
4位−カテコールエストロゲン生成阻害剤
【課題】正常なホルモン作用及びカテコールエストロゲンの解毒作用を阻害することなく、4位-カテコールエストロゲンの生成を阻害する優れた乳がん発生予防因子の提供。
【解決手段】下記一般式で表されるメトキシフラボノイド化合物を有効成分とする、4位-カテコールエストロゲン生成阻害剤。

(R1及びR2は水酸基又はメトキシ基、R3は水素原子又は水酸基、R4は水酸基又はメトキシ基を表し、nは0〜4。)
【解決手段】下記一般式で表されるメトキシフラボノイド化合物を有効成分とする、4位-カテコールエストロゲン生成阻害剤。
(R1及びR2は水酸基又はメトキシ基、R3は水素原子又は水酸基、R4は水酸基又はメトキシ基を表し、nは0〜4。)
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、メトキシフラボノイド化合物を有効成分とする4位−カテコールエストロゲン生成阻害剤に関する。詳しくは、本発明は、正常なホルモン作用及びカテコールエストロゲンの解毒作用を阻害することなく、4位−カテコールエストロゲン生成に関与する代謝酵素(CYP1B1)を特異的に阻害しうる薬剤に関するものである。
【背景技術】
【0002】
(1)乳がんの罹患率、死亡率の増加は、近年際だっている。乳がんはホルモン依存性腫瘍であり、そのリスク要因としては、遺伝的要因の他にカロリーが高く脂肪の多い食事、初潮年齢、閉経年齢、出産経験の有無などがあるが、女性ホルモンであるエストロゲンに暴露される時間が長いほどリスクが高くなると考えられている。
エストロゲン(エストロン、エストラジオール(E2))は、乳腺組織内で薬物代謝酵素の一つであるチトクロムP450(CYP1A1、CYP1B1)により、それぞれ2位及び4位のカテコール体(2−OHE2、4−OHE2)へ代謝される。
【0003】
しかしながら、カテコール体はキノン体へ変換する場合があり、特に4位のカテコール体はキノン体(E2−3,4Q)となって活性酸素を生じたり、DNAや染色体チューブリンに付加体を形成したりして、遺伝子突然変異や染色体不安定性を引き起こし、発がん性を示すことが明らかにされた(非特許文献1参照)
【0004】
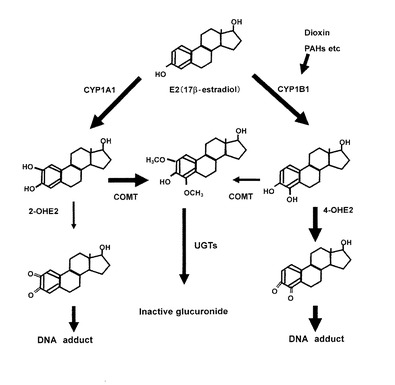
正常な乳腺組織内では、カテコール体はメチル化酵素であるカテコールメチルトランスフェラーゼ(COMT)によりメチル化されて安定なメトキシ体となった後、UDP-glucuronosyltransferase(UGTs)によって不活性のグルクロニド(inactive glucuronide)に抱合化され、組織外へ排出される。しかしながら、乳がん組織内ではこの解毒機構が十分に機能せず、正常組織内よりもCYP1B1の遺伝子発現が亢進し、4位のカテコール体の生成が亢進していることが報告されている(非特許文献2参照)。図1にエストロゲン代謝のメカニズムを表す概略図を示す。
【0005】
(2)また、ホルモン補充療法剤プレマリンに含まれる馬尿より抽出したエキリンやエキレニンも、E2と同様に代謝されて4位のカテコール体(4−OHEQ、4−OHEN)となり、DNA付加体を形成する。ホルモン補充療法の副作用として乳がんや子宮がんの発生が問題となっているが、これらの4位のカテコール体がその原因のひとつとして考えられている。
【0006】
(3)CYP1B1の発現には、ダイオキシンや多環芳香族炭化水素(PAHs)が関与していると考えられている。正常乳腺細胞がダイオキシンやPAHsなどに暴露されると、それらは脂溶性に富むため、細胞膜を容易に通過し核内レセプターであるaryl hydrocarbon receptor(AhR)と結合する。AhRはbHLH−PAS型転写因子の構造を持つ受容体型転写因子であり、リガンドに結合することで核内に移行する。核内でAhR nuclear translocator(Arnt)とヘテロダイマーを形成し、標的遺伝子上流の5'-CACGC-3'をコア配列とするXRE(xenobiotic responsive element)と結合することで標的遺伝子であるCYP1A1/1B1、グルタチオン−S−トランスフェラーゼ(GST)、UGTsが誘導される。
【0007】
薬物代謝型CYPの多くが肝臓で発現しているのに対し、CYP1B1は肝臓では発現しておらず、肺、腎、乳腺や精巣、卵巣、子宮などの生殖器で発現が認められている(非特許文献3参照)。したがって、ダイオキシンやPAHsなどによる暴露によってこれらの臓器でCYP1B1の発現が誘導されると、カテコールエストロゲン生成が亢進され、それがこれらの臓器における発がんのリスク因子となると考えられる。
よって、CYP1B1の働きを特異的に阻害し、4位−カテコールエストロゲンの生成を抑制する物質が見いだされれば、有望な乳がん発生予防因子になりうると考えられる。
【0008】
(4)ところで、フラボノイドはC6−C3−C6の基本骨格を持つポリフェノール化合物であり、野菜、ナッツ類、果物やコーヒー、お茶、赤ワインなどの飲料中に存在する。フラボノイドの機能性については多くの報告があるが、生体内でのフラボノイド健康促進機能として、ヒドロキシラジカル、スーパーオキサイドアニオン、過酸化ラジカルの消去活性があり、膜、蛋白質、DNAへの酸化的ダメージが関与する病気の予防に対し重要な働きをする。
【0009】
フラボノイドは、冠状動脈性心疾患、肝臓病、多くの癌の発生を減少させ予防することがヒトの疫学調査や動物実験により示されている。また、フラボノイドは、CYPによる発がん物質(BaPなど)の代謝活性化抑制作用や生成した活性種(フリーラジカル)などを消去することも報告されている(非特許文献4)。また、メトキシフラボノイドのジメトキシフラボンやスチルベンのレスベラトロル(resveratrol)は、CYP1B1の酵素活性や遺伝子発現を抑制することが報告されている(非特許文献5、6参照)。
【0010】
さらに、フラボンの3’にメトキシ基がつき、4’にアミノ基などが置換しているものにAhRへのアンタゴニスト作用があることも報告されている(非特許文献7参照)。しかし一方で、お茶のカテキンである(-)-epigallocatechin-3-gallate(EGCG)は、カテコールエストロゲンの重要な解毒酵素であるCOMTを競合的に阻害する作用があることがNagaiらによって示された。EGCGなどは、カテコール構造を有するためだと考えられる(非特許文献7)。
【0011】
CYP1B1を特異的に阻害する化合物として2,4,3’,5’−テトラメチルスチルベン(TMS)が報告されているが、この化合物の阻害様式は競合阻害であった(非特許文献8、9)。
Chaudharyらの報告によると、タマリキセチン(tamarixetin)の親化合物のケルセチン(qurcetin)やケムフェロール(kaempherol)、ミリセチン(myricetin)は、10μMになるとCYP1A1とCYP1B1の酵素活性を抑制した(非特許文献10)。MCF10細胞を用いたresveratrolの効果についての報告では、20μM以上でCYP1B1のmRNAの発現を抑制した。E2を基質とした酵素活性も濃度依存的に阻害が見られていた。しかしながら、CYP1A1のmRNAの発現、CYP1A1による酵素活性も同時に抑制していた(非特許文献6)。
乳腺組織ではCYP1A1も働いており、2位のカテコール体を経てCOMTによりメトキシ体になった2−MeOE2には、抗がん作用があることが報告されている。また、2−MeOE2は細胞の増殖、チュウブリンの重合、血管新生作用を阻害する効果がある。従って、CYP1A1も抑制してしまうとE2の解毒代謝機構の一つである2位水酸化とそれに伴うCOMTによるメトキシ化も抑制してしまい、E2のホメオスタシスが崩れてしまう。
【0012】
Doostdarらの報告によると、アカセチン(acacetin)及びジオスメチン(diosmetin)はCYP1A及びCYP1B1に対する阻害作用を示し、ホモエリオディクチオール(homoeriodictyol)はCYP1B1に対し特異的に阻害作用を示した。しかし、ホモエリオディクチオールは、0.1μM以上の比較的高濃度でしか選択的阻害作用を示さない(非特許文献11、12)。
【0013】
(5)ある種のフラボノイド系化合物がヒトのエストロゲン受容体への結合能を有しエストロゲン活性発現抑制効果を有することも報告されている(特許文献1参照)。フラボノイド系化合物はその他にも、紫外線誘発プロスタグランジンE2産生抑制作用(特許文献2参照)、TNF−α産生抑制作用(特許文献3参照)など、種々の生理活性を有することが知られている。
【0014】
乳がん発生予防因子として有用であるためには、それ自身にエストロゲン作用(アゴニスト活性)及びCYP1B1を誘導する作用がないことが必要である。しかしながら、ホルモン補充療法時には、エストロゲン活性を抑制してしまうと、ホルモン補充療法の効果が得られなくなる。
【0015】
(6)そこで、CYP1B1を誘導することなくその作用を特異的に阻害して4位のカテコール体(4位−カテコールエストロゲン)の生成を阻害する作用を低濃度で示し、且つエストロゲンやホルモン補充療法剤等の正常なホルモン作用を阻害せず、またCYP1A1の酵素活性やCOMTの解毒作用をも阻害しない優れた乳がん発生予防因子の開発が望まれているが、未だ見いだされていなかった。
【0016】
【特許文献1】特開2003−2830号公報
【特許文献2】特開2003−192588号公報
【特許文献3】特開2005−213178号公報
【非特許文献1】Tsuchiya Y, Nakajima M, Yokoi T.: Cytochrom P450-mediated metabolism of estrogens and its regulation in human.: Cancer letter 227(2005)
【非特許文献2】Eleanor G.R, Alaa F.B, Prabu D.D, Jane L.M, James A.E, William W.W, Sheila M.H and Ercole L.C: Relative imbalances in estrogen metabolism and conjugation in breast tissue of women with carcinoma: potential biomarkers of susceptibility to cancer.: Carcinogenesis 24 697-702(2003)
【非特許文献3】土屋裕樹、中島美紀、横井毅. ヒトCYP1B1の転写調節機構:発癌とエストロゲンのホメオスタシス:生化学、76、1565−1569
【非特許文献4】吉川敏一編集、フラボノイドの医学、講談社、東京 1998
【非特許文献5】Wen X, Walle U.Kristina and Walle T: 5,7-Dimethoxyflavone downregulates CYP1A1 expression and Benzo[a]Pyrene-induced DNA binding in Hep G2 cell. Carcinogenesis, 26, 803-809(2005)
【非特許文献6】Chen Z-H, Hurch Y-J, Na H-K, Kim H-H, Chun Y-J, Kim D-H, Kang K-S, Cho M-H and Sur Y-J: Resveratol inhibits TCDD-induced expression of CYP1A1 and CYP1B1 and catechol estrogen-mediated oxidative DNA damage in cultured human mammary epithelial cells. Carcinogenesis, 25(10), 2005-2013(2004)
【非特許文献7】Nagai M, Conney AH, Zhu BT.: Strong inhibitory effects of common tea catechins and bioflavonoids on the O-methylation of catechol estrogens catalyzed by human liver cytosolic catechol-O-methyltransferase. DMD. 32, 497-504(2004)
【非特許文献8】Chun YJ, Ryu SY, Jeong TC, Kim MY.: Mechanism-based inhibition of human cytochrome P450 1A1 by rhapontigenin. DMD. 29 389-93. 2001
【非特許文献9】Chun YJ, Kim S, Kim D, Lee SK, Guengerich FP.: A new selective and potent inhibitor of human cytochrome P450 1B1 and its application to antimutagenesis. Cancer Res. 61 8164-70 (2001)
【非特許文献10】Chaudhary A and Willett K. L.: Inhibition of human cytochrome CYP1 enzymes by flavonoids of St. John's wort. Toxicology, 217, 194-205(2006)
【非特許文献11】Doostdar H, Burke MD, Mayer RT: Bioflavonoids: selective substrates and inhibitors for cytochrome P450 CYP1A and CYP1B1. Toxicology, 144, 31-38(2000)
【非特許文献12】Chun YJ, Kim S: Discovery of Cytochrome P450 1B1 Inhibitors as New Promising Anti-Cancer Agents. Medicinal Research Reviews, Vol. 23, No. 6, 657-668(2003)
