
1型ウシヘルペスウイルスの欠失突然変異体、それに基づくワクチンおよび1型ウシヘルペスウイルスを検出するためのキット
【課題】牛を1型ウシヘルペスウイルス(BHV−1)から守る為の弱毒性生ワクチン及び不活化ワクチンを提供する。
【解決手段】糖蛋白質gE−遺伝子中に欠失を有する1型ウシヘルペスウイルスの欠失突然変異株。突然変異株は、さらにチミジンキナーゼ遺伝子および/または糖蛋白質gI遺伝子の欠失を有し、非対称遺伝子の挿入を有する。gE−遺伝子またはその一部から成る再結合核酸。糖蛋白質gE、その上に基礎をおくペプチド、糖蛋白質gEとgIとの複合物、それらに対する抗体。これらの物質のいずれか1つから成るワクチンと診断キット。
【解決手段】糖蛋白質gE−遺伝子中に欠失を有する1型ウシヘルペスウイルスの欠失突然変異株。突然変異株は、さらにチミジンキナーゼ遺伝子および/または糖蛋白質gI遺伝子の欠失を有し、非対称遺伝子の挿入を有する。gE−遺伝子またはその一部から成る再結合核酸。糖蛋白質gE、その上に基礎をおくペプチド、糖蛋白質gEとgIとの複合物、それらに対する抗体。これらの物質のいずれか1つから成るワクチンと診断キット。
【発明の詳細な説明】
【技術分野】
【0001】
発明の分野
本発明は、病原体によって引き起こされる疾病に関係したワクチン注射および診断法の分野に関するもので、弱毒性ワクチンまたは不活化ワクチンを生成する古典的な方法とDNA組換技術に基づく近代的方法両方の使用に関係している。
より詳細には、本発明は、動物、特に牛を1型ウシヘルペスウイルス(BHV-1)から守るための弱毒性生ワクチンおよび不活化ワクチンに関するもので、これらのワクチンは、安全でかつ効能があるばかりでなく、ワクチン接種をうけた群における、非感染動物と感染動物とを識別することも可能にする。
ワクチン接種群における非感染動物と感染動物の識別のための試験に使用が可能である診断キットも本発明の一部である。
【背景技術】
【0002】
発明の背景
牛の伝染性鼻腔気管炎ウイルス(IBRV)および伝染性濃外陰部腔炎ウイルス(IPVV)を含むBHV-1は、牛の呼吸器疾患、生殖能力、障害の解明に重要な役割を果たす。BHV-1は、急性感染後、潜伏した形で宿主内にそのまま残っていることが多い。潜伏ウイルスはとりわけ、ストレス(これは臨床現象を伴うことも、伴わないこともある)の影響下で再び活性化し、続いて排泄されることがあり得る。結果として、感染した牛は終生BHV-1を蔓延するものとみなす必要がある。BHV-1はオランダの養牛場の推定75%において風土病として発生している。特に、高齢牛の血清反応が陽性である。
【0003】
BHV-1感染予防接種に利用される多くの不活化("死んだ")ワクチンおよび弱毒("生きた")ワクチンがある。不活化ワクチンは、たとえば熱処理や照射、またはエタノールやホルマリン処理によって、BHV-1ウイルスを死滅させて作られる。しかしながら、これらによっては不充分な予防しか行えないことが多い。弱毒ワクチンは、同種(ウシ)細胞またはブタの細胞やイヌの細胞のような異種細胞への多数回にわたる継代によって作られ、その後ウイルスは物理的または化学的にも処理されることもある。このようにして、未知の突然変異/欠失がウイルスゲノム中に発達し、これがしばしばウイルスの疾病誘発特性を低下させる。弱毒ワクチンは不活化ワクチンよりも優れた予防を行うが、これは特に宿主の免疫系に、より多くのウイルス性抗原を与えるからである。生きたワクチンの重要な他の利点として、それらは鼻腔内投与が可能であるということがあり、すなわち、ここは感染後に野性型ウイルスの増殖が初めて起こるところである。生きたワクチンはまだ改良の余地がある。いくつかの生きたワクチンはさらに、流産を引き起こす可能性を有しているように思われ、これは特に筋肉内投与後に顕著になる。さらにまた、生きたワクチンはすべて、ワクチンを接種された牛に潜伏して残っていると思われる。また、該ワクチンが野性型ウイルスとほとんど相違するところがなければ、病原性への復帰が起こる可能性もある。しかし、重要な問題の1つは、BHV-1ワクチンは、野性型ウイルスによる感染を予防することができないということである。その結果、ワクチン接種を受けた牛も野性型BHV-1を蔓延させる可能性がある。
【0004】
適切なBHV-1抑制計画のためには、野性型ウイルスと識別することができる効果的かつ安全なワクチンを自由に使えることが必要である。何故ならば、効果的ワクチンの利用によりBHV-1の蔓延をかなりの程度低下させることができ、ワクチンと野性型ウイルスの識別が可能な試験によって、ワクチン接種を受けた牛の群より感染した牛を探りあて、それを取り除くことが可能となるからである。
【0005】
ところで、従来のワクチンよりも一層安全と思われる野性型ウイルスとの識別が可能なBHV-1ワクチンの開発が行われてきた。流産を引き起こす程度もより低く、潜伏の頻度も低く、再活性化され得ることもないチミジンキナーゼ欠失変異株が単離された。さらに、組換えDNA技術を用いて、(血清学的技術により野性型BHV-1とワクチンの区別を可能にする)糖蛋白質gIIIのための遺伝子に欠失を有するBHV-1ワクチンが構成された。しかしながら、これらのワクチンに対してもさらに幾つかの異論がある。一方では、チミジンキナーゼ遺伝子はウイルス複製に関与しており、複製の程度が低い程、より程度の低い防御につながる可能性がある。他方、該糖蛋白質gIIIは防御抗体の生成に重要であり、この防御抗体は、gIII欠失ワクチンの効果を低下させる。実際上の問題として、一般には最高の防御が得られる、組換えワクチンの鼻腔内投与が幾つかの国では認可されていないという問題がある。したがって、有効かつ安全で、さらに野性型BHV-1と区別することが可能なワクチンが求められており、さらにまたこれは、かかるワクチンの少なくとも1つは、組換えDNA技術によって構成されたウイルスよりも、むしろ従来の方法で弱毒化されたウイルスに基づいたものであることが望ましい。
【0006】
ところで、細胞培養における継代によって、糖蛋白質gEのための遺伝子を欠くBHV-1菌株が得られた。我々が行った研究の最初の結果は、この遺伝子は、野性型BHV-1に関する血清学的識別に非常に役立ち、病原性の発原に関与していることを示している。したがって、その欠失は安全性に寄与しており、これによってチミジンキナーゼ欠失の使用は不要となるかもしれない。糖蛋白質gEはく防御の誘導にとって糖蛋白質gEほど重要ではないと思われる。従来の方法で弱毒化された野性型ウイルスと血清学的に識別することが可能な、従来の方法で弱毒化されたBHV-1菌株は独自のものである。ここで初めて述べた、gE-遺伝子の位置とDNA配列は、これまでは知られていなかった。また、これから誘導することができるオリゴヌタレオチドポリペプチドおよびオリゴペプチドも知られていなかった。また、gE-遺伝子に基づく血清学的識別を行うための試験も独自のものである。
【0007】
この"従来の"gE欠失変異株の重要な利点("従来の"とは従来の弱毒ウイルス単離方法を使用することを意味している)は、親換えワクチンに関する限りでは、鼻腔内投与が禁止されている国々において、この"従来の"gE欠失変異株を鼻腔内投与することが可能になるであろうという点である。しかしながら、安全に関して別の観点からよく考慮して、この従来のgE欠失ワクチンの他に、明確に限定された組換えの型も構成された。これらの組換えワクチンもまた欠失を有しており、-また、チミジンキナーゼ遺伝子の欠失も有していてもよく、または有していなくてもよい-および、これらの組換えワクチンは、異種の遺伝子の発現のためのベクタとして用いることもできる。これらの組換えワクチンは全て、同じgE特有試験によって、野性型ウイルスと区別することができる。一組の異なるワクチンに対して標準的な試験を利用することは、国際的な努力としてのBHV-1の戦いにおいて強力な利点となり得る。かかる方法がBHV-1ワクチンの分野において述べられたことはない。
【0008】
牛における抗BHV-1反応の血清分析によって、抗gE抗体の重要部分は、糖蛋白質gEおよび他のBHV-1糖蛋白質である糖蛋白質gIによって形成される複合物に対して向けられていることが示された。したがって、かかる複合物特有の抗体の存在を示すこともできる血清試験は、抗-gE抗体を検出することができるだけの試験よりも高感受性であると思われる。gE単欠失変異株によりワクチン接種を受けた牛は、抗-gI/gE抗体の検出を妨げることが有り得る抗-gI抗体を作る可能性がある。したがって、この発明はgI/gE二重欠失を有するワクチンを含む。
【発明の開示】
【0009】
発明の要約
本発明は、糖蛋白質gE-遺伝子に欠失を有するBHV-1の欠失変異株を提供する。"〜に欠失を"という語は、全体としての遺伝子の欠失を含むことを意図している。本発明の好適な実施例は、以下に述べる欠失変異株Difivac-1のような、弱毒処置によって引き起こされた糖蛋白質gE-遺伝子における欠失を有するBHV-1の欠失変異株によって構成される。
【0010】
本発明の他の好適な実施例は、以下に述べる欠失変異株1B7または1B8のような組換えDNA技術によって作られた糖蛋白質gE-遺伝子に欠失を有しているBHV-1の欠失変異株から成る。
【0011】
本発明の他の好適な実施例は、以下に述べるgI、/gE二重欠失変異株Difivac-IEのような、糖蛋白質gE-遺伝子および糖蛋白質gI一遺伝子に各々欠失を有するBHV-1の二重欠失変異株から成る。
【0012】
さらにまた、最高の安全性を期して、本発明によれば糖蛋白質gE-遺伝子およびチミジンキナーゼ遺伝子のそれぞれに欠失を有するBHV-1の欠失変異株が好適である。また本発明は、糖蛋自質gE-遺伝子、糖蛋白質gI-遺伝子、チミジンキナーゼ遺伝子に欠失を有するBHV-1の欠失変異株も含んでいる。
【0013】
本発明は、動物を、特に哺乳動物を、さらに特にウシ科の動物を上記において明確に述べたBHV-1欠失変異株を有するBHV-1から守るために、これらにワクチン接種をするためのワクチンの組成、適切な担体またはアジュバントを提供する。前記構成は、生きたワクチン組成でも不活化ワクチン組成でもよい。
【0014】
本発明はさらに、糖蛋白質gE-遺伝子に欠失を有し、組換えDNA技術によって導かれた異種遺伝子を含むBHV-1の変異株において実施されている。好適には、糖蛋白質gE-遺伝子の位置に、組換えDNA技術によって導かれた異種遺伝子を含むBHV-1の変異株に関するもので、前記異種遺伝子は、gE-遺伝子の調節配列の管理下にあり、シグナルペプチドに関してコード化するgE-遺伝子の一部に任意に付加される。前記異種遺伝子は、BHV-1の異なるプロモータの管理下、または異種プロモータの管理下にあることも可能である。BHV-1の変異株が、糖蛋白質gE-遺伝子における欠失に加えて、さらに、チミジンキナーゼ遺伝子における欠失、および/または糖蛋白質gI-遺伝子における欠失などの欠失を有している場合には、前記異種遺伝子を、この付加欠失の位置に挿入してもよい。複数の挿入も別の選択肢であり、1つの欠失の位置にまとめて行うか、幾つかの欠失の位置に分けて行うかされる。
【0015】
好適に導かれた異種遺伝子は、別の病原体の免疫原性蛋白質またはペプチドについて、または免疫反応を促進するサイトカインについてコード化する。適切なサイトカインの例としてはインターロイキンー2、αインターフェロン、γインターフェロンがある。
【0016】
本発明はまた、動物を、特に哺乳動物を、さらに特にウシ科の動物を(異なる)病原体より守るため、これらにワクチンを接種するための、(生きた、または不活化)ワクチンの組成を提供するもので、該ワクチンの組成には、免疫原性蛋白質または他の病原体ペプチドについてコード化している異種遺伝子を有しているBHV-1突然変異株およびアジュバントの適切な担体が含まれる。勿論、この防御は1つ以上の病原体に関するものでよく、すなわち、変異株が複数の異種遺伝子を含む多価ワクチンに関するものでよい。
【0017】
さらにまた本発明は、BHV-1の糖蛋白質gE-遺伝子、この糖蛋白質gE-遺伝子の一部、またはこの糖蛋白質gE-遺伝子から誘導されるヌタレオチド配列を含む組換え核酸を含む組成に関する。この組成は、BHV-1の糖蛋白質gE-遺伝子、この糖蛋白質gE-遺伝子の一部、この糖蛋白質gE-遺伝子から誘導されたヌクレオチド配列を含む組換え核酸の挿入を有するクローニングベクタまたは発現ベクタを含むことができる。
【0018】
また本発明は、BHV-1の糖蛋白質gE、この糖蛋白質gEの一部、この糖蛋白質gEから誘導されたペプチド、または糖蛋白質gEおよびgIの複合物を含む組成、およびBHV-1の糖蛋白質gE、この糖蛋白質gEの一部、この糖蛋白質gEから誘導されたペプチド、または糖蛋白質gEとgIの複合物に特有の抗体を有する組成を含む。"抗体"とは、ポリクローナル抗体調製物と、大半の用途に好適なモノクローナル抗体の両方を意昧していると解釈される。"糖蛋白質gEの一部"および"糖蛋白質gEから誘導されるペプチド"という用語は、一般に少なくとも約8アミノ酸の長さを有するであろうgE特有のアミノ酸配列を意味していると解釈される。
【0019】
さらに本発明は、試料中の、特に、動物、特に哺乳動物、さらに特にウシ科の動物の、血液または血清、血液細胞、乳、涙などの体液、肺洗浄液、鼻水、精液、特に精漿、唾液、痰、または組織、特に神経組織のような生物学的試料中の、BHV-1の核酸を検出するための、BHV-1の糖蛋白質gE-遺伝子から誘導されたヌクレオチド配列を有する核酸プローブまたは核酸プライマを含む診断用キット、および核酸検出試験に適した検出手段に関する。
【0020】
さらに本発明は、試料中の、特に、動物、特に哺乳動物、さらに特にウシ科の動物の血液または血清、血液細胞、乳、涙のような体液、肺洗浄液、鼻水、精液、特に精漿、唾液、痰、または組織のような生物学的試料中の、BHV-1に特有の抗体を検出するための、BHV-1の糖蛋白質gE、この糖蛋白質の一部、この糖蛋白質gEから誘導されたペプチド、または糖蛋白質gEおよびgIの複合物を含む診断用キット、および抗体検出試験に適した検出手段に関する。該診断用キットは、さらに、BHV-1の糖蛋白質gEに特有の、またはBHV-1の糖蛋白質のgEとgIの複合物に特有の、1つまたはそれ以上の抗体を含めてもよい。
【0021】
また、本発明は、試料中の、特に、動物、特に哺乳動物、さらに特にウシ科の動物の血液または血清、血液細胞、乳、涙などの体液、肺洗浄液、鼻水、精液、特に精漿、唾液、痰、または組織、特に神経組織のような生物学的試料中の、BHV-1の蛋臼質を検出するための、BHV-1の糖蛋白質gEに特有の、またはBHV-1の糖蛋白質gEとgIの複合物に特有の1つまたはそれ以上の抗体を含む診断用キット、および蛋白質検出試験に適した検出手段に関する。
【0022】
さらに本発明は、BHV-1の糖蛋白質gE-遺伝子を含む核酸の存在、BHV-1の糖蛋白質gE、またはBHV-1の糖蛋白質gEとgIの複合物の存在、またはBHV-1の糖蛋白質gEに特有の、またはBHV-1の糖蛋白質gEとgIの複合物に特有の抗体の存在について、動物の、特に哺乳動物の、さらに特にウシ科の動物のBHV-1感染を判断するための、前記動物の試料の、特に、血液または血清、血液細胞、精液、特に精漿、唾液、痰、涙などの体液、肺洗浄液、鼻水、乳、または組織、特に神経組織のような生物学的試料の試験を含む方法を提供する。試験を行う試料は、本発明に従った組成のワクチンをそれまでに接種されたことのない動物のもの、または本発明に従って調製されたワクチンをそれまでに接種されたことのある動物のものとすることができる。
【0023】
発明の詳細な説明
本発明は、生ワクチンおよび不活化ワクチン両方から成る一組のBHV-1ワクチンに関するもので、これらはどちらも糖蛋白質gE-遺伝子の全部または一部を欠いている。この組は、自然gE欠失変異株も、チミジンキナーゼ遺伝子および/または糖蛋白質gI-遺伝子の欠失を含むことも含まないこともある構成gE欠失変異株も、および異種遺伝子ベクタとして使用される構成gE欠失変異株も含む。さらに本発明は、BHV-1糖蛋白質gE-遺伝子をコード化するヌクレオチド配列、これらの配列から誘導されるオリゴヌタレオチド、該糖蛋白質gEそれ自身、そこから誘導されるペプチド、および該gE糖蛋白質とそこから誘導されるペプチドに対して向けられる(モノクローナルまたはポリクローナル)抗体に関する。また本発明は、BHV-1の糖蛋白質gEとgIの複合物、およびかかる複合物に対して向けられる抗体に関する。
本発明に従ったこれらの物質は、以下のために使用することができる。
【0024】
1)BHV-1によって引き起こされる疾病を予防するための牛のワクチン接種。
これはBHV-1に感染した動物と、ワクチン接種した動物の間の区別が可能な従来のワクチンおよび構成ワクチンを一緒に使用することが可能。
【0025】
2)BHV-1による疾病および防御抗原に対するコード化配列をBHV-1欠失変異株の中に組込むことができる他の病原体により引き起こされる疾病を予防するための牛のワクチン接種。
【0026】
3)牛の血液、血清、乳、他の体液について、それらが野性型BHV-1によって感染しているか、gE欠失変異株でワクチン接種されているかを、血清学的に、または核酸検出技術(たとえばPCR)により判断するためのそれらの試験。
【0027】
オリゴペプチド、ポリペプチド、およびBHV-1の糖蛋白質g-遺伝子と糖蛋白質gI-遺伝子のコード化配列より誘導される糖蛋白質の合成。
糖蛋白質gE-遺伝子(図3A)およびこの遺伝子についてコード化する単離DNAフラグメントについての、例に記載のDNA配列分析の結果により、標準的な分子生物学的処置を用いて、gE蛋白質のペプチド(オリゴまたはポリペプチド)を合成すること、および原核的方法(バクテリア中)または真核的方法(たとえば、ネズミの細胞において)により、gE蛋白質の全体を、またはその一部を表すことが共に可能となる。これらの方法によって、たとえばgE特有のモノクローナル抗体の生成に役立つことができるgE特有の抗原を得ることができる。さらに、gE特有の抗原(および、gE特有のモノクローナル抗体)を血清試験において使用することができ、BHV-1 gE欠失ワクチンの接種をした動物と野性型BHV-1ウイルスに感染した動物の間の区別が可能となる。
【0028】
例に記載の、糖蛋白質gI-遺伝子、およびこの遺伝子についてコード化する単離DNAフラグメントについて、糖蛋白質gEを表す真核細胞と共に、部分的にDNA配列を分析した結果、真核細胞中のgI/gE複合物を表すことができる(図13および図14参照)。この糖蛋白質複合物は、gI/gE特有のモノクローナル抗体を作るために使用することができる。また、このgI/gE複合物は、単gE BHV-1欠失変異株、または二重gI/gE BHV-1欠失変異株によるワクチン接種を受けた牛と野性型BHV-1ウイルスに感染した牛の区別をする血清試験における抗原としても使用することができる。
【0029】
gE特有ペプチド
既知の蛋白質コード化配列に基づき、自動合成装置により、約40〜50のアミノ酸のポリペプチドを作ることができる。BHV-1菌株LamのgE糖蛋白質の蛋白質コード化配列が解明されたので(図3A)、このBHV-1 gE糖蛋臼質のポリペプチドの合成が可能となる。かかるポリペプチドを用いて、標準的な方法に従って、マウスやウサギなどの実験動物を免疫化し、gE特有抗体を作ることができる。さらに、これらのgE特有ペプチドを用いて、抗gE抗体がgE蛋白質(エピトープ)と反応する位置を、たとえばPEPSCAN法(Geysen et a1.,1984,Proc.Natl.Acad,Sci.USA 81,3998〜4002)により、さらに特定することができる。gE特有のオリゴペプチドも抗gE抗体を実証する血清試験において使用することができる。
【0030】
gEの原核発現
細菌中にてgE蛋白質を合成するため(すなわち、gEの原核発現)、糖蛋白質gEについて、またはその一部についてコード化するDNAフラグメントを、原核発現ベクタ中にタローン化しなければならない。原核発現ベクタは、環状DNA分子で、これは別個に複製してゆく分子(プラスミド)として、細菌中で自らを維持することができる。これらの発現ベクタは、抗性物質抵抗についてコード化を行い、したがって発現ベクタを伴う細菌の選択を可能にする。1つ、またはそれ以上の標識遺伝子を含む。さらに、発現ベタタは、(制御可能であることも多い)プロモータ領域を有しており、該領域の背後にDNAフラグメントを結合することができ、これらのフラグメントは、プロモータの影響により発現される。現行の多くの原核発現ベクタにおいては、いわゆる担体蛋白質に融合し、所望の蛋白質が発現している。そのために、ベクタ中ではプロモータの後に、担体蛋白質についてのコード化配列が位置しており、所望のDNAフラグメントがこれに直接隣接して結合されている。融合蛋白質は、多くの場合より安定で、認識および/または単離もより簡単である。一定の細菌株にて、特定の融合蛋白が達成することができる定常状態レベルは融合によって、および菌株によって異なっている。種々の組合わせを試みることが慣例になっている。
【0031】
糖蛋白質gE-遺伝子の真核発現
蛋白質の原核発現は幾つかの利点をもたらしてくれるが、蛋白質は真核細胞で起こるグリコシル化のような修飾変更を欠いている。その結果、真核発現蛋白質は多くの場合より適切な抗原である。マウスの細胞のような真核細胞における蛋白質の異種発現の場合、真核発現ベクタが使用される。これらのベクタは、プラスミドで大腸菌細胞内で増殖させることが可能であるばかりでなく、真核細胞中にて安定した状態で生存することもできる。これらのベクタは、原核選択マーカの他に、真核選択マーカも含む。原核発現ベクタに似て、真核発現ベクタは、所望の遺伝子が後に結合することができるプロモータ領域を含んでいる。しかしながら、真核ベクタにおけるプロモータ配列は、真核細胞に特有である。また、真核ベクタにおいては、担体蛋白質に対する融合が利用されることは滅多にない。これらのベクタは、標準的なトランス・フェタション法(F.L.Graham and A. J. van der Eb. 1973, Virology 52,456〜467)により真核細胞中に導かれる。真核プラスミドベクタの他に、ウィルスベクタもあり、この場合には、異種遺伝子がウィルス(たとえば、レトロウィルス、ヘルペスウィルス、ワクシニアウィルス)のゲノム中に導かれる。次いで、真核細胞に組換えウィルスを感染させることができる。
一般に、個々の遺伝子産物にとって、ベクタや細胞はどういったタイプのものが最適であるかということを予測することはできない。大抵の場合、幾つかの組合せが試みられている。
【0032】
糖蛋白質gEおよび糖蛋自質gI両方の真核発現蛋白質が得る最終構造は、蛋白質の一次アミノ酸配列、折たたみ、翻訳後修飾等に依存している。蛋白質の構造に寄与する重要なファクターは、1つまたはそれ以上の他の蛋白質との相互作用である。BHV-1糖蛋白質gEは、少なくとも1つの他の糖蛋白質、すなわちBHV-1糖蛋白質gIとの複合物を形成することを我々は発見した。かかる複合物を初めて示したものは、候補的抗-gEモノクローナル抗体1,51,67,75,78による我々の結果であった(表2参照)。これらのモノクローナル抗体は、Difivac-1とも、またLam gE-とも反応しなかったばかりでなく、糖蛋白質gE発現3T3細胞を認識することもできなかった。しかしながら、これらのモノクローナル抗体は、Difivac-1に感染後はgE-発現3T3細胞とは反応せず、これらのモノクローナル抗体のための適切な抗原コンホメーションを糖蛋白質gEに与えるために、因子を補足することが必要であることを示した。我々が行ったモノクローナル抗体81との放射線免疫沈澱試験の幾つかにおいて、見かけ上の分子量63KDの蛋臼質が共沈することが判った。ヘルペス単ウィルス糖蛋白質gEは、これに匹敵する分子量の蛋白質(HSV1糖蛋白質gI)と複合物を形成するという事実に鑑み、我々はBHV-1糖蛋白質gEは、糖蛋白質gIのBHV-1同族体と複合物を形成するものと推定した。このBHV-1 gE/gI複合物を研究するため、および適切な抗原構造を有するgE抗原を作るため、これらの両蛋白質を1つの真核細胞中で発現させた。このために、糖蛋白質gEだけの真核発現のために記載したものと同じ処置を適用した。ただし、前提条件として、選択可能な種々の真核標識を有する発現ベクタの使用を1つだけ追加した。
【0033】
血清試験
gEに対する抗体に基づき、Difivac-1のワクチン接種をされた牛と、野性型BHV-1に感染した牛を区別するための血清学的方法は、好適にはgEに対して向けられたモノクローナル抗体の使用に基づく。これらの抗体は、以下の方法で用いることができる。
【0034】
a)Van Oirschot et al(Journal of Virological Metbods 22,191〜206,1988)により述べられた原則による。Aujeszky病のウイルスに対するgI抗体検出のための、この酵素結合イムノソルベント検定法(ELISA)では、抗体は、gI上に2つの異なったエピトープを有する2つのモノクローナル抗体の反応に対するブロック効果によって実証される。この試験は、以下のように実施される。マイタロタイタープレートを37℃で1晩、モノクローナル抗体1で被覆し、その後、たとえば4℃または-20℃にて保存する。試験される血清は、別の未被覆マイクロタイタープレートにおいて、37℃にてたとえば2時間、抗原と共に前保温される。モノクローナル抗体1が被覆されたプレートを、たとえば5回洗浄し、その後セイヨウワサビペルオキシダーゼ(HRPO)と結合したモノクローナル抗体2をこれらのプレートに添加する。次に、前保温された血清-抗原混合物を、2つのモノクローナル抗体が置かれているプレートに転移させ、37℃にてたとえば1時問保温する。これらのプレートを洗浄し、各ウェルに基質を添加する。室温にて約2時間経過させた後、これらのプレートについてスペクトル光度計による読取りを行う。各プレートには、4つの陰性対照血清および陽性血清の4つの連続希釈物が含まれる。光学濃度が同じプレートで試験された4つの陰性対照血清の平均光学濃度の50%を下回る血清は陽性とみなされる。
【0035】
b)間接2重抗体サンドイッチ(IDAS)法による。
この場合は、マイクロタイタープレートは、gE蛋白質に対して向けられたモノクローナルまたはポリクローナル血清で被覆する。gE抗原調製物との保温の結果、被覆にgEが結合する。試験される牛の血清中のgEに対して特別に向けられた抗体は続いてgEに結合する。これらの結合抗体は、抗-ウシ免疫グロブリン複合物によって認識される。この複合物中の抗体は、ペルオキシダーゼ酵素に共有結合する。最後に、この結合複合物は、色素原基質の添加により、可視のものにされる。この反応の特性は、gE-抗原調製物の代わりに、gE-陰性対照調製物により同じ処置を実施して調べる。各マイクロタイタープレート上には、陽性および陰性の対照血清が含まれる。この試験は、陽性の血清が一定の希釈度において陽性を示す場合に有効である。光学濃度が標準陰性対照血清よりも0.2高い場合には、血清は陽性である。
【0036】
c)2において述べたIDAS法では、ただし試験される血清を保温した後であるが、抗-ウシ免疫グロブリン複合物の代わりに、抗-gEモノクローナル抗体/HRPOが使用される。抗-gEペプチド血清または抗-gEポリクローナル血清を抗-gEモノタローナル抗体の代わりに使用してもよい。これらのプレートを洗浄し、各々のウエルに色素原基質を添加する。室温にてたとえば、約2時間経過させた後、これらのプレートについてスペクトル光度計による読取りを行う。各プレートには、4つの陰性対照血清および陽性血清の4つの連続希釈物が含まれる。光学濃度が同じプレートで試験された4つの陰性対照血清の平均光学濃度の50%を下回る血清は陽性とみなされる。
【0037】
d)精製、未精製を問わないウイルス抗原がマイクロタイタープレートに一晩被覆されるブロックELISA法による。これらのプレートにおいて、試験される血清は、37℃にてたとえば1時問または1時間以上保温される。洗浄処置をした後、これらのプレートに抗-gEモノクローナル抗体を添加し、37℃にてたとえぱ1時間保温する。抗-gEペプチド血清または抗-gEポリクローナル血清を抗-gEモノクローナル抗体の代わりに使用してもよい。これらのプレートを洗浄し、各ウエルに、色素原基質を添加する。室温にて、たとえば2時間経過させた後、これらのプレートについて、スペクトル光度計による読取りを行う。4つの陰性対照血清および陽性血清4つの連続希釈物が含まれる。光学濃度が、同じプレートで試験された4つの陰性対照血清の平均光学濃度の50%を下回る血清は陽性とみなされる。
【0038】
上記の方法のいずれにおいても、gEを含む、従来の方法で成長させたウイルス抗原を使用することができるが、原核また真核を介して発現するgE-抗原も使用することができる。あるいはまた、BHV-lgE配列に基づくオリゴペプチドを従来の抗原の代わりに、上記診断試験において使用することができるであろう。さらにまた、かかるオリゴペプチドを、Kemp et al.による記事、Science 241,1352〜1354,1988に記載の方法に従った、いわゆる"Cow-side"試験の開発に使用することができるであろう。かかる試験は、感染動物中に存在しているgEに対して向けられている抗体によるオリゴペプチド抗原配列の連結に基づいたものになるであろう。このような試験の場合、オリゴペプチドを、牛の赤血球に対して向けられたモノクローナル抗体に連結させる必要があるであろう。
【0039】
ポリメラーゼ連鎖反応を使用する核酸分析
たとえば、オリゴヌクレオチド(プローブおよびプライマ)をポリメラーゼ連鎖反応に用いて、ワクチン接触を受けた動物と感染した動物を区別することができる。ポリメラーゼ連鎖(PCR)は、病原体の核酸を短時間に数十億倍に増殖させることができる技術である(Depolymerase kettingreactie. P.F,Hilderink, J.A.Wagenaar,J.W.B.van der Giessen and B.A.M. van der Zeijst,1990,Tijdschrift voor Diergeneeskunde deel 115,1111〜1117)。gEオリゴヌタレオチドは、gE陽性ゲノムにおいて、gE陰性ゲノムにおけるものとは異なる生成物が形成されるように選択することができる。これは、gE欠失ワクチンをワクチン接種された動物は、RCR試験において陽性信号を出すという利点がある。しかしながら、この方法は、試験される動物から採取した試料中、たとえば血液中のウイルスの核酸の存在に依存している。
BHV-1急性感染後は、血液中にBHV-1特有の核酸を実証することができる見込みは充分にあるが、しかしながら、BHV-1核酸も、潜伏時に血清中に実証することができるかどうかはまだ確められていない。
【0040】
ベクタとしてのBHV-1の使用
BHV-1ゲノム中の異種遣伝子の発現には、その異種遺伝子が挿入されるべき領域についての正確な情報を自由にできることが必要である。必須配列についての妨害はいかなるものもあってはならず、また異種遺伝子の発現に調節配列が利用できなければならない。gE-遺伝子は、必須ではなく、したがって、異種遺伝子gE遺伝子を交換しても問題はない。結果として、この異種遺伝子を、gE遺伝子が調節配列の影響下にあるように配置することができる。しかしながら、gE遺伝子の調節配列を使用する必要はない。異種遺伝子の発現は、代わりに、他のもの、たとえば種々の遺伝子のより強い調節配列により制御することができる。また、gE遺伝子の(輸出)信号ペプチドに異種遺伝子を連結することも可能であり、したがって異種遺伝子生成物の分泌に影響させることができる。gE遺伝子およびgE蛋白質についての詳細な知識によって、非常に適切に調整された方法で、ベクタとしてBHV-1を用いる可能性が与えられることは明らかである。発生するベクタは、さらに野性型と血清学的に区別することができる。異種遺伝子を発現するBHV-1変異株の構成は、例に示したgE欠失変異株の構成と同じ方法で実施することができる。しかしながら、次に、欠失フラグメントは、異種遺伝子が欠失位置に配置されているフラグメントと交換すべきである。
【0041】
例
1)自然gE欠失変異株の単離および同定
a)自然変異株の単離
ゲノムDNAを、従来法により弱毒化された多くのワクチンから標準的方法に従って単離し、制限酵素を用いて分析した。特に、野性型BHV-1ウイルスとの区別を可能とするに適切と思われるゲノム偏差について研究した。
特に、BHV-1ゲノムのUs領域に注意した。これは、その領域に-ヘルペス単ウイルス1との類似により-非必須糖蛋白質に対する多くの遺伝子コーディングが配置されているからである〔Identification of a herpes simplex virus1 glycoprotein gene within a gene cluster dispensable for growth in cell culture, R. Longnecker,S.Chatterjec,R.J. Whitley and B.Roizwan(1987)Proc、Natl. Acad. Sci,84,4303〜4307〕。
ユーゴスラビア、サグレフ大学より入手したBHV-1ワクチンバッチ(Lugovic et a1.,Veterinarski Arhiv 55,241〜245,1985)は、ウシの胚腎臓細胞およびウシの胚気管細胞(Ebtr)上で多くの継代を行った後に、正常なUs領域の他に、偏差のあるUs領域を有していることを証明した。さらにこのワクチンは、Ebtr細胞に大きなプラークと小さなプラークの両方を形成するように思われた。この混合群より、それぞれの国ごとに選択された小さなプラークで偏差のあるUs領域を有するウイルスを、3つの限界希釈ステップによって単離した。この経路を介して単離されたウイルスをさらに試験し、Difivac-1を希釈した。これは、1992年5月27日Institut Pasteur,Paris,Franceに寄託番号1-1213として寄託された。
【0042】
b)Difivac-1におけるgE遺伝子の欠失の同定
US領域におけるこの偏差をさらに分析するために、Difivac-1のゲノムDNAを標準的な方法で単離し、サザンブロット分析を行った(図1A)。32Pのラベルを貼った野性型HindIIIKフラグメントを有するこのブロットのハイブリッド形成により、Us領域の中心に位置しているこのフラグメントは、Difivac-1においては、1,0キロベース(kb)程度短い。さらに、この分析によって、欠けている部分の位置をおおよそ推定することができた(図1B)。この欠失をさらに分析するために、野性型BHV-1菌株LamのUs領域を単離し、原核ベクトル中にクローン化した。そのために、標準的な方法に従って、Lam菌株(図2A)のゲノムDNAを単離し、ベクタ、pUC18、pACYCおよびpBR322中にクローン化した(図2B)。欠失の推定位置周辺の領域の物理的地図を作成した(Fig.2c)。この物理的地図からスタートして、この領域のヌクレオチド配列の決定に適したサブクローンをベクタpKUN19とpUC18で構成した(図2D)。これらのサブクローンを用いて、全領域の2つの鎖のヌクレオチド配列(図2Cに指示)をサンガー法を用いて決定した。このヌクレオチド配列(SEQ.ID.NO.1)をPc、/Geneプログラムを用いて分析した。概念翻訳から、ヌクレオチド(nt)168〜nt1893が575のアミノ酸のオープンリーディングフレームのための遺伝暗号を指定する(図3A)ものと推定された。さらに分析の結果、このアミノ酸配列は図3Bに示されているように、貫膜糖蛋白質の特性を有していることが示された。実は、最初の26のアミノ酸(aa)は、典型的真核性の輸出信号として認識され、aa423とaa450の間の領域は、貫膜領域として認識される。さらに、3つのポテンシャルN-結合、糖鎖形式部位がこの配列において発生する。この推定アミノ酸配列は、ヘルペス単ウイルス(HSV)の糖『蛋白質gE-遺伝子に明らかに類似したところを示している。(図4Aおよび図4B参照)。これらおよびその他の類似点により、見つけられた遺伝子は、BHV-1のgE同族体であるという結論を正当化するものである。この理由から、遺伝子はgEと呼ばれる。このBHV-1 gE-遺伝子がDifivac-1においてどの程度欠失しているのかを判断するために、p318フラグメントを単離した。p318フラグメントは、要求されるBHV-gEがリーディングルームを開く前に、Alul部位55ntで開始し、リーディングルームの後の133ntを終了する。ゲノムDifivac-1DNAを、サザンブロットハイブリッド形成を用いて、このp318フラグメントで分析した。この結果、Difivac-1は検出可能なp318配列を全く含んでいないことが明らかとなった(図5)。この試験によりDifivac-1は欠失を有し、この欠失はgE遺伝子全体に広がっていることが明らかに実証されていることが確認された。
【0043】
欠失領域の大きさと位置を決定するために、Difivac-1のUs領域を包括しているゲノム配列を原核ベクタ中にクローン化した(図11Cを参照)。14.5kb EcoRIフラグメントをpACYCベクタ中にクローン化し、P775と命名した。7.4kb HindIIIフラグメントを独自にpUC18ベクタ中にクローン化し、p728と命名した。クローンp728より、2つのサブクローンを単離した。すなわち、クローンp737中の1.4kbPst1フラグメントとクローンp754中の350bp AluI-Pst1フラグメントである。これらのクローンの制限酵素分析とサザンブロット分析(データは示されていない)により、Difivac-1のgE欠失の長さは2.7kbであり、gE遺伝子から丁度5'でスタートし、Us領域の境界で終了することが実証された。これらの2.7kbは、反復領域の逸脱拡張として、gE遺伝子に対向したUs領域に位置した重複1kbセグメントによって置きかえられた(図11bを参照)。この分析の結果を確認するために、および正確な組換え点を決定するために、クローンp754の大半の挿入についてのヌクレオチド配列を決定し、野性型配列を比較した(図12参照)。この分析により組換え点は、gE遺伝子の出発コドンから77bp上流に位置していることが判った。
【0044】
c)Difivac-1の安全性と効能についての評価
7週齢のBHV-1血清反応陰性の特有の病原体をもっていない牛でDifivac-1を試験した。8体の牛に2ml,105TCID50の鼻腔内ワクチン接種を行った。2ml中の1mlは各々の鼻孔にスプレーした。各々別々の独立した区画に収容されていた、BHV-1血清反応陰性の特有病原体を持っていない7週齢の8体の牛に2mlの培養基を鼻腔内投与し、非ワクチン接種対照として用いた。ワクチン接種から5週後に、ワクチン接種牛と非ワクチン接種牛に107TCID50、の強毒性BHV-1菌株、Iowaを鼻腔内接種した。接種から6週後に推定される潜伏ウイルスを再活化させるため、全牛に5日間デキサメタゾンにより筋肉内処理を行った。臨床微候、直腸温、発育を監視した。布で拭き取った鼻水からウイルスの単離を行い、血清中の中和抗体力価を求めた。
【0045】
ワクチン接種後の、牛の挙動、食欲、直腸温、発育率は正常のままであったが、ワクチンを接種した牛は、いくらか漿液性鼻水の分泌と唾液の分泌過多があった。Difivac-1は、ワクチン接種後、布で拭き取った鼻水から排泄された(図17)。ワクチン接種牛は全て、BHV-1に対する中和抗体を生成した。
菌株接種後、14日までに、非ワクチン接種牛は全て、無感情、食欲の喪失、目ヤニおよび鼻水の分泌、下あごの歯肉の赤らみ、鼻の粘膜の強い傷害が見られ、また接種後4日で発育が停止した。ワクチンを接種した牛の鼻の粘膜の傷害は小さく、速やかに治癒するもので、発育が止まることはなかった。菌株接種後の1日毎の臨床評価、直腸温、発育の推移を図18,図19および図20に示した。菌種接種後、全牛とも鼻からウイルスを排泄したが、ウイルス排泄の量と期間は、ワクチン接種牛は著しく低かった(図21)。ワクチン接種牛においては、二次抗体反応が見られ、非ワクチン接種牛は全て、菌種接種後抗体を生成した。再活化後、接種ウイルスを1体のワクチン接種牛および5体の非ワクチン接種牛から単離した。Difivac-1を再活化することはできなかった。
【0046】
上記の結果により、Difivac-1は、若年牛においては殆ど病気の微候を引き起こさず、再活化されないことが実証された。Difivac-1は、菌種接種後病気の程度とウイルスの排泄量を著しく低下させた。
結論として、Difivac-1は、BHV-1感染予防のために牛に使用される、安全かつ効能のあるワクチンである。
【0047】
2)BHV-1の組換えgE欠失変異株の構成
Difivac-1よりも分子的により明確で、かつ所望されるならば、gE-遺伝子の欠失に加えて、たとえばチミジンキナーゼ遺伝子に欠失を有した識別可能なBHV-1ワクチンと自由に利用できるようにするために、Difivac-1の他に、組換えgE欠失変異株を構成した。糖蛋白質gE-遺伝子の決められた位置からスタートし、gE遺伝子を側面に配置するクローン化されたDNAフラグメントを用いて、gE欠失フラグメントを構成することができた。標準的な技術(F.L.Graham and A.J.van der Eb 1973,Virology52,456〜467)を用いて、,この欠失フラグメントを野性型BHV-1菌株のゲノム中で組換えることができ、その結果gE欠失変異株となった。
【0048】
a)gE欠失フラグメントの構成
【0049】
gEフラグメントの構成のために、一方においては、全gE配列を欠き、他方野性型ゲノムと組み換えることができる十分なフランキング配列を含むフラグメントが目標とされた。5'(上流)側では、gE遺伝子の開始コドンの前に18ntを終える1.2kb PstI-AsuIIフラグメントが選択された。3'(下流)フラグメントについては、gE遺伝子の停止ユドンの前に2ntを開始する、1.2kb EcoNI-DraIフラグメントが選択された(図6)。
【0050】
gE欠失フラグメントの構成については、gE遺伝子の5'側に位置した、BHV-1菌株Lamの8.4kb HindIIIKフラグメントからの1.4kb PstI-SmaIフラグメントが、プラスミドpUC18のSmaIおよびPstI中にサブクローンされた。このクローンはp515と命名した。gEの3'側に位置し、4.1kb HindIII-EcoRIクローンからくるEcoNI-SmaIフラグメントはp515の特異なAsuII部位中にクローンされた。このように、gE欠失フラグメントの構成は完結し、このようにして構成されたクローンをp519と命名した。原則的にはp519の全PstI-SmaI挿入をgE欠失フラグメントとして利用することができるが、これは望ましくない。実際は、PstI-SmaIは、Us領域をフランクする反復配列中に、約100〜150塩基対(bp)の拡張を行う。この100〜150bpの部分は、gE遺伝子が位置しておらず、したがって所望されていない組換生成物を作り出すことができないUs領域の他方の反復配列と組換えることができた。このため、PstI-DraIフラグメントが組換え実験のために選択され、したがって100bpの反復が除去された。
【0051】
b)野性型BHV-1のゲノムによるgE欠失フラグメントの組換え
構成gE欠失フラグメントと野性型BHV-1のゲノムの間の組換えを効果的なものとするために、2つのDNA分子のマイクログラム量を、F.L.Graham and A.J.van der Eb(1973、Virology52,456〜467)の標準的な方法に従って、ウシの胚気管(Ebtr)細胞に同時トランスフェクションする。細胞の組換えメカニズムは、細胞によって組み込まれた少しの割合のDNA分子(2〜4%)の組換えを導く。組換えられたgE欠失変異株の選択のために、トランスフェクション後に形成されるウィルス混合物を、新鮮なEbtr細胞培養物上に播種する。大抵の場合、これによって発達する、別々のウィルス群(プラク)が、1つのウィルスから生じる。BHV-1菌株LamのgE欠失変異株の単離のために、これらのプラクの内の230を単離し、DifivaC-1に感染した細胞と反応しない、BHV-1特有のモノクロナール抗体による標準的免疫法により試験を行った。これらのモノクロナール抗体を、糖蛋白質gEに対して向ける。230のプラークの内5つがこれらのモノクロナール抗体と反応しなかった。これらの5つのプラークのDNAについてさらに研究を行った。
【0052】
c)BHV-1菌株Lamの構成gE欠失変異
株のDNA分析上述の5つの候補gE欠失変異株の内3つ(1B7,1B8および2H10)のDNA調製物について、標準的なサザンブロット分析技術(Sambrook et al.1989)を用いて試験した。ゲル電気泳動と、プローブとして2.3kb PstI-DraI欠失フラグメントによるサザンブロットハイブリット化がその後に続く、PstIとDraIによるこれらのDNAの2重の消化は、ウィルス群1B7と1B8のゲノムのgE遺伝子は、所望の方法で正確に取り除かれたことを示している。図7Aおよび図7B参照。群2H10は、偏差PstI-DraIフラグメントを有している。gE特有プローブによるサザンブロットハイブリット化により、3つのDNA調製物のいずれにもgE配列は置かれていないことを示している(結果については示されていない)。BHV-1ウィルス群1B7および1B8は、故意の組換gE欠失変異株である。BHV-1ウィルス群1B7はワクチン特性について試験を行った。
【0053】
d)チミジンキナーゼ/gE二重欠失変異種の構成
わずか1つの遺伝子に欠失を有するBHV-1組換欠失変異株は、十分に毒性が低下したものではないかもしれないので、BHV-1菌株LamおよびHarberinkのチミジンキナーゼ(CTK)遺伝子にも欠失を与えた。これらの変異株は、上記のgE欠失変異種に対して使用される方法と類似の方法で構成された(結果は示されていない)。これらのTK欠失変異株はTK/gE二重欠失変異株の構成に使用した。
【0054】
e)糖蛋白質gI/糖蛋白質gE二重欠失変異
株の構成gE単欠失変異株のワクチン接種をうけた牛は、抗gI/gE抗体の検出を妨害するおそれのある抗-gI抗体を生成する可能性があるため(以下に考察)、gI/gE2重欠失を有するワクチンについても発明した。このようなgI/gE二重欠失変異株は、gE単欠失変異種の構成に使用されるのと同じ処置を用いて構成することができる。gE遺伝子の5'端部を包括する、1.8kb PstIフラグメントの上流側端部のヌクレオチド配列の部分的分析により、他のヘルペスウィルスに見られるgI相同物に対して有意な相同関係を有するオープンリーディングフレームが明らかになった。図13および図14を参照のこと。gE遺伝子の下流に位置した、gI遺伝子およびEcoNI-SmaIフラグメントの推定5'端部を包囲している350bp SmaI-PstIフラグメントを用いて、gI/gE欠失フラグメントを構成することができた。このフラグメントは野性型ゲノムと組換え、BHV-lgI/gE欠失変異株をもたらすことができた。図16を参照のこと。理論的にはさらにまだ生成することができる80〜90アミノ酸は、抗-gI/gE抗体の欠失を防げることができる抗体を導き出すことはできないであろう。さらに、gI遺伝子の配列分析により、完全なgIコード化領域を包括するgI欠失の構成が可能となるであろう。このgI/gE二重欠失変異種をDifivac-IEと命名した。
【0055】
f)Lam gE-およびLam gE-,TK-変異株の安全性と効能の評価
Lam gE-およびLam gE-,TK-,BHV-1変異株菌株のワクチン特性を7週齢の、BHV-1血清反応陰性の特有病原体を持たない牛において試験した。各々の変異株を6体の牛に鼻腔内スプレーした。各牛の全投与量は、2ml培養基において105TCID50となり、2m1中1mlは各々の鼻孔にスプレーした。他の6体の牛はウィルスを含まない培養基を鼻腔内投与し非ワクチン投与対照として用いた。ワクチン投与から5週後に、ワクチン投与および非ワクチン投与いずれの牛も、すべての牛に強毒性BHV-1菌株lowa107TCID50を鼻腔内接種した。ワクチン接種および菌株接種後、臨床徴候、直腸温、体重を監視した。鼻のウィルスの放出日数の決定のために布で拭きとった鼻水を採取した。
ワクチン接種後の、牛の挙動、食欲、直腸温、発育率は正常を維持した。漿液性鼻水の分泌および鼻の粘膜の小さな傷害が、ワクチン接種牛すべてに観察された。ワクチン接種の牛の鼻から約7日間ウィルスを単離することができた(表1)。
【0056】
菌接種後、非ワクチン接種の対照牛すべてが、無感情、食欲の喪失、目や鼻からの分泌、下あごの歯肉の赤らみ、鼻の粘膜の強度の傷害を示し、鼻水率が減少したLam gE-,TK-をワクチン接種した牛はすべて、鼻水の分泌が幾分増加し、鼻の粘膜の小さな傷害がいくらか見られた。Lam gE-をワクチン接種した牛のすべてが、鼻水の分泌または鼻の粘膜の傷害が進行したわけではなかった。無感情、食欲の喪失、疾病についてのその他の徴候は、ワクチン接種牛では観察されなかった。菌接種後の直腸温、発育、臨床評価は図22,23および24に示されている。非ワクチン接種牛はワクチン接種牛の2倍の長さにわたってウィルスを鼻から放出した(表1)。
【0057】
上記の結果は、Lam gE-およびLam gE-,TK-BHV-1突然変異菌株は、若年の牛においては、病気の臨床徴候をほとんど誘発していないことを示している。これらの突然変異菌株はどちらも菌接種後の病気を予防し、鼻のウィルス放出期間を50%短縮させた。
Lam gE-およびLam gE-,TK-B、HV-1突然変異菌株は安全で、BHV-1感染予防のワクチンとして牛に用いて効能を有するものである。
【0058】
3)gEの原核発現
BHV-1糖蛋白質gE-遺伝子の原核発現の場合、これまでのところpGEX発原ベクタ(D.B.Smith and K.S.Johnson,Gene 67(1988)31〜40)が使用されてきた。イソプロピルチオガラクトシド(IPTG)によって発現に誘導されることができる、tacプロモータの影響下にある日本住血吸虫からの担体蛋白質グルタチオンS-転移酵素(GST)についてpGEXベタタはコード化する。GST-gE融合蛋白質の例は構成pGEX-2T600s3の生成物である(図8A)。この構成において、標準的な分子生物学的技術を用いて(Sambrook et al. 1989)、gE蛋白質の200アミノ酸のN末端領域についてコード化する、600bp SmaIフラグメントがGST遺伝子の後に連結された。この構成は3つの部分から成る設計とされ、600bpフラグメントの異なったリーディングフレームがGSTに連結された。これらの構成はすべてIPTGと共に誘発された大腸菌株DH5α内に導入され、形成された蛋白質はウェスタンブロッテングによるポリアクリルアミドゲル電気泳動後、ニトロセルロースに転移させた。抗-GSTによる免疫性検出により、gE蛋白質領域についてコード化する適切なリーディングフレーム(No.3)だけが、27k(GST)t20k(gE)=47kの推定サイズの顕著な融合蛋白質の発現を導くことが実証された。Difivac-1と反応しない我々が単離した3つのモノクロナール抗体は、ウェスタンブロット中の47k D GST-gE融合蛋白質を認識する。図8B参照。
【0059】
4)糖蛋白質gE-遺伝子の真核発現
糖蛋白質gE-遺伝子の真核発現については、これまでは、とりわけ、ベクタ、pEVHisが選択されてきた。pEVHisベクタは、真核遺伝標識として、ヒスチジノールデヒドロギナーゼについてのHisD遺伝子コードを有しており〔EC1.1.1.23〕(C.Hartmann and R.Mulligan,1988,Proc,Natl.Acad Sci.USA 85,8047〜8051)、このヒスナジノールデヒドロギナーゼが、細胞の2.5mMヒスチジノールの毒性濃度を長びかせる原因となっている。さらに、このベクタは、ヒトのサイトメガにウィルス(HCMV)の即時型遺伝子のプロモータ領域を、該遺伝子の後に位置している特異な制限酵素部位と共に有している。pEVHis/gE発現ベクタの構成のためには、糖蛋白質gE-遺伝子の全コード領域を有するフラグメントが使用された。gEの従来のオープンリーティングフレームの前のAluI部位55でスタートし、その後の133bpで終了する。この領域がpEVHisベクタのHCMVプロモータの後でクローン化され、これによって構成pEVHis/gEが形成された(図9)。このpEVHis/gEを大腸菌DH5の細胞中にて増殖させ、塩化セシウム勾配法によって精製した(Sambrook et al.,1989)。この精製DNAを、Graham and Vander Ebの方法に従って、Balb/c-3T3細胞にトランスフェクトした。形質転換細胞をヒスチジノールと共に選択した結果、20の耐ヒスチジノールコロニーを単離することができた。これらのコロニーを免疫ペロキシダーゼ単分子層試験(IPMA)によって、モノクロナール抗体81によって試験した。4つのコロニ一によって、gE蛋白質の発現が証明された。これらの4つのコロニーの内、3T3gEクローン9を、高度のgE発現を有するサブクローンの単離に使用した。この方法によって単離されたクローン(3T3gE9.5と命名)を候補抗gEモノクロナール抗体の特性づけのために用いた。
【0060】
5)同一細胞におけるBHV-1糖蛋白質gEおよびBHV-1糖蛋白質gI双方の真核発現
BHV-1糖蛋白質gEと同一の細胞においてBHV-1糖蛋白質gIを発現させるため、まず最初にBHV-lgI遺伝子の推定位置を決めた。ヘルペス単ウィルス糖蛋白質gI遺伝子は、糖蛋白質gE遺伝子のまさに上流に位置しているので、BHV-1 gI遺伝子は、対応の位置に位置しているものと推定された。これを試験するため、BHV-1 gE位置が開始したところより約1kb上流に位置した、283ヌクレオチドの領域の配列を決定した。この領域の概念翻訳により、第2リーディングフレームは、ヘルペス単ウィルス糖蛋白質gIと相同の94アミノ酸配列についてコード化する(図13および図14参照)。相同セグメントは、スタートコドンから約80のアミノ酸であるので、BHV-1 gI遺伝子のオープンリーディングフレームの推定開始は、配列領域から約250nt上流と見積もられる。これより、配列領域から400nt上流でスタートし、gE遺伝子内で終了する、1.7kbSmalフラグメントは、BHV-lgIl遺伝子の完全なコード化領域を含んでいるべきであることが推定された。この1.7kbSmalフラグメントが真核ベクタMSV-neo内にクローン化された(図15参照)。このベクタは、強いマウス肉腫ウィルスプロモータと、抗生物質G-418硫酸ゲネティシンに対する耐性についてコード化するセレクター遺伝子neoを含んでいる。結果として生じる構成MSVneoGIが、大腸菌DH5α細胞中で増殖され、Graha and Van der Ebの方法を用いて、3T3gE9.5細胞中にトランスフェクトされた。トランスフェクトされた細胞は、400mg、ゲネティシン/ml培養基で選択され、耐性コロニーを単離し、3T3gE9.5細胞と反応しなかった候補抗-gEモノクロナール抗体で試験した。これによって、たとえば、野性型BHV-1だけでなく、モノクロナール抗体66と反応した3T3gE/gIR 20クローンを選択した。
【0061】
6)候補抗-gEモノクロナール抗体の特性づけ
野性型BHV-1に対してモノクロナール抗体を生成し、Difivac-1に感染したEbtr細胞との反応不能性について選択した。これらのモノクロナール抗体を下記との反応性について試験する。
a)Lam gE-欠失変異株
b)ウェスタンブロットにおける、上記の原核発現生成物
c)上記のgE発現Balb/c-3T3細胞
d)c)に記載のDifivac-1に感染した細胞
e)gE/gI複合物を発現するBa1b/c-3T3細胞
a,c,dおよびeとの反応性試験については、免疫ペロキシダーゼ単分子層試験(IPMA)を用いた。表2の結果により、我々が生成したものは、gEに対して向けられているモノクローナル抗体(2,3,4,52,66,68,72,81)およびgE/gI複合物上の立体配座抗原ドメインに対して向けられている可能性のあるモノクローナル抗体(NO.1,51,53,67,75,78)であることが判る。種々のモノクローナル抗体によって認識される抗原ドメインの地図を作るための競含IPMAにより、少なくとも4つの抗原ドメインが糖蛋白質gE上に存在すること、およびおそらく1つのドメインがgE/gI複合物により形成されることが示された(表2)。
【0062】
BHV-1に感染した牛の抗-gE抗体の検出
感染した牛の血清中にgE抗体が存在しているか否かを調べるために、間接ブロッキングIPMAを16の候補gE-モノクローナル抗体に実施し、以下の8血清を選択した:
−Difivac-1のワクチン接種を受け、毒性Iowa菌株が接種された牛の菌接種14日後に採取された2血清。
−試験のためBHV-1サブタイプ1ウイルスに感染させた牛の、感染20ケ月後に採取された2血清。これらの牛のうちの一体は接触感染によって感染させた。
−試験のためBHV-1サブタイプ2bウイルスに感染させた牛の、感染20ケ月後に採取された2血清。これらの牛のうちの一体は接触感染によって感染させた。
−ts変異株ワクチンの接種を行い、3週後にBHV-1サブタイプ2bウイルスを接種させた特有病原体のない牛の、菌接種7週後に採取された1血清。
−ts異変株ワクチンの接種を行い、3週後にBHV-1サブタイプ2bウイルスを接種させた、ノトバイオートの牛の、菌接種7週後に採取された1血清。
【0063】
表2により、これらの血清はどれも、gE上の抗原性ドメインIIIおよびIVに対する、およびgE/gI複合物上に位置していると思われる抗原性ドメイン1に対する抗体を含んでいることが判る。gEは、明らかにBHV-1-感染とワクチン接種された牛を区別するための適切な血清学的遺伝標識であると結論づけることができる。
【0064】
7)BHV-l gE-特有プライマを用いるPCR処置によるBHV-1核酸の検出
BHV-lgE遺伝子の決定されたヌクレオチド配列からスタートして、PCRに適切なプライマの対を、Lowe et alによるプライマ選択プログラム(T. Lowe, J.Sharefkin, S, Qi Yang and C. W .Dieffenback 1990, Nucleic Acids Res, 18, 1757〜1761)を用いて選択した。これらのプライマをP3、P4と命名し、図に示した。これらのプライマは159nt離れて位置しており、200ntのフラグメントの増殖を導く。プライマP3、P4と単離されたBHV-1 DNAを用いて、PCR処置の条件を最適化した。これは特に、MgCl2の濃度の変更、グリセロール濃度、サイクル条件に関するものである。BHV-lDNAの増殖のためにP3、P4を使用するため最適と考えられる緩衝液は、10mM Tris pH 8.0, 50mM Kca1, 0.01%ゼラチン, 2.6mM MgC12および20%グリセロールである。最適のサイクル条件と考えられるものは(Perkin Elber Cetus DNA Thermal Cycler),サイクル1〜51については、1分98℃, 30秒96℃, 30秒55℃, 45秒72℃である。PCR増殖後、得られた200ntDNAフラグメントを2%アガロースゲル上で電気泳動にかけ、ニトロセルロースにブロットし、続いてサザンブロット分析を行った。サザンブロット分析に使用された32P dCTPのラベルが貼られたプローブが、プライマ結合部位間に位置している137bp TagIフラグメントである(図10)。ハイブリッドフィルターの自動X線撮影により、200bpバンドを観察することができた。さらに、このルートを介して、10BHV-1ゲノムだけの増殖(約1.5×10-15μgDNA)が検出可能と思われる信号を導く(結果は示されていない)。同等の方法で、BHV-1糖蛋白質gIIIのコード化配列(D.R.Fitrpatrick, L.A. Babiuk and T.Zamb, 1989,, Virolagy 173, 46〜57)に基づくプライマを用いてPCR処置を行った。野生型BHV-1 DNAとgE欠失変異株ワクチンの区別を可能とするために、DNA試料にgE-特有PCR分析およびgIII-特有PCR分析の両分析を行った。このような試験において、Difivac-1DNA調製物はgIII陽性およびgE陰性であることが判った。ウシの精液中のBHV-lDNAの検出は、BHV-1特有PCR処置の重要な用途であると思われるので、BHV-1に感染したウシの精液にgE-特有PCRの実施を試みた。しかしながら、精液中の未知の成分がポリメラーゼ連鎖反応に強い抑制効果を有している。したがって、ウシの精液からBHV-1 DNAを単離するプロトコールが展開された。ウシの精液からDNAを単離するため、30μ1の精液を、1mg/m1プロティナーゼK(pk)とともに、60℃にて、全量300μ1の0.15M NaC1, 0.5%Na-サルコシル, 40mM DTT中にて培養する。1時間後、試料を室温にまで冷却し、300μl 6M NaIを添加し、5分間培養する。この混合物より、標準クロロホルム/イソアミルエタノール抽出によりDNAを単離し、1容量のイソプロパノールで沈殿させた。沈殿物を2.5M NH4AC/70%エタノールで洗浄し、60℃にて1時間10mM Tris pH7.4,1mM EDTA,0.5%Tween80および0.1mg/mlpk中で、第2培養のために再懸濁する。このDNA調製物は直接ポリメラーゼ連鎖反応下に置くことができる。
【0065】
Lam gE-またはLam gE-/TK-を接種後および毒性の強いBHV-1菌株を投与後、これらの接種されまたは無接種の牛から垂れる鼻水ウイルス
【表1】

