後に得られる殺菌活性を比較することにより、さらなる精製が達成された。表IIを再度参照すると、多くのタンパク質が陰イオン交換樹脂に吸着された一方、カラムにより吸着されなかったタンパク質はより多くの異種殺菌抗体を引き出した。
【0137】
【表3】
NT:試験せず
*臨床分離株539は、同じ発生から単離された8529の相同菌株である。
【0138】
図1Aに示されるように、SDS−PAGEにより確認されるように、非吸着画分中に2つの主要タンパク質が存在した。これらのタンパク質を同定するため、2種類の分析を実行した。一方の分析は、限定タンパク質分解(図1Aおよび図1B参照)を実行し、続いてペプチドの単離および直接的タンパク質配列決定を行うものであった。他方の分析は、SDS−PAGEを実行し、続いてゲル切り出し、タンパク質分解消化、およびLS−MS/MS(液体クロマトグラフィータンデム質量分析)を行い、(図3参照)対象となる調製物の成分についての質量スペクトル情報を得ることであった。(後に本節中で説明されるペプチドマッピングおよび配列決定方法を参照)
N.meningitidis A Sangerゲノム配列は、ZaugurskyおよびRussel,2001,BioTechniques,31:636−659に記載される方法およびアルゴリズムを用いて分析した。このマイニング分析により、12,000以上の考え得るオープンリーディングフレーム(ORF)がもたらされた。上述の直接配列データおよび質量スペクトルデータは、非吸着画分の主要成分が、Sangerデータベースの分析中に存在するいくつかのORFの生成物であることを示した。この方法論により同定された3つの主なタンパク質は、ORF4431、5163および2086に対応する(図1Bおよび3参照)。
【0139】
ORF4431は、画分中で同定された最も支配的なタンパク質であったが、組換え脂質化4431に対するマウス抗体は殺菌性ではなく、動物モデルにおいて防御応答をもたらさなかった。ORF5163の付加的な分析は進行中である。
【0140】
本明細書中で記載される調製物の2番目に支配的な成分は、ORF2086の生成物に対応する。
免疫原性方法:
抗血清の調製:
言及される場合を除き、タンパク質組成物/ワクチンは、25μgの総タンパク質で製剤し、20μgQS−21をアジュバントとした。0.2mLの用量を皮下(臀部)注射により、6〜8週齢雌スイスウェブスターマウスに0週目および4週目に投与した。0週目および4週目に血液を採取し、最終瀉血を6週目に実行した。
殺菌アッセイ:
殺菌アッセイは、本質的に記載されたように実行した(Mountzouros and Howell,2000,J.Clin.Microbiol.38(8):2878−2884参照)。SBAについての補体媒介抗体依存性の殺菌力価は、アッセイに導入された標的分子の≧50%を致死させる試験血清の最高希釈の逆数として表した(BC50力価)。
2086タンパク質の同定に用いた方法:
臭化シアン開裂およびフラグメントの直接的配列決定:
陰イオン交換非吸着画分(AEUF)の臭化シアン開裂。AEUFを90%冷エタノールにより沈降させ、70%蟻酸中の10mg/mL臭化シアンを用いて1mg/mLのタンパク質濃度に可溶化した。反応は、暗所において室温で夜通し実行した。開裂生成物を高速真空によって乾燥させ、ペレットを、HE/0.1%還元TX−100で可溶化した。SDS−PAGE、続いてN末端アミノ酸配列決定を用いて、この画分の成分を同定した。
成分を同定するためのプロテアーゼ消化/逆相/N末端配列決定:
AEUFを、GluC(V8)、LysCまたはArgCで消化した。タンパク質と酵素の比は、30μgタンパク質と1μg酵素であった。消化は夜通し37℃にて実行した。消化されたタンパク質混合物(30μg)を、7ミクロンAquapore RF−300カラムに通し、0.1%トリフルオロ酢酸中の10〜95%アセトニトリルの勾配で溶出し、ピークを手作業で収集した。無タンパク質ブランクも走らせ、これからのピークを試料クロマトグラムから減じた。試料ランにおいてのみ生じるピークを質量分析器により分析し、明確な質量を与える試料をN末端アミノ酸配列決定について分析した。
N末端アミノ酸配列決定:
ブロットから切り出されたバンドの場合、タンパク質試料を、SDSゲルからPVDF膜に移し、アミドブラック(脱イオン水中の10%酢酸、0.1%アミドブラック)で染色し、10%酢酸中で脱色した。次に所望のタンパク質バンドを、メタノールで洗浄した円刃刀またはミニ−Exactoナイフを用いて全10レーンから切り出し、Applied Biosystems477Aタンパク質シークエンサーの反応カートリッジに入れた。溶液中の試料の直接配列決定の場合、Prosorbカートリッジを集め、PVDFを60μLのメタノールで湿らせた。PVDFを50μLの脱イオン水で洗浄し、試料(50μL)をPVDFに置いた。50μLの脱イオン水を使用して試料を洗浄した後、Prosorb PVDFを押し抜き、乾燥し、Applied Biosystems477Aタンパク質シークエンサーの反応カートリッジに入れた。両方の方法について、Applied Biosystems N末端シークエンサーを、最適ブロット条件下で12サイクル以上(1サイクル ブランク、1サイクル 標準、および所望の残基同定のための10サイクル以上)走らせ、PTH−アミノ酸検出をApplied Biosystems 120A PTHアナライザーで行った。サイクルは、アナログチャートレコーダーおよび計測器ソフトウェアを介してデジタル式の両方で収集した。アミノ酸の帰属は、アナログおよびデジタルデータの両方を使用して、PTH−アミノ酸の標準セットとアナライザー上でのそれら個々の保持時間の比較により行った(システイン残基は変換中に破壊され、検出されなかった)。複数の配列情報を単一の残基から得ることが可能であり、一次対二次帰属はシグナル強度に基づいて行われた。
LC−MS/MS
IEFにより精製されたタンパク質試料を、SDSポリアクリルアミドゲル電気泳動によりさらに分析した。タンパク質は、クーマシーブルー染色により視覚化し、対象となるバンドを手動で切り出し、その後還元し、アルキル化し、そして自動in−gelトリプシン消化ロボット(1)を用いてin situでトリプシンにより消化(Promega,Madison,WI)した。消化後、ペプチド抽出物を、Savant Speed Vac Concentrator(ThermoQuest,Holdbrook,NY)を使用して、最終容積10〜20μLに濃縮した。
【0141】
ペプチド抽出物を、自動マイクロエレクトロスプレイ逆相HPLCで分析した。簡単に言えば、マイクロエレクトロスプレイインターフェースは、10umC18逆相ビーズ(YMC,Wilmington,NC)で長さ10cmに充填した、Picofrit石英ガラススプレイニードル、長さ50cm、ID75um、8umオリフィス直径(New Objective,Cambridge MA)で構成された。Picofritニードルは、質量分析器の前に位置する手製基部に保持された光ファイバーホルダー(Melles Griot,Irvine,CA)に取り付けた。カラムの後部を、チタンユニオンを介して配管し、エレクトロスプレイインターフェースに電気的接続を供給した。ユニオンを、ある長さの石英ガラスキャピラリー(FSC)管と共に、HPLC溶媒ポンプ(ABI 140C,Perkin−Elmer,Norwalk,CT)に連結されたFAMOSオートサンプラー(LC−Packings,San Francisco,CA)に連結した。HPLC溶媒ポンプは、50μL/分の流量を送達し、これをPEEKマイクロタイトスプリッティングティー(Upchurch Scientific,Oak Harbor,WA)を使用して250nL/分に低下させ、FSC移送ラインを用いてオートサンプラーに送った。LCポンプおよびオートサンプラーは各々、それらの内部ユーザープログラムを使用して制御された。試料をプラスチック製オートサンプラーバイアルに入れ、密封し、5μL試料ループを使用して注入した。
マイクロキャピラリーHPLC−質量分析:
in−gel消化物由来の抽出ペプチドを、マイクロエレクトロスプレイHPLCシステムにより0〜50%溶媒Bの50分間勾配(A:0.1M HOAc、B:90% MeCN/0.1M HOAc)を用いて分離した。ペプチド分析は、スプレイ電位1.5kVで作動し、加熱キャピラリー温度150℃を使用したFinnigan LCQイオントラップ質量分析器(ThermoQuest,San Jose,CA)で行った。データは、機器と共に提供されたデータ取得ソフトウェアを使用して、自動MS/MSモードで得た。取得方法は、1MSスキャン(375〜1200m/z)、続いてMSスキャン中の最も豊富な3種のイオンのMS/MSスキャンを包含した。動的排除関数および同位体排除関数を使用して分析されるペプチドイオンの数を増加させた(設定:3amu=排除幅、3分=排除期間、30秒=排除前期間、3amu=同位体排除幅)。MS/MSデータの自動分析は、N.meningitidisの完全ゲノム由来のタンパク質データベース(Sangerより)を使用して、Finnigan Bioworksデータ解析パッケージに含まれたSEQUESTコンピュータアルゴリズム(ThermoQuest,San Jose,CA)を使用して実行した。研究の結果を図3に示す。
(実施例2)
組換え脂質化P2086(RLP2086)のクローニング:
A.)天然のリーダー配列:
供給源物質:
ORF2086遺伝子を、8529と呼ばれるセログループB Neisseria meningitidis株の臨床分離菌からPCRにより増幅させた。この菌株のセログループ、セロタイプおよびセロサブタイプは、括弧内に示される;8529(B:15、P1:7b、3)。この髄膜炎菌株は、The RIVM,Bilthoven,The Netherlandsから得た。
PCR増幅およびクローニング戦略:
ORF2086の目視検査は、この遺伝子が潜在的なリポタンパク質シグナル配列を有することを示した。特許権によって保護されている隠れマルコフモデルリポタンパク質アルゴリズム(Hidden Markov Model Lipoprotein algorithm)を使用した付加的な解析は、ORF2086がリポタンパク質シグナル配列を含むことを確認した。より自然に近い立体配座でP2086を組換え発現させるために、オリゴヌクレオチドプライマーは、リポタンパク質シグナル配列を持つ全長遺伝子を増幅するように設計し、そしてN.meningitidis A ORF2086のSanger配列の分析に基づいた。2086遺伝子を、ポリメラーゼ連鎖反応(PCR)[ABI2400サーマルサイクラー,Applied Biosystems,Foster City,CA]によりN.meningitidis菌株8529から増幅した。正しいサイズの増幅産物を結合し、pCR2.1−TOPO(Invitrogen)にクローニングした。プラスミドDNAを、NdeIおよびBamHIで制限消化し、ゲル精製し、pET−27b(+)ベクター(Novagen)に結合した。
【0142】
本明細書に記載のオリゴヌクレオチドプライマーを、β−シアノエチルホスホルアミダイト・ケミストリー(Applied Biosystems,Foster City CA)を使用して、PerSeptive Biosystemsオリゴヌクレオチドシンセサイザー(Applied Biosystems,Foster City CA)で合成した。
天然のリーダー配列を使用したrLP2086タンパク質発現:
図5を参照すると、プラスミドpPX7340をBLR(DE3)pLysS宿主細胞(Life Sciences)に形質転換/形質移入、または感染させた。1つの形質転換体を選択し、2%グルコース、カナマイシン(30μg/mL)、クロラムフェニコール(30μg/mL)、およびテトラサイクリン(12μg/mL)を含有するテリフィックブロス(Terrific Broth)50mLに接種した。一晩培養した培養物のOD600は6.0であった。一晩培養した培養物を、1%グリセロールおよび同じ抗生物質を含有するテリフィックブロス1リットルで希釈した。出発OD600は、0.4であった。2時間後、OD600は1.6であり、誘導前試料を採取した。OD600=1に等価の細胞は遠心分離し、上澄みを除去した。全細胞ペレットを150μLのTris−EDTAバッファーおよび150μLの2xSDS−PAGE試料バッファーに再懸濁した。IPTGを添加し、最終濃度1mMとした。3.5時間後、記載されるように誘導後試料を採取し、SDS−PAGEで分析した(図4参照)。
rLP2086の精製:
rLP2086を、ディファレンシャル界面活性剤抽出後にE.coliから可溶化した。天然環境にあるP2086とは異なり、rLP2086は、Triton X−100またはZwittergent 3−12によって著しく可溶化されなかった。rLP2086のバルクをサルコシルで可溶化し、それはN.meningitidisにおける相互作用とは異なってE.coliの外膜成分と相互作用することを示唆した。ひとたび可溶化されると、rLP2086は天然タンパク質と同様に精製され、その中に混在する多くのE.coliタンパク質はアニオン交換樹脂にpH=8で吸着させることにより除去した。その理論的pIより2分の1pH単位上であるにもかかわらず、rLP2086はpH8で吸着されないままであった。それ以上の精製は、pH4.5におけるカチオン交換樹脂へのrLP2086の吸着により達成された。