【発明の開示】
【発明が解決しようとする課題】
【0017】
本発明は、乳がんの発生予防因子の探索を目的とし、正常なホルモン作用及びカテコールエストロゲンの解毒作用を阻害することなく、4位−カテコールエストロゲン生成に関与する代謝酵素(CYP1B1)を特異的に阻害することによって、乳がん発生の重要なリスク因子である4位−カテコールエストロゲンの生成を阻害しうる物質を提供することを課題とする。
【課題を解決するための手段】
【0018】
本発明者は鋭意検討した結果、特定構造を有するメトキシフラボノイド化合物が、上記課題を解決しうることを見いだし、本発明を完成するに至った。
【0019】
すなわち、本発明は、以下の(1)〜(4)に示す4位−カテコールエストロゲン生成阻害剤及び(5)〜(6)に示すCYP1B1活性阻害剤に関する。
(1)下記一般式(1)で表されるメトキシフラボノイド化合物を有効成分とする、4位−カテコールエストロゲン生成阻害剤。
【0020】
【化4】
【0021】
(一般式(1)中、R1及びR2はそれぞれ独立して水酸基又はメトキシ基を表し、R1とR2の少なくとも一方はメトキシ基である。R3は水素原子又は水酸基を表す。R4は水酸基又はメトキシ基を表し、nは0〜4である。)
【0022】
(2)前記メトキシフラボノイド化合物が、一般式(2)で表されることを特徴とする、(1)記載の4位−カテコールエストロゲン生成阻害剤。
【0023】
【化5】
【0024】
(一般式(2)中、R1及びR2はそれぞれ独立して水酸基又はメトキシ基を表し、R1とR2の少なくとも一方はメトキシ基である。R3は水素原子又は水酸基を表す。)
【0025】
(3)前記メトキシフラボノイド化合物が、クリソエリオール、タマリキセチン、及びイソラムネチンからなる群から選択されるものであることを特徴とする、(2)記載の4位−カテコールエストロゲン生成阻害剤。
(4)前記メトキシフラボノイド化合物がクリソエリオールであることを特徴とする、(3)記載の4位−カテコールエストロゲン生成阻害剤。
(5)下記一般式(1)で表されるメトキシフラボノイド化合物を有効成分とする、CYP1B1活性阻害剤。
【0026】
【化6】
【0027】
(一般式(1)中、R1及びR2はそれぞれ独立して水酸基又はメトキシ基を表し、R1とR2の少なくとも一方はメトキシ基である。R3は水素原子又は水酸基を表す。R4は水酸基又はメトキシ基を表し、nは0〜4である。)
(6)前記メトキシフラボノイド化合物がクリソエリオールであることを特徴とする、(5)記載のCYP1B1活性阻害剤。
【発明の効果】
【0028】
本発明の一般式(1)で表される特定のメトキシフラボノイド化合物は、CYP1B1の酵素活性を特異的に阻害する作用及びそれによって乳がん発生のリスク要因となる4位のカテコールエストロゲンの生成を阻害する作用を有するものであり、以下の作用的特徴を備える。
【0029】
(1)カテコール構造を有しないフラボノイド化合物であり、CYP1B1を特異的に阻害しCOMT及びCYP1A1の活性を阻害しないため、2位のカテコール体生成及びそれによる細胞内の解毒代謝機構の正常な働きを保つことができ、エストロゲンの細胞内でのホメオスタシスに影響を与えない。
【0030】
(2)エストロゲンに対するアゴニスト活性は弱く、1μM以下の低濃度ではアンタゴニスト活性を有しないので、エストロゲンの正常なホルモン作用を阻害しない。したがって、ホルモン補充療法においてもホルモン投与による効果を犠牲にすることなく該療法の副作用である乳がん、子宮がんの発生を抑制する可能性がある。
【0031】
(3)それ自身でCYP1B1を誘導する作用はほとんどない。
(4)植物成分であるため、安全性に優れている。
よって、安全で且つ極めて優れた乳がん発生予防因子として有用であり、またCYP1B1を特異的に阻害するので酵素阻害の試薬として利用することもできる。
【発明を実施するための最良の形態】
【0032】
以下に、本発明を実施するための最良の形態を説明する。
(1)メトキシフラボノイド化合物
本発明のカテコールエストロゲン生成阻害剤及びCYP1B1活性阻害剤は、下記一般式(1)で表されるメトキシフラボノイド化合物を有効成分とする。
【0033】
【化7】
【0034】
上記一般式(1)中、置換基R1及びR2はそれぞれ独立して水酸基又はメトキシ基を表し、R1とR2の少なくとも一方はメトキシ基である。好ましくは、R1とR2の一方は水酸基で他方はメトキシ基である。より好ましくは、R1がメトキシ基であり、R2が水酸基である。すなわち、フラボノイド骨格のB環の3’位がメトキシ基に置換しているフラボノイド化合物が好ましい。
【0035】
R3は水素原子又は水酸基を表し、好ましくは水素原子である。置換基R4は水酸基又はメトキシ基を表し、nは0〜4である。好ましくは、R4は水酸基であり、nは2である。最も好ましくは、R4は水酸基であってA環の5位と7位に置換しているものである。
本発明のメトキシフラボノイド化合物の例を以下に化学式で示す。
【0036】
【化8】
【0037】
これらのうち本発明のより好ましいメトキシフラボノイド化合物は、以下の一般式(2)で表されるメトキシフラボノイド化合物である。
【0038】
【化9】
【0039】
上記一般式(2)で表されるメトキシフラボノイド化合物の具体例としては、上述した化学式で表される化合物のうち、フラボノイド骨格のB環の3’位にメトキシ基が置換しているフラボノイド化合物であるクリソエリオール(Chrysoeriol)とタマリキセチン(Tamarixetin)、及びフラボノイド骨格のB環の4’位にメトキシ基が置換しているフラボノイド化合物であるイソラムネチン(Isorhamnetin)とジオスメチン(Diosmetin)が挙げられる。そのうち、より好ましいものはフラボノイド骨格のB環の3’位がメトキシ基のクリソエリオールとタマリキセチンであり、特に好ましいものはクリソエリオールである。クリソエリオールは特異的なCYP1B1阻害作用及び4位のカテコールエストロゲン生成阻害作用を低濃度で発揮することができ、しかも低濃度(例えば0.1μM以下程度)ではAhRに結合せずCYP1B1の遺伝子発現を誘導する可能性が低いことから、本発明のメトキシフラボノイド化合物のなかでも特に好ましい。
【0040】
(2)CYP1B1活性阻害作用及び4位のカテコールエストロゲン生成阻害作用
図1に示すように、エストロゲン(17竈-estradiol;E2)は、CYP1A1及びCYP1B1により、2位及び4位のカテコール体(2−OHE2、4−OHE2)へ代謝されるが、正常な乳腺組織内ではこれらのカテコール体はCOMTの解毒作用によりメトキシ化された後、UGTsにより抱合化され組織外へ排出される(図1参照)。
【0041】
しかしながら、ダイオキシンやPAHsなどによる暴露によって、CYP1B1の遺伝子発現が亢進されると、4位のカテコールエストロゲンの生成が亢進する。生成した4位のカテコールエストロゲンは、セミキノン体やキノン体(E2−3,4Q)への酸化反応を介してフリーラジカルを生成し、DNAを損傷したり、DNA付加体を形成したりして、乳がんや子宮がんを誘発すると考えられている。
【0042】
薬物代謝酵素であるCYPは、脂溶性低分子化合物の酸化反応に関わるヘムタンパク質性モノオキシゲナーゼである。CYPはPAHs、ヘテロサイクリックアミン、芳香族アミン、ニトロ多環芳香族炭化水素等の代謝的活性化を触媒する。PAHsそのものは変異原性を示さないが、CYP1B1によって代謝活性化され、生成したエポキシ体がDNAに結合して遺伝子の突然変異を惹起し、発がんを誘発することが知られている。
【0043】
CYP1B1は、正常組織に比べて腫瘍組織で高発現しており、発がんとの関連が示唆されているが、PAHs等によりAhRを介して誘導が起こる。またE2によっても誘導がかかることが報告されており、これもまた発がんを亢進させる結果となる。ヒト乳がん組織や子宮内膜がん組織では正常と比較して4−OHE2が多く検出されている。かくしてPAHsによる化学発がんでも、ホルモン依存性の癌においても、CYP1B1は癌化を促進する方向へ機能している。
【0044】
本発明者は、CYP1B1を特異的に阻害して乳がん発生予防因子となりうるものを、いくつかのフラボノイド化合物から探索した結果、上記一般式(1)で表されるメトキシフラボノイド化合物を見いだしたものである。
【0045】
本発明のカテコールエストロゲン生成阻害剤は、エストロゲンから4位のカテコール体(4位−カテコールエストロゲン)への変換において鍵となる酵素であるCYP1B1の働きを阻害することにより、4位のカテコールエストロゲンの生成を抑制する作用を有する。
【0046】
また、本発明のカテコールエストロゲン生成阻害剤は、CYP1B1の阻害作用に加えて、次のような作用的特徴を有する。すなわち、CYP1B1を特異的に阻害しCOMT及びCYP1A1の活性を阻害しないため、抗ガン作用を有する2位のカテコール体生成及びそれによる細胞内の解毒代謝機構の正常な働きを保つことができ、エストロゲンの細胞内でのホメオスタシスに影響を与えない。また、肝臓に多く発現するCYP3A4の活性も阻害しない。
【0047】
また、1μM以下の低濃度ではエストロゲンに対するアンタゴニスト活性を有しないので、エストロゲンの正常なホルモン作用を阻害しない。したがって、ホルモン補充療法においてはホルモン投与による効果を犠牲にすることなく該療法の副作用である乳がん、子宮がんの発生を抑制する可能性がある。また、乳がん発生のリスク要因であるCYP1B1を誘導することもない。よって、目的とするCYP1B1のみを選択的に阻害して乳がん発生のリスク要因である4位−カテコールエストロゲンの生成を抑制することができ、極めて優れた乳がん発生抑制因子となりうる。
【0048】
(3)濃度
本発明のメトキシフラボノイド化合物の上述した優れた特異的CYP1B1活性阻害作用及び4位−カテコールエストロゲン生成阻害作用は、比較的低濃度で顕著に発揮される。顕著な効果を示す濃度の制限は特にないが、好ましい濃度(血中濃度)の目安としては、0.1μM以下である。低濃度でCYP1B1に対して特異的に阻害作用を示すので、酵素阻害の試薬としても有用である。特にクリソエリオールが効果を発揮する好ましい濃度範囲は0.01〜0.1μMである。クリソエリオールは0.1μM以下、より好ましくは0.05μM以下の低濃度で優れた特異的CYP1B1活性阻害作用及び4位−カテコールエストロゲン生成阻害作用を示す。
【0049】
(4)入手方法
本発明のカテコールエストロゲン生成阻害剤及びCYP1B1活性阻害剤の有効成分である上記一般式(1)で表される特定のメトキシフラボノイド化合物は、試薬として市販されているものを用いることができる。また、エゴマ中に多く含まれるルテオリンのメチル化体であるから、エゴマからルテオリンを抽出し、これをメチル化剤を用いてメチル化することによっても入手可能である。
【実施例】
【0050】
以下に、実施例を挙げて本発明を具体的に説明するが、本発明はこれらの実施例にのみ限定されるものではない。
【0051】
<実施例1>
エトキシレゾルフィン(ER)はCYP1A1/1B1のO−脱アルキル化反応(EROD)により蛍光物質であるレゾルフィンへと代謝される。この代謝反応を利用してレゾルフィンを励起させてその蛍光強度を測定することによりCYP1A1/1B1の酵素活性を測定することができる(ERODアッセイ)。
また、CYP3A4は肝臓で最も発現しているCYPであるが、このCYP3A4の酵素活性は、ルシフェリン−6’−ベンジルエーテル(luciferin 6' benzyl ether)を基質として、生成したluciferinを測定することにより測定することができる。
本実施例では、メトキシフラボノイド化合物としてクリソエリオール、タマリキセチン、及びイソラムネチンを用い、CYP1A1/1B1/3A4の各酵素と上述した基質を用いてメトキシフラボノイド化合物の酵素阻害活性を測定した。
【0052】
1.実験方法
[CYP1A1/1B1酵素阻害活性;ERODアッセイ]
(1)試薬調製
(i)1Mリン酸バッファー(pH7.4)
NaCl[特級/和光純薬工業];8.0g
KCl[特級/和光純薬工業];0.2g
KH2PO4[特級/和光純薬工業];0.2g
NaHPO4・12H2O[特級/和光純薬工業];2.48g
上記の試薬を蒸留水に溶解し、最終容量を1L(リットル)とした。
【0053】
(ii)酵素調製液(1ウェル(well)あたり)
0.1Mリン酸バッファー;56μl
1M・MgCl2溶液(wako)(97.2mgを100mlの純水に溶解した);4μl
1MG−6−P溶液(オリエンタル酵母)(30.4mgのG−6−Pを1mlの純水に溶解した);4μl
0.1MNADPH溶液(オリエンタル酵母)(88.7mgのNADPHを1mlの純水に溶解した);8μl
0,1MNADH溶液(オリエンタル酵母)(70.9mgのNADHを1mlの純水に溶解した);8μl
【0054】
(iii)2.5mMエトキシレゾルフィン溶液(SIGMA)
1gを2.64mlDMSOで溶解した。
(iv)15μMエトキシレゾルフィン溶液
上記(iii)30μlに対してリン酸バッファー4970μlを加えた;40μl
【0055】
(v)CYP1A1/1B1マイクロゾーム溶液(第一化学)
RecombinantCYP1A1/1B1supersome
(14.25/12.9μg/0.1Mリン酸バッファー1ml);40μl
(vi)フラボノイド溶液(フナコシ)
DMSOに溶解した最終濃度の100倍の濃さにしたフラボノイド2μlをリン酸バッファー40μlに溶解した;40μl
【0056】
(2)検量線用レゾルフィン溶液(TCI)
レゾルフィンをDMSOに溶解し、最終濃度0〜0.2μMの範囲になるようにリン酸バッファーに添加し、レゾルフィン溶液を作成して96ウェル(穴)プレートを用いて蛍光プレートリーダーで測定した。
【0057】
(3)酵素活性測定方法(ERODアッセイ)
96ウェルプレートにフラボノイド溶液を40μl入れ、酵素調製液を80μl加えた。次にマイクロゾーム溶液を40μl加えて5分間37℃でインキュベートした後、そこに15μMエトキシレゾルフィンを加えて5分間インキュベート後、蛍光プレートリーダーで測定をした(励起530nm、発光590nm)。