【0066】
【表2】
【図面の簡単な説明】
【0067】
【図1】BHV-1菌株Difivac-1およびlowaのサザンブロヅト分析 Aは、Difivac-1とlowaとのゲノムDNAのサザンブロットの自動放射線図である。1と3レーン中で、Difivac-1DNAは、HindIIIとPstIとによる制限酵素消化後に、用いられた。2と4レーン中で、lowa DNAは、HindIIIとPst1とによる制限酵素消化後に用いられた。フラグメントの大きさは、キロベース(kb)で示される。 Beckman L5-65超遠心分離のSW27ロータ中で20,000回/分で、pH7.4のトリス10mMと150mM NaClと1mM EDTAとの中で、25%(w/w)庶糖クッションを用いて2時間、ウイルスで汚染されたEbtr細胞と共に培養液(約450cm2ローラボトルあたり70ml)を遠心分離することによって、ウイルスDNAが単離された。得られたウイルスベレットから、DNAが標準の手法で単離された(J.Sambrook,E,F.Fritsch and T.Manfatis,1989,Molecular cloning:a laboratory manual,2nd ed.Cold Spring Harbor Laboratory Press,NewYork)。このDNA上で制限酵素消化は、製造者によって供給されるSuRE、/Cut緩衝液中でBoehrnger Mannheimからの酵素で行われた。 0.7%アガロゲルを水平電気泳動して分離し、ニトロセルローズフィルタ(Schleicher & Schuell,lnc)でブロットした後、フィルタは6時間42℃で50%ホルムアルデヒド中で3×SSC(1×SSC=0.15MのNaClと0.015Mのクエン酸ナトリウム,pH7.4)と、50μlの変成サケ精液DNA(Sigma)/mlと、0.02%フィコールと、0.1%ドデシルと硫酸ナトリウム(SDS)との中に予備混合された。さらに混合物は、同じ溶液を加えることによって、32PdCTP(Amersham)で標識を付されたHindIII Kフラグメントを完成させた(HindIII Kフラグメントの選択は、1型ウシヘルペスウイルスからのDNAのクローニングと分裂位置のマッピングに基づく(Cooper菌種)John F.Mayfield,Peter J.Good,HollyJ .VanOort,Alphonso R、Campbell and David A.Reed;Journel of Virology (1983) 259〜264)。混合の12〜14時間後、フィルタは2時間、0,1%SDSと0.1×SSCとの中で60℃で洗浄された。HindIII Kフラグメントは、標準のクローニング手法に従って、pUC18ベクタ中でクローン化された(J,Sambrook, E,F,Fritsh and T、Maniatis, 1989, Molecular cloning la labovatony manua1,2nd ed.Cold Spring Harbor labovatort Press,NewYork)。pUC8.4HindIII KクローンのHindIIIによる消化後、pUC18ベクタは、0.7%の低融点アガロース(BRL、Life Techovlogies,Inc.)ゲルで電気泳動によって再び8.4kb HindIIIにフラグメントから分離され、基準のフェノール抽出とエタノール沈澱とによってアガロースから分離された。分離されたHindIII Kフラグメントは、Boehringer Mannheimからのラドン感知DNA標識装置1OO4.760で標識を付された。混合化フィルターの自動放射線図は、-70℃でKodak XARフィルムの36時間露出で、反射スクリーンを使って行われた。 BはIowaとDifivac-1の7.4kb HindIIIフラグメントとの8.4kb HindKフラグメントの物理的マップである。6kb PstIフラグメントの共泳動とDifivac-1中の1.8kb PstIフラグメントの存在を考えて、欠失はハッチを付された面積で求められる。
【図2】Difivac-1中に欠出領域を包んだ野性型BHV-1フラグメントのサブクローニング A中でBHV-1ゲノムの成分は、長ユニーク(UL)領域、短ユニーク(US)領域および2つの反復物(lrとTr)として示される。このマップは、Cooper菌種の公表された分析に基づいている(John F.Rlayfield, Peter J.Good, Holly J VanOort, Alphonso R. Campbell and David A. Reed: Journal of Virology (1983) 259〜264)。 Bの中でフラグメントは、細胞核のないベクタ、すなわちpACYC中の15.2kb EcoRIフラグメントと、pUC18中の8.4kb HindIIIフラグメントと、pBR322中の4.1kb EcoRI-HindIIIフラグメントとの中ヘクローンされたUS領域で示される。ウイルスDNAフラグメントの単離は、図1Aの表示中に述べられている手法に従って実施された。種々のベクタ中へこれらのフラグメントをクローンすることは、基準の手法で行われた(J.Sambrook, E.F.Fritsch and T.Maniatis, 1989, Molecular cloning:a loboratory mannal,2nd ed.Cold Spring Harbor Laboratony press, NewYork)。 cの中で物理マップは、Difivac-1中で要求される欠失が当てはめられる領域を示す。 Dの中で、この領域のサブクローンが分析のために用いられることを示す。2つのPstlフラグメントはpKUN19の中へ、そして残ったフラグメントはpUC18の中ヘクローンされた。
【図3】Aは、〔右端にあるAluIの確認位置から左端にあるHincIIの確認位置に至る〕図2C中で示されるようなDifivac-1中に欠失された要求される位置を囲むBHV-1のLam種のUS領域からの2027個のヌクレオチドのヌクレオチド配列である。図2Dで示されるサブクローンの挿入をしたヌクレオチド配列は、Sangerらのジデオキシ配列法を用いた2つの基準で分析することによって決定された(F.Sanger, S.Nicklen and A.R.Coulson, 1977, Proc.Nat1.Acad.Sci.USA 74,5463〜5467)。端末に対して、PharmaciaのT7配列装置が、製造者によって指定される手法に従って用いられた。放射線標識のために、〔35S〕dATP(Amersham)が用いられた。人工産物と比較して、GLの多い領域の配列分析は、Pharmacia装置の7-デアザアデニン-dGTP変形物で複生された。ヌタレオチド配列の下に示されたものは、3文字のコードで、575アミノ酸残基のオープンリーディングフレームのアミノ酸配列である。それは、ヌクレオチドの概念上の転移後に見付かった。この転移は、一般的なコードに基づきPC遺伝子コンピュータプログラムを用いて決定された(PC遺伝子翻訳バージョン 1987年11月)。この575アミノ酸のオープンリーディングフレームは、ヌクレオチド168のメチオニンで始まり、ヌクレオチド1893の停止コドンで終わる。 575アミノ酸残基のオープンリーディングフレームの構造分析は、PC遺伝子コンピュータプログラムでまた実行される。真核の輸出信号からの最初の26アミノ酸が、「シグナルペプチド」によって図中に示される。6.2の点でこの配列を分裂する信号が、アミノ酸26とアミノ酸27との間で起こされる。アミノ酸575の配列は、アミノ酸残基の線によって指示される。3つの可能なN一結合のグリコレーシヨン位置(NXT/S)を有する。RaoとArgosの方法に従えば、貫膜螺旋による図で示されるアミノ酸423とアミノ酸450との間に貫膜領域がある。制限酵素AsuII, SmaI, HindIIIおよびEcoNIのための確認配列(位置)は下線が付される。このポリペプチドの計算された分子量は61212である。 Bは、上で述べた575アミノ酸オープンリーディングフレームの構造上の特性を模式的に表したものである。
【図4】BHV-1 gE遺伝子のアミノ酸配列と、ヘルペス単体ウイルス(HSV)gE遺伝子および他のgE均質遺伝子〔疑似狂犬病ウイルス(PRV)gIと帯状庖疹ウイルス(VZV)gpl〕のアミノ酸配列との比較 この比較のために用いた配列は、次の公表されたものである。 HSV:1型ヘルペス単体ウイルスのゲノム中のUS領域の配列決定と遺伝子内容(D.J.McGeoch, A.Dolan, S.Donald and F.J.Rixon (1985) Journal Mol.Biol.181,1〜13) VZV:帯状庖疹ウイルス遺伝子のU。成分のDNA配列(A.J.Davidson (1983), EMBO Journal 2, 2203〜2209) PRV:ヘルペス単体ウイルスと帯状庖疹ウイルス糖蛋白とに同族体である2つの疑似狂犬病ウイルス糖蛋白のための単離遺伝子にλgtllの使用(E.A.PetroVsKis, J.G.Timmins and L.E.Post (1986) Journal of Virology 60,185〜193) これらの配列は配列分析プログラムMultalinを使って比較された(F.Corpet (1988), Nucl. Acids Res.16,10881〜10890)。 Aの中で図は、4つのアミノ酸配列が模式的に示されている。ここに確認された貫膜部(TM)が各々に示される。確立された輸出信号配列(SP)と可能なN-結合グリコシレーション位置(I)に加えて、2つの保護される領域が示される。その中で、システン残基の相対位置(CCC)は不変である。 Bの中で結果は4つのgE翻訳の中央に位置するシスティンの多い領域のMultalin比較が示される。アステリスクは、一致したアミノ酸と類似アミノ酸クローンを示す。
【図5】Difivac-1とIowaとのサザンブロット分析で得られた写真図 パネルAは、BstI(1,2),EcoRI(3,4)およびHindIII(5,6)を有するDifivac-1とIowa消化制限酵素のゲノムDNAである。これは、0.7%アガロースゲルで分離され、ニトロセルローズでブロットされ、図1Aの説明で述べられた手法に従ってBHV-1菌株Lamの32Pで標識を付されたHindIIIKフラグメントで混合される。 パネルBは、Aと同様に同じゲルのニトロセルローズブロットであり、BHV-1 gE-種プローブp318で混合された。プローブは、図2C中で示したAluI-HincII中で比較された。
【図6】BHV-1の欠失のあるgEフラグメントの構成 Aの中でgEの位置と用いられたクローンとが示される。BHV-1遺伝子の成分は、UL領域、Us領域および2つの反復(lrとTr)として示される gE遺伝子の5’側に位置する領域を得るために、BHV-1菌種Lamの8,4kbHindIIIKからの1.4kbPstI-SmaIフラグメントが、プラスミドpUC18のSmaIとPstI位置の中ヘサブクローンされた。このクローンは、p515と呼ばれ、B中に示される。gEの3'側に位置するEcoNI-SmaIフラグメントは、4.lkbHindIII−EcoRIクローンに由来し、p515の普通のAsuIIヘクローンされる。AsuIIの休止にEcoNIの休止を結びつけるために、クローンp515はAsoIIで消化され、その後クレノウ酵素(Boehringer Mlannheim)と、基準の手法に従ってAsuII休止中へ1つのシトシン残基を供給すべきdCTPとで処理した(Sambrook et al,1989)。この付加シトシンは、D中でアステリスクで示される。p515は、またSmaI酵素で消化され、その後EcoNIフラグメントは、このベクタ中へ結合される。このようにして構成されたクローンは、p519と呼ばれる。
【図7】Aは、1B7,1B8および2H10のDNA調整品のサザンブロット分析で得られた写真である。DNA単離物、消化制限酵素、ブロットおよび混合物は、図1Aの説明で述べられた手法に従って作られた。DNA調整品1B7,1B8および2H10のPstI-DraIによる二重消化後、フラグメントは0.7%アカロースで分離され、直後にニトロセルローズフィルタがブロットされた。このフィルタは、プローブとして、32PdCTPで標識を付された2.3kbPstI- DraI欠失フラグメントと混合された。レーン1〜3中で試料1B7,1B8および2HlOがそれぞれ分離された。レーン中でLam菌株の野性型BHV-1 DNAが、レーン15中で2.3kb欠失フラグメントが示される。 Bは、BHV-1菌種Lamの15.2kb EcoRIフラグメントの物理マップである。マップは、PstI,DraIおよびHindIIIの確認位置と7A中で述べた混合プローブの位置を示す。
【図8】BHV-1 gEの無核表現 BHV-1 gEの無核細胞を表現するために、gE遺伝子の600bpSmaIフラグメントは、ベクタpGEX-2T中で、日本住血吸虫のグルダチオン-S-転移酵素のコード化領域に3つのリーデングフレーム中で融合された(D.B,Smith and K.S.Johnson,Gene 67(1988)31〜40)。SmaIフラグメントの方位確認性(syn)を有する再結合分子は、基準の手法を用いて、制限酵素によって同定された。この融合構造を有するE.coliDH5αクローンは、pGEX-2T600sl,pGEX-2T600s2およびpGEX-2T600s3と呼ばれた。 Aは、pGEX-2T600sの構成の1つの図である。GST-gE融合物のためにコード化する領域のNH2側にあるものは、タフプロモータ領域を導入しうるイソプロピルチオガラタトシド(IPTG)である。 Bは、pGEX-2T600sで転移されたDH5αの細胞の全蛋白質調整物のウエスタンブロット分析で得られた写真である。pGEX-2T600s1,pGEX-2T600s2およびpGEX-2T600s3構成で転移されたDH5α細胞の徹夜の培養物は、50μg/mlアンピシリンを加え、Luria-Bertani(LB)媒中で1/10継続され、1時間後培養物はIPTGに5時問導入された。 これらの導入された培養物は、5分間6,000Gで遠心分離し、1×ラウエル混合物(2%SDS、10%グリセロール、5%メルカプトエタノールおよび0.01%ブロムフェノールブルー)中に混合〔1.5ml培養物は、500μlラウエル混合物中に混ぜる〕され、95℃に5分間加熱された。そして、レーンあたり50μlが、基準の手法に従って、12.5%ポリアクリルアミドゲルで水平的に分離され、引続いて製造者によって明確にされた条件で、LKB一ムルチフールII型Navaブロットシステムを用いて、ニトロセルローズフイタが半乾燥ブロットされた。 レーンM中で、予め印を付された蛋白質が用いられ、(BRL Life Technologies,lnc.236K,112K,71K,44K,28K,18Kおよび15K)、レーン1,2,3中で、DH5α細胞の全蛋白調整物は、pGEX-2T600sl,pGEX-2T600s2およびpGEX-2T600s3の各々3つのフレームで転移される。 パネルA中で、結果は、抗−GST血清によってウエスタンブロット分析で見られる。フィルタは、基準の手法(E.Harlow and D.Lane(1986)、Antibodies:a laboratory manual,Cold Spring Harbor Loboratory,New York)にしたがって、ウサギ血清ポリクロナール抗GSTを加えた阻止緩衝液(PBS+2%粉乳および0.05%トゥイーン20)中で培養された。そして、フィルタは洗浄され、ヤギ対ウサギ免疫グロブリン血清を接合した西洋ワサビパーオキシターゼ(HRPO)を加えて培養された。その後、結合されたヤギ抗体は、色素体(ヂアゾベンジジン、クロロナフトールおよびH2O2)で免疫化学的に検出された。矢印によって指示されるGST融合物は、フレーム3中でのみ約47kの確認された大きさを有する。 パネルBにおいて、結果は、モノクロナール抗体Mab4で、ウエスタンブロット分析で見られる。パネルA中のようにデュプロフィルタは阻止され、Mabで培養され、ウサギ対マウス血清を接合したHRPOで培養される。その後結合ウサギ抗体は、色素体で免疫化学的に検出される。レーン3(フレーム3)で見られるバンドは、47Kの大きさで、矢印によって指示される。
【図9】BHV-1 gE遺伝子の無核表現のためのpEVHis gEプラスミドの構成 gE遺伝子の無核表現のために、コード化すべき領域の全gEは、基準の手法を用いて表現ベクタpEVHisのHCMV促進領域の背後で方向性をもってクローンされる(Sambrook et a1,1989)。gEのオープンリーディングフレームの前に55bpで出発する394bpA1UIフラグメントは、pUC18の中ヘクローンされ、p201と呼ばれる。そして、p201をHincIIが消化した後、1740bp HincIIフラグメントは、gE遺伝子の大部分から成り、p201中へクローンされる。これは、pUC18のボリリンカ中で、gEの出発コードの前にあるA1Ul位置55bpからgEの停止コードの後にあるHincH位置133bp)までの全gEコード化領域から成るプラスミドp318中に生じた。ベタタのポリリンカ中で制限酵素位置を用いて、このフラグメントは、酵素BamHIとSphiでp318から切られた。最初p318はSphIで消化され、その後SphI位置は、クレノウポリメラーゼとdNTPポリメラーゼとを使って満たされた。BamHIでの消化後、1.9kb挿入物は、pUCl8ベクタから低融点アガロース中で分離され、端末をBamHIとEcoRVで消化されたpEVHis中で結ばれた。このように形成されたプラスミドは、pEVHis/gEと呼ばれた。
【図10】BHV-1DNAを検出するためにPCR手順で得られるgE-種プライマとプローブの位置 図中には、ヌクレオチド1272から2027に至るBHV-1糖蛋白質の核酸配列が示される〔配列は図3から得られる〕。gE-種PCR手法のために用いられるプライマは、P3とP4と呼ばれる。P3とP4の位置に結合したプライマは、下線が付される。P3のヌクレオチド配列は、5'-ACG-TGG-TGG-TGC-CAG-TTA-GC-3'である(配列標別番号:2)。P4のヌクレオチド配列は、(上述の配列に結合するプライマを補足して)5'-ACC-AAA-CTT-TGA-ACC-CAG-AGC-G-3'である(配列標別番号:3)。DNAで拡大されたPCRの検出のためにサザンブロット混合を用いたプローブは、プライマが結合する位置の間に位置する137bp TaqIフラグメントであり、そのフラグメントの)端末が指示される。図3と比較のために、HindIIIとEcoNIの位置がまた示される。
【図11】Difivac-1のgE欠失のマッピング Aは、野性型BHV-1菌種Lamの15.5kb EcoRIフラグメントの物理マップを示す。Bは、Difivac-1の14.5kb EcoRIフラグメントの物理マップを示す。2つのEcoRIフラグメントは、各々のウイルスのゲノムのUPS領域を完全に覆う。gE遺伝子の位置とgI遺伝子の推定上の位置は、白四角で指示される。マップAとBとは、このような方法で位置決めされ、各マップ中の6kb PstIフラグメントは、一直線にされる。両方のマップ中で、内部反復と端部反復との配列は、斜線を付された四角で示される。反復の下方の矢印は、これらの配列の方向を示す。 Aの中でUS領域の部分は、Difivac-1菌種中で失敗があることを示す。 Cは、gE欠失をマップ化し、B中に示される物理マップを得るために用いられるクローン化Difivac-1フラグメントの位置を示す。クローンp728,p737およびp754の挿入の下の矢印は、再結合点を決めるために配列化された領域を示す。 略語: A=A1UI,E=EcoRI,P=PstI,H=HindIII,γ=再結合点,IR=中間反復,TR=端末反復
【図12】Difivac-1のUS領域における正確な再結合位置の決定 Difivac-1菌種中に見付かったgE欠失の正確な境界を決定するために、クローンp754ならびにp728およびp737とが配列された。これらのクローンの挿入は、図11中に指示される。用いられる配列決定の手法は、図3の説明中に述べられている。 Aにおいて、A1UI-PstIフラグメントの最大の配列が示される。この配列は、gE遺伝子のプロモータ領域で出発する。推定のTATAボックスは、下線が付されている。点γ(=再結一合点)において、このプロモータ領域は、Us領域の反対の位置で見付かった裏返しの反復と呼ばれる配列に接合される。正確な再結合点は、US領域の反対位置で見付けられた反復のコピーを有するgEプロモータ位置で見付けられた反復を含むことによって決定される。この配列が分岐する点は、B(I)の中でγで示される。類似の比較は、Difivac-1中で見付けられるgEプロモータ配列と野性型菌種Lam中で見付けられたgEプロモータとで行われる。この配列が分岐する点は、B(II)の中で示され、またγで示される。見付けられた再結合点は同じである。
【図13】BHV-1 gI遺伝子の部分配列分析 BHV-1 gIとgEとの双方の遺伝子(図11に示す)中に達するBHV-1菌種Lamの1.8kb PstIクローンを用いてBHV-1 gIのコード化領域の範囲内で、284ヌクレオチドの配列が決定される。配列決定に用いられる手法は、図3の説明の中で述べられている。配列は、PC/遺伝子コンピュータプログラムバージョン1.03(1987年11月)による汎用コードに基づいて転位された。第2リーディングフレームによってコード化されたアミノ酸配列は、ヌクレオチド配列の後に1つの書かれたコードとして与えられる。このアミノ酸配列は、他のヘルペスウイルスgI同族体(図14に示す)のコード化領域に均等である。
【図14】推定のBHV-1 gI遺伝子の部分アミノ酸配列と、ヘルペス単体ウイルス(HSV1)gI遺伝子、疑似狂犬病ウイルス(PRV)gp63遺伝子および帯状庖疹ウイルス(VZV)gpIV遺伝子のコード化領域の相当部分とのアミノ酸比較 PRV配列はそのコード化領域のアミノ酸82で出発し、HSV1配列はそのコード化領域のアミノ酸で出発し、VZVはそのコード化領域のアミノ酸76で出発する。用いられる配列は、図4の説明で述べられた報告書で公表されている。比較は、Multalinコンピュータプログラムを使って行われた。アステリスクは、同一のアミノ酸を示し、コロンは類似のアミノ酸を示す。
【図15】BHV-1 gI-遺伝子の無核表現のためのMSVneoGIプラスミドの構成 BHV-1 gI-遺伝子の部分配列のアミノ酸配列の比較に基づいて、BHV-1 gI-遺伝子の推定位置は評価される。この評価に基づいて、1.7kb SmaIフラグメントは、BHV-1 gE遺伝子の完全コード化領域を含むことが推論される。この1.7kb SmaIフラグメントの位置は、A中に示される。この1.7kb SmaIフラグメントの丸くなった端末に、BamHIリンカが基準の手法を用いて結び合わされる。この結果生じた物は、BamHIで消化され、無核表現ベクタMSV-neo中で結び合えわされる。MSV-neoベタタは、強いプロモータ性を有するMSV-LTRの背後の一般的なBamHI位置を有する。このベクタは、Rijsewijk et al.,1987EMBO J.6,127〜131中で述べられている。
【図16】BHV-1 gI/gE二重欠失フラグメントの構成 BHV-1のUS領域中における糖蛋白質gE遺伝子の位置と糖蛋白質gI遺伝子の推定位置は、図A中に表現される。斜線を付された部分は、US領域を境界する反復を示す。Bは、双方の遺伝子の位置を考慮して、本質的な制限酵素位置の物理マップを示す。gI/gE欠失フラグメントを構成に関して、gI遺伝子を包み込む1.7kb SmaIフラグメントを含むクローンP1.7-SmaI/0がPstIで消化される。残留350bp SmaI-PstI挿入のPstIで消化される。残留350bp SmaI-PstI挿入のPstI位置は、基準の分子生物学的手法を用いて、丸くなった端末を作る。EcoNI-SmaIフラグメント(図6B参照)は、図6A中で記載された4.1kb HindIII-EcoRIフラグメントから単離され、丸くなった端末に作られ、修正PstI位置に結び合わされる。これは、CとD中に図示される。クローンP△IEの結果から、1.4kb SmaI-DraIフラグメントは、野性型BHV-1DNAと再結合して単離されうる。 略号 E=EcoRI,H=HindIII,S=SmaI,P=PstI,ENI=EcoNI,D=DraI,kb=キロベース,US=短ユニーク
【図17】接種後の牛から垂れた平均鼻水のウイルス ●=Difivac-1を接種、○=接種なし
【図18】毒性の強いBHV-1菌株投与後の平均発病牛数、記号は図17に同じ
【図19】毒性の強いBHV-1菌株を投与した牛の平均体温、記号は図17に同じ
【図20】毒性の強いBHV-1菌株投与後の牛の平均成育量、記号は図17に同じ
【図21】毒性の強いBHV-1菌株投与後の牛から垂れた平均鼻水、記号は図17に同じ
【図22】毒性の強いBHV-1菌株投与後の牛の平均体温 ●=Lam gE-を接種、○=Lam gE-/TK-を接種、×=無接種
【図23】毒性の強いBHV-1菌株投与後の牛の平均成育量、記号は図22と同じ
【図24】毒性の強いBHV-1菌株投与後の平均発病牛数、記号は図22と同じ
【図1A】
【図1B】
【発明の詳細な説明】
【技術分野】
【0001】
発明の分野
本発明は、病原体によって引き起こされる疾病に関係したワクチン注射および診断法の分野に関するもので、弱毒性ワクチンまたは不活化ワクチンを生成する古典的な方法とDNA組換技術に基づく近代的方法両方の使用に関係している。
より詳細には、本発明は、動物、特に牛を1型ウシヘルペスウイルス(BHV-1)から守るための弱毒性生ワクチンおよび不活化ワクチンに関するもので、これらのワクチンは、安全でかつ効能があるばかりでなく、ワクチン接種をうけた群における、非感染動物と感染動物とを識別することも可能にする。
ワクチン接種群における非感染動物と感染動物の識別のための試験に使用が可能である診断キットも本発明の一部である。
【背景技術】
【0002】
発明の背景
牛の伝染性鼻腔気管炎ウイルス(IBRV)および伝染性濃疱外陰部腔炎ウイルス(IPVV)を含むBHV-1は、牛の呼吸器疾患、生殖能力障害の解明に重要な役割を果たす。BHV-1は、急性感染後、潜伏した形で宿主内にそのまま残っていることが多い。潜伏ウイルスはとりわけ、ストレス(これは臨床現象を伴うことも、伴わないこともある)の影響下で再び活性化し、続いて排泄されることがあり得る。結果として、感染した牛は終生BHV-1を蔓延するものとみなす必要がある。BHV-1はオランダの養牛場の推定75%において風土病として発生している。特に、高齢牛の血清反応が陽性である。
【0003】
BHV-1感染予防接種に利用される多くの不活化("死んだ")ワクチンおよび弱毒("生きた")ワクチンがある。不活化ワクチンは、たとえば熱処理や照射、またはエタノールやホルマリン処理によって、BHV-1ウイルスを死滅させて作られる。しかしながら、これらによっては不充分な予防しか行えないことが多い。弱毒ワクチンは、同種(ウシ)細胞またはブタの細胞やイヌの細胞のような異種細胞への多数回にわたる継代によって作られ、その後ウイルスは物理的または化学的にも処理されることもある。このようにして、未知の突然変異/欠失がウイルスゲノム中に発達し、これがしばしばウイルスの疾病誘発特性を低下させる。弱毒ワクチンは不活化ワクチンよりも優れた予防を行うが、これは特に宿主の免疫系に、より多くのウイルス性抗原を与えるからである。生きたワクチンの重要な他の利点として、それらは鼻腔内投与が可能であるということがあり、すなわち、ここは感染後に野生型ウイルスの増殖が初めて起こるところである。生きたワクチンはまだ改良の余地がある。いくつかの生きたワクチンはさらに、流産を引き起こす可能性を有しているように思われ、これは特に筋肉内投与後に顕著になる。さらにまた、生きたワクチンはすべて、ワクチンを接種された牛に潜伏して残っていると思われる。また、該ワクチンが野生型ウイルスとほとんど相違するところがなければ、病原性への復帰が起こる可能性もある。しかし、重要な問題の1つは、BHV-1ワクチンは、野生型ウイルスによる感染を予防することができないということである。その結果、ワクチン接種を受けた牛も野生型BHV-1を蔓延させる可能性がある。
【0004】
適切なBHV-1抑制計画のためには、野生型ウイルスと識別することができる効果的かつ安全なワクチンを自由に使えることが必要である。何故ならば、効果的ワクチンの利用によりBHV-1の蔓延をかなりの程度低下させることができ、ワクチンと野生型ウイルスの識別が可能な試験によって、ワクチン接種を受けた牛の群より感染した牛を探りあて、それを取り除くことが可能となるからである。
【0005】
ところで、従来のワクチンよりも一層安全と思われる野生型ウイルスとの識別が可能なBHV-1ワクチンの開発が行われてきた。流産を引き起こす程度もより低く、潜伏の頻度も低く、再活性化され得ることもないチミジンキナーゼ欠失変異体が単離された。さらに、組換えDNA技術を用いて、(血清学的技術により野生型BHV-1とワクチンの区別を可能にする)糖蛋白質gIIIのための遺伝子に欠失を有するBHV-1ワクチンが構築された。しかしながら、これらのワクチンに対してもさらに幾つかの異論がある。一方では、チミジンキナーゼ遺伝子はウイルス複製に関与しており、複製の程度が低い程、より程度の低い防御につながる可能性がある。他方、該糖蛋白質gIIIは防御抗体の生成に重要であり、この防御抗体は、gIII欠失ワクチンの効果を低下させる。実際上の問題として、一般には最高の防御が得られる、組換えワクチンの鼻腔内投与が幾つかの国では認可されていないという問題がある。したがって、有効かつ安全で、さらに野生型BHV-1と区別することが可能なワクチンが求められており、さらにまたこれは、かかるワクチンの少なくとも1つは、組換えDNA技術によって構築されたウイルスよりも、むしろ従来の方法で弱毒化されたウイルスに基づいたものであることが望ましい。
【0006】
ところで、細胞培養における継代によって、糖蛋白質gEのための遺伝子を欠くBHV-1菌株が得られた。我々が行った研究の最初の結果は、この遺伝子は、野生型BHV-1に関する血清学的識別に非常に役立ち、病原性の発現に関与していることを示している。したがって、その欠失は安全性に寄与しており、これによってチミジンキナーゼ欠失の使用は不要となるかもしれない。糖蛋白質gEは、防御の誘導にとって糖蛋白質gIIIほど重要ではないと思われる。従来の方法で弱毒化された野生型ウイルスと血清学的に識別することが可能な、従来の方法で弱毒化されたBHV-1菌株は独自のものである。ここで初めて述べた、gE-遺伝子の位置とDNA配列は、これまでは知られていなかった。また、これらに由来することができるオリゴヌクレオチドポリペプチドおよびオリゴペプチドも知られていなかった。また、gE-遺伝子に基づく血清学的識別を行うための試験も独自のものである。
【0007】
この"従来の"gE欠失変異体の重要な利点("従来の"とは従来の弱毒ウイルス単離方法を使用することを意味している)は、親換えワクチンに関する限りでは、鼻腔内投与が禁止されている国々において、この"従来の"gE欠失変異体を鼻腔内投与することが可能になるであろうという点である。しかしながら、安全に関して別の観点からよく考慮して、この従来のgE欠失ワクチンの他に、明確に限定された組換えの型も構築された。これらの組換えワクチンもまた欠失を有しており、-また、チミジンキナーゼ遺伝子の欠失も有していてもよく、または有していなくてもよい-および、これらの組換えワクチンは、異種の遺伝子の発現のためのベクタとして用いることもできる。これらの組換えワクチンは全て、同じgEに特異的な試験によって、野生型ウイルスと区別することができる。一組の異なるワクチンに対して標準的な試験を利用することは、国際的な努力としてのBHV-1の戦いにおいて強力な利点となり得る。かかる方法がBHV-1ワクチンの分野において述べられたことはない。
【0008】
牛における抗BHV-1反応の血清分析によって、抗gE抗体の重要部分は、糖蛋白質gEおよび他のBHV-1糖蛋白質である糖蛋白質gIによって形成される複合物に対して向けられていることが示された。したがって、かかる複合物に特異的な抗体の存在を示すこともできる血清試験は、抗-gE抗体を検出することができるだけの試験よりも高感受性であると思われる。gE単欠失変異体によりワクチン接種を受けた牛は、抗-gI/gE抗体の検出を妨げることが有り得る抗-gI抗体を作る可能性がある。したがって、この発明はgI/gE二重欠失を有するワクチンを含む。
【発明の開示】
【0009】
発明の要約
本発明は、糖蛋白質gE-遺伝子に欠失を有するBHV-1の欠失変異体を提供する。"〜に欠失を"という語は、全体としての遺伝子の欠失を含むことを意図している。本発明の好適な実施例は、以下に述べる欠失変異体Difivac-1のような、弱毒処置によって引き起こされた糖蛋白質gE-遺伝子における欠失を有するBHV-1の欠失変異体によって構成される。
【0010】
本発明の他の好適な実施例は、以下に述べる欠失変異体1B7または1B8のような組換えDNA技術によって作られた糖蛋白質gE-遺伝子に欠失を有しているBHV-1の欠失変異体から成る。
【0011】
本発明の他の好適な実施例は、以下に述べるgI、/gE二重欠失変異体Difivac-IEのような、糖蛋白質gE-遺伝子および糖蛋白質gI一遺伝子に各々欠失を有するBHV-1の二重欠失変異体から成る。
【0012】
さらにまた、最高の安全性を期して、本発明によれば糖蛋白質gE-遺伝子およびチミジンキナーゼ遺伝子のそれぞれに欠失を有するBHV-1の欠失変異体が好適である。また本発明は、糖蛋自質gE-遺伝子、糖蛋白質gI-遺伝子、チミジンキナーゼ遺伝子に欠失を有するBHV-1の欠失変異体も含んでいる。
【0013】
本発明は、動物を、特に哺乳動物を、さらに特にウシ科の動物を上記において明確に述べたBHV-1欠失変異体を有するBHV-1から守るために、これらにワクチン接種をするためのワクチンの組成、適切な担体またはアジュバントを提供する。前記構成は、生きたワクチン組成でも不活化ワクチン組成でもよい。
【0014】
本発明はさらに、糖蛋白質gE-遺伝子に欠失を有し、組換えDNA技術によって導入された異種遺伝子を含むBHV-1の変異体において実施されている。好適には、糖蛋白質gE-遺伝子の位置に、組換えDNA技術によって導入された異種遺伝子を含むBHV-1の変異体に関するもので、前記異種遺伝子は、gE-遺伝子の調節配列の管理下にあり、シグナルペプチドをコードするgE-遺伝子の一部に任意に付加される。前記異種遺伝子は、BHV-1の異なるプロモータの管理下、または異種プロモータの管理下にあることも可能である。BHV-1の変異体が、糖蛋白質gE-遺伝子における欠失に加えて、さらに、チミジンキナーゼ遺伝子における欠失、および/または糖蛋白質gI-遺伝子における欠失などの欠失を有している場合には、前記異種遺伝子を、この付加欠失の位置に挿入してもよい。複数の挿入も別の選択肢であり、1つの欠失の位置にまとめて行うか、幾つかの欠失の位置に分けて行うかされる。
【0015】
好適に導入された異種遺伝子は、別の病原体の免疫原性蛋白質またはペプチドについて、または免疫反応を促進するサイトカインをコードする。適切なサイトカインの例としてはインターロイキンー2、αインターフェロン、γインターフェロンがある。
【0016】
本発明はまた、動物を、特に哺乳動物を、さらに特にウシ科の動物を(異なる)病原体より守るため、これらにワクチンを接種するための、(生きた、または不活化)ワクチンの組成を提供するもので、該ワクチンの組成には、免疫原性蛋白質または他の病原体ペプチドをコードしている異種遺伝子を有しているBHV-1突然変異体およびアジュバントの適切な担体が含まれる。勿論、この防御は1つ以上の病原体に関するものでよく、すなわち、変異体が複数の異種遺伝子を含む多価ワクチンに関するものでよい。
【0017】
さらにまた本発明は、BHV-1の糖蛋白質gE-遺伝子、この糖蛋白質gE-遺伝子の一部、またはこの糖蛋白質gE-遺伝子に由来するヌクレオチド配列を含む組換え核酸を含む組成物に関する。この組成は、BHV-1の糖蛋白質gE-遺伝子、この糖蛋白質gE-遺伝子の一部、この糖蛋白質gE-遺伝子に由来するヌクレオチド配列を含む組換え核酸の挿入を有するクローニングベクタまたは発現ベクタを含むことができる。
【0018】
また本発明は、BHV-1の糖蛋白質gE、この糖蛋白質gEの一部、この糖蛋白質gEに由来するペプチド、または糖蛋白質gEおよびgIの複合物を含む組成物、およびBHV-1の糖蛋白質gE、この糖蛋白質gEの一部、この糖蛋白質gEに由来するペプチド、または糖蛋白質gEとgIの複合物に特異的な抗体を有する組成物を含む。"抗体"とは、大半の用途に好適な、ポリクローナル抗体調製物と、大半の用途に好適なモノクローナル抗体の両方を意昧していると解釈される。"糖蛋白質gEの一部"および"糖蛋白質gEに由来するペプチド"という用語は、一般に少なくとも約8アミノ酸の長さを有するであろうgEに特異的なアミノ酸配列を意味していると解釈される。
【0019】
さらに本発明は、試料中の、特に、動物、特に哺乳動物、さらに特にウシ科の動物の、血液または血清、血液細胞、乳、涙などの体液、肺洗浄液、鼻水、精液、特に精漿、唾液、痰、または組織、特に神経組織のような生物学的試料中の、BHV-1の核酸を検出するための、BHV-1の糖蛋白質gE-遺伝子に由来するヌクレオチド配列を有する核酸プローブまたはプライマ、および核酸検出試験に適した検出手段を含む診断用キットに関する。
【0020】
さらに本発明は、試料中の、特に、動物、特に哺乳動物、さらに特にウシ科の動物の血液または血清、血液細胞、乳、涙のような体液、肺洗浄液、鼻水、精液、特に精漿、唾液、痰、または組織のような生物学的試料中の、BHV-1に特異的な抗体を検出するための、BHV-1の糖蛋白質gE、この糖蛋白質の一部、この糖蛋白質gEに由来するペプチド、または糖蛋白質gEおよびgIの複合物、および抗体検出試験に適した検出手段を含む診断用キットに関する。該診断用キットは、さらに、BHV-1の糖蛋白質gEに特異的な、またはBHV-1の糖蛋白質のgEとgIの複合物に特異的な1つまたはそれ以上の抗体を含めてもよい。
【0021】
また、本発明は、試料中の、特に、動物、特に哺乳動物、さらに特にウシ科の動物の血液または血清、血液細胞、乳、涙などの体液、肺洗浄液、鼻水、精液、特に精漿、唾液、痰、または組織、特に神経細胞のような生物学的試料中の、BHV-1の蛋白質を検出するための、BHV-1の糖蛋白質gEに特異的な、またはBHV-1の糖蛋白質gEとgIの複合物に特異的な1つまたはそれ以上の抗体および蛋白質検出試験に適した検出手段を含む診断用キットに関する。
【0022】
さらに本発明は、BHV-1の糖蛋白質gE-遺伝子を含む核酸の存在、BHV-1の糖蛋白質gE、またはBHV-1の糖蛋白質gEとgIの複合物の存在、またはBHV-1の糖蛋白質gEに特異的な、またはBHV-1の糖蛋白質gEとgIの複合物に特異的な抗体の存在について、動物の特に哺乳動物の、さらに特にウシ科の動物のBHV-1感染を判断するための、前記動物の試料の、特に、血液または血清、血液細胞、精液、特に精漿、唾液、痰、涙などの体液、肺洗浄液、鼻水、乳、または組織、特に神経組織のような生物学的試料の試験を含む方法を提供する。試験を行う試料は、本発明に従った組成のワクチンをそれまでに接種されたことのない動物のもの、または本発明に従って調製されたワクチンをそれまでに接種されたことのある動物のものとすることができる。
【0023】
発明の詳細な説明
本発明は、生ワクチンおよび不活化ワクチン両方から成る一組のBHV-1ワクチンに関するもので、これらはどちらも糖蛋白質gE-遺伝子の全部または一部を欠いている。この組は、チミジンキナーゼ遺伝子および/または糖蛋白質gI-遺伝子の欠失を含むことも含まないこともある自然gE欠失変異体および構築されたgE欠失変異体も含み、さらに異種遺伝子ベクタとして使用される構築されたgE欠失変異体も含む。さらに本発明は、BHV-1糖蛋白質gE-遺伝子をコードするヌクレオチド配列、これらの配列に由来するオリゴヌクレオチド、該糖蛋白質gEそれ自身、それらに由来するペプチド、および該gE糖蛋白質とそれらに由来するペプチドに対して誘導される(モノクローナルまたはポリクローナル)抗体に関する。また本発明は、BHV-1の糖蛋白質gEとgIの複合物、およびかかる複合物に対して誘導される抗体に関する。
本発明に従ったこれらの物質は、以下のために使用することができる。
【0024】
1)BHV-1によって引き起こされる疾病を予防するための牛のワクチン接種。
これはBHV-1に感染した動物と、ワクチン接種した動物の間の区別が可能で、従来のワクチンおよび構築されたワクチンを一緒に使用することが可能なものである。
【0025】
2)BHV-1による疾病および防御抗原に対するコード配列をBHV-1欠失変異体の中に組込むことができる他の病原体により引き起こされる疾病を予防するための牛のワクチン接種。
【0026】
3)牛の血液、血清、乳、他の体液について、それらが野生型BHV-1によって感染しているか、gE欠失変異体でワクチン接種されているかを、血清学的に、または核酸検出技術(たとえばPCR)により判断するためのそれらの試験。
【0027】
オリゴペプチド、ポリペプチド、およびBHV-1の糖蛋白質g-遺伝子と糖蛋白質gI-遺伝子のコード配列に由来する糖蛋白質の合成。
糖蛋白質gE-遺伝子(図3A)およびこの遺伝子をコードする単離DNAフラグメントについての、例に記載のDNA配列分析の結果により、標準的な分子生物学的処置を用いて、gE蛋白質のペプチド(オリゴまたはポリペプチド)を合成すること、および原核的方法(バクテリア中)または真核的方法(たとえば、ネズミの細胞において)により、gE蛋白質の全体を、またはその一部を発現させることが共に可能となる。これらの方法によって、たとえばgEに特異的なモノクローナル抗体の生成に役立つことができるgEに特異的な抗原を得ることができる。さらに、gEに特異的な抗原(および、gEに特異的なモノクローナル抗体)を血清試験において使用することができ、BHV-1 gE欠失ワクチンの接種をした動物と野生型BHV-1ウイルスに感染した動物の間の区別が可能となる。
【0028】
例に記載の、糖蛋白質gI-遺伝子、およびこの遺伝子をコードする単離DNAフラグメントについて、糖蛋白質gEを発現する真核細胞と共に、部分的にDNA配列を分析した結果、真核細胞中のgI/gE複合物を発現させることができる(図13および図14参照)。この糖蛋白質複合物は、gI/gEに特異的なモノクローナル抗体を作るために使用することができる。また、このgI/gE複合物は、単gE BHV-1欠失変異体、または二重gI/gE BHV-1欠失変異体によるワクチン接種を受けた牛と野生型BHV-1ウイルスに感染した牛の区別をする血清試験における抗原としても使用することができる。
【0029】
gEに特異的なペプチド
既知の蛋白質コード配列に基づき、自動合成装置により、約40〜50のアミノ酸のポリペプチドを作ることができる。BHV-1菌株LamのgE糖蛋白質の蛋白質コード配列が解明されたので(図3A)、このBHV-1 gE糖蛋白質のポリペプチドの合成が可能となる。かかるポリペプチドを用いて、標準的な方法に従って、マウスやウサギなどの実験動物を免疫化し、gEに特異的な抗体を作ることができる。さらに、これらのgEに特異的なペプチドを用いて、抗gE抗体がgE蛋白質(エピトープ)と反応する位置を、たとえばPEPSCAN法(Geysen et a1.,1984,Proc.Natl.Acad,Sci.USA 81,3998〜4002)により、さらに特定することができる。gEに特異的なオリゴペプチドも抗gE抗体を実証する血清試験において使用することができる。
【0030】
gEの原核発現
細菌中にてgE蛋白質を合成するため(すなわち、gEの原核発現)、糖蛋白質gEを、またはその一部をコードするDNAフラグメントを、原核発現ベクタ中にタローン化しなければならない。原核発現ベクタは、環状DNA分子で、これは別個に複製してゆく分子(プラスミド)として、細菌中で自らを維持することができる。これらの発現ベクタは、抗生物質抵抗についてコードする1つまたはそれ以上の標識遺伝子を含み、したがって発現ベクタを伴う細菌の選択を可能にする。さらに、発現ベクタは、(制御可能であることも多い)プロモータ領域を有しており、該領域の背後にDNAフラグメントを結合することができ、これらのフラグメントは、プロモータの影響により発現される。現行の多くの原核発現ベクタにおいては、いわゆる担体蛋白質に融合し、所望の蛋白質が発現している。そのために、ベクタ中ではプロモータの後に、担体蛋白質についてのコード配列が位置しており、所望のDNAフラグメントがこれに直接隣接して結合されている。融合蛋白質は、多くの場合より安定で、認識および/または単離もより簡単である。一定の細菌株にて、特定の融合蛋白が達成することができる定常状態レベルは融合によって、および菌株によって異なっている。種々の組合わせを試みることが慣例になっている。
【0031】
糖蛋白質gE-遺伝子の真核発現
蛋白質の原核発現は幾つかの利点をもたらしてくれるが、蛋白質は真核細胞で起こるグリコシル化のような修飾変更を欠いている。その結果、真核発現蛋白質は多くの場合より適切な抗原である。マウスの細胞のような真核細胞における蛋白質の異種発現の場合、真核発現ベクタが使用される。これらのベクタは、プラスミドで大腸菌細胞内で増殖させることが可能であるばかりでなく、真核細胞中にて安定した状態で生存することもできる。これらのベクタは、原核選択マーカの他に、真核選択マーカも含む。原核発現ベクタに似て、真核発現ベクタは、所望の遺伝子が後に結合することができるプロモータ領域を含んでいる。しかしながら、真核ベクタにおけるプロモータ配列は、真核細胞に特有である。また、真核ベクタにおいては、担体蛋白質に対する融合が利用されることは滅多にない。これらのベクタは、標準的なトランス・フェタション法(F.L.Graham and A. J. van der Eb. 1973, Virology 52,456〜467)により真核細胞中に導入される。真核プラスミドベクタの他に、ウィルスベクタもあり、この場合には、異種遺伝子がウィルス(たとえば、レトロウィルス、ヘルペスウィルス、ワクシニアウィルス)のゲノム中に導入される。次いで、真核細胞に組換えウィルスを感染させることができる。
一般に、個々の遺伝子産物にとって、ベクタや細胞はどういったタイプのものが最適であるかということを予測することはできない。大抵の場合、幾つかの組合せが試みられている。
【0032】
糖蛋白質gEおよび糖蛋自質gI両方の真核発現
蛋白質が得る最終構造は、蛋白質の一次アミノ酸配列、折たたみ、翻訳後修飾等に依存している。蛋白質の構造に寄与する重要なファクタは、1つまたはそれ以上の他の蛋白質との相互作用である。BHV-1糖蛋白質gEは、少なくとも1つの他の糖蛋白質、すなわちBHV-1糖蛋白質gIとの複合物を形成することを我々は発見した。かかる複合物を初めて示したものは、候補的抗-gEモノクローナル抗体1,51,67,75,78による我々の結果であった(表2参照)。これらのモノクローナル抗体は、Difivac-1とも、またLam gE-とも反応しなかったばかりでなく、糖蛋白質gE発現3T3細胞を認識することもできなかった。しかしながら、これらのモノクローナル抗体は、Difivac-1に感染後はgE-発現3T3細胞とは反応せず、これらのモノクローナル抗体のための適切な抗原コンホメーションを糖蛋白質gEに与えるために、因子を補足することが必要であることを示した。我々が行ったモノクローナル抗体81との放射線免疫沈澱試験の幾つかにおいて、見かけ上の分子量63KDの蛋白質が共沈することが判った。ヘルペス単ウィルス糖蛋白質gEは、これに匹敵する分子量の蛋白質(HSV1糖蛋白質gI)と複合物を形成するという事実に鑑み、我々はBHV-1糖蛋白質gEは、糖蛋白質gIのBHV-1相同体と複合物を形成するものと推定した。このBHV-1 gE/gI複合物を研究するため、および適切な抗原構造を有するgE抗原を作るため、これらの両蛋白質を1つの真核細胞中で発現させた。このために、糖蛋白質gEだけの真核発現のために記載したものと同じ処置を適用した。ただし、前提条件として、選択可能な種々の真核標識を有する発現ベクタの使用を1つだけ追加した。
【0033】
血清試験
gEに対する抗体に基づき、Difivac-1のワクチン接種をされた牛と、野生型BHV-1に感染した牛を区別するための血清学的方法は、好適にはgEに対して誘導されたモノクローナル抗体の使用に基づく。これらの抗体は、以下の方法で用いることができる。
【0034】
a)Van Oirschot et al(Journal of Virological Methods 22,191〜206,1988)により述べられた原則による。Aujeszky病のウイルスに対するgI抗体検出のための、この酵素結合イムノソルベント検定法(ELISA)では、抗体は、gI上に2つの異なったエピトープを有する2つのモノクローナル抗体の反応に対するブロック効果によって実証される。この試験は、以下のように実施される。マイタロタイタープレートを37℃で1晩、モノクローナル抗体1で被覆し、その後、たとえば4℃または-20℃にて保存する。試験される血清は、別の未被覆マイクロタイタープレートにおいて、37℃にてたとえば2時間、抗原と共に前保温される。モノクローナル抗体1が被覆されたプレートを、たとえば5回洗浄し、その後セイヨウワサビペルオキシダーゼ(HRPO)と結合したモノクローナル抗体2をこれらのプレートに添加する。次に、前保温された血清-抗原混合物を、2つのモノクローナル抗体が置かれているプレートに転移させ、37℃にてたとえば1時問保温する。これらのプレートを洗浄し、各ウェルに基質を添加する。室温にて約2時間経過させた後、これらのプレートについてスペクトル光度計による読取りを行う。各プレートには、4つの陰性対照血清および陽性血清の4つの連続希釈物が含まれる。光学濃度が同じプレートで試験された4つの陰性対照血清の平均光学濃度の50%を下回る血清は陽性とみなされる。
【0035】
b)間接2重抗体サンドイッチ(IDAS)法による。
この場合は、マイクロタイタープレートは、gE蛋白質に対して誘導されたモノクローナル抗体またはポリクローナル血清で被覆する。gE抗原調製物との保温の結果、被覆にgEが結合する。試験される牛の血清中のgEに対して特別に誘導された抗体は続いてgEに結合する。これらの結合抗体は、抗-ウシ免疫グロブリン複合物によって認識される。この複合物中の抗体は、ペルオキシダーゼ酵素に共有結合する。最後に、この結合複合物は、色素原基質の添加により、可視のものにされる。この反応の特性は、gE-抗原調製物の代わりに、gE-陰性対照調製物により同じ処置を実施して調べる。各マイクロタイタープレート上には、陽性および陰性の対照血清が含まれる。この試験は、陽性の血清が一定の希釈度において陽性を示す場合に有効である。光学濃度が標準陰性対照血清よりも0.2高い場合には、血清は陽性である。
【0036】
c)2において述べたIDAS法では、ただし試験される血清を保温した後であるが、抗-ウシ免疫グロブリン複合物の代わりに、抗-gEモノクローナル抗体/HRPOが使用される。抗-gEペプチド血清または抗-gEポリクローナル血清を抗-gEモノクローナル抗体の代わりに使用してもよい。これらのプレートを洗浄し、各々のウエルに色素原基質を添加する。室温にてたとえば、約2時間経過させた後、これらのプレートについてスペクトル光度計による読取りを行う。各プレートには、4つの陰性対照血清および陽性血清の4つの連続希釈物が含まれる。光学濃度が同じプレートで試験された4つの陰性対照血清の平均光学濃度の50%を下回る血清は陽性とみなされる。
【0037】
d)精製、未精製を問わないウイルス抗原がマイクロタイタープレートに一晩被覆されるブロックELISA法に従って、これらのプレートにおいて、試験される血清は、37℃にてたとえば1時問または1時間以上保温される。洗浄処置をした後、これらのプレートに抗-gEモノクローナル抗体を添加し、37℃にてたとえぱ1時間保温する。抗-gEペプチド血清または抗-gEポリクローナル血清を抗-gEモノクローナル抗体の代わりに使用してもよい。これらのプレートを洗浄し、各ウエルに、色素原基質を添加する。室温にて、たとえば2時間経過させた後、これらのプレートについて、スペクトル光度計による読取りを行う。4つの陰性対照血清および陽性血清4つの連続希釈物が含まれる。光学濃度が、同じプレートで試験された4つの陰性対照血清の平均光学濃度の50%を下回る血清は陽性とみなされる。
