
(R)−ヒドロキシニトリルリアーゼの製造方法
【課題】 (R)−ヒドロキシニトリルリアーゼの異種発現において、(R)−ヒドロキシニトリルリアーゼの産生効率を高め、光学活性シアノヒドリンの工業的な製造方法を提供する。
【解決手段】 バラ科植物由来(R)−ヒドロキシニトリルリアーゼをコードする遺伝子の塩基配列が改変されてなるDNAであって、該改変が、該DNAが宿主細胞に導入されたときの(R)−ヒドロキシニトリルリアーゼの発現量を改変されていないDNAを導入した場合と比べて増大させるものである、該DNA。
【解決手段】 バラ科植物由来(R)−ヒドロキシニトリルリアーゼをコードする遺伝子の塩基配列が改変されてなるDNAであって、該改変が、該DNAが宿主細胞に導入されたときの(R)−ヒドロキシニトリルリアーゼの発現量を改変されていないDNAを導入した場合と比べて増大させるものである、該DNA。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、(R)−ヒドロキシニトリルリアーゼをコードする遺伝子の塩基配列が改変されてなるDNA、及び該DNAを用いて光学活性シアノヒドリンを製造する方法に関する。
【背景技術】
【0002】
生体触媒を用いる合成プロセスは化学工業にとってきわめて重要になっている。また、生体触媒を使った化学反応の実施は、キラル又はプロキラルな成分との化学反応で複数のエナンチオマーのうちの一方を選択的に変換又は生成するという、基質特異性の高い生体触媒の性質を利用することが可能な応用分野では、とりわけ興味深い。酵素が持つこの有利な性質を利用するには、その酵素を容易にかつ十分な量で入手できることが必須条件になる。
【0003】
光学活性シアノヒドリンは特に興味深い光学活性化合物群であり、医薬活性物質、ビタミン又はピレスロイド化合物などの生物活性物質の製造に使用されるα−ヒドロキシ酸、α−ヒドロキシケトン、β−アミノアルコールの合成等に重要である。シアノヒドリンは、カルボニル化合物のカルボニル基へのシアン化水素酸の付加によって製造される。例えば(S)−シアノヒドリンなどの光学活性化合物の工業的生産はパラゴムノキ(Heveabrasiliensis)由来の酵素(S)−ヒドロキシニトリルリアーゼを利用することによって可能になっており、例えば特許文献1〜4などに記載されている。
【0004】
また、非特許文献1などには、サクラ属植物が(R)−ヒドロキシニトリルリアーゼのアイソザイムを含有しうることが開示されている。これらのアイソザイムは当該植物の様々な組織に異なるレベルで発現される。アーモンドと近縁関係にあるアメリカザクラには、現在までに5種類の異なるアイソザイムが同定され、それらの遺伝子が配列決定されている。アーモンド由来のアイソザイムは今までのところ1つだけが非特許文献2に記載されており、このアイソザイム遺伝子は花組織にもっとも強く発現される。前記(R)−ヒドロキシニトリルリアーゼのアイソザイムの遺伝子は既に単離され、そのcDNAが配列決定されている。しかし、(R)−ヒドロキシニトリルリアーゼはこれらの植物の組織に微量に含まれているに過ぎず、工業的に利用することは困難であった。
【0005】
(R)−ヒドロキシニトリルリアーゼ遺伝子の異種発現については、真核細胞である酵母(サッカロミセス属又はピキア属)を宿主として、アーモンド由来のヒドロキシニトリルリアーゼを遺伝子組換えにより製造した例がある(特許文献5)。しかし、ヒドロキシニトリルリアーゼの異種発現において、酵母以外を宿主として(R)−ヒドロキシニトリルリアーゼの製造に成功した例はなく、また該酵素の発現量を増大させることについても報告されていない。
【0006】
【特許文献1】WO97/03204
【特許文献2】EP 0 951 561
【特許文献3】EP 0 927 766
【特許文献4】特表平11−508775
【特許文献5】特開2002−330791
【非特許文献1】Plant Physiology,April(1999)119:1535〜1546
【非特許文献2】Planta(1998)206:388〜393
【発明の開示】
【発明が解決しようとする課題】
【0007】
本発明の課題は、(R)−ヒドロキシニトリルリアーゼの異種発現において、(R)−ヒドロキシニトリルリアーゼの産生効率を高め、光学活性シアノヒドリンの工業的な製造方法を提供することである。
【課題を解決するための手段】
【0008】
本発明者らは鋭意検討を行った結果、バラ科植物由来(R)−ヒドロキシニトリルリアーゼをコードする遺伝子の塩基配列に対し、特定の改変を加え、これを宿主細胞で発現させることにより、(R)−ヒドロキシニトリルリアーゼを効率的に産生できることを見いだし、本発明を完成するに至った。
【0009】
すなわち、本発明は以下の発明を包含する。
(1)バラ科植物由来(R)−ヒドロキシニトリルリアーゼをコードする遺伝子の塩基配列が改変されてなるDNAであって、該改変が、該DNAが宿主細胞に導入されたときの(R)−ヒドロキシニトリルリアーゼの発現量を改変されていないDNAを導入した場合と比べて増大させるものである、該DNA。
(2)改変が、以下の(a)〜(c)の工程:
(a)バラ科植物由来(R)−ヒドロキシニトリルリアーゼをコードする遺伝子の塩基配列に含まれるシグナルペプチドをコードする塩基配列を除去すること、
(b)バラ科植物由来(R)−ヒドロキシニトリルリアーゼをコードする遺伝子の塩基配列中のコドンを、そのアミノ酸配列を変化させることなく、宿主細胞におけるアミノ酸ごとのコドン使用頻度が10%以上となるように改変すること、及び
(c)バラ科植物由来(R)−ヒドロキシニトリルリアーゼをコードする遺伝子の塩基配列にタグ配列を付加すること
からなる群から選択される少なくとも1つの工程によって実施されるものである、(1)記載のDNA。
(3)配列番号8で表される塩基配列を含み、塩基長が4000塩基以下のDNA。
(4)(1)〜(3)のいずれかに記載のDNAを含む組換えベクター。
(5)(4)記載の組換えベクターを含む形質転換体。
(6)(1)〜(3)のいずれかに記載のDNAを発現させ、得られた(R)−ヒドロキシニトリルリアーゼとカルボニル化合物及びシアノ基供与体とを反応させ、反応液から光学活性シアノヒドリンを回収することを含む、光学活性シアノヒドリンの製造方法。
【発明の効果】
【0010】
本発明により、(R)−ヒドロキシニトリルリアーゼの異種発現における発現量を増大することができ、(R)−ヒドロキシニトリルリアーゼを効率的に産生することができる。それにより光学活性シアノヒドリンの工業的な製造が可能になる。
【発明を実施するための最良の形態】
【0011】
本発明において、(R)−ヒドロキシニトリルリアーゼとは、当技術分野において通常用いられる意味を有し、シアノ基供与体の存在下、カルボニル化合物を基質としてシアノヒドリンを合成する反応を触媒する活性を有する酵素を意味する。
【0012】
本発明は、バラ科植物由来の(R)−ヒドロキシニトリルリアーゼを対象とする。バラ科植物としては、特に限定されないが、例えば、サクラ属、キンミズヒキ属、アプリコット属、セラサス属、リンゴ属、メスピラス属、ペルシカ属、ピルス属、ヤマザクラ属、バラ属、キイチゴ属から選択される属に属する植物が挙げられ、より具体的には、アーモンド、セイヨウヤマザクラ、カリン、サンザシ、セイヨウサンザシ、マルメロ、ビワ、イチゴ、リンゴ、アメリカンレッドプラム、ホンアンズ、アンズ、セイヨウミザクラ、スミノミザクラ、セイヨウスモモ、ハイブリッド、ウメ、シナミザクラ、モモ、サントウ、コモンピーチ、ネクタリン、スモモ、ユスラウメ、セイヨウナシ、ニホンナシ、チュウゴクナシ、ノイバラ、ハマナシ、ノバラ、ハイブッシュブラックベリー、ヨーロッパキイチゴ、エバーグリーンブラックベリー、サーモンベリー、パシフィックブラックベリー等が挙げられる。本発明においては、好ましくはサクラ属(Prunus属)に属する植物、より好ましくはアーモンド(Prunus amygdalus)を使用する。
【0013】
本発明において、バラ科植物由来(R)−ヒドロキシニトリルリアーゼをコードする遺伝子とは、バラ科植物のゲノムDNA上の(R)−ヒドロキシニトリルリアーゼ遺伝子DNA、及び(R)−ヒドロキシニトリルリアーゼをコードするcDNAを包含するものとする。本発明において、バラ科植物由来(R)−ヒドロキシニトリルリアーゼをコードする遺伝子の塩基配列は、好ましくは配列番号1で表される塩基配列である。該遺伝子の塩基配列が改変されてなるDNAは、上記ゲノムDNA上の(R)−ヒドロキシニトリルリアーゼ遺伝子の塩基配列、又は(R)−ヒドロキシニトリルリアーゼcDNAの塩基配列、好ましくは(R)−ヒドロキシニトリルリアーゼcDNAの塩基配列に対して、改変するための処理を行った結果得られるDNAを意味する。
【0014】
上記塩基配列の改変は、好ましくは、該DNAを宿主細胞に導入したときの、(R)−ヒドロキシニトリルリアーゼの発現量を、改変しない場合と比べて10%以上、好ましくは30%以上、より好ましくは50%以上増大させるものである。すなわち、改変された塩基配列からなるDNAを宿主細胞に導入して発現させた場合に発現される(R)−ヒドロキシニトリルリアーゼが、改変していない塩基配列からなるDNAを導入した場合と比較して、10%以上増大する。
【0015】
本発明において、発現量を、改変していないDNAを導入した場合と比べて増大させるとは、改変していないDNAを導入した場合に全く発現しなかったものが、発現するようになる場合も包含する。
【0016】
ここで発現量の比較は遺伝子配列を改変したものとしないものでそれぞれ誘導発現した菌体をSDS−PAGEに供する際にOD660nmの吸光度が10になるように濃縮または希釈してから、SDS−PAGEに供し、得られた画像データからバンドの濃さをデンシトグラムで計算して目的バンドの濃さを比較して算出するものと定義する。
【0017】
(R)−ヒドロキシニトリルリアーゼの発現量を増大するような塩基配列の改変方法としては、当技術分野で通常用いられるものを使用することができ、特に制限されない。好ましくは、以下の(a)〜(c)の工程:(a)バラ科植物由来(R)−ヒドロキシニトリルリアーゼをコードする遺伝子の塩基配列に含まれるシグナルペプチドをコードする塩基配列を除去すること、(b)バラ科植物由来(R)−ヒドロキシニトリルリアーゼをコードする遺伝子の塩基配列中のコドンを、そのアミノ酸配列を変化させることなく、宿主細胞におけるアミノ酸ごとのコドン使用頻度が10%以上となるように改変すること、及び(c)バラ科植物由来(R)−ヒドロキシニトリルリアーゼをコードする遺伝子の塩基配列にタグ配列を付加すること、からなる群から選択される少なくとも1つの工程によって実施できる。(a)〜(c)の工程のうち1つのみを実施してもよいし、2つを実施してもよい。複数の工程を実施する場合、実施する工程の順序は特に制限されない。本発明においては、少なくとも(a)の工程を実施することが好ましい。上記(a)〜(c)の工程について以下に説明する。
【0018】
(a)シグナルペプチドをコードする塩基配列(シグナル配列)の除去
シグナル配列の除去方法は、当技術分野で通常用いられる方法を使用でき、特に限定されないが、例えば、既知の(R)−ヒドロキシニトリルリアーゼ遺伝子を鋳型としてシグナル配列が除去された配列を増幅するように設計された適当なプライマーを用いてPCRを行う方法、DNA合成手法による全合成法、既知の(R)−ヒドロキシニトリルリアーゼ遺伝子のサイトダイレクティッドミュータジェネシス(M.J.Zollerら、Methods in Enzymology(1983)100:468−500)による方法、又は既知の(R)−ヒドロキシニトリルリアーゼ遺伝子の一部を合成DNAフラグメントと差し換える方法などが挙げられる。
【0019】
例えば、配列番号4で表される塩基配列からなる(R)−ヒドロキシニトリルリアーゼ遺伝子において、シグナル配列は1〜81番塩基で表される(Swiss−Prot:アクセッションナンバーO24243参照)ので、配列番号4で表される遺伝子を鋳型として、配列番号4の82番塩基以降の塩基配列を増幅するように設計されたプライマーを用いてPCRを実施することにより、シグナル配列の除去されたDNAを得ることができる。
【0020】
(b)コドン使用頻度の調節
本改変方法は、そのアミノ酸配列を変化させることなく、宿主におけるアミノ酸ごとのコドン使用頻度が10%以上となるように改変することで、(R)−ヒドロキシニトリルリアーゼの発現量を増加させるものである。本発明において、コドン使用頻度とは、宿主細胞内で発現する全ての遺伝子に含まれる全コドン中の、同種アミノ酸をコードするコドンの使用回数に対する1つのコドンの使用回数の百分率をいう。
【0021】
ここで、改変した遺伝子のコドンのうち、宿主細胞における使用頻度が最も低いコドンの使用割合が10%以上、より好ましくは12%以上、さらに好ましくは14%となるようにする。10%を下回ると発現量の増加が少ない場合があるためである。各宿主におけるコドン使用頻度は、データベースとして公開されているものを使用できる。
【0022】
宿主として大腸菌を使用する場合、Escherichia coli K−12 MG1655(XanaGenome(Microbial Genome Database)参照)のコドン使用頻度データを利用し、表1に示すようにコドンを調節することができる。例として具体的には、表1を参照すると、Argをコードする使用頻度が2.2%であるコドンAGGを、使用頻度が37.9%であるコドンCGTへ改変する。
【0023】
【表1】

【0024】
例えば、配列番号4で表される塩基配列からなる(R)−ヒドロキシニトリルリアーゼ遺伝子のコドン使用頻度を上記手順に従って調節することにより、配列番号5で表される塩基配列からなるDNAが得られる。従って、本発明の改変DNAには、配列番号5で表される塩基配列からなるDNAが包含される。このようにして得られたDNAを、大腸菌、特にBL21株に導入して発現させることにより、(R)−ヒドロキシニトリルリアーゼの発現量を増大させることができる。
【0025】
本発明においては、(R)−ヒドロキシニトリルリアーゼ遺伝子の塩基配列において、コドン使用頻度を調節した後で、さらにシグナル配列を除去することにより改変を実施するのが好ましい。例えば、配列番号4で表される塩基配列からなる(R)−ヒドロキシニトリルリアーゼ遺伝子のコドン使用頻度を調節することにより得られる配列番号5で表される塩基配列において、シグナル配列に相当する1〜81番塩基を除去することにより、改変を実施することができる。このようにして得られるDNAとして、配列番号8で表される塩基配列からなるDNAが挙げられる。
