
13Cメチル基標識したタンパク質を合成する方法
【課題】大腸菌、酵母のタンパク質合成系等では、安定同位体を標識したタンパク質自体が合成できない又はリフォールディングしていない状態の安定同位体を標識したタンパク質が発現されることが多数報告されている。
【解決手段】13C標識したα-ケトイソ吉草酸又はα-ケト酪酸を含む培地を用いて本発明の合成系でタンパク質を発現させたところ、他の合成系では困難であった立体構造を維持した13Cメチル基標識したバリン、ロイシン及び/又はイソロイシンを含むタンパク質を合成できることを新規に見出し、本発明を完成した。
【解決手段】13C標識したα-ケトイソ吉草酸又はα-ケト酪酸を含む培地を用いて本発明の合成系でタンパク質を発現させたところ、他の合成系では困難であった立体構造を維持した13Cメチル基標識したバリン、ロイシン及び/又はイソロイシンを含むタンパク質を合成できることを新規に見出し、本発明を完成した。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、立体構造が維持された13Cメチル基標識したバリン、ロイシン及び/又はイソロイシンを含むタンパク質を合成する方法に関する。
【背景技術】
【0002】
{核磁気共鳴(NMR)について}
原子レベルの解像度でのタンパク質の構造、動力学、相互作用、立体構造変化等の研究には、NMRが使用されている。1980年代末まで、1H−NMR実験を使用するタンパク質の構造決定は小分子(<10kDa)に限定されていた。しかし、最近では、均一な13C及び/又は15N標識法、並びに多次元的NMR技法が、20〜25kDaの分子量も解析することができ、特に、NMRでの大型のタンパク質及びそれらの複合体(>25kDa)の構造及び動力学を研究するための、2H、13C、および15Nを使用する高性能で安定した同位体標識法が提案されている。
【0003】
初めての三重共鳴三次元NMR実験の証明の試料として、大腸菌系で発現する、均一に13C−及び15Nで二重標識したCa2+−結合タンパク質カルモジュリン(CaM)が使用された。それ以来、新規なNMR実験の開発と並行して、大腸菌を使用した様々な改変改善された試料調製法が提案されている。
【0004】
大腸菌を使用したタンパク質合成法(大腸菌合成系)は、NMR試料調製には非常に一般的となった。しかし、大腸菌合成系により合成できるタンパク質は制限されている。特に、大腸菌合成系で発現する多くの真核生物タンパク質は、正確に折り畳まれず、本来の翻訳後修飾ができない、さらには大腸菌合成系では真核生物タンパク質自体が合成できないという問題があった。
【0005】
(タバコBY−2懸濁液培養細胞)
タバコBY−2懸濁液培養細胞は、基礎および応用植物科学で使用される最も人気がある植物細胞系の一つである。タバコBY−2懸濁液培養細胞の特性は、幅広く活用されており、NMR試料の調製に適している。BY−2用Linsmaier-Skoog培地は調製が容易であり、動物細胞および無細胞抽出物に使用される培地よりもはるかに経済的である。
しかし、タバコBY−2を使用する既知のタンパク質発現方法は、タンパク質の生産性に関して重大な問題があった。タバコBY−2細胞によって得られる外来タンパク質の発現レベルは低く、NMR試料への使用は制限されている。
【0006】
本発明者の1人である森正之博士らは、ウイルスベクターを使用することによって、懸濁液培養BY−2細胞のタンパク質の生産性を著しく改善することに成功した(参照:非特許文献1)。この改善したタンパク質合成系では、煩雑なウイルス接種手順は不要である。エストラジオールを該系に添加することで、効率良くウイルスRNAの蓄積が誘導される。さらに、該誘導に続いて、タバコBY−2懸濁液培養細胞で組換えタンパク質が効率良く大量に合成される(参照:特許文献1〜4)。
【0007】
また、本発明者らは、上記タバコBY−2懸濁液培養細胞を使用したタンパク質合成系を用いて15N標識タンパク質を合成できたことを報告している(参照:非特許文献2)。 しかし、該文献は、「13Cメチル基標識したバリン、ロイシン及び/又はイソロイシンを含むタンパク質を合成する方法」の開示又は示唆がない。
【先行技術文献】
【特許文献】
【0008】
【特許文献1】特開2011−83232
【特許文献2】特開2005−245228
【特許文献3】WO2008/136253
【特許文献4】特開2005−237301
【特許文献5】特開2005−102652
【0009】
【非特許文献1】Dohi K. et al(2006) Arch Virol 151:1075-1084
【非特許文献2】Ohki S. etal(2008) J Biomol NMR 42:271-277
【非特許文献3】Amy M. etal(2010) J Biomol NMR 46:75-87
【発明の概要】
【発明が解決しようとする課題】
【0010】
多数のタンパク質合成系が開発されているが、NMR試料を調製する普遍的方法は、現在まで確立されていない。また、大腸菌、酵母のタンパク質合成系等では、安定同位体を標識したタンパク質自体が合成できない又はリフォールディングしていない状態の(正しい立体構造を維持していない)安定同位体を標識したタンパク質が発現されることが多数報告されている。
【課題を解決するための手段】
【0011】
本発明者らは、上記課題を解決するために、13C標識したα-ケトイソ吉草酸又はα-ケト酪酸を含む培地を用いて本発明の合成系でタンパク質を発現させたところ、他の合成系では困難であった立体構造を維持した13Cメチル基標識したバリン、ロイシン及び/又はイソロイシンを含むタンパク質を合成できることを新規に見出し、本発明を完成した。
【0012】
すなわち、本発明は以下の通りである。
「1.以下の工程を含む13Cメチル基標識したバリン、ロイシン及び/又はイソロイシンを含むタンパク質の合成方法:
(1)ホルモンによって活性化される性質を有する転写因子をコードする遺伝子と、転写因子発現用プロモーターとが連結されてなる転写因子発現用DNA断片を宿主細胞に導入する第一形質転換体を作成する工程;
(2)RNAを遺伝子とするウイルスに標的タンパク質をコードする遺伝子を挿入したウイルスベクターのcDNAと、該転写因子で転写誘導される転写誘導型プロモーターとが連結されてなる目的タンパク質発現用DNA断片を、該第一形質転換体に導入して第二形質転換体を作成する工程;
(3)該第二形質転換体をα-ケトイソ吉草酸及び/又はα-ケト酪酸を含む培地で培養して、13C標識タンパク質を得る工程、ここで、該ホルモンは培養前及び/又は培養中に該培地に添加されている。
2.前記α-ケトイソ吉草酸及び/又はα-ケト酪酸を含む培地において、α-ケトイソ吉草酸及びα-ケト酪酸の濃度が、それぞれ、100mg〜1000mg/Lである前項1の合成方法。
3.前記α-ケトイソ吉草酸及び/又はα-ケト酪酸を含む培地において、α-ケトイソ吉草酸及びα-ケト酪酸の濃度が、それぞれ、150mg〜350mg/Lである前項1又は2の合成方法。
4.前記標的タンパク質が、分子内にジスルフィド結合を有する前項1〜3のいずれか1の合成法。
5.分子内にジスルフィド結合を有するタンパク質が、以下のいずれか1から選ばれる前項4の合成方法。
(1)単鎖抗体
(2)分泌タンパク質
(3)膜タンパク質
6.前記標的タンパク質が、以下のいずれか1から選ばれる前項1〜3のいずれか1の合成法。
(1)ウシ膵臓トリプシン阻害薬(BPTI)
(2)ストマジェン(stomagen)
(3)EPF2(EPIDERMAL PATTERNING FACTOR 2)
(4)ストレプトマイセス・スブチリシン阻害剤(SSI)
(5)AFP1(alpha-fetoprotein)
7.前記ホルモンが、エストロジェンまたはステロイドホルモンである前項1〜6のいずれか1の合成法。
8.前記エストロジェンで活性化される性質を有する転写因子としてLexA−VP16−hERを用い、上記転写誘導型プロモーターとしてOLexA−46を用いる前項7の合成法。
9.前記ウイルスベクターが、植物ウイルス由来である前項8の合成法。
10.前記宿主細胞および形質転換体が、植物体又は植物由来培養細胞である前項1〜9のいずれか1の合成法。
11.前記ホルモンがエストロジェンであり、
前記宿主細胞がタバコBY2細胞であり、
前記転写因子がLexA−VP16−hERであり、
前記転写誘導型プロモーターがOLexA−46であり、
前記ウイルスベクターがトバモウイルス属に属するウイルス由来である、ことを特徴とする、前項1〜10のいずれか1の合成法。
12.以下を含む13Cメチル基標識したバリン、ロイシン及び/又はイソロイシンを含むタンパク質の合成用キット。
(1)形質転換したBY2細胞
(2)α-ケトイソ吉草酸及び/又はα-ケト酪酸」
【発明の効果】
【0013】
本発明の13C標識タンパク質合成法では、他の合成系では困難であった立体構造を維持したタンパク質を合成することができ、さらにバリン、ロイシン、イソロイシンのメチル基を明瞭に13C標識することができる。
【図面の簡単な説明】
【0014】
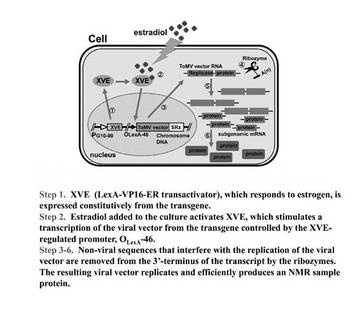
【図1】本発明のタンパク質の合成系の概要
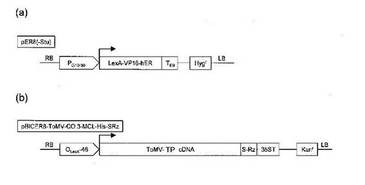
【図2】(a)転写因子発現用DNAの一例、(b)目的タンパク質発現用DNA断片の一例
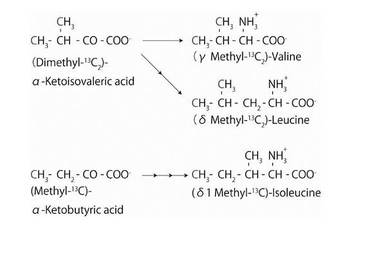
【図3】培地に添加される13C標識α-ケトイソ吉草酸又はα-ケト酪酸並びに生成される13C標識アミノ酸
【図4】均一13C標識したBPTIのNMRスペクトル
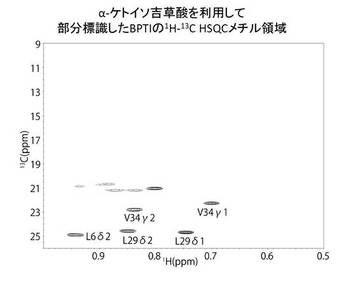
【図5】13Cメチル基標識したバリン及びロイシンを含むBPTIのNMRスペクトル
【図6】13Cメチル基標識したイソロイシンを含むBPTIのNMRスペクトル
【図7】各αケトイソ吉草酸又はαケト酪酸濃度で合成したBPTIの合成量、なお「Con」は13C標識したα-ケトイソ吉草酸又はα-ケト酪酸を含まない培地で合成したBPTIの合成量
【発明を実施するための形態】
【0015】
(本発明のタンパク質合成系)
本発明の13C標識タンパク質を合成する方法で使用するタンパク質合成系は、特許文献1〜5及び非特許文献1、2に記載の合成系を利用することができる。また、図1のタンパク質の合成系の概要図を参照することができる。
本合成系の一例について下記で具体的に説明する。
【0016】
(A)13C標識タンパク質生産用形質転換体の生産方法
