
高酸化還元電位型ラッカーゼの生産方法
【課題】
白色腐朽菌の中から液体培養においてラッカーゼ分泌が盛んな種を選抜し、様々なラッカーゼ誘導成分を用いて誘導性の高い成分と成分に対しての応答性に優れた菌株を選抜し酸化還元電位を持つラッカーゼを効率よく生産すること、より具体的には、精製工程を必要としない純度で、大量の高酸化還元電位型ラッカーゼの分泌生産系の確立をすること。
【解決手段】
オツネンタケモドキ、特に野生株オツネンタケモドキIBRC05015株(寄託番号 NITE P-822)から高酸化還元電位型ラッカーゼを分泌生産する方法であって、銅添加培地を使用することを特徴とする。
白色腐朽菌の中から液体培養においてラッカーゼ分泌が盛んな種を選抜し、様々なラッカーゼ誘導成分を用いて誘導性の高い成分と成分に対しての応答性に優れた菌株を選抜し酸化還元電位を持つラッカーゼを効率よく生産すること、より具体的には、精製工程を必要としない純度で、大量の高酸化還元電位型ラッカーゼの分泌生産系の確立をすること。
【解決手段】
オツネンタケモドキ、特に野生株オツネンタケモドキIBRC05015株(寄託番号 NITE P-822)から高酸化還元電位型ラッカーゼを分泌生産する方法であって、銅添加培地を使用することを特徴とする。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、高酸化還元電位型ラッカーゼの分泌生産方法に関し、特に、銅イオンを添加した培地でオツネンタケモドキIBRC05015株を培養することにより、大量かつ高純度のラッカーゼを培地中へ分泌生産させる方法に関する。
【背景技術】
【0002】
ラッカーゼは微生物から植物に至る生物において広く保有されている酵素である。その役割は幅広く、植物では細胞壁のリグニン合成、菌類ではリグニン分解や子実体形成、メラニン合成など多岐に渡る(非特許文献1、2)。
【0003】
ラッカーゼはその反応の多様性から産業的価値が高く、様々な形で利用化研究がなされている。多くのラッカーゼの中でも、担子菌由来のラッカーゼは他の生物に比べ非常に高い酸化還元電位を持つため、基質の酸化能力に優れている。そのため、産業上の利用価値が特に高いと考えられている。
【0004】
製紙工程でのリグニン処理、洗剤における染料の転写の阻止、繊維の脱色、フェノール樹脂の製造及び廃水処理などである。種々のラッカーゼが触媒する反応は類似しているが、酸化能力、すなわち高い酸化還元電位を持つことが、種々の工業利用において必須と考えられている(非特許文献1、2)。すなわち、担子菌由来の高酸化還元電位型ラッカーゼを利用する事で、様々な産業用途への適用が進むと考えられる。
【0005】
現時点で商業生産されている担子菌ラッカーゼは、ホウロクタケ属のラッカーゼに限られるが(特許文献1、2)、ラッカーゼ単一の酵素標品として使用するためには、精製工程等を必要とする。また同属のTrametes vesicolorのラッカーゼの効率的生産にはリグノセルロース性基質を利用するなどの方法が取られるが(特許文献3)、このときマンガンペルオキシダーゼの同時発現が起きるため、ラッカーゼ標品を製造するためには精製工程を必要とすることに変わりない。
【0006】
近年、燃料電池用の電極素材用にラッカーゼを用いる研究も進んでおり(特許文献4)酸化還元電位が高く、高純度なラッカーゼを安価に効率よく生産する方法が求められている。
【先行技術文献】
【特許文献】
【0007】
【特許文献1】特開平8−70861号公報
【特許文献2】特開平9−56378号公報
【特許文献3】特開2008−538914号公報
【特許文献4】特開2007534115号公報
【非特許文献】
【0008】
【非特許文献1】Riva, S. Trends in Biotechnology Vol.24 No.5: 219-226 (2006).
【非特許文献2】Baldrian, P. FEMS Microbiol. Rev. Vol.30:215-242 (2006).
【非特許文献3】Ryu SH, Lee AY, Kim M.(2008) Molecular characteristics of two laccase from the basidiomycete fungus Polyporus brumalis. J Microbiol. 46:62-9. 非特許文献3については後で述べる。
【発明の概要】
【発明が解決しようとする課題】
【0009】
本発明は、酸化還元電位が高いことが知られている白色腐朽菌の中から液体培養においてラッカーゼ分泌が盛んな種を選抜し、さらに同菌種から生産性の高い株を選抜すること、様々なラッカーゼ誘導成分を用いて誘導性の高い成分と成分に対しての応答性に優れた菌株を選抜し酸化還元電位を持つラッカーゼを効率よく生産することを課題とする。より具体的には、精製工程を必要としない純度で、大量の高酸化還元電位型ラッカーゼの分泌生産系の確立を課題とする。
【課題を解決するための手段】
【0010】
本発明者らは上記課題を解決すべく鋭意研究した結果、オツネンタケモドキIBRC05015株を用いてラッカーゼの高純度、高分泌生産系の確立に成功した。さらに生産されたラッカーゼが、白色腐朽菌ラッカーゼの中でも、最も高いレベルの酸化還元電位を持ち、様々な基質を酸化可能であることを確認した。
【0011】
すなわち本発明は、寄託したIBRC05015株を用いた液体培養によって提供されるラッカーゼの高効率生産法であって、以下の技術を基礎とする。
【0012】
オツネンタケモドキIBRC05015株(寄託番号 NITE P-822)から高酸化還元電位型ラッカーゼを分泌生産する方法であって、銅添加培地を使用することを特徴とする、大量かつ高純度の高酸化還元電位型ラッカーゼを分泌生産する方法。
【発明の効果】
【0013】
本発明の方法によれば、酸素を基質として様々なフェノール系物質を酸化可能である、産業上有用な酵素、ラッカーゼのうち、担子菌抗酸化還元電位型酵素であるラッカーゼを、簡便かつ大量、高純度で分泌生産することができる。
【図面の簡単な説明】
【0014】
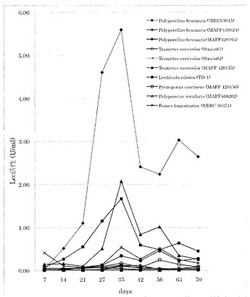
【図1】図1は、各菌糸培養液中のラッカーゼ活性(Polyporellus brumalis; IBRC05015)を示した図である。
【図2】図2は、本発明の酵素の複数の基質に対する反応性を示した図である。
【図3】図3は、IBRC05015株と基準購入株とのCu2+添加によるラッカーゼ活性誘導の比較を示した図である。
【図4】図4は、IBRC05015株でラッカーゼ活性誘導に最適なCu2+添加濃度を示した図である。
【図5】図5は、Cu2+添加により誘導される培地中のタンパク質成分の変化を示した図である。
【図6】図6Aは、Cu2+添加の有無とラッカーゼ活性変動の経時比較で、図6BはCu2+添加によるpblac1遺伝子のmRNA量の変化を示した図である。
【発明を実施するための形態】
【0015】
本発明による高酸化還元電位型ラッカーゼを分泌生産する方法は、オツネンタケモドキIBRC05015株と銅添加培地を使用することを特徴とする。
【0016】
IBRC01015株の培養液は通常の担子菌培養に用いられる培地であればかまわないが、ここに0.1-0.5mM程度の銅イオンを添加することにより、分泌されるタンパク質のほとんどをラッカーゼとし、かつラッカーゼの生産量を飛躍的に引き上げることが可能である。
【0017】