【0038】
上記の方法のいずれにおいても、gEを含む、従来の方法で成長させたウイルス抗原を使用することができるが、原核または真核を介して発現するgE-抗原も使用することができる。あるいはまた、BHV-1 gE配列に基づくオリゴペプチドを従来の抗原の代わりに、上記診断試験において使用することができるであろう。さらにまた、かかるオリゴペプチドを、Kemp et al.による記事、Science 241,1352〜1354,1988に記載の方法に従った、いわゆる"Cow-side"試験の開発に使用することができるであろう。かかる試験は、感染動物中に存在しているgEに対して誘導された抗体によるオリゴペプチド抗原配列の連結に基づいたものになるであろう。このような試験の場合、オリゴペプチドを、牛の赤血球に対して誘導されたモノクローナル抗体に連結させる必要があるであろう。
【0039】
ポリメラーゼ連鎖反応を使用する核酸分析
たとえば、オリゴヌクレオチド(プローブおよびプライマ)をポリメラーゼ連鎖反応に用いて、ワクチン接触を受けた動物と感染した動物を区別することができる。ポリメラーゼ連鎖(PCR)は、病原体の核酸を短時間に数十億倍に増殖させることができる技術である(Depolymerase kettingreactie, P.F.Hilderink, J.A.Wagenaar,J.W.B.van der Giessen and B.A.M. van der Zeijst,1990,Tijdschrift voor Diergeneeskunde deel 115,1111〜1117)。gEオリゴヌクレオチドは、gE陽性ゲノムにおいて、gE陰性ゲノムにおけるものとは異なる生成物が形成されるように選択することができる。これは、gE欠失ワクチンをワクチン接種された動物は、PCR試験において陽性シグナルを出すという利点がある。しかしながら、この方法は、試験される動物から採取した試料中、たとえば血液中のウイルスの核酸の存在に依存している。
BHV-1急性感染後は、血液中にBHV-1特有の核酸を実証することができる見込みは充分にあるが、しかしながら、BHV-1核酸も、潜伏時に血清中に実証することができるかどうかはまだ確められていない。
【0040】
ベクタとしてのBHV-1の使用
BHV-1ゲノム中の異種遣伝子の発現には、その異種遺伝子が挿入されるべき領域についての正確な情報を自由にできることが必要である。必須配列についての妨害はいかなるものもあってはならず、また異種遺伝子の発現に調節配列が利用できなければならない。gE-遺伝子は、必須ではなく、したがって、gE遺伝子を異種遺伝子で置き換えても問題はない。結果として、この異種遺伝子を、gE遺伝子が調節配列の影響下にあるように配置することができる。しかしながら、gE遺伝子の調節配列を使用する必要はない。異種遺伝子の発現は、代わりに、他のもの、たとえば種々の遺伝子のより強い調節配列により制御することができる。また、gE遺伝子の(移行)シグナルペプチドに異種遺伝子を連結することも可能であり、したがって異種遺伝子生成物の分泌に影響させることができる。gE遺伝子およびgE蛋白質についての詳細な知識によって、非常に適切に調整された方法で、ベクタとしてBHV-1を用いる可能性が与えられることは明らかである。発生するベクタは、さらに野生型と血清学的に区別することができる。異種遺伝子を発現するBHV-1変異体の構築は、例に示したgE欠失変異体の構築と同じ方法で実施することができる。しかしながら、そのときは、欠失フラグメントは、異種遺伝子が欠失位置に配置されているフラグメントと置き換えられるべきである。
【0041】
例
1)自然gE欠失変異体の単離および同定
a)自然変異体の単離
ゲノムDNAを、従来法により弱毒化された多くのワクチンから標準的方法に従って単離し、制限酵素を用いて分析した。特に、野生型BHV-1ウイルスとの区別を可能とするに適切と思われるゲノムの逸脱(genome deviation)について調べた。
特に、BHV-1ゲノムのUs領域に注意した。これは、その領域に-ヘルペス単ウイルス1との類似により-非必須糖蛋白質をコードする多くの遺伝子が恐らく配置されているからである〔Identification of a herpes simplex virus1 glycoprotein gene within a gene cluster dispensable for growth in cell culture, R. Longnecker,S.Chatterjec,R.J. Whitley and B.Roizwan(1987)Proc、Natl. Acad. Sci,84,4303〜4307〕。
ユーゴスラビア、サグレフ大学より入手したBHV-1ワクチンバッチ(Lugovic et a1.,Veterinarski Arhiv 55,241〜245,1985)は、ウシの胚腎臓細胞およびウシの胚気管細胞(Ebtr)上で多くの継代を行った後に、正常なUs領域の他に、逸脱のあるUs領域を有していることを証明した。さらにこのワクチンは、Ebtr細胞に大きなプラークと小さなプラークの両方を形成するように思われた。この混合群より、それぞれの国ごとに選択された小さなプラークで偏差のあるUs領域を有するウイルスを、3つの限界希釈ステップによって単離した。この経路を介して単離されたウイルスをさらに試験し、Difivac-1を希釈した。これは、1992年5月27日Institut Pasteur,Paris,Franceに寄託番号1-1213として寄託された。
【0042】
b)Difivac-1におけるgE遺伝子の欠失の同定
US領域におけるこの逸脱をさらに分析するために、Difivac-1のゲノムDNAを標準的な方法で単離し、サザンブロット分析を行った(図1A)。このブロットを32Pでラベルされた野生型HindIIIKフラグメントとハイブリッド形成させることにより、Us領域の中心に位置しているこのフラグメントは、Difivac-1においては、1.0キロベース(kb)程度短いことが確認された。さらに、この分析によって、欠けている部分の位置をおおよそ推定することができた(図1B)。この欠失をさらに分析するために、野生型BHV-1菌株LamのUs領域を単離し、原核ベクトル中にクローン化した。そのために、標準的な方法に従って、Lam菌株(図2A)のゲノムDNAを単離し、ベクタ、pUC18、pACYCおよびpBR322中にクローン化した(図2B)。欠失の推定位置周辺の領域の物理的地図を作成した(図2C)。この物理的地図からスタートして、この領域のヌクレオチド配列の決定に適したサブクローンをベクタpKUN19とpUC18で構築した(図2D)。これらのサブクローンを用いて、全領域の2つの鎖のヌクレオチド配列(図2Cに指示)をサンガー法を用いて決定した。このヌクレオチド配列(SEQ.ID.NO.1)をPc、/Geneプログラムを用いて分析した。概念翻訳から、ヌクレオチド(nt)168〜nt1893が575のアミノ酸のオープンリーディングフレームのための遺伝暗号を指定する(図3A)ものと推定された。さらに分析の結果、このアミノ酸配列は図3Bに示されているように、貫膜糖蛋白質の特性を有していることが示された。実は、最初の26のアミノ酸(aa)は、典型的真核性の移行シグナルとして認識され、aa423とaa450の間の領域は、貫膜領域として認識される。さらに、3つのポテンシャルN-結合、糖鎖形式部位がこの配列において発生する。この推定アミノ酸配列は、ヘルペス単ウイルス(HSV)の糖蛋白質gE-遺伝子に明らかに類似したところを示している。(図4Aおよび図4B参照)。これらおよびその他の類似点により、見つけられた遺伝子は、BHV-1のgE相同体であるという結論を正当化するものである。この理由から、遺伝子はgEと呼ばれる。このBHV-1 gE-遺伝子がDifivac-1においてどの程度欠失しているのかを判断するために、p318フラグメントを単離した。p318フラグメントは、要求されるBHV-1 gEオープンリーディングルーム上流のAluI部位55ntで開始し、オープンリーディングルーム後の133ntで終了する。ゲノムDifivac-1 DNAを、サザンブロットハイブリッド形成を用いて、このp318フラグメントで分析した。この結果、Difivac-1は検出可能なp318配列を全く含んでいないことが明らかとなった(図5)。この試験によりDifivac-1は欠失を有し、この欠失はgE遺伝子全体に広がっていることが明らかに実証されていることが確認された。
【0043】
欠失領域の大きさと位置を決定するために、Difivac-1のUs領域を包括しているゲノム配列を原核ベクタ中にクローン化した(図11Cを参照)。14.5kb EcoRIフラグメントをpACYCベクタ中にクローン化し、P775と命名した。7.4kb HindIIIフラグメントを独自にpUC18ベクタ中にクローン化し、p728と命名した。クローンp728より、2つのサブクローンを単離した。すなわち、クローンp737中の1.4kb PstIフラグメントとクローンp754中の350bp AluI-PstIフラグメントである。これらのクローンの制限酵素分析とサザンブロット分析(データは示されていない)により、Difivac-1のgE欠失の長さは2.7kbであり、gE遺伝子から丁度5'でスタートし、Us領域の境界で終了することが実証された。これらの2.7kbは、反復領域の異常伸長物として、gE遺伝子に対向したUs領域に位置した重複1kbセグメントによって置き換えられた(図11bを参照)。この分析の結果を確認するために、および正確な組換え点を決定するために、クローンp754の大半の挿入物についてのヌクレオチド配列を決定し、野生型配列を比較した(図12参照)。この分析により組換え点は、gE遺伝子の開始コドンから77bp上流に位置していることが判った。
【0044】
c)Difivac-1の安全性と効能についての評価
7週齢のBHV-1血清反応陰性の特定の病原体をもっていない牛でDifivac-1を試験した。8体の牛に2ml,105TCID50の鼻腔内ワクチン接種を行った。2ml中の1mlは各々の鼻孔にスプレーした。各々別々の独立した区画に収容されていた、BHV-1血清反応陰性の特定病原体を持っていない7週齢の8体の牛に2mlの培養基を鼻腔内投与し、非ワクチン接種対照として用いた。ワクチン接種から5週後に、ワクチン接種牛と非ワクチン接種牛に107TCID50、の強毒性BHV-1菌株、Iowaを鼻腔内接種した。接種から6週後に推定される潜伏ウイルスを再活化させるため、全牛に5日間デキサメタゾンにより筋肉内処理を行った。臨床微候、直腸温、発育を監視した。布で拭き取った鼻水からウイルスの単離を行い、血清中の中和抗体力価を求めた。
【0045】
ワクチン接種後の、牛の挙動、食欲、直腸温、発育率は正常のままであったが、ワクチンを接種した牛は、いくらか漿液性鼻水の分泌と唾液の分泌過多があった。Difivac-1は、ワクチン接種後、布で拭き取った鼻水から排泄された(図17)。ワクチン接種牛は全て、BHV-1に対する中和抗体を生成した。
菌株接種後、14日までに、非ワクチン接種牛は全て、無感情、食欲の喪失、目ヤニおよび鼻水の分泌、下あごの歯肉の赤らみ、鼻の粘膜の強い傷害が見られ、また接種後4日で発育が停止した。ワクチンを接種した牛の鼻の粘膜の傷害は小さく、速やかに治癒するもので、発育が止まることはなかった。菌株接種後の1日毎の臨床評価、直腸温、発育の推移を図18,図19および図20に示した。菌種接種後、全牛とも鼻からウイルスを排泄したが、ウイルス排泄の量と期間は、ワクチン接種牛は著しく低かった(図21)。ワクチン接種牛においては、二次抗体反応が見られ、非ワクチン接種牛は全て、菌種接種後抗体を生成した。
再活化後、接種ウイルスを1体のワクチン接種牛および5体の非ワクチン接種牛から単離した。Difivac-1を再活化することはできなかった。
【0046】
上記の結果により、Difivac-1は、若年牛においては殆ど病気の微候を引き起こさず、再活化されないことが実証された。Difivac-1は、菌種接種後病気の程度とウイルスの排泄量を著しく低下させた。
結論として、Difivac-1は、BHV-1感染予防のために牛に使用される、安全かつ効能のあるワクチンである。
【0047】
2)BHV-1の組換えgE欠失変異体の構築
Difivac-1よりも分子的により明確で、かつ所望されるならば、gE-遺伝子の欠失に加えて、たとえばチミジンキナーゼ遺伝子に欠失を有した識別可能なBHV-1ワクチンと自由に利用できるようにするために、Difivac-1の他に、組換えgE欠失変異体を構築した。糖蛋白質gE-遺伝子の決められた位置からスタートし、gE遺伝子を側面に配置するクローン化されたDNAフラグメントを用いて、gE欠失フラグメントを構築することができた。標準的な技術(F.L.Graham and A.J.van der Eb 1973,Virology52,456〜467)を用いて、,この欠失フラグメントを野生型BHV-1菌株のゲノム中で組換えることができ、その結果gE欠失変異体となった。
【0048】
a)gE欠失フラグメントの構築
【0049】
gEフラグメントの構築のために、一方においては、全gE配列を欠き、他方野生型ゲノムと組み換えることができる十分なフランキング配列を含むフラグメントが目標とされた。5'(上流)側では、gE遺伝子の開始コドン18nt上流で終了する1.2kb PstI-AsuIIフラグメントが選択された。3'(下流)フラグメントについては、gE遺伝子の停止コドンの2nt上流で開始する、1.2kb EcoNI-DraIフラグメントが選択された(図6)。
【0050】
gE欠失フラグメントの構築については、gE遺伝子の5'側に位置した、BHV-1菌株Lamの8.4kb HindIIIKフラグメントからの1.4kb PstI-SmaIフラグメントが、プラスミドpUC18のSmaIおよびPstI中にサブクローンされた。このクローンはp515と命名した。gEの3'側に位置し、4.1kb HindIII-EcoRIクローンからくるEcoNI-SmaIフラグメントはp515の特異なAsuII部位中にクローンされた。このように、gE欠失フラグメントの構築は完結し、このようにして構築されたクローンをp519と命名した。原則的にはp519の全PstI-SmaI挿入をgE欠失フラグメントとして利用することができるが、これは望ましくない。実際は、PstI-SmaIは、Us領域をフランクする反復配列中に、約100〜150塩基対(bp)の伸長を行う。この100〜150bpの部分は、gE遺伝子が位置しておらず、したがって所望されていない組換え生成物を作り出すことができないUs領域の他方の反復配列と組換えることができた。このため、PstI-DraIフラグメントが組換え実験のために選択され、したがって100bpの反復が除去された。
【0051】
b)野生型BHV-1のゲノムによるgE欠失フラグメントの組換え
構築されたgE欠失フラグメントと野生型BHV-1のゲノムの間の組換えを効果的なものとするために、2つのDNA分子のマイクログラム量を、F.L.Graham and A.J.van der Eb(1973、Virology52,456〜467)の標準的な方法に従って、ウシの胚気管(Ebtr)細胞に同時トランスフェクションする。細胞の組換えメカニズムは、細胞によって組み込まれた少しの割合のDNA分子(2〜4%)の組換えを導く。組換えられたgE欠失変異体の選択のために、トランスフェクション後に形成されるウィルス混合物を、新鮮なEbtr細胞培養物上に播種する。大抵の場合、これによって発達する、別々のウィルス群(プラーク)が、1つのウィルスから生じる。BHV-1菌株LamのgE欠失変異体の単離のために、これらの230のプラークを単離し、DifivaC-1に感染した細胞とは反応しない、BHV-1に特異的なモノクローナル抗体による標準的な免疫法により試験を行った。これらのモノクローナル抗体は、糖蛋白質gEに対して誘導される。230のプラークの内5つがこれらのモノクローナル抗体と反応しなかった。これらの5つのプラークのDNAをさらに調べた。
【0052】
c)BHV-1菌株Lamの構築されたgE欠失変異体のDNA分析
上述の5つの候補gE欠失変異体の内3つ(1B7,1B8および2H10)のDNA調製物について、標準的なサザンブロット分析技術(Sambrook et al.1989)を用いて試験した。PstIとDraIによってこれらのDNA調製物を2重消化し、続いてゲル電気泳動と、プローブとして2.3kb PstI-DraI欠失フラグメントを用いてサザンブロットハイブリット化によって、ウィルス群1B7と1B8のゲノムのgE遺伝子は、所望の方法で正確に取り除かれたことが判る(図7Aおよび図7B参照)。群2H10は、逸脱したPstI-DraIフラグメントを有している。gEに特異的なプローブによるサザンブロットハイブリット化により、3つのDNA調製物のいずれにもgE配列は置かれていないことを示している(結果については示されていない)。BHV-1ウィルス群1B7および1B8は、故意の組換えgE欠失変異体である。BHV-1ウィルス群1B7はワクチン特性について試験を行った。
【0053】
d)チミジンキナーゼ/gE二重欠失変異体の構築
わずか1つの遺伝子に欠失を有するBHV-1組換え欠失変異体は、十分に毒性が低下したものではないかもしれないので、BHV-1菌株LamおよびHarberinkのチミジンキナーゼ(CTK)遺伝子にも欠失を与えた。これらの変異体は、上記のgE欠失変異体に対して使用される方法と類似の方法で構築された(結果は示されていない)。これらのTK欠失変異体はTK/gE二重欠失変異体の構築に使用した。
【0054】
e)糖蛋白質gI/糖蛋白質gE二重欠失変異体の構築
gE単欠失変異体のワクチン接種をうけた牛は、抗gI/gE抗体の検出を妨害するおそれのある抗-gI抗体を生成する可能性があるため(以下に考察)、gI/gE2重欠失を有するワクチンについても発明した。このようなgI/gE2重欠失変異体は、gE単欠失変異体の構築に使用されるのと同じ処置を用いて構築することができる。gE遺伝子の5'側端部をカバーする、1.8kb PstIフラグメントの上流側端部のヌクレオチド配列の部分的分析により、他のヘルペスウィルスに見られるgI相同物に対して有意な相同関係を有するオープンリーディングフレームが明らかになった(図13および図14を参照のこと)。gI遺伝子の推定5'側端部を包む350bp SmaI-PstIフラグメントおよびgE遺伝子の下流に位置したEcoNI-SmaIフラグメントを用いて、gI/gE欠失フラグメントを構築することができる。このフラグメントは野生型ゲノムと組換え、BHV-1 gI/gE欠失変異体をもたらすことができる。図16を参照のこと。理論的にはさらにまだ生成され得る80〜90のアミノ酸は、抗-gI/gE抗体の欠失を防げることができる抗体を導き出すことはできないであろう。さらに、gI遺伝子の配列分析により、全gIコード領域にわたるgI欠失の構築が可能となるであろう。このgI/gE二重欠失変異体をDifivac-IEと命名した。
【0055】
f)Lam gE-およびLam gE-,TK-変異体の安全性と効能の評価
Lam gE-およびLam gE-,TK-,BHV-1変異菌株のワクチン特性を7週齢の、BHV-1血清反応陰性の特有病原体を持たない牛において試験した。各々の変異体を6体の牛に鼻腔内スプレーした。各牛の全投与量は、2ml培養基において105TCID50となり、2ml中1mlは各々の鼻孔にスプレーした。他の6体の牛はウィルスを含まない培養基を鼻腔内投与し、非ワクチン投与対照として用いた。ワクチン投与から5週後に、ワクチン投与および非ワクチン投与いずれの牛も、すべての牛に強毒性BHV-1菌株Iowa107TCID50を鼻腔内接種した。ワクチン接種および菌株接種後、臨床徴候、直腸温、体重を監視した。鼻のウィルスの放出日数の決定のために布で拭きとった鼻水を採取した。
ワクチン接種後の、牛の挙動、食欲、直腸温、発育率は正常を維持した。漿液性鼻水の分泌および鼻の粘膜の小さな傷害が、ワクチン接種牛すべてに観察された。ワクチン接種の牛の鼻から約7日間ウィルスを単離することができた(表1)。
【0056】
菌接種後、非ワクチン接種の対照牛すべてが、無感情、食欲の喪失、目や鼻からの分泌、下あごの歯肉の赤らみ、鼻の粘膜の強度の傷害を示し、発育が抑制された。Lam gE-,TK-をワクチン接種した牛はすべて、鼻水の分泌が幾分増加し、鼻の粘膜の小さな傷害がいくらか見られた。Lam gE-をワクチン接種した牛のすべてが、鼻水の分泌または鼻の粘膜の傷害が進行したわけではなかった。無感情、食欲の喪失、疾病についてのその他の徴候は、ワクチン接種牛では観察されなかった。菌接種後の直腸温、発育、臨床評価は図22,図23および図24に示されている。非ワクチン接種牛はワクチン接種牛の2倍の長さにわたってウィルスを鼻から放出した(表1)。
【0057】
上記の結果は、Lam gE-およびLam gE-,TK-BHV-1突然変異菌株は、若年の牛においては、病気の臨床徴候をほとんど誘発していないことを示している。これらの突然変異菌株はどちらも菌接種後の病気を予防し、鼻のウィルス放出期間を50%短縮させた。
Lam gE-およびLam gE-,TK- BHV-1突然変異菌株は安全で、BHV-1感染予防のワクチンとして牛に用いて効能を有するものである。
【0058】
3)gEの原核発現
BHV-1糖蛋白質gE-遺伝子の原核発現の場合、これまでのところpGEX発現ベクタ(D.B.Smith and K.S.Johnson,Gene 67(1988)31〜40)が使用されてきた。イソプロピルチオガラクトシド(IPTG)によって発現を誘導されることができ、tacプロモータの影響下にある日本住血吸虫由来の担体蛋白質グルタチオンS-転移酵素(GST)を、pGEXベクタはコードする。GST-gE融合蛋白質の例としては構築されたpGEX-2T600s3の生成物がある(図8A)。この構築物において、標準的な分子生物学的技術を用いて(Sambrook et al. 1989)、gE蛋白質の200アミノ酸のN末端領域をコードする。600bp SmaIフラグメントがGST遺伝子の後に連結された。この構築物は3つの部分から成る設計とされ、600bpフラグメントの異なったリーディングフレームがGSTに連結された。これらの構築物はすべてIPTGと共に誘発された大腸菌株DH5α内に導入され、形成された蛋白質はウェスタンブロッテングによるポリアクリルアミドゲル電気泳動後、ニトロセルロースに転移された。抗-GSTによる免疫性検出により、gE蛋白質領域をコードする適切なリーディングフレーム(No.3)だけが、27k(GST)t20k(gE)=47kの推定サイズの顕著な融合蛋白質の発現を導くことが実証された。Difivac-1と反応しない我々が単離した3つのモノクローナル抗体は、ウェスタンブロット中の47kD GST-gE融合蛋白質を認識する。図8B参照。
【0059】
4)糖蛋白質gE-遺伝子の真核発現
糖蛋白質gE-遺伝子の真核発現については、これまでは、とりわけ、ベクタ、pEVHisが選択されてきた。pEVHisベクタは、真核遺伝標識として、ヒスチジノールデヒドロゲナーゼについてのHisD遺伝子コードを有しており〔EC1.1.1.23〕(C.Hartmann and R.Mulligan,1988,Proc,Natl.Acad Sci.USA 85,8047〜8051)、このヒスチジノールデヒドロゲナーゼが、細胞の2.5mMヒスチジノールの毒性濃度を長びかせる原因となっている。さらに、このベクタは、ヒトのサイトメガロウィルス(HCMV)の極初期遺伝子のプロモータ領域を、該遺伝子の後に位置している特異な制限酵素部位と共に有している。pEVHis/gE発現ベクタの構築のためには、糖蛋白質gE-遺伝子の全コード領域を有するフラグメントが使用された。gEの従来のオープンリーティングフレームの上流のAluI部位55でスタートし、その下流の133bpで終了する。この領域がpEVHisベクタのHCMVプロモータの後方でクローン化され、これによって構築物pEVHis/gEが形成された(図9)。このpEVHis/gEを大腸菌株DH5αの細胞中にて増殖させ、塩化セシウム勾配法によって精製した(Sambrook et al.,1989)。この精製DNAを、Graham and Van der Ebの方法に従って、Balb/c-3T3細胞にトランスフェクトした。形質転換細胞をヒスチジノールを用いて選抜した結果、20の耐ヒスチジノールコロニーを単離することができた。これらのコロニーを免疫ペルオキシダーゼ単分子層試験(IPMA)によって、モノクローナル抗体81によって試験した。4つのコロニ一によって、gE蛋白質の発現が証明された。これらの4つのコロニーの内、3T3gEクローン9を、高度のgE発現を有するサブクローンの単離に使用した。この方法によって単離されたクローン(3T3gE9.5と命名)を候補抗gEモノクローナル抗体の特性づけのために用いた。
【0060】
5)同一細胞におけるBHV-1糖蛋白質gEおよびBHV-1糖蛋白質gI双方の真核発現
BHV-1糖蛋白質gEと同一の細胞においてBHV-1糖蛋白質gIを発現させるため、まず最初にBHV-1 gI遺伝子の推定位置を決めた。ヘルペス単ウィルス糖蛋白質gI遺伝子は、糖蛋白質gE遺伝子のまさに上流に位置しているので、BHV-1 gI遺伝子は、対応の位置に位置しているものと推定された。これを試験するため、BHV-1 gE位置の開始点より約1kb上流に位置した、283ヌクレオチドの領域の配列を決定した。この領域の概念翻訳により、第2リーディングフレームは、ヘルペス単ウィルス糖蛋白質gIと相同の94アミノ酸配列をコードする(図13および図14参照)ことが判った。相同セグメントは、スタートコドンから約80のアミノ酸であるので、BHV-1 gI遺伝子のオープンリーディングフレームの推定開始点は、配列領域から約250nt上流と見積もられる。これより、配列領域から400nt上流でスタートし、gE遺伝子内で終了する、1.7kb SmaIフラグメントは、BHV-1 gI遺伝子の完全なコード領域を含んでいるべきであることが推定された。この1.7kb SmaIフラグメントが真核ベクタMSV-neo内にクローン化された(図15参照)。このベクタは、強いマウス肉腫ウィルスプロモータと、抗生物質G-418硫酸ゲネティシンに対する耐性についてコードするセレクター遺伝子neoを含んでいる。結果として生じる構築物MSVneoGIが、大腸菌DH5α細胞中で増殖され、Graham and Van der Ebの方法を用いて、3T3gE9.5細胞中にトランスフェクトされた。トランスフェクトされた細胞は、400μgゲネティシン/ml培養基で選抜され、耐性コロニーを単離し、3T3gE9.5細胞と反応しなかった候補抗-gEモノクローナル抗体で試験した。これによって、たとえば、野生型BHV-1だけでなく、モノクローナル抗体66と反応した3T3gE/g IR20クローンを選択した。
【0061】
6)候補抗-gEモノクローナル抗体の特性づけ
野生型BHV-1に対してモノクローナル抗体を生成し、Difivac-1に感染したEbtr細胞との反応不能性について選択した。これらのモノクローナル抗体を下記との反応性について試験する。
a)Lam gE-欠失変異体
b)ウェスタンブロットにおける、上記の原核発現生成物
c)上記のgE発現Balb/c-3T3細胞
d)c)に記載のDifivac-1に感染した細胞
e)gE/gI複合物を発現するBalb/c-3T3細胞
a,c,dおよびeとの反応性試験については、免疫ペルオキシダーゼ単分子層試験(IPMA)を用いた。表2の結果により、我々が生成したものは、gEに対して誘導されるモノクローナル抗体(2,3,4,52,66,68,72,81)およびgE/gI複合物上の立体配座抗原ドメインに対して誘導され得る可能性のあるモノクローナル抗体(NO.1,51,53,67,75,78)であることが判る。種々のモノクローナル抗体によって認識される抗原ドメインの地図を作るための競合IPMAにより、少なくとも4つの抗原ドメインが糖蛋白質gE上に存在すること、およびおそらく1つのドメインがgE/gI複合物により形成されることが示された(表2)。
【0062】
BHV-1に感染した牛の抗-gE抗体の検出
感染した牛の血清中にgE抗体が存在しているか否かを調べるために、間接ブロッキングIPMAを16の候補gE-モノクローナル抗体に実施し、以下の8血清を選択した:
−Difivac-1のワクチン接種を受け、毒性Iowa菌株が接種された牛の菌接種14日後に採取された2血清。
−試験のためBHV-1サブタイプ1ウイルスに感染させた牛の、感染20ケ月後に採取された2血清。これらの牛のうちの一体は接触感染によって感染させた。
−試験のためBHV-1サブタイプ2bウイルスに感染させた牛の、感染20ケ月後に採取された2血清。これらの牛のうちの一体は接触感染によって感染させた。
−ts変異体ワクチンの接種を行い、3週後にBHV-1サブタイプ2bウイルスを接種させた特有病原体のない牛の、菌接種7週後に採取された1血清。
−ts変異体ワクチンの接種を行い、3週後にBHV-1サブタイプ2bウイルスを接種させた、ノトバイオートの牛の、菌接種7週後に採取された1血清。
【0063】
表2により、これらの血清はどれも、gE上の抗原性ドメインIIIおよびIVに対する、およびgE/gI複合物上に位置していると思われる抗原性ドメインIに対する抗体を含んでいることが判る。gEは、明らかにBHV-1-感染とワクチン接種された牛を区別するための適切な血清学的遺伝標識であると結論づけることができる。
【0064】
7)BHV-1 gE-特異型プライマを用いるPCR処置によるBHV-1核酸の検出
BHV-1 gE遺伝子の決定されたヌクレオチド配列からスタートして、PCRに適切なプライマの対を、Lowe et alによるプライマ選択プログラム(T. Lowe, J.Sharefkin, S, Qi Yang and C. W .Dieffenback 1990, Nucleic Acids Res, 18, 1757〜1761)を用いて選択した。これらのプライマをP3、P4と命名し、図10に示した。これらのプライマは159nt離れて位置しており、200ntのフラグメントの増殖を導く。プライマP3、P4と単離されたBHV-1 DNAを用いて、PCR処置の条件を最適化した。これは特に、MgCl2の濃度の変更、グリセロール濃度、サイクル条件に関するものである。BHV-1 DNAの増殖のためにP3、P4を使用するため最適と考えられる緩衝液は、10mM Tris pH 8.0, 50mM KCl, 0.01%ゼラチン, 2.6mM MgC12および20%グリセロールである。最適のサイクル条件と考えられるものは(Perkin Elber Cetus DNA Thermal Cycler),サイクル1〜51については、1分98℃, 30秒96℃, 30秒55℃, 45秒72℃である。PCR増殖後、得られた200nt DNAフラグメントを2%アガロースゲル上で電気泳動にかけ、ニトロセルロースにブロットし、続いてサザンブロット分析を行った。サザンブロット分析に使用された32P dCTPのラベルされたプローブが、プライマ結合部位間に位置している137bp TagIフラグメントである(図10)。ハイブリッドフィルターの自動X線撮影により、200bpバンドを観察することができた。さらに、このルートを介して、10BHV-1ゲノムだけの増殖(約1.5×10-15μgDNA)が検出可能と思われるシグナルを導く(結果は示されていない)。同等の方法で、BHV-1糖蛋白質gIIIのコード配列(D.R.Fitrpatrick, L.A. Babiuk and T.Zamb, 1989, Virolagy 173, 46〜57)に基づくプライマを用いてPCR処置を行った。野生型BHV-1 DNAとgE欠失変異体ワクチンの区別を可能とするために、DNA試料にgE-特異型PCR分析およびgIII-特異型PCR分析の両分析を行った。このような試験において、Difivac-1 DNA調製物はgIII陽性およびgE陰性であることが判った。
ウシの精液中のBHV-1 DNAの検出は、BHV-1特異型PCR処置の重要な用途であると思われるので、BHV-1に感染したウシの精液にgE-特異型PCRの実施を試みた。しかしながら、精液中の未知の成分がポリメラーゼ連鎖反応に強い抑制効果を有している。したがって、ウシの精液からBHV-1 DNAを単離するプロトコールが展開された。ウシの精液からDNAを単離するため、30μlの精液を、1mg/mlプロティナーゼK(pk)とともに、60℃にて、全量300μlの0.15M NaCl, 0.5%Na-サルコシル, 40mM DTT中にて培養する。1時間後、試料を室温にまで冷却し、300μl 6M NaIを添加し、5分間培養する。この混合物より、標準クロロホルム/イソアミルエタノール抽出によりDNAを単離し、1容量のイソプロパノールで沈殿させた。沈殿物を2.5M NH4AC/70%エタノールで洗浄し、60℃にて1時間10mM Tris pH7.4,1mM EDTA,0.5%Tween80および0.1mg/ml pk中で、第2培養のために再懸濁する。このDNA調製物は直接ポリメラーゼ連鎖反応下に置くことができる。
【0065】
Lam gE-またはLam gE-/TK-を接種後および毒性の強いBHV-1菌株を投与後、これらの接種されまたは無接種の牛のウイルスを含む鼻水の垂れ
【表1】
【0066】
gE-Mabsの特性
【表2】
+:間接ブロッキングIPMAにおいてテストされた8つの全血清のブロッキングした割合が50%よりも大きい場合
±:±50%の場合
−:50%未満の場合
【図面の簡単な説明】
【0067】
【図1】BHV-1菌株Difivac-1およびIowaのサザンブロット分析 Aは、Difivac-1とIowaとのゲノムDNAのサザンブロットのオートラジオグラムである。1と3レーン中で、Difivac-1 DNAは、HindIIIとPstIとによる制限酵素消化後に、用いられた。2と4レーン中で、Iowa DNAは、HindIIIとPstIとによる制限酵素消化後に用いられた。フラグメントの大きさは、キロベース(kb)で示される。 Beckman L5-65超遠心分離のSW27ロータ中で20,000回/分でpH7.4のトリス10mMと150mM NaClと1mM EDTAとの中で、25%(w/w)庶糖クッションを用いて2時間、ウイルスで汚染されたEbtr細胞と共に培養液(約450cm2ローラボトルあたり70ml)を遠心分離することによって、ウイルスDNAが単離された。得られたウイルスペレットから、DNAが標準の手法で単離された(J.Sambrook,E.F.Fritsch and T.Maniatis,1989,Molecular cloning:a laboratory manual,2nd ed.Cold Spring Harbor Laboratory Press, New York)。このDNA上で制限酵素消化は、Boehrnger Mannheim社によって供給されるSuRE/cut緩衝液中で酵素を用いて行われた。 0.7%アガロースゲルを水平電気泳動して分離し、ニトロセルロースフィルタ(Schleicher & Schuell,Inc)でブロットした後、フィルタは6時間42℃で50%ホルムアルデヒド中で3×SSC(1×SSC=0.15MのNaClと0.015Mのクエン酸ナトリウム,pH7.4)と、50μlの変成サケ精液DNA(Sigma)/mlと、0.02%フィコールと、0.1%ドデシル硫酸ナトリウム(SDS)との中に予備ハイブリダイズされた。さらに同溶液に32PdCTP(Amersham)でラベルされたHindIII Kフラグメントを加えることによってハイブリダイゼーションが行われた(HindIII Kフラグメントの選択は、1型ウシヘルペスウイルスからのDNAのクローニングと分裂位置のマッピングに基づく(Cooper菌種)John F.Mayfield, Peter J.Good, Holly J. VanOort, Alphonso R. Campbell and David A.Reed;Journel of Virology (1983) 259〜264)。ハイブリダイゼーションの12〜14時間後、フィルタは2時間、0.1%SDSと0.1×SSCとの中で60℃で洗浄された。HindIII Kフラグメントは、標準のクローニング手法に従って、pUC18ベクタ中でクローン化された(J.Sambrook, E.F.Fritsh and T.Maniatis, 1989, Molecular cloning la laboratory manual,2nd ed.Cold Spring Harbor labovatort Press, New York)。pUC8.4HindIII KクローンのHindIIIによる消化後、pUC18ベクタは、0.7%の低融点アガロース(BRL、Life Technologies,Inc.)ゲルで再び電気泳動によって8.4kb HindIIIにフラグメントから分離され、基準のフェノール抽出とエタノール沈澱とによってアガロースから分離された。分離されたHindIII Kフラグメントは、Boehringer Mannheim社製のランダムプライムドDNA標識キット1OO4.760で標識を付された。ハイブリダイズされたフィルターのオートラジオグラムは、-70℃でKodak XARフィルムの36時間露出で、反射スクリーンを使って行われた。 BはIowaとDifivac-1の7.4kb HindIIIフラグメントとの8.4kb HindIII Kフラグメントの物理的マップである。6kb PstIフラグメントの共泳動とDifivac-1中の1.8kb PstIフラグメントの欠如を考えて、欠失はハッチングされた面積領域で起こったと推定される。
【図2】Difivac-1中の欠失領域周囲の野生型BHV-1フラグメントのサブクローニング Aにおいて、BHV-1ゲノムの成分は、長ユニーク(UL)領域、短ユニーク(US)領域および2つの反復物(IrとTr)として示される。このマップは、Cooper菌種の公表された分析に基づいている(John F.Mayfield, Peter J.Good, Holly J. VanOort, Alphonso R. Campbell and David A. Reed: Journal of Virology (1983) 259〜264)。 Bにおいて、フラグメントは、原核ベクタ中へクローンされたUS領域で示される。すなわちpACYC中の15.2kb EcoRIフラグメント、pUC18中の8.4kb HindIIIフラグメント、およびpBR322中の4.1kb EcoRI-HindIIIフラグメントである。ウイルスDNAフラグメントの単離は、図1Aの表示中に述べられている手法に従って実施された。種々のベクタ中へこれらのフラグメントをクローン化することは、標準の手法で行われた(J.Sambrook, E.F.Fritsch and T.Maniatis, 1989, Molecular cloning:a loboratory manual,2nd ed.Cold Spring Harbor Laboratory press, New York)。 Cにおいて、物理マップは、Difivac-1中で要求される欠失が位置する領域を示す。 Dにおいて、さらなる分析のために用いられるこの領域のサブクローンが示されている。2つのPstIフラグメントはpKUN19の中で、そして残ったフラグメントはpUC18の中でクローンされた。
【図3】Aは、〔左端にあるAluIの認識サイトから右端にあるHincIIの認識サイトに至る〕図2C中で示されるようなDifivac-1中において欠失された要求される位置周囲の、BHV-1のLam種の、US領域の2027個のヌクレオチドのヌクレオチド配列である。図2Dで示されるサブクローンの挿入をしたヌクレオチド配列は、Sangerらのジデオキシ配列法を用いた2つの基準で分析することによって決定された(F.Sanger, S.Nicklen and A.R.Coulson, 1977, Proc.Natl.Acad.Sci.USA 74,5463〜5467)。この目的に対して、Pharmacia製のT7配列キットが、同製造者によって指定される手法に従って用いられた。放射線標識のために、〔35S〕dATP(Amersham製)が用いられた。コンプレッション現象の影響を併うGLの多い領域の配列分析は、Pharmacia製キットの7-デアザアデニン-dGTP変形物を用いて繰り返された。ヌクレオチド配列の下に3文字のコードで示されたものは、ヌクレオチドの概念上の翻訳後に見つかった575アミノ酸残基のオープンリーディングフレームのアミノ酸配列である。この翻訳は、一般的なコードに基づきPC遺伝子コンピュータプログラムを用いて決定された(PC遺伝子翻訳バージョン1.03 1987年11月)。この575アミノ酸のオープンリーディングフレームは、ヌクレオチド168のメチオニンで始まり、ヌクレオチド1893の停止コドンで終わる。 575アミノ酸残基のオープンリーディングフレームの構造分析は、PC遺伝子コンピュータプログラムで実行される。最初の26アミノ酸が、真核性の移行シグナルを形成し、「シグナルペプチド」で図中に示される。6.2の点で、このシグナル配列の切断が、アミノ酸26とアミノ酸27との間で予想される。アミノ酸575の配列は、アミノ酸残基の線によって指示される。3つの可能なN結合のグリコシレーション位置(NXT/S)を有する。RaoとArgosの方法に従えば、「貫膜螺旋」で図中に示される貫膜領域がアミノ酸423とアミノ酸450との間にある。制限酵素AsuII, SmaI, HindIIIおよびEcoNIのための認識配列(サイト)は下線が付される。このポリペプチドの計算された分子量は61212である。 Bは、上で述べた575アミノ酸オープンリーディングフレームの構造上の特性を模式的に表したものである。
【図4】BHV-1 gE遺伝子のアミノ酸配列と、ヘルペス単体ウイルス(HSV)gE遺伝子および他のgE相同遺伝子〔疑似狂犬病ウイルス(PRV)gIと帯状疱疹ウイルス(VZV)gpI〕のアミノ酸配列との比較 この比較のために用いた配列は、次の公表されたものである。 HSV:1型ヘルペス単体ウイルスのゲノム中のUS領域の配列決定と遺伝子内容(D.J.McGeoch, A.Dolan, S.Donald and F.J.Rixon (1985) Journal Mol.Biol.181,1〜13) VZV:帯状疱疹ウイルス遺伝子のUs成分のDNA配列(A.J.Davidson (1983), EMBO Journal 2, 2203〜2209) PRV:ヘルペス単体ウイルスと帯状疱疹ウイルス糖蛋白とに相同性を有する2つの疑似狂犬病ウイルス糖蛋白のための単離遺伝子にλgt11の使用(E.A.Petrovskis, J.G.Timmins and L.E.Post (1986) Journal of Virology 60,185〜193) これらの配列は配列分析プログラムMultalinを使って比較された(F.Corpet (1988), Nucl. Acids Res.16,10881〜10890)。 Aの中で、4つのアミノ酸配列の模式的図が示されている。ここに予測された貫膜部(TM)が各々に示される。予測された移行シグナル配列(SP)と考えられるN-結合グリコシレーション位置(I)に加えて、2つの保存される領域が示される。その中で、システン残基の相対位置(CCC)は不変である。 Bの中で結果は4つのgE翻訳の中央に位置するシスティンの多い領域のMultalin比較が示される。アステリスクは、一致したアミノ酸と類似アミノ酸クローンを示す。
【図5】Difivac-1とIowaとのサザンブロット分析で得られた写真図 パネルAは、BstI(1,2),EcoRI(3,4)およびHindIII(5,6)を有するDifivac-1とIowa消化制限酵素のゲノムDNAである。これは、0.7%アガロースゲルで分離され、ニトロセルロースでブロットされ、図1Aの説明で述べられた手法に従ってBHV-1菌株Lamの32PでラベルされたHindIIIKフラグメントでハイブリダイズされる。 パネルBは、Aと同様に同じゲルのニトロセルロースブロットであり、BHV-1 gE-特異型プローブp318でハイブリダイズされた。プローブは、図2C中で示したAluI-HincII全領域を含む。
【図6】gEフラグメント欠失BHV-1の構築 Aの中でgEの位置と用いられたクローンとが示される。BHV-1ゲノムの成分は、UL領域、Us領域および2つの反復領域(IrとTr)として示される。gE遺伝子の5’側末端に位置する領域を得るために、BHV-1菌種Lamの8.4kb HindIIIK由来の1.4kb PstI-SmaIフラグメントが、プラスミドpUC18のSmaIとPstIサイトの中ヘサブクローンされた。このクローンは、p515と呼ばれ、図6B中に示される。gEの3'末端側に位置するEcoNI-SmaIフラグメントは、4.1kb HindIII−EcoRIクローンに由来し、p515の普通のAsuIIヘクローンされる。AsuIIの末端にEcoNIの末端を結合するために、クローンp515はAsuIIで消化され、その後クレノウ酵素(Boehringer Mannheim)と、標準的手法に従ってAsuIIの末端へ1つのシトシン残基を供給するdCTPとで処理した(Sambrook et al,1989)。この付加シトシンは、D中でアステリスクで示される。p515は、またSmaI酵素で消化され、その後EcoNIフラグメントは、このベクタ中へ結合される。このようにして構築されたクローンは、p519と呼ばれる。
【図7】Aは、1B7,1B8および2H10のDNA調製品のサザンブロット分析で得られた写真である。DNA単離物、消化制限酵素、ブロットおよびハイブリダイゼーションは、図1Aの説明で述べられた手法に従って作られた。DNA調製品1B7,1B8および2H10のPstI-DraIによる二重消化後、フラグメントは0.7%アガロースで分離され、直後にニトロセルロースフィルタにブロットされた。このフィルタは、プローブとして、32PdCTPでラベルされた2.3kb PstI-DraI欠失フラグメントとハイブリダイズされた。レーン1〜3中で試料1B7,1B8および2H10がそれぞれ分離された。レーン中でLam菌株の野生型BHV-1 DNAが、レーン5中で2.3kb欠失フラグメントが示される。 Bは、BHV-1菌種Lamの15.2kb EcoRIフラグメントの物理マップである。マップは、PstI,DraIおよびHindIIIの認識サイトと7A中で述べたハイブリダイゼーションプローブのサイトを示す。
【図8】BHV-1 gEの原核発現 BHV-1 gEの原核発現のために、gE遺伝子の600bpSmaIフラグメントは、ベクタpGEX-2T中で、日本住血吸虫のグルダチオン-S-転移酵素のコード領域に3つのリーデングフレーム中で融合された(D.B.Smith and K.S.Johnson,Gene 67(1988)31〜40)。SmaIフラグメントの適切な(syn)方向性を有する組換え分子は、標準の手法を用いて、制限酵素分析法によって同定された。この融合構造を有するE.coli DH5αクローンは、pGEX-2T600s1,pGEX-2T600s2およびpGEX-2T600s3と呼ばれた。 Aは、pGEX-2T600s構築物の1つの図である。GST-gE融合物をコードする領域のNH2側にあるものは、イソプロピルチオガラクトシド(IPTG)で誘導し得るtacプロモータ領域である。 Bは、pGEX-2T600sでトランスフェクトされたDH5αの細胞の全蛋白質調製物のウエスタンブロット分析で得られた写真である。pGEX-2T600s1,pGEX-2T600s2およびpGEX-2T600s3構築物でトランスフェクトされたDH5α細胞のオーバーナイトの培養物は、50μg/mlアンピシリンを加えたLuria-Bertani(LB)培地中で1/10の割合で残っており、1時間後培養物はIPTGによって5時問誘導された。これらの誘導された培養物は、5分間6,000Gで遠心分離し、1×ラウエル混合物(2%SDS、10%グリセロール、5%メルカプトエタノールおよび0.01%ブロムフェノールブルー)中に混合〔1.5ml培養物を、500μlラウエル混合物中に混ぜる〕され、95℃に5分間加熱された。そして、1レーンあたり50μlで、標準の手法に従って、12.5%ポリアクリルアミドゲルで水平的に分離され、引続いて製造者によって指定された条件で、LKB-ムルチフールII型Navaブロットシステムを用いて、ニトロセルロースフィルタがセミドライ式でブロットされた。 レーンM中で、予め印を付されたマーカー蛋白質が用いられ、(BRL Life Technologies,Inc.236K,112K,71K,44K,28K,18Kおよび15K)、レーン1,2,3中では、pGEX-2T600s1,pGEX-2T600s2およびpGEX-2T600s3各々3つのフレームでトランスフェクトされたDH5α細胞の全蛋白調製物はが用いられた。 パネルA中で、結果は、抗-GST血清を用いるウエスタンブロット分析で見ることができる。フィルタは、標準の手法(E.Harlow and D.Lane(1986)、Antibodies:a laboratory manual,Cold Spring Harbor Loboratory,New York)にしたがって、ウサギ血清ポリクローナル抗GSTを加えたブロッキング緩衝液(PBS+2%粉乳および0.05%トゥイーン20)中でインキュベートされた。そして、フィルタは洗浄され、ヤギ抗ウサギ免疫グロブリン血清を結合した西洋ワサビパーオキシターゼ(HRPO)を加えてインキュベートされた。その後、結合されたヤギ抗体は、色素体(ヂアゾベンジジン、クロロナフトールおよびH2O2)で免疫化学的に検出された。矢印によって指示されるGST融合物は、フレーム3中でのみ約47kの確認された大きさを有する。 パネルBにおいて、結果は、モノクローナル抗体Mab4を用いる、ウエスタンブロット分析で見ることができる。この目的に対しパネルA中のようにデュプロフィルタはブロッキングされ、Mabでインキュベートされ、ウサギ抗マウス血清を結合したHRPOでインキュベートされる。その後結合ウサギ抗体は、色素体で免疫化学的に検出される。レーン3(フレーム3)で見られるバンドは、47Kの大きさで、矢印によって指示される。
【図9】BHV-1 gE遺伝子の原核発現のためのpEVHis gEプラスミドの構築 gE遺伝子の原核発現のために、gEコード領域は、標準の手法を用いて発現ベクタpEVHisのHCMVプロモータ領域の背後の適切な位置においてクローン化される(Sambrook et a1,1989)。gEのオープンリーディングフレーム55bp上流で開始する394bpA1UIフラグメントは、pUC18の中ヘクローンされ、p201と呼ばれる。そして、p201をHincIIが消化した後、1740bp HincIIフラグメントは、gE遺伝子の大部分から成り、p201中へクローンされる。これは、pUC18のポリリンカ中で、gEの開始コドンの上流にあるAluI位置55bpからgEの停止コドンの下流にあるHincII位置133bpまでの全gEコード領域を含むプラスミドp318になる。ベクタのポリリンカにおける制限酵素位置を用いて、このフラグメントは、酵素BamHIとSphIでp318から切られた。最初p318はSphIで消化され、その後SphIサイトは、クレノウポリメラーゼとdNTPポリメラーゼとを使って満たされた。BamHIでの消化後、1.9kb挿入物は、pUC18ベクタから低融点アガロース中で分離され、BamHIとEcoRVで消化されたpEVHis中で結合された。このように形成されたプラスミドは、pEVHis/gEと呼ばれた。
