
麻疹−マラリア混合ワクチン
本発明は、いくつかの熱帯熱マラリア原虫抗原をコードする異種核酸を含む、異なる弱毒化組み換え麻疹-マラリアベクターを含有する麻疹-マラリア混合ワクチンに関する。好ましくは、本発明は、熱帯熱マラリア原虫のスポロゾイト周囲(CS)タンパク質、熱帯熱マラリア原虫のメロゾイト表面タンパク質1(MSP-1)およびその誘導体(p-42; p-83-30-38)をそのグリコシル化型および分泌型でコードする核酸、ならびに熱帯熱マラリア原虫頂端膜抗原1(AMA1)をそのアンカー型および分泌型でコードする核酸を含むウイルスベクターに関する。ウイルスベクターは、ワクチンとして使用され、対象となる遺伝子の送達において効率的であり、関連免疫細胞に効率的に結合し感染する株に基づく、弱毒化麻疹ウイルスに由来する。好ましい実施形態において、CS、MSP1およびAMA1タンパク質は、これらが哺乳動物、好ましくはヒトにおいて強力な免疫反応を引き起こすように該ウイルスから産生され、このタンパク質の発現は、ヒトコドンへの最適化により上昇する。さらに、本発明は、マラリアの予防的治療における組み換えワクチンの使用に関する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、いくつかの熱帯熱マラリア原虫(Plasmodium falciparum)抗原をコードする異種核酸を含む、異なる弱毒化組み換え麻疹-マラリアベクターを含有する麻疹-マラリア混合ワクチンに関する。好ましくは、本発明は、熱帯熱マラリア原虫のスポロゾイト周囲(CS)タンパク質、熱帯熱マラリア原虫のメロゾイト表面タンパク質1(MSP-1)およびその誘導体(p-42; p-83-30-38)をそのグリコシル化型および分泌型でコードする核酸、ならびに熱帯熱マラリア原虫頂端膜抗原1(AMA1)をそのアンカー型および分泌型でコードする核酸を含むウイルスベクターに関する。ウイルスベクターは、ワクチンとして使用され、対象となる遺伝子の送達において効率的であり、関連免疫細胞に効率的に結合し感染する株に基づく、弱毒化麻疹ウイルスに由来する。好ましい実施形態において、CS、MSP1およびAMA1タンパク質は、これらが哺乳動物、好ましくはヒトにおいて強力な免疫反応を引き起こすように該ウイルスから産生され、これらのタンパク質の発現は、ヒトコドンへの最適化により上昇する。さらに、本発明は、マラリアの予防的治療における組み換えワクチンの使用に関する。
【背景技術】
【0002】
麻疹ウイルス
本発明は、熱帯熱マラリア原虫(Pf)の抗原を発現する弱毒化組み換え麻疹ウイルスを含有するワクチンに関し、麻疹およびマラリア抗原の両方に対する免疫をもたらす、組み換え麻疹-マラリアワクチンの調製のためのそれらの使用に関する。
【0003】
麻疹ウイルス(MV)は、モノネガウイルス(Mononegavirales)目、すなわち、非分節型マイナス鎖RNAゲノムを有するウイルスのメンバーである。MVの非分節型ゲノムは、アンチメッセージ・ポラリティ(antimessage polarity)を有し、したがってこのゲノムRNAは、in vivoまたはin vitroのどちらでも翻訳されない。さらにMVの非分節型ゲノムは、リボ核タンパク質(RNP)複合体(以下を参照されたい)の形態の3種のウイルスタンパク質と特異的に結合した場合にのみ生物学的に活性である。非分節型(-)鎖RNAウイルスの転写および複製およびウイルス粒子としてのそれらの集合は、広範囲にわたって再考されている(1)。麻疹ウイルスの転写および複製は、感染細胞の核に関与するのではなく、むしろ感染細胞の細胞質において起こる。麻疹ウイルスのゲノムは、6種の遺伝子(N、P、M、F、HおよびLと称される)由来の6種の主要な構造タンパク質をコードする遺伝子、ならびに構成的免疫反応の相殺に関与し、転写/複写の制御に関与するP遺伝子のCおよびVに由来するさらなる2種の非構造タンパク質をコードする遺伝子を含む。遺伝子の順番は、3'N、P(CおよびVを含む)、M、F、HおよびL 5'である。さらに、3'-末端領域から、約50ヌクレオチドの短いリーダーRNAが転写される。言及した遺伝子は、ウイルスのリボヌクレオカプシド(RNP)のタンパク質、すなわち、核タンパク質(N)、リンタンパク質(P)および大型のポリメラーゼ/レプリカーゼタンパク質(L)をそれぞれコードし、これらはゲノムRNAに非常に堅く結合し、RNPを形成する。他の遺伝子は、ヘマグルチニン(H)タンパク質、融合(F)タンパク質およびマトリクス(M)タンパク質を含むウイルスエンベロープのタンパク質をコードする。MV遺伝子の転写は漸減勾配に従い、ポリメラーゼがゲノムの鋳型に作用するとき、上流遺伝子から作られるRNAを、下流遺伝子から作られるRNAより多く合成する。この不連続な転写様式において、mRNAはキャップされ、ポリアデニル化される。逆に、この複写様式において、Lタンパク質は完全長のアンチゲノムRNAおよびゲノムRNAを作製し、完全長のアンチゲノムRNAおよびゲノムRNAは直ちにN、PおよびLタンパク質にカバーされ、感染性子孫RNPを形成する。
【0004】
麻疹ウイルスは、1954年に単離され、EndersおよびPeeblesは、麻疹に冒された小児であるDavid Edmostonの血液を初代ヒト腎臓細胞に接種し、得られたMVのEdmoston株(2)を、その後さまざまな細胞系における成長に適合させた。ニワトリ胚、ニワトリ胚線維芽細胞(CEF)および/またはイヌ腎臓細胞ならびにヒト二倍体細胞に対する適合により、弱毒化Edmonston AおよびB(3)、Zagreb(EZ)ならびにAIK-Cシードが作製された。Edmonston Bは、最初のMVワクチンとして、1963年に認可された。CEFに対するEdmonston AおよびBのさらなる継代は、より弱毒化されたSchwarzおよびMoratenウイルスを産生し(3)、それらの配列が同一であることが最近示された(4; 5)。Edmonston Bワクチンは反応原性(reactogenic)なので、1975年に放棄され、Schwarz/ Moratenワクチンに取って代わられた。いくつかの他のワクチン株:日本ではAIK-C、Schwarz F88、CAM70、TD97、ロシアではLeningrad-16、ならびにEdmonston Zagrebもまた使用される。CAM70およびTD97中国株は、Edmonstonに由来しない。Schwarz/ MoratenおよびAIK-Cワクチンは、CEFに対して作製された。Zagrebワクチンは、ヒト二倍体(WI-38)に対して作製された。今日、Schwarz/Moraten、AIK-CおよびEZワクチンは、一般的に使用されるが(6)、原理的には、すべてが1つの特有のMV血清型であるこれらの弱毒化ワクチン株の任意の1つは、安全であり、長期持続性免疫反応を誘導することが証明されており、本発明の目的のために使用できる。
【0005】
MVワクチンは、単回または2回の低用量注射後に生涯免疫を誘導する。麻疹に対する保護は、抗体によって、およびCD4およびCD8のT細胞によって媒介される。MV特異的抗体およびCD8細胞の持続は、ワクチン接種後25年ほどの間示されている(7)。
【0006】
MVワクチンは、大部分の国において大規模な生産が容易であり、低コストで分配できる。MVゲノムの弱毒化は多数の突然変異の有利な組み合わせによりもたらされるので、ワクチンは非常に安定であり、病原性への復帰変異は今までにまったく観察されていない(6)。
【0007】
安全性に関しては、MVは、細胞質において排他的に複製するので、宿主DNA内への統合の可能性はない。これらの特徴から、生きた弱毒化MVワクチンを、多価ワクチン接種ベクターとして使用するための魅力的な候補である。このようなワクチンは、ベクターウイルスそれ自体に対するように、他の病原性物質に対する、長期持続性免疫保護の誘発において効率的であることを証明できる。
【0008】
Martin Billeterおよび同僚は、Edmonston MVのアンチゲノムに対応するcDNAをクローニングし、国際特許出願WO97/06270に記載のように、ウイルスをレスキューする初めてかつ効率的な逆遺伝学手順を確立した(8)。組み換え麻疹ウイルスは、ヘルパー細胞系293-3-46から回収され、安定してトランスフェクトされ、MV NおよびPタンパク質ならびにバクテリオファージT7 RNAポリメラーゼを発現する。任意の変異体または組み換えMVのレスキューのために、次いで、ヘルパー細胞系は、Lタンパク質をコードする発現するプラスミドを一時的にトランスフェクトされ、最も重要には、子孫MVを発生させるために適合性のある、任意の突然変異された、または組み換えのアンチゲノムRNAを得るために適切に構築された任意のアンチゲノムプラスミドを一時的にトランスフェクトされる。一時的トランスフェクトステップは、好ましくは内在するT7 RNAポリメラーゼによって、まず転写に導かれる。得られたアンチゲノムRNAは、直ちに(発生したときの状態で)ウイルスN、PおよびLタンパク質によってカバーされ、ゲノムRNPが産生されるアンチゲノムRNPを得る。第2に、ゲノムRNPは結合したLによって転写され、すべてのウイルスmRNAおよびそれぞれのタンパク質を得る。最終的に、ゲノムRNPおよびアンチゲノムRNPの両方が複製よって増幅される。
【0009】
この手順のわずかな変形において、安定にトランスフェクトされた293-3-46ヘルパー細胞を使用するのではなく、原特許明細書に詳細に説明された5種のプラスミド:N、PおよびT7ポリメラーゼ(ヘルパー細胞系を作り出すために以前使用した)をコードするプラスミドならびにLをコードするプラスミドおよびアンチゲノムプラスミドのすべてを同時に使用して、市販の293T細胞を一時的にトランスフェクトした。「完全な一時的トランスフェクション」手順において、変異体発現プラスミドの使用が可能であり、T7 RNAポリメラーゼ全体の使用を避け、代わりに内在RNAポリメラーゼIIを利用してLタンパク質およびアンチゲノムをさらに発現させることが可能であることに留意されたい(9)。
【0010】
個々の組み換えMVをレスキューするために、利用するアンチゲノムプラスミドは、対象となる異種抗原をコードするヌクレオチド配列(異種ヌクレオチド配列)を組み込まれ、MV-特異的転写の開始配列および終止配列に隣接し、したがって追加転写ユニット(ATU)を形成する、麻疹ウイルスの完全長アンチゲノム(+)RNAをコードするcDNAを含む。このMV Edmonston株のベクターは、最初のMVレスキューの発明者によって、外来遺伝子の発現のために開発され(10)、その挿入能力の大きさ(5kb程度)およびトランス遺伝子、例えばB型肝炎ウイルス(Hepatitis B virus)表面抗原、サルまたはヒトの免疫不全ウイルス(SIVまたはHIV)、ムンプスウイルスおよびヒトIL-12などの発現における高い安定性(11;12)が実証されている。特に、初期のころに、B型肝炎ウイルスの表面抗原およびコア抗原を、個別にまたは組み合わせのいずれかで発現する組み換え麻疹ウイルスが作製され、遺伝子改変マウスにおいて液性免疫反応を誘導することが示された。
【0011】
麻疹ウイルスの特性および特にin vivoで高力価の中和抗体を誘発するその能力および長期持続性細胞免疫反応の強力な誘導因子であるその特性の観察から、本発明者らは、麻疹ウイルスが、好ましくは動物において、より好ましくはヒト宿主において、少なくともある程度の保護を得るために適切であり得る前記マラリア原虫に対する中和抗体を誘導する、熱帯熱マラリア原虫由来の抗原を発現する組み換えウイルスの作製のための優れた候補であり得ることを提唱する。
【0012】
特に、本発明においてMV株および特にワクチン株は、幼児集団はMV免疫を有していないので、曝露された幼児集団において設計された組み換えMV中でその成分が発現される、麻疹ウイルスおよび熱帯熱マラリア原虫の両方に対する免疫を誘導するための候補ベクターとして選択される。
【0013】
既にMV免疫化された個人であっても、本発明の組み換え体であるMVウイルスの再投与により、抗MV抗体の強化がもたらされるので、成人集団はMV組み換え体による免疫化からさらに利益を得ることができる(13)。
【0014】
本発明は、熱帯熱マラリア原虫由来の異種遺伝子を担持する組み換え麻疹ウイルスの調製に特に関する。
【0015】
本発明に従った、組み換え麻疹ウイルスの有利な免疫学的特性は、麻疹ウイルスに感受性の動物の中から選択された動物モデルにおいて示すことができ、異種抗原に対する、および/または麻疹ウイルスに対する液性免疫反応および/または細胞性免疫反応を決定する。免疫反応の特徴づけのためのモデルとしての使用に適切なこのような動物の中で、当業者は、特に、MVの特異的受容体の1つであるCD46を発現するトランスジェニックマウスを使用できる。最も有望な組み換え体は、その後サルにおいて試験され得る。
【0016】
組み換え麻疹ウイルスのヌクレオチド配列は、6の倍数であるヌクレオチドの総数を含まなくてはならない。このいわゆる「6の規則」に対する固守は、MVだけでなく、パラミクソウイルス亜科(Paramyxovirinae)のサブファミリーに属するすべてのウイルスに関する絶対的必要条件である。明らかに、それぞれが6ヌクレオチドと接触するNタンパク質分子は、正確に5'末端から3'末端までのゲノムRNAおよびアンチゲノムRNAをカバーしなければならない。
【0017】
ATUの位置は、アンチゲノムcDNAに沿って変え得ることに留意されたい。したがって、上記のMVのmRNAの天然発現の勾配を利用して、挿入されたATUの発現レベルを適切なレベルに変えることができる。ATUの好ましい位置は、L遺伝子の上流、M遺伝子から上流およびN遺伝子の上流であり、それぞれ、異種タンパク質の低い、中程度のおよび強い発現をもたらす。
【0018】
マラリア原虫
マラリアは、現在、世界中、特に熱帯および亜熱帯地域において最も流行している感染性疾患の1つを表す。1年に、マラリア感染症は、世界中で1億人に重症な病気をもたらし、発展途上国および新興国において主に年少幼児が100万から300万人死亡する。マラリアの広範な発生および発生率の上昇は、DDTの広範な禁止ならびに薬剤耐性寄生虫および殺虫剤耐性寄生虫ベクターの数の増加の結果である。他の因子は、環境変化および気候の変化、騒乱ならびに人口の流動性の増加を含む。
【0019】
マラリアは、アピコンプレックス(Apicomplexa)門由来のプラスモディウム(Plasmodium)属に属する、蚊によって運ばれる血液寄生原生動物(hematoprotozoan parasites)によって引き起こされる。プラスモディウム属の4つの種:四日熱マラリアに関与する四日熱マラリア原虫(P. malariae)、両方とも三日熱マラリアを起こす三日熱マラリア原虫(P. vivax)および卵型マラリア原虫(P. ovale)ならびに熱帯熱マラリアの病原体であり、ほとんどすべての致死性感染症に関与する熱帯熱マラリア原虫はヒトに感染する。他にも多くが動物において疾患の原因となり、例えば、プラスモディウム・ヨエリ(P. yoelii)およびプラスモディウム・ベルゲイ(P. berghei)はマウスにおいて疾患の原因となる。
【0020】
マラリア原虫は、いくつかの段階からなるライフサイクルを有する。各段階は、対応する発生段階特異的抗原に対する特異的免疫反応を誘導できる。マラリア原虫は、雌のハマダラカのいくつかの種によって人間に伝播する。感染した蚊は、哺乳動物の血流中にマラリア原虫の「スポロゾイト」型を注入する。スポロゾイトは循環中で数分維持され、その後肝細胞を侵す。この段階において、寄生虫は細胞外環境に位置し、主にスポロゾイト表面の主要成分である「スポロゾイト周囲」(CS)タンパク質を対象として、抗体の攻撃に曝露される。肝臓においてひとたび寄生虫が複製すると、いわゆる「ジゾント」に発達する。これらのジゾントは、最大20,000/感染細胞の比率で発生する。寄生虫のこの細胞内段階の間、宿主免疫反応の主役はTリンパ球、特にCD8+ Tリンパ球である。肝臓感染の約1週間後、数千のいわゆる「メロゾイト」が血流中に放出される。頂端膜抗原1(AMA1)およびメロゾイト表面タンパク質1(MSP1)は、両方とも、感染肝臓細胞から出現するメロゾイト上に存在し、それらは赤血球の侵入に関与する無生殖血液期のメロゾイトの必須成分である。それらがひとたび赤血球に進入すると、抗体媒介免疫反応およびT細胞により分泌されるサイトカインの標的となる。赤血球の侵入後、メロゾイトはいくつかの複写段階を経て、いわゆる「トロホゾイト」ならびにシゾントおよびメロゾイトをもたらし、これらは新たな赤血球に感染できる。限られた量のトロホゾイトは、寄生虫の有性世代を構成する「ガメトサイト」に進化できる。感受性の蚊が赤血球を摂取した場合、赤血球からガメトサイトが放出され、いくつかの雄のガメトサイトおよび1つの雌のガメトサイトがもたらされる。これらのガメートの受精は、接合体の形成なたびにその後のオーキネートへの転換、次いでオーシストへの転換および最終的な唾液腺スポロゾイトへの転換をもたらす。ガメトサイト期特異的表面抗原に対する標的化抗体は、蚊の中腸内でこのサイクルを遮断できる。このような抗体は、哺乳動物宿主を保護しないが、感染した蚊の数およびそれらの寄生負荷を減少することによって、マラリアの伝播を減少させる。
【0021】
MSP-1は、190〜200kDa (d-190)の前駆体として合成され、これは、シゾゴニーの間に83、30、38および42kDa(d-42)の断片にタンパク質分解的にプロセシングされる(14)。赤血球侵入の時点で、42kDaは、残りの複合体と一緒に取り除かれる33kDaの断片および2つの表皮成長因子(EGF)様ドメインを含有し、侵入の間にメロゾイト膜に結合し続ける19kDaの断片とにさらに切断される。この第2の切断は、赤血球侵入が成功するための前提条件である(15)。
【0022】
MSP-1は、K1およびMAD20のプロトタイプによって特徴付けられる二形成対立遺伝子内に高度な保存を示す、基本的二形成タンパク質である。
【0023】
AMA-1(16)は、構造的に保存されたI型内在性膜タンパク質であり、熱帯熱マラリア原虫中に622aa(PfAMA-1)を含み、細胞質領域(50aa)、膜貫通領域ならびにN末端のプロ配列としてフォールディングされたectoドメインおよび3つのドメイン(DI、DII、DIII)に組織化されている。タンパク質の発現は、後期シゾゴニーにおいて最大であり、AMA-1(83kDa)の前駆体がタンパク質分解的にプロセシングされ、プロ配列が切り離され、タンパク質が66kDaの形態に転換され、これによってメロゾイトの再局在化が可能になる。抗体は主にDIおよびDIIを認識し、いくつかの対立遺伝子変異体と同等によく反応すると思われる。DIIIとの抗体反応は一般的に低く、そのレベルは成人において増加する(17、18)。
【0024】
PfAMA-1は64の多形位置を含有し(プロ配列中に9、ectoドメイン中に52、細胞質領域中に3)、それらの大部分が二形成性であり、これらは宿主免疫反応の重要なエピトープである。PfAMA-1に基づくワクチンを開発するために、多形をカバーすることが重要なはずである: Diversity Covering(DiCo1、2および3)PfAMA-1は、PfAMA1 ectoドメインの天然発生多形を最大限に表す人工配列である。それらが、多様なPfAMA1対立遺伝子を担持するさまざまな寄生虫に対して機能性である免疫反応を誘導することが示されている。この取り組みは、PfAMA1を標的とするワクチンを、広範囲の天然発生PfAMA1対立遺伝子に対する強力かつ機能的な保護を誘導可能に作製できる手段を提示できる(19)。
【0025】
CSタンパク質(CSP)は約420aaおよび58kDaの分子量を有する。スポロゾイトの主要表面タンパク質を表し、その機能は、オーシスト(oocystis)由来のスポロゾイトの突然変異および正に帯電したアミノ酸の保存されたモチーフから媒介される肝細胞の侵入のために基本的である。CSPは、5'末端および3'末端の2つの非反復領域ならびに4残基長のモチーフの複数の反復からなり、CSP内の主要なエピトープを表す可変の種特異的中心領域で組織化される。CSPは、シゾゴニーの少なくとも最初の3日間検出可能であり続けるので、細胞外スポロゾイトを対象とする抗体媒介免疫反応およびシゾントを対象とする細胞媒介免疫反応の両方のために魅力的なワクチン標的であると考えられる(20)。
【0026】
マラリアワクチン開発の現在の取り組みは、上記のように、寄生虫が存在できる異なる段階に従って分類できる。
【0027】
ワクチン候補は、3つの型に識別できる:i)スポロゾイトおよび/またはシゾント感染細胞を対象とする前赤内期ワクチン。これらの型のワクチンは主にCSに基づき、液性免疫反応および細胞性免疫反応によって媒介されて理想的には無菌免疫をもたらしマラリア感染を予防するはずである;ii)メロゾイト感染細胞を対象とする無性生殖血液期ワクチン:MSP1およびAMA1は、臨床的重症度を最小にするように設計されたマラリアワクチン候補をもたらす。これらのワクチンは、罹患率および死亡率を低下させるはずであり、寄生虫が赤血球へ進入すること、および/または赤血球中において発達することを防止することを意味する;iii)寄生虫が蚊宿主中で発達することを妨害するように設計された伝播阻止ワクチン。この型のワクチンは、集団規模のマラリア感染率の減少を支持するはずである。これらのワクチンとは別に、寄生虫のライフサイクルの複数の段階を標的とするマラリアワクチンの開発の実現可能性が、いわゆる多成分ワクチンおよび/または多段階ワクチンにおいて追求されている。
【0028】
今日の世界規模のマラリアワクチンのポートフォリオは、47種の新しいワクチン候補が有望であり、31種が前臨床開発、16種が臨床試験に絞り込まれている。これらの1つである、GSK BiologicalsおよびPATH-MVIにより開発されたRTS,Sワクチンは2008年に最終のフェーズIII臨床試験に入っているはずである(21)。他の興味深いワクチン候補は、ベクターとして使用される生の組み換えウイルス、例えば、国際特許出願US2006127413に記載のModified Vaccinia Ankara(MVA)、ポックスウイルス(US6214353、AU7060294、AU1668197、WO9428930およびUS5756101)、アデノウイルス(US2007071726、US2005265974、US2007088156およびCA2507915)、寒冷適用弱毒化インフルエンザウイルスに基づくワクチン、または酵母、例えばメタノール資化性酵母(Pichia pastoris)およびサッカロミセス属種(Saccharomyces spp)に基づくワクチン、または細菌発現系、例えば、サルモネラ属種(Salmonella spp.)(US5112749)および大腸菌(Escherichia coli)(EB0191748)に基づくワクチンである(22)。
【先行技術文献】
【特許文献】
【0029】
【特許文献1】国際特許出願WO97/06270
【特許文献2】国際特許出願US2006127413
【特許文献3】US6214353
【特許文献4】AU7060294
【特許文献5】AU1668197
【特許文献6】WO9428930
【特許文献7】US5756101
【特許文献8】US2007071726
【特許文献9】US2005265974
【特許文献10】US2007088156
【特許文献11】CA2507915
【特許文献12】US5112749
【特許文献13】EB0191748
【発明の概要】
【発明が解決しようとする課題】
【0030】
現在、マラリアに対して利用可能な市販のワクチンはないが、マラリアに対するワクチンの開発は30年以上前に既に開始されている。多くの因子がマラリアワクチンの開発を困難かつ挑戦的にしている。第1に、寄生虫のサイズおよび遺伝的複雑性は、各感染症が、ヒト免疫系に対して数千の抗原を提示することを意味する。これらのうちのどれがワクチン開発のための有用な標的であり得るかを理解することが複雑であり、今日までに、少なくとも40の異なる有望な抗原が特定されている。第2に、寄生虫は、ヒト宿主においてさえいくつかのライフステージの間に変化し、ライフサイクルの各段階において、免疫系に対する異なるサブセットの分子を提示する。第3に、寄生虫は、ヒト免疫系を混乱させ、潜伏し、誤った方向に導くことが可能な一連の戦略を進化させる。最終的に、異なる種だけではなく、異なる株の多重マラリア感染症も同時に有する可能性がある。
【0031】
したがって、本発明は、いくつかの熱帯熱マラリア原虫抗原をコードする異種核酸を含む、異なる弱毒化組み換え麻疹-マラリアベクターを含有する麻疹-マラリア混合ワクチンを提供することによって、先行技術が長期にわたって感じていた必要性を満たす。
【課題を解決するための手段】
【0032】
本発明の一実施形態において、麻疹およびマラリアの両方に対する免疫反応および保護を誘発できるマラリア抗原を発現する、組み換え麻疹ワクチンウイルスを含む麻疹-マラリア混合ワクチンを提供する。
【0033】
別の実施形態において、本発明は、MSP1マラリア抗原を発現するヌクレオチド配列を有する組み換え麻疹ワクチンウイルスを提供する。好ましい実施形態において、組み換え麻疹ワクチンウイルスは、3D7株およびFCB1株由来のアンカー型または分泌型の両方においてマラリア抗原d190またはd83-30-38またはd42を発現するヌクレオチド配列を有する。
【0034】
さらに別の実施形態において、本発明は、Diversity Covering (DiCo) AMA1マラリア抗原を発現するヌクレオチド配列を有する、組み換え麻疹ワクチンウイルスを提供する。
【0035】
さらに別の実施形態において、本発明は、CSマラリア抗原を発現するヌクレオチド配列を有する、組み換え麻疹ワクチンウイルスを提供する。
【図面の簡単な説明】
【0036】
【図1】麻疹ウイルスのアンチゲノムDNA p(+)MV-EZの略図である。p(+)MV-EZは、T7 RNAポリメラーゼプロモーター(T7)の調節下で、それぞれ1の位置(麻疹ウイルスのN遺伝子の前)、2の位置(麻疹ウイルスのP遺伝子とM遺伝子との間)および3の位置(麻疹ウイルスのH遺伝子およびL遺伝子の間)に3つのATUを含有し、肝炎のデルタリボザイムおよびT7 RNAポリメラーゼターミネーター(δT7t)によって正確に終止される、麻疹ウイルス(Edmoston Zagreb)の完全配列を含有するpBluescript由来のプラスミドである。プラスミドのサイズは18941bpである。
【図2】3D7株由来のMSP-1合成遺伝子(d-190)を表す図である。d-190遺伝子断片(d-83-30-38およびd-42)の隣接領域に関するコーディングヌクレオチドおよび対応するアミノ酸を示す。クローニング手順のために加えた特有の制限部位は色で示す; SP:シグナルペプチド;GPI:膜アンカー型領域のためにコードされたグリコシル-ホスファチジル-イノシトール配列。
