
高効率細胞融合法
【課題】融合細胞を高効率で産生できる細胞融合法を提供する。
【解決手段】免疫細胞に対して、インビトロでオリグデオキシヌクレオチド・サイトカインを含む培養液で抗体産生細胞を培養する培養工程を設ける免疫工程を施した後、電気的細胞融合を行う、細胞融合方法。
【解決手段】免疫細胞に対して、インビトロでオリグデオキシヌクレオチド・サイトカインを含む培養液で抗体産生細胞を培養する培養工程を設ける免疫工程を施した後、電気的細胞融合を行う、細胞融合方法。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、動物細胞同士の細胞融合法及び該方法に用いる細胞融合促進剤に関する。
【背景技術】
【0002】
抗体産生機構
抗体(免疫グロブリン)は、脊椎動物の感染防御機構において重要な役割を担うものであり、免疫細胞(リンパ球)のうちでB細胞により産生される。ヒト等哺乳動物の体内では、それぞれ異なる抗体を作る能力を備えたB細胞が、それぞれの細胞表面に数千万種以上とも数億種以上ともいわれる抗体を提示して抗原を待ち受け、あらゆる抗原に対応する体制を整えている。抗原刺激により抗体産生機能が活性化された特定のB細胞に対して、体内では、速やかに抗体産生活性化を促進する機構にスイッチが入り、抗体産生細胞の分化を経て抗原特異的な抗体の産生に至る。
【0003】
抗体の機能
生体内で産生された抗体は抗原の様々な部位に特異的に結合することにより毒素の中和や、オプソニン化による食細胞への貪食の促進、補体を活性化して細菌の溶菌誘導等の効果を発揮し、感染防御機構において抗原の除去を補助している。抗体の効果は各アイソタイプで異なっており、IgG、IgA、IgM、IgE、IgD、IgEに分類される。特にIgGは血中や細胞外組織液中では主要なアイソタイプであり、抗原との親和性、抗原中和、オプソニン化、補体の活性化能が高い場合が多い。単一の抗体産生細胞は1種類のアイソタイプで抗原の特定の1箇所に結合する抗体を産生しており、その抗体産生細胞をクローン化し得られる抗体は、モノクローナル抗体として利用されている。
【0004】
モノクローナル抗体の利用
生化学やタンパク質科学研究の分野においてモノクローナル抗体はその特異的認識能から標的物質の標識や選別等、幅広く使用されており、バイオツールとして必須であり、抗体の品質は研究の進展に大きく影響する。また、医療の分野においては、妊娠、成人病、感染症等の臨床診断薬として多く使われている。多種多様な抗原に対応可能な生体分子であることから、テーラーメイド医療の実現や副作用の軽減が期待されており、癌等の治療薬としても注目されている。モノクローナル抗体は研究から医療まで応用範囲が急速に拡大しており、有用な抗体を確実かつ効率的に作製する技術の確立が求められている。
【0005】
モノクローナル抗体作製過程の技術的問題点
モノクローナル抗体の作製は主に、1.動物への免疫、2.抗体産生細胞と癌細胞の融合、3.融合細胞(ハイブリドーマ)の選別、と3つの工程がある。工程3は抗体産生細胞と癌細胞の両方の機能を獲得した細胞のみを選別する試薬を用いることにより、既に確立されている。しかし、工程1、2には問題点があり、有用な抗体を確実に効率的に得るには課題を多く残している。
【0006】
問題点1(免疫)
実験動物を十分な抗原量で免疫にすれば、抗原特異的抗体産生細胞が取得できる場合が多いが、免疫原性の低い、すなわち免疫系の活性化能の低い場合は、抗体産生細胞や特異性が高い抗体を取得することは難しい。そのため十分な免疫応答を引き起こし、十分量の抗原特異的抗体産生細胞を取得するには、実験動物に対して、免疫活性化能を持つ抗原とアジュバントとの同時投与や、1回の投与量を多くし多数回にわたっての投与、多数の動物に免疫を行う必要がある。これらの処置にはコストと時間が掛かることに加えて、動物に対する副作用や処置自体による苦痛が大きく、動物保護の観点からも問題がある。したがって、インビボ免疫法において、抗原特異性の高い抗体を高効率で産生できる技術の開発が必要とされている。
【0007】
問題点2(細胞融合)
抗体産生細胞から大量の抗体を得るためには、抗体産生細胞とガン細胞であるミエローマ細胞とを融合させ、不死化させる必要がある。モノクローナル抗体の作製が困難な理由の一つに、この融合効率がきわめて低いことがある。低融合効率が、陽性クローンを多数取れない主要因であり、そのため認識能の高い抗体や、アゴニスト活性、アンタゴニスト活性等の機能を持った抗体の取得が大変難しくなっている。この細胞融合を用いずに抗体様の物質を得るために、ファージを使った作製方法等があるが、今のところ抗体ほどに特異性・親和性の高いものを得るまでにはなっていない。そのため、高効率の細胞融合法の開発は、抗体作製において非常に重要な技術となっている。
【0008】
細胞融合(PEG)の問題点
しかしながら、抗体産生細胞とミエローマ細胞の融合は難しく、その融合効率はきわめて低く、10-5程度である。ポリエチレングリコール(PEG)を用いた融合法は専用機器を必要とせず、試薬が安価であるため、主に用いられている。この方法は技術的に非常に難しく、薬剤自体に毒性があることもあり、効率が低いだけでなく、再現性も低いのが実情である。一方電気的パルスを用いた細胞融合法は、PEG法よりも融合効率が一桁程度高くなると言われている。
【0009】
電気的細胞融合法
電気的細胞融合法は交流電圧を印加することで電極間の抗体産生細胞とミエローマ細胞を接触させ、その後の直流パルス電圧を印加させることで細胞膜に穿孔が開き、接触している細胞同士の融合を引き起こす。この原理によりポリエチレングリコールよりも高い融合効率を実現できる。
しかし、抗体産生細胞は刺激されていない状態では融合効率が著しく低下することや、抗体産生細胞とミエローマ細胞の直径が異なっており、各々の細胞の穿孔する最適な電圧が異なること等が問題となっている。これらの原因により再現性が低いことがネックとなっており、システムとして確立されているとは言えない。また、融合装置を必要とすることもあり、その大きな有用性に注目されながらも未だに広く利用されていない。したがって、電気的細胞融合法において、確実に高効率に細胞融合を実現する技術の開発が強く望まれている。
【先行技術文献】
【特許文献】
【0010】
【特許文献1】特開2009−148296号公報
【特許文献2】特表2010−504750号公報
【非特許文献】
【0011】
【非特許文献1】Yu et al.J Immunol Methods.2008 Jul 31;336(2):142-51
【非特許文献2】Schmidt et al.J Immunol Methods.2001 Sep 1;255(1-2):93-102.
【非特許文献3】B.R.Lentz,Eur Biophys J.2007 Apr;36(4-5):315-26.
【非特許文献4】Li et al.Proc Natl Acad Sci USA.2006 Mar 7;103(10):3557-62.
【非特許文献5】Crowe Vaccine.2009 Dec 30;27 Suppl 6:G47-51.
【非特許文献6】Kwekkeboom et al.J Immunol Methods.1993 Mar 15;160(1):117-27.
【発明の概要】
【発明が解決しようとする課題】
【0012】
本発明の課題は、上記した電気的細胞融合法の問題点を解消し、確実に高い効率で細胞融合させることが可能となる手段を新たに提供することにある。
【課題を解決するための手段】
【0013】
そこで、本発明者等は、従来の電気的細胞融合法を検討したところ、融合効率が低く、陽性クローン数が少ないという欠点のいずれもが抗体産生細胞の活性化不足に起因するのではないか、という推論をたて、免疫系の有効な活性化法について鋭意研究を重ねた結果、従来の電気的細胞融合法を飛躍的に改善する基本プロトコールを開発するに至った。
すなわち、本発明は、細胞の調整、培養方法および最適な免疫系刺激因子に関する基本プロトコールに係るものであり、具体的には、細胞融合に際して、体外に取り出した免疫細胞を免疫賦活物質を含有する培地で培養する工程を加えることにより、目的抗原を認識するB細胞がさらに特異的に活性化されるだけでなく、ミエローマ細胞との融合効率の上昇をもたらすことが可能であるとの知見を得るとともに、さらに研究を進め、免疫刺激物質として特にCpGモチーフを含むオリゴヌクレオチドを用いた場合には、細胞融合効率が、他の免疫賦活物質と比較して全く予想外に飛躍的に高まることを見いだした。そして、これにより、当該抗原特異的抗体産生細胞を片親としたハイブリドーマから抗原特異的モノクローナル抗体をインビトロで効率的に生産可能であることを確認し、本発明を完成させた。
本発明は以下のとおりである。
【0014】
(1)免疫細胞とミエローマ細胞とを電気的細胞融合法で融合するに際して、予め、CpGモチーフを有するオリゴヌクレオチドを含有する培地で免疫細胞を培養することを特徴とする、細胞融合方法。
(2)CpGモチーフを有するオリゴヌクレオチドが、以下の塩基配列を有することを特徴とする上記(1)に記載の細胞融合方法。
【化1】

(ただし、上記塩基配列中、N1N2のうち少なくとも一つはプリン塩基であり、N3N4のうち少なくとも一つはピリミジン塩基である。)
(3)CpGモチーフを有するオリゴヌクレオチドが、5’−GACGTT−3’、5’−TGGCGT−3’、5’−GACGCT−3’、5’−AGCGTT−3’又は5’−GAGCTT−3’からなる配列を有することを特徴とする上記(2)に記載の細胞融合方法。
(4)CpGモチーフを有するオリゴヌクレオチドが、
5'-tcctGACGTTcctgacgtt-3'又は5'-tccatGACGTTcctgacgtt-3'、5'-tAGCGTT-3'、5'-tGACGCT-3'、又は5'-tccatGAGCTTcctgacgtt-3'であることを特徴とする上記(3)に記載の細胞融合方法。
(5)培地が、さらにサイトカインを含有することを特徴とする、上記(1)〜(4)のいずれかに記載の細胞融合方法。
(6)CpGモチーフを有するオリゴヌクレオチドからなる電気的細胞融合促進剤。
(7)CpGモチーフを有するオリゴヌクレオチドが、以下の塩基配列を有することを特徴とする上記(6)に記載の電気的細胞融合促進剤。
【化1】
(ただし、上記塩基配列中、N1N2のうち少なくとも一つはプリン塩基であり、N3N4のうち少なくとも一つはピリミジン塩基である。)
(8)CpGモチーフを有するオリゴヌクレオチドが、5’−GACGTT−3’、5’−TGGCGT−3’、5’−GACGCT−3’、5’−AGCGTT−3’又は5’−GAGCTT−3’からなる配列を有することを特徴とする上記(7)に記載の電気的細胞融合促進剤。
(9)CpGモチーフを有するオリゴヌクレオチドが、
5'-tcctGACGTTcctgacgtt-3'、5'-tcctTGGCGTcctgacgtt-3'、5'-tAGCGTT-3'、5'-tGACGCT-3'、又は5'-tccatGAGCTTcctgacgtt-3'であることを特徴とする上記(8)に記載の電気的細胞融合促進剤。
(10)上記(6)〜(9)のいずれかに記載の電気的細胞融合促進剤とサイトカインを含有することを特徴とする、電気的細胞融合試薬キット。
(11)抗体産生細胞とミエローマ細胞とを電気的細胞融合法で融合するに際して、予め、CpGモチーフを有するオリゴヌクレオチドを含有する培地で抗体細胞を培養することを特徴とする、モノクローナル抗体産生ハイブリドーマの製造方法。
