
高機能化HVJ−Eの開発
【課題】抗腫瘍効果がより改善された改良型センダイウイルスエンベロープ(HVJ−E)を有効成分とする抗癌剤を提供する。
【解決手段】C末端側に機能性ポリペプチドが連結した融合タンパク質、該融合タンパク質を含む複合体タンパク質、ならびに、該融合タンパク質または該タンパク質複合体をエンベロープに含有する改良型センダイウイルスエンベロープ(HVJ−E)を有効成分とする抗癌剤。
【解決手段】C末端側に機能性ポリペプチドが連結した融合タンパク質、該融合タンパク質を含む複合体タンパク質、ならびに、該融合タンパク質または該タンパク質複合体をエンベロープに含有する改良型センダイウイルスエンベロープ(HVJ−E)を有効成分とする抗癌剤。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、センダイウイルス(Hemagglutinating Virus of Japan,以下HVJと表記することもある)を構成するHNタンパク質のN末端側部分アミノ酸配列からなる改変HNタンパク質、該改変HNタンパク質のC末端側に機能性ポリペプチドが連結した融合タンパク質、または、該融合タンパク質を含むタンパク質複合体、さらに、該融合タンパク質または該タンパク質複合体をエンベロープに含有する改良型センダイウイルスエンベロープ(以下HVJ−Eと表記することもある)を有効成分とする抗癌剤等に関する。
【背景技術】
【0002】
センダイウイルス(HVJ)のゲノムを不活化して増殖能が喪失された、HVJエンベロープ(HVJ−E)は、他の分子をその中に封入することができることから、生体機能分子のベクター(運び屋)として開発されてきた(特許文献1、特許文献2など)。そして、HVJ−Eに抗癌剤を封入して、またはHVJ−Eと抗癌剤を併用して投与することで、抗癌効果が増強されることも分かってきた(特許文献3、非特許文献1、非特許文献2)。このように、HVJ−Eは、抗癌剤のためのベクターとして、またそれ自体が抗癌剤の補助剤として研究開発が進められている(非特許文献3)。さらには、HVJ−Eのみを投与した場合でも抗癌作用を示すことが報告されている(特許文献3、非特許文献4)。HVJ−Eは、樹状細胞や腫瘍細胞からInterferon−α、−βやIL−6などのサイトカイン、CCL−2、3、4、5、CXCL−10などのケモカインの分泌を促し、NK細胞の浸潤の活性化、CD4+、CD8+T cellの浸潤と活性化、それによる腫瘍に対するCTL誘導を引きおこす(非特許文献4)。しかしながら、HVJ−EにはInterferon−γ(IFN−γ)の産生促進能が認められておらず、Effector T cellには直接作用することはできないと考えられる。IFN−γ産生を誘導するサイトカインとしては種々の分子が知られているが、代表的な分子としてはIL−12が挙げられる。IL−12がIFN−γの産生を誘導することで抗腫瘍活性を示すことは、当業者には十分に公知の事実である(例えば、非特許文献5、非特許文献6など)。
【先行技術文献】
【特許文献】
【0003】
【特許文献1】WO01/57204
【特許文献2】WO2004/035779
【特許文献3】WO2005/094878
【非特許文献】
【0004】
【非特許文献1】Biochem Biophys Res Commun. 2007 Dec 21;364(3):423−8. Epub 2007 Oct 12
【非特許文献2】Cancer Chemother Pharmacol. 2008 May;61(6):973−8. Epub 2007 Jul 26
【非特許文献3】Expert Opin Drug Deliv. 2008 Feb;5(2):221−33. Review
【非特許文献4】Int J Cancer. 2009 May 15;124(10):2478−87
【非特許文献5】J. Immunol. 1994, 153, 1697− 706
【非特許文献6】J. Leukoc. Biol. 55:280−288; 1994
【発明の概要】
【発明が解決しようとする課題】
【0005】
本発明の課題は、HVJ−Eにより、IL−12等の抗腫瘍性ポリペプチドの抗腫瘍効果を飛躍的に増大させる新たな技術を開発し、これに基づき優れた抗癌剤を提供することである。また、本発明は、この新たな技術に有用な改変タンパク質や、改良型HVJ−Eを提供することを課題とする。
【課題を解決するための手段】
【0006】
本発明者らは上記課題を解決すべく、鋭意検討の結果、HVJ−Eを構成するHNタンパク質の特定の領域のC末端側に所望のポリペプチドを連結させることにより改変HNタンパク質と所望のタンパク質との融合タンパク質を作成し、この融合タンパク質を用いてHVJ−Eを構成すると、所望のタンパク質がHVJ−Eの外側表面に発現することを見出した。この所望のタンパク質として、一本鎖IL−12を用いると、一本鎖IL−12がHVJ−E粒子の表面に発現した、改変HVJ−Eを得ることができた。この改変HVJ−Eは、HVJ−EとIL−12とを単に共投与した場合に比べて、はるかに高い抗腫瘍効果を示したことから、HVJ−Eの粒子表面にIL−12等のIFN−γ分泌促進能を有するポリペプチドを結合させると、極めて高い抗腫瘍効果を得ることが出来ることを見出し、本発明を完成させるに到った。すなわち、本発明は、
[1]HVJのHNタンパク質のN末端部分アミノ酸配列と同一または実質的に同一のアミノ酸配列からなる改変HNタンパク質であって、該部分アミノ酸配列が111〜209アミノ酸の長さを有する、改変HNタンパク質;
[2]該部分アミノ酸配列が140〜180アミノ酸の長さを有する、上記[1]記載の改変HNタンパク質;
[3]下記(1)または(2)のポリペプチドである、上記[1]記載の改変HNタンパク質:
(1)配列番号6の第1〜160アミノ酸に該当するアミノ酸配列からなるポリペプチド、
(2)配列番号6の第1〜160アミノ酸に該当するアミノ酸配列において1または複数の残基が置換、欠失又は付加されたアミノ酸配列を含み、且つHVJに発現された場合に粒子表面に局在しうる特徴を有するポリペプチド;
[4]上記[1]〜[3]のいずれか1つに記載の改変HNタンパク質の残基及び第1の機能性ポリペプチド残基を含む融合タンパク質であって、該改変HNタンパク質残基のC末端側に該機能性ポリペプチド残基が機能可能に連結されている、融合タンパク質;
[5]機能性ポリペプチドがIFN−γ分泌促進能を有するポリペプチドである、上記[4]記載の融合タンパク質;
[6]IFN−γ分泌促進能を有するポリペプチドが一本鎖IL−12である、上記[5]記載の融合タンパク質;
[7]一本鎖IL−12が、下記(1)または(2)のポリペプチドである、上記[6]記載の融合タンパク質:
(1)配列番号2の第103〜625アミノ酸に該当するアミノ酸配列を含むポリペプチド、
(2)配列番号2の第103〜625アミノ酸に該当するアミノ酸配列において1または複数の残基が置換、欠失又は付加されたアミノ酸配列を含み、且つIFN−γ分泌促進能を有するポリペプチド;
[8]機能性ポリペプチドが抗体の定常領域(Fc)のCH2−CH3ドメインである、上記[4]記載の融合タンパク質;
[9]抗体の定常領域(Fc)のCH2−CH3ドメインがIgG2aに由来するものである、上記[8]記載の融合タンパク質;
[10]抗体の定常領域のCH2−CH3ドメインが、下記(1)または(2)のポリペプチドである、上記[9]記載の融合タンパク質:
(1)配列番号4の第171〜389アミノ酸に該当するアミノ酸配列を含むポリペプチド、
(2)配列番号4の第171〜389アミノ酸に該当するアミノ酸配列において1または複数の残基が置換、欠失又は付加されたアミノ酸配列を含み、且つプロテインAとの結合活性を有するポリペプチド;
[11]上記[1]〜[3]のいずれか1つに記載の改変HNタンパク質、又は上記[4]〜[10]のいずれか1つに記載の融合タンパク質をコードする核酸;
[12]上記[8]〜[10]のいずれか1つに記載の第1の融合タンパク質、並びに
プロテインAのZZドメイン及び第2の機能性ポリペプチド残基を含む第2の融合タンパク質を含む、タンパク質複合体であって、第1の融合タンパク質と第2の融合タンパク質とが、第1の融合タンパク質に含まれるCH2−CH3ドメインと第2の融合タンパク質に含まれるZZドメインとの間の結合を介して複合体を形成している、タンパク質複合体;
[13]第2の機能性ポリペプチドが、一本鎖IL−12である、上記[12]記載のタンパク質複合体;
[14]一本鎖IL−12が、下記(1)または(2)のポリペプチドであり、且つZZドメインが、下記(3)または(4)のポリペプチドである、上記[13]記載のタンパク質複合体:
(1)配列番号2の第103〜625アミノ酸に該当するアミノ酸配列を含むポリペプチド、
(2)配列番号2の第103〜625アミノ酸に該当するアミノ酸配列において1または複数の残基が置換、欠失又は付加されたアミノ酸配列を含み、且つIFN−γ分泌促進能を有するポリペプチド、
(3)配列番号2の第42〜99アミノ酸に該当するアミノ酸配列を含むポリペプチド、
(4)配列番号2の第42〜99アミノ酸に該当するアミノ酸配列において1または複数の残基が欠失、置換または付加されたアミノ酸配列を含み、且つ抗体分子の定常領域(Fc)への結合活性を有するポリペプチド;
[15]上記[1]〜[3]のいずれか1つに記載の改変HNタンパク質、上記[4]〜[10]のいずれか1つに記載の融合タンパク質、または上記[12]〜[14]のいずれか1つに記載のタンパク質複合体をエンベロープに含有する、HVJ−E;
[16]上記[6]もしくは[7]に記載の融合タンパク質または上記[13]もしくは[14]に記載のタンパク質複合体をエンベロープに含有する、上記[15]記載のHVJ−E;
[17]IFN−γ分泌促進能を有するポリペプチドをエンベロープ外側に提示する形で担持する、HVJ−E;
[18]IFN−γ分泌促進能を有するポリペプチドが、IL−12である、上記[17]記載のHVJ−E;
[19]IL−12が一本鎖IL−12である、上記[18]記載のHVJ−E;
[20]上記[6]もしくは[7]に記載の融合タンパク質または上記[13]もしくは[14]に記載のタンパク質複合体をエンベロープに含有する、上記[18]記載のHVJ−E;
[21]上記[17]〜[20]のいずれか1つに記載のHVJ−Eを有効成分として含む、抗癌剤;
[22]メラノーマ、前立腺癌、膀胱癌および大腸癌からなる群より選択される癌の治療または予防に用いられる、上記[21]記載の抗癌剤;
[23]1回当たりの投与量が、HVJ−Eとして1000HAU以下であり、かつ該HVJ−Eに含まれるIL−12として100pg以下であることを特徴とする、上記[21]記載のHVJ−Eを有効成分として含む抗癌剤;
[24]上記[21]記載のHVJ−Eの治療上有効量を癌患者に投与することを特徴とする、癌の治療方法;
[25]1回当たりの投与量が、HVJ−Eとして1000HAU以下であり、かつ該HVJ−Eに含まれるIL−12として100pg以下であることを特徴とする、上記[24]記載の癌の治療方法;
を提供する。
【発明の効果】
【0007】
本発明の改変HNタンパク質や融合タンパク質を用いれば、HVJ−Eの表面に所望のポリペプチドを発現させることが可能となる。
例えば、改変HNタンパク質と一本鎖IL−12等のIFN−γ分泌促進能を有するポリペプチドとの融合タンパク質を用いることにより、エンベロープの外側に該ポリペプチドを提示する形で担持するHVJ−Eを得ることが可能である。
エンベロープの外側にIFN−γ分泌促進能を有するポリペプチドを提示する形で担持するHVJ−Eは、高い抗腫瘍効果を有しており、抗腫瘍剤として有用である。
癌の治療においては、外科的切除法、放射線治療法、化学療法などが存在するが、その根治は依然大きな課題である。本発明は、細胞実験および生体内実験を通じて、本発明の改良型HVJ−Eの癌への直接投与で、IFN−γの産生を誘導し、癌を著しく退縮させることを示した。
【図面の簡単な説明】
【0008】
【図1】1本鎖IL−12のZZドメインがHVJ−E表面の抗体の定常領域(Fc)に会合することにより抗腫瘍効果を向上させたHVJ−Eの作製スキームを示した図である。
【図2】融合タンパク質を発現させ、細胞膜上に局在させた細胞にネイティブなHVJを感染させることにより、エンベロープに融合タンパク質を組み込んだ組換えHVJの作製の過程を示した図である。
【図3】(a)HNタンパク質の細胞外ドメインを段階的に欠失させた各融合タンパク質の構造を示す図である。(b)(a)に示した各融合タンパク質の細胞膜での局在を示す免疫染色図である。(c)(a)に示した各融合タンパク質の細胞内発現をウエスタンブロットで確認した図である(d)(a)に示した各融合タンパク質の細胞膜局在をウエスタンブロットで確認した図である。
【図4】(a)HNタンパク質の細胞外ドメインを細胞外ドメインから50アミノ酸残基、80アミノ酸残基、100アミノ酸残基、120アミノ酸残基、150アミノ酸残基だけ残した各融合タンパク質の構造を示す図である。(b)(a)に示した各融合タンパク質の細胞膜での局在を示す免疫染色図である。(c)(a)に示した各融合タンパク質の細胞内発現をウエスタンブロットで確認した図である(d)(a)に示した各融合タンパク質の細胞膜局在をウエスタンブロットで確認した図である。
【図5】(a)HNタンパク質の細胞外ドメインを細胞外ドメインから100アミノ酸残基、だけ残した融合タンパク質にマウス由来IgG2aのFcのCH2−CH3ドメインを付加したFc−HNタンパク質の構造を示す図である。(b)(a)に示したFc−HNタンパク質の細胞内発現をウエスタンブロットで確認した図である。(c)(a)に示したFc−HNタンパク質の細胞膜での局在を示す免疫染色図である。(d)(a)に示したFc−HNタンパク質の細胞膜局在をウエスタンブロットで確認した図である。(e)(a)に示したFc−HNタンパク質のプロテインAセファロースとの共沈を示す図である。
【図6】(a)プロテインAのZZドメインと一本鎖IL−12とを融合させて作製したZZ−IL−12タンパク質の構造を示す図である。(b)各ZZ−IL−12タンパク質のインターフェロンγ(IFNγ)の分泌刺激効果を示す図である。
【図7】ZZ−scIL12とFc−HN−HVJとの結合能を示す図である。
【図8】ZZ−scIL12とFc−HN−HVJとの24時間インキュベーション後の結合保持能を示す図である。
【図9】ZZ−scIL12結合型Fc−HVJ−Eの樹状細胞(DC)および脾臓細胞への作用を示す図である。図9左図は、各HVJ−Eの発現を示すウエスタンブロット図である。図9中図は、ZZ−scIL12結合型Fc−HVJ−Eの樹状細胞(DC)からのIFN−γ分泌促進能を示す図である。図9右図は、ZZ−scIL12結合型Fc−HVJ−Eの樹状細胞(DC)からのIL−6分泌促進能を示す図である。
【図10】ZZ−scIL12結合型Fc−HVJ−EとZZ−scIL12単独投与との脾臓細胞へのIFN−γ分泌促進作用の比較を示す図である。
【図11】ZZ−scIL12結合型Fc−HVJ−Eのin vivoでの抗腫瘍作用を示す図である。
【図12】ZZ−scIL12結合型Fc−HVJ−Eのin vivoでの免疫活性化能を示す図である。図12左図は、ZZ−scIL12結合型Fc−HVJ−EのIFN−γ分泌誘導能を示すELISPOTアッセイ図である。図12右図は、ZZ−scIL12結合型Fc−HVJ−EのCTL活性を示すCr51リリーシングCTLアッセイ図である。
【図13】ZZ−scIL12結合型Fc−HVJ−Eと、非結合scIL12およびHVJ−Eの併用投与との抗癌作用の比較を示す図である。
【図14】ZZ−scIL12結合型Fc−HVJ−Eと、非結合scIL12およびHVJ−Eの併用との脾細胞からのIFN−γ分泌促進能の比較を示す図である。
【発明を実施するための形態】
【0009】
以下に、本発明を詳細に説明する。
本発明は、センダイウイルス(HVJ)のHNタンパク質のN末端部分アミノ酸配列と同一または実質的に同一のアミノ酸配列からなる改変HNタンパク質であって、該部分アミノ酸配列が111〜209アミノ酸の長さを有する、改変HNタンパク質を提供する。
好ましくは、該部分アミノ酸配列が140〜180アミノ酸の長さを有する。
さらに好ましくは、本発明の改変タンパク質は、下記(1)または(2)のポリペプチドである:
(1)配列番号6の第1〜160アミノ酸に該当するアミノ酸配列からなるポリペプチド、
(2)配列番号6の第1〜160アミノ酸に該当するアミノ酸配列において1または複数の残基が置換、欠失又は付加されたアミノ酸配列からなり、且つHVJに発現された場合に粒子表面に局在しうる特徴を有するポリペプチド。
【0010】
本発明で用いられるセンダイウイルス(HVJ)は、例えばVR−105、VR−907等をAmerican Type Culture Collection(ATCC:P.O.Box 1549,Manassas,VA 20108 USA)より購入することができる。
【0011】
本発明で用いられるHNタンパク質とは、センダイウイルス(HVJ)のエンベロープを構成する公知のコンポーネントの一つであり、宿主細胞への結合に関わる赤血球凝集素/ノイラミニダーゼ(HN)遺伝子によってコードされる。HNタンパク質のアミノ酸配列も公知である。本明細書においては、HNタンパク質は、例えば配列番号6に示されるアミノ酸配列からなるタンパク質を意味する。
【0012】
発明者らは、後述する実施例において確認するように、HNタンパク質の種々の欠失変異体を作成し、HNタンパク質が、細胞膜に局在し、ウイルス粒子形成の際にHVJのエンベロープを構成するためのHNタンパク質中の必要領域を検討し、その範囲を見出した。その結果、HNタンパク質の翻訳開始メチオニン残基から始まり、細胞内ドメイン、膜貫通ドメインに加えて細胞外ドメインの一部をコードする部分アミノ酸配列(以下、N末端部分アミノ酸配列とも表記してもよい)が必須であることを見出した。そして、該部分アミノ酸配列が、111アミノ酸以上、好ましくは140アミノ酸以上の長さを有していれば、C末端側の領域を欠失していても、該変異体HNタンパク質は、安定してHVJのエンベロープを構成し得ることを見出した。
【0013】
更にこのHNタンパク質のN末端部分ポリペプチドのC末端側に任意の機能性ポリペプチドを連結した融合タンパク質を調製し、この機能性ポリペプチドがHVJのエンベロープの外側に提示されるように、該融合タンパク質を安定してHVJ−E上に発現させる方法について検討する過程で、該融合タンパク質に含まれるHNタンパク質のN末端部分アミノ酸配列の長さが、209アミノ酸(好ましくは180アミノ酸)よりも長いと、かえって該融合タンパク質が安定してHVJ−E上に発現され難いことを見出した。
【0014】
即ち、HNタンパク質のN末端部分アミノ酸配列の長さは、通常、111〜209アミノ酸、好ましくは、140〜180アミノ酸、最も好ましくは、160アミノ酸である。
【0015】
尚、HNタンパク質のN末端部分アミノ酸配列における、第1のアミノ酸は、HNタンパク質のN末端の第1メチオニン(翻訳開始メチオニン残基)である。
【0016】
本発明の改変HNタンパク質は、哺乳動物細胞(例えばサル腎臓細胞LLCMK2)において発現したときに、該細胞の細胞膜上に局在する(これを、細胞膜へ局在する活性という)。「局在」とは、細胞内の他の領域よりも細胞膜における発現量が高いことを意味する。改変HNタンパク質が細胞膜上へ局在するか否かは、該改変HNタンパク質を発現することが出来る発現ベクターにより、哺乳動物細胞(例えばサル腎臓細胞LLCMK2)をトランスフェクトし、得られたトランスフェクタントの細胞膜や各オルガネラにおける改変HNタンパク質の発現を、改変HNタンパク質を特異的に認識する抗体を用いたウェスタンブロットにより解析することにより、確認することが出来る。
【0017】
また、本発明の改変HNタンパク質は、HVJのウイルス粒子形成の際に、エンベロープへ取り込まれることにより、該エンベロープを構成し得る(これを、HVJ−Eを構成する活性という)。即ち、本発明の改変HNタンパク質を哺乳動物細胞において発現させ、該哺乳動物細胞にHVJを感染させると、HVJ感染細胞から生じるHVJのエンベロープに、該改変タンパク質が取り込まれ、該エンベロープを構成する。改変HNタンパク質が、かかる特徴を有するか否かは、該改変HNタンパク質を発現することが出来る発現ベクターにより、哺乳動物細胞(例えばサル腎臓細胞LLCMK2)をトランスフェクトし、得られたトランスフェクタントにHVJを感染させ、感染したトランスフェクタントから生じたHVJを回収し、該HVJからエンベロープを単離し、該エンベロープにおける改変HNタンパク質の発現を、改変HNタンパク質を特異的に認識する抗体を用いたウェスタンブロットにより解析することにより、確認することが出来る。
【0018】
HVJのHNタンパク質のN末端部分アミノ酸配列と同一のアミノ酸配列とは、配列番号6に示されるアミノ酸配列のアミノ酸番号1番のメチオニンから始まり、アミノ酸番号111番のアミノ酸乃至アミノ酸番号209番のアミノ酸で終了する、配列番号6に示されるアミノ酸配列の部分アミノ酸配列(好ましくは配列番号6に示されるアミノ酸配列のアミノ酸番号1番のメチオニンから始まり、アミノ酸番号140番のアミノ酸乃至アミノ酸番号180番のアミノ酸で終了する、配列番号6に示されるアミノ酸配列の部分アミノ酸配列、より好ましくは、配列番号6に示されるアミノ酸配列のアミノ酸番号1番のメチオニンから始まり、アミノ酸番号160番のアミノ酸で終了する、配列番号6に示されるアミノ酸配列の部分アミノ酸配列)と完全同一のアミノ酸配列が挙げられる。
【0019】
HVJのHNタンパク質のN末端部分アミノ酸配列と実質的に同一のアミノ酸配列とは、上述のHVJのHNタンパク質のN末端部分アミノ酸配列と同一のアミノ酸配列と約80%以上、好ましくは約90%以上、さらに好ましくは約95%以上、特に好ましくは約97%以上の相同性を有するアミノ酸配列などが挙げられる。ここで「相同性」とは、当該技術分野において公知の数学的アルゴリズムを用いて2つのアミノ酸配列をアラインさせた場合の、最適なアラインメント(好ましくは、該アルゴリズムは最適なアラインメントのために配列の一方もしくは両方へのギャップの導入を考慮し得るものである)における、オーバーラップする全アミノ酸残基に対する同一アミノ酸および類似アミノ酸残基の割合(%)を意味する。「類似アミノ酸」とは物理化学的性質において類似したアミノ酸を意味し、例えば、芳香族アミノ酸(Phe、Trp、Tyr)、脂肪族アミノ酸(Ala、Leu、Ile、Val)、極性アミノ酸(Gln、Asn)、塩基性アミノ酸(Lys、Arg、His)、酸性アミノ酸(Glu、Asp)、水酸基を有するアミノ酸(Ser、Thr)、側鎖の小さいアミノ酸(Gly、Ala、Ser、Thr、Met)などの同じグループに分類されるアミノ酸同士が挙げられる。このような類似アミノ酸による置換は蛋白質の表現型に変化をもたらさない(即ち、保存的アミノ酸置換である)ことが予測される。保存的アミノ酸置換の具体例は当該技術分野で周知であり、種々の文献に記載されている(例えば、Bowieら,Science, 247: 1306−1310 (1990)を参照)。アミノ酸(配列)が置換、欠失または挿入されている場合、その置換、欠失または挿入の位置は、上述の、細胞膜へ局在する活性、及びHVJ−Eを構成する活性が実質的に保持される限り、特に限定されない。「実質的に保持される」とは、少なくともネイティブなHNタンパク質(全長HNタンパク質)と比較して、有意に高い細胞膜へ局在する活性を保持していることを意味する。
【0020】
本明細書におけるアミノ酸配列の相同性は、相同性計算アルゴリズムNCBI BLAST(National Center for Biotechnology Information Basic Local Alignment Search Tool)を用い、以下の条件(期待値=10;ギャップを許す;マトリクス=BLOSUM62;フィルタリング=OFF)にて計算することができる。アミノ酸配列の相同性を決定するための他のアルゴリズムとしては、例えば、Karlinら, Proc. Natl. Acad. Sci. USA, 90: 5873−5877 (1993)に記載のアルゴリズム[該アルゴリズムはNBLASTおよびXBLASTプログラム(version 2.0)に組み込まれている(Altschulら, Nucleic Acids Res., 25: 3389−3402 (1997))]、Needlemanら, J. Mol. Biol., 48: 444−453 (1970)に記載のアルゴリズム[該アルゴリズムはGCGソフトウェアパッケージ中のGAPプログラムに組み込まれている]、MyersおよびMiller, CABIOS, 4: 11−17 (1988)に記載のアルゴリズム[該アルゴリズムはCGC配列アラインメントソフトウェアパッケージの一部であるALIGNプログラム(version 2.0)に組み込まれている]、Pearsonら, Proc. Natl. Acad. Sci. USA, 85: 2444−2448 (1988)に記載のアルゴリズム[該アルゴリズムはGCGソフトウェアパッケージ中のFASTAプログラムに組み込まれている]等が挙げられ、それらも同様に好ましく用いられ得る。