【図3】FCB1株由来のMSP-1合成遺伝子(d-190)を表す図である。d-190遺伝子断片(d-83-30-38およびd-42)の隣接領域に関するコーディングヌクレオチドおよび対応するアミノ酸を示す。クローニング手順のために加えた特有の制限部位は色で示す; SP:シグナルペプチド;GPI:膜結合領域のためにコードされたグリコシル-ホスファチジル-イノシトール配列。SPおよびGPI領域は3D7株由来である。
【図4】組み換え麻疹-マラリアプラスミドのp(+)MV2-EZ-d190-3D7の略図である。それは、SgrAI-BssHII消化によって麻疹ゲノムの2の位置にクローニングされる、タンパク質のGPIアンカー型をコードする5253bpのd-190マラリア遺伝子(3D7株)を含有する、p(+)MV-EZに由来するプラスミドである。組み換えプラスミドのサイズは24323bpである。
【図5】組み換え麻疹-マラリアプラスミドのp(+)MV2-EZ-d190*-3D7の略図である。それは、SgrAI-BssHII消化によって麻疹ゲノムの2の位置にクローニングされる、タンパク質の分泌型をコードする5160bpのd-190*マラリア遺伝子(3D7株)を含有する、p(+)MV-EZに由来するプラスミドである。組み換えプラスミドのサイズは24227bpである。
【図6】組み換え麻疹-マラリアプラスミドのp(+)MV3-EZ-d190-3D7またはp(+)MV3-EZ-dl90*-3D7の略図である。それは、SgrAI-BssHII消化によって麻疹ゲノムの3の位置にクローニングされる、タンパク質のGPIアンカー型をコードする5253bpのd-190マラリア遺伝子(3D7株)またはタンパク質の分泌型をコードする5160bpのd-190*マラリア遺伝子(3D7株)を含有する、p(+)MV-EZに由来するプラスミドである。組み換えプラスミドp(+)MV3-EZ-d190は24323bpであり、p(+)MV3-EZ-d190*は24227bpである。
【図7】組み換え麻疹-マラリアプラスミドのp(+)MV2-EZ-d83-30-8-3D7の略図である。それは、SgrAI-BssHII消化によって麻疹ゲノムの2の位置にクローニングされる、タンパク質のGPIアンカー型をコードする4122bpのd-83-30-38マラリア遺伝子(3D7株)を含有する、p(+)MV-EZに由来するプラスミドである。組み換えプラスミドのサイズは23195bpである。
【図8】組み換え麻疹-マラリアプラスミドのp(+)MV2-EZ-d83-30-38*-3D7の略図である。それは、SgrAI-BssHII消化によって麻疹ゲノムの2の位置にクローニングされる、タンパク質の分泌型をコードする4029bpのd-83-30-38*遺伝子(3D7株)を含有する、p(+)MV-EZに由来するプラスミドである。組み換えプラスミドのサイズは23105bpである。
【図9】組み換え麻疹-マラリアプラスミドのp(+)MV3-EZ-d83-30-38-3D7またはp(+)MV3-EZ-d83-30-38*-3D7の略図である。それは、SgrAI-BssHII消化によって麻疹ゲノムの3の位置にクローニングされる、タンパク質のGPIアンカー型をコードする4122bpのd-83-30-38マラリア遺伝子(3D7株)またはタンパク質の分泌型をコードする4029bpのd-83-30-38*遺伝子(3D7株)を含有する、p(+)MV-EZに由来するプラスミドである。組み換えプラスミドp(+)MV3-EZ-d83-30-38は23195bpであり、p(+)MV3-EZ-d83-30-38*は23105bpである。
【図10】組み換え麻疹-マラリアプラスミドのp(+)MV2-EZ-d42-3D7の略図である。それは、SgrAI-BssHII消化によって麻疹ゲノムの2の位置にクローニングされる、タンパク質のGPIアンカー型をコードする1347bpのd-42マラリア遺伝子(3D7株)を含有する、p(+)MV-EZに由来するプラスミドである。組み換えプラスミドのサイズは20417bpである。
【図11】組み換え麻疹-マラリアプラスミドのp(+)MV2-EZ-d42*-3D7の略図である。それは、SgrAI-BssHII消化によって麻疹ゲノムの2の位置にクローニングされる、タンパク質の分泌型をコードする1254bpのd-42*マラリア遺伝子(3D7株)を含有する、p(+)MV-EZに由来するプラスミドである。組み換えプラスミドのサイズは20345bpである。
【図12】組み換え麻疹-マラリアプラスミドのp(+)MV3-EZ-d42-3D7またはp(+)MV3-EZ-d42*-3D7の略図である。それは、SgrAI-BssHII消化によって麻疹ゲノムの3の位置にクローニングされる、タンパク質のGPIアンカー型をコードする1347bpのd-42マラリア遺伝子(3D7株)またはタンパク質の分泌型をコードする1254bpのd-42*マラリア遺伝子(3D7株)を含有する、p(+)MV-EZに由来するプラスミドである。組み換えプラスミドp(+)MV3-EZ-d42のサイズは20417bpであり、p(+)MV3-EZ-d42*は20345bpである。
【図13】組み換え麻疹-マラリアプラスミドのp(+)MV2-EZ-d190-FCB1の略図である。それは、SgrAI-BssHII消化によって麻疹ゲノムの2の位置にクローニングされる、タンパク質のGPIアンカー型をコードする5013bpのd-190マラリア遺伝子(FCB1株)を含有する、p(+)MV-EZに由来するプラスミドである。組み換えプラスミドのサイズは24083bpである。
【図14】組み換え麻疹-マラリアプラスミドのp(+)MV3-EZ-d190-FCB1の略図である。それは、SgrAI-BssHII消化によって麻疹ゲノムの3の位置にクローニングされる、タンパク質のGPIアンカー型をコードする5013bpのd-190マラリア遺伝子(FCB1株)を含有する、p(+)MV-EZに由来するプラスミドである。組み換えプラスミドp(+)MV3-EZ-d190のサイズは24083bpである。
【図15】CS合成遺伝子を表す図である。CS遺伝子の隣接領域に関するコーディングヌクレオチドおよび対応するアミノ酸を示す。クローニング手順のために加えた特有の制限部位は色で示す。
【図16】組み換え麻疹-マラリアプラスミドのp(+)MV2-EZ-CSの略図である。それは、SgrAI-BssHII消化によって麻疹ゲノムの2の位置にクローニングされる、1119bpのCS遺伝子を含有する、p(+)MV-EZに由来するプラスミドである。組み換えプラスミドのサイズは20219bpである。
【図17】組み換え麻疹-マラリアプラスミドのp(+)MV3-EZ-CSの略図である。それは、SgrAI-BssHII消化によって麻疹ゲノムの3の位置にクローニングされる、1119bpのCS遺伝子を含有する、p(+)MV-EZに由来するプラスミドである。組み換えプラスミドのサイズは20219bpである。
【図18】DiCo-1完全合成遺伝子を表す図である。DiCo-1完全遺伝子ドメイン(ectoおよびtrans-cyto)の隣接領域に関するコーディングヌクレオチドおよび対応するアミノ酸を示す。クローニング手順のために加えた特有の制限部位は色で示す; SP:シグナルペプチド(ヒトコドンに最適化されている)。
【図19】DiCo-1 ecto合成遺伝子を表す図である。DiCo-1 ectoドメインの隣接領域に関するコーディングヌクレオチドおよび対応するアミノ酸を示す。クローニング手順のために加えた特有の制限部位は色で示す; SP:シグナルペプチド(ヒトコドンに最適化されている)。
【図20】組み換え麻疹-マラリアプラスミドのp(+)MV2-EZ-DiCo1-完全の略図である。それは、SgrAI-BssHII消化によって麻疹ゲノムの2の位置にクローニングされる、タンパク質の膜貫通型をコードする1689bpのDiCo1完全遺伝子を含有する、p(+)MV-EZに由来するプラスミドである。組み換えプラスミドのサイズは20753bpである。
【図21】組み換え麻疹-マラリアプラスミドのp(+)MV3-EZ-DiCo1-完全の略図である。それは、SgrAI-BssHII消化によって麻疹ゲノムの3の位置にクローニングされる、タンパク質の膜貫通型をコードする1689bpのDiCo1完全遺伝子を含有する、p(+)MV-EZに由来するプラスミドである。組み換えプラスミドのサイズは20753bpである。
【図22】組み換え麻疹-マラリアプラスミドのp(+)MV2-EZ -DiCo1- ectoの略図である。それは、SgrAI-BssHII消化によって麻疹ゲノムの2の位置にクローニングされる、タンパク質の分泌型をコードする1458bpのDiCo1 ecto遺伝子を含有する、p(+)MV-EZに由来するプラスミドである。組み換えプラスミドのサイズは20525bpである。
【図23】組み換え麻疹-マラリアプラスミドのp(+)MV3-EZ -DiCo1- ectoの略図である。それは、SgrAI-BssHII消化によって麻疹ゲノムの3の位置にクローニングされる、タンパク質の分泌型をコードする1458bpのDiCo1 ecto遺伝子を含有する、p(+)MV-EZに由来するプラスミドである。組み換えプラスミドのサイズは20525bpである。
【図24】p(+)MV2EZ-GFPの完全ヌクレオチド配列の図である。この配列は、ヌクレオチドの位置に関して以下のように説明できる:- 592-608 T7プロモーター- 609-17354 MV Edmoston Zagrebアンチゲノム- 4049-4054 MluI制限部位- 4060-4067 SgrAI制限部位- 4079-4084 BssHII制限部位- 4085-4801 緑色蛍光タンパク質(GFP) ORF- 4805-4810 BssHII制限部位- 4817-4822 AatII制限部位- 17355-17580 HDVリボザイムおよびT7ターミネーター
【図25】p(+)MV3EZ-GFPの完全ヌクレオチド配列の図である。この配列は、ヌクレオチドの位置に関して以下のように説明できる:- 592-608 T7プロモーター- 609-17359 MV Edmoston Zagrebアンチゲノム- 9851-9856 MluI制限部位- 9862-9869 SgrAI制限部位- 9886-9891 BssHII制限部位- 9892-10608 緑色蛍光タンパク質(GFP) ORF- 10612-10617 BssHII制限部位- 10624-10629 AatII制限部位- 17360-17585 HDVリボザイムおよびT7ターミネーター
【図26】AN101TEの図であり、これは、本発明者らによってクローニングされたMSP1 d-190 3D7配列のORFである。この配列は、ヌクレオチドの位置に関して以下のように説明できる:- 1-3 開始コドン- 4-99 d-1903D7シグナルペプチド- 100-105 BamHI制限部位- 4014-4020 BstEII制限部位- 5152-5157 AclI制限部位- 5158-5250 GPI配列- 5251-5253 終止コドン
【図27】AN102TEの図であり、これは、本発明者らによってクローニングされたMSP1 d-190* 3D7配列のORFである。この配列は、ヌクレオチドの位置に関して以下のように説明できる:- 1-3 開始コドン- 4-99 d- 190*3D7シグナルペプチド- 100-105 BamHI制限部位- 4014-4020 BstEII制限部位- 5152-5157 AclI制限部位- 5158-5160 終止コドン
【図28】AN103TEの図であり、これは、本発明者らによってクローニングされたMSP1 d-83-30-38 3D7配列のORFである。この配列は、ヌクレオチドの位置に関して以下のように説明できる:- 1-3 開始コドン- 4-99 d-83-30-38 3D7シグナルペプチド- 100-105 BamHI制限部位- 4014-4020 BstEII制限部位- 4021-4026 AclI制限部位- 4027-4119 GPI配列- 4120-4122 終止コドン
【図29】AN104TEの図であり、これは、本発明者らによってクローニングされたMSP1 d-83-30-38* 3D7配列のORFである。この配列は、ヌクレオチドの位置に関して以下のように説明できる:- 1-3 開始コドン- 4-99 d-83-30-38* 3D7シグナルペプチド- 100-105 BamHI制限部位- 4014-4020 BstEII制限部位- 4027-4029 終止コドン
【図30】AN105TEの図であり、これは、本発明者らによってクローニングされたMSP1 d-42 3D7配列のORFである。この配列は、ヌクレオチドの位置に関して以下のように説明できる:- 1-3 開始コドン- 4-99 d-42 3D7シグナルペプチド- 100-105 BamHI制限部位- 108-114 BstEII制限部位- 1246-1251 AclI制限部位s- 1252-1344 GPI配列- 1345-1347 終止コドン
【図31】AN106TEの図であり、これは、本発明者らによってクローニングされたMSP1 d-42* 3D7配列のORFである。この配列は、ヌクレオチドの位置に関して以下のように説明できる:- 1-3 開始コドン- 4-99 d-42* 3D7シグナルペプチド- 100-105 BamHI制限部位- 108-114 BstEII制限部位- 1246-1251 AclI制限部位s- 1252-1254 終止コドン
【図32】AN107TEの図であり、これは、本発明者らによってクローニングされたMSP1 d-190 FCB1配列のORFである。この配列は、ヌクレオチドの位置に関して以下のように説明できる:- 1-3 開始コドン- 4-99 d-190 FCB1シグナルペプチド- 100-105 BamHI制限部位- 146-151 HindIII制限部位- 3825-3831 BstEII制限部位- 4912-4917 AclI制限部位s- 4918-5010 GPI配列- 5011-5013 終止コドン
【図33】AN108TEの図であり、これは、本発明者らによってクローニングされたCS配列のORFである。この配列は、ヌクレオチドの位置に関して以下のように説明できる:- 1-3 開始コドン- 4-1116 CS配列- 1117-1119 終止コドン
【図34】AN109TEの図であり、これは、本発明者らによってクローニングされたDiCo 1完全配列のORFである。この配列は、ヌクレオチドの位置に関して以下のように説明できる:- 1-3 開始コドン- 4-99 DiCo 1完全シグナルペプチド- 100-105 BamHI制限部位- 106-1686 DiCo 1完全配列ORF- 1687-1689 終止コドン
【図35】AN110TEの図であり、これは、本発明者らによってクローニングされたDiCo 1 ecto配列のORFである。この配列は、ヌクレオチドの位置に関して以下のように説明できる:- 1-3 開始コドン- 4-99 DiCo 1 ecto完全シグナルペプチド- 100-105 BamHI制限部位- 106-1455 DiCo 1 ecto完全配列ORF- 1456-1458 終止コドン
【図36】組み換え麻疹-p-42マラリアウイルスのMVウイルスワクチンに感染後のVero細胞において生み出された、比較可能な細胞変性効果を表す図である。
【図37】麻疹ベクター(MV3EZ-d-42 SgrAI)の3の位置に挿入されたd-42 3D7トランス遺伝子の発現の図である。1、5および10継代由来の細胞溶解物を、空の麻疹ベクター(MVEZ)および陰性対照(MV3L1、組み換えMV-パピローマウイルス)に対してウェスタンブロットにより分析した。
【図38】免疫蛍光法によって分析された、麻疹ベクター(MV3EZ-d-42 SgrAI)の3の位置に挿入されたトランス遺伝子d-42 3D7の発現を、空の麻疹ベクター(MVEZ)および陰性対照(MV2EZL1、組み換えMV-パピローマウイルス)と比較した図である。それらは免疫染色の前後に光学顕微鏡を使用して見られたので、矢印は同じシンシチウムを指す。
【図39】MVウイルスワクチンの成長動態曲線と比較した、組み換え麻疹-p-42マラリアウイルスの成長動態曲線の図である。
【図40】麻疹ベクター(MV2-3EZ-d-190 SgrAI FCB1)の2の位置および3の位置に挿入されたd-190 FCB1トランス遺伝子の発現の図である。細胞溶解物を、空の麻疹ベクター(MVEZ)および陰性対照(MV2EZL1、組み換えMV-パピローマウイルス)に対してウェスタンブロットにより分析した。
【図41】MVウイルスワクチンの成長動態曲線と比較した、組み換え麻疹- p-190-FCB1マラリアウイルスの成長動態曲線の図である。
【発明を実施するための形態】
【0037】
本発明の目的は、熱帯熱マラリア原虫のCS、MSP-1またはその一部およびAMA-1またはその一部を分泌型または表面アンカー型でコードする、安定に統合されたDNA配列を含有できる組み換え麻疹ベクターから麻疹-マラリア混合ワクチンを作製することである。
【0038】
本発明は、感受性のトランスジェニックマウス、サルおよびヒト宿主において、PfCS、PfMSP-1およびPfAMA-1の抗原を感染、複写および発現できる組み換えMV-マラリアウイルスのレスキューをさらに含むものである。
【0039】
さらに、本発明は、2種の異なる抗原が同じベクター内で同時にクローニングされ、発現し、それらの両方に対する免疫をもたらす、多価組み換え麻疹-マラリアベクターの構築を含むことを意図する。
【0040】
さらに、本発明は、それぞれが、宿主において免疫反応を誘発する様式で異なる遺伝子を担持し、寄生虫のライフサイクルの異なる段階を対象とする異なる抗原を発現する、3種の異なる組み換え麻疹-マラリアウイルスの組み合わせに関する。
【0041】
さらに、本発明は、欠陥干渉粒子(DI)を欠く組み換え麻疹-マラリアウイルスの作製プロセスを含む。DIは、任意の作製系においてウイルスの成長を有意に阻害し、ヒト個体において免疫反応の抑制に成功することが知られている。
【0042】
さらに、本発明はこのような組み換えウイルスを含有するワクチンの作製方法を含む。
【0043】
以下の実施例は、本発明を実施する好ましい様式を記載する。これらの実施例は例示のために提供されると理解されるべきであり、本発明の範囲を決して限定しないものとして解釈されるべきである。
【実施例1】
【0044】
組み換えMV-PfMSP-1プラスミドの構築
すべてのクローニング手順は、Sambrookら(1989年)に記載の技術により実施した。
【0045】
すべての制限酵素は、New England BioLabsから入手し、オリゴヌクレオチドPCRプライマーおよびDNAポリリンカーはInvitrogenから入手した。
【0046】
分泌型およびアンカー型のPfMSP1およびその断片(d-83-30-38およびd-42)は、いずれも化学的に合成し、ヒトコドンに最適化した。それらをpZE21MV中間ベクター内にクローニングし、SgrAIクローニング部位を5'末端に付加することによってわずかに改変し、その後Kozak配列(TCATCA)に最適化した。これらの改変は、MWG Biotechにおいて配列決定により確認されている。
【0047】
MAD20プロトタイプに属する3D7株およびK1プロトタイプに属するFCB1株由来の組み換えMV-PfMSP-1プラスミド、GPIアンカー型および分泌型(*)のリスト:
p(+)MV2EZ-d-190-SgrAI (3D7)
p(+)MV3EZ-d-190-SgrAI (3D7)
p(+)MV2EZ-d-83-30-38-SgrAI (3D7)
p(+)MV3EZ-d-83-30-38-SgrAI (3D7)
p(+)MV2EZ-d-42-SgrAI (3D7)
p(+)MV3EZ-d-42-SgrAI (3D7)
p(+)MV2EZ-d-190*-SgrAI (3D7)
p(+)MV3EZ-d-190*-SgrAI (3D7)
p(+)MV2EZ-d-83-30-38*-SgrAI (3D7)
p(+)MV3EZ-d-83-30-38*-SgrAI (3D7)
p(+)MV2EZ-d-42*-SgrAI (3D7)
p(+)MV3EZ-d-42*-SgrAI (3D7)
p(+)MV2EZ-d-190-SgrAI (FCBl)
p(+)MV3EZ-d-190-SgrAI (FCBl)
【0048】
1a)p(+)MV2EZ-d-190-SgrAI (3D7、24323bp)およびp(+)MV3EZ-d-190-SgrAI(3D7、24323bp)の構築
緑色蛍光タンパク質(GFP)を含有する1μgのMVプラスミドDNA(p(+)MV2-3EZ-GFP Berna株、19774bp:図24および25)を、1ユニットのSgrAIおよびBssHII制限酵素両方を用いて、それらの最適温度において2時間、最終容量50μlで消化した。すべての消化されたDNAを1%アガロースゲルに添加し、80ボルトで約2時間泳動させた。次いで、固有のバンド(19048bp)をゲルから取り出し、QIAEXゲル精製によって精製し、260nmにおける吸光度によりDNA濃度を計算し、1μg/mlに対して調整した。
【0049】
中間プラスミド内に挿入された1μgのd-190遺伝子(pZE21 MV-d- 190 SgrAI、7564bp)を、SgrAI-BssHII消化(各酵素1ユニット)によって、それらの最適温度において2時間、最終容量50μlで取り出した。すべての消化されたDNAを1%アガロースゲルに添加し、80ボルトで約2時間泳動させた。次いで、固有のバンド(5275bp)をゲルから取り出し、QIAEXゲル精製キットによって精製し、260nmにおける吸光度によりDNA濃度を計算し、1μg/mlに調整した。
【0050】
このようにして、ベクター(MV DNA:図1)およびインサート(d-190 DNA:図2)を、等モル比で16℃において一晩、1ユニットのT4 DNAリガーゼおよびその独自の反応バッファーを使用して、最終容量10μlでライゲーションした。
【0051】
次いで、XL10 Gold化学的コンピテントセルを、全ライゲーション容量を用いて、標準的形質転換プロトコル(Sambrookら、1989年)に従って形質転換し、LB-Agarプレートにプレーティングし、アンピシリン耐性に関して選択した。コロニーを、DNAプラスミド調製(QIAGEN、ミニ、ミディおよびマキシキット)および制限酵素消化によってスクリーニングした。正しいクローンを、配列決定のためにMWGに送り、DNA Striderソフトウェアを使用して、仮定した配列とアライメントしたこの配列は、100%の同一性を示した。
【0052】
MVベクターの2の位置(SgrAI、4060位およびBssHII、9335位)に挿入されたd-190-3D7遺伝子を図4に表し、そのオープンリーディングフレーム(ORF)を図26に載せた。
【0053】
MVベクターの3の位置(SgrAI、9862位およびBssHII、15137位)に挿入されたd-190-3D7遺伝子を図6に表した。
【0054】
組み換え麻疹-マラリアプラスミドのゲノムの長さ(609位のACCから開始し、21884位のGGTまで)は6の倍数であり、組み換えMV2-3-d-190-3D7ウイルスのレスキューが可能であった。
【0055】
1b)p(+)MV2EZ-d-83-30-38-SgrAI(3D7、23195bp)およびp(+)MV3EZ-d-83-30-38-SgrAI(3D7、23195bp)の構築。
麻疹ベクターを、実施例3aに詳細に記載のように調製した。
【0056】
pZE21MV-d- 190 SgrAIをBstEII-AclIで消化し、d-42断片を切り出し、接着性のBstEII末端およびAclI末端を有するポリリンカーをライゲーションし、中間プラスミドのpZE21MV-d-83-30-38-SgrAI(6436bp)を得た。
【0057】
ポリリンカーの配列は5'-GTCACCAGCGGCCGCAA-3'であった。
【0058】
1μgのpZE21MV-d-83-30-38 SgrAIを、それらの最適温度において約2時間、最終容量50μlでSgrAI-BssHH (各酵素1ユニット)で消化した。すべての消化されたDNAを1%アガロースゲルに添加し、80ボルトで約2時間泳動させた。次いで、固有のバンド(4147bp)をゲルから取り出し、QIAEXゲル精製キットによって精製し、260nmにおける吸光度によりDNA濃度を計算し、1μg/mlに調製した。
【0059】
このようにして、ベクター(MV DNA:図1)およびインサート(d-83-30-38DNA:図2)を、等モル比で16℃において一晩、1ユニットのT4 DNAリガーゼおよびその独自の反応バッファーを使用して、最終容量10μlでライゲーションした。
【0060】
次いで、XL10 Gold化学的コンピテントセルを、全ライゲーション容量を用いて、標準的形質転換プロトコル(Sambrookら、1989年)に従って形質転換し、LB-Agarプレートにプレーティングし、アンピシリン耐性に関して選択した。コロニーを、DNAプラスミド調製(QIAGEN、ミニ、ミディおよびマキシキット)および制限酵素消化によってスクリーニングした。正しいクローンを、配列決定のためにMWGに送り、この配列を、DNA Striderソフトウェアを使用して、仮定した配列とアライメントした。
【0061】
正しいクローンを、配列決定のためにMWGに送り、DNA Striderソフトウェアを使用して、仮定した配列とアライメントしたこの配列は、100%の同一性を示した。
【0062】
MVベクターの2の位置(SgrAI、4060位およびBssHII、8207位)に挿入されたd-83-30-38-3D7遺伝子を図7に表し、そのオープンリーディングフレーム(ORF)を図28に載せた。
【0063】
MVベクターの3の位置(SgrAI、9862位およびBssHII、14006位)に挿入されたd-83-30-38-3D7遺伝子を図9に表した。
【0064】
組み換え麻疹-マラリアプラスミドのゲノムの長さ(609位のACCから開始し、20756位のGGTまで)は6の倍数であり、組み換えMV2-3-d-83-30-38-3D7ウイルスのレスキューが可能であった。
【0065】
1c)p(+)MV2EZ-d-42-SgrAI (3D7、20417bp)およびp(+)MV3EZ-d-42-SgrAI (3D7、20417bp)の構築。
麻疹ベクターを、実施例3aに詳細に記載のように調製した。
【0066】
