
高粘度キサンタンポリマー調製物
【課題】キサンタンポリマーの分子長が増加すると、キサンタン組成物の粘度が高くなる比粘度特性を有するキサンタンを提供する。
【解決手段】Xanthomonas campestris培養物中で、orfXではなく、gumD〜gumGからなる群より選択される遺伝子でもないgumBおよびgumCの遺伝子産物の量を選択的に増加させることからなる。増加させる方法は、gumBおよびgumCの1つまたは複数の追加コピーを導入することからなる。
【解決手段】Xanthomonas campestris培養物中で、orfXではなく、gumD〜gumGからなる群より選択される遺伝子でもないgumBおよびgumCの遺伝子産物の量を選択的に増加させることからなる。増加させる方法は、gumBおよびgumCの1つまたは複数の追加コピーを導入することからなる。
【発明の詳細な説明】
【技術分野】
【0001】
発明の分野
本発明は微生物生産物の分野に関する。特に、様々な工業目的のために改善された特性を有する微生物生産物に関する。
【0002】
本出願は2003年3月21日に出願された米国特許仮出願第60/456,245号の恩典を主張する。
【0003】
本特許文書の開示内容の一部は、著作権保護を受ける材料を含む。著作権所有者は、特許庁特許ファイルまたは記録中に現れる場合、特許文書または特許開示のいずれかによる複写に全く異存はない。そうでなければ、何であれ、著作権全てを留保する。
【背景技術】
【0004】
発明の背景
キサンタンの化学構造は、主鎖の交互のグルコース残基に付着した、マンノース、グルクロン酸およびマンノースからなる三糖側鎖を有する直線セルロース(1→4)-β-D-グルコースポリマーから構成される。(Milas and Rinaudo、Carbohydrate Research、76、189-196、1979(非特許文献1))。このように、キサンタンは五糖繰り返しユニットを有する分枝鎖ポリマーとして記述することができ;通常のキサンタンは典型的には、2000〜3000の五糖繰り返しユニットを有する。キサンタンポリマーは典型的は、マンノース残基のアセチル化およびピルビル化(pyruvylation)により修飾される。
【0005】
キサントモナス(Xanthomonas)細菌の作用により生合成水溶性多糖キサンタンガムが生成する糖発酵は、よく知られている。最先の研究は米国農務省により実施されており、米国特許第3,000,790号(特許文献1)において記述されている。キサントモナス親水性コロイド(「キサンタン」)は細胞外ヘテロ多糖である。
【0006】
キサンタンはキサントモナス属の細菌の好気性液内発酵により生成する。発酵培地は典型的には、炭水化物(例えば糖)、微量元素および他の栄養素を含む。発酵が完了すると直ちに、得られた発酵ブロス(溶液)は典型的には加熱処理される。キサンタン発酵ブロスおよび溶液の加熱処理により、転移温度(TM)またはそれ以上の温度で未変性キサンタンの構造変化が起こり、高粘度キサンタンが生成することが確立している。熱処理はまた、キサンタン中の生存微生物および望ましくない酵素活性を破壊するという有益な効果を有する。熱処理後、キサンタンをアルコール沈澱により回収する。しかしながら、キサンタン発酵ブロスの熱処理には欠点もあり、例えば、キサンタンの熱分解である。キサンタン溶液またはブロスを、TMを超える温度で加熱する、またはTMを超える温度で数秒を超える時間維持すると、キサンタンの熱分解が起こる。キサンタンの分解により不可逆的に粘度が減少する。したがって、熱処理はキサンタンの品質および粘稠性を制御する重要な技術である。
【0007】
キサンタンの品質は、主として2つの粘度試験により決定される:水道水中での低せん断速度粘度(「LSRV」)および高塩溶液中での海水粘度(「SWV」)。TMまたはそれを超える温度でのキサンタン発酵ブロスを殺菌(pasteurization)することによって、より高いLSRVおよびSWV値により示されるように、より高粘度のキサンタンが得られることがわかっている。
【0008】
キサンタンポリマーは多くの状況で使用される。キサンタンは様々な産業用途を有し、例えば、油井掘削泥における使用、水攻法による石油の二次回収における粘度制御添加剤、食物中の増粘剤、安定化剤、ならびに乳化、懸濁およびサイズ剤としての使用が挙げられる(Encyclopedia of Polymer Science and Engineering、2nd Edition、Editors John Wiley & Sons、901-918、1989(非特許文献2))。キサンタンはまた、化粧料調製物、薬学的媒質、および同様の組成物中で使用することができる。
【0009】
当技術分野では、未殺菌(unpasteurized)状態で高い比粘度特性を有するキサンタンポリマーを生成する必要がある。そのような比粘度の高いキサンタンポリマーは、例えば、食品、工業および油田用途に対し、同等のキサンタン濃度でより高い粘度を提供することができる。
【先行技術文献】
【特許文献】
【0010】
【特許文献1】米国特許第3,000,790号
【非特許文献】
【0011】
【非特許文献1】Milas and Rinaudo、Carbohydrate Research、76、189-196、1979
【非特許文献2】Encyclopedia of Polymer Science and Engineering、2nd Edition、Editors John Wiley & Sons、901-918、1989
【発明の概要】
【0012】
第1の態様では、未殺菌キサンタン組成物を提供する。組成物は、gumBおよびgumCを過剰発現する細胞により提供することができ、gumBおよびgumCを過剰発現しない対応株よりも少なくとも20%高い固有粘度を有する。
【0013】
第2の態様では、キサンタン組成物を提供する。キサンタン組成物は、一定範囲の分子長を有するキサンタン分子集団を含む。集団の少なくとも1%が原子間力顕微鏡観察法で測定すると、3umを超える長さを有する。
【0014】
本発明の第3の態様では、野生型株により産生されるものに比べ粘度が増加したキサンタンポリマー調製物を産生するための方法を提供する。gumBおよびgumCの遺伝子産物の量をキサントモナス・カンペストリス(Xanthomonas campestris)培養物中で選択的に増加させる。orfXの遺伝子産物の量は選択的に増加させない。gumD〜gumGからなる群より選択される遺伝子産物の量も選択的に増加されない。このように、より粘度の高いキサンタンポリマー調製物を培養物により産生させる。
【0015】
本発明の第4の態様では、野生型株により産生されるものに比べ粘度が増加したキサンタンポリマー調製物を産生するための方法を提供する。キサントモナス・カンペストリス株を、培地中、キサンタンポリマーを産生する条件下で培養する。株は選択的に、野生型株に比べ、orfXの遺伝子産物でもgumD〜gumGからなる群より選択される遺伝子でもなく、gumBおよびgumCの遺伝子産物をより多く産生する。
【0016】
本発明の第5の態様では、未殺菌キサンタン組成物を提供する。組成物はgumBおよびgumCを過剰発現する細胞により作成される。組成物は、gumBおよびgumCを過剰発現しない対応株由来のキサンタンよりも少なくとも10%多い海水粘度を有する。
【0017】
このように、本発明は対応する野生型株により同様に産生されるものに比べ粘度が増加したキサンタン組成物を用いる技術を提供する。
【図面の簡単な説明】
【0018】
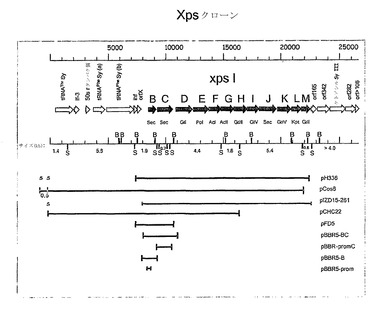
【図1】xpsB-M(キサンタン多糖合成)オペロンとしても知られている、gumB-Mオペロンの遺伝子地図に対する遺伝子構築物を示す図である。
【図2】図2Aおよび2Bは、それぞれgumBおよびgumCタンパク質産物発現のウエスタンブロット分析を示す図である。
【図3】キサンタンガム試料に対する固有粘度プロットを示す図であり、そのうちの1つは遺伝子の余分なコピーを保有するプラスミドの存在により、gumBおよびgumCを過剰発現する。
【発明を実施するための形態】
【0019】
発明の詳細な説明
オペロンにおいて他の遺伝子に比べgumBおよびgumC遺伝子産物が過剰発現すると、1重量を基本に、より高い粘度を有するキサンタン産物が得られることは本発明者らの発見である。出願人はいずれの特別な動作理論にも縛られることは望まないが、一定の遺伝子産物の比率がシフトするとキサンタンポリマー分子のサイズ分布がシフトすると考えられる。キサンタンが野生型細胞により産生される場合に比べ、かなりの数の分子の分子長が長くなる。これらのより長い分子により、集団または調製物の粘度が高くなる。
【0020】
当技術分野では、キサンタン調製物を殺菌することにより粘度を増加させることができることが知られている。Talashekら、米国特許第6,391,596号を参照のこと。しかしながら、gumBおよびgumCの過剰発現の結果見られる粘度の増加は、殺菌をしなくても観察される。それでもなお、本発明の産物をその後に殺菌するとより高粘度の調製物が得られる。
【0021】
粘度の増加を達成するには、gumBおよびgumCの両方の過剰発現が必要であると考えられる。どちらかの遺伝子を単独で試験すると、増加が観察されなかった。gumBおよびgumCの過剰発現は、gumB〜Mオペロンの他の遺伝子に対して評価することができる。それらの遺伝子のうちのいずれに対しても過剰発現が効果を達成するのに十分である場合、orfXおよびgumDに対する過剰発現が特に重要である可能性がある。OrfXは、gumBのすぐ上流の、gumAと呼ばれるゲノムのセグメンとして以前公表された小さな翻訳領域である。最近では2つの翻訳領域が前者gumA領域で区別されており、ihfおよびorfXである。遺伝子gumD〜gumMの全てに対して過剰発現であることが望ましい場合がある。
【0022】
所望の遺伝子産物の過剰発現は、当技術分野で周知の任意の手段により達成してもよい。そのような手段としては、例えば、キサンタンを生成するキサントモナス・カンペストリス細胞または他の細菌に所望の遺伝子産物をコードする遺伝子の追加のコピーを導入すること、および例えば誘導プロモータを用いて所望の遺伝子産物を誘導することが挙げられるが、これらに限定されない。キサンタンを生成する他の細菌としては、キサンタン生合成遺伝子を含むように遺伝子操作されたものが挙げられる。gumBおよびgumC遺伝子は、1つまたは複数のベクター上に、すなわち、組み合わせてまたは別個に導入することができる。
【0023】
本発明により使用することができる誘導プロモータとしては、当技術分野で周知の全てのもの、例えば、lacプロモータ、araプロモータ、tetプロモータ、およびtacプロモータが挙げられる。これらのプロモータに対する天然および人工の誘導物質は当技術分野で周知であり、いずれを使用しても都合がよい。遺伝子の追加のコピーを、例えば、プラスミドまたはウイルスベクターに導入することができる。所望の遺伝子の追加のコピーは染色体外で維持することができ、またはゲノムに組み込むことができる。
【0024】
培養ブロスからのキサンタンの回収は典型的には、1つまたは複数のプロセッシング(processing)段階を含む。キサンタンは加熱処理してもよい。キサンタンはアルコール、例えばイソプロピルアルコール、エチルアルコール、またはプロピルアルコールで沈澱させてもよい。典型的には、細胞は培養ブロスから特異的に除去されない。
【0025】
生合成により生成されるキサンタン分子は典型的には一定のサイズ分布を有する。本発明の粘度増加は、平均長よりもずっと長い分子のその数を増加させることにより、または平均長より幾分長い分子のその数を著しく増加させることにより達成してもよい。長さが増大した分子のその数は莫大である必要はない。少なくとも長さが1、3、5、7、9または11%増大した分子で十分である。長さが増大した分子は、原子間力顕微鏡で測定すると、3、4、5、6、7、8または9umを超えていてもよい。3、4、5、6、7、8または9umよりも長い分子により提供される総キサンタン集団の質量のパーセンテージは、その集団内のそれらの数の割合よりも大きくなる。このように、キサンタン分子の総質量の少なくとも1、3、5、10、15、20または25%が、3umを超える長さを有する分子により提供されていてもよい。
【0026】
