
7Sグロブリンαサブユニット欠失ダイズの検出方法
【課題】ダイズ貯蔵タンパク質の一つである7Sグロブリンαサブユニット欠失ダイズの高感度な検出方法の提供。
【解決手段】ダイズ由来の核酸試料について、CG−2遺伝子(7Sα遺伝子)の第1エキソンの第215位〜第217位の配列AAA中への4個のアデニン塩基の挿入を検出することを特徴とする、7Sグロブリンαサブユニット欠失ダイズの検出方法。更に、上記検出方法のための、特定な配列からなるプライマーセットの提供。
【解決手段】ダイズ由来の核酸試料について、CG−2遺伝子(7Sα遺伝子)の第1エキソンの第215位〜第217位の配列AAA中への4個のアデニン塩基の挿入を検出することを特徴とする、7Sグロブリンαサブユニット欠失ダイズの検出方法。更に、上記検出方法のための、特定な配列からなるプライマーセットの提供。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、7Sグロブリンαサブユニット欠失ダイズを検出する方法、及びその方法に用いるプライマーに関する。
【背景技術】
【0002】
ダイズ貯蔵タンパク質のひとつである7Sグロブリン(別名:β-コングリシニン)は、α’、α及びβの3種類のサブユニットで構成されている。そのうち、7Sグロブリンのαサブユニット(以下、7Sαとも略記する)は、ダイズ陽性のアトピー性皮膚炎患者のうち25%の患者の血清によって検出された主要アレルゲンである。
【0003】
ダイズ貯蔵タンパク質には、7Sグロブリンの他、2Sグロブリン、11Sグロブリン、及び15Sグロブリンが知られている。7Sグロブリンは、アレルゲンであるだけでなく、11Sグロブリンに比べて栄養価や加工適性が劣るとされる。ダイズの7Sグロブリン含量は11Sグロブリンの含量との間で負の相関が認められることから、7Sグロブリンαサブユニット及びα’サブユニットを欠失させることによって相対的に11Sグロブリンの含量を高めることができる。そのため、7Sグロブリンαサブユニット及びα’サブユニットを欠失させて機能性を強化したダイズ品種の開発が進められている。
【0004】
7Sグロブリンのサブユニット変異体として、7Sグロブリンα’サブユニット(7Sα’)を欠失した「毛振(Keburi)」、7Sグロブリンαサブユニット及びβサブユニットの生成量が低下した「秣食豆公503(Moshidou Gong 503)」、秣食豆公503の7Sα及びβの低下性と毛振の7Sα’欠失が導入された「刈系434号(Kari-kei 434)」、刈系434号のγ線照射により作出された7Sα欠失品種「ゆめみのり(Yumeminori)」などを含む多様なダイズ品種・系統が知られている。これらのダイズ品種・系統の研究から、Region Cと呼ばれるダイズ染色体領域内のCG-1遺伝子が7Sグロブリンα’サブユニットをコードする遺伝子であること、及びRegion Aと呼ばれるダイズ染色体領域内のCG-2遺伝子及びCG-3遺伝子が7Sグロブリンαサブユニットをコードする遺伝子であることが明らかになった(非特許文献1及び2)。「毛振」ではCG-1遺伝子及びCG-2遺伝子が欠失していることが示されており(非特許文献2及び3)、「秣食豆公503」の7Sα低下性がCG-3遺伝子の欠失に起因することも報告されている(非特許文献3)。このようにダイズ7Sグロブリンサブユニットに関する研究は確実に進んできている。
【0005】
しかしながら、7Sグロブリンサブユニット変異の検出には、現在でも、手間を要するタンパク質レベルの手法が使用されている。そのような手法では、分析試料として、ダイズ種子粉末あるいは未分解のタンパク質を含むダイズ加工物の調製が必要である。また例えばタンパク質の検出の際に用いられるSDS-PAGEはゲルの作成に熟練を要するし、SDS-PAGEに続いて行うCBB法による検出は、感度が低いため異品種の僅かな混入などを検出できない場合がある。一方、SDS-PAGEに続いて行う銀染色法は、感度は高いが操作が煩雑であるため多数サンプルの分析が困難である。
【0006】
以上のような背景から、7Sグロブリンαサブユニット及びα’サブユニット欠失ダイズ品種の効率的な育成やそれらダイズ品種に関する異品種混入検査などに利用できる高感度な簡易判定技術の開発が求められている。
【0007】
【非特許文献1】Harada J.J. et al., Plant Cell (1989) 1: p.415-425
【非特許文献2】Ladin B.F. et al., J. Mol. Appl. Genet. (1984) 2: p.372-380
【非特許文献3】Yoshino et al., Breed Sci. (2002) 52: p.285-292
【発明の開示】
【発明が解決しようとする課題】
【0008】
本発明は、7Sグロブリンαサブユニット欠失ダイズの高感度な検出方法を提供することを課題とする。
【課題を解決するための手段】
【0009】
