
高収率のセルロース産生活性がある新規なグルコンアセトバクター属菌株

【課題】高収率のセルロース産生活性がある新規なグルコンアセトバクター属菌株を提供する。
【解決手段】セルロース産生活性がある新規なグルコンアセトバクター属菌株であって、ナノ構造のセルロースを非常に高効率にて産生しうる新規な菌株。該菌株は、セルロース産生活性に非常に優れており、産生されたセルロースは熱特性に非常に優れたナノ構造のバクテリアセルロースであって、バイオナノ繊維に有効に用いることができる。特に、樹脂と含浸してセルロース系樹脂を製造する上で用いることができる他、前記セルロース系樹脂は液晶表示装置の基板用に用いて好適である。
【解決手段】セルロース産生活性がある新規なグルコンアセトバクター属菌株であって、ナノ構造のセルロースを非常に高効率にて産生しうる新規な菌株。該菌株は、セルロース産生活性に非常に優れており、産生されたセルロースは熱特性に非常に優れたナノ構造のバクテリアセルロースであって、バイオナノ繊維に有効に用いることができる。特に、樹脂と含浸してセルロース系樹脂を製造する上で用いることができる他、前記セルロース系樹脂は液晶表示装置の基板用に用いて好適である。
【発明の詳細な説明】
【技術分野】
【0001】
本発明はセルロースを産生しうる新規なグルコンアセトバクター属菌株に係り、さらに詳しくは、ナノ構造のセルロースを非常に高収率にて産生しうる新規なグルコンアセトバクター属菌株に関する。
【背景技術】
【0002】
セルロースは、地球上で、最も多く存在する天然高分子の一種であり、略1011トンのセルロースが生合成されていると推算されており、ほとんどは高等植物の細胞壁繊維質成分を合成する。
【0003】
セルロースの強靭性は全体構造に起因するが、その分子は14,000単位に至る鎖や微細繊維を形成し、これらが水素結合によって維持される捩れた綱状束をなしている。このようなセルロースの豊富な物量と他の天然高分子とは異なる物性により、様々な産業分野において再生資源として活用するための研究が盛んになされている。
【0004】
1886年にブラウン(A.J.Brown)によって酢酸菌がセルロースを産生するということが報告されて以来、微生物によって産生されるセルロース(Bacterial Cellulose;以下「BC」と称する)は新素材として絶えず研究対象となっている。特に、BCはリボン状繊維(ribbin−like bundles)で構成されるのに対し、植物由来のセルロースは微細繊維(microfibrils)の束(bundles)状に形成される。
【0005】
リボン状繊維で構成されるBCは、微細繊維の束状に形成される植物由来のセルロースとは異なり、リグニン及びヘミセルロースが全く含まれていない純粋状態で産生され、高い機械的強度、高保湿性、高結晶性、生分解性などのBCならではの長所のため、様々な産業分野において様々な用途に開発されている。
【0006】
一方、液晶表示装置(LCD)の基板素材としては、熱膨張係数が低くて透過率が高く、しかも、微細な太さを有する必要があり、現在は、高価なs−glassが用いられている。
しかし、ナノ構造のセルロースを用いることにより、s−glassよりも熱特性が向上することが報告されている。ナノ構造のセルロースは微生物によって産生しうることが知られている(Schraamm, M. and S. Hestrin. 1054. Factors affecting production of cellulose at the air/liquid interface of a culture of Acetobacter xylinum.J.Gen. Microbiol. 11:123−129)。このため、このようなセルロース産生活性がある微生物を開発することは、経済的に非常に有利である。
【発明の概要】
【発明が解決しようとする課題】
【0007】
しかし、BCを産生する代表的な微生物として、アセトバクター・キシリナム(acetobacter xylinum)が知られているが、セルロースの産生効率が低く、これに代替することができる、新たな微生物の開発が切望されている。
上述した目的を達成するために、本発明は、グルコンアセトバクター属菌株:該属菌株の培養物:前記グルコンアセトバクター属菌株、前記菌株の培養物、前記菌株または培養物の濃縮物およびこれらの乾燥物からなる群より選ばれた1種以上を含むセルロース産生用組成物:セルロースの製造方法及びセルロース系樹脂の製造方法を提供することを目的とする。
【課題を解決するための手段】
【0008】
本発明者らは、セルロース産生効率に非常に優れた新たな微生物を開発するために鋭意研究した結果、グルコンアセトバクター属に属する新規な菌株を開発し、前記菌株が非常に有効にセルロースを産生しうるということを見出し、本発明を完成するに至った。
【0009】
本発明は、セルロース産生活性があるグルコンアセトバクター(gluconacetobacter)属菌株であって、前記菌株は、グルコンアセトバクター属(gluconacetobacter sp.)D44菌株(寄託番号KCCM11078P)またはグルコンアセトバクター・インターメディウス(Gluconacetobacter intermedius)D47菌株(寄託番号KCCM11079P)であることを特徴とするグルコンアセトバクター属菌株を提供する。
また、本発明は、前記グルコンアセトバクター属菌株の培養物を提供する。
さらに、本発明は、前記グルコンアセトバクター属菌株、前記菌株の培養物、前記菌株または培養物の濃縮物およびこれらの乾燥物からなる群より選ばれたいずれか1種以上を含むセルロース産生用組成物を提供する。
また、本発明は、前記グルコンアセトバクター属菌株を培養する工程と、前記培養物からセルロースを得る工程とを含むセルロースの製造方法を提供する。
さらに、本発明は、前記グルコンアセトバクター属菌株によって産生されたセルロースを準備する工程と、前記セルロースを樹脂と含浸させる工程とを含むセルロース系樹脂の製造方法を提供する。
【図面の簡単な説明】
【0010】
【図1】本発明のD44菌株の形態学的な観察結果を示すものであり、(a)は、光学顕微鏡による観察結果を示し、(b)は、走査電子顕微鏡(SEM)による観察結果を示す。
【図2A】本発明のD44菌株の16s rDNA配列をグルコンアセトバクター・ラエティカスの16s rDNA配列と比較して類似度を測定した結果を示すものである(*:塩基配列が一致する部分)。
【図2B】本発明のD44菌株の16s rDNA配列をグルコンアセトバクター・ラエティカスの16s rDNA配列と比較して類似度を測定した結果を示すものである(*:塩基配列が一致する部分)。
【図2C】本発明のD44菌株の16s rDNA配列をグルコンアセトバクター・ラエティカスの16s rDNA配列と比較して類似度を測定した結果を示すものである(*:塩基配列が一致する部分)。
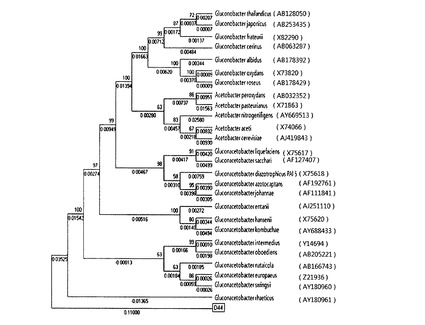
【図3】本発明のD44菌株の酢酸菌科に属するグルコンアセトバクター属、グルコノバクター属およびアセトバクター属の菌に対する系統樹を示すものである。
【図4】本発明のD44菌株のセルロースの生成有無を確認した結果である。
【図5】本発明のD47菌株の形態学的な観察結果を示すものであり、(a)は、光学顕微鏡による観察結果であり、(b)は、走査電子顕微鏡(SEM)による観察結果である。
【図6A】本発明のD47菌株の16s rDNA配列をグルコンアセトバクター・インターメディウスの16s rDNA配列と比較して類似度を測定した結果を示すものである(*:塩基配列が一致する部分)。
【図6B】本発明のD47菌株の16s rDNA配列をグルコンアセトバクター・インターメディウスの16s rDNA配列と比較して類似度を測定した結果を示すものである(*:塩基配列が一致する部分)。
【図6C】本発明のD47菌株の16s rDNA配列をグルコンアセトバクター・インターメディウスの16s rDNA配列と比較して類似度を測定した結果を示すものである(*:塩基配列が一致する部分)。
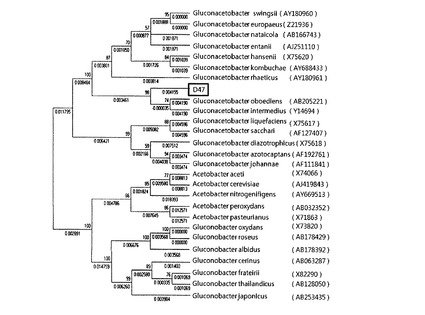
【図7】本発明のD47菌株の酢酸菌科に属するグルコンアセトバクター属、グルコノバクターおよびアセトバクター属の菌に対する系統樹を示すものである。
【図8】本発明のD47菌株のセルロース生成有無を確認した結果である。
【図9A】本発明のD47菌株をH.S培地で培養して産生されたセルロースを示すものである。
【図9B】本発明のD47菌株をココナツ混合培地(2)で培養して産生されたセルロースを示すものである。
【発明を実施するための形態】
【0011】
本発明のグルコンアセトバクター属菌株、具体的に、グルコンアセトバクター属D44菌株(寄託番号KCCM11078P)またはグルコンアセトバクター・インターメディウスD47菌株(寄託番号KCCM11079P)は、セルロース産生能に非常に優れた新規な菌株である。
【0012】
本発明の菌株は酢酸菌(acetobacteraceae)の一種であり、これに限定されないが、酢類から分離することができ、好ましくは、柿酢、より好ましくは、柿酢の表膜から分離することができる。
本発明の一実施の形態においては、柿酢の表膜から高収率のセルロース産生活性がある本発明の菌株を分離し、前記セルロース産生活性があることは公知の方法により確認することができる。例えば、本発明の菌株をH.S培地(下記表1参照)に接種して培地に表膜を形成するかどうか、または、Carr培地(下記表1参照)に接種して色変化を引き起こすかどうかなどにより確認することができるが、これに限定されない(<実施例1>参照)。
また、本発明の菌株を同定するために形態学的特徴、生化学的特徴を分析したところ、本発明の菌株は下記表2に示す形態学的特徴を有し、下記表3(D44)および表4(D47)に示す生化学的特徴を有するグルコンアセトバクター属菌株である(<実施例2>参照)。
【0013】
さらに、本発明の菌株がグルコンアセトバクター属に属する菌株であるかどうかをそれぞれ配列番号1の塩基配列で示される16s rDNA配列(D44)および配列番号2の塩基配列で示される16s rDNA配列(D47)を得て分析した結果、D44菌株の場合、グルコンアセトバクター・ラエティカスの16s rDNA配列に最も類似しているが、類似度は87.215%に過ぎないことが分析された。本発明の菌株はグルコンアセトバクター属に属する新規な菌株であることが確認された。D47菌株はグルコンアセトバクター・インターメディウスの16s rDNA配列に最も類似しており、類似度は99.776%であることが分析された。本発明の菌株はグルコンアセトバクター属に属する菌株であることが確認された(<実施例2>参照)。
本発明者らは上記のようにして分離および同定された本発明の菌株をそれぞれグルコンアセトバクター属菌株D44およびグルコンアセトバクター・インターメディウスD47と命名し、前記菌株を2009年11月12日付けで韓国微生物保存センター(KCCM)寄託機関にそれぞれKFCC11470P寄託番号(D44)およびKFCC11471P寄託番号(D47)で寄託した。この後、2010年3月26日付けで再び韓国微生物保存センター(KCCM)寄託機関にそれぞれKCCM11078P寄託番号(D44)およびKCCM11079P寄託番号(D47)で国際寄託した。
【0014】
本発明のグルコンアセトバクター属菌株の培養物は、前記本発明の菌株を微生物の培養に用いられる公知の培地および培養方法を用いて製造することができる。
本発明のグルコンアセトバクター属菌株を培養するために用いられる培地は、通常、微生物の培養に用いられる培地が挙げられ、好ましくは、酢酸菌の培養に用いられる培地、より好ましくは、下記表1または表7に示す培地等が挙げられる。さらに好ましくは、セルロース産生のためのものであり、具体的に、下記表1に示すH.S培地または下記表7に示すココナツ混合培地(2)が挙げられる。これらを用いることにより、セルロースを一層高収率にて産生することができる(<実施例4>参照)。
【0015】
