
高親和性HIVT細胞レセプター
【課題】SLYNTVATL−HLA-A*0201複合体に対して、1μMより少ないか等しい親和性(KD)及び/又は1×10-3 S-1か若しくはそれより遅い解離速度(koff)を有するTCRを提供する。
【解決手段】特定のアミノ酸配列を有するTCRα鎖可変領域及び特定のアミノ酸配列を有するTCRβ鎖可変領域を含んでなるT細胞レセプター(TCR)。TCRが細胞により提示され、特定のアミノ酸配列を含んでなるときは、該細胞は天然型T細胞ではない。
【解決手段】特定のアミノ酸配列を有するTCRα鎖可変領域及び特定のアミノ酸配列を有するTCRβ鎖可変領域を含んでなるT細胞レセプター(TCR)。TCRが細胞により提示され、特定のアミノ酸配列を含んでなるときは、該細胞は天然型T細胞ではない。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、HIV Gagポリペプチド由来SLYNTVATL−HLA-A*0201への結合特性を有するT細胞レセプター(TCR)に関する。このTCRは、少なくとも1つのTCRα鎖可変ドメイン及び/又は少なくとも1つのTCRβ鎖可変ドメインを含んでなり、1μMより少ないか若しくは等しいSLYNTVATL−HLA-A*0201複合体に対するKD及び/又は1×10-3 S-1か若しくはそれより遅いSLYNTVATL−HLA-A*0201複合体に対する解離速度(off-rate)(koff)を有する。
【背景技術】
【0002】
(発明の背景)
ヒト免疫不全ウイルス(HIV)は、後天性免疫不全症候群(AIDS)の原因因子である。このウイルスは、レンチウイルス群に属するエンベロープを有するレトロウイルスである。SLYNTVATL(配列番号16)ペプチドは、ヒト免疫不全ウイルス-1(HIV-1)を作り上げている9つの遺伝子の1つであるGag遺伝子のg17遺伝子産物に由来する。このペプチドはHLA-A*0201に搭載(load)されてHIV感染細胞の表面に提示される。したがって、SLYNTVATL−HLA-A2*0201複合体は、例えば細胞毒性又は免疫刺激性薬剤を感染細胞に送達する目的で、TCRが標的することができるHIVマーカーを提供する。しかし、この目的には、TCRがペプチド−HLA複合体に対して高い親和性及び/又は遅い解離速度を有することが望ましい。
【発明の概要】
【課題を解決するための手段】
【0003】
(発明の簡単な説明)
本発明は、SLYNTVATL−HLA-A*0201複合体に対し、1μMより少ないか若しくはこれと等しい親和性(KD)及び/又は1×10-3 S-1か若しくはそれより遅い解離速度(koff)を有するTCR(但し、該TCRが細胞により提示され且つ配列番号1及び2を含んでなる場合には、該細胞は天然型T細胞ではない)を初めて利用可能にする。このTCRは、単独で又は治療剤との組合せで、前記複合体を提示するHIV感染細胞を標的するに有用である。
【図面の簡単な説明】
【0004】
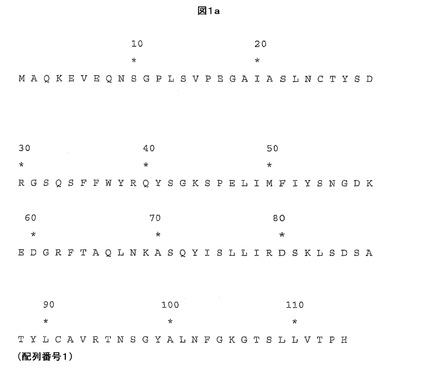
【図1a】親HIV Gag TCRのα鎖可変ドメインアミノ酸配列を示す。
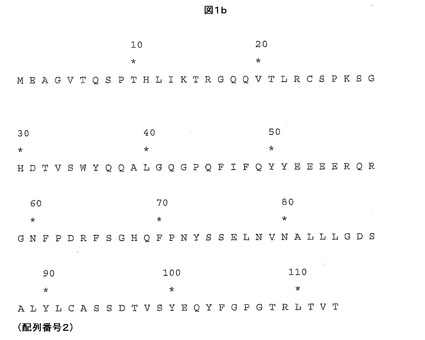
【図1b】親HIV Gag TCRのβ鎖可変ドメインアミノ酸配列を示す。
【図2】可溶性型の親HIV Gag TCRα鎖(a)及びβ鎖(b)のDNA配列を示す。
【図3】図2のDNA配列から産生されるHIV Gag TCRα鎖(a)及びβ鎖(b)の細胞外アミノ酸配列を示す。
【0005】
【図4】非天然型ジスルフィド結合を形成するための追加のシステイン残基をコードするように変異した可溶性型のHIV Gag TCRα鎖(a)及びβ鎖(b)のDNA配列を示す。
【図5】図4のDNA配列から産生されるHIV Gag TCRα鎖(a)及びβ鎖(b)の細胞外アミノ酸配列を示す。
【図6】高親和性HIV Gag TCR変形体のα鎖可変ドメインアミノ酸配列を示す。
【図7】高親和性HIV Gag TCR変形体のβ鎖可変ドメインアミノ酸配列を示す。
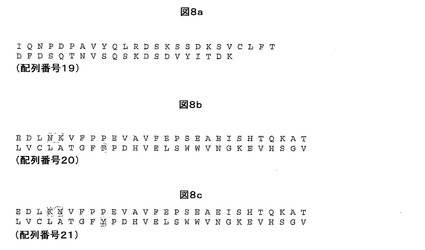
【図8】TRAC(a)、TRBC1(b)及びTRBC2(c)の可溶性部分のアミノ酸配列を示す。
【0006】
【図9−1】pEX954プラスミドのDNA配列を示す。
【図9−2】図9−1の続きである。
【図10−1】pEX821プラスミドのDNA配列を示す。
【図10−2】図10−1の続きである。
【0007】
【図11】ペプチドリンカーを介して野生型ヒトIL-2に融合した可溶性の親HIV Gag TCR変形体のβ鎖アミノ酸配列を示す。
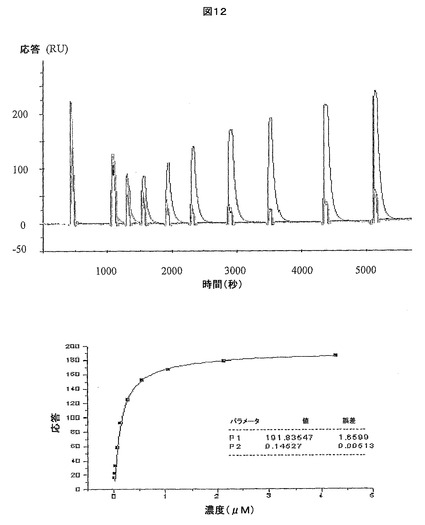
【図12】ジスルフィド連結した可溶性の親HIV Gag TCRとSLYNTVATL−HLA-A*0201複合体との相互作用について作成したBiacore応答曲線を示す。
【図13】pEX954プラスミドのプラスミドマップを示す。
【図14】pEX821プラスミドのプラスミドマップを示す。
【0008】
【図15】ヒトT細胞中での発現について最適化された親HIV Gag TCRα鎖(a)及びβ鎖(b)の全長DNA配列を示す。
【図16】a:親HIV Gag TCRα鎖の全長アミノ酸配列を示す。b:ヒトT細胞中での発現について最適化された親HIV Gag TCRβ鎖の全長アミノ酸配列を示す。
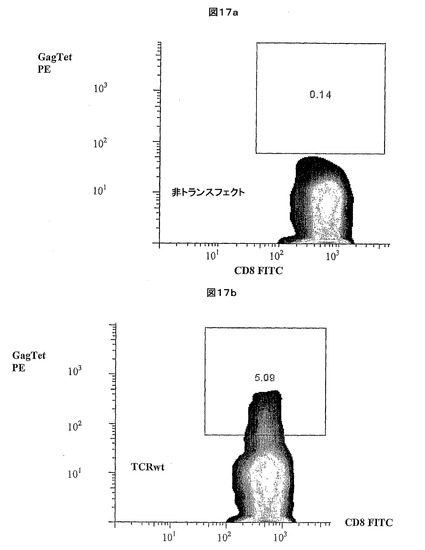
【図17】a:コントロールの非形質転換CD8+T細胞についてのFACS分析データを示す。b:形質転換CD8+T細胞の表面上での親HIV Gag TCRの発現を示すFACS分析データを示す。
【0009】
【図18】可溶性のジスルフィド連結高親和性c11c6 HIV Gag TCRα鎖(a)及びβ鎖(b)のアミノ酸配列を示す。
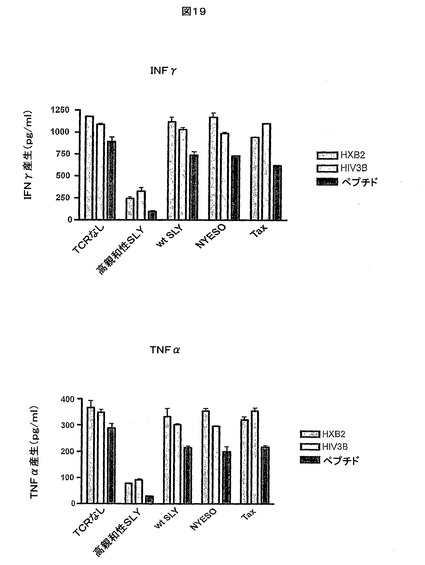
【図19】IFN-γ産生及びTNF-α産生により測定したときの、可溶性のジスルフィド連結高親和性c11c6 HIV Gag TCRがHIVに感染したTo細胞の存在下でSLYNTVATL−HLA-A*0201反応性OX84ポリクローナルT細胞株の活性化を阻害する能力を示す。
【0010】
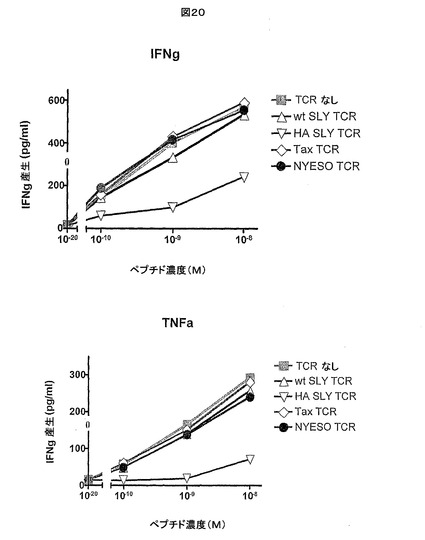
【図20】IFN-γ産生及びTNF-α産生により測定したときの、可溶性のジスルフィド連結高親和性c11c6 HIV Gag TCRがSLYNTVATLペプチドでパルスした非感染To細胞の存在下でSLYNTVATL−HLA-A*0201反応性OX84ポリクローナルT細胞株の活性化を阻害する能力を示す。
【図21】可溶性のジスルフィド連結高親和性c11c6 HIV Gag TCRがSLYNTVATLペプチドでパルスしたT2細胞を染色する能力を示す。
【発明を実施するための形態】
【0011】
(発明の詳細な説明)
本発明は、1μMより少ないか等しいSLYNTVATL−HLA-A*0201複合体に対するKD及び/又は1×10-3 S-1かそれより遅いSLYNTVATL−HLA-A*0201複合体に対する解離速度(koff)を有し、但し細胞により提示され且つ配列番号1及び2を含んでなる場合には該細胞は天然型T細胞ではないことを特徴とする、SLYNTVATL−HLA-A*0201への結合特性を有し且つ少なくとも1つのTCRα鎖可変ドメイン及び/又は少なくとも1つのTCRβ鎖可変ドメインを含んでなるT細胞レセプター(TCR)を提供する。KD及び/又は(koff)の測定は、公知の方法のいずれによっても行うことができる。好ましい方法は、実施例4の表面プラズモン共鳴(Biacore)法である。
【0012】
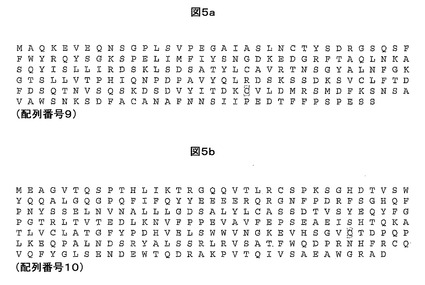
比較のために、親のHIV gag TCR(TCRα鎖については配列番号9を、TCRβ鎖については配列番号10を参照)のジスルフィド連結した可溶性変形体とSLYNTVATL−HLA-A*0201複合体との相互作用は、実施例4のBiacoreに基づく方法により測定すると、約85nMのKD及び2.21×10-2 S-1の解離速度(koff)を有する。
SLYNTVATL−HLA-A*0201複合体に特異的な親HIV Gag TCRは、以下のVα鎖遺伝子及びVβ鎖遺伝子使用(usage)を有する:
α鎖 − TRAV12.2
β鎖:− TRBV 5.6
【0013】
親HIV Gag TCRは、SLYNTVATL−HLA-A*0201複合体との間の相互作用に対して高い親和性及び/又は遅い解離速度を有する本発明の他のTCRを製造できる鋳型として使用することができる。したがって、本発明は、少なくとも1つの相補性決定領域(CDR)及び/又は可変ドメインフレームワーク領域において、親HIV Gag TCRのα鎖可変ドメイン(図1a及び配列番号1を参照)及び/又はβ鎖可変ドメイン(図1b及び配列番号2を参照)に対して変異したTCRを包含する。また、本発明のTCRの可変ドメイン中の他の超可変領域、例えば超可変4(HV4)領域が、高親和性変異体TCR内で変異していてもよいと企図される。
【0014】
ファージディスプレイは、TCR変形体のライブラリを作製し得る1つの手段を提供する。各々が非天然型ジスルフィド鎖間結合を含有するTCR変形体のライブラリのファージディスプレイ及びその後のスクリーニングに適切な方法は、Liら,(2005) Nature Biotech 23(3):349-354及びWO 2004/04404に詳述されている。
天然型TCRはへテロ二量体αβ又はγδ形態で存在する。しかし、単一のTCRα鎖又はTCRβ鎖からなる組換えTCRが、ペプチドMHC分子に結合することが以前に示されている。
1つの実施形態において、本発明のTCRは、α鎖可変ドメイン及びTCRβ鎖可変ドメインを両方含んでなる。
【0015】
当業者には自明であろうが、TCRα鎖配列及び/又はTCRβ鎖配列中の変異は、1又はそれより多い置換、欠失又は挿入であり得る。これら変異は、任意の適切な方法(ポリメラーゼ連鎖反応(PCR)に基づくもの、制限酵素ベースのクローニング、又はライゲーション非依存性クローニング(LIC)手法を包含するがこれらに限定されない)を使用して実行することができる。これら方法は、多くの標準的な分子生物学の教科書に詳述されている。ポリメラーゼ連鎖反応(PCR)変異誘発及び制限酵素ベースのクローニングに関する更なる詳細に関しては、Sambrook & Russell,(2001) Molecular Cloning - A Laboratory Manual (第3版) CSHL Pressを参照。LIC手順についての更なる情報は、Rashtchian(1995) Curr Opin Biotechnol 6(1):30-6に見出すことができる。
【0016】
HIV Gag TCRのものに類似するVα遺伝子及びVβ遺伝子使用、したがって類似するアミノ酸配列を含んでなる任意のαβTCRは、簡便な鋳型TCRを作り得ることに留意すべきである。次いで、鋳型αβTCRの可変ドメインの一方又は両方をコードするDNA中に、本発明の変異した高親和性TCRを作製するに必要な変更を導入することができる。当業者には自明であろうが、必要な変異は、多くの方法(例えば、部位特異的変異誘発)により導入し得る。
【0017】
本発明のTCRは、下記に列挙するものに対応するTCRα鎖可変ドメインアミノ酸の1又はそれより多くが、図1a及び配列番号1中の親HIV Gag TCRα鎖可変ドメインを提供する配列のこれら位置に存在するアミノ酸に対して変異しているTCRを包含する。
【0018】
特に反する記載がない限り、本明細書中のTCRアミノ酸配列は、一般には、N末端メチオニン(Met又はM)残基を含んで提供される。当業者には公知であろうが、この残基は、組換えタンパク質の産生の間に除去され得る。これもまた当業者には自明であろうが、TCRのpMHC結合特性に実質的に影響することなく、C末端及び/又はN末端に提供される配列を1、2、3、4、5又はそれより多くの残基だけ短縮することもまた可能である。全てのそのような平凡な(trivial)変形体は、本発明に包含される。
【0019】
本明細書中で使用する場合、用語「可変領域」は、所定のTCRの、TCRα鎖に関してはTRAC遺伝子により、TCRβ鎖に関してはTRBC1遺伝子又はTRBC2遺伝子のいずれかによりコードされる定常ドメイン内に含まれない全てのアミノ酸を包含すると理解される(T cell receptor Factsbook(2001) LeFranc and LeFranc,Academic Press,ISBN 0-12-441352-8)
。
本明細書で使用する場合、用語「可変ドメイン」は、所定のTCRの、TCRα鎖に関してはTRAV遺伝子により、TCRβ鎖に関してはTRBV遺伝子によりコードされる全てのアミノ酸を包含すると理解される(T cell receptor Factsbook(2001) LeFranc and LeFranc,Academic Press,ISBN 0-12-441352-8)。
【0020】
当業者には公知であるが、TCRレパートリの多様性の一部は、(本明細書中で定義するとおりの)可変領域と定常ドメインとの間の境界のコドンによりコードされるアミノ酸に生じる変化に起因する。例えば、親HIV Gag TCR配列のこの境界に存在するコドンは、可変領域配列のC末端でのヒスチジン(H)残基の存在を生じる。このヒスチジンは、図8aに示されるTRAC遺伝子によりコードされるN末端アスパラギン(N)残基に置き換わる。
【0021】
本発明の実施形態は、95T、96N、97S、98G及び100Aに相当するα鎖可変領域アミノ酸のうちの1又はそれより多くの変異、例えばアミノ酸:
95S又はG
96A
97H
98D
100S
を含んでなる変異したTCRを包含する。
上記で使用した番号付けは、図1a及び配列番号1に示したものと同じである。
【0022】
本発明の実施形態はまた、下記に列挙するものに対応するTCRβ鎖可変領域アミノ酸の1又はそれより多くの変異を含んでなるTCRを包含し、それらは図1b及び配列番号2の天然型HIV Gag TCRβ鎖の天然型HIV Gag TCRα鎖可変領域のために提供される配列中のこれら位置に存在するアミノ酸に関して変異している。変異してもよいとされるアミノ酸は、51Y、52E、53E及び54Eであり、例えば:
51V又はA
52R又はL
53G
54V
上記で使用した番号付けは、図1b及び配列番号2で示したものと同じである。
【0023】
本発明の更なる好ましい実施形態は、図6に示される変異したα鎖可変領域アミノ酸配列(配列番号11〜13)の1つを含んでなるTCRにより提供される。このようなTCRの表現型上サイレントな変形体もまた、本発明の一部を形成する。
本発明の追加の好ましい実施形態は、図7に示される変異したβ鎖可変領域アミノ酸配列(配列番号14及び15)の1つを含んでなるTCRにより提供される。このようなTCRの表現型上サイレントな変形体もまた、本発明の一部を形成する。
【0024】
天然型TCRはへテロ二量体αβ又はγδ形態で存在する。しかし、αα又はββのホモ二量体からなる組換えTCRが、ペプチドMHC分子に結合することが以前に示されている。したがって、本発明の1つの実施形態は、TCRαα又はTCRββのホモ二量体により提供される。
更に好ましい実施形態は、下記に列挙するα鎖可変領域アミノ酸配列とβ鎖可変領域アミノ酸配列との組合せを含んでなる本発明のTCRにより提供され、このようなTCRの表現型上サイレントな変形体もまた、本発明の一部を形成する:
【0025】
【表1】

【0026】
別の好ましい実施形態では、上記で詳述した可変領域の組合せを含んでなる本発明のTCRは、図8aに示されるα鎖定常ドメインアミノ酸配列(配列番号19)並びに図8b及び8cに示されるβ鎖アミノ酸定常ドメイン配列(配列番号20及び21)の一方、又はそれらの表現型上サイレントな変形体を更に含んでなる。
【0027】
本明細書中で使用される場合、用語「表現型上サイレントな変形体」は、SLYNTVATL−HLA-A*0201複合体に対して1μMより少ないか若しくはこれと等しいKDを有し及び/又は1×10-3 S-1か若しくはそれより遅い解離速度(koff)を有するTCRをいうと理解される。例えば、当業者には公知であるが、SLYNTVATL−HLA-A*0201複合体との相互作用に関する親和性及び/又は解離速度を変化させることなく、上記で詳述したものと比較して軽微な変化が定常ドメイン及び/又は可変領域に組み込まれているTCRを作製することは可能であり得る。このような平凡な変形体は本発明の範囲に包含される。1又はそれより多い保存的置換がなされたTCRもまた、本発明の一部を形成する。
【0028】
1つの広い観点では、本発明のTCRは、WO 04/033685及びWO 03/020763に記載されるような単鎖TCR(scTCR)又は二量体TCR(dTCR)のいずれかの形態である。
【0029】
適切なscTCR形態は、TCRα鎖可変領域に相当するアミノ酸配列により構成される第1のセグメントと、TCRβ鎖定常ドメイン細胞外配列に相当するアミノ酸配列のN末端に融合したTCRβ鎖可変領域配列に相当するアミノ酸配列により構成される第2のセグメントと、第1のセグメントのC末端を第2のセグメントのN末端に連結するリンカー配列とを含んでなる。
【0030】
或いは、第1のセグメントは、TCRβ鎖可変領域に相当するアミノ酸配列により構成されてもよく、第2のセグメントは、TCRα鎖定常ドメイン細胞外配列に相当するアミノ酸配列のN末端に融合したTCRα鎖可変領域に相当するアミノ酸配列により構成されてもよい。
【0031】
上記scTCRは、第1の鎖と第2の鎖との間のジスルフィド結合を更に含んでなってもよく、このジスルフィド結合は、天然型αβT細胞レセプター中に等価物がないものであり、リンカー配列の長さ及びジスルフィド結合の位置は、第1及び第2のセグメントの可変ドメイン配列が天然型αβT細胞レセプター中と実質的に同様に互いに配向するような長さ及び位置である。
【0032】
より具体的には、第1のセグメントは、TCRα鎖定常ドメイン細胞外配列に相当するアミノ酸配列のN末端に融合したTCRα鎖可変領域に相当するアミノ酸配列により構成されていてもよく、第2のセグメントは、TCRβ鎖定常ドメイン細胞外配列に相当するアミノ酸配列のN末端に融合したTCRβ鎖可変領域に相当するアミノ酸配列により構成されていてもよく、ジスルフィド結合は、第1の鎖と第2の鎖との間に提供されてもよく、このジスルフィド結合は、天然型αβT細胞レセプター中に等価物がないものである。
【0033】
上記scTCR形態では、リンカー配列は、第1のセグメントのC末端を第2のセグメントのN末端に連結してもよく、式-PGGG-(SGGGG)n-P-(式中、nは5又は6であり、Pはプロリンであり、Gはグリシンであり、Sはセリンである)を有していてもよい。
-PGGG-SGGGGSGGGGSGGGGSGGGGSGGGG-P (配列番号17)
-PGGG-SGGGGSGGGGSGGGGSGGGGSGGGGSGGGG-P (配列番号18)
【0034】
本発明のTCRの適切なdTCR形態は、TCRα鎖可変領域配列に相当する配列がTCRα鎖定常ドメイン細胞外配列に相当する配列のN末端に融合している第1のポリペプチドと、TCRβ鎖可変領域配列に相当する配列がTCRβ鎖定常ドメイン細胞外配列に相当する配列のN末端に融合している第2のポリペプチドとを含んでなり、第1及び第2のポリペプチドが天然型αβT細胞レセプター中に等価物がないジスルフィド結合により連結している。