本発明者らは、上記課題を解決すべく鋭意研究を行った。その結果、7Sグロブリンαサブユニット(7Sα)を欠失したダイズ品種「ゆめみのり」において7Sα遺伝子であるCG-2遺伝子の第1エキソンに4個のアデニン塩基が挿入された変異が生じていることを見出した。「ゆめみのり」ではその4個のアデニン塩基の挿入によってCG-2遺伝子にフレームシフトが生じ、ごく早期に終止コドンが現れるようになっていることから、この挿入変異によって7Sグロブリンαサブユニットが生成されなくなったと考えられた。これらの知見に基づき、本発明者らは、「ゆめみのり」において見出されたCG-2遺伝子の第1エキソン中の4塩基のアデニンの挿入をダイズ核酸について検出することによって、7Sグロブリンαサブユニット欠失ダイズを検出する手法を開発し、本発明を完成するに至った。
【0010】
すなわち本発明は以下の通りである。
【0011】
[1] ダイズ由来の核酸試料について、CG−2遺伝子の第1エキソンの第215位〜第217位の配列AAA中への4個のアデニン塩基の挿入を検出する工程を特徴とする、7Sグロブリンαサブユニット欠失ダイズの検出方法。
この方法では、CG−2遺伝子の第1エキソンは、配列番号1で示す塩基配列からなるものであることが好ましい。
またこの方法の前記検出工程は、好ましくは、配列番号3で示す塩基配列上の第215〜第221位の配列AAAAAAAを含む少なくとも15塩基の連続した配列からなるDNAをプライマー又はプローブとして用いて検出する工程である。
【0012】
さらにその検出工程は、配列番号3で示す塩基配列上の第215〜第221位の配列AAAAAAAを含む15〜50塩基の連続した配列又はそれに相補的な配列からなるポリヌクレオチドプライマーを用いたPCR増幅により検出する工程であることが好ましい。
そのようなプライマーとしては、配列番号12で示す塩基配列からなるポリヌクレオチドプライマーが好適である。
さらに本発明では、配列番号12で示す塩基配列からなるポリヌクレオチドプライマーとともに、配列番号14で示す塩基配列からなるポリヌクレオチドプライマーをPCR増幅に用いることも好ましい。
この方法で検出される7Sグロブリンαサブユニット欠失ダイズは、好ましくは「ゆめみのり」、「ゆめみのり」の兄弟系統、又はそれらの子孫系統である。
【0013】
[2] 上記[1]に記載の方法において、前記プライマーとともに、CG−1遺伝子特異的プライマーセットをPCR増幅に用いることを特徴とする、ダイズの7Sグロブリンαサブユニット欠失性及び7Sグロブリンα’サブユニット欠失性の同時判定方法。
この方法で用いるCG−1遺伝子特異的プライマーセットとしては、配列番号15で示す塩基配列からなるポリヌクレオチドプライマーと、配列番号16で示す塩基配列からなるポリヌクレオチドプライマーとからなる組み合わせが好適である。
【0014】
[3] 上記[2]に記載の方法で得られた判定結果に基づいて、異系統のダイズの混在の有無を判定することを特徴とする、ダイズの混入検査法。
【0015】
[4] 配列番号3で示す塩基配列上の第215〜第221位の配列AAAAAAAを含む少なくとも15塩基の連続した配列からなるDNA。
【0016】
[5] 配列番号3で示す塩基配列上の第215〜第221位の配列AAAAAAAを含む15〜50塩基の連続した配列又はそれに相補的な配列からなるポリヌクレオチドプライマー。
【0017】
[6] 配列番号12で示す塩基配列からなるポリヌクレオチドプライマーと、配列番号14で示す塩基配列からなるポリヌクレオチドプライマーとからなる、プライマーセット。
【0018】
[7] 配列番号12〜14で示す塩基配列の各々からなる3種のポリヌクレオチドプライマーを含むキット。このキットは、好適には、「ゆめみのり」、「ゆめみのり」の兄弟系統、又はそれらの子孫系統である7Sグロブリンαサブユニット欠失ダイズを検出するためのキットである。
【0019】
[8] 配列番号12〜16で示す塩基配列の各々からなる5種のポリヌクレオチドプライマーを含むキット。このキットは、配列番号17及び18で示す塩基配列の各々からなる2種のポリヌクレオチドプライマーをさらに含むことが好ましい。このキットは、好適には、「ゆめみのり」、「ゆめみのり」の兄弟系統、又はそれらの子孫系統である7Sグロブリンαサブユニット欠失ダイズを検出するためのキットである。
【発明の効果】
【0020】
本発明の方法は、「ゆめみのり」、「ゆめみのり」の兄弟系統、又はそれらの子孫系統などの7Sグロブリンαサブユニットを欠失したダイズ系統を、高感度にかつ簡便に検出することができる。
【発明を実施するための最良の形態】
【0021】
1.7Sグロブリンαサブユニット欠失ダイズの検出
1)検出すべき変異
本発明の方法において検出すべき変異、すなわちダイズCG-2遺伝子の第1エキソン中への4個のアデニン塩基の挿入変異は、最初に、ダイズ品種「ゆめみのり」において見出されたものである。
【0022】
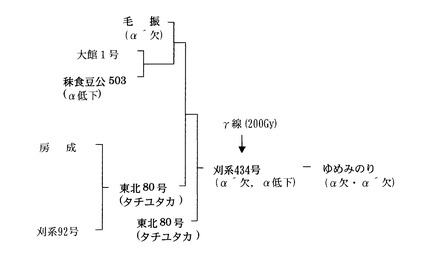