【0021】
より好ましくは、HVJのHNタンパク質のN末端部分アミノ酸配列と実質的に同一のアミノ酸配列とは、上述のHVJのHNタンパク質のN末端部分アミノ酸配列と同一のアミノ酸配列と約80%以上、好ましくは約90%以上、さらに好ましくは約95%以上、特に好ましくは約97%以上の同一性を有するアミノ酸配列である。
【0022】
HVJのHNタンパク質のN末端部分アミノ酸配列と実質的に同一のアミノ酸配列からなる改変HNタンパク質は、前記の配列番号6に示されるアミノ酸配列のアミノ酸番号1番のメチオニンから始まり、アミノ酸番号111番のアミノ酸乃至アミノ酸番号209番のアミノ酸で終了する、配列番号6に示されるアミノ酸配列の部分アミノ酸配列(好ましくは配列番号6に示されるアミノ酸配列のアミノ酸番号1番のメチオニンから始まり、アミノ酸番号140番のアミノ酸乃至アミノ酸番号180番のアミノ酸で終了する、配列番号6に示されるアミノ酸配列の部分アミノ酸配列、より好ましくは、配列番号6に示されるアミノ酸配列のアミノ酸番号1番のメチオニンから始まり、アミノ酸番号160番のアミノ酸で終了する、配列番号6に示されるアミノ酸配列の部分アミノ酸配列)と実質的に同一のアミノ酸配列を含み、且つ配列番号6の第1〜160アミノ酸に該当するアミノ酸配列からなるポリペプチドと実質的に同質の活性を有する蛋白質を意味する。
「実質的に同質の活性」における「活性」は、上述の「細胞膜へ局在する活性」、及び「HVJ−Eを構成する活性」を意味する。「実質的に同質」とは、それらの活性が定性的に同じであることを意味する。したがって、上記の各活性は同等(例えば、約0.5〜約2倍)であることが好ましいが、これらの活性の程度などの量的要素は異なっていてもよい。
【0023】
また、HVJのHNタンパク質のN末端部分アミノ酸配列と実質的に同一のアミノ酸配列には、例えば、(1)上述のHVJのHNタンパク質のN末端部分アミノ酸配列と同一のアミノ酸配列のうち1または2個以上(好ましくは1〜30個程度、より好ましくは1〜10個程度、いっそう好ましくは1〜数(2、3、4もしくは5)個)のアミノ酸が欠失したアミノ酸配列、(2)上述のHVJのHNタンパク質のN末端部分アミノ酸配列と同一のアミノ酸配列に1または2個以上(好ましくは1〜30個程度、より好ましくは1〜10個程度、いっそう好ましくは1〜数(2、3、4もしくは5)個)のアミノ酸が付加したアミノ酸配列、(3)上述のHVJのHNタンパク質のN末端部分アミノ酸配列と同一のアミノ酸配列に1または2個以上(好ましくは1〜30個程度、より好ましくは1〜10個程度、いっそう好ましくは1〜数(2、3、4もしくは5)個)のアミノ酸が挿入されたアミノ酸配列、(4)上述のHVJのHNタンパク質のN末端部分アミノ酸配列と同一のアミノ酸配列のうち1または2個以上(好ましくは1〜30個程度、より好ましくは1〜10個程度、いっそう好ましくは1〜数(2、3、4もしくは5)個)のアミノ酸が他のアミノ酸で置換されたアミノ酸配列、または(5)それらを組み合わせたアミノ酸配列を含有する蛋白質なども含まれる。
【0024】
本発明の最も好ましい改変HNタンパク質は、下記(1)または(2)のポリペプチドである:
(1)配列番号6の第1〜160アミノ酸に該当するアミノ酸配列からなるポリペプチド、
(2)配列番号6の第1〜160アミノ酸に該当するアミノ酸配列において1または複数の残基が置換、欠失又は付加されたアミノ酸配列を含み、且つHVJに発現された場合に粒子表面に局在しうる特徴を有するポリペプチド。
【0025】
本発明の改変HNタンパク質は、例えば下記の融合タンパク質の構築や、下記の融合タンパク質の機能評価におけるネガティブコントロール等として有用である。
【0026】
本発明はさらに、前記改変HNタンパク質の残基及び第1の機能性分子を含む改変HNタンパク質であって、該改変HNタンパク質残基のC末端側に該機能性分子が機能可能に連結されている、改変HNタンパク質を提供する。ここで、「連結」とは、機能性分子が、改変HNタンパク質残基のC末端側に繋がっている状態を達成するものであれば、いかなる結合様式であっても特に限定されないが、好ましくはペプチド結合によって、改変HNタンパク質残基と機能性分子とが連結している。また、「機能可能に連結されている」とは、機能性分子がその機能を発揮できるように連結されていることを意味する。従って、機能性分子がその機能を発揮できる限りは、例えば、連結部位にリンカーやタグを介して改変HNタンパク質の残基と機能性分子が連結されていてもよい。本明細書における機能性分子とは、何らかの生物学的機能または活性を有している、ポリペプチド(例えば、タンパク質、ペプチド)、低分子化合物も含む、あらゆる分子を包含しうる。好ましくは、ポリペプチド(例えば、タンパク質、ペプチド)またはその類似体である。従って好ましくは、本発明は、前記改変HNタンパク質の残基及び第1の機能性ポリペプチド残基を含む融合タンパク質であって、該改変HNタンパク質残基のC末端側に該機能性ポリペプチド残基が機能可能に連結されている、融合タンパク質を提供する。該融合タンパク質においては、前記改変HNタンパク質の残基と第1の機能性ポリペプチド残基とが、結合手又はペプチドリンカーを介してペプチド結合により結合し、1本のポリペプチドを構成している。
【0027】
ペプチドリンカーのアミノ酸配列は、改変HNタンパク質の残基と第1の機能性ポリペプチド残基との機能的な連結を可能にする限り特に限定されない。ペプチドリンカーの長さは、改変HNタンパク質の残基と第1の機能性ポリペプチド残基との機能的な連結を可能にする限り特に限定されないが、通常1〜30アミノ酸、好ましくは1〜10アミノ酸である。
【0028】
本発明の融合タンパク質は、哺乳動物細胞(例えばサル腎臓細胞LLCMK2)において発現したときに、該細胞の細胞膜上に局在する(これを、細胞膜へ局在する活性という)。「局在」とは、細胞内の他の領域よりも細胞膜における発現量が高いことを意味する。融合タンパク質が細胞膜上へ局在するか否かは、該融合タンパク質を発現することが出来る発現ベクターにより、哺乳動物細胞(例えばサル腎臓細胞LLCMK2)をトランスフェクトし、得られたトランスフェクタントの細胞膜や各オルガネラにおける融合タンパク質の発現を、融合タンパク質を特異的に認識する抗体を用いたウェスタンブロットにより解析することにより、確認することが出来る。
【0029】
更に、本発明の融合タンパク質を哺乳動物細胞において発現すると、該融合タンパク質に含まれる機能性ポリペプチド領域が、細胞の外側に提示する形で、該哺乳動物細胞の細胞膜上に局在する。機能性ポリペプチド領域が、細胞の外側に提示されているか否かは、該機能性ポリペプチドを特異的に認識する抗体を用いて、フローサイトメーターや免疫組織染色により確認することが出来る。
【0030】
また、本発明の融合タンパク質は、HVJのウイルス粒子形成の際に、エンベロープへ取り込まれることにより、該エンベロープを構成し得る(これを、HVJ−Eを構成する活性という)。即ち、本発明の融合タンパク質を哺乳動物細胞において発現させ、該哺乳動物細胞にHVJを感染させると、HVJ感染細胞から生じるHVJのエンベロープに、該融合タンパク質が取り込まれ、該エンベロープを構成する。融合タンパク質が、かかる特徴を有するか否かは、該融合タンパク質を発現することが出来る発現ベクターにより、哺乳動物細胞(例えばサル腎臓細胞LLCMK2)をトランスフェクトし、得られたトランスフェクタントにHVJを感染させ、感染したトランスフェクタントから生じたHVJを回収し、該HVJからエンベロープを単離し、該エンベロープにおける融合タンパク質の発現を、融合タンパク質を特異的に認識する抗体を用いたウェスタンブロットにより解析することにより、確認することが出来る。
【0031】
更に、本発明の融合タンパク質が、HVJ−Eへ取り込まれると、該HVJ−Eは、該融合タンパク質に含まれる機能性ポリペプチド領域を、HVJ−Eの外側に提示する形で、該融合タンパク質を担持することとなる。機能性ポリペプチド領域が、HVJ−Eの外側に提示されているか否かは、該機能性ポリペプチドを特異的に認識する抗体を用いて、フローサイトメーターや免疫組織染色により確認することが出来る。
【0032】
本明細書における機能性ポリペプチドとは、何らかの生物学的機能または活性を有しているポリペプチドを言う。好ましくは、タンパク質、ペプチドまたはその類似体である。例えば、何らかの抗体またはその抗原結合部位、各種のサイトカイン、増殖因子、ホルモン、受容体または細胞膜タンパク質に対するリガンドなどが挙げられる。例えば、この機能性ポリペプチドとして細胞特異的表面抗原に対する抗体を用いた場合、本発明の融合タンパク質をHVJ−Eに取り込ませると、該HVJ−Eの外側に、該抗体が提示されるので、その抗原を表面に発現する細胞に選択的にHVJ−Eをターゲティングさせることが可能となる。また、機能性ポリペプチドとして何らかのサイトカインを用いた場合は、本発明の融合タンパク質をHVJ−Eに取り込ませると、該HVJ−Eの外側に、該サイトカインが提示されるので、そのサイトカイン受容体を細胞表面に発現する細胞特異的にHVJ−Eを選択的に作用させるとともに、該サイトカインで該細胞を刺激することも可能である。例えば、抗腫瘍活性を有するタンパク質(例えば、IL−12等のIFN−γ分泌促進能を有するポリペプチド)を機能性ポリペプチドとして用いた場合、本発明の融合タンパク質をHVJ−Eに取り込ませると、該HVJ−Eの外側に、抗腫瘍活性を有するタンパク質が提示されるので、HVJ−Eによる抗腫瘍活性に加えて、この機能性ポリペプチドの有する抗腫瘍活性も発揮されることで、より強力な抗腫瘍効果を得ることが可能である。
【0033】
本発明で用いる機能性ポリペプチドの好ましい一実施態様としては、IFN−γ分泌促進能を有する分子が挙げられる。このIFN−γ分泌促進能は、当業者に公知の方法および実施例に記載の方法で測定することが可能であるとともに、これらの方法に当業者が日常的な活動の範囲内にある改変を加えて測定してもよい。IFN−γ分泌促進能を有するポリペプチドとしては、哺乳動物(例、ヒト)の脾細胞または樹状細胞からのIFN−γ分泌促進能を有していればどのような分子であってもよいが、例えば、哺乳動物のIL−12が挙げられる。IL−12は、2つの異なるサブユニット(p35およびp45)から構成されるヘテロダイマーであってもよく、該2つのサブユニットが機能可能に連結された一本鎖IL−12であってもよい。好ましくは、一本鎖IL−12である。一本鎖IL−12を構築する技術は、例えばGraham J. Lieschke, et al., Nature Biotechnology. 15. 35−40 (1997)等に記載されており、当業者であればこれを容易に製造することが出来る。本発明において用いられる一本鎖IL−12は、IL−12の2つのサブユニットp35およびp40を一本のポリペプチドとして連結させたもので、好ましくはp40のC末端側にp35が機能可能に連結されている。p35とp40との間にはペプチドリンカーを含んでいても良く、好ましくは、Gly−Gly−Gly−Serからなるリンカーを1〜5単位、より好ましくは3単位含んでいる。そのような一本鎖IL−12としては、具体的には、下記(1)または(2)のポリペプチド:
(1)配列番号2の第103〜625アミノ酸に該当するアミノ酸配列を含むポリペプチド、
(2)配列番号2の第103〜625アミノ酸に該当するアミノ酸配列において1または複数の残基が置換、欠失又は付加されたアミノ酸配列を含み、且つIFN−γ分泌促進能を有するポリペプチドがあげられる。
【0034】
(2)の一本鎖IL−12のポリペプチドには、(1)配列番号2の第103〜625アミノ酸に該当するアミノ酸配列のうち1または2個以上(好ましくは1〜30個程度、より好ましくは1〜10個程度、いっそう好ましくは1〜数(2、3、4もしくは5)個)のアミノ酸が欠失したアミノ酸配列、(2)配列番号2の第103〜625アミノ酸に該当するアミノ酸配列に1または2個以上(好ましくは1〜30個程度、より好ましくは1〜10個程度、いっそう好ましくは1〜数(2、3、4もしくは5)個)のアミノ酸が付加したアミノ酸配列、(3)配列番号2の第103〜625アミノ酸に該当するアミノ酸配列に1または2個以上(好ましくは1〜30個程度、より好ましくは1〜10個程度、いっそう好ましくは1〜数(2、3、4もしくは5)個)のアミノ酸が挿入されたアミノ酸配列、(4)配列番号2の第103〜625アミノ酸に該当するアミノ酸配列のうち1または2個以上(好ましくは1〜30個程度、より好ましくは1〜10個程度、いっそう好ましくは1〜数(2、3、4もしくは5)個)のアミノ酸が他のアミノ酸で置換されたアミノ酸配列、または(5)それらを組み合わせたアミノ酸配列を含み、且つ哺乳動物(例、ヒト)の脾細胞または樹状細胞からのIFN−γ分泌促進能を有するポリペプチドが含まれる。
【0035】
尚、本発明の配列番号2で表されるアミノ酸配列は、アミノ酸番号1〜24:シグナルペプチド(p40)、アミノ酸番号25〜41:Flag−tag HA−tag、アミノ酸番号42〜99:ZZ−ドメイン、アミノ酸番号100〜102:リンカー、アミノ酸番号103〜417:IL−12−p40サブユニット、アミノ酸番号418〜432:リンカー、アミノ酸番号433〜625:IL−12−p35サブユニットで構成されている。
【0036】
本発明で用いる機能性ポリペプチドの別の好ましい一実施態様としては、抗体の定常領域(Fc)のCH2−CH3ドメインが挙げられる。このCH2−CH3ドメインは、プロテインAのZZドメインとの結合活性を有する限り、どのイムノグロブリンクラス(例えば、IgG、IgM、IgA、IgD、IgE)のものに由来してもよく、さらにどのサブクラス(例えば、IgG1、IgG2、IgG3、IgG4、IgA1、IgA2)のものに由来してもよく、また各CH2−CH3ドメインのアミノ酸配列に1又は複数個のアミノ酸が欠失・置換・付加されたものであってよい。また、CH2−CH3ドメインがプロテインAのZZドメインとの結合活性を有する限り、当該イムノグロブリンはどの動物種(マウス、ラット、ヒト等の哺乳動物、鳥類等)に由来してもよい。好ましい例としては、IgG2(たとえば、マウスIgG2)のCH2−CH3ドメインが挙げられる。そのような抗体の定常領域(Fc)のCH2−CH3ドメインとしては、具体的には、下記(1)または(2)のポリペプチドが挙げられる:
(1)配列番号4の第171〜389アミノ酸に該当するアミノ酸配列を含むポリペプチド、
(2)配列番号4の第171〜389アミノ酸に該当するアミノ酸配列において1または複数の残基が置換、欠失又は付加されたアミノ酸配列を含み、且つプロテインAのZZドメインとの結合活性を有するポリペプチド。
【0037】
(2)の一本鎖IL−12のポリペプチドには、(1) 配列番号4の第171〜389アミノ酸に該当するアミノ酸配列のうち1または2個以上(好ましくは1〜30個程度、より好ましくは1〜10個程度、いっそう好ましくは1〜数(2、3、4もしくは5)個)のアミノ酸が欠失したアミノ酸配列、(2) 配列番号4の第171〜389アミノ酸に該当するアミノ酸配列に1または2個以上(好ましくは1〜30個程度、より好ましくは1〜10個程度、いっそう好ましくは1〜数(2、3、4もしくは5)個)のアミノ酸が付加したアミノ酸配列、(3) 配列番号4の第171〜389アミノ酸に該当するアミノ酸配列に1または2個以上(好ましくは1〜30個程度、より好ましくは1〜10個程度、いっそう好ましくは1〜数(2、3、4もしくは5)個)のアミノ酸が挿入されたアミノ酸配列、(4) 配列番号4の第171〜389アミノ酸に該当するアミノ酸配列のうち1または2個以上(好ましくは1〜30個程度、より好ましくは1〜10個程度、いっそう好ましくは1〜数(2、3、4もしくは5)個)のアミノ酸が他のアミノ酸で置換されたアミノ酸配列、または(5)それらを組み合わせたアミノ酸配列を含み、且つプロテインAのZZドメインとの結合活性を有するポリペプチドが含まれる。
【0038】
尚、本発明の配列番号4で表されるアミノ酸配列は、アミノ酸番号1〜160:配列番号6の第1〜160アミノ酸配列、アミノ酸番号161〜170:Myc tag、アミノ酸番号171〜389:IgG2a−CH2−CH3ドメインで構成されている。
【0039】
ここで、上記の抗体の定常領域(Fc)のCH2−CH3ドメインは、プロテインAのZZドメインと結合することができる。従って、CH2−CH3ドメインを含む本発明の融合タンパク質(「第1の融合タンパク質」と表記することもある)を発現することが出来る発現ベクターにより、哺乳動物細胞(例えばサル腎臓細胞LLCMK2)をトランスフェクトし、得られたトランスフェクタントにHVJを感染させ、感染したトランスフェクタントから生じたHVJを回収し、該HVJからエンベロープを単離することにより、CH2−CH3ドメインをHVJのエンベロープの外側に提示する形で担持するHVJ−Eを得ることが出来る。更に、ZZドメインと機能可能に連結された前記の機能性ポリペプチドを含む融合タンパク質(「第2の融合タンパク質」と表記することもある)を、HVJ−E上の第1の融合タンパク質へZZドメインを介して結合することにより、所望の機能性ポリペプチドをHVJのエンベロープの外側に提示する形で担持するHVJ−Eを提供することができる。IgG2由来のCH2−CH3ドメインを用いた場合、第2の融合タンパク質が結合したHVJ−Eの血中安定性が高いため、体内に投与した場合でも血中に第2の融合タンパク質が結合したHVJ−Eが長時間存在し、第2の融合タンパク質に含まれる機能性ポリペプチドの機能を発揮するのに有利である。ただし、この他のサブクラスのイムノグロブリンに由来するCH2−CH3ドメインもまた、HVJ−Eと機能性ポリペプチドとの複合体の使用目的に応じて、当業者が選択することができる。
【0040】
従って、本発明は、上記の第1の融合タンパク質、及びプロテインAのZZドメイン及び第2の機能性ポリペプチド残基を含む第2の融合タンパク質を含む、タンパク質複合体であって、第1の融合タンパク質と第2の融合タンパク質とが、第1の融合タンパク質に含まれるCH2−CH3ドメインと第2の融合タンパク質に含まれるZZドメインとの間の結合を介して複合体を形成している、タンパク質複合体を提供する。
【0041】
本発明の第2の融合タンパク質に含まれる機能性ポリペプチドの好ましい一実施態様としては、前記のIFN−γ分泌促進能を有する分子が挙げられる。従って、そのような分子としては、IL−12が挙げられ、好ましくは、一本鎖IL−12が挙げられる。本発明のタンパク質複合体としては、具体的には、一本鎖IL−12が、下記(1)または(2)のポリペプチドであり、且つZZドメインが、下記(3)または(4)のポリペプチドである、タンパク質複合体を挙げることができる:
(1)配列番号2の第103〜625アミノ酸に該当するアミノ酸配列を含むポリペプチド、
(2)配列番号2の第103〜625アミノ酸に該当するアミノ酸配列において1または複数の残基が置換、欠失又は付加されたアミノ酸配列を含み、且つIFN−γ分泌促進能を有するポリペプチド、
(3)配列番号2の第42〜99アミノ酸に該当するアミノ酸配列を含むポリペプチド、
(4)配列番号2の第42〜99アミノ酸に該当するアミノ酸配列において1または複数の残基が欠失、置換または付加されたアミノ酸配列を含み、且つ抗体分子のFc領域への結合活性を有するポリペプチド。
【0042】
さらに、本発明は上記の本発明の改変HNタンパク質又は融合タンパク質をコードする核酸を提供する。該核酸は、本発明の改変HNタンパク質や融合タンパク質を遺伝子工学的手法により製造するために有用である。核酸はDNA又はRNAである。また、核酸は1本鎖又は2本鎖のいずれであってもよい。
【0043】
好ましい一実施態様としては、本発明の改変HNタンパク質をコードする核酸として、配列番号5に示される塩基配列の最も短い範囲で1番目〜333番目の塩基配列であって、最も長い範囲で1番目〜627番目の塩基配列を含有するDNA、または配列番号5に示される塩基配列の最も短い範囲で1番目〜333番目の塩基配列であって、最も長い範囲で1番目〜627番目の塩基配列の相補鎖配列とストリンジェントな条件下でハイブリダイズする塩基配列を含有し、前記したHNタンパク質と実質的に同質の活性を有する蛋白質をコードするDNAなどが挙げられる。より好ましくは、本発明の改変HNタンパク質をコードする核酸として、配列番号5に示される塩基配列の最も短い範囲で1番目〜420番目の塩基配列であって、最も長い範囲で1番目〜540番目の塩基配列を含有するDNA、または配列番号5に示される塩基配列の最も短い範囲で1番目〜420番目の塩基配列であって、最も長い範囲で1番目〜540番目の塩基配列の相補鎖配列とストリンジェントな条件下でハイブリダイズする塩基配列を含有し、前記したHNタンパク質と実質的に同質の活性を有する蛋白質をコードするDNA、最も好ましくは、配列番号5に示される塩基配列の1〜480までの塩基配列を含有するDNA、または配列番号5に示される塩基配列の1〜480までの塩基配列の相補鎖配列とストリンジェントな条件下でハイブリダイズする塩基配列を含有し、前記したHNタンパク質と実質的に同質の活性を有する蛋白質をコードするDNAである。
【0044】
別の好ましい一実施態様としては、配列番号6に表されるアミノ酸配列の配列番号1〜160のポリペプチドに一本鎖IL−12ポリペプチドが機能可能に連結されたタンパク質をコードする核酸を提供する。配列番号6に表されるアミノ酸配列の配列番号1〜160のポリペプチドをコードするDNAとしては、例えば、配列番号5に示される塩基配列の1〜480までの塩基配列を含有するDNA、または配列番号5に示される塩基配列の1〜480までの塩基配列の相補鎖配列とストリンジェントな条件下でハイブリダイズする塩基配列を含有し、前記したHNタンパク質と実質的に同質の活性を有する蛋白質をコードするDNAなどが挙げられる。また、一本鎖IL−12ポリペプチドをコードするDNAとしては、例えば、配列番号1に示される塩基配列の307〜1875までの塩基配列を含有するDNA、または配列番号1に示される塩基配列の307〜1875までの塩基配列の相補鎖配列とストリンジェントな条件下でハイブリダイズする塩基配列を含有し、前記したIL−12タンパク質と実質的に同質の活性を有する蛋白質をコードするDNAなどが挙げられる。
別の好ましい一実施態様としては、配列番号6に表されるアミノ酸配列の配列番号1〜160のポリペプチドにIgG2aのCH2−CH3ドメインのポリペプチドが機能可能に連結されたタンパク質をコードする核酸を提供する。配列番号6に表されるアミノ酸配列の配列番号1〜160のポリペプチドをコードするDNAとしては前記の通りである。また、IgG2aのCH2−CH3ドメインポリペプチドをコードするDNAとしては、例えば、配列番号3に示される塩基配列の511〜1167までの塩基配列を含有するDNA、または配列番号3に示される塩基配列の511〜1167までの塩基配列の相補鎖配列とストリンジェントな条件下でハイブリダイズする塩基配列を含有し、前記したCH2−CH3ドメインと実質的に同質の活性を有する蛋白質をコードするDNAなどが挙げられる。
前記の塩基配列とストリンジェントな条件下でハイブリダイズできるDNAとしては、例えば、配列番号5(または配列番号1または3)に示される塩基配列と約70%以上、好ましくは約80%以上、さらに好ましくは約90%以上、特に好ましくは約95%以上の相同性を有する塩基配列を含有するDNAなどが用いられる。
本明細書における塩基配列の相同性は、相同性計算アルゴリズムNCBI BLAST(National Center for Biotechnology Information Basic Local Alignment Search Tool)を用い、以下の条件(期待値=10;ギャップを許す;フィルタリング=ON;マッチスコア=1;ミスマッチスコア=−3)にて計算することができる。塩基配列の相同性を決定するための他のアルゴリズムとしては、上記したアミノ酸配列の相同性計算アルゴリズムが同様に好ましく例示される。