【図10】BHV-1 DNAを検出するためにPCR手法のためのgE-特異型プライマとプローブの位置 図中には、ヌクレオチド1272から2027に至るBHV-1糖蛋白質の核酸配列が示される〔配列は図3から得られる〕。gE-特異型PCR手法のために用いられるプライマは、P3とP4と呼ばれる。P3とP4のプライマ結合サイトは、下線が付される。P3のヌクレオチド配列は、5'-ACG-TGG-TGG-TGC-CAG-TTA-GC-3'である(配列標別番号:2)。P4のヌクレオチド配列は、(上述のプライマ結合配列に相補的である)5'-ACC-AAA-CTT-TGA-ACC-CAG-AGC-G-3'である(配列標別番号:3)。PCRで増幅されたDNAの検出のためのサザンブロットハイブリダイゼーションで用いられたプローブは、プライマが結合するサイトの間に位置する137bp TaqIフラグメントであり、そのフラグメントの末端が示される。図3と比較のために、HindIIIとEcoNIサイトがまた示される。
【図11】Difivac-1のgE欠失のマッピング Aは、野生型BHV-1菌種Lamの15.5kb EcoRIフラグメントの物理マップを示す。Bは、Difivac-1の14.5kb EcoRIフラグメントの物理マップを示す。2つのEcoRIフラグメントは、各々のウイルスのゲノムのUS領域を完全にカバーする。gE遺伝子の位置とgI遺伝子の推定上の位置は、白ヌキの四角で指示される。マップAとBとは、各マップ中の6kb PstIフラグメントが、一列に並べられるように位置決めされる。両方のマップ中で、内部反復領域と末端反復領域との配列は、斜線を付された四角で示される。反復の下方の矢印は、これらの配列の方向を示す。 Aにおいて、Difivac-1菌種中で欠失があるUS領域の部分が示された。 Cは、gE欠失をマップ化し、B中に示される物理マップを得るために用いられるクローン化Difivac-1フラグメントの位置を示す。クローンp728,p737およびp754の挿入物の下の矢印は、組換え点を決めるために配列決定された領域を示す。 略語: A=A1UI,E=EcoRI,P=PstI,H=HindIII,γ=組換え点,IR=中間反復領域,TR=末端反復領域
【図12】Difivac-1のUS領域における正確な組換え位置の決定 Difivac-1菌種中に見付かったgE欠失の正確な境界を決定するために、クローンp754ならびにp728およびp737とが配列決定された。これらのクローンの挿入は、図11中に指示される。用いられる配列決定の手法は、図3の説明中に述べられている。 Aにおいて、A1UI-PstIフラグメントの大部分の配列が示される。この配列は、gE遺伝子のプロモータ領域で開始する。推定のTATAボックスは、下線が付されている。点γ(=組換え点)において、このプロモータ領域は、Us領域の反対の位置で見付かった逆方向反復と呼ばれる配列に接合される。正確な組換え点は、US領域の反対位置で見付けられた反復のコピーとgEプロモータ領域で見付けられた反復を比較することによって決定される。この配列が分岐する点は、B(I)の中でγで示される。類似の比較は、Difivac-1中で見付けられるgEプロモータ配列と野生型菌種Lam中で見付けられたgEプロモータとで行われる。この配列が分岐する点は、B(II)の中で示され、またγで示される。見付けられた組換え点は同じである。
【図13】BHV-1 gI遺伝子の部分配列分析 BHV-1 gIとgEとの双方の遺伝子(図11に示す)中に達するBHV-1菌種Lamの1.8kb PstIクローンを用いてBHV-1 gIのコード領域の範囲内の、284ヌクレオチドの配列が決定された。配列決定に用いられる手法は、図3の説明の中で述べられている。配列は、PC/遺伝子コンピュータプログラムバージョン1.03(1987年11月)による汎用コードに基づいて翻訳された。第2リーディングフレームによってコードされるアミノ酸配列は、ヌクレオチド配列の下にワンレターコードとして与えられる。このアミノ酸配列は、他のヘルペスウイルスのgI相同体(図14に示す)のコード領域に相同である。
【図14】推定のBHV-1 gI遺伝子の部分アミノ酸配列と、ヘルペス単体ウイルス(HSV1)gI遺伝子、疑似狂犬病ウイルス(PRV)gp63遺伝子および帯状疱疹ウイルス(VZV)gpIV遺伝子のコード領域の相当部分とのアミノ酸比較 PRV配列はそのコード領域のアミノ酸82で開始し、HSV1配列はそのコード領域のアミノ酸で開始し、VZVはそのコード領域のアミノ酸76で開始する。用いられる配列は、図4の説明で述べられた報告書で公表されている。比較は、Multalinコンピュータプログラムを使って行われた。アステリスクは、同一のアミノ酸を示し、コロンは類似のアミノ酸を示す。
【図15】BHV-1 gI-遺伝子の原核発現のためのMSVneoGIプラスミドの構築 BHV-1 gI-遺伝子の部分配列のアミノ酸配列の比較に基づいて、BHV-1 gI-遺伝子の推定位置は推測される。この推測に基づいて、1.7kb SmaIフラグメントは、BHV-1 gE遺伝子の全コード領域を含むことが推論される。この1.7kb SmaIフラグメントの位置は、A中に示される。この1.7kb SmaIフラグメントの丸くなった平滑末端に、BamHIリンカが標準の手法を用いて結び合わされる。この結果生じた物は、BamHIで消化され、原核発現ベクタMSV-neo中で結び合わされる。MSV-neoベクタは、強いプロモータ性を有するMSV-LTRの背後にユニークなBamHIサイトを有する。このベクタは、Rijsewijk et al.,1987EMBO J.6,127〜131中で述べられている。
【図16】BHV-1 gI/gE二重欠失フラグメントの構築 BHV-1のUS領域中における糖蛋白質gE遺伝子の位置と糖蛋白質gI遺伝子の推定位置は、図A中に表現される。斜線を付された部分は、US領域の境界を成す反復物を示す。Bは、双方の遺伝子の位置を考慮して、必然的な制限酵素サイトの物理マップを示す。gI/gE欠失フラグメントを構築するために、gI遺伝子を包み込む1.7kb SmaIフラグメントを含むクローンp1.7-SmaI/oがPstIで消化される。残留350bp SmaI-PstI挿入物のPstIサイトは、標準の分子生物学的手法を用いて、平滑末端を作る。EcoNI-SmaIフラグメント(図6B参照)は、図6A中で記載された4.1kb HindIII-EcoRIフラグメントから単離され、平滑末端を作られ、上述の修正されたPstIサイトに結合される。これは、CとD中に図示される。結果としてできたクローンPΔIEから、1.4kb SmaI-DraIフラグメントは、野生型BHV-1 DNAと組換えるために単離される。 略号 E=EcoRI,H=HindIII,S=SmaI,P=PstI,ENI=EcoNI,D=DraI,kb=キロベース,US=ユニークショート
【図17】接種後の牛から垂れた鼻水の平均ウイルス数 ●=Difivac-1を接種、○=接種なし
【図18】毒性の強いBHV-1菌株投与後の平均発病牛数、記号は図17に同じ
【図19】毒性の強いBHV-1菌株を投与した牛の平均体温、記号は図17に同じ
【図20】毒性の強いBHV-1菌株投与後の牛の平均成育量、記号は図17に同じ
【図21】毒性の強いBHV-1菌株投与後の牛から垂れた鼻水の平均ウイルス数、記号は図17に同じ
【図22】毒性の強いBHV-1菌株投与後の牛の平均体温 ●=Lam gE-を接種、○=Lam gE-/TK-を接種、×=無接種
【図23】毒性の強いBHV-1菌株投与後の牛の平均成育量、記号は図22と同じ
【図24】毒性の強いBHV-1菌株投与後の平均発病牛数、記号は図22と同じ
【技術分野】
【0001】
発明の分野
本発明は、病原体によって引き起こされる疾病に関係したワクチン注射および診断法の分野に関するもので、弱毒性ワクチンまたは不活化ワクチンを生成する古典的な方法とDNA組換技術に基づく近代的方法両方の使用に関係している。
より詳細には、本発明は、動物、特に牛を1型ウシヘルペスウイルス(BHV-1)から守るための弱毒性生ワクチンおよび不活化ワクチンに関するもので、これらのワクチンは、安全でかつ効能があるばかりでなく、ワクチン接種をうけた群における、非感染動物と感染動物とを識別することも可能にする。
ワクチン接種群における非感染動物と感染動物の識別のための試験に使用が可能である診断キットも本発明の一部である。
【背景技術】
【0002】
発明の背景
牛の伝染性鼻腔気管炎ウイルス(IBRV)および伝染性濃外陰部腔炎ウイルス(IPVV)を含むBHV-1は、牛の呼吸器疾患、生殖能力、障害の解明に重要な役割を果たす。BHV-1は、急性感染後、潜伏した形で宿主内にそのまま残っていることが多い。潜伏ウイルスはとりわけ、ストレス(これは臨床現象を伴うことも、伴わないこともある)の影響下で再び活性化し、続いて排泄されることがあり得る。結果として、感染した牛は終生BHV-1を蔓延するものとみなす必要がある。BHV-1はオランダの養牛場の推定75%において風土病として発生している。特に、高齢牛の血清反応が陽性である。
【0003】
BHV-1感染予防接種に利用される多くの不活化("死んだ")ワクチンおよび弱毒("生きた")ワクチンがある。不活化ワクチンは、たとえば熱処理や照射、またはエタノールやホルマリン処理によって、BHV-1ウイルスを死滅させて作られる。しかしながら、これらによっては不充分な予防しか行えないことが多い。弱毒ワクチンは、同種(ウシ)細胞またはブタの細胞やイヌの細胞のような異種細胞への多数回にわたる継代によって作られ、その後ウイルスは物理的または化学的にも処理されることもある。このようにして、未知の突然変異/欠失がウイルスゲノム中に発達し、これがしばしばウイルスの疾病誘発特性を低下させる。弱毒ワクチンは不活化ワクチンよりも優れた予防を行うが、これは特に宿主の免疫系に、より多くのウイルス性抗原を与えるからである。生きたワクチンの重要な他の利点として、それらは鼻腔内投与が可能であるということがあり、すなわち、ここは感染後に野性型ウイルスの増殖が初めて起こるところである。生きたワクチンはまだ改良の余地がある。いくつかの生きたワクチンはさらに、流産を引き起こす可能性を有しているように思われ、これは特に筋肉内投与後に顕著になる。さらにまた、生きたワクチンはすべて、ワクチンを接種された牛に潜伏して残っていると思われる。また、該ワクチンが野性型ウイルスとほとんど相違するところがなければ、病原性への復帰が起こる可能性もある。しかし、重要な問題の1つは、BHV-1ワクチンは、野性型ウイルスによる感染を予防することができないということである。その結果、ワクチン接種を受けた牛も野性型BHV-1を蔓延させる可能性がある。
【0004】
適切なBHV-1抑制計画のためには、野性型ウイルスと識別することができる効果的かつ安全なワクチンを自由に使えることが必要である。何故ならば、効果的ワクチンの利用によりBHV-1の蔓延をかなりの程度低下させることができ、ワクチンと野性型ウイルスの識別が可能な試験によって、ワクチン接種を受けた牛の群より感染した牛を探りあて、それを取り除くことが可能となるからである。
【0005】
ところで、従来のワクチンよりも一層安全と思われる野性型ウイルスとの識別が可能なBHV-1ワクチンの開発が行われてきた。流産を引き起こす程度もより低く、潜伏の頻度も低く、再活性化され得ることもないチミジンキナーゼ欠失変異株が単離された。さらに、組換えDNA技術を用いて、(血清学的技術により野性型BHV-1とワクチンの区別を可能にする)糖蛋白質gIIIのための遺伝子に欠失を有するBHV-1ワクチンが構成された。しかしながら、これらのワクチンに対してもさらに幾つかの異論がある。一方では、チミジンキナーゼ遺伝子はウイルス複製に関与しており、複製の程度が低い程、より程度の低い防御につながる可能性がある。他方、該糖蛋白質gIIIは防御抗体の生成に重要であり、この防御抗体は、gIII欠失ワクチンの効果を低下させる。実際上の問題として、一般には最高の防御が得られる、組換えワクチンの鼻腔内投与が幾つかの国では認可されていないという問題がある。したがって、有効かつ安全で、さらに野性型BHV-1と区別することが可能なワクチンが求められており、さらにまたこれは、かかるワクチンの少なくとも1つは、組換えDNA技術によって構成されたウイルスよりも、むしろ従来の方法で弱毒化されたウイルスに基づいたものであることが望ましい。
【0006】
ところで、細胞培養における継代によって、糖蛋白質gEのための遺伝子を欠くBHV-1菌株が得られた。我々が行った研究の最初の結果は、この遺伝子は、野性型BHV-1に関する血清学的識別に非常に役立ち、病原性の発原に関与していることを示している。したがって、その欠失は安全性に寄与しており、これによってチミジンキナーゼ欠失の使用は不要となるかもしれない。糖蛋白質gEはく防御の誘導にとって糖蛋白質gEほど重要ではないと思われる。従来の方法で弱毒化された野性型ウイルスと血清学的に識別することが可能な、従来の方法で弱毒化されたBHV-1菌株は独自のものである。ここで初めて述べた、gE-遺伝子の位置とDNA配列は、これまでは知られていなかった。また、これから誘導することができるオリゴヌタレオチドポリペプチドおよびオリゴペプチドも知られていなかった。また、gE-遺伝子に基づく血清学的識別を行うための試験も独自のものである。
【0007】
この"従来の"gE欠失変異株の重要な利点("従来の"とは従来の弱毒ウイルス単離方法を使用することを意味している)は、親換えワクチンに関する限りでは、鼻腔内投与が禁止されている国々において、この"従来の"gE欠失変異株を鼻腔内投与することが可能になるであろうという点である。しかしながら、安全に関して別の観点からよく考慮して、この従来のgE欠失ワクチンの他に、明確に限定された組換えの型も構成された。これらの組換えワクチンもまた欠失を有しており、-また、チミジンキナーゼ遺伝子の欠失も有していてもよく、または有していなくてもよい-および、これらの組換えワクチンは、異種の遺伝子の発現のためのベクタとして用いることもできる。これらの組換えワクチンは全て、同じgE特有試験によって、野性型ウイルスと区別することができる。一組の異なるワクチンに対して標準的な試験を利用することは、国際的な努力としてのBHV-1の戦いにおいて強力な利点となり得る。かかる方法がBHV-1ワクチンの分野において述べられたことはない。
【0008】
牛における抗BHV-1反応の血清分析によって、抗gE抗体の重要部分は、糖蛋白質gEおよび他のBHV-1糖蛋白質である糖蛋白質gIによって形成される複合物に対して向けられていることが示された。したがって、かかる複合物特有の抗体の存在を示すこともできる血清試験は、抗-gE抗体を検出することができるだけの試験よりも高感受性であると思われる。gE単欠失変異株によりワクチン接種を受けた牛は、抗-gI/gE抗体の検出を妨げることが有り得る抗-gI抗体を作る可能性がある。したがって、この発明はgI/gE二重欠失を有するワクチンを含む。
【発明の開示】
【0009】
発明の要約
本発明は、糖蛋白質gE-遺伝子に欠失を有するBHV-1の欠失変異株を提供する。"〜に欠失を"という語は、全体としての遺伝子の欠失を含むことを意図している。本発明の好適な実施例は、以下に述べる欠失変異株Difivac-1のような、弱毒処置によって引き起こされた糖蛋白質gE-遺伝子における欠失を有するBHV-1の欠失変異株によって構成される。
【0010】
本発明の他の好適な実施例は、以下に述べる欠失変異株1B7または1B8のような組換えDNA技術によって作られた糖蛋白質gE-遺伝子に欠失を有しているBHV-1の欠失変異株から成る。
【0011】
本発明の他の好適な実施例は、以下に述べるgI、/gE二重欠失変異株Difivac-IEのような、糖蛋白質gE-遺伝子および糖蛋白質gI一遺伝子に各々欠失を有するBHV-1の二重欠失変異株から成る。
【0012】
さらにまた、最高の安全性を期して、本発明によれば糖蛋白質gE-遺伝子およびチミジンキナーゼ遺伝子のそれぞれに欠失を有するBHV-1の欠失変異株が好適である。また本発明は、糖蛋自質gE-遺伝子、糖蛋白質gI-遺伝子、チミジンキナーゼ遺伝子に欠失を有するBHV-1の欠失変異株も含んでいる。
【0013】
本発明は、動物を、特に哺乳動物を、さらに特にウシ科の動物を上記において明確に述べたBHV-1欠失変異株を有するBHV-1から守るために、これらにワクチン接種をするためのワクチンの組成、適切な担体またはアジュバントを提供する。前記構成は、生きたワクチン組成でも不活化ワクチン組成でもよい。
【0014】
本発明はさらに、糖蛋白質gE-遺伝子に欠失を有し、組換えDNA技術によって導かれた異種遺伝子を含むBHV-1の変異株において実施されている。好適には、糖蛋白質gE-遺伝子の位置に、組換えDNA技術によって導かれた異種遺伝子を含むBHV-1の変異株に関するもので、前記異種遺伝子は、gE-遺伝子の調節配列の管理下にあり、シグナルペプチドに関してコード化するgE-遺伝子の一部に任意に付加される。前記異種遺伝子は、BHV-1の異なるプロモータの管理下、または異種プロモータの管理下にあることも可能である。BHV-1の変異株が、糖蛋白質gE-遺伝子における欠失に加えて、さらに、チミジンキナーゼ遺伝子における欠失、および/または糖蛋白質gI-遺伝子における欠失などの欠失を有している場合には、前記異種遺伝子を、この付加欠失の位置に挿入してもよい。複数の挿入も別の選択肢であり、1つの欠失の位置にまとめて行うか、幾つかの欠失の位置に分けて行うかされる。
【0015】
好適に導かれた異種遺伝子は、別の病原体の免疫原性蛋白質またはペプチドについて、または免疫反応を促進するサイトカインについてコード化する。適切なサイトカインの例としてはインターロイキンー2、αインターフェロン、γインターフェロンがある。
【0016】
本発明はまた、動物を、特に哺乳動物を、さらに特にウシ科の動物を(異なる)病原体より守るため、これらにワクチンを接種するための、(生きた、または不活化)ワクチンの組成を提供するもので、該ワクチンの組成には、免疫原性蛋白質または他の病原体ペプチドについてコード化している異種遺伝子を有しているBHV-1突然変異株およびアジュバントの適切な担体が含まれる。勿論、この防御は1つ以上の病原体に関するものでよく、すなわち、変異株が複数の異種遺伝子を含む多価ワクチンに関するものでよい。
【0017】
さらにまた本発明は、BHV-1の糖蛋白質gE-遺伝子、この糖蛋白質gE-遺伝子の一部、またはこの糖蛋白質gE-遺伝子から誘導されるヌタレオチド配列を含む組換え核酸を含む組成に関する。この組成は、BHV-1の糖蛋白質gE-遺伝子、この糖蛋白質gE-遺伝子の一部、この糖蛋白質gE-遺伝子から誘導されたヌクレオチド配列を含む組換え核酸の挿入を有するクローニングベクタまたは発現ベクタを含むことができる。
【0018】
また本発明は、BHV-1の糖蛋白質gE、この糖蛋白質gEの一部、この糖蛋白質gEから誘導されたペプチド、または糖蛋白質gEおよびgIの複合物を含む組成、およびBHV-1の糖蛋白質gE、この糖蛋白質gEの一部、この糖蛋白質gEから誘導されたペプチド、または糖蛋白質gEとgIの複合物に特有の抗体を有する組成を含む。"抗体"とは、ポリクローナル抗体調製物と、大半の用途に好適なモノクローナル抗体の両方を意昧していると解釈される。"糖蛋白質gEの一部"および"糖蛋白質gEから誘導されるペプチド"という用語は、一般に少なくとも約8アミノ酸の長さを有するであろうgE特有のアミノ酸配列を意味していると解釈される。
【0019】
さらに本発明は、試料中の、特に、動物、特に哺乳動物、さらに特にウシ科の動物の、血液または血清、血液細胞、乳、涙などの体液、肺洗浄液、鼻水、精液、特に精漿、唾液、痰、または組織、特に神経組織のような生物学的試料中の、BHV-1の核酸を検出するための、BHV-1の糖蛋白質gE-遺伝子から誘導されたヌクレオチド配列を有する核酸プローブまたは核酸プライマを含む診断用キット、および核酸検出試験に適した検出手段に関する。
【0020】
さらに本発明は、試料中の、特に、動物、特に哺乳動物、さらに特にウシ科の動物の血液または血清、血液細胞、乳、涙のような体液、肺洗浄液、鼻水、精液、特に精漿、唾液、痰、または組織のような生物学的試料中の、BHV-1に特有の抗体を検出するための、BHV-1の糖蛋白質gE、この糖蛋白質の一部、この糖蛋白質gEから誘導されたペプチド、または糖蛋白質gEおよびgIの複合物を含む診断用キット、および抗体検出試験に適した検出手段に関する。該診断用キットは、さらに、BHV-1の糖蛋白質gEに特有の、またはBHV-1の糖蛋白質のgEとgIの複合物に特有の、1つまたはそれ以上の抗体を含めてもよい。
【0021】
また、本発明は、試料中の、特に、動物、特に哺乳動物、さらに特にウシ科の動物の血液または血清、血液細胞、乳、涙などの体液、肺洗浄液、鼻水、精液、特に精漿、唾液、痰、または組織、特に神経組織のような生物学的試料中の、BHV-1の蛋臼質を検出するための、BHV-1の糖蛋白質gEに特有の、またはBHV-1の糖蛋白質gEとgIの複合物に特有の1つまたはそれ以上の抗体を含む診断用キット、および蛋白質検出試験に適した検出手段に関する。
【0022】
さらに本発明は、BHV-1の糖蛋白質gE-遺伝子を含む核酸の存在、BHV-1の糖蛋白質gE、またはBHV-1の糖蛋白質gEとgIの複合物の存在、またはBHV-1の糖蛋白質gEに特有の、またはBHV-1の糖蛋白質gEとgIの複合物に特有の抗体の存在について、動物の、特に哺乳動物の、さらに特にウシ科の動物のBHV-1感染を判断するための、前記動物の試料の、特に、血液または血清、血液細胞、精液、特に精漿、唾液、痰、涙などの体液、肺洗浄液、鼻水、乳、または組織、特に神経組織のような生物学的試料の試験を含む方法を提供する。試験を行う試料は、本発明に従った組成のワクチンをそれまでに接種されたことのない動物のもの、または本発明に従って調製されたワクチンをそれまでに接種されたことのある動物のものとすることができる。
【0023】
発明の詳細な説明
本発明は、生ワクチンおよび不活化ワクチン両方から成る一組のBHV-1ワクチンに関するもので、これらはどちらも糖蛋白質gE-遺伝子の全部または一部を欠いている。この組は、自然gE欠失変異株も、チミジンキナーゼ遺伝子および/または糖蛋白質gI-遺伝子の欠失を含むことも含まないこともある構成gE欠失変異株も、および異種遺伝子ベクタとして使用される構成gE欠失変異株も含む。さらに本発明は、BHV-1糖蛋白質gE-遺伝子をコード化するヌクレオチド配列、これらの配列から誘導されるオリゴヌタレオチド、該糖蛋白質gEそれ自身、そこから誘導されるペプチド、および該gE糖蛋白質とそこから誘導されるペプチドに対して向けられる(モノクローナルまたはポリクローナル)抗体に関する。また本発明は、BHV-1の糖蛋白質gEとgIの複合物、およびかかる複合物に対して向けられる抗体に関する。
本発明に従ったこれらの物質は、以下のために使用することができる。
【0024】
1)BHV-1によって引き起こされる疾病を予防するための牛のワクチン接種。
これはBHV-1に感染した動物と、ワクチン接種した動物の間の区別が可能な従来のワクチンおよび構成ワクチンを一緒に使用することが可能。
【0025】
2)BHV-1による疾病および防御抗原に対するコード化配列をBHV-1欠失変異株の中に組込むことができる他の病原体により引き起こされる疾病を予防するための牛のワクチン接種。
【0026】
3)牛の血液、血清、乳、他の体液について、それらが野性型BHV-1によって感染しているか、gE欠失変異株でワクチン接種されているかを、血清学的に、または核酸検出技術(たとえばPCR)により判断するためのそれらの試験。
【0027】
オリゴペプチド、ポリペプチド、およびBHV-1の糖蛋白質g-遺伝子と糖蛋白質gI-遺伝子のコード化配列より誘導される糖蛋白質の合成。
糖蛋白質gE-遺伝子(図3A)およびこの遺伝子についてコード化する単離DNAフラグメントについての、例に記載のDNA配列分析の結果により、標準的な分子生物学的処置を用いて、gE蛋白質のペプチド(オリゴまたはポリペプチド)を合成すること、および原核的方法(バクテリア中)または真核的方法(たとえば、ネズミの細胞において)により、gE蛋白質の全体を、またはその一部を表すことが共に可能となる。これらの方法によって、たとえばgE特有のモノクローナル抗体の生成に役立つことができるgE特有の抗原を得ることができる。さらに、gE特有の抗原(および、gE特有のモノクローナル抗体)を血清試験において使用することができ、BHV-1 gE欠失ワクチンの接種をした動物と野性型BHV-1ウイルスに感染した動物の間の区別が可能となる。
【0028】
例に記載の、糖蛋白質gI-遺伝子、およびこの遺伝子についてコード化する単離DNAフラグメントについて、糖蛋白質gEを表す真核細胞と共に、部分的にDNA配列を分析した結果、真核細胞中のgI/gE複合物を表すことができる(図13および図14参照)。この糖蛋白質複合物は、gI/gE特有のモノクローナル抗体を作るために使用することができる。また、このgI/gE複合物は、単gE BHV-1欠失変異株、または二重gI/gE BHV-1欠失変異株によるワクチン接種を受けた牛と野性型BHV-1ウイルスに感染した牛の区別をする血清試験における抗原としても使用することができる。
【0029】
gE特有ペプチド
既知の蛋白質コード化配列に基づき、自動合成装置により、約40〜50のアミノ酸のポリペプチドを作ることができる。BHV-1菌株LamのgE糖蛋白質の蛋白質コード化配列が解明されたので(図3A)、このBHV-1 gE糖蛋臼質のポリペプチドの合成が可能となる。かかるポリペプチドを用いて、標準的な方法に従って、マウスやウサギなどの実験動物を免疫化し、gE特有抗体を作ることができる。さらに、これらのgE特有ペプチドを用いて、抗gE抗体がgE蛋白質(エピトープ)と反応する位置を、たとえばPEPSCAN法(Geysen et a1.,1984,Proc.Natl.Acad,Sci.USA 81,3998〜4002)により、さらに特定することができる。gE特有のオリゴペプチドも抗gE抗体を実証する血清試験において使用することができる。
【0030】
gEの原核発現
細菌中にてgE蛋白質を合成するため(すなわち、gEの原核発現)、糖蛋白質gEについて、またはその一部についてコード化するDNAフラグメントを、原核発現ベクタ中にタローン化しなければならない。原核発現ベクタは、環状DNA分子で、これは別個に複製してゆく分子(プラスミド)として、細菌中で自らを維持することができる。これらの発現ベクタは、抗性物質抵抗についてコード化を行い、したがって発現ベクタを伴う細菌の選択を可能にする。1つ、またはそれ以上の標識遺伝子を含む。さらに、発現ベタタは、(制御可能であることも多い)プロモータ領域を有しており、該領域の背後にDNAフラグメントを結合することができ、これらのフラグメントは、プロモータの影響により発現される。現行の多くの原核発現ベクタにおいては、いわゆる担体蛋白質に融合し、所望の蛋白質が発現している。そのために、ベクタ中ではプロモータの後に、担体蛋白質についてのコード化配列が位置しており、所望のDNAフラグメントがこれに直接隣接して結合されている。融合蛋白質は、多くの場合より安定で、認識および/または単離もより簡単である。一定の細菌株にて、特定の融合蛋白が達成することができる定常状態レベルは融合によって、および菌株によって異なっている。種々の組合わせを試みることが慣例になっている。
【0031】
糖蛋白質gE-遺伝子の真核発現
蛋白質の原核発現は幾つかの利点をもたらしてくれるが、蛋白質は真核細胞で起こるグリコシル化のような修飾変更を欠いている。その結果、真核発現蛋白質は多くの場合より適切な抗原である。マウスの細胞のような真核細胞における蛋白質の異種発現の場合、真核発現ベクタが使用される。これらのベクタは、プラスミドで大腸菌細胞内で増殖させることが可能であるばかりでなく、真核細胞中にて安定した状態で生存することもできる。これらのベクタは、原核選択マーカの他に、真核選択マーカも含む。原核発現ベクタに似て、真核発現ベクタは、所望の遺伝子が後に結合することができるプロモータ領域を含んでいる。しかしながら、真核ベクタにおけるプロモータ配列は、真核細胞に特有である。また、真核ベクタにおいては、担体蛋白質に対する融合が利用されることは滅多にない。これらのベクタは、標準的なトランス・フェタション法(F.L.Graham and A. J. van der Eb. 1973, Virology 52,456〜467)により真核細胞中に導かれる。真核プラスミドベクタの他に、ウィルスベクタもあり、この場合には、異種遺伝子がウィルス(たとえば、レトロウィルス、ヘルペスウィルス、ワクシニアウィルス)のゲノム中に導かれる。次いで、真核細胞に組換えウィルスを感染させることができる。
一般に、個々の遺伝子産物にとって、ベクタや細胞はどういったタイプのものが最適であるかということを予測することはできない。大抵の場合、幾つかの組合せが試みられている。
【0032】
糖蛋白質gEおよび糖蛋自質gI両方の真核発現蛋白質が得る最終構造は、蛋白質の一次アミノ酸配列、折たたみ、翻訳後修飾等に依存している。蛋白質の構造に寄与する重要なファクターは、1つまたはそれ以上の他の蛋白質との相互作用である。BHV-1糖蛋白質gEは、少なくとも1つの他の糖蛋白質、すなわちBHV-1糖蛋白質gIとの複合物を形成することを我々は発見した。かかる複合物を初めて示したものは、候補的抗-gEモノクローナル抗体1,51,67,75,78による我々の結果であった(表2参照)。これらのモノクローナル抗体は、Difivac-1とも、またLam gE-とも反応しなかったばかりでなく、糖蛋白質gE発現3T3細胞を認識することもできなかった。しかしながら、これらのモノクローナル抗体は、Difivac-1に感染後はgE-発現3T3細胞とは反応せず、これらのモノクローナル抗体のための適切な抗原コンホメーションを糖蛋白質gEに与えるために、因子を補足することが必要であることを示した。我々が行ったモノクローナル抗体81との放射線免疫沈澱試験の幾つかにおいて、見かけ上の分子量63KDの蛋臼質が共沈することが判った。ヘルペス単ウィルス糖蛋白質gEは、これに匹敵する分子量の蛋白質(HSV1糖蛋白質gI)と複合物を形成するという事実に鑑み、我々はBHV-1糖蛋白質gEは、糖蛋白質gIのBHV-1同族体と複合物を形成するものと推定した。このBHV-1 gE/gI複合物を研究するため、および適切な抗原構造を有するgE抗原を作るため、これらの両蛋白質を1つの真核細胞中で発現させた。このために、糖蛋白質gEだけの真核発現のために記載したものと同じ処置を適用した。ただし、前提条件として、選択可能な種々の真核標識を有する発現ベクタの使用を1つだけ追加した。
【0033】
血清試験
gEに対する抗体に基づき、Difivac-1のワクチン接種をされた牛と、野性型BHV-1に感染した牛を区別するための血清学的方法は、好適にはgEに対して向けられたモノクローナル抗体の使用に基づく。これらの抗体は、以下の方法で用いることができる。
【0034】
a)Van Oirschot et al(Journal of Virological Metbods 22,191〜206,1988)により述べられた原則による。Aujeszky病のウイルスに対するgI抗体検出のための、この酵素結合イムノソルベント検定法(ELISA)では、抗体は、gI上に2つの異なったエピトープを有する2つのモノクローナル抗体の反応に対するブロック効果によって実証される。この試験は、以下のように実施される。マイタロタイタープレートを37℃で1晩、モノクローナル抗体1で被覆し、その後、たとえば4℃または-20℃にて保存する。試験される血清は、別の未被覆マイクロタイタープレートにおいて、37℃にてたとえば2時間、抗原と共に前保温される。モノクローナル抗体1が被覆されたプレートを、たとえば5回洗浄し、その後セイヨウワサビペルオキシダーゼ(HRPO)と結合したモノクローナル抗体2をこれらのプレートに添加する。次に、前保温された血清-抗原混合物を、2つのモノクローナル抗体が置かれているプレートに転移させ、37℃にてたとえば1時問保温する。これらのプレートを洗浄し、各ウェルに基質を添加する。室温にて約2時間経過させた後、これらのプレートについてスペクトル光度計による読取りを行う。各プレートには、4つの陰性対照血清および陽性血清の4つの連続希釈物が含まれる。光学濃度が同じプレートで試験された4つの陰性対照血清の平均光学濃度の50%を下回る血清は陽性とみなされる。
【0035】
b)間接2重抗体サンドイッチ(IDAS)法による。
この場合は、マイクロタイタープレートは、gE蛋白質に対して向けられたモノクローナルまたはポリクローナル血清で被覆する。gE抗原調製物との保温の結果、被覆にgEが結合する。試験される牛の血清中のgEに対して特別に向けられた抗体は続いてgEに結合する。これらの結合抗体は、抗-ウシ免疫グロブリン複合物によって認識される。この複合物中の抗体は、ペルオキシダーゼ酵素に共有結合する。最後に、この結合複合物は、色素原基質の添加により、可視のものにされる。この反応の特性は、gE-抗原調製物の代わりに、gE-陰性対照調製物により同じ処置を実施して調べる。各マイクロタイタープレート上には、陽性および陰性の対照血清が含まれる。この試験は、陽性の血清が一定の希釈度において陽性を示す場合に有効である。光学濃度が標準陰性対照血清よりも0.2高い場合には、血清は陽性である。
【0036】
c)2において述べたIDAS法では、ただし試験される血清を保温した後であるが、抗-ウシ免疫グロブリン複合物の代わりに、抗-gEモノクローナル抗体/HRPOが使用される。抗-gEペプチド血清または抗-gEポリクローナル血清を抗-gEモノタローナル抗体の代わりに使用してもよい。これらのプレートを洗浄し、各々のウエルに色素原基質を添加する。室温にてたとえば、約2時間経過させた後、これらのプレートについてスペクトル光度計による読取りを行う。各プレートには、4つの陰性対照血清および陽性血清の4つの連続希釈物が含まれる。光学濃度が同じプレートで試験された4つの陰性対照血清の平均光学濃度の50%を下回る血清は陽性とみなされる。
【0037】
d)精製、未精製を問わないウイルス抗原がマイクロタイタープレートに一晩被覆されるブロックELISA法による。これらのプレートにおいて、試験される血清は、37℃にてたとえば1時問または1時間以上保温される。洗浄処置をした後、これらのプレートに抗-gEモノクローナル抗体を添加し、37℃にてたとえぱ1時間保温する。抗-gEペプチド血清または抗-gEポリクローナル血清を抗-gEモノクローナル抗体の代わりに使用してもよい。これらのプレートを洗浄し、各ウエルに、色素原基質を添加する。室温にて、たとえば2時間経過させた後、これらのプレートについて、スペクトル光度計による読取りを行う。4つの陰性対照血清および陽性血清4つの連続希釈物が含まれる。光学濃度が、同じプレートで試験された4つの陰性対照血清の平均光学濃度の50%を下回る血清は陽性とみなされる。
【0038】
上記の方法のいずれにおいても、gEを含む、従来の方法で成長させたウイルス抗原を使用することができるが、原核また真核を介して発現するgE-抗原も使用することができる。あるいはまた、BHV-lgE配列に基づくオリゴペプチドを従来の抗原の代わりに、上記診断試験において使用することができるであろう。さらにまた、かかるオリゴペプチドを、Kemp et al.による記事、Science 241,1352〜1354,1988に記載の方法に従った、いわゆる"Cow-side"試験の開発に使用することができるであろう。かかる試験は、感染動物中に存在しているgEに対して向けられている抗体によるオリゴペプチド抗原配列の連結に基づいたものになるであろう。このような試験の場合、オリゴペプチドを、牛の赤血球に対して向けられたモノクローナル抗体に連結させる必要があるであろう。
【0039】
ポリメラーゼ連鎖反応を使用する核酸分析
たとえば、オリゴヌクレオチド(プローブおよびプライマ)をポリメラーゼ連鎖反応に用いて、ワクチン接触を受けた動物と感染した動物を区別することができる。ポリメラーゼ連鎖(PCR)は、病原体の核酸を短時間に数十億倍に増殖させることができる技術である(Depolymerase kettingreactie. P.F,Hilderink, J.A.Wagenaar,J.W.B.van der Giessen and B.A.M. van der Zeijst,1990,Tijdschrift voor Diergeneeskunde deel 115,1111〜1117)。gEオリゴヌタレオチドは、gE陽性ゲノムにおいて、gE陰性ゲノムにおけるものとは異なる生成物が形成されるように選択することができる。これは、gE欠失ワクチンをワクチン接種された動物は、RCR試験において陽性信号を出すという利点がある。しかしながら、この方法は、試験される動物から採取した試料中、たとえば血液中のウイルスの核酸の存在に依存している。
BHV-1急性感染後は、血液中にBHV-1特有の核酸を実証することができる見込みは充分にあるが、しかしながら、BHV-1核酸も、潜伏時に血清中に実証することができるかどうかはまだ確められていない。
【0040】
ベクタとしてのBHV-1の使用
BHV-1ゲノム中の異種遣伝子の発現には、その異種遺伝子が挿入されるべき領域についての正確な情報を自由にできることが必要である。必須配列についての妨害はいかなるものもあってはならず、また異種遺伝子の発現に調節配列が利用できなければならない。gE-遺伝子は、必須ではなく、したがって、異種遺伝子gE遺伝子を交換しても問題はない。結果として、この異種遺伝子を、gE遺伝子が調節配列の影響下にあるように配置することができる。しかしながら、gE遺伝子の調節配列を使用する必要はない。異種遺伝子の発現は、代わりに、他のもの、たとえば種々の遺伝子のより強い調節配列により制御することができる。また、gE遺伝子の(輸出)信号ペプチドに異種遺伝子を連結することも可能であり、したがって異種遺伝子生成物の分泌に影響させることができる。gE遺伝子およびgE蛋白質についての詳細な知識によって、非常に適切に調整された方法で、ベクタとしてBHV-1を用いる可能性が与えられることは明らかである。発生するベクタは、さらに野性型と血清学的に区別することができる。異種遺伝子を発現するBHV-1変異株の構成は、例に示したgE欠失変異株の構成と同じ方法で実施することができる。しかしながら、次に、欠失フラグメントは、異種遺伝子が欠失位置に配置されているフラグメントと交換すべきである。
【0041】
例
1)自然gE欠失変異株の単離および同定
a)自然変異株の単離
ゲノムDNAを、従来法により弱毒化された多くのワクチンから標準的方法に従って単離し、制限酵素を用いて分析した。特に、野性型BHV-1ウイルスとの区別を可能とするに適切と思われるゲノム偏差について研究した。
特に、BHV-1ゲノムのUs領域に注意した。これは、その領域に-ヘルペス単ウイルス1との類似により-非必須糖蛋白質に対する多くの遺伝子コーディングが配置されているからである〔Identification of a herpes simplex virus1 glycoprotein gene within a gene cluster dispensable for growth in cell culture, R. Longnecker,S.Chatterjec,R.J. Whitley and B.Roizwan(1987)Proc、Natl. Acad. Sci,84,4303〜4307〕。
ユーゴスラビア、サグレフ大学より入手したBHV-1ワクチンバッチ(Lugovic et a1.,Veterinarski Arhiv 55,241〜245,1985)は、ウシの胚腎臓細胞およびウシの胚気管細胞(Ebtr)上で多くの継代を行った後に、正常なUs領域の他に、偏差のあるUs領域を有していることを証明した。さらにこのワクチンは、Ebtr細胞に大きなプラークと小さなプラークの両方を形成するように思われた。この混合群より、それぞれの国ごとに選択された小さなプラークで偏差のあるUs領域を有するウイルスを、3つの限界希釈ステップによって単離した。この経路を介して単離されたウイルスをさらに試験し、Difivac-1を希釈した。これは、1992年5月27日Institut Pasteur,Paris,Franceに寄託番号1-1213として寄託された。
【0042】
b)Difivac-1におけるgE遺伝子の欠失の同定
US領域におけるこの偏差をさらに分析するために、Difivac-1のゲノムDNAを標準的な方法で単離し、サザンブロット分析を行った(図1A)。32Pのラベルを貼った野性型HindIIIKフラグメントを有するこのブロットのハイブリッド形成により、Us領域の中心に位置しているこのフラグメントは、Difivac-1においては、1,0キロベース(kb)程度短い。さらに、この分析によって、欠けている部分の位置をおおよそ推定することができた(図1B)。この欠失をさらに分析するために、野性型BHV-1菌株LamのUs領域を単離し、原核ベクトル中にクローン化した。そのために、標準的な方法に従って、Lam菌株(図2A)のゲノムDNAを単離し、ベクタ、pUC18、pACYCおよびpBR322中にクローン化した(図2B)。欠失の推定位置周辺の領域の物理的地図を作成した(Fig.2c)。この物理的地図からスタートして、この領域のヌクレオチド配列の決定に適したサブクローンをベクタpKUN19とpUC18で構成した(図2D)。これらのサブクローンを用いて、全領域の2つの鎖のヌクレオチド配列(図2Cに指示)をサンガー法を用いて決定した。このヌクレオチド配列(SEQ.ID.NO.1)をPc、/Geneプログラムを用いて分析した。概念翻訳から、ヌクレオチド(nt)168〜nt1893が575のアミノ酸のオープンリーディングフレームのための遺伝暗号を指定する(図3A)ものと推定された。さらに分析の結果、このアミノ酸配列は図3Bに示されているように、貫膜糖蛋白質の特性を有していることが示された。実は、最初の26のアミノ酸(aa)は、典型的真核性の輸出信号として認識され、aa423とaa450の間の領域は、貫膜領域として認識される。さらに、3つのポテンシャルN-結合、糖鎖形式部位がこの配列において発生する。この推定アミノ酸配列は、ヘルペス単ウイルス(HSV)の糖『蛋白質gE-遺伝子に明らかに類似したところを示している。(図4Aおよび図4B参照)。これらおよびその他の類似点により、見つけられた遺伝子は、BHV-1のgE同族体であるという結論を正当化するものである。この理由から、遺伝子はgEと呼ばれる。このBHV-1 gE-遺伝子がDifivac-1においてどの程度欠失しているのかを判断するために、p318フラグメントを単離した。p318フラグメントは、要求されるBHV-gEがリーディングルームを開く前に、Alul部位55ntで開始し、リーディングルームの後の133ntを終了する。ゲノムDifivac-1DNAを、サザンブロットハイブリッド形成を用いて、このp318フラグメントで分析した。この結果、Difivac-1は検出可能なp318配列を全く含んでいないことが明らかとなった(図5)。この試験によりDifivac-1は欠失を有し、この欠失はgE遺伝子全体に広がっていることが明らかに実証されていることが確認された。
【0043】
欠失領域の大きさと位置を決定するために、Difivac-1のUs領域を包括しているゲノム配列を原核ベクタ中にクローン化した(図11Cを参照)。14.5kb EcoRIフラグメントをpACYCベクタ中にクローン化し、P775と命名した。7.4kb HindIIIフラグメントを独自にpUC18ベクタ中にクローン化し、p728と命名した。クローンp728より、2つのサブクローンを単離した。すなわち、クローンp737中の1.4kbPst1フラグメントとクローンp754中の350bp AluI-Pst1フラグメントである。これらのクローンの制限酵素分析とサザンブロット分析(データは示されていない)により、Difivac-1のgE欠失の長さは2.7kbであり、gE遺伝子から丁度5'でスタートし、Us領域の境界で終了することが実証された。これらの2.7kbは、反復領域の逸脱拡張として、gE遺伝子に対向したUs領域に位置した重複1kbセグメントによって置きかえられた(図11bを参照)。この分析の結果を確認するために、および正確な組換え点を決定するために、クローンp754の大半の挿入についてのヌクレオチド配列を決定し、野性型配列を比較した(図12参照)。この分析により組換え点は、gE遺伝子の出発コドンから77bp上流に位置していることが判った。
【0044】
c)Difivac-1の安全性と効能についての評価
7週齢のBHV-1血清反応陰性の特有の病原体をもっていない牛でDifivac-1を試験した。8体の牛に2ml,105TCID50の鼻腔内ワクチン接種を行った。2ml中の1mlは各々の鼻孔にスプレーした。各々別々の独立した区画に収容されていた、BHV-1血清反応陰性の特有病原体を持っていない7週齢の8体の牛に2mlの培養基を鼻腔内投与し、非ワクチン接種対照として用いた。ワクチン接種から5週後に、ワクチン接種牛と非ワクチン接種牛に107TCID50、の強毒性BHV-1菌株、Iowaを鼻腔内接種した。接種から6週後に推定される潜伏ウイルスを再活化させるため、全牛に5日間デキサメタゾンにより筋肉内処理を行った。臨床微候、直腸温、発育を監視した。布で拭き取った鼻水からウイルスの単離を行い、血清中の中和抗体力価を求めた。
【0045】
ワクチン接種後の、牛の挙動、食欲、直腸温、発育率は正常のままであったが、ワクチンを接種した牛は、いくらか漿液性鼻水の分泌と唾液の分泌過多があった。Difivac-1は、ワクチン接種後、布で拭き取った鼻水から排泄された(図17)。ワクチン接種牛は全て、BHV-1に対する中和抗体を生成した。
菌株接種後、14日までに、非ワクチン接種牛は全て、無感情、食欲の喪失、目ヤニおよび鼻水の分泌、下あごの歯肉の赤らみ、鼻の粘膜の強い傷害が見られ、また接種後4日で発育が停止した。ワクチンを接種した牛の鼻の粘膜の傷害は小さく、速やかに治癒するもので、発育が止まることはなかった。菌株接種後の1日毎の臨床評価、直腸温、発育の推移を図18,図19および図20に示した。菌種接種後、全牛とも鼻からウイルスを排泄したが、ウイルス排泄の量と期間は、ワクチン接種牛は著しく低かった(図21)。ワクチン接種牛においては、二次抗体反応が見られ、非ワクチン接種牛は全て、菌種接種後抗体を生成した。再活化後、接種ウイルスを1体のワクチン接種牛および5体の非ワクチン接種牛から単離した。Difivac-1を再活化することはできなかった。
【0046】
上記の結果により、Difivac-1は、若年牛においては殆ど病気の微候を引き起こさず、再活化されないことが実証された。Difivac-1は、菌種接種後病気の程度とウイルスの排泄量を著しく低下させた。
結論として、Difivac-1は、BHV-1感染予防のために牛に使用される、安全かつ効能のあるワクチンである。
【0047】
2)BHV-1の組換えgE欠失変異株の構成
Difivac-1よりも分子的により明確で、かつ所望されるならば、gE-遺伝子の欠失に加えて、たとえばチミジンキナーゼ遺伝子に欠失を有した識別可能なBHV-1ワクチンと自由に利用できるようにするために、Difivac-1の他に、組換えgE欠失変異株を構成した。糖蛋白質gE-遺伝子の決められた位置からスタートし、gE遺伝子を側面に配置するクローン化されたDNAフラグメントを用いて、gE欠失フラグメントを構成することができた。標準的な技術(F.L.Graham and A.J.van der Eb 1973,Virology52,456〜467)を用いて、,この欠失フラグメントを野性型BHV-1菌株のゲノム中で組換えることができ、その結果gE欠失変異株となった。
【0048】
a)gE欠失フラグメントの構成
【0049】
gEフラグメントの構成のために、一方においては、全gE配列を欠き、他方野性型ゲノムと組み換えることができる十分なフランキング配列を含むフラグメントが目標とされた。5'(上流)側では、gE遺伝子の開始コドンの前に18ntを終える1.2kb PstI-AsuIIフラグメントが選択された。3'(下流)フラグメントについては、gE遺伝子の停止ユドンの前に2ntを開始する、1.2kb EcoNI-DraIフラグメントが選択された(図6)。
【0050】
gE欠失フラグメントの構成については、gE遺伝子の5'側に位置した、BHV-1菌株Lamの8.4kb HindIIIKフラグメントからの1.4kb PstI-SmaIフラグメントが、プラスミドpUC18のSmaIおよびPstI中にサブクローンされた。このクローンはp515と命名した。gEの3'側に位置し、4.1kb HindIII-EcoRIクローンからくるEcoNI-SmaIフラグメントはp515の特異なAsuII部位中にクローンされた。このように、gE欠失フラグメントの構成は完結し、このようにして構成されたクローンをp519と命名した。原則的にはp519の全PstI-SmaI挿入をgE欠失フラグメントとして利用することができるが、これは望ましくない。実際は、PstI-SmaIは、Us領域をフランクする反復配列中に、約100〜150塩基対(bp)の拡張を行う。この100〜150bpの部分は、gE遺伝子が位置しておらず、したがって所望されていない組換生成物を作り出すことができないUs領域の他方の反復配列と組換えることができた。このため、PstI-DraIフラグメントが組換え実験のために選択され、したがって100bpの反復が除去された。
【0051】
b)野性型BHV-1のゲノムによるgE欠失フラグメントの組換え
構成gE欠失フラグメントと野性型BHV-1のゲノムの間の組換えを効果的なものとするために、2つのDNA分子のマイクログラム量を、F.L.Graham and A.J.van der Eb(1973、Virology52,456〜467)の標準的な方法に従って、ウシの胚気管(Ebtr)細胞に同時トランスフェクションする。細胞の組換えメカニズムは、細胞によって組み込まれた少しの割合のDNA分子(2〜4%)の組換えを導く。組換えられたgE欠失変異株の選択のために、トランスフェクション後に形成されるウィルス混合物を、新鮮なEbtr細胞培養物上に播種する。大抵の場合、これによって発達する、別々のウィルス群(プラク)が、1つのウィルスから生じる。BHV-1菌株LamのgE欠失変異株の単離のために、これらのプラクの内の230を単離し、DifivaC-1に感染した細胞と反応しない、BHV-1特有のモノクロナール抗体による標準的免疫法により試験を行った。これらのモノクロナール抗体を、糖蛋白質gEに対して向ける。230のプラークの内5つがこれらのモノクロナール抗体と反応しなかった。これらの5つのプラークのDNAについてさらに研究を行った。
【0052】
c)BHV-1菌株Lamの構成gE欠失変異
株のDNA分析上述の5つの候補gE欠失変異株の内3つ(1B7,1B8および2H10)のDNA調製物について、標準的なサザンブロット分析技術(Sambrook et al.1989)を用いて試験した。ゲル電気泳動と、プローブとして2.3kb PstI-DraI欠失フラグメントによるサザンブロットハイブリット化がその後に続く、PstIとDraIによるこれらのDNAの2重の消化は、ウィルス群1B7と1B8のゲノムのgE遺伝子は、所望の方法で正確に取り除かれたことを示している。図7Aおよび図7B参照。群2H10は、偏差PstI-DraIフラグメントを有している。gE特有プローブによるサザンブロットハイブリット化により、3つのDNA調製物のいずれにもgE配列は置かれていないことを示している(結果については示されていない)。BHV-1ウィルス群1B7および1B8は、故意の組換gE欠失変異株である。BHV-1ウィルス群1B7はワクチン特性について試験を行った。
【0053】
d)チミジンキナーゼ/gE二重欠失変異種の構成
わずか1つの遺伝子に欠失を有するBHV-1組換欠失変異株は、十分に毒性が低下したものではないかもしれないので、BHV-1菌株LamおよびHarberinkのチミジンキナーゼ(CTK)遺伝子にも欠失を与えた。これらの変異株は、上記のgE欠失変異種に対して使用される方法と類似の方法で構成された(結果は示されていない)。これらのTK欠失変異株はTK/gE二重欠失変異株の構成に使用した。
【0054】
e)糖蛋白質gI/糖蛋白質gE二重欠失変異