前記D44菌株は、好ましくは、グルコース20g/L、酵母抽出物5g/L、ペプトン5g/L、Na2HPO42.7g/Lおよびクエン酸ナトリウム1.15g/Lを含むH.S培地またはココナツ原液100%(w/v)のココナツ液体培地で培養することができる。前記酵母抽出物(CAS番号:8013−01−02)(ヨーロッパ既存化学物質名録:232−387−9)の酵母は40%以下の蛋白質を含有しており、1g当たりに塩酸ジアミン0.12mg以下、リボフラビン0.04mg以下、ニコチン酸0.25mg以下である無発酵性の黄白色の板状または粉末であり、ユニークな臭いの物理化学的特性を有している。
前記D47菌株は、好ましくは、前記H.S培地、前記ココナツ液体培地、またはココナツ原液、砂糖および硫安を含み、好ましくは、前記ココナツ原液85〜94.9重量部、砂糖5〜15重量部、および硫安0.1〜1重量部を含むココナツ混合培地で培養することができる。
前記硫安とは硫酸アンモニウムの略称であり、硫酸とアンモニアとを反応させて得られる白い結晶を意味し、好ましくは、純度10〜99.9%、より好ましくは、純度10〜30%、最も好ましくは、純度15〜25%の硫酸アンモニウムを使用することができる。
【0016】
本発明の菌株の培養物は、前記培地に本発明の菌株を接種し、当該分野で公知の微生物の培養方法(例えば、静置培養、攪拌培養)により、好ましくは、酢酸菌の培養方法により、より好ましくは、1重量%(培地の総重量に対して)にて接種し、28〜30℃で4〜7日間培養する方法により製造することができる。より好ましくは、本発明の菌株を1重量%(培地の総重量に対して)にて培地に接種し、30℃で培養することにより製造する。
【0017】
本発明の培養物は、これに限定されないが、好ましくは、本発明の菌株によって産生されたセルロースを含む。
一方、本発明のセルロース産生用組成物は、前記本発明のグルコンアセトバクター属菌株、前記菌株の培養物、前記菌株または培養物の濃縮物およびこれらの乾燥物からなる群より選ばれた1種以上を含む。
本発明の菌株は、セルロースを非常に有効に産生することができるので、前記菌株、前記菌株の培養物、前記菌株または培養物の濃縮物およびこれらの乾燥物はセルロースを産生する組成物として用いて好適である。
前記菌株の培養物の製造方法については上述した通りであり、これに限定されないが、好ましくは、本発明の菌株をセルロース産生用培地、下記表1に示すH.S培地または下記表7に示すココナツ混合培地(2)に接種し、これに限定されないが、好ましくは、静置培養または攪拌培養、好ましくは静置培養して培養物を製造する方法が挙げられる。前記菌株または培養物の濃縮物およびこれらの乾燥物は、当該分野に公知の微生物または培養物の濃縮または乾燥方法により容易に製造することができる。
前記本発明のグルコンアセトバクター属菌株、前記菌株の培養物、前記菌株または培養物の濃縮物およびこれらの乾燥物は、これに限定されないが、好ましくは、本発明のセルロース産生用組成物に1重量%にて含有される。
【0018】
本発明のセルロースの製造方法は、前記グルコンアセトバクター属菌株を培養する工程と、前記培養物からセルロースを得る工程とを含む。
前記菌株の培養方法は上述した通りであり、本発明のセルロースを産生するためには、これに限定されないが、本発明の菌株の培養によりセルロースの膜が形成されたものを水酸化ナトリウム溶液に浸漬させた後、蒸溜水により洗浄して純粋なセルロースを産生する方法が挙げられる。このようにして産生されたセルロースは、これに限定されないが、好ましくは、膜状である(<実施例3>参照)。
【0019】
本発明の菌株によって産生されたセルロースは滑らかであり、堅固性および弾力性に優れており、厚さが約0.3mm〜約12mmであり、既存のバクテリアセルロースよりも厚さが顕著に向上した、熱特性に非常に優れたナノ構造のセルロースである(<実施例3>および<実施例4>参照)。
さらに、本発明の菌株は、セルロース産生活性に非常に優れており、特に、BCを産生する代表的な微生物であるアセトバクター・キシリナム(acetobacter xylinum)と比較して、最少約3倍、最大約97.5倍の顕著に高い効率にてセルロースを産生することができる(<実施例3>および<実施例4>参照)。
このため、本発明の菌株によって産生されたセルロースは熱特性に非常に優れたナノ構造のバクテリアセルロースであり、バイオナノ繊維として利用するのに好適である。
【0020】
本発明のセルロース系樹脂の製造方法は、前記本発明のグルコンアセトバクター属菌株によって産生されたセルロースを準備する工程と、前記セルロースを樹脂と含浸させる工程とを含む。
前記セルロース系樹脂は、本発明のグルコンアセトバクター属菌株によって産生されたセルロースを用いたものであれば、当該分野に公知の製造方法により製造することができる。例えば、本発明のグルコンアセトバクター属菌株によって産生されたセルロースに熱可塑性樹脂および結合剤を混入し、これを圧出成形して製造することができる。
【0021】
前記熱可塑性樹脂は、ポリプロピレン、ポリエチレン、ポリスチレンおよびポリ塩化ビニールからなる群より選ばれたものであることが好ましい。
前記結合剤は、マレイン化エチレン、マレイン化プロピレンおよびシランよりなる群から選ばれる1種以上であることが好ましい。
これらセルロース、熱可塑性樹脂及び結合剤からなる混合物には、安定剤、顔料、染料、無機物または有機物添加剤、電磁波遮断剤またはこれらの混合物の中から選ばれた1種以上の添加剤をさらに添加してもよい。
【0022】
このようにして製造されたセルロース樹脂は熱特性に非常に優れたナノ構造のバクテリアセルロースが含まれているため、液晶表示装置の基板用に用いることができる。
したがって、本発明は、前記セルロース系樹脂の製造方法であって、セルロース系樹脂を製造する工程と、前記製造されたセルロース系樹脂を用いて液晶表示装置の基板を製造する工程とを含む液晶表示装置の基板の製造方法を提供する(Masaya Nogi and Hiroyuki Yano, Transparent Nanocomposites Based on Cellulose Produced by Bacteria Offer Potential Innovation in the Electronics Device Industry, Advanced materials 2008, pp1849-1852)。
また、本発明は、前記セルロース系樹脂を用いて製造された液晶表示装置の基板を提供する。
【0023】
セルロースを産生する菌において、セルロースは、UV光の致死効果から細胞を防御し、菌の集落を形成して栄養源として同じ基質を使用する競争者から防御効果をもたらすと共に、細胞に湿気を提供して乾燥を防ぐことが知られている。
また、本発明のように微生物によって産生されたセルロースは、繊維の結晶化度、吸湿性などが高く、スピーカの振動板などへの産業的な応用可能性が高く、セルロース膜はゲル状態で肌触り性が良く、身体の表面に馴染み易く、さらに、医薬品や化粧材料成分の補水能が高く、皮膚の表面を保湿状態に維持することができて、医療用パッドや化粧パッド(例えば、マスクパック)、人工皮膚などに応用することができ、現在、セルロースを用いたマスクパックは商用化工程まで至っている。
さらに、セルロース膜を5%のNaOHまたは0.5%のNaClにより処理してセルロース成分以外の物質を除去した後にこれを圧着すれば高弾性膜になることが判明し、日本のソニー社ではスピーカの音量振動板に応用して高性能ヘッドホンとして市販している。
【0024】
また、本発明のように微生物によって産生されたセルロースを用いて紙を製紙すれば、紙の強度や弾性率が対照群に比べて2.5倍も増加する。合成高分子繊維、芳香剤ポリアミドやポリアミド単繊維に5〜20%添加してシートを製作すれば、合成繊維に比べて、強度、熱安定性、形態安定性などに優れたシートが得られる。合成繊維だけではなく、ガラス繊維にセルロースを混入してシート状にすれば、空気ろ過用フィルター、プリント配線基板、床材料などに応用することが可能となる。
このような本発明の微生物由来のセルロースは、産業用素材、食品素材など多岐に亘って活用することができ、特に、環境にやさしい素材であるという点は無限の開発可能性と多様性を有している(Jeong.,Y.J.,Lee.,I.S.. A view of utilizing cellulose produced by acetobacter bacteria. Food Industry and Nutrition.,V ol.5.,25〜29.,2000.参照)。
【0025】
本発明の菌株はセルロース産生活性に非常に優れており、本発明の菌株によって産生されたセルロースは熱特性に非常に優れたナノ構造のバクテリアセルロースであって、バイオナノ繊維として有効に用いることができる。特に、樹脂と含浸してセルロース系樹脂を製造する上で用いることができ、しかも、前記セルロース系樹脂は液晶表示装置の基板用に有効に用いることができる。
【0026】
以下、実施例を挙げて本発明による高収率のセルロース産生活性がある新規なグルコンアセトバクター属菌株について詳述する。
但し、下記の実施例は本発明を例示するものに過ぎず、本発明の内容が下記の実施例に限定されることはない。
【0027】
<参考例1>
実験に供された培地および培養条件
本発明の新規な菌株を分離および同定するために、下記表1に示す培地を使用した。なお、下記の実施例において特別に言及がない限り、前記培地を用いて本発明のセルロースの高収率産生菌株を30℃で7日間培養して実験を行った。
【0028】
【表1】

【0029】
<実施例1>
セルロースを高収率にて産生する新規な菌株の分離
<実施例1−1>
グルコンアセトバクター属D44菌株の分離
本実験に供された菌株を在来式柿酢(全羅北道完州郡鳳東邑栗所里)の表膜から分離した。具体的に、柿酢の複数の部分を直接的に採取し、採取された部分によってA、B、C、Dと区別して命名し、これらのうちDは柿酢の表膜から採取した試料である。
前記柿酢A、B、C、D原液を1%ずつH.S培地100mに接種して7日間30℃で培養した。前記培養後、H.S培地の表膜にセルロースを生成するD試料で単一菌株を10−7まで希釈して固体H.S培地に塗抹した後、ランダムでコロニーを取って再び液体H.S培地に接種し、これらのうち培地に表膜を形成する菌株を分離した。
【0030】
前記柿酢のD部分から分離された菌株を変色範囲がpH5.2〜6.8であり、酸性で黄色、塩基性で紫色を帯びるブロモクレゾールパープル含有Carr培地に接種し、黄色から紫色へと、さらに紫色から黄色へと色変化があることを確認することにより、前記分離された菌株がエタノールを酢酸に酸化させ、二酸化炭素と水に再酸化させる酸化経路を有していることを改めて確認することができた。これにより、前記菌株がセルロース産生菌株の特性を有していることが分かる(Jang. O.Y., Joo. K.H., Lee. J.H., Baik. C.G. 2003. Growth characteristics and production of cellulose of microorganisms in static culture vinegar. Korean J. Food SCI. tech. vol. 35, No. 6, pp. 1150〜1154)。
本発明者らは、前記セルロース生成活性がある菌株を「D44」と命名した。
【0031】
<実施例1−2>
グルコンアセトバクター・インターメディウス(Gluconacetobacter intermedius)D47菌株の分離
本実験に供された菌株を在来式柿酢(全羅北道完州郡鳳東邑栗所里)の表膜から分離した。具体的に、在来式柿酢を製造するときに生成される被膜を農協から得て、滅菌済み解剖用はさみで十分に切断した後、少量をH.S培地に接種して7日間30℃で培養した。前記培養後、H.S培地の表膜にセルロースが産生された試料で単一菌株を10−7まで希釈して固体H.S培地に塗抹した後、ランダムでコロニーを取って液体H.S培地に再接種し、これらのうち培地に表膜を形成する菌株を分離した。
前記柿酢の表膜部分から分離された菌株を、変色範囲がpH5.2〜6.8であり、酸性で黄色、塩基性で紫色を帯びるブロモクレゾールパープル含有Carr培地に接種し、黄色から紫色へと、さらに紫色から黄色へと色変化があることを確認することにより、前記分離された菌株がセルロース生成活性があることを最終的に確認した。