本発明において生産されるラッカーゼのアミノ酸配列は配列番号1に示されるようにPolyporellus brumalis(学名:ポリポレルスブルマリス、日本名:オツネンタケモドキ)の分泌型ラッカーゼであり、このことは分泌タンパク質のN末端アミノ酸配列のシークエンスと、銅イオン添加によって発現誘導が確認されるラッカーゼ遺伝子の配列からも確認できる。
【0018】
以下、実施例に基づいて本発明を詳細に説明する。なお、本発明は実施例に限定されるものではない。
【実施例】
【0019】
多数の担子菌株を収集し、液体培養に供した時にラッカーゼ生産量(培地中へ分泌されるラッカーゼ活性)が多いものを選抜した。用いた株は独立行政法人農業生物資源ジーンバンクより入手した基準培養菌株Polyporellus(Polyporus) brumalis (オツネンタケモドキ) MAFF;420024、MAFF420194、Polyporus arcularis (アミスギタケ) MAFF 460202、Polyporus squamosus (アミヒラタケ) MAFF 460203、 Polyporellus badius(アシグロタケ) MAFF 420217、 Polyporellus varius (キアシグロタケ) MAFF 420751、Trametes versicolor (カワラタケ) MAFF 420155、 Polyporellus tuberaster (タマチョレイタケ) MAFF 420261、Ganoderma lucidum (霊芝) MAFF 420210、Ganoderma tsugae MAFF 420118、Pycnoporus coccineus (ヒイロタケ) MAFF 420150、Pycnoporus cinnabarinus (シュタケ) MAFF 420151及び独立行政法人製品評価基盤機構 生物遺伝資源部門(NBRC)(H14年財団法人発酵研究所(IFO)から委託業務移管)より入手した基準培養株Polyporellus(Polyporus) brumalis (オツネンタケモドキ)IFO;30741 Formes fomentarius (ツリガネタケ)IFO;30371を含む。
【0020】
北東北の山中から採取した野生きのこ[シイタケ、キアシグロタケ、オツネンタケモドキ、カワラタケ、ニクウチワタケ、ナメコ、クリタケ、ツキヨタケ、ツガサルノコシカケ、コフキサルノコシカケ (推定名称) を含む。]から菌糸塊を0.25×MYPG寒天培地(0.25% Malt extract、0.1% Yrast extract、0.1% Pepton、0.5% Glucose、2% Agar)に植菌し、生育を確認した、25℃で5〜10日生育させた後、スクリーニングのため上記培地へ植え継ぎを繰りかえして単離したTD-1(シイタケ野生種)、キアシグロタケ、オツネンタケモドキ、カワラタケ、ニクウチワタケ、ナメコ、クリタケ、ツキヨタケ、ツガサルノコシカケ、コフキサルノコシカケ (推定名称) を含む。菌糸単離後は種菌プレートから成長点の約5mm内側を内径10mmのコルクボーラーでくり抜き、これを0.5×MYPG培地 (0.5% Malt extract、0.2% Yeast extract、0.2% pepton、1% Glucose) 100ml入り300ml三角フラスコへ5個づつ植菌し、25℃で4週間振とう培養を行った。
【0021】
ラッカーゼ活性測定には基質には2,2’-Azino-bis (3-etylbenzothiazoline-6-sulfonic acid) diammonium salt [ABTS; SIGMAモル吸光係数ε; 36,000 (M-1cm-1)]を基質として用いた。酵素液含むpH4.0のMcIlvaine Buffer100μl中で、30℃で10分間プレインキュベート後、終濃度1mMのABTSを加え反応を開始した。30℃10分間反応させた後、反応液と等量の5%TCAを加え反応を停止し、420nmの吸光度で測定した。1Uは、1分間に1μmolのABTSを触媒する酵素量と定義した。
【0022】
ラッカーゼ活性の算出は、以下に示す式に従った。
[ラッカーゼ活性算出式]:
U/ml= 吸光度(OD420nm)×1000(μmol/ml) ×0.2(ml) ×1000(U/ml)
36000(M-1cm-1)×10(min) ×反応酵素液量(μl)
【0023】
粗酵素液のタンパク質の定量には Advanced protein assay reagent (5×concentarate Cytoskeleton社, USA) を用いたBradford法により検出し、標準タンパク質として牛血清アルブミン (BSA) を使用して、595nmの吸光度を測定した。
【0024】
各菌糸培養液中のラッカーゼ活性を図1に示す。
図に示した結果から、最も高いラッカーゼ生産量を示し、タンパク質分泌量、単位タンパク質当たりのラッカーゼ比活性も高い結果を示した。ITS領域の配列より本株をPolyporellus brumalis (オツネンタケモドキ)と同定した。寄託センターより入手したオツネンタケモドキ株では、野生株ほどの高いラッカーゼ生産が見られなかった。
【0025】
以下、オツネンタケモドキ野生株をIBRC05015として、本株が生産するラッカーゼについて詳細に解析を行った。
【0026】
<オツネンタケモドキIBRC05015からの酵素精製>
オツネンタケモドキIBRC05015は1.5%寒天を含む0.25×MYPG培地で9cmシャーレを用いて1週間25℃で培養し、液体培養用の種菌とした。シャーレ一枚分の表面菌糸を掻きとり、0.5×MYPG液体培地に接種した。液体培養は、25℃で振とう培養で、24日間培養を行い、培養ろ液からラッカーゼを精製した。培養ろ液に80%飽和の硫酸アンモニウムを加えてタンパク質を塩析し、沈殿を10mMリン酸バッファー(pH7.0) に溶解した。得られたタンパク質溶液に30%飽和の硫酸アンモニウムを加え、上清を疎水クロマトグラフィー用のタンパク質サンプルとし、HiLoad Phenylカラム(GE healthcare)に供した。
【0027】
HiLoad Phenylカラムに吸着したタンパク質は10 mMリン酸バッファー(pH7.0)と30%飽和硫酸アンモニウムを含む同バッファーによる直線的濃度勾配(30~0%)による溶出を行った。ラッカーゼ活性のあるフラクションを集めてAmicon Ultra-4 (5000MWCO、Millipore社) にて限外濾過を行い25mlに濃縮した。濃縮したタンパク質サンプルはゲルろ過カラムPD-10 (GE helthcare) に供した後、陰イオン交換カラム(MonoQ 5/50 GL、GE Healthcare社)に吸着させ、10 mMリン酸バッファー(pH7.0)と0.5M NaClを含む同バッファーによる直線的濃度勾配(0-0.5M)による溶出を行った。流速については、0.5ml/minとし、各フラクションを0.5mlとした。ラッカーゼ活性を有するフラクションをAmicon Ultra-4 (Millipore 5000MWCO) 限外濾過により濃縮し、ゲル濾過を行った。ゲルろ過はSuperdex 75 10/30カラム(Millipore社)を用い、10mMリン酸バッファー(pH 6.5)、流速0.25ml/minで行った。64.3kDaの精製ラッカーゼが得られこれをpbLac1とした。
以下精製ラッカーゼ(PbLac1)を用いて諸性質の解析を行った。
【0028】
<活性等測定法>
1) 酵素の活性に及ぼすpHと温度の影響
緩衝液はKCl-HCl buffer(pH1.0)、Glycine-HCl buffer(pH2.0〜3.0)、McIlvaine buffer(pH4.0〜6.0)、Tris-HCl buffer(pH7.0〜9.0)、CAPS buffer (pH10.0、CAPS; DOJINDO) を使用した。基質はABTSを用い、至適pHと至適温度の測定時間は10分間とした。
