
4−ケト−D−リボン酸、4−ケト−D−リボース及びそれらの製造方法
【課題】新規化合物である4−ケト−D−リボン酸やその製造方法、及び4−ケト−D−リボースやその製造方法を提供すること。
【解決手段】4−ケト−D−リボン酸生産菌や4−ケト−D−リボース生産菌、例えば4−ケト−D−リボン酸や4−ケト−D−リボース生産能を有するグルコノバクター・サブオキシダンスの生体菌を、フレンチプレスを用いて破砕処理した後、遠心操作により得られた沈殿物を細胞膜画分として単離し、その後かかる細胞膜画分とD−リボースとを反応させると、代謝産物として4−ケト−D−リボースに加えて、4−ケト−D−リボン酸が生成される。
【解決手段】4−ケト−D−リボン酸生産菌や4−ケト−D−リボース生産菌、例えば4−ケト−D−リボン酸や4−ケト−D−リボース生産能を有するグルコノバクター・サブオキシダンスの生体菌を、フレンチプレスを用いて破砕処理した後、遠心操作により得られた沈殿物を細胞膜画分として単離し、その後かかる細胞膜画分とD−リボースとを反応させると、代謝産物として4−ケト−D−リボースに加えて、4−ケト−D−リボン酸が生成される。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、4−ケト−D−リボン酸若しくはその塩又はそれらの誘導体や、4−ケト−D−リボース又はその誘導体や、4−ケト−D−リボン酸や4−ケト−D−リボースの製造方法に関する。
【背景技術】
【0002】
キノプロテインD−グルコースデヒドロゲナーゼ(quinoprotein D-glucose dehydrogenase;GDH)がD−グルコースを酸化することによりD−グルコン酸(GA)を生成し、またフラボプロテインGAデヒドロゲナーゼ(flavoprotein GA dehydrogenase;GADH)がGAを酸化することにより2−ケト−D−グルコン酸(2KGA)を生成し、さらに2KGAデヒドロゲナーゼ(2KGA dehydrogenase;2KGDH)が2KGAを酸化することにより2,5−ジケト−D−グルコン酸(25DKA)を生成する、D−グルコースを多段階的に酸化する経路が酢酸菌にあることが知られている(例えば非特許文献1参照)。また、キノプロテイングリセロールデヒドロゲナーゼがGAを酸化することにより5−ケト−D−グルコン酸(5KGA)を生成することも知られている(例えば非特許文献2参照)。グルコース酸化反応に関与するこれらの酵素は、すべて細胞膜タンパク質であり、かかる酵素が有する酵素活性領域は細胞質膜周辺に露出していることが知られている。上記D−グルコースの酸化産物、すなわちD−グルコースの代謝産物は、L−アスコルビン酸(ビタミンC)と同様に様々な化学製品及び製薬品を製造する過程において、広く利用されている重要な化合物であることが知られている。また、ライヒシュタイン法によるビタミンCの工業的生産において、ビタミンCの合成原料として2−ケト−L−ギュロン酸が使用されるが、かかる2−ケト−L−ギュロン酸は、NADPH依存的な25DKAリダクターゼ(NADPH-dependent 25DKA reductase)が立体構造特異的に25DKAを還元することにより生成されることが知られている。
【0003】
他方、25DKA以降の代謝産物に関する研究は、様々な菌株を用いて盛んに行われているものの、十分な知見又は成果が得られていないのが現状である。例えば、酢酸菌としてアセトバクター・メラノゲナム(Acetobacter melanogenum;A.Melanogenum)を用いた場合、D−グルコースの代謝産物としてD−リキシウロン酸(D-lyxuronic acid)が生成されることが報告されており(例えば非特許文献3)、かかる非特許文献3においてD−リキシウロン酸の生成過程における25DKAの代謝産物として、4−ケト−D−アラボン酸が示唆されているが、4−ケト−D−アラボン酸は精製又は単離はおろか、その存在すら確認できていない。すなわち、4−ケト−D−アラボン酸の存在についてはこれまで知られていなかった。
【0004】
本発明者らは、長年の間、酢酸菌を用いた糖類の生成反応について、鋭意研究を続けている。その研究過程で、アルゼンチン産のケフィア水から単離した酢酸菌であるグルコンアセトバクター・リケファシエンス(Gluconacetobacter liquefaciens:Ga.liquefaciens)株を、D−グルコース、D−グルコン酸又は2−ケト−D−グルコン酸を含有する培地で培養を行った結果、対数増殖期において高効率で2,5−ジケト−D−グルコン酸(25DKA)を代謝産物として生成する能力があることを報告している(例えば、非特許文献4参照)。また、かかるグルコンアセトバクター・リケファシエンス株の細胞膜画分と25DKAとを反応させると、新規物質として4−ケト−D−アラボン酸や4−ケト−D−アラビノースが生成されることも報告している(例えば、非特許文献4参照)。
【先行技術文献】
【非特許文献】
【0005】
【非特許文献1】Matsushita et al. Academic Press, London, pp. 247-301 (1994).
【非特許文献2】Matsushita et al. Appl. Environ. Microbiol., 69, 1959-1966 (2003).
【非特許文献3】Ameyama M and Kondo K, Bull. Agr. Chem. Soc. Jpn., 22, 271-272, 380-386, (1958).
【非特許文献4】Adachi et al., Biosci. Biotechnol. Biochem., 74 (12), 2555-2558, 2010.
