
鱗翅目の電位依存性カルシウムチャネル
【課題】鱗翅目のカルシウムチャネルサブユニットをコード化する新しい核酸配列、ならびに、組換え型体の発現および同様のものからなる宿主細胞を提供する。
【解決手段】単離した鱗翅目のカルシウムチャネルサブユニット、鱗翅目のカルシウムチャネルサブユニットを発現している宿主細胞、鱗翅目のカルシウムチャネルサブユニットの作製し、鱗翅目のカルシウムチャネルサブユニットに特異的な抗体を作製し、モジュレーター識別法および/または鱗翅目のカルシウムチャネルのインヒビターをも作製する。
【解決手段】単離した鱗翅目のカルシウムチャネルサブユニット、鱗翅目のカルシウムチャネルサブユニットを発現している宿主細胞、鱗翅目のカルシウムチャネルサブユニットの作製し、鱗翅目のカルシウムチャネルサブユニットに特異的な抗体を作製し、モジュレーター識別法および/または鱗翅目のカルシウムチャネルのインヒビターをも作製する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、農芸化学、獣医学または薬学の分野で有用な核酸配列に関する。本発明は特に、ポリペプチドと、農薬または医薬品として有効な化合物の識別または開発で有用となるポリペプチドをコード化する核酸配列に関する。
【背景技術】
【0002】
電位依存性カルシウムチャネルは、生理学的応答の調節で重要な役割を果たしている。すなわち、電位依存性カルシウムチャネルの脱分極により、チャネル孔を通じてのカルシウムの通過が生じ、これによって、筋収縮および神経伝達物質の分泌などのさまざまな生理学的応答が生じると考えられている(非特許文献1);(非特許文献2);(非特許文献3);(非特許文献4);(非特許文献5);(非特許文献6);および(非特許文献7)。同様に、殺虫剤などの生物学的に活性な化合物を識別するための手段としてこのチャネルを標的とするための方法を開発しようとの要求が存在する(非特許文献8);(非特許文献9);(非特許文献10);(非特許文献11)。
【0003】
【非特許文献1】Jeziorskiら、J.Exper.Bio.,203、841−856ページ(2000)
【非特許文献2】Edwin W.McCleskey、Current Opinion in Neurobiology、4,304−312ページ(1994)
【非特許文献3】Herman Moreno Davila,Ananals New York Academy of Science,102−117ページ
【非特許文献4】Stephen W.Jones,J.of Bioenergetics and Biomembranes,30巻、4号、299−312ページ(1998)
【非特許文献5】Hofmannら、Anna.Rev.Neurosci.,17、399−418ページ(1994)
【非特許文献6】Edward Perez−Reyes and Toni Schneider, Kidney International、48巻、1111−1124ページ(1995)
【非特許文献7】AstraZeneca Pharmaceuticals Presentation,第28回National Medicinal Chemistry Symposium of the American Chemical Society,(2002年6月)
【非特許文献8】Mintzら、Nature、355巻、827−829ページ(1992)
【非特許文献9】Wangら、Nature Structural Biology、7巻、6号、505−512ページ(2000)
【非特許文献10】Wangら、Eur.J.Biochem,264,488−494ページ(1999)
【非特許文献11】Fletcherら、Nature Structural Biology、4巻、7号、559−566ページ(1997)
【0004】
電位依存性カルシウムチャネルは、ヒト、ウサギ、ラット、マウス、シビレエイ(Discopyge Ommata)などの脊椎動物で発現し、これらの動物からクローン化されている(非特許文献12);(非特許文献13);(非特許文献14);(非特許文献15);(非特許文献16);(非特許文献17);(非特許文献18);(非特許文献19);(非特許文献20)。電位依存性カルシウムチャネルは、線虫(Caenorhabditis elegans)、Styllphora pistilla、Bdellourea candida、キタユウレイクラゲ(Cyanea capillata)、ヤリイカ(Loligo bleeker)、ゾウアメフラシ(Aplysia californica)、ショウジョウバエ(Drosophila melanogaster)、イエバエ(Musca domestica)、チャバネゴキブリ(Blatella germanica)およびマボヤ(Halocynthia roretz)などの無脊椎動物でも発現し、これらの動物からクローン化されている(非特許文献21)。
【0005】
【非特許文献12】Velicelebeら、Methods in Enzymology、294巻、20−47ページ(1999)
【非特許文献13】Herman Moreno Davila、Annals New York Academy of Sciences、102−117ページ
【非特許文献14】Hofmannら、Anna.Rev.Neurosci,17,399−418ページ(1994)
【非特許文献15】Edward Perez−Reyes and Toni Schneider, Kidney International、48巻、1111−1124ページ(1995)
【非特許文献16】Astra Zeneca Pharmaceuticals Presentation,第28回National Medicinal Chemistry Symposium of the American Chemical Society,(2002年6月)
【非特許文献17】Horneら、Proc.Natl.Acad.Sci.,90巻,3787−3791ページ(1993)
【非特許文献18】Roussetら、J.of Physiology,532,3,583−593ページ(2001)
【非特許文献19】Neelandsら、J.Physiology,2933−2944ページ(2000)
【非特許文献20】Brustら、Neuropharmacology、32巻、11号、1089−1102ページ(1993)
【非特許文献21】Jeziorskiら、J.Exper.Bio.,203,841−856ページ(2000))
【発明の開示】
【発明が解決しようとする課題】
【0006】
本発明は、農芸化学、獣医学または薬学の分野で有用な核酸配列に関する。本発明は特に、ポリペプチドと、農薬または医薬品として有効な化合物の識別または開発で有用となるポリペプチドをコード化する核酸配列に関する。
【課題を解決するための手段】
【0007】
本発明の一実施形態は、農薬または医薬品としての(潜在的な)活性を有する化合物の識別または開発で有用なアミノ酸配列をコード化する核酸配列、または、これらのアミノ酸配列の発現に利用することの可能な核酸配列に関連するものである。以下に詳述するこれらの核酸配列は、その突然変異体およびフラグメントを含めて、ここでは「本発明の核酸配列」とも呼ぶ。
【0008】
本発明の別の実施形態は、本発明の核酸配列によってコード化されたり、本発明の核酸配列の適切な発現によって得られる可能性のある、蛋白質またはポリペプチドなどのアミノ酸配列に関連するものである。以下に詳述するこれらのアミノ酸配列は、その突然変異体およびフラグメントを含めて、ここでは「本発明のアミノ酸配列」とも呼ぶ。
【0009】
本発明のさらに別の実施形態は、例えば本発明のアミノ酸配列の発現についてなど、宿主細胞または宿主生物のトランスフォーメーションにおいて、好ましくは後述のような適切な遺伝子構成の形態での、本発明の核酸配列の利用に関連するものである。本発明はさらに、本発明の核酸配列でトランスフォームしたり、本発明のアミノ酸配列を発現することのできる宿主細胞または宿主生物とも関連する。
【0010】
さらにまた別の実施形態では本発明は、上述の核酸配列、アミノ酸配列、遺伝子構成、宿主細胞または宿主生物を使用しての、本発明のアミノ酸配列の生物学的活性を変調または阻害することが可能な化合物の識別または開発のための方法に関連するものである。通常は測定またはスクリーニングで利用されることになるこうした方法については、以下に詳述する。
【0011】
さらに別の実施形態では、本発明は、in vitroで、または、好ましくはin vivoで(も)、本発明のアミノ酸配列(の生物学的活性)を変調する化合物または、本発明のアミノ酸配列と相互作用する化合物と関連する。本発明はさらに、こうした化合物を含有する合成物ならびに、これらの合成物の調製および害虫の制御でのこうした化合物の利用とも関連する。
【0012】
今回の発明の核酸はここでは一括して「本発明の核酸」と呼ぶ。さらに、以下の発明の詳細な記述の本文では適切な場合には、「本発明の核酸配列」および「本発明の核酸」という語は、基本的に同等であり、基本的に相互変換が可能であると考えても差し支えない。
【0013】
さらに、今回の発明の目的では、核酸とは、通常は結合している別の核酸分子および配列の少なくとも一つから分離されている場合には、−例えば、本来の生物学的起源などから−「基本的に分離された(形態のもの)」と考える。同様に、ポリペプチドとは、通常は結合している別のポリペプチド分子から効率的に分離されている場合には、−例えば、本来の生物学的起源などから−「基本的に分離された(形態のもの)」と考える。核酸またはポリペプチドは、特に、少なくとも2倍、特に少なくとも10倍、さらに特に少なくとも100倍、および、最高1000倍以上に精製されている場合には、「基本的に分離されている」と考える。
【発明を実施するための最良の形態】
【0014】
今回の発明は、化合物およびその他の因子とのin vitroまたはin vivoでの相互作用において、本発明のアミノ酸配列が(潜在的な)「標的」として利用可能であるという所見から確立された(「標的」という語は、本明細書に援用して組み込まれる(特許文献1)での定義のように、当該技術での通常の意味を持つものである)。その結果、(以下に記述された方法などにより)本発明のアミノ酸配列と相互作用するとされている化合物または因子は、農芸化学、獣医学または薬学の分野で活性物質として有用なものとなる可能性がある。
【0015】
【特許文献1】国際公開第98/06737号
【0016】
一実施形態では、本発明は、本発明の核酸配列を含む核酸に関し、好ましくは(基本的に)分離された形態で、特に配列番号1という核酸配列と関連する。配列番号1の核酸配列は、後述の実施例の項で詳述された方法で、オオタバコガ(Heliothis virescens)の生体に由来またはこれから分離されたものである。この配列は、オオタバコガ(Heliothis virescens)の電位依存性カルシウムチャネルサブユニットα1についての不完全なコード化配列を示す。
【0017】
別の実施形態では、本発明は、本発明の核酸配列を含む核酸に関し、好ましくは(基本的に)分離された形態で、特に配列番号5という核酸配列と関連する。配列番号5の核酸配列はキメラ配列であり、これでは配列番号1のオオタバコガ(Heliothis virescens)の電位依存性カルシウムチャネルサブユニットα1が、ワタアブラムシcDNAライブラリ(実施例1)に由来する配列番号3で列挙した核酸1から429までのストップコード化ンと共に示されている。このアブラムシ由来の配列は、アブラムシの電位依存性カルシウムチャネルサブユニットα1に由来する。この配列の構成については、後述の実施例の項で詳述する。この配列は、機能的キメラコード化配列を示す。
【0018】
別の実施形態では、本発明は、本発明の核酸配列を含む核酸に関し、好ましくは(基本的に)分離された形態で、特に配列番号7、配列番号9または配列番号11という核酸配列と関連する。配列番号7、配列番号9または配列番号11という核酸配列は、後述の実施例の項で詳述された方法などで、オオタバコガ(Heliothis virescens)の生体に由来またはこれから分離されたものである。これらの配列はそれぞれ、オオタバコガ(Heliothis virescens)の電位依存性カルシウムチャネルサブユニットβ-1、β-2、β-3についての完全なコード化配列を示している。
【0019】
別の実施形態では、本発明は、本発明の核酸配列を含む核酸に関し、好ましくは(基本的に)分離された形態で、特に配列番号13という核酸配列と関連する。配列番号13という核酸配列は、後述の実施例の項で詳述された方法で、ワタアブラムシ(Aphis gossypii)の生体に由来またはこれから分離されたものである。この配列は、ワタアブラムシ(Aphis gossypii)の電位依存性カルシウムチャネルサブユニットα2δについての完全なコード化配列を示している。
【0020】
別の実施形態では、本発明は、本発明の核酸配列を含む核酸に関し、好ましくは(基本的に)分離された形態で、特に配列番号15または配列番号17という核酸配列と関連する。配列番号15または配列番号17という核酸配列は、後述の実施例の項で詳述された方法などで、オオタバコガ(Heliothis virescens)の生体に由来またはこれから分離されたものである。これらの配列はそれぞれ、オオタバコガ(Heliothis virescens)の電位依存性カルシウムチャネルサブユニットα2δ−1およびα2δ−2についての完全なコード化配列を示している。
【0021】
一般的に、本発明の核酸配列は、核酸の形態をとっている場合には、DNAまたはRNAであると思われ、一本鎖または二本鎖となることがある。例えば、本発明の核酸配列がゲノムDNA、cDNAまたは合成DNAとなることがある(企図した宿主細胞または宿主生物で発現するように特に適応させたコード化ンを利用しているDNAなどで、例えば、Vector NTI(InforMax,Inc.,メリーランド州ベセズダ)のBackTranslate解析ツールなどの適切なコンピュータプログラムを用いてデザインされたものなどであることがある)。従って、本発明の核酸配列は、イントロン配列を含むことがあり、一般的に種々のスプライシング変異体からなることもある。
【0022】
これとは別の実施形態として、本発明の核酸配列に向けての二本鎖RNA分子に関連するものがある(うち一本の鎖は通常、少なくとも本発明の核酸配列の一部からなる)。こうした二本鎖RNA分子は、遺伝子機能のRNA干渉の研究で特に有用である(Zamoreら、Cell 101:25−33(2000))。本発明はさらに、こうした二本鎖RNA分子を提供するために利用することが可能な遺伝子構造とも関連する(E.coliなどの菌株での例など、宿主細胞または宿主生物での適切な発現によるものなど)。こうした構造については、(非特許文献22)といった参考文献がある。
【0023】
【非特許文献22】Maniatisら、Molecular Cloning, a Laboratory Manual(Cold Spring Harbor Press,1989)
【0024】
広い意味では、「本発明の核酸配列」という語は以下のものから構成される:
−配列番号1、配列番号5、配列番号7、配列番号9、配列番号11、配列番号13、配列番号15および配列番号17の核酸配列の一部またはフラグメント;
−下記に詳述するような、配列番号1、配列番号5、配列番号7、配列番号9、配列番号11、配列番号13、配列番号15および配列番号17の核酸配列の(天然または合成による)突然変異体、変異体、対立遺伝子、類縁体、オルソログ(本明細書では以下、総称して「突然変異体」と呼ぶ);
−こうした(天然または合成による)突然変異体の一部またはフラグメント;
−少なくとも一つのさらに別の核酸配列と、配列番号1、配列番号5、配列番号7、配列番号9、配列番号11、配列番号13、配列番号15および配列番号17(またはこれらの一部またはフラグメント)の核酸配列の核酸融合物;
−少なくとも一つのさらに別の核酸配列と、(天然または合成による)突然変異体(またはその一部もしくはフラグメント)の核酸融合物;
こうした突然変異体、一部、フラグメントまたは融合物は、主として以下に記述するようなものである。
【0025】
本発明は、上述の核酸配列の種々のスプライシング変異体からも構成されている。
【0026】
主として、本発明の核酸配列は少なくとも500個の長さのヌクレオチドからなり、好ましいものとしては最低1,000個のヌクレオチド、さらに好ましいものとしては最低2,000個のヌクレオチド、最高ではほぼ8,000個のヌクレオチド、望ましくは多くの場合7,500個のヌクレオチド、さらに好ましい場合には多くの場合7,000個のヌクレオチドとなる。
【0027】
配列番号1の核酸配列の一部もしくはフラグメント、またはその(天然または突然変異による)突然変異体の一部もしくはフラグメントの例には、5’または3’切断核酸配列または、フレームスタートコード化ンまたはストップコード化ンの導入を受けた核酸配列が含まれるが、これに限定されるものではない。さらに、本発明の核酸配列の一つまたは複数についての二つまたはそれ以上のこうした部分またはフラグメントが、適切に組み合わされて(フレーム中のリガンドなど)、さらに本発明の核酸配列となることがある。
【0028】
望ましくは、こうした一部またはフラグメントは、配列番号1、配列番号5、配列番号7、配列番号9、配列番号11、配列番号13、配列番号15および配列番号17の核酸配列からの少なくとも100個、望ましくは250個、さらに望ましくは少なくとも500個、さらに1,000個以上となることもあるヌクレオチドからなる少なくとも一つの連続的伸展から構成されるものとなる。配列番号5の場合には、本発明によるフラグメントは、配列番号5からの少なくとも5個、望ましくは10個、さらに望ましくは20個以上のヌクレオチドからなる。
【0029】
さらに、当業者によれば、本明細書の公開に基づいて(別の個体の)同一生物種(例えば、異なる菌株または系列の個体からなどのもので、突然変異体を含むがそれに限定しない)からの配列番号1、配列番号5、配列番号7、配列番号9、配列番号11、配列番号13、配列番号15および配列番号17の核酸配列の天然の「突然変異体」(上述のもの)を識別したり、入手したり、分離したりすることが可能となると予想される。また、当業者によれば、本明細書の公開に基づいて配列番号1の核酸配列の合成突然変異体(本明細書上述で定義したもの)を提示したり、提供することができるようになると予想される。
【0030】
具体的な実施形態の一つとしては、配列番号1、配列番号5、配列番号7、配列番号9、配列番号11、配列番号13、配列番号15および配列番号17またはその一部またはそのフラグメントの核酸配列をコード化する突然変異体がある。
本明細書で記述した突然変異体は、配列番号1、配列番号5、配列番号7、配列番号9、配列番号11、配列番号13、配列番号15および配列番号17の核酸配列についての一つまたは複数、望ましくはすべての構造的特徴を有しているか、または、以下に記述した特徴を保存していることが好ましい。
【0031】
特に、本明細書で記述したあらゆる突然変異体、一部またはフラグメントは、少なくとも、本発明の対応するアミノ酸配列の活性部位または触媒部位および本発明の対応するアミノ酸の結合ドメインをコード化しているものとすることができる。
【0032】
さらに、本明細書で記述したあらゆる突然変異体、一部またはフラグメントは、好ましくはヌクレオチドレベルで、少なくとも75%、望ましくは最低80%、さらに望ましくは少なくとも85%、特に90%以上、および最高95%以上にわたり、配列番号1、配列番号5、配列番号7、配列番号9、配列番号11、配列番号13、配列番号15および配列番号17の核酸配列とある程度の「配列の同一性」を有することになる。
【0033】
さらに、主として、本発明の核酸配列のあらゆる突然変異体、一部またはフラグメントは、アミノ酸レベルで、少なくとも80%、特に90%以上、および最高95%以上にわたり、配列番号1、配列番号5、配列番号7、配列番号9、配列番号11、配列番号13、配列番号15および配列番号17のアミノ酸配列とある程度の「配列の同一性」を有することになるが、この「配列同一性」の割合については、以下で算定する。
【0034】
こうした目的では、配列番号1、配列番号5、配列番号7、配列番号9、配列番号11、配列番号13、配列番号15または配列番号17の配列と比較して、ヌクレオチドの欠失、挿入、置換または付加を、単独ヌクレオチド位での相違と考え、該当する核酸配列において、配列番号1、配列番号5、配列番号7、配列番号9、配列番号11、配列番号13、配列番号15または配列番号17の核酸配列での対応する位置でヌクレオチドが同一となっているヌクレオチドの数を、該当する核酸配列での総ヌクレオチド数で割って100%を掛けることにより、該当する核酸配列と配列番号1、配列番号5、配列番号7、配列番号9、配列番号11、配列番号13、配列番号15または配列番号17の核酸配列との間の「配列同一性」を算定することができる。
【0035】
全体的な配列アラインメントを行い、配列同一性を決定する好ましいコンピュータプログラムとしてはclustalWがあり(Higginsら、Nucl.Acids Res.22:4673−4680(1994))、これは、種々のコンピュータプラットフォームで利用可能であると発表されている。蛋白質配列アラインメントについてのclustalWプログラムの好ましいパラメータは、ktuple=1,diagonals=5,windows=5,gap=3,Score=PERCENTAGE,matrix=BLOSUM、open pnalty=10.0、および、extension penalty=0.5である。
【0036】
さらに、好ましい意図としては、本明細書で記述したあらゆる突然変異体、一部またはフラグメントは、下記の標準的な測定法により測定した場合、最低50%、好ましくは最低75%、最高90%といった範囲で、配列番号1、配列番号5、配列番号7、配列番号9、配列番号11、配列番号13、配列番号15または配列番号17の配列について上で記載されている生物学的活性に基本的に類似した生物学的活性を有する蛋白質またはポリペプチドをコード化することになる。
【0037】
本明細書で記述したあらゆる突然変異体、一部およびフラグメントは、配列番号1、配列番号5、配列番号7、配列番号9、配列番号11、配列番号13、配列番号15または配列番号17の核酸配列と、「きわめて厳密な」条件下で、ハイブリダイゼーションすることが可能であることが好ましい。当業者にとってはこうした条件は、本明細書に援用して組み入れられている上述のSambrookら、およびAusubelらなどの標準的なハンドブック、および、(特許文献2)、(特許文献3)または(特許文献4)などから、明確なものとなるはずである。
【0038】
【特許文献2】欧州特許第0967284号
【特許文献3】欧州特許第1085089号
【特許文献4】国際公開第00/55318号
【0039】
本発明の核酸配列を一つまたは複数の別の核酸配列と融合させたものを利用することも、(上述のように)本発明の意図の範囲内であり、こうした配列には、コード化配列、非コード化配列または制御配列が含まれるが、これらに限定されるわけではない。望ましくは、こうした融合においては、一つまたは複数の別の核酸配列が操作可能な形で、本発明の核酸配列と結合している(下記参照)(例えば、別の核酸配列がコード化配列である場合には、ヌクレオチド融合物は下記のように蛋白質融合物をコード化する)。
【0040】
別の実施形態は、本発明は、本発明の核酸配列に対するアンチセンス分子に関連する。
【0041】
本発明の核酸は、やはり下記に記述するように、遺伝子構成物の形を取ることがある。本発明の遺伝子構成物は一般的に少なくとも一つの本発明の核酸配列から構成され、下記に記述するように、それ自体で知られている遺伝子構成物の一つまたは複数の成分と連結していることがある。こうした遺伝子構成物はDNAまたはRNAであることがあり、好ましくは二本鎖DNAである。構成物は、企図する宿主細胞または宿主生物のトランスフォーメーションに適した形態、企図する宿主細胞のゲノムDNAへの組み込みに適した形態または、企図する宿主生物での独立した複製、維持および伝達に適した形態で存在することもある。遺伝子構成物は例えば、プラスミド、コスミド、酵母人工染色体(「YAC」)、ウイルスベクターまたはトランスポゾンなどのベクターの形態で存在することがある。特に、ベクターは発現ベクター、すなわち、in vitroまたはin vivo(下記のような適切な宿主細胞または宿主生物において)で発現させることの可能なベクターである。本発明の核酸配列からなる発現ベクターは、本明細書では組換え発現ベクターとも呼ばれている。これらの構成物は本明細書では「本発明の遺伝子構成物」とも呼ばれることになる。
【0042】
好ましい実施形態では、組換え発現ベクターでのこうした構成物は、以下のものから構成される:
a)本発明の核酸配列で、以下のものに操作可能な形で結合したもの:
b)プロモーターおよび任意の適切なターミネーター等の一つまたは複数の調節成分;
および、さらに任意により:
c)それ自体で知られている一つまたは複数の遺伝子構成物の別の成分;ここで、「調節成分」、「プロモーター」、「ターミネーター」、「別の成分」および「操作可能な形での結合」という語は、以下に示す意味を有している。
【0043】
上述の一つまたは複数の「別の成分」として、本発明の遺伝子構成物は一般的に、一つまたは複数の適切な調節成分(適切なプロモーター、エンハンサーまたはターミネーター、3’−または5’−非翻訳領域(「UTR」)配列、リーダー配列、選択マーカー、発現マーカーもしくはレポーター遺伝子、または、トランスフォーメーションもしくは組み込み(の効率)を促進若しくは増大させる可能性のある成分など)を含有する。こうした遺伝子構成物に対してのこれらおよびその他の適切な成分は、当業者にとっては明白なものであり、場合によっては使用する構成物のタイプ、企図された宿主細胞または宿主生物、対象となる発明の核酸配列が発現される方法(構成的発現、一過性の発現または誘発性の発現)、および、使用されるトランスフォーメーション技術に依存することがある。
【0044】
本発明の遺伝子構成物では、一つまたは複数の成分が、本発明の核酸配列または相互に「操作可能な形で」結合していることが望ましく、これは一般的に、互いに機能的な関連で存在することを意味する。例えば、プロモーターがコード化配列の転写または発現を開始または制御または調節することができる場合には、このプロモーターはコード化配列と「操作可能な形で」結合していると考えられる(この場合、このコード化配列は、このプロモーターの「制御下にある」と理解しなければならない)。
【0045】
一般的に、二つの核酸配列が操作可能な形で結合している場合には、これらは同一の方向を向いており、通常は同一のリーディングフレーム内に存在する。さらにこれらは通常、基本的に隣接しているが、必ずしもこれが必要とされるわけでもない。
【0046】
好ましくは、本発明で使用される遺伝子構成物の任意のその他の成分は、企図された宿主細胞または宿主生物内で企図された生物学的機能を示せる状態であることが好ましい。
【0047】
例えば、プロモーター、エンハンサーまたはターミネーターが企図された宿主細胞または宿主生物内で「操作可能」でなければならず、これは、(例えば)上述のプロモーターが、それが操作可能な形で結合した核酸配列、すなわちコード化配列の転写の開始、制御または調節をできるものでなければならないことを意味する。
【0048】
こうしたプロモーターは、構成的プロモーターまたは誘発性プロモーターであることがあり、さらに、宿主細胞または宿主生物の特定の発達段階で発現を提示したり(するのみであったり)、特定の細胞、組織、臓器または多細胞宿主生物の一部で発現を提示したり(するのみであったり)することもある。
【0049】
一部の特に好ましいプロモーターには、サイトメガロウイルス(「CMV」)、ラウス肉腫ウイルス(「RSV」)、pSVL SV40 Late Promoter Expression Vector(Pharmacia Biotech Inc., ニュージャージー州Piscataway)などのシミアンウイルス40(「SV40」)、または単純疱疹ウイルス(「HSV」)といった哺乳動物細胞での発現のための構成的プロモーターや、Jarvisらによって記述され(非特許文献23)、NovagenからpIEベクターとして販売されている(Novagen, Inc.ウィスコンシン州マディソン)即時型バキュロウイルスプロモーターなどの昆虫型構成的プロモーター、または、Bunchらによって記述され(非特許文献24)、Invitrogenからベクターとして販売されている(Invitrogen Corporation、カリフォルニア州カールズバッド)Drosophila metallothioneinプロモーターなどの昆虫型誘発性プロモーターが含まれるが、これに限定されるわけではない。
【0050】
【非特許文献23】Methods in Molecular Biology、39巻、Baculovirus Expression Protocols, ed.C.Richardson,. Hamana Press Inc., Totowa, NJ(1995)
【非特許文献24】Nucleic Acids Research、6巻、3号、1043−106(1988)
【0051】
選択マーカーは、適切な選択条件下で、本発明の核酸配列により(成功裏に)トランスフォームを受けた宿主細胞または宿主生物を、(成功裏に)トランスフォームを受けていない宿主細胞または生物と区別できるものでなければならない。こうしたマーカーの好ましい例としては、抗生物質(ジェネチシンもしくはG−418(GIBCO−BRL、ニューヨーク州グランドアイランド)、カナマイシン、または、アンピシリンなど)に対する耐性を付与する遺伝子、温度耐性を付与する遺伝子、または、トランスフォームをしていない細胞または生物の生存に必要不可欠なある種の因子、化合物または(栄養)成分が培地内に存在しなくても宿主細胞または宿主生物が生存できるようにする遺伝子などがあるが、これに限定されるものではない。
【0052】
リーダー配列は、企図した宿主細胞または宿主生物で、翻訳後に希望の変形を行ったり、転写されたmRNAをシグナルペプチドなどの細胞の希望の場所または細胞小器官に方向付けるようにしたりするものでなければならない。リーダー配列は、対象となる細胞から発現産物を分泌させることもある。このような場合、リーダー配列は、ピコルナウイルスリーダー、ポティウイルスリーダー、ヒト免疫グロブリン重鎖結合蛋白質(「BiP」)、タバコモザイクウイルスリーダー(「TMV」)およびトウモロコシクロロテック斑紋ウイルスリーダー(「MCMV」)などの宿主細胞または宿主生物内で操作可能なプロ配列、プレ配列またはプレプロ配列となることがあるが、これらに限定されるわけではない。
【0053】
発現マーカーおよびレポーター遺伝子は、宿主細胞または宿主生物内で遺伝子構造(上に存在する遺伝子または核酸配列)の発現を検出するものでなければならない。発現マーカーではさらに、任意により、細胞の特定の部位もしくは細胞小器官、または、特定の細胞、組織、臓器または多細胞生物の一部などで、発現産物の位置を確認できることがある。こうしたレポーター遺伝子は、本発明のアミノ酸配列との融合蛋白質として発現することもある。好ましいものの一部として、GFPなどの蛍光蛋白質、ベクター内で得られるV5エピトープもしくはポリヒスチジンなどの抗体認識蛋白質およびInvitrogenから供給されている抗体、または、ニッケルカラムでの精製を可能にするポリヒスチジンやメトトレキセートカラムでの精製を可能にするジヒドロ葉酸レダクターゼなどの精製アフィニティ・ハンドル、または、E.coliのβガラクトシダーゼ遺伝子などの遺伝子を発現している細胞を選別できるマーカーなどがあるが、これらに限定されるわけではない。
【0054】
プロモーター、選択マーカー、リーダー配列、発現マーカーおよび、本発明の遺伝子構成物中に存在するまたはそこで利用される可能性のあるその他の成分−ターミネーター、転写エンハンサーもしくは翻訳エンハンサーまたは組み込み因子−についての限定的ではない例の一部としては、上述のSambrookらおよびAusubelらなどの一般的なハンドブック、W.B.Woodらの”The nematode Caenorhabditis elegans”,Cold Spring Harbor Laboratory Press(1988)、D.L.Riddleらの“C.ELEGANS II” Cold Spring Harbor Laboratory Press(1997)、さらに、(特許文献5)、(特許文献6)、(特許文献7)、(特許文献8)、(特許文献9)、(特許文献10)、(特許文献11)、(特許文献12)、(特許文献13)、(特許文献14)および(特許文献15)などに言及されており、これらはそれぞれ、本明細書に援用して組み込まれている。当業者にとっては、その他の実施例は、明確なものである。
【0055】
【特許文献5】国際公開第95/07463号
【特許文献6】国際公開第96/23810号
【特許文献7】国際公開第95/07463号
【特許文献8】国際公開第95/21191号
【特許文献9】国際公開第97/11094号
【特許文献10】国際公開第97/42320号
【特許文献11】国際公開第98/06737号
【特許文献12】国際公開第98/21355号
【特許文献13】米国特許第6,207,410号
【特許文献14】米国特許第5,693,492号
【特許文献15】欧州特許第1085089号
【0056】
本発明の別の実施形態は、トランスフォームを受けたり、本発明での一つまたは複数の核酸配列、一つまたは複数の核酸もしくは一つまたは複数の遺伝子構成物を含む宿主細胞または宿主生物に関連するものである。本発明は、本発明のアミノ酸配列の一つまたは複数を(適切な条件下などで)発現する、または、(少なくとも)発現することのできる宿主細胞または宿主生物とも関連する。一部の実施形態では、宿主細胞が、機能性フラグメントおよびキメラなどを含む、キメラとしての電位依存性カルシウムチャネルのα1、TBW β−1および/またはβ−2および/またはβ−3およびアブラムシα2δおよび/またはTBW α2δ−1および/またはα2δ−2サブユニットを発現する組換えベクターから構成され、α1、βおよびα2δサブユニットのうちの少なくとも一つが、本発明の、望ましくは配列番号5、配列番号7、配列番号9、配列番号11、配列番号13、配列番号15および/または配列番号17といった、α1、TBW β−1および/またはβ−2および/またはβ−3ならびにTBW α2δ−1および/またはα2δ−2および/またはアブラムシα2δサブユニットのキメラとなる、機能性の電位依存性カルシウムチャネルを形成することがある。一部の好ましい実施形態では、宿主細胞が、機能性フラグメントおよびキメラなどを含む、電位依存性カルシウムチャネルのα1、βおよびα2δサブユニットを発現する組換えベクターから構成され、望ましくは配列番号5、配列番号7、配列番号9、配列番号11、配列番号13、配列番号15、および/または配列番号17といった、本発明のα1、TBW β−1および/またはβ−2および/またはβ−3ならびにTBW α2δ−1および/またはα2δ−2サブユニットのうちの少なくとも二つのキメラとなる、機能性の電位依存性カルシウムチャネルを形成する。一部の好ましい実施形態では、宿主細胞が、本発明の核酸分子であるコード化配列からのα1、βおよびα2δサブユニットを発現する組換えベクターから構成され、望ましくは、配列番号5と、配列番号7、配列番号9、配列番号11、配列番号13のうちの一つまたは複数と、配列番号15および/または配列番号17のうちの一つまたは複数といった、本発明のα1と、TBW β−1および/またはβ−2および/またはβ−3と、TBW α2δ−1および/またはα2δ−2および/またはアブラムシα2δサブユニットのキメラとなる。こうした宿主細胞または宿主生物は本明細書では集合的に、「本発明の宿主細胞または宿主生物」と呼ばれることもある。
【0057】
以下に例示するように、宿主細胞は、適切な細胞(真菌、原核細胞または真核細胞)または細胞系列となることがある:
−E.coli、Bacillus、StreptomycesおよびPseudomontasなどの菌株、ただしこれらに限定されない;
−AspergillusおよびTrichodermaなどの真菌細胞種、ただしこれらに限定されない;
−KluyveromycesまたはSaccharomycesなどの酵母細胞種、ただしこれらに限定されない;
−アフリカツメガエル(Xenopus)の卵母細胞などの両生類の細胞または細胞系列。
【0058】
殺虫性化合物の発見および開発で本発明の核酸配列を使用(しようと)する場合に特に有用であると思われる一つの具体的な実施形態では、宿主細胞が、以下のような昆虫由来の細胞または細胞系列となることがある:
−Spodoptera SF9およびSf21細胞などの鱗翅目に由来する細胞または細胞系列、ただしこれらに限定されない;
−Aphisに由来する細胞または細胞系列;
−シュナイダー細胞およびKc細胞など、Drosohilaに由来する細胞または細胞系列;および
−オオタバコガ(Heliothis virescens)など、対象となる有害生物種(後述のもの)に由来する細胞または細胞系列。