中間プラスミド(pZE21MV-d-42 SgrAI、3658bp)を挿入された1μgのd-42遺伝子を、それらの最適温度において約2時間、最終容量50μlでSgrAI-BssHII (各酵素1ユニット)消化によって取り出した。すべての消化されたDNAを1%アガロースゲルに添加し、80ボルトで約2時間泳動させた。次いで、固有のバンド(1369bp)をゲルから取り出し、QIAEXゲル精製キットによって精製し、260nmにおける吸光度によりDNA濃度を計算し、1μg/mlに調製した。
【0067】
このようにして、ベクター(MV DNA:図1)およびインサート(d-42DNA:図2)を、等モル比で16℃において一晩、1ユニットのT4 DNAリガーゼおよびその独自の反応バッファーを使用して、最終容量10μlでライゲーションした。
【0068】
次いで、XL10 Gold化学的コンピテントセルを、全ライゲーション容量を用いて、標準的形質転換プロトコル(Sambrookら、1989年)に従って形質転換し、LB-Agarプレートにプレーティングし、アンピシリン耐性に関して選択した。コロニーを、DNAプラスミド調製(QIAGEN、ミニ、ミディおよびマキシキット)および制限酵素消化によってスクリーニングした。正しいクローンを、配列決定のためにMWGに送り、DNA Striderソフトウェアを使用して、仮定した配列とアライメントしたこの配列は、100%の同一性を示した。
【0069】
MVベクターの2の位置(SgrAI、4060位およびBssHII、5429位)に挿入されたd-42-3D7遺伝子を図10に表し、そのオープンリーディングフレーム(ORF)を図30に載せた。
【0070】
MVベクターの3の位置(SgrAI、9862位およびBssHII、11231位)に挿入されたd-42-3D7遺伝子を図12に表した。
【0071】
組み換え麻疹-マラリアプラスミドのゲノムの長さ(609位のACCから開始し、17978位のGGTまで)は6の倍数であり、組み換えMV2-3- d-42-3D7ウイルスのレスキューが可能であった。
【0072】
組み換え麻疹-p-42マラリアウイルスおよびMVワクチンは、同様の細胞変性効果を誘導した(図36)。トランス遺伝子はむしろ安定して発現され、その発現は、ヒト2倍体細胞MRC5におけるウイルスの連続する10継代後に、単一の最初にレスキューされたクローンに由来する、分析したすべての子孫クローンにおいて完全に維持されていた(図37〜38)。
【0073】
組み換えMV-マラリアウイルスおよびMVワクチンの成長曲線は、同じ動態を示した(図39)。
【0074】
1d)p(+)MV2EZ-d- 190*-SgrAI(3D7、24227bp)およびp(+)MV3EZ-d-190*- SgrAI(3D7、24227bp)の構築。
麻疹ベクターを、実施例3aに詳細に記載のように調製した。
【0075】
中間ベクターpZE2 IMVd- 190-SgrAIを鋳型として使用して、PCR反応を実施し、AclI(5434位)部位とClaI(5536位)部位との間に位置するGPIアンカー領域を欠失させた。
【0076】
PCR増幅を、プルーフリーディングPfu DNAポリメラーゼ(Stratagene)を使用して実施した。合成オリゴヌクレオチドプライマーのDNA配列を、MVヌクレオチドに関しては小文字で、非MVヌクレオチドに関しては大文字で示し、関連制限エンドヌクレアーゼ認識部位の配列には下線を引いた。以下のオリゴヌクレオチドプライマーを使用した: For-C ClaI、5'-CCAATAAACGTTTAAT AGatcgattacgcgcgctctagc -3'およびRev-AvrII、5'-gcctttgagtgagctgatacc-3'。
【0077】
For-ClaIは、ClaIおよびBssHII部位のレベルにおいて鋳型と相同であり、オーバーハング(大文字)および2つの終止コドン(TAATAG)、AclI部位(AACGTT)ならびにAclIのための6bpの長い保護部位(CCAATA)を含有する。いわゆるPCR-GPIにおいて、および最終構築体d-190*において、AclIはClaIに近くなる。
【0078】
Rev-Avrllは鋳型と相同である(5704位から5724まで)。
【0079】
PCR産物は207bp長であり、:AclI + AvrIIを用いたその消化およびAclI +AvrIIで予備消化された中間ベクターpZE21MVd-190-SgrAIとのライゲーションにより、pZE21 MVd- 190*- SgrAIを作製した。
【0080】
詳細には、AclI + AvrIIを用いたベクターの消化により、7318bpおよび246bp (欠失されたGPI領域を含有する)の2つのバンドが生じ、7.3kb-断片が、QIAEX II精製キット(Qiagen)を使用することによってアガロースゲルから精製され、消化されたAclI+Avrll PCR(インサート)とライゲーションされ、pZE21 MVd- l90*-SgrAIが得られた。
【0081】
陽性クローンをスクリーニングするために、NcoI消化を実施し、d-190*中間ベクターから7kbの単一バンドならびに元のGPI-アンカー構築体から1.3および5.7kbの2つのバンドが生じた。
【0082】
確定した組み換えp(+)MeV2EZ-d190*およびp(+)MeV3EZ-d190*(図5および図6)を構築するために、「6の規則」に従って、MeVベクターおよび中間プラスミドを、SgrAI+BssHIIを用いて消化し、その後互いをライゲーションした。
【0083】
詳細には、SgrAI+BssHIIで消化されたpZE21 MVd- 190*- SgrAIは、5.2kb +1.3kb + 900bpの3つのバンドを生じた。D-190*配列は5.2kbの断片に含有され、この断片を切断し、精製し、SgrAI+BssHIIで消化されたMeV2EZおよびMeV3EZベクター(19Kb長)と、等モル比で16℃において一晩、1ユニットのT4 DNAリガーゼを使用してライゲーションした。
【0084】
次いで、XL10 Gold化学的コンピテントセルを、全ライゲーション容量を用いて、標準的形質転換プロトコル(Sambrookら、1989年)に従って形質転換し、LB-Agarプレートにプレーティングし、アンピシリン耐性に関して選択した。コロニーを、DNAプラスミド調製(QIAGEN、ミニ、ミディおよびマキシキット)および制限酵素消化によってスクリーニングした。正しいクローンを、配列決定のためにMWGに送り、DNA Striderソフトウェアを使用して、仮定した配列とアライメントしたこの配列は、100%の同一性を示した。
【0085】
MVベクターの2の位置(SgrAI、4060位およびBssHII、9239位)に挿入されたd-190*- 3D7遺伝子を図5に表し、そのオープンリーディングフレーム(ORF)を図27に載せた。
【0086】
MVベクターの3の位置(SgrAI、9862位およびBssHII、15041位)に挿入されたd-190*- 3D7遺伝子を図6に表した。
【0087】
組み換え麻疹-マラリアプラスミドのゲノムの長さ(609位のACCから開始し、21788位のGGTまで)は6の倍数であり、組み換えMV2-3- d-190*-3D7ウイルスのレスキューが可能であった。
【0088】
1e)p(+)MV2EZ-d-83-30-38*-SgrAI(3D7、23105bp)およびp(+)MV3EZ-d-83-30-38*- SgrAI (3D7、23105bp)の構築。
麻疹ベクターを、実施例3aに詳細に記載のように調製した。
【0089】
中間ベクターのpZE 21MV-d-190SgrAIをBstEII-ClaIで消化し、d-42断片およびAclI部位(5434位)とおよびClaI(5536位)部位との間に位置するGPI領域を切り出し、接着性のBstEII 末端およびClaI末端を有するポリリンカーをライゲーションし、中間プラスミドのpZE21MV- d-83-30-38*-SgrAI(6436bp)を得た。
【0090】
ポリリンカーの配列は5'-GTCACCGGGGAATAATAGCGCAT-3'であった。
【0091】
合成オリゴヌクレオチドポリリンカーのDNA配列を、非MVヌクレオチドに関して大文字で示し、関連制限エンドヌクレアーゼ認識部位の配列には下線を引いた。
【0092】
ポリリンカーはBstEII(GTCACC)およびClaI(AT)付着末端、2つの終止コドン (TAATAG)および6の規則を守るためのトリプレット(GCG)を含有する。
【0093】
1μgのpZE21MV-d-83-30-38* SgrAIを、それらの最適温度において約2時間、最終容量50μlでSgrAI-BssHII(各酵素1ユニット)消化した。すべての消化されたDNAを1%アガロースゲルに添加し、80ボルトで約2時間泳動させた。次いで、固有のバンド(4057bp)をゲルから取り出し、QIAEXゲル精製キットによって精製し、260nmにおける吸光度によりDNA濃度を計算し、1μg/mlに調製した。
【0094】
このようにして、ベクター(MV DNA:図1)およびインサート(d-83-30-38* DNA:図2)を、等モル比で16℃において一晩、1ユニットのT4 DNAリガーゼおよびその独自の反応バッファーを使用して、最終容量10μlでライゲーションした。
【0095】
次いで、XL10 Gold化学的コンピテントセルを、全ライゲーション容量を用いて、標準的形質転換プロトコル(Sambrookら、1989年)に従って形質転換し、LB-Agarプレートにプレーティングし、アンピシリン耐性に関して選択した。コロニーを、DNAプラスミド調製(QIAGEN、ミニ、ミディおよびマキシキット)および制限酵素消化によってスクリーニングした。正しいクローンを配列決定のためにMWGに送り、この配列を、DNA Striderソフトウェアを使用して、仮定した配列とアライメントした。正しいクローンを配列決定のためにMWGに送り、DNA Striderソフトウェアを使用して、仮定した配列とアライメントしたこの配列は、100%の同一性を示した。
【0096】
MVベクターの2の位置(SgrAI、4060位およびBssHII、8117位)に挿入されたd-83-30-38*-3D7遺伝子を図8に表し、そのオープンリーディングフレーム(ORF)を、図29に載せた。
【0097】
MVベクターの3の位置(SgrAI、9862位およびBssHII、13919位)に挿入されたd-83-30-38*- 3D7遺伝子を図9に表した。
【0098】
組み換え麻疹-マラリアプラスミドのゲノムの長さ(609位のACCから開始し、20666位のGGTまで)は6の倍数であり、組み換えMV2-3- d-83-30-38*-3D7ウイルスのレスキューが可能であった。
【0099】
1f)p(+)MV2EZ-d-42*-SgrAI(3D7、20345bp)およびp(+)MV3EZ-d-42*-SgrAI(3D7、20345bp)の構築。
麻疹ベクターを、実施例3aに詳細に記載のように調製した。
【0100】
中間ベクターpZE21MVd-42-SgrAI(3658bp)を鋳型として使用して、PCR反応を実施し、AclI(1528位)部位とClaI(1630位)部位との間に位置するGPIアンカー領域を欠失させた。
【0101】
PCR増幅を、プルーフリーディングPfu DNAポリメラーゼ(Stratagene)を使用して実施した。合成オリゴヌクレオチドプライマーのDNA配列を、MVヌクレオチドに関しては小文字で、非MVヌクレオチドに関しては大文字で示し、関連制限エンドヌクレアーゼ認識部位の配列には下線を引いた。以下のオリゴヌクレオチドプライマーを使用した: For-C ClaI、5'-CCAATAAACGTTTAAT AGatcgattacgcgcgctctagc -3'およびRev-AvrII、5'-gcctttgagtgagctgatacc-3'。
【0102】
For-ClaIは、ClaI(1630位)部位およびBssHII(1639位)部位のレベルにおいて鋳型と相同であり、オーバーハング(大文字)および2つの終止コドン(TAATAG)、AclI部位(AACGTT)ならびにAclIのための6bpの長い保護部位(CCAATA)を含有する。いわゆるPCR-GPIにおいて、および最終構築体d-42*において、AclIはClaIに近くなる。
Rev-Avrllは鋳型と相同である(1798位から1818位まで)。
【0103】
PCR産物は207bp長であり、:AclI + AvrIIを用いたその消化およびAclI +AvrIIで予備消化された中間ベクターpZE21MV d-42-SgrAIとのライゲーションにより、pZE21 MV d-42*- SgrAIを作製した。
【0104】
詳細には、AclI + Avrllを用いたベクターの消化により、3412bpおよび246bp (欠失されたGPI領域を含有する)の2つのバンドが生じ、3.4kb-断片が、QIAEX II精製キット(Qiagen)を使用することによってアガロースゲルから精製され、消化されたAclI+AvrII PCR(インサート)とライゲーションされ、pZE21 MVd-42*-SgrAIが得られた。
【0105】
陽性クローンをスクリーニングするために、NcoI消化を実施し、d-42*中間ベクターから3.4kbの単一バンドならびに元のGPI-アンカー構築体から1.3および2.3kbの2つのバンドが生じた。
【0106】
確定した組み換えp(+)MeV2EZ- d42*およびp(+)MeV3EZ- d42*を構築するために、「6の規則」に従って、MeVベクターおよび中間プラスミドを、SgrAI+BssHIIを用いて消化し、その後互いをライゲーションした。
【0107】
詳細には、SgrAI+BssHII+ SpeIで消化されたpZE21 MV d-42*- SgrAIは、1.3kb + 936bp + 800bp + 400bpの4つのバンドを生じた。d-42*配列は1.3kbの断片に含有され、この断片を切断し、精製し、SgrAI+BssHIIにより消化されたMeV2EZおよびMeV3EZベクター(19Kb長)と、等モル比で16℃において一晩、1ユニットのT4 DNAリガーゼを使用してライゲーションした。
【0108】
次いで、XL10 Gold化学的コンピテントセルを、全ライゲーション容量を用いて、標準的形質転換プロトコル(Sambrookら、1989年)に従って形質転換し、LB-Agarプレートにプレーティングし、アンピシリン耐性に関して選択した。コロニーを、DNAプラスミド調製(QIAGEN、ミニ、ミディおよびマキシキット)および制限酵素消化によってスクリーニングした。正しいクローンを、配列決定のためにMWGに送り、DNA Striderソフトウェアを使用して、仮定した配列とアライメントしたこの配列は、100%の同一性を示した。
【0109】
MVベクターの2の位置(SgrAI、4060位およびBssHII、5357位)に挿入されたd-42*- 3D7遺伝子を図11に表し、そのオープンリーディングフレーム(ORF)を図31に載せた。
【0110】
MVベクターの3の位置(SgrAI、9862位およびBssHII、11159位)に挿入されたd-42*-3D7遺伝子を図12に表した。
【0111】
組み換え麻疹-マラリアプラスミドのゲノムの長さ(609位のACCから開始し、17906位のGGTまで)は6の倍数であり、組み換えMV2-3- d-42*-3D7ウイルスのレスキューが可能であった。
【0112】
1g)p(+)MV2EZ-d-190- SgrAI(FCB1、24083bp)およびp(+)MV3EZ-d-190-SgrAI(FCB1、24083bp)の構築。
まず第1に、FCB 1株のMSP-1のための合成遺伝子の、中間プラスミドpZE21MV-SgrAIへのクローニングを実施し、3D7株のMSP-1由来のシグナルペプチドおよびGPI-アンカー領域を保った。D-190遺伝子(FCB1)を、pZE23f-GX-190Hと呼ばれる中間ベクターから以下のように段階的に得た:
i).1μgのプラスミドpZE21MV-d-190-SgrAI(3D7)を、それらの最適温度において2時間、最終容量50μlで、HindIII + AclI制限酵素を用いて消化した。すべての消化されたDNAを1%アガロースゲルに添加し、80ボルトで約2時間泳動させた。次いで、ベクターに対応する固有のバンド(2558 bp)をゲルから取り出し、QIAEXゲル精製によって精製し、260nmにおける吸光度によりDNA濃度を計算した。
【0113】
ii).MSP-1/FCB1のd-42部分を増幅し、回収するために、pZE23f-GX-190Hを鋳型として使用してPCR反応を実施した。PCR増幅を、プルーフリーディングPfu DNAポリメラーゼ(Stratagene)を使用して実施した。合成オリゴヌクレオチドプライマーのDNA配列を、MVヌクレオチドに関しては小文字で、非MVヌクレオチドに関しては大文字で示し、関連制限エンドヌクレアーゼ認識部位の配列には下線を引いた。
【0114】
以下のオリゴヌクレオチドプライマーを使用し、ρZE23f-GX-190H配列を設計した: For-1 FCB1、5'-CCCAAGCTTccaggtggtcaccggAgagctgtcactcc -3'およびRev-1 FCB1, 5'- GCCTGCaacgttGCTagagctggagcaGaaGatcccgtcg -3'。
【0115】
For-1 FCB1は、4509位から4538位の鋳型に相同であり、BstEII部位(ggtcacc)を含む。A(大文字)は、鋳型においてはtであり、SgrAI部位を除去するために改変されている。For-1 FCB1は、3bpの長い保護部位(CCC)の後ろにHindIII部位(AAGCTT)およびオーバーハング(大文字)を含有する。
【0116】
Rev-1 FCB1はAclI部位(aacgtt)を含有し、その前に6-bpの保護部位(GCCTGC)が付く。Rev-1 FCB1は、セリンをコードするトリプレットのGCTが導入され、6の規則に従っており、ポリ(A)部位を回避するために2つのaがGに改変されている。
【0117】
得られたPCR-HindIII-AclI(1.1kb)をHindIII + AclI消化し、等モルで16℃において一晩、HindIII + AdIIを用いて予備消化されたpZE21MV-d- 190- SgrAI(ステップi)とライゲーションし、pZE21MV-d-42-SgrAI-FCB1(3657bp)を得た。次いで、XL10 Gold化学的コンピテントセルを、全ライゲーション容量を用いて、標準的形質転換プロトコル(Sambrookら、1989年)に従って形質転換し、LB-Agarプレートにプレーティングし、アンピシリン耐性に関して選択した。コロニーを、DNAプラスミド調製(QIAGEN、ミニ、ミディおよびマキシキット)およびHindIII+AclIを用いた制限酵素消化によってスクリーニングした(期待される断片2558bp + 1099bp)。
【0118】
iii).ステップiiに記載のように得られたpZE21MV-d-42- SgrAI-FCB1を、HindIII + BstEII(HindIII、428位およびBstEII、440位)で消化し、開いたベクターに対応する特有のバンド(3645bp)を1%アガロースゲルに添加し、ゲルから取り出し、QIAEXゲル精製によって精製し、260nmにおける吸光度によりDNA濃度を計算した。
【0119】
iv).pZE23f-GX-190HをHindIII + BstEIIで消化し、d-83-30-38/FCB1断片に対応する、3679bpの特有のバンド(インサート)を、前記のようにゲルから精製した。
【0120】
v).pZE21MV-d-42-SgrAI-FCB1から得られた、HindIII + BstEIIにより消化された3657bpの断片(ベクター)を、pZE23f-GX- 190Hから消化によって得た、d-83-30-38/FCB1を含有する3679bpのHindIII + BstEII断片(インサート)とライゲーションした。ライゲーションを、等モルで16℃において一晩、1ユニットのT4 DNAリガーゼを使用して実施し、pZE21MV-d-190-SgrAI-FCB1(7324bp)を得た。その後、XL10 Gold化学的コンピテントセルを、全ライゲーション容量を用いて、標準的形質転換プロトコル(Sambrookら、1989年)に従って形質転換し、LB-Agarプレートにプレーティングし、アンピシリン耐性に関して選択した。コロニーを、DNAプラスミド調製(QIAGEN、ミニ、ミディおよびマキシキット)および制限酵素消化によってスクリーニングした。
【0121】
p(+)MV2EZ-d-190-SgrAI-FCB1およびp(+)MV3EZ-d-190-SgrAI-FCB1を構築するために、麻疹ベクターを、実施例3aに詳細に記載のように調製した。
【0122】
中間プラスミド内に挿入された1μgのd-190/FCB1遺伝子(pZE21MV-d-190 SgrAI-FCB1、7324bp)を、SgrAI-BssHII消化(各酵素1ユニット)によって、それらの最適温度において2時間、最終容量50μlで取り出した。すべての消化されたDNAを1%アガロースゲルに添加し、80ボルトで約2時間泳動させた。次いで、固有のバンド(5035bp)をゲルから取り出し、QIAEXゲル精製キットによって精製し、260nmにおける吸光度によりDNA濃度を計算し、1μg/mlに対して調整した。
【0123】
このようにして、ベクター(MV DNA:図1)およびインサート(d-190/FCB1 DNA:図3)を、等モル比で16℃において一晩、1ユニットのT4 DNAリガーゼおよびその独自の反応バッファーを使用して、最終容量10μlでライゲーションした。
【0124】
次いで、XL10 Gold化学的コンピテントセルを、全ライゲーション容量を用いて、標準的形質転換プロトコル(Sambrookら、1989年)に従って形質転換し、LB-Agarプレートにプレーティングし、アンピシリン耐性に関して選択した。コロニーを、DNAプラスミド調製(QIAGEN、ミニ、ミディおよびマキシキット)および制限酵素消化によってスクリーニングした。正しいクローンを、配列決定のためにMWGに送り、DNA Striderソフトウェアを使用して、仮定した配列とアライメントしたこの配列は、100%の同一性を示した。
【0125】
MVベクターの2の位置(SgrAI、4060位およびBssHII、9095位)に挿入されたd-190- FCB1遺伝子を図13に表し、そのオープンリーディングフレーム(ORF)を図32に載せた。
【0126】
MVベクターの3の位置(SgrAI、9862位およびBssHII、14897位)に挿入されたd-190- FCB1遺伝子を図14に表した。
【0127】
組み換え麻疹-マラリアプラスミドのゲノムの長さ(609位のACCから開始し、21884位のGGTまで)は6の倍数であり、組み換えMV2-3- d-190-FCB1ウイルスのレスキューが可能であった。
【0128】
トランス遺伝子はむしろ安定して発現され、その発現は、ヒト2倍体細胞MRC5におけるウイルスの連続する10継代後に、単一の最初にレスキューされたクローンに由来する、分析したすべての子孫クローンにおいて完全に維持されていた(図40)。
【0129】
組み換えMV-マラリアウイルスおよびMVワクチンの成長曲線は、同じ動態を示した(図41)。
【実施例2】
【0130】
DiCo1核酸配列の設計
アミノ酸DiCo1配列(ecto、トランスおよび細胞質ドメイン: aa97〜622)から開始し、DNA Strider softwareを使用して、DiCo1 DNA変性配列と、BLASTアライメント後にDiCo1に最も類似した配列を表す、選択されたPfAMA1遺伝子(受託番号AAG141.1)とを比較して、対応する核酸配列を設計した。
【0131】
5'末端に、適切な特有の制限部位を、クローニング部位として付加し(MluIおよびSgrAI)、その後ろに最適KOZAC配列およびヒト最適化シグナルペプチド(SP)を付加した。3'末端に、2つの終止コドンおよびBssHIIクローニング部位を付加した。このスキームに従って、本発明者らは、(麻疹ベクター内におけるさらなる発現のために「6の規則」を尊重して) DiCo1タンパク質のアンカー型および分泌型をコードする2種のヌクレオチド配列を設計した:第1の遺伝子は外形質(ectoplasmasmic)、膜貫通および細胞質ドメインを含み(図18)、一方、第2の遺伝子はectoドメイン単独に対応する(図19)。この2種の配列は、AT%含有量を減少させ、ポリ(A)配列およびRNAの不安定モチーフを回避するために、GENEARTによってヒトコドンに最適化されている。
【0132】
DiCo1の完全ORFおよびDiCo1のectoドメインORFを、それぞれ図34および35に載せる。
【実施例3】
【0133】
組み換えMV-PfΑMA-1プラスミドの構築
すべてのクローニング手順は、Sambrookら(1989年)に記載の技術により実施した。
【0134】
PfΑMA-1、特に分泌型およびアンカー型のDiversity Covering配列1(DiCo1)は、いずれも化学的に合成し、ヒトコドンに最適化した。
【0135】
最適化されたコドンのDiCo1分泌型およびアンカー型を、SgrAI+BssHIIで消化し、予備消化したMeV2EZおよびMeV3EZベクター(19Kb長)と、等モル比で16℃において一晩、1ユニットのT4 DNAリガーゼを使用してライゲーションし、以下の組み換えMV-PfΑMA-1プラスミド:p(+)MV2EZ-DiCo1-完全(図20)、p(+)MV3EZ-DiCo1-完全(図21)、p(+) MV2EZ-DiCo1-ecto(図22)およびp(+)MV3EZ-DiCo1-ecto(図23)を得た。
【実施例4】
【0136】
組み換えMV-PfCSプラスミドの構築
p(+)MV2EZ-CS-SgrAI(20219bp)およびp(+)MV3EZ- CS-SgrAI (20219bp)の構築
すべてのクローニング手順は、Sambrookら(1989年)に記載の技術により実施した。
【0137】
中間ベクターpAdApt35Bsu.CS.Pfalc.aa-sub.gccにクローングされたPfCS1をPCRによって増幅し、確定的MVベクターに直接クローニングし、2種の組み換えMV-PfCSプラスミド: p(+) MV2EZ-CSおよびp(+)MV3EZ-CSを得た。
【0138】
詳細には、CS遺伝子(図15)を増幅し、回収するために、pAdApt35Bsu.CS.Pfalc.aa-sub.gccを鋳型として使用してPCR反応を実施した。PCR増幅は、プルーフリーディングPfu DNAポリメラーゼ(Stratagene)を使用して実施した。合成オリゴヌクレオチドプライマーのDNA配列を、MVヌクレオチドに関しては小文字で、非MVヌクレオチドに関しては大文字で示し、関連制限エンドヌクレアーゼ認識部位の配列には下線を引いた。
【0139】