【発明の概要】
【発明が解決しようとする課題】
【0006】
本発明の課題は、4−ケト−D−リボン酸やその製造方法、及び4−ケト−D−リボースやその製造方法を提供することにある。
【課題を解決するための手段】
【0007】
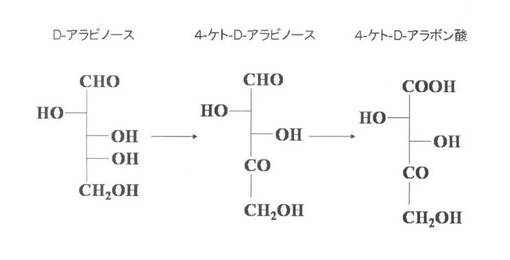
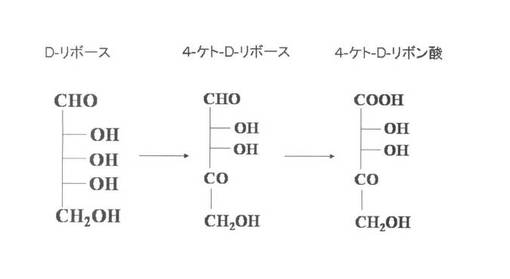
本発明者らは、4−ケト−D−アラボン酸や4−ケト−D−アラビノースが、D−アラビノースからも生成されることを見いだしている(図1参照)。D−アラビノースにおける4位の炭素の水酸基が脱水素化されることにより、D−アラビノースから4−ケト−D−アラビノースが生成されると推測した。また、D−リボースは、D−アラビノースの2位の炭素の立体配置が異なる異性体であり、両者は異なる化合物であるが、3位以下の炭素の構造が同一であることに着目し、D−リボースも同様に脱水素化されるのではないかと考えた。そこで、グルコノバクター・サブオキシダンスの細胞膜画分を単離した後、かかる細胞膜画分とD−リボースとを反応させた結果、4−ケト−D−リボースに加えて、4−ケト−D−リボン酸が生成されることを見いだし(図2参照)、本発明を完成するに至った。
【0008】
すなわち本発明は、(1)以下の式[1]で表される4−ケト−D−リボン酸若しくはその塩、又はそれらの誘導体に関する。
【0009】
【化1】

【0010】
また本発明は、(2)4−ケト−D−リボン酸生産菌又はその細胞膜画分と、D−リボースとを接触させることを特徴とする式[1]で表される4−ケト−D−リボン酸の製造方法や、(3)4−ケト−D−リボン酸生産菌が、グルコノバクター・サブオキシダンス(G.suboxydans)であることを特徴とする上記(2)に記載の製造方法に関する。
【0011】
さらに本発明は、(4)以下の式[2]で表される4−ケト−D−リボース又はその誘導体に関する。
【0012】
【化2】
【0013】
本発明はまた、(5)4−ケト−D−リボース生産菌又はその細胞膜画分と、D−リボースとを接触させることを特徴とする式[2]で表される4−ケト−D−リボースの製造方法や、(6)4−ケト−D−リボース生産菌が、グルコノバクター・サブオキシダンスであることを特徴とする上記(5)に記載の製造方法に関する。
【発明の効果】
【0014】
本発明によると、新規物質である4−ケト−D−リボン酸や4−ケト−D−リボースを提供することができる。また、4−ケト−D−リボン酸や4−ケト−D−リボースは、D−リボース代謝における新規化合物であることから、他の糖類代謝化合物と同様、試薬としての利用や様々な化学製品及び製薬品開発における合成原料としての利用や、4位の炭素が反応性豊かなケトンである点を考慮すると、種々な置換基の導入が可能であり、容易に創薬ブロックとして利用できるほか、易生物分解性の高分子合成素材となることが期待される。すなわち、4−ケト−D−リボン酸や4−ケト−D−リボースは、ケトンを持たない通常の五炭糖と異なって、4位の炭素に反応性が豊かなケトンを有していることからさまざまな置換基を導入することができるので、アデニンアラビノシド等の従来の五炭糖の応用例よりも、遥かに拡大した応用範囲を提供できる。また、ケトンをもつカルボン酸であるピルビン酸と同様に、4−ケト−D−リボン酸や4−ケト−D−リボースを原料とするポリマー合成への展開が期待できる。さらに、近年抗生物質耐性菌が病院等で重篤な問題とされていることから、これらの問題を解決する医薬中間体原料、すなわち新しい抗生物質創製のための側鎖材料への展開も期待される。
【図面の簡単な説明】
【0015】
【図1】4−ケト−D−アラボン酸生産菌におけるD−アラビノース代謝経路を示す図である。
【図2】4−ケト−D−リボン酸及び4−ケト−D−リボース生産菌におけるD−リボース代謝経路を示す図である。
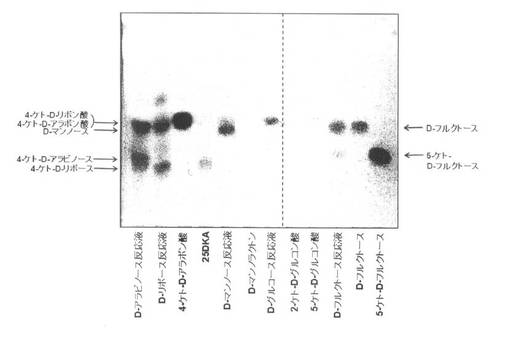
【図3】各種糖類(D−アラビノース、D−リボース、D−マンノース、D−グルコース及びD−フルクトース)と、4−ケト−D−リボン酸及び4−ケト−D−リボース生産菌の細胞膜画分とを反応させた反応産物を薄層クロマトグラフィーで検出した結果を示す図である。4−ケト−D−アラボン酸、25DKA、D−マンノラクトン、2−ケト−D−グルコン酸、5−ケト−D−グルコン酸、D−フルクトース及び5−ケト−D−フルクトースは、コントロールとして用いている。
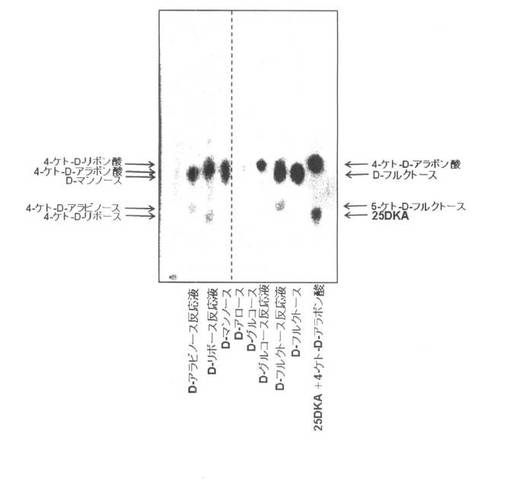
【図4】各種糖類(D−アラビノース、D−リボース、D−グルコース及びD−フルクトース)と、4−ケト−D−リボン酸生産菌や4−ケト−D−リボース生産菌の細胞膜画分とを反応させた反応産物を薄層クロマトグラフィーで検出した結果を示す図である。4−ケトーD−アラボン酸、D−マンノース、D−アロース、D−グルコース、D−フルクトース及び25DKA+4−ケト−D−アラボン酸は、コントロールとして用いている。
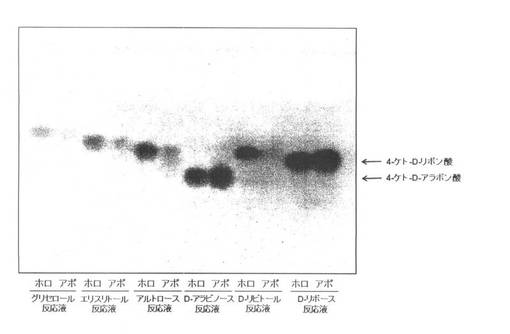
【図5】各種糖類(グリセロール、エリスリトール、アルトロース、D−アラビノース、D−リビトール及びD−リボース)と、4−ケト−D−リボン酸及び4−ケト−D−リボース生産菌の細胞膜画分(ホロ酵素;ホロ)及びEDTAを用いてアポ化処理した細胞膜画分(アポ酵素;アポ)とをそれぞれ反応させた反応産物を薄層クロマトグラフィーで検出した結果を示す図である。
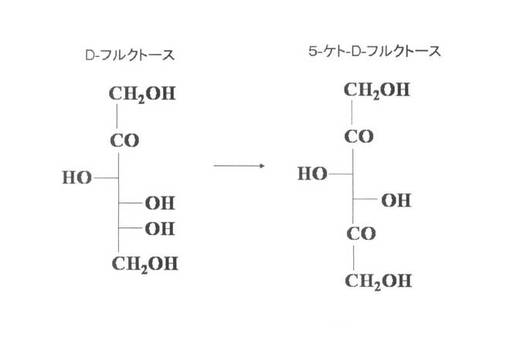
【図6】4−ケト−D−リボン酸及び4−ケト−D−リボース生産菌におけるD−フルクトース代謝経路を示す図である。
【発明を実施するための形態】
【0016】
本発明の4−ケト−D−リボン酸は、4−ケト−D−リボン酸生産菌又はその細胞膜画分と、D−リボースとを接触させることにより製造することができる。また、本発明の4−ケト−D−リボースは、4−ケト−D−リボース生産菌又はその細胞膜画分と、D−リボースとを接触させることにより製造することができる。