【0143】
rLP2086の均一性をSDS−PAGE後の図2に示す。rLP2086の質量はMALDI−TOF質量スペクトル分析により27,836であると確認された。この質量は理論的質量の27,100と736異なり、これは細菌リポタンパク質に共通なN末端脂質修飾の質量に近似する。天然およびrLP2086は共に外膜リポタンパク質であると考えられる。N末端配列決定の試みは妨げられ、このことは末端修飾と一致する。
精製方法:
P2086を発現するBLR DE3 pLysS細胞の凍結ペレットを、20mL/g細胞湿重量で10mM HEPES−NaOH/1mM EDTA/1μg/mL Pefabloc SCプロテアーゼ阻害剤(Roche)pH7.4(HEP)に再懸濁し、マイクロフルイダイザー(Microfluidics Corporation Model 110Y)により溶解した。細胞ライゼートは150,000×gで1時間遠心分離した。ペレットはHEPで2回洗浄し、2回遠心分離し、得られた膜ペレットは一晩凍結した。該ペレットは10mM HEPES−NaOH/1mM MgCl2/1%TX−100 pH7.4で30分間可溶化し、続いて150,000xgで30分間遠心分離した。これを3回繰り返した。膜ペレットは上記のように50mM Tris−HCl/5mM EDTA/1%Zwittergent3−12 pH8で2回洗浄し、続いて50mM Tris−HCl/5mM EDTA/1%Zwittergent3−14 pH8および50mM Tris−HCl/5mM EDTA/1%Zwittergent3−14/0.5M NaCl pH8のそれぞれで2回洗浄した。
【0144】
次にrLP2086を、50mM Tris−HCl/5mM EDTA/1%サルコシル pH8で可溶化した。このサルコシル抽出物を1%Zwittergent3−14(Z3−14)に調整し、30倍過剰の50mM Tris−HCl/5mM EDTA/1%Z3−14に対して2回透析した。透析したrLP2086抽出物は90%エタノールで沈殿させ、残存するサルコシルを除去し、50mM Tris−HCl/5mM EDTA/1%Z3−14 pH8(TEZ)で可溶化した。不溶な物質は遠心分離により除去し、上澄みは陰イオン交換クロマトグラフィーカラムを通し、rLP2086を非結合画分に集めた。次に非結合画分を30倍過剰の25mM NaAc/1%Z3−14pH4.5に対して2回透析し、カチオン交換クロマトグラフィーカラムを通した。rLP2086は0〜0.3M NaCl勾配で溶出し、SDS−PAGE(クーマシー染色)により分析した。rLP2086プールはレーザーデンシトメトリーにより84%純粋であることが確認された。
rLP2086サブファミリーBに対する抗血清の表面反応性および殺菌活性:
表VIIを参照すると、サブファミリーB菌株8526由来の精製rLP2086に対する抗血清は、全細胞ELISAにより試験した10種の2086サブファミリーB菌株すべてに対して表面反応性を示した。殺菌活性は異種セロサブタイプ抗原、PorAを発現する10種の2086サブファミリーB菌株のうち9種に対して検出された。これらの菌株は西ヨーロッパ、アメリカ、オーストラリア、およびニュージーランド全体にわたってセログループB髄膜炎菌性疾患を引き起こす代表的な菌株である。殺菌アッセイで致死されなかった唯一の菌株、870227は、全細胞ELISAにより抗rLP2086(サブファミリーB)血清と強く反応し、この菌株がP2086に共通のエピトープを持つタンパク質を発現することを示した。
【0145】
表VIIに挙げた2086サブファミリーA菌株も同様に、全細胞ELISAにより表面反応性を試験した。これらの菌株3種の中の2種は非常に低レベルの反応性を有することが分かり、いくつかの2086サブファミリーA菌株はrLP2086サブファミリーBに対して産生された抗体と交差反応性ではない可能性があることを示した。菌株8529から2086サブファミリーB遺伝子を同定するために使用されるPCR増幅手順もまた、菌株870446、NMBおよび6557に対して実行した。2086サブファミリーBのPCR増幅産物は全く検出されなかった。
免疫原性方法:
抗血清の調製:
ワクチンを、前に実施例1に記載されるように製剤した。しかしながら、10μg投与量を使用した。
全細胞酵素結合免疫吸着アッセイ(ELISA):
N.meningitidis全細胞懸濁液を、無菌0.01M ホスフェート、0.137M NaCl、0.002M KCl(PBS)中、620nmにおける吸光度0.1に希釈した。この懸濁液から、0.1mLをNunc Bac T96ウェルプレート(Cat# 2−69620)の各々のウェルに添加した。細胞は室温で3日間、プレート上で乾燥し、その後覆い、反転させ、4℃で保存した。プレートは洗浄バッファー(0.01M Tris−HCl、0.139M NaCl/KCl、0.1%ドデシルポリ(オキシエチレレングリコールエーテル)n n=23(Brij−35(登録商標)、ICI Americas,Inc.,Wilmington,Delawareから入手可能)、pH7.0〜7.4)で3回洗浄した。抗血清の希釈物を、PBS、0.05%Tween−20/アジドで調製し、そして0.1mLを被覆したプレートに移した。プレートを2時間37℃でインキュベートした。プレートは洗浄バッファーで3回洗浄した。ヤギ−抗マウスIgG AP(Southern Biotech)をPBS/0.05%Tween−20で1:1500に希釈し、0.1mLを各々のウェルに添加し、プレートを37℃で2時間インキュベートした。プレートは(上記のように)洗浄した。基質溶液は、1M ジエタノールアミン/0.5mM MgCl2中にp−ニトロフェニルホスフェート(Sigma)を1mg/mLに溶解することにより調製した。基質はウェルあたり0.1mLでプレートに添加し、室温で1時間インキュベートした。反応は50μL/ウェルの3N NaOHで停止させ、プレートは、690nmを基準として、405nmで読み取った。
B.)P4リーダー配列:
PCR増幅およびクローニング法:
rLP2086発現を最適化するために、2086遺伝子を型分類不能なHaemophilus influenzaeのP4シグナル配列の後にクローニングした(Greenら、1991)。リポタンパク質クローニングに使用されたプライマーが表IVに列挙してあり、化合物番号:5658、5660、6473、6543および6385によって識別される。ORF2086は、以下の化合物番号5658および5660を持つプライマーを使用してN.meningitidisセログループB菌株8529から増幅された。ORF2086は、以下の化合物番号6385および5660を持つプライマーを使用してN.meningitidisセログループB菌株CDC1573から増幅された。ORF2086は、以下の化合物番号6473および6543を持つプライマーを使用してN.meningitidisセログループB菌株2996から増幅された。N末端(5’)プライマーは、2086遺伝子の成熟領域(システインのすぐ下流のアミノ酸位置3のセリン残基において開始)に相同であるように設計した。制限部位BamHI(GGATTC)はそれぞれのN−末端プライマーの5’末端に組み込まれ、アミノ酸位置2において成熟タンパク質にグリシン残基が挿入された。C末端(3’)プライマーは2086遺伝子のC−末端に相同であるように設計され、終止コドンおよびクローニングのためにSphI部位を含有した。各N.meningitidis B菌株から増幅されたフラグメントを、中間ベクターにクローニングし、配列分析によってスクリーニングした。
【0146】
正しいクローン由来のプラスミドDNAを、BamHIおよびSphI制限酵素(New England Biolabs,(NEB))で消化した。pLP339という名称のベクター(出願者の譲受人により供給された)を発現ベクターとして選択した。このベクターは、pBAD18−Cmバックボーン(Beckwithら、1995)を使用し、P4リポタンパク質シグナル配列および型分類不能なヘモフィルス インフルエンザのP4遺伝子(Greenら、1991)を含有する。pLP339ベクターは、制限酵素BamHIにより部分的に消化され、その後SphIで消化された。増幅された2086フラグメント(BamHI/SphI)は各々別個にpLP339ベクター(部分的BamHI/SphI)に結合された。このクローニング戦略は、成熟2086遺伝子をP4リポタンパク質シグナル配列の後に配置する。BamHI部位はP4シグナル配列と2086遺伝子間のクローニング接合部に残る(図7に示したプラスミド構築物を参照されたい)。以下のものはBamHIクローニング接合部における配列の例である:
[P4シグナル配列]−TGT GGA TCC−[残存2086成熟核酸配列]
[P4シグナル配列]−Cys Gly Ser−[残存2086成熟アミノ酸配列]
図7を参照すると、各増幅されたフラグメントはP4リーダー配列を含有する改変pBAD18−Cmベクターにクローニングされた。発酵はrP4LP2086(組換えP4脂質化2086)を発現する組換えE.coli BLR pPX7343で行い、付加的グルコースの添加により細胞密度増大を試みた。発酵装置は、1%グルコースを補足したSambrookの完全M9最小培地10Lで満たした。
【0147】
発酵装置中のグルコースの初期濃度は45g/Lであった。初期ODが約0.25になるように発酵装置に菌を接種した。約OD25で、付加的な20g/Lグルコースを添加した。培養物はOD63.4、グルコース枯渇において1%アラビノースで誘導した。発酵は、誘導後3時間まで継続した。試料は誘導後t=0、1、2、3に保存し、タンパク質はBSAを用いて定量した。t=3において、タンパク質収量は約0.35g/L、そして7%総細胞タンパク質であった。約10Lの培養物から全部で895グラムの湿細胞ペーストが採取された。
【0148】
rP4LP2086の精製は、上記の実施例2、節Aに記載のものと同じ方法を使用して行った。
【0149】
本明細書に記載のオリゴヌクレオチドプライマーを、−シアノエチルホスホルアミダイト・ケミストリー(Applied Biosystems,Foster City CA)を使用して、PerSeptive Biosystemsオリゴヌクレオチドシンセサイザー(Applied Biosystems,Foster City CA)で合成した。ORF2086遺伝子ファミリーのPCR増幅に使用されるプライマーは表IVに挙げられ、それは本発明のプライマーの非限定的な例を示す。
【0150】
【表4】
(実施例3)
非脂質化成熟2086タンパク質の発生遺伝学:
2086タンパク質の免疫原性をさらに評価するため、P2086の非脂質化型のクローニングおよび発現を行った。
ORF2086のPCR遺伝子増幅:
様々な染色由来の2086遺伝子が、参照により本明細書に組み込まれる国際出願PCT/US02/32369号(2003年8月7日、国際公開第03/063766号として公開)、および国際出願PCT/US04/11901号(2004年11月4日、国際公開第04/094596号として公開)に記載されるように、プライマーで増幅され得る。
【0151】
これらのプライマーの特徴としては、各プライマーにおける合成BglII制限部位、化合物番号6406および6474における合成NdeI制限部位が含まれ、そして3種すべてのリーディングフレームの停止コドンは化合物番号5135および6605に存在する。プライマー番号6406および6474は、第2のアミノ末端コドン(ACG)に融合したATG(Met)を持つ2086遺伝子を増幅し、成熟2086ポリペプチドの単一アミノ酸置換(TGC Cysを置換)を表す。
【0152】
PCRクローニングベクターは、TOPO−PCR2.1、Invitrogen,Valencia,CAであった。
【0153】
非脂質化2086タンパク質を発現するために使用されたベクターはNovagen,Madison,WIのpET9aであった。
【0154】
E.coliクローニング菌株は、Top10,Invitrogen,Carlsbad,CAであった。
【0155】
E.coli発現菌株は、BLR(DE3)pLysS,Novagen,Madison,WIであった。
【0156】
クローニング目的のための培地は、グリセロールの代わりの1%無菌グルコース、および適切な抗生物質(アンピシリンまたはカナマイシン)を含む、Sambrookら、によるテリフィックブロス液または寒天培地であった。
【0157】
プラスミド精製は、Qiagen Spin Miniprepキット(Valencia,CA)により行った。
非脂質化2086発現のための産生菌株または細胞系統の調製:
