
高親和性TCRタンパク質および方法
【課題】野生型TCRよりもリガンドに対して高い親和性を有するT細胞レセプター(TCR)を提供することを、本発明の課題とする。
【解決手段】これら高親和性TCRは、T細胞レセプターコード配列を変異誘発して、T細胞レセプタータンパク質コード配列の変異体の種々の集団を作製する工程;このT細胞レセプター変異体コード配列を酵母細胞に形質変換する工程;この酵母細胞の表面上にこのT細胞レセプター変異体コード配列の発現を誘導する工程;および野生型T細胞レセプタータンパク質よりもペプチド/MHCリガンドに対してより高い親和性を有するT細胞レセプター変異物を発現する細胞を選択する工程によって取得された。この高親和性TCRは、抗体または単鎖抗体の代わりに使用され得る。
【解決手段】これら高親和性TCRは、T細胞レセプターコード配列を変異誘発して、T細胞レセプタータンパク質コード配列の変異体の種々の集団を作製する工程;このT細胞レセプター変異体コード配列を酵母細胞に形質変換する工程;この酵母細胞の表面上にこのT細胞レセプター変異体コード配列の発現を誘導する工程;および野生型T細胞レセプタータンパク質よりもペプチド/MHCリガンドに対してより高い親和性を有するT細胞レセプター変異物を発現する細胞を選択する工程によって取得された。この高親和性TCRは、抗体または単鎖抗体の代わりに使用され得る。
【発明の詳細な説明】
【技術分野】
【0001】
(関連出願の相互参照)
本願は、米国特許第09/009,388号(1998年1月20日出願)の一部継続である。本願は、米国予備出願第60/169,179号(1999年12月6日出願)の利益を主張する。
【0002】
(連邦政府の研究支援の承認)
本発明は、少なくとも一部分、国立衛生研究所からの財政的支援でなされた。従って、米国政府は、本発明に特定の権利を有する。
【背景技術】
【0003】
(発明の背景)
本発明の分野は、分子生物学であり、詳細には、組換え宿主細胞の細胞表面上に提示された免疫細胞レセプターのコンビナトリアルライブラリーの場合に関する。より詳細には、本発明は、組換え酵母細胞の表面上に提示された高い親和性のT細胞レセプタータンパク質のライブラリー、可溶性の高い親和性のTCRレセプタータンパク質、特定のペプチド/MHC対への高い親和性の結合に関して選択された高い親和性のTCRタンパク質、MHC決定基の非存在下における特定の抗原への結合に関して選択された高い親和性のTCRタンパク質、ならびに数ある適用の中で診断方法および画像化アッセイにおける選択された高い親和性のTCR誘導体の使用に関する。
【0004】
T細胞レセプター(TCR)および抗体は、異なるクラスのリガンドを認識するために開発されている。抗体は、可溶性抗原に結合する膜結合かつ可溶性タンパク質として機能し、一方、天然において、TCRは、細胞関連ペプチド/MHC抗原に結合する膜結合分子としてのみ機能する。抗体:抗原相互作用の全エネルギーは、外来抗原にのみ集中しているが、TCR:ペプチド/MHC相互作用のエネルギーのうちの実質的な画分は、自己MHC分子に指向されるようである[Manningら(1998)Immunity 8:413:425]。さらに、抗体は、体細胞変異および親和性成熟のプロセスに大きく起因して、TCRのものよりも高い位の大きさであるリガンド−結合親和性を有し得る。その正常な細胞の状況において、TCRは、体細胞変異を引起さず、そして胸腺選択のプロセスは、親和性の狭いウィンドウを維持することによって操作されるようである[Alamら(1996)Nature 381:616−620]。細胞表面におけるTCRのアクセサリー分子CD4またはCD8との関連はまた、TCRの機能的な親和性に影響を及ぼし得る[Garciaら(1996)Nature 384:577−581]。これらの差異にもかかわらず、2つのタンパク質の3次元構造は、顕著に類似し、抗原と接触する分子の単一表面上に超可変領域形成ループを有する。
【0005】
これらの構造類似性に基づいて、異種発現系においてTCRおよび抗体の細胞外ドメインの、可溶性かつ表面提示形態の産生を成功させることに有意な差異が存在したことは、いくらか驚くべきである。多くの抗体は、現在、インタクトもしくはFabフラグメント形態としてか、または単鎖(sc)フラグメント−可変(Fv)タンパク質としてかのいずれかで、高収率かつ可溶性で発現されている。さらに、新規に単離され、そして/またはファージディスプレイ技術(より最近では、酵母−ディスプレイ技術)の使用を介して改善された、多数の抗原結合Fvフラグメントが存在する[BoderおよびWittrup(1997)Nat.Biotechnol.15:553−557;Kiekeら(1997)Prot.Eng.10:1303−1310]。抗体フラグメントに関するこれらの発現系は、構造研究ならびに診断抗体および治療抗体の設計に重要である。
【0006】
対照的に、数個のTCR分子の3次元構造は、可溶性で相当折り畳まれたTCRの発現に対するかなりの努力のあとにのみ決定された[BentleyおよびMariuzza(1996)Ann.Rev.Immunol.14:563−590]。FabとTCRとの間の基礎を探求することにおける困難のうちの1つは、抗体における大量の配列多様性であり、そしてTCR可変(V)領域は、どの特徴のV領域が抗原結合以外の機能(例えば、V領域の対形成および結合動力学、可溶性、ならびに折り畳み)に重要であるかを識別するための努力を複雑にする。これらの性質に関して、TCRのV領域および抗体を比較する研究は、相対的にほとんどない。
【0007】
それにもかかわらず、マウスT細胞クローン2C由来のTCRは、現在、Escherichia coliにおいてscVαVβ(scTCR)として[Soo Hooら(1992)Proc.Natl.Acad.Sci.USA 89:4759−4763]、骨髄腫細胞から脂質連結VαCαVβCβ二量体として[SlanetzおよびBothwell(1991)Eur.J.Immunol.21:179−183]、および昆虫細胞から分泌VαCαVβCβ二量体として[Garciaら(1996)Science 274:209−219]発現されている。2C scTCRは、ほとんどのscFvと比較して、相対的に低い可溶性を有するが、その可溶性は、アミノ末端のチオレドキシンへの融合によって約10倍増加する[Schodinら(1996)Molec.Immunol.33:819−829]。可溶性で相当折り畳まれたVαVβドメインを産生することにおける困難は、他のTCRに対して拡大されている[Udakaら(1993)前出;Sykulevら(1994)前出;Manningら(1998)前出]。可溶性または表面提示能力のいずれかにおけるTCRとFvとの間の明らかな差異に関する分子説明は、適切に探査されていない。2CscTCRが、Vα/Vβ界面またはCD3εシグナル伝達サブユニットと相互作用することが疑われるVβフレームワークの領域における特定の単一部位変異の、ランダムなライブラリーからの選択後に、酵母表面ディスプレイシステムで発現され得ることが示された。これらの変異(これらのうちのいくつかは、抗体V領域に天然に見出される)は、TCR中のこれらの位置の重要性を反映し、TCR結合特性のさらなる操作に関する基礎を提供する。
【発明の概要】
【課題を解決するための手段】
【0008】
(発明の要旨)
本発明は、組換え宿主細胞、例えば、酵母細胞(望ましくはSaccharomyces cerevisiae)の表面上に提示された免疫T細胞レセプターポリペプチドのコンビナトリアルライブラリーを提供する。このようなライブラリーから、高い親和性のTCRポリペプチド(同族のリガンド:主要組織適合遺伝子複合体のタンパク質(pMHC)に結合するペプチドの複合体に関して、野生型よりも高い親和性を示すもの)が単離され得る。望ましくは、pMHCに対するTCRペプチドの親和性は、例えば、当該分野で公知の方法によって測定される場合、約107〜約1010の解離定数に反映される。可溶性の高い親和性のTCRをコードする核酸を含むDNAライブラリーが提供され、ここで、このTCRは、変異体TCRコード配列を作製するためのTCRを変異誘発すること;変異体TCRの変異体TCRコード配列を含むDNAを酵母細胞に形質転換すること;この変異体TCRが酵母細胞の表面上に提示されるようにこの変異体TCRコード配列の発現を誘導すること;この酵母細胞を、ペプチド/MHCリガンドを結合する蛍光標識と接触して、選択された酵母細胞を産生すること;および、最も高い蛍光を示す酵母細胞を単離する、方法によって作製される。野生型T細胞レセプタータンパク質よりもペプチド/MHCリガンドに対して高い親和性を有する、酵母細胞の表面上に提示されたT細胞レセプタータンパク質のライブラリーがまた、提供され、ここで、このライブラリーは、T細胞レセプタータンパク質コード配列を変異誘発して変化に富むT細胞レセプタータンパク質コード配列変異体の集団を作製すること;このT細胞レセプター変異体コード配列を、酵母細胞に形質転換すること;酵母細胞の表面上でこのT細胞レセプター変異体コード配列の発現を誘導すること;そして、野生型T細胞レセプタータンパク質よりもペプチド/MHCリガンドに対して高い親和性を有する、T細胞レセプター変異体を発現する細胞が選択することによって形成される。
【0009】
本発明はさらに、同族リガンドに対して高い親和性の結合を示すTCRタンパク質(細胞結合形態でかまたは可溶性形態で)を提供する。本発明において、TCRタンパク質が結合するリガンドは、ペプチド/MHC複合体であるか、または選択プロセス(望ましくは、繰り返された選択プロセス)に起因して、このリガンドは、MHC成分(例えば、スーパー抗原)を含まないリガンドであり得る。このリガンドは、とりわけ、ペプチド、タンパク質、炭水化物部分、または脂質部分であり得る。これらの可溶性の高い親和性のTCRは、変異体TCRコード配列を作製するためにTCRを変異誘発する工程;変異体TCRのための変異体TCRコード配列を含むDNAを酵母細胞に形質転換する工程;この変異体TCRが酵母細胞の表面上に提示されるように、この変異体TCRコード配列の発現を誘導する工程;この酵母細胞を、ペプチド/MHCリガンドに結合する蛍光標識と接触して、選択された酵母細胞を生成する工程;および、最も高い蛍光を示す酵母細胞を単離する工程を包含する方法によって作製され得る。この可溶性の高い親和性のTCRは、好ましくは酵母ディスプレイによって単離される。
【0010】
本発明はさらに、高い親和性のTCRタンパク質の同族リガンドを検出するための方法を提供し、この方法は、高い親和性のTCRタンパク質を同族リガンドと結合させる工程を包含し、ここで、この高い親和性のTCRタンパク質が検出可能に標識化されるか、または添加される二次的な検出可能なタンパク質(例えば、同族リガンドに結合する領域以外の領域においてTCRに特異的な抗体)が存在する。リガンドを同定するために高い親和性のTCRを用いるための好ましい方法は:高い親和性のTCRを検出可能な標識で標識化する工程;この標識化TCRをリガンドと接触する工程;標識化TCRが結合するリガンドを同定する工程を包含する。好ましくは、このリガンドは、野生型TCRよりも高い親和性でTCRに結合するペプチド/MHCリガンドである。このリガンドを同定する方法は、当業者に公知である。直接または間接的に、TCRタンパク質の検出を可能にする適切な標識としては、蛍光化合物、化学発光化合物、放射性同位体、発色団などが挙げられるが、これらに限定されない。
【0011】
高い親和性のTCRタンパク質は、医学、獣医学もしくは植物の診断設定における標的リガンドの定性測定および定量測定のための手段としてか、または組織材料もしくは植物材料の同定のために、実験室で使用され得る。同様に、本発明の高い親和性のTCRは、標的材料またはリガンドの検出および/または定量のための試薬として使用され得る。選択されたペプチド/MHCリガンドに結合する高い親和性のTCRを用いる方法がまた、提供され、この方法は、この高い親和性のTCRをこの選択されたペプチド/MHCリガンドに結合する標識で標識化する工程;この標識化された高い親和性のTCRをMHC分子を含む細胞と接触させる工程を包含する。本発明の高い親和性のタンパク質(ここで、このタンパク質は、高い親和性および特異性で腫瘍細胞抗原に特異的に結合する)は、特定の癌の型の診断試験に使用され得るか、または生物(好ましくは、哺乳動物、そしてまた好ましくは、ヒト)において、腫瘍の位置を確認するためならびに/または腫瘍のサイズおよび数を概算するための画像化試験の際に生物で使用され得る。診断試験のためにpMHCに結合する高い親和性のTCRを用いるための方法が提供され、この方法は、高い親和性のTCRを検出可能な標識で標識化する工程;この高い親和性のTCRが高い親和性を有するリガンドを含む細胞と、この高い親和性のTCRを接触させる工程;およびこの標識を検出する工程を包含する。この方法において、標識は、特定のペプチド/MHCリガンドに結合するために選択され得、これによって、特定のペプチド/MHCリガンドを発現する細胞が、標的化される。細胞表面上で特定のペプチド/MHC分子に対する診断プローブとして高い親和性のTCRを用いるための好ましい方法は:高い親和性のTCRを、特定のペプチド/MHCリガンドに結合する検出可能な標識で標識化する工程;このTCRを細胞と接触する工程;およびこの標識を検出する工程を包含する。特定の使用に依存した使用のために選択された検出可能な標識、および適切なの選択は、関連分野の当業者の十分に範囲内である。一般的に、目的のリガンドに対する高い親和性結合に関して選択されたTCRタンパク質は、このリガンドに特異的な抗体が使用され得る方法(TCR 対 抗体タンパク質に対して手順の改変がなされ、このような改変は、当該分野で公知である)に使用され得る。
【0012】
高い親和性のTCR、望ましくは可溶性単鎖(sc)TCRは、自己免疫疾患における細胞または組織の自己免疫破壊をブロックするために使用され得、ここで、標的細胞の表面上で細胞傷害性リンパ球によって認識される部位は、高い親和性のTCRによって結合される部位と同じである。細胞の自己免疫破壊をブロックするための好ましい方法は、標的細胞の表面上でTリンパ球によって認識される部位に高い親和性を有するTCRを、細胞と接触させる工程を包含し、これによって、細胞の自己免疫破壊がブロックされる。
【0013】
可溶性の、高い親和性のscTCRは、治療化合物(例えば、制癌化合物、治療放射性核種または細胞傷害性タンパク質)に結合され得、ここで、scTCRの同族のリガンドは、新生物形成細胞表面マーカーである。あるいは、高い親和性の可溶性scTCRの結合特異性は、病原体感染された標的細胞であり得(例えば、ウイルス、細菌、または原生動物感染された標的細胞)、そして、標的細胞が感染因子のさらなる複製無しに排除され得るように毒性分子が結合され得る。不活性病原体に対して高い親和性のTCRを使用する提供された方法は:この病原体に毒性である分子を、高い親和性のTCRに結合させる工程;およびこの病原体を発現する細胞と、このTCRを接触させる工程を包含する。「毒性」は、病原体が病原体の複製を回避または阻害することを意味する。
【0014】
疾患を処置するために高い親和性のTCRを使用するための方法がまた提供され、この方法は:新生物形成細胞表面マーカーに対して高い親和性を有するTCRを、治療化合物と結合させる工程:およびこのTCRを細胞と接触させる工程を包含する。TCRと結合され得る、疾患の進行を遅延する際に有用である任意の治療化合物が、使用され得る。治療化合物をTCRと結合する方法は、当該分野で公知である。
【0015】
高い親和性のTCR変異体に対する遺伝子を、T細胞の表面上で変異体の発現を可能にするシステムにクローニングするための方法がまた、提供され、この方法は:高い親和性のTCR変異体を作製するためにTCRを変異させる工程;このTCR変異体をベクターにクローニングする工程;このベクターをT細胞にトランスフェクトする工程;高い親和性のTCR変異体をT細胞の表面上に発現させる工程を包含する。この方法は、さらに:ペプチド/MHCリガンドによって他のT細胞よりも大きな程度に活性化されるT細胞を選択する工程を包含する。トランスフェクト/感染されたT細胞は、選択されたペプチド保有MHC細胞の認識に有用であり得る。これらのトランスフェクト/感染されたT細胞は、患者の疾患を処置する際に有用であり、ここで、患者由来のT細胞は、取り出され、そして高い親和性のTCR変異体を発現するベクターで形質転換され、そしてこの患者に戻される(ここで、このT細胞は、患者の野生型T細胞よりも、ペプチド/MHCリガンドによって大きな程度に活性化される)。
【0016】
可溶性の高い親和性のTCR分子は、関連分野の当業者に容易に明らかであるように、ほとんどの適用に対して抗体または単鎖抗体の代わりに使用され得る。
本願発明は、例えば、以下を提供する:
(項目1) リガンドを同定するために高親和性TCRを使用するための方法であって、以下:
高親和性TCRを標識化する工程;
該標識化TCRをリガンドと接触させる工程;
該標識化TCRと結合する該リガンドを同定する工程、
を包含する、方法。
(項目2) 前記標識が、蛍光化合物、化学発光化合物、放射性同位元素および発色団からなる群より選択される、項目1に記載の方法。
(項目3) 前記リガンドが、ペプチド/MHCリガンドである、項目1に記載の方法。(項目4) 選択されたペプチド/MHCリガンドに結合するために高親和性TCRを使用するための方法であって、以下:
該高親和性TCRを、該選択されたペプチド/MHCリガンドに結合する標識で標識化する工程;
該標識化高親和性TCRを、MHC分子を含む細胞と接触させる工程、
を包含する、方法。
(項目5) 前記標識が、蛍光化合物、化学発光化合物、放射性同位元素および発色団からなる群より選択される、項目4に記載の方法。
(項目6) 細胞の表面上の特定のペプチド/MHC分子に対する診断用プローブとして高親和性TCRを使用するための方法であって、以下:
高親和性TCRを、特定のペプチド/MHCリガンドに結合する標識で標識化する工程;
該TCRを細胞と接触させる工程;
該標識を検出する工程、
を包含する、方法。
(項目7) 診断試験のためにpMHCに結合する高親和性TCRを使用するための方法であって、以下:
該高親和性TCRを検出可能な標識で標識化する工程;
該高親和性TCRを細胞と接触させる工程;
該標識を検出する工程、
を包含する、方法。
(項目8) 前記存在する標識の数が検出される、項目7に記載の方法。
(項目9) 前記標識の位置が、生物体において検出される、項目7に記載の方法。
(項目10) 前記標識が、特定のペプチド/MHCリガンドに結合し、それによって、特定のペプチド/MHCリガンドを発現する細胞が標的化される、項目7に記載の方法。(項目11) 細胞の自己免疫破壊をブロックするための方法であって、標的細胞の表面上のTリンパ球によって認識される部位に対して高い親和性を有するTCRを、細胞と接触させる工程であって、それによって、細胞の自己免疫破壊がブロックされる、工程、
を包含する、方法。
(項目12) 疾患を処置するために高親和性TCRを使用するための方法であって、以下:
新生物細胞表面マーカーに対する高い親和性を有するTCRを、治療化合物と結合させる工程;および、
該TCRを細胞と接触させる工程、
を包含する、方法。
(項目13) 病原体を不活性化するために高親和性TCRを使用する方法であって、以下:
病原体に対して毒性のある分子を、高親和性TCRに結合する工程;および、
該TCRを、該病原体を発現する細胞と接触させる工程、
を包含する、方法。
(項目14) 前記病原体が、ウイルス、細菌および原生動物からなる群より選択される、項目13に記載の方法。
(項目15) 野生型TCRよりもリガンドに対して高い親和性を有する、可溶性T細胞レセプター(TCR)。
(項目16) 前記リガンドが、ペプチド/MHCリガンドである、項目15に記載の可溶性高親和性TCR。
(項目17) 項目15に記載の可溶性高親和性TCRであって、該高親和性TCRが、以下:
TCRを変異誘発して、変異体TCRコード配列を作製する工程;
変異体TCRについての該変異体TCRコード配列を含むDNAを、酵母細胞に形質転換する工程;
該変異体TCRコード配列の発現を誘導する工程であって、その結果、該変異体TCRが、該酵母細胞の表面上に提示される、工程;
該酵母細胞を、蛍光標識と接触させる工程であって、該蛍光標識は、ペプチド/MHCリガンドに結合して、選択された酵母細胞を産生する、工程;および
最も高い蛍光を示す酵母細胞を単離する工程、
を包含する方法によって作製される、可溶性高親和性TCR。
(項目18) 酵母ディスプレイによって単離された、項目15に記載の可溶性高親和性TCR。
(項目19) 可溶性高親和性TCRをコードする核酸を含む、DNAライブラリーであって、該TCRが、以下:
TCRを変異誘発して、変異体TCRコード配列を作製する工程;
変異体TCRについての該変異体TCRコード配列を含むDNAを、酵母細胞に形質転換する工程;
該変異体TCRコード配列の発現を誘導する工程であって、その結果、該変異体TCRが、該酵母細胞の表面上に提示される工程;
該酵母細胞を、蛍光標識と接触させる工程であって、該蛍光標識は、ペプチド/MHCリガンドに結合して、選択された酵母細胞を産生する、工程;および
最も高い蛍光を示す酵母細胞を単離する工程、
を包含する方法によって作製される、
DNAライブラリー。
(項目20) 野生型T細胞レセプタータンパク質よりもペプチド/MHCリガンドに対して高い親和性を有する、酵母細胞の表面上に提示されたT細胞レセプタータンパク質のライブラリーであって、ここで、該ライブラリーは、
T細胞レセプタータンパク質コード配列を変異誘発して、T細胞レセプタータンパク質コード配列の変異体の多様な集団を生成する工程;
該T細胞レセプター変異体コード配列を、酵母細胞に形質転換する工程;
酵母細胞の表面上での該T細胞レセプター変異体コード配列の発現を誘導する工程;および、
野生型T細胞レセプタータンパク質よりもペプチド/MHCリガンドに対して高い親和性を有するT細胞レセプター変異体を発現する細胞を選択する工程、
によって形成される、
ライブラリー。
(項目21) T細胞の表面上に高親和性TCR変異体の発現を可能にする系に、該高親和性TCR変異体についての遺伝子をクローニングする方法であって、以下:
TCRを変異誘発して、高親和性TCR変異体を作製する工程;
該TCR変異体を、ベクターにクローニングする工程;
該ベクターを、T細胞にトランスフェクトする工程;
該T細胞の表面上に該高親和性TCR変異体を発現させる工程、
を包含する、方法。
(項目22) 野生型よりもペプチド/MHCリガンドによって活性化されるT細胞を選択する工程をさらに包含する、項目21に記載の方法。
(項目23) 前記トランスフェクト/感染されたT細胞が、選択されたペプチド保有MHC細胞の認識のために使用される、項目21に記載の方法。
(項目24) 項目21の方法によって作製された、T細胞。
【図面の簡単な説明】
【0017】
【図1】図1は、野生型および変異型2C TCRをそれらの表面に発現する酵母細胞のフローサイトメトリー分析を示す。野生型(T7)および変異型(qL2、qL7)scTCRを示す酵母細胞は、抗Vβ8抗体F23.2(120nM)、特異的な同種抗原ペプチドのMHC、QL9/Ld/Ig(40nM)、または無ペプチドMCMV(配列番号1)/Ld/Ig(40nM)で染色した。結合をFITC結合体化ヤギ抗マウスIgG F(ab’)2によって検出し、そして、フローサイトメトリーによって分析した。ネガティブ集団(例えば、F23.2染色でみられるような)は、すべての酵母のディスプレイされたタンパク質について観察され、そして、表面融合タンパク質を発現し得ない、増殖または誘導の段階の細胞に起因すると考えられている(Kiekeら、(1999)Proc.Natl.Acad.Sci.USA 96:5651−5656;BoderおよびWittrup(1997)Nat.Biotech.15:553−557;Kiekeら、(1997)Protein Engineering 10:1303−1310)。
【図2】図2は、Ldに結合された異なったQL9改変体ペプチドに対する変異体scTCRの結合の優れた特異性分析を示す。本来のT細胞クローン2Cおよび種々の酵母クローンを、野生型QL9(P5F)、QL9の位置5の改変体(P5Y、P5H、P5E)またはMCMV(配列番号1)でロードしたLd/Ig二量体への結合について、フローサイトメトリーによって分析した。結合をFITC標識化したヤギ抗マウスIgGを用いて検出した。相対的な蛍光を2つの異なったアプローチによって測定した。T細胞クローン2Cについては、種々のペプチド/LdIg二量体の結合を、QL9/Ld染色(MFUpMHC/MFUQL9−Ld)に対して調整した。酵母細胞について、各ペプチド/Ld二量体の結合を、抗Vβ8抗体F23.2(MFUpMHC/MFUF23.2)による結合に対して調整した。後者は、異なった変異体が、野生型QL9/Ldに対する結合についてお互いに比較されるのを可能とした。
【図3】図3は、可溶性scTCRによるQL9/Ld結合を示す。QL9を用いてロードしたT2−Ld細胞を125I標識化した抗LdFabフラグメント(30−5−7)および種々の濃度の未標識化のFab(黒菱形)、scTCR−T7(黒四角)、または変異体scTCR−qL2(黒丸)と共にインキュベートした。結合したおよび未結合の125I 30−5−7 Fabフラグメントを、オリーブ油/ヂブチルフタレートを介して、遠心分離によって分離した。コントロールペプチドMCMV(配列番号1)を用いてロードしたT2−Ld細胞に対する125I標識化した抗LdFabフラグメントの結合は、scTCRの最も高い濃度においてさえ阻害されなかった(データは示されていない)。
【図4A】図4Aは、細胞表面ペプチド/MHCへのscTCR/ビオチンの結合のフローサイトメトリー分析を示す。ペプチド−ロードしたT2−Ld細胞を、ビオチン標識化したqL2scTCR(約0.3μM)またはT7scTCR(約1.6μM)scTCRと共にインキュベートし、その後、ストレプトアビジン−PEと共にインキュベートし、フローサイトメトリーによって分析した。図4A:QL9(影なし)、p2Ca(軽い影付き)、またはMCMV(配列番号1)(暗い影付き)を用いてロードし、そしてqL2scTCR/ビオチンで染色したT2−Ld細胞のフローサイトメトリーヒストグラム。図4B:QL9、p2Ca、またはMCMV(配列番号1)を用いてロードし、そして第2のSA−PEのみ、T7scTCR/ビオチン+SA−PE、またはqL2scTCR/ビオチン+SA−PEのいずれかで染色したT2−Ld細胞の平均蛍光単位(MFU)。図4C:昆虫細胞から発現された変異体qL2の可溶性の高い親和性形態は、非常に低い濃度のペプチド−MHC複合体を検出し得る。Ld複合体は、37℃で1.5時間、種々の濃度のQL9ペプチドを用いてインキュベーションによってT2−Ld細胞(3×106/ml)の表面上で上方制御された。約2×105細胞をトランスフェクトされたDrosophila melanogaster(昆虫)SC2細胞由来のTCRを用いて、30分間氷上で染色した(Garcia,K.C.ら、(1997)Proc Natl Acad Sci USA 94(25)、13838−13843)。次いで、細胞を洗浄し、そしてストレプトアビジン−PEが後に続く、ビオチン標識化した抗Vb IgG(F23.1)で染色し、そしてフローサイトメトリーによって分析した。
【図4B】図4Bは、細胞表面ペプチド/MHCへのscTCR/ビオチンの結合のフローサイトメトリー分析を示す。ペプチド−ロードしたT2−Ld細胞を、ビオチン標識化したqL2scTCR(約0.3μM)またはT7scTCR(約1.6μM)scTCRと共にインキュベートし、その後、ストレプトアビジン−PEと共にインキュベートし、フローサイトメトリーによって分析した。図4A:QL9(影なし)、p2Ca(軽い影付き)、またはMCMV(配列番号1)(暗い影付き)を用いてロードし、そしてqL2scTCR/ビオチンで染色したT2−Ld細胞のフローサイトメトリーヒストグラム。図4B:QL9、p2Ca、またはMCMV(配列番号1)を用いてロードし、そして第2のSA−PEのみ、T7scTCR/ビオチン+SA−PE、またはqL2scTCR/ビオチン+SA−PEのいずれかで染色したT2−Ld細胞の平均蛍光単位(MFU)。図4C:昆虫細胞から発現された変異体qL2の可溶性の高い親和性形態は、非常に低い濃度のペプチド−MHC複合体を検出し得る。Ld複合体は、37℃で1.5時間、種々の濃度のQL9ペプチドを用いてインキュベーションによってT2−Ld細胞(3×106/ml)の表面上で上方制御された。約2×105細胞をトランスフェクトされたDrosophila melanogaster(昆虫)SC2細胞由来のTCRを用いて、30分間氷上で染色した(Garcia,K.C.ら、(1997)Proc Natl Acad Sci USA 94(25)、13838−13843)。次いで、細胞を洗浄し、そしてストレプトアビジン−PEが後に続く、ビオチン標識化した抗Vb IgG(F23.1)で染色し、そしてフローサイトメトリーによって分析した。