本発明者らは、前記セルロース生成活性がある菌株を「D47」と命名した。
【0032】
<実施例2>
セルロースを高収率にて産生する新規な菌株の同定および寄託
<2−1>形態学的特徴
前記<実施例1>において分離されたD44菌株およびD47菌株のそれぞれの大きさ、形状、グラム染色およびコロニーの形状、色などを観察し、具体的に、GYC培地でD44菌株およびD47菌株それぞれのコロニーを観察し、D44菌株およびD47菌株がそれぞれ生成したセルロースを菌が除去されていない状態で光学顕微鏡および走査転写顕微鏡(SEM)により観察した。
より具体的に、光学顕微鏡(Nikon ECLIPSE E600、日本)を用いてD44菌株およびD47菌株の形状とグラム染色を、走査電子顕微鏡(SEM)によりD44菌株およびD47菌の大きさと菌が産生したセルロース繊維を検鏡した。特に、電子顕微鏡はChaeの方法(Chae. C.M.1977. Maceration methods for SEM obserbation ’A legal medicine lecture room’. Kyungpook National University. p 40−58)を変形して利用し、具体的に、D44菌株およびD47菌体を2%(v/v)グルタルアルデヒド (Glutaraldehyde)により2時間前固定し、2%(w/v)のOsO4により2時間後固定してエタノールにより順次に脱水した後、イソアミルアセタート(isoamyl acetate)を処理して55℃で乾燥し、金コートして検鏡した。D44菌株およびD47菌株の形態学的特徴
についての観察結果を表2と、図1および図5に示す。
【0033】
【表2】
【0034】
表2と、図1および図5に示すように、D44菌株およびD47菌株はグラム陰性であり、桿状であり、胞子を形成せず、単一(single)または対(pair)で配列されて存在する。特に、図1および図5に示すように、D44菌株およびD47によって生成されたセルロース繊維内に前記菌株が内在されている。
【0035】
<2−2>生化学的特徴
前記D44菌株およびD47菌株の生化学的特徴を調べるために、D44菌株およびD47菌株をそれぞれH.S液体培地に培養した後に採取した菌液を新たな培地に継代培養して使用した。
具体的に、本発明の新規なD44菌株およびD47菌株をそれぞれFrateur培地に接種して培養した後、培地中のCaCO3が中和するかどうかを確認してD44菌株およびD47菌株のエタノール酸化経路を確認した。
また、グリセロールでケトン体を形成するかどうかを確認するために、YEG固体培地に前記各菌株をそれぞれ塗抹して5日間30℃で培養した後、フェーリング液をコロニーの上に数滴滴下した後、オレンジ色への変色有無により確認した。
さらに、エタノールと塩化ナトリウムに対して菌株の生育とセルロースの生成有無を確認するために、SM培地に塩化ナトリウム0.5、1.0、1.5、2.0%(w/v)とエタノール10%(v/v)をそれぞれ添加した培地に、D44菌株およびD47菌株をそれぞれ接種して7日間30℃で静置培養した後に観察した。
【0036】
加えて、GYC固体培地にD44菌株およびD47菌株をそれぞれ塗抹して7日間30℃で培養した後に褐色色素の形成有無を確認し、γ−ピロンの生成有無を確認するために、葡萄糖50g/L、果糖50g/Lをそれぞれ含有するYPC培地に菌を塗抹して7日間30℃で培養した後に塩化第二鉄(ferric chloride)5%(w/v)溶液を滴下して確認した。このとき、紫色に変色すれば、菌株が葡萄糖と果糖を用いてγ−ピロンを生成することができると決めた(Jang. O.Y., Joo. K.H., Lee. J.H., Baik. C.G. 2003. Growth characteristics and production of cellulose of microorganisms in static culture vinegar. Korean J. Food SCI. tech. vol. 35, No. 6, pp. 1150〜1154)。
【0037】
さらに、D44菌株およびD47菌株の酢酸に対する抵抗性を調べるために、酢酸の濃度を2%(v/v)〜10%(v/v)に調節したGYP培地にD44菌株およびD47菌株をそれぞれ接種し、次いで、7日間30℃で培養後に調査した。
これらに加えて、生化学的実験はAPI 20NEキット(Bio−Merieux、フランス)を用いて行い、導出された結果をバージーズ・マニュアル・オブ・システマティック・バクテリオロジー(Bergy’s manual of systematic bacteriology)に準じて同定した(Krig, N.R., J.G. Holt. 1984. Bergey’s Manual of Systematic Bacteriology Vol. 1. The William and Wilkins Co.U.S.A.)。
前記実験結果を下記表3および表4に示し、その結果をアセトバクター属菌株、グルコノバクター属菌株およびグルコンアセトバクター属菌株と比較した。前記グルコンアセトバクター属菌は、Lee. O.S.文献(Lee. O.S., Jang. S.Y.,Jeong. Y.J. 2002. Effect of Ethanol on the Production of Cellulose and Acetic Acid by Gluconacetobacter persimmonensis KJ145. J .Korean Soc. Food sci. Nutr. 31(4).572−577.)に記載のものによる。
【0038】
【表3】
1)+:陽性、2)−:陰性、3)ND:未決定
【0039】
前記表3に示すように、本発明の新規なD44菌株をFrateur培地に接種して培養した結果、培地中のCaCO3が中和して透明環が形成されることを観察することができ、培養12日後にCaCO3が再沈殿されることを確認することができる。要するに、D44菌株はエタノールを酢酸に、酢酸をCO2とH2Oに酸化することが分かる(Son. H.J., Kim. K.K., Kim. H.S., Kim.Y.G. 1999. Isolation and identification of cellulose−producing bacteria. J. Agri. Tech & Dev. Inst.)。
【0040】
また、本発明のD44菌株は5%(w/v)葡萄糖と果糖を用いてγ−ピロンを生成することができず、グリセロールからのケトン体形成反応で陽性を示す。さらに、前記D44菌株はγ−ガラクトシダーゼ陽性、硝酸(nitrate)から亜硝酸(nitrite)に還元させ、アセタート、果糖、葡萄糖、マンニトルを酸化させることはできるものの、乳酸(lactate)は酸化させることができない。
さらに、本発明のD44菌株は、インドール産生、ウレアーゼ、ゼラチン液化反応で陰性を示し、0.5、1.0、1.5、2.0%(w/v)の塩化ナトリウムに生育可能であり、特に、0.5%(w/v)の塩化ナトリウムでセルロースも生成することができる。しかし、2%(v/v)〜10%(v/v)のアセタート濃度でD44は生育することができないため耐性がないことが観察され、褐色色素を生成することができずに透明で且つ白いコロニーを形成する。
前記結果から、本発明のD44菌株はグルコンアセトバクター(Gluconacetobacter)属菌株であることが分かる。
【0041】
【表4】
1)+:陽性、2)−:陰性、3)ND:未決定
【0042】
前記表4に示すように、本発明のD47菌株をFrateur培地に接種して培養した結果、培地中のCaCO3が中和して透明環が形成されることを観察することができ、培養8日後、CaCO3が再沈殿されることを確認することができる。要するに、D47菌株は、エタノールを酢酸に、酢酸をCO2とH2Oに酸化することが分かる(Son. H.J., Kim. K.K., Kim. H.S., Kim.Y.G. 1999. Isolation and identification of cellulose−producing bacteria. J. Agri. Tech & Dev. Inst.)。
また、本発明のD47菌株は、5%葡萄糖(w/v)と果糖を用いてγ−ピロンを生成することができず、前記D44菌株とは異なり、グリセロールからのケトン体形成反応で陰性を示す。なお、前記D47菌株は、前記D44菌株とは異なり、β−ガラクトシダーゼ陰性であり、硝酸(nitrate)から亜硝酸(nitrite)に還元させることができなかった。しかし、前記D44菌株と同様に、アセタート、果糖、葡萄糖、マンニトルを酸化させることはできるものの、乳酸は酸化させることができない。
【0043】
さらに、本発明のD47菌株は、インドール産生、ウレアーゼ、ゼラチン液化反応で陰性を示し、0.5、1.0、1.5、2.0%(w/v)の塩化ナトリウムに生育可能であり、特に、0.5%の塩化ナトリウムでセルロースも生成することができる。しかしながら、2%(v/v)〜10%(v/v)のアセタート濃度でD47は生育することができずに耐性がないことが観察され、前記D44菌株とは異なり、薄い褐色のコロニーを形成する。
前記結果から、本発明のD47菌株は、グルコンアセトバクター(Gluconacetobacter)属菌株であることが分かる。
【0044】
<2−3>16s rDNA分析
前記<実施例1>において分離されたD44菌株およびD47菌株のgDNAを抽出するために、まず、H.S培地の表膜に生成されたセルロースを除去するために、セルラーゼ(Celluclast、nonozyme)を1%(v/v)添加して37℃で1時間攪拌して培養し、DNeasy Blood & Tissueキット(QIAGEN、Germany)を用いて16s rDNAを抽出および増幅し、16s rDNAシーケンシングを通して配列番号1のD44菌株の16s rDNAの全体配列および配列番号2のD47菌株の16s rDNAの全体配列をそれぞれ得た。菌株の同定のために候補菌株からゲノムDNAをDNeasy(R) Blood & Tissueキット(QIAGEN, Germany)を用いて分離した。DNAの精製は、製造社のガイドラインに準じて行った。
【0045】
前記D44菌株およびD47菌株の16s rDNAのクローニングのために、プライマーとしては、配列番号3の5’−AGAGTTTGATCMT GGCTCAG−3’(順方向)と、配列番号4の5’−ACGGGCGGGTGTGTRC−3’(逆方向)を用いてPCR増幅を行った。具体的に、PCR増幅は、100ng鋳型DNA、0.5μMのプライマーDNA、0.2mM dNTPs、10×Ex Taq緩衝液およびTaq重合酵素(TaKaRa bio Inc., Japan)0.025U/μlを用いて95℃で30秒変性、58℃で30秒の結合、72℃で1分30秒の伸張とする35サイクルをBiometra社製サーモサイクラー(Tampa、USA)により増幅させた。PCR産物を1%のアガロースゲル電気泳動を行うことで確認した。確認されたPCR産物をPCR精製キット(Bioneer、Korea)を用いて精製した後、T4リガーゼ(ligase)(Promega、USA)を用いてpGEM T−easyベクター(Promega、USA)に連結し、16s rDNAシーケンシングを通して前記各菌株の塩基配列情報を得た。
【0046】
前記得られたD44菌株およびD47菌株の16s rDNA配列からリボソーム遺伝子銀行からDNAの相同性を調査して同定に参考し、Bioeditプログラム(
1298607716195_0.html
)、Clustal Xプログラム、Megaプログラム(http://www.megasoftware.net/)を用いて類似同定された微生物との系統樹(phylogenetic tree)を描いて類似種間の関係を観察した。
【0047】