本発明に係る13C標識タンパク質生産用第一形質転換体の生産方法は、(1)ホルモンによって活性化される性質を有する転写因子をコードする遺伝子と転写因子発現用プロモーターとが連結されてなる転写因子発現用DNA断片を細胞に導入する第一形質転換体作成工程と、必要に応じて(2)該第一形質転換体の中から該転写因子を発現する形質転換体を選抜する選抜工程と、(3)RNAを遺伝子とするウイルスに目的タンパク質をコードする遺伝子を挿入したウイルスベクターのcDNAと、該転写因子で転写誘導される転写誘導型プロモーターとが連結されてなる目的タンパク質発現用DNA断片を、第一形質転換体に導入して第二形質転換体を作成する工程と、を含む。以下、工程ごとに説明する。
【0017】
(A−1)第一形質転換体作成工程
第一形質転換体作成工程は、ホルモンによって活性化される性質を有する転写因子をコードする遺伝子と、該転写因子の発現を担うプロモーター(転写因子発現用プロモーター)とを連結して構築したDNA断片(転写因子発現用DNA断片)を、適当な宿主細胞に導入する工程である。
該工程によって、後述するウイルスベクター発現用プロモーターの転写誘導を行なう転写因子を安定的に高発現する形質転換体(細胞)の生産(作出)を行なう。すなわち宿主細胞の転写因子の発現に最適な染色体上の位置に、該転写因子をコードする遺伝子が組み込まれた形質転換体(細胞)を生産(作出)する工程である。
【0018】
<転写因子発現用DNA断片>
転写因子発現用DNA断片は、転写因子発現用プロモーターの下流に転写因子をコードする遺伝子が連結されている。また、該遺伝子の他、ベクター配列、ターミネーター、薬剤耐性マーカー等のDNAセグメントが含まれていてもよい。かかる転写因子発現用DNA断片の構築方法は、通常の遺伝子工学的手法を用いて行なえばよい。
【0019】
なお、転写因子発現用プロモーターは、転写因子を発現することが可能なものであれば特に限定されるものではなく、また恒常的にプロモーター活性を有するもの(以下、恒常的プロモーターと称する)であってもよいし、さらに転写因子によってプロモーター活性が誘導されるものであってもよい。ただし、転写因子の発現をさらに別の転写因子で制御することは、目的タンパク質の発現系自体が複雑化すること、コスト面で不利等の理由により、上記転写発現用プロモーターとしては、恒常的プロモーターの方がより好ましいといえる。恒常的プロモーターの例としては、カリフラワーモザイクウイルス35Sプロモーター、PG10−90(Ishige, F., Takaichi, M., Foster, R.,Chua, N. H. and Oeda, K.(1999)A G-box motif(GCCACGTGCC) tetramer confers high-level constitutive expressionin dicot and monocot plants. Plant J. 20, 127-133. 参照)、ユビキチンプロモーター、アクチンプロモーター等が挙げられる。
【0020】
一方、使用する転写因子は、ホルモンによって活性化される性質を有する転写因子であって、後述する第二形質転換体作成工程において導入する目的タンパク質発現用DNA断片中に含まれるプロモーターを転写誘導するものであれば特に限定されない。転写因子を活性化するホルモンとしては、エストロジェン(好ましくはエストラジオール、より好ましくはβエストラジオール)、ステロイドホルモン、エクジソン等が挙げられる。かかる転写因子は、ホルモンが存在しない状態においては不活性型でありプロモーターの転写誘導を行なうことができないが、ホルモンが存在することによって活性型に変化しプロモーターの転写誘導を行なうことができる。この性質を利用すれば目的タンパク質の生産をより綿密に制御できるため、より安全に目的タンパク質の生産を行なうことができるといえる。すなわち、目的タンパク質生産を行なう必要がないとき、あるいは目的タンパク質生産を行なってはいけないときには、合成系にホルモンを添加しなければよい。
【0021】
ホルモンによって活性化される性質を有する転写因子、および該転写因子によって転写誘導されるプロモーターの組み合わせとしては、例えば、ステロイドホルモンで活性化される転写因子であるGVGと該転写因子によって転写誘導されるプロモーターである6xUASgal4の組み合わせ、エストロジェンで活性化される転写因子LexA−VP16−hERと該転写因子によって転写誘導可能なプロモーターであるOLexA−46(Zuo J, Niu QW, Chua NH. "An estrogen receptor-basedtransactivator XVE mediates highly inducible gene exprssion in transgenicplants." Plant J.2000, 24: 265-273 参照)の組み合わせ、エクジソンで活性化される転写因子であるエクジソンレセプターGR Act and DBDとヘルペスウイルストランスアクチベーションドメインHecR LBDのキメラタンパク質と該転写因子によって転写誘導可能なプロモーターGREの組み合わせ等が挙げられる。ただし、宿主として植物体および植物細胞を利用する場合においては、活性化に用いるホルモンによる宿主への悪影響が少ないという理由からエストロジェンで活性化される転写因子LexA−VP16−hERと該転写因子によって転写誘導可能なプロモーターであるOLexA−46の組み合わせが好ましい。
【0022】
図2(a)に転写因子発現用DNAの一例を示す。図2(a)は、形質転換用ベクターであるTiプラスミドpER8(−Stu)の一部分を示している。同図中左から転写因子発現用の恒常的プロモーターPG10−90、その下流にエストロジェンレセプターを含む融合転写因子LexA−VP16−hER、その3'末端にターミネーター配列TE9、またその下流に薬剤耐性マーカーとしてハイグロマイシン耐性遺伝子Hygrがある。
【0023】
<転写因子発現用DNA断片を導入する宿主細胞およびDNA断片導入方法>
上記転写因子発現用DNA断片が導入される宿主細胞は特に限定されるものではなく、動物由来細胞であっても植物由来細胞であってもよい。ただし、植物由来細胞は、動物由来細胞に比して、増殖速度が速くコンタミネーションのリスクが少ない点、培地作成費用が非常に安価であるという点において、植物由来細胞がより好ましい。なお、動物由来細胞および植物由来細胞とは、細胞、組織、並びに器官も含む意味である。特に液体培地等で培養可能な細胞(培養細胞)が好ましい。
【0024】
また由来となる動物としては、特に限定されるものでないが、ヒト、サル、イヌ、ヒツジ、ヤギ、ウサギ、マウス、ラット、モルモット、チャイニーズハムスター、ウシ、ウマ、ブタ、メダカやゼブラフィッシュ等の魚類、カイコ、夜蛾(Spodoptera frugiperda)等が挙げられる。一方由来となる植物としては、特に限定されるものではないが、イネ、シロイナズナ、オオムギ、コムギ、タバコ、トマト、キュウリ、ダイズ、ジャガイモ、トウモロコシ、ニチニチソウ、シロイヌナズナ、アルファルファが挙げられる。その他、枯草菌や乳酸菌などの菌類、酵母など単細胞生物が宿主細胞として利用できる。
【0025】
また動物由来細胞の例としては特に限定されるものではないが、HeLa細胞、CHO細胞、メラノーマ細胞、マウス3T3細胞が挙げられる。また植物由来細胞の例としては、タバコBY2細胞、ジャガイモ由来、イネ由来、サツマイモ由来、ダイズ由来、パセリ由来、シロイヌナズナ由来、コムギ由来、トウモロコシ由来細胞、ニチニチソウ由来細胞が挙げられる。
【0026】
後述する実施例において、タバコBY2細胞を宿主として用いている。タバコBY2細胞(Toshiyuki nagata, Yasuyuki Nemoto, and Seiichiro Hasezawa"Tobacco BY-2 Cell Line as the "Hela" Cell in the Cell Biologyof Higher Plants" International Review of cytology, vol.132, p.p. 1-30(1992) 、およびhttp://www.riken.go.jp/r-world/info/release/press/2003/030620/ 参照)は、植物培養細胞株としては、世界中で最も広く用いられているものであり、最も増殖速度が速いこと、遺伝子操作が容易なこと、大量培養を容易に行うことができるという理由から好ましい。
【0027】
また、転写因子発現用DNA断片を宿主細胞へ導入する形質転換方法は特に限定されるものではなく、宿主細胞の種類に応じた適切な形質転換方法を用いればよい。例えば、特別なベクター配列を有しないDNA断片を導入する場合には、エレクトロポレーション法、パーティクルガン法、リン酸カルシウム法等を用いることができる。また植物由来細胞への一般的な形質転換法としては、アグロバクテリウムを用いた形質転換法(アグロバクテリウム法)を挙げることができ、本発明でもアグロバクテリウム法を好適に用いることができる。ただしアグロバクテリウム法を用いて形質転換する場合には、本発明にかかるDNA断片を含むTiプラスミドを構築する必要がある。またその他プロトプラスト/スフェロプラスト法、リン酸カルシウム法、リポソーム法、DEAEデキストラン法等の従来公知の方法を好適に用いることができる。
【0028】
転写因子発現用DNA断片は、宿主細胞のゲノムに組み込まれることが好ましい。転写因子発現用DNA断片がゲノムに組み込まれることにより、細胞分裂後の娘細胞にもベクターの構成に含まれる遺伝子を確実に伝達することが可能となり、転写因子(タンパク質)の生産効率を維持することが可能となるからである。ゲノムは染色体(核ゲノム)に限定されるものではなく、ミトコンドリアゲノムや葉緑体ゲノムも含まれる。
【0029】
また、転写因子発現用DNA断片が宿主細胞に導入されたか否かを確認する方法は、特に限定されるものではなく、公知の各種の方法を用いることができる。具体的には、各種マーカーを用いればよい。例えば、宿主細胞中で欠失している遺伝子をマーカーとして用い、このマーカーと組み換え植物ウイルス遺伝子とを含むプラスミド等を発現ベクターとして宿主細胞に導入する。これによってマーカー遺伝子の発現から本発明の遺伝子の導入を確認することができる。例えば、薬剤耐性マーカー(ハイグロマイシン耐性遺伝子、Hygr)を用いれば、ハイグロマイシンを含有する培地中で、形質転換候補株を培養することにより、生育してきた細胞株を形質転換体として選抜することが可能となる。
その他のマーカーとしては、ビアラホス耐性マーカー、カナマイシン耐性マーカー等が植物細胞の選抜に有効であり、ピューロマイシン耐性マーカー、ブレオマイシン耐性マーカー、XGPRT遺伝子、DHFR遺伝子、チミジンキナーゼ遺伝子等が動物細胞の選抜には有効である。
【0030】
(A−2)選抜工程
本形質転換体の生産方法において、必要に応じて、選抜工程を行う。該選抜工程では、上記第一形質転換体(細胞)の中から前記転写因子を安定的に高発現する形質転換体(細胞)の選抜を行なう。すなわち転写因子の発現に最適な宿主細胞の染色体上の位置に、該転写因子をコードする遺伝子が組み込まれた形質転換体(細胞)を選抜(スクリーニング)する工程である。