【図4C】図4Cは、細胞表面ペプチド/MHCへのscTCR/ビオチンの結合のフローサイトメトリー分析を示す。ペプチド−ロードしたT2−Ld細胞を、ビオチン標識化したqL2scTCR(約0.3μM)またはT7scTCR(約1.6μM)scTCRと共にインキュベートし、その後、ストレプトアビジン−PEと共にインキュベートし、フローサイトメトリーによって分析した。図4A:QL9(影なし)、p2Ca(軽い影付き)、またはMCMV(配列番号1)(暗い影付き)を用いてロードし、そしてqL2scTCR/ビオチンで染色したT2−Ld細胞のフローサイトメトリーヒストグラム。図4B:QL9、p2Ca、またはMCMV(配列番号1)を用いてロードし、そして第2のSA−PEのみ、T7scTCR/ビオチン+SA−PE、またはqL2scTCR/ビオチン+SA−PEのいずれかで染色したT2−Ld細胞の平均蛍光単位(MFU)。図4C:昆虫細胞から発現された変異体qL2の可溶性の高い親和性形態は、非常に低い濃度のペプチド−MHC複合体を検出し得る。Ld複合体は、37℃で1.5時間、種々の濃度のQL9ペプチドを用いてインキュベーションによってT2−Ld細胞(3×106/ml)の表面上で上方制御された。約2×105細胞をトランスフェクトされたDrosophila melanogaster(昆虫)SC2細胞由来のTCRを用いて、30分間氷上で染色した(Garcia,K.C.ら、(1997)Proc Natl Acad Sci USA 94(25)、13838−13843)。次いで、細胞を洗浄し、そしてストレプトアビジン−PEが後に続く、ビオチン標識化した抗Vb IgG(F23.1)で染色し、そしてフローサイトメトリーによって分析した。
【図5】図5は、ストレプトアビジン−PEが続く、ビオチン標識化したペプチド/MHC複合体、OVA/Kb、dEV8/KbまたはSIYR(配列番号2)/Kbを用いて染色した変異体scTCR(3SQ2と呼ばれる)を示す酵母のフローサイトメトリーヒストグラムを示す。scTCRの存在についてのポジティブコントロールとして、酵母を、FITCヤギ抗マウスF(ab’)2が続く、Vβ特異的Ig、F23.2で染色した。
【図6】図6は、T2−Kb腫瘍細胞を特定のペプチド(OVA、dEV8またはSIYR(配列番号2))と共にインキュベートし、そしてストレプトアビジン−PEが続く、ビオチン標識化した可溶性scTCR、3SQ2で染色し、フローサイトメトリーによって分析したことを示す。Kb細胞の存在についてのポジティブコントロールとして、T2−Kb細胞をビオチン標識化した抗体B8.24.3(これは、結合されたペプチドに関わらずKbを認識する)を用いて染色した。
【図7】図7は、dEV8/Kbを用いる複数回の仕分け後、酵母VαCDR3ライブラリーを、ストレプトアビジン−PEが続く、ビオチン標識化したdEV8/Kbで染色し、そしてフローサイトメトリーによって分析したことを示す。
【図8】図8は、ビオチン標識化したストレプトアビジン−PEを用いる染色が続く、ビオチン標識化したペプチド/MHC複合体、OVA/KbまたはdEV8/Kbで染色した変異体scTCR(4d1と呼ばれる)を示す酵母のフローサイトメトリーヒストグラムを示す。ポジティブコントロールとして、酵母を、F23,2Igを用いてscTCR Vβの存在について分析し、そして抗6His抗体または抗HA Ig、12CA5を用いてエピトープタグについて分析した。
【図9】図9は、変異体T細胞レセプターqL2を用いてトランスフェクトしたT細胞は、低濃度でペプチドMHCを発現する標的細胞を認識し、そしてそれによって刺激され得ることを示す。T細胞ハイブリドーマ細胞株58−/−(Letourneur,F.,およびB.Malissen.(1989)Eur J Immunol 19(12)、2269−74)を野生型(2C)または変異型(qL2)TCRを用いてトランスフェクトし、そして1ウェルあたり7.5×104個のトランスフェクトされた細胞をQL9ペプチドの存在下で、T2−Ld細胞(7.5×104ウェル)と共に37℃でインキュベートした。約30時間後、上清を回収し、そして、T細胞によって放出されたIL−2についてアッセイした:上清をIL−2依存性細胞株、HT2(5×103/ウェル)と共に37℃で18時間インキュベートした。HT2細胞の増殖を3[H]チミジンの取り込みによって測定した。平均CPMは、3連ウェルの平均を示す。QL9ペプチドの非存在下では、IL−2は全く放出されなかった(データは示されていない)。
【発明を実施するための形態】
【0018】
(好ましい実施形態の詳細な説明)
明細書および特許請求の範囲(このような用語に対して与えられる範囲を含む)の明瞭なそして一貫した理解を提供するために、以下の定義が提供される。
【0019】
コード配列は、タンパク質のアミノ酸配列についてコードするか、またはtRNAまたはrRNAのような機能的なRNAについてコードする遺伝子またはcDNAの一部である。
【0020】
相補体または相補的な配列は、ワトソン−クリック塩基対形成法則に従う別の配列のヌクレオチドと水素結合した二重鎖を形成するヌクレオチドの配列を意味する。例えば、5’−AAGGCT−3’についての相補的な塩基配列は、3’−TTCCGA−5’である。
【0021】
下流は、DNAまたはRNAにおける任意の部位の3’側を意味する。
【0022】
発現は、構造的なRNA(rRNA、tRNA)またはメッセンジャーRNA(mRNA)への遺伝子の転写および引き続くmRNAのタンパク質への翻訳をいう。
【0023】
具体的に例示されたTCR配列に機能的に等価であるアミノ酸配列は、単一または複数のアミノ酸置換によって、アミノ酸の付加および/または欠失によって、改変されたアミノ酸配列をいい、ここで、1つ以上のアミノ酸は化学的に改変されているが、それにも関わらず、本発明の細胞結合または可溶性TCRタンパク質の結合特異性および高い親和性結合活性を保持する。機能的に等価なヌクレオチド配列は、具体的に例示された細胞結合または可溶性TCRタンパク質と同一の生物学的活性を実質的に有するポリペプチドをコードするヌクレオチド配列である。本発明の状況において、可溶性TCRタンパク質は、ネイティブな細胞結合TCRの一部を欠き、そして、溶液中で安定である(すなわち、それは、本明細書中で記載されるように処理される場合、そしてタンパク質溶液についての標準的な条件下で、一般的に溶液中で凝集しない)。
【0024】
2つの核酸配列は、これらの配列が別々の生物に由来する場合、このような生物が異なった種であろうとなかろうと、これらの配列が、同一の生物において同一の配列中で天然に存在しない限り、お互いに異種である。
【0025】
相同性は、2つのヌクレオチド間またはアミノ酸配列間の同一性の程度をいう。
【0026】
単離されたは、天然の状態からヒトの手によって変更されることを意味する。「単離された」組成物または物質が天然に存在する場合、それは、その本来の環境から変化されるかまたは取り出されるか、あるいはその両方である。例えば、生きている動物に天然に存在するポリヌクレオチドまたはポリペプチドは単離されていないが、その用語が本明細書中で使用される場合、天然の状態で共存する物質から分離された同一のポリヌクレオチドまたはポリペプチドは単離されている。
【0027】
リンカー領域は、タンパク質の2つの機能的または構造的ドメインを作動可能に連結するアミノ酸配列である。
【0028】
核酸構築物は、天然に存在する遺伝子から単離されたか、他に天然に存在しない様式で組み合わされそして並列された核酸のセグメントを含むように改変された核酸分子である。
【0029】
核酸分子は、3’−5’−リン酸ジエステル結合によって結合されたデオキシリボヌクレオチドまたはリボヌクレオチドのいずれかを含む単一または二重鎖の直鎖状ポリヌクレオチドを意味する。
【0030】
2つのDNA配列は、連結の性質が、それらの配列がお互いに対してそれらの正常な機能をもたらす能力を妨げない場合、作動可能に連結されている。例えば、プロモーター領域は、このプロモーターがコード配列の転写をもたらし得る場合、コード配列に作動可能に連結されている。
【0031】
ポリペプチドは、ペプチド結合によって連結されているアミノ酸の直鎖状ポリマーである。
【0032】
プロモーターは、シス作用DNA配列を意味し、一般的に80〜120塩基対長であり、そして遺伝子の開始部位の上流に位置し、これに対してRNAポリメラーゼは、結合し得そして正確な転写を開始する。転写調節の開始/終了調節を提供しそして/または下流のコード配列の発現を増強する(増加する)さらなる転写調節配列に関連し得る。
【0033】
組換え核酸分子(例えば、組換えDNA分子)は、2つ以上の非相同性DNA分子の連結を介してインビトロで形成された新規の核酸配列である(例えば、少なくとも1つのブローニング部位にクローン化された外来のDNAの1つ以上のインサートを含む組換えプラスミド)。
【0034】
形質転換は、異なった遺伝子型の別の細胞由来の精製された組換えDNAの外部適用によってゲノムまたは細胞の方向付けられた改変を意味し、被験体細胞のゲノム中へのその取り込みおよび組み込みを生じる。細菌においては、この組換えられたDNAは、代表的には細菌の染色体には組み込まれないが、その代わりに、プラスミドとして自律的に複製する。
【0035】
上流は、DNAまたはRNA中の任意の部位の5’側を意味する。
【0036】
ベクターは、宿主細胞中で自律的に複製可能でありそして外来DNAを受け入れ可能である核酸分子である。ベクターは、複製の自分自身の起源、外来DNAの挿入について使用され得る制限エンドヌクレアーゼについての1つ以上の独特の制限部位、そして通常抗生物質耐性をコードする遺伝子のような選択マーカー、およびしばしば挿入されたDNAの発現のための認識配列(例えば、プロモーター)を保有する。一般的なベクターとしては、プラスミドベクターおよびファージベクターが挙げられる。
【0037】
高い親和性T細胞レセプター(TCR)は、野生型TCRよりもより強力に標的リガンドへ結合する操作されたTCRを意味する。
【0038】
T細胞は、αβヘテロ二量体T細胞レセプター(TCR)を介したMHC産物に結合された外来ペプチドを認識する。TCRレパートリーは、抗体重鎖および軽鎖遺伝子において使用される同一の遺伝子再配列機構によって作製された広範な多様性を有する[Tonegawa,S.(1988)Biosci.Rep.8:3−26]。多様性の大部分は、αおよびβ鎖の相補性決定領域3(CDR3)をコードする可変(V)および連結(J)(または、多様性、D)領域の接合部において作成される[DavisおよびBjorkman(1988)Nature 334:395−402]。しかし、TCRは、抗体が体細胞点変異を生じるようには、おそらく同時には、体細胞点変異を起さない。TCRはまた、抗体と同一の程度の親和性変異を起こさない。TCRは、天然に存在するように、105から106M−1まで変化する親和性を有するようであり、これに対して、抗体は代表的に、105から109M−1まで変化する親和性を有する[Davisら、(1998)Annu.Rev.Immunol.16:523−544;Eisenら、(1996)Adv.Protein Chem.49:1−56]。TCRにおける体細胞変異の非存在は、より低い親和性に関連し得るが、TCRがより高い親和性を有する選択的な利点が存在しないことがまた議論されている。実際、T細胞活性化の連続的に引き起こす[Valituttiら(1995)Nature 375:148−151]および速度論的校正[Rabinowitzら(1996)Proc.Natl.Acad.USA 93:1401−1405]モデルは共に、より長いオフレート(off−rate)(より高い親和性と関連した)はシグナル伝達プロセスに有害であることを示唆する。より高い親和性TCRは、T細胞応答について必要なペプチド特異性を維持し得ないこともまたありうる。例えば、MHCの溝内に結合されたペプチドは、制限されたアクセス可能な表面を示し[Bjorkman,P.J.(1997)Cell 89:167−170]、これは、代わりに、相互作用おいて生成され得るエネルギーの量を制限し得る。他方、MHCへリックスに対してエネルギーを向けることによって、TCRの親和性を上昇することは、おそらくネガティブな選択の間の胸腺欠失を生じる[Bevan,M.J.(1997)Immunity 7:175−178]。
【0039】
我々は、T細胞レセプタータンパク質のより高い親和性に対する固有な構造特性または遺伝的制限は存在しないことを示す。より高い親和性TCR変異体は、酵母ディスプレイコンビナトリアル技術およびTCR変異体(例えば、VαおよびVβCDR3変異体)を使用することによって、インビボ選択圧力の非存在下で、生成される。標的リガンドに対する比較的強力な結合について選択された変異体(特定のp/MHC複合体)は、100倍以上高い親和性(すなわち、p/MHCについて約10nMのKd)を有し得、そして、これらの変異体は、高度のペプチド特異性を保持した。プロリンまたはグリシンが豊富な保存されたCDR3モチーフを有するTCRタンパク質についての強力な優先性がまた明らかである。高い親和性TCRの可溶性単量体形態は、抗原提示細胞上のp/MHC複合体を直接検出可能であった。これらの発見は、TCRの親和性成熟が、少なくともインビトロで可能であることを証明する。従って、操作されたTCRタンパク質は、特定のリガンドを標的化するために使用され得、これらとしては、MHC成分の非存在下において特定のp/MHC複合体およびペプチド、タンパク質または他のリガンドが挙げられる。
【0040】
ペプチド特異性を保持するより高い親和性のTCRを作製することが可能か否かを試験するために、本発明者らは、特徴付けられたTCRを、指向性インビトロ進化のプロセスに供した。ファージディスプレイ技術[Clacksonら(1991)Nature 352:624−628]は、抗体とTCR V領域との間の広範囲な構造的類似性にもかかわらず、単鎖TCR(scTCR、Vβ−リンカー−Vα)の操作における成功はまだ証明されていない。しかし、本発明者らは、scTCRが、抗体操作において首尾良いことが証明された系[BoderおよびWittrup(1997)(前出);Kiekeら(1997)前出]において、酵母の表面上に提示され得る[Kiekeら(1999)Proc.Natl.Acad.Sci.USA 96:5651−5656]ということを近年示した。CTLクローン2C由来のscTCRの温度安定化改変体(T7と
呼ばれる)[Shustaら(1999)J. Mol.Biol.292:949−956]を、本研究において使用した。CTLクローン2Cは、酵素2−オキソグルタレートデヒドロゲナーゼ由来の[Udakaら(1993)Proc.Natl.Acad.Sci.USA 90:11272−11276]、p2Caと呼ばれる結合した八量体ペプチドを有する同種異系抗原Ldを認識する。九量体改変体QL9はまた、CTL 2Cにより認識されるが、2C TCRによる10倍高い親和性を有する[Sykulevら(1994)Proc.Natl.Acad.Sci.USA 91:11487−11491]。構造的研究は、2C TCRのCDR3αがペプチドに近く、そしてpMHC複合体に付随するためにコンホメーション変化を受けることが示されたが[Garcia(1998)Science 279: 1166−1172]、アラニンスキャニング変異誘発は、CDR3αループが、結合相互作用に対して最少エネルギーの寄与をすることを示す[Manning(1998)前出]。従って、本発明者らは、変異誘発の試みを、CDR3αの先端を形成する5つの残基に集中させた。
【0041】
105の独立したTCR−CDR3α酵母変異体のライブラリーを、蛍光で標識化されたQL9/Ldリガンドを用いるフローサイトメトリーによる選択にかけた[Dal P
ortoら(1993)Proc.Natl.Acad.Sci.USA 90:6671−6675]。4回の選別および増殖の後、15個の異なる酵母コロニーを、リガンドに結合する能力について、scTCR改変体T7(これは、wt CRD3α配列を保有する)に比較して試験した(図1)。抗Vβ8.2抗体F23.2(これは、タンパク質のCDR1領域およびCDR2領域における残基を認識する)を、コントロールとして使用して、wt scTCR−T7変異体およびscTCR変異体(図1におけるqL2およびqL7ならびにその他)が、各々scTCRのほぼ等しい表面レベルを有することが示された(図1)。対照的に、可溶性QL9/Ldリガンドは、各変異体酵母クローンに非常によく結合するが、wt scTCR−T7には結合しなかった。MCMV(配列番号1)/Ld複合体(これは、CTLクローン2Cにより認識されない)は、scTCR変異体にも、wt scTCR−T7にも結合せず、このことは、scTCR変異体がペプチド特異性を保持していることを示した。変異体TCRの相対的親和性はまた、一定の濃度でQL9/Ldリガンドで観察されるシグナルにおける差異に基づいて、クローンによって変化するようであった。
【0042】
15の変異体のCDR3α配列は、全て、出発した2C TCR配列(表1)と異なっていた。BLASTアラインメントアルゴリズムによる比較は、2つのモチーフへと配列を整列させた。あるモチーフは、残基ストレッチの中間にグリシンを含んでいたが、別のモチーフは、3つの直列プロリンを含んでいた。3つのプロリン全てが、最も高い親和性部位を生成させる際に重要であるという証拠が、変異体q3rを用いた結果により示唆される。変異体q3rは、3つのプロリンのうち2つのみを含んでおり、そして三重プロリン変異体と比較して減少した結合を示した。グリシン含有変異体は、カルボキシ側に対する2つの残基のうち正に荷電した残基を選択し(7/9)、そしてアミノ側に対する2つの残基のうち芳香族のかつ/または正に荷電した残基を選択する(4/9および5/9)ようであった。理論に拘束されることは望まないが、モチーフにおける102位のグリシン残基についての選択は、CDR3αループが増加した親和性を達成するためにこの残基の周りでのコンホメーションの柔軟性を必要とすることを示していると考えられる。このことは、リガンドが配位したTCRのCDR3αループとリガンドが配位していないTCRのCDR3αループとの間で観察された、大きな(6Å)のコンホメーション差異と一致する[Garciaら(1998)前出]。グリシンが抗体のV(D)J接合部における最も一般的な残基であり、そしてグリシンの存在は、近年、(4−ヒドロキシ−3−ニトロフェニル)アセチルハプテンに応じて増加した親和性と関連付けられたことに留意することもまた興味深い[Furukawaら(1999)Immunity 11:329−338]。
【0043】
グリシンを含有する単離物と対照的に、CDR3αループの先端におけるプロリンリッチ配列についての選択は、どんな理論にも束縛されることを望まないが、これらのTCR分子がより高い親和性を付与するより堅固なコンホメーションを示すということを示すと考えられる。その親和性成熟誘導体に比較して低い親和性の生殖細胞系抗体のX線結晶解析構造は、高い親和性の状態が、ハプテンを付随する配置における抗体の安定化に起因し得たことを示した[Wedemayerら(1997)Science 276:1665−1669]。同様に、生殖細胞系抗体に類似し得るscTCRのNMR溶液構造は、CDR3αおよびβループが、両方とも有意な運動性を示したことを示した[Hareら(1999)Nat.Struct.Biol.6:574−581]。TCR:pMHC相互作用の最近の熱力学的研究はまた、結合におけるコンホメーション変化の重要性を示唆した[Willcoxら(1999)Immunity 10:357−365;Bonifaceら(1999)Proc.Natl.Acad.Sci.USA 96:11446−11451]。本明細書中で考察されるTCR変異体の構造研究および熱力学的研究により、本発明者らは、2つのCDR3αモチーフ(Gly対Proリッチ)がより高い親和性を付与する機構において異なるか否かを調べることが可能になった。
【0044】
scTCR変異体は、無効な(null)(無関連な)ペプチド/Ld複合体MCMV(配列番号1)/Ldに結合しなかったが、親和性の増加が、微細な(fine)特異性における変化と関連付けられたことは可能なままであった。この問題を調べるために、本発明者らは、以前にそのwt2C TCRについての結合親和性における有意な差異を示すことが支援された、QL9の5位(Phe)ペプチド改変体を使用した[Schlueter(1996)J.Immunol.157:4478−4485]。酵母表面上の種々のTCR変異体およびクローン2Cに対するこれらのpMHCの結合を、フローサイトメトリーにより測定した。図2に示されるように、2C上のネイティブTCRは、チロシンまたはヒスチジンのいずれかを5位に含むがグルタミン酸は含まないQL9改変体に結合し得る。より高い親和性のTCR変異体の各々は、保存的チロシン置換ペプチドを認識する能力を保持し、そしてこれらは、グルタミン酸置換ペプチドを認識できないようであった。しかし、TCR変異体のうちのいくつか(qL2、qL5、およびqL7)は、ヒスチジン置換ペプチドに(異なる程度であるが)結合したが、その他の変異体(qL1、qL3、およびqL8)は、このペプチドに結合しなかった(このアッセイの検出限界内)。従って、CDR3αループは、ペプチドの微細な特異性の認識に影響し得るが、TCRが含まれる唯一の領域ではない。CDR3に指向された選択に関する初期の研究[Sant’Angeloら(1996)Immunity 4:367−376;Jorgensenら(1992)Nature 355:224−230]から示唆されたように、ペプチド特異性に対する効果は、改変体ペプチドとのCDR3α残基の直接の相互作用に起因し得る。あるいは、結合エネルギーは、Ld分子自体におけるペプチドに誘導される変化に向けられ得る。後者の可能性は、2C TCR:QL9/Ld相互作用の場合におけるよりもおそらく起こりやすい。なぜならば、QL9の5位は、Ld溝に向いていると推測されたからである[Schlueterら(1996)前出,Speirら(1998)Immunity 8:553−562]。微細特異性分析はまた、同族のペプチドについて増加したか、または少なくとも変更された特異性を有するTCRを操作することが可能であることを示す。従って、単一のTCRの短い領域(CDR3α)のみの指向された進化は、所望のペプチド結合特異性および/または増加した結合親和性を有する多くのTCR改変体の単離を可能にする。
【0045】
選択されたCDR3α変異体に関連する親和性の増加の大きさを決定するために、野生型T7scTCRおよびqL2scTCRは、酵母分泌系において可溶性形態として発現された。精製されたscTCR調製物を、T2−Ld細胞の表面上でLd上に負荷されたQL9またはMCMV(配列番号1)に対する、125I標識化抗LdFabフラグメントの結合を遮断する能力について比較した[Manning(1998)前出、Sykulevら(1994)Immunity 1:15−22]。予測されたように、T7もqL2scTCRも、MCMV(配列番号1)ペプチドでアップレギュレートされたT2−Ld細胞に対する125I−Fabフラグメントの結合を阻害することができなかった。しかし、T7およびqL2の両方が、QL9/Ldに対する抗LdFabフラグメントの結合を阻害し得た(図3)。qL2scTCR改変体は、結合の阻害において標識化されていないFabフラグメントと同程度の効力であったが、T7scTCRは、160倍も小さい効力(4回の独立した適定の中で、平均して140倍の差)であった。QL9/LdについてのscTCRのKD値を、その阻害曲線から、T7について1.5μMおよびqL2について9.0nMと算出した。T7についての値は、2CscTCRについて以前に報告された3.2μMのKDと密接に一致した[Manningら(1999)J:Exp.Med.189:461−470]。これらの発見は、CDR3α指向性変異誘発と組み合わされた酵母系が、特定のpMHCリガンドについての、少なくとも約100倍高い固有の結合親和性を有する変異体の選択を可能にするということを示す。
【0046】
可溶性scTCRがそのpMHCリガンドに関して高い親和性を有する場合、抗体のように、細胞表面結合抗原についての特異的プローブとして有用である。このことを確かめるために、この可溶性T7およびqL2scTCRを、ビオチン化し、そして標識化されたscTCR分子を、QL9、p2Ca、またはMCMV(配列番号1)を負荷されたT2−Ld細胞とインキュベートした。qL2scTCR(T7 scTCRではない)は、QL9またはp2CaとインキュベートされたT2細胞の容易に検出可能な染色を生じた(図4A〜4B)。p2Caアップレギュレートされた細胞がまた、qL2scTCRにより容易に検出されたことは重要である。なぜならば、p2Caは、同種反応性(alloreactive)クローン2Cにより認識されたペプチドの天然で処理された形態であり、そしてこれは、2C TCRについてQL9/Ld複合体よりも低い親和性でさえ有するからである[Sykulevら(1994)前出]。
【0047】
本発明者らの研究において記載される高親和性レセプターは、VJ接合部における改変により誘導された(T細胞における遺伝子再配列によりインビボで非常に有効に作用する同じプロセス)(DavisおよびBjorkman(1988)Nature 334:395−402)。本発明者らがインビトロで高親和性TCRの広範なセットを容易に単離し得たという事実は、高親和性レセプターに対する遺伝的制限も構造的制限も存在しないということを示す。このことは、インビボで見出されるTCRの固有の低い親和性が、より高い親和性についての選択の欠如、およびおそらくより低い親和性の選択に起因するという見解を支持する(Sykulevら(1995)Proc.Natl.Acad.Sci.USA 92:11990−11992;Valituttiら(1995)Nature 375:148−151;Rabinowitzら(1996)Proc.Natl.Acad.Sci.USA 93:1401−1405)。この点において、本発明のより高い親和性のTCRはここで、T細胞応答に対する親和性の効果についての仮定を直接的に試験するための試薬を提供する((Davisら(1998)Ann.Rev.Immunol.16:523−544;Sykulevら(1995)(前出);Valituttiら(1995)(前出);Rabinowitzら1996)前出)。
【0048】
要約すると、本発明者らは、T細胞レセプター(抗体と同程度広範なタンパク質のクラスを代表する)が、抗体と同様に操作されて、高い親和性の、抗原特異的プローブを与え得ることを示した。さらに、高親和性レセプターの可溶性変形は、細胞上の特異的ペプチド/MHC複合体を直接検出し得る。従って、これらの操作されたタンパク質は、例えば、腫瘍細胞についての診断剤として有用である。高親和性TCRの可溶性誘導体は、高親和性の抗原特異的プローブとして有用であるか、またはさらに操作され得る。適切に標識化された場合(またはこの可溶性TCRに検出可能なリガンドにより結合される場合)の可溶性TCR誘導体は、細胞(例えば、腫瘍細胞もしくはその他の新形成細胞の誘導された表面、またはウイルス感染細胞もしくはその他の疾患細胞の診断的抗原)上の特異的ペプチド/MCHC複合体についてのプローブとして作用し得る。高親和性TCR細胞に結合したタンパク質または可溶性誘導体についての他の適用としては、特定の自己免疫疾患の診断または研究における使用が挙げられる。特徴的なペプチド/MHCまたはその他のマーカー表面抗原が公知であるかまたは同定され得る場合、高親和性の可溶性TCRは、その細胞表面部分への特異的結合のために単離され得、そして診断または治療において使用され得る。破壊されることが所望される細胞が、高親和性TCR(望ましくは可溶性TCRタンパク質)により認識される特徴的かつ特異的細胞表面部分を有する場合、高親和性TCRタンパク質(望ましくは可溶性誘導体)は、癌の処置または他の障害における治療剤として、細胞傷害性因子に結合して使用され得る。同様に、本明細書中に記載される可溶性高親和性TCRは、画像化剤にカップリングされ得、そして腫瘍細胞が存在する身体内の部位(ここでTCRが、高親和性および特異性を有する腫瘍細胞マーカーに特異的に結合する)を同定するために使用され得る。不適切に自己免疫破壊の標的にされてきた細胞または組織の表面に結合した高親和性TCRは、標的細胞または組織の細胞表面への結合について細胞傷害性リンパ球と競合することにより、これらの細胞傷害性リンパ球による自己免疫組織破壊を低減させ得る。
【0049】
これらの結果はまた、T細胞応答における重要で、基本的な疑問の状況において考慮され得る。T細胞レセプターについて以前に観察された低い親和性は、体細胞変異の非存在に起因するのか、またはT細胞に対して作用するインビボでの選択的圧力に起因するのか?本発明者らの研究において記載される高親和性レセプターは、VJ接合部でのバリエーションに由来した(T細胞において非常に有効に作用する同じプロセス)[DavisおよびBjorkman(1988)前出]。本発明者らが、インビトロで高親和性TCRの広範なセットを容易に単離し得たという事実は、高親和性レセプターへの構造的制限または遺伝的制限が存在しないことを示す。このことは、インビボで見出される元々低い親和性のTCRが、より高い親和性の選択の欠如およびおそらくより低い親和性の選択の欠如に起因すると言う見解を支持する[Sykulevら(1995)Proc.Natl.Acad.Sci.USA 92:11990−11992;Rabinowitzら(1996)前出]。この点において、ここでより高い親和性のTCRは、T細胞応答に対する親和性の効果についての仮説を直接試験するための試薬を提供する[Davisら(1998)(前出);Sykulevら(1995)(前出);Valituttiら(1995)(前出);Rabinowitzら(1996)前出]。
【0050】