【0035】
第1のポリペプチドは、TCRα鎖定常ドメイン細胞外配列に相当する配列のN末端に融合したTCRα鎖可変領域配列を含んでなってもよく、第2のポリペプチドでは、TCRβ鎖可変領域配列に相当する配列がTCRβ鎖定常ドメイン細胞外配列に相当する配列のN末端に融合し、第1及び第2のポリペプチドは、TRAC*01のエキソン1のThr 48及びTRBC1*01若しくはTRBC2*01のエキソン1のSer 57又はこれらの非ヒト等価物から置換したシステイン残基同士間のジスルフィド結合により連結されている(本明細書中の「TRAC」などの命名法は、T cell receptor Factsbook(2001) LeFranc and LeFranc,Academic Press,ISBN 0-12-441352-8による)。
【0036】
本発明のTCRのdTCR形態又はscTCR形態は、ヒトαβTCRの細胞外定常ドメイン及び可変領域の配列に相当するアミノ酸配列を有していてもよく、ジスルフィド結合は、前記定常ドメイン配列のアミノ酸残基同士を連結してもよく、このジスルフィド結合は、天然型TCR中には等価物がない。ジスルフィド結合は、β炭素原子が天然型TCR中で0.6nm未満離れているアミノ酸残基に相当するシステイン残基同士間、例えばTRAC*01のエキソン1のThr 48及びTRBC1*01若しくはTRBC2*01のエキソン1のSer 57又はこれらの非ヒト等価物から置換されたシステイン残基同士間にある。システインが導入されてジスルフィド結合を形成することができる他の部位は、TCRα鎖についてはTRAC*01のエキソン1中の、TCRβ鎖についてはTRBC1*01又はTRBC2*01のエキソン1中の以下の残基である:
【0037】
【表2】
【0038】
上記で言及した非天然型ジスルフィド結合に加えて、本発明のTCRのdTCR形態又はscTCR形態は、天然型TCR中のジスルフィド結合により連結される残基に相当する残基同士間のジスルフィド結合を含んでもよい。
本発明のTCRのdTCR形態又はscTCR形態は、好ましくは、天然型TCRの膜貫通配列に相当する配列も細胞質配列に相当する配列も含有しない。
【0039】
本発明のTCRは、SLYNTVATL−HLA-A2*0201に強く結合する。これらTCRはまた、HLA-A*0201が搭載する場合にはHIV Gag由来SLYNTVATLの天然に存在する変形体に、変化した程度ではあるが未だ有用な程度で結合する。AIDs患者から単離されたSLYNTVATL変形体には以下が含まれる(Sewellら,(1997) Eur J Immunol. 27:2323-2329):
【0040】
SLFNTVATL
SLFNTVAVL
SLSNTVATL
SSFNTVATL
SLLNTVATL
SLYNTIATL
SLYNTIAVL
SLFNTIATL
SLFNTIAVL
SLFNFVATL
変異アミノ酸には下線を付する。
【0041】
PEG化TCR単量体
1つの特定の実施形態において、本発明のTCRは、少なくとも1つのポリアルキレングリコール鎖と結合している。この結合は、当業者に公知の多くの方法で引き起こされ得る。好ましい実施形態では、ポリアルキレン鎖は、TCRと共有結合している。更なる実施形態では、本発明のこの観点のポリエチレングリコール鎖は、少なくとも2つのポリエチレン反復単位を含んでなる。
【0042】
多価TCR複合体
本発明の1つの観点は、少なくとも2つの本発明のTCRを含んでなる多価TCR複合体を提供する。この観点の1つの実施形態では、少なくとも2つのTCR分子は、リンカー部分を介して連結して多価複合体を形成する。好ましくは、この複合体は水溶性であるので、リンカー部分はそのように選択されるべきである。更に、リンカー部分は、形成される複合体の構造的多様性が最小となるように、TCR分子の規定位置に付着することができるべきであることが好ましい。この観点の1つの実施形態は、ポリマー鎖又はペプチド性リンカー配列が各TCRの可変領域配列中に位置しないアミノ酸残基同士の間に伸びる本発明のTCR複合体により提供される。
【0043】
本発明の複合体は医薬で使用され得るので、リンカー部分は、薬学的適合性(例えば免疫原性)に当然払うべき注意をもって選択されるべきである。
上記の望ましい基準を満たすリンカー部分の例は、当該分野(例えば抗体フラグメントを連結する技術分野)で公知である。
【0044】
本発明の多価TCR分子の製造での使用に好ましい2つのクラスのリンカーが存在する。TCRがポリアルキレングリコール鎖により連結されている本発明のTCR複合体は、この観点の1つの実施形態を提供する。
【0045】
第1は、親水性ポリマー、例えばポリアルキレングリコールである。最も通常に使用されるこのクラスのものは、ポリエチレングリコールすなわちPEGをベースにする。その構造を下記に示す。
HOCH2CH2O(CH2CH2O)n−CH2CH2OH
(式中、nは2より大きい)。しかし、他の適切な(任意に置換されていてもよい)ポリアルキレングリコールをベースにするものもある。ポリプロピレングリコー及びエチレングリコールとプロピレングリコールとのコポリマーが挙げられる。
【0046】
このようなポリマーは、治療剤、特にポリペプチド治療剤又はタンパク質治療剤を処理するか又はこれに接合するかして、該治療剤のPKプロフィールに有益な変化(例えば、減少した腎クリアランス、向上した血漿半減期、減少した免疫原性及び向上した溶解性)を達成するために使用され得る。PEG−治療剤接合体のPKプロフィールのこのような改善は、PEG分子が該治療剤の周囲に、免疫系との反応を立体的に障害し、タンパク質分解性分解を減少させる「殻」を形成することに起因すると考えられる(Caseyら(2000) Tumor Targeting 4 235-244)。使用する親水性ポリマーのサイズは、具体的には、TCR複合体の意図する治療用途に基づいて選択し得る。医薬製剤におけるPEG及び類似分子の使用を詳述する多くの総説論文及び本が存在する。例えば、Harris(1992) Polyethylene Glycol Chemistry - Biotechnical and Biomedical Applications,Plenum,New York,NY.、又はHarris & Zalipsky(1997) Chemistry and Biological Applications of Polyethylene Glycol ACS Books,Washington,D.C.を参照。
【0047】
使用するポリマーは、直鎖状又は分枝状の構造を有することができる。分枝状PEG分子又はその誘導体は、グリセロール及びグリセロールオリゴマー、ペンタエリスリトール、ソルビトール並びにリジンを含む分枝成分の付加により誘導することができる。
【0048】
通常、ポリマーは、該ポリマーがTCR中の標的部位へ連結することが可能となるように、その構造中、例えば一方又は両方の端部及び/又は主鎖からの分枝上に、化学反応性の基を有する。下記に示すように、この化学反応性の基は親水性ポリマーに直接結合させてもよいし、又は親水性ポリマーと反応性化学成分との間にスペーサ基/成分が存在していてもよい:
反応性化学成分−親水性ポリマー−反応性化学成分
反応性化学成分−スペーサ−親水性ポリマー−スペーサ−反応性化学成分
【0049】
上記で概説したタイプの構築物の形成に使用するスペーサは、非反応性の化学的に安定な鎖である任意の有機成分であり得る。このようなスペーサには、以下が含まれるがそれらに限定されない:
-(CH2)n- (式中、n=2〜5)
-(CH2)3NHCO(CH2)2
【0050】
二価アルキレンスペーサ基がポリアルキレングリコール鎖とTCRへのその結合点との間に位置する本発明のTCR複合体は、この観点の更なる実施形態を提供する。
ポリアルキレングリコール鎖が少なくとも2つのポリエチレングリコール反復単位を含んでなる本発明のTCR複合体は、この観点の更なる実施形態を提供する。
【0051】
本発明において有用であり得る反応性化学成分に直接か又はスペーサを介して連結する親水性ポリマーの商業的供給業者は多く存在する。これら供給業者としては、Nektar Therapeutics(CA、米国)、NOF Corporation(日本)、Sunbio(韓国)及びEnzon Pharmaceuticals(NJ、米国)が挙げられる。
【0052】
本発明において有用であり得る反応性化学成分に直接か又はスペーサを介して連結する市販の親水性ポリマーには、以下のものが含まれるがそれらに限定されない。
【表3】
【0053】
種々のカップリング化学成分を使用して、ポリマー分子をタンパク質治療薬やペプチド治療薬とカップリングすることができる。最も適切なカップリング化学成分の選択は、所望するカップリング部位に大きく依存する。例えば、以下のカップリング化学成分が、PEG分子の1又はそれ以上の末端に付着されるために使用されてきた(出典:Nektar Molecular Engineering Catalogue 2003):
【0054】
N−マレイミド
ビニルスルホン
炭酸ベンゾトリアゾール
スクシンイミジルプロピオネート
スクシンイミジルブタノエート(succinimidyl butanoate)
チオエステル
アセトアルデヒド
アクリラート
ビオチン
一級アミン
【0055】
上記のように、非PEGベースのポリマーもまた、本発明のTCRを多量化するために適切なリンカーを提供する。例えば、脂肪族鎖により連結されたマレイミド末端を含有する成分、例えばBMH及びBMOE(Pierce,製品番号22330及び22323)が使用できる。
【0056】
ペプチド性リンカーが他方の好ましいクラスのTCRリンカーである。これらリンカーは、アミノ酸の鎖から構成され、単純なリンカー又はTCR分子を付着させることができる多量体化ドメインを作製するために機能する。ビオチン/ストレプトアビジン系は、以前に、インビトロ結合研究用のTCR四量体を作製するために使用された(WO/99/60119を参照)。しかし、ストレプトアビジンは微生物由来のポリペプチドであり、よって治療剤における使用に理想的には適していない。
【0057】
TCRがヒト多量体化ドメインに由来するペプチド性リンカーにより連結されている本発明のTCR複合体は、この観点の更なる実施形態を提供する。
【0058】
多価TCR複合体の作製に使用できる多量体化ドメインを含有する多くのヒトタンパク質が存在する。例えば、p53の四量体化ドメインは、単量体scFvフラグメントと比較して、増大した血清残存率及び有意に減少した解離速度を示すscFv抗体フラグメント四量体を作製するために利用されている(Willudaら(2001) J. Biol. Chem. 276 (17) 14385-14392)。ヘモグロビンもまた、この種の適用におそらく使用できる四量体化ドメインを有する。
【0059】
少なくとも一方が治療剤に結合されている少なくとも2つのTCRを含んでなる本発明の多価TRC複合体はこの観点の最後の実施形態を提供する。
1つの観点では、本発明のTCR(又はその多価複合体)は、択一的又は追加的に、そのα鎖又はβ鎖のC末端又はN末端に反応性のシステインを含んでなり得る。
【0060】
診断及び治療用途
1つの観点では、本発明のTCRは治療剤又は検出可能な成分と結合させてもよい。例えば、治療剤又は検出可能な成分はTCRに共有結合させてもよい。
本発明の1つの実施形態において、治療剤又は検出可能な成分は、一方又は両方のTCR鎖のC末端に共有結合している。
【0061】
1つの観点において、本発明のscTCR又はdTCRの一方若しくは両方の鎖が検出可能な成分、例えば診断目的に適切な標識で標識されてもよい。このような標識TCRは、TCRリガンドと該TCRリガンドに特異的なTCR(又は多量体高親和性TCR複合体)とを接触させ、TCRリガンドへの結合を検出することを含んでなる、SLYNTVATL−HLA-A*0201複合体を検出する方法において有用である。例えばビオチン化ヘテロ二量体を用いて形成された四量体TCR複合体では、蛍光ストレプトアビジンを使用し、検出可能な標識を提供することができる。このような蛍光標識TCR四量体は、例えばこれら高親和性TCRが特異的であるSLYNTVATL−HLA-A*0201複合体を有する抗原提示細胞を検出するために、FACS分析で使用するに適切である。
【0062】
本発明の可溶性TCRが検出され得る別の様式は、TCR特異的抗体、特にモノクローナル抗体の使用による。多くの市販の抗TCR抗体、例えばαF1及びβF1(それぞれα鎖及びβ鎖の定常領域を認識する)が存在する。
【0063】
更なる観点において、本発明のTCR(又はその多価複合体)は、択一的又は追加的に、例えば細胞殺傷に使用するための毒性成分又は免疫エフェクター分子(例えばインターロイキン又はサイトカイン)であり得る治療剤と結合(例えば、共有結合又はその他の結合)していてもよい。本発明の多価TCR複合体は、非多量体の野生型又は本発明のT細胞レセプターへテロ二量体と比べて、TCRリガンドに関して増強した結合能力を有し得る。したがって、本発明による多価TCR複合体は、SLYNTVATL−HLA-A*0201複合体を提示する細胞をインビトロ又はインビボで追跡又は標的するために特に有用であり、またそのような用途を有する更なる多価TCR複合体の製造のための中間体としても有用である。したがって、これらTCR又は多価TCR複合体は、インビボでの使用のために医薬的に許容される製剤で提供され得る。
【0064】
本発明はまた、標的細胞に治療剤を送達するための方法を提供する。この方法は、可能性のある標的細胞を本発明によるTCR又は多価TCR複合体と、標的細胞へのTCR又は多価TCR複合体の付着を可能にする条件下で接触させることを含んでなり、当該TCR又は多価TCR複合体は、SLYNTVATL−HLA-A*0201複合体に特異的であり、これに治療剤が結合している。
【0065】
特に、本発明の可溶性TCR又は多価TCR複合体を使用して、特定の抗原を提示する細胞の位置に治療剤を送達することができる。このことは、多くの状況で、特にHIV感染細胞に対して有用である。治療剤は、その効果を局所的にではあるが、該治療剤が結合する細胞に限らず発揮するように送達され得る。したがって、1つの特定の戦略は、SLYNTVATL−HLA-A*0201複合体に特異的な本発明のTCR又は多価TCR複合体に連結した細胞毒性又は免疫刺激性の分子を考案する。
【0066】
多くの治療剤、例えば放射活性化合物、酵素(例えばパーフォリン)又は化学療法剤(例えばシスプラチン)がこの用途に用いられ得る。確実に所望の位置で毒性効果が発揮されるために、毒素は、それがゆっくりと放出されるように、ストレプトアビジンに連結したリポソーム内にあり得る。このことにより、体内での輸送の間の損傷効果が防止され、TCRと該当する抗原提示細胞との結合後に毒素が最大効果を有することが確実になる。
【0067】
他の適切な治療剤として、以下のものが挙げられる:
・小分子細胞毒性物質、すなわち、哺乳動物細胞を殺傷する能力を有する、分子量700ダルトン未満の化合物。このような化合物はまた、細胞毒性効果を有することができる毒性金属を含有し得る。更に、これら小分子細胞毒性物質にはまた、プロドラッグ、すなわち、生理学的条件下で崩壊又は変換して細胞毒性物質を放出する化合物が含まれると理解される。このような物質の例には、シスプラチン、メイタンシン(maytansine)誘導体、ラケルマイシン(rachelmycin)、カリケアマイシン(calicheamicin)、ドセタキセル、エトポシド、ゲムシタビン、イホスファミド、イリノテカン、メルファラン、ミトキサントロン、ソルフィマーソディウムホトフィリンII(sorfimer sodiumphotofrin II)、テモゾロマイド(temozolmide)、トポテカン、トリメトレキサート(trimetreate)、グルクロナート、オーリスタチンE(auristatin E)、ビンクリスチン及びドキソルビシンが含まれる;
【0068】
・ペプチド細胞毒素、すなわち、哺乳動物細胞を殺傷する能力を有するタンパク質又はそのフラグメント。例には、リシン、ジフテリア毒素、シュードモナス細菌外毒素A、DNAアーゼ及びRNAアーゼが含まれる;
・放射性核種、すなわち、1以上のα粒子若しくはβ粒子又はγ線の同時放射を伴って崩壊する元素の不安定同位体。例には、ヨウ素131、レニウム186、インジウム111、イットリウム90、ビスマス210及び213、アクチニウム225及びアスタチン213が含まれるがこれらに限定されない;キレート化剤が、高親和性TCR又はその多量体へのこれら放射性核種の結合を促進するために使用されてもよい;
・プロドラッグ、抗体を指向する酵素プロドラッグを含むがこれに限定されない;
【0069】
・免疫刺激剤、すなわち、免疫応答を刺激する成分。サイトカイン(例えば、IL-2及びIFN)、スーパー抗原及びその変異体、TCR-HLA融合体及びケモカイン(例えばIL-8)、血小板第4因子、メラノーマ増殖刺激タンパク質など、抗体又はそのフラグメント、補体活性因子、異種タンパク質ドメイン、同種タンパク質ドメイン、ウイルス性/細菌性タンパク質ドメイン、ウイルス性/細菌性ペプチド及び抗T細胞決定基抗体(例えば、抗CD3又は抗CD28)又は抗体アナログ(例えば、ナノボディ(商標)(Nanobodies)及びアフィボディ(商標)(Affydodies))を含むがこれらに限定されない。
【0070】
本発明の可溶性TCR又は多価TCR複合体は、プロドラッグを薬物に変換し得る酵素に連結されてもよい。これにより、プロドラッグが薬物を必要とする(すなわち、sTCRにより標的にされる)部位でのみ薬物に変換することが可能になる。
【0071】
本明細書中で開示される高親和性SLYNTVATL(配列番号16)−HLA-A*0201特異的TCRは、AIDSの診断方法及び治療方法に使用され得ると考えられる。
治療には、HIV感染(CD4+)細胞の近傍への治療剤の局在化が、毒素又は免疫刺激剤の効果を増強する。ワクチン送達には、ワクチン抗原を抗原提示細胞の近傍に局在させ、該抗原の効力を増強させ得る。この方法はまた造影目的にも適用できる。
【0072】
1つの実施形態は、本発明のTCRを含んでなる膜調製物により提供される。この膜調製物は、細胞から調製されてもよいし、又は人工膜を含んでなってもよい。
別の実施形態は、本発明のTCRをコードする核酸を含んでなる発現ベクターを有する細胞により提供される。例えば、この細胞はT細胞であり得る。
【0073】
本発明の更なる実施形態は、以下:
医薬的に許容されるキャリアと共に、本発明のTCR若しくは多価TCR複合体(任意に治療剤と結合していてもよい)、又は本発明のTCRを含んでなる膜調製物、又は本発明のTCRをコードする核酸を含んでなる発現ベクターを有する複数の細胞
を含んでなる医薬組成物により提供される。
【0074】