株の構成gE単欠失変異株のワクチン接種をうけた牛は、抗gI/gE抗体の検出を妨害するおそれのある抗-gI抗体を生成する可能性があるため(以下に考察)、gI/gE2重欠失を有するワクチンについても発明した。このようなgI/gE二重欠失変異株は、gE単欠失変異種の構成に使用されるのと同じ処置を用いて構成することができる。gE遺伝子の5'端部を包括する、1.8kb PstIフラグメントの上流側端部のヌクレオチド配列の部分的分析により、他のヘルペスウィルスに見られるgI相同物に対して有意な相同関係を有するオープンリーディングフレームが明らかになった。図13および図14を参照のこと。gE遺伝子の下流に位置した、gI遺伝子およびEcoNI-SmaIフラグメントの推定5'端部を包囲している350bp SmaI-PstIフラグメントを用いて、gI/gE欠失フラグメントを構成することができた。このフラグメントは野性型ゲノムと組換え、BHV-lgI/gE欠失変異株をもたらすことができた。図16を参照のこと。理論的にはさらにまだ生成することができる80〜90アミノ酸は、抗-gI/gE抗体の欠失を防げることができる抗体を導き出すことはできないであろう。さらに、gI遺伝子の配列分析により、完全なgIコード化領域を包括するgI欠失の構成が可能となるであろう。このgI/gE二重欠失変異種をDifivac-IEと命名した。
【0055】
f)Lam gE-およびLam gE-,TK-変異株の安全性と効能の評価
Lam gE-およびLam gE-,TK-,BHV-1変異株菌株のワクチン特性を7週齢の、BHV-1血清反応陰性の特有病原体を持たない牛において試験した。各々の変異株を6体の牛に鼻腔内スプレーした。各牛の全投与量は、2ml培養基において105TCID50となり、2m1中1mlは各々の鼻孔にスプレーした。他の6体の牛はウィルスを含まない培養基を鼻腔内投与し非ワクチン投与対照として用いた。ワクチン投与から5週後に、ワクチン投与および非ワクチン投与いずれの牛も、すべての牛に強毒性BHV-1菌株lowa107TCID50を鼻腔内接種した。ワクチン接種および菌株接種後、臨床徴候、直腸温、体重を監視した。鼻のウィルスの放出日数の決定のために布で拭きとった鼻水を採取した。
ワクチン接種後の、牛の挙動、食欲、直腸温、発育率は正常を維持した。漿液性鼻水の分泌および鼻の粘膜の小さな傷害が、ワクチン接種牛すべてに観察された。ワクチン接種の牛の鼻から約7日間ウィルスを単離することができた(表1)。
【0056】
菌接種後、非ワクチン接種の対照牛すべてが、無感情、食欲の喪失、目や鼻からの分泌、下あごの歯肉の赤らみ、鼻の粘膜の強度の傷害を示し、鼻水率が減少したLam gE-,TK-をワクチン接種した牛はすべて、鼻水の分泌が幾分増加し、鼻の粘膜の小さな傷害がいくらか見られた。Lam gE-をワクチン接種した牛のすべてが、鼻水の分泌または鼻の粘膜の傷害が進行したわけではなかった。無感情、食欲の喪失、疾病についてのその他の徴候は、ワクチン接種牛では観察されなかった。菌接種後の直腸温、発育、臨床評価は図22,23および24に示されている。非ワクチン接種牛はワクチン接種牛の2倍の長さにわたってウィルスを鼻から放出した(表1)。
【0057】
上記の結果は、Lam gE-およびLam gE-,TK-BHV-1突然変異菌株は、若年の牛においては、病気の臨床徴候をほとんど誘発していないことを示している。これらの突然変異菌株はどちらも菌接種後の病気を予防し、鼻のウィルス放出期間を50%短縮させた。
Lam gE-およびLam gE-,TK-B、HV-1突然変異菌株は安全で、BHV-1感染予防のワクチンとして牛に用いて効能を有するものである。
【0058】
3)gEの原核発現
BHV-1糖蛋白質gE-遺伝子の原核発現の場合、これまでのところpGEX発原ベクタ(D.B.Smith and K.S.Johnson,Gene 67(1988)31〜40)が使用されてきた。イソプロピルチオガラクトシド(IPTG)によって発現に誘導されることができる、tacプロモータの影響下にある日本住血吸虫からの担体蛋白質グルタチオンS-転移酵素(GST)についてpGEXベタタはコード化する。GST-gE融合蛋白質の例は構成pGEX-2T600s3の生成物である(図8A)。この構成において、標準的な分子生物学的技術を用いて(Sambrook et al. 1989)、gE蛋白質の200アミノ酸のN末端領域についてコード化する、600bp SmaIフラグメントがGST遺伝子の後に連結された。この構成は3つの部分から成る設計とされ、600bpフラグメントの異なったリーディングフレームがGSTに連結された。これらの構成はすべてIPTGと共に誘発された大腸菌株DH5α内に導入され、形成された蛋白質はウェスタンブロッテングによるポリアクリルアミドゲル電気泳動後、ニトロセルロースに転移させた。抗-GSTによる免疫性検出により、gE蛋白質領域についてコード化する適切なリーディングフレーム(No.3)だけが、27k(GST)t20k(gE)=47kの推定サイズの顕著な融合蛋白質の発現を導くことが実証された。Difivac-1と反応しない我々が単離した3つのモノクロナール抗体は、ウェスタンブロット中の47k D GST-gE融合蛋白質を認識する。図8B参照。
【0059】
4)糖蛋白質gE-遺伝子の真核発現
糖蛋白質gE-遺伝子の真核発現については、これまでは、とりわけ、ベクタ、pEVHisが選択されてきた。pEVHisベクタは、真核遺伝標識として、ヒスチジノールデヒドロギナーゼについてのHisD遺伝子コードを有しており〔EC1.1.1.23〕(C.Hartmann and R.Mulligan,1988,Proc,Natl.Acad Sci.USA 85,8047〜8051)、このヒスナジノールデヒドロギナーゼが、細胞の2.5mMヒスチジノールの毒性濃度を長びかせる原因となっている。さらに、このベクタは、ヒトのサイトメガにウィルス(HCMV)の即時型遺伝子のプロモータ領域を、該遺伝子の後に位置している特異な制限酵素部位と共に有している。pEVHis/gE発現ベクタの構成のためには、糖蛋白質gE-遺伝子の全コード領域を有するフラグメントが使用された。gEの従来のオープンリーティングフレームの前のAluI部位55でスタートし、その後の133bpで終了する。この領域がpEVHisベクタのHCMVプロモータの後でクローン化され、これによって構成pEVHis/gEが形成された(図9)。このpEVHis/gEを大腸菌DH5の細胞中にて増殖させ、塩化セシウム勾配法によって精製した(Sambrook et al.,1989)。この精製DNAを、Graham and Vander Ebの方法に従って、Balb/c-3T3細胞にトランスフェクトした。形質転換細胞をヒスチジノールと共に選択した結果、20の耐ヒスチジノールコロニーを単離することができた。これらのコロニーを免疫ペロキシダーゼ単分子層試験(IPMA)によって、モノクロナール抗体81によって試験した。4つのコロニ一によって、gE蛋白質の発現が証明された。これらの4つのコロニーの内、3T3gEクローン9を、高度のgE発現を有するサブクローンの単離に使用した。この方法によって単離されたクローン(3T3gE9.5と命名)を候補抗gEモノクロナール抗体の特性づけのために用いた。
【0060】
5)同一細胞におけるBHV-1糖蛋白質gEおよびBHV-1糖蛋白質gI双方の真核発現
BHV-1糖蛋白質gEと同一の細胞においてBHV-1糖蛋白質gIを発現させるため、まず最初にBHV-lgI遺伝子の推定位置を決めた。ヘルペス単ウィルス糖蛋白質gI遺伝子は、糖蛋白質gE遺伝子のまさに上流に位置しているので、BHV-1 gI遺伝子は、対応の位置に位置しているものと推定された。これを試験するため、BHV-1 gE位置が開始したところより約1kb上流に位置した、283ヌクレオチドの領域の配列を決定した。この領域の概念翻訳により、第2リーディングフレームは、ヘルペス単ウィルス糖蛋白質gIと相同の94アミノ酸配列についてコード化する(図13および図14参照)。相同セグメントは、スタートコドンから約80のアミノ酸であるので、BHV-1 gI遺伝子のオープンリーディングフレームの推定開始は、配列領域から約250nt上流と見積もられる。これより、配列領域から400nt上流でスタートし、gE遺伝子内で終了する、1.7kbSmalフラグメントは、BHV-lgIl遺伝子の完全なコード化領域を含んでいるべきであることが推定された。この1.7kbSmalフラグメントが真核ベクタMSV-neo内にクローン化された(図15参照)。このベクタは、強いマウス肉腫ウィルスプロモータと、抗生物質G-418硫酸ゲネティシンに対する耐性についてコード化するセレクター遺伝子neoを含んでいる。結果として生じる構成MSVneoGIが、大腸菌DH5α細胞中で増殖され、Graha and Van der Ebの方法を用いて、3T3gE9.5細胞中にトランスフェクトされた。トランスフェクトされた細胞は、400mg、ゲネティシン/ml培養基で選択され、耐性コロニーを単離し、3T3gE9.5細胞と反応しなかった候補抗-gEモノクロナール抗体で試験した。これによって、たとえば、野性型BHV-1だけでなく、モノクロナール抗体66と反応した3T3gE/gIR 20クローンを選択した。
【0061】
6)候補抗-gEモノクロナール抗体の特性づけ
野性型BHV-1に対してモノクロナール抗体を生成し、Difivac-1に感染したEbtr細胞との反応不能性について選択した。これらのモノクロナール抗体を下記との反応性について試験する。
a)Lam gE-欠失変異株
b)ウェスタンブロットにおける、上記の原核発現生成物
c)上記のgE発現Balb/c-3T3細胞
d)c)に記載のDifivac-1に感染した細胞
e)gE/gI複合物を発現するBa1b/c-3T3細胞
a,c,dおよびeとの反応性試験については、免疫ペロキシダーゼ単分子層試験(IPMA)を用いた。表2の結果により、我々が生成したものは、gEに対して向けられているモノクローナル抗体(2,3,4,52,66,68,72,81)およびgE/gI複合物上の立体配座抗原ドメインに対して向けられている可能性のあるモノクローナル抗体(NO.1,51,53,67,75,78)であることが判る。種々のモノクローナル抗体によって認識される抗原ドメインの地図を作るための競含IPMAにより、少なくとも4つの抗原ドメインが糖蛋白質gE上に存在すること、およびおそらく1つのドメインがgE/gI複合物により形成されることが示された(表2)。
【0062】
BHV-1に感染した牛の抗-gE抗体の検出
感染した牛の血清中にgE抗体が存在しているか否かを調べるために、間接ブロッキングIPMAを16の候補gE-モノクローナル抗体に実施し、以下の8血清を選択した:
−Difivac-1のワクチン接種を受け、毒性Iowa菌株が接種された牛の菌接種14日後に採取された2血清。
−試験のためBHV-1サブタイプ1ウイルスに感染させた牛の、感染20ケ月後に採取された2血清。これらの牛のうちの一体は接触感染によって感染させた。
−試験のためBHV-1サブタイプ2bウイルスに感染させた牛の、感染20ケ月後に採取された2血清。これらの牛のうちの一体は接触感染によって感染させた。
−ts変異株ワクチンの接種を行い、3週後にBHV-1サブタイプ2bウイルスを接種させた特有病原体のない牛の、菌接種7週後に採取された1血清。
−ts異変株ワクチンの接種を行い、3週後にBHV-1サブタイプ2bウイルスを接種させた、ノトバイオートの牛の、菌接種7週後に採取された1血清。
【0063】
表2により、これらの血清はどれも、gE上の抗原性ドメインIIIおよびIVに対する、およびgE/gI複合物上に位置していると思われる抗原性ドメイン1に対する抗体を含んでいることが判る。gEは、明らかにBHV-1-感染とワクチン接種された牛を区別するための適切な血清学的遺伝標識であると結論づけることができる。
【0064】
7)BHV-l gE-特有プライマを用いるPCR処置によるBHV-1核酸の検出
BHV-lgE遺伝子の決定されたヌクレオチド配列からスタートして、PCRに適切なプライマの対を、Lowe et alによるプライマ選択プログラム(T. Lowe, J.Sharefkin, S, Qi Yang and C. W .Dieffenback 1990, Nucleic Acids Res, 18, 1757〜1761)を用いて選択した。これらのプライマをP3、P4と命名し、図に示した。これらのプライマは159nt離れて位置しており、200ntのフラグメントの増殖を導く。プライマP3、P4と単離されたBHV-1 DNAを用いて、PCR処置の条件を最適化した。これは特に、MgCl2の濃度の変更、グリセロール濃度、サイクル条件に関するものである。BHV-lDNAの増殖のためにP3、P4を使用するため最適と考えられる緩衝液は、10mM Tris pH 8.0, 50mM Kca1, 0.01%ゼラチン, 2.6mM MgC12および20%グリセロールである。最適のサイクル条件と考えられるものは(Perkin Elber Cetus DNA Thermal Cycler),サイクル1〜51については、1分98℃, 30秒96℃, 30秒55℃, 45秒72℃である。PCR増殖後、得られた200ntDNAフラグメントを2%アガロースゲル上で電気泳動にかけ、ニトロセルロースにブロットし、続いてサザンブロット分析を行った。サザンブロット分析に使用された32P dCTPのラベルが貼られたプローブが、プライマ結合部位間に位置している137bp TagIフラグメントである(図10)。ハイブリッドフィルターの自動X線撮影により、200bpバンドを観察することができた。さらに、このルートを介して、10BHV-1ゲノムだけの増殖(約1.5×10-15μgDNA)が検出可能と思われる信号を導く(結果は示されていない)。同等の方法で、BHV-1糖蛋白質gIIIのコード化配列(D.R.Fitrpatrick, L.A. Babiuk and T.Zamb, 1989,, Virolagy 173, 46〜57)に基づくプライマを用いてPCR処置を行った。野生型BHV-1 DNAとgE欠失変異株ワクチンの区別を可能とするために、DNA試料にgE-特有PCR分析およびgIII-特有PCR分析の両分析を行った。このような試験において、Difivac-1DNA調製物はgIII陽性およびgE陰性であることが判った。ウシの精液中のBHV-lDNAの検出は、BHV-1特有PCR処置の重要な用途であると思われるので、BHV-1に感染したウシの精液にgE-特有PCRの実施を試みた。しかしながら、精液中の未知の成分がポリメラーゼ連鎖反応に強い抑制効果を有している。したがって、ウシの精液からBHV-1 DNAを単離するプロトコールが展開された。ウシの精液からDNAを単離するため、30μ1の精液を、1mg/m1プロティナーゼK(pk)とともに、60℃にて、全量300μ1の0.15M NaC1, 0.5%Na-サルコシル, 40mM DTT中にて培養する。1時間後、試料を室温にまで冷却し、300μl 6M NaIを添加し、5分間培養する。この混合物より、標準クロロホルム/イソアミルエタノール抽出によりDNAを単離し、1容量のイソプロパノールで沈殿させた。沈殿物を2.5M NH4AC/70%エタノールで洗浄し、60℃にて1時間10mM Tris pH7.4,1mM EDTA,0.5%Tween80および0.1mg/mlpk中で、第2培養のために再懸濁する。このDNA調製物は直接ポリメラーゼ連鎖反応下に置くことができる。
【0065】
Lam gE-またはLam gE-/TK-を接種後および毒性の強いBHV-1菌株を投与後、これらの接種されまたは無接種の牛から垂れる鼻水ウイルス
【表1】
【0066】
【表2】
【図面の簡単な説明】
【0067】
【図1】BHV-1菌株Difivac-1およびlowaのサザンブロヅト分析 Aは、Difivac-1とlowaとのゲノムDNAのサザンブロットの自動放射線図である。1と3レーン中で、Difivac-1DNAは、HindIIIとPstIとによる制限酵素消化後に、用いられた。2と4レーン中で、lowa DNAは、HindIIIとPst1とによる制限酵素消化後に用いられた。フラグメントの大きさは、キロベース(kb)で示される。 Beckman L5-65超遠心分離のSW27ロータ中で20,000回/分で、pH7.4のトリス10mMと150mM NaClと1mM EDTAとの中で、25%(w/w)庶糖クッションを用いて2時間、ウイルスで汚染されたEbtr細胞と共に培養液(約450cm2ローラボトルあたり70ml)を遠心分離することによって、ウイルスDNAが単離された。得られたウイルスベレットから、DNAが標準の手法で単離された(J.Sambrook,E,F.Fritsch and T.Manfatis,1989,Molecular cloning:a laboratory manual,2nd ed.Cold Spring Harbor Laboratory Press,NewYork)。このDNA上で制限酵素消化は、製造者によって供給されるSuRE、/Cut緩衝液中でBoehrnger Mannheimからの酵素で行われた。 0.7%アガロゲルを水平電気泳動して分離し、ニトロセルローズフィルタ(Schleicher & Schuell,lnc)でブロットした後、フィルタは6時間42℃で50%ホルムアルデヒド中で3×SSC(1×SSC=0.15MのNaClと0.015Mのクエン酸ナトリウム,pH7.4)と、50μlの変成サケ精液DNA(Sigma)/mlと、0.02%フィコールと、0.1%ドデシルと硫酸ナトリウム(SDS)との中に予備混合された。さらに混合物は、同じ溶液を加えることによって、32PdCTP(Amersham)で標識を付されたHindIII Kフラグメントを完成させた(HindIII Kフラグメントの選択は、1型ウシヘルペスウイルスからのDNAのクローニングと分裂位置のマッピングに基づく(Cooper菌種)John F.Mayfield,Peter J.Good,HollyJ .VanOort,Alphonso R、Campbell and David A.Reed;Journel of Virology (1983) 259〜264)。混合の12〜14時間後、フィルタは2時間、0,1%SDSと0.1×SSCとの中で60℃で洗浄された。HindIII Kフラグメントは、標準のクローニング手法に従って、pUC18ベクタ中でクローン化された(J,Sambrook, E,F,Fritsh and T、Maniatis, 1989, Molecular cloning la labovatony manua1,2nd ed.Cold Spring Harbor labovatort Press,NewYork)。pUC8.4HindIII KクローンのHindIIIによる消化後、pUC18ベクタは、0.7%の低融点アガロース(BRL、Life Techovlogies,Inc.)ゲルで電気泳動によって再び8.4kb HindIIIにフラグメントから分離され、基準のフェノール抽出とエタノール沈澱とによってアガロースから分離された。分離されたHindIII Kフラグメントは、Boehringer Mannheimからのラドン感知DNA標識装置1OO4.760で標識を付された。混合化フィルターの自動放射線図は、-70℃でKodak XARフィルムの36時間露出で、反射スクリーンを使って行われた。 BはIowaとDifivac-1の7.4kb HindIIIフラグメントとの8.4kb HindKフラグメントの物理的マップである。6kb PstIフラグメントの共泳動とDifivac-1中の1.8kb PstIフラグメントの存在を考えて、欠失はハッチを付された面積で求められる。
【図2】Difivac-1中に欠出領域を包んだ野性型BHV-1フラグメントのサブクローニング A中でBHV-1ゲノムの成分は、長ユニーク(UL)領域、短ユニーク(US)領域および2つの反復物(lrとTr)として示される。このマップは、Cooper菌種の公表された分析に基づいている(John F.Rlayfield, Peter J.Good, Holly J VanOort, Alphonso R. Campbell and David A. Reed: Journal of Virology (1983) 259〜264)。 Bの中でフラグメントは、細胞核のないベクタ、すなわちpACYC中の15.2kb EcoRIフラグメントと、pUC18中の8.4kb HindIIIフラグメントと、pBR322中の4.1kb EcoRI-HindIIIフラグメントとの中ヘクローンされたUS領域で示される。ウイルスDNAフラグメントの単離は、図1Aの表示中に述べられている手法に従って実施された。種々のベクタ中へこれらのフラグメントをクローンすることは、基準の手法で行われた(J.Sambrook, E.F.Fritsch and T.Maniatis, 1989, Molecular cloning:a loboratory mannal,2nd ed.Cold Spring Harbor Laboratony press, NewYork)。 cの中で物理マップは、Difivac-1中で要求される欠失が当てはめられる領域を示す。 Dの中で、この領域のサブクローンが分析のために用いられることを示す。2つのPstlフラグメントはpKUN19の中へ、そして残ったフラグメントはpUC18の中ヘクローンされた。
【図3】Aは、〔右端にあるAluIの確認位置から左端にあるHincIIの確認位置に至る〕図2C中で示されるようなDifivac-1中に欠失された要求される位置を囲むBHV-1のLam種のUS領域からの2027個のヌクレオチドのヌクレオチド配列である。図2Dで示されるサブクローンの挿入をしたヌクレオチド配列は、Sangerらのジデオキシ配列法を用いた2つの基準で分析することによって決定された(F.Sanger, S.Nicklen and A.R.Coulson, 1977, Proc.Nat1.Acad.Sci.USA 74,5463〜5467)。端末に対して、PharmaciaのT7配列装置が、製造者によって指定される手法に従って用いられた。放射線標識のために、〔35S〕dATP(Amersham)が用いられた。人工産物と比較して、GLの多い領域の配列分析は、Pharmacia装置の7-デアザアデニン-dGTP変形物で複生された。ヌタレオチド配列の下に示されたものは、3文字のコードで、575アミノ酸残基のオープンリーディングフレームのアミノ酸配列である。それは、ヌクレオチドの概念上の転移後に見付かった。この転移は、一般的なコードに基づきPC遺伝子コンピュータプログラムを用いて決定された(PC遺伝子翻訳バージョン 1987年11月)。この575アミノ酸のオープンリーディングフレームは、ヌクレオチド168のメチオニンで始まり、ヌクレオチド1893の停止コドンで終わる。 575アミノ酸残基のオープンリーディングフレームの構造分析は、PC遺伝子コンピュータプログラムでまた実行される。真核の輸出信号からの最初の26アミノ酸が、「シグナルペプチド」によって図中に示される。6.2の点でこの配列を分裂する信号が、アミノ酸26とアミノ酸27との間で起こされる。アミノ酸575の配列は、アミノ酸残基の線によって指示される。3つの可能なN一結合のグリコレーシヨン位置(NXT/S)を有する。RaoとArgosの方法に従えば、貫膜螺旋による図で示されるアミノ酸423とアミノ酸450との間に貫膜領域がある。制限酵素AsuII, SmaI, HindIIIおよびEcoNIのための確認配列(位置)は下線が付される。このポリペプチドの計算された分子量は61212である。 Bは、上で述べた575アミノ酸オープンリーディングフレームの構造上の特性を模式的に表したものである。
【図4】BHV-1 gE遺伝子のアミノ酸配列と、ヘルペス単体ウイルス(HSV)gE遺伝子および他のgE均質遺伝子〔疑似狂犬病ウイルス(PRV)gIと帯状庖疹ウイルス(VZV)gpl〕のアミノ酸配列との比較 この比較のために用いた配列は、次の公表されたものである。 HSV:1型ヘルペス単体ウイルスのゲノム中のUS領域の配列決定と遺伝子内容(D.J.McGeoch, A.Dolan, S.Donald and F.J.Rixon (1985) Journal Mol.Biol.181,1〜13) VZV:帯状庖疹ウイルス遺伝子のU。成分のDNA配列(A.J.Davidson (1983), EMBO Journal 2, 2203〜2209) PRV:ヘルペス単体ウイルスと帯状庖疹ウイルス糖蛋白とに同族体である2つの疑似狂犬病ウイルス糖蛋白のための単離遺伝子にλgtllの使用(E.A.PetroVsKis, J.G.Timmins and L.E.Post (1986) Journal of Virology 60,185〜193) これらの配列は配列分析プログラムMultalinを使って比較された(F.Corpet (1988), Nucl. Acids Res.16,10881〜10890)。 Aの中で図は、4つのアミノ酸配列が模式的に示されている。ここに確認された貫膜部(TM)が各々に示される。確立された輸出信号配列(SP)と可能なN-結合グリコシレーション位置(I)に加えて、2つの保護される領域が示される。その中で、システン残基の相対位置(CCC)は不変である。 Bの中で結果は4つのgE翻訳の中央に位置するシスティンの多い領域のMultalin比較が示される。アステリスクは、一致したアミノ酸と類似アミノ酸クローンを示す。
【図5】Difivac-1とIowaとのサザンブロット分析で得られた写真図 パネルAは、BstI(1,2),EcoRI(3,4)およびHindIII(5,6)を有するDifivac-1とIowa消化制限酵素のゲノムDNAである。これは、0.7%アガロースゲルで分離され、ニトロセルローズでブロットされ、図1Aの説明で述べられた手法に従ってBHV-1菌株Lamの32Pで標識を付されたHindIIIKフラグメントで混合される。 パネルBは、Aと同様に同じゲルのニトロセルローズブロットであり、BHV-1 gE-種プローブp318で混合された。プローブは、図2C中で示したAluI-HincII中で比較された。
【図6】BHV-1の欠失のあるgEフラグメントの構成 Aの中でgEの位置と用いられたクローンとが示される。BHV-1遺伝子の成分は、UL領域、Us領域および2つの反復(lrとTr)として示される gE遺伝子の5’側に位置する領域を得るために、BHV-1菌種Lamの8,4kbHindIIIKからの1.4kbPstI-SmaIフラグメントが、プラスミドpUC18のSmaIとPstI位置の中ヘサブクローンされた。このクローンは、p515と呼ばれ、B中に示される。gEの3'側に位置するEcoNI-SmaIフラグメントは、4.lkbHindIII−EcoRIクローンに由来し、p515の普通のAsuIIヘクローンされる。AsuIIの休止にEcoNIの休止を結びつけるために、クローンp515はAsoIIで消化され、その後クレノウ酵素(Boehringer Mlannheim)と、基準の手法に従ってAsuII休止中へ1つのシトシン残基を供給すべきdCTPとで処理した(Sambrook et al,1989)。この付加シトシンは、D中でアステリスクで示される。p515は、またSmaI酵素で消化され、その後EcoNIフラグメントは、このベクタ中へ結合される。このようにして構成されたクローンは、p519と呼ばれる。
【図7】Aは、1B7,1B8および2H10のDNA調整品のサザンブロット分析で得られた写真である。DNA単離物、消化制限酵素、ブロットおよび混合物は、図1Aの説明で述べられた手法に従って作られた。DNA調整品1B7,1B8および2H10のPstI-DraIによる二重消化後、フラグメントは0.7%アカロースで分離され、直後にニトロセルローズフィルタがブロットされた。このフィルタは、プローブとして、32PdCTPで標識を付された2.3kbPstI- DraI欠失フラグメントと混合された。レーン1〜3中で試料1B7,1B8および2HlOがそれぞれ分離された。レーン中でLam菌株の野性型BHV-1 DNAが、レーン15中で2.3kb欠失フラグメントが示される。 Bは、BHV-1菌種Lamの15.2kb EcoRIフラグメントの物理マップである。マップは、PstI,DraIおよびHindIIIの確認位置と7A中で述べた混合プローブの位置を示す。
【図8】BHV-1 gEの無核表現 BHV-1 gEの無核細胞を表現するために、gE遺伝子の600bpSmaIフラグメントは、ベクタpGEX-2T中で、日本住血吸虫のグルダチオン-S-転移酵素のコード化領域に3つのリーデングフレーム中で融合された(D.B,Smith and K.S.Johnson,Gene 67(1988)31〜40)。SmaIフラグメントの方位確認性(syn)を有する再結合分子は、基準の手法を用いて、制限酵素によって同定された。この融合構造を有するE.coliDH5αクローンは、pGEX-2T600sl,pGEX-2T600s2およびpGEX-2T600s3と呼ばれた。 Aは、pGEX-2T600sの構成の1つの図である。GST-gE融合物のためにコード化する領域のNH2側にあるものは、タフプロモータ領域を導入しうるイソプロピルチオガラタトシド(IPTG)である。 Bは、pGEX-2T600sで転移されたDH5αの細胞の全蛋白質調整物のウエスタンブロット分析で得られた写真である。pGEX-2T600s1,pGEX-2T600s2およびpGEX-2T600s3構成で転移されたDH5α細胞の徹夜の培養物は、50μg/mlアンピシリンを加え、Luria-Bertani(LB)媒中で1/10継続され、1時間後培養物はIPTGに5時問導入された。 これらの導入された培養物は、5分間6,000Gで遠心分離し、1×ラウエル混合物(2%SDS、10%グリセロール、5%メルカプトエタノールおよび0.01%ブロムフェノールブルー)中に混合〔1.5ml培養物は、500μlラウエル混合物中に混ぜる〕され、95℃に5分間加熱された。そして、レーンあたり50μlが、基準の手法に従って、12.5%ポリアクリルアミドゲルで水平的に分離され、引続いて製造者によって明確にされた条件で、LKB一ムルチフールII型Navaブロットシステムを用いて、ニトロセルローズフイタが半乾燥ブロットされた。 レーンM中で、予め印を付された蛋白質が用いられ、(BRL Life Technologies,lnc.236K,112K,71K,44K,28K,18Kおよび15K)、レーン1,2,3中で、DH5α細胞の全蛋白調整物は、pGEX-2T600sl,pGEX-2T600s2およびpGEX-2T600s3の各々3つのフレームで転移される。 パネルA中で、結果は、抗−GST血清によってウエスタンブロット分析で見られる。フィルタは、基準の手法(E.Harlow and D.Lane(1986)、Antibodies:a laboratory manual,Cold Spring Harbor Loboratory,New York)にしたがって、ウサギ血清ポリクロナール抗GSTを加えた阻止緩衝液(PBS+2%粉乳および0.05%トゥイーン20)中で培養された。そして、フィルタは洗浄され、ヤギ対ウサギ免疫グロブリン血清を接合した西洋ワサビパーオキシターゼ(HRPO)を加えて培養された。その後、結合されたヤギ抗体は、色素体(ヂアゾベンジジン、クロロナフトールおよびH2O2)で免疫化学的に検出された。矢印によって指示されるGST融合物は、フレーム3中でのみ約47kの確認された大きさを有する。 パネルBにおいて、結果は、モノクロナール抗体Mab4で、ウエスタンブロット分析で見られる。パネルA中のようにデュプロフィルタは阻止され、Mabで培養され、ウサギ対マウス血清を接合したHRPOで培養される。その後結合ウサギ抗体は、色素体で免疫化学的に検出される。レーン3(フレーム3)で見られるバンドは、47Kの大きさで、矢印によって指示される。
【図9】BHV-1 gE遺伝子の無核表現のためのpEVHis gEプラスミドの構成 gE遺伝子の無核表現のために、コード化すべき領域の全gEは、基準の手法を用いて表現ベクタpEVHisのHCMV促進領域の背後で方向性をもってクローンされる(Sambrook et a1,1989)。gEのオープンリーディングフレームの前に55bpで出発する394bpA1UIフラグメントは、pUC18の中ヘクローンされ、p201と呼ばれる。そして、p201をHincIIが消化した後、1740bp HincIIフラグメントは、gE遺伝子の大部分から成り、p201中へクローンされる。これは、pUC18のボリリンカ中で、gEの出発コードの前にあるA1Ul位置55bpからgEの停止コードの後にあるHincH位置133bp)までの全gEコード化領域から成るプラスミドp318中に生じた。ベタタのポリリンカ中で制限酵素位置を用いて、このフラグメントは、酵素BamHIとSphiでp318から切られた。最初p318はSphIで消化され、その後SphI位置は、クレノウポリメラーゼとdNTPポリメラーゼとを使って満たされた。BamHIでの消化後、1.9kb挿入物は、pUCl8ベクタから低融点アガロース中で分離され、端末をBamHIとEcoRVで消化されたpEVHis中で結ばれた。このように形成されたプラスミドは、pEVHis/gEと呼ばれた。
【図10】BHV-1DNAを検出するためにPCR手順で得られるgE-種プライマとプローブの位置 図中には、ヌクレオチド1272から2027に至るBHV-1糖蛋白質の核酸配列が示される〔配列は図3から得られる〕。gE-種PCR手法のために用いられるプライマは、P3とP4と呼ばれる。P3とP4の位置に結合したプライマは、下線が付される。P3のヌクレオチド配列は、5'-ACG-TGG-TGG-TGC-CAG-TTA-GC-3'である(配列標別番号:2)。P4のヌクレオチド配列は、(上述の配列に結合するプライマを補足して)5'-ACC-AAA-CTT-TGA-ACC-CAG-AGC-G-3'である(配列標別番号:3)。DNAで拡大されたPCRの検出のためにサザンブロット混合を用いたプローブは、プライマが結合する位置の間に位置する137bp TaqIフラグメントであり、そのフラグメントの)端末が指示される。図3と比較のために、HindIIIとEcoNIの位置がまた示される。
【図11】Difivac-1のgE欠失のマッピング Aは、野性型BHV-1菌種Lamの15.5kb EcoRIフラグメントの物理マップを示す。Bは、Difivac-1の14.5kb EcoRIフラグメントの物理マップを示す。2つのEcoRIフラグメントは、各々のウイルスのゲノムのUPS領域を完全に覆う。gE遺伝子の位置とgI遺伝子の推定上の位置は、白四角で指示される。マップAとBとは、このような方法で位置決めされ、各マップ中の6kb PstIフラグメントは、一直線にされる。両方のマップ中で、内部反復と端部反復との配列は、斜線を付された四角で示される。反復の下方の矢印は、これらの配列の方向を示す。 Aの中でUS領域の部分は、Difivac-1菌種中で失敗があることを示す。 Cは、gE欠失をマップ化し、B中に示される物理マップを得るために用いられるクローン化Difivac-1フラグメントの位置を示す。クローンp728,p737およびp754の挿入の下の矢印は、再結合点を決めるために配列化された領域を示す。 略語: A=A1UI,E=EcoRI,P=PstI,H=HindIII,γ=再結合点,IR=中間反復,TR=端末反復
【図12】Difivac-1のUS領域における正確な再結合位置の決定 Difivac-1菌種中に見付かったgE欠失の正確な境界を決定するために、クローンp754ならびにp728およびp737とが配列された。これらのクローンの挿入は、図11中に指示される。用いられる配列決定の手法は、図3の説明中に述べられている。 Aにおいて、A1UI-PstIフラグメントの最大の配列が示される。この配列は、gE遺伝子のプロモータ領域で出発する。推定のTATAボックスは、下線が付されている。点γ(=再結一合点)において、このプロモータ領域は、Us領域の反対の位置で見付かった裏返しの反復と呼ばれる配列に接合される。正確な再結合点は、US領域の反対位置で見付けられた反復のコピーを有するgEプロモータ位置で見付けられた反復を含むことによって決定される。この配列が分岐する点は、B(I)の中でγで示される。類似の比較は、Difivac-1中で見付けられるgEプロモータ配列と野性型菌種Lam中で見付けられたgEプロモータとで行われる。この配列が分岐する点は、B(II)の中で示され、またγで示される。見付けられた再結合点は同じである。
【図13】BHV-1 gI遺伝子の部分配列分析 BHV-1 gIとgEとの双方の遺伝子(図11に示す)中に達するBHV-1菌種Lamの1.8kb PstIクローンを用いてBHV-1 gIのコード化領域の範囲内で、284ヌクレオチドの配列が決定される。配列決定に用いられる手法は、図3の説明の中で述べられている。配列は、PC/遺伝子コンピュータプログラムバージョン1.03(1987年11月)による汎用コードに基づいて転位された。第2リーディングフレームによってコード化されたアミノ酸配列は、ヌクレオチド配列の後に1つの書かれたコードとして与えられる。このアミノ酸配列は、他のヘルペスウイルスgI同族体(図14に示す)のコード化領域に均等である。
【図14】推定のBHV-1 gI遺伝子の部分アミノ酸配列と、ヘルペス単体ウイルス(HSV1)gI遺伝子、疑似狂犬病ウイルス(PRV)gp63遺伝子および帯状庖疹ウイルス(VZV)gpIV遺伝子のコード化領域の相当部分とのアミノ酸比較 PRV配列はそのコード化領域のアミノ酸82で出発し、HSV1配列はそのコード化領域のアミノ酸で出発し、VZVはそのコード化領域のアミノ酸76で出発する。用いられる配列は、図4の説明で述べられた報告書で公表されている。比較は、Multalinコンピュータプログラムを使って行われた。アステリスクは、同一のアミノ酸を示し、コロンは類似のアミノ酸を示す。
【図15】BHV-1 gI-遺伝子の無核表現のためのMSVneoGIプラスミドの構成 BHV-1 gI-遺伝子の部分配列のアミノ酸配列の比較に基づいて、BHV-1 gI-遺伝子の推定位置は評価される。この評価に基づいて、1.7kb SmaIフラグメントは、BHV-1 gE遺伝子の完全コード化領域を含むことが推論される。この1.7kb SmaIフラグメントの位置は、A中に示される。この1.7kb SmaIフラグメントの丸くなった端末に、BamHIリンカが基準の手法を用いて結び合わされる。この結果生じた物は、BamHIで消化され、無核表現ベクタMSV-neo中で結び合えわされる。MSV-neoベタタは、強いプロモータ性を有するMSV-LTRの背後の一般的なBamHI位置を有する。このベクタは、Rijsewijk et al.,1987EMBO J.6,127〜131中で述べられている。
【図16】BHV-1 gI/gE二重欠失フラグメントの構成 BHV-1のUS領域中における糖蛋白質gE遺伝子の位置と糖蛋白質gI遺伝子の推定位置は、図A中に表現される。斜線を付された部分は、US領域を境界する反復を示す。Bは、双方の遺伝子の位置を考慮して、本質的な制限酵素位置の物理マップを示す。gI/gE欠失フラグメントを構成に関して、gI遺伝子を包み込む1.7kb SmaIフラグメントを含むクローンP1.7-SmaI/0がPstIで消化される。残留350bp SmaI-PstI挿入のPstIで消化される。残留350bp SmaI-PstI挿入のPstI位置は、基準の分子生物学的手法を用いて、丸くなった端末を作る。EcoNI-SmaIフラグメント(図6B参照)は、図6A中で記載された4.1kb HindIII-EcoRIフラグメントから単離され、丸くなった端末に作られ、修正PstI位置に結び合わされる。これは、CとD中に図示される。クローンP△IEの結果から、1.4kb SmaI-DraIフラグメントは、野性型BHV-1DNAと再結合して単離されうる。 略号 E=EcoRI,H=HindIII,S=SmaI,P=PstI,ENI=EcoNI,D=DraI,kb=キロベース,US=短ユニーク
【図17】接種後の牛から垂れた平均鼻水のウイルス ●=Difivac-1を接種、○=接種なし
【図18】毒性の強いBHV-1菌株投与後の平均発病牛数、記号は図17に同じ
【図19】毒性の強いBHV-1菌株を投与した牛の平均体温、記号は図17に同じ
【図20】毒性の強いBHV-1菌株投与後の牛の平均成育量、記号は図17に同じ
【図21】毒性の強いBHV-1菌株投与後の牛から垂れた平均鼻水、記号は図17に同じ
【図22】毒性の強いBHV-1菌株投与後の牛の平均体温 ●=Lam gE-を接種、○=Lam gE-/TK-を接種、×=無接種
【図23】毒性の強いBHV-1菌株投与後の牛の平均成育量、記号は図22と同じ
【図24】毒性の強いBHV-1菌株投与後の平均発病牛数、記号は図22と同じ
【図1A】
【図1B】
【発明の詳細な説明】
【技術分野】
【0001】
発明の分野
本発明は、病原体によって引き起こされる疾病に関係したワクチン注射および診断法の分野に関するもので、弱毒性ワクチンまたは不活化ワクチンを生成する古典的な方法とDNA組換技術に基づく近代的方法両方の使用に関係している。
より詳細には、本発明は、動物、特に牛を1型ウシヘルペスウイルス(BHV-1)から守るための弱毒性生ワクチンおよび不活化ワクチンに関するもので、これらのワクチンは、安全でかつ効能があるばかりでなく、ワクチン接種をうけた群における、非感染動物と感染動物とを識別することも可能にする。
ワクチン接種群における非感染動物と感染動物の識別のための試験に使用が可能である診断キットも本発明の一部である。
【背景技術】
【0002】
発明の背景
牛の伝染性鼻腔気管炎ウイルス(IBRV)および伝染性濃疱外陰部腔炎ウイルス(IPVV)を含むBHV-1は、牛の呼吸器疾患、生殖能力障害の解明に重要な役割を果たす。BHV-1は、急性感染後、潜伏した形で宿主内にそのまま残っていることが多い。潜伏ウイルスはとりわけ、ストレス(これは臨床現象を伴うことも、伴わないこともある)の影響下で再び活性化し、続いて排泄されることがあり得る。結果として、感染した牛は終生BHV-1を蔓延するものとみなす必要がある。BHV-1はオランダの養牛場の推定75%において風土病として発生している。特に、高齢牛の血清反応が陽性である。
【0003】
BHV-1感染予防接種に利用される多くの不活化("死んだ")ワクチンおよび弱毒("生きた")ワクチンがある。不活化ワクチンは、たとえば熱処理や照射、またはエタノールやホルマリン処理によって、BHV-1ウイルスを死滅させて作られる。しかしながら、これらによっては不充分な予防しか行えないことが多い。弱毒ワクチンは、同種(ウシ)細胞またはブタの細胞やイヌの細胞のような異種細胞への多数回にわたる継代によって作られ、その後ウイルスは物理的または化学的にも処理されることもある。このようにして、未知の突然変異/欠失がウイルスゲノム中に発達し、これがしばしばウイルスの疾病誘発特性を低下させる。弱毒ワクチンは不活化ワクチンよりも優れた予防を行うが、これは特に宿主の免疫系に、より多くのウイルス性抗原を与えるからである。生きたワクチンの重要な他の利点として、それらは鼻腔内投与が可能であるということがあり、すなわち、ここは感染後に野生型ウイルスの増殖が初めて起こるところである。生きたワクチンはまだ改良の余地がある。いくつかの生きたワクチンはさらに、流産を引き起こす可能性を有しているように思われ、これは特に筋肉内投与後に顕著になる。さらにまた、生きたワクチンはすべて、ワクチンを接種された牛に潜伏して残っていると思われる。また、該ワクチンが野生型ウイルスとほとんど相違するところがなければ、病原性への復帰が起こる可能性もある。しかし、重要な問題の1つは、BHV-1ワクチンは、野生型ウイルスによる感染を予防することができないということである。その結果、ワクチン接種を受けた牛も野生型BHV-1を蔓延させる可能性がある。
【0004】
適切なBHV-1抑制計画のためには、野生型ウイルスと識別することができる効果的かつ安全なワクチンを自由に使えることが必要である。何故ならば、効果的ワクチンの利用によりBHV-1の蔓延をかなりの程度低下させることができ、ワクチンと野生型ウイルスの識別が可能な試験によって、ワクチン接種を受けた牛の群より感染した牛を探りあて、それを取り除くことが可能となるからである。
【0005】
ところで、従来のワクチンよりも一層安全と思われる野生型ウイルスとの識別が可能なBHV-1ワクチンの開発が行われてきた。流産を引き起こす程度もより低く、潜伏の頻度も低く、再活性化され得ることもないチミジンキナーゼ欠失変異体が単離された。さらに、組換えDNA技術を用いて、(血清学的技術により野生型BHV-1とワクチンの区別を可能にする)糖蛋白質gIIIのための遺伝子に欠失を有するBHV-1ワクチンが構築された。しかしながら、これらのワクチンに対してもさらに幾つかの異論がある。一方では、チミジンキナーゼ遺伝子はウイルス複製に関与しており、複製の程度が低い程、より程度の低い防御につながる可能性がある。他方、該糖蛋白質gIIIは防御抗体の生成に重要であり、この防御抗体は、gIII欠失ワクチンの効果を低下させる。実際上の問題として、一般には最高の防御が得られる、組換えワクチンの鼻腔内投与が幾つかの国では認可されていないという問題がある。したがって、有効かつ安全で、さらに野生型BHV-1と区別することが可能なワクチンが求められており、さらにまたこれは、かかるワクチンの少なくとも1つは、組換えDNA技術によって構築されたウイルスよりも、むしろ従来の方法で弱毒化されたウイルスに基づいたものであることが望ましい。
【0006】
ところで、細胞培養における継代によって、糖蛋白質gEのための遺伝子を欠くBHV-1菌株が得られた。我々が行った研究の最初の結果は、この遺伝子は、野生型BHV-1に関する血清学的識別に非常に役立ち、病原性の発現に関与していることを示している。したがって、その欠失は安全性に寄与しており、これによってチミジンキナーゼ欠失の使用は不要となるかもしれない。糖蛋白質gEは、防御の誘導にとって糖蛋白質gIIIほど重要ではないと思われる。従来の方法で弱毒化された野生型ウイルスと血清学的に識別することが可能な、従来の方法で弱毒化されたBHV-1菌株は独自のものである。ここで初めて述べた、gE-遺伝子の位置とDNA配列は、これまでは知られていなかった。また、これらに由来することができるオリゴヌクレオチドポリペプチドおよびオリゴペプチドも知られていなかった。また、gE-遺伝子に基づく血清学的識別を行うための試験も独自のものである。
【0007】
この"従来の"gE欠失変異体の重要な利点("従来の"とは従来の弱毒ウイルス単離方法を使用することを意味している)は、親換えワクチンに関する限りでは、鼻腔内投与が禁止されている国々において、この"従来の"gE欠失変異体を鼻腔内投与することが可能になるであろうという点である。しかしながら、安全に関して別の観点からよく考慮して、この従来のgE欠失ワクチンの他に、明確に限定された組換えの型も構築された。これらの組換えワクチンもまた欠失を有しており、-また、チミジンキナーゼ遺伝子の欠失も有していてもよく、または有していなくてもよい-および、これらの組換えワクチンは、異種の遺伝子の発現のためのベクタとして用いることもできる。これらの組換えワクチンは全て、同じgEに特異的な試験によって、野生型ウイルスと区別することができる。一組の異なるワクチンに対して標準的な試験を利用することは、国際的な努力としてのBHV-1の戦いにおいて強力な利点となり得る。かかる方法がBHV-1ワクチンの分野において述べられたことはない。
【0008】
牛における抗BHV-1反応の血清分析によって、抗gE抗体の重要部分は、糖蛋白質gEおよび他のBHV-1糖蛋白質である糖蛋白質gIによって形成される複合物に対して向けられていることが示された。したがって、かかる複合物に特異的な抗体の存在を示すこともできる血清試験は、抗-gE抗体を検出することができるだけの試験よりも高感受性であると思われる。gE単欠失変異体によりワクチン接種を受けた牛は、抗-gI/gE抗体の検出を妨げることが有り得る抗-gI抗体を作る可能性がある。したがって、この発明はgI/gE二重欠失を有するワクチンを含む。
【発明の開示】
【0009】
発明の要約
本発明は、糖蛋白質gE-遺伝子に欠失を有するBHV-1の欠失変異体を提供する。"〜に欠失を"という語は、全体としての遺伝子の欠失を含むことを意図している。本発明の好適な実施例は、以下に述べる欠失変異体Difivac-1のような、弱毒処置によって引き起こされた糖蛋白質gE-遺伝子における欠失を有するBHV-1の欠失変異体によって構成される。
【0010】
本発明の他の好適な実施例は、以下に述べる欠失変異体1B7または1B8のような組換えDNA技術によって作られた糖蛋白質gE-遺伝子に欠失を有しているBHV-1の欠失変異体から成る。
【0011】
本発明の他の好適な実施例は、以下に述べるgI、/gE二重欠失変異体Difivac-IEのような、糖蛋白質gE-遺伝子および糖蛋白質gI一遺伝子に各々欠失を有するBHV-1の二重欠失変異体から成る。
【0012】
さらにまた、最高の安全性を期して、本発明によれば糖蛋白質gE-遺伝子およびチミジンキナーゼ遺伝子のそれぞれに欠失を有するBHV-1の欠失変異体が好適である。また本発明は、糖蛋自質gE-遺伝子、糖蛋白質gI-遺伝子、チミジンキナーゼ遺伝子に欠失を有するBHV-1の欠失変異体も含んでいる。
【0013】
本発明は、動物を、特に哺乳動物を、さらに特にウシ科の動物を上記において明確に述べたBHV-1欠失変異体を有するBHV-1から守るために、これらにワクチン接種をするためのワクチンの組成、適切な担体またはアジュバントを提供する。前記構成は、生きたワクチン組成でも不活化ワクチン組成でもよい。
【0014】
本発明はさらに、糖蛋白質gE-遺伝子に欠失を有し、組換えDNA技術によって導入された異種遺伝子を含むBHV-1の変異体において実施されている。好適には、糖蛋白質gE-遺伝子の位置に、組換えDNA技術によって導入された異種遺伝子を含むBHV-1の変異体に関するもので、前記異種遺伝子は、gE-遺伝子の調節配列の管理下にあり、シグナルペプチドをコードするgE-遺伝子の一部に任意に付加される。前記異種遺伝子は、BHV-1の異なるプロモータの管理下、または異種プロモータの管理下にあることも可能である。BHV-1の変異体が、糖蛋白質gE-遺伝子における欠失に加えて、さらに、チミジンキナーゼ遺伝子における欠失、および/または糖蛋白質gI-遺伝子における欠失などの欠失を有している場合には、前記異種遺伝子を、この付加欠失の位置に挿入してもよい。複数の挿入も別の選択肢であり、1つの欠失の位置にまとめて行うか、幾つかの欠失の位置に分けて行うかされる。
【0015】
好適に導入された異種遺伝子は、別の病原体の免疫原性蛋白質またはペプチドについて、または免疫反応を促進するサイトカインをコードする。適切なサイトカインの例としてはインターロイキンー2、αインターフェロン、γインターフェロンがある。
【0016】
本発明はまた、動物を、特に哺乳動物を、さらに特にウシ科の動物を(異なる)病原体より守るため、これらにワクチンを接種するための、(生きた、または不活化)ワクチンの組成を提供するもので、該ワクチンの組成には、免疫原性蛋白質または他の病原体ペプチドをコードしている異種遺伝子を有しているBHV-1突然変異体およびアジュバントの適切な担体が含まれる。勿論、この防御は1つ以上の病原体に関するものでよく、すなわち、変異体が複数の異種遺伝子を含む多価ワクチンに関するものでよい。
【0017】
さらにまた本発明は、BHV-1の糖蛋白質gE-遺伝子、この糖蛋白質gE-遺伝子の一部、またはこの糖蛋白質gE-遺伝子に由来するヌクレオチド配列を含む組換え核酸を含む組成物に関する。この組成は、BHV-1の糖蛋白質gE-遺伝子、この糖蛋白質gE-遺伝子の一部、この糖蛋白質gE-遺伝子に由来するヌクレオチド配列を含む組換え核酸の挿入を有するクローニングベクタまたは発現ベクタを含むことができる。
【0018】
また本発明は、BHV-1の糖蛋白質gE、この糖蛋白質gEの一部、この糖蛋白質gEに由来するペプチド、または糖蛋白質gEおよびgIの複合物を含む組成物、およびBHV-1の糖蛋白質gE、この糖蛋白質gEの一部、この糖蛋白質gEに由来するペプチド、または糖蛋白質gEとgIの複合物に特異的な抗体を有する組成物を含む。"抗体"とは、大半の用途に好適な、ポリクローナル抗体調製物と、大半の用途に好適なモノクローナル抗体の両方を意昧していると解釈される。"糖蛋白質gEの一部"および"糖蛋白質gEに由来するペプチド"という用語は、一般に少なくとも約8アミノ酸の長さを有するであろうgEに特異的なアミノ酸配列を意味していると解釈される。
【0019】