(12)培地がさらにサイトカインを含有することを特徴とする、上記(11)に記載のハイブリドーマの製造方法。
(13)上記(11)又は(12)に記載の方法により得られたモノクローナル抗体産生ハイブリドーマ。
(14)上記(13)に記載のハイブリドーマを培地に培養し、モノクローナル抗体を採取することを特徴とする、モノクローナル抗体の製造方法。
【発明の効果】
【0015】
本発明の細胞融合法によれば、従来困難であった極めて高効率の細胞融合が可能となる。また、用いた抗体産生細胞に対する、目的とする機能を有するモノクローナル抗体産生ハイブリドーマの出現率は極めて高く、細胞融合に際し、抗体産生細胞を従来のように大量に必要とせず、動物を免疫する抗原量も少量ですむ。また、従来法によれば、例えば、一次スクリーニングで抗原と結合するハイブリドーマの選別、2次スクリーニングで該ハイブリドーマの目的とする機能性の評価をそれぞれ行っていたが、本発明の融合法によれば、このような選別及び評価は、一回のスクリーニングで行うことが可能となる。したがって、本発明の細胞融合法によれば、有用なハイブリドーマあるいは目的とするモノクローナル抗体を極めて効率的、かつ短時間、安価に提供でき、これらを通じて医薬あるいは化粧品その他の分野において大いに貢献できる。
【図面の簡単な説明】
【0016】
【図1】各種免疫刺激物質と刺激日数による融合効率の変化を示す図である。
【図2】CpG ODN刺激免疫細胞の融合電圧による融合効率の変化を示す図である。
【図3】CpG ODN刺激時の細胞培養濃度による融合効率の変化を示す図である。
【図4】各種免疫刺激物質刺激2日後の免疫細胞の直径を測定した結果を示す図である。
【図5】CpG ODN刺激の有無によるB細胞の肥大の変化を測定した結果を示す図である。
【図6】CpG ODN刺激の有無による形質細胞の肥大の変化を測定した結果を示す図である。
【図7】各種免疫刺激物質の刺激により、得られる抗原特異的IgG産生細胞数の差違を示す図である。
【図8】CpG ODNの配列バリエーションによる融合効率を測定した結果を示す図である。
【図9】本発明の細胞融合法により得られたハイブリドーマの培養上澄みを使用した検出試験により、Gaussia luciferaseの発光阻害機能抗体を産生するハイブリドーマが取得可能であることを示す図である。
【図10】上記発光阻害機能抗体の発光阻害機能評価の結果を示す図である。
【発明を実施するための形態】
【0017】
本発明は、細胞融合に先立ち、予め免疫細胞をCpGモチーフを含むオリゴヌクレオリド(以下、CpG ODNということがある。)あるいはさらにサイトカインを含有する培地に培養して、インビトロで免疫刺激し、次いで培養した免疫細胞とミエローマ細胞とを電気的細胞融合法により融合させて、ハイブリドーマを得るものである。
【0018】
本発明において「免疫細胞」とは、動物を抗原で免疫した後、該動物の末梢血、脾臓、リンパ節、扁桃、パイエル板、虫垂等の2次リンパ組織から採取した抗体産生細胞を含むリンパ球をいう。採取源の生物種はどの哺乳動物由来であってもよいが、治療用に用いる抗原特異的モノクローナル抗体を得るためには、同じ生物種由来、例えばヒト用であればヒト由来のリンパ球が好ましい。
本発明において、インビトロでの免疫刺激に先立ち、あらかじめ赤血球除去工程を設けることが好ましいが、設けることなく幼若化工程に付してもよい。
本発明で使用するCpG ODN、CpG motif(5'-CG-3')を持つオリゴヌクレオチドで、このmotifが免疫細胞活性化の活性中心であり、特にCの非メチル化が活性に必須である。
本発明で使用するCpG ODNのより具体的な例は、 5’−N1N2CGN3N4−3’(配列番号1)の塩基配列を有するものである。(ただし、上記塩基配列中、N1N2のうち少なくとも一つはプリン塩基であり、N3XN4のうち少なくとも一つはピリミジン塩基である。)
【0019】
このようなCpG ODNは、免疫刺激物質として、それ自体公知である。
CG前後の配列はCGを中心に回文構造の場合が多く、マウス等のげっ歯類では5'-GACGTT-3'(配列番号2)を、ヒトでは5'-GTCGTT-3'(配列番号3)を含むオリゴヌクレオチドが免疫活性化能を持つことが報告されている。
また、その他のCpG ODNとしては、例えば5’−TGGCGT−3’(配列番号4)、5’−GACGCT−3’(配列番号5)、5’−AGCGTT−3’(配列番号6)又は5’−GAGCTT−3’(配列番号12)の配列を有するオリゴヌクレオチドが挙げられる。
また、CG前後の配列のパターンによって作用が異なり、作用別にA-type、B-type、C-typeの3つのグループに分類される。A-typeは形質細胞様樹状細胞のインターフェロンαの産生を誘導し、B-typeはB細胞を活性化しIgM、インターロイキン6、インターロイキン10の産生を誘導し、C-typeはA-type、B-type双方の作用を有するが、本発明においてはそのいずれもが使用できる。
【0020】
電気的細胞融合において、このようなCpG ODNを使用する場合、他の免疫刺激物質、例えば、LPS(リポポリサッカライド)、PWM(pokeweed mitogen)、PHA(フィトヘムアグルチニン)、EBV(Epstein-Barrウィルス)等に比べて格段に高い確率で融合細胞が得られ、極めて顕著に細胞融合効率を上昇させる。したがって、本発明のCpG ODNは細胞融合促進剤として有用である。
また、サイトカインの使用はさらに融合効率を上昇させ、CpG ODNとサイトカインは電気的細胞融合試薬キットとして有用である。本発明において使用するサイトカインとしては、例えばインターロイキン2、インターロイキン4、インターロイキン5、インターロイキン6、インターロイキン13、インターロイキン21等が挙げられる。
【0021】
本発明において免疫細胞を培養する際の培地としては、通常の細胞培養用培地、例えばRPMI培地、DMEM培地、ハム−F12培地、RDF培地、ERDF培地、又はこれらの混合培地を用いることができるが、これらの中では、RPMI培地が特に好ましい。35〜38℃、好ましくは37℃の温度条件下で培養するが、その際に1%〜10%、好ましくは約5%のCO2環境下でインキュベートすることが好ましい。培地中の免疫細胞の細胞密度は、0.5×106〜1×107 cells/ml範囲内に、好ましくは2×106 cells/mlで用いる。当該培地は、免疫細胞を採取後懸濁液とする際の懸濁液としても、また当該懸濁液から遠心分離により免疫細胞を濃縮分離する際の洗浄液としても用いることができ、ハイブリドーマの培養液としても用いることができる。
【0022】
本発明においては細胞融合手段として、電気的細胞融合法を用いるが、電気的細胞融合法は、電極を配した細胞懸濁液収容部材に、融合する細胞を懸濁し、所定の浸透圧に調整した溶液を収容し、電極から細胞を接着する(パールチェーンの形成)ための交流、及び細胞融合させるための直流パルスを順次与えて細胞を融合させる手段である。この手段自体はよく知られている。
【0023】
本発明においては上記の各手段を組み合わせて細胞融合を行うが、その概要は図1に示され、そのプロセスは以下のとおりである。
まず、マウス等の動物に抗原を投与し、その抗原に対する抗体産生細胞を誘導し、免疫組織(脾臓、リンパ節)から、免疫細胞を取り出す。次いで、免疫細胞を上記した培地で培養して幼若化するが、このとき培地に上記したCpG ODNを添加する。添加量は、培養液に対し0.05〜100uMであり、好ましくは0.2〜10uMである。
本発明においては、CpG ODNとともに上記サイトカインを併用することが好ましく、さらに上記したLPS等の他の免疫刺激物質を組み合わせて添加してもよい。
【0024】
CpG ODN含有培地での培養時間については、後記する実施例の実験によれば、細胞融合効率は1日でも極めて高いが、2日が最大となり、3日以上では急速に効率が低下する。また、抗体産生能は1日で最大である。したがって、培養は2日以内で終了することが好ましい。
このようなCpG ODN含有培地での培養により、抗体産生細胞は顕著に活性化され、活発に増殖・巨大化する。
次いで、このように活性化された抗体産生細胞を含む免疫細胞は、培地から取り出して洗浄した後、所定の浸透圧に調整された緩衝液においてミエローマ細胞と1対1となるよう混合され、1〜10×106cells/ml、好ましくは2〜4×106cells/mlで、電極を配した細胞懸濁液収容部材に投入し、電極から交流及び直流パルスを印加して、電気的細胞融合を行う。
【0025】
上記緩衝液の浸透圧は、グルコース、スクロース、トレハロース等の糖類、イノシトール、マンニトール等の糖アルコール類、あるいは塩化マグネシウム、塩化カルシウム、リン酸カリウム等により調整するが、浸透圧は590〜90mOsm/kg、好ましくは280〜90mOsm/kgに調整する。
また、電極間のギャップが2mmの場合、上記交流印加は、10〜50Vで10〜60秒、好ましくは25〜40Vで10〜30秒である。上記直流パルスの印加は、150〜450Vで5〜50マイクロ秒を1〜4サイクル。好ましくは200〜300Vで10〜40マイクロ秒を2〜3サイクルである。本発明においては、上記直流パルス印加後、細胞融合を促進させ安定化させるために、さらに交流電圧を印加することが好ましい、10〜50Vで10〜60秒、好ましくは25〜40Vで10〜30秒である。
【0026】
電気的細胞融合後、フェノールレッド不含のRPMIで培養後、常法により、HAT培地でハイブリドーマを選別し、さらに限界希釈法により得たハイブリドーマの各クローンを培養し、得られた各クローンの培養上澄みについて、例えば、上記動物に免疫した抗原を使用した酵素免疫測定法(ELISA)により、発色強度を測定することによって、該抗原との特異的反応性が高いものを検出し、また、さらに必要により、増殖性の高いクローンを選定することにより、目的とするIgGモノクローナル抗体を産生するハイブリドーマクローンを得ることができる。
特に本発明の電気的細胞融合法による場合、用いた抗体産生細胞に対する、目的とする機能を有するモノクローナル抗体産生ハイブリドーマの出現率は極めて高く、細胞融合に際し、抗体産生細胞を従来のように大量に必要としない。また、従来法のように一次、二次の2回のスクリーニングを行うことなく、1回のスクリーニングで目的とするモノクローナル抗体産生ハイブリドーマを取得することが可能となる。
【実施例】
【0027】
実施例1
免疫マウスの脾臓を用いた各種免疫刺激物質による融合効率の変化
(1-1)KLHの免疫
抗原KLHとFREUND’S ADJUVANT Complete(Sigma社)を混合し、BALB/cマウス(♀)の腹腔とフットパッドにKLH 100μg/匹となるよう投与し、1次免疫を1週間行った。続いて、抗原KLHとFREUND’S ADJUVANT In Complete(Sigma社)を混合し、1次免疫と同様に2次免疫を行った。
(1-2)免疫細胞の調整
免疫を行ったマウスより脾臓を摘出し、RPMI培養液(Sigma社)中に分散させ、セルストレーナー(BD社)を通し夾雑物を取り除いた。次いで、Lympholyte-M(CEDARLANE社)上に細胞懸濁液を重層し、23℃ 1000g 20分間遠心し、赤血球を取り除き、培養液で2回洗浄した。
【0028】
(1-3)免疫細胞の幼若化