本発明はまた、AIDSを患う対象に、本発明のTCR若しくは多価TCR複合体又は本発明のTCRを含んでなる膜調製物又は本発明のTCRをコードする核酸を含んでなる発現ベクターを有する複数の細胞の有効量を投与することを含んでなる、AIDSの治療法を提供する。関連する実施形態では、本発明は、AIDS治療用組成物の製造における、本発明のTCR若しくは多価TCR複合体又は本発明のTCRを含んでなる膜調製物又は本発明のTCRをコードする核酸を含んでなる発現ベクターを有する複数の細胞の使用を提供する。本発明のこれら使用及び方法の更なる具体的実施形態が提供され、そこでは、本発明のTCR若しくは多価TCR複合体又は本発明のTCRを含んでなる膜調製物は、治療剤と結合した形態で投与される。他の好ましい実施形態では、本発明のTCRをコードする核酸を含んでなる発現ベクターを有する細胞はCD8+T細胞である。
【0075】
本発明による治療用又は造影用TCRは、通常、滅菌の、一般には薬学的に許容されるキャリアを含む医薬組成物の一部として供給される。この医薬組成物は(所望される患者への投与法に依存して)任意の適切な形態であり得る。この医薬組成物は、単位剤形で提供されてもよく、一般には密封容器中で提供され、キットの一部として提供されてもよい。そのようなキットは、通常(必ずというわけではないが)、使用のための指示書を含む。キットは、前記の単位剤形を複数含み得る。
【0076】
理論により制限されることは望まないが、本発明のTCRは、治療剤(例えば免疫刺激剤及び/又は細胞傷害剤)をHIV感染(CD4+)細胞に送達することができる効果的な標的化剤(targeting agent)を提供すると考えられる。詳細には、従来の抗レトロウイルス薬治療及び/又はIL-2治療との組合せで、免疫刺激剤及び/又は細胞傷害剤と結合させると本発明のTCRの投与は、HIV感染細胞を標的することができると考えられる。
【0077】
以下は、現在米国で使用が承認されている抗レトロウイルスの一覧である:
アジェネラーゼ(アムプレナビル)−プロテアーゼ阻害剤
コンビビル−レトロビル(300mg)とエピビル(150mg)の組合せ
クリキシバン(インジナビル)−プロテアーゼ阻害剤
エピビル(3tc/ラミブジン)−ヌクレオシドアナログ逆転写酵素阻害剤
エプジコム(2つのヌクレオシド逆転写酵素阻害剤の組合せ(同じ丸剤中のNRTI;600mgのザイアジェン(アバカビル)及び300mgのエピビル(3TC)
【0078】
エムトリバ[エムトリシタビン(FTC)]
フォートベイス(サキナビル)−プロテアーゼ阻害剤
フューゼオン(Fuzeon)(エンフュービルタイド(enfuvirtide))−融合阻害剤
ハイビッド(ddc/ザルシタビン)−ヌクレオシドアナログ逆転写酵素阻害剤
インビラーゼ(サキナビル)−プロテアーゼ阻害剤
【0079】
カレトラ(ロピナビル)−プロテアーゼ阻害剤
レクシヴァ(ホスアンプレナビル)−2003年10月20日承認のプロテアーゼ阻害剤
ノービア(リトナビル)−プロテアーゼ阻害剤
レスクリプター(デラビルジン)−非ヌクレオシドアナログ逆転写酵素阻害剤
レトロビル、AZT(ジドブジン)−ヌクレオシドアナログ逆転写酵素阻害剤
【0080】
レイアタッツ(アタザナビル;BMS-232632)−プロテアーゼ阻害剤
サスティバ(エファビレンツ)−非ヌクレオシドアナログ逆転写酵素阻害剤
トリジビル(1錠剤中3つの非ヌクレオシド;アバカビル+ジドブジン+ラミブジン)
ツルバダ(エムトリシタビン+テノホビルDF)
ヴァイデックス(ddl/ジダノシン)−ヌクレオシドアナログ逆転写酵素阻害剤
ヴァイデックスEC;(ddl/ジダノシン)−ヌクレオシドアナログ逆転写酵素阻害剤;
【0081】
ビラセプト(ネルフィナビル)−プロテアーゼ阻害剤
ビラミューン(ネビラピン)−非ヌクレオシドアナログ逆転写酵素阻害剤
ビリアード(フマル酸テノホビルジソプロキシル)−ヌクレオチド逆転写酵素阻害剤(アデノシンクラス)
ゼリット(d4t/スタブジン)−ヌクレオシドアナログ逆転写酵素阻害剤
ザイアジェン(アバカビル)−ヌクレオシドアナログ逆転写酵素阻害剤
【0082】
医薬組成物は、任意の適切な経路、例えば、非経口経路、経皮経路又は吸入を介する経路(好ましくは非経口経路(皮下、筋肉内又は(最も好ましい)静脈内を含む))による投与に適合され得る。このような組成物は、医薬の技術分野において公知の任意の方法(例えば、滅菌条件下で活性成分をキャリア又は賦形剤と混合すること)により製造され得る。
【0083】
本発明の物質の投薬量は、治療すべき疾患又は障害、治療すべき個体の年齢及び状態などに依存して、広範囲に変わり得る。最終的には、医師が使用すべき適切な投薬量を決定する。
【0084】
追加の観点
本発明のscTCR又はdTCR(好ましくは、ヒト配列に対応する定常配列及び可変配列により構成される)は、実質的に純粋な形態で、又は精製若しくは単離された調製物として提供されてもよい。例えば、本発明のscTCR又はdTCRは、他のタンパク質を実質的に含まない形態で提供されてもよい。
【0085】
本発明のTCRをコードする核酸の配列は、宿主細胞で得られる発現レベルを最適化するように変更されてもよい。宿主細胞は任意の適切な原核細胞又は真核細胞であり得る。例えば、宿主細胞はE.coli細胞又はヒトT細胞であってもよい。これら遺伝子配列に対してなされる変更はサイレントである。すなわち、コードされるアミノ酸配列を変更しない。そのような発現最適化サービスを提供する会社は、GeneArt(ドイツ)を始めとして数多く存在する。
【0086】
本発明はまた、(i)少なくとも1つのTCRα鎖可変ドメイン及び/又は少なくとも1つのTCRβ鎖可変ドメインを含んでなり、且つ(ii)1μMより少ないか若しくは等しいSLYNTVATL−HLA-A*0201複合体に対するKD及び/又は1×10-3か若しくはそれより遅いSLYNTVATL−HLA-A*0201複合体に対する解離速度(koff)を有することを特徴とするSLYNTVATL−HLA-A*0201への結合特性を有する高親和性TCRを作製する方法を提供する。この方法は、
(a)親HIV Gag TCRのα鎖可変ドメイン及びβ鎖可変ドメインを含んでなり、そのα鎖可変ドメイン及びβ鎖可変ドメインの一方又は両方が請求項7及び8で特定したアミノ酸の1又はそれ以上において変異を含んでなるTCRを作製し;
(b)この変異TCRをSLYNTVATL−HLA-A*0201と、該TCRのSLYNTVATL−HLA-A*0201への結合を可能にするに適切な条件下で接触させ、
その相互作用のKD及び/又はkoffを測定する
ことを含んでなる。
【0087】
本発明の各観点の好ましい特徴は、必要な変更を加えてではあるが、他の観点の各々についても同様である。本明細書中で言及する先行技術文献は法が許す最大限の範囲で本明細書に援用する。
【実施例】
【0088】
実施例
以下の実施例で本発明を更に説明する。以下の実施例は、如何なる態様でも本発明の範囲を制限しない。
以下、添付の図面に言及する:
【0089】
図1a及び1bはそれぞれ、親HIV Gag TCRのα鎖可変ドメインアミノ酸配列及びβ鎖可変ドメインアミノ酸配列を詳述する。
図2a及び2bはそれぞれ、可溶性型の親HIV Gag TCRα鎖及びβ鎖のDNA配列を示す。
図3a及び3bはそれぞれ、図2a及び2bのDNA配列から産生されるHIV Gag TCRα鎖及びβ鎖の細胞外アミノ酸配列を示す。
【0090】
図4a及び4bはそれぞれ、非天然型ジスルフィド結合を形成するための追加のシステイン残基をコードするように変異した可溶性型のHIV Gag TCRα鎖及びβ鎖のDNA配列を示す。変異コドンを影付きで示し、導入した制限酵素認識部位に下線を付す。
図5a及び5bはそれぞれ、図4a及び4bのDNA配列から産生されるHIV Gag TCRα鎖及びβ鎖の細胞外アミノ酸配列を示す。各鎖における導入システインを影付きで示す。
【0091】
図6は、高親和性HIV Gag TCR変形体のα鎖可変ドメインアミノ酸配列を詳述する。
図7は、高親和性HIV Gag TCR変形体のβ鎖可変ドメインアミノ酸配列を詳述する。
図8aは、TRACの可溶性部分のアミノ酸配列を詳述する。
図8bは、TRBC1の可溶性部分のアミノ酸配列を詳述する。
図8cは、TRBC2の可溶性部分のアミノ酸配列を詳述する。
【0092】
図9は、pEX954プラスミドのDNA配列を詳述する。
図10は、pEX821プラスミドのDNA配列を詳述する。
図11は、ペプチドリンカーを介して野生型ヒトIL-2に融合した可溶性の親HIV Gag TCR
変形体のβ鎖アミノ酸配列を詳述する。リンカー及びIL-2のアミノ酸をイタリック体で示す。
図12は、ジスルフィド連結した可溶性の親HIV Gag TCRとSLYNTVATL−HLA-A*0201複合体との相互作用について作成したBiacore応答曲線を提供する。
【0093】
図13は、pEX954プラスミドのプラスミドマップを提供する。
図14は、pEX821プラスミドのプラスミドマップを提供する。
図15aは、ヒトT細胞中での発現について最適化された親HIV Gag TCRα鎖の全長DNA配列を提供する。
図15bは、ヒトT細胞中での発現について最適化された親HIV Gag TCRβ鎖の全長DNA配列を提供する。
【0094】
図16aは、親HIV Gag TCRα鎖の全長アミノ酸配列を提供する。
図16bは、ヒトT細胞中での発現について最適化された親HIV Gag TCRβ鎖の全長アミノ酸配列を提供する。
図17aは、コントロールの非形質転換CD8+T細胞についてのFACS分析データを提供する。
図17bは、形質転換CD8+T細胞の表面上での親HIV Gag TCRの発現を示すFACS分析データを提供する。
【0095】
図18a及び18bはそれぞれ、可溶性のジスルフィド連結高親和性c11c6 HIV Gag TCRのα鎖及びβ鎖アミノ酸配列を提供する。
図19は、IFN-γ産生及びTNF-α産生により測定したときの、可溶性のジスルフィド連結高親和性c11c6 HIV Gag TCRがHIVに感染したTo細胞の存在下でSLYNTVATL−HLA-A*0201反応性OX84ポリクローナルT細胞株の活性化を阻害する能力を示す。
【0096】
図20は、IFN-γ産生及びTNF-α産生により測定したときの、可溶性のジスルフィド連結高親和性c11c6 HIV Gag TCRがSLYNTVATLペプチドでパルスした非感染To細胞の存在下でSLYNTVATL−HLA-A*0201反応性OX84ポリクローナルT細胞株の活性化を阻害する能力を示す。
図21は、可溶性のジスルフィド連結高親和性c11c6 HIV Gag TCRがSLYNTVATLペプチドでパルスしたT2細胞を染色する能力を示す。
【0097】
実施例1 − 親HIV Gag TCR可変領域を含んでなる可溶性のジスルフィド連結TCRの作製
図4a及び4bは、SLYNTVATL−HLA-A*0201複合体に特異的な親TCRの可溶性ジスルフィド連結αβ鎖のDNA配列を提供する。これらDNA配列は、多くの受託研究会社、例えばGeneArt(ドイツ)により新たに合成され得る。また、これらDNA配列のpGMT7ベースの発現プラスミド(これは、E.coli株BL21-DE3における高レベル発現用のT7プロモーターを含有する(pLysS)(Panら,Biotechniques(2000) 29 (6):1234-8))中へのライゲーションを容易にするために、これらDNA配列に制限酵素認識部位を付加することもできる。
【0098】
TCRα鎖配列は導入したClaI及びSalII制限酵素認識部位を含有する。この配列を、ClaI及びXhoIで切断したpEX954(図9及び13を参照)中にライゲートした。
TCRβ鎖配列は導入したAseI及びAgeI制限酵素認識部位を含有する。この配列を、NdeI/AgeIで切断したpEX821(図10及び14を参照)中にライゲートした。
【0099】
TCR鎖をコードするDNA中に導入した制限酵素認識部位
ClaI - ATCGAT
SalII - GTCGAC
AseI - ATTAAT
AgeI - ACCGGT
【0100】
ライゲーション
ラピッドDNAライゲーションキット(Roche)を製造業者の指示書に従って用い、切断したTCRα鎖及びβ鎖のDNAと切断したベクターとをライゲートした。
ライゲートしたプラスミドをコンピテントE.coli株XL1-blue細胞に形質転換し、100mg/mlアンピシリン含有LB/寒天プレート上に播種した。37℃で一晩のインキュベーション後、1つのコロニーを採取し、100mg/mlアンピシリンを含有する10mlのLB中で振盪させながら37℃で一晩増殖させた。クローン化プラスミドを、Miniprepキット(Qiagen)を用いて精製し、挿入物を自動DNAシーケンサ(Lark Technologies)を用いて配列決定した。
【0101】
図5a及び5bはそれぞれ、図4a及び4bのDNA配列から作製した、可溶性のジスルフィド連結した親HIV gag TCRのα鎖及びβ鎖の細胞外アミノ酸配列を示す。
【0102】
実施例2 − 可溶性のジスルフィド連結HIV Gag TCRの高親和性変形体の作製
実施例1に記載されるように作製した可溶性のジスルフィド連結天然型HIV Gag TCRは、SLYNTVATL(配列番号16)−HLA-A*0201複合体に対して増大した親和性を有する本発明のTCRを作製するための鋳型として用いることができる。
ファージディスプレイは、高親和性変異体を同定するためにHIV Gag TCR変形体のライブラリを作成することができる1つの手段である。例えば、(Liら(2005) Nature Biotech
23 (3):349-354)に記載されるTCRファージディスプレイ及びスクリーニング法は、HIV Gag TCRに適合させて適用することができる。
【0103】
適切なTCR鎖と組み合せるとSLYNTVATL−HLA-A*0201複合体に対して高親和性を示す変異TCRα鎖及びβ鎖可変ドメインのアミノ酸配列を、それぞれ図6及び7に列挙する(それぞれ配列番号11〜13及び14〜15)。当業者に公知であるように、これらの変異鎖を作製するために要する必要なコドン変更は、これらの鎖をコードするDNA中に、部位特異的変異誘発により導入することができる(StratageneのQuickChange(商標) Site-Directed Mutagenesis Kit)。
【0104】
簡潔には、これは、所望のコドン変更が組み込まれたプライマーと、変異誘発の鋳型として該当するTCR鎖DNAを含有するプラスミドとを用いることにより達成される。
変異誘発は次の条件を用いて行った:全容量50μl中、50ngのプラスミド鋳型、1μlの10mM dNTP、5μlの製造業者により供給された10×Pfu DNAポリメラーゼ緩衝液、25pmol
のfwdプライマー、25pmolのrevプライマー、1μlのpfu DNAポリメラーゼ。95℃で2分間の最初の変性工程後、反応物を、変性(95℃、10秒)、アニーリング(55℃、10秒)及び伸長(72℃、8分)の25サイクルに供した。得られた生成物をDpnI制限酵素で消化して鋳型プラスミドを除き、E.coli株XL1-blueに形質転換した。変異誘発を配列決定により確証した。
【0105】
実施例3 − 可溶性TCRの発現、リフォールディング及び精製
それぞれ実施例1又は2のように作製した変異α鎖及びβ鎖を含む発現プラスミドを、E.coli株BL21pLysSに別々に形質転換し、1つのアンピシリン耐性コロニーを37℃にてTYP(アンピシリン100μg/ml)培地中で0.4のOD600まで増殖させた後、0.5mM IPTGでタンパク質発現を誘導した。誘導の3時間後に、Beckman J-6B中で4000rpmにて30分間の遠心分離により細胞を回収した。細胞ペレットを、50mMのTris-HCl、25%(w/v)のスクロース、1mMのNaEDTA、0.1%(w/v)のNaAzide、10mMのDTTを含有する緩衝液(pH8.0)中に再懸濁した。一晩の凍結−解凍工程の後、再懸濁細胞を、Milsonix XL2020ソニケータ中での標準の12mm径プローブを使用する1分間のバーストで合計約10分間の超音波処理に付した。封入体ペレットを、Beckman J2-21遠心機における13000rpmにて30分間の遠心分離により回収した。次いで、3回の界面活性剤での洗浄を行い、細胞残渣及び膜成分を除去した。各回、封入体ペレットをTriton緩衝液(50mMのTris-HCl、0.5%のTriton-X100、200mMのNaCl、10mMのNaEDTA、0.1%(w/v)のNaAzide、2mMのDTT、pH8.0)中でホモジナイズした後、Beckman J2-21における13000rpmにて15分間の遠心分離によりペレット化した。次いで、以下の緩衝液中での同様な洗浄により界面活性剤及び塩を除去した:50mMのTris-HCl、1mMのNaEDTA、0.1%(w/v)のNaAzide、2mMのDTT、pH8.0。最後に、封入体を30mgずつ小分けし、−70℃にて凍結させた。封入体タンパク質の収率を、6Mグアニジン-HClで可溶化し、Bradford色素結合アッセイ(PerBio)で測定することにより定量した。
【0106】
約30mgのTCRβ鎖及び60mgのTCRα鎖の可溶化封入体を凍結ストックから解凍し、次いでサンプルを混合し、混合物を15mlのグアニジン溶液(6M塩酸グアニジン、10mM酢酸ナトリウム、10mM EDTA)中に希釈して、完全な鎖変性を確実に行った。次いで、十分に還元され変性したTCR鎖を含有するグアニジン溶液を、以下の1リットルのリフォールディング緩衝液中に注入した:100mM Tris pH8.5、400mM L-アルギニン、2mM EDTA、5mM還元型グルタチオン、0.5mM酸化型グルタチオン、5M尿素、0.2mM PMSF。酸化還元対(2-メルカプトエチルアミン及びシスタミン(それぞれ最終濃度6.6mM及び3.7mMまで))を加え、約5分後に変性TCR鎖を加えた。この溶液を5時間±15分間放置した。リフォールディングしたTCRを、Spectrapor 1メンブレン(Spectrum;製品番号132670)中で、10Lの10mM Tris(pH8.1)に対し、5℃±3℃にて18〜20時間透析した。この時間の後、透析緩衝液を新鮮な10mM Tris(pH8.1)(10L)に変え、透析を5℃±3℃にて更に20〜22時間継続した。
【0107】
透析したリフォールディング物をPOROS 50HQアニオン交換カラムにロードし、Akta Purifier(Pharmacia)を用いて50カラム容量の0〜500mM NaClグラジエントで結合タンパク質を溶出することにより、sTCRを分解産物及び不純物から分離した。ピーク画分を4℃にて貯蔵し、クーマシー染色SDS-PAGEで分析した後、プールして濃縮した。最後に、sTCRを精製して、HBS-EP緩衝液(10mM HEPES pH7.4、150mM NaCl、3.5mM EDTA、0.05% Nonidet p40)で予め平衡化したSuperdex 200HRゲル濾過カラムを用いて特徴付けを行った。約50kDaの相対分子量で溶出するピークをプールし、濃縮した後、BIAcore表面プラズモン共鳴分析により特徴付けを行った。
【0108】
実施例4 − 特異pMHCへのsTCR結合のBiacore表面プラズモン共鳴特徴付け
表面プラズモン共鳴バイオセンサ(BIAcore 3000TM)を使用して、ペプチド−MHCリガンドへのsTCRの結合を分析した。これは、半配向様式(semi-oriented fashion)でストレプトアビジン被覆結合表面に固定した単一pMHC複合体(下記で説明)を作製し、同時に4つまでの異なるpMHC(別個のフローセルに固定)への可溶性T細胞レセプターの結合の効率的な試験を可能にすることにより容易となった。HLA複合体の手動での注入により、正確なレベルの固定化クラスI分子を容易に操作することが可能となった。
