
高親和性のタンパク質間相互作用検出・スクリーニング方法
【課題】親和性の高いタンパク質間相互作用を効率よく検出またはスクリーニングする方法を提供すること。
【解決手段】本発明の酵母を用いるタンパク質間相互作用検出方法は、(1)第1のタンパク質とGタンパク質γサブユニット細胞膜非結合変異型との融合タンパク質を発現する第1の発現カセットを調製する工程、(2)第2のタンパク質と細胞膜局在化シグナル配列との融合タンパク質を発現する第2の発現カセットを調製する工程、(3)第3のタンパク質を細胞質に発現する第3の発現カセットを調製する工程、(4)該工程(1)、(2)および(3)でそれぞれ得られた該第1、2および3の発現カセットを1倍体酵母に導入して形質転換酵母を調製する工程、および(5)該工程(4)で得られた該形質転換酵母におけるフェロモンシグナル伝達を検出する、またはフェロモンシグナル伝達を示す形質転換酵母をスクリーニングする工程を含む。
【解決手段】本発明の酵母を用いるタンパク質間相互作用検出方法は、(1)第1のタンパク質とGタンパク質γサブユニット細胞膜非結合変異型との融合タンパク質を発現する第1の発現カセットを調製する工程、(2)第2のタンパク質と細胞膜局在化シグナル配列との融合タンパク質を発現する第2の発現カセットを調製する工程、(3)第3のタンパク質を細胞質に発現する第3の発現カセットを調製する工程、(4)該工程(1)、(2)および(3)でそれぞれ得られた該第1、2および3の発現カセットを1倍体酵母に導入して形質転換酵母を調製する工程、および(5)該工程(4)で得られた該形質転換酵母におけるフェロモンシグナル伝達を検出する、またはフェロモンシグナル伝達を示す形質転換酵母をスクリーニングする工程を含む。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、高親和性のタンパク質間相互作用検出・スクリーニング方法に関する。特に、競合タンパク質を用いるタンパク質間相互作用検出・スクリーニング方法に関する。
【背景技術】
【0002】
タンパク質間相互作用検出方法として酵母ツーハイブリッド法が知られている。この方法は、遺伝子の発現調節に関与する転写因子が、遺伝子(DNA)上の特定の塩基配列を認識して結合するドメイン(DNA結合ドメイン:BD)と、遺伝子の転写に必須の基本因子に作用して遺伝子の転写を活性化するドメイン(転写活性化ドメイン:AD)とから構成されており、それぞれのドメインが分離できることに基づく。すなわち、タンパク質間相互作用を検出したい1対のタンパク質(XとY)のうち、一方(X)を転写因子のDNA結合ドメインと融合させて1つのハイブリッドタンパク質(X−BD)を形成し、他方(Y)を転写因子の転写活性化ドメインと融合させてもう1つのハイブリッドタンパク質(Y−AD)を形成すると、X−Y間に相互作用が存在する場合のみ、遺伝子の転写が活性化される。この遺伝子の転写活性化は、BDが結合する特定の塩基配列の下流にレポーター遺伝子を連結することによって、レポーター遺伝子の発現により検出することができる。酵母ツーハイブリッド法に利用される転写因子としては、Gal4、LexA−B42、LexA−VP16などが挙げられる。
【0003】
酵母は、単細胞性の真核生物であり、古くから食品加工などに用いられている。酵母は、真核生物の中で最も研究されており、酵母の遺伝子組換え技術も発達している。酵母は、真核生物という点では、人類など哺乳類をはじめとする高等真核生物と共通するため、高等真核生物のタンパク質を解析するために、これらのタンパク質の発現用宿主として汎用されている。真核生物のタンパク質間相互作用検出方法として酵母ツーハイブリッド法は有用である。
【0004】
しかし、細胞核内の転写因子を用いる酵母ツーハイブリッド法は、核移行が困難なタンパク質、および元来転写活性化ドメインを有するタンパク質(例えば、転写因子)に適用することが難しいという問題がある。
【0005】
そこで、近年、タンパク質の局在性や機能に応じて酵母ツーハイブリッド法の変法が考案されている。非特許文献1および2には、酵母のRasシグナル伝達経路を利用する方法が記載されている。この方法では、Rasの細胞膜結合能をタンパク質間相互作用で代替することで、酵母の増殖を指標としてタンパク質間相互作用を検出する。Ras伝達経路のシグナル増幅効果を利用することにより高感度にタンパク質間相互作用を検出することができる。すなわち、タンパク質間相互作用を検出したい1対のタンパク質(XとY)のうち、一方(X)を細胞膜に固定し、他方(Y)をヒト由来グアニンヌクレオチド交換因子(hSOS)または細胞膜結合能を欠損させたRas恒常活性変異型(mRas)と融合させてハイブリッドタンパク質(それぞれY−hSOSまたはY−mRas)を形成すると、X−Y間に相互作用が存在する場合のみ、hSOSまたはmRasが細胞膜に局在化し、Rasのシグナル伝達経路が活性化される。このシグナル伝達経路の活性化は、酵母の増殖を指標として検出することができる。
【0006】
しかしながら、この方法では、酵母の温度感受性変異株(cdc25−2)を使用する必要があり、酵母の増殖に温度制限が課せられる。すなわち、タンパク質のライブラリの構築を感受性温度でない25℃で行い、タンパク質間相互作用の検出またはスクリーニングを感受性温度である36℃で行う必要があるため、酵母の最適生育温度である30℃に比べて酵母の増殖速度が大きく低下し、迅速なタンパク質間相互作用の検出またはスクリーニングが困難であるという問題がある。
【0007】
非特許文献3には、Gタンパク質による接合シグナル(フェロモンシグナル)伝達経路を利用したタンパク質間相互作用の検出方法が記載されている。1倍体酵母は対となる接合型の1倍体酵母に対してフェロモンを分泌し、相手のフェロモンを感知すると接合を促すシグナルを細胞内に伝達する。細胞膜上にはフェロモンと結合する受容体が存在しており、この受容体は細胞内のGタンパク質と呼ばれる3量体のタンパク質と共役している。フェロモンが受容体に結合すると、Gタンパク質はαサブユニットとβγ複合体に解離する。αサブユニットとγサブユニットとはそれぞれ細胞膜に結合しているため、Gタンパク質の解離後においてもαサブユニットとβγ複合体は細胞膜に局在化する。βγ複合体のβサブユニットが細胞膜を介してエフェクターに作用することでシグナルが下流に伝達される。一方、γサブユニットの細胞膜結合能を欠損させた変異型(Gγcyto)では、Gタンパク質の解離によってβγ複合体(Gβγcyto複合体)が細胞質中に遊離してしまい、エフェクターに作用できなくなるため、シグナルが下流に伝達されなくなる。非特許文献3では、このGβγcyto複合体を細胞膜に局在化させるために、タンパク質間相互作用を利用している。まず、タンパク質(A)をGγcytoと融合させた融合タンパク質(A−Gγcyto)を発現させる。次に、タンパク質(A)と相互作用するタンパク質(B)を細胞膜に発現させる(Bmem)。タンパク質(A)とタンパク質(B)とが相互作用すると、A−GγcytoからなるGβγcyto複合体が細胞膜に局在化し接合シグナル伝達が回復する。したがって、接合シグナル伝達を検出することで、タンパク質(A)とタンパク質(B)との間の相互作用を検出できる。フェロモン刺激を受けた酵母では、Gタンパク質による接合シグナル伝達を介して様々な応答が起こり、様々な遺伝子の転写が活性化される。そこで、フェロモン応答的なプロモーターの下流にレポーター遺伝子を組み込み、接合シグナル伝達をレポーター遺伝子の発現として検出している。非特許文献3では、タンパク質(A)として、ヒトIgGのFc部分を用い、タンパク質(B)として、スタフィロコッカス・アウレウス(Staphylococcus aureus)のプロテインA由来のZドメインを用い、そしてレポーター遺伝子として、緑色蛍光タンパク質EGFPを用いて、レポーターの発する蛍光強度をフローサイトメーターで解析することにより、タンパク質間相互作用を定量的に評価している。その結果、結合定数8.0×103M-1という比較的弱い相互作用の検出にも成功している。
【0008】
しかしながら、非特許文献1〜3の方法では、一定の親和性を有するタンパク質間相互作用がもれなく検出またはスクリーニングされ、親和性の高いタンパク質間相互作用を選択的に検出またはスクリーニングすることが困難である。
【先行技術文献】
【非特許文献】
【0009】
【非特許文献1】A. Aronheimら、「Membrane targeting of the nucleotide exchange factor Sos is sufficient for activating the Ras signaling pathway」、Cell、1994年、第78巻、pp.949-961
【非特許文献2】Y.C. Broderら、「The ras recruitment system, a novel approach to the study of protein-protein interactions」、Curr. Biol.、1998年、第8巻、pp.1121-1124
【非特許文献3】N. Fukudaら、「Construction of a novel detection system for protein-protein interactions using yeast G-protein signaling」、FEBS J.、 2009年、第276巻、pp.2636-2644
【発明の概要】
【発明が解決しようとする課題】
【0010】
本発明は、親和性の高いタンパク質間相互作用を効率よく検出またはスクリーニングする方法を提供することを目的とする。
【課題を解決するための手段】
【0011】
本発明者らは、上記課題を解決するために鋭意研究を重ねた結果、酵母のGタンパク質による接合シグナル(フェロモンシグナル)伝達経路を利用したタンパク質間相互作用の検出方法において、酵母の細胞質に競合タンパク質を発現させることによって、競合タンパク質よりも高い親和性を有するタンパク質を効率よく検出またはスクリーニングできることを見出し、本発明を完成した。
【0012】
