
高温登熟性イネ
【課題】イネ登熟期の気温上昇及びCO2濃度上昇にさらされても、米生産性の低下を抑制することができ、高品質な良食味米を供給することができる、高温登熟性に優れた高温登熟性イネの提供。
【解決手段】本発明の高温登熟性イネは、イネ種子をカルス培養することによりカルス培養細胞を得る工程と、前記カルス培養細胞を懸濁培養することによりイネゲノムの変異が生じた懸濁培養細胞を得る工程と、前記懸濁培養細胞を再分化することにより再分化個体を得る工程と、前記再分化個体から高温登熟性イネを選別する工程と、を含む育種方法により育種される。
【解決手段】本発明の高温登熟性イネは、イネ種子をカルス培養することによりカルス培養細胞を得る工程と、前記カルス培養細胞を懸濁培養することによりイネゲノムの変異が生じた懸濁培養細胞を得る工程と、前記懸濁培養細胞を再分化することにより再分化個体を得る工程と、前記再分化個体から高温登熟性イネを選別する工程と、を含む育種方法により育種される。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、高温登熟性イネに関する。
【背景技術】
【0002】
近年、地球温暖化に伴うイネ登熟期の気温上昇及びCO2濃度上昇は、デンプン集積不良による米生産性の低下、品質の低下等の問題を引き起こしている。特に、新潟県などの良食味米・高品質米の生産地では、登熟期の気温上昇等によるイネの品質低下が、米生産農家の収入に直接影響するだけでなく、産地のブランドイメージを壊すことにもなりかねず、極めて深刻な問題となっており、高温登熟性に優れたイネが求められている。
【0003】
このような問題を解決するために、例えば、イネ細胞を培養して突然変異を生じさせ、イネの品質を改良する方法が提案されている(例えば、非特許文献1及び2参照)。しかしながら、これらの提案では、早生化、短桿化、短穂化、穂数型化、小粒化、味度低下等のイネが得られたにすぎず、高温登熟性に優れたイネを提供するには至っていない。
【0004】
したがって、イネ登熟期の気温上昇及びCO2濃度上昇にさらされても、米生産性の低下を抑制することができ、高品質な良食味米を供給することができる、高温登熟性に優れた高温登熟性イネの開発が強く求められているのが現状である。
【先行技術文献】
【非特許文献】
【0005】
【非特許文献1】Shigemune A.,Tamura Y.,Aoki H.,Liang Z.−W.,Miyao A.,Hirochika H.,and Yatou O.Screening of grain mutants in the Tos17 mutant panel of rice.Hokuriku Crop Sci.38:50−52(2003).
【非特許文献2】Tateyama M.,Kobayashi W.,Mikami T.,Nakahori T.,and Tsugawa H.Characteristics of progenies derived from rice suspension cultured cells.Tohoku Agric.Res.51:7−8(1998).
【発明の概要】
【発明が解決しようとする課題】
【0006】
本発明は、前記従来における諸問題を解決し、以下の目的を達成することを課題とする。即ち、本発明は、イネ登熟期の気温上昇及びCO2濃度上昇にさらされても、米生産性の低下を抑制することができ、高品質な良食味米を供給することができる、高温登熟性に優れた高温登熟性イネを提供することを目的とする。
【課題を解決するための手段】
【0007】
前記課題を解決するため、本発明者らは鋭意検討した結果、以下のような知見を得た。即ち、イネ種子をカルス培養することによりカルス培養細胞を得る工程と、前記カルス培養細胞を懸濁培養することによりイネゲノムの変異が生じた懸濁培養細胞を得る工程と、前記懸濁培養細胞を再分化することにより再分化個体を得る工程と、前記再分化個体から高温登熟性イネを選別する工程と、を含む育種方法により、イネ登熟期の気温上昇及びCO2濃度上昇にさらされても、米生産性の低下を抑制することができ、高品質な良食味米を供給することができる、高温登熟性に優れた高温登熟性イネを提供することができることを知見した。
【0008】
本発明は、本発明者らによる前記知見に基づくものであり、前記課題を解決するための手段としては、以下の通りである。即ち、
<1> イネ種子をカルス培養することによりカルス培養細胞を得る工程と、前記カルス培養細胞を懸濁培養することによりイネゲノムの変異が生じた懸濁培養細胞を得る工程と、前記懸濁培養細胞を再分化することにより再分化個体を得る工程と、前記再分化個体から高温登熟性イネを選別する工程と、を含む育種方法により育種されたことを特徴とする高温登熟性イネである。
<2> イネゲノムが、レトロトランスポゾンの活性化により変異して、高温登熟性及び高CO2登熟性を獲得する前記<1>に記載の高温登熟性イネである。
<3> 登熟期平均気温28℃以上35℃以下で生育させた時に得られる米の正常粒率が、40%以上である前記<1>から<2>のいずれかに記載の高温登熟性イネである。
<4> 登熟期平均CO2濃度550ppm以上650ppm以下で生育させた時に得られる米の正常粒率が、40%以上である前記<1>から<2>のいずれかに記載の高温登熟性イネである。
<5> 高温登熟性イネの品種が、コシヒカリである前記<1>から<4>のいずれかに記載の高温登熟性イネである。
<6> イネ種子をカルス培養することによりカルス培養細胞を得る工程と、前記カルス培養細胞を懸濁培養することによりイネゲノムの変異が生じた懸濁培養細胞を得る工程と、前記懸濁培養細胞を再分化することにより再分化個体を得る工程と、前記再分化個体から高温登熟性イネを選別する工程と、を含むことを特徴とする高温登熟性イネの育種方法である。
<7> カルス培養細胞を得る工程が、イネ種子(玄米)をDKN培地上でカルス培養することによりカルス培養細胞を得る工程である前記<6>に記載の高温登熟性イネの育種方法である。
【発明の効果】
【0009】
本発明によると、従来における諸問題を解決することができ、イネ登熟期の気温上昇及びCO2濃度上昇にさらされても、米生産性の低下を抑制することができ、高品質な良食味米を供給することができる、高温登熟性に優れた高温登熟性イネを提供することができる。
【図面の簡単な説明】
【0010】
【図1】図1は、本発明の高温登熟性イネの育種方法の工程の一例を示す図である。
【図2A】図2Aは、本発明の高温登熟性イネの一例を示す図である。
【図2B】図2Bは、粒数の低下が見られた系統のイネの一例を示す図である。
【図2C】図2Cは、品質低下が見られた系統のイネの一例を示す図である。
【図2D】図2Dは、青未熟粒が多発した系統のイネの一例を示す図である。
【図2E】図2Eは、不稔が生じた系統のイネの一例を示す図である。
【図2F】図2Fは、野生型のイネの一例を示す図である。
【図3A】図3Aは、本発明の高温登熟性イネの評価結果を示す図である。
【図3B】図3Bは、本発明の高温登熟性イネの評価結果を示す図である。
【図3C】図3Cは、本発明の高温登熟性イネの評価結果を示す図である。
【図3D】図3Dは、本発明の高温登熟性イネの評価結果を示す図である。
【図4A】図4Aは、野生型のイネの玄米品質の一例を示す図である。
【図4B】図4Bは、野生型のイネの玄米品質の一例を示す図である。
【図4C】図4Cは、本発明の高温登熟性イネの玄米品質の一例を示す図である。
【図4D】図4Dは、本発明の高温登熟性イネの玄米品質の一例を示す図である。
【発明を実施するための形態】
【0011】
(高温登熟性イネ)
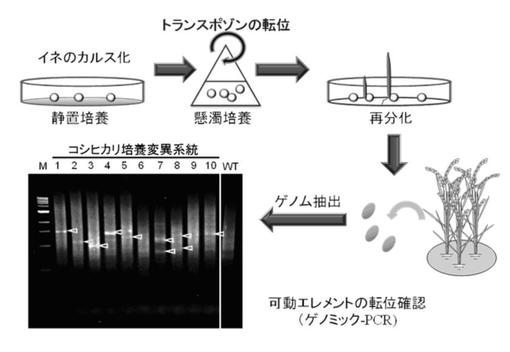
本発明の高温登熟性イネは、高温耐性イネと称することもでき、イネ種子をカルス培養することによりカルス培養細胞を得る工程(カルス培養工程)と、前記カルス培養細胞を懸濁培養することによりイネゲノムの変異が生じた懸濁培養細胞を得る工程(懸濁培養工程)と、前記懸濁培養細胞を再分化することにより再分化個体を得る工程(再分化工程)と、前記再分化個体から高温登熟性イネを選別する工程(高温登熟性イネ選別工程)と、を含む育種方法により育種される。
【0012】
<カルス培養工程>
前記カルス培養工程は、イネ種子をカルス培養することによりカルス培養細胞を得る工程である。
【0013】