上記(1-2)で調整した免疫細胞をRPMI培養液に懸濁し、培養プレートに10mlずつ分注した。各プレートには、添加無し、0.25μM CpG ODN(北海道システム・サイエンス株式会社に委託して作成)、10μg/ml LPS(Sigma社)、0.5μg/ml抗CD40抗体(R&D Systems社)と10ng/ml IL-2(PEPROTECH社)、2.4μg/ml PWM(Sigma社)を添加し、37℃、5% CO2インキュベータ内で1日から5日間培養した。なお、上記CpG ODNの塩基配列は下記のとおりである。
5’−tccatgacgttcatgacgtt−3’(配列番号7)
(1-4)ミエローマ細胞の調整
エレクトロフュージョン3日前にミエローマ細胞を起眠し、1×106/mlの濃度を超えないようRPMI培養液に懸濁し37℃、5% CO2インキュベータ内で培養した。
【0029】
(1-5)エレクトロフュージョン
幼若化した免疫細胞とミエローマ細胞を各々180g 5分間遠心し、290mOsm Inositol、0.5mM塩化マグネシウム、0.1mM塩化カルシウム、1mMリン酸カリウム、pH7.2バッファーに懸濁し、2回洗浄した。洗浄した細胞を5×106/mlの濃度になるよう185mOsm Inositolバッファーに懸濁し、免疫細胞とミエローマ細胞を1:1で混合した。細胞混合液80μlを2mmギャップ電極のエレクトロフュージョンチャンバーへ添加し、185mOsm Inositolバッファーに懸濁してから2分後に電気的細胞融合装置LF100(ベックス社)を用いて、アラインメント35V 30sec、パルス300V 20μsec 2cycle、ポストアラインメント35V 30secのパルスを与え、10分間静置し、エレクトロフュージョンを行った。その後、細胞を20%FCS(HyClone社)、5% Briclone(QED BIOSCIENCE社)含有のフェノールレッド不含RPMI培養液10mlに懸濁し96ウェルプレートマイクロに100μl/ウェルで分注し、37℃、5% CO2インキュベータ内で培養した。
【0030】
(1-6)細胞融合効率の測定
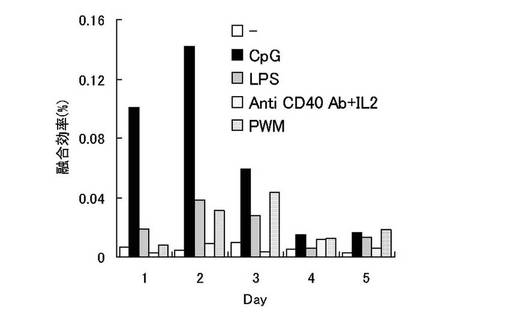
各免疫刺激物質で幼若化した免疫細胞は刺激開始から1日毎に5日間回収し、各々エレクトロフュージョンを行った。エレクトロフュージョンの翌日に細胞を播種した96穴プレートに20%FCS、5% Briclone、2×HAT(Sigma社)含有のRPMI培養液を100μl/ウェルで分注した。ハイブリドーマのコロニーが観察出来るまで、7日から9日間37℃ 5% CO2インキュベータ内で培養し、顕微鏡下でコロニー数を計測し、各免疫刺激物質による幼若化1〜5日間の細胞融合効率を算出した。結果を(図1)に示す。
各免疫刺激物質は幼若化開始から2〜3日で融合効率が高くなり、特にCpG ODNを用いた高効率融合法の場合、刺激開始から2日目に融合効率の上昇がピークとなり、形成されるコロニー数が圧倒的に多いことがわかる。
【0031】
実施例2
各種免疫刺激物質による刺激後のエレクトロフュージョン融合電圧による融合効率の変化
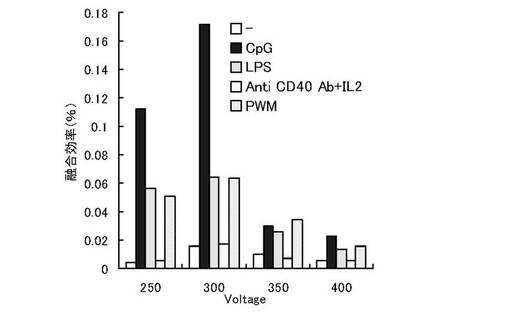
エレクトロフュージョンのパルス条件を除いて実施例1と同様に実験を実施した。エレクトロフュージョンはパルスを250V,300V,350V,400Vの4条件で実施し、実施例1と同様に細胞融合効率を測定した。結果を(図2)に示す。各免疫刺激物質いずれも300Vで最も融合効率が高く、特にCpG ODNによって幼若化された場合、融合効率が上昇し、コロニー形成に優位であることが示された。
【0032】
実施例3
CpG ODN刺激時の細胞培養濃度による融合効率の変化
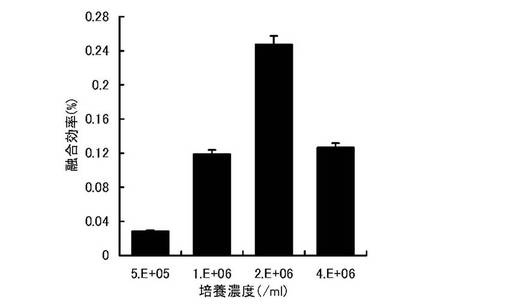
免疫細胞の幼若化の細胞濃度を除いて実施例1と同様に実験を実施した。免疫細胞の幼若化のステップでは0.5×106/ml,1×106/ml,2×106/ml,4×106/mlの細胞濃度となるよう各々RPMI培養液に懸濁し、培養プレートに10mlずつ分注した。各プレートには、CpG ODNを0.25μMとなるよう添加し、37℃、5% CO2インキュベータ内で2日間培養した。その後、実施例1と同様に細胞融合効率を測定した。結果を(図3)に示す。幼若化のステップの培養濃度は融合効率に大きく影響を及ぼしており、2×106/mlで培養した時に最も融合効率が上昇する。
【0033】
実施例4
各種免疫刺激物質刺激2日後の細胞の直径
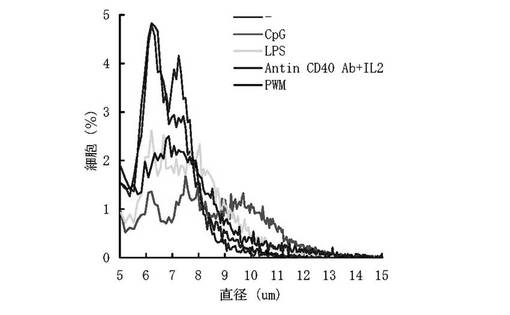
実施例1と同様にKLHの免疫、免疫細胞の調整、各種免疫刺激物質による免疫細胞の幼若化を行い、2日後に細胞カウントバッファーで100倍に希釈し、コールターカウンター(ベックマンコールター社)で細胞の直径を測定した。結果を(図4)に示す。他の刺激物質と比較してCpG ODNの実験区ではより免疫細胞の直径が拡大しており、平均直径の拡大も突出している。エレクトロフュージョンにおいて細胞の直径は融合電圧を決定する要因の1つである。ミエローマ細胞と免疫細胞の直径は大きく異なっており、2つの細胞間の直径差を減少させることは細胞融合効率に大きく影響を及ぼす。CpG ODNによる幼若化は免疫細胞の活性化と共に細胞をより肥大させ、ミエローマ細胞の直径に近づけることで融合効率を上昇させていると考えられる。
【0034】
実施例5
CpG ODN刺激2日後のB細胞の肥大
(5-1)B細胞の染色
実施例1と同様にKLHの免疫、免疫細胞の調整、CpG ODNによる免疫細胞の幼若化を行い、刺激後2日の免疫細胞を300gで5分間遠心し、0.5%BSA,2mM EDTA/PBS pH7.6バッファーに懸濁し、1回洗浄した。洗浄した細胞1×106/ml 100μlにFITCラベルした抗マウスCD45R抗体(Sigma社)4μlを加え、4℃で20分間、暗所でB細胞を染色した。染色後、1mlの0.5%BSA,2mM EDTA/PBS pH7.6バッファーを加え、300gで5分間遠心し、上清を除き、1mlの0.5%BSA,2mM EDTA/PBS pH7.6バッファーを加えた。その後、細胞懸濁液をセルストレーナー(日本ベクトン・ディッキン社)に通し測定用の細胞を調整した。
(5-2)B細胞肥大の測定
調整した細胞懸濁液をフローサイトメーター(日本ベクトン・ディッキン社)でFITCの蛍光強度を測定することによりB細胞を検出し、その前方散乱光により細胞の大きさを測定した。結果を(図5)に示す。CpG ODNで2日間培養し幼若化した実験区ではCD45R陽性のB細胞中の70%以上がURの領域内に存在し、細胞が肥大しており、細胞融合に有利であることが示された。CpG ODNによる幼若化前あるいはCpG ODN無添加で2日間培養した実験区ではB細胞の肥大はほとんど見られなかった。
【0035】
実施例6
CpG ODN刺激2日後の形質細胞の肥大
(5-1)形質細胞の染色
実施例1と同様にKLHの免疫、免疫細胞の調整、CpG ODNによる免疫細胞の幼若化後2日の細胞を300gで5分間遠心し、0.5%BSA,2mM EDTA/PBS pH7.6バッファーに懸濁し、1回洗浄した。洗浄した細胞1×106/ml 100μlにPEラベルした抗マウスCD138抗体(BD Phamingen社)2μlを加え、4℃で20分間、暗所で抗体産生細胞である形質細胞を染色した。染色後、1mlの0.5%BSA,2mM EDTA/PBS pH7.6バッファーを加え、300gで5分間遠心し、上清を除き、1mlの0.5%BSA,2mM EDTA/PBS pH7.6バッファーを加えた。その後、細胞懸濁液をセルストレーナー(日本ベクトン・ディッキン社)に通し測定用の細胞を調整した。
【0036】
(6-2)形質細胞肥大の測定
調整した細胞懸濁液をフローサイトメーター(日本ベクトン・ディッキン社)でPEの蛍光強度を測定することにより形質細胞を検出し、その前方散乱光により細胞の大きさを測定した。結果を(図6)に示す。CpG ODNで2日間培養し幼若化した実験区ではCD45R陽性のB細胞と同様に形質細胞中の70%以上がURの領域内に存在し、細胞が肥大しており、細胞融合に有利であることが示された。CpG ODNによる幼若化前あるいはCpG ODN無添加で2日間培養した実験区で形質細胞の肥大はほとんど見られなかった。
【0037】
実施例7
抗原特異的IgG産生細胞数の増大
(7-1)免疫細胞の刺激と調整
実施例1と同様にKLHの免疫、免疫細胞の調整、各種免疫刺激物質により1日から5日間刺激し、1日毎に細胞を回収し、300gで5分間遠心後、RPMIで1回洗浄した。洗浄した細胞は細胞数をカウントし、5×105/mlとなるようRPMI培地に懸濁した。
(7-2)Elispot
コーティングバッファーで10μg/mlに希釈したKLHを、ニトロセルロースメンブレンのマルチスクリーン96ウェルマイクロプレート(ミリポア社)に100μl/ウェルで分注し4℃ 1晩コーティングした。その後、コーティングバッファーを除き、純水で2回洗浄を行い、10%FBS/RPMIを250μl/ウェルで分注し37℃、2時間、5% CO2インキュベータ内でブロッキングした。10%FBS/RPMIを除き、7-1で調整した細胞懸濁液を100μl/ウェルで分注し37℃ 24時間 5% CO2インキュベータ内で培養した。培養後、細胞懸濁液を除き、0.05% Tween20/PBSで6回洗浄し、0.5%BSA/PBSで4000倍に希釈したアルカリフォスファターゼ標識の抗マウスIgG抗体(サザンバイオテック社)を50μl/ウェルで分注し37℃、1時間、5% CO2インキュベータ内で反応させた。反応液を除き、0.05% Tween20/PBSで3回洗浄し、続いてPBSで3回洗浄後、アルカリフォスファターゼ発色液(Sigma社)を100μl/ウェルで分注し、数分間発色させ、スポットの確認後、流水ですすぎ反応を停止した。ニトロセルロースメンブレンを暗所で乾燥し、スポットを計測した。
【0038】
結果を(図7)に示す。刺激物質無添加の実験区と比較して、CpG ODN,LPS,PWMで刺激を加え幼若化された実験区では抗原特異的IgG産生細胞数が増大しているのが分かる。特にCpG ODN刺激1日から2日では顕著な増大を示しており、これはCpG ODNがTLR9を介してB細胞を活性化していることに由来すると考えられる。TLR9はB細胞の分化が進み、最終分化形態である形質細胞に近づくに連れて発現が上昇する。従って、CpG ODNによりB細胞集団のTLR9の発現が高い形質細胞では刺激が増強されることにより、TLR9の発現が低いB細胞、即ち分化が進んでいないB細胞よりも優先的に、細胞の活性化や細胞肥大等の幼若化が誘導されると考えられる。このことは他の免疫刺激物質に比べCpG ODNによる幼若化が抗体産生B細胞の細胞融合に有利に働くことを示していると考えられる。
【0039】
実施例8
CpG ODNの配列バリエーションによる融合効率の変化
CpG ODNによる免疫細胞の幼若化の条件を除いて実施例1と同様に実験を実施した。免疫細胞の幼若化はCpG無添加と各CpG ODN 1826(配列番号8)、CpG ODN AG(配列番号9)、CpG ODN CT(配列番号10)、CpG control(配列番号11)を0.25μMとなるよう添加し、37℃、5% CO2インキュベータ内で2日間培養した。使用した各CpG ODNの塩基配列は以下のとおりである。
1)tccatGACGTTcctgacgtt(配列番号8)