以下のオリゴヌクレオチドプライマーを使用し、: pAdApt35Bsu.CS.Pfalc.aa-sub.gcc配列上に設計した: For-SgrAI、5'- ACTTCTCACCGGTGTggaagcttgccaccatgat-3'およびRev-BssHII-CS、5'- TAGCGCGCtctagaggatccttatcagc -3'。
【0140】
For-SgrAIはHindIII部位(aagctt)を含む1356位から1375位までが鋳型と相同である。それは、6-bpの長い保護部位(ACTTCT)の後ろにオーバーハング(大文字)およびSgrAI制限部位(CACCGGTG)を含有する。
【0141】
Rev-BssHII-CSは、オーバーハング(大文字)およびBssHII制限部位(GCGCGC)を含有し、Rev-BssHII-CSは、PCR-CS(1187bp)においてXbaI(tctaga)に近い。得られたPCR-CSをSgrAI + BssHIIで消化して、SgrAI+BssHIIで予備消化したMeV2EZおよびMeV3EZベクター(19Kb長)と、等モル比で16℃において一晩、1ユニットのT4 DNAリガーゼを使用してライゲーションし、それぞれp(+)MV2EZ-CS-SgrAI(20219bp、図16)およびp(+)MV3EZ- CS-SgrAI(20219bp、図17)を得た。CSのORFを図33に載せる。
【0142】
次いで、XL10 Gold化学的コンピテントセルを、全ライゲーション容量を用いて、標準的形質転換プロトコル(Sambrookら、1989年)に従って形質転換し、LB-Agarプレートにプレーティングし、アンピシリン耐性に関して選択した。コロニーを、DNAプラスミド調製(QIAGEN、ミニ、ミディおよびマキシキット)および制限酵素消化によってスクリーニングした。正しいクローンを、配列決定のためにMWGに送り、DNA Striderソフトウェアを使用して、仮定した配列とアライメントしたこの配列は、100%の同一性を示した。
【実施例5】
【0143】
細胞およびウイルス
細胞を、Vero細胞(アフリカミドリザルの腎臓)のために5%(FCS)を補充したダルベッコ変法イーグル培地(DMEM)、293T細胞(ヒト胎児腎臓)のために10% FCSおよび1%ペニシリン/ストレプトマイシン(P/S)を補充したダルベッコ変法イーグル培地、MRC-5(ヒト胎児線維芽細胞)のためにGlutamax(F12)および10% FCSを補充したDMEM、293-3-46のために10% FCSおよび1.2mg/mlのG 418を補充したDMEMにおいて単層で維持した。
【0144】
MVウイルスストックを、約107pfu/mlの力価に達するまで成長させるため、組み換えウイルスおよびワクチンのEdmoston Zagreb株をMRC-5細胞において増殖させ、プラーク精製を、シンシチウムを35mmのMRC-5細胞培養液に移し、まず10cmの皿、その後175cmのフラスコに増殖させることによって実施した。シンシチウムが約90%形成されたと判断されたとき、ウイルスストックを175cm2の培養から作製した。いわゆる「無細胞ウイルス分画」に対応する培地を回収し、凍結および解凍を3回行い、細胞のデブリを回避するために遠沈させた。次いでこの培地を-80℃で保存した。いわゆる「細胞結合型ウイルス分画」に対応する細胞を、3mlのOPTIMEM(Gibco BRL)に削り取り、その後3回凍結および解凍を行い、遠沈させ、上清(surnatant)を-80℃で保存した。
【実施例6】
【0145】
プラスミドのトランスフェクションおよびMVウイルスのレスキュー
トランスフェクトされた場合、約50〜70%の集密に達するように、293T細胞を35mmのウェルに播種した。トランスフェクションの4時間前、培地を10%FCSを含有する3mlのDMEMに取り替えた。すべての組み換えプラスミドを、QIAGENプラスミド調製キットに従って調製した。DNAのリン酸Ca2+共沈法のキットはInvitrogenによる。
【0146】
細胞に、以下の最終濃度でプラスミドを共トランスフェクトした:pCA-L 0.5μg、pCA-N 0.5μg、pCA-P 0.1μg、pCA T7 1μgおよび組み換え麻疹-マラリアプラスミド4μg。H2Oで希釈された5種すべてのプラスミドを、2MのCaCl2を含有するエッペンドルフチューブに加え、この混合液を、HEPESバッファーを含有する別のエッペンドルフチューブに振とう状態下で加え、室温(RT)において30分間インキュベートした。このようにして、共沈物を培養液に滴下し、37℃、5%CO2において約18時間トランスフェクションを実施した。その後、トランスフェクション培地を、10% FCSを含有する3mlのDMEMに取り替えた。
【0147】
麻疹のNタンパク質およびPタンパク質を安定に発現する293-3-46ヘルパー細胞(ヒト胎児腎臓細胞)、ならびにT7 RNAポリメラーゼを使用する、組み換え麻疹-マラリアワクチンウイルスを得るための別の方法を、これ以降に記載する。ウイルス性RNAポリメラーゼ(巨大タンパク質、L)を、細胞を、15ngのプラスミドpeMCLaを用いて共トランスフェクトすることによって発現させた。トランスフェクション効率を改善するために、300ngのpSC6-T7 Neoを加えた。カルシウムリン酸法を、トランスフェクションのために使用した。
【0148】
細胞がまだ半集密のとき、トランスフェクションの3〜4日後に最初のシンシチウムが現れた。シンシチウムの形成がより容易に進行するように、各35mmウェルのほぼ集密の細胞単層を、10cmの皿に移した。個々のシンシチウムを、300μlのトランスフェクション培地にとり、700μlのOPTIMEMを含有する滅菌エッペンドルフチューブに入れ、凍結および解凍を3ラウンド行い、-80℃で保存した。
【実施例7】
【0149】
プラークアッセイによるウイルスの滴定
ウイルス調製液の10倍段階希釈を、OPTIMEMを使用して、最終容量0.5mlまで実施した。個々の希釈液を35mmのVero細胞培養物に加えた。ウイルス吸着の1時間後、接種材料を取り除き、感染細胞に、5% FCSおよび1%の低融点アガロース(LMPアガロース)を含有する2mlのDMEMを重ねた。37℃および5% CO2におけるインキュベーション5日後、培養物を1mlの10% TCAに1時間固定し、次いで、30分間UV架橋した。アガロースの重ねを除去後、細胞単層を、4%エタノールに溶解したクリスタルバイオレットで染色し、水で洗浄し、倒立顕微鏡下でプラークをカウントした。
【実施例8】
【0150】
組み換えウイルスのMRC-5におけるウイルス連続継代
レスキューされたウイルスを、MRC5細胞において10回連続継代させ、直径10cmのプレートに播種し、これらを標準的MVウイルスおよび組み換えMVウイルスに、0.01 PFU/細胞のMOIで感染させた。単層を完全に感染させた後、各培養液の1%上清を使用して、その後のMRC5細胞単層を感染させた。トランス遺伝子の発現および安定性を試験するために、継代1、5および10由来のウイルスを、ウェスタンブロットおよび免疫蛍光法による、発現のさらなる特徴付けに使用した。
【実施例9】
【0151】
ウェスタンブロットおよび免疫蛍光法
MVおよびマラリア両方の発現を分析するために、ウェスタンブロットおよび免疫蛍光法を実施した。
【0152】
ウェスタンブロットのために、35mmの皿(1〜5×105)に播種したVero細胞を、90%の集密に関して翌日モニターし、対照としてMVEZを含む、細胞結合型ウイルス分画由来のクリアなウイルス懸濁液を、0.1 MOI(Multiplicity Of Infection)を使用して感染させた。約80%のシンシチウム形成が観察されたとき、細胞をまずPBSを用いて洗浄し、次いで1mlのPBSに削り取り、エッペンドルフチューブに回収し、2000 RPM/4分において、遠心分離にかけた。その後、細胞を、プロテアーゼ阻害剤カクテル(Complete Mini、Roche、1 836 153)を補充した70μlの溶解バッファー(1% NP-40、50mM Tris pH8、150mM NaCl)を用いて、5分/RTで溶解した。上清を13000 RPM/5分において遠心分離にかけることによってクリアにし、新しいチューブに移し、30μlの4×ローディングバッファー(Invitrogen)を加え、試料を混合し、95℃/2分で煮沸し、遠沈させ、-20℃で保存した。
【0153】
SDS-PAGE泳動を実施し、還元条件において1×ランニングバッファーを使用し、200V(開時100〜125mA、終了時60〜80mA)において50分間、NuPAGE 12% Bis-アクリルアミドゲル中を泳動させた。
【0154】
次いで、セミドライ法を使用して、分離された細胞タンパク質をニトロセルロース膜に14V/1h30において転写した。
【0155】
一次抗体として、少なくとも1:30000にPBSTで希釈されたMSP1-p-83に対するウサギポリクローナル抗体および少なくとも1:50000に希釈された、MSP1- p-42,*に対するウサギポリクローナル抗体を使用した。二次抗体は、化学発光増強キット(enhanced chemiluminescence kit)(ECL(商標)、Amersham LifeScience)によってバンドの可視化が可能な、ホースラディッシュペルオキシダーゼで標識されたブタ抗ウサギ抗体であった。
【0156】
免疫蛍光法のために、Vero細胞を、35mmのウェル中の24mm×24mmのガラスカバースリップに播種し、一晩培養し、レスキューされた組み換えウイルスに感染させた。感染3日後に、カバースリップ上の細胞を、PBS中3.7%のパラホルムアルデヒドを用いて固定させ、0.1% TX-100を用いて透過処理し、ブロッキング溶液(1% BSAを含有するPBS)を用いて1時間洗浄し、特異的抗体を用いて染色した。p-42のp-19部分においてEGF様ドメインを認識する、マウスハイブリドーマ上清mAb 5.2を1: 100希釈で使用し、次いで1:250に希釈したFITCH標識ヤギ抗マウス血清を使用した。
【実施例10】
【0157】
成長動態曲線
35mm皿に播種したMRC5細胞(1〜5×105)を、90%の集密に関してモニターし、対照としてMVEZを含む、細胞結合型ウイルス分画由来のクリアなウイルス懸濁液を、0.1 MOIを使用して感染させた。いわゆる「無細胞ウイルス分画」および「細胞結合型ウイルス分画」に対応する試料を、1週間の間毎日回収し、滴定した。
【実施例11】
【0158】
マウスの免疫化
上記のレスキューされた組み換えMV-マラリアウイルスの免疫原性力を、MV感染に対して感受性であるトランスジェニックマウスIFNAR/CD46において免疫化試験を実施することによって証明した。動物を衛生的状況下で飼育し、6〜8週齢において免疫化した。以下に、2種の組み換え麻疹-マラリアウイルス:MeV2EZ-d-p42-SgrAI(GPIアンカー型)およびMeV2EZ-d-p42*(分泌型)によるマウス免疫化の例を提供する。免疫化を、各組み換えMV-マラリアの105 PFUを使用する筋肉注射で、0、4および8週において3回の注射で実施した。組み換え-空麻疹(rMVEZ13-空クローン)を用いて免疫化されたマウスは、陰性対照として機能する。UV不活性化rMVを対照として使用して、免疫反応の活性化に関するウイルス複写の効果を決定した。MVベクター化抗原の免疫反応を、精製d-42タンパク質(0.5mg/ml)と比較して試験した:マウスを、不完全フロイントアジュバント中の20μgのタンパク質を用いて皮下に免疫化した。
【0159】
免疫化IFNAR/CD46マウス(6匹/試験群および3匹/対照群)由来の血清中のMV-特異的抗体の存在を、IgG抗体検出のために、麻疹ウイルスEIA bulk(ATCC VR-24)でコーティングされた96-マイクロウェルプレート使用するELISAによって決定した。タンパク質を、0.05Mの炭酸バッファー(pH 9.4)を用いて0.6μg/mlに希釈し、100μl/ウェルを96ウェルマイクロタイタープレートに加えた。このプレートを、4℃において一晩インキュベートし、PBS/0.05% Tween 20(PT)(ph7.4)を用いて洗浄し、PT(0.1ml/ウェル)-10%BSAとともに37℃において60分間インキュベートし、PTを用いて再度洗浄した。被験血清の2倍段階希釈液を加え(100μl/ウェル)、プレートを37℃において60分間インキュベートした。このプレートをPTを用いて洗浄し、PTで1:2000に希釈した100μlのヤギ抗マウスIgG HRPとともに37℃において30分間インキュベートした。このプレートをPTを用いて洗浄し、100μlのOPD(o-フェニレンジアミン(Phenylendiamin)、Fluka 78411)とともにインキュベートした。反応を、3〜4分後に停止させた。プレートを、MicroElisa Readerで、波長490nmにおいて読み取った。陰性対照より3倍高い示度を、陽性の反応として判定した。
【0160】
免疫化CD46マウス(少なくとも10匹/試験群)の血清中のMV-マラリア-特異的抗体の存在を、ELISAアッセイによって決定した。簡潔には、96マイクロウェルプレートを、炭酸バッファーpH9.4を用いて希釈した50ng/ウェルのMSP-1-d42 3D7株でコーティングした。プレートを、4℃において一晩インキュベートし、PBS/0.05% Tween 20(PT)を用いて洗浄した。その後、非特異的相互作用を、PTに溶解した10%脱脂乳を用いて、37℃において1時間ブロックし、ウェルを、PTを用いて再度洗浄した。プレートを、マウス血清のさまざまな希釈液(1:200から出発して、その後2倍段階希釈)、ペルオキシダーゼ標識ヤギ抗マウスIgGおよびOPD基質とともに連続してインキュベートした。光学密度値を490nmで測定した。カットオフ・バックグラウンドレベル(MV免疫化マウス由来血清の平均値×係数2.1)を上回る値を陽性と考えた。力価は、限界の希釈倍率の逆数として表した。
【0161】
麻疹に対する液性免疫反応を図Aに示す。マラリアp42に対する液性免疫反応を図Bに示す。
【0162】
【化1】

【0163】
【化2】
【実施例12】
【0164】
プラーク精製による、欠陥干渉粒子(DI)からのマラリア抗原を発現する組み換え麻疹ウイルスの精製。
パラミクソウイルス(Paramyxoviruses)、特に麻疹ウイルスによる一定数の継代後、欠陥干渉粒子(DI)の蓄積が起こるであろうことが文献により知られている(23、24)。これらのDIがさまざまな欠陥:ワクチンの安全性に関する負の影響、生産におけるウイルス収率に関する負の影響、ゲノムの不安定性およびワクチン接種後の免疫反応の抑制を発生させることが記載されている。本発明者らの新しい組み換えウイルスに伴うこのようなDIを回避するために、本発明者らは、本発明者らが293T細胞の代わりにMRC5細胞を使用することを除いて、実施例6に記載のようにプラーク精製の方法を適用した。クリアな十分確定されたシンシチウムの形成後、新鮮なMRC5組織培養におけるさらなる継代のために、本発明者らは、顕微鏡下でマイクロピペットを用いてこのような材料を吸引した。
【実施例13】
【0165】
限界希釈による、欠陥干渉粒子(DI)からのマラリア抗原を発現する組み換え麻疹ウイルスの精製。
限界希釈技術を、マイクロプレートにおいて適用した。すべてのウェルにおいて、MRC5細胞の新鮮な単層を発生させた。組み換え麻疹-マラリアウイルスを含有するウイルス懸濁液を、2倍希釈で調製した。シンシチウムが検出された最新の単層のウェルから、上清をピペットで吸引した。上清を、MRC5細胞を含有する懸濁液と混合した。この混合液を、4℃において1時間インキュベートした。最終的に、それを小型のコースターフラスコ(Costar flask)に移し、35℃/5%CO2においてインキュベートし、10日後に組み換え麻疹-マラリアウイルスを精製するために収集した。
【実施例14】
【0166】
麻疹-マラリア混合ワクチンの作製
上記の組み換え麻疹-マラリアウイルスのワーキングシードを、1750cm2のローラーボトル中のMRC5細胞の単層上で、35℃において10日間インキュベートした。細胞を、健康状態および集密に関して毎日モニターした。シンシチウム形成が最も高いレベルである10日目に、上清を液体窒素中の保存用鋼制シリンダーにポンプで汲み上げた。同じ手順を、2日後に繰り返した。すべての試験(ウイルス力価、ゲノムの安定性、ウイルスの安全性、細胞の安全性、化学分析、無菌性など)を実施後、収集物を解凍し、ゼラチン、ソルビトール、アミノ酸および他の糖を含有する安定剤と、最終希釈の105になるように混合した。自動充填機を用いて、小型の凍結乾燥ボトル(F3)に0.5mlずつ接種した。特別に計算した凍結乾燥プログラムを使用して、凍結乾燥プロセスの間の生成物の最大生存を保証した。
(参考文献)
【技術分野】
【0001】
本発明は、いくつかの熱帯熱マラリア原虫(Plasmodium falciparum)抗原をコードする異種核酸を含む、異なる弱毒化組み換え麻疹-マラリアベクターを含有する麻疹-マラリア混合ワクチンに関する。好ましくは、本発明は、熱帯熱マラリア原虫のスポロゾイト周囲(CS)タンパク質、熱帯熱マラリア原虫のメロゾイト表面タンパク質1(MSP-1)およびその誘導体(p-42; p-83-30-38)をそのグリコシル化型および分泌型でコードする核酸、ならびに熱帯熱マラリア原虫頂端膜抗原1(AMA1)をそのアンカー型および分泌型でコードする核酸を含むウイルスベクターに関する。ウイルスベクターは、ワクチンとして使用され、対象となる遺伝子の送達において効率的であり、関連免疫細胞に効率的に結合し感染する株に基づく、弱毒化麻疹ウイルスに由来する。好ましい実施形態において、CS、MSP1およびAMA1タンパク質は、これらが哺乳動物、好ましくはヒトにおいて強力な免疫反応を引き起こすように該ウイルスから産生され、これらのタンパク質の発現は、ヒトコドンへの最適化により上昇する。さらに、本発明は、マラリアの予防的治療における組み換えワクチンの使用に関する。
【背景技術】
【0002】
麻疹ウイルス
本発明は、熱帯熱マラリア原虫(Pf)の抗原を発現する弱毒化組み換え麻疹ウイルスを含有するワクチンに関し、麻疹およびマラリア抗原の両方に対する免疫をもたらす、組み換え麻疹-マラリアワクチンの調製のためのそれらの使用に関する。
【0003】
麻疹ウイルス(MV)は、モノネガウイルス(Mononegavirales)目、すなわち、非分節型マイナス鎖RNAゲノムを有するウイルスのメンバーである。MVの非分節型ゲノムは、アンチメッセージ・ポラリティ(antimessage polarity)を有し、したがってこのゲノムRNAは、in vivoまたはin vitroのどちらでも翻訳されない。さらにMVの非分節型ゲノムは、リボ核タンパク質(RNP)複合体(以下を参照されたい)の形態の3種のウイルスタンパク質と特異的に結合した場合にのみ生物学的に活性である。非分節型(-)鎖RNAウイルスの転写および複製およびウイルス粒子としてのそれらの集合は、広範囲にわたって再考されている(1)。麻疹ウイルスの転写および複製は、感染細胞の核に関与するのではなく、むしろ感染細胞の細胞質において起こる。麻疹ウイルスのゲノムは、6種の遺伝子(N、P、M、F、HおよびLと称される)由来の6種の主要な構造タンパク質をコードする遺伝子、ならびに構成的免疫反応の相殺に関与し、転写/複写の制御に関与するP遺伝子のCおよびVに由来するさらなる2種の非構造タンパク質をコードする遺伝子を含む。遺伝子の順番は、3'N、P(CおよびVを含む)、M、F、HおよびL 5'である。さらに、3'-末端領域から、約50ヌクレオチドの短いリーダーRNAが転写される。言及した遺伝子は、ウイルスのリボヌクレオカプシド(RNP)のタンパク質、すなわち、核タンパク質(N)、リンタンパク質(P)および大型のポリメラーゼ/レプリカーゼタンパク質(L)をそれぞれコードし、これらはゲノムRNAに非常に堅く結合し、RNPを形成する。他の遺伝子は、ヘマグルチニン(H)タンパク質、融合(F)タンパク質およびマトリクス(M)タンパク質を含むウイルスエンベロープのタンパク質をコードする。MV遺伝子の転写は漸減勾配に従い、ポリメラーゼがゲノムの鋳型に作用するとき、上流遺伝子から作られるRNAを、下流遺伝子から作られるRNAより多く合成する。この不連続な転写様式において、mRNAはキャップされ、ポリアデニル化される。逆に、この複写様式において、Lタンパク質は完全長のアンチゲノムRNAおよびゲノムRNAを作製し、完全長のアンチゲノムRNAおよびゲノムRNAは直ちにN、PおよびLタンパク質にカバーされ、感染性子孫RNPを形成する。
【0004】
麻疹ウイルスは、1954年に単離され、EndersおよびPeeblesは、麻疹に冒された小児であるDavid Edmostonの血液を初代ヒト腎臓細胞に接種し、得られたMVのEdmoston株(2)を、その後さまざまな細胞系における成長に適合させた。ニワトリ胚、ニワトリ胚線維芽細胞(CEF)および/またはイヌ腎臓細胞ならびにヒト二倍体細胞に対する適合により、弱毒化Edmonston AおよびB(3)、Zagreb(EZ)ならびにAIK-Cシードが作製された。Edmonston Bは、最初のMVワクチンとして、1963年に認可された。CEFに対するEdmonston AおよびBのさらなる継代は、より弱毒化されたSchwarzおよびMoratenウイルスを産生し(3)、それらの配列が同一であることが最近示された(4; 5)。Edmonston Bワクチンは反応原性(reactogenic)なので、1975年に放棄され、Schwarz/ Moratenワクチンに取って代わられた。いくつかの他のワクチン株:日本ではAIK-C、Schwarz F88、CAM70、TD97、ロシアではLeningrad-16、ならびにEdmonston Zagrebもまた使用される。CAM70およびTD97中国株は、Edmonstonに由来しない。Schwarz/ MoratenおよびAIK-Cワクチンは、CEFに対して作製された。Zagrebワクチンは、ヒト二倍体(WI-38)に対して作製された。今日、Schwarz/Moraten、AIK-CおよびEZワクチンは、一般的に使用されるが(6)、原理的には、すべてが1つの特有のMV血清型であるこれらの弱毒化ワクチン株の任意の1つは、安全であり、長期持続性免疫反応を誘導することが証明されており、本発明の目的のために使用できる。
【0005】
MVワクチンは、単回または2回の低用量注射後に生涯免疫を誘導する。麻疹に対する保護は、抗体によって、およびCD4およびCD8のT細胞によって媒介される。MV特異的抗体およびCD8細胞の持続は、ワクチン接種後25年ほどの間示されている(7)。
【0006】
MVワクチンは、大部分の国において大規模な生産が容易であり、低コストで分配できる。MVゲノムの弱毒化は多数の突然変異の有利な組み合わせによりもたらされるので、ワクチンは非常に安定であり、病原性への復帰変異は今までにまったく観察されていない(6)。
【0007】
安全性に関しては、MVは、細胞質において排他的に複製するので、宿主DNA内への統合の可能性はない。これらの特徴から、生きた弱毒化MVワクチンを、多価ワクチン接種ベクターとして使用するための魅力的な候補である。このようなワクチンは、ベクターウイルスそれ自体に対するように、他の病原性物質に対する、長期持続性免疫保護の誘発において効率的であることを証明できる。
【0008】
Martin Billeterおよび同僚は、Edmonston MVのアンチゲノムに対応するcDNAをクローニングし、国際特許出願WO97/06270に記載のように、ウイルスをレスキューする初めてかつ効率的な逆遺伝学手順を確立した(8)。組み換え麻疹ウイルスは、ヘルパー細胞系293-3-46から回収され、安定してトランスフェクトされ、MV NおよびPタンパク質ならびにバクテリオファージT7 RNAポリメラーゼを発現する。任意の変異体または組み換えMVのレスキューのために、次いで、ヘルパー細胞系は、Lタンパク質をコードする発現するプラスミドを一時的にトランスフェクトされ、最も重要には、子孫MVを発生させるために適合性のある、任意の突然変異された、または組み換えのアンチゲノムRNAを得るために適切に構築された任意のアンチゲノムプラスミドを一時的にトランスフェクトされる。一時的トランスフェクトステップは、好ましくは内在するT7 RNAポリメラーゼによって、まず転写に導かれる。得られたアンチゲノムRNAは、直ちに(発生したときの状態で)ウイルスN、PおよびLタンパク質によってカバーされ、ゲノムRNPが産生されるアンチゲノムRNPを得る。第2に、ゲノムRNPは結合したLによって転写され、すべてのウイルスmRNAおよびそれぞれのタンパク質を得る。最終的に、ゲノムRNPおよびアンチゲノムRNPの両方が複製よって増幅される。
【0009】
この手順のわずかな変形において、安定にトランスフェクトされた293-3-46ヘルパー細胞を使用するのではなく、原特許明細書に詳細に説明された5種のプラスミド:N、PおよびT7ポリメラーゼ(ヘルパー細胞系を作り出すために以前使用した)をコードするプラスミドならびにLをコードするプラスミドおよびアンチゲノムプラスミドのすべてを同時に使用して、市販の293T細胞を一時的にトランスフェクトした。「完全な一時的トランスフェクション」手順において、変異体発現プラスミドの使用が可能であり、T7 RNAポリメラーゼ全体の使用を避け、代わりに内在RNAポリメラーゼIIを利用してLタンパク質およびアンチゲノムをさらに発現させることが可能であることに留意されたい(9)。
【0010】
個々の組み換えMVをレスキューするために、利用するアンチゲノムプラスミドは、対象となる異種抗原をコードするヌクレオチド配列(異種ヌクレオチド配列)を組み込まれ、MV-特異的転写の開始配列および終止配列に隣接し、したがって追加転写ユニット(ATU)を形成する、麻疹ウイルスの完全長アンチゲノム(+)RNAをコードするcDNAを含む。このMV Edmonston株のベクターは、最初のMVレスキューの発明者によって、外来遺伝子の発現のために開発され(10)、その挿入能力の大きさ(5kb程度)およびトランス遺伝子、例えばB型肝炎ウイルス(Hepatitis B virus)表面抗原、サルまたはヒトの免疫不全ウイルス(SIVまたはHIV)、ムンプスウイルスおよびヒトIL-12などの発現における高い安定性(11;12)が実証されている。特に、初期のころに、B型肝炎ウイルスの表面抗原およびコア抗原を、個別にまたは組み合わせのいずれかで発現する組み換え麻疹ウイルスが作製され、遺伝子改変マウスにおいて液性免疫反応を誘導することが示された。
【0011】