まず、前記D44菌株の16s rDNA配列とグルコンアセトバクター・ラエティカスの16s rDNA配列(AY180961)との類似度を比較した結果、類似度が87.215%に過ぎなかった。普通、菌株同定の類似度が約98〜99%でなければ類似菌として判定することができないが、D44菌株とグルコンアセトバクター・ラエティカスとの類似度は87.215%に過ぎず、本発明のD44菌株は新規なものであることが分かる。
Clustal Xプログラムを用いて本発明のD44とグルコンアセトバクター・ラエティカスとの類似度を再確認し、その結果を図2Aから図2Cに示した。前記図2Aから図2Cに示すように、D44とグルコンアセトバクター・ラエティカスとの類似度は87.215%に過ぎない。
【0048】
前記類似度結果を参考として、同じ科に属する微生物を基にD44との系統を調べるために、Bioeditプログラム、Clustal Xプログラム、Megaプログラムを用いた。具体的に、酢酸菌(acetobacteraceae)科に属するグルコンアセトバクター(Gluconacetobacter)属とグルコノバクター(Gluconobacter)属、アセトバクター(acetobacter)属の類似菌に対して近隣結合法により作成した系統樹を図3に示した。
【0049】
次に、前記D47菌株の16s rDNA配列とグルコンアセトバクター・インターメディウスの16s rDNA配列(Y14694)との類似度を比較した結果、99.774%の類似度を示した。このため、前記D47菌株はグルコンアセトバクター・インターメディウスの類似菌であると判定され、グルコンアセトバクター・インターメディウスD47と命名した。
さらに、Clustal Xプログラムを用いて本発明のD47とグルコンアセトバクター・インターメディウスとの類似度を再確認し、その結果を図6Aから図6Cに示した。前記図6Aから図6Cに示すように、D47とグルコンアセトバクター・インターメディウスが99%類似していることが分かる。
但し、D47菌株は、前記<実施例2−2>の表4に示すように、従来のグルコンアセトバクター属菌株とは異なり、グリセロール(glycerol)でジヒドロキシアセトン(dihydroxyacetone)を形成するケトン体生成(ketogenesis)反応をし、乳酸(lactate)を過酸化させることができず、0.5%(w/v)のNaClをH.S培地に添加して培養するときに培地の表面にセルロースを形成しうるなど生化学的特性が従来のグルコンアセトバクター属菌株とは全く異なる新規な菌株であることを確認することができ、セルロース産生能においても、後述するように、従来のグルコンアセトバクター属菌株に比べて顕著に高い産生活性を有していることを確認した。
【0050】
前記類似度結果を参考にして、同じ科に属する微生物を基にD47の系統を調べるために、Bioeditプログラム、Clustal Xプログラム、Megaプログラムを用いた。具体的に、酢酸菌科に属するグルコンアセトバクター属とグルコノバクター属、アセトバクター属の類似菌に対して近隣結合法により作成した系統樹を図7に示した。
【0051】
<2−4>本発明の菌株の寄託
本発明者らは、上記のようにして分離および同定された本発明のグルコンアセトバクター属菌株を2010年3月26日付けで韓国微生物保存センター(KCCM)国際寄託機関にそれぞれKCCM11078P寄託番号(D44)およびKCCM11079P寄託番号(D47)で寄託した。
【0052】
<実施例3>
本発明の菌株のセルロース産生活性
<3−1>本発明の菌株のセルロース産生活性
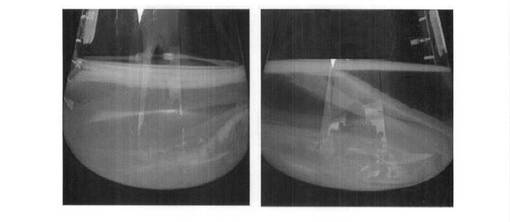
前記D44菌株およびD47菌株をそれぞれH.S培地に接種してセルロースの生成有無を確認した結果を図4および図8に示す。
前記図4および図8に示すように、本発明のD44菌株およびD47菌株は培養してから2〜3日経過後に表膜にセルロースを形成し、培養期間が長くなるにつれてD47によって生成されたセルロース膜はさらに厚くなって多重に形成される。
また、前記H.S培地にD44菌株およびD47菌株をそれぞれ1重量%接種(培地の総重量に対して)し、30℃で7日間静置培養した後、産生されたセルロースを得て乾燥重量を測定し、その結果を表5および図9Aに示す。具体的に、前記静置培養後、H.S培地の表膜に形成されたセルロースを回収して水道水により洗浄し、次いで、少量の0.1N NaOH溶液に入れて1時間沸騰して溶菌させた。前記回収されたセルロースを水道水によりさらに洗浄した後、少量の蒸溜水に一日中入れて中和させた。中和されたセルロースをろ過して90℃で抗量になるまで乾燥した後、乾燥されたセルロースをデシケーターで冷却した後に乾燥重量を測定した。
【0053】
【表5】
【0054】
前記本発明のD44菌株を用いて産生されたセルロース膜は、表面が滑らかであり、堅固性および弾力性があり、厚さは約0.3mmであった。これに対し、前記本発明のD47菌株を用いて産生されたセルロース膜は、表面が滑らかであり、堅固性及び弾力性があり、厚さは約4mmであり、遥かに高いセルロース活性を示している。さらに、後述するように、ココナツ混合培地(2)でD47菌株を用いて産生されたセルロース膜の厚さは約12mmであって、顕著に高いセルロース活性を示している。
【0055】
<3−2>本発明の菌株のセルロース産生活性の比較
本発明のD44菌株およびD47菌株のセルロース産生活性をセルロース産生に用いられる公知の菌株であるアセトバクター・キシリナム菌株(KCCM40274)と比較した。
前記本発明のD44菌株とD47菌株、およびアセトバクター・キシリナム CCM40274菌株をそれぞれH.S培地に1重量%接種(培地の総重量に対して)した後に30℃で7日間静置培養し、次いで、前記実施例<3−1>により産生されたセルロースの乾燥重量を測定し、その結果を下記表6に示す。
【0056】
【表6】
【0057】
前記表6に示すように、本発明のD44菌株およびD47菌株は、アセトバクター・キシリナムと比較して、D44菌株の場合に約3倍、そしてD47菌株の場合に約4.34倍のセルロースを産生することができ、本発明の菌株の中でも、特にD47菌株は、D44菌株に比べて約1.27倍さらに多量のセルロースを産生することができ、セルロースを非常に高収率にて産生することができるということが分かる。
【0058】
<参考例2>
培地の組成によるセルロース産生量の評価に供された培地および培養条件
本発明の菌株がH.S培地で産生するセルロースの量を一層増大させるための要因の一つとして、培地の組成による影響を確認するために、下記表7に示す組成で製造された培地に菌を培養した。
菌培養液は1%(v/v)にし、温度は30℃に維持し、7日間培養後に乾燥重量を測定し、反復実験を通して平均値として使用した。
【0059】
【表7】
【0060】
1)ココナツ原液:100%原液であり、www.onedrinks.com(One world enterproses,LLC, USA. mede in brazil)で輸入
2)硫安:Sigma(USA)社製純度99%の硫化アンモニウム
3)硫安:Chia Tai Co., Ltd.(Thailand)社製純度21%の硫化アンモニウム
【0061】
<実施例4>
培地の組成による本発明の菌株のセルロース産生活性の増進試験
前記表7に示すように、培地組成を異ならせて、本発明のD44菌株およびD47菌株が産生したセルロースの乾燥重量および厚さを測定し、その結果を下記表8に示す。測定結果は、前記実施例3−2で用いられたアセトバクター・キシリナム菌株(KCCM40274)と比較した。
【0062】
【表8】
【0063】
前記表8に示すように、本発明のD44菌株およびD47菌株は、セルロースを産生する代表菌株であるアセトバクター・キシリナム菌株に比べて、セルロース産生量が全般的に遥かに高く現れ、特に、D47菌株の場合、全ての培地で前記菌株のうち最も多量のセルロースを産生した。
具体的に、H.S培地およびココナツ液体培地でD44菌株が産生したセルロースの乾燥重量は、アセトバクター・キシリナム菌株に比べて、それぞれ19倍、50倍に達し、D47菌株はそれぞれ22倍、97.5倍に達することから、本発明の菌株がセルロースを遥かに高い収率にて産生することを確認した。
一方、ココナツ混合培地(1)、(2)でのセルロースの産生量を調べてみると、代表菌株であるアセトバクター・キシリナム菌株と本発明のD44菌株との間にはセルロースの産生量に差がなかったが、本発明のD47菌株の場合、産生したセルロースの乾燥重量がココナツ混合培地(1)で0.80g/Lであり、ココナツ混合培地(2)で5.10g/Lであることが判明し、それぞれ4倍、25.5倍に達するセルロースの産生量を示して、アセトバクター・キシリナム菌株だけでなく、本発明のD44菌株よりも顕著に高い収率にてセルロースを産生することができるということを確認した。
【0064】
結論的に、本発明の菌株は、セルロースを産生する代表菌株であるアセトバクター・キシリナム菌株に比べて、遥かに高い収率にてセルロースを産生することができ、このようなセルロースの産生効率は培地の組成によって異なってくる。
すなわち、D44菌株は、ココナツ混合培地よりもH.S合成培地で約9.5倍セルロースをさらに多く産生し、D47菌株はH.S合成培地よりもココナツ原液に砂糖と硫安を添加して製造したココナツ混合培地(2)でセルロースを約2.32倍さらに多く産生し、ココナツ混合培地(1)に比べては約6.4倍セルロースをさらに多く産生することが分かる。なお、D47菌株がココナツ混合培地(2)で産生したセルロースの厚さを測定した結果、H.S合成培地で測定された4mmに比べて、約3倍増加された約12mmの厚さを示し、厚さが顕著に増加されていることが分かる(図9Aおよび図9B)。
【産業上の利用可能性】
【0065】
本発明の菌株が産生したセルロースは、液晶表示装置の基板、スピーカの振動板、医療用パッドや化粧パッド、人工皮膚、空気ろ過用フィルター、プリント配線基板、床材料等の種々の産業用素材、食品素材など多岐にわたって利用することができる。
【技術分野】
【0001】
本発明はセルロースを産生しうる新規なグルコンアセトバクター属菌株に係り、さらに詳しくは、ナノ構造のセルロースを非常に高収率にて産生しうる新規なグルコンアセトバクター属菌株に関する。
【背景技術】
【0002】
セルロースは、地球上で、最も多く存在する天然高分子の一種であり、略1011トンのセルロースが生合成されていると推算されており、ほとんどは高等植物の細胞壁繊維質成分を合成する。
【0003】
セルロースの強靭性は全体構造に起因するが、その分子は14,000単位に至る鎖や微細繊維を形成し、これらが水素結合によって維持される捩れた綱状束をなしている。このようなセルロースの豊富な物量と他の天然高分子とは異なる物性により、様々な産業分野において再生資源として活用するための研究が盛んになされている。
【0004】
1886年にブラウン(A.J.Brown)によって酢酸菌がセルロースを産生するということが報告されて以来、微生物によって産生されるセルロース(Bacterial Cellulose;以下「BC」と称する)は新素材として絶えず研究対象となっている。特に、BCはリボン状繊維(ribbin−like bundles)で構成されるのに対し、植物由来のセルロースは微細繊維(microfibrils)の束(bundles)状に形成される。
【0005】
リボン状繊維で構成されるBCは、微細繊維の束状に形成される植物由来のセルロースとは異なり、リグニン及びヘミセルロースが全く含まれていない純粋状態で産生され、高い機械的強度、高保湿性、高結晶性、生分解性などのBCならではの長所のため、様々な産業分野において様々な用途に開発されている。
【0006】
一方、液晶表示装置(LCD)の基板素材としては、熱膨張係数が低くて透過率が高く、しかも、微細な太さを有する必要があり、現在は、高価なs−glassが用いられている。
しかし、ナノ構造のセルロースを用いることにより、s−glassよりも熱特性が向上することが報告されている。ナノ構造のセルロースは微生物によって産生しうることが知られている(Schraamm, M. and S. Hestrin. 1054. Factors affecting production of cellulose at the air/liquid interface of a culture of Acetobacter xylinum.J.Gen. Microbiol. 11:123−129)。このため、このようなセルロース産生活性がある微生物を開発することは、経済的に非常に有利である。
