
魚の癌モデル
本発明は、プロモーターに作動可能に連結した発癌性の核酸がゲノムに組み込まれた魚を対象とする。その魚を産生する方法およびそれらの使用法も提供する。その魚は、癌遺伝子媒介性の腫瘍性もしくは過形成性形質転換を調節する、または化学療法もしくは放射線治療に対する感受性を調節する薬物もしくは物質をスクリーニングする方法に有利に利用され得る。不死の腫瘍細胞株、不死の腫瘍細胞株を産生する方法およびそれらの使用法も提供する。

【発明の詳細な説明】
【技術分野】
【0001】
[発明の背景]
本発明は、発癌性の核酸を含む魚、魚の腫瘍形成モデル、不死の腫瘍細胞株および抗癌剤のスクリーニングを対象とする。
【背景技術】
【0002】
本発明の背景を明らかにするために本明細書にて用いる出版物や他の資料、また具体的には実施に関するさらなる詳細を提供する事例は、参照により組み込まれ、また便宜のために以下の文章中に著者および日付により引用し、さらに補足した参考文献一覧に著者のアルファベット順に列挙する。
【0003】
疾患状態の動物モデルは、特定の疾患の根底にある生化学的メカニズムを同定し、ならびに疾患を根絶するまたはその症状を軽減するための治療剤を発見するのに重要な役割を果たす。例えば、家族性高コレステロール血症のウサギモデル、非インスリン依存性糖尿病のラットモデル、癌のマウスモデルおよび突発性心房血栓症(spontaneous atrial thrombosis)のハムスターモデルが知られている。さらに、マウス、ネコおよびイヌなどのさまざまな種において遺伝子疾患の動物モデルが発症的に生じている。そのような大きな動物を用いる研究にはいくつかの欠点がある。例えば、そのようなモデルに用いる多くの動物は比較的大きな脊椎動物であり、かなり広い研究スペースを取り、餌代もしくは維持費が高く、生殖周期が遅く、一度にもうける子孫が比較的少なく、所望する全ての疾患状態を効率的に模倣することができない。
【0004】
形質転換魚は疾患モデルの開発に最近利用されている。広範な魚がこの目的に利用され得る。典型的な魚には、硬骨魚、例えば、ゼブラフィッシュ(ダニオ・レリオ(Danio rerio))、メダカ(オリジアス・ラティペス(Oryzias latipes))、マミチョグ(フンドゥルス・ヘテロクリトゥス(Fundulus heteroclitus))、カダヤシ(フンドゥルス属)、ナマズ(イクタルルス(Ictalurus)属)(アメリカナマズなど);コイ(キプリヌス(Cyprinus)属)、例えば、一般的なコイ;およびマスまたはサケ(例えば、イワナ(サルベニルス(Salvelinus))属、大西洋サケ(サルモ(Salmo))属および太平洋サケ(オンコリンクス(Oncorhynchus))属)などがある。ゼブラフィッシュは発生、生理学および疾患を様々な角度から調べるための、確立されたモデルとなっている。
【0005】
ゼブラフィッシュは小さく、子宮外で発生し、また世代時間が短いので、特に有利である。ゼブラフィッシュは実験室環境で維持できるほど経済的であり、また繁殖力が高い;一匹のメスは一週間につき何百もの子孫をもうけることができる。それぞれ5日齢になった魚は、心臓、脳、血液および膵臓など哺乳類に存在する臓器系のほとんどを備え、自由に泳ぎ/餌を採る生命体である。ゼブラフィッシュの胚は外部で発生し、透明であるため、インビボで進行する細胞および組織の発生過程を直接目視でき、これにより遺伝子および小分子薬の大規模なスクリーニングが容易になる。過去数年間、多数の出版物において細胞型に制限的な(cell-type restricted)発現パターンで緑色蛍光タンパク質(GFP)を発現する形質転換魚系が報告されている(Gong et al., 2001; Kennedy et al., 2001; Long et al., 1997; Moss et al., 1996; Motoike et al., 2000; Park et al., 2000)。これまで、蛍光形質転換ゼブラフィッシュを用いた研究では、主に細胞や組織の発生過程を撮像することに焦点が置かれていた。そのような形質転換ゼブラフィッシュ系により、組織の形態形成の発生的研究が促進されることに加え、別々の細胞集団を高解像度で撮像することが可能なため、遺伝的および薬理学的スクリーニングも容易となる。
【0006】
癌を誘導する多くの潜在メカニズムは、これから完全に解明しなければならない。これらの疾患において変異した遺伝子を同定することで、癌全体への新たな見識が得られるであろう。さらに、標的化学療法の開発において新しい標的薬物を同定するためには、疾患の進行を抑制または遅延する、または化学療法もしくは放射線誘導性プログラム細胞死に対する感受性を阻害する、遺伝的または化学的抑制因子が同定できる脊椎動物モデル系を用いることが必要であろう。例えば、特異的標的の事前の知識を必要としないモデル系が必要である。調節性標的薬物または物質の安全性および有効性が実証された後に標的の解明を行うと、薬物を発見する過程での時間と費用の両方を節約できる。
【0007】
癌などの様々な疾患状態の細胞および分子遺伝学的特性をさらに理解する必要がある。適切な動物モデルは、癌をより理解し、また癌を処置または予防するためのより有効な薬物の開発を行うために非常に有益である。本発明はこれらの必要性を解決することを目的とする。
【発明の開示】
【0008】
[発明の概要]
本発明は発癌性の核酸を含む魚、魚の腫瘍形成モデル、不死の腫瘍細胞株および抗癌剤のスクリーニングを対象とする。
【0009】
従って、第1の局面において、本発明は、プロモーターに作動可能に連結した、本明細書において癌遺伝子とも称する発癌性の核酸を含むコンストラクトを提供する。このコンストラクトは、好ましくはマーカーをさらに含む。このコンストラクトはベクターの一部ともなり得る。好ましくは、このプロモーターは器官もしくは組織特異的プロモーターまたは最小プロモーターである。当分野で知られるように、癌遺伝子は、発現すると細胞増殖の調節を改変するか、またはプログラム細胞死を阻止する遺伝子である。広範な癌遺伝子が本明細書に記載の核酸コンストラクトに利用できる。癌遺伝子はウイルスまたは細胞起源であり得る。細胞起源の癌遺伝子には内在性癌遺伝子が含まれる。そのような癌遺伝子が発現すると、細胞に腫瘍性または過形成性形質転換が起こる。その癌遺伝子は、その癌遺伝子の完全配列、好ましくはその癌遺伝子の発癌性形態であり得る。またはその癌遺伝子はその癌遺伝子の発癌能を維持するその癌遺伝子のフラグメントであり得る。
【0010】
第2の局面において、本発明は、上記のコンストラクトをゲノムに含む、またはいくつかの細胞においてのみその癌遺伝子を発現する(ランダム/モザイク発現)形質転換魚、特に形質転換ゼブラフィッシュ(ダニオ・レリオ)を提供する。
【0011】
第3の局面において、本発明は、不死の腫瘍細胞株および不死の腫瘍細胞株を産生する方法を提供する。一実施態様において、不死の腫瘍細胞株は、魚において癌遺伝子を発現させて腫瘍を作り出し、腫瘍細胞を単離し、単離した腫瘍細胞を培養して不死の腫瘍細胞株を産生することによって、得られる。
【0012】
第4の局面において、本発明は、癌遺伝子に関連する癌を処置するのに有用な薬物を同定するために、化合物をスクリーニングする方法を提供する。
【0013】
[発明の詳細な説明]
本発明は発癌性の核酸を含む魚、魚の腫瘍形成モデル、不死の腫瘍細胞株および抗癌剤のスクリーニングを記載する。
【0014】
一実施態様において、本発明は、プロモーターに作動可能に連結した、本明細書において癌遺伝子とも称する発癌性の核酸を含むコンストラクトを対象とする。このコンストラクトは、好ましくはマーカーをさらに含む。このコンストラクトはベクターの一部ともなり得る。好ましくは、そのプロモーターは器官特異的プロモーター、組織特異的プロモーターまたは最小プロモーターである。「最小プロモーター」なる語は本明細書において、そのプロモーターが機能性プロモーターを含む最小の配列から構成されることを意味する。最小プロモーターを同定する技術は当分野でよく知られている(Baliga, 2001)。当分野で知られるように、癌遺伝子は、発現すると細胞増殖の調節を改変するか、またはプログラム細胞死を阻止する遺伝子である。広範な癌遺伝子が本明細書に記載の核酸コンストラクトに利用できる。癌遺伝子はウイルスまたは細胞起源であり得る。細胞起源の癌遺伝子には内在性癌遺伝子が含まれる。そのような癌遺伝子が発現すると、細胞に腫瘍性または過形成性形質転換が起こる。癌遺伝子は、その癌遺伝子の完全配列、好ましくはその癌遺伝子の発癌性形態であり得る。またはその癌遺伝子は、その癌遺伝子の発癌能を維持するその癌遺伝子のフラグメントであり得る。
【0015】
典型的な癌遺伝子には、活性化RAS、MYC、SRC、FOS、JUN、MYB、RAS、ABL、BCL2、HOX11、HOX11L2、TAL1/SCL、LMO1、LMO2、EGFR、MYCN、MDM2、CDK4、GLI1、IGF2、活性化EGFR、変異型遺伝子(FLT3-ITDなど)、変異型および活性型のTP53、PAX3、PAX7、BCR/ABL、HER2/NEU、FLT3R、FLT3-ITD、SRC、RAS、ABL、TAN1、PTC、B-RAF、PML-RARアルファ、E2A-PBX1およびNPM-ALK、ならびにPAXおよびFKHR遺伝子ファミリーのメンバーの融合などがある。
【0016】
他の典型的な癌遺伝子は当分野でよく知られており、そのような例のいくつかは、例えば、「The Genetic Basis of Human Cancer」(Vogelstein, B.およびKinzler, K. W.編 McGraw-Hill, New York, N.Y., 1998)に記載されている。そのような遺伝子の相同体も用いることができる。そのような遺伝子の哺乳類相同体は、内在性の魚の遺伝子と区別できるので、好ましい。さらに好ましいのは、そのような遺伝子のヒト相同体である。そのような癌遺伝子に対応する配列は、その癌遺伝子のヒト相同体を含めて周知であり、例えばGenBankデータベースから見付けることができる。
【0017】
癌遺伝子は形質転換魚に発症することを望む癌の形態に基づいて選択する。例えば、RASファミリー(K-、H-またはN-RAS)の変異型または活性化遺伝子は、腎臓、膵臓または結腸癌など幅広い種類の癌の誘導に用いることができ、HOX11およびTAL1はT細胞癌の誘導などに用いることができる。本発明は特定の癌遺伝子配列に限定されない。例えば、その癌遺伝子の形質転換能が増大または低下した、その癌遺伝子ヌクレオチド配列の改変型または本明細書に記載の他の癌遺伝子ヌクレオチド配列も、包含される。
【0018】
本発明の一実施態様において、本発明に利用される癌遺伝子は、発癌性のrasポリペプチド配列をコードする。全Rasタンパク質のうち164のN末端アミノ酸は高度に保存されている(図1B)。残りの25のC末端残基のみがK-、H-およびN-Rasの間でかなり多様である。主にコドン12、13または61に起こる発癌性変異は、GTP加水分解の触媒部位に影響する。Rasの変異型はGTPに結合したままであり、細胞増殖のためのシグナルを構成的に伝達する。ras遺伝子の活性化変異はヒトの癌の約30%に関係している。しかしながら、異なる種類のヒトの癌には異なるRasタンパク質の発癌性形態が見られる。例えば、K-Rasの変異は、膵臓、結腸および腺癌に共通しているが(Bos, 1989)、血液悪性腫瘍は主にN-Rasに変異がある(Ahuja et al., 1990)。
【0019】
別の実施態様において、本発明に利用する癌遺伝子は、発癌性のK-rasポリペプチドをコードする。さらなる実施態様において、本発明に利用する癌遺伝子は、ゼブラフィッシュの発癌性のzK-rasBポリペプチドをコードする(配列番号8)。さらなる実施態様において、本発明に利用する癌遺伝子は、最適にアライメントした場合に、本明細書に記載の発癌性のrasヌクレオチド配列または他の癌遺伝子ヌクレオチド配列のヌクレオチド配列に対して、(適切なヌクレオチド挿入または置換を含んで)少なくとも約60%、好ましくは少なくとも約70%、より好ましくは少なくとも約80%、最も好ましくは少なくとも約90%の同一性を有し得る。別の実施態様において、本発明の発癌性のrasポリペプチドは、配列番号8のポリペプチド、ならびに配列番号8のポリペプチドに対して少なくとも65%の類似性(好ましくは少なくとも65%の同一性)または少なくとも70%の類似性(好ましくは少なくとも70%の同一性)または少なくとも75%の類似性(好ましくは少なくとも75%の同一性)または少なくとも80%の類似性(好ましくは少なくとも80%の同一性)または少なくとも85%の類似性(好ましくは少なくとも85%の同一性)または少なくとも90%の類似性(好ましくは少なくとも90%の同一性)または少なくとも95%の類似性(好ましくは95%の同一性)を有するポリペプチドを含む。当分野で知られるように2ポリペプチド間の「類似性」は、あるポリペプチドのアミノ酸配列とその保存的アミノ酸置換を、第2のポリペプチド配列と比較して決定する。
【0020】
同一性とは、2つのポリペプチドまたは2つのポリヌクレオチド配列鎖間のマッチの同一性により測定される、2配列の関連性の度合を意味する。同一性は容易に計算できる。2つのポリヌクレオチドまたはポリペプチド配列間の同一性を測定する方法は多数存在するものの、「同一性」なる用語は、当業者に周知である(「Computational Molecular Biology」、Lesk, A. M., 編、Oxford University Press, New York, 1988;「Biocomputing: Informatics and Genome Projects」、Smith, D. W., 編、Academic Press, New York, 1993;「Computer Analysis of Sequence Data」、Part I, Griffin, A. M.およびGriffin, H. G., 編、Humana Press, New Jersey, 1994;「Sequence Analysis in Molecular Biology」、von Heinje, G., Academic Press, 1987; および「Sequence Analysis Primer」、Gribskov, M.およびDevereux, J., 編、M Stockton Press, New York, 1991)。2つの配列間の同一性を決定するのに一般に用いられる方法には、「Guide to Huge Computers」、Martin J. Bishop編、Academic Press、San Diego、1994およびCarillo, H.およびLipman, D.,「SIAM J Applied Math.」48:1073 (1988)などに記載の方法があるがこれらに限定されない。同一性を測定するための好ましい方法は、2つの被験配列間に最大のマッチが生じるように構成されている。そのような方法はコンピュータープログラムに体系化されている。2配列間の同一性を決定する好ましいコンピュータープログラム法には、GCG (Genetics Computer Group, Madison Wis.)プログラムパッケージ(Devereux, J., et al., Nucleic Acids Research 12:387 (1984))、BLASTP、BLASTN、FASTA (Altschul et al. (1990); Altschul et al. (1997))などがあるがこれらに限定されない。よく知られているSmith Watermanアルゴリズムも同一性の決定に用いることができる。
【0021】
例として、参照ヌクレオチド配列に対して少なくとも、例えば95%の「同一性」を有するヌクレオチド配列を有するポリヌクレオチドとは、そのポリヌクレオチドのヌクレオチド配列が、参照ヌクレオチド配列の100ヌクレオチドにつき点変異を5つまで含み得る以外は、参照配列に対して同一であることを意図している。言い換えると、参照ヌクレオチド配列に対して少なくとも95%同一なヌクレオチド配列を有するポリヌクレオチドが生じるには、参照配列中の5%までのヌクレオチドが欠失する、または別のヌクレオチドと置換される、あるいは参照配列の全ヌクレオチドの5%までの数のヌクレオチドが、その参照配列に挿入される場合がある。参照配列のこれらの変異は、参照ヌクレオチド配列の5または3末端部位またはそれらの末端部位の間のどこかに生じ、参照配列のヌクレオチド内に個別に分散するか、または参照配列内に1以上の連続した群として分散している可能性がある。同様の考え方が2つのポリペプチド配列間の同一性を決定する場合にも当てはまる。
【0022】
さらに、癌遺伝子は、発癌性rasヌクレオチド配列、または発癌性K-rasヌクレオチド配列、または配列番号8に示すポリペプチドをコードするゼブラフィッシュの発癌性のzK-rasB、または本明細書に記載の他の癌遺伝子ヌクレオチド配列に対して実質的類似性を有するヌクレオチド配列を含み得る。「実質的類似性」なる語は、本明細書において、そのヌクレオチド配列が参照ヌクレオチド配列と十分に類似し、中程度のストリンジェントな条件下において参照ヌクレオチド配列にハイブリダイズするであろうことを意味する。類似性を決定するこの方法は、本発明が属する分野において周知である。手短に説明すると、中程度のストリンジェントな条件は、Sambrook et al. (「Molecular Cloning: A Laboratory Manual」、第2版、1巻、101〜104頁、Cold Spring Harbor Laboratory Press (1989))に、前洗浄溶液に5 X SSC(塩化ナトリウム/クエン酸ナトリウム溶液)、0.5%ドデシル硫酸ナトリウム(SDS)、1.0mMエチレンジアミン四酢酸(EDTA) (pH 8.0)を用いること、ハイブリダイゼーションおよび55℃、5 X SSCの洗浄条件を含むと規定されている。癌遺伝子のヌクレオチド配列にさらに必要なことは、細胞に腫瘍性形質転換を施す能力を有するタンパク質をコードすることである。つまり、そのタンパク質は正常細胞(すなわち非癌性細胞)を癌性細胞(すなわち腫瘍)へと転換する能力を有する。
【0023】
本明細書においては導入遺伝子(本明細書に記載のゲノムに導入する遺伝子)でもある癌遺伝子は、インビトロで合成しても、または生物学的起源から単離してもよい。そのような合成および単離方法は当業者に周知である。本明細書に用いる、「導入遺伝子」または「導入遺伝子(transgene)コンストラクト」または「遺伝子導入(transgenic)コンストラクト」または「遺伝子導入(transgenic)DNA配列」なる用語は、互換的に用いられ、一般に遺伝子産物の発現を開始し、また調節する能力を持つ調節領域(例えばプロモーターおよびエンハンサー配列)を含む核酸分子を意味するが、これに限定されない。遺伝子導入コンストラクトは、関連遺伝子産物の発現および/または安定性を調節するための、ポリアデニル化配列などの相互適合性の他のあらゆるDNA要素も含み得る。導入遺伝子コンストラクトはまた、作動可能に結合したDNA配列の、ゼブラフィッシュのゲノムへの統合を促進するよう機能する他のDNA配列、および組換え核酸配列の増殖、選択、操作および/または移動に用いるあらゆる核酸系(例えばプラスミド発現ベクター)に含まれるあらゆる関連DNA要素を含み得る。
【0024】
導入遺伝子コンストラクトは、魚に導入して形質転換魚を産生するための遺伝子素材である。本明細書において用いられる「形質転換(transgenic)」なる用語は、人工的に導入されたDNA分子を含む生物およびその子孫を意味する。導入方法および、しばしば導入遺伝子コンストラクトの構造によっては、導入遺伝子コンストラクトは外来コンストラクトとなる。導入遺伝子コンストラクトはあらゆる核酸配列からなるものであってもよいが、開示する形質転換魚に用いるためには導入遺伝子コンストラクトは、発現産物をコードする配列に発現用配列(expression sequences)を作動可能に連結させて組み合わせるのが好ましい。導入遺伝子コンストラクトは好ましくは、発現または安定性のための、またはそのコンストラクトを魚ゲノムへ組み込むことを補助するための、他の構成要素をも含む。本明細書に用いられる、作動可能に(operablyまたはoperatively)連結されたと称される、導入遺伝子コンストラクトの各構成要素とは、それらに望まれる目的のために共に機能できるよう連結されている各構成要素を意味する。例えば、プロモーターとコード領域は、プロモーターが機能してコード領域の転写が起こるならば、作動可能に連結している。
【0025】
発現用配列は、開示する導入遺伝子コンストラクトにおいて、コンストラクトにコードされた発現産物の発現を媒介させるために用いられる。本明細書において用いられる、発現用配列には、プロモーター、上流エレメント、エンハンサーおよび応答エレメントが含まれる。開示するコンストラクトに用いる発現用配列は、相同発現用配列であるのが好ましい。開示する形質転換魚に用いる導入遺伝子コンストラクトの構成要素に関して、本明細書に用いる相同とは、その構成要素が用いる魚の種(species)または種類(type)の野生型であるまたはそれに由来することを示す。逆に、非相同とはその構成要素が用いる魚の種または種類の野生型でもなくそれに由来もしないことを示す。
【0026】
本明細書において用いる発現用配列は、2つの主要なクラス、つまりプロモーターおよびエンハンサーに分けられる。プロモーターは一般に、転写開始部位に対して比較的固定された位置において機能するDNA配列(複数の場合もある)である。プロモーターは、RNAポリメラーゼと転写因子の基本的な相互作用に必要なコアエレメントを含み、また上流エレメントおよび応答エレメントを含み得る。エンハンサーは一般に、転写開始部位からの距離が不定の位置において機能する、いずれかの配向性であり得るDNA配列を意味する。エンハンサーは近くのプロモーターから始まる転写を増大するように機能する。エンハンサーはまた、転写の調節を媒介する応答エレメントをしばしば含む。プロモーターも、転写の調節を媒介する応答エレメントを含み得る。
【0027】
エンハンサーは遺伝子の発現を調節する。この作用は、プロモーターに作動可能に連結したレポーター遺伝子を含有する導入コンストラクトがエンハンサーのドメインに挿入された時にだけ発現する、いわゆるエンハンサートラップ法において観察され、利用されている(O’Kane et al., 1987; Allen et al., 1988; Kothary et al., 1988; Gossler et al., 1989; Parinov et al., 2004)。そのような場合、コンストラクトの発現は新たに関連づけられたエンハンサーの様式に従って調節される。開示する形質転換魚に、最小または短いプロモーターのみを有する遺伝子導入コンストラクトを用いて、様々な組織に対して癌遺伝子の発現を駆動することができる。エンハンサートラップ法の利点は、同じコンストラクトを用いて様々な組織に腫瘍を誘導できることである。従って、そのようなコンストラクトを用いて形質転換した形質転換集団をスクリーニングして、組織学的に異なるタイプの腫瘍を有する動物または動物系を産出する。
【0028】
コードされたペプチドまたはタンパク質が発現するためには、導入遺伝子コンストラクトに、RNAに転写されるとコードされた発現産物の翻訳を媒介する配列も必要である。そのような配列は一般に、転写RNAの5'非翻訳領域に見られる。この領域は、そのコンストラクト上の転写開始部位と翻訳開始部位(すなわち開始コドン)間の領域に対応する。コンストラクトの5'非翻訳領域は、そのコンストラクトに用いたプロモーターに正常に関連づけられた5'非翻訳領域、発現産物をコードする配列に正常に関連づけられた5'非翻訳領域、そのプロモーターもしくは発現産物をコードする配列と無関係の遺伝子の5'非翻訳領域、またはこれらの5'非翻訳領域の混成体に由来し得る。好ましくは、5'非翻訳領域は、そのコンストラクトが導入されるべき魚と相同である。好ましくは、5'非翻訳領域は用いたプロモーターと正常に関連づけられたものである。
【0029】
開示する形質転換魚に用いる導入遺伝子コンストラクトは、(発現の検出および定量化のための)レポータータンパク質をコードする。本明細書において用いられるレポータータンパク質は、発現すると特異的に検出することができるあらゆるタンパク質である。レポータータンパク質は、発現用配列からの発現を検出または定量化するのに有用である。例えば、レポータータンパク質をコードするヌクレオチド配列を組織特異的な発現用配列に作動可能に連結すると、系統発生を綿密に研究することができる。そのような研究において、レポータータンパク質は、細胞遊走など、発生過程をモニタリングするためのマーカーとして働く。多くのレポータータンパク質が知られており、他の生物において同様の目的に用いられている。これらには、特異的に検出可能な産物を産生することができる、βガラクトシダーゼ、ルシフェラーゼおよびアルカリフォスファタ−ゼなどの酵素、および直接検出が可能なタンパク質などがある。実質的には、あらゆるタンパク質は、例えばそのタンパク質に特異的な抗体を用いて直接検出することができる。
【0030】
外来因子を加える必要なく直接検出できるレポータータンパク質を用いることは、ゼブラフィッシュの胚発生過程での遺伝子発現を検出または評価するのに好ましい。レポータータンパク質および組織特異的発現用配列をコードするコンストラクトを含む形質転換ゼブラフィッシュ胚によって、発生的に調節された遺伝子の空間的および時間的な発現パターンを解析するための高速リアルタイムインビボ系(rapid real time in vivo system)を提供することができる。
【0031】
開示する導入遺伝子コンストラクトは好ましくは、コンストラクトからの発現または安定性を改善する他の配列を含む。例えば、タンパク質をコードするコンストラクト上にポリアデニル化シグナルを含むと、導入遺伝子からの転写物が確実に加工されmRNAとして輸送される。発現コンストラクトにおけるポリアデニル化シグナルの同定および利用は確立されている。相同ポリアデニル化シグナルを導入遺伝子コンストラクトに用いるのが好ましい。
【0032】
上記の指針によって、癌遺伝子はプロモーターに作動可能に連結している。好ましくは、プロモーターは器官または組織(細胞を含む)特異的プロモーターである。最も好ましくは、プロモーターは上皮細胞に特異的なケラチン8 (krt8)プロモーターである(Gong et al., 2002)。好ましくは、プロモーターは、エンハンサートラップとして用いることができる短縮されたkrt8 プロモーター(Parinov et al., 2004)または最小プロモーターである。プロモーターの他の例には、組換え活性化遺伝子(RAG)(RAG1およびRAG2を含む);T細胞に特異的な、非レセプター型チロシンキナーゼをコードするLCK;IgMエンハンサーエレメントおよびCD2のプロモーターが含まれる。組織制限的(tissue-ristricted)発現を導くいくつかのプロモーターが同定されており、例えば、リンパ系組織のためのゼブラフィッシュRAG1(Jessen et al., 1999)およびゼブラフィッシュRAG2(Jessen et al., 2001)、および神経特異的発現のためのIslet-1(Motoike et al., 2000)、膵臓のためのPDX-1およびインスリン(Huang et al., 2001)などがある。本明細書に記載の組織特異的プロモーターのヌクレオチド配列に対して、少なくとも約70%の同一性、少なくとも約80%の同一性、さらに少なくとも約90%の同一性を有するプロモーターも包含されるが、ただし作動可能に連結した癌遺伝子の転写を促進することが条件である。ほとんどの哺乳類プロモーターは魚においてはあまり働かないことが確認されているので、ゼブラフィッシュ、フグまたは他種の魚のゲノムの調節配列をしばしば目的のコード配列の上流、内部および下流に特異的にクローニングしなければならず、これは当業者に習慣的な方法によって行うことができる。
【0033】
本明細書にて規定したように、ヌクレオチド配列は、別のヌクレオチド配列と機能的関係にある時、別のヌクレオチド配列と「作動可能に連結し」ている。例えば、コード配列がプロモーター配列に作動可能に連結している場合、このことは一般にそのプロモーターがコード配列の転写を促進し得ることを意味する。作動可能に連結しているということは、連結しているDNA配列が、典型的には近接していること、また2つのタンパク質コード領域をつなぐ必要があれば、近接しかつリーティングフレーム内にあることを意味する。エンハンサーはプロモーターから数キロベース離れていても機能し得ること、またイントロン配列は様々な長さであり得ることから、いくつかのヌクレオチド配列は作動可能に連結するが近接しないこともあり得る。
【0034】
