
高い中鎖長モノマー含有量を有するポリヒドロキシルアルカノアートコポリマーを製造する方法
本発明は、高い3−ヒドロキシへキサのアートモノマー含有量を有するポリヒドロキシアルカノアートコポリマーの、組換え遺伝子発現を通じた製造に関する。

【発明の詳細な説明】
【技術分野】
【0001】
関連出願
本出願は、米国仮出願番号61/260,164、タイトル「高い中鎖長モノマー含有量を有するポリヒドロキシルアルカノアートコポリマーを製造する方法(Methods for Producing Polyhydroxyalkanoate Copolymer with High Medium Chain Length Monomer Content)」、出願日2009年11月11日、の35U.S.C.120条に基づく優先権を主張し、その全開示は、全体として本明細書に参照として組み込まれる。
【0002】
技術分野
本発明は、組換え体による遺伝子発現を通じた高い中鎖長モノマー含有量を有するポリヒドロキシアルカノアートコポリマーの製造に関する。
【背景技術】
【0003】
商業的に有用な複合バイオポリマーであるポリヒドロキシルブチラートは、多数の細菌によって産生される細胞間物質である。ポリヒドロキシブチラート(PHB)は、ポリマーの化学的および物理的特性両方に基づく有用な生物材料である。PHBは、生分解性/熱可塑性材料として、ある抗生物質の有機合成のためのキラル中心の給源として、および薬剤送達および骨置換のためのマトリクスとしての有用性を含む、様々な潜在的応用性を有する。in vivoにおいて、該ポリマーは内部的に分解されて、ヒトの血液の通常の構成物質であるヒドロキシブチラートになる。ポリヒドロキシアルカノアート(PHA、PHBを含む)およびコポリマーの製造の様々な側面が、例えば米国特許第5,534,432号、米国特許第5,663,063号、米国特許第5,798,235号および米国特許第7,202,064号などに記載されている。
【0004】
例として、細菌Ralstonia eutrophaは、高レベルのポリヒドロキシアルカノアート(PHA)生体プラスチックを蓄積することでよく知られている。この生物の野生型は、典型的にはポリヒドロキシブチラートのホモポリマーを産生する。このポリマーは、β−ケトチオラーゼ(PhaA)、アセトアセチル−CoAリダクターゼ(PhaB)およびポリヒドロキシアルカノアートシンターゼ(PhaC)の作用によりアセチル−CoAから産生される。PHBはもろく、その分解温度に近い融点を有するため、有用な生体プラスチックではない。
【0005】
3−ヒドロキシへキサノアートモノマーをポリマー鎖に導入することで、PHBよりも頑強で低い融点を有する材料、したがってPHBよりプラスチックの性能を増大させ、PHBより多くの適用に適するようにしたものが獲得されることが、以前に実証された。
【0006】
米国特許第7,235,621号には、非常に特定の条件下における3−ヒドロキシブチラートと3−ヒドロキシへキサのアートとのコポリマーの製造が記載されている。米国特許第7,235,621号には、高いラウリン酸含有量を有する特定の植物油が、コポリマーを産生する微生物の炭素給源として要求されることが記載されている。これらの要求される油は、ヤシ、ダイズおよびナタネなどのより普通の油中に見出されるものより短鎖の脂肪酸を有している。かかる制限的な炭素給源の要求があっても、米国特許第7,235,621号に開示されたコポリマー中の3−ヒドロキシヘキサノアートの最高量は、13.8mol%である。
【0007】
米国特許公開公報2009/0130731号は、3−ヒドロキシブチラートと3−ヒドロキシへキサのアートとのコポリマーの、PHAシンターゼ遺伝子(phaC)および3−ケトアシル−ACPリダクターゼ遺伝子(fabG)を組換えにより発現する細菌中での製造を記載する。しかしながら、米国特許公開公報2009/0130731号に開示されたコポリマー中の3−ヒドロキシヘキサノアートの最高量は、4mol%である。
【発明の概要】
【0008】
本発明は、高い中鎖長モノマー含有量を有するポリヒドロキシアルカノアートコポリマーの、組換え遺伝子発現を通じた製造に関する。
いくつかの側面において、任意の植物油を炭素給源として使用して、少なくとも約4mol%または5重量%の中鎖長モノマー含有量を有するポリヒドロキシアルカノアートコポリマーを産生する細胞または生物が提供される。いくつかの態様において、細胞または生物は、任意の植物油を炭素源として使用して、少なくとも約4mol%または5重量%のHHx含有量を有する3−ヒドロキシルブチラートと3−ヒドロキシルヘキサノアートとのコポリマー(ポリ(HB−co−HHx))を産生する。いくつかの態様において、細胞中の3−ヒドロキシルブチラートの正常な合成は破壊されている。いくつかの態様において、アセトアセチル−CoAリダクターゼをコードする遺伝子は欠損している。いくつかの態様において、細胞が、Ralstonia eutropha細胞であり、1または2以上のphaB1、phaB2およびphaB3遺伝子は破壊されている。いくつかの態様において、phaB3遺伝子は破壊されている。
【0009】
いくつかの態様において、細胞は、非内在性のPHAシンターゼ遺伝子を組換えにより発現する。いくつかの態様において、非内在性PHAシンターゼ遺伝子は、Aeromonas caviaePHAシンターゼ遺伝子またはRhodococcus aetherivoransPHAシンターゼ遺伝子である。いくつかの態様において、Rhodococcus aetherivoransPHAシンターゼ遺伝子は、配列番号4をコードするRhodococcus aetherivorans I24 D12 PHAシンターゼ遺伝子または配列番号2をコードするRhodococcus aetherivorans I24 C09 PHAシンターゼ遺伝子である。いくつかの態様において、Rhodococcus aetherivorans I24 D12 PHAシンターゼ遺伝子は、配列番号3を含むかまたは配列番号3からなる、および/あるいはRhodococcus aetherivorans I24 C09 PHAシンターゼ遺伝子は、配列番号1もしくは配列番号1の開始コドンがTTGからATGに変えられたものを含むかまたは配列番号1もしくは配列番号1の開始コドンがTTGからATGに変えられたものからなる。
【0010】
いくつかの態様において、細胞は、エノイル−CoAヒドラターゼ遺伝子を組換えにより発現する。いくつかの態様において、エノイル−CoAヒドラターゼ遺伝子は、Aeromonas caviaeエノイル−CoAヒドラターゼ遺伝子またはPseudomonas aeruginosaエノイル−CoAヒドラターゼ遺伝子である。いくつかの態様において、Pseudomonas aeruginosaエノイル−CoAヒドラターゼ遺伝子は、Pseudomonas aeruginosa phaJ1遺伝子(遺伝子PA3302)またはPseudomonas aeruginosa phaJ2遺伝子(遺伝子PA1018)である。
【0011】
いくつかの態様において、非内在性PHAシンターゼ遺伝子および/またはエノイル−CoAヒドラターゼ遺伝子は、増幅される。いくつかの態様において、モノマー含有量は、少なくとも約5mol%、少なくとも約6mol%、少なくとも約7mol%、少なくとも約10mol%、少なくとも約15mol%、または少なくとも約20mol%、またはそれ以上である
【0012】
いくつかの態様において、モノマー含有量は、少なくとも約6重量%、少なくとも約8重量%、少なくとも約10重量%、少なくとも約15重量%、少なくとも約10重量%、または少なくとも約25重量%、またはそれ以上である。
【0013】
いくつかの態様において、細胞または生物は、細菌細胞、真菌細胞(酵母細胞を含む)、植物細胞、昆虫細胞または動物細胞である。いくつかの態様において、細胞は、Ralstonia菌種、Aeromonas菌種、Rhizobium菌種、Alcaligenis菌種またはPseudomonas菌種の細胞である。いくつかの態様において、細胞は、Ralstonia eutropha、Aeromonas caviae、Rhizobium japonicum、Alcaligenis eutrophusまたはPseudomonas oleovorans細胞である。いくつかの好ましい態様において、細胞は、Ralstonia eutropha細胞である。
【0014】
いくつかの態様において、非内在性PHAシンターゼ遺伝子および/またはエノイル−CoAヒドラターゼ遺伝子は、プラスミドから発現される。
いくつかの態様において、非内在性PHAシンターゼ遺伝子および/またはエノイル−CoAヒドラターゼ遺伝子は、細胞のゲノムに組み込まれている。
【0015】
他の側面において、高い中鎖長モノマー含有量を有するポリヒドロキシアルカノアートコポリマーを製造する方法が提供される。本方法は、前述の細胞または生物を培養し、少なくとも約4mol%または5重量%の中鎖長モノマー含有量を有するコポリマーを製造することを含む。いくつかの態様において、本方法は、前述の細胞を培養し、少なくとも約4mol%または5重量%のHHx含有量を有する(ポリ(HB−co−HHx))を製造することを含み、ここで3−ヒドロキシルブチラートと3−ヒドロキシルヘキサノアートとのコポリマー(ポリ(HB−co−HHx))が製造される。いくつかの態様において、本方法は、コポリマーを細胞から回収することをさらに含む。
【0016】
いくつかの態様において、製造されるコポリマーの量は、細胞の乾燥重量の少なくとも約20%、細胞の乾燥重量の少なくとも約30%、細胞の乾燥重量の少なくとも約40%、細胞の乾燥重量の少なくとも約50%、細胞の乾燥重量の少なくとも約60%、またはそれ以上である。
【0017】
他の側面において、少なくとも約4mol%または5重量%の中鎖長モノマー含有量を有するポリヒドロキシルアルカノアートコポリマーを産生する細胞を製造する方法が提供される。本方法は、細胞中に少なくとも1つのRhodococcus aetherivoransPHAシンターゼ遺伝子を組み換えにより発現することを含む。いくつかの態様において、細胞は、少なくとも約4mol%または5重量%のHHx含有量を有する3−ヒドロキシルブチラートと3−ヒドロキシルヘキサノアートとのコポリマー(ポリ(HB−co−HHx))を産生する。いくつかの態様において、Rhodococcus aetherivoransPHAシンターゼ遺伝子は、配列番号4をコードするRhodococcus aetherivorans I24 D12 PHAシンターゼ遺伝子または配列番号2をコードするRhodococcus aetherivorans I24 C09 PHAシンターゼ遺伝子である。いくつかの態様において、Rhodococcus aetherivorans I24 D12 PHAシンターゼ遺伝子は、配列番号3を含むかまたは配列番号3からなる、および/あるいはRhodococcus aetherivorans I24 C09 PHAシンターゼ遺伝子は、配列番号1もしくは配列番号1の開始コドンがTTGからATGに変えられたものを含むかまたは配列番号1もしくは配列番号1の開始コドンがTTGからATGに変えられたものからなる。
【0018】
いくつかの態様において、本方法は、エノイル−CoAヒドラターゼ遺伝子を組換えにより発現することをさらに含む。いくつかの態様において、エノイル−CoAヒドラターゼ遺伝子は、Aeromonas caviaeエノイル−CoAヒドラターゼ遺伝子またはPseudomonas aeruginosaエノイル−CoAヒドラターゼ遺伝子である。いくつかの態様において、Pseudomonas aeruginosaエノイル−CoAヒドラターゼ遺伝子は、Pseudomonas aeruginosa phaJ1遺伝子(遺伝子PA3302)またはPseudomonas aeruginosa phaJ2遺伝子(遺伝子PA1018)である。
【0019】
いくつかの態様において、非内在性PHAシンターゼ遺伝子および/またはエノイル−CoAヒドラターゼ遺伝子は、増幅される。
いくつかの態様において、細胞は、細菌細胞、真菌細胞(酵母細胞を含む)、植物細胞、昆虫細胞または動物細胞である。いくつかの態様において、細胞は、Ralstonia菌種、Aeromonas菌種、Rhizobium菌種、Alcaligenis菌種またはPseudomonas菌種の細胞である。いくつかの態様において、細胞は、Ralstonia eutropha、Aeromonas caviae、Rhizobium japonicum、Alcaligenis eutrophusまたはPseudomonas oleovoransの細胞である。いくつかの好ましい態様において、細胞は、Ralstonia eutrophaの細胞である。
【0020】
いくつかの態様において、非内在性PHAシンターゼ遺伝子および/またはエノイル−CoAヒドラターゼ遺伝子は、プラスミドから発現される。
いくつかの態様において、非内在性PHAシンターゼ遺伝子および/またはエノイル−CoAヒドラターゼ遺伝子は、細胞のゲノムに組み込まれている。
【0021】
いくつかの側面において、1または2以上の、少なくとも約4mol%または5重量%の中鎖長モノマー含有量を有するコポリマーを製造する方法が提供される。本方法は、前述の方法により細胞を製造すること、および該細胞の集団を培養することを含む。いくつかの態様において、1または2以上のコポリマーは、3−ヒドロキシルブチラートと3−ヒドロキシルヘキサノアートとのコポリマー(ポリ(HB−co−HHx))である。いくつかの態様において、本方法は、細胞集団から1または2以上のコポリマーを収集することをさらに含む。
【0022】
いくつかの態様において、モノマー含有量は、少なくとも約5mol%、少なくとも約6mol%、少なくとも約7mol%、少なくとも約10mol%、少なくとも約15mol%、少なくとも約20mol%、またはそれ以上である。
いくつかの態様において、モノマー含有量は、少なくとも約6重量%、少なくとも約8重量%、少なくとも約10重量%、少なくとも約15重量%、少なくとも約20重量%、少なくとも約25重量%、またはそれ以上である。
【0023】
いくつかの態様において、製造されるコポリマーの量は、細胞の乾燥重量の少なくとも約20%、細胞の乾燥重量の少なくとも約30%、細胞の乾燥重量の少なくとも約40%、細胞の乾燥重量の少なくとも約50%、細胞の乾燥重量の少なくとも約60%、またはそれ以上である。
【0024】
他の側面において、配列番号2または配列番号4をコードする、単離された核酸分子が提供される。いくつかの態様において、該単離された核酸分子は、配列番号1または配列番号3に記載のヌクレオチド配列を含む。いくつかの態様において、単離された核酸分子は、配列番号1または配列番号3に記載のヌクレオチド配列と、少なくとも80%の同一性、少なくとも90%の同一性、少なくとも95%の同一性、少なくとも98%の同一性、またはそれ以上の同一性を有する。
【0025】
いくつかの態様において、前述の核酸分子によりコードされる、単離されたポリペプチドが提供される。
いくつかの態様において、前述の単離された核酸分子を含むベクターが提供される。
いくつかの態様において、前述の単離された核酸分子を組換えにより発現する細胞が提供される。いくつかの態様において、核酸分子は、ベクターから発現される。いくつかの態様において、核酸分子は、細胞のゲノムに組み込まれている。
本発明のこれらのおよび他の側面、ならびにそれらの様々な態様は、図面および本発明の詳細な説明を参照することにより、より明白となるであろう。
添付の図面はスケール通りに描かれていることを意図していない。本図面において、様々な図中に描かれている各同一のまたはほぼ同一の構成要素は、同様の番号で表される。明瞭さのため、全ての構成要素が全ての図面において名前を付けられているわけではないだろう。図面において:
【図面の簡単な説明】
【0026】
【図1】図1はPHAポリマーの構造および特性を示す。上、ポリ(HB);下、ポリ(HB−co−HHx)
【0027】
【図2】図2はPhaA、PhaBおよびPhaCの、PHAコポリマー合成の経路への関与を示す。
【図3】図3は、脂肪酸β−酸化とPHAコポリマー合成の経路との相互作用、およびモノマーが脂肪酸代謝の副産物として作られることを概略的に示す。
【図4】図4は、PHAコポリマー合成の経路におけるPhaAおよびPhaBのブロッキングの効果、およびモノマーが脂肪酸代謝の副産物として作られることを概略的に示す。
【0028】
【図5】図5は、phaB1、phaB2および/またはphaB3欠損を有する数菌株による、フルクトース限定培地中におけるPHB合成を示す。野生型、および1変異、2変異および3変異株の結果が示されている。
【図6】図6は、phaB1、phaB2および/またはphaB3欠損を有する数菌株における、フルクトース限定培地中におけるNADPHとのリダクターゼ活性を示す。野生型、および1変異、2変異および3変異株の結果が示されている。
【0029】
【図7】図7は、phaB1、phaB2および/またはphaB3欠損を有する数菌株における、フルクトース限定培地中におけるNADHとのリダクターゼ活性を示す。野生型、および1変異、2変異および3変異株の結果が示されている。
【図8】図8は、phaB欠損を有する菌株における相補的リダクターゼ変異の効果を示す。phaB1、phaB2およびphaB3遺伝子は独立して菌株Re2115(ΔphaB123)に戻された。別の株は、Re2115に戻したfabGを有していた。野生型、変異型および相補化された菌株におけるPHB合成の結果が示されている。
【0030】
【図9】図9は、図8に記載されている菌株のPhaA活性を示す。
【図10】図10は、図8に記載されている菌株によって産生されたPHBポリマーの分子量を示す。
【図11】図11は、PhaBの基質特異性を示す。
【図12】図12は、PhaJの基質特異性を示す。
【図13】図13は、用いたPHAシンターゼ、炭素源、PHA含有量、およびmol%HHxを示し、文献からの数菌株を列挙する。
【0031】
【図14】図14は、遺伝子型(用いたPHAシンターゼを含む)、PHA含有量(細胞乾燥重量の%として)、および重量%HHxを示して、構築された菌株およびかかる菌株からの結果を列挙する。
【図15】図15は、構築されたPHAオペロン(phaCD12−phaA−phaJ1Pa)が増幅されおよびプラスミドにクローニングされ、それが菌株Re2133に形質転換されるという菌株構築の手法を示す。
【発明を実施するための形態】
【0032】
発明の詳細な説明
本発明は、一部において、高い中鎖長モノマー含有量を有するポリヒドロキシアルカノアートコポリマーなどのバイオプラスチックを製造する方法、および細菌発酵および他の細胞および生物においてバイオプラスチックの産生を増大させる方法に関する。コポリマー化してポリヒドロキシアルカノアートコポリマーを製造するモノマーは、3−ヒドロキシブチラートおよび炭素鎖長が5以上である3−ヒドロキシアルカン酸(例えば、米国特許第7,341,856号およびそこに引用された参考文献を参照のこと、その開示のそれぞれは、それらの教示として参照により組み込まれる)を含む。この一例として、3−ヒドロキシブチラートと3−ヒドロキシヘキサノアートとのコポリマー(ポリ(HB−co−HHx))および特に高いHHx含有量を有するコポリマーがその物理的特性により有用である。本発明はまた、一部において、菌株が脂肪酸または任意の植物油を炭素源として用いて生育されたとき、ポリ(3−ヒドロキシブチラート−co−3−ヒドロキシヘキサノアート)などの高い中鎖長モノマー含有量を有するポリヒドロキシアルカノアートコポリマーを高度に蓄積することができる、Ralstonia eutropha菌株などの細胞に関する。
【0033】
かかる細胞により作られるコポリマーの3−ヒドロキシルヘキサノアート含有量は、文献に記載されている、植物油から製造される任意の材料に匹敵するかまたはそれを超えるものである。対照的に、米国特許第7,235,621号に記載の方法は、許容可能なHHx含有量のコポリマーを製造するために、構成脂肪酸中にラウリン酸を有する特定の植物油の使用が求められる。それであっても、米国特許第7,235,621号に記載の方法を用いて製造されるコポリマーのHHx含有量は、本明細書に記載される方法を用いて製造されたコポリマーのHHx含有量よりも劣るものである。
【0034】
以下の特徴は、独立しておよび組み合わせとして、本発明に含まれる。
いくつかの態様において、細胞中のPHBの正常な合成が破壊されている。例えば、下記実施例で示されるとおり、PHBは、Ralstonia eutrophaにおいてアセトアセチル−CoAリダクターゼをコードする遺伝子を欠損させることにより、破壊されている。本発明の特定の態様において、phaB3が破壊されており;phaB3は、従前文献において特徴付けられておらず、この特定の目的に用いられてこなかった遺伝子である。しかし本発明はかかる態様に限定されず、したがって、アセトアセチル−CoAリダクターゼの発現を低減させることを含む、正常なPHB生合成を破壊する他の方法も含まれる。
【0035】
いくつかの態様において、内在性PHAシンターゼ遺伝子が破壊され、新たなPHAシンターゼに置き換えられている。しかし本発明はかかる態様に限定されず、したがって、内在性PHAシンターゼの発現を低減させることを含む、正常な内在性PHAシンターゼ活性を破壊する他の方法を含む。例えば、下記実施例に示されるように、Ralstonia eutropha中の野生型PHAシンターゼ遺伝子が欠損され、新たなシンターゼ遺伝子が菌株に付与された。新たなPHAシンターゼは、高い割合の3−ヒドロキシヘキサノアートモノマーを組み込むことができる。いくつかの態様において、新たなシンターゼ遺伝子はRhodococcus aetherivorans I24由来(本明細書において「RhodococcusCO9シンターゼ」(配列番号1および2)または「RhodococcusD12シンターゼ」(配列番号3および4)ともいう)である。しかし本発明はかかる態様に限定されず、したがって、高い割合の中鎖長モノマー、特に3−ヒドロキシヘキサノアートモノマーを組み込む類似の能力を有する他のPHAシンターゼ遺伝子もまた、類似のやり方で用いることができる。
【0036】
いくつかの態様において、特定のエノイル−CoAヒドラターゼ遺伝子が細胞に導入され、コポリマーに組み込まれるモノマーの産生を増大する。例えば、下記実施例において示されるとおり、Pserudomonas aeruginosa PAO1からのphaJ1(遺伝子PA3302)と呼ばれる(R)−特異的エノイルCoAヒドラターゼ遺伝子が、Ralstonia eutropha菌株に導入された。酵素PhaJ1は、3−ヒドロキシブチリル−CoAおよび3−ヒドロキシヘキサノイル−CoAモノマーを産生し、これらはその後PHAシンターゼによりポリマー化される。Pseudomonas aeruginosa PAO1のphaJ2遺伝子(遺伝子PA1018)もまた用いることができる。Pseudomonas aeruginosa PAO1ゲノムのGenBank受託番号は、AE004091である。しかし本発明はかかる態様に限定されず、したがって、3−ヒドロキシブチリル−CoAおよび3−ヒドロキシヘキサノイル−CoAモノマーを産生する類似の能力を有する1または2以上の他のエノイル−CoA遺伝子もまた、類似のやり方で用いることができる。