【0059】
宿主細胞は、CHO細胞、BHK細胞およびヒト細胞またはHEK,HeLaおよびCOSなどの細胞系列といった哺乳動物細胞または細胞系列となることもあるが、これらの限定されるわけではない。
【0060】
宿主生物は、以下のような適切な多細胞生物(脊椎動物または無脊椎動物)となることがあるが、これらに限定されるわけではない:
−C.elegansなどのCaenorhabditis属に由来する線形動物、ただしこれに限定されない;
−Aphis,Drosophila,Heliothisまたは、対象となる有害生物種(上述のもの)などの昆虫、ただしこれらに限定されない;
−ゼブラフィッシュなど、その他のよく知られているモデル生物;
−ラットまたはマウスなどの哺乳動物;
前述のハンドブックおよび特許出願からの例のように、当業者にとってはその他の適切な宿主細胞または宿主生物は明白なものである。
【0061】
本発明の核酸配列が多細胞生物で発現した場合には、当該生物の全体で発現する可能性も、該当する細胞、組織、臓器または一部に特異的なプロモーターの制御下での発現などの場合には、一つまたは複数の特殊な細胞、組織、臓器またはその一部のみで発現する可能性もあることに留意しなければならない。
【0062】
核酸配列は、やはり発達または生活環での該当する段階に特異的なプロモーターの制御下での発現などの場合には、宿主細胞または宿主生物の発達または生活環の特定の段階のみで発現することもある。さらに、すでに上で述べたように、該当する発現は、構成的なものであったり、一過性のものであったり、誘発性のものであったりする可能性がある。
【0063】
これらの宿主細胞または宿主生物は、(さらに、宿主生物の場合には:少なくともその一つの細胞、一部、組織または臓器で)本発明のアミノ酸配列を(適切な条件下などで)発現する、または、(少なくとも)発現することのできるものであることが好ましい。本発明にはさらに、本発明の宿主細胞または宿主生物のその後の世代、後代および子孫が対象となり、これらは例えば、細胞分裂または有性生殖もしくは無性生殖で得られたものとなることがある。
【0064】
別の態様では、本発明は、好ましくは(基本的に)分離された形態での核酸と関連しており、この核酸は、(本明細書に定義するような)本発明のアミノ酸配列、特に、配列番号2、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16および配列番号18のアミノ酸をコード化しているか、これら本発明のアミノ酸配列の発現に利用することが可能なものである。
【0065】
配列番号2、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16および配列番号18のアミノ酸配列は、当業者に知られている、蛋白質の分離および精製のための技術を用いて、上述の生物種から分離されることがある。これとは別に、配列番号2、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16および配列番号18のアミノ酸配列は、以下に詳述するように、適切な宿主細胞または宿主生物で、適切な核酸配列−それぞれ、配列番号1、配列番号5、配列番号7、配列番号9、配列番号11、配列番号13、配列番号15および配列番号17の核酸配列および、その適切な突然変異体など−を適切に発現させることによって得られることがある。
【0066】
別の態様では、本発明は、好ましくは(基本的に)分離された形態での蛋白質またはポリペプチドに関連しており、該当する蛋白質またはポリペプチドは、(上述に定義するような)本発明のアミノ酸配列、特に配列番号2、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16および配列番号18のアミノ酸配列からなっている。
【0067】
広い意味では、「本発明のアミノ酸配列」という語は以下のものから構成される:
−配列番号2、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16および配列番号18のアミノ酸配列の一部またはフラグメント;
−配列番号2、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16および配列番号18のアミノ酸配列の(天然または合成による)突然変異体、変異体、対立遺伝子、類縁体、オルソログ(本明細書では以下、総称して「類縁体」と呼ぶ);
−こうした類縁体の一部またはフラグメント;
−配列番号2、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16および配列番号18のアミノ酸配列(またはその一部もしくはフラグメント)と、少なくとも一つの別のアミノ酸残基またはアミノ酸配列との融合物;
−類縁体(またはその一部若しくはフラグメント)のアミノ酸と、少なくとも一つのさらに別のアミノ酸残基またはアミノ酸配列との融合物;
こうした突然変異体、一部、フラグメントまたは融合物は、以下に記述するようなものであることが好ましい。
【0068】
「本発明のアミノ酸配列」という語は、プレフォーム、プロフォームもしくはプレプロフォームまたは適切なリーダー配列との融合体など、上述のアミノ酸配列の「不完全な」形態をも包含する。さらに、本発明のアミノ酸配列は、該当のアミノ酸配列の発現または産生に使用した宿主細胞または宿主生物によっては、翻訳後プロセシングまたは適切なグリコシル化を受けたり;またはその他の変形(該当技術ではそれ自体が知られている化学的手法などにより)を受けたりすることがある。
【0069】
配列番号2、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16および配列番号18のアミノ酸配列の一部もしくはフラグメント、または、その突然変異体を含む、その(天然または合成による)類縁体の一部若しくはフラグメントの例には、N末端およびC末端切断アミノ酸配列などがあるが、これらに限定されない。さらに、本発明のアミノ酸配列の一つまたは複数の一部またはフラグメントの二つまたはそれ以上が適切に統合して、本発明のアミノ酸配列となることがある。
【0070】
本発明のアミノ酸配列は、少なくとも100個、好ましくは最低250個、さらに望ましくは最低350個の長さのアミノ酸、最大では2,500個、好ましくは多くの場合に2,000個、さらに望ましくは少なくとも1,750個のアミノ酸を有していることが好ましい。
【0071】
こうした一部またはフラグメントは、配列番号2、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16および配列番号18のアミノ酸配列からの、少なくとも5個、望ましくは最低10個、さらに望ましくは最低20個、さらに好ましいものとしては最低30個以上のアミノ酸からなる、少なくとも一つの連続的伸展から構成されるものとなる。配列番号6のフラグメントの場合には、フラグメントは、配列番号2からの少なくとも4個、望ましくは8個、さらに望ましくは15個以上のアミノ酸を含んでいなければならない。
【0072】
特に、本明細書で記述した一部またはフラグメントは、(少なくとも)本発明での活性部位若しくは触媒部位に対応するアミノ酸配列、または、本発明での結合ドメインに対応するアミノ酸配列から構成されるものである。当業者にとっては明確であるように、こうした一部またはフラグメントには、(下記で全般的に記述するように)測定技術およびスクリーニング技術、ならびに、(該当する部位またはフラグメントが結晶の形で提供される場合には)X線結晶解析で、特別な利用法が見出される可能性がある。
【0073】
さらに、当業者によれば、本明細書の公開に基づいて、配列番号2、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16または配列番号18のアミノ酸配列の天然の「類縁体」(上述のもの)を識別したり、入手したり、分離したりすることが可能となると予想される。こうした突然変異体は、(別の個体の)同一生物種(例えば、異なる菌株または細胞系列からのもので、突然変異株または細胞系列を含むがそれに限定しない)から、または、別の生物種(の個体)から得ることができた。例えば、こうした類縁体は、上述の昆虫種から得ることができた。
【0074】
さらに、当業者によれば、本明細書の公開に基づいて、配列番号2、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16または配列番号18のアミノ酸配列の合成「類縁体」(上述で定義したもの)を提示したり、提供することができるようになると予想される。
【0075】
本明細書で記述した突然変異体は、配列番号2、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16または配列番号18のアミノ酸配列についての一つまたは複数、望ましくはすべての構造的特徴を有しているか、または、以下に記述した特徴を保存しているものであることが好ましい。
【0076】
本明細書で記述したあらゆる類縁体、一部またはフラグメントは、アミノ酸レベルで、少なくとも80%、特に90%以上および最高95%以上の度合いで、配列番号2、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16または配列番号18のアミノ酸と、「配列の同一性」を有するものであることが好ましい。
【0077】
こうした目的では、−配列番号2、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16または配列番号18のアミノ酸配列と比較して−アミノ酸残基の欠失、挿入、置換または付加を、単独アミノ酸(位)での相違と考え、該当するアミノ酸配列において配列番号2、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16または配列番号18のアミノ酸配列での対応する位置でアミノ酸残基が同一となっているアミノ酸残基の数を、該当するアミノ酸配列での総アミノ酸数で割って100%を掛けることにより、該当するアミノ酸配列と配列番号2、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16または配列番号18のアミノ酸配列との間の「配列同一性」を算定することができる。上述のように、こうした算定で配列アラインメントの対比を実施するために適した方法は、clustalWプログラムである。
【0078】
さらに、望ましくは、本明細書で記述したあらゆる類縁体、一部またはフラグメントは、下記の標準的な測定法により測定した場合、最低10%、好ましくは最低50%、さらに望ましくは最低75%および、最高90%といった範囲で、配列番号2、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16および配列番号18の配列について、上で記載されている生物学的活性と基本的に類似した生物学的活性を有することになる。
【0079】
蛋白質融合の提供などのように、本発明のアミノ酸配列を一つまたは複数の別のアミノ酸配列と融合させたものを利用することも、(上述のように)本発明の意図の範囲内である。以下に詳述するように、一般的に、こうした融合物は、適切な宿主細胞または宿主生物で−本発明の核酸配列を、一つまたは複数の別のコード化配列と適切に融合させるなど−本発明の適切な核酸配列の適切な発現によって得られる。
【0080】
具体的な実施形態の一つでは、こうした融合物が、グルタチオン−S−トランスフェラーゼ(「GST」)、緑色蛍光蛋白質(「GFP」)、ルシフェラーゼまたはその他の蛍光蛋白質などのような、レポーター蛋白質と融合した本発明のアミノ酸配列から構成されることがある。当業者にとっては明らかであるように、こうした融合物が、発現解析および同様の方法で特に有用とされることがある。
【0081】
別の実施形態では、該当するアミノ酸または残基に対するアフィニティ法を用いる場合など、融合相手が、発現したアミノ酸配列の精製で利用される可能性のあるアミノ酸配列またはアミノ酸残基となることがある。その後、該当する配列または残基は除去され(化学的切断または酵素的切断などにより)、本発明の核酸配列が得られる(この目的のためには、アミノ酸配列または残基は、切断可能なリンカー配列を介して、本発明のアミノ酸配列に任意で結合することができる)。こうした残基の好ましい例としては、複数のヒスチジン残基およびグルタチオン残基があるが、これらに限定されない。
【0082】
好ましいものではあるが限定的ではない態様では、こうした融合物は、下記の標準的な測定法により測定した場合、最低10%、好ましくは最低50%、さらに望ましくは最低75%および、最高90%といった範囲で、配列番号2、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16または配列番号18の配列について、上で記載されている生物学的活性と基本的に類似した生物学的活性を有することになる。
【0083】
本発明の核酸配列およびアミノ酸配列は、一般的に、以下の構造的特徴または保存された特徴のうちの一つまたは複数の存在によって、特徴付けられると思われる。
【0084】
Heliothis verescensの遺伝子については、配列番号1は、オープンリーディングフレームを取り囲むcDNA配列であり;配列番号2は、配列番号1によってコード化された蛋白質である。HeliothisのN型電位依存性カルシウムチャネルの蛋白質配列は、以下の表に示すように、他のN型電位依存性カルシウムチャネルの蛋白質と関連しており、関連性の値は、clustalWおよび上述に示したパラメータを用いて決定された。
【0085】
【表1】

【0086】
1 Geneband Accession No.AAC47406,(非特許文献25)
2 Geneband Accession No.AAC36868,(非特許文献26)
【0087】
【非特許文献25】Smith,L.A.ら、J.Neurosci.16(24)、7868−7879、1996
【非特許文献26】Waterston,R.,Science 282(5396)、2012−2018、1998
【0088】
他のカルシウムチャネルから類推されるように、機能性蛋白質は単量体である可能性が高い。(非特許文献27)などを参照されたい。
【0089】
【非特許文献27】Hannan and Hall,In Comparative Molecular Neurobiology,Y.Pichon,1993、Birkhuase Verlag(スイス・バーゼル)
【0090】
上述に基づき、本発明は特定の説明または機序に特に限定されていないが、核酸配列およびアミノ酸配列は、電位依存性カルシウムチャネルとしての(生物学的な)活性を有する。特に、今回の発明は、穿孔虫、根切り虫、アワヨトウ、オオタバコガ、fruit worm、アオムシ、ガおよびlopperなど、特にオオタバコガの鱗翅目の族の昆虫と、穿孔および吸引に適した口器を有する半翅類の昆虫から、電位依存性カルシウムチャネルとして活性が示されている。
【0091】
当該技術で知られているように、こうした種類の生物学的活性は、標識された既知のリガンドとの結合部位での完全な競合;蛍光共鳴エネルギー移動などの蛋白質相互作用に基づく蛍光測定法によるカルシウム蛍光色素を用いたカルシウム流動性の測定(Velicelebeらを参照)による、経時的な蛍光の消失または蛍光法もしくは比色法によるレポーターアッセイ;電位依存性カルシウムチャネルの測定に適したあらゆる技術など、標準的な測定法を用いて測定することができる。生物学的活性は、カルシウム蛍光色素を用いたカルシウム流動性を測定することによって測定することが好ましい。
【0092】
本発明の別の実施形態は、ある程度厳密な条件下、好ましくは厳密性が高いものおよび特に厳密な条件下(いずれも、上述のもの)で、本発明の核酸配列とハイブリダイゼーションすることが可能な核酸プローブに関連する。こうした核酸プローブは、例えば、本発明の核酸配列の検出もしくは分離で利用されたり、本発明の核酸配列の増幅でプライマーとして利用されることがある;いずれも、それ自体で知られている技術を用いており、やはり、上述のSambrookらおよびAusubelらなどの一般的なハンドブックに記述されている。
【0093】
本発明の別の核酸配列を検出または分離する際に利用する場合には、こうした核酸プローブは、通常は15個から100個、好ましくは20個から80個の間の長さのヌクレオチドを有することが好ましい。増幅のプライマーとして使用する場合には、こうした核酸プローブは、25個から75個、好ましくは20個から40個の間の長さのヌクレオチドを有することになる。
【0094】
一般的に、こうしたプローブは、当業者が、適切なコンピュータプログラムを任意に使用しながら、本発明の核酸配列またはアミノ酸配列−特に、配列番号1または配列番号2、配列番号5または配列番号6、配列番号7または配列番号8、配列番号9または配列番号10、配列番号11または配列番号12、配列番号13または配列番号14、配列番号15または配列番号16および配列番号17まはた配列番号18から開始することによってデザインすることができる。さらに、当業者にとっては明白であるように、こうしたプローブは変性プローブとすることもできる。
【0095】
別の態様では、本発明は、今回の発明の核酸配列の突然変異体および遺伝子構成物を調製するための方法と関連する。
【0096】
今回の発明の核酸配列の天然の突然変異体は、実施例で記載されている方法と基本的に類似した方法、または、以下のような代替法で、得ることができる:
−適切な発現ベクター系での、対象となる生物者からのDNAライブラリの構築と、それに続く突然変異配列の直接発現;
−適切な発現ベクター系での、対象となる生物者からのDNAライブラリの構築と、それに続く、本発明のプローブ(下記のもの)または本発明の核酸配列を用いての、該当するライブラリのスクリーニング;
−対象となる生物者からの、突然変異配列をコード化するmRNAの分離と、それに続く、逆転写酵素を用いてのcDNAの合成;
または、それ自体で知られている何らかの他の適切な方法または技術により得るが、これについては、例えば、Sambrookら、”Molecular Cloning:A Laboratory Manual”(第2版)、1−3巻、Cold Spring Harbor Laboratory Press(1989)およびF.Ausubelら、”Current protocols in molecular biology”,Green Publishing and Wiley Interscience、ニューヨーク(1987)などの標準的なハンドブックで述べられている。
【0097】
今回の発明の核酸配列のこうした合成配列を作製するための技術は、当業者にとっては明らかであり、例えば、自動化DNA合成法;部位特異的突然変異誘発;天然に存在する一つまたは複数の配列の二つまたはそれ以上の部分を結合しての、切断された発現産物の発現を導く突然変異の誘発;一つまたは複数の制限酵素認識部位の誘発(適切な制限酵素を用いた場合に容易に切断または結合されると思われる部位または領域を作製するなど)および、例えば天然に存在する電位依存性カルシウムチャネルの配列をテンプレートとして利用して、一つまたは複数の「不適正」プライマーを用いてのPCR法による突然変異の誘発といったものがあるが、これらに限定されるわけではない。これらおよびその他の手法は、当業者にとっては明らかなものであり、やはり、上述のような、SambroodらおよびAusubelらなどの標準的なハンドブックに記載されている。
【0098】
本発明の遺伝子構成物は一般的に、上述の、SambrookらおよびAusubelらなどの一般的なハンドブックに記載されている技術を用いるなどして、本発明の核酸配列を、上述の一つまたは複数の別の成分と適切に結合させることによって得られる。
【0099】
本発明の遺伝子構成物は、それ自体で知られている適切な(発現)ベクターに、本発明の核酸配列を挿入することによって得られることが多くなる。限定的なものではないが好ましい適切な発現ベクターの実施例の一部に、以下のようなものがある:
−哺乳動物細胞での発現のためのベクター:pSVL SV40(Pharmacia),pMAMneo(Clontech),pcDNA3(Invitrogen),pMC1neo(Stratagene),pSG5(Stratagene),pMSG(Pharmacia),pIND(Invitrogen),EBO−pSV−neo(ATCC37593)、pBPV−1(8−2)(ATCC37110),pdBPV−MMTneo(342−12)(ATCC37224),pRSV gpt(ATCC37119),pRSVneo(ATCC37198),pSV2−dhfr(ATCC37146),pUCTag(ATCC37460)および1ZD35(ATCC37565);
−細菌での発現のためのベクター:pETベクター(Novagen)およびpQEベクター(Qiagen);
−酵母またはその他の真菌細胞での発現のためのベクター:pYES32(Invitrigen)およびPichia発現ベクター(Invitrogen);
−昆虫細胞での発現のためのベクター:pBlueBacII(Invitroge),pEI1(Novagen),pMT/V5His(Invitrogen)。
【0100】
別の態様では、本発明は、本発明の核酸配列、本発明の核酸または本発明の遺伝子構成物による宿主細胞または宿主生物のトランスフォーミングのための方法に関連する。本発明はさらに、宿主細胞または宿主生物をトランスフォームする本発明の核酸配列、本発明の核酸または本発明の遺伝子構成物の利用とも関連する。
【0101】
具体的な実施形態では、宿主細胞または宿主生物で本発明の核酸配列の発現が、元の(すなわち天然の)宿主細胞または宿主生物に比べて低下することがある。これは、例えば、技術領域でよく知られているアンチセンス技術またはRNA干渉技術を用いた一時的な方法、もしくは、本発明の核酸配列のランダムな、部位特異的または化学的な突然変異誘発を用いた構成的な方法で、行った場合などに生ずることがある。
【0102】
適切なトランスフォーメーション技術は、当業者には明白であり、企図されている宿主細胞または宿主生物もしくは、利用予定の遺伝子構成物に依存することがある。限定的なものではないが好ましい適切な技術の例には、粒子射撃法によるトランスフォーメーション、(マイクロ)インジェクション、トランスフェクション(適切なトランスポゾンを使用したものなど)、電気穿孔法およびリポフェクチン法などがある。これらの技術およびその他の適切な技術については、やはり、上述のハンドブックおよび特許出願に記載されている。
【0103】
トランスフォーメーション後には、本発明の核酸配列または遺伝子構成物によってトランスフォーメーションが成功した宿主細胞または宿主生物の検出および選別の手順がとられることがある。これは、例えば、本発明の遺伝子構成物中に存在する選択可能なマーカーに基づく選択手順、または、特異的な抗体などを用いての本発明のアミノ酸配列の検出によるステップなどとなることがある。
【0104】
トランスフォーメーションを受けた宿主細胞(安定な細胞系列の形をとることがある)または宿主生物(安定な突然変異系列または株の形をとることがある)は、今回の発明の別の態様を形成する。
【0105】
さらに別の態様では、本発明は、本発明のアミノ酸配列を産生するための方法に関連する。
【0106】
本発明のアミノ酸配列の発現を生じさせたり、得たりするためには、一般的に、本発明の(希望の)アミノ酸配列が発現または産生するような条件下で、トランスフォーメーションを受けた宿主細胞またはトランスフォーメーションを受けた宿主生物を、保持したり、維持したり、培養したりすることになる。適切な条件は、当業者にとっては明らかであり、通常、使用した宿主細胞または宿主生物、ならびに、本発明の(関連する)核酸配列の発現を制御する制御因子に依存する。これについても、上の、本発明の遺伝子構成物についてのパラグラフで述べた、ハンドブックおよび特許出願に記載されている。
【0107】
一般的に、適切な条件には、適切な培地の使用、適切な栄養源または適切な栄養分の存在、適切な温度の利用、ならびに、任意により、適切な誘発因子または化合物の存在(本発明の核酸配列が、誘発性プロモーターの制御下にある場合など)が含まれることがある;これらはいずれも、当業者によって選択される。この場合にも、こうした条件下で、本発明のアミノ酸配列は、構成的な方法、一過性の方法または、適切な誘発が行われた場合にのみ、発現できる。
【0108】
本発明のアミノ酸配列が(最初は)不完全な形で生じ(上述のように)、その後、使用した宿主細胞または宿主生物に応じて、トランスフォーメーション後の変形を受けることがあるということも、当業者には明白である。さらに、本発明のアミノ酸配列は、使用した宿主細胞または宿主生物に応じて、グリコシル化を受けることがある。
【0109】
本発明のアミノ酸配列はその後、宿主細胞または宿主生物、もしくは、該当する宿主細胞または宿主生物が培養された培地から、(分離用)クロマトグラフィー法および電気泳動法、沈降分離法、アフィニティ法(本発明のアミノ酸配列と融合した、特異的な切断可能なアミノ酸配列を使用するなど)および、調製用の免疫学的手法(分離予定のアミノ酸配列に対する抗体を使用するなど)などの、それ自体が知られている蛋白質分離精製技術を用いて、分離される。
【0110】
実施形態の一つでは、該当する配列もしくは、その抗原部位またはエピトープに対して特異的な抗体を得るために、こうして得られたアミノ酸配列を利用することもある。
【0111】
実施形態の一つでは、今回の発明は、今回の発明のアミノ酸配列、望ましくは、配列番号2、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16または配列番号18、もしくは、これらの類縁体、変異体、対立遺伝子、オルソログ、一部、フラグメントまたは、配列番号6に特異的な抗体が、配列番号4などのアブラムシα1と交差反応しない場合には、エピトープに対して特異的に作製された、モノクローナル抗体およびポリクローナル抗体などの抗体に関連する。
【0112】
本発明の別の態様となるこうした抗体は、(特許文献16)、(特許文献17)、(特許文献18)、(特許文献19)、(特許文献20)、(特許文献21)および(特許文献22)などに記述されている、それ自体が知られた方法で作製される。排他的ではないものの、こうした方法は、全般的に、該当するアミノ酸配列に対しての抗体が生ずるような方法に従って、免疫応答性の宿主を本発明の妥当なアミノ酸配列または免疫原性を有するその一部(特定のエピトープなど)で免疫し、その後、該当の宿主より得られた血液または血清などから、生じた抗体を回収することによるものであることが多い。
【0113】
【特許文献16】英国特許出願公開第2357768号
【特許文献17】米国特許第5,693,492号
【特許文献18】国際公開第95/32734号
【特許文献19】国際公開第96/23882号
【特許文献20】国際公開第98/02456号
【特許文献21】国際公開第98/41633号
【特許文献22】国際公開第98/49306号
【0114】
例えば、ポリクローナル抗体は、ヤギ、ウサギ、ヒツジ、ラット、ブタまたはマウスなどの適切な宿主を、任意により、免疫原性のキャリアー(ウシ血清アルブミンまたはキーホールリンペットヘモシニアンなど)、もしくは、フロイントのアジュバント、サポニン、水酸化アルミニウムまたは同様の界面活性剤などのアジュバントを用いて、本発明のアミノ酸配列(のエピトープ)で免疫することによって、得ることができる。適切な免疫応答が生じた後に(通常は1〜7日以内)、やはり(特許文献23)などに言及されている、それ自体が知られている方法により、免疫した動物から採取した血液または血清から抗体を分離するが、任意によって、既知の免疫測定法技術を用いての、希望する性質(特異性など)を有する抗体についてのスクリーニング段階が含まれることがある。
【0115】
【特許文献23】国際公開第96/23882号
【0116】
例えば、モノクローナル抗体は、ハイブリドーマをベースとしたものおよび同様の方法など、培養物での連続的な細胞系列を用いて産生させることができ、これについても基本的には上述で引用した参考文献に記載されている。従って、本発明のアミノ酸配列に対するモノクローナル抗体を産生する細胞または細胞系列は、本発明のアミノ酸配列に対する抗体産生法と同様、本発明の別の態様を形成し、こうした方法では一般的に、こうした細胞の培養と、培養物または倍地の抗体の分離が関与し、やはり、それ自体が知られている技術が用いられる。
【0117】
さらに、本発明のアミノ酸配列に対するFabフラグメント(F(ab)2,Fab’およびFabフラグメント)は、それぞれ、ペプシンまたは別のプロテアーゼによる抗体の消化、ジスルフィド結合の開裂、ならびに、パパインおよび還元剤による処理によって得ることができる。Fab−発現ライブラリは、Huseらの方法、Science 245:1275−1281等によって得ることができる。
【0118】
別の実施形態では、本発明のアミノ酸配列、または、こうしたアミノ酸配列をコード化し、発現する組換え発現ベクターからなる宿主細胞もしくは宿主生物を用いて、本発明のアミノ酸配列の(生物学的)活性を変調したりその他の点でこれと相互作用をすることのできる化合物またはその他の因子を、識別したり開発したりすることもでき、こうした利用法により、本発明の別の態様が形成される。当業者には明白なように、こうした状況では、本発明のアミノ酸配列は、こうした化合物または因子との相互作用の標的となる。
【0119】
こうした状況では、「変調する」、「変調」、「モジュレーター」および「標的」という語は、当該技術でのその通常の意味を有し、とりわけ、国際公開第98/06737号で定義が示されている。一般的に、モジュレーターは、生物学的活性または過程(例えば、本発明のアミノ酸配列の生物学的活性など)の機能的性質を、促進、阻害または低減またはその他の変更、感化または影響(「変調」と総称する)することのできる化合物または因子である。
【0120】
こうした状況では、本発明のアミノ酸配列は、in vitroでの変調(測定またはスクリーニングの一部として)もしくは、in vivoでの変調(標的を変調することが知られている化合物または因子による変調で、こうした化合物または因子は、例えば、農芸化学、獣医学または薬学での利用で活性化合物として利用されることがある)の標的となることがある。
【0121】
例えば、本発明のアミノ酸配列、宿主細胞または宿主生物は、一次的スクリーニング(標的に関して未知の活性を有する被験化合物の一連の群またはライブラリから、標的のモジュレーターを識別するために利用されるスクリーニング法など)または二次的測定(一次的スクリーニングで該当したものを確認したり、該当するものを導くための化学の一部などとして、該当する分子の最適化に使用される測定法)など、本発明のアミノ酸配列のモジュレーターを識別したり開発したりするために利用することのできる測定法またはスクリーニング法の一部として利用されることがある
【0122】
例えば、こうした測定法またはスクリーニング法は、in vitro測定法またはスクリーニング法として構成されることがあり、これは一般的に、標的に対する潜在的モジュレーターとして検討対象となる化合物または因子(本明細書では以下、「被験化合物」と呼ぶ)を標的と結合させ、該当する結合を生じた信号を測定するというものになる。こうしたin vitroスクリーニングに適した技術は、当業者には明白なものとなり、(非特許文献28)および(非特許文献29)などに記載されている。例えば、こうした測定法またはスクリーニング法が、結合測定法またはスクリーニング法として構成されることがあり、この場合、被験化合物は、標的からの検出可能なリガンドを置換するために用いられ(放射性リガンドまたは蛍光リガンドなど)、これによって、モジュレーターによって標的から置換されたリガンドの量が決定される。
【0123】
【非特許文献28】Eldegrawiら(1987).FASEB J.,1巻、262−271ページ
【非特許文献29】Rauhら(1990),Trends in Pharmacol.Sci.,11巻、325−329ページ
【0124】
こうした測定法またはスクリーニング法は、細胞をベースとした測定法またはスクリーニング法として構成されることもあり、この場合、本発明の宿主細胞は、被験化合物と結合したりこれにさらされ、これに基づいて、宿主細胞による少なくとも一つの生物学的応答が測定される。
【0125】
さらに、こうした測定法またはスクリーニング法は、動物一個体でのスクリーニングとして構成されることがあり、この場合、本発明の宿主生物は、被験化合物と接触したりこれにさらされ、これに基づいて、宿主生物による少なくとも一つの生物学的応答(表現型の変化、挙動の変化または、麻痺または死亡を含むがこれに限定されない生理学的変化など)が測定される。
【0126】
従って、一般的に、上述の測定法およびスクリーニング法は、特に、被験化合物による標的の変調を示す信号が発生するような方法で、被験化合物が標的(または、標的を発現している宿主細胞または宿主生物)と接触するための少なくとも一つの段階から構成される。ついで、その後の段階で、該当する信号が測定される。
【0127】
従って、ある態様では、本発明は、本発明の該当するアミノ酸配列と被験化合物との相互作用を示すような信号を生じさせるための方法に関連しており、該当する方法は少なくとも以下の手順からなっている:
a)本発明のアミノ酸配列もしくは、アミノ酸配列を含有または発現する宿主細胞または宿主生物を、該当する被験化合物と該当するアミノ酸配列との間の相互作用を示す信号が発生するような方法で、該当する被験化合物と接触させる手順;および、任意により、
b)これによって生ずる信号を検出する手順。
【0128】
別の態様では、本発明は、本発明のアミノ酸配列のモジュレーターおよび/またはインヒビターを識別するための方法に関連しており、
該当する方法は少なくとも以下の手順からなっている:
a)本発明のアミノ酸配列もしくは、アミノ酸配列を含有または発現する宿主細胞または宿主生物を、該当する被験化合物と該当する標的との間の相互作用を示す信号が発生するような方法で、該当する被験化合物と接触させる手順;および、任意により、
b)これによって生ずる信号を検出し、該当する信号が該当するアミノ酸配列のモジュレーターおよび/またはインヒビターを識別するような手順。
【0129】
従って、今回の発明は、鱗翅目のカルシウムチャネル蛋白質活性のモジュレーターを識別する方法を示すものである。この方法は、核酸配列の少なくとも一つが本発明の核酸配列で、被験化合物の存在下でカルシウムチャネルを発現するような、機能性カルシウムチャネルをコード化する拡散配列を含む組換え発現ベクターから構成される被験細胞をカルシウムを含有する溶液と接触させ、被験細胞での細胞内カルシウム量を検出することによっての、試験方法の実施段階からなっている。好ましい実施形態では、機能性カルシウムチャネルは、本発明の二つまたはそれ以上のアミノ酸配列から作られている。一部の実施形態では、機能性カルシウムチャネルが、望ましくは、配列番号6と、配列番号8、配列番号10または配列番号12のうちの一つ、ならびに、配列番号14、配列番号16または配列番号18のうちの一つという、本発明の三つのアミノ酸配列から作られている。この方法は、鱗翅目のカルシウムチャネルをコード化し、鱗翅目のカルシウムチャネルを発現する核酸配列を含有する組換え発現ベクターからなる陰性対照細胞を、被験化合物の非存在下でカルシウムを含有する溶液と接触させ、陰性対照細胞での細胞内カルシウムの量を検出することによって陰性対照測定を実施する手順からも構成されている。被験細胞での細胞内カルシウムの量を、陰性対照細胞での細胞内カルシウムの量と比較する。陰性対照細胞での細胞内カルシウムの量と比較した場合の被験細胞での細胞内カルシウムの量の変化は、被験化合物が鱗翅目のカルシウムチャネル蛋白質活性のモジュレーターであることを示すものである。好ましい実施形態の一部では、被験細胞がCHOまたはHEKとなる。好ましい実施形態の一部では、該当する細胞内部の色素が細胞内カルシウムと相互作用することによって生じた蛍光を測定するという測定法を用いることにより、細胞内カルシウムが検出される。一部の好ましい実施形態では、この方法はさらに、鱗翅目のカルシウムチャネルをコード化し、鱗翅目のカルシウムチャネルを発現する核酸配列を含有する組換え発現ベクターからなる陽性対照細胞を、被験化合物の非存在下および鱗翅目のカルシウムチャネル作動物質の存在下でカルシウムを含有する溶液と接触させ、該当する陽性対照細胞での細胞内カルシウム量を検出することにより、陽性対照測定を行うというものである。一部の実施形態では、この方法はさらに、鱗翅目のカルシウムチャネルを発現しない鱗翅目のカルシウムチャネル陰性細胞を、被験化合物の非存在下でカルシウムを含有する溶液と接触させて、該当する鱗翅目のカルシウムチャネル陰性対照細胞に取り込まれたカルシウムの量を検出することによって、二番目のタイプの陰性対照測定を実施するか;および/または、鱗翅目のカルシウムチャネルを発現しない鱗翅目のカルシウムチャネル陰性細胞を、被験化合物の非存在下および鱗翅目のカルシウムチャネル作動物質の存在下でカルシウムを含有する溶液と接触させて、該当する鱗翅目のカルシウムチャネル陰性対照細胞に取り込まれたカルシウムの量を検出することによって、三番目のタイプの陰性対照測定を実施するかの、いずれかの段階からなっている。好ましい実施形態では、この方法で利用される鱗翅目のカルシウムチャネル蛋白質は、配列番号2または配列番号6、その突然変異体、そのフラグメントからなる群から選択されたアミノ酸配列を有しており、これは、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16および配列番号18の群、その突然変異体、そのフラグメントからなる群から選択されたさらに別のサブユニットの一つまたは複数と組み合わされることがある。一部の実施形態では、鱗翅目のカルシウムチャネルをコード化する核酸配列は、配列番号1または配列番号5で、補助的なサブユニットが存在する場合には、これは、配列番号7、配列番号9、配列番号11、配列番号13、配列番号15または配列番号17をコード化するものとなる。