【0017】
本発明の4−ケト−D−リボン酸の塩としては、例えばナトリウム塩、カリウム塩、リチウム塩等のアルカリ金属塩や、カルシウム塩、マグネシウム塩等のアルカリ土類金属塩や、アルミニウム塩、鉄塩、亜鉛塩、銅塩、ニッケル塩、コバルト塩等の金属塩や、アンモニウム塩等の無機塩や、t−オクチルアミン塩、ジベンジルアミン塩、モルホリン塩、グルコサミン塩、フェニルグリシンアルキルエステル塩、エチレンジアミン塩、N−メチルグルカミン塩等の有機アミン塩や、グリシン塩、リジン塩、アルギニン塩、オルニチン塩、アスパラギン塩等のアミノ酸塩を挙げることができ、これらの中でも取り扱いが容易なことから、カルシウム塩を好適に例示することができる。また本発明の4−ケト−D−リボン酸やその塩の誘導体としては、4−ケト−D−リボン酸のカルボキシル基がエステルに置換したものや、水酸基がエーテルに置換した誘導体を例示することができる。また本発明の4−ケト−D−リボースの誘導体としては、水酸基がエーテルに置換した誘導体を例示することができる。
【0018】
上記4−ケト−D−リボン酸生産菌としては、グルコノバクター属に属する4−ケト−D−リボン酸生産菌を例示することができ、グルコノバクター属に属する4−ケト−D−リボン酸生産菌としては、4−ケト−D−リボン酸生産能を有するグルコノバクター・サブオキシダンスを例示することができる。また、上記4−ケト−D−リボース生産菌としては、グルコノバクター属に属する4−ケト−D−リボース生産菌を例示することができ、グルコノバクター属に属する4−ケト−D−リボース生産菌としては、4−ケト−D−リボース生産能を有するグルコノバクター・サブオキシダンスを例示することができる。より具体的には、4−ケト−D−リボン酸及び4−ケト−D−リボース生産能を有するグルコノバクター・サブオキシダンス(寄託番号:IFO12528[NBRC12528])を挙げることができる。上記D−リボースとしては、市販のものを購入して用いてもよいし、自ら調製したものを用いてもよい。
【0019】
上記4−ケト−D−リボン酸及び/又は4−ケト−D−リボース生産菌の生菌体をD−リボースと接触させる場合は、菌体を回収、洗浄した洗浄細胞を有利に用いることができる。また、4−ケト−D−リボン酸及び/又は4−ケト−D−リボース生産菌の細胞膜画分を、D−リボースと接触させる場合は、D−リボース代謝系に関与する細胞膜タンパク質酵素活性が維持された菌体処理物を有利に用いることができる。かかる菌体処理物を得るためには、細胞膜タンパク質酵素活性を有した状態で細胞膜と一体化したものとして回収し、濃縮することが好ましい。例えば、フレンチプレス、ビーズホモジナイザー、超音波破砕機やガラスビーズ等を用いて物理的に4−ケト−D−リボン酸及び/又は4−ケト−D−リボース生産菌を破砕処理した後、細胞質成分を含んだ細胞質画分を遠心操作などにより除去することにより、上記細胞膜画分を有利に調製することができる。
【0020】
上記生菌体や細胞膜画分と、D−リボースとを接触させる方法は、両者を溶液等の生化学的な反応が可能な条件下で接触させる方法がよく、溶液、pH、時間、温度等の反応条件は、あらかじめペーパークロマトグラフィー法等を用いて4−ケト−D−リボン酸や4−ケト−D−リボースの生成量を確認することで最適な条件を適宜選択することができる。例えば、10〜100mg/mlの膜濃度に調製した細胞膜画分を、pH3.0〜pH7.0の酢酸緩衝液(10〜100mM)中で、終濃度10〜500mMになるようにD−リボースを加え、数時間〜二夜、15〜35℃で150回転の振盪培養器で反応させることができる。
【0021】
以下に、実施例等を挙げて本発明を具体的に説明するが、本発明の技術的範囲は、これら実施例等により限定されるものではない。
【実施例1】
【0022】
[単離した細胞膜画分を用いた4−ケト−D−リボン酸及び4−ケト−D−リボースの製造]
グルコノバクター・サブオキシダンス(IFO12528;NBRC12528)を、基礎培地(0.3%グリセロール、0.5%酵母エキス、0.2%ポリペプトン)を用いて、36時間(定常増殖期の初期に相当)30℃で振盪培養を行った。Klett濁度計の値が300〜350を示した細胞培養液を、遠心分離により集菌した後、0.1M酢酸緩衝液(pH6.0)で洗浄し、その後再度0.1M酢酸緩衝液(pH6.0)に縣濁した細胞を、フレンチプレス(アミンコ[AMINCO]社製、加圧力1000kg/cm2)を用いて破砕した。得られた細胞破砕液から遠心操作(2000rpm、10分、4℃)により上清を回収した後、再度遠心操作(50000rpm、60分超、4℃)により得られた沈殿物を、細胞膜画分として単離した。細胞膜画分を再度遠心操作(50000rpm、60分超、4℃)することにより、細胞質由来の成分を完全に除去した後、0.1M酢酸緩衝液(pH6.0)に縣濁し、その後ガラスホモゲナイザー(米国Wheaton社製;55ml容)で均一な縣濁液にした細胞膜画分を、膜濃度が30mg/mlになるように調整した。調整した細胞膜画分を、以下の実験に使用した。
【0023】
0.5mlの細胞膜画分と、基質として、0.5mlの0.5M各種糖類(D−アラビノース、D−リボース、D−マンノース、D−グルコース及びD−フルクトース)溶液とを混合した後、25℃で150回転の振盪培養器で一夜(16時間程度)反応させた。その後、10μlの100%TCA溶液により膜画分を変性沈殿させ、反応産物を含む上清を、t−ブタノール:蟻酸:水=4:2:1.5の展開溶媒を用いた薄層クロマトグラフィー(TLC)処理を室温で行い、2,3,5−トリフェニルテトラゾリウムクロライド(TTC)溶液で染色した(図3)。TTC染色は、無色のTTCが還元されると、赤色を呈するトリフェニルホルマザン(TPF)に変換されるため、ケトン基等の強い還元作用を有する化合物が存在する場合に、発色が検出される。また同様に、1mlの0.5M各種標準(4−ケト−D−アラボン酸、25DKA、D−マンノラクトン、2−ケト−D−グルコン酸、5−ケト−D−グルコン酸、D−フルクトース及び5−ケト−D−フルクトース)を用いてコントロールとした。