当該選抜工程における具体的選抜方法は上記第一形質転換工程で得られた形質転換体(細胞)の中から目的とする転写因子を安定的に高発現する形質転換体を選抜する方法であれば特に限定されるものではない。
【0031】
(A−3)第二形質転換体作成工程
第二形質転換工程は、第一形質転換体、好ましくは、上記選抜工程で得られた転写因子を高発現する形質転換体(細胞){以下、転写因子高発現形質転換体(細胞)と称する}に、RNAを遺伝子とするウイルスに目的タンパク質をコードする遺伝子を挿入したウイルスベクターのcDNAと、上記転写因子高発現形質転換体が発現する転写因子で転写誘導される転写誘導型プロモーターとが連結されてなる発現用DNA断片を導入する工程である。あらかじめ選抜された、転写因子を高発現する形質転換体に、目的タンパク質をコードする遺伝子が挿入されたウイルスベクターのcDNAを導入することにより、目的タンパク質を高生産する形質転換体(細胞)を生産することができる。
【0032】
<目的タンパク質発現用DNA断片>
目的タンパク質発現用DNA断片は、形質転換体(細胞)に目的タンパク質を生産させるために使用するDNA断片であり、少なくともRNAを遺伝子とするウイルスに目的タンパク質をコードする遺伝子を挿入したウイルスベクターのcDNAと、上記転写因子高発現形質転換体(細胞)が発現する転写因子で転写誘導される転写誘導型プロモーターと、を含む。
【0033】
<ウイルスベクター>
ウイルスベクターとしては、RNAを遺伝子とするウイルス由来のウイルスベクターであれば特に限定されるものではなく、二本鎖RNAウイルス、一本鎖マイナス鎖RNAウイルス、一本鎖プラス鎖RNAウイルス由来のウイルスベクターを用いることができる。中でも細胞内でcDNAから転写されたRNA自体がmRNAとして機能するという理由から、一本鎖プラス鎖RNAを遺伝子とするウイルス由来のウイルスベクターであることが特に好ましい。
【0034】
また、ウイルスベクターは植物ウイルス由来のウイルスベクターに限定されるものではなく、動物ウイルス、ファージを含むあらゆるRNAウイルス由来のウイルスベクターを用いることが可能である。例えば、ポティ属(Potyvirus属)ウイルス、ククモウイルス属(Cucumovirus属)ウイルス(例えばキュウリモザイクウイルス(CMV))、ポテックスウイルス属(Potexvirus属)ウイルス(例えばジャガイモXウイルス(PVX))、トンブスウイルス属(Tombusvirus属)ウイルス(例えばトマトブッシースタントウイルス(TBSV)、Cymbidiumu ringspot virus (CymRSV))、カルモウイルス属(Carmovirus属)ウイルス(例えば、Turnip crinkle virus (TCV))、トバモウイルス属(Tobamovirus属)ウイルス(例えば、タバコモザイクウイルス(TMV)、トマトモザイクウイルス(ToMV))が挙げられる。
【0035】
また、上記ウイルスベクターのcDNAの3'末端には、リボザイム配列が結合していることが好ましい。形質転換体の細胞内でcDNAから転写されるウイルスRNAの3'末端に付加されウイルスの複製能力低下の原因となるターミネーター由来の配列やポリA配列を切断することができるからである。その結果ウイルスの複製能力の低下を防止することができ、目的タンパク質の高生産が可能となる。結合されるリボザイム配列は、上記3'末端に付加された余分な配列を切断できるものであればよく、特に限定されるものではない。例えば、肝炎デルタウイルスのリボザイム配列(GenBank accession No. X77627他)またはサテライトタバコリングスポットウイルスのリボザイム配列(GenBank accession No. M17439)を用いることができる。
【0036】
図2(b)に目的タンパク質発現用DNA断片の一例を示す。図2(b)は、形質転換用ベクターであるTiプラスミドpBICER8−ToMV−CO.3−TP−His−SRZの一部分を示している。同図中左からエストロジェンによって活性化される融合転写因子LexA−VP16−hERによって転写誘導されるプロモーターOLexA−46、その下流に目的タンパク遺伝子(以下、TP遺伝子と称する)を組み込んだトマトモザイクウイルスベクターToMV−TPcDNA、その3'末端にサテライトタバコリングスポットウイルスリボザイム配列S−Rz、さらに35Sターミネーター配列35ST、またその下流に薬剤耐性マーカーとしてカナマイシン耐性遺伝子Kanrがある。
【0037】
本形質転換体(細胞)の生産方法の特に好ましい実施形態としては、
宿主細胞がタバコBY2細胞であり、
転写因子がLexA−VP16−hERであり、
転写誘導型プロモーターがOLexA−46であり、
ウイルスベクターがトバモウイルス属に属するウイルス由来であり、
ウイルスベクターがウイルスの移行タンパク質をコードする遺伝子が削除されており、 ウイルスの外被タンパク質の開始コドンが無効化されており、かつ、目的タンパク質をコードする遺伝子が、ウイルスの外被タンパク質をコードする遺伝子に置換されて挿入されているウイルスベクターであり、
ウイルスベクターのcDNAの3'末端に、リボザイム配列が結合しており、かつ
転写因子発現用DNA断片、および目的タンパク質発現用DNA断片が、アグロバクテリウム法により導入される、
ことである。この実施形態によれば、目的タンパク質を、天然物中と同様の活性を維持しつつ、効率的に生産することが可能である。
【0038】
(B)13C標識タンパク質生産用形質転換体を用いた13C標識タンパク質の合成方法
本発明の13C標識タンパク質の合成方法は、上記第二形質転換体をα-ケトイソ吉草酸及び/又はα-ケト酪酸を含む培地で培養して、13Cメチル基標識したバリン、ロイシン及び/又はイソロイシンを含むタンパク質を得ることを特徴としている。
また、本発明の13C標識タンパク質の合成方法は、好ましくは、目的タンパク質をコードする遺伝子が導入された第二形質転換体から目的タンパク質を回収することにより行なう。
しかしながら、宿主細胞(植物細胞)の中の全てのタンパク質がαケトイソ吉草酸とαケト酪酸でメチル基標識されている。よって、目的タンパク質を大量発現しなくても、形質転換されていない細胞自体からタンパク質を精製することも可能である。
加えて、目的タンパク質を大量に取得するためには、該形質転換体の数を培養・栽培・育成等によって増加させ、該増加させた第二形質転換体から目的タンパク質を回収することにより行なうことが好ましい。
第二形質転換体の培養・栽培・育成条件等は、α-ケトイソ吉草酸及び/又はα-ケト酪酸を含む培地を使用する限りは特に限定されるものではなく、該形質転換体に好適な条件を適宜選択して用いればよい。
植物由来培養細胞を培養する際の培地としては特に限定されるものではないが、無機塩類、炭素源、ビタミン類、アミノ酸が加えられている場合がある。さらに、ココナツミルクや酵母エキスを加えて成長を促進させる場合がある。その他、オーキシンとサイトカイニン、ジベレリン、アブシジン酸、エチレン等の植物ホルモンを添加する場合がある。また培養条件であるが、光、温度、通気の有無等を培養する細胞に応じて最適なものを採用すればよい。
【0039】
例えば、タバコBY2細胞を培養条件の一例としては、50〜1000mg/lα-ケトイソ吉草酸及び/又はα-ケト酪酸、370mg/lリン酸二水素カリウム、1mg/lチアミン塩酸、3%スクロース、0.2mg/l2,4−Dを含むMS培地を用い、暗所、26℃、135回転/分で旋回振盪培養後、1/100量を一週間ごとに継代することが挙げられる。
本発明者らは、実施例3により、αケト酪酸又はαケトイソ吉草酸の培地中の最適な濃度を確認している。本発明のαケト酪酸又はαケトイソ吉草酸の培地中の最適な濃度は、大腸菌のタンパク質合成系で使用する濃度と比較して、非常に高い。
培地中のα-ケトイソ吉草酸及び/又はα-ケト酪酸は、50〜1000mg/Lであり、好ましくは100〜800mg/Lであり、より好ましくは150〜350mg/Lであり、最も好ましくは約250mg/mLである。
【0040】
加えて、本形質転換体の種類としては特に限定されるものではなく、植物・動物の個体であっても、植物・動物由来培養細胞であってもよい。ただし、迅速に大量に目的タンパク質を生産するためには、形質転換体は培養可能な細胞(培養細胞)であることが好ましい。さらには、細胞の取り扱いのしやすさ・培地が安価である等の理由から、植物由来培養細胞であることが好ましい。上記植物由来培養細胞としては、例えばタバコ由来BY2細胞が挙げられる。
【0041】
(目的タンパク質)
本発明の13C標識タンパク質の合成方法において、合成する目的タンパク質は特に限定されないが、好ましくは、他のタンパク質合成系(特に、大腸菌合成系、酵母合成系)では、合成できないタンパク質又は正しくリフォールディングできない(正しい立体構造を維持できない)タンパク質を対象とする。
例えば、分子内にジスルフィド結合を有するタンパク質、特に単鎖抗体、分泌タンパク質、膜タンパク質が挙げられる。
より詳しくは、ウシ膵臓トリプシン阻害薬(BPT1)、ストマジェン(stomagen)、EPF2(EPIDERMAL PATTERNING FACTOR 2)、ストレプトマイセス・スブチリシン阻害剤(SSI)、AFP1(alpha-fetoprotein)等が挙げられる。これらのタンパク質は、他のタンパク質合成系(特に、大腸菌系、酵母系)では合成できない又はリフォールディングできないタンパク質である。
【0042】
(目的タンパク質の収率向上)
本発明では、目的タンパク質の収率向上及び標識率向上のために、糖類(例えば、シュークロース,グルコース,フルクトース)並びにアミノ酸、アミノ酸代謝に関与する酵素を阻害する化合物を添加することができる。
【0043】
(NMR測定)
上記本発明の13C標識タンパク質の合成方法で得られた13C標識タンパク質は、NMR測定法により、タンパク質全体の情報から、13Cメチル基標識したバリン、ロイシン及び/又はイソロイシンの情報だけを抽出して観測することができる。
ここで用いられるNMR測定法としては、NMRに用いられ得る方法であれば溶液、固体にかかわらず如何なる方法も用いることができる。具体的には、同種核多次元NMR測定法または異種核多次元NMR測定法のいずれでもよく、例えば溶液同種核多次元NMR測定法としては、COSY、TOCSY、NOESY、ROESY等が挙げられ、異種核多次元NMR測定法としては、HSQC、HMQC、CH-COSY、CBCANH、CBCA(CO)NH、HNCO等が挙げられるが特に限定されない。
【0044】
(13Cメチル基標識したバリン、ロイシン及び/又はイソロイシンを含むタンパク質の合成用キット)
本発明の13Cメチル基標識したバリン、ロイシン及び/又はイソロイシンを含むタンパク質の合成用キットは、少なくとも以下を含む。
(1)形質転換したBY2細胞
(2)α-ケトイソ吉草酸及び/又はα-ケト酪酸