麻疹ウイルスの特性および特にin vivoで高力価の中和抗体を誘発するその能力および長期持続性細胞免疫反応の強力な誘導因子であるその特性の観察から、本発明者らは、麻疹ウイルスが、好ましくは動物において、より好ましくはヒト宿主において、少なくともある程度の保護を得るために適切であり得る前記マラリア原虫に対する中和抗体を誘導する、熱帯熱マラリア原虫由来の抗原を発現する組み換えウイルスの作製のための優れた候補であり得ることを提唱する。
【0012】
特に、本発明においてMV株および特にワクチン株は、幼児集団はMV免疫を有していないので、曝露された幼児集団において設計された組み換えMV中でその成分が発現される、麻疹ウイルスおよび熱帯熱マラリア原虫の両方に対する免疫を誘導するための候補ベクターとして選択される。
【0013】
既にMV免疫化された個人であっても、本発明の組み換え体であるMVウイルスの再投与により、抗MV抗体の強化がもたらされるので、成人集団はMV組み換え体による免疫化からさらに利益を得ることができる(13)。
【0014】
本発明は、熱帯熱マラリア原虫由来の異種遺伝子を担持する組み換え麻疹ウイルスの調製に特に関する。
【0015】
本発明に従った、組み換え麻疹ウイルスの有利な免疫学的特性は、麻疹ウイルスに感受性の動物の中から選択された動物モデルにおいて示すことができ、異種抗原に対する、および/または麻疹ウイルスに対する液性免疫反応および/または細胞性免疫反応を決定する。免疫反応の特徴づけのためのモデルとしての使用に適切なこのような動物の中で、当業者は、特に、MVの特異的受容体の1つであるCD46を発現するトランスジェニックマウスを使用できる。最も有望な組み換え体は、その後サルにおいて試験され得る。
【0016】
組み換え麻疹ウイルスのヌクレオチド配列は、6の倍数であるヌクレオチドの総数を含まなくてはならない。このいわゆる「6の規則」に対する固守は、MVだけでなく、パラミクソウイルス亜科(Paramyxovirinae)のサブファミリーに属するすべてのウイルスに関する絶対的必要条件である。明らかに、それぞれが6ヌクレオチドと接触するNタンパク質分子は、正確に5'末端から3'末端までのゲノムRNAおよびアンチゲノムRNAをカバーしなければならない。
【0017】
ATUの位置は、アンチゲノムcDNAに沿って変え得ることに留意されたい。したがって、上記のMVのmRNAの天然発現の勾配を利用して、挿入されたATUの発現レベルを適切なレベルに変えることができる。ATUの好ましい位置は、L遺伝子の上流、M遺伝子から上流およびN遺伝子の上流であり、それぞれ、異種タンパク質の低い、中程度のおよび強い発現をもたらす。
【0018】
マラリア原虫
マラリアは、現在、世界中、特に熱帯および亜熱帯地域において最も流行している感染性疾患の1つを表す。1年に、マラリア感染症は、世界中で1億人に重症な病気をもたらし、発展途上国および新興国において主に年少幼児が100万から300万人死亡する。マラリアの広範な発生および発生率の上昇は、DDTの広範な禁止ならびに薬剤耐性寄生虫および殺虫剤耐性寄生虫ベクターの数の増加の結果である。他の因子は、環境変化および気候の変化、騒乱ならびに人口の流動性の増加を含む。
【0019】
マラリアは、アピコンプレックス(Apicomplexa)門由来のプラスモディウム(Plasmodium)属に属する、蚊によって運ばれる血液寄生原生動物(hematoprotozoan parasites)によって引き起こされる。プラスモディウム属の4つの種:四日熱マラリアに関与する四日熱マラリア原虫(P. malariae)、両方とも三日熱マラリアを起こす三日熱マラリア原虫(P. vivax)および卵型マラリア原虫(P. ovale)ならびに熱帯熱マラリアの病原体であり、ほとんどすべての致死性感染症に関与する熱帯熱マラリア原虫はヒトに感染する。他にも多くが動物において疾患の原因となり、例えば、プラスモディウム・ヨエリ(P. yoelii)およびプラスモディウム・ベルゲイ(P. berghei)はマウスにおいて疾患の原因となる。
【0020】
マラリア原虫は、いくつかの段階からなるライフサイクルを有する。各段階は、対応する発生段階特異的抗原に対する特異的免疫反応を誘導できる。マラリア原虫は、雌のハマダラカのいくつかの種によって人間に伝播する。感染した蚊は、哺乳動物の血流中にマラリア原虫の「スポロゾイト」型を注入する。スポロゾイトは循環中で数分維持され、その後肝細胞を侵す。この段階において、寄生虫は細胞外環境に位置し、主にスポロゾイト表面の主要成分である「スポロゾイト周囲」(CS)タンパク質を対象として、抗体の攻撃に曝露される。肝臓においてひとたび寄生虫が複製すると、いわゆる「ジゾント」に発達する。これらのジゾントは、最大20,000/感染細胞の比率で発生する。寄生虫のこの細胞内段階の間、宿主免疫反応の主役はTリンパ球、特にCD8+ Tリンパ球である。肝臓感染の約1週間後、数千のいわゆる「メロゾイト」が血流中に放出される。頂端膜抗原1(AMA1)およびメロゾイト表面タンパク質1(MSP1)は、両方とも、感染肝臓細胞から出現するメロゾイト上に存在し、それらは赤血球の侵入に関与する無生殖血液期のメロゾイトの必須成分である。それらがひとたび赤血球に進入すると、抗体媒介免疫反応およびT細胞により分泌されるサイトカインの標的となる。赤血球の侵入後、メロゾイトはいくつかの複写段階を経て、いわゆる「トロホゾイト」ならびにシゾントおよびメロゾイトをもたらし、これらは新たな赤血球に感染できる。限られた量のトロホゾイトは、寄生虫の有性世代を構成する「ガメトサイト」に進化できる。感受性の蚊が赤血球を摂取した場合、赤血球からガメトサイトが放出され、いくつかの雄のガメトサイトおよび1つの雌のガメトサイトがもたらされる。これらのガメートの受精は、接合体の形成なたびにその後のオーキネートへの転換、次いでオーシストへの転換および最終的な唾液腺スポロゾイトへの転換をもたらす。ガメトサイト期特異的表面抗原に対する標的化抗体は、蚊の中腸内でこのサイクルを遮断できる。このような抗体は、哺乳動物宿主を保護しないが、感染した蚊の数およびそれらの寄生負荷を減少することによって、マラリアの伝播を減少させる。
【0021】
MSP-1は、190〜200kDa (d-190)の前駆体として合成され、これは、シゾゴニーの間に83、30、38および42kDa(d-42)の断片にタンパク質分解的にプロセシングされる(14)。赤血球侵入の時点で、42kDaは、残りの複合体と一緒に取り除かれる33kDaの断片および2つの表皮成長因子(EGF)様ドメインを含有し、侵入の間にメロゾイト膜に結合し続ける19kDaの断片とにさらに切断される。この第2の切断は、赤血球侵入が成功するための前提条件である(15)。
【0022】
MSP-1は、K1およびMAD20のプロトタイプによって特徴付けられる二形成対立遺伝子内に高度な保存を示す、基本的二形成タンパク質である。
【0023】
AMA-1(16)は、構造的に保存されたI型内在性膜タンパク質であり、熱帯熱マラリア原虫中に622aa(PfAMA-1)を含み、細胞質領域(50aa)、膜貫通領域ならびにN末端のプロ配列としてフォールディングされたectoドメインおよび3つのドメイン(DI、DII、DIII)に組織化されている。タンパク質の発現は、後期シゾゴニーにおいて最大であり、AMA-1(83kDa)の前駆体がタンパク質分解的にプロセシングされ、プロ配列が切り離され、タンパク質が66kDaの形態に転換され、これによってメロゾイトの再局在化が可能になる。抗体は主にDIおよびDIIを認識し、いくつかの対立遺伝子変異体と同等によく反応すると思われる。DIIIとの抗体反応は一般的に低く、そのレベルは成人において増加する(17、18)。
【0024】
PfAMA-1は64の多形位置を含有し(プロ配列中に9、ectoドメイン中に52、細胞質領域中に3)、それらの大部分が二形成性であり、これらは宿主免疫反応の重要なエピトープである。PfAMA-1に基づくワクチンを開発するために、多形をカバーすることが重要なはずである: Diversity Covering(DiCo1、2および3)PfAMA-1は、PfAMA1 ectoドメインの天然発生多形を最大限に表す人工配列である。それらが、多様なPfAMA1対立遺伝子を担持するさまざまな寄生虫に対して機能性である免疫反応を誘導することが示されている。この取り組みは、PfAMA1を標的とするワクチンを、広範囲の天然発生PfAMA1対立遺伝子に対する強力かつ機能的な保護を誘導可能に作製できる手段を提示できる(19)。
【0025】
CSタンパク質(CSP)は約420aaおよび58kDaの分子量を有する。スポロゾイトの主要表面タンパク質を表し、その機能は、オーシスト(oocystis)由来のスポロゾイトの突然変異および正に帯電したアミノ酸の保存されたモチーフから媒介される肝細胞の侵入のために基本的である。CSPは、5'末端および3'末端の2つの非反復領域ならびに4残基長のモチーフの複数の反復からなり、CSP内の主要なエピトープを表す可変の種特異的中心領域で組織化される。CSPは、シゾゴニーの少なくとも最初の3日間検出可能であり続けるので、細胞外スポロゾイトを対象とする抗体媒介免疫反応およびシゾントを対象とする細胞媒介免疫反応の両方のために魅力的なワクチン標的であると考えられる(20)。
【0026】
マラリアワクチン開発の現在の取り組みは、上記のように、寄生虫が存在できる異なる段階に従って分類できる。
【0027】
ワクチン候補は、3つの型に識別できる:i)スポロゾイトおよび/またはシゾント感染細胞を対象とする前赤内期ワクチン。これらの型のワクチンは主にCSに基づき、液性免疫反応および細胞性免疫反応によって媒介されて理想的には無菌免疫をもたらしマラリア感染を予防するはずである;ii)メロゾイト感染細胞を対象とする無性生殖血液期ワクチン:MSP1およびAMA1は、臨床的重症度を最小にするように設計されたマラリアワクチン候補をもたらす。これらのワクチンは、罹患率および死亡率を低下させるはずであり、寄生虫が赤血球へ進入すること、および/または赤血球中において発達することを防止することを意味する;iii)寄生虫が蚊宿主中で発達することを妨害するように設計された伝播阻止ワクチン。この型のワクチンは、集団規模のマラリア感染率の減少を支持するはずである。これらのワクチンとは別に、寄生虫のライフサイクルの複数の段階を標的とするマラリアワクチンの開発の実現可能性が、いわゆる多成分ワクチンおよび/または多段階ワクチンにおいて追求されている。
【0028】
今日の世界規模のマラリアワクチンのポートフォリオは、47種の新しいワクチン候補が有望であり、31種が前臨床開発、16種が臨床試験に絞り込まれている。これらの1つである、GSK BiologicalsおよびPATH-MVIにより開発されたRTS,Sワクチンは2008年に最終のフェーズIII臨床試験に入っているはずである(21)。他の興味深いワクチン候補は、ベクターとして使用される生の組み換えウイルス、例えば、国際特許出願US2006127413に記載のModified Vaccinia Ankara(MVA)、ポックスウイルス(US6214353、AU7060294、AU1668197、WO9428930およびUS5756101)、アデノウイルス(US2007071726、US2005265974、US2007088156およびCA2507915)、寒冷適用弱毒化インフルエンザウイルスに基づくワクチン、または酵母、例えばメタノール資化性酵母(Pichia pastoris)およびサッカロミセス属種(Saccharomyces spp)に基づくワクチン、または細菌発現系、例えば、サルモネラ属種(Salmonella spp.)(US5112749)および大腸菌(Escherichia coli)(EB0191748)に基づくワクチンである(22)。
【先行技術文献】
【特許文献】
【0029】
【特許文献1】国際特許出願WO97/06270
【特許文献2】国際特許出願US2006127413
【特許文献3】US6214353
【特許文献4】AU7060294
【特許文献5】AU1668197
【特許文献6】WO9428930
【特許文献7】US5756101
【特許文献8】US2007071726
【特許文献9】US2005265974
【特許文献10】US2007088156
【特許文献11】CA2507915
【特許文献12】US5112749
【特許文献13】EB0191748
【発明の概要】
【発明が解決しようとする課題】
【0030】
現在、マラリアに対して利用可能な市販のワクチンはないが、マラリアに対するワクチンの開発は30年以上前に既に開始されている。多くの因子がマラリアワクチンの開発を困難かつ挑戦的にしている。第1に、寄生虫のサイズおよび遺伝的複雑性は、各感染症が、ヒト免疫系に対して数千の抗原を提示することを意味する。これらのうちのどれがワクチン開発のための有用な標的であり得るかを理解することが複雑であり、今日までに、少なくとも40の異なる有望な抗原が特定されている。第2に、寄生虫は、ヒト宿主においてさえいくつかのライフステージの間に変化し、ライフサイクルの各段階において、免疫系に対する異なるサブセットの分子を提示する。第3に、寄生虫は、ヒト免疫系を混乱させ、潜伏し、誤った方向に導くことが可能な一連の戦略を進化させる。最終的に、異なる種だけではなく、異なる株の多重マラリア感染症も同時に有する可能性がある。
【0031】
したがって、本発明は、いくつかの熱帯熱マラリア原虫抗原をコードする異種核酸を含む、異なる弱毒化組み換え麻疹-マラリアベクターを含有する麻疹-マラリア混合ワクチンを提供することによって、先行技術が長期にわたって感じていた必要性を満たす。
【課題を解決するための手段】
【0032】
本発明の一実施形態において、麻疹およびマラリアの両方に対する免疫反応および保護を誘発できるマラリア抗原を発現する、組み換え麻疹ワクチンウイルスを含む麻疹-マラリア混合ワクチンを提供する。
【0033】
別の実施形態において、本発明は、MSP1マラリア抗原を発現するヌクレオチド配列を有する組み換え麻疹ワクチンウイルスを提供する。好ましい実施形態において、組み換え麻疹ワクチンウイルスは、3D7株およびFCB1株由来のアンカー型または分泌型の両方においてマラリア抗原d190またはd83-30-38またはd42を発現するヌクレオチド配列を有する。
【0034】
さらに別の実施形態において、本発明は、Diversity Covering (DiCo) AMA1マラリア抗原を発現するヌクレオチド配列を有する、組み換え麻疹ワクチンウイルスを提供する。
【0035】
さらに別の実施形態において、本発明は、CSマラリア抗原を発現するヌクレオチド配列を有する、組み換え麻疹ワクチンウイルスを提供する。
【図面の簡単な説明】
【0036】
【図1】麻疹ウイルスのアンチゲノムDNA p(+)MV-EZの略図である。p(+)MV-EZは、T7 RNAポリメラーゼプロモーター(T7)の調節下で、それぞれ1の位置(麻疹ウイルスのN遺伝子の前)、2の位置(麻疹ウイルスのP遺伝子とM遺伝子との間)および3の位置(麻疹ウイルスのH遺伝子およびL遺伝子の間)に3つのATUを含有し、肝炎のデルタリボザイムおよびT7 RNAポリメラーゼターミネーター(δT7t)によって正確に終止される、麻疹ウイルス(Edmoston Zagreb)の完全配列を含有するpBluescript由来のプラスミドである。プラスミドのサイズは18941bpである。
【図2】3D7株由来のMSP-1合成遺伝子(d-190)を表す図である。d-190遺伝子断片(d-83-30-38およびd-42)の隣接領域に関するコーディングヌクレオチドおよび対応するアミノ酸を示す。クローニング手順のために加えた特有の制限部位は色で示す; SP:シグナルペプチド;GPI:膜アンカー型領域のためにコードされたグリコシル-ホスファチジル-イノシトール配列。
【図3】FCB1株由来のMSP-1合成遺伝子(d-190)を表す図である。d-190遺伝子断片(d-83-30-38およびd-42)の隣接領域に関するコーディングヌクレオチドおよび対応するアミノ酸を示す。クローニング手順のために加えた特有の制限部位は色で示す; SP:シグナルペプチド;GPI:膜結合領域のためにコードされたグリコシル-ホスファチジル-イノシトール配列。SPおよびGPI領域は3D7株由来である。
【図4】組み換え麻疹-マラリアプラスミドのp(+)MV2-EZ-d190-3D7の略図である。それは、SgrAI-BssHII消化によって麻疹ゲノムの2の位置にクローニングされる、タンパク質のGPIアンカー型をコードする5253bpのd-190マラリア遺伝子(3D7株)を含有する、p(+)MV-EZに由来するプラスミドである。組み換えプラスミドのサイズは24323bpである。
【図5】組み換え麻疹-マラリアプラスミドのp(+)MV2-EZ-d190*-3D7の略図である。それは、SgrAI-BssHII消化によって麻疹ゲノムの2の位置にクローニングされる、タンパク質の分泌型をコードする5160bpのd-190*マラリア遺伝子(3D7株)を含有する、p(+)MV-EZに由来するプラスミドである。組み換えプラスミドのサイズは24227bpである。
【図6】組み換え麻疹-マラリアプラスミドのp(+)MV3-EZ-d190-3D7またはp(+)MV3-EZ-dl90*-3D7の略図である。それは、SgrAI-BssHII消化によって麻疹ゲノムの3の位置にクローニングされる、タンパク質のGPIアンカー型をコードする5253bpのd-190マラリア遺伝子(3D7株)またはタンパク質の分泌型をコードする5160bpのd-190*マラリア遺伝子(3D7株)を含有する、p(+)MV-EZに由来するプラスミドである。組み換えプラスミドp(+)MV3-EZ-d190は24323bpであり、p(+)MV3-EZ-d190*は24227bpである。
【図7】組み換え麻疹-マラリアプラスミドのp(+)MV2-EZ-d83-30-8-3D7の略図である。それは、SgrAI-BssHII消化によって麻疹ゲノムの2の位置にクローニングされる、タンパク質のGPIアンカー型をコードする4122bpのd-83-30-38マラリア遺伝子(3D7株)を含有する、p(+)MV-EZに由来するプラスミドである。組み換えプラスミドのサイズは23195bpである。
【図8】組み換え麻疹-マラリアプラスミドのp(+)MV2-EZ-d83-30-38*-3D7の略図である。それは、SgrAI-BssHII消化によって麻疹ゲノムの2の位置にクローニングされる、タンパク質の分泌型をコードする4029bpのd-83-30-38*遺伝子(3D7株)を含有する、p(+)MV-EZに由来するプラスミドである。組み換えプラスミドのサイズは23105bpである。
【図9】組み換え麻疹-マラリアプラスミドのp(+)MV3-EZ-d83-30-38-3D7またはp(+)MV3-EZ-d83-30-38*-3D7の略図である。それは、SgrAI-BssHII消化によって麻疹ゲノムの3の位置にクローニングされる、タンパク質のGPIアンカー型をコードする4122bpのd-83-30-38マラリア遺伝子(3D7株)またはタンパク質の分泌型をコードする4029bpのd-83-30-38*遺伝子(3D7株)を含有する、p(+)MV-EZに由来するプラスミドである。組み換えプラスミドp(+)MV3-EZ-d83-30-38は23195bpであり、p(+)MV3-EZ-d83-30-38*は23105bpである。
【図10】組み換え麻疹-マラリアプラスミドのp(+)MV2-EZ-d42-3D7の略図である。それは、SgrAI-BssHII消化によって麻疹ゲノムの2の位置にクローニングされる、タンパク質のGPIアンカー型をコードする1347bpのd-42マラリア遺伝子(3D7株)を含有する、p(+)MV-EZに由来するプラスミドである。組み換えプラスミドのサイズは20417bpである。
【図11】組み換え麻疹-マラリアプラスミドのp(+)MV2-EZ-d42*-3D7の略図である。それは、SgrAI-BssHII消化によって麻疹ゲノムの2の位置にクローニングされる、タンパク質の分泌型をコードする1254bpのd-42*マラリア遺伝子(3D7株)を含有する、p(+)MV-EZに由来するプラスミドである。組み換えプラスミドのサイズは20345bpである。
【図12】組み換え麻疹-マラリアプラスミドのp(+)MV3-EZ-d42-3D7またはp(+)MV3-EZ-d42*-3D7の略図である。それは、SgrAI-BssHII消化によって麻疹ゲノムの3の位置にクローニングされる、タンパク質のGPIアンカー型をコードする1347bpのd-42マラリア遺伝子(3D7株)またはタンパク質の分泌型をコードする1254bpのd-42*マラリア遺伝子(3D7株)を含有する、p(+)MV-EZに由来するプラスミドである。組み換えプラスミドp(+)MV3-EZ-d42のサイズは20417bpであり、p(+)MV3-EZ-d42*は20345bpである。
【図13】組み換え麻疹-マラリアプラスミドのp(+)MV2-EZ-d190-FCB1の略図である。それは、SgrAI-BssHII消化によって麻疹ゲノムの2の位置にクローニングされる、タンパク質のGPIアンカー型をコードする5013bpのd-190マラリア遺伝子(FCB1株)を含有する、p(+)MV-EZに由来するプラスミドである。組み換えプラスミドのサイズは24083bpである。
【図14】組み換え麻疹-マラリアプラスミドのp(+)MV3-EZ-d190-FCB1の略図である。それは、SgrAI-BssHII消化によって麻疹ゲノムの3の位置にクローニングされる、タンパク質のGPIアンカー型をコードする5013bpのd-190マラリア遺伝子(FCB1株)を含有する、p(+)MV-EZに由来するプラスミドである。組み換えプラスミドp(+)MV3-EZ-d190のサイズは24083bpである。
【図15】CS合成遺伝子を表す図である。CS遺伝子の隣接領域に関するコーディングヌクレオチドおよび対応するアミノ酸を示す。クローニング手順のために加えた特有の制限部位は色で示す。
【図16】組み換え麻疹-マラリアプラスミドのp(+)MV2-EZ-CSの略図である。それは、SgrAI-BssHII消化によって麻疹ゲノムの2の位置にクローニングされる、1119bpのCS遺伝子を含有する、p(+)MV-EZに由来するプラスミドである。組み換えプラスミドのサイズは20219bpである。
【図17】組み換え麻疹-マラリアプラスミドのp(+)MV3-EZ-CSの略図である。それは、SgrAI-BssHII消化によって麻疹ゲノムの3の位置にクローニングされる、1119bpのCS遺伝子を含有する、p(+)MV-EZに由来するプラスミドである。組み換えプラスミドのサイズは20219bpである。
【図18】DiCo-1完全合成遺伝子を表す図である。DiCo-1完全遺伝子ドメイン(ectoおよびtrans-cyto)の隣接領域に関するコーディングヌクレオチドおよび対応するアミノ酸を示す。クローニング手順のために加えた特有の制限部位は色で示す; SP:シグナルペプチド(ヒトコドンに最適化されている)。
【図19】DiCo-1 ecto合成遺伝子を表す図である。DiCo-1 ectoドメインの隣接領域に関するコーディングヌクレオチドおよび対応するアミノ酸を示す。クローニング手順のために加えた特有の制限部位は色で示す; SP:シグナルペプチド(ヒトコドンに最適化されている)。
【図20】組み換え麻疹-マラリアプラスミドのp(+)MV2-EZ-DiCo1-完全の略図である。それは、SgrAI-BssHII消化によって麻疹ゲノムの2の位置にクローニングされる、タンパク質の膜貫通型をコードする1689bpのDiCo1完全遺伝子を含有する、p(+)MV-EZに由来するプラスミドである。組み換えプラスミドのサイズは20753bpである。
【図21】組み換え麻疹-マラリアプラスミドのp(+)MV3-EZ-DiCo1-完全の略図である。それは、SgrAI-BssHII消化によって麻疹ゲノムの3の位置にクローニングされる、タンパク質の膜貫通型をコードする1689bpのDiCo1完全遺伝子を含有する、p(+)MV-EZに由来するプラスミドである。組み換えプラスミドのサイズは20753bpである。
【図22】組み換え麻疹-マラリアプラスミドのp(+)MV2-EZ -DiCo1- ectoの略図である。それは、SgrAI-BssHII消化によって麻疹ゲノムの2の位置にクローニングされる、タンパク質の分泌型をコードする1458bpのDiCo1 ecto遺伝子を含有する、p(+)MV-EZに由来するプラスミドである。組み換えプラスミドのサイズは20525bpである。
【図23】組み換え麻疹-マラリアプラスミドのp(+)MV3-EZ -DiCo1- ectoの略図である。それは、SgrAI-BssHII消化によって麻疹ゲノムの3の位置にクローニングされる、タンパク質の分泌型をコードする1458bpのDiCo1 ecto遺伝子を含有する、p(+)MV-EZに由来するプラスミドである。組み換えプラスミドのサイズは20525bpである。
【図24】p(+)MV2EZ-GFPの完全ヌクレオチド配列の図である。この配列は、ヌクレオチドの位置に関して以下のように説明できる:- 592-608 T7プロモーター- 609-17354 MV Edmoston Zagrebアンチゲノム- 4049-4054 MluI制限部位- 4060-4067 SgrAI制限部位- 4079-4084 BssHII制限部位- 4085-4801 緑色蛍光タンパク質(GFP) ORF- 4805-4810 BssHII制限部位- 4817-4822 AatII制限部位- 17355-17580 HDVリボザイムおよびT7ターミネーター
【図25】p(+)MV3EZ-GFPの完全ヌクレオチド配列の図である。この配列は、ヌクレオチドの位置に関して以下のように説明できる:- 592-608 T7プロモーター- 609-17359 MV Edmoston Zagrebアンチゲノム- 9851-9856 MluI制限部位- 9862-9869 SgrAI制限部位- 9886-9891 BssHII制限部位- 9892-10608 緑色蛍光タンパク質(GFP) ORF- 10612-10617 BssHII制限部位- 10624-10629 AatII制限部位- 17360-17585 HDVリボザイムおよびT7ターミネーター
【図26】AN101TEの図であり、これは、本発明者らによってクローニングされたMSP1 d-190 3D7配列のORFである。この配列は、ヌクレオチドの位置に関して以下のように説明できる:- 1-3 開始コドン- 4-99 d-1903D7シグナルペプチド- 100-105 BamHI制限部位- 4014-4020 BstEII制限部位- 5152-5157 AclI制限部位- 5158-5250 GPI配列- 5251-5253 終止コドン