【0026】
従って、本発明は、以下の(a)又は(b)のDNAを包含する:
(a)配列番号8で表される塩基配列を含み、塩基長が4000塩基以下、好ましくは2500塩基以下、より好ましくは1600塩基以下のDNA、
(b)配列番号8で表される塩基配列と少なくとも80%、好ましくは少なくとも90%、より好ましくは少なくとも95%の相同性を有し、(R)−ヒドロキシニトリルリアーゼ活性を有するタンパク質をコードする塩基配列を含み、塩基長が4000塩基以下、好ましくは2500塩基以下、より好ましくは1600塩基以下のDNA。
【0027】
(c)タグ配列の付加
本改変方法は、バラ科植物由来(R)−ヒドロキシニトリルリアーゼをコードする遺伝子の塩基配列にタグ配列を付加することによって、該タンパク質の発現量を増大させるものである。
【0028】
付加するタグ配列としては、(R)−ヒドロキシニトリルリアーゼの宿主細胞における発現を増大するものであれば特に制限されない。例えば、タンパク質を可溶化する機能を有するタグ配列、具体的には、Trxタグ配列(チオレドキシンをコードする配列)、及びNusタグ配列(大腸菌データベース上で最も優れた可溶性ポテンシャルを有すると同定された配列)、ならびにタンパク質のフォールディングを促進する機能を有するタグ配列、具体的にはDSbタグ配列(ジスルフィド結合の形成と異性化を触媒する酵素をコードする配列)が挙げられる。例えば、Trxタグ配列がコードするチオレドキシンと(R)−ヒドロキシニトリルリアーゼとが融合して発現するように、Trgタグ配列を付加することにより、(R)−ヒドロキシニトリルリアーゼが宿主細胞の細胞質中に可溶性画分として発現されやすくなる。
【0029】
タグ配列の付加は、例えば、対象とするタグ配列を含む発現ベクターに、目的とするDNAを連結することにより実施することができる。Trxタグ配列を含む発現ベクターとしては、pET32a(+)、pET32b(+)、pET32c(+)(Novagen社製)等が挙げられる。DSbタグ配列を含む発現ベクターとしては、pET39b(+)、pET40b(+)、(Novagen社製)等が挙げられる。Nusタグ配列を含む発現ベクターとしては、pET43.1a(+)、pET43.1b(+)、pET43.1c(+)、pET44a(+)、pET44b(+)、pET44c(+)(Novagen社製)等が挙げられる。
【0030】
上記のようにして改変された塩基配列からなるDNAをベクター中に連結し、得られた組換えベクターを用いて宿主細胞を形質転換し、該形質転換体を培養することにより、または改変された塩基配列からなるDNAを無細胞タンパク質合成系で発現させることにより、(R)−ヒドロキシニトリルリアーゼを産生することができる。従って、本発明はまた、上記のように改変されたDNAを含む組換えベクター及び該組換えベクターを含む形質転換体に関する。
【0031】
ベクターには、宿主細胞で自律的に増殖し得るファージ、コスミド、人工染色体又はプラスミドが使用される。プラスミドとしては、大腸菌由来のプラスミド(例えばpET21a(+)、pET32a(+)、pET39b(+)、pET40b(+)、pET43.1a(+)、pET44a(+)、pKK223−3、pGEX4T、pUC118、pUC119、pUC18、pUC19等)、枯草菌由来のプラスミド(例えばpUB110、pTP5等)、酵母由来のプラスミド(例えばYEp13、YEp24、YCp50等)などが挙げられ、ファージDNAとしてはλファージ(λgt11、λZAP等)が挙げられる。さらに、ワクシニアウイルスなどの動物ウイルス、バキュロウイルスなどの昆虫ウイルスベクターを用いることもできる。(R)−ヒドロキシニトリルリアーゼの生産性向上のため、より強力なプロモーターを有するベクターを用いるのが好ましく、大腸菌由来のベクターとしては、pET21a、pKK223−3等が好ましい。
【0032】
ベクターに本発明の改変されたDNAを挿入するには、まず、精製された該DNAを適当な制限酵素で切断し、適当なベクターDNAの制限酵素部位又はマルチクローニングサイトに挿入してベクターに連結する方法などが採用される。例えば本発明の改変されたDNAは、通常知られている方法により合成することができ、ベクターに組み込むため、適当な制限酵素の切断部位を両末端に含むように、プライマーを用いてPCR法により増幅してもよい。PCR反応の条件は、当業者が適宜決定することができるが、例えば98℃で30秒間の変性、57℃で15秒間のアニーリング、74℃で30秒間の重合を1サイクルとして30サイクル反応させ、増幅された遺伝子を得ることができる。
【0033】
その他、哺乳動物細胞において用いられる組換えベクターには、プロモーター、本発明のDNAのほか、所望によりエンハンサーなどのシスエレメント、スプライシングシグナル、ポリA付加シグナル、選択マーカー、リボソーム結合配列(SD配列)などが連結されていてもよい。選択マーカーの例としては、カナマイシン、アンピシリン、テトラサイクリン、クロラムフェニコールなどの抗生物質が挙げられるがこれに限定されない。
【0034】
DNA断片とベクター断片とを連結させるには、公知のDNAリガーゼを用いる。そして、DNA断片とベクター断片とをアニーリングさせた後連結させ、組換えベクターを作成する。好ましくは市販のライゲーションキット、例えばライゲーションhigh(東洋紡株式会社製)を用いて、規定の条件にてライゲーション反応を行なうことにより組換えベクターを得ることができる。
【0035】
また、これらのベクターを、必要であればボイル法、アルカリSDS法などの精製手段により精製し、さらに例えばエタノール沈殿法、ポリエチレングリコール沈殿法などの濃縮手段により濃縮することができる。
【0036】
形質転換に使用する宿主としては、(R)−ヒドロキシニトリルリアーゼを発現できるものであれば特に限定されるものではない。例えば、細菌(大腸菌、枯草菌等)、酵母、動物細胞(COS細胞、CHO細胞等)、昆虫細胞が挙げられる。本発明においては、細菌、特に大腸菌を使用するのが好ましく、DH5α細胞、BL21(DE3)細胞、BL21(DE3)pLysS細胞、OrigamiB2細胞等を用いるのがより好ましく、中でも生産性に優れる点でBL21(DE3)細胞を用いるのが最も好ましい。
【0037】
細菌を宿主とする場合は、組換えベクターが該細菌中で自律複製可能であると同時に、プロモーター、リボゾーム結合配列(SD配列)、本発明の改変されたDNA、転写終結配列を含むことが好ましい。所望によりエンハンサーなどのシスエレメント、選択マーカー、オペレーター、プロモーターを制御する遺伝子が含まれていてもよい。プロモーターは、大腸菌等の宿主細胞中で発現できるものであればいずれを用いてもよい。大腸菌の場合、具体的には、T7プロモーター、Lacプロモーター、Tacプロモーターなどが挙げられる。組換えベクターの導入方法は、細菌にDNAを導入する方法であれば特に限定されるものではない。例えばカルシウムイオンを用いる方法、エレクトロポレーション法等が挙げられる。
【0038】
本発明のDNAを含む形質転換体は、その組換えベクターが有するマーカー遺伝子により、例えば、アンピシリン、カナマイシンなどの抗生物質を含むLB培地寒天プレート上でコロニーを形成することにより選抜することができるが、クローニングされた宿主細胞が本発明の組換えベクターにより形質転換されたものかどうかを確認するため、一部を用いて、PCR法によるインサートの増幅確認、又はシーケンサーを用いたダイデオキシ法による配列解析をしてもよい。
【0039】
本発明はまた、本発明の改変DNAを発現させ、得られた(R)−ヒドロキシニトリルリアーゼとカルボニル化合物及びシアノ基供与体とを反応させ、反応液から光学活性シアノヒドリンを回収することを含む、光学活性シアノヒドリンの製造方法に関する。
【0040】
このような光学活性シアノヒドリンの製造方法としては、上記形質転換体を培養し、得られた培養物とカルボニル化合物及びシアノ基供与体とを反応させ、反応液から光学活性シアノヒドリンを回収することを含む方法が挙げられる。あるいは、無細胞タンパク質合成系を用いる方法が挙げられる。無細胞タンパク質合成系は、細胞抽出液を用いて試験管内でタンパク質を合成する系である。「無細胞タンパク質合成系」は、mRNAの情報を読み取ってリボソーム上でタンパク質を合成する無細胞翻訳系とDNAを鋳型としてRNAを合成する無細胞転写系との両者を含む。無細胞タンパク質合成系は、系を容易に改変することができるため、目的のタンパク質に適した発現系を構築しやすいという利点がある。なお、無細胞タンパク質合成系の詳細については、特開2000−175695号などに記載されている。
【0041】
従って、(R)−ヒドロキシニトリルリアーゼとカルボニル化合物及びシアノ基供与体とを反応させることには、精製された(R)−ヒドロキシニトリルリアーゼとカルボニル化合物及びシアノ基供与体とを反応させることだけでなく、形質転換体を培養した培養物とカルボニル化合物及びシアノ基供与体とを反応させること、ならびに無細胞タンパク質合成系から得られる反応液とカルボニル化合物及びシアノ基供与体とを反応させることも含まれる。
【0042】
ここで「培養物」とは、培養上清、培養細胞、培養菌体、細胞又は菌体の破砕物、及びこれらから精製された(R)−ヒドロキシニトリルリアーゼタンパク質自体のいずれをも意味するものである。本発明においては、好ましくは精製(R)−ヒドロキシニトリルリアーゼを用いる。
【0043】
上記形質転換体を培地で培養する方法は、宿主の培養に用いられる通常の方法に従って行われる。大腸菌や酵母菌等の微生物を宿主として得られた形質転換体を培養する培地としては、微生物が資化し得る炭素源、窒素源、無機塩類等を含有し、形質転換体の培養を効率的に行うことができる培地であれば、天然培地、合成培地のいずれを用いてもよい。
【0044】
炭素源としては資化可能な炭素化合物であればよく、例えば、グリセリンなどのポリオール類、又はピルビン酸、コハク酸もしくはクエン酸等の有機酸類を使用することができる。また、窒素源としては利用可能な窒素化合物であればよく、例えば、ペプトン、肉エキス、酵母エキス、カゼイン加水分解物、大豆粕アルカリ抽出物、又はアンモニアもしくはその塩などを使用することができる。その他、リン酸塩、炭酸塩、硫酸塩、マグネシウム、カルシウム、カリウム、鉄、マンガン、亜鉛などの塩類、特定のアミノ酸、特定のビタミン、消泡剤なども必要に応じて使用してもよい。また、イソプロピル−β−D−チオガラクトピラノシドなどのタンパク質発現誘導剤を必要に応じて培地に添加してもよい。
【0045】
培養は、通常、振盪培養又は通気攪拌培養などの好気的条件下、好ましくは0〜40℃、より好ましくは10〜37℃、特に好ましくは15〜37℃で培養を行う。培養期間中、培地のpHは宿主の発育が可能で、生産された(R)−ヒドロキシニトリルリアーゼの活性が損なわれない範囲で適宜変更することができるが、好ましくはpH4〜8程度の範囲である。pHの調整は、無機又は有機酸、アルカリ溶液等を用いて行う。培養中は必要に応じてアンピシリンやテトラサイクリン等の抗生物質を培地に添加してもよい。
【0046】
培養後、(R)−ヒドロキシニトリルリアーゼが菌体内又は細胞内に生産される場合には、通常知られている方法、例えば、菌体又は細胞を機械的方法、リゾチームなどを用いた酵素的方法又は界面活性剤などを用いた化学的処理によって破壊することにより当該酵素を抽出できる。また、当該酵素が菌体外又は細胞外に生産される場合には、培養液をそのまま使用するか、遠心分離等により菌体又は細胞を除去する。
【0047】
(R)−ヒドロキシニトリルリアーゼが封入体として得られる場合には、好ましくはさらにリフォールディングを行うことにより、活性型でかつ可溶性の(R)−ヒドロキシニトリルリアーゼを得ることができる。リフォールディングは、当技術分野で通常用いられる方法に従って実施することができ、例えば、Applied and Environmental Microbiology(1998)64:1405−1411に記載された方法に従って、実施することができる。
【0048】
具体的には、培養後、遠心分離等の方法により集菌した後、細胞を破砕し、封入体を変性剤を用いて可溶化することによって、(R)−ヒドロキシニトリルリアーゼ類を抽出することができる。細胞の破砕は、常法で、たとえば超音波により実施できる。懸濁媒体としてはpH5.0〜8.0のpH値に調整した好適な緩衝液(例えばクエン酸緩衝液やリン酸緩衝液等)を用いることが好ましい。この際、細胞の破砕を促進させるためEDTAを添加してもよい。このようにして細胞を破砕した後に、不溶性成分(封入体)を任意の方法で、遠心分離するか、濾過することにより分取する。異種のタンパク質をできる限り除去するため、たとえば水、リン酸緩衝液を用いて洗浄することが好ましいが、場合により界面活性剤や4M程度の尿素で洗浄してもよい。得られた沈殿(ペレット)を変性剤を用いて可溶化するが、変性剤としては、公知の変性剤、特にグアニジンまたは尿素を使用することができる。この可溶化に当っての変性剤の濃度は、グアニジンでは4〜8モル/L、好ましくは約6モル/L、尿素では5〜9モル/L、好ましくは約8モル/Lである。グアニジンは通常グアニジン塩酸塩等のグアニジンの酸付加塩として用いられる。
【0049】
上記のようにして封入体の可溶化を行った後、遠心分離等で不純物を除去し、得られた上澄液を必要により精製工程に付した後、リフォールディング(活性化、再生化)を行うことができる。リフォールディングは、精製した(R)−ヒドロキシニトリルリアーゼ類にレドックスバッファーを添加するか、あるいは(R)−ヒドロキシニトリルリアーゼ類を含有する上澄液をレドックスバッファーで希釈することにより行われる。(R)−ヒドロキシニトリルリアーゼ類を含有する上澄液をレドックスバッファーで希釈する場合、変性剤の濃度を活性化に適した中性pHにおいて不作用濃度まで希釈することが必要であり、グアジニンでは0〜2.0モル/L、好ましくは約1モル/L以下まで、尿素では0〜4.0モル/L、好ましくは約2モル/L以下まで希釈する。
【0050】
レドックスバッファーとしては、酸化型グルタチオン(GSSG)および還元型グルタチオン(GSH)、システインおよびシスチン、またはシステアミンおよびシスタミンを含有する緩衝液(例、リン酸緩衝液、酢酸緩衝液、クエン酸緩衝液、Tris/HCl緩衝液)が好ましいものとして挙げられ、中でもGSSGおよびGSHを含有するTris/HCl緩衝液が好ましい。GSSGおよびGSHを用いる場合、GSSGの濃度は0.1〜10ミリモル/L、好ましくは0.1〜1.0ミリモル/L、GSHの濃度は0.1〜10ミリモル/L、好ましくは0.1〜1.0ミリモル/Lである。