【0130】
一部の実施形態によると、鱗翅目のカルシウムチャネル蛋白質活性のインヒビターを識別する方法は、鱗翅目のカルシウムチャネルをコード化し、鱗翅目のカルシウムチャネルを発現する核酸配列を含有する組換え発現ベクターからなる被験細胞を、被験化合物の存在下でカルシウムを含有する溶液および鱗翅目のカルシウムチャネル作動物質と接触させて、該当する被験細胞での細胞内カルシウムの量を検出することにより、検査測定法を実施するという段階から構成される。対照測定も、鱗翅目のカルシウムチャネルをコード化し、鱗翅目のカルシウムチャネルを発現する核酸配列を含有する組換え発現ベクターからなる陰性対照細胞を、被験化合物の非存在下でカルシウムを含有する溶液および鱗翅目のカルシウムチャネル作動物質と接触させて、該当する陰性対照細胞での細胞内カルシウムの量を検出することにより実施される。被験細胞での細胞内カルシウムの量を、対照細胞での細胞内カルシウムの量と比較する。対照細胞での細胞内カルシウムの量と比較した場合の被験細胞での細胞内カルシウムの量の減少は、被験化合物が鱗翅目のカルシウムチャネル蛋白質活性のインヒビターであることを示すものである。一部の好ましい実施形態では、被験細胞は、CHOまたはHEKである。一部の好ましい実施形態では、細胞内カルシウムは、細胞内部の色素が細胞内カルシウムと相互作用することによって生じた蛍光を測定するという測定法を用いることによって検出される。一部の好ましい実施形態では、鱗翅目のカルシウムチャネル蛋白質は、配列番号2または配列番号6、その突然変異体、そのフラグメントからなる群から選択されたアミノ酸配列を有しており、これは、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16および配列番号18の群、その突然変異体、そのフラグメントからなる群から選択されたさらに別のサブユニットの一つまたは複数と組み合わされることがある。一部の実施形態では、該当する鱗翅目のカルシウムチャネルをコード化する核酸配列は、配列番号1または配列番号5で、補助的なサブユニットが存在する場合には、これは、配列番号7、配列番号9、配列番号11、配列番号13、配列番号15または配列番号17をコード化するものとなる。
【0131】
被験化合物は、化合物の一連の群またはライブラリの一部となることがあり、当業者には明らかなように、これは、多種多様な一連の群またはライブラリ、もしくは、対象となる一連の群またはライブラリとなることがある。こうしたスクリーニングに使用されるライブラリは、技術で知られている化学的過程、または、化学合成での従来の手段を組み合わせて用いることで、調製が可能である。
本発明の測定法およびスクリーニング法は、例えば、適切なロボットを使用した自動化法などで、中程度のスループットからハイスループットで実施されることがある。特に、この実施形態では、本発明の方法は、標準的な24穴、96穴、384穴、1536穴または3456穴プレートなどのマルチウェルプレートのウェル内で標的を被験化合物と接触させることによって実施されることがある。
【0132】
通常は、本発明の測定法およびスクリーニング法では、各々の測定について、標的または宿主細胞または宿主生物を、単独の被験化合物のみと接触させる。しかし、例えば、該当する併用によって相乗効果が生ずるか否かを検討するなどで、標的を二つまたはそれ以上の被験化合物と、同時にまたは連続的に接触させることも、本発明の意図の範囲内である。
【0133】
被験化合物が本発明のアミノ酸配列に対してモジュレーターおよび/またはインヒビターとして識別された場合には(本明細書上述のようなスクリーニングまたは測定によって)、それ自体が、本発明の関連アミノ酸、望ましくは、配列番号2、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16または配列番号18、その突然変異体、そのフラグメント、さらに望ましくは、配列番号2、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16または配列番号18のモジュレーターおよび/またはインヒビターとして利用されたり(農芸化学、獣医学または薬学での利用を目的とした作用物質として)、または、例えば、溶解度、吸収性、バイオアベイラビリティー、毒性、安定性、持続性、環境への影響などの性質を改善するために、任意によりさらに最適化されることがある。当業者にとっては、例えば二番目の測定法(の一部)などとして、本発明の核酸配列、望ましくは、配列番号1、配列番号5、配列番号7、配列番号9、配列番号11、配列番号13、配列番号15または配列番号17、アミノ酸配列、宿主細胞または宿主生物および方法が、最適化法などで、さらに利用される可能性があることは明白である。
【0134】
本発明は、モジュレーターおよび/またはインヒビター(被験化合物、化合物または因子)が、標的の変調、阻害または標的との相互作用を行う(in vivoまたはin vitroで)際の特定の方法または機序を、特に限定するものではない。例えば、モジュレーターおよび/またはインヒビターは、作動物質、拮抗物質、逆作動物質、部分作動物質、競合的インヒビター、非競合的インヒビター、補助因子、アロステリックインヒビターまたは標的に対するその他のアロステリック因子であることがあったり、別の蛋白質またはポリペプチド、受容体、もしくは、細胞の細胞小器官の一部のように、他の生物学的化合物への標的の結合を促進または低下させる化合物または因子であったりする。同様に、モジュレーターおよび/またはインヒビターは、(活性部位、アロステリック部位、結合ドメイン、もしくは、共有結合または水素結合により標的の別の部位で)標的と結合したり、標的の活性部位を(可逆的、不可逆的または競合的に)遮断および/または阻害したり、標的の結合ドメインを(可逆的、不可逆的または競合的に)遮断および/または阻害したり、標的の構造に影響または変更を与えたりすることがある。
【0135】
同様に、被験化合物、モジュレーターおよび/またはインヒビターは、例えば、以下のようなものであることがある:
−標的の既知の基質の類縁体;
−2〜20個、望ましくは3〜15個のアミノ酸残基からなるオリゴペプチド;
−アンチセンスまたは二本鎖RNA分子;
−蛋白質、ポリペプチド;
−補助因子または補助因子の類縁体。
【0136】
被験化合物、モジュレーターおよび/またはインヒビターが、参照化合物または因子であることもあり、これは、標的を変調、阻害したり、標的と相互作用することが知られている化合物(標的に対する既知の基質またはインヒビターなど)であったり、標的が属する一般的なクラスの他のものを、変調、阻害したり、これらと相互作用することが一般的に知られている化合物または因子(該当するクラスの既知の基質またはインヒビターなど)であったりすることがある。
【0137】
しかし、被験化合物、モジュレーターおよび/またはインヒビターは小分子であることが望ましく、これの意味することは、分子量が1500未満、望ましくは1000未満のものということである。これは例えば、有機分子、無機分子または有機金属分子などであり、可溶性塩類などの適切な塩の形態をとっていることもある。「小分子」という語は、(総)分子量が上述の範囲である限りは、複合体、キレートおよび同様の分子を網羅するものでもある。
【0138】
すでに上で述べたように、本発明のアミノ酸配列、望ましくは、配列番号2、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16または配列番号18のモジュレーターおよび/またはインヒビターとして識別または開発されている化合物または因子(およびこれらの化合物の前駆物質)は、農芸化学、獣医学または薬学での成分調製など、農芸化学、獣医学または薬学の領域で、作用物質として有用なものとなる可能性があり、こうしたモジュレーターとこれらを含有する成分はどちらも、本発明の別の態様である。
【0139】
例えば、農芸化学の領域では、本発明のモジュレーターおよび/またはインヒビターは、有害生物(の侵襲)の予防または制御などで、接触剤および植物浸透剤の双方として、殺虫剤、殺線虫剤、軟体動物駆除剤、殺蠕虫剤、殺ダニ剤またはその他のタイプの農薬もしくは殺生物剤として利用されることがある。同様に、モジュレーターおよび/またはインヒビターは、例えば、作物保護剤、家庭内用殺虫剤、または、有害生物によって生ずる損傷を予防または処理(種子、木材もしくは、貯蔵した作物または果物の保護など)するための薬剤として利用されることがある。本発明のモジュレーターおよび/またはインヒビターは、殺虫剤として利用されることが好ましい。こうした適用では、企図する最終的な使用法に適した処方品とするために、本発明の一つまたは複数のモジュレーターおよび/またはインヒビターが、一つまたは複数の農学的に許容されている担体、アジュバントまたは希釈剤と−さらに、それ自体で(例えば)植物保護剤(作用スペクトルを広げ、任意により相乗作用を生ずるため)、除草剤、肥料または植物成長調整剤としての活性を有する一つまたは複数の別の化合物も任意により併用して−適切に組み合わせられることがある。こうした処方品は、例えば、溶液、乳剤、散布剤、濃縮剤、エアロゾル、スプレイ、粉末、流動物、粒子、顆粒、錠剤、燻蒸剤、餌もしくはその他の適切な固体、半固体または液体といった処方品の形であることがあり、さらに、任意により、適切な溶剤、乳化剤、安定剤、界面活性剤、消泡剤、湿潤剤、展着剤、固着剤、誘引剤または(餌については)食物成分を含有することもある。”Pesticidal Formulation Research”、ACS発行物(1969)および”Pesticide Formulation”,Wade van Valkenburg Ed,Marcel Dekker発行物(1973)などの標準的なマニュアルで言及されている。
【0140】
こうした成分は一般的に、本発明の一つまたは複数のモジュレーターおよび/またはインヒビターを適切な含量で含有しており、これは一般的に、総成分の重量の0.1〜99%、特に10〜50%範囲である。
【0141】
本発明のモジュレーターおよび/またはインヒビターならびに成分は、特に、例えば、以下の目の望ましくないまたは有害な昆虫(成体および、幼虫などの未成熟体)を駆除または制御するための殺虫剤として有用である:
−Pissodes strobi、Diabrotica undecimpunctata howardiおよびLeptinotarsa decemlineataなどの甲虫目;
−Rhagoletis pomonella,Mayetiola destructorおよびLiriomyza huidobrensisなどの双翅目;
−Neodiprion taedae tsugae、Camponotus pennsylvanicusおよびSolenopsis wagneriなどの膜翅目;
−Pseudatomoscelis seriatus,Lygus lineolaris(Palisot de beauvois)およびAcrosternum hilareなどの半翅目;
−Aphis gossypiiなどの同翅亜目およびアブラムシ;ならびに
−オオタバコガ(Heliothis verescens)などの鱗翅目。
【0142】
有害または望ましくない生物を制御するために使用する際には、これらの生物が、生物の制御(死滅または麻痺など)に適した量で、本発明のモジュレーター、インヒビターまたは成分に直接接触することになる。こうした量は、当業者により(制御対象となる生物種に対して化合物を検討することにより)容易に決定することが可能で、通常は、約10〜500g/ha程度、特に100〜250g/haとなる。
【0143】
本発明のモジュレーター、インヒビターまたは成分は、系統的に適用されることもあり(制御対象となる生物の環境または土壌中などに)、植物、種子、果物などを保護するために、やはり、当業者によって決定することの可能な適切な量で、適用されることもある。本発明のモジュレーターおよび/またはインヒビターは、例えば、こうした組成で通常に使用される他の殺虫化合物と置き換えるために、−添加剤などとして−それ自体が知られている成分に組み込まれることがある。
【0144】
具体的な実施形態の一つでは、本発明のモジュレーターおよび/またはインヒビターならびに成分は、農芸化学、獣医学またはヒトの健康の領域で、寄生虫、特に以下のような寄生性節足動物、線形動物および蠕虫によって生ずる感染症または損傷または不快感を予防または治療するために使用されることがある:
−マダニ、ダニ、ノミ、シラミ、サシバエ、ツノサシバエ、アオバエおよび、その他、咬み傷を生じたり吸血性の外部寄生虫などの、外寄生性節足動物;
−蠕虫などの内部寄生虫;
さらに、こうした寄生虫によって引き起こされたり伝播する疾患の予防または治療にも使用される。こうした目的のため、本発明のモジュレーターおよび/またはインヒビターは、例えば、錠剤、経口溶液または乳剤、注射用溶液または乳剤、ローション、エアロゾル、スプレイ、粉末、浸漬剤または濃縮剤として処方されることがある。
【0145】
動物およびヒトの健康の領域では、本発明のモジュレーター、インヒビターおよび成分は、本発明のアミノ酸配列が標的として関与する疾患または障害の予防または治療にも使用されることがある。この目的のためには、本発明のモジュレーターおよび/またはインヒビターは、当業者にとっては明白な、薬学または獣医学での利用が許容されている一つまたは複数の添加剤、担体または希釈剤と共に処方されることがある。
【0146】
このように、さらに別の態様で、本発明は、本明細書上述のような、農芸化学、獣医学または薬学での利用を目的とした成分の調製での本発明のモジュレーターおよび/またはインヒビターの利用に関連している。本発明は、やはり上述のように、有害生物の制御および有害生物によって引き起こされる感染症または損傷の予防での、本発明のモジュレーター、インヒビターおよび成分の利用に関連する。
【0147】
本発明についてはここで、以下の非限定的な実施例により、さらに例証する。
(実施例)
【0148】
実施例1−cDNAライブラリの構成および配列データベース
【0149】
【表2】
【0150】
全長クローンの豊富なcDNAライブラリが、(a)混合ライフステージのワタアブラムシ(ワタアブラムシcDNAライブラリ)および、(b)虫齢の高いHeliothis virescenesの幼虫から切断した頭部(TBW頭部cDNAライブラリ)から精製された、ポリ(A)含有RNAより、Invitrogen Corporation(カルフォルニア州カールスバッド)によって構築された。これらのライブラリはいずれも、プラスミドベクターpCMV−SPORT6.1で構成され、E.coli DH10B(TonA)にトランスフォームされた。これらのライブラリは、さらに増幅されることなく、グリセロール培養物として、−80℃で貯蔵された。Invitrogen Corporationにより、ワタアブラムシcDNAライブラリの一部が増幅され、標準化されて、384穴プレートに並べられた。並べられたクローンについては、クローン化されたcDNAの5’末端および3’末端に隣接するベクタープライマーを用いて、自動化配列決定分析が行われた(Genome Therapeutics、マサチューセッツ州ウォルサム)。配列読み取り結果は、質およびベクターの混入についてトリミングが行われ、コンティグに構築されて、BLASTコンパチブルデータベースに入力された。検索ツールのBasic Local Alignment Sequence Tool(BLAST)スートおよびこれらのツールの使用法ならびに、BLASTコンパチブルデータベースへのアクセスは、米国立予防衛生研究所(メリーランド州ベセズダ)の全米バイオテクノロジー情報センターから行うことができる。
【0151】
さらに、虫齢の高いHeliothis virescenesの幼虫の腹側神経索および頭部から分離したポリ(A)含有RNAより、cDNAライブラリが構成された(TBW神経索ライブラリ)。約195個の神経索および頭部を、第5虫齢初期のTBW幼虫から切断採取し、ドライアイス上で急速に冷凍した。この組織は、室温にて3mLのTRIZOLTM試薬(Invitrogen Corporation)でホモジナイズし、製造業者のプロトコルに従って、全RNAを抽出した。RNAの質は、UV吸収分光光度法(320nm〜220nm)および、少量のグリコシル化試料の1×NorthernMaxTM−Gly緩衝液を含有する1%アガロースゲルでの電気泳動により確認した(Ambion、テキサス州オースチン)。ポリ(A)含有RNAは、製造業者の記述に従って、Oligotex(登録商標)mRNA分離キット(QIAGEN Inc.、カリフォルニア州バレンシア)を用いて、アフィニティクロマトグラフィーにより全RNAから選択した。(非特許文献30)の方法の改良法により、cDNAを合成した。要約すると、6.2μgのポリ(A)含有RNAを、11.2μlの80%(v/v)グリセロールおよび400pmolのSacdTプライマー(表I)と混合し、全量を24μlとした。プライマー/RNA混合物は、サーモサイクラーで65℃にて10分加熱した後に45℃まで冷却し、やはり45℃に予温した76μlの合成緩衝液と混合した。ファーストストランドcDNA合成反応の最終組成は:50mMのTris−HCl(25℃でpH8.3)、75mMのKCl、3mMのMg2Cl、10mMのジチオスレイトール(DTT)、0.6mMのdGTP、0.6mMのdATP、0.6mMのTTP、0.6mMの5−メチル−dCTP、50μg/mlのウシ血清アルブミン、2000単位/mlのRNaseOUTTMリボヌクレアーゼインヒビター(Invitrogen Corporation)および20,000単位/mlのSuperScriptTM逆転写酵素(Invitrogen Corporation)であった。反応は、45℃での2分間のインキュベート、2分間をかけての35℃への冷却、35度での2分間のインキュベート、50℃への上昇と5分間のインキュベート、最終的に55℃への上昇と60分間のインキュベートによるものであった。ファーストストランド反応は、EDTAを10mMへ、ナトリウムN−ラウロイルサルコシンを0.1%(w/v)、プロテイナーゼK(Invitrogen Corporation)を0.2mg/mlへと加えることにより、終了させた。混合物を45℃で15分間インキュベートした後、フェノール:クロロホルム:イソアミルアルコール(25:24:1)で1回抽出した。水相を除去し、100μlの5MのNH4OAcと500μlのエタノールを加えて、cDNAを−20度で60分間沈降させた。その後、遠心分離によりcDNAを採集し、70%エタノールで1回洗浄した後、500μlの0.1×TE(1mMのTris−HCl(pH8.0)−0.1mMのEDTA)に溶解して、氷浴上で200μlのセカンドストランド合成緩衝液とあわせた。セカンドストランド反応の最終組成は、20mMのTris−HCl(pH8.3)、130mMのKCl、10mMの(NH4)2SO4、5mMのMgCl2、10mMのDTT、0.6mMのdATP、0.6mMのdGTP、0.6mMのdCTP、0.6mMのTTP、10Mmのb−NAD、50μg/mlのBSA、3単位のE.coli RNaseH(Invitrogen Corporation)、2.5単位のE.coli DNAリガーゼ(Invitrogen Corporation)および25単位のE.coli DNAポリメラーゼ(Invitrogen Corporation)であった。セカンドストランド合成は、15℃にて150分間行った。酵素混合物は70℃での10分間の加熱により不活化し、その後、氷浴上に放置した。1μl(5単位)のT4 DNAポリメラーゼ(Invitrogen Corporation)を加え、15℃での15分間のインキュベートにより、二本鎖cDNAを平滑末端とした。方向性のクローニングを促進するために、平滑末端としたcDNAに二本鎖Xhoアダプターを加え、その後、ファーストストランド合成のためのプライマーで独特の切断を行うSacIによる切断によって、3’末端から除去した。アダプターは、それぞれ2nmolのオリゴヌクレオチドXhoAd4とXhoAd5(表I)を40μlの20mMNaClであわせ、混合物を70℃で3分間加熱してから、10分間をかけて25℃にゆっくりと冷却することによって、形成させた。アニールしたXhoアダプター(450pmol)は、50mMのTris−HCl(pH7.5)、10mMのMgCl2、1mMのATP、10mMのDTT、25μg/mlのBSAおよび600単位のT4 DNAリガーゼ(New England Biolabs、マサチューセッツ州ビバリー)を含有する30μlの反応溶液中で、1.7μgの二本鎖dDNAと8℃にて一晩結合させた。上述の、プロテアーゼKによる切断、フェノール:クロロホルムによる抽出およびエタノールによる沈降で再度cDNAを精製した。その後、1μgのcDNAあたり25単位の酵素の割合で、SacIによりcDNAを切断し、10mMのTris−HCl(pH8.0)、1mMのEDTA、0.1%(w/v)のSDSおよび0.1MのNaClで平衡化した2mlのSepharose CL−2B(Sigma Chemical Co.,ミズーリ州セントルイス)ゲル濾過カラムでサイズ分画を行った。ピーク分画(730ngのcDNA)をプールし、エタノール沈降法によって濃縮して、10μlの0.1XTE(pH8.0)に再溶解した。クローニングのために、57mMのTris−HCl(pH7.5)、11.4mMのMgCl2、1.1mMのATP、11.4mMのDTTおよび28μg/mlのBSAを含有する4.4μlの溶液中で、50ngのcDNAを、バクテリオファージλgtGreen1Sから調製した、500ngのXhoI/SacIファージアームとあわせた(cDNAとベクターの分子比は2:1)。0.3μl(3単位)のT4ポリヌクレオチドキナーゼ(New England Biolabs)を加え、混合液を37℃で15分間インキュベートして、cDNA上のXhoアダプターをリン酸化した。その後、0.3μl(120単位)のT4 DNAリガーゼ(New England Biolabs)を加えてcDNAをベクターと結合させ、混合液を15℃にて一晩インキュベートした。バクテリオファージλgtGreen1Sはλgt11からの派生物であり(非特許文献31)、これでは、(a)野生型バクテリオファージλゲノムのSalI−Xhoフラグメント架橋座標33245−33449が除去され、(b)E.coli lacZ遺伝子を含有する独特のSacI−EcoRIフラグメントが除去されて、直線型のpGreen1S(+)Lox2(Xba)プラスミドに置き換えられている(図1A)。pGreen1S(+)Lox2(Xba)はpBluescript SK(−)の派生物であり(Stratagene、カリフォルニア州ラホヤ)、これでは、(a)f1の起点とアンピシリン耐性遺伝子とのあいだにpZL1のincA遺伝子(Invitrogen Corporation)が挿入され、(b)colE1の起点が除去されて、pACYC184の低コピー数のp15Aの起点に置き換えられ(ATCC数は37033、American Type Culture Collection、バージニア州マナッサス)、(c)タンデム反復配列をしている二つのloxP部位(非特許文献32)が独特のMfeIにより分離されて、PmeI切断酵素認識部位がp15Aの起点とアンピシリン耐性遺伝子とのあいだに挿入されている。MfeIおよびPmeIによるpGreen1S(+)Lox2(Xba)の直線化により、λgtGreen1Sに挿入される形のプラスミドが産生される(図1B)。製造業者によって記述されているように、結合反応の一部は、Gigapack III Goldパッキングエキス(Stratagene)を用いて、in vitroでファージにパッケージした。組換えプラスミドの調製には、パッケージしたファージをE.coliのDH10B(Zip)(Invitrogen Corporation)株上に撒き込み、アンピシリンまたはカルベニシリンのいずれかを含有するLB平板上での増殖により、コロニーを選択した。E.coliのDH10B(Zip)株はcreリコンビナーゼを発現し、バクテリオファージλの複製を抑制するため、creを介する組換えによって、プロプラスミドに隣接するloxP部位で、ファージベクターからプラスミドを切除し、これを独自に複製するプラスミドとして確立することができる。
【0152】
【非特許文献30】CarninciおよびHayashizaki(Methods in Enzymology、303巻、19−44ページ(1999))
【非特許文献31】Young and Davis, Proc.Natl.Acad.USA,80巻、1194−1198ページ(1983)
【非特許文献32】Sternberg and Hamilton,J.Mol.Biol.,150巻、467−86ページ(1981)
【0153】
実施例2−オオタバコガ(「TBW」)電位依存性カルシウムチャネルα1サブユニット配列の識別
材料および方法
TBW頭部ポリA RNAの分離。0.1%のジエチルピロカルボネート水溶液(「DEPC」;Aldrich Chemical Co.,Inc.、ウィスコンシン州ミルウォーキー)を37℃にて約16時間インキュベートした後、オートクレーブで60分間処理した。ガラス器具はすべて、250℃で4時間加熱し、ボトルの蓋はすべて、0.1%のDEPC溶液で浸漬した。Braunホモジナイザーのマイクロプローブ(B.Braun Biotech International、ペンシルバニア州アレンタウン)は50mlの100%エタノール(「EtOH」)に浸漬した後、25mlのRNAzolB(CINNA−BIOTECX Labs,Inc.、テキサス州ヒューストンから入手可能な塩酸グアニジニウム標品)で稼動させた。TBWの頭部は、第4虫齢のオオタバコガ幼虫から切断採取し、氷浴上の風袋を測定した遠心管中に入れた。約0.5gのTBW頭部材料を入手した後、TBW頭部は使用時まで−70℃にて凍結した。その後、頭部を4mlのTRIZOLTM試薬に入れ、室温にて最大速度で30秒間ホモジナイズした。ホモジナイズが終了した時点で、2mlのクロロホルムを加え、生じた混合液をSS34ローター(Sorvall Products,L.P.、ノースカロライナ州アッシュビル)中で室温にて12000gで15分間遠心分離した。この手順の完了後、上部水層を15mlの新しいコニカルチューブに入れ、等量のイソプロパノールを加えた。生じた混合液をSS34ローター中で室温にて12000gで10分間遠心分離した。その後、生じた沈塊を10mlの75%エタノール水溶液で洗浄し、SS34ローター中で室温にて7500gで5分間遠心分離した。上清を流去し、生じた沈殿塊を室温で5分間、風乾させた。その後、生じた沈殿塊を、1mlの上述の0.1%DEPC溶液に溶解した。全RNAの濃度は、UV分光法で測定した(Amersham BiosciencesのGeneQuant、ニュージャージー州Piscataway)。500μgの全RNAを用い、Micro−FastTrackTM 2.0キット(Invitrogen Corporation)のプロトコルに従って、ポリA mRNAを分離した。ポリA mRNAの濃度は、UV分光法により測定した。ポリA mRNA溶液は、その後の利用のために、−70℃で保存した。
【0154】
ファーストストランドcDNAの合成。逆転写は、1.0μgのテンプレートRNAに鳥類骨髄芽球症ウイルス(「AMV」)逆転写酵素(Invitrogen Corporation)を加えることによって開始した。逆転写反応には、Invitrogen cDNA Cycleキットに含まれている以下の試薬も関与した:4μlの5X逆転写緩衝液、1μlのリボヌクレアーゼインヒビター、1μlの100mM dNTP、1μlの80mMリン酸ナトリウムおよび、ランダムプライマー、オリゴ(dT)プライマーまたは遺伝子特異的プライマーのうちのいずれか。5’および3’でのRACE反応では、製造業者の指示に従い、オリゴ(dT)プライマーまたはランダムプライマーまたは遺伝子特異的プライマーにより、SMARTTM RACE cDNA増幅キット(BD Biosciences Clontech、カリフォルニア州パロアルト)を用いて、mRNAの逆転写を行った。反応は、GeneAmp(登録商標)9700サーマルサイクラー(Applied Biosystems、カリフォルニア州フォスターシティー)上で行い、42℃にて60分間維持した。その後、95℃にて5分間加熱した後に4℃で5分間放置することによって、AMV RTを不活化した。Ambion(テキサス州オースチン)FirstChoiceTM RLM−RACEキットを用いて、製造業者の指示に従って、5’−RACE反応を行った。
【0155】
PCRによる増幅。製造業者の指示に従い、Advantage 2 PCRキット(BD Biosciences Clontech)で供給された緩衝液とdNTPを用いて、50μlのcDNA反応液を作製した。変性プライマーを利用した増幅では通常、50〜60℃の範囲のアニーリング温度を利用し、アイソフォーム特異的プライマーを用いるものでは65〜70℃の範囲のアニーリング温度を利用した。3’および5’RACE反応は、SMARTTM RACE cDNA Amplificationキット(BD Biosciences Clontech)により供給されたプライマーとプロトコルを利用して、製造業者の指示に従って行った。生じたPCR産物は、アガロースゲル電気泳動法により性質検討した。上述の方法で二次的な「ネステッド」増幅を行った後、NuSieve(登録商標)ゲル(FMC Corp.、ペンシルバニア州フィラデルフィア)からバンドを切り出し、Quiagenゲル抽出キットを用いて、製造業者の指示に従って精製した。
【0156】
プライマーの合成およびデザイン。オリゴヌクレオチドは、Seq Wright Inc.(テキサス州ヒューストン)が合成し、使用前に蒸留水で溶解する凍結乾燥品として提供された。PCRプライマーおよびプローブがデザインされ、InforMax, Inc.(メリーランド州ベセズダ)からのVectorNTI suite 6.0ソフトウェアと、Roseら(Nucleic Acids Research、26巻、1628−1635ページ(1998))からのConsensus−Degenerate Hybrid Oligonucleotide Primers Software(「CODEHOP」)を用いて、アニーリング温度が推定された。
【0157】
サブクローニングおよび配列決定。精製PCRフラグメントは、TOPO TA Cloningキット(Invitrogen Corp.)を用いて、製造業者の指示に従って、pCR2.1−TOPOベクターにサブクローンされた。生じたpCR2.1−TOPOベクターの配列を決定し、その後、VectorNTI suite6.0ソフトウェアを用いて解析された。
【0158】
プライマー。使用したプライマーは、表IIに示すようなものであった:
【0159】
【表3】
【0160】
【表4】
【0161】
【表5】
【0162】
上述のオリゴヌクレオチドの中で、変性プライマーは、N(A、G、CまたはT)、H(A、CまたはT)、S(CまたはG)、Y(CまたはT)、W(AまたはT)、D(A、GまたはT)またはR(AまたはG)の記号の入った位置に、単量体の統計学的混合物を組み込んでいる[IUPAC総会と一致]。
【0163】
オオタバコガ(Heliothis virescens)のα1サブユニット配列の増幅
オオタバコガ(Heliothis virescens)のα1サブユニットについての増幅の記述はいずれも、配列番号1で示された配列位置を指定している。
【0164】
RT−PCR反応では、プライマー1および2は、オオタバコガ(Heliothis virescens)の配列での3674位から4113位のヌクレオチドからのフラグメントを増幅するために使用され、これについてのクローン化およびサブクローン化が行われた。
【0165】
Heliothisの配列は、標準化していないTBW cDNAライブラリプラスミドDNAを用いたPCRにより(実施例1)、プライマー3および5を用いて、上流方向に伸展され、その後、プライマー4および5を用いてネステッドPCRが行われた。生じたPCR産物からは、2621位から3770位のヌクレオチドからの配列情報が得られ、これについてのクローン化およびサブクローン化が行われた。プライマー5は、ライブラリベクター配列からのSP6である。
Heliothisの配列は、標準化していないTBW cDNAライブラリプラスミドDNAを用いたPCRにより(実施例1)、プライマー6および8を用いて、下流方向に伸展され、その後、プライマー7および8を用いてネステッドPCRが行われた。生じたPCR産物からは、4029位から5047位のヌクレオチドからの配列情報が得られ、これについてのクローン化およびサブクローン化が行われた。プライマー8は、ライブラリベクター配列からのT7である。
【0166】
プライマー9および11を用いての3’−RACEによる3’−末端cDNAの回復と、プライマー10および12を用いてのその後のネステッドPCRの試みでは、驚くべきことに、生じたPCR産物から、3’−末端cDNA配列の代わりに、1589位から2149位のヌクレオチドからの配列情報が得られた。このPCR産物のクローン化およびサブクローン化が行われた。その後、2149位から2621位のヌクレオチドからのHeliothis配列が、SMARTTM RACE cDNA増幅キット(BD Biosciences Clontech)からのランダム六量体を用いた逆転写により得られ、ついで、プライマー13および15を用いてタッチダウンPCRが行われた。その後、生じたcDNAは、ネステッドプライマー14および16を用いて増幅され、2056位から2685位のヌクレオチドからのアンプリマーが得られ、これについてのクローン化およびサブクローン化が行われた。
【0167】
Heliothisの配列は、さらに、SMARTTM RACE cDNA増幅キットからのランダム六量体を用いた逆転写によって上流方向に伸展され、ついで、プライマー17および18を用いたタッチダウンPCRが行われた。その後、生じたcDNAは、ネステッドプライマー17および19を用いて増幅された。生じたPCR産物からは、524位から1645位のヌクレオチドからの配列情報が得られ、これについてのクローン化およびサブクローン化が行われた。
Heliothisの配列は、さらに、SMARTTM RACE cDNA増幅キットからのランダム六量体を用いた逆転写によって上流方向に伸展され、ついで、プライマー11および20を用いたタッチダウンPCRが行われた。ついで、この反応で生じたcDNAを、ネステッドプライマー12および21を用いて増幅した。生じたPCR産物からは、181位から613位のヌクレオチドからの配列情報が得られ、これについてのクローン化およびサブクローン化が行われた。
【0168】
その後、FirstChoiceTM RLM−RACEキット(Ambion)からのランダム六量体を用いて、逆転写によりHeliothisの配列の5’−末端cDNAが回復され、プライマー22および24を用いてタッチダウンPCRが行われた。この反応によって生じたcDNAは、ネステッドプライマー23および25を用いて増幅した。生じたPCR産物からは、1位から252位のヌクレオチドからの配列情報が得られ、これについてのクローン化およびサブクローン化が行われた。
【0169】
全長キメラα1サブユニットの構成
重複しているDNAセグメントで制限酵素認識部位を介して上述PCRフラグメントをつなぎあわせることにより、配列番号1についての全長クローンが構成された。翻訳停止コード化ンを加えるために、結合されたクローンをNotIで切断し、その後、ワタアブラムシライブラリ(実施例1)から得られた配列番号3に列挙されている1位から429位のヌクレオチドからの停止コード化ンを有するアブラムシα1遺伝子の3’−末端を融合させて、発現についての全長キメラクローンを作製した。キメラDNAおよび蛋白質配列は、配列番号5および6のように示されている。全長キメラα1サブユニットは、発現ベクターpCDNA3.1(Invitrogen Corp)に挿入された。
【0170】
実施例3−オオタバコガ(「TBW」)電位依存性カルシウムチャネルβサブユニット配列の識別
材料および方法
実施例2と同様。
【0171】
プライマー。使用したプライマーは、表IIIに示すようなものであった:
【0172】
【表6】
【0173】