【0037】
いくつかの態様において、本明細書に記載された遺伝子のコピー数を調節して、細胞によって蓄積されるPHAの量を変化させることができる。例えば、下記実施例において示されるとおり、最初にRhodococcus D12シンターゼおよびPseudomonas aeruginosaphaJ1遺伝子をRalstonia eutrophaゲノムに組み込んだ。その後遺伝子のコピー数を、遺伝子をプラスミドにクローニングすることにより増大させ、該プラスミドを、アセトアセチル−CoAリダクターゼ遺伝子および天然のPHAシンターゼ遺伝子がゲノムから欠損したRalstonia eutropha菌株に導入した。プラスミドを保有する菌株は、遺伝子がゲノムにのみ存在する菌株よりも顕著に多くのポリマーを産生した。しかし本発明はかかる態様に限定されず、したがって、遺伝子のコピー数を調節(好ましくは増大)する他の方法も含まれる。
【0038】
本発明の側面は、高いHHx含有量を有する3−ヒドロキシルブチラートと3−ヒドロキシルヘキサノアートとのコポリマー(ポリ(HB−co−HHx))などの高い中鎖長モノマー含有量を有するポリヒドロキシルアルカノアートコポリマーを、細胞中での組換え遺伝子発現を通じて製造する方法および組成物に関する。1つの態様において、本明細書に記載されるのは、植物油を炭素源とした、非内在性PHAシンターゼおよびエノイル−CoAヒドラターゼ遺伝子を組み換えにより発現し、アセトアセチル−CoAリダクターゼをコードする遺伝子が欠損している細胞における、5重量%よりも多い(4mol%よりも多い)HHx含有量を有するポリ(HB−co−HHx)の製造である。このシステムは、広く様々な適用を有する分子である、高いHHx含有量を有するポリ(HB−co−HHx)を製造する効果的で新たな方法を表す。
【0039】
本発明の側面によれば、1または2以上の酵素を組換えにより発現する細胞および、HHx含有量を有する(ポリ(HB−co−HHx))などの、高い中鎖長モノマー含有量を有するポリヒドロキシルアルカノアートコポリマーの製造におけるかかる細胞の使用が提供される。当然のことながら、PHAシンターゼ遺伝子を含む、かかる酵素をコードする遺伝子を様々な給源から得ることができる。当業者ならば気づくであろうように、これらの酵素の相同遺伝子を他の種から得ることができ、例えばNCBIのインターネットサイト(www.ncbi.nlm.nih.gov)において利用できるタンパク質BLASTN調査などの相同性調査により同定することができる。かかる遺伝子は、その特定の遺伝子を含む任意の種のDNAからのDNAからPCR増幅することができる。いくつかの態様において、遺伝子配列は合成であるおよび/または導入される細胞に対してコドン最適化されている。本明細書に記載されるような、酵素をコードする遺伝子を得るための任意の方法が、本発明と適合する。
【0040】
タンパク質発現の最適化は、例えば細菌細胞における発現のためのコドン最適化を通してなどにより、酵素をコードする遺伝子が細胞に導入する前に修飾されるいくつかの態様において求められ得る。様々な生物のコドン使用頻度はCodon Usage Database(www.kazusa.or.jp/codon/)においてアクセスできる。
本明細書に記載された方法、酵素、細胞および生物は、高い中鎖長モノマー含有量を有するポリヒドロキシルアルカノアートコポリマーの製造を提供する。モノマー含有量はしばしばモルパーセント(mol%)で表現される。いくつかの態様において、コポリマーは、少なくとも4mol%、5mol%、6mol%、7mol%、8mol%、9mol%、10mol%、11mol%、12mol%、13mol%、14mol%、15mol%、16mol%、17mol%、18mol%、19mol%、20mol%、21mol%、22mol%、23mol%、24mol%、25mol%、26mol%、27mol%、28mol%、29mol%、30mol%、31mol%、32mol%、33mol%、34mol%、35mol%、またはそれ以上の、3−ヒドロキシヘキサノアートなどの中鎖長モノマー含有量で製造される。かかる中鎖長モノマー含有量を有するコポリマーは、例えば任意の脂肪酸または油(例えば植物油)を炭素源として用いて製造することができる。
【0041】
モノマー含有量を表現する別の方法は、重量パーセント(重量%)である。モルパーセントは、重量パーセントの値に0.8を掛けることにより、重量パーセントから概算することができる;同様に、重量パーセントは、モルパーセントの値に1.25を掛けることにより、モルパーセントから概算することができる。したがって、コポリマーは、少なくとも5重量%、6重量%、7重量%、8重量%、9重量%、10重量%、11重量%、12重量%、13重量%、14重量%、15重量%、16重量%、17重量%、18重量%、19重量%、20重量%、21重量%、22重量%、23重量%、24重量%、25重量%、26重量%、27重量%、28重量%、29重量%、30重量%、31重量%、32重量%、33重量%、34重量%、35重量%、またはそれ以上の、3−ヒドロキシヘキサノアートなどの中鎖長モノマー含有量で製造される。かかる中鎖長モノマー含有量を有するコポリマーは、例えば任意の脂肪酸または油(例えば植物油)を生物または細胞の炭素源として用いて製造することができる。
【0042】
いくつかの態様において、細胞または生物により製造されるコポリマーの量は、細胞または生物の乾燥重量(細胞乾燥重量)の少なくとも20%、22%、24%、26%、28%、30%、35%、40%、45%、50%、55%、60%、またはそれ以上である。
上述したように米国特許第7,235,621号に記載の方法には、許容可能なHHx含有量のコポリマーを製造するために、構成脂肪酸中にある量のラウリン酸を有する特定の植物油の使用が求められる。米国特許第7,235,621号で報告されているコポリマーの最大HHx含有量は、13.8%である。本明細書に記載された方法、酵素、細胞および生物は、米国特許第7,235,621号で要求されるようなラウリン酸含有量を構成脂肪酸中に有する油または脂肪である炭素源を使用することなく、HHx含有量がより多いコポリマーを製造するのに向いている。
【0043】
有用なポリマーの分子量は、約10,000と400万ダルトンとの間、および好ましくは50,000ダルトンと150万ダルトンとの間など、10,000ダルトンより多いものを含む。
本発明によれば、高い中鎖長モノマー含有量を有するポリヒドロキシルアルカノアートコポリマーが製造される。モノマー単位は当該技術分野において知られており、ヒドロキシブチラート、ヒドロキシバレラート、ヒドロキシヘキサノアート、ヒドロキシヘプタノアート、ヒドロキシオクタノアート、ヒドロキシノナノアート、ヒドロキシデカノアート、ヒドロキシウンデカノアート、およびヒドロキシドデカノアート単位を含む。いくつかの態様において、製造されるコポリマーは、ポリ(3−ヒドロキシブチラート−co−3−ヒドロキシヘキサノアート)など、ヒドロキシブチラートおよびヒドロキシヘキサノアートモノマーを含む。
【0044】
本発明は、原核細胞および真核細胞を含む、PHAシンターゼによってポリマー化されるモノマーを作るための遺伝子を発現する任意のタイプの細胞を包含する。いくつかの態様において、細胞は最近細胞である。いくつかの態様において、細菌細胞はRalstonia属、Aeromonas属、Rhizobium属、Alcaligenes属またはPseudomonas属細胞である。いくつかの態様において、細胞は、Ralstonia eutropha、Aeromonas caviae、Rhizobium japonicum、Alcaligenes eutrophusまたはPseudomonas oleovorans細胞である。1つの好ましい態様において、細胞は、例えばSaccharomyces属、Schizosaccharomyces属、Pichia属、Phaffia属、Hansenula属、Kluyveromyces属、Candida属、Talaromyces属、Brettanomyces属、Schwanniomyces属、Pachysolen属、Debaryomyces属、Yarrowia属または工業的倍数体酵母株などの酵母細胞などの真菌細胞である。コポリマーの製造に有用な酵母種および株の例は、米国特許第7,083,972号に記載されており、その開示はその教示についての参照により組み込まれる。真菌の他の例は、Aspergillus属、Pennicilium属、Fusarium属、Rhizopus属、Acremonium属、Neurospora属、Sordaria属、Magnaporthe属、Allomyces属、Ustilago属、Botrytis属、およびTrichoderma属を含む。他の態様において、細胞は藻類細胞、哺乳類細胞または植物細胞である。
【0045】
当然のことながら、本発明に適合するいくつかの細胞は、本発明に付随する1または2以上の遺伝子の内在性コピーを、組換えコピーと共に発現してもよい。いくつかの態様において、細胞が本発明に付随する1または2以上の遺伝子の内在性コピーを有する場合、本方法は内在的に発現する遺伝子の組換えコピーを付加することを必ずしも要求しない。いくつかの態様において、細胞は、本明細書に記載の経路からの1または2以上の酵素を内在的に発現してもよく、本明細書に記載の経路からの1または2以上の酵素を組換えにより発現してもよい。
【0046】
当然のことながら、本発明に適合するいくつかの細胞は、内在性生化学的経路を、1または2以上の組換え遺伝子のその経路、類似の経路、または相補的な経路とともに発現してもよい。例えば、R. eutrophaはモノマー産生のための遺伝子を発現する。実施例において示されているように、これらの細胞においてphaJが付加的に発現されて、PHAシンターゼによってポリマー化されるモノマー産生が増強された。
組織優先型プロモーター(tissue-preferred promoter)の使用を含む、モノマーを製造および/またはポリマー化するための関連する酵素の植物における発現によって、PHAポリマーおよびコポリマーを製造する方法は、例えば米国特許第5,534,432号および米国特許第7,341,856号に記載されており、いずれもこの教示について参照により本明細書に組み込まれる。
【0047】
興味のある生物または細胞のゲノムに存在する遺伝子からの発現および/または、例えばプラスミドなど、ベクターなどの1または2以上の染色体外核酸にある遺伝子からの発現を含む、生物および細胞における遺伝子の発現は当該技術分野においてよく知られている。修飾ゲノム、染色体外核酸およびかかる修飾ゲノムまたは染色体外核酸を含む生物または細胞を構築するための方法、材料および技術は、当該技術分野において周知であり、そのうちいくつかの方法、材料および技術は本明細書の他の場所に記載されている。
【0048】
いくつかの態様において、ポリ(HB−co−HHx)コポリマーなどの、高い中鎖長モノマー含有量を有する1または2以上のポリヒドロキシルアルカノアートコポリマーの製造のために最適化された細胞の使用が有利であり得る。例えば、生化学的経路の1または2以上の構成を変異して競合する経路を除去し、ポリ(HB−co−HHx)コポリマーなどの、高い中鎖長モノマー含有量を有する1または2以上のポリヒドロキシルアルカノアートコポリマーをより多く産生させることが最適であり得る。例えば、いくつかの態様において、3HB−CoA合成に寄与する1または2以上の遺伝子を辺または欠損させることにより、3HB−CoA合成を低減させる。いくつかの態様において、このことはアセトアセチル−CoAリダクターゼ活性の低減により達成することができる。例えばR. eutrophaにおいて、アセトアセチル−CoAリダクターゼ活性(PhaB)は、例えば1または2以上のphaB遺伝子を変異させることまたは(一部をまたは完全に)欠損させることによって、phaB発現を低減させることにより、低減させることができる。本明細書で例示されるように、これはphaB1、phaB2およびphaB3遺伝子をR. eutrophaゲノムから欠損させることにより達成できた。
【0049】
いくつかの態様において、ポリ(HB−co−HHx)コポリマーなどの、高い中鎖長モノマー含有量を有する1または2以上のポリヒドロキシルアルカノアートコポリマーの増強された産生に導く変異のスクリーニングが、ランダム変異原性スクリーニングまたは既知の変異のスクリーニングを通して実施されてもよい。いくつかの態様において、ポリ(HB−co−HHx)コポリマーなどの、高い中鎖長モノマー含有量を有する1または2以上のポリヒドロキシルアルカノアートコポリマーの増大した産生のためのこれらの断片を有する細胞または生物のスクリーニングを通して、ポリ(HB−co−HHx)コポリマーなどの、高い中鎖長モノマー含有量を有する1または2以上のポリヒドロキシルアルカノアートコポリマーの産生の増加に導くゲノム領域同定のためにゲノム断片のショットガンクローニングを用いることができる。いくつかの場合において、同一の細胞または生物中で1または2以上の変異が組み合わされてもよい。
【0050】
本明細書に記載された様々な細胞を生育するための培地は、当業者に周知である。例えば、米国特許第5,534,432号、これらの教示は参照として本明細書に組み込まれる、は、細菌細胞の生育に適したかかる培地の例を提供する。
当該技術分野における通常の技術を有するものには理解できるように、1または2以上のヒドロキシ酸およびそのコポリマーの産生のための最適な培養条件は、細胞のタイプ、生育培地および生育条件を含む多くの要因に影響され得る。培養温度は生物または細胞が生育可能な温度であり、好ましくは20℃から40℃である。例えば摂氏20、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35、36、37、38、39または40度、あるいはその間の任意の値であってよい。培養時間は特に限定されないが、約1から10日であってよい。
【0051】
ポリ(HB−co−HHx)コポリマーなどの、高い中鎖長モノマー含有量を有する1または2以上のポリヒドロキシルアルカノアートコポリマーの産生を最適化するために日常的な実験を通して変化し得る他の限定されない要因は、用いられる特定の炭素源、培養培地のタイプ、培養培地のpH、およびポリ(HB−co−HHx)を収穫する前に細胞を培養する時間量を含む。いくつかの態様において、3〜4日など、細胞を数日間培養した後に最適な産生が達成される。しかし当然のことながら、日常的な実験で、上記パラメータおよび他の類似のパラメータを変化および最適化できる。
【0052】
生物または細胞は、コポリマーの製造を可能にする炭素源を含む培養培地中で培養される。いくつかの態様において、任意の植物油、脂肪酸または脂肪酸誘導体あるいはその組み合わせなどの、任意の油および/または脂肪酸が炭素源として用いられる。植物油の例は、パーム油、大豆油、ナタネ油、コーン油、綿実油、ピーナッツ油、ココナッツ油、およびベニバナ油を含み、さらなる油または脂肪酸は当該技術分野において周知である。コポリマーの最適な製造のための炭素源組成物は、用いられる生物または細胞の特定の株に依存し得る。培養培地の他の構成成分は、当業者には知られているであろうように、いくつかの態様において窒素源、無機塩、(抗生物質などの)選択的成分、または他の栄養源などの様々な組み合わせを含む。
【0053】
本発明に関連する細胞を生育するのに用いる液体培養は、当該技術分野において知られ、および用いられる任意の培養容器に格納されることができる。いくつかの態様において、大量の、本発明に関連するポリ(HB−co−HHx)コポリマーなどの高い中鎖長モノマー含有量を有する1または2以上のポリヒドロキシルアルカノアートコポリマーの製造に、攪拌槽型反応器などの通気反応容器内での大規模製造を用いることができる。
【0054】
PHAポリマーおよびコポリマーを継続的に製造する方法もまた当業者に知られており、かかる方法は例えば米国特許第5,534,432号、これはこの教示について参照により本明細書に組み込まれる、などに記載されている。
PHAコポリマーは、当該技術分野において知られた方法で製造された生物、細胞または培養物から単離することができる。この一例は下記の実施例に示されているものであり、ポリマーは凍結乾燥された細胞から、クロロホルム中48時間で抽出されている。例えば、米国特許第5,942,597号、第5,918,747号、第5,899,339号、第5,849,854号および第5,821,299号、欧州特許公開公報第859858号、国際公開公報第97/07229号、第WO97/07230号および第WO97/15681号などもまた参照のこと、これらのそれぞれはこれらの教示について本明細書に参照として組み込まれる。
【0055】
1つの非限定的な例として、米国特許出願2009/0130731号に記載された方法を採用し得る。培養後、細菌細胞を遠心分離機などにより培養培地から分離し、細菌細胞を蒸留水およびメタノールなどで洗浄し、乾燥させる。乾燥させた細菌細胞から、クロロホルムなどの有機溶媒を用いてポリエステルを抽出する。細菌細胞構成成分は、このポリエステルを含む有機溶媒溶液からろ過などにより除去され、メタノールまたはヘキサンなどの貧溶媒をろ液に添加し、ポリエステルを沈殿させる。さらに、ろ過または遠心分離により上清を除去し、乾燥させる。他の方法は当業者に周知であろう。
【0056】
本発明によって製造されたコポリマーは、当該技術分野において知られた任意の多くの用途に用いることができる。例えば、米国特許第7,455,999号には、PHAポリマーの大量の用途およびポリマーの物性が「組成物の適用」と標題されたセクションおよびそこに引用された参照に記載されており、これらの開示のそれぞれは、その教示について参照として組み込まれる。
【0057】
いくつかの態様において、本発明は単離されたまたは実質的に精製されたPHAシンターゼ核酸またはポリペプチド、かかる核酸またはポリペプチドを含むまたは発現する構築物およびベクター、およびかかる核酸、ポリペプチド、構築物またはベクターを含む細胞または生物を包含する。本明細書に開示されるように、高濃度の3−ヒドロキシヘキサノアートモノマーを組み込むことができるRhodococcus aetherivorans I24からのPHAシンターゼが同定され、分子クローニング手法により単離された。単離されたPHAシンターゼはまた、本明細書において「Rhodococcus CO2シンターゼ」(配列番号1および2)または「Rhodococcus D12シンターゼ」(配列番号3および4)ともいう。
【0058】
「単離された」または「精製された」核酸分子またはタンパク質、またはその生物学的に活性な一部分は、実質的に他の細胞物質または組換え技術により製造された場合には培養培地を含まず、または化学的に合成された場合は実質的に化学物質前駆体または他の化学物質を含まない。好ましくは、「単離された」核酸は、核酸が由来する生物のゲノムDNAにおいては天然に隣接する核酸配列(好ましくはタンパク質コード配列)(すなわち核酸の5’および3’末端に存在する配列)を含まない。例えば、さまざまな態様において、単離された核酸分子は5kb、4kb、3kb、2kb、1kb、0.5kbまたは0.1kb未満の核酸が由来する生物のゲノムDNAにおいては天然に隣接する核酸配列を含み得る。細胞物質を実質的に含まないポリペプチド(タンパク質ともいう)は、(乾燥重量で)約30%、20%、10%、5%の不純物ポリペプチドを含むポリペプチドの調製物を含む。ポリペプチドまたはその生物学的に活性な一部分が組換えにより製造される場合、好ましくは培養培地が、(乾燥重量で)約30%、20%、10%または5%未満の化学物質前駆体またはポリペプチドではない他の構成物質に相当する。
【0059】
開示された核酸分子の断片および変異体およびそれによりコードされるポリペプチドもまた、本発明に包含される。「断片」により、ヌクレオチド配列の一部分およびそれによりコードされるアミノ酸配列の一部分が意図される。ヌクレオチド配列の断片は、天然ポリペプチドの生物学的活性を維持したポリペプチド断片をコードし得る。代替的に、ハイブリダイゼーションプローブとして有用なヌクレオチド配列の断片は、一般的に天然ポリペプチドの生物学的活性を維持したポリペプチド断片をコードしない。したがって、ヌクレオチド配列の断片は、少なくとも約20ヌクレオチド、約50ヌクレオチド、約100ヌクレオチド、および本明細書に開示のポリペプチドをコードする全長ヌクレオチドまでの範囲であってよい。
【0060】
ポリペプチドの生物学的に活性な一部分をコードする本発明のヌクレオチド配列の断片は、少なくとも15、25、30、50、100、150、200、250、300、350、400、450、500または550の隣接したアミノ酸または全長ポリペプチド中に存在する全アミノ酸数(例えば配列番号2については562アミノ酸または配列番号1については561アミノ酸)までをコードすることができる。ポリペプチドの生物学的に活性な一部分は、本明細書に開示されたヌクレオチド配列の1つの一部分を単離し、(例えばin vitroの組換え発現により)ポリペプチドのコードされた一部分を発現させ、ポリペプチドのコードされた一部分の活性を評価することにより調製できる。本明細書に開示のヌクレオチド配列の断片である核酸分子(ポリペプチドの生物学的に活性な断片をコードするかどうかにかかわらず)は、少なくとも15、20、30、45、60、75、100、150、200、250、300、350、400、450、500、550、600、650、700、800、900、1000、1100、1200、1300、1400、1500または1600ヌクレオチド、または本明細書に開示の全長ヌクレオチド配列に存在するヌクレオチド数(例えば配列番号1については1689ヌクレオチドまたは配列番号3については1686ヌクレオチド)までを含む。
【0061】
「変異体」により、本明細書に記載の配列と実質的に同様の配列を意図する。ヌクレオチド配列について、保存的変異体は、遺伝情報の縮退により、本発明のコポリマー合成に関与するポリペプチドまたは他の酵素の1つのアミノ酸配列をコードするそれらの配列を含む。これらのような天然に存在する対立遺伝子多型は、例えばポリメラーゼ連鎖反応(PCR)および以下に概説するハイブリダイゼーション技術などの周知の分子生物学の技術を用いて同定することができる。変異体ヌクレオチド配列はまた、例えば部位特異的変異導入(site-directed mutagenesis)などにより発生するが依然として本明細書に開示されたポリペプチドをコードするものなど、合成的に誘導されたヌクレオチド配列を含む。一般的に、本発明の特定のヌクレオチド配列の変異体は、本明細書の多の部分で記載された配列アラインメントプログラムにより、初期設定のパラメータを用いて決定された特定のヌクレオチド配列と、少なくとも50%、60%、65%、70%、75%、80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%またはそれ以上の配列同一性を有するだろう。
【0062】