【0051】
また、レドックスバッファーには、通常、グリセリン等の添加剤を添加してもよい。添加量としてグリセリンは1〜50%、好ましくは5〜20%の濃度で添加するのがよい。また、当該酵素は補酵素としてFADが必要なので、これを添加してもよい。添加量としては1〜1000μM、好ましくは10〜100μMの濃度で添加するのがよい。
【0052】
該リフォールディングに当っての温度は0〜30℃、好ましくは0〜15℃、pHは7〜9、好ましくはpH7.5〜8.5である。リフォールディングに要する時間は通常1時間〜10週間、好ましくは3日〜1週間である。
【0053】
その後、タンパク質の単離精製に用いられる一般的な生化学的方法、例えば硫安分画法、有機溶媒沈殿法、イオン交換体などによる吸着処理法、イオン交換クロマトグラフィー、疎水クロマトグラフィー、ゲルろ過クロマトグラフィー、アフィニティークロマトグラフィー、電気泳動法等を単独で又は適宜組み合わせて用いることにより、前記培養物中から(R)−ヒドロキシニトリルリアーゼを単離精製することができる。(R)−ヒドロキシニトリルリアーゼが得られたか否かは、SDS−ポリアクリルアミドゲル電気泳動等により確認することができる。
【0054】
本発明の形質転換体の上記培養物、すなわち、培養上清、培養細胞、培養菌体、細胞若しくは菌体の破砕物、又は精製(R)−ヒドロキシニトリルリアーゼタンパク質と、カルボニル化合物及びシアノ基供与基とを反応させることにより、光学活性シアノヒドリンを製造する。シアノヒドリンを合成する酵素反応の形態については制限されず、水・有機溶媒混合系、有機溶媒系、有機溶媒水二相系、固定化酵素を使う反応系などのいずれの反応系においても効果的に実施することができる。
【0055】
光学活性シアノヒドリンとは、一方の鏡像異性体が他方の鏡像異性体より多く含まれているシアノヒドリン、又は、いずれか一方の鏡像異性体のみからなるシアノヒドリンをいう。(R)−ヒドロキシニトリルリアーゼを用いることにより、用いた基質に応じて(R)−シアノヒドリン及び(S)−シアノヒドリンの双方を製造することができる。
【0056】
ここでカルボニル化合物とは、アルデヒド又はケトンをいい、具体的には、次式(I):
R1−CO−R2 (I)
(式中、R1及びR2は、互いに同一でも異なっていてもよく、それぞれ水素原子又は炭素数22以下の1価の炭化水素基を表し、前記炭化水素基中、−CH2−並びに−CH3のCH2はカルボニル基、スルホニル基、−O−又は−S−で置き換えられていてもよく、=CH2は=O又は=Sで置き換えられていてもよく、また−CH2−のC−H、−CH3のC−H、>CH−のC−H、=CH−のC−H並びに=CH2のC−Hは、N又はC−ハロゲンで置き換えられていてもよく、また、R1及びR2は、共同して2価の基を表してもよい。)で示される。
【0057】
前記式(I)において、炭素数22以下の1価の炭化水素基とは、直鎖状又は分岐状の鎖状炭化水素基、側鎖のない又は側鎖のある単環式炭化水素基、側鎖のない又は側鎖のある多環式炭化水素基、側鎖のない又は側鎖のあるスピロ炭化水素基、側鎖のない又は側鎖のある環集合構造の炭化水素基、あるいは、前記の環式炭化水素基が置換した鎖状炭化水素基のいずれをも含む。また、飽和な炭化水素基並びに不飽和な炭化水素基のいずれをも含むが、不飽和な炭化水素基において、C=C=Cのアレン構造を含む基は除く。
【0058】
なお、以下においては、側鎖のない芳香族基、側鎖のある芳香族基、並びに、フェニルフェニル基又は側鎖のあるフェニルフェニル基などを併せて、アリール基といい、このアリール基で置換された直鎖状又は分岐状のアルキル基をアラルキル基という。他の環式炭化水素基に関しても、特に明記しない場合、環上に側鎖のないものとあるものを併せて指す場合には、単にシクロアルキル基等の名称を用いる。鎖状炭化水素基についても、直鎖状のものと分岐状のものを併せて指す場合には、単にアルキル基等の名称を用いる。
【0059】
前記炭化水素基中、−CH2−がカルボニル基、スルホニル基、−O−又は−S−で置き換えられると、それぞれケトン、スルホン、エーテル又はチオエーテルの構造が導入され、−CH3の−CH2−がカルボニル基、−O−又は−S−で置き換わると、それぞれホルミル基(アルデヒド)、水酸基又はメルカプト基に変わり、あるいは、末端の=CH2が=O又は=Sに置き換わると、ケトン、チオケトンの構造が導入されることを意味し、また、−CH2−のC−HがNに変わると、−NH−となり、>CH−のC−HがNに変わると、>N−となり、=CH−のC−HがNに変わると、=N−となり、末端の−CH3のC−HがNに変わると、−NH2が導入され、=CH2のC−HがNに変わると、=NHとなる。また、−CH3、−CH2−、=CH−、≡CH又は>CH−のC−HがC−ハロゲンで置き換えられると、当該炭素上へハロゲン原子を置換することになる。なお、炭素鎖中における−O−、−S−、Nへの置き換えは、当該炭化水素基に対する、それぞれオキサ置換、チア置換、アザ置換に当たり、例えば、炭化水素環の環の骨格炭素で起こると、炭化水素環のそれぞれ含酸素複素環、含硫黄複素環、含窒素複素環への変換となる。該炭化水素基中、CH2並びにC−Hにおける置き換えは、それぞれ独立に行われてよく、加えて、前記の置き換えを行った後、なお当該炭素上にCH2又はC−Hが残存する際には、更に置き換えがなされてもよい。更には、前記の置き換えにより、−CH2−CH3の−CO−O−H;カルボン酸構造への変換などもなされる。
【0060】
本明細書において、ハロゲン原子とは、フッ素原子、塩素原子、臭素原子、ヨウ素原子を指すが、フッ素原子、塩素原子、臭素原子が好ましい。
【0061】
従って、前記炭化水素基としては、鎖状炭化水素基並びに環式炭化水素基など環構造を有する炭化水素基のいずれをも選択でき、例えば、飽和鎖状炭化水素基である直鎖状又は分岐状のアルキル基、不飽和鎖状炭化水素基である直鎖状又は分岐状のアルケニル基、直鎖状又は分岐状のアルキニル基、直鎖状又は分岐状のアルカジエニル基など、飽和な環式炭化水素基であるシクロアルキル基、不飽和な環式炭化水素基であるシクロアルケニル基、シクロアルキニル基、シクロアルカジエニル基など、芳香族炭化水素基であるアリール基、アラルキル基、アリールアルケニル基などが挙げられる。
【0062】
更に詳しくいえば、直鎖状又は分岐状のアルキル基としては、例えばメチル基、エチル基、プロピル基、イソプロピル基、ブチル基、1−メチルプロピル基、ペンチル基、1−メチルブチル基、ヘキシル基、1−メチルペンチル基、ヘプチル基、1−メチルヘキシル基、1−エチルペンチル基、オクチル基、ノニル基、デシル基、ウンデシル基、ドデシル基、トリデシル基、テトラデシル基、2−メチルプロピル基、2−メチルブチル基、3−メチルブチル基、2−メチルペンチル基、3−メチルペンチル基、4−メチルペンチル基、メチルヘキシル基、メチルヘプチル基、メチルオクチル基、メチルノニル基、1,1−ジメチルエチル基、1,1−ジメチルプロピル基、2,6−ジメチルヘプチル基、3,7−ジメチルオクチル基、2−エチルヘキシル基など、シクロアルキルアルキル基としては、シクロペンチルメチル基、シクロヘキシルメチル基など、シクロアルキル基としては、シクロプロピル基、シクロブチル基、シクロペンチル基、メチルシクロペンチル基、シクロヘキシル基、メチルシクロヘキシル基、シクロヘプチル基、シクロオクチル基など、ビシクロアルキル基としては、ノルボルニル基、ビシクロ[2.2.2]オクチル基、アダマンチル基などが挙げられる。直鎖状又は分岐状のアルケニル基としては、例えばビニル基、アリル基、クロチル基(2−ブテニル基)、イソプロペニル基(1−メチルビニル基)など、シクロアルケニル基又はシクロアルカジエニル基としては、シクロペンテニル基、シクロペンタジエニル基、シクロヘキセニル基、シクロヘキサンジエニル基などが挙げられる。直鎖状又は分岐状のアルキニル基としては、例えばエチニル基、プロピニル基、ブチニル基などが挙げられる。アリール基としては、例えばフェニル基、1−ナフチル基、2−ナフチル基、2−フェニルフェニル基、3−フェニルフェニル基、4−フェニルフェニル基、9−アントリル基、メチルフェニル基、ジメチルフェニル基、トリメチルフェニル基、エチルフェニル基、メチルエチルフェニル基、ジエチルフェニル基、プロピルフェニル基、ブチルフェニル基などが挙げられる。アラルキル基としては、例えばベンジル基、1−ナフチルメチル基、2−ナフチルメチル基、フェネチル基(2−フェニルエチル基)、1−フェニルエチル基、フェニルプロピル基、フェニルブチル基、フェニルペンチル基、フェニルヘキシル基、メチルベンジル基、メチルフェネチル基、ジメチルベンジル基、ジメチルフェネチル基、トリメチルベンジル基、エチルベンジル基、ジエチルベンジル基などが挙げられる。アリールアルケニル基としては、例えばスチリル基、メチルスチリル基、エチルスチリル基、ジメチルスチリル基、3−フェニル−2−プロペニル基などが挙げられる。
【0063】
前記炭化水素基中のCH2がカルボニル基、スルホニル基、O又はSで、又はC−HがN又はC−ハロゲンで置き換えられた基としては、ケトン、アルデヒド、カルボン酸、スルホン、エーテル、チオエーテル、アミン、アルコール、チオール、ハロゲン、複素環(例えば、含酸素複素環、含硫黄複素環、含窒素複素環)などの構造を一つ以上含む基が挙げられる。なお、含酸素複素環、含硫黄複素環、含窒素複素環とは、環式炭化水素基の環骨格の炭素がそれぞれ酸素、硫黄、窒素で置き換わるものを意味し、更には、これらヘテロ原子置換が二種以上ある複素環であってもよい。前記の置換を有する炭化水素基としては、例えば、ケトン構造のアセチルメチル基、アセチルフェニル基;スルホン構造のメタンスルホニルメチル基;エーテル構造のメトキシメチル基、メトキシエチル基、エトキシエチル基、メトキシプロピル基、ブトキシエチル基、エトキシエトキシエチル基、メトキシフェニル基、ジメトキシフェニル基、フェノキシメチル基;チオエーテル構造のメチルチオメチル基、メチルチオフェニル基;アミン構造のアミノメチル基、2−アミノエチル基、2−アミノプロピル基、3−アミノプロピル基、2,3−ジアミノプロピル基、2−アミノブチル基、3−アミノブチル基、4−アミノブチル基、2,3−ジアミノブチル基、2,4−ジアミノブチル基、3,4−ジアミノブチル基、2,3,4−トリアミノブチル基、メチルアミノメチル基、ジメチルアミノメチル基、メチルアミノエチル基、プロピルアミノメチル基、シクロペンチルアミノメチル基、アミノフェニル基、ジアミノフェニル基、アミノメチルフェニル基;含酸素複素環のテトラヒドロフラニル基、テトラヒドロピラニル基、モルホリルエチル基;含酸素複素芳香環のフリル基、フルフリル基、ベンゾフリル基、ベンゾフルフリル基;含硫黄複素芳香環のチエニル基;含窒素複素芳香環のピロリル基、イミダゾリル基、オキサゾリル基、チアジアゾリル基、ピリジル基、ピリミジニル基、ピリダジニル基、ピラジニル基、テトラジニル基、キノリル基、イソキノリル基、ピリジルメチル基;アルコール構造の2−ヒドロキシエチル基、2−ヒドロキシプロピル基、3−ヒドロキシプロピル基、2,3−ジヒドロキシプロピル基、2−ヒドロキシブチル基、3−ヒドロキシブチル基、4−ヒドロキシブチル基、2,3−ジヒドロキシブチル基、2,4−ジヒドロキシブチル基、3,4−ジヒドロキシブチル基、2,3,4−トリヒドロキシブチル基、ヒドロキシフェニル基、ジヒドロキシフェニル基、ヒドロキシメチルフェニル基、ヒドロキシエチルフェニル基;チオール構造の2−メルカプトエチル基、2−メルカプトプロピル基、3−メルカプトプロピル基、2,3−ジメルカプトプロピル基、2−メルカプトブチル基、3−メルカプトブチル基、4−メルカプトブチル基、メルカプトフェニル基;ハロゲン化炭化水素基である2−クロロエチル基、2−クロロプロピル基、3−クロロプロピル基、2−クロロブチル基、3−クロロブチル基、4−クロロブチル基、フルオロフェニル基、クロロフェニル基、ブロモフェニル基、ジフルオロフェニル基、ジクロロフェニル基、ジブロモフェニル基、クロロフルオロフェニル基、トリフルオロフェニル基、トリクロロフェニル基、フルオロメチルフェニル基、トリフルオロメチルフェニル基;アミン構造とアルコール構造を有する2−アミノ−3−ヒドロキシプロピル基、3−アミノ−2−ヒドロキシプロピル基、2−アミノ−3−ヒドロキシブチル基、3−アミノ−2−ヒドロキシブチル基、2−アミノ−4−ヒドロキシブチル基、4−アミノ−2−ヒドロキシブチル基、3−アミノ−4−ヒドロキシブチル基、4−アミノ−3−ヒドロキシブチル基、2,4−ジアミノ−3−ヒドロキシブチル基、3−アミノ−2,4−ジヒドロキシブチル基、2,3−ジアミノ−4−ヒドロキシブチル基、4−アミノ−2,3−ジヒドロキシブチル基、3,4−ジアミノ−2−ヒドロキシブチル基、2−アミノ−3,4−ジヒドロキシブチル基、アミノヒドロキシフェニル基;ハロゲンと水酸基で置換された炭化水素基であるフルオロヒドロキシフェニル基、クロロヒドロキシフェニル基;カルボン構造のカルボキシフェニル基などが挙げられる。
【0064】
R1及びR2で表される非対称の2価の基としては、特に制限はなく、例えば、ノルボルナン−2−イリデン、2−ノルボルネン−5−イリデンが挙げられる。
【0065】
前記式(I)で示されるカルボニル化合物としては、例えば、ベンズアルデヒド、m−フェノキシベンズアルデヒド、p−メチルベンズアルデヒド、o−クロロベンズアルデヒド、m−クロロベンズアルデヒド、p−クロロベンズアルデヒド、m−ニトロベンズアルデヒド、3,4−メチレンジオキシベンズアルデヒド、2,3−メチレンジオキシベンズアルデヒド、フェニルアセトアルデヒド、フルフラール等の芳香族アルデヒド;アセトアルデヒド、ブチルアルデヒド、イソブチルアルデヒド、バレルアルデヒド、シクロヘキサンアルデヒド等の脂肪族アルデヒド;エチルメチルケトン、ブチルメチルケトン、メチルプロピルケトン、イソプロピルメチルケトン、メチルペンチルケトン、メチル(2−メチルプロピル)ケトン、メチル(3−メチルブチル)ケトン等の飽和脂肪族ケトン;メチル(2−プロペニル)ケトン、(3−ブテニル)メチルケトン等の不飽和脂肪族ケトン;(3−クロロプロピル)メチルケトン等のアルキル(ハロアルキル)ケトン;2−(アルコキシカルボニルアミノ)−3−シクロヘキシルプロピオンアルデヒド等の2−(保護アミノ)アルデヒド;3−メチルチオプロピオンアルデヒド等のアルキルチオ脂肪族アルデヒドが挙げられる。
【0066】