前記カルス培養する方法としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、イネ種子(玄米)を培地上でカルス培養する方法などが挙げられ、具体的には、殺菌したイネ種子(玄米)をカルス誘導培地(固体培地)に置床して暗所で培養してカルス誘導を行った後、カルス誘導した種子から芽と根を除去し、芽と根を除去したカルスをカルス選抜培地(固体培地)にて明所で培養する方法などが挙げられ、“Daigen M.,Kawakami O.,and Nagasawa Y.Efficient anther culture method of the Japonica rice cultivar Koshihikari.Breed.Sci.50:197−202(2000).”及び“Ogawa T.,Fukuoka H.,Yano H.,and Ohkawa Y.Relationships between nitrite reductase activity and genotype−dependent callus growth in rice cell cultures.Plant Cell Rep.18:576−581(1999).”等の文献を参照して培養することができる。
【0014】
前記カルス培養する際の温度としては、特に制限はなく、目的に応じて適宜選択することができるが、高温登熟性イネの育種に好適なカルス培養細胞を得ることができる点で、25℃〜30℃が好ましい。
【0015】
前記カルス培養する際の光量としては、特に制限はなく、目的に応じて適宜選択することができるが、高温登熟性イネの育種に好適なカルス培養細胞を得ることができる点で、カルス誘導する際の光量として、15,000ルクス〜30,000ルクスが好ましく、カルス選抜する際の光量として、15,000ルクス〜30,000ルクスが好ましい。
【0016】
前記カルス培養する際の培地としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、KSP培地、DKN培地などが挙げられるが、高温登熟性イネの育種に好適なカルス培養細胞を得ることができる点で、DKN培地が好ましい。
【0017】
前記カルス培養する際の培地のpHとしては、特に制限はなく、目的に応じて適宜選択することができるが、高温登熟性イネの育種に好適な点で、pH5〜pH6が好ましい。
【0018】
前記カルス培養する際の培地に添加するイネ種子としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、JA等の市販品を用いてもよく、(独)農業生物資源研究所ゲノムリソースセンター等から入手してもよい。
【0019】
前記カルス培養する際の培地に対するイネ種子の添加量としては、特に制限はなく、目的に応じて適宜選択することができるが、高温登熟性イネの育種に好適なカルス培養細胞を得ることができる点で、固体培地プレート1枚(直径90mm×20mm)あたり、15個〜20個が好ましい。
【0020】
前記カルス培養する際の培養期間としては、特に制限はなく、目的に応じて適宜選択することができるが、高温登熟性イネの育種に好適なカルス培養細胞を得ることができる点で、カルス誘導する際の培養期間としては、5日間〜10日間が好ましく、カルス選抜する際の培養期間としては、10日間〜65日間が好ましい。
【0021】
前記カルス培養に用いられる培養装置としては、培養温度及び光量を調節することができる装置であれば、特に制限はなく、目的に応じて適宜選択することができ、例えば、グロースチャンバー(MLR−350T、SANYO社製)などが挙げられ、培養容器としては、固体培地を形成することができる培養容器であれば、特に制限はなく、目的に応じて適宜選択することができ、例えば、シャーレ、三角フラスコなどが挙げられる。
【0022】
<懸濁培養工程>
前記懸濁培養工程は、前記カルス培養細胞を懸濁培養することによりイネゲノムの変異が生じた懸濁培養細胞を得る工程である。
【0023】
前記懸濁培養する方法としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、前記カルス培養で用いたカルス選抜培地(液体培地)を用いて前記カルス培養細胞を撹拌又は振盪しながら培養する方法などが挙げられ、“Daigen M.,Kawakami O.,and Nagasawa Y.Efficient anther culture method of the Japonica rice cultivar Koshihikari.Breed.Sci.50:197−202(2000).”及び“Ogawa T.,Fukuoka H.,Yano H.,and Ohkawa Y.Relationships between nitrite reductase activity and genotype−dependent callus growth in rice cell cultures.Plant Cell Rep.18:576−581(1999).”等の文献を参照して培養することができる。
【0024】
前記懸濁培養する際の温度としては、特に制限はなく、目的に応じて適宜選択することができるが、高温登熟性イネの育種に好適な培養変異を生じさせることができる点で、25℃〜30℃が好ましい。
【0025】
前記懸濁培養する際の光量としては、特に制限はなく、目的に応じて適宜選択することができるが、高温登熟性イネの育種に好適な懸濁培養細胞を得ることができる点で、15,000ルクス〜30,000ルクスが好ましい。
【0026】
前記懸濁培養する際の振盪条件としては、特に制限はなく、目的に応じて適宜選択することができるが、高温登熟性イネの育種に好適な培養変異を生じさせることができる点で、60rpm〜80rpmが好ましい。
【0027】
前記懸濁培養する際の培地としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、KSP培地、DKN培地などが挙げられるが、高温登熟性イネの育種に好適な培養変異を生じさせることができる点で、DKN培地が好ましい。
【0028】
前記懸濁培養する際の培地のpHとしては、特に制限はなく、目的に応じて適宜選択することができるが、高温登熟性イネの育種に好適な培養変異を生じさせることができる点で、pH5〜pH6が好ましい。
【0029】
前記懸濁培養する際の培地に対する前記カルス培養細胞の播種量としては、特に制限はなく、目的に応じて適宜選択することができるが、高温登熟性イネの育種に好適な培養変異を生じさせることができる点で、1.0×107cells/cm2〜1.0×108cells/cm2が好ましい。
【0030】
前記懸濁培養する際の培養期間としては、特に制限はなく、目的に応じて適宜選択することができるが、高温登熟性イネの育種に好適な培養変異を生じさせることができる点で、50日間〜130日間が好ましい。
【0031】
前記懸濁培養に用いられる培養装置としては、培養温度及び光量を調節することができる装置であれば、特に制限はなく、目的に応じて適宜選択することができ、例えば、グロースチャンバー(MLR−350T、SANYO社製)などが挙げられ、培養容器としては、液体培地を振盪することができる培養容器であれば、特に制限はなく、目的に応じて適宜選択することができ、例えば、三角フラスコなどが挙げられる。
【0032】
前記懸濁培養することにより変異するイネゲノムとしては、特に制限はなく、目的に応じて適宜選択することができるが、高温登熟性イネの育種に好適な変異を生じさせることができる点で、イネゲノムのレトロトランスポゾン(逆転写酵素を使って転位するタイプのトランスポゾン(可動遺伝因子))が好ましく、これらの中でも、Tos17、Karma、Lullabyがより好ましい。なお、前記変異が生じたことを確認する方法としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、後述する高温登熟性イネのゲノム及び比較のために野生型のイネのゲノムの各ゲノムを抽出して、ゲノミックPCRにより各ゲノムを増幅させたものを、AFLP(Amplified Fragment Length Polymorphism、増幅断片長多型)解析することにより野生型のイネゲノムの変異が生じていることを確認する方法などが挙げられる。
【0033】
前記懸濁培養することによりカルス培養細胞のイネゲノムに内在するレトロトランスポゾンが活性化される。前記レトロトランスポゾンが活性化されることにより、前記レトロトランスポゾンが変異する。本発明の高温登熟性イネは、このレトロトランスポゾンが変異したことにより得られることが示唆されている。
【0034】
<再分化工程>
前記再分化工程は、前記懸濁培養細胞を再分化することにより再分化個体を得る工程である。
【0035】
前記再分化する方法としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、前記懸濁培養細胞を前記再分化個体形成培地(固体培地)に置床して明所で培養する方法などが挙げられ、“Daigen M.,Kawakami O.,and Nagasawa Y.Efficient anther culture method of the Japonica rice cultivar Koshihikari.Breed.Sci.50:197−202(2000).”及び“Ogawa T.,Fukuoka H.,Yano H.,and Ohkawa Y.Relationships between nitrite reductase activity and genotype−dependent callus growth in rice cell cultures.Plant Cell Rep.18:576−581(1999).”等の文献を参照して培養することができる。