【0045】
ハイブリダイゼーションは、自体公知の方法あるいはそれに準じる方法、例えば、モレキュラー・クローニング(Molecular Cloning)第2版(J. Sambrook et al., Cold Spring Harbor Lab. Press, 1989)に記載の方法などに従って行なうことができる。また、市販のライブラリーを使用する場合、ハイブリダイゼーションは、添付の使用説明書に記載の方法に従って行なうことができる。ハイブリダイゼーションは、好ましくは、ストリンジェントな条件に従って行なうことができる。
ハイストリンジェントな条件としては、例えば、6×SSC(sodium chloride/sodium citrate)中45℃でのハイブリダイゼーション反応の後、0.2×SSC/0.1% SDS中65℃での一回以上の洗浄などが挙げられる。当業者は、ハイブリダイゼーション溶液の塩濃度、ハイブリダゼーション反応の温度、プローブ濃度、プローブの長さ、ミスマッチの数、ハイブリダイゼーション反応の時間、洗浄液の塩濃度、洗浄の温度等を適宜変更することにより、所望のストリンジェンシーに容易に調節することができる。
【0046】
配列番号6に表されるアミノ酸配列の配列番号1〜160のポリペプチド、一本鎖IL−12ポリペプチドおよびIgG2aのCH2−CH3ドメインのポリペプチドをコードするDNAは、化学的にDNA鎖を合成するか、もしくは合成した一部オーバーラップするオリゴDNA短鎖を、PCR法を利用して接続することにより、配列番号6に表されるアミノ酸配列の配列番号1〜160のポリペプチドおよび一本鎖IL−12ポリペプチドの全長をコードするDNAまたは配列番号6に表されるアミノ酸配列の配列番号1〜160のポリペプチドおよびIgG2aのCH2−CH3ドメインのポリペプチドの全長をコードするDNAを構築することも可能である。
【0047】
上記のようにしてクローン化されたDNAは、目的によりそのまま、または所望により制限酵素で消化するか、リンカーを付加した後に、使用することができる。配列番号6に表されるアミノ酸配列の配列番号1〜160のポリペプチドをコードするDNA、一本鎖IL−12ポリペプチドをコードするDNA、IgG2aのCH2−CH3ドメインのポリペプチドをコードするDNAを、別個にクローン化した場合には、適当な制限酵素、リンカー、リガーゼ等を用いて、それらを配列番号6に表されるアミノ酸配列の配列番号1〜160のポリペプチド、一本鎖IL−12ポリペプチドの順序または配列番号6に表されるアミノ酸配列の配列番号1〜160のポリペプチド、IgG2aのCH2−CH3ドメインのポリペプチドの順序で連結する。
【0048】
次いで、得られた前記DNAは、必要に応じて、その5’末端側に翻訳開始コドンとしてのATGを、また3’末端側には翻訳終止コドンとしてのTAA、TGAまたはTAGをそれぞれ導入し、制限酵素およびDNAリガーゼを用いて、適当な発現ベクター中のプロモーターの下流に連結される。翻訳開始コドンや翻訳終止コドンは、適当な合成DNAアダプターを用いて付加することができる。
【0049】
本発明の核酸を、発現ベクター内のプロモーターの制御下に組み込むことにより、本発明の改変HNタンパク質又は融合タンパク質を発現し得る発現ベクター(本発明の発現ベクター)を製造することが出来る。
【0050】
発現ベクターとしては、動物細胞発現プラスミド(例:pCAGGS 、pA1−11、pXT1、pRc/CMV、pRc/RSV、pcDNAI/Neo);レトロウイルス、ワクシニアウイルス、アデノウイルスなどの動物ウイルスベクターなどが用いられる。
プロモーターとしては、遺伝子の発現に用いる宿主の細胞内で機能可能なプロモーターであれば、いかなるものでもよい。
例えば、宿主が動物細胞である場合、CAGプロモーター、β−アクチンプロモーター、SRαプロモーター、SV40プロモーター、LTRプロモーター、CMV(サイトメガロウイルス)プロモーター、RSV(ラウス肉腫ウイルス)プロモーター、MoMuLV(モロニーマウス白血病ウイルス)LTR、HSV−TK(単純ヘルペスウイルスチミジンキナーゼ)プロモーターなどが用いられる。
【0051】
発現ベクターとしては、上記の他に、所望によりエンハンサー、スプライシングシグナル、ポリA付加シグナル、選択マーカー、SV40複製起点(以下、SV40 oriと略称する場合がある)などを含有しているものを用いることができる。選択マーカーとしては、例えば、ジヒドロ葉酸還元酵素遺伝子(以下、dhfrと略称する場合がある、メソトレキセート(MTX)耐性)、アンピシリン耐性遺伝子(以下、amprと略称する場合がある)、ネオマイシン耐性遺伝子(以下、neorと略称する場合がある、G418耐性)等が挙げられる。特に、dhfr遺伝子欠損チャイニーズハムスター細胞を用い、dhfr遺伝子を選択マーカーとして使用する場合、チミジンを含まない培地によって目的遺伝子を選択することもできる。
【0052】
本発明の発現ベクターで宿主を形質転換し、得られる形質転換体を培養することによって、本発明の改変タンパク質又は本発明の融合タンパク質を製造することができる。
宿主としては、例えば、動物細胞、動物などの分泌発現に適したものが用いられる。
【0053】
動物細胞としては、例えば、COS−7、Vero、CHO、CHO(dhfr−)、CHO−K1、L、AtT−20、GH3、FL、HEK293、NIH3T3、Balb3T3、FM3A、L929、SP2/0、P3U1、B16、P388、LLCMK2などの細胞が用いられる。
動物としては、トランスジェニック系の確立した哺乳動物(例えば、マウス、ラット、ウサギ、ヒツジ、ブタ、ウシ)もしくはニワトリなどが挙げられる。
【0054】
形質転換は、宿主の種類に応じ、公知の方法に従って実施することができる。
動物細胞は、例えば、細胞工学別冊8 新細胞工学実験プロトコール,263−267 (1995)(秀潤社発行)、Virology,52巻,456 (1973)に記載の方法に従って形質転換することができる。
動物は、例えば、トランスジェニック動物の開発(シーエムシー出版),(2001)に記載の方法に従って形質転換することができる。
【0055】
形質転換体の培養は、宿主の種類に応じ、公知の方法に従って実施することができる。
宿主が動物細胞である形質転換体を培養する場合の培地としては、例えば、約5〜約20%の胎児ウシ血清を含む最小必須培地(MEM),ダルベッコ改変イーグル培地(DMEM),RPMI 1640培地,199培地などが用いられる。培地のpHは、好ましくは約6〜約8である。培養は、通常約30〜約40℃で、約15〜約60時間行なわれる。必要に応じて通気や撹拌を行ってもよい。
宿主が動物である場合、遺伝子導入した受精卵から、常法に従ってトランスジェニック動物を得、通常の飼育条件下で飼育、哺乳動物の乳やニワトリの卵を採取すればよい。
【0056】
このようにして得られた本発明の改変タンパク質又は融合タンパク質を発現する細胞から、適切なタンパク質精製法(例えば、HPLC、アフィニティクロマトグラフィ等)を用いて、本発明の改変タンパク質又は融合タンパク質を精製することにより、単離又は精製された本発明の改変タンパク質又は融合タンパク質を得ることが出来る。単離又は精製とは、目的とする成分以外の成分を除去する操作がなされていることを意味する。哺乳動物細胞中では、本発明の改変タンパク質又は融合タンパク質は、細胞膜へ局在するので、該細胞を界面活性剤で処理し、この溶解液を精製操作に付すことにより、単離又は精製された本発明の改変タンパク質又は融合タンパク質を得ることが出来る。単離又は精製された本発明の改変タンパク質又は融合タンパク質の純度は、タンパク質純度として、通常50%(w/w)以上、好ましくは80%(w/w)以上、より好ましくは90%(w/w)以上、最も好ましくは実質的に100%である。
【0057】
上記のようにして得られた、本発明の改変HNタンパク質又は融合タンパク質を発現するトランスフェクタントへHVJを感染させ、感染したトランスフェクタントから生じたHVJを回収し、該HVJからエンベロープを単離することにより、本発明の改変HNタンパク質又は融合タンパク質をエンベロープに含有する、HVJを製造することが出来る。ここで、本発明の融合タンパク質が、HVJのエンベロープへ取り込まれると、該HVJは、該融合タンパク質に含まれる機能性ポリペプチド領域を、HVJのエンベロープの外側に提示する形で、該融合タンパク質を担持する。更に、該HVJを紫外線等に曝露することによってゲノムRNAで不活性化することで、HVJ−Eを製造することが出来る。従って、本発明は、本発明の改変HNタンパク質又は融合タンパク質をエンベロープに含有する、HVJ−Eを提供する。
【0058】
本明細書で使用される場合、「HVJ−E」とは、HVJの最も外側に位置する膜状の構造物を意味する。HVJにおいては、エンベロープは、通常、ウイルスゲノム及びカプシドタンパク質を覆っている。HVJ−Eは、通常、その内腔に種々の物質を担持することが可能であり、且つ細胞膜へ融合し、その内腔に含まれる物質を細胞内へ移入する活性を有する。そのため、HVJ−Eの内腔に、所望の物質(遺伝子(例えば、DNA、RNAなど)、タンパク質(またはペプチド)、化合物(抗癌剤、抗菌剤、免疫促進剤など)を封入し、これを個体へ投与することにより、種々の組織や細胞の内部へ、所望の物質を送達することが可能である。すなわち、HVJ−Eは種々の物質のベクターとして使用可能である。本発明において使用されるHVJ−Eは、その内腔に、生体内や細胞内への導入が意図される物資を封入していても、封入していなくてもよい。
【0059】
HVJ−Eについてより詳しくは、例えば、特開2001−286282号公報(WO01/57204号公報)、特開2002−065278号公報、WO03/014338号公報等に記載されており、具体的には例えば特開2001−286282号公報の実施例8などに従って調製することができる。
【0060】
本発明に用いられるHVJ−Eは、好ましくは、単離且つ精製されたものである。「単離且つ精製」とは、目的物以外の成分(ウイルスゲノム、カプシドタンパク質等)を除去する操作がなされていることを意味する。
【0061】
本発明は、また、本発明のタンパク質複合体をエンベロープに含有する、HVJ−Eを提供する。ここで、本発明のタンパク質複合体に含有される、本発明の融合タンパク質(第1の融合タンパク質)は、HVJ−Eへ取り込まれており、該融合タンパク質に含まれる機能性ポリペプチド領域(CH2−CH3ドメイン)が、HVJ−Eの外側に提示されている。ここへ、本発明のタンパク質複合体に含有される第2の融合タンパク質が結合することより、結果として、第2の融合タンパク質に含まれる第2の機能性ポリペプチドが、HVJ−Eの外側に提示されることとなる。
【0062】
本発明のタンパク質複合体をHVJのエンベロープに含有させるためには、抗体の定常領域(Fc)のCH2−CH3ドメインを機能性ポリペプチドとして含有する本発明の融合タンパク質(第一の融合タンパク質)をHVJのエンベロープに組み込ませたHVJ−Eを作製し、その後、該HVJ−Eと別途調製したZZドメインに機能可能に連結された機能性ポリペプチドを含む第2の融合タンパク質とを結合させて複合体を形成させる。複合体形成のためには、両分子を含む溶液を調製し、5分以上、好ましくは15分以上、例えば30分または1時間、ゆるやかに攪拌または静置するとよい。この溶液は、生理的緩衝液および生理的食塩水など、タンパク質の変性をもたらさない緩衝液中で行うことが好ましい。複合体の形成は、常法の免疫学的手法で確認することもでき、例えば機能性ポリペプチドが一本鎖IL−12である場合は、ショ糖密度勾配遠心に供してHVJ−EとZZドメインに連結された一本鎖IL−12が同じショ糖密度の画分に回収されることを、免疫学的手法またはIL−12活性の測定により確認することもできる。
【0063】
本発明は、またIFN−γ分泌促進能を有するポリペプチドをエンベロープの外側に提示する形で担持する、HVJ−Eを提供する。IFN−γ分泌促進能を有するポリペプチドの定義は上述の通りであり、該ポリペプチドとしては、哺乳動物のIL−12を挙げることができる。IL−12は、2つの異なるサブユニット(p35およびp45)から構成されるヘテロダイマーであってもよく、該2つのサブユニットが機能可能に連結された一本鎖IL−12であってもよい。好ましくは、一本鎖IL−12である。一本鎖IL−12としては、配列番号1の第103〜625アミノ酸に該当するアミノ酸配列を含むポリペプチドを挙げることが出来る。
【0064】
IFN−γ分泌促進能を有するポリペプチドをエンベロープの外側に提示する形で担持させる方法としては、上述のように、機能性ポリペプチドとしてIFN−γ分泌促進能を有するポリペプチドを含む本発明の融合ポリペプチドや、第2の機能性ポリペプチドとしてIFN−γ分泌促進能を有するポリペプチドを含む本発明のタンパク質複合体を、エンベロープに含有させる方法が好適なものとして挙げられる。但し、これらの方法に特に限定されるものではなく、当業者に公知の方法でIFN−γ分泌促進能を有するポリペプチドをエンベロープの外側に提示する形で担持させることができる。例えば、HVJ−Eの外側をビオチン化し、ストレプトアビジンをコンジュゲートしたIFN−γ分泌促進能を有するポリペプチドを、ビオチン化したHVJ−Eへ結合させる方法等を挙げることができる。
【0065】
本態様のHVJ−Eに含有されるIFN−γ分泌促進能を有するポリペプチドの量は、特に限定されないが、IFN−γ分泌促進能を有するポリペプチドとしてIL−12を用いる場合には、その含有量は、HVJ−E 100HAUあたり、1〜10000pg、好ましくは10〜1000pg、より好ましくは100〜500pg(例えば、250pg)である。本明細書においてHAU(Hemagglutinating unit)とは赤血球を凝集させる活性の単位であり、HVJ−Eの1HAUとはニワトリの赤血球と混合した場合に凝集を起こす最小単位のことをいう。例えば1000HAUとは、目的の検体の1000分の1量をニワトリの赤血球と混合した場合には凝集を起こすが、それ未満では凝集を起こさない量をいう。実際の測定は、公知の方法で行うことが可能であるが、例えば、96ウェルの試験プレートのウェルに2倍希釈系列でHVJ−Eを50μlずつ入れ、そこにニワトリの赤血球懸濁液を50μL添加して計100μLとし、溶液の凝集の有無を目視で確認し、その希釈率を基に計算することができる。
【0066】
IL−12等のIFN−γ分泌促進能を有するポリペプチド自体は抗腫瘍活性を有しており、また、HVJ−Eも抗腫瘍活性を有しているが、IFN−γ分泌促進能を有するポリペプチドをエンベロープの外側に提示する形で担持させることにより、抗腫瘍活性が飛躍的に上昇する。
【0067】
HVJ−Eは、化学療法剤、治療用外来遺伝子等の治療物質をその中に含有することで、標的となる細胞に高効率で治療物質を導入することができることは当業者にとっては公知である。また、本発明者らは、HVJ−E自体に抗腫瘍活性があることを見出している。本発明のHVJ−Eにおいては、機能性分子をその粒子表面に露出されることが可能であり、この機能性分子の作用により、従来公知であった抗腫瘍活性を増強させたり、HVJ−Eを特定の腫瘍へ標的化し得ることを見出した。従って、本発明は、前記本発明のHVJ−Eを有効成分として含む抗癌剤と提供する。本発明の抗癌剤は、あらゆる種類の癌(固形癌、細胞癌など)に有効である。消化器系、泌尿器科系、神経系、結合組織における癌、皮膚癌等あらゆる種類の癌に適用可能であるが、一例として、メラノーマ、前立腺癌、大腸癌などが挙げられる。本発明のHVJ−Eは低毒性であり、そのまま液剤として、または適当な剤型の医薬組成物として、ヒトまたは他の哺乳動物(例、マウス、ラット、ウサギ、ヒツジ、ブタ、ウシ、ネコ、イヌ、サルなど)に対して経口的または非経口的(例、血管内投与、皮下投与など)に投与することができる。
【0068】
本発明のHVJ−Eを癌の治療剤などの医薬として使用する場合、自体公知の方法に従って製剤化し、投与することができる。本発明のHVJ−Eは常套手段に従って製剤化することができる。該HVJ−Eは、そのままで、あるいは摂取促進のための補助剤とともに、遺伝子銃やハイドロゲルカテーテルのようなカテーテルによって投与することができる。あるいは、エアロゾル化して吸入剤として気管内に局所投与することもできる。
【0069】
本発明のHVJ−Eは、それ自体を投与してもよいし、または適当な医薬組成物として投与してもよい。投与に用いられる医薬組成物としては、本発明のHVJ−Eと薬理学的に許容され得る担体、希釈剤もしくは賦形剤とを含むものであってよい。このような医薬組成物は、経口または非経口投与に適する剤形として提供される。
【0070】
非経口投与のための組成物としては、例えば、注射剤、坐剤等が用いられ、注射剤は静脈注射剤、皮下注射剤、皮内注射剤、筋肉注射剤、点滴注射剤等の剤形を包含しても良い。このような注射剤は、公知の方法に従って調製できる。注射剤の調製方法としては、例えば、上記本発明のHVJ−Eを通常注射剤に用いられる無菌の水性液、または油性液に溶解、懸濁または乳化することによって調製できる。注射用の水性液としては、例えば、生理食塩水、ブドウ糖やその他の補助薬を含む等張液等が用いられ、適当な溶解補助剤、例えば、アルコール(例、エタノール)、ポリアルコール(例、プロピレングリコール、ポリエチレングリコール)、非イオン界面活性剤〔例、ポリソルベート80、HCO−50(polyoxyethylene(50mol)adduct of hydrogenated castor oil)〕等と併用してもよい。油性液としては、例えば、ゴマ油、大豆油等が用いられ、溶解補助剤として安息香酸ベンジル、ベンジルアルコール等を併用してもよい。調製された注射液は、適当なアンプルに充填されることが好ましい。直腸投与に用いられる坐剤は、上記HVJ−Eを通常の坐薬用基剤に混合することによって調製されてもよい。
【0071】
経口投与のための組成物としては、固体または液体の剤形、具体的には錠剤(糖衣錠、フィルムコーティング錠を含む)、丸剤、顆粒剤、散剤、カプセル剤(ソフトカプセル剤を含む)、シロップ剤、乳剤、懸濁剤等が挙げられる。このような組成物は公知の方法によって製造され、製剤分野において通常用いられる担体、希釈剤もしくは賦形剤を含有していても良い。錠剤用の担体、賦形剤としては、例えば、乳糖、でんぷん、蔗糖、ステアリン酸マグネシウムが用いられる。
【0072】
本発明のHVJ−Eを注射により投与する場合、腫瘍部位、またはその周辺部に、HVJ−Eを直接注射により投与することができる。投与量は、腫瘍の大きさ、患者の年齢、体重、状態等を考慮して、医者または医療従事者が適宜決定することができる。腫瘍体積が1000mm3以下(例えば、200mm3程度など)の場合、1腫瘍部位あたり1回当たりのHVJ−Eの投与量を10〜10000HAU、好ましくは10〜5000HAU、さらに好ましくは50〜1000HAU、例えば100HAUとすることができる。あるいはまた、体重1kgあたり、300000HAU以下であることが好ましい。
【0073】
本発明のHVJ−Eとして、IL−12をエンベロープの外側に提示する形で担持する、HVJ−Eを用いた場合における、IL−12としての投与量は、腫瘍体積が1000mm3以下(例えば、200mm3程度など)の場合、1腫瘍部位あたりの1回あたりのIL−12の投与量を0.1pg〜1000pg、1〜500pg、さらに好ましくは10〜50pg(例えば25pg)とすることができる。HVJ−E自体の抗腫瘍効果とIL−12の抗腫瘍効果とが相まって、極めて強力な抗腫瘍活性を達成することが可能であるため、HVJ−EやIL−12を、それぞれ単独で投与する場合や、HVJ−EとIL−12とを単に混合して投与する場合と比較して、HVJ−E及びIL−12それぞれの投与量を減らすことが可能であり、安全性の観点から有利である。例えば、1腫瘍部位あたり1回当たりの投与量を、HVJ−Eとして、1000HAU以下、好ましくは50〜1000HAU、より好ましくは100HAUとすることができる。また、同じ場合、該投与量を、IL−12として、1腫瘍部位あたり100pg以下、例えば10〜100pg、好ましくは10〜50pg(例えば25pg)とすることができる。このような低い投与量は、特に、腫瘍体積が1000mm3以下の場合に、有効である。また、HVJ−EとIL−12 がそれぞれ所望の量含まれるように、抗体の定常領域(Fc)のCH2−CH3ドメインが機能可能に連結された第1の融合タンパク質をエンベロープに含むHVJ−Eと第2の融合タンパク質とを混合する際にその比率を適宜変更することができる。また腫瘍体積が1000mm3以上の場合であっても、例えばHVJ−Eが1腫瘍部位あたりの1回投与量として10〜100000HAU、IL−12が1pg〜10000pgの範囲から、患者の状態に応じて投与量を決定することができる。同様に投与回数も腫瘍の大きさ、患者の年齢、体重、状態等を考慮して、医者または医療従事者が適宜決定することができる。
【0074】
なお、前記した各組成物は、本発明のHVJ−Eとの配合により好ましくない相互作用を生じない限り他の活性成分を含有してもよい。
【実施例】
【0075】
改変HVJ−Eの作製スキーム
本実施例に用いる1本鎖IL−12(以下、scIL−12とも表す)との会合により抗腫瘍効果を向上させた改変HVJ−Eの作製スキームを図1に示す。図1では、この機能性分子がIL−12の場合を例として記載しているが、細胞表面の受容体に結合する各種の分子等、所望の機能性分子をこの方法でHVJ−Eの表面に提示させることができることは、当業者であれば容易に理解可能である。
すなわち、機能性ポリペプチドを提示するHVJ−Eには、イムノグロブリン(Ig)分子の定常領域(Fc)のCH2−CH3ドメインをウイルス表面に提示するFc−HNタンパク質と、所望の機能性ポリペプチドとプロテインAのZZドメインとを融合させたZZ融合タンパク質との、2種の融合タンパク質が用いられた。
まず、天然型HVJ−Eのウイルス粒子表面に提示されたHNタンパク質の粒子外ドメインを抗体の定常領域(Fc)に置換したHN−Fc融合タンパク質を作製し、これをHVJ−EのHNタンパク質の代わりにウイルス粒子に提示するFc−HN−HVJ−Eを作製した(図1a)。
より詳細には、HNタンパク質の粒子外ドメインを抗体の定常領域(Fc)に置換したHN−Fc融合タンパク質をコードする遺伝子を細胞に導入して該細胞でHN−Fc融合タンパク質を発現させた。この細胞に天然型HNタンパク質を持つ天然型HVJ−Eを感染させて増幅させた。すると、このウイルス増幅の過程で、細胞内で発現され細胞外にFcを提示していたHN−Fc融合タンパク質を粒子外被に取り込んでFc部を粒子表面に提示したウイルス(Fc−HN−HVJ−E)が細胞外に放出された(図2)。
次いで、一本鎖IL−12にプロテインAのZZドメイン(該ドメインがFcと結合する)を付加したscIL−12−ZZ融合タンパク質(ZZ−IL12)を作製した(図1b)。
Fc−HN−HVJ−EとZZ−IL12とを混合することで、Fc−HN−HVJ−Eの粒子表面のFcにZZ−IL12のZZドメインが結合して、ZZ−IL12とFc−HN−HVJ−Eとが会合し、HVJ−Eの粒子表面にIL−12が提示されたZZ−IL12−Fc−HN−HVJ−Eが得られた(図1a+b)。
より具体的に本実施例に用いたZZ−IL12−Fc−HN−HVJ−Eの作製方法を以下に記載するが、この記載により本発明の態様がこれに限定されるものではない。
【0076】
1.Fc−HN−HVJの作製
(1)ウイルス粒子に取り込まれるために必要なHNタンパク質内の領域の特定
まず、ウイルス粒子に取り込まれ得るHN−Fc融合タンパク質を作製するにあたり、HNタンパク質のどの領域がウイルス粒子に取り込まれるために必須であるのかを特定した。
HNタンパク質中のウイルス粒子の外に提示される領域(細胞外ドメイン)のC末端側から欠失させて、6種類のリコンビナントHN発現ベクター(Full length HN、ecto−400aa−HN、ecto−300aa−HN、ecto−200aa−HN、ecto−100aa−HN、ecto−0aa−HN)を作製した(図3(a))。これらのリコンビナントHNは、免疫学的検出を可能とするために、便宜上、C末端側にMycタグを融合させた。このリコンビナントHN発現ベクターをサル腎臓細胞LLCMK2に導入し、その発現をウエスタンブロットにより確認したところ、所望の分子量の位置に各リコンビナントHNが発現されていた(図3(c))。このウエスタンブロットで検出される発現量がかなり少なかったecto−0aa−HN以外のリコンビナントHNについて、細胞内局在を免疫染色法で確認した。Triton存在下では、いずれも細胞質に検出されたが、Toriton非存在下で本来の細胞内局を見たところ、Full length HN 及びecto−100aa−HNのみが細胞膜表面に局在している事が明らかとなった(図3(b))。さらに、これらのリコンビナント HN 発現細胞に HVJ を感染させると、ecto−100aa−HN のみが感染細胞由来 HVJ に取り込まれていることが確認された(図3(d))。