上述のオリゴヌクレオチドの中で、変性プライマーは、N(A、G、CまたはT)、H(A、CまたはT)、S(CまたはG)、Y(CまたはT)、W(AまたはT)、D(A、GまたはT)またはR(AまたはG)の記号の入った位置に、単量体の統計学的混合物を組み込んでいる[IUPAC総会と一致]。
【0174】
オオタバコガ(Heliothis virescens)のβサブユニット配列の増幅
オオタバコガ(Heliothis virescens)についての増幅の記述はいずれも、配列番号7で示された配列位置を指定している。
【0175】
RT−PCR反応では、プライマー1および2は、オオタバコガ(Heliothis virescens)の配列での874位から1140位のヌクレオチドからのフラグメントを増幅するために使用され、これについてのクローン化およびサブクローン化が行われた。
【0176】
このフラグメントから、プライマー3および4ならびに、RecActiveTM Gene Enrichmentキット(Active Motif、カリフォルニア州カールズバッド)からのビオチン−dNTP混合物を含有するPCR反応によって、ビオチル化プローブ(900位から1113位のヌクレオチドより)が合成された。このビオチル化プローブはその後、ゲル精製し、標準化されていないTBW cDNAライブラリ(実施例1)からrecAを介する遺伝子選択により、Active Motif社(カリフォルニア州カールズバッド)のRecActiveTM Gene Enrichmentキットにより、全長TBW βサブユニットcDNAをクローンするために利用された。3種類の全長クローンの配列が決定され、これは、配列番号7、8および9で示されている。配列番号7、8および9からそれぞれ翻訳された蛋白質配列に対応する配列番号10、11および12に示すように、複数のアイソフォームが認められた。これらのcDNAは最初に発現ベクターpCMV−SPORT6.1(Invitrogen Corp.)でクローン化され、発現ベクターへの追加サブクローニングを必要としなかった。
【0177】
実施例4−ワタアブラムシ電位依存性カルシウムチャネルα2δサブユニットの配列の識別
【0178】
【表7】
【0179】
蛋白質配列CG12295−PBを質問内容として用いて、ワタアブラムシコンティグデータベースのTBLASTN検索を実施することにより(実施例1)、ワタアブラムシカルシウムチャネルα2δの配列が識別された。CG12295−PBは、遺伝子CG12295について予測された翻訳産物であり、Drosophila melanogasterゲノムで識別された3種類のカルシウムチャネルα2δ遺伝子のうちの一つである。ワタアブラムシα2δコンティグそれぞれを用いたDrosophila蛋白質データベースのバージョン3のBLASTXによる相互検索から、明らかにCG12295のワタアブラムシオルソログに属するコンティグの一つ(CAg12001、1489bp)が識別された。
【0180】
コンティグCAg12001に対応する全長cDNAが、RecActiveTMキットおよびActive Motif社(カリフォルニア州カールズバッド)からのプロトコルを用いて、RecAを介する遺伝子増強により、ワタアブラムシcDNAライブラリから分離された。要約すると、約106個の一次トランスフォーマントからなる一定量のワタアブラムシcDNAライブラリから、プラスミドDNAが調製された。表示時の偏りは、50μg/mlのカルベニシリンを添加した1LのTerrific Brothで細胞を30℃にて16時間増殖させることにより、最小限にした。プローブ合成のテンプレートは、CAg12001特異的なプライマーであるAph040984U531およびAph040984L959(表IV)を用いたPCRにより、ライブラリDNAから合成し、Oligo 6プライマー解析ソフトウェア(Molecular Biology Insights,Inc.,コロラド州カスケード)を用いて命名した。PCR反応では、最終容積15μl中に、100ngのライブラリDNA、5pmolのAph040984U531およびAph040984L959プライマー、0.3μlの10mM dNTP混合液(それぞれ10mMのdATP,dGTP,dCTPおよびTTP)、1.5μlの10×TitaniumTM Taq緩衝液、0.3μlのTitaniumTM Taq DNAポリメラーゼ(BD Biosciences Clontech、カリフォルニア州パロアルト)が含まれていた。反応では、94℃での2分間の加熱の後に、94℃での30秒間の加熱変性、55℃での30秒間のアニーリングおよび、72℃での1.5分間のプライマー伸展を1サイクルとして、35サイクルのDNA合成を行った。反応産物は、低融点アガロースゲル(SealPlaque(登録商標)GTG(登録商標)アガロース、BioWhittaker Molecular Applications、メイン州ロックランド)上で分離し、ゲルから450bpと予測されるバンドを抽出して、PicoGreen(登録商標)dsDNA定量試薬(Molecular Probes、オレゴン州ユージーン)による蛍光染色により定量した。精製フラグメントをテンプレートとして用いて、100pgのテンプレートフラグメント、10pmolのAph040984U531およびAph040984L959プライマー、7μlのビオチン−dNTP混合液(Active Motif)、5μlの10×TitaniumTM Taq緩衝液および1μlのTitaniumTM Taq DNAポリメラーゼを含有する50μlのPCR反応液中で、ビオチル化DNAの合成を行った。ビオチル化プローブは、1%の低融点アガロースゲル上で精製し、PicoGreen(登録商標)による蛍光染色によって定量した。ついで、50ngのプローブフラグメントを用いて、RecActiveTMキットにより提供されたプロトコルを利用して、5μgのワタアブラムシライブラリDNAからα2δクローンを選択した。最初の選択により約105個のコロニーがプールされ、プラスミドDNAが調製された。プールDNAの定量的PCR分析から、標的とするα2δ配列が開始時のライブラリと比較して約60倍に増大していることが示された。単独コロニースクリーニングで十分な量を得るために、50ngのビオチル化プローブを用いて、最初に得られたプラスミドDNAのうちの3μgを再度選択した。2回目の選択で得られたDNAを用いてE.coli DH10B細胞をトランスフォームした後、ビオチル化プローブ合成で使用した領域に隣接するCAg12001特異的プライマー(Aph040984U451およびAph040984L1104プライマー、)を用いたPCRにより、個々のコロニーのスクリーニングを行った。2回の選択の後、スクリーニングされたコロニー36個のうち34個がCAg12001配列に対して陽性だった。各々の陽性コロニーからプラスミドDNAを調製し、HindIIIおよびKpnIによる切断によって、cDNAインサートの大きさを決定した。一つのクローン(RA1.11)からのcDNAインサートは、完全に配列決定されている。RA1.11は、4383bpのインサート(配列番号13)を有しており、1245個のアミノ酸オープンリーディングフレームを含み(配列番号14)、これは、CG12295−PBと、1239残基にわたって41%のアミノ酸配列を共有していた。さらに別の15個の全長クローンについての制限酵素解析および部分配列決定から、少なくともcDNAの二つの異なる領域が関与した複雑な別のスプライシングのパターンが示された。別のスプライシングのパターンの一つでは、配列番号14の805−815位の残基によって示される11個のアミノ酸セグメントでの相違の存在が関与している。このセグメントは、Drosophila CG12295−PB α2δ蛋白質配列には存在しないが、配列番号16および18に示されるTBW α2δ蛋白質配列では、同一の位置に局在しており、存在状態の異なる11個のアミノ酸配列で部分的な相同性を有している(実施例5など)。
【0181】
実施例5−オオタバコガ(「TBW」)電位依存性カルシウムチャネルα2δサブユニット配列の識別
【0182】
【表8】
【0183】
最初に、TBW頭部cDNAライブラリ(実施例1)から、以下の3種類の昆虫α2δオルソログで保存された蛋白質配列ブロックよりデザインされたコンセンサス変性ハイブリッドオリゴヌクレオチドプライマー(CODEHOPプライマー)を用いたPCRにより、TBW α2δ CG12295オルソログの5’末端から得られたフラグメントを合成した:Drosophila melanogasterからのCG12295−PB、Anopheles gambiaeからのENSANGP00000021218および、ワタアブラムシ(Aphis gossypii)からのRA1.11(配列番号14、実施例4)。Block Maker(Henikoff. and Henikoff,Nucleic Acid Research、19巻、6565−6572ページ(1991))を用いてこれら3種類の配列で保存された蛋白質ブロックのデータベースを構築し、CODEHOPプライマーデザインプログラムの入力値として使用した(Roseら、Nucleic Acid Research、26巻、1628−1635ページ(1998))。変性正方向プライマーのHvCaA2D1F(表V)は、配列番号81の5’側でもっとも保存されている蛋白質ブロックのアミノ酸配列VQTWAEKLから得られ、一方、変性逆方向プライマーのHvCaA2D1R(表V)は、配列番号82でN末端から蛋白質側方向へのおおむね1/3の位置に局在する保存配列CNQAIMIVSの相補鎖から得られた。TBW頭部cDNAライブラリ(実施例1)から分離された100ngのプラスミドDNAを用いてタッチダウンPCRを行い、アガロースゲル電気泳動により900bpのフラグメントを分離して、pBluescript SK(−)(Stratagene)にクローン化した。DNA配列解析から、バンドはDrosophila遺伝子CG12295のTBWオルソログに由来するものであることが確認された。この900bpのフラグメントの配列から、2種類の非変性遺伝子特異的プライマー(HvCaA2DU388およびHvCaA2DL775、表V)をデザインし、PCR反応のアニーリング温度を55℃ではなく62℃とした以外は、実施例4に記載されているのと同様にして、このプライマーを用いてビオチン化プローブを調製した。このビオチン化プローブは、TBW神経索ライブラリからの5μgのプラスミドDNAから開始した、2回のrecAを介した遺伝子増幅(実施例4など)で用いた。2回目の選択後にPCRの検討対象となった24個のコロニーのうち3個は、TBW α2δ配列について陽性であった。これら3つのプラスミドはいずれも、〜5kbのインサートを含有していた。クローンのうちの二つ(クローン2およびクローン7)は、全体についての配列決定が行われた。クローン2(5028bp、配列番号15)は、1271個のアミノ酸(配列番号16)によるオープンリーディングフレームを有しており、クローン7(4892bp、配列番号17)は、1258個のアミノ酸(配列番号18)によるオープンリーディングフレーム蛋白質を有していた。クローン2に存在する5’UTRセグメントは、クローン7で認められたものに比べて98bp長かったが、二つの5’UTR配列はその他の点では同一である。しかし、二つのcDNAのコード化領域で69個の単独ヌクレオチドに多型性が存在し、3’UTRには少数の挿入/欠失が存在することから、二つのクローンは、ライブラリの構築に使用した非近交系TBW集団で、異なる対立遺伝子に由来すると思われる。さらに、クローン2には、クローン7と比較した場合、コード化領域に以下の二つの小さな挿入を含有している:490−410位の残基でのアミノ酸2個の挿入(VK)および、842−852位の残基でのアミノ酸11個の挿入(PLTKVIGLLPR、配列番号83)。コード化領域には多数の単独ヌクレオチドの多型性が含まれているが、蛋白質配列の相違を引き起こすのはこの二つのみであり、どちらの変化も、保存的なアミノ酸置換となっている。
【0184】
実施例6−アフリカツメガエル卵母細胞でのオオタバコガ(「TBW」)電位依存性カルシウムチャネルの機能性の発現
材料および方法
ポリ(A)末端を有するキャップ構造mRNAの構築。pCDNA3.1でのキメラα1サブユニットは、NsiIにより直線状にした;pCMV−SPORT6.1 1でのTBW βサブユニットは、DrdIにより直線状にした;pCMV−SPORT6.1のアブラムシα2δは、NotIにより直線状にした;pGreen1S(+)Lox2(Xba)のTBW□2□は、NheIにより直線状にした。ついで、直線状にしたDNAを、フェノール−クロロホルム抽出と、エタノール沈降により精製した。このDNAを用い、Ambion社からのmMESSAGE mMACHINETM高収量キャップ構造mRNA転写キットのマニュアルに従って、キャップ構造mRNAを作製した。転写反応の後、試料をDNaseIで処理して、テンプレートDNAを除去し、その後、Ambion社のポリ(A)ポリメラーゼを用いて、RNAにポリ(A)末端を付加した。ポリ(A)末端を有するキャップ構造mRNAには、フェノール−クロロホルム抽出と、エタノール沈降を行った。ポリ(A)末端を有するキャップ構造mRNAの濃度は、UV分光法により測定した。mRNA溶液は、その後の利用のために、−20℃で保存した。
【0185】
卵母細胞発現およびパッチクランプ法:アフリカツメガエル卵母細胞遺伝子発現で使用した方法は、技術で十分に確立されているものである(非特許文献33)および(非特許文献34)。この手法を以下に要約する。0.15%のTricaine(3−アミノ安息香酸エチルエステル)に沈めて麻酔したアフリカツメガエル卵母細胞(Xenopus laevis)(NASCO、ウィスコンシン州Fort Atkinson)から、未受精の卵母細胞を外科的に摘出した。卵母細胞は、カルシウムを含まない生理食塩液(OR2:82.5mMのNaCl,2mMのKCl,1mMのMgCl2,5mMのHEPES,pH7.4)で洗浄し、その後、4mg/mlのコラゲナーゼを含有するOR2中で、オービタルシェイカー上にて17℃で2時間インキュベートした。ついで、ピンセットを用いて卵母細胞に機械的な濾胞除去を行った後、通常のカルシウム含有生理食塩液(ND96:96mMのNaCl,2mMのKCl,1mMのMgCl2,1.8mMのCaCl2,5mMのHEPES,pH7.4)に入れた。ピストン式のマイクロインジェクター(Drummond Nanoject,Drummond Scientific Company、ペンシルバニア州Broomall)を用いて、希望する遺伝子または遺伝子群からのRNA転写物を、各々の卵母細胞にインジェクションした。通常の実験では、4〜6ngの各々のサブユニットのRNAを、23nLの水に入れて導入した。インジェクションを受けた卵母細胞を、オービタルシェイカー上にて17℃でインキュベートし、2〜4日でインジェクションしたチャネルの機能的な発現を検討した。
【0186】
【非特許文献33】Methods in Enzymology、207巻、Rudy,B.
【非特許文献34】Iverson,L.E編著、セクションII、A.Expression of Ion channels in Xenopus oocytes、225−390ページ)
【0187】
卵母細胞は、二電極電圧固定装置(AxoClamp 2B,Axon Instrument、カリフォルニア州ユニオンシティー)を用いて、イオンチャネルの機能的な発現について検討した。標準的な電圧固定法での電流および記録法を用いた。記録電極は、Narishigeピペットプラー(PP830型、Narishige International USA、ニューヨーク州ロングアイランド)を用いて組み立て、4mMの酢酸カリウムを充填した場合の入力抵抗が0.8〜3メガオームとなるように調節した。データは、アナログ−デジタルインターフェース(Digidata 1322,Axon Instruments)を備えたPentium(登録商標)クラスのコンピュータを用いて、pClamp8ソフトウェア(Axon Instruments)により統合した。カルシウムチャネルの性能を評価するための記録は、内因性のカルシウム活性化電流を最小限にするために処方されたNS生理食塩液中(40mMのBa(OH)2,50mMのD−グルコン酸ナトリウム、1mMのKOH,0.1mMのEDTA,10mMのHEPES,0.1mMのニフルム酸、pH7.4)で行った。
【0188】
結果:3種類の無脊椎動物カルシウムチャネルサブユニットをさまざまな形で組み合わせたものについて検討を行った。配列番号6で示されるα1サブユニット、配列番号10で示されるβ−2サブユニット、配列番号18で示されるα2δサブユニットは、オオタバコガ(Heliothis virescens)に由来するものであった。ワタアブラムシからの配列番号14で示されるα2δサブユニットについても、このサブユニットが天然のH.virescens α2δと置換できるか否かを調べるために、検討を行った。
【0189】
各々のサブユニットのRNAを、6ng/卵の用量でインジェクションした。各々のバッチの回収卵からの卵にインジェクションした対照電位依存性チャネルは、大量の電流を生じたという事実(データ未発表)にもかかわらず、α1サブユニット単独では電位依存性の電流を得ることはできなかった。α1サブユニットとβ−2サブユニットが共に発現した場合には、図2に示すように通常は10〜20nAの範囲で、中程度の電流を生じた。しかし、電流の発現は不安定であり、卵ごとに一定していなかった。α1サブユニットおよびβ−2サブユニットにアブラムシα2δサブユニットを加えると、わずかではあるが明確な、電位感受性N型カルシウム電流が生じた(図3)。アブラムシα2δをTBW α2δ−2サブユニットに置換した場合にも、明確な電位感受性N型カルシウム電流が生じた(図4)が、観察された最大電流の大きさは、40nA程度の大きさまで増大した。
【0190】
上述の方法で集められたデータから、固定電圧の場合に電位段階に応じて発生した電流は、培地で増殖させたニューロンから生ずる、全細胞パッチクランプ記録で観察されるN型カルシウムチャネルに類似していると結論付けられる(非特許文献35)。試験条件下でのクローン化されたチャネルと、培養ニューロンで観察されたものはどちらも、−30mV付近で活性化ポイントを示し、見かけ上の逆転電位は+50mV付近である。卵母細胞で観察された電流は減少性であるが、その動態は、培養昆虫ニューロンで観察される、緩慢に不活化する電流を暗示するものである。さらに、配列番号6で示されるキメラα1サブユニットは、適切な昆虫βおよびα2δカルシウムチャネルサブユニットの存在下でN型カルシウムチャネルを暗示する、機能性電位感受性チャネルを形成することができると結論付けられる。
【0191】
【非特許文献35】Hayashi and Levine, Exp.Biol.、171巻、15−42ページ(1992)
【図面の簡単な説明】
【0192】
【図1.A】コピー数の少ないプラスミドベクターpGreen1S(+)Lox2(Xba)を図示したものである。複数のクローニング部位は、KpnIおよびSacI制限酵素切断部位の間の小さな領域に局在しており、T3およびT7 RNAポリメラーゼに対するプロモーターによって隣接している。p15Aの矢印は、DNA複製でコピー数の少ないp15Aプラスミドの起点部の位置を示している。loxPの矢印は、Creリコンビナーゼによる部位特異的組換えで使用される34bpの反復配列の部位を示している。AmpRの矢印は、抗生物質のアンピシリンおよびカルベニシリンに対する耐性を付与するβ−ラクタマーゼ遺伝子の位置および方向を示している。incAは、その存在によってE.coliのDH10B(ZIP)株でのCreリコンビナーゼを発現するP1プラスミドの反復を遮断するDNAセグメントの位置を示している。AmpRの矢印は、抗生物質のアンピシリンおよびカルベニシリンに対する耐性を付与するβ−ラクタマーゼ遺伝子の位置および方向を示している。incAは、その存在によってE.coliのDH10B(ZIP)株でのCreリコンビナーゼを発現するP1プラスミドの反復を遮断するDNAセグメントの位置を示している。f1(+)originの矢印は、繊維状ファージf1のローリングサイクル複製の起点の位置と方向を示している。図1A:バクテリオファージλgt11の誘導体に挿入を行う前のプラスミドの環状構造を示す。pGreen1S(+)Lox2(Xba)は、制限酵素MfeIおよびPmeIによって切断され、バクテリオファージλgt11の誘導体の(破壊された)SacI部位とEco RI部位のあいだに巨大な直線フラグメントが挿入された。λgtGreen1SのXhoIおよびSacIによる切断により、ファージに二つのアームが生ずるが、cDNAライブラリの構成ではこれらを利用することが可能である。
【図1.B】コピー数の少ないプラスミドベクターpGreen1S(+)Lox2(Xba)を図示したものである。複数のクローニング部位は、KpnIおよびSacI制限酵素切断部位の間の小さな領域に局在しており、T3およびT7 RNAポリメラーゼに対するプロモーターによって隣接している。p15Aの矢印は、DNA複製でコピー数の少ないp15Aプラスミドの起点部の位置を示している。loxPの矢印は、Creリコンビナーゼによる部位特異的組換えで使用される34bpの反復配列の部位を示している。AmpRの矢印は、抗生物質のアンピシリンおよびカルベニシリンに対する耐性を付与するβ−ラクタマーゼ遺伝子の位置および方向を示している。incAは、その存在によってE.coliのDH10B(ZIP)株でのCreリコンビナーゼを発現するP1プラスミドの反復を遮断するDNAセグメントの位置を示している。f1(+)originの矢印は、繊維状ファージf1のローリングサイクル複製の起点の位置と方向を示している。図1B:バクテリオファージクローニングベクターλgtGreen1Sに存在する形での、直線型のプラスミドの構造を示す。pGreen1S(+)Lox2(Xba)は、制限酵素MfeIおよびPmeIによって切断され、バクテリオファージλgt11の誘導体の(破壊された)SacI部位とEco RI部位のあいだに巨大な直線フラグメントが挿入された。λgtGreen1SのXhoIおよびSacIによる切断により、ファージに二つのアームが生ずるが、cDNAライブラリの構成ではこれらを利用することが可能である。
【図2】昆虫α1サブユニットとTBW β−2サブユニットを注入したアフリカツメガエル(Xenopus)卵母細胞での電位ステップに応じて生じた電流を示す。最大電流と電位との間の相関関係図である。最大電流は、マーカー1とマーカー2(逆三角形)とのあいだで決定された。
【図3】昆虫α1サブユニット、TBW β−2サブユニットおよびワタアブラムシα2δサブユニットを注入したアフリカツメガエル(Xenopus)卵母細胞での電位ステップに応じて生じた電流を示す。最大電流と電位との間の相関関係図である。最大電流は、マーカー1とマーカー2(逆三角形)とのあいだで決定された。
【図4】昆虫α1サブユニット、TBW β−2サブユニットおよびTBW α2δ−2サブユニットを注入したアフリカツメガエル(Xenopus)卵母細胞での電位ステップに応じて生じた電流を示す。最大電流と電位との間の相関関係図である。最大電流は、マーカー1とマーカー2(逆三角形)とのあいだで決定された。
【配列表】
【技術分野】
【0001】
本発明は、農芸化学、獣医学または薬学の分野で有用な核酸配列に関する。本発明は特に、ポリペプチドと、農薬または医薬品として有効な化合物の識別または開発で有用となるポリペプチドをコード化する核酸配列に関する。
【背景技術】
【0002】
電位依存性カルシウムチャネルは、生理学的応答の調節で重要な役割を果たしている。すなわち、電位依存性カルシウムチャネルの脱分極により、チャネル孔を通じてのカルシウムの通過が生じ、これによって、筋収縮および神経伝達物質の分泌などのさまざまな生理学的応答が生じると考えられている(非特許文献1);(非特許文献2);(非特許文献3);(非特許文献4);(非特許文献5);(非特許文献6);および(非特許文献7)。同様に、殺虫剤などの生物学的に活性な化合物を識別するための手段としてこのチャネルを標的とするための方法を開発しようとの要求が存在する(非特許文献8);(非特許文献9);(非特許文献10);(非特許文献11)。
【0003】
【非特許文献1】Jeziorskiら、J.Exper.Bio.,203、841−856ページ(2000)
【非特許文献2】Edwin W.McCleskey、Current Opinion in Neurobiology、4,304−312ページ(1994)
【非特許文献3】Herman Moreno Davila,Ananals New York Academy of Science,102−117ページ
【非特許文献4】Stephen W.Jones,J.of Bioenergetics and Biomembranes,30巻、4号、299−312ページ(1998)
【非特許文献5】Hofmannら、Anna.Rev.Neurosci.,17、399−418ページ(1994)
【非特許文献6】Edward Perez−Reyes and Toni Schneider, Kidney International、48巻、1111−1124ページ(1995)
【非特許文献7】AstraZeneca Pharmaceuticals Presentation,第28回National Medicinal Chemistry Symposium of the American Chemical Society,(2002年6月)
【非特許文献8】Mintzら、Nature、355巻、827−829ページ(1992)
【非特許文献9】Wangら、Nature Structural Biology、7巻、6号、505−512ページ(2000)
【非特許文献10】Wangら、Eur.J.Biochem,264,488−494ページ(1999)
【非特許文献11】Fletcherら、Nature Structural Biology、4巻、7号、559−566ページ(1997)
【0004】
電位依存性カルシウムチャネルは、ヒト、ウサギ、ラット、マウス、シビレエイ(Discopyge Ommata)などの脊椎動物で発現し、これらの動物からクローン化されている(非特許文献12);(非特許文献13);(非特許文献14);(非特許文献15);(非特許文献16);(非特許文献17);(非特許文献18);(非特許文献19);(非特許文献20)。電位依存性カルシウムチャネルは、線虫(Caenorhabditis elegans)、Styllphora pistilla、Bdellourea candida、キタユウレイクラゲ(Cyanea capillata)、ヤリイカ(Loligo bleeker)、ゾウアメフラシ(Aplysia californica)、ショウジョウバエ(Drosophila melanogaster)、イエバエ(Musca domestica)、チャバネゴキブリ(Blatella germanica)およびマボヤ(Halocynthia roretz)などの無脊椎動物でも発現し、これらの動物からクローン化されている(非特許文献21)。
【0005】
【非特許文献12】Velicelebeら、Methods in Enzymology、294巻、20−47ページ(1999)
【非特許文献13】Herman Moreno Davila、Annals New York Academy of Sciences、102−117ページ
【非特許文献14】Hofmannら、Anna.Rev.Neurosci,17,399−418ページ(1994)
【非特許文献15】Edward Perez−Reyes and Toni Schneider, Kidney International、48巻、1111−1124ページ(1995)
【非特許文献16】Astra Zeneca Pharmaceuticals Presentation,第28回National Medicinal Chemistry Symposium of the American Chemical Society,(2002年6月)
【非特許文献17】Horneら、Proc.Natl.Acad.Sci.,90巻,3787−3791ページ(1993)
【非特許文献18】Roussetら、J.of Physiology,532,3,583−593ページ(2001)
【非特許文献19】Neelandsら、J.Physiology,2933−2944ページ(2000)
【非特許文献20】Brustら、Neuropharmacology、32巻、11号、1089−1102ページ(1993)
【非特許文献21】Jeziorskiら、J.Exper.Bio.,203,841−856ページ(2000))
【発明の開示】
【発明が解決しようとする課題】
【0006】
本発明は、農芸化学、獣医学または薬学の分野で有用な核酸配列に関する。本発明は特に、ポリペプチドと、農薬または医薬品として有効な化合物の識別または開発で有用となるポリペプチドをコード化する核酸配列に関する。
【課題を解決するための手段】
【0007】
本発明の一実施形態は、農薬または医薬品としての(潜在的な)活性を有する化合物の識別または開発で有用なアミノ酸配列をコード化する核酸配列、または、これらのアミノ酸配列の発現に利用することの可能な核酸配列に関連するものである。以下に詳述するこれらの核酸配列は、その突然変異体およびフラグメントを含めて、ここでは「本発明の核酸配列」とも呼ぶ。
【0008】
本発明の別の実施形態は、本発明の核酸配列によってコード化されたり、本発明の核酸配列の適切な発現によって得られる可能性のある、蛋白質またはポリペプチドなどのアミノ酸配列に関連するものである。以下に詳述するこれらのアミノ酸配列は、その突然変異体およびフラグメントを含めて、ここでは「本発明のアミノ酸配列」とも呼ぶ。
【0009】
本発明のさらに別の実施形態は、例えば本発明のアミノ酸配列の発現についてなど、宿主細胞または宿主生物のトランスフォーメーションにおいて、好ましくは後述のような適切な遺伝子構成の形態での、本発明の核酸配列の利用に関連するものである。本発明はさらに、本発明の核酸配列でトランスフォームしたり、本発明のアミノ酸配列を発現することのできる宿主細胞または宿主生物とも関連する。
【0010】
さらにまた別の実施形態では本発明は、上述の核酸配列、アミノ酸配列、遺伝子構成、宿主細胞または宿主生物を使用しての、本発明のアミノ酸配列の生物学的活性を変調または阻害することが可能な化合物の識別または開発のための方法に関連するものである。通常は測定またはスクリーニングで利用されることになるこうした方法については、以下に詳述する。
【0011】
さらに別の実施形態では、本発明は、in vitroで、または、好ましくはin vivoで(も)、本発明のアミノ酸配列(の生物学的活性)を変調する化合物または、本発明のアミノ酸配列と相互作用する化合物と関連する。本発明はさらに、こうした化合物を含有する合成物ならびに、これらの合成物の調製および害虫の制御でのこうした化合物の利用とも関連する。
【0012】
今回の発明の核酸はここでは一括して「本発明の核酸」と呼ぶ。さらに、以下の発明の詳細な記述の本文では適切な場合には、「本発明の核酸配列」および「本発明の核酸」という語は、基本的に同等であり、基本的に相互変換が可能であると考えても差し支えない。
【0013】
さらに、今回の発明の目的では、核酸とは、通常は結合している別の核酸分子および配列の少なくとも一つから分離されている場合には、−例えば、本来の生物学的起源などから−「基本的に分離された(形態のもの)」と考える。同様に、ポリペプチドとは、通常は結合している別のポリペプチド分子から効率的に分離されている場合には、−例えば、本来の生物学的起源などから−「基本的に分離された(形態のもの)」と考える。核酸またはポリペプチドは、特に、少なくとも2倍、特に少なくとも10倍、さらに特に少なくとも100倍、および、最高1000倍以上に精製されている場合には、「基本的に分離されている」と考える。
【発明を実施するための最良の形態】
【0014】
今回の発明は、化合物およびその他の因子とのin vitroまたはin vivoでの相互作用において、本発明のアミノ酸配列が(潜在的な)「標的」として利用可能であるという所見から確立された(「標的」という語は、本明細書に援用して組み込まれる(特許文献1)での定義のように、当該技術での通常の意味を持つものである)。その結果、(以下に記述された方法などにより)本発明のアミノ酸配列と相互作用するとされている化合物または因子は、農芸化学、獣医学または薬学の分野で活性物質として有用なものとなる可能性がある。
【0015】
【特許文献1】国際公開第98/06737号
【0016】
一実施形態では、本発明は、本発明の核酸配列を含む核酸に関し、好ましくは(基本的に)分離された形態で、特に配列番号1という核酸配列と関連する。配列番号1の核酸配列は、後述の実施例の項で詳述された方法で、オオタバコガ(Heliothis virescens)の生体に由来またはこれから分離されたものである。この配列は、オオタバコガ(Heliothis virescens)の電位依存性カルシウムチャネルサブユニットα1についての不完全なコード化配列を示す。
【0017】
別の実施形態では、本発明は、本発明の核酸配列を含む核酸に関し、好ましくは(基本的に)分離された形態で、特に配列番号5という核酸配列と関連する。配列番号5の核酸配列はキメラ配列であり、これでは配列番号1のオオタバコガ(Heliothis virescens)の電位依存性カルシウムチャネルサブユニットα1が、ワタアブラムシcDNAライブラリ(実施例1)に由来する配列番号3で列挙した核酸1から429までのストップコード化ンと共に示されている。このアブラムシ由来の配列は、アブラムシの電位依存性カルシウムチャネルサブユニットα1に由来する。この配列の構成については、後述の実施例の項で詳述する。この配列は、機能的キメラコード化配列を示す。
【0018】
別の実施形態では、本発明は、本発明の核酸配列を含む核酸に関し、好ましくは(基本的に)分離された形態で、特に配列番号7、配列番号9または配列番号11という核酸配列と関連する。配列番号7、配列番号9または配列番号11という核酸配列は、後述の実施例の項で詳述された方法などで、オオタバコガ(Heliothis virescens)の生体に由来またはこれから分離されたものである。これらの配列はそれぞれ、オオタバコガ(Heliothis virescens)の電位依存性カルシウムチャネルサブユニットβ-1、β-2、β-3についての完全なコード化配列を示している。
【0019】
別の実施形態では、本発明は、本発明の核酸配列を含む核酸に関し、好ましくは(基本的に)分離された形態で、特に配列番号13という核酸配列と関連する。配列番号13という核酸配列は、後述の実施例の項で詳述された方法で、ワタアブラムシ(Aphis gossypii)の生体に由来またはこれから分離されたものである。この配列は、ワタアブラムシ(Aphis gossypii)の電位依存性カルシウムチャネルサブユニットα2δについての完全なコード化配列を示している。
【0020】
別の実施形態では、本発明は、本発明の核酸配列を含む核酸に関し、好ましくは(基本的に)分離された形態で、特に配列番号15または配列番号17という核酸配列と関連する。配列番号15または配列番号17という核酸配列は、後述の実施例の項で詳述された方法などで、オオタバコガ(Heliothis virescens)の生体に由来またはこれから分離されたものである。これらの配列はそれぞれ、オオタバコガ(Heliothis virescens)の電位依存性カルシウムチャネルサブユニットα2δ−1およびα2δ−2についての完全なコード化配列を示している。
【0021】
一般的に、本発明の核酸配列は、核酸の形態をとっている場合には、DNAまたはRNAであると思われ、一本鎖または二本鎖となることがある。例えば、本発明の核酸配列がゲノムDNA、cDNAまたは合成DNAとなることがある(企図した宿主細胞または宿主生物で発現するように特に適応させたコード化ンを利用しているDNAなどで、例えば、Vector NTI(InforMax,Inc.,メリーランド州ベセズダ)のBackTranslate解析ツールなどの適切なコンピュータプログラムを用いてデザインされたものなどであることがある)。従って、本発明の核酸配列は、イントロン配列を含むことがあり、一般的に種々のスプライシング変異体からなることもある。
【0022】
これとは別の実施形態として、本発明の核酸配列に向けての二本鎖RNA分子に関連するものがある(うち一本の鎖は通常、少なくとも本発明の核酸配列の一部からなる)。こうした二本鎖RNA分子は、遺伝子機能のRNA干渉の研究で特に有用である(Zamoreら、Cell 101:25−33(2000))。本発明はさらに、こうした二本鎖RNA分子を提供するために利用することが可能な遺伝子構造とも関連する(E.coliなどの菌株での例など、宿主細胞または宿主生物での適切な発現によるものなど)。こうした構造については、(非特許文献22)といった参考文献がある。
【0023】
【非特許文献22】Maniatisら、Molecular Cloning, a Laboratory Manual(Cold Spring Harbor Press,1989)