結果、pbLac1の至適pHはABTSを基質とした場合pH4.0であり至適温度は40℃であった。
【0029】
2) 酵素の安定性に及ぼすpHと温度の影響
実験に用いた緩衝液は5 1)に示した。本実験では終濃度50μg/mlの牛胎児血清アルブミンを加えた緩衝液を使用し、基質はABTSを用いた。pH安定性試験は各pHにて30℃、12時間インキュベートを行なった。温度安定性試験ではpH 6.0で各温度での加温を行い、30分後に残存活性を測定した。
結果、pH3.0-6.0の範囲で40%以上のラッカーゼ活性を保持し安定であった。さらにpH6.0 のMcIlvaine緩衝液中では30分の加温処理で60℃まで安定であった。
【0030】
3) 阻害剤の影響
これまでラッカーゼの阻害剤として報告のある金属キレーター剤を中心に(EDTA、p-Cumaric acid 、Kojic acid、Tropolon、L-Cysteine、 NaN3、DTT)阻害剤の影響について検討を行なった。反応条件はMcIlvaine buffer (pH4.0)で、ABTSを基質として用い、30℃で10分間反応を行なった。阻害剤無添加時の活性に対する阻害剤添加時のラッカーゼ活性の相対値(%)を算出した。
結果、1mMのL-Cysteine、NaN3、DTTによってpbLac1の活性はほぼ完全に阻害された。また、金属イオンのキレーターであるEDTAは10mMでラッカーゼ活性を50%阻害し、p-cumaric acid、Kojic acidはどちらも1mMで約70%ラッカーゼ活性を阻害した。
【0031】
4) 金属イオンの影響
金属イオン(Ca2+、Cd2+、Co2+、Cu2+、K+、Mg2+、Mn2+、Na2+、Sn2+、Zn2+)無添加時の活性に対する金属イオン添加時のラッカーゼ活性の相対値(%)を算出した。反応条件は阻害剤実験と同様の条件で行なった。
結果、10mMのCu2+を添加した時に1.38倍の活性上昇が見られた。しかし、その他の金属イオンに関しては10mMの各金属イオンを添加すると70%程度に活性が低下し、Cd2+1mMで活性が48.8%まで低下した。一番顕著に阻害が見られたのは1mMのSn2+を添加した時で、ラッカーゼ活性が4.6%にまで低下した。
【0032】
5)基質特異性の検討
本酵素の基質特異性を比較するために表1(Table.1)に示したフェノール化合物を基質としてMcIlvaine緩衝液中で10分間反応させ、反応後の吸光度を測定し、1分間当たりに酸化される基質量を算出し、Lineweaver-Burk plot (Lineweaver and Burk 1934)を作成した。各基質で測定した吸光度の波長とモル吸光度係数(ε)を、表1 に示した。
【0033】
【表1】

【0034】
解析の結果、PbLac1は分泌型ラッカーゼには珍しくDOPAを基質にし、幅広い基質特異性を示した。
【0035】
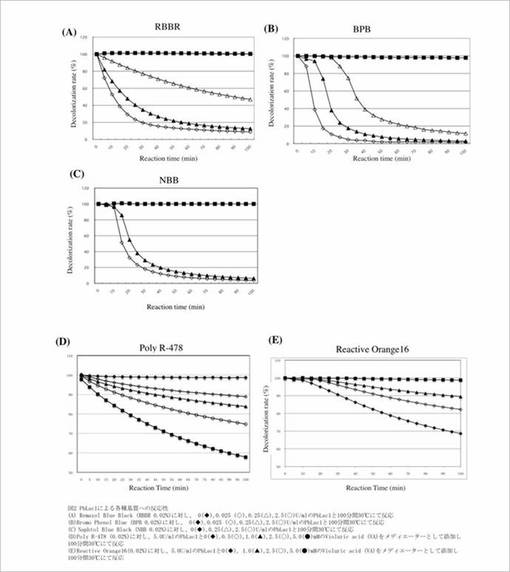
またメディエーターの非存在下においてアントラキノン系色素であるRemazol Blue Black (RBBR)、ジアゾ色素であるNaphtol Blue Black (NBB)の脱色が可能であり、メディエーターの存在で高分子色素Poly R-478、モノアゾ色素 Reactive Orange 16も脱色できたことから、難分解性環境汚染物質の分解に有効である可能性を示唆した(図2)。
【0036】
<pblac1のcDNAおよびゲノムDNAのクローニング>
オツネンタケモドキよりRNAを精製し、オツネンタケモドキラッカーゼのcDNA,ゲノムDNAの配列を以下の手順により決定した。
【0037】
[方法]
得られたタンパク質のN-末端のアミノ酸配列をもとにプライマー(LccO-3;ATHWSNTGYGAYCCNAA, LccO-4L;SNTGYGAYCCNAAYCA) を設計し、3’RACEを行った。RACE用のRNAは酵素精製用に培養した菌糸より抽出し、抽出はMasterPureTM Yeast RNA抽出キット(EPICENTRERBiotechnologies社、USA) を用いて行った。また、3' RACE用のテンプレートcDNAは、SMART RACEキット(BD Biosciences)を用いて行った。また、得られた配列をもとに5’RACE を行った5’RACE用のcDNAはGeneRACERキット(Invitrogen) を用いて合成した。cDNAの合成は各キットのマニュアルに従った。また、PCRの条件は98℃ for 30sec,55℃ for 30sec,and 72℃ for 3min,(30cycles)で行った。
【0038】
各遺伝子の3’及び5’RACEを行う際に用いたプライマーを下記に示す。
LccO-375U-RACE CGCGCTCCCGTCCAACTCGTCCATC for RACE-PCR
LccO-243U-RACE GGCGTTCAACTTCAATGGCTCAGAC for RACE-PCR
LccO-149L-RACE AGGTCAGTCTCCACAAGCAGGTTCG for RACE-PCR
LccO-264L-RACE GGAGGCGCCGTTGATGAAAAAGTCT for RACE-PCR
【0039】
[結果]
得られた配列は非特許文献3に記載のあるオツネンタケモドキのラッカーゼと同一のものであった(配列番号1)。
【0040】
この配列をもとに、アミノ酸配列を推定し、精製ラッカーゼの配列と比較を行ったところ同一のものであることが確認できた(配列番号2)。
【0041】
<誘導剤がオツネンタケモドキIBRC05015株のラッカーゼ活性へ与える影響>
[方法]
菌糸は種菌プレートから成長点の約5mm内側を内径10mmのコルクボーラーでくり抜き、これを7個ずつ0.5×MYPG培地 (0.5% Malt extract、0.2% Yeast extract、0.2% pepton、1% Glucose) 50ml (容量200ml用の三角フラスコ) に植菌した。誘導剤はTable.4に示した濃度で植菌時に液体培地へ添加し、25℃で4週間振とう培養を行い、7日毎にラッカーゼ活性の測定を行った。菌糸は種菌プレートから成長点の約5mm内側を内径10mmのコルクボーラーでくり抜き、これを5個ずつ0.5×MYPG培地 (0.5% Malt extract、0.2% Yeast extract、0.2% pepton、1% Glucose) 50ml (容量200ml用の三角フラスコ) に入れた。
【0042】
[結果]
ラッカーゼの誘導剤として表2に示す誘導剤を用いて比較を行った。そのうち最もラッカーゼの誘導能が高かったのは0.25mMのCu2+を培地へ添加した条件下で、28日目でラッカーゼ活性が34.64U/mlに達した。この値はCu2+無添加時の最大値の21倍に相当し、試したInducerの中では最も誘導率が高かった。また、Cu2+を含む金属イオン添加時には乾燥菌糸体重量が増加する傾向が見られた。
【0043】
【表2】
【0044】
<IBRC05015株と基準購入株のCu2+誘導によるラッカーゼ活性変動の比較>
オツネンタケモドキIBRC05015株はCu2+添加でラッカーゼ活性の上昇が見られたことから、基準購入株とラッカーゼ活性誘導の比較を行った。
[方法]
植菌方法及び、培養法、活性測定法は誘導剤の検討と同様の条件で行った。Cu2+誘導は植菌時に0.5mMのCuSO4を添加した。