本コンストラクトはさらに好ましくは、マーカーまたはレポーター遺伝子を含む。好ましい実施態様において、蛍光タンパク質遺伝子(例えば、EGFP、GFP、RFP、BFP、YFPまたはdsRED2)またはルシフェラーゼタンパク質遺伝子のようなレポーター遺伝子が癌遺伝子に先行する。最も好ましい実施態様において、マーカーは高感度緑色蛍光タンパク質(EGFP)である(Zhang et al., 1996)。EGFPは感度の高いレポータータンパク質なので好ましい。好ましい実施態様において、マーカーと癌遺伝子の融合物は、その融合遺伝子がプロモーターの制御下になるように調製する。マーカーは融合タンパク質のN末端を構成し、癌遺伝子産物は融合タンパク質のC末端を構成するのが好ましい。この好ましい実施態様において、本コンストラクトは、プロモーター−マーカー−癌遺伝子を含むキメラ導入遺伝子を含む。
【0035】
本明細書において具体的なマーカーの使用を開示し論じたが、具体的に開示したマーカーに本発明が限定されるということは決してない。さらに多くのレポータータンパク質が知られており、同様の目的に使用されている。これらには、特異的に検出可能な産物を産生することができる、βガラクトシダーゼ、ルシフェラーゼ、クロラムフェニコール・アシルトランスフェラーゼ(acytransferase)、βグルクロニダーゼおよびアルカリフォスファタ−ゼなどの酵素、および直接検出可能なタンパク質などがある。実質的には、あらゆるタンパク質が、例えばそのタンパク質に特異的な抗体を用いて直接検出することができる。容易に検出できるレポーターであればいずれも、EGFPの代わりに用いてよい。真核生物細胞の陽性または陰性選択に適するさらなるマーカー(および関連する抗生物質)は、とりわけ、SambrookとRussell (2001)、「Molecular Cloning」、第3版、Cold Spring Harbor Laboratory Press、Cold Spring Harbor、New York、および、Ausubelら(1992)、「Current Protocols in Molecular Biology」、John Wiley & Sons(定期的更新を含む)に開示されている。報告されているあらゆるマーカー並びに当分野で知られる他の物は、本発明の実施に用い得る。
【0036】
コンストラクトは、遺伝子コード領域の上流にプロモーター配列を用いて作製することができる。まず、ゲノムのプロモーター配列をマーカー(例えばEGFP)の上流にクローニングし、それらの配列が発生過程でこの蛍光マーカーの発現を組織特異的に駆動できるかどうかを確認し、そうであれば同じ配列をK-rasおよび他の癌遺伝子の発現を駆動するために用いる。当業者に周知の、他の(例えばゼブラフィッシュの)、ドーパミン作動性の(dopaminanergic)神経系のチロシンヒドロキシラーゼ、筋肉系のMYO-D、骨髄系のMPOまたはPU.1をコードする遺伝子からクローニングされたプロモーターのような、器官および組織特異的プロモーターの構築に、同様の方法が利用できる。
【0037】
好ましい実施態様において、本発明のコンストラクトは、ゼブラフィッシュのkrt8プロモーター、マーカーとしてのEGFPおよびzK-rasなどの癌遺伝子を含む。ゼブラフィッシュのkrt8プロモーターは、皮膚の上皮組織において、またエンハンサートラップ効果によって他の様々な組織に対して、導入遺伝子つまり癌遺伝子の発現を駆動するので、用いると有利である(Parinov et al., 2004)。別のプロモーターを利用する場合、そのプロモーターもエンハンサートラップ効果を示すことが好ましい。エンハンサートラップが特定のプロモーターと正常には結びつかない場合、エンハンサートラップ系もコンストラクトの一部にしてもよい。この局面において、組み込み部位に作用する調節領域(例えばエンハンサーおよび/またはリプレッサー)の特性に依拠する無作為なパターンの発現を促進する「エンハンサートラップ」方策を適用し、そのコンストラクトには、対応する活性化配列(例えばUAS(Gal4に特異的な上流活性化配列))の制御下、所定のDNA結合タンパク質(例えばGal4-VP 16融合物)およびレポーター遺伝子産物に作動可能に連結した最小プロモーターエレメントを含むエンハンサートラップ系をさらに含ませ、あとはその遺伝子導入コンストラクトが魚のゲノムに無作為に挿入および/または転位(transpose)されればよい。例えば、エンハンサートラップ系は、転位因子(Sleeping Beauty、Tc1/mariner-likeファミリー(Grabher et al., 2003; Ivics et al., 1999; Parinov et al., 2004)およびトウモロコシAc/Ds(2005年5月17日出願の米国仮特許出願第60/681,447号を参照。参照により本明細書に組み込まれる。)などがあるがこれらに限定されない)を用いてゼブラフィッシュ内に創出でき、また所望の発現パターンを示す魚を繁殖させ、他の形質転換魚と全く同じように利用できる。
【0038】
導入遺伝子は輸送ベクターに含めることができる。本明細書に用いられる、当分野で周知のベクターとは、標的細胞の形質転換(すなわち、外来DNAを個々の細胞のゲノムに組み込むことによってその細胞の遺伝物質を改変する工程)を導くように設計された遺伝子素材を含む核酸コンストラクトを意味する。ベクターは、位置的および順次的に配向された複数の遺伝子要素を含み得る(つまり、そのベクターを微量注入した単細胞受精胚において、カセット内のその核酸が、転写、および必要に応じて翻訳されるように、他の必要または所望の要素が作動可能に連結している)。
【0039】
組換え発現ベクターは、当業者に周知の方法によって、また例えば本明細書の引用文献に記載されているように、上記のヌクレオチド配列をベクターに組み込むことによって構築することができる。本発明に有用な多種多様なベクターが知られている。適切なベクターには、プラスミドベクター、レトロウイルスベクター(例えばMiller et al., 1993を参照)、アデノウイルスベクター(例えばErzurum et al., 1993; Zabner et al., 1994; Davidson et al., 1993を参照)、アデノ随伴ウイルスベクター(例えばFlotte et al., 1993を参照)、ヘルペスウイルスベクター(例えばAnderson et al., 1993を参照)およびレンチウイルスベクター(例えば、Lever, 2000を参照)を含むウイルスベクターなどがある。遺伝子組換え効果を増大させるために、ベクターにトランスポゾン(Davidson et al., 2003; Fadool et al., 1998; Kawakami et al., 2000; Parinov et al., 2004;米国仮特許出願第60/681,447号明細書)またはSce-Iメガヌクレアーゼ部位(Thermes et al., 2002)を含ませることができる。
【0040】
本発明のさらなる局面は、形質転換魚を産生するためのコンストラクトまたはベクターの使用である。本明細書で用いられる形質転換魚とは、外来コンストラクトを導入した魚または魚の子孫を意味する。コンストラクトを導入した魚には、コンストラクトを導入した胚細胞から発生した魚も含まれる。本明細書に用いられる外来コンストラクトとは、動物に、人工的に導入される、または最初に人工的に導入された核酸である。人工的導入なる用語は、正常な生殖または遺伝的交雑によるコンストラクトの導入を除外することを意図している。つまり、交雑育種による、動物の系統(lineまたはstrain)への遺伝子または形質の生来の導入は除くことを意図している。しかしながら、外来コンストラクト(つまり、元々は人工的に導入されたコンストラクト)を持つ魚からそのコンストラクトが通常の繁殖によって移入されて生じた魚は、外来コンストラクトを持つと見なされる。そのような魚は、外来コンストラクトを導入した魚の子孫である。本明細書において用いる魚の子孫とは、有性生殖またはクローニングによってその魚から由来し、その魚から遺伝物質を受け継いだあらゆる魚である。この文脈において、クローニングとは、その魚のDNA、細胞または細胞群から遺伝的に同一の魚を産生することを意味する。別の魚がその魚から由来する魚を祖先または創始者の魚と称する。本明細書において用いる、細胞または細胞群(例えば胚細胞群)からの魚の発生、または細胞または細胞群の魚への発生とは、受精卵または胚細胞(およびそれらの子孫)が成長し、分裂し、分化して成魚を形成する発生過程を意味する。
【0041】
開示するコンストラクトおよび方法はあらゆるタイプの魚に用いることができる。本明細書において用いる魚とは、まとめて魚鋼と称される分類群のあらゆるメンバーを意味する。市販のまたは科学的に関心のある魚の種および変種に属する魚を用いるのが好ましい。そのような魚には、サケ、マス、マグロ、オヒョウ(halibut)、ナマズ、ゼブラフィッシュ、メダカ、コイ、ティラピア、キンギョおよびドジョウなどがあるが、これらに限定されない。
【0042】
開示するコンストラクトおよび方法に用いるために最も好ましい魚は、ゼブラフィッシュ、ダニオ・レリオである。ゼブラフィッシュは一般的な無脊椎実験生物の利点を多く有し、脊椎動物としての利点もさらに含むので、ますます人気の出ている実験動物である。ゼブラフィッシュの別の重要な利点は、線虫類のように大部分が透明であることである(Kimmel, 1989)。一般的なゼブラフィッシュの飼育および維持の仕方はStreisinger (1984)およびWesterfield (2000)に記載されている。
【0043】
ゼブラフィッシュの胚は容易に得られほぼ透明である。これらの特性により、レポータータンパク質および組織特異的発現用配列をコードするコンストラクトを含む形質転換ゼブラフィッシュ胚によって、発生的に調節された遺伝子の空間的および時間的な発現パターンを解析するための高速リアルタイムインビボ系を提供することができる。さらに、ゼブラフィッシュの胚発生は極めて速い。胚は、24時間以内に、機能性の心臓や循環血球を含む全ての主要器官の原基を発生させる(Kimmel, 1989)。同じ望ましい特性のいくつかまたは全てを有する他の魚も好ましい。
【0044】
形質転換魚は本明細書に記載のコンストラクトを用いて調製する。一実施態様において、方法は、核酸、つまり本明細書に記載のコンストラクトまたはベクターを、魚の受精卵(つまり魚の胚など)または魚の未受精卵の核酸に導入することを含む。魚の受精卵を用いる場合、その方法はその魚の胚を形質転換魚に成長させることを含む。核酸を未受精卵に導入する場合、その方法は卵を受精させること、およびその魚の胚を形質転換魚に成長させることを含む。当分野に既知の、機械的方法、化学的方法、親油性方法、レトロウイルス感染法およびエレクトロポレーションなどの様々な方法によって、その核酸を卵に導入することができる。典型的な機械的方法には、例えば微量注入などがある。典型的な化学的方法には、例えばリン酸カルシウムまたはDEAE-デキストラン(DEAE-Dextran)の使用などがある。典型的な親油性の方法には、脂質媒介性トランスフェクションのためのリポソームおよび他のカチオン性物質の使用などがある。そのような方法は一般に当分野でよく知られており、多くのそのような方法が、例えば「Gene Transfer Methods」:「Introducing DNA into Living Cells and Organisms」(P. A. NortonおよびL. F. Steel, 編、Biotechniques Press, 2000);および「Current Protocols in Molecular Biology」(Ausubel et al., 編、John Wiley & Sons)に記載されている。魚に関する微量注入技術は、例えばChen and Powers (1990)およびFletcher and Davis (1991)にさらにより完全に記載されている。魚に関するエレクトロポレーション技術は、例えばPowers et al. (1992)およびLu et al. (1992)にさらにより完全に記載されている。レトロウイルスベクター(汎親和性(pantropic)レトロウイルスベクターなど)を感染させてDNAを魚の卵または胚へ導入する技術は、例えばBurns et al. (1993)にさらに記載されている。
【0045】
導入遺伝子を含むベクターまたは他の核酸は、所望の発生段階で未受精卵または受精卵に導入すればよい。本明細書に記載の異なる導入遺伝子をそれぞれコードする複数のベクターを用いてもよい。受精卵または胚を用いる場合、核酸は胚(つまり発生の1細胞の段階)へ導入するのが好ましい。しかしながら、2細胞の段階、4細胞の段階など、より後の発生段階でも核酸を投与してもよい。従って、核酸を桑実胚、胞胚などに導入してもよい。上記の遺伝子導入コンストラクトが組み込まれた少なくとも1つの単離核酸分子を受精卵(zygote)へ導入する。さらに、より後の発生段階の卵へ核酸を導入する場合、上記の遺伝子導入コンストラクトが組み込まれた少なくとも1つの単離核酸分子を、例えば桑実胚、胞胚などの少なくとも1つの細胞に導入する。
【0046】
魚の卵は適切な魚から標準法により得ることができる。多くの魚は例えばペットショップから購入できる。受精卵は当分野で既知の方法によって得ることができる。例えば、所望の数の、約3から約12月齢のような適切な年齢の魚を、所望のメスとオスの割合(例えば約2:1)で、適切なサイズの水槽などの容器に入れるとよい。例えば交尾の後、適切な時間(例えば約10から60分間)水槽の繁殖部屋(nuptial chamber)に魚をおいて、卵を採取してもよい。そのような方法は例えばCulp et al. (1991)に記載されている。あるいは、当業者に周知の方法によって魚の卵を人工的に受精させてもよい。当業者はそのような魚の受精卵を得る他の方法にも精通している。
【0047】
核酸コンストラクトを魚の卵または胚へ導入した後、成魚への成長を促す環境にその魚の卵または胚をおく。そのような環境は、例えばE3卵海水(egg water)において28.5℃で15日間成長させ、次いで循環系の水に16日目まで入れておくことを含み得る(Westerfield, 2000)。
【0048】
導入遺伝子を持つ魚は、あらゆる適切な手段によって同定することができる。ゼブラフィッシュの胚発生段階で遺伝子発現を検出または評価するには、さらなる外来因子を要さずに直接検出できるEGFPのようなレポータータンパク質を用いるのが好ましい。レポータータンパク質をコードするコンストラクトを含む形質転換ゼブラフィッシュ胚によって、発生的に調節された遺伝子の空間的および時間的な発現パターンを解析するための高速リアルタイムインビボ系を提供することができる。あるいは、形質転換魚である可能性がある魚のゲノムをコンストラクト配列の存在について探索することができる。実際に導入遺伝子を発現する形質転換魚を同定するために、発現産物の存在をアッセイすることもできる。そのような同定のためのいくつかの技術が知られていて、形質転換動物に用いられており、ほとんどは形質転換魚に適用することができる。導入遺伝子コンストラクトに存在するまたはそのコンストラクトに特徴的な核酸配列についての、形質転換魚である可能性がある魚または実際の形質転換魚の探索は、好ましくはサザンまたはノーザンブロッティングによって行う。ポリメラーゼ連鎖反応(PCR)または他の配列特異的核酸増幅技術を用いて検出するのも好ましい。形質転換ゼブラフィッシュを同定するための好ましい技術を実施例に記載する。
【0049】
開示する形質転換魚における発現パターンの同定は、種々の組織の(組織特異的発現)、発生の様々な時点の(発生的に調節された発現または発生段階に特異的な発現)、種々の細胞系列の(細胞系列に特異的な発現)導入遺伝子の発現を測定または同定することによって行うことができる。これらの評価は、例えば発生過程の細胞系列における発現を測定する(および変化があれば観察する)ことと組み合わせることもできる。検出すべき発現産物の性質が、これらの分析のうちのいくつかの好適性に影響する可能性がある。ある面では、魚の種々の組織を解剖し、個々の組織サンプルにおける発現をアッセイすることができる。そのような評価は、ほとんど全ての発現産物について行うことができる。この技術は形質転換動物に共通して用いられ、組織特異的発現の評価に有用である。発現は、生化学的に、酵素学的に、表現型的にまたはモデル魚において測定することができる。
【0050】
この技術はまた、種々の発生段階において発現産物をアッセイして、発生過程における経時的発現を評価するのにも用いることができる。発現産物の検出に、サンプルの固定、または成長する胚または魚を破壊するまたは殺す他の処置が必要な場合、複数の胚を使わなければならない。これは、それぞれの胚の発現パターンが同じかまたは類似すると予測される場合にのみ実用的である。これは、安定で予測可能な発現が起こる本発明の形質転換魚を用いる場合に相当する。発生過程で導入遺伝子の発現パターンを評価するより好ましい方法は、生きた胚および動物において検出可能な発現産物を用いることである。
【0051】
ゼブラフィッシュにおいて、神経系および他の器官の原基は受精後24時間以内に現れる。ほぼ透明のゼブラフィッシュ胚が母親の外側で発生するため、細胞系列の祖先細胞の起源および遊走を、形質転換魚における発現産物の発現を追跡することによりモニターすることができる。さらに、特定遺伝子の調節をこれらの魚において研究することができる。
【0052】
本発明のさらなる局面は、形質転換魚に誘導された腫瘍から不死の腫瘍細胞株を産生すること、およびそうして産生された不死の腫瘍細胞株である。一実施態様において、不死の腫瘍細胞株は、魚において癌遺伝子を発現させて腫瘍を作り出し、腫瘍から細胞を単離し、単離した腫瘍細胞を培養して不死の腫瘍細胞株を産生することによって、得られる。魚の腫瘍から単離した腫瘍細胞から不死の腫瘍細胞株を産生する工程は、当業者に周知の従来技術を用いて行う。適切な技術は、「Culture of Animal Cells」:「A Manual of Basic Techniques」、第5版 (R.I. Freshney, Wiley-Liss, Inc., New York, 2005)、「Animal Cell Culture」:「A Practical Approach」第3版 (J. Masters (編), Oxford University Press, Oxford, 2000)および「Animal Cell Culture」(J.W. PollardおよびJ.M. Walker, Humana Press, Totowa, New Jersey, USA, 1990)に記載されている。一実施態様において、腫瘍細胞は、1細胞段階にPkrt8-EGFP-K-rasB(V12)コンストラクトを注射した魚において生じた腫瘍から単離する。単離された細胞は、培養中で長期間維持でき、不死である。例えば、1つの細胞株は、約24から36時間毎に1細胞分裂という急速な増殖速度を維持しながら、20を超える連続的継代を経ることができた。この腫瘍細胞株は、培養中で多数の継代を通して、そして亜致死的に放射線照射した宿主に移植した後も、EGFP発現を維持した。移植した魚の様々な器官に腫瘍が生じ、その腫瘍細胞株は悪性のものと確認された。このように、Pkrt8-EGFP-K-rasB(V12)誘導性腫瘍の細胞は、培養条件において長期間の増殖能がある。亜致死的にγ線を照射した宿主に培養細胞を移植すると腫瘍が形成され広がっていく。
【0053】
本発明によって産生された腫瘍細胞株は、腫瘍進行および転移のモデルとして有用である。腫瘍細胞株は、異なるコンストラクトによってさらに形質転換でき、または修飾でき、宿主に移植して移植および移植細胞と宿主間の様々な相互作用の研究に用いることができる。さらに、移植した不死の腫瘍細胞株を有する宿主は、腫瘍の進行または転移を阻害する化合物をスクリーニングするのに有用である。また、不死の腫瘍細胞株は腫瘍細胞増殖を阻害する化合物をスクリーニングするのに有用である。移植した宿主または不死の腫瘍細胞株を用いる化合物のスクリーニングは、形質転換魚を用いる化合物のスクリーニングについて本明細書に記載した方法と同様の技術を含む、当業者に周知の技術を用いて行うことができる。
【0054】
本発明の形質転換魚(または不死の腫瘍細胞株を移植した宿主または不死の腫瘍細胞株)は、目的の遺伝子によってコードされたタンパク質に対する効果を調節する化合物をスクリーニングするのに有用である。この実施態様において、癌遺伝子媒介性あるいは誘導性の腫瘍性もしくは過形成性形質転換を調節する、または、放射線もしくは化学療法による処置に対する形質転換細胞の感受性を調節する薬物もしくは物質をスクリーニングする方法を提供する。方法は以下を含む:(a)本明細書に記載の形質転換魚(例えば、成体の形質転換魚または形質転換魚の胚)を被験薬物もしくは物質と接触させる、あるいは曝すこと(ここで、形質転換魚のゲノムには、プロモーターに作動可能に連結した癌遺伝子を含む核酸が安定に取り込まれている);(b)被験薬物もしくは物質が、癌遺伝子媒介性の腫瘍性もしくは過形成性形質転換を抑制するもしくは促進するかどうか、または、放射線もしくは化学療法による処置に対する形質転換細胞の感受性を調節するかどうかを判定する;および(c)その被験薬物もしくは物質が癌遺伝子媒介性の腫瘍性もしくは過形成性形質転換を抑制もしくは促進する、または、放射線もしくは化学療法による処置に対する形質転換細胞の感受性を調節する場合、その被験薬物もしくは物質を癌遺伝子媒介性の腫瘍性もしくは過形成性形質転換を調節する、または、放射線もしくは化学療法による形質転換細胞の処置に対する感受性を調節する薬物もしくは物質として分類すること。本明細書に記載のように、調節には、癌遺伝子媒介性の腫瘍性もしくは過形成性形質転換の速度を含む癌遺伝子媒介性の腫瘍性もしくは過形成性形質転換、または、放射線もしくは化学療法による処置に対する形質転換細胞の感受性を、抑制もしくは減少または促進もしくは刺激することが含まれ得る。被験化合物は、本コンストラクトを含む形質転換魚に投与できる。あるいは好ましくは、形質転換魚が存在する水中にその化合物を投薬して、魚が接触面(例えばえら、皮膚および腸)を通して薬物を取得するようにできる。
【0055】
本発明によって様々な被験化合物が評価できる。特定の実施態様において、試験する化合物はライブラリーに由来する(つまり化合物ライブラリーのメンバーである)。ペプチドライブラリーの使用は当分野において確立されているが、ベンゾジアゼピン(Bunin and Ellman, 1992; DeWitt et al., 1993)、ペプトイド(Zuckermann, 1994)、オリゴカルバメート(Cho et al., 1993)およびヒダントイン(DeWitt et al., 1993)のような他の化合物の混合物が合成できる新しい技術が開発されている。104〜105の多様性を有する生体小分子の分子ライブラリーを合成する手法が報告されている(Carell et al., 1994a; Carell et al., 1994b)。
【0056】
本発明の化合物は、当分野に既知のコンビナトリアル(combinatorial)ライブラリー法の非常に多数の手段のいずれかを用いて得ることができ、それには生物学的ライブラリー;空間的にアドレス可能なパラレル固相または溶液相(spatially addressable parallel solid phase or solution phase)ライブラリー;デコンボリューション(deconvolution)が必要な合成ライブラリー法、「1ビーズ1化合物(one-bead one-compound)」ライブラリー法およびアフィニティークロマトグラフィー選別を用いる合成ライブラリー法などがある。生物学的ライブラリー法はペプチドライブラリーに限定されるが、他の4つの手法はペプチド、非ペプチドオリゴマーまたは小分子の化合物ライブラリーに適用できる(Lam, 1997)。分子ライブラリー合成のための他の典型的な方法は当分野において、例えばErb et al. (1994)、Horwell et al. (1996)およびGallop et al. (1994)に見ることができる。
【0057】
化合物ライブラリーは、溶液中(例えばHoughten et al., 1992)またはビーズ上(Lam et al., 1991)、チップ(Fodor et al., 1993)、細菌(米国特許第5,223,409号明細書)、胞子(米国特許第5,223,409号明細書)、プラスミド(Cull et al., 1992)またはファージ上(Scott and Smith, 1990; Devlin et al., 1990; Cwirla et al., 1990; Felici et al., 1991)に存在し得る。さらに別の実施態様において、コンビナトリアルポリペプチドはcDNAライブラリーから合成する。
【0058】
活性によってスクリーニングできる典型的な化合物には、ペプチド、核酸、炭水化物、生体小分子および天然物抽出物ライブラリーなどがあるが、これらに限定されない。
【0059】
被験薬物もしくは物質は一般には、癌遺伝子媒介性の腫瘍性もしくは過形成性形質転換を調節する、または放射線もしくは化学療法による処置に対する形質転換細胞の感受性を調節する能力を有すると想定される薬物もしくは物質を同定するための、大規模なロボット操作による何千もの化合物のスクリーニングから同定される。そのようなスクリーニングは慣習的であり、これらの、および他のスクリーニング法は当業者によく知られている。被験薬物または物質は、癌遺伝子RNAおよび/または癌遺伝子タンパク質産物の発現、または発癌性の形質転換過程に関する他の遺伝子のRNAまたはタンパク質発現を、抑制あるいは改変、または促進し得る。さらに、被験薬物もしくは物質は、腫瘍性/過形成性の形質転換過程、または放射線もしくは化学療法による処置に対する形質転換細胞の感受性に、直接または間接的に関与する他の分子の活性を、阻害または刺激し得る。広範な薬物もしくは物質(本明細書に記載したものや他のものなど)が本発明のスクリーニング法において試験できる。
【0060】
被験薬物もしくは物質は一般には、癌遺伝子媒介性の腫瘍性または過形成性形質転換を抑制あるいは改変または促進するのに必要な量および期間で投与する。そのような量および期間は当業者が既知の標準的手順によって決定すればよい。形質転換魚は一般に所望の期間、被験薬物もしくは物質に接触させる。
【0061】
本発明の一実施態様において、被験薬物または物質が、癌遺伝子媒介性の腫瘍性もしくは過形成性形質転換を抑制あるいは改変または促進するかどうかの判定は、魚に形成された腫瘍の量および/または大きさを測定する、および/または腫瘍形成の発症率(rate of onset)を測定することによって行ってもよい。癌遺伝子媒介性あるいは誘導性の腫瘍性もしくは過形成性形質転換、または放射線もしくは化学療法による処置に対する形質転換細胞の感受性の調節の他の指標を測定することもできる。例えば、レポーター遺伝子−癌遺伝子融合コントラクトを用いて、当業者に周知の方法および本明細書に記載の方法によって、レポーター遺伝子の発現を測定することができる。例えば、EGFP-癌遺伝子融合コンストラクトに作動可能に連結した組織特異的プロモーターを利用して、測定すべき特定組織において特異的に発現されたタンパク質からEGFP 蛍光を放射させることができるであろう。転移性腫瘍についてのさらなる可視的なまたはその他のスクリーニングを用いることもできる。
【0062】
本発明のさらに別の形態において、癌遺伝子媒介性あるいは誘導性の悪性腫瘍を含む腫瘍性もしくは過形成性形質転換(腫瘍性もしくは過形成性増殖の発症率など)を調節する(つまり促進、抑制あるいは改変する)変異を同定する方法を提供する。一形態において、方法は、遺伝子修飾因子(genetic modifier)スクリーニングの使用を含む。そのようなスクリーニングは本明細書に記載の形質転換魚の順遺伝学的適応性(forward genetic capability)を利用する。悪性腫瘍の発症率を促進する変異は、例えば腫瘍抑制遺伝子、癌遺伝子または腫瘍性もしくは過形成性過程に関与する他の遺伝子で観察されうる。ゲノムの安定性を改変する他の変異も悪性腫瘍の発症率を促進しうる。腫瘍性もしくは過形成性増殖の発症率を抑制する変異には、例えば、未だ同定されていないタンパク質を含め、悪性表現型に必要なタンパク質などがある。そのような変異を同定する方法は、米国特許出願公開第2004/0117867 A1号明細書に記載されており、参照により本明細書に組み込まれる。
【0063】