【0109】
ビオチン化クラスI HLA-A*0201分子を、構成サブユニットタンパク質及び合成ペプチドを含有する細菌発現封入体から、インビトロでリフォールディングさせ、続いて精製し、インビトロにて酵素でビオチン化した(O'Callaghanら(1999) Anal. Biochem. 266:9-15)。そのタンパク質の膜貫通ドメイン及び細胞質ドメインと置換したC末端ビオチン化タグを有するHLA-A*0201-重鎖を適切な構築物中で発現させた。〜75mg/リットル細菌培養物の封入体発現レベルが得られた。MHC軽鎖又はβ2-ミクログロブリンもまた、E.coli中で適切な構築物から封入体として〜500mg/リットル細菌培養物のレベルで発現させた。
【0110】
E. coli細胞を溶解し、封入体を約80%純度まで精製した。封入体からのタンパク質を6Mのグアニジン-HCl、50mMのTris(pH8.1)、100mMのNaCl、10mMのDTT、10mMのEDTA中で変性させ、5℃より低いリフォールディング緩衝液中に変性タンパク質の単一パルス(single pulse of denatured protein)を添加することにより、0.4MのL-アルギニン-HCl、100mMのTris(pH8.1)、3.7mMのシスタミン、6.6mMのβ-システアミン、HLA-A*0201分子が搭載するために必要な4mg/mlのSLYNTVATLペプチド中で30mg/リットルの重鎖、30mg/リットルのβ2mの濃度にてリフォールディングさせた。リフォールディングは、4℃にて少なくとも1時間で完了に到達させた。
【0111】
緩衝液を、10容量の10mM Tris(pH8.1)での透析により交換した。溶液のイオン強度を十分に減少させるために、2回の緩衝液交換が必要であった。次いで、タンパク質溶液を1.5μm酢酸セルロースフィルターに通して濾過し、POROS 50HQアニオン交換カラム(8ml床容量)に充填した。タンパク質を0〜500mMのNaCl線形勾配で溶出させた。HLA-A*0201−ペプチド複合体は約250mM NaClで溶出した。ピーク画分を収集し、プロテアーゼ阻害剤のカクテル(Calbiochem)を加え、画分を氷上で冷却した。
【0112】
ビオチン化タグを付したpMHC分子を、10mM Tris(pH8.1)、5mM NaCl中に、同じ緩衝液中で平衡化したPharmacia迅速脱塩カラムを使用して移して緩衝液を交換した。溶出に際して即座に、タンパク質含有画分を氷上で冷却し、プロテアーゼ阻害剤カクテル(Calbiochem)を加えた。次いで、ビオチン化試薬を加えた:1mMビオチン、5mM ATP(pH8に緩衝化)、7.5mM MgCl2及び5μg/ml BirA酵素(O’Callaghanら(1999)Anal.Biochem.266:9-15に従って精製)。次いで、混合物を室温にて一晩インキュベートした。
【0113】
ゲル濾過クロマトグラフィーを使用してビオチン化pHLA-A*0201分子を精製した。Pharmacia Superdex 75 HR 10/30カラムを濾過PBSで予め平衡化し、1mlのビオチン化反応混合物を充填し、カラムをPBSで0.5ml/分にて展開した。ビオチン化pHLA-A*0201分子は、約15mlで単一ピークとして溶出した。タンパク質を含む画分をプールし、氷上で冷却し、プロテアーゼ阻害剤カクテルを加えた。クーマシー結合アッセイ(PerBio)を使用してタンパク質濃度を測定し、ビオチン化pHLA-A*0201分子のアリコートを−20℃で凍結保存した。標準的なアミンカップリング法によりストレプトアビジンを固定化した。
【0114】
この固定化複合体はT細胞レセプター及びコレセプターCD8αα(共に可溶性相中に注入され得る)の両方に結合することができる。低濃度(少なくとも40μg/ml)でさえTCRの特異的結合が得られる。このことは、このTCRが比較的安定であることを示す。sTCRのpMHC結合特性は、sTCRが可溶性相又は固定化相のいずれかで使用される場合、質的にも量的にも同様であることが観察される。これは、可溶性種の部分的な活性についての重要なコントロールであり、また、ビオチン化pMHC複合体が非ビオチン化複合体と生物学的に同程度に活性であることを示唆する。
【0115】
新規鎖間結合を含有するHIV Gag sTCRとそのリガンド/MHC複合体又は無関係のHLA−ペプチド組合せ(これらの製造は上記で説明)との間の相互作用を、Biacore 3000TM表面プラズモン共鳴(SPR)バイオセンサで分析した。SPRは、レセプターリガンド相互作用を検出しその親和性及び動力学的パラメータを分析するために使用することができる原理である、小さなフローセル内のセンサ表面近くでの屈折率の変化(応答単位(RU)で表示)を測定する。β2mに架橋したビオチンとフローセルの活性化表面に化学的に架橋されているストレプトアビジンとの間の結合を介して、個々のHLA−ペプチド複合体を別個のフローセルに固定化することにより、プローブフローセルを準備した。次いで、異なるフローセルの表面上にsTCRを一定流速で通過させ、そうしている間のSPR応答を測定することにより、アッセイを実施した。
【0116】
平衡結合定数の測定
親の又は変異したHIV Gag sTCRの系列希釈物を調製し、5μl/分の一定流速で2つの異なる表面(一方は〜1000RUの特異SLYNTVATL−HLA-A*0201複合体を被覆し、2つめは〜1000RUの非特異HLA-A2-ペプチド複合体を被覆)に注入した。応答は、コントロールセルの測定値を用いて各濃度について規格化した。規格化データ応答を、TCRサンプルの濃度に対してプロットし、平衡結合定数KDを算出するために双曲線にフィットさせた(Price & Dwek,Principles and Problems in Physical Chemistry for Biochemists (第2版) 1979,Clarendon Press,Oxford)。
【0117】
動力学パラメータの測定
高親和性TCRについて、解離速度定数kd及び結合速度定数kaを実験的に測定することによりKDを決定した。平衡定数KDはkd/kaとして算出した。
1つは〜300RUの特異HLA-A2-nyesoペプチド複合体で被覆し、2つめは〜300RUの非特異HLA-A2−ペプチド複合体で被覆した2つの異なるセル上にTCRを注入した。流速を50μl/分に設定した。代表的には、250μlのTCRを〜3μM濃度で注入した。次いで、反応がベースラインに戻るまで緩衝液を流した。Biaevaluationソフトウェアを用いて動力学パラメータを算出した。また、半減期の算出を可能とするために、解離相を一次指数関数減衰式にフィットさせた。
【0118】
結果
可溶性のジスルフィド連結した天然型HIV Gag TCR(それぞれ配列番号9及び10に詳述されるαTCR鎖及びβTCR鎖からなる)とSLYNTVATL−HLA-A*0201複合体との間の相互作用を、上記の方法を用いて分析し、85nMのKD及び2.21×10-2 S-1の解離速度(koff)が示された(Biacore応答曲線については図12を参照)。
以下の表で特定されるTCRは、1μMより少ないか若しくは等しいKD及び/又は1×10-3 S-1か若しくはそれより遅いkoffを有する。
【0119】
【表4】
【0120】
実施例5 − 可溶性の高親和性HIV Gag TCR−WTヒトIL-2融合タンパク質の作製
実質的に実施例1〜3に記載の方法を使用して、可溶性の高親和性HIV Gag TCR−WTヒトIL-2融合タンパク質を作製することができる。簡潔には、所望のリンカー及びWTヒトIL-2をコードするDNAを、可溶性のジスルフィド連結親HIV Gag TCRβ鎖のDNA配列の3'末端にTAA(「ストップ」)コドンの直前で付加する。図11は、WTヒトIL-2にリンカー配列を介して融合したジスルフィド連結親HIV Gag TCRβ鎖を含んでなる融合タンパク質のアミノ酸配列を提供する(配列番号24)。この融合タンパク質のリンカー及びIL-2部分はイタリック体で示されている。次いで、この構築物をコードするDNAは、pEX821中にライゲートすることができる。次いで、可溶性の親HIV Gag TCR−IL-2融合タンパク質は、実質的に実施例3に記載された方法を使用して、このβ鎖融合タンパク質を図5aに詳述される可溶性のジスルフィド連結親HIV Gag α鎖TCR鎖(配列番号9)と組み合せることにより、発現させることができる。
【0121】
実施例6 − T細胞表面での親HIV Gag TCRの組換え発現
親HIV Gag TCR鎖のシグナル配列、細胞外ドメイン、膜貫通ドメイン及び細胞内ドメインをコードするDNA構築物を合成した(GeneArt、ドイツ)。これらTCRα鎖及びTCRβ鎖のDNA配列(それぞれ図15a及び15bに提供)は、親HIV Gag TCR DNA配列から、天然型アミノ酸配列を維持しつつ、ヒトT細胞中での該コードTCR鎖の発現レベルを増大させるように変更する。図16a及び16bはそれぞれ、図15a及び15bのDNA配列によりコードされる全長アミノ酸配列を提供する。
【0122】
次いで、TCRα鎖及びTCRβ鎖のDNA配列を、レンチウイルス発現ベクター中に一緒に挿入した。このベクターは、親HIV Gag TCRα鎖及びβ鎖の両方をコードするDNAを、該TCR鎖を分離する手足口病ウイルス(FMDV)2A切断因子アミノ酸配列(LLNFDLLKLAGDVESNPG(配列番号31))とインフレームで単一オープンリーディングフレームとして含有する(de Felipeら,Genet Vaccines Ther (2004) 2 (1):13)。mRNA翻訳に際して、TCRα鎖はそのC末端に2Aペプチド配列を有して作製され、TCRβ鎖は別個のポリペプチドとして作製される。
【0123】
T細胞を上記レンチウイルスベクターを用いて形質導入した。簡潔には、抗CD3/抗CD28ビーズを使用して初代T細胞を24時間刺激した。次いで、TCR遺伝子を発現する濃縮レンチウイルス上清を、刺激したT細胞と共にインキュベートしてウイルス形質導入した。次いで、抗CD3/抗CD28ビーズを除去し、200〜300fLの「静止容量(resting volume)」を達成するまで形質導入T細胞を培養した。
形質導入細胞表面上での親HIV Gag TCRの提示は、HLA-A*0201−SLYNTVALT PEテトラマーを使用するFACS分析及び抗CD8モノクローナル抗体FITC同時染色により確証した。
【0124】
結果
図17bは、形質導入CD8+細胞表面上での親HIV Gag TCR発現の成功を示すFACS分析データを提供する。図17aは、コントロールの非形質導入T細胞を使用して作成したFACS分析データを提供する。
【0125】
実施例7 − 可溶性の高親和性HIV Gag TCRによるCTL活性化の阻害
以下のアッセイを行い、可溶性の高親和性c11c6 HIV Gag TCRがSLYNTVATL−HLA-A*0201反応性ポリクローナルT細胞株の活性化を阻害し得ることを示した。
【0126】
HIV感染細胞の存在下でのOX84 SLYNTVATL−HLA-A*0201反応性ポリクローナルT細胞株活性化の阻害
この実験に使用した可溶性のc11c6高親和性HIV Gag TCRは、それぞれ図6c(配列番号13)及び図7b(配列番号15)に示すTCRα鎖可変ドメイン及びTCRβ鎖可変領域を含んでいた。この可溶性TCRのTCRα鎖及びβ鎖の完全アミノ酸配列はそれぞれ、図18a(配列番号29)及び図18b(配列番号30)に提供される。
IFN-γ産生及びTNF-α産生をCTL活性化についての読み取り値として使用した。
【0127】
試薬
R10アッセイ培地:10% FCS(熱不活化,Gibco,cat# 10108-165)、88% RPMI 1640(Gibco,cat# 42401-018)、1%グルタミン(Gibco,cat# 25030-024)及び1%ペニシリン/ストレプトマイシン(Gibco,cat# 15070-063)。
ペプチド:(種々の供給源から入手)最初にDMSO(Sigma,cat# D2650)中に4mg/mlで溶解し、凍結。
BDTM Cytometric Bead Array Kit、ヒトTh1/Th2 cytokine Kit II (BD Biosciences,San Diego,US)はアッセイに必要な全ての試薬を含む。
【0128】
T細胞活性化アッセイ
慢性HIV感染To標的細胞(HXB2及びHIV3B HIV Lab株)を洗浄し、R10培地中に再懸濁した。コントロールとして、非感染To標的細胞を1nMのSLYNTVATLペプチドで、37℃、5%CO2にて30分間パルスした。
【0129】
試験サンプル:
96ウェルU底プレートのウェル当たりR10培地中25,000のHIV感染To標的細胞。
ウェル当たりR10培地中2×10-7Mの高親和性c11c6 HIV Gag TCR又は親HIV Gag TCR。
ウェル当たりR10培地中5000のOX84ポリクローナルエフェクターT細胞株。
コントロール:
無関係の可溶性TCR(HLA-A*0201−Tax特異的TCR及びHLA-A*0201−NY-ESO特異的TCR)又は高親和性 HIV Gag TCRに代えた上記と同じもの。
【0130】
次いで、プレートを37℃、5%CO2にて4時間インキュベートした。培養上清を除去し、以下の方法を用いて存在するIFN-γ及びTNF-αのレベルを測定した。
【0131】
IFN-γアッセイ及びTNF-αアッセイ
(a)抗IFNγ捕捉抗体及び(b)抗TNFα捕捉抗体で被覆したBDTM Cytometric Beadsを製造業者の指示書に従い調製した。
次いで、以下の添加物を含むアッセイチューブを調製した:
BDアッセイ希釈剤中の50μlの抗IFNγ及び抗TNFα混合BDTM Cytometric Beads
50μlのPE検出試薬
【0132】
続いて、いずれか:
T細胞活性化アッセイウェルから採取した50μlの捕捉上清(試験サンプル)、
又は
ストック標準の系列希釈により一連の濃度で調製した50μlのIFNγ及びTNFα混合標準(較正標準)
【0133】
次いで、チューブを暗所で3時間インキュベートした後、1mlのBD洗浄緩衝液で洗浄し、遠心分離した。最後に、ビーズを300μlの洗浄緩衝液中に再懸濁し、存在するIFNγ及びTNFαのレベルを、製造業者の指示書に従ってフローサイトメトリにより決定した。
【0134】
非感染SLYNTVATLペプチドパルスTo細胞の存在下でのSLYNTVATL−HLA-A*0201特異的OX84ポリクローナルT株の阻害
上記CTL活性化アッセイに使用した同じ試薬及び方法を使用した。但し、2000のOX84ポリクローナルエフェクターT細胞を各T細胞活性化アッセイに使用し、非感染Toリンパ芽球様細胞(10-10〜10-8MのSLYNTVATLペプチドでパルス)を標的細胞として使用した。
【0135】
結果
可溶性の高親和性c11c6 HIV Gag TCRは、IFN-γ産生及びTNF-α産生により測定したところ、HIV感染To細胞の存在下で、SLYNTVATL−HLA-A*0201反応性OX84ポリクローナルT細胞株の活性化を強力に阻害した(図19を参照)。
可溶性の高親和性c11c6 HIV Gag TCRは、IFN-γ産生及びTNF-α産生により測定したところ、SLYNTVATLでパルスした非感染To細胞の存在下で、SLYNTVATL−HLA-A*0201反応性OX84ポリクローナルT細胞株の活性化を強力に阻害した(図20を参照)。
【0136】
実施例8 − 高親和性c11c6 HIV Gag TCRを使用する蛍光顕微鏡法による、ペプチドでパルスしたT2細胞上の細胞表面SLYNTVATL−HLA-A*0201抗原の定量
ペプチドパルスしたT2リンパ芽球様細胞上のSLYNTVATL−HLA-A*0201抗原の数を、可溶性の高親和性c11c6 HIV Gag TCRを用いる単一分子蛍光顕微鏡法により決定した(1つの蛍光シグナルは、標的細胞表面上の同族pMHCリガンドに結合した単一の標識TCRに関連するとの仮定に基づく)。このことは、ビオチン化TCRを用いて抗原発現ガン細胞を標的し、その後、細胞結合TCRをストレプトアビジン−Rフィコエリトリン(PE)コンジュゲートにより標識することによって容易となった。次いで、個々のPE分子を、三次元蛍光顕微鏡法により撮像した。
【0137】
T2リンパ芽球様細胞を、一連の濃度(10-5〜10-10M)で90分間37℃にてHIV Gag由来SLYNTVATLペプチド又は無関係のペプチド(SLLMWITQC)でパルスした。パルス後、細胞を500μlのPBSで2回洗浄した。細胞を200μlのTCR溶液(0.5% BSAアルブミン含有PBS中100nMの高親和性c11c6 HIV Gag TCR)中で30分間室温にてインキュベートした。TCR溶液を除去し、細胞を500μlのPBSで3回洗浄した。細胞を、200μlのストレプトアビジン−PE溶液(0.5% BSA含有PBS中5μg ml-1のストレプトアビジン−PE)中で室温にて暗所で20分間インキュベートした。ストレプトアビジン−PE溶液を除去し、細胞を500μlのPBSで3回洗浄した。洗浄媒体を除去し、細胞を、フェノールレッドを含まない400μlのR10に維持した後、蛍光顕微鏡法により撮像した。
【0138】
蛍光顕微鏡法。蛍光顕微鏡法は、63×油浸対物レンズ(Zeiss)を備えるAxiovert 200M(Zeiss)顕微鏡を用いて行った。300Wキセノンアークランプ(Sutter)を含むラムダLS光源を照明に用い、光路内に0.3及び0.6の中性フィルタを配置することにより最適レベルまで光度を下げた。励起光スペクトル及び発光スペクトルを、TRITC/DiIフィルタセット(Chroma)を用いて分離した。細胞を、z-スタック取り込みにより三次元で撮像した(21面、1μm間隔)。画像の取得及び分析は、Metamorphソフトウェア(Universal Imaging)を、記載されたようにして行った(Irvineら,Nature 419:p845-9、及びPurbhooら,Nature Immunology 5:p524〜30)。
【0139】
結果
図21に示されるように、上記方法を使用して、ペプチドパルスしたT2細胞の表面上のSLYNTVATL−HLA-A*0201抗原に結合した高親和性c11c6 HIV Gag TCRの撮像に成功した。これら結果は、高親和性c6c11 HIV Gag TCRを使用する、SLYNTVATLペプチドでパルスした細胞上のエピトープを計数するための閾値が約10-9Mペプチドであることを示す。
【技術分野】
【0001】
本発明は、HIV Gagポリペプチド由来SLYNTVATL−HLA-A*0201への結合特性を有するT細胞レセプター(TCR)に関する。このTCRは、少なくとも1つのTCRα鎖可変ドメイン及び/又は少なくとも1つのTCRβ鎖可変ドメインを含んでなり、1μMより少ないか若しくは等しいSLYNTVATL−HLA-A*0201複合体に対するKD及び/又は1×10-3 S-1か若しくはそれより遅いSLYNTVATL−HLA-A*0201複合体に対する解離速度(off-rate)(koff)を有する。
【背景技術】
【0002】
(発明の背景)