遺伝子コードの縮重に起因して、多数の機能的に等価なヌクレオチド配列が、同じアミノ酸配列をコードするということが、当業者に理解される。
【0051】
さらに、当業者は、標準的な変異誘発技術により、本明細書中に記載される抗原決定活性アッセイを組み合わせて、変更されたTCR配列を得、そしてそれらを特定の結合活性を有するポリペプチドの発現について試験し得る。当該分野で公知の有用な変異誘発技術としては、限定せずに、オリゴヌクレオチド指向性変異誘発、領域特異的変異誘発、リンカースキャニング変異誘発、およびPCRによる部位指向性変異誘発が挙げられる[例えば、Sambrookら(1989)およびAusubelら(1999)を参照のこと]。
【0052】
改変体TCRコード配列を得る際に、当業者は、TCR由来のタンパク質が、生物学的活性の損失も減少もなしに、特定のアミノ酸置換、付加、欠失および翻訳後修飾により改変され得ることを認識する。特に、保存的アミノ酸置換、すなわち、1つのアミノ酸の、類似のサイズ、電荷、極性およびコンホメーションの別のアミノ酸との置換が、タンパク質の機能を有意に変更しないようであることは周知である。タンパク質を構成する20種の標準的なアミノ酸は、以下のような保存的アミノ酸の4つのグループに広く分類され得る:非極性(疎水性)グループは、アラニン、イソロイシン、ロイシン、メチオニン、フェニルアラニン、プロリン、トリプトファンおよびバリンを含む;極性(非荷電、中性)グループは、アスパラギン、システイン、グルタミン、グリシン、セリン、トレオニンおよびチロシンを含む;正に荷電した(塩基性)グループは、アルギニン、ヒスチジンおよびリシンを含む;そして負に荷電した(酸性)グループは、アスパラギン酸およびグルタミン酸を含む。タンパク質中の1つのアミノ酸を同じグループ内の別のアミノ酸と置換することは、そのタンパク質の生物学的活性に不利な影響を有さないようである。
【0053】
ヌクレオチド配列間の相同性は、DNAハイブリダイゼーション分析によって決定され得、ここでは、二本鎖DNAハイブリッドの安定性は、生じる塩基対形成の程度に依存する。高温および/または低塩含量の条件は、ハイブリッドの安定性を減少させ、そして選択された程度より少ない相同性を有する配列のアニール化を妨げるために変更させ得る。例えば、約55%のG−C含量を有する配列に対して、40〜50℃、6×SSC(塩化ナトリウム/クエン酸ナトリウム緩衝液)および0.1% SDS(ドデシル硫酸ナトリウム)というハイブリダイゼーションおよび洗浄条件は、約60〜70%の相同性を示し、50〜65℃、1×SSCおよび0.1% SDSのハイブリダイゼーションおよび洗浄条件は、約82〜97%の相同性を示し、そして52℃、0.1×SSCおよび0.1% SDSのハイブリダイゼーションおよび洗浄条件は、約99〜100%の相同性を示す。ヌクレオチド配列およびアミノ酸配列を比較するため(そして相同性の程度を測定するため)の広範囲のコンピュータプログラムはまた、利用可能であり、そして市販されるソフトウエアとフリーソフトウエアの両方の供給源を提供する表は、Ausubelら(1999)において見出される。すぐに利用可能な配列比較および複数の配列整列アルゴリズムは、それぞれ、Basic Local Alignment Search Tool(BLAST)(Altschulら、1997)およびClustalWプログラムである。BLASTは、http://www.ncbi.nlm.nih.govでインターネット上で利用可能であり、そしてClustalWのバージョンは、http://www2.ebi.ac.ukで利用可能である。
【0054】
微生物の工業的株(例えば、Aspergillus niger、Aspergillus ficuum、Aspergillus awamori、Aspergillus oryzae、Trichoderma reesei、Mucor miehei、Kluyveromyces lactis、Pichia pastoris、Saccharomyces cerevisiae、Escherichia coli、Bacillus subtilisまたはBacillus licheniformis)あるいは植物種(例えば、カノーラ、ダイズ、コーン、トマト、オオムギ、ライムギ、コムギ)は、TCRペプチドの組換え産生のための宿主細胞として使用され得る。高親和性TCRタンパク質または可溶なタンパク質の異種発現における第1工程として、発現構築物は、TCRまたは可溶なTCRのコード配列、ならびに制御配列(例えば、プロモーター、エンハンサーおよびターミネーター)を含むように構築される。シグナル配列および選択マーカーのような他の配列はまた、含まれ得る。scTCRの細胞外発現を達成するために、発現構築物は、分泌性シグナル配列を含み得る。シグナル配列は、細胞質発現が所望される場合、発現構築物上に含まれない。プロモーターおよびシグナル配列は、宿主細胞において機能的であり、そしてTCRまたは可溶なTCRタンパク質の発現および分泌を提供する。転写ターミネーターは、効果的な転写を保証するために含まれる。発現またはタンパク質精製を増強する補助的配列はまた、発現構築物に含まれ得る。
【0055】
種々のプロモーター(転写開始調節領域)は、本発明に従って使用され得る。適切なプロモーターの選択は、提案される発現宿主に依存する。異種供給源からのプロモーターは、それらが選択された宿主において機能的である限り、使用され得る。
【0056】
プロモーター選択はまた、所望の有効性およびペプチドまたはタンパク質の産生のレベルに依存する。誘導プロモーター(例えば、tac)は、しばしば、E.coliにおけるタンパク質発現のレベルを劇的に増加させるために利用される。タンパク質の過剰発現は、宿主細胞に有害であり得る。その結果、宿主細胞の増殖は制限される。誘導プロモーター系の使用によって、宿主細胞は、遺伝子発現の誘導前に、受容可能な密度まで培養されることが可能となり、それによって、より高い産生物収量を容易にする。
【0057】
種々のシグナル配列が、本発明に従って使用され得る。TCRコード配列と同種のシグナル配列が、使用され得る。あるいは、効果的な分泌および発現宿主におけるプロセシングのために選択されるかまたは設計されたシグナル配列はまた、使用され得る。例えば、適切なシグナル配列/宿主細胞の組としては、B.subtilisにおける分泌のためのB.subtilis sacBシグナル配列、およびSaccharomyces cerevisiae α接合因子またはP.pastoris分泌のためのP.pastoris酸性ホスファターゼphoIシグナル配列が挙げられる。シグナル配列は、タンパク質コード配列に対するシグナルペプチダーゼ切断部位をコードする配列によって、または通常10より少ないコドンからなる短いヌクレオチドブリッジ(bridge)(ここで、このブリッジは、下流のTCR配列の正しい読み取り枠を保証する)によって、直接連結され得る。
【0058】
転写および翻訳を増強するためのエレメントは、真核生物のタンパク質発現系について同定されている。例えば、カリフラワーモザイクウイルス(CaMV)プロモーター1000bpを異種プロモーターのいずれかの端に配置することは、植物細胞において転写レベルを10〜400倍増強させ得る。発現構築物はまた、適切な翻訳開始配列を含む。適切な翻訳開始のためのKozakコンセンサス配列を含むための発現構築物の改変は、翻訳のレベルを10倍増大させ得る。
【0059】
選択マーカーがしばしば利用され(これは、発現構築物の一部分であっても、または発現構築物と離れて(例えば、発現ベクターによって運ばれる)いてもよい)、その結果、マーカーは、目的の遺伝子と異なる部位で統合され得る。例としては、抗生物質に耐性を与える(例えば、blaは、E.coli宿主細胞に対してアンピシリン耐性を与え、nptIIは、広範囲の原核生物細胞および真核生物細胞に対してカナマイシン耐性を与える)マーカー、または宿主を最少培地で増殖させるマーカー(例えば、HIS4は、P.pastorisまたはHis−S.cerevisiaeがヒスチジンの非存在下で増殖することを可能にする)が挙げられる。選択可能なマーカーは、それ自身の転写および翻訳の開始ならびに停止の調節領域を有し、マーカーの独立した発現を可能にする。抗生物質耐性がマーカーとして利用される場合、選択のための抗生物質濃度は、その抗生物質に依存して変化し、通常、抗生物質10〜600μg/培地1mLの範囲である。
【0060】
発現構築物は、公知の組換えDNA技術(Sambrookら、1989;Ausubelら、1999)を利用することによって構築される。制限酵素消化および連結は、DNAの2つのフラグメントを連結させるために利用される基本的工程である。DNAフラグメントの末端は、連結前に改変を必要とし得、そしてこれは、突出を埋めること、ヌクレアーゼ(例えば、ExoIII)でフラグメント(単数または複数)の末端部分を欠失すること、部位を指向された突然変異誘発によって、またはPCRによって新しい塩基対を添加することによって達成され得る。ポリリンカーおよびアダプターは、選択されたフラグメントの連結を容易にするために利用され得る。発現構築物は、代表的に、E.coliの制限、連結および形質転換の数ラウンド(round)を利用する段階で構築される。発現構築のために適切な多くのクローニングベクターは、当該分野において公知である(λZAPおよびpBLUESCRIPT SK−1、Stratagene、LaJolla、CA;pET、Novagen Inc.Madison、WI−Ausubelら、1999において引用される)、そして特定の選択が、本発明には重要ではない。クローニングベクターの選択は、宿主細胞への発現構築物の導入のために選択される遺伝子転移系によって影響される。各段階の終わりに、得られた構築物は、制限、DNA配列、ハイブリダイゼーションおよびPCR分析によって分析され得る。
【0061】
発現構築物は、クローニングベクター構築物(線状か環状かのいずれか)として宿主に形質転換され得るか、あるいはクローニングベクターから除去され得、そして送達ベクターとして使用され得るか、または送達ベクター内に導入され得る。送達ベクターは、選択された宿主細胞型において、発現構築物の導入および保持を容易にする。発現構築物は、多くの公知の遺伝子導入系(例えば、天然の能力(competence)、化学的に媒介される形質転換、プロトプラスト形質転換、エレクトロポレーション、微粒子銃形質転換、トランスフェクション、または結合)(Ausubelら、1999;Sambrookら、1989)のいずれかによって宿主細胞に導入される。選択される遺伝子転移系は、使用される宿主細胞およびベクター系に依存する。
【0062】
例えば、発現構築物は、プロトプラスト形質転換またはエレクトロポレーションによってS.cerevisiae細胞に導入され得る。S.cerevisiaeのエレクトロポレーションは、容易に達成され、そしてスフェロプラスト形質転換に匹敵する形質転換効率をもたらす。
【0063】
リガンド結合部位以外の部位でTCRタンパク質によって特異的に反応するモノクローナルまたはポリクローナル(好ましくは、モノクローナル)抗体は、当該分野において公知である方法によって作製され得る。例えば、HarlowおよびLane(1988)Antibodies:A Laboratory Manual、Cold Spring Harbor Laboratories;Goding(1986)Monocolonal Antibodies:Principles and Practice 第2版、Academic Press、New York;およびAusubelら(1999)Molecular Biology、John Wiley & Sons,Inc.、New YorkのCurrent Protocolsを参照のこと。
【0064】
特定のpMHCに特異的な細胞結合形態または可溶形態の高親和性のTCRタンパク質は、例えば、生物学的サンプル(例えば、細胞、組織サンプル、生検材料、体液など)のスクリーニングのため、または試験サンプルにおける同族pMHCの存在を検出するための診断プローブとして有用である。しばしば、高親和性のTCRタンパク質は、検出可能なシグナルを提供する基質を連結させる(共有結合か非共有結合かのいずれか)ことによって標識化される。適切な標識としては、放射性核種、酵素、基質、補助因子、インヒビター、蛍光試薬、化学発光試薬、磁気粒子などが挙げられるが、これらに限定されない。さらに、TCRタンパク質は、第2の結合分子に対するリガンドに結合し得:例えば、TCRタンパク質は、ビオチン標識化され得る。従って、標的細胞または分子に結合したTCRの検出は、検出可能なストレプトアビジン(蛍光分子、放射活性分子、化学発光分子、または他の検出可能な分子が付着されるか、または、発色団基質が利用可能である酵素が付着される、ストレプトアビジン)の結合によって達成され得る。scTCRタンパク質に共有結合するべきこのような標識物および/または有毒な化合物の使用を記載する米国特許のとしては、以下の号が挙げられるが、これらに限定されない:
【0065】
【数1】

標識化されたTCRタンパク質は、使用される標識に適切なモニタリングデバイスまたは方法を用いて検出され得る。蛍光顕微鏡または蛍光活性化セルソーティングは、標識が蛍光部分である場合に使用され得、そして標識が、放射性核種である場合、γカウンティング、オートラジオグラフィまたは液体シンチレーションカウンティングが、例えば、その方法が分析されているサンプルおよび使用される放射性核種に適切であるという条件で使用され得る。さらに、本明細書中に述べられるMHC成分の非存在下で同族pHMCリガ
ンドまたは他のリガンドに対する結合部位の一部でないTCRタンパク質の部分を認識する検出可能な分子または粒子がある場合、第2の検出分子または粒子が利用され得る。当業者は、インサイチュでの診断画像処理に有用な化合物を知っている;例えば、米国特許第5,101,827号、同第5,059,413号を参照のこと。インビボの治療および/または画像処理に有用な放射性核種はとしては、111インジウム、97ルビジウム、125ヨウ素、131ヨウ素、123ヨウ素、67ガリウム、99テクネチウムが挙げられる。毒素としては、一旦、TCR−毒素複合体が細胞に結合される場合、毒性部分が内部移行し、その結果、その細胞傷害性効果を及ぼし得るという条件では、とりわけ、ジフテリア毒素、リシンおよびトウゴマ毒素が挙げられる。免疫毒性テクノロジーは、当該分野において周知であり、そして適切な毒性分子としては、以下が挙げられるが、これらに限定されない;化学療法薬剤(例えば、ビンデシン)、葉酸代謝拮抗財(例えば、メトトレキセート、シスプラチン、マイトマイシン)、アンスロサイクリン(anthrocyclines)(例えば、ダウノマイシン、ダウノルビシンまたはアドリアマイシン)およびリボソーム不活性化タンパク質のような細胞傷害性タンパク質(例えば、ジフテリア毒素、ヤマゴボウ抗ウイルスタンパク質、アブリン、リシン、シュードモナス属体外毒素Aまたはこれらの組換え誘導体)。一般的に、例えば、OlsnesおよびPihl(1982)Pharmac.Ther.25:355−381およびMonoclonal Antibodies for
Cancer Detection and Therapy、BaldwinおよびByers編、159頁−179頁、Academic Press、1985を参照のこと。
【0066】
特定のpMHCリガンドに特異的な高親和性TCRタンパク質は、特定のpMHCに関連する疾患に罹患すると考えられる動物(ヒトを含む)を診断において有用である。本発明のscTCR分子は、本質的に、任意の抗原を基本的に検出するのに有用であり、新生物状態、異常なタンパク質、あるいは細菌、真菌、ウイルス、原虫、酵母、線虫または他の寄生生物による感染またはインフェステーションに関連するものが挙げられるが、これらに限定されない。高親和性のscTCRタンパク質はまた、生産された異常なタンパク質が存在する特定の遺伝子疾患の診断の際に使用され得る。これらの高親和性タンパク質の例示的な適用は、自己免疫疾患の処置においてであり、その中には、既知のpMHCがある。I型糖尿病は、免疫破壊を誘発する自己抗原に関して比較的よく特徴付けられる。多発性硬化症、セリアック病、炎症性腸疾患、クローン病および慢性関節リウマチは、このような適用に対するさらなる候補疾患である。自己免疫破壊を不適切に標的化する、細胞または組織の表面上のp/MHC複合体に対する結合特異性を有する高親和性のTCR(可溶性)タンパク質は、細胞傷害性リンパ球による標的細胞への結合について競合することによって、自己免疫破壊のアンタゴニストとして役立ち得る。対照的に、高親和性TCRタンパク質(望ましくは、可溶性の一本鎖TCRタンパク質)(これは、破壊が有益である細胞表面上の抗原またはp/MHCに特異的に結合する)は、有毒な化合物(例えば、毒素または放射性核種)に結合し得、その結果、標的細胞への結合は、細胞傷害性リンパ球による引き続いての結合および破壊を引き起こす。破壊のために標識化された細胞は、新生物細胞(例えば、腫瘍細胞)、ウイルス、細菌、または原虫で感染された細胞、あるいは他の疾患原因因子もしくは寄生生物であり得、すなわち、それは、細菌、酵母、真菌、原虫または所望されない細胞であり得る。このような高親和性のscTCRタンパク質は、本明細書中に記載される方法によって得ることができ、そして続いて、目的の特定のリガンドについてスクリーニングするために使用され得る。
【0067】
高親和性のTCR組成物は、当該分野において公知の任意の方法によって処方され得る。これらは、代表的に、特に経静脈投与、経腹膜投与、または滑液投与(特定の疾患によって決定される経路で)のための注射剤として、または、経鼻または経口投与のための処方物(液体溶液か懸濁液かのいずれか)として調製され得る。注射または他の投与前の液体中の、溶液または懸濁液に適切な固体の形態はまた、調製され得る。この調製物はまた、例えば、乳化され得るか、またはリポソーム中にカプセル化されたタンパク質(単数および複数)/ペプチド(単数および複数)であり得る。
【0068】
活性な成分はしばしば、賦形剤またはキャリアと混合され、これらは、薬学的に受容可能であり、そして活性な成分と共存可能である。適切な賦形剤としては、水、生理的食塩水、ブドウ糖、グリセロール、エタノールなど、およびそれらの組合せが挙げられるが、これらに限定されない。注射可能な、エアロゾルまたは経鼻処方物の形態の高親和性のTCRタンパク質の濃度は、通常、0.05〜5mg/mlの範囲にある。特定の有効な用量の選択は、公知であり、そして当業者によって過度の実験なしに行われる。同じ用量が、他の粘膜表面に投与され得る。
【0069】
さらに、所望される場合、ワクチンは、少量の補助的物質(例えば、湿潤剤または乳化剤、pH緩衝剤、および/またはワクチンの効果を増強するアジュバント)を含み得る。有効であり得るアジュバントの例としては、以下が挙げられるが、これらの限定されない:水酸化アルミニウム;N−アセチル−ムラミル−L−スレオニル−D−イソグルタミン(thr−MDP);N−アセチル−nor−ムラミル−L−アラニル−D−イソグルタミン(CGP 11637、nor−MDPと呼ばれる);N−アセチルムラミル−L−アラニル−D−イソグルタミニル−L−アラニン−2−(1’−2’−ジパルミトイル−sn−グリセロ−3ヒドロキシホスホリル)−エチルアミン(CGP 19835A、MTP−PEと呼ばれる)およびRIBI(これは、細菌から抽出された3つの成分を含む:2%のスクアレン/Tween 80乳濁液中のモノホスホリルリピドA、トレハロースジマイコレート(dimycolate)および細胞壁骨格(MPL+TDM+CWS)。当該分野において公知であるこのようなさらなる処方物および投与の形態はまた、使用され得る。
【0070】
本発明の高親和性TCRタンパク質、および/または高親和性TCRタンパク質に対して類似(90%より大きい同一性)の一次構造を有するpMHC結合フラグメント(同族のリガンドについて高親和性を維持する)は、中性形態または塩形態としてワクチン中に処方され得る。薬学的に受容可能な塩としては、酸付加塩(ペプチドの遊離アミノ酸基で形成された)が挙げられるがこれらに限定されない。この塩は無機酸(例えば、塩酸またはリン酸)および有機酸(例えば、酢酸、シュウ酸、酒石酸、またはマレイン酸)で形成される。遊離のカルボキシル基で形成された塩はまた、無機塩基(例えば、水酸化ナトリウム、水酸化カリウム、水酸化アンモニウム、水酸化カルシウム、または水酸化第二鉄)、および有機塩基(例えば、イソプロピルアミン、トリメチルアミン、2−エチルアミノ−エタノール、ヒスチジン、およびプロカイン)に由来し得る。
【0071】
治療用途のための高親和性TCRタンパク質(例えば、細胞毒性化合物に結合体化されたタンパク質)を、当該分野で公知の方法に従って、投与処方物に適切な様式で、そして予防的におよび/または治療的に効果的であるような量および様式で、投与する。投与されるべき量は、一般に1用量あたり約100〜20,000μgタンパク質の範囲、より一般的には、1用量あたり約1000〜10,000μgの範囲である。同様の組成物が、画像化における用途のために標識化した高親和性TCRタンパク質を用いて類似の様式で投与され、例えば、自己免疫攻撃下でかつ同属のpMHCを含む組織を検出するか、またはその表面に同属のpMHCを保有する癌細胞を検出し得る。投与されるのに必要な活性成分の正確な量は、医師または獣医師の判定に依存し得、そして各個体について特有であり得るが、このような決定は、このような医師の当該分野の範囲内である。
【0072】
ワクチンまたは他の免疫原性組成物は、単回用量;2用量スケジュール(例えば、2〜8週間間隔);または複数用量スケジュールで与えられ得る。複数用量スケジュールは、ワクチン接種の一次コースが1〜10以上の別の用量、続いて、免疫応答を維持するおよび/または強化するために必要な場合、引き続く時間間隔で投与される他の用量(例えば、第二用量は1〜4カ月)、そして必要な場合、数ヶ月後に引き続く用量を含み得るスケジュールである。本発明のレトロウイルス様粒子で免疫したヒト(または他の動物)は、同属のレトロウイルスによる感染から防御される。
【0073】
クローニング、DNA単離、増幅および精製のため、DNAリガーゼ、DNAポリメラーゼ、制限エンドヌクレアーゼなどを含む酵素反応のための標準的技術、ならびに種々の別の技術は、公知の技術であり、そして当業者によって通常使用される。多数の標準的技術が、以下に記載されている:Sambrookら(1989)Molecular Cloning:第二版、Cold Spring Harbor Laboratory,Plainview,New York;Maniatisら(1982)Molecular Cloning、Cold Spring Harbor Laboratory,Plainview,New York;Wu(編)(1993)Meth.Enzymol.218,PartI;Wu(編)(1979)Meth Enzymol.68:Wuら(編)(1983)Meth Enzymol.100および101;GrossmanおよびMoldave(編)Meth.Enzymol.65;Miller(編)(1972)Experiments in Molecular
Genetics,Cold Spring Harbor Laboratory,Cold Spring Harbor,New York;Old and Primrose(1981)Principles of Gene
Manipulation,University of California Press,Berkeley;SchleifおよびWensink(1982)Practical Methods in Molecular Biology;Glover(編)(1985)DNA Cloning第IおよびII号、IRL Press,Oxford,UK;Hames and
Higgins(編)(1985)Nucleic Acid Hybridization,IRL Press,Oxford,UK;ならびにSetlowおよびHollaender(1979)Genetic Engineering:Principles and Methods、1〜4号、Plenum Press,New York。ここで使用した略号および命名法は、この分野で標準的とみなされ、そしてここで引用される文献のような専門の文献中に通常用いられている。
【0074】
本出願に引用される全ての参考文献は、本開示と矛盾しない程度まで、本明細書に提供される開示および実験手順を補充するために、本明細書において参考として援用される。
【0075】
以下の実施例は、例示的な目的のために提供されるものであり、そして本明細書に記載される本発明の特許請求の範囲を限定する意図ではない。当業者に対して生じる、例示された記事および/または方法における任意のバリエーションは、本発明の範囲内におさまることが意図される。
【0076】
(実施例)
高い親和性のTCRを作成するための一般的方法は、1998年1月20日出願、米国特許出願番号09/009,388号、および1999年1月20日出願WO99/36569(これらは、本明細書の開示と矛盾しない程度まで、参考として援用される)に与えられる。
【0077】
(実施例1.ライブラリーの構築)
指向性進化のための足場として用いた2C scTCR(T7)は、TCRの安定性を増大させるがpMHC結合を可能にすることが示されている、6つの変異体(βG17E、βG42E βL81S,αL43P、αW82R、およびαI118N)を含んだ[例えば、Shusta,E.V.ら(1999)J.Mol.Biol.292,949〜956]。
【0078】
AGA2−特異的上流プライマー(GGCAGCCCCATAAACACACAGTAT(配列番号3))および変性下流プライマーCTTTTGTGCCGGATCCAAATGTCAG(SNN)5GCTCACAGCACAGAAGTACACGGCCGAGTCGCTC(配列番号4)を用いて、T7 scTCR VαCDR3の変異性PCRを、実施した。
下線を付した塩基(配列番号4の最初から11番目のG、終わりから13番目のG)は、特有のBamHIおよびEagI制限部位を誘導するサイレント変異の位置を示す。精製したPCR産物を、NdeIおよびBamHIを用いて消化し、そしてNdeI−BamHIで消化したT7/pCT302に連結した[BoderおよびWittrup(1997)前出;Kiekeら(1999)前出;Shustaら(1999)前出]。連結混合物をDH10Bエレクトロ−コンピテントなE.coli(Gibco BRI,Gaithersburg,MD)中に形質転換し、そして形質転換体を、アンピシリン100μg/ml補充した250mLのLB中にプールし、そして37℃で一晩増殖した。プラスミドDNAを、GietzおよびSchiestl[Geitzら(1995)Yeast 11:355〜360]の方法によって、酵母(Saccharomyces
cerevisiae)株EBY100中に形質転換した。
【0079】
(実施例2.細胞ソーティング)
酵母ライブラリー[Shustaら(1999)Curr.Opin.Biotechnol.10:117〜122]を、SD−CAA(2%デキストロース、0.67%酵母窒素ベース、1%カザミノ酸(Difco、Livonia,MI))中で、30℃で、OD600=4.0まで増殖した。表面scTCR発現を誘導するため、酵母を遠心分離によってペレット化し、SG−CAA(2%ガラクトース、0.67%酵母窒素ベース、1%カザミノ酸)中で、OD600=1.0まで再懸濁し、そして20℃で24時間インキュベートした。一般に、約107細胞/チューブを、50μlのQL9/Ld/IgGダイマー[Dal Portoら(1993)前出](これは0.5mg/mlBSA(PBS−BSA)を補充したリン酸緩衝化生理食塩水(pH7.4)中で希釈してある)とともに、1時間氷上でインキュベートした。インキュベーション後、細胞を洗浄し、そしてFITC結合体化ヤギ抗マウスIgG F(ab7)2(Kirkegaard&Perry,Gaithersburg,MD)含有PBS−BSAを用いて30分間標識した。次いで、ソーティングの直前に、PBS−BSA中で酵母を洗浄し、そして再懸濁した。コールター753ベンチを用いたFACSソーティングによって最高の蛍光を示す細胞を、単離した。単離後、ソーティングした細胞をSD−CAA中で増殖し、そして以降の回の選択のためにSG−CAA中で誘導した。引き続く4回の総ソーティングを実施した。染色に用いたQL9/Ld/IgGダイマーの濃度は、ソート1〜3については50μg/mlであり、そして最終のソートについては0.5μg/mlであった。各ソートから単離した総細胞のパーセンテージは、それぞれ5.55%、2.68%、2.56%および0.58%であった。ソート3および4のアリコートをSD−CAA上にプレートし、個々のクローンを単離した。クローンは、コールターEpicsXL装置を用いたフローサイトメトリーにより分析した。
【0080】
(実施例3.可溶性のscTCR産生)