【0058】
なお、本実験では酵素反応の阻害形式を解析するためにラインウィーバー・バークプロット(Lineweaber-Burk plot)をとったが、これに関してはインキュベート時間を30分間とした。すなわち、上記ERODアッセイにおいて15μMエトキシレゾルフィン溶液を加えた後、30分間インキュベートした。なお、使用したエトキシレゾルフィンの濃度は0、0.186、0.374、0.75、1.5、3及び6μMであった。
【0059】
[CYP3A4酵素阻害活性]
(1)試薬調製(1wellあたり)
(i)4×CYP3A4/リン酸バッファー/Substrate Reaction Mixture
Recombinant CYP3A4 supersome;5.8μl
1Mリン酸バッファー; 2.5μl
Substrate Reaction Mixture; 0.5μl
純水; 3.7μl
Total; 12.5μl
【0060】
(ii)クリソエリオール溶液;12.5μl
(iii)2×NADPH
65mM NADPH; 1 μl
1mM G−6−P; 1.65μl
1mM MgCl2; 1.65μl
1Mリン酸バッファー; 10 μl
純水; 10.7 μl
Total; 25 μl
【0061】
(iv)LDR溶液
Luciferin Detection Reagent+P450-Glo Buffer;50μl
【0062】
(2)CYP3A4の酵素測定法
(i)と(ii)を12.5μlずつルミノスキャン用ホワイト96wellプレートに入れて10分37℃でインキュベートした。その後、(iii)の溶液を25μl添加後、30分37℃でインキュベートした。反応後(iv)の溶液を添加した後、ルミノスキャンで検出した。
【0063】
2.実験結果
(1)CYP1A1/1B1酵素阻害活性の測定結果
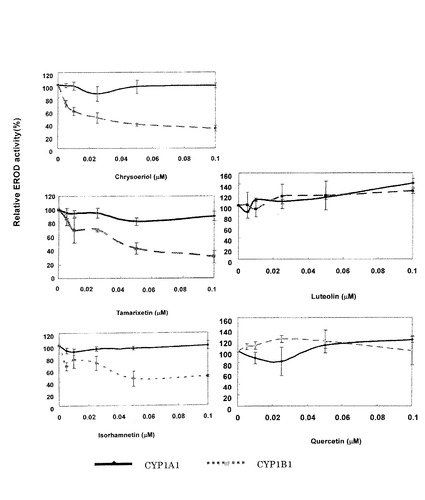
CYP1A1/1B1(14.25/12.9μg)に対してメトキシフラボノイド化合物を0〜0.1μM作用させ、ERODアッセイによる阻害活性を測定した結果を図2に示した。図2よりクリソエリオール、タマリキセチン及びイソラムネチンにCYP1B1に対する特異的な酵素阻害作用が見られ、IC50(酵素活性を50%阻害する濃度)は、それぞれ0.028μM、0.045μM及び0.048μMであった。一方、メチル化されていないケルセチン(quercetin)とルテオリン(luteolin)には、CYP1A1/1B1いずれに対しても阻害効果は見られなかった。
【0064】
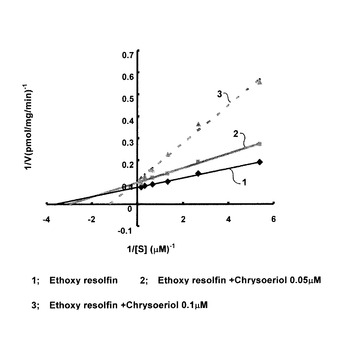
クリソエリオールの阻害様式を確認するためにラインウィーバー・バークプロットをとったところ、図3に示したようにクリソエリオールのCYP1B1に対する阻害様式は非競合阻害だと考えられる。
【0065】
図2からフラボノイド骨格のB環の3’位がメトキシ基に置換しているクリソエリオールとタマリキセチンに、CYP1B1の酵素活性を特異的に低濃度で阻害する作用が顕著に見られた。フラボノイド骨格のB環がメトキシ基に置換していること、特にB環の3’位がメトキシ基に置換していることがCYP1B1の酵素活性を特異的に阻害するのに重要であると考えられる。
【0066】
(2)CYP3A4酵素阻害活性の測定結果
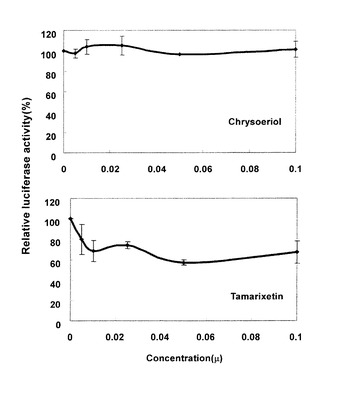
図4にCYP3A4酵素阻害活性の測定結果を示した。それによれば、クリソエリオールによるCYP3A4酵素阻害作用は見られなかった。タマリキセチンによる阻害作用もわずかであった。CYP3A4は肝臓でよく発現しており、本発明のメトキシフラボノイド化合物がCYP3A4酵素阻害作用を示さないことから、肝臓における副作用は少ない可能性が示された。
【0067】
<実施例2>
本実施例では、本発明のメトキシフラボノイド化合物のうち、特に実施例1でCYP1B1の酵素活性を特異的に阻害したクリソエリオールを用いて、E2から2−OHE2及び4−OHE2への代謝に対し阻害が起こるかどうかをGC/MS分析により検討した。
【0068】
1.実験方法
(1)試薬調製
(i)1Mリン酸バッファー(pH7.4)
実施例1の(1)試薬調製(i)に準じて調製した。
【0069】
(ii)酵素調製液(1チューブ(tube)あたり)
0.1Mリン酸バッファー;112μl
1M・MgCl2溶液(wako)(97.2mgを100mlの純水に溶解した);8μl
1MG−6−P溶液(オリエンタル酵母)(30.4mgのG−6−Pを1mlの純水に溶解した);8μl
0.1MNADPH溶液(オリエンタル酵母)(88.7mgのNADPHを1mlの純水に溶解した);16μl
0,1MNADH溶液(オリエンタル酵母)(70.9mgのNADHを1mlの純水に溶解した);16μl
【0070】
(iii)クリソエリオール溶液(フナコシ)
メタノールで調製したクリソエリオールをリン酸バッファーに溶解した;80μl
(iv)E2溶液
E2をメタノールで調整し最終濃度20μMになるようにリン酸バッファーに添加した;80μl
(v)CYP1A1/1B1マイクロゾーム溶液(第一化学)
CYP1A1/1B1(18.4/38.4μg/800μlリン酸バッファー);80μl
【0071】
(2)酵素反応・2−OHE2及び4−OHE2の抽出
15mlチューブにクリソエリオール、酵素調製液、マイクロゾーム溶液を混合し、5分37℃でプレインキュベートした。5分後、混合液の中にE2溶液を入れ40分インキュベートした。40分後反応を止めるためにジクロロメタン5ml添加した。よく振った後、上層をとり、下層のジクロロメタン層に硫酸ナトリウムを添加し、脱水した。ジクロロメタン層の水分が硫酸ナトリウムにより除去され、さらに添加した硫酸ナトリウムがさらさらになったところで溶液をフィルターに通して内部標準物質であるequiline(1μM)を50μl加えた。
【0072】
(3)GC/MS分析
脱水したジクロロメタン抽出液を窒素で濃縮し、それにTMSを50μlずつ添加後、1時間室温で放置し、GC/MSで測定した(TMS化後は、1日間−10℃冷凍保存可能)。
【0073】
(4)GC/MS条件
GC/MS Agilent.6890
注入口:300℃
オーブン:100℃で5分加熱後、15℃/分で300℃まで昇温し、300℃で20分保持した。
SIM:
内部標準−TMS:340(m/Z);22.42min
2OHE2−TMS:504(m/Z);23.95min
4OHE2−TMS:504(m/Z);24.59min
検出器:300℃
キャリアー:He,1ml/min
カラム:DB−5(Agilent)
60×0.32mm×0.25mm
【0074】
(5)統計処理
分散分析を行った後、対照群として指定した1群と多群を比較したDunnett法を用いて検定を行った。
【0075】
2.実験結果
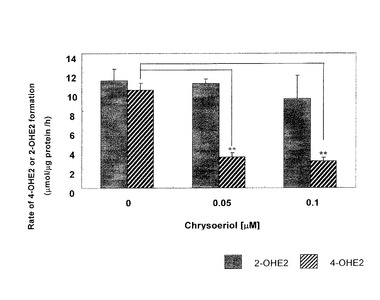
GC/MS分析の結果を図5に示す。クリソエリオールは、CYP1B1による4−OHE2の生成を0.05μMで69%抑制したが、CYP1A1による2−OHE2の生成に対しては抑制作用を示さなかった。この結果は実施例1の結果と同様に、クリソエリオールがCYP1B1を特異的に阻害することを示している。
【0076】
<実施例3>
エストロゲンのカテコール体である4−OHE2および2−OHE2は、COMTによりメチル化され無毒化する。肝サイトゾール中にはCOMTが存在する。本実験では、肝サイトゾール及びドナー基質としてトリチウムラベルしたS-adenosyl-L-methionine(SAM)を用いて、4−OHE2からメチル化された4−MeOE2を検出するメチレーションアッセイを行い、クリソエリオールがCOMTの阻害をしないかどうかを確認した。
【0077】
1.実験方法
(1)試料調製
(i)Tris−HCl(pH7.4)
TRIZMA・BASE(Sigma)60.55gを400mlの純水に溶解し、塩酸でpHを7.4に調節して500mlまでメスアップした。
(ii)24mM・MgCl2
MgCl2(Wako)228.5mgを100mlの純水に溶解した。
(iii)20mM・DTT
DTT(DL-dithiothreitol)60mgを10mlの純水に溶解した。
(iv)1mM cold・SAM([methyl]-S-adenosyl-L-methionine)
32mMのSAM溶液を1mMになるようにエタノールで希釈して、−20℃で保存した。
【0078】
(2)メチレーションアッセイ
(i)反応液の組成
下記表1に示す組成で反応液を調製した。
【0079】
【表1】
【0080】
(ii)反応方法
MgCl2、DTT、cold・SAMを混合したものにTris−HCl(pH7.4)と基質(4−OHE2)を添加してHot・SAMを入れた(ネガコンはここまで)。次に、試料を添加したものとしないものに肝サイトゾールを加え37℃で20分反応を行った。反応後、DDW500μlと反応を止めるためのn−ヘプタン3mlを加え、vortexをかけてから遠心を2000rpmで10分行った。その後、上澄を1ml分注し、シンチバースを4ml加え、vortexをかけた後に液体シンチレーションカウンターで測定した。
【0081】
2.実験結果
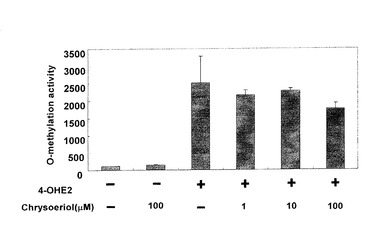
メチレーションアッセイを行った結果、図6に示すようにクリソエリオールにCOMT阻害効果は見られなかった。また4−OHE2を入れずに反応を行ったところ、クリソエリオール自身がCOMTによりメトキシ化されることはなかった。
【0082】
<実施例4>
MCF7細胞においては、CYP1A1よりもCYP1B1が発現しており、細胞内でE2は代謝され、最終的に2−MeOE2及び4−MeOE2の抱合体となって細胞外へ排泄される。そこで、クリソエリオールの存在下、非存在下で細胞を培養し、CYP1B1が細胞内でも阻害されるかどうか、培地中に排泄された2−MeOE2及び4−MeOE2抱合体量を、竈-glucuronidase/sulfataseによる加水分解後、クーロアレイHPLC法により測定した。
【0083】
1.実験方法
(1)使用した細胞
MCF−7:乳癌細胞[乳腺上皮:転移性腫瘍:胸水・腺癌](69 years)
静岡県立大学大学院生活健康科学研究科生体防御研究室、久留戸涼子先生(Dr. Hanspaul Hagenmaier, Institute of Organic Chemistry, University of Tuebingen, Germany)から分与頂いたものを使用。
【0084】
(2)使用した培地(培地1Lあたり)
(i)粉末培地(phenol red)
DMEM(Dulbecco's modified Eagle's medium)[日水製薬];9.5g
炭酸水素ナトリウム[特級/和光純薬];1.5g
L-glutamine[SIGMA];0.58g
カナマイシン[ICN];0.1g
アンピシリン[ICN];0.1g
非働化牛胎児血清(FBS)[ICN];110ml
【0085】
(ii)液体培地(phenol red free medium)
以下に示す組成の培地を調製後、pH7.6に合わせたのち、組織培養フィルター[半径75mm/ポアサイズ0.2μm/NALGENE]で滅菌後使用した。
DMEM(phenol red free)[Gibco];500ml
L-glutamine[SIGMA];0.29g
カナマイシン[ICN];0.05g
アンピシリン[ICN];0.05g
Charcoal/Dextran treated FBS[Hyclone];55ml
【0086】
(ii)1M酢酸アンモニウム(pH4.4)溶液
酢酸アンモニウム(Wako)53.96gを700mlの純水に溶解させた。
(iii)A液
アセトニトリル(関東化学HPLC用);450ml
メタノール(関東化学HPLC用);150ml
純水;2100ml
1M酢酸アンモニウム;300ml
(iv)B液
アセトニトリル(関東化学HPLC用);1500ml
メタノール(関東化学HPLC用);600ml
純水;600ml
1M酢酸アンモニウム;300ml
【0087】
(v)スタンダード原液
20mM E2;5.44mg
20mM 4OHE2;5.76mg
20mM 2MeOE2;6.04mg
20mM 4MeOE2;6.04mg
それぞれをDMSO1mlに溶解した。
(vi)スタンダード溶液
上記(v)で調製したスタンダード原液をB液で20倍希釈→100倍希釈→100倍希釈の順で最終濃度を100nMに調製した。
【0088】
(3)細胞培養
MCF7を1×104cell/cm2になるようにLPディッシュに播き、5日間培養した。5日後、培地をDMEM培地からE2フリーのDMEM培地に変え、0.05及び0.1μMのクリソエリオールを添加し、その15分後に1μMのE2を添加して24時間培養した。培地を採取し、遠心を行った後、上清を液体窒素で凍結し、−20℃で保存した。
【0089】
(4)抽出
培地2mlに対して2N酢酸アンモニウムを0.5ml加えて酸性とし、竈-glucuronidase/sulfatase(SIGMA)を10μl加えて遮光して37℃で17時間酵素処理を行った。その後、ジクロロメタンを2.5ml加えてボルテックスを30秒かけ抽出し、3000rpmで遠心を3層になるまで行った。上層を別の試験管に移し、下層(ジクロロメタン層)をパスツールで取って目盛付試験管に移し、濃縮遠心エバポレーターにより濃縮した。上層を同様に再抽出し、ジクロロメタン層を最初のジクロロメタン層に加え、完全に乾固した。HPLC移動層B液を100μl加えボルテックスをかけて溶解した後、3000rpmで15分遠心し、上清をクーロアレイ用バイアルに入れてクーロアレイHPLC分析を行った。
【0090】
(5)定量分析
定量分析は(6)に示すHPLC−ECD条件下で、ピーク面積を測定することにより行った。
【0091】
(6)HPLC−ECD条件