【0045】
以下、実施例を挙げて本発明を詳細に説明するが、本発明の範囲はこれらの実施例により限定されるものではない。
【実施例1】
【0046】
(13C標識タンパク質の合成)
以下の方法及び材料により、目的タンパク質である13C標識BPTIを合成した。詳細は、以下の通りである。
【0047】
1.Tiプラスミドの構築
3個のジスルフィド結合を含むBPTIを目的タンパク質とした。BPTIには、ER標的シグナル配列の1つである19のアミノ酸配列(Lys−Thr−Asn−Leu−Phe−Leu−Phe−Leu−Ile−Phe−Ser−Leu−Leu−Leu−Ser−Leu−Ser−Ser−Ala)、His6配列、続いて因子Xa消化配列をN末端に結合させた。
pBICER8−ToMVerG3(SF3)SrzをベースとするTiプラスミドに、BPTIをコードするDNAフラグメントを挿入した{参照:Dohi K. et al (2006) Arch Virol 151:1075-1084}。
【0048】
2.BY−2細胞に使用した培養条件
窒素源として未標識KNO3および未標識NH4NO3を含む改変Linsmaier-Skoog培地{参照:Nagata T. et al(1992)Int Rev Cytol132:1-30}中、暗所26℃、135rpmで継続的に振盪しながら、タバコBY−2懸濁液培養細胞を増殖させた。該細胞は、7日毎に1:100に希釈し新鮮培地で継代培養し維持した。
【0049】
3.BY−2細胞の形質転換
Tiプラスミドを含むアグロバクテリウムツメファシエンス(Agrobacterium tumefaciens)株LBA4404を使用することによって、BY−2細胞を形質転換した。100mg/Lカナマイシンと500mg/Lカルベニシリンを含む寒天培地で遺伝子導入したBY−2系統を選択した。一次スクリーニング中、6ウェル培養プレート内の3mLの液体培地で懸濁培養細胞を増殖し、次いで135rpmで継続的に振盪させた100mLフラスコ内の30mLの液体培地に移した。2〜3週間の初期培養後、抗生物質を加えずに懸濁液細胞を維持した。
【0050】
4.13C標識タンパク質の合成
13C標識されているメチル基を含むバリン及びロイシンを含むBPTI(参照:図3)を合成するために、安定同位体標識した濃度250mg/Lのα-ケトイソ吉草酸を上記培地に溶解した。
13C標識されているメチル基を含むイソロイシンを含むBPTI(参照:図3)を合成するために、安定同位体標識した濃度250mg/Lのα-ケト酪酸を上記培地に溶解した。
なお、コントロールとして、均一13C標識されているBPTIを合成するために、安定同位体標識した濃度3000mg/Lの13C標識スクロースを培地に溶解している。
30mlの上記それぞれの培地中で目的タンパク質をコードする遺伝子導入BY−2細胞を7日間プレインキュベートし、続いて50〜100mLの新鮮培地中で1:20の割合で再度継代培養した。再継代培養から48時間後、タンパク質合成を開始させるために、培地に終濃度10μMの17β−エストラジオールを加えた。
次いで、目的タンパク質を合成するために、該培養培地を26℃でインキュベートし、135rpmで振盪維持した。最後に、誘導から72時間後にBY−2細胞を回収した。
【0051】
5.13C標識タンパク質の精製
TSE緩衝液{50 mM Tris-HCl(pH 8.0),100 mM NaCl,および1 mM EDTA}に、沈殿した上記回収したBY−2細胞を再懸濁し、超音波処理により破砕した。さらに、ライセートを15,000gで10分間遠心分離し破片を除去した。加えて、Ni2+アフィニティカラム及び因子Xaをそれぞれ精製、タグ配列の消化に使用した。
SDS−PAGEによって13C標識タンパク質が合成されていることを確認した。
以下の実施例2のNMR測定の内部ロック用に10%D2Oを含む緩衝液{10 mM Tris-HCl (pH 7.0),100 mM KCl,および0.02 % NaN3)}で13C標識タンパク質を含む試料を濃縮した。次いで、該試料は、NMR測定用のShigemiマイクロチューブ(株式会社シゲミ、日本国東京都)に移した。
【実施例2】
【0052】
(NMR測定)
上記実施例1で合成した13C標識されているメチル基を含むバリン及びロイシンを含むBPTI、13C標識されているメチル基を含むイソロイシンを含むBPTI、及び均一13C標識されているBPTIをNMR測定した。詳細は、以下の通りである。
【0053】
Bruker社製AVANCE3で1H共鳴周波数800MHzにより実施した。z軸PFGコイルを備えた三重共鳴プローブを使用し、NMR測定中の試料温度は25℃に維持した。1H−13HのHSQC実験で得られた全FIDデータを512×128(t2×t1)複素マトリックスとして記録し、加工し、NMRPipeおよびNMRDrawを使用し、リナックスPC上に表示した。
【0054】
(NMR測定結果)
均一13C標識されているBPTIのNMRスペクトルを図4に示した。
13C標識されているメチル基を含むバリン及びロイシンを含むBPTIのNMRスペクトルを図5に示した。
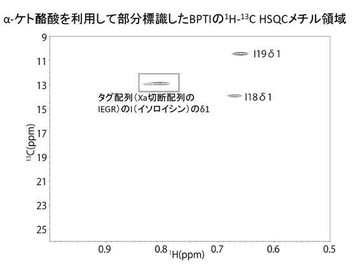
13C標識されているメチル基を含むイソロイシンを含むBPTIのNMRスペクトルを図6に示した。
図4〜図6のNMRスペクトルから明らかなように、本発明の13C標識タンパク質合成方法で得られた13C標識タンパク質は、明瞭なNMRスペクトルを示した。特に、図4のNMRスペクトルを、従来のBPTIのNMRスペクトルと比較すると、BPTIが立体構造を維持して発現していることを確認できた。
さらに、図5のNMRスペクトルを、図4のNMRスペクトルと比較したことにより、バリン及びロイシンのみが明瞭に標識されていることを確認した。
加えて、図6のNMRスペクトルを、図4のNMRスペクトルと比較したことにより、イソロイシンのみが明瞭に標識されていることを確認した。
以上により、本発明の方法で合成したBPTIタンパク質は、他の合成系では困難であった立体構造を維持しており、さらにバリン、ロイシン、イソロイシンのメチル基を明瞭に13C標識できることを確認した。
【実施例3】
【0055】
(αケト酪酸又はαケトイソ吉草酸の培地中の最適な濃度)
αケト酪酸又はαケトイソ吉草酸の培地中の最適な濃度を設定した。詳細は、以下の通りである。
【0056】
上記実施例1の方法に従い、培地に添加するαケトイソ吉草酸を100mg/L、250mg/L又は500mg/Lで合成した13C標識されているメチル基を含むバリン及びロイシンを含むBPTI並びに培地に添加するαケト酪酸を100mg/L、250mg/L又は500mg/Lで合成した13C標識されているメチル基を含むイソロイシンを含むBPTIを得た。
各αケトイソ吉草酸又はαケト酪酸濃度で合成したBPTIの合成量は、吸光度値を基にして算出した。
【0057】
(αケト酪酸又はαケトイソ吉草酸の培地中の最適な濃度の測定結果)
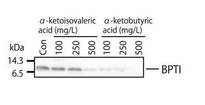
各αケトイソ吉草酸又はαケト酪酸濃度で合成したBPTIの合成量を図7に示した。
図7で示された合成量から明らかなように、αケト酪酸又はαケトイソ吉草酸の培地中の最適な濃度は約250mg/Lであることを確認した。
さらに、本発明のαケト酪酸又はαケトイソ吉草酸の培地中の最適な濃度は、大腸菌のタンパク質合成系で使用する濃度と比較して、非常に高い。
【0058】
(総論)
本発明の13C標識タンパク質合成法では、他の合成系では困難であった立体構造を維持したタンパク質を合成することでき、さらにバリン、ロイシン、イソロイシンのメチル基を明瞭に13C標識することができた。
【産業上の利用可能性】
【0059】
本発明では、他の合成系では困難であった立体構造を維持したタンパク質を合成することでき、さらにバリン、ロイシン、イソロイシンのメチル基を明瞭に13C標識することができる方法を提供することができる。
【技術分野】
【0001】
本発明は、立体構造が維持された13Cメチル基標識したバリン、ロイシン及び/又はイソロイシンを含むタンパク質を合成する方法に関する。
【背景技術】
【0002】
{核磁気共鳴(NMR)について}
原子レベルの解像度でのタンパク質の構造、動力学、相互作用、立体構造変化等の研究には、NMRが使用されている。1980年代末まで、1H−NMR実験を使用するタンパク質の構造決定は小分子(<10kDa)に限定されていた。しかし、最近では、均一な13C及び/又は15N標識法、並びに多次元的NMR技法が、20〜25kDaの分子量も解析することができ、特に、NMRでの大型のタンパク質及びそれらの複合体(>25kDa)の構造及び動力学を研究するための、2H、13C、および15Nを使用する高性能で安定した同位体標識法が提案されている。
【0003】
初めての三重共鳴三次元NMR実験の証明の試料として、大腸菌系で発現する、均一に13C−及び15Nで二重標識したCa2+−結合タンパク質カルモジュリン(CaM)が使用された。それ以来、新規なNMR実験の開発と並行して、大腸菌を使用した様々な改変改善された試料調製法が提案されている。
【0004】
大腸菌を使用したタンパク質合成法(大腸菌合成系)は、NMR試料調製には非常に一般的となった。しかし、大腸菌合成系により合成できるタンパク質は制限されている。特に、大腸菌合成系で発現する多くの真核生物タンパク質は、正確に折り畳まれず、本来の翻訳後修飾ができない、さらには大腸菌合成系では真核生物タンパク質自体が合成できないという問題があった。
【0005】
(タバコBY−2懸濁液培養細胞)
タバコBY−2懸濁液培養細胞は、基礎および応用植物科学で使用される最も人気がある植物細胞系の一つである。タバコBY−2懸濁液培養細胞の特性は、幅広く活用されており、NMR試料の調製に適している。BY−2用Linsmaier-Skoog培地は調製が容易であり、動物細胞および無細胞抽出物に使用される培地よりもはるかに経済的である。
しかし、タバコBY−2を使用する既知のタンパク質発現方法は、タンパク質の生産性に関して重大な問題があった。タバコBY−2細胞によって得られる外来タンパク質の発現レベルは低く、NMR試料への使用は制限されている。
【0006】
本発明者の1人である森正之博士らは、ウイルスベクターを使用することによって、懸濁液培養BY−2細胞のタンパク質の生産性を著しく改善することに成功した(参照:非特許文献1)。この改善したタンパク質合成系では、煩雑なウイルス接種手順は不要である。エストラジオールを該系に添加することで、効率良くウイルスRNAの蓄積が誘導される。さらに、該誘導に続いて、タバコBY−2懸濁液培養細胞で組換えタンパク質が効率良く大量に合成される(参照:特許文献1〜4)。
【0007】