T7およびqL2のオープンリーディングフレームを、pCT302 NheI−XhoIから切り出し、そしてNheI−XhoIで消化したpRSGALT(酵母発現プラスミド)に連結した[Shustaら(1999)前出]。ライゲーション(連結)したプラスミドをDH10Bエレクトロ−コンピテントなE.coli(Gibco BRL)中に形質転換した。プラスミドDNAを細菌培養物から単離し、そしてSaccharomyces cerevisiae BJ5464(α ura3−52 trp1 leu2 1 his3 200 pep4::HIS3 prb1 1.6R can1GAL)中に形質転換した[Shustaら(1999)前出]。酵母クローンを30℃で48時間1リットルのSD−CAA/Trp(20mg/mL トリプトファン)中で増殖した。scTCR分泌を誘導するため、細胞を4000×gでの遠心分離によってペレット化し、1mg/mlBSAを補充した1リットルのSG−CAA/Trp中に再懸濁し、そして20℃で72時間インキュベートした。培養上清を4000×gでの遠心分離によって回収し、約50mlに濃縮し、そしてPBS、pH8.0に対して透析した。6His−タグ化scTCRを、ネイティブなニッケルアフィニティークロマトグラフィー(Ni−NTA Superflow,Qiagen,Valencia,CA;5mMおよび20mM イミダゾール、pH8.0洗浄;250mMイミダゾール溶出)によって精製した[Shustaら(1999)前出]。
【0081】
(実施例4.細胞結合アッセイ)
QL9/Ldに対する可溶性scTCRの結合を、以前に記載した競合様式でモニターした[Manningら(1998)前出;Sykulevら(1994)前出]。ペプチド−上方制御(アップレギュレート)T2−Ld細胞(3×105/ウェル)を、125I標識化抗LdFabs(30−5−7)および種々の濃度のscTCRの存在下で、氷上で1時間インキュベートした。結合および未結合の125I30−5−7Fabをオリーブオイル/ジブチルフタラートによる遠心分離によって分離した。阻害曲線を作成し、最大阻害の50%を生じるインヒビター濃度を決定した。解離定数をChengおよびPursoffの式を用いて算出した[Cheng(1973)Biochem.Pharm.22:3099〜3108]。細胞結合pMHCに対するscTCRの直接結合をモニターするため、ペプチド−アップレギュレートT2−Ld細胞(5×105/チューブ)を、ビオチン化可溶性scTCRとともに、氷上で40分間インキュベートし、続いてストレプトアビジン−フィコエリトリン(PharMingen,San
Diego,CA)を用いて30分間染色した。フローサイトメトリーによって細胞蛍光を検出した。
【0082】
(実施例5.異なるペプチドおよび異なるMHC分子(Kb)について特異的である高親和性TCRの同定)
TCRのCDR3α領域の酵母ディスプレイ変異体の同じライブラリーを用いて、異なるMHC分子に対して結合した異なるペプチドになお特異的である、より親和性の高いTCRについて選択することが可能であった。この場合、SIYRと呼ばれるペプチド(SIYRYYGL(配列番号5))を、Kbと呼ばれるMHC分子に結合し、そしてこのリガンド複合体を、蛍光形態中で用いて、フローサイトメトリーにより選択した。高い親和性のTCRを発現する16のクローンを、配列決定した。各々のクローンは、CDR3α領域において異なる配列を示した(表2)。
【0083】
これらのTCRの特異性の例として、変異3SQ2を、二次試薬単独(SA:PE)、抗−Vβ抗体F23.2および3つのペプチド/Kb複合体(OVA/Kb,dEV8/Kb、およびSIYR/Kb)を含む、種々の因子で染色した。図5に示すように、元来の選択に用いたpMHC(SIYR/Kb)のみが、変異TCRに対して結合するのに十分な親和性を有した。野生型TCRは、試験したいずれの濃度でもSIYR/Kbリガンドに結合しなかった(データ示さず)。
【0084】
変異TCR3SQ2はまた、酵母分泌系における可溶性タンパク質として発現され、そして、それが腫瘍細胞の表面上のpMHCに直接結合する能力についてビオチン化後に試験された。図6に示すように、標識化した3SQ2は、適切なペプチドSIYRのみを発現した腫瘍細胞に非常によく結合した。染色は、ペプチドの存在にかかわらず、任意のKb分子に結合する(図6)高親和性抗Kbモノクローナル抗体B8.24.3とほぼ同じ強度であった。
【0085】
(実施例6.同じMHC分子(Kb)に結合した異なるペプチドについて特異的である高親和性TCRの同定)
同じTCR足場が、同じMHCに対して結合した異なるペプチドに対しても、より高い親和性の形態を単離するために用いられ得るか否かを決定するため、本発明者らは、Kbに結合した、dEV8(EQYKFYSV(配列番号6))と呼ばれるペプチドを用いてTCR CDR3αライブラリーをスクリーニングした。ビオチン化dEV8/Kbリガンド(その後、フィコエリトリン−ストレプトビアジン,PE−SA)を用いたフローサイトメトリーによるいくつかのソート後、dEV8/Kbに結合した酵母細胞の有意な富化が存在した(図7においてPEレベルによって示されるように)。
【0086】
dEV8/Kbでの選択によって単離されたクローンのうち6つを配列決定したところ、CDR3配列は全て異なっていた(表3)。これらの配列は類似であったが、SIYR/Kbを用いた選択によって単離されたクローンの配列とは異なっていた(2つの例である、3SQ2および3SQ5をまた、表3に示す)。同じTCRライブラリーの変異体を用いてさえ、異なる抗原に対して、より高い親和性のTCRを単離することが可能であると結論できる。
【0087】
単離されたクローンの抗原特異性を証明するため、dEV8/Kb選択クローンのうちの1つ(4d1)を、異なる抗体およびリガンドのパネルを用いて試験した(図6)。予期された通り、このTCRは、3つの適切な抗体(抗−Vβ8抗体 F23.2,抗HA
タグ 抗体、および抗His タグ抗体)、およびdEV8/Kb抗原とは反応するが、他の抗原、OVA/Kbとは反応しない。野生型TCRは、ペプチドKb複合体のいずれ
とも結合しない(データ示さず)。このように、高い親和性のTCRは、選択された抗原に特異的であった。
【0088】
(実施例7.TCRの異なる領域(β鎖の相補性決定領域3)においてランダムなTCRライブラリーを作成することによる、高親和性TCRの同定)
上記の本実施例は、CDR3と呼ばれるα鎖の領域内で変異したTCRのライブラリーを用いた。TCRの他の領域がまた、より高い親和性のTCRを生成するように変異され得ることを示すため、β鎖のCDR3領域の5連続アミノ酸残基内のランダム変異のライブラリーを、出発物質としてqL2 TCR変異体を用いて、生成した。次いで、このライブラリーを、qL2変異体で検出した濃度以下のQL9/Ldリガンドで選択した。5つの酵母クローン(フローサイトメトリーによって選択した)を配列決定したところ、各々は、異なるヌクレオチドおよびアミノ酸配列を示した(図8)。変異された5つのアミノ酸領域内には配列の顕著な保存が存在しており、このことは、この配列モチーフが高い親和性について最適であったことを示唆していた。本発明者らは、特定のpMHCに対して、より高い親和性を有する誘導体を生じるためにTCRの異なる領域を変異することが可能であると結論付ける。
【0089】
上記の詳細な説明は、多くの特殊性を含み、これらは、本発明の範囲を限定するように解釈されるべきではなく、単に、本発明の現在の好適な実施形態のいくつかの例示を提供するだけである。例えば、特別に例示されたリガンド以外のリガンドが用いられてもよい。従って、本発明の範囲は、示された実施例によってではなく、添付の特許請求の範囲およびその法的な等価物によって決定されるべきである。本明細書に引用される全ての参考文献は、本明細書における開示と矛盾しない程度まで援用される。
【0090】
【表1】
【0091】
【表2】
【0092】
【表3】
【0093】
【表4】
【0094】
(配列表)
【0095】
【0096】
【0097】
【0098】
【0099】
【0100】
【0101】
【0102】
【0103】
(配列表)
SEQUENCE LISTING
<110> BOARD OF TRUSTEES OF THE UNIVERSITY OF ILLINOIS
<120> HIGH AFFINITY TCR PROTEINS AND METHODS
<130> F111502CDU
<150> US 60/169,179
<151> 1999-12-06
<160> 53
<170> PatentIn version 3.0
<210> 1
<211> 4
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 1
Met Cys Met Val
1
<210> 2
<211> 4
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 2
Ser Ile Tyr Arg
1
<210> 3
<211> 24
<212> PRT
<213> ARTIFICAL SEQUENCE
<400> 3
Gly Gly CysAlaGly Cys Cys Cys CysAla Thr Ala Ala Ala Cys Ala
1 5 10 15
Cys Ala Cys AlaGly Thr Ala Thr
20
<210> 4
<211> 74
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 4
Cys Thr ThrThrThr Gly Thr Gly CysCys Gly GlyAla Thr Cys Cys
1 5 10 15
Ala Ala Ala ThrGly Thr CysAla Gly Ser Asn Asn Ser Asn AsnSer
20 25 30
Asn Asn Ser AsnAsn Ser Asn Asn Gly CysThr Cys Ala Cys Ala Gly
35 40 45
Cys Ala Cys AlaGly Ala Ala Gly Thr Ala CysAla Cys Gly Gly Cys
50 55 60
Cys Gly Ala GlyThr Cys GlyCys Thr Cys
65 70
<210> 5
<211> 8
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 5
Ser Ile Tyr ArgTyr Tyr GlyLeu
1 5
<210> 6
<211> 8
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 6
Glu Gln TyrLysPhe Tyr Ser Val
1 5
<210> 7
<211> 7
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 7
Ser Gly Phe Ala Ser Ala Leu
1 5
<210> 8
<211> 7
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 8
Ser Ser Tyr GlyAsn Tyr Leu
1 5
<210> 9
<211> 7
<212> PRT
<213> ARTIFICAL SEQUENCE
<400> 9
Ser Arg Arg GlyHis Ala Leu
1 5
<210> 10
<211> 7
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 10
Ser Ser Arg Gly Thr Ala Leu
1 5
<210> 11
<211> 7
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 11
Ser His Phe GlyThr Arg Leu
1 5
<210> 12
<211> 7
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 12
Ser Met Phe GlyThr Arg Leu
1 5
<210> 13
<211> 7
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 13
Ser His Gln GlyArg Tyr Leu
1 5
<210> 14
<211> 7
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 14
Ser Tyr Leu GlyLeu Arg Leu
1 5
<210> 15
<211> 7
<212> PRT
<213> ARTIFICAL SEQUENCE
<400> 15
Ser Lys His GlyIle His Leu
1 5
<210> 16
<211> 7
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 16
Ser Leu Thr GlyArg Tyr Leu
1 5
<210> 17
<211> 7
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 17
Ser Leu Pro ProPro Leu Leu
1 5
<210> 18
<211> 7
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 18
Ser Ile Pro ThrPro Ser Leu
1 5
<210> 19
<211> 7
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 19
Ser Asn Pro ProPro Leu Leu
1 5
<210> 20
<211> 7
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 20
Ser Asp Pro ProPro Leu Leu
1 5
<210> 21
<211> 7
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 21
Ser Ser Pro ProPro Arg Leu
1 5
<210> 22
<211> 7
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 22
Ser Ala Pro ProPro Ile Leu
1 5
<210> 23
<211> 7
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 23
Ser Gly Thr HisPro Phe Leu
1 5
<210> 24
<211> 7
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 24
Ser Gly His LeuPro Phe Leu
1 5
<210> 25
<211> 7
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 25
Ser Asp Ser LysPro Phe Leu
1 5
<210> 26
<211> 7
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 26
Ser Ser Asp Arg Pro Tyr Leu
1 5
<210> 27
<211> 7
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 27
Ser Leu Glu ArgPro Tyr Leu
1 5
<210> 28
<211> 7
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 28
Ser Arg Glu AlaPro Tyr Leu
1 5
<210> 29
<211> 7
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 29
Ser Leu His ArgPro Ala Leu
1 5
<210> 30
<211> 7
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 30
Ser Leu His ArgPro Ala Leu
1 5
<210> 31
<211> 7
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 31
Ser Ser Asn ArgPro Ala Leu
1 5
<210> 32
<211> 7
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 32
Ser Thr Asp ArgPro Ser Leu
1 5
<210> 33
<211> 7
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 33
Ser Gly Ser ArgPro Thr Leu
1 5
<210> 34
<211> 7
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 34
Ser Leu Val ThrPro Ala Leu
1 5
<210> 35
<211> 7
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 35
Ser Ala Thr SerPro Ala Leu
1 5
<210> 36
<211> 7
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 36
Ser Ser Ile AsnPro Ala Leu
1 5
<210> 37
<211> 7
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 37
Ser Ala Ser TyrPro Ser Leu
1 5
<210> 38
<211> 7
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 38
Ser Arg Trp Thr Ser Gly Leu
1 5
<210> 39
<211> 7
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 39
Ser Gly Ser ArgPro Ala Leu
1 5
<210> 40
<211> 7
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 40
Ser Leu Thr HisHis Phe Leu
1 5
<210> 41
<211> 7
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 41
Ser Met Thr HisHis Phe Leu
1 5
<210> 42
<211> 7
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 42
Ser Leu Ser ArgPro Tyr Leu
1 5
<210> 43
<211> 7
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 43
Ser Leu Thr Arg Pro Tyr Leu
1 5
<210> 44
<211> 7
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 44
Ser Thr Tyr ArgHis Tyr Leu
1 5
<210> 45
<211> 7
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 45
Ser Gly Leu AlaArg Pro Leu
1 5
<210> 46
<211> 7
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 46
Ser Leu His ArgPro Ala Leu
1 5
<210> 47
<211> 7
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 47
Ser Gly Thr HisPro Phe Leu
1 5
<210> 48
<211> 7
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 48
Gly Gly GlyGlyThr Leu Tyr
1 5
<210> 49
<211> 7
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 49
Gly Gly GlyGlyVal Leu Tyr
1 5
<210> 50
<211> 7
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 50
Gly Leu GlyGlyIle Leu Tyr
1 5
<210> 51
<211> 7
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 51
Gly Gln GlyGlyVal Leu Tyr
1 5
<210> 52
<211> 7
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 52
Gly Ser Gly GlyIle Ile Tyr
1 5
<210> 53
<211> 7
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 53
Gly Gly GlyGlyIle Leu Tyr
1 5
【技術分野】
【0001】
(関連出願の相互参照)
本願は、米国特許第09/009,388号(1998年1月20日出願)の一部継続である。本願は、米国予備出願第60/169,179号(1999年12月6日出願)の利益を主張する。
【0002】
(連邦政府の研究支援の承認)
本発明は、少なくとも一部分、国立衛生研究所からの財政的支援でなされた。従って、米国政府は、本発明に特定の権利を有する。
【背景技術】
【0003】
(発明の背景)
本発明の分野は、分子生物学であり、詳細には、組換え宿主細胞の細胞表面上に提示された免疫細胞レセプターのコンビナトリアルライブラリーの場合に関する。より詳細には、本発明は、組換え酵母細胞の表面上に提示された高い親和性のT細胞レセプタータンパク質のライブラリー、可溶性の高い親和性のTCRレセプタータンパク質、特定のペプチド/MHC対への高い親和性の結合に関して選択された高い親和性のTCRタンパク質、MHC決定基の非存在下における特定の抗原への結合に関して選択された高い親和性のTCRタンパク質、ならびに数ある適用の中で診断方法および画像化アッセイにおける選択された高い親和性のTCR誘導体の使用に関する。
【0004】
T細胞レセプター(TCR)および抗体は、異なるクラスのリガンドを認識するために開発されている。抗体は、可溶性抗原に結合する膜結合かつ可溶性タンパク質として機能し、一方、天然において、TCRは、細胞関連ペプチド/MHC抗原に結合する膜結合分子としてのみ機能する。抗体:抗原相互作用の全エネルギーは、外来抗原にのみ集中しているが、TCR:ペプチド/MHC相互作用のエネルギーのうちの実質的な画分は、自己MHC分子に指向されるようである[Manningら(1998)Immunity 8:413:425]。さらに、抗体は、体細胞変異および親和性成熟のプロセスに大きく起因して、TCRのものよりも高い位の大きさであるリガンド−結合親和性を有し得る。その正常な細胞の状況において、TCRは、体細胞変異を引起さず、そして胸腺選択のプロセスは、親和性の狭いウィンドウを維持することによって操作されるようである[Alamら(1996)Nature 381:616−620]。細胞表面におけるTCRのアクセサリー分子CD4またはCD8との関連はまた、TCRの機能的な親和性に影響を及ぼし得る[Garciaら(1996)Nature 384:577−581]。これらの差異にもかかわらず、2つのタンパク質の3次元構造は、顕著に類似し、抗原と接触する分子の単一表面上に超可変領域形成ループを有する。
【0005】
これらの構造類似性に基づいて、異種発現系においてTCRおよび抗体の細胞外ドメインの、可溶性かつ表面提示形態の産生を成功させることに有意な差異が存在したことは、いくらか驚くべきである。多くの抗体は、現在、インタクトもしくはFabフラグメント形態としてか、または単鎖(sc)フラグメント−可変(Fv)タンパク質としてかのいずれかで、高収率かつ可溶性で発現されている。さらに、新規に単離され、そして/またはファージディスプレイ技術(より最近では、酵母−ディスプレイ技術)の使用を介して改善された、多数の抗原結合Fvフラグメントが存在する[BoderおよびWittrup(1997)Nat.Biotechnol.15:553−557;Kiekeら(1997)Prot.Eng.10:1303−1310]。抗体フラグメントに関するこれらの発現系は、構造研究ならびに診断抗体および治療抗体の設計に重要である。
【0006】
対照的に、数個のTCR分子の3次元構造は、可溶性で相当折り畳まれたTCRの発現に対するかなりの努力のあとにのみ決定された[BentleyおよびMariuzza(1996)Ann.Rev.Immunol.14:563−590]。FabとTCRとの間の基礎を探求することにおける困難のうちの1つは、抗体における大量の配列多様性であり、そしてTCR可変(V)領域は、どの特徴のV領域が抗原結合以外の機能(例えば、V領域の対形成および結合動力学、可溶性、ならびに折り畳み)に重要であるかを識別するための努力を複雑にする。これらの性質に関して、TCRのV領域および抗体を比較する研究は、相対的にほとんどない。
【0007】
それにもかかわらず、マウスT細胞クローン2C由来のTCRは、現在、Escherichia coliにおいてscVαVβ(scTCR)として[Soo Hooら(1992)Proc.Natl.Acad.Sci.USA 89:4759−4763]、骨髄腫細胞から脂質連結VαCαVβCβ二量体として[SlanetzおよびBothwell(1991)Eur.J.Immunol.21:179−183]、および昆虫細胞から分泌VαCαVβCβ二量体として[Garciaら(1996)Science 274:209−219]発現されている。2C scTCRは、ほとんどのscFvと比較して、相対的に低い可溶性を有するが、その可溶性は、アミノ末端のチオレドキシンへの融合によって約10倍増加する[Schodinら(1996)Molec.Immunol.33:819−829]。可溶性で相当折り畳まれたVαVβドメインを産生することにおける困難は、他のTCRに対して拡大されている[Udakaら(1993)前出;Sykulevら(1994)前出;Manningら(1998)前出]。可溶性または表面提示能力のいずれかにおけるTCRとFvとの間の明らかな差異に関する分子説明は、適切に探査されていない。2CscTCRが、Vα/Vβ界面またはCD3εシグナル伝達サブユニットと相互作用することが疑われるVβフレームワークの領域における特定の単一部位変異の、ランダムなライブラリーからの選択後に、酵母表面ディスプレイシステムで発現され得ることが示された。これらの変異(これらのうちのいくつかは、抗体V領域に天然に見出される)は、TCR中のこれらの位置の重要性を反映し、TCR結合特性のさらなる操作に関する基礎を提供する。
【発明の概要】
【課題を解決するための手段】
【0008】
(発明の要旨)
本発明は、組換え宿主細胞、例えば、酵母細胞(望ましくはSaccharomyces cerevisiae)の表面上に提示された免疫T細胞レセプターポリペプチドのコンビナトリアルライブラリーを提供する。このようなライブラリーから、高い親和性のTCRポリペプチド(同族のリガンド:主要組織適合遺伝子複合体のタンパク質(pMHC)に結合するペプチドの複合体に関して、野生型よりも高い親和性を示すもの)が単離され得る。望ましくは、pMHCに対するTCRペプチドの親和性は、例えば、当該分野で公知の方法によって測定される場合、約107〜約1010の解離定数に反映される。可溶性の高い親和性のTCRをコードする核酸を含むDNAライブラリーが提供され、ここで、このTCRは、変異体TCRコード配列を作製するためのTCRを変異誘発すること;変異体TCRの変異体TCRコード配列を含むDNAを酵母細胞に形質転換すること;この変異体TCRが酵母細胞の表面上に提示されるようにこの変異体TCRコード配列の発現を誘導すること;この酵母細胞を、ペプチド/MHCリガンドを結合する蛍光標識と接触して、選択された酵母細胞を産生すること;および、最も高い蛍光を示す酵母細胞を単離する、方法によって作製される。野生型T細胞レセプタータンパク質よりもペプチド/MHCリガンドに対して高い親和性を有する、酵母細胞の表面上に提示されたT細胞レセプタータンパク質のライブラリーがまた、提供され、ここで、このライブラリーは、T細胞レセプタータンパク質コード配列を変異誘発して変化に富むT細胞レセプタータンパク質コード配列変異体の集団を作製すること;このT細胞レセプター変異体コード配列を、酵母細胞に形質転換すること;酵母細胞の表面上でこのT細胞レセプター変異体コード配列の発現を誘導すること;そして、野生型T細胞レセプタータンパク質よりもペプチド/MHCリガンドに対して高い親和性を有する、T細胞レセプター変異体を発現する細胞が選択することによって形成される。
【0009】
本発明はさらに、同族リガンドに対して高い親和性の結合を示すTCRタンパク質(細胞結合形態でかまたは可溶性形態で)を提供する。本発明において、TCRタンパク質が結合するリガンドは、ペプチド/MHC複合体であるか、または選択プロセス(望ましくは、繰り返された選択プロセス)に起因して、このリガンドは、MHC成分(例えば、スーパー抗原)を含まないリガンドであり得る。このリガンドは、とりわけ、ペプチド、タンパク質、炭水化物部分、または脂質部分であり得る。これらの可溶性の高い親和性のTCRは、変異体TCRコード配列を作製するためにTCRを変異誘発する工程;変異体TCRのための変異体TCRコード配列を含むDNAを酵母細胞に形質転換する工程;この変異体TCRが酵母細胞の表面上に提示されるように、この変異体TCRコード配列の発現を誘導する工程;この酵母細胞を、ペプチド/MHCリガンドに結合する蛍光標識と接触して、選択された酵母細胞を生成する工程;および、最も高い蛍光を示す酵母細胞を単離する工程を包含する方法によって作製され得る。この可溶性の高い親和性のTCRは、好ましくは酵母ディスプレイによって単離される。