固有粘度測定は、本発明の調製物を特徴づけるさらに別の方法である。この型の測定を用いて観察される増加は、野生型株により産生されるものよりも、5、10、15、20、25、30、または35%を超えて大きい場合がある。比較のための適当な対照は、試験される株に最も密接に関係する対応株である。このように、gumBおよびgumCの追加のコピーを有する株を試験する場合、最もよい対照は、gumBおよびgumCの追加のコピーの存在を除き、同じ遺伝的相補を有する。より多くのgumBおよびgumC遺伝子産物を産生させる誘導物質により誘導されている培養物を試験する場合、最良の対照は、誘導されていない同じ株の培養物である。本発明の調製物を特徴づけるために海水粘度もまた使用することができる。この型の測定を用いて観察される増加は、野生型により産生されるものよりも、5、10、15、20、25、30、または35%を超えて大きい場合がある。
【0027】
キサンタンを、特性を改善するために多くの製品中の成分として使用する。特性としては、2〜3例を挙げると、粘度、粒子の懸濁、口当たり(mouth feeling)、バルクが挙げられる。他の特性としては、水の結合、増粘剤、エマルジョン安定化、起泡増強、およびせん断-希薄化(shear-thinning)が挙げられる。そのような製品としては、食品、例えばサラダドレッシング、シロップ、ジュース飲料、および冷菓が挙げられる。そのような製品としてはまた、印刷染料、油掘削流体(oil drilling fluid)、セラミック釉薬(ceramic glaze)、および薬学的組成物が挙げられる。後者の場合、キサンタンは担体として、または放出制御基質として使用することができる。キサンタンを使用することができる他の製品としては、洗浄液、塗料およびインク、壁紙接着剤、農薬、磨歯剤、ならびに酵素および細胞固定化剤が挙げられる。
【0028】
本発明について、本発明を実施するのに現在好ましい様式を含む特定の実施例に関し説明してきたが、当業者であれば、添付の特許請求の範囲で記述した本発明の精神および範囲の中にある、上記システムおよび技術について多くの変更および置換が存在することは理解するであろう。
【実施例】
【0029】
実施例1 株の構築
X.カンペストリスの完全なgum遺伝子領域を保有する断片を単離するために、野生型X.カンペストリス株、NRRL B-1459(1)のゲノムライブラリを、Sau3AIにより部分的に消化した総DNAのクローニングにより、宿主範囲の広いコスミドベクターpRK311(2)を用いて構築した。このライブラリは、全体として、大腸菌(E.coli)S17-1(3)〜Gum- X.カンペストリス突然変異体2895(4)に一致した。pIZD15-261(5)と呼ばれるいくつかのムコイド接合完了体(mucoid exconjugant)から単離したコスミドの1つは、完全gum領域を包含する16kbの断片を含む。グラフ表示の図1、およびオペロンの遺伝子のリストの表1を参照のこと。
【0030】
(表1)キサンタン多糖合成をコードする染色体領域中の遺伝子名称リスト

* 遺伝子位置は、da Silva、A.C.R.ら(Nature、Vol. 417、pg.459-463、2002)により記述されているように、X.カンペストリスpv.カンペストリスATCC33913(GenBank寄託:AE008922)の遺伝子配列に従う
【0031】
pBBR5-BCプラスミドを構築するこために、SpeI-BglIIで消化したpIZD15-261由来の4026bp断片をpKmob19(8)のXbaI部位とBamHI部位の間にクローニングし、pGum02-19S(5)を得た。SphIでの消化によりプラスミドpGum02-19Sから2855bp断片を放出させた。この断片を、あらかじめSphIを用いて消化したpUC18(9)にクローニングし、pUC18-BCASを形成させた。
【0032】
gumプロモータならびにgumBおよびgumC遺伝子を含む、HindIII-XbaI断片を、HindIII-XbaI消化pBBRR1-MCS5(10)(GenBank アクセッション番号U25061)にクローニングすることにより最終プラスミド(pBBR5-BC)を構築した。
【0033】
得られたpBBR5-BCプラスミドのヌクレオチド配列をSEQ ID NO:1に示す。(gumBおよびgumCの予想アミノ酸配列はそれぞれ、SEQ ID NO:2およびSEQ ID NO:3に示す)。この宿主範囲の広い、コピー数が中程度のプラスミドは長さが7.1kbであり、IncP、IncQおよびIncW群プラスミド、ならびにCo1E1系およびP15a系レプリコンと適合する。RK2伝達機能がトランスで提供されると、伝達起点(mobRK2)の存在により、広範囲の細菌内に接合により伝達することが可能となる。このプラスミドは、ゲンタマイシン耐性遺伝子も保有し、かつLacZαペプチドをコードする遺伝子内に位置するpBluescript II KS多重クローニング部位を含む(Stratagene、La Jolla、Ca、USAからのpBluescript II KS)。
【0034】
pBBR5-BCからのGumBおよびGumCタンパク質の発現を証明するために、プラスミドをX.カンペストリス変異体1231に導入した。変異体では、全gum(xps)遺伝子クラスターが欠失した。どちらのタンパク質も変異体株におけるウエスタンブロットにより検出した。
【0035】
(表2)本研究で使用または構築した細菌株およびプラスミド
a 数はgum領域のヌクレオチド配列の位置に対応する(GenBank、アクセッション番号 U222511)
【0036】
細菌株、プラスミド、および増殖条件
この研究で使用した株およびプラスミドを表2に示す。大腸菌(E.coli)株は37℃のLuria-Betani培地で増殖させた。X.カンペストリス株はTY(トリプトン5g、酵母抽出物3g、およびH2O 1リットルにつきCaCl2 0.7g)またはYM培地(12)、28℃で増殖させた。Sigma(St.Louis、Mo.)からの抗生物質を、必要に応じて下記濃度(μg/mL)で補った:X.カンペストリスでは、ゲンタマイシン、30;およびテトラサイクリン、10;大腸菌では、ゲンタマイシン、10;カナマイシン、30;アンピシリン、100;およびテトラサイクリン、10。
【0037】
DNA生物化学
大腸菌およびX.カンペストリス由来のプラスミドDNAを、QIAprep Spin Miniprep Kit(QIAGEN、ヒルデン、ドイツ)を用いて調製した。DNA制限、アガロースゲル電気泳動およびクローニング手順を、確立したプロトコル(13)に従い実施した。全ての構築物をDNA配列決定により検証した。プラスミドDNAを、Bio-Rad(リッチモンド、カリフォルニア州)により指示されているようにエレクトロポレーションにより大腸菌およびX.カンペストリス細胞に導入した(使用パラメータ:大腸菌:200Ω、25μF、2500V、およびX.カンペストリス:1000Ω、25μF、2500V)。
【0038】
ヌクレオチドおよびタンパク質配列の分析
ヌクレオチドおよびアミノ酸配列を、MacVector Sequence Analysis Software(Oxford Molecular Limited、ケンブリッジ、英国)を用いて分析した。
【0039】
実施例2 gumBおよびgumC発現のウエスタン分析
ウエスタン分析により、gumBおよびgumC遺伝子産物が、gumBおよびgumCの余分なコピーを有するX.カンペストリス株で過剰発現されることが確認された。図2を参照のこと。
【0040】
実施例3 固有粘度決定
gumBおよびgumC遺伝子の多重、プラスミドコードコピーを有する(XWCM1/pBBR5BC)、ならびに有さない(XWCM1)X.カンペストリス株から調製したキサンタン試料を比較した。炭素源としてブドウ糖を使用したフラスコ発酵物を振盪し、これらの株からキサンタンを得た。
【0041】
精製および非精製キサンタン試料について粘度を測定することにより固有粘度を決定した。gumBおよびgumCの多重コピーを有するX.カンペストリス株由来のキサンタンでは固有粘度の増加が観察された。固有粘度は、同一の溶媒および温度条件下で測定すると、一定のポリマー型では分子量に比例する。そのため、gumBおよびgumCの多重コピーを有するX.カンペストリス株由来のキサンタンは対照株由来のキサンタンよりも高い分子量を有する。
【0042】
方法:2種のブロスについてそれぞれ5つの振盪フラスコで試験を行った。各型のブロスを合わせ、総体積を測定した。その後、ブロスをイソプロピルアルコールに沈澱させた。(注:ブロスが約3%のガムを含むと推定した。総ブロス体積を測定し、3%をかけると、おおよその乾燥ガム重量が得られた。この概算を使用して、約0.5%のガム溶液を作製するのに必要とされる水の量を計算した)。その後、沈澱の湿性繊維を0.01M NaCl中で混合することにより直ちに再水和させ、約0.5%のガム溶液を生成させた。繊維を良好なせん断で3時間、3ブレードの直径2インチのプロペラ撹拌機を用いて混合し、その後、一晩放置した。下記手順を使用して、固有粘度測定のための試料を調製した。
【0043】
上記で調製した〜0.5%ガム溶液を、Gelman Science 293mm圧力濾過ユニットを用いて濾過する。溶液は最初に、20μMagnaナイロンフィルター(N22SP29325)を通して濾過する。フィルターを〜60psiまで加圧し、溶液を清浄なビーカー中に回収する(注:流速が〜5ドリップ/分まで減少するとフィルターを交換する)。
【0044】
第1の濾過段階に続き、上記濾過ユニットを用い試料をさらに2回濾過する。第1に、Millipore 8.0μフィルター(SCWP293 25)を通し、その後、Gelman Versapor(登録商標)293mm 1.2μフィルター(66397)を通す。濾過した試料を各濾過段階後、洗浄なビーカー中に回収する。
【0045】
濾過後、〜600mlのガム溶液をSpectra/Por(登録商標)透析管、直径28.6mmのSpectrum#S732706(MWCO 12,000〜14,000)に入れる。管を〜18から20インチの長さに切断し、一端で結び目を作る。溶液を管に追加し、端から〜2インチ以内になるまで満たす。管内にトラップされる空気ができるだけ少なくなるように管で第2の結び目を作る。全てのガム溶液が透析管に入れられるまで続ける。
【0046】
ガム溶液を含む管の外側を〜1分間、脱イオン水ですすぎ、その後、管を0.01M NaClの容器内に入れる。塩溶液が透析管を完全に覆うようにしなければならない。
【0047】
管を0.01M NaCl溶液中に4日間静置し、毎日NaCl溶液を交換する。4日後、管の一端を切り開き、注意深くガム溶液を清浄なビーカーに移す。
【0048】
下記手順を用いて、濾過した透析溶液について固体分析を実施する。
【0049】
±0.0002gまで秤量できる化学天秤を用いて、清浄なアルミニウム製の秤量皿VWR Cat#25433-008の質量を秤量し、記録する。(A)
【0050】
清浄なピペットを使用して、約10mlのガム溶液をアルミニウムパンに添加し、パンとガム溶液を合わせた正確な質量を記録する。(B)
【0051】
溶液を有するパンを105℃の乾燥オーブン内に入れ、24時間放置する。
【0052】
24時間後、オーブンからパンを取り出し、冷却し、再び秤量する。パンと残った乾燥ガムの質量を記録する。(C)
【0053】
パン+ガム溶液の質量(B)からアルミニウムパンの質量(A)を引く。乾燥ガム+パンの質量(C)からアルミニウムパン(A)の質量を引く。最初の値(B-A)で第2の値(C-A)を割る。この値に100をかけると固体の割合(%)が得られる・
【0054】
注:固体分析を、上記手順を用いて各濾過した透析溶液に対し、3連で実施した。計算した固体の割合(%)を各試料に対し平均化し、その平均値を使用した。
【0055】
各溶液に対する固体決定に基づき、0.01M NaClを用い試料を0.25%総ガム濃度まで希釈する。
【0056】
固有粘度測定を、Vilastic Viscoelasticity Analyzer(Vilastic Scientific, Inc.、オースチン、TX、半径0.0537cm×長さ6.137cmの管が取り付けられている)を用いて実施した。測定する前に機器を水で較正し、測定完了後確認した。周波数2.0Hz、定歪1.0、積分時間10秒に設定した、機器TIMETソフトウエアプロトコルを用いて測定を実施した。温度は23.5℃に維持した。0.25%ガム溶液を希釈することにより試料を調製した。各希釈液を20分間混合し、測定する前、一晩冷却して放置した。各希釈液に対し、6回測定し、平均した。下記表3は希釈液および各調製試料に対して得られた平均粘度を示す。