また、上記と同様に、ecto−50aa−HN、ecto−80aa−HN、ecto−120aa−HNおよびecto−150aa−HNを作製して(図4(a))、同様に細胞内発現(図4(c))、細胞内局在(図4(b))およびウイルス粒子への取り込み(図4(d))を確認したところ、ecto−50aa−HNは、細胞内発現が低く、細胞膜表面への局在も確認できなかったが、ecto−80aa−HN、ecto−120aa−HNおよびecto−150aa−HNについては、十分に発現され細胞膜表面に局在することが確認された。ウイルス粒子への取り込みについては、ecto−80aa−HNおよびecto−120aa−HNにおいて、認められた(図4(d))。
以上より、C末端切断型のHNタンパク質がウイルス粒子に取り込まれるためには、HNタンパク質の細胞外ドメインのN末端側の少なくとも51残基以上、好ましくは80残基以上(N末端から111残基以上、好ましくは140残基以上)が必要であることがわかった。また、N末端部分アミノ酸配列の長さが209アミノ酸(好ましくは180アミノ酸)よりも長いと、かえってC末端切断型のHNタンパク質はウイルス粒子に取り込まれなくなることが分かった。
(2)Fc−HN−HVJの作製
次いで、上記ecto−100aa−HNを利用してFc−HN−HVJを作製した。まず、ecto−100aa−HNのMycタグのC末端側にマウスIgG2aFcドメインのCH2−CH3ドメインをさらに付加したFc融合ecto−100aa−HN(HN−Fc)を作製した(図5(a))。このHN−Fcについても、上記と同様の方法で、細胞内発現(図5(b))、細胞内局在(図5(c))、ウイルス粒子への取り込み(図5(d))を確認できた。さらに、このecto−100aa−HNのプロテインA結合能を調べるために、Fc−HNとプロテインAセファロースとの共沈実験を行った。ecto−100aa−HN発現LLCMK2およびFc−HN発現LLCMK2をそれぞれに溶解させてプロテインAセファロースと混合後、プロテインAセファロースを沈殿させた。ecto−100aa−HNはプロテインAセファロースと共沈せず、全て細胞溶解液(lysate)中に残ったが、Fc−HNはプロテインAセファロースと共沈し、プロテインAとの結合が可能である事が確認された(図5(e))。
【0077】
2.ZZドメイン融合型機能性ポリペプチドの作製(機能性ポリペプチドが一本鎖IL−12である場合)
プロテインAのZZドメインと一本鎖IL−12とを融合させて、ZZ−IL−12を作製した。まず、Lieschkeらの報告(Nature Biotechnology, 15, 35−40 (1997))にならって一本鎖IL−12(scIL12)を作製した。具体的には、IL−12の第一のサブユニットp40と第二のサブユニットp35との間にGly−Gly−Gly−Serからなるリンカーが3つ連結して挟まれ、さらにp40のN末端側にシグナルペプチドを連結したものである(図6(a))。本実施例においては、免疫学的検出の便宜のために、シグナルペプチドC端とp40サブユニットのN端との間にFlagおよびヘマグルチニン(HA)のタグを挿入し(Tag−scIL12)、さらに、HAタグのC端とp40サブユニットの間にプロテインAのZZドメインを挿入したものを遺伝子工学的常法により作製した(ZZ−scIL12)。また、ネガティブコントロールとしてp35サブユニットを欠損したものも作製した(ZZ−p40)。なお、このシグナルペプチドはタンパク質生成後切断される。これらが、IL−12として機能しうるかどうかを確認するために、Tag−scIL12、ZZ−scIL12およびZZ−p40について、脾細胞からのインターフェロンγ(IFNγ)の分泌刺激効果を測定した。マウス大腿骨から骨髄細胞を採取し、DC−Medium(RPMI(10%FBS、100unit/ml penicillin、0.1mg/ml streptomycin、10ng/ml GM−CSF、4nl/ml 2−mercaptethanole))で6日間培養後、浮遊細胞を樹状細胞(DC)として回収し、1×105cells/100μl/wellで96 well plateにまいた。
Tag−scIL12、ZZ−scIL12およびZZ−p40のそれぞれを、10 ng/mlの濃度となるよう各wellに加え(total 200 μl)、24h 37℃でインキュベーションし、96well plateを遠心(440×g、5 min)することで、上清を回収し、上清中のIFNγをELISAで測定した。その結果、ZZ−p40はIFNγ分泌誘導を示さなかったが、Tag−scIL12およびZZ−scIL12は共に、IFNγ分泌誘導を示し(図6(b))、ここで作製したZZ−scIL12がIL−12として作用することが確認された。
【0078】
3.ZZ−scIL12とFc−HN−HVJとの結合能
ZZ−scIL12が、Fc−HNを粒子表面に提示したHVJ(Fc−HN−HVJ)と結合して複合体を形成しうるかどうかを調べた。
上記Fc−HNの発現ベクターをLLCMK2に導入後、HVJを該細胞に1.5MOIにて感染させて増幅させたHVJを回収することにより、Fc−HNを粒子表面に提示したFc−HN−HVJを得た。このFc−HN−HVJ液(TFBバッファー:20mM Tris−HCl(pH7.0)、150mM NaCl、1mM MgCl2に懸濁)とZZ−scIL−12液(ZZ−scIL12発現ベクター導入CHO−K1培養上清(血清不含HAM’S F−12培地))とを、体積比1:20で混合し(4℃、一晩、転倒混和)、その後、ショ糖密度勾配による遠心分離(25−50%スクロース、100,000×g、11h)後、scIL12をHAタグに対する抗体で、HVJはHVJに発現しているMタンパク質に対する抗体で、それぞれショ糖濃度画分のウェスタンブロットにより検出した。Fc−HNを提示していないHVJとZZ−scIL12とを混合した場合は、ZZ−scIL12は低ショ糖濃度の画分に、HVJは高ショ糖濃度の画分に、それぞれ分離される(図7右上図)のに対し、Fc−HN−HVJと混合した場合には、ZZ−scIL12はFc−HN−HVJと同じ高ショ糖濃度の画分にも検出され、混合液中のZZ−scIL12はFc−HN−HVJと複合体を形成していることが確認された。なお、ZZドメインを有さないscIL−12(Tag−scIL12)はFc−HN−HVJと混合した場合でも、Fc−HN−HVJと同じ高ショ糖濃度の画分には検出されなかった(図7左下図)。したがって、Fc−HN−HVJとZZ−scIL−12とは、Fc部とZZドメインの結合を介して会合し複合体を形成するものと考える。
【0079】
4.ZZ−scIL12とHVJ−Eとの複合体の血清中での結合安定性
前述のようにして得られたFc−HN−HVJを、公知の方法(WO2006/011580など参照のこと)に従って精製し、Fc−HVJ−Eを得た。このFc−HVJ−EとZZ−scIL12とを上記と同様に混合し、血清中で37℃で0〜24時間インキュベートした後、上記同様にウエスタンブロットに供した。その結果、24時間のインキュベーションにおいても、ZZ−scIL12とFc−HVJ−Eの結合が維持されている事が確認された(図8)。
【0080】
5.ZZ−scIL12結合型Fc−HVJ−Eの樹状細胞(DC)および脾臓細胞への作用
Fc−HVJ−Eを樹状細胞(DC)に対するZZ−scIL12結合型Fc−HVJ−Eの作用を検討した。DCの調製は、マウス大腿骨から骨髄細胞を採取し、DC−Medium(RPMI(10%FBS、100unit/ml penicillin、0.1mg/ml streptomycin、10ng/ml GM−CSF、4nl/ml 2−mercaptethanole))で6日間培養した後、浮遊細胞(DC)を回収し、1×105cells/100μl/wellで96well plateにまいた。その後、天然型HVJ−E、Fc−HN−HVJ−EおよびZZscIL12と複合体を形成したZZ−scIL12結合型Fc−HN−HVJ−EHVJ−Eを、それぞれ10、50または100HAU(100μl培地中)を各ウェルに加え、24時間、37℃でインキュベーションした。この96well plateを440×g、5minで遠心し、上清を回収し、上清中のIFN−γおよびIL−6の量をELISAで測定した。その結果を図9に示す。ZZ−scIL12結合型Fc−HVJ−Eは、DCからのIFN−γ分泌を促進した(図9中図)。天然型HVJ−EおよびFc−HN−HVJ−EもIFN−γ分泌促進を誘導したが、ZZ−scIL12結合型Fc−HVJ−Eが最もIFN−γ分泌促進能に優れていた。また、DCからのIL−6分泌促進能については、天然型HVJ−Eよりは低いものの、Fc−HN−HVJ−EおよびZZ−scIL12結合型Fc−HVJ−Eとも、IL−6の分泌も促進させることが確認された(図9右図)。
さらにまた、脾臓細胞におけるIFN−γ分泌促進についても検討した。脾臓細胞は、マウス脾臓から脾細胞を回収後、塩化アンモニウム−トリス緩衝液を加えて赤血球を溶血させた後、細胞をsplenocyte−Medium(RPMI(10%FBS、100unit/ml penicillin、0.1mg/ml streptomycin、4nl/ml 2−mercaptethanole))に懸濁し、2×105cells/100μl/wellで96well plateにまいた。これに上記と同様に、天然型HVJ−E、Fc−HN−HVJ−EおよびZZ−scIL12 結合型Fc−HN−HVJ−EHVJ−Eを、それぞれ10、50または100HAU(100μl培地中)を、ならびにIL−12を25pg、50pgおよび250pg(100μl培地中)をそれぞれ、各ウェルに加え24h 37℃でインキュベーションした。この96well plateを440×g、5minで遠心し、上清を回収し、上清中のIFN−γの量をELISAで測定した。その結果を図10に示す。天然型HVJ−EおよびFc−HN−HVJ−Eは、脾臓細胞のIFN−γ分泌を誘導せず、IL−12も殆どIFN−γ分泌を誘導しなかったが、ZZ−scIL12結合型Fc−HVJ−EはIFN−γ分泌を誘導した(図10)。すなわち、ZZ−scIL12結合型Fc−HVJ−Eの脾細胞からのIFN−γ分泌促進に関しては、IL−12とHVJ−Eとの相加効果を上回るものである。
【0081】
6.ZZ−scIL12結合型Fc−HVJ−Eのin vivoでの抗腫瘍作用
マウスの背にF10メラノーマをの皮内注入し、その腫瘍径が2〜4mmになったところでZZ−scIL12結合型Fc−HVJ−E(HVJ−E:100HAU、IL−12:250pg)を2日おきに3回腫瘍内注射をし、その後の腫瘍径や免疫系活性化能を測定した。
ZZ−scIL12結合型Fc−HVJ−E投与後の腫瘍体積の増大はほとんど認められず、天然型HVJ−Eと比較しても明らかに強い腫瘍増殖抑制が認められた(図11左図)。なお、本発明者らは、同様の方法で、通常のHVJ−Eが前立腺癌の腫瘍体積の増加をほぼ90%以上抑制するという事実を発表しているが(WO2005/094878)、これに用いたHVJ−Eは5000HAUという高用量であり、Fc−HN−HVJ−E+ZZ−scIL12が、HVJ−Eの力価として100HAUという低用量でメラノーマに対して同レベルの効果を呈するということは驚くべき事実であった。また、ZZ−scIL12結合型Fc−HVJ−E投与群の4個体中、2個体は完全に寛解した。このことから、ZZ−scIL12結合型Fc−HVJ−Eは、100HAUという低容量でも強力な抗腫瘍作用を示すことがわかった。
【0082】
7.ZZ−scIL12結合型Fc−HVJ−Eのin vivoでの免疫活性化能の検討
マウス背部の皮内に5×105 cellsのF10メラノーマを注入し、腫瘍が約3〜5mmになった時点で、天然型HVJ−E100HAUまたはZZ−scIL12結合型Fc−HVJ−E(HVJ−E:100HAU、IL−12:250pg)を腫瘍内に2日おきに3回投与した。最終投与から10日後に各マウスの脾細胞を採取した。
マイトマイシンCで処理(15μg/ml、45min、37℃)したF10メラノーマを、採取した脾細胞に10:1の割合で混合し(脾細胞:F10=2×107:2×106(5ml))、37℃で48h インキュベートした後、脾細胞を回収し、Mouse IFN−γ Development Module(R&D)のプロトコールに従ってELISPOTアッセイを行った。なお、ELISPOTアッセイにおいては、メンブレン上での脾細胞とマイトマイシンC処理F10メラノーマとの比は、脾細胞:F10=3×105:0.2×104(100μl/well)で行い、インキュベーションの時間は24時間とした。
天然型HVJ−Eに比べてZZ−scIL12結合型Fc−HVJ−Eは、約5倍以上のIFN−γ分泌誘導効果を示した(図12左図)。
また、Cr51リリーシングCTLアッセイにより、ZZ−scIL12結合型Fc−HVJ−E刺激脾細胞による免疫系活性化を調べた。Cr51リリーシングCTLアッセイは、常法にしたがい、下記の要領で実施した。F10メラノーマをマイトマイシンCで処理(15μg/ml、45min、37℃)後、上記採取した脾細胞とメラノーマを10:1割合で混合し(脾細胞:F10=5×107:5×106(20ml))、37℃で7日間インキュベーションした。この際、培地にはIL−2(final 10ng/ml)を添加しておき、インキュベーション4日後には10mlの培地(5ng/ml IL−2含有)を再度添加した。脾細胞を回収し、96well plateに2×106、1×106、5×105、2.5×105、1.25×105、0.625×105 cells/100μl/wellの2倍希釈系列を作製する。また、ポジティブコントロールとして1%NP−40溶液、ネガティブコントロールとしてRPMI培地を使用した。F10メラノーマを5×106 cells/0.2mlに調整し、1.85MBqの51Crを加え、37℃、45minインキュベーションし、F10メラノーマを洗浄して2×104cells/100μl/wellで脾細胞をまいたプレートに加え、4hインキュベーションした。そのプレートを1000rpm、5minで遠心後、各wellの上清を100μl回収し、51Crを測定した。
その結果、やはりZZ−scIL12結合型Fc−HVJ−Eは、天然型HVJ−Eに比べてかなり強いCTL活性を示した(図12右図)。
【0083】
8.ZZ−scIL12結合型Fc−HVJ−Eと、非結合scIL12およびHVJ−Eの併用投与との抗癌作用の比較
ZZ−scIL12結合型Fc−HVJ−E(HVJ−E:100HAU、IL−12:250pg)、またはZZドメインを有していないためFc−HVJ−Eとは結合しないTag−scIL12(IL−12:250pg)とHVJ−E100HAUとの混合液とを、上記同様F10メラノーマを注入し腫瘍が約2〜4mmになったマウスに腫瘍内に2日おきに3回投与した。回投与から14日目において、各マウスの腫瘍を観察し、完全に腫瘍が消失したマウスの割合を図13に示す。ZZ−scIL12結合型Fc−HVJ−Eを投与した群では30%以上で腫瘍が全く消失し、それ以後再発することなく90日後でも生存していた。一方、Fc−HN−HVJ−Eと結合しないTag−scIL12とFc−HN−HVJ−Eとを共投与した群では、5%程度しか腫瘍寛解が認められなかった(図13)。このことから、IL−12とHVJ−Eとが結合して複合体を形成することで強い抗腫瘍活性を示すことがわかった。
このことは、ZZ−scIL12結合型Fc−HVJ−Eの抗癌作用は、IL−12とHVJ−Eとが結合して複合体を形成した状態で癌細胞に作用することによって、IL−12とHVJ−Eというそれぞれに抗癌作用を奏するものの単なる組合せによる効果の増大を大きく上回るということを示している。
【0084】
9.ZZ−scIL12結合型Fc−HVJ−Eと、非結合scIL12およびHVJ−Eの併用との脾細胞からのIFN−γ分泌促進能の比較
上記と同様、ZZ−scIL12結合型Fc−HVJ−Eを投与した場合と、同量のscIL12とHVJ−Eとを結合していない状態で併用投与した場合とにおける、脾細胞からのIFN−γ分泌促進刺激を調べた。F10メラノーマを注入し腫瘍が約3〜5mmになったマウスに腫瘍内に、HVJ−E単独、Fc−HVJ−E+ZZscIL12複合体(250pg scIL12/100HAU HVJ−E、またはFc−HVJ−EとTagscIL−12との混合液(250pg scIL12/100HAU HVJ−E)を、10、50または100HAU HVJ−Eを、2日おきに3回投与した。最終投与から10日後に各マウスの脾細胞を採取し、脾細胞から分泌されたIFN−γを定量した(図14)。その結果、高濃度投与時においては結合型、非結合型共に同程度に高いIFN−γ分泌を誘導するが、低濃度投与時においては結合型の方が非結合型と比較して高いIFN−γ分泌誘導を引き起こすことが明らかとなった。
【産業上の利用可能性】
【0085】
本発明の一本鎖IL−12がHVJ−Eの表面に連結した改良型HVJ−E、さらに該改良型HVJ−Eを有効成分とする抗癌剤は、従来公知のHVJ−E単独の抗癌剤よりも低用量かつ著しい抗腫瘍効果を発揮し、新規な癌治療剤として有用である。
【技術分野】
【0001】
本発明は、センダイウイルス(Hemagglutinating Virus of Japan,以下HVJと表記することもある)を構成するHNタンパク質のN末端側部分アミノ酸配列からなる改変HNタンパク質、該改変HNタンパク質のC末端側に機能性ポリペプチドが連結した融合タンパク質、または、該融合タンパク質を含むタンパク質複合体、さらに、該融合タンパク質または該タンパク質複合体をエンベロープに含有する改良型センダイウイルスエンベロープ(以下HVJ−Eと表記することもある)を有効成分とする抗癌剤等に関する。
【背景技術】
【0002】
センダイウイルス(HVJ)のゲノムを不活化して増殖能が喪失された、HVJエンベロープ(HVJ−E)は、他の分子をその中に封入することができることから、生体機能分子のベクター(運び屋)として開発されてきた(特許文献1、特許文献2など)。そして、HVJ−Eに抗癌剤を封入して、またはHVJ−Eと抗癌剤を併用して投与することで、抗癌効果が増強されることも分かってきた(特許文献3、非特許文献1、非特許文献2)。このように、HVJ−Eは、抗癌剤のためのベクターとして、またそれ自体が抗癌剤の補助剤として研究開発が進められている(非特許文献3)。さらには、HVJ−Eのみを投与した場合でも抗癌作用を示すことが報告されている(特許文献3、非特許文献4)。HVJ−Eは、樹状細胞や腫瘍細胞からInterferon−α、−βやIL−6などのサイトカイン、CCL−2、3、4、5、CXCL−10などのケモカインの分泌を促し、NK細胞の浸潤の活性化、CD4+、CD8+T cellの浸潤と活性化、それによる腫瘍に対するCTL誘導を引きおこす(非特許文献4)。しかしながら、HVJ−EにはInterferon−γ(IFN−γ)の産生促進能が認められておらず、Effector T cellには直接作用することはできないと考えられる。IFN−γ産生を誘導するサイトカインとしては種々の分子が知られているが、代表的な分子としてはIL−12が挙げられる。IL−12がIFN−γの産生を誘導することで抗腫瘍活性を示すことは、当業者には十分に公知の事実である(例えば、非特許文献5、非特許文献6など)。
【先行技術文献】
【特許文献】
【0003】
【特許文献1】WO01/57204
【特許文献2】WO2004/035779
【特許文献3】WO2005/094878
【非特許文献】
【0004】
【非特許文献1】Biochem Biophys Res Commun. 2007 Dec 21;364(3):423−8. Epub 2007 Oct 12
【非特許文献2】Cancer Chemother Pharmacol. 2008 May;61(6):973−8. Epub 2007 Jul 26
【非特許文献3】Expert Opin Drug Deliv. 2008 Feb;5(2):221−33. Review
【非特許文献4】Int J Cancer. 2009 May 15;124(10):2478−87
【非特許文献5】J. Immunol. 1994, 153, 1697− 706
【非特許文献6】J. Leukoc. Biol. 55:280−288; 1994
【発明の概要】
【発明が解決しようとする課題】
【0005】
本発明の課題は、HVJ−Eにより、IL−12等の抗腫瘍性ポリペプチドの抗腫瘍効果を飛躍的に増大させる新たな技術を開発し、これに基づき優れた抗癌剤を提供することである。また、本発明は、この新たな技術に有用な改変タンパク質や、改良型HVJ−Eを提供することを課題とする。
【課題を解決するための手段】
【0006】
本発明者らは上記課題を解決すべく、鋭意検討の結果、HVJ−Eを構成するHNタンパク質の特定の領域のC末端側に所望のポリペプチドを連結させることにより改変HNタンパク質と所望のタンパク質との融合タンパク質を作成し、この融合タンパク質を用いてHVJ−Eを構成すると、所望のタンパク質がHVJ−Eの外側表面に発現することを見出した。この所望のタンパク質として、一本鎖IL−12を用いると、一本鎖IL−12がHVJ−E粒子の表面に発現した、改変HVJ−Eを得ることができた。この改変HVJ−Eは、HVJ−EとIL−12とを単に共投与した場合に比べて、はるかに高い抗腫瘍効果を示したことから、HVJ−Eの粒子表面にIL−12等のIFN−γ分泌促進能を有するポリペプチドを結合させると、極めて高い抗腫瘍効果を得ることが出来ることを見出し、本発明を完成させるに到った。すなわち、本発明は、
[1]HVJのHNタンパク質のN末端部分アミノ酸配列と同一または実質的に同一のアミノ酸配列からなる改変HNタンパク質であって、該部分アミノ酸配列が111〜209アミノ酸の長さを有する、改変HNタンパク質;
[2]該部分アミノ酸配列が140〜180アミノ酸の長さを有する、上記[1]記載の改変HNタンパク質;
[3]下記(1)または(2)のポリペプチドである、上記[1]記載の改変HNタンパク質:
(1)配列番号6の第1〜160アミノ酸に該当するアミノ酸配列からなるポリペプチド、
(2)配列番号6の第1〜160アミノ酸に該当するアミノ酸配列において1または複数の残基が置換、欠失又は付加されたアミノ酸配列を含み、且つHVJに発現された場合に粒子表面に局在しうる特徴を有するポリペプチド;
[4]上記[1]〜[3]のいずれか1つに記載の改変HNタンパク質の残基及び第1の機能性ポリペプチド残基を含む融合タンパク質であって、該改変HNタンパク質残基のC末端側に該機能性ポリペプチド残基が機能可能に連結されている、融合タンパク質;
[5]機能性ポリペプチドがIFN−γ分泌促進能を有するポリペプチドである、上記[4]記載の融合タンパク質;
[6]IFN−γ分泌促進能を有するポリペプチドが一本鎖IL−12である、上記[5]記載の融合タンパク質;
[7]一本鎖IL−12が、下記(1)または(2)のポリペプチドである、上記[6]記載の融合タンパク質:
(1)配列番号2の第103〜625アミノ酸に該当するアミノ酸配列を含むポリペプチド、
(2)配列番号2の第103〜625アミノ酸に該当するアミノ酸配列において1または複数の残基が置換、欠失又は付加されたアミノ酸配列を含み、且つIFN−γ分泌促進能を有するポリペプチド;
[8]機能性ポリペプチドが抗体の定常領域(Fc)のCH2−CH3ドメインである、上記[4]記載の融合タンパク質;
[9]抗体の定常領域(Fc)のCH2−CH3ドメインがIgG2aに由来するものである、上記[8]記載の融合タンパク質;
[10]抗体の定常領域のCH2−CH3ドメインが、下記(1)または(2)のポリペプチドである、上記[9]記載の融合タンパク質:
(1)配列番号4の第171〜389アミノ酸に該当するアミノ酸配列を含むポリペプチド、
(2)配列番号4の第171〜389アミノ酸に該当するアミノ酸配列において1または複数の残基が置換、欠失又は付加されたアミノ酸配列を含み、且つプロテインAとの結合活性を有するポリペプチド;
[11]上記[1]〜[3]のいずれか1つに記載の改変HNタンパク質、又は上記[4]〜[10]のいずれか1つに記載の融合タンパク質をコードする核酸;