2086遺伝子は、髄膜炎菌株8529に由来する染色体DNAからポリメラーゼ連鎖反応(PCR)[AmpliTaq and ABI2400サーマルサイクラー,Applied Biosystems,Foster City,CA]により増幅した。2086遺伝子のPCR増幅は、それぞれの反応において化合物番号6474および5135によって同定された2種のオリゴヌレオチドプライマーを使用した。増幅された2086PCR産物はTOPO−PCR2.1クローニングベクターに直接クローニングし、100μg/mlアンピシリンおよび20μg/ml X−Galを補ったテリフィックブロス寒天培地で選択した。白色コロニーを選択し、培養した。プラスミドDNAは、Qiagen miniprepキットを使用して調製し、該プラスミドをPCRフラグメントインサートに関してスクリーニングした。PCRインサートプラスミドを、DNA配列決定にかけた(ABI377シークエンサーでのBig Dye化学,Applied Biosystems,Foster City,CA)。
【0158】
適切なDNA配列を示すプラスミドを、BglII制限酵素で消化し、BglIIフラグメントを、GeneClean II精製キット(Bio101、Carlsbad,CA)を使用してゲル精製した。精製BglIIフラグメントを、発現ベクターpET9aのBamHI部位にクローニングした。pET9a/2086クローンは30μg/mlのカナマイシンを補ったテリフィックブロスプレート上で選択した。カナマイシン耐性クローンを培養し、miniprepプラスミドDNAを調製した。プラスミドはBamHI部位における2086遺伝子の適切な配向に関してスクリーニングした。適切に配置されたプラスミドは、2086遺伝子のアミノ末端へのT7−抗原の融合物(rP2086T7)を表す。これらのrP2086T7遺伝子融合物を、BLR(DE3)pLysSに形質転換し、テリフィックブロス/Kanプレート上で選択し、テリフィックブロスで培養し、rP2086T7融合タンパク質を発現するように1mM IPTG(イソプロピルβ−D−チオガラクトピラノシド)により誘導した。rP2086T7融合タンパク質は高レベルで発現された。
【0159】
次に、これらの融合プラスミドをNdeI制限消化にかけ、それによりT7抗原を欠失し、ベクターにより提供されたATGスタートに成熟2086遺伝子を直接連結する。これらのNdeI欠失プラスミドを、Top10細胞に形質転換し、テリフィックブロス/Kanプレート上で選択した。候補クローンを培養し、ミニプレッププラスミドDNAを調製した。プラスミドDNAをDNA配列決定にかけ、欠失および2086遺伝子配列の完全性を確認した。これらのプラスミドはpPX7328と呼ぶプラスミドマップによって表される(図6)。適切なDNA配列を表すプラスミドを、BLR(DE3)pLysSに形質転換し、テリフィックブロス/Kanプレート上で選択し、テリフィックブロスで培養し、IPTGにより2086タンパク質を発現するように誘導した。pET9aベクターは、T7タグが除去されると、菌株BLR(DE3)pLysSに成熟2086タンパク質を発現できなかった。
非脂質化2086タンパク質の産生:
精製プラスミドDNAを用い、発現菌株BLR(DE3)pLysSを形質転換した。プラスミドを持つBLR(DE3)pLysS細胞は、カナマイシン耐性で、1mM IPTGの添加により高レベルのPorAタンパク質を発現するように誘導することができる。rP2086T7融合タンパク質は、E.coli細胞系統BLR(DE3)pLysS中に不溶性封入体として、全タンパク質の約40%において発現させることができる。この精製融合タンパク質は、マウスを免疫するために用いられ、異種髄膜炎菌株に対する有意レベルの殺菌抗体を生成した(表V参照)。
2086非脂質化遺伝子変異誘発:
PCRプライマー変異誘発は2086遺伝子の5’末端で行われた。成熟rP2086T7の高発現レベルを示しつつT7−タグを除去できるかどうかを確かめるための発現研究が進行中である。
非脂質化rP2086T7の精製:
非脂質化rP2086T7を発現するE.coliBLR(DE3)pLysS細胞を、10mM HEPES−NaOH/5mM EDTA/1mM Pefabloc SC pH7.4中で、マイクロフルイダイザーにより溶解した。細胞ライゼートを18,000xgで30分間遠心分離した。封入体ペレットを50mM Tris−HCl/5mM EDTA/1%Triton X−100 pH8で3回洗浄し、都度24,000xgで30分間遠心分離した。次に封入体ペレットを50mM Tris−HCl/5mM EDTA/1%Zwittergent3−14 pH8で2回洗浄し、都度24,000xgで15分間遠心分離した。続いて封入体ペレットを50mM Tris−HCl/5mM EDTA/4M 尿素 pH8で2時間可溶化し、その後不溶性物質を遠心分離により除去した。上澄み(可溶化rP2086T7)を4つの等しい試料に分けた。保存溶液を使用して、1つは50mM Tris−HCl/5mM EDTA/250mM NaCl/2M 尿素 pH8(界面活性剤を含まない)に調整し、1つは50mM Tris−HCl/5mM EDTA/250mM NaCl/2M 尿素/1%水素化Triton X−100 pH8(TX−100)に調整し、1つは50mM Tris−HCl/5mM EDTA/250mM NaCl/2M 尿素/1%Zwittergent3−12 pH8(Z3−12)に調整し、そして1つは50mM Tris−HCl/5mM EDTA/250mM NaCl/2M 尿素/1%Zwittergent3−14 pH8(Z3−14)に調整した。尿素を除去するために、尿素を含まないそれぞれのバッファーに対して試料を透析した。次に尿素を含まずに60mM NaClを含むそれぞれのバッファーに対して試料を透析し、NaCl濃度を減少させた。不溶性物質は2,000xgで15分間遠心分離することにより除去し、得られた上澄み(再生rP2086T7)をその後の実験に使用した。rP2086T7の均一性は、クーマシー染色したSDS−PAGEおよびレーザーデンシトメトリーを使用して測定し、91〜95%であることが見出された。
【0160】
免疫原性手順−実施例2に記載の通り
この精製融合タンパク質を使用してマウスを免疫化し、異種髄膜炎菌株に対する有意レベルの殺菌抗体を生成した。(以下の表V参照):
【0161】
【表5】
(* rLP2086によるマウスの免疫化によって生成された陽性対照血清)
(実施例4)
ORF2086のキメラクローンの開発
菌株CDC−1573由来の2086タンパク質N末端領域は、菌株8529および2996由来の2086遺伝子には存在しない反復部分を含有する(図8参照)。この反復部分は、E.coliベースの2つの発現系(pETおよびpBAD)由来の高レベルの組換え2086タンパク質発現に関与すると考えられる。pETおよびpBAD発現系におけるCDC−1573 2086遺伝子由来の組換えタンパク質発現レベルは、同じ系を使用した菌株8529および2996による2086遺伝子由来の組換え体発現レベルに比較して有意に良好であった。3種すべての菌株に由来する2086遺伝子のN末端領域は、この反復部分以外は比較的相同である。したがって、pETおよびpBAD系を使用した場合、菌株8529および2996由来の2086遺伝子にCDC−1573N末端を融合することにより、これらの遺伝子から発現される組換え2086タンパク質レベルが増大すると考えることは妥当である。
材料および方法:
菌株8529および2996由来の染色体DNAを精製し、キメラ2086遺伝子のPCR増幅のための鋳型として使用した。化合物番号6721および5135を持つPCRプライマーを使用して菌株8529由来のキメラ2086遺伝子を増幅し、化合物番号6721および6605を持つPCRプライマーを使用して、菌株2996由来のキメラ2086遺伝子を増幅した。PCR産物は、InvitrogenのPCR2.1 TOPOベクターに直接クローニングし、次にDNA配列分析によりスクリーニングして、無傷のキメラ2086遺伝子を同定した。次にその遺伝子をBglIIによりPCR2.1ベクターから開裂し、BglIIフラグメントをpET9aプラスミドのBamHI部位に挿入した。プラスミドインサートが適切な配向であることをスクリーニングした後、NdeI消化にかけた。線状NdeIフラグメントは自己連結し、pET9aベクターによって与えられたT7−タグ配列を含有する小NdeIフラグメントの欠失が達成された。この欠失は、キメラ2086遺伝子の5’末端にT7プロモーターを直接結合する。NdeI欠失プラスミドをE.coli菌株BL21(DE3)に形質転換し、カナマイシン耐性コロニーをIPTG誘導によるキメラ2086タンパク質発現に関してスクリーニングした。
【0162】
初期の研究は、pET9a系で発現される場合、菌株2996由来のキメラ2086遺伝子は、天然の2996/2086遺伝子に比較して2倍の組換えタンパク質を発現することを示す。pBAD系は現在のところ試験されていない。
【0163】
ただ1回の実験が行われただけであるが、データは、キメラ2086遺伝子に由来する、増大した用途があることを示唆する。菌株8529および2996由来2086遺伝子に対するCDC−1573N−末端融合物の生成により、増大した組換え2086タンパク質発現がもたらされる。
(実施例5)
N.meningitidis菌株の2086PCRスクリーニング:
臨床分離菌間の2086遺伝子の保存を確かめるために、88のN.meningitidis菌株にPCR増幅を行った。
【0164】
ORF2086の当初のPCR同定は、化合物番号:4623、4624および4625により同定された表IV(先の実施例2参照)に列挙されるプライマーを使用した。これらのプライマーは、SangerのN.meningitidisセログループA配列に基づいて設計された。多数の菌株のスクリーニングを容易にするため、内部プライマーが2086遺伝子に関して設計された。全部で88のN.meningitidis菌株を、化合物番号5005および5007により同定された、新規に設計された内部2086プライマーを用いて、PCRによりスクリーニングした。これらのプライマーを用い、本出願人らは、88のN.meningitidis菌株の中の63(約70%)を同定することができた(表VI−A参照)。
【0165】
SangerのN.meningitidisセログループA配列およびTIGRのN.meningitidisセログループB配列中の2086遺伝子を囲む伸長領域を調べ、並べた。プライマーは、2086遺伝子の上流および下流領域に対応するように設計した。目的は、配列比較のためにこれらのプライマーを使用して、多様なN.meningitidis菌株から全長2086遺伝子より大きいものを増幅することであった。化合物No.6470および6472を使用した1種の菌株(6577)のPCR増幅は、低収率の産物という結果になった。菌株6557増幅産物をクローニングし、プラスミドDNAを配列分析にかけた。結果は、以前見られたよりも配列可変性の大きい新型の2086遺伝子を示した。菌株6557由来の2086遺伝子は、配列決定された他の菌株に対してアミノ酸レベルで約75%同一であった。興味深いことに、菌株6557は、上述の2086PCRスクリーニングにより以前陰性とされた菌株30%の中の1種であった。
【0166】
菌株6557内のC末端可変領域に特異的な内部プライマーが設計された。これらのプライマーを使用して、2086PCRスクリーニングによる試験で以前陰性とされた約30%の菌株中のより可変性の2086遺伝子をスクリーニングした。これらの新規に同定された内部2086プライマー(化合物番号6495および6496により識別)を使用して、入手可能なすべてのN.meningitidis菌株(n=88)をPCRによりスクリーニングした。2086に関してPCRにより以前陰性とされた約30%のN.meningitidis菌株だけがこのスクリーニングでPCR陽性であった。以前のPCR陰性(約30%)菌株から増幅された遺伝子の組は、新型2086遺伝子または2086遺伝子の第2ファミリーを表すべきであり、本明細書では2086サブファミリーAと呼ぶ。8529由来のプライマーにより約70%の菌株から増幅された2086遺伝子の組は、本明細書ではサブファミリーBと呼ぶ。
【0167】
PCR増幅研究に使用されるN.meningitidis菌株は、以下の表、表VI Aおよび表VI Bから選択された。表に列挙された菌株は、前に本明細書中で開示されたものに加えて、N.meningitidis菌株の例として提供されるが、それらに限定されない。表VI Aに列挙された菌株は、2086タンパク質サブファミリーAに分類され、表VI Bに列挙された菌株は、2086タンパク質サブファミリーBに分類される。各表中に列挙された菌株は、セロサブタイプにより分類される。菌株は表中に示すような以下の4つの供与源から入手可能である:MPHL−Manchester Public Health Laboratory,Manchester,UK;RIVM,Bilthoven,The Netherlands;University of Iowa,College of Medicine,Department of Microbiology,Iowa City,IA;およびWalter Reed Army Institute of Research,Washington,D.C.