「ゆめみのり」は、ダイズの主要アレルゲンである7Sグロブリンαサブユニット(7Sα)が欠失しており、栄養価の高い11Sグロブリンの含有量が向上した、低アレルゲン性のダイズ品種である。「ゆめみのり」は、「毛振」の7Sα’欠失形質(すなわち、7Sα’が生成されない)と「秣食豆公503」の7Sα低下形質(すなわち、7Sαの生成量が低下する)とを導入した「刈系434号」に、γ線照射(200Gy)することによって得られた7Sα欠失変異体から確立した1系統に基づいて、品種登録された。図1に、ダイズ品種「ゆめみのり」の系譜を示す。「ゆめみのり」は、7Sα欠失形質(7Sαが生成されない)と7Sα’欠失形質(7Sα’が生成されない)の両方を示す。なおダイズ品種「ゆめみのり」は、独立行政法人 農業・生物系特定産業技術研究機構 東北農業研究センターから入手することができる。
【0023】
本発明者らは、「ゆめみのり」において7Sα遺伝子の1つであるCG-2遺伝子の第1エキソン中に4個のアデニン塩基の挿入が生じていること、その結果CG-2遺伝子にフレームシフトが起き、さらに終止コドンが早期に生じていることを見出した。「ゆめみのり」のCG-2遺伝子の第1エキソン配列を原品種「刈系434号」と比較すると、この4個のアデニン塩基の挿入箇所は、「刈系434号」のCG-2遺伝子第1エキソンの第215位〜第217位に位置する3アデニン配列(配列AAA)内に相当する。「ゆめみのり」においてはその3アデニン配列が、4個のアデニン塩基が挿入されたことにより、7アデニン配列(配列AAAAAAA)となっている(「ゆめみのり」のCG-2遺伝子の第1エキソンの第215位〜第221位)。「ゆめみのり」はもう1つの7Sα遺伝子であるCG-3遺伝子を完全欠失(遺伝子構造自体を完全に喪失)しているので、「ゆめみのり」の7Sα欠失形質が、CG-2遺伝子の第1エキソン中に4個のアデニン塩基が挿入された結果として引き起こされたことは明らかである。本発明では、第1エキソンの第215位〜第217位に位置する3アデニン配列(配列AAA)内に4個のアデニン塩基が挿入されたCG-2遺伝子を、便宜的に、欠失型CG-2遺伝子と呼ぶ。
【0024】
「ゆめみのり」のCG-2遺伝子の第1エキソン中への4個のアデニン塩基の挿入は、もともとは、「ゆめみのり」の育種過程で「刈系434号」にγ線照射(200Gy)することによって得られた7Sα欠失変異体に生じた変異である。この7Sα欠失変異体からは「ゆめみのり」と同様の形質をもった系統が他にも作成されており、それらの系統でもそのようなCG-2遺伝子の第1エキソン中への4個のアデニン塩基の挿入が認められる。本発明では、7Sα欠失変異体から作成したこれらの系統を便宜的に『「ゆめみのり」の兄弟系統』と呼ぶ。
【0025】
CG-2遺伝子の第1エキソン中への4個のアデニン塩基の挿入は、「ゆめみのり」又は「ゆめみのり」の兄弟系統の子孫系統にも受け継がれる。本発明において「ゆめみのり」又は「ゆめみのり」の兄弟系統の子孫系統とは、「ゆめみのり」又は「ゆめみのり」の兄弟系統を育種の母本として用いて、例えば突然変異誘発処理や他のダイズ系統・品種との交配によって作製された、後代のダイズを言う。「ゆめみのり」又は「ゆめみのり」の兄弟系統の子孫系統は、CG-2遺伝子の第1エキソン中への上記の4個のアデニン塩基の挿入をヘテロ接合又はホモ接合で有し得る。
【0026】
本発明においては、限定するものではないが上述の「ゆめみのり」、「ゆめみのり」の兄弟系統、及びそれらの子孫系統のように、CG-2遺伝子の第1エキソンの第215位〜第217位に位置する3アデニン配列内への4個のアデニン塩基の挿入によってそのCG-2遺伝子の7Sグロブリンαサブユニット産生機能が失われたダイズを、「7Sグロブリンαサブユニット欠失ダイズ(7Sα欠失ダイズ)」と呼ぶ。7Sα欠失ダイズは、そのCG-2遺伝子中への4個のアデニン塩基の挿入変異をヘテロ接合又はホモ接合で有していてもよい。CG-2遺伝子とCG-3遺伝子は密接に連鎖していることから、この7Sグロブリンαサブユニット欠失ダイズは、通常は、「ゆめみのり」と同様にCG-3遺伝子を完全欠失している。
【0027】
多くのダイズ品種が、7Sグロブリンαサブユニット遺伝子としてCG-2およびCG-3の2つを保持している。しかし、「毛振」はCG-2遺伝子を完全欠失しており、7Sα遺伝子としてはCG-3遺伝子を保持する。一方、「秣食豆公503」及び「刈系434号」は、CG-3遺伝子を完全欠失しており、7Sα遺伝子としてはCG-2遺伝子を保持する。後述の実施例に示されるように、「毛振」のCG-3遺伝子と「刈系434号」のCG-2遺伝子とは、少なくとも開始コドンから終止コドンまでのゲノム配列領域内では同一の塩基配列を有しており、「ゆめみのり」のようなCG-2遺伝子における4個のアデニン塩基の挿入は含まない。本発明では、「ゆめみのり」で認められた上記の4個のアデニン塩基の挿入を含まない7Sα遺伝子、例えば「毛振」のCG-3遺伝子及び「刈系434号」のCG-2遺伝子などを、便宜的に、野生型7Sα遺伝子と呼ぶ。
【0028】
従って本発明では、ダイズ由来の核酸試料について、その核酸中に含まれるダイズ7Sα遺伝子であるCG-2遺伝子の第1エキソンの第215位〜第217位に位置する3アデニン塩基配列(配列AAA)中の4個のアデニン塩基の挿入を検出することにより、7Sグロブリンαサブユニット欠失ダイズを検出することができる。
【0029】