【0057】
(表3)
【0058】
ガム濃度に対し減少した比粘度(ηsp/c)をプロットすることにより固有粘度を決定した。ηsp/c=((ηc-η0)/η0)、式中、ηc=ガムの粘度である。切片から固有粘度が得られる。図3を参照のこと。
【0059】
XWCM1/pBBR5-BC変異体では固有粘度が増加するのは、分子量が増加するためだと考えられる。この実験を実施した場合と同一の溶媒および温度条件下で測定すると、一定のポリマー型では固有粘度は分子量に比例する。[η]と分子量の間の関係は、Mark-Houwinkの式[η]=kMaにより与えられる。式中、kおよびaは特定の温度の特定の溶媒中の特定のポリマー型に対する定数である。定数「a」は正の数であるので、[η]の増加は、Mark-Houwinkの式に従わない異なる分子配座を有しなければ、分子量(M)の増加によってのみ得られる。
【0060】
実施例4 手順−低せん断速度粘度測定
精製キサンタン試料について低せん断速度粘度測定を実施した。LSRVを測定するのに使用した手順の詳細を以下に示す。対照株由来のキサンタンに比べ、gumBおよびgumCの複数コピーを有する株由来のキサンタンに対する粘度の増加が観察された。データから、鎖長の増加にはgumBおよびgumC両方の過剰発現が必要であること;gumBまたはgumCいずれかの単独の過剰発現では鎖長増加には不十分であることが示唆される。
【0061】
材料および装置:
1. 標準(合成)生水(1000ppmのNaClおよび40ppmのCa++または147ppmのCaCl2.2H2Oを含む水):適した容器に入れた20リットルの蒸留水に試薬グレードのNaCl 20gおよび試薬グレードのCaCl2.2H2O 2.94mgを溶解することにより調製する。
2. 0.01gmまで正確に測定することができる天秤。
3. Brookfield LV粘度計、スピンドル#1、およびスピンドルGuard。
4. 標準の実験室用ガラス器具。
5. 標準の実験室用撹拌ベンチ。RAE撹拌モーター(C25U)および3ブレードプロペラを備えた撹拌シャフト(5/16'')は置換してもよい。
【0062】
手順:
1. 600mlのBerzelius(背の高い形態)ビーカーに秤量した合成生水299mlに、生成物0.75gm(0.001gmの位まで秤量)を、800ppmで撹拌しながら、徐々に添加する。
2. 800rpmで4時間撹拌した後、溶液を撹拌ベンチから取り出し、30分間放置する。
3. 温度を室温に調節し、3rpmでNo.1スピンドルを備えたBrookfield LV粘度計を用いて粘度を測定する。スピンドルを3分間回転させた後、粘度を記録する。
【0063】
実施例5 タンパク質発現の定量
細胞溶解物をウエスタンブロットおよび免疫検出分析にかけ、プラスミドによりコードされたGumBおよびGumCレベルを明確にした。4つの独立したブロットを分析した。同じ試料に対する絶対値は各定量において再現性がなかったが、試料間の相対量は全ての測定において同じままであった。
【0064】
GumBおよびGumCに対し産生された抗体の調製
GumCタンパク質のアミノ酸残基53-447をコードする1184bpのDNA断片を、PCR増幅により作製した。下記プライマーを使用した。
PCR産物をNdeIおよびBamHIを用いて消化し、pET22b(+)にサブクローニングし、得られたプラスミド(pET-C)を大腸菌株BL21(DE3)に導入した。
【0065】
OD6000.6まで、50μg/mlのカルベニシリンを含むL-ブロス中で増殖させた大腸菌BL21(pET-C)を、1mM IPTGで3時間誘導した。総細胞溶解物を、37℃で30分間、1mgのリゾチームを含む溶解緩衝液(50mM Tris/HCl pH8、1mM EDTA pH8、100mM NaCl、1mM PMSF、0.1mg/ml DNase、0.5% Triton X-100)で処理し、その後氷上で超音波処理することにより調製した。低速遠心分離(Eppendrof、4000xg、5分間)により細胞片を除去し、上清を、10分間の1400xgの遠心分離により、可溶性画分とペレット(封入体)画分に分画した。元の細胞溶解物と同じ体積の溶解緩衝液で2度、2mg/mlのDOCを含む溶解緩衝液で1度、続いて、水で3度、ペレット画分を洗浄した。処理後、タンパク質をSDS-PAGEにより分離し、過剰産生したGumCタンパク質を含む主バンドを切断し、ウサギを免疫化するために溶離した。
【0066】
OD6000.6まで、50μgのカルベニシリン、25μg/mlのカナマイシンを含むL-ブロス中で増殖させた大腸菌JM109(pQE-Xps#6、pREP4)を、1mM IPTGで3時間誘導した。総細胞溶解物を、37℃で30分間、1mg/mlのリゾチームを含む溶解緩衝液(50mM Tris/HCl pH8、1mM EDTA pH8、100mM NaCl、1mM PMSF、0.1mg/ml DNase、0.5% Triton X-100)で処理し、その後氷上で超音波処理することにより調製した。低速遠心分離(Eppendrof、4000xg、5分間)により細胞片を除去し、上清を、10分間の14000xgの遠心分離により、可溶性画分とペレット(封入体)画分に分画した。ペレット画分を溶解緩衝液で2度洗浄し、6Mの塩酸グアニジンを含む100mMのリン酸緩衝液(pH7)、5mM DTTおよび5mM EDTAに再懸濁させ、封入体を、緩衝液D(4M GdnHCl、50mM リン酸緩衝液(pH7)、150mM NaCl)で予め平衡化したFPLC Superdex HR200(Pharmacia Biotech)上でクロマトグラフにかけた。GumBを含む画分をプールし、マウスを免疫化するのに使用した。
【0067】
プラスミドpFD5、pBBR-promC、およびpBBR5-Bの構築
gumBおよびgumC遺伝子を含む3141bp断片を、pIZD15-261のBamHI(#318および#3459)を用いた部分消化により得、BamHIで消化したpRK404にクローニングし、プラスミドpFD5を得た。pGum02-19をEcoRI(#1979)およびBamHI(#3459)で消化して1480bp断片を単離し、同じ酵素であらかじめ消化したpBBR1MCS-5にクローニングし、pBBR-promCを得た。MCS内のHindIIIおよびEcoRI(#1979)でpGum02-19を消化して1233bp断片を得、これをpBBR1MCS-5にクローニングし、プラスミドpBBR5-Bを得た。
【0068】
上記のように調製したGumCを用い、雌のニュージーランドホワイトウサギを免疫化した。500μgのタンパク質をフロイント完全アジュバントと共にウサギに最初に注入し、続いて、隔週に、250μgのタンパク質を不完全アジュバントと共に3回注入した。BALB/c雌マウスを上記のように調製したGumBを用いて免疫化した。100μgのタンパク質をフロイント完全アジュバントと共にマウスに最初に注入し、続いて、1週間に1度、50μgのタンパク質を不完全アジュバントと共に3回注入した。ポリクローナル抗体を、Harlow & Lane((1999)「Using antibodies:a laboratory manual」、Cold Spring Harbor Laboratory Press、Cold Spring Harbor、N.Y.)により記述されているように調製し、抗血清を-70℃で保存した。血清を大腸菌BL21(pET22b+)およびXc1231アセトン粉末の両方に吸着させてGumC特異的抗体を得た(Harlow & Lane、上記)。
【0069】
タンパク質抽出物
プラスミドをエレクトロポレーションにより親株PRM-1に導入した。得られた株を28℃、YM培地、250rpmで中間対数期まで増殖させた。遠心分離により細胞を収集し、生体重を決定した。ペレットを10mM Tris/HCl、10mM EDTA(pH 8.0)で2度洗浄しエキソポリサッカライドを除去し、同じ緩衝液中に100mg/mlの濃度で再懸濁させた。50μlの各試料に100μlの緩衝液A(10mM Tris/HCl、10mM EDTA(pH8.0)、1.5% SDS)を添加した後、混合物を室温で10分間、その後、100℃で12分間インキュベートした。細胞溶解物を14000xg(Eppendorf 5415C)で5分間遠心分離にかけ、収集した上清を総タンパク質抽出物と名付けた。各溶解物のタンパク質濃度を、SDSの存在下、標準としてBSAを使用し、Markwell法((1978)「A modification of the Lowry procedure to simplify protein determination in membrane and lipoprotein samples.」Anal Biochem 87(1)、206-10)により決定した。
【0070】
SDS-PAGEおよび免疫検出
細胞溶解物(30μg/レーン)を試料緩衝液(125mM Tris/HCl、pH 6.8;4% SDS、20mM DTT、0.05% ブロモフェノールブルー、20% グリセロール)と混合し、2分間沸騰させた。Schaggerおよびvon Jagowの方法((1987)「Tricine-sodium dodecyl sulfate-polyacrylamide gel electrophresis for the separation of proteins in the range from 1 to 100kDa.」Analytical Biochemistry 166(2)、368-79)に従い、SDS-10%ポリアクリルアミドゲルによりタンパク質を分離した。Immobilon-P膜(PVDF、Millipore)への半乾燥転写システム(Hoefer Semiphor unit)を用いエレクトロブロッティングを実施した。転写は10mM CAPS(pH11)、10%(v/v)メタノールを含む緩衝液中で、30分間、2.5mA/cm2のゲル表面積で実施した。エレクトロトランスファー(electrotransfer)の完了後直ちに、ブロットを0.5% Ponceau-Sレッドで染色し、トランスファーの質を評価し、Milli-Q(登録商標)-グレード水で洗浄した。ブロットを一晩中、4℃で、5%の脱脂乳粉末を含むTBST(150mM NaCl、10mM Tris/HCl pH8、0.05% Tween-20)(Harlow & Lane、上記)でブロックし、その後、抗GumB(1:3000)または抗GumC(1:5000)抗体と共に3%の脱脂乳粉末を含むTBST中、室温で3時間インキュベートした。アルカリホスファターゼ結合ヤギ抗マウスIgGまたは抗ウサギIgG(Sigma)をそれぞれ、製造者により記述されているように、検出に使用した。ブロットを3度TBSTで洗浄し、ニトロブルーテトラゾリウム-5-ブロモ-4-クロロ-3-インドリルホスフェート(NBT/BCIP、Promega)を含む溶液中で展開させた。市販のタンパク質マーカーMW-SDS-70L(Sigma)を使用してSDS-PAGEの較正を実施した。
【0071】
ブロット定量
GumBおよびGumCタンパク質バンドの強度を、NBT/BCIP発現フィルターをUVPデンシトメーター(Ultra Violet Products)で走査することにより決定し、GelWorks 1D Analysis software(NonLinear Dynamics Ltd)を用いて定量化した。各フィルターは、野生型細胞における染色体にコードされたGumBおよびGumCのレベルを確立するためにPRM-1(pBBR-prom)抽出物の基準レーンを含んだ。相対量のGumBおよびGumCを観察した。図2Aおよび2Bを参照のこと。
【0072】
実施例6 手順−分子間力顕微鏡観察法を使用した分子長または分子量決定
分子間力顕微鏡観察法(AFM)または走査型プローブ顕微鏡観察法(SPM)と呼ばれる直接可視化技術を使用して、gumBおよびgumCの多重コピーを有する(XWCM1/pBBR5-BC)、有さない(XWCM1)X.カンペストリス株由来のキサンタン分子の長さを画像化した。AFMを実施するために使用した手順の詳細を以下に示す。本発明者らは、gumBおよびgumCの多重コピーを有する株により産生されたキサンタン分子の平均分子輪郭長(contour length)が、親株のものよりずっと長いことを観察した。
【0073】