【0045】
[結果]
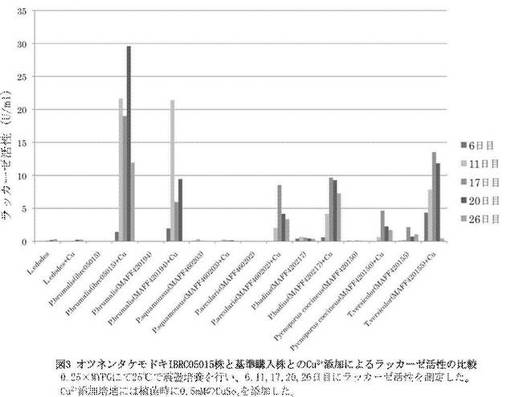
オツネンタケモドキIBRC05015株は基準購入株P. brumalis (MAFF420194)と比較してもCu2+に対する応答性が高く、培養11日目以降常時高いラッカーゼ活性を示すことが分かった。またP. arcularis (MAFF 460202)、P. badius (MAFF 420217)、P. coccineus (MAFF 420150) も同様にCu2+に対するラッカーゼ活性の上昇が見られたが、オツネンタケモドキには及ばなかった。また、T. versicolor (MAFF 420155)と比較してもオツネンタケモドキIBRC05015株のラッカーゼ誘導率は高かった(図3)。
【0046】
<Cu2+濃度がオツネンタケモドキIBRC05015株ラッカーゼ活性へ与える影響>
[方法]
菌糸は種菌プレートから成長点の約5mm内側を内径10mmのコルクボーラーでくり抜き、これを5個ずつ0.5×MYPG培地 (0.5% Malt extract、0.2% Yeast extract、0.2% pepton、1% Glucose) 50ml (容量200ml用の三角フラスコ) に入れた。CuSO4は植菌時に終濃度0、0.0005、0.005、0.05、0.1、0.25、0.5、1.0、2.5、5.0mMの濃度で0.5×MYPG液体培地中へ添加し、7日毎にラッカーゼ活性測定を行った。
【0047】
[結果]
Cu2+によってラッカーゼ活性が上昇することから、最適Cu2+添加濃度について検討を行った。0、0.005、0.05、0.50、0.1、0.25、0.5、1.0、2.5、5.0mMのCu2+を植菌時に添加し、ラッカーゼ活性を測定した。
【0048】
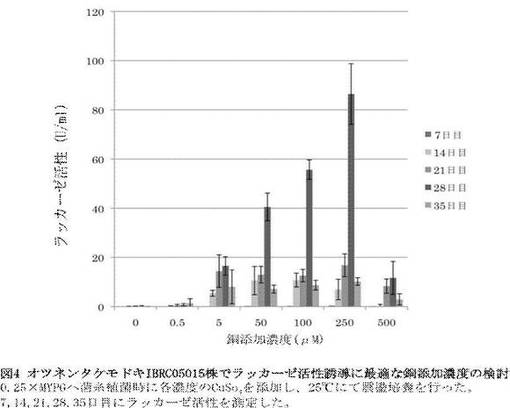
結果0.25mMのCu2+添加時に28日目で最も高いラッカーゼ活性を示すことが分かった(86.4U/ml)、次いで0.1mMのCu2+添加時(55.7U/ml)、0.5mM Cu2+添加時(40.5μM)、 0.005mM Cu2+添加時(16.6U/ml)の順でラッカーゼ活性が高い傾向が見られ、0.5mMでは急激にラッカーゼ活性の低下が見られることが分かった(3.0U/ml)。このことから、最適なCu2+添加濃度は0.25mMであることが示された(図4)。
【0049】
<Cu2+添加時に誘導されるラッカーゼについて>
Cu2+添加時にLac活性の急激な上昇が見られるが、Cu2+添加により誘導される蛋白質の有無を調べた。Cu2+添加時のオツネンタケモドキ培養ろ液を電気泳動し、CBB染色及びラッカーゼ活性染色を行った (図5)。
【0050】
[方法]
粗酵素液に、等量のサンプルバッファー (トリスSDSβMERサンプル処理液; 第一化学薬品) を添加し、5分間ボイルした。これを泳動ゲル [パジェルNPU-10L; ATTO (株)]にアプライし、1×Tris-Glycine-SDS Buffer (BioRad社) を用いて定電流20mAで泳動を行った。CBB染色は常法に従い、Commassie Brilliant Blue R250 (CBB) (PAGE Blue83;第一化学) で行った。
【0051】
また分子量マーカーにはプレシジョンplusスタンダード (BioRad) を用いた。活性染色のNative-PAGEは非還元条件下で行った。ラッカーゼ活性のユニット(U)を等しく調整した各粗酵素液に、等量のサンプルバッファー(Integrated Separation Systems;第一化学薬品)を添加して、泳動ゲル(パジェルNPU-10L) (ATTO (株))にアプライした。泳動は1×Tris-Glycine Buffer (BioRad)を用いて定電流20mAにて泳動を行った。
【0052】
泳動後、pH4.0のMcIlvaine 緩衝液にてpHを平衡化した後、終濃度1mMのABTSを含むMcIlvaine 緩衝液(pH4.0)と30℃で反応させた。N末端側アミノ酸配列決定は、粗精製酵素はSDS-PAGE後PVDF膜にブロットし、クマシーブリリアントブルーR250で染色した。目的バンド付近を切り取り、50%メタノールで脱色し、蒸留水で2時間洗浄して酢酸を除去し風乾したPVDF膜をサンプルとした。N末端アミノ酸配列の解析は (株)ニッピバイオマトリックス研究所に依頼した。なお、同社ではプロテインシークエンサーにABI procise 491 HT (ABI) を使用している。
【0053】
[結果]
Cu2+添加時にラッカーゼ活性を示す、70kDa付近の誘導バンドが存在することが明らかとなった。本バンドは全分泌蛋白質に占める割合が高く、Cu2+添加時には特異的に誘導されるラッカーゼであることが考えられた。そこで、本蛋白質の精製を試み、最終的に単一バンドになるまで精製を行った。精製されたラッカーゼはCu非添加時のラッカーゼと比較すると若干分子量が大きかったが、N末端配列を決定したところ、PbLac1(Rye et al. 2008) と完全に一致することが分かり、Cu2+添加によって誘導されているラッカーゼはPbLac1であることが明らかとなった(図5)。
【0054】
<Cu2+添加時のpblac1の誘導>
[方法]
Cu2+添加時にpblac1が誘導されている可能性が示唆されたため、リアルタイムPCRを用いてCu2+添加によるpblac1遺伝子のmRNA量の変化を調べた(図7)。Cu2+は植菌時に添加した。Cu2+添加から4日目〜35日目にかけて顕著なラッカーゼ活性上昇が見られたため(図6A)、その時期の菌糸を回収し、リアルタイムPCRを行った(図6B)。
【0055】
結果、10日目のCu2+添加画分でpblac1のmRNAレベルの上昇が見られ、14日目にmRNAレベルがピークに達し、16日には若干の減少が見られたが、Cu2+非添加と比較すると已然としてpblac1のmRNAレベルは高かった。
【0056】
<酸化還元電位測定>
ラッカーゼの酸化能力の指標の一つである酸化還元電位を以下の方法で測定した。
PbLac1の酸化還元電位測定にはサイクリックボルタモグラム法と無隔膜バルグ全電解法を用いた。ボルタモグラム法での測定で、大まかな電位の概算を行い、無隔膜バルグ法でより正確な電位の数値を計算した。サイクリックボルタモグラム法で最もクリアな結果が出たのはパージを行ったアルゴン飽和条件で、以前からの報告(Kamitaka et al. 2007)にあるように、Multicopper oxidaseのDETタイプの生物電気的反応を示し、触媒波はtype1Cu部位のE0辺りから上昇してきた。
【0057】
その結果、PbLac1のE0値は797 ± 10mV(標準水素電極: NHE)、無隔膜バルグ全電解法では4回の測定の後、平均して811mV ± 5mV vs 標準水素電極: NHE)だった(表3)。この値はサイクリックボルタモグラム法で測定した電位とほぼ同じ値を示し、より正確な数値が得られた。この値はTrametes hirsute (ThL; 0.78V)やTrametes versicolor (TvL; 0.78V)のようなTypeI Cu部位の酸化還元電位が高いラッカーゼと比較しても、同等かそれ以上であった。
【0058】
【表3】
【産業上の利用可能性】
【0059】
本発明によるラッカーゼは、非常に高い酸化還元電位を持つため、基質の酸化能力に優れており、産業上の利用価値が高い。
製紙工程でのリグニン処理、洗剤における染料の転写の阻止、繊維の脱色、フェノール樹脂の製造及び廃水処理など、また、燃料電池用の電極素材への適用など、種々の工業利用にきわめて有用である。