本明細書に記載のヌクレオチド配列は、他の生物、特に他の細菌であるが本明細書に記載された他の生物または細胞も、から対応する配列を単離するのに用いることができる。このやり方において、PCR、ハイブリダイゼーション等の方法を、本明細書に記載の配列とのそれらの配列ホモロジーに基づき、かかる配列の同定に用いることができる。本明細書に記載された全ヌクレオチド配列またはその断片との配列同一性に基づいて単離された配列は、本発明に包含される。かかる配列は、開示された配列のオルソログである配列を含む。「オルソログ」により、種分化の結果、共通の祖先遺伝子に由来し異なる種において見出される遺伝子を意図する。異なる種において見出される遺伝子は、本明細書の他の場所で定義されるように、そのヌクレオチド配列および/またはそのコードするポリペプチド配列が実質的同一性を共有する場合に、オルソログであると考えられる。
【0063】
PCRベースの取り組みにおいて、オリゴヌクレオチドプライマーは、PCR反応において使用して、対応するDNA配列を任意の興味のある生物から抽出したcDNAまたはゲノムDNAから増幅するために設計できる。PCRプライマーの設計およびPCRクローニングのための方法は一般的に当該技術分野に知られており、例えばSambrook et al. (1989) Molecular Cloning: A Laboratory Manual(2d ed., Cold Spring Harbor Laboratory Press, Plainview, N.Y.)などに開示されている。Innis et al., eds.(1990)PCR Protocols: A Guide to Methods and Applications(Academic Press, New York)、Innis and Gelfand, eds.(1995)PCR Strategies(Academic Press, New York)、およびInnis and Gelfand, eds.(1999)PCR Methods Manual(Academic Press, New York)も参照のこと。PCRの既知の方法は、これに限定するものではないが、ペアプライマー、ネステッドプライマー、単一の特異的プライマー、縮退プライマー、遺伝子特異的プライマーベクター特異的プライマー部分的ミスマッチプライマーなどを用いる方法を含む。
【0064】
ハイブリダイゼーション技術において、基地のヌクレオチド配列の全部または一部が、選ばれた生物または細胞からクローンされたゲノムDNA断片またはcDNA断片の集団(ゲノムまたはcDNAライブラリーなど)の中に存在する他の対応するヌクレオチド配列と選択的にハイブリダイズするプローブとして用いられる。ハイブリダイゼーションプローブは、ゲノムDNA断片、cDNA断片、RNA断片または他のオリゴヌクレオチドであってよく、当該技術分野において周知の任意の検出可能なマーカーで標識されていてよい。したがって、例えば、ハイブリダイゼーションのためのプローブは、本明細書に開示されたヌクレオチド配列またはその縮退物に基づいて、標識合成オリゴヌクレオチドにより作られ得る。ハイブリダイゼーションのためのおよびcDNAおよびゲノムライブラリーの構築のためのプローブを調製するための方法は当該技術分野において知られており、例えばSambrook et al. (1989) Molecular Cloning: A Laboratory Manual(2d ed., Cold Spring Harbor Laboratory Press, Plainview, N.Y.)などに開示されている。
【0065】
かかる配列のハイブリダイゼーションは、ストリンジェントな条件化で行われてよい。「ストリンジェントな条件」または「ストリンジェントなハイブリダイゼーション条件」により、プローブがその標的配列に、他の配列に比べて検出可能に多量な程度(例えば、少なくともバックグラウンドの2倍以上)にハイブリダイズするであろう条件を意図する。ストリンジェントな条件は配列依存であり、異なる状況において異なるであろう。ハイブリダイゼーションおよび/または洗浄条件のストリンジェンシーのコントロールにより、プローブに100%相補的な標的配列を同定できる(相同的プロービング(homologous probing))。代替的に、ストリンジェンシー条件を調整して、配列におけるいくつかのミスマッチを許容し、より低い程度の類似性を検出することもできる(異種性プロービング(heterologous probing))。一般的に、プローブは約1000ヌクレオチド長未満であり、好ましくは500ヌクレオチド長未満である。ストリンジェントなハイブリダイゼーション条件下でのハイブリダイゼーションにおいて用いられるさまざまな技術、方法、条件および組成物は、当該技術分野において周知であり、Tijssen(1993)Laboratory Techniques in Biochemistry and Molecular Biology--Hybridization with Nucleic Acid Probes, Part I, Chapter 2(Elsevier, N.Y.)、およびAusubel et al., eds.(1995)Current Protocols in Molecular Biology, Chapter 2(Greene Publishing and Wiley-Interscience, New York)などの参照文献中に見出すことができる。Sambrook et al. (1989) Molecular Cloning: A Laboratory Manual(2d ed., Cold Spring Harbor Laboratory Press, Plainview, N.Y.)参照のこと。
【0066】
本明細書に開示されたPHAシンターゼ配列にストリンジェントな条件下でハイブリダイズする単離された配列またはその断片は、本発明に包含される。いくつかの態様において、かかる配列は、開示された配列と少なくとも約50%、60%、70%、75%、80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%またはそれ以上の相同性であるだろう。つまり、配列の配列同一性は、開示された配列と少なくとも約50%、60%、70%、75%、80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%またはそれ以上の配列同一性を共有してよい。
【0067】
比較のための配列のアラインメントの方法は、当該技術分野において周知である。任意の2配列間のパーセント配列同一性の決定は、数学的アルゴリズムを用いて達成することができる。かかる数学的アルゴリズムの非限定的な例は、Myers and Miller (1988) CABIOS 4:11-17のアルゴリズム、Smith et al. (1981) Adv. Appl. Math. 2:482のローカルホモロジーアルゴリズム、Needleman and Wunsch (1970) J. Mol. Biol. 48:443-453のホモロジーアラインメントアルゴリズム、Pearson and Lipman (1988) Proc. Natl. Acad. Sci. 85:2444-2448の類似性検索法(search-for-similarity-method)、Karlin and Altschul (1990) Proc. Natl. Acad. Sci. USA 87: 2264-2268のアルゴリズム、Karlin and Altschul (1993) Proc. Natl. Acad. Sci. USA 90:5873-5877のように改変したものなどである。
【0068】
これらの数学的アルゴリズムのコンピュータによる実現は、配列を比較して配列同一性を決定するのに利用できる。かかる実現は、これに限定するものではないが、PC/遺伝子プログラムにおけるCLUSTAL(Intelligenetics, Mountain View, Calif.から購入可能)、ALIGNプログラム(バージョン2.0)およびWisconsin Genetics Software Package, Version 8中のGAP、BESTFIT、BLAST、FASTAおよびTFASTA(Genetics Computer Group(GCG), 575 Science Drive, Madison, Wis., USAから購入可能)を含む。BLASTおよび関連するプログラムもまた(他の配列比較プログラムと同様)インターネットを介して、国立生物工学情報センター(NCBI:例えばwww.ncbi.nlm.nih.gov or blast.ncbi.nlm.nih.gov/Blast.cgi参照のこと)のウェブサイトで利用可能である。このプログラムを用いたアラインメントは、初期設定のパラメータを用いて実行することができる。Altschul et al (1990) J. Mol. Biol. 215:403のBLASTプログラムは、Karlin and Altschul (1990) のアルゴリズムに基づいている。BLASTヌクレオチド検索は、例えばBLASTNプログラムで、スコア=100、文字長=12で実行し、本発明の明細書に記載のポリペプチドをコードするヌクレオチド配列と相同なヌクレオチド配列を得ることができる。BLASTタンパク質検索は、BLASTXプログラムで、スコア=50、文字長=3で実行し、本発明の明細書に記載のポリペプチドのアミノ酸配列と相同なアミノ酸配列を得ることができる。比較の目的でギャップアラインメントを得るため、Altschul et al. (1997) Nucleic Acids Res. 25:3389-3402に記載されているように、Gapped BLAST(BLAST2.0において)を用いることができる。代替的に、PSI-BLAST(BLAST2.0において)を用いて分子間の距離関係を検出する反復検索を実行することができる。Altschul et al. (1997)参照のこと。BLAST、Gapped BLAST、PSI-BLASTを用いる場合、それぞれのプログラム(例えばヌクレオチド配列についてBLASTN、タンパク質についてBLASTXなど)の初期設定パラメータを用いることができる。
【0069】
いくつかの態様において、本明細書に記載の1または2以上の遺伝子は、1または2以上の組換え発現ベクターにおいて発現する。かかるベクターは、オペロン配置においてなど、独立してまたは組み合わせて遺伝子を含み得る。本明細書で用いる場合、「ベクター」は、異なる遺伝環境間の輸送のためまたは宿主細胞における発現のため、所望の配列または配列(複数)が制限およびライゲーションにより挿入され得る、任意の数の核酸であり得る。ベクターは、RNAベクターもまた利用可能であるものの、典型的にはDNAからできている。ベクターはこれに限定するものではないが、プラスミド、フォスミド、ファージミド、ウイルスゲノムおよび人工染色体を含む。
【0070】
クローニングベクターは、自発的に複製することができるまたは宿主細胞のゲノムに統合されることができるものであり、さらにベクターが決定可能なやり方で切断され得、新たな組換えベクターがその宿主細胞における複製能力を維持するように所望のDNA配列がライゲーションされ得る、1または2以上のエンドヌクレアーゼ制限部位により定義されるものである。プラスミドの場合、所望の配列の複製は、プラスミドが宿主細菌などの宿主細胞内でコピー数を増加させるにつれ何度も起こり得、または有糸分裂により宿主が増殖する前に宿主ごとに一度だけ起こり得る。ファージの場合、複製は溶解相の間能動的に起こり得、または溶原性相の間受動的に起こり得る。
【0071】
発現ベクターは、所望のDNA配列が、制限およびライゲーションにより作動可能に制御配列に加わるように挿入され得、RNA転写物として発現し得るものである。ベクターはさらに、該ベクターにより変換または形質転換されたかまたはされていない細胞の同定に用いるのに適した1または2以上のマーカー配列を含んでもよい。マーカーは、例えば、抗生物質又は他の化合物に対する抵抗性または感受性を増加するまたは低減するタンパク質をコードする遺伝子、当該技術分野において知られた標準的なアッセイによってその活性を検出可能な酵素をコードする遺伝子(例えばβ−ガラクトシダーゼ、ルシフェラーゼまたはアルカリホスファターゼ)、および変換または形質転換された細胞、宿主、コロニーまたはプラークのフェノタイプに視覚的に影響する遺伝子(例えば緑色蛍光タンパク質)を含む。好ましいベクターは、自立した複製および作動可能に加わったDNAセグメントに存在する構造的遺伝子産物の発現が可能なものである。
【0072】
本明細書で使用される場合、コード配列および制御配列は、制御配列の影響下または制御下でコード配列が発現または転写される状態に置かれるように共有結合された場合、「作動可能に(operably)」加えられると言われる。コード配列が機能的タンパク質に翻訳されることが所望される場合、5’制御配列におけるプロモーターの導入がコード配列の転写の結果となり、2つのDNA間の結合の性質が(1)フレームシフト変異の導入の結果とならず、(2)プロモーター領域がコード配列の転写を指示する能力を妨害せず、または(3)対応するNA転写物がタンパク質に翻訳される能力を妨害しない場合に、2つのDNA配列が作動可能に加えられると言われる。したがって、得られた転写物が所望のタンパク質またはポリペプチドに翻訳され得るようにプロモーター領域がDNA配列の転写に作用できる場合、プロモーター領域はコード配列に作動可能に加えられるということになる。
【0073】
クレームされた発明の任意の酵素をコードする核酸分子が細胞内で発現された場合、その発現を指示するのに様々な転写制御配列(例えばプロモーター/エンハンサー配列など)を用いることができる。プロモーターは、遺伝子の発現の正常な制御を提供する、天然のプロモーター、すなわちその内在性の構成成分である遺伝子のプロモーター、であり得る。いくつかの態様において、プロモーターは構成的、すなわちその関連する遺伝子の連続的な転写を許容するように制御されていない、であり得る。分子の存在または比存在によりコントロールされるプロモーターなど、様々な条件的プロモーターもまた用いられ得る。
【0074】
遺伝子発現に必要な制御配列の正確な性質は、種または細胞型の間で変化してもよいが、一般的に、必要に応じて、TATAボックス、キャッピング配列、CAAT配列など、それぞれ転写および翻訳の開始に関与する5’非転写および5’非翻訳配列を含む。特に、かかる5’非転写制御配列は、作動可能に加えられた遺伝子の転写コントロールのためのプロモーター領域を含むだろう。制御配列はまた、記載されているように、エンハンサー配列または上流のアクチベーター配列を含んでよい。本発明のベクターは、任意に、5’リーダーまたはシグナル配列を含んでよい。適切なベクターの選択および設計は、当業者の能力および裁量の範囲内である。
【0075】
発現に必要な全ての要素を含む発現ベクターは、商業的に入手可能であり、当業者に知られている。例えばSambrook et al., Molecular Cloning: A Laboratory Manual, Second Edition, Cold Spring Harbor Laboratory Press, 1989を参照のこと。細胞は、異種性DNA(RNA)の細胞内への導入により、遺伝子操作されている。その異種性DNA(RNA)は、宿主細胞において該異種性DNAを発現できるように、転写要素の作動可能なコントロール下に置かれている。高い中鎖長モノマー含有量を有するポリヒドロキシアルカノアートコポリマー、特に高いHHx含有量を有するポリ(HB−co−HHx)、の産生を促進する遺伝子配列の異種性発現は、実施例のセクションにおいて実証されている。高いHHx含有量を有するポリ(HB−co−HHx)などの、高い中鎖長モノマー含有量を有するポリヒドロキシアルカノアートコポリマーを製造するための新規な方法もまた、他の細菌細胞、古細菌、真菌(酵母を含む)、哺乳類細胞、植物細胞などにおいて実施できる。
【0076】
本明細書に記載された酵素をコードする核酸分子は、当該技術分野において標準的な方法および技術を用いて、細胞または細胞(複数)に導入することができる。例えば核酸分子は、化学的形質転換およびエレクトロポレーション、形質導入、粒子衝突などを含む形質転換などの標準的なプロトコルにより導入できる。クレームされた発明の酵素をコードする1または2以上の核酸分子の発現はまた、核酸分子を細胞のゲノム内に組み込むことによっても達成され得る。
【実施例】
【0077】
例1:PhaB活性が低減した菌株
モノマー3−ヒドロキシブチリル−CoA(3HB−CoA)は、β−ケトチオラーゼ(PhaA)およびアセトアセチル−CoAリダクターゼ(PhaB)により、アセチル−CoAから合成される。Ralstonia eutrophaゲノムにおけるタンパク質配列の分析は、このタンパク質のよく研究されたバージョン(PhaAはH16_A1438、PhaB1はH16_A1439)に対する多くの潜在的ホモログを予測する。3HB−CoA合成を防ぐため、我々は遺伝子phaB1、phaB2およびphaB3をR. eutrophaゲノムからきれいに欠損させた。
【0078】
マーカー無しの欠損は、York[1]出典の方法を用いて行った。興味有る遺伝子の上流および下流のDNA配列をPCRで増幅した。この配列を、オーバーラップPCRを介して組み合わせて単一の隣接した一続きのDNAとした。この手法の間用いられたプライマーは、BamHI部位がDNAの終端に付加され、SwaI部位が上流および下流領域の間に挿入されるように設計された。この構築物はpGY46の骨格のBamHI部位にクローニングされ、R. eutrophaにおける興味有る遺伝子のマーカー無しの欠損を作るのに用いられるプラスミドを作製した[2]。プラスミドはE. coli S17-1に形質転換され、接合を介してR. eutrophaに導入した。欠損は、興味有る遺伝子の上流および下流にハイブリダイズする診断プライマーを用いたPCRにより確認された。
【0079】
全てのphaB遺伝子が、独立しておよび組み合わせて欠損した菌株のシリーズを構築した(下記表1参照のこと)。これらの菌株は、フルクトースを唯一の炭素源とする限定培地で生育された。PHB蓄積は窒素を限定することにより誘導した。
【表1】
【0080】
これらの培養物からのサンプルを様々な時点において取り、ポリ(3−ヒドロキシブチラート)(PHB)含有量について、クロトン酸アッセイ[3]によりアッセイした(図5)。phaB1およびphaB3の両方を欠失する菌株(すなわちRe2113およびRe2115)は、野生型菌株よりも顕著に少ないPHBを産生した。NADPHおよびNADH依存性アセトアセチル−CoAリダクターゼ活性は、確立された方法[4]により計測した(図6、7)。我々は、低量のPHBを作る菌株は、顕著に低減したNADPH依存性リダクターゼレベルを有し、このことはPHB蓄積における減少が実際に、PhaB活性の排除に起因する3HB−CoA合成の欠失に起因することを提示する。
【0081】
我々は、phaB欠損を有する菌株における相補的リダクターゼ変異の効果を検討した。phaB1、phaB2およびphaB3遺伝子を独立して菌株Re2115(ΔphaB123)のゲノムのphaB1遺伝子座に付加して戻し、菌株Re2139、Re2140およびRe2143を作製した。これらの実験の途中で、phaB3の開始コドンが誤って注釈されていることが発見され、正しく注釈された配列を用いた(表2においてphaB3correctと表記されている)。脂肪酸合成に関与するリダクターゼをコードするfabG遺伝子もまた、菌株Re2115(ΔphaB123)のphaB1遺伝子座に付加され、菌株Re2142を作製した。全ての挿入された遺伝子は同一のリボソーム結合部位を有する。
【0082】
【表2】
これらの培養物からのサンプルを様々な時点において取り、ポリ(3−ヒドロキシブチラート)(PHB)含有量について、クロトン酸アッセイ[3]によりアッセイした(図8)。
図9および図10にそれぞれ示されているように、PhaA活性およびこれらの菌株により産生されたPHBポリマーの分子量もまた計測された。PhaA活性は[6]に記載の方法により計測された。
【0083】
コポリマーの分子量は、ポリスチレン標準を用いたゲル浸透クロマトグラフィによって計測した。PHBを凍結乾燥した細胞から、クロロホルム中48時間で抽出した。PHB溶液を3mg/mLで調製した。抽出の後、溶液をろ過して不溶性のバイオマスを除去した。溶解したポリマーを、PLgel Olexisガードカラム(Polymer Laboratoriesパーツ番号PL1110-1400)および2つの一連のPLgel Olexis分析カラム(Polymer Laboratoriesパーツ番号PL1110-6400)を装備したAgilent 1100 HPLCを用いて分析した。100μLの各溶液を注入し、カラムから溶出される際に屈折率検出器によりポリマーを検出した。分子量を、Agilent GPC分析ソフトウェアを用いて、得られたクロマトグラムから決定した。システムを、1,110から13,155,000g/molまでのサイズ範囲の一連のポリスチレン標準(Polymer Laboratoriesパーツ番号PL2010-0104)を用いて校正した。イソプロパノールを全ての校正標準および実験サンプルに内部標準として含ませた。
【0084】
例2:ポリ(3−ヒドロキシブチラート−co−3−ヒドロキシヘキサノアート)製造のための菌株
我々は、限定された3HB−CoA合成能力を有する菌株は、高いHHx含有量を有する3−ヒドロキシブチラートと3−ヒドロキシヘキサノアートとのコポリマー(ポリ(HB−co−HHx))の合成が可能な菌株の設計においてよい出発点になるだろうと予測した。この予測は、シンターゼが3HB−CoAおよび3HHx−CoAをポリマー化可能であったとしても、野生型R. eutrophaにおける3HB−CoAの高い細胞内濃度が、ポリマーへのHHx組み込みを限定するだろうという我々の信念に基づいていた。加えて、R. eutrophaシンターゼは3HB−CoAと3HV−CoAとをポリマー化するのみである。PhaB基質特異性は図11に示されている(参考文献[4]も参照のこと)。
【0085】
我々の予測を試験するため、我々は最初に、pGY46を用いて天然のPHAシンターゼ(phaC1)をRe2115から欠損させ[2]、菌株Re2133を作製した。我々はそこでAeromonas caviaeからのおよびRhodococcus aetherivorans I24(D12およびC09シンターゼ)からの他のシンターゼを試験した。例えば、Rhodococcus aetherivorans I24からのD12シンターゼを、[2]出典の手法を用いて、Re2133ゲノムのphaC1遺伝子座に挿入した。これは、最初にpGY46中のDNAの上流および下流領域の間に部位特異的変異導入を介してSwaI部位を挿入し、次にD12シンターゼ遺伝子をSwaI部位にクローニングして入れることで、菌株Re2135を作製した。
【0086】
菌株Re2135を、パーム油を炭素源として用いて、フラスコの中で生育した。我々は3HHx−CoAモノマーが脂肪酸代謝の副産物として作られ(図4参照)、したがって油/脂肪酸を炭素源として使用することがコポリマー産生における不可欠であると信じていた。これらの培養物におけるPHA蓄積は、窒素限定により誘導された。細胞およびポリマーの組成におけるポリマー含有量を、標準的なメタノリシス手法[5]により計測した(図14)。この方法は、PHAモノマーを、関連するメチルエステルに変換し、それをガスクロマトグラフィーにより分離および定量する。我々は、Re2135が少量のPHAを蓄積するが、このPHAは高レベルのHHxモノマーを含むことを見出した。