【0024】
広い意味では、「本発明の核酸配列」という語は以下のものから構成される:
−配列番号1、配列番号5、配列番号7、配列番号9、配列番号11、配列番号13、配列番号15および配列番号17の核酸配列の一部またはフラグメント;
−下記に詳述するような、配列番号1、配列番号5、配列番号7、配列番号9、配列番号11、配列番号13、配列番号15および配列番号17の核酸配列の(天然または合成による)突然変異体、変異体、対立遺伝子、類縁体、オルソログ(本明細書では以下、総称して「突然変異体」と呼ぶ);
−こうした(天然または合成による)突然変異体の一部またはフラグメント;
−少なくとも一つのさらに別の核酸配列と、配列番号1、配列番号5、配列番号7、配列番号9、配列番号11、配列番号13、配列番号15および配列番号17(またはこれらの一部またはフラグメント)の核酸配列の核酸融合物;
−少なくとも一つのさらに別の核酸配列と、(天然または合成による)突然変異体(またはその一部もしくはフラグメント)の核酸融合物;
こうした突然変異体、一部、フラグメントまたは融合物は、主として以下に記述するようなものである。
【0025】
本発明は、上述の核酸配列の種々のスプライシング変異体からも構成されている。
【0026】
主として、本発明の核酸配列は少なくとも500個の長さのヌクレオチドからなり、好ましいものとしては最低1,000個のヌクレオチド、さらに好ましいものとしては最低2,000個のヌクレオチド、最高ではほぼ8,000個のヌクレオチド、望ましくは多くの場合7,500個のヌクレオチド、さらに好ましい場合には多くの場合7,000個のヌクレオチドとなる。
【0027】
配列番号1の核酸配列の一部もしくはフラグメント、またはその(天然または突然変異による)突然変異体の一部もしくはフラグメントの例には、5’または3’切断核酸配列または、フレームスタートコード化ンまたはストップコード化ンの導入を受けた核酸配列が含まれるが、これに限定されるものではない。さらに、本発明の核酸配列の一つまたは複数についての二つまたはそれ以上のこうした部分またはフラグメントが、適切に組み合わされて(フレーム中のリガンドなど)、さらに本発明の核酸配列となることがある。
【0028】
望ましくは、こうした一部またはフラグメントは、配列番号1、配列番号5、配列番号7、配列番号9、配列番号11、配列番号13、配列番号15および配列番号17の核酸配列からの少なくとも100個、望ましくは250個、さらに望ましくは少なくとも500個、さらに1,000個以上となることもあるヌクレオチドからなる少なくとも一つの連続的伸展から構成されるものとなる。配列番号5の場合には、本発明によるフラグメントは、配列番号5からの少なくとも5個、望ましくは10個、さらに望ましくは20個以上のヌクレオチドからなる。
【0029】
さらに、当業者によれば、本明細書の公開に基づいて(別の個体の)同一生物種(例えば、異なる菌株または系列の個体からなどのもので、突然変異体を含むがそれに限定しない)からの配列番号1、配列番号5、配列番号7、配列番号9、配列番号11、配列番号13、配列番号15および配列番号17の核酸配列の天然の「突然変異体」(上述のもの)を識別したり、入手したり、分離したりすることが可能となると予想される。また、当業者によれば、本明細書の公開に基づいて配列番号1の核酸配列の合成突然変異体(本明細書上述で定義したもの)を提示したり、提供することができるようになると予想される。
【0030】
具体的な実施形態の一つとしては、配列番号1、配列番号5、配列番号7、配列番号9、配列番号11、配列番号13、配列番号15および配列番号17またはその一部またはそのフラグメントの核酸配列をコード化する突然変異体がある。
本明細書で記述した突然変異体は、配列番号1、配列番号5、配列番号7、配列番号9、配列番号11、配列番号13、配列番号15および配列番号17の核酸配列についての一つまたは複数、望ましくはすべての構造的特徴を有しているか、または、以下に記述した特徴を保存していることが好ましい。
【0031】
特に、本明細書で記述したあらゆる突然変異体、一部またはフラグメントは、少なくとも、本発明の対応するアミノ酸配列の活性部位または触媒部位および本発明の対応するアミノ酸の結合ドメインをコード化しているものとすることができる。
【0032】
さらに、本明細書で記述したあらゆる突然変異体、一部またはフラグメントは、好ましくはヌクレオチドレベルで、少なくとも75%、望ましくは最低80%、さらに望ましくは少なくとも85%、特に90%以上、および最高95%以上にわたり、配列番号1、配列番号5、配列番号7、配列番号9、配列番号11、配列番号13、配列番号15および配列番号17の核酸配列とある程度の「配列の同一性」を有することになる。
【0033】
さらに、主として、本発明の核酸配列のあらゆる突然変異体、一部またはフラグメントは、アミノ酸レベルで、少なくとも80%、特に90%以上、および最高95%以上にわたり、配列番号1、配列番号5、配列番号7、配列番号9、配列番号11、配列番号13、配列番号15および配列番号17のアミノ酸配列とある程度の「配列の同一性」を有することになるが、この「配列同一性」の割合については、以下で算定する。
【0034】
こうした目的では、配列番号1、配列番号5、配列番号7、配列番号9、配列番号11、配列番号13、配列番号15または配列番号17の配列と比較して、ヌクレオチドの欠失、挿入、置換または付加を、単独ヌクレオチド位での相違と考え、該当する核酸配列において、配列番号1、配列番号5、配列番号7、配列番号9、配列番号11、配列番号13、配列番号15または配列番号17の核酸配列での対応する位置でヌクレオチドが同一となっているヌクレオチドの数を、該当する核酸配列での総ヌクレオチド数で割って100%を掛けることにより、該当する核酸配列と配列番号1、配列番号5、配列番号7、配列番号9、配列番号11、配列番号13、配列番号15または配列番号17の核酸配列との間の「配列同一性」を算定することができる。
【0035】
全体的な配列アラインメントを行い、配列同一性を決定する好ましいコンピュータプログラムとしてはclustalWがあり(Higginsら、Nucl.Acids Res.22:4673−4680(1994))、これは、種々のコンピュータプラットフォームで利用可能であると発表されている。蛋白質配列アラインメントについてのclustalWプログラムの好ましいパラメータは、ktuple=1,diagonals=5,windows=5,gap=3,Score=PERCENTAGE,matrix=BLOSUM、open pnalty=10.0、および、extension penalty=0.5である。
【0036】
さらに、好ましい意図としては、本明細書で記述したあらゆる突然変異体、一部またはフラグメントは、下記の標準的な測定法により測定した場合、最低50%、好ましくは最低75%、最高90%といった範囲で、配列番号1、配列番号5、配列番号7、配列番号9、配列番号11、配列番号13、配列番号15または配列番号17の配列について上で記載されている生物学的活性に基本的に類似した生物学的活性を有する蛋白質またはポリペプチドをコード化することになる。
【0037】
本明細書で記述したあらゆる突然変異体、一部およびフラグメントは、配列番号1、配列番号5、配列番号7、配列番号9、配列番号11、配列番号13、配列番号15または配列番号17の核酸配列と、「きわめて厳密な」条件下で、ハイブリダイゼーションすることが可能であることが好ましい。当業者にとってはこうした条件は、本明細書に援用して組み入れられている上述のSambrookら、およびAusubelらなどの標準的なハンドブック、および、(特許文献2)、(特許文献3)または(特許文献4)などから、明確なものとなるはずである。
【0038】
【特許文献2】欧州特許第0967284号
【特許文献3】欧州特許第1085089号
【特許文献4】国際公開第00/55318号
【0039】
本発明の核酸配列を一つまたは複数の別の核酸配列と融合させたものを利用することも、(上述のように)本発明の意図の範囲内であり、こうした配列には、コード化配列、非コード化配列または制御配列が含まれるが、これらに限定されるわけではない。望ましくは、こうした融合においては、一つまたは複数の別の核酸配列が操作可能な形で、本発明の核酸配列と結合している(下記参照)(例えば、別の核酸配列がコード化配列である場合には、ヌクレオチド融合物は下記のように蛋白質融合物をコード化する)。
【0040】
別の実施形態は、本発明は、本発明の核酸配列に対するアンチセンス分子に関連する。
【0041】
本発明の核酸は、やはり下記に記述するように、遺伝子構成物の形を取ることがある。本発明の遺伝子構成物は一般的に少なくとも一つの本発明の核酸配列から構成され、下記に記述するように、それ自体で知られている遺伝子構成物の一つまたは複数の成分と連結していることがある。こうした遺伝子構成物はDNAまたはRNAであることがあり、好ましくは二本鎖DNAである。構成物は、企図する宿主細胞または宿主生物のトランスフォーメーションに適した形態、企図する宿主細胞のゲノムDNAへの組み込みに適した形態または、企図する宿主生物での独立した複製、維持および伝達に適した形態で存在することもある。遺伝子構成物は例えば、プラスミド、コスミド、酵母人工染色体(「YAC」)、ウイルスベクターまたはトランスポゾンなどのベクターの形態で存在することがある。特に、ベクターは発現ベクター、すなわち、in vitroまたはin vivo(下記のような適切な宿主細胞または宿主生物において)で発現させることの可能なベクターである。本発明の核酸配列からなる発現ベクターは、本明細書では組換え発現ベクターとも呼ばれている。これらの構成物は本明細書では「本発明の遺伝子構成物」とも呼ばれることになる。
【0042】
好ましい実施形態では、組換え発現ベクターでのこうした構成物は、以下のものから構成される:
a)本発明の核酸配列で、以下のものに操作可能な形で結合したもの:
b)プロモーターおよび任意の適切なターミネーター等の一つまたは複数の調節成分;
および、さらに任意により:
c)それ自体で知られている一つまたは複数の遺伝子構成物の別の成分;ここで、「調節成分」、「プロモーター」、「ターミネーター」、「別の成分」および「操作可能な形での結合」という語は、以下に示す意味を有している。
【0043】
上述の一つまたは複数の「別の成分」として、本発明の遺伝子構成物は一般的に、一つまたは複数の適切な調節成分(適切なプロモーター、エンハンサーまたはターミネーター、3’−または5’−非翻訳領域(「UTR」)配列、リーダー配列、選択マーカー、発現マーカーもしくはレポーター遺伝子、または、トランスフォーメーションもしくは組み込み(の効率)を促進若しくは増大させる可能性のある成分など)を含有する。こうした遺伝子構成物に対してのこれらおよびその他の適切な成分は、当業者にとっては明白なものであり、場合によっては使用する構成物のタイプ、企図された宿主細胞または宿主生物、対象となる発明の核酸配列が発現される方法(構成的発現、一過性の発現または誘発性の発現)、および、使用されるトランスフォーメーション技術に依存することがある。
【0044】
本発明の遺伝子構成物では、一つまたは複数の成分が、本発明の核酸配列または相互に「操作可能な形で」結合していることが望ましく、これは一般的に、互いに機能的な関連で存在することを意味する。例えば、プロモーターがコード化配列の転写または発現を開始または制御または調節することができる場合には、このプロモーターはコード化配列と「操作可能な形で」結合していると考えられる(この場合、このコード化配列は、このプロモーターの「制御下にある」と理解しなければならない)。
【0045】
一般的に、二つの核酸配列が操作可能な形で結合している場合には、これらは同一の方向を向いており、通常は同一のリーディングフレーム内に存在する。さらにこれらは通常、基本的に隣接しているが、必ずしもこれが必要とされるわけでもない。
【0046】
好ましくは、本発明で使用される遺伝子構成物の任意のその他の成分は、企図された宿主細胞または宿主生物内で企図された生物学的機能を示せる状態であることが好ましい。
【0047】
例えば、プロモーター、エンハンサーまたはターミネーターが企図された宿主細胞または宿主生物内で「操作可能」でなければならず、これは、(例えば)上述のプロモーターが、それが操作可能な形で結合した核酸配列、すなわちコード化配列の転写の開始、制御または調節をできるものでなければならないことを意味する。
【0048】
こうしたプロモーターは、構成的プロモーターまたは誘発性プロモーターであることがあり、さらに、宿主細胞または宿主生物の特定の発達段階で発現を提示したり(するのみであったり)、特定の細胞、組織、臓器または多細胞宿主生物の一部で発現を提示したり(するのみであったり)することもある。
【0049】
一部の特に好ましいプロモーターには、サイトメガロウイルス(「CMV」)、ラウス肉腫ウイルス(「RSV」)、pSVL SV40 Late Promoter Expression Vector(Pharmacia Biotech Inc., ニュージャージー州Piscataway)などのシミアンウイルス40(「SV40」)、または単純疱疹ウイルス(「HSV」)といった哺乳動物細胞での発現のための構成的プロモーターや、Jarvisらによって記述され(非特許文献23)、NovagenからpIEベクターとして販売されている(Novagen, Inc.ウィスコンシン州マディソン)即時型バキュロウイルスプロモーターなどの昆虫型構成的プロモーター、または、Bunchらによって記述され(非特許文献24)、Invitrogenからベクターとして販売されている(Invitrogen Corporation、カリフォルニア州カールズバッド)Drosophila metallothioneinプロモーターなどの昆虫型誘発性プロモーターが含まれるが、これに限定されるわけではない。
【0050】
【非特許文献23】Methods in Molecular Biology、39巻、Baculovirus Expression Protocols, ed.C.Richardson,. Hamana Press Inc., Totowa, NJ(1995)
【非特許文献24】Nucleic Acids Research、6巻、3号、1043−106(1988)
【0051】
選択マーカーは、適切な選択条件下で、本発明の核酸配列により(成功裏に)トランスフォームを受けた宿主細胞または宿主生物を、(成功裏に)トランスフォームを受けていない宿主細胞または生物と区別できるものでなければならない。こうしたマーカーの好ましい例としては、抗生物質(ジェネチシンもしくはG−418(GIBCO−BRL、ニューヨーク州グランドアイランド)、カナマイシン、または、アンピシリンなど)に対する耐性を付与する遺伝子、温度耐性を付与する遺伝子、または、トランスフォームをしていない細胞または生物の生存に必要不可欠なある種の因子、化合物または(栄養)成分が培地内に存在しなくても宿主細胞または宿主生物が生存できるようにする遺伝子などがあるが、これに限定されるものではない。
【0052】
リーダー配列は、企図した宿主細胞または宿主生物で、翻訳後に希望の変形を行ったり、転写されたmRNAをシグナルペプチドなどの細胞の希望の場所または細胞小器官に方向付けるようにしたりするものでなければならない。リーダー配列は、対象となる細胞から発現産物を分泌させることもある。このような場合、リーダー配列は、ピコルナウイルスリーダー、ポティウイルスリーダー、ヒト免疫グロブリン重鎖結合蛋白質(「BiP」)、タバコモザイクウイルスリーダー(「TMV」)およびトウモロコシクロロテック斑紋ウイルスリーダー(「MCMV」)などの宿主細胞または宿主生物内で操作可能なプロ配列、プレ配列またはプレプロ配列となることがあるが、これらに限定されるわけではない。
【0053】
発現マーカーおよびレポーター遺伝子は、宿主細胞または宿主生物内で遺伝子構造(上に存在する遺伝子または核酸配列)の発現を検出するものでなければならない。発現マーカーではさらに、任意により、細胞の特定の部位もしくは細胞小器官、または、特定の細胞、組織、臓器または多細胞生物の一部などで、発現産物の位置を確認できることがある。こうしたレポーター遺伝子は、本発明のアミノ酸配列との融合蛋白質として発現することもある。好ましいものの一部として、GFPなどの蛍光蛋白質、ベクター内で得られるV5エピトープもしくはポリヒスチジンなどの抗体認識蛋白質およびInvitrogenから供給されている抗体、または、ニッケルカラムでの精製を可能にするポリヒスチジンやメトトレキセートカラムでの精製を可能にするジヒドロ葉酸レダクターゼなどの精製アフィニティ・ハンドル、または、E.coliのβガラクトシダーゼ遺伝子などの遺伝子を発現している細胞を選別できるマーカーなどがあるが、これらに限定されるわけではない。
【0054】
プロモーター、選択マーカー、リーダー配列、発現マーカーおよび、本発明の遺伝子構成物中に存在するまたはそこで利用される可能性のあるその他の成分−ターミネーター、転写エンハンサーもしくは翻訳エンハンサーまたは組み込み因子−についての限定的ではない例の一部としては、上述のSambrookらおよびAusubelらなどの一般的なハンドブック、W.B.Woodらの”The nematode Caenorhabditis elegans”,Cold Spring Harbor Laboratory Press(1988)、D.L.Riddleらの“C.ELEGANS II” Cold Spring Harbor Laboratory Press(1997)、さらに、(特許文献5)、(特許文献6)、(特許文献7)、(特許文献8)、(特許文献9)、(特許文献10)、(特許文献11)、(特許文献12)、(特許文献13)、(特許文献14)および(特許文献15)などに言及されており、これらはそれぞれ、本明細書に援用して組み込まれている。当業者にとっては、その他の実施例は、明確なものである。
【0055】
【特許文献5】国際公開第95/07463号
【特許文献6】国際公開第96/23810号
【特許文献7】国際公開第95/07463号
【特許文献8】国際公開第95/21191号
【特許文献9】国際公開第97/11094号
【特許文献10】国際公開第97/42320号
【特許文献11】国際公開第98/06737号
【特許文献12】国際公開第98/21355号
【特許文献13】米国特許第6,207,410号
【特許文献14】米国特許第5,693,492号
【特許文献15】欧州特許第1085089号
【0056】
本発明の別の実施形態は、トランスフォームを受けたり、本発明での一つまたは複数の核酸配列、一つまたは複数の核酸もしくは一つまたは複数の遺伝子構成物を含む宿主細胞または宿主生物に関連するものである。本発明は、本発明のアミノ酸配列の一つまたは複数を(適切な条件下などで)発現する、または、(少なくとも)発現することのできる宿主細胞または宿主生物とも関連する。一部の実施形態では、宿主細胞が、機能性フラグメントおよびキメラなどを含む、キメラとしての電位依存性カルシウムチャネルのα1、TBW β−1および/またはβ−2および/またはβ−3およびアブラムシα2δおよび/またはTBW α2δ−1および/またはα2δ−2サブユニットを発現する組換えベクターから構成され、α1、βおよびα2δサブユニットのうちの少なくとも一つが、本発明の、望ましくは配列番号5、配列番号7、配列番号9、配列番号11、配列番号13、配列番号15および/または配列番号17といった、α1、TBW β−1および/またはβ−2および/またはβ−3ならびにTBW α2δ−1および/またはα2δ−2および/またはアブラムシα2δサブユニットのキメラとなる、機能性の電位依存性カルシウムチャネルを形成することがある。一部の好ましい実施形態では、宿主細胞が、機能性フラグメントおよびキメラなどを含む、電位依存性カルシウムチャネルのα1、βおよびα2δサブユニットを発現する組換えベクターから構成され、望ましくは配列番号5、配列番号7、配列番号9、配列番号11、配列番号13、配列番号15、および/または配列番号17といった、本発明のα1、TBW β−1および/またはβ−2および/またはβ−3ならびにTBW α2δ−1および/またはα2δ−2サブユニットのうちの少なくとも二つのキメラとなる、機能性の電位依存性カルシウムチャネルを形成する。一部の好ましい実施形態では、宿主細胞が、本発明の核酸分子であるコード化配列からのα1、βおよびα2δサブユニットを発現する組換えベクターから構成され、望ましくは、配列番号5と、配列番号7、配列番号9、配列番号11、配列番号13のうちの一つまたは複数と、配列番号15および/または配列番号17のうちの一つまたは複数といった、本発明のα1と、TBW β−1および/またはβ−2および/またはβ−3と、TBW α2δ−1および/またはα2δ−2および/またはアブラムシα2δサブユニットのキメラとなる。こうした宿主細胞または宿主生物は本明細書では集合的に、「本発明の宿主細胞または宿主生物」と呼ばれることもある。
【0057】
以下に例示するように、宿主細胞は、適切な細胞(真菌、原核細胞または真核細胞)または細胞系列となることがある:
−E.coli、Bacillus、StreptomycesおよびPseudomontasなどの菌株、ただしこれらに限定されない;
−AspergillusおよびTrichodermaなどの真菌細胞種、ただしこれらに限定されない;
−KluyveromycesまたはSaccharomycesなどの酵母細胞種、ただしこれらに限定されない;
−アフリカツメガエル(Xenopus)の卵母細胞などの両生類の細胞または細胞系列。
【0058】
殺虫性化合物の発見および開発で本発明の核酸配列を使用(しようと)する場合に特に有用であると思われる一つの具体的な実施形態では、宿主細胞が、以下のような昆虫由来の細胞または細胞系列となることがある:
−Spodoptera SF9およびSf21細胞などの鱗翅目に由来する細胞または細胞系列、ただしこれらに限定されない;
−Aphisに由来する細胞または細胞系列;
−シュナイダー細胞およびKc細胞など、Drosohilaに由来する細胞または細胞系列;および
−オオタバコガ(Heliothis virescens)など、対象となる有害生物種(後述のもの)に由来する細胞または細胞系列。
【0059】
宿主細胞は、CHO細胞、BHK細胞およびヒト細胞またはHEK,HeLaおよびCOSなどの細胞系列といった哺乳動物細胞または細胞系列となることもあるが、これらの限定されるわけではない。
【0060】
宿主生物は、以下のような適切な多細胞生物(脊椎動物または無脊椎動物)となることがあるが、これらに限定されるわけではない:
−C.elegansなどのCaenorhabditis属に由来する線形動物、ただしこれに限定されない;
−Aphis,Drosophila,Heliothisまたは、対象となる有害生物種(上述のもの)などの昆虫、ただしこれらに限定されない;
−ゼブラフィッシュなど、その他のよく知られているモデル生物;
−ラットまたはマウスなどの哺乳動物;
前述のハンドブックおよび特許出願からの例のように、当業者にとってはその他の適切な宿主細胞または宿主生物は明白なものである。
【0061】
本発明の核酸配列が多細胞生物で発現した場合には、当該生物の全体で発現する可能性も、該当する細胞、組織、臓器または一部に特異的なプロモーターの制御下での発現などの場合には、一つまたは複数の特殊な細胞、組織、臓器またはその一部のみで発現する可能性もあることに留意しなければならない。
【0062】
核酸配列は、やはり発達または生活環での該当する段階に特異的なプロモーターの制御下での発現などの場合には、宿主細胞または宿主生物の発達または生活環の特定の段階のみで発現することもある。さらに、すでに上で述べたように、該当する発現は、構成的なものであったり、一過性のものであったり、誘発性のものであったりする可能性がある。
【0063】
これらの宿主細胞または宿主生物は、(さらに、宿主生物の場合には:少なくともその一つの細胞、一部、組織または臓器で)本発明のアミノ酸配列を(適切な条件下などで)発現する、または、(少なくとも)発現することのできるものであることが好ましい。本発明にはさらに、本発明の宿主細胞または宿主生物のその後の世代、後代および子孫が対象となり、これらは例えば、細胞分裂または有性生殖もしくは無性生殖で得られたものとなることがある。
【0064】
別の態様では、本発明は、好ましくは(基本的に)分離された形態での核酸と関連しており、この核酸は、(本明細書に定義するような)本発明のアミノ酸配列、特に、配列番号2、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16および配列番号18のアミノ酸をコード化しているか、これら本発明のアミノ酸配列の発現に利用することが可能なものである。
【0065】
配列番号2、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16および配列番号18のアミノ酸配列は、当業者に知られている、蛋白質の分離および精製のための技術を用いて、上述の生物種から分離されることがある。これとは別に、配列番号2、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16および配列番号18のアミノ酸配列は、以下に詳述するように、適切な宿主細胞または宿主生物で、適切な核酸配列−それぞれ、配列番号1、配列番号5、配列番号7、配列番号9、配列番号11、配列番号13、配列番号15および配列番号17の核酸配列および、その適切な突然変異体など−を適切に発現させることによって得られることがある。
【0066】
別の態様では、本発明は、好ましくは(基本的に)分離された形態での蛋白質またはポリペプチドに関連しており、該当する蛋白質またはポリペプチドは、(上述に定義するような)本発明のアミノ酸配列、特に配列番号2、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16および配列番号18のアミノ酸配列からなっている。
【0067】
広い意味では、「本発明のアミノ酸配列」という語は以下のものから構成される:
−配列番号2、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16および配列番号18のアミノ酸配列の一部またはフラグメント;
−配列番号2、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16および配列番号18のアミノ酸配列の(天然または合成による)突然変異体、変異体、対立遺伝子、類縁体、オルソログ(本明細書では以下、総称して「類縁体」と呼ぶ);
−こうした類縁体の一部またはフラグメント;
−配列番号2、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16および配列番号18のアミノ酸配列(またはその一部もしくはフラグメント)と、少なくとも一つの別のアミノ酸残基またはアミノ酸配列との融合物;
−類縁体(またはその一部若しくはフラグメント)のアミノ酸と、少なくとも一つのさらに別のアミノ酸残基またはアミノ酸配列との融合物;
こうした突然変異体、一部、フラグメントまたは融合物は、以下に記述するようなものであることが好ましい。
【0068】
「本発明のアミノ酸配列」という語は、プレフォーム、プロフォームもしくはプレプロフォームまたは適切なリーダー配列との融合体など、上述のアミノ酸配列の「不完全な」形態をも包含する。さらに、本発明のアミノ酸配列は、該当のアミノ酸配列の発現または産生に使用した宿主細胞または宿主生物によっては、翻訳後プロセシングまたは適切なグリコシル化を受けたり;またはその他の変形(該当技術ではそれ自体が知られている化学的手法などにより)を受けたりすることがある。
【0069】
配列番号2、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16および配列番号18のアミノ酸配列の一部もしくはフラグメント、または、その突然変異体を含む、その(天然または合成による)類縁体の一部若しくはフラグメントの例には、N末端およびC末端切断アミノ酸配列などがあるが、これらに限定されない。さらに、本発明のアミノ酸配列の一つまたは複数の一部またはフラグメントの二つまたはそれ以上が適切に統合して、本発明のアミノ酸配列となることがある。
【0070】
本発明のアミノ酸配列は、少なくとも100個、好ましくは最低250個、さらに望ましくは最低350個の長さのアミノ酸、最大では2,500個、好ましくは多くの場合に2,000個、さらに望ましくは少なくとも1,750個のアミノ酸を有していることが好ましい。
【0071】
こうした一部またはフラグメントは、配列番号2、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16および配列番号18のアミノ酸配列からの、少なくとも5個、望ましくは最低10個、さらに望ましくは最低20個、さらに好ましいものとしては最低30個以上のアミノ酸からなる、少なくとも一つの連続的伸展から構成されるものとなる。配列番号6のフラグメントの場合には、フラグメントは、配列番号2からの少なくとも4個、望ましくは8個、さらに望ましくは15個以上のアミノ酸を含んでいなければならない。
【0072】
特に、本明細書で記述した一部またはフラグメントは、(少なくとも)本発明での活性部位若しくは触媒部位に対応するアミノ酸配列、または、本発明での結合ドメインに対応するアミノ酸配列から構成されるものである。当業者にとっては明確であるように、こうした一部またはフラグメントには、(下記で全般的に記述するように)測定技術およびスクリーニング技術、ならびに、(該当する部位またはフラグメントが結晶の形で提供される場合には)X線結晶解析で、特別な利用法が見出される可能性がある。
【0073】
さらに、当業者によれば、本明細書の公開に基づいて、配列番号2、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16または配列番号18のアミノ酸配列の天然の「類縁体」(上述のもの)を識別したり、入手したり、分離したりすることが可能となると予想される。こうした突然変異体は、(別の個体の)同一生物種(例えば、異なる菌株または細胞系列からのもので、突然変異株または細胞系列を含むがそれに限定しない)から、または、別の生物種(の個体)から得ることができた。例えば、こうした類縁体は、上述の昆虫種から得ることができた。
【0074】
さらに、当業者によれば、本明細書の公開に基づいて、配列番号2、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16または配列番号18のアミノ酸配列の合成「類縁体」(上述で定義したもの)を提示したり、提供することができるようになると予想される。
【0075】
本明細書で記述した突然変異体は、配列番号2、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16または配列番号18のアミノ酸配列についての一つまたは複数、望ましくはすべての構造的特徴を有しているか、または、以下に記述した特徴を保存しているものであることが好ましい。
【0076】
本明細書で記述したあらゆる類縁体、一部またはフラグメントは、アミノ酸レベルで、少なくとも80%、特に90%以上および最高95%以上の度合いで、配列番号2、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16または配列番号18のアミノ酸と、「配列の同一性」を有するものであることが好ましい。
【0077】
こうした目的では、−配列番号2、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16または配列番号18のアミノ酸配列と比較して−アミノ酸残基の欠失、挿入、置換または付加を、単独アミノ酸(位)での相違と考え、該当するアミノ酸配列において配列番号2、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16または配列番号18のアミノ酸配列での対応する位置でアミノ酸残基が同一となっているアミノ酸残基の数を、該当するアミノ酸配列での総アミノ酸数で割って100%を掛けることにより、該当するアミノ酸配列と配列番号2、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16または配列番号18のアミノ酸配列との間の「配列同一性」を算定することができる。上述のように、こうした算定で配列アラインメントの対比を実施するために適した方法は、clustalWプログラムである。
【0078】
さらに、望ましくは、本明細書で記述したあらゆる類縁体、一部またはフラグメントは、下記の標準的な測定法により測定した場合、最低10%、好ましくは最低50%、さらに望ましくは最低75%および、最高90%といった範囲で、配列番号2、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16および配列番号18の配列について、上で記載されている生物学的活性と基本的に類似した生物学的活性を有することになる。
【0079】
蛋白質融合の提供などのように、本発明のアミノ酸配列を一つまたは複数の別のアミノ酸配列と融合させたものを利用することも、(上述のように)本発明の意図の範囲内である。以下に詳述するように、一般的に、こうした融合物は、適切な宿主細胞または宿主生物で−本発明の核酸配列を、一つまたは複数の別のコード化配列と適切に融合させるなど−本発明の適切な核酸配列の適切な発現によって得られる。
【0080】
具体的な実施形態の一つでは、こうした融合物が、グルタチオン−S−トランスフェラーゼ(「GST」)、緑色蛍光蛋白質(「GFP」)、ルシフェラーゼまたはその他の蛍光蛋白質などのような、レポーター蛋白質と融合した本発明のアミノ酸配列から構成されることがある。当業者にとっては明らかであるように、こうした融合物が、発現解析および同様の方法で特に有用とされることがある。
【0081】
別の実施形態では、該当するアミノ酸または残基に対するアフィニティ法を用いる場合など、融合相手が、発現したアミノ酸配列の精製で利用される可能性のあるアミノ酸配列またはアミノ酸残基となることがある。その後、該当する配列または残基は除去され(化学的切断または酵素的切断などにより)、本発明の核酸配列が得られる(この目的のためには、アミノ酸配列または残基は、切断可能なリンカー配列を介して、本発明のアミノ酸配列に任意で結合することができる)。こうした残基の好ましい例としては、複数のヒスチジン残基およびグルタチオン残基があるが、これらに限定されない。
【0082】
好ましいものではあるが限定的ではない態様では、こうした融合物は、下記の標準的な測定法により測定した場合、最低10%、好ましくは最低50%、さらに望ましくは最低75%および、最高90%といった範囲で、配列番号2、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16または配列番号18の配列について、上で記載されている生物学的活性と基本的に類似した生物学的活性を有することになる。
【0083】
本発明の核酸配列およびアミノ酸配列は、一般的に、以下の構造的特徴または保存された特徴のうちの一つまたは複数の存在によって、特徴付けられると思われる。
【0084】
Heliothis verescensの遺伝子については、配列番号1は、オープンリーディングフレームを取り囲むcDNA配列であり;配列番号2は、配列番号1によってコード化された蛋白質である。HeliothisのN型電位依存性カルシウムチャネルの蛋白質配列は、以下の表に示すように、他のN型電位依存性カルシウムチャネルの蛋白質と関連しており、関連性の値は、clustalWおよび上述に示したパラメータを用いて決定された。
【0085】
【表1】
【0086】
1 Geneband Accession No.AAC47406,(非特許文献25)
2 Geneband Accession No.AAC36868,(非特許文献26)
【0087】
【非特許文献25】Smith,L.A.ら、J.Neurosci.16(24)、7868−7879、1996
【非特許文献26】Waterston,R.,Science 282(5396)、2012−2018、1998
【0088】
他のカルシウムチャネルから類推されるように、機能性蛋白質は単量体である可能性が高い。(非特許文献27)などを参照されたい。
【0089】
【非特許文献27】Hannan and Hall,In Comparative Molecular Neurobiology,Y.Pichon,1993、Birkhuase Verlag(スイス・バーゼル)
【0090】
上述に基づき、本発明は特定の説明または機序に特に限定されていないが、核酸配列およびアミノ酸配列は、電位依存性カルシウムチャネルとしての(生物学的な)活性を有する。特に、今回の発明は、穿孔虫、根切り虫、アワヨトウ、オオタバコガ、fruit worm、アオムシ、ガおよびlopperなど、特にオオタバコガの鱗翅目の族の昆虫と、穿孔および吸引に適した口器を有する半翅類の昆虫から、電位依存性カルシウムチャネルとして活性が示されている。