【技術分野】
【0001】
本発明は、高酸化還元電位型ラッカーゼの分泌生産方法に関し、特に、銅イオンを添加した培地でオツネンタケモドキIBRC05015株を培養することにより、大量かつ高純度のラッカーゼを培地中へ分泌生産させる方法に関する。
【背景技術】
【0002】
ラッカーゼは微生物から植物に至る生物において広く保有されている酵素である。その役割は幅広く、植物では細胞壁のリグニン合成、菌類ではリグニン分解や子実体形成、メラニン合成など多岐に渡る(非特許文献1、2)。
【0003】
ラッカーゼはその反応の多様性から産業的価値が高く、様々な形で利用化研究がなされている。多くのラッカーゼの中でも、担子菌由来のラッカーゼは他の生物に比べ非常に高い酸化還元電位を持つため、基質の酸化能力に優れている。そのため、産業上の利用価値が特に高いと考えられている。
【0004】
製紙工程でのリグニン処理、洗剤における染料の転写の阻止、繊維の脱色、フェノール樹脂の製造及び廃水処理などである。種々のラッカーゼが触媒する反応は類似しているが、酸化能力、すなわち高い酸化還元電位を持つことが、種々の工業利用において必須と考えられている(非特許文献1、2)。すなわち、担子菌由来の高酸化還元電位型ラッカーゼを利用する事で、様々な産業用途への適用が進むと考えられる。
【0005】
現時点で商業生産されている担子菌ラッカーゼは、ホウロクタケ属のラッカーゼに限られるが(特許文献1、2)、ラッカーゼ単一の酵素標品として使用するためには、精製工程等を必要とする。また同属のTrametes vesicolorのラッカーゼの効率的生産にはリグノセルロース性基質を利用するなどの方法が取られるが(特許文献3)、このときマンガンペルオキシダーゼの同時発現が起きるため、ラッカーゼ標品を製造するためには精製工程を必要とすることに変わりない。
【0006】
近年、燃料電池用の電極素材用にラッカーゼを用いる研究も進んでおり(特許文献4)酸化還元電位が高く、高純度なラッカーゼを安価に効率よく生産する方法が求められている。
【先行技術文献】
【特許文献】
【0007】
【特許文献1】特開平8−70861号公報
【特許文献2】特開平9−56378号公報
【特許文献3】特開2008−538914号公報
【特許文献4】特開2007534115号公報
【非特許文献】
【0008】
【非特許文献1】Riva, S. Trends in Biotechnology Vol.24 No.5: 219-226 (2006).
【非特許文献2】Baldrian, P. FEMS Microbiol. Rev. Vol.30:215-242 (2006).
【非特許文献3】Ryu SH, Lee AY, Kim M.(2008) Molecular characteristics of two laccase from the basidiomycete fungus Polyporus brumalis. J Microbiol. 46:62-9. 非特許文献3については後で述べる。
【発明の概要】
【発明が解決しようとする課題】
【0009】
本発明は、酸化還元電位が高いことが知られている白色腐朽菌の中から液体培養においてラッカーゼ分泌が盛んな種を選抜し、さらに同菌種から生産性の高い株を選抜すること、様々なラッカーゼ誘導成分を用いて誘導性の高い成分と成分に対しての応答性に優れた菌株を選抜し酸化還元電位を持つラッカーゼを効率よく生産することを課題とする。より具体的には、精製工程を必要としない純度で、大量の高酸化還元電位型ラッカーゼの分泌生産系の確立を課題とする。
【課題を解決するための手段】
【0010】
本発明者らは上記課題を解決すべく鋭意研究した結果、オツネンタケモドキIBRC05015株を用いてラッカーゼの高純度、高分泌生産系の確立に成功した。さらに生産されたラッカーゼが、白色腐朽菌ラッカーゼの中でも、最も高いレベルの酸化還元電位を持ち、様々な基質を酸化可能であることを確認した。
【0011】
すなわち本発明は、寄託したIBRC05015株を用いた液体培養によって提供されるラッカーゼの高効率生産法であって、以下の技術を基礎とする。
【0012】
オツネンタケモドキIBRC05015株(寄託番号 NITE P-822)から高酸化還元電位型ラッカーゼを分泌生産する方法であって、銅添加培地を使用することを特徴とする、大量かつ高純度の高酸化還元電位型ラッカーゼを分泌生産する方法。
【発明の効果】
【0013】
本発明の方法によれば、酸素を基質として様々なフェノール系物質を酸化可能である、産業上有用な酵素、ラッカーゼのうち、担子菌抗酸化還元電位型酵素であるラッカーゼを、簡便かつ大量、高純度で分泌生産することができる。
【図面の簡単な説明】
【0014】
【図1】図1は、各菌糸培養液中のラッカーゼ活性(Polyporellus brumalis; IBRC05015)を示した図である。
【図2】図2は、本発明の酵素の複数の基質に対する反応性を示した図である。
【図3】図3は、IBRC05015株と基準購入株とのCu2+添加によるラッカーゼ活性誘導の比較を示した図である。
【図4】図4は、IBRC05015株でラッカーゼ活性誘導に最適なCu2+添加濃度を示した図である。
【図5】図5は、Cu2+添加により誘導される培地中のタンパク質成分の変化を示した図である。
【図6】図6Aは、Cu2+添加の有無とラッカーゼ活性変動の経時比較で、図6BはCu2+添加によるpblac1遺伝子のmRNA量の変化を示した図である。
【発明を実施するための形態】
【0015】
本発明による高酸化還元電位型ラッカーゼを分泌生産する方法は、オツネンタケモドキIBRC05015株と銅添加培地を使用することを特徴とする。
【0016】
IBRC01015株の培養液は通常の担子菌培養に用いられる培地であればかまわないが、ここに0.1-0.5mM程度の銅イオンを添加することにより、分泌されるタンパク質のほとんどをラッカーゼとし、かつラッカーゼの生産量を飛躍的に引き上げることが可能である。
【0017】
本発明において生産されるラッカーゼのアミノ酸配列は配列番号1に示されるようにPolyporellus brumalis(学名:ポリポレルスブルマリス、日本名:オツネンタケモドキ)の分泌型ラッカーゼであり、このことは分泌タンパク質のN末端アミノ酸配列のシークエンスと、銅イオン添加によって発現誘導が確認されるラッカーゼ遺伝子の配列からも確認できる。
【0018】
以下、実施例に基づいて本発明を詳細に説明する。なお、本発明は実施例に限定されるものではない。
【実施例】
【0019】
多数の担子菌株を収集し、液体培養に供した時にラッカーゼ生産量(培地中へ分泌されるラッカーゼ活性)が多いものを選抜した。用いた株は独立行政法人農業生物資源ジーンバンクより入手した基準培養菌株Polyporellus(Polyporus) brumalis (オツネンタケモドキ) MAFF;420024、MAFF420194、Polyporus arcularis (アミスギタケ) MAFF 460202、Polyporus squamosus (アミヒラタケ) MAFF 460203、 Polyporellus badius(アシグロタケ) MAFF 420217、 Polyporellus varius (キアシグロタケ) MAFF 420751、Trametes versicolor (カワラタケ) MAFF 420155、 Polyporellus tuberaster (タマチョレイタケ) MAFF 420261、Ganoderma lucidum (霊芝) MAFF 420210、Ganoderma tsugae MAFF 420118、Pycnoporus coccineus (ヒイロタケ) MAFF 420150、Pycnoporus cinnabarinus (シュタケ) MAFF 420151及び独立行政法人製品評価基盤機構 生物遺伝資源部門(NBRC)(H14年財団法人発酵研究所(IFO)から委託業務移管)より入手した基準培養株Polyporellus(Polyporus) brumalis (オツネンタケモドキ)IFO;30741 Formes fomentarius (ツリガネタケ)IFO;30371を含む。