【図27】AN102TEの図であり、これは、本発明者らによってクローニングされたMSP1 d-190* 3D7配列のORFである。この配列は、ヌクレオチドの位置に関して以下のように説明できる:- 1-3 開始コドン- 4-99 d- 190*3D7シグナルペプチド- 100-105 BamHI制限部位- 4014-4020 BstEII制限部位- 5152-5157 AclI制限部位- 5158-5160 終止コドン
【図28】AN103TEの図であり、これは、本発明者らによってクローニングされたMSP1 d-83-30-38 3D7配列のORFである。この配列は、ヌクレオチドの位置に関して以下のように説明できる:- 1-3 開始コドン- 4-99 d-83-30-38 3D7シグナルペプチド- 100-105 BamHI制限部位- 4014-4020 BstEII制限部位- 4021-4026 AclI制限部位- 4027-4119 GPI配列- 4120-4122 終止コドン
【図29】AN104TEの図であり、これは、本発明者らによってクローニングされたMSP1 d-83-30-38* 3D7配列のORFである。この配列は、ヌクレオチドの位置に関して以下のように説明できる:- 1-3 開始コドン- 4-99 d-83-30-38* 3D7シグナルペプチド- 100-105 BamHI制限部位- 4014-4020 BstEII制限部位- 4027-4029 終止コドン
【図30】AN105TEの図であり、これは、本発明者らによってクローニングされたMSP1 d-42 3D7配列のORFである。この配列は、ヌクレオチドの位置に関して以下のように説明できる:- 1-3 開始コドン- 4-99 d-42 3D7シグナルペプチド- 100-105 BamHI制限部位- 108-114 BstEII制限部位- 1246-1251 AclI制限部位s- 1252-1344 GPI配列- 1345-1347 終止コドン
【図31】AN106TEの図であり、これは、本発明者らによってクローニングされたMSP1 d-42* 3D7配列のORFである。この配列は、ヌクレオチドの位置に関して以下のように説明できる:- 1-3 開始コドン- 4-99 d-42* 3D7シグナルペプチド- 100-105 BamHI制限部位- 108-114 BstEII制限部位- 1246-1251 AclI制限部位s- 1252-1254 終止コドン
【図32】AN107TEの図であり、これは、本発明者らによってクローニングされたMSP1 d-190 FCB1配列のORFである。この配列は、ヌクレオチドの位置に関して以下のように説明できる:- 1-3 開始コドン- 4-99 d-190 FCB1シグナルペプチド- 100-105 BamHI制限部位- 146-151 HindIII制限部位- 3825-3831 BstEII制限部位- 4912-4917 AclI制限部位s- 4918-5010 GPI配列- 5011-5013 終止コドン
【図33】AN108TEの図であり、これは、本発明者らによってクローニングされたCS配列のORFである。この配列は、ヌクレオチドの位置に関して以下のように説明できる:- 1-3 開始コドン- 4-1116 CS配列- 1117-1119 終止コドン
【図34】AN109TEの図であり、これは、本発明者らによってクローニングされたDiCo 1完全配列のORFである。この配列は、ヌクレオチドの位置に関して以下のように説明できる:- 1-3 開始コドン- 4-99 DiCo 1完全シグナルペプチド- 100-105 BamHI制限部位- 106-1686 DiCo 1完全配列ORF- 1687-1689 終止コドン
【図35】AN110TEの図であり、これは、本発明者らによってクローニングされたDiCo 1 ecto配列のORFである。この配列は、ヌクレオチドの位置に関して以下のように説明できる:- 1-3 開始コドン- 4-99 DiCo 1 ecto完全シグナルペプチド- 100-105 BamHI制限部位- 106-1455 DiCo 1 ecto完全配列ORF- 1456-1458 終止コドン
【図36】組み換え麻疹-p-42マラリアウイルスのMVウイルスワクチンに感染後のVero細胞において生み出された、比較可能な細胞変性効果を表す図である。
【図37】麻疹ベクター(MV3EZ-d-42 SgrAI)の3の位置に挿入されたd-42 3D7トランス遺伝子の発現の図である。1、5および10継代由来の細胞溶解物を、空の麻疹ベクター(MVEZ)および陰性対照(MV3L1、組み換えMV-パピローマウイルス)に対してウェスタンブロットにより分析した。
【図38】免疫蛍光法によって分析された、麻疹ベクター(MV3EZ-d-42 SgrAI)の3の位置に挿入されたトランス遺伝子d-42 3D7の発現を、空の麻疹ベクター(MVEZ)および陰性対照(MV2EZL1、組み換えMV-パピローマウイルス)と比較した図である。それらは免疫染色の前後に光学顕微鏡を使用して見られたので、矢印は同じシンシチウムを指す。
【図39】MVウイルスワクチンの成長動態曲線と比較した、組み換え麻疹-p-42マラリアウイルスの成長動態曲線の図である。
【図40】麻疹ベクター(MV2-3EZ-d-190 SgrAI FCB1)の2の位置および3の位置に挿入されたd-190 FCB1トランス遺伝子の発現の図である。細胞溶解物を、空の麻疹ベクター(MVEZ)および陰性対照(MV2EZL1、組み換えMV-パピローマウイルス)に対してウェスタンブロットにより分析した。
【図41】MVウイルスワクチンの成長動態曲線と比較した、組み換え麻疹- p-190-FCB1マラリアウイルスの成長動態曲線の図である。
【発明を実施するための形態】
【0037】
本発明の目的は、熱帯熱マラリア原虫のCS、MSP-1またはその一部およびAMA-1またはその一部を分泌型または表面アンカー型でコードする、安定に統合されたDNA配列を含有できる組み換え麻疹ベクターから麻疹-マラリア混合ワクチンを作製することである。
【0038】
本発明は、感受性のトランスジェニックマウス、サルおよびヒト宿主において、PfCS、PfMSP-1およびPfAMA-1の抗原を感染、複写および発現できる組み換えMV-マラリアウイルスのレスキューをさらに含むものである。
【0039】
さらに、本発明は、2種の異なる抗原が同じベクター内で同時にクローニングされ、発現し、それらの両方に対する免疫をもたらす、多価組み換え麻疹-マラリアベクターの構築を含むことを意図する。
【0040】
さらに、本発明は、それぞれが、宿主において免疫反応を誘発する様式で異なる遺伝子を担持し、寄生虫のライフサイクルの異なる段階を対象とする異なる抗原を発現する、3種の異なる組み換え麻疹-マラリアウイルスの組み合わせに関する。
【0041】
さらに、本発明は、欠陥干渉粒子(DI)を欠く組み換え麻疹-マラリアウイルスの作製プロセスを含む。DIは、任意の作製系においてウイルスの成長を有意に阻害し、ヒト個体において免疫反応の抑制に成功することが知られている。
【0042】
さらに、本発明はこのような組み換えウイルスを含有するワクチンの作製方法を含む。
【0043】
以下の実施例は、本発明を実施する好ましい様式を記載する。これらの実施例は例示のために提供されると理解されるべきであり、本発明の範囲を決して限定しないものとして解釈されるべきである。
【実施例1】
【0044】
組み換えMV-PfMSP-1プラスミドの構築
すべてのクローニング手順は、Sambrookら(1989年)に記載の技術により実施した。
【0045】
すべての制限酵素は、New England BioLabsから入手し、オリゴヌクレオチドPCRプライマーおよびDNAポリリンカーはInvitrogenから入手した。
【0046】
分泌型およびアンカー型のPfMSP1およびその断片(d-83-30-38およびd-42)は、いずれも化学的に合成し、ヒトコドンに最適化した。それらをpZE21MV中間ベクター内にクローニングし、SgrAIクローニング部位を5'末端に付加することによってわずかに改変し、その後Kozak配列(TCATCA)に最適化した。これらの改変は、MWG Biotechにおいて配列決定により確認されている。
【0047】
MAD20プロトタイプに属する3D7株およびK1プロトタイプに属するFCB1株由来の組み換えMV-PfMSP-1プラスミド、GPIアンカー型および分泌型(*)のリスト:
p(+)MV2EZ-d-190-SgrAI (3D7)
p(+)MV3EZ-d-190-SgrAI (3D7)
p(+)MV2EZ-d-83-30-38-SgrAI (3D7)
p(+)MV3EZ-d-83-30-38-SgrAI (3D7)
p(+)MV2EZ-d-42-SgrAI (3D7)
p(+)MV3EZ-d-42-SgrAI (3D7)
p(+)MV2EZ-d-190*-SgrAI (3D7)
p(+)MV3EZ-d-190*-SgrAI (3D7)
p(+)MV2EZ-d-83-30-38*-SgrAI (3D7)
p(+)MV3EZ-d-83-30-38*-SgrAI (3D7)
p(+)MV2EZ-d-42*-SgrAI (3D7)
p(+)MV3EZ-d-42*-SgrAI (3D7)
p(+)MV2EZ-d-190-SgrAI (FCBl)
p(+)MV3EZ-d-190-SgrAI (FCBl)
【0048】
1a)p(+)MV2EZ-d-190-SgrAI (3D7、24323bp)およびp(+)MV3EZ-d-190-SgrAI(3D7、24323bp)の構築
緑色蛍光タンパク質(GFP)を含有する1μgのMVプラスミドDNA(p(+)MV2-3EZ-GFP Berna株、19774bp:図24および25)を、1ユニットのSgrAIおよびBssHII制限酵素両方を用いて、それらの最適温度において2時間、最終容量50μlで消化した。すべての消化されたDNAを1%アガロースゲルに添加し、80ボルトで約2時間泳動させた。次いで、固有のバンド(19048bp)をゲルから取り出し、QIAEXゲル精製によって精製し、260nmにおける吸光度によりDNA濃度を計算し、1μg/mlに対して調整した。
【0049】
中間プラスミド内に挿入された1μgのd-190遺伝子(pZE21 MV-d- 190 SgrAI、7564bp)を、SgrAI-BssHII消化(各酵素1ユニット)によって、それらの最適温度において2時間、最終容量50μlで取り出した。すべての消化されたDNAを1%アガロースゲルに添加し、80ボルトで約2時間泳動させた。次いで、固有のバンド(5275bp)をゲルから取り出し、QIAEXゲル精製キットによって精製し、260nmにおける吸光度によりDNA濃度を計算し、1μg/mlに調整した。
【0050】
このようにして、ベクター(MV DNA:図1)およびインサート(d-190 DNA:図2)を、等モル比で16℃において一晩、1ユニットのT4 DNAリガーゼおよびその独自の反応バッファーを使用して、最終容量10μlでライゲーションした。
【0051】
次いで、XL10 Gold化学的コンピテントセルを、全ライゲーション容量を用いて、標準的形質転換プロトコル(Sambrookら、1989年)に従って形質転換し、LB-Agarプレートにプレーティングし、アンピシリン耐性に関して選択した。コロニーを、DNAプラスミド調製(QIAGEN、ミニ、ミディおよびマキシキット)および制限酵素消化によってスクリーニングした。正しいクローンを、配列決定のためにMWGに送り、DNA Striderソフトウェアを使用して、仮定した配列とアライメントしたこの配列は、100%の同一性を示した。
【0052】
MVベクターの2の位置(SgrAI、4060位およびBssHII、9335位)に挿入されたd-190-3D7遺伝子を図4に表し、そのオープンリーディングフレーム(ORF)を図26に載せた。
【0053】
MVベクターの3の位置(SgrAI、9862位およびBssHII、15137位)に挿入されたd-190-3D7遺伝子を図6に表した。
【0054】
組み換え麻疹-マラリアプラスミドのゲノムの長さ(609位のACCから開始し、21884位のGGTまで)は6の倍数であり、組み換えMV2-3-d-190-3D7ウイルスのレスキューが可能であった。
【0055】
1b)p(+)MV2EZ-d-83-30-38-SgrAI(3D7、23195bp)およびp(+)MV3EZ-d-83-30-38-SgrAI(3D7、23195bp)の構築。
麻疹ベクターを、実施例3aに詳細に記載のように調製した。
【0056】
pZE21MV-d- 190 SgrAIをBstEII-AclIで消化し、d-42断片を切り出し、接着性のBstEII末端およびAclI末端を有するポリリンカーをライゲーションし、中間プラスミドのpZE21MV-d-83-30-38-SgrAI(6436bp)を得た。
【0057】
ポリリンカーの配列は5'-GTCACCAGCGGCCGCAA-3'であった。
【0058】
1μgのpZE21MV-d-83-30-38 SgrAIを、それらの最適温度において約2時間、最終容量50μlでSgrAI-BssHH (各酵素1ユニット)で消化した。すべての消化されたDNAを1%アガロースゲルに添加し、80ボルトで約2時間泳動させた。次いで、固有のバンド(4147bp)をゲルから取り出し、QIAEXゲル精製キットによって精製し、260nmにおける吸光度によりDNA濃度を計算し、1μg/mlに調製した。
【0059】
このようにして、ベクター(MV DNA:図1)およびインサート(d-83-30-38DNA:図2)を、等モル比で16℃において一晩、1ユニットのT4 DNAリガーゼおよびその独自の反応バッファーを使用して、最終容量10μlでライゲーションした。
【0060】
次いで、XL10 Gold化学的コンピテントセルを、全ライゲーション容量を用いて、標準的形質転換プロトコル(Sambrookら、1989年)に従って形質転換し、LB-Agarプレートにプレーティングし、アンピシリン耐性に関して選択した。コロニーを、DNAプラスミド調製(QIAGEN、ミニ、ミディおよびマキシキット)および制限酵素消化によってスクリーニングした。正しいクローンを、配列決定のためにMWGに送り、この配列を、DNA Striderソフトウェアを使用して、仮定した配列とアライメントした。
【0061】
正しいクローンを、配列決定のためにMWGに送り、DNA Striderソフトウェアを使用して、仮定した配列とアライメントしたこの配列は、100%の同一性を示した。
【0062】
MVベクターの2の位置(SgrAI、4060位およびBssHII、8207位)に挿入されたd-83-30-38-3D7遺伝子を図7に表し、そのオープンリーディングフレーム(ORF)を図28に載せた。
【0063】
MVベクターの3の位置(SgrAI、9862位およびBssHII、14006位)に挿入されたd-83-30-38-3D7遺伝子を図9に表した。
【0064】
組み換え麻疹-マラリアプラスミドのゲノムの長さ(609位のACCから開始し、20756位のGGTまで)は6の倍数であり、組み換えMV2-3-d-83-30-38-3D7ウイルスのレスキューが可能であった。
【0065】
1c)p(+)MV2EZ-d-42-SgrAI (3D7、20417bp)およびp(+)MV3EZ-d-42-SgrAI (3D7、20417bp)の構築。
麻疹ベクターを、実施例3aに詳細に記載のように調製した。
【0066】
中間プラスミド(pZE21MV-d-42 SgrAI、3658bp)を挿入された1μgのd-42遺伝子を、それらの最適温度において約2時間、最終容量50μlでSgrAI-BssHII (各酵素1ユニット)消化によって取り出した。すべての消化されたDNAを1%アガロースゲルに添加し、80ボルトで約2時間泳動させた。次いで、固有のバンド(1369bp)をゲルから取り出し、QIAEXゲル精製キットによって精製し、260nmにおける吸光度によりDNA濃度を計算し、1μg/mlに調製した。
【0067】
このようにして、ベクター(MV DNA:図1)およびインサート(d-42DNA:図2)を、等モル比で16℃において一晩、1ユニットのT4 DNAリガーゼおよびその独自の反応バッファーを使用して、最終容量10μlでライゲーションした。
【0068】
次いで、XL10 Gold化学的コンピテントセルを、全ライゲーション容量を用いて、標準的形質転換プロトコル(Sambrookら、1989年)に従って形質転換し、LB-Agarプレートにプレーティングし、アンピシリン耐性に関して選択した。コロニーを、DNAプラスミド調製(QIAGEN、ミニ、ミディおよびマキシキット)および制限酵素消化によってスクリーニングした。正しいクローンを、配列決定のためにMWGに送り、DNA Striderソフトウェアを使用して、仮定した配列とアライメントしたこの配列は、100%の同一性を示した。
【0069】
MVベクターの2の位置(SgrAI、4060位およびBssHII、5429位)に挿入されたd-42-3D7遺伝子を図10に表し、そのオープンリーディングフレーム(ORF)を図30に載せた。
【0070】
MVベクターの3の位置(SgrAI、9862位およびBssHII、11231位)に挿入されたd-42-3D7遺伝子を図12に表した。
【0071】
組み換え麻疹-マラリアプラスミドのゲノムの長さ(609位のACCから開始し、17978位のGGTまで)は6の倍数であり、組み換えMV2-3- d-42-3D7ウイルスのレスキューが可能であった。
【0072】
組み換え麻疹-p-42マラリアウイルスおよびMVワクチンは、同様の細胞変性効果を誘導した(図36)。トランス遺伝子はむしろ安定して発現され、その発現は、ヒト2倍体細胞MRC5におけるウイルスの連続する10継代後に、単一の最初にレスキューされたクローンに由来する、分析したすべての子孫クローンにおいて完全に維持されていた(図37〜38)。
【0073】
組み換えMV-マラリアウイルスおよびMVワクチンの成長曲線は、同じ動態を示した(図39)。
【0074】
1d)p(+)MV2EZ-d- 190*-SgrAI(3D7、24227bp)およびp(+)MV3EZ-d-190*- SgrAI(3D7、24227bp)の構築。
麻疹ベクターを、実施例3aに詳細に記載のように調製した。
【0075】
中間ベクターpZE2 IMVd- 190-SgrAIを鋳型として使用して、PCR反応を実施し、AclI(5434位)部位とClaI(5536位)部位との間に位置するGPIアンカー領域を欠失させた。
【0076】
PCR増幅を、プルーフリーディングPfu DNAポリメラーゼ(Stratagene)を使用して実施した。合成オリゴヌクレオチドプライマーのDNA配列を、MVヌクレオチドに関しては小文字で、非MVヌクレオチドに関しては大文字で示し、関連制限エンドヌクレアーゼ認識部位の配列には下線を引いた。以下のオリゴヌクレオチドプライマーを使用した: For-C ClaI、5'-CCAATAAACGTTTAAT AGatcgattacgcgcgctctagc -3'およびRev-AvrII、5'-gcctttgagtgagctgatacc-3'。
【0077】
For-ClaIは、ClaIおよびBssHII部位のレベルにおいて鋳型と相同であり、オーバーハング(大文字)および2つの終止コドン(TAATAG)、AclI部位(AACGTT)ならびにAclIのための6bpの長い保護部位(CCAATA)を含有する。いわゆるPCR-GPIにおいて、および最終構築体d-190*において、AclIはClaIに近くなる。
【0078】
Rev-Avrllは鋳型と相同である(5704位から5724まで)。
【0079】
PCR産物は207bp長であり、:AclI + AvrIIを用いたその消化およびAclI +AvrIIで予備消化された中間ベクターpZE21MVd-190-SgrAIとのライゲーションにより、pZE21 MVd- 190*- SgrAIを作製した。
【0080】
詳細には、AclI + AvrIIを用いたベクターの消化により、7318bpおよび246bp (欠失されたGPI領域を含有する)の2つのバンドが生じ、7.3kb-断片が、QIAEX II精製キット(Qiagen)を使用することによってアガロースゲルから精製され、消化されたAclI+Avrll PCR(インサート)とライゲーションされ、pZE21 MVd- l90*-SgrAIが得られた。
【0081】
陽性クローンをスクリーニングするために、NcoI消化を実施し、d-190*中間ベクターから7kbの単一バンドならびに元のGPI-アンカー構築体から1.3および5.7kbの2つのバンドが生じた。
【0082】
確定した組み換えp(+)MeV2EZ-d190*およびp(+)MeV3EZ-d190*(図5および図6)を構築するために、「6の規則」に従って、MeVベクターおよび中間プラスミドを、SgrAI+BssHIIを用いて消化し、その後互いをライゲーションした。
【0083】
詳細には、SgrAI+BssHIIで消化されたpZE21 MVd- 190*- SgrAIは、5.2kb +1.3kb + 900bpの3つのバンドを生じた。D-190*配列は5.2kbの断片に含有され、この断片を切断し、精製し、SgrAI+BssHIIで消化されたMeV2EZおよびMeV3EZベクター(19Kb長)と、等モル比で16℃において一晩、1ユニットのT4 DNAリガーゼを使用してライゲーションした。
【0084】
次いで、XL10 Gold化学的コンピテントセルを、全ライゲーション容量を用いて、標準的形質転換プロトコル(Sambrookら、1989年)に従って形質転換し、LB-Agarプレートにプレーティングし、アンピシリン耐性に関して選択した。コロニーを、DNAプラスミド調製(QIAGEN、ミニ、ミディおよびマキシキット)および制限酵素消化によってスクリーニングした。正しいクローンを、配列決定のためにMWGに送り、DNA Striderソフトウェアを使用して、仮定した配列とアライメントしたこの配列は、100%の同一性を示した。
【0085】
MVベクターの2の位置(SgrAI、4060位およびBssHII、9239位)に挿入されたd-190*- 3D7遺伝子を図5に表し、そのオープンリーディングフレーム(ORF)を図27に載せた。
【0086】
MVベクターの3の位置(SgrAI、9862位およびBssHII、15041位)に挿入されたd-190*- 3D7遺伝子を図6に表した。
【0087】
組み換え麻疹-マラリアプラスミドのゲノムの長さ(609位のACCから開始し、21788位のGGTまで)は6の倍数であり、組み換えMV2-3- d-190*-3D7ウイルスのレスキューが可能であった。
【0088】
1e)p(+)MV2EZ-d-83-30-38*-SgrAI(3D7、23105bp)およびp(+)MV3EZ-d-83-30-38*- SgrAI (3D7、23105bp)の構築。
麻疹ベクターを、実施例3aに詳細に記載のように調製した。
【0089】
中間ベクターのpZE 21MV-d-190SgrAIをBstEII-ClaIで消化し、d-42断片およびAclI部位(5434位)とおよびClaI(5536位)部位との間に位置するGPI領域を切り出し、接着性のBstEII 末端およびClaI末端を有するポリリンカーをライゲーションし、中間プラスミドのpZE21MV- d-83-30-38*-SgrAI(6436bp)を得た。
【0090】
ポリリンカーの配列は5'-GTCACCGGGGAATAATAGCGCAT-3'であった。
【0091】
合成オリゴヌクレオチドポリリンカーのDNA配列を、非MVヌクレオチドに関して大文字で示し、関連制限エンドヌクレアーゼ認識部位の配列には下線を引いた。
【0092】
ポリリンカーはBstEII(GTCACC)およびClaI(AT)付着末端、2つの終止コドン (TAATAG)および6の規則を守るためのトリプレット(GCG)を含有する。
【0093】
1μgのpZE21MV-d-83-30-38* SgrAIを、それらの最適温度において約2時間、最終容量50μlでSgrAI-BssHII(各酵素1ユニット)消化した。すべての消化されたDNAを1%アガロースゲルに添加し、80ボルトで約2時間泳動させた。次いで、固有のバンド(4057bp)をゲルから取り出し、QIAEXゲル精製キットによって精製し、260nmにおける吸光度によりDNA濃度を計算し、1μg/mlに調製した。
【0094】
このようにして、ベクター(MV DNA:図1)およびインサート(d-83-30-38* DNA:図2)を、等モル比で16℃において一晩、1ユニットのT4 DNAリガーゼおよびその独自の反応バッファーを使用して、最終容量10μlでライゲーションした。
【0095】
次いで、XL10 Gold化学的コンピテントセルを、全ライゲーション容量を用いて、標準的形質転換プロトコル(Sambrookら、1989年)に従って形質転換し、LB-Agarプレートにプレーティングし、アンピシリン耐性に関して選択した。コロニーを、DNAプラスミド調製(QIAGEN、ミニ、ミディおよびマキシキット)および制限酵素消化によってスクリーニングした。正しいクローンを配列決定のためにMWGに送り、この配列を、DNA Striderソフトウェアを使用して、仮定した配列とアライメントした。正しいクローンを配列決定のためにMWGに送り、DNA Striderソフトウェアを使用して、仮定した配列とアライメントしたこの配列は、100%の同一性を示した。
【0096】
MVベクターの2の位置(SgrAI、4060位およびBssHII、8117位)に挿入されたd-83-30-38*-3D7遺伝子を図8に表し、そのオープンリーディングフレーム(ORF)を、図29に載せた。
【0097】
MVベクターの3の位置(SgrAI、9862位およびBssHII、13919位)に挿入されたd-83-30-38*- 3D7遺伝子を図9に表した。
【0098】
組み換え麻疹-マラリアプラスミドのゲノムの長さ(609位のACCから開始し、20666位のGGTまで)は6の倍数であり、組み換えMV2-3- d-83-30-38*-3D7ウイルスのレスキューが可能であった。
【0099】
1f)p(+)MV2EZ-d-42*-SgrAI(3D7、20345bp)およびp(+)MV3EZ-d-42*-SgrAI(3D7、20345bp)の構築。
麻疹ベクターを、実施例3aに詳細に記載のように調製した。
【0100】
中間ベクターpZE21MVd-42-SgrAI(3658bp)を鋳型として使用して、PCR反応を実施し、AclI(1528位)部位とClaI(1630位)部位との間に位置するGPIアンカー領域を欠失させた。
【0101】