Column: ESA ODS(4.6×250mm)
Injection Volume:50・l
Temperature:25℃
Detector Potentials:-10,80,170,260,350,440,530,
Gradient solvent: A; acetnitrile, methanol, water, ammonium acetate/ B; acetnitrile, methanol, water, ammonium acetate
Flow rate:1.0ml/min
【0092】
(7)統計処理
分散分析を行った後、対照群として指定した1群と多群を比較したDunnett法を用いて検定を行った。
【0093】
2.実験結果
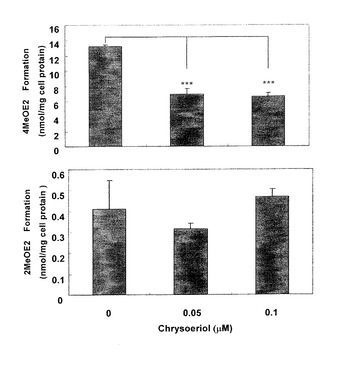
クリソエリオールがCYP1B1を細胞内でも選択的に阻害するかどうかについて、培地中に排泄された2−MeOE2及び4−MeOE2抱合体量を、竈-glucuronidase/sulfataseで加水分解後、クーロアレイHPLC法により測定して確認した。結果を図7に示した。クリソエリオールは、4−MeOE2の生成を0.05μM、0.1μMで有意に抑制したが、2−MeOE2の生成に対しては抑制作用を示さなかった。実施例3によりクリソエリオールがE2の各カテコール体のメチル化を阻害しなかったので、この結果は実施例1〜2の結果と同じであり、本結果はクリソエリオールが細胞内においてもCYP1B1を低濃度で特異的に阻害したことを示している。
【0094】
<実施例5>
本実施例では、本発明のメトキシフラボノイド化合物がそれ自身でエストロゲン作用やCYP1B1を誘導する作用がないかどうかを確認した。すなわち、本発明のメトキシフラボノイド化合物のエストロゲンレセプター結合性を検討し、エストロゲンレセプターへのアンタゴニスト又はアゴニスト活性の有無を検討した。
【0095】
1.実験方法
(1)使用した細胞
ERE−GFP−MCF−7;静岡県立大学大学院生活健康科学研究科生体防御研究室、久留戸涼子先生の開発によるものを用いた。ERE−GFP−MCF7細胞はMCF7細胞にgreen-fluorescent-protein reportor vectorが組み込まれており、EREにE2又はE2様物質が結合すると蛍光を発する。
【0096】
(2)試薬調製
(i)液体培地(phenol red free medium)
DMEM(phenol red free)[Gibco];500ml
L-glutamine[SIGMA];0.29g
カナマイシン[ICN];0.05g
アンピシリン[ICN];0.05g
Charcoal/Dextran treated FBS [Hyclone];55ml
(ii)テストサンプル
クリソエリオールをDMSOに溶解した。
【0097】
(3)培養・測定方法
ERE−GFP−MCF−7細胞を96ウェル(well)プレートに1×104/well播き、細胞が接着するまで4時間インキュベートした。4時間後、10-8Mエストロゲンと種々の濃度のクリソエリオール又はクリソエリオールのみを細胞に添加した。添加してから3日目に細胞にCell counting kitを10μlずつ添加し、2時間後に細胞数を測定した(450nm, Reference 630nm)。細胞数を測定後、wellの溶液を取り除き、蛍光をFluorImager SIで測定をした。
【0098】
2.実験結果
蛍光測定結果を図8に示す。クリソエリオールは、10-8〜10-5の濃度でエストロゲンレセプターへの結合活性が上昇していたが、アンタゴニスト作用は見られなかった。
【0099】
<実施例6>
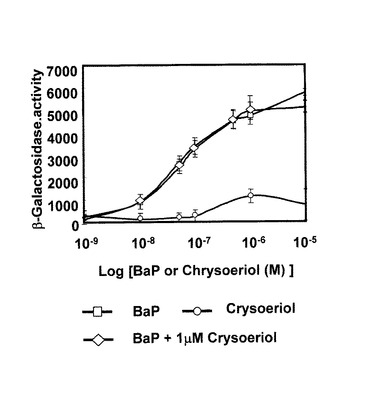
本実施例では、lacZ reporter plasmidならびにヒトAhRとAhR核内輸送酵素をコードする遺伝子を含む遺伝子組み換え酵母Saccharomyces cerevisiae YCM3を用いたレポータージーンアッセイを行った。BaPなどのリガンドがAhRに結合するとβ−ガラクトシダーゼ(竈-galactosidase)が誘導され、本酵素の活性を測定することによりAhRへの結合性が評価できる。本実験では本発明のメトキシフラボノイド化合物のAhRへの結合性を検討し、アゴニスト作用やアンタゴニスト作用がないかを確認した。
【0100】
1.実験方法
(1)使用した酵母
LacZ reporter plasmidならびにヒトAhRとAhR核内輸送酵素をコードする遺伝子を含む遺伝子組み換え酵母Saccharomyces cerevisiae YCM3株
【0101】
(2)試薬等の調製方法
(i)酵母前培養培地
D(+)グルコース;2g
Yeast Nitrogen Base w/0 Amino Acid(BD);0.67g
アデニン塩酸塩(Wako);0.01g
L−ヒスチジン塩酸塩−水和物(Wako);0.01g
L−ロイシン(Wako);0.01g
L(+)−リシン(Wako);0.01g
これらを100mlの純水に溶解した。
【0102】
(ii)酵母本培養培地
ガラクトース;2g
Yeast Nitrogen Base w/0 Amino Acid(BD);0.67g
アデニン塩酸塩(Wako);0.01g
L−ヒスチジン塩酸塩−水和物(Wako);0.01g
L−ロイシン(Wako);0.01g
L(+)−リシン(Wako);0.01g
これらを100mlの純水に溶解した。
【0103】
(iii)Z−Buffer
NaH2PO4・12H2O[特級/和光純薬工業];4.3002g
NaH2PO4・2H2O;1.2438g
KCl;0.1519g
MgSO4・7H2O;0.0492g
これらを200mlの蒸留水に溶解した。
【0104】
(iv)溶菌剤
N-Laurosyl cosine solution(SIGMA);30mg
メルカプトエタノール(Wako);81μl
Z−Buffer;30ml
【0105】
(v)反応液
Z−buffer;15ml
メルカプトエタノール(Wako);45μl
2−ニトロフェニル−β−D−ガラクトピラノシド(Wako);60mg
(vi)クリソエリオール10〜0.001μM(フナコシ)
各種フラボノイドを10mMになるようにDMSOに溶解し、−20℃保存した。それを10〜0.001μM濃度に調製した。
【0106】
(3)測定方法
96ウェル(Well)プレートにBaPとメトキシフラボノイド0.001〜10μMの範囲で添加した。次にガラクトースScに培地を変えた酵母を200μlずつ加えて1晩置いた。翌日、培養した酵母の濁度を630nmで測定した。そこから30μl取り、別のプレートに溶液を移し、そこに溶菌剤を120μl添加して13分インキュベートした。その後、反応液を50μlずつ添加し、数分置いた後(色が出てきたら)450nmで測定した。
【0107】
2.実験結果
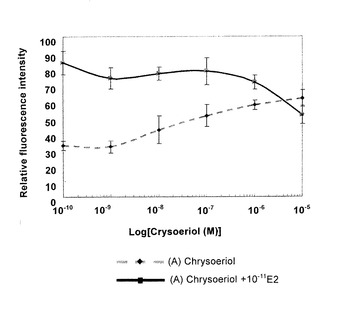
図9に示すように、クリソエリオールのAhR結合活性は非常に弱く、0.1μM以下ではアゴニスト作用は全く見られず、1μMでアンタゴニスト作用も見られなかった。
【0108】
上記実施例1〜5の結果から以下のことが明らかとなった。
(1)実施例1で示されたように、本発明のメトキシフラボノイド化合物は、CYP1B1の酵素活性を低濃度で特異的に阻害し、CYP1A1及びCYP3A4の酵素活性を阻害しない。
【0109】
(2)実施例2において、本発明のメトキシフラボノイド化合物がCYP1A1/1B1によるE2から2−OHE2及び4−OHE2への代謝に対し阻害作用を示すかどうかをGC/MS分析により検討した結果、CYP1B1についてはエトキシレゾルフィンを用いた実施例1の結果と同様の阻害作用が見られ、0.05μMで有意に4−OHE2の生成を69%抑制した。CYP1A1については、ほとんど影響が見られなかった。なお、CYP1B1を特異的に阻害する化合物として2,3,4,5−テトラメチルスチルベン(TMS)が報告されているが、TMSのCYP1B1による4‐OHE2生成阻害活性は、IC50が0.25μMであり、本発明のカテコールエストロゲン生成阻害剤のほうが低濃度で阻害活性を示しており、有効であると考えられる。
【0110】
(3)カテコールエストロゲンの重要な解毒酵素であるCOMTは、カテコールエストロゲンをメチル化するが、クリソエリオールのCOMTへの影響について、肝サイトゾールを用いたメチレーションアッセイを行った実施例3の結果から、クリソエリオールはCOMTを阻害せず、同時にクリソエリオール自身がメチル化されることもなかった。カテキン、エピカテキン、EGCG、ケルセチン、フィセチン(fisetin)のようなカテコール構造を有するフラボノイドは、COMTによるカテコールエストロゲンのメチル化を阻害するが、クリソエリオールをはじめとする本発明のメトキシフラボノイド化合物はカテコール構造を持っておらずCOMTを阻害しないことから、カテコールエストロゲンのメチル化には影響がなくCYP1A1により生成する2−OHE2の代謝を阻害しないため、エストロゲンの細胞内でのホメオスタシスに影響を与えないと思われる。
【0111】
(4)実施例4において、MCF−7細胞を用いて培地中のカテコールエストロゲン代謝物の解析を行い、クリソエリオールがCYP1B1に対して細胞内においても特異的に阻害するかどうかを検討した。その結果、クリソエリオール0.05μM及び0.1μMをそれぞれ作用させた場合、培地中に排泄された4−MeOE2量が低下したが、2−MeOE2量の低下は見られなかった。このことからクリソエリオールは細胞内においてもCYP1B1の酵素に対して特異的に阻害し、細胞内で4−OHE2の生成のみを抑制しうると思われる。従って、クリソエリオールをはじめとする本発明のメトキシフラボノイド化合物はE2が4位の水酸基の酸化、キノン体の生成を経てDNA付加体の形成を促進するのを抑制すると思われる。
【0112】
(5)実施例5に示すように、クリソエリオールは1μM以下ではエストロゲンレセプターに対してアンタゴニスト作用を示さない。高濃度になるにつれてE2へのアゴニスト作用をわずかではあるが示した、すなわちわずかにエストロゲン活性が見られた。しかし、E2に対するアンタゴニスト作用、すなわちエストロゲン活性抑制作用は見られなかった。一方、タマリキセチンにはエストロゲンレセプターへの結合性もE2に対するアンタゴニスト作用も見られなかった。従って、本発明のメトキシフラボノイド化合物は弱いエストロゲン様を示すが、エストロゲンレセプターへのE2の結合作用に対して低濃度ではアンタゴニスト的に働く可能性は低いと考えられる。
【0113】
(6)実施例6に示すように、クリソエリオールはAhRにも低濃度では結合しないことから、クリソエリオール自身はCYPを誘導しないことが示唆された。従ってクリソエリオールはCYP1B1によるE2から4−OHE2の生成を特異的に阻害する因子として有用であると思われる。
【産業上の利用可能性】
【0114】
本発明のメトキシフラボノイド化合物は、低濃度でCYP1B1の酵素活性を特異的に阻害し、乳がんの原因とされる4−OHE2の生成を抑制する作用を有する。しかも、CYP1A1及びCOMTを阻害しないため細胞内の解毒作用が正常に働き、エストロゲンの細胞内でのホメオスタシスに影響を与えない。また、CYP3A4の酵素活性を阻害しないため、肝臓における副作用が少ない可能性がある。また、エストロゲンに対するアンタゴニスト活性を有しないので、エストロゲンの正常なホルモン作用を阻害しない。
【0115】
したがって、ホルモン補充療法においてもホルモン投与による効果を犠牲にすることなく該療法の副作用である乳がん、子宮がんの発生を抑制することができる。よって、安全で且つ極めて優れた乳がん発生予防因子として有用であり、がん予防・治療の分野への貢献が期待される。また、CYP1B1の酵素阻害試薬として利用することもできる。
【図面の簡単な説明】
【0116】
【図1】エストロゲン代謝のメカニズムを表す概略図である。
【図2】実施例1のCYP1A1/1B1酵素阻害活性(ERODアッセイ)の結果を示すグラフである。
【図3】実施例1のラインウィーバー・バークプロットをとった結果を示すグラフである。
【図4】実施例1のCYP3A4酵素阻害活性の測定結果を示すグラフである。
【図5】実施例2のGC/MS分析の結果を示すグラフである。
【図6】実施例3のメチレーションアッセイの結果を示すグラフである。
【図7】実施例4のクーロアレイHPLC法による測定結果を示すグラフである。
【図8】実施例5のERE−GFP−MCF−7細胞の蛍光測定結果を示すグラフである。
【図9】実施例6のレポータージーンアッセイの結果を示すグラフである。
【符号の説明】
【0117】
1 エトキシレゾルフィン
2 エトキシレゾルフィン+クリソエリオール0.05μM
3 エトキシレゾルフィン+クリソエリオール0.1μM
【技術分野】
【0001】
本発明は、メトキシフラボノイド化合物を有効成分とする4位−カテコールエストロゲン生成阻害剤に関する。詳しくは、本発明は、正常なホルモン作用及びカテコールエストロゲンの解毒作用を阻害することなく、4位−カテコールエストロゲン生成に関与する代謝酵素(CYP1B1)を特異的に阻害しうる薬剤に関するものである。
【背景技術】
【0002】
(1)乳がんの罹患率、死亡率の増加は、近年際だっている。乳がんはホルモン依存性腫瘍であり、そのリスク要因としては、遺伝的要因の他にカロリーが高く脂肪の多い食事、初潮年齢、閉経年齢、出産経験の有無などがあるが、女性ホルモンであるエストロゲンに暴露される時間が長いほどリスクが高くなると考えられている。
エストロゲン(エストロン、エストラジオール(E2))は、乳腺組織内で薬物代謝酵素の一つであるチトクロムP450(CYP1A1、CYP1B1)により、それぞれ2位及び4位のカテコール体(2−OHE2、4−OHE2)へ代謝される。
【0003】
しかしながら、カテコール体はキノン体へ変換する場合があり、特に4位のカテコール体はキノン体(E2−3,4Q)となって活性酸素を生じたり、DNAや染色体チューブリンに付加体を形成したりして、遺伝子突然変異や染色体不安定性を引き起こし、発がん性を示すことが明らかにされた(非特許文献1参照)