本発明において、基質としてのカルボニル化合物をシアノヒドリンに変換するためには、シアノ基供与体の存在下で酵素反応を実施する。本明細書中、シアノ基供与体とは、反応系へ添加することによって、シアニド、すなわちシアン化物イオン(CN−)を生じる物質を意味し、例えば、シアン化水素、青酸(シアン化水素酸)、シアン化ナトリウムやシアン化カリウムなどのシアン化水素の塩、又は、アセトンシアノヒドリン等のシアノヒドリン類が挙げられる。特に回収リサイクルが容易な青酸(シアン化水素酸)を用いるのが好ましい。シアノ基供与体の供給方法としては常法により液体として供給する方法、又は常法により気体として供給する方法のいずれをも採用することができる。
【0067】
本発明においては、反応原料の濃度を高め、生産性を高めるために、反応溶媒として、水と実質的に混和しない有機溶媒を用いる。ここで、「水と実質的に混和しない有機溶媒」とは、水に任意の割合で溶解する溶媒を除く有機溶媒を意味する。有機溶媒としては、水と実質的に混和せず、基質及び生成物を充分に溶解し、酵素反応に悪影響を与えないものであれば特に制限なく用いることができる。このような有機溶媒は、原料のアルデヒド又はケトンの物性、生成物であるシアノヒドリンの物性に応じて適宜選択することができる。
【0068】
水と実質的に混和しない有機溶媒としては、具体的には、ハロゲン化されていてもよい炭化水素系溶媒(例えば、直鎖状、分岐状又は環状の飽和又は不飽和脂肪族炭化水素、芳香族炭化水素)、例えば、ペンタン、ヘキサン、シクロヘキサン、ベンゼン、トルエン、キシレン、塩化メチレン、クロロホルムなど;ハロゲン化されていてもよいアルコール系溶媒(例えば、直鎖状、分岐状又は環状の飽和又は不飽和脂肪族アルコール、アラルキルアルコール)、例えば、n−ブタノール、イソブタノール、t−ブタノール、ヘキサノール、シクロヘキサノール、n−アミルアルコールなど;ハロゲン化されていてもよいエーテル系溶媒(例えば、直鎖状、分岐状又は環状の飽和又は不飽和脂肪族エーテル、芳香族エーテル)、例えば、ジエチルエーテル、ジプロピルエーテル、ジイソプロピルエーテル、ジブチルエーテル、t−ブチルメチルエーテル、ジメトキシエタンなど;ハロゲン化されていてもよいエステル系溶媒(例えば、直鎖状、分岐状又は環状の飽和又は不飽和脂肪族エステル、芳香族エステル)、例えば、ギ酸メチル、酢酸メチル、酢酸エチル、酢酸ブチル、プロピオン酸メチル等が挙げられ、これらを単独で用いてもまた2種以上を混合して用いてもよい。特に、ジイソプロピルエーテル、ジブチルエーテル、t-ブチルメチルエーテル、酢酸エチルを用いるのが好ましい。
【0069】
前記有機溶媒は、水又は水性緩衝液で飽和されているのが好ましい。ここで水性緩衝液としては、特に制限はないが、酵素活性の最適pH(pH4〜7)の付近において緩衝能を発揮する緩衝液、例えば、リン酸、クエン酸、グルタル酸、リンゴ酸、マロン酸、o−フタル酸、コハク酸などの塩等によって構成される緩衝液等が好ましく用いられる。
【0070】
本発明において、酵素、基質及びシアノ基供与体の使用量、反応温度は、用いる基質に応じて適宜決定される。通常、酵素の使用量は基質であるカルボニル化合物50ミリモルに対して250〜100,000単位、好ましくは500〜50,000単位である。本明細書において、当該酵素(R)−ヒドロキシニトリルリアーゼの活性1単位は、DL−マンデロニトリルを基質に反応させて1分間にベンズアルデヒド1μモルを生成する活性と定義する。測定は0.2Mクエン酸−NaOH緩衝液(pH5.5)7mLにDLマンデロニトリルを0.931μL添加した液を分光光度計のあらかじめ20℃に調温した対照セル、測定セルに3mLずつ量りとり、酵素液を添加したときのベンズアルデヒドの生成速度を波長249.6nmの吸光度変化を測定することによって算出する。カルボニル化合物の濃度は通常0.1〜10モル/Lの範囲に設定し、シアノ基供与体は用いるカルボニル化合物に対して1〜5倍モル、好ましくは1.1〜3倍モルの濃度で添加する。本反応は基質濃度によって酵素活性及び反応速度が変化するので、用いるカルボニル化合物の種類に応じて基質濃度を適宜決定する。反応時間は、基質であるカルボニル化合物の転換率が80%以上、好ましくは90%以上に達するまでの時間が適当であるが、これに限定されない。反応温度は酵素の活性が十分発揮される温度であればよく、通常0〜40℃、好ましくは4〜30℃である。
【0071】
反応系において、回分式で反応を行う場合には、撹拌などにより、酵素が反応系内に分散するようにする。カラムなどに固定化酵素を充填して反応を行う場合には、基質を含む溶液を適当な流速でカラムに流入させ、流出液を採取することで実施できる。回分反応の場合には、反応が完結した時点で混合を止め、生産物が溶解している有機相を常法により取り出すことで生産物を回収できる。これらの酵素は初回と同じ方法で調製した基質を含む溶液と混合することによって再使用することができる。
【0072】
酵素反応終了後のシアノヒドリンを含む反応液からの反応溶媒及び未反応のシアノ基供与体の回収は、蒸留等の操作をすることにより行うことができる。蒸留は、シアノヒドリンが高温では不安定であるため、常圧高温下で実施するよりも、比較的低い温度で減圧下で実施することが好ましい。また、この蒸留操作においては、公知のシアノヒドリンの安定化剤を添加することもできる。安定化剤としては蒸留ボトムを酸性に維持できるものであればよく、p-トルエンスルホン酸、酢酸などの有機酸、硫酸などの無機酸などをシアノヒドリンに対して1/200〜1/10モル添加することで実施できる。
【実施例】
【0073】
実施例1 アーモンド葉からのcDNAの調製及びアーモンド葉由来(R)−ヒドロキシニトリルリアーゼ遺伝子の調製
アーモンドの葉を材料として、全mRNAの抽出を行った。回収したmRNAを鋳型として、cDNA合成を行い、アーモンドのcDNAライブラリーを作成した。GeneBank Accession No.Y08211のアーモンド花由来の(R)−ヒドロキシニトリルリアーゼ遺伝子の配列情報(配列番号1)を参考にして、下記PCRプライマーを合成した。
センスプライマー:5’−GGCATCGATGGAGAAATCAACAACAATG−3’(配列番号2)
アンチセンスプライマー:5’−GCGTCGACTCACATGGACTCTTG−3’(配列番号3)
【0074】
これらのプライマーを使用し、上記のcDNAを鋳型としてPCRを行ったところ、上記GeneBank Accession No.Y08211のアーモンド花由来の(R)−ヒドロキシニトリルリアーゼ遺伝子と相同性の高い(99.5%)(R)−ヒドロキシニトリルリアーゼ遺伝子(以下RHNL−KR1と称する)を獲得した。その塩基配列を配列番号4に示す。
【0075】
実施例2 (R)−ヒドロキシニトリルリアーゼ遺伝子の塩基配列を改変したDNAの作成
配列番号4で表される塩基配列を有する(R)−ヒドロキシニトリルリアーゼを大腸菌で発現させるためにコドン使用頻度を検討し、上記表1に従って、アミノ酸配列を変化させることなくコドン使用頻度の低いものをコドン使用頻度の高いものに変更した配列(以下、RHNL−KR2と称する)を作成した。RHNL−KR2の塩基配列を配列番号5に示す。得られた配列に基づき(R)−ヒドロキシニトリルリアーゼ遺伝子を改変したDNAを合成した。
【0076】
さらに配列番号5で表される塩基配列においてシグナル配列(配列番号5の1〜81番塩基)を除去した遺伝子を得るために下記のPCRプライマーを合成した。
センスプライマー:5’−GGGCATATGGCCAACACTTCTGCTCACGATTTCAGCTAC−3’(配列番号6)
アンチセンスプライマー:5’−CCCGGATCCTTACATGGATTCCTGAATGTTGTGAATAGC−3’(配列番号7)
【0077】
これらのプライマーを使用し、上記の配列番号5で表される塩基配列からなるDNAを鋳型としてPCRを行い、シグナル配列を除去したDNA(RHNL−KR2−2と称する)を獲得した。その塩基配列を配列番号8に示す。
【0078】
実施例3 アーモンド葉由来(R)−ヒドロキシニトリルリアーゼ遺伝子の塩基配列を改変したDNAの大腸菌における発現
配列番号8で表される塩基配列からなるDNAを、大腸菌用ベクターpET21a(+)(Novagen社製)のマルチクローニングサイトのNdeIサイトとBamHIサイトの間に導入して組換えベクターpET21a(+)/RHNL−KR2−2を調製した。このプラスミドベクターを大腸菌株BL21(DE3)(Novagen社製)に形質転換により導入した。また、ここで得られた遺伝子組換え大腸菌をBL21(DE3)/pET21a(+)/RHNL−KR2−2と称する。
【0079】
上記で構築された組換え大腸菌株(BL21(DE3)/pET21a(+)/RHNL−KR2−2)を以下の方法により培養し、組換えタンパク質を得た。
【0080】
まず、15mL試験管中でLB培地5mLに菌株1コロニー分を植菌し、培養温度37℃、振盪速度170rpmで12時間、前培養を行った。続いて、500mL坂口フラスコ中でLB培地100mLにそれぞれの前培養液2mLを植菌し、培養温度17℃、振盪速度130rpmで培養を開始し、培養液の細胞密度の測定を、660nmにおける吸光度(OD660nmと称する)を測定することにより行い、OD660nmが0.4〜0.7になるまで培養後、タンパク質発現誘導剤であるイソプロピル−β−D−チオガラクトピラノシド(IPTG)を最終濃度が1mMになるように添加した。その後培養を継続し、OD660nmが3を越えるまで約24時間、本培養を行った。
【0081】
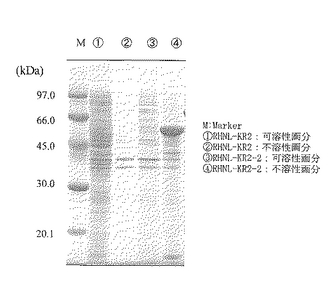
上記培養後、培養液を回収し、遠心分離(15000rpm、5分間)して菌体細胞を得た。得られた細胞はクエン酸緩衝液中で超音波処理により破砕した。破砕処理液は遠心分離(15000rpm、30分間)により可溶性上清と不溶性沈殿に分離した。このそれぞれの画分をSDS−PAGEに供したところ、目的のタンパク質と思われる57kDaのバンドが不溶性沈殿として菌体内全タンパク質の20%以上を占める量で発現されていた。SDS−PAGEの結果を図1の3レーン及び4レーンに示す。3レーンは可溶性画分の結果を表し、4レーンは不溶性画分の結果を表す。
【0082】
SDS−PAGEは通常の方法で行い、ゲルにはATTO社製PAGEL NPU−12.5Lを用い、電流を20mAで80分間流した後、クマシーブリリアントブルーで染色した。目的のタンパク質の発現量は画像処理ソフトウェアのScion Image Release Beta 4.0.2(Scion Corporation製)を用いてSDS−PAGEのイメージ画像のデンシトグラムを作成することにより、算出した。
【0083】
実施例4 アーモンド葉由来(R)−ヒドロキシニトリルリアーゼ遺伝子の塩基配列を改変したDNAを使用した光学活性シアノヒドリンの合成
実施例3に準じて調製した培養液1Lから遠心分離により、3gの菌体細胞を得た。この菌体を0.2Mクエン酸緩衝液(pH5.5)30mLに懸濁後、超音波破砕し、遠心分離(15000rpm、30分)した。得られたペレットを100mM Tris/HCl、4% Triton X−100、2M 尿素(pH8.0)で懸濁し、30℃で30分インキュベートし、遠心分離(15000rpm、30分)した。次に得られたペレットを蒸留水で懸濁し、遠心分離(15000rpm、30分)した。得られたペレットを凍結乾燥して粗精製封入体120mgを得た。
【0084】
この粗精製封入体50mgを20mM Tris/HCl、8M 尿素、30mM ジチオスレイトール(pH8.0)10mLに溶解し、遠心分離(15000rpm、30分)後、上澄み液を20mM Tris/HCl、10%グリセリン、50μM FAD、1mM 酸化型グルタチオン、1mM 還元型グルタチオン(pH8.0)で100倍に希釈し、4℃で、1週間リフォールディングを行った。
【0085】
リフォールディングの終了した再生液を限外ろ過で濃縮して10U/mLの容量活性の酵素液を作成した。この酵素液1mLに対して1gの固定化担体(多孔性シリカゲル、MB−5D、富士シリシア化学製)を混合して固定化酵素を作成した。t−ブチルメチルエーテルを溶媒に用い、青酸0.15M、ベンズアルデヒド0.1Mの濃度で上述の固定化酵素を触媒として反応を行ったところ、光学純度90%ee以上のR−マンデロニトリルが得られた。
【0086】
比較例1 アーモンド葉由来(R)−ヒドロキシニトリルリアーゼ遺伝子の塩基配列においてシグナルペプチド除去しないDNAの大腸菌での発現
実施例2で記載した配列番号5で表される塩基配列からなるRHNL−KR2について、実施例3に準じて遺伝子組換え大腸菌BL21(DE3)/pET21a(+)/RHNL−KR2を作成し、実施例3に準じて培養を行い、菌体を回収し、超音波破砕し、可溶性上清、不溶性沈殿をSDS−PAGEに供した。結果を図1の1レーン及び2レーンに示す。1レーンは可溶性画分の結果を示し、2レーンは不溶性画分の結果を示す。目的のRHNL−KR2と思われるバンド(61kDa)は検出されなかった。
【0087】
以上から、本発明により(R)−ヒドロキシニトリルリアーゼの発現量が増大されることが明らかとなった。
【図面の簡単な説明】
【0088】
【図1】実施例3及び比較例1におけるSDS−PAGEの結果を示す。
【配列表フリーテキスト】
【0089】
配列番号2〜8:合成DNA
【技術分野】
【0001】
本発明は、(R)−ヒドロキシニトリルリアーゼをコードする遺伝子の塩基配列が改変されてなるDNA、及び該DNAを用いて光学活性シアノヒドリンを製造する方法に関する。
【背景技術】
【0002】
生体触媒を用いる合成プロセスは化学工業にとってきわめて重要になっている。また、生体触媒を使った化学反応の実施は、キラル又はプロキラルな成分との化学反応で複数のエナンチオマーのうちの一方を選択的に変換又は生成するという、基質特異性の高い生体触媒の性質を利用することが可能な応用分野では、とりわけ興味深い。酵素が持つこの有利な性質を利用するには、その酵素を容易にかつ十分な量で入手できることが必須条件になる。
【0003】
光学活性シアノヒドリンは特に興味深い光学活性化合物群であり、医薬活性物質、ビタミン又はピレスロイド化合物などの生物活性物質の製造に使用されるα−ヒドロキシ酸、α−ヒドロキシケトン、β−アミノアルコールの合成等に重要である。シアノヒドリンは、カルボニル化合物のカルボニル基へのシアン化水素酸の付加によって製造される。例えば(S)−シアノヒドリンなどの光学活性化合物の工業的生産はパラゴムノキ(Heveabrasiliensis)由来の酵素(S)−ヒドロキシニトリルリアーゼを利用することによって可能になっており、例えば特許文献1〜4などに記載されている。
【0004】