【0010】
本発明はさらに、高い親和性のTCRタンパク質の同族リガンドを検出するための方法を提供し、この方法は、高い親和性のTCRタンパク質を同族リガンドと結合させる工程を包含し、ここで、この高い親和性のTCRタンパク質が検出可能に標識化されるか、または添加される二次的な検出可能なタンパク質(例えば、同族リガンドに結合する領域以外の領域においてTCRに特異的な抗体)が存在する。リガンドを同定するために高い親和性のTCRを用いるための好ましい方法は:高い親和性のTCRを検出可能な標識で標識化する工程;この標識化TCRをリガンドと接触する工程;標識化TCRが結合するリガンドを同定する工程を包含する。好ましくは、このリガンドは、野生型TCRよりも高い親和性でTCRに結合するペプチド/MHCリガンドである。このリガンドを同定する方法は、当業者に公知である。直接または間接的に、TCRタンパク質の検出を可能にする適切な標識としては、蛍光化合物、化学発光化合物、放射性同位体、発色団などが挙げられるが、これらに限定されない。
【0011】
高い親和性のTCRタンパク質は、医学、獣医学もしくは植物の診断設定における標的リガンドの定性測定および定量測定のための手段としてか、または組織材料もしくは植物材料の同定のために、実験室で使用され得る。同様に、本発明の高い親和性のTCRは、標的材料またはリガンドの検出および/または定量のための試薬として使用され得る。選択されたペプチド/MHCリガンドに結合する高い親和性のTCRを用いる方法がまた、提供され、この方法は、この高い親和性のTCRをこの選択されたペプチド/MHCリガンドに結合する標識で標識化する工程;この標識化された高い親和性のTCRをMHC分子を含む細胞と接触させる工程を包含する。本発明の高い親和性のタンパク質(ここで、このタンパク質は、高い親和性および特異性で腫瘍細胞抗原に特異的に結合する)は、特定の癌の型の診断試験に使用され得るか、または生物(好ましくは、哺乳動物、そしてまた好ましくは、ヒト)において、腫瘍の位置を確認するためならびに/または腫瘍のサイズおよび数を概算するための画像化試験の際に生物で使用され得る。診断試験のためにpMHCに結合する高い親和性のTCRを用いるための方法が提供され、この方法は、高い親和性のTCRを検出可能な標識で標識化する工程;この高い親和性のTCRが高い親和性を有するリガンドを含む細胞と、この高い親和性のTCRを接触させる工程;およびこの標識を検出する工程を包含する。この方法において、標識は、特定のペプチド/MHCリガンドに結合するために選択され得、これによって、特定のペプチド/MHCリガンドを発現する細胞が、標的化される。細胞表面上で特定のペプチド/MHC分子に対する診断プローブとして高い親和性のTCRを用いるための好ましい方法は:高い親和性のTCRを、特定のペプチド/MHCリガンドに結合する検出可能な標識で標識化する工程;このTCRを細胞と接触する工程;およびこの標識を検出する工程を包含する。特定の使用に依存した使用のために選択された検出可能な標識、および適切なの選択は、関連分野の当業者の十分に範囲内である。一般的に、目的のリガンドに対する高い親和性結合に関して選択されたTCRタンパク質は、このリガンドに特異的な抗体が使用され得る方法(TCR 対 抗体タンパク質に対して手順の改変がなされ、このような改変は、当該分野で公知である)に使用され得る。
【0012】
高い親和性のTCR、望ましくは可溶性単鎖(sc)TCRは、自己免疫疾患における細胞または組織の自己免疫破壊をブロックするために使用され得、ここで、標的細胞の表面上で細胞傷害性リンパ球によって認識される部位は、高い親和性のTCRによって結合される部位と同じである。細胞の自己免疫破壊をブロックするための好ましい方法は、標的細胞の表面上でTリンパ球によって認識される部位に高い親和性を有するTCRを、細胞と接触させる工程を包含し、これによって、細胞の自己免疫破壊がブロックされる。
【0013】
可溶性の、高い親和性のscTCRは、治療化合物(例えば、制癌化合物、治療放射性核種または細胞傷害性タンパク質)に結合され得、ここで、scTCRの同族のリガンドは、新生物形成細胞表面マーカーである。あるいは、高い親和性の可溶性scTCRの結合特異性は、病原体感染された標的細胞であり得(例えば、ウイルス、細菌、または原生動物感染された標的細胞)、そして、標的細胞が感染因子のさらなる複製無しに排除され得るように毒性分子が結合され得る。不活性病原体に対して高い親和性のTCRを使用する提供された方法は:この病原体に毒性である分子を、高い親和性のTCRに結合させる工程;およびこの病原体を発現する細胞と、このTCRを接触させる工程を包含する。「毒性」は、病原体が病原体の複製を回避または阻害することを意味する。
【0014】
疾患を処置するために高い親和性のTCRを使用するための方法がまた提供され、この方法は:新生物形成細胞表面マーカーに対して高い親和性を有するTCRを、治療化合物と結合させる工程:およびこのTCRを細胞と接触させる工程を包含する。TCRと結合され得る、疾患の進行を遅延する際に有用である任意の治療化合物が、使用され得る。治療化合物をTCRと結合する方法は、当該分野で公知である。
【0015】
高い親和性のTCR変異体に対する遺伝子を、T細胞の表面上で変異体の発現を可能にするシステムにクローニングするための方法がまた、提供され、この方法は:高い親和性のTCR変異体を作製するためにTCRを変異させる工程;このTCR変異体をベクターにクローニングする工程;このベクターをT細胞にトランスフェクトする工程;高い親和性のTCR変異体をT細胞の表面上に発現させる工程を包含する。この方法は、さらに:ペプチド/MHCリガンドによって他のT細胞よりも大きな程度に活性化されるT細胞を選択する工程を包含する。トランスフェクト/感染されたT細胞は、選択されたペプチド保有MHC細胞の認識に有用であり得る。これらのトランスフェクト/感染されたT細胞は、患者の疾患を処置する際に有用であり、ここで、患者由来のT細胞は、取り出され、そして高い親和性のTCR変異体を発現するベクターで形質転換され、そしてこの患者に戻される(ここで、このT細胞は、患者の野生型T細胞よりも、ペプチド/MHCリガンドによって大きな程度に活性化される)。
【0016】
可溶性の高い親和性のTCR分子は、関連分野の当業者に容易に明らかであるように、ほとんどの適用に対して抗体または単鎖抗体の代わりに使用され得る。
本願発明は、例えば、以下を提供する:
(項目1) リガンドを同定するために高親和性TCRを使用するための方法であって、以下:
高親和性TCRを標識化する工程;
該標識化TCRをリガンドと接触させる工程;
該標識化TCRと結合する該リガンドを同定する工程、
を包含する、方法。
(項目2) 前記標識が、蛍光化合物、化学発光化合物、放射性同位元素および発色団からなる群より選択される、項目1に記載の方法。
(項目3) 前記リガンドが、ペプチド/MHCリガンドである、項目1に記載の方法。(項目4) 選択されたペプチド/MHCリガンドに結合するために高親和性TCRを使用するための方法であって、以下:
該高親和性TCRを、該選択されたペプチド/MHCリガンドに結合する標識で標識化する工程;
該標識化高親和性TCRを、MHC分子を含む細胞と接触させる工程、
を包含する、方法。
(項目5) 前記標識が、蛍光化合物、化学発光化合物、放射性同位元素および発色団からなる群より選択される、項目4に記載の方法。
(項目6) 細胞の表面上の特定のペプチド/MHC分子に対する診断用プローブとして高親和性TCRを使用するための方法であって、以下:
高親和性TCRを、特定のペプチド/MHCリガンドに結合する標識で標識化する工程;
該TCRを細胞と接触させる工程;
該標識を検出する工程、
を包含する、方法。
(項目7) 診断試験のためにpMHCに結合する高親和性TCRを使用するための方法であって、以下:
該高親和性TCRを検出可能な標識で標識化する工程;
該高親和性TCRを細胞と接触させる工程;
該標識を検出する工程、
を包含する、方法。
(項目8) 前記存在する標識の数が検出される、項目7に記載の方法。
(項目9) 前記標識の位置が、生物体において検出される、項目7に記載の方法。
(項目10) 前記標識が、特定のペプチド/MHCリガンドに結合し、それによって、特定のペプチド/MHCリガンドを発現する細胞が標的化される、項目7に記載の方法。(項目11) 細胞の自己免疫破壊をブロックするための方法であって、標的細胞の表面上のTリンパ球によって認識される部位に対して高い親和性を有するTCRを、細胞と接触させる工程であって、それによって、細胞の自己免疫破壊がブロックされる、工程、
を包含する、方法。
(項目12) 疾患を処置するために高親和性TCRを使用するための方法であって、以下:
新生物細胞表面マーカーに対する高い親和性を有するTCRを、治療化合物と結合させる工程;および、
該TCRを細胞と接触させる工程、
を包含する、方法。
(項目13) 病原体を不活性化するために高親和性TCRを使用する方法であって、以下:
病原体に対して毒性のある分子を、高親和性TCRに結合する工程;および、
該TCRを、該病原体を発現する細胞と接触させる工程、
を包含する、方法。
(項目14) 前記病原体が、ウイルス、細菌および原生動物からなる群より選択される、項目13に記載の方法。
(項目15) 野生型TCRよりもリガンドに対して高い親和性を有する、可溶性T細胞レセプター(TCR)。
(項目16) 前記リガンドが、ペプチド/MHCリガンドである、項目15に記載の可溶性高親和性TCR。
(項目17) 項目15に記載の可溶性高親和性TCRであって、該高親和性TCRが、以下:
TCRを変異誘発して、変異体TCRコード配列を作製する工程;
変異体TCRについての該変異体TCRコード配列を含むDNAを、酵母細胞に形質転換する工程;
該変異体TCRコード配列の発現を誘導する工程であって、その結果、該変異体TCRが、該酵母細胞の表面上に提示される、工程;
該酵母細胞を、蛍光標識と接触させる工程であって、該蛍光標識は、ペプチド/MHCリガンドに結合して、選択された酵母細胞を産生する、工程;および
最も高い蛍光を示す酵母細胞を単離する工程、
を包含する方法によって作製される、可溶性高親和性TCR。
(項目18) 酵母ディスプレイによって単離された、項目15に記載の可溶性高親和性TCR。
(項目19) 可溶性高親和性TCRをコードする核酸を含む、DNAライブラリーであって、該TCRが、以下:
TCRを変異誘発して、変異体TCRコード配列を作製する工程;
変異体TCRについての該変異体TCRコード配列を含むDNAを、酵母細胞に形質転換する工程;
該変異体TCRコード配列の発現を誘導する工程であって、その結果、該変異体TCRが、該酵母細胞の表面上に提示される工程;
該酵母細胞を、蛍光標識と接触させる工程であって、該蛍光標識は、ペプチド/MHCリガンドに結合して、選択された酵母細胞を産生する、工程;および
最も高い蛍光を示す酵母細胞を単離する工程、
を包含する方法によって作製される、
DNAライブラリー。
(項目20) 野生型T細胞レセプタータンパク質よりもペプチド/MHCリガンドに対して高い親和性を有する、酵母細胞の表面上に提示されたT細胞レセプタータンパク質のライブラリーであって、ここで、該ライブラリーは、
T細胞レセプタータンパク質コード配列を変異誘発して、T細胞レセプタータンパク質コード配列の変異体の多様な集団を生成する工程;
該T細胞レセプター変異体コード配列を、酵母細胞に形質転換する工程;
酵母細胞の表面上での該T細胞レセプター変異体コード配列の発現を誘導する工程;および、
野生型T細胞レセプタータンパク質よりもペプチド/MHCリガンドに対して高い親和性を有するT細胞レセプター変異体を発現する細胞を選択する工程、
によって形成される、
ライブラリー。
(項目21) T細胞の表面上に高親和性TCR変異体の発現を可能にする系に、該高親和性TCR変異体についての遺伝子をクローニングする方法であって、以下:
TCRを変異誘発して、高親和性TCR変異体を作製する工程;
該TCR変異体を、ベクターにクローニングする工程;
該ベクターを、T細胞にトランスフェクトする工程;
該T細胞の表面上に該高親和性TCR変異体を発現させる工程、
を包含する、方法。
(項目22) 野生型よりもペプチド/MHCリガンドによって活性化されるT細胞を選択する工程をさらに包含する、項目21に記載の方法。
(項目23) 前記トランスフェクト/感染されたT細胞が、選択されたペプチド保有MHC細胞の認識のために使用される、項目21に記載の方法。
(項目24) 項目21の方法によって作製された、T細胞。
【図面の簡単な説明】
【0017】
【図1】図1は、野生型および変異型2C TCRをそれらの表面に発現する酵母細胞のフローサイトメトリー分析を示す。野生型(T7)および変異型(qL2、qL7)scTCRを示す酵母細胞は、抗Vβ8抗体F23.2(120nM)、特異的な同種抗原ペプチドのMHC、QL9/Ld/Ig(40nM)、または無ペプチドMCMV(配列番号1)/Ld/Ig(40nM)で染色した。結合をFITC結合体化ヤギ抗マウスIgG F(ab’)2によって検出し、そして、フローサイトメトリーによって分析した。ネガティブ集団(例えば、F23.2染色でみられるような)は、すべての酵母のディスプレイされたタンパク質について観察され、そして、表面融合タンパク質を発現し得ない、増殖または誘導の段階の細胞に起因すると考えられている(Kiekeら、(1999)Proc.Natl.Acad.Sci.USA 96:5651−5656;BoderおよびWittrup(1997)Nat.Biotech.15:553−557;Kiekeら、(1997)Protein Engineering 10:1303−1310)。
【図2】図2は、Ldに結合された異なったQL9改変体ペプチドに対する変異体scTCRの結合の優れた特異性分析を示す。本来のT細胞クローン2Cおよび種々の酵母クローンを、野生型QL9(P5F)、QL9の位置5の改変体(P5Y、P5H、P5E)またはMCMV(配列番号1)でロードしたLd/Ig二量体への結合について、フローサイトメトリーによって分析した。結合をFITC標識化したヤギ抗マウスIgGを用いて検出した。相対的な蛍光を2つの異なったアプローチによって測定した。T細胞クローン2Cについては、種々のペプチド/LdIg二量体の結合を、QL9/Ld染色(MFUpMHC/MFUQL9−Ld)に対して調整した。酵母細胞について、各ペプチド/Ld二量体の結合を、抗Vβ8抗体F23.2(MFUpMHC/MFUF23.2)による結合に対して調整した。後者は、異なった変異体が、野生型QL9/Ldに対する結合についてお互いに比較されるのを可能とした。
【図3】図3は、可溶性scTCRによるQL9/Ld結合を示す。QL9を用いてロードしたT2−Ld細胞を125I標識化した抗LdFabフラグメント(30−5−7)および種々の濃度の未標識化のFab(黒菱形)、scTCR−T7(黒四角)、または変異体scTCR−qL2(黒丸)と共にインキュベートした。結合したおよび未結合の125I 30−5−7 Fabフラグメントを、オリーブ油/ヂブチルフタレートを介して、遠心分離によって分離した。コントロールペプチドMCMV(配列番号1)を用いてロードしたT2−Ld細胞に対する125I標識化した抗LdFabフラグメントの結合は、scTCRの最も高い濃度においてさえ阻害されなかった(データは示されていない)。
【図4A】図4Aは、細胞表面ペプチド/MHCへのscTCR/ビオチンの結合のフローサイトメトリー分析を示す。ペプチド−ロードしたT2−Ld細胞を、ビオチン標識化したqL2scTCR(約0.3μM)またはT7scTCR(約1.6μM)scTCRと共にインキュベートし、その後、ストレプトアビジン−PEと共にインキュベートし、フローサイトメトリーによって分析した。図4A:QL9(影なし)、p2Ca(軽い影付き)、またはMCMV(配列番号1)(暗い影付き)を用いてロードし、そしてqL2scTCR/ビオチンで染色したT2−Ld細胞のフローサイトメトリーヒストグラム。図4B:QL9、p2Ca、またはMCMV(配列番号1)を用いてロードし、そして第2のSA−PEのみ、T7scTCR/ビオチン+SA−PE、またはqL2scTCR/ビオチン+SA−PEのいずれかで染色したT2−Ld細胞の平均蛍光単位(MFU)。図4C:昆虫細胞から発現された変異体qL2の可溶性の高い親和性形態は、非常に低い濃度のペプチド−MHC複合体を検出し得る。Ld複合体は、37℃で1.5時間、種々の濃度のQL9ペプチドを用いてインキュベーションによってT2−Ld細胞(3×106/ml)の表面上で上方制御された。約2×105細胞をトランスフェクトされたDrosophila melanogaster(昆虫)SC2細胞由来のTCRを用いて、30分間氷上で染色した(Garcia,K.C.ら、(1997)Proc Natl Acad Sci USA 94(25)、13838−13843)。次いで、細胞を洗浄し、そしてストレプトアビジン−PEが後に続く、ビオチン標識化した抗Vb IgG(F23.1)で染色し、そしてフローサイトメトリーによって分析した。
【図4B】図4Bは、細胞表面ペプチド/MHCへのscTCR/ビオチンの結合のフローサイトメトリー分析を示す。ペプチド−ロードしたT2−Ld細胞を、ビオチン標識化したqL2scTCR(約0.3μM)またはT7scTCR(約1.6μM)scTCRと共にインキュベートし、その後、ストレプトアビジン−PEと共にインキュベートし、フローサイトメトリーによって分析した。図4A:QL9(影なし)、p2Ca(軽い影付き)、またはMCMV(配列番号1)(暗い影付き)を用いてロードし、そしてqL2scTCR/ビオチンで染色したT2−Ld細胞のフローサイトメトリーヒストグラム。図4B:QL9、p2Ca、またはMCMV(配列番号1)を用いてロードし、そして第2のSA−PEのみ、T7scTCR/ビオチン+SA−PE、またはqL2scTCR/ビオチン+SA−PEのいずれかで染色したT2−Ld細胞の平均蛍光単位(MFU)。図4C:昆虫細胞から発現された変異体qL2の可溶性の高い親和性形態は、非常に低い濃度のペプチド−MHC複合体を検出し得る。Ld複合体は、37℃で1.5時間、種々の濃度のQL9ペプチドを用いてインキュベーションによってT2−Ld細胞(3×106/ml)の表面上で上方制御された。約2×105細胞をトランスフェクトされたDrosophila melanogaster(昆虫)SC2細胞由来のTCRを用いて、30分間氷上で染色した(Garcia,K.C.ら、(1997)Proc Natl Acad Sci USA 94(25)、13838−13843)。次いで、細胞を洗浄し、そしてストレプトアビジン−PEが後に続く、ビオチン標識化した抗Vb IgG(F23.1)で染色し、そしてフローサイトメトリーによって分析した。
【図4C】図4Cは、細胞表面ペプチド/MHCへのscTCR/ビオチンの結合のフローサイトメトリー分析を示す。ペプチド−ロードしたT2−Ld細胞を、ビオチン標識化したqL2scTCR(約0.3μM)またはT7scTCR(約1.6μM)scTCRと共にインキュベートし、その後、ストレプトアビジン−PEと共にインキュベートし、フローサイトメトリーによって分析した。図4A:QL9(影なし)、p2Ca(軽い影付き)、またはMCMV(配列番号1)(暗い影付き)を用いてロードし、そしてqL2scTCR/ビオチンで染色したT2−Ld細胞のフローサイトメトリーヒストグラム。図4B:QL9、p2Ca、またはMCMV(配列番号1)を用いてロードし、そして第2のSA−PEのみ、T7scTCR/ビオチン+SA−PE、またはqL2scTCR/ビオチン+SA−PEのいずれかで染色したT2−Ld細胞の平均蛍光単位(MFU)。図4C:昆虫細胞から発現された変異体qL2の可溶性の高い親和性形態は、非常に低い濃度のペプチド−MHC複合体を検出し得る。Ld複合体は、37℃で1.5時間、種々の濃度のQL9ペプチドを用いてインキュベーションによってT2−Ld細胞(3×106/ml)の表面上で上方制御された。約2×105細胞をトランスフェクトされたDrosophila melanogaster(昆虫)SC2細胞由来のTCRを用いて、30分間氷上で染色した(Garcia,K.C.ら、(1997)Proc Natl Acad Sci USA 94(25)、13838−13843)。次いで、細胞を洗浄し、そしてストレプトアビジン−PEが後に続く、ビオチン標識化した抗Vb IgG(F23.1)で染色し、そしてフローサイトメトリーによって分析した。
【図5】図5は、ストレプトアビジン−PEが続く、ビオチン標識化したペプチド/MHC複合体、OVA/Kb、dEV8/KbまたはSIYR(配列番号2)/Kbを用いて染色した変異体scTCR(3SQ2と呼ばれる)を示す酵母のフローサイトメトリーヒストグラムを示す。scTCRの存在についてのポジティブコントロールとして、酵母を、FITCヤギ抗マウスF(ab’)2が続く、Vβ特異的Ig、F23.2で染色した。
【図6】図6は、T2−Kb腫瘍細胞を特定のペプチド(OVA、dEV8またはSIYR(配列番号2))と共にインキュベートし、そしてストレプトアビジン−PEが続く、ビオチン標識化した可溶性scTCR、3SQ2で染色し、フローサイトメトリーによって分析したことを示す。Kb細胞の存在についてのポジティブコントロールとして、T2−Kb細胞をビオチン標識化した抗体B8.24.3(これは、結合されたペプチドに関わらずKbを認識する)を用いて染色した。
【図7】図7は、dEV8/Kbを用いる複数回の仕分け後、酵母VαCDR3ライブラリーを、ストレプトアビジン−PEが続く、ビオチン標識化したdEV8/Kbで染色し、そしてフローサイトメトリーによって分析したことを示す。
【図8】図8は、ビオチン標識化したストレプトアビジン−PEを用いる染色が続く、ビオチン標識化したペプチド/MHC複合体、OVA/KbまたはdEV8/Kbで染色した変異体scTCR(4d1と呼ばれる)を示す酵母のフローサイトメトリーヒストグラムを示す。ポジティブコントロールとして、酵母を、F23,2Igを用いてscTCR Vβの存在について分析し、そして抗6His抗体または抗HA Ig、12CA5を用いてエピトープタグについて分析した。
【図9】図9は、変異体T細胞レセプターqL2を用いてトランスフェクトしたT細胞は、低濃度でペプチドMHCを発現する標的細胞を認識し、そしてそれによって刺激され得ることを示す。T細胞ハイブリドーマ細胞株58−/−(Letourneur,F.,およびB.Malissen.(1989)Eur J Immunol 19(12)、2269−74)を野生型(2C)または変異型(qL2)TCRを用いてトランスフェクトし、そして1ウェルあたり7.5×104個のトランスフェクトされた細胞をQL9ペプチドの存在下で、T2−Ld細胞(7.5×104ウェル)と共に37℃でインキュベートした。約30時間後、上清を回収し、そして、T細胞によって放出されたIL−2についてアッセイした:上清をIL−2依存性細胞株、HT2(5×103/ウェル)と共に37℃で18時間インキュベートした。HT2細胞の増殖を3[H]チミジンの取り込みによって測定した。平均CPMは、3連ウェルの平均を示す。QL9ペプチドの非存在下では、IL−2は全く放出されなかった(データは示されていない)。
【発明を実施するための形態】
【0018】
(好ましい実施形態の詳細な説明)
明細書および特許請求の範囲(このような用語に対して与えられる範囲を含む)の明瞭なそして一貫した理解を提供するために、以下の定義が提供される。
【0019】
コード配列は、タンパク質のアミノ酸配列についてコードするか、またはtRNAまたはrRNAのような機能的なRNAについてコードする遺伝子またはcDNAの一部である。
【0020】
相補体または相補的な配列は、ワトソン−クリック塩基対形成法則に従う別の配列のヌクレオチドと水素結合した二重鎖を形成するヌクレオチドの配列を意味する。例えば、5’−AAGGCT−3’についての相補的な塩基配列は、3’−TTCCGA−5’である。
【0021】
下流は、DNAまたはRNAにおける任意の部位の3’側を意味する。
【0022】
発現は、構造的なRNA(rRNA、tRNA)またはメッセンジャーRNA(mRNA)への遺伝子の転写および引き続くmRNAのタンパク質への翻訳をいう。
【0023】
具体的に例示されたTCR配列に機能的に等価であるアミノ酸配列は、単一または複数のアミノ酸置換によって、アミノ酸の付加および/または欠失によって、改変されたアミノ酸配列をいい、ここで、1つ以上のアミノ酸は化学的に改変されているが、それにも関わらず、本発明の細胞結合または可溶性TCRタンパク質の結合特異性および高い親和性結合活性を保持する。機能的に等価なヌクレオチド配列は、具体的に例示された細胞結合または可溶性TCRタンパク質と同一の生物学的活性を実質的に有するポリペプチドをコードするヌクレオチド配列である。本発明の状況において、可溶性TCRタンパク質は、ネイティブな細胞結合TCRの一部を欠き、そして、溶液中で安定である(すなわち、それは、本明細書中で記載されるように処理される場合、そしてタンパク質溶液についての標準的な条件下で、一般的に溶液中で凝集しない)。
【0024】
2つの核酸配列は、これらの配列が別々の生物に由来する場合、このような生物が異なった種であろうとなかろうと、これらの配列が、同一の生物において同一の配列中で天然に存在しない限り、お互いに異種である。
【0025】
相同性は、2つのヌクレオチド間またはアミノ酸配列間の同一性の程度をいう。
【0026】
単離されたは、天然の状態からヒトの手によって変更されることを意味する。「単離された」組成物または物質が天然に存在する場合、それは、その本来の環境から変化されるかまたは取り出されるか、あるいはその両方である。例えば、生きている動物に天然に存在するポリヌクレオチドまたはポリペプチドは単離されていないが、その用語が本明細書中で使用される場合、天然の状態で共存する物質から分離された同一のポリヌクレオチドまたはポリペプチドは単離されている。
【0027】
リンカー領域は、タンパク質の2つの機能的または構造的ドメインを作動可能に連結するアミノ酸配列である。
【0028】
核酸構築物は、天然に存在する遺伝子から単離されたか、他に天然に存在しない様式で組み合わされそして並列された核酸のセグメントを含むように改変された核酸分子である。
【0029】
核酸分子は、3’−5’−リン酸ジエステル結合によって結合されたデオキシリボヌクレオチドまたはリボヌクレオチドのいずれかを含む単一または二重鎖の直鎖状ポリヌクレオチドを意味する。
【0030】
2つのDNA配列は、連結の性質が、それらの配列がお互いに対してそれらの正常な機能をもたらす能力を妨げない場合、作動可能に連結されている。例えば、プロモーター領域は、このプロモーターがコード配列の転写をもたらし得る場合、コード配列に作動可能に連結されている。
【0031】
ポリペプチドは、ペプチド結合によって連結されているアミノ酸の直鎖状ポリマーである。
【0032】
プロモーターは、シス作用DNA配列を意味し、一般的に80〜120塩基対長であり、そして遺伝子の開始部位の上流に位置し、これに対してRNAポリメラーゼは、結合し得そして正確な転写を開始する。転写調節の開始/終了調節を提供しそして/または下流のコード配列の発現を増強する(増加する)さらなる転写調節配列に関連し得る。
【0033】
組換え核酸分子(例えば、組換えDNA分子)は、2つ以上の非相同性DNA分子の連結を介してインビトロで形成された新規の核酸配列である(例えば、少なくとも1つのブローニング部位にクローン化された外来のDNAの1つ以上のインサートを含む組換えプラスミド)。
【0034】
形質転換は、異なった遺伝子型の別の細胞由来の精製された組換えDNAの外部適用によってゲノムまたは細胞の方向付けられた改変を意味し、被験体細胞のゲノム中へのその取り込みおよび組み込みを生じる。細菌においては、この組換えられたDNAは、代表的には細菌の染色体には組み込まれないが、その代わりに、プラスミドとして自律的に複製する。
【0035】
上流は、DNAまたはRNA中の任意の部位の5’側を意味する。
【0036】
ベクターは、宿主細胞中で自律的に複製可能でありそして外来DNAを受け入れ可能である核酸分子である。ベクターは、複製の自分自身の起源、外来DNAの挿入について使用され得る制限エンドヌクレアーゼについての1つ以上の独特の制限部位、そして通常抗生物質耐性をコードする遺伝子のような選択マーカー、およびしばしば挿入されたDNAの発現のための認識配列(例えば、プロモーター)を保有する。一般的なベクターとしては、プラスミドベクターおよびファージベクターが挙げられる。
【0037】
高い親和性T細胞レセプター(TCR)は、野生型TCRよりもより強力に標的リガンドへ結合する操作されたTCRを意味する。
【0038】