【0091】
当該技術で知られているように、こうした種類の生物学的活性は、標識された既知のリガンドとの結合部位での完全な競合;蛍光共鳴エネルギー移動などの蛋白質相互作用に基づく蛍光測定法によるカルシウム蛍光色素を用いたカルシウム流動性の測定(Velicelebeらを参照)による、経時的な蛍光の消失または蛍光法もしくは比色法によるレポーターアッセイ;電位依存性カルシウムチャネルの測定に適したあらゆる技術など、標準的な測定法を用いて測定することができる。生物学的活性は、カルシウム蛍光色素を用いたカルシウム流動性を測定することによって測定することが好ましい。
【0092】
本発明の別の実施形態は、ある程度厳密な条件下、好ましくは厳密性が高いものおよび特に厳密な条件下(いずれも、上述のもの)で、本発明の核酸配列とハイブリダイゼーションすることが可能な核酸プローブに関連する。こうした核酸プローブは、例えば、本発明の核酸配列の検出もしくは分離で利用されたり、本発明の核酸配列の増幅でプライマーとして利用されることがある;いずれも、それ自体で知られている技術を用いており、やはり、上述のSambrookらおよびAusubelらなどの一般的なハンドブックに記述されている。
【0093】
本発明の別の核酸配列を検出または分離する際に利用する場合には、こうした核酸プローブは、通常は15個から100個、好ましくは20個から80個の間の長さのヌクレオチドを有することが好ましい。増幅のプライマーとして使用する場合には、こうした核酸プローブは、25個から75個、好ましくは20個から40個の間の長さのヌクレオチドを有することになる。
【0094】
一般的に、こうしたプローブは、当業者が、適切なコンピュータプログラムを任意に使用しながら、本発明の核酸配列またはアミノ酸配列−特に、配列番号1または配列番号2、配列番号5または配列番号6、配列番号7または配列番号8、配列番号9または配列番号10、配列番号11または配列番号12、配列番号13または配列番号14、配列番号15または配列番号16および配列番号17まはた配列番号18から開始することによってデザインすることができる。さらに、当業者にとっては明白であるように、こうしたプローブは変性プローブとすることもできる。
【0095】
別の態様では、本発明は、今回の発明の核酸配列の突然変異体および遺伝子構成物を調製するための方法と関連する。
【0096】
今回の発明の核酸配列の天然の突然変異体は、実施例で記載されている方法と基本的に類似した方法、または、以下のような代替法で、得ることができる:
−適切な発現ベクター系での、対象となる生物者からのDNAライブラリの構築と、それに続く突然変異配列の直接発現;
−適切な発現ベクター系での、対象となる生物者からのDNAライブラリの構築と、それに続く、本発明のプローブ(下記のもの)または本発明の核酸配列を用いての、該当するライブラリのスクリーニング;
−対象となる生物者からの、突然変異配列をコード化するmRNAの分離と、それに続く、逆転写酵素を用いてのcDNAの合成;
または、それ自体で知られている何らかの他の適切な方法または技術により得るが、これについては、例えば、Sambrookら、”Molecular Cloning:A Laboratory Manual”(第2版)、1−3巻、Cold Spring Harbor Laboratory Press(1989)およびF.Ausubelら、”Current protocols in molecular biology”,Green Publishing and Wiley Interscience、ニューヨーク(1987)などの標準的なハンドブックで述べられている。
【0097】
今回の発明の核酸配列のこうした合成配列を作製するための技術は、当業者にとっては明らかであり、例えば、自動化DNA合成法;部位特異的突然変異誘発;天然に存在する一つまたは複数の配列の二つまたはそれ以上の部分を結合しての、切断された発現産物の発現を導く突然変異の誘発;一つまたは複数の制限酵素認識部位の誘発(適切な制限酵素を用いた場合に容易に切断または結合されると思われる部位または領域を作製するなど)および、例えば天然に存在する電位依存性カルシウムチャネルの配列をテンプレートとして利用して、一つまたは複数の「不適正」プライマーを用いてのPCR法による突然変異の誘発といったものがあるが、これらに限定されるわけではない。これらおよびその他の手法は、当業者にとっては明らかなものであり、やはり、上述のような、SambroodらおよびAusubelらなどの標準的なハンドブックに記載されている。
【0098】
本発明の遺伝子構成物は一般的に、上述の、SambrookらおよびAusubelらなどの一般的なハンドブックに記載されている技術を用いるなどして、本発明の核酸配列を、上述の一つまたは複数の別の成分と適切に結合させることによって得られる。
【0099】
本発明の遺伝子構成物は、それ自体で知られている適切な(発現)ベクターに、本発明の核酸配列を挿入することによって得られることが多くなる。限定的なものではないが好ましい適切な発現ベクターの実施例の一部に、以下のようなものがある:
−哺乳動物細胞での発現のためのベクター:pSVL SV40(Pharmacia),pMAMneo(Clontech),pcDNA3(Invitrogen),pMC1neo(Stratagene),pSG5(Stratagene),pMSG(Pharmacia),pIND(Invitrogen),EBO−pSV−neo(ATCC37593)、pBPV−1(8−2)(ATCC37110),pdBPV−MMTneo(342−12)(ATCC37224),pRSV gpt(ATCC37119),pRSVneo(ATCC37198),pSV2−dhfr(ATCC37146),pUCTag(ATCC37460)および1ZD35(ATCC37565);
−細菌での発現のためのベクター:pETベクター(Novagen)およびpQEベクター(Qiagen);
−酵母またはその他の真菌細胞での発現のためのベクター:pYES32(Invitrigen)およびPichia発現ベクター(Invitrogen);
−昆虫細胞での発現のためのベクター:pBlueBacII(Invitroge),pEI1(Novagen),pMT/V5His(Invitrogen)。
【0100】
別の態様では、本発明は、本発明の核酸配列、本発明の核酸または本発明の遺伝子構成物による宿主細胞または宿主生物のトランスフォーミングのための方法に関連する。本発明はさらに、宿主細胞または宿主生物をトランスフォームする本発明の核酸配列、本発明の核酸または本発明の遺伝子構成物の利用とも関連する。
【0101】
具体的な実施形態では、宿主細胞または宿主生物で本発明の核酸配列の発現が、元の(すなわち天然の)宿主細胞または宿主生物に比べて低下することがある。これは、例えば、技術領域でよく知られているアンチセンス技術またはRNA干渉技術を用いた一時的な方法、もしくは、本発明の核酸配列のランダムな、部位特異的または化学的な突然変異誘発を用いた構成的な方法で、行った場合などに生ずることがある。
【0102】
適切なトランスフォーメーション技術は、当業者には明白であり、企図されている宿主細胞または宿主生物もしくは、利用予定の遺伝子構成物に依存することがある。限定的なものではないが好ましい適切な技術の例には、粒子射撃法によるトランスフォーメーション、(マイクロ)インジェクション、トランスフェクション(適切なトランスポゾンを使用したものなど)、電気穿孔法およびリポフェクチン法などがある。これらの技術およびその他の適切な技術については、やはり、上述のハンドブックおよび特許出願に記載されている。
【0103】
トランスフォーメーション後には、本発明の核酸配列または遺伝子構成物によってトランスフォーメーションが成功した宿主細胞または宿主生物の検出および選別の手順がとられることがある。これは、例えば、本発明の遺伝子構成物中に存在する選択可能なマーカーに基づく選択手順、または、特異的な抗体などを用いての本発明のアミノ酸配列の検出によるステップなどとなることがある。
【0104】
トランスフォーメーションを受けた宿主細胞(安定な細胞系列の形をとることがある)または宿主生物(安定な突然変異系列または株の形をとることがある)は、今回の発明の別の態様を形成する。
【0105】
さらに別の態様では、本発明は、本発明のアミノ酸配列を産生するための方法に関連する。
【0106】
本発明のアミノ酸配列の発現を生じさせたり、得たりするためには、一般的に、本発明の(希望の)アミノ酸配列が発現または産生するような条件下で、トランスフォーメーションを受けた宿主細胞またはトランスフォーメーションを受けた宿主生物を、保持したり、維持したり、培養したりすることになる。適切な条件は、当業者にとっては明らかであり、通常、使用した宿主細胞または宿主生物、ならびに、本発明の(関連する)核酸配列の発現を制御する制御因子に依存する。これについても、上の、本発明の遺伝子構成物についてのパラグラフで述べた、ハンドブックおよび特許出願に記載されている。
【0107】
一般的に、適切な条件には、適切な培地の使用、適切な栄養源または適切な栄養分の存在、適切な温度の利用、ならびに、任意により、適切な誘発因子または化合物の存在(本発明の核酸配列が、誘発性プロモーターの制御下にある場合など)が含まれることがある;これらはいずれも、当業者によって選択される。この場合にも、こうした条件下で、本発明のアミノ酸配列は、構成的な方法、一過性の方法または、適切な誘発が行われた場合にのみ、発現できる。
【0108】
本発明のアミノ酸配列が(最初は)不完全な形で生じ(上述のように)、その後、使用した宿主細胞または宿主生物に応じて、トランスフォーメーション後の変形を受けることがあるということも、当業者には明白である。さらに、本発明のアミノ酸配列は、使用した宿主細胞または宿主生物に応じて、グリコシル化を受けることがある。
【0109】
本発明のアミノ酸配列はその後、宿主細胞または宿主生物、もしくは、該当する宿主細胞または宿主生物が培養された培地から、(分離用)クロマトグラフィー法および電気泳動法、沈降分離法、アフィニティ法(本発明のアミノ酸配列と融合した、特異的な切断可能なアミノ酸配列を使用するなど)および、調製用の免疫学的手法(分離予定のアミノ酸配列に対する抗体を使用するなど)などの、それ自体が知られている蛋白質分離精製技術を用いて、分離される。
【0110】
実施形態の一つでは、該当する配列もしくは、その抗原部位またはエピトープに対して特異的な抗体を得るために、こうして得られたアミノ酸配列を利用することもある。
【0111】
実施形態の一つでは、今回の発明は、今回の発明のアミノ酸配列、望ましくは、配列番号2、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16または配列番号18、もしくは、これらの類縁体、変異体、対立遺伝子、オルソログ、一部、フラグメントまたは、配列番号6に特異的な抗体が、配列番号4などのアブラムシα1と交差反応しない場合には、エピトープに対して特異的に作製された、モノクローナル抗体およびポリクローナル抗体などの抗体に関連する。
【0112】
本発明の別の態様となるこうした抗体は、(特許文献16)、(特許文献17)、(特許文献18)、(特許文献19)、(特許文献20)、(特許文献21)および(特許文献22)などに記述されている、それ自体が知られた方法で作製される。排他的ではないものの、こうした方法は、全般的に、該当するアミノ酸配列に対しての抗体が生ずるような方法に従って、免疫応答性の宿主を本発明の妥当なアミノ酸配列または免疫原性を有するその一部(特定のエピトープなど)で免疫し、その後、該当の宿主より得られた血液または血清などから、生じた抗体を回収することによるものであることが多い。
【0113】
【特許文献16】英国特許出願公開第2357768号
【特許文献17】米国特許第5,693,492号
【特許文献18】国際公開第95/32734号
【特許文献19】国際公開第96/23882号
【特許文献20】国際公開第98/02456号
【特許文献21】国際公開第98/41633号
【特許文献22】国際公開第98/49306号
【0114】
例えば、ポリクローナル抗体は、ヤギ、ウサギ、ヒツジ、ラット、ブタまたはマウスなどの適切な宿主を、任意により、免疫原性のキャリアー(ウシ血清アルブミンまたはキーホールリンペットヘモシニアンなど)、もしくは、フロイントのアジュバント、サポニン、水酸化アルミニウムまたは同様の界面活性剤などのアジュバントを用いて、本発明のアミノ酸配列(のエピトープ)で免疫することによって、得ることができる。適切な免疫応答が生じた後に(通常は1〜7日以内)、やはり(特許文献23)などに言及されている、それ自体が知られている方法により、免疫した動物から採取した血液または血清から抗体を分離するが、任意によって、既知の免疫測定法技術を用いての、希望する性質(特異性など)を有する抗体についてのスクリーニング段階が含まれることがある。
【0115】
【特許文献23】国際公開第96/23882号
【0116】
例えば、モノクローナル抗体は、ハイブリドーマをベースとしたものおよび同様の方法など、培養物での連続的な細胞系列を用いて産生させることができ、これについても基本的には上述で引用した参考文献に記載されている。従って、本発明のアミノ酸配列に対するモノクローナル抗体を産生する細胞または細胞系列は、本発明のアミノ酸配列に対する抗体産生法と同様、本発明の別の態様を形成し、こうした方法では一般的に、こうした細胞の培養と、培養物または倍地の抗体の分離が関与し、やはり、それ自体が知られている技術が用いられる。
【0117】
さらに、本発明のアミノ酸配列に対するFabフラグメント(F(ab)2,Fab’およびFabフラグメント)は、それぞれ、ペプシンまたは別のプロテアーゼによる抗体の消化、ジスルフィド結合の開裂、ならびに、パパインおよび還元剤による処理によって得ることができる。Fab−発現ライブラリは、Huseらの方法、Science 245:1275−1281等によって得ることができる。
【0118】
別の実施形態では、本発明のアミノ酸配列、または、こうしたアミノ酸配列をコード化し、発現する組換え発現ベクターからなる宿主細胞もしくは宿主生物を用いて、本発明のアミノ酸配列の(生物学的)活性を変調したりその他の点でこれと相互作用をすることのできる化合物またはその他の因子を、識別したり開発したりすることもでき、こうした利用法により、本発明の別の態様が形成される。当業者には明白なように、こうした状況では、本発明のアミノ酸配列は、こうした化合物または因子との相互作用の標的となる。
【0119】
こうした状況では、「変調する」、「変調」、「モジュレーター」および「標的」という語は、当該技術でのその通常の意味を有し、とりわけ、国際公開第98/06737号で定義が示されている。一般的に、モジュレーターは、生物学的活性または過程(例えば、本発明のアミノ酸配列の生物学的活性など)の機能的性質を、促進、阻害または低減またはその他の変更、感化または影響(「変調」と総称する)することのできる化合物または因子である。
【0120】
こうした状況では、本発明のアミノ酸配列は、in vitroでの変調(測定またはスクリーニングの一部として)もしくは、in vivoでの変調(標的を変調することが知られている化合物または因子による変調で、こうした化合物または因子は、例えば、農芸化学、獣医学または薬学での利用で活性化合物として利用されることがある)の標的となることがある。
【0121】
例えば、本発明のアミノ酸配列、宿主細胞または宿主生物は、一次的スクリーニング(標的に関して未知の活性を有する被験化合物の一連の群またはライブラリから、標的のモジュレーターを識別するために利用されるスクリーニング法など)または二次的測定(一次的スクリーニングで該当したものを確認したり、該当するものを導くための化学の一部などとして、該当する分子の最適化に使用される測定法)など、本発明のアミノ酸配列のモジュレーターを識別したり開発したりするために利用することのできる測定法またはスクリーニング法の一部として利用されることがある
【0122】
例えば、こうした測定法またはスクリーニング法は、in vitro測定法またはスクリーニング法として構成されることがあり、これは一般的に、標的に対する潜在的モジュレーターとして検討対象となる化合物または因子(本明細書では以下、「被験化合物」と呼ぶ)を標的と結合させ、該当する結合を生じた信号を測定するというものになる。こうしたin vitroスクリーニングに適した技術は、当業者には明白なものとなり、(非特許文献28)および(非特許文献29)などに記載されている。例えば、こうした測定法またはスクリーニング法が、結合測定法またはスクリーニング法として構成されることがあり、この場合、被験化合物は、標的からの検出可能なリガンドを置換するために用いられ(放射性リガンドまたは蛍光リガンドなど)、これによって、モジュレーターによって標的から置換されたリガンドの量が決定される。
【0123】
【非特許文献28】Eldegrawiら(1987).FASEB J.,1巻、262−271ページ
【非特許文献29】Rauhら(1990),Trends in Pharmacol.Sci.,11巻、325−329ページ
【0124】
こうした測定法またはスクリーニング法は、細胞をベースとした測定法またはスクリーニング法として構成されることもあり、この場合、本発明の宿主細胞は、被験化合物と結合したりこれにさらされ、これに基づいて、宿主細胞による少なくとも一つの生物学的応答が測定される。
【0125】
さらに、こうした測定法またはスクリーニング法は、動物一個体でのスクリーニングとして構成されることがあり、この場合、本発明の宿主生物は、被験化合物と接触したりこれにさらされ、これに基づいて、宿主生物による少なくとも一つの生物学的応答(表現型の変化、挙動の変化または、麻痺または死亡を含むがこれに限定されない生理学的変化など)が測定される。
【0126】
従って、一般的に、上述の測定法およびスクリーニング法は、特に、被験化合物による標的の変調を示す信号が発生するような方法で、被験化合物が標的(または、標的を発現している宿主細胞または宿主生物)と接触するための少なくとも一つの段階から構成される。ついで、その後の段階で、該当する信号が測定される。
【0127】
従って、ある態様では、本発明は、本発明の該当するアミノ酸配列と被験化合物との相互作用を示すような信号を生じさせるための方法に関連しており、該当する方法は少なくとも以下の手順からなっている:
a)本発明のアミノ酸配列もしくは、アミノ酸配列を含有または発現する宿主細胞または宿主生物を、該当する被験化合物と該当するアミノ酸配列との間の相互作用を示す信号が発生するような方法で、該当する被験化合物と接触させる手順;および、任意により、
b)これによって生ずる信号を検出する手順。
【0128】
別の態様では、本発明は、本発明のアミノ酸配列のモジュレーターおよび/またはインヒビターを識別するための方法に関連しており、
該当する方法は少なくとも以下の手順からなっている:
a)本発明のアミノ酸配列もしくは、アミノ酸配列を含有または発現する宿主細胞または宿主生物を、該当する被験化合物と該当する標的との間の相互作用を示す信号が発生するような方法で、該当する被験化合物と接触させる手順;および、任意により、
b)これによって生ずる信号を検出し、該当する信号が該当するアミノ酸配列のモジュレーターおよび/またはインヒビターを識別するような手順。
【0129】
従って、今回の発明は、鱗翅目のカルシウムチャネル蛋白質活性のモジュレーターを識別する方法を示すものである。この方法は、核酸配列の少なくとも一つが本発明の核酸配列で、被験化合物の存在下でカルシウムチャネルを発現するような、機能性カルシウムチャネルをコード化する拡散配列を含む組換え発現ベクターから構成される被験細胞をカルシウムを含有する溶液と接触させ、被験細胞での細胞内カルシウム量を検出することによっての、試験方法の実施段階からなっている。好ましい実施形態では、機能性カルシウムチャネルは、本発明の二つまたはそれ以上のアミノ酸配列から作られている。一部の実施形態では、機能性カルシウムチャネルが、望ましくは、配列番号6と、配列番号8、配列番号10または配列番号12のうちの一つ、ならびに、配列番号14、配列番号16または配列番号18のうちの一つという、本発明の三つのアミノ酸配列から作られている。この方法は、鱗翅目のカルシウムチャネルをコード化し、鱗翅目のカルシウムチャネルを発現する核酸配列を含有する組換え発現ベクターからなる陰性対照細胞を、被験化合物の非存在下でカルシウムを含有する溶液と接触させ、陰性対照細胞での細胞内カルシウムの量を検出することによって陰性対照測定を実施する手順からも構成されている。被験細胞での細胞内カルシウムの量を、陰性対照細胞での細胞内カルシウムの量と比較する。陰性対照細胞での細胞内カルシウムの量と比較した場合の被験細胞での細胞内カルシウムの量の変化は、被験化合物が鱗翅目のカルシウムチャネル蛋白質活性のモジュレーターであることを示すものである。好ましい実施形態の一部では、被験細胞がCHOまたはHEKとなる。好ましい実施形態の一部では、該当する細胞内部の色素が細胞内カルシウムと相互作用することによって生じた蛍光を測定するという測定法を用いることにより、細胞内カルシウムが検出される。一部の好ましい実施形態では、この方法はさらに、鱗翅目のカルシウムチャネルをコード化し、鱗翅目のカルシウムチャネルを発現する核酸配列を含有する組換え発現ベクターからなる陽性対照細胞を、被験化合物の非存在下および鱗翅目のカルシウムチャネル作動物質の存在下でカルシウムを含有する溶液と接触させ、該当する陽性対照細胞での細胞内カルシウム量を検出することにより、陽性対照測定を行うというものである。一部の実施形態では、この方法はさらに、鱗翅目のカルシウムチャネルを発現しない鱗翅目のカルシウムチャネル陰性細胞を、被験化合物の非存在下でカルシウムを含有する溶液と接触させて、該当する鱗翅目のカルシウムチャネル陰性対照細胞に取り込まれたカルシウムの量を検出することによって、二番目のタイプの陰性対照測定を実施するか;および/または、鱗翅目のカルシウムチャネルを発現しない鱗翅目のカルシウムチャネル陰性細胞を、被験化合物の非存在下および鱗翅目のカルシウムチャネル作動物質の存在下でカルシウムを含有する溶液と接触させて、該当する鱗翅目のカルシウムチャネル陰性対照細胞に取り込まれたカルシウムの量を検出することによって、三番目のタイプの陰性対照測定を実施するかの、いずれかの段階からなっている。好ましい実施形態では、この方法で利用される鱗翅目のカルシウムチャネル蛋白質は、配列番号2または配列番号6、その突然変異体、そのフラグメントからなる群から選択されたアミノ酸配列を有しており、これは、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16および配列番号18の群、その突然変異体、そのフラグメントからなる群から選択されたさらに別のサブユニットの一つまたは複数と組み合わされることがある。一部の実施形態では、鱗翅目のカルシウムチャネルをコード化する核酸配列は、配列番号1または配列番号5で、補助的なサブユニットが存在する場合には、これは、配列番号7、配列番号9、配列番号11、配列番号13、配列番号15または配列番号17をコード化するものとなる。
【0130】
一部の実施形態によると、鱗翅目のカルシウムチャネル蛋白質活性のインヒビターを識別する方法は、鱗翅目のカルシウムチャネルをコード化し、鱗翅目のカルシウムチャネルを発現する核酸配列を含有する組換え発現ベクターからなる被験細胞を、被験化合物の存在下でカルシウムを含有する溶液および鱗翅目のカルシウムチャネル作動物質と接触させて、該当する被験細胞での細胞内カルシウムの量を検出することにより、検査測定法を実施するという段階から構成される。対照測定も、鱗翅目のカルシウムチャネルをコード化し、鱗翅目のカルシウムチャネルを発現する核酸配列を含有する組換え発現ベクターからなる陰性対照細胞を、被験化合物の非存在下でカルシウムを含有する溶液および鱗翅目のカルシウムチャネル作動物質と接触させて、該当する陰性対照細胞での細胞内カルシウムの量を検出することにより実施される。被験細胞での細胞内カルシウムの量を、対照細胞での細胞内カルシウムの量と比較する。対照細胞での細胞内カルシウムの量と比較した場合の被験細胞での細胞内カルシウムの量の減少は、被験化合物が鱗翅目のカルシウムチャネル蛋白質活性のインヒビターであることを示すものである。一部の好ましい実施形態では、被験細胞は、CHOまたはHEKである。一部の好ましい実施形態では、細胞内カルシウムは、細胞内部の色素が細胞内カルシウムと相互作用することによって生じた蛍光を測定するという測定法を用いることによって検出される。一部の好ましい実施形態では、鱗翅目のカルシウムチャネル蛋白質は、配列番号2または配列番号6、その突然変異体、そのフラグメントからなる群から選択されたアミノ酸配列を有しており、これは、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16および配列番号18の群、その突然変異体、そのフラグメントからなる群から選択されたさらに別のサブユニットの一つまたは複数と組み合わされることがある。一部の実施形態では、該当する鱗翅目のカルシウムチャネルをコード化する核酸配列は、配列番号1または配列番号5で、補助的なサブユニットが存在する場合には、これは、配列番号7、配列番号9、配列番号11、配列番号13、配列番号15または配列番号17をコード化するものとなる。
【0131】
被験化合物は、化合物の一連の群またはライブラリの一部となることがあり、当業者には明らかなように、これは、多種多様な一連の群またはライブラリ、もしくは、対象となる一連の群またはライブラリとなることがある。こうしたスクリーニングに使用されるライブラリは、技術で知られている化学的過程、または、化学合成での従来の手段を組み合わせて用いることで、調製が可能である。
本発明の測定法およびスクリーニング法は、例えば、適切なロボットを使用した自動化法などで、中程度のスループットからハイスループットで実施されることがある。特に、この実施形態では、本発明の方法は、標準的な24穴、96穴、384穴、1536穴または3456穴プレートなどのマルチウェルプレートのウェル内で標的を被験化合物と接触させることによって実施されることがある。
【0132】
通常は、本発明の測定法およびスクリーニング法では、各々の測定について、標的または宿主細胞または宿主生物を、単独の被験化合物のみと接触させる。しかし、例えば、該当する併用によって相乗効果が生ずるか否かを検討するなどで、標的を二つまたはそれ以上の被験化合物と、同時にまたは連続的に接触させることも、本発明の意図の範囲内である。
【0133】
被験化合物が本発明のアミノ酸配列に対してモジュレーターおよび/またはインヒビターとして識別された場合には(本明細書上述のようなスクリーニングまたは測定によって)、それ自体が、本発明の関連アミノ酸、望ましくは、配列番号2、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16または配列番号18、その突然変異体、そのフラグメント、さらに望ましくは、配列番号2、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16または配列番号18のモジュレーターおよび/またはインヒビターとして利用されたり(農芸化学、獣医学または薬学での利用を目的とした作用物質として)、または、例えば、溶解度、吸収性、バイオアベイラビリティー、毒性、安定性、持続性、環境への影響などの性質を改善するために、任意によりさらに最適化されることがある。当業者にとっては、例えば二番目の測定法(の一部)などとして、本発明の核酸配列、望ましくは、配列番号1、配列番号5、配列番号7、配列番号9、配列番号11、配列番号13、配列番号15または配列番号17、アミノ酸配列、宿主細胞または宿主生物および方法が、最適化法などで、さらに利用される可能性があることは明白である。
【0134】
本発明は、モジュレーターおよび/またはインヒビター(被験化合物、化合物または因子)が、標的の変調、阻害または標的との相互作用を行う(in vivoまたはin vitroで)際の特定の方法または機序を、特に限定するものではない。例えば、モジュレーターおよび/またはインヒビターは、作動物質、拮抗物質、逆作動物質、部分作動物質、競合的インヒビター、非競合的インヒビター、補助因子、アロステリックインヒビターまたは標的に対するその他のアロステリック因子であることがあったり、別の蛋白質またはポリペプチド、受容体、もしくは、細胞の細胞小器官の一部のように、他の生物学的化合物への標的の結合を促進または低下させる化合物または因子であったりする。同様に、モジュレーターおよび/またはインヒビターは、(活性部位、アロステリック部位、結合ドメイン、もしくは、共有結合または水素結合により標的の別の部位で)標的と結合したり、標的の活性部位を(可逆的、不可逆的または競合的に)遮断および/または阻害したり、標的の結合ドメインを(可逆的、不可逆的または競合的に)遮断および/または阻害したり、標的の構造に影響または変更を与えたりすることがある。
【0135】
同様に、被験化合物、モジュレーターおよび/またはインヒビターは、例えば、以下のようなものであることがある:
−標的の既知の基質の類縁体;
−2〜20個、望ましくは3〜15個のアミノ酸残基からなるオリゴペプチド;
−アンチセンスまたは二本鎖RNA分子;
−蛋白質、ポリペプチド;
−補助因子または補助因子の類縁体。
【0136】
被験化合物、モジュレーターおよび/またはインヒビターが、参照化合物または因子であることもあり、これは、標的を変調、阻害したり、標的と相互作用することが知られている化合物(標的に対する既知の基質またはインヒビターなど)であったり、標的が属する一般的なクラスの他のものを、変調、阻害したり、これらと相互作用することが一般的に知られている化合物または因子(該当するクラスの既知の基質またはインヒビターなど)であったりすることがある。
【0137】
しかし、被験化合物、モジュレーターおよび/またはインヒビターは小分子であることが望ましく、これの意味することは、分子量が1500未満、望ましくは1000未満のものということである。これは例えば、有機分子、無機分子または有機金属分子などであり、可溶性塩類などの適切な塩の形態をとっていることもある。「小分子」という語は、(総)分子量が上述の範囲である限りは、複合体、キレートおよび同様の分子を網羅するものでもある。
【0138】
すでに上で述べたように、本発明のアミノ酸配列、望ましくは、配列番号2、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16または配列番号18のモジュレーターおよび/またはインヒビターとして識別または開発されている化合物または因子(およびこれらの化合物の前駆物質)は、農芸化学、獣医学または薬学での成分調製など、農芸化学、獣医学または薬学の領域で、作用物質として有用なものとなる可能性があり、こうしたモジュレーターとこれらを含有する成分はどちらも、本発明の別の態様である。
【0139】
例えば、農芸化学の領域では、本発明のモジュレーターおよび/またはインヒビターは、有害生物(の侵襲)の予防または制御などで、接触剤および植物浸透剤の双方として、殺虫剤、殺線虫剤、軟体動物駆除剤、殺蠕虫剤、殺ダニ剤またはその他のタイプの農薬もしくは殺生物剤として利用されることがある。同様に、モジュレーターおよび/またはインヒビターは、例えば、作物保護剤、家庭内用殺虫剤、または、有害生物によって生ずる損傷を予防または処理(種子、木材もしくは、貯蔵した作物または果物の保護など)するための薬剤として利用されることがある。本発明のモジュレーターおよび/またはインヒビターは、殺虫剤として利用されることが好ましい。こうした適用では、企図する最終的な使用法に適した処方品とするために、本発明の一つまたは複数のモジュレーターおよび/またはインヒビターが、一つまたは複数の農学的に許容されている担体、アジュバントまたは希釈剤と−さらに、それ自体で(例えば)植物保護剤(作用スペクトルを広げ、任意により相乗作用を生ずるため)、除草剤、肥料または植物成長調整剤としての活性を有する一つまたは複数の別の化合物も任意により併用して−適切に組み合わせられることがある。こうした処方品は、例えば、溶液、乳剤、散布剤、濃縮剤、エアロゾル、スプレイ、粉末、流動物、粒子、顆粒、錠剤、燻蒸剤、餌もしくはその他の適切な固体、半固体または液体といった処方品の形であることがあり、さらに、任意により、適切な溶剤、乳化剤、安定剤、界面活性剤、消泡剤、湿潤剤、展着剤、固着剤、誘引剤または(餌については)食物成分を含有することもある。”Pesticidal Formulation Research”、ACS発行物(1969)および”Pesticide Formulation”,Wade van Valkenburg Ed,Marcel Dekker発行物(1973)などの標準的なマニュアルで言及されている。
【0140】
こうした成分は一般的に、本発明の一つまたは複数のモジュレーターおよび/またはインヒビターを適切な含量で含有しており、これは一般的に、総成分の重量の0.1〜99%、特に10〜50%範囲である。
【0141】
本発明のモジュレーターおよび/またはインヒビターならびに成分は、特に、例えば、以下の目の望ましくないまたは有害な昆虫(成体および、幼虫などの未成熟体)を駆除または制御するための殺虫剤として有用である:
−Pissodes strobi、Diabrotica undecimpunctata howardiおよびLeptinotarsa decemlineataなどの甲虫目;
−Rhagoletis pomonella,Mayetiola destructorおよびLiriomyza huidobrensisなどの双翅目;
−Neodiprion taedae tsugae、Camponotus pennsylvanicusおよびSolenopsis wagneriなどの膜翅目;
−Pseudatomoscelis seriatus,Lygus lineolaris(Palisot de beauvois)およびAcrosternum hilareなどの半翅目;
−Aphis gossypiiなどの同翅亜目およびアブラムシ;ならびに
−オオタバコガ(Heliothis verescens)などの鱗翅目。
【0142】
有害または望ましくない生物を制御するために使用する際には、これらの生物が、生物の制御(死滅または麻痺など)に適した量で、本発明のモジュレーター、インヒビターまたは成分に直接接触することになる。こうした量は、当業者により(制御対象となる生物種に対して化合物を検討することにより)容易に決定することが可能で、通常は、約10〜500g/ha程度、特に100〜250g/haとなる。
【0143】
本発明のモジュレーター、インヒビターまたは成分は、系統的に適用されることもあり(制御対象となる生物の環境または土壌中などに)、植物、種子、果物などを保護するために、やはり、当業者によって決定することの可能な適切な量で、適用されることもある。本発明のモジュレーターおよび/またはインヒビターは、例えば、こうした組成で通常に使用される他の殺虫化合物と置き換えるために、−添加剤などとして−それ自体が知られている成分に組み込まれることがある。
【0144】
具体的な実施形態の一つでは、本発明のモジュレーターおよび/またはインヒビターならびに成分は、農芸化学、獣医学またはヒトの健康の領域で、寄生虫、特に以下のような寄生性節足動物、線形動物および蠕虫によって生ずる感染症または損傷または不快感を予防または治療するために使用されることがある:
−マダニ、ダニ、ノミ、シラミ、サシバエ、ツノサシバエ、アオバエおよび、その他、咬み傷を生じたり吸血性の外部寄生虫などの、外寄生性節足動物;
−蠕虫などの内部寄生虫;
さらに、こうした寄生虫によって引き起こされたり伝播する疾患の予防または治療にも使用される。こうした目的のため、本発明のモジュレーターおよび/またはインヒビターは、例えば、錠剤、経口溶液または乳剤、注射用溶液または乳剤、ローション、エアロゾル、スプレイ、粉末、浸漬剤または濃縮剤として処方されることがある。
【0145】
動物およびヒトの健康の領域では、本発明のモジュレーター、インヒビターおよび成分は、本発明のアミノ酸配列が標的として関与する疾患または障害の予防または治療にも使用されることがある。この目的のためには、本発明のモジュレーターおよび/またはインヒビターは、当業者にとっては明白な、薬学または獣医学での利用が許容されている一つまたは複数の添加剤、担体または希釈剤と共に処方されることがある。
【0146】
このように、さらに別の態様で、本発明は、本明細書上述のような、農芸化学、獣医学または薬学での利用を目的とした成分の調製での本発明のモジュレーターおよび/またはインヒビターの利用に関連している。本発明は、やはり上述のように、有害生物の制御および有害生物によって引き起こされる感染症または損傷の予防での、本発明のモジュレーター、インヒビターおよび成分の利用に関連する。
【0147】
本発明についてはここで、以下の非限定的な実施例により、さらに例証する。
(実施例)
【0148】
実施例1−cDNAライブラリの構成および配列データベース
【0149】
【表2】
【0150】