本発明におけるダイズのCG-2遺伝子は、限定するものではないが、例えばGenBank登録番号AB051865(ダイズ品種「Forrest」のゲノム配列;配列番号5)、同登録番号X17698(mRNA配列)、同登録番号AB030839(mRNA配列)に示された配列を有するものでありうる。本発明のダイズCG-2遺伝子は、便宜的には、登録番号AB051865に記載された塩基配列(配列番号5)の開始コドンから終止コドンまでの配列(第490位〜第2855位)を含むゲノムDNA領域、又は登録番号X17698(mRNA配列)の開始コドンから終止コドンまでの配列(第1位〜第1818位)を含むmRNA又はcDNA、あるいはそれらの塩基配列と非常に高い相同性(好ましくは98%以上、より好ましくは99%以上)を有する配列からなるゲノムDNA、mRNA又はcDNAとみなすこともできる。
【0030】
ダイズCG-2遺伝子は、通常はゲノム上で第1エキソン〜第6エキソンを含む。その第1エキソン(上記4アデニン塩基が挿入される)は、ダイズCG-2遺伝子中、登録番号AB051865に示す塩基配列(配列番号5)の第490位〜第1273位の領域に相当する。登録番号AB051865に示す塩基配列の第490位〜第1273位の領域は、「毛振」のCG-3遺伝子や「刈系434号」のCG-2遺伝子とも完全に一致していたことから、その領域を「野生型7Sα第1エキソン」と呼び、その野生型7Sα第1エキソンの塩基配列を配列番号1に示す。これに対し、「ゆめみのり」の、上記4アデニン塩基挿入部位を含む、CG-2遺伝子中の野生型7Sα第1エキソンに相当する領域(便宜的に、「挿入変異型7Sα第1エキソン」と呼ぶ)の塩基配列を、配列番号3に示す。
【0031】
2)挿入変異の検出
本発明では、7Sグロブリンαサブユニット欠失ダイズを検出するため、上記1)で述べたダイズCG-2遺伝子における挿入変異(CG-2遺伝子の第1エキソン中への4個のアデニン塩基の挿入)を、ダイズ由来の核酸を被験試料として検出する。
【0032】
この検出に供するダイズ(Glycine max Merrill)は、任意の品種又は系統のものであってよく、例えば、以下に限定するものではないが、「タチユタカ(Glycine max cv. Tachiyutaka)」、「毛振(Glycine max cv. Keburi)」、「秣食豆公503(Glycine max cv. Moshidou Gong 503」、「ゆめみのり(Glycine max cv. Yumeminori)」、「刈系434号(Glycine max, line Kari-kei434)」などが挙げられる。上記検出に供するダイズは、ダイズのゲノムを少なくとも1セット含む限り、例えばダイズと他の植物とのハイブリッド植物や遺伝子組換え植物などであってもよい。
【0033】
本発明において「ダイズ由来の核酸試料」とは、上記のような供試ダイズの植物体の一部(葉など)又は種子等から調製されたダイズ核酸(DNA又はRNA)を意味する。核酸試料を調製するためのダイズ試料の形態は特に限定されず、一般的に用いられている種子粉末等のタンパク質調製に適した試料だけでなく、ダイズから採取した葉(幼葉)、茎、花などの核酸抽出可能な任意の試料を用いることができる。本発明では、ダイズ由来の核酸試料として、ダイズから抽出されたゲノムDNA又はゲノムDNAをクローン化したゲノムライブラリーを用いることが好ましい。あるいは本発明におけるダイズ由来の核酸試料として、ダイズから抽出したmRNAから逆転写PCRによって合成されたcDNA、又はそのcDNAを組み込んだcDNAライブラリーを用いてもよい。本発明におけるダイズ由来の核酸試料は、PCR法などによって増幅した核酸増幅断片であってもよい。ダイズからの核酸の抽出、精製、調製などの実験手順は、分子生物学の分野で慣用されている手法に従えばよい。
【0034】
CG-2遺伝子中の挿入変異の検出は、SNP、塩基多型又は塩基変異を検出するために当業者が利用可能な公知の様々な方法を行うことができる(Orita et al., Proc. Natl. Acad. Sci., 86: 2766-2770等を参照されたい)。
【0035】
そのような変異検出法としては、より具体的には、例えば、dCAPS法〔Plant J 14, 381-385 (1998)〕、CAPS(Cleaved Amplified Polymorphic Sequence)法〔「モデル植物の実験プロトコール」細胞工学別冊 植物細胞工学シリーズ15、2001年4月2日発行、秀潤社、p.69-76〕、PCRダイレクトシークエンス法〔Biotechniques, 11, 246-249 (1991)〕、AP-PCR(Arbitrarily Primed-PCR)法〔Nucl. Acids Res., 18, 7213-7218 (1990)〕、PCR-SSCP(一本鎖DNA高次構造多型)法〔Biotechniques, 16,296-297 (1994), Biotechniques, 21, 510-514 (1996)〕、ASO(Allele Specific Oligonucleotide)ハイブリダイゼーション法〔Clin. Chim. Acta, 