T細胞は、αβヘテロ二量体T細胞レセプター(TCR)を介したMHC産物に結合された外来ペプチドを認識する。TCRレパートリーは、抗体重鎖および軽鎖遺伝子において使用される同一の遺伝子再配列機構によって作製された広範な多様性を有する[Tonegawa,S.(1988)Biosci.Rep.8:3−26]。多様性の大部分は、αおよびβ鎖の相補性決定領域3(CDR3)をコードする可変(V)および連結(J)(または、多様性、D)領域の接合部において作成される[DavisおよびBjorkman(1988)Nature 334:395−402]。しかし、TCRは、抗体が体細胞点変異を生じるようには、おそらく同時には、体細胞点変異を起さない。TCRはまた、抗体と同一の程度の親和性変異を起こさない。TCRは、天然に存在するように、105から106M−1まで変化する親和性を有するようであり、これに対して、抗体は代表的に、105から109M−1まで変化する親和性を有する[Davisら、(1998)Annu.Rev.Immunol.16:523−544;Eisenら、(1996)Adv.Protein Chem.49:1−56]。TCRにおける体細胞変異の非存在は、より低い親和性に関連し得るが、TCRがより高い親和性を有する選択的な利点が存在しないことがまた議論されている。実際、T細胞活性化の連続的に引き起こす[Valituttiら(1995)Nature 375:148−151]および速度論的校正[Rabinowitzら(1996)Proc.Natl.Acad.USA 93:1401−1405]モデルは共に、より長いオフレート(off−rate)(より高い親和性と関連した)はシグナル伝達プロセスに有害であることを示唆する。より高い親和性TCRは、T細胞応答について必要なペプチド特異性を維持し得ないこともまたありうる。例えば、MHCの溝内に結合されたペプチドは、制限されたアクセス可能な表面を示し[Bjorkman,P.J.(1997)Cell 89:167−170]、これは、代わりに、相互作用おいて生成され得るエネルギーの量を制限し得る。他方、MHCへリックスに対してエネルギーを向けることによって、TCRの親和性を上昇することは、おそらくネガティブな選択の間の胸腺欠失を生じる[Bevan,M.J.(1997)Immunity 7:175−178]。
【0039】
我々は、T細胞レセプタータンパク質のより高い親和性に対する固有な構造特性または遺伝的制限は存在しないことを示す。より高い親和性TCR変異体は、酵母ディスプレイコンビナトリアル技術およびTCR変異体(例えば、VαおよびVβCDR3変異体)を使用することによって、インビボ選択圧力の非存在下で、生成される。標的リガンドに対する比較的強力な結合について選択された変異体(特定のp/MHC複合体)は、100倍以上高い親和性(すなわち、p/MHCについて約10nMのKd)を有し得、そして、これらの変異体は、高度のペプチド特異性を保持した。プロリンまたはグリシンが豊富な保存されたCDR3モチーフを有するTCRタンパク質についての強力な優先性がまた明らかである。高い親和性TCRの可溶性単量体形態は、抗原提示細胞上のp/MHC複合体を直接検出可能であった。これらの発見は、TCRの親和性成熟が、少なくともインビトロで可能であることを証明する。従って、操作されたTCRタンパク質は、特定のリガンドを標的化するために使用され得、これらとしては、MHC成分の非存在下において特定のp/MHC複合体およびペプチド、タンパク質または他のリガンドが挙げられる。
【0040】
ペプチド特異性を保持するより高い親和性のTCRを作製することが可能か否かを試験するために、本発明者らは、特徴付けられたTCRを、指向性インビトロ進化のプロセスに供した。ファージディスプレイ技術[Clacksonら(1991)Nature 352:624−628]は、抗体とTCR V領域との間の広範囲な構造的類似性にもかかわらず、単鎖TCR(scTCR、Vβ−リンカー−Vα)の操作における成功はまだ証明されていない。しかし、本発明者らは、scTCRが、抗体操作において首尾良いことが証明された系[BoderおよびWittrup(1997)(前出);Kiekeら(1997)前出]において、酵母の表面上に提示され得る[Kiekeら(1999)Proc.Natl.Acad.Sci.USA 96:5651−5656]ということを近年示した。CTLクローン2C由来のscTCRの温度安定化改変体(T7と
呼ばれる)[Shustaら(1999)J. Mol.Biol.292:949−956]を、本研究において使用した。CTLクローン2Cは、酵素2−オキソグルタレートデヒドロゲナーゼ由来の[Udakaら(1993)Proc.Natl.Acad.Sci.USA 90:11272−11276]、p2Caと呼ばれる結合した八量体ペプチドを有する同種異系抗原Ldを認識する。九量体改変体QL9はまた、CTL 2Cにより認識されるが、2C TCRによる10倍高い親和性を有する[Sykulevら(1994)Proc.Natl.Acad.Sci.USA 91:11487−11491]。構造的研究は、2C TCRのCDR3αがペプチドに近く、そしてpMHC複合体に付随するためにコンホメーション変化を受けることが示されたが[Garcia(1998)Science 279: 1166−1172]、アラニンスキャニング変異誘発は、CDR3αループが、結合相互作用に対して最少エネルギーの寄与をすることを示す[Manning(1998)前出]。従って、本発明者らは、変異誘発の試みを、CDR3αの先端を形成する5つの残基に集中させた。
【0041】
105の独立したTCR−CDR3α酵母変異体のライブラリーを、蛍光で標識化されたQL9/Ldリガンドを用いるフローサイトメトリーによる選択にかけた[Dal P
ortoら(1993)Proc.Natl.Acad.Sci.USA 90:6671−6675]。4回の選別および増殖の後、15個の異なる酵母コロニーを、リガンドに結合する能力について、scTCR改変体T7(これは、wt CRD3α配列を保有する)に比較して試験した(図1)。抗Vβ8.2抗体F23.2(これは、タンパク質のCDR1領域およびCDR2領域における残基を認識する)を、コントロールとして使用して、wt scTCR−T7変異体およびscTCR変異体(図1におけるqL2およびqL7ならびにその他)が、各々scTCRのほぼ等しい表面レベルを有することが示された(図1)。対照的に、可溶性QL9/Ldリガンドは、各変異体酵母クローンに非常によく結合するが、wt scTCR−T7には結合しなかった。MCMV(配列番号1)/Ld複合体(これは、CTLクローン2Cにより認識されない)は、scTCR変異体にも、wt scTCR−T7にも結合せず、このことは、scTCR変異体がペプチド特異性を保持していることを示した。変異体TCRの相対的親和性はまた、一定の濃度でQL9/Ldリガンドで観察されるシグナルにおける差異に基づいて、クローンによって変化するようであった。
【0042】
15の変異体のCDR3α配列は、全て、出発した2C TCR配列(表1)と異なっていた。BLASTアラインメントアルゴリズムによる比較は、2つのモチーフへと配列を整列させた。あるモチーフは、残基ストレッチの中間にグリシンを含んでいたが、別のモチーフは、3つの直列プロリンを含んでいた。3つのプロリン全てが、最も高い親和性部位を生成させる際に重要であるという証拠が、変異体q3rを用いた結果により示唆される。変異体q3rは、3つのプロリンのうち2つのみを含んでおり、そして三重プロリン変異体と比較して減少した結合を示した。グリシン含有変異体は、カルボキシ側に対する2つの残基のうち正に荷電した残基を選択し(7/9)、そしてアミノ側に対する2つの残基のうち芳香族のかつ/または正に荷電した残基を選択する(4/9および5/9)ようであった。理論に拘束されることは望まないが、モチーフにおける102位のグリシン残基についての選択は、CDR3αループが増加した親和性を達成するためにこの残基の周りでのコンホメーションの柔軟性を必要とすることを示していると考えられる。このことは、リガンドが配位したTCRのCDR3αループとリガンドが配位していないTCRのCDR3αループとの間で観察された、大きな(6Å)のコンホメーション差異と一致する[Garciaら(1998)前出]。グリシンが抗体のV(D)J接合部における最も一般的な残基であり、そしてグリシンの存在は、近年、(4−ヒドロキシ−3−ニトロフェニル)アセチルハプテンに応じて増加した親和性と関連付けられたことに留意することもまた興味深い[Furukawaら(1999)Immunity 11:329−338]。
【0043】
グリシンを含有する単離物と対照的に、CDR3αループの先端におけるプロリンリッチ配列についての選択は、どんな理論にも束縛されることを望まないが、これらのTCR分子がより高い親和性を付与するより堅固なコンホメーションを示すということを示すと考えられる。その親和性成熟誘導体に比較して低い親和性の生殖細胞系抗体のX線結晶解析構造は、高い親和性の状態が、ハプテンを付随する配置における抗体の安定化に起因し得たことを示した[Wedemayerら(1997)Science 276:1665−1669]。同様に、生殖細胞系抗体に類似し得るscTCRのNMR溶液構造は、CDR3αおよびβループが、両方とも有意な運動性を示したことを示した[Hareら(1999)Nat.Struct.Biol.6:574−581]。TCR:pMHC相互作用の最近の熱力学的研究はまた、結合におけるコンホメーション変化の重要性を示唆した[Willcoxら(1999)Immunity 10:357−365;Bonifaceら(1999)Proc.Natl.Acad.Sci.USA 96:11446−11451]。本明細書中で考察されるTCR変異体の構造研究および熱力学的研究により、本発明者らは、2つのCDR3αモチーフ(Gly対Proリッチ)がより高い親和性を付与する機構において異なるか否かを調べることが可能になった。
【0044】
scTCR変異体は、無効な(null)(無関連な)ペプチド/Ld複合体MCMV(配列番号1)/Ldに結合しなかったが、親和性の増加が、微細な(fine)特異性における変化と関連付けられたことは可能なままであった。この問題を調べるために、本発明者らは、以前にそのwt2C TCRについての結合親和性における有意な差異を示すことが支援された、QL9の5位(Phe)ペプチド改変体を使用した[Schlueter(1996)J.Immunol.157:4478−4485]。酵母表面上の種々のTCR変異体およびクローン2Cに対するこれらのpMHCの結合を、フローサイトメトリーにより測定した。図2に示されるように、2C上のネイティブTCRは、チロシンまたはヒスチジンのいずれかを5位に含むがグルタミン酸は含まないQL9改変体に結合し得る。より高い親和性のTCR変異体の各々は、保存的チロシン置換ペプチドを認識する能力を保持し、そしてこれらは、グルタミン酸置換ペプチドを認識できないようであった。しかし、TCR変異体のうちのいくつか(qL2、qL5、およびqL7)は、ヒスチジン置換ペプチドに(異なる程度であるが)結合したが、その他の変異体(qL1、qL3、およびqL8)は、このペプチドに結合しなかった(このアッセイの検出限界内)。従って、CDR3αループは、ペプチドの微細な特異性の認識に影響し得るが、TCRが含まれる唯一の領域ではない。CDR3に指向された選択に関する初期の研究[Sant’Angeloら(1996)Immunity 4:367−376;Jorgensenら(1992)Nature 355:224−230]から示唆されたように、ペプチド特異性に対する効果は、改変体ペプチドとのCDR3α残基の直接の相互作用に起因し得る。あるいは、結合エネルギーは、Ld分子自体におけるペプチドに誘導される変化に向けられ得る。後者の可能性は、2C TCR:QL9/Ld相互作用の場合におけるよりもおそらく起こりやすい。なぜならば、QL9の5位は、Ld溝に向いていると推測されたからである[Schlueterら(1996)前出,Speirら(1998)Immunity 8:553−562]。微細特異性分析はまた、同族のペプチドについて増加したか、または少なくとも変更された特異性を有するTCRを操作することが可能であることを示す。従って、単一のTCRの短い領域(CDR3α)のみの指向された進化は、所望のペプチド結合特異性および/または増加した結合親和性を有する多くのTCR改変体の単離を可能にする。
【0045】
選択されたCDR3α変異体に関連する親和性の増加の大きさを決定するために、野生型T7scTCRおよびqL2scTCRは、酵母分泌系において可溶性形態として発現された。精製されたscTCR調製物を、T2−Ld細胞の表面上でLd上に負荷されたQL9またはMCMV(配列番号1)に対する、125I標識化抗LdFabフラグメントの結合を遮断する能力について比較した[Manning(1998)前出、Sykulevら(1994)Immunity 1:15−22]。予測されたように、T7もqL2scTCRも、MCMV(配列番号1)ペプチドでアップレギュレートされたT2−Ld細胞に対する125I−Fabフラグメントの結合を阻害することができなかった。しかし、T7およびqL2の両方が、QL9/Ldに対する抗LdFabフラグメントの結合を阻害し得た(図3)。qL2scTCR改変体は、結合の阻害において標識化されていないFabフラグメントと同程度の効力であったが、T7scTCRは、160倍も小さい効力(4回の独立した適定の中で、平均して140倍の差)であった。QL9/LdについてのscTCRのKD値を、その阻害曲線から、T7について1.5μMおよびqL2について9.0nMと算出した。T7についての値は、2CscTCRについて以前に報告された3.2μMのKDと密接に一致した[Manningら(1999)J:Exp.Med.189:461−470]。これらの発見は、CDR3α指向性変異誘発と組み合わされた酵母系が、特定のpMHCリガンドについての、少なくとも約100倍高い固有の結合親和性を有する変異体の選択を可能にするということを示す。
【0046】
可溶性scTCRがそのpMHCリガンドに関して高い親和性を有する場合、抗体のように、細胞表面結合抗原についての特異的プローブとして有用である。このことを確かめるために、この可溶性T7およびqL2scTCRを、ビオチン化し、そして標識化されたscTCR分子を、QL9、p2Ca、またはMCMV(配列番号1)を負荷されたT2−Ld細胞とインキュベートした。qL2scTCR(T7 scTCRではない)は、QL9またはp2CaとインキュベートされたT2細胞の容易に検出可能な染色を生じた(図4A〜4B)。p2Caアップレギュレートされた細胞がまた、qL2scTCRにより容易に検出されたことは重要である。なぜならば、p2Caは、同種反応性(alloreactive)クローン2Cにより認識されたペプチドの天然で処理された形態であり、そしてこれは、2C TCRについてQL9/Ld複合体よりも低い親和性でさえ有するからである[Sykulevら(1994)前出]。
【0047】
本発明者らの研究において記載される高親和性レセプターは、VJ接合部における改変により誘導された(T細胞における遺伝子再配列によりインビボで非常に有効に作用する同じプロセス)(DavisおよびBjorkman(1988)Nature 334:395−402)。本発明者らがインビトロで高親和性TCRの広範なセットを容易に単離し得たという事実は、高親和性レセプターに対する遺伝的制限も構造的制限も存在しないということを示す。このことは、インビボで見出されるTCRの固有の低い親和性が、より高い親和性についての選択の欠如、およびおそらくより低い親和性の選択に起因するという見解を支持する(Sykulevら(1995)Proc.Natl.Acad.Sci.USA 92:11990−11992;Valituttiら(1995)Nature 375:148−151;Rabinowitzら(1996)Proc.Natl.Acad.Sci.USA 93:1401−1405)。この点において、本発明のより高い親和性のTCRはここで、T細胞応答に対する親和性の効果についての仮定を直接的に試験するための試薬を提供する((Davisら(1998)Ann.Rev.Immunol.16:523−544;Sykulevら(1995)(前出);Valituttiら(1995)(前出);Rabinowitzら1996)前出)。
【0048】
要約すると、本発明者らは、T細胞レセプター(抗体と同程度広範なタンパク質のクラスを代表する)が、抗体と同様に操作されて、高い親和性の、抗原特異的プローブを与え得ることを示した。さらに、高親和性レセプターの可溶性変形は、細胞上の特異的ペプチド/MHC複合体を直接検出し得る。従って、これらの操作されたタンパク質は、例えば、腫瘍細胞についての診断剤として有用である。高親和性TCRの可溶性誘導体は、高親和性の抗原特異的プローブとして有用であるか、またはさらに操作され得る。適切に標識化された場合(またはこの可溶性TCRに検出可能なリガンドにより結合される場合)の可溶性TCR誘導体は、細胞(例えば、腫瘍細胞もしくはその他の新形成細胞の誘導された表面、またはウイルス感染細胞もしくはその他の疾患細胞の診断的抗原)上の特異的ペプチド/MCHC複合体についてのプローブとして作用し得る。高親和性TCR細胞に結合したタンパク質または可溶性誘導体についての他の適用としては、特定の自己免疫疾患の診断または研究における使用が挙げられる。特徴的なペプチド/MHCまたはその他のマーカー表面抗原が公知であるかまたは同定され得る場合、高親和性の可溶性TCRは、その細胞表面部分への特異的結合のために単離され得、そして診断または治療において使用され得る。破壊されることが所望される細胞が、高親和性TCR(望ましくは可溶性TCRタンパク質)により認識される特徴的かつ特異的細胞表面部分を有する場合、高親和性TCRタンパク質(望ましくは可溶性誘導体)は、癌の処置または他の障害における治療剤として、細胞傷害性因子に結合して使用され得る。同様に、本明細書中に記載される可溶性高親和性TCRは、画像化剤にカップリングされ得、そして腫瘍細胞が存在する身体内の部位(ここでTCRが、高親和性および特異性を有する腫瘍細胞マーカーに特異的に結合する)を同定するために使用され得る。不適切に自己免疫破壊の標的にされてきた細胞または組織の表面に結合した高親和性TCRは、標的細胞または組織の細胞表面への結合について細胞傷害性リンパ球と競合することにより、これらの細胞傷害性リンパ球による自己免疫組織破壊を低減させ得る。
【0049】
これらの結果はまた、T細胞応答における重要で、基本的な疑問の状況において考慮され得る。T細胞レセプターについて以前に観察された低い親和性は、体細胞変異の非存在に起因するのか、またはT細胞に対して作用するインビボでの選択的圧力に起因するのか?本発明者らの研究において記載される高親和性レセプターは、VJ接合部でのバリエーションに由来した(T細胞において非常に有効に作用する同じプロセス)[DavisおよびBjorkman(1988)前出]。本発明者らが、インビトロで高親和性TCRの広範なセットを容易に単離し得たという事実は、高親和性レセプターへの構造的制限または遺伝的制限が存在しないことを示す。このことは、インビボで見出される元々低い親和性のTCRが、より高い親和性の選択の欠如およびおそらくより低い親和性の選択の欠如に起因すると言う見解を支持する[Sykulevら(1995)Proc.Natl.Acad.Sci.USA 92:11990−11992;Rabinowitzら(1996)前出]。この点において、ここでより高い親和性のTCRは、T細胞応答に対する親和性の効果についての仮説を直接試験するための試薬を提供する[Davisら(1998)(前出);Sykulevら(1995)(前出);Valituttiら(1995)(前出);Rabinowitzら(1996)前出]。
【0050】
遺伝子コードの縮重に起因して、多数の機能的に等価なヌクレオチド配列が、同じアミノ酸配列をコードするということが、当業者に理解される。
【0051】
さらに、当業者は、標準的な変異誘発技術により、本明細書中に記載される抗原決定活性アッセイを組み合わせて、変更されたTCR配列を得、そしてそれらを特定の結合活性を有するポリペプチドの発現について試験し得る。当該分野で公知の有用な変異誘発技術としては、限定せずに、オリゴヌクレオチド指向性変異誘発、領域特異的変異誘発、リンカースキャニング変異誘発、およびPCRによる部位指向性変異誘発が挙げられる[例えば、Sambrookら(1989)およびAusubelら(1999)を参照のこと]。
【0052】
改変体TCRコード配列を得る際に、当業者は、TCR由来のタンパク質が、生物学的活性の損失も減少もなしに、特定のアミノ酸置換、付加、欠失および翻訳後修飾により改変され得ることを認識する。特に、保存的アミノ酸置換、すなわち、1つのアミノ酸の、類似のサイズ、電荷、極性およびコンホメーションの別のアミノ酸との置換が、タンパク質の機能を有意に変更しないようであることは周知である。タンパク質を構成する20種の標準的なアミノ酸は、以下のような保存的アミノ酸の4つのグループに広く分類され得る:非極性(疎水性)グループは、アラニン、イソロイシン、ロイシン、メチオニン、フェニルアラニン、プロリン、トリプトファンおよびバリンを含む;極性(非荷電、中性)グループは、アスパラギン、システイン、グルタミン、グリシン、セリン、トレオニンおよびチロシンを含む;正に荷電した(塩基性)グループは、アルギニン、ヒスチジンおよびリシンを含む;そして負に荷電した(酸性)グループは、アスパラギン酸およびグルタミン酸を含む。タンパク質中の1つのアミノ酸を同じグループ内の別のアミノ酸と置換することは、そのタンパク質の生物学的活性に不利な影響を有さないようである。
【0053】
ヌクレオチド配列間の相同性は、DNAハイブリダイゼーション分析によって決定され得、ここでは、二本鎖DNAハイブリッドの安定性は、生じる塩基対形成の程度に依存する。高温および/または低塩含量の条件は、ハイブリッドの安定性を減少させ、そして選択された程度より少ない相同性を有する配列のアニール化を妨げるために変更させ得る。例えば、約55%のG−C含量を有する配列に対して、40〜50℃、6×SSC(塩化ナトリウム/クエン酸ナトリウム緩衝液)および0.1% SDS(ドデシル硫酸ナトリウム)というハイブリダイゼーションおよび洗浄条件は、約60〜70%の相同性を示し、50〜65℃、1×SSCおよび0.1% SDSのハイブリダイゼーションおよび洗浄条件は、約82〜97%の相同性を示し、そして52℃、0.1×SSCおよび0.1% SDSのハイブリダイゼーションおよび洗浄条件は、約99〜100%の相同性を示す。ヌクレオチド配列およびアミノ酸配列を比較するため(そして相同性の程度を測定するため)の広範囲のコンピュータプログラムはまた、利用可能であり、そして市販されるソフトウエアとフリーソフトウエアの両方の供給源を提供する表は、Ausubelら(1999)において見出される。すぐに利用可能な配列比較および複数の配列整列アルゴリズムは、それぞれ、Basic Local Alignment Search Tool(BLAST)(Altschulら、1997)およびClustalWプログラムである。BLASTは、http://www.ncbi.nlm.nih.govでインターネット上で利用可能であり、そしてClustalWのバージョンは、http://www2.ebi.ac.ukで利用可能である。
【0054】
微生物の工業的株(例えば、Aspergillus niger、Aspergillus ficuum、Aspergillus awamori、Aspergillus oryzae、Trichoderma reesei、Mucor miehei、Kluyveromyces lactis、Pichia pastoris、Saccharomyces cerevisiae、Escherichia coli、Bacillus subtilisまたはBacillus licheniformis)あるいは植物種(例えば、カノーラ、ダイズ、コーン、トマト、オオムギ、ライムギ、コムギ)は、TCRペプチドの組換え産生のための宿主細胞として使用され得る。高親和性TCRタンパク質または可溶なタンパク質の異種発現における第1工程として、発現構築物は、TCRまたは可溶なTCRのコード配列、ならびに制御配列(例えば、プロモーター、エンハンサーおよびターミネーター)を含むように構築される。シグナル配列および選択マーカーのような他の配列はまた、含まれ得る。scTCRの細胞外発現を達成するために、発現構築物は、分泌性シグナル配列を含み得る。シグナル配列は、細胞質発現が所望される場合、発現構築物上に含まれない。プロモーターおよびシグナル配列は、宿主細胞において機能的であり、そしてTCRまたは可溶なTCRタンパク質の発現および分泌を提供する。転写ターミネーターは、効果的な転写を保証するために含まれる。発現またはタンパク質精製を増強する補助的配列はまた、発現構築物に含まれ得る。
【0055】
種々のプロモーター(転写開始調節領域)は、本発明に従って使用され得る。適切なプロモーターの選択は、提案される発現宿主に依存する。異種供給源からのプロモーターは、それらが選択された宿主において機能的である限り、使用され得る。
【0056】
プロモーター選択はまた、所望の有効性およびペプチドまたはタンパク質の産生のレベルに依存する。誘導プロモーター(例えば、tac)は、しばしば、E.coliにおけるタンパク質発現のレベルを劇的に増加させるために利用される。タンパク質の過剰発現は、宿主細胞に有害であり得る。その結果、宿主細胞の増殖は制限される。誘導プロモーター系の使用によって、宿主細胞は、遺伝子発現の誘導前に、受容可能な密度まで培養されることが可能となり、それによって、より高い産生物収量を容易にする。
【0057】
種々のシグナル配列が、本発明に従って使用され得る。TCRコード配列と同種のシグナル配列が、使用され得る。あるいは、効果的な分泌および発現宿主におけるプロセシングのために選択されるかまたは設計されたシグナル配列はまた、使用され得る。例えば、適切なシグナル配列/宿主細胞の組としては、B.subtilisにおける分泌のためのB.subtilis sacBシグナル配列、およびSaccharomyces cerevisiae α接合因子またはP.pastoris分泌のためのP.pastoris酸性ホスファターゼphoIシグナル配列が挙げられる。シグナル配列は、タンパク質コード配列に対するシグナルペプチダーゼ切断部位をコードする配列によって、または通常10より少ないコドンからなる短いヌクレオチドブリッジ(bridge)(ここで、このブリッジは、下流のTCR配列の正しい読み取り枠を保証する)によって、直接連結され得る。
【0058】
転写および翻訳を増強するためのエレメントは、真核生物のタンパク質発現系について同定されている。例えば、カリフラワーモザイクウイルス(CaMV)プロモーター1000bpを異種プロモーターのいずれかの端に配置することは、植物細胞において転写レベルを10〜400倍増強させ得る。発現構築物はまた、適切な翻訳開始配列を含む。適切な翻訳開始のためのKozakコンセンサス配列を含むための発現構築物の改変は、翻訳のレベルを10倍増大させ得る。
【0059】
選択マーカーがしばしば利用され(これは、発現構築物の一部分であっても、または発現構築物と離れて(例えば、発現ベクターによって運ばれる)いてもよい)、その結果、マーカーは、目的の遺伝子と異なる部位で統合され得る。例としては、抗生物質に耐性を与える(例えば、blaは、E.coli宿主細胞に対してアンピシリン耐性を与え、nptIIは、広範囲の原核生物細胞および真核生物細胞に対してカナマイシン耐性を与える)マーカー、または宿主を最少培地で増殖させるマーカー(例えば、HIS4は、P.pastorisまたはHis−S.cerevisiaeがヒスチジンの非存在下で増殖することを可能にする)が挙げられる。選択可能なマーカーは、それ自身の転写および翻訳の開始ならびに停止の調節領域を有し、マーカーの独立した発現を可能にする。抗生物質耐性がマーカーとして利用される場合、選択のための抗生物質濃度は、その抗生物質に依存して変化し、通常、抗生物質10〜600μg/培地1mLの範囲である。
【0060】
発現構築物は、公知の組換えDNA技術(Sambrookら、1989;Ausubelら、1999)を利用することによって構築される。制限酵素消化および連結は、DNAの2つのフラグメントを連結させるために利用される基本的工程である。DNAフラグメントの末端は、連結前に改変を必要とし得、そしてこれは、突出を埋めること、ヌクレアーゼ(例えば、ExoIII)でフラグメント(単数または複数)の末端部分を欠失すること、部位を指向された突然変異誘発によって、またはPCRによって新しい塩基対を添加することによって達成され得る。ポリリンカーおよびアダプターは、選択されたフラグメントの連結を容易にするために利用され得る。発現構築物は、代表的に、E.coliの制限、連結および形質転換の数ラウンド(round)を利用する段階で構築される。発現構築のために適切な多くのクローニングベクターは、当該分野において公知である(λZAPおよびpBLUESCRIPT SK−1、Stratagene、LaJolla、CA;pET、Novagen Inc.Madison、WI−Ausubelら、1999において引用される)、そして特定の選択が、本発明には重要ではない。クローニングベクターの選択は、宿主細胞への発現構築物の導入のために選択される遺伝子転移系によって影響される。各段階の終わりに、得られた構築物は、制限、DNA配列、ハイブリダイゼーションおよびPCR分析によって分析され得る。
【0061】