【0004】
正常な乳腺組織内では、カテコール体はメチル化酵素であるカテコールメチルトランスフェラーゼ(COMT)によりメチル化されて安定なメトキシ体となった後、UDP-glucuronosyltransferase(UGTs)によって不活性のグルクロニド(inactive glucuronide)に抱合化され、組織外へ排出される。しかしながら、乳がん組織内ではこの解毒機構が十分に機能せず、正常組織内よりもCYP1B1の遺伝子発現が亢進し、4位のカテコール体の生成が亢進していることが報告されている(非特許文献2参照)。図1にエストロゲン代謝のメカニズムを表す概略図を示す。
【0005】
(2)また、ホルモン補充療法剤プレマリンに含まれる馬尿より抽出したエキリンやエキレニンも、E2と同様に代謝されて4位のカテコール体(4−OHEQ、4−OHEN)となり、DNA付加体を形成する。ホルモン補充療法の副作用として乳がんや子宮がんの発生が問題となっているが、これらの4位のカテコール体がその原因のひとつとして考えられている。
【0006】
(3)CYP1B1の発現には、ダイオキシンや多環芳香族炭化水素(PAHs)が関与していると考えられている。正常乳腺細胞がダイオキシンやPAHsなどに暴露されると、それらは脂溶性に富むため、細胞膜を容易に通過し核内レセプターであるaryl hydrocarbon receptor(AhR)と結合する。AhRはbHLH−PAS型転写因子の構造を持つ受容体型転写因子であり、リガンドに結合することで核内に移行する。核内でAhR nuclear translocator(Arnt)とヘテロダイマーを形成し、標的遺伝子上流の5'-CACGC-3'をコア配列とするXRE(xenobiotic responsive element)と結合することで標的遺伝子であるCYP1A1/1B1、グルタチオン−S−トランスフェラーゼ(GST)、UGTsが誘導される。
【0007】
薬物代謝型CYPの多くが肝臓で発現しているのに対し、CYP1B1は肝臓では発現しておらず、肺、腎、乳腺や精巣、卵巣、子宮などの生殖器で発現が認められている(非特許文献3参照)。したがって、ダイオキシンやPAHsなどによる暴露によってこれらの臓器でCYP1B1の発現が誘導されると、カテコールエストロゲン生成が亢進され、それがこれらの臓器における発がんのリスク因子となると考えられる。
よって、CYP1B1の働きを特異的に阻害し、4位−カテコールエストロゲンの生成を抑制する物質が見いだされれば、有望な乳がん発生予防因子になりうると考えられる。
【0008】
(4)ところで、フラボノイドはC6−C3−C6の基本骨格を持つポリフェノール化合物であり、野菜、ナッツ類、果物やコーヒー、お茶、赤ワインなどの飲料中に存在する。フラボノイドの機能性については多くの報告があるが、生体内でのフラボノイド健康促進機能として、ヒドロキシラジカル、スーパーオキサイドアニオン、過酸化ラジカルの消去活性があり、膜、蛋白質、DNAへの酸化的ダメージが関与する病気の予防に対し重要な働きをする。
【0009】
フラボノイドは、冠状動脈性心疾患、肝臓病、多くの癌の発生を減少させ予防することがヒトの疫学調査や動物実験により示されている。また、フラボノイドは、CYPによる発がん物質(BaPなど)の代謝活性化抑制作用や生成した活性種(フリーラジカル)などを消去することも報告されている(非特許文献4)。また、メトキシフラボノイドのジメトキシフラボンやスチルベンのレスベラトロル(resveratrol)は、CYP1B1の酵素活性や遺伝子発現を抑制することが報告されている(非特許文献5、6参照)。
【0010】
さらに、フラボンの3’にメトキシ基がつき、4’にアミノ基などが置換しているものにAhRへのアンタゴニスト作用があることも報告されている(非特許文献7参照)。しかし一方で、お茶のカテキンである(-)-epigallocatechin-3-gallate(EGCG)は、カテコールエストロゲンの重要な解毒酵素であるCOMTを競合的に阻害する作用があることがNagaiらによって示された。EGCGなどは、カテコール構造を有するためだと考えられる(非特許文献7)。
【0011】
CYP1B1を特異的に阻害する化合物として2,4,3’,5’−テトラメチルスチルベン(TMS)が報告されているが、この化合物の阻害様式は競合阻害であった(非特許文献8、9)。
Chaudharyらの報告によると、タマリキセチン(tamarixetin)の親化合物のケルセチン(qurcetin)やケムフェロール(kaempherol)、ミリセチン(myricetin)は、10μMになるとCYP1A1とCYP1B1の酵素活性を抑制した(非特許文献10)。MCF10細胞を用いたresveratrolの効果についての報告では、20μM以上でCYP1B1のmRNAの発現を抑制した。E2を基質とした酵素活性も濃度依存的に阻害が見られていた。しかしながら、CYP1A1のmRNAの発現、CYP1A1による酵素活性も同時に抑制していた(非特許文献6)。
乳腺組織ではCYP1A1も働いており、2位のカテコール体を経てCOMTによりメトキシ体になった2−MeOE2には、抗がん作用があることが報告されている。また、2−MeOE2は細胞の増殖、チュウブリンの重合、血管新生作用を阻害する効果がある。従って、CYP1A1も抑制してしまうとE2の解毒代謝機構の一つである2位水酸化とそれに伴うCOMTによるメトキシ化も抑制してしまい、E2のホメオスタシスが崩れてしまう。
【0012】
Doostdarらの報告によると、アカセチン(acacetin)及びジオスメチン(diosmetin)はCYP1A及びCYP1B1に対する阻害作用を示し、ホモエリオディクチオール(homoeriodictyol)はCYP1B1に対し特異的に阻害作用を示した。しかし、ホモエリオディクチオールは、0.1μM以上の比較的高濃度でしか選択的阻害作用を示さない(非特許文献11、12)。
【0013】
(5)ある種のフラボノイド系化合物がヒトのエストロゲン受容体への結合能を有しエストロゲン活性発現抑制効果を有することも報告されている(特許文献1参照)。フラボノイド系化合物はその他にも、紫外線誘発プロスタグランジンE2産生抑制作用(特許文献2参照)、TNF−α産生抑制作用(特許文献3参照)など、種々の生理活性を有することが知られている。
【0014】
乳がん発生予防因子として有用であるためには、それ自身にエストロゲン作用(アゴニスト活性)及びCYP1B1を誘導する作用がないことが必要である。しかしながら、ホルモン補充療法時には、エストロゲン活性を抑制してしまうと、ホルモン補充療法の効果が得られなくなる。
【0015】
(6)そこで、CYP1B1を誘導することなくその作用を特異的に阻害して4位のカテコール体(4位−カテコールエストロゲン)の生成を阻害する作用を低濃度で示し、且つエストロゲンやホルモン補充療法剤等の正常なホルモン作用を阻害せず、またCYP1A1の酵素活性やCOMTの解毒作用をも阻害しない優れた乳がん発生予防因子の開発が望まれているが、未だ見いだされていなかった。
【0016】
【特許文献1】特開2003−2830号公報
【特許文献2】特開2003−192588号公報
【特許文献3】特開2005−213178号公報
【非特許文献1】Tsuchiya Y, Nakajima M, Yokoi T.: Cytochrom P450-mediated metabolism of estrogens and its regulation in human.: Cancer letter 227(2005)
【非特許文献2】Eleanor G.R, Alaa F.B, Prabu D.D, Jane L.M, James A.E, William W.W, Sheila M.H and Ercole L.C: Relative imbalances in estrogen metabolism and conjugation in breast tissue of women with carcinoma: potential biomarkers of susceptibility to cancer.: Carcinogenesis 24 697-702(2003)
【非特許文献3】土屋裕樹、中島美紀、横井毅. ヒトCYP1B1の転写調節機構:発癌とエストロゲンのホメオスタシス:生化学、76、1565−1569
【非特許文献4】吉川敏一編集、フラボノイドの医学、講談社、東京 1998
【非特許文献5】Wen X, Walle U.Kristina and Walle T: 5,7-Dimethoxyflavone downregulates CYP1A1 expression and Benzo[a]Pyrene-induced DNA binding in Hep G2 cell. Carcinogenesis, 26, 803-809(2005)
【非特許文献6】Chen Z-H, Hurch Y-J, Na H-K, Kim H-H, Chun Y-J, Kim D-H, Kang K-S, Cho M-H and Sur Y-J: Resveratol inhibits TCDD-induced expression of CYP1A1 and CYP1B1 and catechol estrogen-mediated oxidative DNA damage in cultured human mammary epithelial cells. Carcinogenesis, 25(10), 2005-2013(2004)
【非特許文献7】Nagai M, Conney AH, Zhu BT.: Strong inhibitory effects of common tea catechins and bioflavonoids on the O-methylation of catechol estrogens catalyzed by human liver cytosolic catechol-O-methyltransferase. DMD. 32, 497-504(2004)
【非特許文献8】Chun YJ, Ryu SY, Jeong TC, Kim MY.: Mechanism-based inhibition of human cytochrome P450 1A1 by rhapontigenin. DMD. 29 389-93. 2001
【非特許文献9】Chun YJ, Kim S, Kim D, Lee SK, Guengerich FP.: A new selective and potent inhibitor of human cytochrome P450 1B1 and its application to antimutagenesis. Cancer Res. 61 8164-70 (2001)
【非特許文献10】Chaudhary A and Willett K. L.: Inhibition of human cytochrome CYP1 enzymes by flavonoids of St. John's wort. Toxicology, 217, 194-205(2006)
【非特許文献11】Doostdar H, Burke MD, Mayer RT: Bioflavonoids: selective substrates and inhibitors for cytochrome P450 CYP1A and CYP1B1. Toxicology, 144, 31-38(2000)
【非特許文献12】Chun YJ, Kim S: Discovery of Cytochrome P450 1B1 Inhibitors as New Promising Anti-Cancer Agents. Medicinal Research Reviews, Vol. 23, No. 6, 657-668(2003)
【発明の開示】
【発明が解決しようとする課題】
【0017】
本発明は、乳がんの発生予防因子の探索を目的とし、正常なホルモン作用及びカテコールエストロゲンの解毒作用を阻害することなく、4位−カテコールエストロゲン生成に関与する代謝酵素(CYP1B1)を特異的に阻害することによって、乳がん発生の重要なリスク因子である4位−カテコールエストロゲンの生成を阻害しうる物質を提供することを課題とする。
【課題を解決するための手段】
【0018】
本発明者は鋭意検討した結果、特定構造を有するメトキシフラボノイド化合物が、上記課題を解決しうることを見いだし、本発明を完成するに至った。
【0019】
すなわち、本発明は、以下の(1)〜(4)に示す4位−カテコールエストロゲン生成阻害剤及び(5)〜(6)に示すCYP1B1活性阻害剤に関する。
(1)下記一般式(1)で表されるメトキシフラボノイド化合物を有効成分とする、4位−カテコールエストロゲン生成阻害剤。
【0020】
【化4】
【0021】
(一般式(1)中、R1及びR2はそれぞれ独立して水酸基又はメトキシ基を表し、R1とR2の少なくとも一方はメトキシ基である。R3は水素原子又は水酸基を表す。R4は水酸基又はメトキシ基を表し、nは0〜4である。)
【0022】
(2)前記メトキシフラボノイド化合物が、一般式(2)で表されることを特徴とする、(1)記載の4位−カテコールエストロゲン生成阻害剤。
【0023】
【化5】
【0024】
(一般式(2)中、R1及びR2はそれぞれ独立して水酸基又はメトキシ基を表し、R1とR2の少なくとも一方はメトキシ基である。R3は水素原子又は水酸基を表す。)
【0025】
(3)前記メトキシフラボノイド化合物が、クリソエリオール、タマリキセチン、及びイソラムネチンからなる群から選択されるものであることを特徴とする、(2)記載の4位−カテコールエストロゲン生成阻害剤。
(4)前記メトキシフラボノイド化合物がクリソエリオールであることを特徴とする、(3)記載の4位−カテコールエストロゲン生成阻害剤。
(5)下記一般式(1)で表されるメトキシフラボノイド化合物を有効成分とする、CYP1B1活性阻害剤。