2)tAGCGTT(配列番号9)
3)tGACGCT(配列番号10)
4)tccatGAGCTTcctgacgtt(配列番号11)
細胞融合効率を測定した結果を(図8)に示す。CpG無添加と活性中心を改変したCpG controlではほとんど細胞は融合しないが、CpG ODN 1826及びCpG ODN AG、CpG ODN CTはいずれも同様の融合効率が得られた。
【0040】
実施例9
Gaussia
luciferaseの発光阻害機能抗体の取得
(9-1)Gaussia luciferaseの免疫
抗原Gaussia luciferaseとFREUND Complete ADJUVANT(Sigma社)を混合し、BALB/cマウス(♀)の腹腔にGaussia luciferase 100μg/匹となるよう投与し、1次免疫を2週間行った。続いて、抗原KLHとFREUND Incomplete ADJUVANT(Sigma社)を混合し、1次免疫と同様に2回免疫を行った。最後の免疫から2週間後にマウスの腹腔に抗原100μgを投与しブーストした。
【0041】
(9-2)Gaussia luciferase免疫細胞の幼若化
実施例1と同様に免疫細胞の調整を行い、0.25μM CpG ODNを添加したRPMI培養液に2×106/mlとなるよう懸濁し、培養プレートに10ml分注後、37℃、5% CO2インキュベータ内で2日間培養し、細胞を幼若化した。
(9-3)エレクトロフュージョン
幼若化したGaussia luciferase免疫細胞と実施例1と同様に調整したミエローマ細胞を各々180g 5分間遠心し、280mOsm Inositolバッファーに懸濁し、2回洗浄した。洗浄した細胞を5×106/mlの濃度になるよう150mOsm Inositolバッファーに懸濁し、免疫細胞とミエローマ細胞を1:1で混合した。細胞混合液80μlを2mmギャップ電極のエレクトロフュージョンチャンバーへ添加し、150mOsm Inositolバッファーに懸濁してから2分後にLF100(ベックス社)を用いて、アラインメント35V 30sec、パルス300V 20μsec 2cycle、ポストアラインメント35V 30secのパルスを与え、10分間静置し、エレクトロフュージョンを行った。その後、細胞を20%FCS(HyClone社)、5% Briclone(QED BIOSCIENCE社)含有のフェノールレッド不含RPMI培養液10mlに懸濁し96ウェルマイクロプレートに100μl/ウェルで分注し、37℃、5% CO2インキュベータ内で1日培養し、翌日に20%FCS、5% Briclone、2×HAT含有のRPMI培養液を100μl/ウェルで分注した。7日間37℃ 5% CO2インキュベータ内で培養した。
【0042】
(9-4)抗体産生ハイブリドーマの発光阻害スクリーニング
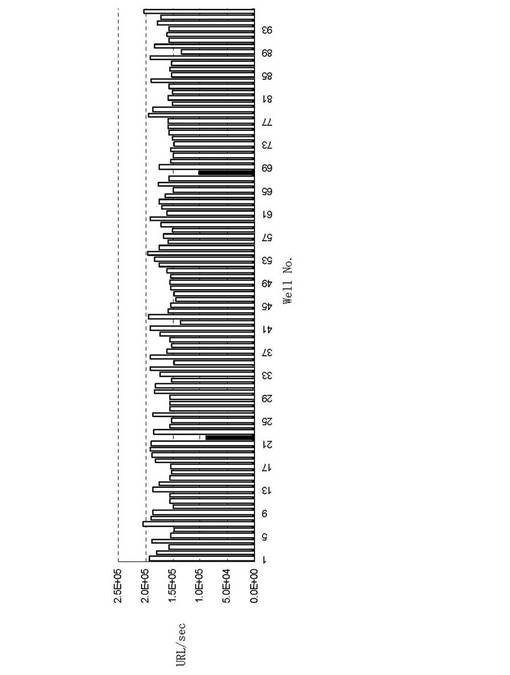
Gaussia luciferaseを12.5μg/mlとなるよう発光バッファー(200mM NaCl,50mM Tris,0.08% Triton X-100 pH7.6)で希釈し、遮光96ウェルマイクロプレートに40μl/ウェルで分注した。上記で7日間培養したハイブリドーマの培養上清50μlを各ウェルに分注し、室温30分間反応した。発光基質のCoelenterazineを0.5μMとなるよう発光バッファーで希釈し、40μl/ウェルで分注し10分後にルミノメーター、Centro LB960(ベルトールド社)で発光を測定した。結果を(図8)に示す。96ウェル中のNo.22とNo.68の2ウェルのハイブリドーマ培養上清に発光阻害が認められ、1次スクリーニングのみによって機能性抗体の取得に成功した。細胞融合は一般的に専用機器を必要としないPEG法が広く使用されている。PEG法の場合、コロニー出現効率は使用する免疫細胞の0.001%以下と細胞融合の効率が低い。また、PEG法よりも高効率である従来法によるエレクトロポレーションによる細胞融合効率はPEG法の3〜10倍と言われている。
【0043】
(図7)で示されているように、免疫細胞中の目的IgG抗体産生クローンは0.1%以下である。さらに、目的IgG抗体産生クローン中の機能性抗体産生クローン取得の割合は減少し、発光阻害機能を有する抗Gaussia lusifrase抗体の場合10%以下となる。これらの要因により、従来法では、高価値の抗体を得るためには多くのマウスに抗原の免疫を行ない、大量の免疫細胞の細胞融合を行う必要があった。加えて、大量の免疫細胞を用するため、数10〜数100枚の96ウェルマイクロプレートにハイブリドーマを播種し検体が多くなる。そのため、1次スクリーニングで抗原に結合する抗体産生クローンを選抜し、2次スクリーニングで機能性を評価する手順が必要となり、抗体取得のためにコスト・時間・労力等を多く費やしている。本高効率融合法の場合、細胞融合効率は約0.1%以上とPEG法と比較すると100倍以上の効率で細胞融合が可能となった。また、2×105の免疫細胞から開始し2ウェルから発光阻害抗体を検出したことから、従来法と比べて圧倒的な効率で機能性抗体を取得できたことが分かる。このことは、実施例7で前述したようにCpG ODNが抗体産生細胞の細胞融合に有利な活性化を引き起こしていることを支持しており、抗体産生細胞のハイブリドーマを効率的に得られた結果、1次スクリーニングで機能性の高い高価値抗体の取得に成功したと考えられる。
【0044】
実施例10
抗ルシフェラーゼ抗体の発光阻害機能評価
(10-1)ハイブリドーマのクローニング
実施例9で取得した発光阻害陽性ウェルのハイブリドーマを2.5×102/mlとなるよう20%FCS、5% Briclone、1×HAT含有のRPMI培養液に懸濁し、96ウェルマイクロプレートに200μl/ウェルで分注した。その後、37℃ 5% CO2インキュベータ内で培養した。培養7日後に顕微鏡下でシングルクローンのみが存在するウェルを確認し、培養上清を回収し、(9-4)と同様に発光阻害活性を確認した。発光阻害抗体陽性の2ウェルから各々クローニングを行い、ハイブリドーマクローン11-6と61-2を得た。
(10-2)腹水の作製
11-6と61-2のクローンをそれぞれ2×106/500μlとなるようPBSに懸濁し、マウスの腹腔内に投与した。10日後に腹腔から腹水を回収し、室温で1時間静置後、4℃、3000r.p.m 15分間遠心し、中間層の腹水を回収した。
【0045】
(10-3)腹水からのIgGの精製
腹水3mlを15mlの結合バッファー(PBS pH7.4)で希釈し、1ml Pritein Gカラム(GE社)へ1分間に0.2mlの流速で送液しIgG抗体をカラムに結合する。次いで5mlの結合バッファーを送液しカラムを洗浄後、5mlの溶出バッファー(0.1M Glycine-HCl pH2.7)を送液し抗体を溶出した。溶出した抗体溶液は500μl毎に中和バッファー(1M Tris-HCl pH9.0)を50μlを分注したチューブに回収し、直ちに中和後、各フラクションをOD280で測定し、抗体の濃度を算出した。11-6は2.2mg/mlの濃度で2.2ml、61-2は3.2mg/mlの濃度で1.6mlを回収した。
【0046】
(10-4)取得抗体の発光阻害機能評価
Gaussia luciferaseを12.5μg/mlとなるよう発光バッファーで希釈し、遮光96ウェルマイクロプレートに40μl/ウェルで分注した。精製した抗Gaussia lusiferase抗体の各クローン(20-3,11-6,61-2)を0.2mg/mlから0.032mg/mlまで発光バッファーで段階的に希釈し、50μlを各ウェルに分注し、室温30分間反応した。反応後にCoelenterazineを0.5μMとなるよう発光バッファーで希釈し、40μl/ウェルで分注し、10分後にルミノメーター、Centro LB960(ベルトールド社)で発光を測定した。結果を(図10)に示す。発光阻害機能の無い20-3は抗体濃度に関わらず、一定の発光であったのに対して、11-6と61-2は抗体濃度依存的にGaussia luciferaseの発光を阻害した。これにより、1次スクリーニングのみで得られたクローンから機能性抗体の取得が可能であることを証明した。本高効率融合法により高価値の抗体の取得に強力なツールの提供が可能となった。
【技術分野】
【0001】
本発明は、動物細胞同士の細胞融合法及び該方法に用いる細胞融合促進剤に関する。
【背景技術】
【0002】
抗体産生機構
抗体(免疫グロブリン)は、脊椎動物の感染防御機構において重要な役割を担うものであり、免疫細胞(リンパ球)のうちでB細胞により産生される。ヒト等哺乳動物の体内では、それぞれ異なる抗体を作る能力を備えたB細胞が、それぞれの細胞表面に数千万種以上とも数億種以上ともいわれる抗体を提示して抗原を待ち受け、あらゆる抗原に対応する体制を整えている。抗原刺激により抗体産生機能が活性化された特定のB細胞に対して、体内では、速やかに抗体産生活性化を促進する機構にスイッチが入り、抗体産生細胞の分化を経て抗原特異的な抗体の産生に至る。
【0003】
抗体の機能
生体内で産生された抗体は抗原の様々な部位に特異的に結合することにより毒素の中和や、オプソニン化による食細胞への貪食の促進、補体を活性化して細菌の溶菌誘導等の効果を発揮し、感染防御機構において抗原の除去を補助している。抗体の効果は各アイソタイプで異なっており、IgG、IgA、IgM、IgE、IgD、IgEに分類される。特にIgGは血中や細胞外組織液中では主要なアイソタイプであり、抗原との親和性、抗原中和、オプソニン化、補体の活性化能が高い場合が多い。単一の抗体産生細胞は1種類のアイソタイプで抗原の特定の1箇所に結合する抗体を産生しており、その抗体産生細胞をクローン化し得られる抗体は、モノクローナル抗体として利用されている。
【0004】
モノクローナル抗体の利用
生化学やタンパク質科学研究の分野においてモノクローナル抗体はその特異的認識能から標的物質の標識や選別等、幅広く使用されており、バイオツールとして必須であり、抗体の品質は研究の進展に大きく影響する。また、医療の分野においては、妊娠、成人病、感染症等の臨床診断薬として多く使われている。多種多様な抗原に対応可能な生体分子であることから、テーラーメイド医療の実現や副作用の軽減が期待されており、癌等の治療薬としても注目されている。モノクローナル抗体は研究から医療まで応用範囲が急速に拡大しており、有用な抗体を確実かつ効率的に作製する技術の確立が求められている。
【0005】
モノクローナル抗体作製過程の技術的問題点
モノクローナル抗体の作製は主に、1.動物への免疫、2.抗体産生細胞と癌細胞の融合、3.融合細胞(ハイブリドーマ)の選別、と3つの工程がある。工程3は抗体産生細胞と癌細胞の両方の機能を獲得した細胞のみを選別する試薬を用いることにより、既に確立されている。しかし、工程1、2には問題点があり、有用な抗体を確実に効率的に得るには課題を多く残している。
【0006】
問題点1(免疫)
実験動物を十分な抗原量で免疫にすれば、抗原特異的抗体産生細胞が取得できる場合が多いが、免疫原性の低い、すなわち免疫系の活性化能の低い場合は、抗体産生細胞や特異性が高い抗体を取得することは難しい。そのため十分な免疫応答を引き起こし、十分量の抗原特異的抗体産生細胞を取得するには、実験動物に対して、免疫活性化能を持つ抗原とアジュバントとの同時投与や、1回の投与量を多くし多数回にわたっての投与、多数の動物に免疫を行う必要がある。これらの処置にはコストと時間が掛かることに加えて、動物に対する副作用や処置自体による苦痛が大きく、動物保護の観点からも問題がある。したがって、インビボ免疫法において、抗原特異性の高い抗体を高効率で産生できる技術の開発が必要とされている。