【0168】
【表6A】
【0169】
【表6B−1】
【0170】
【表6B−2】
その他の菌株は、感染した個体から分離菌として容易に入手可能である。
(実施例6)
髄膜炎菌株に対するrLP2086抗血清の反応性:
以下の表、表VIIは、上述のrLP2086の交差反応性および交差保護能力を示す。表に示すように、rLP2086は、全細胞ELISA(WCE)力価、殺菌アッセイ(BCA)および幼若ラット(IR)アッセイを含む、多様な手法を使用して処理および分析して、2086タンパク質に対して引き起こされたポリクローナル抗体の細菌表面反応性を確認した。
【0171】
【表7】
+ 菌血病において10分の1減少より大
− 菌血病において10分の1減少より小
(実施例7)
ORF2086タンパク質を発現させるための種々の構築物を調製した。以下の表、表VIIIは、例を示し、本発明の実施を例示するために提供されるr2086構築物表であるが、それらに限定されない。
【0172】
【表8】
(実施例8)
LOS除去外膜タンパク質を用いたさらなる研究から、異種セロサブタイプを発現する菌株に殺菌抗体を誘導することができる、PorA以外の外膜タンパク質(単数または複数)を産生する付加的な菌株を同定した。以下に、本発明の一実施態様による付加的なタンパク質、具体的には髄膜炎菌免疫原性組成物に必要なタンパク質の数を減少させることができる外膜リポタンパク質を同定するためのさらなる研究を記載する。これらのさらなる研究は、以前の実施例に記載された研究を補足する。
【0173】
複数菌株に対する免疫および殺菌アッセイと共に、細胞下分画、ディファレンシャル界面活性剤抽出、等電点分離法、およびイオン交換クロマトグラフィーを使用して、対象となる小グループのタンパク質を同定した。主成分の直接配列決定は、N末端が遮断されていることを示した。内部タンパク質配列は、化学的およびタンパク質分解消化に由来するポリペプチドの直接配列決定により得た。グループA髄膜炎菌株のゲノム配列は、Sanger Centerからダウンロードし、既存および本出願者らが所有するアルゴリズムを使用して本出願者らのバイオインフォマティックグループが分析し、検索データベースを調製した。ペプチド配列データは、ORF2086が関心のあるものであることを示した。このorfに基づいたプライマーを使用して、菌株8529からP2086遺伝子をPCRした。遺伝子配列の分析、N末端が遮断されているという事実、およびその細胞下局在は、P2086が脂質化外膜タンパク質(LP2086)であることを示した。rLP2086−8529および別の髄膜炎菌株由来の変異体は、H.インフルエンザP4シグナル配列を使用してE.coli中でリポタンパク質として組換え発現された。これらの組換えタンパク質を、ディファレンシャル界面活性剤抽出によりE.coli膜から単離し、イオン交換クロマトグラフィーを使用して精製し、マウスを免疫するために使用した。マウス抗LP2086血清は、N.meningitidisのいくつかの異なるセロサブタイプ株に対する殺菌活性の促進が可能であった。多くのN.meningitidis菌株由来のP2086遺伝子のさらなる分析は、これらの配列がサブファミリーAおよびサブファミリーBと呼ばれる2種のグループに分類されることを示した。(図12参照)。サブファミリーBタンパク質に対して誘導された抗血清はサブファミリーBタンパク質を発現する9種の菌株、およびサブファミリーAタンパク質を発現する1種の菌株に対して殺菌性であった。サブファミリーA抗血清はサブファミリーA系統に対して殺菌性であった。1種のrPorAと1種のrLP2086の混合物は、いずれかのタンパク質だけによって誘導されたもの以上の広いワクチン適用範囲を持つ相補的抗体を誘導した。
【0174】
これらの観察は以下の結論に至る。rLP2086抗原は、異種PorAおよび異種P2086タンパク質を発現する髄膜炎菌株に対して殺菌抗体を誘導することができる。抗原のP2086ファミリーは、単独または別の髄膜炎菌抗原との組み合わせのいずれかで、有用なワクチンであるか、または免疫原性であり得る。
【0175】
以下、先の研究について詳細に説明する。可溶性外膜タンパク質(sOMPs)の複合体混合物は、異種PorAタンパク質を発現する菌株に対してPorA非依存的殺菌抗体を誘導することが見出された。ディファレンシャル界面活性剤抽出、等電点分離法およびイオン交換クロマトグラフィー、続くマウス免疫化の工程を使用して免疫学的に活性な成分を追跡した。
【0176】
各ステップで、髄膜炎菌性疾患の世界的疫学を代表するセロサブタイプ抗原を含有するいくつかの菌株に対して、血清は表面反応性および殺菌活性についてアッセイされた。
【0177】
この分離および免疫化の工程を使用して、グループB N.meningitidisに関する新規な交差反応性免疫原性候補を同定した。
【0178】
PorA欠乏菌株の調製−porA染色体座位を、菌株2996由来のプラスミドpPX7016にクローニングした。プラスミド内では、porAプロモーター、S/Dボックスおよび初めの38のN末端コドンが欠失し、自己含有KanR発現カセットにより置換されている。該プラスミドは制限酵素で線状にされ、セロサブタイプ菌株PI:5,2;PI:9;PI:7,16;PI:15;PI:4;PI:3&PI:10に自然に形質転換された。カナマイシン耐性形質転換体を選択し、ELISAにおいてセロサブタイプ特異的モノクローナル抗体によりPorAの損失をスクリーニングした。
【0179】
殺菌アッセイ:Mountzouros,K.T.and Howell,A.P.Detection of Complement−Medicated Antibody−Dependent Bactericidal Activity in a Fluorescence−Based Serum Bactericidal Assay for Group b Neisseria meningitidis.J Clin Microbiol.2000;38:2878−2884を参照されたい。
【0180】
全細胞酵素結合免疫吸着アッセイ(ELISA):N.meningitidis全細胞懸濁液を、滅菌0.01Mホスフェート、0.137M NaCl、0.002M KCl(PBS)中で、620nmにおける光学密度度0.1に希釈した。この懸濁液から、Nunc Bac T96ウェルプレート(Cat#2−69620)の各ウェルに0.1mL添加した。細胞を37℃で一晩、プレート上で乾燥させ、その後被覆し、反転させ、4℃で保存した。プレートは、洗浄バッファー(0.01M Tris−HCl、0.139M NaCl/KCl、0.1% Brij−35、pH7.0〜7.4)で3回洗浄した。抗血清の希釈物は、PBS、0.05%Tween−20/アジドで調製し、0.1mLを被覆プレートに移し、37℃で2時間インキュベートした。プレートは洗浄バッファー中で3回洗浄した。ヤギ抗マウスIgG AP(Southern Biotech)をPBS/0.05%Tween−20で1:1500に希釈し、0.1mLを各ウェルに添加し、プレートを37℃で2時間インキュベートした。プレートを(上記の通り)洗浄した。基質溶液は、ジエタノールアミンで、p−ニトロフェニルホスフェート(Sigma)を1mg/mlに希釈することにより調製した。ウェルあたり0.1mLの基質をプレートに添加し、室温で1時間インキュベートした。反応は50ul/ウェルの3N NaOHで停止し、690nmを参照として405nmでプレートを読んだ。
【0181】
組換えPorA誘導:BLR(DE3)/pET9a菌株を、Kan−30および2%グルコースを補ったハイソイ(HySoy)ブロス(Sheffield Products)中で、37℃で一晩培養した。翌朝、O/N培養物をハイソイブロス、Kan−30および1%グリセロールで希釈し、37℃で1時間培養した。これらの培養物にIPTGを最終濃度1mMで添加することにより誘導した。培養物はさらに2〜3時間培養し、その後採取した。
【0182】
組換えPorA精製:rPorAを、E.coli封入体から8M尿素で可溶化し、尿素を全く含まないバッファーに対して透析して再生した。再生されたrPorAを透析ろ過により濃縮し、バッファーはG25カラムでNaPO4 pH6に交換した。次に、透析したrPorAをカチオン交換カラム(S Fractogel)に流し、1M NaClで溶出した。
【0183】
菌株8529(P1.7−2,3)のsOMPsは、異種セロサブタイプを発現する菌株に対し、マウスにおいてPorA非依存性殺菌活性を誘導する。以下の表、表IXは、調べた菌株における殺菌活性を示す。
【0184】
【表9−1】
【0185】
【表9−2】
sOMPsの調製:N.meningitidis膜を、TX−100、Zwittergent3−14,およびZwittergent3−14+0.5N NaClで抽出した。上記のsOMPを、Zwittergent3−14/0.5M NaCl抽出物中で可溶化した。抽出は当業者に周知の技術、例えば、参照により本明細書に組み込まれる米国特許第6,355,253号を使用して実行する。
【0186】
免疫原性:雌スイスウェブスターマウスを、0週目および4週目に20ug QS−21をアジュバントとして添加した25ug総タンパク質量で免疫化した。瀉血およびデータ分析を6週目に行った。
【0187】
1 殺菌性(BC50)力価(生細胞数を50%減少させる抗血清の希釈度の逆数として表される)。0週目の正常マウス血清のBC50力価は<25であった。
【0188】
2 NST=セロタイプ分類不能
以下の表、表Xはおよび表XIは、サブファミリーAおよびサブファミリーB両方の組換え脂質化P2086(rLP2086)の精製および特徴の概要を示す。
サブファミリーA rLP2086精製
【0189】
【表10】
サブファミリーB rLP2086精製
【0190】
【表11】
精製法:すべての変異体は、TX−100によりE.coli膜から可溶化された(サルコシルまたは尿素で可溶化されたrLP2086−8529を除く)。さらなる精製は、Tris−HClまたはNaPO4バッファー中でアニオン交換(TMAE)、サイズ排除および/またはカチオン交換(S Fractogel)クロマトグラフィーの組み合わせにより行った。
【0191】
1 菌株8529由来のP2086と比較したアミノ酸相同性
2 SDS−PAGEおよびコロイド状クーマシー染色バンド(Simply Blue染色)のレーザーデンシトメトリーにより確認した純度
同種および異種菌株に対して試験された、サブファミリーBメンバー、rLP2086−8529の免疫原性。
【0192】
以下の表XIIは、同種および異種菌株に対して試験された、サブファミリーBメンバー、rLP2086−8529の免疫原性を示す。
【0193】
【表12】