【0087】
我々は次に、この菌株においてモノマー合成を増大させる遺伝子について調査した。文献からのいくつかの菌株が図13に記載してあり、これには用いられたPHAシンターゼ、炭素源、PHA含有量およびHHxのmol%が示されている。
我々はエノイル−CoAヒドラターゼをコードする遺伝子(phaJ遺伝子)に焦点を当てた。いくつかのphaJがRe2135ゲノムのphaB1遺伝子座に挿入された:Aeromonas caviaeおよびPseudomonas aeruginosa(phaJ1およびphaJ2)からのエノイル−CoAヒドラターゼ。PhaJ基質特異性は図12に示されている([7]も参照のこと)。
【0088】
遺伝子は、当初はB1欠損ベクターであったものの、上流および下流DNA領域の間のSwaI部位にクローニングされた。遺伝子は上記の手法を用いてゲノムに挿入され、パーム油を唯一の炭素源とする限定培地中で生育した。我々は、Pseudomonas aeruginosa PAO1からの遺伝子phaJ1が、依然として高いHHx含有量を含みながら、より多量のポリマーの産生に導くことを見出した。phaJ1をゲノムに含む菌株をRe2152と名付けた。
【0089】
菌株構築および結果は、遺伝子型(用いたPHAシンターゼを含む)、PHA含有量(細胞乾燥重量の%として)、およびHHxの重量%を示す、図14に記載されている。mol%HHxとして計測されたHHx含有量は、常に重量%として計測されたHHx含有量よりも小さい数字であった。mol%は、重量%の値に0.8を掛けることにより、重量%から概算できる。例えば、25重量%のHHxは20mol%のHHxに相当し、30重量%のHHxは24mol%のHHxに相当する。
【0090】
我々は、我々の新たに開発したPHAオペロン(phaCD12−phaA−phaJ1Pa)における酵素的ステップの1つが、PHA産生を限定しているだろうと仮説を立てた。遺伝子発現を増大させるため、我々はこのオペロンをRe2152からPCRによって増幅し、プラスミドpBBR1MCS−2のKpnIとHindIIIとの間にクローニングした。この新たなプラスミド(pCB81)をRe2133に形質転換し、得られた菌株を、300μg/mLのカナマイシンを含有するパーム油限定培地で生育した。手法は図15に記載されている。このように増幅された遺伝子発現がPHA産生または得られたPHAのモノマー組成にネガティブに影響するかを決定するため、我々は得られた菌株によって産生されたポリマーにおけるPHAおよびHHX含有量を分析した。予想外にも、培養物の分析は、細胞が高いPHA含有量および高レベルのHHxをポリマー内に有するコポリマーを産生したことを示した。
我々は、共通して、25〜30重量%のHHx、これは20〜24mol%のHHxに相当する、を有するコポリマーを得た。33重量%(27mol%)までのHHx値を有するコポリマーが、菌株をパーム油で生育した場合に得られた。
【0091】
例3:ポリマー特性評価
新たに開発したPHAオペロン(phaCD12−phaA−phaJ1Pa)を含む菌株により産生されたPHAコポリマーの特性を決定した。PHAコポリマーの分子量は、ゲル浸透クロマトグラフィにより、ポリスチレン標準と比較して120,000〜150,000g/molであることが見出された。
【0092】
OHAコポリマーの熱的特性を、示差走査熱量測定を用いて計測した。サンプルをアルミニウムの小皿に乗せ、Perkin ElmerのPyris 1 DSCを用いて分析した。用いた温度プログラムは、(1)50℃で1分間維持、(2)20℃/分で−40℃まで冷却、(3)−40℃で3分維持、(4)20℃/分で200℃まで加熱、(5)200℃で1分維持、(6)20℃/分で−40℃まで冷却、(7)−40℃で3分維持、(8)20℃/分で50℃まで加熱、であった。ガラス転移温度を、吸熱勾配に変化が起こる温度として同定した。融点を、吸熱の最高ピークとして同定した。DSC分析は、上記の菌株により産生された、27mol%のHHxを含有するコポリマーは−4℃のガラス転移温度を有することを明らかにした。
【0093】
例4:Rhodococcus aetherivorans I24のPHAシンターゼについての配列データ
潜在的なシンターゼ遺伝子を、既知のシンターゼペプチドの配列に対してRhodococcus aetherivorans I24ゲノム配列BLASTNすることにより同定した。Rhodococcus aetherivorans I24のゲノムにおける予測されるタンパク質配列の分析に基づき、2つのPHAシンターゼがゲノムからクローニングされ、それはまた本明細書においてCO9シンターゼおよびD12シンターゼと言われる。両方のPHAシンターゼとも、R.eutrophaにおいて(上記の通り)発現したときに、活性であると決定された。
【0094】
BLASTN配列分析は、クローニングされたPHAシンターゼの両方の遺伝子が、よく研究されたシンターゼのうちシュードモナスからのシンターゼに最も近いことを示した。これらシュードモナスのシンターゼは非常に広範な基質特異性を有することが知られている。CO9シンターゼおよびD12シンターゼはC7〜C8までの基質をポリマー化し、これはR. eutrophaからのシンターゼよりも広い基質特異性である。
【0095】
【表3】
【0096】
【表4】
【0097】
【表5】
【0098】
【表6】
【0099】
【表7】
【0100】
当業者であれば、本明細書に記載された発明の特定の態様に対する多くの均等物を認識する、または日常的な実験を超えない程度の実験を用いて突き止められるだろう。かかる均等物は、特許請求の範囲に包含されることを意図する。
本明細書に開示された全ての参考文献は、本明細書に引用された目的のために、その全開示が参照により組み込まれる。
【技術分野】
【0001】
関連出願
本出願は、米国仮出願番号61/260,164、タイトル「高い中鎖長モノマー含有量を有するポリヒドロキシルアルカノアートコポリマーを製造する方法(Methods for Producing Polyhydroxyalkanoate Copolymer with High Medium Chain Length Monomer Content)」、出願日2009年11月11日、の35U.S.C.120条に基づく優先権を主張し、その全開示は、全体として本明細書に参照として組み込まれる。
【0002】
技術分野
本発明は、組換え体による遺伝子発現を通じた高い中鎖長モノマー含有量を有するポリヒドロキシアルカノアートコポリマーの製造に関する。
【背景技術】
【0003】
商業的に有用な複合バイオポリマーであるポリヒドロキシルブチラートは、多数の細菌によって産生される細胞間物質である。ポリヒドロキシブチラート(PHB)は、ポリマーの化学的および物理的特性両方に基づく有用な生物材料である。PHBは、生分解性/熱可塑性材料として、ある抗生物質の有機合成のためのキラル中心の給源として、および薬剤送達および骨置換のためのマトリクスとしての有用性を含む、様々な潜在的応用性を有する。in vivoにおいて、該ポリマーは内部的に分解されて、ヒトの血液の通常の構成物質であるヒドロキシブチラートになる。ポリヒドロキシアルカノアート(PHA、PHBを含む)およびコポリマーの製造の様々な側面が、例えば米国特許第5,534,432号、米国特許第5,663,063号、米国特許第5,798,235号および米国特許第7,202,064号などに記載されている。
【0004】
例として、細菌Ralstonia eutrophaは、高レベルのポリヒドロキシアルカノアート(PHA)生体プラスチックを蓄積することでよく知られている。この生物の野生型は、典型的にはポリヒドロキシブチラートのホモポリマーを産生する。このポリマーは、β−ケトチオラーゼ(PhaA)、アセトアセチル−CoAリダクターゼ(PhaB)およびポリヒドロキシアルカノアートシンターゼ(PhaC)の作用によりアセチル−CoAから産生される。PHBはもろく、その分解温度に近い融点を有するため、有用な生体プラスチックではない。
【0005】
3−ヒドロキシへキサノアートモノマーをポリマー鎖に導入することで、PHBよりも頑強で低い融点を有する材料、したがってPHBよりプラスチックの性能を増大させ、PHBより多くの適用に適するようにしたものが獲得されることが、以前に実証された。
【0006】
米国特許第7,235,621号には、非常に特定の条件下における3−ヒドロキシブチラートと3−ヒドロキシへキサのアートとのコポリマーの製造が記載されている。米国特許第7,235,621号には、高いラウリン酸含有量を有する特定の植物油が、コポリマーを産生する微生物の炭素給源として要求されることが記載されている。これらの要求される油は、ヤシ、ダイズおよびナタネなどのより普通の油中に見出されるものより短鎖の脂肪酸を有している。かかる制限的な炭素給源の要求があっても、米国特許第7,235,621号に開示されたコポリマー中の3−ヒドロキシヘキサノアートの最高量は、13.8mol%である。
【0007】
米国特許公開公報2009/0130731号は、3−ヒドロキシブチラートと3−ヒドロキシへキサのアートとのコポリマーの、PHAシンターゼ遺伝子(phaC)および3−ケトアシル−ACPリダクターゼ遺伝子(fabG)を組換えにより発現する細菌中での製造を記載する。しかしながら、米国特許公開公報2009/0130731号に開示されたコポリマー中の3−ヒドロキシヘキサノアートの最高量は、4mol%である。
【発明の概要】
【0008】
本発明は、高い中鎖長モノマー含有量を有するポリヒドロキシアルカノアートコポリマーの、組換え遺伝子発現を通じた製造に関する。
いくつかの側面において、任意の植物油を炭素給源として使用して、少なくとも約4mol%または5重量%の中鎖長モノマー含有量を有するポリヒドロキシアルカノアートコポリマーを産生する細胞または生物が提供される。いくつかの態様において、細胞または生物は、任意の植物油を炭素源として使用して、少なくとも約4mol%または5重量%のHHx含有量を有する3−ヒドロキシルブチラートと3−ヒドロキシルヘキサノアートとのコポリマー(ポリ(HB−co−HHx))を産生する。いくつかの態様において、細胞中の3−ヒドロキシルブチラートの正常な合成は破壊されている。いくつかの態様において、アセトアセチル−CoAリダクターゼをコードする遺伝子は欠損している。いくつかの態様において、細胞が、Ralstonia eutropha細胞であり、1または2以上のphaB1、phaB2およびphaB3遺伝子は破壊されている。いくつかの態様において、phaB3遺伝子は破壊されている。
【0009】
いくつかの態様において、細胞は、非内在性のPHAシンターゼ遺伝子を組換えにより発現する。いくつかの態様において、非内在性PHAシンターゼ遺伝子は、Aeromonas caviaePHAシンターゼ遺伝子またはRhodococcus aetherivoransPHAシンターゼ遺伝子である。いくつかの態様において、Rhodococcus aetherivoransPHAシンターゼ遺伝子は、配列番号4をコードするRhodococcus aetherivorans I24 D12 PHAシンターゼ遺伝子または配列番号2をコードするRhodococcus aetherivorans I24 C09 PHAシンターゼ遺伝子である。いくつかの態様において、Rhodococcus aetherivorans I24 D12 PHAシンターゼ遺伝子は、配列番号3を含むかまたは配列番号3からなる、および/あるいはRhodococcus aetherivorans I24 C09 PHAシンターゼ遺伝子は、配列番号1もしくは配列番号1の開始コドンがTTGからATGに変えられたものを含むかまたは配列番号1もしくは配列番号1の開始コドンがTTGからATGに変えられたものからなる。
【0010】
いくつかの態様において、細胞は、エノイル−CoAヒドラターゼ遺伝子を組換えにより発現する。いくつかの態様において、エノイル−CoAヒドラターゼ遺伝子は、Aeromonas caviaeエノイル−CoAヒドラターゼ遺伝子またはPseudomonas aeruginosaエノイル−CoAヒドラターゼ遺伝子である。いくつかの態様において、Pseudomonas aeruginosaエノイル−CoAヒドラターゼ遺伝子は、Pseudomonas aeruginosa phaJ1遺伝子(遺伝子PA3302)またはPseudomonas aeruginosa phaJ2遺伝子(遺伝子PA1018)である。
【0011】
いくつかの態様において、非内在性PHAシンターゼ遺伝子および/またはエノイル−CoAヒドラターゼ遺伝子は、増幅される。いくつかの態様において、モノマー含有量は、少なくとも約5mol%、少なくとも約6mol%、少なくとも約7mol%、少なくとも約10mol%、少なくとも約15mol%、または少なくとも約20mol%、またはそれ以上である
【0012】
いくつかの態様において、モノマー含有量は、少なくとも約6重量%、少なくとも約8重量%、少なくとも約10重量%、少なくとも約15重量%、少なくとも約10重量%、または少なくとも約25重量%、またはそれ以上である。
【0013】
いくつかの態様において、細胞または生物は、細菌細胞、真菌細胞(酵母細胞を含む)、植物細胞、昆虫細胞または動物細胞である。いくつかの態様において、細胞は、Ralstonia菌種、Aeromonas菌種、Rhizobium菌種、Alcaligenis菌種またはPseudomonas菌種の細胞である。いくつかの態様において、細胞は、Ralstonia eutropha、Aeromonas caviae、Rhizobium japonicum、Alcaligenis eutrophusまたはPseudomonas oleovorans細胞である。いくつかの好ましい態様において、細胞は、Ralstonia eutropha細胞である。
【0014】
いくつかの態様において、非内在性PHAシンターゼ遺伝子および/またはエノイル−CoAヒドラターゼ遺伝子は、プラスミドから発現される。
いくつかの態様において、非内在性PHAシンターゼ遺伝子および/またはエノイル−CoAヒドラターゼ遺伝子は、細胞のゲノムに組み込まれている。
【0015】
他の側面において、高い中鎖長モノマー含有量を有するポリヒドロキシアルカノアートコポリマーを製造する方法が提供される。本方法は、前述の細胞または生物を培養し、少なくとも約4mol%または5重量%の中鎖長モノマー含有量を有するコポリマーを製造することを含む。いくつかの態様において、本方法は、前述の細胞を培養し、少なくとも約4mol%または5重量%のHHx含有量を有する(ポリ(HB−co−HHx))を製造することを含み、ここで3−ヒドロキシルブチラートと3−ヒドロキシルヘキサノアートとのコポリマー(ポリ(HB−co−HHx))が製造される。いくつかの態様において、本方法は、コポリマーを細胞から回収することをさらに含む。
【0016】
いくつかの態様において、製造されるコポリマーの量は、細胞の乾燥重量の少なくとも約20%、細胞の乾燥重量の少なくとも約30%、細胞の乾燥重量の少なくとも約40%、細胞の乾燥重量の少なくとも約50%、細胞の乾燥重量の少なくとも約60%、またはそれ以上である。
【0017】
他の側面において、少なくとも約4mol%または5重量%の中鎖長モノマー含有量を有するポリヒドロキシルアルカノアートコポリマーを産生する細胞を製造する方法が提供される。本方法は、細胞中に少なくとも1つのRhodococcus aetherivoransPHAシンターゼ遺伝子を組み換えにより発現することを含む。いくつかの態様において、細胞は、少なくとも約4mol%または5重量%のHHx含有量を有する3−ヒドロキシルブチラートと3−ヒドロキシルヘキサノアートとのコポリマー(ポリ(HB−co−HHx))を産生する。いくつかの態様において、Rhodococcus aetherivoransPHAシンターゼ遺伝子は、配列番号4をコードするRhodococcus aetherivorans I24 D12 PHAシンターゼ遺伝子または配列番号2をコードするRhodococcus aetherivorans I24 C09 PHAシンターゼ遺伝子である。いくつかの態様において、Rhodococcus aetherivorans I24 D12 PHAシンターゼ遺伝子は、配列番号3を含むかまたは配列番号3からなる、および/あるいはRhodococcus aetherivorans I24 C09 PHAシンターゼ遺伝子は、配列番号1もしくは配列番号1の開始コドンがTTGからATGに変えられたものを含むかまたは配列番号1もしくは配列番号1の開始コドンがTTGからATGに変えられたものからなる。
【0018】
いくつかの態様において、本方法は、エノイル−CoAヒドラターゼ遺伝子を組換えにより発現することをさらに含む。いくつかの態様において、エノイル−CoAヒドラターゼ遺伝子は、Aeromonas caviaeエノイル−CoAヒドラターゼ遺伝子またはPseudomonas aeruginosaエノイル−CoAヒドラターゼ遺伝子である。いくつかの態様において、Pseudomonas aeruginosaエノイル−CoAヒドラターゼ遺伝子は、Pseudomonas aeruginosa phaJ1遺伝子(遺伝子PA3302)またはPseudomonas aeruginosa phaJ2遺伝子(遺伝子PA1018)である。
【0019】
いくつかの態様において、非内在性PHAシンターゼ遺伝子および/またはエノイル−CoAヒドラターゼ遺伝子は、増幅される。
いくつかの態様において、細胞は、細菌細胞、真菌細胞(酵母細胞を含む)、植物細胞、昆虫細胞または動物細胞である。いくつかの態様において、細胞は、Ralstonia菌種、Aeromonas菌種、Rhizobium菌種、Alcaligenis菌種またはPseudomonas菌種の細胞である。いくつかの態様において、細胞は、Ralstonia eutropha、Aeromonas caviae、Rhizobium japonicum、Alcaligenis eutrophusまたはPseudomonas oleovoransの細胞である。いくつかの好ましい態様において、細胞は、Ralstonia eutrophaの細胞である。
【0020】
いくつかの態様において、非内在性PHAシンターゼ遺伝子および/またはエノイル−CoAヒドラターゼ遺伝子は、プラスミドから発現される。
いくつかの態様において、非内在性PHAシンターゼ遺伝子および/またはエノイル−CoAヒドラターゼ遺伝子は、細胞のゲノムに組み込まれている。
【0021】
いくつかの側面において、1または2以上の、少なくとも約4mol%または5重量%の中鎖長モノマー含有量を有するコポリマーを製造する方法が提供される。本方法は、前述の方法により細胞を製造すること、および該細胞の集団を培養することを含む。いくつかの態様において、1または2以上のコポリマーは、3−ヒドロキシルブチラートと3−ヒドロキシルヘキサノアートとのコポリマー(ポリ(HB−co−HHx))である。いくつかの態様において、本方法は、細胞集団から1または2以上のコポリマーを収集することをさらに含む。
【0022】
いくつかの態様において、モノマー含有量は、少なくとも約5mol%、少なくとも約6mol%、少なくとも約7mol%、少なくとも約10mol%、少なくとも約15mol%、少なくとも約20mol%、またはそれ以上である。
いくつかの態様において、モノマー含有量は、少なくとも約6重量%、少なくとも約8重量%、少なくとも約10重量%、少なくとも約15重量%、少なくとも約20重量%、少なくとも約25重量%、またはそれ以上である。
【0023】
いくつかの態様において、製造されるコポリマーの量は、細胞の乾燥重量の少なくとも約20%、細胞の乾燥重量の少なくとも約30%、細胞の乾燥重量の少なくとも約40%、細胞の乾燥重量の少なくとも約50%、細胞の乾燥重量の少なくとも約60%、またはそれ以上である。
【0024】
他の側面において、配列番号2または配列番号4をコードする、単離された核酸分子が提供される。いくつかの態様において、該単離された核酸分子は、配列番号1または配列番号3に記載のヌクレオチド配列を含む。いくつかの態様において、単離された核酸分子は、配列番号1または配列番号3に記載のヌクレオチド配列と、少なくとも80%の同一性、少なくとも90%の同一性、少なくとも95%の同一性、少なくとも98%の同一性、またはそれ以上の同一性を有する。
【0025】
いくつかの態様において、前述の核酸分子によりコードされる、単離されたポリペプチドが提供される。
いくつかの態様において、前述の単離された核酸分子を含むベクターが提供される。
いくつかの態様において、前述の単離された核酸分子を組換えにより発現する細胞が提供される。いくつかの態様において、核酸分子は、ベクターから発現される。いくつかの態様において、核酸分子は、細胞のゲノムに組み込まれている。
本発明のこれらのおよび他の側面、ならびにそれらの様々な態様は、図面および本発明の詳細な説明を参照することにより、より明白となるであろう。
添付の図面はスケール通りに描かれていることを意図していない。本図面において、様々な図中に描かれている各同一のまたはほぼ同一の構成要素は、同様の番号で表される。明瞭さのため、全ての構成要素が全ての図面において名前を付けられているわけではないだろう。図面において:
【図面の簡単な説明】
【0026】
【図1】図1はPHAポリマーの構造および特性を示す。上、ポリ(HB);下、ポリ(HB−co−HHx)
【0027】
【図2】図2はPhaA、PhaBおよびPhaCの、PHAコポリマー合成の経路への関与を示す。
【図3】図3は、脂肪酸β−酸化とPHAコポリマー合成の経路との相互作用、およびモノマーが脂肪酸代謝の副産物として作られることを概略的に示す。
【図4】図4は、PHAコポリマー合成の経路におけるPhaAおよびPhaBのブロッキングの効果、およびモノマーが脂肪酸代謝の副産物として作られることを概略的に示す。
【0028】
【図5】図5は、phaB1、phaB2および/またはphaB3欠損を有する数菌株による、フルクトース限定培地中におけるPHB合成を示す。野生型、および1変異、2変異および3変異株の結果が示されている。
【図6】図6は、phaB1、phaB2および/またはphaB3欠損を有する数菌株における、フルクトース限定培地中におけるNADPHとのリダクターゼ活性を示す。野生型、および1変異、2変異および3変異株の結果が示されている。
【0029】
【図7】図7は、phaB1、phaB2および/またはphaB3欠損を有する数菌株における、フルクトース限定培地中におけるNADHとのリダクターゼ活性を示す。野生型、および1変異、2変異および3変異株の結果が示されている。
【図8】図8は、phaB欠損を有する菌株における相補的リダクターゼ変異の効果を示す。phaB1、phaB2およびphaB3遺伝子は独立して菌株Re2115(ΔphaB123)に戻された。別の株は、Re2115に戻したfabGを有していた。野生型、変異型および相補化された菌株におけるPHB合成の結果が示されている。
【0030】