【0036】
前記再分化する際の温度としては、特に制限はなく、目的に応じて適宜選択することができるが、高温登熟性イネの育種に好適な再分化個体を得ることができる点で、25℃〜30℃が好ましい。
【0037】
前記再分化する際の光量としては、特に制限はなく、目的に応じて適宜選択することができるが、高温登熟性イネの育種に好適な再分化個体を得ることができる点で、15,000ルクス〜30,000ルクスが好ましい。
【0038】
前記再分化する際の培地としては、特に制限はなく、目的に応じて適宜選択することができるが、高温登熟性イネの育種に好適な再分化個体を得ることができる点で、KSP培地、DKN培地等にカイネチン、アブシシン酸、カザミノ酸、ソルビトール等を添加した培地、KSP培地、DKN培地などに、インドール酢酸、カザミノ酸、6−ベンジルアミノプリン等を添加した培地などが挙げられるが、高温登熟性イネの育種に好適な培養変異を生じさせることができる点で、DKN培地を利用した培地が好ましい。
【0039】
前記再分化する際の培地のpHとしては、特に制限はなく、目的に応じて適宜選択することができるが、高温登熟性イネの育種に好適な再分化個体を得ることができる点で、pH5〜pH6が好ましい。
【0040】
前記再分化する際の前記懸濁培養細胞の播種量としては、特に制限はなく、目的に応じて適宜選択することができるが、高温登熟性イネの育種に好適な再分化個体を得ることができる点で、1.0×107cells/cm2〜1.0×108cells/cm2が好ましい。
【0041】
前記再分化する際の培養期間としては、特に制限はなく、目的に応じて適宜選択することができるが、高温登熟性イネの育種に好適な再分化個体を得ることができる点で、30日間〜50日間が好ましい。
【0042】
前記再分化する際に用いられる培養装置としては、培養温度及び光量を調節することができる装置であれば、特に制限はなく、目的に応じて適宜選択することができ、例えば、グロースチャンバー(MLR−350T、SANYO社製)などが挙げられ、培養容器としては、固体培地を形成することができる培養容器であれば、特に制限はなく、目的に応じて適宜選択することができ、例えば、シャーレ、三角フラスコなどが挙げられる。
【0043】
<高温登熟性イネ選別工程>
前記高温登熟性イネ選別工程は、前記再分化個体から高温登熟性イネを選別する工程である。
【0044】
前記高温登熟性イネを選別する方法としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、登熟期平均気温を一定温度以上にしてイネを栽培し、得られた米の正常粒率を測定することにより選別する方法、登熟期平均CO2濃度を一定濃度以上にしてイネを栽培し、得られた米の正常粒率を測定することにより選別する方法などが挙げられる。
【0045】
前記高温登熟性イネを選別する際の登熟期平均気温としては、特に制限はなく、目的に応じて適宜選択することができるが、目的とする高温登熟性イネの選別に好適な点で、28℃以上35℃以下が好ましい。
【0046】
前記高温登熟性イネを選別する際の登熟期平均CO2濃度としては、特に制限はなく、目的に応じて適宜選択することができるが、目的とする高温登熟性イネの選別に好適な点で、550ppm以上650ppm以下が好ましい。
【0047】
前記正常粒率は、玄米全体に占める正常粒の百分率をいう。前記正常粒(整粒)は、豊満で左右乃至上下均整の取れた形状であり、側面の縦溝が浅く、全体が透明で、表面が光沢を有する。前記整粒判定は、目視あるいは市販の穀粒判別器(例えば、RGQI 10B、(株)サタケ製)により行うことができる。
【0048】
前記正常粒率としては、特に制限はなく、目的に応じて適宜選択することができるが、目的とする高温登熟性イネの選別に好適な点で、40%以上が好ましく、60%以上がより好ましい。
【0049】
前記高温登熟性イネを選別する際の培養土としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、通常イネに用いられる培養土などが挙げられる。
【0050】
<高温登熟性イネの性質>
本発明の高温登熟性イネは、前記高温登熟性イネの育種方法により好適に育種される。
本発明の高温登熟性イネは、登熟期平均気温28℃以上35℃以下で生育させた時に得られる米の正常粒率が、40%以上であることが好ましい。また、本発明の高温登熟性イネは、登熟期平均CO2濃度550ppm以上650ppm以下で生育させた時に得られる米の正常粒率が、40%以上であることが好ましい。換言すれば、高温乃至高CO2濃度でも良好に生育可能な高温耐熱性及び高CO2濃度耐性に優れたイネである。前記高温登熟性イネは、通常知られているイネと同様、日本では、花期が7〜8月、結実期は9月〜10月であり、花の大きさが6mm程度、丈が50cm〜90cm程度である。
【0051】
前記高温登熟性イネの生育期は、通常知られているイネの生育期と同様、栄養生長期(発芽から穂の基となるもの(穂の原基)ができるまでの期間)、生殖生長期(穂の原基ができてから出穂乃至開花までの期間)、登熟期(出穂乃至開花から成熟までの期間)に分類される。高温登熟性能を有さないイネでは、前記登熟期にイネを高温にさらすと、籾(イネの果実)のデンプン合成能力等が低下することが報告されている。これは、デンプン合成系酵素の発現が高温によって低下し、デンプンの分解酵素(α−アミラーゼ)の活性が高温によって上昇するためである。一方、本発明の高温登熟性イネは、登熟期に高温にさらされても、デンプン合成能力が低下せず、前記デンプンの分解酵素の活性が抑制される性質を有する。
【0052】
前記高温登熟性イネの品種としては、特に制限はなく、目的に応じて適宜選択することができるが、例えば、コシヒカリ、ひとめぼれ、あきたこまち、きらら397、はえぬき、ほしのゆめ、つがるロマン、ななつぼし、などが好適に挙げられる。これらの中でも、本発明の高温登熟性イネの育種方法により好適に育種することができる点で、コシヒカリが特に好ましい。
【0053】
前記高温登熟性イネは、高温登熟性に優れるため、登熟期の異常高温や高CO2濃度にさらされても、米生産性の低下を抑制することができ、高品質な良食味米を供給することができるため、地球温暖化に伴うイネの高温登熟障害の被害対策として好適に活用することができる。
【実施例】
【0054】
以下に本発明の実施例を説明するが、本発明は、これらの実施例に何ら限定されるものではない。
【0055】
(実験例1)
<高温登熟性イネの製造>
−カルス培養工程−
イネをカルス培養することによりカルス培養細胞を得た。
イネのカルス培養は、以下のようにして行った。
籾摺り器を用いて、新潟県農業総合研究所から入手したコシヒカリ完熟種子の籾殻を剥き、1本の50mLファルコンチューブに、イネ種子(玄米)200粒を入れた。このチューブに、70質量%エタノールを10mL入れ、1分間表面を殺菌し、その後滅菌蒸留水で洗浄した。更に、このチューブに1質量%の次亜塩素酸ナトリウム溶液を加え、振盪器を用いて10分間穏やかに振盪した後、溶液を捨て、これを3回繰り返した。そして、約30mLの滅菌蒸留水で10分間振盪し、この洗浄を3回繰り返した。表1に記載のDKNカルス誘導培地(固体培地:90mm×20mmシャーレ使用)に、洗浄後の玄米を埋めた。このとき、胚は上向きにし、培地から胚が3mm程度はみ出るように埋めた。また、培地プレート(90mm×20mmシャーレ使用)1枚あたり、20個となるように埋めた。これを29℃、明所(20,000ルクス)で1週間培養してコシヒカリのカルスを誘導させた。次に、クリーンベンチ内で、ピンセットを用い、上述のカルス誘導した種子から芽と根を除去した。このカルス誘導した玄米を、ピンセットを用いて表1に記載のDKNカルス選抜培地へ移し替えた後、29℃の明所(20,000ルクス)で2カ月、1カ月、2週間の各期間で培養して、カルス培養細胞を得た。
【0056】
【表1】

【0057】
−懸濁培養工程−
前記カルス培養細胞を懸濁培養することにより懸濁培養細胞を得た。
カルス培養細胞の懸濁培養は、以下のようにして行った。
2カ月、1カ月、2週間培養した各カルス培養細胞を、表1に記載のDKNカルス選抜培地の液体培地(三角フラスコ使用)(300mL)を用いて、撹拌又は振盪しながら培養した。また、前記カルス培養細胞の前記液体培地に対する播種量が、2.7×107cells/cm2、培養の振盪条件が、60rpmとなるように懸濁培養した。そして、29℃、明所(20,000ルクス)で60日間培養してコシヒカリの懸濁培養細胞を得た。