また、非特許文献1などには、サクラ属植物が(R)−ヒドロキシニトリルリアーゼのアイソザイムを含有しうることが開示されている。これらのアイソザイムは当該植物の様々な組織に異なるレベルで発現される。アーモンドと近縁関係にあるアメリカザクラには、現在までに5種類の異なるアイソザイムが同定され、それらの遺伝子が配列決定されている。アーモンド由来のアイソザイムは今までのところ1つだけが非特許文献2に記載されており、このアイソザイム遺伝子は花組織にもっとも強く発現される。前記(R)−ヒドロキシニトリルリアーゼのアイソザイムの遺伝子は既に単離され、そのcDNAが配列決定されている。しかし、(R)−ヒドロキシニトリルリアーゼはこれらの植物の組織に微量に含まれているに過ぎず、工業的に利用することは困難であった。
【0005】
(R)−ヒドロキシニトリルリアーゼ遺伝子の異種発現については、真核細胞である酵母(サッカロミセス属又はピキア属)を宿主として、アーモンド由来のヒドロキシニトリルリアーゼを遺伝子組換えにより製造した例がある(特許文献5)。しかし、ヒドロキシニトリルリアーゼの異種発現において、酵母以外を宿主として(R)−ヒドロキシニトリルリアーゼの製造に成功した例はなく、また該酵素の発現量を増大させることについても報告されていない。
【0006】
【特許文献1】WO97/03204
【特許文献2】EP 0 951 561
【特許文献3】EP 0 927 766
【特許文献4】特表平11−508775
【特許文献5】特開2002−330791
【非特許文献1】Plant Physiology,April(1999)119:1535〜1546
【非特許文献2】Planta(1998)206:388〜393
【発明の開示】
【発明が解決しようとする課題】
【0007】
本発明の課題は、(R)−ヒドロキシニトリルリアーゼの異種発現において、(R)−ヒドロキシニトリルリアーゼの産生効率を高め、光学活性シアノヒドリンの工業的な製造方法を提供することである。
【課題を解決するための手段】
【0008】
本発明者らは鋭意検討を行った結果、バラ科植物由来(R)−ヒドロキシニトリルリアーゼをコードする遺伝子の塩基配列に対し、特定の改変を加え、これを宿主細胞で発現させることにより、(R)−ヒドロキシニトリルリアーゼを効率的に産生できることを見いだし、本発明を完成するに至った。
【0009】
すなわち、本発明は以下の発明を包含する。
(1)バラ科植物由来(R)−ヒドロキシニトリルリアーゼをコードする遺伝子の塩基配列が改変されてなるDNAであって、該改変が、該DNAが宿主細胞に導入されたときの(R)−ヒドロキシニトリルリアーゼの発現量を改変されていないDNAを導入した場合と比べて増大させるものである、該DNA。
(2)改変が、以下の(a)〜(c)の工程:
(a)バラ科植物由来(R)−ヒドロキシニトリルリアーゼをコードする遺伝子の塩基配列に含まれるシグナルペプチドをコードする塩基配列を除去すること、
(b)バラ科植物由来(R)−ヒドロキシニトリルリアーゼをコードする遺伝子の塩基配列中のコドンを、そのアミノ酸配列を変化させることなく、宿主細胞におけるアミノ酸ごとのコドン使用頻度が10%以上となるように改変すること、及び
(c)バラ科植物由来(R)−ヒドロキシニトリルリアーゼをコードする遺伝子の塩基配列にタグ配列を付加すること
からなる群から選択される少なくとも1つの工程によって実施されるものである、(1)記載のDNA。
(3)配列番号8で表される塩基配列を含み、塩基長が4000塩基以下のDNA。
(4)(1)〜(3)のいずれかに記載のDNAを含む組換えベクター。
(5)(4)記載の組換えベクターを含む形質転換体。
(6)(1)〜(3)のいずれかに記載のDNAを発現させ、得られた(R)−ヒドロキシニトリルリアーゼとカルボニル化合物及びシアノ基供与体とを反応させ、反応液から光学活性シアノヒドリンを回収することを含む、光学活性シアノヒドリンの製造方法。
【発明の効果】
【0010】
本発明により、(R)−ヒドロキシニトリルリアーゼの異種発現における発現量を増大することができ、(R)−ヒドロキシニトリルリアーゼを効率的に産生することができる。それにより光学活性シアノヒドリンの工業的な製造が可能になる。
【発明を実施するための最良の形態】
【0011】
本発明において、(R)−ヒドロキシニトリルリアーゼとは、当技術分野において通常用いられる意味を有し、シアノ基供与体の存在下、カルボニル化合物を基質としてシアノヒドリンを合成する反応を触媒する活性を有する酵素を意味する。
【0012】
本発明は、バラ科植物由来の(R)−ヒドロキシニトリルリアーゼを対象とする。バラ科植物としては、特に限定されないが、例えば、サクラ属、キンミズヒキ属、アプリコット属、セラサス属、リンゴ属、メスピラス属、ペルシカ属、ピルス属、ヤマザクラ属、バラ属、キイチゴ属から選択される属に属する植物が挙げられ、より具体的には、アーモンド、セイヨウヤマザクラ、カリン、サンザシ、セイヨウサンザシ、マルメロ、ビワ、イチゴ、リンゴ、アメリカンレッドプラム、ホンアンズ、アンズ、セイヨウミザクラ、スミノミザクラ、セイヨウスモモ、ハイブリッド、ウメ、シナミザクラ、モモ、サントウ、コモンピーチ、ネクタリン、スモモ、ユスラウメ、セイヨウナシ、ニホンナシ、チュウゴクナシ、ノイバラ、ハマナシ、ノバラ、ハイブッシュブラックベリー、ヨーロッパキイチゴ、エバーグリーンブラックベリー、サーモンベリー、パシフィックブラックベリー等が挙げられる。本発明においては、好ましくはサクラ属(Prunus属)に属する植物、より好ましくはアーモンド(Prunus amygdalus)を使用する。
【0013】
本発明において、バラ科植物由来(R)−ヒドロキシニトリルリアーゼをコードする遺伝子とは、バラ科植物のゲノムDNA上の(R)−ヒドロキシニトリルリアーゼ遺伝子DNA、及び(R)−ヒドロキシニトリルリアーゼをコードするcDNAを包含するものとする。本発明において、バラ科植物由来(R)−ヒドロキシニトリルリアーゼをコードする遺伝子の塩基配列は、好ましくは配列番号1で表される塩基配列である。該遺伝子の塩基配列が改変されてなるDNAは、上記ゲノムDNA上の(R)−ヒドロキシニトリルリアーゼ遺伝子の塩基配列、又は(R)−ヒドロキシニトリルリアーゼcDNAの塩基配列、好ましくは(R)−ヒドロキシニトリルリアーゼcDNAの塩基配列に対して、改変するための処理を行った結果得られるDNAを意味する。
【0014】
上記塩基配列の改変は、好ましくは、該DNAを宿主細胞に導入したときの、(R)−ヒドロキシニトリルリアーゼの発現量を、改変しない場合と比べて10%以上、好ましくは30%以上、より好ましくは50%以上増大させるものである。すなわち、改変された塩基配列からなるDNAを宿主細胞に導入して発現させた場合に発現される(R)−ヒドロキシニトリルリアーゼが、改変していない塩基配列からなるDNAを導入した場合と比較して、10%以上増大する。
【0015】
本発明において、発現量を、改変していないDNAを導入した場合と比べて増大させるとは、改変していないDNAを導入した場合に全く発現しなかったものが、発現するようになる場合も包含する。
【0016】
ここで発現量の比較は遺伝子配列を改変したものとしないものでそれぞれ誘導発現した菌体をSDS−PAGEに供する際にOD660nmの吸光度が10になるように濃縮または希釈してから、SDS−PAGEに供し、得られた画像データからバンドの濃さをデンシトグラムで計算して目的バンドの濃さを比較して算出するものと定義する。
【0017】
(R)−ヒドロキシニトリルリアーゼの発現量を増大するような塩基配列の改変方法としては、当技術分野で通常用いられるものを使用することができ、特に制限されない。好ましくは、以下の(a)〜(c)の工程:(a)バラ科植物由来(R)−ヒドロキシニトリルリアーゼをコードする遺伝子の塩基配列に含まれるシグナルペプチドをコードする塩基配列を除去すること、(b)バラ科植物由来(R)−ヒドロキシニトリルリアーゼをコードする遺伝子の塩基配列中のコドンを、そのアミノ酸配列を変化させることなく、宿主細胞におけるアミノ酸ごとのコドン使用頻度が10%以上となるように改変すること、及び(c)バラ科植物由来(R)−ヒドロキシニトリルリアーゼをコードする遺伝子の塩基配列にタグ配列を付加すること、からなる群から選択される少なくとも1つの工程によって実施できる。(a)〜(c)の工程のうち1つのみを実施してもよいし、2つを実施してもよい。複数の工程を実施する場合、実施する工程の順序は特に制限されない。本発明においては、少なくとも(a)の工程を実施することが好ましい。上記(a)〜(c)の工程について以下に説明する。
【0018】
(a)シグナルペプチドをコードする塩基配列(シグナル配列)の除去
シグナル配列の除去方法は、当技術分野で通常用いられる方法を使用でき、特に限定されないが、例えば、既知の(R)−ヒドロキシニトリルリアーゼ遺伝子を鋳型としてシグナル配列が除去された配列を増幅するように設計された適当なプライマーを用いてPCRを行う方法、DNA合成手法による全合成法、既知の(R)−ヒドロキシニトリルリアーゼ遺伝子のサイトダイレクティッドミュータジェネシス(M.J.Zollerら、Methods in Enzymology(1983)100:468−500)による方法、又は既知の(R)−ヒドロキシニトリルリアーゼ遺伝子の一部を合成DNAフラグメントと差し換える方法などが挙げられる。
【0019】
例えば、配列番号4で表される塩基配列からなる(R)−ヒドロキシニトリルリアーゼ遺伝子において、シグナル配列は1〜81番塩基で表される(Swiss−Prot:アクセッションナンバーO24243参照)ので、配列番号4で表される遺伝子を鋳型として、配列番号4の82番塩基以降の塩基配列を増幅するように設計されたプライマーを用いてPCRを実施することにより、シグナル配列の除去されたDNAを得ることができる。
【0020】
(b)コドン使用頻度の調節
本改変方法は、そのアミノ酸配列を変化させることなく、宿主におけるアミノ酸ごとのコドン使用頻度が10%以上となるように改変することで、(R)−ヒドロキシニトリルリアーゼの発現量を増加させるものである。本発明において、コドン使用頻度とは、宿主細胞内で発現する全ての遺伝子に含まれる全コドン中の、同種アミノ酸をコードするコドンの使用回数に対する1つのコドンの使用回数の百分率をいう。
【0021】
ここで、改変した遺伝子のコドンのうち、宿主細胞における使用頻度が最も低いコドンの使用割合が10%以上、より好ましくは12%以上、さらに好ましくは14%となるようにする。10%を下回ると発現量の増加が少ない場合があるためである。各宿主におけるコドン使用頻度は、データベースとして公開されているものを使用できる。
【0022】
宿主として大腸菌を使用する場合、Escherichia coli K−12 MG1655(XanaGenome(Microbial Genome Database)参照)のコドン使用頻度データを利用し、表1に示すようにコドンを調節することができる。例として具体的には、表1を参照すると、Argをコードする使用頻度が2.2%であるコドンAGGを、使用頻度が37.9%であるコドンCGTへ改変する。
【0023】
【表1】
【0024】
例えば、配列番号4で表される塩基配列からなる(R)−ヒドロキシニトリルリアーゼ遺伝子のコドン使用頻度を上記手順に従って調節することにより、配列番号5で表される塩基配列からなるDNAが得られる。従って、本発明の改変DNAには、配列番号5で表される塩基配列からなるDNAが包含される。このようにして得られたDNAを、大腸菌、特にBL21株に導入して発現させることにより、(R)−ヒドロキシニトリルリアーゼの発現量を増大させることができる。
【0025】
本発明においては、(R)−ヒドロキシニトリルリアーゼ遺伝子の塩基配列において、コドン使用頻度を調節した後で、さらにシグナル配列を除去することにより改変を実施するのが好ましい。例えば、配列番号4で表される塩基配列からなる(R)−ヒドロキシニトリルリアーゼ遺伝子のコドン使用頻度を調節することにより得られる配列番号5で表される塩基配列において、シグナル配列に相当する1〜81番塩基を除去することにより、改変を実施することができる。このようにして得られるDNAとして、配列番号8で表される塩基配列からなるDNAが挙げられる。
【0026】
従って、本発明は、以下の(a)又は(b)のDNAを包含する:
(a)配列番号8で表される塩基配列を含み、塩基長が4000塩基以下、好ましくは2500塩基以下、より好ましくは1600塩基以下のDNA、
(b)配列番号8で表される塩基配列と少なくとも80%、好ましくは少なくとも90%、より好ましくは少なくとも95%の相同性を有し、(R)−ヒドロキシニトリルリアーゼ活性を有するタンパク質をコードする塩基配列を含み、塩基長が4000塩基以下、好ましくは2500塩基以下、より好ましくは1600塩基以下のDNA。
【0027】
(c)タグ配列の付加
本改変方法は、バラ科植物由来(R)−ヒドロキシニトリルリアーゼをコードする遺伝子の塩基配列にタグ配列を付加することによって、該タンパク質の発現量を増大させるものである。
【0028】
付加するタグ配列としては、(R)−ヒドロキシニトリルリアーゼの宿主細胞における発現を増大するものであれば特に制限されない。例えば、タンパク質を可溶化する機能を有するタグ配列、具体的には、Trxタグ配列(チオレドキシンをコードする配列)、及びNusタグ配列(大腸菌データベース上で最も優れた可溶性ポテンシャルを有すると同定された配列)、ならびにタンパク質のフォールディングを促進する機能を有するタグ配列、具体的にはDSbタグ配列(ジスルフィド結合の形成と異性化を触媒する酵素をコードする配列)が挙げられる。例えば、Trxタグ配列がコードするチオレドキシンと(R)−ヒドロキシニトリルリアーゼとが融合して発現するように、Trgタグ配列を付加することにより、(R)−ヒドロキシニトリルリアーゼが宿主細胞の細胞質中に可溶性画分として発現されやすくなる。
【0029】
タグ配列の付加は、例えば、対象とするタグ配列を含む発現ベクターに、目的とするDNAを連結することにより実施することができる。Trxタグ配列を含む発現ベクターとしては、pET32a(+)、pET32b(+)、pET32c(+)(Novagen社製)等が挙げられる。DSbタグ配列を含む発現ベクターとしては、pET39b(+)、pET40b(+)、(Novagen社製)等が挙げられる。Nusタグ配列を含む発現ベクターとしては、pET43.1a(+)、pET43.1b(+)、pET43.1c(+)、pET44a(+)、pET44b(+)、pET44c(+)(Novagen社製)等が挙げられる。
【0030】