さらに本発明は、試料中の、特に、動物、特に哺乳動物、さらに特にウシ科の動物の、血液または血清、血液細胞、乳、涙などの体液、肺洗浄液、鼻水、精液、特に精漿、唾液、痰、または組織、特に神経組織のような生物学的試料中の、BHV-1の核酸を検出するための、BHV-1の糖蛋白質gE-遺伝子に由来するヌクレオチド配列を有する核酸プローブまたはプライマ、および核酸検出試験に適した検出手段を含む診断用キットに関する。
【0020】
さらに本発明は、試料中の、特に、動物、特に哺乳動物、さらに特にウシ科の動物の血液または血清、血液細胞、乳、涙のような体液、肺洗浄液、鼻水、精液、特に精漿、唾液、痰、または組織のような生物学的試料中の、BHV-1に特異的な抗体を検出するための、BHV-1の糖蛋白質gE、この糖蛋白質の一部、この糖蛋白質gEに由来するペプチド、または糖蛋白質gEおよびgIの複合物、および抗体検出試験に適した検出手段を含む診断用キットに関する。該診断用キットは、さらに、BHV-1の糖蛋白質gEに特異的な、またはBHV-1の糖蛋白質のgEとgIの複合物に特異的な1つまたはそれ以上の抗体を含めてもよい。
【0021】
また、本発明は、試料中の、特に、動物、特に哺乳動物、さらに特にウシ科の動物の血液または血清、血液細胞、乳、涙などの体液、肺洗浄液、鼻水、精液、特に精漿、唾液、痰、または組織、特に神経細胞のような生物学的試料中の、BHV-1の蛋白質を検出するための、BHV-1の糖蛋白質gEに特異的な、またはBHV-1の糖蛋白質gEとgIの複合物に特異的な1つまたはそれ以上の抗体および蛋白質検出試験に適した検出手段を含む診断用キットに関する。
【0022】
さらに本発明は、BHV-1の糖蛋白質gE-遺伝子を含む核酸の存在、BHV-1の糖蛋白質gE、またはBHV-1の糖蛋白質gEとgIの複合物の存在、またはBHV-1の糖蛋白質gEに特異的な、またはBHV-1の糖蛋白質gEとgIの複合物に特異的な抗体の存在について、動物の特に哺乳動物の、さらに特にウシ科の動物のBHV-1感染を判断するための、前記動物の試料の、特に、血液または血清、血液細胞、精液、特に精漿、唾液、痰、涙などの体液、肺洗浄液、鼻水、乳、または組織、特に神経組織のような生物学的試料の試験を含む方法を提供する。試験を行う試料は、本発明に従った組成のワクチンをそれまでに接種されたことのない動物のもの、または本発明に従って調製されたワクチンをそれまでに接種されたことのある動物のものとすることができる。
【0023】
発明の詳細な説明
本発明は、生ワクチンおよび不活化ワクチン両方から成る一組のBHV-1ワクチンに関するもので、これらはどちらも糖蛋白質gE-遺伝子の全部または一部を欠いている。この組は、チミジンキナーゼ遺伝子および/または糖蛋白質gI-遺伝子の欠失を含むことも含まないこともある自然gE欠失変異体および構築されたgE欠失変異体も含み、さらに異種遺伝子ベクタとして使用される構築されたgE欠失変異体も含む。さらに本発明は、BHV-1糖蛋白質gE-遺伝子をコードするヌクレオチド配列、これらの配列に由来するオリゴヌクレオチド、該糖蛋白質gEそれ自身、それらに由来するペプチド、および該gE糖蛋白質とそれらに由来するペプチドに対して誘導される(モノクローナルまたはポリクローナル)抗体に関する。また本発明は、BHV-1の糖蛋白質gEとgIの複合物、およびかかる複合物に対して誘導される抗体に関する。
本発明に従ったこれらの物質は、以下のために使用することができる。
【0024】
1)BHV-1によって引き起こされる疾病を予防するための牛のワクチン接種。
これはBHV-1に感染した動物と、ワクチン接種した動物の間の区別が可能で、従来のワクチンおよび構築されたワクチンを一緒に使用することが可能なものである。
【0025】
2)BHV-1による疾病および防御抗原に対するコード配列をBHV-1欠失変異体の中に組込むことができる他の病原体により引き起こされる疾病を予防するための牛のワクチン接種。
【0026】
3)牛の血液、血清、乳、他の体液について、それらが野生型BHV-1によって感染しているか、gE欠失変異体でワクチン接種されているかを、血清学的に、または核酸検出技術(たとえばPCR)により判断するためのそれらの試験。
【0027】
オリゴペプチド、ポリペプチド、およびBHV-1の糖蛋白質g-遺伝子と糖蛋白質gI-遺伝子のコード配列に由来する糖蛋白質の合成。
糖蛋白質gE-遺伝子(図3A)およびこの遺伝子をコードする単離DNAフラグメントについての、例に記載のDNA配列分析の結果により、標準的な分子生物学的処置を用いて、gE蛋白質のペプチド(オリゴまたはポリペプチド)を合成すること、および原核的方法(バクテリア中)または真核的方法(たとえば、ネズミの細胞において)により、gE蛋白質の全体を、またはその一部を発現させることが共に可能となる。これらの方法によって、たとえばgEに特異的なモノクローナル抗体の生成に役立つことができるgEに特異的な抗原を得ることができる。さらに、gEに特異的な抗原(および、gEに特異的なモノクローナル抗体)を血清試験において使用することができ、BHV-1 gE欠失ワクチンの接種をした動物と野生型BHV-1ウイルスに感染した動物の間の区別が可能となる。
【0028】
例に記載の、糖蛋白質gI-遺伝子、およびこの遺伝子をコードする単離DNAフラグメントについて、糖蛋白質gEを発現する真核細胞と共に、部分的にDNA配列を分析した結果、真核細胞中のgI/gE複合物を発現させることができる(図13および図14参照)。この糖蛋白質複合物は、gI/gEに特異的なモノクローナル抗体を作るために使用することができる。また、このgI/gE複合物は、単gE BHV-1欠失変異体、または二重gI/gE BHV-1欠失変異体によるワクチン接種を受けた牛と野生型BHV-1ウイルスに感染した牛の区別をする血清試験における抗原としても使用することができる。
【0029】
gEに特異的なペプチド
既知の蛋白質コード配列に基づき、自動合成装置により、約40〜50のアミノ酸のポリペプチドを作ることができる。BHV-1菌株LamのgE糖蛋白質の蛋白質コード配列が解明されたので(図3A)、このBHV-1 gE糖蛋白質のポリペプチドの合成が可能となる。かかるポリペプチドを用いて、標準的な方法に従って、マウスやウサギなどの実験動物を免疫化し、gEに特異的な抗体を作ることができる。さらに、これらのgEに特異的なペプチドを用いて、抗gE抗体がgE蛋白質(エピトープ)と反応する位置を、たとえばPEPSCAN法(Geysen et a1.,1984,Proc.Natl.Acad,Sci.USA 81,3998〜4002)により、さらに特定することができる。gEに特異的なオリゴペプチドも抗gE抗体を実証する血清試験において使用することができる。
【0030】
gEの原核発現
細菌中にてgE蛋白質を合成するため(すなわち、gEの原核発現)、糖蛋白質gEを、またはその一部をコードするDNAフラグメントを、原核発現ベクタ中にタローン化しなければならない。原核発現ベクタは、環状DNA分子で、これは別個に複製してゆく分子(プラスミド)として、細菌中で自らを維持することができる。これらの発現ベクタは、抗生物質抵抗についてコードする1つまたはそれ以上の標識遺伝子を含み、したがって発現ベクタを伴う細菌の選択を可能にする。さらに、発現ベクタは、(制御可能であることも多い)プロモータ領域を有しており、該領域の背後にDNAフラグメントを結合することができ、これらのフラグメントは、プロモータの影響により発現される。現行の多くの原核発現ベクタにおいては、いわゆる担体蛋白質に融合し、所望の蛋白質が発現している。そのために、ベクタ中ではプロモータの後に、担体蛋白質についてのコード配列が位置しており、所望のDNAフラグメントがこれに直接隣接して結合されている。融合蛋白質は、多くの場合より安定で、認識および/または単離もより簡単である。一定の細菌株にて、特定の融合蛋白が達成することができる定常状態レベルは融合によって、および菌株によって異なっている。種々の組合わせを試みることが慣例になっている。
【0031】
糖蛋白質gE-遺伝子の真核発現
蛋白質の原核発現は幾つかの利点をもたらしてくれるが、蛋白質は真核細胞で起こるグリコシル化のような修飾変更を欠いている。その結果、真核発現蛋白質は多くの場合より適切な抗原である。マウスの細胞のような真核細胞における蛋白質の異種発現の場合、真核発現ベクタが使用される。これらのベクタは、プラスミドで大腸菌細胞内で増殖させることが可能であるばかりでなく、真核細胞中にて安定した状態で生存することもできる。これらのベクタは、原核選択マーカの他に、真核選択マーカも含む。原核発現ベクタに似て、真核発現ベクタは、所望の遺伝子が後に結合することができるプロモータ領域を含んでいる。しかしながら、真核ベクタにおけるプロモータ配列は、真核細胞に特有である。また、真核ベクタにおいては、担体蛋白質に対する融合が利用されることは滅多にない。これらのベクタは、標準的なトランス・フェタション法(F.L.Graham and A. J. van der Eb. 1973, Virology 52,456〜467)により真核細胞中に導入される。真核プラスミドベクタの他に、ウィルスベクタもあり、この場合には、異種遺伝子がウィルス(たとえば、レトロウィルス、ヘルペスウィルス、ワクシニアウィルス)のゲノム中に導入される。次いで、真核細胞に組換えウィルスを感染させることができる。
一般に、個々の遺伝子産物にとって、ベクタや細胞はどういったタイプのものが最適であるかということを予測することはできない。大抵の場合、幾つかの組合せが試みられている。
【0032】
糖蛋白質gEおよび糖蛋自質gI両方の真核発現
蛋白質が得る最終構造は、蛋白質の一次アミノ酸配列、折たたみ、翻訳後修飾等に依存している。蛋白質の構造に寄与する重要なファクタは、1つまたはそれ以上の他の蛋白質との相互作用である。BHV-1糖蛋白質gEは、少なくとも1つの他の糖蛋白質、すなわちBHV-1糖蛋白質gIとの複合物を形成することを我々は発見した。かかる複合物を初めて示したものは、候補的抗-gEモノクローナル抗体1,51,67,75,78による我々の結果であった(表2参照)。これらのモノクローナル抗体は、Difivac-1とも、またLam gE-とも反応しなかったばかりでなく、糖蛋白質gE発現3T3細胞を認識することもできなかった。しかしながら、これらのモノクローナル抗体は、Difivac-1に感染後はgE-発現3T3細胞とは反応せず、これらのモノクローナル抗体のための適切な抗原コンホメーションを糖蛋白質gEに与えるために、因子を補足することが必要であることを示した。我々が行ったモノクローナル抗体81との放射線免疫沈澱試験の幾つかにおいて、見かけ上の分子量63KDの蛋白質が共沈することが判った。ヘルペス単ウィルス糖蛋白質gEは、これに匹敵する分子量の蛋白質(HSV1糖蛋白質gI)と複合物を形成するという事実に鑑み、我々はBHV-1糖蛋白質gEは、糖蛋白質gIのBHV-1相同体と複合物を形成するものと推定した。このBHV-1 gE/gI複合物を研究するため、および適切な抗原構造を有するgE抗原を作るため、これらの両蛋白質を1つの真核細胞中で発現させた。このために、糖蛋白質gEだけの真核発現のために記載したものと同じ処置を適用した。ただし、前提条件として、選択可能な種々の真核標識を有する発現ベクタの使用を1つだけ追加した。
【0033】
血清試験
gEに対する抗体に基づき、Difivac-1のワクチン接種をされた牛と、野生型BHV-1に感染した牛を区別するための血清学的方法は、好適にはgEに対して誘導されたモノクローナル抗体の使用に基づく。これらの抗体は、以下の方法で用いることができる。
【0034】
a)Van Oirschot et al(Journal of Virological Methods 22,191〜206,1988)により述べられた原則による。Aujeszky病のウイルスに対するgI抗体検出のための、この酵素結合イムノソルベント検定法(ELISA)では、抗体は、gI上に2つの異なったエピトープを有する2つのモノクローナル抗体の反応に対するブロック効果によって実証される。この試験は、以下のように実施される。マイタロタイタープレートを37℃で1晩、モノクローナル抗体1で被覆し、その後、たとえば4℃または-20℃にて保存する。試験される血清は、別の未被覆マイクロタイタープレートにおいて、37℃にてたとえば2時間、抗原と共に前保温される。モノクローナル抗体1が被覆されたプレートを、たとえば5回洗浄し、その後セイヨウワサビペルオキシダーゼ(HRPO)と結合したモノクローナル抗体2をこれらのプレートに添加する。次に、前保温された血清-抗原混合物を、2つのモノクローナル抗体が置かれているプレートに転移させ、37℃にてたとえば1時問保温する。これらのプレートを洗浄し、各ウェルに基質を添加する。室温にて約2時間経過させた後、これらのプレートについてスペクトル光度計による読取りを行う。各プレートには、4つの陰性対照血清および陽性血清の4つの連続希釈物が含まれる。光学濃度が同じプレートで試験された4つの陰性対照血清の平均光学濃度の50%を下回る血清は陽性とみなされる。
【0035】
b)間接2重抗体サンドイッチ(IDAS)法による。
この場合は、マイクロタイタープレートは、gE蛋白質に対して誘導されたモノクローナル抗体またはポリクローナル血清で被覆する。gE抗原調製物との保温の結果、被覆にgEが結合する。試験される牛の血清中のgEに対して特別に誘導された抗体は続いてgEに結合する。これらの結合抗体は、抗-ウシ免疫グロブリン複合物によって認識される。この複合物中の抗体は、ペルオキシダーゼ酵素に共有結合する。最後に、この結合複合物は、色素原基質の添加により、可視のものにされる。この反応の特性は、gE-抗原調製物の代わりに、gE-陰性対照調製物により同じ処置を実施して調べる。各マイクロタイタープレート上には、陽性および陰性の対照血清が含まれる。この試験は、陽性の血清が一定の希釈度において陽性を示す場合に有効である。光学濃度が標準陰性対照血清よりも0.2高い場合には、血清は陽性である。
【0036】
c)2において述べたIDAS法では、ただし試験される血清を保温した後であるが、抗-ウシ免疫グロブリン複合物の代わりに、抗-gEモノクローナル抗体/HRPOが使用される。抗-gEペプチド血清または抗-gEポリクローナル血清を抗-gEモノクローナル抗体の代わりに使用してもよい。これらのプレートを洗浄し、各々のウエルに色素原基質を添加する。室温にてたとえば、約2時間経過させた後、これらのプレートについてスペクトル光度計による読取りを行う。各プレートには、4つの陰性対照血清および陽性血清の4つの連続希釈物が含まれる。光学濃度が同じプレートで試験された4つの陰性対照血清の平均光学濃度の50%を下回る血清は陽性とみなされる。
【0037】
d)精製、未精製を問わないウイルス抗原がマイクロタイタープレートに一晩被覆されるブロックELISA法に従って、これらのプレートにおいて、試験される血清は、37℃にてたとえば1時問または1時間以上保温される。洗浄処置をした後、これらのプレートに抗-gEモノクローナル抗体を添加し、37℃にてたとえぱ1時間保温する。抗-gEペプチド血清または抗-gEポリクローナル血清を抗-gEモノクローナル抗体の代わりに使用してもよい。これらのプレートを洗浄し、各ウエルに、色素原基質を添加する。室温にて、たとえば2時間経過させた後、これらのプレートについて、スペクトル光度計による読取りを行う。4つの陰性対照血清および陽性血清4つの連続希釈物が含まれる。光学濃度が、同じプレートで試験された4つの陰性対照血清の平均光学濃度の50%を下回る血清は陽性とみなされる。
【0038】
上記の方法のいずれにおいても、gEを含む、従来の方法で成長させたウイルス抗原を使用することができるが、原核または真核を介して発現するgE-抗原も使用することができる。あるいはまた、BHV-1 gE配列に基づくオリゴペプチドを従来の抗原の代わりに、上記診断試験において使用することができるであろう。さらにまた、かかるオリゴペプチドを、Kemp et al.による記事、Science 241,1352〜1354,1988に記載の方法に従った、いわゆる"Cow-side"試験の開発に使用することができるであろう。かかる試験は、感染動物中に存在しているgEに対して誘導された抗体によるオリゴペプチド抗原配列の連結に基づいたものになるであろう。このような試験の場合、オリゴペプチドを、牛の赤血球に対して誘導されたモノクローナル抗体に連結させる必要があるであろう。
【0039】
ポリメラーゼ連鎖反応を使用する核酸分析
たとえば、オリゴヌクレオチド(プローブおよびプライマ)をポリメラーゼ連鎖反応に用いて、ワクチン接触を受けた動物と感染した動物を区別することができる。ポリメラーゼ連鎖(PCR)は、病原体の核酸を短時間に数十億倍に増殖させることができる技術である(Depolymerase kettingreactie, P.F.Hilderink, J.A.Wagenaar,J.W.B.van der Giessen and B.A.M. van der Zeijst,1990,Tijdschrift voor Diergeneeskunde deel 115,1111〜1117)。gEオリゴヌクレオチドは、gE陽性ゲノムにおいて、gE陰性ゲノムにおけるものとは異なる生成物が形成されるように選択することができる。これは、gE欠失ワクチンをワクチン接種された動物は、PCR試験において陽性シグナルを出すという利点がある。しかしながら、この方法は、試験される動物から採取した試料中、たとえば血液中のウイルスの核酸の存在に依存している。
BHV-1急性感染後は、血液中にBHV-1特有の核酸を実証することができる見込みは充分にあるが、しかしながら、BHV-1核酸も、潜伏時に血清中に実証することができるかどうかはまだ確められていない。
【0040】
ベクタとしてのBHV-1の使用
BHV-1ゲノム中の異種遣伝子の発現には、その異種遺伝子が挿入されるべき領域についての正確な情報を自由にできることが必要である。必須配列についての妨害はいかなるものもあってはならず、また異種遺伝子の発現に調節配列が利用できなければならない。gE-遺伝子は、必須ではなく、したがって、gE遺伝子を異種遺伝子で置き換えても問題はない。結果として、この異種遺伝子を、gE遺伝子が調節配列の影響下にあるように配置することができる。しかしながら、gE遺伝子の調節配列を使用する必要はない。異種遺伝子の発現は、代わりに、他のもの、たとえば種々の遺伝子のより強い調節配列により制御することができる。また、gE遺伝子の(移行)シグナルペプチドに異種遺伝子を連結することも可能であり、したがって異種遺伝子生成物の分泌に影響させることができる。gE遺伝子およびgE蛋白質についての詳細な知識によって、非常に適切に調整された方法で、ベクタとしてBHV-1を用いる可能性が与えられることは明らかである。発生するベクタは、さらに野生型と血清学的に区別することができる。異種遺伝子を発現するBHV-1変異体の構築は、例に示したgE欠失変異体の構築と同じ方法で実施することができる。しかしながら、そのときは、欠失フラグメントは、異種遺伝子が欠失位置に配置されているフラグメントと置き換えられるべきである。
【0041】
例
1)自然gE欠失変異体の単離および同定
a)自然変異体の単離
ゲノムDNAを、従来法により弱毒化された多くのワクチンから標準的方法に従って単離し、制限酵素を用いて分析した。特に、野生型BHV-1ウイルスとの区別を可能とするに適切と思われるゲノムの逸脱(genome deviation)について調べた。
特に、BHV-1ゲノムのUs領域に注意した。これは、その領域に-ヘルペス単ウイルス1との類似により-非必須糖蛋白質をコードする多くの遺伝子が恐らく配置されているからである〔Identification of a herpes simplex virus1 glycoprotein gene within a gene cluster dispensable for growth in cell culture, R. Longnecker,S.Chatterjec,R.J. Whitley and B.Roizwan(1987)Proc、Natl. Acad. Sci,84,4303〜4307〕。
ユーゴスラビア、サグレフ大学より入手したBHV-1ワクチンバッチ(Lugovic et a1.,Veterinarski Arhiv 55,241〜245,1985)は、ウシの胚腎臓細胞およびウシの胚気管細胞(Ebtr)上で多くの継代を行った後に、正常なUs領域の他に、逸脱のあるUs領域を有していることを証明した。さらにこのワクチンは、Ebtr細胞に大きなプラークと小さなプラークの両方を形成するように思われた。この混合群より、それぞれの国ごとに選択された小さなプラークで偏差のあるUs領域を有するウイルスを、3つの限界希釈ステップによって単離した。この経路を介して単離されたウイルスをさらに試験し、Difivac-1を希釈した。これは、1992年5月27日Institut Pasteur,Paris,Franceに寄託番号1-1213として寄託された。
【0042】
b)Difivac-1におけるgE遺伝子の欠失の同定
US領域におけるこの逸脱をさらに分析するために、Difivac-1のゲノムDNAを標準的な方法で単離し、サザンブロット分析を行った(図1A)。このブロットを32Pでラベルされた野生型HindIIIKフラグメントとハイブリッド形成させることにより、Us領域の中心に位置しているこのフラグメントは、Difivac-1においては、1.0キロベース(kb)程度短いことが確認された。さらに、この分析によって、欠けている部分の位置をおおよそ推定することができた(図1B)。この欠失をさらに分析するために、野生型BHV-1菌株LamのUs領域を単離し、原核ベクトル中にクローン化した。そのために、標準的な方法に従って、Lam菌株(図2A)のゲノムDNAを単離し、ベクタ、pUC18、pACYCおよびpBR322中にクローン化した(図2B)。欠失の推定位置周辺の領域の物理的地図を作成した(図2C)。この物理的地図からスタートして、この領域のヌクレオチド配列の決定に適したサブクローンをベクタpKUN19とpUC18で構築した(図2D)。これらのサブクローンを用いて、全領域の2つの鎖のヌクレオチド配列(図2Cに指示)をサンガー法を用いて決定した。このヌクレオチド配列(SEQ.ID.NO.1)をPc、/Geneプログラムを用いて分析した。概念翻訳から、ヌクレオチド(nt)168〜nt1893が575のアミノ酸のオープンリーディングフレームのための遺伝暗号を指定する(図3A)ものと推定された。さらに分析の結果、このアミノ酸配列は図3Bに示されているように、貫膜糖蛋白質の特性を有していることが示された。実は、最初の26のアミノ酸(aa)は、典型的真核性の移行シグナルとして認識され、aa423とaa450の間の領域は、貫膜領域として認識される。さらに、3つのポテンシャルN-結合、糖鎖形式部位がこの配列において発生する。この推定アミノ酸配列は、ヘルペス単ウイルス(HSV)の糖蛋白質gE-遺伝子に明らかに類似したところを示している。(図4Aおよび図4B参照)。これらおよびその他の類似点により、見つけられた遺伝子は、BHV-1のgE相同体であるという結論を正当化するものである。この理由から、遺伝子はgEと呼ばれる。このBHV-1 gE-遺伝子がDifivac-1においてどの程度欠失しているのかを判断するために、p318フラグメントを単離した。p318フラグメントは、要求されるBHV-1 gEオープンリーディングルーム上流のAluI部位55ntで開始し、オープンリーディングルーム後の133ntで終了する。ゲノムDifivac-1 DNAを、サザンブロットハイブリッド形成を用いて、このp318フラグメントで分析した。この結果、Difivac-1は検出可能なp318配列を全く含んでいないことが明らかとなった(図5)。この試験によりDifivac-1は欠失を有し、この欠失はgE遺伝子全体に広がっていることが明らかに実証されていることが確認された。
【0043】
欠失領域の大きさと位置を決定するために、Difivac-1のUs領域を包括しているゲノム配列を原核ベクタ中にクローン化した(図11Cを参照)。14.5kb EcoRIフラグメントをpACYCベクタ中にクローン化し、P775と命名した。7.4kb HindIIIフラグメントを独自にpUC18ベクタ中にクローン化し、p728と命名した。クローンp728より、2つのサブクローンを単離した。すなわち、クローンp737中の1.4kb PstIフラグメントとクローンp754中の350bp AluI-PstIフラグメントである。これらのクローンの制限酵素分析とサザンブロット分析(データは示されていない)により、Difivac-1のgE欠失の長さは2.7kbであり、gE遺伝子から丁度5'でスタートし、Us領域の境界で終了することが実証された。これらの2.7kbは、反復領域の異常伸長物として、gE遺伝子に対向したUs領域に位置した重複1kbセグメントによって置き換えられた(図11bを参照)。この分析の結果を確認するために、および正確な組換え点を決定するために、クローンp754の大半の挿入物についてのヌクレオチド配列を決定し、野生型配列を比較した(図12参照)。この分析により組換え点は、gE遺伝子の開始コドンから77bp上流に位置していることが判った。
【0044】
c)Difivac-1の安全性と効能についての評価
7週齢のBHV-1血清反応陰性の特定の病原体をもっていない牛でDifivac-1を試験した。8体の牛に2ml,105TCID50の鼻腔内ワクチン接種を行った。2ml中の1mlは各々の鼻孔にスプレーした。各々別々の独立した区画に収容されていた、BHV-1血清反応陰性の特定病原体を持っていない7週齢の8体の牛に2mlの培養基を鼻腔内投与し、非ワクチン接種対照として用いた。ワクチン接種から5週後に、ワクチン接種牛と非ワクチン接種牛に107TCID50、の強毒性BHV-1菌株、Iowaを鼻腔内接種した。接種から6週後に推定される潜伏ウイルスを再活化させるため、全牛に5日間デキサメタゾンにより筋肉内処理を行った。臨床微候、直腸温、発育を監視した。布で拭き取った鼻水からウイルスの単離を行い、血清中の中和抗体力価を求めた。
【0045】
ワクチン接種後の、牛の挙動、食欲、直腸温、発育率は正常のままであったが、ワクチンを接種した牛は、いくらか漿液性鼻水の分泌と唾液の分泌過多があった。Difivac-1は、ワクチン接種後、布で拭き取った鼻水から排泄された(図17)。ワクチン接種牛は全て、BHV-1に対する中和抗体を生成した。
菌株接種後、14日までに、非ワクチン接種牛は全て、無感情、食欲の喪失、目ヤニおよび鼻水の分泌、下あごの歯肉の赤らみ、鼻の粘膜の強い傷害が見られ、また接種後4日で発育が停止した。ワクチンを接種した牛の鼻の粘膜の傷害は小さく、速やかに治癒するもので、発育が止まることはなかった。菌株接種後の1日毎の臨床評価、直腸温、発育の推移を図18,図19および図20に示した。菌種接種後、全牛とも鼻からウイルスを排泄したが、ウイルス排泄の量と期間は、ワクチン接種牛は著しく低かった(図21)。ワクチン接種牛においては、二次抗体反応が見られ、非ワクチン接種牛は全て、菌種接種後抗体を生成した。
再活化後、接種ウイルスを1体のワクチン接種牛および5体の非ワクチン接種牛から単離した。Difivac-1を再活化することはできなかった。
【0046】
上記の結果により、Difivac-1は、若年牛においては殆ど病気の微候を引き起こさず、再活化されないことが実証された。Difivac-1は、菌種接種後病気の程度とウイルスの排泄量を著しく低下させた。
結論として、Difivac-1は、BHV-1感染予防のために牛に使用される、安全かつ効能のあるワクチンである。
【0047】
2)BHV-1の組換えgE欠失変異体の構築
Difivac-1よりも分子的により明確で、かつ所望されるならば、gE-遺伝子の欠失に加えて、たとえばチミジンキナーゼ遺伝子に欠失を有した識別可能なBHV-1ワクチンと自由に利用できるようにするために、Difivac-1の他に、組換えgE欠失変異体を構築した。糖蛋白質gE-遺伝子の決められた位置からスタートし、gE遺伝子を側面に配置するクローン化されたDNAフラグメントを用いて、gE欠失フラグメントを構築することができた。標準的な技術(F.L.Graham and A.J.van der Eb 1973,Virology52,456〜467)を用いて、,この欠失フラグメントを野生型BHV-1菌株のゲノム中で組換えることができ、その結果gE欠失変異体となった。
【0048】
a)gE欠失フラグメントの構築
【0049】
gEフラグメントの構築のために、一方においては、全gE配列を欠き、他方野生型ゲノムと組み換えることができる十分なフランキング配列を含むフラグメントが目標とされた。5'(上流)側では、gE遺伝子の開始コドン18nt上流で終了する1.2kb PstI-AsuIIフラグメントが選択された。3'(下流)フラグメントについては、gE遺伝子の停止コドンの2nt上流で開始する、1.2kb EcoNI-DraIフラグメントが選択された(図6)。
【0050】
gE欠失フラグメントの構築については、gE遺伝子の5'側に位置した、BHV-1菌株Lamの8.4kb HindIIIKフラグメントからの1.4kb PstI-SmaIフラグメントが、プラスミドpUC18のSmaIおよびPstI中にサブクローンされた。このクローンはp515と命名した。gEの3'側に位置し、4.1kb HindIII-EcoRIクローンからくるEcoNI-SmaIフラグメントはp515の特異なAsuII部位中にクローンされた。このように、gE欠失フラグメントの構築は完結し、このようにして構築されたクローンをp519と命名した。原則的にはp519の全PstI-SmaI挿入をgE欠失フラグメントとして利用することができるが、これは望ましくない。実際は、PstI-SmaIは、Us領域をフランクする反復配列中に、約100〜150塩基対(bp)の伸長を行う。この100〜150bpの部分は、gE遺伝子が位置しておらず、したがって所望されていない組換え生成物を作り出すことができないUs領域の他方の反復配列と組換えることができた。このため、PstI-DraIフラグメントが組換え実験のために選択され、したがって100bpの反復が除去された。
【0051】
b)野生型BHV-1のゲノムによるgE欠失フラグメントの組換え
構築されたgE欠失フラグメントと野生型BHV-1のゲノムの間の組換えを効果的なものとするために、2つのDNA分子のマイクログラム量を、F.L.Graham and A.J.van der Eb(1973、Virology52,456〜467)の標準的な方法に従って、ウシの胚気管(Ebtr)細胞に同時トランスフェクションする。細胞の組換えメカニズムは、細胞によって組み込まれた少しの割合のDNA分子(2〜4%)の組換えを導く。組換えられたgE欠失変異体の選択のために、トランスフェクション後に形成されるウィルス混合物を、新鮮なEbtr細胞培養物上に播種する。大抵の場合、これによって発達する、別々のウィルス群(プラーク)が、1つのウィルスから生じる。BHV-1菌株LamのgE欠失変異体の単離のために、これらの230のプラークを単離し、DifivaC-1に感染した細胞とは反応しない、BHV-1に特異的なモノクローナル抗体による標準的な免疫法により試験を行った。これらのモノクローナル抗体は、糖蛋白質gEに対して誘導される。230のプラークの内5つがこれらのモノクローナル抗体と反応しなかった。これらの5つのプラークのDNAをさらに調べた。
【0052】
c)BHV-1菌株Lamの構築されたgE欠失変異体のDNA分析
上述の5つの候補gE欠失変異体の内3つ(1B7,1B8および2H10)のDNA調製物について、標準的なサザンブロット分析技術(Sambrook et al.1989)を用いて試験した。PstIとDraIによってこれらのDNA調製物を2重消化し、続いてゲル電気泳動と、プローブとして2.3kb PstI-DraI欠失フラグメントを用いてサザンブロットハイブリット化によって、ウィルス群1B7と1B8のゲノムのgE遺伝子は、所望の方法で正確に取り除かれたことが判る(図7Aおよび図7B参照)。群2H10は、逸脱したPstI-DraIフラグメントを有している。gEに特異的なプローブによるサザンブロットハイブリット化により、3つのDNA調製物のいずれにもgE配列は置かれていないことを示している(結果については示されていない)。BHV-1ウィルス群1B7および1B8は、故意の組換えgE欠失変異体である。BHV-1ウィルス群1B7はワクチン特性について試験を行った。
【0053】
d)チミジンキナーゼ/gE二重欠失変異体の構築
わずか1つの遺伝子に欠失を有するBHV-1組換え欠失変異体は、十分に毒性が低下したものではないかもしれないので、BHV-1菌株LamおよびHarberinkのチミジンキナーゼ(CTK)遺伝子にも欠失を与えた。これらの変異体は、上記のgE欠失変異体に対して使用される方法と類似の方法で構築された(結果は示されていない)。これらのTK欠失変異体はTK/gE二重欠失変異体の構築に使用した。
【0054】
e)糖蛋白質gI/糖蛋白質gE二重欠失変異体の構築
gE単欠失変異体のワクチン接種をうけた牛は、抗gI/gE抗体の検出を妨害するおそれのある抗-gI抗体を生成する可能性があるため(以下に考察)、gI/gE2重欠失を有するワクチンについても発明した。このようなgI/gE2重欠失変異体は、gE単欠失変異体の構築に使用されるのと同じ処置を用いて構築することができる。gE遺伝子の5'側端部をカバーする、1.8kb PstIフラグメントの上流側端部のヌクレオチド配列の部分的分析により、他のヘルペスウィルスに見られるgI相同物に対して有意な相同関係を有するオープンリーディングフレームが明らかになった(図13および図14を参照のこと)。gI遺伝子の推定5'側端部を包む350bp SmaI-PstIフラグメントおよびgE遺伝子の下流に位置したEcoNI-SmaIフラグメントを用いて、gI/gE欠失フラグメントを構築することができる。このフラグメントは野生型ゲノムと組換え、BHV-1 gI/gE欠失変異体をもたらすことができる。図16を参照のこと。理論的にはさらにまだ生成され得る80〜90のアミノ酸は、抗-gI/gE抗体の欠失を防げることができる抗体を導き出すことはできないであろう。さらに、gI遺伝子の配列分析により、全gIコード領域にわたるgI欠失の構築が可能となるであろう。このgI/gE二重欠失変異体をDifivac-IEと命名した。
【0055】
f)Lam gE-およびLam gE-,TK-変異体の安全性と効能の評価
Lam gE-およびLam gE-,TK-,BHV-1変異菌株のワクチン特性を7週齢の、BHV-1血清反応陰性の特有病原体を持たない牛において試験した。各々の変異体を6体の牛に鼻腔内スプレーした。各牛の全投与量は、2ml培養基において105TCID50となり、2ml中1mlは各々の鼻孔にスプレーした。他の6体の牛はウィルスを含まない培養基を鼻腔内投与し、非ワクチン投与対照として用いた。ワクチン投与から5週後に、ワクチン投与および非ワクチン投与いずれの牛も、すべての牛に強毒性BHV-1菌株Iowa107TCID50を鼻腔内接種した。ワクチン接種および菌株接種後、臨床徴候、直腸温、体重を監視した。鼻のウィルスの放出日数の決定のために布で拭きとった鼻水を採取した。
ワクチン接種後の、牛の挙動、食欲、直腸温、発育率は正常を維持した。漿液性鼻水の分泌および鼻の粘膜の小さな傷害が、ワクチン接種牛すべてに観察された。ワクチン接種の牛の鼻から約7日間ウィルスを単離することができた(表1)。
【0056】
菌接種後、非ワクチン接種の対照牛すべてが、無感情、食欲の喪失、目や鼻からの分泌、下あごの歯肉の赤らみ、鼻の粘膜の強度の傷害を示し、発育が抑制された。Lam gE-,TK-をワクチン接種した牛はすべて、鼻水の分泌が幾分増加し、鼻の粘膜の小さな傷害がいくらか見られた。Lam gE-をワクチン接種した牛のすべてが、鼻水の分泌または鼻の粘膜の傷害が進行したわけではなかった。無感情、食欲の喪失、疾病についてのその他の徴候は、ワクチン接種牛では観察されなかった。菌接種後の直腸温、発育、臨床評価は図22,図23および図24に示されている。非ワクチン接種牛はワクチン接種牛の2倍の長さにわたってウィルスを鼻から放出した(表1)。
【0057】
上記の結果は、Lam gE-およびLam gE-,TK-BHV-1突然変異菌株は、若年の牛においては、病気の臨床徴候をほとんど誘発していないことを示している。これらの突然変異菌株はどちらも菌接種後の病気を予防し、鼻のウィルス放出期間を50%短縮させた。
Lam gE-およびLam gE-,TK- BHV-1突然変異菌株は安全で、BHV-1感染予防のワクチンとして牛に用いて効能を有するものである。
【0058】
3)gEの原核発現
BHV-1糖蛋白質gE-遺伝子の原核発現の場合、これまでのところpGEX発現ベクタ(D.B.Smith and K.S.Johnson,Gene 67(1988)31〜40)が使用されてきた。イソプロピルチオガラクトシド(IPTG)によって発現を誘導されることができ、tacプロモータの影響下にある日本住血吸虫由来の担体蛋白質グルタチオンS-転移酵素(GST)を、pGEXベクタはコードする。GST-gE融合蛋白質の例としては構築されたpGEX-2T600s3の生成物がある(図8A)。この構築物において、標準的な分子生物学的技術を用いて(Sambrook et al. 1989)、gE蛋白質の200アミノ酸のN末端領域をコードする。600bp SmaIフラグメントがGST遺伝子の後に連結された。この構築物は3つの部分から成る設計とされ、600bpフラグメントの異なったリーディングフレームがGSTに連結された。これらの構築物はすべてIPTGと共に誘発された大腸菌株DH5α内に導入され、形成された蛋白質はウェスタンブロッテングによるポリアクリルアミドゲル電気泳動後、ニトロセルロースに転移された。抗-GSTによる免疫性検出により、gE蛋白質領域をコードする適切なリーディングフレーム(No.3)だけが、27k(GST)t20k(gE)=47kの推定サイズの顕著な融合蛋白質の発現を導くことが実証された。Difivac-1と反応しない我々が単離した3つのモノクローナル抗体は、ウェスタンブロット中の47kD GST-gE融合蛋白質を認識する。図8B参照。
【0059】
4)糖蛋白質gE-遺伝子の真核発現
糖蛋白質gE-遺伝子の真核発現については、これまでは、とりわけ、ベクタ、pEVHisが選択されてきた。pEVHisベクタは、真核遺伝標識として、ヒスチジノールデヒドロゲナーゼについてのHisD遺伝子コードを有しており〔EC1.1.1.23〕(C.Hartmann and R.Mulligan,1988,Proc,Natl.Acad Sci.USA 85,8047〜8051)、このヒスチジノールデヒドロゲナーゼが、細胞の2.5mMヒスチジノールの毒性濃度を長びかせる原因となっている。さらに、このベクタは、ヒトのサイトメガロウィルス(HCMV)の極初期遺伝子のプロモータ領域を、該遺伝子の後に位置している特異な制限酵素部位と共に有している。pEVHis/gE発現ベクタの構築のためには、糖蛋白質gE-遺伝子の全コード領域を有するフラグメントが使用された。gEの従来のオープンリーティングフレームの上流のAluI部位55でスタートし、その下流の133bpで終了する。この領域がpEVHisベクタのHCMVプロモータの後方でクローン化され、これによって構築物pEVHis/gEが形成された(図9)。このpEVHis/gEを大腸菌株DH5αの細胞中にて増殖させ、塩化セシウム勾配法によって精製した(Sambrook et al.,1989)。この精製DNAを、Graham and Van der Ebの方法に従って、Balb/c-3T3細胞にトランスフェクトした。形質転換細胞をヒスチジノールを用いて選抜した結果、20の耐ヒスチジノールコロニーを単離することができた。これらのコロニーを免疫ペルオキシダーゼ単分子層試験(IPMA)によって、モノクローナル抗体81によって試験した。4つのコロニ一によって、gE蛋白質の発現が証明された。これらの4つのコロニーの内、3T3gEクローン9を、高度のgE発現を有するサブクローンの単離に使用した。この方法によって単離されたクローン(3T3gE9.5と命名)を候補抗gEモノクローナル抗体の特性づけのために用いた。
【0060】
5)同一細胞におけるBHV-1糖蛋白質gEおよびBHV-1糖蛋白質gI双方の真核発現
BHV-1糖蛋白質gEと同一の細胞においてBHV-1糖蛋白質gIを発現させるため、まず最初にBHV-1 gI遺伝子の推定位置を決めた。ヘルペス単ウィルス糖蛋白質gI遺伝子は、糖蛋白質gE遺伝子のまさに上流に位置しているので、BHV-1 gI遺伝子は、対応の位置に位置しているものと推定された。これを試験するため、BHV-1 gE位置の開始点より約1kb上流に位置した、283ヌクレオチドの領域の配列を決定した。この領域の概念翻訳により、第2リーディングフレームは、ヘルペス単ウィルス糖蛋白質gIと相同の94アミノ酸配列をコードする(図13および図14参照)ことが判った。相同セグメントは、スタートコドンから約80のアミノ酸であるので、BHV-1 gI遺伝子のオープンリーディングフレームの推定開始点は、配列領域から約250nt上流と見積もられる。これより、配列領域から400nt上流でスタートし、gE遺伝子内で終了する、1.7kb SmaIフラグメントは、BHV-1 gI遺伝子の完全なコード領域を含んでいるべきであることが推定された。この1.7kb SmaIフラグメントが真核ベクタMSV-neo内にクローン化された(図15参照)。このベクタは、強いマウス肉腫ウィルスプロモータと、抗生物質G-418硫酸ゲネティシンに対する耐性についてコードするセレクター遺伝子neoを含んでいる。結果として生じる構築物MSVneoGIが、大腸菌DH5α細胞中で増殖され、Graham and Van der Ebの方法を用いて、3T3gE9.5細胞中にトランスフェクトされた。トランスフェクトされた細胞は、400μgゲネティシン/ml培養基で選抜され、耐性コロニーを単離し、3T3gE9.5細胞と反応しなかった候補抗-gEモノクローナル抗体で試験した。これによって、たとえば、野生型BHV-1だけでなく、モノクローナル抗体66と反応した3T3gE/g IR20クローンを選択した。
【0061】
6)候補抗-gEモノクローナル抗体の特性づけ
野生型BHV-1に対してモノクローナル抗体を生成し、Difivac-1に感染したEbtr細胞との反応不能性について選択した。これらのモノクローナル抗体を下記との反応性について試験する。
a)Lam gE-欠失変異体
b)ウェスタンブロットにおける、上記の原核発現生成物
c)上記のgE発現Balb/c-3T3細胞
d)c)に記載のDifivac-1に感染した細胞
e)gE/gI複合物を発現するBalb/c-3T3細胞
a,c,dおよびeとの反応性試験については、免疫ペルオキシダーゼ単分子層試験(IPMA)を用いた。表2の結果により、我々が生成したものは、gEに対して誘導されるモノクローナル抗体(2,3,4,52,66,68,72,81)およびgE/gI複合物上の立体配座抗原ドメインに対して誘導され得る可能性のあるモノクローナル抗体(NO.1,51,53,67,75,78)であることが判る。種々のモノクローナル抗体によって認識される抗原ドメインの地図を作るための競合IPMAにより、少なくとも4つの抗原ドメインが糖蛋白質gE上に存在すること、およびおそらく1つのドメインがgE/gI複合物により形成されることが示された(表2)。
【0062】
BHV-1に感染した牛の抗-gE抗体の検出
感染した牛の血清中にgE抗体が存在しているか否かを調べるために、間接ブロッキングIPMAを16の候補gE-モノクローナル抗体に実施し、以下の8血清を選択した:
−Difivac-1のワクチン接種を受け、毒性Iowa菌株が接種された牛の菌接種14日後に採取された2血清。
−試験のためBHV-1サブタイプ1ウイルスに感染させた牛の、感染20ケ月後に採取された2血清。これらの牛のうちの一体は接触感染によって感染させた。
−試験のためBHV-1サブタイプ2bウイルスに感染させた牛の、感染20ケ月後に採取された2血清。これらの牛のうちの一体は接触感染によって感染させた。
−ts変異体ワクチンの接種を行い、3週後にBHV-1サブタイプ2bウイルスを接種させた特有病原体のない牛の、菌接種7週後に採取された1血清。
−ts変異体ワクチンの接種を行い、3週後にBHV-1サブタイプ2bウイルスを接種させた、ノトバイオートの牛の、菌接種7週後に採取された1血清。
【0063】
表2により、これらの血清はどれも、gE上の抗原性ドメインIIIおよびIVに対する、およびgE/gI複合物上に位置していると思われる抗原性ドメインIに対する抗体を含んでいることが判る。gEは、明らかにBHV-1-感染とワクチン接種された牛を区別するための適切な血清学的遺伝標識であると結論づけることができる。
【0064】
7)BHV-1 gE-特異型プライマを用いるPCR処置によるBHV-1核酸の検出
BHV-1 gE遺伝子の決定されたヌクレオチド配列からスタートして、PCRに適切なプライマの対を、Lowe et alによるプライマ選択プログラム(T. Lowe, J.Sharefkin, S, Qi Yang and C. W .Dieffenback 1990, Nucleic Acids Res, 18, 1757〜1761)を用いて選択した。これらのプライマをP3、P4と命名し、図10に示した。これらのプライマは159nt離れて位置しており、200ntのフラグメントの増殖を導く。プライマP3、P4と単離されたBHV-1 DNAを用いて、PCR処置の条件を最適化した。これは特に、MgCl2の濃度の変更、グリセロール濃度、サイクル条件に関するものである。BHV-1 DNAの増殖のためにP3、P4を使用するため最適と考えられる緩衝液は、10mM Tris pH 8.0, 50mM KCl, 0.01%ゼラチン, 2.6mM MgC12および20%グリセロールである。最適のサイクル条件と考えられるものは(Perkin Elber Cetus DNA Thermal Cycler),サイクル1〜51については、1分98℃, 30秒96℃, 30秒55℃, 45秒72℃である。PCR増殖後、得られた200nt DNAフラグメントを2%アガロースゲル上で電気泳動にかけ、ニトロセルロースにブロットし、続いてサザンブロット分析を行った。サザンブロット分析に使用された32P dCTPのラベルされたプローブが、プライマ結合部位間に位置している137bp TagIフラグメントである(図10)。ハイブリッドフィルターの自動X線撮影により、200bpバンドを観察することができた。さらに、このルートを介して、10BHV-1ゲノムだけの増殖(約1.5×10-15μgDNA)が検出可能と思われるシグナルを導く(結果は示されていない)。同等の方法で、BHV-1糖蛋白質gIIIのコード配列(D.R.Fitrpatrick, L.A. Babiuk and T.Zamb, 1989, Virolagy 173, 46〜57)に基づくプライマを用いてPCR処置を行った。野生型BHV-1 DNAとgE欠失変異体ワクチンの区別を可能とするために、DNA試料にgE-特異型PCR分析およびgIII-特異型PCR分析の両分析を行った。このような試験において、Difivac-1 DNA調製物はgIII陽性およびgE陰性であることが判った。
ウシの精液中のBHV-1 DNAの検出は、BHV-1特異型PCR処置の重要な用途であると思われるので、BHV-1に感染したウシの精液にgE-特異型PCRの実施を試みた。しかしながら、精液中の未知の成分がポリメラーゼ連鎖反応に強い抑制効果を有している。したがって、ウシの精液からBHV-1 DNAを単離するプロトコールが展開された。ウシの精液からDNAを単離するため、30μlの精液を、1mg/mlプロティナーゼK(pk)とともに、60℃にて、全量300μlの0.15M NaCl, 0.5%Na-サルコシル, 40mM DTT中にて培養する。1時間後、試料を室温にまで冷却し、300μl 6M NaIを添加し、5分間培養する。この混合物より、標準クロロホルム/イソアミルエタノール抽出によりDNAを単離し、1容量のイソプロパノールで沈殿させた。沈殿物を2.5M NH4AC/70%エタノールで洗浄し、60℃にて1時間10mM Tris pH7.4,1mM EDTA,0.5%Tween80および0.1mg/ml pk中で、第2培養のために再懸濁する。このDNA調製物は直接ポリメラーゼ連鎖反応下に置くことができる。
【0065】
Lam gE-またはLam gE-/TK-を接種後および毒性の強いBHV-1菌株を投与後、これらの接種されまたは無接種の牛のウイルスを含む鼻水の垂れ
【表1】
【0066】
gE-Mabsの特性
【表2】
+:間接ブロッキングIPMAにおいてテストされた8つの全血清のブロッキングした割合が50%よりも大きい場合
±:±50%の場合
−:50%未満の場合
【図面の簡単な説明】
【0067】
【図1】BHV-1菌株Difivac-1およびIowaのサザンブロット分析 Aは、Difivac-1とIowaとのゲノムDNAのサザンブロットのオートラジオグラムである。1と3レーン中で、Difivac-1 DNAは、HindIIIとPstIとによる制限酵素消化後に、用いられた。2と4レーン中で、Iowa DNAは、HindIIIとPstIとによる制限酵素消化後に用いられた。フラグメントの大きさは、キロベース(kb)で示される。 Beckman L5-65超遠心分離のSW27ロータ中で20,000回/分でpH7.4のトリス10mMと150mM NaClと1mM EDTAとの中で、25%(w/w)庶糖クッションを用いて2時間、ウイルスで汚染されたEbtr細胞と共に培養液(約450cm2ローラボトルあたり70ml)を遠心分離することによって、ウイルスDNAが単離された。得られたウイルスペレットから、DNAが標準の手法で単離された(J.Sambrook,E.F.Fritsch and T.Maniatis,1989,Molecular cloning:a laboratory manual,2nd ed.Cold Spring Harbor Laboratory Press, New York)。このDNA上で制限酵素消化は、Boehrnger Mannheim社によって供給されるSuRE/cut緩衝液中で酵素を用いて行われた。 0.7%アガロースゲルを水平電気泳動して分離し、ニトロセルロースフィルタ(Schleicher & Schuell,Inc)でブロットした後、フィルタは6時間42℃で50%ホルムアルデヒド中で3×SSC(1×SSC=0.15MのNaClと0.015Mのクエン酸ナトリウム,pH7.4)と、50μlの変成サケ精液DNA(Sigma)/mlと、0.02%フィコールと、0.1%ドデシル硫酸ナトリウム(SDS)との中に予備ハイブリダイズされた。さらに同溶液に32PdCTP(Amersham)でラベルされたHindIII Kフラグメントを加えることによってハイブリダイゼーションが行われた(HindIII Kフラグメントの選択は、1型ウシヘルペスウイルスからのDNAのクローニングと分裂位置のマッピングに基づく(Cooper菌種)John F.Mayfield, Peter J.Good, Holly J. VanOort, Alphonso R. Campbell and David A.Reed;Journel of Virology (1983) 259〜264)。ハイブリダイゼーションの12〜14時間後、フィルタは2時間、0.1%SDSと0.1×SSCとの中で60℃で洗浄された。HindIII Kフラグメントは、標準のクローニング手法に従って、pUC18ベクタ中でクローン化された(J.Sambrook, E.F.Fritsh and T.Maniatis, 1989, Molecular cloning la laboratory manual,2nd ed.Cold Spring Harbor labovatort Press, New York)。pUC8.4HindIII KクローンのHindIIIによる消化後、pUC18ベクタは、0.7%の低融点アガロース(BRL、Life Technologies,Inc.)ゲルで再び電気泳動によって8.4kb HindIIIにフラグメントから分離され、基準のフェノール抽出とエタノール沈澱とによってアガロースから分離された。分離されたHindIII Kフラグメントは、Boehringer Mannheim社製のランダムプライムドDNA標識キット1OO4.760で標識を付された。ハイブリダイズされたフィルターのオートラジオグラムは、-70℃でKodak XARフィルムの36時間露出で、反射スクリーンを使って行われた。 BはIowaとDifivac-1の7.4kb HindIIIフラグメントとの8.4kb HindIII Kフラグメントの物理的マップである。6kb PstIフラグメントの共泳動とDifivac-1中の1.8kb PstIフラグメントの欠如を考えて、欠失はハッチングされた面積領域で起こったと推定される。