【発明の概要】
【発明が解決しようとする課題】
【0007】
しかし、BCを産生する代表的な微生物として、アセトバクター・キシリナム(acetobacter xylinum)が知られているが、セルロースの産生効率が低く、これに代替することができる、新たな微生物の開発が切望されている。
上述した目的を達成するために、本発明は、グルコンアセトバクター属菌株:該属菌株の培養物:前記グルコンアセトバクター属菌株、前記菌株の培養物、前記菌株または培養物の濃縮物およびこれらの乾燥物からなる群より選ばれた1種以上を含むセルロース産生用組成物:セルロースの製造方法及びセルロース系樹脂の製造方法を提供することを目的とする。
【課題を解決するための手段】
【0008】
本発明者らは、セルロース産生効率に非常に優れた新たな微生物を開発するために鋭意研究した結果、グルコンアセトバクター属に属する新規な菌株を開発し、前記菌株が非常に有効にセルロースを産生しうるということを見出し、本発明を完成するに至った。
【0009】
本発明は、セルロース産生活性があるグルコンアセトバクター(gluconacetobacter)属菌株であって、前記菌株は、グルコンアセトバクター属(gluconacetobacter sp.)D44菌株(寄託番号KCCM11078P)またはグルコンアセトバクター・インターメディウス(Gluconacetobacter intermedius)D47菌株(寄託番号KCCM11079P)であることを特徴とするグルコンアセトバクター属菌株を提供する。
また、本発明は、前記グルコンアセトバクター属菌株の培養物を提供する。
さらに、本発明は、前記グルコンアセトバクター属菌株、前記菌株の培養物、前記菌株または培養物の濃縮物およびこれらの乾燥物からなる群より選ばれたいずれか1種以上を含むセルロース産生用組成物を提供する。
また、本発明は、前記グルコンアセトバクター属菌株を培養する工程と、前記培養物からセルロースを得る工程とを含むセルロースの製造方法を提供する。
さらに、本発明は、前記グルコンアセトバクター属菌株によって産生されたセルロースを準備する工程と、前記セルロースを樹脂と含浸させる工程とを含むセルロース系樹脂の製造方法を提供する。
【図面の簡単な説明】
【0010】
【図1】本発明のD44菌株の形態学的な観察結果を示すものであり、(a)は、光学顕微鏡による観察結果を示し、(b)は、走査電子顕微鏡(SEM)による観察結果を示す。
【図2A】本発明のD44菌株の16s rDNA配列をグルコンアセトバクター・ラエティカスの16s rDNA配列と比較して類似度を測定した結果を示すものである(*:塩基配列が一致する部分)。
【図2B】本発明のD44菌株の16s rDNA配列をグルコンアセトバクター・ラエティカスの16s rDNA配列と比較して類似度を測定した結果を示すものである(*:塩基配列が一致する部分)。
【図2C】本発明のD44菌株の16s rDNA配列をグルコンアセトバクター・ラエティカスの16s rDNA配列と比較して類似度を測定した結果を示すものである(*:塩基配列が一致する部分)。
【図3】本発明のD44菌株の酢酸菌科に属するグルコンアセトバクター属、グルコノバクター属およびアセトバクター属の菌に対する系統樹を示すものである。
【図4】本発明のD44菌株のセルロースの生成有無を確認した結果である。
【図5】本発明のD47菌株の形態学的な観察結果を示すものであり、(a)は、光学顕微鏡による観察結果であり、(b)は、走査電子顕微鏡(SEM)による観察結果である。
【図6A】本発明のD47菌株の16s rDNA配列をグルコンアセトバクター・インターメディウスの16s rDNA配列と比較して類似度を測定した結果を示すものである(*:塩基配列が一致する部分)。
【図6B】本発明のD47菌株の16s rDNA配列をグルコンアセトバクター・インターメディウスの16s rDNA配列と比較して類似度を測定した結果を示すものである(*:塩基配列が一致する部分)。
【図6C】本発明のD47菌株の16s rDNA配列をグルコンアセトバクター・インターメディウスの16s rDNA配列と比較して類似度を測定した結果を示すものである(*:塩基配列が一致する部分)。
【図7】本発明のD47菌株の酢酸菌科に属するグルコンアセトバクター属、グルコノバクターおよびアセトバクター属の菌に対する系統樹を示すものである。
【図8】本発明のD47菌株のセルロース生成有無を確認した結果である。
【図9A】本発明のD47菌株をH.S培地で培養して産生されたセルロースを示すものである。
【図9B】本発明のD47菌株をココナツ混合培地(2)で培養して産生されたセルロースを示すものである。
【発明を実施するための形態】
【0011】
本発明のグルコンアセトバクター属菌株、具体的に、グルコンアセトバクター属D44菌株(寄託番号KCCM11078P)またはグルコンアセトバクター・インターメディウスD47菌株(寄託番号KCCM11079P)は、セルロース産生能に非常に優れた新規な菌株である。
【0012】
本発明の菌株は酢酸菌(acetobacteraceae)の一種であり、これに限定されないが、酢類から分離することができ、好ましくは、柿酢、より好ましくは、柿酢の表膜から分離することができる。
本発明の一実施の形態においては、柿酢の表膜から高収率のセルロース産生活性がある本発明の菌株を分離し、前記セルロース産生活性があることは公知の方法により確認することができる。例えば、本発明の菌株をH.S培地(下記表1参照)に接種して培地に表膜を形成するかどうか、または、Carr培地(下記表1参照)に接種して色変化を引き起こすかどうかなどにより確認することができるが、これに限定されない(<実施例1>参照)。
また、本発明の菌株を同定するために形態学的特徴、生化学的特徴を分析したところ、本発明の菌株は下記表2に示す形態学的特徴を有し、下記表3(D44)および表4(D47)に示す生化学的特徴を有するグルコンアセトバクター属菌株である(<実施例2>参照)。
【0013】
さらに、本発明の菌株がグルコンアセトバクター属に属する菌株であるかどうかをそれぞれ配列番号1の塩基配列で示される16s rDNA配列(D44)および配列番号2の塩基配列で示される16s rDNA配列(D47)を得て分析した結果、D44菌株の場合、グルコンアセトバクター・ラエティカスの16s rDNA配列に最も類似しているが、類似度は87.215%に過ぎないことが分析された。本発明の菌株はグルコンアセトバクター属に属する新規な菌株であることが確認された。D47菌株はグルコンアセトバクター・インターメディウスの16s rDNA配列に最も類似しており、類似度は99.776%であることが分析された。本発明の菌株はグルコンアセトバクター属に属する菌株であることが確認された(<実施例2>参照)。
本発明者らは上記のようにして分離および同定された本発明の菌株をそれぞれグルコンアセトバクター属菌株D44およびグルコンアセトバクター・インターメディウスD47と命名し、前記菌株を2009年11月12日付けで韓国微生物保存センター(KCCM)寄託機関にそれぞれKFCC11470P寄託番号(D44)およびKFCC11471P寄託番号(D47)で寄託した。この後、2010年3月26日付けで再び韓国微生物保存センター(KCCM)寄託機関にそれぞれKCCM11078P寄託番号(D44)およびKCCM11079P寄託番号(D47)で国際寄託した。
【0014】
本発明のグルコンアセトバクター属菌株の培養物は、前記本発明の菌株を微生物の培養に用いられる公知の培地および培養方法を用いて製造することができる。
本発明のグルコンアセトバクター属菌株を培養するために用いられる培地は、通常、微生物の培養に用いられる培地が挙げられ、好ましくは、酢酸菌の培養に用いられる培地、より好ましくは、下記表1または表7に示す培地等が挙げられる。さらに好ましくは、セルロース産生のためのものであり、具体的に、下記表1に示すH.S培地または下記表7に示すココナツ混合培地(2)が挙げられる。これらを用いることにより、セルロースを一層高収率にて産生することができる(<実施例4>参照)。
【0015】
前記D44菌株は、好ましくは、グルコース20g/L、酵母抽出物5g/L、ペプトン5g/L、Na2HPO42.7g/Lおよびクエン酸ナトリウム1.15g/Lを含むH.S培地またはココナツ原液100%(w/v)のココナツ液体培地で培養することができる。前記酵母抽出物(CAS番号:8013−01−02)(ヨーロッパ既存化学物質名録:232−387−9)の酵母は40%以下の蛋白質を含有しており、1g当たりに塩酸ジアミン0.12mg以下、リボフラビン0.04mg以下、ニコチン酸0.25mg以下である無発酵性の黄白色の板状または粉末であり、ユニークな臭いの物理化学的特性を有している。
前記D47菌株は、好ましくは、前記H.S培地、前記ココナツ液体培地、またはココナツ原液、砂糖および硫安を含み、好ましくは、前記ココナツ原液85〜94.9重量部、砂糖5〜15重量部、および硫安0.1〜1重量部を含むココナツ混合培地で培養することができる。
前記硫安とは硫酸アンモニウムの略称であり、硫酸とアンモニアとを反応させて得られる白い結晶を意味し、好ましくは、純度10〜99.9%、より好ましくは、純度10〜30%、最も好ましくは、純度15〜25%の硫酸アンモニウムを使用することができる。
【0016】
本発明の菌株の培養物は、前記培地に本発明の菌株を接種し、当該分野で公知の微生物の培養方法(例えば、静置培養、攪拌培養)により、好ましくは、酢酸菌の培養方法により、より好ましくは、1重量%(培地の総重量に対して)にて接種し、28〜30℃で4〜7日間培養する方法により製造することができる。より好ましくは、本発明の菌株を1重量%(培地の総重量に対して)にて培地に接種し、30℃で培養することにより製造する。
【0017】
本発明の培養物は、これに限定されないが、好ましくは、本発明の菌株によって産生されたセルロースを含む。
一方、本発明のセルロース産生用組成物は、前記本発明のグルコンアセトバクター属菌株、前記菌株の培養物、前記菌株または培養物の濃縮物およびこれらの乾燥物からなる群より選ばれた1種以上を含む。
本発明の菌株は、セルロースを非常に有効に産生することができるので、前記菌株、前記菌株の培養物、前記菌株または培養物の濃縮物およびこれらの乾燥物はセルロースを産生する組成物として用いて好適である。
前記菌株の培養物の製造方法については上述した通りであり、これに限定されないが、好ましくは、本発明の菌株をセルロース産生用培地、下記表1に示すH.S培地または下記表7に示すココナツ混合培地(2)に接種し、これに限定されないが、好ましくは、静置培養または攪拌培養、好ましくは静置培養して培養物を製造する方法が挙げられる。前記菌株または培養物の濃縮物およびこれらの乾燥物は、当該分野に公知の微生物または培養物の濃縮または乾燥方法により容易に製造することができる。
前記本発明のグルコンアセトバクター属菌株、前記菌株の培養物、前記菌株または培養物の濃縮物およびこれらの乾燥物は、これに限定されないが、好ましくは、本発明のセルロース産生用組成物に1重量%にて含有される。
【0018】
本発明のセルロースの製造方法は、前記グルコンアセトバクター属菌株を培養する工程と、前記培養物からセルロースを得る工程とを含む。
前記菌株の培養方法は上述した通りであり、本発明のセルロースを産生するためには、これに限定されないが、本発明の菌株の培養によりセルロースの膜が形成されたものを水酸化ナトリウム溶液に浸漬させた後、蒸溜水により洗浄して純粋なセルロースを産生する方法が挙げられる。このようにして産生されたセルロースは、これに限定されないが、好ましくは、膜状である(<実施例3>参照)。
【0019】
本発明の菌株によって産生されたセルロースは滑らかであり、堅固性および弾力性に優れており、厚さが約0.3mm〜約12mmであり、既存のバクテリアセルロースよりも厚さが顕著に向上した、熱特性に非常に優れたナノ構造のセルロースである(<実施例3>および<実施例4>参照)。