【0020】
北東北の山中から採取した野生きのこ[シイタケ、キアシグロタケ、オツネンタケモドキ、カワラタケ、ニクウチワタケ、ナメコ、クリタケ、ツキヨタケ、ツガサルノコシカケ、コフキサルノコシカケ (推定名称) を含む。]から菌糸塊を0.25×MYPG寒天培地(0.25% Malt extract、0.1% Yrast extract、0.1% Pepton、0.5% Glucose、2% Agar)に植菌し、生育を確認した、25℃で5〜10日生育させた後、スクリーニングのため上記培地へ植え継ぎを繰りかえして単離したTD-1(シイタケ野生種)、キアシグロタケ、オツネンタケモドキ、カワラタケ、ニクウチワタケ、ナメコ、クリタケ、ツキヨタケ、ツガサルノコシカケ、コフキサルノコシカケ (推定名称) を含む。菌糸単離後は種菌プレートから成長点の約5mm内側を内径10mmのコルクボーラーでくり抜き、これを0.5×MYPG培地 (0.5% Malt extract、0.2% Yeast extract、0.2% pepton、1% Glucose) 100ml入り300ml三角フラスコへ5個づつ植菌し、25℃で4週間振とう培養を行った。
【0021】
ラッカーゼ活性測定には基質には2,2’-Azino-bis (3-etylbenzothiazoline-6-sulfonic acid) diammonium salt [ABTS; SIGMAモル吸光係数ε; 36,000 (M-1cm-1)]を基質として用いた。酵素液含むpH4.0のMcIlvaine Buffer100μl中で、30℃で10分間プレインキュベート後、終濃度1mMのABTSを加え反応を開始した。30℃10分間反応させた後、反応液と等量の5%TCAを加え反応を停止し、420nmの吸光度で測定した。1Uは、1分間に1μmolのABTSを触媒する酵素量と定義した。
【0022】
ラッカーゼ活性の算出は、以下に示す式に従った。
[ラッカーゼ活性算出式]:
U/ml= 吸光度(OD420nm)×1000(μmol/ml) ×0.2(ml) ×1000(U/ml)
36000(M-1cm-1)×10(min) ×反応酵素液量(μl)
【0023】
粗酵素液のタンパク質の定量には Advanced protein assay reagent (5×concentarate Cytoskeleton社, USA) を用いたBradford法により検出し、標準タンパク質として牛血清アルブミン (BSA) を使用して、595nmの吸光度を測定した。
【0024】
各菌糸培養液中のラッカーゼ活性を図1に示す。
図に示した結果から、最も高いラッカーゼ生産量を示し、タンパク質分泌量、単位タンパク質当たりのラッカーゼ比活性も高い結果を示した。ITS領域の配列より本株をPolyporellus brumalis (オツネンタケモドキ)と同定した。寄託センターより入手したオツネンタケモドキ株では、野生株ほどの高いラッカーゼ生産が見られなかった。
【0025】
以下、オツネンタケモドキ野生株をIBRC05015として、本株が生産するラッカーゼについて詳細に解析を行った。
【0026】
<オツネンタケモドキIBRC05015からの酵素精製>
オツネンタケモドキIBRC05015は1.5%寒天を含む0.25×MYPG培地で9cmシャーレを用いて1週間25℃で培養し、液体培養用の種菌とした。シャーレ一枚分の表面菌糸を掻きとり、0.5×MYPG液体培地に接種した。液体培養は、25℃で振とう培養で、24日間培養を行い、培養ろ液からラッカーゼを精製した。培養ろ液に80%飽和の硫酸アンモニウムを加えてタンパク質を塩析し、沈殿を10mMリン酸バッファー(pH7.0) に溶解した。得られたタンパク質溶液に30%飽和の硫酸アンモニウムを加え、上清を疎水クロマトグラフィー用のタンパク質サンプルとし、HiLoad Phenylカラム(GE healthcare)に供した。
【0027】
HiLoad Phenylカラムに吸着したタンパク質は10 mMリン酸バッファー(pH7.0)と30%飽和硫酸アンモニウムを含む同バッファーによる直線的濃度勾配(30~0%)による溶出を行った。ラッカーゼ活性のあるフラクションを集めてAmicon Ultra-4 (5000MWCO、Millipore社) にて限外濾過を行い25mlに濃縮した。濃縮したタンパク質サンプルはゲルろ過カラムPD-10 (GE helthcare) に供した後、陰イオン交換カラム(MonoQ 5/50 GL、GE Healthcare社)に吸着させ、10 mMリン酸バッファー(pH7.0)と0.5M NaClを含む同バッファーによる直線的濃度勾配(0-0.5M)による溶出を行った。流速については、0.5ml/minとし、各フラクションを0.5mlとした。ラッカーゼ活性を有するフラクションをAmicon Ultra-4 (Millipore 5000MWCO) 限外濾過により濃縮し、ゲル濾過を行った。ゲルろ過はSuperdex 75 10/30カラム(Millipore社)を用い、10mMリン酸バッファー(pH 6.5)、流速0.25ml/minで行った。64.3kDaの精製ラッカーゼが得られこれをpbLac1とした。
以下精製ラッカーゼ(PbLac1)を用いて諸性質の解析を行った。
【0028】
<活性等測定法>
1) 酵素の活性に及ぼすpHと温度の影響
緩衝液はKCl-HCl buffer(pH1.0)、Glycine-HCl buffer(pH2.0〜3.0)、McIlvaine buffer(pH4.0〜6.0)、Tris-HCl buffer(pH7.0〜9.0)、CAPS buffer (pH10.0、CAPS; DOJINDO) を使用した。基質はABTSを用い、至適pHと至適温度の測定時間は10分間とした。
結果、pbLac1の至適pHはABTSを基質とした場合pH4.0であり至適温度は40℃であった。
【0029】
2) 酵素の安定性に及ぼすpHと温度の影響
実験に用いた緩衝液は5 1)に示した。本実験では終濃度50μg/mlの牛胎児血清アルブミンを加えた緩衝液を使用し、基質はABTSを用いた。pH安定性試験は各pHにて30℃、12時間インキュベートを行なった。温度安定性試験ではpH 6.0で各温度での加温を行い、30分後に残存活性を測定した。
結果、pH3.0-6.0の範囲で40%以上のラッカーゼ活性を保持し安定であった。さらにpH6.0 のMcIlvaine緩衝液中では30分の加温処理で60℃まで安定であった。
【0030】
3) 阻害剤の影響
これまでラッカーゼの阻害剤として報告のある金属キレーター剤を中心に(EDTA、p-Cumaric acid 、Kojic acid、Tropolon、L-Cysteine、 NaN3、DTT)阻害剤の影響について検討を行なった。反応条件はMcIlvaine buffer (pH4.0)で、ABTSを基質として用い、30℃で10分間反応を行なった。阻害剤無添加時の活性に対する阻害剤添加時のラッカーゼ活性の相対値(%)を算出した。