189, 153-157 (1990)〕、ARMS(Amplification Refracting Mutation System)法〔Nuc. Acids. Res., 19, 3561-3567 (1991), Nuc. Acids. Res., 20,4831-4837 (1992)〕、変性剤濃度勾配ゲル電気泳動(Denaturing Gradient Gel Electrophoresis; DGGE)法〔Biotechniqus, 27, 1016-1018 (1999)〕、RNaseA切断法〔DNA Cell. Biol., 14, 87-94 (1995)〕、化学切断法〔Biotechniques, 21, 216-218 (1996)〕、DOL(Dye-labeled Oligonucleotide Ligation)法〔Genome Res., 8, 549-556 (1998)〕、MALDI-TOF/MS(Matrix Assisted Laser Desorption-time of Flight/Mass Spectrometry)法〔Genome Res., 7, 378-388 (1997), Eur. J. Clin. Chem. Clin. Biochem., 35, 545-548 (1997)〕、TDI(Template-directed Dye-terminator Incorporation)法〔Proc. Natl. Acad. Sci. USA, 94, 10756-10761 (1997)〕、パドロック・プローブ(Padlock Probe)法〔Nat. Genet., 3, p225-232 (1998)、遺伝子医学, 4, p50-51 (2000)〕、モレキュラー・ビーコン(Molecular Beacons)法〔Nat. Biotechnol.,1, p49-53 (1998)、遺伝子医学、4, p46-48(2000)〕、TaqMan PCR法〔Genet. Anal., 14, 143-149 (1999), J. Clin. Microbiol., 34, 2933-2936 (1996)〕、インベーダー法〔Science, 5109, 778-783(1993), J. Biol. Chem., 30, 21387-21394 (1999), Nat. Biotechnol., 17, 292-296 (1999)〕、ダイナミック・アレル−スペシフィック・ハイブリダイゼーション(Dynamic Allele-Specific Hybridization (DASH))法〔Nat. Biotechnol.,1, p87-88, (1999)、遺伝子医学, 4, p47-48 (2000)〕、UCAN法〔タカラ酒造株式会社ホームページ(http://www.takara.co.jp)参照〕、及びDNAチップまたはDNAマイクロアレイを用いる方法〔Genomics 4, (1989), Drmanae, R., Labat, I., Brukner, I. and Crkvenjakov, R., p114-128、Bio Industry Vol.17 No.4, 「DNAチップ技術」 p5-11 (2000)〕等が挙げられるが、これらの方法に限定されるものではない。
【0036】
これらの方法において好適に用いられる本発明に係る挿入変異を検出するためのプライマー又はプローブは、上記挿入変異を含むCG-2遺伝子と、変異を含まない野生型7Sα遺伝子とを特異的に識別できるように、野生型7Sα第1エキソンの配列(配列番号1)及び「ゆめみのり」の挿入変異型7Sα第1エキソンの配列(配列番号3)に基づいて任意に設計することができる。プライマー又はプライマー対の設計は、当業者には公知の方法に従って行えばよい。プライマーの設計には、例えば、オンラインソフトウェアNetPrimer(PREMIER Biosoft International、http://www.premierbiosoft.com/)を利用することもできる。さらに本発明では、上記挿入変異を含む配列に特異的にハイブリダイズするプライマー又はプローブを設計して用いることがより好ましい。例えば、配列番号3で示す塩基配列上の第215〜第221位の配列AAAAAAAを含む少なくとも15塩基(15塩基〜783塩基)、例えば15塩基〜50塩基(プライマー用など)又は30塩基〜500塩基(プローブ用など)の連続した配列からなるDNAを、プライマー又はプローブとして好適に用いることができる。また、本発明の方法をより簡便な判定法とするためには、配列番号3で示す塩基配列上の第215〜第221位の配列AAAAAAAを含む15〜50塩基の連続した配列又はそれに相補的な配列からなるポリヌクレオチドプライマーを用いたPCR増幅を利用してその増幅断片の有無に基づいて検出することも好ましい。
【0037】
本発明においてPCR増幅を用いた検出法を用いる場合、配列番号3で示す塩基配列上の第215〜第221位の配列AAAAAAAを3'末端付近に含む、15〜50塩基、より好ましくは18〜40塩基の連続した配列又はそれに相補的な配列からなるポリヌクレオチドプライマーを用いることが特に好ましい。このようなプライマーは、CG-2遺伝子において上記挿入変異を有するダイズ核酸のみを高感度に増幅することができる。