本発明は、酵母を用いるタンパク質間相互作用検出方法を提供し、該方法は、(1)第1のタンパク質とGタンパク質γサブユニット細胞膜非結合変異型との融合タンパク質を発現する第1の発現カセットを調製する工程、(2)第2のタンパク質と細胞膜局在化シグナル配列との融合タンパク質を発現する第2の発現カセットを調製する工程、(3)第3のタンパク質を細胞質に発現する第3の発現カセットを調製する工程、(4)該工程(1)、(2)および(3)でそれぞれ得られた該第1、2および3の発現カセットを酵母に導入して形質転換酵母を調製する工程、および(5)該工程(4)で得られた該形質転換酵母におけるフェロモンシグナル伝達を検出する、またはフェロモンシグナル伝達を示す形質転換酵母をスクリーニングする工程を含む。
【0013】
本発明はまた、酵母を用いるタンパク質間相互作用検出方法のさらなる実施態様を提供し、該方法は、(1)第1のタンパク質とGタンパク質γサブユニット細胞膜非結合変異型との融合タンパク質を発現する第1の発現カセットを調製する工程、(2)第2のタンパク質と細胞膜局在化シグナル配列との融合タンパク質を発現する第2の発現カセットを調製する工程、(3)第3のタンパク質を細胞質に発現する第3の発現カセットを調製する工程、(4)該工程(1)、(2)および(3)でそれぞれ得られた該第1、2および3の発現カセットを1倍体酵母に導入して形質転換酵母を調製する工程、(5)該工程(4)で得られた該形質転換酵母を、該形質転換酵母と接合可能な1倍体酵母と接合させて2倍体酵母を調製する工程、および(6)該工程(5)で得られた該2倍体酵母を選択する工程を含む。
【0014】
1つの実施態様では、上記第3のタンパク質は、第1のタンパク質または第2のタンパク質の競合タンパク質である。
【発明の効果】
【0015】
本発明によれば、親和性の高いタンパク質間相互作用を効率よく検出またはスクリーニングすることができる。
【図面の簡単な説明】
【0016】
【図1】本発明のタンパク質間相互作用検出方法の原理の概略を示す模式図である。
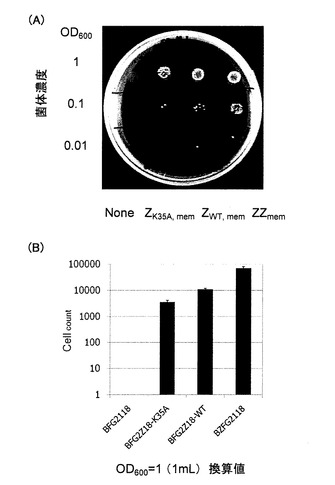
【図2】競合タンパク質を用いない場合における、2倍体選択用培地上で生成した2倍体酵母の生育を示す写真(A)、および菌体濃度OD600=1.0に調整した1mLの酵母懸濁液が生成し得る2倍体酵母のコロニー数を示すグラフ(B)である。
【図3】競合タンパク質としてZK35Aを用いた場合(A)およびZWTを用いた場合(B)における、5μMのαファクター存在下での1倍体酵母での緑色蛍光蛋白質(GFP)レポーター遺伝子の発現を示すグラフである。
【図4】競合タンパク質としてZK35Aを用いた場合(AおよびC)およびZWTを用いた場合(BおよびD)における、2倍体選択用培地上で生成した2倍体酵母の生育を示す写真(AおよびB)、および菌体濃度OD600=1.0に調整した1mLの酵母懸濁液が生成し得る2倍体酵母のコロニー数を示すグラフ(CおよびD)である。
【発明を実施するための形態】
【0017】
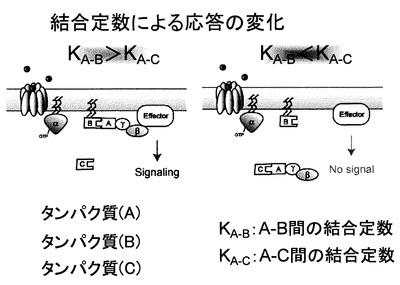
本発明のタンパク質間相互作用検出方法の原理の概略を図1に示す。本発明の方法を、図1を参照して説明する。例えば、タンパク質(A)は、標的タンパク質であり、タンパク質(B)は、標的タンパク質に対する結合タンパク質であり、そしてタンパク質(C)は、タンパク質(A)との相互作用において、タンパク質(B)の競合タンパク質である。すなわち、タンパク質(A)とタンパク質(B)とが相互作用し、タンパク質(A)とタンパク質(C)とが相互作用する。この場合に、競合タンパク質とは、タンパク質(B)とタンパク質(C)との関係において互いのタンパク質をいう。ここでタンパク質(A)−(B)間の結合定数をKA−B、タンパク質(A)−(C)間の結合定数をKA−Cとすると、KA−BがKA−Cより小さいとき(図1の右図)、Gβγ複合体は細胞質にトラップされて接合シグナルの伝達が阻害される。一方、KA−BがKA−Cより大きいとき(図1の左図)、Gβγ複合体は細胞膜に結合できるため、接合シグナルが伝達される。このため、接合シグナル伝達により、タンパク質(A)−タンパク質(B)間の相互作用を検出することによって、標的タンパク質(A)に対して、競合タンパク質(C)と比べて、より親和性の高い(結合定数の大きい)結合タンパク質(B)を検出またはスクリーニングすることが可能となる。
【0018】
ここで、タンパク質間相互作用とは、タンパク質とタンパク質との間の結合または親和性をいう。
【0019】
本発明の酵母を用いるタンパク質間相互作用検出方法は、第1の実施態様では、(1)第1のタンパク質とGタンパク質γサブユニット細胞膜非結合変異型との融合タンパク質を発現する第1の発現カセットを調製する工程、(2)第2のタンパク質と細胞膜局在化シグナル配列との融合タンパク質を発現する第2の発現カセットを調製する工程、(3)第3のタンパク質を細胞質に発現する第3の発現カセットを調製する工程、(4)該工程(1)、(2)および(3)でそれぞれ得られた該第1、2および3の発現カセットを酵母に導入して形質転換酵母を調製する工程、および、(5)該工程(4)で得られた該形質転換酵母におけるフェロモンシグナル伝達を検出する、またはフェロモンシグナル伝達を示す形質転換酵母をスクリーニングする工程を含む。
【0020】
上記の方法において得られた形質転換酵母は、シグナル伝達能を評価できるよう活性化する必要がある。活性化の方法は、Gタンパク質の上流側を活性化するものであれば、特に制限されない。例えば、酵母内在性のGタンパク質共役型受容体に対するリガンド(アゴニスト)を加える方法が挙げられる。a接合型細胞の場合、例えば、配列番号1に示される塩基配列を有する遺伝子(GenBankアクセッション番号:NC001148)にコードされ、例えば、配列番号2に示されるアミノ酸配列(GenBankアクセッション番号:NP015137)を有する前駆体であるMF(alpha)1から生産される13アミノ酸のペプチドであるαファクターを加える方法が挙げられる。
【0021】
フェロモンシグナル伝達を検出する方法またはフェロモンシグナル伝達を示す形質転換酵母をスクリーニングする方法は特に制限されない。好ましくは、フェロモンシグナル応答性のプロモーター(例えば、FUS1、FIG1、AGA1など)の制御下でレポーター遺伝子の発現が誘導されるよう修飾する。レポーター遺伝子としては、例えば、蛍光レポーター遺伝子、生育レポーター遺伝子、発色レポーター遺伝子、発光レポーター遺伝子などが挙げられる。蛍光レポーター遺伝子としては、例えば、緑色蛍光蛋白質(GFP)、赤色蛍光蛋白質(DsRed)、シアン色蛍光蛋白質(CFP)などが挙げられる。生育レポーター遺伝子としては、例えば、HIS3、ADE2、URA3、CAN1などが挙げられる。発色レポーター遺伝子としては、例えば、βガラクトシダーゼ(lacZ)などが挙げられる。発光レポーター遺伝子としては、例えば、ルシフェラーゼ(luc)などが挙げられる。また、フェロモンシグナル伝達によって誘導される細胞周期抑制を利用してもよい。シグナルによって細胞周期が抑制された酵母は増殖が阻害されるため、固体培地上でのハロ形成や液体培地中での菌体密度を測定することにより、シグナル伝達能を検出することまたはシグナル伝達能を有する形質転換酵母をスクリーニングすることができる。
【0022】
本発明の酵母を用いるタンパク質間相互作用検出方法の第2の実施態様は、(1)第1のタンパク質とGタンパク質γサブユニット細胞膜非結合変異型との融合タンパク質を発現する第1の発現カセットを調製する工程、(2)第2のタンパク質と細胞膜局在化シグナル配列との融合タンパク質を発現する第2の発現カセットを調製する工程、(3)第3のタンパク質を細胞質に発現する第3の発現カセットを調製する工程、(4)該工程(1)、(2)および(3)でそれぞれ得られた該第1、2および3の発現カセットを1倍体酵母に導入して形質転換酵母を調製する工程、(5)該工程(4)で得られた該形質転換酵母を、該形質転換酵母と接合可能な1倍体酵母と接合させて2倍体酵母を調製する工程、および(6)該工程(5)で得られた該2倍体酵母を選択する工程を含む。
【0023】
上記の第2の実施態様において、形質転換に用いる1倍体酵母は、接合能を有する必要がある。好ましくはa/α接合型のa型またはα型の1倍体酵母である。いずれも、接合により生成される2倍体酵母を1倍体酵母と区別して選択できるように、適切な選択マーカーを有する必要がある。また、内在性のGタンパク質γサブユニットをコードする遺伝子が欠損している酵母が好ましい。内在性のGタンパク質γサブユニットをコードする遺伝子が欠損していない酵母を用いる場合には、上記第1の発現カセットを1倍体酵母に導入して形質転換酵母を調製する工程において、相同組換えにより内在性Gタンパク質γサブユニットと第1の発現カセットとを置き換える必要がある。酵母の種としては、例えば、サッカロミセス・セレビシエ(Saccharomyces cerevisiae)、シゾサッカロミセス・ポンベ(Schizosaccharomyces pombe)が挙げられる。
【0024】
選択マーカーとしては、例えば、栄養要求性マーカー、薬剤耐性マーカーが挙げられる。栄養要求性マーカーとしては、例えば、各種アミノ酸要求性マーカー、各種核酸要求性マーカーなどが挙げられる。アミノ酸要求性マーカーとしては、例えば、リジン要求性マーカー、メチオニン要求性マーカー、ヒスチジン要求性マーカー、トリプトファン要求性マーカー、ロイシン要求性マーカーなどが挙げられる。核酸要求性マーカーとしては、例えば、ウラシル要求性マーカー、アデニン要求性マーカーなどが挙げられる。薬剤耐性マーカーとしては、例えば、ジェネティシン(Geneticin:G418)耐性マーカー、オーレオバシジンA(Aureobasidin A)耐性マーカーなどが挙げられる。接合する1対の1倍体酵母は、接合により生成される2倍体酵母を1倍体酵母と区別して選択できるように、互いに異なる選択マーカーを有する必要がある。
【0025】
ここで、1倍体とは、ゲノムの1セットを有する酵母の個体をいい、半数体ともいう。本発明では、第1のタンパク質(A)、第2のタンパク質(B)および第3のタンパク質(C)の遺伝子を1倍体の酵母に導入する。
【0026】