【0026】
【化6】
【0027】
(一般式(1)中、R1及びR2はそれぞれ独立して水酸基又はメトキシ基を表し、R1とR2の少なくとも一方はメトキシ基である。R3は水素原子又は水酸基を表す。R4は水酸基又はメトキシ基を表し、nは0〜4である。)
(6)前記メトキシフラボノイド化合物がクリソエリオールであることを特徴とする、(5)記載のCYP1B1活性阻害剤。
【発明の効果】
【0028】
本発明の一般式(1)で表される特定のメトキシフラボノイド化合物は、CYP1B1の酵素活性を特異的に阻害する作用及びそれによって乳がん発生のリスク要因となる4位のカテコールエストロゲンの生成を阻害する作用を有するものであり、以下の作用的特徴を備える。
【0029】
(1)カテコール構造を有しないフラボノイド化合物であり、CYP1B1を特異的に阻害しCOMT及びCYP1A1の活性を阻害しないため、2位のカテコール体生成及びそれによる細胞内の解毒代謝機構の正常な働きを保つことができ、エストロゲンの細胞内でのホメオスタシスに影響を与えない。
【0030】
(2)エストロゲンに対するアゴニスト活性は弱く、1μM以下の低濃度ではアンタゴニスト活性を有しないので、エストロゲンの正常なホルモン作用を阻害しない。したがって、ホルモン補充療法においてもホルモン投与による効果を犠牲にすることなく該療法の副作用である乳がん、子宮がんの発生を抑制する可能性がある。
【0031】
(3)それ自身でCYP1B1を誘導する作用はほとんどない。
(4)植物成分であるため、安全性に優れている。
よって、安全で且つ極めて優れた乳がん発生予防因子として有用であり、またCYP1B1を特異的に阻害するので酵素阻害の試薬として利用することもできる。
【発明を実施するための最良の形態】
【0032】
以下に、本発明を実施するための最良の形態を説明する。
(1)メトキシフラボノイド化合物
本発明のカテコールエストロゲン生成阻害剤及びCYP1B1活性阻害剤は、下記一般式(1)で表されるメトキシフラボノイド化合物を有効成分とする。
【0033】
【化7】
【0034】
上記一般式(1)中、置換基R1及びR2はそれぞれ独立して水酸基又はメトキシ基を表し、R1とR2の少なくとも一方はメトキシ基である。好ましくは、R1とR2の一方は水酸基で他方はメトキシ基である。より好ましくは、R1がメトキシ基であり、R2が水酸基である。すなわち、フラボノイド骨格のB環の3’位がメトキシ基に置換しているフラボノイド化合物が好ましい。
【0035】
R3は水素原子又は水酸基を表し、好ましくは水素原子である。置換基R4は水酸基又はメトキシ基を表し、nは0〜4である。好ましくは、R4は水酸基であり、nは2である。最も好ましくは、R4は水酸基であってA環の5位と7位に置換しているものである。
本発明のメトキシフラボノイド化合物の例を以下に化学式で示す。
【0036】
【化8】
【0037】
これらのうち本発明のより好ましいメトキシフラボノイド化合物は、以下の一般式(2)で表されるメトキシフラボノイド化合物である。
【0038】
【化9】
【0039】
上記一般式(2)で表されるメトキシフラボノイド化合物の具体例としては、上述した化学式で表される化合物のうち、フラボノイド骨格のB環の3’位にメトキシ基が置換しているフラボノイド化合物であるクリソエリオール(Chrysoeriol)とタマリキセチン(Tamarixetin)、及びフラボノイド骨格のB環の4’位にメトキシ基が置換しているフラボノイド化合物であるイソラムネチン(Isorhamnetin)とジオスメチン(Diosmetin)が挙げられる。そのうち、より好ましいものはフラボノイド骨格のB環の3’位がメトキシ基のクリソエリオールとタマリキセチンであり、特に好ましいものはクリソエリオールである。クリソエリオールは特異的なCYP1B1阻害作用及び4位のカテコールエストロゲン生成阻害作用を低濃度で発揮することができ、しかも低濃度(例えば0.1μM以下程度)ではAhRに結合せずCYP1B1の遺伝子発現を誘導する可能性が低いことから、本発明のメトキシフラボノイド化合物のなかでも特に好ましい。
【0040】
(2)CYP1B1活性阻害作用及び4位のカテコールエストロゲン生成阻害作用
図1に示すように、エストロゲン(17竈-estradiol;E2)は、CYP1A1及びCYP1B1により、2位及び4位のカテコール体(2−OHE2、4−OHE2)へ代謝されるが、正常な乳腺組織内ではこれらのカテコール体はCOMTの解毒作用によりメトキシ化された後、UGTsにより抱合化され組織外へ排出される(図1参照)。
【0041】
しかしながら、ダイオキシンやPAHsなどによる暴露によって、CYP1B1の遺伝子発現が亢進されると、4位のカテコールエストロゲンの生成が亢進する。生成した4位のカテコールエストロゲンは、セミキノン体やキノン体(E2−3,4Q)への酸化反応を介してフリーラジカルを生成し、DNAを損傷したり、DNA付加体を形成したりして、乳がんや子宮がんを誘発すると考えられている。
【0042】
薬物代謝酵素であるCYPは、脂溶性低分子化合物の酸化反応に関わるヘムタンパク質性モノオキシゲナーゼである。CYPはPAHs、ヘテロサイクリックアミン、芳香族アミン、ニトロ多環芳香族炭化水素等の代謝的活性化を触媒する。PAHsそのものは変異原性を示さないが、CYP1B1によって代謝活性化され、生成したエポキシ体がDNAに結合して遺伝子の突然変異を惹起し、発がんを誘発することが知られている。
【0043】
CYP1B1は、正常組織に比べて腫瘍組織で高発現しており、発がんとの関連が示唆されているが、PAHs等によりAhRを介して誘導が起こる。またE2によっても誘導がかかることが報告されており、これもまた発がんを亢進させる結果となる。ヒト乳がん組織や子宮内膜がん組織では正常と比較して4−OHE2が多く検出されている。かくしてPAHsによる化学発がんでも、ホルモン依存性の癌においても、CYP1B1は癌化を促進する方向へ機能している。
【0044】
本発明者は、CYP1B1を特異的に阻害して乳がん発生予防因子となりうるものを、いくつかのフラボノイド化合物から探索した結果、上記一般式(1)で表されるメトキシフラボノイド化合物を見いだしたものである。
【0045】
本発明のカテコールエストロゲン生成阻害剤は、エストロゲンから4位のカテコール体(4位−カテコールエストロゲン)への変換において鍵となる酵素であるCYP1B1の働きを阻害することにより、4位のカテコールエストロゲンの生成を抑制する作用を有する。
【0046】
また、本発明のカテコールエストロゲン生成阻害剤は、CYP1B1の阻害作用に加えて、次のような作用的特徴を有する。すなわち、CYP1B1を特異的に阻害しCOMT及びCYP1A1の活性を阻害しないため、抗ガン作用を有する2位のカテコール体生成及びそれによる細胞内の解毒代謝機構の正常な働きを保つことができ、エストロゲンの細胞内でのホメオスタシスに影響を与えない。また、肝臓に多く発現するCYP3A4の活性も阻害しない。
【0047】
また、1μM以下の低濃度ではエストロゲンに対するアンタゴニスト活性を有しないので、エストロゲンの正常なホルモン作用を阻害しない。したがって、ホルモン補充療法においてはホルモン投与による効果を犠牲にすることなく該療法の副作用である乳がん、子宮がんの発生を抑制する可能性がある。また、乳がん発生のリスク要因であるCYP1B1を誘導することもない。よって、目的とするCYP1B1のみを選択的に阻害して乳がん発生のリスク要因である4位−カテコールエストロゲンの生成を抑制することができ、極めて優れた乳がん発生抑制因子となりうる。
【0048】
(3)濃度
本発明のメトキシフラボノイド化合物の上述した優れた特異的CYP1B1活性阻害作用及び4位−カテコールエストロゲン生成阻害作用は、比較的低濃度で顕著に発揮される。顕著な効果を示す濃度の制限は特にないが、好ましい濃度(血中濃度)の目安としては、0.1μM以下である。低濃度でCYP1B1に対して特異的に阻害作用を示すので、酵素阻害の試薬としても有用である。特にクリソエリオールが効果を発揮する好ましい濃度範囲は0.01〜0.1μMである。クリソエリオールは0.1μM以下、より好ましくは0.05μM以下の低濃度で優れた特異的CYP1B1活性阻害作用及び4位−カテコールエストロゲン生成阻害作用を示す。
【0049】
(4)入手方法
本発明のカテコールエストロゲン生成阻害剤及びCYP1B1活性阻害剤の有効成分である上記一般式(1)で表される特定のメトキシフラボノイド化合物は、試薬として市販されているものを用いることができる。また、エゴマ中に多く含まれるルテオリンのメチル化体であるから、エゴマからルテオリンを抽出し、これをメチル化剤を用いてメチル化することによっても入手可能である。
【実施例】
【0050】
以下に、実施例を挙げて本発明を具体的に説明するが、本発明はこれらの実施例にのみ限定されるものではない。
【0051】
<実施例1>
エトキシレゾルフィン(ER)はCYP1A1/1B1のO−脱アルキル化反応(EROD)により蛍光物質であるレゾルフィンへと代謝される。この代謝反応を利用してレゾルフィンを励起させてその蛍光強度を測定することによりCYP1A1/1B1の酵素活性を測定することができる(ERODアッセイ)。
また、CYP3A4は肝臓で最も発現しているCYPであるが、このCYP3A4の酵素活性は、ルシフェリン−6’−ベンジルエーテル(luciferin 6' benzyl ether)を基質として、生成したluciferinを測定することにより測定することができる。
本実施例では、メトキシフラボノイド化合物としてクリソエリオール、タマリキセチン、及びイソラムネチンを用い、CYP1A1/1B1/3A4の各酵素と上述した基質を用いてメトキシフラボノイド化合物の酵素阻害活性を測定した。
【0052】
1.実験方法
[CYP1A1/1B1酵素阻害活性;ERODアッセイ]
(1)試薬調製
(i)1Mリン酸バッファー(pH7.4)
NaCl[特級/和光純薬工業];8.0g
KCl[特級/和光純薬工業];0.2g
KH2PO4[特級/和光純薬工業];0.2g
NaHPO4・12H2O[特級/和光純薬工業];2.48g
上記の試薬を蒸留水に溶解し、最終容量を1L(リットル)とした。
【0053】
(ii)酵素調製液(1ウェル(well)あたり)
0.1Mリン酸バッファー;56μl
1M・MgCl2溶液(wako)(97.2mgを100mlの純水に溶解した);4μl
1MG−6−P溶液(オリエンタル酵母)(30.4mgのG−6−Pを1mlの純水に溶解した);4μl
0.1MNADPH溶液(オリエンタル酵母)(88.7mgのNADPHを1mlの純水に溶解した);8μl
0,1MNADH溶液(オリエンタル酵母)(70.9mgのNADHを1mlの純水に溶解した);8μl
【0054】
(iii)2.5mMエトキシレゾルフィン溶液(SIGMA)
1gを2.64mlDMSOで溶解した。
(iv)15μMエトキシレゾルフィン溶液
上記(iii)30μlに対してリン酸バッファー4970μlを加えた;40μl
【0055】
(v)CYP1A1/1B1マイクロゾーム溶液(第一化学)
RecombinantCYP1A1/1B1supersome
(14.25/12.9μg/0.1Mリン酸バッファー1ml);40μl
(vi)フラボノイド溶液(フナコシ)
DMSOに溶解した最終濃度の100倍の濃さにしたフラボノイド2μlをリン酸バッファー40μlに溶解した;40μl
【0056】
(2)検量線用レゾルフィン溶液(TCI)
レゾルフィンをDMSOに溶解し、最終濃度0〜0.2μMの範囲になるようにリン酸バッファーに添加し、レゾルフィン溶液を作成して96ウェル(穴)プレートを用いて蛍光プレートリーダーで測定した。
【0057】
(3)酵素活性測定方法(ERODアッセイ)
96ウェルプレートにフラボノイド溶液を40μl入れ、酵素調製液を80μl加えた。次にマイクロゾーム溶液を40μl加えて5分間37℃でインキュベートした後、そこに15μMエトキシレゾルフィンを加えて5分間インキュベート後、蛍光プレートリーダーで測定をした(励起530nm、発光590nm)。
【0058】
なお、本実験では酵素反応の阻害形式を解析するためにラインウィーバー・バークプロット(Lineweaber-Burk plot)をとったが、これに関してはインキュベート時間を30分間とした。すなわち、上記ERODアッセイにおいて15μMエトキシレゾルフィン溶液を加えた後、30分間インキュベートした。なお、使用したエトキシレゾルフィンの濃度は0、0.186、0.374、0.75、1.5、3及び6μMであった。
【0059】
[CYP3A4酵素阻害活性]
(1)試薬調製(1wellあたり)
(i)4×CYP3A4/リン酸バッファー/Substrate Reaction Mixture
Recombinant CYP3A4 supersome;5.8μl
1Mリン酸バッファー; 2.5μl
Substrate Reaction Mixture; 0.5μl
純水; 3.7μl
Total; 12.5μl
【0060】
(ii)クリソエリオール溶液;12.5μl
(iii)2×NADPH
65mM NADPH; 1 μl
1mM G−6−P; 1.65μl
1mM MgCl2; 1.65μl
1Mリン酸バッファー; 10 μl
純水; 10.7 μl
Total; 25 μl
【0061】
(iv)LDR溶液
Luciferin Detection Reagent+P450-Glo Buffer;50μl
【0062】
(2)CYP3A4の酵素測定法
(i)と(ii)を12.5μlずつルミノスキャン用ホワイト96wellプレートに入れて10分37℃でインキュベートした。その後、(iii)の溶液を25μl添加後、30分37℃でインキュベートした。反応後(iv)の溶液を添加した後、ルミノスキャンで検出した。
【0063】
2.実験結果
(1)CYP1A1/1B1酵素阻害活性の測定結果