【0058】
−再分化工程−
前記懸濁培養細胞を再分化させることにより再分化個体を得た。
懸濁培養細胞の再分化培養は、以下のようにして行った。
2カ月、1カ月、2週間培養した各カルス培養細胞から得られた各懸濁培養細胞を表2に記載の再分化I培地(固体培地:90mm×20mmシャーレ使用)に置床した。その後、29℃の明所(20,000ルクス)で培養した。次に、新しい再分化I培地への植継ぎを3回ほど繰返し、3cm程度に成長した個体を再分化II培地へ移して、29℃の明所(20,000ルクス)で培養し、再分化個体を得た。その後、実験温室にて再分化個体を培養土(商品名:合成培土3号、製造会社名:JA社製)へ植替え、生育させた。
【0059】
【表2】
【0060】
−高温登熟性イネ選別工程−
前記再分化個体を生育させたイネから高温登熟性イネを選別した。
高温登熟性イネの選別方法は、以下のようにして行った。
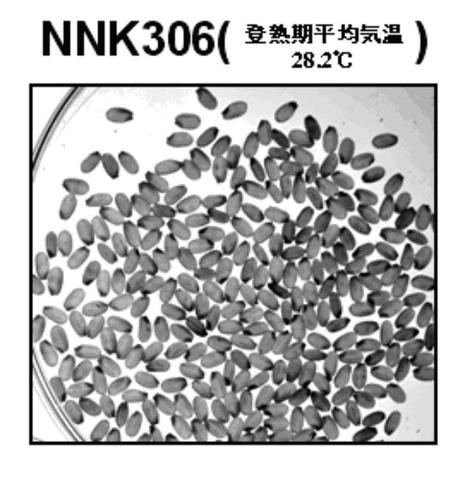
前記培養変異が確認された培養変異系統479系統)について、実験温室水田(登熟期平均気温26.6℃)で生育させ、玄米形質を調査した(図2A〜F参照)。これらの系統のうち、図2Aのように、粒数及び品質の低下が生じず、青未熟粒が多発せず、不稔が生じなかったイネを、コシヒカリ原種よりも品質が良い系統として19系統を得た。なお、図2Bは、粒数の低下が見られた系統のイネ、図2Cは、品質低下が見られた系統のイネ、図2Dは、青未熟粒が多発した系統のイネ、図2Eは、不稔が生じた系統のイネを示し、図2Fは、野生型のイネを示す。これらのコシヒカリ原種よりも品質が良かった19系統の中から、更に品質の良い系統を、3系統(NNK286系統、NNK306系統、及びNNK506系統)選択し、下記試験を行った。
【0061】
<高温登熟性イネ試験>
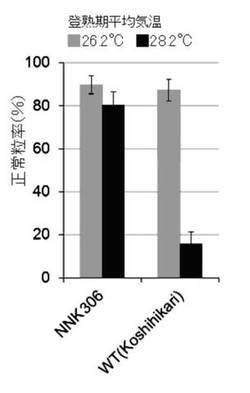
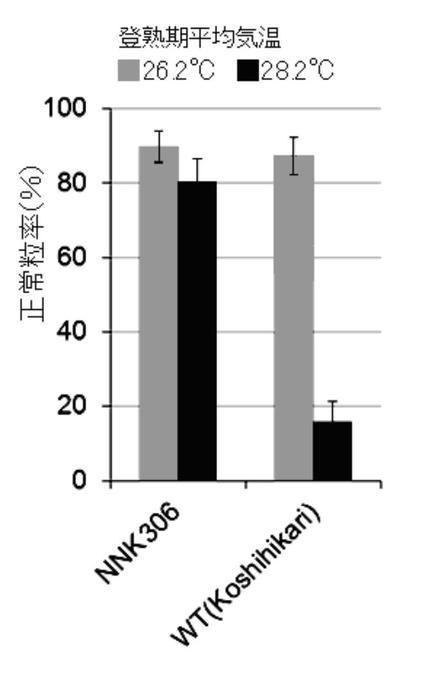
5月上旬及び7月上旬に、鹿児島県農業開発総合センターの圃場において、コシヒカリ培養変異系統(NNK306系統)及び野生(WT)型の苗(n=5)を栽培した。これらの正常粒率を調査した結果を、図3A及び図4A〜図4Dに示す。なお、圃場の登熟期の平均気温は、それぞれ28.2℃、26.2℃であった。
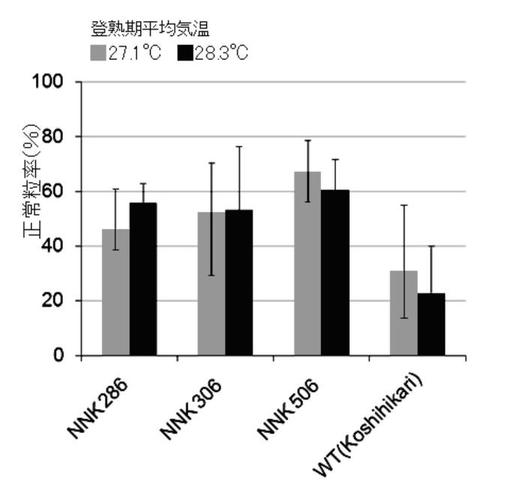
新潟県総合農業研究所作物研究センターの温水掛流し圃場及び一般圃場において、コシヒカリ培養変異系統(NNK286系統、NNK306系統、NNK506系統)及び野生(WT)型の苗(n=4)を栽培した。これらの正常粒率を調査した結果を図3Bに示す。なお、各圃場の登熟期平均気温は28.3℃、27.1℃であった。
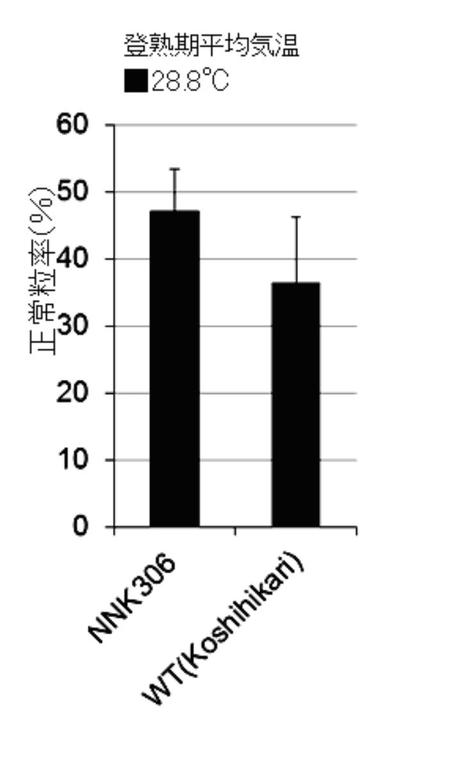
福岡県農業総合試験場の温水掛流し圃場において、コシヒカリ培養変異系統(NNK306系統)及び野生(WT)型の苗(n=5)を栽培した。これらの正常粒率を調査した結果を図3Cに示す。なお、圃場の登熟期平均気温は28.8℃であった。
【0062】
<高CO2登熟性イネ試験>
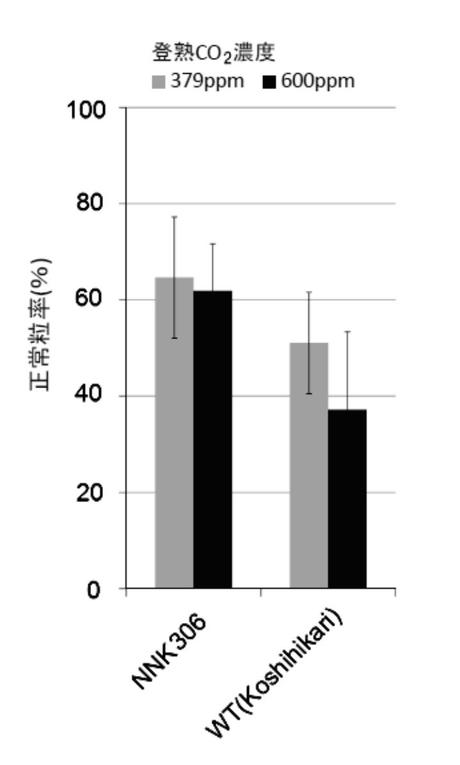
独立行政法人農業環境技術研究所のFACE(Free Air CO2 Enrichment)実験圃場において、コシヒカリ培養変異系統(NNK306系統)及び野生(WT)型の苗(n=12)を栽培した。これらの正常粒率を調査した結果を図3Dに示す。なお、圃場の登熟期平均気温は25.9℃であり、FACE実験圃場の平均CO2濃度(600ppm)は、一般実験圃場の平均CO2濃度(379ppm)の約1.6倍であった。
【0063】
<結果>
本発明の高温登熟性イネの育種方法により育種された3系統(NNK286系統、NNK306系統、NNK506系統)は、イネの登熟期に平均気温28℃以上に曝して栽培しても、正常粒率が40%以上となることがわかった(図3A〜C参照)。また、図4A〜Dより、本発明の高温登熟性イネは、玄米品質の低下も見られず、イネの高温登熟性が改善されたことがわかった。また、本発明の高温登熟性イネの育種方法により育種されたNNK306系統は、イネの登熟期の平均CO2濃度を一般実験圃場の平均CO2濃度の約1.5倍に曝して栽培しても、品質低下が軽減され、正常粒率が60%以上となることがわかった(図3D参照)。
【0064】
最も結果の良かったNNK306系統については、ゲノム解析を行った。
ゲノム解析は、生育させた苗を、CTAB処理、フェノール/クロロフォルム/イソアミルアルコール抽出、エタノール沈殿することにより、NNK306系統のイネゲノム及び野生型のイネゲノムの各ゲノムDNAを抽出して、AFLP(Amplified Fragment Length Polymorphism、増幅断片長多型)解析することにより行った。
まず、各種制限酵素(DraI、HindIII、MspI、タカラバイオ社製)により切断したゲノムDNA断片に2種のオリゴマー(Adapter1、Adapter2−Msp1、Adaprer2−Hind3、Adapter2−blunt end、北海道システムサイエンス社に作製を依頼)をアニールさせることで作製したアダプターを結合させ、アダプター特異的にアニールするプライマー(AP1、北海道システムサイエンス社に作製を依頼)と、トランスポゾン特異的にアニールするプライマー(LTR1、北海道システムサイエンス社に作製を依頼)を用いて、PCR(PCR条件;変性:94℃、30秒間、アニーリング:69℃、30秒間、伸長反応:72℃、4分間、上記反応サイクル数7回、その後変性:94℃、30秒間、アニーリング:61℃、30秒間、伸長反応:72℃、4分間、上記反応サイクル数32回、PCR装置;TAKARA PCR Thermal Cycler Dice TP650、タカラバイオ社製)によりDNA断片を増幅させた。
次に、得られたPCR産物を鋳型にして、nested−PCR(PCR条件;変性:94℃、30秒間、アニーリング・伸長反応:72℃、4分間、上記反応サイクル数5回、その後変性:94℃、30秒間、アニーリング:69℃、30秒間、伸長反応:72℃、4分間、上記反応サイクル数32回、プライマー(LTR2、AP2、北海道システムサイエンス社に作製を依頼)、PCR装置;TAKARA PCR Thermal Cycler Dice TP650、タカラバイオ社製)を行い、増幅断片を得た。なお、用いたオリゴマー、プライマー等の塩基配列を表3に示す。
【0065】
【表3】
【0066】
AFLP解析により、NNK306系統については、レトロトランスポゾンのイネゲノム上の位置が、変異したことがわかった(図1参照)。なお、図1は、DraIで切断したDNAをテンプレートに、アダプター特異的なAP2プライマーとTos17特異的なLTR2プライマーを用いたPCR(nasted−PCR)の電気泳動写真を含み、この電気泳動写真におけるレーン4が、NNK306系統に該当する。これらの結果から、NNK306系統は、カルス培養細胞の懸濁培養により、イネゲノムのレトロトランスポゾンが活性化され、高温登熟性及び高CO2登熟性を獲得したことが示唆された。
【0067】
以上より、本発明の高温登熟性イネは、イネ自身の遺伝子により高温登熟性を獲得し、イネ登熟期の気温上昇及びCO2濃度上昇にさらされても、米生産性の低下を抑制することができ、高品質な良食味米を供給することができ、高温登熟性、及び高CO2登熟性に優れることがわかった。
【産業上の利用可能性】
【0068】
本発明は、イネ登熟期の気温上昇及びCO2濃度上昇にさらされても、米生産性の低下を抑制することができ、高品質な良食味米を供給することができる、高温登熟性に優れた高温登熟性イネを提供することができるため、地球温暖化に伴うイネの高温登熟障害、及び高CO2登熟障害の被害対策として好適に活用することができる。
【技術分野】
【0001】
本発明は、高温登熟性イネに関する。
【背景技術】
【0002】
近年、地球温暖化に伴うイネ登熟期の気温上昇及びCO2濃度上昇は、デンプン集積不良による米生産性の低下、品質の低下等の問題を引き起こしている。特に、新潟県などの良食味米・高品質米の生産地では、登熟期の気温上昇等によるイネの品質低下が、米生産農家の収入に直接影響するだけでなく、産地のブランドイメージを壊すことにもなりかねず、極めて深刻な問題となっており、高温登熟性に優れたイネが求められている。
【0003】
このような問題を解決するために、例えば、イネ細胞を培養して突然変異を生じさせ、イネの品質を改良する方法が提案されている(例えば、非特許文献1及び2参照)。しかしながら、これらの提案では、早生化、短桿化、短穂化、穂数型化、小粒化、味度低下等のイネが得られたにすぎず、高温登熟性に優れたイネを提供するには至っていない。