ヒト免疫不全ウイルス(HIV)は、後天性免疫不全症候群(AIDS)の原因因子である。このウイルスは、レンチウイルス群に属するエンベロープを有するレトロウイルスである。SLYNTVATL(配列番号16)ペプチドは、ヒト免疫不全ウイルス-1(HIV-1)を作り上げている9つの遺伝子の1つであるGag遺伝子のg17遺伝子産物に由来する。このペプチドはHLA-A*0201に搭載(load)されてHIV感染細胞の表面に提示される。したがって、SLYNTVATL−HLA-A2*0201複合体は、例えば細胞毒性又は免疫刺激性薬剤を感染細胞に送達する目的で、TCRが標的することができるHIVマーカーを提供する。しかし、この目的には、TCRがペプチド−HLA複合体に対して高い親和性及び/又は遅い解離速度を有することが望ましい。
【発明の概要】
【課題を解決するための手段】
【0003】
(発明の簡単な説明)
本発明は、SLYNTVATL−HLA-A*0201複合体に対し、1μMより少ないか若しくはこれと等しい親和性(KD)及び/又は1×10-3 S-1か若しくはそれより遅い解離速度(koff)を有するTCR(但し、該TCRが細胞により提示され且つ配列番号1及び2を含んでなる場合には、該細胞は天然型T細胞ではない)を初めて利用可能にする。このTCRは、単独で又は治療剤との組合せで、前記複合体を提示するHIV感染細胞を標的するに有用である。
【図面の簡単な説明】
【0004】
【図1a】親HIV Gag TCRのα鎖可変ドメインアミノ酸配列を示す。
【図1b】親HIV Gag TCRのβ鎖可変ドメインアミノ酸配列を示す。
【図2】可溶性型の親HIV Gag TCRα鎖(a)及びβ鎖(b)のDNA配列を示す。
【図3】図2のDNA配列から産生されるHIV Gag TCRα鎖(a)及びβ鎖(b)の細胞外アミノ酸配列を示す。
【0005】
【図4】非天然型ジスルフィド結合を形成するための追加のシステイン残基をコードするように変異した可溶性型のHIV Gag TCRα鎖(a)及びβ鎖(b)のDNA配列を示す。
【図5】図4のDNA配列から産生されるHIV Gag TCRα鎖(a)及びβ鎖(b)の細胞外アミノ酸配列を示す。
【図6】高親和性HIV Gag TCR変形体のα鎖可変ドメインアミノ酸配列を示す。
【図7】高親和性HIV Gag TCR変形体のβ鎖可変ドメインアミノ酸配列を示す。
【図8】TRAC(a)、TRBC1(b)及びTRBC2(c)の可溶性部分のアミノ酸配列を示す。
【0006】
【図9−1】pEX954プラスミドのDNA配列を示す。
【図9−2】図9−1の続きである。
【図10−1】pEX821プラスミドのDNA配列を示す。
【図10−2】図10−1の続きである。
【0007】
【図11】ペプチドリンカーを介して野生型ヒトIL-2に融合した可溶性の親HIV Gag TCR変形体のβ鎖アミノ酸配列を示す。
【図12】ジスルフィド連結した可溶性の親HIV Gag TCRとSLYNTVATL−HLA-A*0201複合体との相互作用について作成したBiacore応答曲線を示す。
【図13】pEX954プラスミドのプラスミドマップを示す。
【図14】pEX821プラスミドのプラスミドマップを示す。
【0008】
【図15】ヒトT細胞中での発現について最適化された親HIV Gag TCRα鎖(a)及びβ鎖(b)の全長DNA配列を示す。
【図16】a:親HIV Gag TCRα鎖の全長アミノ酸配列を示す。b:ヒトT細胞中での発現について最適化された親HIV Gag TCRβ鎖の全長アミノ酸配列を示す。
【図17】a:コントロールの非形質転換CD8+T細胞についてのFACS分析データを示す。b:形質転換CD8+T細胞の表面上での親HIV Gag TCRの発現を示すFACS分析データを示す。
【0009】
【図18】可溶性のジスルフィド連結高親和性c11c6 HIV Gag TCRα鎖(a)及びβ鎖(b)のアミノ酸配列を示す。
【図19】IFN-γ産生及びTNF-α産生により測定したときの、可溶性のジスルフィド連結高親和性c11c6 HIV Gag TCRがHIVに感染したTo細胞の存在下でSLYNTVATL−HLA-A*0201反応性OX84ポリクローナルT細胞株の活性化を阻害する能力を示す。
【0010】
【図20】IFN-γ産生及びTNF-α産生により測定したときの、可溶性のジスルフィド連結高親和性c11c6 HIV Gag TCRがSLYNTVATLペプチドでパルスした非感染To細胞の存在下でSLYNTVATL−HLA-A*0201反応性OX84ポリクローナルT細胞株の活性化を阻害する能力を示す。
【図21】可溶性のジスルフィド連結高親和性c11c6 HIV Gag TCRがSLYNTVATLペプチドでパルスしたT2細胞を染色する能力を示す。
【発明を実施するための形態】
【0011】
(発明の詳細な説明)
本発明は、1μMより少ないか等しいSLYNTVATL−HLA-A*0201複合体に対するKD及び/又は1×10-3 S-1かそれより遅いSLYNTVATL−HLA-A*0201複合体に対する解離速度(koff)を有し、但し細胞により提示され且つ配列番号1及び2を含んでなる場合には該細胞は天然型T細胞ではないことを特徴とする、SLYNTVATL−HLA-A*0201への結合特性を有し且つ少なくとも1つのTCRα鎖可変ドメイン及び/又は少なくとも1つのTCRβ鎖可変ドメインを含んでなるT細胞レセプター(TCR)を提供する。KD及び/又は(koff)の測定は、公知の方法のいずれによっても行うことができる。好ましい方法は、実施例4の表面プラズモン共鳴(Biacore)法である。
【0012】
比較のために、親のHIV gag TCR(TCRα鎖については配列番号9を、TCRβ鎖については配列番号10を参照)のジスルフィド連結した可溶性変形体とSLYNTVATL−HLA-A*0201複合体との相互作用は、実施例4のBiacoreに基づく方法により測定すると、約85nMのKD及び2.21×10-2 S-1の解離速度(koff)を有する。
SLYNTVATL−HLA-A*0201複合体に特異的な親HIV Gag TCRは、以下のVα鎖遺伝子及びVβ鎖遺伝子使用(usage)を有する:
α鎖 − TRAV12.2
β鎖:− TRBV 5.6
【0013】
親HIV Gag TCRは、SLYNTVATL−HLA-A*0201複合体との間の相互作用に対して高い親和性及び/又は遅い解離速度を有する本発明の他のTCRを製造できる鋳型として使用することができる。したがって、本発明は、少なくとも1つの相補性決定領域(CDR)及び/又は可変ドメインフレームワーク領域において、親HIV Gag TCRのα鎖可変ドメイン(図1a及び配列番号1を参照)及び/又はβ鎖可変ドメイン(図1b及び配列番号2を参照)に対して変異したTCRを包含する。また、本発明のTCRの可変ドメイン中の他の超可変領域、例えば超可変4(HV4)領域が、高親和性変異体TCR内で変異していてもよいと企図される。
【0014】
ファージディスプレイは、TCR変形体のライブラリを作製し得る1つの手段を提供する。各々が非天然型ジスルフィド鎖間結合を含有するTCR変形体のライブラリのファージディスプレイ及びその後のスクリーニングに適切な方法は、Liら,(2005) Nature Biotech 23(3):349-354及びWO 2004/04404に詳述されている。
天然型TCRはへテロ二量体αβ又はγδ形態で存在する。しかし、単一のTCRα鎖又はTCRβ鎖からなる組換えTCRが、ペプチドMHC分子に結合することが以前に示されている。
1つの実施形態において、本発明のTCRは、α鎖可変ドメイン及びTCRβ鎖可変ドメインを両方含んでなる。
【0015】
当業者には自明であろうが、TCRα鎖配列及び/又はTCRβ鎖配列中の変異は、1又はそれより多い置換、欠失又は挿入であり得る。これら変異は、任意の適切な方法(ポリメラーゼ連鎖反応(PCR)に基づくもの、制限酵素ベースのクローニング、又はライゲーション非依存性クローニング(LIC)手法を包含するがこれらに限定されない)を使用して実行することができる。これら方法は、多くの標準的な分子生物学の教科書に詳述されている。ポリメラーゼ連鎖反応(PCR)変異誘発及び制限酵素ベースのクローニングに関する更なる詳細に関しては、Sambrook & Russell,(2001) Molecular Cloning - A Laboratory Manual (第3版) CSHL Pressを参照。LIC手順についての更なる情報は、Rashtchian(1995) Curr Opin Biotechnol 6(1):30-6に見出すことができる。
【0016】
HIV Gag TCRのものに類似するVα遺伝子及びVβ遺伝子使用、したがって類似するアミノ酸配列を含んでなる任意のαβTCRは、簡便な鋳型TCRを作り得ることに留意すべきである。次いで、鋳型αβTCRの可変ドメインの一方又は両方をコードするDNA中に、本発明の変異した高親和性TCRを作製するに必要な変更を導入することができる。当業者には自明であろうが、必要な変異は、多くの方法(例えば、部位特異的変異誘発)により導入し得る。
【0017】
本発明のTCRは、下記に列挙するものに対応するTCRα鎖可変ドメインアミノ酸の1又はそれより多くが、図1a及び配列番号1中の親HIV Gag TCRα鎖可変ドメインを提供する配列のこれら位置に存在するアミノ酸に対して変異しているTCRを包含する。
【0018】
特に反する記載がない限り、本明細書中のTCRアミノ酸配列は、一般には、N末端メチオニン(Met又はM)残基を含んで提供される。当業者には公知であろうが、この残基は、組換えタンパク質の産生の間に除去され得る。これもまた当業者には自明であろうが、TCRのpMHC結合特性に実質的に影響することなく、C末端及び/又はN末端に提供される配列を1、2、3、4、5又はそれより多くの残基だけ短縮することもまた可能である。全てのそのような平凡な(trivial)変形体は、本発明に包含される。
【0019】
本明細書中で使用する場合、用語「可変領域」は、所定のTCRの、TCRα鎖に関してはTRAC遺伝子により、TCRβ鎖に関してはTRBC1遺伝子又はTRBC2遺伝子のいずれかによりコードされる定常ドメイン内に含まれない全てのアミノ酸を包含すると理解される(T cell receptor Factsbook(2001) LeFranc and LeFranc,Academic Press,ISBN 0-12-441352-8)
。
本明細書で使用する場合、用語「可変ドメイン」は、所定のTCRの、TCRα鎖に関してはTRAV遺伝子により、TCRβ鎖に関してはTRBV遺伝子によりコードされる全てのアミノ酸を包含すると理解される(T cell receptor Factsbook(2001) LeFranc and LeFranc,Academic Press,ISBN 0-12-441352-8)。
【0020】
当業者には公知であるが、TCRレパートリの多様性の一部は、(本明細書中で定義するとおりの)可変領域と定常ドメインとの間の境界のコドンによりコードされるアミノ酸に生じる変化に起因する。例えば、親HIV Gag TCR配列のこの境界に存在するコドンは、可変領域配列のC末端でのヒスチジン(H)残基の存在を生じる。このヒスチジンは、図8aに示されるTRAC遺伝子によりコードされるN末端アスパラギン(N)残基に置き換わる。
【0021】
本発明の実施形態は、95T、96N、97S、98G及び100Aに相当するα鎖可変領域アミノ酸のうちの1又はそれより多くの変異、例えばアミノ酸:
95S又はG
96A
97H
98D
100S
を含んでなる変異したTCRを包含する。
上記で使用した番号付けは、図1a及び配列番号1に示したものと同じである。
【0022】
本発明の実施形態はまた、下記に列挙するものに対応するTCRβ鎖可変領域アミノ酸の1又はそれより多くの変異を含んでなるTCRを包含し、それらは図1b及び配列番号2の天然型HIV Gag TCRβ鎖の天然型HIV Gag TCRα鎖可変領域のために提供される配列中のこれら位置に存在するアミノ酸に関して変異している。変異してもよいとされるアミノ酸は、51Y、52E、53E及び54Eであり、例えば:
51V又はA
52R又はL
53G
54V
上記で使用した番号付けは、図1b及び配列番号2で示したものと同じである。
【0023】
本発明の更なる好ましい実施形態は、図6に示される変異したα鎖可変領域アミノ酸配列(配列番号11〜13)の1つを含んでなるTCRにより提供される。このようなTCRの表現型上サイレントな変形体もまた、本発明の一部を形成する。
本発明の追加の好ましい実施形態は、図7に示される変異したβ鎖可変領域アミノ酸配列(配列番号14及び15)の1つを含んでなるTCRにより提供される。このようなTCRの表現型上サイレントな変形体もまた、本発明の一部を形成する。
【0024】
天然型TCRはへテロ二量体αβ又はγδ形態で存在する。しかし、αα又はββのホモ二量体からなる組換えTCRが、ペプチドMHC分子に結合することが以前に示されている。したがって、本発明の1つの実施形態は、TCRαα又はTCRββのホモ二量体により提供される。
更に好ましい実施形態は、下記に列挙するα鎖可変領域アミノ酸配列とβ鎖可変領域アミノ酸配列との組合せを含んでなる本発明のTCRにより提供され、このようなTCRの表現型上サイレントな変形体もまた、本発明の一部を形成する:
【0025】
【表1】
【0026】
別の好ましい実施形態では、上記で詳述した可変領域の組合せを含んでなる本発明のTCRは、図8aに示されるα鎖定常ドメインアミノ酸配列(配列番号19)並びに図8b及び8cに示されるβ鎖アミノ酸定常ドメイン配列(配列番号20及び21)の一方、又はそれらの表現型上サイレントな変形体を更に含んでなる。
【0027】
本明細書中で使用される場合、用語「表現型上サイレントな変形体」は、SLYNTVATL−HLA-A*0201複合体に対して1μMより少ないか若しくはこれと等しいKDを有し及び/又は1×10-3 S-1か若しくはそれより遅い解離速度(koff)を有するTCRをいうと理解される。例えば、当業者には公知であるが、SLYNTVATL−HLA-A*0201複合体との相互作用に関する親和性及び/又は解離速度を変化させることなく、上記で詳述したものと比較して軽微な変化が定常ドメイン及び/又は可変領域に組み込まれているTCRを作製することは可能であり得る。このような平凡な変形体は本発明の範囲に包含される。1又はそれより多い保存的置換がなされたTCRもまた、本発明の一部を形成する。
【0028】
1つの広い観点では、本発明のTCRは、WO 04/033685及びWO 03/020763に記載されるような単鎖TCR(scTCR)又は二量体TCR(dTCR)のいずれかの形態である。
【0029】
適切なscTCR形態は、TCRα鎖可変領域に相当するアミノ酸配列により構成される第1のセグメントと、TCRβ鎖定常ドメイン細胞外配列に相当するアミノ酸配列のN末端に融合したTCRβ鎖可変領域配列に相当するアミノ酸配列により構成される第2のセグメントと、第1のセグメントのC末端を第2のセグメントのN末端に連結するリンカー配列とを含んでなる。
【0030】
或いは、第1のセグメントは、TCRβ鎖可変領域に相当するアミノ酸配列により構成されてもよく、第2のセグメントは、TCRα鎖定常ドメイン細胞外配列に相当するアミノ酸配列のN末端に融合したTCRα鎖可変領域に相当するアミノ酸配列により構成されてもよい。
【0031】
上記scTCRは、第1の鎖と第2の鎖との間のジスルフィド結合を更に含んでなってもよく、このジスルフィド結合は、天然型αβT細胞レセプター中に等価物がないものであり、リンカー配列の長さ及びジスルフィド結合の位置は、第1及び第2のセグメントの可変ドメイン配列が天然型αβT細胞レセプター中と実質的に同様に互いに配向するような長さ及び位置である。
【0032】
より具体的には、第1のセグメントは、TCRα鎖定常ドメイン細胞外配列に相当するアミノ酸配列のN末端に融合したTCRα鎖可変領域に相当するアミノ酸配列により構成されていてもよく、第2のセグメントは、TCRβ鎖定常ドメイン細胞外配列に相当するアミノ酸配列のN末端に融合したTCRβ鎖可変領域に相当するアミノ酸配列により構成されていてもよく、ジスルフィド結合は、第1の鎖と第2の鎖との間に提供されてもよく、このジスルフィド結合は、天然型αβT細胞レセプター中に等価物がないものである。
【0033】
上記scTCR形態では、リンカー配列は、第1のセグメントのC末端を第2のセグメントのN末端に連結してもよく、式-PGGG-(SGGGG)n-P-(式中、nは5又は6であり、Pはプロリンであり、Gはグリシンであり、Sはセリンである)を有していてもよい。
-PGGG-SGGGGSGGGGSGGGGSGGGGSGGGG-P (配列番号17)
-PGGG-SGGGGSGGGGSGGGGSGGGGSGGGGSGGGG-P (配列番号18)
【0034】
本発明のTCRの適切なdTCR形態は、TCRα鎖可変領域配列に相当する配列がTCRα鎖定常ドメイン細胞外配列に相当する配列のN末端に融合している第1のポリペプチドと、TCRβ鎖可変領域配列に相当する配列がTCRβ鎖定常ドメイン細胞外配列に相当する配列のN末端に融合している第2のポリペプチドとを含んでなり、第1及び第2のポリペプチドが天然型αβT細胞レセプター中に等価物がないジスルフィド結合により連結している。
【0035】
第1のポリペプチドは、TCRα鎖定常ドメイン細胞外配列に相当する配列のN末端に融合したTCRα鎖可変領域配列を含んでなってもよく、第2のポリペプチドでは、TCRβ鎖可変領域配列に相当する配列がTCRβ鎖定常ドメイン細胞外配列に相当する配列のN末端に融合し、第1及び第2のポリペプチドは、TRAC*01のエキソン1のThr 48及びTRBC1*01若しくはTRBC2*01のエキソン1のSer 57又はこれらの非ヒト等価物から置換したシステイン残基同士間のジスルフィド結合により連結されている(本明細書中の「TRAC」などの命名法は、T cell receptor Factsbook(2001) LeFranc and LeFranc,Academic Press,ISBN 0-12-441352-8による)。
【0036】
本発明のTCRのdTCR形態又はscTCR形態は、ヒトαβTCRの細胞外定常ドメイン及び可変領域の配列に相当するアミノ酸配列を有していてもよく、ジスルフィド結合は、前記定常ドメイン配列のアミノ酸残基同士を連結してもよく、このジスルフィド結合は、天然型TCR中には等価物がない。ジスルフィド結合は、β炭素原子が天然型TCR中で0.6nm未満離れているアミノ酸残基に相当するシステイン残基同士間、例えばTRAC*01のエキソン1のThr 48及びTRBC1*01若しくはTRBC2*01のエキソン1のSer 57又はこれらの非ヒト等価物から置換されたシステイン残基同士間にある。システインが導入されてジスルフィド結合を形成することができる他の部位は、TCRα鎖についてはTRAC*01のエキソン1中の、TCRβ鎖についてはTRBC1*01又はTRBC2*01のエキソン1中の以下の残基である:
【0037】
【表2】
【0038】
上記で言及した非天然型ジスルフィド結合に加えて、本発明のTCRのdTCR形態又はscTCR形態は、天然型TCR中のジスルフィド結合により連結される残基に相当する残基同士間のジスルフィド結合を含んでもよい。
本発明のTCRのdTCR形態又はscTCR形態は、好ましくは、天然型TCRの膜貫通配列に相当する配列も細胞質配列に相当する配列も含有しない。
【0039】