2倍体とは、ゲノムの2セットを有する酵母の個体をいい、倍数体ともいう。1倍体どうしの接合により生成する。本発明では、2倍体の生成効率により、タンパク質間相互作用を検出またはスクリーニングする。
【0027】
本発明で用いるGタンパク質は、αサブユニット、βサブユニットおよびγサブユニットが形成する複合体からなるヘテロ三量体Gタンパク質のことをいい、Gタンパク質とはグアニンヌクレオチド結合タンパク質の略称である。αサブユニットとγサブユニットとはそれぞれ細胞膜に結合している。フェロモンが受容体に結合すると、Gタンパク質はαサブユニットとβγ複合体に解離する。Gタンパク質の解離後においてもαサブユニットとβγ複合体は細胞膜に局在化する。βγ複合体のβサブユニットが細胞膜を介してエフェクターに作用することでシグナルが下流に伝達される。Gタンパク質の由来としては、接合シグナル伝達に関与する限り、特に制限されない。
【0028】
本発明で用いるGタンパク質のγサブユニットとしては、例えば、配列番号3に示される塩基配列を有する遺伝子(GenBankアクセッション番号:AY557888)にコードされ、例えば、配列番号4に示されるアミノ酸配列(GenBankアクセッション番号:CAA89613)を有するタンパク質であるSte18が挙げられる。γサブユニット細胞膜非結合変異型(Gγcyto)は、細胞膜結合能を欠損している限り、特に制限されない。Gγcytoとしては、例えば、γサブユニットSte18の配列番号4に示されるアミノ酸配列の第106位〜第110位が欠失している変異型が挙げられる。
【0029】
本発明で第1のタンパク質(A)とGγcytoとを融合させる方法は、Gγcytoの機能が発揮できる限り、特に制限されない。例えば、γサブユニットSte18の配列番号4に示されるアミノ酸配列の第105位に、タンパク質(A)のN末端を連結する。
【0030】
本発明で用いる細胞膜局在化シグナル配列は、第2のタンパク質(B)を細胞膜に局在化することができる限り、特に制限されない。好ましくは、Gタンパク質のγサブユニットの細胞膜局在化シグナル配列である。例えば、このシグナルは、γサブユニットSte18の配列番号4に示されるアミノ酸配列の第102位〜第110位に相当する。
【0031】
本発明で第2のタンパク質(B)と細胞膜局在化シグナル配列とを融合させる方法は、細胞膜局在化シグナル配列の機能が発揮できる限り、特に制限されない。例えば、γサブユニットSte18の配列番号4に示されるアミノ酸配列の第102位〜第110位からなる断片をタンパク質(B)のC末端に連結する。
【0032】
本発明でタンパク質(C)を細胞質に発現させる方法は、特に制限されない。
【0033】
タンパク質(A)−Gγcyto融合タンパク質、細胞膜結合性タンパク質(B)および細胞質性タンパク質(C)を酵母で発現させるための発現カセットは、通常、これらのタンパク質のコード領域のほかに、酵母で機能するプロモーター、ターミネーターなどの遺伝子発現調節領域、発現カセットが導入された形質転換酵母を選択するための選択マーカーなどを有する。プロモーターとしては、例えば、PGK1プロモーター、TDH3プロモーター、STE18プロモーターが挙げられる。ターミネーターとしては、例えば、PGK1ターミネーター、TDH3ターミネーター、CYC1ターミネーターが挙げられる。選択マーカーとしては、例えば、ロイシン要求性マーカー、ウラシル要求性マーカー、ジェネティシン(Geneticin:G418)耐性マーカーが挙げられる。
【0034】
発現カセットは、通常、プラスミドとして作製される。プラスミドは、発現カセットのほかに、発現カセットを相同組換えにより酵母染色体に組み込むために必要な断片、またはプラスミドを酵母内で自律複製させるために必要な複製起点を有する。相同組換えに必要な断片としては、組み込みたい染色体上の位置に存在する遺伝子の断片が好ましい。発現カセットの相同組換えによる酵母染色体への組み込みは、当業者が通常用いる方法により行われる。酵母内複製起点としては、例えば、2μ(マルチコピー型)、CEN/ARS(シングルコピー型)が挙げられる。プラスミドの酵母内での自律複製は、当業者が通常用いる方法により行われる。プラスミドは、当業者が通常用いる方法により、大腸菌などの適切な宿主内で増幅され、調製される。プラスミドを調製するための宿主は特に制限されない。好ましくは、大腸菌である。プラスミドはまた、通常、プラスミドの調製を容易にするために、プラスミドが導入された形質転換大腸菌などを選択するための選択マーカーを有する。選択マーカーとしては、例えば、アンピシリン耐性、カナマイシン耐性などの抗生物質耐性が挙げられる。
【0035】
発現カセットは、プラスミドの形状で酵母に導入されてもよく、発現カセットのみが酵母に導入されてもよい。発現カセットは、相同組換えにより、効率的に酵母染色体に組み込むことができる。プラスミドは、選択マーカーに対応する選択条件下で形質転換酵母の培養を継続することにより、酵母内で自律複製させることができる。
【0036】
発現カセットが導入された形質転換酵母は、選択マーカーに対応する選択条件下、すなわち選択マーカーに対応するアミノ酸もしくは核酸の非存在下の培養により、または薬剤存在下の培養により得ることができる。
【0037】
本発明の方法の第2の実施態様では、発現カセットが導入された形質転換酵母と、対応する接合型の1倍体酵母、好ましくは野生型の1倍体酵母とを接合させる。ここで用いる1倍体酵母は、選択マーカーを有し、かつ接合可能であれば、特に制限されない。例えば、発現カセットが導入された形質転換酵母がa型由来であれば、これを対応するa/α接合型のα型の野生酵母と接合させる。1倍体野生酵母は、形質転換酵母との接合により生成する2倍体酵母のみを選択できる選択マーカーを有し、形質転換酵母との接合能を有する限り、特に制限されない。接合により生成する2倍体酵母は、上記選択マーカーを利用することにより、確認・選択することができる。
【0038】
本発明では、接合効率を適宜定量化することもできる。例えば、接合させた酵母の懸濁液を適宜希釈して選択用固体培地上にコロニーを形成させて、コロニー数を計測することにより定量化できる。
【0039】
本発明では、第1のタンパク質(A)、第2のタンパク質(B)および第3のタンパク質(C)は、特定のタンパク質であってもよく、不特定のタンパク質であってもよい。不特定のタンパク質とは、ライブラリを構成するタンパク質などをいう。第3のタンパク質(C)として、第1のタンパク質(A)または第2のタンパク質(B)の競合タンパク質を用いる点が特徴である。この特徴により、特定の第1のタンパク質(A)を用い、ライブラリを構成する第2のタンパク質(B)を用いる場合、第1のタンパク質(A)に対して、競合タンパク質である第3のタンパク質(C)と比べて、より親和性の高い(結合定数の大きい)第2のタンパク質(B)をライブラリの中から検出またはスクリーニングすることが可能となる。
【実施例】
【0040】
以下に、実施例を示して本発明をさらに具体的に説明するが、本発明はこれらに限定されない。
【0041】
(培地)
1%(w/v)酵母エキストラクト、2%(w/v)ペプトンおよび2%(w/v)グルコースを含むYPD培地、または0.67%(w/v)アミノ酸不含酵母ニトロゲンベース(Becton Dickinson社製)および2%(w/v)グルコースを含むSD培地で酵母を培養した。SD培地には選択マーカーに対応するアミノ酸および核酸を適宜添加した。2%(w/v)寒天をこれらの培地に添加してYPDおよびSDの固体培地を調製した。
【0042】
(酵母)
酵母サッカロミセス・セレビシエ(Saccharomyces cerevisiae)の1倍体株BY4741(メチオニン要求性a細胞)およびBY4742(リジン要求性α細胞)、ならびに以下の実施例で用いた酵母株および作製した酵母株の遺伝子型を表1に、そして各株において発現するタンパク質を表2に示す。a型のBY4741を親株として遺伝子組換えにより作製された形質転換株MC−F1は、フェロモン誘導性FIG1プロモーターの制御下においてEGFPレポーター遺伝子を発現する。MC−F1を親株として遺伝子組換えにより作製された形質転換株BFG2Z18−K35A、BFG2Z18−WTおよびBZFG2118はいずれも、タンパク質(A)として、ヒトIgGのFc部分をGタンパク質γサブユニットの細胞膜非結合変異型(Gγcyto)との融合タンパク質(Gγcyto−Fc)として細胞質に発現する。BFG2Z18−K35A、BFG2Z18−WTおよびBZFG2118は、タンパク質(B)として、ヒトIgGのFc部分に対する異なる親和性を有するスタフィロコッカス・アウレウス(Staphylococcus aureus)のプロテインA由来のZドメインを細胞膜に結合して発現する。BFG2Z18−K35Aは、結合定数4.6×106M-1のZドメイン変異型(ZK35A,mem)を発現し、BFG2Z18−WTは、結合定数5.9×107M-1のZドメイン野生型(ZWT,mem)を発現し、そしてBZFG2118は、結合定数6.8×108M-1のZドメイン2量体(ZZmem)を発現する。BFG2118は、ネガティブコントロールであり、Gγcyto−Fcのみを発現する。
【0043】
【表1】

【0044】
【表2】
【0045】
(参考例)
(接合能を指標とするタンパク質間相互作用検出方法)
BFG2Z18−K35A、BFG2Z18−WT、BZFG2118およびBFG2118の各酵母株を初期菌体濃度OD600=0.1で5mLのYPD培地に播種し、無処置の接合相手BY4742と30℃にて3時間共培養した。培養後、酵母を回収し、洗浄後、蒸留水に再懸濁した。各株の接合能を評価するために、酵母懸濁液のOD600=1.0、0.1および0.001の希釈系列を調製し、メチオニンおよびリジンを含まず、20mg/Lのヒスチジン、30mg/Lのロイシンおよび20mg/Lウラシルを含む2倍体選択用SD固体培地上(SD−Met、Lysプレート)にスポットした。この結果、BFG2118は増殖しなかったが、他の3つの株は増殖した(図2A)。
【0046】