【0007】
問題点2(細胞融合)
抗体産生細胞から大量の抗体を得るためには、抗体産生細胞とガン細胞であるミエローマ細胞とを融合させ、不死化させる必要がある。モノクローナル抗体の作製が困難な理由の一つに、この融合効率がきわめて低いことがある。低融合効率が、陽性クローンを多数取れない主要因であり、そのため認識能の高い抗体や、アゴニスト活性、アンタゴニスト活性等の機能を持った抗体の取得が大変難しくなっている。この細胞融合を用いずに抗体様の物質を得るために、ファージを使った作製方法等があるが、今のところ抗体ほどに特異性・親和性の高いものを得るまでにはなっていない。そのため、高効率の細胞融合法の開発は、抗体作製において非常に重要な技術となっている。
【0008】
細胞融合(PEG)の問題点
しかしながら、抗体産生細胞とミエローマ細胞の融合は難しく、その融合効率はきわめて低く、10-5程度である。ポリエチレングリコール(PEG)を用いた融合法は専用機器を必要とせず、試薬が安価であるため、主に用いられている。この方法は技術的に非常に難しく、薬剤自体に毒性があることもあり、効率が低いだけでなく、再現性も低いのが実情である。一方電気的パルスを用いた細胞融合法は、PEG法よりも融合効率が一桁程度高くなると言われている。
【0009】
電気的細胞融合法
電気的細胞融合法は交流電圧を印加することで電極間の抗体産生細胞とミエローマ細胞を接触させ、その後の直流パルス電圧を印加させることで細胞膜に穿孔が開き、接触している細胞同士の融合を引き起こす。この原理によりポリエチレングリコールよりも高い融合効率を実現できる。
しかし、抗体産生細胞は刺激されていない状態では融合効率が著しく低下することや、抗体産生細胞とミエローマ細胞の直径が異なっており、各々の細胞の穿孔する最適な電圧が異なること等が問題となっている。これらの原因により再現性が低いことがネックとなっており、システムとして確立されているとは言えない。また、融合装置を必要とすることもあり、その大きな有用性に注目されながらも未だに広く利用されていない。したがって、電気的細胞融合法において、確実に高効率に細胞融合を実現する技術の開発が強く望まれている。
【先行技術文献】
【特許文献】
【0010】
【特許文献1】特開2009−148296号公報
【特許文献2】特表2010−504750号公報
【非特許文献】
【0011】
【非特許文献1】Yu et al.J Immunol Methods.2008 Jul 31;336(2):142-51
【非特許文献2】Schmidt et al.J Immunol Methods.2001 Sep 1;255(1-2):93-102.
【非特許文献3】B.R.Lentz,Eur Biophys J.2007 Apr;36(4-5):315-26.
【非特許文献4】Li et al.Proc Natl Acad Sci USA.2006 Mar 7;103(10):3557-62.
【非特許文献5】Crowe Vaccine.2009 Dec 30;27 Suppl 6:G47-51.
【非特許文献6】Kwekkeboom et al.J Immunol Methods.1993 Mar 15;160(1):117-27.
【発明の概要】
【発明が解決しようとする課題】
【0012】
本発明の課題は、上記した電気的細胞融合法の問題点を解消し、確実に高い効率で細胞融合させることが可能となる手段を新たに提供することにある。
【課題を解決するための手段】
【0013】
そこで、本発明者等は、従来の電気的細胞融合法を検討したところ、融合効率が低く、陽性クローン数が少ないという欠点のいずれもが抗体産生細胞の活性化不足に起因するのではないか、という推論をたて、免疫系の有効な活性化法について鋭意研究を重ねた結果、従来の電気的細胞融合法を飛躍的に改善する基本プロトコールを開発するに至った。
すなわち、本発明は、細胞の調整、培養方法および最適な免疫系刺激因子に関する基本プロトコールに係るものであり、具体的には、細胞融合に際して、体外に取り出した免疫細胞を免疫賦活物質を含有する培地で培養する工程を加えることにより、目的抗原を認識するB細胞がさらに特異的に活性化されるだけでなく、ミエローマ細胞との融合効率の上昇をもたらすことが可能であるとの知見を得るとともに、さらに研究を進め、免疫刺激物質として特にCpGモチーフを含むオリゴヌクレオチドを用いた場合には、細胞融合効率が、他の免疫賦活物質と比較して全く予想外に飛躍的に高まることを見いだした。そして、これにより、当該抗原特異的抗体産生細胞を片親としたハイブリドーマから抗原特異的モノクローナル抗体をインビトロで効率的に生産可能であることを確認し、本発明を完成させた。
本発明は以下のとおりである。
【0014】
(1)免疫細胞とミエローマ細胞とを電気的細胞融合法で融合するに際して、予め、CpGモチーフを有するオリゴヌクレオチドを含有する培地で免疫細胞を培養することを特徴とする、細胞融合方法。
(2)CpGモチーフを有するオリゴヌクレオチドが、以下の塩基配列を有することを特徴とする上記(1)に記載の細胞融合方法。
【化1】
(ただし、上記塩基配列中、N1N2のうち少なくとも一つはプリン塩基であり、N3N4のうち少なくとも一つはピリミジン塩基である。)
(3)CpGモチーフを有するオリゴヌクレオチドが、5’−GACGTT−3’、5’−TGGCGT−3’、5’−GACGCT−3’、5’−AGCGTT−3’又は5’−GAGCTT−3’からなる配列を有することを特徴とする上記(2)に記載の細胞融合方法。
(4)CpGモチーフを有するオリゴヌクレオチドが、
5'-tcctGACGTTcctgacgtt-3'又は5'-tccatGACGTTcctgacgtt-3'、5'-tAGCGTT-3'、5'-tGACGCT-3'、又は5'-tccatGAGCTTcctgacgtt-3'であることを特徴とする上記(3)に記載の細胞融合方法。
(5)培地が、さらにサイトカインを含有することを特徴とする、上記(1)〜(4)のいずれかに記載の細胞融合方法。
(6)CpGモチーフを有するオリゴヌクレオチドからなる電気的細胞融合促進剤。
(7)CpGモチーフを有するオリゴヌクレオチドが、以下の塩基配列を有することを特徴とする上記(6)に記載の電気的細胞融合促進剤。
【化1】
(ただし、上記塩基配列中、N1N2のうち少なくとも一つはプリン塩基であり、N3N4のうち少なくとも一つはピリミジン塩基である。)
(8)CpGモチーフを有するオリゴヌクレオチドが、5’−GACGTT−3’、5’−TGGCGT−3’、5’−GACGCT−3’、5’−AGCGTT−3’又は5’−GAGCTT−3’からなる配列を有することを特徴とする上記(7)に記載の電気的細胞融合促進剤。
(9)CpGモチーフを有するオリゴヌクレオチドが、
5'-tcctGACGTTcctgacgtt-3'、5'-tcctTGGCGTcctgacgtt-3'、5'-tAGCGTT-3'、5'-tGACGCT-3'、又は5'-tccatGAGCTTcctgacgtt-3'であることを特徴とする上記(8)に記載の電気的細胞融合促進剤。
(10)上記(6)〜(9)のいずれかに記載の電気的細胞融合促進剤とサイトカインを含有することを特徴とする、電気的細胞融合試薬キット。
(11)抗体産生細胞とミエローマ細胞とを電気的細胞融合法で融合するに際して、予め、CpGモチーフを有するオリゴヌクレオチドを含有する培地で抗体細胞を培養することを特徴とする、モノクローナル抗体産生ハイブリドーマの製造方法。
(12)培地がさらにサイトカインを含有することを特徴とする、上記(11)に記載のハイブリドーマの製造方法。
(13)上記(11)又は(12)に記載の方法により得られたモノクローナル抗体産生ハイブリドーマ。
(14)上記(13)に記載のハイブリドーマを培地に培養し、モノクローナル抗体を採取することを特徴とする、モノクローナル抗体の製造方法。
【発明の効果】
【0015】
本発明の細胞融合法によれば、従来困難であった極めて高効率の細胞融合が可能となる。また、用いた抗体産生細胞に対する、目的とする機能を有するモノクローナル抗体産生ハイブリドーマの出現率は極めて高く、細胞融合に際し、抗体産生細胞を従来のように大量に必要とせず、動物を免疫する抗原量も少量ですむ。また、従来法によれば、例えば、一次スクリーニングで抗原と結合するハイブリドーマの選別、2次スクリーニングで該ハイブリドーマの目的とする機能性の評価をそれぞれ行っていたが、本発明の融合法によれば、このような選別及び評価は、一回のスクリーニングで行うことが可能となる。したがって、本発明の細胞融合法によれば、有用なハイブリドーマあるいは目的とするモノクローナル抗体を極めて効率的、かつ短時間、安価に提供でき、これらを通じて医薬あるいは化粧品その他の分野において大いに貢献できる。
【図面の簡単な説明】
【0016】
【図1】各種免疫刺激物質と刺激日数による融合効率の変化を示す図である。
【図2】CpG ODN刺激免疫細胞の融合電圧による融合効率の変化を示す図である。
【図3】CpG ODN刺激時の細胞培養濃度による融合効率の変化を示す図である。
【図4】各種免疫刺激物質刺激2日後の免疫細胞の直径を測定した結果を示す図である。
【図5】CpG ODN刺激の有無によるB細胞の肥大の変化を測定した結果を示す図である。
【図6】CpG ODN刺激の有無による形質細胞の肥大の変化を測定した結果を示す図である。
【図7】各種免疫刺激物質の刺激により、得られる抗原特異的IgG産生細胞数の差違を示す図である。
【図8】CpG ODNの配列バリエーションによる融合効率を測定した結果を示す図である。
【図9】本発明の細胞融合法により得られたハイブリドーマの培養上澄みを使用した検出試験により、Gaussia luciferaseの発光阻害機能抗体を産生するハイブリドーマが取得可能であることを示す図である。
【図10】上記発光阻害機能抗体の発光阻害機能評価の結果を示す図である。
【発明を実施するための形態】
【0017】
本発明は、細胞融合に先立ち、予め免疫細胞をCpGモチーフを含むオリゴヌクレオリド(以下、CpG ODNということがある。)あるいはさらにサイトカインを含有する培地に培養して、インビトロで免疫刺激し、次いで培養した免疫細胞とミエローマ細胞とを電気的細胞融合法により融合させて、ハイブリドーマを得るものである。
【0018】
本発明において「免疫細胞」とは、動物を抗原で免疫した後、該動物の末梢血、脾臓、リンパ節、扁桃、パイエル板、虫垂等の2次リンパ組織から採取した抗体産生細胞を含むリンパ球をいう。採取源の生物種はどの哺乳動物由来であってもよいが、治療用に用いる抗原特異的モノクローナル抗体を得るためには、同じ生物種由来、例えばヒト用であればヒト由来のリンパ球が好ましい。
本発明において、インビトロでの免疫刺激に先立ち、あらかじめ赤血球除去工程を設けることが好ましいが、設けることなく幼若化工程に付してもよい。
本発明で使用するCpG ODN、CpG motif(5'-CG-3')を持つオリゴヌクレオチドで、このmotifが免疫細胞活性化の活性中心であり、特にCの非メチル化が活性に必須である。
本発明で使用するCpG ODNのより具体的な例は、 5’−N1N2CGN3N4−3’(配列番号1)の塩基配列を有するものである。(ただし、上記塩基配列中、N1N2のうち少なくとも一つはプリン塩基であり、N3XN4のうち少なくとも一つはピリミジン塩基である。)