本発明のTCRは、SLYNTVATL−HLA-A2*0201に強く結合する。これらTCRはまた、HLA-A*0201が搭載する場合にはHIV Gag由来SLYNTVATLの天然に存在する変形体に、変化した程度ではあるが未だ有用な程度で結合する。AIDs患者から単離されたSLYNTVATL変形体には以下が含まれる(Sewellら,(1997) Eur J Immunol. 27:2323-2329):
【0040】
SLFNTVATL
SLFNTVAVL
SLSNTVATL
SSFNTVATL
SLLNTVATL
SLYNTIATL
SLYNTIAVL
SLFNTIATL
SLFNTIAVL
SLFNFVATL
変異アミノ酸には下線を付する。
【0041】
PEG化TCR単量体
1つの特定の実施形態において、本発明のTCRは、少なくとも1つのポリアルキレングリコール鎖と結合している。この結合は、当業者に公知の多くの方法で引き起こされ得る。好ましい実施形態では、ポリアルキレン鎖は、TCRと共有結合している。更なる実施形態では、本発明のこの観点のポリエチレングリコール鎖は、少なくとも2つのポリエチレン反復単位を含んでなる。
【0042】
多価TCR複合体
本発明の1つの観点は、少なくとも2つの本発明のTCRを含んでなる多価TCR複合体を提供する。この観点の1つの実施形態では、少なくとも2つのTCR分子は、リンカー部分を介して連結して多価複合体を形成する。好ましくは、この複合体は水溶性であるので、リンカー部分はそのように選択されるべきである。更に、リンカー部分は、形成される複合体の構造的多様性が最小となるように、TCR分子の規定位置に付着することができるべきであることが好ましい。この観点の1つの実施形態は、ポリマー鎖又はペプチド性リンカー配列が各TCRの可変領域配列中に位置しないアミノ酸残基同士の間に伸びる本発明のTCR複合体により提供される。
【0043】
本発明の複合体は医薬で使用され得るので、リンカー部分は、薬学的適合性(例えば免疫原性)に当然払うべき注意をもって選択されるべきである。
上記の望ましい基準を満たすリンカー部分の例は、当該分野(例えば抗体フラグメントを連結する技術分野)で公知である。
【0044】
本発明の多価TCR分子の製造での使用に好ましい2つのクラスのリンカーが存在する。TCRがポリアルキレングリコール鎖により連結されている本発明のTCR複合体は、この観点の1つの実施形態を提供する。
【0045】
第1は、親水性ポリマー、例えばポリアルキレングリコールである。最も通常に使用されるこのクラスのものは、ポリエチレングリコールすなわちPEGをベースにする。その構造を下記に示す。
HOCH2CH2O(CH2CH2O)n−CH2CH2OH
(式中、nは2より大きい)。しかし、他の適切な(任意に置換されていてもよい)ポリアルキレングリコールをベースにするものもある。ポリプロピレングリコー及びエチレングリコールとプロピレングリコールとのコポリマーが挙げられる。
【0046】
このようなポリマーは、治療剤、特にポリペプチド治療剤又はタンパク質治療剤を処理するか又はこれに接合するかして、該治療剤のPKプロフィールに有益な変化(例えば、減少した腎クリアランス、向上した血漿半減期、減少した免疫原性及び向上した溶解性)を達成するために使用され得る。PEG−治療剤接合体のPKプロフィールのこのような改善は、PEG分子が該治療剤の周囲に、免疫系との反応を立体的に障害し、タンパク質分解性分解を減少させる「殻」を形成することに起因すると考えられる(Caseyら(2000) Tumor Targeting 4 235-244)。使用する親水性ポリマーのサイズは、具体的には、TCR複合体の意図する治療用途に基づいて選択し得る。医薬製剤におけるPEG及び類似分子の使用を詳述する多くの総説論文及び本が存在する。例えば、Harris(1992) Polyethylene Glycol Chemistry - Biotechnical and Biomedical Applications,Plenum,New York,NY.、又はHarris & Zalipsky(1997) Chemistry and Biological Applications of Polyethylene Glycol ACS Books,Washington,D.C.を参照。
【0047】
使用するポリマーは、直鎖状又は分枝状の構造を有することができる。分枝状PEG分子又はその誘導体は、グリセロール及びグリセロールオリゴマー、ペンタエリスリトール、ソルビトール並びにリジンを含む分枝成分の付加により誘導することができる。
【0048】
通常、ポリマーは、該ポリマーがTCR中の標的部位へ連結することが可能となるように、その構造中、例えば一方又は両方の端部及び/又は主鎖からの分枝上に、化学反応性の基を有する。下記に示すように、この化学反応性の基は親水性ポリマーに直接結合させてもよいし、又は親水性ポリマーと反応性化学成分との間にスペーサ基/成分が存在していてもよい:
反応性化学成分−親水性ポリマー−反応性化学成分
反応性化学成分−スペーサ−親水性ポリマー−スペーサ−反応性化学成分
【0049】
上記で概説したタイプの構築物の形成に使用するスペーサは、非反応性の化学的に安定な鎖である任意の有機成分であり得る。このようなスペーサには、以下が含まれるがそれらに限定されない:
-(CH2)n- (式中、n=2〜5)
-(CH2)3NHCO(CH2)2
【0050】
二価アルキレンスペーサ基がポリアルキレングリコール鎖とTCRへのその結合点との間に位置する本発明のTCR複合体は、この観点の更なる実施形態を提供する。
ポリアルキレングリコール鎖が少なくとも2つのポリエチレングリコール反復単位を含んでなる本発明のTCR複合体は、この観点の更なる実施形態を提供する。
【0051】
本発明において有用であり得る反応性化学成分に直接か又はスペーサを介して連結する親水性ポリマーの商業的供給業者は多く存在する。これら供給業者としては、Nektar Therapeutics(CA、米国)、NOF Corporation(日本)、Sunbio(韓国)及びEnzon Pharmaceuticals(NJ、米国)が挙げられる。
【0052】
本発明において有用であり得る反応性化学成分に直接か又はスペーサを介して連結する市販の親水性ポリマーには、以下のものが含まれるがそれらに限定されない。
【表3】
【0053】
種々のカップリング化学成分を使用して、ポリマー分子をタンパク質治療薬やペプチド治療薬とカップリングすることができる。最も適切なカップリング化学成分の選択は、所望するカップリング部位に大きく依存する。例えば、以下のカップリング化学成分が、PEG分子の1又はそれ以上の末端に付着されるために使用されてきた(出典:Nektar Molecular Engineering Catalogue 2003):
【0054】
N−マレイミド
ビニルスルホン
炭酸ベンゾトリアゾール
スクシンイミジルプロピオネート
スクシンイミジルブタノエート(succinimidyl butanoate)
チオエステル
アセトアルデヒド
アクリラート
ビオチン
一級アミン
【0055】
上記のように、非PEGベースのポリマーもまた、本発明のTCRを多量化するために適切なリンカーを提供する。例えば、脂肪族鎖により連結されたマレイミド末端を含有する成分、例えばBMH及びBMOE(Pierce,製品番号22330及び22323)が使用できる。
【0056】
ペプチド性リンカーが他方の好ましいクラスのTCRリンカーである。これらリンカーは、アミノ酸の鎖から構成され、単純なリンカー又はTCR分子を付着させることができる多量体化ドメインを作製するために機能する。ビオチン/ストレプトアビジン系は、以前に、インビトロ結合研究用のTCR四量体を作製するために使用された(WO/99/60119を参照)。しかし、ストレプトアビジンは微生物由来のポリペプチドであり、よって治療剤における使用に理想的には適していない。
【0057】
TCRがヒト多量体化ドメインに由来するペプチド性リンカーにより連結されている本発明のTCR複合体は、この観点の更なる実施形態を提供する。
【0058】
多価TCR複合体の作製に使用できる多量体化ドメインを含有する多くのヒトタンパク質が存在する。例えば、p53の四量体化ドメインは、単量体scFvフラグメントと比較して、増大した血清残存率及び有意に減少した解離速度を示すscFv抗体フラグメント四量体を作製するために利用されている(Willudaら(2001) J. Biol. Chem. 276 (17) 14385-14392)。ヘモグロビンもまた、この種の適用におそらく使用できる四量体化ドメインを有する。
【0059】
少なくとも一方が治療剤に結合されている少なくとも2つのTCRを含んでなる本発明の多価TRC複合体はこの観点の最後の実施形態を提供する。
1つの観点では、本発明のTCR(又はその多価複合体)は、択一的又は追加的に、そのα鎖又はβ鎖のC末端又はN末端に反応性のシステインを含んでなり得る。
【0060】
診断及び治療用途
1つの観点では、本発明のTCRは治療剤又は検出可能な成分と結合させてもよい。例えば、治療剤又は検出可能な成分はTCRに共有結合させてもよい。
本発明の1つの実施形態において、治療剤又は検出可能な成分は、一方又は両方のTCR鎖のC末端に共有結合している。
【0061】
1つの観点において、本発明のscTCR又はdTCRの一方若しくは両方の鎖が検出可能な成分、例えば診断目的に適切な標識で標識されてもよい。このような標識TCRは、TCRリガンドと該TCRリガンドに特異的なTCR(又は多量体高親和性TCR複合体)とを接触させ、TCRリガンドへの結合を検出することを含んでなる、SLYNTVATL−HLA-A*0201複合体を検出する方法において有用である。例えばビオチン化ヘテロ二量体を用いて形成された四量体TCR複合体では、蛍光ストレプトアビジンを使用し、検出可能な標識を提供することができる。このような蛍光標識TCR四量体は、例えばこれら高親和性TCRが特異的であるSLYNTVATL−HLA-A*0201複合体を有する抗原提示細胞を検出するために、FACS分析で使用するに適切である。
【0062】
本発明の可溶性TCRが検出され得る別の様式は、TCR特異的抗体、特にモノクローナル抗体の使用による。多くの市販の抗TCR抗体、例えばαF1及びβF1(それぞれα鎖及びβ鎖の定常領域を認識する)が存在する。
【0063】
更なる観点において、本発明のTCR(又はその多価複合体)は、択一的又は追加的に、例えば細胞殺傷に使用するための毒性成分又は免疫エフェクター分子(例えばインターロイキン又はサイトカイン)であり得る治療剤と結合(例えば、共有結合又はその他の結合)していてもよい。本発明の多価TCR複合体は、非多量体の野生型又は本発明のT細胞レセプターへテロ二量体と比べて、TCRリガンドに関して増強した結合能力を有し得る。したがって、本発明による多価TCR複合体は、SLYNTVATL−HLA-A*0201複合体を提示する細胞をインビトロ又はインビボで追跡又は標的するために特に有用であり、またそのような用途を有する更なる多価TCR複合体の製造のための中間体としても有用である。したがって、これらTCR又は多価TCR複合体は、インビボでの使用のために医薬的に許容される製剤で提供され得る。
【0064】
本発明はまた、標的細胞に治療剤を送達するための方法を提供する。この方法は、可能性のある標的細胞を本発明によるTCR又は多価TCR複合体と、標的細胞へのTCR又は多価TCR複合体の付着を可能にする条件下で接触させることを含んでなり、当該TCR又は多価TCR複合体は、SLYNTVATL−HLA-A*0201複合体に特異的であり、これに治療剤が結合している。
【0065】
特に、本発明の可溶性TCR又は多価TCR複合体を使用して、特定の抗原を提示する細胞の位置に治療剤を送達することができる。このことは、多くの状況で、特にHIV感染細胞に対して有用である。治療剤は、その効果を局所的にではあるが、該治療剤が結合する細胞に限らず発揮するように送達され得る。したがって、1つの特定の戦略は、SLYNTVATL−HLA-A*0201複合体に特異的な本発明のTCR又は多価TCR複合体に連結した細胞毒性又は免疫刺激性の分子を考案する。
【0066】
多くの治療剤、例えば放射活性化合物、酵素(例えばパーフォリン)又は化学療法剤(例えばシスプラチン)がこの用途に用いられ得る。確実に所望の位置で毒性効果が発揮されるために、毒素は、それがゆっくりと放出されるように、ストレプトアビジンに連結したリポソーム内にあり得る。このことにより、体内での輸送の間の損傷効果が防止され、TCRと該当する抗原提示細胞との結合後に毒素が最大効果を有することが確実になる。
【0067】
他の適切な治療剤として、以下のものが挙げられる:
・小分子細胞毒性物質、すなわち、哺乳動物細胞を殺傷する能力を有する、分子量700ダルトン未満の化合物。このような化合物はまた、細胞毒性効果を有することができる毒性金属を含有し得る。更に、これら小分子細胞毒性物質にはまた、プロドラッグ、すなわち、生理学的条件下で崩壊又は変換して細胞毒性物質を放出する化合物が含まれると理解される。このような物質の例には、シスプラチン、メイタンシン(maytansine)誘導体、ラケルマイシン(rachelmycin)、カリケアマイシン(calicheamicin)、ドセタキセル、エトポシド、ゲムシタビン、イホスファミド、イリノテカン、メルファラン、ミトキサントロン、ソルフィマーソディウムホトフィリンII(sorfimer sodiumphotofrin II)、テモゾロマイド(temozolmide)、トポテカン、トリメトレキサート(trimetreate)、グルクロナート、オーリスタチンE(auristatin E)、ビンクリスチン及びドキソルビシンが含まれる;
【0068】
・ペプチド細胞毒素、すなわち、哺乳動物細胞を殺傷する能力を有するタンパク質又はそのフラグメント。例には、リシン、ジフテリア毒素、シュードモナス細菌外毒素A、DNAアーゼ及びRNAアーゼが含まれる;
・放射性核種、すなわち、1以上のα粒子若しくはβ粒子又はγ線の同時放射を伴って崩壊する元素の不安定同位体。例には、ヨウ素131、レニウム186、インジウム111、イットリウム90、ビスマス210及び213、アクチニウム225及びアスタチン213が含まれるがこれらに限定されない;キレート化剤が、高親和性TCR又はその多量体へのこれら放射性核種の結合を促進するために使用されてもよい;
・プロドラッグ、抗体を指向する酵素プロドラッグを含むがこれに限定されない;
【0069】
・免疫刺激剤、すなわち、免疫応答を刺激する成分。サイトカイン(例えば、IL-2及びIFN)、スーパー抗原及びその変異体、TCR-HLA融合体及びケモカイン(例えばIL-8)、血小板第4因子、メラノーマ増殖刺激タンパク質など、抗体又はそのフラグメント、補体活性因子、異種タンパク質ドメイン、同種タンパク質ドメイン、ウイルス性/細菌性タンパク質ドメイン、ウイルス性/細菌性ペプチド及び抗T細胞決定基抗体(例えば、抗CD3又は抗CD28)又は抗体アナログ(例えば、ナノボディ(商標)(Nanobodies)及びアフィボディ(商標)(Affydodies))を含むがこれらに限定されない。
【0070】
本発明の可溶性TCR又は多価TCR複合体は、プロドラッグを薬物に変換し得る酵素に連結されてもよい。これにより、プロドラッグが薬物を必要とする(すなわち、sTCRにより標的にされる)部位でのみ薬物に変換することが可能になる。
【0071】
本明細書中で開示される高親和性SLYNTVATL(配列番号16)−HLA-A*0201特異的TCRは、AIDSの診断方法及び治療方法に使用され得ると考えられる。
治療には、HIV感染(CD4+)細胞の近傍への治療剤の局在化が、毒素又は免疫刺激剤の効果を増強する。ワクチン送達には、ワクチン抗原を抗原提示細胞の近傍に局在させ、該抗原の効力を増強させ得る。この方法はまた造影目的にも適用できる。
【0072】
1つの実施形態は、本発明のTCRを含んでなる膜調製物により提供される。この膜調製物は、細胞から調製されてもよいし、又は人工膜を含んでなってもよい。
別の実施形態は、本発明のTCRをコードする核酸を含んでなる発現ベクターを有する細胞により提供される。例えば、この細胞はT細胞であり得る。
【0073】
本発明の更なる実施形態は、以下:
医薬的に許容されるキャリアと共に、本発明のTCR若しくは多価TCR複合体(任意に治療剤と結合していてもよい)、又は本発明のTCRを含んでなる膜調製物、又は本発明のTCRをコードする核酸を含んでなる発現ベクターを有する複数の細胞
を含んでなる医薬組成物により提供される。
【0074】
本発明はまた、AIDSを患う対象に、本発明のTCR若しくは多価TCR複合体又は本発明のTCRを含んでなる膜調製物又は本発明のTCRをコードする核酸を含んでなる発現ベクターを有する複数の細胞の有効量を投与することを含んでなる、AIDSの治療法を提供する。関連する実施形態では、本発明は、AIDS治療用組成物の製造における、本発明のTCR若しくは多価TCR複合体又は本発明のTCRを含んでなる膜調製物又は本発明のTCRをコードする核酸を含んでなる発現ベクターを有する複数の細胞の使用を提供する。本発明のこれら使用及び方法の更なる具体的実施形態が提供され、そこでは、本発明のTCR若しくは多価TCR複合体又は本発明のTCRを含んでなる膜調製物は、治療剤と結合した形態で投与される。他の好ましい実施形態では、本発明のTCRをコードする核酸を含んでなる発現ベクターを有する細胞はCD8+T細胞である。
【0075】
本発明による治療用又は造影用TCRは、通常、滅菌の、一般には薬学的に許容されるキャリアを含む医薬組成物の一部として供給される。この医薬組成物は(所望される患者への投与法に依存して)任意の適切な形態であり得る。この医薬組成物は、単位剤形で提供されてもよく、一般には密封容器中で提供され、キットの一部として提供されてもよい。そのようなキットは、通常(必ずというわけではないが)、使用のための指示書を含む。キットは、前記の単位剤形を複数含み得る。
【0076】
理論により制限されることは望まないが、本発明のTCRは、治療剤(例えば免疫刺激剤及び/又は細胞傷害剤)をHIV感染(CD4+)細胞に送達することができる効果的な標的化剤(targeting agent)を提供すると考えられる。詳細には、従来の抗レトロウイルス薬治療及び/又はIL-2治療との組合せで、免疫刺激剤及び/又は細胞傷害剤と結合させると本発明のTCRの投与は、HIV感染細胞を標的することができると考えられる。
【0077】
以下は、現在米国で使用が承認されている抗レトロウイルスの一覧である:
アジェネラーゼ(アムプレナビル)−プロテアーゼ阻害剤
コンビビル−レトロビル(300mg)とエピビル(150mg)の組合せ
クリキシバン(インジナビル)−プロテアーゼ阻害剤
エピビル(3tc/ラミブジン)−ヌクレオシドアナログ逆転写酵素阻害剤
エプジコム(2つのヌクレオシド逆転写酵素阻害剤の組合せ(同じ丸剤中のNRTI;600mgのザイアジェン(アバカビル)及び300mgのエピビル(3TC)
【0078】
エムトリバ[エムトリシタビン(FTC)]
フォートベイス(サキナビル)−プロテアーゼ阻害剤
フューゼオン(Fuzeon)(エンフュービルタイド(enfuvirtide))−融合阻害剤
ハイビッド(ddc/ザルシタビン)−ヌクレオシドアナログ逆転写酵素阻害剤
インビラーゼ(サキナビル)−プロテアーゼ阻害剤
【0079】
カレトラ(ロピナビル)−プロテアーゼ阻害剤
レクシヴァ(ホスアンプレナビル)−2003年10月20日承認のプロテアーゼ阻害剤