【0038】
配列AAAAAAAを3'末端付近に含むこのようなプライマーを用いた好適なプライマーセットとして、実施例に記載した欠失型CG-2特異的プライマーセット(プライマーCG2MF[配列番号12]及びCG23R[配列番号14])が挙げられる。この欠失型CG-2特異的プライマーセットを用いた場合、上記挿入変異を含むCG-2遺伝子のみが増幅され、上記挿入変異を含まないCG-2遺伝子やCG-3遺伝子(野生型7Sα遺伝子)は増幅されない。従って、この欠失型CG-2特異的プライマーセットを用いれば、CG-2遺伝子の上記挿入変異のみを特異的に検出することができる。さらにこのような検出において、挿入変異を含まない野生型7Sα遺伝子を特異的に検出できる、実施例に記載の野生型7Sα特異的プライマーセット(プライマーCG23F[配列番号13]及びCG23R[配列番号14])をコントロール試薬として用いることも好ましい。
【0039】
PCR増幅を用いた本発明の好適な検出法の具体例は、後述の実施例に記載している。
【0040】
本発明において用いるゲノムDNA及びmRNAの抽出及び精製、cDNAの作製(RT-PCR)、PCR、ベクター中へのライゲーション、細胞の形質転換及びライブラリーの作製、DNAの塩基配列決定、プライマーの合成、突然変異誘発、タンパク質の抽出などの分子生物学的・生化学的実験法は、基本的には、通常の実験書の記載に従って行うことができる。そのような実験書として、例えば、SambrookらのMolecular Cloning, A laboratory manual, 2001, Eds., Sambrook, J. & Russell, DW. Cold Spring Harbor Laboratory Pressを挙げることができる。
【0041】
3)7Sα欠失ダイズの検出及び識別
本発明の方法においては、以上のようにしてCG-2遺伝子の第1エキソン中への上記4アデニン塩基の挿入(挿入変異)を検出した結果から、ダイズの7Sα欠失性が示される。すなわち、CG-2遺伝子の第1エキソン中への上記4アデニン塩基の挿入が確認された場合、試験したダイズでは、少なくとも1つのCG-2遺伝子がフレームシフトを起こして7Sαタンパク質産生能を失っており、そのため7Sαタンパク質を(少なくとも部分的に)欠失していることが示される。
【0042】
従って、本発明の方法において、単一の品種又は系統のダイズ由来の核酸試料について、CG-2遺伝子の第1エキソン中への上記4アデニン塩基の挿入(挿入変異)が確認された場合には、そのダイズは7Sαタンパク質欠失ダイズであると判断することができる。
【0043】
また、本発明の方法において、複数の品種又は系統のダイズ由来の核酸を含む核酸試料について、CG-2遺伝子の第1エキソン中への上記4アデニン塩基の挿入(挿入変異)が確認された場合には、それらのダイズの一部が、7Sαタンパク質欠失ダイズであると判断することができる。
【0044】
本発明の方法においては、野生型7Sα遺伝子の特異的検出を並行して行うことにより、ダイズの7Sα欠失性をさらに詳細に判定することもできる。例えば、上記4アデニン塩基の挿入変異の存在が示され、かつ野生型7Sα遺伝子の存在が示されない場合は、供試ダイズは7Sαタンパク質を完全に欠失していると判定される。またこの場合、野生型7Sα遺伝子の存在が示されないことから、供試ダイズはCG-2遺伝子中の挿入変異についてホモ接合性であると判断できる。さらに、例えば「ゆめみのり」を交配親として作製した「ゆめみのり」の子孫系統について、上記の4アデニン塩基の挿入変異が検出され、かつ野生型7Sα遺伝子の存在が示されない場合には、その子孫系統は、7Sαを完全に欠失しており、「ゆめみのり」の7Sα欠失性の導入に成功したものと判断することができる。
【0045】
また、上記4アデニン塩基の挿入変異の存在が示される一方、野生型7Sα遺伝子の存在も示される場合には、7Sαタンパク質は部分的に欠失している。この場合、通常は、供試ダイズはCG-2遺伝子の挿入変異についてヘテロ接合性であると判定される。
【0046】
このように本発明では、ダイズの7Sα欠失性を判定した結果に基づいて、7Sグロブリンαサブユニット欠失ダイズを識別することもできる。本発明の方法を用いることにより、「ゆめみのり」、「ゆめみのり」の兄弟系統、又はそれらの子孫系統を用いて例えば7Sα欠失ダイズを育種しようとする際に、7Sα欠失形質を有するダイズを早期に選抜することもできる。そのため本発明は、7Sα欠失形質を有するダイズの効率的育種方法にも有用である。
【0047】
本発明において、7Sαの「欠失性」は、生成される7Sαタンパク質自体の欠失の有無だけでなく、7Sαを生成できない7Sα遺伝子、特に、「ゆめみのり」において認められる、第1エキソン中に4アデニン塩基の挿入変異を有し7Sαを産生できないCG-2遺伝子(「欠失型CG-2遺伝子」と呼ぶ)の有無として、理解してもよい。また、後述の7Sα’の「欠失性」も、同様に、生成される7Sα’タンパク質自体の欠失の有無だけでなく、7Sα’を生成できない7Sα’遺伝子、特に、その遺伝子構造全体が欠失しているために7Sα’を産生できないCG-1遺伝子(「欠失型7Sα’遺伝子」と呼ぶ)の有無として、理解することもできる。