また、本発明者らは、上記タバコBY−2懸濁液培養細胞を使用したタンパク質合成系を用いて15N標識タンパク質を合成できたことを報告している(参照:非特許文献2)。 しかし、該文献は、「13Cメチル基標識したバリン、ロイシン及び/又はイソロイシンを含むタンパク質を合成する方法」の開示又は示唆がない。
【先行技術文献】
【特許文献】
【0008】
【特許文献1】特開2011−83232
【特許文献2】特開2005−245228
【特許文献3】WO2008/136253
【特許文献4】特開2005−237301
【特許文献5】特開2005−102652
【0009】
【非特許文献1】Dohi K. et al(2006) Arch Virol 151:1075-1084
【非特許文献2】Ohki S. etal(2008) J Biomol NMR 42:271-277
【非特許文献3】Amy M. etal(2010) J Biomol NMR 46:75-87
【発明の概要】
【発明が解決しようとする課題】
【0010】
多数のタンパク質合成系が開発されているが、NMR試料を調製する普遍的方法は、現在まで確立されていない。また、大腸菌、酵母のタンパク質合成系等では、安定同位体を標識したタンパク質自体が合成できない又はリフォールディングしていない状態の(正しい立体構造を維持していない)安定同位体を標識したタンパク質が発現されることが多数報告されている。
【課題を解決するための手段】
【0011】
本発明者らは、上記課題を解決するために、13C標識したα-ケトイソ吉草酸又はα-ケト酪酸を含む培地を用いて本発明の合成系でタンパク質を発現させたところ、他の合成系では困難であった立体構造を維持した13Cメチル基標識したバリン、ロイシン及び/又はイソロイシンを含むタンパク質を合成できることを新規に見出し、本発明を完成した。
【0012】
すなわち、本発明は以下の通りである。
「1.以下の工程を含む13Cメチル基標識したバリン、ロイシン及び/又はイソロイシンを含むタンパク質の合成方法:
(1)ホルモンによって活性化される性質を有する転写因子をコードする遺伝子と、転写因子発現用プロモーターとが連結されてなる転写因子発現用DNA断片を宿主細胞に導入する第一形質転換体を作成する工程;
(2)RNAを遺伝子とするウイルスに標的タンパク質をコードする遺伝子を挿入したウイルスベクターのcDNAと、該転写因子で転写誘導される転写誘導型プロモーターとが連結されてなる目的タンパク質発現用DNA断片を、該第一形質転換体に導入して第二形質転換体を作成する工程;
(3)該第二形質転換体をα-ケトイソ吉草酸及び/又はα-ケト酪酸を含む培地で培養して、13C標識タンパク質を得る工程、ここで、該ホルモンは培養前及び/又は培養中に該培地に添加されている。
2.前記α-ケトイソ吉草酸及び/又はα-ケト酪酸を含む培地において、α-ケトイソ吉草酸及びα-ケト酪酸の濃度が、それぞれ、100mg〜1000mg/Lである前項1の合成方法。
3.前記α-ケトイソ吉草酸及び/又はα-ケト酪酸を含む培地において、α-ケトイソ吉草酸及びα-ケト酪酸の濃度が、それぞれ、150mg〜350mg/Lである前項1又は2の合成方法。
4.前記標的タンパク質が、分子内にジスルフィド結合を有する前項1〜3のいずれか1の合成法。
5.分子内にジスルフィド結合を有するタンパク質が、以下のいずれか1から選ばれる前項4の合成方法。
(1)単鎖抗体
(2)分泌タンパク質
(3)膜タンパク質
6.前記標的タンパク質が、以下のいずれか1から選ばれる前項1〜3のいずれか1の合成法。
(1)ウシ膵臓トリプシン阻害薬(BPTI)
(2)ストマジェン(stomagen)
(3)EPF2(EPIDERMAL PATTERNING FACTOR 2)
(4)ストレプトマイセス・スブチリシン阻害剤(SSI)
(5)AFP1(alpha-fetoprotein)
7.前記ホルモンが、エストロジェンまたはステロイドホルモンである前項1〜6のいずれか1の合成法。
8.前記エストロジェンで活性化される性質を有する転写因子としてLexA−VP16−hERを用い、上記転写誘導型プロモーターとしてOLexA−46を用いる前項7の合成法。
9.前記ウイルスベクターが、植物ウイルス由来である前項8の合成法。
10.前記宿主細胞および形質転換体が、植物体又は植物由来培養細胞である前項1〜9のいずれか1の合成法。
11.前記ホルモンがエストロジェンであり、
前記宿主細胞がタバコBY2細胞であり、
前記転写因子がLexA−VP16−hERであり、
前記転写誘導型プロモーターがOLexA−46であり、
前記ウイルスベクターがトバモウイルス属に属するウイルス由来である、ことを特徴とする、前項1〜10のいずれか1の合成法。
12.以下を含む13Cメチル基標識したバリン、ロイシン及び/又はイソロイシンを含むタンパク質の合成用キット。
(1)形質転換したBY2細胞
(2)α-ケトイソ吉草酸及び/又はα-ケト酪酸」
【発明の効果】
【0013】
本発明の13C標識タンパク質合成法では、他の合成系では困難であった立体構造を維持したタンパク質を合成することができ、さらにバリン、ロイシン、イソロイシンのメチル基を明瞭に13C標識することができる。
【図面の簡単な説明】
【0014】
【図1】本発明のタンパク質の合成系の概要
【図2】(a)転写因子発現用DNAの一例、(b)目的タンパク質発現用DNA断片の一例
【図3】培地に添加される13C標識α-ケトイソ吉草酸又はα-ケト酪酸並びに生成される13C標識アミノ酸
【図4】均一13C標識したBPTIのNMRスペクトル
【図5】13Cメチル基標識したバリン及びロイシンを含むBPTIのNMRスペクトル
【図6】13Cメチル基標識したイソロイシンを含むBPTIのNMRスペクトル
【図7】各αケトイソ吉草酸又はαケト酪酸濃度で合成したBPTIの合成量、なお「Con」は13C標識したα-ケトイソ吉草酸又はα-ケト酪酸を含まない培地で合成したBPTIの合成量
【発明を実施するための形態】
【0015】
(本発明のタンパク質合成系)
本発明の13C標識タンパク質を合成する方法で使用するタンパク質合成系は、特許文献1〜5及び非特許文献1、2に記載の合成系を利用することができる。また、図1のタンパク質の合成系の概要図を参照することができる。
本合成系の一例について下記で具体的に説明する。
【0016】
(A)13C標識タンパク質生産用形質転換体の生産方法
本発明に係る13C標識タンパク質生産用第一形質転換体の生産方法は、(1)ホルモンによって活性化される性質を有する転写因子をコードする遺伝子と転写因子発現用プロモーターとが連結されてなる転写因子発現用DNA断片を細胞に導入する第一形質転換体作成工程と、必要に応じて(2)該第一形質転換体の中から該転写因子を発現する形質転換体を選抜する選抜工程と、(3)RNAを遺伝子とするウイルスに目的タンパク質をコードする遺伝子を挿入したウイルスベクターのcDNAと、該転写因子で転写誘導される転写誘導型プロモーターとが連結されてなる目的タンパク質発現用DNA断片を、第一形質転換体に導入して第二形質転換体を作成する工程と、を含む。以下、工程ごとに説明する。
【0017】
(A−1)第一形質転換体作成工程
第一形質転換体作成工程は、ホルモンによって活性化される性質を有する転写因子をコードする遺伝子と、該転写因子の発現を担うプロモーター(転写因子発現用プロモーター)とを連結して構築したDNA断片(転写因子発現用DNA断片)を、適当な宿主細胞に導入する工程である。
該工程によって、後述するウイルスベクター発現用プロモーターの転写誘導を行なう転写因子を安定的に高発現する形質転換体(細胞)の生産(作出)を行なう。すなわち宿主細胞の転写因子の発現に最適な染色体上の位置に、該転写因子をコードする遺伝子が組み込まれた形質転換体(細胞)を生産(作出)する工程である。
【0018】
<転写因子発現用DNA断片>
転写因子発現用DNA断片は、転写因子発現用プロモーターの下流に転写因子をコードする遺伝子が連結されている。また、該遺伝子の他、ベクター配列、ターミネーター、薬剤耐性マーカー等のDNAセグメントが含まれていてもよい。かかる転写因子発現用DNA断片の構築方法は、通常の遺伝子工学的手法を用いて行なえばよい。
【0019】
なお、転写因子発現用プロモーターは、転写因子を発現することが可能なものであれば特に限定されるものではなく、また恒常的にプロモーター活性を有するもの(以下、恒常的プロモーターと称する)であってもよいし、さらに転写因子によってプロモーター活性が誘導されるものであってもよい。ただし、転写因子の発現をさらに別の転写因子で制御することは、目的タンパク質の発現系自体が複雑化すること、コスト面で不利等の理由により、上記転写発現用プロモーターとしては、恒常的プロモーターの方がより好ましいといえる。恒常的プロモーターの例としては、カリフラワーモザイクウイルス35Sプロモーター、PG10−90(Ishige, F., Takaichi, M., Foster, R.,Chua, N. H. and Oeda, K.(1999)A G-box motif(GCCACGTGCC) tetramer confers high-level constitutive expressionin dicot and monocot plants. Plant J. 20, 127-133. 参照)、ユビキチンプロモーター、アクチンプロモーター等が挙げられる。
【0020】
一方、使用する転写因子は、ホルモンによって活性化される性質を有する転写因子であって、後述する第二形質転換体作成工程において導入する目的タンパク質発現用DNA断片中に含まれるプロモーターを転写誘導するものであれば特に限定されない。転写因子を活性化するホルモンとしては、エストロジェン(好ましくはエストラジオール、より好ましくはβエストラジオール)、ステロイドホルモン、エクジソン等が挙げられる。かかる転写因子は、ホルモンが存在しない状態においては不活性型でありプロモーターの転写誘導を行なうことができないが、ホルモンが存在することによって活性型に変化しプロモーターの転写誘導を行なうことができる。この性質を利用すれば目的タンパク質の生産をより綿密に制御できるため、より安全に目的タンパク質の生産を行なうことができるといえる。すなわち、目的タンパク質生産を行なう必要がないとき、あるいは目的タンパク質生産を行なってはいけないときには、合成系にホルモンを添加しなければよい。
【0021】