[12]上記[8]〜[10]のいずれか1つに記載の第1の融合タンパク質、並びに
プロテインAのZZドメイン及び第2の機能性ポリペプチド残基を含む第2の融合タンパク質を含む、タンパク質複合体であって、第1の融合タンパク質と第2の融合タンパク質とが、第1の融合タンパク質に含まれるCH2−CH3ドメインと第2の融合タンパク質に含まれるZZドメインとの間の結合を介して複合体を形成している、タンパク質複合体;
[13]第2の機能性ポリペプチドが、一本鎖IL−12である、上記[12]記載のタンパク質複合体;
[14]一本鎖IL−12が、下記(1)または(2)のポリペプチドであり、且つZZドメインが、下記(3)または(4)のポリペプチドである、上記[13]記載のタンパク質複合体:
(1)配列番号2の第103〜625アミノ酸に該当するアミノ酸配列を含むポリペプチド、
(2)配列番号2の第103〜625アミノ酸に該当するアミノ酸配列において1または複数の残基が置換、欠失又は付加されたアミノ酸配列を含み、且つIFN−γ分泌促進能を有するポリペプチド、
(3)配列番号2の第42〜99アミノ酸に該当するアミノ酸配列を含むポリペプチド、
(4)配列番号2の第42〜99アミノ酸に該当するアミノ酸配列において1または複数の残基が欠失、置換または付加されたアミノ酸配列を含み、且つ抗体分子の定常領域(Fc)への結合活性を有するポリペプチド;
[15]上記[1]〜[3]のいずれか1つに記載の改変HNタンパク質、上記[4]〜[10]のいずれか1つに記載の融合タンパク質、または上記[12]〜[14]のいずれか1つに記載のタンパク質複合体をエンベロープに含有する、HVJ−E;
[16]上記[6]もしくは[7]に記載の融合タンパク質または上記[13]もしくは[14]に記載のタンパク質複合体をエンベロープに含有する、上記[15]記載のHVJ−E;
[17]IFN−γ分泌促進能を有するポリペプチドをエンベロープ外側に提示する形で担持する、HVJ−E;
[18]IFN−γ分泌促進能を有するポリペプチドが、IL−12である、上記[17]記載のHVJ−E;
[19]IL−12が一本鎖IL−12である、上記[18]記載のHVJ−E;
[20]上記[6]もしくは[7]に記載の融合タンパク質または上記[13]もしくは[14]に記載のタンパク質複合体をエンベロープに含有する、上記[18]記載のHVJ−E;
[21]上記[17]〜[20]のいずれか1つに記載のHVJ−Eを有効成分として含む、抗癌剤;
[22]メラノーマ、前立腺癌、膀胱癌および大腸癌からなる群より選択される癌の治療または予防に用いられる、上記[21]記載の抗癌剤;
[23]1回当たりの投与量が、HVJ−Eとして1000HAU以下であり、かつ該HVJ−Eに含まれるIL−12として100pg以下であることを特徴とする、上記[21]記載のHVJ−Eを有効成分として含む抗癌剤;
[24]上記[21]記載のHVJ−Eの治療上有効量を癌患者に投与することを特徴とする、癌の治療方法;
[25]1回当たりの投与量が、HVJ−Eとして1000HAU以下であり、かつ該HVJ−Eに含まれるIL−12として100pg以下であることを特徴とする、上記[24]記載の癌の治療方法;
を提供する。
【発明の効果】
【0007】
本発明の改変HNタンパク質や融合タンパク質を用いれば、HVJ−Eの表面に所望のポリペプチドを発現させることが可能となる。
例えば、改変HNタンパク質と一本鎖IL−12等のIFN−γ分泌促進能を有するポリペプチドとの融合タンパク質を用いることにより、エンベロープの外側に該ポリペプチドを提示する形で担持するHVJ−Eを得ることが可能である。
エンベロープの外側にIFN−γ分泌促進能を有するポリペプチドを提示する形で担持するHVJ−Eは、高い抗腫瘍効果を有しており、抗腫瘍剤として有用である。
癌の治療においては、外科的切除法、放射線治療法、化学療法などが存在するが、その根治は依然大きな課題である。本発明は、細胞実験および生体内実験を通じて、本発明の改良型HVJ−Eの癌への直接投与で、IFN−γの産生を誘導し、癌を著しく退縮させることを示した。
【図面の簡単な説明】
【0008】
【図1】1本鎖IL−12のZZドメインがHVJ−E表面の抗体の定常領域(Fc)に会合することにより抗腫瘍効果を向上させたHVJ−Eの作製スキームを示した図である。
【図2】融合タンパク質を発現させ、細胞膜上に局在させた細胞にネイティブなHVJを感染させることにより、エンベロープに融合タンパク質を組み込んだ組換えHVJの作製の過程を示した図である。
【図3】(a)HNタンパク質の細胞外ドメインを段階的に欠失させた各融合タンパク質の構造を示す図である。(b)(a)に示した各融合タンパク質の細胞膜での局在を示す免疫染色図である。(c)(a)に示した各融合タンパク質の細胞内発現をウエスタンブロットで確認した図である(d)(a)に示した各融合タンパク質の細胞膜局在をウエスタンブロットで確認した図である。
【図4】(a)HNタンパク質の細胞外ドメインを細胞外ドメインから50アミノ酸残基、80アミノ酸残基、100アミノ酸残基、120アミノ酸残基、150アミノ酸残基だけ残した各融合タンパク質の構造を示す図である。(b)(a)に示した各融合タンパク質の細胞膜での局在を示す免疫染色図である。(c)(a)に示した各融合タンパク質の細胞内発現をウエスタンブロットで確認した図である(d)(a)に示した各融合タンパク質の細胞膜局在をウエスタンブロットで確認した図である。
【図5】(a)HNタンパク質の細胞外ドメインを細胞外ドメインから100アミノ酸残基、だけ残した融合タンパク質にマウス由来IgG2aのFcのCH2−CH3ドメインを付加したFc−HNタンパク質の構造を示す図である。(b)(a)に示したFc−HNタンパク質の細胞内発現をウエスタンブロットで確認した図である。(c)(a)に示したFc−HNタンパク質の細胞膜での局在を示す免疫染色図である。(d)(a)に示したFc−HNタンパク質の細胞膜局在をウエスタンブロットで確認した図である。(e)(a)に示したFc−HNタンパク質のプロテインAセファロースとの共沈を示す図である。
【図6】(a)プロテインAのZZドメインと一本鎖IL−12とを融合させて作製したZZ−IL−12タンパク質の構造を示す図である。(b)各ZZ−IL−12タンパク質のインターフェロンγ(IFNγ)の分泌刺激効果を示す図である。
【図7】ZZ−scIL12とFc−HN−HVJとの結合能を示す図である。
【図8】ZZ−scIL12とFc−HN−HVJとの24時間インキュベーション後の結合保持能を示す図である。
【図9】ZZ−scIL12結合型Fc−HVJ−Eの樹状細胞(DC)および脾臓細胞への作用を示す図である。図9左図は、各HVJ−Eの発現を示すウエスタンブロット図である。図9中図は、ZZ−scIL12結合型Fc−HVJ−Eの樹状細胞(DC)からのIFN−γ分泌促進能を示す図である。図9右図は、ZZ−scIL12結合型Fc−HVJ−Eの樹状細胞(DC)からのIL−6分泌促進能を示す図である。
【図10】ZZ−scIL12結合型Fc−HVJ−EとZZ−scIL12単独投与との脾臓細胞へのIFN−γ分泌促進作用の比較を示す図である。
【図11】ZZ−scIL12結合型Fc−HVJ−Eのin vivoでの抗腫瘍作用を示す図である。
【図12】ZZ−scIL12結合型Fc−HVJ−Eのin vivoでの免疫活性化能を示す図である。図12左図は、ZZ−scIL12結合型Fc−HVJ−EのIFN−γ分泌誘導能を示すELISPOTアッセイ図である。図12右図は、ZZ−scIL12結合型Fc−HVJ−EのCTL活性を示すCr51リリーシングCTLアッセイ図である。
【図13】ZZ−scIL12結合型Fc−HVJ−Eと、非結合scIL12およびHVJ−Eの併用投与との抗癌作用の比較を示す図である。
【図14】ZZ−scIL12結合型Fc−HVJ−Eと、非結合scIL12およびHVJ−Eの併用との脾細胞からのIFN−γ分泌促進能の比較を示す図である。
【発明を実施するための形態】
【0009】
以下に、本発明を詳細に説明する。
本発明は、センダイウイルス(HVJ)のHNタンパク質のN末端部分アミノ酸配列と同一または実質的に同一のアミノ酸配列からなる改変HNタンパク質であって、該部分アミノ酸配列が111〜209アミノ酸の長さを有する、改変HNタンパク質を提供する。
好ましくは、該部分アミノ酸配列が140〜180アミノ酸の長さを有する。
さらに好ましくは、本発明の改変タンパク質は、下記(1)または(2)のポリペプチドである:
(1)配列番号6の第1〜160アミノ酸に該当するアミノ酸配列からなるポリペプチド、
(2)配列番号6の第1〜160アミノ酸に該当するアミノ酸配列において1または複数の残基が置換、欠失又は付加されたアミノ酸配列からなり、且つHVJに発現された場合に粒子表面に局在しうる特徴を有するポリペプチド。
【0010】
本発明で用いられるセンダイウイルス(HVJ)は、例えばVR−105、VR−907等をAmerican Type Culture Collection(ATCC:P.O.Box 1549,Manassas,VA 20108 USA)より購入することができる。
【0011】
本発明で用いられるHNタンパク質とは、センダイウイルス(HVJ)のエンベロープを構成する公知のコンポーネントの一つであり、宿主細胞への結合に関わる赤血球凝集素/ノイラミニダーゼ(HN)遺伝子によってコードされる。HNタンパク質のアミノ酸配列も公知である。本明細書においては、HNタンパク質は、例えば配列番号6に示されるアミノ酸配列からなるタンパク質を意味する。
【0012】
発明者らは、後述する実施例において確認するように、HNタンパク質の種々の欠失変異体を作成し、HNタンパク質が、細胞膜に局在し、ウイルス粒子形成の際にHVJのエンベロープを構成するためのHNタンパク質中の必要領域を検討し、その範囲を見出した。その結果、HNタンパク質の翻訳開始メチオニン残基から始まり、細胞内ドメイン、膜貫通ドメインに加えて細胞外ドメインの一部をコードする部分アミノ酸配列(以下、N末端部分アミノ酸配列とも表記してもよい)が必須であることを見出した。そして、該部分アミノ酸配列が、111アミノ酸以上、好ましくは140アミノ酸以上の長さを有していれば、C末端側の領域を欠失していても、該変異体HNタンパク質は、安定してHVJのエンベロープを構成し得ることを見出した。
【0013】
更にこのHNタンパク質のN末端部分ポリペプチドのC末端側に任意の機能性ポリペプチドを連結した融合タンパク質を調製し、この機能性ポリペプチドがHVJのエンベロープの外側に提示されるように、該融合タンパク質を安定してHVJ−E上に発現させる方法について検討する過程で、該融合タンパク質に含まれるHNタンパク質のN末端部分アミノ酸配列の長さが、209アミノ酸(好ましくは180アミノ酸)よりも長いと、かえって該融合タンパク質が安定してHVJ−E上に発現され難いことを見出した。
【0014】
即ち、HNタンパク質のN末端部分アミノ酸配列の長さは、通常、111〜209アミノ酸、好ましくは、140〜180アミノ酸、最も好ましくは、160アミノ酸である。
【0015】
尚、HNタンパク質のN末端部分アミノ酸配列における、第1のアミノ酸は、HNタンパク質のN末端の第1メチオニン(翻訳開始メチオニン残基)である。
【0016】
本発明の改変HNタンパク質は、哺乳動物細胞(例えばサル腎臓細胞LLCMK2)において発現したときに、該細胞の細胞膜上に局在する(これを、細胞膜へ局在する活性という)。「局在」とは、細胞内の他の領域よりも細胞膜における発現量が高いことを意味する。改変HNタンパク質が細胞膜上へ局在するか否かは、該改変HNタンパク質を発現することが出来る発現ベクターにより、哺乳動物細胞(例えばサル腎臓細胞LLCMK2)をトランスフェクトし、得られたトランスフェクタントの細胞膜や各オルガネラにおける改変HNタンパク質の発現を、改変HNタンパク質を特異的に認識する抗体を用いたウェスタンブロットにより解析することにより、確認することが出来る。
【0017】
また、本発明の改変HNタンパク質は、HVJのウイルス粒子形成の際に、エンベロープへ取り込まれることにより、該エンベロープを構成し得る(これを、HVJ−Eを構成する活性という)。即ち、本発明の改変HNタンパク質を哺乳動物細胞において発現させ、該哺乳動物細胞にHVJを感染させると、HVJ感染細胞から生じるHVJのエンベロープに、該改変タンパク質が取り込まれ、該エンベロープを構成する。改変HNタンパク質が、かかる特徴を有するか否かは、該改変HNタンパク質を発現することが出来る発現ベクターにより、哺乳動物細胞(例えばサル腎臓細胞LLCMK2)をトランスフェクトし、得られたトランスフェクタントにHVJを感染させ、感染したトランスフェクタントから生じたHVJを回収し、該HVJからエンベロープを単離し、該エンベロープにおける改変HNタンパク質の発現を、改変HNタンパク質を特異的に認識する抗体を用いたウェスタンブロットにより解析することにより、確認することが出来る。
【0018】
HVJのHNタンパク質のN末端部分アミノ酸配列と同一のアミノ酸配列とは、配列番号6に示されるアミノ酸配列のアミノ酸番号1番のメチオニンから始まり、アミノ酸番号111番のアミノ酸乃至アミノ酸番号209番のアミノ酸で終了する、配列番号6に示されるアミノ酸配列の部分アミノ酸配列(好ましくは配列番号6に示されるアミノ酸配列のアミノ酸番号1番のメチオニンから始まり、アミノ酸番号140番のアミノ酸乃至アミノ酸番号180番のアミノ酸で終了する、配列番号6に示されるアミノ酸配列の部分アミノ酸配列、より好ましくは、配列番号6に示されるアミノ酸配列のアミノ酸番号1番のメチオニンから始まり、アミノ酸番号160番のアミノ酸で終了する、配列番号6に示されるアミノ酸配列の部分アミノ酸配列)と完全同一のアミノ酸配列が挙げられる。
【0019】
HVJのHNタンパク質のN末端部分アミノ酸配列と実質的に同一のアミノ酸配列とは、上述のHVJのHNタンパク質のN末端部分アミノ酸配列と同一のアミノ酸配列と約80%以上、好ましくは約90%以上、さらに好ましくは約95%以上、特に好ましくは約97%以上の相同性を有するアミノ酸配列などが挙げられる。ここで「相同性」とは、当該技術分野において公知の数学的アルゴリズムを用いて2つのアミノ酸配列をアラインさせた場合の、最適なアラインメント(好ましくは、該アルゴリズムは最適なアラインメントのために配列の一方もしくは両方へのギャップの導入を考慮し得るものである)における、オーバーラップする全アミノ酸残基に対する同一アミノ酸および類似アミノ酸残基の割合(%)を意味する。「類似アミノ酸」とは物理化学的性質において類似したアミノ酸を意味し、例えば、芳香族アミノ酸(Phe、Trp、Tyr)、脂肪族アミノ酸(Ala、Leu、Ile、Val)、極性アミノ酸(Gln、Asn)、塩基性アミノ酸(Lys、Arg、His)、酸性アミノ酸(Glu、Asp)、水酸基を有するアミノ酸(Ser、Thr)、側鎖の小さいアミノ酸(Gly、Ala、Ser、Thr、Met)などの同じグループに分類されるアミノ酸同士が挙げられる。このような類似アミノ酸による置換は蛋白質の表現型に変化をもたらさない(即ち、保存的アミノ酸置換である)ことが予測される。保存的アミノ酸置換の具体例は当該技術分野で周知であり、種々の文献に記載されている(例えば、Bowieら,Science, 247: 1306−1310 (1990)を参照)。アミノ酸(配列)が置換、欠失または挿入されている場合、その置換、欠失または挿入の位置は、上述の、細胞膜へ局在する活性、及びHVJ−Eを構成する活性が実質的に保持される限り、特に限定されない。「実質的に保持される」とは、少なくともネイティブなHNタンパク質(全長HNタンパク質)と比較して、有意に高い細胞膜へ局在する活性を保持していることを意味する。
【0020】
本明細書におけるアミノ酸配列の相同性は、相同性計算アルゴリズムNCBI BLAST(National Center for Biotechnology Information Basic Local Alignment Search Tool)を用い、以下の条件(期待値=10;ギャップを許す;マトリクス=BLOSUM62;フィルタリング=OFF)にて計算することができる。アミノ酸配列の相同性を決定するための他のアルゴリズムとしては、例えば、Karlinら, Proc. Natl. Acad. Sci. USA, 90: 5873−5877 (1993)に記載のアルゴリズム[該アルゴリズムはNBLASTおよびXBLASTプログラム(version 2.0)に組み込まれている(Altschulら, Nucleic Acids Res., 25: 3389−3402 (1997))]、Needlemanら, J. Mol. Biol., 48: 444−453 (1970)に記載のアルゴリズム[該アルゴリズムはGCGソフトウェアパッケージ中のGAPプログラムに組み込まれている]、MyersおよびMiller, CABIOS, 4: 11−17 (1988)に記載のアルゴリズム[該アルゴリズムはCGC配列アラインメントソフトウェアパッケージの一部であるALIGNプログラム(version 2.0)に組み込まれている]、Pearsonら, Proc. Natl. Acad. Sci. USA, 85: 2444−2448 (1988)に記載のアルゴリズム[該アルゴリズムはGCGソフトウェアパッケージ中のFASTAプログラムに組み込まれている]等が挙げられ、それらも同様に好ましく用いられ得る。
【0021】
より好ましくは、HVJのHNタンパク質のN末端部分アミノ酸配列と実質的に同一のアミノ酸配列とは、上述のHVJのHNタンパク質のN末端部分アミノ酸配列と同一のアミノ酸配列と約80%以上、好ましくは約90%以上、さらに好ましくは約95%以上、特に好ましくは約97%以上の同一性を有するアミノ酸配列である。
【0022】
HVJのHNタンパク質のN末端部分アミノ酸配列と実質的に同一のアミノ酸配列からなる改変HNタンパク質は、前記の配列番号6に示されるアミノ酸配列のアミノ酸番号1番のメチオニンから始まり、アミノ酸番号111番のアミノ酸乃至アミノ酸番号209番のアミノ酸で終了する、配列番号6に示されるアミノ酸配列の部分アミノ酸配列(好ましくは配列番号6に示されるアミノ酸配列のアミノ酸番号1番のメチオニンから始まり、アミノ酸番号140番のアミノ酸乃至アミノ酸番号180番のアミノ酸で終了する、配列番号6に示されるアミノ酸配列の部分アミノ酸配列、より好ましくは、配列番号6に示されるアミノ酸配列のアミノ酸番号1番のメチオニンから始まり、アミノ酸番号160番のアミノ酸で終了する、配列番号6に示されるアミノ酸配列の部分アミノ酸配列)と実質的に同一のアミノ酸配列を含み、且つ配列番号6の第1〜160アミノ酸に該当するアミノ酸配列からなるポリペプチドと実質的に同質の活性を有する蛋白質を意味する。
「実質的に同質の活性」における「活性」は、上述の「細胞膜へ局在する活性」、及び「HVJ−Eを構成する活性」を意味する。「実質的に同質」とは、それらの活性が定性的に同じであることを意味する。したがって、上記の各活性は同等(例えば、約0.5〜約2倍)であることが好ましいが、これらの活性の程度などの量的要素は異なっていてもよい。
【0023】
また、HVJのHNタンパク質のN末端部分アミノ酸配列と実質的に同一のアミノ酸配列には、例えば、(1)上述のHVJのHNタンパク質のN末端部分アミノ酸配列と同一のアミノ酸配列のうち1または2個以上(好ましくは1〜30個程度、より好ましくは1〜10個程度、いっそう好ましくは1〜数(2、3、4もしくは5)個)のアミノ酸が欠失したアミノ酸配列、(2)上述のHVJのHNタンパク質のN末端部分アミノ酸配列と同一のアミノ酸配列に1または2個以上(好ましくは1〜30個程度、より好ましくは1〜10個程度、いっそう好ましくは1〜数(2、3、4もしくは5)個)のアミノ酸が付加したアミノ酸配列、(3)上述のHVJのHNタンパク質のN末端部分アミノ酸配列と同一のアミノ酸配列に1または2個以上(好ましくは1〜30個程度、より好ましくは1〜10個程度、いっそう好ましくは1〜数(2、3、4もしくは5)個)のアミノ酸が挿入されたアミノ酸配列、(4)上述のHVJのHNタンパク質のN末端部分アミノ酸配列と同一のアミノ酸配列のうち1または2個以上(好ましくは1〜30個程度、より好ましくは1〜10個程度、いっそう好ましくは1〜数(2、3、4もしくは5)個)のアミノ酸が他のアミノ酸で置換されたアミノ酸配列、または(5)それらを組み合わせたアミノ酸配列を含有する蛋白質なども含まれる。
【0024】
本発明の最も好ましい改変HNタンパク質は、下記(1)または(2)のポリペプチドである:
(1)配列番号6の第1〜160アミノ酸に該当するアミノ酸配列からなるポリペプチド、
(2)配列番号6の第1〜160アミノ酸に該当するアミノ酸配列において1または複数の残基が置換、欠失又は付加されたアミノ酸配列を含み、且つHVJに発現された場合に粒子表面に局在しうる特徴を有するポリペプチド。
【0025】
本発明の改変HNタンパク質は、例えば下記の融合タンパク質の構築や、下記の融合タンパク質の機能評価におけるネガティブコントロール等として有用である。
【0026】
本発明はさらに、前記改変HNタンパク質の残基及び第1の機能性分子を含む改変HNタンパク質であって、該改変HNタンパク質残基のC末端側に該機能性分子が機能可能に連結されている、改変HNタンパク質を提供する。ここで、「連結」とは、機能性分子が、改変HNタンパク質残基のC末端側に繋がっている状態を達成するものであれば、いかなる結合様式であっても特に限定されないが、好ましくはペプチド結合によって、改変HNタンパク質残基と機能性分子とが連結している。また、「機能可能に連結されている」とは、機能性分子がその機能を発揮できるように連結されていることを意味する。従って、機能性分子がその機能を発揮できる限りは、例えば、連結部位にリンカーやタグを介して改変HNタンパク質の残基と機能性分子が連結されていてもよい。本明細書における機能性分子とは、何らかの生物学的機能または活性を有している、ポリペプチド(例えば、タンパク質、ペプチド)、低分子化合物も含む、あらゆる分子を包含しうる。好ましくは、ポリペプチド(例えば、タンパク質、ペプチド)またはその類似体である。従って好ましくは、本発明は、前記改変HNタンパク質の残基及び第1の機能性ポリペプチド残基を含む融合タンパク質であって、該改変HNタンパク質残基のC末端側に該機能性ポリペプチド残基が機能可能に連結されている、融合タンパク質を提供する。該融合タンパク質においては、前記改変HNタンパク質の残基と第1の機能性ポリペプチド残基とが、結合手又はペプチドリンカーを介してペプチド結合により結合し、1本のポリペプチドを構成している。
【0027】
ペプチドリンカーのアミノ酸配列は、改変HNタンパク質の残基と第1の機能性ポリペプチド残基との機能的な連結を可能にする限り特に限定されない。ペプチドリンカーの長さは、改変HNタンパク質の残基と第1の機能性ポリペプチド残基との機能的な連結を可能にする限り特に限定されないが、通常1〜30アミノ酸、好ましくは1〜10アミノ酸である。
【0028】
本発明の融合タンパク質は、哺乳動物細胞(例えばサル腎臓細胞LLCMK2)において発現したときに、該細胞の細胞膜上に局在する(これを、細胞膜へ局在する活性という)。「局在」とは、細胞内の他の領域よりも細胞膜における発現量が高いことを意味する。融合タンパク質が細胞膜上へ局在するか否かは、該融合タンパク質を発現することが出来る発現ベクターにより、哺乳動物細胞(例えばサル腎臓細胞LLCMK2)をトランスフェクトし、得られたトランスフェクタントの細胞膜や各オルガネラにおける融合タンパク質の発現を、融合タンパク質を特異的に認識する抗体を用いたウェスタンブロットにより解析することにより、確認することが出来る。
【0029】
更に、本発明の融合タンパク質を哺乳動物細胞において発現すると、該融合タンパク質に含まれる機能性ポリペプチド領域が、細胞の外側に提示する形で、該哺乳動物細胞の細胞膜上に局在する。機能性ポリペプチド領域が、細胞の外側に提示されているか否かは、該機能性ポリペプチドを特異的に認識する抗体を用いて、フローサイトメーターや免疫組織染色により確認することが出来る。
【0030】