増殖した3つの株では、タンパク質(A)とタンパク質(B)との間には相互作用が存在するため、Gタンパク質を介するフェロモンシグナル伝達によるa型とα型との接合が生じたものと思われる。一方、BFG2118では、タンパク質(B)を欠くため、このような相互作用は存在せず、増殖が見られなかった。このように、1倍体酵母のa型とα型とのフェロモンシグナル伝達を介する接合能は、タンパク質(A)とタンパク質(B)との相互作用の有無を検出する方法として有効であることが示された。
【0047】
次に、増殖した3つの株の接合能を定量的に評価するために、菌体濃度OD600=1.0に調整した1mLの酵母懸濁液を、プレート上に100〜1000コロニーが成長するように、適宜希釈してSD−Met、Lysプレートに塗布した。コロニー数を計測し、計測したコロニー数に希釈率を乗じて、菌体濃度OD600=1.0に調整した1mLの酵母懸濁液が生成し得る2倍体酵母のコロニー数を求めた。結果を図2Bに示す。図2B中のエラーバーは、3度の独立した実験結果における標準偏差を示す。
【0048】
図2Bからわかるように、タンパク質(A)とタンパク質(B)との間の結合定数に対応して生成し得る2倍体酵母のコロニー数に差異があった。このように、1倍体酵母のa型とα型とのフェロモンシグナル伝達を介する接合能は、タンパク質(A)とタンパク質(B)との相互作用を定量的に評価する方法としても有効であることが示された。
【0049】
(実施例1)
(細胞質で競合タンパク質(C)を発現する形質転換株の作製)
細胞質での競合タンパク質(C)の発現がGタンパク質を介する接合シグナル伝達の回復を阻害するかどうか調べるために、BFG2Z18−K35A、BFG2Z18−WTおよびBZFG2118を遺伝子組換えし、競合タンパク質(C)として、Zドメイン野生型(ZWT)またはZドメイン変異型(ZK35A)を細胞質に発現する形質転換株を作製した。
【0050】
細胞質で競合タンパク質として発現させるZドメイン野生型遺伝子(ZWT)およびZドメイン変異型遺伝子(ZK35A)を酵母染色体上のHOP2遺伝子の上流(PHOP2:HOP2プロモーター領域)に相同組換えにより組み込むため、次のプラスミドを構築した。ZWTおよびZK35Aを、プラスミドpUMZ−WTおよびpUMZ−K35Aからプライマー1(配列番号5)およびプライマー2(配列番号6)を用いてPCRにより増幅し、プラスミドpGK425のSalI−BamHI部位間に挿入し、それぞれプラスミドpLMZ−WTおよびpLMZ−K35Aとした。HOP2プロモーター領域での相同組換えに用いるDNA断片を、MC−F1のゲノムDNAからプライマー3(配列番号7)およびプライマー4(配列番号8)を用いてPCRにより増幅し、プラスミドpLMZ−WTおよびpLMZ−K35AのNotI−SacI部位間にそれぞれ挿入し、それぞれプラスミドpLMZ−WT−HおよびpLMZ−K35A−Hとした。
【0051】
次いで、LEU2−PGK5’−Z−PGK3’−PHOP2(PGK5’:PGK1プロモーター;PGK3’:PGK1ターミネーター)を含むDNA断片をpLMZ−WT−HおよびpLMZ−K35A−HからPHOP2のすぐ上流領域に相同な領域を含む50塩基のプライマー5(配列番号9)およびプライマー6(配列番号10)を用いてPCRにより増幅した。増幅したDNA断片をBGF2Z18−K35A、BFG2Z18−WTおよびBZFG2118の形質転換に用いた。形質転換では、酢酸リチウム法を用いた。形質転換体は、ウラシルおよびロイシンを含まず、20mg/Lのヒスチジンおよび30mg/Lのメチオニンを含むSD培地(SD−Ura、Leuプレート)で選択し、FC1−1、FC2−1、FC3−1、FC1−2、FC2−2およびFC3−2株を得た(表1)。
【0052】
(実施例2)
(細胞質で競合タンパク質(C)を発現する形質転換株における蛍光レポーター遺伝子を指標とするタンパク質間相互作用検出方法)
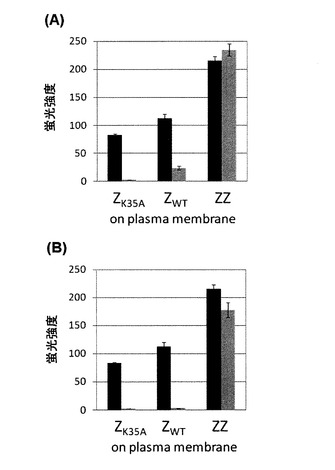
実施例1で得られた形質転換株の蛍光レポーター遺伝子の発現を評価した。競合タンパク質を発現しない例として、BFG2Z18−K35A、BFG2Z18−WTおよびBZFG2118を用いた。競合タンパク質としてZK35Aを発現する例として、FC1−1、FC2−1およびFC3−1を用いた。競合タンパク質としてZWTを発現する例として、FC1−2、FC2−2およびFC3−2を用いた。各酵母株を初期菌体濃度OD600=0.1で5mLのYPD培地に播種した後、最終濃度5μMとなるようαファクターを添加し、30℃にて6時間培養した。各株の蛍光レポーター遺伝子発現を評価するために、培養後、酵母を回収し、フローサイトメーターを用いて細胞の蛍光強度を評価した。結果を図3に示す。図3のグラフにおいて並立する2本カラムのうち、左(濃色)のカラムは競合タンパク質を導入していない酵母株、および右(淡色)のカラムは競合タンパク質を導入した酵母株の結果を示す。図3中のエラーバーは、3度の独立した実験結果における標準偏差を示す。
【0053】
この結果、競合タンパク質(C)を発現しない酵母株BFG2Z18−K35A、BFG2Z18−WTおよびBZFG2118はすべて蛍光を示した。一方で、競合タンパク質(C)ZK35Aを発現する酵母株においては、結合定数の小さい第2のタンパク質(B)を発現しているFC1−1は全く蛍光を示さず、より結合定数の大きい第2のタンパク質(B)を発現しているFC2−1およびFC3−1のみ蛍光を示した(図3A)。同様に、競合タンパク質(C)ZWTを発現する酵母株においては、結合定数の小さい第2のタンパク質(B)を発現しているFC1−2およびFC2−2は全く蛍光を示さず、より結合定数の大きい第2のタンパク質(B)を発現しているFC3−2のみ蛍光を示した(図3B)。
【0054】
(実施例3)
(細胞質で競合タンパク質(C)を発現する形質転換株における接合能を指標とするタンパク質間相互作用検出方法)
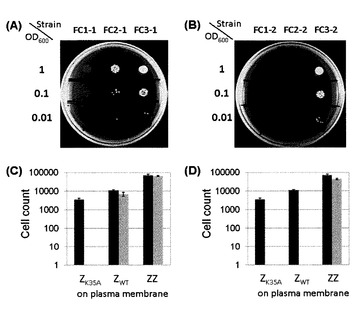
実施例1で得られた形質転換株の接合能を参考例と同様に評価した。結果を図4に示す。図4のグラフにおいて並立する2本カラムのうち、左(濃色)のカラムは競合タンパク質を導入していない酵母株、および右(淡色)のカラムは競合タンパク質を導入した酵母株の結果を示す。図4中のエラーバーは、3度の独立した実験結果における標準偏差を示す。
【0055】
この結果、競合タンパク質ZK35Aを発現するFC2−1およびFC3−1は、2倍体選択用培地で増殖した(図4A)。FC2−1およびFC3−1の接合能は、競合タンパク質(C)を発現しない酵母株とほとんど同等であった(図4C)。同様の結果が、競合タンパク質(C)ZWTを発現する酵母株についても得られた。FC1−2およびFC2−2は、接合能を示さなかったが、FC3−2のみ、競合タンパク質(C)を発現しない酵母株とほとんど同等の接合能を示した(図4Bおよび4D)。
【0056】
(実施例4)
(モデルライブラリからの標的細胞のスクリーニング)
細胞質で競合タンパク質(C)を発現する形質転換株における接合能を指標とするタンパク質間相互作用検出方法が有効かどうかを、モデルライブラリを用いて検証した。モデルライブラリとして、標的細胞(FC3−2またはBZFG2118)とコントロール細胞(FC2−2またはBFGZ18−WT)とが表3に示す初期割合で混合された2つのライブラリを調製した。一方は、競合タンパク質(C)を発現していない酵母株からなり、ZZを発現する少量のBZFG2118とZWTを発現する過剰量のBFG2Z18−WTとを含み、他方は、競合タンパク質(C)を発現している酵母株からなり、ZZを発現する少量のFC3−2とZWTを発現する過剰量のFC2−2とを含む。これらのライブラリは、各1倍体の初期菌体濃度をOD600=0.1に設定して、接合相手のBY4742と30℃にて3時間10mLのYPD培地で共培養した。培養後、酵母を回収し、洗浄後、SD−Met、Lysプレートに塗布し、30℃にて2日間培養した。10コロニーをピックアップし、YPD培地中で一晩別々に増殖させた。ゲノムを抽出し、タンパク質(B)のコード領域をプライマー7(配列番号11)およびプライマー8(配列番号12)を用いてPCRにより増幅した。増幅した断片がZZまたはZWTのいずれに由来するかは、断片の長さで判断した。10コロニー中、ZZの断片が得られたコロニーの割合を、標的細胞の最終割合とした。標的細胞の最終割合を初期割合で除した値をスクリーニング効率として算出し、評価した。結果を表3に示す。
【0057】
【表3】
【0058】
表3に示すように、競合タンパク質(C)を用いるタンパク質間相互作用検出方法は、競合タンパク質を用いない従来の方法よりも明らかに優れたスクリーニング効率を示し、最大で7000倍に達した。
【0059】
これらの結果は、競合タンパク質(C)を用いるタンパク質間相互作用検出方法は、標的タンパク質(A)に対して、もとの結合タンパク質(B)と比べて、より高い親和性を有する結合タンパク質(B)を検出するのに適していることを示唆する。
【産業上の利用可能性】
【0060】
本発明によれば、親和性の高いタンパク質間相互作用を効率よく検出またはスクリーニングすることができる。
【技術分野】
【0001】
本発明は、高親和性のタンパク質間相互作用検出・スクリーニング方法に関する。特に、競合タンパク質を用いるタンパク質間相互作用検出・スクリーニング方法に関する。
【背景技術】
【0002】
タンパク質間相互作用検出方法として酵母ツーハイブリッド法が知られている。この方法は、遺伝子の発現調節に関与する転写因子が、遺伝子(DNA)上の特定の塩基配列を認識して結合するドメイン(DNA結合ドメイン:BD)と、遺伝子の転写に必須の基本因子に作用して遺伝子の転写を活性化するドメイン(転写活性化ドメイン:AD)とから構成されており、それぞれのドメインが分離できることに基づく。すなわち、タンパク質間相互作用を検出したい1対のタンパク質(XとY)のうち、一方(X)を転写因子のDNA結合ドメインと融合させて1つのハイブリッドタンパク質(X−BD)を形成し、他方(Y)を転写因子の転写活性化ドメインと融合させてもう1つのハイブリッドタンパク質(Y−AD)を形成すると、X−Y間に相互作用が存在する場合のみ、遺伝子の転写が活性化される。この遺伝子の転写活性化は、BDが結合する特定の塩基配列の下流にレポーター遺伝子を連結することによって、レポーター遺伝子の発現により検出することができる。酵母ツーハイブリッド法に利用される転写因子としては、Gal4、LexA−B42、LexA−VP16などが挙げられる。