ホルモンによって活性化される性質を有する転写因子、および該転写因子によって転写誘導されるプロモーターの組み合わせとしては、例えば、ステロイドホルモンで活性化される転写因子であるGVGと該転写因子によって転写誘導されるプロモーターである6xUASgal4の組み合わせ、エストロジェンで活性化される転写因子LexA−VP16−hERと該転写因子によって転写誘導可能なプロモーターであるOLexA−46(Zuo J, Niu QW, Chua NH. "An estrogen receptor-basedtransactivator XVE mediates highly inducible gene exprssion in transgenicplants." Plant J.2000, 24: 265-273 参照)の組み合わせ、エクジソンで活性化される転写因子であるエクジソンレセプターGR Act and DBDとヘルペスウイルストランスアクチベーションドメインHecR LBDのキメラタンパク質と該転写因子によって転写誘導可能なプロモーターGREの組み合わせ等が挙げられる。ただし、宿主として植物体および植物細胞を利用する場合においては、活性化に用いるホルモンによる宿主への悪影響が少ないという理由からエストロジェンで活性化される転写因子LexA−VP16−hERと該転写因子によって転写誘導可能なプロモーターであるOLexA−46の組み合わせが好ましい。
【0022】
図2(a)に転写因子発現用DNAの一例を示す。図2(a)は、形質転換用ベクターであるTiプラスミドpER8(−Stu)の一部分を示している。同図中左から転写因子発現用の恒常的プロモーターPG10−90、その下流にエストロジェンレセプターを含む融合転写因子LexA−VP16−hER、その3'末端にターミネーター配列TE9、またその下流に薬剤耐性マーカーとしてハイグロマイシン耐性遺伝子Hygrがある。
【0023】
<転写因子発現用DNA断片を導入する宿主細胞およびDNA断片導入方法>
上記転写因子発現用DNA断片が導入される宿主細胞は特に限定されるものではなく、動物由来細胞であっても植物由来細胞であってもよい。ただし、植物由来細胞は、動物由来細胞に比して、増殖速度が速くコンタミネーションのリスクが少ない点、培地作成費用が非常に安価であるという点において、植物由来細胞がより好ましい。なお、動物由来細胞および植物由来細胞とは、細胞、組織、並びに器官も含む意味である。特に液体培地等で培養可能な細胞(培養細胞)が好ましい。
【0024】
また由来となる動物としては、特に限定されるものでないが、ヒト、サル、イヌ、ヒツジ、ヤギ、ウサギ、マウス、ラット、モルモット、チャイニーズハムスター、ウシ、ウマ、ブタ、メダカやゼブラフィッシュ等の魚類、カイコ、夜蛾(Spodoptera frugiperda)等が挙げられる。一方由来となる植物としては、特に限定されるものではないが、イネ、シロイナズナ、オオムギ、コムギ、タバコ、トマト、キュウリ、ダイズ、ジャガイモ、トウモロコシ、ニチニチソウ、シロイヌナズナ、アルファルファが挙げられる。その他、枯草菌や乳酸菌などの菌類、酵母など単細胞生物が宿主細胞として利用できる。
【0025】
また動物由来細胞の例としては特に限定されるものではないが、HeLa細胞、CHO細胞、メラノーマ細胞、マウス3T3細胞が挙げられる。また植物由来細胞の例としては、タバコBY2細胞、ジャガイモ由来、イネ由来、サツマイモ由来、ダイズ由来、パセリ由来、シロイヌナズナ由来、コムギ由来、トウモロコシ由来細胞、ニチニチソウ由来細胞が挙げられる。
【0026】
後述する実施例において、タバコBY2細胞を宿主として用いている。タバコBY2細胞(Toshiyuki nagata, Yasuyuki Nemoto, and Seiichiro Hasezawa"Tobacco BY-2 Cell Line as the "Hela" Cell in the Cell Biologyof Higher Plants" International Review of cytology, vol.132, p.p. 1-30(1992) 、およびhttp://www.riken.go.jp/r-world/info/release/press/2003/030620/ 参照)は、植物培養細胞株としては、世界中で最も広く用いられているものであり、最も増殖速度が速いこと、遺伝子操作が容易なこと、大量培養を容易に行うことができるという理由から好ましい。
【0027】
また、転写因子発現用DNA断片を宿主細胞へ導入する形質転換方法は特に限定されるものではなく、宿主細胞の種類に応じた適切な形質転換方法を用いればよい。例えば、特別なベクター配列を有しないDNA断片を導入する場合には、エレクトロポレーション法、パーティクルガン法、リン酸カルシウム法等を用いることができる。また植物由来細胞への一般的な形質転換法としては、アグロバクテリウムを用いた形質転換法(アグロバクテリウム法)を挙げることができ、本発明でもアグロバクテリウム法を好適に用いることができる。ただしアグロバクテリウム法を用いて形質転換する場合には、本発明にかかるDNA断片を含むTiプラスミドを構築する必要がある。またその他プロトプラスト/スフェロプラスト法、リン酸カルシウム法、リポソーム法、DEAEデキストラン法等の従来公知の方法を好適に用いることができる。
【0028】
転写因子発現用DNA断片は、宿主細胞のゲノムに組み込まれることが好ましい。転写因子発現用DNA断片がゲノムに組み込まれることにより、細胞分裂後の娘細胞にもベクターの構成に含まれる遺伝子を確実に伝達することが可能となり、転写因子(タンパク質)の生産効率を維持することが可能となるからである。ゲノムは染色体(核ゲノム)に限定されるものではなく、ミトコンドリアゲノムや葉緑体ゲノムも含まれる。
【0029】
また、転写因子発現用DNA断片が宿主細胞に導入されたか否かを確認する方法は、特に限定されるものではなく、公知の各種の方法を用いることができる。具体的には、各種マーカーを用いればよい。例えば、宿主細胞中で欠失している遺伝子をマーカーとして用い、このマーカーと組み換え植物ウイルス遺伝子とを含むプラスミド等を発現ベクターとして宿主細胞に導入する。これによってマーカー遺伝子の発現から本発明の遺伝子の導入を確認することができる。例えば、薬剤耐性マーカー(ハイグロマイシン耐性遺伝子、Hygr)を用いれば、ハイグロマイシンを含有する培地中で、形質転換候補株を培養することにより、生育してきた細胞株を形質転換体として選抜することが可能となる。
その他のマーカーとしては、ビアラホス耐性マーカー、カナマイシン耐性マーカー等が植物細胞の選抜に有効であり、ピューロマイシン耐性マーカー、ブレオマイシン耐性マーカー、XGPRT遺伝子、DHFR遺伝子、チミジンキナーゼ遺伝子等が動物細胞の選抜には有効である。
【0030】
(A−2)選抜工程
本形質転換体の生産方法において、必要に応じて、選抜工程を行う。該選抜工程では、上記第一形質転換体(細胞)の中から前記転写因子を安定的に高発現する形質転換体(細胞)の選抜を行なう。すなわち転写因子の発現に最適な宿主細胞の染色体上の位置に、該転写因子をコードする遺伝子が組み込まれた形質転換体(細胞)を選抜(スクリーニング)する工程である。
当該選抜工程における具体的選抜方法は上記第一形質転換工程で得られた形質転換体(細胞)の中から目的とする転写因子を安定的に高発現する形質転換体を選抜する方法であれば特に限定されるものではない。
【0031】
(A−3)第二形質転換体作成工程
第二形質転換工程は、第一形質転換体、好ましくは、上記選抜工程で得られた転写因子を高発現する形質転換体(細胞){以下、転写因子高発現形質転換体(細胞)と称する}に、RNAを遺伝子とするウイルスに目的タンパク質をコードする遺伝子を挿入したウイルスベクターのcDNAと、上記転写因子高発現形質転換体が発現する転写因子で転写誘導される転写誘導型プロモーターとが連結されてなる発現用DNA断片を導入する工程である。あらかじめ選抜された、転写因子を高発現する形質転換体に、目的タンパク質をコードする遺伝子が挿入されたウイルスベクターのcDNAを導入することにより、目的タンパク質を高生産する形質転換体(細胞)を生産することができる。
【0032】
<目的タンパク質発現用DNA断片>
目的タンパク質発現用DNA断片は、形質転換体(細胞)に目的タンパク質を生産させるために使用するDNA断片であり、少なくともRNAを遺伝子とするウイルスに目的タンパク質をコードする遺伝子を挿入したウイルスベクターのcDNAと、上記転写因子高発現形質転換体(細胞)が発現する転写因子で転写誘導される転写誘導型プロモーターと、を含む。
【0033】
<ウイルスベクター>
ウイルスベクターとしては、RNAを遺伝子とするウイルス由来のウイルスベクターであれば特に限定されるものではなく、二本鎖RNAウイルス、一本鎖マイナス鎖RNAウイルス、一本鎖プラス鎖RNAウイルス由来のウイルスベクターを用いることができる。中でも細胞内でcDNAから転写されたRNA自体がmRNAとして機能するという理由から、一本鎖プラス鎖RNAを遺伝子とするウイルス由来のウイルスベクターであることが特に好ましい。
【0034】
また、ウイルスベクターは植物ウイルス由来のウイルスベクターに限定されるものではなく、動物ウイルス、ファージを含むあらゆるRNAウイルス由来のウイルスベクターを用いることが可能である。例えば、ポティ属(Potyvirus属)ウイルス、ククモウイルス属(Cucumovirus属)ウイルス(例えばキュウリモザイクウイルス(CMV))、ポテックスウイルス属(Potexvirus属)ウイルス(例えばジャガイモXウイルス(PVX))、トンブスウイルス属(Tombusvirus属)ウイルス(例えばトマトブッシースタントウイルス(TBSV)、Cymbidiumu ringspot virus (CymRSV))、カルモウイルス属(Carmovirus属)ウイルス(例えば、Turnip crinkle virus (TCV))、トバモウイルス属(Tobamovirus属)ウイルス(例えば、タバコモザイクウイルス(TMV)、トマトモザイクウイルス(ToMV))が挙げられる。
【0035】
また、上記ウイルスベクターのcDNAの3'末端には、リボザイム配列が結合していることが好ましい。形質転換体の細胞内でcDNAから転写されるウイルスRNAの3'末端に付加されウイルスの複製能力低下の原因となるターミネーター由来の配列やポリA配列を切断することができるからである。その結果ウイルスの複製能力の低下を防止することができ、目的タンパク質の高生産が可能となる。結合されるリボザイム配列は、上記3'末端に付加された余分な配列を切断できるものであればよく、特に限定されるものではない。例えば、肝炎デルタウイルスのリボザイム配列(GenBank accession No. X77627他)またはサテライトタバコリングスポットウイルスのリボザイム配列(GenBank accession No. M17439)を用いることができる。
【0036】
図2(b)に目的タンパク質発現用DNA断片の一例を示す。図2(b)は、形質転換用ベクターであるTiプラスミドpBICER8−ToMV−CO.3−TP−His−SRZの一部分を示している。同図中左からエストロジェンによって活性化される融合転写因子LexA−VP16−hERによって転写誘導されるプロモーターOLexA−46、その下流に目的タンパク遺伝子(以下、TP遺伝子と称する)を組み込んだトマトモザイクウイルスベクターToMV−TPcDNA、その3'末端にサテライトタバコリングスポットウイルスリボザイム配列S−Rz、さらに35Sターミネーター配列35ST、またその下流に薬剤耐性マーカーとしてカナマイシン耐性遺伝子Kanrがある。