【0024】
TTC染色により、D−リボースとの反応液からは、新規に4−ケト−D−リボースと4−ケト−D−リボン酸が生成されていた(図3)。一方、D−アラビノースとの反応液からは、4−ケト−D−アラビノース及び4−ケト−D−アラボン酸が生成されていた。4−ケト−D−リボースの相対移動距離(Rf)値は、4−ケト−D−アラビノースのRf値よりも常に小さく、両者は別の化合物であることを示している。また、D−フルクトースとの反応液からは、未反応のD−フルクトースと、標準の5−ケト−D−フルクトースと同じ位置に5−ケト−D−フルクトースの生成が検出された(図3)。この結果は、D−フルクトースの2位以下の炭素の構造がD−アラビノースやD−リボースの構想と似ているため、D−アラビノースやD−リボースの4位の炭素が酸化されたのと同様に、D−フルクトースの5位の炭素も酸化されて、5−ケト−D−フルクトースが生成されたことを示している(図6)。フルクトースから5−ケトフルクトースを生成する酵素として、フルクトース脱水素酵素(FDH)が報告されているが(M. Ameyama, E. Shinagawa, K. Matsushita, & O. Adachi. D-Fructose dehydrogenase of Gluconobacter industrius: Purification, characterization and application to enzymatic micro-determination of D-fructose. J. Bacteriol., 145, 814-823 (1981))、かかる報告では、グルコノバクター・サブオキシダンスのFDH活性は、全て陰性であることが示されている。
【0025】
上記の結果から、D−アラビノースから4−ケト−D−アラビノースが生成される反応と同じ酵素の働きによって、D−リボースから4−ケト−D−リボースが生成されたと考えることができる。また、かかる新規4−ケト−D−アルドース(4−ケト−D−アラビノースや4−ケト−D−リボース)を生成する酵素は、これまで確認されていない新規酵素であることが予想される。さらに、上記新規4−ケト−D−アルドースとD−フルクトースとが構造類似なものであることから、D−フルクトースから5−ケト−D−フルクトースを生成される反応にも、かかる新規酵素が関与していることを示唆している。また、D−マンノースの1位の炭素のアルデヒド基が酸化されると、D−マンノラクトンが生成されることが知られている。D−マンノラクトンには還元作用がないため、TTCによる強い発色は検出されないのに対し、D−マンノースとの反応液からは、TTCによる発色が検出された(図3)。すなわち、D−マンノースとの反応液からは、D−マンノラクトンは生成されていないことを示している。これらの結果は、細胞膜画分に含まれる新規酵素は、すべての糖類を基質とするのではなく、D−アラビノース、D−リボース、D−フルクトース等の糖類を特異的に基質としていることを示している。
【0026】
上記図3で示した実験では、4−ケト−D−リボン酸と4−ケト−D−アラボン酸とを異なるRf値を示すものとして区別できなかったので、再度実験を行った。室温を除く条件は同じであったが、4−ケト−D−アラボン酸のRf値よりも4−ケト−D−リボン酸のRf値の方がやや大きい結果が得られた。この結果は両者が別の化合物であることを示している(図4)。また、図3の結果と同様に、4−ケト−D−リボースのRf値は、4−ケト−D−アラビノースのRf値よりも小さいこと、また、D−フルクトースの反応液から5−ケト−D−フルクトースが生成されることを示していた。なお、グルコースは分子中にケトン基がないため、TTCによる強い発色はわずかしか検出されなかった。
【実施例2】
【0027】
[4−ケト−D−アラビノース及び4−ケト−D−アラボン酸並びに4−ケト−D−リボース及び4−ケト−D−リボン酸の生成に関与する新規酵素]
D−グルコース等の様々な糖類を酸化する酵素として、グリセリン脱水素酵素(K. Matsushita, K. Fujii, Y. Ano, H. Toyama, M. Shinjoh, N. Tomiyama, T. Miyazaki, T. Hoshino & O. Adachi. 5-Keto-D-gluconate production is catalyzed by a quinoprotein glycerol dehydrogenase, major polyol dehydrogenase, in Gluconobacter sp. Appl. Environ. Microbiol., 69, 1959-1966 (2003))が報告されている。かかるグリセリン脱水素酵素のようなポリオール脱水素酵素は、ピロロキノリンキノン(PQQ)を補酵素とする典型的なキノプロテインであり、EDTAで処理すると、容易に酵素活性が失われることが知られている。そこで、4−ケト−D−アラビノース及び4−ケト−D−アラボン酸並びに4−ケト−D−リボース及び4−ケト−D−リボン酸の生成過程に、上記酵素が関与しているかどうかを検証した。細胞膜画分を、20mMのEDTAを含む溶液(20mMトリス−塩酸緩衝液、pH8.0)中に一夜放置することによりPQQの解離を促し、補酵素を失った酵素、すなわち、アポ酵素を含む細胞膜画分を、アポ膜として調製した。かかるアポ膜及びEDTA未処理の細胞膜画分(ホロ酵素を含む)各0.5mlの細胞膜画分と、基質として0.5mlの0.5M各種基質(グリセロール、エリスリトール、アルトロース、D−アラビノース、D−リビトール及びD−リボース)溶液とを混合した後、25℃で150回転の振盪培養器で2日間反応させた。その後、10μlの100%TCA溶液により膜画分を変性沈殿させ、反応産物を含む上清を、t−ブタノール:蟻酸:水=4:2:1.5の展開溶媒を用いた薄層クロマトグラフィー(TLC)処理を、上記図4の実験と同じ室温で行い、2,3,5−トリフェニルテトラゾリウムクロライド(TTC)溶液で染色した(図5)。