【0003】
酵母は、単細胞性の真核生物であり、古くから食品加工などに用いられている。酵母は、真核生物の中で最も研究されており、酵母の遺伝子組換え技術も発達している。酵母は、真核生物という点では、人類など哺乳類をはじめとする高等真核生物と共通するため、高等真核生物のタンパク質を解析するために、これらのタンパク質の発現用宿主として汎用されている。真核生物のタンパク質間相互作用検出方法として酵母ツーハイブリッド法は有用である。
【0004】
しかし、細胞核内の転写因子を用いる酵母ツーハイブリッド法は、核移行が困難なタンパク質、および元来転写活性化ドメインを有するタンパク質(例えば、転写因子)に適用することが難しいという問題がある。
【0005】
そこで、近年、タンパク質の局在性や機能に応じて酵母ツーハイブリッド法の変法が考案されている。非特許文献1および2には、酵母のRasシグナル伝達経路を利用する方法が記載されている。この方法では、Rasの細胞膜結合能をタンパク質間相互作用で代替することで、酵母の増殖を指標としてタンパク質間相互作用を検出する。Ras伝達経路のシグナル増幅効果を利用することにより高感度にタンパク質間相互作用を検出することができる。すなわち、タンパク質間相互作用を検出したい1対のタンパク質(XとY)のうち、一方(X)を細胞膜に固定し、他方(Y)をヒト由来グアニンヌクレオチド交換因子(hSOS)または細胞膜結合能を欠損させたRas恒常活性変異型(mRas)と融合させてハイブリッドタンパク質(それぞれY−hSOSまたはY−mRas)を形成すると、X−Y間に相互作用が存在する場合のみ、hSOSまたはmRasが細胞膜に局在化し、Rasのシグナル伝達経路が活性化される。このシグナル伝達経路の活性化は、酵母の増殖を指標として検出することができる。
【0006】
しかしながら、この方法では、酵母の温度感受性変異株(cdc25−2)を使用する必要があり、酵母の増殖に温度制限が課せられる。すなわち、タンパク質のライブラリの構築を感受性温度でない25℃で行い、タンパク質間相互作用の検出またはスクリーニングを感受性温度である36℃で行う必要があるため、酵母の最適生育温度である30℃に比べて酵母の増殖速度が大きく低下し、迅速なタンパク質間相互作用の検出またはスクリーニングが困難であるという問題がある。
【0007】
非特許文献3には、Gタンパク質による接合シグナル(フェロモンシグナル)伝達経路を利用したタンパク質間相互作用の検出方法が記載されている。1倍体酵母は対となる接合型の1倍体酵母に対してフェロモンを分泌し、相手のフェロモンを感知すると接合を促すシグナルを細胞内に伝達する。細胞膜上にはフェロモンと結合する受容体が存在しており、この受容体は細胞内のGタンパク質と呼ばれる3量体のタンパク質と共役している。フェロモンが受容体に結合すると、Gタンパク質はαサブユニットとβγ複合体に解離する。αサブユニットとγサブユニットとはそれぞれ細胞膜に結合しているため、Gタンパク質の解離後においてもαサブユニットとβγ複合体は細胞膜に局在化する。βγ複合体のβサブユニットが細胞膜を介してエフェクターに作用することでシグナルが下流に伝達される。一方、γサブユニットの細胞膜結合能を欠損させた変異型(Gγcyto)では、Gタンパク質の解離によってβγ複合体(Gβγcyto複合体)が細胞質中に遊離してしまい、エフェクターに作用できなくなるため、シグナルが下流に伝達されなくなる。非特許文献3では、このGβγcyto複合体を細胞膜に局在化させるために、タンパク質間相互作用を利用している。まず、タンパク質(A)をGγcytoと融合させた融合タンパク質(A−Gγcyto)を発現させる。次に、タンパク質(A)と相互作用するタンパク質(B)を細胞膜に発現させる(Bmem)。タンパク質(A)とタンパク質(B)とが相互作用すると、A−GγcytoからなるGβγcyto複合体が細胞膜に局在化し接合シグナル伝達が回復する。したがって、接合シグナル伝達を検出することで、タンパク質(A)とタンパク質(B)との間の相互作用を検出できる。フェロモン刺激を受けた酵母では、Gタンパク質による接合シグナル伝達を介して様々な応答が起こり、様々な遺伝子の転写が活性化される。そこで、フェロモン応答的なプロモーターの下流にレポーター遺伝子を組み込み、接合シグナル伝達をレポーター遺伝子の発現として検出している。非特許文献3では、タンパク質(A)として、ヒトIgGのFc部分を用い、タンパク質(B)として、スタフィロコッカス・アウレウス(Staphylococcus aureus)のプロテインA由来のZドメインを用い、そしてレポーター遺伝子として、緑色蛍光タンパク質EGFPを用いて、レポーターの発する蛍光強度をフローサイトメーターで解析することにより、タンパク質間相互作用を定量的に評価している。その結果、結合定数8.0×103M-1という比較的弱い相互作用の検出にも成功している。
【0008】
しかしながら、非特許文献1〜3の方法では、一定の親和性を有するタンパク質間相互作用がもれなく検出またはスクリーニングされ、親和性の高いタンパク質間相互作用を選択的に検出またはスクリーニングすることが困難である。
【先行技術文献】
【非特許文献】
【0009】
【非特許文献1】A. Aronheimら、「Membrane targeting of the nucleotide exchange factor Sos is sufficient for activating the Ras signaling pathway」、Cell、1994年、第78巻、pp.949-961
【非特許文献2】Y.C. Broderら、「The ras recruitment system, a novel approach to the study of protein-protein interactions」、Curr. Biol.、1998年、第8巻、pp.1121-1124
【非特許文献3】N. Fukudaら、「Construction of a novel detection system for protein-protein interactions using yeast G-protein signaling」、FEBS J.、 2009年、第276巻、pp.2636-2644
【発明の概要】
【発明が解決しようとする課題】
【0010】
本発明は、親和性の高いタンパク質間相互作用を効率よく検出またはスクリーニングする方法を提供することを目的とする。
【課題を解決するための手段】
【0011】
本発明者らは、上記課題を解決するために鋭意研究を重ねた結果、酵母のGタンパク質による接合シグナル(フェロモンシグナル)伝達経路を利用したタンパク質間相互作用の検出方法において、酵母の細胞質に競合タンパク質を発現させることによって、競合タンパク質よりも高い親和性を有するタンパク質を効率よく検出またはスクリーニングできることを見出し、本発明を完成した。
【0012】
本発明は、酵母を用いるタンパク質間相互作用検出方法を提供し、該方法は、(1)第1のタンパク質とGタンパク質γサブユニット細胞膜非結合変異型との融合タンパク質を発現する第1の発現カセットを調製する工程、(2)第2のタンパク質と細胞膜局在化シグナル配列との融合タンパク質を発現する第2の発現カセットを調製する工程、(3)第3のタンパク質を細胞質に発現する第3の発現カセットを調製する工程、(4)該工程(1)、(2)および(3)でそれぞれ得られた該第1、2および3の発現カセットを酵母に導入して形質転換酵母を調製する工程、および(5)該工程(4)で得られた該形質転換酵母におけるフェロモンシグナル伝達を検出する、またはフェロモンシグナル伝達を示す形質転換酵母をスクリーニングする工程を含む。
【0013】
本発明はまた、酵母を用いるタンパク質間相互作用検出方法のさらなる実施態様を提供し、該方法は、(1)第1のタンパク質とGタンパク質γサブユニット細胞膜非結合変異型との融合タンパク質を発現する第1の発現カセットを調製する工程、(2)第2のタンパク質と細胞膜局在化シグナル配列との融合タンパク質を発現する第2の発現カセットを調製する工程、(3)第3のタンパク質を細胞質に発現する第3の発現カセットを調製する工程、(4)該工程(1)、(2)および(3)でそれぞれ得られた該第1、2および3の発現カセットを1倍体酵母に導入して形質転換酵母を調製する工程、(5)該工程(4)で得られた該形質転換酵母を、該形質転換酵母と接合可能な1倍体酵母と接合させて2倍体酵母を調製する工程、および(6)該工程(5)で得られた該2倍体酵母を選択する工程を含む。
【0014】
1つの実施態様では、上記第3のタンパク質は、第1のタンパク質または第2のタンパク質の競合タンパク質である。
【発明の効果】
【0015】
本発明によれば、親和性の高いタンパク質間相互作用を効率よく検出またはスクリーニングすることができる。
【図面の簡単な説明】
【0016】
【図1】本発明のタンパク質間相互作用検出方法の原理の概略を示す模式図である。
【図2】競合タンパク質を用いない場合における、2倍体選択用培地上で生成した2倍体酵母の生育を示す写真(A)、および菌体濃度OD600=1.0に調整した1mLの酵母懸濁液が生成し得る2倍体酵母のコロニー数を示すグラフ(B)である。
【図3】競合タンパク質としてZK35Aを用いた場合(A)およびZWTを用いた場合(B)における、5μMのαファクター存在下での1倍体酵母での緑色蛍光蛋白質(GFP)レポーター遺伝子の発現を示すグラフである。
【図4】競合タンパク質としてZK35Aを用いた場合(AおよびC)およびZWTを用いた場合(BおよびD)における、2倍体選択用培地上で生成した2倍体酵母の生育を示す写真(AおよびB)、および菌体濃度OD600=1.0に調整した1mLの酵母懸濁液が生成し得る2倍体酵母のコロニー数を示すグラフ(CおよびD)である。
【発明を実施するための形態】
【0017】