ワクチン接種手順:6〜8週齢雌スイスウェブスターマウスを、0週目および4週目に10ug rLP2086−8529+20ug QS−21で免疫化した。データ分析は6週目の瀉血により行った。
【0194】
a rLP2086−8529と比較したP2086のアミノ酸相同性
b 吸光度=0.1における希釈度の逆数として表したエンドポイント力価
c 生細胞数を50%減少させる抗血清の希釈度の逆数として表したBC50力価。0週目の正常マウス血清のBC50力価は<10であった。
【0195】
表XIIIは、同種および異種菌株に対して試験した、サブファミリーBメンバー、rLP2086−2996の免疫原性を示す。
【0196】
【表13】
ワクチン接種手順:6〜8週齢雌スイスウェブスターマウスを、0週目および4週目に10ug rLP2086−2996+20ug QS−21で免疫化した。データ分析は6週目の瀉血で行った。
【0197】
a rLP2086−2996と比較したP2086のアミノ酸相同性
b 吸光度=0.1における希釈度の逆数として表したエンドポイント力価
c 生細胞数を50%減少させる抗血清の希釈度の逆数として表した殺菌性(BC50)力価。0週目の正常マウス血清のBC50力価は<10であった。
【0198】
以下の表XIVは、rLP2086およびrPorAに対する抗血清は、混合され殺菌活性をアッセイした場合に相補的であることを示す。
【0199】
【表14】
ワクチン接種手順:6〜8週齢雌スイスウェブスターマウスを、0週目および4週目に10ug rLP2086−8529/20ug QS−21か、15ug rPorA/100ug MPLのいずれかで免疫化した。データ分析は、6週目の瀉血に関して実行した。
【0200】
a 生細胞数を50%減少させる抗血清の希釈度の逆数として表した殺菌性(BC50)力価。0週目の正常マウス血清のBC50力価は<10であった。
【0201】
以下の表XVは、rLP2086サブファミリーと2種のrPorAとの混合物がマウスに殺菌抗体を誘導することを示す。
【0202】
【表15】
ワクチン接種手順:6〜8週齢雌スイスウェブスターマウスを、0週目および4週目に各タンパク質10ug+20ug QS−21で免疫化した。データ分析は6週目の瀉血で行った。
【0203】
a 生細胞数を50%減少させる抗血清の希釈度の逆数として表した殺菌性(BC50)力価。0週目の正常マウス血清のBC50力価は<10であった。
【0204】
b SfA−サブファミリーA、SfB−サブファミリーB
c 適切な1価対照:rLP2086−8529、rLP2086−2996、rP1.5−1,2−2またはrP1.22−1,14−1抗血清
以下に上記の研究の結果を概説する。抗rLP2086抗血清は、13/16の試験菌株に対して殺菌性である。異なるセロサブタイプを発現する11種の菌株は抗P2086血清により致死する。抗rLP2086血清の殺菌活性は抗rPorA血清に相補的である。P2086とPorAとの混合物は、マウスに相補的殺菌抗体を誘導する。ディファレンシャル界面活性剤抽出、精製および免疫、ならびに多くの菌株に対する機能的抗体アッセイを使用して、新規ワクチン候補を同定することができる。P2086は、P2086およびrPorAの両方において異種の菌株に対し殺菌抗体を誘導するワクチン候補として同定されている。したがって、タンパク質の2086ファミリーは、単独で、または別の髄膜炎菌抗原と組み合わせて有用なワクチンであり得る。
(実施例9)
変化するセログループの髄膜炎菌株を、ORF2086遺伝子の存在についてPCRによりスクリーニングした。結局、100種以上の髄膜炎菌株をスクリーニングした。
【0205】
C末端可変領域に特異的な2組の内部PCRプライマーを利用して、サブファミリーAおよびB遺伝子配列を識別した。約350bpのPCR増幅産物の存在は、2086遺伝子配列が染色体上に存在することを示した。すべての菌株は、予想されたサイズの単一のPCR産物を生じた。全長ORF2086遺伝子のヌクレオチド配列を決定し、整列させ(DNAStar MegAlign)、系統樹を調製するために使用した。
【0206】
2086遺伝子は、pBADアラビノース誘導性プロモーター系においてrLP2086リポタンパク質として組換え発現され、またはIPTG誘導性pET系においてrLP2086非脂質化タンパク質として組換え発現された。これらの組換えタンパク質は、E.coli Bにおいて発現させた。精製組換えタンパク質を使用してマウスを免疫化し、マウス抗血清を、その血清IgG力価および多様な髄膜炎菌株に対する殺菌活性についてアッセイした。
【0207】
ORF2086を、以下のものうちの1つからPCRにより増幅した:全髄膜炎菌細胞、精製染色体DNAまたはプラスミドDNA鋳型。
【0208】
ORF2086遺伝子をベクターpLP339にクローニングし、これはヘモフィルスP4リーダー配列をORF2086遺伝子の5’末端に融合する。E.coli菌株BLRは、pBAD/ORF2086クローン由来のrP2086の脂質化型の組換え発現のための宿主菌株として使用した。(図10A参照)。pBADアラビノース誘導性プロモーターは、P4シグナル/ORF2086融合タンパク質の発現を操作して、rP2086の脂質化型を発現させる。シグナル配列を欠くP2086遺伝子を、高活性T7ファージプロモーターの後にpET9aベクターにクローニングされた。E.coli菌株BL21(DE3)は、pET9a/ORF2086クローン由来のORF2086の非脂質化型の組換え発現のための宿主菌株として使用した。E.coli菌株BL21におけるDE3溶原菌は、lacUV5プロモーターの制御下で、IPTGの添加によりに誘導されてT7RNAポリメラーゼを発現することができる。WCE;FEMS Micro.Lett.,48(1987)367−371およびBCA;J.Clin.Microbiol.,38(2000)2878−2884を参照されたい。
【0209】
遺伝子、ORF2086は、異なるN.meningitidis菌株からクローニングおよび配列決定された。ヌクレオチド配列を整列させ(DNAStar MegAlign)、系統樹調製に使用した。この樹は、ORF2086遺伝子ヌクレオチド配列の2種の異なるサブファミリーを明らかにする。2つの遺伝子サブファミリーは、それらの5’末端において類似するが、それらの3’末端近くではかなりの変化を含む。かなりの変動性があると考えられるが、遺伝子のある種の重要な領域は、異なる菌株間で高度に相同性である。理論により拘束されることを意図することなく、これらの保存された領域は、タンパク質に対して機能的連続性を提供することができ、そしてワクチン標的として利用されることになる交差防御エピトープを表し得る。
【0210】
2086遺伝子は、いくつかのセログループB髄膜炎菌株からクローニングされ、脂質化シグナル配列を伴って、またはそれを伴わずに発現された。T7−タグに融合された非脂質化型は、最高レベルで発現した。T7−タグ配列は、mRNAに安定性を提供し、翻訳されたポリペプチドのレベルを有意に向上し得る。この融合タンパク質は、封入体内に沈着すると考えられ、公知のプロトコルにより容易に精製および再生できる。P2086の脂質化型および非脂質化型は、全細胞タンパク質の約5〜8%で発現され、全タンパク質の約50%としてrP2086を発現するT7−タグ融合物を例外とする。タンパク質の非脂質化型は、可溶性で細胞質中に局在すると考えられる。タンパク質の脂質化型は、膜画分に結合すると考えられ、界面活性剤により可溶化される。天然の脂質化型のタンパク質は、抗原提示に関して優れた三次構造を有することができ、および/または付着した脂質は、より強い免疫原性反応を刺激するアジュバントとして作用し得る。
【0211】
試験したすべてのN.meningitidisB菌株は、1つ2086様遺伝子を有すると考えられる。2086遺伝子の少なくとも2種のファミリーが提示される。2086ホモログは、PCRスクリーニングにより以下のものに同定されている:
N.meningitidisA、B、C,W135、Y
N.lactamica
N.gonorrhoeaeFA1090
いくつかのORF2086遺伝子は、クローニングされ、組換え発現されている
P2086の脂質化バージョンは、種々の髄膜炎菌株から発現された。
【0212】
これらの組換えタンパク質は精製され、マウスに予防接種するために使用されている。
【0213】
結果として生じる抗血清は殺菌性である。
【0214】
P2086の非脂質化バージョンは、様々な菌株から発現された。
【0215】
rLP2086は、rP2086より強い免疫反応を一貫して誘発する。
【0216】
rLP2086はまた、同種および異種髄膜炎菌株の両方に対して高められた殺菌活性を示す。
(実施例11)
【0217】
以下は、P2086が髄膜炎菌株において発現され、そしていくつかの菌株においてP2086発現の付加的な具体例を提供することをさらに示す。
【0218】
細胞ライゼートを、SDS試料バッファーに再懸濁されたプレート培養物由来の細胞で調製し、98℃で4分間加熱し。試料を、ウェルあたり約30〜50ug総タンパク質で、10〜20%プレキャストゲル(ICN)に負荷し、175Vで流した。ゲルをニトロセルロース膜に移し、次にそれを30分間、Tris緩衝生理食塩水(Blotto)中の5%粉末ミルクでブロッキングした。使用した一次抗体は、マウスにおいて個々のrLP2086変異体に対して誘導されたポリクローナル抗血清のプールであった。
【0219】
図17および18を参照すると、ウェスタンブロットは、P2086サブファミリーAおよびB全細胞ライゼートに対するrLP2086マウス抗血清の反応性を示す。サブファミリーA細胞ライゼートブロットの場合、使用される抗血清は、rLP2086−2996、−870446および−250771に対して誘導され、rLP2086−250771はBlottoで1/500に希釈され、他はBlottoで1/1000に希釈された。サブファミリーB細胞ライゼートブロットの場合、使用される抗血清は、rLP2086−8529(Blottoで1/1000に希釈)、−CDC1573、−M982および−880049(これら3種はBlottoで1/500に希釈)に対して誘導された。一次抗血清およびブロットを、4℃で一晩インキュベートした。ブロットを洗浄し、ヤギ抗マウスAP二次をBlottoで1/500に希釈して添加し、ブロットを30分間室温でインキュベートした。洗浄後、ブロットをBCIP/NBT膜ホスファターゼ基質系(KPL)を使用して展開した。
参考文献一覧
本明細書中で参照した参考文献が以下に記載され、それらは、参照によりその全体が本明細書に組み込まれる:
【0220】
【化2】
【0221】
【化3】
【0222】
【化4】
【0223】
【化5】
【0224】
【化6】
【0225】
【化7】
【0226】
【化8】
【0227】
【化9】
【0228】
【化10】
【0229】
【化11】
【0230】
【化12】