0.1gのガムを含む100gの蒸留水を〜3時間混合することにより、0.1% wt%のガム溶液を調製した。1ppmストック溶液を、0.1wt%溶液20μlを20gの0.1M 酢酸アンモニウム溶液中に希釈することにより調製した。20μlの1ppmストック溶液を新たに劈開させたマイカディスク(〜1cm2)上に噴霧した。これらのマイカ試料ディスクを、その後、加熱(〜60℃)真空チャンバー中に〜1時間入れ過剰の水を除去した。その後乾燥マイカディスクをTappingモードのAFMを用いて走査した。全てのAFM像の分子輪郭長をDigital Instrumentsにより提供されたソフトウエアを用いて測定した。
【0074】
キサンタン分子の集団の輪郭長を測定した。この研究の結果を表4に示す。(各サイズクラスの分子は、示した長さより小さい、またはそれと等しい;1つのサイズクラスで示した分子の数は、より小さなサイズクラスで計数した分子を含まない)。これらの結果から、gumBおよびgumCの多重コピーを有するX.カンペストリス株由来のキサンタン分子は、対照株由来のキサンタン分子よりもかなり大きいことが証明された。原子間力顕微鏡観察法(AFM)または走査型電子顕微鏡観察法(SPM)を、市販の機器(Nanoscope IIIa、Digital Instruments、サンタバーバラ、CA)により、窒化珪素カンチレバーチップを使用して実施した。
【0075】
(表4)キサンタン分子輪郭長のAFM測定
【0076】
実施例7 海水粘度の評価
XWCM1/pBBR5-BC株により産生されたキサンタンを、市販のキサンタン製品(Xanvis(商標))と比較し、海水粘度(SWV)に対し評価した。Xanvis(商標)キサンタン製品に対する典型的なSWVは18〜22の範囲である。
【0077】
海水粘度は下記手順を用いて決定した。41.95gの海塩(ASTM D1141-52、Lake Products Co., Inc.製、メリーランドハイツ、ミズーリ州)を1リットルの脱イオン水に溶解することにより調製した。300mlの海水溶液を、Hamilton-Beach 936-2ミキサーに取り付けた混合カップに移した(Hamilton-Beach Div.、ワシントンD.C.)。ミキサー速度制御を低に設定し、単一の溝付ディスクを混合軸に取り付けた。低速設定では、混合軸は約4,000〜6,000rpmで回転する。0.86gのバイオガム製品を15〜30秒にわたり徐々に混合カップに添加し、5分間混合させた。ミキサー速度制御を高(11,000±1,000rpm)に設定し、試験溶液を約5分間混合させた。混合物は、バイオガム製品添加時間から始めて、計45分間混合させた。45分の混合時間の終わりに、2〜3滴のBara Defoam(NL Baroid/NL industries, Inc.、ヒューストン、TX)を添加し、さらに30秒間撹拌し続けた。
【0078】
混合カップをミキサーから取り外し、冷水に浸漬させ、流体温度を25±0.5℃まで降下させた。均質な溶液を確保するために、溶液を冷却後5秒間11,000±1,000rpmで再混合した。溶液を混合カップから400mlのパイレックスビーカーに移し、Fann粘度(Fann粘度計、モデル35A)を測定した。これは低速(約3rpm)で混合することにより達成した。この読み取りにより安定化させ、その後、せん断応力値をダイヤルから読み取り、3rpmでのSWV値として記録した。
【0079】
(表5)XWCM1/pBBR5-BCキサンタンおよびXanvis(商標)キサンタンの品質
a ダイヤル目盛り
【0080】
参考文献
【技術分野】
【0001】
発明の分野
本発明は微生物生産物の分野に関する。特に、様々な工業目的のために改善された特性を有する微生物生産物に関する。
【0002】
本出願は2003年3月21日に出願された米国特許仮出願第60/456,245号の恩典を主張する。
【0003】
本特許文書の開示内容の一部は、著作権保護を受ける材料を含む。著作権所有者は、特許庁特許ファイルまたは記録中に現れる場合、特許文書または特許開示のいずれかによる複写に全く異存はない。そうでなければ、何であれ、著作権全てを留保する。
【背景技術】
【0004】
発明の背景
キサンタンの化学構造は、主鎖の交互のグルコース残基に付着した、マンノース、グルクロン酸およびマンノースからなる三糖側鎖を有する直線セルロース(1→4)-β-D-グルコースポリマーから構成される。(Milas and Rinaudo、Carbohydrate Research、76、189-196、1979(非特許文献1))。このように、キサンタンは五糖繰り返しユニットを有する分枝鎖ポリマーとして記述することができ;通常のキサンタンは典型的には、2000〜3000の五糖繰り返しユニットを有する。キサンタンポリマーは典型的は、マンノース残基のアセチル化およびピルビル化(pyruvylation)により修飾される。
【0005】
キサントモナス(Xanthomonas)細菌の作用により生合成水溶性多糖キサンタンガムが生成する糖発酵は、よく知られている。最先の研究は米国農務省により実施されており、米国特許第3,000,790号(特許文献1)において記述されている。キサントモナス親水性コロイド(「キサンタン」)は細胞外ヘテロ多糖である。
【0006】
キサンタンはキサントモナス属の細菌の好気性液内発酵により生成する。発酵培地は典型的には、炭水化物(例えば糖)、微量元素および他の栄養素を含む。発酵が完了すると直ちに、得られた発酵ブロス(溶液)は典型的には加熱処理される。キサンタン発酵ブロスおよび溶液の加熱処理により、転移温度(TM)またはそれ以上の温度で未変性キサンタンの構造変化が起こり、高粘度キサンタンが生成することが確立している。熱処理はまた、キサンタン中の生存微生物および望ましくない酵素活性を破壊するという有益な効果を有する。熱処理後、キサンタンをアルコール沈澱により回収する。しかしながら、キサンタン発酵ブロスの熱処理には欠点もあり、例えば、キサンタンの熱分解である。キサンタン溶液またはブロスを、TMを超える温度で加熱する、またはTMを超える温度で数秒を超える時間維持すると、キサンタンの熱分解が起こる。キサンタンの分解により不可逆的に粘度が減少する。したがって、熱処理はキサンタンの品質および粘稠性を制御する重要な技術である。
【0007】
キサンタンの品質は、主として2つの粘度試験により決定される:水道水中での低せん断速度粘度(「LSRV」)および高塩溶液中での海水粘度(「SWV」)。TMまたはそれを超える温度でのキサンタン発酵ブロスを殺菌(pasteurization)することによって、より高いLSRVおよびSWV値により示されるように、より高粘度のキサンタンが得られることがわかっている。
【0008】
キサンタンポリマーは多くの状況で使用される。キサンタンは様々な産業用途を有し、例えば、油井掘削泥における使用、水攻法による石油の二次回収における粘度制御添加剤、食物中の増粘剤、安定化剤、ならびに乳化、懸濁およびサイズ剤としての使用が挙げられる(Encyclopedia of Polymer Science and Engineering、2nd Edition、Editors John Wiley & Sons、901-918、1989(非特許文献2))。キサンタンはまた、化粧料調製物、薬学的媒質、および同様の組成物中で使用することができる。
【0009】
当技術分野では、未殺菌(unpasteurized)状態で高い比粘度特性を有するキサンタンポリマーを生成する必要がある。そのような比粘度の高いキサンタンポリマーは、例えば、食品、工業および油田用途に対し、同等のキサンタン濃度でより高い粘度を提供することができる。
【先行技術文献】
【特許文献】
【0010】
【特許文献1】米国特許第3,000,790号
【非特許文献】
【0011】
【非特許文献1】Milas and Rinaudo、Carbohydrate Research、76、189-196、1979
【非特許文献2】Encyclopedia of Polymer Science and Engineering、2nd Edition、Editors John Wiley & Sons、901-918、1989
【発明の概要】
【0012】
第1の態様では、未殺菌キサンタン組成物を提供する。組成物は、gumBおよびgumCを過剰発現する細胞により提供することができ、gumBおよびgumCを過剰発現しない対応株よりも少なくとも20%高い固有粘度を有する。
【0013】
第2の態様では、キサンタン組成物を提供する。キサンタン組成物は、一定範囲の分子長を有するキサンタン分子集団を含む。集団の少なくとも1%が原子間力顕微鏡観察法で測定すると、3umを超える長さを有する。
【0014】
本発明の第3の態様では、野生型株により産生されるものに比べ粘度が増加したキサンタンポリマー調製物を産生するための方法を提供する。gumBおよびgumCの遺伝子産物の量をキサントモナス・カンペストリス(Xanthomonas campestris)培養物中で選択的に増加させる。orfXの遺伝子産物の量は選択的に増加させない。gumD〜gumGからなる群より選択される遺伝子産物の量も選択的に増加されない。このように、より粘度の高いキサンタンポリマー調製物を培養物により産生させる。
【0015】
本発明の第4の態様では、野生型株により産生されるものに比べ粘度が増加したキサンタンポリマー調製物を産生するための方法を提供する。キサントモナス・カンペストリス株を、培地中、キサンタンポリマーを産生する条件下で培養する。株は選択的に、野生型株に比べ、orfXの遺伝子産物でもgumD〜gumGからなる群より選択される遺伝子でもなく、gumBおよびgumCの遺伝子産物をより多く産生する。
【0016】
本発明の第5の態様では、未殺菌キサンタン組成物を提供する。組成物はgumBおよびgumCを過剰発現する細胞により作成される。組成物は、gumBおよびgumCを過剰発現しない対応株由来のキサンタンよりも少なくとも10%多い海水粘度を有する。
【0017】
このように、本発明は対応する野生型株により同様に産生されるものに比べ粘度が増加したキサンタン組成物を用いる技術を提供する。
【図面の簡単な説明】
【0018】
【図1】xpsB-M(キサンタン多糖合成)オペロンとしても知られている、gumB-Mオペロンの遺伝子地図に対する遺伝子構築物を示す図である。
【図2】図2Aおよび2Bは、それぞれgumBおよびgumCタンパク質産物発現のウエスタンブロット分析を示す図である。
【図3】キサンタンガム試料に対する固有粘度プロットを示す図であり、そのうちの1つは遺伝子の余分なコピーを保有するプラスミドの存在により、gumBおよびgumCを過剰発現する。
【発明を実施するための形態】
【0019】
発明の詳細な説明
オペロンにおいて他の遺伝子に比べgumBおよびgumC遺伝子産物が過剰発現すると、1重量を基本に、より高い粘度を有するキサンタン産物が得られることは本発明者らの発見である。出願人はいずれの特別な動作理論にも縛られることは望まないが、一定の遺伝子産物の比率がシフトするとキサンタンポリマー分子のサイズ分布がシフトすると考えられる。キサンタンが野生型細胞により産生される場合に比べ、かなりの数の分子の分子長が長くなる。これらのより長い分子により、集団または調製物の粘度が高くなる。
【0020】
当技術分野では、キサンタン調製物を殺菌することにより粘度を増加させることができることが知られている。Talashekら、米国特許第6,391,596号を参照のこと。しかしながら、gumBおよびgumCの過剰発現の結果見られる粘度の増加は、殺菌をしなくても観察される。それでもなお、本発明の産物をその後に殺菌するとより高粘度の調製物が得られる。
【0021】
粘度の増加を達成するには、gumBおよびgumCの両方の過剰発現が必要であると考えられる。どちらかの遺伝子を単独で試験すると、増加が観察されなかった。gumBおよびgumCの過剰発現は、gumB〜Mオペロンの他の遺伝子に対して評価することができる。それらの遺伝子のうちのいずれに対しても過剰発現が効果を達成するのに十分である場合、orfXおよびgumDに対する過剰発現が特に重要である可能性がある。OrfXは、gumBのすぐ上流の、gumAと呼ばれるゲノムのセグメンとして以前公表された小さな翻訳領域である。最近では2つの翻訳領域が前者gumA領域で区別されており、ihfおよびorfXである。遺伝子gumD〜gumMの全てに対して過剰発現であることが望ましい場合がある。
【0022】