本発明のタンパク質間相互作用検出方法の原理の概略を図1に示す。本発明の方法を、図1を参照して説明する。例えば、タンパク質(A)は、標的タンパク質であり、タンパク質(B)は、標的タンパク質に対する結合タンパク質であり、そしてタンパク質(C)は、タンパク質(A)との相互作用において、タンパク質(B)の競合タンパク質である。すなわち、タンパク質(A)とタンパク質(B)とが相互作用し、タンパク質(A)とタンパク質(C)とが相互作用する。この場合に、競合タンパク質とは、タンパク質(B)とタンパク質(C)との関係において互いのタンパク質をいう。ここでタンパク質(A)−(B)間の結合定数をKA−B、タンパク質(A)−(C)間の結合定数をKA−Cとすると、KA−BがKA−Cより小さいとき(図1の右図)、Gβγ複合体は細胞質にトラップされて接合シグナルの伝達が阻害される。一方、KA−BがKA−Cより大きいとき(図1の左図)、Gβγ複合体は細胞膜に結合できるため、接合シグナルが伝達される。このため、接合シグナル伝達により、タンパク質(A)−タンパク質(B)間の相互作用を検出することによって、標的タンパク質(A)に対して、競合タンパク質(C)と比べて、より親和性の高い(結合定数の大きい)結合タンパク質(B)を検出またはスクリーニングすることが可能となる。
【0018】
ここで、タンパク質間相互作用とは、タンパク質とタンパク質との間の結合または親和性をいう。
【0019】
本発明の酵母を用いるタンパク質間相互作用検出方法は、第1の実施態様では、(1)第1のタンパク質とGタンパク質γサブユニット細胞膜非結合変異型との融合タンパク質を発現する第1の発現カセットを調製する工程、(2)第2のタンパク質と細胞膜局在化シグナル配列との融合タンパク質を発現する第2の発現カセットを調製する工程、(3)第3のタンパク質を細胞質に発現する第3の発現カセットを調製する工程、(4)該工程(1)、(2)および(3)でそれぞれ得られた該第1、2および3の発現カセットを酵母に導入して形質転換酵母を調製する工程、および、(5)該工程(4)で得られた該形質転換酵母におけるフェロモンシグナル伝達を検出する、またはフェロモンシグナル伝達を示す形質転換酵母をスクリーニングする工程を含む。
【0020】
上記の方法において得られた形質転換酵母は、シグナル伝達能を評価できるよう活性化する必要がある。活性化の方法は、Gタンパク質の上流側を活性化するものであれば、特に制限されない。例えば、酵母内在性のGタンパク質共役型受容体に対するリガンド(アゴニスト)を加える方法が挙げられる。a接合型細胞の場合、例えば、配列番号1に示される塩基配列を有する遺伝子(GenBankアクセッション番号:NC001148)にコードされ、例えば、配列番号2に示されるアミノ酸配列(GenBankアクセッション番号:NP015137)を有する前駆体であるMF(alpha)1から生産される13アミノ酸のペプチドであるαファクターを加える方法が挙げられる。
【0021】
フェロモンシグナル伝達を検出する方法またはフェロモンシグナル伝達を示す形質転換酵母をスクリーニングする方法は特に制限されない。好ましくは、フェロモンシグナル応答性のプロモーター(例えば、FUS1、FIG1、AGA1など)の制御下でレポーター遺伝子の発現が誘導されるよう修飾する。レポーター遺伝子としては、例えば、蛍光レポーター遺伝子、生育レポーター遺伝子、発色レポーター遺伝子、発光レポーター遺伝子などが挙げられる。蛍光レポーター遺伝子としては、例えば、緑色蛍光蛋白質(GFP)、赤色蛍光蛋白質(DsRed)、シアン色蛍光蛋白質(CFP)などが挙げられる。生育レポーター遺伝子としては、例えば、HIS3、ADE2、URA3、CAN1などが挙げられる。発色レポーター遺伝子としては、例えば、βガラクトシダーゼ(lacZ)などが挙げられる。発光レポーター遺伝子としては、例えば、ルシフェラーゼ(luc)などが挙げられる。また、フェロモンシグナル伝達によって誘導される細胞周期抑制を利用してもよい。シグナルによって細胞周期が抑制された酵母は増殖が阻害されるため、固体培地上でのハロ形成や液体培地中での菌体密度を測定することにより、シグナル伝達能を検出することまたはシグナル伝達能を有する形質転換酵母をスクリーニングすることができる。
【0022】
本発明の酵母を用いるタンパク質間相互作用検出方法の第2の実施態様は、(1)第1のタンパク質とGタンパク質γサブユニット細胞膜非結合変異型との融合タンパク質を発現する第1の発現カセットを調製する工程、(2)第2のタンパク質と細胞膜局在化シグナル配列との融合タンパク質を発現する第2の発現カセットを調製する工程、(3)第3のタンパク質を細胞質に発現する第3の発現カセットを調製する工程、(4)該工程(1)、(2)および(3)でそれぞれ得られた該第1、2および3の発現カセットを1倍体酵母に導入して形質転換酵母を調製する工程、(5)該工程(4)で得られた該形質転換酵母を、該形質転換酵母と接合可能な1倍体酵母と接合させて2倍体酵母を調製する工程、および(6)該工程(5)で得られた該2倍体酵母を選択する工程を含む。
【0023】
上記の第2の実施態様において、形質転換に用いる1倍体酵母は、接合能を有する必要がある。好ましくはa/α接合型のa型またはα型の1倍体酵母である。いずれも、接合により生成される2倍体酵母を1倍体酵母と区別して選択できるように、適切な選択マーカーを有する必要がある。また、内在性のGタンパク質γサブユニットをコードする遺伝子が欠損している酵母が好ましい。内在性のGタンパク質γサブユニットをコードする遺伝子が欠損していない酵母を用いる場合には、上記第1の発現カセットを1倍体酵母に導入して形質転換酵母を調製する工程において、相同組換えにより内在性Gタンパク質γサブユニットと第1の発現カセットとを置き換える必要がある。酵母の種としては、例えば、サッカロミセス・セレビシエ(Saccharomyces cerevisiae)、シゾサッカロミセス・ポンベ(Schizosaccharomyces pombe)が挙げられる。
【0024】
選択マーカーとしては、例えば、栄養要求性マーカー、薬剤耐性マーカーが挙げられる。栄養要求性マーカーとしては、例えば、各種アミノ酸要求性マーカー、各種核酸要求性マーカーなどが挙げられる。アミノ酸要求性マーカーとしては、例えば、リジン要求性マーカー、メチオニン要求性マーカー、ヒスチジン要求性マーカー、トリプトファン要求性マーカー、ロイシン要求性マーカーなどが挙げられる。核酸要求性マーカーとしては、例えば、ウラシル要求性マーカー、アデニン要求性マーカーなどが挙げられる。薬剤耐性マーカーとしては、例えば、ジェネティシン(Geneticin:G418)耐性マーカー、オーレオバシジンA(Aureobasidin A)耐性マーカーなどが挙げられる。接合する1対の1倍体酵母は、接合により生成される2倍体酵母を1倍体酵母と区別して選択できるように、互いに異なる選択マーカーを有する必要がある。
【0025】
ここで、1倍体とは、ゲノムの1セットを有する酵母の個体をいい、半数体ともいう。本発明では、第1のタンパク質(A)、第2のタンパク質(B)および第3のタンパク質(C)の遺伝子を1倍体の酵母に導入する。
【0026】
2倍体とは、ゲノムの2セットを有する酵母の個体をいい、倍数体ともいう。1倍体どうしの接合により生成する。本発明では、2倍体の生成効率により、タンパク質間相互作用を検出またはスクリーニングする。
【0027】
本発明で用いるGタンパク質は、αサブユニット、βサブユニットおよびγサブユニットが形成する複合体からなるヘテロ三量体Gタンパク質のことをいい、Gタンパク質とはグアニンヌクレオチド結合タンパク質の略称である。αサブユニットとγサブユニットとはそれぞれ細胞膜に結合している。フェロモンが受容体に結合すると、Gタンパク質はαサブユニットとβγ複合体に解離する。Gタンパク質の解離後においてもαサブユニットとβγ複合体は細胞膜に局在化する。βγ複合体のβサブユニットが細胞膜を介してエフェクターに作用することでシグナルが下流に伝達される。Gタンパク質の由来としては、接合シグナル伝達に関与する限り、特に制限されない。
【0028】
本発明で用いるGタンパク質のγサブユニットとしては、例えば、配列番号3に示される塩基配列を有する遺伝子(GenBankアクセッション番号:AY557888)にコードされ、例えば、配列番号4に示されるアミノ酸配列(GenBankアクセッション番号:CAA89613)を有するタンパク質であるSte18が挙げられる。γサブユニット細胞膜非結合変異型(Gγcyto)は、細胞膜結合能を欠損している限り、特に制限されない。Gγcytoとしては、例えば、γサブユニットSte18の配列番号4に示されるアミノ酸配列の第106位〜第110位が欠失している変異型が挙げられる。
【0029】
本発明で第1のタンパク質(A)とGγcytoとを融合させる方法は、Gγcytoの機能が発揮できる限り、特に制限されない。例えば、γサブユニットSte18の配列番号4に示されるアミノ酸配列の第105位に、タンパク質(A)のN末端を連結する。
【0030】
本発明で用いる細胞膜局在化シグナル配列は、第2のタンパク質(B)を細胞膜に局在化することができる限り、特に制限されない。好ましくは、Gタンパク質のγサブユニットの細胞膜局在化シグナル配列である。例えば、このシグナルは、γサブユニットSte18の配列番号4に示されるアミノ酸配列の第102位〜第110位に相当する。
【0031】
本発明で第2のタンパク質(B)と細胞膜局在化シグナル配列とを融合させる方法は、細胞膜局在化シグナル配列の機能が発揮できる限り、特に制限されない。例えば、γサブユニットSte18の配列番号4に示されるアミノ酸配列の第102位〜第110位からなる断片をタンパク質(B)のC末端に連結する。
【0032】
本発明でタンパク質(C)を細胞質に発現させる方法は、特に制限されない。
【0033】