ノービア(リトナビル)−プロテアーゼ阻害剤
レスクリプター(デラビルジン)−非ヌクレオシドアナログ逆転写酵素阻害剤
レトロビル、AZT(ジドブジン)−ヌクレオシドアナログ逆転写酵素阻害剤
【0080】
レイアタッツ(アタザナビル;BMS-232632)−プロテアーゼ阻害剤
サスティバ(エファビレンツ)−非ヌクレオシドアナログ逆転写酵素阻害剤
トリジビル(1錠剤中3つの非ヌクレオシド;アバカビル+ジドブジン+ラミブジン)
ツルバダ(エムトリシタビン+テノホビルDF)
ヴァイデックス(ddl/ジダノシン)−ヌクレオシドアナログ逆転写酵素阻害剤
ヴァイデックスEC;(ddl/ジダノシン)−ヌクレオシドアナログ逆転写酵素阻害剤;
【0081】
ビラセプト(ネルフィナビル)−プロテアーゼ阻害剤
ビラミューン(ネビラピン)−非ヌクレオシドアナログ逆転写酵素阻害剤
ビリアード(フマル酸テノホビルジソプロキシル)−ヌクレオチド逆転写酵素阻害剤(アデノシンクラス)
ゼリット(d4t/スタブジン)−ヌクレオシドアナログ逆転写酵素阻害剤
ザイアジェン(アバカビル)−ヌクレオシドアナログ逆転写酵素阻害剤
【0082】
医薬組成物は、任意の適切な経路、例えば、非経口経路、経皮経路又は吸入を介する経路(好ましくは非経口経路(皮下、筋肉内又は(最も好ましい)静脈内を含む))による投与に適合され得る。このような組成物は、医薬の技術分野において公知の任意の方法(例えば、滅菌条件下で活性成分をキャリア又は賦形剤と混合すること)により製造され得る。
【0083】
本発明の物質の投薬量は、治療すべき疾患又は障害、治療すべき個体の年齢及び状態などに依存して、広範囲に変わり得る。最終的には、医師が使用すべき適切な投薬量を決定する。
【0084】
追加の観点
本発明のscTCR又はdTCR(好ましくは、ヒト配列に対応する定常配列及び可変配列により構成される)は、実質的に純粋な形態で、又は精製若しくは単離された調製物として提供されてもよい。例えば、本発明のscTCR又はdTCRは、他のタンパク質を実質的に含まない形態で提供されてもよい。
【0085】
本発明のTCRをコードする核酸の配列は、宿主細胞で得られる発現レベルを最適化するように変更されてもよい。宿主細胞は任意の適切な原核細胞又は真核細胞であり得る。例えば、宿主細胞はE.coli細胞又はヒトT細胞であってもよい。これら遺伝子配列に対してなされる変更はサイレントである。すなわち、コードされるアミノ酸配列を変更しない。そのような発現最適化サービスを提供する会社は、GeneArt(ドイツ)を始めとして数多く存在する。
【0086】
本発明はまた、(i)少なくとも1つのTCRα鎖可変ドメイン及び/又は少なくとも1つのTCRβ鎖可変ドメインを含んでなり、且つ(ii)1μMより少ないか若しくは等しいSLYNTVATL−HLA-A*0201複合体に対するKD及び/又は1×10-3か若しくはそれより遅いSLYNTVATL−HLA-A*0201複合体に対する解離速度(koff)を有することを特徴とするSLYNTVATL−HLA-A*0201への結合特性を有する高親和性TCRを作製する方法を提供する。この方法は、
(a)親HIV Gag TCRのα鎖可変ドメイン及びβ鎖可変ドメインを含んでなり、そのα鎖可変ドメイン及びβ鎖可変ドメインの一方又は両方が請求項7及び8で特定したアミノ酸の1又はそれ以上において変異を含んでなるTCRを作製し;
(b)この変異TCRをSLYNTVATL−HLA-A*0201と、該TCRのSLYNTVATL−HLA-A*0201への結合を可能にするに適切な条件下で接触させ、
その相互作用のKD及び/又はkoffを測定する
ことを含んでなる。
【0087】
本発明の各観点の好ましい特徴は、必要な変更を加えてではあるが、他の観点の各々についても同様である。本明細書中で言及する先行技術文献は法が許す最大限の範囲で本明細書に援用する。
【実施例】
【0088】
実施例
以下の実施例で本発明を更に説明する。以下の実施例は、如何なる態様でも本発明の範囲を制限しない。
以下、添付の図面に言及する:
【0089】
図1a及び1bはそれぞれ、親HIV Gag TCRのα鎖可変ドメインアミノ酸配列及びβ鎖可変ドメインアミノ酸配列を詳述する。
図2a及び2bはそれぞれ、可溶性型の親HIV Gag TCRα鎖及びβ鎖のDNA配列を示す。
図3a及び3bはそれぞれ、図2a及び2bのDNA配列から産生されるHIV Gag TCRα鎖及びβ鎖の細胞外アミノ酸配列を示す。
【0090】
図4a及び4bはそれぞれ、非天然型ジスルフィド結合を形成するための追加のシステイン残基をコードするように変異した可溶性型のHIV Gag TCRα鎖及びβ鎖のDNA配列を示す。変異コドンを影付きで示し、導入した制限酵素認識部位に下線を付す。
図5a及び5bはそれぞれ、図4a及び4bのDNA配列から産生されるHIV Gag TCRα鎖及びβ鎖の細胞外アミノ酸配列を示す。各鎖における導入システインを影付きで示す。
【0091】
図6は、高親和性HIV Gag TCR変形体のα鎖可変ドメインアミノ酸配列を詳述する。
図7は、高親和性HIV Gag TCR変形体のβ鎖可変ドメインアミノ酸配列を詳述する。
図8aは、TRACの可溶性部分のアミノ酸配列を詳述する。
図8bは、TRBC1の可溶性部分のアミノ酸配列を詳述する。
図8cは、TRBC2の可溶性部分のアミノ酸配列を詳述する。
【0092】
図9は、pEX954プラスミドのDNA配列を詳述する。
図10は、pEX821プラスミドのDNA配列を詳述する。
図11は、ペプチドリンカーを介して野生型ヒトIL-2に融合した可溶性の親HIV Gag TCR
変形体のβ鎖アミノ酸配列を詳述する。リンカー及びIL-2のアミノ酸をイタリック体で示す。
図12は、ジスルフィド連結した可溶性の親HIV Gag TCRとSLYNTVATL−HLA-A*0201複合体との相互作用について作成したBiacore応答曲線を提供する。
【0093】
図13は、pEX954プラスミドのプラスミドマップを提供する。
図14は、pEX821プラスミドのプラスミドマップを提供する。
図15aは、ヒトT細胞中での発現について最適化された親HIV Gag TCRα鎖の全長DNA配列を提供する。
図15bは、ヒトT細胞中での発現について最適化された親HIV Gag TCRβ鎖の全長DNA配列を提供する。
【0094】
図16aは、親HIV Gag TCRα鎖の全長アミノ酸配列を提供する。
図16bは、ヒトT細胞中での発現について最適化された親HIV Gag TCRβ鎖の全長アミノ酸配列を提供する。
図17aは、コントロールの非形質転換CD8+T細胞についてのFACS分析データを提供する。
図17bは、形質転換CD8+T細胞の表面上での親HIV Gag TCRの発現を示すFACS分析データを提供する。
【0095】
図18a及び18bはそれぞれ、可溶性のジスルフィド連結高親和性c11c6 HIV Gag TCRのα鎖及びβ鎖アミノ酸配列を提供する。
図19は、IFN-γ産生及びTNF-α産生により測定したときの、可溶性のジスルフィド連結高親和性c11c6 HIV Gag TCRがHIVに感染したTo細胞の存在下でSLYNTVATL−HLA-A*0201反応性OX84ポリクローナルT細胞株の活性化を阻害する能力を示す。
【0096】
図20は、IFN-γ産生及びTNF-α産生により測定したときの、可溶性のジスルフィド連結高親和性c11c6 HIV Gag TCRがSLYNTVATLペプチドでパルスした非感染To細胞の存在下でSLYNTVATL−HLA-A*0201反応性OX84ポリクローナルT細胞株の活性化を阻害する能力を示す。
図21は、可溶性のジスルフィド連結高親和性c11c6 HIV Gag TCRがSLYNTVATLペプチドでパルスしたT2細胞を染色する能力を示す。
【0097】
実施例1 − 親HIV Gag TCR可変領域を含んでなる可溶性のジスルフィド連結TCRの作製
図4a及び4bは、SLYNTVATL−HLA-A*0201複合体に特異的な親TCRの可溶性ジスルフィド連結αβ鎖のDNA配列を提供する。これらDNA配列は、多くの受託研究会社、例えばGeneArt(ドイツ)により新たに合成され得る。また、これらDNA配列のpGMT7ベースの発現プラスミド(これは、E.coli株BL21-DE3における高レベル発現用のT7プロモーターを含有する(pLysS)(Panら,Biotechniques(2000) 29 (6):1234-8))中へのライゲーションを容易にするために、これらDNA配列に制限酵素認識部位を付加することもできる。
【0098】
TCRα鎖配列は導入したClaI及びSalII制限酵素認識部位を含有する。この配列を、ClaI及びXhoIで切断したpEX954(図9及び13を参照)中にライゲートした。
TCRβ鎖配列は導入したAseI及びAgeI制限酵素認識部位を含有する。この配列を、NdeI/AgeIで切断したpEX821(図10及び14を参照)中にライゲートした。
【0099】
TCR鎖をコードするDNA中に導入した制限酵素認識部位
ClaI - ATCGAT
SalII - GTCGAC
AseI - ATTAAT
AgeI - ACCGGT
【0100】
ライゲーション
ラピッドDNAライゲーションキット(Roche)を製造業者の指示書に従って用い、切断したTCRα鎖及びβ鎖のDNAと切断したベクターとをライゲートした。
ライゲートしたプラスミドをコンピテントE.coli株XL1-blue細胞に形質転換し、100mg/mlアンピシリン含有LB/寒天プレート上に播種した。37℃で一晩のインキュベーション後、1つのコロニーを採取し、100mg/mlアンピシリンを含有する10mlのLB中で振盪させながら37℃で一晩増殖させた。クローン化プラスミドを、Miniprepキット(Qiagen)を用いて精製し、挿入物を自動DNAシーケンサ(Lark Technologies)を用いて配列決定した。
【0101】
図5a及び5bはそれぞれ、図4a及び4bのDNA配列から作製した、可溶性のジスルフィド連結した親HIV gag TCRのα鎖及びβ鎖の細胞外アミノ酸配列を示す。
【0102】
実施例2 − 可溶性のジスルフィド連結HIV Gag TCRの高親和性変形体の作製
実施例1に記載されるように作製した可溶性のジスルフィド連結天然型HIV Gag TCRは、SLYNTVATL(配列番号16)−HLA-A*0201複合体に対して増大した親和性を有する本発明のTCRを作製するための鋳型として用いることができる。
ファージディスプレイは、高親和性変異体を同定するためにHIV Gag TCR変形体のライブラリを作成することができる1つの手段である。例えば、(Liら(2005) Nature Biotech
23 (3):349-354)に記載されるTCRファージディスプレイ及びスクリーニング法は、HIV Gag TCRに適合させて適用することができる。
【0103】
適切なTCR鎖と組み合せるとSLYNTVATL−HLA-A*0201複合体に対して高親和性を示す変異TCRα鎖及びβ鎖可変ドメインのアミノ酸配列を、それぞれ図6及び7に列挙する(それぞれ配列番号11〜13及び14〜15)。当業者に公知であるように、これらの変異鎖を作製するために要する必要なコドン変更は、これらの鎖をコードするDNA中に、部位特異的変異誘発により導入することができる(StratageneのQuickChange(商標) Site-Directed Mutagenesis Kit)。
【0104】
簡潔には、これは、所望のコドン変更が組み込まれたプライマーと、変異誘発の鋳型として該当するTCR鎖DNAを含有するプラスミドとを用いることにより達成される。
変異誘発は次の条件を用いて行った:全容量50μl中、50ngのプラスミド鋳型、1μlの10mM dNTP、5μlの製造業者により供給された10×Pfu DNAポリメラーゼ緩衝液、25pmol
のfwdプライマー、25pmolのrevプライマー、1μlのpfu DNAポリメラーゼ。95℃で2分間の最初の変性工程後、反応物を、変性(95℃、10秒)、アニーリング(55℃、10秒)及び伸長(72℃、8分)の25サイクルに供した。得られた生成物をDpnI制限酵素で消化して鋳型プラスミドを除き、E.coli株XL1-blueに形質転換した。変異誘発を配列決定により確証した。
【0105】
実施例3 − 可溶性TCRの発現、リフォールディング及び精製
それぞれ実施例1又は2のように作製した変異α鎖及びβ鎖を含む発現プラスミドを、E.coli株BL21pLysSに別々に形質転換し、1つのアンピシリン耐性コロニーを37℃にてTYP(アンピシリン100μg/ml)培地中で0.4のOD600まで増殖させた後、0.5mM IPTGでタンパク質発現を誘導した。誘導の3時間後に、Beckman J-6B中で4000rpmにて30分間の遠心分離により細胞を回収した。細胞ペレットを、50mMのTris-HCl、25%(w/v)のスクロース、1mMのNaEDTA、0.1%(w/v)のNaAzide、10mMのDTTを含有する緩衝液(pH8.0)中に再懸濁した。一晩の凍結−解凍工程の後、再懸濁細胞を、Milsonix XL2020ソニケータ中での標準の12mm径プローブを使用する1分間のバーストで合計約10分間の超音波処理に付した。封入体ペレットを、Beckman J2-21遠心機における13000rpmにて30分間の遠心分離により回収した。次いで、3回の界面活性剤での洗浄を行い、細胞残渣及び膜成分を除去した。各回、封入体ペレットをTriton緩衝液(50mMのTris-HCl、0.5%のTriton-X100、200mMのNaCl、10mMのNaEDTA、0.1%(w/v)のNaAzide、2mMのDTT、pH8.0)中でホモジナイズした後、Beckman J2-21における13000rpmにて15分間の遠心分離によりペレット化した。次いで、以下の緩衝液中での同様な洗浄により界面活性剤及び塩を除去した:50mMのTris-HCl、1mMのNaEDTA、0.1%(w/v)のNaAzide、2mMのDTT、pH8.0。最後に、封入体を30mgずつ小分けし、−70℃にて凍結させた。封入体タンパク質の収率を、6Mグアニジン-HClで可溶化し、Bradford色素結合アッセイ(PerBio)で測定することにより定量した。
【0106】
約30mgのTCRβ鎖及び60mgのTCRα鎖の可溶化封入体を凍結ストックから解凍し、次いでサンプルを混合し、混合物を15mlのグアニジン溶液(6M塩酸グアニジン、10mM酢酸ナトリウム、10mM EDTA)中に希釈して、完全な鎖変性を確実に行った。次いで、十分に還元され変性したTCR鎖を含有するグアニジン溶液を、以下の1リットルのリフォールディング緩衝液中に注入した:100mM Tris pH8.5、400mM L-アルギニン、2mM EDTA、5mM還元型グルタチオン、0.5mM酸化型グルタチオン、5M尿素、0.2mM PMSF。酸化還元対(2-メルカプトエチルアミン及びシスタミン(それぞれ最終濃度6.6mM及び3.7mMまで))を加え、約5分後に変性TCR鎖を加えた。この溶液を5時間±15分間放置した。リフォールディングしたTCRを、Spectrapor 1メンブレン(Spectrum;製品番号132670)中で、10Lの10mM Tris(pH8.1)に対し、5℃±3℃にて18〜20時間透析した。この時間の後、透析緩衝液を新鮮な10mM Tris(pH8.1)(10L)に変え、透析を5℃±3℃にて更に20〜22時間継続した。
【0107】
透析したリフォールディング物をPOROS 50HQアニオン交換カラムにロードし、Akta Purifier(Pharmacia)を用いて50カラム容量の0〜500mM NaClグラジエントで結合タンパク質を溶出することにより、sTCRを分解産物及び不純物から分離した。ピーク画分を4℃にて貯蔵し、クーマシー染色SDS-PAGEで分析した後、プールして濃縮した。最後に、sTCRを精製して、HBS-EP緩衝液(10mM HEPES pH7.4、150mM NaCl、3.5mM EDTA、0.05% Nonidet p40)で予め平衡化したSuperdex 200HRゲル濾過カラムを用いて特徴付けを行った。約50kDaの相対分子量で溶出するピークをプールし、濃縮した後、BIAcore表面プラズモン共鳴分析により特徴付けを行った。
【0108】
実施例4 − 特異pMHCへのsTCR結合のBiacore表面プラズモン共鳴特徴付け
表面プラズモン共鳴バイオセンサ(BIAcore 3000TM)を使用して、ペプチド−MHCリガンドへのsTCRの結合を分析した。これは、半配向様式(semi-oriented fashion)でストレプトアビジン被覆結合表面に固定した単一pMHC複合体(下記で説明)を作製し、同時に4つまでの異なるpMHC(別個のフローセルに固定)への可溶性T細胞レセプターの結合の効率的な試験を可能にすることにより容易となった。HLA複合体の手動での注入により、正確なレベルの固定化クラスI分子を容易に操作することが可能となった。
【0109】
ビオチン化クラスI HLA-A*0201分子を、構成サブユニットタンパク質及び合成ペプチドを含有する細菌発現封入体から、インビトロでリフォールディングさせ、続いて精製し、インビトロにて酵素でビオチン化した(O'Callaghanら(1999) Anal. Biochem. 266:9-15)。そのタンパク質の膜貫通ドメイン及び細胞質ドメインと置換したC末端ビオチン化タグを有するHLA-A*0201-重鎖を適切な構築物中で発現させた。〜75mg/リットル細菌培養物の封入体発現レベルが得られた。MHC軽鎖又はβ2-ミクログロブリンもまた、E.coli中で適切な構築物から封入体として〜500mg/リットル細菌培養物のレベルで発現させた。
【0110】
E. coli細胞を溶解し、封入体を約80%純度まで精製した。封入体からのタンパク質を6Mのグアニジン-HCl、50mMのTris(pH8.1)、100mMのNaCl、10mMのDTT、10mMのEDTA中で変性させ、5℃より低いリフォールディング緩衝液中に変性タンパク質の単一パルス(single pulse of denatured protein)を添加することにより、0.4MのL-アルギニン-HCl、100mMのTris(pH8.1)、3.7mMのシスタミン、6.6mMのβ-システアミン、HLA-A*0201分子が搭載するために必要な4mg/mlのSLYNTVATLペプチド中で30mg/リットルの重鎖、30mg/リットルのβ2mの濃度にてリフォールディングさせた。リフォールディングは、4℃にて少なくとも1時間で完了に到達させた。
【0111】
緩衝液を、10容量の10mM Tris(pH8.1)での透析により交換した。溶液のイオン強度を十分に減少させるために、2回の緩衝液交換が必要であった。次いで、タンパク質溶液を1.5μm酢酸セルロースフィルターに通して濾過し、POROS 50HQアニオン交換カラム(8ml床容量)に充填した。タンパク質を0〜500mMのNaCl線形勾配で溶出させた。HLA-A*0201−ペプチド複合体は約250mM NaClで溶出した。ピーク画分を収集し、プロテアーゼ阻害剤のカクテル(Calbiochem)を加え、画分を氷上で冷却した。
【0112】