PCR増幅を、プルーフリーディングPfu DNAポリメラーゼ(Stratagene)を使用して実施した。合成オリゴヌクレオチドプライマーのDNA配列を、MVヌクレオチドに関しては小文字で、非MVヌクレオチドに関しては大文字で示し、関連制限エンドヌクレアーゼ認識部位の配列には下線を引いた。以下のオリゴヌクレオチドプライマーを使用した: For-C ClaI、5'-CCAATAAACGTTTAAT AGatcgattacgcgcgctctagc -3'およびRev-AvrII、5'-gcctttgagtgagctgatacc-3'。
【0102】
For-ClaIは、ClaI(1630位)部位およびBssHII(1639位)部位のレベルにおいて鋳型と相同であり、オーバーハング(大文字)および2つの終止コドン(TAATAG)、AclI部位(AACGTT)ならびにAclIのための6bpの長い保護部位(CCAATA)を含有する。いわゆるPCR-GPIにおいて、および最終構築体d-42*において、AclIはClaIに近くなる。
Rev-Avrllは鋳型と相同である(1798位から1818位まで)。
【0103】
PCR産物は207bp長であり、:AclI + AvrIIを用いたその消化およびAclI +AvrIIで予備消化された中間ベクターpZE21MV d-42-SgrAIとのライゲーションにより、pZE21 MV d-42*- SgrAIを作製した。
【0104】
詳細には、AclI + Avrllを用いたベクターの消化により、3412bpおよび246bp (欠失されたGPI領域を含有する)の2つのバンドが生じ、3.4kb-断片が、QIAEX II精製キット(Qiagen)を使用することによってアガロースゲルから精製され、消化されたAclI+AvrII PCR(インサート)とライゲーションされ、pZE21 MVd-42*-SgrAIが得られた。
【0105】
陽性クローンをスクリーニングするために、NcoI消化を実施し、d-42*中間ベクターから3.4kbの単一バンドならびに元のGPI-アンカー構築体から1.3および2.3kbの2つのバンドが生じた。
【0106】
確定した組み換えp(+)MeV2EZ- d42*およびp(+)MeV3EZ- d42*を構築するために、「6の規則」に従って、MeVベクターおよび中間プラスミドを、SgrAI+BssHIIを用いて消化し、その後互いをライゲーションした。
【0107】
詳細には、SgrAI+BssHII+ SpeIで消化されたpZE21 MV d-42*- SgrAIは、1.3kb + 936bp + 800bp + 400bpの4つのバンドを生じた。d-42*配列は1.3kbの断片に含有され、この断片を切断し、精製し、SgrAI+BssHIIにより消化されたMeV2EZおよびMeV3EZベクター(19Kb長)と、等モル比で16℃において一晩、1ユニットのT4 DNAリガーゼを使用してライゲーションした。
【0108】
次いで、XL10 Gold化学的コンピテントセルを、全ライゲーション容量を用いて、標準的形質転換プロトコル(Sambrookら、1989年)に従って形質転換し、LB-Agarプレートにプレーティングし、アンピシリン耐性に関して選択した。コロニーを、DNAプラスミド調製(QIAGEN、ミニ、ミディおよびマキシキット)および制限酵素消化によってスクリーニングした。正しいクローンを、配列決定のためにMWGに送り、DNA Striderソフトウェアを使用して、仮定した配列とアライメントしたこの配列は、100%の同一性を示した。
【0109】
MVベクターの2の位置(SgrAI、4060位およびBssHII、5357位)に挿入されたd-42*- 3D7遺伝子を図11に表し、そのオープンリーディングフレーム(ORF)を図31に載せた。
【0110】
MVベクターの3の位置(SgrAI、9862位およびBssHII、11159位)に挿入されたd-42*-3D7遺伝子を図12に表した。
【0111】
組み換え麻疹-マラリアプラスミドのゲノムの長さ(609位のACCから開始し、17906位のGGTまで)は6の倍数であり、組み換えMV2-3- d-42*-3D7ウイルスのレスキューが可能であった。
【0112】
1g)p(+)MV2EZ-d-190- SgrAI(FCB1、24083bp)およびp(+)MV3EZ-d-190-SgrAI(FCB1、24083bp)の構築。
まず第1に、FCB 1株のMSP-1のための合成遺伝子の、中間プラスミドpZE21MV-SgrAIへのクローニングを実施し、3D7株のMSP-1由来のシグナルペプチドおよびGPI-アンカー領域を保った。D-190遺伝子(FCB1)を、pZE23f-GX-190Hと呼ばれる中間ベクターから以下のように段階的に得た:
i).1μgのプラスミドpZE21MV-d-190-SgrAI(3D7)を、それらの最適温度において2時間、最終容量50μlで、HindIII + AclI制限酵素を用いて消化した。すべての消化されたDNAを1%アガロースゲルに添加し、80ボルトで約2時間泳動させた。次いで、ベクターに対応する固有のバンド(2558 bp)をゲルから取り出し、QIAEXゲル精製によって精製し、260nmにおける吸光度によりDNA濃度を計算した。
【0113】
ii).MSP-1/FCB1のd-42部分を増幅し、回収するために、pZE23f-GX-190Hを鋳型として使用してPCR反応を実施した。PCR増幅を、プルーフリーディングPfu DNAポリメラーゼ(Stratagene)を使用して実施した。合成オリゴヌクレオチドプライマーのDNA配列を、MVヌクレオチドに関しては小文字で、非MVヌクレオチドに関しては大文字で示し、関連制限エンドヌクレアーゼ認識部位の配列には下線を引いた。
【0114】
以下のオリゴヌクレオチドプライマーを使用し、ρZE23f-GX-190H配列を設計した: For-1 FCB1、5'-CCCAAGCTTccaggtggtcaccggAgagctgtcactcc -3'およびRev-1 FCB1, 5'- GCCTGCaacgttGCTagagctggagcaGaaGatcccgtcg -3'。
【0115】
For-1 FCB1は、4509位から4538位の鋳型に相同であり、BstEII部位(ggtcacc)を含む。A(大文字)は、鋳型においてはtであり、SgrAI部位を除去するために改変されている。For-1 FCB1は、3bpの長い保護部位(CCC)の後ろにHindIII部位(AAGCTT)およびオーバーハング(大文字)を含有する。
【0116】
Rev-1 FCB1はAclI部位(aacgtt)を含有し、その前に6-bpの保護部位(GCCTGC)が付く。Rev-1 FCB1は、セリンをコードするトリプレットのGCTが導入され、6の規則に従っており、ポリ(A)部位を回避するために2つのaがGに改変されている。
【0117】
得られたPCR-HindIII-AclI(1.1kb)をHindIII + AclI消化し、等モルで16℃において一晩、HindIII + AdIIを用いて予備消化されたpZE21MV-d- 190- SgrAI(ステップi)とライゲーションし、pZE21MV-d-42-SgrAI-FCB1(3657bp)を得た。次いで、XL10 Gold化学的コンピテントセルを、全ライゲーション容量を用いて、標準的形質転換プロトコル(Sambrookら、1989年)に従って形質転換し、LB-Agarプレートにプレーティングし、アンピシリン耐性に関して選択した。コロニーを、DNAプラスミド調製(QIAGEN、ミニ、ミディおよびマキシキット)およびHindIII+AclIを用いた制限酵素消化によってスクリーニングした(期待される断片2558bp + 1099bp)。
【0118】
iii).ステップiiに記載のように得られたpZE21MV-d-42- SgrAI-FCB1を、HindIII + BstEII(HindIII、428位およびBstEII、440位)で消化し、開いたベクターに対応する特有のバンド(3645bp)を1%アガロースゲルに添加し、ゲルから取り出し、QIAEXゲル精製によって精製し、260nmにおける吸光度によりDNA濃度を計算した。
【0119】
iv).pZE23f-GX-190HをHindIII + BstEIIで消化し、d-83-30-38/FCB1断片に対応する、3679bpの特有のバンド(インサート)を、前記のようにゲルから精製した。
【0120】
v).pZE21MV-d-42-SgrAI-FCB1から得られた、HindIII + BstEIIにより消化された3657bpの断片(ベクター)を、pZE23f-GX- 190Hから消化によって得た、d-83-30-38/FCB1を含有する3679bpのHindIII + BstEII断片(インサート)とライゲーションした。ライゲーションを、等モルで16℃において一晩、1ユニットのT4 DNAリガーゼを使用して実施し、pZE21MV-d-190-SgrAI-FCB1(7324bp)を得た。その後、XL10 Gold化学的コンピテントセルを、全ライゲーション容量を用いて、標準的形質転換プロトコル(Sambrookら、1989年)に従って形質転換し、LB-Agarプレートにプレーティングし、アンピシリン耐性に関して選択した。コロニーを、DNAプラスミド調製(QIAGEN、ミニ、ミディおよびマキシキット)および制限酵素消化によってスクリーニングした。
【0121】
p(+)MV2EZ-d-190-SgrAI-FCB1およびp(+)MV3EZ-d-190-SgrAI-FCB1を構築するために、麻疹ベクターを、実施例3aに詳細に記載のように調製した。
【0122】
中間プラスミド内に挿入された1μgのd-190/FCB1遺伝子(pZE21MV-d-190 SgrAI-FCB1、7324bp)を、SgrAI-BssHII消化(各酵素1ユニット)によって、それらの最適温度において2時間、最終容量50μlで取り出した。すべての消化されたDNAを1%アガロースゲルに添加し、80ボルトで約2時間泳動させた。次いで、固有のバンド(5035bp)をゲルから取り出し、QIAEXゲル精製キットによって精製し、260nmにおける吸光度によりDNA濃度を計算し、1μg/mlに対して調整した。
【0123】
このようにして、ベクター(MV DNA:図1)およびインサート(d-190/FCB1 DNA:図3)を、等モル比で16℃において一晩、1ユニットのT4 DNAリガーゼおよびその独自の反応バッファーを使用して、最終容量10μlでライゲーションした。
【0124】
次いで、XL10 Gold化学的コンピテントセルを、全ライゲーション容量を用いて、標準的形質転換プロトコル(Sambrookら、1989年)に従って形質転換し、LB-Agarプレートにプレーティングし、アンピシリン耐性に関して選択した。コロニーを、DNAプラスミド調製(QIAGEN、ミニ、ミディおよびマキシキット)および制限酵素消化によってスクリーニングした。正しいクローンを、配列決定のためにMWGに送り、DNA Striderソフトウェアを使用して、仮定した配列とアライメントしたこの配列は、100%の同一性を示した。
【0125】
MVベクターの2の位置(SgrAI、4060位およびBssHII、9095位)に挿入されたd-190- FCB1遺伝子を図13に表し、そのオープンリーディングフレーム(ORF)を図32に載せた。
【0126】
MVベクターの3の位置(SgrAI、9862位およびBssHII、14897位)に挿入されたd-190- FCB1遺伝子を図14に表した。
【0127】
組み換え麻疹-マラリアプラスミドのゲノムの長さ(609位のACCから開始し、21884位のGGTまで)は6の倍数であり、組み換えMV2-3- d-190-FCB1ウイルスのレスキューが可能であった。
【0128】
トランス遺伝子はむしろ安定して発現され、その発現は、ヒト2倍体細胞MRC5におけるウイルスの連続する10継代後に、単一の最初にレスキューされたクローンに由来する、分析したすべての子孫クローンにおいて完全に維持されていた(図40)。
【0129】
組み換えMV-マラリアウイルスおよびMVワクチンの成長曲線は、同じ動態を示した(図41)。
【実施例2】
【0130】
DiCo1核酸配列の設計
アミノ酸DiCo1配列(ecto、トランスおよび細胞質ドメイン: aa97〜622)から開始し、DNA Strider softwareを使用して、DiCo1 DNA変性配列と、BLASTアライメント後にDiCo1に最も類似した配列を表す、選択されたPfAMA1遺伝子(受託番号AAG141.1)とを比較して、対応する核酸配列を設計した。
【0131】
5'末端に、適切な特有の制限部位を、クローニング部位として付加し(MluIおよびSgrAI)、その後ろに最適KOZAC配列およびヒト最適化シグナルペプチド(SP)を付加した。3'末端に、2つの終止コドンおよびBssHIIクローニング部位を付加した。このスキームに従って、本発明者らは、(麻疹ベクター内におけるさらなる発現のために「6の規則」を尊重して) DiCo1タンパク質のアンカー型および分泌型をコードする2種のヌクレオチド配列を設計した:第1の遺伝子は外形質(ectoplasmasmic)、膜貫通および細胞質ドメインを含み(図18)、一方、第2の遺伝子はectoドメイン単独に対応する(図19)。この2種の配列は、AT%含有量を減少させ、ポリ(A)配列およびRNAの不安定モチーフを回避するために、GENEARTによってヒトコドンに最適化されている。
【0132】
DiCo1の完全ORFおよびDiCo1のectoドメインORFを、それぞれ図34および35に載せる。
【実施例3】
【0133】
組み換えMV-PfΑMA-1プラスミドの構築
すべてのクローニング手順は、Sambrookら(1989年)に記載の技術により実施した。
【0134】
PfΑMA-1、特に分泌型およびアンカー型のDiversity Covering配列1(DiCo1)は、いずれも化学的に合成し、ヒトコドンに最適化した。
【0135】
最適化されたコドンのDiCo1分泌型およびアンカー型を、SgrAI+BssHIIで消化し、予備消化したMeV2EZおよびMeV3EZベクター(19Kb長)と、等モル比で16℃において一晩、1ユニットのT4 DNAリガーゼを使用してライゲーションし、以下の組み換えMV-PfΑMA-1プラスミド:p(+)MV2EZ-DiCo1-完全(図20)、p(+)MV3EZ-DiCo1-完全(図21)、p(+) MV2EZ-DiCo1-ecto(図22)およびp(+)MV3EZ-DiCo1-ecto(図23)を得た。
【実施例4】
【0136】
組み換えMV-PfCSプラスミドの構築
p(+)MV2EZ-CS-SgrAI(20219bp)およびp(+)MV3EZ- CS-SgrAI (20219bp)の構築
すべてのクローニング手順は、Sambrookら(1989年)に記載の技術により実施した。
【0137】
中間ベクターpAdApt35Bsu.CS.Pfalc.aa-sub.gccにクローングされたPfCS1をPCRによって増幅し、確定的MVベクターに直接クローニングし、2種の組み換えMV-PfCSプラスミド: p(+) MV2EZ-CSおよびp(+)MV3EZ-CSを得た。
【0138】
詳細には、CS遺伝子(図15)を増幅し、回収するために、pAdApt35Bsu.CS.Pfalc.aa-sub.gccを鋳型として使用してPCR反応を実施した。PCR増幅は、プルーフリーディングPfu DNAポリメラーゼ(Stratagene)を使用して実施した。合成オリゴヌクレオチドプライマーのDNA配列を、MVヌクレオチドに関しては小文字で、非MVヌクレオチドに関しては大文字で示し、関連制限エンドヌクレアーゼ認識部位の配列には下線を引いた。
【0139】
以下のオリゴヌクレオチドプライマーを使用し、: pAdApt35Bsu.CS.Pfalc.aa-sub.gcc配列上に設計した: For-SgrAI、5'- ACTTCTCACCGGTGTggaagcttgccaccatgat-3'およびRev-BssHII-CS、5'- TAGCGCGCtctagaggatccttatcagc -3'。
【0140】
For-SgrAIはHindIII部位(aagctt)を含む1356位から1375位までが鋳型と相同である。それは、6-bpの長い保護部位(ACTTCT)の後ろにオーバーハング(大文字)およびSgrAI制限部位(CACCGGTG)を含有する。
【0141】
Rev-BssHII-CSは、オーバーハング(大文字)およびBssHII制限部位(GCGCGC)を含有し、Rev-BssHII-CSは、PCR-CS(1187bp)においてXbaI(tctaga)に近い。得られたPCR-CSをSgrAI + BssHIIで消化して、SgrAI+BssHIIで予備消化したMeV2EZおよびMeV3EZベクター(19Kb長)と、等モル比で16℃において一晩、1ユニットのT4 DNAリガーゼを使用してライゲーションし、それぞれp(+)MV2EZ-CS-SgrAI(20219bp、図16)およびp(+)MV3EZ- CS-SgrAI(20219bp、図17)を得た。CSのORFを図33に載せる。
【0142】
次いで、XL10 Gold化学的コンピテントセルを、全ライゲーション容量を用いて、標準的形質転換プロトコル(Sambrookら、1989年)に従って形質転換し、LB-Agarプレートにプレーティングし、アンピシリン耐性に関して選択した。コロニーを、DNAプラスミド調製(QIAGEN、ミニ、ミディおよびマキシキット)および制限酵素消化によってスクリーニングした。正しいクローンを、配列決定のためにMWGに送り、DNA Striderソフトウェアを使用して、仮定した配列とアライメントしたこの配列は、100%の同一性を示した。
【実施例5】
【0143】
細胞およびウイルス
細胞を、Vero細胞(アフリカミドリザルの腎臓)のために5%(FCS)を補充したダルベッコ変法イーグル培地(DMEM)、293T細胞(ヒト胎児腎臓)のために10% FCSおよび1%ペニシリン/ストレプトマイシン(P/S)を補充したダルベッコ変法イーグル培地、MRC-5(ヒト胎児線維芽細胞)のためにGlutamax(F12)および10% FCSを補充したDMEM、293-3-46のために10% FCSおよび1.2mg/mlのG 418を補充したDMEMにおいて単層で維持した。
【0144】
MVウイルスストックを、約107pfu/mlの力価に達するまで成長させるため、組み換えウイルスおよびワクチンのEdmoston Zagreb株をMRC-5細胞において増殖させ、プラーク精製を、シンシチウムを35mmのMRC-5細胞培養液に移し、まず10cmの皿、その後175cmのフラスコに増殖させることによって実施した。シンシチウムが約90%形成されたと判断されたとき、ウイルスストックを175cm2の培養から作製した。いわゆる「無細胞ウイルス分画」に対応する培地を回収し、凍結および解凍を3回行い、細胞のデブリを回避するために遠沈させた。次いでこの培地を-80℃で保存した。いわゆる「細胞結合型ウイルス分画」に対応する細胞を、3mlのOPTIMEM(Gibco BRL)に削り取り、その後3回凍結および解凍を行い、遠沈させ、上清(surnatant)を-80℃で保存した。
【実施例6】
【0145】
プラスミドのトランスフェクションおよびMVウイルスのレスキュー
トランスフェクトされた場合、約50〜70%の集密に達するように、293T細胞を35mmのウェルに播種した。トランスフェクションの4時間前、培地を10%FCSを含有する3mlのDMEMに取り替えた。すべての組み換えプラスミドを、QIAGENプラスミド調製キットに従って調製した。DNAのリン酸Ca2+共沈法のキットはInvitrogenによる。
【0146】
細胞に、以下の最終濃度でプラスミドを共トランスフェクトした:pCA-L 0.5μg、pCA-N 0.5μg、pCA-P 0.1μg、pCA T7 1μgおよび組み換え麻疹-マラリアプラスミド4μg。H2Oで希釈された5種すべてのプラスミドを、2MのCaCl2を含有するエッペンドルフチューブに加え、この混合液を、HEPESバッファーを含有する別のエッペンドルフチューブに振とう状態下で加え、室温(RT)において30分間インキュベートした。このようにして、共沈物を培養液に滴下し、37℃、5%CO2において約18時間トランスフェクションを実施した。その後、トランスフェクション培地を、10% FCSを含有する3mlのDMEMに取り替えた。
【0147】
麻疹のNタンパク質およびPタンパク質を安定に発現する293-3-46ヘルパー細胞(ヒト胎児腎臓細胞)、ならびにT7 RNAポリメラーゼを使用する、組み換え麻疹-マラリアワクチンウイルスを得るための別の方法を、これ以降に記載する。ウイルス性RNAポリメラーゼ(巨大タンパク質、L)を、細胞を、15ngのプラスミドpeMCLaを用いて共トランスフェクトすることによって発現させた。トランスフェクション効率を改善するために、300ngのpSC6-T7 Neoを加えた。カルシウムリン酸法を、トランスフェクションのために使用した。
【0148】
細胞がまだ半集密のとき、トランスフェクションの3〜4日後に最初のシンシチウムが現れた。シンシチウムの形成がより容易に進行するように、各35mmウェルのほぼ集密の細胞単層を、10cmの皿に移した。個々のシンシチウムを、300μlのトランスフェクション培地にとり、700μlのOPTIMEMを含有する滅菌エッペンドルフチューブに入れ、凍結および解凍を3ラウンド行い、-80℃で保存した。
【実施例7】
【0149】
プラークアッセイによるウイルスの滴定
ウイルス調製液の10倍段階希釈を、OPTIMEMを使用して、最終容量0.5mlまで実施した。個々の希釈液を35mmのVero細胞培養物に加えた。ウイルス吸着の1時間後、接種材料を取り除き、感染細胞に、5% FCSおよび1%の低融点アガロース(LMPアガロース)を含有する2mlのDMEMを重ねた。37℃および5% CO2におけるインキュベーション5日後、培養物を1mlの10% TCAに1時間固定し、次いで、30分間UV架橋した。アガロースの重ねを除去後、細胞単層を、4%エタノールに溶解したクリスタルバイオレットで染色し、水で洗浄し、倒立顕微鏡下でプラークをカウントした。
【実施例8】
【0150】
組み換えウイルスのMRC-5におけるウイルス連続継代
レスキューされたウイルスを、MRC5細胞において10回連続継代させ、直径10cmのプレートに播種し、これらを標準的MVウイルスおよび組み換えMVウイルスに、0.01 PFU/細胞のMOIで感染させた。単層を完全に感染させた後、各培養液の1%上清を使用して、その後のMRC5細胞単層を感染させた。トランス遺伝子の発現および安定性を試験するために、継代1、5および10由来のウイルスを、ウェスタンブロットおよび免疫蛍光法による、発現のさらなる特徴付けに使用した。
【実施例9】
【0151】
ウェスタンブロットおよび免疫蛍光法
MVおよびマラリア両方の発現を分析するために、ウェスタンブロットおよび免疫蛍光法を実施した。
【0152】
ウェスタンブロットのために、35mmの皿(1〜5×105)に播種したVero細胞を、90%の集密に関して翌日モニターし、対照としてMVEZを含む、細胞結合型ウイルス分画由来のクリアなウイルス懸濁液を、0.1 MOI(Multiplicity Of Infection)を使用して感染させた。約80%のシンシチウム形成が観察されたとき、細胞をまずPBSを用いて洗浄し、次いで1mlのPBSに削り取り、エッペンドルフチューブに回収し、2000 RPM/4分において、遠心分離にかけた。その後、細胞を、プロテアーゼ阻害剤カクテル(Complete Mini、Roche、1 836 153)を補充した70μlの溶解バッファー(1% NP-40、50mM Tris pH8、150mM NaCl)を用いて、5分/RTで溶解した。上清を13000 RPM/5分において遠心分離にかけることによってクリアにし、新しいチューブに移し、30μlの4×ローディングバッファー(Invitrogen)を加え、試料を混合し、95℃/2分で煮沸し、遠沈させ、-20℃で保存した。
【0153】
SDS-PAGE泳動を実施し、還元条件において1×ランニングバッファーを使用し、200V(開時100〜125mA、終了時60〜80mA)において50分間、NuPAGE 12% Bis-アクリルアミドゲル中を泳動させた。
【0154】
次いで、セミドライ法を使用して、分離された細胞タンパク質をニトロセルロース膜に14V/1h30において転写した。
【0155】
一次抗体として、少なくとも1:30000にPBSTで希釈されたMSP1-p-83に対するウサギポリクローナル抗体および少なくとも1:50000に希釈された、MSP1- p-42,*に対するウサギポリクローナル抗体を使用した。二次抗体は、化学発光増強キット(enhanced chemiluminescence kit)(ECL(商標)、Amersham LifeScience)によってバンドの可視化が可能な、ホースラディッシュペルオキシダーゼで標識されたブタ抗ウサギ抗体であった。
【0156】
免疫蛍光法のために、Vero細胞を、35mmのウェル中の24mm×24mmのガラスカバースリップに播種し、一晩培養し、レスキューされた組み換えウイルスに感染させた。感染3日後に、カバースリップ上の細胞を、PBS中3.7%のパラホルムアルデヒドを用いて固定させ、0.1% TX-100を用いて透過処理し、ブロッキング溶液(1% BSAを含有するPBS)を用いて1時間洗浄し、特異的抗体を用いて染色した。p-42のp-19部分においてEGF様ドメインを認識する、マウスハイブリドーマ上清mAb 5.2を1: 100希釈で使用し、次いで1:250に希釈したFITCH標識ヤギ抗マウス血清を使用した。
【実施例10】
【0157】
成長動態曲線
35mm皿に播種したMRC5細胞(1〜5×105)を、90%の集密に関してモニターし、対照としてMVEZを含む、細胞結合型ウイルス分画由来のクリアなウイルス懸濁液を、0.1 MOIを使用して感染させた。いわゆる「無細胞ウイルス分画」および「細胞結合型ウイルス分画」に対応する試料を、1週間の間毎日回収し、滴定した。
【実施例11】
【0158】
マウスの免疫化
上記のレスキューされた組み換えMV-マラリアウイルスの免疫原性力を、MV感染に対して感受性であるトランスジェニックマウスIFNAR/CD46において免疫化試験を実施することによって証明した。動物を衛生的状況下で飼育し、6〜8週齢において免疫化した。以下に、2種の組み換え麻疹-マラリアウイルス:MeV2EZ-d-p42-SgrAI(GPIアンカー型)およびMeV2EZ-d-p42*(分泌型)によるマウス免疫化の例を提供する。免疫化を、各組み換えMV-マラリアの105 PFUを使用する筋肉注射で、0、4および8週において3回の注射で実施した。組み換え-空麻疹(rMVEZ13-空クローン)を用いて免疫化されたマウスは、陰性対照として機能する。UV不活性化rMVを対照として使用して、免疫反応の活性化に関するウイルス複写の効果を決定した。MVベクター化抗原の免疫反応を、精製d-42タンパク質(0.5mg/ml)と比較して試験した:マウスを、不完全フロイントアジュバント中の20μgのタンパク質を用いて皮下に免疫化した。