【0004】
したがって、イネ登熟期の気温上昇及びCO2濃度上昇にさらされても、米生産性の低下を抑制することができ、高品質な良食味米を供給することができる、高温登熟性に優れた高温登熟性イネの開発が強く求められているのが現状である。
【先行技術文献】
【非特許文献】
【0005】
【非特許文献1】Shigemune A.,Tamura Y.,Aoki H.,Liang Z.−W.,Miyao A.,Hirochika H.,and Yatou O.Screening of grain mutants in the Tos17 mutant panel of rice.Hokuriku Crop Sci.38:50−52(2003).
【非特許文献2】Tateyama M.,Kobayashi W.,Mikami T.,Nakahori T.,and Tsugawa H.Characteristics of progenies derived from rice suspension cultured cells.Tohoku Agric.Res.51:7−8(1998).
【発明の概要】
【発明が解決しようとする課題】
【0006】
本発明は、前記従来における諸問題を解決し、以下の目的を達成することを課題とする。即ち、本発明は、イネ登熟期の気温上昇及びCO2濃度上昇にさらされても、米生産性の低下を抑制することができ、高品質な良食味米を供給することができる、高温登熟性に優れた高温登熟性イネを提供することを目的とする。
【課題を解決するための手段】
【0007】
前記課題を解決するため、本発明者らは鋭意検討した結果、以下のような知見を得た。即ち、イネ種子をカルス培養することによりカルス培養細胞を得る工程と、前記カルス培養細胞を懸濁培養することによりイネゲノムの変異が生じた懸濁培養細胞を得る工程と、前記懸濁培養細胞を再分化することにより再分化個体を得る工程と、前記再分化個体から高温登熟性イネを選別する工程と、を含む育種方法により、イネ登熟期の気温上昇及びCO2濃度上昇にさらされても、米生産性の低下を抑制することができ、高品質な良食味米を供給することができる、高温登熟性に優れた高温登熟性イネを提供することができることを知見した。
【0008】
本発明は、本発明者らによる前記知見に基づくものであり、前記課題を解決するための手段としては、以下の通りである。即ち、
<1> イネ種子をカルス培養することによりカルス培養細胞を得る工程と、前記カルス培養細胞を懸濁培養することによりイネゲノムの変異が生じた懸濁培養細胞を得る工程と、前記懸濁培養細胞を再分化することにより再分化個体を得る工程と、前記再分化個体から高温登熟性イネを選別する工程と、を含む育種方法により育種されたことを特徴とする高温登熟性イネである。
<2> イネゲノムが、レトロトランスポゾンの活性化により変異して、高温登熟性及び高CO2登熟性を獲得する前記<1>に記載の高温登熟性イネである。
<3> 登熟期平均気温28℃以上35℃以下で生育させた時に得られる米の正常粒率が、40%以上である前記<1>から<2>のいずれかに記載の高温登熟性イネである。
<4> 登熟期平均CO2濃度550ppm以上650ppm以下で生育させた時に得られる米の正常粒率が、40%以上である前記<1>から<2>のいずれかに記載の高温登熟性イネである。
<5> 高温登熟性イネの品種が、コシヒカリである前記<1>から<4>のいずれかに記載の高温登熟性イネである。
<6> イネ種子をカルス培養することによりカルス培養細胞を得る工程と、前記カルス培養細胞を懸濁培養することによりイネゲノムの変異が生じた懸濁培養細胞を得る工程と、前記懸濁培養細胞を再分化することにより再分化個体を得る工程と、前記再分化個体から高温登熟性イネを選別する工程と、を含むことを特徴とする高温登熟性イネの育種方法である。
<7> カルス培養細胞を得る工程が、イネ種子(玄米)をDKN培地上でカルス培養することによりカルス培養細胞を得る工程である前記<6>に記載の高温登熟性イネの育種方法である。
【発明の効果】
【0009】
本発明によると、従来における諸問題を解決することができ、イネ登熟期の気温上昇及びCO2濃度上昇にさらされても、米生産性の低下を抑制することができ、高品質な良食味米を供給することができる、高温登熟性に優れた高温登熟性イネを提供することができる。
【図面の簡単な説明】
【0010】
【図1】図1は、本発明の高温登熟性イネの育種方法の工程の一例を示す図である。
【図2A】図2Aは、本発明の高温登熟性イネの一例を示す図である。
【図2B】図2Bは、粒数の低下が見られた系統のイネの一例を示す図である。
【図2C】図2Cは、品質低下が見られた系統のイネの一例を示す図である。
【図2D】図2Dは、青未熟粒が多発した系統のイネの一例を示す図である。
【図2E】図2Eは、不稔が生じた系統のイネの一例を示す図である。
【図2F】図2Fは、野生型のイネの一例を示す図である。
【図3A】図3Aは、本発明の高温登熟性イネの評価結果を示す図である。
【図3B】図3Bは、本発明の高温登熟性イネの評価結果を示す図である。
【図3C】図3Cは、本発明の高温登熟性イネの評価結果を示す図である。
【図3D】図3Dは、本発明の高温登熟性イネの評価結果を示す図である。
【図4A】図4Aは、野生型のイネの玄米品質の一例を示す図である。
【図4B】図4Bは、野生型のイネの玄米品質の一例を示す図である。
【図4C】図4Cは、本発明の高温登熟性イネの玄米品質の一例を示す図である。
【図4D】図4Dは、本発明の高温登熟性イネの玄米品質の一例を示す図である。
【発明を実施するための形態】
【0011】
(高温登熟性イネ)
本発明の高温登熟性イネは、高温耐性イネと称することもでき、イネ種子をカルス培養することによりカルス培養細胞を得る工程(カルス培養工程)と、前記カルス培養細胞を懸濁培養することによりイネゲノムの変異が生じた懸濁培養細胞を得る工程(懸濁培養工程)と、前記懸濁培養細胞を再分化することにより再分化個体を得る工程(再分化工程)と、前記再分化個体から高温登熟性イネを選別する工程(高温登熟性イネ選別工程)と、を含む育種方法により育種される。
【0012】
<カルス培養工程>
前記カルス培養工程は、イネ種子をカルス培養することによりカルス培養細胞を得る工程である。
【0013】
前記カルス培養する方法としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、イネ種子(玄米)を培地上でカルス培養する方法などが挙げられ、具体的には、殺菌したイネ種子(玄米)をカルス誘導培地(固体培地)に置床して暗所で培養してカルス誘導を行った後、カルス誘導した種子から芽と根を除去し、芽と根を除去したカルスをカルス選抜培地(固体培地)にて明所で培養する方法などが挙げられ、“Daigen M.,Kawakami O.,and Nagasawa Y.Efficient anther culture method of the Japonica rice cultivar Koshihikari.Breed.Sci.50:197−202(2000).”及び“Ogawa T.,Fukuoka H.,Yano H.,and Ohkawa Y.Relationships between nitrite reductase activity and genotype−dependent callus growth in rice cell cultures.Plant Cell Rep.18:576−581(1999).”等の文献を参照して培養することができる。
【0014】
前記カルス培養する際の温度としては、特に制限はなく、目的に応じて適宜選択することができるが、高温登熟性イネの育種に好適なカルス培養細胞を得ることができる点で、25℃〜30℃が好ましい。
【0015】
前記カルス培養する際の光量としては、特に制限はなく、目的に応じて適宜選択することができるが、高温登熟性イネの育種に好適なカルス培養細胞を得ることができる点で、カルス誘導する際の光量として、15,000ルクス〜30,000ルクスが好ましく、カルス選抜する際の光量として、15,000ルクス〜30,000ルクスが好ましい。
【0016】
前記カルス培養する際の培地としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、KSP培地、DKN培地などが挙げられるが、高温登熟性イネの育種に好適なカルス培養細胞を得ることができる点で、DKN培地が好ましい。
【0017】
前記カルス培養する際の培地のpHとしては、特に制限はなく、目的に応じて適宜選択することができるが、高温登熟性イネの育種に好適な点で、pH5〜pH6が好ましい。
【0018】