【図2】Difivac-1中の欠失領域周囲の野生型BHV-1フラグメントのサブクローニング Aにおいて、BHV-1ゲノムの成分は、長ユニーク(UL)領域、短ユニーク(US)領域および2つの反復物(IrとTr)として示される。このマップは、Cooper菌種の公表された分析に基づいている(John F.Mayfield, Peter J.Good, Holly J. VanOort, Alphonso R. Campbell and David A. Reed: Journal of Virology (1983) 259〜264)。 Bにおいて、フラグメントは、原核ベクタ中へクローンされたUS領域で示される。すなわちpACYC中の15.2kb EcoRIフラグメント、pUC18中の8.4kb HindIIIフラグメント、およびpBR322中の4.1kb EcoRI-HindIIIフラグメントである。ウイルスDNAフラグメントの単離は、図1Aの表示中に述べられている手法に従って実施された。種々のベクタ中へこれらのフラグメントをクローン化することは、標準の手法で行われた(J.Sambrook, E.F.Fritsch and T.Maniatis, 1989, Molecular cloning:a loboratory manual,2nd ed.Cold Spring Harbor Laboratory press, New York)。 Cにおいて、物理マップは、Difivac-1中で要求される欠失が位置する領域を示す。 Dにおいて、さらなる分析のために用いられるこの領域のサブクローンが示されている。2つのPstIフラグメントはpKUN19の中で、そして残ったフラグメントはpUC18の中でクローンされた。
【図3】Aは、〔左端にあるAluIの認識サイトから右端にあるHincIIの認識サイトに至る〕図2C中で示されるようなDifivac-1中において欠失された要求される位置周囲の、BHV-1のLam種の、US領域の2027個のヌクレオチドのヌクレオチド配列である。図2Dで示されるサブクローンの挿入をしたヌクレオチド配列は、Sangerらのジデオキシ配列法を用いた2つの基準で分析することによって決定された(F.Sanger, S.Nicklen and A.R.Coulson, 1977, Proc.Natl.Acad.Sci.USA 74,5463〜5467)。この目的に対して、Pharmacia製のT7配列キットが、同製造者によって指定される手法に従って用いられた。放射線標識のために、〔35S〕dATP(Amersham製)が用いられた。コンプレッション現象の影響を併うGLの多い領域の配列分析は、Pharmacia製キットの7-デアザアデニン-dGTP変形物を用いて繰り返された。ヌクレオチド配列の下に3文字のコードで示されたものは、ヌクレオチドの概念上の翻訳後に見つかった575アミノ酸残基のオープンリーディングフレームのアミノ酸配列である。この翻訳は、一般的なコードに基づきPC遺伝子コンピュータプログラムを用いて決定された(PC遺伝子翻訳バージョン1.03 1987年11月)。この575アミノ酸のオープンリーディングフレームは、ヌクレオチド168のメチオニンで始まり、ヌクレオチド1893の停止コドンで終わる。 575アミノ酸残基のオープンリーディングフレームの構造分析は、PC遺伝子コンピュータプログラムで実行される。最初の26アミノ酸が、真核性の移行シグナルを形成し、「シグナルペプチド」で図中に示される。6.2の点で、このシグナル配列の切断が、アミノ酸26とアミノ酸27との間で予想される。アミノ酸575の配列は、アミノ酸残基の線によって指示される。3つの可能なN結合のグリコシレーション位置(NXT/S)を有する。RaoとArgosの方法に従えば、「貫膜螺旋」で図中に示される貫膜領域がアミノ酸423とアミノ酸450との間にある。制限酵素AsuII, SmaI, HindIIIおよびEcoNIのための認識配列(サイト)は下線が付される。このポリペプチドの計算された分子量は61212である。 Bは、上で述べた575アミノ酸オープンリーディングフレームの構造上の特性を模式的に表したものである。
【図4】BHV-1 gE遺伝子のアミノ酸配列と、ヘルペス単体ウイルス(HSV)gE遺伝子および他のgE相同遺伝子〔疑似狂犬病ウイルス(PRV)gIと帯状疱疹ウイルス(VZV)gpI〕のアミノ酸配列との比較 この比較のために用いた配列は、次の公表されたものである。 HSV:1型ヘルペス単体ウイルスのゲノム中のUS領域の配列決定と遺伝子内容(D.J.McGeoch, A.Dolan, S.Donald and F.J.Rixon (1985) Journal Mol.Biol.181,1〜13) VZV:帯状疱疹ウイルス遺伝子のUs成分のDNA配列(A.J.Davidson (1983), EMBO Journal 2, 2203〜2209) PRV:ヘルペス単体ウイルスと帯状疱疹ウイルス糖蛋白とに相同性を有する2つの疑似狂犬病ウイルス糖蛋白のための単離遺伝子にλgt11の使用(E.A.Petrovskis, J.G.Timmins and L.E.Post (1986) Journal of Virology 60,185〜193) これらの配列は配列分析プログラムMultalinを使って比較された(F.Corpet (1988), Nucl. Acids Res.16,10881〜10890)。 Aの中で、4つのアミノ酸配列の模式的図が示されている。ここに予測された貫膜部(TM)が各々に示される。予測された移行シグナル配列(SP)と考えられるN-結合グリコシレーション位置(I)に加えて、2つの保存される領域が示される。その中で、システン残基の相対位置(CCC)は不変である。 Bの中で結果は4つのgE翻訳の中央に位置するシスティンの多い領域のMultalin比較が示される。アステリスクは、一致したアミノ酸と類似アミノ酸クローンを示す。
【図5】Difivac-1とIowaとのサザンブロット分析で得られた写真図 パネルAは、BstI(1,2),EcoRI(3,4)およびHindIII(5,6)を有するDifivac-1とIowa消化制限酵素のゲノムDNAである。これは、0.7%アガロースゲルで分離され、ニトロセルロースでブロットされ、図1Aの説明で述べられた手法に従ってBHV-1菌株Lamの32PでラベルされたHindIIIKフラグメントでハイブリダイズされる。 パネルBは、Aと同様に同じゲルのニトロセルロースブロットであり、BHV-1 gE-特異型プローブp318でハイブリダイズされた。プローブは、図2C中で示したAluI-HincII全領域を含む。
【図6】gEフラグメント欠失BHV-1の構築 Aの中でgEの位置と用いられたクローンとが示される。BHV-1ゲノムの成分は、UL領域、Us領域および2つの反復領域(IrとTr)として示される。gE遺伝子の5’側末端に位置する領域を得るために、BHV-1菌種Lamの8.4kb HindIIIK由来の1.4kb PstI-SmaIフラグメントが、プラスミドpUC18のSmaIとPstIサイトの中ヘサブクローンされた。このクローンは、p515と呼ばれ、図6B中に示される。gEの3'末端側に位置するEcoNI-SmaIフラグメントは、4.1kb HindIII−EcoRIクローンに由来し、p515の普通のAsuIIヘクローンされる。AsuIIの末端にEcoNIの末端を結合するために、クローンp515はAsuIIで消化され、その後クレノウ酵素(Boehringer Mannheim)と、標準的手法に従ってAsuIIの末端へ1つのシトシン残基を供給するdCTPとで処理した(Sambrook et al,1989)。この付加シトシンは、D中でアステリスクで示される。p515は、またSmaI酵素で消化され、その後EcoNIフラグメントは、このベクタ中へ結合される。このようにして構築されたクローンは、p519と呼ばれる。
【図7】Aは、1B7,1B8および2H10のDNA調製品のサザンブロット分析で得られた写真である。DNA単離物、消化制限酵素、ブロットおよびハイブリダイゼーションは、図1Aの説明で述べられた手法に従って作られた。DNA調製品1B7,1B8および2H10のPstI-DraIによる二重消化後、フラグメントは0.7%アガロースで分離され、直後にニトロセルロースフィルタにブロットされた。このフィルタは、プローブとして、32PdCTPでラベルされた2.3kb PstI-DraI欠失フラグメントとハイブリダイズされた。レーン1〜3中で試料1B7,1B8および2H10がそれぞれ分離された。レーン中でLam菌株の野生型BHV-1 DNAが、レーン5中で2.3kb欠失フラグメントが示される。 Bは、BHV-1菌種Lamの15.2kb EcoRIフラグメントの物理マップである。マップは、PstI,DraIおよびHindIIIの認識サイトと7A中で述べたハイブリダイゼーションプローブのサイトを示す。
【図8】BHV-1 gEの原核発現 BHV-1 gEの原核発現のために、gE遺伝子の600bpSmaIフラグメントは、ベクタpGEX-2T中で、日本住血吸虫のグルダチオン-S-転移酵素のコード領域に3つのリーデングフレーム中で融合された(D.B.Smith and K.S.Johnson,Gene 67(1988)31〜40)。SmaIフラグメントの適切な(syn)方向性を有する組換え分子は、標準の手法を用いて、制限酵素分析法によって同定された。この融合構造を有するE.coli DH5αクローンは、pGEX-2T600s1,pGEX-2T600s2およびpGEX-2T600s3と呼ばれた。 Aは、pGEX-2T600s構築物の1つの図である。GST-gE融合物をコードする領域のNH2側にあるものは、イソプロピルチオガラクトシド(IPTG)で誘導し得るtacプロモータ領域である。 Bは、pGEX-2T600sでトランスフェクトされたDH5αの細胞の全蛋白質調製物のウエスタンブロット分析で得られた写真である。pGEX-2T600s1,pGEX-2T600s2およびpGEX-2T600s3構築物でトランスフェクトされたDH5α細胞のオーバーナイトの培養物は、50μg/mlアンピシリンを加えたLuria-Bertani(LB)培地中で1/10の割合で残っており、1時間後培養物はIPTGによって5時問誘導された。これらの誘導された培養物は、5分間6,000Gで遠心分離し、1×ラウエル混合物(2%SDS、10%グリセロール、5%メルカプトエタノールおよび0.01%ブロムフェノールブルー)中に混合〔1.5ml培養物を、500μlラウエル混合物中に混ぜる〕され、95℃に5分間加熱された。そして、1レーンあたり50μlで、標準の手法に従って、12.5%ポリアクリルアミドゲルで水平的に分離され、引続いて製造者によって指定された条件で、LKB-ムルチフールII型Navaブロットシステムを用いて、ニトロセルロースフィルタがセミドライ式でブロットされた。 レーンM中で、予め印を付されたマーカー蛋白質が用いられ、(BRL Life Technologies,Inc.236K,112K,71K,44K,28K,18Kおよび15K)、レーン1,2,3中では、pGEX-2T600s1,pGEX-2T600s2およびpGEX-2T600s3各々3つのフレームでトランスフェクトされたDH5α細胞の全蛋白調製物はが用いられた。 パネルA中で、結果は、抗-GST血清を用いるウエスタンブロット分析で見ることができる。フィルタは、標準の手法(E.Harlow and D.Lane(1986)、Antibodies:a laboratory manual,Cold Spring Harbor Loboratory,New York)にしたがって、ウサギ血清ポリクローナル抗GSTを加えたブロッキング緩衝液(PBS+2%粉乳および0.05%トゥイーン20)中でインキュベートされた。そして、フィルタは洗浄され、ヤギ抗ウサギ免疫グロブリン血清を結合した西洋ワサビパーオキシターゼ(HRPO)を加えてインキュベートされた。その後、結合されたヤギ抗体は、色素体(ヂアゾベンジジン、クロロナフトールおよびH2O2)で免疫化学的に検出された。矢印によって指示されるGST融合物は、フレーム3中でのみ約47kの確認された大きさを有する。 パネルBにおいて、結果は、モノクローナル抗体Mab4を用いる、ウエスタンブロット分析で見ることができる。この目的に対しパネルA中のようにデュプロフィルタはブロッキングされ、Mabでインキュベートされ、ウサギ抗マウス血清を結合したHRPOでインキュベートされる。その後結合ウサギ抗体は、色素体で免疫化学的に検出される。レーン3(フレーム3)で見られるバンドは、47Kの大きさで、矢印によって指示される。
【図9】BHV-1 gE遺伝子の原核発現のためのpEVHis gEプラスミドの構築 gE遺伝子の原核発現のために、gEコード領域は、標準の手法を用いて発現ベクタpEVHisのHCMVプロモータ領域の背後の適切な位置においてクローン化される(Sambrook et a1,1989)。gEのオープンリーディングフレーム55bp上流で開始する394bpA1UIフラグメントは、pUC18の中ヘクローンされ、p201と呼ばれる。そして、p201をHincIIが消化した後、1740bp HincIIフラグメントは、gE遺伝子の大部分から成り、p201中へクローンされる。これは、pUC18のポリリンカ中で、gEの開始コドンの上流にあるAluI位置55bpからgEの停止コドンの下流にあるHincII位置133bpまでの全gEコード領域を含むプラスミドp318になる。ベクタのポリリンカにおける制限酵素位置を用いて、このフラグメントは、酵素BamHIとSphIでp318から切られた。最初p318はSphIで消化され、その後SphIサイトは、クレノウポリメラーゼとdNTPポリメラーゼとを使って満たされた。BamHIでの消化後、1.9kb挿入物は、pUC18ベクタから低融点アガロース中で分離され、BamHIとEcoRVで消化されたpEVHis中で結合された。このように形成されたプラスミドは、pEVHis/gEと呼ばれた。
【図10】BHV-1 DNAを検出するためにPCR手法のためのgE-特異型プライマとプローブの位置 図中には、ヌクレオチド1272から2027に至るBHV-1糖蛋白質の核酸配列が示される〔配列は図3から得られる〕。gE-特異型PCR手法のために用いられるプライマは、P3とP4と呼ばれる。P3とP4のプライマ結合サイトは、下線が付される。P3のヌクレオチド配列は、5'-ACG-TGG-TGG-TGC-CAG-TTA-GC-3'である(配列標別番号:2)。P4のヌクレオチド配列は、(上述のプライマ結合配列に相補的である)5'-ACC-AAA-CTT-TGA-ACC-CAG-AGC-G-3'である(配列標別番号:3)。PCRで増幅されたDNAの検出のためのサザンブロットハイブリダイゼーションで用いられたプローブは、プライマが結合するサイトの間に位置する137bp TaqIフラグメントであり、そのフラグメントの末端が示される。図3と比較のために、HindIIIとEcoNIサイトがまた示される。
【図11】Difivac-1のgE欠失のマッピング Aは、野生型BHV-1菌種Lamの15.5kb EcoRIフラグメントの物理マップを示す。Bは、Difivac-1の14.5kb EcoRIフラグメントの物理マップを示す。2つのEcoRIフラグメントは、各々のウイルスのゲノムのUS領域を完全にカバーする。gE遺伝子の位置とgI遺伝子の推定上の位置は、白ヌキの四角で指示される。マップAとBとは、各マップ中の6kb PstIフラグメントが、一列に並べられるように位置決めされる。両方のマップ中で、内部反復領域と末端反復領域との配列は、斜線を付された四角で示される。反復の下方の矢印は、これらの配列の方向を示す。 Aにおいて、Difivac-1菌種中で欠失があるUS領域の部分が示された。 Cは、gE欠失をマップ化し、B中に示される物理マップを得るために用いられるクローン化Difivac-1フラグメントの位置を示す。クローンp728,p737およびp754の挿入物の下の矢印は、組換え点を決めるために配列決定された領域を示す。 略語: A=A1UI,E=EcoRI,P=PstI,H=HindIII,γ=組換え点,IR=中間反復領域,TR=末端反復領域
【図12】Difivac-1のUS領域における正確な組換え位置の決定 Difivac-1菌種中に見付かったgE欠失の正確な境界を決定するために、クローンp754ならびにp728およびp737とが配列決定された。これらのクローンの挿入は、図11中に指示される。用いられる配列決定の手法は、図3の説明中に述べられている。 Aにおいて、A1UI-PstIフラグメントの大部分の配列が示される。この配列は、gE遺伝子のプロモータ領域で開始する。推定のTATAボックスは、下線が付されている。点γ(=組換え点)において、このプロモータ領域は、Us領域の反対の位置で見付かった逆方向反復と呼ばれる配列に接合される。正確な組換え点は、US領域の反対位置で見付けられた反復のコピーとgEプロモータ領域で見付けられた反復を比較することによって決定される。この配列が分岐する点は、B(I)の中でγで示される。類似の比較は、Difivac-1中で見付けられるgEプロモータ配列と野生型菌種Lam中で見付けられたgEプロモータとで行われる。この配列が分岐する点は、B(II)の中で示され、またγで示される。見付けられた組換え点は同じである。
【図13】BHV-1 gI遺伝子の部分配列分析 BHV-1 gIとgEとの双方の遺伝子(図11に示す)中に達するBHV-1菌種Lamの1.8kb PstIクローンを用いてBHV-1 gIのコード領域の範囲内の、284ヌクレオチドの配列が決定された。配列決定に用いられる手法は、図3の説明の中で述べられている。配列は、PC/遺伝子コンピュータプログラムバージョン1.03(1987年11月)による汎用コードに基づいて翻訳された。第2リーディングフレームによってコードされるアミノ酸配列は、ヌクレオチド配列の下にワンレターコードとして与えられる。このアミノ酸配列は、他のヘルペスウイルスのgI相同体(図14に示す)のコード領域に相同である。
【図14】推定のBHV-1 gI遺伝子の部分アミノ酸配列と、ヘルペス単体ウイルス(HSV1)gI遺伝子、疑似狂犬病ウイルス(PRV)gp63遺伝子および帯状疱疹ウイルス(VZV)gpIV遺伝子のコード領域の相当部分とのアミノ酸比較 PRV配列はそのコード領域のアミノ酸82で開始し、HSV1配列はそのコード領域のアミノ酸で開始し、VZVはそのコード領域のアミノ酸76で開始する。用いられる配列は、図4の説明で述べられた報告書で公表されている。比較は、Multalinコンピュータプログラムを使って行われた。アステリスクは、同一のアミノ酸を示し、コロンは類似のアミノ酸を示す。
【図15】BHV-1 gI-遺伝子の原核発現のためのMSVneoGIプラスミドの構築 BHV-1 gI-遺伝子の部分配列のアミノ酸配列の比較に基づいて、BHV-1 gI-遺伝子の推定位置は推測される。この推測に基づいて、1.7kb SmaIフラグメントは、BHV-1 gE遺伝子の全コード領域を含むことが推論される。この1.7kb SmaIフラグメントの位置は、A中に示される。この1.7kb SmaIフラグメントの丸くなった平滑末端に、BamHIリンカが標準の手法を用いて結び合わされる。この結果生じた物は、BamHIで消化され、原核発現ベクタMSV-neo中で結び合わされる。MSV-neoベクタは、強いプロモータ性を有するMSV-LTRの背後にユニークなBamHIサイトを有する。このベクタは、Rijsewijk et al.,1987EMBO J.6,127〜131中で述べられている。
【図16】BHV-1 gI/gE二重欠失フラグメントの構築 BHV-1のUS領域中における糖蛋白質gE遺伝子の位置と糖蛋白質gI遺伝子の推定位置は、図A中に表現される。斜線を付された部分は、US領域の境界を成す反復物を示す。Bは、双方の遺伝子の位置を考慮して、必然的な制限酵素サイトの物理マップを示す。gI/gE欠失フラグメントを構築するために、gI遺伝子を包み込む1.7kb SmaIフラグメントを含むクローンp1.7-SmaI/oがPstIで消化される。残留350bp SmaI-PstI挿入物のPstIサイトは、標準の分子生物学的手法を用いて、平滑末端を作る。EcoNI-SmaIフラグメント(図6B参照)は、図6A中で記載された4.1kb HindIII-EcoRIフラグメントから単離され、平滑末端を作られ、上述の修正されたPstIサイトに結合される。これは、CとD中に図示される。結果としてできたクローンPΔIEから、1.4kb SmaI-DraIフラグメントは、野生型BHV-1 DNAと組換えるために単離される。 略号 E=EcoRI,H=HindIII,S=SmaI,P=PstI,ENI=EcoNI,D=DraI,kb=キロベース,US=ユニークショート
【図17】接種後の牛から垂れた鼻水の平均ウイルス数 ●=Difivac-1を接種、○=接種なし
【図18】毒性の強いBHV-1菌株投与後の平均発病牛数、記号は図17に同じ
【図19】毒性の強いBHV-1菌株を投与した牛の平均体温、記号は図17に同じ
【図20】毒性の強いBHV-1菌株投与後の牛の平均成育量、記号は図17に同じ
【図21】毒性の強いBHV-1菌株投与後の牛から垂れた鼻水の平均ウイルス数、記号は図17に同じ
【図22】毒性の強いBHV-1菌株投与後の牛の平均体温 ●=Lam gE-を接種、○=Lam gE-/TK-を接種、×=無接種
【図23】毒性の強いBHV-1菌株投与後の牛の平均成育量、記号は図22と同じ
【図24】毒性の強いBHV-1菌株投与後の平均発病牛数、記号は図22と同じ
【特許請求の範囲】
【請求項1】
糖蛋白質gE-遺伝子中に欠失を有する1型ウシヘルペスウイルスの欠失突然変異株。
【請求項2】
糖蛋白質gE-遺伝子中に減衰手段によって起こる欠失を有する1型ウシヘルペスウイルスの欠失突然変異株。
【請求項3】
糖蛋白質gE-遺伝子中に減衰手段によって起こり、Difivac-1である欠失を有する1型ウシヘルペスウイルスの欠失突然変異株。
【請求項4】
糖蛋白質gE-遺伝子中に再結合DNA技術によって起こる欠失を有する1型ウシヘルペスウイルスの欠失突然変異株。
【請求項5】
糖蛋白質gE-遺伝子中に再結合DNA技術によって起こり、IB7または1B8である欠失を有する1型ウシヘルペスウイルスの欠失突然変異株。
【請求項6】
糖蛋白質gE-遺伝子中と、チミジンキナーゼ遺伝子中とに欠失を有する1型ウシヘルペスウイルスの欠失突然変異株。
【請求項7】
糖蛋白質gE-遺伝子中と、同じくgI-遺伝子中とに欠失を有する1型ウシヘルペスウイルスの欠失突然変異株。
【請求項8】
糖蛋白質gE-遺伝子中と、チミジンキナーゼ遺伝子中と、糖蛋白質gI-遺伝子中とに欠失を有する1型ウシヘルペスウイルスの欠失突然変異株。
【請求項9】
糖蛋白質gE-遺伝子中に欠失を有し、再結合DNA技術によって誘導される非対称遺伝子を含む1型ウシヘルペスウイルスの突然変異株。
【請求項10】
糖蛋白質gE-遺伝子の位置に、再結合DNA技術によって誘導され、調節した配列、たとえばgE-遺伝子または非対称遺伝子の制御下にあり、ペプチド信号のためコード化されたgE-遺伝子の部分に随意に接触させられる非対称遺伝子を含む1型ウシヘルペスウイルスの突然変異株。
【請求項11】
糖蛋白質gE-遺伝子中にある欠失に加えて、チミジンキナーゼ遺伝子中と、糖蛋白質gI-遺伝子中との一方または双方に欠失を有し、再結合DNA技術によって、前記欠失の少なくとも1つの位置に誘導される非対称遺伝子を含む1型ウシヘルペスウイルスの突然変異株。
【請求項12】
糖蛋白質gE-遺伝子中に欠失を有し、再結合DNA技術で誘導され、他の病原菌の免疫原蛋白質のためと、サイドカインのためとにコード化された非対称遺伝子を含む1型ウシヘルペスウイルスの突然変異株。
【請求項13】
1型ウシヘルペスウイルスの糖蛋白質gE-遺伝子を含む再結合核酸、この糖蛋白質gE-遺伝子の一部またはこの糖蛋白質gE-遺伝子から誘導されるヌタレオチド配列から成る組成物。
【請求項14】
1型ウシヘルペスウイルスの糖蛋白質gE-遺伝子を含む再結合核酸、この糖蛋白質gE-遺伝子の一部または、この糖蛋白質gE-遺伝子から誘導されるヌクレオチド配列を挿入するクローンまたは表現ベタタから成る組成物。
【請求項15】
1型ウシヘルペスウイルスの糖蛋白質gE、この糖蛋白質gEの一部、この糖蛋白質gEから誘導されるペプチドまたは1型ウシヘルペスウイルス糖蛋白質gEと同gIとの複合物から成る組成物。
【請求項16】
1型ウシヘルペスウイルスの糖蛋白質、この糖蛋白質gEの一部、この糖蛋白質gEから誘導されたペプチドまたは1型ウシヘルペスウイルス糖蛋白質gEおよび同gIの複合物に特有の組成物。
【請求項17】
1型ウシヘルペスウイルスの糖蛋白質、この糖蛋白質gEの一部、この糖蛋白質gEから誘導されたペプチドまたは1型ウシヘルペスウイルス糖蛋白質gEおよび同gIの複合物に特有のモノクロナール抗体から成る組成物。
【請求項18】
1型ウシヘルペスウイルスの糖蛋白質、この糖蛋白質gEの一部、この糖蛋白質gEから誘導されたペプチドまたは1型ウシヘルペスウイルス糖蛋白質gEおよび同gIの複合物に特有のボリクロナール抗体から成る組成物。
【請求項19】
1型ウシヘルペスウイルスに対して動物を保護するために、動物、好ましくは哺乳動物、さらに好ましくはウシ科の動物に接種するための請求項1〜12のいずれか1項に記載の1型ウシヘルペスウイルスの欠失から成るワクチン組成物とそれに適する担体と免疫助成剤。
【請求項20】
病原菌に対して動物を保護するために、動物、好ましくは哺乳動物、さらに好ましくはウシ科の動物に接種するための糖蛋白質gE-遺伝子中に欠失を有し、再結合DNA技術によって誘導され、病原菌の免疫原蛋白質またはペプチドのためにコード化された非対称遺伝子を含む1型ウシヘルペスウイルスの欠失から成るワクチン組成物とそれに適する担体と免疫助成剤。
【請求項21】
動物、好ましくは哺乳動物、さらに好ましくはウシ科の動物から得られる試料、特に血液、血清、血液細胞、乳、涙のような体液、肺洗浄液、鼻水、精液、特に精漿、痰、または、組織、特に神経組織のような生体試料の中で、1型ウシヘルペスウイルスの糖蛋白質gE-遺伝子から誘導されたヌタレオチド配列を有する核酸プローブまたはプライマから成り、1型ウシヘルペスウイルスの核酸を検出するための診断キットおよび核酸を分析するために適する検出方法。
【請求項22】
動物、好ましくは哺乳動物、さらに好ましくはウシ科の動物から得られる試料、特に血液、血清、唾液、痰、涙のような体液、肺洗浄液、鼻水、乳または組織のような生体試料の中で、BHV-1の糖蛋白質gE、この糖蛋白質gEの一部、この糖蛋白質gEから誘導されるペプチド、または1型ウシヘルペスウイルス糖蛋白質gEおよび同gIの複合物から成り、1型ウシヘルペスウイルスに特有の抗体を検出するための診断キットおよび抗体を分析するために適する検出方法。
【請求項23】
1型ウシヘルペスウイルス糖蛋白質gEまたは1型ウシヘルペスウイルス糖蛋白質gEおよび同gIの複合物に特有の1またはそれ以上の抗体から成る請求項22記載の診断キット。
【請求項24】
動物、好ましくは哺乳動物、さらに好ましくはウシ科の動物から得られる試料、特に血液、血清、血液細胞、乳、涙のような体液、肺洗浄液、鼻水、精液、特に精漿、唾液、痰、または組織、特に神経組織のような生体試料の中で、1型ウシヘルペスウイルスの糖蛋白質gE、または1型ウシヘルペスウイルスgEおよび同gIの複合物に特有の抗体から成り、1型ウシヘルペスウイルスの蛋白質を検出するための診断キットおよび蛋白質を分析するために適する検出方法。
【請求項25】
動物由来の試料、特に血液、血清、血液細胞、精液、特に精漿、唾液、痰、涙のような体液、肺洗浄液、鼻水、乳、組織、特に神経組織のような生体試料を試験することと、1型ウシヘルペスウイルスの糖蛋白質gE-遺伝子から成る核酸の存在、または1型ウシヘルペスウイルス糖蛋白質gEもしくは1型ウシヘルペスウイルス糖蛋白質gEと同gIとの複合物の存在、または1型ウシヘルペスウイルスの糖蛋白質もしくは1型ウシヘルペスウイルス糖蛋白質gEと同gIの複合物のための特有の抗体の存在を試験することとから成る1型ウシヘルペスウイルスに感染した動物、好ましくは哺乳動物、さらに好ましくはウシ科の動物を決める方法。
【請求項26】
動物由来の試料、特に血液、血清、血液細胞、精液、特に精漿、唾液、痰、涙のような体液、肺洗浄液、鼻水、乳、組織、特に神経組織のような生体試料を試験することと、1型ウシヘルペスウイルスの糖蛋白質gE-遺伝子から成る核酸の存在、または1型ウシヘルペスウイルス糖蛋白質gEもしくは1型ウシヘルペスウイルス糖蛋白質gEと同gIとの複合物の存在、または1型ウシヘルペスウイルスの糖蛋白質もしくは1型ウシヘルペスウイルス糖蛋白質gEと同gIの複合物に特有の抗体の存在を試験することと、請求項19によって準備されたワクチンを接種した動物から得られた試料を試験することとから成る1型ウシヘルペスウイルスに感染した動物、好ましくは哺乳動物、さらに好ましくはウシ科の動物を決める方法。
【特許請求の範囲】
【請求項1】
糖蛋白質gE-遺伝子中に欠失を有する、1型ウシヘルペスウイルスの欠失突然変異体。
【請求項2】
糖蛋白質gE-遺伝子中に弱毒処置によって起こる欠失を有する、1型ウシヘルペスウイルスの欠失突然変異体。
【請求項3】
糖蛋白質gE-遺伝子中に弱毒処置によって起こる欠失を有する、1型ウシヘルペスウイルスの欠失突然変異体 Difivac-1。
【請求項4】
糖蛋白質gE-遺伝子中に組換えDNA技術によって構築された欠失を有する、1型ウシヘルペスウイルスの欠失突然変異体。
【請求項5】
糖蛋白質gE-遺伝子中に組換えDNA技術によって構築された欠失を有する、1型ウシヘルペスウイルスの欠失突然変異体 1B7または1B8。
【請求項6】
糖蛋白質gE-遺伝子中と、チミジンキナーゼ遺伝子中とに欠失を有する、1型ウシヘルペスウイルスの欠失突然変異体。
【請求項7】
糖蛋白質gE-遺伝子中と、糖蛋白質gI-遺伝子中とに欠失を有する、1型ウシヘルペスウイルスの欠失突然変異体。
【請求項8】
糖蛋白質gE-遺伝子中と、チミジンキナーゼ遺伝子中と、糖蛋白質gI-遺伝子中とに欠失を有する、1型ウシヘルペスウイルスの欠失突然変異体。
【請求項9】
糖蛋白質gE-遺伝子中に欠失を有し、且つ組換えDNA技術によって導入される異種遺伝子を含む、1型ウシヘルペスウイルスの突然変異体。
【請求項10】
糖蛋白質gE-遺伝子の位置に、組換えDNA技術によって導入された異種遺伝子を含む、1型ウシヘルペスウイルスの突然変異体であって、該異種遺伝子が、調節配列、たとえばgE-遺伝子または異種遺伝子の調節配列の制御下にあり、シグナルペプチドをコードするgE-遺伝子の部分に任意で付着される、1型ウシヘルペスウイルスの突然変異体。
【請求項11】
糖蛋白質gE-遺伝子中にある欠失に加えて、チミジンキナーゼ遺伝子中と、糖蛋白質gI-遺伝子中との一方または双方に欠失を有し、且つ組換えDNA技術によって、前記欠失の少なくとも1つの位置に導入される異種遺伝子を含む、1型ウシヘルペスウイルスの突然変異体。
【請求項12】
糖蛋白質gE-遺伝子中に欠失を有し、且つ組換えDNA技術で導入され、他の病原菌の免疫原蛋白質もしくはペプチドをコードする、またはサイトカインをコードする異種遺伝子を含む、1型ウシヘルペスウイルスの突然変異体。
【請求項13】
1型ウシヘルペスウイルスの糖蛋白質gE-遺伝子、該糖蛋白質gE-遺伝子の一部、または該糖蛋白質gE-遺伝子に由来するヌクレオチド配列を含む組換え核酸を含む、組成物。
【請求項14】
1型ウシヘルペスウイルスの糖蛋白質gE-遺伝子、該糖蛋白質gE-遺伝子の一部、または該糖蛋白質gE-遺伝子に由来するヌクレオチド配列を含む組換え核酸を挿入するクローニングベクタまたは発現ベクタを含む、組成物。
【請求項15】
1型ウシヘルペスウイルスの糖蛋白質gE、該糖蛋白質gEの一部、該糖蛋白質gEに由来するペプチド、または1型ウシヘルペスウイルス糖蛋白質のgEとgIの複合物を含む、組成物。
【請求項16】
1型ウシヘルペスウイルスの糖蛋白質gE、該糖蛋白質gEの一部、該糖蛋白質gEに由来するペプチド、または1型ウシヘルペスウイルス糖蛋白質のgEとgIの複合物に特異的な抗体を含む、組成物。
【請求項17】
1型ウシヘルペスウイルスの糖蛋白質gE、該糖蛋白質gEの一部、該糖蛋白質gEに由来するペプチド、または1型ウシヘルペスウイルス糖蛋白質のgEとgIの複合物に特異的なモノクローナル抗体を含む、組成物。
【請求項18】
1型ウシヘルペスウイルスの糖蛋白質gE、該糖蛋白質gEの一部、該糖蛋白質gEに由来するペプチド、または1型ウシヘルペスウイルス糖蛋白質のgEとgIの複合物に特異的なポリクローナル抗体を含む、組成物。
【請求項19】
1型ウシヘルペスウイルスに対して動物を保護するために、動物、特に哺乳動物、さらに特にウシ科の動物に接種するためのワクチン組成物において、請求項1〜12のいずれか1項に記載の1型ウシヘルペスウイルス変異体とそれに適する担体またはアジュバントとを含む、ワクチン組成物。
【請求項20】
病原菌に対して動物を保護するために、動物、特に哺乳動物、さらに特にウシ科の動物に接種するためのワクチン組成物において、糖蛋白質gE-遺伝子中に欠失を有し、且つ組換えDNA技術によって導入され、病原菌の免疫原蛋白質またはペプチドをコードする異種遺伝子を含む1型ウシヘルペスウイルス変異体とそれに適する担体またはアジュバントとを含む、ワクチン組成物。
【請求項21】
動物、特に哺乳動物、さらに特にウシ科の動物から得られる試料、特に血液、血清、血液細胞、乳、涙のような体液、肺洗浄液、鼻水、精液、特に精漿、唾液、痰、または組織、特に神経組織のような生体試料の中で、1型ウシヘルペスウイルスの核酸を検出するための診断キットにおいて、1型ウシヘルペスウイルスの糖蛋白質gE-遺伝子に由来するヌクレオチド配列を有する核酸プローブまたはプライマ、および核酸検出分析に適する検出手段を含む、診断キット。
【請求項22】
動物、特に哺乳動物、さらに特にウシ科の動物から得られる試料、特に血液、血清、唾液、痰、涙のような体液、肺洗浄液、鼻水、乳または組織のような生体試料の中で、1型ウシヘルペスウイルスに特異的な抗体を検出するための診断キットにおいて、1型ウシヘルペスウイルスの糖蛋白質gE、該糖蛋白質gEの一部、該糖蛋白質gEに由来するペプチド、または1型ウシヘルペスウイルス糖蛋白質のgEとgIの複合物、および抗体検出分析に適する検出手段を含む、診断キット。
【請求項23】
1型ウシヘルペスウイルス糖蛋白質gE、または1型ウシヘルペスウイルス糖蛋白質のgEとgIの複合物に特異的な1またはそれ以上の抗体を含む、請求項22記載の診断キット。
【請求項24】
動物、特に哺乳動物、さらに特にウシ科の動物から得られる試料、特に血液、血清、血液細胞、乳、涙のような体液、肺洗浄液、鼻水、精液、特に精漿、唾液、痰、または組織、特に神経組織のような生体試料の中で、1型ウシヘルペスウイルスの蛋白質を検出するための診断キットにおいて、1型ウシヘルペスウイルスの糖蛋白質gE、または1型ウシヘルペスウイルス糖蛋白質のgEとgIの複合物に特異的な抗体、および蛋白質検出分析に適する検出手段を含む、診断キット。
【請求項25】
動物、特に哺乳動物、さらに特にウシ科の動物から得られる試料、特に血液、血清、血液細胞、精液、特に精漿、唾液、痰、涙のような体液、肺洗浄液、鼻水、乳、または組織、特に神経組織のような生体試料について、1型ウシヘルペスウイルスの糖蛋白質gE-遺伝子を含む核酸の存在、または1型ウシヘルペスウイルス糖蛋白質gEもしくは1型ウシヘルペスウイルス糖蛋白質のgEとgIの複合物の存在、または1型ウシヘルペスウイルスの糖蛋白質gEもしくは1型ウシヘルペスウイルス糖蛋白質のgEとgIの複合物に特異的な抗体の存在を試験する工程を含む、動物、特に哺乳動物、さらに特にウシ科の動物における1型ウシヘルペスウイルスの感染を決定する方法。
【請求項26】
動物、特に哺乳動物、さらに特にウシ科の動物から得られる試料、特に血液、血清、血液細胞、精液、特に精漿、唾液、痰、涙のような体液、肺洗浄液、鼻水、乳、または組織、特に神経組織のような生体試料について、1型ウシヘルペスウイルスの糖蛋白質gE-遺伝子を含む核酸の存在、または1型ウシヘルペスウイルス糖蛋白質gEもしくは1型ウシヘルペスウイルス糖蛋白質のgEとgIの複合物の存在、または1型ウシヘルペスウイルスの糖蛋白質gEもしくは1型ウシヘルペスウイルス糖蛋白質のgEとgIの複合物に特異的な抗体の存在を試験する工程を含む、動物、特に哺乳動物、さらに特にウシ科の動物における1型ウシヘルペスウイルスの感染を決定する方法であって、該試料が請求項19に記載のワクチン調製物を接種した動物から得られる方法。
【請求項1】
糖蛋白質gE-遺伝子中に欠失を有する1型ウシヘルペスウイルスの欠失突然変異株。
【請求項2】
糖蛋白質gE-遺伝子中に減衰手段によって起こる欠失を有する1型ウシヘルペスウイルスの欠失突然変異株。
【請求項3】
糖蛋白質gE-遺伝子中に減衰手段によって起こり、Difivac-1である欠失を有する1型ウシヘルペスウイルスの欠失突然変異株。
【請求項4】
糖蛋白質gE-遺伝子中に再結合DNA技術によって起こる欠失を有する1型ウシヘルペスウイルスの欠失突然変異株。
【請求項5】
糖蛋白質gE-遺伝子中に再結合DNA技術によって起こり、IB7または1B8である欠失を有する1型ウシヘルペスウイルスの欠失突然変異株。
【請求項6】
糖蛋白質gE-遺伝子中と、チミジンキナーゼ遺伝子中とに欠失を有する1型ウシヘルペスウイルスの欠失突然変異株。
【請求項7】
糖蛋白質gE-遺伝子中と、同じくgI-遺伝子中とに欠失を有する1型ウシヘルペスウイルスの欠失突然変異株。
【請求項8】
糖蛋白質gE-遺伝子中と、チミジンキナーゼ遺伝子中と、糖蛋白質gI-遺伝子中とに欠失を有する1型ウシヘルペスウイルスの欠失突然変異株。
【請求項9】
糖蛋白質gE-遺伝子中に欠失を有し、再結合DNA技術によって誘導される非対称遺伝子を含む1型ウシヘルペスウイルスの突然変異株。
【請求項10】
糖蛋白質gE-遺伝子の位置に、再結合DNA技術によって誘導され、調節した配列、たとえばgE-遺伝子または非対称遺伝子の制御下にあり、ペプチド信号のためコード化されたgE-遺伝子の部分に随意に接触させられる非対称遺伝子を含む1型ウシヘルペスウイルスの突然変異株。
【請求項11】
糖蛋白質gE-遺伝子中にある欠失に加えて、チミジンキナーゼ遺伝子中と、糖蛋白質gI-遺伝子中との一方または双方に欠失を有し、再結合DNA技術によって、前記欠失の少なくとも1つの位置に誘導される非対称遺伝子を含む1型ウシヘルペスウイルスの突然変異株。
【請求項12】
糖蛋白質gE-遺伝子中に欠失を有し、再結合DNA技術で誘導され、他の病原菌の免疫原蛋白質のためと、サイドカインのためとにコード化された非対称遺伝子を含む1型ウシヘルペスウイルスの突然変異株。
【請求項13】
1型ウシヘルペスウイルスの糖蛋白質gE-遺伝子を含む再結合核酸、この糖蛋白質gE-遺伝子の一部またはこの糖蛋白質gE-遺伝子から誘導されるヌタレオチド配列から成る組成物。
【請求項14】
1型ウシヘルペスウイルスの糖蛋白質gE-遺伝子を含む再結合核酸、この糖蛋白質gE-遺伝子の一部または、この糖蛋白質gE-遺伝子から誘導されるヌクレオチド配列を挿入するクローンまたは表現ベタタから成る組成物。
【請求項15】
1型ウシヘルペスウイルスの糖蛋白質gE、この糖蛋白質gEの一部、この糖蛋白質gEから誘導されるペプチドまたは1型ウシヘルペスウイルス糖蛋白質gEと同gIとの複合物から成る組成物。
【請求項16】
1型ウシヘルペスウイルスの糖蛋白質、この糖蛋白質gEの一部、この糖蛋白質gEから誘導されたペプチドまたは1型ウシヘルペスウイルス糖蛋白質gEおよび同gIの複合物に特有の組成物。
【請求項17】
1型ウシヘルペスウイルスの糖蛋白質、この糖蛋白質gEの一部、この糖蛋白質gEから誘導されたペプチドまたは1型ウシヘルペスウイルス糖蛋白質gEおよび同gIの複合物に特有のモノクロナール抗体から成る組成物。
【請求項18】
1型ウシヘルペスウイルスの糖蛋白質、この糖蛋白質gEの一部、この糖蛋白質gEから誘導されたペプチドまたは1型ウシヘルペスウイルス糖蛋白質gEおよび同gIの複合物に特有のボリクロナール抗体から成る組成物。
【請求項19】
1型ウシヘルペスウイルスに対して動物を保護するために、動物、好ましくは哺乳動物、さらに好ましくはウシ科の動物に接種するための請求項1〜12のいずれか1項に記載の1型ウシヘルペスウイルスの欠失から成るワクチン組成物とそれに適する担体と免疫助成剤。
【請求項20】
病原菌に対して動物を保護するために、動物、好ましくは哺乳動物、さらに好ましくはウシ科の動物に接種するための糖蛋白質gE-遺伝子中に欠失を有し、再結合DNA技術によって誘導され、病原菌の免疫原蛋白質またはペプチドのためにコード化された非対称遺伝子を含む1型ウシヘルペスウイルスの欠失から成るワクチン組成物とそれに適する担体と免疫助成剤。
【請求項21】
動物、好ましくは哺乳動物、さらに好ましくはウシ科の動物から得られる試料、特に血液、血清、血液細胞、乳、涙のような体液、肺洗浄液、鼻水、精液、特に精漿、痰、または、組織、特に神経組織のような生体試料の中で、1型ウシヘルペスウイルスの糖蛋白質gE-遺伝子から誘導されたヌタレオチド配列を有する核酸プローブまたはプライマから成り、1型ウシヘルペスウイルスの核酸を検出するための診断キットおよび核酸を分析するために適する検出方法。
【請求項22】
動物、好ましくは哺乳動物、さらに好ましくはウシ科の動物から得られる試料、特に血液、血清、唾液、痰、涙のような体液、肺洗浄液、鼻水、乳または組織のような生体試料の中で、BHV-1の糖蛋白質gE、この糖蛋白質gEの一部、この糖蛋白質gEから誘導されるペプチド、または1型ウシヘルペスウイルス糖蛋白質gEおよび同gIの複合物から成り、1型ウシヘルペスウイルスに特有の抗体を検出するための診断キットおよび抗体を分析するために適する検出方法。
【請求項23】
1型ウシヘルペスウイルス糖蛋白質gEまたは1型ウシヘルペスウイルス糖蛋白質gEおよび同gIの複合物に特有の1またはそれ以上の抗体から成る請求項22記載の診断キット。
【請求項24】
動物、好ましくは哺乳動物、さらに好ましくはウシ科の動物から得られる試料、特に血液、血清、血液細胞、乳、涙のような体液、肺洗浄液、鼻水、精液、特に精漿、唾液、痰、または組織、特に神経組織のような生体試料の中で、1型ウシヘルペスウイルスの糖蛋白質gE、または1型ウシヘルペスウイルスgEおよび同gIの複合物に特有の抗体から成り、1型ウシヘルペスウイルスの蛋白質を検出するための診断キットおよび蛋白質を分析するために適する検出方法。
【請求項25】
動物由来の試料、特に血液、血清、血液細胞、精液、特に精漿、唾液、痰、涙のような体液、肺洗浄液、鼻水、乳、組織、特に神経組織のような生体試料を試験することと、1型ウシヘルペスウイルスの糖蛋白質gE-遺伝子から成る核酸の存在、または1型ウシヘルペスウイルス糖蛋白質gEもしくは1型ウシヘルペスウイルス糖蛋白質gEと同gIとの複合物の存在、または1型ウシヘルペスウイルスの糖蛋白質もしくは1型ウシヘルペスウイルス糖蛋白質gEと同gIの複合物のための特有の抗体の存在を試験することとから成る1型ウシヘルペスウイルスに感染した動物、好ましくは哺乳動物、さらに好ましくはウシ科の動物を決める方法。
【請求項26】
動物由来の試料、特に血液、血清、血液細胞、精液、特に精漿、唾液、痰、涙のような体液、肺洗浄液、鼻水、乳、組織、特に神経組織のような生体試料を試験することと、1型ウシヘルペスウイルスの糖蛋白質gE-遺伝子から成る核酸の存在、または1型ウシヘルペスウイルス糖蛋白質gEもしくは1型ウシヘルペスウイルス糖蛋白質gEと同gIとの複合物の存在、または1型ウシヘルペスウイルスの糖蛋白質もしくは1型ウシヘルペスウイルス糖蛋白質gEと同gIの複合物に特有の抗体の存在を試験することと、請求項19によって準備されたワクチンを接種した動物から得られた試料を試験することとから成る1型ウシヘルペスウイルスに感染した動物、好ましくは哺乳動物、さらに好ましくはウシ科の動物を決める方法。
【特許請求の範囲】
【請求項1】
糖蛋白質gE-遺伝子中に欠失を有する、1型ウシヘルペスウイルスの欠失突然変異体。
【請求項2】
糖蛋白質gE-遺伝子中に弱毒処置によって起こる欠失を有する、1型ウシヘルペスウイルスの欠失突然変異体。
【請求項3】
糖蛋白質gE-遺伝子中に弱毒処置によって起こる欠失を有する、1型ウシヘルペスウイルスの欠失突然変異体 Difivac-1。
【請求項4】
糖蛋白質gE-遺伝子中に組換えDNA技術によって構築された欠失を有する、1型ウシヘルペスウイルスの欠失突然変異体。
【請求項5】
糖蛋白質gE-遺伝子中に組換えDNA技術によって構築された欠失を有する、1型ウシヘルペスウイルスの欠失突然変異体 1B7または1B8。
【請求項6】
糖蛋白質gE-遺伝子中と、チミジンキナーゼ遺伝子中とに欠失を有する、1型ウシヘルペスウイルスの欠失突然変異体。
【請求項7】
糖蛋白質gE-遺伝子中と、糖蛋白質gI-遺伝子中とに欠失を有する、1型ウシヘルペスウイルスの欠失突然変異体。
【請求項8】
糖蛋白質gE-遺伝子中と、チミジンキナーゼ遺伝子中と、糖蛋白質gI-遺伝子中とに欠失を有する、1型ウシヘルペスウイルスの欠失突然変異体。
【請求項9】
糖蛋白質gE-遺伝子中に欠失を有し、且つ組換えDNA技術によって導入される異種遺伝子を含む、1型ウシヘルペスウイルスの突然変異体。
【請求項10】
糖蛋白質gE-遺伝子の位置に、組換えDNA技術によって導入された異種遺伝子を含む、1型ウシヘルペスウイルスの突然変異体であって、該異種遺伝子が、調節配列、たとえばgE-遺伝子または異種遺伝子の調節配列の制御下にあり、シグナルペプチドをコードするgE-遺伝子の部分に任意で付着される、1型ウシヘルペスウイルスの突然変異体。
【請求項11】
糖蛋白質gE-遺伝子中にある欠失に加えて、チミジンキナーゼ遺伝子中と、糖蛋白質gI-遺伝子中との一方または双方に欠失を有し、且つ組換えDNA技術によって、前記欠失の少なくとも1つの位置に導入される異種遺伝子を含む、1型ウシヘルペスウイルスの突然変異体。
【請求項12】
糖蛋白質gE-遺伝子中に欠失を有し、且つ組換えDNA技術で導入され、他の病原菌の免疫原蛋白質もしくはペプチドをコードする、またはサイトカインをコードする異種遺伝子を含む、1型ウシヘルペスウイルスの突然変異体。
【請求項13】
1型ウシヘルペスウイルスの糖蛋白質gE-遺伝子、該糖蛋白質gE-遺伝子の一部、または該糖蛋白質gE-遺伝子に由来するヌクレオチド配列を含む組換え核酸を含む、組成物。
【請求項14】
1型ウシヘルペスウイルスの糖蛋白質gE-遺伝子、該糖蛋白質gE-遺伝子の一部、または該糖蛋白質gE-遺伝子に由来するヌクレオチド配列を含む組換え核酸を挿入するクローニングベクタまたは発現ベクタを含む、組成物。
【請求項15】
1型ウシヘルペスウイルスの糖蛋白質gE、該糖蛋白質gEの一部、該糖蛋白質gEに由来するペプチド、または1型ウシヘルペスウイルス糖蛋白質のgEとgIの複合物を含む、組成物。
【請求項16】
1型ウシヘルペスウイルスの糖蛋白質gE、該糖蛋白質gEの一部、該糖蛋白質gEに由来するペプチド、または1型ウシヘルペスウイルス糖蛋白質のgEとgIの複合物に特異的な抗体を含む、組成物。
【請求項17】
1型ウシヘルペスウイルスの糖蛋白質gE、該糖蛋白質gEの一部、該糖蛋白質gEに由来するペプチド、または1型ウシヘルペスウイルス糖蛋白質のgEとgIの複合物に特異的なモノクローナル抗体を含む、組成物。
【請求項18】
1型ウシヘルペスウイルスの糖蛋白質gE、該糖蛋白質gEの一部、該糖蛋白質gEに由来するペプチド、または1型ウシヘルペスウイルス糖蛋白質のgEとgIの複合物に特異的なポリクローナル抗体を含む、組成物。
【請求項19】
1型ウシヘルペスウイルスに対して動物を保護するために、動物、特に哺乳動物、さらに特にウシ科の動物に接種するためのワクチン組成物において、請求項1〜12のいずれか1項に記載の1型ウシヘルペスウイルス変異体とそれに適する担体またはアジュバントとを含む、ワクチン組成物。
【請求項20】
病原菌に対して動物を保護するために、動物、特に哺乳動物、さらに特にウシ科の動物に接種するためのワクチン組成物において、糖蛋白質gE-遺伝子中に欠失を有し、且つ組換えDNA技術によって導入され、病原菌の免疫原蛋白質またはペプチドをコードする異種遺伝子を含む1型ウシヘルペスウイルス変異体とそれに適する担体またはアジュバントとを含む、ワクチン組成物。
【請求項21】
動物、特に哺乳動物、さらに特にウシ科の動物から得られる試料、特に血液、血清、血液細胞、乳、涙のような体液、肺洗浄液、鼻水、精液、特に精漿、唾液、痰、または組織、特に神経組織のような生体試料の中で、1型ウシヘルペスウイルスの核酸を検出するための診断キットにおいて、1型ウシヘルペスウイルスの糖蛋白質gE-遺伝子に由来するヌクレオチド配列を有する核酸プローブまたはプライマ、および核酸検出分析に適する検出手段を含む、診断キット。
【請求項22】
動物、特に哺乳動物、さらに特にウシ科の動物から得られる試料、特に血液、血清、唾液、痰、涙のような体液、肺洗浄液、鼻水、乳または組織のような生体試料の中で、1型ウシヘルペスウイルスに特異的な抗体を検出するための診断キットにおいて、1型ウシヘルペスウイルスの糖蛋白質gE、該糖蛋白質gEの一部、該糖蛋白質gEに由来するペプチド、または1型ウシヘルペスウイルス糖蛋白質のgEとgIの複合物、および抗体検出分析に適する検出手段を含む、診断キット。
【請求項23】
1型ウシヘルペスウイルス糖蛋白質gE、または1型ウシヘルペスウイルス糖蛋白質のgEとgIの複合物に特異的な1またはそれ以上の抗体を含む、請求項22記載の診断キット。
【請求項24】
動物、特に哺乳動物、さらに特にウシ科の動物から得られる試料、特に血液、血清、血液細胞、乳、涙のような体液、肺洗浄液、鼻水、精液、特に精漿、唾液、痰、または組織、特に神経組織のような生体試料の中で、1型ウシヘルペスウイルスの蛋白質を検出するための診断キットにおいて、1型ウシヘルペスウイルスの糖蛋白質gE、または1型ウシヘルペスウイルス糖蛋白質のgEとgIの複合物に特異的な抗体、および蛋白質検出分析に適する検出手段を含む、診断キット。
【請求項25】
動物、特に哺乳動物、さらに特にウシ科の動物から得られる試料、特に血液、血清、血液細胞、精液、特に精漿、唾液、痰、涙のような体液、肺洗浄液、鼻水、乳、または組織、特に神経組織のような生体試料について、1型ウシヘルペスウイルスの糖蛋白質gE-遺伝子を含む核酸の存在、または1型ウシヘルペスウイルス糖蛋白質gEもしくは1型ウシヘルペスウイルス糖蛋白質のgEとgIの複合物の存在、または1型ウシヘルペスウイルスの糖蛋白質gEもしくは1型ウシヘルペスウイルス糖蛋白質のgEとgIの複合物に特異的な抗体の存在を試験する工程を含む、動物、特に哺乳動物、さらに特にウシ科の動物における1型ウシヘルペスウイルスの感染を決定する方法。
【請求項26】
動物、特に哺乳動物、さらに特にウシ科の動物から得られる試料、特に血液、血清、血液細胞、精液、特に精漿、唾液、痰、涙のような体液、肺洗浄液、鼻水、乳、または組織、特に神経組織のような生体試料について、1型ウシヘルペスウイルスの糖蛋白質gE-遺伝子を含む核酸の存在、または1型ウシヘルペスウイルス糖蛋白質gEもしくは1型ウシヘルペスウイルス糖蛋白質のgEとgIの複合物の存在、または1型ウシヘルペスウイルスの糖蛋白質gEもしくは1型ウシヘルペスウイルス糖蛋白質のgEとgIの複合物に特異的な抗体の存在を試験する工程を含む、動物、特に哺乳動物、さらに特にウシ科の動物における1型ウシヘルペスウイルスの感染を決定する方法であって、該試料が請求項19に記載のワクチン調製物を接種した動物から得られる方法。
【図2】