発現構築物は、クローニングベクター構築物(線状か環状かのいずれか)として宿主に形質転換され得るか、あるいはクローニングベクターから除去され得、そして送達ベクターとして使用され得るか、または送達ベクター内に導入され得る。送達ベクターは、選択された宿主細胞型において、発現構築物の導入および保持を容易にする。発現構築物は、多くの公知の遺伝子導入系(例えば、天然の能力(competence)、化学的に媒介される形質転換、プロトプラスト形質転換、エレクトロポレーション、微粒子銃形質転換、トランスフェクション、または結合)(Ausubelら、1999;Sambrookら、1989)のいずれかによって宿主細胞に導入される。選択される遺伝子転移系は、使用される宿主細胞およびベクター系に依存する。
【0062】
例えば、発現構築物は、プロトプラスト形質転換またはエレクトロポレーションによってS.cerevisiae細胞に導入され得る。S.cerevisiaeのエレクトロポレーションは、容易に達成され、そしてスフェロプラスト形質転換に匹敵する形質転換効率をもたらす。
【0063】
リガンド結合部位以外の部位でTCRタンパク質によって特異的に反応するモノクローナルまたはポリクローナル(好ましくは、モノクローナル)抗体は、当該分野において公知である方法によって作製され得る。例えば、HarlowおよびLane(1988)Antibodies:A Laboratory Manual、Cold Spring Harbor Laboratories;Goding(1986)Monocolonal Antibodies:Principles and Practice 第2版、Academic Press、New York;およびAusubelら(1999)Molecular Biology、John Wiley & Sons,Inc.、New YorkのCurrent Protocolsを参照のこと。
【0064】
特定のpMHCに特異的な細胞結合形態または可溶形態の高親和性のTCRタンパク質は、例えば、生物学的サンプル(例えば、細胞、組織サンプル、生検材料、体液など)のスクリーニングのため、または試験サンプルにおける同族pMHCの存在を検出するための診断プローブとして有用である。しばしば、高親和性のTCRタンパク質は、検出可能なシグナルを提供する基質を連結させる(共有結合か非共有結合かのいずれか)ことによって標識化される。適切な標識としては、放射性核種、酵素、基質、補助因子、インヒビター、蛍光試薬、化学発光試薬、磁気粒子などが挙げられるが、これらに限定されない。さらに、TCRタンパク質は、第2の結合分子に対するリガンドに結合し得:例えば、TCRタンパク質は、ビオチン標識化され得る。従って、標的細胞または分子に結合したTCRの検出は、検出可能なストレプトアビジン(蛍光分子、放射活性分子、化学発光分子、または他の検出可能な分子が付着されるか、または、発色団基質が利用可能である酵素が付着される、ストレプトアビジン)の結合によって達成され得る。scTCRタンパク質に共有結合するべきこのような標識物および/または有毒な化合物の使用を記載する米国特許のとしては、以下の号が挙げられるが、これらに限定されない:
【0065】
【数1】
標識化されたTCRタンパク質は、使用される標識に適切なモニタリングデバイスまたは方法を用いて検出され得る。蛍光顕微鏡または蛍光活性化セルソーティングは、標識が蛍光部分である場合に使用され得、そして標識が、放射性核種である場合、γカウンティング、オートラジオグラフィまたは液体シンチレーションカウンティングが、例えば、その方法が分析されているサンプルおよび使用される放射性核種に適切であるという条件で使用され得る。さらに、本明細書中に述べられるMHC成分の非存在下で同族pHMCリガ
ンドまたは他のリガンドに対する結合部位の一部でないTCRタンパク質の部分を認識する検出可能な分子または粒子がある場合、第2の検出分子または粒子が利用され得る。当業者は、インサイチュでの診断画像処理に有用な化合物を知っている;例えば、米国特許第5,101,827号、同第5,059,413号を参照のこと。インビボの治療および/または画像処理に有用な放射性核種はとしては、111インジウム、97ルビジウム、125ヨウ素、131ヨウ素、123ヨウ素、67ガリウム、99テクネチウムが挙げられる。毒素としては、一旦、TCR−毒素複合体が細胞に結合される場合、毒性部分が内部移行し、その結果、その細胞傷害性効果を及ぼし得るという条件では、とりわけ、ジフテリア毒素、リシンおよびトウゴマ毒素が挙げられる。免疫毒性テクノロジーは、当該分野において周知であり、そして適切な毒性分子としては、以下が挙げられるが、これらに限定されない;化学療法薬剤(例えば、ビンデシン)、葉酸代謝拮抗財(例えば、メトトレキセート、シスプラチン、マイトマイシン)、アンスロサイクリン(anthrocyclines)(例えば、ダウノマイシン、ダウノルビシンまたはアドリアマイシン)およびリボソーム不活性化タンパク質のような細胞傷害性タンパク質(例えば、ジフテリア毒素、ヤマゴボウ抗ウイルスタンパク質、アブリン、リシン、シュードモナス属体外毒素Aまたはこれらの組換え誘導体)。一般的に、例えば、OlsnesおよびPihl(1982)Pharmac.Ther.25:355−381およびMonoclonal Antibodies for
Cancer Detection and Therapy、BaldwinおよびByers編、159頁−179頁、Academic Press、1985を参照のこと。
【0066】
特定のpMHCリガンドに特異的な高親和性TCRタンパク質は、特定のpMHCに関連する疾患に罹患すると考えられる動物(ヒトを含む)を診断において有用である。本発明のscTCR分子は、本質的に、任意の抗原を基本的に検出するのに有用であり、新生物状態、異常なタンパク質、あるいは細菌、真菌、ウイルス、原虫、酵母、線虫または他の寄生生物による感染またはインフェステーションに関連するものが挙げられるが、これらに限定されない。高親和性のscTCRタンパク質はまた、生産された異常なタンパク質が存在する特定の遺伝子疾患の診断の際に使用され得る。これらの高親和性タンパク質の例示的な適用は、自己免疫疾患の処置においてであり、その中には、既知のpMHCがある。I型糖尿病は、免疫破壊を誘発する自己抗原に関して比較的よく特徴付けられる。多発性硬化症、セリアック病、炎症性腸疾患、クローン病および慢性関節リウマチは、このような適用に対するさらなる候補疾患である。自己免疫破壊を不適切に標的化する、細胞または組織の表面上のp/MHC複合体に対する結合特異性を有する高親和性のTCR(可溶性)タンパク質は、細胞傷害性リンパ球による標的細胞への結合について競合することによって、自己免疫破壊のアンタゴニストとして役立ち得る。対照的に、高親和性TCRタンパク質(望ましくは、可溶性の一本鎖TCRタンパク質)(これは、破壊が有益である細胞表面上の抗原またはp/MHCに特異的に結合する)は、有毒な化合物(例えば、毒素または放射性核種)に結合し得、その結果、標的細胞への結合は、細胞傷害性リンパ球による引き続いての結合および破壊を引き起こす。破壊のために標識化された細胞は、新生物細胞(例えば、腫瘍細胞)、ウイルス、細菌、または原虫で感染された細胞、あるいは他の疾患原因因子もしくは寄生生物であり得、すなわち、それは、細菌、酵母、真菌、原虫または所望されない細胞であり得る。このような高親和性のscTCRタンパク質は、本明細書中に記載される方法によって得ることができ、そして続いて、目的の特定のリガンドについてスクリーニングするために使用され得る。
【0067】
高親和性のTCR組成物は、当該分野において公知の任意の方法によって処方され得る。これらは、代表的に、特に経静脈投与、経腹膜投与、または滑液投与(特定の疾患によって決定される経路で)のための注射剤として、または、経鼻または経口投与のための処方物(液体溶液か懸濁液かのいずれか)として調製され得る。注射または他の投与前の液体中の、溶液または懸濁液に適切な固体の形態はまた、調製され得る。この調製物はまた、例えば、乳化され得るか、またはリポソーム中にカプセル化されたタンパク質(単数および複数)/ペプチド(単数および複数)であり得る。
【0068】
活性な成分はしばしば、賦形剤またはキャリアと混合され、これらは、薬学的に受容可能であり、そして活性な成分と共存可能である。適切な賦形剤としては、水、生理的食塩水、ブドウ糖、グリセロール、エタノールなど、およびそれらの組合せが挙げられるが、これらに限定されない。注射可能な、エアロゾルまたは経鼻処方物の形態の高親和性のTCRタンパク質の濃度は、通常、0.05〜5mg/mlの範囲にある。特定の有効な用量の選択は、公知であり、そして当業者によって過度の実験なしに行われる。同じ用量が、他の粘膜表面に投与され得る。
【0069】
さらに、所望される場合、ワクチンは、少量の補助的物質(例えば、湿潤剤または乳化剤、pH緩衝剤、および/またはワクチンの効果を増強するアジュバント)を含み得る。有効であり得るアジュバントの例としては、以下が挙げられるが、これらの限定されない:水酸化アルミニウム;N−アセチル−ムラミル−L−スレオニル−D−イソグルタミン(thr−MDP);N−アセチル−nor−ムラミル−L−アラニル−D−イソグルタミン(CGP 11637、nor−MDPと呼ばれる);N−アセチルムラミル−L−アラニル−D−イソグルタミニル−L−アラニン−2−(1’−2’−ジパルミトイル−sn−グリセロ−3ヒドロキシホスホリル)−エチルアミン(CGP 19835A、MTP−PEと呼ばれる)およびRIBI(これは、細菌から抽出された3つの成分を含む:2%のスクアレン/Tween 80乳濁液中のモノホスホリルリピドA、トレハロースジマイコレート(dimycolate)および細胞壁骨格(MPL+TDM+CWS)。当該分野において公知であるこのようなさらなる処方物および投与の形態はまた、使用され得る。
【0070】
本発明の高親和性TCRタンパク質、および/または高親和性TCRタンパク質に対して類似(90%より大きい同一性)の一次構造を有するpMHC結合フラグメント(同族のリガンドについて高親和性を維持する)は、中性形態または塩形態としてワクチン中に処方され得る。薬学的に受容可能な塩としては、酸付加塩(ペプチドの遊離アミノ酸基で形成された)が挙げられるがこれらに限定されない。この塩は無機酸(例えば、塩酸またはリン酸)および有機酸(例えば、酢酸、シュウ酸、酒石酸、またはマレイン酸)で形成される。遊離のカルボキシル基で形成された塩はまた、無機塩基(例えば、水酸化ナトリウム、水酸化カリウム、水酸化アンモニウム、水酸化カルシウム、または水酸化第二鉄)、および有機塩基(例えば、イソプロピルアミン、トリメチルアミン、2−エチルアミノ−エタノール、ヒスチジン、およびプロカイン)に由来し得る。
【0071】
治療用途のための高親和性TCRタンパク質(例えば、細胞毒性化合物に結合体化されたタンパク質)を、当該分野で公知の方法に従って、投与処方物に適切な様式で、そして予防的におよび/または治療的に効果的であるような量および様式で、投与する。投与されるべき量は、一般に1用量あたり約100〜20,000μgタンパク質の範囲、より一般的には、1用量あたり約1000〜10,000μgの範囲である。同様の組成物が、画像化における用途のために標識化した高親和性TCRタンパク質を用いて類似の様式で投与され、例えば、自己免疫攻撃下でかつ同属のpMHCを含む組織を検出するか、またはその表面に同属のpMHCを保有する癌細胞を検出し得る。投与されるのに必要な活性成分の正確な量は、医師または獣医師の判定に依存し得、そして各個体について特有であり得るが、このような決定は、このような医師の当該分野の範囲内である。
【0072】
ワクチンまたは他の免疫原性組成物は、単回用量;2用量スケジュール(例えば、2〜8週間間隔);または複数用量スケジュールで与えられ得る。複数用量スケジュールは、ワクチン接種の一次コースが1〜10以上の別の用量、続いて、免疫応答を維持するおよび/または強化するために必要な場合、引き続く時間間隔で投与される他の用量(例えば、第二用量は1〜4カ月)、そして必要な場合、数ヶ月後に引き続く用量を含み得るスケジュールである。本発明のレトロウイルス様粒子で免疫したヒト(または他の動物)は、同属のレトロウイルスによる感染から防御される。
【0073】
クローニング、DNA単離、増幅および精製のため、DNAリガーゼ、DNAポリメラーゼ、制限エンドヌクレアーゼなどを含む酵素反応のための標準的技術、ならびに種々の別の技術は、公知の技術であり、そして当業者によって通常使用される。多数の標準的技術が、以下に記載されている:Sambrookら(1989)Molecular Cloning:第二版、Cold Spring Harbor Laboratory,Plainview,New York;Maniatisら(1982)Molecular Cloning、Cold Spring Harbor Laboratory,Plainview,New York;Wu(編)(1993)Meth.Enzymol.218,PartI;Wu(編)(1979)Meth Enzymol.68:Wuら(編)(1983)Meth Enzymol.100および101;GrossmanおよびMoldave(編)Meth.Enzymol.65;Miller(編)(1972)Experiments in Molecular
Genetics,Cold Spring Harbor Laboratory,Cold Spring Harbor,New York;Old and Primrose(1981)Principles of Gene
Manipulation,University of California Press,Berkeley;SchleifおよびWensink(1982)Practical Methods in Molecular Biology;Glover(編)(1985)DNA Cloning第IおよびII号、IRL Press,Oxford,UK;Hames and
Higgins(編)(1985)Nucleic Acid Hybridization,IRL Press,Oxford,UK;ならびにSetlowおよびHollaender(1979)Genetic Engineering:Principles and Methods、1〜4号、Plenum Press,New York。ここで使用した略号および命名法は、この分野で標準的とみなされ、そしてここで引用される文献のような専門の文献中に通常用いられている。
【0074】
本出願に引用される全ての参考文献は、本開示と矛盾しない程度まで、本明細書に提供される開示および実験手順を補充するために、本明細書において参考として援用される。
【0075】
以下の実施例は、例示的な目的のために提供されるものであり、そして本明細書に記載される本発明の特許請求の範囲を限定する意図ではない。当業者に対して生じる、例示された記事および/または方法における任意のバリエーションは、本発明の範囲内におさまることが意図される。
【0076】
(実施例)
高い親和性のTCRを作成するための一般的方法は、1998年1月20日出願、米国特許出願番号09/009,388号、および1999年1月20日出願WO99/36569(これらは、本明細書の開示と矛盾しない程度まで、参考として援用される)に与えられる。
【0077】
(実施例1.ライブラリーの構築)
指向性進化のための足場として用いた2C scTCR(T7)は、TCRの安定性を増大させるがpMHC結合を可能にすることが示されている、6つの変異体(βG17E、βG42E βL81S,αL43P、αW82R、およびαI118N)を含んだ[例えば、Shusta,E.V.ら(1999)J.Mol.Biol.292,949〜956]。
【0078】
AGA2−特異的上流プライマー(GGCAGCCCCATAAACACACAGTAT(配列番号3))および変性下流プライマーCTTTTGTGCCGGATCCAAATGTCAG(SNN)5GCTCACAGCACAGAAGTACACGGCCGAGTCGCTC(配列番号4)を用いて、T7 scTCR VαCDR3の変異性PCRを、実施した。
下線を付した塩基(配列番号4の最初から11番目のG、終わりから13番目のG)は、特有のBamHIおよびEagI制限部位を誘導するサイレント変異の位置を示す。精製したPCR産物を、NdeIおよびBamHIを用いて消化し、そしてNdeI−BamHIで消化したT7/pCT302に連結した[BoderおよびWittrup(1997)前出;Kiekeら(1999)前出;Shustaら(1999)前出]。連結混合物をDH10Bエレクトロ−コンピテントなE.coli(Gibco BRI,Gaithersburg,MD)中に形質転換し、そして形質転換体を、アンピシリン100μg/ml補充した250mLのLB中にプールし、そして37℃で一晩増殖した。プラスミドDNAを、GietzおよびSchiestl[Geitzら(1995)Yeast 11:355〜360]の方法によって、酵母(Saccharomyces
cerevisiae)株EBY100中に形質転換した。
【0079】
(実施例2.細胞ソーティング)
酵母ライブラリー[Shustaら(1999)Curr.Opin.Biotechnol.10:117〜122]を、SD−CAA(2%デキストロース、0.67%酵母窒素ベース、1%カザミノ酸(Difco、Livonia,MI))中で、30℃で、OD600=4.0まで増殖した。表面scTCR発現を誘導するため、酵母を遠心分離によってペレット化し、SG−CAA(2%ガラクトース、0.67%酵母窒素ベース、1%カザミノ酸)中で、OD600=1.0まで再懸濁し、そして20℃で24時間インキュベートした。一般に、約107細胞/チューブを、50μlのQL9/Ld/IgGダイマー[Dal Portoら(1993)前出](これは0.5mg/mlBSA(PBS−BSA)を補充したリン酸緩衝化生理食塩水(pH7.4)中で希釈してある)とともに、1時間氷上でインキュベートした。インキュベーション後、細胞を洗浄し、そしてFITC結合体化ヤギ抗マウスIgG F(ab7)2(Kirkegaard&Perry,Gaithersburg,MD)含有PBS−BSAを用いて30分間標識した。次いで、ソーティングの直前に、PBS−BSA中で酵母を洗浄し、そして再懸濁した。コールター753ベンチを用いたFACSソーティングによって最高の蛍光を示す細胞を、単離した。単離後、ソーティングした細胞をSD−CAA中で増殖し、そして以降の回の選択のためにSG−CAA中で誘導した。引き続く4回の総ソーティングを実施した。染色に用いたQL9/Ld/IgGダイマーの濃度は、ソート1〜3については50μg/mlであり、そして最終のソートについては0.5μg/mlであった。各ソートから単離した総細胞のパーセンテージは、それぞれ5.55%、2.68%、2.56%および0.58%であった。ソート3および4のアリコートをSD−CAA上にプレートし、個々のクローンを単離した。クローンは、コールターEpicsXL装置を用いたフローサイトメトリーにより分析した。
【0080】
(実施例3.可溶性のscTCR産生)
T7およびqL2のオープンリーディングフレームを、pCT302 NheI−XhoIから切り出し、そしてNheI−XhoIで消化したpRSGALT(酵母発現プラスミド)に連結した[Shustaら(1999)前出]。ライゲーション(連結)したプラスミドをDH10Bエレクトロ−コンピテントなE.coli(Gibco BRL)中に形質転換した。プラスミドDNAを細菌培養物から単離し、そしてSaccharomyces cerevisiae BJ5464(α ura3−52 trp1 leu2 1 his3 200 pep4::HIS3 prb1 1.6R can1GAL)中に形質転換した[Shustaら(1999)前出]。酵母クローンを30℃で48時間1リットルのSD−CAA/Trp(20mg/mL トリプトファン)中で増殖した。scTCR分泌を誘導するため、細胞を4000×gでの遠心分離によってペレット化し、1mg/mlBSAを補充した1リットルのSG−CAA/Trp中に再懸濁し、そして20℃で72時間インキュベートした。培養上清を4000×gでの遠心分離によって回収し、約50mlに濃縮し、そしてPBS、pH8.0に対して透析した。6His−タグ化scTCRを、ネイティブなニッケルアフィニティークロマトグラフィー(Ni−NTA Superflow,Qiagen,Valencia,CA;5mMおよび20mM イミダゾール、pH8.0洗浄;250mMイミダゾール溶出)によって精製した[Shustaら(1999)前出]。
【0081】
(実施例4.細胞結合アッセイ)
QL9/Ldに対する可溶性scTCRの結合を、以前に記載した競合様式でモニターした[Manningら(1998)前出;Sykulevら(1994)前出]。ペプチド−上方制御(アップレギュレート)T2−Ld細胞(3×105/ウェル)を、125I標識化抗LdFabs(30−5−7)および種々の濃度のscTCRの存在下で、氷上で1時間インキュベートした。結合および未結合の125I30−5−7Fabをオリーブオイル/ジブチルフタラートによる遠心分離によって分離した。阻害曲線を作成し、最大阻害の50%を生じるインヒビター濃度を決定した。解離定数をChengおよびPursoffの式を用いて算出した[Cheng(1973)Biochem.Pharm.22:3099〜3108]。細胞結合pMHCに対するscTCRの直接結合をモニターするため、ペプチド−アップレギュレートT2−Ld細胞(5×105/チューブ)を、ビオチン化可溶性scTCRとともに、氷上で40分間インキュベートし、続いてストレプトアビジン−フィコエリトリン(PharMingen,San
Diego,CA)を用いて30分間染色した。フローサイトメトリーによって細胞蛍光を検出した。
【0082】
(実施例5.異なるペプチドおよび異なるMHC分子(Kb)について特異的である高親和性TCRの同定)
TCRのCDR3α領域の酵母ディスプレイ変異体の同じライブラリーを用いて、異なるMHC分子に対して結合した異なるペプチドになお特異的である、より親和性の高いTCRについて選択することが可能であった。この場合、SIYRと呼ばれるペプチド(SIYRYYGL(配列番号5))を、Kbと呼ばれるMHC分子に結合し、そしてこのリガンド複合体を、蛍光形態中で用いて、フローサイトメトリーにより選択した。高い親和性のTCRを発現する16のクローンを、配列決定した。各々のクローンは、CDR3α領域において異なる配列を示した(表2)。
【0083】
これらのTCRの特異性の例として、変異3SQ2を、二次試薬単独(SA:PE)、抗−Vβ抗体F23.2および3つのペプチド/Kb複合体(OVA/Kb,dEV8/Kb、およびSIYR/Kb)を含む、種々の因子で染色した。図5に示すように、元来の選択に用いたpMHC(SIYR/Kb)のみが、変異TCRに対して結合するのに十分な親和性を有した。野生型TCRは、試験したいずれの濃度でもSIYR/Kbリガンドに結合しなかった(データ示さず)。
【0084】
変異TCR3SQ2はまた、酵母分泌系における可溶性タンパク質として発現され、そして、それが腫瘍細胞の表面上のpMHCに直接結合する能力についてビオチン化後に試験された。図6に示すように、標識化した3SQ2は、適切なペプチドSIYRのみを発現した腫瘍細胞に非常によく結合した。染色は、ペプチドの存在にかかわらず、任意のKb分子に結合する(図6)高親和性抗Kbモノクローナル抗体B8.24.3とほぼ同じ強度であった。
【0085】
(実施例6.同じMHC分子(Kb)に結合した異なるペプチドについて特異的である高親和性TCRの同定)
同じTCR足場が、同じMHCに対して結合した異なるペプチドに対しても、より高い親和性の形態を単離するために用いられ得るか否かを決定するため、本発明者らは、Kbに結合した、dEV8(EQYKFYSV(配列番号6))と呼ばれるペプチドを用いてTCR CDR3αライブラリーをスクリーニングした。ビオチン化dEV8/Kbリガンド(その後、フィコエリトリン−ストレプトビアジン,PE−SA)を用いたフローサイトメトリーによるいくつかのソート後、dEV8/Kbに結合した酵母細胞の有意な富化が存在した(図7においてPEレベルによって示されるように)。
【0086】
dEV8/Kbでの選択によって単離されたクローンのうち6つを配列決定したところ、CDR3配列は全て異なっていた(表3)。これらの配列は類似であったが、SIYR/Kbを用いた選択によって単離されたクローンの配列とは異なっていた(2つの例である、3SQ2および3SQ5をまた、表3に示す)。同じTCRライブラリーの変異体を用いてさえ、異なる抗原に対して、より高い親和性のTCRを単離することが可能であると結論できる。
【0087】
単離されたクローンの抗原特異性を証明するため、dEV8/Kb選択クローンのうちの1つ(4d1)を、異なる抗体およびリガンドのパネルを用いて試験した(図6)。予期された通り、このTCRは、3つの適切な抗体(抗−Vβ8抗体 F23.2,抗HA
タグ 抗体、および抗His タグ抗体)、およびdEV8/Kb抗原とは反応するが、他の抗原、OVA/Kbとは反応しない。野生型TCRは、ペプチドKb複合体のいずれ
とも結合しない(データ示さず)。このように、高い親和性のTCRは、選択された抗原に特異的であった。
【0088】
(実施例7.TCRの異なる領域(β鎖の相補性決定領域3)においてランダムなTCRライブラリーを作成することによる、高親和性TCRの同定)
上記の本実施例は、CDR3と呼ばれるα鎖の領域内で変異したTCRのライブラリーを用いた。TCRの他の領域がまた、より高い親和性のTCRを生成するように変異され得ることを示すため、β鎖のCDR3領域の5連続アミノ酸残基内のランダム変異のライブラリーを、出発物質としてqL2 TCR変異体を用いて、生成した。次いで、このライブラリーを、qL2変異体で検出した濃度以下のQL9/Ldリガンドで選択した。5つの酵母クローン(フローサイトメトリーによって選択した)を配列決定したところ、各々は、異なるヌクレオチドおよびアミノ酸配列を示した(図8)。変異された5つのアミノ酸領域内には配列の顕著な保存が存在しており、このことは、この配列モチーフが高い親和性について最適であったことを示唆していた。本発明者らは、特定のpMHCに対して、より高い親和性を有する誘導体を生じるためにTCRの異なる領域を変異することが可能であると結論付ける。
【0089】
上記の詳細な説明は、多くの特殊性を含み、これらは、本発明の範囲を限定するように解釈されるべきではなく、単に、本発明の現在の好適な実施形態のいくつかの例示を提供するだけである。例えば、特別に例示されたリガンド以外のリガンドが用いられてもよい。従って、本発明の範囲は、示された実施例によってではなく、添付の特許請求の範囲およびその法的な等価物によって決定されるべきである。本明細書に引用される全ての参考文献は、本明細書における開示と矛盾しない程度まで援用される。
【0090】
【表1】
【0091】
【表2】
【0092】
【表3】
【0093】
【表4】
【0094】
(配列表)
【0095】
【0096】
【0097】
【0098】
【0099】
【0100】
【0101】
【0102】
【0103】
(配列表)
SEQUENCE LISTING
<110> BOARD OF TRUSTEES OF THE UNIVERSITY OF ILLINOIS
<120> HIGH AFFINITY TCR PROTEINS AND METHODS
<130> F111502CDU
<150> US 60/169,179
<151> 1999-12-06
<160> 53
<170> PatentIn version 3.0
<210> 1
<211> 4
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 1
Met Cys Met Val
1
<210> 2
<211> 4
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 2
Ser Ile Tyr Arg
1
<210> 3
<211> 24
<212> PRT
<213> ARTIFICAL SEQUENCE
<400> 3
Gly Gly CysAlaGly Cys Cys Cys CysAla Thr Ala Ala Ala Cys Ala
1 5 10 15
Cys Ala Cys AlaGly Thr Ala Thr
20
<210> 4
<211> 74
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 4
Cys Thr ThrThrThr Gly Thr Gly CysCys Gly GlyAla Thr Cys Cys
1 5 10 15
Ala Ala Ala ThrGly Thr CysAla Gly Ser Asn Asn Ser Asn AsnSer
20 25 30
Asn Asn Ser AsnAsn Ser Asn Asn Gly CysThr Cys Ala Cys Ala Gly
35 40 45
Cys Ala Cys AlaGly Ala Ala Gly Thr Ala CysAla Cys Gly Gly Cys
50 55 60
Cys Gly Ala GlyThr Cys GlyCys Thr Cys
65 70
<210> 5
<211> 8
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 5
Ser Ile Tyr ArgTyr Tyr GlyLeu