CYP1A1/1B1(14.25/12.9μg)に対してメトキシフラボノイド化合物を0〜0.1μM作用させ、ERODアッセイによる阻害活性を測定した結果を図2に示した。図2よりクリソエリオール、タマリキセチン及びイソラムネチンにCYP1B1に対する特異的な酵素阻害作用が見られ、IC50(酵素活性を50%阻害する濃度)は、それぞれ0.028μM、0.045μM及び0.048μMであった。一方、メチル化されていないケルセチン(quercetin)とルテオリン(luteolin)には、CYP1A1/1B1いずれに対しても阻害効果は見られなかった。
【0064】
クリソエリオールの阻害様式を確認するためにラインウィーバー・バークプロットをとったところ、図3に示したようにクリソエリオールのCYP1B1に対する阻害様式は非競合阻害だと考えられる。
【0065】
図2からフラボノイド骨格のB環の3’位がメトキシ基に置換しているクリソエリオールとタマリキセチンに、CYP1B1の酵素活性を特異的に低濃度で阻害する作用が顕著に見られた。フラボノイド骨格のB環がメトキシ基に置換していること、特にB環の3’位がメトキシ基に置換していることがCYP1B1の酵素活性を特異的に阻害するのに重要であると考えられる。
【0066】
(2)CYP3A4酵素阻害活性の測定結果
図4にCYP3A4酵素阻害活性の測定結果を示した。それによれば、クリソエリオールによるCYP3A4酵素阻害作用は見られなかった。タマリキセチンによる阻害作用もわずかであった。CYP3A4は肝臓でよく発現しており、本発明のメトキシフラボノイド化合物がCYP3A4酵素阻害作用を示さないことから、肝臓における副作用は少ない可能性が示された。
【0067】
<実施例2>
本実施例では、本発明のメトキシフラボノイド化合物のうち、特に実施例1でCYP1B1の酵素活性を特異的に阻害したクリソエリオールを用いて、E2から2−OHE2及び4−OHE2への代謝に対し阻害が起こるかどうかをGC/MS分析により検討した。
【0068】
1.実験方法
(1)試薬調製
(i)1Mリン酸バッファー(pH7.4)
実施例1の(1)試薬調製(i)に準じて調製した。
【0069】
(ii)酵素調製液(1チューブ(tube)あたり)
0.1Mリン酸バッファー;112μl
1M・MgCl2溶液(wako)(97.2mgを100mlの純水に溶解した);8μl
1MG−6−P溶液(オリエンタル酵母)(30.4mgのG−6−Pを1mlの純水に溶解した);8μl
0.1MNADPH溶液(オリエンタル酵母)(88.7mgのNADPHを1mlの純水に溶解した);16μl
0,1MNADH溶液(オリエンタル酵母)(70.9mgのNADHを1mlの純水に溶解した);16μl
【0070】
(iii)クリソエリオール溶液(フナコシ)
メタノールで調製したクリソエリオールをリン酸バッファーに溶解した;80μl
(iv)E2溶液
E2をメタノールで調整し最終濃度20μMになるようにリン酸バッファーに添加した;80μl
(v)CYP1A1/1B1マイクロゾーム溶液(第一化学)
CYP1A1/1B1(18.4/38.4μg/800μlリン酸バッファー);80μl
【0071】
(2)酵素反応・2−OHE2及び4−OHE2の抽出
15mlチューブにクリソエリオール、酵素調製液、マイクロゾーム溶液を混合し、5分37℃でプレインキュベートした。5分後、混合液の中にE2溶液を入れ40分インキュベートした。40分後反応を止めるためにジクロロメタン5ml添加した。よく振った後、上層をとり、下層のジクロロメタン層に硫酸ナトリウムを添加し、脱水した。ジクロロメタン層の水分が硫酸ナトリウムにより除去され、さらに添加した硫酸ナトリウムがさらさらになったところで溶液をフィルターに通して内部標準物質であるequiline(1μM)を50μl加えた。
【0072】
(3)GC/MS分析
脱水したジクロロメタン抽出液を窒素で濃縮し、それにTMSを50μlずつ添加後、1時間室温で放置し、GC/MSで測定した(TMS化後は、1日間−10℃冷凍保存可能)。
【0073】
(4)GC/MS条件
GC/MS Agilent.6890
注入口:300℃
オーブン:100℃で5分加熱後、15℃/分で300℃まで昇温し、300℃で20分保持した。
SIM:
内部標準−TMS:340(m/Z);22.42min
2OHE2−TMS:504(m/Z);23.95min
4OHE2−TMS:504(m/Z);24.59min
検出器:300℃
キャリアー:He,1ml/min
カラム:DB−5(Agilent)
60×0.32mm×0.25mm
【0074】
(5)統計処理
分散分析を行った後、対照群として指定した1群と多群を比較したDunnett法を用いて検定を行った。
【0075】
2.実験結果
GC/MS分析の結果を図5に示す。クリソエリオールは、CYP1B1による4−OHE2の生成を0.05μMで69%抑制したが、CYP1A1による2−OHE2の生成に対しては抑制作用を示さなかった。この結果は実施例1の結果と同様に、クリソエリオールがCYP1B1を特異的に阻害することを示している。
【0076】
<実施例3>
エストロゲンのカテコール体である4−OHE2および2−OHE2は、COMTによりメチル化され無毒化する。肝サイトゾール中にはCOMTが存在する。本実験では、肝サイトゾール及びドナー基質としてトリチウムラベルしたS-adenosyl-L-methionine(SAM)を用いて、4−OHE2からメチル化された4−MeOE2を検出するメチレーションアッセイを行い、クリソエリオールがCOMTの阻害をしないかどうかを確認した。
【0077】
1.実験方法
(1)試料調製
(i)Tris−HCl(pH7.4)
TRIZMA・BASE(Sigma)60.55gを400mlの純水に溶解し、塩酸でpHを7.4に調節して500mlまでメスアップした。
(ii)24mM・MgCl2
MgCl2(Wako)228.5mgを100mlの純水に溶解した。
(iii)20mM・DTT
DTT(DL-dithiothreitol)60mgを10mlの純水に溶解した。
(iv)1mM cold・SAM([methyl]-S-adenosyl-L-methionine)
32mMのSAM溶液を1mMになるようにエタノールで希釈して、−20℃で保存した。
【0078】
(2)メチレーションアッセイ
(i)反応液の組成
下記表1に示す組成で反応液を調製した。
【0079】
【表1】
【0080】
(ii)反応方法
MgCl2、DTT、cold・SAMを混合したものにTris−HCl(pH7.4)と基質(4−OHE2)を添加してHot・SAMを入れた(ネガコンはここまで)。次に、試料を添加したものとしないものに肝サイトゾールを加え37℃で20分反応を行った。反応後、DDW500μlと反応を止めるためのn−ヘプタン3mlを加え、vortexをかけてから遠心を2000rpmで10分行った。その後、上澄を1ml分注し、シンチバースを4ml加え、vortexをかけた後に液体シンチレーションカウンターで測定した。
【0081】
2.実験結果
メチレーションアッセイを行った結果、図6に示すようにクリソエリオールにCOMT阻害効果は見られなかった。また4−OHE2を入れずに反応を行ったところ、クリソエリオール自身がCOMTによりメトキシ化されることはなかった。
【0082】
<実施例4>
MCF7細胞においては、CYP1A1よりもCYP1B1が発現しており、細胞内でE2は代謝され、最終的に2−MeOE2及び4−MeOE2の抱合体となって細胞外へ排泄される。そこで、クリソエリオールの存在下、非存在下で細胞を培養し、CYP1B1が細胞内でも阻害されるかどうか、培地中に排泄された2−MeOE2及び4−MeOE2抱合体量を、竈-glucuronidase/sulfataseによる加水分解後、クーロアレイHPLC法により測定した。
【0083】
1.実験方法
(1)使用した細胞
MCF−7:乳癌細胞[乳腺上皮:転移性腫瘍:胸水・腺癌](69 years)
静岡県立大学大学院生活健康科学研究科生体防御研究室、久留戸涼子先生(Dr. Hanspaul Hagenmaier, Institute of Organic Chemistry, University of Tuebingen, Germany)から分与頂いたものを使用。
【0084】
(2)使用した培地(培地1Lあたり)
(i)粉末培地(phenol red)
DMEM(Dulbecco's modified Eagle's medium)[日水製薬];9.5g
炭酸水素ナトリウム[特級/和光純薬];1.5g
L-glutamine[SIGMA];0.58g
カナマイシン[ICN];0.1g
アンピシリン[ICN];0.1g
非働化牛胎児血清(FBS)[ICN];110ml
【0085】
(ii)液体培地(phenol red free medium)
以下に示す組成の培地を調製後、pH7.6に合わせたのち、組織培養フィルター[半径75mm/ポアサイズ0.2μm/NALGENE]で滅菌後使用した。
DMEM(phenol red free)[Gibco];500ml
L-glutamine[SIGMA];0.29g
カナマイシン[ICN];0.05g
アンピシリン[ICN];0.05g
Charcoal/Dextran treated FBS[Hyclone];55ml
【0086】
(ii)1M酢酸アンモニウム(pH4.4)溶液
酢酸アンモニウム(Wako)53.96gを700mlの純水に溶解させた。
(iii)A液
アセトニトリル(関東化学HPLC用);450ml
メタノール(関東化学HPLC用);150ml
純水;2100ml
1M酢酸アンモニウム;300ml
(iv)B液
アセトニトリル(関東化学HPLC用);1500ml
メタノール(関東化学HPLC用);600ml
純水;600ml
1M酢酸アンモニウム;300ml
【0087】
(v)スタンダード原液
20mM E2;5.44mg
20mM 4OHE2;5.76mg
20mM 2MeOE2;6.04mg
20mM 4MeOE2;6.04mg
それぞれをDMSO1mlに溶解した。
(vi)スタンダード溶液
上記(v)で調製したスタンダード原液をB液で20倍希釈→100倍希釈→100倍希釈の順で最終濃度を100nMに調製した。
【0088】
(3)細胞培養
MCF7を1×104cell/cm2になるようにLPディッシュに播き、5日間培養した。5日後、培地をDMEM培地からE2フリーのDMEM培地に変え、0.05及び0.1μMのクリソエリオールを添加し、その15分後に1μMのE2を添加して24時間培養した。培地を採取し、遠心を行った後、上清を液体窒素で凍結し、−20℃で保存した。
【0089】
(4)抽出
培地2mlに対して2N酢酸アンモニウムを0.5ml加えて酸性とし、竈-glucuronidase/sulfatase(SIGMA)を10μl加えて遮光して37℃で17時間酵素処理を行った。その後、ジクロロメタンを2.5ml加えてボルテックスを30秒かけ抽出し、3000rpmで遠心を3層になるまで行った。上層を別の試験管に移し、下層(ジクロロメタン層)をパスツールで取って目盛付試験管に移し、濃縮遠心エバポレーターにより濃縮した。上層を同様に再抽出し、ジクロロメタン層を最初のジクロロメタン層に加え、完全に乾固した。HPLC移動層B液を100μl加えボルテックスをかけて溶解した後、3000rpmで15分遠心し、上清をクーロアレイ用バイアルに入れてクーロアレイHPLC分析を行った。
【0090】
(5)定量分析
定量分析は(6)に示すHPLC−ECD条件下で、ピーク面積を測定することにより行った。
【0091】
(6)HPLC−ECD条件
Column: ESA ODS(4.6×250mm)
Injection Volume:50・l
Temperature:25℃
Detector Potentials:-10,80,170,260,350,440,530,
Gradient solvent: A; acetnitrile, methanol, water, ammonium acetate/ B; acetnitrile, methanol, water, ammonium acetate
Flow rate:1.0ml/min
【0092】
(7)統計処理
分散分析を行った後、対照群として指定した1群と多群を比較したDunnett法を用いて検定を行った。
【0093】
2.実験結果
クリソエリオールがCYP1B1を細胞内でも選択的に阻害するかどうかについて、培地中に排泄された2−MeOE2及び4−MeOE2抱合体量を、竈-glucuronidase/sulfataseで加水分解後、クーロアレイHPLC法により測定して確認した。結果を図7に示した。クリソエリオールは、4−MeOE2の生成を0.05μM、0.1μMで有意に抑制したが、2−MeOE2の生成に対しては抑制作用を示さなかった。実施例3によりクリソエリオールがE2の各カテコール体のメチル化を阻害しなかったので、この結果は実施例1〜2の結果と同じであり、本結果はクリソエリオールが細胞内においてもCYP1B1を低濃度で特異的に阻害したことを示している。
【0094】
<実施例5>
本実施例では、本発明のメトキシフラボノイド化合物がそれ自身でエストロゲン作用やCYP1B1を誘導する作用がないかどうかを確認した。すなわち、本発明のメトキシフラボノイド化合物のエストロゲンレセプター結合性を検討し、エストロゲンレセプターへのアンタゴニスト又はアゴニスト活性の有無を検討した。
【0095】
1.実験方法
(1)使用した細胞
ERE−GFP−MCF−7;静岡県立大学大学院生活健康科学研究科生体防御研究室、久留戸涼子先生の開発によるものを用いた。ERE−GFP−MCF7細胞はMCF7細胞にgreen-fluorescent-protein reportor vectorが組み込まれており、EREにE2又はE2様物質が結合すると蛍光を発する。
【0096】
(2)試薬調製
(i)液体培地(phenol red free medium)
DMEM(phenol red free)[Gibco];500ml
L-glutamine[SIGMA];0.29g
カナマイシン[ICN];0.05g
アンピシリン[ICN];0.05g
Charcoal/Dextran treated FBS [Hyclone];55ml