上記のようにして改変された塩基配列からなるDNAをベクター中に連結し、得られた組換えベクターを用いて宿主細胞を形質転換し、該形質転換体を培養することにより、または改変された塩基配列からなるDNAを無細胞タンパク質合成系で発現させることにより、(R)−ヒドロキシニトリルリアーゼを産生することができる。従って、本発明はまた、上記のように改変されたDNAを含む組換えベクター及び該組換えベクターを含む形質転換体に関する。
【0031】
ベクターには、宿主細胞で自律的に増殖し得るファージ、コスミド、人工染色体又はプラスミドが使用される。プラスミドとしては、大腸菌由来のプラスミド(例えばpET21a(+)、pET32a(+)、pET39b(+)、pET40b(+)、pET43.1a(+)、pET44a(+)、pKK223−3、pGEX4T、pUC118、pUC119、pUC18、pUC19等)、枯草菌由来のプラスミド(例えばpUB110、pTP5等)、酵母由来のプラスミド(例えばYEp13、YEp24、YCp50等)などが挙げられ、ファージDNAとしてはλファージ(λgt11、λZAP等)が挙げられる。さらに、ワクシニアウイルスなどの動物ウイルス、バキュロウイルスなどの昆虫ウイルスベクターを用いることもできる。(R)−ヒドロキシニトリルリアーゼの生産性向上のため、より強力なプロモーターを有するベクターを用いるのが好ましく、大腸菌由来のベクターとしては、pET21a、pKK223−3等が好ましい。
【0032】
ベクターに本発明の改変されたDNAを挿入するには、まず、精製された該DNAを適当な制限酵素で切断し、適当なベクターDNAの制限酵素部位又はマルチクローニングサイトに挿入してベクターに連結する方法などが採用される。例えば本発明の改変されたDNAは、通常知られている方法により合成することができ、ベクターに組み込むため、適当な制限酵素の切断部位を両末端に含むように、プライマーを用いてPCR法により増幅してもよい。PCR反応の条件は、当業者が適宜決定することができるが、例えば98℃で30秒間の変性、57℃で15秒間のアニーリング、74℃で30秒間の重合を1サイクルとして30サイクル反応させ、増幅された遺伝子を得ることができる。
【0033】
その他、哺乳動物細胞において用いられる組換えベクターには、プロモーター、本発明のDNAのほか、所望によりエンハンサーなどのシスエレメント、スプライシングシグナル、ポリA付加シグナル、選択マーカー、リボソーム結合配列(SD配列)などが連結されていてもよい。選択マーカーの例としては、カナマイシン、アンピシリン、テトラサイクリン、クロラムフェニコールなどの抗生物質が挙げられるがこれに限定されない。
【0034】
DNA断片とベクター断片とを連結させるには、公知のDNAリガーゼを用いる。そして、DNA断片とベクター断片とをアニーリングさせた後連結させ、組換えベクターを作成する。好ましくは市販のライゲーションキット、例えばライゲーションhigh(東洋紡株式会社製)を用いて、規定の条件にてライゲーション反応を行なうことにより組換えベクターを得ることができる。
【0035】
また、これらのベクターを、必要であればボイル法、アルカリSDS法などの精製手段により精製し、さらに例えばエタノール沈殿法、ポリエチレングリコール沈殿法などの濃縮手段により濃縮することができる。
【0036】
形質転換に使用する宿主としては、(R)−ヒドロキシニトリルリアーゼを発現できるものであれば特に限定されるものではない。例えば、細菌(大腸菌、枯草菌等)、酵母、動物細胞(COS細胞、CHO細胞等)、昆虫細胞が挙げられる。本発明においては、細菌、特に大腸菌を使用するのが好ましく、DH5α細胞、BL21(DE3)細胞、BL21(DE3)pLysS細胞、OrigamiB2細胞等を用いるのがより好ましく、中でも生産性に優れる点でBL21(DE3)細胞を用いるのが最も好ましい。
【0037】
細菌を宿主とする場合は、組換えベクターが該細菌中で自律複製可能であると同時に、プロモーター、リボゾーム結合配列(SD配列)、本発明の改変されたDNA、転写終結配列を含むことが好ましい。所望によりエンハンサーなどのシスエレメント、選択マーカー、オペレーター、プロモーターを制御する遺伝子が含まれていてもよい。プロモーターは、大腸菌等の宿主細胞中で発現できるものであればいずれを用いてもよい。大腸菌の場合、具体的には、T7プロモーター、Lacプロモーター、Tacプロモーターなどが挙げられる。組換えベクターの導入方法は、細菌にDNAを導入する方法であれば特に限定されるものではない。例えばカルシウムイオンを用いる方法、エレクトロポレーション法等が挙げられる。
【0038】
本発明のDNAを含む形質転換体は、その組換えベクターが有するマーカー遺伝子により、例えば、アンピシリン、カナマイシンなどの抗生物質を含むLB培地寒天プレート上でコロニーを形成することにより選抜することができるが、クローニングされた宿主細胞が本発明の組換えベクターにより形質転換されたものかどうかを確認するため、一部を用いて、PCR法によるインサートの増幅確認、又はシーケンサーを用いたダイデオキシ法による配列解析をしてもよい。
【0039】
本発明はまた、本発明の改変DNAを発現させ、得られた(R)−ヒドロキシニトリルリアーゼとカルボニル化合物及びシアノ基供与体とを反応させ、反応液から光学活性シアノヒドリンを回収することを含む、光学活性シアノヒドリンの製造方法に関する。
【0040】
このような光学活性シアノヒドリンの製造方法としては、上記形質転換体を培養し、得られた培養物とカルボニル化合物及びシアノ基供与体とを反応させ、反応液から光学活性シアノヒドリンを回収することを含む方法が挙げられる。あるいは、無細胞タンパク質合成系を用いる方法が挙げられる。無細胞タンパク質合成系は、細胞抽出液を用いて試験管内でタンパク質を合成する系である。「無細胞タンパク質合成系」は、mRNAの情報を読み取ってリボソーム上でタンパク質を合成する無細胞翻訳系とDNAを鋳型としてRNAを合成する無細胞転写系との両者を含む。無細胞タンパク質合成系は、系を容易に改変することができるため、目的のタンパク質に適した発現系を構築しやすいという利点がある。なお、無細胞タンパク質合成系の詳細については、特開2000−175695号などに記載されている。
【0041】
従って、(R)−ヒドロキシニトリルリアーゼとカルボニル化合物及びシアノ基供与体とを反応させることには、精製された(R)−ヒドロキシニトリルリアーゼとカルボニル化合物及びシアノ基供与体とを反応させることだけでなく、形質転換体を培養した培養物とカルボニル化合物及びシアノ基供与体とを反応させること、ならびに無細胞タンパク質合成系から得られる反応液とカルボニル化合物及びシアノ基供与体とを反応させることも含まれる。
【0042】
ここで「培養物」とは、培養上清、培養細胞、培養菌体、細胞又は菌体の破砕物、及びこれらから精製された(R)−ヒドロキシニトリルリアーゼタンパク質自体のいずれをも意味するものである。本発明においては、好ましくは精製(R)−ヒドロキシニトリルリアーゼを用いる。
【0043】
上記形質転換体を培地で培養する方法は、宿主の培養に用いられる通常の方法に従って行われる。大腸菌や酵母菌等の微生物を宿主として得られた形質転換体を培養する培地としては、微生物が資化し得る炭素源、窒素源、無機塩類等を含有し、形質転換体の培養を効率的に行うことができる培地であれば、天然培地、合成培地のいずれを用いてもよい。
【0044】
炭素源としては資化可能な炭素化合物であればよく、例えば、グリセリンなどのポリオール類、又はピルビン酸、コハク酸もしくはクエン酸等の有機酸類を使用することができる。また、窒素源としては利用可能な窒素化合物であればよく、例えば、ペプトン、肉エキス、酵母エキス、カゼイン加水分解物、大豆粕アルカリ抽出物、又はアンモニアもしくはその塩などを使用することができる。その他、リン酸塩、炭酸塩、硫酸塩、マグネシウム、カルシウム、カリウム、鉄、マンガン、亜鉛などの塩類、特定のアミノ酸、特定のビタミン、消泡剤なども必要に応じて使用してもよい。また、イソプロピル−β−D−チオガラクトピラノシドなどのタンパク質発現誘導剤を必要に応じて培地に添加してもよい。
【0045】
培養は、通常、振盪培養又は通気攪拌培養などの好気的条件下、好ましくは0〜40℃、より好ましくは10〜37℃、特に好ましくは15〜37℃で培養を行う。培養期間中、培地のpHは宿主の発育が可能で、生産された(R)−ヒドロキシニトリルリアーゼの活性が損なわれない範囲で適宜変更することができるが、好ましくはpH4〜8程度の範囲である。pHの調整は、無機又は有機酸、アルカリ溶液等を用いて行う。培養中は必要に応じてアンピシリンやテトラサイクリン等の抗生物質を培地に添加してもよい。
【0046】
培養後、(R)−ヒドロキシニトリルリアーゼが菌体内又は細胞内に生産される場合には、通常知られている方法、例えば、菌体又は細胞を機械的方法、リゾチームなどを用いた酵素的方法又は界面活性剤などを用いた化学的処理によって破壊することにより当該酵素を抽出できる。また、当該酵素が菌体外又は細胞外に生産される場合には、培養液をそのまま使用するか、遠心分離等により菌体又は細胞を除去する。
【0047】
(R)−ヒドロキシニトリルリアーゼが封入体として得られる場合には、好ましくはさらにリフォールディングを行うことにより、活性型でかつ可溶性の(R)−ヒドロキシニトリルリアーゼを得ることができる。リフォールディングは、当技術分野で通常用いられる方法に従って実施することができ、例えば、Applied and Environmental Microbiology(1998)64:1405−1411に記載された方法に従って、実施することができる。
【0048】
具体的には、培養後、遠心分離等の方法により集菌した後、細胞を破砕し、封入体を変性剤を用いて可溶化することによって、(R)−ヒドロキシニトリルリアーゼ類を抽出することができる。細胞の破砕は、常法で、たとえば超音波により実施できる。懸濁媒体としてはpH5.0〜8.0のpH値に調整した好適な緩衝液(例えばクエン酸緩衝液やリン酸緩衝液等)を用いることが好ましい。この際、細胞の破砕を促進させるためEDTAを添加してもよい。このようにして細胞を破砕した後に、不溶性成分(封入体)を任意の方法で、遠心分離するか、濾過することにより分取する。異種のタンパク質をできる限り除去するため、たとえば水、リン酸緩衝液を用いて洗浄することが好ましいが、場合により界面活性剤や4M程度の尿素で洗浄してもよい。得られた沈殿(ペレット)を変性剤を用いて可溶化するが、変性剤としては、公知の変性剤、特にグアニジンまたは尿素を使用することができる。この可溶化に当っての変性剤の濃度は、グアニジンでは4〜8モル/L、好ましくは約6モル/L、尿素では5〜9モル/L、好ましくは約8モル/Lである。グアニジンは通常グアニジン塩酸塩等のグアニジンの酸付加塩として用いられる。
【0049】
上記のようにして封入体の可溶化を行った後、遠心分離等で不純物を除去し、得られた上澄液を必要により精製工程に付した後、リフォールディング(活性化、再生化)を行うことができる。リフォールディングは、精製した(R)−ヒドロキシニトリルリアーゼ類にレドックスバッファーを添加するか、あるいは(R)−ヒドロキシニトリルリアーゼ類を含有する上澄液をレドックスバッファーで希釈することにより行われる。(R)−ヒドロキシニトリルリアーゼ類を含有する上澄液をレドックスバッファーで希釈する場合、変性剤の濃度を活性化に適した中性pHにおいて不作用濃度まで希釈することが必要であり、グアジニンでは0〜2.0モル/L、好ましくは約1モル/L以下まで、尿素では0〜4.0モル/L、好ましくは約2モル/L以下まで希釈する。
【0050】
レドックスバッファーとしては、酸化型グルタチオン(GSSG)および還元型グルタチオン(GSH)、システインおよびシスチン、またはシステアミンおよびシスタミンを含有する緩衝液(例、リン酸緩衝液、酢酸緩衝液、クエン酸緩衝液、Tris/HCl緩衝液)が好ましいものとして挙げられ、中でもGSSGおよびGSHを含有するTris/HCl緩衝液が好ましい。GSSGおよびGSHを用いる場合、GSSGの濃度は0.1〜10ミリモル/L、好ましくは0.1〜1.0ミリモル/L、GSHの濃度は0.1〜10ミリモル/L、好ましくは0.1〜1.0ミリモル/Lである。
【0051】
また、レドックスバッファーには、通常、グリセリン等の添加剤を添加してもよい。添加量としてグリセリンは1〜50%、好ましくは5〜20%の濃度で添加するのがよい。また、当該酵素は補酵素としてFADが必要なので、これを添加してもよい。添加量としては1〜1000μM、好ましくは10〜100μMの濃度で添加するのがよい。
【0052】
該リフォールディングに当っての温度は0〜30℃、好ましくは0〜15℃、pHは7〜9、好ましくはpH7.5〜8.5である。リフォールディングに要する時間は通常1時間〜10週間、好ましくは3日〜1週間である。
【0053】
その後、タンパク質の単離精製に用いられる一般的な生化学的方法、例えば硫安分画法、有機溶媒沈殿法、イオン交換体などによる吸着処理法、イオン交換クロマトグラフィー、疎水クロマトグラフィー、ゲルろ過クロマトグラフィー、アフィニティークロマトグラフィー、電気泳動法等を単独で又は適宜組み合わせて用いることにより、前記培養物中から(R)−ヒドロキシニトリルリアーゼを単離精製することができる。(R)−ヒドロキシニトリルリアーゼが得られたか否かは、SDS−ポリアクリルアミドゲル電気泳動等により確認することができる。
【0054】
本発明の形質転換体の上記培養物、すなわち、培養上清、培養細胞、培養菌体、細胞若しくは菌体の破砕物、又は精製(R)−ヒドロキシニトリルリアーゼタンパク質と、カルボニル化合物及びシアノ基供与基とを反応させることにより、光学活性シアノヒドリンを製造する。シアノヒドリンを合成する酵素反応の形態については制限されず、水・有機溶媒混合系、有機溶媒系、有機溶媒水二相系、固定化酵素を使う反応系などのいずれの反応系においても効果的に実施することができる。
【0055】
光学活性シアノヒドリンとは、一方の鏡像異性体が他方の鏡像異性体より多く含まれているシアノヒドリン、又は、いずれか一方の鏡像異性体のみからなるシアノヒドリンをいう。(R)−ヒドロキシニトリルリアーゼを用いることにより、用いた基質に応じて(R)−シアノヒドリン及び(S)−シアノヒドリンの双方を製造することができる。
【0056】
ここでカルボニル化合物とは、アルデヒド又はケトンをいい、具体的には、次式(I):
R1−CO−R2 (I)