タンパク質(A)−Gγcyto融合タンパク質、細胞膜結合性タンパク質(B)および細胞質性タンパク質(C)を酵母で発現させるための発現カセットは、通常、これらのタンパク質のコード領域のほかに、酵母で機能するプロモーター、ターミネーターなどの遺伝子発現調節領域、発現カセットが導入された形質転換酵母を選択するための選択マーカーなどを有する。プロモーターとしては、例えば、PGK1プロモーター、TDH3プロモーター、STE18プロモーターが挙げられる。ターミネーターとしては、例えば、PGK1ターミネーター、TDH3ターミネーター、CYC1ターミネーターが挙げられる。選択マーカーとしては、例えば、ロイシン要求性マーカー、ウラシル要求性マーカー、ジェネティシン(Geneticin:G418)耐性マーカーが挙げられる。
【0034】
発現カセットは、通常、プラスミドとして作製される。プラスミドは、発現カセットのほかに、発現カセットを相同組換えにより酵母染色体に組み込むために必要な断片、またはプラスミドを酵母内で自律複製させるために必要な複製起点を有する。相同組換えに必要な断片としては、組み込みたい染色体上の位置に存在する遺伝子の断片が好ましい。発現カセットの相同組換えによる酵母染色体への組み込みは、当業者が通常用いる方法により行われる。酵母内複製起点としては、例えば、2μ(マルチコピー型)、CEN/ARS(シングルコピー型)が挙げられる。プラスミドの酵母内での自律複製は、当業者が通常用いる方法により行われる。プラスミドは、当業者が通常用いる方法により、大腸菌などの適切な宿主内で増幅され、調製される。プラスミドを調製するための宿主は特に制限されない。好ましくは、大腸菌である。プラスミドはまた、通常、プラスミドの調製を容易にするために、プラスミドが導入された形質転換大腸菌などを選択するための選択マーカーを有する。選択マーカーとしては、例えば、アンピシリン耐性、カナマイシン耐性などの抗生物質耐性が挙げられる。
【0035】
発現カセットは、プラスミドの形状で酵母に導入されてもよく、発現カセットのみが酵母に導入されてもよい。発現カセットは、相同組換えにより、効率的に酵母染色体に組み込むことができる。プラスミドは、選択マーカーに対応する選択条件下で形質転換酵母の培養を継続することにより、酵母内で自律複製させることができる。
【0036】
発現カセットが導入された形質転換酵母は、選択マーカーに対応する選択条件下、すなわち選択マーカーに対応するアミノ酸もしくは核酸の非存在下の培養により、または薬剤存在下の培養により得ることができる。
【0037】
本発明の方法の第2の実施態様では、発現カセットが導入された形質転換酵母と、対応する接合型の1倍体酵母、好ましくは野生型の1倍体酵母とを接合させる。ここで用いる1倍体酵母は、選択マーカーを有し、かつ接合可能であれば、特に制限されない。例えば、発現カセットが導入された形質転換酵母がa型由来であれば、これを対応するa/α接合型のα型の野生酵母と接合させる。1倍体野生酵母は、形質転換酵母との接合により生成する2倍体酵母のみを選択できる選択マーカーを有し、形質転換酵母との接合能を有する限り、特に制限されない。接合により生成する2倍体酵母は、上記選択マーカーを利用することにより、確認・選択することができる。
【0038】
本発明では、接合効率を適宜定量化することもできる。例えば、接合させた酵母の懸濁液を適宜希釈して選択用固体培地上にコロニーを形成させて、コロニー数を計測することにより定量化できる。
【0039】
本発明では、第1のタンパク質(A)、第2のタンパク質(B)および第3のタンパク質(C)は、特定のタンパク質であってもよく、不特定のタンパク質であってもよい。不特定のタンパク質とは、ライブラリを構成するタンパク質などをいう。第3のタンパク質(C)として、第1のタンパク質(A)または第2のタンパク質(B)の競合タンパク質を用いる点が特徴である。この特徴により、特定の第1のタンパク質(A)を用い、ライブラリを構成する第2のタンパク質(B)を用いる場合、第1のタンパク質(A)に対して、競合タンパク質である第3のタンパク質(C)と比べて、より親和性の高い(結合定数の大きい)第2のタンパク質(B)をライブラリの中から検出またはスクリーニングすることが可能となる。
【実施例】
【0040】
以下に、実施例を示して本発明をさらに具体的に説明するが、本発明はこれらに限定されない。
【0041】
(培地)
1%(w/v)酵母エキストラクト、2%(w/v)ペプトンおよび2%(w/v)グルコースを含むYPD培地、または0.67%(w/v)アミノ酸不含酵母ニトロゲンベース(Becton Dickinson社製)および2%(w/v)グルコースを含むSD培地で酵母を培養した。SD培地には選択マーカーに対応するアミノ酸および核酸を適宜添加した。2%(w/v)寒天をこれらの培地に添加してYPDおよびSDの固体培地を調製した。
【0042】
(酵母)
酵母サッカロミセス・セレビシエ(Saccharomyces cerevisiae)の1倍体株BY4741(メチオニン要求性a細胞)およびBY4742(リジン要求性α細胞)、ならびに以下の実施例で用いた酵母株および作製した酵母株の遺伝子型を表1に、そして各株において発現するタンパク質を表2に示す。a型のBY4741を親株として遺伝子組換えにより作製された形質転換株MC−F1は、フェロモン誘導性FIG1プロモーターの制御下においてEGFPレポーター遺伝子を発現する。MC−F1を親株として遺伝子組換えにより作製された形質転換株BFG2Z18−K35A、BFG2Z18−WTおよびBZFG2118はいずれも、タンパク質(A)として、ヒトIgGのFc部分をGタンパク質γサブユニットの細胞膜非結合変異型(Gγcyto)との融合タンパク質(Gγcyto−Fc)として細胞質に発現する。BFG2Z18−K35A、BFG2Z18−WTおよびBZFG2118は、タンパク質(B)として、ヒトIgGのFc部分に対する異なる親和性を有するスタフィロコッカス・アウレウス(Staphylococcus aureus)のプロテインA由来のZドメインを細胞膜に結合して発現する。BFG2Z18−K35Aは、結合定数4.6×106M-1のZドメイン変異型(ZK35A,mem)を発現し、BFG2Z18−WTは、結合定数5.9×107M-1のZドメイン野生型(ZWT,mem)を発現し、そしてBZFG2118は、結合定数6.8×108M-1のZドメイン2量体(ZZmem)を発現する。BFG2118は、ネガティブコントロールであり、Gγcyto−Fcのみを発現する。
【0043】
【表1】
【0044】
【表2】
【0045】
(参考例)
(接合能を指標とするタンパク質間相互作用検出方法)
BFG2Z18−K35A、BFG2Z18−WT、BZFG2118およびBFG2118の各酵母株を初期菌体濃度OD600=0.1で5mLのYPD培地に播種し、無処置の接合相手BY4742と30℃にて3時間共培養した。培養後、酵母を回収し、洗浄後、蒸留水に再懸濁した。各株の接合能を評価するために、酵母懸濁液のOD600=1.0、0.1および0.001の希釈系列を調製し、メチオニンおよびリジンを含まず、20mg/Lのヒスチジン、30mg/Lのロイシンおよび20mg/Lウラシルを含む2倍体選択用SD固体培地上(SD−Met、Lysプレート)にスポットした。この結果、BFG2118は増殖しなかったが、他の3つの株は増殖した(図2A)。
【0046】
増殖した3つの株では、タンパク質(A)とタンパク質(B)との間には相互作用が存在するため、Gタンパク質を介するフェロモンシグナル伝達によるa型とα型との接合が生じたものと思われる。一方、BFG2118では、タンパク質(B)を欠くため、このような相互作用は存在せず、増殖が見られなかった。このように、1倍体酵母のa型とα型とのフェロモンシグナル伝達を介する接合能は、タンパク質(A)とタンパク質(B)との相互作用の有無を検出する方法として有効であることが示された。
【0047】
次に、増殖した3つの株の接合能を定量的に評価するために、菌体濃度OD600=1.0に調整した1mLの酵母懸濁液を、プレート上に100〜1000コロニーが成長するように、適宜希釈してSD−Met、Lysプレートに塗布した。コロニー数を計測し、計測したコロニー数に希釈率を乗じて、菌体濃度OD600=1.0に調整した1mLの酵母懸濁液が生成し得る2倍体酵母のコロニー数を求めた。結果を図2Bに示す。図2B中のエラーバーは、3度の独立した実験結果における標準偏差を示す。
【0048】
図2Bからわかるように、タンパク質(A)とタンパク質(B)との間の結合定数に対応して生成し得る2倍体酵母のコロニー数に差異があった。このように、1倍体酵母のa型とα型とのフェロモンシグナル伝達を介する接合能は、タンパク質(A)とタンパク質(B)との相互作用を定量的に評価する方法としても有効であることが示された。
【0049】
(実施例1)
(細胞質で競合タンパク質(C)を発現する形質転換株の作製)
細胞質での競合タンパク質(C)の発現がGタンパク質を介する接合シグナル伝達の回復を阻害するかどうか調べるために、BFG2Z18−K35A、BFG2Z18−WTおよびBZFG2118を遺伝子組換えし、競合タンパク質(C)として、Zドメイン野生型(ZWT)またはZドメイン変異型(ZK35A)を細胞質に発現する形質転換株を作製した。
【0050】
細胞質で競合タンパク質として発現させるZドメイン野生型遺伝子(ZWT)およびZドメイン変異型遺伝子(ZK35A)を酵母染色体上のHOP2遺伝子の上流(PHOP2:HOP2プロモーター領域)に相同組換えにより組み込むため、次のプラスミドを構築した。ZWTおよびZK35Aを、プラスミドpUMZ−WTおよびpUMZ−K35Aからプライマー1(配列番号5)およびプライマー2(配列番号6)を用いてPCRにより増幅し、プラスミドpGK425のSalI−BamHI部位間に挿入し、それぞれプラスミドpLMZ−WTおよびpLMZ−K35Aとした。HOP2プロモーター領域での相同組換えに用いるDNA断片を、MC−F1のゲノムDNAからプライマー3(配列番号7)およびプライマー4(配列番号8)を用いてPCRにより増幅し、プラスミドpLMZ−WTおよびpLMZ−K35AのNotI−SacI部位間にそれぞれ挿入し、それぞれプラスミドpLMZ−WT−HおよびpLMZ−K35A−Hとした。
【0051】