合理的薬物設計の目的は、目的の生物学的に活性なポリペプチドまたはそれらと相互作用する小分子(例えば作動薬、拮抗薬、阻害薬)の構造類似体を産生し、例えばそのポリペプチドの、より活性なまたは安定な形態である薬物、または例えばインビボでポリペプチドの機能を促進または干渉する薬物を作ることである。合理的薬物設計に用いるいくつかの手法には、3次元構造の解析、アラニンスキャニング、分子モデリングおよび抗イディオタイプ抗体の使用などがある。これらの技術は当業者に周知である。そのような技術は、第1のポリペプチドと第2のポリペプチドにより形成されたタンパク質複合体の3次元構造を規定する原子座標を提供すること、その原子座標に基づいて第1のポリペプチドと第2のポリペプチド間の相互作用を干渉することができる化合物を設計または選択することを含み得る。
【0064】
ポリペプチド活性を調節するまたはその活性に作用する物質を同定した後、その物質をさらに調査してもよい。さらに、その物質を製造してもよく、および/または医薬、医薬組成物もしくは薬物のような組成物の調製(つまり製造または製剤化)に用いてもよい。これらを個体に投与してもよい。
【0065】
ポリペプチド機能のモジュレーターとして同定された物質は、本来、ペプチドまたは非ペプチドであり得る。インビボでの多くの医薬的用途において、非ペプチドの「小分子」はしばしば好ましい。従って、医薬的用途のためにその物質(特にペプチドの場合)の模倣体(mimeticまたはmimic)を設計してもよい。
【0066】
既知の医薬的に活性な化合物の模倣体を設計することは、「リード」化合物に基づく医薬品の開発で知られる手法である。活性化合物の合成が困難である、または高額の費用がかかる場合、または活性化合物が特定の投与方法に適さない場合、例えば、純粋なペプチドが消化管でプロテアーゼによってすぐに分解される傾向があるため経口組成物には適さない活性物質である場合に、この手法は望ましいであろう。模倣体の設計、合成および試験は一般に、大量の分子を目的の特性について無作為にスクリーニングするのを避けるために行う。
【0067】
ファルマコフォアが見付かれば、例えば分光学的技術、x線回折データおよびNMRといった様々な情報源からのデータを用いて、例えば立体化学、結合性、大きさおよび/または電荷といった物理学的特性に従って、その構造をモデル化する。計算機解析(Computational analysis)である、類似性マッピング(原子間の結合性よりも、ファルマコフォアの電荷および/または体積をモデル化する)および他の技術を、このモデル化工程に用いることができる。そのような技術には米国特許第 6,080,576号明細書に開示されている技術などがある。
【0068】
次いで、ファルマコフォアを模倣する化学基を付けることができるテンプレート分子を選択する。生じる模倣体が、リード化合物の生物活性を維持しつつ、簡単に合成でき、薬理学的に許容される可能性が高く、インビボで分解しないように、テンプレート分子およびその上に付ける化学基を便宜的に選択することができる。あるいは、その模倣体がペプチドベースである場合、そのペプチドを環化させ、硬性を増大させて安定性をさらに増すことができる。次いで、この手法で見出された1つまたは複数の模倣体を、それらが目的の特性を有するかどうかを確かめるために、またはそれがどの程度示されるかについて、スクリーニングすることができる。次いで、さらなる最適化または修飾を行って、インビボまたは臨床試験のための1以上の最終模倣体を得る。
【0069】
治療介入に関しては、インビボにおいてあらゆる遺伝子の所与の表現型または発現のあらゆる側面を逆転させ、インビトロにおいてタンパク質活性または結合パートナーとの結合を調節するあらゆる化合物を、ヒトにおけるさらなる開発または潜在的な利用の候補として考慮するべきである。被験物質の用量は、当分野に周知の方法を用いて用量反応曲線に基づいて決定できる。
【0070】
本発明にはいくつかの利点がある。始めに、既存の癌モデルとは異なり、本発明は、成魚における腫瘍形成および抑制の動態に焦点を当てる代わりに、発生のかなり早い段階に形質転換細胞に起こる表現型変化についてスクリーニングすることができる。そのような手法には以下の利点がある:(1)約100%の形質転換細胞が初期に同時に腫瘍形成を起こすこと(検出が1度でよく、再現性ある明確な結果が得られることを意味する)および(2)成魚の代わりに胚を用いること(胚が透明で便利であり、実験時間が短く、必要な空間および薬物量がかなり減少し大規模スクリーニングに有利である)。一実施態様において、癌遺伝子発現を駆動する皮膚特異的プロモーターの選択に成功すると、早期致死が起こらず、大集団の安定な導入遺伝子保有者が産生できる(図5を参照)。
【0071】
魚の胚は一般に用いられるが、癌の症状が起こった成魚も様々な用途に用いることができる。成魚の腫瘍はヒトの腫瘍を、より緊密に反映する。成魚における侵襲性で非同期性の腫瘍形成は、誘導可能な発現系(inducible expression systems)(例えばLangenau et al., 2005に記載のCre/Lox)および例えばガラス魚(glass fish)の成魚のような透明の体を持つ種の魚を用いて取り扱うことができる。
【0072】
安定に組み込まれた導入遺伝子を含む魚を用いることに加え、発癌性コンストラクトを注射した魚も用いることができる。微量注入は扱いやすい技術であり、大規模用の媒体に適用できる。この手法には1つ利点がある:特徴的な形状を持つ個々の標識化細胞(例えば個々のGFP標識化細胞)が観察できる。発癌性コンストラクトの注射によっても、発生の始めの3ヵ月以内の成魚の10%以上において様々な癌が誘導される(図3および4を参照)。これらの腫瘍のいくつかは、保有者の早期致死のために、誘導可能な発現系がなくては安定な導入遺伝子として維持するのが困難な可能性がある。
【0073】
エンハンサートラップ法を用いることにより、異なる組織で同じ導入遺伝子の発現を駆動するために、様々な組織または器官特異的プロモーターを有する膨大なコンストラクトを作製する必要がなくなる(そのようなコンストラクトは必要であれば作製できるが)。ゼブラフィッシュにおいて効果的なエンハンサートラップ法がParinov et al. (2004)に記載されている。通常、エンハンサートラップのパターンはタグ化遺伝子と比較して狭く、細胞の特定のサブセットに対してのみ影響する。最も重要なことは、実質的に無制限の変形パターンを有するエンハンサートラップ系を同じDNAコンストラクトを用いて容易に創出できることである。本発明は、魚の様々な組織に対して腫瘍形成(tumorogenesis)のコンビナトリアルな標的化を行うために、エンハンサートラップ法(の効果)を採用する。例えば、本明細書に記載の皮膚特異的プロモーターを用いるにもかかわらず、同じkrt8-GFP-rasコンストラクトを注射した個体の約15%に、3月齢までに様々な内部器官に組織学的に異なる腫瘍が生じる。生じた腫瘍の種類が多いため、解析は複雑になる可能性がある。とはいうものの、安定な導入遺伝子を含み異なる発現パターンを示す形質転換魚を産生することはできる。そのような安定な形質転換魚の早期致死は誘導可能な系を用いると防ぐことができる。
【0074】
形質転換魚において発癌性のrasが発現すると、多くの場合、発生過程において早期致死の原因となり、安定な形質転換腫瘍産生系を作り出し何代にもわたって維持していく上で問題となるであろう。これらの困難は、誘導可能な系(例えばLangenau et al., 2005に記載のCre/Lox、ならびに当業者に周知の他の誘導可能な系)を用いて克服することができる。そのような場合、癌遺伝子ではなくレポーター(例えばGFP)を発現する形質転換系をスクリーニングで選択することができ、成魚へ成長させ、子孫を得ることができる。必要な時に、全胚への(pan-embryonically)注射もしくは感染によるか、またはCre遺伝子をゲノムに含み発現する形質転換魚と交雑することによって、Creリコンビナーゼを供給することにより、癌遺伝子発現を誘導することができる。
【0075】
別段の記述がない限り、本発明の実施には、当分野の技術範囲である化学、分子生物学、微生物学、組換えDNA、遺伝学、免疫学、細胞生物学、細胞培養および形質転換生物学といった従来技術を使用する。例えば、Maniatisら、「Molecular Cloning」(Cold Spring Harbor Laboratory Press、Cold Spring Harbor、New York、1982);Sambrookら、「Molecular Cloning」第2版(Cold Spring Harbor Laboratory Press、Cold Spring Harbor、New York、1989);SambrookおよびRussell、「Molecular Cloning」第3版(Cold Spring Harbor Laboratory Press、Cold Spring Harbor、New York、2001);Ausubelら、「Current Protocols in Molecular Biology」(John Wiley & Sons、2005現在で最新のもの);Glover、「DNA Cloning」(IRL Press、Oxford、1985);Anand、「Techniques for the Analysis of Complex Genomes」(Academic Press、New York、1992);GuthrieおよびFink、「Guide to Yeast Genetics and Molecular Biology」(Academic Press、New York、1991);HarlowおよびLane、「Antibodys」(Cold Spring Harbor Laboratory Press、Cold Spring Harbor、New York、1998);JakobyおよびPastan、1979;「Nucleic Acid Hybridization」(B. D. Hames & S. J. Higgins編1984);「Transcription And Translation」(B. D. Hames & S. J. Higgins編1984);「Culture Of Animal Cells」(R. I. Freshney、Alan R. Liss、Inc.、1987);「Immobilized Cells And Enzymes」(IRL Press、1986);B. Perbal、「A Practical Guide To Molecular Cloning」(1984);専門書(treatise)「Methos In Enzymology」(Academic Press、Inc.、N.Y.);「Gene Transfer Vectors For Mammalian Cells」(J. H. MillerおよびM. P. Calos編、1987、Cold Spring Harbor Laboratory);「Immunochemical Methods In Cell And Molecular Biology」(MayerおよびWalker、編、Academic Press、London、1987);「Handbook Of Experimental Immunology」、I-IV巻 (D. M. WeirおよびC. C. Blackwell、編、1986);Riott、「Essential Immunology」、第6版、(Blackwell Scientific Publications、Oxford、1988);Hoganら、「Manipulating the Mouse Embryo」、(Cold Spring Harbor Laboratory Press、Cold Spring Harbor、N.Y.、1986);Westerfield、M.、「The zebrafish book. A guide for the laboratory use of zebrafish (Danio rerio)」、第4版、(Univ. of Oregon Press、Eugene、Oregon、2000)を参照のこと。
【実施例】
【0076】
本発明を以下の実施例を参照して説明するが、それらは例示のために示すのであって、決して本発明を限定することを意図するものではない。当分野で周知の標準的技術または以下に具体的に記載する技術を利用した。
【0077】
[実施例1]
材料と方法
コンストラクトおよび微量注入
プライマーKras1:ggagccaaGCGG CCGCatgaccgaatataagcttgtg(配列番号9)およびKras2:ggaaggaaGCGGCCGCtcacattaatgcacatt ttgttttg(配列番号10)を用いてRT PCRによってゼブラフィッシュのK-rasBを増幅し、NotI制限酵素で切断し、480 bpのkrt8プロモーター下にEGFP遺伝子を含むETコンストラクト(Parinov et al 2004)のNotI部位にクローニングした(P-krt8-EGFP-TAA(終止)-K-rasB)。オリゴヌクレオチド:ctgtacaagtTaagcggcGgcatgaccgaatataagcttgtggtc gtgggagctgTaggcg(配列番号11)およびKRR:cgcctAcagctcccacgaccacaagcttatattcggtcatgcCg ccgcttAacttgtacag(配列番号12)を用いて、TAA終止コドンを除去し、K-rasの12aaをV(発癌型)に変えるための部位特異的変異誘発を行った。その結果、部分krt8プロモーターから転写されたmRNA産物は、EGFPとK-RasB-V12の融合タンパク質(P-krt8-EGFP-K-rasB-V12)である。
【0078】
5〜10pgのプラスミドDNAを1〜2細胞段階のゼブラフィッシュの胚に注射した。実際のDNA濃度を、胚の生存率が50%になるように経験的に調整した。
【0079】
薬物試験
BIOMOL(登録商標)のICCB Known Bioactives Library(カタログ番号2840)からの薬物を薬物試験に用いた。5 mg/mlのDMSOストック溶液からの標準的な魚用の水(fish water)に薬物を希釈した。注射した胚を受精後8時間目(hpf)に分別し、正常に発生しているものを薬物処理のために選択した。24ウェルマイクロタイタープレートの各ウェルに10から15の胚を蒔き、10 hpfに1.5 mlの薬物溶液で表面を覆った。胚を暗下で28℃でインキュベートした。受精後5日(dpf)まで毎日状態を観察した。
【0080】
画像解析
落射蛍光用付属品(epi-fluorescence attachment)およびDXM1200Fデジタルカメラ(ニコン)を装備したオリジナルソフトウェアのSMZ1500実体拡大顕微鏡(ニコン)を画像解析に用いた。
【0081】
[実施例2]
ゼブラフィッシュK-rasBコード配列
ゼブラフィッシュK-rasをzgc:85725 cDNA配列(GenBank登録番号BC078646; GI:50925043)(Strausberg et al., 2002)に基づいて設計したPCRプライマーを用いて実施例1に記載したように増幅した。クローニングした配列は公表されたGenBank配列に対して7の単一ヌクレオチド置換を含み、いずれもアミノ酸置換は起こさなかった。188のアミノ酸をコードする増幅産物は、ヒトK-ras2タンパク質のイソ型bに対して96%同一(181/188 aa)で、99.5%相同である(図1)。
【0082】
[実施例3]
K-rasB EGFPの融合コンストラクト
N末端の高感度緑色蛍光タンパク質(EGFP)およびC末端の発癌性(V12) zK-rasB間の融合コンストラクトを作製した。その発現は480bpゼブラフィッシュkrt8プロモーター(Gong et al., 2002; Parinov et al., 2004)を用いて制御した。このプロモーターは初期にはEVLの細胞において、後には皮膚上皮において発現を正常に駆動する。加えて、このプロモーターはエンハンサートラップ法(Parinov et al., 2004)に成功裡に用いることができた。この後者の特性は腫瘍形成を様々な組織へ標的化するのに利用する。核酸配列を表1に示し、タンパク質配列を表2に示す。EGFP配列には下線を引き、zK-rasB配列は太字にしている。修飾されたRasのコドン12および対応するアミノ酸(V12)に下線を引いている。活性コンストラクトにおいてはegfpの終止コドンを変異させ(二重下線)、対照においては変異させずにおいた。コンストラクトを1細胞段階のゼブラフィッシュ胚に微量注入した。
【0083】
【表1】
【0084】
【表2】
【0085】
[実施例4]
P-krt8-EGFP-K-rasB(V12)導入遺伝子を発現する細胞は、早期に腫瘍性の形質転換が起こる。
【0086】
導入遺伝子によって創出された効果の解析では、主にGFP陽性の皮膚上皮細胞(krt8プロモーターからの特異的発現)の形状や動態の変化に焦点を当てた。EGFPおよびK-rasコード配列間に終止コドンを含む、対照コンストラクトP-krt8-EGFP-TAA(終止)-K-rasB(V12)を注射した胚から、正常に見える皮膚上皮細胞が生じた(図2a)(Gong et al., 2002; Parinov et al., 2004)。対照とは反対に、P-krt8-EGFP-K-rasB(V12)コンストラクトを注射した、胚のEGFP陽性細胞は、24 hpfまでに異常な発生が観察された。48 hpf時点では、異常がより明らかになった:ほとんどの皮膚上皮細胞は識別可能な丸い形となり(図2b)、多くは大きくなり、いくつかは胚との接触を失い始め、分離された。72 hpf時点で、それらの細胞はほとんどが脱離し、絨毛膜袋内に留まっている胚の周りに残っていた(図2c)。実質的に、全ての形質転換皮膚細胞が5 dpfまでに脱落し、いくつかの体内細胞のみがGFP陽性のままであった。(これらの細胞における導入遺伝子の異所的発現はエンハンサートラップ効果に誘発された可能性が高い。)対照においては正常に狭く繊維様であった筋肉細胞は、星状の構造となりより大きく成長した。神経細胞は、より大きく成長したほかに、異常な突起を形成した。脊索細胞はわずかにより大きくなり、対照と解剖学的に類似しているようであった。形質転換速度は導入遺伝子発現レベルと相関していた。一方では、早期の形質転換にはより強いGFP発現が伴った。他方では、ほとんど検出できないレベルのEGFPを発現する、正常に見える少数の皮膚上皮細胞が、3〜5 dpf時点に依然として観察できた。
【0087】
注射した魚において初めの2週間に高い死亡率が確認された。注射後生き残った魚の約15%に、1〜3月齢までに様々な内部器官において腫瘍が生じた(図3)。これらの腫瘍は導入遺伝子の発現と強く相関している。
【0088】
[実施例5]
F0の魚における腫瘍形成
Ras-EGFPコンストラクトを注射した魚の15%に、1〜3月齢で様々な組織において腫瘍が生じる(図3)。罹患した魚は出血によって容易に同定できた。これらの腫瘍は極めて悪性であり、様々な組織や器官への広範な多発転移を示す(図4A、4B、4C)。F0の魚に生じる癌は様々な組織学的起源のものである。転移のせいで、元の腫瘍部位を同定するのは困難である。しかしながら、それぞれの魚が持つ癌細胞は一種類のみであるので(解剖による)、これらの腫瘍が別個の起源を持つのではなく、元の部位から転移により広がった結果である可能性が高い。腫瘍には常にGFP発現が伴っている。他方で、明らかな腫瘍を持たない組織において強いGFP発現が確認されることがある。それでもやはり、Ras-EGFP導入遺伝子を高レベルで発現する組織は全て、常に解剖学的に異常である。例えば、脳における発現(図3C、3D)は異常な発生を引き起こすが、明らかな腫瘍形成は観察されない。
【0089】
上記をまとめると、魚におけるP-krt8-EGFP-zK-RasB(V12)の発現は、様々な組織学的起源の腫瘍を誘導するのに用いることができる。魚におけるRas誘導性の癌は侵襲性の成長速度および転移を特徴とする。従って、様々な組織特異的プロモーターまたはエンハンサートラップ法を用いるP-krt8-EGFP-zK-RasB(V12)発現は、魚の癌モデル化において有望である。これらの腫瘍が早期発生性および侵襲性であることは、誘導可能な発現系(例えばCre/lox (Langenau et al., 2005))を適用する必要があることを示唆している。
【0090】
[実施例6]
ゲノムにP-krt8-EGFP-zk-RasB(V12)コンストラクトを含む安定な形質転換F1系は、皮膚細胞に腫瘍性形質転換を示し、次の世代を生み出すことができる。
P-krt8-EGFP-zK-RasB(V12)コンストラクトを注射した魚(F0創始者)を用意し、その子孫(F1)をGFP蛍光について解析した。安定なkrt8-EGFP-zK-RasB(V12)導入遺伝子を含む少数のF1ファミリーを単離した。これらの安定な形質転換魚(図5)のGFP陽性の皮膚細胞は、そのコンストラクトを注射した魚のその細胞と似ている(図2b、c):細胞はkrt8-GFP細胞に典型的な平たい形態を失い、平らな層を形成する代わりにクラスターを形成し、無制限に(uncontrollably)分裂し、体表との接触を失う。4〜5 dpfまでに全てのGFP陽性の皮膚細胞が脱落する、本コンストラクトを注射した魚とは違って、今回の安定な形質転換魚は、新しい細胞が脱落細胞に置き換わるように産生される(それらは5日後においても体表に見られる)。皮膚細胞においてのみEGFP-zK-RasB(V12)を発現する形質転換魚は成体まで成長でき、形質転換性の子孫を生み出すことができる。従って、大規模スクリーニングのために十分な数の胚を産生することが可能である。
【0091】
[実施例7]
既知の腫瘍抑制薬を用いたモデル評価
モデルの創薬実現可能性を評価するために、BIOMOL(登録商標)のICCB Known Bioactives Libraryから選択した化合物を用いてミニスクリーニングを行った。EGFP-zK-rasB(V12)コンストラクトを注射した胚を24ウェルマイクロタイタープレート上で様々な薬物で処理した。10 hpf時点に標準的な魚用の水への薬物の投与を開始した。発生における毒性について予めスクリーニングしたデータに基づいて薬物の濃度を計画した。この予備スクリーニングにおける、これらの細胞に対する薬物の効果を表3に示す。
【0092】
【表3】
【0093】
2つのSrcファミリーキナーゼ阻害薬、PP1およびPP2は同様の効果を示した(図2dおよび2e)。形質転換細胞は平らな形状を維持したが成長し続けた。これは、肥大した星状形態の細胞を生じ、胚に接触し続けた。この効果は、胚発生にとって毒性である3μg/mlの濃度で観察された。1μg/ml濃度の毒性は耐容できたが、しかしながら、EGFP陽性細胞に対する特異的効果もかなり減少した。PP2によって、Eカドヘリン媒介性の細胞付着系が回復し癌の転移的拡大が減少することが、先に実証されている(Nam et al., 2002)。これは、その効果がゼブラフィッシュモデルに現れるメカニズムの1つであり得る。
【0094】
マイトジェン活性化プロテインキナーゼキナーゼ(MEK) PD98059 (Alessi et al., 1995; Dudley et al., 1995; Guyton et al., 1996)の阻害薬は3μg/mlの濃度で、EGFP陽性細胞の成長を強く障害する。72 hpfまでに細胞はより小さくなり、平らな形状を保ち、胚に付着し続ける(図2fおよび2g)。発生に対するこの薬物の毒性作用は、10μg/mlにおいてのみ重篤となった。
【0095】
[実施例8]
ゼブラフィッシュ腫瘍細胞株の確立
1細胞段階にPkrt8-EGFP-K-rasB(V12)コンストラクトを注射した魚の1つは、2月齢までに眼に大きな腫瘍を生じた(図6A)。その腫瘍を剪刀で単離して解剖し、エタノールで滅菌し、滅菌した1Xハンクス液(Hank’s balanced salt solution)および1%抗生物質−抗真菌剤ミックス中で数回洗浄し、カミソリの刃で粉砕した。次いでサンプルを0.14ユニット/mlのLiberase Blendzyme 3を加えた1 mlの1Xハンクス液中、28℃で30分間インキュベートした。完全に解離した後、細胞を1Xハンクス液で洗浄し、10% FBSを含む8 mlの成長培地LFD (50% L-15/15% F-12/35%DMEM)および1%の抗生物質−抗真菌剤ミックスを加え、その混合物を培養フラスコ中28℃でインキュベートした。2日後、細胞を成長培地で洗浄して死んで付着していない細胞を除去し、新しい培地に置き換えた。単層のコンフルエンスができるまで細胞を成長させ、FBSを含まないLFDに剥離させ、培養培地に再懸濁し、106細胞を10 mlフラスコに播種した。
【0096】
以下のように、一連の継代を通してこの細胞株を継代した:2〜4x107の集団に達した後、細胞を剥離させ、新しい培地に再懸濁し(1:20)、約106細胞を5〜6日毎に(2〜4x107細胞)10 mlフラスコに継代した。そのような24継代を通して36時間以内に1細胞分裂という速い増殖速度を維持しながら、細胞を成功裡に継代させた。5、8、10および24継代後、細胞株を液体窒素中でLFD/10%FBS/10%DMSO中に凍結させた。
【0097】
[実施例9]
放射線照射した魚への腫瘍細胞株の移植
移植の2日前に、野生型の成魚に24ガンマ線を照射した。7x107細胞(21継代目)をFBSを含まないLFDに剥離させ、1Xハンクス液で2回洗浄し、1Xハンクス液に105細胞/μlになるまで再懸濁した。亜致死的に放射線照射を受けてから2日後の野生型のレシピエントに25μlハミルトンシリンジを用いて10μlの濃縮細胞懸濁液(106細胞)を腹腔内または筋肉内注射した。亜致死的に放射線照射を受けた他の野生型レシピエントに、10μlの1Xハンクス液を注射した(対照集団)。GFP陽性の腫瘍形成が、注射部位だけでなく注射部位から遠く離れた位置にも観察され、腫瘍細胞が活発に伝播していることを示唆している。さらに、腫瘍細胞および腫瘍病巣が同じ宿主の種々の組織および器官に観察された。
【0098】
本発明を記載する文脈における(特に添付の特許請求の範囲の文脈における)「a」や「an」および「本、その(the)」なる用語の使用、および同様の指示体(referents)は、本明細書に特に別に示したり、または文脈上明らかに矛盾したりしない限り、単数および複数の両方を包含すると解釈されたい。「含む(comprising)」、「有する、持つ(having)」、「含む(including)」および「含む、含有する(containing)」なる用語は、特に別に示さない限りオープンエンドの用語(つまり「含むが、限定されない」ことを意味する)として解釈されたい。本明細書において詳述する値の範囲は、本明細書において特に別に示さない限り、単に、その範囲にあるそれぞれ別個の値を個別に示す簡単明瞭な方法として機能することを意図したもので、それぞれ別個の値は、あたかも本明細書に個別に列挙されているように本明細書に包含される。本明細書に記載の全ての方法は、本明細書に特に別に示したり、または文脈上明らかに矛盾したりしない限り、任意の適切な順序(order)で行うことができる。本明細書に記載の任意のおよび全ての実施例、または典型的な例を示す言葉(例えば「例えば、など、のような(such as)」)の使用は、単に本発明をより明らかにすることを意図したもので、特に別に請求しない限り本発明の範囲を限定するものではない。本明細書におけるどの言葉も、請求の範囲に記載していないあらゆる要素を本発明の実施に不可欠なものとして示していると解釈してはいけない。
【0099】
本発明の方法、魚、不死の腫瘍細胞株および組成物は、本明細書に少しだけ開示している実施態様の様々な形態に組み込まれ得ると理解されたい。本発明の実施態様は、本発明者らに既知の本発明を実施するための最良の形態を含めて、本明細書に記載されている。それらの実施態様の様々な形態は、当業者が前記の説明を読めば明らかになるであろう。本発明者らは当業者がそのような様々な形態を必要に応じて用いることを予想し、また本発明者らは本発明が本明細書に具体的に記載されている以外のようにも実施されることを意図している。従って、本発明は、適用法により認められているように特許請求の範囲に列挙した対象の全ての修飾物および等価物を含む。さらに、本明細書に特に別に示したり、または文脈上明らかに矛盾したりしない限り、上記の要素の全ての潜在的な種々の形態におけるあらゆる組み合わせが本発明に包含される。
【0100】
参考文献一覧
Ahuja, H.G. et al. (1990).「The pattern of mutational involvement of RAS genes in human hematologic malignancies determined by DNA amplification and direct sequencing.」Blood 75:1684-1690.
Alessi, D.R. et al. (1995).「PD098059 is a specific inhibitor of the activation of mitogen-activated protein kinase in vitro and in vivo.」J Biol Chem 270:27489-27494.