【0048】
なお本発明において、ダイズの「系統」とは、品種登録されている純系のダイズ系統、及び品種登録されていない純系のダイズ系統を含むだけでなく、まだ純系として確立されていない育種過程にあるダイズ個体をも含む。
【0049】
2.7Sα欠失性及び7Sα’欠失性の同時判定、並びにダイズの混入検査法
本発明は、CG-2遺伝子の第1エキソン中への4アデニン塩基の挿入を検出する際に、7Sα’遺伝子の完全欠失(遺伝子構造自体の完全な喪失)の検出も同時に行う方法にも関する。7Sα’遺伝子の完全欠失の検出法としては、上記のCG-2遺伝子における挿入変異の検出と同様の検出手法を利用すればよいが、PCR増幅を利用して行う場合には、7Sα’遺伝子中の領域を特異的に増幅するプライマーセットを用いることによって行うことができる。例えば、7Sα’遺伝子の完全欠失は、7Sα’遺伝子を特異的に増幅可能なプライマー又はプローブを公知の7Sα’遺伝子(CG-1遺伝子)の塩基配列(例えばGenBank登録番号AB030838;配列番号11)に基づいて設計し、それを、CG-2遺伝子の第1エキソン中への4アデニン塩基の挿入を検出する工程において、その挿入変異を特異的に検出するためのプライマー又はプローブとともに(好ましくは混合して)使用して、検出することができる。
【0050】
7Sα’遺伝子を特異的に増幅可能なプライマーを含む特に好適なプライマーセットとしては、後述の実施例3に示したCG-1遺伝子特異的プライマーセット(CG1F[配列番号15]及びCG1R[配列番号16])が挙げられる。
【0051】
限定するものではないが、7Sα欠失性及び7Sα’欠失性の同時判定には、実施例3及び図7に記載したAセット及びBセットのプライマーセットが特に有用である。これらのプライマーセットを用いた同時判定法の手順は、実施例に詳述されている。
【0052】
7Sα欠失性及び7Sα’欠失性の同時判定法を用いれば、7Sα欠失性及び7Sα’欠失性の点で異なる性質を持つダイズ系統を、容易に識別することができる。このことは、7Sαの欠失による低アレルゲン性、及び7Sαと7Sα’の欠失による栄養価の高い11Sグロブリン含量の増加という有用な性質に関して、望ましい性質を持つダイズ系統を識別できることを意味する。
【0053】
さらに本発明は、7Sα及び7Sα’の欠失性が異なるダイズ系統の混入を、上記の同時判定試験を利用して検査する方法にも関する。ダイズ由来の核酸試料について、7Sα欠失性及び7Sα’欠失性の判定結果が、目的のダイズに関する結果と異なっていた場合には、その核酸試料の由来するダイズに、7Sα及び7Sα’の欠失性が異なる系統のダイズが混入していることが分かる。本発明のダイズの混入検査法は、検出限界が低く、高感度である。この混入検査法における検出限界は、核酸試料におけるDNA重量に換算した場合、混入率0.1%以上、好ましくは0.5%以上、好ましくは1%以上である。
【0054】
3.その他の実施形態
本発明は、CG-2遺伝子の第1エキソン中への4アデニン塩基の挿入変異を特異的に検出可能なDNA断片にも関する。これらのDNA断片は、プライマー又はプローブとして有用に用いることができる。
【0055】
具体的には、そのようなDNA断片は、「ゆめみのり」に見出されたCG-2遺伝子の挿入変異を含むDNAであって、配列番号3で示す塩基配列上の第215〜第221位の配列AAAAAAAを含む少なくとも15塩基(15塩基〜788塩基)、例えば15塩基〜50塩基(プライマー用など)又は30塩基〜500塩基(プローブ用など)の連続した配列からなるDNAであってよい。
【0056】
本発明は、「ゆめみのり」に見出されたCG-2遺伝子の挿入変異を含むプライマーであって、配列番号3で示す塩基配列上の第215〜第221位の配列AAAAAAAを含む15〜50塩基の連続した配列又はそれに相補的な配列からなるポリヌクレオチドプライマー、特に、配列番号3で示す塩基配列上の第215〜第221位の配列AAAAAAAを3'末端に含む、15〜50塩基、より好ましくは18〜40塩基の連続した配列又はそれに相補的な配列からなるポリヌクレオチドプライマーにも関する。この配列AAAAAAAを3'末端付近(3'末端又はその数塩基5'側に当たる3'末端近傍)に含むプライマーは、上記挿入変異を有するダイズ核酸のみを高感度に増幅することができるため、とりわけ有用である。このようなプライマーを用いた、欠失型CG-2特異的プライマーセット(プライマーCG2MF[配列番号12]及びCG23R[配列番号14])も本発明に含まれる。
【0057】