所望の遺伝子産物の過剰発現は、当技術分野で周知の任意の手段により達成してもよい。そのような手段としては、例えば、キサンタンを生成するキサントモナス・カンペストリス細胞または他の細菌に所望の遺伝子産物をコードする遺伝子の追加のコピーを導入すること、および例えば誘導プロモータを用いて所望の遺伝子産物を誘導することが挙げられるが、これらに限定されない。キサンタンを生成する他の細菌としては、キサンタン生合成遺伝子を含むように遺伝子操作されたものが挙げられる。gumBおよびgumC遺伝子は、1つまたは複数のベクター上に、すなわち、組み合わせてまたは別個に導入することができる。
【0023】
本発明により使用することができる誘導プロモータとしては、当技術分野で周知の全てのもの、例えば、lacプロモータ、araプロモータ、tetプロモータ、およびtacプロモータが挙げられる。これらのプロモータに対する天然および人工の誘導物質は当技術分野で周知であり、いずれを使用しても都合がよい。遺伝子の追加のコピーを、例えば、プラスミドまたはウイルスベクターに導入することができる。所望の遺伝子の追加のコピーは染色体外で維持することができ、またはゲノムに組み込むことができる。
【0024】
培養ブロスからのキサンタンの回収は典型的には、1つまたは複数のプロセッシング(processing)段階を含む。キサンタンは加熱処理してもよい。キサンタンはアルコール、例えばイソプロピルアルコール、エチルアルコール、またはプロピルアルコールで沈澱させてもよい。典型的には、細胞は培養ブロスから特異的に除去されない。
【0025】
生合成により生成されるキサンタン分子は典型的には一定のサイズ分布を有する。本発明の粘度増加は、平均長よりもずっと長い分子のその数を増加させることにより、または平均長より幾分長い分子のその数を著しく増加させることにより達成してもよい。長さが増大した分子のその数は莫大である必要はない。少なくとも長さが1、3、5、7、9または11%増大した分子で十分である。長さが増大した分子は、原子間力顕微鏡で測定すると、3、4、5、6、7、8または9umを超えていてもよい。3、4、5、6、7、8または9umよりも長い分子により提供される総キサンタン集団の質量のパーセンテージは、その集団内のそれらの数の割合よりも大きくなる。このように、キサンタン分子の総質量の少なくとも1、3、5、10、15、20または25%が、3umを超える長さを有する分子により提供されていてもよい。
【0026】
固有粘度測定は、本発明の調製物を特徴づけるさらに別の方法である。この型の測定を用いて観察される増加は、野生型株により産生されるものよりも、5、10、15、20、25、30、または35%を超えて大きい場合がある。比較のための適当な対照は、試験される株に最も密接に関係する対応株である。このように、gumBおよびgumCの追加のコピーを有する株を試験する場合、最もよい対照は、gumBおよびgumCの追加のコピーの存在を除き、同じ遺伝的相補を有する。より多くのgumBおよびgumC遺伝子産物を産生させる誘導物質により誘導されている培養物を試験する場合、最良の対照は、誘導されていない同じ株の培養物である。本発明の調製物を特徴づけるために海水粘度もまた使用することができる。この型の測定を用いて観察される増加は、野生型により産生されるものよりも、5、10、15、20、25、30、または35%を超えて大きい場合がある。
【0027】
キサンタンを、特性を改善するために多くの製品中の成分として使用する。特性としては、2〜3例を挙げると、粘度、粒子の懸濁、口当たり(mouth feeling)、バルクが挙げられる。他の特性としては、水の結合、増粘剤、エマルジョン安定化、起泡増強、およびせん断-希薄化(shear-thinning)が挙げられる。そのような製品としては、食品、例えばサラダドレッシング、シロップ、ジュース飲料、および冷菓が挙げられる。そのような製品としてはまた、印刷染料、油掘削流体(oil drilling fluid)、セラミック釉薬(ceramic glaze)、および薬学的組成物が挙げられる。後者の場合、キサンタンは担体として、または放出制御基質として使用することができる。キサンタンを使用することができる他の製品としては、洗浄液、塗料およびインク、壁紙接着剤、農薬、磨歯剤、ならびに酵素および細胞固定化剤が挙げられる。
【0028】
本発明について、本発明を実施するのに現在好ましい様式を含む特定の実施例に関し説明してきたが、当業者であれば、添付の特許請求の範囲で記述した本発明の精神および範囲の中にある、上記システムおよび技術について多くの変更および置換が存在することは理解するであろう。
【実施例】
【0029】
実施例1 株の構築
X.カンペストリスの完全なgum遺伝子領域を保有する断片を単離するために、野生型X.カンペストリス株、NRRL B-1459(1)のゲノムライブラリを、Sau3AIにより部分的に消化した総DNAのクローニングにより、宿主範囲の広いコスミドベクターpRK311(2)を用いて構築した。このライブラリは、全体として、大腸菌(E.coli)S17-1(3)〜Gum- X.カンペストリス突然変異体2895(4)に一致した。pIZD15-261(5)と呼ばれるいくつかのムコイド接合完了体(mucoid exconjugant)から単離したコスミドの1つは、完全gum領域を包含する16kbの断片を含む。グラフ表示の図1、およびオペロンの遺伝子のリストの表1を参照のこと。
【0030】
(表1)キサンタン多糖合成をコードする染色体領域中の遺伝子名称リスト
* 遺伝子位置は、da Silva、A.C.R.ら(Nature、Vol. 417、pg.459-463、2002)により記述されているように、X.カンペストリスpv.カンペストリスATCC33913(GenBank寄託:AE008922)の遺伝子配列に従う
【0031】
pBBR5-BCプラスミドを構築するこために、SpeI-BglIIで消化したpIZD15-261由来の4026bp断片をpKmob19(8)のXbaI部位とBamHI部位の間にクローニングし、pGum02-19S(5)を得た。SphIでの消化によりプラスミドpGum02-19Sから2855bp断片を放出させた。この断片を、あらかじめSphIを用いて消化したpUC18(9)にクローニングし、pUC18-BCASを形成させた。
【0032】
gumプロモータならびにgumBおよびgumC遺伝子を含む、HindIII-XbaI断片を、HindIII-XbaI消化pBBRR1-MCS5(10)(GenBank アクセッション番号U25061)にクローニングすることにより最終プラスミド(pBBR5-BC)を構築した。
【0033】
得られたpBBR5-BCプラスミドのヌクレオチド配列をSEQ ID NO:1に示す。(gumBおよびgumCの予想アミノ酸配列はそれぞれ、SEQ ID NO:2およびSEQ ID NO:3に示す)。この宿主範囲の広い、コピー数が中程度のプラスミドは長さが7.1kbであり、IncP、IncQおよびIncW群プラスミド、ならびにCo1E1系およびP15a系レプリコンと適合する。RK2伝達機能がトランスで提供されると、伝達起点(mobRK2)の存在により、広範囲の細菌内に接合により伝達することが可能となる。このプラスミドは、ゲンタマイシン耐性遺伝子も保有し、かつLacZαペプチドをコードする遺伝子内に位置するpBluescript II KS多重クローニング部位を含む(Stratagene、La Jolla、Ca、USAからのpBluescript II KS)。
【0034】
pBBR5-BCからのGumBおよびGumCタンパク質の発現を証明するために、プラスミドをX.カンペストリス変異体1231に導入した。変異体では、全gum(xps)遺伝子クラスターが欠失した。どちらのタンパク質も変異体株におけるウエスタンブロットにより検出した。
【0035】
(表2)本研究で使用または構築した細菌株およびプラスミド
a 数はgum領域のヌクレオチド配列の位置に対応する(GenBank、アクセッション番号 U222511)
【0036】
細菌株、プラスミド、および増殖条件
この研究で使用した株およびプラスミドを表2に示す。大腸菌(E.coli)株は37℃のLuria-Betani培地で増殖させた。X.カンペストリス株はTY(トリプトン5g、酵母抽出物3g、およびH2O 1リットルにつきCaCl2 0.7g)またはYM培地(12)、28℃で増殖させた。Sigma(St.Louis、Mo.)からの抗生物質を、必要に応じて下記濃度(μg/mL)で補った:X.カンペストリスでは、ゲンタマイシン、30;およびテトラサイクリン、10;大腸菌では、ゲンタマイシン、10;カナマイシン、30;アンピシリン、100;およびテトラサイクリン、10。
【0037】
DNA生物化学
大腸菌およびX.カンペストリス由来のプラスミドDNAを、QIAprep Spin Miniprep Kit(QIAGEN、ヒルデン、ドイツ)を用いて調製した。DNA制限、アガロースゲル電気泳動およびクローニング手順を、確立したプロトコル(13)に従い実施した。全ての構築物をDNA配列決定により検証した。プラスミドDNAを、Bio-Rad(リッチモンド、カリフォルニア州)により指示されているようにエレクトロポレーションにより大腸菌およびX.カンペストリス細胞に導入した(使用パラメータ:大腸菌:200Ω、25μF、2500V、およびX.カンペストリス:1000Ω、25μF、2500V)。
【0038】
ヌクレオチドおよびタンパク質配列の分析
ヌクレオチドおよびアミノ酸配列を、MacVector Sequence Analysis Software(Oxford Molecular Limited、ケンブリッジ、英国)を用いて分析した。
【0039】
実施例2 gumBおよびgumC発現のウエスタン分析
ウエスタン分析により、gumBおよびgumC遺伝子産物が、gumBおよびgumCの余分なコピーを有するX.カンペストリス株で過剰発現されることが確認された。図2を参照のこと。
【0040】
実施例3 固有粘度決定
gumBおよびgumC遺伝子の多重、プラスミドコードコピーを有する(XWCM1/pBBR5BC)、ならびに有さない(XWCM1)X.カンペストリス株から調製したキサンタン試料を比較した。炭素源としてブドウ糖を使用したフラスコ発酵物を振盪し、これらの株からキサンタンを得た。
【0041】
精製および非精製キサンタン試料について粘度を測定することにより固有粘度を決定した。gumBおよびgumCの多重コピーを有するX.カンペストリス株由来のキサンタンでは固有粘度の増加が観察された。固有粘度は、同一の溶媒および温度条件下で測定すると、一定のポリマー型では分子量に比例する。そのため、gumBおよびgumCの多重コピーを有するX.カンペストリス株由来のキサンタンは対照株由来のキサンタンよりも高い分子量を有する。
【0042】
方法:2種のブロスについてそれぞれ5つの振盪フラスコで試験を行った。各型のブロスを合わせ、総体積を測定した。その後、ブロスをイソプロピルアルコールに沈澱させた。(注:ブロスが約3%のガムを含むと推定した。総ブロス体積を測定し、3%をかけると、おおよその乾燥ガム重量が得られた。この概算を使用して、約0.5%のガム溶液を作製するのに必要とされる水の量を計算した)。その後、沈澱の湿性繊維を0.01M NaCl中で混合することにより直ちに再水和させ、約0.5%のガム溶液を生成させた。繊維を良好なせん断で3時間、3ブレードの直径2インチのプロペラ撹拌機を用いて混合し、その後、一晩放置した。下記手順を使用して、固有粘度測定のための試料を調製した。
【0043】
上記で調製した〜0.5%ガム溶液を、Gelman Science 293mm圧力濾過ユニットを用いて濾過する。溶液は最初に、20μMagnaナイロンフィルター(N22SP29325)を通して濾過する。フィルターを〜60psiまで加圧し、溶液を清浄なビーカー中に回収する(注:流速が〜5ドリップ/分まで減少するとフィルターを交換する)。
【0044】
第1の濾過段階に続き、上記濾過ユニットを用い試料をさらに2回濾過する。第1に、Millipore 8.0μフィルター(SCWP293 25)を通し、その後、Gelman Versapor(登録商標)293mm 1.2μフィルター(66397)を通す。濾過した試料を各濾過段階後、洗浄なビーカー中に回収する。
【0045】