Allen et al. (1988).「Transgene as probes for active chromosomal domains in mouse development.」Nature 333:852-855.
Altschul, S.F. et al., (1990).「Basic local alignment search tool.」J Mol Biol 215:403-410.
Altschul, S.F. et al. (1997).「Gapped BLAST and PSI-BLAST: a new generation of protein database search programs.」Nucleic Acids Res 25:3389-3402.
Andersen, J.K. et al. (1993).「Herpesvirus-mediated gene delivery into the rat brain: specificity and efficiency of the neuron-specific enolase promoter.」Cell Mol
Neurobiol 13:503-515.
Baliga, S,N. (2001).「Promoter analysis by saturation mutagenesis.」Biol. Proced. Online 3:64-69.
Bos, J.L. (1989).「Ras oncogenes in human cancer: a review.」Cancer Res 49:4682-4689.
Bunin, B.A. and Ellman, J.A. et al. (1992).「A general and expedient method for the solid-phase synthesis of 1,4-benzodiazepine derivatives.」J. Am. Chem. Soc. 114:10997-10998.
Burns, J.C. et al.. (1993).「Vesicular stomatitis virus G glycoprotein pseudotyped retroviral vectors: concentration to very high titer and efficient gene transfer into mammalian and nonmammalian cells.」Proc Natl Acad Sci USA 90:8033-8037.
Carell, E. et al. (1994a).「A Novel Procedure for the Synthesis of Libraries Containing Small Organic Molecules.」Angew. Chem Int. Ed. Engl. 33:2059-2061.
Carell, E. et al. (1994b).「A Solution-Phase Screening Procedure for the Isolation of Active Compounds from a Library of Molecules.」Angew. Chem. Int. Ed. Engl. 33:2061-2064.
Chen, T.T. and Powers, D.A. (1990).「Transgenic fish.」Trends Biotechnol 8:209-215.
Cho, C.Y. et al. (1993).「An Unnatural biopolymer.」Science. 261:1303-1305.
Cull, M.G. et al. (1992).「Screening for Receptor Ligands Using Large Libraries of Peptides Linked to the C Terminus of the lac Repressor.」Proc Natl Acad Sci USA 89:1865-1869.
Culp, P. et al. (1991).「High-frequency germ-line transmission of plasmid DNA sequences injected into fertilized zebrafish eggs.」Proc Natl Acad.Sci USA 88:7953-7957.
Cwirla, S.E. et al. (1990).「Peptides on Phage: A Vast Library of Peptides for Identifying Ligands.」Proc. Natl. Acad. Sci. USA 87:6378-6382.
Davidson, A.E. et al. (2003).「Efficient gene delivery and gene expression in zebrafish using the Sleeping Beauty transposon.」Dev Biol 263:191-202.
Davidson, B.L. et al. (1993).「A model system for in vivo gene transfer into the central nervous system using an adenoviral vector.」Nature Genetics 3:219-223.
Devlin, J.L. et al. (1990).「Random Peptide Libraries: A Source of Specific Protein Binding Molecules.」Science 249:404-406.
DeWitt, S.H. et al. (1993).「Diversomers": An Approach to Nonpeptide, Nonoligomeric Chemical Diversity.」Proc. Natl. Acad. Sci. USA 90:6909-6913.
Dudley, D.T. et al. (1995).「A synthetic inhibitor of the mitogen-activated protein kinase cascade.」Proc Natl Acad Sci USA 92:7686-7689.
Erb, E. et al. (1994).「Recursive Deconvolution of Combinatorial Chemical Libraries.」Proc. Natl. Acad. Sci. USA 91:11422-11426.
Erzurum, S.C. et al. (1993).「Protection of human endothelial cells from oxidant injury by adenovirus-mediated transfer of the human catalase cDNA.」Nucleic Acids Res 21:1607-1612.
Fadool, J.M. et al. (1998).「Transposition of the mariner element from Drosophila mauritiana in zebrafish.」Proc Natl Acad Sci USA 95:5182-5186.
Felici, F. et al. (1991).「Selection of antibody ligands from a large library of oligopeptides expressed on a multivalent exposition vector.」J Mol. Biol. 222:301-310.
Fletcher, G.L., and Davis, P.L. (1991).「Transgenic fish for aquaculture.」In Genetic Engineering, Setlow, J.K., ed., Plenum Press.
Flotte, T.R. et al. (1993).「Stable in vivo expression of the cystic fibrosis transmembrane conductance regulator with an adeno-associated virus vector.」Proc Natl Acad Sci USA 90:10613-10617.
Fodor, S. et al. (1993).「Multiplexed biochemical assays with biological chips.」Nature 364:555-556.
Gallop, M.A. et al. (1994).「Applications of combinatorial technologies to drug discovery. 1. Background and peptide combinatorial libraries.」J. Med. Chem. 37:1233-1251.
Gong, Z. et al. (2001).「Green fluorescent protein (GFP) transgenic fish and their applications.」Genetica 111:213-225.
Gong, Z. et al. (2002).「Green fluorescent protein expression in germ-line transmitted transgenic zebrafish under a stratified epithelial promoter from keratin8.」Dev Dyn 223:204-215.
Gossler et al. (1989).「Mouse Embryonic Stem Cells and Reporter Constructs to Detect Developmentally Regulated Genes.」Science 244:463-465.
Grabher, C. et al. (2003).「Transposon-mediated enhancer trapping in medaka.」Gene 322:57-66.
Guyton, K.Z. et al. (1996).「Mitogen-activated protein kinase (MAPK) activation by butylated hydroxytoluene hydroperoxide: implications for cellular survival and tumor promotion.」Cancer Res 36:3480-3465.
Horwell, D. et al (1996).「'Targeted' molecular diversity: design and development of non-peptide antagonists for cholecystokinin and tachykinin receptors.」Immunopharmacology 33:68-72.
Houghten, R.A. et al. (1992).「The use of synthetic peptide combinatorial libraries for the identification of bioactive peptides.」Biotechniques 13:412-421.
Huang, H. et al. (2001).「Pdx-1 knockdown reduces insulin promoter activity in zebrafish.」Genesis 30:134-136.
Ivics, Z. et al. (1999).「Genetic applications of transposons and other repetitive elements in zebrafish.」Methods in Cell Biology 60:99-131.
Jessen, J.R. et al. (1999).「Artificial chromosome transgenesis reveals long-distance negative regulation of rag1 in zebrafish.」Nat Genet 23:15-16.
Jessen, J.R. et al. (2001).「Concurrent expression of recombination activating genes 1 and 2 in zebrafish olfactory sensory neurons.」Genesis 29:156-162.
Johnson, L. et al. (2001).「Somatic activation of the K-ras oncogene causes early onset lung cancer in mice.」Nature, 410:1111-1115.
Kaeakami, K. et al. (2000).「Identification of a functional transposase of the Tol2 element, an Ac-like element from the Japanese medaka fish, and its transposition in the zebrafish germ lineage.」Proc Natl Acad Sci USA 97:11403-11408.
Kennedy, B.N. et al. (2001).「Isolation of a zebrafish rod opsin promoter to generate a transgenic zebrafish line expressing enhanced green fluorescent protein in rod photoreceptors.」J Biol Chem 276:14037-14043.
Kimmel, C.B. (1989).「Genetics and Early Development of Zebrafish.」Trends Genet 5:283-288.
Kothary et al. (1988).「A transgene containing lacZ inserted into the dystonia locus is expressed in neural tube.」Nature 335:435-437.
Lam, K.S. (1997).「Application of combinatorial library methods in cancer research and drug discovery.」Anticancer Drug Des. 12:145-167.
Lam, K.S. et al. (1991).「A new type of synthetic peptide library for identifying ligand-binding activity.」Nature 354:82-84.
Langenau, D.M. et al. (2005).「Cre/lox-regulated transgenic zebrafish model with conditional myc-induced T cell acute lymphoblastic leukemia.」Proc Natl Acad Sci USA 102:7369-7374.
Lever, A.M. (2000).「Lentiviral vectors: progress and potential.」Curr Opin Mol Ther 2:488-496.
Long, Q. et al. (1997).「GATA-1 expression pattern can be recapitulated in living transgenic zebrafish using GFP reporter gene.」Development 124:4105-11.
Lu, J.K. et al. (1992).「Integration, expression and germ-line transmission of foreign growth hormone genes in medaka (Oryzias latipes).」Molec Mar Biol Biotechnol 1:366-375.
Miller, A.D. et al., (1993).「Use of retroviral vectors for gene transfer and expression.」Methods of Enzymology 217:581-599.
Moss, J.B. et al. (1996).「Green Fluorescent Protein Marks Skeletal Muscle in Murine Cell Lines and Zebrafish.」Gene 173:89-98.
Motoike, T. et al. (2000).「Universal GFP reporter for the study of vascular development.」Genesis 28:75-81.
Nam, J.S. et al. (2002).「Src family kinase inhibitor PP2 restores the E-cadherin/catenin cell adhesion system in human cancer cells and reduces cancer metastasis.」Clin Cancer Res. 8:2430-2436.
O'Kane, et al. (1987).「Detection in situ of Genomic Regulatory Elements in Drosophila.」Proc. Natl. Acad. Sci. USA 84:9123-9127.
Parinov, S. et al. (2004).「Enhancer trap transposable element as a tool for identification of developmentally regulated genes in zebrafish in vivo.」Dev Dyn. 231:449-459.
Park, H.C. et al. (2000).「Analysis of upstream elements in the HuC promoter leads to the establishment of transgenic zebrafish with fluorescent neurons.」Dev Biol 227:279-293.
Powers, D.A. et al. (1992).「Electroporation: a method for transferring genes into the gametes of zebrafish (Brachydanio rerio), channel catfish (Ictalurus punctatus), and common carp (Cyprinus carpio).」Molec Mar Biol Biotechnol 1:301-308.
Scott, J.K. and J.P. Smith (1990).「Searching for Peptide Ligands with an Epitope Library.」Science 249:386-390.
Strausberg et al. 2002. Generation and initial analysis of more than 15,000 full-length human and mouse cDNA sequences. Proc. Natl. Acad. Sci. U.S.A. 99:16899-16903.
Streisinger (1984). Attainment of Minimal Biological Variability and Measurements of Genotoxicity: Production of Homozygous Diploid Zebra Fish. Natl. Cancer Inst. Monogr. 65:53-58.
Thermes, V. et al. (2002).「I-SecI meganuclease mediates highly efficient transgenesis in fish.」Mech Dev 118:91-98.
Westerfield, M. (2000). The Zebrafish Book: A guide for the laboratory use of Zebrafish (Danio rerio), 4th ed., University of Oregon Press, Eugene.
Zabner, J. et al. (1994).「Safety and efficacy of repetitive adenovirus-mediated transfer of CFTR cDNA to airway epithelia of primates and cotton rats.」Nature Genetics 6:75-83.
Zhang, G. et al. (1996).「An enhanced green fluorescent protein allows sensitive detection of gene transfer in mammalian cells.」Biochem Biophys Res Commun 227:707-711.
Zuckermann, R.N. (1994).「Discovery of nanomolar ligands for 7-transmembrane G-protein-coupled receptors from a diverse N-(substituted)glycine peptoid library.」J. Med. Chem. 37:2678-2685.
【図面の簡単な説明】
【0101】
【図1A】図1Aは、ヒト(配列番号1)およびゼブラフィッシュ(配列番号2)のK-rasタンパク質配列のアライメントを示す。ヒトc-K-ras2b:GenBank登録番号NP_004976;バージョンNP_004976.2;GI:15718761。zgc:85725 mRNA (cDNAクローンMGC:85725 IMAGE:6968999) GenBank登録番号BC078646;GI:50925043 (Strausberg et al., 2002)によってコードされたダニオ・レリオZgc:85725:GenBank登録番号NP_001003744;バージョンNP_001003744.1;GI:51230608。「l」は保存アミノ酸残基を表し、「*」はアミノ酸の保存的変化を表す。
【0102】
【図1B】図1Bは、Ras発癌性タンパク質ファミリーメンバーのアライメントを示す。全Rasタンパク質のうち164のN末端アミノ酸は高度に保存されている。残りの25のC末端残基のみがかなり多様である(高頻度可変領域)。ダニオ・レリオZgc:85725:GenBank登録番号NP_001003744;バージョンNP_001003744.1;GI:51230608(配列番号2)。カーステン(Kirsten)マウス肉腫ウイルス癌原遺伝子タンパク質RNA:GenBank 登録番号Z23152;バージョンZ23152.1;GI:939929(配列番号3)。ヒトc-K-ras2タンパク質イソ型b:GenBank登録番号NP_004976;バージョンNP_004976.2;GI:15718761(配列番号1)。ヒトc-K-ras2タンパク質イソ型a:GenBank登録番号NP_203524;バージョンNP_203524.1;GI:15718763(配列番号4)。ヒト形質転換タンパク質p21/H-Ras-1:GenBank登録番号P01112;バージョンP01112;GI:131869(配列番号5)。ヒト形質転換タンパク質N-Ras:GenBank登録番号P01111;バージョンP01111;GI:131883(配列番号6)。ダニオ・レリオp21 N-ras癌遺伝子:GenBank登録番号AAB40625;バージョンAAB40625.1;GI:1778053(配列番号7)。
【0103】
【図2】図2はEGFP-Rasコンストラクトを注射したゼブラフィッシュの細胞の腫瘍性形質転換、および種々の薬物の効果を示す。図2aはK-RasB(V12)の発現を防ぐTAA終止コドンを有するP-krt8-EGFP-TAA(終止)-K-rasB(V12)コンストラクトを注射した対照魚を示す。この対照において、EGFP陽性細胞はP-krt8-EGFPの発現を特徴とする正常な形態を持つ。図2bおよび2cは、EGFP-K-rasB(V12)発現により誘導された腫瘍性の表現型を示す。図2dから2gは、PP2(図2dおよび2e)およびPD98059(図2fおよび2g)の薬物の効果を示す。
【0104】
【図3】図3はP-krt8-EGFP-zK-rasB導入遺伝子を注射したF0成魚における腫瘍形成を示す。図3A(3B)、3C(3D)、3F(3E)および3G(3H)は、4種のF0魚の腫瘍の例と、対応するGFP画像を示す。図3A(3B)、3F(3E)および3G(3H)は体内に腫瘍を有する魚を示す。図3C(3D)は、導入遺伝子発現に伴い脳に重篤な異常を現した魚を示す。全ての魚の腫瘍成長部位に広範な出血が広がっている。図3Eは、腫瘍成長が導入遺伝子の発現と密に関係していることを示す。図3A(3B)、3C(3D)および3G(3H)の魚は2月齢である。図3F(3E)の魚は3月齢の繁殖力のあるメスである。図3G(3H)の魚は1月齢で目に見える症状(広範な出血)までに進行した。
【0105】
【図4】図4は、P-krt8-EGFP-zK-rasB導入遺伝子を注射したF0成魚における腫瘍形成の組織化学的分析を示す。パラフィン包埋切片をヘマトキシリン/エオシンで染色した。図4Aおよび4Bは魚1(図3G、3H)を示す:起源が不明な転移性の癌が様々な組織へ広がっている。図4Cは魚2(図3F、3E)を示す:転移性の線維肉腫が様々な組織へ広がっている。図4Dは健康な魚、すなわち対照を示す。V−椎骨;M−筋肉;K−腎臓;T−腫瘍。
【0106】
【図5】図5は、安定なP-krt8-EGFP-zK-rasB導入遺伝子を含むF1世代の魚を示す。正常な形態のP-krt8-EGFP陽性細胞を示すP-krt8-EGFP対照(図5A)。zK-ras(V12)誘導性の表現型(図5B)。
【0107】
【図6】図6は、Pkrt8-EGFP-K-rasB(V12)誘導性の眼の腫瘍から得た腫瘍細胞株の例の起源および特性を示す。図6Aは、この腫瘍細胞株を産生するのに用いた、腫瘍を単離する前の魚を示す。1細胞の段階で胚にPkrt8-EGFP-K-rasB(V12)コンストラクトを注射すると、2月齢までに眼に大きな腫瘍が成長した。腫瘍はGFP陽性であった。この腫瘍を単離し、細胞を培養中で成長させた。図6Bは、新しい継代が急速な増殖を示してから4日目および6日目の培養細胞の例を示す。図6Cは細胞株における接触阻害の喪失を示す。細胞は、病巣が明らかになるまで、成長しコンフルエンスになった(継代後8日目)。この例に示した形成病巣は、明るいEGFP蛍光を示している。培養細胞は弱いGFP蛍光を放射し、その蛍光は、病巣が非常に明るいほど、それを取り巻く単層においては観察が困難になる。図6Dは培養細胞を注射した魚の腫瘍を示す(注射後12日目)。GFP陽性腫瘍の形成は筋肉において明らかである。図6Eは、図6Dに示した魚における腫瘍形成の組織学的分析を示す。ヘマトキシリン/エオシンで染色したパラフィン包埋切片。v−椎骨;m−筋肉;t−腫瘍。
【技術分野】
【0001】
[発明の背景]
本発明は、発癌性の核酸を含む魚、魚の腫瘍形成モデル、不死の腫瘍細胞株および抗癌剤のスクリーニングを対象とする。
【背景技術】
【0002】
本発明の背景を明らかにするために本明細書にて用いる出版物や他の資料、また具体的には実施に関するさらなる詳細を提供する事例は、参照により組み込まれ、また便宜のために以下の文章中に著者および日付により引用し、さらに補足した参考文献一覧に著者のアルファベット順に列挙する。
【0003】
疾患状態の動物モデルは、特定の疾患の根底にある生化学的メカニズムを同定し、ならびに疾患を根絶するまたはその症状を軽減するための治療剤を発見するのに重要な役割を果たす。例えば、家族性高コレステロール血症のウサギモデル、非インスリン依存性糖尿病のラットモデル、癌のマウスモデルおよび突発性心房血栓症(spontaneous atrial thrombosis)のハムスターモデルが知られている。さらに、マウス、ネコおよびイヌなどのさまざまな種において遺伝子疾患の動物モデルが発症的に生じている。そのような大きな動物を用いる研究にはいくつかの欠点がある。例えば、そのようなモデルに用いる多くの動物は比較的大きな脊椎動物であり、かなり広い研究スペースを取り、餌代もしくは維持費が高く、生殖周期が遅く、一度にもうける子孫が比較的少なく、所望する全ての疾患状態を効率的に模倣することができない。
【0004】
形質転換魚は疾患モデルの開発に最近利用されている。広範な魚がこの目的に利用され得る。典型的な魚には、硬骨魚、例えば、ゼブラフィッシュ(ダニオ・レリオ(Danio rerio))、メダカ(オリジアス・ラティペス(Oryzias latipes))、マミチョグ(フンドゥルス・ヘテロクリトゥス(Fundulus heteroclitus))、カダヤシ(フンドゥルス属)、ナマズ(イクタルルス(Ictalurus)属)(アメリカナマズなど);コイ(キプリヌス(Cyprinus)属)、例えば、一般的なコイ;およびマスまたはサケ(例えば、イワナ(サルベニルス(Salvelinus))属、大西洋サケ(サルモ(Salmo))属および太平洋サケ(オンコリンクス(Oncorhynchus))属)などがある。ゼブラフィッシュは発生、生理学および疾患を様々な角度から調べるための、確立されたモデルとなっている。
【0005】
ゼブラフィッシュは小さく、子宮外で発生し、また世代時間が短いので、特に有利である。ゼブラフィッシュは実験室環境で維持できるほど経済的であり、また繁殖力が高い;一匹のメスは一週間につき何百もの子孫をもうけることができる。それぞれ5日齢になった魚は、心臓、脳、血液および膵臓など哺乳類に存在する臓器系のほとんどを備え、自由に泳ぎ/餌を採る生命体である。ゼブラフィッシュの胚は外部で発生し、透明であるため、インビボで進行する細胞および組織の発生過程を直接目視でき、これにより遺伝子および小分子薬の大規模なスクリーニングが容易になる。過去数年間、多数の出版物において細胞型に制限的な(cell-type restricted)発現パターンで緑色蛍光タンパク質(GFP)を発現する形質転換魚系が報告されている(Gong et al., 2001; Kennedy et al., 2001; Long et al., 1997; Moss et al., 1996; Motoike et al., 2000; Park et al., 2000)。これまで、蛍光形質転換ゼブラフィッシュを用いた研究では、主に細胞や組織の発生過程を撮像することに焦点が置かれていた。そのような形質転換ゼブラフィッシュ系により、組織の形態形成の発生的研究が促進されることに加え、別々の細胞集団を高解像度で撮像することが可能なため、遺伝的および薬理学的スクリーニングも容易となる。
【0006】
癌を誘導する多くの潜在メカニズムは、これから完全に解明しなければならない。これらの疾患において変異した遺伝子を同定することで、癌全体への新たな見識が得られるであろう。さらに、標的化学療法の開発において新しい標的薬物を同定するためには、疾患の進行を抑制または遅延する、または化学療法もしくは放射線誘導性プログラム細胞死に対する感受性を阻害する、遺伝的または化学的抑制因子が同定できる脊椎動物モデル系を用いることが必要であろう。例えば、特異的標的の事前の知識を必要としないモデル系が必要である。調節性標的薬物または物質の安全性および有効性が実証された後に標的の解明を行うと、薬物を発見する過程での時間と費用の両方を節約できる。
【0007】
癌などの様々な疾患状態の細胞および分子遺伝学的特性をさらに理解する必要がある。適切な動物モデルは、癌をより理解し、また癌を処置または予防するためのより有効な薬物の開発を行うために非常に有益である。本発明はこれらの必要性を解決することを目的とする。
【発明の開示】
【0008】
[発明の概要]
本発明は発癌性の核酸を含む魚、魚の腫瘍形成モデル、不死の腫瘍細胞株および抗癌剤のスクリーニングを対象とする。
【0009】
従って、第1の局面において、本発明は、プロモーターに作動可能に連結した、本明細書において癌遺伝子とも称する発癌性の核酸を含むコンストラクトを提供する。このコンストラクトは、好ましくはマーカーをさらに含む。このコンストラクトはベクターの一部ともなり得る。好ましくは、このプロモーターは器官もしくは組織特異的プロモーターまたは最小プロモーターである。当分野で知られるように、癌遺伝子は、発現すると細胞増殖の調節を改変するか、またはプログラム細胞死を阻止する遺伝子である。広範な癌遺伝子が本明細書に記載の核酸コンストラクトに利用できる。癌遺伝子はウイルスまたは細胞起源であり得る。細胞起源の癌遺伝子には内在性癌遺伝子が含まれる。そのような癌遺伝子が発現すると、細胞に腫瘍性または過形成性形質転換が起こる。その癌遺伝子は、その癌遺伝子の完全配列、好ましくはその癌遺伝子の発癌性形態であり得る。またはその癌遺伝子はその癌遺伝子の発癌能を維持するその癌遺伝子のフラグメントであり得る。
【0010】
第2の局面において、本発明は、上記のコンストラクトをゲノムに含む、またはいくつかの細胞においてのみその癌遺伝子を発現する(ランダム/モザイク発現)形質転換魚、特に形質転換ゼブラフィッシュ(ダニオ・レリオ)を提供する。