(式中、R1及びR2は、互いに同一でも異なっていてもよく、それぞれ水素原子又は炭素数22以下の1価の炭化水素基を表し、前記炭化水素基中、−CH2−並びに−CH3のCH2はカルボニル基、スルホニル基、−O−又は−S−で置き換えられていてもよく、=CH2は=O又は=Sで置き換えられていてもよく、また−CH2−のC−H、−CH3のC−H、>CH−のC−H、=CH−のC−H並びに=CH2のC−Hは、N又はC−ハロゲンで置き換えられていてもよく、また、R1及びR2は、共同して2価の基を表してもよい。)で示される。
【0057】
前記式(I)において、炭素数22以下の1価の炭化水素基とは、直鎖状又は分岐状の鎖状炭化水素基、側鎖のない又は側鎖のある単環式炭化水素基、側鎖のない又は側鎖のある多環式炭化水素基、側鎖のない又は側鎖のあるスピロ炭化水素基、側鎖のない又は側鎖のある環集合構造の炭化水素基、あるいは、前記の環式炭化水素基が置換した鎖状炭化水素基のいずれをも含む。また、飽和な炭化水素基並びに不飽和な炭化水素基のいずれをも含むが、不飽和な炭化水素基において、C=C=Cのアレン構造を含む基は除く。
【0058】
なお、以下においては、側鎖のない芳香族基、側鎖のある芳香族基、並びに、フェニルフェニル基又は側鎖のあるフェニルフェニル基などを併せて、アリール基といい、このアリール基で置換された直鎖状又は分岐状のアルキル基をアラルキル基という。他の環式炭化水素基に関しても、特に明記しない場合、環上に側鎖のないものとあるものを併せて指す場合には、単にシクロアルキル基等の名称を用いる。鎖状炭化水素基についても、直鎖状のものと分岐状のものを併せて指す場合には、単にアルキル基等の名称を用いる。
【0059】
前記炭化水素基中、−CH2−がカルボニル基、スルホニル基、−O−又は−S−で置き換えられると、それぞれケトン、スルホン、エーテル又はチオエーテルの構造が導入され、−CH3の−CH2−がカルボニル基、−O−又は−S−で置き換わると、それぞれホルミル基(アルデヒド)、水酸基又はメルカプト基に変わり、あるいは、末端の=CH2が=O又は=Sに置き換わると、ケトン、チオケトンの構造が導入されることを意味し、また、−CH2−のC−HがNに変わると、−NH−となり、>CH−のC−HがNに変わると、>N−となり、=CH−のC−HがNに変わると、=N−となり、末端の−CH3のC−HがNに変わると、−NH2が導入され、=CH2のC−HがNに変わると、=NHとなる。また、−CH3、−CH2−、=CH−、≡CH又は>CH−のC−HがC−ハロゲンで置き換えられると、当該炭素上へハロゲン原子を置換することになる。なお、炭素鎖中における−O−、−S−、Nへの置き換えは、当該炭化水素基に対する、それぞれオキサ置換、チア置換、アザ置換に当たり、例えば、炭化水素環の環の骨格炭素で起こると、炭化水素環のそれぞれ含酸素複素環、含硫黄複素環、含窒素複素環への変換となる。該炭化水素基中、CH2並びにC−Hにおける置き換えは、それぞれ独立に行われてよく、加えて、前記の置き換えを行った後、なお当該炭素上にCH2又はC−Hが残存する際には、更に置き換えがなされてもよい。更には、前記の置き換えにより、−CH2−CH3の−CO−O−H;カルボン酸構造への変換などもなされる。
【0060】
本明細書において、ハロゲン原子とは、フッ素原子、塩素原子、臭素原子、ヨウ素原子を指すが、フッ素原子、塩素原子、臭素原子が好ましい。
【0061】
従って、前記炭化水素基としては、鎖状炭化水素基並びに環式炭化水素基など環構造を有する炭化水素基のいずれをも選択でき、例えば、飽和鎖状炭化水素基である直鎖状又は分岐状のアルキル基、不飽和鎖状炭化水素基である直鎖状又は分岐状のアルケニル基、直鎖状又は分岐状のアルキニル基、直鎖状又は分岐状のアルカジエニル基など、飽和な環式炭化水素基であるシクロアルキル基、不飽和な環式炭化水素基であるシクロアルケニル基、シクロアルキニル基、シクロアルカジエニル基など、芳香族炭化水素基であるアリール基、アラルキル基、アリールアルケニル基などが挙げられる。
【0062】
更に詳しくいえば、直鎖状又は分岐状のアルキル基としては、例えばメチル基、エチル基、プロピル基、イソプロピル基、ブチル基、1−メチルプロピル基、ペンチル基、1−メチルブチル基、ヘキシル基、1−メチルペンチル基、ヘプチル基、1−メチルヘキシル基、1−エチルペンチル基、オクチル基、ノニル基、デシル基、ウンデシル基、ドデシル基、トリデシル基、テトラデシル基、2−メチルプロピル基、2−メチルブチル基、3−メチルブチル基、2−メチルペンチル基、3−メチルペンチル基、4−メチルペンチル基、メチルヘキシル基、メチルヘプチル基、メチルオクチル基、メチルノニル基、1,1−ジメチルエチル基、1,1−ジメチルプロピル基、2,6−ジメチルヘプチル基、3,7−ジメチルオクチル基、2−エチルヘキシル基など、シクロアルキルアルキル基としては、シクロペンチルメチル基、シクロヘキシルメチル基など、シクロアルキル基としては、シクロプロピル基、シクロブチル基、シクロペンチル基、メチルシクロペンチル基、シクロヘキシル基、メチルシクロヘキシル基、シクロヘプチル基、シクロオクチル基など、ビシクロアルキル基としては、ノルボルニル基、ビシクロ[2.2.2]オクチル基、アダマンチル基などが挙げられる。直鎖状又は分岐状のアルケニル基としては、例えばビニル基、アリル基、クロチル基(2−ブテニル基)、イソプロペニル基(1−メチルビニル基)など、シクロアルケニル基又はシクロアルカジエニル基としては、シクロペンテニル基、シクロペンタジエニル基、シクロヘキセニル基、シクロヘキサンジエニル基などが挙げられる。直鎖状又は分岐状のアルキニル基としては、例えばエチニル基、プロピニル基、ブチニル基などが挙げられる。アリール基としては、例えばフェニル基、1−ナフチル基、2−ナフチル基、2−フェニルフェニル基、3−フェニルフェニル基、4−フェニルフェニル基、9−アントリル基、メチルフェニル基、ジメチルフェニル基、トリメチルフェニル基、エチルフェニル基、メチルエチルフェニル基、ジエチルフェニル基、プロピルフェニル基、ブチルフェニル基などが挙げられる。アラルキル基としては、例えばベンジル基、1−ナフチルメチル基、2−ナフチルメチル基、フェネチル基(2−フェニルエチル基)、1−フェニルエチル基、フェニルプロピル基、フェニルブチル基、フェニルペンチル基、フェニルヘキシル基、メチルベンジル基、メチルフェネチル基、ジメチルベンジル基、ジメチルフェネチル基、トリメチルベンジル基、エチルベンジル基、ジエチルベンジル基などが挙げられる。アリールアルケニル基としては、例えばスチリル基、メチルスチリル基、エチルスチリル基、ジメチルスチリル基、3−フェニル−2−プロペニル基などが挙げられる。
【0063】
前記炭化水素基中のCH2がカルボニル基、スルホニル基、O又はSで、又はC−HがN又はC−ハロゲンで置き換えられた基としては、ケトン、アルデヒド、カルボン酸、スルホン、エーテル、チオエーテル、アミン、アルコール、チオール、ハロゲン、複素環(例えば、含酸素複素環、含硫黄複素環、含窒素複素環)などの構造を一つ以上含む基が挙げられる。なお、含酸素複素環、含硫黄複素環、含窒素複素環とは、環式炭化水素基の環骨格の炭素がそれぞれ酸素、硫黄、窒素で置き換わるものを意味し、更には、これらヘテロ原子置換が二種以上ある複素環であってもよい。前記の置換を有する炭化水素基としては、例えば、ケトン構造のアセチルメチル基、アセチルフェニル基;スルホン構造のメタンスルホニルメチル基;エーテル構造のメトキシメチル基、メトキシエチル基、エトキシエチル基、メトキシプロピル基、ブトキシエチル基、エトキシエトキシエチル基、メトキシフェニル基、ジメトキシフェニル基、フェノキシメチル基;チオエーテル構造のメチルチオメチル基、メチルチオフェニル基;アミン構造のアミノメチル基、2−アミノエチル基、2−アミノプロピル基、3−アミノプロピル基、2,3−ジアミノプロピル基、2−アミノブチル基、3−アミノブチル基、4−アミノブチル基、2,3−ジアミノブチル基、2,4−ジアミノブチル基、3,4−ジアミノブチル基、2,3,4−トリアミノブチル基、メチルアミノメチル基、ジメチルアミノメチル基、メチルアミノエチル基、プロピルアミノメチル基、シクロペンチルアミノメチル基、アミノフェニル基、ジアミノフェニル基、アミノメチルフェニル基;含酸素複素環のテトラヒドロフラニル基、テトラヒドロピラニル基、モルホリルエチル基;含酸素複素芳香環のフリル基、フルフリル基、ベンゾフリル基、ベンゾフルフリル基;含硫黄複素芳香環のチエニル基;含窒素複素芳香環のピロリル基、イミダゾリル基、オキサゾリル基、チアジアゾリル基、ピリジル基、ピリミジニル基、ピリダジニル基、ピラジニル基、テトラジニル基、キノリル基、イソキノリル基、ピリジルメチル基;アルコール構造の2−ヒドロキシエチル基、2−ヒドロキシプロピル基、3−ヒドロキシプロピル基、2,3−ジヒドロキシプロピル基、2−ヒドロキシブチル基、3−ヒドロキシブチル基、4−ヒドロキシブチル基、2,3−ジヒドロキシブチル基、2,4−ジヒドロキシブチル基、3,4−ジヒドロキシブチル基、2,3,4−トリヒドロキシブチル基、ヒドロキシフェニル基、ジヒドロキシフェニル基、ヒドロキシメチルフェニル基、ヒドロキシエチルフェニル基;チオール構造の2−メルカプトエチル基、2−メルカプトプロピル基、3−メルカプトプロピル基、2,3−ジメルカプトプロピル基、2−メルカプトブチル基、3−メルカプトブチル基、4−メルカプトブチル基、メルカプトフェニル基;ハロゲン化炭化水素基である2−クロロエチル基、2−クロロプロピル基、3−クロロプロピル基、2−クロロブチル基、3−クロロブチル基、4−クロロブチル基、フルオロフェニル基、クロロフェニル基、ブロモフェニル基、ジフルオロフェニル基、ジクロロフェニル基、ジブロモフェニル基、クロロフルオロフェニル基、トリフルオロフェニル基、トリクロロフェニル基、フルオロメチルフェニル基、トリフルオロメチルフェニル基;アミン構造とアルコール構造を有する2−アミノ−3−ヒドロキシプロピル基、3−アミノ−2−ヒドロキシプロピル基、2−アミノ−3−ヒドロキシブチル基、3−アミノ−2−ヒドロキシブチル基、2−アミノ−4−ヒドロキシブチル基、4−アミノ−2−ヒドロキシブチル基、3−アミノ−4−ヒドロキシブチル基、4−アミノ−3−ヒドロキシブチル基、2,4−ジアミノ−3−ヒドロキシブチル基、3−アミノ−2,4−ジヒドロキシブチル基、2,3−ジアミノ−4−ヒドロキシブチル基、4−アミノ−2,3−ジヒドロキシブチル基、3,4−ジアミノ−2−ヒドロキシブチル基、2−アミノ−3,4−ジヒドロキシブチル基、アミノヒドロキシフェニル基;ハロゲンと水酸基で置換された炭化水素基であるフルオロヒドロキシフェニル基、クロロヒドロキシフェニル基;カルボン構造のカルボキシフェニル基などが挙げられる。
【0064】
R1及びR2で表される非対称の2価の基としては、特に制限はなく、例えば、ノルボルナン−2−イリデン、2−ノルボルネン−5−イリデンが挙げられる。
【0065】
前記式(I)で示されるカルボニル化合物としては、例えば、ベンズアルデヒド、m−フェノキシベンズアルデヒド、p−メチルベンズアルデヒド、o−クロロベンズアルデヒド、m−クロロベンズアルデヒド、p−クロロベンズアルデヒド、m−ニトロベンズアルデヒド、3,4−メチレンジオキシベンズアルデヒド、2,3−メチレンジオキシベンズアルデヒド、フェニルアセトアルデヒド、フルフラール等の芳香族アルデヒド;アセトアルデヒド、ブチルアルデヒド、イソブチルアルデヒド、バレルアルデヒド、シクロヘキサンアルデヒド等の脂肪族アルデヒド;エチルメチルケトン、ブチルメチルケトン、メチルプロピルケトン、イソプロピルメチルケトン、メチルペンチルケトン、メチル(2−メチルプロピル)ケトン、メチル(3−メチルブチル)ケトン等の飽和脂肪族ケトン;メチル(2−プロペニル)ケトン、(3−ブテニル)メチルケトン等の不飽和脂肪族ケトン;(3−クロロプロピル)メチルケトン等のアルキル(ハロアルキル)ケトン;2−(アルコキシカルボニルアミノ)−3−シクロヘキシルプロピオンアルデヒド等の2−(保護アミノ)アルデヒド;3−メチルチオプロピオンアルデヒド等のアルキルチオ脂肪族アルデヒドが挙げられる。
【0066】
本発明において、基質としてのカルボニル化合物をシアノヒドリンに変換するためには、シアノ基供与体の存在下で酵素反応を実施する。本明細書中、シアノ基供与体とは、反応系へ添加することによって、シアニド、すなわちシアン化物イオン(CN−)を生じる物質を意味し、例えば、シアン化水素、青酸(シアン化水素酸)、シアン化ナトリウムやシアン化カリウムなどのシアン化水素の塩、又は、アセトンシアノヒドリン等のシアノヒドリン類が挙げられる。特に回収リサイクルが容易な青酸(シアン化水素酸)を用いるのが好ましい。シアノ基供与体の供給方法としては常法により液体として供給する方法、又は常法により気体として供給する方法のいずれをも採用することができる。
【0067】
本発明においては、反応原料の濃度を高め、生産性を高めるために、反応溶媒として、水と実質的に混和しない有機溶媒を用いる。ここで、「水と実質的に混和しない有機溶媒」とは、水に任意の割合で溶解する溶媒を除く有機溶媒を意味する。有機溶媒としては、水と実質的に混和せず、基質及び生成物を充分に溶解し、酵素反応に悪影響を与えないものであれば特に制限なく用いることができる。このような有機溶媒は、原料のアルデヒド又はケトンの物性、生成物であるシアノヒドリンの物性に応じて適宜選択することができる。
【0068】
水と実質的に混和しない有機溶媒としては、具体的には、ハロゲン化されていてもよい炭化水素系溶媒(例えば、直鎖状、分岐状又は環状の飽和又は不飽和脂肪族炭化水素、芳香族炭化水素)、例えば、ペンタン、ヘキサン、シクロヘキサン、ベンゼン、トルエン、キシレン、塩化メチレン、クロロホルムなど;ハロゲン化されていてもよいアルコール系溶媒(例えば、直鎖状、分岐状又は環状の飽和又は不飽和脂肪族アルコール、アラルキルアルコール)、例えば、n−ブタノール、イソブタノール、t−ブタノール、ヘキサノール、シクロヘキサノール、n−アミルアルコールなど;ハロゲン化されていてもよいエーテル系溶媒(例えば、直鎖状、分岐状又は環状の飽和又は不飽和脂肪族エーテル、芳香族エーテル)、例えば、ジエチルエーテル、ジプロピルエーテル、ジイソプロピルエーテル、ジブチルエーテル、t−ブチルメチルエーテル、ジメトキシエタンなど;ハロゲン化されていてもよいエステル系溶媒(例えば、直鎖状、分岐状又は環状の飽和又は不飽和脂肪族エステル、芳香族エステル)、例えば、ギ酸メチル、酢酸メチル、酢酸エチル、酢酸ブチル、プロピオン酸メチル等が挙げられ、これらを単独で用いてもまた2種以上を混合して用いてもよい。特に、ジイソプロピルエーテル、ジブチルエーテル、t-ブチルメチルエーテル、酢酸エチルを用いるのが好ましい。
【0069】