ビオチン化タグを付したpMHC分子を、10mM Tris(pH8.1)、5mM NaCl中に、同じ緩衝液中で平衡化したPharmacia迅速脱塩カラムを使用して移して緩衝液を交換した。溶出に際して即座に、タンパク質含有画分を氷上で冷却し、プロテアーゼ阻害剤カクテル(Calbiochem)を加えた。次いで、ビオチン化試薬を加えた:1mMビオチン、5mM ATP(pH8に緩衝化)、7.5mM MgCl2及び5μg/ml BirA酵素(O’Callaghanら(1999)Anal.Biochem.266:9-15に従って精製)。次いで、混合物を室温にて一晩インキュベートした。
【0113】
ゲル濾過クロマトグラフィーを使用してビオチン化pHLA-A*0201分子を精製した。Pharmacia Superdex 75 HR 10/30カラムを濾過PBSで予め平衡化し、1mlのビオチン化反応混合物を充填し、カラムをPBSで0.5ml/分にて展開した。ビオチン化pHLA-A*0201分子は、約15mlで単一ピークとして溶出した。タンパク質を含む画分をプールし、氷上で冷却し、プロテアーゼ阻害剤カクテルを加えた。クーマシー結合アッセイ(PerBio)を使用してタンパク質濃度を測定し、ビオチン化pHLA-A*0201分子のアリコートを−20℃で凍結保存した。標準的なアミンカップリング法によりストレプトアビジンを固定化した。
【0114】
この固定化複合体はT細胞レセプター及びコレセプターCD8αα(共に可溶性相中に注入され得る)の両方に結合することができる。低濃度(少なくとも40μg/ml)でさえTCRの特異的結合が得られる。このことは、このTCRが比較的安定であることを示す。sTCRのpMHC結合特性は、sTCRが可溶性相又は固定化相のいずれかで使用される場合、質的にも量的にも同様であることが観察される。これは、可溶性種の部分的な活性についての重要なコントロールであり、また、ビオチン化pMHC複合体が非ビオチン化複合体と生物学的に同程度に活性であることを示唆する。
【0115】
新規鎖間結合を含有するHIV Gag sTCRとそのリガンド/MHC複合体又は無関係のHLA−ペプチド組合せ(これらの製造は上記で説明)との間の相互作用を、Biacore 3000TM表面プラズモン共鳴(SPR)バイオセンサで分析した。SPRは、レセプターリガンド相互作用を検出しその親和性及び動力学的パラメータを分析するために使用することができる原理である、小さなフローセル内のセンサ表面近くでの屈折率の変化(応答単位(RU)で表示)を測定する。β2mに架橋したビオチンとフローセルの活性化表面に化学的に架橋されているストレプトアビジンとの間の結合を介して、個々のHLA−ペプチド複合体を別個のフローセルに固定化することにより、プローブフローセルを準備した。次いで、異なるフローセルの表面上にsTCRを一定流速で通過させ、そうしている間のSPR応答を測定することにより、アッセイを実施した。
【0116】
平衡結合定数の測定
親の又は変異したHIV Gag sTCRの系列希釈物を調製し、5μl/分の一定流速で2つの異なる表面(一方は〜1000RUの特異SLYNTVATL−HLA-A*0201複合体を被覆し、2つめは〜1000RUの非特異HLA-A2-ペプチド複合体を被覆)に注入した。応答は、コントロールセルの測定値を用いて各濃度について規格化した。規格化データ応答を、TCRサンプルの濃度に対してプロットし、平衡結合定数KDを算出するために双曲線にフィットさせた(Price & Dwek,Principles and Problems in Physical Chemistry for Biochemists (第2版) 1979,Clarendon Press,Oxford)。
【0117】
動力学パラメータの測定
高親和性TCRについて、解離速度定数kd及び結合速度定数kaを実験的に測定することによりKDを決定した。平衡定数KDはkd/kaとして算出した。
1つは〜300RUの特異HLA-A2-nyesoペプチド複合体で被覆し、2つめは〜300RUの非特異HLA-A2−ペプチド複合体で被覆した2つの異なるセル上にTCRを注入した。流速を50μl/分に設定した。代表的には、250μlのTCRを〜3μM濃度で注入した。次いで、反応がベースラインに戻るまで緩衝液を流した。Biaevaluationソフトウェアを用いて動力学パラメータを算出した。また、半減期の算出を可能とするために、解離相を一次指数関数減衰式にフィットさせた。
【0118】
結果
可溶性のジスルフィド連結した天然型HIV Gag TCR(それぞれ配列番号9及び10に詳述されるαTCR鎖及びβTCR鎖からなる)とSLYNTVATL−HLA-A*0201複合体との間の相互作用を、上記の方法を用いて分析し、85nMのKD及び2.21×10-2 S-1の解離速度(koff)が示された(Biacore応答曲線については図12を参照)。
以下の表で特定されるTCRは、1μMより少ないか若しくは等しいKD及び/又は1×10-3 S-1か若しくはそれより遅いkoffを有する。
【0119】
【表4】
【0120】
実施例5 − 可溶性の高親和性HIV Gag TCR−WTヒトIL-2融合タンパク質の作製
実質的に実施例1〜3に記載の方法を使用して、可溶性の高親和性HIV Gag TCR−WTヒトIL-2融合タンパク質を作製することができる。簡潔には、所望のリンカー及びWTヒトIL-2をコードするDNAを、可溶性のジスルフィド連結親HIV Gag TCRβ鎖のDNA配列の3'末端にTAA(「ストップ」)コドンの直前で付加する。図11は、WTヒトIL-2にリンカー配列を介して融合したジスルフィド連結親HIV Gag TCRβ鎖を含んでなる融合タンパク質のアミノ酸配列を提供する(配列番号24)。この融合タンパク質のリンカー及びIL-2部分はイタリック体で示されている。次いで、この構築物をコードするDNAは、pEX821中にライゲートすることができる。次いで、可溶性の親HIV Gag TCR−IL-2融合タンパク質は、実質的に実施例3に記載された方法を使用して、このβ鎖融合タンパク質を図5aに詳述される可溶性のジスルフィド連結親HIV Gag α鎖TCR鎖(配列番号9)と組み合せることにより、発現させることができる。
【0121】
実施例6 − T細胞表面での親HIV Gag TCRの組換え発現
親HIV Gag TCR鎖のシグナル配列、細胞外ドメイン、膜貫通ドメイン及び細胞内ドメインをコードするDNA構築物を合成した(GeneArt、ドイツ)。これらTCRα鎖及びTCRβ鎖のDNA配列(それぞれ図15a及び15bに提供)は、親HIV Gag TCR DNA配列から、天然型アミノ酸配列を維持しつつ、ヒトT細胞中での該コードTCR鎖の発現レベルを増大させるように変更する。図16a及び16bはそれぞれ、図15a及び15bのDNA配列によりコードされる全長アミノ酸配列を提供する。
【0122】
次いで、TCRα鎖及びTCRβ鎖のDNA配列を、レンチウイルス発現ベクター中に一緒に挿入した。このベクターは、親HIV Gag TCRα鎖及びβ鎖の両方をコードするDNAを、該TCR鎖を分離する手足口病ウイルス(FMDV)2A切断因子アミノ酸配列(LLNFDLLKLAGDVESNPG(配列番号31))とインフレームで単一オープンリーディングフレームとして含有する(de Felipeら,Genet Vaccines Ther (2004) 2 (1):13)。mRNA翻訳に際して、TCRα鎖はそのC末端に2Aペプチド配列を有して作製され、TCRβ鎖は別個のポリペプチドとして作製される。
【0123】
T細胞を上記レンチウイルスベクターを用いて形質導入した。簡潔には、抗CD3/抗CD28ビーズを使用して初代T細胞を24時間刺激した。次いで、TCR遺伝子を発現する濃縮レンチウイルス上清を、刺激したT細胞と共にインキュベートしてウイルス形質導入した。次いで、抗CD3/抗CD28ビーズを除去し、200〜300fLの「静止容量(resting volume)」を達成するまで形質導入T細胞を培養した。
形質導入細胞表面上での親HIV Gag TCRの提示は、HLA-A*0201−SLYNTVALT PEテトラマーを使用するFACS分析及び抗CD8モノクローナル抗体FITC同時染色により確証した。
【0124】
結果
図17bは、形質導入CD8+細胞表面上での親HIV Gag TCR発現の成功を示すFACS分析データを提供する。図17aは、コントロールの非形質導入T細胞を使用して作成したFACS分析データを提供する。
【0125】
実施例7 − 可溶性の高親和性HIV Gag TCRによるCTL活性化の阻害
以下のアッセイを行い、可溶性の高親和性c11c6 HIV Gag TCRがSLYNTVATL−HLA-A*0201反応性ポリクローナルT細胞株の活性化を阻害し得ることを示した。
【0126】
HIV感染細胞の存在下でのOX84 SLYNTVATL−HLA-A*0201反応性ポリクローナルT細胞株活性化の阻害
この実験に使用した可溶性のc11c6高親和性HIV Gag TCRは、それぞれ図6c(配列番号13)及び図7b(配列番号15)に示すTCRα鎖可変ドメイン及びTCRβ鎖可変領域を含んでいた。この可溶性TCRのTCRα鎖及びβ鎖の完全アミノ酸配列はそれぞれ、図18a(配列番号29)及び図18b(配列番号30)に提供される。
IFN-γ産生及びTNF-α産生をCTL活性化についての読み取り値として使用した。
【0127】
試薬
R10アッセイ培地:10% FCS(熱不活化,Gibco,cat# 10108-165)、88% RPMI 1640(Gibco,cat# 42401-018)、1%グルタミン(Gibco,cat# 25030-024)及び1%ペニシリン/ストレプトマイシン(Gibco,cat# 15070-063)。
ペプチド:(種々の供給源から入手)最初にDMSO(Sigma,cat# D2650)中に4mg/mlで溶解し、凍結。
BDTM Cytometric Bead Array Kit、ヒトTh1/Th2 cytokine Kit II (BD Biosciences,San Diego,US)はアッセイに必要な全ての試薬を含む。
【0128】
T細胞活性化アッセイ
慢性HIV感染To標的細胞(HXB2及びHIV3B HIV Lab株)を洗浄し、R10培地中に再懸濁した。コントロールとして、非感染To標的細胞を1nMのSLYNTVATLペプチドで、37℃、5%CO2にて30分間パルスした。
【0129】
試験サンプル:
96ウェルU底プレートのウェル当たりR10培地中25,000のHIV感染To標的細胞。
ウェル当たりR10培地中2×10-7Mの高親和性c11c6 HIV Gag TCR又は親HIV Gag TCR。
ウェル当たりR10培地中5000のOX84ポリクローナルエフェクターT細胞株。
コントロール:
無関係の可溶性TCR(HLA-A*0201−Tax特異的TCR及びHLA-A*0201−NY-ESO特異的TCR)又は高親和性 HIV Gag TCRに代えた上記と同じもの。
【0130】
次いで、プレートを37℃、5%CO2にて4時間インキュベートした。培養上清を除去し、以下の方法を用いて存在するIFN-γ及びTNF-αのレベルを測定した。
【0131】
IFN-γアッセイ及びTNF-αアッセイ
(a)抗IFNγ捕捉抗体及び(b)抗TNFα捕捉抗体で被覆したBDTM Cytometric Beadsを製造業者の指示書に従い調製した。
次いで、以下の添加物を含むアッセイチューブを調製した:
BDアッセイ希釈剤中の50μlの抗IFNγ及び抗TNFα混合BDTM Cytometric Beads
50μlのPE検出試薬
【0132】
続いて、いずれか:
T細胞活性化アッセイウェルから採取した50μlの捕捉上清(試験サンプル)、
又は
ストック標準の系列希釈により一連の濃度で調製した50μlのIFNγ及びTNFα混合標準(較正標準)
【0133】
次いで、チューブを暗所で3時間インキュベートした後、1mlのBD洗浄緩衝液で洗浄し、遠心分離した。最後に、ビーズを300μlの洗浄緩衝液中に再懸濁し、存在するIFNγ及びTNFαのレベルを、製造業者の指示書に従ってフローサイトメトリにより決定した。
【0134】
非感染SLYNTVATLペプチドパルスTo細胞の存在下でのSLYNTVATL−HLA-A*0201特異的OX84ポリクローナルT株の阻害
上記CTL活性化アッセイに使用した同じ試薬及び方法を使用した。但し、2000のOX84ポリクローナルエフェクターT細胞を各T細胞活性化アッセイに使用し、非感染Toリンパ芽球様細胞(10-10〜10-8MのSLYNTVATLペプチドでパルス)を標的細胞として使用した。
【0135】
結果
可溶性の高親和性c11c6 HIV Gag TCRは、IFN-γ産生及びTNF-α産生により測定したところ、HIV感染To細胞の存在下で、SLYNTVATL−HLA-A*0201反応性OX84ポリクローナルT細胞株の活性化を強力に阻害した(図19を参照)。
可溶性の高親和性c11c6 HIV Gag TCRは、IFN-γ産生及びTNF-α産生により測定したところ、SLYNTVATLでパルスした非感染To細胞の存在下で、SLYNTVATL−HLA-A*0201反応性OX84ポリクローナルT細胞株の活性化を強力に阻害した(図20を参照)。
【0136】
実施例8 − 高親和性c11c6 HIV Gag TCRを使用する蛍光顕微鏡法による、ペプチドでパルスしたT2細胞上の細胞表面SLYNTVATL−HLA-A*0201抗原の定量
ペプチドパルスしたT2リンパ芽球様細胞上のSLYNTVATL−HLA-A*0201抗原の数を、可溶性の高親和性c11c6 HIV Gag TCRを用いる単一分子蛍光顕微鏡法により決定した(1つの蛍光シグナルは、標的細胞表面上の同族pMHCリガンドに結合した単一の標識TCRに関連するとの仮定に基づく)。このことは、ビオチン化TCRを用いて抗原発現ガン細胞を標的し、その後、細胞結合TCRをストレプトアビジン−Rフィコエリトリン(PE)コンジュゲートにより標識することによって容易となった。次いで、個々のPE分子を、三次元蛍光顕微鏡法により撮像した。
【0137】
T2リンパ芽球様細胞を、一連の濃度(10-5〜10-10M)で90分間37℃にてHIV Gag由来SLYNTVATLペプチド又は無関係のペプチド(SLLMWITQC)でパルスした。パルス後、細胞を500μlのPBSで2回洗浄した。細胞を200μlのTCR溶液(0.5% BSAアルブミン含有PBS中100nMの高親和性c11c6 HIV Gag TCR)中で30分間室温にてインキュベートした。TCR溶液を除去し、細胞を500μlのPBSで3回洗浄した。細胞を、200μlのストレプトアビジン−PE溶液(0.5% BSA含有PBS中5μg ml-1のストレプトアビジン−PE)中で室温にて暗所で20分間インキュベートした。ストレプトアビジン−PE溶液を除去し、細胞を500μlのPBSで3回洗浄した。洗浄媒体を除去し、細胞を、フェノールレッドを含まない400μlのR10に維持した後、蛍光顕微鏡法により撮像した。
【0138】
蛍光顕微鏡法。蛍光顕微鏡法は、63×油浸対物レンズ(Zeiss)を備えるAxiovert 200M(Zeiss)顕微鏡を用いて行った。300Wキセノンアークランプ(Sutter)を含むラムダLS光源を照明に用い、光路内に0.3及び0.6の中性フィルタを配置することにより最適レベルまで光度を下げた。励起光スペクトル及び発光スペクトルを、TRITC/DiIフィルタセット(Chroma)を用いて分離した。細胞を、z-スタック取り込みにより三次元で撮像した(21面、1μm間隔)。画像の取得及び分析は、Metamorphソフトウェア(Universal Imaging)を、記載されたようにして行った(Irvineら,Nature 419:p845-9、及びPurbhooら,Nature Immunology 5:p524〜30)。
【0139】
結果
図21に示されるように、上記方法を使用して、ペプチドパルスしたT2細胞の表面上のSLYNTVATL−HLA-A*0201抗原に結合した高親和性c11c6 HIV Gag TCRの撮像に成功した。これら結果は、高親和性c6c11 HIV Gag TCRを使用する、SLYNTVATLペプチドでパルスした細胞上のエピトープを計数するための閾値が約10-9Mペプチドであることを示す。
【特許請求の範囲】
【請求項1】
配列番号1のアミノ酸配列を有するTCRα鎖可変領域及び配列番号2のアミノ酸配列を有するTCRβ鎖可変領域を含んでなるT細胞レセプター(TCR)。
【請求項2】
配列番号5のアミノ酸配列を有するα鎖及び配列番号6のアミノ酸配列を有するβ鎖を含んでなる、請求項1に記載のTCR。
【請求項3】
配列番号9のアミノ酸配列を有するα鎖及び配列番号10のアミノ酸配列を有するβ鎖を含んでなる、請求項1に記載のTCR。
【請求項4】
膜貫通領域及び細胞質領域を欠いている請求項1〜3のいずれか1項に記載のTCR。
【請求項5】
配列番号27のアミノ酸配列を有するTCRα鎖及び配列番号28のアミノ酸配列を有するTCRβ鎖をコードする核酸を含んでなるTCR発現ベクターを保有する細胞。
【請求項6】
請求項1〜3のいずれか1項に記載のTCRをその表面に提示している非天然型T細胞。
【請求項1】
配列番号1のアミノ酸配列を有するTCRα鎖可変領域及び配列番号2のアミノ酸配列を有するTCRβ鎖可変領域を含んでなるT細胞レセプター(TCR)。
【請求項2】
配列番号5のアミノ酸配列を有するα鎖及び配列番号6のアミノ酸配列を有するβ鎖を含んでなる、請求項1に記載のTCR。
【請求項3】
配列番号9のアミノ酸配列を有するα鎖及び配列番号10のアミノ酸配列を有するβ鎖を含んでなる、請求項1に記載のTCR。
【請求項4】
膜貫通領域及び細胞質領域を欠いている請求項1〜3のいずれか1項に記載のTCR。
【請求項5】
配列番号27のアミノ酸配列を有するTCRα鎖及び配列番号28のアミノ酸配列を有するTCRβ鎖をコードする核酸を含んでなるTCR発現ベクターを保有する細胞。
【請求項6】
請求項1〜3のいずれか1項に記載のTCRをその表面に提示している非天然型T細胞。
【図1a】

【図1b】
【図2】
【図3】

【図4】

【図5】
【図6】

【図7】

【図8】
【図9−1】

【図9−2】

【図10−1】

【図10−2】

【図11】

【図12】
【図15】

【図16】

【図17】
【図18】

【図19】
【図20】
【図21】
【図13】
【図14】
【図1b】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9−1】
【図9−2】
【図10−1】
【図10−2】
【図11】
【図12】
【図15】
【図16】
【図17】
【図18】
【図19】
【図20】
【図21】
【図13】
【図14】
【公開番号】特開2012−144543(P2012−144543A)
【公開日】平成24年8月2日(2012.8.2)
【国際特許分類】
【出願番号】特願2012−49174(P2012−49174)
【出願日】平成24年3月6日(2012.3.6)
【分割の表示】特願2008−503585(P2008−503585)の分割
【原出願日】平成18年3月29日(2006.3.29)
【出願人】(510019129)イムノコア リミテッド (3)
【Fターム(参考)】
【公開日】平成24年8月2日(2012.8.2)
【国際特許分類】
【出願日】平成24年3月6日(2012.3.6)
【分割の表示】特願2008−503585(P2008−503585)の分割
【原出願日】平成18年3月29日(2006.3.29)
【出願人】(510019129)イムノコア リミテッド (3)
【Fターム(参考)】
[ Back to top ]