さらに、本発明の菌株は、セルロース産生活性に非常に優れており、特に、BCを産生する代表的な微生物であるアセトバクター・キシリナム(acetobacter xylinum)と比較して、最少約3倍、最大約97.5倍の顕著に高い効率にてセルロースを産生することができる(<実施例3>および<実施例4>参照)。
このため、本発明の菌株によって産生されたセルロースは熱特性に非常に優れたナノ構造のバクテリアセルロースであり、バイオナノ繊維として利用するのに好適である。
【0020】
本発明のセルロース系樹脂の製造方法は、前記本発明のグルコンアセトバクター属菌株によって産生されたセルロースを準備する工程と、前記セルロースを樹脂と含浸させる工程とを含む。
前記セルロース系樹脂は、本発明のグルコンアセトバクター属菌株によって産生されたセルロースを用いたものであれば、当該分野に公知の製造方法により製造することができる。例えば、本発明のグルコンアセトバクター属菌株によって産生されたセルロースに熱可塑性樹脂および結合剤を混入し、これを圧出成形して製造することができる。
【0021】
前記熱可塑性樹脂は、ポリプロピレン、ポリエチレン、ポリスチレンおよびポリ塩化ビニールからなる群より選ばれたものであることが好ましい。
前記結合剤は、マレイン化エチレン、マレイン化プロピレンおよびシランよりなる群から選ばれる1種以上であることが好ましい。
これらセルロース、熱可塑性樹脂及び結合剤からなる混合物には、安定剤、顔料、染料、無機物または有機物添加剤、電磁波遮断剤またはこれらの混合物の中から選ばれた1種以上の添加剤をさらに添加してもよい。
【0022】
このようにして製造されたセルロース樹脂は熱特性に非常に優れたナノ構造のバクテリアセルロースが含まれているため、液晶表示装置の基板用に用いることができる。
したがって、本発明は、前記セルロース系樹脂の製造方法であって、セルロース系樹脂を製造する工程と、前記製造されたセルロース系樹脂を用いて液晶表示装置の基板を製造する工程とを含む液晶表示装置の基板の製造方法を提供する(Masaya Nogi and Hiroyuki Yano, Transparent Nanocomposites Based on Cellulose Produced by Bacteria Offer Potential Innovation in the Electronics Device Industry, Advanced materials 2008, pp1849-1852)。
また、本発明は、前記セルロース系樹脂を用いて製造された液晶表示装置の基板を提供する。
【0023】
セルロースを産生する菌において、セルロースは、UV光の致死効果から細胞を防御し、菌の集落を形成して栄養源として同じ基質を使用する競争者から防御効果をもたらすと共に、細胞に湿気を提供して乾燥を防ぐことが知られている。
また、本発明のように微生物によって産生されたセルロースは、繊維の結晶化度、吸湿性などが高く、スピーカの振動板などへの産業的な応用可能性が高く、セルロース膜はゲル状態で肌触り性が良く、身体の表面に馴染み易く、さらに、医薬品や化粧材料成分の補水能が高く、皮膚の表面を保湿状態に維持することができて、医療用パッドや化粧パッド(例えば、マスクパック)、人工皮膚などに応用することができ、現在、セルロースを用いたマスクパックは商用化工程まで至っている。
さらに、セルロース膜を5%のNaOHまたは0.5%のNaClにより処理してセルロース成分以外の物質を除去した後にこれを圧着すれば高弾性膜になることが判明し、日本のソニー社ではスピーカの音量振動板に応用して高性能ヘッドホンとして市販している。
【0024】
また、本発明のように微生物によって産生されたセルロースを用いて紙を製紙すれば、紙の強度や弾性率が対照群に比べて2.5倍も増加する。合成高分子繊維、芳香剤ポリアミドやポリアミド単繊維に5〜20%添加してシートを製作すれば、合成繊維に比べて、強度、熱安定性、形態安定性などに優れたシートが得られる。合成繊維だけではなく、ガラス繊維にセルロースを混入してシート状にすれば、空気ろ過用フィルター、プリント配線基板、床材料などに応用することが可能となる。
このような本発明の微生物由来のセルロースは、産業用素材、食品素材など多岐に亘って活用することができ、特に、環境にやさしい素材であるという点は無限の開発可能性と多様性を有している(Jeong.,Y.J.,Lee.,I.S.. A view of utilizing cellulose produced by acetobacter bacteria. Food Industry and Nutrition.,V ol.5.,25〜29.,2000.参照)。
【0025】
本発明の菌株はセルロース産生活性に非常に優れており、本発明の菌株によって産生されたセルロースは熱特性に非常に優れたナノ構造のバクテリアセルロースであって、バイオナノ繊維として有効に用いることができる。特に、樹脂と含浸してセルロース系樹脂を製造する上で用いることができ、しかも、前記セルロース系樹脂は液晶表示装置の基板用に有効に用いることができる。
【0026】
以下、実施例を挙げて本発明による高収率のセルロース産生活性がある新規なグルコンアセトバクター属菌株について詳述する。
但し、下記の実施例は本発明を例示するものに過ぎず、本発明の内容が下記の実施例に限定されることはない。
【0027】
<参考例1>
実験に供された培地および培養条件
本発明の新規な菌株を分離および同定するために、下記表1に示す培地を使用した。なお、下記の実施例において特別に言及がない限り、前記培地を用いて本発明のセルロースの高収率産生菌株を30℃で7日間培養して実験を行った。
【0028】
【表1】
【0029】
<実施例1>
セルロースを高収率にて産生する新規な菌株の分離
<実施例1−1>
グルコンアセトバクター属D44菌株の分離
本実験に供された菌株を在来式柿酢(全羅北道完州郡鳳東邑栗所里)の表膜から分離した。具体的に、柿酢の複数の部分を直接的に採取し、採取された部分によってA、B、C、Dと区別して命名し、これらのうちDは柿酢の表膜から採取した試料である。
前記柿酢A、B、C、D原液を1%ずつH.S培地100mに接種して7日間30℃で培養した。前記培養後、H.S培地の表膜にセルロースを生成するD試料で単一菌株を10−7まで希釈して固体H.S培地に塗抹した後、ランダムでコロニーを取って再び液体H.S培地に接種し、これらのうち培地に表膜を形成する菌株を分離した。
【0030】
前記柿酢のD部分から分離された菌株を変色範囲がpH5.2〜6.8であり、酸性で黄色、塩基性で紫色を帯びるブロモクレゾールパープル含有Carr培地に接種し、黄色から紫色へと、さらに紫色から黄色へと色変化があることを確認することにより、前記分離された菌株がエタノールを酢酸に酸化させ、二酸化炭素と水に再酸化させる酸化経路を有していることを改めて確認することができた。これにより、前記菌株がセルロース産生菌株の特性を有していることが分かる(Jang. O.Y., Joo. K.H., Lee. J.H., Baik. C.G. 2003. Growth characteristics and production of cellulose of microorganisms in static culture vinegar. Korean J. Food SCI. tech. vol. 35, No. 6, pp. 1150〜1154)。
本発明者らは、前記セルロース生成活性がある菌株を「D44」と命名した。
【0031】
<実施例1−2>
グルコンアセトバクター・インターメディウス(Gluconacetobacter intermedius)D47菌株の分離
本実験に供された菌株を在来式柿酢(全羅北道完州郡鳳東邑栗所里)の表膜から分離した。具体的に、在来式柿酢を製造するときに生成される被膜を農協から得て、滅菌済み解剖用はさみで十分に切断した後、少量をH.S培地に接種して7日間30℃で培養した。前記培養後、H.S培地の表膜にセルロースが産生された試料で単一菌株を10−7まで希釈して固体H.S培地に塗抹した後、ランダムでコロニーを取って液体H.S培地に再接種し、これらのうち培地に表膜を形成する菌株を分離した。
前記柿酢の表膜部分から分離された菌株を、変色範囲がpH5.2〜6.8であり、酸性で黄色、塩基性で紫色を帯びるブロモクレゾールパープル含有Carr培地に接種し、黄色から紫色へと、さらに紫色から黄色へと色変化があることを確認することにより、前記分離された菌株がセルロース生成活性があることを最終的に確認した。
本発明者らは、前記セルロース生成活性がある菌株を「D47」と命名した。
【0032】
<実施例2>
セルロースを高収率にて産生する新規な菌株の同定および寄託
<2−1>形態学的特徴
前記<実施例1>において分離されたD44菌株およびD47菌株のそれぞれの大きさ、形状、グラム染色およびコロニーの形状、色などを観察し、具体的に、GYC培地でD44菌株およびD47菌株それぞれのコロニーを観察し、D44菌株およびD47菌株がそれぞれ生成したセルロースを菌が除去されていない状態で光学顕微鏡および走査転写顕微鏡(SEM)により観察した。
より具体的に、光学顕微鏡(Nikon ECLIPSE E600、日本)を用いてD44菌株およびD47菌株の形状とグラム染色を、走査電子顕微鏡(SEM)によりD44菌株およびD47菌の大きさと菌が産生したセルロース繊維を検鏡した。特に、電子顕微鏡はChaeの方法(Chae. C.M.1977. Maceration methods for SEM obserbation ’A legal medicine lecture room’. Kyungpook National University. p 40−58)を変形して利用し、具体的に、D44菌株およびD47菌体を2%(v/v)グルタルアルデヒド (Glutaraldehyde)により2時間前固定し、2%(w/v)のOsO4により2時間後固定してエタノールにより順次に脱水した後、イソアミルアセタート(isoamyl acetate)を処理して55℃で乾燥し、金コートして検鏡した。D44菌株およびD47菌株の形態学的特徴
についての観察結果を表2と、図1および図5に示す。
【0033】
【表2】
【0034】
表2と、図1および図5に示すように、D44菌株およびD47菌株はグラム陰性であり、桿状であり、胞子を形成せず、単一(single)または対(pair)で配列されて存在する。特に、図1および図5に示すように、D44菌株およびD47によって生成されたセルロース繊維内に前記菌株が内在されている。
【0035】
<2−2>生化学的特徴
前記D44菌株およびD47菌株の生化学的特徴を調べるために、D44菌株およびD47菌株をそれぞれH.S液体培地に培養した後に採取した菌液を新たな培地に継代培養して使用した。
具体的に、本発明の新規なD44菌株およびD47菌株をそれぞれFrateur培地に接種して培養した後、培地中のCaCO3が中和するかどうかを確認してD44菌株およびD47菌株のエタノール酸化経路を確認した。
また、グリセロールでケトン体を形成するかどうかを確認するために、YEG固体培地に前記各菌株をそれぞれ塗抹して5日間30℃で培養した後、フェーリング液をコロニーの上に数滴滴下した後、オレンジ色への変色有無により確認した。
さらに、エタノールと塩化ナトリウムに対して菌株の生育とセルロースの生成有無を確認するために、SM培地に塩化ナトリウム0.5、1.0、1.5、2.0%(w/v)とエタノール10%(v/v)をそれぞれ添加した培地に、D44菌株およびD47菌株をそれぞれ接種して7日間30℃で静置培養した後に観察した。
【0036】
加えて、GYC固体培地にD44菌株およびD47菌株をそれぞれ塗抹して7日間30℃で培養した後に褐色色素の形成有無を確認し、γ−ピロンの生成有無を確認するために、葡萄糖50g/L、果糖50g/Lをそれぞれ含有するYPC培地に菌を塗抹して7日間30℃で培養した後に塩化第二鉄(ferric chloride)5%(w/v)溶液を滴下して確認した。このとき、紫色に変色すれば、菌株が葡萄糖と果糖を用いてγ−ピロンを生成することができると決めた(Jang. O.Y., Joo. K.H., Lee. J.H., Baik. C.G. 2003. Growth characteristics and production of cellulose of microorganisms in static culture vinegar. Korean J. Food SCI. tech. vol. 35, No. 6, pp. 1150〜1154)。