また、本発明の融合タンパク質は、HVJのウイルス粒子形成の際に、エンベロープへ取り込まれることにより、該エンベロープを構成し得る(これを、HVJ−Eを構成する活性という)。即ち、本発明の融合タンパク質を哺乳動物細胞において発現させ、該哺乳動物細胞にHVJを感染させると、HVJ感染細胞から生じるHVJのエンベロープに、該融合タンパク質が取り込まれ、該エンベロープを構成する。融合タンパク質が、かかる特徴を有するか否かは、該融合タンパク質を発現することが出来る発現ベクターにより、哺乳動物細胞(例えばサル腎臓細胞LLCMK2)をトランスフェクトし、得られたトランスフェクタントにHVJを感染させ、感染したトランスフェクタントから生じたHVJを回収し、該HVJからエンベロープを単離し、該エンベロープにおける融合タンパク質の発現を、融合タンパク質を特異的に認識する抗体を用いたウェスタンブロットにより解析することにより、確認することが出来る。
【0031】
更に、本発明の融合タンパク質が、HVJ−Eへ取り込まれると、該HVJ−Eは、該融合タンパク質に含まれる機能性ポリペプチド領域を、HVJ−Eの外側に提示する形で、該融合タンパク質を担持することとなる。機能性ポリペプチド領域が、HVJ−Eの外側に提示されているか否かは、該機能性ポリペプチドを特異的に認識する抗体を用いて、フローサイトメーターや免疫組織染色により確認することが出来る。
【0032】
本明細書における機能性ポリペプチドとは、何らかの生物学的機能または活性を有しているポリペプチドを言う。好ましくは、タンパク質、ペプチドまたはその類似体である。例えば、何らかの抗体またはその抗原結合部位、各種のサイトカイン、増殖因子、ホルモン、受容体または細胞膜タンパク質に対するリガンドなどが挙げられる。例えば、この機能性ポリペプチドとして細胞特異的表面抗原に対する抗体を用いた場合、本発明の融合タンパク質をHVJ−Eに取り込ませると、該HVJ−Eの外側に、該抗体が提示されるので、その抗原を表面に発現する細胞に選択的にHVJ−Eをターゲティングさせることが可能となる。また、機能性ポリペプチドとして何らかのサイトカインを用いた場合は、本発明の融合タンパク質をHVJ−Eに取り込ませると、該HVJ−Eの外側に、該サイトカインが提示されるので、そのサイトカイン受容体を細胞表面に発現する細胞特異的にHVJ−Eを選択的に作用させるとともに、該サイトカインで該細胞を刺激することも可能である。例えば、抗腫瘍活性を有するタンパク質(例えば、IL−12等のIFN−γ分泌促進能を有するポリペプチド)を機能性ポリペプチドとして用いた場合、本発明の融合タンパク質をHVJ−Eに取り込ませると、該HVJ−Eの外側に、抗腫瘍活性を有するタンパク質が提示されるので、HVJ−Eによる抗腫瘍活性に加えて、この機能性ポリペプチドの有する抗腫瘍活性も発揮されることで、より強力な抗腫瘍効果を得ることが可能である。
【0033】
本発明で用いる機能性ポリペプチドの好ましい一実施態様としては、IFN−γ分泌促進能を有する分子が挙げられる。このIFN−γ分泌促進能は、当業者に公知の方法および実施例に記載の方法で測定することが可能であるとともに、これらの方法に当業者が日常的な活動の範囲内にある改変を加えて測定してもよい。IFN−γ分泌促進能を有するポリペプチドとしては、哺乳動物(例、ヒト)の脾細胞または樹状細胞からのIFN−γ分泌促進能を有していればどのような分子であってもよいが、例えば、哺乳動物のIL−12が挙げられる。IL−12は、2つの異なるサブユニット(p35およびp45)から構成されるヘテロダイマーであってもよく、該2つのサブユニットが機能可能に連結された一本鎖IL−12であってもよい。好ましくは、一本鎖IL−12である。一本鎖IL−12を構築する技術は、例えばGraham J. Lieschke, et al., Nature Biotechnology. 15. 35−40 (1997)等に記載されており、当業者であればこれを容易に製造することが出来る。本発明において用いられる一本鎖IL−12は、IL−12の2つのサブユニットp35およびp40を一本のポリペプチドとして連結させたもので、好ましくはp40のC末端側にp35が機能可能に連結されている。p35とp40との間にはペプチドリンカーを含んでいても良く、好ましくは、Gly−Gly−Gly−Serからなるリンカーを1〜5単位、より好ましくは3単位含んでいる。そのような一本鎖IL−12としては、具体的には、下記(1)または(2)のポリペプチド:
(1)配列番号2の第103〜625アミノ酸に該当するアミノ酸配列を含むポリペプチド、
(2)配列番号2の第103〜625アミノ酸に該当するアミノ酸配列において1または複数の残基が置換、欠失又は付加されたアミノ酸配列を含み、且つIFN−γ分泌促進能を有するポリペプチドがあげられる。
【0034】
(2)の一本鎖IL−12のポリペプチドには、(1)配列番号2の第103〜625アミノ酸に該当するアミノ酸配列のうち1または2個以上(好ましくは1〜30個程度、より好ましくは1〜10個程度、いっそう好ましくは1〜数(2、3、4もしくは5)個)のアミノ酸が欠失したアミノ酸配列、(2)配列番号2の第103〜625アミノ酸に該当するアミノ酸配列に1または2個以上(好ましくは1〜30個程度、より好ましくは1〜10個程度、いっそう好ましくは1〜数(2、3、4もしくは5)個)のアミノ酸が付加したアミノ酸配列、(3)配列番号2の第103〜625アミノ酸に該当するアミノ酸配列に1または2個以上(好ましくは1〜30個程度、より好ましくは1〜10個程度、いっそう好ましくは1〜数(2、3、4もしくは5)個)のアミノ酸が挿入されたアミノ酸配列、(4)配列番号2の第103〜625アミノ酸に該当するアミノ酸配列のうち1または2個以上(好ましくは1〜30個程度、より好ましくは1〜10個程度、いっそう好ましくは1〜数(2、3、4もしくは5)個)のアミノ酸が他のアミノ酸で置換されたアミノ酸配列、または(5)それらを組み合わせたアミノ酸配列を含み、且つ哺乳動物(例、ヒト)の脾細胞または樹状細胞からのIFN−γ分泌促進能を有するポリペプチドが含まれる。
【0035】
尚、本発明の配列番号2で表されるアミノ酸配列は、アミノ酸番号1〜24:シグナルペプチド(p40)、アミノ酸番号25〜41:Flag−tag HA−tag、アミノ酸番号42〜99:ZZ−ドメイン、アミノ酸番号100〜102:リンカー、アミノ酸番号103〜417:IL−12−p40サブユニット、アミノ酸番号418〜432:リンカー、アミノ酸番号433〜625:IL−12−p35サブユニットで構成されている。
【0036】
本発明で用いる機能性ポリペプチドの別の好ましい一実施態様としては、抗体の定常領域(Fc)のCH2−CH3ドメインが挙げられる。このCH2−CH3ドメインは、プロテインAのZZドメインとの結合活性を有する限り、どのイムノグロブリンクラス(例えば、IgG、IgM、IgA、IgD、IgE)のものに由来してもよく、さらにどのサブクラス(例えば、IgG1、IgG2、IgG3、IgG4、IgA1、IgA2)のものに由来してもよく、また各CH2−CH3ドメインのアミノ酸配列に1又は複数個のアミノ酸が欠失・置換・付加されたものであってよい。また、CH2−CH3ドメインがプロテインAのZZドメインとの結合活性を有する限り、当該イムノグロブリンはどの動物種(マウス、ラット、ヒト等の哺乳動物、鳥類等)に由来してもよい。好ましい例としては、IgG2(たとえば、マウスIgG2)のCH2−CH3ドメインが挙げられる。そのような抗体の定常領域(Fc)のCH2−CH3ドメインとしては、具体的には、下記(1)または(2)のポリペプチドが挙げられる:
(1)配列番号4の第171〜389アミノ酸に該当するアミノ酸配列を含むポリペプチド、
(2)配列番号4の第171〜389アミノ酸に該当するアミノ酸配列において1または複数の残基が置換、欠失又は付加されたアミノ酸配列を含み、且つプロテインAのZZドメインとの結合活性を有するポリペプチド。
【0037】
(2)の一本鎖IL−12のポリペプチドには、(1) 配列番号4の第171〜389アミノ酸に該当するアミノ酸配列のうち1または2個以上(好ましくは1〜30個程度、より好ましくは1〜10個程度、いっそう好ましくは1〜数(2、3、4もしくは5)個)のアミノ酸が欠失したアミノ酸配列、(2) 配列番号4の第171〜389アミノ酸に該当するアミノ酸配列に1または2個以上(好ましくは1〜30個程度、より好ましくは1〜10個程度、いっそう好ましくは1〜数(2、3、4もしくは5)個)のアミノ酸が付加したアミノ酸配列、(3) 配列番号4の第171〜389アミノ酸に該当するアミノ酸配列に1または2個以上(好ましくは1〜30個程度、より好ましくは1〜10個程度、いっそう好ましくは1〜数(2、3、4もしくは5)個)のアミノ酸が挿入されたアミノ酸配列、(4) 配列番号4の第171〜389アミノ酸に該当するアミノ酸配列のうち1または2個以上(好ましくは1〜30個程度、より好ましくは1〜10個程度、いっそう好ましくは1〜数(2、3、4もしくは5)個)のアミノ酸が他のアミノ酸で置換されたアミノ酸配列、または(5)それらを組み合わせたアミノ酸配列を含み、且つプロテインAのZZドメインとの結合活性を有するポリペプチドが含まれる。
【0038】
尚、本発明の配列番号4で表されるアミノ酸配列は、アミノ酸番号1〜160:配列番号6の第1〜160アミノ酸配列、アミノ酸番号161〜170:Myc tag、アミノ酸番号171〜389:IgG2a−CH2−CH3ドメインで構成されている。
【0039】
ここで、上記の抗体の定常領域(Fc)のCH2−CH3ドメインは、プロテインAのZZドメインと結合することができる。従って、CH2−CH3ドメインを含む本発明の融合タンパク質(「第1の融合タンパク質」と表記することもある)を発現することが出来る発現ベクターにより、哺乳動物細胞(例えばサル腎臓細胞LLCMK2)をトランスフェクトし、得られたトランスフェクタントにHVJを感染させ、感染したトランスフェクタントから生じたHVJを回収し、該HVJからエンベロープを単離することにより、CH2−CH3ドメインをHVJのエンベロープの外側に提示する形で担持するHVJ−Eを得ることが出来る。更に、ZZドメインと機能可能に連結された前記の機能性ポリペプチドを含む融合タンパク質(「第2の融合タンパク質」と表記することもある)を、HVJ−E上の第1の融合タンパク質へZZドメインを介して結合することにより、所望の機能性ポリペプチドをHVJのエンベロープの外側に提示する形で担持するHVJ−Eを提供することができる。IgG2由来のCH2−CH3ドメインを用いた場合、第2の融合タンパク質が結合したHVJ−Eの血中安定性が高いため、体内に投与した場合でも血中に第2の融合タンパク質が結合したHVJ−Eが長時間存在し、第2の融合タンパク質に含まれる機能性ポリペプチドの機能を発揮するのに有利である。ただし、この他のサブクラスのイムノグロブリンに由来するCH2−CH3ドメインもまた、HVJ−Eと機能性ポリペプチドとの複合体の使用目的に応じて、当業者が選択することができる。
【0040】
従って、本発明は、上記の第1の融合タンパク質、及びプロテインAのZZドメイン及び第2の機能性ポリペプチド残基を含む第2の融合タンパク質を含む、タンパク質複合体であって、第1の融合タンパク質と第2の融合タンパク質とが、第1の融合タンパク質に含まれるCH2−CH3ドメインと第2の融合タンパク質に含まれるZZドメインとの間の結合を介して複合体を形成している、タンパク質複合体を提供する。
【0041】
本発明の第2の融合タンパク質に含まれる機能性ポリペプチドの好ましい一実施態様としては、前記のIFN−γ分泌促進能を有する分子が挙げられる。従って、そのような分子としては、IL−12が挙げられ、好ましくは、一本鎖IL−12が挙げられる。本発明のタンパク質複合体としては、具体的には、一本鎖IL−12が、下記(1)または(2)のポリペプチドであり、且つZZドメインが、下記(3)または(4)のポリペプチドである、タンパク質複合体を挙げることができる:
(1)配列番号2の第103〜625アミノ酸に該当するアミノ酸配列を含むポリペプチド、
(2)配列番号2の第103〜625アミノ酸に該当するアミノ酸配列において1または複数の残基が置換、欠失又は付加されたアミノ酸配列を含み、且つIFN−γ分泌促進能を有するポリペプチド、
(3)配列番号2の第42〜99アミノ酸に該当するアミノ酸配列を含むポリペプチド、
(4)配列番号2の第42〜99アミノ酸に該当するアミノ酸配列において1または複数の残基が欠失、置換または付加されたアミノ酸配列を含み、且つ抗体分子のFc領域への結合活性を有するポリペプチド。
【0042】
さらに、本発明は上記の本発明の改変HNタンパク質又は融合タンパク質をコードする核酸を提供する。該核酸は、本発明の改変HNタンパク質や融合タンパク質を遺伝子工学的手法により製造するために有用である。核酸はDNA又はRNAである。また、核酸は1本鎖又は2本鎖のいずれであってもよい。
【0043】
好ましい一実施態様としては、本発明の改変HNタンパク質をコードする核酸として、配列番号5に示される塩基配列の最も短い範囲で1番目〜333番目の塩基配列であって、最も長い範囲で1番目〜627番目の塩基配列を含有するDNA、または配列番号5に示される塩基配列の最も短い範囲で1番目〜333番目の塩基配列であって、最も長い範囲で1番目〜627番目の塩基配列の相補鎖配列とストリンジェントな条件下でハイブリダイズする塩基配列を含有し、前記したHNタンパク質と実質的に同質の活性を有する蛋白質をコードするDNAなどが挙げられる。より好ましくは、本発明の改変HNタンパク質をコードする核酸として、配列番号5に示される塩基配列の最も短い範囲で1番目〜420番目の塩基配列であって、最も長い範囲で1番目〜540番目の塩基配列を含有するDNA、または配列番号5に示される塩基配列の最も短い範囲で1番目〜420番目の塩基配列であって、最も長い範囲で1番目〜540番目の塩基配列の相補鎖配列とストリンジェントな条件下でハイブリダイズする塩基配列を含有し、前記したHNタンパク質と実質的に同質の活性を有する蛋白質をコードするDNA、最も好ましくは、配列番号5に示される塩基配列の1〜480までの塩基配列を含有するDNA、または配列番号5に示される塩基配列の1〜480までの塩基配列の相補鎖配列とストリンジェントな条件下でハイブリダイズする塩基配列を含有し、前記したHNタンパク質と実質的に同質の活性を有する蛋白質をコードするDNAである。
【0044】
別の好ましい一実施態様としては、配列番号6に表されるアミノ酸配列の配列番号1〜160のポリペプチドに一本鎖IL−12ポリペプチドが機能可能に連結されたタンパク質をコードする核酸を提供する。配列番号6に表されるアミノ酸配列の配列番号1〜160のポリペプチドをコードするDNAとしては、例えば、配列番号5に示される塩基配列の1〜480までの塩基配列を含有するDNA、または配列番号5に示される塩基配列の1〜480までの塩基配列の相補鎖配列とストリンジェントな条件下でハイブリダイズする塩基配列を含有し、前記したHNタンパク質と実質的に同質の活性を有する蛋白質をコードするDNAなどが挙げられる。また、一本鎖IL−12ポリペプチドをコードするDNAとしては、例えば、配列番号1に示される塩基配列の307〜1875までの塩基配列を含有するDNA、または配列番号1に示される塩基配列の307〜1875までの塩基配列の相補鎖配列とストリンジェントな条件下でハイブリダイズする塩基配列を含有し、前記したIL−12タンパク質と実質的に同質の活性を有する蛋白質をコードするDNAなどが挙げられる。
別の好ましい一実施態様としては、配列番号6に表されるアミノ酸配列の配列番号1〜160のポリペプチドにIgG2aのCH2−CH3ドメインのポリペプチドが機能可能に連結されたタンパク質をコードする核酸を提供する。配列番号6に表されるアミノ酸配列の配列番号1〜160のポリペプチドをコードするDNAとしては前記の通りである。また、IgG2aのCH2−CH3ドメインポリペプチドをコードするDNAとしては、例えば、配列番号3に示される塩基配列の511〜1167までの塩基配列を含有するDNA、または配列番号3に示される塩基配列の511〜1167までの塩基配列の相補鎖配列とストリンジェントな条件下でハイブリダイズする塩基配列を含有し、前記したCH2−CH3ドメインと実質的に同質の活性を有する蛋白質をコードするDNAなどが挙げられる。
前記の塩基配列とストリンジェントな条件下でハイブリダイズできるDNAとしては、例えば、配列番号5(または配列番号1または3)に示される塩基配列と約70%以上、好ましくは約80%以上、さらに好ましくは約90%以上、特に好ましくは約95%以上の相同性を有する塩基配列を含有するDNAなどが用いられる。
本明細書における塩基配列の相同性は、相同性計算アルゴリズムNCBI BLAST(National Center for Biotechnology Information Basic Local Alignment Search Tool)を用い、以下の条件(期待値=10;ギャップを許す;フィルタリング=ON;マッチスコア=1;ミスマッチスコア=−3)にて計算することができる。塩基配列の相同性を決定するための他のアルゴリズムとしては、上記したアミノ酸配列の相同性計算アルゴリズムが同様に好ましく例示される。
【0045】
ハイブリダイゼーションは、自体公知の方法あるいはそれに準じる方法、例えば、モレキュラー・クローニング(Molecular Cloning)第2版(J. Sambrook et al., Cold Spring Harbor Lab. Press, 1989)に記載の方法などに従って行なうことができる。また、市販のライブラリーを使用する場合、ハイブリダイゼーションは、添付の使用説明書に記載の方法に従って行なうことができる。ハイブリダイゼーションは、好ましくは、ストリンジェントな条件に従って行なうことができる。
ハイストリンジェントな条件としては、例えば、6×SSC(sodium chloride/sodium citrate)中45℃でのハイブリダイゼーション反応の後、0.2×SSC/0.1% SDS中65℃での一回以上の洗浄などが挙げられる。当業者は、ハイブリダイゼーション溶液の塩濃度、ハイブリダゼーション反応の温度、プローブ濃度、プローブの長さ、ミスマッチの数、ハイブリダイゼーション反応の時間、洗浄液の塩濃度、洗浄の温度等を適宜変更することにより、所望のストリンジェンシーに容易に調節することができる。
【0046】
配列番号6に表されるアミノ酸配列の配列番号1〜160のポリペプチド、一本鎖IL−12ポリペプチドおよびIgG2aのCH2−CH3ドメインのポリペプチドをコードするDNAは、化学的にDNA鎖を合成するか、もしくは合成した一部オーバーラップするオリゴDNA短鎖を、PCR法を利用して接続することにより、配列番号6に表されるアミノ酸配列の配列番号1〜160のポリペプチドおよび一本鎖IL−12ポリペプチドの全長をコードするDNAまたは配列番号6に表されるアミノ酸配列の配列番号1〜160のポリペプチドおよびIgG2aのCH2−CH3ドメインのポリペプチドの全長をコードするDNAを構築することも可能である。
【0047】
上記のようにしてクローン化されたDNAは、目的によりそのまま、または所望により制限酵素で消化するか、リンカーを付加した後に、使用することができる。配列番号6に表されるアミノ酸配列の配列番号1〜160のポリペプチドをコードするDNA、一本鎖IL−12ポリペプチドをコードするDNA、IgG2aのCH2−CH3ドメインのポリペプチドをコードするDNAを、別個にクローン化した場合には、適当な制限酵素、リンカー、リガーゼ等を用いて、それらを配列番号6に表されるアミノ酸配列の配列番号1〜160のポリペプチド、一本鎖IL−12ポリペプチドの順序または配列番号6に表されるアミノ酸配列の配列番号1〜160のポリペプチド、IgG2aのCH2−CH3ドメインのポリペプチドの順序で連結する。
【0048】
次いで、得られた前記DNAは、必要に応じて、その5’末端側に翻訳開始コドンとしてのATGを、また3’末端側には翻訳終止コドンとしてのTAA、TGAまたはTAGをそれぞれ導入し、制限酵素およびDNAリガーゼを用いて、適当な発現ベクター中のプロモーターの下流に連結される。翻訳開始コドンや翻訳終止コドンは、適当な合成DNAアダプターを用いて付加することができる。
【0049】
本発明の核酸を、発現ベクター内のプロモーターの制御下に組み込むことにより、本発明の改変HNタンパク質又は融合タンパク質を発現し得る発現ベクター(本発明の発現ベクター)を製造することが出来る。
【0050】
発現ベクターとしては、動物細胞発現プラスミド(例:pCAGGS 、pA1−11、pXT1、pRc/CMV、pRc/RSV、pcDNAI/Neo);レトロウイルス、ワクシニアウイルス、アデノウイルスなどの動物ウイルスベクターなどが用いられる。
プロモーターとしては、遺伝子の発現に用いる宿主の細胞内で機能可能なプロモーターであれば、いかなるものでもよい。
例えば、宿主が動物細胞である場合、CAGプロモーター、β−アクチンプロモーター、SRαプロモーター、SV40プロモーター、LTRプロモーター、CMV(サイトメガロウイルス)プロモーター、RSV(ラウス肉腫ウイルス)プロモーター、MoMuLV(モロニーマウス白血病ウイルス)LTR、HSV−TK(単純ヘルペスウイルスチミジンキナーゼ)プロモーターなどが用いられる。
【0051】
発現ベクターとしては、上記の他に、所望によりエンハンサー、スプライシングシグナル、ポリA付加シグナル、選択マーカー、SV40複製起点(以下、SV40 oriと略称する場合がある)などを含有しているものを用いることができる。選択マーカーとしては、例えば、ジヒドロ葉酸還元酵素遺伝子(以下、dhfrと略称する場合がある、メソトレキセート(MTX)耐性)、アンピシリン耐性遺伝子(以下、amprと略称する場合がある)、ネオマイシン耐性遺伝子(以下、neorと略称する場合がある、G418耐性)等が挙げられる。特に、dhfr遺伝子欠損チャイニーズハムスター細胞を用い、dhfr遺伝子を選択マーカーとして使用する場合、チミジンを含まない培地によって目的遺伝子を選択することもできる。
【0052】
本発明の発現ベクターで宿主を形質転換し、得られる形質転換体を培養することによって、本発明の改変タンパク質又は本発明の融合タンパク質を製造することができる。
宿主としては、例えば、動物細胞、動物などの分泌発現に適したものが用いられる。
【0053】
動物細胞としては、例えば、COS−7、Vero、CHO、CHO(dhfr−)、CHO−K1、L、AtT−20、GH3、FL、HEK293、NIH3T3、Balb3T3、FM3A、L929、SP2/0、P3U1、B16、P388、LLCMK2などの細胞が用いられる。
動物としては、トランスジェニック系の確立した哺乳動物(例えば、マウス、ラット、ウサギ、ヒツジ、ブタ、ウシ)もしくはニワトリなどが挙げられる。
【0054】
形質転換は、宿主の種類に応じ、公知の方法に従って実施することができる。
動物細胞は、例えば、細胞工学別冊8 新細胞工学実験プロトコール,263−267 (1995)(秀潤社発行)、Virology,52巻,456 (1973)に記載の方法に従って形質転換することができる。
動物は、例えば、トランスジェニック動物の開発(シーエムシー出版),(2001)に記載の方法に従って形質転換することができる。
【0055】
形質転換体の培養は、宿主の種類に応じ、公知の方法に従って実施することができる。
宿主が動物細胞である形質転換体を培養する場合の培地としては、例えば、約5〜約20%の胎児ウシ血清を含む最小必須培地(MEM),ダルベッコ改変イーグル培地(DMEM),RPMI 1640培地,199培地などが用いられる。培地のpHは、好ましくは約6〜約8である。培養は、通常約30〜約40℃で、約15〜約60時間行なわれる。必要に応じて通気や撹拌を行ってもよい。
宿主が動物である場合、遺伝子導入した受精卵から、常法に従ってトランスジェニック動物を得、通常の飼育条件下で飼育、哺乳動物の乳やニワトリの卵を採取すればよい。
【0056】
このようにして得られた本発明の改変タンパク質又は融合タンパク質を発現する細胞から、適切なタンパク質精製法(例えば、HPLC、アフィニティクロマトグラフィ等)を用いて、本発明の改変タンパク質又は融合タンパク質を精製することにより、単離又は精製された本発明の改変タンパク質又は融合タンパク質を得ることが出来る。単離又は精製とは、目的とする成分以外の成分を除去する操作がなされていることを意味する。哺乳動物細胞中では、本発明の改変タンパク質又は融合タンパク質は、細胞膜へ局在するので、該細胞を界面活性剤で処理し、この溶解液を精製操作に付すことにより、単離又は精製された本発明の改変タンパク質又は融合タンパク質を得ることが出来る。単離又は精製された本発明の改変タンパク質又は融合タンパク質の純度は、タンパク質純度として、通常50%(w/w)以上、好ましくは80%(w/w)以上、より好ましくは90%(w/w)以上、最も好ましくは実質的に100%である。
【0057】