【0011】
第3の局面において、本発明は、不死の腫瘍細胞株および不死の腫瘍細胞株を産生する方法を提供する。一実施態様において、不死の腫瘍細胞株は、魚において癌遺伝子を発現させて腫瘍を作り出し、腫瘍細胞を単離し、単離した腫瘍細胞を培養して不死の腫瘍細胞株を産生することによって、得られる。
【0012】
第4の局面において、本発明は、癌遺伝子に関連する癌を処置するのに有用な薬物を同定するために、化合物をスクリーニングする方法を提供する。
【0013】
[発明の詳細な説明]
本発明は発癌性の核酸を含む魚、魚の腫瘍形成モデル、不死の腫瘍細胞株および抗癌剤のスクリーニングを記載する。
【0014】
一実施態様において、本発明は、プロモーターに作動可能に連結した、本明細書において癌遺伝子とも称する発癌性の核酸を含むコンストラクトを対象とする。このコンストラクトは、好ましくはマーカーをさらに含む。このコンストラクトはベクターの一部ともなり得る。好ましくは、そのプロモーターは器官特異的プロモーター、組織特異的プロモーターまたは最小プロモーターである。「最小プロモーター」なる語は本明細書において、そのプロモーターが機能性プロモーターを含む最小の配列から構成されることを意味する。最小プロモーターを同定する技術は当分野でよく知られている(Baliga, 2001)。当分野で知られるように、癌遺伝子は、発現すると細胞増殖の調節を改変するか、またはプログラム細胞死を阻止する遺伝子である。広範な癌遺伝子が本明細書に記載の核酸コンストラクトに利用できる。癌遺伝子はウイルスまたは細胞起源であり得る。細胞起源の癌遺伝子には内在性癌遺伝子が含まれる。そのような癌遺伝子が発現すると、細胞に腫瘍性または過形成性形質転換が起こる。癌遺伝子は、その癌遺伝子の完全配列、好ましくはその癌遺伝子の発癌性形態であり得る。またはその癌遺伝子は、その癌遺伝子の発癌能を維持するその癌遺伝子のフラグメントであり得る。
【0015】
典型的な癌遺伝子には、活性化RAS、MYC、SRC、FOS、JUN、MYB、RAS、ABL、BCL2、HOX11、HOX11L2、TAL1/SCL、LMO1、LMO2、EGFR、MYCN、MDM2、CDK4、GLI1、IGF2、活性化EGFR、変異型遺伝子(FLT3-ITDなど)、変異型および活性型のTP53、PAX3、PAX7、BCR/ABL、HER2/NEU、FLT3R、FLT3-ITD、SRC、RAS、ABL、TAN1、PTC、B-RAF、PML-RARアルファ、E2A-PBX1およびNPM-ALK、ならびにPAXおよびFKHR遺伝子ファミリーのメンバーの融合などがある。
【0016】
他の典型的な癌遺伝子は当分野でよく知られており、そのような例のいくつかは、例えば、「The Genetic Basis of Human Cancer」(Vogelstein, B.およびKinzler, K. W.編 McGraw-Hill, New York, N.Y., 1998)に記載されている。そのような遺伝子の相同体も用いることができる。そのような遺伝子の哺乳類相同体は、内在性の魚の遺伝子と区別できるので、好ましい。さらに好ましいのは、そのような遺伝子のヒト相同体である。そのような癌遺伝子に対応する配列は、その癌遺伝子のヒト相同体を含めて周知であり、例えばGenBankデータベースから見付けることができる。
【0017】
癌遺伝子は形質転換魚に発症することを望む癌の形態に基づいて選択する。例えば、RASファミリー(K-、H-またはN-RAS)の変異型または活性化遺伝子は、腎臓、膵臓または結腸癌など幅広い種類の癌の誘導に用いることができ、HOX11およびTAL1はT細胞癌の誘導などに用いることができる。本発明は特定の癌遺伝子配列に限定されない。例えば、その癌遺伝子の形質転換能が増大または低下した、その癌遺伝子ヌクレオチド配列の改変型または本明細書に記載の他の癌遺伝子ヌクレオチド配列も、包含される。
【0018】
本発明の一実施態様において、本発明に利用される癌遺伝子は、発癌性のrasポリペプチド配列をコードする。全Rasタンパク質のうち164のN末端アミノ酸は高度に保存されている(図1B)。残りの25のC末端残基のみがK-、H-およびN-Rasの間でかなり多様である。主にコドン12、13または61に起こる発癌性変異は、GTP加水分解の触媒部位に影響する。Rasの変異型はGTPに結合したままであり、細胞増殖のためのシグナルを構成的に伝達する。ras遺伝子の活性化変異はヒトの癌の約30%に関係している。しかしながら、異なる種類のヒトの癌には異なるRasタンパク質の発癌性形態が見られる。例えば、K-Rasの変異は、膵臓、結腸および腺癌に共通しているが(Bos, 1989)、血液悪性腫瘍は主にN-Rasに変異がある(Ahuja et al., 1990)。
【0019】
別の実施態様において、本発明に利用する癌遺伝子は、発癌性のK-rasポリペプチドをコードする。さらなる実施態様において、本発明に利用する癌遺伝子は、ゼブラフィッシュの発癌性のzK-rasBポリペプチドをコードする(配列番号8)。さらなる実施態様において、本発明に利用する癌遺伝子は、最適にアライメントした場合に、本明細書に記載の発癌性のrasヌクレオチド配列または他の癌遺伝子ヌクレオチド配列のヌクレオチド配列に対して、(適切なヌクレオチド挿入または置換を含んで)少なくとも約60%、好ましくは少なくとも約70%、より好ましくは少なくとも約80%、最も好ましくは少なくとも約90%の同一性を有し得る。別の実施態様において、本発明の発癌性のrasポリペプチドは、配列番号8のポリペプチド、ならびに配列番号8のポリペプチドに対して少なくとも65%の類似性(好ましくは少なくとも65%の同一性)または少なくとも70%の類似性(好ましくは少なくとも70%の同一性)または少なくとも75%の類似性(好ましくは少なくとも75%の同一性)または少なくとも80%の類似性(好ましくは少なくとも80%の同一性)または少なくとも85%の類似性(好ましくは少なくとも85%の同一性)または少なくとも90%の類似性(好ましくは少なくとも90%の同一性)または少なくとも95%の類似性(好ましくは95%の同一性)を有するポリペプチドを含む。当分野で知られるように2ポリペプチド間の「類似性」は、あるポリペプチドのアミノ酸配列とその保存的アミノ酸置換を、第2のポリペプチド配列と比較して決定する。
【0020】
同一性とは、2つのポリペプチドまたは2つのポリヌクレオチド配列鎖間のマッチの同一性により測定される、2配列の関連性の度合を意味する。同一性は容易に計算できる。2つのポリヌクレオチドまたはポリペプチド配列間の同一性を測定する方法は多数存在するものの、「同一性」なる用語は、当業者に周知である(「Computational Molecular Biology」、Lesk, A. M., 編、Oxford University Press, New York, 1988;「Biocomputing: Informatics and Genome Projects」、Smith, D. W., 編、Academic Press, New York, 1993;「Computer Analysis of Sequence Data」、Part I, Griffin, A. M.およびGriffin, H. G., 編、Humana Press, New Jersey, 1994;「Sequence Analysis in Molecular Biology」、von Heinje, G., Academic Press, 1987; および「Sequence Analysis Primer」、Gribskov, M.およびDevereux, J., 編、M Stockton Press, New York, 1991)。2つの配列間の同一性を決定するのに一般に用いられる方法には、「Guide to Huge Computers」、Martin J. Bishop編、Academic Press、San Diego、1994およびCarillo, H.およびLipman, D.,「SIAM J Applied Math.」48:1073 (1988)などに記載の方法があるがこれらに限定されない。同一性を測定するための好ましい方法は、2つの被験配列間に最大のマッチが生じるように構成されている。そのような方法はコンピュータープログラムに体系化されている。2配列間の同一性を決定する好ましいコンピュータープログラム法には、GCG (Genetics Computer Group, Madison Wis.)プログラムパッケージ(Devereux, J., et al., Nucleic Acids Research 12:387 (1984))、BLASTP、BLASTN、FASTA (Altschul et al. (1990); Altschul et al. (1997))などがあるがこれらに限定されない。よく知られているSmith Watermanアルゴリズムも同一性の決定に用いることができる。
【0021】
例として、参照ヌクレオチド配列に対して少なくとも、例えば95%の「同一性」を有するヌクレオチド配列を有するポリヌクレオチドとは、そのポリヌクレオチドのヌクレオチド配列が、参照ヌクレオチド配列の100ヌクレオチドにつき点変異を5つまで含み得る以外は、参照配列に対して同一であることを意図している。言い換えると、参照ヌクレオチド配列に対して少なくとも95%同一なヌクレオチド配列を有するポリヌクレオチドが生じるには、参照配列中の5%までのヌクレオチドが欠失する、または別のヌクレオチドと置換される、あるいは参照配列の全ヌクレオチドの5%までの数のヌクレオチドが、その参照配列に挿入される場合がある。参照配列のこれらの変異は、参照ヌクレオチド配列の5または3末端部位またはそれらの末端部位の間のどこかに生じ、参照配列のヌクレオチド内に個別に分散するか、または参照配列内に1以上の連続した群として分散している可能性がある。同様の考え方が2つのポリペプチド配列間の同一性を決定する場合にも当てはまる。
【0022】
さらに、癌遺伝子は、発癌性rasヌクレオチド配列、または発癌性K-rasヌクレオチド配列、または配列番号8に示すポリペプチドをコードするゼブラフィッシュの発癌性のzK-rasB、または本明細書に記載の他の癌遺伝子ヌクレオチド配列に対して実質的類似性を有するヌクレオチド配列を含み得る。「実質的類似性」なる語は、本明細書において、そのヌクレオチド配列が参照ヌクレオチド配列と十分に類似し、中程度のストリンジェントな条件下において参照ヌクレオチド配列にハイブリダイズするであろうことを意味する。類似性を決定するこの方法は、本発明が属する分野において周知である。手短に説明すると、中程度のストリンジェントな条件は、Sambrook et al. (「Molecular Cloning: A Laboratory Manual」、第2版、1巻、101〜104頁、Cold Spring Harbor Laboratory Press (1989))に、前洗浄溶液に5 X SSC(塩化ナトリウム/クエン酸ナトリウム溶液)、0.5%ドデシル硫酸ナトリウム(SDS)、1.0mMエチレンジアミン四酢酸(EDTA) (pH 8.0)を用いること、ハイブリダイゼーションおよび55℃、5 X SSCの洗浄条件を含むと規定されている。癌遺伝子のヌクレオチド配列にさらに必要なことは、細胞に腫瘍性形質転換を施す能力を有するタンパク質をコードすることである。つまり、そのタンパク質は正常細胞(すなわち非癌性細胞)を癌性細胞(すなわち腫瘍)へと転換する能力を有する。
【0023】
本明細書においては導入遺伝子(本明細書に記載のゲノムに導入する遺伝子)でもある癌遺伝子は、インビトロで合成しても、または生物学的起源から単離してもよい。そのような合成および単離方法は当業者に周知である。本明細書に用いる、「導入遺伝子」または「導入遺伝子(transgene)コンストラクト」または「遺伝子導入(transgenic)コンストラクト」または「遺伝子導入(transgenic)DNA配列」なる用語は、互換的に用いられ、一般に遺伝子産物の発現を開始し、また調節する能力を持つ調節領域(例えばプロモーターおよびエンハンサー配列)を含む核酸分子を意味するが、これに限定されない。遺伝子導入コンストラクトは、関連遺伝子産物の発現および/または安定性を調節するための、ポリアデニル化配列などの相互適合性の他のあらゆるDNA要素も含み得る。導入遺伝子コンストラクトはまた、作動可能に結合したDNA配列の、ゼブラフィッシュのゲノムへの統合を促進するよう機能する他のDNA配列、および組換え核酸配列の増殖、選択、操作および/または移動に用いるあらゆる核酸系(例えばプラスミド発現ベクター)に含まれるあらゆる関連DNA要素を含み得る。
【0024】
導入遺伝子コンストラクトは、魚に導入して形質転換魚を産生するための遺伝子素材である。本明細書において用いられる「形質転換(transgenic)」なる用語は、人工的に導入されたDNA分子を含む生物およびその子孫を意味する。導入方法および、しばしば導入遺伝子コンストラクトの構造によっては、導入遺伝子コンストラクトは外来コンストラクトとなる。導入遺伝子コンストラクトはあらゆる核酸配列からなるものであってもよいが、開示する形質転換魚に用いるためには導入遺伝子コンストラクトは、発現産物をコードする配列に発現用配列(expression sequences)を作動可能に連結させて組み合わせるのが好ましい。導入遺伝子コンストラクトは好ましくは、発現または安定性のための、またはそのコンストラクトを魚ゲノムへ組み込むことを補助するための、他の構成要素をも含む。本明細書に用いられる、作動可能に(operablyまたはoperatively)連結されたと称される、導入遺伝子コンストラクトの各構成要素とは、それらに望まれる目的のために共に機能できるよう連結されている各構成要素を意味する。例えば、プロモーターとコード領域は、プロモーターが機能してコード領域の転写が起こるならば、作動可能に連結している。
【0025】
発現用配列は、開示する導入遺伝子コンストラクトにおいて、コンストラクトにコードされた発現産物の発現を媒介させるために用いられる。本明細書において用いられる、発現用配列には、プロモーター、上流エレメント、エンハンサーおよび応答エレメントが含まれる。開示するコンストラクトに用いる発現用配列は、相同発現用配列であるのが好ましい。開示する形質転換魚に用いる導入遺伝子コンストラクトの構成要素に関して、本明細書に用いる相同とは、その構成要素が用いる魚の種(species)または種類(type)の野生型であるまたはそれに由来することを示す。逆に、非相同とはその構成要素が用いる魚の種または種類の野生型でもなくそれに由来もしないことを示す。
【0026】
本明細書において用いる発現用配列は、2つの主要なクラス、つまりプロモーターおよびエンハンサーに分けられる。プロモーターは一般に、転写開始部位に対して比較的固定された位置において機能するDNA配列(複数の場合もある)である。プロモーターは、RNAポリメラーゼと転写因子の基本的な相互作用に必要なコアエレメントを含み、また上流エレメントおよび応答エレメントを含み得る。エンハンサーは一般に、転写開始部位からの距離が不定の位置において機能する、いずれかの配向性であり得るDNA配列を意味する。エンハンサーは近くのプロモーターから始まる転写を増大するように機能する。エンハンサーはまた、転写の調節を媒介する応答エレメントをしばしば含む。プロモーターも、転写の調節を媒介する応答エレメントを含み得る。
【0027】
エンハンサーは遺伝子の発現を調節する。この作用は、プロモーターに作動可能に連結したレポーター遺伝子を含有する導入コンストラクトがエンハンサーのドメインに挿入された時にだけ発現する、いわゆるエンハンサートラップ法において観察され、利用されている(O’Kane et al., 1987; Allen et al., 1988; Kothary et al., 1988; Gossler et al., 1989; Parinov et al., 2004)。そのような場合、コンストラクトの発現は新たに関連づけられたエンハンサーの様式に従って調節される。開示する形質転換魚に、最小または短いプロモーターのみを有する遺伝子導入コンストラクトを用いて、様々な組織に対して癌遺伝子の発現を駆動することができる。エンハンサートラップ法の利点は、同じコンストラクトを用いて様々な組織に腫瘍を誘導できることである。従って、そのようなコンストラクトを用いて形質転換した形質転換集団をスクリーニングして、組織学的に異なるタイプの腫瘍を有する動物または動物系を産出する。
【0028】
コードされたペプチドまたはタンパク質が発現するためには、導入遺伝子コンストラクトに、RNAに転写されるとコードされた発現産物の翻訳を媒介する配列も必要である。そのような配列は一般に、転写RNAの5'非翻訳領域に見られる。この領域は、そのコンストラクト上の転写開始部位と翻訳開始部位(すなわち開始コドン)間の領域に対応する。コンストラクトの5'非翻訳領域は、そのコンストラクトに用いたプロモーターに正常に関連づけられた5'非翻訳領域、発現産物をコードする配列に正常に関連づけられた5'非翻訳領域、そのプロモーターもしくは発現産物をコードする配列と無関係の遺伝子の5'非翻訳領域、またはこれらの5'非翻訳領域の混成体に由来し得る。好ましくは、5'非翻訳領域は、そのコンストラクトが導入されるべき魚と相同である。好ましくは、5'非翻訳領域は用いたプロモーターと正常に関連づけられたものである。
【0029】
開示する形質転換魚に用いる導入遺伝子コンストラクトは、(発現の検出および定量化のための)レポータータンパク質をコードする。本明細書において用いられるレポータータンパク質は、発現すると特異的に検出することができるあらゆるタンパク質である。レポータータンパク質は、発現用配列からの発現を検出または定量化するのに有用である。例えば、レポータータンパク質をコードするヌクレオチド配列を組織特異的な発現用配列に作動可能に連結すると、系統発生を綿密に研究することができる。そのような研究において、レポータータンパク質は、細胞遊走など、発生過程をモニタリングするためのマーカーとして働く。多くのレポータータンパク質が知られており、他の生物において同様の目的に用いられている。これらには、特異的に検出可能な産物を産生することができる、βガラクトシダーゼ、ルシフェラーゼおよびアルカリフォスファタ−ゼなどの酵素、および直接検出が可能なタンパク質などがある。実質的には、あらゆるタンパク質は、例えばそのタンパク質に特異的な抗体を用いて直接検出することができる。
【0030】
外来因子を加える必要なく直接検出できるレポータータンパク質を用いることは、ゼブラフィッシュの胚発生過程での遺伝子発現を検出または評価するのに好ましい。レポータータンパク質および組織特異的発現用配列をコードするコンストラクトを含む形質転換ゼブラフィッシュ胚によって、発生的に調節された遺伝子の空間的および時間的な発現パターンを解析するための高速リアルタイムインビボ系(rapid real time in vivo system)を提供することができる。
【0031】
開示する導入遺伝子コンストラクトは好ましくは、コンストラクトからの発現または安定性を改善する他の配列を含む。例えば、タンパク質をコードするコンストラクト上にポリアデニル化シグナルを含むと、導入遺伝子からの転写物が確実に加工されmRNAとして輸送される。発現コンストラクトにおけるポリアデニル化シグナルの同定および利用は確立されている。相同ポリアデニル化シグナルを導入遺伝子コンストラクトに用いるのが好ましい。
【0032】
上記の指針によって、癌遺伝子はプロモーターに作動可能に連結している。好ましくは、プロモーターは器官または組織(細胞を含む)特異的プロモーターである。最も好ましくは、プロモーターは上皮細胞に特異的なケラチン8 (krt8)プロモーターである(Gong et al., 2002)。好ましくは、プロモーターは、エンハンサートラップとして用いることができる短縮されたkrt8 プロモーター(Parinov et al., 2004)または最小プロモーターである。プロモーターの他の例には、組換え活性化遺伝子(RAG)(RAG1およびRAG2を含む);T細胞に特異的な、非レセプター型チロシンキナーゼをコードするLCK;IgMエンハンサーエレメントおよびCD2のプロモーターが含まれる。組織制限的(tissue-ristricted)発現を導くいくつかのプロモーターが同定されており、例えば、リンパ系組織のためのゼブラフィッシュRAG1(Jessen et al., 1999)およびゼブラフィッシュRAG2(Jessen et al., 2001)、および神経特異的発現のためのIslet-1(Motoike et al., 2000)、膵臓のためのPDX-1およびインスリン(Huang et al., 2001)などがある。本明細書に記載の組織特異的プロモーターのヌクレオチド配列に対して、少なくとも約70%の同一性、少なくとも約80%の同一性、さらに少なくとも約90%の同一性を有するプロモーターも包含されるが、ただし作動可能に連結した癌遺伝子の転写を促進することが条件である。ほとんどの哺乳類プロモーターは魚においてはあまり働かないことが確認されているので、ゼブラフィッシュ、フグまたは他種の魚のゲノムの調節配列をしばしば目的のコード配列の上流、内部および下流に特異的にクローニングしなければならず、これは当業者に習慣的な方法によって行うことができる。
【0033】
本明細書にて規定したように、ヌクレオチド配列は、別のヌクレオチド配列と機能的関係にある時、別のヌクレオチド配列と「作動可能に連結し」ている。例えば、コード配列がプロモーター配列に作動可能に連結している場合、このことは一般にそのプロモーターがコード配列の転写を促進し得ることを意味する。作動可能に連結しているということは、連結しているDNA配列が、典型的には近接していること、また2つのタンパク質コード領域をつなぐ必要があれば、近接しかつリーティングフレーム内にあることを意味する。エンハンサーはプロモーターから数キロベース離れていても機能し得ること、またイントロン配列は様々な長さであり得ることから、いくつかのヌクレオチド配列は作動可能に連結するが近接しないこともあり得る。
【0034】
本コンストラクトはさらに好ましくは、マーカーまたはレポーター遺伝子を含む。好ましい実施態様において、蛍光タンパク質遺伝子(例えば、EGFP、GFP、RFP、BFP、YFPまたはdsRED2)またはルシフェラーゼタンパク質遺伝子のようなレポーター遺伝子が癌遺伝子に先行する。最も好ましい実施態様において、マーカーは高感度緑色蛍光タンパク質(EGFP)である(Zhang et al., 1996)。EGFPは感度の高いレポータータンパク質なので好ましい。好ましい実施態様において、マーカーと癌遺伝子の融合物は、その融合遺伝子がプロモーターの制御下になるように調製する。マーカーは融合タンパク質のN末端を構成し、癌遺伝子産物は融合タンパク質のC末端を構成するのが好ましい。この好ましい実施態様において、本コンストラクトは、プロモーター−マーカー−癌遺伝子を含むキメラ導入遺伝子を含む。
【0035】
本明細書において具体的なマーカーの使用を開示し論じたが、具体的に開示したマーカーに本発明が限定されるということは決してない。さらに多くのレポータータンパク質が知られており、同様の目的に使用されている。これらには、特異的に検出可能な産物を産生することができる、βガラクトシダーゼ、ルシフェラーゼ、クロラムフェニコール・アシルトランスフェラーゼ(acytransferase)、βグルクロニダーゼおよびアルカリフォスファタ−ゼなどの酵素、および直接検出可能なタンパク質などがある。実質的には、あらゆるタンパク質が、例えばそのタンパク質に特異的な抗体を用いて直接検出することができる。容易に検出できるレポーターであればいずれも、EGFPの代わりに用いてよい。真核生物細胞の陽性または陰性選択に適するさらなるマーカー(および関連する抗生物質)は、とりわけ、SambrookとRussell (2001)、「Molecular Cloning」、第3版、Cold Spring Harbor Laboratory Press、Cold Spring Harbor、New York、および、Ausubelら(1992)、「Current Protocols in Molecular Biology」、John Wiley & Sons(定期的更新を含む)に開示されている。報告されているあらゆるマーカー並びに当分野で知られる他の物は、本発明の実施に用い得る。
【0036】
コンストラクトは、遺伝子コード領域の上流にプロモーター配列を用いて作製することができる。まず、ゲノムのプロモーター配列をマーカー(例えばEGFP)の上流にクローニングし、それらの配列が発生過程でこの蛍光マーカーの発現を組織特異的に駆動できるかどうかを確認し、そうであれば同じ配列をK-rasおよび他の癌遺伝子の発現を駆動するために用いる。当業者に周知の、他の(例えばゼブラフィッシュの)、ドーパミン作動性の(dopaminanergic)神経系のチロシンヒドロキシラーゼ、筋肉系のMYO-D、骨髄系のMPOまたはPU.1をコードする遺伝子からクローニングされたプロモーターのような、器官および組織特異的プロモーターの構築に、同様の方法が利用できる。
【0037】
好ましい実施態様において、本発明のコンストラクトは、ゼブラフィッシュのkrt8プロモーター、マーカーとしてのEGFPおよびzK-rasなどの癌遺伝子を含む。ゼブラフィッシュのkrt8プロモーターは、皮膚の上皮組織において、またエンハンサートラップ効果によって他の様々な組織に対して、導入遺伝子つまり癌遺伝子の発現を駆動するので、用いると有利である(Parinov et al., 2004)。別のプロモーターを利用する場合、そのプロモーターもエンハンサートラップ効果を示すことが好ましい。エンハンサートラップが特定のプロモーターと正常には結びつかない場合、エンハンサートラップ系もコンストラクトの一部にしてもよい。この局面において、組み込み部位に作用する調節領域(例えばエンハンサーおよび/またはリプレッサー)の特性に依拠する無作為なパターンの発現を促進する「エンハンサートラップ」方策を適用し、そのコンストラクトには、対応する活性化配列(例えばUAS(Gal4に特異的な上流活性化配列))の制御下、所定のDNA結合タンパク質(例えばGal4-VP 16融合物)およびレポーター遺伝子産物に作動可能に連結した最小プロモーターエレメントを含むエンハンサートラップ系をさらに含ませ、あとはその遺伝子導入コンストラクトが魚のゲノムに無作為に挿入および/または転位(transpose)されればよい。例えば、エンハンサートラップ系は、転位因子(Sleeping Beauty、Tc1/mariner-likeファミリー(Grabher et al., 2003; Ivics et al., 1999; Parinov et al., 2004)およびトウモロコシAc/Ds(2005年5月17日出願の米国仮特許出願第60/681,447号を参照。参照により本明細書に組み込まれる。)などがあるがこれらに限定されない)を用いてゼブラフィッシュ内に創出でき、また所望の発現パターンを示す魚を繁殖させ、他の形質転換魚と全く同じように利用できる。
【0038】
導入遺伝子は輸送ベクターに含めることができる。本明細書に用いられる、当分野で周知のベクターとは、標的細胞の形質転換(すなわち、外来DNAを個々の細胞のゲノムに組み込むことによってその細胞の遺伝物質を改変する工程)を導くように設計された遺伝子素材を含む核酸コンストラクトを意味する。ベクターは、位置的および順次的に配向された複数の遺伝子要素を含み得る(つまり、そのベクターを微量注入した単細胞受精胚において、カセット内のその核酸が、転写、および必要に応じて翻訳されるように、他の必要または所望の要素が作動可能に連結している)。
【0039】
組換え発現ベクターは、当業者に周知の方法によって、また例えば本明細書の引用文献に記載されているように、上記のヌクレオチド配列をベクターに組み込むことによって構築することができる。本発明に有用な多種多様なベクターが知られている。適切なベクターには、プラスミドベクター、レトロウイルスベクター(例えばMiller et al., 1993を参照)、アデノウイルスベクター(例えばErzurum et al., 1993; Zabner et al., 1994; Davidson et al., 1993を参照)、アデノ随伴ウイルスベクター(例えばFlotte et al., 1993を参照)、ヘルペスウイルスベクター(例えばAnderson et al., 1993を参照)およびレンチウイルスベクター(例えば、Lever, 2000を参照)を含むウイルスベクターなどがある。遺伝子組換え効果を増大させるために、ベクターにトランスポゾン(Davidson et al., 2003; Fadool et al., 1998; Kawakami et al., 2000; Parinov et al., 2004;米国仮特許出願第60/681,447号明細書)またはSce-Iメガヌクレアーゼ部位(Thermes et al., 2002)を含ませることができる。
【0040】
本発明のさらなる局面は、形質転換魚を産生するためのコンストラクトまたはベクターの使用である。本明細書で用いられる形質転換魚とは、外来コンストラクトを導入した魚または魚の子孫を意味する。コンストラクトを導入した魚には、コンストラクトを導入した胚細胞から発生した魚も含まれる。本明細書に用いられる外来コンストラクトとは、動物に、人工的に導入される、または最初に人工的に導入された核酸である。人工的導入なる用語は、正常な生殖または遺伝的交雑によるコンストラクトの導入を除外することを意図している。つまり、交雑育種による、動物の系統(lineまたはstrain)への遺伝子または形質の生来の導入は除くことを意図している。しかしながら、外来コンストラクト(つまり、元々は人工的に導入されたコンストラクト)を持つ魚からそのコンストラクトが通常の繁殖によって移入されて生じた魚は、外来コンストラクトを持つと見なされる。そのような魚は、外来コンストラクトを導入した魚の子孫である。本明細書において用いる魚の子孫とは、有性生殖またはクローニングによってその魚から由来し、その魚から遺伝物質を受け継いだあらゆる魚である。この文脈において、クローニングとは、その魚のDNA、細胞または細胞群から遺伝的に同一の魚を産生することを意味する。別の魚がその魚から由来する魚を祖先または創始者の魚と称する。本明細書において用いる、細胞または細胞群(例えば胚細胞群)からの魚の発生、または細胞または細胞群の魚への発生とは、受精卵または胚細胞(およびそれらの子孫)が成長し、分裂し、分化して成魚を形成する発生過程を意味する。
【0041】
開示するコンストラクトおよび方法はあらゆるタイプの魚に用いることができる。本明細書において用いる魚とは、まとめて魚鋼と称される分類群のあらゆるメンバーを意味する。市販のまたは科学的に関心のある魚の種および変種に属する魚を用いるのが好ましい。そのような魚には、サケ、マス、マグロ、オヒョウ(halibut)、ナマズ、ゼブラフィッシュ、メダカ、コイ、ティラピア、キンギョおよびドジョウなどがあるが、これらに限定されない。
【0042】
開示するコンストラクトおよび方法に用いるために最も好ましい魚は、ゼブラフィッシュ、ダニオ・レリオである。ゼブラフィッシュは一般的な無脊椎実験生物の利点を多く有し、脊椎動物としての利点もさらに含むので、ますます人気の出ている実験動物である。ゼブラフィッシュの別の重要な利点は、線虫類のように大部分が透明であることである(Kimmel, 1989)。一般的なゼブラフィッシュの飼育および維持の仕方はStreisinger (1984)およびWesterfield (2000)に記載されている。
【0043】
ゼブラフィッシュの胚は容易に得られほぼ透明である。これらの特性により、レポータータンパク質および組織特異的発現用配列をコードするコンストラクトを含む形質転換ゼブラフィッシュ胚によって、発生的に調節された遺伝子の空間的および時間的な発現パターンを解析するための高速リアルタイムインビボ系を提供することができる。さらに、ゼブラフィッシュの胚発生は極めて速い。胚は、24時間以内に、機能性の心臓や循環血球を含む全ての主要器官の原基を発生させる(Kimmel, 1989)。同じ望ましい特性のいくつかまたは全てを有する他の魚も好ましい。
【0044】
形質転換魚は本明細書に記載のコンストラクトを用いて調製する。一実施態様において、方法は、核酸、つまり本明細書に記載のコンストラクトまたはベクターを、魚の受精卵(つまり魚の胚など)または魚の未受精卵の核酸に導入することを含む。魚の受精卵を用いる場合、その方法はその魚の胚を形質転換魚に成長させることを含む。核酸を未受精卵に導入する場合、その方法は卵を受精させること、およびその魚の胚を形質転換魚に成長させることを含む。当分野に既知の、機械的方法、化学的方法、親油性方法、レトロウイルス感染法およびエレクトロポレーションなどの様々な方法によって、その核酸を卵に導入することができる。典型的な機械的方法には、例えば微量注入などがある。典型的な化学的方法には、例えばリン酸カルシウムまたはDEAE-デキストラン(DEAE-Dextran)の使用などがある。典型的な親油性の方法には、脂質媒介性トランスフェクションのためのリポソームおよび他のカチオン性物質の使用などがある。そのような方法は一般に当分野でよく知られており、多くのそのような方法が、例えば「Gene Transfer Methods」:「Introducing DNA into Living Cells and Organisms」(P. A. NortonおよびL. F. Steel, 編、Biotechniques Press, 2000);および「Current Protocols in Molecular Biology」(Ausubel et al., 編、John Wiley & Sons)に記載されている。魚に関する微量注入技術は、例えばChen and Powers (1990)およびFletcher and Davis (1991)にさらにより完全に記載されている。魚に関するエレクトロポレーション技術は、例えばPowers et al. (1992)およびLu et al. (1992)にさらにより完全に記載されている。レトロウイルスベクター(汎親和性(pantropic)レトロウイルスベクターなど)を感染させてDNAを魚の卵または胚へ導入する技術は、例えばBurns et al. (1993)にさらに記載されている。