濾過後、〜600mlのガム溶液をSpectra/Por(登録商標)透析管、直径28.6mmのSpectrum#S732706(MWCO 12,000〜14,000)に入れる。管を〜18から20インチの長さに切断し、一端で結び目を作る。溶液を管に追加し、端から〜2インチ以内になるまで満たす。管内にトラップされる空気ができるだけ少なくなるように管で第2の結び目を作る。全てのガム溶液が透析管に入れられるまで続ける。
【0046】
ガム溶液を含む管の外側を〜1分間、脱イオン水ですすぎ、その後、管を0.01M NaClの容器内に入れる。塩溶液が透析管を完全に覆うようにしなければならない。
【0047】
管を0.01M NaCl溶液中に4日間静置し、毎日NaCl溶液を交換する。4日後、管の一端を切り開き、注意深くガム溶液を清浄なビーカーに移す。
【0048】
下記手順を用いて、濾過した透析溶液について固体分析を実施する。
【0049】
±0.0002gまで秤量できる化学天秤を用いて、清浄なアルミニウム製の秤量皿VWR Cat#25433-008の質量を秤量し、記録する。(A)
【0050】
清浄なピペットを使用して、約10mlのガム溶液をアルミニウムパンに添加し、パンとガム溶液を合わせた正確な質量を記録する。(B)
【0051】
溶液を有するパンを105℃の乾燥オーブン内に入れ、24時間放置する。
【0052】
24時間後、オーブンからパンを取り出し、冷却し、再び秤量する。パンと残った乾燥ガムの質量を記録する。(C)
【0053】
パン+ガム溶液の質量(B)からアルミニウムパンの質量(A)を引く。乾燥ガム+パンの質量(C)からアルミニウムパン(A)の質量を引く。最初の値(B-A)で第2の値(C-A)を割る。この値に100をかけると固体の割合(%)が得られる・
【0054】
注:固体分析を、上記手順を用いて各濾過した透析溶液に対し、3連で実施した。計算した固体の割合(%)を各試料に対し平均化し、その平均値を使用した。
【0055】
各溶液に対する固体決定に基づき、0.01M NaClを用い試料を0.25%総ガム濃度まで希釈する。
【0056】
固有粘度測定を、Vilastic Viscoelasticity Analyzer(Vilastic Scientific, Inc.、オースチン、TX、半径0.0537cm×長さ6.137cmの管が取り付けられている)を用いて実施した。測定する前に機器を水で較正し、測定完了後確認した。周波数2.0Hz、定歪1.0、積分時間10秒に設定した、機器TIMETソフトウエアプロトコルを用いて測定を実施した。温度は23.5℃に維持した。0.25%ガム溶液を希釈することにより試料を調製した。各希釈液を20分間混合し、測定する前、一晩冷却して放置した。各希釈液に対し、6回測定し、平均した。下記表3は希釈液および各調製試料に対して得られた平均粘度を示す。
【0057】
(表3)
【0058】
ガム濃度に対し減少した比粘度(ηsp/c)をプロットすることにより固有粘度を決定した。ηsp/c=((ηc-η0)/η0)、式中、ηc=ガムの粘度である。切片から固有粘度が得られる。図3を参照のこと。
【0059】
XWCM1/pBBR5-BC変異体では固有粘度が増加するのは、分子量が増加するためだと考えられる。この実験を実施した場合と同一の溶媒および温度条件下で測定すると、一定のポリマー型では固有粘度は分子量に比例する。[η]と分子量の間の関係は、Mark-Houwinkの式[η]=kMaにより与えられる。式中、kおよびaは特定の温度の特定の溶媒中の特定のポリマー型に対する定数である。定数「a」は正の数であるので、[η]の増加は、Mark-Houwinkの式に従わない異なる分子配座を有しなければ、分子量(M)の増加によってのみ得られる。
【0060】
実施例4 手順−低せん断速度粘度測定
精製キサンタン試料について低せん断速度粘度測定を実施した。LSRVを測定するのに使用した手順の詳細を以下に示す。対照株由来のキサンタンに比べ、gumBおよびgumCの複数コピーを有する株由来のキサンタンに対する粘度の増加が観察された。データから、鎖長の増加にはgumBおよびgumC両方の過剰発現が必要であること;gumBまたはgumCいずれかの単独の過剰発現では鎖長増加には不十分であることが示唆される。
【0061】
材料および装置:
1. 標準(合成)生水(1000ppmのNaClおよび40ppmのCa++または147ppmのCaCl2.2H2Oを含む水):適した容器に入れた20リットルの蒸留水に試薬グレードのNaCl 20gおよび試薬グレードのCaCl2.2H2O 2.94mgを溶解することにより調製する。
2. 0.01gmまで正確に測定することができる天秤。
3. Brookfield LV粘度計、スピンドル#1、およびスピンドルGuard。
4. 標準の実験室用ガラス器具。
5. 標準の実験室用撹拌ベンチ。RAE撹拌モーター(C25U)および3ブレードプロペラを備えた撹拌シャフト(5/16'')は置換してもよい。
【0062】
手順:
1. 600mlのBerzelius(背の高い形態)ビーカーに秤量した合成生水299mlに、生成物0.75gm(0.001gmの位まで秤量)を、800ppmで撹拌しながら、徐々に添加する。
2. 800rpmで4時間撹拌した後、溶液を撹拌ベンチから取り出し、30分間放置する。
3. 温度を室温に調節し、3rpmでNo.1スピンドルを備えたBrookfield LV粘度計を用いて粘度を測定する。スピンドルを3分間回転させた後、粘度を記録する。
【0063】
実施例5 タンパク質発現の定量
細胞溶解物をウエスタンブロットおよび免疫検出分析にかけ、プラスミドによりコードされたGumBおよびGumCレベルを明確にした。4つの独立したブロットを分析した。同じ試料に対する絶対値は各定量において再現性がなかったが、試料間の相対量は全ての測定において同じままであった。
【0064】
GumBおよびGumCに対し産生された抗体の調製
GumCタンパク質のアミノ酸残基53-447をコードする1184bpのDNA断片を、PCR増幅により作製した。下記プライマーを使用した。
PCR産物をNdeIおよびBamHIを用いて消化し、pET22b(+)にサブクローニングし、得られたプラスミド(pET-C)を大腸菌株BL21(DE3)に導入した。
【0065】
OD6000.6まで、50μg/mlのカルベニシリンを含むL-ブロス中で増殖させた大腸菌BL21(pET-C)を、1mM IPTGで3時間誘導した。総細胞溶解物を、37℃で30分間、1mgのリゾチームを含む溶解緩衝液(50mM Tris/HCl pH8、1mM EDTA pH8、100mM NaCl、1mM PMSF、0.1mg/ml DNase、0.5% Triton X-100)で処理し、その後氷上で超音波処理することにより調製した。低速遠心分離(Eppendrof、4000xg、5分間)により細胞片を除去し、上清を、10分間の1400xgの遠心分離により、可溶性画分とペレット(封入体)画分に分画した。元の細胞溶解物と同じ体積の溶解緩衝液で2度、2mg/mlのDOCを含む溶解緩衝液で1度、続いて、水で3度、ペレット画分を洗浄した。処理後、タンパク質をSDS-PAGEにより分離し、過剰産生したGumCタンパク質を含む主バンドを切断し、ウサギを免疫化するために溶離した。
【0066】
OD6000.6まで、50μgのカルベニシリン、25μg/mlのカナマイシンを含むL-ブロス中で増殖させた大腸菌JM109(pQE-Xps#6、pREP4)を、1mM IPTGで3時間誘導した。総細胞溶解物を、37℃で30分間、1mg/mlのリゾチームを含む溶解緩衝液(50mM Tris/HCl pH8、1mM EDTA pH8、100mM NaCl、1mM PMSF、0.1mg/ml DNase、0.5% Triton X-100)で処理し、その後氷上で超音波処理することにより調製した。低速遠心分離(Eppendrof、4000xg、5分間)により細胞片を除去し、上清を、10分間の14000xgの遠心分離により、可溶性画分とペレット(封入体)画分に分画した。ペレット画分を溶解緩衝液で2度洗浄し、6Mの塩酸グアニジンを含む100mMのリン酸緩衝液(pH7)、5mM DTTおよび5mM EDTAに再懸濁させ、封入体を、緩衝液D(4M GdnHCl、50mM リン酸緩衝液(pH7)、150mM NaCl)で予め平衡化したFPLC Superdex HR200(Pharmacia Biotech)上でクロマトグラフにかけた。GumBを含む画分をプールし、マウスを免疫化するのに使用した。
【0067】
プラスミドpFD5、pBBR-promC、およびpBBR5-Bの構築
gumBおよびgumC遺伝子を含む3141bp断片を、pIZD15-261のBamHI(#318および#3459)を用いた部分消化により得、BamHIで消化したpRK404にクローニングし、プラスミドpFD5を得た。pGum02-19をEcoRI(#1979)およびBamHI(#3459)で消化して1480bp断片を単離し、同じ酵素であらかじめ消化したpBBR1MCS-5にクローニングし、pBBR-promCを得た。MCS内のHindIIIおよびEcoRI(#1979)でpGum02-19を消化して1233bp断片を得、これをpBBR1MCS-5にクローニングし、プラスミドpBBR5-Bを得た。
【0068】
上記のように調製したGumCを用い、雌のニュージーランドホワイトウサギを免疫化した。500μgのタンパク質をフロイント完全アジュバントと共にウサギに最初に注入し、続いて、隔週に、250μgのタンパク質を不完全アジュバントと共に3回注入した。BALB/c雌マウスを上記のように調製したGumBを用いて免疫化した。100μgのタンパク質をフロイント完全アジュバントと共にマウスに最初に注入し、続いて、1週間に1度、50μgのタンパク質を不完全アジュバントと共に3回注入した。ポリクローナル抗体を、Harlow & Lane((1999)「Using antibodies:a laboratory manual」、Cold Spring Harbor Laboratory Press、Cold Spring Harbor、N.Y.)により記述されているように調製し、抗血清を-70℃で保存した。血清を大腸菌BL21(pET22b+)およびXc1231アセトン粉末の両方に吸着させてGumC特異的抗体を得た(Harlow & Lane、上記)。
【0069】
タンパク質抽出物
プラスミドをエレクトロポレーションにより親株PRM-1に導入した。得られた株を28℃、YM培地、250rpmで中間対数期まで増殖させた。遠心分離により細胞を収集し、生体重を決定した。ペレットを10mM Tris/HCl、10mM EDTA(pH 8.0)で2度洗浄しエキソポリサッカライドを除去し、同じ緩衝液中に100mg/mlの濃度で再懸濁させた。50μlの各試料に100μlの緩衝液A(10mM Tris/HCl、10mM EDTA(pH8.0)、1.5% SDS)を添加した後、混合物を室温で10分間、その後、100℃で12分間インキュベートした。細胞溶解物を14000xg(Eppendorf 5415C)で5分間遠心分離にかけ、収集した上清を総タンパク質抽出物と名付けた。各溶解物のタンパク質濃度を、SDSの存在下、標準としてBSAを使用し、Markwell法((1978)「A modification of the Lowry procedure to simplify protein determination in membrane and lipoprotein samples.」Anal Biochem 87(1)、206-10)により決定した。
【0070】
SDS-PAGEおよび免疫検出