次いで、LEU2−PGK5’−Z−PGK3’−PHOP2(PGK5’:PGK1プロモーター;PGK3’:PGK1ターミネーター)を含むDNA断片をpLMZ−WT−HおよびpLMZ−K35A−HからPHOP2のすぐ上流領域に相同な領域を含む50塩基のプライマー5(配列番号9)およびプライマー6(配列番号10)を用いてPCRにより増幅した。増幅したDNA断片をBGF2Z18−K35A、BFG2Z18−WTおよびBZFG2118の形質転換に用いた。形質転換では、酢酸リチウム法を用いた。形質転換体は、ウラシルおよびロイシンを含まず、20mg/Lのヒスチジンおよび30mg/Lのメチオニンを含むSD培地(SD−Ura、Leuプレート)で選択し、FC1−1、FC2−1、FC3−1、FC1−2、FC2−2およびFC3−2株を得た(表1)。
【0052】
(実施例2)
(細胞質で競合タンパク質(C)を発現する形質転換株における蛍光レポーター遺伝子を指標とするタンパク質間相互作用検出方法)
実施例1で得られた形質転換株の蛍光レポーター遺伝子の発現を評価した。競合タンパク質を発現しない例として、BFG2Z18−K35A、BFG2Z18−WTおよびBZFG2118を用いた。競合タンパク質としてZK35Aを発現する例として、FC1−1、FC2−1およびFC3−1を用いた。競合タンパク質としてZWTを発現する例として、FC1−2、FC2−2およびFC3−2を用いた。各酵母株を初期菌体濃度OD600=0.1で5mLのYPD培地に播種した後、最終濃度5μMとなるようαファクターを添加し、30℃にて6時間培養した。各株の蛍光レポーター遺伝子発現を評価するために、培養後、酵母を回収し、フローサイトメーターを用いて細胞の蛍光強度を評価した。結果を図3に示す。図3のグラフにおいて並立する2本カラムのうち、左(濃色)のカラムは競合タンパク質を導入していない酵母株、および右(淡色)のカラムは競合タンパク質を導入した酵母株の結果を示す。図3中のエラーバーは、3度の独立した実験結果における標準偏差を示す。
【0053】
この結果、競合タンパク質(C)を発現しない酵母株BFG2Z18−K35A、BFG2Z18−WTおよびBZFG2118はすべて蛍光を示した。一方で、競合タンパク質(C)ZK35Aを発現する酵母株においては、結合定数の小さい第2のタンパク質(B)を発現しているFC1−1は全く蛍光を示さず、より結合定数の大きい第2のタンパク質(B)を発現しているFC2−1およびFC3−1のみ蛍光を示した(図3A)。同様に、競合タンパク質(C)ZWTを発現する酵母株においては、結合定数の小さい第2のタンパク質(B)を発現しているFC1−2およびFC2−2は全く蛍光を示さず、より結合定数の大きい第2のタンパク質(B)を発現しているFC3−2のみ蛍光を示した(図3B)。
【0054】
(実施例3)
(細胞質で競合タンパク質(C)を発現する形質転換株における接合能を指標とするタンパク質間相互作用検出方法)
実施例1で得られた形質転換株の接合能を参考例と同様に評価した。結果を図4に示す。図4のグラフにおいて並立する2本カラムのうち、左(濃色)のカラムは競合タンパク質を導入していない酵母株、および右(淡色)のカラムは競合タンパク質を導入した酵母株の結果を示す。図4中のエラーバーは、3度の独立した実験結果における標準偏差を示す。
【0055】
この結果、競合タンパク質ZK35Aを発現するFC2−1およびFC3−1は、2倍体選択用培地で増殖した(図4A)。FC2−1およびFC3−1の接合能は、競合タンパク質(C)を発現しない酵母株とほとんど同等であった(図4C)。同様の結果が、競合タンパク質(C)ZWTを発現する酵母株についても得られた。FC1−2およびFC2−2は、接合能を示さなかったが、FC3−2のみ、競合タンパク質(C)を発現しない酵母株とほとんど同等の接合能を示した(図4Bおよび4D)。
【0056】
(実施例4)
(モデルライブラリからの標的細胞のスクリーニング)
細胞質で競合タンパク質(C)を発現する形質転換株における接合能を指標とするタンパク質間相互作用検出方法が有効かどうかを、モデルライブラリを用いて検証した。モデルライブラリとして、標的細胞(FC3−2またはBZFG2118)とコントロール細胞(FC2−2またはBFGZ18−WT)とが表3に示す初期割合で混合された2つのライブラリを調製した。一方は、競合タンパク質(C)を発現していない酵母株からなり、ZZを発現する少量のBZFG2118とZWTを発現する過剰量のBFG2Z18−WTとを含み、他方は、競合タンパク質(C)を発現している酵母株からなり、ZZを発現する少量のFC3−2とZWTを発現する過剰量のFC2−2とを含む。これらのライブラリは、各1倍体の初期菌体濃度をOD600=0.1に設定して、接合相手のBY4742と30℃にて3時間10mLのYPD培地で共培養した。培養後、酵母を回収し、洗浄後、SD−Met、Lysプレートに塗布し、30℃にて2日間培養した。10コロニーをピックアップし、YPD培地中で一晩別々に増殖させた。ゲノムを抽出し、タンパク質(B)のコード領域をプライマー7(配列番号11)およびプライマー8(配列番号12)を用いてPCRにより増幅した。増幅した断片がZZまたはZWTのいずれに由来するかは、断片の長さで判断した。10コロニー中、ZZの断片が得られたコロニーの割合を、標的細胞の最終割合とした。標的細胞の最終割合を初期割合で除した値をスクリーニング効率として算出し、評価した。結果を表3に示す。
【0057】
【表3】
【0058】
表3に示すように、競合タンパク質(C)を用いるタンパク質間相互作用検出方法は、競合タンパク質を用いない従来の方法よりも明らかに優れたスクリーニング効率を示し、最大で7000倍に達した。
【0059】
これらの結果は、競合タンパク質(C)を用いるタンパク質間相互作用検出方法は、標的タンパク質(A)に対して、もとの結合タンパク質(B)と比べて、より高い親和性を有する結合タンパク質(B)を検出するのに適していることを示唆する。
【産業上の利用可能性】
【0060】
本発明によれば、親和性の高いタンパク質間相互作用を効率よく検出またはスクリーニングすることができる。
【特許請求の範囲】
【請求項1】
酵母を用いるタンパク質間相互作用検出方法であって、
(1)第1のタンパク質とGタンパク質γサブユニット細胞膜非結合変異型との融合タンパク質を発現する第1の発現カセットを調製する工程、
(2)第2のタンパク質と細胞膜局在化シグナル配列との融合タンパク質を発現する第2の発現カセットを調製する工程、
(3)第3のタンパク質を細胞質に発現する第3の発現カセットを調製する工程、
(4)該工程(1)、(2)および(3)でそれぞれ得られた該第1、2および3の発現カセットを酵母に導入して形質転換酵母を調製する工程、および
(5)該工程(4)で得られた該形質転換酵母におけるフェロモンシグナル伝達を検出する、またはフェロモンシグナル伝達を示す形質転換酵母をスクリーニングする工程
を含む、方法。
【請求項2】
酵母を用いるタンパク質間相互作用検出方法であって、
(1)第1のタンパク質とGタンパク質γサブユニット細胞膜非結合変異型との融合タンパク質を発現する第1の発現カセットを調製する工程、
(2)第2のタンパク質と細胞膜局在化シグナル配列との融合タンパク質を発現する第2の発現カセットを調製する工程、
(3)第3のタンパク質を細胞質に発現する第3の発現カセットを調製する工程、
(4)該工程(1)、(2)および(3)でそれぞれ得られた該第1、2および3の発現カセットを1倍体酵母に導入して形質転換酵母を調製する工程、
(5)該工程(4)で得られた該形質転換酵母を、該形質転換酵母と接合可能な1倍体酵母と接合させて2倍体酵母を調製する工程、および
(6)該工程(5)で得られた該2倍体酵母を選択する工程
を含む、方法。
【請求項3】
前記第3のタンパク質が、第1のタンパク質または第2のタンパク質の競合タンパク質である、請求項1または2に記載の方法。
【請求項1】
酵母を用いるタンパク質間相互作用検出方法であって、
(1)第1のタンパク質とGタンパク質γサブユニット細胞膜非結合変異型との融合タンパク質を発現する第1の発現カセットを調製する工程、
(2)第2のタンパク質と細胞膜局在化シグナル配列との融合タンパク質を発現する第2の発現カセットを調製する工程、
(3)第3のタンパク質を細胞質に発現する第3の発現カセットを調製する工程、
(4)該工程(1)、(2)および(3)でそれぞれ得られた該第1、2および3の発現カセットを酵母に導入して形質転換酵母を調製する工程、および
(5)該工程(4)で得られた該形質転換酵母におけるフェロモンシグナル伝達を検出する、またはフェロモンシグナル伝達を示す形質転換酵母をスクリーニングする工程
を含む、方法。
【請求項2】
酵母を用いるタンパク質間相互作用検出方法であって、
(1)第1のタンパク質とGタンパク質γサブユニット細胞膜非結合変異型との融合タンパク質を発現する第1の発現カセットを調製する工程、
(2)第2のタンパク質と細胞膜局在化シグナル配列との融合タンパク質を発現する第2の発現カセットを調製する工程、
(3)第3のタンパク質を細胞質に発現する第3の発現カセットを調製する工程、
(4)該工程(1)、(2)および(3)でそれぞれ得られた該第1、2および3の発現カセットを1倍体酵母に導入して形質転換酵母を調製する工程、
(5)該工程(4)で得られた該形質転換酵母を、該形質転換酵母と接合可能な1倍体酵母と接合させて2倍体酵母を調製する工程、および
(6)該工程(5)で得られた該2倍体酵母を選択する工程
を含む、方法。
【請求項3】
前記第3のタンパク質が、第1のタンパク質または第2のタンパク質の競合タンパク質である、請求項1または2に記載の方法。
【図1】

【図2】
【図3】
【図4】
【図2】
【図3】
【図4】
【公開番号】特開2011−155839(P2011−155839A)
【公開日】平成23年8月18日(2011.8.18)
【国際特許分類】
【出願番号】特願2010−17509(P2010−17509)
【出願日】平成22年1月28日(2010.1.28)
【出願人】(504150450)国立大学法人神戸大学 (421)
【Fターム(参考)】
【公開日】平成23年8月18日(2011.8.18)
【国際特許分類】
【出願日】平成22年1月28日(2010.1.28)
【出願人】(504150450)国立大学法人神戸大学 (421)
【Fターム(参考)】
[ Back to top ]