前記カルス培養する際の培地に添加するイネ種子としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、JA等の市販品を用いてもよく、(独)農業生物資源研究所ゲノムリソースセンター等から入手してもよい。
【0019】
前記カルス培養する際の培地に対するイネ種子の添加量としては、特に制限はなく、目的に応じて適宜選択することができるが、高温登熟性イネの育種に好適なカルス培養細胞を得ることができる点で、固体培地プレート1枚(直径90mm×20mm)あたり、15個〜20個が好ましい。
【0020】
前記カルス培養する際の培養期間としては、特に制限はなく、目的に応じて適宜選択することができるが、高温登熟性イネの育種に好適なカルス培養細胞を得ることができる点で、カルス誘導する際の培養期間としては、5日間〜10日間が好ましく、カルス選抜する際の培養期間としては、10日間〜65日間が好ましい。
【0021】
前記カルス培養に用いられる培養装置としては、培養温度及び光量を調節することができる装置であれば、特に制限はなく、目的に応じて適宜選択することができ、例えば、グロースチャンバー(MLR−350T、SANYO社製)などが挙げられ、培養容器としては、固体培地を形成することができる培養容器であれば、特に制限はなく、目的に応じて適宜選択することができ、例えば、シャーレ、三角フラスコなどが挙げられる。
【0022】
<懸濁培養工程>
前記懸濁培養工程は、前記カルス培養細胞を懸濁培養することによりイネゲノムの変異が生じた懸濁培養細胞を得る工程である。
【0023】
前記懸濁培養する方法としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、前記カルス培養で用いたカルス選抜培地(液体培地)を用いて前記カルス培養細胞を撹拌又は振盪しながら培養する方法などが挙げられ、“Daigen M.,Kawakami O.,and Nagasawa Y.Efficient anther culture method of the Japonica rice cultivar Koshihikari.Breed.Sci.50:197−202(2000).”及び“Ogawa T.,Fukuoka H.,Yano H.,and Ohkawa Y.Relationships between nitrite reductase activity and genotype−dependent callus growth in rice cell cultures.Plant Cell Rep.18:576−581(1999).”等の文献を参照して培養することができる。
【0024】
前記懸濁培養する際の温度としては、特に制限はなく、目的に応じて適宜選択することができるが、高温登熟性イネの育種に好適な培養変異を生じさせることができる点で、25℃〜30℃が好ましい。
【0025】
前記懸濁培養する際の光量としては、特に制限はなく、目的に応じて適宜選択することができるが、高温登熟性イネの育種に好適な懸濁培養細胞を得ることができる点で、15,000ルクス〜30,000ルクスが好ましい。
【0026】
前記懸濁培養する際の振盪条件としては、特に制限はなく、目的に応じて適宜選択することができるが、高温登熟性イネの育種に好適な培養変異を生じさせることができる点で、60rpm〜80rpmが好ましい。
【0027】
前記懸濁培養する際の培地としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、KSP培地、DKN培地などが挙げられるが、高温登熟性イネの育種に好適な培養変異を生じさせることができる点で、DKN培地が好ましい。
【0028】
前記懸濁培養する際の培地のpHとしては、特に制限はなく、目的に応じて適宜選択することができるが、高温登熟性イネの育種に好適な培養変異を生じさせることができる点で、pH5〜pH6が好ましい。
【0029】
前記懸濁培養する際の培地に対する前記カルス培養細胞の播種量としては、特に制限はなく、目的に応じて適宜選択することができるが、高温登熟性イネの育種に好適な培養変異を生じさせることができる点で、1.0×107cells/cm2〜1.0×108cells/cm2が好ましい。
【0030】
前記懸濁培養する際の培養期間としては、特に制限はなく、目的に応じて適宜選択することができるが、高温登熟性イネの育種に好適な培養変異を生じさせることができる点で、50日間〜130日間が好ましい。
【0031】
前記懸濁培養に用いられる培養装置としては、培養温度及び光量を調節することができる装置であれば、特に制限はなく、目的に応じて適宜選択することができ、例えば、グロースチャンバー(MLR−350T、SANYO社製)などが挙げられ、培養容器としては、液体培地を振盪することができる培養容器であれば、特に制限はなく、目的に応じて適宜選択することができ、例えば、三角フラスコなどが挙げられる。
【0032】
前記懸濁培養することにより変異するイネゲノムとしては、特に制限はなく、目的に応じて適宜選択することができるが、高温登熟性イネの育種に好適な変異を生じさせることができる点で、イネゲノムのレトロトランスポゾン(逆転写酵素を使って転位するタイプのトランスポゾン(可動遺伝因子))が好ましく、これらの中でも、Tos17、Karma、Lullabyがより好ましい。なお、前記変異が生じたことを確認する方法としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、後述する高温登熟性イネのゲノム及び比較のために野生型のイネのゲノムの各ゲノムを抽出して、ゲノミックPCRにより各ゲノムを増幅させたものを、AFLP(Amplified Fragment Length Polymorphism、増幅断片長多型)解析することにより野生型のイネゲノムの変異が生じていることを確認する方法などが挙げられる。
【0033】
前記懸濁培養することによりカルス培養細胞のイネゲノムに内在するレトロトランスポゾンが活性化される。前記レトロトランスポゾンが活性化されることにより、前記レトロトランスポゾンが変異する。本発明の高温登熟性イネは、このレトロトランスポゾンが変異したことにより得られることが示唆されている。
【0034】
<再分化工程>
前記再分化工程は、前記懸濁培養細胞を再分化することにより再分化個体を得る工程である。
【0035】
前記再分化する方法としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、前記懸濁培養細胞を前記再分化個体形成培地(固体培地)に置床して明所で培養する方法などが挙げられ、“Daigen M.,Kawakami O.,and Nagasawa Y.Efficient anther culture method of the Japonica rice cultivar Koshihikari.Breed.Sci.50:197−202(2000).”及び“Ogawa T.,Fukuoka H.,Yano H.,and Ohkawa Y.Relationships between nitrite reductase activity and genotype−dependent callus growth in rice cell cultures.Plant Cell Rep.18:576−581(1999).”等の文献を参照して培養することができる。
【0036】
前記再分化する際の温度としては、特に制限はなく、目的に応じて適宜選択することができるが、高温登熟性イネの育種に好適な再分化個体を得ることができる点で、25℃〜30℃が好ましい。
【0037】
前記再分化する際の光量としては、特に制限はなく、目的に応じて適宜選択することができるが、高温登熟性イネの育種に好適な再分化個体を得ることができる点で、15,000ルクス〜30,000ルクスが好ましい。
【0038】
前記再分化する際の培地としては、特に制限はなく、目的に応じて適宜選択することができるが、高温登熟性イネの育種に好適な再分化個体を得ることができる点で、KSP培地、DKN培地等にカイネチン、アブシシン酸、カザミノ酸、ソルビトール等を添加した培地、KSP培地、DKN培地などに、インドール酢酸、カザミノ酸、6−ベンジルアミノプリン等を添加した培地などが挙げられるが、高温登熟性イネの育種に好適な培養変異を生じさせることができる点で、DKN培地を利用した培地が好ましい。
【0039】
前記再分化する際の培地のpHとしては、特に制限はなく、目的に応じて適宜選択することができるが、高温登熟性イネの育種に好適な再分化個体を得ることができる点で、pH5〜pH6が好ましい。
【0040】
前記再分化する際の前記懸濁培養細胞の播種量としては、特に制限はなく、目的に応じて適宜選択することができるが、高温登熟性イネの育種に好適な再分化個体を得ることができる点で、1.0×107cells/cm2〜1.0×108cells/cm2が好ましい。