結果、1mMのL-Cysteine、NaN3、DTTによってpbLac1の活性はほぼ完全に阻害された。また、金属イオンのキレーターであるEDTAは10mMでラッカーゼ活性を50%阻害し、p-cumaric acid、Kojic acidはどちらも1mMで約70%ラッカーゼ活性を阻害した。
【0031】
4) 金属イオンの影響
金属イオン(Ca2+、Cd2+、Co2+、Cu2+、K+、Mg2+、Mn2+、Na2+、Sn2+、Zn2+)無添加時の活性に対する金属イオン添加時のラッカーゼ活性の相対値(%)を算出した。反応条件は阻害剤実験と同様の条件で行なった。
結果、10mMのCu2+を添加した時に1.38倍の活性上昇が見られた。しかし、その他の金属イオンに関しては10mMの各金属イオンを添加すると70%程度に活性が低下し、Cd2+1mMで活性が48.8%まで低下した。一番顕著に阻害が見られたのは1mMのSn2+を添加した時で、ラッカーゼ活性が4.6%にまで低下した。
【0032】
5)基質特異性の検討
本酵素の基質特異性を比較するために表1(Table.1)に示したフェノール化合物を基質としてMcIlvaine緩衝液中で10分間反応させ、反応後の吸光度を測定し、1分間当たりに酸化される基質量を算出し、Lineweaver-Burk plot (Lineweaver and Burk 1934)を作成した。各基質で測定した吸光度の波長とモル吸光度係数(ε)を、表1 に示した。
【0033】
【表1】
【0034】
解析の結果、PbLac1は分泌型ラッカーゼには珍しくDOPAを基質にし、幅広い基質特異性を示した。
【0035】
またメディエーターの非存在下においてアントラキノン系色素であるRemazol Blue Black (RBBR)、ジアゾ色素であるNaphtol Blue Black (NBB)の脱色が可能であり、メディエーターの存在で高分子色素Poly R-478、モノアゾ色素 Reactive Orange 16も脱色できたことから、難分解性環境汚染物質の分解に有効である可能性を示唆した(図2)。
【0036】
<pblac1のcDNAおよびゲノムDNAのクローニング>
オツネンタケモドキよりRNAを精製し、オツネンタケモドキラッカーゼのcDNA,ゲノムDNAの配列を以下の手順により決定した。
【0037】
[方法]
得られたタンパク質のN-末端のアミノ酸配列をもとにプライマー(LccO-3;ATHWSNTGYGAYCCNAA, LccO-4L;SNTGYGAYCCNAAYCA) を設計し、3’RACEを行った。RACE用のRNAは酵素精製用に培養した菌糸より抽出し、抽出はMasterPureTM Yeast RNA抽出キット(EPICENTRERBiotechnologies社、USA) を用いて行った。また、3' RACE用のテンプレートcDNAは、SMART RACEキット(BD Biosciences)を用いて行った。また、得られた配列をもとに5’RACE を行った5’RACE用のcDNAはGeneRACERキット(Invitrogen) を用いて合成した。cDNAの合成は各キットのマニュアルに従った。また、PCRの条件は98℃ for 30sec,55℃ for 30sec,and 72℃ for 3min,(30cycles)で行った。
【0038】
各遺伝子の3’及び5’RACEを行う際に用いたプライマーを下記に示す。
LccO-375U-RACE CGCGCTCCCGTCCAACTCGTCCATC for RACE-PCR
LccO-243U-RACE GGCGTTCAACTTCAATGGCTCAGAC for RACE-PCR
LccO-149L-RACE AGGTCAGTCTCCACAAGCAGGTTCG for RACE-PCR
LccO-264L-RACE GGAGGCGCCGTTGATGAAAAAGTCT for RACE-PCR
【0039】
[結果]
得られた配列は非特許文献3に記載のあるオツネンタケモドキのラッカーゼと同一のものであった(配列番号1)。
【0040】
この配列をもとに、アミノ酸配列を推定し、精製ラッカーゼの配列と比較を行ったところ同一のものであることが確認できた(配列番号2)。
【0041】
<誘導剤がオツネンタケモドキIBRC05015株のラッカーゼ活性へ与える影響>
[方法]
菌糸は種菌プレートから成長点の約5mm内側を内径10mmのコルクボーラーでくり抜き、これを7個ずつ0.5×MYPG培地 (0.5% Malt extract、0.2% Yeast extract、0.2% pepton、1% Glucose) 50ml (容量200ml用の三角フラスコ) に植菌した。誘導剤はTable.4に示した濃度で植菌時に液体培地へ添加し、25℃で4週間振とう培養を行い、7日毎にラッカーゼ活性の測定を行った。菌糸は種菌プレートから成長点の約5mm内側を内径10mmのコルクボーラーでくり抜き、これを5個ずつ0.5×MYPG培地 (0.5% Malt extract、0.2% Yeast extract、0.2% pepton、1% Glucose) 50ml (容量200ml用の三角フラスコ) に入れた。
【0042】
[結果]
ラッカーゼの誘導剤として表2に示す誘導剤を用いて比較を行った。そのうち最もラッカーゼの誘導能が高かったのは0.25mMのCu2+を培地へ添加した条件下で、28日目でラッカーゼ活性が34.64U/mlに達した。この値はCu2+無添加時の最大値の21倍に相当し、試したInducerの中では最も誘導率が高かった。また、Cu2+を含む金属イオン添加時には乾燥菌糸体重量が増加する傾向が見られた。
【0043】
【表2】
【0044】
<IBRC05015株と基準購入株のCu2+誘導によるラッカーゼ活性変動の比較>
オツネンタケモドキIBRC05015株はCu2+添加でラッカーゼ活性の上昇が見られたことから、基準購入株とラッカーゼ活性誘導の比較を行った。
[方法]
植菌方法及び、培養法、活性測定法は誘導剤の検討と同様の条件で行った。Cu2+誘導は植菌時に0.5mMのCuSO4を添加した。
【0045】
[結果]
オツネンタケモドキIBRC05015株は基準購入株P. brumalis (MAFF420194)と比較してもCu2+に対する応答性が高く、培養11日目以降常時高いラッカーゼ活性を示すことが分かった。またP. arcularis (MAFF 460202)、P. badius (MAFF 420217)、P. coccineus (MAFF 420150) も同様にCu2+に対するラッカーゼ活性の上昇が見られたが、オツネンタケモドキには及ばなかった。また、T. versicolor (MAFF 420155)と比較してもオツネンタケモドキIBRC05015株のラッカーゼ誘導率は高かった(図3)。
【0046】
<Cu2+濃度がオツネンタケモドキIBRC05015株ラッカーゼ活性へ与える影響>
[方法]
菌糸は種菌プレートから成長点の約5mm内側を内径10mmのコルクボーラーでくり抜き、これを5個ずつ0.5×MYPG培地 (0.5% Malt extract、0.2% Yeast extract、0.2% pepton、1% Glucose) 50ml (容量200ml用の三角フラスコ) に入れた。CuSO4は植菌時に終濃度0、0.0005、0.005、0.05、0.1、0.25、0.5、1.0、2.5、5.0mMの濃度で0.5×MYPG液体培地中へ添加し、7日毎にラッカーゼ活性測定を行った。
【0047】
[結果]