【0045】
導入遺伝子を含むベクターまたは他の核酸は、所望の発生段階で未受精卵または受精卵に導入すればよい。本明細書に記載の異なる導入遺伝子をそれぞれコードする複数のベクターを用いてもよい。受精卵または胚を用いる場合、核酸は胚(つまり発生の1細胞の段階)へ導入するのが好ましい。しかしながら、2細胞の段階、4細胞の段階など、より後の発生段階でも核酸を投与してもよい。従って、核酸を桑実胚、胞胚などに導入してもよい。上記の遺伝子導入コンストラクトが組み込まれた少なくとも1つの単離核酸分子を受精卵(zygote)へ導入する。さらに、より後の発生段階の卵へ核酸を導入する場合、上記の遺伝子導入コンストラクトが組み込まれた少なくとも1つの単離核酸分子を、例えば桑実胚、胞胚などの少なくとも1つの細胞に導入する。
【0046】
魚の卵は適切な魚から標準法により得ることができる。多くの魚は例えばペットショップから購入できる。受精卵は当分野で既知の方法によって得ることができる。例えば、所望の数の、約3から約12月齢のような適切な年齢の魚を、所望のメスとオスの割合(例えば約2:1)で、適切なサイズの水槽などの容器に入れるとよい。例えば交尾の後、適切な時間(例えば約10から60分間)水槽の繁殖部屋(nuptial chamber)に魚をおいて、卵を採取してもよい。そのような方法は例えばCulp et al. (1991)に記載されている。あるいは、当業者に周知の方法によって魚の卵を人工的に受精させてもよい。当業者はそのような魚の受精卵を得る他の方法にも精通している。
【0047】
核酸コンストラクトを魚の卵または胚へ導入した後、成魚への成長を促す環境にその魚の卵または胚をおく。そのような環境は、例えばE3卵海水(egg water)において28.5℃で15日間成長させ、次いで循環系の水に16日目まで入れておくことを含み得る(Westerfield, 2000)。
【0048】
導入遺伝子を持つ魚は、あらゆる適切な手段によって同定することができる。ゼブラフィッシュの胚発生段階で遺伝子発現を検出または評価するには、さらなる外来因子を要さずに直接検出できるEGFPのようなレポータータンパク質を用いるのが好ましい。レポータータンパク質をコードするコンストラクトを含む形質転換ゼブラフィッシュ胚によって、発生的に調節された遺伝子の空間的および時間的な発現パターンを解析するための高速リアルタイムインビボ系を提供することができる。あるいは、形質転換魚である可能性がある魚のゲノムをコンストラクト配列の存在について探索することができる。実際に導入遺伝子を発現する形質転換魚を同定するために、発現産物の存在をアッセイすることもできる。そのような同定のためのいくつかの技術が知られていて、形質転換動物に用いられており、ほとんどは形質転換魚に適用することができる。導入遺伝子コンストラクトに存在するまたはそのコンストラクトに特徴的な核酸配列についての、形質転換魚である可能性がある魚または実際の形質転換魚の探索は、好ましくはサザンまたはノーザンブロッティングによって行う。ポリメラーゼ連鎖反応(PCR)または他の配列特異的核酸増幅技術を用いて検出するのも好ましい。形質転換ゼブラフィッシュを同定するための好ましい技術を実施例に記載する。
【0049】
開示する形質転換魚における発現パターンの同定は、種々の組織の(組織特異的発現)、発生の様々な時点の(発生的に調節された発現または発生段階に特異的な発現)、種々の細胞系列の(細胞系列に特異的な発現)導入遺伝子の発現を測定または同定することによって行うことができる。これらの評価は、例えば発生過程の細胞系列における発現を測定する(および変化があれば観察する)ことと組み合わせることもできる。検出すべき発現産物の性質が、これらの分析のうちのいくつかの好適性に影響する可能性がある。ある面では、魚の種々の組織を解剖し、個々の組織サンプルにおける発現をアッセイすることができる。そのような評価は、ほとんど全ての発現産物について行うことができる。この技術は形質転換動物に共通して用いられ、組織特異的発現の評価に有用である。発現は、生化学的に、酵素学的に、表現型的にまたはモデル魚において測定することができる。
【0050】
この技術はまた、種々の発生段階において発現産物をアッセイして、発生過程における経時的発現を評価するのにも用いることができる。発現産物の検出に、サンプルの固定、または成長する胚または魚を破壊するまたは殺す他の処置が必要な場合、複数の胚を使わなければならない。これは、それぞれの胚の発現パターンが同じかまたは類似すると予測される場合にのみ実用的である。これは、安定で予測可能な発現が起こる本発明の形質転換魚を用いる場合に相当する。発生過程で導入遺伝子の発現パターンを評価するより好ましい方法は、生きた胚および動物において検出可能な発現産物を用いることである。
【0051】
ゼブラフィッシュにおいて、神経系および他の器官の原基は受精後24時間以内に現れる。ほぼ透明のゼブラフィッシュ胚が母親の外側で発生するため、細胞系列の祖先細胞の起源および遊走を、形質転換魚における発現産物の発現を追跡することによりモニターすることができる。さらに、特定遺伝子の調節をこれらの魚において研究することができる。
【0052】
本発明のさらなる局面は、形質転換魚に誘導された腫瘍から不死の腫瘍細胞株を産生すること、およびそうして産生された不死の腫瘍細胞株である。一実施態様において、不死の腫瘍細胞株は、魚において癌遺伝子を発現させて腫瘍を作り出し、腫瘍から細胞を単離し、単離した腫瘍細胞を培養して不死の腫瘍細胞株を産生することによって、得られる。魚の腫瘍から単離した腫瘍細胞から不死の腫瘍細胞株を産生する工程は、当業者に周知の従来技術を用いて行う。適切な技術は、「Culture of Animal Cells」:「A Manual of Basic Techniques」、第5版 (R.I. Freshney, Wiley-Liss, Inc., New York, 2005)、「Animal Cell Culture」:「A Practical Approach」第3版 (J. Masters (編), Oxford University Press, Oxford, 2000)および「Animal Cell Culture」(J.W. PollardおよびJ.M. Walker, Humana Press, Totowa, New Jersey, USA, 1990)に記載されている。一実施態様において、腫瘍細胞は、1細胞段階にPkrt8-EGFP-K-rasB(V12)コンストラクトを注射した魚において生じた腫瘍から単離する。単離された細胞は、培養中で長期間維持でき、不死である。例えば、1つの細胞株は、約24から36時間毎に1細胞分裂という急速な増殖速度を維持しながら、20を超える連続的継代を経ることができた。この腫瘍細胞株は、培養中で多数の継代を通して、そして亜致死的に放射線照射した宿主に移植した後も、EGFP発現を維持した。移植した魚の様々な器官に腫瘍が生じ、その腫瘍細胞株は悪性のものと確認された。このように、Pkrt8-EGFP-K-rasB(V12)誘導性腫瘍の細胞は、培養条件において長期間の増殖能がある。亜致死的にγ線を照射した宿主に培養細胞を移植すると腫瘍が形成され広がっていく。
【0053】
本発明によって産生された腫瘍細胞株は、腫瘍進行および転移のモデルとして有用である。腫瘍細胞株は、異なるコンストラクトによってさらに形質転換でき、または修飾でき、宿主に移植して移植および移植細胞と宿主間の様々な相互作用の研究に用いることができる。さらに、移植した不死の腫瘍細胞株を有する宿主は、腫瘍の進行または転移を阻害する化合物をスクリーニングするのに有用である。また、不死の腫瘍細胞株は腫瘍細胞増殖を阻害する化合物をスクリーニングするのに有用である。移植した宿主または不死の腫瘍細胞株を用いる化合物のスクリーニングは、形質転換魚を用いる化合物のスクリーニングについて本明細書に記載した方法と同様の技術を含む、当業者に周知の技術を用いて行うことができる。
【0054】
本発明の形質転換魚(または不死の腫瘍細胞株を移植した宿主または不死の腫瘍細胞株)は、目的の遺伝子によってコードされたタンパク質に対する効果を調節する化合物をスクリーニングするのに有用である。この実施態様において、癌遺伝子媒介性あるいは誘導性の腫瘍性もしくは過形成性形質転換を調節する、または、放射線もしくは化学療法による処置に対する形質転換細胞の感受性を調節する薬物もしくは物質をスクリーニングする方法を提供する。方法は以下を含む:(a)本明細書に記載の形質転換魚(例えば、成体の形質転換魚または形質転換魚の胚)を被験薬物もしくは物質と接触させる、あるいは曝すこと(ここで、形質転換魚のゲノムには、プロモーターに作動可能に連結した癌遺伝子を含む核酸が安定に取り込まれている);(b)被験薬物もしくは物質が、癌遺伝子媒介性の腫瘍性もしくは過形成性形質転換を抑制するもしくは促進するかどうか、または、放射線もしくは化学療法による処置に対する形質転換細胞の感受性を調節するかどうかを判定する;および(c)その被験薬物もしくは物質が癌遺伝子媒介性の腫瘍性もしくは過形成性形質転換を抑制もしくは促進する、または、放射線もしくは化学療法による処置に対する形質転換細胞の感受性を調節する場合、その被験薬物もしくは物質を癌遺伝子媒介性の腫瘍性もしくは過形成性形質転換を調節する、または、放射線もしくは化学療法による形質転換細胞の処置に対する感受性を調節する薬物もしくは物質として分類すること。本明細書に記載のように、調節には、癌遺伝子媒介性の腫瘍性もしくは過形成性形質転換の速度を含む癌遺伝子媒介性の腫瘍性もしくは過形成性形質転換、または、放射線もしくは化学療法による処置に対する形質転換細胞の感受性を、抑制もしくは減少または促進もしくは刺激することが含まれ得る。被験化合物は、本コンストラクトを含む形質転換魚に投与できる。あるいは好ましくは、形質転換魚が存在する水中にその化合物を投薬して、魚が接触面(例えばえら、皮膚および腸)を通して薬物を取得するようにできる。
【0055】
本発明によって様々な被験化合物が評価できる。特定の実施態様において、試験する化合物はライブラリーに由来する(つまり化合物ライブラリーのメンバーである)。ペプチドライブラリーの使用は当分野において確立されているが、ベンゾジアゼピン(Bunin and Ellman, 1992; DeWitt et al., 1993)、ペプトイド(Zuckermann, 1994)、オリゴカルバメート(Cho et al., 1993)およびヒダントイン(DeWitt et al., 1993)のような他の化合物の混合物が合成できる新しい技術が開発されている。104〜105の多様性を有する生体小分子の分子ライブラリーを合成する手法が報告されている(Carell et al., 1994a; Carell et al., 1994b)。
【0056】
本発明の化合物は、当分野に既知のコンビナトリアル(combinatorial)ライブラリー法の非常に多数の手段のいずれかを用いて得ることができ、それには生物学的ライブラリー;空間的にアドレス可能なパラレル固相または溶液相(spatially addressable parallel solid phase or solution phase)ライブラリー;デコンボリューション(deconvolution)が必要な合成ライブラリー法、「1ビーズ1化合物(one-bead one-compound)」ライブラリー法およびアフィニティークロマトグラフィー選別を用いる合成ライブラリー法などがある。生物学的ライブラリー法はペプチドライブラリーに限定されるが、他の4つの手法はペプチド、非ペプチドオリゴマーまたは小分子の化合物ライブラリーに適用できる(Lam, 1997)。分子ライブラリー合成のための他の典型的な方法は当分野において、例えばErb et al. (1994)、Horwell et al. (1996)およびGallop et al. (1994)に見ることができる。
【0057】
化合物ライブラリーは、溶液中(例えばHoughten et al., 1992)またはビーズ上(Lam et al., 1991)、チップ(Fodor et al., 1993)、細菌(米国特許第5,223,409号明細書)、胞子(米国特許第5,223,409号明細書)、プラスミド(Cull et al., 1992)またはファージ上(Scott and Smith, 1990; Devlin et al., 1990; Cwirla et al., 1990; Felici et al., 1991)に存在し得る。さらに別の実施態様において、コンビナトリアルポリペプチドはcDNAライブラリーから合成する。
【0058】
活性によってスクリーニングできる典型的な化合物には、ペプチド、核酸、炭水化物、生体小分子および天然物抽出物ライブラリーなどがあるが、これらに限定されない。
【0059】
被験薬物もしくは物質は一般には、癌遺伝子媒介性の腫瘍性もしくは過形成性形質転換を調節する、または放射線もしくは化学療法による処置に対する形質転換細胞の感受性を調節する能力を有すると想定される薬物もしくは物質を同定するための、大規模なロボット操作による何千もの化合物のスクリーニングから同定される。そのようなスクリーニングは慣習的であり、これらの、および他のスクリーニング法は当業者によく知られている。被験薬物または物質は、癌遺伝子RNAおよび/または癌遺伝子タンパク質産物の発現、または発癌性の形質転換過程に関する他の遺伝子のRNAまたはタンパク質発現を、抑制あるいは改変、または促進し得る。さらに、被験薬物もしくは物質は、腫瘍性/過形成性の形質転換過程、または放射線もしくは化学療法による処置に対する形質転換細胞の感受性に、直接または間接的に関与する他の分子の活性を、阻害または刺激し得る。広範な薬物もしくは物質(本明細書に記載したものや他のものなど)が本発明のスクリーニング法において試験できる。
【0060】
被験薬物もしくは物質は一般には、癌遺伝子媒介性の腫瘍性または過形成性形質転換を抑制あるいは改変または促進するのに必要な量および期間で投与する。そのような量および期間は当業者が既知の標準的手順によって決定すればよい。形質転換魚は一般に所望の期間、被験薬物もしくは物質に接触させる。
【0061】
本発明の一実施態様において、被験薬物または物質が、癌遺伝子媒介性の腫瘍性もしくは過形成性形質転換を抑制あるいは改変または促進するかどうかの判定は、魚に形成された腫瘍の量および/または大きさを測定する、および/または腫瘍形成の発症率(rate of onset)を測定することによって行ってもよい。癌遺伝子媒介性あるいは誘導性の腫瘍性もしくは過形成性形質転換、または放射線もしくは化学療法による処置に対する形質転換細胞の感受性の調節の他の指標を測定することもできる。例えば、レポーター遺伝子−癌遺伝子融合コントラクトを用いて、当業者に周知の方法および本明細書に記載の方法によって、レポーター遺伝子の発現を測定することができる。例えば、EGFP-癌遺伝子融合コンストラクトに作動可能に連結した組織特異的プロモーターを利用して、測定すべき特定組織において特異的に発現されたタンパク質からEGFP 蛍光を放射させることができるであろう。転移性腫瘍についてのさらなる可視的なまたはその他のスクリーニングを用いることもできる。
【0062】
本発明のさらに別の形態において、癌遺伝子媒介性あるいは誘導性の悪性腫瘍を含む腫瘍性もしくは過形成性形質転換(腫瘍性もしくは過形成性増殖の発症率など)を調節する(つまり促進、抑制あるいは改変する)変異を同定する方法を提供する。一形態において、方法は、遺伝子修飾因子(genetic modifier)スクリーニングの使用を含む。そのようなスクリーニングは本明細書に記載の形質転換魚の順遺伝学的適応性(forward genetic capability)を利用する。悪性腫瘍の発症率を促進する変異は、例えば腫瘍抑制遺伝子、癌遺伝子または腫瘍性もしくは過形成性過程に関与する他の遺伝子で観察されうる。ゲノムの安定性を改変する他の変異も悪性腫瘍の発症率を促進しうる。腫瘍性もしくは過形成性増殖の発症率を抑制する変異には、例えば、未だ同定されていないタンパク質を含め、悪性表現型に必要なタンパク質などがある。そのような変異を同定する方法は、米国特許出願公開第2004/0117867 A1号明細書に記載されており、参照により本明細書に組み込まれる。
【0063】
合理的薬物設計の目的は、目的の生物学的に活性なポリペプチドまたはそれらと相互作用する小分子(例えば作動薬、拮抗薬、阻害薬)の構造類似体を産生し、例えばそのポリペプチドの、より活性なまたは安定な形態である薬物、または例えばインビボでポリペプチドの機能を促進または干渉する薬物を作ることである。合理的薬物設計に用いるいくつかの手法には、3次元構造の解析、アラニンスキャニング、分子モデリングおよび抗イディオタイプ抗体の使用などがある。これらの技術は当業者に周知である。そのような技術は、第1のポリペプチドと第2のポリペプチドにより形成されたタンパク質複合体の3次元構造を規定する原子座標を提供すること、その原子座標に基づいて第1のポリペプチドと第2のポリペプチド間の相互作用を干渉することができる化合物を設計または選択することを含み得る。
【0064】
ポリペプチド活性を調節するまたはその活性に作用する物質を同定した後、その物質をさらに調査してもよい。さらに、その物質を製造してもよく、および/または医薬、医薬組成物もしくは薬物のような組成物の調製(つまり製造または製剤化)に用いてもよい。これらを個体に投与してもよい。
【0065】
ポリペプチド機能のモジュレーターとして同定された物質は、本来、ペプチドまたは非ペプチドであり得る。インビボでの多くの医薬的用途において、非ペプチドの「小分子」はしばしば好ましい。従って、医薬的用途のためにその物質(特にペプチドの場合)の模倣体(mimeticまたはmimic)を設計してもよい。
【0066】
既知の医薬的に活性な化合物の模倣体を設計することは、「リード」化合物に基づく医薬品の開発で知られる手法である。活性化合物の合成が困難である、または高額の費用がかかる場合、または活性化合物が特定の投与方法に適さない場合、例えば、純粋なペプチドが消化管でプロテアーゼによってすぐに分解される傾向があるため経口組成物には適さない活性物質である場合に、この手法は望ましいであろう。模倣体の設計、合成および試験は一般に、大量の分子を目的の特性について無作為にスクリーニングするのを避けるために行う。
【0067】
ファルマコフォアが見付かれば、例えば分光学的技術、x線回折データおよびNMRといった様々な情報源からのデータを用いて、例えば立体化学、結合性、大きさおよび/または電荷といった物理学的特性に従って、その構造をモデル化する。計算機解析(Computational analysis)である、類似性マッピング(原子間の結合性よりも、ファルマコフォアの電荷および/または体積をモデル化する)および他の技術を、このモデル化工程に用いることができる。そのような技術には米国特許第 6,080,576号明細書に開示されている技術などがある。
【0068】
次いで、ファルマコフォアを模倣する化学基を付けることができるテンプレート分子を選択する。生じる模倣体が、リード化合物の生物活性を維持しつつ、簡単に合成でき、薬理学的に許容される可能性が高く、インビボで分解しないように、テンプレート分子およびその上に付ける化学基を便宜的に選択することができる。あるいは、その模倣体がペプチドベースである場合、そのペプチドを環化させ、硬性を増大させて安定性をさらに増すことができる。次いで、この手法で見出された1つまたは複数の模倣体を、それらが目的の特性を有するかどうかを確かめるために、またはそれがどの程度示されるかについて、スクリーニングすることができる。次いで、さらなる最適化または修飾を行って、インビボまたは臨床試験のための1以上の最終模倣体を得る。
【0069】
治療介入に関しては、インビボにおいてあらゆる遺伝子の所与の表現型または発現のあらゆる側面を逆転させ、インビトロにおいてタンパク質活性または結合パートナーとの結合を調節するあらゆる化合物を、ヒトにおけるさらなる開発または潜在的な利用の候補として考慮するべきである。被験物質の用量は、当分野に周知の方法を用いて用量反応曲線に基づいて決定できる。
【0070】
本発明にはいくつかの利点がある。始めに、既存の癌モデルとは異なり、本発明は、成魚における腫瘍形成および抑制の動態に焦点を当てる代わりに、発生のかなり早い段階に形質転換細胞に起こる表現型変化についてスクリーニングすることができる。そのような手法には以下の利点がある:(1)約100%の形質転換細胞が初期に同時に腫瘍形成を起こすこと(検出が1度でよく、再現性ある明確な結果が得られることを意味する)および(2)成魚の代わりに胚を用いること(胚が透明で便利であり、実験時間が短く、必要な空間および薬物量がかなり減少し大規模スクリーニングに有利である)。一実施態様において、癌遺伝子発現を駆動する皮膚特異的プロモーターの選択に成功すると、早期致死が起こらず、大集団の安定な導入遺伝子保有者が産生できる(図5を参照)。
【0071】
魚の胚は一般に用いられるが、癌の症状が起こった成魚も様々な用途に用いることができる。成魚の腫瘍はヒトの腫瘍を、より緊密に反映する。成魚における侵襲性で非同期性の腫瘍形成は、誘導可能な発現系(inducible expression systems)(例えばLangenau et al., 2005に記載のCre/Lox)および例えばガラス魚(glass fish)の成魚のような透明の体を持つ種の魚を用いて取り扱うことができる。
【0072】
安定に組み込まれた導入遺伝子を含む魚を用いることに加え、発癌性コンストラクトを注射した魚も用いることができる。微量注入は扱いやすい技術であり、大規模用の媒体に適用できる。この手法には1つ利点がある:特徴的な形状を持つ個々の標識化細胞(例えば個々のGFP標識化細胞)が観察できる。発癌性コンストラクトの注射によっても、発生の始めの3ヵ月以内の成魚の10%以上において様々な癌が誘導される(図3および4を参照)。これらの腫瘍のいくつかは、保有者の早期致死のために、誘導可能な発現系がなくては安定な導入遺伝子として維持するのが困難な可能性がある。
【0073】
エンハンサートラップ法を用いることにより、異なる組織で同じ導入遺伝子の発現を駆動するために、様々な組織または器官特異的プロモーターを有する膨大なコンストラクトを作製する必要がなくなる(そのようなコンストラクトは必要であれば作製できるが)。ゼブラフィッシュにおいて効果的なエンハンサートラップ法がParinov et al. (2004)に記載されている。通常、エンハンサートラップのパターンはタグ化遺伝子と比較して狭く、細胞の特定のサブセットに対してのみ影響する。最も重要なことは、実質的に無制限の変形パターンを有するエンハンサートラップ系を同じDNAコンストラクトを用いて容易に創出できることである。本発明は、魚の様々な組織に対して腫瘍形成(tumorogenesis)のコンビナトリアルな標的化を行うために、エンハンサートラップ法(の効果)を採用する。例えば、本明細書に記載の皮膚特異的プロモーターを用いるにもかかわらず、同じkrt8-GFP-rasコンストラクトを注射した個体の約15%に、3月齢までに様々な内部器官に組織学的に異なる腫瘍が生じる。生じた腫瘍の種類が多いため、解析は複雑になる可能性がある。とはいうものの、安定な導入遺伝子を含み異なる発現パターンを示す形質転換魚を産生することはできる。そのような安定な形質転換魚の早期致死は誘導可能な系を用いると防ぐことができる。
【0074】
形質転換魚において発癌性のrasが発現すると、多くの場合、発生過程において早期致死の原因となり、安定な形質転換腫瘍産生系を作り出し何代にもわたって維持していく上で問題となるであろう。これらの困難は、誘導可能な系(例えばLangenau et al., 2005に記載のCre/Lox、ならびに当業者に周知の他の誘導可能な系)を用いて克服することができる。そのような場合、癌遺伝子ではなくレポーター(例えばGFP)を発現する形質転換系をスクリーニングで選択することができ、成魚へ成長させ、子孫を得ることができる。必要な時に、全胚への(pan-embryonically)注射もしくは感染によるか、またはCre遺伝子をゲノムに含み発現する形質転換魚と交雑することによって、Creリコンビナーゼを供給することにより、癌遺伝子発現を誘導することができる。
【0075】
別段の記述がない限り、本発明の実施には、当分野の技術範囲である化学、分子生物学、微生物学、組換えDNA、遺伝学、免疫学、細胞生物学、細胞培養および形質転換生物学といった従来技術を使用する。例えば、Maniatisら、「Molecular Cloning」(Cold Spring Harbor Laboratory Press、Cold Spring Harbor、New York、1982);Sambrookら、「Molecular Cloning」第2版(Cold Spring Harbor Laboratory Press、Cold Spring Harbor、New York、1989);SambrookおよびRussell、「Molecular Cloning」第3版(Cold Spring Harbor Laboratory Press、Cold Spring Harbor、New York、2001);Ausubelら、「Current Protocols in Molecular Biology」(John Wiley & Sons、2005現在で最新のもの);Glover、「DNA Cloning」(IRL Press、Oxford、1985);Anand、「Techniques for the Analysis of Complex Genomes」(Academic Press、New York、1992);GuthrieおよびFink、「Guide to Yeast Genetics and Molecular Biology」(Academic Press、New York、1991);HarlowおよびLane、「Antibodys」(Cold Spring Harbor Laboratory Press、Cold Spring Harbor、New York、1998);JakobyおよびPastan、1979;「Nucleic Acid Hybridization」(B. D. Hames & S. J. Higgins編1984);「Transcription And Translation」(B. D. Hames & S. J. Higgins編1984);「Culture Of Animal Cells」(R. I. Freshney、Alan R. Liss、Inc.、1987);「Immobilized Cells And Enzymes」(IRL Press、1986);B. Perbal、「A Practical Guide To Molecular Cloning」(1984);専門書(treatise)「Methos In Enzymology」(Academic Press、Inc.、N.Y.);「Gene Transfer Vectors For Mammalian Cells」(J. H. MillerおよびM. P. Calos編、1987、Cold Spring Harbor Laboratory);「Immunochemical Methods In Cell And Molecular Biology」(MayerおよびWalker、編、Academic Press、London、1987);「Handbook Of Experimental Immunology」、I-IV巻 (D. M. WeirおよびC. C. Blackwell、編、1986);Riott、「Essential Immunology」、第6版、(Blackwell Scientific Publications、Oxford、1988);Hoganら、「Manipulating the Mouse Embryo」、(Cold Spring Harbor Laboratory Press、Cold Spring Harbor、N.Y.、1986);Westerfield、M.、「The zebrafish book. A guide for the laboratory use of zebrafish (Danio rerio)」、第4版、(Univ. of Oregon Press、Eugene、Oregon、2000)を参照のこと。
【実施例】
【0076】
本発明を以下の実施例を参照して説明するが、それらは例示のために示すのであって、決して本発明を限定することを意図するものではない。当分野で周知の標準的技術または以下に具体的に記載する技術を利用した。
【0077】
[実施例1]
材料と方法
コンストラクトおよび微量注入
プライマーKras1:ggagccaaGCGG CCGCatgaccgaatataagcttgtg(配列番号9)およびKras2:ggaaggaaGCGGCCGCtcacattaatgcacatt ttgttttg(配列番号10)を用いてRT PCRによってゼブラフィッシュのK-rasBを増幅し、NotI制限酵素で切断し、480 bpのkrt8プロモーター下にEGFP遺伝子を含むETコンストラクト(Parinov et al 2004)のNotI部位にクローニングした(P-krt8-EGFP-TAA(終止)-K-rasB)。オリゴヌクレオチド:ctgtacaagtTaagcggcGgcatgaccgaatataagcttgtggtc gtgggagctgTaggcg(配列番号11)およびKRR:cgcctAcagctcccacgaccacaagcttatattcggtcatgcCg ccgcttAacttgtacag(配列番号12)を用いて、TAA終止コドンを除去し、K-rasの12aaをV(発癌型)に変えるための部位特異的変異誘発を行った。その結果、部分krt8プロモーターから転写されたmRNA産物は、EGFPとK-RasB-V12の融合タンパク質(P-krt8-EGFP-K-rasB-V12)である。
【0078】
5〜10pgのプラスミドDNAを1〜2細胞段階のゼブラフィッシュの胚に注射した。実際のDNA濃度を、胚の生存率が50%になるように経験的に調整した。
【0079】
薬物試験
BIOMOL(登録商標)のICCB Known Bioactives Library(カタログ番号2840)からの薬物を薬物試験に用いた。5 mg/mlのDMSOストック溶液からの標準的な魚用の水(fish water)に薬物を希釈した。注射した胚を受精後8時間目(hpf)に分別し、正常に発生しているものを薬物処理のために選択した。24ウェルマイクロタイタープレートの各ウェルに10から15の胚を蒔き、10 hpfに1.5 mlの薬物溶液で表面を覆った。胚を暗下で28℃でインキュベートした。受精後5日(dpf)まで毎日状態を観察した。
【0080】
画像解析
落射蛍光用付属品(epi-fluorescence attachment)およびDXM1200Fデジタルカメラ(ニコン)を装備したオリジナルソフトウェアのSMZ1500実体拡大顕微鏡(ニコン)を画像解析に用いた。
【0081】
[実施例2]
ゼブラフィッシュK-rasBコード配列
ゼブラフィッシュK-rasをzgc:85725 cDNA配列(GenBank登録番号BC078646; GI:50925043)(Strausberg et al., 2002)に基づいて設計したPCRプライマーを用いて実施例1に記載したように増幅した。クローニングした配列は公表されたGenBank配列に対して7の単一ヌクレオチド置換を含み、いずれもアミノ酸置換は起こさなかった。188のアミノ酸をコードする増幅産物は、ヒトK-ras2タンパク質のイソ型bに対して96%同一(181/188 aa)で、99.5%相同である(図1)。
【0082】
[実施例3]
K-rasB EGFPの融合コンストラクト
N末端の高感度緑色蛍光タンパク質(EGFP)およびC末端の発癌性(V12) zK-rasB間の融合コンストラクトを作製した。その発現は480bpゼブラフィッシュkrt8プロモーター(Gong et al., 2002; Parinov et al., 2004)を用いて制御した。このプロモーターは初期にはEVLの細胞において、後には皮膚上皮において発現を正常に駆動する。加えて、このプロモーターはエンハンサートラップ法(Parinov et al., 2004)に成功裡に用いることができた。この後者の特性は腫瘍形成を様々な組織へ標的化するのに利用する。核酸配列を表1に示し、タンパク質配列を表2に示す。EGFP配列には下線を引き、zK-rasB配列は太字にしている。修飾されたRasのコドン12および対応するアミノ酸(V12)に下線を引いている。活性コンストラクトにおいてはegfpの終止コドンを変異させ(二重下線)、対照においては変異させずにおいた。コンストラクトを1細胞段階のゼブラフィッシュ胚に微量注入した。
【0083】
【表1】
【0084】
【表2】
【0085】
[実施例4]
P-krt8-EGFP-K-rasB(V12)導入遺伝子を発現する細胞は、早期に腫瘍性の形質転換が起こる。