【0037】
さらに、D44菌株およびD47菌株の酢酸に対する抵抗性を調べるために、酢酸の濃度を2%(v/v)〜10%(v/v)に調節したGYP培地にD44菌株およびD47菌株をそれぞれ接種し、次いで、7日間30℃で培養後に調査した。
これらに加えて、生化学的実験はAPI 20NEキット(Bio−Merieux、フランス)を用いて行い、導出された結果をバージーズ・マニュアル・オブ・システマティック・バクテリオロジー(Bergy’s manual of systematic bacteriology)に準じて同定した(Krig, N.R., J.G. Holt. 1984. Bergey’s Manual of Systematic Bacteriology Vol. 1. The William and Wilkins Co.U.S.A.)。
前記実験結果を下記表3および表4に示し、その結果をアセトバクター属菌株、グルコノバクター属菌株およびグルコンアセトバクター属菌株と比較した。前記グルコンアセトバクター属菌は、Lee. O.S.文献(Lee. O.S., Jang. S.Y.,Jeong. Y.J. 2002. Effect of Ethanol on the Production of Cellulose and Acetic Acid by Gluconacetobacter persimmonensis KJ145. J .Korean Soc. Food sci. Nutr. 31(4).572−577.)に記載のものによる。
【0038】
【表3】
1)+:陽性、2)−:陰性、3)ND:未決定
【0039】
前記表3に示すように、本発明の新規なD44菌株をFrateur培地に接種して培養した結果、培地中のCaCO3が中和して透明環が形成されることを観察することができ、培養12日後にCaCO3が再沈殿されることを確認することができる。要するに、D44菌株はエタノールを酢酸に、酢酸をCO2とH2Oに酸化することが分かる(Son. H.J., Kim. K.K., Kim. H.S., Kim.Y.G. 1999. Isolation and identification of cellulose−producing bacteria. J. Agri. Tech & Dev. Inst.)。
【0040】
また、本発明のD44菌株は5%(w/v)葡萄糖と果糖を用いてγ−ピロンを生成することができず、グリセロールからのケトン体形成反応で陽性を示す。さらに、前記D44菌株はγ−ガラクトシダーゼ陽性、硝酸(nitrate)から亜硝酸(nitrite)に還元させ、アセタート、果糖、葡萄糖、マンニトルを酸化させることはできるものの、乳酸(lactate)は酸化させることができない。
さらに、本発明のD44菌株は、インドール産生、ウレアーゼ、ゼラチン液化反応で陰性を示し、0.5、1.0、1.5、2.0%(w/v)の塩化ナトリウムに生育可能であり、特に、0.5%(w/v)の塩化ナトリウムでセルロースも生成することができる。しかし、2%(v/v)〜10%(v/v)のアセタート濃度でD44は生育することができないため耐性がないことが観察され、褐色色素を生成することができずに透明で且つ白いコロニーを形成する。
前記結果から、本発明のD44菌株はグルコンアセトバクター(Gluconacetobacter)属菌株であることが分かる。
【0041】
【表4】
1)+:陽性、2)−:陰性、3)ND:未決定
【0042】
前記表4に示すように、本発明のD47菌株をFrateur培地に接種して培養した結果、培地中のCaCO3が中和して透明環が形成されることを観察することができ、培養8日後、CaCO3が再沈殿されることを確認することができる。要するに、D47菌株は、エタノールを酢酸に、酢酸をCO2とH2Oに酸化することが分かる(Son. H.J., Kim. K.K., Kim. H.S., Kim.Y.G. 1999. Isolation and identification of cellulose−producing bacteria. J. Agri. Tech & Dev. Inst.)。
また、本発明のD47菌株は、5%葡萄糖(w/v)と果糖を用いてγ−ピロンを生成することができず、前記D44菌株とは異なり、グリセロールからのケトン体形成反応で陰性を示す。なお、前記D47菌株は、前記D44菌株とは異なり、β−ガラクトシダーゼ陰性であり、硝酸(nitrate)から亜硝酸(nitrite)に還元させることができなかった。しかし、前記D44菌株と同様に、アセタート、果糖、葡萄糖、マンニトルを酸化させることはできるものの、乳酸は酸化させることができない。
【0043】
さらに、本発明のD47菌株は、インドール産生、ウレアーゼ、ゼラチン液化反応で陰性を示し、0.5、1.0、1.5、2.0%(w/v)の塩化ナトリウムに生育可能であり、特に、0.5%の塩化ナトリウムでセルロースも生成することができる。しかしながら、2%(v/v)〜10%(v/v)のアセタート濃度でD47は生育することができずに耐性がないことが観察され、前記D44菌株とは異なり、薄い褐色のコロニーを形成する。
前記結果から、本発明のD47菌株は、グルコンアセトバクター(Gluconacetobacter)属菌株であることが分かる。
【0044】
<2−3>16s rDNA分析
前記<実施例1>において分離されたD44菌株およびD47菌株のgDNAを抽出するために、まず、H.S培地の表膜に生成されたセルロースを除去するために、セルラーゼ(Celluclast、nonozyme)を1%(v/v)添加して37℃で1時間攪拌して培養し、DNeasy Blood & Tissueキット(QIAGEN、Germany)を用いて16s rDNAを抽出および増幅し、16s rDNAシーケンシングを通して配列番号1のD44菌株の16s rDNAの全体配列および配列番号2のD47菌株の16s rDNAの全体配列をそれぞれ得た。菌株の同定のために候補菌株からゲノムDNAをDNeasy(R) Blood & Tissueキット(QIAGEN, Germany)を用いて分離した。DNAの精製は、製造社のガイドラインに準じて行った。
【0045】
前記D44菌株およびD47菌株の16s rDNAのクローニングのために、プライマーとしては、配列番号3の5’−AGAGTTTGATCMT GGCTCAG−3’(順方向)と、配列番号4の5’−ACGGGCGGGTGTGTRC−3’(逆方向)を用いてPCR増幅を行った。具体的に、PCR増幅は、100ng鋳型DNA、0.5μMのプライマーDNA、0.2mM dNTPs、10×Ex Taq緩衝液およびTaq重合酵素(TaKaRa bio Inc., Japan)0.025U/μlを用いて95℃で30秒変性、58℃で30秒の結合、72℃で1分30秒の伸張とする35サイクルをBiometra社製サーモサイクラー(Tampa、USA)により増幅させた。PCR産物を1%のアガロースゲル電気泳動を行うことで確認した。確認されたPCR産物をPCR精製キット(Bioneer、Korea)を用いて精製した後、T4リガーゼ(ligase)(Promega、USA)を用いてpGEM T−easyベクター(Promega、USA)に連結し、16s rDNAシーケンシングを通して前記各菌株の塩基配列情報を得た。
【0046】
前記得られたD44菌株およびD47菌株の16s rDNA配列からリボソーム遺伝子銀行からDNAの相同性を調査して同定に参考し、Bioeditプログラム(
1298607716195_0.html
)、Clustal Xプログラム、Megaプログラム(http://www.megasoftware.net/)を用いて類似同定された微生物との系統樹(phylogenetic tree)を描いて類似種間の関係を観察した。
【0047】
まず、前記D44菌株の16s rDNA配列とグルコンアセトバクター・ラエティカスの16s rDNA配列(AY180961)との類似度を比較した結果、類似度が87.215%に過ぎなかった。普通、菌株同定の類似度が約98〜99%でなければ類似菌として判定することができないが、D44菌株とグルコンアセトバクター・ラエティカスとの類似度は87.215%に過ぎず、本発明のD44菌株は新規なものであることが分かる。
Clustal Xプログラムを用いて本発明のD44とグルコンアセトバクター・ラエティカスとの類似度を再確認し、その結果を図2Aから図2Cに示した。前記図2Aから図2Cに示すように、D44とグルコンアセトバクター・ラエティカスとの類似度は87.215%に過ぎない。
【0048】
前記類似度結果を参考として、同じ科に属する微生物を基にD44との系統を調べるために、Bioeditプログラム、Clustal Xプログラム、Megaプログラムを用いた。具体的に、酢酸菌(acetobacteraceae)科に属するグルコンアセトバクター(Gluconacetobacter)属とグルコノバクター(Gluconobacter)属、アセトバクター(acetobacter)属の類似菌に対して近隣結合法により作成した系統樹を図3に示した。
【0049】
次に、前記D47菌株の16s rDNA配列とグルコンアセトバクター・インターメディウスの16s rDNA配列(Y14694)との類似度を比較した結果、99.774%の類似度を示した。このため、前記D47菌株はグルコンアセトバクター・インターメディウスの類似菌であると判定され、グルコンアセトバクター・インターメディウスD47と命名した。
さらに、Clustal Xプログラムを用いて本発明のD47とグルコンアセトバクター・インターメディウスとの類似度を再確認し、その結果を図6Aから図6Cに示した。前記図6Aから図6Cに示すように、D47とグルコンアセトバクター・インターメディウスが99%類似していることが分かる。
但し、D47菌株は、前記<実施例2−2>の表4に示すように、従来のグルコンアセトバクター属菌株とは異なり、グリセロール(glycerol)でジヒドロキシアセトン(dihydroxyacetone)を形成するケトン体生成(ketogenesis)反応をし、乳酸(lactate)を過酸化させることができず、0.5%(w/v)のNaClをH.S培地に添加して培養するときに培地の表面にセルロースを形成しうるなど生化学的特性が従来のグルコンアセトバクター属菌株とは全く異なる新規な菌株であることを確認することができ、セルロース産生能においても、後述するように、従来のグルコンアセトバクター属菌株に比べて顕著に高い産生活性を有していることを確認した。
【0050】
前記類似度結果を参考にして、同じ科に属する微生物を基にD47の系統を調べるために、Bioeditプログラム、Clustal Xプログラム、Megaプログラムを用いた。具体的に、酢酸菌科に属するグルコンアセトバクター属とグルコノバクター属、アセトバクター属の類似菌に対して近隣結合法により作成した系統樹を図7に示した。
【0051】
<2−4>本発明の菌株の寄託
本発明者らは、上記のようにして分離および同定された本発明のグルコンアセトバクター属菌株を2010年3月26日付けで韓国微生物保存センター(KCCM)国際寄託機関にそれぞれKCCM11078P寄託番号(D44)およびKCCM11079P寄託番号(D47)で寄託した。
【0052】
<実施例3>
本発明の菌株のセルロース産生活性
<3−1>本発明の菌株のセルロース産生活性
前記D44菌株およびD47菌株をそれぞれH.S培地に接種してセルロースの生成有無を確認した結果を図4および図8に示す。
前記図4および図8に示すように、本発明のD44菌株およびD47菌株は培養してから2〜3日経過後に表膜にセルロースを形成し、培養期間が長くなるにつれてD47によって生成されたセルロース膜はさらに厚くなって多重に形成される。