【0028】
グリセロース、エリスリトール、アルトロース及びD−リビトールを基質として用いた場合、ホロ酵素として含まれる細胞膜画分では、それぞれが対応するケトン化合物(グリセロールからジオキシアセトン、エリスリトールからエリスルロース、アルトロースから5−ケト−アルトロース、及びD−リビトールからD−リブロース)が検出されたのに対し、アポ酵素として含まれる細胞膜画分では、大幅に発色レベルが減退していた(図5)。一方、D−アラビノース及びD−リボースを基質として用いたとき、アポ酵素として含まれる細胞膜画分を用いた場合に検出される、4−ケト−D−アラボン酸及び4−ケト−D−リボン酸のそれぞれの発色レベルは、ホロ酵素として含まれる細胞膜画分を用いた場合と同じであった(図5)。すなわち、D−アラビノース及びD−リボースそれぞれの4−ケトD−アラビノース及び4−ケト−D−リボースへの反応には、従来知られているグリセリン脱水素酵素のようなポリオール脱水素酵素が関与しているのではなく、新規酵素が関わっている可能性を示している。
【産業上の利用可能性】
【0029】
本発明の4−ケト−D−リボン酸や4−ケト−D−リボースは、D−リボース代謝における新規化合物であることから、他の糖質代謝化合物と同様、様々な化学製品及び製薬品開発における合成原料として有用されることや、4位の炭素がケトンである点を考慮すると、さまざまな置換基を容易に導入できるので、創薬ブロックとしての用途や易生物分解性の高分子合成素材となることが期待される。
【技術分野】
【0001】
本発明は、4−ケト−D−リボン酸若しくはその塩又はそれらの誘導体や、4−ケト−D−リボース又はその誘導体や、4−ケト−D−リボン酸や4−ケト−D−リボースの製造方法に関する。
【背景技術】
【0002】
キノプロテインD−グルコースデヒドロゲナーゼ(quinoprotein D-glucose dehydrogenase;GDH)がD−グルコースを酸化することによりD−グルコン酸(GA)を生成し、またフラボプロテインGAデヒドロゲナーゼ(flavoprotein GA dehydrogenase;GADH)がGAを酸化することにより2−ケト−D−グルコン酸(2KGA)を生成し、さらに2KGAデヒドロゲナーゼ(2KGA dehydrogenase;2KGDH)が2KGAを酸化することにより2,5−ジケト−D−グルコン酸(25DKA)を生成する、D−グルコースを多段階的に酸化する経路が酢酸菌にあることが知られている(例えば非特許文献1参照)。また、キノプロテイングリセロールデヒドロゲナーゼがGAを酸化することにより5−ケト−D−グルコン酸(5KGA)を生成することも知られている(例えば非特許文献2参照)。グルコース酸化反応に関与するこれらの酵素は、すべて細胞膜タンパク質であり、かかる酵素が有する酵素活性領域は細胞質膜周辺に露出していることが知られている。上記D−グルコースの酸化産物、すなわちD−グルコースの代謝産物は、L−アスコルビン酸(ビタミンC)と同様に様々な化学製品及び製薬品を製造する過程において、広く利用されている重要な化合物であることが知られている。また、ライヒシュタイン法によるビタミンCの工業的生産において、ビタミンCの合成原料として2−ケト−L−ギュロン酸が使用されるが、かかる2−ケト−L−ギュロン酸は、NADPH依存的な25DKAリダクターゼ(NADPH-dependent 25DKA reductase)が立体構造特異的に25DKAを還元することにより生成されることが知られている。
【0003】
他方、25DKA以降の代謝産物に関する研究は、様々な菌株を用いて盛んに行われているものの、十分な知見又は成果が得られていないのが現状である。例えば、酢酸菌としてアセトバクター・メラノゲナム(Acetobacter melanogenum;A.Melanogenum)を用いた場合、D−グルコースの代謝産物としてD−リキシウロン酸(D-lyxuronic acid)が生成されることが報告されており(例えば非特許文献3)、かかる非特許文献3においてD−リキシウロン酸の生成過程における25DKAの代謝産物として、4−ケト−D−アラボン酸が示唆されているが、4−ケト−D−アラボン酸は精製又は単離はおろか、その存在すら確認できていない。すなわち、4−ケト−D−アラボン酸の存在についてはこれまで知られていなかった。
【0004】
本発明者らは、長年の間、酢酸菌を用いた糖類の生成反応について、鋭意研究を続けている。その研究過程で、アルゼンチン産のケフィア水から単離した酢酸菌であるグルコンアセトバクター・リケファシエンス(Gluconacetobacter liquefaciens:Ga.liquefaciens)株を、D−グルコース、D−グルコン酸又は2−ケト−D−グルコン酸を含有する培地で培養を行った結果、対数増殖期において高効率で2,5−ジケト−D−グルコン酸(25DKA)を代謝産物として生成する能力があることを報告している(例えば、非特許文献4参照)。また、かかるグルコンアセトバクター・リケファシエンス株の細胞膜画分と25DKAとを反応させると、新規物質として4−ケト−D−アラボン酸や4−ケト−D−アラビノースが生成されることも報告している(例えば、非特許文献4参照)。
【先行技術文献】
【非特許文献】
【0005】
【非特許文献1】Matsushita et al. Academic Press, London, pp. 247-301 (1994).
【非特許文献2】Matsushita et al. Appl. Environ. Microbiol., 69, 1959-1966 (2003).
【非特許文献3】Ameyama M and Kondo K, Bull. Agr. Chem. Soc. Jpn., 22, 271-272, 380-386, (1958).
【非特許文献4】Adachi et al., Biosci. Biotechnol. Biochem., 74 (12), 2555-2558, 2010.