【0086】
導入遺伝子によって創出された効果の解析では、主にGFP陽性の皮膚上皮細胞(krt8プロモーターからの特異的発現)の形状や動態の変化に焦点を当てた。EGFPおよびK-rasコード配列間に終止コドンを含む、対照コンストラクトP-krt8-EGFP-TAA(終止)-K-rasB(V12)を注射した胚から、正常に見える皮膚上皮細胞が生じた(図2a)(Gong et al., 2002; Parinov et al., 2004)。対照とは反対に、P-krt8-EGFP-K-rasB(V12)コンストラクトを注射した、胚のEGFP陽性細胞は、24 hpfまでに異常な発生が観察された。48 hpf時点では、異常がより明らかになった:ほとんどの皮膚上皮細胞は識別可能な丸い形となり(図2b)、多くは大きくなり、いくつかは胚との接触を失い始め、分離された。72 hpf時点で、それらの細胞はほとんどが脱離し、絨毛膜袋内に留まっている胚の周りに残っていた(図2c)。実質的に、全ての形質転換皮膚細胞が5 dpfまでに脱落し、いくつかの体内細胞のみがGFP陽性のままであった。(これらの細胞における導入遺伝子の異所的発現はエンハンサートラップ効果に誘発された可能性が高い。)対照においては正常に狭く繊維様であった筋肉細胞は、星状の構造となりより大きく成長した。神経細胞は、より大きく成長したほかに、異常な突起を形成した。脊索細胞はわずかにより大きくなり、対照と解剖学的に類似しているようであった。形質転換速度は導入遺伝子発現レベルと相関していた。一方では、早期の形質転換にはより強いGFP発現が伴った。他方では、ほとんど検出できないレベルのEGFPを発現する、正常に見える少数の皮膚上皮細胞が、3〜5 dpf時点に依然として観察できた。
【0087】
注射した魚において初めの2週間に高い死亡率が確認された。注射後生き残った魚の約15%に、1〜3月齢までに様々な内部器官において腫瘍が生じた(図3)。これらの腫瘍は導入遺伝子の発現と強く相関している。
【0088】
[実施例5]
F0の魚における腫瘍形成
Ras-EGFPコンストラクトを注射した魚の15%に、1〜3月齢で様々な組織において腫瘍が生じる(図3)。罹患した魚は出血によって容易に同定できた。これらの腫瘍は極めて悪性であり、様々な組織や器官への広範な多発転移を示す(図4A、4B、4C)。F0の魚に生じる癌は様々な組織学的起源のものである。転移のせいで、元の腫瘍部位を同定するのは困難である。しかしながら、それぞれの魚が持つ癌細胞は一種類のみであるので(解剖による)、これらの腫瘍が別個の起源を持つのではなく、元の部位から転移により広がった結果である可能性が高い。腫瘍には常にGFP発現が伴っている。他方で、明らかな腫瘍を持たない組織において強いGFP発現が確認されることがある。それでもやはり、Ras-EGFP導入遺伝子を高レベルで発現する組織は全て、常に解剖学的に異常である。例えば、脳における発現(図3C、3D)は異常な発生を引き起こすが、明らかな腫瘍形成は観察されない。
【0089】
上記をまとめると、魚におけるP-krt8-EGFP-zK-RasB(V12)の発現は、様々な組織学的起源の腫瘍を誘導するのに用いることができる。魚におけるRas誘導性の癌は侵襲性の成長速度および転移を特徴とする。従って、様々な組織特異的プロモーターまたはエンハンサートラップ法を用いるP-krt8-EGFP-zK-RasB(V12)発現は、魚の癌モデル化において有望である。これらの腫瘍が早期発生性および侵襲性であることは、誘導可能な発現系(例えばCre/lox (Langenau et al., 2005))を適用する必要があることを示唆している。
【0090】
[実施例6]
ゲノムにP-krt8-EGFP-zk-RasB(V12)コンストラクトを含む安定な形質転換F1系は、皮膚細胞に腫瘍性形質転換を示し、次の世代を生み出すことができる。
P-krt8-EGFP-zK-RasB(V12)コンストラクトを注射した魚(F0創始者)を用意し、その子孫(F1)をGFP蛍光について解析した。安定なkrt8-EGFP-zK-RasB(V12)導入遺伝子を含む少数のF1ファミリーを単離した。これらの安定な形質転換魚(図5)のGFP陽性の皮膚細胞は、そのコンストラクトを注射した魚のその細胞と似ている(図2b、c):細胞はkrt8-GFP細胞に典型的な平たい形態を失い、平らな層を形成する代わりにクラスターを形成し、無制限に(uncontrollably)分裂し、体表との接触を失う。4〜5 dpfまでに全てのGFP陽性の皮膚細胞が脱落する、本コンストラクトを注射した魚とは違って、今回の安定な形質転換魚は、新しい細胞が脱落細胞に置き換わるように産生される(それらは5日後においても体表に見られる)。皮膚細胞においてのみEGFP-zK-RasB(V12)を発現する形質転換魚は成体まで成長でき、形質転換性の子孫を生み出すことができる。従って、大規模スクリーニングのために十分な数の胚を産生することが可能である。
【0091】
[実施例7]
既知の腫瘍抑制薬を用いたモデル評価
モデルの創薬実現可能性を評価するために、BIOMOL(登録商標)のICCB Known Bioactives Libraryから選択した化合物を用いてミニスクリーニングを行った。EGFP-zK-rasB(V12)コンストラクトを注射した胚を24ウェルマイクロタイタープレート上で様々な薬物で処理した。10 hpf時点に標準的な魚用の水への薬物の投与を開始した。発生における毒性について予めスクリーニングしたデータに基づいて薬物の濃度を計画した。この予備スクリーニングにおける、これらの細胞に対する薬物の効果を表3に示す。
【0092】
【表3】
【0093】
2つのSrcファミリーキナーゼ阻害薬、PP1およびPP2は同様の効果を示した(図2dおよび2e)。形質転換細胞は平らな形状を維持したが成長し続けた。これは、肥大した星状形態の細胞を生じ、胚に接触し続けた。この効果は、胚発生にとって毒性である3μg/mlの濃度で観察された。1μg/ml濃度の毒性は耐容できたが、しかしながら、EGFP陽性細胞に対する特異的効果もかなり減少した。PP2によって、Eカドヘリン媒介性の細胞付着系が回復し癌の転移的拡大が減少することが、先に実証されている(Nam et al., 2002)。これは、その効果がゼブラフィッシュモデルに現れるメカニズムの1つであり得る。
【0094】
マイトジェン活性化プロテインキナーゼキナーゼ(MEK) PD98059 (Alessi et al., 1995; Dudley et al., 1995; Guyton et al., 1996)の阻害薬は3μg/mlの濃度で、EGFP陽性細胞の成長を強く障害する。72 hpfまでに細胞はより小さくなり、平らな形状を保ち、胚に付着し続ける(図2fおよび2g)。発生に対するこの薬物の毒性作用は、10μg/mlにおいてのみ重篤となった。
【0095】
[実施例8]
ゼブラフィッシュ腫瘍細胞株の確立
1細胞段階にPkrt8-EGFP-K-rasB(V12)コンストラクトを注射した魚の1つは、2月齢までに眼に大きな腫瘍を生じた(図6A)。その腫瘍を剪刀で単離して解剖し、エタノールで滅菌し、滅菌した1Xハンクス液(Hank’s balanced salt solution)および1%抗生物質−抗真菌剤ミックス中で数回洗浄し、カミソリの刃で粉砕した。次いでサンプルを0.14ユニット/mlのLiberase Blendzyme 3を加えた1 mlの1Xハンクス液中、28℃で30分間インキュベートした。完全に解離した後、細胞を1Xハンクス液で洗浄し、10% FBSを含む8 mlの成長培地LFD (50% L-15/15% F-12/35%DMEM)および1%の抗生物質−抗真菌剤ミックスを加え、その混合物を培養フラスコ中28℃でインキュベートした。2日後、細胞を成長培地で洗浄して死んで付着していない細胞を除去し、新しい培地に置き換えた。単層のコンフルエンスができるまで細胞を成長させ、FBSを含まないLFDに剥離させ、培養培地に再懸濁し、106細胞を10 mlフラスコに播種した。
【0096】
以下のように、一連の継代を通してこの細胞株を継代した:2〜4x107の集団に達した後、細胞を剥離させ、新しい培地に再懸濁し(1:20)、約106細胞を5〜6日毎に(2〜4x107細胞)10 mlフラスコに継代した。そのような24継代を通して36時間以内に1細胞分裂という速い増殖速度を維持しながら、細胞を成功裡に継代させた。5、8、10および24継代後、細胞株を液体窒素中でLFD/10%FBS/10%DMSO中に凍結させた。
【0097】
[実施例9]
放射線照射した魚への腫瘍細胞株の移植
移植の2日前に、野生型の成魚に24ガンマ線を照射した。7x107細胞(21継代目)をFBSを含まないLFDに剥離させ、1Xハンクス液で2回洗浄し、1Xハンクス液に105細胞/μlになるまで再懸濁した。亜致死的に放射線照射を受けてから2日後の野生型のレシピエントに25μlハミルトンシリンジを用いて10μlの濃縮細胞懸濁液(106細胞)を腹腔内または筋肉内注射した。亜致死的に放射線照射を受けた他の野生型レシピエントに、10μlの1Xハンクス液を注射した(対照集団)。GFP陽性の腫瘍形成が、注射部位だけでなく注射部位から遠く離れた位置にも観察され、腫瘍細胞が活発に伝播していることを示唆している。さらに、腫瘍細胞および腫瘍病巣が同じ宿主の種々の組織および器官に観察された。
【0098】
本発明を記載する文脈における(特に添付の特許請求の範囲の文脈における)「a」や「an」および「本、その(the)」なる用語の使用、および同様の指示体(referents)は、本明細書に特に別に示したり、または文脈上明らかに矛盾したりしない限り、単数および複数の両方を包含すると解釈されたい。「含む(comprising)」、「有する、持つ(having)」、「含む(including)」および「含む、含有する(containing)」なる用語は、特に別に示さない限りオープンエンドの用語(つまり「含むが、限定されない」ことを意味する)として解釈されたい。本明細書において詳述する値の範囲は、本明細書において特に別に示さない限り、単に、その範囲にあるそれぞれ別個の値を個別に示す簡単明瞭な方法として機能することを意図したもので、それぞれ別個の値は、あたかも本明細書に個別に列挙されているように本明細書に包含される。本明細書に記載の全ての方法は、本明細書に特に別に示したり、または文脈上明らかに矛盾したりしない限り、任意の適切な順序(order)で行うことができる。本明細書に記載の任意のおよび全ての実施例、または典型的な例を示す言葉(例えば「例えば、など、のような(such as)」)の使用は、単に本発明をより明らかにすることを意図したもので、特に別に請求しない限り本発明の範囲を限定するものではない。本明細書におけるどの言葉も、請求の範囲に記載していないあらゆる要素を本発明の実施に不可欠なものとして示していると解釈してはいけない。
【0099】
本発明の方法、魚、不死の腫瘍細胞株および組成物は、本明細書に少しだけ開示している実施態様の様々な形態に組み込まれ得ると理解されたい。本発明の実施態様は、本発明者らに既知の本発明を実施するための最良の形態を含めて、本明細書に記載されている。それらの実施態様の様々な形態は、当業者が前記の説明を読めば明らかになるであろう。本発明者らは当業者がそのような様々な形態を必要に応じて用いることを予想し、また本発明者らは本発明が本明細書に具体的に記載されている以外のようにも実施されることを意図している。従って、本発明は、適用法により認められているように特許請求の範囲に列挙した対象の全ての修飾物および等価物を含む。さらに、本明細書に特に別に示したり、または文脈上明らかに矛盾したりしない限り、上記の要素の全ての潜在的な種々の形態におけるあらゆる組み合わせが本発明に包含される。
【0100】
参考文献一覧
Ahuja, H.G. et al. (1990).「The pattern of mutational involvement of RAS genes in human hematologic malignancies determined by DNA amplification and direct sequencing.」Blood 75:1684-1690.
Alessi, D.R. et al. (1995).「PD098059 is a specific inhibitor of the activation of mitogen-activated protein kinase in vitro and in vivo.」J Biol Chem 270:27489-27494.
Allen et al. (1988).「Transgene as probes for active chromosomal domains in mouse development.」Nature 333:852-855.
Altschul, S.F. et al., (1990).「Basic local alignment search tool.」J Mol Biol 215:403-410.
Altschul, S.F. et al. (1997).「Gapped BLAST and PSI-BLAST: a new generation of protein database search programs.」Nucleic Acids Res 25:3389-3402.
Andersen, J.K. et al. (1993).「Herpesvirus-mediated gene delivery into the rat brain: specificity and efficiency of the neuron-specific enolase promoter.」Cell Mol
Neurobiol 13:503-515.
Baliga, S,N. (2001).「Promoter analysis by saturation mutagenesis.」Biol. Proced. Online 3:64-69.
Bos, J.L. (1989).「Ras oncogenes in human cancer: a review.」Cancer Res 49:4682-4689.
Bunin, B.A. and Ellman, J.A. et al. (1992).「A general and expedient method for the solid-phase synthesis of 1,4-benzodiazepine derivatives.」J. Am. Chem. Soc. 114:10997-10998.
Burns, J.C. et al.. (1993).「Vesicular stomatitis virus G glycoprotein pseudotyped retroviral vectors: concentration to very high titer and efficient gene transfer into mammalian and nonmammalian cells.」Proc Natl Acad Sci USA 90:8033-8037.
Carell, E. et al. (1994a).「A Novel Procedure for the Synthesis of Libraries Containing Small Organic Molecules.」Angew. Chem Int. Ed. Engl. 33:2059-2061.
Carell, E. et al. (1994b).「A Solution-Phase Screening Procedure for the Isolation of Active Compounds from a Library of Molecules.」Angew. Chem. Int. Ed. Engl. 33:2061-2064.
Chen, T.T. and Powers, D.A. (1990).「Transgenic fish.」Trends Biotechnol 8:209-215.
Cho, C.Y. et al. (1993).「An Unnatural biopolymer.」Science. 261:1303-1305.
Cull, M.G. et al. (1992).「Screening for Receptor Ligands Using Large Libraries of Peptides Linked to the C Terminus of the lac Repressor.」Proc Natl Acad Sci USA 89:1865-1869.
Culp, P. et al. (1991).「High-frequency germ-line transmission of plasmid DNA sequences injected into fertilized zebrafish eggs.」Proc Natl Acad.Sci USA 88:7953-7957.
Cwirla, S.E. et al. (1990).「Peptides on Phage: A Vast Library of Peptides for Identifying Ligands.」Proc. Natl. Acad. Sci. USA 87:6378-6382.
Davidson, A.E. et al. (2003).「Efficient gene delivery and gene expression in zebrafish using the Sleeping Beauty transposon.」Dev Biol 263:191-202.
Davidson, B.L. et al. (1993).「A model system for in vivo gene transfer into the central nervous system using an adenoviral vector.」Nature Genetics 3:219-223.
Devlin, J.L. et al. (1990).「Random Peptide Libraries: A Source of Specific Protein Binding Molecules.」Science 249:404-406.
DeWitt, S.H. et al. (1993).「Diversomers": An Approach to Nonpeptide, Nonoligomeric Chemical Diversity.」Proc. Natl. Acad. Sci. USA 90:6909-6913.
Dudley, D.T. et al. (1995).「A synthetic inhibitor of the mitogen-activated protein kinase cascade.」Proc Natl Acad Sci USA 92:7686-7689.
Erb, E. et al. (1994).「Recursive Deconvolution of Combinatorial Chemical Libraries.」Proc. Natl. Acad. Sci. USA 91:11422-11426.
Erzurum, S.C. et al. (1993).「Protection of human endothelial cells from oxidant injury by adenovirus-mediated transfer of the human catalase cDNA.」Nucleic Acids Res 21:1607-1612.
Fadool, J.M. et al. (1998).「Transposition of the mariner element from Drosophila mauritiana in zebrafish.」Proc Natl Acad Sci USA 95:5182-5186.
Felici, F. et al. (1991).「Selection of antibody ligands from a large library of oligopeptides expressed on a multivalent exposition vector.」J Mol. Biol. 222:301-310.
Fletcher, G.L., and Davis, P.L. (1991).「Transgenic fish for aquaculture.」In Genetic Engineering, Setlow, J.K., ed., Plenum Press.
Flotte, T.R. et al. (1993).「Stable in vivo expression of the cystic fibrosis transmembrane conductance regulator with an adeno-associated virus vector.」Proc Natl Acad Sci USA 90:10613-10617.
Fodor, S. et al. (1993).「Multiplexed biochemical assays with biological chips.」Nature 364:555-556.
Gallop, M.A. et al. (1994).「Applications of combinatorial technologies to drug discovery. 1. Background and peptide combinatorial libraries.」J. Med. Chem. 37:1233-1251.
Gong, Z. et al. (2001).「Green fluorescent protein (GFP) transgenic fish and their applications.」Genetica 111:213-225.
Gong, Z. et al. (2002).「Green fluorescent protein expression in germ-line transmitted transgenic zebrafish under a stratified epithelial promoter from keratin8.」Dev Dyn 223:204-215.
Gossler et al. (1989).「Mouse Embryonic Stem Cells and Reporter Constructs to Detect Developmentally Regulated Genes.」Science 244:463-465.
Grabher, C. et al. (2003).「Transposon-mediated enhancer trapping in medaka.」Gene 322:57-66.
Guyton, K.Z. et al. (1996).「Mitogen-activated protein kinase (MAPK) activation by butylated hydroxytoluene hydroperoxide: implications for cellular survival and tumor promotion.」Cancer Res 36:3480-3465.
Horwell, D. et al (1996).「'Targeted' molecular diversity: design and development of non-peptide antagonists for cholecystokinin and tachykinin receptors.」Immunopharmacology 33:68-72.
Houghten, R.A. et al. (1992).「The use of synthetic peptide combinatorial libraries for the identification of bioactive peptides.」Biotechniques 13:412-421.
Huang, H. et al. (2001).「Pdx-1 knockdown reduces insulin promoter activity in zebrafish.」Genesis 30:134-136.
Ivics, Z. et al. (1999).「Genetic applications of transposons and other repetitive elements in zebrafish.」Methods in Cell Biology 60:99-131.
Jessen, J.R. et al. (1999).「Artificial chromosome transgenesis reveals long-distance negative regulation of rag1 in zebrafish.」Nat Genet 23:15-16.
Jessen, J.R. et al. (2001).「Concurrent expression of recombination activating genes 1 and 2 in zebrafish olfactory sensory neurons.」Genesis 29:156-162.
Johnson, L. et al. (2001).「Somatic activation of the K-ras oncogene causes early onset lung cancer in mice.」Nature, 410:1111-1115.
Kaeakami, K. et al. (2000).「Identification of a functional transposase of the Tol2 element, an Ac-like element from the Japanese medaka fish, and its transposition in the zebrafish germ lineage.」Proc Natl Acad Sci USA 97:11403-11408.
Kennedy, B.N. et al. (2001).「Isolation of a zebrafish rod opsin promoter to generate a transgenic zebrafish line expressing enhanced green fluorescent protein in rod photoreceptors.」J Biol Chem 276:14037-14043.
Kimmel, C.B. (1989).「Genetics and Early Development of Zebrafish.」Trends Genet 5:283-288.
Kothary et al. (1988).「A transgene containing lacZ inserted into the dystonia locus is expressed in neural tube.」Nature 335:435-437.
Lam, K.S. (1997).「Application of combinatorial library methods in cancer research and drug discovery.」Anticancer Drug Des. 12:145-167.
Lam, K.S. et al. (1991).「A new type of synthetic peptide library for identifying ligand-binding activity.」Nature 354:82-84.
Langenau, D.M. et al. (2005).「Cre/lox-regulated transgenic zebrafish model with conditional myc-induced T cell acute lymphoblastic leukemia.」Proc Natl Acad Sci USA 102:7369-7374.
Lever, A.M. (2000).「Lentiviral vectors: progress and potential.」Curr Opin Mol Ther 2:488-496.
Long, Q. et al. (1997).「GATA-1 expression pattern can be recapitulated in living transgenic zebrafish using GFP reporter gene.」Development 124:4105-11.
Lu, J.K. et al. (1992).「Integration, expression and germ-line transmission of foreign growth hormone genes in medaka (Oryzias latipes).」Molec Mar Biol Biotechnol 1:366-375.
Miller, A.D. et al., (1993).「Use of retroviral vectors for gene transfer and expression.」Methods of Enzymology 217:581-599.
Moss, J.B. et al. (1996).「Green Fluorescent Protein Marks Skeletal Muscle in Murine Cell Lines and Zebrafish.」Gene 173:89-98.
Motoike, T. et al. (2000).「Universal GFP reporter for the study of vascular development.」Genesis 28:75-81.
Nam, J.S. et al. (2002).「Src family kinase inhibitor PP2 restores the E-cadherin/catenin cell adhesion system in human cancer cells and reduces cancer metastasis.」Clin Cancer Res. 8:2430-2436.
O'Kane, et al. (1987).「Detection in situ of Genomic Regulatory Elements in Drosophila.」Proc. Natl. Acad. Sci. USA 84:9123-9127.
Parinov, S. et al. (2004).「Enhancer trap transposable element as a tool for identification of developmentally regulated genes in zebrafish in vivo.」Dev Dyn. 231:449-459.
Park, H.C. et al. (2000).「Analysis of upstream elements in the HuC promoter leads to the establishment of transgenic zebrafish with fluorescent neurons.」Dev Biol 227:279-293.
Powers, D.A. et al. (1992).「Electroporation: a method for transferring genes into the gametes of zebrafish (Brachydanio rerio), channel catfish (Ictalurus punctatus), and common carp (Cyprinus carpio).」Molec Mar Biol Biotechnol 1:301-308.
Scott, J.K. and J.P. Smith (1990).「Searching for Peptide Ligands with an Epitope Library.」Science 249:386-390.
Strausberg et al. 2002. Generation and initial analysis of more than 15,000 full-length human and mouse cDNA sequences. Proc. Natl. Acad. Sci. U.S.A. 99:16899-16903.
Streisinger (1984). Attainment of Minimal Biological Variability and Measurements of Genotoxicity: Production of Homozygous Diploid Zebra Fish. Natl. Cancer Inst. Monogr. 65:53-58.
Thermes, V. et al. (2002).「I-SecI meganuclease mediates highly efficient transgenesis in fish.」Mech Dev 118:91-98.
Westerfield, M. (2000). The Zebrafish Book: A guide for the laboratory use of Zebrafish (Danio rerio), 4th ed., University of Oregon Press, Eugene.
Zabner, J. et al. (1994).「Safety and efficacy of repetitive adenovirus-mediated transfer of CFTR cDNA to airway epithelia of primates and cotton rats.」Nature Genetics 6:75-83.