また、前記H.S培地にD44菌株およびD47菌株をそれぞれ1重量%接種(培地の総重量に対して)し、30℃で7日間静置培養した後、産生されたセルロースを得て乾燥重量を測定し、その結果を表5および図9Aに示す。具体的に、前記静置培養後、H.S培地の表膜に形成されたセルロースを回収して水道水により洗浄し、次いで、少量の0.1N NaOH溶液に入れて1時間沸騰して溶菌させた。前記回収されたセルロースを水道水によりさらに洗浄した後、少量の蒸溜水に一日中入れて中和させた。中和されたセルロースをろ過して90℃で抗量になるまで乾燥した後、乾燥されたセルロースをデシケーターで冷却した後に乾燥重量を測定した。
【0053】
【表5】
【0054】
前記本発明のD44菌株を用いて産生されたセルロース膜は、表面が滑らかであり、堅固性および弾力性があり、厚さは約0.3mmであった。これに対し、前記本発明のD47菌株を用いて産生されたセルロース膜は、表面が滑らかであり、堅固性及び弾力性があり、厚さは約4mmであり、遥かに高いセルロース活性を示している。さらに、後述するように、ココナツ混合培地(2)でD47菌株を用いて産生されたセルロース膜の厚さは約12mmであって、顕著に高いセルロース活性を示している。
【0055】
<3−2>本発明の菌株のセルロース産生活性の比較
本発明のD44菌株およびD47菌株のセルロース産生活性をセルロース産生に用いられる公知の菌株であるアセトバクター・キシリナム菌株(KCCM40274)と比較した。
前記本発明のD44菌株とD47菌株、およびアセトバクター・キシリナム CCM40274菌株をそれぞれH.S培地に1重量%接種(培地の総重量に対して)した後に30℃で7日間静置培養し、次いで、前記実施例<3−1>により産生されたセルロースの乾燥重量を測定し、その結果を下記表6に示す。
【0056】
【表6】
【0057】
前記表6に示すように、本発明のD44菌株およびD47菌株は、アセトバクター・キシリナムと比較して、D44菌株の場合に約3倍、そしてD47菌株の場合に約4.34倍のセルロースを産生することができ、本発明の菌株の中でも、特にD47菌株は、D44菌株に比べて約1.27倍さらに多量のセルロースを産生することができ、セルロースを非常に高収率にて産生することができるということが分かる。
【0058】
<参考例2>
培地の組成によるセルロース産生量の評価に供された培地および培養条件
本発明の菌株がH.S培地で産生するセルロースの量を一層増大させるための要因の一つとして、培地の組成による影響を確認するために、下記表7に示す組成で製造された培地に菌を培養した。
菌培養液は1%(v/v)にし、温度は30℃に維持し、7日間培養後に乾燥重量を測定し、反復実験を通して平均値として使用した。
【0059】
【表7】
【0060】
1)ココナツ原液:100%原液であり、www.onedrinks.com(One world enterproses,LLC, USA. mede in brazil)で輸入
2)硫安:Sigma(USA)社製純度99%の硫化アンモニウム
3)硫安:Chia Tai Co., Ltd.(Thailand)社製純度21%の硫化アンモニウム
【0061】
<実施例4>
培地の組成による本発明の菌株のセルロース産生活性の増進試験
前記表7に示すように、培地組成を異ならせて、本発明のD44菌株およびD47菌株が産生したセルロースの乾燥重量および厚さを測定し、その結果を下記表8に示す。測定結果は、前記実施例3−2で用いられたアセトバクター・キシリナム菌株(KCCM40274)と比較した。
【0062】
【表8】
【0063】
前記表8に示すように、本発明のD44菌株およびD47菌株は、セルロースを産生する代表菌株であるアセトバクター・キシリナム菌株に比べて、セルロース産生量が全般的に遥かに高く現れ、特に、D47菌株の場合、全ての培地で前記菌株のうち最も多量のセルロースを産生した。
具体的に、H.S培地およびココナツ液体培地でD44菌株が産生したセルロースの乾燥重量は、アセトバクター・キシリナム菌株に比べて、それぞれ19倍、50倍に達し、D47菌株はそれぞれ22倍、97.5倍に達することから、本発明の菌株がセルロースを遥かに高い収率にて産生することを確認した。
一方、ココナツ混合培地(1)、(2)でのセルロースの産生量を調べてみると、代表菌株であるアセトバクター・キシリナム菌株と本発明のD44菌株との間にはセルロースの産生量に差がなかったが、本発明のD47菌株の場合、産生したセルロースの乾燥重量がココナツ混合培地(1)で0.80g/Lであり、ココナツ混合培地(2)で5.10g/Lであることが判明し、それぞれ4倍、25.5倍に達するセルロースの産生量を示して、アセトバクター・キシリナム菌株だけでなく、本発明のD44菌株よりも顕著に高い収率にてセルロースを産生することができるということを確認した。
【0064】
結論的に、本発明の菌株は、セルロースを産生する代表菌株であるアセトバクター・キシリナム菌株に比べて、遥かに高い収率にてセルロースを産生することができ、このようなセルロースの産生効率は培地の組成によって異なってくる。
すなわち、D44菌株は、ココナツ混合培地よりもH.S合成培地で約9.5倍セルロースをさらに多く産生し、D47菌株はH.S合成培地よりもココナツ原液に砂糖と硫安を添加して製造したココナツ混合培地(2)でセルロースを約2.32倍さらに多く産生し、ココナツ混合培地(1)に比べては約6.4倍セルロースをさらに多く産生することが分かる。なお、D47菌株がココナツ混合培地(2)で産生したセルロースの厚さを測定した結果、H.S合成培地で測定された4mmに比べて、約3倍増加された約12mmの厚さを示し、厚さが顕著に増加されていることが分かる(図9Aおよび図9B)。
【産業上の利用可能性】
【0065】
本発明の菌株が産生したセルロースは、液晶表示装置の基板、スピーカの振動板、医療用パッドや化粧パッド、人工皮膚、空気ろ過用フィルター、プリント配線基板、床材料等の種々の産業用素材、食品素材など多岐にわたって利用することができる。
【特許請求の範囲】
【請求項1】
セルロース産生活性があるグルコンアセトバクター(gluconacetobacter)属菌株であって、
前記菌株は、グルコンアセトバクター属(gluconacetobacter sp.)D44菌株(寄託番号KCCM11078P)またはグルコンアセトバクター・インターメディウス(Gluconacetobacter intermedius)D47菌株(寄託番号KCCM11079P)であることを特徴とするグルコンアセトバクター属菌株。
【請求項2】
前記D44菌株は、配列番号1の16s rDNA塩基配列を有する請求項1に記載のグルコンアセトバクター属菌株。
【請求項3】
前記D47菌株は、配列番号2の16s rDNA塩基配列を有する請求項1に記載のグルコンアセトバクター属菌株。
【請求項4】
請求項1のグルコンアセトバクター属菌株の培養物。
【請求項5】
前記培養物はセルロースを含む請求項4に記載の培養物。
【請求項6】
請求項1のグルコンアセトバクター属菌株、前記菌株の培養物、前記菌株または培養物の濃縮物およびこれらの乾燥物からなる群より選ばれた1種以上を含むセルロース産生用組成物。
【請求項7】
請求項1のグルコンアセトバクター属菌株を培養する工程及び
前記培養物からセルロースを得る工程を含むセルロースの製造方法。
【請求項8】
前記培養工程においては、
グルコンアセトバクター属D44菌株(寄託番号KCCM11078P)を、
グルコース20g/L、酵母抽出物5g/L、ペプトン5g/L、Na2HPO42.7g/Lおよびクエン酸ナトリウム1.15g/Lを含むH.S培地で培養する請求項7に記載のセルロースの製造方法。
【請求項9】
前記培養工程においては、
グルコンアセトバクター・インターメディウスD47菌株(寄託番号KCCM11079P)を、
ココナツ原液85〜94.9重量部、砂糖5〜15重量部および硫安0.1〜1重量部を含む培地で培養する請求項7に記載のセルロースの製造方法。
【請求項10】
請求項1のグルコンアセトバクター属菌株によって産生されたセルロースを準備する工程及び
前記セルロースを樹脂と含浸させる工程を含むセルロース系樹脂の製造方法。
【請求項11】
前記セルロース系樹脂は、液晶表示装置の基板用として用いられるものである請求項10に記載のセルロース系樹脂の製造方法。
【請求項1】
セルロース産生活性があるグルコンアセトバクター(gluconacetobacter)属菌株であって、
前記菌株は、グルコンアセトバクター属(gluconacetobacter sp.)D44菌株(寄託番号KCCM11078P)またはグルコンアセトバクター・インターメディウス(Gluconacetobacter intermedius)D47菌株(寄託番号KCCM11079P)であることを特徴とするグルコンアセトバクター属菌株。
【請求項2】
前記D44菌株は、配列番号1の16s rDNA塩基配列を有する請求項1に記載のグルコンアセトバクター属菌株。
【請求項3】
前記D47菌株は、配列番号2の16s rDNA塩基配列を有する請求項1に記載のグルコンアセトバクター属菌株。
【請求項4】
請求項1のグルコンアセトバクター属菌株の培養物。
【請求項5】
前記培養物はセルロースを含む請求項4に記載の培養物。
【請求項6】
請求項1のグルコンアセトバクター属菌株、前記菌株の培養物、前記菌株または培養物の濃縮物およびこれらの乾燥物からなる群より選ばれた1種以上を含むセルロース産生用組成物。
【請求項7】
請求項1のグルコンアセトバクター属菌株を培養する工程及び
前記培養物からセルロースを得る工程を含むセルロースの製造方法。
【請求項8】
前記培養工程においては、
グルコンアセトバクター属D44菌株(寄託番号KCCM11078P)を、
グルコース20g/L、酵母抽出物5g/L、ペプトン5g/L、Na2HPO42.7g/Lおよびクエン酸ナトリウム1.15g/Lを含むH.S培地で培養する請求項7に記載のセルロースの製造方法。
【請求項9】
前記培養工程においては、
グルコンアセトバクター・インターメディウスD47菌株(寄託番号KCCM11079P)を、
ココナツ原液85〜94.9重量部、砂糖5〜15重量部および硫安0.1〜1重量部を含む培地で培養する請求項7に記載のセルロースの製造方法。
【請求項10】
請求項1のグルコンアセトバクター属菌株によって産生されたセルロースを準備する工程及び
前記セルロースを樹脂と含浸させる工程を含むセルロース系樹脂の製造方法。
【請求項11】
前記セルロース系樹脂は、液晶表示装置の基板用として用いられるものである請求項10に記載のセルロース系樹脂の製造方法。
【図2A】


【図3】
【図6A】

【図6B】

【図6C】

【図7】
【図1】

【図2B】

【図2C】

【図4】
【図5】

【図8】

【図9A】

【図9B】

【図3】
【図6A】
【図6B】
【図6C】
【図7】
【図1】
【図2B】
【図2C】
【図4】
【図5】
【図8】
【図9A】
【図9B】
【公開番号】特開2011−177172(P2011−177172A)
【公開日】平成23年9月15日(2011.9.15)
【国際特許分類】
【出願番号】特願2011−41619(P2011−41619)
【出願日】平成23年2月28日(2011.2.28)
【出願人】(390019839)三星電子株式会社 (8,520)
【氏名又は名称原語表記】Samsung Electronics Co.,Ltd.
【住所又は居所原語表記】416,Maetan−dong,Yeongtong−gu,Suwon−si,Gyeonggi−do,Republic of Korea
【Fターム(参考)】
【公開日】平成23年9月15日(2011.9.15)
【国際特許分類】
【出願日】平成23年2月28日(2011.2.28)
【出願人】(390019839)三星電子株式会社 (8,520)
【氏名又は名称原語表記】Samsung Electronics Co.,Ltd.
【住所又は居所原語表記】416,Maetan−dong,Yeongtong−gu,Suwon−si,Gyeonggi−do,Republic of Korea
【Fターム(参考)】
[ Back to top ]