1 5
<210> 6
<211> 8
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 6
Glu Gln TyrLysPhe Tyr Ser Val
1 5
<210> 7
<211> 7
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 7
Ser Gly Phe Ala Ser Ala Leu
1 5
<210> 8
<211> 7
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 8
Ser Ser Tyr GlyAsn Tyr Leu
1 5
<210> 9
<211> 7
<212> PRT
<213> ARTIFICAL SEQUENCE
<400> 9
Ser Arg Arg GlyHis Ala Leu
1 5
<210> 10
<211> 7
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 10
Ser Ser Arg Gly Thr Ala Leu
1 5
<210> 11
<211> 7
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 11
Ser His Phe GlyThr Arg Leu
1 5
<210> 12
<211> 7
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 12
Ser Met Phe GlyThr Arg Leu
1 5
<210> 13
<211> 7
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 13
Ser His Gln GlyArg Tyr Leu
1 5
<210> 14
<211> 7
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 14
Ser Tyr Leu GlyLeu Arg Leu
1 5
<210> 15
<211> 7
<212> PRT
<213> ARTIFICAL SEQUENCE
<400> 15
Ser Lys His GlyIle His Leu
1 5
<210> 16
<211> 7
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 16
Ser Leu Thr GlyArg Tyr Leu
1 5
<210> 17
<211> 7
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 17
Ser Leu Pro ProPro Leu Leu
1 5
<210> 18
<211> 7
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 18
Ser Ile Pro ThrPro Ser Leu
1 5
<210> 19
<211> 7
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 19
Ser Asn Pro ProPro Leu Leu
1 5
<210> 20
<211> 7
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 20
Ser Asp Pro ProPro Leu Leu
1 5
<210> 21
<211> 7
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 21
Ser Ser Pro ProPro Arg Leu
1 5
<210> 22
<211> 7
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 22
Ser Ala Pro ProPro Ile Leu
1 5
<210> 23
<211> 7
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 23
Ser Gly Thr HisPro Phe Leu
1 5
<210> 24
<211> 7
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 24
Ser Gly His LeuPro Phe Leu
1 5
<210> 25
<211> 7
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 25
Ser Asp Ser LysPro Phe Leu
1 5
<210> 26
<211> 7
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 26
Ser Ser Asp Arg Pro Tyr Leu
1 5
<210> 27
<211> 7
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 27
Ser Leu Glu ArgPro Tyr Leu
1 5
<210> 28
<211> 7
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 28
Ser Arg Glu AlaPro Tyr Leu
1 5
<210> 29
<211> 7
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 29
Ser Leu His ArgPro Ala Leu
1 5
<210> 30
<211> 7
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 30
Ser Leu His ArgPro Ala Leu
1 5
<210> 31
<211> 7
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 31
Ser Ser Asn ArgPro Ala Leu
1 5
<210> 32
<211> 7
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 32
Ser Thr Asp ArgPro Ser Leu
1 5
<210> 33
<211> 7
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 33
Ser Gly Ser ArgPro Thr Leu
1 5
<210> 34
<211> 7
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 34
Ser Leu Val ThrPro Ala Leu
1 5
<210> 35
<211> 7
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 35
Ser Ala Thr SerPro Ala Leu
1 5
<210> 36
<211> 7
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 36
Ser Ser Ile AsnPro Ala Leu
1 5
<210> 37
<211> 7
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 37
Ser Ala Ser TyrPro Ser Leu
1 5
<210> 38
<211> 7
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 38
Ser Arg Trp Thr Ser Gly Leu
1 5
<210> 39
<211> 7
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 39
Ser Gly Ser ArgPro Ala Leu
1 5
<210> 40
<211> 7
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 40
Ser Leu Thr HisHis Phe Leu
1 5
<210> 41
<211> 7
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 41
Ser Met Thr HisHis Phe Leu
1 5
<210> 42
<211> 7
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 42
Ser Leu Ser ArgPro Tyr Leu
1 5
<210> 43
<211> 7
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 43
Ser Leu Thr Arg Pro Tyr Leu
1 5
<210> 44
<211> 7
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 44
Ser Thr Tyr ArgHis Tyr Leu
1 5
<210> 45
<211> 7
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 45
Ser Gly Leu AlaArg Pro Leu
1 5
<210> 46
<211> 7
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 46
Ser Leu His ArgPro Ala Leu
1 5
<210> 47
<211> 7
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 47
Ser Gly Thr HisPro Phe Leu
1 5
<210> 48
<211> 7
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 48
Gly Gly GlyGlyThr Leu Tyr
1 5
<210> 49
<211> 7
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 49
Gly Gly GlyGlyVal Leu Tyr
1 5
<210> 50
<211> 7
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 50
Gly Leu GlyGlyIle Leu Tyr
1 5
<210> 51
<211> 7
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 51
Gly Gln GlyGlyVal Leu Tyr
1 5
<210> 52
<211> 7
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 52
Gly Ser Gly GlyIle Ile Tyr
1 5
<210> 53
<211> 7
<212> PRT
<213> ARTIFICIAL SEQUENCE
<400> 53
Gly Gly GlyGlyIle Leu Tyr
1 5
【特許請求の範囲】
【請求項1】
リガンドに対して高い親和性を有し、該リガンドに対して107M−1より大きい解離定数を示す、可溶性T細胞レセプター(TCR)であって、ここで、該可溶性TCRがCDR中に1つ以上の変異を含む、可溶性TCR。
【請求項2】
前記TCRが前記リガンドに対して107M−1〜1010M−1の解離定数を示す、請求項1に記載の可溶性TCR。
【請求項3】
前記リガンドがペプチド/MHCリガンドである、請求項1に記載の可溶性TCR。
【請求項4】
前記リガンドがスーパー抗原である、請求項1に記載の可溶性TCR。
【請求項5】
前記1つ以上の変異がCDR3αまたはCDR3βにおいて存在する、請求項1に記載の可溶性TCR。
【請求項6】
選択されたリガンドに対して107M−1より大きい解離定数を示す高親和性TCRをT細胞表面上で発現する、T細胞であって、ここで、該高親和性TCRがCDR中に1つ以上の変異を含む、T細胞。
【請求項7】
前記TCRが前記リガンドに対して107M−1〜1010M−1の解離定数を示す、請求項6に記載のT細胞。
【請求項8】
前記リガンドがペプチド/MHCリガンドである、請求項6に記載のT細胞。
【請求項9】
前記リガンドがスーパー抗原である、請求項6に記載のT細胞。
【請求項10】
前記高親和性TCRがCDR3αまたはCDR3βにおいて1つ以上の変異を有する変異体である、請求項6に記載のT細胞。
【請求項11】
標的細胞の自己免疫破壊をブロックする方法であって、該方法が、該標的細胞の表面上のTリンパ球によって認識される部位に対して高い親和性を有する可溶性変異体TCRと、該標的細胞とを接触させ、それにより該標的細胞の自己免疫破壊がブロックされ、ここで該可溶性、高親和性変異体TCRが該標的細胞の表面上のTリンパ球によって認識される部位に対して107M−1より大きい解離定数を示す、工程を包含する、方法。
【請求項12】
前記高親和性変異体TCRが前記該標的細胞の表面上のTリンパ球によって認識される部位に対して107M−1〜1010M−1の解離定数を示す、請求項11に記載の方法。
【請求項13】
前記高親和性TCRがCDRにおいて1つ以上の変異を有する変異体である、請求項11に記載の方法。
【請求項14】
前記高親和性TCRがCDR3αまたはCDR3βにおいて1つ以上の変異を有する変異体である、請求項13に記載の方法。
【請求項15】
所望でない細胞を殺傷するための薬学的組成物であって、該組成物が高親和性TCRを含み、ここで該TCRが該細胞の細胞表面マーカーに対して高い親和性を有し、そして治療化合物と結合されて、治療TCR誘導体を形成し、ここで該高親和性TCRが該所望でない細胞の細胞表面マーカーに対して107M−1より大きい解離定数を示し、ここで、該高親和性TCRがCDR中に1つ以上の変異を含む、組成物。
【請求項16】
前記高親和性TCRが前記所望でない細胞の細胞表面マーカーに対して107M−1〜1010M−1の解離定数を示す、請求項15に記載の組成物。
【請求項17】
前記高親和性TCRがCDR3αまたはCDR3βにおいて1つ以上の変異を有する変異体である、請求項15に記載の組成物。
【請求項18】
高親和性TCRを細胞表面上の選択されたペプチド/MHCリガンドを有する該細胞に結合する方法であって、該方法が、以下:
該選択されたペプチド/MHCリガンドに対して107M−1より大きい解離定数を示す変異体TCRを提供する工程;
該高親和性TCRを標識化する工程;および、
該標識化高親和性TCRを細胞表面上に1つ以上のペプチド/MHCリガンドを有する該細胞を含むサンプルと接触させ、該高親和性TCRを該サンプル中に存在する選択されたペプチド/MHCリガンドに結合する工程、
を包含する、方法。
【請求項19】
前記変異体TCRが前記選択されたペプチド/MHCリガンドに対して107M−1〜1010M−1の解離定数を示す、請求項18に記載の方法。
【請求項20】
前記変異体TCRがCDRにおいて1つ以上の変異を有する、請求項18に記載の方法。
【請求項21】
前記変異体TCRがCDR3αまたはCDR3βにおいて1つ以上の変異を有する、請求項20に記載の方法。
【請求項22】
T細胞の表面上に高親和性TCR変異体の発現を可能にする系に、該高親和性TCR変異体についての遺伝子をクローニングする方法であって、以下:
TCRを変異させて、TCRの同族リガンドに対して少なくとも107M−1の解離定数を示す高親和性TCR変異体を作製する工程;
該TCR変異体をベクターにクローニングする工程;
該ベクターをT細胞にトランスフェクトする工程;および
該T細胞の表面上に該高親和性TCR変異体を発現する工程、
を包含する、方法。
【請求項23】
前記トランスフェクトされたT細胞が選択されたペプチド保有MHC細胞の認識のために使用される、請求項22に記載の方法。
【請求項24】
前記高親和性TCR変異体がCDRにおいて1つ以上の変異を有する、請求項22に記載の方法。
【請求項25】
前記高親和性TCR変異体がCDR3αまたはCDR3βにおいて1つ以上の変異を有する、請求項24に記載の方法。
【請求項26】
請求項22の方法によって作製される、T細胞。
【請求項27】
変異体高親和性TCRの同族リガンドに対して107M−1より大きい解離定数を示す該変異体高親和性TCRをコードする、DNA配列であって、ここで、該変異体高親和性TCRがCDR中に1つ以上の変異を含む、DNA配列。
【請求項28】
前記TCR変異体が該TCR変異体同族リガンドに対して107M−1〜1010M−1の解離定数を示す、請求項27に記載のDNA配列。
【請求項29】
前記TCR変異体がCDR3αまたはCDR3βにおいて1つ以上の変異を有する、請求項27に記載のDNA配列。
【請求項30】
治療化合物に結合された可溶性の高親和性単鎖TCRを含む、治療TCR誘導体であって、ここで、該可溶性の高親和性単鎖TCRが、その同族のリガンドに対して107M−1より大きい解離定数を示し、ここで、該可溶性の高親和性TCRがCDR中に1つ以上の変異を含む、治療TCR誘導体。
【請求項31】
前記治療化合物が抗癌剤、治療放射性核種または細胞傷害性タンパク質である、請求項30に記載の治療TCR誘導体。
【請求項32】
前記可溶性の高親和性TCRが病原体に感染した細胞に特異的に結合する、請求項30に記載の治療TCR誘導体。
【請求項33】
前記治療化合物が、病原体に対して有毒である分子である、請求項32に記載の治療TCR誘導体。
【請求項34】
高親和性TCR変異体を発現するベクターで形質転換されたT細胞であって、ここで該形質転換されたT細胞が野生型T細胞よりもより大きな程度で活性化され得、ここで、該高親和性TCR変異体が選択されたリガンドに対して107M−1より大きい解離定数を示し、ここで、該高親和性TCR変異体がCDR中に1つ以上の変異を含む、T細胞。
【請求項35】
細胞の表面上の特異的ペプチド/MHCリガンドを検出する診断プローブであって、該プローブが該特異的ペプチド/MHCリガンドに結合する検出可能に標識化された高親和性TCRを含み、ここで、該検出可能に標識化された高親和性TCRが特異的ペプチド/MHCリガンドに対して107M−1より大きい解離定数を示し、ここで、該検出可能に標識化された高親和性TCRがCDR中に1つ以上の変異を含む、プローブ。
【請求項36】
病原体を不活化するための高親和性TCRを含む薬学的組成物であって、該高親和性TCRが該病原体に対して有毒である分子に結合され、ここで、該高親和性TCRが該分子に対して107M−1より大きい解離定数を示し、ここで、該高親和性TCRがCDR中に1つ以上の変異を含む、薬学的組成物。
【請求項37】
前記病原体が、以下:
ウイルス、細菌および原生生物
からなる群より選択される、請求項36に記載の組成物。
【請求項38】
薬学的キャリア中に高親和性TCRを含む、薬学的組成物であって、ここで、該高親和性TCRがリガンドに対して107M−1より大きい解離定数を示し、ここで、該高親和性TCRがCDR中に1つ以上の変異を含む、薬学的組成物。
【請求項1】
リガンドに対して高い親和性を有し、該リガンドに対して107M−1より大きい解離定数を示す、可溶性T細胞レセプター(TCR)であって、ここで、該可溶性TCRがCDR中に1つ以上の変異を含む、可溶性TCR。
【請求項2】
前記TCRが前記リガンドに対して107M−1〜1010M−1の解離定数を示す、請求項1に記載の可溶性TCR。
【請求項3】
前記リガンドがペプチド/MHCリガンドである、請求項1に記載の可溶性TCR。
【請求項4】
前記リガンドがスーパー抗原である、請求項1に記載の可溶性TCR。
【請求項5】
前記1つ以上の変異がCDR3αまたはCDR3βにおいて存在する、請求項1に記載の可溶性TCR。
【請求項6】
選択されたリガンドに対して107M−1より大きい解離定数を示す高親和性TCRをT細胞表面上で発現する、T細胞であって、ここで、該高親和性TCRがCDR中に1つ以上の変異を含む、T細胞。
【請求項7】
前記TCRが前記リガンドに対して107M−1〜1010M−1の解離定数を示す、請求項6に記載のT細胞。
【請求項8】
前記リガンドがペプチド/MHCリガンドである、請求項6に記載のT細胞。
【請求項9】
前記リガンドがスーパー抗原である、請求項6に記載のT細胞。
【請求項10】
前記高親和性TCRがCDR3αまたはCDR3βにおいて1つ以上の変異を有する変異体である、請求項6に記載のT細胞。
【請求項11】
標的細胞の自己免疫破壊をブロックする方法であって、該方法が、該標的細胞の表面上のTリンパ球によって認識される部位に対して高い親和性を有する可溶性変異体TCRと、該標的細胞とを接触させ、それにより該標的細胞の自己免疫破壊がブロックされ、ここで該可溶性、高親和性変異体TCRが該標的細胞の表面上のTリンパ球によって認識される部位に対して107M−1より大きい解離定数を示す、工程を包含する、方法。
【請求項12】
前記高親和性変異体TCRが前記該標的細胞の表面上のTリンパ球によって認識される部位に対して107M−1〜1010M−1の解離定数を示す、請求項11に記載の方法。
【請求項13】
前記高親和性TCRがCDRにおいて1つ以上の変異を有する変異体である、請求項11に記載の方法。
【請求項14】
前記高親和性TCRがCDR3αまたはCDR3βにおいて1つ以上の変異を有する変異体である、請求項13に記載の方法。
【請求項15】
所望でない細胞を殺傷するための薬学的組成物であって、該組成物が高親和性TCRを含み、ここで該TCRが該細胞の細胞表面マーカーに対して高い親和性を有し、そして治療化合物と結合されて、治療TCR誘導体を形成し、ここで該高親和性TCRが該所望でない細胞の細胞表面マーカーに対して107M−1より大きい解離定数を示し、ここで、該高親和性TCRがCDR中に1つ以上の変異を含む、組成物。
【請求項16】
前記高親和性TCRが前記所望でない細胞の細胞表面マーカーに対して107M−1〜1010M−1の解離定数を示す、請求項15に記載の組成物。
【請求項17】
前記高親和性TCRがCDR3αまたはCDR3βにおいて1つ以上の変異を有する変異体である、請求項15に記載の組成物。
【請求項18】
高親和性TCRを細胞表面上の選択されたペプチド/MHCリガンドを有する該細胞に結合する方法であって、該方法が、以下:
該選択されたペプチド/MHCリガンドに対して107M−1より大きい解離定数を示す変異体TCRを提供する工程;
該高親和性TCRを標識化する工程;および、
該標識化高親和性TCRを細胞表面上に1つ以上のペプチド/MHCリガンドを有する該細胞を含むサンプルと接触させ、該高親和性TCRを該サンプル中に存在する選択されたペプチド/MHCリガンドに結合する工程、
を包含する、方法。
【請求項19】
前記変異体TCRが前記選択されたペプチド/MHCリガンドに対して107M−1〜1010M−1の解離定数を示す、請求項18に記載の方法。
【請求項20】
前記変異体TCRがCDRにおいて1つ以上の変異を有する、請求項18に記載の方法。
【請求項21】
前記変異体TCRがCDR3αまたはCDR3βにおいて1つ以上の変異を有する、請求項20に記載の方法。
【請求項22】
T細胞の表面上に高親和性TCR変異体の発現を可能にする系に、該高親和性TCR変異体についての遺伝子をクローニングする方法であって、以下:
TCRを変異させて、TCRの同族リガンドに対して少なくとも107M−1の解離定数を示す高親和性TCR変異体を作製する工程;
該TCR変異体をベクターにクローニングする工程;
該ベクターをT細胞にトランスフェクトする工程;および
該T細胞の表面上に該高親和性TCR変異体を発現する工程、
を包含する、方法。
【請求項23】
前記トランスフェクトされたT細胞が選択されたペプチド保有MHC細胞の認識のために使用される、請求項22に記載の方法。
【請求項24】
前記高親和性TCR変異体がCDRにおいて1つ以上の変異を有する、請求項22に記載の方法。
【請求項25】
前記高親和性TCR変異体がCDR3αまたはCDR3βにおいて1つ以上の変異を有する、請求項24に記載の方法。
【請求項26】
請求項22の方法によって作製される、T細胞。
【請求項27】
変異体高親和性TCRの同族リガンドに対して107M−1より大きい解離定数を示す該変異体高親和性TCRをコードする、DNA配列であって、ここで、該変異体高親和性TCRがCDR中に1つ以上の変異を含む、DNA配列。
【請求項28】
前記TCR変異体が該TCR変異体同族リガンドに対して107M−1〜1010M−1の解離定数を示す、請求項27に記載のDNA配列。
【請求項29】
前記TCR変異体がCDR3αまたはCDR3βにおいて1つ以上の変異を有する、請求項27に記載のDNA配列。
【請求項30】
治療化合物に結合された可溶性の高親和性単鎖TCRを含む、治療TCR誘導体であって、ここで、該可溶性の高親和性単鎖TCRが、その同族のリガンドに対して107M−1より大きい解離定数を示し、ここで、該可溶性の高親和性TCRがCDR中に1つ以上の変異を含む、治療TCR誘導体。
【請求項31】
前記治療化合物が抗癌剤、治療放射性核種または細胞傷害性タンパク質である、請求項30に記載の治療TCR誘導体。
【請求項32】
前記可溶性の高親和性TCRが病原体に感染した細胞に特異的に結合する、請求項30に記載の治療TCR誘導体。
【請求項33】
前記治療化合物が、病原体に対して有毒である分子である、請求項32に記載の治療TCR誘導体。
【請求項34】
高親和性TCR変異体を発現するベクターで形質転換されたT細胞であって、ここで該形質転換されたT細胞が野生型T細胞よりもより大きな程度で活性化され得、ここで、該高親和性TCR変異体が選択されたリガンドに対して107M−1より大きい解離定数を示し、ここで、該高親和性TCR変異体がCDR中に1つ以上の変異を含む、T細胞。
【請求項35】
細胞の表面上の特異的ペプチド/MHCリガンドを検出する診断プローブであって、該プローブが該特異的ペプチド/MHCリガンドに結合する検出可能に標識化された高親和性TCRを含み、ここで、該検出可能に標識化された高親和性TCRが特異的ペプチド/MHCリガンドに対して107M−1より大きい解離定数を示し、ここで、該検出可能に標識化された高親和性TCRがCDR中に1つ以上の変異を含む、プローブ。
【請求項36】
病原体を不活化するための高親和性TCRを含む薬学的組成物であって、該高親和性TCRが該病原体に対して有毒である分子に結合され、ここで、該高親和性TCRが該分子に対して107M−1より大きい解離定数を示し、ここで、該高親和性TCRがCDR中に1つ以上の変異を含む、薬学的組成物。
【請求項37】
前記病原体が、以下:
ウイルス、細菌および原生生物
からなる群より選択される、請求項36に記載の組成物。
【請求項38】
薬学的キャリア中に高親和性TCRを含む、薬学的組成物であって、ここで、該高親和性TCRがリガンドに対して107M−1より大きい解離定数を示し、ここで、該高親和性TCRがCDR中に1つ以上の変異を含む、薬学的組成物。
【図1】

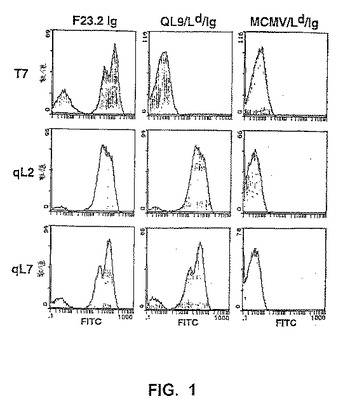
【図2】
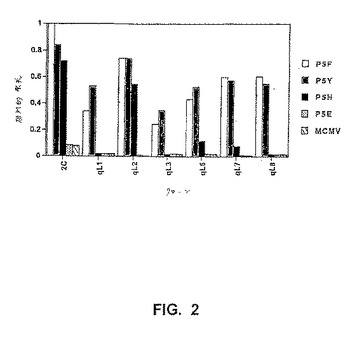
【図3】
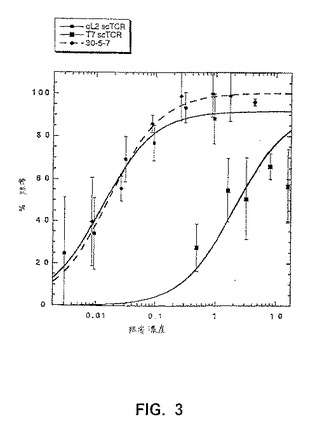
【図4A】
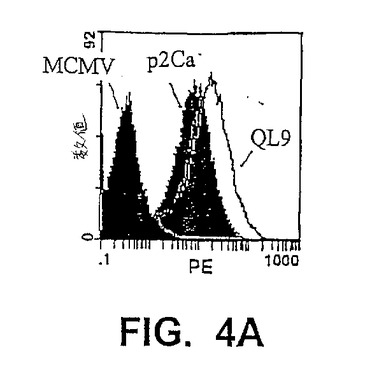
【図4B】
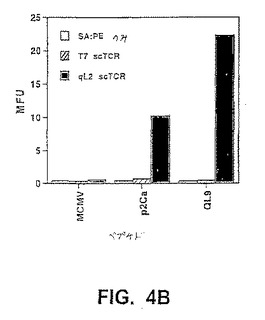
【図4C】
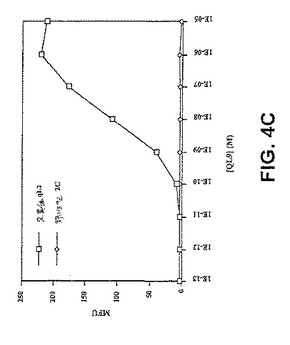
【図5】
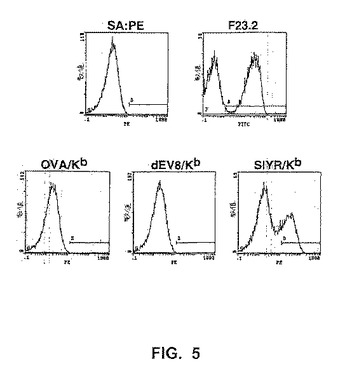
【図6】
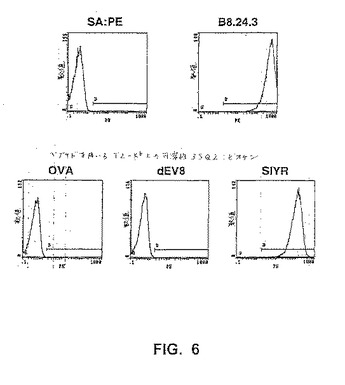
【図7】
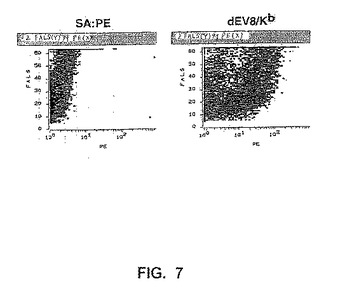
【図8】
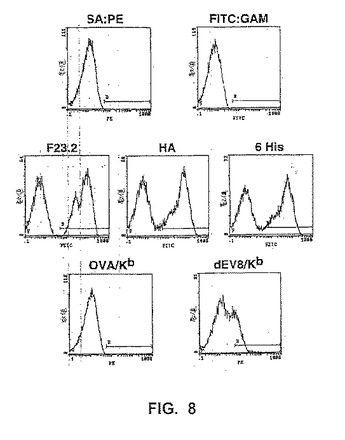
【図9】
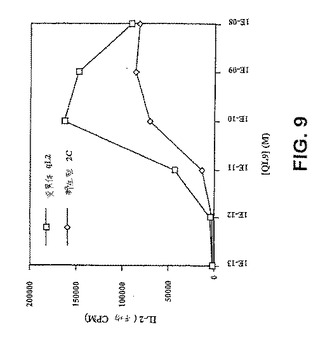
【図2】
【図3】
【図4A】
【図4B】
【図4C】
【図5】
【図6】
【図7】
【図8】
【図9】
【公開番号】特開2012−65672(P2012−65672A)
【公開日】平成24年4月5日(2012.4.5)
【国際特許分類】
【外国語出願】
【出願番号】特願2011−290233(P2011−290233)
【出願日】平成23年12月29日(2011.12.29)
【分割の表示】特願2001−548658(P2001−548658)の分割
【原出願日】平成12年12月6日(2000.12.6)
【出願人】(502201136)ボード オブ トラスティーズ オブ ザ ユニバーシティ オブ イリノイ (2)
【Fターム(参考)】
【公開日】平成24年4月5日(2012.4.5)
【国際特許分類】
【出願番号】特願2011−290233(P2011−290233)
【出願日】平成23年12月29日(2011.12.29)
【分割の表示】特願2001−548658(P2001−548658)の分割
【原出願日】平成12年12月6日(2000.12.6)
【出願人】(502201136)ボード オブ トラスティーズ オブ ザ ユニバーシティ オブ イリノイ (2)
【Fターム(参考)】
[ Back to top ]