【0159】
免疫化IFNAR/CD46マウス(6匹/試験群および3匹/対照群)由来の血清中のMV-特異的抗体の存在を、IgG抗体検出のために、麻疹ウイルスEIA bulk(ATCC VR-24)でコーティングされた96-マイクロウェルプレート使用するELISAによって決定した。タンパク質を、0.05Mの炭酸バッファー(pH 9.4)を用いて0.6μg/mlに希釈し、100μl/ウェルを96ウェルマイクロタイタープレートに加えた。このプレートを、4℃において一晩インキュベートし、PBS/0.05% Tween 20(PT)(ph7.4)を用いて洗浄し、PT(0.1ml/ウェル)-10%BSAとともに37℃において60分間インキュベートし、PTを用いて再度洗浄した。被験血清の2倍段階希釈液を加え(100μl/ウェル)、プレートを37℃において60分間インキュベートした。このプレートをPTを用いて洗浄し、PTで1:2000に希釈した100μlのヤギ抗マウスIgG HRPとともに37℃において30分間インキュベートした。このプレートをPTを用いて洗浄し、100μlのOPD(o-フェニレンジアミン(Phenylendiamin)、Fluka 78411)とともにインキュベートした。反応を、3〜4分後に停止させた。プレートを、MicroElisa Readerで、波長490nmにおいて読み取った。陰性対照より3倍高い示度を、陽性の反応として判定した。
【0160】
免疫化CD46マウス(少なくとも10匹/試験群)の血清中のMV-マラリア-特異的抗体の存在を、ELISAアッセイによって決定した。簡潔には、96マイクロウェルプレートを、炭酸バッファーpH9.4を用いて希釈した50ng/ウェルのMSP-1-d42 3D7株でコーティングした。プレートを、4℃において一晩インキュベートし、PBS/0.05% Tween 20(PT)を用いて洗浄した。その後、非特異的相互作用を、PTに溶解した10%脱脂乳を用いて、37℃において1時間ブロックし、ウェルを、PTを用いて再度洗浄した。プレートを、マウス血清のさまざまな希釈液(1:200から出発して、その後2倍段階希釈)、ペルオキシダーゼ標識ヤギ抗マウスIgGおよびOPD基質とともに連続してインキュベートした。光学密度値を490nmで測定した。カットオフ・バックグラウンドレベル(MV免疫化マウス由来血清の平均値×係数2.1)を上回る値を陽性と考えた。力価は、限界の希釈倍率の逆数として表した。
【0161】
麻疹に対する液性免疫反応を図Aに示す。マラリアp42に対する液性免疫反応を図Bに示す。
【0162】
【化1】
【0163】
【化2】
【実施例12】
【0164】
プラーク精製による、欠陥干渉粒子(DI)からのマラリア抗原を発現する組み換え麻疹ウイルスの精製。
パラミクソウイルス(Paramyxoviruses)、特に麻疹ウイルスによる一定数の継代後、欠陥干渉粒子(DI)の蓄積が起こるであろうことが文献により知られている(23、24)。これらのDIがさまざまな欠陥:ワクチンの安全性に関する負の影響、生産におけるウイルス収率に関する負の影響、ゲノムの不安定性およびワクチン接種後の免疫反応の抑制を発生させることが記載されている。本発明者らの新しい組み換えウイルスに伴うこのようなDIを回避するために、本発明者らは、本発明者らが293T細胞の代わりにMRC5細胞を使用することを除いて、実施例6に記載のようにプラーク精製の方法を適用した。クリアな十分確定されたシンシチウムの形成後、新鮮なMRC5組織培養におけるさらなる継代のために、本発明者らは、顕微鏡下でマイクロピペットを用いてこのような材料を吸引した。
【実施例13】
【0165】
限界希釈による、欠陥干渉粒子(DI)からのマラリア抗原を発現する組み換え麻疹ウイルスの精製。
限界希釈技術を、マイクロプレートにおいて適用した。すべてのウェルにおいて、MRC5細胞の新鮮な単層を発生させた。組み換え麻疹-マラリアウイルスを含有するウイルス懸濁液を、2倍希釈で調製した。シンシチウムが検出された最新の単層のウェルから、上清をピペットで吸引した。上清を、MRC5細胞を含有する懸濁液と混合した。この混合液を、4℃において1時間インキュベートした。最終的に、それを小型のコースターフラスコ(Costar flask)に移し、35℃/5%CO2においてインキュベートし、10日後に組み換え麻疹-マラリアウイルスを精製するために収集した。
【実施例14】
【0166】
麻疹-マラリア混合ワクチンの作製
上記の組み換え麻疹-マラリアウイルスのワーキングシードを、1750cm2のローラーボトル中のMRC5細胞の単層上で、35℃において10日間インキュベートした。細胞を、健康状態および集密に関して毎日モニターした。シンシチウム形成が最も高いレベルである10日目に、上清を液体窒素中の保存用鋼制シリンダーにポンプで汲み上げた。同じ手順を、2日後に繰り返した。すべての試験(ウイルス力価、ゲノムの安定性、ウイルスの安全性、細胞の安全性、化学分析、無菌性など)を実施後、収集物を解凍し、ゼラチン、ソルビトール、アミノ酸および他の糖を含有する安定剤と、最終希釈の105になるように混合した。自動充填機を用いて、小型の凍結乾燥ボトル(F3)に0.5mlずつ接種した。特別に計算した凍結乾燥プログラムを使用して、凍結乾燥プロセスの間の生成物の最大生存を保証した。
(参考文献)
【特許請求の範囲】
【請求項1】
麻疹およびマラリアの両方に対する免疫反応および保護を誘発できるマラリア抗原を発現する、組み換え麻疹ワクチンウイルスを含む麻疹-マラリア混合ワクチン。
【請求項2】
組み換え麻疹ワクチンウイルスが単一または異なるマラリア抗原を発現する、請求項1に記載の麻疹-マラリア混合ワクチン。
【請求項3】
組み換え麻疹ワクチンウイルスがMSP1マラリア抗原を発現する、請求項1に記載の麻疹-マラリア混合ワクチン。
【請求項4】
MSP1マラリア抗原が190〜200KDa(d190)である、請求項1に記載の麻疹-マラリア混合ワクチン。
【請求項5】
MSP1マラリア抗原がd83-30-38である、請求項1に記載の麻疹-マラリア混合ワクチン。
【請求項6】
MSP1マラリア抗原がd42である、請求項1に記載の麻疹-マラリア混合ワクチン。
【請求項7】
組み換え麻疹ワクチンウイルスが、アンカー型および分泌型両方のMSP1マラリア抗原を発現する、請求項1から6のいずれか一項に記載の麻疹-マラリア混合ワクチン。
【請求項8】
組み換え麻疹ワクチンウイルスが、3D7株およびMAD20株、好ましくはFCB1株のアンカー型および分泌型両方のMSP1マラリア抗原を発現する、請求項1から7のいずれか一項に記載の麻疹-マラリア混合ワクチン。
【請求項9】
組み換え麻疹ワクチンウイルスが、FCB1株のアンカー型および分泌型両方のMSP1マラリア抗原を発現する、請求項1から8のいずれか一項に記載の麻疹-マラリア混合ワクチン。
【請求項10】
組み換え麻疹ワクチンウイルスが、Diversity Covering(DiCo)AMA1マラリア抗原を発現する、請求項1に記載の麻疹-マラリア混合ワクチン。
【請求項11】
組み換え麻疹ワクチンウイルスが、AMA1マラリア抗原のDiCo-1を発現する、請求項1に記載の麻疹-マラリア混合ワクチン。
【請求項12】
組み換え麻疹ワクチンウイルスが、AMA1マラリア抗原のDiCo-2を発現する、請求項1に記載の麻疹-マラリア混合ワクチン。
【請求項13】
組み換え麻疹ワクチンウイルスが、AMA1マラリア抗原のDiCo-3を発現する、請求項1に記載の麻疹-マラリア混合ワクチン。
【請求項14】
組み換え麻疹ワクチンウイルスが、AMA1マラリア抗原のDiCo-1、DiCo-2およびDiCo-3を発現する、請求項1に記載の麻疹-マラリア混合ワクチン。
【請求項15】
組み換え麻疹ワクチンウイルスが、膜貫通型および分泌型のAMA1マラリア抗原のDiversity Covering(DiCo)を発現する、請求項10から14のいずれか一項に記載の麻疹-マラリア混合ワクチン。
【請求項16】
組み換え麻疹ワクチンウイルスが、CSマラリア抗原を発現する、請求項1に記載の麻疹-マラリア混合ワクチン。
【請求項17】
マラリア抗原が、組み換え麻疹ワクチンウイルスのPタンパク質とMタンパク質との間またはHタンパク質とLタンパク質との間にクローニングされる、請求項1から16のいずれか一項に記載の麻疹-マラリア混合ワクチン。
【請求項18】
マラリアの抗原のヌクレオチド配列を含む麻疹ワクチンウイルスベクター。
【請求項19】
ヌクレオチド配列が、図26〜図35から選択される、請求項18に記載のベクター。
【請求項20】
図24〜図25から選択されるヌクレオチド配列をさらに含む、請求項18または19に記載のベクター。
【請求項21】
ヌクレオチド配列が、MSP1またはDiversity Covering(DiCo)のAMA1またはCSタンパク質のd83-30-38およびd42およびd190の断片から選択されるマラリア抗原をコードする、請求項18に記載のベクター。
【請求項22】
請求項18に記載のベクターを含む宿主。
【請求項23】
大腸菌(E.coli)または哺乳動物細胞系から選択される、請求項22に記載の宿主。
【請求項24】
組み換え麻疹ウイルスが、Edmoston Zagreb由来のワクチン株を起源とする、請求項1から23のいずれか一項に記載の麻疹-マラリア混合ワクチン。
【請求項25】
組み換え麻疹ワクチンウイルスが、MSP1またはDiversity Covering(DiCo)AMA1またはCSタンパク質のd83-30-38およびd42およびd190の断片から選択される、少なくとも1種のマラリア抗原を発現する、請求項1から24のいずれか一項に記載の麻疹-マラリア混合ワクチン。
【請求項26】
組み換え麻疹ワクチンウイルスが、MSP1もしくはDiversity Covering(DiCo)AMA1もしくはCSタンパク質のd83-30-38およびd42およびd190の断片またはそれらの組み合わせから選択される、2種以上のマラリア抗原を発現する、請求項1から25のいずれか一項に記載の麻疹-マラリア混合ワクチン。
【請求項27】
組み換え麻疹ワクチンウイルスが、以下の配列
MSP-1 d-190-3D7 AN101TE
MSP-1 d-190*-3D7 AN102TE
MSP-1 d-83-30-38-3D7 AN103TE
MSP-1 d-83-30-38*-3D7 AN104TE
MSP-1 d-42-3D7 AN105TE
MSP-1 d-42*-3D7 AN106TE
MSP-1 d-190-FCBl AN107TE
CS AN108TE
DiCo1-完全 AN109TE
DiCo1-ecto AN110TE
を含む、請求項1から26のいずれか一項に記載の麻疹-マラリア混合ワクチン。
【請求項28】
組み換え麻疹ワクチンウイルスが、マラリア抗原に加えてアジュバント特性を有するタンパク質をコードする、請求項1から27のいずれか一項に記載の麻疹-マラリア混合ワクチン。
【請求項29】
マラリア抗原に加えてインターロイキン、好ましくはインターロイキン2をコードする組み換え麻疹ウイルスを含む、請求項1から28のいずれか一項に記載の麻疹-マラリア混合ワクチン。
【請求項30】
記載の組み換え麻疹マラリアウイルスの1種またはこれらのウイルスの2種から数種の混合物を含む、請求項1から29のいずれか一項に記載のワクチン。
【請求項31】
記載の組み換え麻疹マラリアウイルスまたは、これらのウイルスの2種から数種の混合物が欠陥干渉粒子(DI)を欠いている、請求項1から30のいずれか一項に記載のワクチン。
【請求項32】
偶発的に発生したDI粒子がプラーク精製によって除去された、請求項1から31のいずれか一項に記載のワクチン。
【請求項33】
偶発的に発生したDI粒子が限界希釈によって除去された、請求項1から32のいずれか一項に記載のワクチン。
【請求項34】
偶発的に発生したDI粒子が、物理的方法、例えば分画遠心分離によって除去された、請求項1から33のいずれか一項に記載のワクチン。
【請求項35】
混合ワクチンの成分であり、他の成分が、天然に弱毒化された、または組み換えの、風疹、流行性耳下腺炎、水痘または別の生弱毒化ワクチンウイルスを、単独または組み合わせたものである、請求項1から34のいずれか一項に記載のワクチン。
【請求項36】
非経口適用に理想的な、適切な安定剤、例えばゼラチンおよび/または主成分としてヒト血清アルブミンおよびソルビトールを含む、請求項1から35のいずれか一項に記載のワクチン。
【請求項37】
鼻腔内適用に理想的な、適切な安定剤および/またはアジュバントを含む、請求項1から36のいずれか一項に記載のワクチン。
【請求項38】
吸入適用に理想的な、適切な安定剤および/またはアジュバントを含む、請求項1から37のいずれか一項に記載のワクチン。
【請求項39】
経口適用に理想的な、適切な安定剤および/またはアジュバントを含む、請求項1から38のいずれか一項に記載のワクチン。
【請求項40】
経皮適用に理想的な、適切な安定剤および/またはアジュバントを含む、請求項1から39のいずれか一項に記載のワクチン。
【請求項41】
任意の坐薬製剤化に理想的な、適切な安定剤および/またはアジュバントを含む、請求項1から40のいずれか一項に記載のワクチン。
【請求項42】
安定剤および/またはアジュバントを含む、麻疹-マラリア混合ワクチンの組成物。
【請求項1】
麻疹およびマラリアの両方に対する免疫反応および保護を誘発できるマラリア抗原を発現する、組み換え麻疹ワクチンウイルスを含む麻疹-マラリア混合ワクチン。
【請求項2】
組み換え麻疹ワクチンウイルスが単一または異なるマラリア抗原を発現する、請求項1に記載の麻疹-マラリア混合ワクチン。
【請求項3】
組み換え麻疹ワクチンウイルスがMSP1マラリア抗原を発現する、請求項1に記載の麻疹-マラリア混合ワクチン。
【請求項4】
MSP1マラリア抗原が190〜200KDa(d190)である、請求項1に記載の麻疹-マラリア混合ワクチン。
【請求項5】
MSP1マラリア抗原がd83-30-38である、請求項1に記載の麻疹-マラリア混合ワクチン。
【請求項6】
MSP1マラリア抗原がd42である、請求項1に記載の麻疹-マラリア混合ワクチン。
【請求項7】
組み換え麻疹ワクチンウイルスが、アンカー型および分泌型両方のMSP1マラリア抗原を発現する、請求項1から6のいずれか一項に記載の麻疹-マラリア混合ワクチン。
【請求項8】
組み換え麻疹ワクチンウイルスが、3D7株およびMAD20株、好ましくはFCB1株のアンカー型および分泌型両方のMSP1マラリア抗原を発現する、請求項1から7のいずれか一項に記載の麻疹-マラリア混合ワクチン。
【請求項9】
組み換え麻疹ワクチンウイルスが、FCB1株のアンカー型および分泌型両方のMSP1マラリア抗原を発現する、請求項1から8のいずれか一項に記載の麻疹-マラリア混合ワクチン。
【請求項10】
組み換え麻疹ワクチンウイルスが、Diversity Covering(DiCo)AMA1マラリア抗原を発現する、請求項1に記載の麻疹-マラリア混合ワクチン。
【請求項11】
組み換え麻疹ワクチンウイルスが、AMA1マラリア抗原のDiCo-1を発現する、請求項1に記載の麻疹-マラリア混合ワクチン。
【請求項12】
組み換え麻疹ワクチンウイルスが、AMA1マラリア抗原のDiCo-2を発現する、請求項1に記載の麻疹-マラリア混合ワクチン。
【請求項13】
組み換え麻疹ワクチンウイルスが、AMA1マラリア抗原のDiCo-3を発現する、請求項1に記載の麻疹-マラリア混合ワクチン。
【請求項14】
組み換え麻疹ワクチンウイルスが、AMA1マラリア抗原のDiCo-1、DiCo-2およびDiCo-3を発現する、請求項1に記載の麻疹-マラリア混合ワクチン。
【請求項15】
組み換え麻疹ワクチンウイルスが、膜貫通型および分泌型のAMA1マラリア抗原のDiversity Covering(DiCo)を発現する、請求項10から14のいずれか一項に記載の麻疹-マラリア混合ワクチン。
【請求項16】
組み換え麻疹ワクチンウイルスが、CSマラリア抗原を発現する、請求項1に記載の麻疹-マラリア混合ワクチン。
【請求項17】
マラリア抗原が、組み換え麻疹ワクチンウイルスのPタンパク質とMタンパク質との間またはHタンパク質とLタンパク質との間にクローニングされる、請求項1から16のいずれか一項に記載の麻疹-マラリア混合ワクチン。
【請求項18】
マラリアの抗原のヌクレオチド配列を含む麻疹ワクチンウイルスベクター。
【請求項19】
ヌクレオチド配列が、図26〜図35から選択される、請求項18に記載のベクター。
【請求項20】
図24〜図25から選択されるヌクレオチド配列をさらに含む、請求項18または19に記載のベクター。
【請求項21】
ヌクレオチド配列が、MSP1またはDiversity Covering(DiCo)のAMA1またはCSタンパク質のd83-30-38およびd42およびd190の断片から選択されるマラリア抗原をコードする、請求項18に記載のベクター。
【請求項22】
請求項18に記載のベクターを含む宿主。
【請求項23】
大腸菌(E.coli)または哺乳動物細胞系から選択される、請求項22に記載の宿主。
【請求項24】
組み換え麻疹ウイルスが、Edmoston Zagreb由来のワクチン株を起源とする、請求項1から23のいずれか一項に記載の麻疹-マラリア混合ワクチン。
【請求項25】
組み換え麻疹ワクチンウイルスが、MSP1またはDiversity Covering(DiCo)AMA1またはCSタンパク質のd83-30-38およびd42およびd190の断片から選択される、少なくとも1種のマラリア抗原を発現する、請求項1から24のいずれか一項に記載の麻疹-マラリア混合ワクチン。
【請求項26】
組み換え麻疹ワクチンウイルスが、MSP1もしくはDiversity Covering(DiCo)AMA1もしくはCSタンパク質のd83-30-38およびd42およびd190の断片またはそれらの組み合わせから選択される、2種以上のマラリア抗原を発現する、請求項1から25のいずれか一項に記載の麻疹-マラリア混合ワクチン。
【請求項27】
組み換え麻疹ワクチンウイルスが、以下の配列
MSP-1 d-190-3D7 AN101TE
MSP-1 d-190*-3D7 AN102TE
MSP-1 d-83-30-38-3D7 AN103TE
MSP-1 d-83-30-38*-3D7 AN104TE
MSP-1 d-42-3D7 AN105TE
MSP-1 d-42*-3D7 AN106TE
MSP-1 d-190-FCBl AN107TE
CS AN108TE
DiCo1-完全 AN109TE
DiCo1-ecto AN110TE
を含む、請求項1から26のいずれか一項に記載の麻疹-マラリア混合ワクチン。
【請求項28】
組み換え麻疹ワクチンウイルスが、マラリア抗原に加えてアジュバント特性を有するタンパク質をコードする、請求項1から27のいずれか一項に記載の麻疹-マラリア混合ワクチン。
【請求項29】
マラリア抗原に加えてインターロイキン、好ましくはインターロイキン2をコードする組み換え麻疹ウイルスを含む、請求項1から28のいずれか一項に記載の麻疹-マラリア混合ワクチン。
【請求項30】
記載の組み換え麻疹マラリアウイルスの1種またはこれらのウイルスの2種から数種の混合物を含む、請求項1から29のいずれか一項に記載のワクチン。
【請求項31】
記載の組み換え麻疹マラリアウイルスまたは、これらのウイルスの2種から数種の混合物が欠陥干渉粒子(DI)を欠いている、請求項1から30のいずれか一項に記載のワクチン。
【請求項32】
偶発的に発生したDI粒子がプラーク精製によって除去された、請求項1から31のいずれか一項に記載のワクチン。
【請求項33】
偶発的に発生したDI粒子が限界希釈によって除去された、請求項1から32のいずれか一項に記載のワクチン。
【請求項34】
偶発的に発生したDI粒子が、物理的方法、例えば分画遠心分離によって除去された、請求項1から33のいずれか一項に記載のワクチン。
【請求項35】
混合ワクチンの成分であり、他の成分が、天然に弱毒化された、または組み換えの、風疹、流行性耳下腺炎、水痘または別の生弱毒化ワクチンウイルスを、単独または組み合わせたものである、請求項1から34のいずれか一項に記載のワクチン。
【請求項36】
非経口適用に理想的な、適切な安定剤、例えばゼラチンおよび/または主成分としてヒト血清アルブミンおよびソルビトールを含む、請求項1から35のいずれか一項に記載のワクチン。
【請求項37】
鼻腔内適用に理想的な、適切な安定剤および/またはアジュバントを含む、請求項1から36のいずれか一項に記載のワクチン。
【請求項38】
吸入適用に理想的な、適切な安定剤および/またはアジュバントを含む、請求項1から37のいずれか一項に記載のワクチン。
【請求項39】
経口適用に理想的な、適切な安定剤および/またはアジュバントを含む、請求項1から38のいずれか一項に記載のワクチン。
【請求項40】
経皮適用に理想的な、適切な安定剤および/またはアジュバントを含む、請求項1から39のいずれか一項に記載のワクチン。
【請求項41】
任意の坐薬製剤化に理想的な、適切な安定剤および/またはアジュバントを含む、請求項1から40のいずれか一項に記載のワクチン。
【請求項42】
安定剤および/またはアジュバントを含む、麻疹-マラリア混合ワクチンの組成物。
【図1】

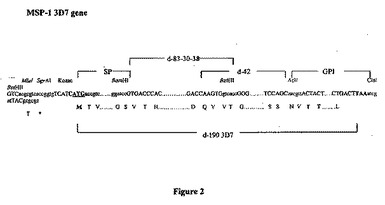
【図3】
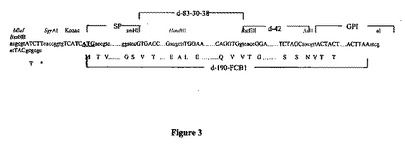
【図4】
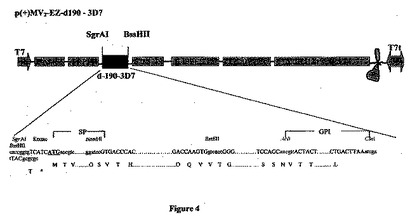
【図5】
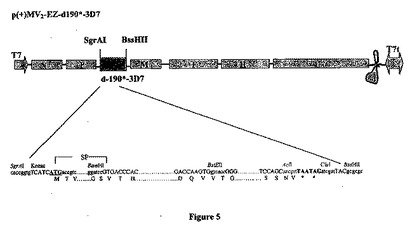
【図6】
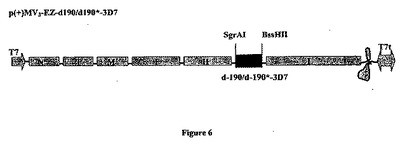
【図7】
【図8】
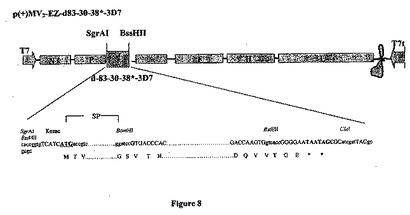
【図9】
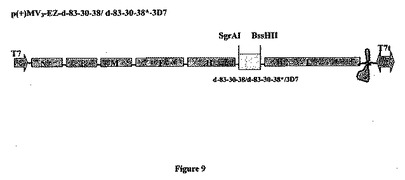
【図10】
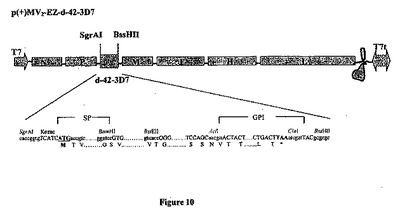
【図11】
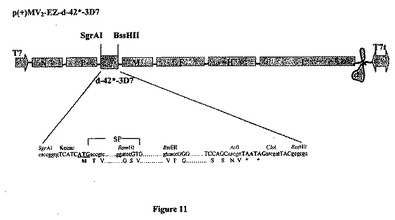
【図12】
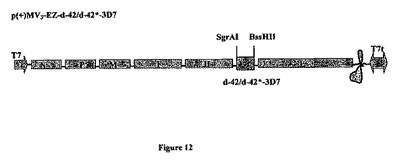
【図13】
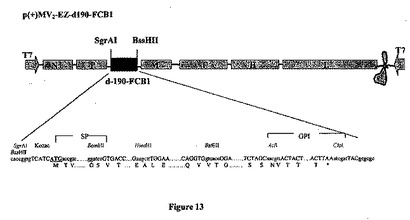
【図14】
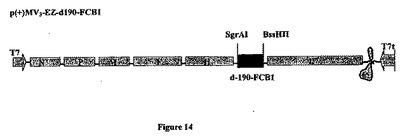

【図16】
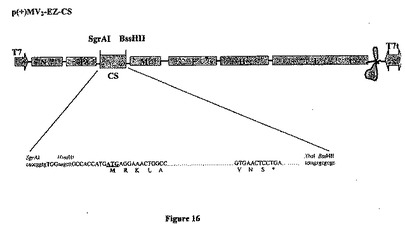
【図17】
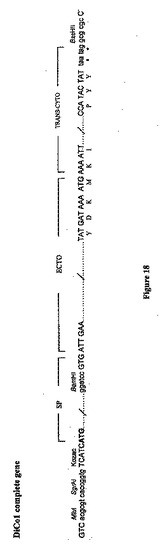

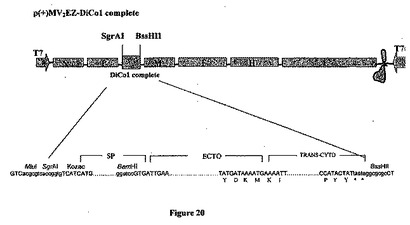
【図22】
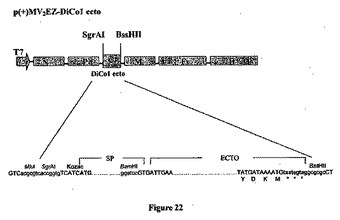
【図23】
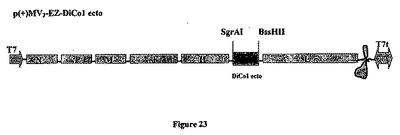
【図24−1】
【図24−2】
【図24−3】
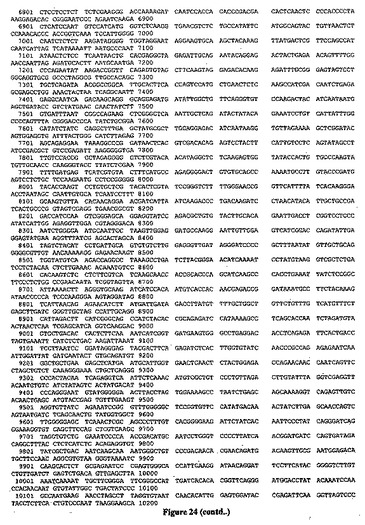
【図24−4】
【図24−5】
【図24−6】
【図25−1】
【図25−2】
【図25−3】
【図25−4】
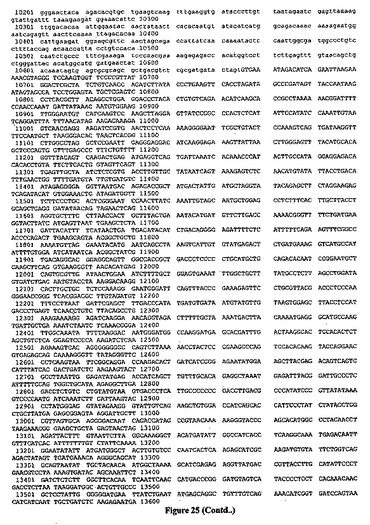
【図25−5】
【図25−6】
【図26−1】
【図26−2】
【図27−1】
【図27−2】
【図28−1】
【図28−2】

【図29−1】
【図29−2】

【図30】

【図31】

【図32−1】
【図32−2】
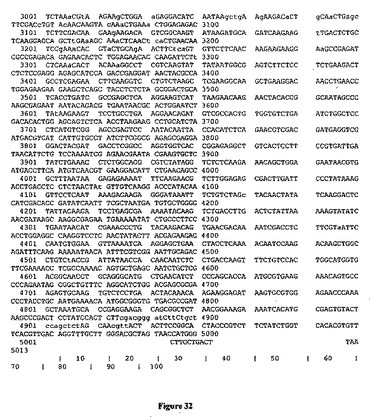
【図33】

【図34】
【図35】
【図36】
【図37】
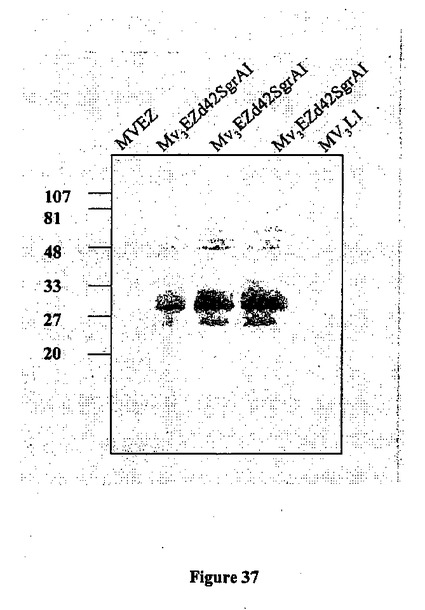
【図38】

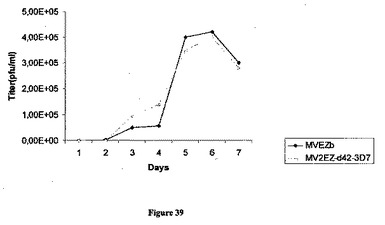
【図40】
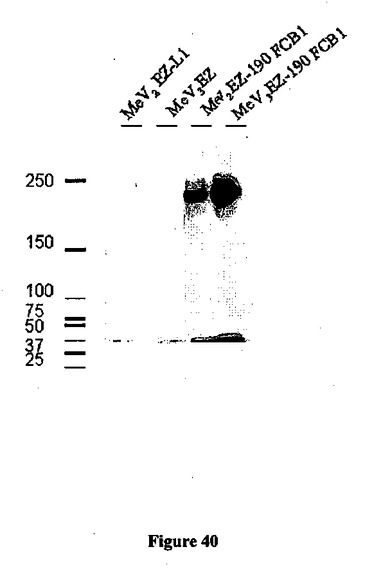
【図2】
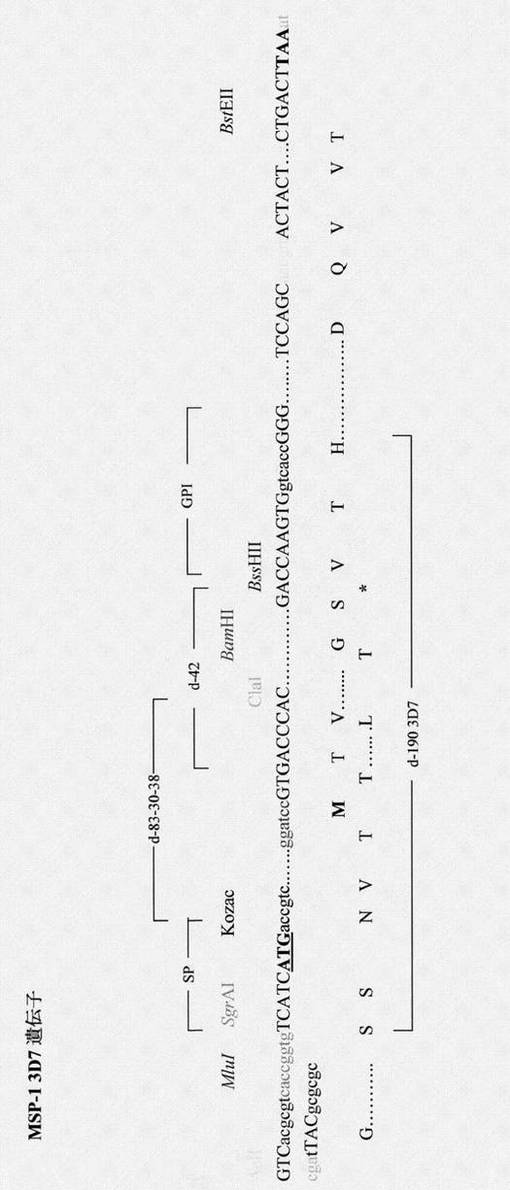
【図15】

【図18】
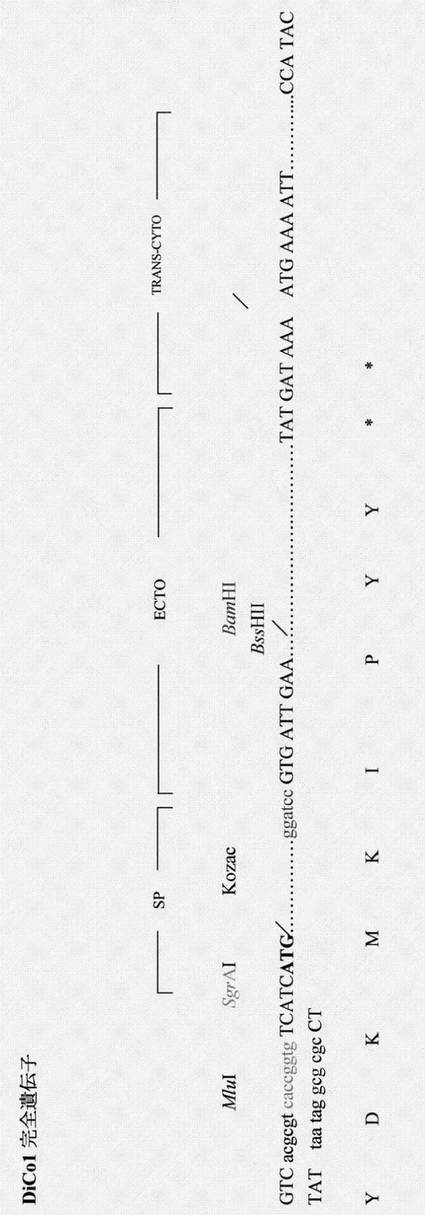
【図19】
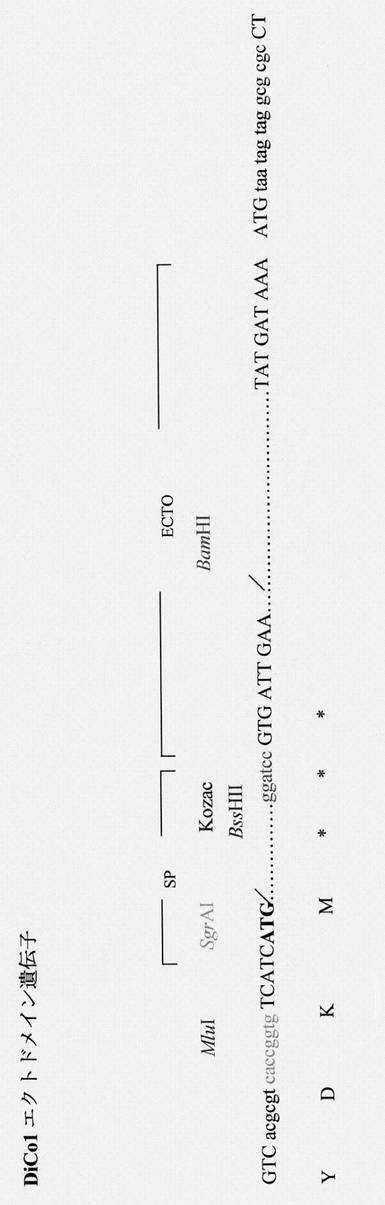
【図20】
【図21】
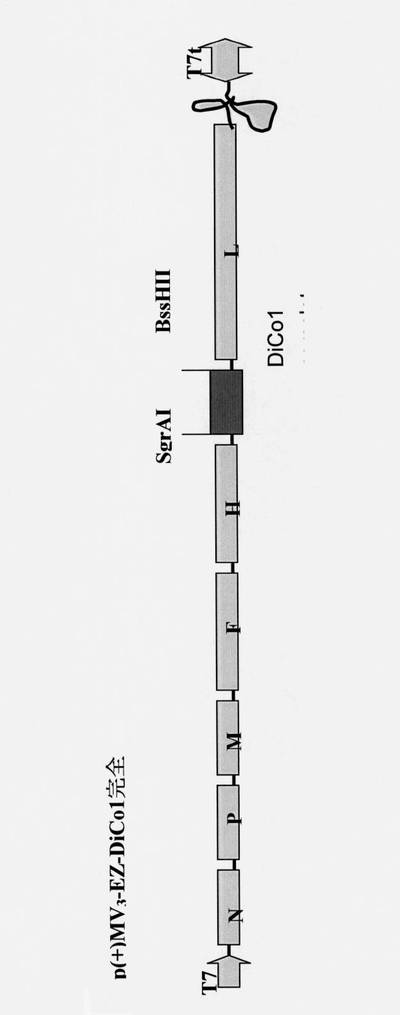
【図39】
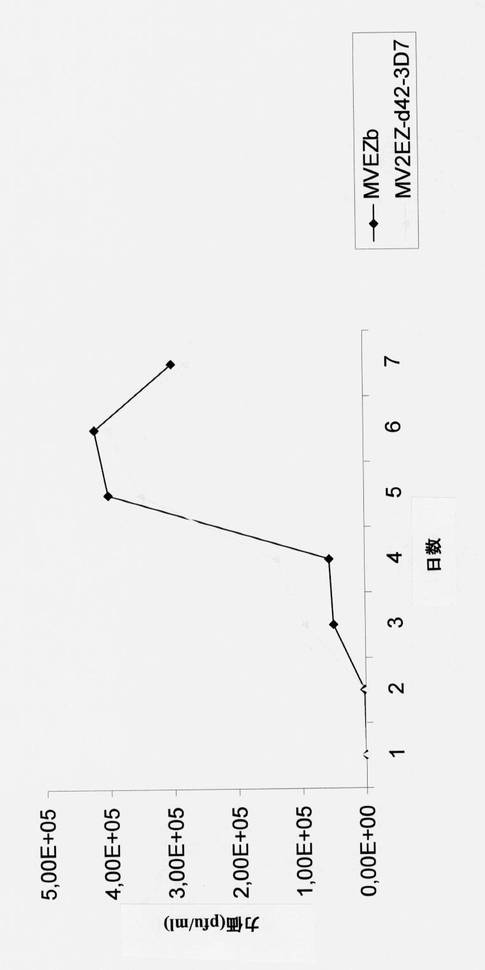
【図41】
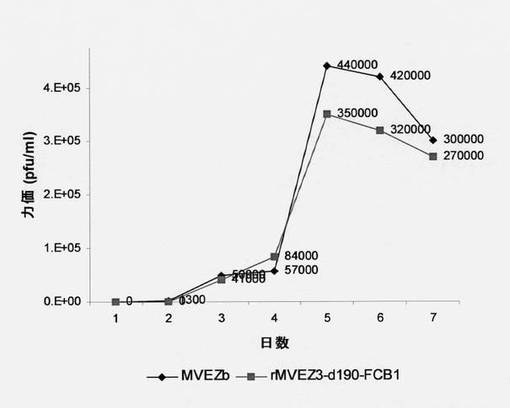
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図16】
【図17】
【図22】
【図23】
【図24−1】
【図24−2】
【図24−3】
【図24−4】
【図24−5】
【図24−6】
【図25−1】
【図25−2】
【図25−3】
【図25−4】
【図25−5】
【図25−6】
【図26−1】
【図26−2】
【図27−1】
【図27−2】
【図28−1】
【図28−2】
【図29−1】
【図29−2】
【図30】
【図31】
【図32−1】
【図32−2】
【図33】
【図34】
【図35】
【図36】
【図37】
【図38】
【図40】
【図2】
【図15】
【図18】
【図19】
【図20】
【図21】
【図39】
【図41】
【公表番号】特表2012−526102(P2012−526102A)
【公表日】平成24年10月25日(2012.10.25)
【国際特許分類】
【出願番号】特願2012−509150(P2012−509150)
【出願日】平成22年5月3日(2010.5.3)
【国際出願番号】PCT/IN2010/000287
【国際公開番号】WO2010/128524
【国際公開日】平成22年11月11日(2010.11.11)
【出願人】(507365927)カディラ・ヘルスケア・リミテッド (26)
【Fターム(参考)】
【公表日】平成24年10月25日(2012.10.25)
【国際特許分類】
【出願日】平成22年5月3日(2010.5.3)
【国際出願番号】PCT/IN2010/000287
【国際公開番号】WO2010/128524
【国際公開日】平成22年11月11日(2010.11.11)
【出願人】(507365927)カディラ・ヘルスケア・リミテッド (26)
【Fターム(参考)】
[ Back to top ]