【0231】
【化13】
本発明は今や十分に説明されているが、本明細書中で示された本発明の趣旨または範囲を逸脱することなく、本発明に対し多くの変更および修正を行い得ることが当業者には明らかであろう。上記は、本発明の好ましい実施態様をいくつかの考え得る代替案と共に説明している。しかしながら、これらの実施態様は、単に例のためであり、本発明はそれらに限定されない。
【特許請求の範囲】
【請求項1】
ポリヌクレオチドであって、
(a) 配列番号1−11の奇数番号配列のいずれかと少なくとも約95%配列同一性を有するヌクレオチド配列、または
(b) 配列番号2−12の偶数番号配列のいずれかのアミノ酸配列と少なくとも約95%配列同一性を有するアミノ酸配列を含むポリペプチドをコードするヌクレオチド、
を含む
ポリヌクレオチド。
【請求項2】
前記ヌクレオチド配列が、配列番号1−11の奇数番号配列のいずれかと少なくとも約95%配列同一性を有する、請求項1に記載のポリヌクレオチド。
【請求項3】
前記ヌクレオチド配列が、配列番号1−11の奇数番号配列のいずれかを含む、請求項1に記載のポリヌクレオチド。
【請求項4】
前記ヌクレオチド配列が、配列番号2−12の偶数番号配列のいずれかのアミノ酸配列と少なくとも約95%配列同一性を有するアミノ酸配列を含むポリペプチドをコードする、請求項1に記載のポリヌクレオチド。
【請求項5】
前記ヌクレオチド配列が、配列番号2−12の偶数番号配列のうちのいずれかを含むポリペプチドをコードする、請求項1に記載のポリヌクレオチド。
【請求項6】
前記ヌクレオチド配列が、配列番号1−5の奇数番号配列のいずれかと少なくとも約95%配列同一性を有する、請求項1に記載のポリヌクレオチド。
【請求項7】
前記ヌクレオチド配列が、配列番号1−5の奇数番号配列のいずれかを含む、請求項1に記載のポリヌクレオチド。
【請求項8】
前記ヌクレオチド配列が、配列番号2−6の偶数番号配列のいずれかのアミノ酸配列と少なくとも約95%配列同一性を有するアミノ酸配列を含むポリペプチドをコードする、請求項1に記載のポリヌクレオチド。
【請求項9】
前記ヌクレオチド配列が、配列番号2−6の偶数番号配列のうちのいずれかを含むポリペプチドをコードする、請求項1に記載のポリヌクレオチド。
【請求項10】
前記ヌクレオチド配列が、配列番号7−11の奇数番号配列のいずれかと少なくとも約95%配列同一性を有する、請求項1に記載のポリヌクレオチド。
【請求項11】
前記ヌクレオチド配列が、配列番号7−11の奇数番号配列のうちのいずれかを含むポリペプチドをコードする、請求項1に記載のポリヌクレオチド。
【請求項12】
前記ヌクレオチド配列が、配列番号8−12の偶数番号配列のいずれかのアミノ酸配列と少なくとも約95%配列同一性を有するアミノ酸配列を含むポリペプチドをコードする、請求項1に記載のポリヌクレオチド。
【請求項13】
前記ヌクレオチド配列が、配列番号8−12の偶数番号配列のいずれかを含むポリペプチドをコードする、請求項1に記載のポリヌクレオチド。
【請求項14】
前記ポリヌクレオチドが、N.meningitidisセログループBに対する抗体を誘導するポリペプチドをコードする、請求項1に記載のポリヌクレオチド。
【請求項15】
前記ポリヌクレオチドが、N.meningitidisセログループBサブファミリーAに対する抗体を誘導するポリペプチドをコードする、請求項1に記載のポリヌクレオチド。
【請求項16】
前記ポリヌクレオチドが、N.meningitidisセログループBサブファミリーBに対する抗体を誘導するポリペプチドをコードする、請求項1に記載のポリヌクレオチド。
【請求項17】
前記ポリヌクレオチドが組換えポリヌクレオチドである、請求項1〜16のいずれかに記載のポリヌクレオチド。
【請求項18】
前記ポリヌクレオチドが天然供給源から単離される、請求項1〜16のいずれかに記載のポリヌクレオチド。
【請求項19】
請求項1〜18のいずれかに記載のポリヌクレオチドを含むベクター。
【請求項20】
前記ベクターがプラスミドである、請求項19に記載のベクター。
【請求項21】
前記ベクターがファージである、請求項19に記載のベクター。
【請求項22】
前記ベクターがバクテリオファージである、請求項19に記載のベクター。
【請求項23】
前記ベクターが穏和ファージ(moderate phage)である、請求項19に記載のベクター。
【請求項24】
請求項19〜23のいずれかに記載のベクターを含む、組換え細胞。
【請求項25】
前記組換え細胞がハイブリドーマである、請求項24に記載の組換え細胞。
【請求項26】
前記組換え細胞がトリオーマである、請求項24に記載の組換え細胞。
【請求項27】
ポリペプチドであって、
(a) 配列番号2−12の偶数番号配列のいずれかと少なくとも95%配列同一性を有するアミノ酸配列、
(b) 配列番号1−11の奇数番号配列のいずれかと少なくとも95%配列同一性を有するヌクレオチド配列によってコードされるアミノ酸配列、
(c) (a)もしくは(b)に記載されたアミノ酸配列の少なくとも1つの免疫原性部分、あるいは
(d) (a)もしくは(b)に記載されたアミノ酸配列または(c)に記載された免疫原性部分の少なくとも1つの生物学的等価物、
を含む
ポリペプチド。
【請求項28】
前記ポリペプチドが、配列番号2−12の偶数番号配列のいずれかと少なくとも95%配列同一性を有するアミノ酸配列を含む、請求項27に記載のポリペプチド。
【請求項29】
前記ポリペプチドが、配列番号2−12の偶数番号配列のいずれかのアミノ酸配列を含む、請求項27に記載のポリペプチド。
【請求項30】
前記ポリペプチドが、配列番号1−11の奇数番号配列のいずれかと少なくとも95%配列同一性を有するヌクレオチド配列によってコードされるアミノ酸配列を含む、請求項27に記載のポリペプチド。
【請求項31】
前記ポリペプチドが、配列番号1−11の奇数番号配列のいずれかの核酸配列によってコードされるアミノ酸配列を含む、請求項27に記載のポリペプチド。
【請求項32】
前記ポリペプチドが、配列番号2−6の偶数番号配列のいずれかと少なくとも95%配列同一性を有するアミノ酸配列を含む、請求項27に記載のポリペプチド。
【請求項33】
前記ポリペプチドが、配列番号2−6の偶数番号配列のいずれかのアミノ酸配列を含む、請求項27に記載のポリペプチド。
【請求項34】
前記ポリペプチドが、配列番号1−5の奇数番号配列のいずれかと少なくとも95%配列同一性を有するヌクレオチド配列によってコードされるアミノ酸配列を含む、請求項27に記載のポリペプチド。
【請求項35】
前記ポリペプチドが、配列番号1−5の奇数番号配列のいずれかの核酸配列によってコードされるアミノ酸配列を含む、請求項27に記載のポリペプチド。
【請求項36】
前記ポリペプチドが、配列番号8−12の偶数番号配列のいずれかと少なくとも95%配列同一性を有するアミノ酸配列を含む、請求項27に記載のポリペプチド:。
【請求項37】
前記ポリペプチドが、配列番号8−12の偶数番号配列のいずれかのアミノ酸配列を含む、請求項27に記載のポリペプチド。
【請求項38】
前記ポリペプチドが、配列番号7−11の奇数番号配列のいずれかと少なくとも95%配列同一性を有するヌクレオチド配列によってコードされるアミノ酸配列を含む、請求項27に記載のポリペプチド。
【請求項39】
前記ポリペプチドが、配列番号7−11の奇数番号配列のいずれかの核酸配列によってコードされるアミノ酸配列を含む、請求項27に記載のポリペプチド。
【請求項40】
少なくとも1種のPorA、PorB、トランスフェリン結合タンパク質、または不透明タンパク質(opacity protein)(Opc)をさらに含む、請求項27に記載のポリペプチド。
【請求項41】
Neisseria speciesの少なくとも1種の付加的な表面抗原をさらに含み、該付加的な表面抗原が、非ORF2086タンパク質である、請求項27に記載のポリペプチド。
【請求項42】
前記ポリペプチドが、質量分析により測定される場合に、約26,000〜約30,000の分子量を有する、請求項27に記載のポリペプチド。
【請求項43】
前記ポリペプチドが、10%−20%SDSポリアクリルアミドゲル上で測定される場合に、約28〜35kDaの分子量を有する、請求項27に記載のポリペプチド。
【請求項44】
前記ポリペプチドが非脂質化タンパク質である、請求項27に記載のポリペプチド。
【請求項45】
前記ポリペプチドが組換えタンパク質である、請求項27に記載のポリペプチド。
【請求項46】
前記ポリペプチドが天然のNeisseria speciesから単離される、請求項27に記載のポリペプチド。
【請求項47】
抗体であって、
(a) 偶数番号の配列番号2−12のいずれかのアミノ酸配列を含むポリペプチドと免疫特異的に結合するポリペプチド、または
(b) (a)に記載されたポリペプチドの少なくとも1つの免疫原性部分、または
(c) (a)に記載されたポリペプチドもしくは(b)に記載された1つの免疫原性部分の少なくとも1つの生物学的等価物、
のいずれかを含む
抗体。
【請求項48】
少なくとも1つの抗体がモノクローナル抗体である、請求項47に記載の抗体。
【請求項49】
ポリヌクレオチド、ベクター、組換え細胞、ポリペプチドまたは請求項1〜48のいずれかに記載の抗体を含む、組成物。
【請求項50】
前記組成物が、薬学的に許容されるバッファー、希釈剤、およびアジュバントまたは担体をさらに含む、請求項49に記載の組成物。
【請求項51】
前記組成物が担体をさらに含む、請求項49に記載の組成物。
【請求項52】
前記組成物がアジュバントをさらに含む、請求項49に記載の組成物。
【請求項53】
前記アジュバントが液体を含む、請求項52に記載の組成物。
【請求項54】
前記ポリペプチドがリポタンパク質である、請求項49に記載の組成物。
【請求項55】
前記組成物が多糖をさらに含む、請求項49に記載の組成物。
【請求項56】
前記組成物が、前記ポリペプチドと結合体を形成する付加的なペプチド、ポリペプチドまたはタンパク質を含む、請求項49に記載の組成物。
【請求項57】
前記付加的なペプチド、ポリペプチドまたはタンパク質が、哺乳動物において2種以上の細菌に対する免疫応答を引き起こす結合体を形成する、請求項56に記載の組成物。
【請求項58】
前記組成物が、付加的なペプチド、ポリペプチドまたはタンパク質をコードする核酸配列をさらに含む、請求項49に記載の組成物。
【請求項59】
組成物であって、
(a) 配列番号1−5の奇数番号配列のいずれかと少なくとも約95%配列同一性または配列番号2−6の偶数番号配列のいずれかのアミノ酸配列をコードするヌクレオチド配列と少なくとも約95%配列同一性を有するヌクレオチド配列を含む第1のポリヌクレオチドと、
(b) 配列番号7−11の奇数番号配列のいずれかと少なくとも約95%配列同一性または配列番号8−12の偶数番号配列のいずれかのアミノ酸配列をコードするヌクレオチド配列と少なくとも約95%配列同一性を有するヌクレオチド配列を含む第2のポリヌクレオチドと、