【0019】
このようなCpG ODNは、免疫刺激物質として、それ自体公知である。
CG前後の配列はCGを中心に回文構造の場合が多く、マウス等のげっ歯類では5'-GACGTT-3'(配列番号2)を、ヒトでは5'-GTCGTT-3'(配列番号3)を含むオリゴヌクレオチドが免疫活性化能を持つことが報告されている。
また、その他のCpG ODNとしては、例えば5’−TGGCGT−3’(配列番号4)、5’−GACGCT−3’(配列番号5)、5’−AGCGTT−3’(配列番号6)又は5’−GAGCTT−3’(配列番号12)の配列を有するオリゴヌクレオチドが挙げられる。
また、CG前後の配列のパターンによって作用が異なり、作用別にA-type、B-type、C-typeの3つのグループに分類される。A-typeは形質細胞様樹状細胞のインターフェロンαの産生を誘導し、B-typeはB細胞を活性化しIgM、インターロイキン6、インターロイキン10の産生を誘導し、C-typeはA-type、B-type双方の作用を有するが、本発明においてはそのいずれもが使用できる。
【0020】
電気的細胞融合において、このようなCpG ODNを使用する場合、他の免疫刺激物質、例えば、LPS(リポポリサッカライド)、PWM(pokeweed mitogen)、PHA(フィトヘムアグルチニン)、EBV(Epstein-Barrウィルス)等に比べて格段に高い確率で融合細胞が得られ、極めて顕著に細胞融合効率を上昇させる。したがって、本発明のCpG ODNは細胞融合促進剤として有用である。
また、サイトカインの使用はさらに融合効率を上昇させ、CpG ODNとサイトカインは電気的細胞融合試薬キットとして有用である。本発明において使用するサイトカインとしては、例えばインターロイキン2、インターロイキン4、インターロイキン5、インターロイキン6、インターロイキン13、インターロイキン21等が挙げられる。
【0021】
本発明において免疫細胞を培養する際の培地としては、通常の細胞培養用培地、例えばRPMI培地、DMEM培地、ハム−F12培地、RDF培地、ERDF培地、又はこれらの混合培地を用いることができるが、これらの中では、RPMI培地が特に好ましい。35〜38℃、好ましくは37℃の温度条件下で培養するが、その際に1%〜10%、好ましくは約5%のCO2環境下でインキュベートすることが好ましい。培地中の免疫細胞の細胞密度は、0.5×106〜1×107 cells/ml範囲内に、好ましくは2×106 cells/mlで用いる。当該培地は、免疫細胞を採取後懸濁液とする際の懸濁液としても、また当該懸濁液から遠心分離により免疫細胞を濃縮分離する際の洗浄液としても用いることができ、ハイブリドーマの培養液としても用いることができる。
【0022】
本発明においては細胞融合手段として、電気的細胞融合法を用いるが、電気的細胞融合法は、電極を配した細胞懸濁液収容部材に、融合する細胞を懸濁し、所定の浸透圧に調整した溶液を収容し、電極から細胞を接着する(パールチェーンの形成)ための交流、及び細胞融合させるための直流パルスを順次与えて細胞を融合させる手段である。この手段自体はよく知られている。
【0023】
本発明においては上記の各手段を組み合わせて細胞融合を行うが、その概要は図1に示され、そのプロセスは以下のとおりである。
まず、マウス等の動物に抗原を投与し、その抗原に対する抗体産生細胞を誘導し、免疫組織(脾臓、リンパ節)から、免疫細胞を取り出す。次いで、免疫細胞を上記した培地で培養して幼若化するが、このとき培地に上記したCpG ODNを添加する。添加量は、培養液に対し0.05〜100uMであり、好ましくは0.2〜10uMである。
本発明においては、CpG ODNとともに上記サイトカインを併用することが好ましく、さらに上記したLPS等の他の免疫刺激物質を組み合わせて添加してもよい。
【0024】
CpG ODN含有培地での培養時間については、後記する実施例の実験によれば、細胞融合効率は1日でも極めて高いが、2日が最大となり、3日以上では急速に効率が低下する。また、抗体産生能は1日で最大である。したがって、培養は2日以内で終了することが好ましい。
このようなCpG ODN含有培地での培養により、抗体産生細胞は顕著に活性化され、活発に増殖・巨大化する。
次いで、このように活性化された抗体産生細胞を含む免疫細胞は、培地から取り出して洗浄した後、所定の浸透圧に調整された緩衝液においてミエローマ細胞と1対1となるよう混合され、1〜10×106cells/ml、好ましくは2〜4×106cells/mlで、電極を配した細胞懸濁液収容部材に投入し、電極から交流及び直流パルスを印加して、電気的細胞融合を行う。
【0025】
上記緩衝液の浸透圧は、グルコース、スクロース、トレハロース等の糖類、イノシトール、マンニトール等の糖アルコール類、あるいは塩化マグネシウム、塩化カルシウム、リン酸カリウム等により調整するが、浸透圧は590〜90mOsm/kg、好ましくは280〜90mOsm/kgに調整する。
また、電極間のギャップが2mmの場合、上記交流印加は、10〜50Vで10〜60秒、好ましくは25〜40Vで10〜30秒である。上記直流パルスの印加は、150〜450Vで5〜50マイクロ秒を1〜4サイクル。好ましくは200〜300Vで10〜40マイクロ秒を2〜3サイクルである。本発明においては、上記直流パルス印加後、細胞融合を促進させ安定化させるために、さらに交流電圧を印加することが好ましい、10〜50Vで10〜60秒、好ましくは25〜40Vで10〜30秒である。
【0026】
電気的細胞融合後、フェノールレッド不含のRPMIで培養後、常法により、HAT培地でハイブリドーマを選別し、さらに限界希釈法により得たハイブリドーマの各クローンを培養し、得られた各クローンの培養上澄みについて、例えば、上記動物に免疫した抗原を使用した酵素免疫測定法(ELISA)により、発色強度を測定することによって、該抗原との特異的反応性が高いものを検出し、また、さらに必要により、増殖性の高いクローンを選定することにより、目的とするIgGモノクローナル抗体を産生するハイブリドーマクローンを得ることができる。
特に本発明の電気的細胞融合法による場合、用いた抗体産生細胞に対する、目的とする機能を有するモノクローナル抗体産生ハイブリドーマの出現率は極めて高く、細胞融合に際し、抗体産生細胞を従来のように大量に必要としない。また、従来法のように一次、二次の2回のスクリーニングを行うことなく、1回のスクリーニングで目的とするモノクローナル抗体産生ハイブリドーマを取得することが可能となる。
【実施例】
【0027】
実施例1
免疫マウスの脾臓を用いた各種免疫刺激物質による融合効率の変化
(1-1)KLHの免疫
抗原KLHとFREUND’S ADJUVANT Complete(Sigma社)を混合し、BALB/cマウス(♀)の腹腔とフットパッドにKLH 100μg/匹となるよう投与し、1次免疫を1週間行った。続いて、抗原KLHとFREUND’S ADJUVANT In Complete(Sigma社)を混合し、1次免疫と同様に2次免疫を行った。
(1-2)免疫細胞の調整
免疫を行ったマウスより脾臓を摘出し、RPMI培養液(Sigma社)中に分散させ、セルストレーナー(BD社)を通し夾雑物を取り除いた。次いで、Lympholyte-M(CEDARLANE社)上に細胞懸濁液を重層し、23℃ 1000g 20分間遠心し、赤血球を取り除き、培養液で2回洗浄した。
【0028】
(1-3)免疫細胞の幼若化
上記(1-2)で調整した免疫細胞をRPMI培養液に懸濁し、培養プレートに10mlずつ分注した。各プレートには、添加無し、0.25μM CpG ODN(北海道システム・サイエンス株式会社に委託して作成)、10μg/ml LPS(Sigma社)、0.5μg/ml抗CD40抗体(R&D Systems社)と10ng/ml IL-2(PEPROTECH社)、2.4μg/ml PWM(Sigma社)を添加し、37℃、5% CO2インキュベータ内で1日から5日間培養した。なお、上記CpG ODNの塩基配列は下記のとおりである。
5’−tccatgacgttcatgacgtt−3’(配列番号7)
(1-4)ミエローマ細胞の調整
エレクトロフュージョン3日前にミエローマ細胞を起眠し、1×106/mlの濃度を超えないようRPMI培養液に懸濁し37℃、5% CO2インキュベータ内で培養した。
【0029】
(1-5)エレクトロフュージョン
幼若化した免疫細胞とミエローマ細胞を各々180g 5分間遠心し、290mOsm Inositol、0.5mM塩化マグネシウム、0.1mM塩化カルシウム、1mMリン酸カリウム、pH7.2バッファーに懸濁し、2回洗浄した。洗浄した細胞を5×106/mlの濃度になるよう185mOsm Inositolバッファーに懸濁し、免疫細胞とミエローマ細胞を1:1で混合した。細胞混合液80μlを2mmギャップ電極のエレクトロフュージョンチャンバーへ添加し、185mOsm Inositolバッファーに懸濁してから2分後に電気的細胞融合装置LF100(ベックス社)を用いて、アラインメント35V 30sec、パルス300V 20μsec 2cycle、ポストアラインメント35V 30secのパルスを与え、10分間静置し、エレクトロフュージョンを行った。その後、細胞を20%FCS(HyClone社)、5% Briclone(QED BIOSCIENCE社)含有のフェノールレッド不含RPMI培養液10mlに懸濁し96ウェルプレートマイクロに100μl/ウェルで分注し、37℃、5% CO2インキュベータ内で培養した。
【0030】
(1-6)細胞融合効率の測定
各免疫刺激物質で幼若化した免疫細胞は刺激開始から1日毎に5日間回収し、各々エレクトロフュージョンを行った。エレクトロフュージョンの翌日に細胞を播種した96穴プレートに20%FCS、5% Briclone、2×HAT(Sigma社)含有のRPMI培養液を100μl/ウェルで分注した。ハイブリドーマのコロニーが観察出来るまで、7日から9日間37℃ 5% CO2インキュベータ内で培養し、顕微鏡下でコロニー数を計測し、各免疫刺激物質による幼若化1〜5日間の細胞融合効率を算出した。結果を(図1)に示す。
各免疫刺激物質は幼若化開始から2〜3日で融合効率が高くなり、特にCpG ODNを用いた高効率融合法の場合、刺激開始から2日目に融合効率の上昇がピークとなり、形成されるコロニー数が圧倒的に多いことがわかる。
【0031】
実施例2
各種免疫刺激物質による刺激後のエレクトロフュージョン融合電圧による融合効率の変化
エレクトロフュージョンのパルス条件を除いて実施例1と同様に実験を実施した。エレクトロフュージョンはパルスを250V,300V,350V,400Vの4条件で実施し、実施例1と同様に細胞融合効率を測定した。結果を(図2)に示す。各免疫刺激物質いずれも300Vで最も融合効率が高く、特にCpG ODNによって幼若化された場合、融合効率が上昇し、コロニー形成に優位であることが示された。
【0032】
実施例3
CpG ODN刺激時の細胞培養濃度による融合効率の変化