細胞溶解物(30μg/レーン)を試料緩衝液(125mM Tris/HCl、pH 6.8;4% SDS、20mM DTT、0.05% ブロモフェノールブルー、20% グリセロール)と混合し、2分間沸騰させた。Schaggerおよびvon Jagowの方法((1987)「Tricine-sodium dodecyl sulfate-polyacrylamide gel electrophresis for the separation of proteins in the range from 1 to 100kDa.」Analytical Biochemistry 166(2)、368-79)に従い、SDS-10%ポリアクリルアミドゲルによりタンパク質を分離した。Immobilon-P膜(PVDF、Millipore)への半乾燥転写システム(Hoefer Semiphor unit)を用いエレクトロブロッティングを実施した。転写は10mM CAPS(pH11)、10%(v/v)メタノールを含む緩衝液中で、30分間、2.5mA/cm2のゲル表面積で実施した。エレクトロトランスファー(electrotransfer)の完了後直ちに、ブロットを0.5% Ponceau-Sレッドで染色し、トランスファーの質を評価し、Milli-Q(登録商標)-グレード水で洗浄した。ブロットを一晩中、4℃で、5%の脱脂乳粉末を含むTBST(150mM NaCl、10mM Tris/HCl pH8、0.05% Tween-20)(Harlow & Lane、上記)でブロックし、その後、抗GumB(1:3000)または抗GumC(1:5000)抗体と共に3%の脱脂乳粉末を含むTBST中、室温で3時間インキュベートした。アルカリホスファターゼ結合ヤギ抗マウスIgGまたは抗ウサギIgG(Sigma)をそれぞれ、製造者により記述されているように、検出に使用した。ブロットを3度TBSTで洗浄し、ニトロブルーテトラゾリウム-5-ブロモ-4-クロロ-3-インドリルホスフェート(NBT/BCIP、Promega)を含む溶液中で展開させた。市販のタンパク質マーカーMW-SDS-70L(Sigma)を使用してSDS-PAGEの較正を実施した。
【0071】
ブロット定量
GumBおよびGumCタンパク質バンドの強度を、NBT/BCIP発現フィルターをUVPデンシトメーター(Ultra Violet Products)で走査することにより決定し、GelWorks 1D Analysis software(NonLinear Dynamics Ltd)を用いて定量化した。各フィルターは、野生型細胞における染色体にコードされたGumBおよびGumCのレベルを確立するためにPRM-1(pBBR-prom)抽出物の基準レーンを含んだ。相対量のGumBおよびGumCを観察した。図2Aおよび2Bを参照のこと。
【0072】
実施例6 手順−分子間力顕微鏡観察法を使用した分子長または分子量決定
分子間力顕微鏡観察法(AFM)または走査型プローブ顕微鏡観察法(SPM)と呼ばれる直接可視化技術を使用して、gumBおよびgumCの多重コピーを有する(XWCM1/pBBR5-BC)、有さない(XWCM1)X.カンペストリス株由来のキサンタン分子の長さを画像化した。AFMを実施するために使用した手順の詳細を以下に示す。本発明者らは、gumBおよびgumCの多重コピーを有する株により産生されたキサンタン分子の平均分子輪郭長(contour length)が、親株のものよりずっと長いことを観察した。
【0073】
0.1gのガムを含む100gの蒸留水を〜3時間混合することにより、0.1% wt%のガム溶液を調製した。1ppmストック溶液を、0.1wt%溶液20μlを20gの0.1M 酢酸アンモニウム溶液中に希釈することにより調製した。20μlの1ppmストック溶液を新たに劈開させたマイカディスク(〜1cm2)上に噴霧した。これらのマイカ試料ディスクを、その後、加熱(〜60℃)真空チャンバー中に〜1時間入れ過剰の水を除去した。その後乾燥マイカディスクをTappingモードのAFMを用いて走査した。全てのAFM像の分子輪郭長をDigital Instrumentsにより提供されたソフトウエアを用いて測定した。
【0074】
キサンタン分子の集団の輪郭長を測定した。この研究の結果を表4に示す。(各サイズクラスの分子は、示した長さより小さい、またはそれと等しい;1つのサイズクラスで示した分子の数は、より小さなサイズクラスで計数した分子を含まない)。これらの結果から、gumBおよびgumCの多重コピーを有するX.カンペストリス株由来のキサンタン分子は、対照株由来のキサンタン分子よりもかなり大きいことが証明された。原子間力顕微鏡観察法(AFM)または走査型電子顕微鏡観察法(SPM)を、市販の機器(Nanoscope IIIa、Digital Instruments、サンタバーバラ、CA)により、窒化珪素カンチレバーチップを使用して実施した。
【0075】
(表4)キサンタン分子輪郭長のAFM測定
【0076】
実施例7 海水粘度の評価
XWCM1/pBBR5-BC株により産生されたキサンタンを、市販のキサンタン製品(Xanvis(商標))と比較し、海水粘度(SWV)に対し評価した。Xanvis(商標)キサンタン製品に対する典型的なSWVは18〜22の範囲である。
【0077】
海水粘度は下記手順を用いて決定した。41.95gの海塩(ASTM D1141-52、Lake Products Co., Inc.製、メリーランドハイツ、ミズーリ州)を1リットルの脱イオン水に溶解することにより調製した。300mlの海水溶液を、Hamilton-Beach 936-2ミキサーに取り付けた混合カップに移した(Hamilton-Beach Div.、ワシントンD.C.)。ミキサー速度制御を低に設定し、単一の溝付ディスクを混合軸に取り付けた。低速設定では、混合軸は約4,000〜6,000rpmで回転する。0.86gのバイオガム製品を15〜30秒にわたり徐々に混合カップに添加し、5分間混合させた。ミキサー速度制御を高(11,000±1,000rpm)に設定し、試験溶液を約5分間混合させた。混合物は、バイオガム製品添加時間から始めて、計45分間混合させた。45分の混合時間の終わりに、2〜3滴のBara Defoam(NL Baroid/NL industries, Inc.、ヒューストン、TX)を添加し、さらに30秒間撹拌し続けた。
【0078】
混合カップをミキサーから取り外し、冷水に浸漬させ、流体温度を25±0.5℃まで降下させた。均質な溶液を確保するために、溶液を冷却後5秒間11,000±1,000rpmで再混合した。溶液を混合カップから400mlのパイレックスビーカーに移し、Fann粘度(Fann粘度計、モデル35A)を測定した。これは低速(約3rpm)で混合することにより達成した。この読み取りにより安定化させ、その後、せん断応力値をダイヤルから読み取り、3rpmでのSWV値として記録した。
【0079】
(表5)XWCM1/pBBR5-BCキサンタンおよびXanvis(商標)キサンタンの品質
a ダイヤル目盛り
【0080】
参考文献
【特許請求の範囲】
【請求項1】
gumBおよびgumCを過剰発現する細胞に由来し、gumBおよびgumCを過剰発現しない対応株由来のキサンタンよりも少なくとも20%高い固有粘度を有する、未殺菌キサンタン組成物。
【請求項2】
対応株由来のキサンタンよりも少なくとも25%高い固有粘度を有する、請求項1記載の未殺菌(unpasteurized)キサンタン組成物。
【請求項3】
対応株由来のキサンタンよりも少なくとも30%高い固有粘度を有する、請求項1記載の未殺菌キサンタン組成物。
【請求項4】
一定範囲の分子長を有するキサンタン分子の集団を含み、その集団の少なくとも1%が、原子間力顕微鏡観察法により測定すると、少なくとも3umの長さを有する、キサンタン組成物。
【請求項5】
集団の少なくとも5%が、原子間力顕微鏡観察法により測定すると、少なくとも3umの長さを有する、請求項4記載の方法。
【請求項6】
一定範囲の分子長を有するキサンタン分子の集団を含み、その集団の少なくとも1%が、原子間力顕微鏡観察法により測定すると、少なくとも4umの長さを有する、キサンタン組成物。
【請求項7】
集団の少なくとも1%が、少なくとも5umの長さを有する、請求項6記載のキサンタン組成物。
【請求項8】
集団の少なくとも1%が、少なくとも7umの長さを有する、請求項6記載のキサンタン組成物。
【請求項9】
一定範囲の分子長を有するキサンタン分子の集団を含むキサンタン組成物であって、その組成物中のキサンタン分子の総質量の少なくとも5%が、原子間力顕微鏡観察法により測定すると、3umを超える分子長を有するキサンタン分子によるものである、組成物。
【請求項10】
組成物中のキサンタン分子の総質量の少なくとも10%が、原子間力顕微鏡観察法により測定すると、3umを超える分子長を有するキサンタン分子によるものである、請求項9記載のキサンタン組成物。
【請求項11】
組成物中のキサンタン分子の総質量の少なくとも15%が、原子間力顕微鏡観察法により測定すると、3umを超える分子長を有するキサンタン分子によるものである、請求項9記載のキサンタン組成物。
【請求項12】
組成物中のキサンタン分子の総質量の少なくとも20%が、原子間力顕微鏡観察法により測定すると、3umを超える分子長を有するキサンタン分子によるものである、請求項9記載のキサンタン組成物。
【請求項13】
請求項1記載のキサンタン組成物を含む食品。
【請求項14】
請求項4記載のキサンタン組成物を含む食品。
【請求項15】
請求項6記載のキサンタン組成物を含む食品。
【請求項16】
請求項9記載のキサンタン組成物を含む食品。
【請求項17】
サラダドレッシング、シロップ、ジュース飲料、および冷菓からなる群より選択される、請求項1、4、6、または9のいずれか一項記載の食品。
【請求項18】
請求項1記載のキサンタン組成物を含む印刷染料。
【請求項19】
請求項4記載のキサンタン組成物を含む印刷染料。
【請求項20】
請求項6記載のキサンタン組成物を含む印刷染料。
【請求項21】
請求項9記載のキサンタン組成物を含む印刷染料。
【請求項22】
請求項1記載のキサンタン組成物を含む油掘削流体。
【請求項23】
請求項4記載のキサンタン組成物を含む油掘削流体。
【請求項24】
請求項6記載のキサンタン組成物を含む油掘削流体。
【請求項25】
請求項9記載のキサンタン組成物を含む油掘削流体。
【請求項26】
請求項1記載のキサンタン組成物を含むセラミック釉薬。
【請求項27】
請求項4記載のキサンタン組成物を含むセラミック釉薬。
【請求項28】
請求項6記載のキサンタン組成物を含むセラミック釉薬。
【請求項29】
請求項9記載のキサンタン組成物を含むセラミック釉薬。
【請求項30】
請求項1記載のキサンタン組成物を含む薬学的組成物。
【請求項31】
請求項4記載のキサンタン組成物を含む薬学的組成物。
【請求項32】
請求項6記載のキサンタン組成物を含む薬学的組成物。
【請求項33】
請求項9記載のキサンタン組成物を含む薬学的組成物。
【請求項34】
放出制御製剤である、請求項30記載の薬学的組成物。
【請求項35】
放出制御製剤である、請求項31記載の薬学的組成物。
【請求項36】
放出制御製剤である、請求項32記載の薬学的組成物。
【請求項37】
放出制御製剤である、請求項33記載の薬学的組成物。
【請求項38】
放出制御製剤である、請求項34記載の薬学的組成物。
【請求項39】
以下の段階を含む、野生型株により産生されるものに比べ粘度が増大しているキサンタンポリマー調製物を産生する方法:
キサントモナス・カンペストリス(Xanthomonas campestris)培養物中で、orfXではなく、gumD〜gumGからなる群より選択される遺伝子でもないgumBおよびgumCの遺伝子産物の量を選択的に増加させる段階であって、高粘度キサンタンポリマー調製物は培養物により産生される段階。
【請求項40】
選択的に増加させる段階が、キサントモナス・カンペストリスに、gumBおよびgumCの1つまたは複数の追加のコピーを導入することにより実施される、請求項39記載の方法。
【請求項41】
選択的に増加させる段階が、キサントモナス・カンペストリスに、gumD〜gumGではないgumBおよびgumCの1つまたは複数の追加のコピーを導入することにより実施される、請求項39記載の方法。
【請求項42】
選択的に増加させる段階が、キサントモナス・カンペストリスに、orfXではなく、gumD〜gumGでもないgumBおよびgumCの1つまたは複数の追加のコピーを導入することにより実施される、請求項39記載の方法。
【請求項43】
追加のコピーが染色体外遺伝因子上にある、請求項40記載の方法。
【請求項44】
染色体外遺伝因子がプラスミドである、請求項43記載の方法。
【請求項45】
プラスミドが宿主範囲の広いプラスミドである、請求項44記載の方法。
【請求項46】
追加のコピーがキサントモナス・カンペストリスのゲノムに組み込まれる、請求項39記載の方法。
【請求項47】
選択的に増加させる段階が、誘導プロモータと、誘導プロモータからの発現を増加させる誘導物質とを使用してgumBおよびgumCの発現を誘導することにより実施される、請求項39記載の方法。
【請求項48】
調製物から、より高粘度のキサンタンポリマーを回収する段階をさらに含む、請求項39記載の方法。
【請求項49】
より高粘度のキサンタンポリマー調製物からキサンタンポリマーを沈澱させる段階をさらに含む、請求項39記載の方法。
【請求項50】