【0041】
前記再分化する際の培養期間としては、特に制限はなく、目的に応じて適宜選択することができるが、高温登熟性イネの育種に好適な再分化個体を得ることができる点で、30日間〜50日間が好ましい。
【0042】
前記再分化する際に用いられる培養装置としては、培養温度及び光量を調節することができる装置であれば、特に制限はなく、目的に応じて適宜選択することができ、例えば、グロースチャンバー(MLR−350T、SANYO社製)などが挙げられ、培養容器としては、固体培地を形成することができる培養容器であれば、特に制限はなく、目的に応じて適宜選択することができ、例えば、シャーレ、三角フラスコなどが挙げられる。
【0043】
<高温登熟性イネ選別工程>
前記高温登熟性イネ選別工程は、前記再分化個体から高温登熟性イネを選別する工程である。
【0044】
前記高温登熟性イネを選別する方法としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、登熟期平均気温を一定温度以上にしてイネを栽培し、得られた米の正常粒率を測定することにより選別する方法、登熟期平均CO2濃度を一定濃度以上にしてイネを栽培し、得られた米の正常粒率を測定することにより選別する方法などが挙げられる。
【0045】
前記高温登熟性イネを選別する際の登熟期平均気温としては、特に制限はなく、目的に応じて適宜選択することができるが、目的とする高温登熟性イネの選別に好適な点で、28℃以上35℃以下が好ましい。
【0046】
前記高温登熟性イネを選別する際の登熟期平均CO2濃度としては、特に制限はなく、目的に応じて適宜選択することができるが、目的とする高温登熟性イネの選別に好適な点で、550ppm以上650ppm以下が好ましい。
【0047】
前記正常粒率は、玄米全体に占める正常粒の百分率をいう。前記正常粒(整粒)は、豊満で左右乃至上下均整の取れた形状であり、側面の縦溝が浅く、全体が透明で、表面が光沢を有する。前記整粒判定は、目視あるいは市販の穀粒判別器(例えば、RGQI 10B、(株)サタケ製)により行うことができる。
【0048】
前記正常粒率としては、特に制限はなく、目的に応じて適宜選択することができるが、目的とする高温登熟性イネの選別に好適な点で、40%以上が好ましく、60%以上がより好ましい。
【0049】
前記高温登熟性イネを選別する際の培養土としては、特に制限はなく、目的に応じて適宜選択することができ、例えば、通常イネに用いられる培養土などが挙げられる。
【0050】
<高温登熟性イネの性質>
本発明の高温登熟性イネは、前記高温登熟性イネの育種方法により好適に育種される。
本発明の高温登熟性イネは、登熟期平均気温28℃以上35℃以下で生育させた時に得られる米の正常粒率が、40%以上であることが好ましい。また、本発明の高温登熟性イネは、登熟期平均CO2濃度550ppm以上650ppm以下で生育させた時に得られる米の正常粒率が、40%以上であることが好ましい。換言すれば、高温乃至高CO2濃度でも良好に生育可能な高温耐熱性及び高CO2濃度耐性に優れたイネである。前記高温登熟性イネは、通常知られているイネと同様、日本では、花期が7〜8月、結実期は9月〜10月であり、花の大きさが6mm程度、丈が50cm〜90cm程度である。
【0051】
前記高温登熟性イネの生育期は、通常知られているイネの生育期と同様、栄養生長期(発芽から穂の基となるもの(穂の原基)ができるまでの期間)、生殖生長期(穂の原基ができてから出穂乃至開花までの期間)、登熟期(出穂乃至開花から成熟までの期間)に分類される。高温登熟性能を有さないイネでは、前記登熟期にイネを高温にさらすと、籾(イネの果実)のデンプン合成能力等が低下することが報告されている。これは、デンプン合成系酵素の発現が高温によって低下し、デンプンの分解酵素(α−アミラーゼ)の活性が高温によって上昇するためである。一方、本発明の高温登熟性イネは、登熟期に高温にさらされても、デンプン合成能力が低下せず、前記デンプンの分解酵素の活性が抑制される性質を有する。
【0052】
前記高温登熟性イネの品種としては、特に制限はなく、目的に応じて適宜選択することができるが、例えば、コシヒカリ、ひとめぼれ、あきたこまち、きらら397、はえぬき、ほしのゆめ、つがるロマン、ななつぼし、などが好適に挙げられる。これらの中でも、本発明の高温登熟性イネの育種方法により好適に育種することができる点で、コシヒカリが特に好ましい。
【0053】
前記高温登熟性イネは、高温登熟性に優れるため、登熟期の異常高温や高CO2濃度にさらされても、米生産性の低下を抑制することができ、高品質な良食味米を供給することができるため、地球温暖化に伴うイネの高温登熟障害の被害対策として好適に活用することができる。
【実施例】
【0054】
以下に本発明の実施例を説明するが、本発明は、これらの実施例に何ら限定されるものではない。
【0055】
(実験例1)
<高温登熟性イネの製造>
−カルス培養工程−
イネをカルス培養することによりカルス培養細胞を得た。
イネのカルス培養は、以下のようにして行った。
籾摺り器を用いて、新潟県農業総合研究所から入手したコシヒカリ完熟種子の籾殻を剥き、1本の50mLファルコンチューブに、イネ種子(玄米)200粒を入れた。このチューブに、70質量%エタノールを10mL入れ、1分間表面を殺菌し、その後滅菌蒸留水で洗浄した。更に、このチューブに1質量%の次亜塩素酸ナトリウム溶液を加え、振盪器を用いて10分間穏やかに振盪した後、溶液を捨て、これを3回繰り返した。そして、約30mLの滅菌蒸留水で10分間振盪し、この洗浄を3回繰り返した。表1に記載のDKNカルス誘導培地(固体培地:90mm×20mmシャーレ使用)に、洗浄後の玄米を埋めた。このとき、胚は上向きにし、培地から胚が3mm程度はみ出るように埋めた。また、培地プレート(90mm×20mmシャーレ使用)1枚あたり、20個となるように埋めた。これを29℃、明所(20,000ルクス)で1週間培養してコシヒカリのカルスを誘導させた。次に、クリーンベンチ内で、ピンセットを用い、上述のカルス誘導した種子から芽と根を除去した。このカルス誘導した玄米を、ピンセットを用いて表1に記載のDKNカルス選抜培地へ移し替えた後、29℃の明所(20,000ルクス)で2カ月、1カ月、2週間の各期間で培養して、カルス培養細胞を得た。
【0056】
【表1】
【0057】
−懸濁培養工程−
前記カルス培養細胞を懸濁培養することにより懸濁培養細胞を得た。
カルス培養細胞の懸濁培養は、以下のようにして行った。
2カ月、1カ月、2週間培養した各カルス培養細胞を、表1に記載のDKNカルス選抜培地の液体培地(三角フラスコ使用)(300mL)を用いて、撹拌又は振盪しながら培養した。また、前記カルス培養細胞の前記液体培地に対する播種量が、2.7×107cells/cm2、培養の振盪条件が、60rpmとなるように懸濁培養した。そして、29℃、明所(20,000ルクス)で60日間培養してコシヒカリの懸濁培養細胞を得た。
【0058】
−再分化工程−
前記懸濁培養細胞を再分化させることにより再分化個体を得た。
懸濁培養細胞の再分化培養は、以下のようにして行った。
2カ月、1カ月、2週間培養した各カルス培養細胞から得られた各懸濁培養細胞を表2に記載の再分化I培地(固体培地:90mm×20mmシャーレ使用)に置床した。その後、29℃の明所(20,000ルクス)で培養した。次に、新しい再分化I培地への植継ぎを3回ほど繰返し、3cm程度に成長した個体を再分化II培地へ移して、29℃の明所(20,000ルクス)で培養し、再分化個体を得た。その後、実験温室にて再分化個体を培養土(商品名:合成培土3号、製造会社名:JA社製)へ植替え、生育させた。
【0059】
【表2】
【0060】
−高温登熟性イネ選別工程−
前記再分化個体を生育させたイネから高温登熟性イネを選別した。
高温登熟性イネの選別方法は、以下のようにして行った。
前記培養変異が確認された培養変異系統479系統)について、実験温室水田(登熟期平均気温26.6℃)で生育させ、玄米形質を調査した(図2A〜F参照)。これらの系統のうち、図2Aのように、粒数及び品質の低下が生じず、青未熟粒が多発せず、不稔が生じなかったイネを、コシヒカリ原種よりも品質が良い系統として19系統を得た。なお、図2Bは、粒数の低下が見られた系統のイネ、図2Cは、品質低下が見られた系統のイネ、図2Dは、青未熟粒が多発した系統のイネ、図2Eは、不稔が生じた系統のイネを示し、図2Fは、野生型のイネを示す。これらのコシヒカリ原種よりも品質が良かった19系統の中から、更に品質の良い系統を、3系統(NNK286系統、NNK306系統、及びNNK506系統)選択し、下記試験を行った。
【0061】
<高温登熟性イネ試験>