全長クローンの豊富なcDNAライブラリが、(a)混合ライフステージのワタアブラムシ(ワタアブラムシcDNAライブラリ)および、(b)虫齢の高いHeliothis virescenesの幼虫から切断した頭部(TBW頭部cDNAライブラリ)から精製された、ポリ(A)含有RNAより、Invitrogen Corporation(カルフォルニア州カールスバッド)によって構築された。これらのライブラリはいずれも、プラスミドベクターpCMV−SPORT6.1で構成され、E.coli DH10B(TonA)にトランスフォームされた。これらのライブラリは、さらに増幅されることなく、グリセロール培養物として、−80℃で貯蔵された。Invitrogen Corporationにより、ワタアブラムシcDNAライブラリの一部が増幅され、標準化されて、384穴プレートに並べられた。並べられたクローンについては、クローン化されたcDNAの5’末端および3’末端に隣接するベクタープライマーを用いて、自動化配列決定分析が行われた(Genome Therapeutics、マサチューセッツ州ウォルサム)。配列読み取り結果は、質およびベクターの混入についてトリミングが行われ、コンティグに構築されて、BLASTコンパチブルデータベースに入力された。検索ツールのBasic Local Alignment Sequence Tool(BLAST)スートおよびこれらのツールの使用法ならびに、BLASTコンパチブルデータベースへのアクセスは、米国立予防衛生研究所(メリーランド州ベセズダ)の全米バイオテクノロジー情報センターから行うことができる。
【0151】
さらに、虫齢の高いHeliothis virescenesの幼虫の腹側神経索および頭部から分離したポリ(A)含有RNAより、cDNAライブラリが構成された(TBW神経索ライブラリ)。約195個の神経索および頭部を、第5虫齢初期のTBW幼虫から切断採取し、ドライアイス上で急速に冷凍した。この組織は、室温にて3mLのTRIZOLTM試薬(Invitrogen Corporation)でホモジナイズし、製造業者のプロトコルに従って、全RNAを抽出した。RNAの質は、UV吸収分光光度法(320nm〜220nm)および、少量のグリコシル化試料の1×NorthernMaxTM−Gly緩衝液を含有する1%アガロースゲルでの電気泳動により確認した(Ambion、テキサス州オースチン)。ポリ(A)含有RNAは、製造業者の記述に従って、Oligotex(登録商標)mRNA分離キット(QIAGEN Inc.、カリフォルニア州バレンシア)を用いて、アフィニティクロマトグラフィーにより全RNAから選択した。(非特許文献30)の方法の改良法により、cDNAを合成した。要約すると、6.2μgのポリ(A)含有RNAを、11.2μlの80%(v/v)グリセロールおよび400pmolのSacdTプライマー(表I)と混合し、全量を24μlとした。プライマー/RNA混合物は、サーモサイクラーで65℃にて10分加熱した後に45℃まで冷却し、やはり45℃に予温した76μlの合成緩衝液と混合した。ファーストストランドcDNA合成反応の最終組成は:50mMのTris−HCl(25℃でpH8.3)、75mMのKCl、3mMのMg2Cl、10mMのジチオスレイトール(DTT)、0.6mMのdGTP、0.6mMのdATP、0.6mMのTTP、0.6mMの5−メチル−dCTP、50μg/mlのウシ血清アルブミン、2000単位/mlのRNaseOUTTMリボヌクレアーゼインヒビター(Invitrogen Corporation)および20,000単位/mlのSuperScriptTM逆転写酵素(Invitrogen Corporation)であった。反応は、45℃での2分間のインキュベート、2分間をかけての35℃への冷却、35度での2分間のインキュベート、50℃への上昇と5分間のインキュベート、最終的に55℃への上昇と60分間のインキュベートによるものであった。ファーストストランド反応は、EDTAを10mMへ、ナトリウムN−ラウロイルサルコシンを0.1%(w/v)、プロテイナーゼK(Invitrogen Corporation)を0.2mg/mlへと加えることにより、終了させた。混合物を45℃で15分間インキュベートした後、フェノール:クロロホルム:イソアミルアルコール(25:24:1)で1回抽出した。水相を除去し、100μlの5MのNH4OAcと500μlのエタノールを加えて、cDNAを−20度で60分間沈降させた。その後、遠心分離によりcDNAを採集し、70%エタノールで1回洗浄した後、500μlの0.1×TE(1mMのTris−HCl(pH8.0)−0.1mMのEDTA)に溶解して、氷浴上で200μlのセカンドストランド合成緩衝液とあわせた。セカンドストランド反応の最終組成は、20mMのTris−HCl(pH8.3)、130mMのKCl、10mMの(NH4)2SO4、5mMのMgCl2、10mMのDTT、0.6mMのdATP、0.6mMのdGTP、0.6mMのdCTP、0.6mMのTTP、10Mmのb−NAD、50μg/mlのBSA、3単位のE.coli RNaseH(Invitrogen Corporation)、2.5単位のE.coli DNAリガーゼ(Invitrogen Corporation)および25単位のE.coli DNAポリメラーゼ(Invitrogen Corporation)であった。セカンドストランド合成は、15℃にて150分間行った。酵素混合物は70℃での10分間の加熱により不活化し、その後、氷浴上に放置した。1μl(5単位)のT4 DNAポリメラーゼ(Invitrogen Corporation)を加え、15℃での15分間のインキュベートにより、二本鎖cDNAを平滑末端とした。方向性のクローニングを促進するために、平滑末端としたcDNAに二本鎖Xhoアダプターを加え、その後、ファーストストランド合成のためのプライマーで独特の切断を行うSacIによる切断によって、3’末端から除去した。アダプターは、それぞれ2nmolのオリゴヌクレオチドXhoAd4とXhoAd5(表I)を40μlの20mMNaClであわせ、混合物を70℃で3分間加熱してから、10分間をかけて25℃にゆっくりと冷却することによって、形成させた。アニールしたXhoアダプター(450pmol)は、50mMのTris−HCl(pH7.5)、10mMのMgCl2、1mMのATP、10mMのDTT、25μg/mlのBSAおよび600単位のT4 DNAリガーゼ(New England Biolabs、マサチューセッツ州ビバリー)を含有する30μlの反応溶液中で、1.7μgの二本鎖dDNAと8℃にて一晩結合させた。上述の、プロテアーゼKによる切断、フェノール:クロロホルムによる抽出およびエタノールによる沈降で再度cDNAを精製した。その後、1μgのcDNAあたり25単位の酵素の割合で、SacIによりcDNAを切断し、10mMのTris−HCl(pH8.0)、1mMのEDTA、0.1%(w/v)のSDSおよび0.1MのNaClで平衡化した2mlのSepharose CL−2B(Sigma Chemical Co.,ミズーリ州セントルイス)ゲル濾過カラムでサイズ分画を行った。ピーク分画(730ngのcDNA)をプールし、エタノール沈降法によって濃縮して、10μlの0.1XTE(pH8.0)に再溶解した。クローニングのために、57mMのTris−HCl(pH7.5)、11.4mMのMgCl2、1.1mMのATP、11.4mMのDTTおよび28μg/mlのBSAを含有する4.4μlの溶液中で、50ngのcDNAを、バクテリオファージλgtGreen1Sから調製した、500ngのXhoI/SacIファージアームとあわせた(cDNAとベクターの分子比は2:1)。0.3μl(3単位)のT4ポリヌクレオチドキナーゼ(New England Biolabs)を加え、混合液を37℃で15分間インキュベートして、cDNA上のXhoアダプターをリン酸化した。その後、0.3μl(120単位)のT4 DNAリガーゼ(New England Biolabs)を加えてcDNAをベクターと結合させ、混合液を15℃にて一晩インキュベートした。バクテリオファージλgtGreen1Sはλgt11からの派生物であり(非特許文献31)、これでは、(a)野生型バクテリオファージλゲノムのSalI−Xhoフラグメント架橋座標33245−33449が除去され、(b)E.coli lacZ遺伝子を含有する独特のSacI−EcoRIフラグメントが除去されて、直線型のpGreen1S(+)Lox2(Xba)プラスミドに置き換えられている(図1A)。pGreen1S(+)Lox2(Xba)はpBluescript SK(−)の派生物であり(Stratagene、カリフォルニア州ラホヤ)、これでは、(a)f1の起点とアンピシリン耐性遺伝子とのあいだにpZL1のincA遺伝子(Invitrogen Corporation)が挿入され、(b)colE1の起点が除去されて、pACYC184の低コピー数のp15Aの起点に置き換えられ(ATCC数は37033、American Type Culture Collection、バージニア州マナッサス)、(c)タンデム反復配列をしている二つのloxP部位(非特許文献32)が独特のMfeIにより分離されて、PmeI切断酵素認識部位がp15Aの起点とアンピシリン耐性遺伝子とのあいだに挿入されている。MfeIおよびPmeIによるpGreen1S(+)Lox2(Xba)の直線化により、λgtGreen1Sに挿入される形のプラスミドが産生される(図1B)。製造業者によって記述されているように、結合反応の一部は、Gigapack III Goldパッキングエキス(Stratagene)を用いて、in vitroでファージにパッケージした。組換えプラスミドの調製には、パッケージしたファージをE.coliのDH10B(Zip)(Invitrogen Corporation)株上に撒き込み、アンピシリンまたはカルベニシリンのいずれかを含有するLB平板上での増殖により、コロニーを選択した。E.coliのDH10B(Zip)株はcreリコンビナーゼを発現し、バクテリオファージλの複製を抑制するため、creを介する組換えによって、プロプラスミドに隣接するloxP部位で、ファージベクターからプラスミドを切除し、これを独自に複製するプラスミドとして確立することができる。
【0152】
【非特許文献30】CarninciおよびHayashizaki(Methods in Enzymology、303巻、19−44ページ(1999))
【非特許文献31】Young and Davis, Proc.Natl.Acad.USA,80巻、1194−1198ページ(1983)
【非特許文献32】Sternberg and Hamilton,J.Mol.Biol.,150巻、467−86ページ(1981)
【0153】
実施例2−オオタバコガ(「TBW」)電位依存性カルシウムチャネルα1サブユニット配列の識別
材料および方法
TBW頭部ポリA RNAの分離。0.1%のジエチルピロカルボネート水溶液(「DEPC」;Aldrich Chemical Co.,Inc.、ウィスコンシン州ミルウォーキー)を37℃にて約16時間インキュベートした後、オートクレーブで60分間処理した。ガラス器具はすべて、250℃で4時間加熱し、ボトルの蓋はすべて、0.1%のDEPC溶液で浸漬した。Braunホモジナイザーのマイクロプローブ(B.Braun Biotech International、ペンシルバニア州アレンタウン)は50mlの100%エタノール(「EtOH」)に浸漬した後、25mlのRNAzolB(CINNA−BIOTECX Labs,Inc.、テキサス州ヒューストンから入手可能な塩酸グアニジニウム標品)で稼動させた。TBWの頭部は、第4虫齢のオオタバコガ幼虫から切断採取し、氷浴上の風袋を測定した遠心管中に入れた。約0.5gのTBW頭部材料を入手した後、TBW頭部は使用時まで−70℃にて凍結した。その後、頭部を4mlのTRIZOLTM試薬に入れ、室温にて最大速度で30秒間ホモジナイズした。ホモジナイズが終了した時点で、2mlのクロロホルムを加え、生じた混合液をSS34ローター(Sorvall Products,L.P.、ノースカロライナ州アッシュビル)中で室温にて12000gで15分間遠心分離した。この手順の完了後、上部水層を15mlの新しいコニカルチューブに入れ、等量のイソプロパノールを加えた。生じた混合液をSS34ローター中で室温にて12000gで10分間遠心分離した。その後、生じた沈塊を10mlの75%エタノール水溶液で洗浄し、SS34ローター中で室温にて7500gで5分間遠心分離した。上清を流去し、生じた沈殿塊を室温で5分間、風乾させた。その後、生じた沈殿塊を、1mlの上述の0.1%DEPC溶液に溶解した。全RNAの濃度は、UV分光法で測定した(Amersham BiosciencesのGeneQuant、ニュージャージー州Piscataway)。500μgの全RNAを用い、Micro−FastTrackTM 2.0キット(Invitrogen Corporation)のプロトコルに従って、ポリA mRNAを分離した。ポリA mRNAの濃度は、UV分光法により測定した。ポリA mRNA溶液は、その後の利用のために、−70℃で保存した。
【0154】
ファーストストランドcDNAの合成。逆転写は、1.0μgのテンプレートRNAに鳥類骨髄芽球症ウイルス(「AMV」)逆転写酵素(Invitrogen Corporation)を加えることによって開始した。逆転写反応には、Invitrogen cDNA Cycleキットに含まれている以下の試薬も関与した:4μlの5X逆転写緩衝液、1μlのリボヌクレアーゼインヒビター、1μlの100mM dNTP、1μlの80mMリン酸ナトリウムおよび、ランダムプライマー、オリゴ(dT)プライマーまたは遺伝子特異的プライマーのうちのいずれか。5’および3’でのRACE反応では、製造業者の指示に従い、オリゴ(dT)プライマーまたはランダムプライマーまたは遺伝子特異的プライマーにより、SMARTTM RACE cDNA増幅キット(BD Biosciences Clontech、カリフォルニア州パロアルト)を用いて、mRNAの逆転写を行った。反応は、GeneAmp(登録商標)9700サーマルサイクラー(Applied Biosystems、カリフォルニア州フォスターシティー)上で行い、42℃にて60分間維持した。その後、95℃にて5分間加熱した後に4℃で5分間放置することによって、AMV RTを不活化した。Ambion(テキサス州オースチン)FirstChoiceTM RLM−RACEキットを用いて、製造業者の指示に従って、5’−RACE反応を行った。
【0155】
PCRによる増幅。製造業者の指示に従い、Advantage 2 PCRキット(BD Biosciences Clontech)で供給された緩衝液とdNTPを用いて、50μlのcDNA反応液を作製した。変性プライマーを利用した増幅では通常、50〜60℃の範囲のアニーリング温度を利用し、アイソフォーム特異的プライマーを用いるものでは65〜70℃の範囲のアニーリング温度を利用した。3’および5’RACE反応は、SMARTTM RACE cDNA Amplificationキット(BD Biosciences Clontech)により供給されたプライマーとプロトコルを利用して、製造業者の指示に従って行った。生じたPCR産物は、アガロースゲル電気泳動法により性質検討した。上述の方法で二次的な「ネステッド」増幅を行った後、NuSieve(登録商標)ゲル(FMC Corp.、ペンシルバニア州フィラデルフィア)からバンドを切り出し、Quiagenゲル抽出キットを用いて、製造業者の指示に従って精製した。
【0156】
プライマーの合成およびデザイン。オリゴヌクレオチドは、Seq Wright Inc.(テキサス州ヒューストン)が合成し、使用前に蒸留水で溶解する凍結乾燥品として提供された。PCRプライマーおよびプローブがデザインされ、InforMax, Inc.(メリーランド州ベセズダ)からのVectorNTI suite 6.0ソフトウェアと、Roseら(Nucleic Acids Research、26巻、1628−1635ページ(1998))からのConsensus−Degenerate Hybrid Oligonucleotide Primers Software(「CODEHOP」)を用いて、アニーリング温度が推定された。
【0157】
サブクローニングおよび配列決定。精製PCRフラグメントは、TOPO TA Cloningキット(Invitrogen Corp.)を用いて、製造業者の指示に従って、pCR2.1−TOPOベクターにサブクローンされた。生じたpCR2.1−TOPOベクターの配列を決定し、その後、VectorNTI suite6.0ソフトウェアを用いて解析された。
【0158】
プライマー。使用したプライマーは、表IIに示すようなものであった:
【0159】
【表3】
【0160】
【表4】
【0161】
【表5】
【0162】
上述のオリゴヌクレオチドの中で、変性プライマーは、N(A、G、CまたはT)、H(A、CまたはT)、S(CまたはG)、Y(CまたはT)、W(AまたはT)、D(A、GまたはT)またはR(AまたはG)の記号の入った位置に、単量体の統計学的混合物を組み込んでいる[IUPAC総会と一致]。
【0163】
オオタバコガ(Heliothis virescens)のα1サブユニット配列の増幅
オオタバコガ(Heliothis virescens)のα1サブユニットについての増幅の記述はいずれも、配列番号1で示された配列位置を指定している。
【0164】
RT−PCR反応では、プライマー1および2は、オオタバコガ(Heliothis virescens)の配列での3674位から4113位のヌクレオチドからのフラグメントを増幅するために使用され、これについてのクローン化およびサブクローン化が行われた。
【0165】
Heliothisの配列は、標準化していないTBW cDNAライブラリプラスミドDNAを用いたPCRにより(実施例1)、プライマー3および5を用いて、上流方向に伸展され、その後、プライマー4および5を用いてネステッドPCRが行われた。生じたPCR産物からは、2621位から3770位のヌクレオチドからの配列情報が得られ、これについてのクローン化およびサブクローン化が行われた。プライマー5は、ライブラリベクター配列からのSP6である。
Heliothisの配列は、標準化していないTBW cDNAライブラリプラスミドDNAを用いたPCRにより(実施例1)、プライマー6および8を用いて、下流方向に伸展され、その後、プライマー7および8を用いてネステッドPCRが行われた。生じたPCR産物からは、4029位から5047位のヌクレオチドからの配列情報が得られ、これについてのクローン化およびサブクローン化が行われた。プライマー8は、ライブラリベクター配列からのT7である。
【0166】
プライマー9および11を用いての3’−RACEによる3’−末端cDNAの回復と、プライマー10および12を用いてのその後のネステッドPCRの試みでは、驚くべきことに、生じたPCR産物から、3’−末端cDNA配列の代わりに、1589位から2149位のヌクレオチドからの配列情報が得られた。このPCR産物のクローン化およびサブクローン化が行われた。その後、2149位から2621位のヌクレオチドからのHeliothis配列が、SMARTTM RACE cDNA増幅キット(BD Biosciences Clontech)からのランダム六量体を用いた逆転写により得られ、ついで、プライマー13および15を用いてタッチダウンPCRが行われた。その後、生じたcDNAは、ネステッドプライマー14および16を用いて増幅され、2056位から2685位のヌクレオチドからのアンプリマーが得られ、これについてのクローン化およびサブクローン化が行われた。
【0167】
Heliothisの配列は、さらに、SMARTTM RACE cDNA増幅キットからのランダム六量体を用いた逆転写によって上流方向に伸展され、ついで、プライマー17および18を用いたタッチダウンPCRが行われた。その後、生じたcDNAは、ネステッドプライマー17および19を用いて増幅された。生じたPCR産物からは、524位から1645位のヌクレオチドからの配列情報が得られ、これについてのクローン化およびサブクローン化が行われた。
Heliothisの配列は、さらに、SMARTTM RACE cDNA増幅キットからのランダム六量体を用いた逆転写によって上流方向に伸展され、ついで、プライマー11および20を用いたタッチダウンPCRが行われた。ついで、この反応で生じたcDNAを、ネステッドプライマー12および21を用いて増幅した。生じたPCR産物からは、181位から613位のヌクレオチドからの配列情報が得られ、これについてのクローン化およびサブクローン化が行われた。
【0168】
その後、FirstChoiceTM RLM−RACEキット(Ambion)からのランダム六量体を用いて、逆転写によりHeliothisの配列の5’−末端cDNAが回復され、プライマー22および24を用いてタッチダウンPCRが行われた。この反応によって生じたcDNAは、ネステッドプライマー23および25を用いて増幅した。生じたPCR産物からは、1位から252位のヌクレオチドからの配列情報が得られ、これについてのクローン化およびサブクローン化が行われた。
【0169】
全長キメラα1サブユニットの構成
重複しているDNAセグメントで制限酵素認識部位を介して上述PCRフラグメントをつなぎあわせることにより、配列番号1についての全長クローンが構成された。翻訳停止コード化ンを加えるために、結合されたクローンをNotIで切断し、その後、ワタアブラムシライブラリ(実施例1)から得られた配列番号3に列挙されている1位から429位のヌクレオチドからの停止コード化ンを有するアブラムシα1遺伝子の3’−末端を融合させて、発現についての全長キメラクローンを作製した。キメラDNAおよび蛋白質配列は、配列番号5および6のように示されている。全長キメラα1サブユニットは、発現ベクターpCDNA3.1(Invitrogen Corp)に挿入された。
【0170】
実施例3−オオタバコガ(「TBW」)電位依存性カルシウムチャネルβサブユニット配列の識別
材料および方法
実施例2と同様。
【0171】
プライマー。使用したプライマーは、表IIIに示すようなものであった:
【0172】
【表6】
【0173】
上述のオリゴヌクレオチドの中で、変性プライマーは、N(A、G、CまたはT)、H(A、CまたはT)、S(CまたはG)、Y(CまたはT)、W(AまたはT)、D(A、GまたはT)またはR(AまたはG)の記号の入った位置に、単量体の統計学的混合物を組み込んでいる[IUPAC総会と一致]。
【0174】
オオタバコガ(Heliothis virescens)のβサブユニット配列の増幅
オオタバコガ(Heliothis virescens)についての増幅の記述はいずれも、配列番号7で示された配列位置を指定している。
【0175】
RT−PCR反応では、プライマー1および2は、オオタバコガ(Heliothis virescens)の配列での874位から1140位のヌクレオチドからのフラグメントを増幅するために使用され、これについてのクローン化およびサブクローン化が行われた。
【0176】
このフラグメントから、プライマー3および4ならびに、RecActiveTM Gene Enrichmentキット(Active Motif、カリフォルニア州カールズバッド)からのビオチン−dNTP混合物を含有するPCR反応によって、ビオチル化プローブ(900位から1113位のヌクレオチドより)が合成された。このビオチル化プローブはその後、ゲル精製し、標準化されていないTBW cDNAライブラリ(実施例1)からrecAを介する遺伝子選択により、Active Motif社(カリフォルニア州カールズバッド)のRecActiveTM Gene Enrichmentキットにより、全長TBW βサブユニットcDNAをクローンするために利用された。3種類の全長クローンの配列が決定され、これは、配列番号7、8および9で示されている。配列番号7、8および9からそれぞれ翻訳された蛋白質配列に対応する配列番号10、11および12に示すように、複数のアイソフォームが認められた。これらのcDNAは最初に発現ベクターpCMV−SPORT6.1(Invitrogen Corp.)でクローン化され、発現ベクターへの追加サブクローニングを必要としなかった。
【0177】
実施例4−ワタアブラムシ電位依存性カルシウムチャネルα2δサブユニットの配列の識別
【0178】
【表7】
【0179】
蛋白質配列CG12295−PBを質問内容として用いて、ワタアブラムシコンティグデータベースのTBLASTN検索を実施することにより(実施例1)、ワタアブラムシカルシウムチャネルα2δの配列が識別された。CG12295−PBは、遺伝子CG12295について予測された翻訳産物であり、Drosophila melanogasterゲノムで識別された3種類のカルシウムチャネルα2δ遺伝子のうちの一つである。ワタアブラムシα2δコンティグそれぞれを用いたDrosophila蛋白質データベースのバージョン3のBLASTXによる相互検索から、明らかにCG12295のワタアブラムシオルソログに属するコンティグの一つ(CAg12001、1489bp)が識別された。
【0180】
コンティグCAg12001に対応する全長cDNAが、RecActiveTMキットおよびActive Motif社(カリフォルニア州カールズバッド)からのプロトコルを用いて、RecAを介する遺伝子増強により、ワタアブラムシcDNAライブラリから分離された。要約すると、約106個の一次トランスフォーマントからなる一定量のワタアブラムシcDNAライブラリから、プラスミドDNAが調製された。表示時の偏りは、50μg/mlのカルベニシリンを添加した1LのTerrific Brothで細胞を30℃にて16時間増殖させることにより、最小限にした。プローブ合成のテンプレートは、CAg12001特異的なプライマーであるAph040984U531およびAph040984L959(表IV)を用いたPCRにより、ライブラリDNAから合成し、Oligo 6プライマー解析ソフトウェア(Molecular Biology Insights,Inc.,コロラド州カスケード)を用いて命名した。PCR反応では、最終容積15μl中に、100ngのライブラリDNA、5pmolのAph040984U531およびAph040984L959プライマー、0.3μlの10mM dNTP混合液(それぞれ10mMのdATP,dGTP,dCTPおよびTTP)、1.5μlの10×TitaniumTM Taq緩衝液、0.3μlのTitaniumTM Taq DNAポリメラーゼ(BD Biosciences Clontech、カリフォルニア州パロアルト)が含まれていた。反応では、94℃での2分間の加熱の後に、94℃での30秒間の加熱変性、55℃での30秒間のアニーリングおよび、72℃での1.5分間のプライマー伸展を1サイクルとして、35サイクルのDNA合成を行った。反応産物は、低融点アガロースゲル(SealPlaque(登録商標)GTG(登録商標)アガロース、BioWhittaker Molecular Applications、メイン州ロックランド)上で分離し、ゲルから450bpと予測されるバンドを抽出して、PicoGreen(登録商標)dsDNA定量試薬(Molecular Probes、オレゴン州ユージーン)による蛍光染色により定量した。精製フラグメントをテンプレートとして用いて、100pgのテンプレートフラグメント、10pmolのAph040984U531およびAph040984L959プライマー、7μlのビオチン−dNTP混合液(Active Motif)、5μlの10×TitaniumTM Taq緩衝液および1μlのTitaniumTM Taq DNAポリメラーゼを含有する50μlのPCR反応液中で、ビオチル化DNAの合成を行った。ビオチル化プローブは、1%の低融点アガロースゲル上で精製し、PicoGreen(登録商標)による蛍光染色によって定量した。ついで、50ngのプローブフラグメントを用いて、RecActiveTMキットにより提供されたプロトコルを利用して、5μgのワタアブラムシライブラリDNAからα2δクローンを選択した。最初の選択により約105個のコロニーがプールされ、プラスミドDNAが調製された。プールDNAの定量的PCR分析から、標的とするα2δ配列が開始時のライブラリと比較して約60倍に増大していることが示された。単独コロニースクリーニングで十分な量を得るために、50ngのビオチル化プローブを用いて、最初に得られたプラスミドDNAのうちの3μgを再度選択した。2回目の選択で得られたDNAを用いてE.coli DH10B細胞をトランスフォームした後、ビオチル化プローブ合成で使用した領域に隣接するCAg12001特異的プライマー(Aph040984U451およびAph040984L1104プライマー、)を用いたPCRにより、個々のコロニーのスクリーニングを行った。2回の選択の後、スクリーニングされたコロニー36個のうち34個がCAg12001配列に対して陽性だった。各々の陽性コロニーからプラスミドDNAを調製し、HindIIIおよびKpnIによる切断によって、cDNAインサートの大きさを決定した。一つのクローン(RA1.11)からのcDNAインサートは、完全に配列決定されている。RA1.11は、4383bpのインサート(配列番号13)を有しており、1245個のアミノ酸オープンリーディングフレームを含み(配列番号14)、これは、CG12295−PBと、1239残基にわたって41%のアミノ酸配列を共有していた。さらに別の15個の全長クローンについての制限酵素解析および部分配列決定から、少なくともcDNAの二つの異なる領域が関与した複雑な別のスプライシングのパターンが示された。別のスプライシングのパターンの一つでは、配列番号14の805−815位の残基によって示される11個のアミノ酸セグメントでの相違の存在が関与している。このセグメントは、Drosophila CG12295−PB α2δ蛋白質配列には存在しないが、配列番号16および18に示されるTBW α2δ蛋白質配列では、同一の位置に局在しており、存在状態の異なる11個のアミノ酸配列で部分的な相同性を有している(実施例5など)。
【0181】
実施例5−オオタバコガ(「TBW」)電位依存性カルシウムチャネルα2δサブユニット配列の識別
【0182】
【表8】
【0183】
最初に、TBW頭部cDNAライブラリ(実施例1)から、以下の3種類の昆虫α2δオルソログで保存された蛋白質配列ブロックよりデザインされたコンセンサス変性ハイブリッドオリゴヌクレオチドプライマー(CODEHOPプライマー)を用いたPCRにより、TBW α2δ CG12295オルソログの5’末端から得られたフラグメントを合成した:Drosophila melanogasterからのCG12295−PB、Anopheles gambiaeからのENSANGP00000021218および、ワタアブラムシ(Aphis gossypii)からのRA1.11(配列番号14、実施例4)。Block Maker(Henikoff. and Henikoff,Nucleic Acid Research、19巻、6565−6572ページ(1991))を用いてこれら3種類の配列で保存された蛋白質ブロックのデータベースを構築し、CODEHOPプライマーデザインプログラムの入力値として使用した(Roseら、Nucleic Acid Research、26巻、1628−1635ページ(1998))。変性正方向プライマーのHvCaA2D1F(表V)は、配列番号81の5’側でもっとも保存されている蛋白質ブロックのアミノ酸配列VQTWAEKLから得られ、一方、変性逆方向プライマーのHvCaA2D1R(表V)は、配列番号82でN末端から蛋白質側方向へのおおむね1/3の位置に局在する保存配列CNQAIMIVSの相補鎖から得られた。TBW頭部cDNAライブラリ(実施例1)から分離された100ngのプラスミドDNAを用いてタッチダウンPCRを行い、アガロースゲル電気泳動により900bpのフラグメントを分離して、pBluescript SK(−)(Stratagene)にクローン化した。DNA配列解析から、バンドはDrosophila遺伝子CG12295のTBWオルソログに由来するものであることが確認された。この900bpのフラグメントの配列から、2種類の非変性遺伝子特異的プライマー(HvCaA2DU388およびHvCaA2DL775、表V)をデザインし、PCR反応のアニーリング温度を55℃ではなく62℃とした以外は、実施例4に記載されているのと同様にして、このプライマーを用いてビオチン化プローブを調製した。このビオチン化プローブは、TBW神経索ライブラリからの5μgのプラスミドDNAから開始した、2回のrecAを介した遺伝子増幅(実施例4など)で用いた。2回目の選択後にPCRの検討対象となった24個のコロニーのうち3個は、TBW α2δ配列について陽性であった。これら3つのプラスミドはいずれも、〜5kbのインサートを含有していた。クローンのうちの二つ(クローン2およびクローン7)は、全体についての配列決定が行われた。クローン2(5028bp、配列番号15)は、1271個のアミノ酸(配列番号16)によるオープンリーディングフレームを有しており、クローン7(4892bp、配列番号17)は、1258個のアミノ酸(配列番号18)によるオープンリーディングフレーム蛋白質を有していた。クローン2に存在する5’UTRセグメントは、クローン7で認められたものに比べて98bp長かったが、二つの5’UTR配列はその他の点では同一である。しかし、二つのcDNAのコード化領域で69個の単独ヌクレオチドに多型性が存在し、3’UTRには少数の挿入/欠失が存在することから、二つのクローンは、ライブラリの構築に使用した非近交系TBW集団で、異なる対立遺伝子に由来すると思われる。さらに、クローン2には、クローン7と比較した場合、コード化領域に以下の二つの小さな挿入を含有している:490−410位の残基でのアミノ酸2個の挿入(VK)および、842−852位の残基でのアミノ酸11個の挿入(PLTKVIGLLPR、配列番号83)。コード化領域には多数の単独ヌクレオチドの多型性が含まれているが、蛋白質配列の相違を引き起こすのはこの二つのみであり、どちらの変化も、保存的なアミノ酸置換となっている。
【0184】
実施例6−アフリカツメガエル卵母細胞でのオオタバコガ(「TBW」)電位依存性カルシウムチャネルの機能性の発現
材料および方法
ポリ(A)末端を有するキャップ構造mRNAの構築。pCDNA3.1でのキメラα1サブユニットは、NsiIにより直線状にした;pCMV−SPORT6.1 1でのTBW βサブユニットは、DrdIにより直線状にした;pCMV−SPORT6.1のアブラムシα2δは、NotIにより直線状にした;pGreen1S(+)Lox2(Xba)のTBW□2□は、NheIにより直線状にした。ついで、直線状にしたDNAを、フェノール−クロロホルム抽出と、エタノール沈降により精製した。このDNAを用い、Ambion社からのmMESSAGE mMACHINETM高収量キャップ構造mRNA転写キットのマニュアルに従って、キャップ構造mRNAを作製した。転写反応の後、試料をDNaseIで処理して、テンプレートDNAを除去し、その後、Ambion社のポリ(A)ポリメラーゼを用いて、RNAにポリ(A)末端を付加した。ポリ(A)末端を有するキャップ構造mRNAには、フェノール−クロロホルム抽出と、エタノール沈降を行った。ポリ(A)末端を有するキャップ構造mRNAの濃度は、UV分光法により測定した。mRNA溶液は、その後の利用のために、−20℃で保存した。
【0185】
卵母細胞発現およびパッチクランプ法:アフリカツメガエル卵母細胞遺伝子発現で使用した方法は、技術で十分に確立されているものである(非特許文献33)および(非特許文献34)。この手法を以下に要約する。0.15%のTricaine(3−アミノ安息香酸エチルエステル)に沈めて麻酔したアフリカツメガエル卵母細胞(Xenopus laevis)(NASCO、ウィスコンシン州Fort Atkinson)から、未受精の卵母細胞を外科的に摘出した。卵母細胞は、カルシウムを含まない生理食塩液(OR2:82.5mMのNaCl,2mMのKCl,1mMのMgCl2,5mMのHEPES,pH7.4)で洗浄し、その後、4mg/mlのコラゲナーゼを含有するOR2中で、オービタルシェイカー上にて17℃で2時間インキュベートした。ついで、ピンセットを用いて卵母細胞に機械的な濾胞除去を行った後、通常のカルシウム含有生理食塩液(ND96:96mMのNaCl,2mMのKCl,1mMのMgCl2,1.8mMのCaCl2,5mMのHEPES,pH7.4)に入れた。ピストン式のマイクロインジェクター(Drummond Nanoject,Drummond Scientific Company、ペンシルバニア州Broomall)を用いて、希望する遺伝子または遺伝子群からのRNA転写物を、各々の卵母細胞にインジェクションした。通常の実験では、4〜6ngの各々のサブユニットのRNAを、23nLの水に入れて導入した。インジェクションを受けた卵母細胞を、オービタルシェイカー上にて17℃でインキュベートし、2〜4日でインジェクションしたチャネルの機能的な発現を検討した。
【0186】
【非特許文献33】Methods in Enzymology、207巻、Rudy,B.