【図9】図9は、図8に記載されている菌株のPhaA活性を示す。
【図10】図10は、図8に記載されている菌株によって産生されたPHBポリマーの分子量を示す。
【図11】図11は、PhaBの基質特異性を示す。
【図12】図12は、PhaJの基質特異性を示す。
【図13】図13は、用いたPHAシンターゼ、炭素源、PHA含有量、およびmol%HHxを示し、文献からの数菌株を列挙する。
【0031】
【図14】図14は、遺伝子型(用いたPHAシンターゼを含む)、PHA含有量(細胞乾燥重量の%として)、および重量%HHxを示して、構築された菌株およびかかる菌株からの結果を列挙する。
【図15】図15は、構築されたPHAオペロン(phaCD12−phaA−phaJ1Pa)が増幅されおよびプラスミドにクローニングされ、それが菌株Re2133に形質転換されるという菌株構築の手法を示す。
【発明を実施するための形態】
【0032】
発明の詳細な説明
本発明は、一部において、高い中鎖長モノマー含有量を有するポリヒドロキシアルカノアートコポリマーなどのバイオプラスチックを製造する方法、および細菌発酵および他の細胞および生物においてバイオプラスチックの産生を増大させる方法に関する。コポリマー化してポリヒドロキシアルカノアートコポリマーを製造するモノマーは、3−ヒドロキシブチラートおよび炭素鎖長が5以上である3−ヒドロキシアルカン酸(例えば、米国特許第7,341,856号およびそこに引用された参考文献を参照のこと、その開示のそれぞれは、それらの教示として参照により組み込まれる)を含む。この一例として、3−ヒドロキシブチラートと3−ヒドロキシヘキサノアートとのコポリマー(ポリ(HB−co−HHx))および特に高いHHx含有量を有するコポリマーがその物理的特性により有用である。本発明はまた、一部において、菌株が脂肪酸または任意の植物油を炭素源として用いて生育されたとき、ポリ(3−ヒドロキシブチラート−co−3−ヒドロキシヘキサノアート)などの高い中鎖長モノマー含有量を有するポリヒドロキシアルカノアートコポリマーを高度に蓄積することができる、Ralstonia eutropha菌株などの細胞に関する。
【0033】
かかる細胞により作られるコポリマーの3−ヒドロキシルヘキサノアート含有量は、文献に記載されている、植物油から製造される任意の材料に匹敵するかまたはそれを超えるものである。対照的に、米国特許第7,235,621号に記載の方法は、許容可能なHHx含有量のコポリマーを製造するために、構成脂肪酸中にラウリン酸を有する特定の植物油の使用が求められる。それであっても、米国特許第7,235,621号に記載の方法を用いて製造されるコポリマーのHHx含有量は、本明細書に記載される方法を用いて製造されたコポリマーのHHx含有量よりも劣るものである。
【0034】
以下の特徴は、独立しておよび組み合わせとして、本発明に含まれる。
いくつかの態様において、細胞中のPHBの正常な合成が破壊されている。例えば、下記実施例で示されるとおり、PHBは、Ralstonia eutrophaにおいてアセトアセチル−CoAリダクターゼをコードする遺伝子を欠損させることにより、破壊されている。本発明の特定の態様において、phaB3が破壊されており;phaB3は、従前文献において特徴付けられておらず、この特定の目的に用いられてこなかった遺伝子である。しかし本発明はかかる態様に限定されず、したがって、アセトアセチル−CoAリダクターゼの発現を低減させることを含む、正常なPHB生合成を破壊する他の方法も含まれる。
【0035】
いくつかの態様において、内在性PHAシンターゼ遺伝子が破壊され、新たなPHAシンターゼに置き換えられている。しかし本発明はかかる態様に限定されず、したがって、内在性PHAシンターゼの発現を低減させることを含む、正常な内在性PHAシンターゼ活性を破壊する他の方法を含む。例えば、下記実施例に示されるように、Ralstonia eutropha中の野生型PHAシンターゼ遺伝子が欠損され、新たなシンターゼ遺伝子が菌株に付与された。新たなPHAシンターゼは、高い割合の3−ヒドロキシヘキサノアートモノマーを組み込むことができる。いくつかの態様において、新たなシンターゼ遺伝子はRhodococcus aetherivorans I24由来(本明細書において「RhodococcusCO9シンターゼ」(配列番号1および2)または「RhodococcusD12シンターゼ」(配列番号3および4)ともいう)である。しかし本発明はかかる態様に限定されず、したがって、高い割合の中鎖長モノマー、特に3−ヒドロキシヘキサノアートモノマーを組み込む類似の能力を有する他のPHAシンターゼ遺伝子もまた、類似のやり方で用いることができる。
【0036】
いくつかの態様において、特定のエノイル−CoAヒドラターゼ遺伝子が細胞に導入され、コポリマーに組み込まれるモノマーの産生を増大する。例えば、下記実施例において示されるとおり、Pserudomonas aeruginosa PAO1からのphaJ1(遺伝子PA3302)と呼ばれる(R)−特異的エノイルCoAヒドラターゼ遺伝子が、Ralstonia eutropha菌株に導入された。酵素PhaJ1は、3−ヒドロキシブチリル−CoAおよび3−ヒドロキシヘキサノイル−CoAモノマーを産生し、これらはその後PHAシンターゼによりポリマー化される。Pseudomonas aeruginosa PAO1のphaJ2遺伝子(遺伝子PA1018)もまた用いることができる。Pseudomonas aeruginosa PAO1ゲノムのGenBank受託番号は、AE004091である。しかし本発明はかかる態様に限定されず、したがって、3−ヒドロキシブチリル−CoAおよび3−ヒドロキシヘキサノイル−CoAモノマーを産生する類似の能力を有する1または2以上の他のエノイル−CoA遺伝子もまた、類似のやり方で用いることができる。
【0037】
いくつかの態様において、本明細書に記載された遺伝子のコピー数を調節して、細胞によって蓄積されるPHAの量を変化させることができる。例えば、下記実施例において示されるとおり、最初にRhodococcus D12シンターゼおよびPseudomonas aeruginosaphaJ1遺伝子をRalstonia eutrophaゲノムに組み込んだ。その後遺伝子のコピー数を、遺伝子をプラスミドにクローニングすることにより増大させ、該プラスミドを、アセトアセチル−CoAリダクターゼ遺伝子および天然のPHAシンターゼ遺伝子がゲノムから欠損したRalstonia eutropha菌株に導入した。プラスミドを保有する菌株は、遺伝子がゲノムにのみ存在する菌株よりも顕著に多くのポリマーを産生した。しかし本発明はかかる態様に限定されず、したがって、遺伝子のコピー数を調節(好ましくは増大)する他の方法も含まれる。
【0038】
本発明の側面は、高いHHx含有量を有する3−ヒドロキシルブチラートと3−ヒドロキシルヘキサノアートとのコポリマー(ポリ(HB−co−HHx))などの高い中鎖長モノマー含有量を有するポリヒドロキシルアルカノアートコポリマーを、細胞中での組換え遺伝子発現を通じて製造する方法および組成物に関する。1つの態様において、本明細書に記載されるのは、植物油を炭素源とした、非内在性PHAシンターゼおよびエノイル−CoAヒドラターゼ遺伝子を組み換えにより発現し、アセトアセチル−CoAリダクターゼをコードする遺伝子が欠損している細胞における、5重量%よりも多い(4mol%よりも多い)HHx含有量を有するポリ(HB−co−HHx)の製造である。このシステムは、広く様々な適用を有する分子である、高いHHx含有量を有するポリ(HB−co−HHx)を製造する効果的で新たな方法を表す。
【0039】
本発明の側面によれば、1または2以上の酵素を組換えにより発現する細胞および、HHx含有量を有する(ポリ(HB−co−HHx))などの、高い中鎖長モノマー含有量を有するポリヒドロキシルアルカノアートコポリマーの製造におけるかかる細胞の使用が提供される。当然のことながら、PHAシンターゼ遺伝子を含む、かかる酵素をコードする遺伝子を様々な給源から得ることができる。当業者ならば気づくであろうように、これらの酵素の相同遺伝子を他の種から得ることができ、例えばNCBIのインターネットサイト(www.ncbi.nlm.nih.gov)において利用できるタンパク質BLASTN調査などの相同性調査により同定することができる。かかる遺伝子は、その特定の遺伝子を含む任意の種のDNAからのDNAからPCR増幅することができる。いくつかの態様において、遺伝子配列は合成であるおよび/または導入される細胞に対してコドン最適化されている。本明細書に記載されるような、酵素をコードする遺伝子を得るための任意の方法が、本発明と適合する。
【0040】
タンパク質発現の最適化は、例えば細菌細胞における発現のためのコドン最適化を通してなどにより、酵素をコードする遺伝子が細胞に導入する前に修飾されるいくつかの態様において求められ得る。様々な生物のコドン使用頻度はCodon Usage Database(www.kazusa.or.jp/codon/)においてアクセスできる。
本明細書に記載された方法、酵素、細胞および生物は、高い中鎖長モノマー含有量を有するポリヒドロキシルアルカノアートコポリマーの製造を提供する。モノマー含有量はしばしばモルパーセント(mol%)で表現される。いくつかの態様において、コポリマーは、少なくとも4mol%、5mol%、6mol%、7mol%、8mol%、9mol%、10mol%、11mol%、12mol%、13mol%、14mol%、15mol%、16mol%、17mol%、18mol%、19mol%、20mol%、21mol%、22mol%、23mol%、24mol%、25mol%、26mol%、27mol%、28mol%、29mol%、30mol%、31mol%、32mol%、33mol%、34mol%、35mol%、またはそれ以上の、3−ヒドロキシヘキサノアートなどの中鎖長モノマー含有量で製造される。かかる中鎖長モノマー含有量を有するコポリマーは、例えば任意の脂肪酸または油(例えば植物油)を炭素源として用いて製造することができる。
【0041】
モノマー含有量を表現する別の方法は、重量パーセント(重量%)である。モルパーセントは、重量パーセントの値に0.8を掛けることにより、重量パーセントから概算することができる;同様に、重量パーセントは、モルパーセントの値に1.25を掛けることにより、モルパーセントから概算することができる。したがって、コポリマーは、少なくとも5重量%、6重量%、7重量%、8重量%、9重量%、10重量%、11重量%、12重量%、13重量%、14重量%、15重量%、16重量%、17重量%、18重量%、19重量%、20重量%、21重量%、22重量%、23重量%、24重量%、25重量%、26重量%、27重量%、28重量%、29重量%、30重量%、31重量%、32重量%、33重量%、34重量%、35重量%、またはそれ以上の、3−ヒドロキシヘキサノアートなどの中鎖長モノマー含有量で製造される。かかる中鎖長モノマー含有量を有するコポリマーは、例えば任意の脂肪酸または油(例えば植物油)を生物または細胞の炭素源として用いて製造することができる。
【0042】
いくつかの態様において、細胞または生物により製造されるコポリマーの量は、細胞または生物の乾燥重量(細胞乾燥重量)の少なくとも20%、22%、24%、26%、28%、30%、35%、40%、45%、50%、55%、60%、またはそれ以上である。
上述したように米国特許第7,235,621号に記載の方法には、許容可能なHHx含有量のコポリマーを製造するために、構成脂肪酸中にある量のラウリン酸を有する特定の植物油の使用が求められる。米国特許第7,235,621号で報告されているコポリマーの最大HHx含有量は、13.8%である。本明細書に記載された方法、酵素、細胞および生物は、米国特許第7,235,621号で要求されるようなラウリン酸含有量を構成脂肪酸中に有する油または脂肪である炭素源を使用することなく、HHx含有量がより多いコポリマーを製造するのに向いている。
【0043】
有用なポリマーの分子量は、約10,000と400万ダルトンとの間、および好ましくは50,000ダルトンと150万ダルトンとの間など、10,000ダルトンより多いものを含む。
本発明によれば、高い中鎖長モノマー含有量を有するポリヒドロキシルアルカノアートコポリマーが製造される。モノマー単位は当該技術分野において知られており、ヒドロキシブチラート、ヒドロキシバレラート、ヒドロキシヘキサノアート、ヒドロキシヘプタノアート、ヒドロキシオクタノアート、ヒドロキシノナノアート、ヒドロキシデカノアート、ヒドロキシウンデカノアート、およびヒドロキシドデカノアート単位を含む。いくつかの態様において、製造されるコポリマーは、ポリ(3−ヒドロキシブチラート−co−3−ヒドロキシヘキサノアート)など、ヒドロキシブチラートおよびヒドロキシヘキサノアートモノマーを含む。
【0044】
本発明は、原核細胞および真核細胞を含む、PHAシンターゼによってポリマー化されるモノマーを作るための遺伝子を発現する任意のタイプの細胞を包含する。いくつかの態様において、細胞は最近細胞である。いくつかの態様において、細菌細胞はRalstonia属、Aeromonas属、Rhizobium属、Alcaligenes属またはPseudomonas属細胞である。いくつかの態様において、細胞は、Ralstonia eutropha、Aeromonas caviae、Rhizobium japonicum、Alcaligenes eutrophusまたはPseudomonas oleovorans細胞である。1つの好ましい態様において、細胞は、例えばSaccharomyces属、Schizosaccharomyces属、Pichia属、Phaffia属、Hansenula属、Kluyveromyces属、Candida属、Talaromyces属、Brettanomyces属、Schwanniomyces属、Pachysolen属、Debaryomyces属、Yarrowia属または工業的倍数体酵母株などの酵母細胞などの真菌細胞である。コポリマーの製造に有用な酵母種および株の例は、米国特許第7,083,972号に記載されており、その開示はその教示についての参照により組み込まれる。真菌の他の例は、Aspergillus属、Pennicilium属、Fusarium属、Rhizopus属、Acremonium属、Neurospora属、Sordaria属、Magnaporthe属、Allomyces属、Ustilago属、Botrytis属、およびTrichoderma属を含む。他の態様において、細胞は藻類細胞、哺乳類細胞または植物細胞である。
【0045】
当然のことながら、本発明に適合するいくつかの細胞は、本発明に付随する1または2以上の遺伝子の内在性コピーを、組換えコピーと共に発現してもよい。いくつかの態様において、細胞が本発明に付随する1または2以上の遺伝子の内在性コピーを有する場合、本方法は内在的に発現する遺伝子の組換えコピーを付加することを必ずしも要求しない。いくつかの態様において、細胞は、本明細書に記載の経路からの1または2以上の酵素を内在的に発現してもよく、本明細書に記載の経路からの1または2以上の酵素を組換えにより発現してもよい。
【0046】
当然のことながら、本発明に適合するいくつかの細胞は、内在性生化学的経路を、1または2以上の組換え遺伝子のその経路、類似の経路、または相補的な経路とともに発現してもよい。例えば、R. eutrophaはモノマー産生のための遺伝子を発現する。実施例において示されているように、これらの細胞においてphaJが付加的に発現されて、PHAシンターゼによってポリマー化されるモノマー産生が増強された。
組織優先型プロモーター(tissue-preferred promoter)の使用を含む、モノマーを製造および/またはポリマー化するための関連する酵素の植物における発現によって、PHAポリマーおよびコポリマーを製造する方法は、例えば米国特許第5,534,432号および米国特許第7,341,856号に記載されており、いずれもこの教示について参照により本明細書に組み込まれる。
【0047】
興味のある生物または細胞のゲノムに存在する遺伝子からの発現および/または、例えばプラスミドなど、ベクターなどの1または2以上の染色体外核酸にある遺伝子からの発現を含む、生物および細胞における遺伝子の発現は当該技術分野においてよく知られている。修飾ゲノム、染色体外核酸およびかかる修飾ゲノムまたは染色体外核酸を含む生物または細胞を構築するための方法、材料および技術は、当該技術分野において周知であり、そのうちいくつかの方法、材料および技術は本明細書の他の場所に記載されている。
【0048】
いくつかの態様において、ポリ(HB−co−HHx)コポリマーなどの、高い中鎖長モノマー含有量を有する1または2以上のポリヒドロキシルアルカノアートコポリマーの製造のために最適化された細胞の使用が有利であり得る。例えば、生化学的経路の1または2以上の構成を変異して競合する経路を除去し、ポリ(HB−co−HHx)コポリマーなどの、高い中鎖長モノマー含有量を有する1または2以上のポリヒドロキシルアルカノアートコポリマーをより多く産生させることが最適であり得る。例えば、いくつかの態様において、3HB−CoA合成に寄与する1または2以上の遺伝子を辺または欠損させることにより、3HB−CoA合成を低減させる。いくつかの態様において、このことはアセトアセチル−CoAリダクターゼ活性の低減により達成することができる。例えばR. eutrophaにおいて、アセトアセチル−CoAリダクターゼ活性(PhaB)は、例えば1または2以上のphaB遺伝子を変異させることまたは(一部をまたは完全に)欠損させることによって、phaB発現を低減させることにより、低減させることができる。本明細書で例示されるように、これはphaB1、phaB2およびphaB3遺伝子をR. eutrophaゲノムから欠損させることにより達成できた。
【0049】
いくつかの態様において、ポリ(HB−co−HHx)コポリマーなどの、高い中鎖長モノマー含有量を有する1または2以上のポリヒドロキシルアルカノアートコポリマーの増強された産生に導く変異のスクリーニングが、ランダム変異原性スクリーニングまたは既知の変異のスクリーニングを通して実施されてもよい。いくつかの態様において、ポリ(HB−co−HHx)コポリマーなどの、高い中鎖長モノマー含有量を有する1または2以上のポリヒドロキシルアルカノアートコポリマーの増大した産生のためのこれらの断片を有する細胞または生物のスクリーニングを通して、ポリ(HB−co−HHx)コポリマーなどの、高い中鎖長モノマー含有量を有する1または2以上のポリヒドロキシルアルカノアートコポリマーの産生の増加に導くゲノム領域同定のためにゲノム断片のショットガンクローニングを用いることができる。いくつかの場合において、同一の細胞または生物中で1または2以上の変異が組み合わされてもよい。
【0050】
本明細書に記載された様々な細胞を生育するための培地は、当業者に周知である。例えば、米国特許第5,534,432号、これらの教示は参照として本明細書に組み込まれる、は、細菌細胞の生育に適したかかる培地の例を提供する。
当該技術分野における通常の技術を有するものには理解できるように、1または2以上のヒドロキシ酸およびそのコポリマーの産生のための最適な培養条件は、細胞のタイプ、生育培地および生育条件を含む多くの要因に影響され得る。培養温度は生物または細胞が生育可能な温度であり、好ましくは20℃から40℃である。例えば摂氏20、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35、36、37、38、39または40度、あるいはその間の任意の値であってよい。培養時間は特に限定されないが、約1から10日であってよい。
【0051】
ポリ(HB−co−HHx)コポリマーなどの、高い中鎖長モノマー含有量を有する1または2以上のポリヒドロキシルアルカノアートコポリマーの産生を最適化するために日常的な実験を通して変化し得る他の限定されない要因は、用いられる特定の炭素源、培養培地のタイプ、培養培地のpH、およびポリ(HB−co−HHx)を収穫する前に細胞を培養する時間量を含む。いくつかの態様において、3〜4日など、細胞を数日間培養した後に最適な産生が達成される。しかし当然のことながら、日常的な実験で、上記パラメータおよび他の類似のパラメータを変化および最適化できる。
【0052】
生物または細胞は、コポリマーの製造を可能にする炭素源を含む培養培地中で培養される。いくつかの態様において、任意の植物油、脂肪酸または脂肪酸誘導体あるいはその組み合わせなどの、任意の油および/または脂肪酸が炭素源として用いられる。植物油の例は、パーム油、大豆油、ナタネ油、コーン油、綿実油、ピーナッツ油、ココナッツ油、およびベニバナ油を含み、さらなる油または脂肪酸は当該技術分野において周知である。コポリマーの最適な製造のための炭素源組成物は、用いられる生物または細胞の特定の株に依存し得る。培養培地の他の構成成分は、当業者には知られているであろうように、いくつかの態様において窒素源、無機塩、(抗生物質などの)選択的成分、または他の栄養源などの様々な組み合わせを含む。
【0053】
本発明に関連する細胞を生育するのに用いる液体培養は、当該技術分野において知られ、および用いられる任意の培養容器に格納されることができる。いくつかの態様において、大量の、本発明に関連するポリ(HB−co−HHx)コポリマーなどの高い中鎖長モノマー含有量を有する1または2以上のポリヒドロキシルアルカノアートコポリマーの製造に、攪拌槽型反応器などの通気反応容器内での大規模製造を用いることができる。
【0054】
PHAポリマーおよびコポリマーを継続的に製造する方法もまた当業者に知られており、かかる方法は例えば米国特許第5,534,432号、これはこの教示について参照により本明細書に組み込まれる、などに記載されている。
PHAコポリマーは、当該技術分野において知られた方法で製造された生物、細胞または培養物から単離することができる。この一例は下記の実施例に示されているものであり、ポリマーは凍結乾燥された細胞から、クロロホルム中48時間で抽出されている。例えば、米国特許第5,942,597号、第5,918,747号、第5,899,339号、第5,849,854号および第5,821,299号、欧州特許公開公報第859858号、国際公開公報第97/07229号、第WO97/07230号および第WO97/15681号などもまた参照のこと、これらのそれぞれはこれらの教示について本明細書に参照として組み込まれる。
【0055】
1つの非限定的な例として、米国特許出願2009/0130731号に記載された方法を採用し得る。培養後、細菌細胞を遠心分離機などにより培養培地から分離し、細菌細胞を蒸留水およびメタノールなどで洗浄し、乾燥させる。乾燥させた細菌細胞から、クロロホルムなどの有機溶媒を用いてポリエステルを抽出する。細菌細胞構成成分は、このポリエステルを含む有機溶媒溶液からろ過などにより除去され、メタノールまたはヘキサンなどの貧溶媒をろ液に添加し、ポリエステルを沈殿させる。さらに、ろ過または遠心分離により上清を除去し、乾燥させる。他の方法は当業者に周知であろう。
【0056】