以下の段階を含む、野生型株により産生されるものに比べ粘度が増大しているキサンタンポリマー調製物を産生する方法:
培地中、キサンタンポリマーを産生する条件下でキサントモナス・カンペストリス株を培養する段階であって、野生型株に比べ、株が、orfXではなく、gumD〜gu
mGからなる群より選択される遺伝子でもないgumBおよびgumCの遺伝子産物を選択的により多く産生する段階。
【請求項51】
株が、gumD 1コピーあたり、1コピーを超えるgumBおよびgumCを有する、請求項50記載の方法。
【請求項52】
株が、gumD〜gumG 1コピーあたり、1コピーを超えるgumBおよびgumCを有する、請求項50記載の方法。
【請求項53】
株が、gumD〜gumGからなる群より選択される遺伝子1コピーあたり、1コピーを超えるgumBおよびgumCを有する、請求項50記載の方法。
【請求項54】
株が、orfX 1コピーあたり、1コピーを超えるgumBおよびgumCを有する、請求項50記載の方法。
【請求項55】
株が、orfXおよびgumD〜gumG 1コピーあたり、1コピーを超えるgumBおよびgumCを有する、請求項50記載の方法。
【請求項56】
株が、凝集して少なくとも1コピーのgumBおよびgumCを保有する1つまたは複数のプラスミドを保有する、請求項50記載の方法。
【請求項57】
培地から、より高粘度のキサンタンポリマーを回収する段階をさらに含む、請求項50記載の方法。
【請求項58】
培地からキサンタンポリマーを沈澱させる段階をさらに含む、請求項50記載の方法。
【請求項59】
gumBおよびgumCを過剰発現する細胞に由来し、gumBおよびgumCを過剰発現しない対応株由来のキサンタンよりも少なくとも10%高い海水粘度を有する、未殺菌キサンタン組成物。
【請求項60】
脱イオン水1リットルあたり海塩41.95g、キサンタン濃度0.86g/lの溶液中で海水粘度を測定すると、DR>25の海水粘度を有する、請求項59記載のキサンタン組成物。
【請求項61】
gumBおよびgumCを過剰発現しない対応株由来のキサンタンよりも少なくとも15%高い海水粘度を有する、請求項59記載のキサンタン組成物。
【請求項62】
請求項59記載のキサンタン組成物を含む油掘削流体。
【請求項63】
請求項61記載のキサンタン組成物を含む油掘削流体。
【請求項1】
gumBおよびgumCを過剰発現する細胞に由来し、gumBおよびgumCを過剰発現しない対応株由来のキサンタンよりも少なくとも20%高い固有粘度を有する、未殺菌キサンタン組成物。
【請求項2】
対応株由来のキサンタンよりも少なくとも25%高い固有粘度を有する、請求項1記載の未殺菌(unpasteurized)キサンタン組成物。
【請求項3】
対応株由来のキサンタンよりも少なくとも30%高い固有粘度を有する、請求項1記載の未殺菌キサンタン組成物。
【請求項4】
一定範囲の分子長を有するキサンタン分子の集団を含み、その集団の少なくとも1%が、原子間力顕微鏡観察法により測定すると、少なくとも3umの長さを有する、キサンタン組成物。
【請求項5】
集団の少なくとも5%が、原子間力顕微鏡観察法により測定すると、少なくとも3umの長さを有する、請求項4記載の方法。
【請求項6】
一定範囲の分子長を有するキサンタン分子の集団を含み、その集団の少なくとも1%が、原子間力顕微鏡観察法により測定すると、少なくとも4umの長さを有する、キサンタン組成物。
【請求項7】
集団の少なくとも1%が、少なくとも5umの長さを有する、請求項6記載のキサンタン組成物。
【請求項8】
集団の少なくとも1%が、少なくとも7umの長さを有する、請求項6記載のキサンタン組成物。
【請求項9】
一定範囲の分子長を有するキサンタン分子の集団を含むキサンタン組成物であって、その組成物中のキサンタン分子の総質量の少なくとも5%が、原子間力顕微鏡観察法により測定すると、3umを超える分子長を有するキサンタン分子によるものである、組成物。
【請求項10】
組成物中のキサンタン分子の総質量の少なくとも10%が、原子間力顕微鏡観察法により測定すると、3umを超える分子長を有するキサンタン分子によるものである、請求項9記載のキサンタン組成物。
【請求項11】
組成物中のキサンタン分子の総質量の少なくとも15%が、原子間力顕微鏡観察法により測定すると、3umを超える分子長を有するキサンタン分子によるものである、請求項9記載のキサンタン組成物。
【請求項12】
組成物中のキサンタン分子の総質量の少なくとも20%が、原子間力顕微鏡観察法により測定すると、3umを超える分子長を有するキサンタン分子によるものである、請求項9記載のキサンタン組成物。
【請求項13】
請求項1記載のキサンタン組成物を含む食品。
【請求項14】
請求項4記載のキサンタン組成物を含む食品。
【請求項15】
請求項6記載のキサンタン組成物を含む食品。
【請求項16】
請求項9記載のキサンタン組成物を含む食品。
【請求項17】
サラダドレッシング、シロップ、ジュース飲料、および冷菓からなる群より選択される、請求項1、4、6、または9のいずれか一項記載の食品。
【請求項18】
請求項1記載のキサンタン組成物を含む印刷染料。
【請求項19】
請求項4記載のキサンタン組成物を含む印刷染料。
【請求項20】
請求項6記載のキサンタン組成物を含む印刷染料。
【請求項21】
請求項9記載のキサンタン組成物を含む印刷染料。
【請求項22】
請求項1記載のキサンタン組成物を含む油掘削流体。
【請求項23】
請求項4記載のキサンタン組成物を含む油掘削流体。
【請求項24】
請求項6記載のキサンタン組成物を含む油掘削流体。
【請求項25】
請求項9記載のキサンタン組成物を含む油掘削流体。
【請求項26】
請求項1記載のキサンタン組成物を含むセラミック釉薬。
【請求項27】
請求項4記載のキサンタン組成物を含むセラミック釉薬。
【請求項28】
請求項6記載のキサンタン組成物を含むセラミック釉薬。
【請求項29】
請求項9記載のキサンタン組成物を含むセラミック釉薬。
【請求項30】
請求項1記載のキサンタン組成物を含む薬学的組成物。
【請求項31】
請求項4記載のキサンタン組成物を含む薬学的組成物。
【請求項32】
請求項6記載のキサンタン組成物を含む薬学的組成物。
【請求項33】
請求項9記載のキサンタン組成物を含む薬学的組成物。
【請求項34】
放出制御製剤である、請求項30記載の薬学的組成物。
【請求項35】
放出制御製剤である、請求項31記載の薬学的組成物。
【請求項36】
放出制御製剤である、請求項32記載の薬学的組成物。
【請求項37】
放出制御製剤である、請求項33記載の薬学的組成物。
【請求項38】
放出制御製剤である、請求項34記載の薬学的組成物。
【請求項39】
以下の段階を含む、野生型株により産生されるものに比べ粘度が増大しているキサンタンポリマー調製物を産生する方法:
キサントモナス・カンペストリス(Xanthomonas campestris)培養物中で、orfXではなく、gumD〜gumGからなる群より選択される遺伝子でもないgumBおよびgumCの遺伝子産物の量を選択的に増加させる段階であって、高粘度キサンタンポリマー調製物は培養物により産生される段階。
【請求項40】
選択的に増加させる段階が、キサントモナス・カンペストリスに、gumBおよびgumCの1つまたは複数の追加のコピーを導入することにより実施される、請求項39記載の方法。
【請求項41】
選択的に増加させる段階が、キサントモナス・カンペストリスに、gumD〜gumGではないgumBおよびgumCの1つまたは複数の追加のコピーを導入することにより実施される、請求項39記載の方法。
【請求項42】
選択的に増加させる段階が、キサントモナス・カンペストリスに、orfXではなく、gumD〜gumGでもないgumBおよびgumCの1つまたは複数の追加のコピーを導入することにより実施される、請求項39記載の方法。
【請求項43】
追加のコピーが染色体外遺伝因子上にある、請求項40記載の方法。
【請求項44】
染色体外遺伝因子がプラスミドである、請求項43記載の方法。
【請求項45】
プラスミドが宿主範囲の広いプラスミドである、請求項44記載の方法。
【請求項46】
追加のコピーがキサントモナス・カンペストリスのゲノムに組み込まれる、請求項39記載の方法。
【請求項47】
選択的に増加させる段階が、誘導プロモータと、誘導プロモータからの発現を増加させる誘導物質とを使用してgumBおよびgumCの発現を誘導することにより実施される、請求項39記載の方法。
【請求項48】
調製物から、より高粘度のキサンタンポリマーを回収する段階をさらに含む、請求項39記載の方法。
【請求項49】
より高粘度のキサンタンポリマー調製物からキサンタンポリマーを沈澱させる段階をさらに含む、請求項39記載の方法。
【請求項50】
以下の段階を含む、野生型株により産生されるものに比べ粘度が増大しているキサンタンポリマー調製物を産生する方法:
培地中、キサンタンポリマーを産生する条件下でキサントモナス・カンペストリス株を培養する段階であって、野生型株に比べ、株が、orfXではなく、gumD〜gu
mGからなる群より選択される遺伝子でもないgumBおよびgumCの遺伝子産物を選択的により多く産生する段階。
【請求項51】
株が、gumD 1コピーあたり、1コピーを超えるgumBおよびgumCを有する、請求項50記載の方法。
【請求項52】
株が、gumD〜gumG 1コピーあたり、1コピーを超えるgumBおよびgumCを有する、請求項50記載の方法。
【請求項53】
株が、gumD〜gumGからなる群より選択される遺伝子1コピーあたり、1コピーを超えるgumBおよびgumCを有する、請求項50記載の方法。
【請求項54】
株が、orfX 1コピーあたり、1コピーを超えるgumBおよびgumCを有する、請求項50記載の方法。
【請求項55】
株が、orfXおよびgumD〜gumG 1コピーあたり、1コピーを超えるgumBおよびgumCを有する、請求項50記載の方法。
【請求項56】
株が、凝集して少なくとも1コピーのgumBおよびgumCを保有する1つまたは複数のプラスミドを保有する、請求項50記載の方法。
【請求項57】
培地から、より高粘度のキサンタンポリマーを回収する段階をさらに含む、請求項50記載の方法。
【請求項58】
培地からキサンタンポリマーを沈澱させる段階をさらに含む、請求項50記載の方法。
【請求項59】
gumBおよびgumCを過剰発現する細胞に由来し、gumBおよびgumCを過剰発現しない対応株由来のキサンタンよりも少なくとも10%高い海水粘度を有する、未殺菌キサンタン組成物。
【請求項60】
脱イオン水1リットルあたり海塩41.95g、キサンタン濃度0.86g/lの溶液中で海水粘度を測定すると、DR>25の海水粘度を有する、請求項59記載のキサンタン組成物。
【請求項61】
gumBおよびgumCを過剰発現しない対応株由来のキサンタンよりも少なくとも15%高い海水粘度を有する、請求項59記載のキサンタン組成物。
【請求項62】
請求項59記載のキサンタン組成物を含む油掘削流体。
【請求項63】
請求項61記載のキサンタン組成物を含む油掘削流体。
【図1】

【図2】

【図3】

【図2】
【図3】
【公開番号】特開2011−67205(P2011−67205A)
【公開日】平成23年4月7日(2011.4.7)
【国際特許分類】
【出願番号】特願2010−209019(P2010−209019)
【出願日】平成22年9月17日(2010.9.17)
【分割の表示】特願2006−507320(P2006−507320)の分割
【原出願日】平成16年3月18日(2004.3.18)
【公序良俗違反の表示】
(特許庁注:以下のものは登録商標)
1.パイレックス
【出願人】(501190918)シーピー ケルコ ユー.エス.インク. (3)
【出願人】(507169417)ファンダシオン インスティテュート ルロワール (1)
【Fターム(参考)】
【公開日】平成23年4月7日(2011.4.7)
【国際特許分類】
【出願日】平成22年9月17日(2010.9.17)
【分割の表示】特願2006−507320(P2006−507320)の分割
【原出願日】平成16年3月18日(2004.3.18)
【公序良俗違反の表示】
(特許庁注:以下のものは登録商標)
1.パイレックス
【出願人】(501190918)シーピー ケルコ ユー.エス.インク. (3)
【出願人】(507169417)ファンダシオン インスティテュート ルロワール (1)
【Fターム(参考)】
[ Back to top ]