免疫細胞の幼若化の細胞濃度を除いて実施例1と同様に実験を実施した。免疫細胞の幼若化のステップでは0.5×106/ml,1×106/ml,2×106/ml,4×106/mlの細胞濃度となるよう各々RPMI培養液に懸濁し、培養プレートに10mlずつ分注した。各プレートには、CpG ODNを0.25μMとなるよう添加し、37℃、5% CO2インキュベータ内で2日間培養した。その後、実施例1と同様に細胞融合効率を測定した。結果を(図3)に示す。幼若化のステップの培養濃度は融合効率に大きく影響を及ぼしており、2×106/mlで培養した時に最も融合効率が上昇する。
【0033】
実施例4
各種免疫刺激物質刺激2日後の細胞の直径
実施例1と同様にKLHの免疫、免疫細胞の調整、各種免疫刺激物質による免疫細胞の幼若化を行い、2日後に細胞カウントバッファーで100倍に希釈し、コールターカウンター(ベックマンコールター社)で細胞の直径を測定した。結果を(図4)に示す。他の刺激物質と比較してCpG ODNの実験区ではより免疫細胞の直径が拡大しており、平均直径の拡大も突出している。エレクトロフュージョンにおいて細胞の直径は融合電圧を決定する要因の1つである。ミエローマ細胞と免疫細胞の直径は大きく異なっており、2つの細胞間の直径差を減少させることは細胞融合効率に大きく影響を及ぼす。CpG ODNによる幼若化は免疫細胞の活性化と共に細胞をより肥大させ、ミエローマ細胞の直径に近づけることで融合効率を上昇させていると考えられる。
【0034】
実施例5
CpG ODN刺激2日後のB細胞の肥大
(5-1)B細胞の染色
実施例1と同様にKLHの免疫、免疫細胞の調整、CpG ODNによる免疫細胞の幼若化を行い、刺激後2日の免疫細胞を300gで5分間遠心し、0.5%BSA,2mM EDTA/PBS pH7.6バッファーに懸濁し、1回洗浄した。洗浄した細胞1×106/ml 100μlにFITCラベルした抗マウスCD45R抗体(Sigma社)4μlを加え、4℃で20分間、暗所でB細胞を染色した。染色後、1mlの0.5%BSA,2mM EDTA/PBS pH7.6バッファーを加え、300gで5分間遠心し、上清を除き、1mlの0.5%BSA,2mM EDTA/PBS pH7.6バッファーを加えた。その後、細胞懸濁液をセルストレーナー(日本ベクトン・ディッキン社)に通し測定用の細胞を調整した。
(5-2)B細胞肥大の測定
調整した細胞懸濁液をフローサイトメーター(日本ベクトン・ディッキン社)でFITCの蛍光強度を測定することによりB細胞を検出し、その前方散乱光により細胞の大きさを測定した。結果を(図5)に示す。CpG ODNで2日間培養し幼若化した実験区ではCD45R陽性のB細胞中の70%以上がURの領域内に存在し、細胞が肥大しており、細胞融合に有利であることが示された。CpG ODNによる幼若化前あるいはCpG ODN無添加で2日間培養した実験区ではB細胞の肥大はほとんど見られなかった。
【0035】
実施例6
CpG ODN刺激2日後の形質細胞の肥大
(5-1)形質細胞の染色
実施例1と同様にKLHの免疫、免疫細胞の調整、CpG ODNによる免疫細胞の幼若化後2日の細胞を300gで5分間遠心し、0.5%BSA,2mM EDTA/PBS pH7.6バッファーに懸濁し、1回洗浄した。洗浄した細胞1×106/ml 100μlにPEラベルした抗マウスCD138抗体(BD Phamingen社)2μlを加え、4℃で20分間、暗所で抗体産生細胞である形質細胞を染色した。染色後、1mlの0.5%BSA,2mM EDTA/PBS pH7.6バッファーを加え、300gで5分間遠心し、上清を除き、1mlの0.5%BSA,2mM EDTA/PBS pH7.6バッファーを加えた。その後、細胞懸濁液をセルストレーナー(日本ベクトン・ディッキン社)に通し測定用の細胞を調整した。
【0036】
(6-2)形質細胞肥大の測定
調整した細胞懸濁液をフローサイトメーター(日本ベクトン・ディッキン社)でPEの蛍光強度を測定することにより形質細胞を検出し、その前方散乱光により細胞の大きさを測定した。結果を(図6)に示す。CpG ODNで2日間培養し幼若化した実験区ではCD45R陽性のB細胞と同様に形質細胞中の70%以上がURの領域内に存在し、細胞が肥大しており、細胞融合に有利であることが示された。CpG ODNによる幼若化前あるいはCpG ODN無添加で2日間培養した実験区で形質細胞の肥大はほとんど見られなかった。
【0037】
実施例7
抗原特異的IgG産生細胞数の増大
(7-1)免疫細胞の刺激と調整
実施例1と同様にKLHの免疫、免疫細胞の調整、各種免疫刺激物質により1日から5日間刺激し、1日毎に細胞を回収し、300gで5分間遠心後、RPMIで1回洗浄した。洗浄した細胞は細胞数をカウントし、5×105/mlとなるようRPMI培地に懸濁した。
(7-2)Elispot
コーティングバッファーで10μg/mlに希釈したKLHを、ニトロセルロースメンブレンのマルチスクリーン96ウェルマイクロプレート(ミリポア社)に100μl/ウェルで分注し4℃ 1晩コーティングした。その後、コーティングバッファーを除き、純水で2回洗浄を行い、10%FBS/RPMIを250μl/ウェルで分注し37℃、2時間、5% CO2インキュベータ内でブロッキングした。10%FBS/RPMIを除き、7-1で調整した細胞懸濁液を100μl/ウェルで分注し37℃ 24時間 5% CO2インキュベータ内で培養した。培養後、細胞懸濁液を除き、0.05% Tween20/PBSで6回洗浄し、0.5%BSA/PBSで4000倍に希釈したアルカリフォスファターゼ標識の抗マウスIgG抗体(サザンバイオテック社)を50μl/ウェルで分注し37℃、1時間、5% CO2インキュベータ内で反応させた。反応液を除き、0.05% Tween20/PBSで3回洗浄し、続いてPBSで3回洗浄後、アルカリフォスファターゼ発色液(Sigma社)を100μl/ウェルで分注し、数分間発色させ、スポットの確認後、流水ですすぎ反応を停止した。ニトロセルロースメンブレンを暗所で乾燥し、スポットを計測した。
【0038】
結果を(図7)に示す。刺激物質無添加の実験区と比較して、CpG ODN,LPS,PWMで刺激を加え幼若化された実験区では抗原特異的IgG産生細胞数が増大しているのが分かる。特にCpG ODN刺激1日から2日では顕著な増大を示しており、これはCpG ODNがTLR9を介してB細胞を活性化していることに由来すると考えられる。TLR9はB細胞の分化が進み、最終分化形態である形質細胞に近づくに連れて発現が上昇する。従って、CpG ODNによりB細胞集団のTLR9の発現が高い形質細胞では刺激が増強されることにより、TLR9の発現が低いB細胞、即ち分化が進んでいないB細胞よりも優先的に、細胞の活性化や細胞肥大等の幼若化が誘導されると考えられる。このことは他の免疫刺激物質に比べCpG ODNによる幼若化が抗体産生B細胞の細胞融合に有利に働くことを示していると考えられる。
【0039】
実施例8
CpG ODNの配列バリエーションによる融合効率の変化
CpG ODNによる免疫細胞の幼若化の条件を除いて実施例1と同様に実験を実施した。免疫細胞の幼若化はCpG無添加と各CpG ODN 1826(配列番号8)、CpG ODN AG(配列番号9)、CpG ODN CT(配列番号10)、CpG control(配列番号11)を0.25μMとなるよう添加し、37℃、5% CO2インキュベータ内で2日間培養した。使用した各CpG ODNの塩基配列は以下のとおりである。
1)tccatGACGTTcctgacgtt(配列番号8)
2)tAGCGTT(配列番号9)
3)tGACGCT(配列番号10)
4)tccatGAGCTTcctgacgtt(配列番号11)
細胞融合効率を測定した結果を(図8)に示す。CpG無添加と活性中心を改変したCpG controlではほとんど細胞は融合しないが、CpG ODN 1826及びCpG ODN AG、CpG ODN CTはいずれも同様の融合効率が得られた。
【0040】
実施例9
Gaussia
luciferaseの発光阻害機能抗体の取得
(9-1)Gaussia luciferaseの免疫
抗原Gaussia luciferaseとFREUND Complete ADJUVANT(Sigma社)を混合し、BALB/cマウス(♀)の腹腔にGaussia luciferase 100μg/匹となるよう投与し、1次免疫を2週間行った。続いて、抗原KLHとFREUND Incomplete ADJUVANT(Sigma社)を混合し、1次免疫と同様に2回免疫を行った。最後の免疫から2週間後にマウスの腹腔に抗原100μgを投与しブーストした。
【0041】
(9-2)Gaussia luciferase免疫細胞の幼若化
実施例1と同様に免疫細胞の調整を行い、0.25μM CpG ODNを添加したRPMI培養液に2×106/mlとなるよう懸濁し、培養プレートに10ml分注後、37℃、5% CO2インキュベータ内で2日間培養し、細胞を幼若化した。
(9-3)エレクトロフュージョン
幼若化したGaussia luciferase免疫細胞と実施例1と同様に調整したミエローマ細胞を各々180g 5分間遠心し、280mOsm Inositolバッファーに懸濁し、2回洗浄した。洗浄した細胞を5×106/mlの濃度になるよう150mOsm Inositolバッファーに懸濁し、免疫細胞とミエローマ細胞を1:1で混合した。細胞混合液80μlを2mmギャップ電極のエレクトロフュージョンチャンバーへ添加し、150mOsm Inositolバッファーに懸濁してから2分後にLF100(ベックス社)を用いて、アラインメント35V 30sec、パルス300V 20μsec 2cycle、ポストアラインメント35V 30secのパルスを与え、10分間静置し、エレクトロフュージョンを行った。その後、細胞を20%FCS(HyClone社)、5% Briclone(QED BIOSCIENCE社)含有のフェノールレッド不含RPMI培養液10mlに懸濁し96ウェルマイクロプレートに100μl/ウェルで分注し、37℃、5% CO2インキュベータ内で1日培養し、翌日に20%FCS、5% Briclone、2×HAT含有のRPMI培養液を100μl/ウェルで分注した。7日間37℃ 5% CO2インキュベータ内で培養した。
【0042】
(9-4)抗体産生ハイブリドーマの発光阻害スクリーニング
Gaussia luciferaseを12.5μg/mlとなるよう発光バッファー(200mM NaCl,50mM Tris,0.08% Triton X-100 pH7.6)で希釈し、遮光96ウェルマイクロプレートに40μl/ウェルで分注した。上記で7日間培養したハイブリドーマの培養上清50μlを各ウェルに分注し、室温30分間反応した。発光基質のCoelenterazineを0.5μMとなるよう発光バッファーで希釈し、40μl/ウェルで分注し10分後にルミノメーター、Centro LB960(ベルトールド社)で発光を測定した。結果を(図8)に示す。96ウェル中のNo.22とNo.68の2ウェルのハイブリドーマ培養上清に発光阻害が認められ、1次スクリーニングのみによって機能性抗体の取得に成功した。細胞融合は一般的に専用機器を必要としないPEG法が広く使用されている。PEG法の場合、コロニー出現効率は使用する免疫細胞の0.001%以下と細胞融合の効率が低い。また、PEG法よりも高効率である従来法によるエレクトロポレーションによる細胞融合効率はPEG法の3〜10倍と言われている。
【0043】