本発明によって製造されたコポリマーは、当該技術分野において知られた任意の多くの用途に用いることができる。例えば、米国特許第7,455,999号には、PHAポリマーの大量の用途およびポリマーの物性が「組成物の適用」と標題されたセクションおよびそこに引用された参照に記載されており、これらの開示のそれぞれは、その教示について参照として組み込まれる。
【0057】
いくつかの態様において、本発明は単離されたまたは実質的に精製されたPHAシンターゼ核酸またはポリペプチド、かかる核酸またはポリペプチドを含むまたは発現する構築物およびベクター、およびかかる核酸、ポリペプチド、構築物またはベクターを含む細胞または生物を包含する。本明細書に開示されるように、高濃度の3−ヒドロキシヘキサノアートモノマーを組み込むことができるRhodococcus aetherivorans I24からのPHAシンターゼが同定され、分子クローニング手法により単離された。単離されたPHAシンターゼはまた、本明細書において「Rhodococcus CO2シンターゼ」(配列番号1および2)または「Rhodococcus D12シンターゼ」(配列番号3および4)ともいう。
【0058】
「単離された」または「精製された」核酸分子またはタンパク質、またはその生物学的に活性な一部分は、実質的に他の細胞物質または組換え技術により製造された場合には培養培地を含まず、または化学的に合成された場合は実質的に化学物質前駆体または他の化学物質を含まない。好ましくは、「単離された」核酸は、核酸が由来する生物のゲノムDNAにおいては天然に隣接する核酸配列(好ましくはタンパク質コード配列)(すなわち核酸の5’および3’末端に存在する配列)を含まない。例えば、さまざまな態様において、単離された核酸分子は5kb、4kb、3kb、2kb、1kb、0.5kbまたは0.1kb未満の核酸が由来する生物のゲノムDNAにおいては天然に隣接する核酸配列を含み得る。細胞物質を実質的に含まないポリペプチド(タンパク質ともいう)は、(乾燥重量で)約30%、20%、10%、5%の不純物ポリペプチドを含むポリペプチドの調製物を含む。ポリペプチドまたはその生物学的に活性な一部分が組換えにより製造される場合、好ましくは培養培地が、(乾燥重量で)約30%、20%、10%または5%未満の化学物質前駆体またはポリペプチドではない他の構成物質に相当する。
【0059】
開示された核酸分子の断片および変異体およびそれによりコードされるポリペプチドもまた、本発明に包含される。「断片」により、ヌクレオチド配列の一部分およびそれによりコードされるアミノ酸配列の一部分が意図される。ヌクレオチド配列の断片は、天然ポリペプチドの生物学的活性を維持したポリペプチド断片をコードし得る。代替的に、ハイブリダイゼーションプローブとして有用なヌクレオチド配列の断片は、一般的に天然ポリペプチドの生物学的活性を維持したポリペプチド断片をコードしない。したがって、ヌクレオチド配列の断片は、少なくとも約20ヌクレオチド、約50ヌクレオチド、約100ヌクレオチド、および本明細書に開示のポリペプチドをコードする全長ヌクレオチドまでの範囲であってよい。
【0060】
ポリペプチドの生物学的に活性な一部分をコードする本発明のヌクレオチド配列の断片は、少なくとも15、25、30、50、100、150、200、250、300、350、400、450、500または550の隣接したアミノ酸または全長ポリペプチド中に存在する全アミノ酸数(例えば配列番号2については562アミノ酸または配列番号1については561アミノ酸)までをコードすることができる。ポリペプチドの生物学的に活性な一部分は、本明細書に開示されたヌクレオチド配列の1つの一部分を単離し、(例えばin vitroの組換え発現により)ポリペプチドのコードされた一部分を発現させ、ポリペプチドのコードされた一部分の活性を評価することにより調製できる。本明細書に開示のヌクレオチド配列の断片である核酸分子(ポリペプチドの生物学的に活性な断片をコードするかどうかにかかわらず)は、少なくとも15、20、30、45、60、75、100、150、200、250、300、350、400、450、500、550、600、650、700、800、900、1000、1100、1200、1300、1400、1500または1600ヌクレオチド、または本明細書に開示の全長ヌクレオチド配列に存在するヌクレオチド数(例えば配列番号1については1689ヌクレオチドまたは配列番号3については1686ヌクレオチド)までを含む。
【0061】
「変異体」により、本明細書に記載の配列と実質的に同様の配列を意図する。ヌクレオチド配列について、保存的変異体は、遺伝情報の縮退により、本発明のコポリマー合成に関与するポリペプチドまたは他の酵素の1つのアミノ酸配列をコードするそれらの配列を含む。これらのような天然に存在する対立遺伝子多型は、例えばポリメラーゼ連鎖反応(PCR)および以下に概説するハイブリダイゼーション技術などの周知の分子生物学の技術を用いて同定することができる。変異体ヌクレオチド配列はまた、例えば部位特異的変異導入(site-directed mutagenesis)などにより発生するが依然として本明細書に開示されたポリペプチドをコードするものなど、合成的に誘導されたヌクレオチド配列を含む。一般的に、本発明の特定のヌクレオチド配列の変異体は、本明細書の多の部分で記載された配列アラインメントプログラムにより、初期設定のパラメータを用いて決定された特定のヌクレオチド配列と、少なくとも50%、60%、65%、70%、75%、80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%またはそれ以上の配列同一性を有するだろう。
【0062】
本明細書に記載のヌクレオチド配列は、他の生物、特に他の細菌であるが本明細書に記載された他の生物または細胞も、から対応する配列を単離するのに用いることができる。このやり方において、PCR、ハイブリダイゼーション等の方法を、本明細書に記載の配列とのそれらの配列ホモロジーに基づき、かかる配列の同定に用いることができる。本明細書に記載された全ヌクレオチド配列またはその断片との配列同一性に基づいて単離された配列は、本発明に包含される。かかる配列は、開示された配列のオルソログである配列を含む。「オルソログ」により、種分化の結果、共通の祖先遺伝子に由来し異なる種において見出される遺伝子を意図する。異なる種において見出される遺伝子は、本明細書の他の場所で定義されるように、そのヌクレオチド配列および/またはそのコードするポリペプチド配列が実質的同一性を共有する場合に、オルソログであると考えられる。
【0063】
PCRベースの取り組みにおいて、オリゴヌクレオチドプライマーは、PCR反応において使用して、対応するDNA配列を任意の興味のある生物から抽出したcDNAまたはゲノムDNAから増幅するために設計できる。PCRプライマーの設計およびPCRクローニングのための方法は一般的に当該技術分野に知られており、例えばSambrook et al. (1989) Molecular Cloning: A Laboratory Manual(2d ed., Cold Spring Harbor Laboratory Press, Plainview, N.Y.)などに開示されている。Innis et al., eds.(1990)PCR Protocols: A Guide to Methods and Applications(Academic Press, New York)、Innis and Gelfand, eds.(1995)PCR Strategies(Academic Press, New York)、およびInnis and Gelfand, eds.(1999)PCR Methods Manual(Academic Press, New York)も参照のこと。PCRの既知の方法は、これに限定するものではないが、ペアプライマー、ネステッドプライマー、単一の特異的プライマー、縮退プライマー、遺伝子特異的プライマーベクター特異的プライマー部分的ミスマッチプライマーなどを用いる方法を含む。
【0064】
ハイブリダイゼーション技術において、基地のヌクレオチド配列の全部または一部が、選ばれた生物または細胞からクローンされたゲノムDNA断片またはcDNA断片の集団(ゲノムまたはcDNAライブラリーなど)の中に存在する他の対応するヌクレオチド配列と選択的にハイブリダイズするプローブとして用いられる。ハイブリダイゼーションプローブは、ゲノムDNA断片、cDNA断片、RNA断片または他のオリゴヌクレオチドであってよく、当該技術分野において周知の任意の検出可能なマーカーで標識されていてよい。したがって、例えば、ハイブリダイゼーションのためのプローブは、本明細書に開示されたヌクレオチド配列またはその縮退物に基づいて、標識合成オリゴヌクレオチドにより作られ得る。ハイブリダイゼーションのためのおよびcDNAおよびゲノムライブラリーの構築のためのプローブを調製するための方法は当該技術分野において知られており、例えばSambrook et al. (1989) Molecular Cloning: A Laboratory Manual(2d ed., Cold Spring Harbor Laboratory Press, Plainview, N.Y.)などに開示されている。
【0065】
かかる配列のハイブリダイゼーションは、ストリンジェントな条件化で行われてよい。「ストリンジェントな条件」または「ストリンジェントなハイブリダイゼーション条件」により、プローブがその標的配列に、他の配列に比べて検出可能に多量な程度(例えば、少なくともバックグラウンドの2倍以上)にハイブリダイズするであろう条件を意図する。ストリンジェントな条件は配列依存であり、異なる状況において異なるであろう。ハイブリダイゼーションおよび/または洗浄条件のストリンジェンシーのコントロールにより、プローブに100%相補的な標的配列を同定できる(相同的プロービング(homologous probing))。代替的に、ストリンジェンシー条件を調整して、配列におけるいくつかのミスマッチを許容し、より低い程度の類似性を検出することもできる(異種性プロービング(heterologous probing))。一般的に、プローブは約1000ヌクレオチド長未満であり、好ましくは500ヌクレオチド長未満である。ストリンジェントなハイブリダイゼーション条件下でのハイブリダイゼーションにおいて用いられるさまざまな技術、方法、条件および組成物は、当該技術分野において周知であり、Tijssen(1993)Laboratory Techniques in Biochemistry and Molecular Biology--Hybridization with Nucleic Acid Probes, Part I, Chapter 2(Elsevier, N.Y.)、およびAusubel et al., eds.(1995)Current Protocols in Molecular Biology, Chapter 2(Greene Publishing and Wiley-Interscience, New York)などの参照文献中に見出すことができる。Sambrook et al. (1989) Molecular Cloning: A Laboratory Manual(2d ed., Cold Spring Harbor Laboratory Press, Plainview, N.Y.)参照のこと。
【0066】
本明細書に開示されたPHAシンターゼ配列にストリンジェントな条件下でハイブリダイズする単離された配列またはその断片は、本発明に包含される。いくつかの態様において、かかる配列は、開示された配列と少なくとも約50%、60%、70%、75%、80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%またはそれ以上の相同性であるだろう。つまり、配列の配列同一性は、開示された配列と少なくとも約50%、60%、70%、75%、80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%またはそれ以上の配列同一性を共有してよい。
【0067】
比較のための配列のアラインメントの方法は、当該技術分野において周知である。任意の2配列間のパーセント配列同一性の決定は、数学的アルゴリズムを用いて達成することができる。かかる数学的アルゴリズムの非限定的な例は、Myers and Miller (1988) CABIOS 4:11-17のアルゴリズム、Smith et al. (1981) Adv. Appl. Math. 2:482のローカルホモロジーアルゴリズム、Needleman and Wunsch (1970) J. Mol. Biol. 48:443-453のホモロジーアラインメントアルゴリズム、Pearson and Lipman (1988) Proc. Natl. Acad. Sci. 85:2444-2448の類似性検索法(search-for-similarity-method)、Karlin and Altschul (1990) Proc. Natl. Acad. Sci. USA 87: 2264-2268のアルゴリズム、Karlin and Altschul (1993) Proc. Natl. Acad. Sci. USA 90:5873-5877のように改変したものなどである。
【0068】
これらの数学的アルゴリズムのコンピュータによる実現は、配列を比較して配列同一性を決定するのに利用できる。かかる実現は、これに限定するものではないが、PC/遺伝子プログラムにおけるCLUSTAL(Intelligenetics, Mountain View, Calif.から購入可能)、ALIGNプログラム(バージョン2.0)およびWisconsin Genetics Software Package, Version 8中のGAP、BESTFIT、BLAST、FASTAおよびTFASTA(Genetics Computer Group(GCG), 575 Science Drive, Madison, Wis., USAから購入可能)を含む。BLASTおよび関連するプログラムもまた(他の配列比較プログラムと同様)インターネットを介して、国立生物工学情報センター(NCBI:例えばwww.ncbi.nlm.nih.gov or blast.ncbi.nlm.nih.gov/Blast.cgi参照のこと)のウェブサイトで利用可能である。このプログラムを用いたアラインメントは、初期設定のパラメータを用いて実行することができる。Altschul et al (1990) J. Mol. Biol. 215:403のBLASTプログラムは、Karlin and Altschul (1990) のアルゴリズムに基づいている。BLASTヌクレオチド検索は、例えばBLASTNプログラムで、スコア=100、文字長=12で実行し、本発明の明細書に記載のポリペプチドをコードするヌクレオチド配列と相同なヌクレオチド配列を得ることができる。BLASTタンパク質検索は、BLASTXプログラムで、スコア=50、文字長=3で実行し、本発明の明細書に記載のポリペプチドのアミノ酸配列と相同なアミノ酸配列を得ることができる。比較の目的でギャップアラインメントを得るため、Altschul et al. (1997) Nucleic Acids Res. 25:3389-3402に記載されているように、Gapped BLAST(BLAST2.0において)を用いることができる。代替的に、PSI-BLAST(BLAST2.0において)を用いて分子間の距離関係を検出する反復検索を実行することができる。Altschul et al. (1997)参照のこと。BLAST、Gapped BLAST、PSI-BLASTを用いる場合、それぞれのプログラム(例えばヌクレオチド配列についてBLASTN、タンパク質についてBLASTXなど)の初期設定パラメータを用いることができる。
【0069】
いくつかの態様において、本明細書に記載の1または2以上の遺伝子は、1または2以上の組換え発現ベクターにおいて発現する。かかるベクターは、オペロン配置においてなど、独立してまたは組み合わせて遺伝子を含み得る。本明細書で用いる場合、「ベクター」は、異なる遺伝環境間の輸送のためまたは宿主細胞における発現のため、所望の配列または配列(複数)が制限およびライゲーションにより挿入され得る、任意の数の核酸であり得る。ベクターは、RNAベクターもまた利用可能であるものの、典型的にはDNAからできている。ベクターはこれに限定するものではないが、プラスミド、フォスミド、ファージミド、ウイルスゲノムおよび人工染色体を含む。
【0070】
クローニングベクターは、自発的に複製することができるまたは宿主細胞のゲノムに統合されることができるものであり、さらにベクターが決定可能なやり方で切断され得、新たな組換えベクターがその宿主細胞における複製能力を維持するように所望のDNA配列がライゲーションされ得る、1または2以上のエンドヌクレアーゼ制限部位により定義されるものである。プラスミドの場合、所望の配列の複製は、プラスミドが宿主細菌などの宿主細胞内でコピー数を増加させるにつれ何度も起こり得、または有糸分裂により宿主が増殖する前に宿主ごとに一度だけ起こり得る。ファージの場合、複製は溶解相の間能動的に起こり得、または溶原性相の間受動的に起こり得る。
【0071】
発現ベクターは、所望のDNA配列が、制限およびライゲーションにより作動可能に制御配列に加わるように挿入され得、RNA転写物として発現し得るものである。ベクターはさらに、該ベクターにより変換または形質転換されたかまたはされていない細胞の同定に用いるのに適した1または2以上のマーカー配列を含んでもよい。マーカーは、例えば、抗生物質又は他の化合物に対する抵抗性または感受性を増加するまたは低減するタンパク質をコードする遺伝子、当該技術分野において知られた標準的なアッセイによってその活性を検出可能な酵素をコードする遺伝子(例えばβ−ガラクトシダーゼ、ルシフェラーゼまたはアルカリホスファターゼ)、および変換または形質転換された細胞、宿主、コロニーまたはプラークのフェノタイプに視覚的に影響する遺伝子(例えば緑色蛍光タンパク質)を含む。好ましいベクターは、自立した複製および作動可能に加わったDNAセグメントに存在する構造的遺伝子産物の発現が可能なものである。
【0072】
本明細書で使用される場合、コード配列および制御配列は、制御配列の影響下または制御下でコード配列が発現または転写される状態に置かれるように共有結合された場合、「作動可能に(operably)」加えられると言われる。コード配列が機能的タンパク質に翻訳されることが所望される場合、5’制御配列におけるプロモーターの導入がコード配列の転写の結果となり、2つのDNA間の結合の性質が(1)フレームシフト変異の導入の結果とならず、(2)プロモーター領域がコード配列の転写を指示する能力を妨害せず、または(3)対応するNA転写物がタンパク質に翻訳される能力を妨害しない場合に、2つのDNA配列が作動可能に加えられると言われる。したがって、得られた転写物が所望のタンパク質またはポリペプチドに翻訳され得るようにプロモーター領域がDNA配列の転写に作用できる場合、プロモーター領域はコード配列に作動可能に加えられるということになる。
【0073】
クレームされた発明の任意の酵素をコードする核酸分子が細胞内で発現された場合、その発現を指示するのに様々な転写制御配列(例えばプロモーター/エンハンサー配列など)を用いることができる。プロモーターは、遺伝子の発現の正常な制御を提供する、天然のプロモーター、すなわちその内在性の構成成分である遺伝子のプロモーター、であり得る。いくつかの態様において、プロモーターは構成的、すなわちその関連する遺伝子の連続的な転写を許容するように制御されていない、であり得る。分子の存在または比存在によりコントロールされるプロモーターなど、様々な条件的プロモーターもまた用いられ得る。
【0074】
遺伝子発現に必要な制御配列の正確な性質は、種または細胞型の間で変化してもよいが、一般的に、必要に応じて、TATAボックス、キャッピング配列、CAAT配列など、それぞれ転写および翻訳の開始に関与する5’非転写および5’非翻訳配列を含む。特に、かかる5’非転写制御配列は、作動可能に加えられた遺伝子の転写コントロールのためのプロモーター領域を含むだろう。制御配列はまた、記載されているように、エンハンサー配列または上流のアクチベーター配列を含んでよい。本発明のベクターは、任意に、5’リーダーまたはシグナル配列を含んでよい。適切なベクターの選択および設計は、当業者の能力および裁量の範囲内である。
【0075】
発現に必要な全ての要素を含む発現ベクターは、商業的に入手可能であり、当業者に知られている。例えばSambrook et al., Molecular Cloning: A Laboratory Manual, Second Edition, Cold Spring Harbor Laboratory Press, 1989を参照のこと。細胞は、異種性DNA(RNA)の細胞内への導入により、遺伝子操作されている。その異種性DNA(RNA)は、宿主細胞において該異種性DNAを発現できるように、転写要素の作動可能なコントロール下に置かれている。高い中鎖長モノマー含有量を有するポリヒドロキシアルカノアートコポリマー、特に高いHHx含有量を有するポリ(HB−co−HHx)、の産生を促進する遺伝子配列の異種性発現は、実施例のセクションにおいて実証されている。高いHHx含有量を有するポリ(HB−co−HHx)などの、高い中鎖長モノマー含有量を有するポリヒドロキシアルカノアートコポリマーを製造するための新規な方法もまた、他の細菌細胞、古細菌、真菌(酵母を含む)、哺乳類細胞、植物細胞などにおいて実施できる。
【0076】
本明細書に記載された酵素をコードする核酸分子は、当該技術分野において標準的な方法および技術を用いて、細胞または細胞(複数)に導入することができる。例えば核酸分子は、化学的形質転換およびエレクトロポレーション、形質導入、粒子衝突などを含む形質転換などの標準的なプロトコルにより導入できる。クレームされた発明の酵素をコードする1または2以上の核酸分子の発現はまた、核酸分子を細胞のゲノム内に組み込むことによっても達成され得る。
【実施例】
【0077】
例1:PhaB活性が低減した菌株
モノマー3−ヒドロキシブチリル−CoA(3HB−CoA)は、β−ケトチオラーゼ(PhaA)およびアセトアセチル−CoAリダクターゼ(PhaB)により、アセチル−CoAから合成される。Ralstonia eutrophaゲノムにおけるタンパク質配列の分析は、このタンパク質のよく研究されたバージョン(PhaAはH16_A1438、PhaB1はH16_A1439)に対する多くの潜在的ホモログを予測する。3HB−CoA合成を防ぐため、我々は遺伝子phaB1、phaB2およびphaB3をR. eutrophaゲノムからきれいに欠損させた。
【0078】
マーカー無しの欠損は、York[1]出典の方法を用いて行った。興味有る遺伝子の上流および下流のDNA配列をPCRで増幅した。この配列を、オーバーラップPCRを介して組み合わせて単一の隣接した一続きのDNAとした。この手法の間用いられたプライマーは、BamHI部位がDNAの終端に付加され、SwaI部位が上流および下流領域の間に挿入されるように設計された。この構築物はpGY46の骨格のBamHI部位にクローニングされ、R. eutrophaにおける興味有る遺伝子のマーカー無しの欠損を作るのに用いられるプラスミドを作製した[2]。プラスミドはE. coli S17-1に形質転換され、接合を介してR. eutrophaに導入した。欠損は、興味有る遺伝子の上流および下流にハイブリダイズする診断プライマーを用いたPCRにより確認された。