【発明の概要】
【発明が解決しようとする課題】
【0006】
本発明の課題は、4−ケト−D−リボン酸やその製造方法、及び4−ケト−D−リボースやその製造方法を提供することにある。
【課題を解決するための手段】
【0007】
本発明者らは、4−ケト−D−アラボン酸や4−ケト−D−アラビノースが、D−アラビノースからも生成されることを見いだしている(図1参照)。D−アラビノースにおける4位の炭素の水酸基が脱水素化されることにより、D−アラビノースから4−ケト−D−アラビノースが生成されると推測した。また、D−リボースは、D−アラビノースの2位の炭素の立体配置が異なる異性体であり、両者は異なる化合物であるが、3位以下の炭素の構造が同一であることに着目し、D−リボースも同様に脱水素化されるのではないかと考えた。そこで、グルコノバクター・サブオキシダンスの細胞膜画分を単離した後、かかる細胞膜画分とD−リボースとを反応させた結果、4−ケト−D−リボースに加えて、4−ケト−D−リボン酸が生成されることを見いだし(図2参照)、本発明を完成するに至った。
【0008】
すなわち本発明は、(1)以下の式[1]で表される4−ケト−D−リボン酸若しくはその塩、又はそれらの誘導体に関する。
【0009】
【化1】
【0010】
また本発明は、(2)4−ケト−D−リボン酸生産菌又はその細胞膜画分と、D−リボースとを接触させることを特徴とする式[1]で表される4−ケト−D−リボン酸の製造方法や、(3)4−ケト−D−リボン酸生産菌が、グルコノバクター・サブオキシダンス(G.suboxydans)であることを特徴とする上記(2)に記載の製造方法に関する。
【0011】
さらに本発明は、(4)以下の式[2]で表される4−ケト−D−リボース又はその誘導体に関する。
【0012】
【化2】
【0013】
本発明はまた、(5)4−ケト−D−リボース生産菌又はその細胞膜画分と、D−リボースとを接触させることを特徴とする式[2]で表される4−ケト−D−リボースの製造方法や、(6)4−ケト−D−リボース生産菌が、グルコノバクター・サブオキシダンスであることを特徴とする上記(5)に記載の製造方法に関する。
【発明の効果】
【0014】
本発明によると、新規物質である4−ケト−D−リボン酸や4−ケト−D−リボースを提供することができる。また、4−ケト−D−リボン酸や4−ケト−D−リボースは、D−リボース代謝における新規化合物であることから、他の糖類代謝化合物と同様、試薬としての利用や様々な化学製品及び製薬品開発における合成原料としての利用や、4位の炭素が反応性豊かなケトンである点を考慮すると、種々な置換基の導入が可能であり、容易に創薬ブロックとして利用できるほか、易生物分解性の高分子合成素材となることが期待される。すなわち、4−ケト−D−リボン酸や4−ケト−D−リボースは、ケトンを持たない通常の五炭糖と異なって、4位の炭素に反応性が豊かなケトンを有していることからさまざまな置換基を導入することができるので、アデニンアラビノシド等の従来の五炭糖の応用例よりも、遥かに拡大した応用範囲を提供できる。また、ケトンをもつカルボン酸であるピルビン酸と同様に、4−ケト−D−リボン酸や4−ケト−D−リボースを原料とするポリマー合成への展開が期待できる。さらに、近年抗生物質耐性菌が病院等で重篤な問題とされていることから、これらの問題を解決する医薬中間体原料、すなわち新しい抗生物質創製のための側鎖材料への展開も期待される。
【図面の簡単な説明】
【0015】
【図1】4−ケト−D−アラボン酸生産菌におけるD−アラビノース代謝経路を示す図である。
【図2】4−ケト−D−リボン酸及び4−ケト−D−リボース生産菌におけるD−リボース代謝経路を示す図である。
【図3】各種糖類(D−アラビノース、D−リボース、D−マンノース、D−グルコース及びD−フルクトース)と、4−ケト−D−リボン酸及び4−ケト−D−リボース生産菌の細胞膜画分とを反応させた反応産物を薄層クロマトグラフィーで検出した結果を示す図である。4−ケト−D−アラボン酸、25DKA、D−マンノラクトン、2−ケト−D−グルコン酸、5−ケト−D−グルコン酸、D−フルクトース及び5−ケト−D−フルクトースは、コントロールとして用いている。
【図4】各種糖類(D−アラビノース、D−リボース、D−グルコース及びD−フルクトース)と、4−ケト−D−リボン酸生産菌や4−ケト−D−リボース生産菌の細胞膜画分とを反応させた反応産物を薄層クロマトグラフィーで検出した結果を示す図である。4−ケトーD−アラボン酸、D−マンノース、D−アロース、D−グルコース、D−フルクトース及び25DKA+4−ケト−D−アラボン酸は、コントロールとして用いている。
【図5】各種糖類(グリセロール、エリスリトール、アルトロース、D−アラビノース、D−リビトール及びD−リボース)と、4−ケト−D−リボン酸及び4−ケト−D−リボース生産菌の細胞膜画分(ホロ酵素;ホロ)及びEDTAを用いてアポ化処理した細胞膜画分(アポ酵素;アポ)とをそれぞれ反応させた反応産物を薄層クロマトグラフィーで検出した結果を示す図である。
【図6】4−ケト−D−リボン酸及び4−ケト−D−リボース生産菌におけるD−フルクトース代謝経路を示す図である。
【発明を実施するための形態】
【0016】
本発明の4−ケト−D−リボン酸は、4−ケト−D−リボン酸生産菌又はその細胞膜画分と、D−リボースとを接触させることにより製造することができる。また、本発明の4−ケト−D−リボースは、4−ケト−D−リボース生産菌又はその細胞膜画分と、D−リボースとを接触させることにより製造することができる。
【0017】
本発明の4−ケト−D−リボン酸の塩としては、例えばナトリウム塩、カリウム塩、リチウム塩等のアルカリ金属塩や、カルシウム塩、マグネシウム塩等のアルカリ土類金属塩や、アルミニウム塩、鉄塩、亜鉛塩、銅塩、ニッケル塩、コバルト塩等の金属塩や、アンモニウム塩等の無機塩や、t−オクチルアミン塩、ジベンジルアミン塩、モルホリン塩、グルコサミン塩、フェニルグリシンアルキルエステル塩、エチレンジアミン塩、N−メチルグルカミン塩等の有機アミン塩や、グリシン塩、リジン塩、アルギニン塩、オルニチン塩、アスパラギン塩等のアミノ酸塩を挙げることができ、これらの中でも取り扱いが容易なことから、カルシウム塩を好適に例示することができる。また本発明の4−ケト−D−リボン酸やその塩の誘導体としては、4−ケト−D−リボン酸のカルボキシル基がエステルに置換したものや、水酸基がエーテルに置換した誘導体を例示することができる。また本発明の4−ケト−D−リボースの誘導体としては、水酸基がエーテルに置換した誘導体を例示することができる。
【0018】
上記4−ケト−D−リボン酸生産菌としては、グルコノバクター属に属する4−ケト−D−リボン酸生産菌を例示することができ、グルコノバクター属に属する4−ケト−D−リボン酸生産菌としては、4−ケト−D−リボン酸生産能を有するグルコノバクター・サブオキシダンスを例示することができる。また、上記4−ケト−D−リボース生産菌としては、グルコノバクター属に属する4−ケト−D−リボース生産菌を例示することができ、グルコノバクター属に属する4−ケト−D−リボース生産菌としては、4−ケト−D−リボース生産能を有するグルコノバクター・サブオキシダンスを例示することができる。より具体的には、4−ケト−D−リボン酸及び4−ケト−D−リボース生産能を有するグルコノバクター・サブオキシダンス(寄託番号:IFO12528[NBRC12528])を挙げることができる。上記D−リボースとしては、市販のものを購入して用いてもよいし、自ら調製したものを用いてもよい。