前記有機溶媒は、水又は水性緩衝液で飽和されているのが好ましい。ここで水性緩衝液としては、特に制限はないが、酵素活性の最適pH(pH4〜7)の付近において緩衝能を発揮する緩衝液、例えば、リン酸、クエン酸、グルタル酸、リンゴ酸、マロン酸、o−フタル酸、コハク酸などの塩等によって構成される緩衝液等が好ましく用いられる。
【0070】
本発明において、酵素、基質及びシアノ基供与体の使用量、反応温度は、用いる基質に応じて適宜決定される。通常、酵素の使用量は基質であるカルボニル化合物50ミリモルに対して250〜100,000単位、好ましくは500〜50,000単位である。本明細書において、当該酵素(R)−ヒドロキシニトリルリアーゼの活性1単位は、DL−マンデロニトリルを基質に反応させて1分間にベンズアルデヒド1μモルを生成する活性と定義する。測定は0.2Mクエン酸−NaOH緩衝液(pH5.5)7mLにDLマンデロニトリルを0.931μL添加した液を分光光度計のあらかじめ20℃に調温した対照セル、測定セルに3mLずつ量りとり、酵素液を添加したときのベンズアルデヒドの生成速度を波長249.6nmの吸光度変化を測定することによって算出する。カルボニル化合物の濃度は通常0.1〜10モル/Lの範囲に設定し、シアノ基供与体は用いるカルボニル化合物に対して1〜5倍モル、好ましくは1.1〜3倍モルの濃度で添加する。本反応は基質濃度によって酵素活性及び反応速度が変化するので、用いるカルボニル化合物の種類に応じて基質濃度を適宜決定する。反応時間は、基質であるカルボニル化合物の転換率が80%以上、好ましくは90%以上に達するまでの時間が適当であるが、これに限定されない。反応温度は酵素の活性が十分発揮される温度であればよく、通常0〜40℃、好ましくは4〜30℃である。
【0071】
反応系において、回分式で反応を行う場合には、撹拌などにより、酵素が反応系内に分散するようにする。カラムなどに固定化酵素を充填して反応を行う場合には、基質を含む溶液を適当な流速でカラムに流入させ、流出液を採取することで実施できる。回分反応の場合には、反応が完結した時点で混合を止め、生産物が溶解している有機相を常法により取り出すことで生産物を回収できる。これらの酵素は初回と同じ方法で調製した基質を含む溶液と混合することによって再使用することができる。
【0072】
酵素反応終了後のシアノヒドリンを含む反応液からの反応溶媒及び未反応のシアノ基供与体の回収は、蒸留等の操作をすることにより行うことができる。蒸留は、シアノヒドリンが高温では不安定であるため、常圧高温下で実施するよりも、比較的低い温度で減圧下で実施することが好ましい。また、この蒸留操作においては、公知のシアノヒドリンの安定化剤を添加することもできる。安定化剤としては蒸留ボトムを酸性に維持できるものであればよく、p-トルエンスルホン酸、酢酸などの有機酸、硫酸などの無機酸などをシアノヒドリンに対して1/200〜1/10モル添加することで実施できる。
【実施例】
【0073】
実施例1 アーモンド葉からのcDNAの調製及びアーモンド葉由来(R)−ヒドロキシニトリルリアーゼ遺伝子の調製
アーモンドの葉を材料として、全mRNAの抽出を行った。回収したmRNAを鋳型として、cDNA合成を行い、アーモンドのcDNAライブラリーを作成した。GeneBank Accession No.Y08211のアーモンド花由来の(R)−ヒドロキシニトリルリアーゼ遺伝子の配列情報(配列番号1)を参考にして、下記PCRプライマーを合成した。
センスプライマー:5’−GGCATCGATGGAGAAATCAACAACAATG−3’(配列番号2)
アンチセンスプライマー:5’−GCGTCGACTCACATGGACTCTTG−3’(配列番号3)
【0074】
これらのプライマーを使用し、上記のcDNAを鋳型としてPCRを行ったところ、上記GeneBank Accession No.Y08211のアーモンド花由来の(R)−ヒドロキシニトリルリアーゼ遺伝子と相同性の高い(99.5%)(R)−ヒドロキシニトリルリアーゼ遺伝子(以下RHNL−KR1と称する)を獲得した。その塩基配列を配列番号4に示す。
【0075】
実施例2 (R)−ヒドロキシニトリルリアーゼ遺伝子の塩基配列を改変したDNAの作成
配列番号4で表される塩基配列を有する(R)−ヒドロキシニトリルリアーゼを大腸菌で発現させるためにコドン使用頻度を検討し、上記表1に従って、アミノ酸配列を変化させることなくコドン使用頻度の低いものをコドン使用頻度の高いものに変更した配列(以下、RHNL−KR2と称する)を作成した。RHNL−KR2の塩基配列を配列番号5に示す。得られた配列に基づき(R)−ヒドロキシニトリルリアーゼ遺伝子を改変したDNAを合成した。
【0076】
さらに配列番号5で表される塩基配列においてシグナル配列(配列番号5の1〜81番塩基)を除去した遺伝子を得るために下記のPCRプライマーを合成した。
センスプライマー:5’−GGGCATATGGCCAACACTTCTGCTCACGATTTCAGCTAC−3’(配列番号6)
アンチセンスプライマー:5’−CCCGGATCCTTACATGGATTCCTGAATGTTGTGAATAGC−3’(配列番号7)
【0077】
これらのプライマーを使用し、上記の配列番号5で表される塩基配列からなるDNAを鋳型としてPCRを行い、シグナル配列を除去したDNA(RHNL−KR2−2と称する)を獲得した。その塩基配列を配列番号8に示す。
【0078】
実施例3 アーモンド葉由来(R)−ヒドロキシニトリルリアーゼ遺伝子の塩基配列を改変したDNAの大腸菌における発現
配列番号8で表される塩基配列からなるDNAを、大腸菌用ベクターpET21a(+)(Novagen社製)のマルチクローニングサイトのNdeIサイトとBamHIサイトの間に導入して組換えベクターpET21a(+)/RHNL−KR2−2を調製した。このプラスミドベクターを大腸菌株BL21(DE3)(Novagen社製)に形質転換により導入した。また、ここで得られた遺伝子組換え大腸菌をBL21(DE3)/pET21a(+)/RHNL−KR2−2と称する。
【0079】
上記で構築された組換え大腸菌株(BL21(DE3)/pET21a(+)/RHNL−KR2−2)を以下の方法により培養し、組換えタンパク質を得た。
【0080】
まず、15mL試験管中でLB培地5mLに菌株1コロニー分を植菌し、培養温度37℃、振盪速度170rpmで12時間、前培養を行った。続いて、500mL坂口フラスコ中でLB培地100mLにそれぞれの前培養液2mLを植菌し、培養温度17℃、振盪速度130rpmで培養を開始し、培養液の細胞密度の測定を、660nmにおける吸光度(OD660nmと称する)を測定することにより行い、OD660nmが0.4〜0.7になるまで培養後、タンパク質発現誘導剤であるイソプロピル−β−D−チオガラクトピラノシド(IPTG)を最終濃度が1mMになるように添加した。その後培養を継続し、OD660nmが3を越えるまで約24時間、本培養を行った。
【0081】
上記培養後、培養液を回収し、遠心分離(15000rpm、5分間)して菌体細胞を得た。得られた細胞はクエン酸緩衝液中で超音波処理により破砕した。破砕処理液は遠心分離(15000rpm、30分間)により可溶性上清と不溶性沈殿に分離した。このそれぞれの画分をSDS−PAGEに供したところ、目的のタンパク質と思われる57kDaのバンドが不溶性沈殿として菌体内全タンパク質の20%以上を占める量で発現されていた。SDS−PAGEの結果を図1の3レーン及び4レーンに示す。3レーンは可溶性画分の結果を表し、4レーンは不溶性画分の結果を表す。
【0082】
SDS−PAGEは通常の方法で行い、ゲルにはATTO社製PAGEL NPU−12.5Lを用い、電流を20mAで80分間流した後、クマシーブリリアントブルーで染色した。目的のタンパク質の発現量は画像処理ソフトウェアのScion Image Release Beta 4.0.2(Scion Corporation製)を用いてSDS−PAGEのイメージ画像のデンシトグラムを作成することにより、算出した。
【0083】
実施例4 アーモンド葉由来(R)−ヒドロキシニトリルリアーゼ遺伝子の塩基配列を改変したDNAを使用した光学活性シアノヒドリンの合成
実施例3に準じて調製した培養液1Lから遠心分離により、3gの菌体細胞を得た。この菌体を0.2Mクエン酸緩衝液(pH5.5)30mLに懸濁後、超音波破砕し、遠心分離(15000rpm、30分)した。得られたペレットを100mM Tris/HCl、4% Triton X−100、2M 尿素(pH8.0)で懸濁し、30℃で30分インキュベートし、遠心分離(15000rpm、30分)した。次に得られたペレットを蒸留水で懸濁し、遠心分離(15000rpm、30分)した。得られたペレットを凍結乾燥して粗精製封入体120mgを得た。
【0084】
この粗精製封入体50mgを20mM Tris/HCl、8M 尿素、30mM ジチオスレイトール(pH8.0)10mLに溶解し、遠心分離(15000rpm、30分)後、上澄み液を20mM Tris/HCl、10%グリセリン、50μM FAD、1mM 酸化型グルタチオン、1mM 還元型グルタチオン(pH8.0)で100倍に希釈し、4℃で、1週間リフォールディングを行った。
【0085】
リフォールディングの終了した再生液を限外ろ過で濃縮して10U/mLの容量活性の酵素液を作成した。この酵素液1mLに対して1gの固定化担体(多孔性シリカゲル、MB−5D、富士シリシア化学製)を混合して固定化酵素を作成した。t−ブチルメチルエーテルを溶媒に用い、青酸0.15M、ベンズアルデヒド0.1Mの濃度で上述の固定化酵素を触媒として反応を行ったところ、光学純度90%ee以上のR−マンデロニトリルが得られた。
【0086】
比較例1 アーモンド葉由来(R)−ヒドロキシニトリルリアーゼ遺伝子の塩基配列においてシグナルペプチド除去しないDNAの大腸菌での発現
実施例2で記載した配列番号5で表される塩基配列からなるRHNL−KR2について、実施例3に準じて遺伝子組換え大腸菌BL21(DE3)/pET21a(+)/RHNL−KR2を作成し、実施例3に準じて培養を行い、菌体を回収し、超音波破砕し、可溶性上清、不溶性沈殿をSDS−PAGEに供した。結果を図1の1レーン及び2レーンに示す。1レーンは可溶性画分の結果を示し、2レーンは不溶性画分の結果を示す。目的のRHNL−KR2と思われるバンド(61kDa)は検出されなかった。
【0087】
以上から、本発明により(R)−ヒドロキシニトリルリアーゼの発現量が増大されることが明らかとなった。
【図面の簡単な説明】
【0088】
【図1】実施例3及び比較例1におけるSDS−PAGEの結果を示す。
【配列表フリーテキスト】
【0089】
配列番号2〜8:合成DNA
【特許請求の範囲】
【請求項1】
バラ科植物由来(R)−ヒドロキシニトリルリアーゼをコードする遺伝子の塩基配列が改変されてなるDNAであって、該改変が、該DNAが宿主細胞に導入されたときの(R)−ヒドロキシニトリルリアーゼの発現量を改変されていないDNAを導入した場合と比べて増大させるものである、該DNA。
【請求項2】
改変が、以下の(a)〜(c)の工程:
(a)バラ科植物由来(R)−ヒドロキシニトリルリアーゼをコードする遺伝子の塩基配列に含まれるシグナルペプチドをコードする塩基配列を除去すること、
(b)バラ科植物由来(R)−ヒドロキシニトリルリアーゼをコードする遺伝子の塩基配列中のコドンを、そのアミノ酸配列を変化させることなく、宿主細胞におけるアミノ酸ごとのコドン使用頻度が10%以上となるように改変すること、及び
(c)バラ科植物由来(R)−ヒドロキシニトリルリアーゼをコードする遺伝子の塩基配列にタグ配列を付加すること
からなる群から選択される少なくとも1つの工程によって実施されるものである、請求項1記載のDNA。
【請求項3】
配列番号8で表される塩基配列を含み、塩基長が4000塩基以下のDNA。
【請求項4】
請求項1〜3のいずれか1項記載のDNAを含む組換えベクター。
【請求項5】
請求項4記載の組換えベクターを含む形質転換体。
【請求項6】
請求項1〜3のいずれか1項記載のDNAを発現させ、得られた(R)−ヒドロキシニトリルリアーゼとカルボニル化合物及びシアノ基供与体とを反応させ、反応液から光学活性シアノヒドリンを回収することを含む、光学活性シアノヒドリンの製造方法。
【請求項1】
バラ科植物由来(R)−ヒドロキシニトリルリアーゼをコードする遺伝子の塩基配列が改変されてなるDNAであって、該改変が、該DNAが宿主細胞に導入されたときの(R)−ヒドロキシニトリルリアーゼの発現量を改変されていないDNAを導入した場合と比べて増大させるものである、該DNA。
【請求項2】
改変が、以下の(a)〜(c)の工程:
(a)バラ科植物由来(R)−ヒドロキシニトリルリアーゼをコードする遺伝子の塩基配列に含まれるシグナルペプチドをコードする塩基配列を除去すること、
(b)バラ科植物由来(R)−ヒドロキシニトリルリアーゼをコードする遺伝子の塩基配列中のコドンを、そのアミノ酸配列を変化させることなく、宿主細胞におけるアミノ酸ごとのコドン使用頻度が10%以上となるように改変すること、及び
(c)バラ科植物由来(R)−ヒドロキシニトリルリアーゼをコードする遺伝子の塩基配列にタグ配列を付加すること
からなる群から選択される少なくとも1つの工程によって実施されるものである、請求項1記載のDNA。
【請求項3】
配列番号8で表される塩基配列を含み、塩基長が4000塩基以下のDNA。
【請求項4】
請求項1〜3のいずれか1項記載のDNAを含む組換えベクター。
【請求項5】
請求項4記載の組換えベクターを含む形質転換体。
【請求項6】
請求項1〜3のいずれか1項記載のDNAを発現させ、得られた(R)−ヒドロキシニトリルリアーゼとカルボニル化合物及びシアノ基供与体とを反応させ、反応液から光学活性シアノヒドリンを回収することを含む、光学活性シアノヒドリンの製造方法。
【図1】

【公開番号】特開2006−141263(P2006−141263A)
【公開日】平成18年6月8日(2006.6.8)
【国際特許分類】
【出願番号】特願2004−334857(P2004−334857)
【出願日】平成16年11月18日(2004.11.18)
【出願人】(504132272)国立大学法人京都大学 (1,269)
【出願人】(000004628)株式会社日本触媒 (2,292)
【Fターム(参考)】
【公開日】平成18年6月8日(2006.6.8)
【国際特許分類】
【出願日】平成16年11月18日(2004.11.18)
【出願人】(504132272)国立大学法人京都大学 (1,269)
【出願人】(000004628)株式会社日本触媒 (2,292)
【Fターム(参考)】
[ Back to top ]