【0037】
本形質転換体(細胞)の生産方法の特に好ましい実施形態としては、
宿主細胞がタバコBY2細胞であり、
転写因子がLexA−VP16−hERであり、
転写誘導型プロモーターがOLexA−46であり、
ウイルスベクターがトバモウイルス属に属するウイルス由来であり、
ウイルスベクターがウイルスの移行タンパク質をコードする遺伝子が削除されており、 ウイルスの外被タンパク質の開始コドンが無効化されており、かつ、目的タンパク質をコードする遺伝子が、ウイルスの外被タンパク質をコードする遺伝子に置換されて挿入されているウイルスベクターであり、
ウイルスベクターのcDNAの3'末端に、リボザイム配列が結合しており、かつ
転写因子発現用DNA断片、および目的タンパク質発現用DNA断片が、アグロバクテリウム法により導入される、
ことである。この実施形態によれば、目的タンパク質を、天然物中と同様の活性を維持しつつ、効率的に生産することが可能である。
【0038】
(B)13C標識タンパク質生産用形質転換体を用いた13C標識タンパク質の合成方法
本発明の13C標識タンパク質の合成方法は、上記第二形質転換体をα-ケトイソ吉草酸及び/又はα-ケト酪酸を含む培地で培養して、13Cメチル基標識したバリン、ロイシン及び/又はイソロイシンを含むタンパク質を得ることを特徴としている。
また、本発明の13C標識タンパク質の合成方法は、好ましくは、目的タンパク質をコードする遺伝子が導入された第二形質転換体から目的タンパク質を回収することにより行なう。
しかしながら、宿主細胞(植物細胞)の中の全てのタンパク質がαケトイソ吉草酸とαケト酪酸でメチル基標識されている。よって、目的タンパク質を大量発現しなくても、形質転換されていない細胞自体からタンパク質を精製することも可能である。
加えて、目的タンパク質を大量に取得するためには、該形質転換体の数を培養・栽培・育成等によって増加させ、該増加させた第二形質転換体から目的タンパク質を回収することにより行なうことが好ましい。
第二形質転換体の培養・栽培・育成条件等は、α-ケトイソ吉草酸及び/又はα-ケト酪酸を含む培地を使用する限りは特に限定されるものではなく、該形質転換体に好適な条件を適宜選択して用いればよい。
植物由来培養細胞を培養する際の培地としては特に限定されるものではないが、無機塩類、炭素源、ビタミン類、アミノ酸が加えられている場合がある。さらに、ココナツミルクや酵母エキスを加えて成長を促進させる場合がある。その他、オーキシンとサイトカイニン、ジベレリン、アブシジン酸、エチレン等の植物ホルモンを添加する場合がある。また培養条件であるが、光、温度、通気の有無等を培養する細胞に応じて最適なものを採用すればよい。
【0039】
例えば、タバコBY2細胞を培養条件の一例としては、50〜1000mg/lα-ケトイソ吉草酸及び/又はα-ケト酪酸、370mg/lリン酸二水素カリウム、1mg/lチアミン塩酸、3%スクロース、0.2mg/l2,4−Dを含むMS培地を用い、暗所、26℃、135回転/分で旋回振盪培養後、1/100量を一週間ごとに継代することが挙げられる。
本発明者らは、実施例3により、αケト酪酸又はαケトイソ吉草酸の培地中の最適な濃度を確認している。本発明のαケト酪酸又はαケトイソ吉草酸の培地中の最適な濃度は、大腸菌のタンパク質合成系で使用する濃度と比較して、非常に高い。
培地中のα-ケトイソ吉草酸及び/又はα-ケト酪酸は、50〜1000mg/Lであり、好ましくは100〜800mg/Lであり、より好ましくは150〜350mg/Lであり、最も好ましくは約250mg/mLである。
【0040】
加えて、本形質転換体の種類としては特に限定されるものではなく、植物・動物の個体であっても、植物・動物由来培養細胞であってもよい。ただし、迅速に大量に目的タンパク質を生産するためには、形質転換体は培養可能な細胞(培養細胞)であることが好ましい。さらには、細胞の取り扱いのしやすさ・培地が安価である等の理由から、植物由来培養細胞であることが好ましい。上記植物由来培養細胞としては、例えばタバコ由来BY2細胞が挙げられる。
【0041】
(目的タンパク質)
本発明の13C標識タンパク質の合成方法において、合成する目的タンパク質は特に限定されないが、好ましくは、他のタンパク質合成系(特に、大腸菌合成系、酵母合成系)では、合成できないタンパク質又は正しくリフォールディングできない(正しい立体構造を維持できない)タンパク質を対象とする。
例えば、分子内にジスルフィド結合を有するタンパク質、特に単鎖抗体、分泌タンパク質、膜タンパク質が挙げられる。
より詳しくは、ウシ膵臓トリプシン阻害薬(BPT1)、ストマジェン(stomagen)、EPF2(EPIDERMAL PATTERNING FACTOR 2)、ストレプトマイセス・スブチリシン阻害剤(SSI)、AFP1(alpha-fetoprotein)等が挙げられる。これらのタンパク質は、他のタンパク質合成系(特に、大腸菌系、酵母系)では合成できない又はリフォールディングできないタンパク質である。
【0042】
(目的タンパク質の収率向上)
本発明では、目的タンパク質の収率向上及び標識率向上のために、糖類(例えば、シュークロース,グルコース,フルクトース)並びにアミノ酸、アミノ酸代謝に関与する酵素を阻害する化合物を添加することができる。
【0043】
(NMR測定)
上記本発明の13C標識タンパク質の合成方法で得られた13C標識タンパク質は、NMR測定法により、タンパク質全体の情報から、13Cメチル基標識したバリン、ロイシン及び/又はイソロイシンの情報だけを抽出して観測することができる。
ここで用いられるNMR測定法としては、NMRに用いられ得る方法であれば溶液、固体にかかわらず如何なる方法も用いることができる。具体的には、同種核多次元NMR測定法または異種核多次元NMR測定法のいずれでもよく、例えば溶液同種核多次元NMR測定法としては、COSY、TOCSY、NOESY、ROESY等が挙げられ、異種核多次元NMR測定法としては、HSQC、HMQC、CH-COSY、CBCANH、CBCA(CO)NH、HNCO等が挙げられるが特に限定されない。
【0044】
(13Cメチル基標識したバリン、ロイシン及び/又はイソロイシンを含むタンパク質の合成用キット)
本発明の13Cメチル基標識したバリン、ロイシン及び/又はイソロイシンを含むタンパク質の合成用キットは、少なくとも以下を含む。
(1)形質転換したBY2細胞
(2)α-ケトイソ吉草酸及び/又はα-ケト酪酸
【0045】
以下、実施例を挙げて本発明を詳細に説明するが、本発明の範囲はこれらの実施例により限定されるものではない。
【実施例1】
【0046】
(13C標識タンパク質の合成)
以下の方法及び材料により、目的タンパク質である13C標識BPTIを合成した。詳細は、以下の通りである。
【0047】
1.Tiプラスミドの構築
3個のジスルフィド結合を含むBPTIを目的タンパク質とした。BPTIには、ER標的シグナル配列の1つである19のアミノ酸配列(Lys−Thr−Asn−Leu−Phe−Leu−Phe−Leu−Ile−Phe−Ser−Leu−Leu−Leu−Ser−Leu−Ser−Ser−Ala)、His6配列、続いて因子Xa消化配列をN末端に結合させた。
pBICER8−ToMVerG3(SF3)SrzをベースとするTiプラスミドに、BPTIをコードするDNAフラグメントを挿入した{参照:Dohi K. et al (2006) Arch Virol 151:1075-1084}。
【0048】
2.BY−2細胞に使用した培養条件
窒素源として未標識KNO3および未標識NH4NO3を含む改変Linsmaier-Skoog培地{参照:Nagata T. et al(1992)Int Rev Cytol132:1-30}中、暗所26℃、135rpmで継続的に振盪しながら、タバコBY−2懸濁液培養細胞を増殖させた。該細胞は、7日毎に1:100に希釈し新鮮培地で継代培養し維持した。
【0049】
3.BY−2細胞の形質転換
Tiプラスミドを含むアグロバクテリウムツメファシエンス(Agrobacterium tumefaciens)株LBA4404を使用することによって、BY−2細胞を形質転換した。100mg/Lカナマイシンと500mg/Lカルベニシリンを含む寒天培地で遺伝子導入したBY−2系統を選択した。一次スクリーニング中、6ウェル培養プレート内の3mLの液体培地で懸濁培養細胞を増殖し、次いで135rpmで継続的に振盪させた100mLフラスコ内の30mLの液体培地に移した。2〜3週間の初期培養後、抗生物質を加えずに懸濁液細胞を維持した。
【0050】
4.13C標識タンパク質の合成
13C標識されているメチル基を含むバリン及びロイシンを含むBPTI(参照:図3)を合成するために、安定同位体標識した濃度250mg/Lのα-ケトイソ吉草酸を上記培地に溶解した。
13C標識されているメチル基を含むイソロイシンを含むBPTI(参照:図3)を合成するために、安定同位体標識した濃度250mg/Lのα-ケト酪酸を上記培地に溶解した。
なお、コントロールとして、均一13C標識されているBPTIを合成するために、安定同位体標識した濃度3000mg/Lの13C標識スクロースを培地に溶解している。
30mlの上記それぞれの培地中で目的タンパク質をコードする遺伝子導入BY−2細胞を7日間プレインキュベートし、続いて50〜100mLの新鮮培地中で1:20の割合で再度継代培養した。再継代培養から48時間後、タンパク質合成を開始させるために、培地に終濃度10μMの17β−エストラジオールを加えた。
次いで、目的タンパク質を合成するために、該培養培地を26℃でインキュベートし、135rpmで振盪維持した。最後に、誘導から72時間後にBY−2細胞を回収した。
【0051】
5.13C標識タンパク質の精製
TSE緩衝液{50 mM Tris-HCl(pH 8.0),100 mM NaCl,および1 mM EDTA}に、沈殿した上記回収したBY−2細胞を再懸濁し、超音波処理により破砕した。さらに、ライセートを15,000gで10分間遠心分離し破片を除去した。加えて、Ni2+アフィニティカラム及び因子Xaをそれぞれ精製、タグ配列の消化に使用した。
SDS−PAGEによって13C標識タンパク質が合成されていることを確認した。
以下の実施例2のNMR測定の内部ロック用に10%D2Oを含む緩衝液{10 mM Tris-HCl (pH 7.0),100 mM KCl,および0.02 % NaN3)}で13C標識タンパク質を含む試料を濃縮した。次いで、該試料は、NMR測定用のShigemiマイクロチューブ(株式会社シゲミ、日本国東京都)に移した。
【実施例2】
【0052】
(NMR測定)