【0019】
上記4−ケト−D−リボン酸及び/又は4−ケト−D−リボース生産菌の生菌体をD−リボースと接触させる場合は、菌体を回収、洗浄した洗浄細胞を有利に用いることができる。また、4−ケト−D−リボン酸及び/又は4−ケト−D−リボース生産菌の細胞膜画分を、D−リボースと接触させる場合は、D−リボース代謝系に関与する細胞膜タンパク質酵素活性が維持された菌体処理物を有利に用いることができる。かかる菌体処理物を得るためには、細胞膜タンパク質酵素活性を有した状態で細胞膜と一体化したものとして回収し、濃縮することが好ましい。例えば、フレンチプレス、ビーズホモジナイザー、超音波破砕機やガラスビーズ等を用いて物理的に4−ケト−D−リボン酸及び/又は4−ケト−D−リボース生産菌を破砕処理した後、細胞質成分を含んだ細胞質画分を遠心操作などにより除去することにより、上記細胞膜画分を有利に調製することができる。
【0020】
上記生菌体や細胞膜画分と、D−リボースとを接触させる方法は、両者を溶液等の生化学的な反応が可能な条件下で接触させる方法がよく、溶液、pH、時間、温度等の反応条件は、あらかじめペーパークロマトグラフィー法等を用いて4−ケト−D−リボン酸や4−ケト−D−リボースの生成量を確認することで最適な条件を適宜選択することができる。例えば、10〜100mg/mlの膜濃度に調製した細胞膜画分を、pH3.0〜pH7.0の酢酸緩衝液(10〜100mM)中で、終濃度10〜500mMになるようにD−リボースを加え、数時間〜二夜、15〜35℃で150回転の振盪培養器で反応させることができる。
【0021】
以下に、実施例等を挙げて本発明を具体的に説明するが、本発明の技術的範囲は、これら実施例等により限定されるものではない。
【実施例1】
【0022】
[単離した細胞膜画分を用いた4−ケト−D−リボン酸及び4−ケト−D−リボースの製造]
グルコノバクター・サブオキシダンス(IFO12528;NBRC12528)を、基礎培地(0.3%グリセロール、0.5%酵母エキス、0.2%ポリペプトン)を用いて、36時間(定常増殖期の初期に相当)30℃で振盪培養を行った。Klett濁度計の値が300〜350を示した細胞培養液を、遠心分離により集菌した後、0.1M酢酸緩衝液(pH6.0)で洗浄し、その後再度0.1M酢酸緩衝液(pH6.0)に縣濁した細胞を、フレンチプレス(アミンコ[AMINCO]社製、加圧力1000kg/cm2)を用いて破砕した。得られた細胞破砕液から遠心操作(2000rpm、10分、4℃)により上清を回収した後、再度遠心操作(50000rpm、60分超、4℃)により得られた沈殿物を、細胞膜画分として単離した。細胞膜画分を再度遠心操作(50000rpm、60分超、4℃)することにより、細胞質由来の成分を完全に除去した後、0.1M酢酸緩衝液(pH6.0)に縣濁し、その後ガラスホモゲナイザー(米国Wheaton社製;55ml容)で均一な縣濁液にした細胞膜画分を、膜濃度が30mg/mlになるように調整した。調整した細胞膜画分を、以下の実験に使用した。
【0023】
0.5mlの細胞膜画分と、基質として、0.5mlの0.5M各種糖類(D−アラビノース、D−リボース、D−マンノース、D−グルコース及びD−フルクトース)溶液とを混合した後、25℃で150回転の振盪培養器で一夜(16時間程度)反応させた。その後、10μlの100%TCA溶液により膜画分を変性沈殿させ、反応産物を含む上清を、t−ブタノール:蟻酸:水=4:2:1.5の展開溶媒を用いた薄層クロマトグラフィー(TLC)処理を室温で行い、2,3,5−トリフェニルテトラゾリウムクロライド(TTC)溶液で染色した(図3)。TTC染色は、無色のTTCが還元されると、赤色を呈するトリフェニルホルマザン(TPF)に変換されるため、ケトン基等の強い還元作用を有する化合物が存在する場合に、発色が検出される。また同様に、1mlの0.5M各種標準(4−ケト−D−アラボン酸、25DKA、D−マンノラクトン、2−ケト−D−グルコン酸、5−ケト−D−グルコン酸、D−フルクトース及び5−ケト−D−フルクトース)を用いてコントロールとした。
【0024】
TTC染色により、D−リボースとの反応液からは、新規に4−ケト−D−リボースと4−ケト−D−リボン酸が生成されていた(図3)。一方、D−アラビノースとの反応液からは、4−ケト−D−アラビノース及び4−ケト−D−アラボン酸が生成されていた。4−ケト−D−リボースの相対移動距離(Rf)値は、4−ケト−D−アラビノースのRf値よりも常に小さく、両者は別の化合物であることを示している。また、D−フルクトースとの反応液からは、未反応のD−フルクトースと、標準の5−ケト−D−フルクトースと同じ位置に5−ケト−D−フルクトースの生成が検出された(図3)。この結果は、D−フルクトースの2位以下の炭素の構造がD−アラビノースやD−リボースの構想と似ているため、D−アラビノースやD−リボースの4位の炭素が酸化されたのと同様に、D−フルクトースの5位の炭素も酸化されて、5−ケト−D−フルクトースが生成されたことを示している(図6)。フルクトースから5−ケトフルクトースを生成する酵素として、フルクトース脱水素酵素(FDH)が報告されているが(M. Ameyama, E. Shinagawa, K. Matsushita, & O. Adachi. D-Fructose dehydrogenase of Gluconobacter industrius: Purification, characterization and application to enzymatic micro-determination of D-fructose. J. Bacteriol., 145, 814-823 (1981))、かかる報告では、グルコノバクター・サブオキシダンスのFDH活性は、全て陰性であることが示されている。
【0025】
上記の結果から、D−アラビノースから4−ケト−D−アラビノースが生成される反応と同じ酵素の働きによって、D−リボースから4−ケト−D−リボースが生成されたと考えることができる。また、かかる新規4−ケト−D−アルドース(4−ケト−D−アラビノースや4−ケト−D−リボース)を生成する酵素は、これまで確認されていない新規酵素であることが予想される。さらに、上記新規4−ケト−D−アルドースとD−フルクトースとが構造類似なものであることから、D−フルクトースから5−ケト−D−フルクトースを生成される反応にも、かかる新規酵素が関与していることを示唆している。また、D−マンノースの1位の炭素のアルデヒド基が酸化されると、D−マンノラクトンが生成されることが知られている。D−マンノラクトンには還元作用がないため、TTCによる強い発色は検出されないのに対し、D−マンノースとの反応液からは、TTCによる発色が検出された(図3)。すなわち、D−マンノースとの反応液からは、D−マンノラクトンは生成されていないことを示している。これらの結果は、細胞膜画分に含まれる新規酵素は、すべての糖類を基質とするのではなく、D−アラビノース、D−リボース、D−フルクトース等の糖類を特異的に基質としていることを示している。