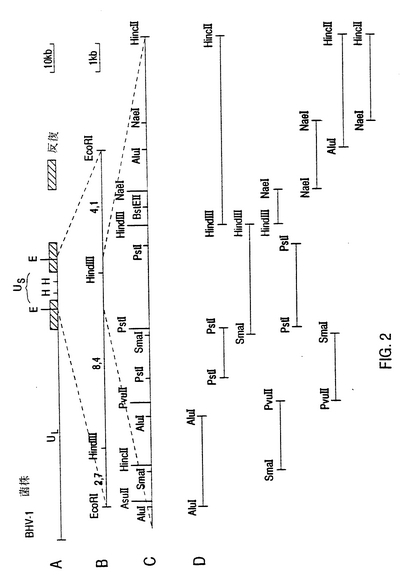
【図3A1】

【図3A2】

【図3A3】

【図3A4】

【図3A5】

【図3A6】

【図3B】
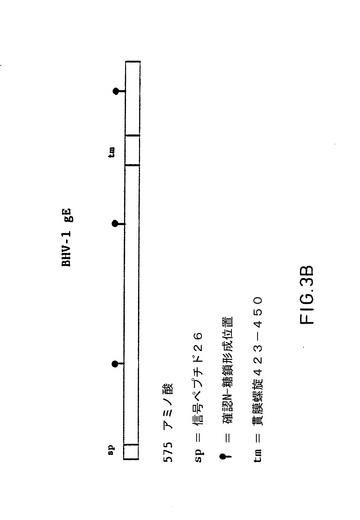
【図4A】
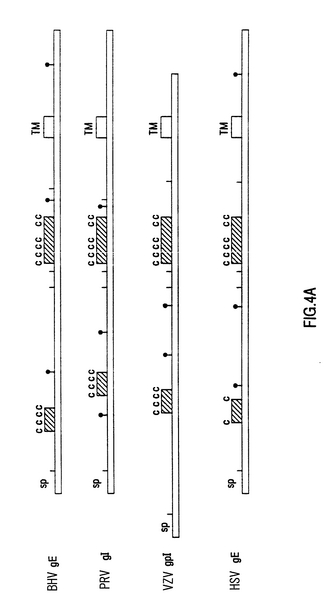
【図4B】

【図5】

【図6】

【図7A】
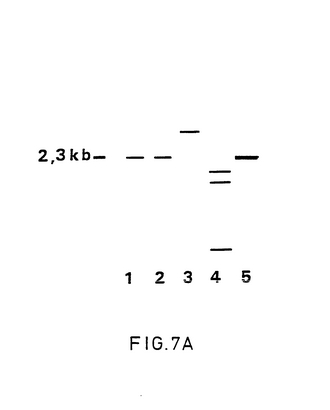
【図7B】

【図8A】
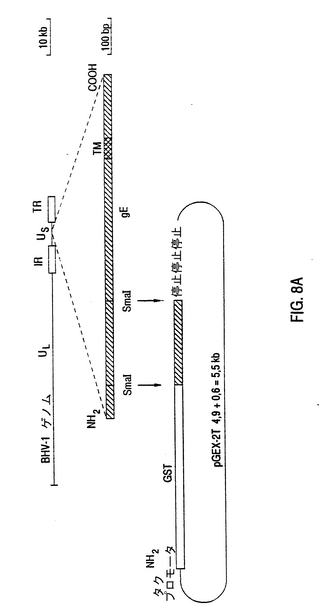
【図8B】

【図9】
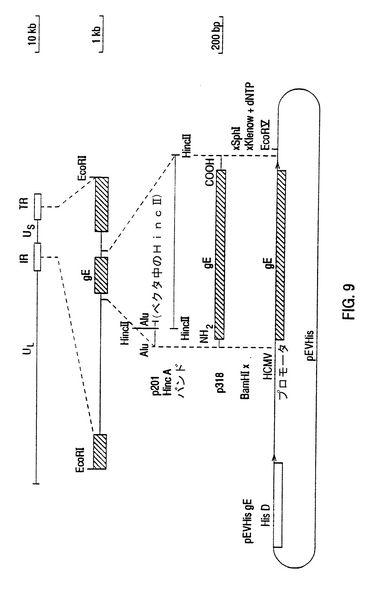
【図10】
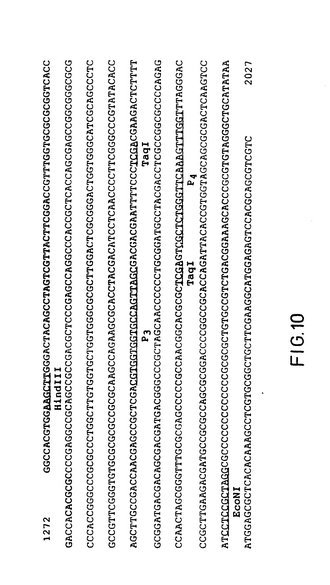
【図11】
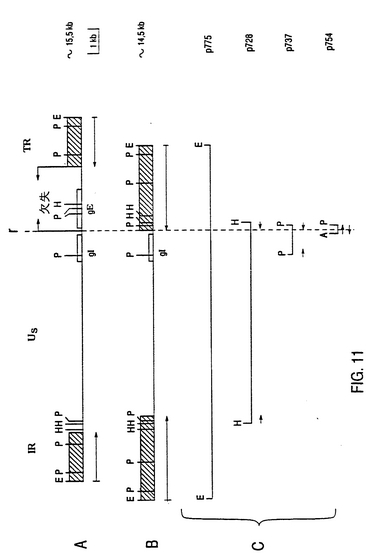
【図12】

【図13】
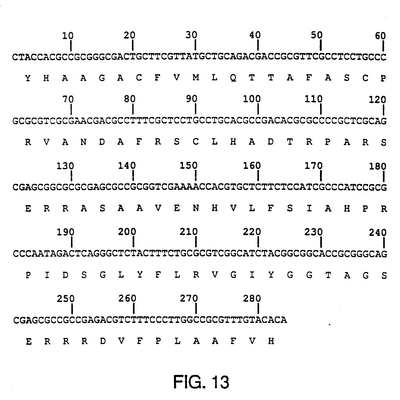
【図14】

【図15】
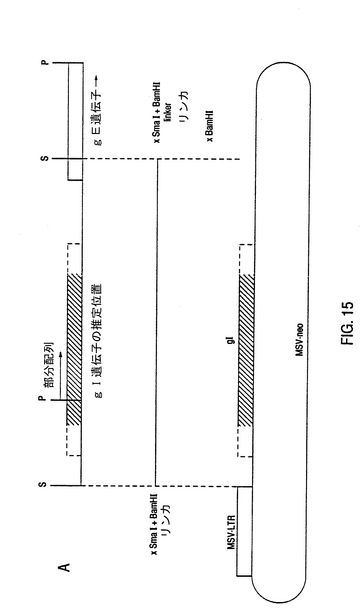
【図16】
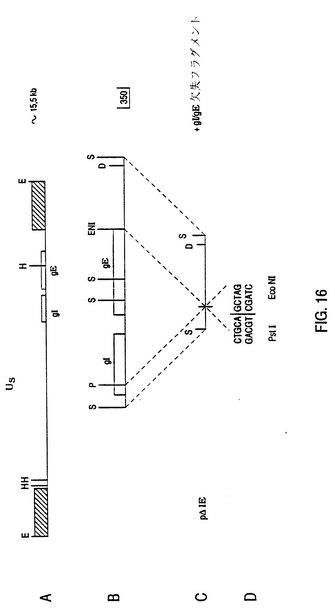
【図17】

【図18】

【図19】

【図20】
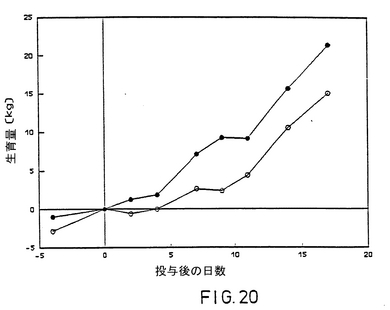
【図21】
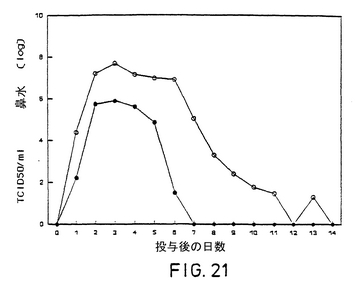
【図22】

【図23】
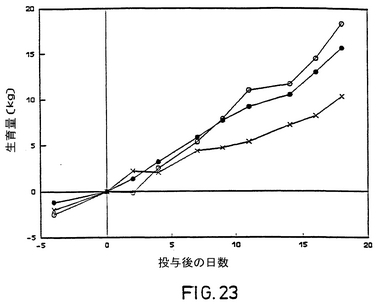
【図24】
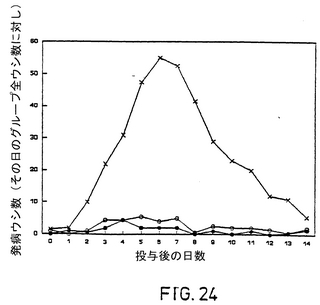
【図3A1】
【図3A2】
【図3A3】
【図3A4】
【図3A5】
【図3A6】
【図3B】
【図4A】
【図4B】
【図5】
【図6】
【図7A】
【図7B】
【図8A】
【図8B】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【図19】
【図20】
【図21】
【図22】
【図23】
【図24】
【公開番号】特開2006−296426(P2006−296426A)
【公開日】平成18年11月2日(2006.11.2)
【国際特許分類】
【出願番号】特願2006−109215(P2006−109215)
【出願日】平成18年4月11日(2006.4.11)
【分割の表示】特願平5−500368の分割
【原出願日】平成4年6月5日(1992.6.5)
【出願人】(500012318)
【Fターム(参考)】
【公開日】平成18年11月2日(2006.11.2)
【国際特許分類】
【出願日】平成18年4月11日(2006.4.11)
【分割の表示】特願平5−500368の分割
【原出願日】平成4年6月5日(1992.6.5)
【出願人】(500012318)
【Fターム(参考)】
[ Back to top ]