【非特許文献34】Iverson,L.E編著、セクションII、A.Expression of Ion channels in Xenopus oocytes、225−390ページ)
【0187】
卵母細胞は、二電極電圧固定装置(AxoClamp 2B,Axon Instrument、カリフォルニア州ユニオンシティー)を用いて、イオンチャネルの機能的な発現について検討した。標準的な電圧固定法での電流および記録法を用いた。記録電極は、Narishigeピペットプラー(PP830型、Narishige International USA、ニューヨーク州ロングアイランド)を用いて組み立て、4mMの酢酸カリウムを充填した場合の入力抵抗が0.8〜3メガオームとなるように調節した。データは、アナログ−デジタルインターフェース(Digidata 1322,Axon Instruments)を備えたPentium(登録商標)クラスのコンピュータを用いて、pClamp8ソフトウェア(Axon Instruments)により統合した。カルシウムチャネルの性能を評価するための記録は、内因性のカルシウム活性化電流を最小限にするために処方されたNS生理食塩液中(40mMのBa(OH)2,50mMのD−グルコン酸ナトリウム、1mMのKOH,0.1mMのEDTA,10mMのHEPES,0.1mMのニフルム酸、pH7.4)で行った。
【0188】
結果:3種類の無脊椎動物カルシウムチャネルサブユニットをさまざまな形で組み合わせたものについて検討を行った。配列番号6で示されるα1サブユニット、配列番号10で示されるβ−2サブユニット、配列番号18で示されるα2δサブユニットは、オオタバコガ(Heliothis virescens)に由来するものであった。ワタアブラムシからの配列番号14で示されるα2δサブユニットについても、このサブユニットが天然のH.virescens α2δと置換できるか否かを調べるために、検討を行った。
【0189】
各々のサブユニットのRNAを、6ng/卵の用量でインジェクションした。各々のバッチの回収卵からの卵にインジェクションした対照電位依存性チャネルは、大量の電流を生じたという事実(データ未発表)にもかかわらず、α1サブユニット単独では電位依存性の電流を得ることはできなかった。α1サブユニットとβ−2サブユニットが共に発現した場合には、図2に示すように通常は10〜20nAの範囲で、中程度の電流を生じた。しかし、電流の発現は不安定であり、卵ごとに一定していなかった。α1サブユニットおよびβ−2サブユニットにアブラムシα2δサブユニットを加えると、わずかではあるが明確な、電位感受性N型カルシウム電流が生じた(図3)。アブラムシα2δをTBW α2δ−2サブユニットに置換した場合にも、明確な電位感受性N型カルシウム電流が生じた(図4)が、観察された最大電流の大きさは、40nA程度の大きさまで増大した。
【0190】
上述の方法で集められたデータから、固定電圧の場合に電位段階に応じて発生した電流は、培地で増殖させたニューロンから生ずる、全細胞パッチクランプ記録で観察されるN型カルシウムチャネルに類似していると結論付けられる(非特許文献35)。試験条件下でのクローン化されたチャネルと、培養ニューロンで観察されたものはどちらも、−30mV付近で活性化ポイントを示し、見かけ上の逆転電位は+50mV付近である。卵母細胞で観察された電流は減少性であるが、その動態は、培養昆虫ニューロンで観察される、緩慢に不活化する電流を暗示するものである。さらに、配列番号6で示されるキメラα1サブユニットは、適切な昆虫βおよびα2δカルシウムチャネルサブユニットの存在下でN型カルシウムチャネルを暗示する、機能性電位感受性チャネルを形成することができると結論付けられる。
【0191】
【非特許文献35】Hayashi and Levine, Exp.Biol.、171巻、15−42ページ(1992)
【図面の簡単な説明】
【0192】
【図1.A】コピー数の少ないプラスミドベクターpGreen1S(+)Lox2(Xba)を図示したものである。複数のクローニング部位は、KpnIおよびSacI制限酵素切断部位の間の小さな領域に局在しており、T3およびT7 RNAポリメラーゼに対するプロモーターによって隣接している。p15Aの矢印は、DNA複製でコピー数の少ないp15Aプラスミドの起点部の位置を示している。loxPの矢印は、Creリコンビナーゼによる部位特異的組換えで使用される34bpの反復配列の部位を示している。AmpRの矢印は、抗生物質のアンピシリンおよびカルベニシリンに対する耐性を付与するβ−ラクタマーゼ遺伝子の位置および方向を示している。incAは、その存在によってE.coliのDH10B(ZIP)株でのCreリコンビナーゼを発現するP1プラスミドの反復を遮断するDNAセグメントの位置を示している。AmpRの矢印は、抗生物質のアンピシリンおよびカルベニシリンに対する耐性を付与するβ−ラクタマーゼ遺伝子の位置および方向を示している。incAは、その存在によってE.coliのDH10B(ZIP)株でのCreリコンビナーゼを発現するP1プラスミドの反復を遮断するDNAセグメントの位置を示している。f1(+)originの矢印は、繊維状ファージf1のローリングサイクル複製の起点の位置と方向を示している。図1A:バクテリオファージλgt11の誘導体に挿入を行う前のプラスミドの環状構造を示す。pGreen1S(+)Lox2(Xba)は、制限酵素MfeIおよびPmeIによって切断され、バクテリオファージλgt11の誘導体の(破壊された)SacI部位とEco RI部位のあいだに巨大な直線フラグメントが挿入された。λgtGreen1SのXhoIおよびSacIによる切断により、ファージに二つのアームが生ずるが、cDNAライブラリの構成ではこれらを利用することが可能である。
【図1.B】コピー数の少ないプラスミドベクターpGreen1S(+)Lox2(Xba)を図示したものである。複数のクローニング部位は、KpnIおよびSacI制限酵素切断部位の間の小さな領域に局在しており、T3およびT7 RNAポリメラーゼに対するプロモーターによって隣接している。p15Aの矢印は、DNA複製でコピー数の少ないp15Aプラスミドの起点部の位置を示している。loxPの矢印は、Creリコンビナーゼによる部位特異的組換えで使用される34bpの反復配列の部位を示している。AmpRの矢印は、抗生物質のアンピシリンおよびカルベニシリンに対する耐性を付与するβ−ラクタマーゼ遺伝子の位置および方向を示している。incAは、その存在によってE.coliのDH10B(ZIP)株でのCreリコンビナーゼを発現するP1プラスミドの反復を遮断するDNAセグメントの位置を示している。f1(+)originの矢印は、繊維状ファージf1のローリングサイクル複製の起点の位置と方向を示している。図1B:バクテリオファージクローニングベクターλgtGreen1Sに存在する形での、直線型のプラスミドの構造を示す。pGreen1S(+)Lox2(Xba)は、制限酵素MfeIおよびPmeIによって切断され、バクテリオファージλgt11の誘導体の(破壊された)SacI部位とEco RI部位のあいだに巨大な直線フラグメントが挿入された。λgtGreen1SのXhoIおよびSacIによる切断により、ファージに二つのアームが生ずるが、cDNAライブラリの構成ではこれらを利用することが可能である。
【図2】昆虫α1サブユニットとTBW β−2サブユニットを注入したアフリカツメガエル(Xenopus)卵母細胞での電位ステップに応じて生じた電流を示す。最大電流と電位との間の相関関係図である。最大電流は、マーカー1とマーカー2(逆三角形)とのあいだで決定された。
【図3】昆虫α1サブユニット、TBW β−2サブユニットおよびワタアブラムシα2δサブユニットを注入したアフリカツメガエル(Xenopus)卵母細胞での電位ステップに応じて生じた電流を示す。最大電流と電位との間の相関関係図である。最大電流は、マーカー1とマーカー2(逆三角形)とのあいだで決定された。
【図4】昆虫α1サブユニット、TBW β−2サブユニットおよびTBW α2δ−2サブユニットを注入したアフリカツメガエル(Xenopus)卵母細胞での電位ステップに応じて生じた電流を示す。最大電流と電位との間の相関関係図である。最大電流は、マーカー1とマーカー2(逆三角形)とのあいだで決定された。
【配列表】
【特許請求の範囲】
【請求項1】
配列番号2、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16および配列番号18、その突然変異体、およびそのフラグメントからなるグループから選択されたアミノ酸配列を有する、実質的に純粋な蛋白質。
【請求項2】
前記蛋白質が、配列番号2、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16または配列番号18のアミノ酸配列を有する、請求項1記載の蛋白質。
【請求項3】
請求項1の蛋白質をコード化する核酸配列から構成される、分離された核酸分子。
【請求項4】
請求項3の核酸分子を含む、組換え発現ベクター。
【請求項5】
請求項4の組換え発現ベクターを含む、宿主細胞。
【請求項6】
請求項2の蛋白質をコード化する核酸配列を含む、分離された核酸分子。
【請求項7】
請求項6の核酸分子を含む、組換え発現ベクター。
【請求項8】
請求項7の組換え発現ベクターを含む、宿主細胞。
【請求項9】
配列番号1、少なくとも10個のヌクレオチドを有するそのフラグメント、配列番号5、少なくとも10個のヌクレオチドを有するそのフラグメント、配列番号7、少なくとも10個のヌクレオチドを有するそのフラグメント、配列番号9、少なくとも10個のヌクレオチドを有するそのフラグメント、配列番号11、少なくとも10個のヌクレオチドを有するそのフラグメント、配列番号13、少なくとも10個のヌクレオチドを有するそのフラグメント、配列番号15、少なくとも10個のヌクレオチドを有するそのフラグメント、配列番号17、および、少なくとも10個のヌクレオチドを有するそのフラグメントからなる、分離された核酸分子であって、配列番号5のフラグメントは、配列番号1のヌクレオチドを少なくとも5個含んでいる、核酸分子。
【請求項10】
配列番号1、配列番号5、配列番号7、配列番号9、配列番号11、配列番号13、配列番号15および配列番号17からなる、請求項9記載の核酸分子。
【請求項11】
請求項10の核酸分子を含む組換え発現ベクター。
【請求項12】
請求項11の組換え発現ベクターを含む、宿主細胞。
【請求項13】
少なくとも10個のヌクレオチドを有する配列番号1のフラグメント、少なくとも10個のヌクレオチドを有する配列番号5のフラグメント、少なくとも10個のヌクレオチドを有する配列番号7のフラグメント、少なくとも10個のヌクレオチドを有する配列番号9のフラグメント、少なくとも10個のヌクレオチドを有する配列番号11のフラグメント、少なくとも10個のヌクレオチドを有する配列番号13のフラグメント、少なくとも10個のヌクレオチドを有する配列番号15のフラグメント、および、少なくとも10個のヌクレオチドを有する配列番号17のフラグメントからなる核酸分子であって、配列番号5のフラグメントは、配列番号1のヌクレオチドを少なくとも5個含んでいる、請求項9記載の核酸分子。
【請求項14】
12個から150個のヌクレオチドを有する配列番号1のフラグメント、12個から150個のヌクレオチドを有する配列番号5のフラグメント、12個から150個のヌクレオチドを有する配列番号7のフラグメント、12個から150個のヌクレオチドを有する配列番号9のフラグメント、12個から150個のヌクレオチドを有する配列番号11のフラグメント、12個から150個のヌクレオチドを有する配列番号13のフラグメント、12個から150個のヌクレオチドを有する配列番号15のフラグメント、および、12個から150個のヌクレオチドを有する配列番号17のフラグメントからなる核酸分子であって、配列番号5のフラグメントは、配列番号1のヌクレオチドを少なくとも5個含んでいる、請求項9記載の核酸分子。
【請求項15】
15個から150個のヌクレオチドを有する配列番号1のフラグメント、15個から150個のヌクレオチドを有する配列番号5のフラグメント、15個から150個のヌクレオチドを有する配列番号7のフラグメント、15個から150個のヌクレオチドを有する配列番号9のフラグメント、15個から150個のヌクレオチドを有する配列番号11のフラグメント、15個から150個のヌクレオチドを有する配列番号13のフラグメント、15個から150個のヌクレオチドを有する配列番号15のフラグメント、および、15個から150個のヌクレオチドを有する配列番号17のフラグメントからなる核酸分子であって、配列番号5のフラグメントは、配列番号1のヌクレオチドを少なくとも5個含んでいる、請求項9記載の核酸分子。
【請求項16】
配列番号2、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16または配列番号18のエピトープに結合する、分離された抗体であって、配列番号6に結合する抗体は、配列番号4と交差反応しない抗体。
【請求項17】
前記抗体がモノクローナル抗体である、請求項16記載の抗体。
【請求項18】
鱗翅目のカルシウムチャネル蛋白質活性のモジュレーターを識別する方法であって、
被験細胞が、キメラTBW電位依存性カルシウムチャネルα1サブユニットをコード化する核酸配列を含む組換え発現ベクター、TBW電位依存性カルシウムチャネルβサブユニットをコード化する核酸配列を含む組換え発現ベクターおよびTBW電位依存性カルシウムチャネルα2δサブユニットをコード化する核酸配列を含む組換え発現ベクターのうちの少なくとも一つを含み、前記被験細胞が機能性カルシウムチャネルを発現する場合に、機能性カルシウムチャネルを含む細胞を、被験化合物の存在下でカルシウムを含有する溶液と接触させ、前記被験細胞での細胞内カルシウムの量を検出することによって行う、被験測定法を実施することと、
前記被験細胞が、キメラTBW電位依存性カルシウムチャネルα1サブユニットをコード化する核酸配列を含む組換え発現ベクター、TBW電位依存性カルシウムチャネルβサブユニットをコード化する核酸配列を含む組換え発現ベクターおよびTBW電位依存性カルシウムチャネルα2δサブユニットをコード化する核酸配列を含む組換え発現ベクターのうちの少なくとも一つを含み、陰性細胞が機能性カルシウムチャネルを発現する場合に、機能性カルシウムチャネルを含む陰性対照細胞を、前記被験化合物の非存在下でカルシウムを含有する溶液と接触させ、前記陰性対照細胞での細胞内カルシウムの量を検出することによって行う、陰性対照測定法を実施することと、
前記被験細胞での細胞内カルシウム量を、陰性対照細胞での細胞内カルシウム量と比較することとを含み、
前記陰性対照細胞での細胞内カルシウム量を、前記被験細胞でのカルシウム量と比較して変化が認められた場合に、前記被験化合物はカルシウムチャネル活性のモジュレーターであることが示される、方法。
【請求項19】
前記被験細胞がアフリカツメガエル卵母細胞、CHO細胞またはHEK細胞である、請求項18記載の方法。
【請求項20】
前記細胞の内部の色素を細胞内カルシウムと接触させることで生ずる蛍光を測定する方法を使用することによって、前記細胞内カルシウムを検出する、請求項18記載の方法。
【請求項21】
機能性カルシウムチャネルを含む陽性対照細胞を、前記被験化合物の非存在下およびカルシウムチャネル作動物質の存在下でカルシウムを含有する溶液と接触させ、前記陽性対照細胞での細胞内カルシウム量を検出することによって、さらに陽性対照測定を実施する、請求項18記載の方法。
【請求項22】
前記カルシウムチャネルが鱗翅目のカルシウムチャネルである、請求項21記載の方法。
【請求項23】
機能性カルシウムチャネルを発現していない陰性対照細胞を、前記被験化合物の非存在下でカルシウムを含有する溶液と接触させ、前記カルシウムチャネル陰性対照細胞によって取り込まれるカルシウムの量を検出することによる、二番目のタイプの陰性対照測定法を実施すること、および/または
機能性カルシウムチャネルを発現していない陰性対照細胞を、前記被験化合物の非存在下およびカルシウムチャネル作動物質の存在下で、カルシウムを含有する溶液と接触させ、前記カルシウムチャネル陰性対照細胞によって取り込まれるカルシウムの量を検出することによる、三番目のタイプの陰性対照測定法を実施することを含む、請求項18記載の方法。
【請求項24】
前記カルシウムチャネルが鱗翅目のカルシウムチャネルである、請求項23記載の方法。
【請求項25】
前記機能性カルシウムチャネルが、キメラTBW電位依存性カルシウムチャネルα1サブユニット、TBW電位依存性カルシウムチャネルβサブユニットおよびTBW電位依存性カルシウムチャネルα2δサブユニットのうち少なくとも一つを含み、前記キメラTBW電位依存性カルシウムチャネルα1サブユニットは、配列番号2、その突然変異体、そのフラグメントからなり、前記TBW電位依存性カルシウムチャネルβサブユニットは配列番号8、配列番号10、配列番号12、その突然変異体およびそのフラグメントからなり、前記TBW電位依存性カルシウムチャネルα2δサブユニットは配列番号16、配列番号18、その突然変異体およびそのフラグメントからなる、請求項18記載の方法。
【請求項26】
前記被験細胞がアフリカツメガエル卵母細胞、CHO細胞またはHEK細胞である、請求項25記載の方法。
【請求項27】
前記細胞の内部の色素を細胞内カルシウムと接触させることで生ずる蛍光を測定する方法を使用することによって、前記細胞内カルシウムを検出する、請求項25記載の方法。
【請求項28】
前記機能性カルシウムチャネルが、配列番号2からなるキメラTBW電位依存性カルシウムチャネルα1サブユニット、配列番号8、配列番号10または配列番号12からなるTBW電位依存性カルシウムチャネルβサブユニット、および、配列番号16または配列番号18からなるTBW電位依存性カルシウムチャネルα2δサブユニットを含んでいる、請求項25記載の方法。
【請求項29】
前記キメラTBW電位依存性カルシウムチャネルα1サブユニットが配列番号6を含んでいる、請求項28記載の方法。
【請求項30】
前記キメラTBW電位依存性カルシウムチャネルα1サブユニットをコード化する前記核酸配列が配列番号1からなり、前記TBW電位依存性カルシウムチャネルβサブユニットをコード化する前記核酸配列が配列番号7、配列番号9または配列番号11からなり、前記TBW電位依存性カルシウムチャネルα2δサブユニットをコード化する前記核酸配列が配列番号15または配列番号17からなる、請求項28記載の方法。
【請求項31】
前記キメラTBW電位依存性カルシウムチャネルα1サブユニットをコード化する核酸配列が配列番号5を含んでいる、請求項30記載の方法。
【請求項32】
鱗翅目のカルシウムチャネル蛋白質活性のインヒビターを識別する方法であって、
被験細胞が、キメラTBW電位依存性カルシウムチャネルα1サブユニットをコード化する核酸配列を含む組換え発現ベクター、TBW電位依存性カルシウムチャネルβサブユニットをコード化する核酸配列を含む組換え発現ベクターおよびTBW電位依存性カルシウムチャネルα2δサブユニットをコード化する核酸配列を含む組換え発現ベクターのうちの少なくとも一つを含み、前記被験細胞が機能性カルシウムチャネルを発現する場合に、機能性カルシウムチャネルを含む前記被験細胞を、被験化合物の存在下で、カルシウムおよびカルシウムチャネル作動物質を含有する溶液と接触させ、前記被験細胞での細胞内カルシウムの量を検出することによって行う、被験測定法を実施することと、
陰性対照細胞が機能性カルシウムチャネルを発現する場合に、機能性カルシウムチャネルを含む前記陰性対照細胞を、前記被験化合物の非存在下でカルシウムおよびカルシウムチャネル作動物質を含有する溶液と接触させ、前記陰性対照細胞での細胞内カルシウム量を検出することによって行う、対照測定法を実施することと、
前記被験細胞での細胞内カルシウム量を、前記対照細胞での細胞内カルシウム量と比較することとを含み、
前記被験細胞での細胞内カルシウム量に、前記対照細胞での細胞内カルシウム量と比較して減少が認められた場合に、被験化合物はカルシウムチャネル活性のインヒビターであることが示される、方法。
【請求項33】
前記被験細胞がアフリカツメガエル卵母細胞、CHO細胞またはHEK細胞である、請求項32記載の方法。
【請求項34】
前記細胞の内部の色素を細胞内カルシウムと接触させることで生ずる蛍光を測定する方法を使用することによって、前記細胞内カルシウムを検出する、請求項32記載の方法。
【請求項35】
前記機能性カルシウムチャネルが、配列番号2、その突然変異体および、そのフラグメントからなるキメラTBW電位依存性カルシウムチャネルα1サブユニットと、配列番号8、配列番号10、配列番号12、その突然変異体および、そのフラグメントからなるTBW電位依存性カルシウムチャネルβサブユニット、ならびに、配列番号16、配列番号18、その突然変異体および、そのフラグメントからなるTBW電位依存性カルシウムチャネルα2δサブユニットのうちの少なくとも一つを含んでいる、請求項38記載の方法。
【請求項36】
前記被験細胞がアフリカツメガエル卵母細胞、CHO細胞またはHEK細胞である、請求項35記載の方法。
【請求項37】
前記細胞の内部の色素を細胞内カルシウムと接触させることで生ずる蛍光を測定する方法を使用することによって、前記細胞内カルシウムを検出する、請求項35記載の方法。
【請求項38】
前記機能性カルシウムチャネルが、配列番号2からなるキメラTBW電位依存性カルシウムチャネルα1サブユニットと配列番号8、配列番号10または配列番号12からなるTBW電位依存性カルシウムチャネルβサブユニット、および、配列番号16または配列番号18からなるTBW電位依存性カルシウムチャネルα2δサブユニットを含んでいる、請求項35記載の方法。
【請求項39】
前記キメラTBW電位依存性カルシウムチャネルα1サブユニットが配列番号6を含んでいる、請求項38記載の方法。
【請求項40】
前記キメラTBW電位依存性カルシウムチャネルα1サブユニットをコード化する前記核酸配列が配列番号1からなり、前記TBW電位依存性カルシウムチャネルβサブユニットをコード化する前記核酸配列が配列番号7、配列番号9または配列番号11からなり、前記TBW電位依存性カルシウムチャネルα2δサブユニットをコード化する前記核酸配列が配列番号15または配列番号17からなる、請求項38記載の方法。
【請求項41】
配列番号2、その突然変異体、およびそのフラグメント、配列番号6、その突然変異体、およびそのフラグメント、配列番号8、その突然変異体、およびそのフラグメント、配列番号10、その突然変異体、およびそのフラグメント、配列番号12、その突然変異体、およびそのフラグメント、配列番号16、その突然変異体、およびそのフラグメント、配列番号18、その突然変異体、およびそのフラグメント;からなるグループから選択された前記アミノ酸配列を有する、分離された蛋白質の調製方法であって、請求項5の宿主細胞から前記蛋白質を分離する方法。
【請求項42】
昆虫を、請求項18に記載の方法で識別されたモジュレーターと接触させることを含む、昆虫制御法。
【請求項43】
昆虫を、請求項32に記載の方法で識別されたインヒビターと接触させることを含む、昆虫制御法。
【請求項1】
配列番号2、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16および配列番号18、その突然変異体、およびそのフラグメントからなるグループから選択されたアミノ酸配列を有する、実質的に純粋な蛋白質。
【請求項2】
前記蛋白質が、配列番号2、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16または配列番号18のアミノ酸配列を有する、請求項1記載の蛋白質。
【請求項3】
請求項1の蛋白質をコード化する核酸配列から構成される、分離された核酸分子。
【請求項4】
請求項3の核酸分子を含む、組換え発現ベクター。
【請求項5】
請求項4の組換え発現ベクターを含む、宿主細胞。
【請求項6】
請求項2の蛋白質をコード化する核酸配列を含む、分離された核酸分子。
【請求項7】
請求項6の核酸分子を含む、組換え発現ベクター。
【請求項8】
請求項7の組換え発現ベクターを含む、宿主細胞。
【請求項9】
配列番号1、少なくとも10個のヌクレオチドを有するそのフラグメント、配列番号5、少なくとも10個のヌクレオチドを有するそのフラグメント、配列番号7、少なくとも10個のヌクレオチドを有するそのフラグメント、配列番号9、少なくとも10個のヌクレオチドを有するそのフラグメント、配列番号11、少なくとも10個のヌクレオチドを有するそのフラグメント、配列番号13、少なくとも10個のヌクレオチドを有するそのフラグメント、配列番号15、少なくとも10個のヌクレオチドを有するそのフラグメント、配列番号17、および、少なくとも10個のヌクレオチドを有するそのフラグメントからなる、分離された核酸分子であって、配列番号5のフラグメントは、配列番号1のヌクレオチドを少なくとも5個含んでいる、核酸分子。
【請求項10】
配列番号1、配列番号5、配列番号7、配列番号9、配列番号11、配列番号13、配列番号15および配列番号17からなる、請求項9記載の核酸分子。
【請求項11】
請求項10の核酸分子を含む組換え発現ベクター。
【請求項12】
請求項11の組換え発現ベクターを含む、宿主細胞。
【請求項13】
少なくとも10個のヌクレオチドを有する配列番号1のフラグメント、少なくとも10個のヌクレオチドを有する配列番号5のフラグメント、少なくとも10個のヌクレオチドを有する配列番号7のフラグメント、少なくとも10個のヌクレオチドを有する配列番号9のフラグメント、少なくとも10個のヌクレオチドを有する配列番号11のフラグメント、少なくとも10個のヌクレオチドを有する配列番号13のフラグメント、少なくとも10個のヌクレオチドを有する配列番号15のフラグメント、および、少なくとも10個のヌクレオチドを有する配列番号17のフラグメントからなる核酸分子であって、配列番号5のフラグメントは、配列番号1のヌクレオチドを少なくとも5個含んでいる、請求項9記載の核酸分子。
【請求項14】
12個から150個のヌクレオチドを有する配列番号1のフラグメント、12個から150個のヌクレオチドを有する配列番号5のフラグメント、12個から150個のヌクレオチドを有する配列番号7のフラグメント、12個から150個のヌクレオチドを有する配列番号9のフラグメント、12個から150個のヌクレオチドを有する配列番号11のフラグメント、12個から150個のヌクレオチドを有する配列番号13のフラグメント、12個から150個のヌクレオチドを有する配列番号15のフラグメント、および、12個から150個のヌクレオチドを有する配列番号17のフラグメントからなる核酸分子であって、配列番号5のフラグメントは、配列番号1のヌクレオチドを少なくとも5個含んでいる、請求項9記載の核酸分子。
【請求項15】
15個から150個のヌクレオチドを有する配列番号1のフラグメント、15個から150個のヌクレオチドを有する配列番号5のフラグメント、15個から150個のヌクレオチドを有する配列番号7のフラグメント、15個から150個のヌクレオチドを有する配列番号9のフラグメント、15個から150個のヌクレオチドを有する配列番号11のフラグメント、15個から150個のヌクレオチドを有する配列番号13のフラグメント、15個から150個のヌクレオチドを有する配列番号15のフラグメント、および、15個から150個のヌクレオチドを有する配列番号17のフラグメントからなる核酸分子であって、配列番号5のフラグメントは、配列番号1のヌクレオチドを少なくとも5個含んでいる、請求項9記載の核酸分子。
【請求項16】
配列番号2、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16または配列番号18のエピトープに結合する、分離された抗体であって、配列番号6に結合する抗体は、配列番号4と交差反応しない抗体。
【請求項17】
前記抗体がモノクローナル抗体である、請求項16記載の抗体。
【請求項18】
鱗翅目のカルシウムチャネル蛋白質活性のモジュレーターを識別する方法であって、
被験細胞が、キメラTBW電位依存性カルシウムチャネルα1サブユニットをコード化する核酸配列を含む組換え発現ベクター、TBW電位依存性カルシウムチャネルβサブユニットをコード化する核酸配列を含む組換え発現ベクターおよびTBW電位依存性カルシウムチャネルα2δサブユニットをコード化する核酸配列を含む組換え発現ベクターのうちの少なくとも一つを含み、前記被験細胞が機能性カルシウムチャネルを発現する場合に、機能性カルシウムチャネルを含む細胞を、被験化合物の存在下でカルシウムを含有する溶液と接触させ、前記被験細胞での細胞内カルシウムの量を検出することによって行う、被験測定法を実施することと、
前記被験細胞が、キメラTBW電位依存性カルシウムチャネルα1サブユニットをコード化する核酸配列を含む組換え発現ベクター、TBW電位依存性カルシウムチャネルβサブユニットをコード化する核酸配列を含む組換え発現ベクターおよびTBW電位依存性カルシウムチャネルα2δサブユニットをコード化する核酸配列を含む組換え発現ベクターのうちの少なくとも一つを含み、陰性細胞が機能性カルシウムチャネルを発現する場合に、機能性カルシウムチャネルを含む陰性対照細胞を、前記被験化合物の非存在下でカルシウムを含有する溶液と接触させ、前記陰性対照細胞での細胞内カルシウムの量を検出することによって行う、陰性対照測定法を実施することと、
前記被験細胞での細胞内カルシウム量を、陰性対照細胞での細胞内カルシウム量と比較することとを含み、
前記陰性対照細胞での細胞内カルシウム量を、前記被験細胞でのカルシウム量と比較して変化が認められた場合に、前記被験化合物はカルシウムチャネル活性のモジュレーターであることが示される、方法。
【請求項19】
前記被験細胞がアフリカツメガエル卵母細胞、CHO細胞またはHEK細胞である、請求項18記載の方法。
【請求項20】
前記細胞の内部の色素を細胞内カルシウムと接触させることで生ずる蛍光を測定する方法を使用することによって、前記細胞内カルシウムを検出する、請求項18記載の方法。
【請求項21】
機能性カルシウムチャネルを含む陽性対照細胞を、前記被験化合物の非存在下およびカルシウムチャネル作動物質の存在下でカルシウムを含有する溶液と接触させ、前記陽性対照細胞での細胞内カルシウム量を検出することによって、さらに陽性対照測定を実施する、請求項18記載の方法。
【請求項22】
前記カルシウムチャネルが鱗翅目のカルシウムチャネルである、請求項21記載の方法。
【請求項23】
機能性カルシウムチャネルを発現していない陰性対照細胞を、前記被験化合物の非存在下でカルシウムを含有する溶液と接触させ、前記カルシウムチャネル陰性対照細胞によって取り込まれるカルシウムの量を検出することによる、二番目のタイプの陰性対照測定法を実施すること、および/または
機能性カルシウムチャネルを発現していない陰性対照細胞を、前記被験化合物の非存在下およびカルシウムチャネル作動物質の存在下で、カルシウムを含有する溶液と接触させ、前記カルシウムチャネル陰性対照細胞によって取り込まれるカルシウムの量を検出することによる、三番目のタイプの陰性対照測定法を実施することを含む、請求項18記載の方法。
【請求項24】
前記カルシウムチャネルが鱗翅目のカルシウムチャネルである、請求項23記載の方法。
【請求項25】
前記機能性カルシウムチャネルが、キメラTBW電位依存性カルシウムチャネルα1サブユニット、TBW電位依存性カルシウムチャネルβサブユニットおよびTBW電位依存性カルシウムチャネルα2δサブユニットのうち少なくとも一つを含み、前記キメラTBW電位依存性カルシウムチャネルα1サブユニットは、配列番号2、その突然変異体、そのフラグメントからなり、前記TBW電位依存性カルシウムチャネルβサブユニットは配列番号8、配列番号10、配列番号12、その突然変異体およびそのフラグメントからなり、前記TBW電位依存性カルシウムチャネルα2δサブユニットは配列番号16、配列番号18、その突然変異体およびそのフラグメントからなる、請求項18記載の方法。
【請求項26】
前記被験細胞がアフリカツメガエル卵母細胞、CHO細胞またはHEK細胞である、請求項25記載の方法。
【請求項27】
前記細胞の内部の色素を細胞内カルシウムと接触させることで生ずる蛍光を測定する方法を使用することによって、前記細胞内カルシウムを検出する、請求項25記載の方法。
【請求項28】
前記機能性カルシウムチャネルが、配列番号2からなるキメラTBW電位依存性カルシウムチャネルα1サブユニット、配列番号8、配列番号10または配列番号12からなるTBW電位依存性カルシウムチャネルβサブユニット、および、配列番号16または配列番号18からなるTBW電位依存性カルシウムチャネルα2δサブユニットを含んでいる、請求項25記載の方法。
【請求項29】
前記キメラTBW電位依存性カルシウムチャネルα1サブユニットが配列番号6を含んでいる、請求項28記載の方法。
【請求項30】
前記キメラTBW電位依存性カルシウムチャネルα1サブユニットをコード化する前記核酸配列が配列番号1からなり、前記TBW電位依存性カルシウムチャネルβサブユニットをコード化する前記核酸配列が配列番号7、配列番号9または配列番号11からなり、前記TBW電位依存性カルシウムチャネルα2δサブユニットをコード化する前記核酸配列が配列番号15または配列番号17からなる、請求項28記載の方法。
【請求項31】
前記キメラTBW電位依存性カルシウムチャネルα1サブユニットをコード化する核酸配列が配列番号5を含んでいる、請求項30記載の方法。
【請求項32】
鱗翅目のカルシウムチャネル蛋白質活性のインヒビターを識別する方法であって、
被験細胞が、キメラTBW電位依存性カルシウムチャネルα1サブユニットをコード化する核酸配列を含む組換え発現ベクター、TBW電位依存性カルシウムチャネルβサブユニットをコード化する核酸配列を含む組換え発現ベクターおよびTBW電位依存性カルシウムチャネルα2δサブユニットをコード化する核酸配列を含む組換え発現ベクターのうちの少なくとも一つを含み、前記被験細胞が機能性カルシウムチャネルを発現する場合に、機能性カルシウムチャネルを含む前記被験細胞を、被験化合物の存在下で、カルシウムおよびカルシウムチャネル作動物質を含有する溶液と接触させ、前記被験細胞での細胞内カルシウムの量を検出することによって行う、被験測定法を実施することと、
陰性対照細胞が機能性カルシウムチャネルを発現する場合に、機能性カルシウムチャネルを含む前記陰性対照細胞を、前記被験化合物の非存在下でカルシウムおよびカルシウムチャネル作動物質を含有する溶液と接触させ、前記陰性対照細胞での細胞内カルシウム量を検出することによって行う、対照測定法を実施することと、
前記被験細胞での細胞内カルシウム量を、前記対照細胞での細胞内カルシウム量と比較することとを含み、
前記被験細胞での細胞内カルシウム量に、前記対照細胞での細胞内カルシウム量と比較して減少が認められた場合に、被験化合物はカルシウムチャネル活性のインヒビターであることが示される、方法。
【請求項33】
前記被験細胞がアフリカツメガエル卵母細胞、CHO細胞またはHEK細胞である、請求項32記載の方法。
【請求項34】
前記細胞の内部の色素を細胞内カルシウムと接触させることで生ずる蛍光を測定する方法を使用することによって、前記細胞内カルシウムを検出する、請求項32記載の方法。
【請求項35】
前記機能性カルシウムチャネルが、配列番号2、その突然変異体および、そのフラグメントからなるキメラTBW電位依存性カルシウムチャネルα1サブユニットと、配列番号8、配列番号10、配列番号12、その突然変異体および、そのフラグメントからなるTBW電位依存性カルシウムチャネルβサブユニット、ならびに、配列番号16、配列番号18、その突然変異体および、そのフラグメントからなるTBW電位依存性カルシウムチャネルα2δサブユニットのうちの少なくとも一つを含んでいる、請求項38記載の方法。
【請求項36】
前記被験細胞がアフリカツメガエル卵母細胞、CHO細胞またはHEK細胞である、請求項35記載の方法。
【請求項37】
前記細胞の内部の色素を細胞内カルシウムと接触させることで生ずる蛍光を測定する方法を使用することによって、前記細胞内カルシウムを検出する、請求項35記載の方法。
【請求項38】
前記機能性カルシウムチャネルが、配列番号2からなるキメラTBW電位依存性カルシウムチャネルα1サブユニットと配列番号8、配列番号10または配列番号12からなるTBW電位依存性カルシウムチャネルβサブユニット、および、配列番号16または配列番号18からなるTBW電位依存性カルシウムチャネルα2δサブユニットを含んでいる、請求項35記載の方法。
【請求項39】
前記キメラTBW電位依存性カルシウムチャネルα1サブユニットが配列番号6を含んでいる、請求項38記載の方法。
【請求項40】
前記キメラTBW電位依存性カルシウムチャネルα1サブユニットをコード化する前記核酸配列が配列番号1からなり、前記TBW電位依存性カルシウムチャネルβサブユニットをコード化する前記核酸配列が配列番号7、配列番号9または配列番号11からなり、前記TBW電位依存性カルシウムチャネルα2δサブユニットをコード化する前記核酸配列が配列番号15または配列番号17からなる、請求項38記載の方法。
【請求項41】
配列番号2、その突然変異体、およびそのフラグメント、配列番号6、その突然変異体、およびそのフラグメント、配列番号8、その突然変異体、およびそのフラグメント、配列番号10、その突然変異体、およびそのフラグメント、配列番号12、その突然変異体、およびそのフラグメント、配列番号16、その突然変異体、およびそのフラグメント、配列番号18、その突然変異体、およびそのフラグメント;からなるグループから選択された前記アミノ酸配列を有する、分離された蛋白質の調製方法であって、請求項5の宿主細胞から前記蛋白質を分離する方法。
【請求項42】
昆虫を、請求項18に記載の方法で識別されたモジュレーターと接触させることを含む、昆虫制御法。
【請求項43】
昆虫を、請求項32に記載の方法で識別されたインヒビターと接触させることを含む、昆虫制御法。
【図1.A】

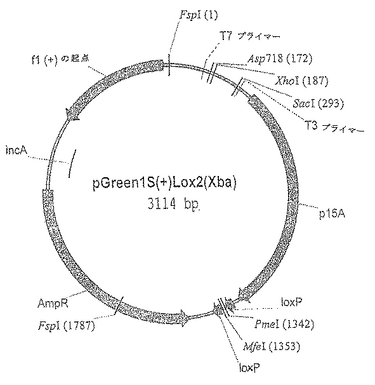
【図1.B】
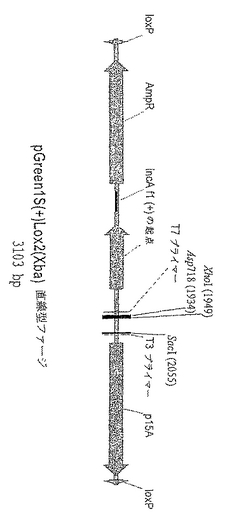
【図2】
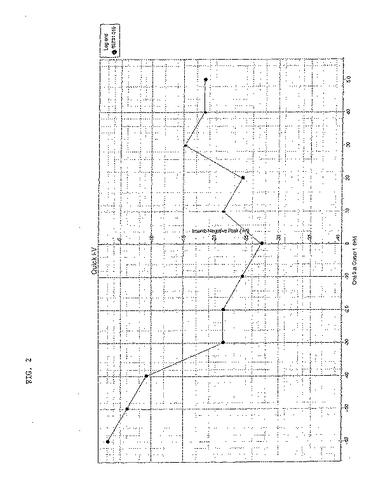
【図3】
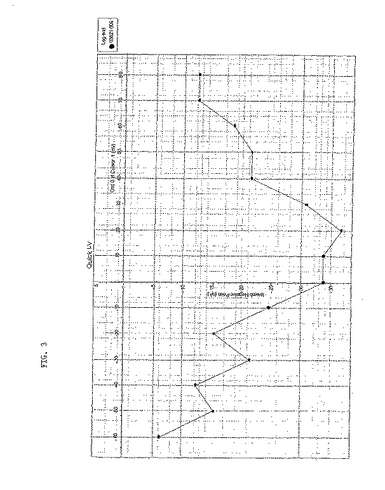
【図4】
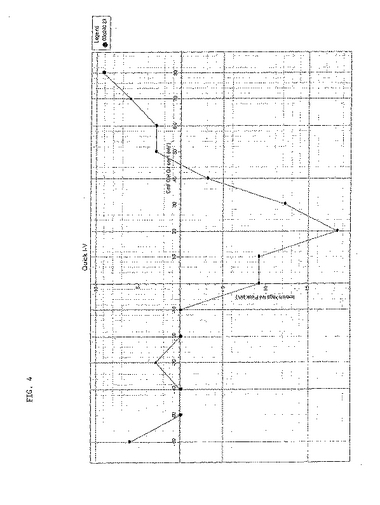
【図1.B】
【図2】
【図3】
【図4】
【公表番号】特表2006−508652(P2006−508652A)
【公表日】平成18年3月16日(2006.3.16)
【国際特許分類】
【出願番号】特願2004−552089(P2004−552089)
【出願日】平成15年11月10日(2003.11.10)
【国際出願番号】PCT/US2003/036001
【国際公開番号】WO2004/044553
【国際公開日】平成16年5月27日(2004.5.27)
【出願人】(391022452)エフ エム シー コーポレーション (74)
【氏名又は名称原語表記】FMC CORPORATION
【Fターム(参考)】
【公表日】平成18年3月16日(2006.3.16)
【国際特許分類】
【出願日】平成15年11月10日(2003.11.10)
【国際出願番号】PCT/US2003/036001
【国際公開番号】WO2004/044553
【国際公開日】平成16年5月27日(2004.5.27)
【出願人】(391022452)エフ エム シー コーポレーション (74)
【氏名又は名称原語表記】FMC CORPORATION
【Fターム(参考)】
[ Back to top ]