上記のようにして得られた、本発明の改変HNタンパク質又は融合タンパク質を発現するトランスフェクタントへHVJを感染させ、感染したトランスフェクタントから生じたHVJを回収し、該HVJからエンベロープを単離することにより、本発明の改変HNタンパク質又は融合タンパク質をエンベロープに含有する、HVJを製造することが出来る。ここで、本発明の融合タンパク質が、HVJのエンベロープへ取り込まれると、該HVJは、該融合タンパク質に含まれる機能性ポリペプチド領域を、HVJのエンベロープの外側に提示する形で、該融合タンパク質を担持する。更に、該HVJを紫外線等に曝露することによってゲノムRNAで不活性化することで、HVJ−Eを製造することが出来る。従って、本発明は、本発明の改変HNタンパク質又は融合タンパク質をエンベロープに含有する、HVJ−Eを提供する。
【0058】
本明細書で使用される場合、「HVJ−E」とは、HVJの最も外側に位置する膜状の構造物を意味する。HVJにおいては、エンベロープは、通常、ウイルスゲノム及びカプシドタンパク質を覆っている。HVJ−Eは、通常、その内腔に種々の物質を担持することが可能であり、且つ細胞膜へ融合し、その内腔に含まれる物質を細胞内へ移入する活性を有する。そのため、HVJ−Eの内腔に、所望の物質(遺伝子(例えば、DNA、RNAなど)、タンパク質(またはペプチド)、化合物(抗癌剤、抗菌剤、免疫促進剤など)を封入し、これを個体へ投与することにより、種々の組織や細胞の内部へ、所望の物質を送達することが可能である。すなわち、HVJ−Eは種々の物質のベクターとして使用可能である。本発明において使用されるHVJ−Eは、その内腔に、生体内や細胞内への導入が意図される物資を封入していても、封入していなくてもよい。
【0059】
HVJ−Eについてより詳しくは、例えば、特開2001−286282号公報(WO01/57204号公報)、特開2002−065278号公報、WO03/014338号公報等に記載されており、具体的には例えば特開2001−286282号公報の実施例8などに従って調製することができる。
【0060】
本発明に用いられるHVJ−Eは、好ましくは、単離且つ精製されたものである。「単離且つ精製」とは、目的物以外の成分(ウイルスゲノム、カプシドタンパク質等)を除去する操作がなされていることを意味する。
【0061】
本発明は、また、本発明のタンパク質複合体をエンベロープに含有する、HVJ−Eを提供する。ここで、本発明のタンパク質複合体に含有される、本発明の融合タンパク質(第1の融合タンパク質)は、HVJ−Eへ取り込まれており、該融合タンパク質に含まれる機能性ポリペプチド領域(CH2−CH3ドメイン)が、HVJ−Eの外側に提示されている。ここへ、本発明のタンパク質複合体に含有される第2の融合タンパク質が結合することより、結果として、第2の融合タンパク質に含まれる第2の機能性ポリペプチドが、HVJ−Eの外側に提示されることとなる。
【0062】
本発明のタンパク質複合体をHVJのエンベロープに含有させるためには、抗体の定常領域(Fc)のCH2−CH3ドメインを機能性ポリペプチドとして含有する本発明の融合タンパク質(第一の融合タンパク質)をHVJのエンベロープに組み込ませたHVJ−Eを作製し、その後、該HVJ−Eと別途調製したZZドメインに機能可能に連結された機能性ポリペプチドを含む第2の融合タンパク質とを結合させて複合体を形成させる。複合体形成のためには、両分子を含む溶液を調製し、5分以上、好ましくは15分以上、例えば30分または1時間、ゆるやかに攪拌または静置するとよい。この溶液は、生理的緩衝液および生理的食塩水など、タンパク質の変性をもたらさない緩衝液中で行うことが好ましい。複合体の形成は、常法の免疫学的手法で確認することもでき、例えば機能性ポリペプチドが一本鎖IL−12である場合は、ショ糖密度勾配遠心に供してHVJ−EとZZドメインに連結された一本鎖IL−12が同じショ糖密度の画分に回収されることを、免疫学的手法またはIL−12活性の測定により確認することもできる。
【0063】
本発明は、またIFN−γ分泌促進能を有するポリペプチドをエンベロープの外側に提示する形で担持する、HVJ−Eを提供する。IFN−γ分泌促進能を有するポリペプチドの定義は上述の通りであり、該ポリペプチドとしては、哺乳動物のIL−12を挙げることができる。IL−12は、2つの異なるサブユニット(p35およびp45)から構成されるヘテロダイマーであってもよく、該2つのサブユニットが機能可能に連結された一本鎖IL−12であってもよい。好ましくは、一本鎖IL−12である。一本鎖IL−12としては、配列番号1の第103〜625アミノ酸に該当するアミノ酸配列を含むポリペプチドを挙げることが出来る。
【0064】
IFN−γ分泌促進能を有するポリペプチドをエンベロープの外側に提示する形で担持させる方法としては、上述のように、機能性ポリペプチドとしてIFN−γ分泌促進能を有するポリペプチドを含む本発明の融合ポリペプチドや、第2の機能性ポリペプチドとしてIFN−γ分泌促進能を有するポリペプチドを含む本発明のタンパク質複合体を、エンベロープに含有させる方法が好適なものとして挙げられる。但し、これらの方法に特に限定されるものではなく、当業者に公知の方法でIFN−γ分泌促進能を有するポリペプチドをエンベロープの外側に提示する形で担持させることができる。例えば、HVJ−Eの外側をビオチン化し、ストレプトアビジンをコンジュゲートしたIFN−γ分泌促進能を有するポリペプチドを、ビオチン化したHVJ−Eへ結合させる方法等を挙げることができる。
【0065】
本態様のHVJ−Eに含有されるIFN−γ分泌促進能を有するポリペプチドの量は、特に限定されないが、IFN−γ分泌促進能を有するポリペプチドとしてIL−12を用いる場合には、その含有量は、HVJ−E 100HAUあたり、1〜10000pg、好ましくは10〜1000pg、より好ましくは100〜500pg(例えば、250pg)である。本明細書においてHAU(Hemagglutinating unit)とは赤血球を凝集させる活性の単位であり、HVJ−Eの1HAUとはニワトリの赤血球と混合した場合に凝集を起こす最小単位のことをいう。例えば1000HAUとは、目的の検体の1000分の1量をニワトリの赤血球と混合した場合には凝集を起こすが、それ未満では凝集を起こさない量をいう。実際の測定は、公知の方法で行うことが可能であるが、例えば、96ウェルの試験プレートのウェルに2倍希釈系列でHVJ−Eを50μlずつ入れ、そこにニワトリの赤血球懸濁液を50μL添加して計100μLとし、溶液の凝集の有無を目視で確認し、その希釈率を基に計算することができる。
【0066】
IL−12等のIFN−γ分泌促進能を有するポリペプチド自体は抗腫瘍活性を有しており、また、HVJ−Eも抗腫瘍活性を有しているが、IFN−γ分泌促進能を有するポリペプチドをエンベロープの外側に提示する形で担持させることにより、抗腫瘍活性が飛躍的に上昇する。
【0067】
HVJ−Eは、化学療法剤、治療用外来遺伝子等の治療物質をその中に含有することで、標的となる細胞に高効率で治療物質を導入することができることは当業者にとっては公知である。また、本発明者らは、HVJ−E自体に抗腫瘍活性があることを見出している。本発明のHVJ−Eにおいては、機能性分子をその粒子表面に露出されることが可能であり、この機能性分子の作用により、従来公知であった抗腫瘍活性を増強させたり、HVJ−Eを特定の腫瘍へ標的化し得ることを見出した。従って、本発明は、前記本発明のHVJ−Eを有効成分として含む抗癌剤と提供する。本発明の抗癌剤は、あらゆる種類の癌(固形癌、細胞癌など)に有効である。消化器系、泌尿器科系、神経系、結合組織における癌、皮膚癌等あらゆる種類の癌に適用可能であるが、一例として、メラノーマ、前立腺癌、大腸癌などが挙げられる。本発明のHVJ−Eは低毒性であり、そのまま液剤として、または適当な剤型の医薬組成物として、ヒトまたは他の哺乳動物(例、マウス、ラット、ウサギ、ヒツジ、ブタ、ウシ、ネコ、イヌ、サルなど)に対して経口的または非経口的(例、血管内投与、皮下投与など)に投与することができる。
【0068】
本発明のHVJ−Eを癌の治療剤などの医薬として使用する場合、自体公知の方法に従って製剤化し、投与することができる。本発明のHVJ−Eは常套手段に従って製剤化することができる。該HVJ−Eは、そのままで、あるいは摂取促進のための補助剤とともに、遺伝子銃やハイドロゲルカテーテルのようなカテーテルによって投与することができる。あるいは、エアロゾル化して吸入剤として気管内に局所投与することもできる。
【0069】
本発明のHVJ−Eは、それ自体を投与してもよいし、または適当な医薬組成物として投与してもよい。投与に用いられる医薬組成物としては、本発明のHVJ−Eと薬理学的に許容され得る担体、希釈剤もしくは賦形剤とを含むものであってよい。このような医薬組成物は、経口または非経口投与に適する剤形として提供される。
【0070】
非経口投与のための組成物としては、例えば、注射剤、坐剤等が用いられ、注射剤は静脈注射剤、皮下注射剤、皮内注射剤、筋肉注射剤、点滴注射剤等の剤形を包含しても良い。このような注射剤は、公知の方法に従って調製できる。注射剤の調製方法としては、例えば、上記本発明のHVJ−Eを通常注射剤に用いられる無菌の水性液、または油性液に溶解、懸濁または乳化することによって調製できる。注射用の水性液としては、例えば、生理食塩水、ブドウ糖やその他の補助薬を含む等張液等が用いられ、適当な溶解補助剤、例えば、アルコール(例、エタノール)、ポリアルコール(例、プロピレングリコール、ポリエチレングリコール)、非イオン界面活性剤〔例、ポリソルベート80、HCO−50(polyoxyethylene(50mol)adduct of hydrogenated castor oil)〕等と併用してもよい。油性液としては、例えば、ゴマ油、大豆油等が用いられ、溶解補助剤として安息香酸ベンジル、ベンジルアルコール等を併用してもよい。調製された注射液は、適当なアンプルに充填されることが好ましい。直腸投与に用いられる坐剤は、上記HVJ−Eを通常の坐薬用基剤に混合することによって調製されてもよい。
【0071】
経口投与のための組成物としては、固体または液体の剤形、具体的には錠剤(糖衣錠、フィルムコーティング錠を含む)、丸剤、顆粒剤、散剤、カプセル剤(ソフトカプセル剤を含む)、シロップ剤、乳剤、懸濁剤等が挙げられる。このような組成物は公知の方法によって製造され、製剤分野において通常用いられる担体、希釈剤もしくは賦形剤を含有していても良い。錠剤用の担体、賦形剤としては、例えば、乳糖、でんぷん、蔗糖、ステアリン酸マグネシウムが用いられる。
【0072】
本発明のHVJ−Eを注射により投与する場合、腫瘍部位、またはその周辺部に、HVJ−Eを直接注射により投与することができる。投与量は、腫瘍の大きさ、患者の年齢、体重、状態等を考慮して、医者または医療従事者が適宜決定することができる。腫瘍体積が1000mm3以下(例えば、200mm3程度など)の場合、1腫瘍部位あたり1回当たりのHVJ−Eの投与量を10〜10000HAU、好ましくは10〜5000HAU、さらに好ましくは50〜1000HAU、例えば100HAUとすることができる。あるいはまた、体重1kgあたり、300000HAU以下であることが好ましい。
【0073】
本発明のHVJ−Eとして、IL−12をエンベロープの外側に提示する形で担持する、HVJ−Eを用いた場合における、IL−12としての投与量は、腫瘍体積が1000mm3以下(例えば、200mm3程度など)の場合、1腫瘍部位あたりの1回あたりのIL−12の投与量を0.1pg〜1000pg、1〜500pg、さらに好ましくは10〜50pg(例えば25pg)とすることができる。HVJ−E自体の抗腫瘍効果とIL−12の抗腫瘍効果とが相まって、極めて強力な抗腫瘍活性を達成することが可能であるため、HVJ−EやIL−12を、それぞれ単独で投与する場合や、HVJ−EとIL−12とを単に混合して投与する場合と比較して、HVJ−E及びIL−12それぞれの投与量を減らすことが可能であり、安全性の観点から有利である。例えば、1腫瘍部位あたり1回当たりの投与量を、HVJ−Eとして、1000HAU以下、好ましくは50〜1000HAU、より好ましくは100HAUとすることができる。また、同じ場合、該投与量を、IL−12として、1腫瘍部位あたり100pg以下、例えば10〜100pg、好ましくは10〜50pg(例えば25pg)とすることができる。このような低い投与量は、特に、腫瘍体積が1000mm3以下の場合に、有効である。また、HVJ−EとIL−12 がそれぞれ所望の量含まれるように、抗体の定常領域(Fc)のCH2−CH3ドメインが機能可能に連結された第1の融合タンパク質をエンベロープに含むHVJ−Eと第2の融合タンパク質とを混合する際にその比率を適宜変更することができる。また腫瘍体積が1000mm3以上の場合であっても、例えばHVJ−Eが1腫瘍部位あたりの1回投与量として10〜100000HAU、IL−12が1pg〜10000pgの範囲から、患者の状態に応じて投与量を決定することができる。同様に投与回数も腫瘍の大きさ、患者の年齢、体重、状態等を考慮して、医者または医療従事者が適宜決定することができる。
【0074】
なお、前記した各組成物は、本発明のHVJ−Eとの配合により好ましくない相互作用を生じない限り他の活性成分を含有してもよい。
【実施例】
【0075】
改変HVJ−Eの作製スキーム
本実施例に用いる1本鎖IL−12(以下、scIL−12とも表す)との会合により抗腫瘍効果を向上させた改変HVJ−Eの作製スキームを図1に示す。図1では、この機能性分子がIL−12の場合を例として記載しているが、細胞表面の受容体に結合する各種の分子等、所望の機能性分子をこの方法でHVJ−Eの表面に提示させることができることは、当業者であれば容易に理解可能である。
すなわち、機能性ポリペプチドを提示するHVJ−Eには、イムノグロブリン(Ig)分子の定常領域(Fc)のCH2−CH3ドメインをウイルス表面に提示するFc−HNタンパク質と、所望の機能性ポリペプチドとプロテインAのZZドメインとを融合させたZZ融合タンパク質との、2種の融合タンパク質が用いられた。
まず、天然型HVJ−Eのウイルス粒子表面に提示されたHNタンパク質の粒子外ドメインを抗体の定常領域(Fc)に置換したHN−Fc融合タンパク質を作製し、これをHVJ−EのHNタンパク質の代わりにウイルス粒子に提示するFc−HN−HVJ−Eを作製した(図1a)。
より詳細には、HNタンパク質の粒子外ドメインを抗体の定常領域(Fc)に置換したHN−Fc融合タンパク質をコードする遺伝子を細胞に導入して該細胞でHN−Fc融合タンパク質を発現させた。この細胞に天然型HNタンパク質を持つ天然型HVJ−Eを感染させて増幅させた。すると、このウイルス増幅の過程で、細胞内で発現され細胞外にFcを提示していたHN−Fc融合タンパク質を粒子外被に取り込んでFc部を粒子表面に提示したウイルス(Fc−HN−HVJ−E)が細胞外に放出された(図2)。
次いで、一本鎖IL−12にプロテインAのZZドメイン(該ドメインがFcと結合する)を付加したscIL−12−ZZ融合タンパク質(ZZ−IL12)を作製した(図1b)。
Fc−HN−HVJ−EとZZ−IL12とを混合することで、Fc−HN−HVJ−Eの粒子表面のFcにZZ−IL12のZZドメインが結合して、ZZ−IL12とFc−HN−HVJ−Eとが会合し、HVJ−Eの粒子表面にIL−12が提示されたZZ−IL12−Fc−HN−HVJ−Eが得られた(図1a+b)。
より具体的に本実施例に用いたZZ−IL12−Fc−HN−HVJ−Eの作製方法を以下に記載するが、この記載により本発明の態様がこれに限定されるものではない。
【0076】
1.Fc−HN−HVJの作製
(1)ウイルス粒子に取り込まれるために必要なHNタンパク質内の領域の特定
まず、ウイルス粒子に取り込まれ得るHN−Fc融合タンパク質を作製するにあたり、HNタンパク質のどの領域がウイルス粒子に取り込まれるために必須であるのかを特定した。
HNタンパク質中のウイルス粒子の外に提示される領域(細胞外ドメイン)のC末端側から欠失させて、6種類のリコンビナントHN発現ベクター(Full length HN、ecto−400aa−HN、ecto−300aa−HN、ecto−200aa−HN、ecto−100aa−HN、ecto−0aa−HN)を作製した(図3(a))。これらのリコンビナントHNは、免疫学的検出を可能とするために、便宜上、C末端側にMycタグを融合させた。このリコンビナントHN発現ベクターをサル腎臓細胞LLCMK2に導入し、その発現をウエスタンブロットにより確認したところ、所望の分子量の位置に各リコンビナントHNが発現されていた(図3(c))。このウエスタンブロットで検出される発現量がかなり少なかったecto−0aa−HN以外のリコンビナントHNについて、細胞内局在を免疫染色法で確認した。Triton存在下では、いずれも細胞質に検出されたが、Toriton非存在下で本来の細胞内局を見たところ、Full length HN 及びecto−100aa−HNのみが細胞膜表面に局在している事が明らかとなった(図3(b))。さらに、これらのリコンビナント HN 発現細胞に HVJ を感染させると、ecto−100aa−HN のみが感染細胞由来 HVJ に取り込まれていることが確認された(図3(d))。
また、上記と同様に、ecto−50aa−HN、ecto−80aa−HN、ecto−120aa−HNおよびecto−150aa−HNを作製して(図4(a))、同様に細胞内発現(図4(c))、細胞内局在(図4(b))およびウイルス粒子への取り込み(図4(d))を確認したところ、ecto−50aa−HNは、細胞内発現が低く、細胞膜表面への局在も確認できなかったが、ecto−80aa−HN、ecto−120aa−HNおよびecto−150aa−HNについては、十分に発現され細胞膜表面に局在することが確認された。ウイルス粒子への取り込みについては、ecto−80aa−HNおよびecto−120aa−HNにおいて、認められた(図4(d))。
以上より、C末端切断型のHNタンパク質がウイルス粒子に取り込まれるためには、HNタンパク質の細胞外ドメインのN末端側の少なくとも51残基以上、好ましくは80残基以上(N末端から111残基以上、好ましくは140残基以上)が必要であることがわかった。また、N末端部分アミノ酸配列の長さが209アミノ酸(好ましくは180アミノ酸)よりも長いと、かえってC末端切断型のHNタンパク質はウイルス粒子に取り込まれなくなることが分かった。
(2)Fc−HN−HVJの作製
次いで、上記ecto−100aa−HNを利用してFc−HN−HVJを作製した。まず、ecto−100aa−HNのMycタグのC末端側にマウスIgG2aFcドメインのCH2−CH3ドメインをさらに付加したFc融合ecto−100aa−HN(HN−Fc)を作製した(図5(a))。このHN−Fcについても、上記と同様の方法で、細胞内発現(図5(b))、細胞内局在(図5(c))、ウイルス粒子への取り込み(図5(d))を確認できた。さらに、このecto−100aa−HNのプロテインA結合能を調べるために、Fc−HNとプロテインAセファロースとの共沈実験を行った。ecto−100aa−HN発現LLCMK2およびFc−HN発現LLCMK2をそれぞれに溶解させてプロテインAセファロースと混合後、プロテインAセファロースを沈殿させた。ecto−100aa−HNはプロテインAセファロースと共沈せず、全て細胞溶解液(lysate)中に残ったが、Fc−HNはプロテインAセファロースと共沈し、プロテインAとの結合が可能である事が確認された(図5(e))。
【0077】
2.ZZドメイン融合型機能性ポリペプチドの作製(機能性ポリペプチドが一本鎖IL−12である場合)
プロテインAのZZドメインと一本鎖IL−12とを融合させて、ZZ−IL−12を作製した。まず、Lieschkeらの報告(Nature Biotechnology, 15, 35−40 (1997))にならって一本鎖IL−12(scIL12)を作製した。具体的には、IL−12の第一のサブユニットp40と第二のサブユニットp35との間にGly−Gly−Gly−Serからなるリンカーが3つ連結して挟まれ、さらにp40のN末端側にシグナルペプチドを連結したものである(図6(a))。本実施例においては、免疫学的検出の便宜のために、シグナルペプチドC端とp40サブユニットのN端との間にFlagおよびヘマグルチニン(HA)のタグを挿入し(Tag−scIL12)、さらに、HAタグのC端とp40サブユニットの間にプロテインAのZZドメインを挿入したものを遺伝子工学的常法により作製した(ZZ−scIL12)。また、ネガティブコントロールとしてp35サブユニットを欠損したものも作製した(ZZ−p40)。なお、このシグナルペプチドはタンパク質生成後切断される。これらが、IL−12として機能しうるかどうかを確認するために、Tag−scIL12、ZZ−scIL12およびZZ−p40について、脾細胞からのインターフェロンγ(IFNγ)の分泌刺激効果を測定した。マウス大腿骨から骨髄細胞を採取し、DC−Medium(RPMI(10%FBS、100unit/ml penicillin、0.1mg/ml streptomycin、10ng/ml GM−CSF、4nl/ml 2−mercaptethanole))で6日間培養後、浮遊細胞を樹状細胞(DC)として回収し、1×105cells/100μl/wellで96 well plateにまいた。
Tag−scIL12、ZZ−scIL12およびZZ−p40のそれぞれを、10 ng/mlの濃度となるよう各wellに加え(total 200 μl)、24h 37℃でインキュベーションし、96well plateを遠心(440×g、5 min)することで、上清を回収し、上清中のIFNγをELISAで測定した。その結果、ZZ−p40はIFNγ分泌誘導を示さなかったが、Tag−scIL12およびZZ−scIL12は共に、IFNγ分泌誘導を示し(図6(b))、ここで作製したZZ−scIL12がIL−12として作用することが確認された。
【0078】
3.ZZ−scIL12とFc−HN−HVJとの結合能
ZZ−scIL12が、Fc−HNを粒子表面に提示したHVJ(Fc−HN−HVJ)と結合して複合体を形成しうるかどうかを調べた。
上記Fc−HNの発現ベクターをLLCMK2に導入後、HVJを該細胞に1.5MOIにて感染させて増幅させたHVJを回収することにより、Fc−HNを粒子表面に提示したFc−HN−HVJを得た。このFc−HN−HVJ液(TFBバッファー:20mM Tris−HCl(pH7.0)、150mM NaCl、1mM MgCl2に懸濁)とZZ−scIL−12液(ZZ−scIL12発現ベクター導入CHO−K1培養上清(血清不含HAM’S F−12培地))とを、体積比1:20で混合し(4℃、一晩、転倒混和)、その後、ショ糖密度勾配による遠心分離(25−50%スクロース、100,000×g、11h)後、scIL12をHAタグに対する抗体で、HVJはHVJに発現しているMタンパク質に対する抗体で、それぞれショ糖濃度画分のウェスタンブロットにより検出した。Fc−HNを提示していないHVJとZZ−scIL12とを混合した場合は、ZZ−scIL12は低ショ糖濃度の画分に、HVJは高ショ糖濃度の画分に、それぞれ分離される(図7右上図)のに対し、Fc−HN−HVJと混合した場合には、ZZ−scIL12はFc−HN−HVJと同じ高ショ糖濃度の画分にも検出され、混合液中のZZ−scIL12はFc−HN−HVJと複合体を形成していることが確認された。なお、ZZドメインを有さないscIL−12(Tag−scIL12)はFc−HN−HVJと混合した場合でも、Fc−HN−HVJと同じ高ショ糖濃度の画分には検出されなかった(図7左下図)。したがって、Fc−HN−HVJとZZ−scIL−12とは、Fc部とZZドメインの結合を介して会合し複合体を形成するものと考える。
【0079】
4.ZZ−scIL12とHVJ−Eとの複合体の血清中での結合安定性
前述のようにして得られたFc−HN−HVJを、公知の方法(WO2006/011580など参照のこと)に従って精製し、Fc−HVJ−Eを得た。このFc−HVJ−EとZZ−scIL12とを上記と同様に混合し、血清中で37℃で0〜24時間インキュベートした後、上記同様にウエスタンブロットに供した。その結果、24時間のインキュベーションにおいても、ZZ−scIL12とFc−HVJ−Eの結合が維持されている事が確認された(図8)。
【0080】
5.ZZ−scIL12結合型Fc−HVJ−Eの樹状細胞(DC)および脾臓細胞への作用