5月上旬及び7月上旬に、鹿児島県農業開発総合センターの圃場において、コシヒカリ培養変異系統(NNK306系統)及び野生(WT)型の苗(n=5)を栽培した。これらの正常粒率を調査した結果を、図3A及び図4A〜図4Dに示す。なお、圃場の登熟期の平均気温は、それぞれ28.2℃、26.2℃であった。
新潟県総合農業研究所作物研究センターの温水掛流し圃場及び一般圃場において、コシヒカリ培養変異系統(NNK286系統、NNK306系統、NNK506系統)及び野生(WT)型の苗(n=4)を栽培した。これらの正常粒率を調査した結果を図3Bに示す。なお、各圃場の登熟期平均気温は28.3℃、27.1℃であった。
福岡県農業総合試験場の温水掛流し圃場において、コシヒカリ培養変異系統(NNK306系統)及び野生(WT)型の苗(n=5)を栽培した。これらの正常粒率を調査した結果を図3Cに示す。なお、圃場の登熟期平均気温は28.8℃であった。
【0062】
<高CO2登熟性イネ試験>
独立行政法人農業環境技術研究所のFACE(Free Air CO2 Enrichment)実験圃場において、コシヒカリ培養変異系統(NNK306系統)及び野生(WT)型の苗(n=12)を栽培した。これらの正常粒率を調査した結果を図3Dに示す。なお、圃場の登熟期平均気温は25.9℃であり、FACE実験圃場の平均CO2濃度(600ppm)は、一般実験圃場の平均CO2濃度(379ppm)の約1.6倍であった。
【0063】
<結果>
本発明の高温登熟性イネの育種方法により育種された3系統(NNK286系統、NNK306系統、NNK506系統)は、イネの登熟期に平均気温28℃以上に曝して栽培しても、正常粒率が40%以上となることがわかった(図3A〜C参照)。また、図4A〜Dより、本発明の高温登熟性イネは、玄米品質の低下も見られず、イネの高温登熟性が改善されたことがわかった。また、本発明の高温登熟性イネの育種方法により育種されたNNK306系統は、イネの登熟期の平均CO2濃度を一般実験圃場の平均CO2濃度の約1.5倍に曝して栽培しても、品質低下が軽減され、正常粒率が60%以上となることがわかった(図3D参照)。
【0064】
最も結果の良かったNNK306系統については、ゲノム解析を行った。
ゲノム解析は、生育させた苗を、CTAB処理、フェノール/クロロフォルム/イソアミルアルコール抽出、エタノール沈殿することにより、NNK306系統のイネゲノム及び野生型のイネゲノムの各ゲノムDNAを抽出して、AFLP(Amplified Fragment Length Polymorphism、増幅断片長多型)解析することにより行った。
まず、各種制限酵素(DraI、HindIII、MspI、タカラバイオ社製)により切断したゲノムDNA断片に2種のオリゴマー(Adapter1、Adapter2−Msp1、Adaprer2−Hind3、Adapter2−blunt end、北海道システムサイエンス社に作製を依頼)をアニールさせることで作製したアダプターを結合させ、アダプター特異的にアニールするプライマー(AP1、北海道システムサイエンス社に作製を依頼)と、トランスポゾン特異的にアニールするプライマー(LTR1、北海道システムサイエンス社に作製を依頼)を用いて、PCR(PCR条件;変性:94℃、30秒間、アニーリング:69℃、30秒間、伸長反応:72℃、4分間、上記反応サイクル数7回、その後変性:94℃、30秒間、アニーリング:61℃、30秒間、伸長反応:72℃、4分間、上記反応サイクル数32回、PCR装置;TAKARA PCR Thermal Cycler Dice TP650、タカラバイオ社製)によりDNA断片を増幅させた。
次に、得られたPCR産物を鋳型にして、nested−PCR(PCR条件;変性:94℃、30秒間、アニーリング・伸長反応:72℃、4分間、上記反応サイクル数5回、その後変性:94℃、30秒間、アニーリング:69℃、30秒間、伸長反応:72℃、4分間、上記反応サイクル数32回、プライマー(LTR2、AP2、北海道システムサイエンス社に作製を依頼)、PCR装置;TAKARA PCR Thermal Cycler Dice TP650、タカラバイオ社製)を行い、増幅断片を得た。なお、用いたオリゴマー、プライマー等の塩基配列を表3に示す。
【0065】
【表3】
【0066】
AFLP解析により、NNK306系統については、レトロトランスポゾンのイネゲノム上の位置が、変異したことがわかった(図1参照)。なお、図1は、DraIで切断したDNAをテンプレートに、アダプター特異的なAP2プライマーとTos17特異的なLTR2プライマーを用いたPCR(nasted−PCR)の電気泳動写真を含み、この電気泳動写真におけるレーン4が、NNK306系統に該当する。これらの結果から、NNK306系統は、カルス培養細胞の懸濁培養により、イネゲノムのレトロトランスポゾンが活性化され、高温登熟性及び高CO2登熟性を獲得したことが示唆された。
【0067】
以上より、本発明の高温登熟性イネは、イネ自身の遺伝子により高温登熟性を獲得し、イネ登熟期の気温上昇及びCO2濃度上昇にさらされても、米生産性の低下を抑制することができ、高品質な良食味米を供給することができ、高温登熟性、及び高CO2登熟性に優れることがわかった。
【産業上の利用可能性】
【0068】
本発明は、イネ登熟期の気温上昇及びCO2濃度上昇にさらされても、米生産性の低下を抑制することができ、高品質な良食味米を供給することができる、高温登熟性に優れた高温登熟性イネを提供することができるため、地球温暖化に伴うイネの高温登熟障害、及び高CO2登熟障害の被害対策として好適に活用することができる。
【特許請求の範囲】
【請求項1】
イネ種子をカルス培養することによりカルス培養細胞を得る工程と、
前記カルス培養細胞を懸濁培養することによりイネゲノムの変異が生じた懸濁培養細胞を得る工程と、
前記懸濁培養細胞を再分化することにより再分化個体を得る工程と、
前記再分化個体から高温登熟性イネを選別する工程と、
を含む育種方法により育種されたことを特徴とする高温登熟性イネ。
【請求項2】
イネゲノムが、レトロトランスポゾンの活性化により変異して、高温登熟性及び高CO2登熟性を獲得する請求項1に記載の高温登熟性イネ。
【請求項3】
登熟期平均気温28℃以上35℃以下で生育させた時に得られる米の正常粒率が、40%以上である請求項1から2のいずれかに記載の高温登熟性イネ。
【請求項4】
登熟期平均CO2濃度550ppm以上650ppm以下で生育させた時に得られる米の正常粒率が、40%以上である請求項1から2のいずれかに記載の高温登熟性イネ。
【請求項1】
イネ種子をカルス培養することによりカルス培養細胞を得る工程と、
前記カルス培養細胞を懸濁培養することによりイネゲノムの変異が生じた懸濁培養細胞を得る工程と、
前記懸濁培養細胞を再分化することにより再分化個体を得る工程と、
前記再分化個体から高温登熟性イネを選別する工程と、
を含む育種方法により育種されたことを特徴とする高温登熟性イネ。
【請求項2】
イネゲノムが、レトロトランスポゾンの活性化により変異して、高温登熟性及び高CO2登熟性を獲得する請求項1に記載の高温登熟性イネ。
【請求項3】
登熟期平均気温28℃以上35℃以下で生育させた時に得られる米の正常粒率が、40%以上である請求項1から2のいずれかに記載の高温登熟性イネ。
【請求項4】
登熟期平均CO2濃度550ppm以上650ppm以下で生育させた時に得られる米の正常粒率が、40%以上である請求項1から2のいずれかに記載の高温登熟性イネ。
【図1】

【図2A】

【図2B】

【図2C】

【図2D】

【図2E】

【図2F】

【図3A】
【図3B】
【図3C】
【図3D】
【図4A】

【図4B】

【図4C】

【図4D】
【図2A】
【図2B】
【図2C】
【図2D】
【図2E】
【図2F】
【図3A】
【図3B】
【図3C】
【図3D】
【図4A】
【図4B】
【図4C】
【図4D】
【公開番号】特開2013−102698(P2013−102698A)
【公開日】平成25年5月30日(2013.5.30)
【国際特許分類】
【出願番号】特願2011−246579(P2011−246579)
【出願日】平成23年11月10日(2011.11.10)
【国等の委託研究の成果に係る記載事項】(出願人による申告)平成23年度農林水産省「新農業展開ゲノムプロジェクト」委託事業、産業技術力強化法第19条の適用を受ける特許出願
【出願人】(304027279)国立大学法人 新潟大学 (310)
【Fターム(参考)】
【公開日】平成25年5月30日(2013.5.30)
【国際特許分類】
【出願日】平成23年11月10日(2011.11.10)
【国等の委託研究の成果に係る記載事項】(出願人による申告)平成23年度農林水産省「新農業展開ゲノムプロジェクト」委託事業、産業技術力強化法第19条の適用を受ける特許出願
【出願人】(304027279)国立大学法人 新潟大学 (310)
【Fターム(参考)】
[ Back to top ]