Zhang, G. et al. (1996).「An enhanced green fluorescent protein allows sensitive detection of gene transfer in mammalian cells.」Biochem Biophys Res Commun 227:707-711.
Zuckermann, R.N. (1994).「Discovery of nanomolar ligands for 7-transmembrane G-protein-coupled receptors from a diverse N-(substituted)glycine peptoid library.」J. Med. Chem. 37:2678-2685.
【図面の簡単な説明】
【0101】
【図1A】図1Aは、ヒト(配列番号1)およびゼブラフィッシュ(配列番号2)のK-rasタンパク質配列のアライメントを示す。ヒトc-K-ras2b:GenBank登録番号NP_004976;バージョンNP_004976.2;GI:15718761。zgc:85725 mRNA (cDNAクローンMGC:85725 IMAGE:6968999) GenBank登録番号BC078646;GI:50925043 (Strausberg et al., 2002)によってコードされたダニオ・レリオZgc:85725:GenBank登録番号NP_001003744;バージョンNP_001003744.1;GI:51230608。「l」は保存アミノ酸残基を表し、「*」はアミノ酸の保存的変化を表す。
【0102】
【図1B】図1Bは、Ras発癌性タンパク質ファミリーメンバーのアライメントを示す。全Rasタンパク質のうち164のN末端アミノ酸は高度に保存されている。残りの25のC末端残基のみがかなり多様である(高頻度可変領域)。ダニオ・レリオZgc:85725:GenBank登録番号NP_001003744;バージョンNP_001003744.1;GI:51230608(配列番号2)。カーステン(Kirsten)マウス肉腫ウイルス癌原遺伝子タンパク質RNA:GenBank 登録番号Z23152;バージョンZ23152.1;GI:939929(配列番号3)。ヒトc-K-ras2タンパク質イソ型b:GenBank登録番号NP_004976;バージョンNP_004976.2;GI:15718761(配列番号1)。ヒトc-K-ras2タンパク質イソ型a:GenBank登録番号NP_203524;バージョンNP_203524.1;GI:15718763(配列番号4)。ヒト形質転換タンパク質p21/H-Ras-1:GenBank登録番号P01112;バージョンP01112;GI:131869(配列番号5)。ヒト形質転換タンパク質N-Ras:GenBank登録番号P01111;バージョンP01111;GI:131883(配列番号6)。ダニオ・レリオp21 N-ras癌遺伝子:GenBank登録番号AAB40625;バージョンAAB40625.1;GI:1778053(配列番号7)。
【0103】
【図2】図2はEGFP-Rasコンストラクトを注射したゼブラフィッシュの細胞の腫瘍性形質転換、および種々の薬物の効果を示す。図2aはK-RasB(V12)の発現を防ぐTAA終止コドンを有するP-krt8-EGFP-TAA(終止)-K-rasB(V12)コンストラクトを注射した対照魚を示す。この対照において、EGFP陽性細胞はP-krt8-EGFPの発現を特徴とする正常な形態を持つ。図2bおよび2cは、EGFP-K-rasB(V12)発現により誘導された腫瘍性の表現型を示す。図2dから2gは、PP2(図2dおよび2e)およびPD98059(図2fおよび2g)の薬物の効果を示す。
【0104】
【図3】図3はP-krt8-EGFP-zK-rasB導入遺伝子を注射したF0成魚における腫瘍形成を示す。図3A(3B)、3C(3D)、3F(3E)および3G(3H)は、4種のF0魚の腫瘍の例と、対応するGFP画像を示す。図3A(3B)、3F(3E)および3G(3H)は体内に腫瘍を有する魚を示す。図3C(3D)は、導入遺伝子発現に伴い脳に重篤な異常を現した魚を示す。全ての魚の腫瘍成長部位に広範な出血が広がっている。図3Eは、腫瘍成長が導入遺伝子の発現と密に関係していることを示す。図3A(3B)、3C(3D)および3G(3H)の魚は2月齢である。図3F(3E)の魚は3月齢の繁殖力のあるメスである。図3G(3H)の魚は1月齢で目に見える症状(広範な出血)までに進行した。
【0105】
【図4】図4は、P-krt8-EGFP-zK-rasB導入遺伝子を注射したF0成魚における腫瘍形成の組織化学的分析を示す。パラフィン包埋切片をヘマトキシリン/エオシンで染色した。図4Aおよび4Bは魚1(図3G、3H)を示す:起源が不明な転移性の癌が様々な組織へ広がっている。図4Cは魚2(図3F、3E)を示す:転移性の線維肉腫が様々な組織へ広がっている。図4Dは健康な魚、すなわち対照を示す。V−椎骨;M−筋肉;K−腎臓;T−腫瘍。
【0106】
【図5】図5は、安定なP-krt8-EGFP-zK-rasB導入遺伝子を含むF1世代の魚を示す。正常な形態のP-krt8-EGFP陽性細胞を示すP-krt8-EGFP対照(図5A)。zK-ras(V12)誘導性の表現型(図5B)。
【0107】
【図6】図6は、Pkrt8-EGFP-K-rasB(V12)誘導性の眼の腫瘍から得た腫瘍細胞株の例の起源および特性を示す。図6Aは、この腫瘍細胞株を産生するのに用いた、腫瘍を単離する前の魚を示す。1細胞の段階で胚にPkrt8-EGFP-K-rasB(V12)コンストラクトを注射すると、2月齢までに眼に大きな腫瘍が成長した。腫瘍はGFP陽性であった。この腫瘍を単離し、細胞を培養中で成長させた。図6Bは、新しい継代が急速な増殖を示してから4日目および6日目の培養細胞の例を示す。図6Cは細胞株における接触阻害の喪失を示す。細胞は、病巣が明らかになるまで、成長しコンフルエンスになった(継代後8日目)。この例に示した形成病巣は、明るいEGFP蛍光を示している。培養細胞は弱いGFP蛍光を放射し、その蛍光は、病巣が非常に明るいほど、それを取り巻く単層においては観察が困難になる。図6Dは培養細胞を注射した魚の腫瘍を示す(注射後12日目)。GFP陽性腫瘍の形成は筋肉において明らかである。図6Eは、図6Dに示した魚における腫瘍形成の組織学的分析を示す。ヘマトキシリン/エオシンで染色したパラフィン包埋切片。v−椎骨;m−筋肉;t−腫瘍。
【特許請求の範囲】
【請求項1】
プロモーターに作動可能に連結した癌遺伝子を含む核酸がゲノムに安定に組み込まれている形質転換魚、および前記核酸であってエンハンサートラップ効果の特性を有する核酸がいくつかの細胞に取り込まれている形質転換魚からなる群から選択される形質転換魚。
【請求項2】
プロモーターが、器官特異的プロモーター、組織特異的プロモーター、最小プロモーターであるか、またはエンハンサートラッププロモーターの特性を有する、請求項1に記載の形質転換魚。
【請求項3】
組織特異的プロモーターがケラチン8(krt8ある、請求項2に記載の形質転換魚。
【請求項4】
癌遺伝子が、RAS、MYC、CYCLIN D1、FOS、JUN、MYB、BCL2、HOX11、HOX11L2、LYL1、TAL1/SCL、LMO1、LMO2、MYCN、MDM2、CDK4、GLI1、IGF2、活性化RAS、活性化EGFR、変異型FLT3-ITD、変異型および活性型のTP53、PAX3、PAX7、BCR/ABL、HER2/NEU、FLT3R、NPM-ALK、SRC、ABL、TAN1、PTC、B-RAF、PML-RARアルファおよびE2A-PBX1からなる群から選択される、請求項1に記載の形質転換魚。
【請求項5】
癌遺伝子がRas癌遺伝子である、請求項4に記載の形質転換魚。
【請求項6】
癌遺伝子が、該癌遺伝子の相同体である、請求項4に記載の形質転換魚。
【請求項7】
相同体が哺乳類の相同体である、請求項6に記載の形質転換魚。
【請求項8】
癌遺伝子が、(i) zK-rasB、(ii) zK-rasB相同体および(iii) zK-rasBと実質的に類似する癌遺伝子、からなる群から選択される、請求項1に記載の形質転換魚。
【請求項9】
癌遺伝子がレポーター遺伝子と融合している、請求項1に記載の形質転換魚。
【請求項10】
レポーター遺伝子が、蛍光タンパク質遺伝子、ルシフェラーゼ、βガラクトシダーゼ、クロラムフェニコール、アシルトランスフェラーゼ、βグルクロニダーゼおよびアルカリフォスファタ−ゼからなる群から選択される、請求項9に記載の形質転換魚。
【請求項11】
蛍光タンパク質遺伝子がEGFPである、請求項10に記載の形質転換魚。
【請求項12】
前記核酸がkrt8プロモーターと作動可能に連結したK-ras癌遺伝子を含み、該K-ras癌遺伝子は高感度緑色蛍光タンパク質遺伝子と融合している、請求項1に記載の形質転換魚。
【請求項13】
K-ras癌遺伝子がzK-rasB(V12)であり、krt8プロモーターがゼブラフィッシュkrt8である、請求項12に記載の形質転換魚。
【請求項14】
癌遺伝子が癌遺伝子媒介性の癌の進行を誘導し、癌が非ホジキンリンパ腫、高悪性度星細胞腫、横紋筋肉腫、神経芽細胞腫、神経内分泌癌、膵癌、卵巣癌、精巣癌、胃癌、結腸癌、腎癌、黒色腫、急性骨髄性白血病、慢性骨髄性白血病およびT細胞急性リンパ性白血病からなる群から選択される、請求項1に記載の形質転換魚。
【請求項15】
癌遺伝子が腫瘍を誘導する、請求項1に記載の形質転換魚。
【請求項16】
不死の腫瘍細胞株を調製する方法であって、請求項15に記載の形質転換魚の腫瘍から細胞を単離すること、および単離した腫瘍細胞を不死の腫瘍細胞株の産生に適した条件下で培養することを含む方法。
【請求項17】
請求項16に記載の方法により調製した不死の腫瘍細胞株を移植した魚を含む、腫瘍の進行および転移に関する動物モデル。
【請求項18】
腫瘍の進行または転移を阻害する薬物をスクリーニングする方法であって、請求項17の動物モデルを被験薬物または物質と接触させる、あるいは曝すこと、および該被験薬物または物質が腫瘍の進行または転移を阻害するかどうかを判定することを含む方法。
【請求項19】
腫瘍細胞の成長を阻害する薬物をスクリーニングする方法であって、請求項16に記載の方法により調製した不死の腫瘍細胞株を被験薬物または物質と接触させること、および該薬物または物質が腫瘍細胞の成長を阻害するかどうかを判定することを含む方法。
【請求項20】
癌遺伝子媒介性の腫瘍性または過形成性形質転換を調節する薬物または物質をスクリーニングする方法であって、以下を含む方法:
形質転換魚を被験薬物または物質と接触させる、あるいは曝す;
該薬物または物質が癌遺伝子媒介性の腫瘍性または過形成性形質転換を調節するかどうかを判定する;および
該薬物または物質が癌遺伝子媒介性の腫瘍性または過形成性形質転換を調節する場合、該薬物または物質を癌遺伝子媒介性の腫瘍性または過形成性形質転換を調節する薬物または物質として分類する、
ここで該形質転換魚は、プロモーターに作動可能に連結した癌遺伝子を含む核酸がゲノムに安定に組み込まれている形質転換魚、および前記核酸であってエンハンサートラップ効果の特性を有する核酸がいくつかの細胞に取り込まれている形質転換魚からなる群から選択される。
【請求項21】
プロモーターが、器官特異的プロモーター、組織特異的プロモーターまたは最小プロモーターである、請求項20に記載の方法。
【請求項22】
組織特異的プロモーターがケラチン8 (krt8)である、請求項21に記載の方法。
【請求項23】
癌遺伝子がRAS、MYC、CYCLIN D1、FOS、JUN、MYB、BCL2、HOX11、HOX11L2、LYL1、TAL1/SCL、LMO1、LMO2、MYCN、MDM2、CDK4、GLI1、IGF2、活性化RAS、活性化EGFR、変異型FLT3-ITD、変異型および活性型のTP53、PAX3、PAX7、BCR/ABL、HER2/NEU、FLT3R、NPM-ALK、SRC、ABL、TAN1、PTC、B-RAF、PML-RARアルファおよびE2A-PBX1からなる群から選択される、請求項20に記載の方法。
【請求項24】
癌遺伝子がRas癌遺伝子である、請求項23に記載の方法。
【請求項25】
癌遺伝子が、該癌遺伝子の相同体である、請求項23に記載の方法。
【請求項26】
相同体が哺乳類の相同体である、請求項25に記載の形質転換魚。
【請求項27】
癌遺伝子が、(i) zK-rasB、(ii) zK-rasB相同体および(iii) zK-rasBと実質的に類似する癌遺伝子、からなる群から選択される、請求項20に記載の方法。
【請求項28】
癌遺伝子がレポーター遺伝子と融合している、請求項20に記載の方法。
【請求項29】
レポーター遺伝子が、蛍光タンパク質遺伝子、ルシフェラーゼ、βガラクトシダーゼ、クロラムフェニコール、アシルトランスフェラーゼ、βグルクロニダーゼおよびアルカリフォスファタ−ゼからなる群から選択される、請求項28に記載の方法。
【請求項30】
蛍光タンパク質遺伝子がEGFPである、請求項29に記載の方法。
【請求項31】
前記核酸がkrt8プロモーターと作動可能に連結したK-ras癌遺伝子を含み、該K-ras癌遺伝子は高感度緑色蛍光タンパク質遺伝子と融合している、請求項20に記載の方法。
【請求項32】
K-ras癌遺伝子がzK-rasB(V12)であり、krt8プロモーターがゼブラフィッシュkrt8である、請求項31に記載の方法。
【請求項33】
癌遺伝子が癌遺伝子媒介性の癌の進行を誘導し、癌が非ホジキンリンパ腫、高悪性度星細胞腫、横紋筋肉腫、神経芽細胞腫、神経内分泌癌、膵癌、卵巣癌、精巣癌、胃癌、結腸癌、腎癌、黒色腫、急性骨髄性白血病、慢性骨髄性白血病およびT細胞急性リンパ性白血病からなる群から選択される、請求項20に記載の方法。
【請求項34】
癌遺伝子媒介性の腫瘍性または過形成性形質転換に起因する腫瘍形成の発症率を測定することをさらに含む、請求項20に記載の方法。
【請求項35】
癌遺伝子媒介性の腫瘍性または過形成性形質転換に起因する腫瘍の量または大きさを測定することをさらに含む、請求項20に記載の方法。
【請求項36】
被験薬物または物質がアンチセンスDNA、アンチセンスRNAまたは低分子干渉RNAである、請求項20に記載の方法。
【請求項37】
形質転換魚が形質転換魚の胚である、請求項20に記載の方法。
【請求項38】
形質転換魚が形質転換ゼブラフィッシュである、請求項20に記載の方法。
【請求項39】
形質転換魚の胚が形質転換ゼブラフィッシュの胚である、請求項38に記載の方法。
【請求項1】
プロモーターに作動可能に連結した癌遺伝子を含む核酸がゲノムに安定に組み込まれている形質転換魚、および前記核酸であってエンハンサートラップ効果の特性を有する核酸がいくつかの細胞に取り込まれている形質転換魚からなる群から選択される形質転換魚。
【請求項2】
プロモーターが、器官特異的プロモーター、組織特異的プロモーター、最小プロモーターであるか、またはエンハンサートラッププロモーターの特性を有する、請求項1に記載の形質転換魚。
【請求項3】
組織特異的プロモーターがケラチン8(krt8ある、請求項2に記載の形質転換魚。
【請求項4】
癌遺伝子が、RAS、MYC、CYCLIN D1、FOS、JUN、MYB、BCL2、HOX11、HOX11L2、LYL1、TAL1/SCL、LMO1、LMO2、MYCN、MDM2、CDK4、GLI1、IGF2、活性化RAS、活性化EGFR、変異型FLT3-ITD、変異型および活性型のTP53、PAX3、PAX7、BCR/ABL、HER2/NEU、FLT3R、NPM-ALK、SRC、ABL、TAN1、PTC、B-RAF、PML-RARアルファおよびE2A-PBX1からなる群から選択される、請求項1に記載の形質転換魚。
【請求項5】
癌遺伝子がRas癌遺伝子である、請求項4に記載の形質転換魚。
【請求項6】
癌遺伝子が、該癌遺伝子の相同体である、請求項4に記載の形質転換魚。
【請求項7】
相同体が哺乳類の相同体である、請求項6に記載の形質転換魚。
【請求項8】
癌遺伝子が、(i) zK-rasB、(ii) zK-rasB相同体および(iii) zK-rasBと実質的に類似する癌遺伝子、からなる群から選択される、請求項1に記載の形質転換魚。
【請求項9】
癌遺伝子がレポーター遺伝子と融合している、請求項1に記載の形質転換魚。
【請求項10】
レポーター遺伝子が、蛍光タンパク質遺伝子、ルシフェラーゼ、βガラクトシダーゼ、クロラムフェニコール、アシルトランスフェラーゼ、βグルクロニダーゼおよびアルカリフォスファタ−ゼからなる群から選択される、請求項9に記載の形質転換魚。
【請求項11】
蛍光タンパク質遺伝子がEGFPである、請求項10に記載の形質転換魚。
【請求項12】
前記核酸がkrt8プロモーターと作動可能に連結したK-ras癌遺伝子を含み、該K-ras癌遺伝子は高感度緑色蛍光タンパク質遺伝子と融合している、請求項1に記載の形質転換魚。
【請求項13】
K-ras癌遺伝子がzK-rasB(V12)であり、krt8プロモーターがゼブラフィッシュkrt8である、請求項12に記載の形質転換魚。
【請求項14】
癌遺伝子が癌遺伝子媒介性の癌の進行を誘導し、癌が非ホジキンリンパ腫、高悪性度星細胞腫、横紋筋肉腫、神経芽細胞腫、神経内分泌癌、膵癌、卵巣癌、精巣癌、胃癌、結腸癌、腎癌、黒色腫、急性骨髄性白血病、慢性骨髄性白血病およびT細胞急性リンパ性白血病からなる群から選択される、請求項1に記載の形質転換魚。
【請求項15】
癌遺伝子が腫瘍を誘導する、請求項1に記載の形質転換魚。
【請求項16】
不死の腫瘍細胞株を調製する方法であって、請求項15に記載の形質転換魚の腫瘍から細胞を単離すること、および単離した腫瘍細胞を不死の腫瘍細胞株の産生に適した条件下で培養することを含む方法。
【請求項17】
請求項16に記載の方法により調製した不死の腫瘍細胞株を移植した魚を含む、腫瘍の進行および転移に関する動物モデル。
【請求項18】
腫瘍の進行または転移を阻害する薬物をスクリーニングする方法であって、請求項17の動物モデルを被験薬物または物質と接触させる、あるいは曝すこと、および該被験薬物または物質が腫瘍の進行または転移を阻害するかどうかを判定することを含む方法。
【請求項19】
腫瘍細胞の成長を阻害する薬物をスクリーニングする方法であって、請求項16に記載の方法により調製した不死の腫瘍細胞株を被験薬物または物質と接触させること、および該薬物または物質が腫瘍細胞の成長を阻害するかどうかを判定することを含む方法。
【請求項20】
癌遺伝子媒介性の腫瘍性または過形成性形質転換を調節する薬物または物質をスクリーニングする方法であって、以下を含む方法:
形質転換魚を被験薬物または物質と接触させる、あるいは曝す;
該薬物または物質が癌遺伝子媒介性の腫瘍性または過形成性形質転換を調節するかどうかを判定する;および
該薬物または物質が癌遺伝子媒介性の腫瘍性または過形成性形質転換を調節する場合、該薬物または物質を癌遺伝子媒介性の腫瘍性または過形成性形質転換を調節する薬物または物質として分類する、
ここで該形質転換魚は、プロモーターに作動可能に連結した癌遺伝子を含む核酸がゲノムに安定に組み込まれている形質転換魚、および前記核酸であってエンハンサートラップ効果の特性を有する核酸がいくつかの細胞に取り込まれている形質転換魚からなる群から選択される。
【請求項21】
プロモーターが、器官特異的プロモーター、組織特異的プロモーターまたは最小プロモーターである、請求項20に記載の方法。
【請求項22】
組織特異的プロモーターがケラチン8 (krt8)である、請求項21に記載の方法。
【請求項23】
癌遺伝子がRAS、MYC、CYCLIN D1、FOS、JUN、MYB、BCL2、HOX11、HOX11L2、LYL1、TAL1/SCL、LMO1、LMO2、MYCN、MDM2、CDK4、GLI1、IGF2、活性化RAS、活性化EGFR、変異型FLT3-ITD、変異型および活性型のTP53、PAX3、PAX7、BCR/ABL、HER2/NEU、FLT3R、NPM-ALK、SRC、ABL、TAN1、PTC、B-RAF、PML-RARアルファおよびE2A-PBX1からなる群から選択される、請求項20に記載の方法。
【請求項24】
癌遺伝子がRas癌遺伝子である、請求項23に記載の方法。
【請求項25】
癌遺伝子が、該癌遺伝子の相同体である、請求項23に記載の方法。
【請求項26】
相同体が哺乳類の相同体である、請求項25に記載の形質転換魚。
【請求項27】
癌遺伝子が、(i) zK-rasB、(ii) zK-rasB相同体および(iii) zK-rasBと実質的に類似する癌遺伝子、からなる群から選択される、請求項20に記載の方法。
【請求項28】
癌遺伝子がレポーター遺伝子と融合している、請求項20に記載の方法。
【請求項29】
レポーター遺伝子が、蛍光タンパク質遺伝子、ルシフェラーゼ、βガラクトシダーゼ、クロラムフェニコール、アシルトランスフェラーゼ、βグルクロニダーゼおよびアルカリフォスファタ−ゼからなる群から選択される、請求項28に記載の方法。
【請求項30】
蛍光タンパク質遺伝子がEGFPである、請求項29に記載の方法。
【請求項31】
前記核酸がkrt8プロモーターと作動可能に連結したK-ras癌遺伝子を含み、該K-ras癌遺伝子は高感度緑色蛍光タンパク質遺伝子と融合している、請求項20に記載の方法。
【請求項32】
K-ras癌遺伝子がzK-rasB(V12)であり、krt8プロモーターがゼブラフィッシュkrt8である、請求項31に記載の方法。
【請求項33】
癌遺伝子が癌遺伝子媒介性の癌の進行を誘導し、癌が非ホジキンリンパ腫、高悪性度星細胞腫、横紋筋肉腫、神経芽細胞腫、神経内分泌癌、膵癌、卵巣癌、精巣癌、胃癌、結腸癌、腎癌、黒色腫、急性骨髄性白血病、慢性骨髄性白血病およびT細胞急性リンパ性白血病からなる群から選択される、請求項20に記載の方法。
【請求項34】
癌遺伝子媒介性の腫瘍性または過形成性形質転換に起因する腫瘍形成の発症率を測定することをさらに含む、請求項20に記載の方法。
【請求項35】
癌遺伝子媒介性の腫瘍性または過形成性形質転換に起因する腫瘍の量または大きさを測定することをさらに含む、請求項20に記載の方法。
【請求項36】
被験薬物または物質がアンチセンスDNA、アンチセンスRNAまたは低分子干渉RNAである、請求項20に記載の方法。
【請求項37】
形質転換魚が形質転換魚の胚である、請求項20に記載の方法。
【請求項38】
形質転換魚が形質転換ゼブラフィッシュである、請求項20に記載の方法。
【請求項39】
形質転換魚の胚が形質転換ゼブラフィッシュの胚である、請求項38に記載の方法。
【図1A】


【図1B】

【図2】
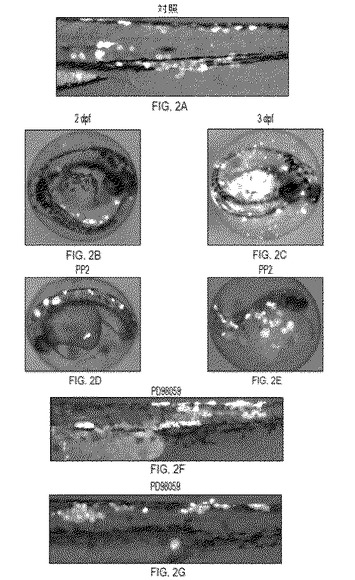
【図3】

【図4】
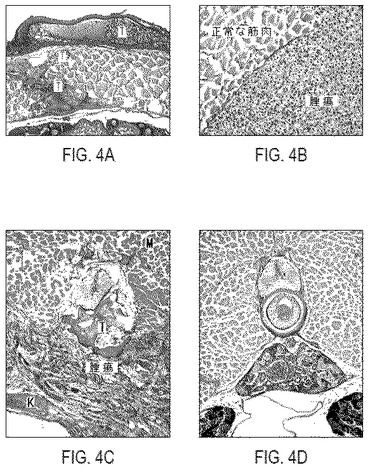
【図5】
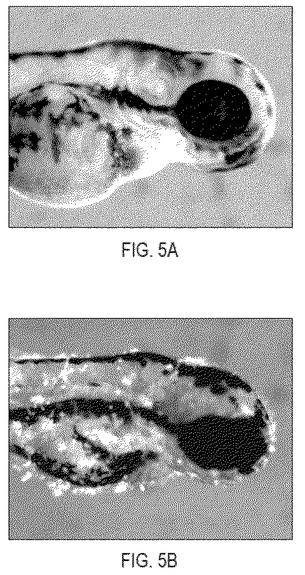
【図6】
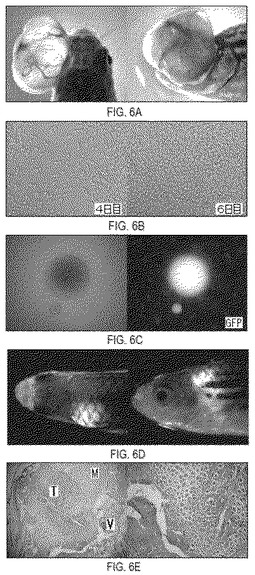
【図1B】
【図2】
【図3】
【図4】
【図5】
【図6】
【公表番号】特表2009−501544(P2009−501544A)
【公表日】平成21年1月22日(2009.1.22)
【国際特許分類】
【出願番号】特願2008−522744(P2008−522744)
【出願日】平成18年7月18日(2006.7.18)
【国際出願番号】PCT/SG2006/000202
【国際公開番号】WO2007/011312
【国際公開日】平成19年1月25日(2007.1.25)
【出願人】(505381426)テマセク・ライフ・サイエンシーズ・ラボラトリー・リミテッド (4)
【氏名又は名称原語表記】Temasek Life Sciences Laboratory Limited
【Fターム(参考)】
【公表日】平成21年1月22日(2009.1.22)
【国際特許分類】
【出願日】平成18年7月18日(2006.7.18)
【国際出願番号】PCT/SG2006/000202
【国際公開番号】WO2007/011312
【国際公開日】平成19年1月25日(2007.1.25)
【出願人】(505381426)テマセク・ライフ・サイエンシーズ・ラボラトリー・リミテッド (4)
【氏名又は名称原語表記】Temasek Life Sciences Laboratory Limited
【Fターム(参考)】
[ Back to top ]