【0026】
上記図3で示した実験では、4−ケト−D−リボン酸と4−ケト−D−アラボン酸とを異なるRf値を示すものとして区別できなかったので、再度実験を行った。室温を除く条件は同じであったが、4−ケト−D−アラボン酸のRf値よりも4−ケト−D−リボン酸のRf値の方がやや大きい結果が得られた。この結果は両者が別の化合物であることを示している(図4)。また、図3の結果と同様に、4−ケト−D−リボースのRf値は、4−ケト−D−アラビノースのRf値よりも小さいこと、また、D−フルクトースの反応液から5−ケト−D−フルクトースが生成されることを示していた。なお、グルコースは分子中にケトン基がないため、TTCによる強い発色はわずかしか検出されなかった。
【実施例2】
【0027】
[4−ケト−D−アラビノース及び4−ケト−D−アラボン酸並びに4−ケト−D−リボース及び4−ケト−D−リボン酸の生成に関与する新規酵素]
D−グルコース等の様々な糖類を酸化する酵素として、グリセリン脱水素酵素(K. Matsushita, K. Fujii, Y. Ano, H. Toyama, M. Shinjoh, N. Tomiyama, T. Miyazaki, T. Hoshino & O. Adachi. 5-Keto-D-gluconate production is catalyzed by a quinoprotein glycerol dehydrogenase, major polyol dehydrogenase, in Gluconobacter sp. Appl. Environ. Microbiol., 69, 1959-1966 (2003))が報告されている。かかるグリセリン脱水素酵素のようなポリオール脱水素酵素は、ピロロキノリンキノン(PQQ)を補酵素とする典型的なキノプロテインであり、EDTAで処理すると、容易に酵素活性が失われることが知られている。そこで、4−ケト−D−アラビノース及び4−ケト−D−アラボン酸並びに4−ケト−D−リボース及び4−ケト−D−リボン酸の生成過程に、上記酵素が関与しているかどうかを検証した。細胞膜画分を、20mMのEDTAを含む溶液(20mMトリス−塩酸緩衝液、pH8.0)中に一夜放置することによりPQQの解離を促し、補酵素を失った酵素、すなわち、アポ酵素を含む細胞膜画分を、アポ膜として調製した。かかるアポ膜及びEDTA未処理の細胞膜画分(ホロ酵素を含む)各0.5mlの細胞膜画分と、基質として0.5mlの0.5M各種基質(グリセロール、エリスリトール、アルトロース、D−アラビノース、D−リビトール及びD−リボース)溶液とを混合した後、25℃で150回転の振盪培養器で2日間反応させた。その後、10μlの100%TCA溶液により膜画分を変性沈殿させ、反応産物を含む上清を、t−ブタノール:蟻酸:水=4:2:1.5の展開溶媒を用いた薄層クロマトグラフィー(TLC)処理を、上記図4の実験と同じ室温で行い、2,3,5−トリフェニルテトラゾリウムクロライド(TTC)溶液で染色した(図5)。
【0028】
グリセロース、エリスリトール、アルトロース及びD−リビトールを基質として用いた場合、ホロ酵素として含まれる細胞膜画分では、それぞれが対応するケトン化合物(グリセロールからジオキシアセトン、エリスリトールからエリスルロース、アルトロースから5−ケト−アルトロース、及びD−リビトールからD−リブロース)が検出されたのに対し、アポ酵素として含まれる細胞膜画分では、大幅に発色レベルが減退していた(図5)。一方、D−アラビノース及びD−リボースを基質として用いたとき、アポ酵素として含まれる細胞膜画分を用いた場合に検出される、4−ケト−D−アラボン酸及び4−ケト−D−リボン酸のそれぞれの発色レベルは、ホロ酵素として含まれる細胞膜画分を用いた場合と同じであった(図5)。すなわち、D−アラビノース及びD−リボースそれぞれの4−ケトD−アラビノース及び4−ケト−D−リボースへの反応には、従来知られているグリセリン脱水素酵素のようなポリオール脱水素酵素が関与しているのではなく、新規酵素が関わっている可能性を示している。
【産業上の利用可能性】
【0029】
本発明の4−ケト−D−リボン酸や4−ケト−D−リボースは、D−リボース代謝における新規化合物であることから、他の糖質代謝化合物と同様、様々な化学製品及び製薬品開発における合成原料として有用されることや、4位の炭素がケトンである点を考慮すると、さまざまな置換基を容易に導入できるので、創薬ブロックとしての用途や易生物分解性の高分子合成素材となることが期待される。
【特許請求の範囲】
【請求項1】
以下の式[1]で表される4−ケト−D−リボン酸若しくはその塩、又はそれらの誘導体。
【化1】
【請求項2】
4−ケト−D−リボン酸生産菌又はその細胞膜画分と、D−リボースとを接触させることを特徴とする以下の式[1]で表される4−ケト−D−リボン酸の製造方法。
【化2】
【請求項3】
4−ケト−D−リボン酸生産菌が、グルコノバクター・サブオキシダンス(G.suboxydans)であることを特徴とする請求項2に記載の製造方法。
【請求項4】
以下の式[2]で表される4−ケト−D−リボース又はその誘導体。
【化3】
【請求項5】
4−ケト−D−リボース生産菌又はその細胞膜画分と、D−リボースとを接触させることを特徴とする以下の式[2]で表される4−ケト−D−リボースの製造方法。
【化4】
【請求項6】
4−ケト−D−リボース生産菌が、グルコノバクター・サブオキシダンス(G.suboxydans)であることを特徴とする請求項5に記載の製造方法。
【請求項1】
以下の式[1]で表される4−ケト−D−リボン酸若しくはその塩、又はそれらの誘導体。
【化1】
【請求項2】
4−ケト−D−リボン酸生産菌又はその細胞膜画分と、D−リボースとを接触させることを特徴とする以下の式[1]で表される4−ケト−D−リボン酸の製造方法。
【化2】
【請求項3】
4−ケト−D−リボン酸生産菌が、グルコノバクター・サブオキシダンス(G.suboxydans)であることを特徴とする請求項2に記載の製造方法。
【請求項4】
以下の式[2]で表される4−ケト−D−リボース又はその誘導体。
【化3】
【請求項5】
4−ケト−D−リボース生産菌又はその細胞膜画分と、D−リボースとを接触させることを特徴とする以下の式[2]で表される4−ケト−D−リボースの製造方法。
【化4】
【請求項6】
4−ケト−D−リボース生産菌が、グルコノバクター・サブオキシダンス(G.suboxydans)であることを特徴とする請求項5に記載の製造方法。
【図1】

【図2】
【図3】
【図4】
【図5】
【図6】
【図2】
【図3】
【図4】
【図5】
【図6】
【公開番号】特開2012−184190(P2012−184190A)
【公開日】平成24年9月27日(2012.9.27)
【国際特許分類】
【出願番号】特願2011−47632(P2011−47632)
【出願日】平成23年3月4日(2011.3.4)
【出願人】(304020177)国立大学法人山口大学 (579)
【Fターム(参考)】
【公開日】平成24年9月27日(2012.9.27)
【国際特許分類】
【出願日】平成23年3月4日(2011.3.4)
【出願人】(304020177)国立大学法人山口大学 (579)
【Fターム(参考)】
[ Back to top ]