Fc−HVJ−Eを樹状細胞(DC)に対するZZ−scIL12結合型Fc−HVJ−Eの作用を検討した。DCの調製は、マウス大腿骨から骨髄細胞を採取し、DC−Medium(RPMI(10%FBS、100unit/ml penicillin、0.1mg/ml streptomycin、10ng/ml GM−CSF、4nl/ml 2−mercaptethanole))で6日間培養した後、浮遊細胞(DC)を回収し、1×105cells/100μl/wellで96well plateにまいた。その後、天然型HVJ−E、Fc−HN−HVJ−EおよびZZscIL12と複合体を形成したZZ−scIL12結合型Fc−HN−HVJ−EHVJ−Eを、それぞれ10、50または100HAU(100μl培地中)を各ウェルに加え、24時間、37℃でインキュベーションした。この96well plateを440×g、5minで遠心し、上清を回収し、上清中のIFN−γおよびIL−6の量をELISAで測定した。その結果を図9に示す。ZZ−scIL12結合型Fc−HVJ−Eは、DCからのIFN−γ分泌を促進した(図9中図)。天然型HVJ−EおよびFc−HN−HVJ−EもIFN−γ分泌促進を誘導したが、ZZ−scIL12結合型Fc−HVJ−Eが最もIFN−γ分泌促進能に優れていた。また、DCからのIL−6分泌促進能については、天然型HVJ−Eよりは低いものの、Fc−HN−HVJ−EおよびZZ−scIL12結合型Fc−HVJ−Eとも、IL−6の分泌も促進させることが確認された(図9右図)。
さらにまた、脾臓細胞におけるIFN−γ分泌促進についても検討した。脾臓細胞は、マウス脾臓から脾細胞を回収後、塩化アンモニウム−トリス緩衝液を加えて赤血球を溶血させた後、細胞をsplenocyte−Medium(RPMI(10%FBS、100unit/ml penicillin、0.1mg/ml streptomycin、4nl/ml 2−mercaptethanole))に懸濁し、2×105cells/100μl/wellで96well plateにまいた。これに上記と同様に、天然型HVJ−E、Fc−HN−HVJ−EおよびZZ−scIL12 結合型Fc−HN−HVJ−EHVJ−Eを、それぞれ10、50または100HAU(100μl培地中)を、ならびにIL−12を25pg、50pgおよび250pg(100μl培地中)をそれぞれ、各ウェルに加え24h 37℃でインキュベーションした。この96well plateを440×g、5minで遠心し、上清を回収し、上清中のIFN−γの量をELISAで測定した。その結果を図10に示す。天然型HVJ−EおよびFc−HN−HVJ−Eは、脾臓細胞のIFN−γ分泌を誘導せず、IL−12も殆どIFN−γ分泌を誘導しなかったが、ZZ−scIL12結合型Fc−HVJ−EはIFN−γ分泌を誘導した(図10)。すなわち、ZZ−scIL12結合型Fc−HVJ−Eの脾細胞からのIFN−γ分泌促進に関しては、IL−12とHVJ−Eとの相加効果を上回るものである。
【0081】
6.ZZ−scIL12結合型Fc−HVJ−Eのin vivoでの抗腫瘍作用
マウスの背にF10メラノーマをの皮内注入し、その腫瘍径が2〜4mmになったところでZZ−scIL12結合型Fc−HVJ−E(HVJ−E:100HAU、IL−12:250pg)を2日おきに3回腫瘍内注射をし、その後の腫瘍径や免疫系活性化能を測定した。
ZZ−scIL12結合型Fc−HVJ−E投与後の腫瘍体積の増大はほとんど認められず、天然型HVJ−Eと比較しても明らかに強い腫瘍増殖抑制が認められた(図11左図)。なお、本発明者らは、同様の方法で、通常のHVJ−Eが前立腺癌の腫瘍体積の増加をほぼ90%以上抑制するという事実を発表しているが(WO2005/094878)、これに用いたHVJ−Eは5000HAUという高用量であり、Fc−HN−HVJ−E+ZZ−scIL12が、HVJ−Eの力価として100HAUという低用量でメラノーマに対して同レベルの効果を呈するということは驚くべき事実であった。また、ZZ−scIL12結合型Fc−HVJ−E投与群の4個体中、2個体は完全に寛解した。このことから、ZZ−scIL12結合型Fc−HVJ−Eは、100HAUという低容量でも強力な抗腫瘍作用を示すことがわかった。
【0082】
7.ZZ−scIL12結合型Fc−HVJ−Eのin vivoでの免疫活性化能の検討
マウス背部の皮内に5×105 cellsのF10メラノーマを注入し、腫瘍が約3〜5mmになった時点で、天然型HVJ−E100HAUまたはZZ−scIL12結合型Fc−HVJ−E(HVJ−E:100HAU、IL−12:250pg)を腫瘍内に2日おきに3回投与した。最終投与から10日後に各マウスの脾細胞を採取した。
マイトマイシンCで処理(15μg/ml、45min、37℃)したF10メラノーマを、採取した脾細胞に10:1の割合で混合し(脾細胞:F10=2×107:2×106(5ml))、37℃で48h インキュベートした後、脾細胞を回収し、Mouse IFN−γ Development Module(R&D)のプロトコールに従ってELISPOTアッセイを行った。なお、ELISPOTアッセイにおいては、メンブレン上での脾細胞とマイトマイシンC処理F10メラノーマとの比は、脾細胞:F10=3×105:0.2×104(100μl/well)で行い、インキュベーションの時間は24時間とした。
天然型HVJ−Eに比べてZZ−scIL12結合型Fc−HVJ−Eは、約5倍以上のIFN−γ分泌誘導効果を示した(図12左図)。
また、Cr51リリーシングCTLアッセイにより、ZZ−scIL12結合型Fc−HVJ−E刺激脾細胞による免疫系活性化を調べた。Cr51リリーシングCTLアッセイは、常法にしたがい、下記の要領で実施した。F10メラノーマをマイトマイシンCで処理(15μg/ml、45min、37℃)後、上記採取した脾細胞とメラノーマを10:1割合で混合し(脾細胞:F10=5×107:5×106(20ml))、37℃で7日間インキュベーションした。この際、培地にはIL−2(final 10ng/ml)を添加しておき、インキュベーション4日後には10mlの培地(5ng/ml IL−2含有)を再度添加した。脾細胞を回収し、96well plateに2×106、1×106、5×105、2.5×105、1.25×105、0.625×105 cells/100μl/wellの2倍希釈系列を作製する。また、ポジティブコントロールとして1%NP−40溶液、ネガティブコントロールとしてRPMI培地を使用した。F10メラノーマを5×106 cells/0.2mlに調整し、1.85MBqの51Crを加え、37℃、45minインキュベーションし、F10メラノーマを洗浄して2×104cells/100μl/wellで脾細胞をまいたプレートに加え、4hインキュベーションした。そのプレートを1000rpm、5minで遠心後、各wellの上清を100μl回収し、51Crを測定した。
その結果、やはりZZ−scIL12結合型Fc−HVJ−Eは、天然型HVJ−Eに比べてかなり強いCTL活性を示した(図12右図)。
【0083】
8.ZZ−scIL12結合型Fc−HVJ−Eと、非結合scIL12およびHVJ−Eの併用投与との抗癌作用の比較
ZZ−scIL12結合型Fc−HVJ−E(HVJ−E:100HAU、IL−12:250pg)、またはZZドメインを有していないためFc−HVJ−Eとは結合しないTag−scIL12(IL−12:250pg)とHVJ−E100HAUとの混合液とを、上記同様F10メラノーマを注入し腫瘍が約2〜4mmになったマウスに腫瘍内に2日おきに3回投与した。回投与から14日目において、各マウスの腫瘍を観察し、完全に腫瘍が消失したマウスの割合を図13に示す。ZZ−scIL12結合型Fc−HVJ−Eを投与した群では30%以上で腫瘍が全く消失し、それ以後再発することなく90日後でも生存していた。一方、Fc−HN−HVJ−Eと結合しないTag−scIL12とFc−HN−HVJ−Eとを共投与した群では、5%程度しか腫瘍寛解が認められなかった(図13)。このことから、IL−12とHVJ−Eとが結合して複合体を形成することで強い抗腫瘍活性を示すことがわかった。
このことは、ZZ−scIL12結合型Fc−HVJ−Eの抗癌作用は、IL−12とHVJ−Eとが結合して複合体を形成した状態で癌細胞に作用することによって、IL−12とHVJ−Eというそれぞれに抗癌作用を奏するものの単なる組合せによる効果の増大を大きく上回るということを示している。
【0084】
9.ZZ−scIL12結合型Fc−HVJ−Eと、非結合scIL12およびHVJ−Eの併用との脾細胞からのIFN−γ分泌促進能の比較
上記と同様、ZZ−scIL12結合型Fc−HVJ−Eを投与した場合と、同量のscIL12とHVJ−Eとを結合していない状態で併用投与した場合とにおける、脾細胞からのIFN−γ分泌促進刺激を調べた。F10メラノーマを注入し腫瘍が約3〜5mmになったマウスに腫瘍内に、HVJ−E単独、Fc−HVJ−E+ZZscIL12複合体(250pg scIL12/100HAU HVJ−E、またはFc−HVJ−EとTagscIL−12との混合液(250pg scIL12/100HAU HVJ−E)を、10、50または100HAU HVJ−Eを、2日おきに3回投与した。最終投与から10日後に各マウスの脾細胞を採取し、脾細胞から分泌されたIFN−γを定量した(図14)。その結果、高濃度投与時においては結合型、非結合型共に同程度に高いIFN−γ分泌を誘導するが、低濃度投与時においては結合型の方が非結合型と比較して高いIFN−γ分泌誘導を引き起こすことが明らかとなった。
【産業上の利用可能性】
【0085】
本発明の一本鎖IL−12がHVJ−Eの表面に連結した改良型HVJ−E、さらに該改良型HVJ−Eを有効成分とする抗癌剤は、従来公知のHVJ−E単独の抗癌剤よりも低用量かつ著しい抗腫瘍効果を発揮し、新規な癌治療剤として有用である。
【特許請求の範囲】
【請求項1】
HVJのHNタンパク質のN末端部分アミノ酸配列と同一または実質的に同一のアミノ酸配列からなる改変HNタンパク質であって、該部分アミノ酸配列が111〜209アミノ酸の長さを有する、改変HNタンパク質。
【請求項2】
該部分アミノ酸配列が140〜180アミノ酸の長さを有する、請求項1記載の改変HNタンパク質。
【請求項3】
下記(1)または(2)のポリペプチドである、請求項1記載の改変HNタンパク質:
(1)配列番号6の第1〜160アミノ酸に該当するアミノ酸配列からなるポリペプチド、
(2)配列番号6の第1〜160アミノ酸に該当するアミノ酸配列において1または複数の残基が置換、欠失又は付加されたアミノ酸配列を含み、且つHVJに発現された場合に粒子表面に局在しうる特徴を有するポリペプチド。
【請求項4】
請求項1〜3のいずれか1項に記載の改変HNタンパク質の残基及び第1の機能性ポリペプチド残基を含む融合タンパク質であって、該改変HNタンパク質残基のC末端側に該機能性ポリペプチド残基が機能可能に連結されている、融合タンパク質。
【請求項5】
機能性ポリペプチドがIFN−γ分泌促進能を有するポリペプチドである、請求項4記載の融合タンパク質。
【請求項6】
IFN−γ分泌促進能を有するポリペプチドが一本鎖IL−12である、請求項5記載の融合タンパク質。
【請求項7】
一本鎖IL−12が、下記(1)または(2)のポリペプチドである、請求項6記載の融合タンパク質:
(1)配列番号2の第103〜625アミノ酸に該当するアミノ酸配列を含むポリペプチド、
(2)配列番号2の第103〜625アミノ酸に該当するアミノ酸配列において1または複数の残基が置換、欠失又は付加されたアミノ酸配列を含み、且つIFN−γ分泌促進能を有するポリペプチド。
【請求項8】
機能性ポリペプチドが抗体の定常領域(Fc)のCH2−CH3ドメインである、請求項4記載の融合タンパク質。
【請求項9】
抗体の定常領域(Fc)のCH2−CH3ドメインがIgG2aに由来するものである、請求項8記載の融合タンパク質。
【請求項10】
抗体の定常領域のCH2−CH3ドメインが、下記(1)または(2)のポリペプチドである、請求項9記載の融合タンパク質:
(1)配列番号4の第171〜389アミノ酸に該当するアミノ酸配列を含むポリペプチド、
(2)配列番号4の第171〜389アミノ酸に該当するアミノ酸配列において1または複数の残基が置換、欠失又は付加されたアミノ酸配列を含み、且つプロテインAとの結合活性を有するポリペプチド。
【請求項11】
請求項1〜3のいずれか1項に記載の改変HNタンパク質、又は請求項4〜10のいずれか1項に記載の融合タンパク質をコードする核酸。
【請求項12】
請求項8〜10のいずれか1に項記載の第1の融合タンパク質、並びにプロテインAのZZドメイン及び第2の機能性ポリペプチド残基を含む第2の融合タンパク質を含む、タンパク質複合体であって、第1の融合タンパク質と第2の融合タンパク質とが、第1の融合タンパク質に含まれるCH2−CH3ドメインと第2の融合タンパク質に含まれるZZドメインとの間の結合を介して複合体を形成している、タンパク質複合体。
【請求項13】
第2の機能性ポリペプチドが、一本鎖IL−12である、請求項12記載のタンパク質複合体。
【請求項14】
一本鎖IL−12が、下記(1)または(2)のポリペプチドであり、且つZZドメインが、下記(3)または(4)のポリペプチドである、請求項13記載のタンパク質複合体:
(1)配列番号2の第103〜625アミノ酸に該当するアミノ酸配列を含むポリペプチド、
(2)配列番号2の第103〜625アミノ酸に該当するアミノ酸配列において1または複数の残基が置換、欠失又は付加されたアミノ酸配列を含み、且つIFN−γ分泌促進能を有するポリペプチド、
(3)配列番号2の第42〜99アミノ酸に該当するアミノ酸配列を含むポリペプチド、
(4)配列番号2の第42〜99アミノ酸に該当するアミノ酸配列において1または複数の残基が欠失、置換または付加されたアミノ酸配列を含み、且つ抗体分子の定常領域(Fc)への結合活性を有するポリペプチド。
【請求項15】
請求項1〜3のいずれか1項に記載の改変HNタンパク質、請求項4〜10のいずれか1項に記載の融合タンパク質、または請求項12〜14のいずれか1項に記載のタンパク質複合体をエンベロープに含有する、HVJ−E。
【請求項16】
請求項6もしくは7に記載の融合タンパク質または請求項13もしくは14に記載のタンパク質複合体をエンベロープに含有する、請求項15記載のHVJ−E。
【請求項17】
IFN−γ分泌促進能を有するポリペプチドをエンベロープ外側に提示する形で担持する、HVJ−E。
【請求項18】
IFN−γ分泌促進能を有するポリペプチドが、IL−12である、請求項17記載のHVJ−E。
【請求項19】
IL−12が一本鎖IL−12である、請求項18記載のHVJ−E。
【請求項20】
請求項6もしくは7に記載の融合タンパク質または請求項13もしくは14に記載のタンパク質複合体をエンベロープに含有する、請求項18記載のHVJ−E。
【請求項21】
請求項17〜20のいずれか1項に記載のHVJ−Eを有効成分として含む、抗癌剤。
【請求項22】
メラノーマ、前立腺癌、膀胱癌および大腸癌からなる群より選択される癌の治療または予防に用いられる、請求項21記載の抗癌剤。
【請求項23】
1回当たりの投与量が、HVJ−Eとして1000HAU以下であり、かつ該HVJ−Eに含まれるIL−12として100pg以下であることを特徴とする、請求項18〜20のいずれか1項に記載のHVJ−Eを有効成分として含む抗癌剤。
【請求項24】
請求項18〜20のいずれか1項に記載のHVJ−Eの治療上有効量を癌患者に投与することを特徴とする、癌の治療方法。
【請求項25】
1回当たりの投与量が、HVJ−Eとして1000HAU以下であり、かつ該HVJ−Eに含まれるIL−12として100pg以下であることを特徴とする、請求項24記載の癌の治療方法。
【請求項1】
HVJのHNタンパク質のN末端部分アミノ酸配列と同一または実質的に同一のアミノ酸配列からなる改変HNタンパク質であって、該部分アミノ酸配列が111〜209アミノ酸の長さを有する、改変HNタンパク質。
【請求項2】
該部分アミノ酸配列が140〜180アミノ酸の長さを有する、請求項1記載の改変HNタンパク質。
【請求項3】
下記(1)または(2)のポリペプチドである、請求項1記載の改変HNタンパク質:
(1)配列番号6の第1〜160アミノ酸に該当するアミノ酸配列からなるポリペプチド、
(2)配列番号6の第1〜160アミノ酸に該当するアミノ酸配列において1または複数の残基が置換、欠失又は付加されたアミノ酸配列を含み、且つHVJに発現された場合に粒子表面に局在しうる特徴を有するポリペプチド。
【請求項4】
請求項1〜3のいずれか1項に記載の改変HNタンパク質の残基及び第1の機能性ポリペプチド残基を含む融合タンパク質であって、該改変HNタンパク質残基のC末端側に該機能性ポリペプチド残基が機能可能に連結されている、融合タンパク質。
【請求項5】
機能性ポリペプチドがIFN−γ分泌促進能を有するポリペプチドである、請求項4記載の融合タンパク質。
【請求項6】
IFN−γ分泌促進能を有するポリペプチドが一本鎖IL−12である、請求項5記載の融合タンパク質。
【請求項7】
一本鎖IL−12が、下記(1)または(2)のポリペプチドである、請求項6記載の融合タンパク質:
(1)配列番号2の第103〜625アミノ酸に該当するアミノ酸配列を含むポリペプチド、
(2)配列番号2の第103〜625アミノ酸に該当するアミノ酸配列において1または複数の残基が置換、欠失又は付加されたアミノ酸配列を含み、且つIFN−γ分泌促進能を有するポリペプチド。
【請求項8】
機能性ポリペプチドが抗体の定常領域(Fc)のCH2−CH3ドメインである、請求項4記載の融合タンパク質。
【請求項9】
抗体の定常領域(Fc)のCH2−CH3ドメインがIgG2aに由来するものである、請求項8記載の融合タンパク質。
【請求項10】
抗体の定常領域のCH2−CH3ドメインが、下記(1)または(2)のポリペプチドである、請求項9記載の融合タンパク質:
(1)配列番号4の第171〜389アミノ酸に該当するアミノ酸配列を含むポリペプチド、
(2)配列番号4の第171〜389アミノ酸に該当するアミノ酸配列において1または複数の残基が置換、欠失又は付加されたアミノ酸配列を含み、且つプロテインAとの結合活性を有するポリペプチド。
【請求項11】
請求項1〜3のいずれか1項に記載の改変HNタンパク質、又は請求項4〜10のいずれか1項に記載の融合タンパク質をコードする核酸。
【請求項12】
請求項8〜10のいずれか1に項記載の第1の融合タンパク質、並びにプロテインAのZZドメイン及び第2の機能性ポリペプチド残基を含む第2の融合タンパク質を含む、タンパク質複合体であって、第1の融合タンパク質と第2の融合タンパク質とが、第1の融合タンパク質に含まれるCH2−CH3ドメインと第2の融合タンパク質に含まれるZZドメインとの間の結合を介して複合体を形成している、タンパク質複合体。
【請求項13】
第2の機能性ポリペプチドが、一本鎖IL−12である、請求項12記載のタンパク質複合体。
【請求項14】
一本鎖IL−12が、下記(1)または(2)のポリペプチドであり、且つZZドメインが、下記(3)または(4)のポリペプチドである、請求項13記載のタンパク質複合体:
(1)配列番号2の第103〜625アミノ酸に該当するアミノ酸配列を含むポリペプチド、
(2)配列番号2の第103〜625アミノ酸に該当するアミノ酸配列において1または複数の残基が置換、欠失又は付加されたアミノ酸配列を含み、且つIFN−γ分泌促進能を有するポリペプチド、
(3)配列番号2の第42〜99アミノ酸に該当するアミノ酸配列を含むポリペプチド、
(4)配列番号2の第42〜99アミノ酸に該当するアミノ酸配列において1または複数の残基が欠失、置換または付加されたアミノ酸配列を含み、且つ抗体分子の定常領域(Fc)への結合活性を有するポリペプチド。
【請求項15】
請求項1〜3のいずれか1項に記載の改変HNタンパク質、請求項4〜10のいずれか1項に記載の融合タンパク質、または請求項12〜14のいずれか1項に記載のタンパク質複合体をエンベロープに含有する、HVJ−E。
【請求項16】
請求項6もしくは7に記載の融合タンパク質または請求項13もしくは14に記載のタンパク質複合体をエンベロープに含有する、請求項15記載のHVJ−E。
【請求項17】
IFN−γ分泌促進能を有するポリペプチドをエンベロープ外側に提示する形で担持する、HVJ−E。
【請求項18】
IFN−γ分泌促進能を有するポリペプチドが、IL−12である、請求項17記載のHVJ−E。
【請求項19】
IL−12が一本鎖IL−12である、請求項18記載のHVJ−E。
【請求項20】
請求項6もしくは7に記載の融合タンパク質または請求項13もしくは14に記載のタンパク質複合体をエンベロープに含有する、請求項18記載のHVJ−E。
【請求項21】
請求項17〜20のいずれか1項に記載のHVJ−Eを有効成分として含む、抗癌剤。
【請求項22】
メラノーマ、前立腺癌、膀胱癌および大腸癌からなる群より選択される癌の治療または予防に用いられる、請求項21記載の抗癌剤。
【請求項23】
1回当たりの投与量が、HVJ−Eとして1000HAU以下であり、かつ該HVJ−Eに含まれるIL−12として100pg以下であることを特徴とする、請求項18〜20のいずれか1項に記載のHVJ−Eを有効成分として含む抗癌剤。
【請求項24】
請求項18〜20のいずれか1項に記載のHVJ−Eの治療上有効量を癌患者に投与することを特徴とする、癌の治療方法。
【請求項25】
1回当たりの投与量が、HVJ−Eとして1000HAU以下であり、かつ該HVJ−Eに含まれるIL−12として100pg以下であることを特徴とする、請求項24記載の癌の治療方法。
【図2】


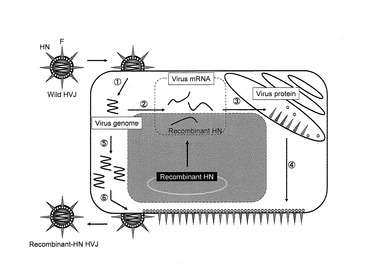
【図9】
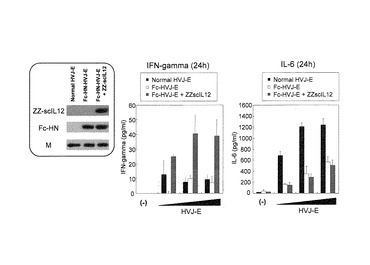
【図10】
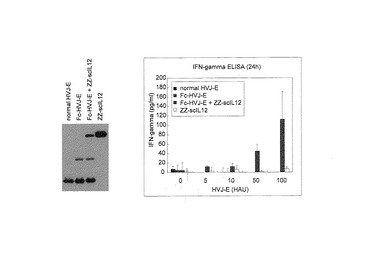
【図11】
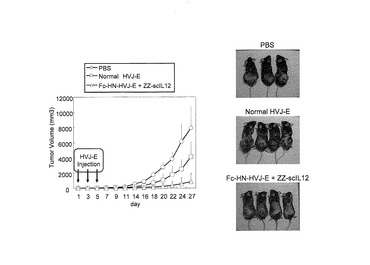
【図12】
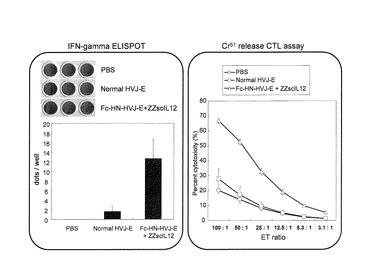
【図1】
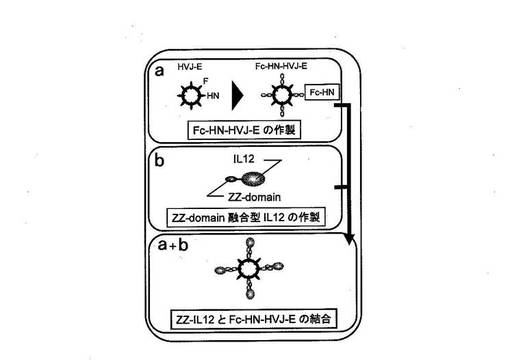
【図3】
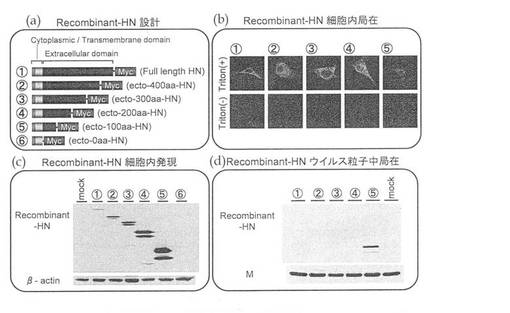
【図4】
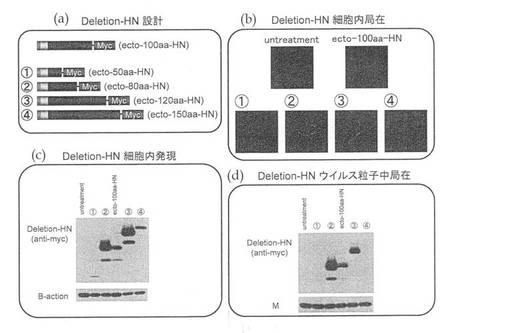
【図5】
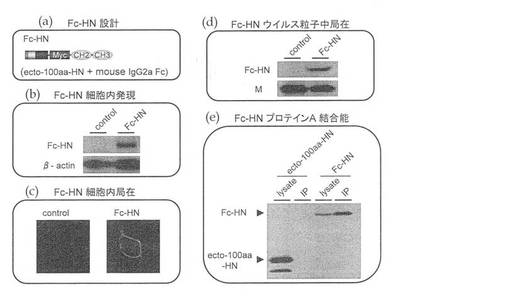
【図6】
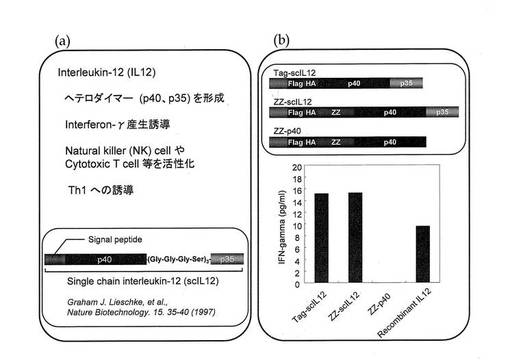
【図7】
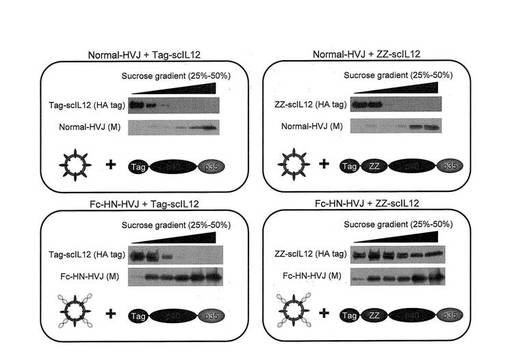
【図8】
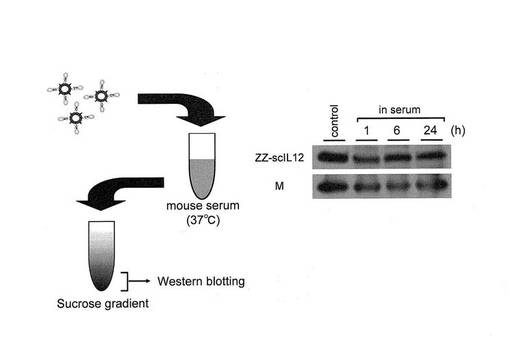
【図13】
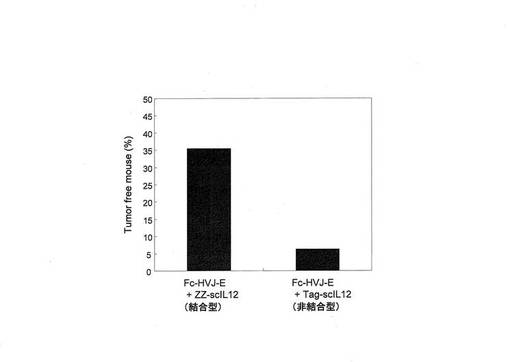
【図14】
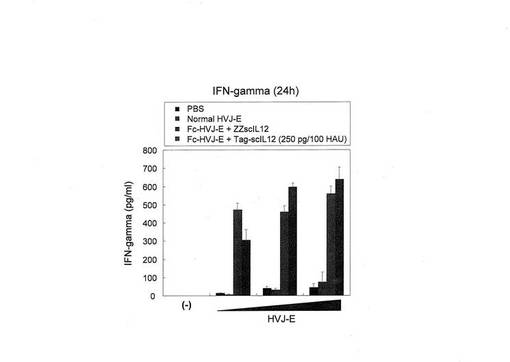
【図9】
【図10】
【図11】
【図12】
【図1】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図13】
【図14】
【公開番号】特開2011−50292(P2011−50292A)
【公開日】平成23年3月17日(2011.3.17)
【国際特許分類】
【出願番号】特願2009−201114(P2009−201114)
【出願日】平成21年8月31日(2009.8.31)
【国等の委託研究の成果に係る記載事項】(出願人による申告)平成21年度、文部科学省、「関西広域バイオメディカルクラスター構想(大阪北部(彩都)地域)に伴う研究委託業務」に係る再委託契約、産業技術力強化法第19条の適用を受ける特許出願
【出願人】(302060281)ジェノミディア株式会社 (7)
【出願人】(504176911)国立大学法人大阪大学 (1,536)
【Fターム(参考)】
【公開日】平成23年3月17日(2011.3.17)
【国際特許分類】
【出願日】平成21年8月31日(2009.8.31)
【国等の委託研究の成果に係る記載事項】(出願人による申告)平成21年度、文部科学省、「関西広域バイオメディカルクラスター構想(大阪北部(彩都)地域)に伴う研究委託業務」に係る再委託契約、産業技術力強化法第19条の適用を受ける特許出願
【出願人】(302060281)ジェノミディア株式会社 (7)
【出願人】(504176911)国立大学法人大阪大学 (1,536)
【Fターム(参考)】
[ Back to top ]