Cu2+によってラッカーゼ活性が上昇することから、最適Cu2+添加濃度について検討を行った。0、0.005、0.05、0.50、0.1、0.25、0.5、1.0、2.5、5.0mMのCu2+を植菌時に添加し、ラッカーゼ活性を測定した。
【0048】
結果0.25mMのCu2+添加時に28日目で最も高いラッカーゼ活性を示すことが分かった(86.4U/ml)、次いで0.1mMのCu2+添加時(55.7U/ml)、0.5mM Cu2+添加時(40.5μM)、 0.005mM Cu2+添加時(16.6U/ml)の順でラッカーゼ活性が高い傾向が見られ、0.5mMでは急激にラッカーゼ活性の低下が見られることが分かった(3.0U/ml)。このことから、最適なCu2+添加濃度は0.25mMであることが示された(図4)。
【0049】
<Cu2+添加時に誘導されるラッカーゼについて>
Cu2+添加時にLac活性の急激な上昇が見られるが、Cu2+添加により誘導される蛋白質の有無を調べた。Cu2+添加時のオツネンタケモドキ培養ろ液を電気泳動し、CBB染色及びラッカーゼ活性染色を行った (図5)。
【0050】
[方法]
粗酵素液に、等量のサンプルバッファー (トリスSDSβMERサンプル処理液; 第一化学薬品) を添加し、5分間ボイルした。これを泳動ゲル [パジェルNPU-10L; ATTO (株)]にアプライし、1×Tris-Glycine-SDS Buffer (BioRad社) を用いて定電流20mAで泳動を行った。CBB染色は常法に従い、Commassie Brilliant Blue R250 (CBB) (PAGE Blue83;第一化学) で行った。
【0051】
また分子量マーカーにはプレシジョンplusスタンダード (BioRad) を用いた。活性染色のNative-PAGEは非還元条件下で行った。ラッカーゼ活性のユニット(U)を等しく調整した各粗酵素液に、等量のサンプルバッファー(Integrated Separation Systems;第一化学薬品)を添加して、泳動ゲル(パジェルNPU-10L) (ATTO (株))にアプライした。泳動は1×Tris-Glycine Buffer (BioRad)を用いて定電流20mAにて泳動を行った。
【0052】
泳動後、pH4.0のMcIlvaine 緩衝液にてpHを平衡化した後、終濃度1mMのABTSを含むMcIlvaine 緩衝液(pH4.0)と30℃で反応させた。N末端側アミノ酸配列決定は、粗精製酵素はSDS-PAGE後PVDF膜にブロットし、クマシーブリリアントブルーR250で染色した。目的バンド付近を切り取り、50%メタノールで脱色し、蒸留水で2時間洗浄して酢酸を除去し風乾したPVDF膜をサンプルとした。N末端アミノ酸配列の解析は (株)ニッピバイオマトリックス研究所に依頼した。なお、同社ではプロテインシークエンサーにABI procise 491 HT (ABI) を使用している。
【0053】
[結果]
Cu2+添加時にラッカーゼ活性を示す、70kDa付近の誘導バンドが存在することが明らかとなった。本バンドは全分泌蛋白質に占める割合が高く、Cu2+添加時には特異的に誘導されるラッカーゼであることが考えられた。そこで、本蛋白質の精製を試み、最終的に単一バンドになるまで精製を行った。精製されたラッカーゼはCu非添加時のラッカーゼと比較すると若干分子量が大きかったが、N末端配列を決定したところ、PbLac1(Rye et al. 2008) と完全に一致することが分かり、Cu2+添加によって誘導されているラッカーゼはPbLac1であることが明らかとなった(図5)。
【0054】
<Cu2+添加時のpblac1の誘導>
[方法]
Cu2+添加時にpblac1が誘導されている可能性が示唆されたため、リアルタイムPCRを用いてCu2+添加によるpblac1遺伝子のmRNA量の変化を調べた(図7)。Cu2+は植菌時に添加した。Cu2+添加から4日目〜35日目にかけて顕著なラッカーゼ活性上昇が見られたため(図6A)、その時期の菌糸を回収し、リアルタイムPCRを行った(図6B)。
【0055】
結果、10日目のCu2+添加画分でpblac1のmRNAレベルの上昇が見られ、14日目にmRNAレベルがピークに達し、16日には若干の減少が見られたが、Cu2+非添加と比較すると已然としてpblac1のmRNAレベルは高かった。
【0056】
<酸化還元電位測定>
ラッカーゼの酸化能力の指標の一つである酸化還元電位を以下の方法で測定した。
PbLac1の酸化還元電位測定にはサイクリックボルタモグラム法と無隔膜バルグ全電解法を用いた。ボルタモグラム法での測定で、大まかな電位の概算を行い、無隔膜バルグ法でより正確な電位の数値を計算した。サイクリックボルタモグラム法で最もクリアな結果が出たのはパージを行ったアルゴン飽和条件で、以前からの報告(Kamitaka et al. 2007)にあるように、Multicopper oxidaseのDETタイプの生物電気的反応を示し、触媒波はtype1Cu部位のE0辺りから上昇してきた。
【0057】
その結果、PbLac1のE0値は797 ± 10mV(標準水素電極: NHE)、無隔膜バルグ全電解法では4回の測定の後、平均して811mV ± 5mV vs 標準水素電極: NHE)だった(表3)。この値はサイクリックボルタモグラム法で測定した電位とほぼ同じ値を示し、より正確な数値が得られた。この値はTrametes hirsute (ThL; 0.78V)やTrametes versicolor (TvL; 0.78V)のようなTypeI Cu部位の酸化還元電位が高いラッカーゼと比較しても、同等かそれ以上であった。
【0058】
【表3】
【産業上の利用可能性】
【0059】
本発明によるラッカーゼは、非常に高い酸化還元電位を持つため、基質の酸化能力に優れており、産業上の利用価値が高い。
製紙工程でのリグニン処理、洗剤における染料の転写の阻止、繊維の脱色、フェノール樹脂の製造及び廃水処理など、また、燃料電池用の電極素材への適用など、種々の工業利用にきわめて有用である。
【特許請求の範囲】
【請求項1】
オツネンタケモドキIBRC05015株(寄託番号 NITE P-822)から高酸化還元電位型ラッカーゼを分泌生産する方法であって、銅添加培地を使用することを特徴とする、大量かつ高純度の高酸化還元電位型ラッカーゼを分泌生産する方法。
【請求項2】
オツネンタケモドキが野生株に由来するものであることを特徴とする請求項1に記載の方法。
【請求項1】
オツネンタケモドキIBRC05015株(寄託番号 NITE P-822)から高酸化還元電位型ラッカーゼを分泌生産する方法であって、銅添加培地を使用することを特徴とする、大量かつ高純度の高酸化還元電位型ラッカーゼを分泌生産する方法。
【請求項2】
オツネンタケモドキが野生株に由来するものであることを特徴とする請求項1に記載の方法。
【図1】


【図2】
【図3】
【図4】
【図5】

【図6】

【図2】
【図3】
【図4】
【図5】
【図6】
【公開番号】特開2011−130732(P2011−130732A)
【公開日】平成23年7月7日(2011.7.7)
【国際特許分類】
【出願番号】特願2009−295057(P2009−295057)
【出願日】平成21年12月25日(2009.12.25)
【出願人】(390025793)岩手県 (38)
【Fターム(参考)】
【公開日】平成23年7月7日(2011.7.7)
【国際特許分類】
【出願日】平成21年12月25日(2009.12.25)
【出願人】(390025793)岩手県 (38)
【Fターム(参考)】
[ Back to top ]