を含む、
組成物。
【請求項60】
前記第1のポリヌクレオチドが、配列番号1−5の奇数番号配列のいずれかを含む、請求項59に記載の組成物。
【請求項61】
前記第2のポリヌクレオチド配列が、配列番号7−11の奇数番号配列のいずれか含む、請求項59に記載の組成物。
【請求項62】
組成物であって、
(a) 配列番号2−6の偶数番号配列のいずれかと少なくとも約95%配列同一性を有するアミノ酸配列を含む第1のポリペプチドと、
(b) 配列番号8−12の偶数番号配列のいずれかと少なくとも約95%配列同一性を有するアミノ酸配列を含む第2のポリペプチドと、
を含む
組成物。
【請求項63】
前記第1のポリペプチドが、配列番号2−6の偶数番号配列のいずれかのアミノ酸配列を含む、請求項62に記載の組成物。
【請求項64】
前記第2のポリペプチドが、配列番号8−12の偶数番号配列のいずれかのアミノ酸配列を含む、請求項62に記載の組成物。
【請求項65】
組成物であって、
(a) 偶数番号の配列番号2−12のいずれかのアミノ酸配列を含むポリペプチド、
(b) 奇数番号の配列番号1−11のいずれかの核酸配列を含むポリヌクレオチドによりコードされたポリペプチド、
(c) (a)または(b)に記載されたポリペプチドの免疫原性部分、あるいは
(d) (a)もしくは(b)に記載されたポリペプチドまたは(c)に記載された免疫原性フラグメントの生物学的等価物
のいずれかを、
Neisseria speciesから単離および精製、または組換えにより調製するステップを含むプロセスにより調製される
組成物。
【請求項66】
哺乳動物において免疫応答を引き起こすための薬剤の調製における、請求項49〜65にいずれかに記載の組成物の使用。
【請求項67】
前記組成物が非経口的に投与される、請求項66に記載の使用。
【請求項68】
前記組成物が粘膜的に投与される、請求項66に記載の使用。
【請求項69】
哺乳動物における細菌性髄膜炎に対して効果的な薬剤における、請求項49〜65のいずれかに記載の組成物の使用。
【請求項70】
前記組成物が非経口的に投与される、請求項69に記載の組成物の使用。
【請求項71】
前記組成物が粘膜的に投与される、請求項69に記載の組成物の使用。
【請求項72】
前記組成物が皮下または筋肉内注射により投与される、請求項69に記載の組成物の使用。
【請求項73】
組成物を調製する方法であって、
本明細書中で記載されるポリペプチドのいずれかをコードする核酸配列を宿主細胞中で発現させるステップを含む、
方法。
【請求項74】
前記核酸配列がin vivoで発現される、請求項73に記載の方法。
【請求項75】
前記核酸配列がin vitroで発現される、請求項73に記載の方法。
【請求項76】
前記ポリペプチドを発現させるためにP4リーダー配列を前記核酸配列と関連付けるステップをさらに含む、請求項73に記載の方法。
【請求項77】
抗体組成物を調製する方法であって、
本明細書中で記載されるポリペプチド、タンパク質、免疫原性部分または生物学的等価物のいずれかを含む組成物を動物に導入した後、該動物から抗体を回収するステップを含む、
方法。
【請求項78】
哺乳動物において免疫応答を引き起こす方法であって、
請求項49〜65に記載の1種以上の組成物の有効量を該哺乳動物に投与するステップを含む、
方法。
【請求項79】
前記組成物が非経口的に投与される、請求項78に記載の方法。
【請求項80】
前記組成物が粘膜的に投与される、前記の請求項78に記載の方法。
【請求項81】
哺乳動物において細菌性髄膜炎を予防または処置する方法であって、
請求項49〜65に記載の1種以上の組成物の有効量を該哺乳動物に投与するステップを含む、
方法。
【請求項82】
前記組成物が非経口的に投与される、請求項81に記載の方法。
【請求項83】
前記組成物が粘膜的に投与される、請求項81に記載の方法。
【請求項84】
前記組成物が皮下または筋肉内注射により投与される、請求項81に記載の方法。
【請求項1】
ポリヌクレオチドであって、
(a) 配列番号1−11の奇数番号配列のいずれかと少なくとも約95%配列同一性を有するヌクレオチド配列、または
(b) 配列番号2−12の偶数番号配列のいずれかのアミノ酸配列と少なくとも約95%配列同一性を有するアミノ酸配列を含むポリペプチドをコードするヌクレオチド、
を含む
ポリヌクレオチド。
【請求項2】
前記ヌクレオチド配列が、配列番号1−11の奇数番号配列のいずれかと少なくとも約95%配列同一性を有する、請求項1に記載のポリヌクレオチド。
【請求項3】
前記ヌクレオチド配列が、配列番号1−11の奇数番号配列のいずれかを含む、請求項1に記載のポリヌクレオチド。
【請求項4】
前記ヌクレオチド配列が、配列番号2−12の偶数番号配列のいずれかのアミノ酸配列と少なくとも約95%配列同一性を有するアミノ酸配列を含むポリペプチドをコードする、請求項1に記載のポリヌクレオチド。
【請求項5】
前記ヌクレオチド配列が、配列番号2−12の偶数番号配列のうちのいずれかを含むポリペプチドをコードする、請求項1に記載のポリヌクレオチド。
【請求項6】
前記ヌクレオチド配列が、配列番号1−5の奇数番号配列のいずれかと少なくとも約95%配列同一性を有する、請求項1に記載のポリヌクレオチド。
【請求項7】
前記ヌクレオチド配列が、配列番号1−5の奇数番号配列のいずれかを含む、請求項1に記載のポリヌクレオチド。
【請求項8】
前記ヌクレオチド配列が、配列番号2−6の偶数番号配列のいずれかのアミノ酸配列と少なくとも約95%配列同一性を有するアミノ酸配列を含むポリペプチドをコードする、請求項1に記載のポリヌクレオチド。
【請求項9】
前記ヌクレオチド配列が、配列番号2−6の偶数番号配列のうちのいずれかを含むポリペプチドをコードする、請求項1に記載のポリヌクレオチド。
【請求項10】
前記ヌクレオチド配列が、配列番号7−11の奇数番号配列のいずれかと少なくとも約95%配列同一性を有する、請求項1に記載のポリヌクレオチド。
【請求項11】
前記ヌクレオチド配列が、配列番号7−11の奇数番号配列のうちのいずれかを含むポリペプチドをコードする、請求項1に記載のポリヌクレオチド。
【請求項12】
前記ヌクレオチド配列が、配列番号8−12の偶数番号配列のいずれかのアミノ酸配列と少なくとも約95%配列同一性を有するアミノ酸配列を含むポリペプチドをコードする、請求項1に記載のポリヌクレオチド。
【請求項13】
前記ヌクレオチド配列が、配列番号8−12の偶数番号配列のいずれかを含むポリペプチドをコードする、請求項1に記載のポリヌクレオチド。
【請求項14】
前記ポリヌクレオチドが、N.meningitidisセログループBに対する抗体を誘導するポリペプチドをコードする、請求項1に記載のポリヌクレオチド。
【請求項15】
前記ポリヌクレオチドが、N.meningitidisセログループBサブファミリーAに対する抗体を誘導するポリペプチドをコードする、請求項1に記載のポリヌクレオチド。
【請求項16】
前記ポリヌクレオチドが、N.meningitidisセログループBサブファミリーBに対する抗体を誘導するポリペプチドをコードする、請求項1に記載のポリヌクレオチド。
【請求項17】
前記ポリヌクレオチドが組換えポリヌクレオチドである、請求項1〜16のいずれかに記載のポリヌクレオチド。
【請求項18】
前記ポリヌクレオチドが天然供給源から単離される、請求項1〜16のいずれかに記載のポリヌクレオチド。
【請求項19】
請求項1〜18のいずれかに記載のポリヌクレオチドを含むベクター。
【請求項20】
前記ベクターがプラスミドである、請求項19に記載のベクター。
【請求項21】
前記ベクターがファージである、請求項19に記載のベクター。
【請求項22】
前記ベクターがバクテリオファージである、請求項19に記載のベクター。
【請求項23】
前記ベクターが穏和ファージ(moderate phage)である、請求項19に記載のベクター。
【請求項24】
請求項19〜23のいずれかに記載のベクターを含む、組換え細胞。
【請求項25】
前記組換え細胞がハイブリドーマである、請求項24に記載の組換え細胞。
【請求項26】
前記組換え細胞がトリオーマである、請求項24に記載の組換え細胞。
【請求項27】
ポリペプチドであって、
(a) 配列番号2−12の偶数番号配列のいずれかと少なくとも95%配列同一性を有するアミノ酸配列、
(b) 配列番号1−11の奇数番号配列のいずれかと少なくとも95%配列同一性を有するヌクレオチド配列によってコードされるアミノ酸配列、
(c) (a)もしくは(b)に記載されたアミノ酸配列の少なくとも1つの免疫原性部分、あるいは
(d) (a)もしくは(b)に記載されたアミノ酸配列または(c)に記載された免疫原性部分の少なくとも1つの生物学的等価物、
を含む
ポリペプチド。
【請求項28】
前記ポリペプチドが、配列番号2−12の偶数番号配列のいずれかと少なくとも95%配列同一性を有するアミノ酸配列を含む、請求項27に記載のポリペプチド。
【請求項29】
前記ポリペプチドが、配列番号2−12の偶数番号配列のいずれかのアミノ酸配列を含む、請求項27に記載のポリペプチド。
【請求項30】
前記ポリペプチドが、配列番号1−11の奇数番号配列のいずれかと少なくとも95%配列同一性を有するヌクレオチド配列によってコードされるアミノ酸配列を含む、請求項27に記載のポリペプチド。
【請求項31】
前記ポリペプチドが、配列番号1−11の奇数番号配列のいずれかの核酸配列によってコードされるアミノ酸配列を含む、請求項27に記載のポリペプチド。
【請求項32】
前記ポリペプチドが、配列番号2−6の偶数番号配列のいずれかと少なくとも95%配列同一性を有するアミノ酸配列を含む、請求項27に記載のポリペプチド。
【請求項33】
前記ポリペプチドが、配列番号2−6の偶数番号配列のいずれかのアミノ酸配列を含む、請求項27に記載のポリペプチド。
【請求項34】
前記ポリペプチドが、配列番号1−5の奇数番号配列のいずれかと少なくとも95%配列同一性を有するヌクレオチド配列によってコードされるアミノ酸配列を含む、請求項27に記載のポリペプチド。
【請求項35】
前記ポリペプチドが、配列番号1−5の奇数番号配列のいずれかの核酸配列によってコードされるアミノ酸配列を含む、請求項27に記載のポリペプチド。
【請求項36】
前記ポリペプチドが、配列番号8−12の偶数番号配列のいずれかと少なくとも95%配列同一性を有するアミノ酸配列を含む、請求項27に記載のポリペプチド:。
【請求項37】
前記ポリペプチドが、配列番号8−12の偶数番号配列のいずれかのアミノ酸配列を含む、請求項27に記載のポリペプチド。
【請求項38】
前記ポリペプチドが、配列番号7−11の奇数番号配列のいずれかと少なくとも95%配列同一性を有するヌクレオチド配列によってコードされるアミノ酸配列を含む、請求項27に記載のポリペプチド。