上記実施例1で合成した13C標識されているメチル基を含むバリン及びロイシンを含むBPTI、13C標識されているメチル基を含むイソロイシンを含むBPTI、及び均一13C標識されているBPTIをNMR測定した。詳細は、以下の通りである。
【0053】
Bruker社製AVANCE3で1H共鳴周波数800MHzにより実施した。z軸PFGコイルを備えた三重共鳴プローブを使用し、NMR測定中の試料温度は25℃に維持した。1H−13HのHSQC実験で得られた全FIDデータを512×128(t2×t1)複素マトリックスとして記録し、加工し、NMRPipeおよびNMRDrawを使用し、リナックスPC上に表示した。
【0054】
(NMR測定結果)
均一13C標識されているBPTIのNMRスペクトルを図4に示した。
13C標識されているメチル基を含むバリン及びロイシンを含むBPTIのNMRスペクトルを図5に示した。
13C標識されているメチル基を含むイソロイシンを含むBPTIのNMRスペクトルを図6に示した。
図4〜図6のNMRスペクトルから明らかなように、本発明の13C標識タンパク質合成方法で得られた13C標識タンパク質は、明瞭なNMRスペクトルを示した。特に、図4のNMRスペクトルを、従来のBPTIのNMRスペクトルと比較すると、BPTIが立体構造を維持して発現していることを確認できた。
さらに、図5のNMRスペクトルを、図4のNMRスペクトルと比較したことにより、バリン及びロイシンのみが明瞭に標識されていることを確認した。
加えて、図6のNMRスペクトルを、図4のNMRスペクトルと比較したことにより、イソロイシンのみが明瞭に標識されていることを確認した。
以上により、本発明の方法で合成したBPTIタンパク質は、他の合成系では困難であった立体構造を維持しており、さらにバリン、ロイシン、イソロイシンのメチル基を明瞭に13C標識できることを確認した。
【実施例3】
【0055】
(αケト酪酸又はαケトイソ吉草酸の培地中の最適な濃度)
αケト酪酸又はαケトイソ吉草酸の培地中の最適な濃度を設定した。詳細は、以下の通りである。
【0056】
上記実施例1の方法に従い、培地に添加するαケトイソ吉草酸を100mg/L、250mg/L又は500mg/Lで合成した13C標識されているメチル基を含むバリン及びロイシンを含むBPTI並びに培地に添加するαケト酪酸を100mg/L、250mg/L又は500mg/Lで合成した13C標識されているメチル基を含むイソロイシンを含むBPTIを得た。
各αケトイソ吉草酸又はαケト酪酸濃度で合成したBPTIの合成量は、吸光度値を基にして算出した。
【0057】
(αケト酪酸又はαケトイソ吉草酸の培地中の最適な濃度の測定結果)
各αケトイソ吉草酸又はαケト酪酸濃度で合成したBPTIの合成量を図7に示した。
図7で示された合成量から明らかなように、αケト酪酸又はαケトイソ吉草酸の培地中の最適な濃度は約250mg/Lであることを確認した。
さらに、本発明のαケト酪酸又はαケトイソ吉草酸の培地中の最適な濃度は、大腸菌のタンパク質合成系で使用する濃度と比較して、非常に高い。
【0058】
(総論)
本発明の13C標識タンパク質合成法では、他の合成系では困難であった立体構造を維持したタンパク質を合成することでき、さらにバリン、ロイシン、イソロイシンのメチル基を明瞭に13C標識することができた。
【産業上の利用可能性】
【0059】
本発明では、他の合成系では困難であった立体構造を維持したタンパク質を合成することでき、さらにバリン、ロイシン、イソロイシンのメチル基を明瞭に13C標識することができる方法を提供することができる。
【特許請求の範囲】
【請求項1】
以下の工程を含む13Cメチル基標識したバリン、ロイシン及び/又はイソロイシンを含むタンパク質の合成方法:
(1)ホルモンによって活性化される性質を有する転写因子をコードする遺伝子と、転写因子発現用プロモーターとが連結されてなる転写因子発現用DNA断片を宿主細胞に導入する第一形質転換体を作成する工程;
(2)RNAを遺伝子とするウイルスに標的タンパク質をコードする遺伝子を挿入したウイルスベクターのcDNAと、該転写因子で転写誘導される転写誘導型プロモーターとが連結されてなる目的タンパク質発現用DNA断片を、該第一形質転換体に導入して第二形質転換体を作成する工程;
(3)該第二形質転換体をα-ケトイソ吉草酸及び/又はα-ケト酪酸を含む培地で培養して、13C標識タンパク質を得る工程、ここで、該ホルモンは培養前及び/又は培養中に該培地に添加されている。
【請求項2】
前記α-ケトイソ吉草酸及び/又はα-ケト酪酸を含む培地において、α-ケトイソ吉草酸及びα-ケト酪酸の濃度が、それぞれ、100mg〜1000mg/Lである請求項1の合成方法。
【請求項3】
前記α-ケトイソ吉草酸及び/又はα-ケト酪酸を含む培地において、α-ケトイソ吉草酸及びα-ケト酪酸の濃度が、それぞれ、150mg〜350mg/Lである請求項1又は2の合成方法。
【請求項4】
前記標的タンパク質が、分子内にジスルフィド結合を有する請求項1〜3のいずれか1の合成法。
【請求項5】
分子内にジスルフィド結合を有するタンパク質が、以下のいずれか1から選ばれる請求項4の合成方法。
(1)単鎖抗体
(2)分泌タンパク質
(3)膜タンパク質
【請求項6】
前記標的タンパク質が、以下のいずれか1から選ばれる請求項1〜3のいずれか1の合成法。
(1)ウシ膵臓トリプシン阻害薬(BPTI)
(2)ストマジェン(stomagen)
(3)EPF2(EPIDERMAL PATTERNING FACTOR 2)
(4)ストレプトマイセス・スブチリシン阻害剤(SSI)
(5)AFP1(alpha-fetoprotein)
【請求項7】
前記ホルモンが、エストロジェンまたはステロイドホルモンである請求項1〜6のいずれか1の合成法。
【請求項8】
前記エストロジェンで活性化される性質を有する転写因子としてLexA−VP16−hERを用い、前記転写誘導型プロモーターとしてOLexA−46を用いる請求項7の合成法。
【請求項9】
前記ウイルスベクターが、植物ウイルス由来である請求項8の合成法。
【請求項10】
前記宿主細胞および形質転換体が、植物体又は植物由来培養細胞である請求項1〜9のいずれか1の合成法。
【請求項11】
前記ホルモンがエストロジェンであり、
前記宿主細胞がタバコBY2細胞であり、
前記転写因子がLexA−VP16−hERであり、
前記転写誘導型プロモーターがOLexA−46であり、
前記ウイルスベクターがトバモウイルス属に属するウイルス由来である、ことを特徴とする、請求項1〜10のいずれか1の合成法。
【請求項12】
以下を含む13Cメチル基標識したバリン、ロイシン及び/又はイソロイシンを含むタンパク質の合成用キット。
(1)形質転換したBY2細胞
(2)α-ケトイソ吉草酸及び/又はα-ケト酪酸
【請求項1】
以下の工程を含む13Cメチル基標識したバリン、ロイシン及び/又はイソロイシンを含むタンパク質の合成方法:
(1)ホルモンによって活性化される性質を有する転写因子をコードする遺伝子と、転写因子発現用プロモーターとが連結されてなる転写因子発現用DNA断片を宿主細胞に導入する第一形質転換体を作成する工程;
(2)RNAを遺伝子とするウイルスに標的タンパク質をコードする遺伝子を挿入したウイルスベクターのcDNAと、該転写因子で転写誘導される転写誘導型プロモーターとが連結されてなる目的タンパク質発現用DNA断片を、該第一形質転換体に導入して第二形質転換体を作成する工程;
(3)該第二形質転換体をα-ケトイソ吉草酸及び/又はα-ケト酪酸を含む培地で培養して、13C標識タンパク質を得る工程、ここで、該ホルモンは培養前及び/又は培養中に該培地に添加されている。
【請求項2】
前記α-ケトイソ吉草酸及び/又はα-ケト酪酸を含む培地において、α-ケトイソ吉草酸及びα-ケト酪酸の濃度が、それぞれ、100mg〜1000mg/Lである請求項1の合成方法。
【請求項3】
前記α-ケトイソ吉草酸及び/又はα-ケト酪酸を含む培地において、α-ケトイソ吉草酸及びα-ケト酪酸の濃度が、それぞれ、150mg〜350mg/Lである請求項1又は2の合成方法。
【請求項4】
前記標的タンパク質が、分子内にジスルフィド結合を有する請求項1〜3のいずれか1の合成法。
【請求項5】
分子内にジスルフィド結合を有するタンパク質が、以下のいずれか1から選ばれる請求項4の合成方法。
(1)単鎖抗体
(2)分泌タンパク質
(3)膜タンパク質
【請求項6】
前記標的タンパク質が、以下のいずれか1から選ばれる請求項1〜3のいずれか1の合成法。
(1)ウシ膵臓トリプシン阻害薬(BPTI)
(2)ストマジェン(stomagen)
(3)EPF2(EPIDERMAL PATTERNING FACTOR 2)
(4)ストレプトマイセス・スブチリシン阻害剤(SSI)
(5)AFP1(alpha-fetoprotein)
【請求項7】
前記ホルモンが、エストロジェンまたはステロイドホルモンである請求項1〜6のいずれか1の合成法。
【請求項8】
前記エストロジェンで活性化される性質を有する転写因子としてLexA−VP16−hERを用い、前記転写誘導型プロモーターとしてOLexA−46を用いる請求項7の合成法。
【請求項9】
前記ウイルスベクターが、植物ウイルス由来である請求項8の合成法。
【請求項10】
前記宿主細胞および形質転換体が、植物体又は植物由来培養細胞である請求項1〜9のいずれか1の合成法。
【請求項11】
前記ホルモンがエストロジェンであり、
前記宿主細胞がタバコBY2細胞であり、
前記転写因子がLexA−VP16−hERであり、
前記転写誘導型プロモーターがOLexA−46であり、
前記ウイルスベクターがトバモウイルス属に属するウイルス由来である、ことを特徴とする、請求項1〜10のいずれか1の合成法。
【請求項12】
以下を含む13Cメチル基標識したバリン、ロイシン及び/又はイソロイシンを含むタンパク質の合成用キット。
(1)形質転換したBY2細胞
(2)α-ケトイソ吉草酸及び/又はα-ケト酪酸
【図1】


【図2】
【図3】
【図4】

【図5】
【図6】
【図7】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【公開番号】特開2013−34475(P2013−34475A)
【公開日】平成25年2月21日(2013.2.21)
【国際特許分類】
【出願番号】特願2012−151942(P2012−151942)
【出願日】平成24年7月6日(2012.7.6)
【公序良俗違反の表示】
(特許庁注:以下のものは登録商標)
1.リナックス
【出願人】(304024430)国立大学法人北陸先端科学技術大学院大学 (169)
【Fターム(参考)】
【公開日】平成25年2月21日(2013.2.21)
【国際特許分類】
【出願日】平成24年7月6日(2012.7.6)
【公序良俗違反の表示】
(特許庁注:以下のものは登録商標)
1.リナックス
【出願人】(304024430)国立大学法人北陸先端科学技術大学院大学 (169)
【Fターム(参考)】
[ Back to top ]