(ii)テストサンプル
クリソエリオールをDMSOに溶解した。
【0097】
(3)培養・測定方法
ERE−GFP−MCF−7細胞を96ウェル(well)プレートに1×104/well播き、細胞が接着するまで4時間インキュベートした。4時間後、10-8Mエストロゲンと種々の濃度のクリソエリオール又はクリソエリオールのみを細胞に添加した。添加してから3日目に細胞にCell counting kitを10μlずつ添加し、2時間後に細胞数を測定した(450nm, Reference 630nm)。細胞数を測定後、wellの溶液を取り除き、蛍光をFluorImager SIで測定をした。
【0098】
2.実験結果
蛍光測定結果を図8に示す。クリソエリオールは、10-8〜10-5の濃度でエストロゲンレセプターへの結合活性が上昇していたが、アンタゴニスト作用は見られなかった。
【0099】
<実施例6>
本実施例では、lacZ reporter plasmidならびにヒトAhRとAhR核内輸送酵素をコードする遺伝子を含む遺伝子組み換え酵母Saccharomyces cerevisiae YCM3を用いたレポータージーンアッセイを行った。BaPなどのリガンドがAhRに結合するとβ−ガラクトシダーゼ(竈-galactosidase)が誘導され、本酵素の活性を測定することによりAhRへの結合性が評価できる。本実験では本発明のメトキシフラボノイド化合物のAhRへの結合性を検討し、アゴニスト作用やアンタゴニスト作用がないかを確認した。
【0100】
1.実験方法
(1)使用した酵母
LacZ reporter plasmidならびにヒトAhRとAhR核内輸送酵素をコードする遺伝子を含む遺伝子組み換え酵母Saccharomyces cerevisiae YCM3株
【0101】
(2)試薬等の調製方法
(i)酵母前培養培地
D(+)グルコース;2g
Yeast Nitrogen Base w/0 Amino Acid(BD);0.67g
アデニン塩酸塩(Wako);0.01g
L−ヒスチジン塩酸塩−水和物(Wako);0.01g
L−ロイシン(Wako);0.01g
L(+)−リシン(Wako);0.01g
これらを100mlの純水に溶解した。
【0102】
(ii)酵母本培養培地
ガラクトース;2g
Yeast Nitrogen Base w/0 Amino Acid(BD);0.67g
アデニン塩酸塩(Wako);0.01g
L−ヒスチジン塩酸塩−水和物(Wako);0.01g
L−ロイシン(Wako);0.01g
L(+)−リシン(Wako);0.01g
これらを100mlの純水に溶解した。
【0103】
(iii)Z−Buffer
NaH2PO4・12H2O[特級/和光純薬工業];4.3002g
NaH2PO4・2H2O;1.2438g
KCl;0.1519g
MgSO4・7H2O;0.0492g
これらを200mlの蒸留水に溶解した。
【0104】
(iv)溶菌剤
N-Laurosyl cosine solution(SIGMA);30mg
メルカプトエタノール(Wako);81μl
Z−Buffer;30ml
【0105】
(v)反応液
Z−buffer;15ml
メルカプトエタノール(Wako);45μl
2−ニトロフェニル−β−D−ガラクトピラノシド(Wako);60mg
(vi)クリソエリオール10〜0.001μM(フナコシ)
各種フラボノイドを10mMになるようにDMSOに溶解し、−20℃保存した。それを10〜0.001μM濃度に調製した。
【0106】
(3)測定方法
96ウェル(Well)プレートにBaPとメトキシフラボノイド0.001〜10μMの範囲で添加した。次にガラクトースScに培地を変えた酵母を200μlずつ加えて1晩置いた。翌日、培養した酵母の濁度を630nmで測定した。そこから30μl取り、別のプレートに溶液を移し、そこに溶菌剤を120μl添加して13分インキュベートした。その後、反応液を50μlずつ添加し、数分置いた後(色が出てきたら)450nmで測定した。
【0107】
2.実験結果
図9に示すように、クリソエリオールのAhR結合活性は非常に弱く、0.1μM以下ではアゴニスト作用は全く見られず、1μMでアンタゴニスト作用も見られなかった。
【0108】
上記実施例1〜5の結果から以下のことが明らかとなった。
(1)実施例1で示されたように、本発明のメトキシフラボノイド化合物は、CYP1B1の酵素活性を低濃度で特異的に阻害し、CYP1A1及びCYP3A4の酵素活性を阻害しない。
【0109】
(2)実施例2において、本発明のメトキシフラボノイド化合物がCYP1A1/1B1によるE2から2−OHE2及び4−OHE2への代謝に対し阻害作用を示すかどうかをGC/MS分析により検討した結果、CYP1B1についてはエトキシレゾルフィンを用いた実施例1の結果と同様の阻害作用が見られ、0.05μMで有意に4−OHE2の生成を69%抑制した。CYP1A1については、ほとんど影響が見られなかった。なお、CYP1B1を特異的に阻害する化合物として2,3,4,5−テトラメチルスチルベン(TMS)が報告されているが、TMSのCYP1B1による4‐OHE2生成阻害活性は、IC50が0.25μMであり、本発明のカテコールエストロゲン生成阻害剤のほうが低濃度で阻害活性を示しており、有効であると考えられる。
【0110】
(3)カテコールエストロゲンの重要な解毒酵素であるCOMTは、カテコールエストロゲンをメチル化するが、クリソエリオールのCOMTへの影響について、肝サイトゾールを用いたメチレーションアッセイを行った実施例3の結果から、クリソエリオールはCOMTを阻害せず、同時にクリソエリオール自身がメチル化されることもなかった。カテキン、エピカテキン、EGCG、ケルセチン、フィセチン(fisetin)のようなカテコール構造を有するフラボノイドは、COMTによるカテコールエストロゲンのメチル化を阻害するが、クリソエリオールをはじめとする本発明のメトキシフラボノイド化合物はカテコール構造を持っておらずCOMTを阻害しないことから、カテコールエストロゲンのメチル化には影響がなくCYP1A1により生成する2−OHE2の代謝を阻害しないため、エストロゲンの細胞内でのホメオスタシスに影響を与えないと思われる。
【0111】
(4)実施例4において、MCF−7細胞を用いて培地中のカテコールエストロゲン代謝物の解析を行い、クリソエリオールがCYP1B1に対して細胞内においても特異的に阻害するかどうかを検討した。その結果、クリソエリオール0.05μM及び0.1μMをそれぞれ作用させた場合、培地中に排泄された4−MeOE2量が低下したが、2−MeOE2量の低下は見られなかった。このことからクリソエリオールは細胞内においてもCYP1B1の酵素に対して特異的に阻害し、細胞内で4−OHE2の生成のみを抑制しうると思われる。従って、クリソエリオールをはじめとする本発明のメトキシフラボノイド化合物はE2が4位の水酸基の酸化、キノン体の生成を経てDNA付加体の形成を促進するのを抑制すると思われる。
【0112】
(5)実施例5に示すように、クリソエリオールは1μM以下ではエストロゲンレセプターに対してアンタゴニスト作用を示さない。高濃度になるにつれてE2へのアゴニスト作用をわずかではあるが示した、すなわちわずかにエストロゲン活性が見られた。しかし、E2に対するアンタゴニスト作用、すなわちエストロゲン活性抑制作用は見られなかった。一方、タマリキセチンにはエストロゲンレセプターへの結合性もE2に対するアンタゴニスト作用も見られなかった。従って、本発明のメトキシフラボノイド化合物は弱いエストロゲン様を示すが、エストロゲンレセプターへのE2の結合作用に対して低濃度ではアンタゴニスト的に働く可能性は低いと考えられる。
【0113】
(6)実施例6に示すように、クリソエリオールはAhRにも低濃度では結合しないことから、クリソエリオール自身はCYPを誘導しないことが示唆された。従ってクリソエリオールはCYP1B1によるE2から4−OHE2の生成を特異的に阻害する因子として有用であると思われる。
【産業上の利用可能性】
【0114】
本発明のメトキシフラボノイド化合物は、低濃度でCYP1B1の酵素活性を特異的に阻害し、乳がんの原因とされる4−OHE2の生成を抑制する作用を有する。しかも、CYP1A1及びCOMTを阻害しないため細胞内の解毒作用が正常に働き、エストロゲンの細胞内でのホメオスタシスに影響を与えない。また、CYP3A4の酵素活性を阻害しないため、肝臓における副作用が少ない可能性がある。また、エストロゲンに対するアンタゴニスト活性を有しないので、エストロゲンの正常なホルモン作用を阻害しない。
【0115】
したがって、ホルモン補充療法においてもホルモン投与による効果を犠牲にすることなく該療法の副作用である乳がん、子宮がんの発生を抑制することができる。よって、安全で且つ極めて優れた乳がん発生予防因子として有用であり、がん予防・治療の分野への貢献が期待される。また、CYP1B1の酵素阻害試薬として利用することもできる。
【図面の簡単な説明】
【0116】
【図1】エストロゲン代謝のメカニズムを表す概略図である。
【図2】実施例1のCYP1A1/1B1酵素阻害活性(ERODアッセイ)の結果を示すグラフである。
【図3】実施例1のラインウィーバー・バークプロットをとった結果を示すグラフである。
【図4】実施例1のCYP3A4酵素阻害活性の測定結果を示すグラフである。
【図5】実施例2のGC/MS分析の結果を示すグラフである。
【図6】実施例3のメチレーションアッセイの結果を示すグラフである。
【図7】実施例4のクーロアレイHPLC法による測定結果を示すグラフである。
【図8】実施例5のERE−GFP−MCF−7細胞の蛍光測定結果を示すグラフである。
【図9】実施例6のレポータージーンアッセイの結果を示すグラフである。
【符号の説明】
【0117】
1 エトキシレゾルフィン
2 エトキシレゾルフィン+クリソエリオール0.05μM
3 エトキシレゾルフィン+クリソエリオール0.1μM
【特許請求の範囲】
【請求項1】
下記一般式(1)で表されるメトキシフラボノイド化合物を有効成分とする、4位−カテコールエストロゲン生成阻害剤。
【化1】
(一般式(1)中、R1及びR2はそれぞれ独立して水酸基又はメトキシ基を表し、R1とR2の少なくとも一方はメトキシ基である。R3は水素原子又は水酸基を表す。R4は水酸基又はメトキシ基を表し、nは0〜4である。)
【請求項2】
前記メトキシフラボノイド化合物が、一般式(2)で表されることを特徴とする、請求項1記載の4位−カテコールエストロゲン生成阻害剤。
【化2】
(一般式(2)中、R1及びR2はそれぞれ独立して水酸基又はメトキシ基を表し、R1とR2の少なくとも一方はメトキシ基である。R3は水素原子又は水酸基を表す。)
【請求項3】
前記メトキシフラボノイド化合物が、クリソエリオール、タマリキセチン、及びイソラムネチンからなる群から選択されるものであることを特徴とする、請求項2記載の4位−カテコールエストロゲン生成阻害剤。
【請求項4】
前記メトキシフラボノイド化合物がクリソエリオールであることを特徴とする、請求項3記載の4位−カテコールエストロゲン生成阻害剤。
【請求項5】
下記一般式(1)で表されるメトキシフラボノイド化合物を有効成分とする、CYP1B1活性阻害剤。
【化3】
(一般式(1)中、R1及びR2はそれぞれ独立して水酸基又はメトキシ基を表し、R1とR2の少なくとも一方はメトキシ基である。R3は水素原子又は水酸基を表す。R4は水酸基又はメトキシ基を表し、nは0〜4である。)
【請求項6】
前記メトキシフラボノイド化合物がクリソエリオールであることを特徴とする、請求項5記載のCYP1B1活性阻害剤。
【請求項1】
下記一般式(1)で表されるメトキシフラボノイド化合物を有効成分とする、4位−カテコールエストロゲン生成阻害剤。
【化1】
(一般式(1)中、R1及びR2はそれぞれ独立して水酸基又はメトキシ基を表し、R1とR2の少なくとも一方はメトキシ基である。R3は水素原子又は水酸基を表す。R4は水酸基又はメトキシ基を表し、nは0〜4である。)
【請求項2】
前記メトキシフラボノイド化合物が、一般式(2)で表されることを特徴とする、請求項1記載の4位−カテコールエストロゲン生成阻害剤。
【化2】
(一般式(2)中、R1及びR2はそれぞれ独立して水酸基又はメトキシ基を表し、R1とR2の少なくとも一方はメトキシ基である。R3は水素原子又は水酸基を表す。)
【請求項3】
前記メトキシフラボノイド化合物が、クリソエリオール、タマリキセチン、及びイソラムネチンからなる群から選択されるものであることを特徴とする、請求項2記載の4位−カテコールエストロゲン生成阻害剤。
【請求項4】
前記メトキシフラボノイド化合物がクリソエリオールであることを特徴とする、請求項3記載の4位−カテコールエストロゲン生成阻害剤。
【請求項5】
下記一般式(1)で表されるメトキシフラボノイド化合物を有効成分とする、CYP1B1活性阻害剤。
【化3】
(一般式(1)中、R1及びR2はそれぞれ独立して水酸基又はメトキシ基を表し、R1とR2の少なくとも一方はメトキシ基である。R3は水素原子又は水酸基を表す。R4は水酸基又はメトキシ基を表し、nは0〜4である。)
【請求項6】
前記メトキシフラボノイド化合物がクリソエリオールであることを特徴とする、請求項5記載のCYP1B1活性阻害剤。
【図1】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【公開番号】特開2007−291040(P2007−291040A)
【公開日】平成19年11月8日(2007.11.8)
【国際特許分類】
【出願番号】特願2006−122998(P2006−122998)
【出願日】平成18年4月27日(2006.4.27)
【出願人】(802000020)財団法人浜松科学技術研究振興会 (63)
【Fターム(参考)】
【公開日】平成19年11月8日(2007.11.8)
【国際特許分類】
【出願日】平成18年4月27日(2006.4.27)
【出願人】(802000020)財団法人浜松科学技術研究振興会 (63)
【Fターム(参考)】
[ Back to top ]