【請求項39】
前記ポリペプチドが、配列番号7−11の奇数番号配列のいずれかの核酸配列によってコードされるアミノ酸配列を含む、請求項27に記載のポリペプチド。
【請求項40】
少なくとも1種のPorA、PorB、トランスフェリン結合タンパク質、または不透明タンパク質(opacity protein)(Opc)をさらに含む、請求項27に記載のポリペプチド。
【請求項41】
Neisseria speciesの少なくとも1種の付加的な表面抗原をさらに含み、該付加的な表面抗原が、非ORF2086タンパク質である、請求項27に記載のポリペプチド。
【請求項42】
前記ポリペプチドが、質量分析により測定される場合に、約26,000〜約30,000の分子量を有する、請求項27に記載のポリペプチド。
【請求項43】
前記ポリペプチドが、10%−20%SDSポリアクリルアミドゲル上で測定される場合に、約28〜35kDaの分子量を有する、請求項27に記載のポリペプチド。
【請求項44】
前記ポリペプチドが非脂質化タンパク質である、請求項27に記載のポリペプチド。
【請求項45】
前記ポリペプチドが組換えタンパク質である、請求項27に記載のポリペプチド。
【請求項46】
前記ポリペプチドが天然のNeisseria speciesから単離される、請求項27に記載のポリペプチド。
【請求項47】
抗体であって、
(a) 偶数番号の配列番号2−12のいずれかのアミノ酸配列を含むポリペプチドと免疫特異的に結合するポリペプチド、または
(b) (a)に記載されたポリペプチドの少なくとも1つの免疫原性部分、または
(c) (a)に記載されたポリペプチドもしくは(b)に記載された1つの免疫原性部分の少なくとも1つの生物学的等価物、
のいずれかを含む
抗体。
【請求項48】
少なくとも1つの抗体がモノクローナル抗体である、請求項47に記載の抗体。
【請求項49】
ポリヌクレオチド、ベクター、組換え細胞、ポリペプチドまたは請求項1〜48のいずれかに記載の抗体を含む、組成物。
【請求項50】
前記組成物が、薬学的に許容されるバッファー、希釈剤、およびアジュバントまたは担体をさらに含む、請求項49に記載の組成物。
【請求項51】
前記組成物が担体をさらに含む、請求項49に記載の組成物。
【請求項52】
前記組成物がアジュバントをさらに含む、請求項49に記載の組成物。
【請求項53】
前記アジュバントが液体を含む、請求項52に記載の組成物。
【請求項54】
前記ポリペプチドがリポタンパク質である、請求項49に記載の組成物。
【請求項55】
前記組成物が多糖をさらに含む、請求項49に記載の組成物。
【請求項56】
前記組成物が、前記ポリペプチドと結合体を形成する付加的なペプチド、ポリペプチドまたはタンパク質を含む、請求項49に記載の組成物。
【請求項57】
前記付加的なペプチド、ポリペプチドまたはタンパク質が、哺乳動物において2種以上の細菌に対する免疫応答を引き起こす結合体を形成する、請求項56に記載の組成物。
【請求項58】
前記組成物が、付加的なペプチド、ポリペプチドまたはタンパク質をコードする核酸配列をさらに含む、請求項49に記載の組成物。
【請求項59】
組成物であって、
(a) 配列番号1−5の奇数番号配列のいずれかと少なくとも約95%配列同一性または配列番号2−6の偶数番号配列のいずれかのアミノ酸配列をコードするヌクレオチド配列と少なくとも約95%配列同一性を有するヌクレオチド配列を含む第1のポリヌクレオチドと、
(b) 配列番号7−11の奇数番号配列のいずれかと少なくとも約95%配列同一性または配列番号8−12の偶数番号配列のいずれかのアミノ酸配列をコードするヌクレオチド配列と少なくとも約95%配列同一性を有するヌクレオチド配列を含む第2のポリヌクレオチドと、
を含む、
組成物。
【請求項60】
前記第1のポリヌクレオチドが、配列番号1−5の奇数番号配列のいずれかを含む、請求項59に記載の組成物。
【請求項61】
前記第2のポリヌクレオチド配列が、配列番号7−11の奇数番号配列のいずれか含む、請求項59に記載の組成物。
【請求項62】
組成物であって、
(a) 配列番号2−6の偶数番号配列のいずれかと少なくとも約95%配列同一性を有するアミノ酸配列を含む第1のポリペプチドと、
(b) 配列番号8−12の偶数番号配列のいずれかと少なくとも約95%配列同一性を有するアミノ酸配列を含む第2のポリペプチドと、
を含む
組成物。
【請求項63】
前記第1のポリペプチドが、配列番号2−6の偶数番号配列のいずれかのアミノ酸配列を含む、請求項62に記載の組成物。
【請求項64】
前記第2のポリペプチドが、配列番号8−12の偶数番号配列のいずれかのアミノ酸配列を含む、請求項62に記載の組成物。
【請求項65】
組成物であって、
(a) 偶数番号の配列番号2−12のいずれかのアミノ酸配列を含むポリペプチド、
(b) 奇数番号の配列番号1−11のいずれかの核酸配列を含むポリヌクレオチドによりコードされたポリペプチド、
(c) (a)または(b)に記載されたポリペプチドの免疫原性部分、あるいは
(d) (a)もしくは(b)に記載されたポリペプチドまたは(c)に記載された免疫原性フラグメントの生物学的等価物
のいずれかを、
Neisseria speciesから単離および精製、または組換えにより調製するステップを含むプロセスにより調製される
組成物。
【請求項66】
哺乳動物において免疫応答を引き起こすための薬剤の調製における、請求項49〜65にいずれかに記載の組成物の使用。
【請求項67】
前記組成物が非経口的に投与される、請求項66に記載の使用。
【請求項68】
前記組成物が粘膜的に投与される、請求項66に記載の使用。
【請求項69】
哺乳動物における細菌性髄膜炎に対して効果的な薬剤における、請求項49〜65のいずれかに記載の組成物の使用。
【請求項70】
前記組成物が非経口的に投与される、請求項69に記載の組成物の使用。
【請求項71】
前記組成物が粘膜的に投与される、請求項69に記載の組成物の使用。
【請求項72】
前記組成物が皮下または筋肉内注射により投与される、請求項69に記載の組成物の使用。
【請求項73】
組成物を調製する方法であって、
本明細書中で記載されるポリペプチドのいずれかをコードする核酸配列を宿主細胞中で発現させるステップを含む、
方法。
【請求項74】
前記核酸配列がin vivoで発現される、請求項73に記載の方法。
【請求項75】
前記核酸配列がin vitroで発現される、請求項73に記載の方法。
【請求項76】
前記ポリペプチドを発現させるためにP4リーダー配列を前記核酸配列と関連付けるステップをさらに含む、請求項73に記載の方法。
【請求項77】
抗体組成物を調製する方法であって、
本明細書中で記載されるポリペプチド、タンパク質、免疫原性部分または生物学的等価物のいずれかを含む組成物を動物に導入した後、該動物から抗体を回収するステップを含む、
方法。
【請求項78】
哺乳動物において免疫応答を引き起こす方法であって、
請求項49〜65に記載の1種以上の組成物の有効量を該哺乳動物に投与するステップを含む、
方法。
【請求項79】
前記組成物が非経口的に投与される、請求項78に記載の方法。
【請求項80】
前記組成物が粘膜的に投与される、前記の請求項78に記載の方法。
【請求項81】
哺乳動物において細菌性髄膜炎を予防または処置する方法であって、
請求項49〜65に記載の1種以上の組成物の有効量を該哺乳動物に投与するステップを含む、
方法。
【請求項82】
前記組成物が非経口的に投与される、請求項81に記載の方法。
【請求項83】
前記組成物が粘膜的に投与される、請求項81に記載の方法。
【請求項84】
前記組成物が皮下または筋肉内注射により投与される、請求項81に記載の方法。
【図1A】

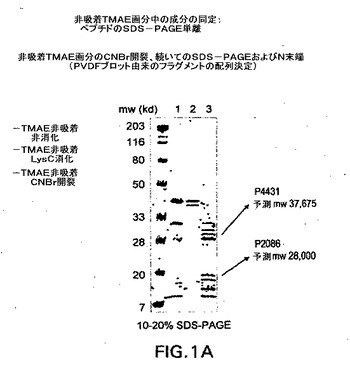
【図1B】
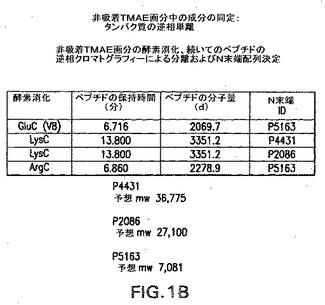
【図2】
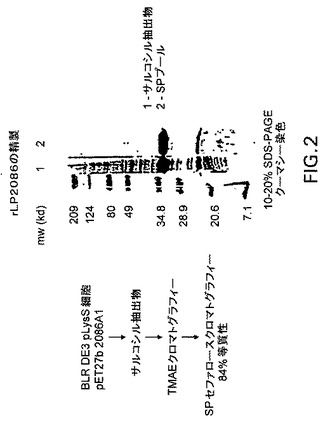
【図3】

【図4】

【図5】
【図6】
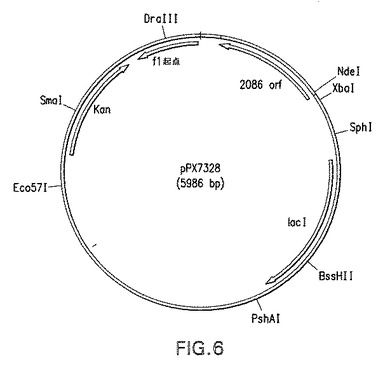
【図7】
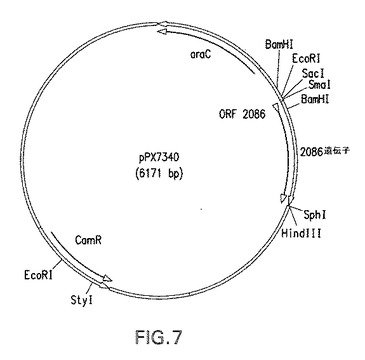
【図8】
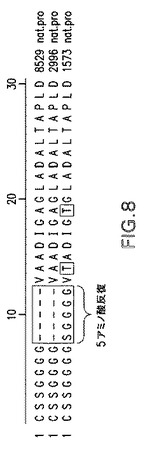
【図9A】
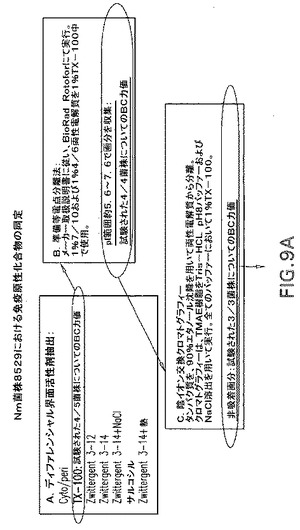
【図9B】
【図10】
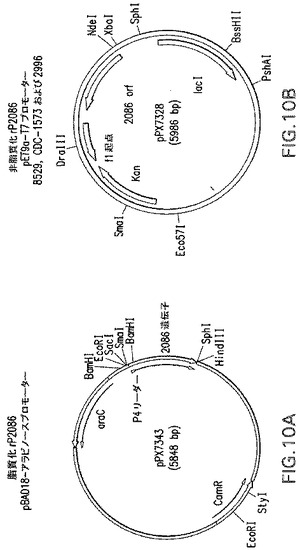
【図11】
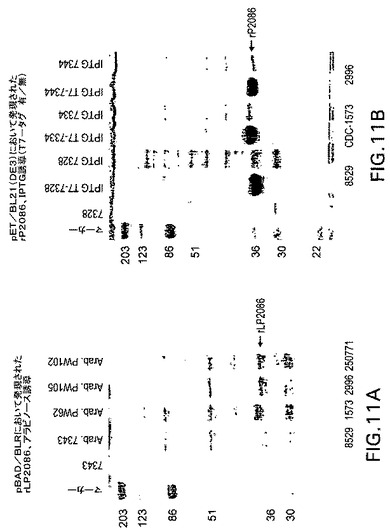
【図12A】
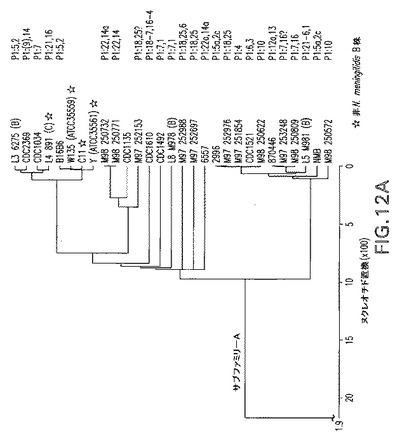
【図12B】

【図13】
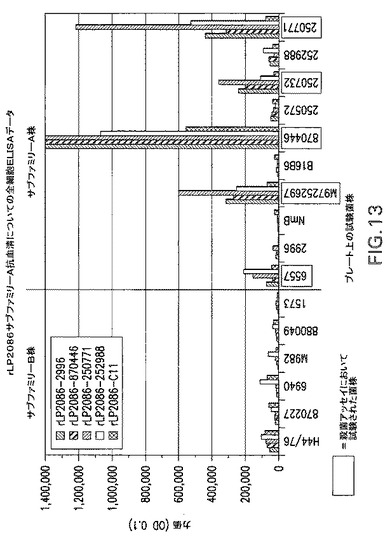
【図14】
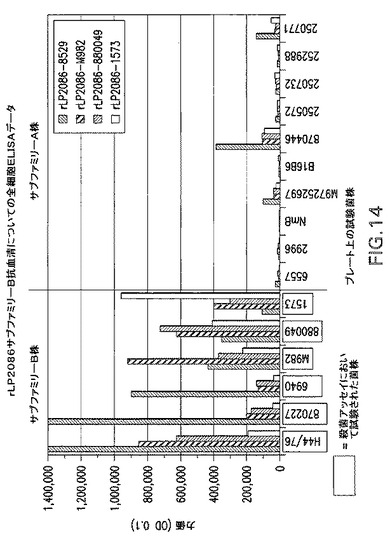
【図15】
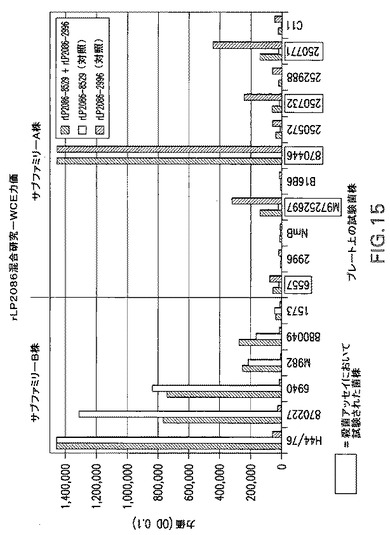
【図16】
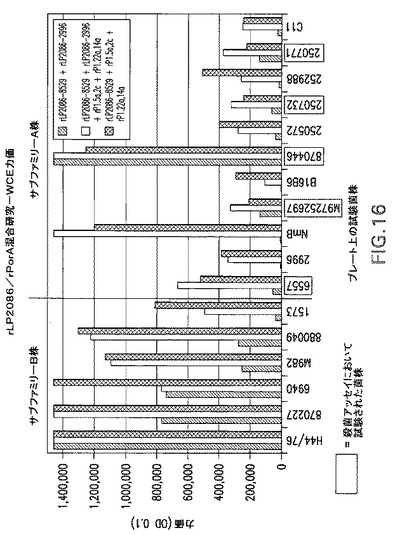
【図17】
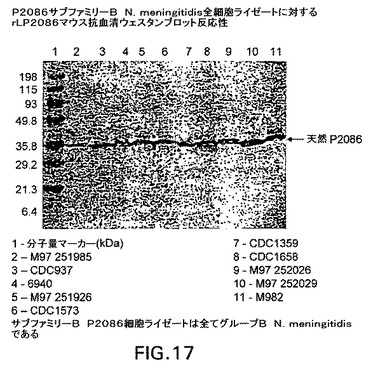
【図18】
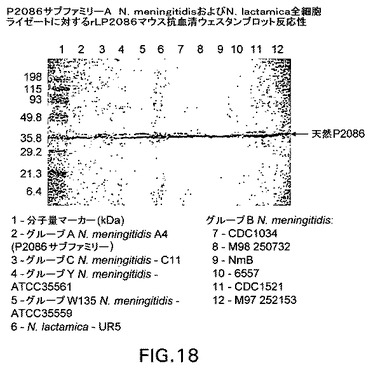
【図1B】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9A】
【図9B】
【図10】
【図11】
【図12A】
【図12B】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【公表番号】特表2010−512792(P2010−512792A)
【公表日】平成22年4月30日(2010.4.30)
【国際特許分類】
【出願番号】特願2009−542957(P2009−542957)
【出願日】平成19年12月21日(2007.12.21)
【国際出願番号】PCT/US2007/026238
【国際公開番号】WO2008/079372
【国際公開日】平成20年7月3日(2008.7.3)
【出願人】(591011502)ワイス エルエルシー (573)
【Fターム(参考)】
【公表日】平成22年4月30日(2010.4.30)
【国際特許分類】
【出願日】平成19年12月21日(2007.12.21)
【国際出願番号】PCT/US2007/026238
【国際公開番号】WO2008/079372
【国際公開日】平成20年7月3日(2008.7.3)
【出願人】(591011502)ワイス エルエルシー (573)
【Fターム(参考)】
[ Back to top ]