【0079】
全てのphaB遺伝子が、独立しておよび組み合わせて欠損した菌株のシリーズを構築した(下記表1参照のこと)。これらの菌株は、フルクトースを唯一の炭素源とする限定培地で生育された。PHB蓄積は窒素を限定することにより誘導した。
【表1】
【0080】
これらの培養物からのサンプルを様々な時点において取り、ポリ(3−ヒドロキシブチラート)(PHB)含有量について、クロトン酸アッセイ[3]によりアッセイした(図5)。phaB1およびphaB3の両方を欠失する菌株(すなわちRe2113およびRe2115)は、野生型菌株よりも顕著に少ないPHBを産生した。NADPHおよびNADH依存性アセトアセチル−CoAリダクターゼ活性は、確立された方法[4]により計測した(図6、7)。我々は、低量のPHBを作る菌株は、顕著に低減したNADPH依存性リダクターゼレベルを有し、このことはPHB蓄積における減少が実際に、PhaB活性の排除に起因する3HB−CoA合成の欠失に起因することを提示する。
【0081】
我々は、phaB欠損を有する菌株における相補的リダクターゼ変異の効果を検討した。phaB1、phaB2およびphaB3遺伝子を独立して菌株Re2115(ΔphaB123)のゲノムのphaB1遺伝子座に付加して戻し、菌株Re2139、Re2140およびRe2143を作製した。これらの実験の途中で、phaB3の開始コドンが誤って注釈されていることが発見され、正しく注釈された配列を用いた(表2においてphaB3correctと表記されている)。脂肪酸合成に関与するリダクターゼをコードするfabG遺伝子もまた、菌株Re2115(ΔphaB123)のphaB1遺伝子座に付加され、菌株Re2142を作製した。全ての挿入された遺伝子は同一のリボソーム結合部位を有する。
【0082】
【表2】
これらの培養物からのサンプルを様々な時点において取り、ポリ(3−ヒドロキシブチラート)(PHB)含有量について、クロトン酸アッセイ[3]によりアッセイした(図8)。
図9および図10にそれぞれ示されているように、PhaA活性およびこれらの菌株により産生されたPHBポリマーの分子量もまた計測された。PhaA活性は[6]に記載の方法により計測された。
【0083】
コポリマーの分子量は、ポリスチレン標準を用いたゲル浸透クロマトグラフィによって計測した。PHBを凍結乾燥した細胞から、クロロホルム中48時間で抽出した。PHB溶液を3mg/mLで調製した。抽出の後、溶液をろ過して不溶性のバイオマスを除去した。溶解したポリマーを、PLgel Olexisガードカラム(Polymer Laboratoriesパーツ番号PL1110-1400)および2つの一連のPLgel Olexis分析カラム(Polymer Laboratoriesパーツ番号PL1110-6400)を装備したAgilent 1100 HPLCを用いて分析した。100μLの各溶液を注入し、カラムから溶出される際に屈折率検出器によりポリマーを検出した。分子量を、Agilent GPC分析ソフトウェアを用いて、得られたクロマトグラムから決定した。システムを、1,110から13,155,000g/molまでのサイズ範囲の一連のポリスチレン標準(Polymer Laboratoriesパーツ番号PL2010-0104)を用いて校正した。イソプロパノールを全ての校正標準および実験サンプルに内部標準として含ませた。
【0084】
例2:ポリ(3−ヒドロキシブチラート−co−3−ヒドロキシヘキサノアート)製造のための菌株
我々は、限定された3HB−CoA合成能力を有する菌株は、高いHHx含有量を有する3−ヒドロキシブチラートと3−ヒドロキシヘキサノアートとのコポリマー(ポリ(HB−co−HHx))の合成が可能な菌株の設計においてよい出発点になるだろうと予測した。この予測は、シンターゼが3HB−CoAおよび3HHx−CoAをポリマー化可能であったとしても、野生型R. eutrophaにおける3HB−CoAの高い細胞内濃度が、ポリマーへのHHx組み込みを限定するだろうという我々の信念に基づいていた。加えて、R. eutrophaシンターゼは3HB−CoAと3HV−CoAとをポリマー化するのみである。PhaB基質特異性は図11に示されている(参考文献[4]も参照のこと)。
【0085】
我々の予測を試験するため、我々は最初に、pGY46を用いて天然のPHAシンターゼ(phaC1)をRe2115から欠損させ[2]、菌株Re2133を作製した。我々はそこでAeromonas caviaeからのおよびRhodococcus aetherivorans I24(D12およびC09シンターゼ)からの他のシンターゼを試験した。例えば、Rhodococcus aetherivorans I24からのD12シンターゼを、[2]出典の手法を用いて、Re2133ゲノムのphaC1遺伝子座に挿入した。これは、最初にpGY46中のDNAの上流および下流領域の間に部位特異的変異導入を介してSwaI部位を挿入し、次にD12シンターゼ遺伝子をSwaI部位にクローニングして入れることで、菌株Re2135を作製した。
【0086】
菌株Re2135を、パーム油を炭素源として用いて、フラスコの中で生育した。我々は3HHx−CoAモノマーが脂肪酸代謝の副産物として作られ(図4参照)、したがって油/脂肪酸を炭素源として使用することがコポリマー産生における不可欠であると信じていた。これらの培養物におけるPHA蓄積は、窒素限定により誘導された。細胞およびポリマーの組成におけるポリマー含有量を、標準的なメタノリシス手法[5]により計測した(図14)。この方法は、PHAモノマーを、関連するメチルエステルに変換し、それをガスクロマトグラフィーにより分離および定量する。我々は、Re2135が少量のPHAを蓄積するが、このPHAは高レベルのHHxモノマーを含むことを見出した。
【0087】
我々は次に、この菌株においてモノマー合成を増大させる遺伝子について調査した。文献からのいくつかの菌株が図13に記載してあり、これには用いられたPHAシンターゼ、炭素源、PHA含有量およびHHxのmol%が示されている。
我々はエノイル−CoAヒドラターゼをコードする遺伝子(phaJ遺伝子)に焦点を当てた。いくつかのphaJがRe2135ゲノムのphaB1遺伝子座に挿入された:Aeromonas caviaeおよびPseudomonas aeruginosa(phaJ1およびphaJ2)からのエノイル−CoAヒドラターゼ。PhaJ基質特異性は図12に示されている([7]も参照のこと)。
【0088】
遺伝子は、当初はB1欠損ベクターであったものの、上流および下流DNA領域の間のSwaI部位にクローニングされた。遺伝子は上記の手法を用いてゲノムに挿入され、パーム油を唯一の炭素源とする限定培地中で生育した。我々は、Pseudomonas aeruginosa PAO1からの遺伝子phaJ1が、依然として高いHHx含有量を含みながら、より多量のポリマーの産生に導くことを見出した。phaJ1をゲノムに含む菌株をRe2152と名付けた。
【0089】
菌株構築および結果は、遺伝子型(用いたPHAシンターゼを含む)、PHA含有量(細胞乾燥重量の%として)、およびHHxの重量%を示す、図14に記載されている。mol%HHxとして計測されたHHx含有量は、常に重量%として計測されたHHx含有量よりも小さい数字であった。mol%は、重量%の値に0.8を掛けることにより、重量%から概算できる。例えば、25重量%のHHxは20mol%のHHxに相当し、30重量%のHHxは24mol%のHHxに相当する。
【0090】
我々は、我々の新たに開発したPHAオペロン(phaCD12−phaA−phaJ1Pa)における酵素的ステップの1つが、PHA産生を限定しているだろうと仮説を立てた。遺伝子発現を増大させるため、我々はこのオペロンをRe2152からPCRによって増幅し、プラスミドpBBR1MCS−2のKpnIとHindIIIとの間にクローニングした。この新たなプラスミド(pCB81)をRe2133に形質転換し、得られた菌株を、300μg/mLのカナマイシンを含有するパーム油限定培地で生育した。手法は図15に記載されている。このように増幅された遺伝子発現がPHA産生または得られたPHAのモノマー組成にネガティブに影響するかを決定するため、我々は得られた菌株によって産生されたポリマーにおけるPHAおよびHHX含有量を分析した。予想外にも、培養物の分析は、細胞が高いPHA含有量および高レベルのHHxをポリマー内に有するコポリマーを産生したことを示した。
我々は、共通して、25〜30重量%のHHx、これは20〜24mol%のHHxに相当する、を有するコポリマーを得た。33重量%(27mol%)までのHHx値を有するコポリマーが、菌株をパーム油で生育した場合に得られた。
【0091】
例3:ポリマー特性評価
新たに開発したPHAオペロン(phaCD12−phaA−phaJ1Pa)を含む菌株により産生されたPHAコポリマーの特性を決定した。PHAコポリマーの分子量は、ゲル浸透クロマトグラフィにより、ポリスチレン標準と比較して120,000〜150,000g/molであることが見出された。
【0092】
OHAコポリマーの熱的特性を、示差走査熱量測定を用いて計測した。サンプルをアルミニウムの小皿に乗せ、Perkin ElmerのPyris 1 DSCを用いて分析した。用いた温度プログラムは、(1)50℃で1分間維持、(2)20℃/分で−40℃まで冷却、(3)−40℃で3分維持、(4)20℃/分で200℃まで加熱、(5)200℃で1分維持、(6)20℃/分で−40℃まで冷却、(7)−40℃で3分維持、(8)20℃/分で50℃まで加熱、であった。ガラス転移温度を、吸熱勾配に変化が起こる温度として同定した。融点を、吸熱の最高ピークとして同定した。DSC分析は、上記の菌株により産生された、27mol%のHHxを含有するコポリマーは−4℃のガラス転移温度を有することを明らかにした。
【0093】
例4:Rhodococcus aetherivorans I24のPHAシンターゼについての配列データ
潜在的なシンターゼ遺伝子を、既知のシンターゼペプチドの配列に対してRhodococcus aetherivorans I24ゲノム配列BLASTNすることにより同定した。Rhodococcus aetherivorans I24のゲノムにおける予測されるタンパク質配列の分析に基づき、2つのPHAシンターゼがゲノムからクローニングされ、それはまた本明細書においてCO9シンターゼおよびD12シンターゼと言われる。両方のPHAシンターゼとも、R.eutrophaにおいて(上記の通り)発現したときに、活性であると決定された。
【0094】
BLASTN配列分析は、クローニングされたPHAシンターゼの両方の遺伝子が、よく研究されたシンターゼのうちシュードモナスからのシンターゼに最も近いことを示した。これらシュードモナスのシンターゼは非常に広範な基質特異性を有することが知られている。CO9シンターゼおよびD12シンターゼはC7〜C8までの基質をポリマー化し、これはR. eutrophaからのシンターゼよりも広い基質特異性である。
【0095】
【表3】
【0096】
【表4】
【0097】
【表5】
【0098】
【表6】
【0099】
【表7】
【0100】
当業者であれば、本明細書に記載された発明の特定の態様に対する多くの均等物を認識する、または日常的な実験を超えない程度の実験を用いて突き止められるだろう。かかる均等物は、特許請求の範囲に包含されることを意図する。
本明細書に開示された全ての参考文献は、本明細書に引用された目的のために、その全開示が参照により組み込まれる。
【特許請求の範囲】
【請求項1】
任意の植物油を炭素源として使用して、少なくとも約4mol%または5重量%の中鎖長モノマー含有量を有するポリヒドロキシルアルカノアートコポリマーを産生する細胞。
【請求項2】
細胞が、任意の植物油を炭素源として使用して、少なくとも約4mol%または5重量%のHHx含有量を有する3−ヒドロキシルブチラートと3−ヒドロキシルヘキサノアートとのコポリマー(ポリ(HB−co−HHx))を産生する、請求項1に記載の細胞。
【請求項3】
細胞中の3−ヒドロキシルブチラートの正常な合成が破壊されている、請求項1または2に記載の細胞。
【請求項4】
アセトアセチル−CoAリダクターゼをコードする遺伝子が欠損している、請求項3に記載の細胞。
【請求項5】
細胞が、Ralstonia eutropha細胞であり、1または2以上のphaB1、phaB2およびphaB3遺伝子が破壊されている、請求項4に記載の細胞。
【請求項6】
phaB3遺伝子が破壊されている、請求項5に記載の細胞。
【請求項7】
細胞が、非内在性のPHAシンターゼ遺伝子を組換えにより発現する、請求項1〜6のいずれか一項に記載の細胞。
【請求項8】
非内在性PHAシンターゼ遺伝子が、Aeromonas caviaePHAシンターゼ遺伝子またはRhodococcus aetherivoransPHAシンターゼ遺伝子である、請求項7に記載の細胞。
【請求項9】
Rhodococcus aetherivoransPHAシンターゼ遺伝子が、配列番号4をコードするRhodococcus aetherivorans I24 D12 PHAシンターゼ遺伝子または配列番号2をコードするRhodococcus aetherivorans I24 C09 PHAシンターゼ遺伝子である、請求項8に記載の細胞。
【請求項10】
Rhodococcus aetherivorans I24 D12 PHAシンターゼ遺伝子が、配列番号3を含むかまたは配列番号3からなる、および/あるいはRhodococcus aetherivorans I24 C09 PHAシンターゼ遺伝子が、配列番号1もしくは配列番号1の開始コドンがTTGからATGに変えられたものを含むかまたは配列番号1もしくは配列番号1の開始コドンがTTGからATGに変えられたものからなる、請求項9に記載の細胞
【請求項11】
細胞が、エノイル−CoAヒドラターゼ遺伝子を組換えにより発現する、請求項1〜10のいずれか一項に記載の細胞。
【請求項12】
エノイル−CoAヒドラターゼ遺伝子が、Aeromonas caviaeエノイル−CoAヒドラターゼ遺伝子またはPseudomonas aeruginosaエノイル−CoAヒドラターゼ遺伝子である、請求項11に記載の細胞。
【請求項13】
Pseudomonas aeruginosaエノイル−CoAヒドラターゼ遺伝子が、Pseudomonas aeruginosa phaJ1遺伝子(遺伝子PA3302)またはPseudomonas aeruginosa phaJ2遺伝子(遺伝子PA1018)である、請求項12に記載の細胞。
【請求項14】
非内在性PHAシンターゼ遺伝子および/またはエノイル−CoAヒドラターゼ遺伝子が、増幅される、請求項7〜13のいずれか一項に記載の細胞。
【請求項15】
モノマー含有量が、少なくとも約5mol%である、請求項1〜14のいずれか一項に記載の細胞。
【請求項16】
モノマー含有量が、少なくとも約6mol%である、請求項15に記載の細胞。
【請求項17】
モノマー含有量が、少なくとも約7mol%である、請求項16に記載の細胞。
【請求項18】
モノマー含有量が、少なくとも約10mol%である、請求項17に記載の細胞。
【請求項19】
モノマー含有量が、少なくとも約15mol%である、請求項18に記載の細胞。
【請求項20】
モノマー含有量が、少なくとも約20mol%である、請求項19に記載の細胞。
【請求項21】
モノマー含有量が、少なくとも約6重量%である、請求項1〜14のいずれか一項に記載の細胞。
【請求項22】
モノマー含有量が、少なくとも約8重量%である、請求項21に記載の細胞。
【請求項23】
モノマー含有量が、少なくとも約10重量%である、請求項22に記載の細胞。
【請求項24】
モノマー含有量が、少なくとも約15重量%である、請求項23に記載の細胞。
【請求項25】
モノマー含有量が、少なくとも約20重量%である、請求項24に記載の細胞。
【請求項26】
モノマー含有量が、少なくとも約25重量%である、請求項25に記載の細胞。
【請求項27】
細胞が、細菌細胞、真菌細胞(酵母細胞を含む)、植物細胞、昆虫細胞または動物細胞である、請求項1〜26のいずれか一項に記載の細胞。
【請求項28】
細胞が、細菌細胞または真菌細胞である、請求項27に記載の細胞。
【請求項29】
細胞が、Ralstonia菌種、Aeromonas菌種、Rhizobium菌種、Alcaligenis菌種またはPseudomonas菌種の細胞である、請求項28に記載の細胞。
【請求項30】
細胞が、Ralstonia eutropha、Aeromonas caviae、Rhizobium japonicum、Alcaligenis eutrophusまたはPseudomonas oleovorans細胞である、請求項29に記載の細胞。
【請求項31】
細胞が、Ralstonia eutropha細胞である、請求項30に記載の細胞。
【請求項32】
非内在性PHAシンターゼ遺伝子および/またはエノイル−CoAヒドラターゼ遺伝子が、プラスミドから発現される、請求項7〜31のいずれか一項に記載の細胞。
【請求項33】
非内在性PHAシンターゼ遺伝子および/またはエノイル−CoAヒドラターゼ遺伝子が、細胞のゲノムに組み込まれている、請求項7〜31のいずれか一項に記載の細胞。
【請求項34】
高い中鎖長モノマー含有量を有するポリヒドロキシルアルカノアートコポリマーを製造する方法であって、該方法が、請求項1〜33のいずれか一項に記載の細胞を培養し、少なくとも約4mol%または5重量%の中鎖長モノマー含有量を有するコポリマーを製造することを含む、前記方法。
【請求項35】
方法が、請求項2〜33のいずれか一項に記載の細胞を培養し、少なくとも約4mol%または5重量%のHHx含有量を有する(ポリ(HB−co−HHx))を製造することを含み、ここで3−ヒドロキシルブチラートと3−ヒドロキシルヘキサノアートとのコポリマー(ポリ(HB−co−HHx))が製造される、請求項34に記載の方法。
【請求項36】
コポリマーを細胞から回収することをさらに含む、請求項34または35に記載の方法。
【請求項37】
製造されるコポリマーの量が、細胞の乾燥重量の少なくとも約20%である、請求項35または36に記載の方法。
【請求項38】
製造されるコポリマーの量が、細胞の乾燥重量の少なくとも約30%である、請求項37に記載の方法。
【請求項39】
製造されるコポリマーの量が、細胞の乾燥重量の少なくとも約40%である、請求項38に記載の方法。
【請求項40】
製造されるコポリマーの量が、細胞の乾燥重量の少なくとも約50%である、請求項39に記載の方法。
【請求項41】
製造されるコポリマーの量が、細胞の乾燥重量の少なくとも約60%である、請求項40に記載の方法。
【請求項42】
少なくとも約4mol%または5重量%の中鎖長モノマー含有量を有するポリヒドロキシルアルカノアートコポリマーを産生する細胞を製造する方法であって、細胞中に少なくとも1つのRhodococcus aetherivoransPHAシンターゼ遺伝子を組み換えにより発現することを含む、前記方法。
【請求項43】
細胞が、少なくとも約4mol%または5重量%のHHx含有量を有する3−ヒドロキシルブチラートと3−ヒドロキシルヘキサノアートとのコポリマー(ポリ(HB−co−HHx))を産生する、請求項42に記載の方法。
【請求項44】
Rhodococcus aetherivoransPHAシンターゼ遺伝子が、配列番号4をコードするRhodococcus aetherivorans I24 D12 PHAシンターゼ遺伝子または配列番号2をコードするRhodococcus aetherivorans I24 C09 PHAシンターゼ遺伝子である、請求項42または43に記載の方法。
【請求項45】
Rhodococcus aetherivorans I24 D12 PHAシンターゼ遺伝子が、配列番号3を含むかまたは配列番号3からなる、および/あるいはRhodococcus aetherivorans I24 C09 PHAシンターゼ遺伝子が、配列番号1もしくは配列番号1の開始コドンがTTGからATGに変えられたものを含むかまたは配列番号1もしくは配列番号1の開始コドンがTTGからATGに変えられたものからなる、請求項42〜44のいずれか一項に記載の方法。
【請求項46】
エノイル−CoAヒドラターゼ遺伝子を組換えにより発現することをさらに含む、請求項42〜45のいずれか一項に記載の方法。
【請求項47】
エノイル−CoAヒドラターゼ遺伝子が、Aeromonas caviaeエノイル−CoAヒドラターゼ遺伝子またはPseudomonas aeruginosaエノイル−CoAヒドラターゼ遺伝子である、請求項46に記載の方法。
【請求項48】
Pseudomonas aeruginosaエノイル−CoAヒドラターゼ遺伝子が、Pseudomonas aeruginosa phaJ1遺伝子(遺伝子PA3302)またはPseudomonas aeruginosa phaJ2遺伝子(遺伝子PA1018)である、請求項47に記載の細胞。
【請求項49】
非内在性PHAシンターゼ遺伝子および/またはエノイル−CoAヒドラターゼ遺伝子が、増幅される、請求項42〜48のいずれか一項に記載の方法。
【請求項50】
細胞が、細菌細胞、真菌細胞(酵母細胞を含む)、植物細胞、昆虫細胞または動物細胞である、請求項42〜49のいずれか一項に記載の方法。
【請求項51】
細胞が、細菌細胞または真菌細胞である、請求項50に記載の方法。
【請求項52】
細胞が、Ralstonia菌種、Aeromonas菌種、Rhizobium菌種、Alcaligenis菌種またはPseudomonas菌種の細胞である、請求項51に記載の方法。
【請求項53】
細胞が、Ralstonia eutropha、Aeromonas caviae、Rhizobium japonicum、Alcaligenis eutrophusまたはPseudomonas oleovoransの細胞である、請求項52に記載の方法。
【請求項54】
細胞が、Ralstonia eutrophaの細胞である、請求項53に記載の方法。
【請求項55】
非内在性PHAシンターゼ遺伝子および/またはエノイル−CoAヒドラターゼ遺伝子が、プラスミドから発現される、請求項42〜54のいずれか一項に記載の方法。
【請求項56】
非内在性PHAシンターゼ遺伝子および/またはエノイル−CoAヒドラターゼ遺伝子が、細胞のゲノムに組み込まれている、請求項42〜54のいずれか一項に記載の方法。
【請求項57】
1または2以上の、少なくとも約4mol%または5重量%の中鎖長モノマー含有量を有するコポリマーを製造する方法であって、該方法が、請求項C1〜C14のいずれか一項に記載の細胞を製造すること、および該細胞の集団を培養することを含む、前記方法。
【請求項58】
1または2以上のコポリマーが、3−ヒドロキシルブチラートと3−ヒドロキシルヘキサノアートとのコポリマー(ポリ(HB−co−HHx))である、請求項57に記載の方法。
【請求項59】
細胞集団から1または2以上のコポリマーを収集することをさらに含む、請求項57または58に記載の方法。
【請求項60】