(図7)で示されているように、免疫細胞中の目的IgG抗体産生クローンは0.1%以下である。さらに、目的IgG抗体産生クローン中の機能性抗体産生クローン取得の割合は減少し、発光阻害機能を有する抗Gaussia lusifrase抗体の場合10%以下となる。これらの要因により、従来法では、高価値の抗体を得るためには多くのマウスに抗原の免疫を行ない、大量の免疫細胞の細胞融合を行う必要があった。加えて、大量の免疫細胞を用するため、数10〜数100枚の96ウェルマイクロプレートにハイブリドーマを播種し検体が多くなる。そのため、1次スクリーニングで抗原に結合する抗体産生クローンを選抜し、2次スクリーニングで機能性を評価する手順が必要となり、抗体取得のためにコスト・時間・労力等を多く費やしている。本高効率融合法の場合、細胞融合効率は約0.1%以上とPEG法と比較すると100倍以上の効率で細胞融合が可能となった。また、2×105の免疫細胞から開始し2ウェルから発光阻害抗体を検出したことから、従来法と比べて圧倒的な効率で機能性抗体を取得できたことが分かる。このことは、実施例7で前述したようにCpG ODNが抗体産生細胞の細胞融合に有利な活性化を引き起こしていることを支持しており、抗体産生細胞のハイブリドーマを効率的に得られた結果、1次スクリーニングで機能性の高い高価値抗体の取得に成功したと考えられる。
【0044】
実施例10
抗ルシフェラーゼ抗体の発光阻害機能評価
(10-1)ハイブリドーマのクローニング
実施例9で取得した発光阻害陽性ウェルのハイブリドーマを2.5×102/mlとなるよう20%FCS、5% Briclone、1×HAT含有のRPMI培養液に懸濁し、96ウェルマイクロプレートに200μl/ウェルで分注した。その後、37℃ 5% CO2インキュベータ内で培養した。培養7日後に顕微鏡下でシングルクローンのみが存在するウェルを確認し、培養上清を回収し、(9-4)と同様に発光阻害活性を確認した。発光阻害抗体陽性の2ウェルから各々クローニングを行い、ハイブリドーマクローン11-6と61-2を得た。
(10-2)腹水の作製
11-6と61-2のクローンをそれぞれ2×106/500μlとなるようPBSに懸濁し、マウスの腹腔内に投与した。10日後に腹腔から腹水を回収し、室温で1時間静置後、4℃、3000r.p.m 15分間遠心し、中間層の腹水を回収した。
【0045】
(10-3)腹水からのIgGの精製
腹水3mlを15mlの結合バッファー(PBS pH7.4)で希釈し、1ml Pritein Gカラム(GE社)へ1分間に0.2mlの流速で送液しIgG抗体をカラムに結合する。次いで5mlの結合バッファーを送液しカラムを洗浄後、5mlの溶出バッファー(0.1M Glycine-HCl pH2.7)を送液し抗体を溶出した。溶出した抗体溶液は500μl毎に中和バッファー(1M Tris-HCl pH9.0)を50μlを分注したチューブに回収し、直ちに中和後、各フラクションをOD280で測定し、抗体の濃度を算出した。11-6は2.2mg/mlの濃度で2.2ml、61-2は3.2mg/mlの濃度で1.6mlを回収した。
【0046】
(10-4)取得抗体の発光阻害機能評価
Gaussia luciferaseを12.5μg/mlとなるよう発光バッファーで希釈し、遮光96ウェルマイクロプレートに40μl/ウェルで分注した。精製した抗Gaussia lusiferase抗体の各クローン(20-3,11-6,61-2)を0.2mg/mlから0.032mg/mlまで発光バッファーで段階的に希釈し、50μlを各ウェルに分注し、室温30分間反応した。反応後にCoelenterazineを0.5μMとなるよう発光バッファーで希釈し、40μl/ウェルで分注し、10分後にルミノメーター、Centro LB960(ベルトールド社)で発光を測定した。結果を(図10)に示す。発光阻害機能の無い20-3は抗体濃度に関わらず、一定の発光であったのに対して、11-6と61-2は抗体濃度依存的にGaussia luciferaseの発光を阻害した。これにより、1次スクリーニングのみで得られたクローンから機能性抗体の取得が可能であることを証明した。本高効率融合法により高価値の抗体の取得に強力なツールの提供が可能となった。
【特許請求の範囲】
【請求項1】
免疫細胞とミエローマ細胞とを電気的細胞融合法で融合するに際して、予め、CpGモチーフを有するオリゴヌクレオチドを含有する培地で免疫細胞を培養することを特徴とする、細胞融合方法。
【請求項2】
CpGモチーフを有するオリゴヌクレオチドが、以下の塩基配列を有することを特徴とする請求項1に記載の細胞融合方法。
【化1】
(ただし、上記塩基配列中、N1N2のうち少なくとも一つはプリン塩基であり、N3N4のうち少なくとも一つはピリミジン塩基である。)
【請求項3】
CpGモチーフを有するオリゴヌクレオチドが、5’−GACGTT−3’、5’−TGGCGT−3’、5’−GACGCT−3’、5’−AGCGTT−3’又は 5’−GAGCTT−3’からなる配列を有することを特徴とする請求項2に記載の細胞融合方法。
【請求項4】
CpGモチーフを有するオリゴヌクレオチドが、
5'-tcctGACGTTcctgacgtt-3'又は5'-tccatGACGTTcctgacgtt-3'、5'-tAGCGTT-3'、5'-tGACGCT-3'、又は5'-tccatGAGCTTcctgacgtt-3'であることを特徴とする請求項3に記載の細胞融合方法。
【請求項5】
培地が、さらにサイトカインを含有することを特徴とする、請求項1〜4のいずれかに記載の細胞融合方法。
【請求項6】
CpGモチーフを有するオリゴヌクレオチドからなる電気的細胞融合促進剤。
【請求項7】
CpGモチーフを有するオリゴヌクレオチドが、以下の塩基配列を有することを特徴とする請求項6に記載の電気的細胞融合促進剤。
【化1】
(ただし、上記塩基配列中、N1N2のうち少なくとも一つはプリン塩基であり、N3N4のうち少なくとも一つはピリミジン塩基である。)
【請求項8】
CpGモチーフを有するオリゴヌクレオチドが、5’−GACGTT−3’、5’−TGGCGT−3’、5’−GACGCT−3’、5’−AGCGTT−3’、又は5’−GAGCTT−3’からなる配列を有することを特徴とする請求項7に記載の電気的細胞融合促進剤。
【請求項9】
CpGモチーフを有するオリゴヌクレオチドが、
5'-tcctGACGTTcctgacgtt-3'、5'-tcctTGGCGTcctgacgtt-3'、5'-tAGCGTT-3'、5'-tGACGCT-3'、又は5'-tccatGAGCTTcctgacgtt-3'であることを特徴とする請求項8に記載の電気的細胞融合促進剤。
【請求項10】
請求項6〜9のいずれかに記載の電気的細胞融合促進剤とサイトカインを含有することを特徴とする、電気的細胞融合試薬キット。
【請求項11】
抗体産生細胞とミエローマ細胞とを電気的細胞融合法で融合するに際して、予め、CpGモチーフを有するオリゴヌクレオチドを含有する培地で抗体細胞を培養することを特徴とする、モノクローナル抗体産生ハイブリドーマの製造方法。
【請求項12】
培地がさらにサイトカインを含有することを特徴とする、請求項11に記載のハイブリドーマの製造方法。
【請求項13】
請求項11又は12に記載の方法により得られたモノクローナル抗体産生ハイブリドーマ。
【請求項14】
請求項13に記載のハイブリドーマを培地に培養し、モノクローナル抗体を採取することを特徴とする、モノクローナル抗体の製造方法。
【請求項1】
免疫細胞とミエローマ細胞とを電気的細胞融合法で融合するに際して、予め、CpGモチーフを有するオリゴヌクレオチドを含有する培地で免疫細胞を培養することを特徴とする、細胞融合方法。
【請求項2】
CpGモチーフを有するオリゴヌクレオチドが、以下の塩基配列を有することを特徴とする請求項1に記載の細胞融合方法。
【化1】
(ただし、上記塩基配列中、N1N2のうち少なくとも一つはプリン塩基であり、N3N4のうち少なくとも一つはピリミジン塩基である。)
【請求項3】
CpGモチーフを有するオリゴヌクレオチドが、5’−GACGTT−3’、5’−TGGCGT−3’、5’−GACGCT−3’、5’−AGCGTT−3’又は 5’−GAGCTT−3’からなる配列を有することを特徴とする請求項2に記載の細胞融合方法。
【請求項4】
CpGモチーフを有するオリゴヌクレオチドが、
5'-tcctGACGTTcctgacgtt-3'又は5'-tccatGACGTTcctgacgtt-3'、5'-tAGCGTT-3'、5'-tGACGCT-3'、又は5'-tccatGAGCTTcctgacgtt-3'であることを特徴とする請求項3に記載の細胞融合方法。
【請求項5】
培地が、さらにサイトカインを含有することを特徴とする、請求項1〜4のいずれかに記載の細胞融合方法。
【請求項6】
CpGモチーフを有するオリゴヌクレオチドからなる電気的細胞融合促進剤。
【請求項7】
CpGモチーフを有するオリゴヌクレオチドが、以下の塩基配列を有することを特徴とする請求項6に記載の電気的細胞融合促進剤。
【化1】
(ただし、上記塩基配列中、N1N2のうち少なくとも一つはプリン塩基であり、N3N4のうち少なくとも一つはピリミジン塩基である。)
【請求項8】
CpGモチーフを有するオリゴヌクレオチドが、5’−GACGTT−3’、5’−TGGCGT−3’、5’−GACGCT−3’、5’−AGCGTT−3’、又は5’−GAGCTT−3’からなる配列を有することを特徴とする請求項7に記載の電気的細胞融合促進剤。
【請求項9】
CpGモチーフを有するオリゴヌクレオチドが、
5'-tcctGACGTTcctgacgtt-3'、5'-tcctTGGCGTcctgacgtt-3'、5'-tAGCGTT-3'、5'-tGACGCT-3'、又は5'-tccatGAGCTTcctgacgtt-3'であることを特徴とする請求項8に記載の電気的細胞融合促進剤。
【請求項10】
請求項6〜9のいずれかに記載の電気的細胞融合促進剤とサイトカインを含有することを特徴とする、電気的細胞融合試薬キット。
【請求項11】
抗体産生細胞とミエローマ細胞とを電気的細胞融合法で融合するに際して、予め、CpGモチーフを有するオリゴヌクレオチドを含有する培地で抗体細胞を培養することを特徴とする、モノクローナル抗体産生ハイブリドーマの製造方法。
【請求項12】
培地がさらにサイトカインを含有することを特徴とする、請求項11に記載のハイブリドーマの製造方法。
【請求項13】
請求項11又は12に記載の方法により得られたモノクローナル抗体産生ハイブリドーマ。
【請求項14】
請求項13に記載のハイブリドーマを培地に培養し、モノクローナル抗体を採取することを特徴とする、モノクローナル抗体の製造方法。
【図1】

【図2】
【図3】
【図4】
【図5】

【図6】

【図7】

【図8】

【図9】
【図10】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【公開番号】特開2012−157312(P2012−157312A)
【公開日】平成24年8月23日(2012.8.23)
【国際特許分類】
【出願番号】特願2011−20472(P2011−20472)
【出願日】平成23年2月2日(2011.2.2)
【出願人】(301021533)独立行政法人産業技術総合研究所 (6,529)
【Fターム(参考)】
【公開日】平成24年8月23日(2012.8.23)
【国際特許分類】
【出願日】平成23年2月2日(2011.2.2)
【出願人】(301021533)独立行政法人産業技術総合研究所 (6,529)
【Fターム(参考)】
[ Back to top ]