モノマー含有量が、少なくとも約5mol%である、請求項57〜59のいずれか一項に記載の細胞。
【請求項61】
モノマー含有量が、少なくとも約6mol%である、請求項60に記載の細胞。
【請求項62】
モノマー含有量が、少なくとも約7mol%である、請求項61に記載の細胞。
【請求項63】
モノマー含有量が、少なくとも約10mol%である、請求項62に記載の細胞。
【請求項64】
モノマー含有量が、少なくとも約15mol%である、請求項63に記載の細胞。
【請求項65】
モノマー含有量が、少なくとも約20mol%である、請求項64に記載の細胞。
【請求項66】
モノマー含有量が、少なくとも約6重量%である、請求項57〜59のいずれか一項に記載の細胞。
【請求項67】
モノマー含有量が、少なくとも約8重量%である、請求項66に記載の細胞。
【請求項68】
モノマー含有量が、少なくとも約10重量%である、請求項67に記載の細胞。
【請求項69】
モノマー含有量が、少なくとも約15重量%である、請求項68に記載の細胞。
【請求項70】
モノマー含有量が、少なくとも約20重量%である、請求項69に記載の細胞。
【請求項71】
モノマー含有量が、少なくとも約25重量%である、請求項70に記載の細胞。
【請求項72】
製造されるコポリマーの量が、細胞の乾燥重量の少なくとも約20%である、請求項57〜71のいずれか一項に記載の方法。
【請求項73】
製造されるコポリマーの量が、細胞の乾燥重量の少なくとも約30%である、請求項72に記載の方法。
【請求項74】
製造されるコポリマーの量が、細胞の乾燥重量の少なくとも約40%である、請求項73に記載の方法。
【請求項75】
製造されるコポリマーの量が、細胞の乾燥重量の少なくとも約50%である、請求項74に記載の方法。
【請求項76】
製造されるコポリマーの量が、細胞の乾燥重量の少なくとも約60%である、請求項75に記載の方法。
【請求項77】
配列番号2をコードする、単離された核酸分子。
【請求項78】
配列番号1もしくは配列番号1の開始コドンがTTGからATGに変えられたものであるヌクレオチド配列を含む、単離された核酸分子。
【請求項79】
配列番号1に記載のヌクレオチド配列と、少なくとも80%の同一性を有する、単離された核酸分子。
【請求項80】
単離された核酸分子が、配列番号1に記載のヌクレオチド配列と少なくとも90%の同一性を有する、請求項79に記載の単離された核酸分子。
【請求項81】
単離された核酸分子が、配列番号1に記載のヌクレオチド配列と少なくとも95%の同一性を有する、請求項80に記載の単離された核酸分子。
【請求項82】
単離された核酸分子が、配列番号1に記載のヌクレオチド配列と少なくとも98%の同一性を有する、請求項81に記載の単離された核酸分子。
【請求項83】
請求項77〜82のいずれか一項に記載の核酸分子によりコードされる、単離されたポリペプチド。
【請求項84】
請求項77〜82のいずれか一項に記載の単離された核酸分子を含むベクター。
【請求項85】
請求項77〜82のいずれか一項に記載の単離された核酸分子を組換えにより発現する細胞。
【請求項86】
核酸分子が、ベクターから発現される、請求項85に記載の細胞。
【請求項87】
核酸分子が、細胞のゲノムに組み込まれている、請求項85に記載の細胞。
【請求項88】
配列番号4をコードする、単離された核酸分子。
【請求項89】
配列番号3に記載のヌクレオチド配列を含む、単離された核酸分子。
【請求項90】
配列番号3に記載のヌクレオチド配列と、少なくとも80%の同一性を有する、単離された核酸分子。
【請求項91】
単離された核酸分子が、配列番号3に記載のヌクレオチド配列と少なくとも90%の同一性を有する、請求項90に記載の単離された核酸分子。
【請求項92】
単離された核酸分子が、配列番号3に記載のヌクレオチド配列と少なくとも95%の同一性を有する、請求項91に記載の単離された核酸分子。
【請求項93】
単離された核酸分子が、配列番号3に記載のヌクレオチド配列と少なくとも98%の同一性を有する、請求項92に記載の単離された核酸分子。
【請求項94】
請求項88〜93のいずれか一項に記載の核酸分子によりコードされる、単離されたポリペプチド。
【請求項95】
請求項88〜93のいずれか一項に記載の単離された核酸分子を含むベクター。
【請求項96】
請求項88〜93のいずれか一項に記載の単離された核酸分子を組換えにより発現する細胞。
【請求項97】
核酸分子が、ベクターから発現される、請求項96に記載の細胞。
【請求項98】
核酸分子が、細胞のゲノムに組み込まれている、請求項96に記載の細胞。
【請求項1】
任意の植物油を炭素源として使用して、少なくとも約4mol%または5重量%の中鎖長モノマー含有量を有するポリヒドロキシルアルカノアートコポリマーを産生する細胞。
【請求項2】
細胞が、任意の植物油を炭素源として使用して、少なくとも約4mol%または5重量%のHHx含有量を有する3−ヒドロキシルブチラートと3−ヒドロキシルヘキサノアートとのコポリマー(ポリ(HB−co−HHx))を産生する、請求項1に記載の細胞。
【請求項3】
細胞中の3−ヒドロキシルブチラートの正常な合成が破壊されている、請求項1または2に記載の細胞。
【請求項4】
アセトアセチル−CoAリダクターゼをコードする遺伝子が欠損している、請求項3に記載の細胞。
【請求項5】
細胞が、Ralstonia eutropha細胞であり、1または2以上のphaB1、phaB2およびphaB3遺伝子が破壊されている、請求項4に記載の細胞。
【請求項6】
phaB3遺伝子が破壊されている、請求項5に記載の細胞。
【請求項7】
細胞が、非内在性のPHAシンターゼ遺伝子を組換えにより発現する、請求項1〜6のいずれか一項に記載の細胞。
【請求項8】
非内在性PHAシンターゼ遺伝子が、Aeromonas caviaePHAシンターゼ遺伝子またはRhodococcus aetherivoransPHAシンターゼ遺伝子である、請求項7に記載の細胞。
【請求項9】
Rhodococcus aetherivoransPHAシンターゼ遺伝子が、配列番号4をコードするRhodococcus aetherivorans I24 D12 PHAシンターゼ遺伝子または配列番号2をコードするRhodococcus aetherivorans I24 C09 PHAシンターゼ遺伝子である、請求項8に記載の細胞。
【請求項10】
Rhodococcus aetherivorans I24 D12 PHAシンターゼ遺伝子が、配列番号3を含むかまたは配列番号3からなる、および/あるいはRhodococcus aetherivorans I24 C09 PHAシンターゼ遺伝子が、配列番号1もしくは配列番号1の開始コドンがTTGからATGに変えられたものを含むかまたは配列番号1もしくは配列番号1の開始コドンがTTGからATGに変えられたものからなる、請求項9に記載の細胞
【請求項11】
細胞が、エノイル−CoAヒドラターゼ遺伝子を組換えにより発現する、請求項1〜10のいずれか一項に記載の細胞。
【請求項12】
エノイル−CoAヒドラターゼ遺伝子が、Aeromonas caviaeエノイル−CoAヒドラターゼ遺伝子またはPseudomonas aeruginosaエノイル−CoAヒドラターゼ遺伝子である、請求項11に記載の細胞。
【請求項13】
Pseudomonas aeruginosaエノイル−CoAヒドラターゼ遺伝子が、Pseudomonas aeruginosa phaJ1遺伝子(遺伝子PA3302)またはPseudomonas aeruginosa phaJ2遺伝子(遺伝子PA1018)である、請求項12に記載の細胞。
【請求項14】
非内在性PHAシンターゼ遺伝子および/またはエノイル−CoAヒドラターゼ遺伝子が、増幅される、請求項7〜13のいずれか一項に記載の細胞。
【請求項15】
モノマー含有量が、少なくとも約5mol%である、請求項1〜14のいずれか一項に記載の細胞。
【請求項16】
モノマー含有量が、少なくとも約6mol%である、請求項15に記載の細胞。
【請求項17】
モノマー含有量が、少なくとも約7mol%である、請求項16に記載の細胞。
【請求項18】
モノマー含有量が、少なくとも約10mol%である、請求項17に記載の細胞。
【請求項19】
モノマー含有量が、少なくとも約15mol%である、請求項18に記載の細胞。
【請求項20】
モノマー含有量が、少なくとも約20mol%である、請求項19に記載の細胞。
【請求項21】
モノマー含有量が、少なくとも約6重量%である、請求項1〜14のいずれか一項に記載の細胞。
【請求項22】
モノマー含有量が、少なくとも約8重量%である、請求項21に記載の細胞。
【請求項23】
モノマー含有量が、少なくとも約10重量%である、請求項22に記載の細胞。
【請求項24】
モノマー含有量が、少なくとも約15重量%である、請求項23に記載の細胞。
【請求項25】
モノマー含有量が、少なくとも約20重量%である、請求項24に記載の細胞。
【請求項26】
モノマー含有量が、少なくとも約25重量%である、請求項25に記載の細胞。
【請求項27】
細胞が、細菌細胞、真菌細胞(酵母細胞を含む)、植物細胞、昆虫細胞または動物細胞である、請求項1〜26のいずれか一項に記載の細胞。
【請求項28】
細胞が、細菌細胞または真菌細胞である、請求項27に記載の細胞。
【請求項29】
細胞が、Ralstonia菌種、Aeromonas菌種、Rhizobium菌種、Alcaligenis菌種またはPseudomonas菌種の細胞である、請求項28に記載の細胞。
【請求項30】
細胞が、Ralstonia eutropha、Aeromonas caviae、Rhizobium japonicum、Alcaligenis eutrophusまたはPseudomonas oleovorans細胞である、請求項29に記載の細胞。
【請求項31】
細胞が、Ralstonia eutropha細胞である、請求項30に記載の細胞。
【請求項32】
非内在性PHAシンターゼ遺伝子および/またはエノイル−CoAヒドラターゼ遺伝子が、プラスミドから発現される、請求項7〜31のいずれか一項に記載の細胞。
【請求項33】
非内在性PHAシンターゼ遺伝子および/またはエノイル−CoAヒドラターゼ遺伝子が、細胞のゲノムに組み込まれている、請求項7〜31のいずれか一項に記載の細胞。
【請求項34】
高い中鎖長モノマー含有量を有するポリヒドロキシルアルカノアートコポリマーを製造する方法であって、該方法が、請求項1〜33のいずれか一項に記載の細胞を培養し、少なくとも約4mol%または5重量%の中鎖長モノマー含有量を有するコポリマーを製造することを含む、前記方法。
【請求項35】
方法が、請求項2〜33のいずれか一項に記載の細胞を培養し、少なくとも約4mol%または5重量%のHHx含有量を有する(ポリ(HB−co−HHx))を製造することを含み、ここで3−ヒドロキシルブチラートと3−ヒドロキシルヘキサノアートとのコポリマー(ポリ(HB−co−HHx))が製造される、請求項34に記載の方法。
【請求項36】
コポリマーを細胞から回収することをさらに含む、請求項34または35に記載の方法。
【請求項37】
製造されるコポリマーの量が、細胞の乾燥重量の少なくとも約20%である、請求項35または36に記載の方法。
【請求項38】
製造されるコポリマーの量が、細胞の乾燥重量の少なくとも約30%である、請求項37に記載の方法。
【請求項39】
製造されるコポリマーの量が、細胞の乾燥重量の少なくとも約40%である、請求項38に記載の方法。
【請求項40】
製造されるコポリマーの量が、細胞の乾燥重量の少なくとも約50%である、請求項39に記載の方法。
【請求項41】
製造されるコポリマーの量が、細胞の乾燥重量の少なくとも約60%である、請求項40に記載の方法。
【請求項42】
少なくとも約4mol%または5重量%の中鎖長モノマー含有量を有するポリヒドロキシルアルカノアートコポリマーを産生する細胞を製造する方法であって、細胞中に少なくとも1つのRhodococcus aetherivoransPHAシンターゼ遺伝子を組み換えにより発現することを含む、前記方法。
【請求項43】
細胞が、少なくとも約4mol%または5重量%のHHx含有量を有する3−ヒドロキシルブチラートと3−ヒドロキシルヘキサノアートとのコポリマー(ポリ(HB−co−HHx))を産生する、請求項42に記載の方法。
【請求項44】
Rhodococcus aetherivoransPHAシンターゼ遺伝子が、配列番号4をコードするRhodococcus aetherivorans I24 D12 PHAシンターゼ遺伝子または配列番号2をコードするRhodococcus aetherivorans I24 C09 PHAシンターゼ遺伝子である、請求項42または43に記載の方法。
【請求項45】
Rhodococcus aetherivorans I24 D12 PHAシンターゼ遺伝子が、配列番号3を含むかまたは配列番号3からなる、および/あるいはRhodococcus aetherivorans I24 C09 PHAシンターゼ遺伝子が、配列番号1もしくは配列番号1の開始コドンがTTGからATGに変えられたものを含むかまたは配列番号1もしくは配列番号1の開始コドンがTTGからATGに変えられたものからなる、請求項42〜44のいずれか一項に記載の方法。
【請求項46】
エノイル−CoAヒドラターゼ遺伝子を組換えにより発現することをさらに含む、請求項42〜45のいずれか一項に記載の方法。
【請求項47】
エノイル−CoAヒドラターゼ遺伝子が、Aeromonas caviaeエノイル−CoAヒドラターゼ遺伝子またはPseudomonas aeruginosaエノイル−CoAヒドラターゼ遺伝子である、請求項46に記載の方法。
【請求項48】
Pseudomonas aeruginosaエノイル−CoAヒドラターゼ遺伝子が、Pseudomonas aeruginosa phaJ1遺伝子(遺伝子PA3302)またはPseudomonas aeruginosa phaJ2遺伝子(遺伝子PA1018)である、請求項47に記載の細胞。
【請求項49】
非内在性PHAシンターゼ遺伝子および/またはエノイル−CoAヒドラターゼ遺伝子が、増幅される、請求項42〜48のいずれか一項に記載の方法。
【請求項50】
細胞が、細菌細胞、真菌細胞(酵母細胞を含む)、植物細胞、昆虫細胞または動物細胞である、請求項42〜49のいずれか一項に記載の方法。
【請求項51】
細胞が、細菌細胞または真菌細胞である、請求項50に記載の方法。
【請求項52】
細胞が、Ralstonia菌種、Aeromonas菌種、Rhizobium菌種、Alcaligenis菌種またはPseudomonas菌種の細胞である、請求項51に記載の方法。
【請求項53】
細胞が、Ralstonia eutropha、Aeromonas caviae、Rhizobium japonicum、Alcaligenis eutrophusまたはPseudomonas oleovoransの細胞である、請求項52に記載の方法。
【請求項54】
細胞が、Ralstonia eutrophaの細胞である、請求項53に記載の方法。
【請求項55】
非内在性PHAシンターゼ遺伝子および/またはエノイル−CoAヒドラターゼ遺伝子が、プラスミドから発現される、請求項42〜54のいずれか一項に記載の方法。
【請求項56】
非内在性PHAシンターゼ遺伝子および/またはエノイル−CoAヒドラターゼ遺伝子が、細胞のゲノムに組み込まれている、請求項42〜54のいずれか一項に記載の方法。
【請求項57】
1または2以上の、少なくとも約4mol%または5重量%の中鎖長モノマー含有量を有するコポリマーを製造する方法であって、該方法が、請求項C1〜C14のいずれか一項に記載の細胞を製造すること、および該細胞の集団を培養することを含む、前記方法。
【請求項58】
1または2以上のコポリマーが、3−ヒドロキシルブチラートと3−ヒドロキシルヘキサノアートとのコポリマー(ポリ(HB−co−HHx))である、請求項57に記載の方法。
【請求項59】
細胞集団から1または2以上のコポリマーを収集することをさらに含む、請求項57または58に記載の方法。
【請求項60】
モノマー含有量が、少なくとも約5mol%である、請求項57〜59のいずれか一項に記載の細胞。
【請求項61】
モノマー含有量が、少なくとも約6mol%である、請求項60に記載の細胞。
【請求項62】
モノマー含有量が、少なくとも約7mol%である、請求項61に記載の細胞。
【請求項63】
モノマー含有量が、少なくとも約10mol%である、請求項62に記載の細胞。
【請求項64】
モノマー含有量が、少なくとも約15mol%である、請求項63に記載の細胞。
【請求項65】
モノマー含有量が、少なくとも約20mol%である、請求項64に記載の細胞。
【請求項66】
モノマー含有量が、少なくとも約6重量%である、請求項57〜59のいずれか一項に記載の細胞。
【請求項67】
モノマー含有量が、少なくとも約8重量%である、請求項66に記載の細胞。
【請求項68】
モノマー含有量が、少なくとも約10重量%である、請求項67に記載の細胞。
【請求項69】
モノマー含有量が、少なくとも約15重量%である、請求項68に記載の細胞。
【請求項70】
モノマー含有量が、少なくとも約20重量%である、請求項69に記載の細胞。
【請求項71】
モノマー含有量が、少なくとも約25重量%である、請求項70に記載の細胞。
【請求項72】
製造されるコポリマーの量が、細胞の乾燥重量の少なくとも約20%である、請求項57〜71のいずれか一項に記載の方法。
【請求項73】
製造されるコポリマーの量が、細胞の乾燥重量の少なくとも約30%である、請求項72に記載の方法。
【請求項74】
製造されるコポリマーの量が、細胞の乾燥重量の少なくとも約40%である、請求項73に記載の方法。
【請求項75】
製造されるコポリマーの量が、細胞の乾燥重量の少なくとも約50%である、請求項74に記載の方法。
【請求項76】
製造されるコポリマーの量が、細胞の乾燥重量の少なくとも約60%である、請求項75に記載の方法。
【請求項77】
配列番号2をコードする、単離された核酸分子。
【請求項78】
配列番号1もしくは配列番号1の開始コドンがTTGからATGに変えられたものであるヌクレオチド配列を含む、単離された核酸分子。
【請求項79】
配列番号1に記載のヌクレオチド配列と、少なくとも80%の同一性を有する、単離された核酸分子。
【請求項80】
単離された核酸分子が、配列番号1に記載のヌクレオチド配列と少なくとも90%の同一性を有する、請求項79に記載の単離された核酸分子。
【請求項81】
単離された核酸分子が、配列番号1に記載のヌクレオチド配列と少なくとも95%の同一性を有する、請求項80に記載の単離された核酸分子。
【請求項82】
単離された核酸分子が、配列番号1に記載のヌクレオチド配列と少なくとも98%の同一性を有する、請求項81に記載の単離された核酸分子。
【請求項83】
請求項77〜82のいずれか一項に記載の核酸分子によりコードされる、単離されたポリペプチド。
【請求項84】
請求項77〜82のいずれか一項に記載の単離された核酸分子を含むベクター。
【請求項85】
請求項77〜82のいずれか一項に記載の単離された核酸分子を組換えにより発現する細胞。
【請求項86】
核酸分子が、ベクターから発現される、請求項85に記載の細胞。
【請求項87】
核酸分子が、細胞のゲノムに組み込まれている、請求項85に記載の細胞。
【請求項88】
配列番号4をコードする、単離された核酸分子。
【請求項89】
配列番号3に記載のヌクレオチド配列を含む、単離された核酸分子。
【請求項90】
配列番号3に記載のヌクレオチド配列と、少なくとも80%の同一性を有する、単離された核酸分子。
【請求項91】
単離された核酸分子が、配列番号3に記載のヌクレオチド配列と少なくとも90%の同一性を有する、請求項90に記載の単離された核酸分子。
【請求項92】
単離された核酸分子が、配列番号3に記載のヌクレオチド配列と少なくとも95%の同一性を有する、請求項91に記載の単離された核酸分子。
【請求項93】
単離された核酸分子が、配列番号3に記載のヌクレオチド配列と少なくとも98%の同一性を有する、請求項92に記載の単離された核酸分子。
【請求項94】
請求項88〜93のいずれか一項に記載の核酸分子によりコードされる、単離されたポリペプチド。
【請求項95】
請求項88〜93のいずれか一項に記載の単離された核酸分子を含むベクター。
【請求項96】
請求項88〜93のいずれか一項に記載の単離された核酸分子を組換えにより発現する細胞。
【請求項97】
核酸分子が、ベクターから発現される、請求項96に記載の細胞。
【請求項98】
核酸分子が、細胞のゲノムに組み込まれている、請求項96に記載の細胞。
【図1】

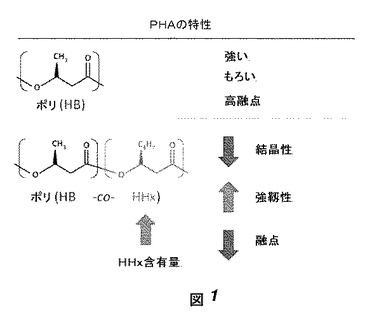
【図2】
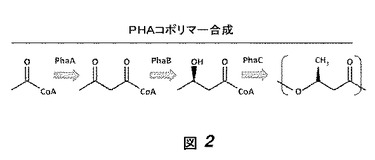
【図3】
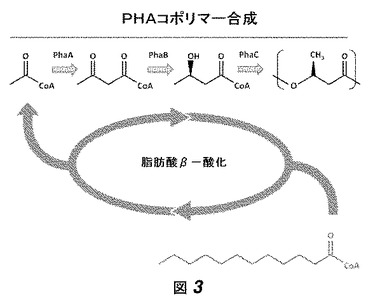
【図4】
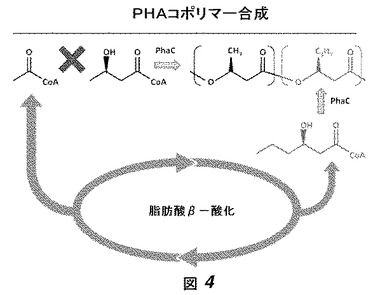
【図5】
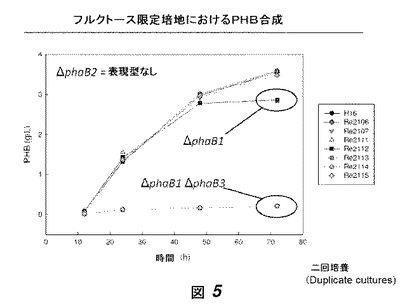
【図6】
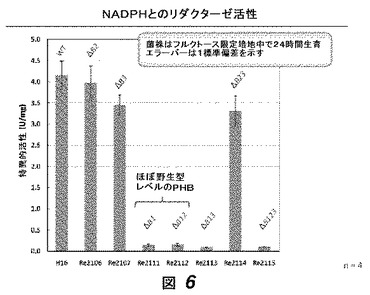
【図7】
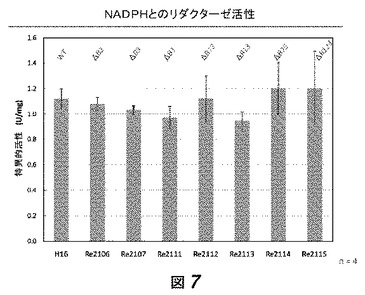
【図8】
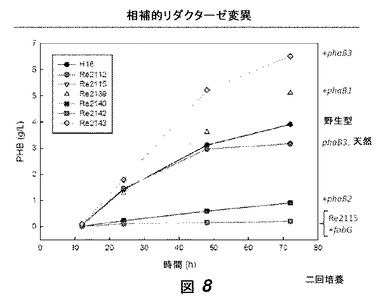
【図9】
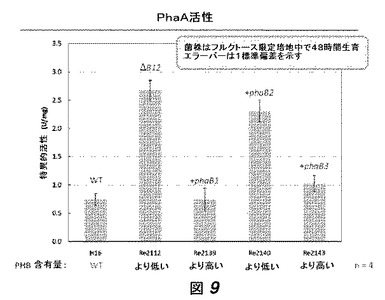
【図10】
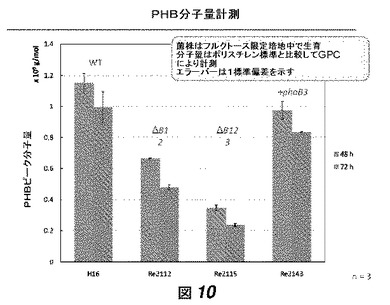
【図11】
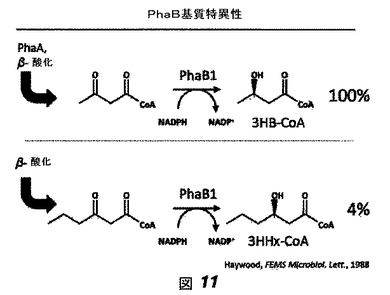
【図12】
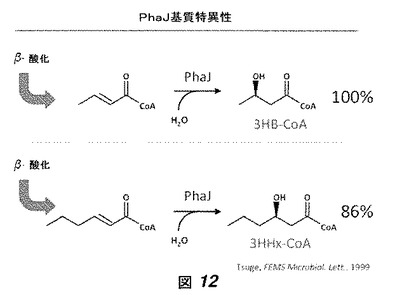
【図13】

【図14】

【図15】
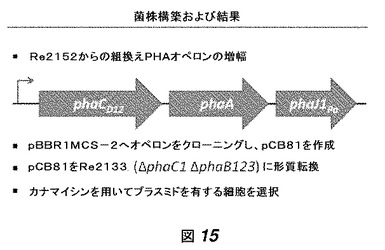
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【公表番号】特表2013−510572(P2013−510572A)
【公表日】平成25年3月28日(2013.3.28)
【国際特許分類】
【出願番号】特願2012−538939(P2012−538939)
【出願日】平成22年11月10日(2010.11.10)
【国際出願番号】PCT/US2010/056196
【国際公開番号】WO2011/060048
【国際公開日】平成23年5月19日(2011.5.19)
【出願人】(596060697)マサチューセッツ インスティテュート オブ テクノロジー (233)
【Fターム(参考)】
【公表日】平成25年3月28日(2013.3.28)
【国際特許分類】
【出願日】平成22年11月10日(2010.11.10)
【国際出願番号】PCT/US2010/056196
【国際公開番号】WO2011/060048
【国際公開日】平成23年5月19日(2011.5.19)
【出願人】(596060697)マサチューセッツ インスティテュート オブ テクノロジー (233)
【Fターム(参考)】
[ Back to top ]