また本発明では、CG-2遺伝子中の挿入変異を特異的に検出できるプライマーに加えてコントロールプライマー等をさらに含む、検出に便利に使用できるプライマーセットにも関する。例えば、実施例3に記載した欠失型CG-2特異的プライマーセットと野生型7Sα特異的プライマーセットは、2回のPCRによって7Sαの有無を判定できる便利な共優性マーカーとして用いることができる。これらのマーカーを用いることによって、育種選抜において重要であるが従来のタンパク質での分析では検出不可能な、上記4アデニン塩基の挿入変異を有し7Sαを産生できないCG-2遺伝子(欠失型CG-2遺伝子)についてヘテロ接合であるダイズ系統を識別することができる。またこれらのマーカーを用いることによって、7Sα欠失ダイズ系統が野生型ダイズ系統に混入した場合の検出なども高感度に行うことができる。
【0058】
本発明は、CG-2遺伝子の上記挿入変異の有無と7Sα’遺伝子の有無をマルチプレックスPCRによって同時に検出することができる、実施例3に記載したCG-1特異的プライマーセットと欠失型CG-2特異的プライマーセットを含むプライマーセットにも関する。このプライマーセットには、実施例3に記載のようにさらにポジティブコントロール用プライマーセットを含んでもよい(実施例3のセットA)。本発明は、この検出のコントロール実験用に有用な、実施例3に記載の野生型7Sα特異的プライマーセットとポジティブコントロール用プライマーセットとを含むプライマーセット(実施例3のセットB)にも関する。これらのプライマーセットを用いて同時検出を行えば、欠失型CG-2遺伝子の有無と7Sα’遺伝子の有無が重要となる、7Sグロブリンαサブユニット変異体を利用した育種やダイズアレルゲン性の分析などにおいて、分析コストや労力を顕著に節減することができる。
【0059】
さらに、これらのプライマーセットは、ダイズの7Sα欠失性や7Sα’欠失性の検出用のキット、ダイズの識別用キット、ダイズの異系統混入検査キットなどに含めて提供することもありうる。このような各種キットも、本発明の範囲に含まれる。
【0060】
なお上記のようなDNA断片、並びにプライマー及びプローブなどは、当業者に公知の核酸調製法、例えば、限定するものではないが、化学合成法(液相法、固相法、カラム法及びバッチ法等)、クロマトグラフィー法による精製抽出法、PCR法、制限酵素消化等を利用して製造することができる。Applied Biosystems 3400 DNA合成機(Applied Biosystems)などの核酸合成装置も市販されている。
【0061】
4.配列の説明
・配列番号1: GenBank登録番号AB051865の配列中の、ダイズ7Sα遺伝子の第1エキソンに相当する塩基配列。野生型7Sα第1エキソンである。
【0062】
・配列番号2: 配列番号1の野生型7Sα第1エキソンにコードされるアミノ酸配列。
【0063】
・配列番号3: 「ゆめみのり」のダイズCG-2遺伝子(7Sα遺伝子)の、第1エキソンに相当する塩基配列。第215位〜第221位に挿入変異を含む配列AAAAAAAが存在し、その結果この第1エキソン領域内に終止コドンが生じている。
【0064】
・配列番号4: 配列番号3の配列にコードされるアミノ酸配列。
【0065】
・配列番号5: GenBank登録番号AB051865の塩基配列。ダイズ7Sα遺伝子を含むゲノム配列である。
【0066】
・配列番号6: 配列番号5の配列にコードされるアミノ酸配列。
【0067】
・配列番号7: GenBank登録番号X17698の塩基配列。野生型ダイズ7Sα遺伝子のmRNA配列である。
【0068】
・配列番号8: 刈系434号及び毛振の7Sα遺伝子を含むゲノム配列。
【0069】
・配列番号9: 刈系434号及び毛振の7Sα遺伝子にコードされるアミノ酸配列。
【0070】
・配列番号10:「ゆめみのり」のCG-2遺伝子(7Sα遺伝子)を含むゲノム配列。
【0071】
・配列番号11: GenBank登録番号AB051838の塩基配列。ダイズ7Sα’遺伝子(CG-1遺伝子)のmRNA配列である。
【0072】
・配列番号12: CG2MFプライマー
・配列番号13: CG23Fプライマー
・配列番号14: CG23Rプライマー
・配列番号15: CG1Fプライマー
・配列番号16: CG1Rプライマー
・配列番号17: 30KFプライマー
・配列番号18: 30KRプライマー
・配列番号19: CG2TFプライマー
・配列番号20: CG2TRプライマー
【実施例】
【0073】
以下、実施例及び図面を参照して本発明を説明するが、本発明はこれらの実施例に限定されるものではない。
【0074】
以下の実施例では、γ線照射によって7Sグロブリンα’サブユニット(7Sα’)および7Sグロブリンαサブユニット(7Sα)を欠失したダイズ品種「ゆめみのり」、「ゆめみのり」の原品種「刈系434号」、「ゆめみのり」の反復親「タチユタカ」、遺伝資源より見出された7Sサブユニットの変異体「毛振」および「秣食豆公503」を供試した。
【0075】
各品種における、7Sα’およびαをコードする遺伝子の有無は次の通りである。
【0076】
【表1】

【0077】
表1中、表現型について「欠失」とは、当該タンパク質の生成量がほぼゼロである完全欠失を意味し、「低下」とは当該タンパク質の生成量が野生型に比べて減少していることを意味し、「正常」とは当該タンパク質の生成量が野生型と同等であることを意味する。
【0078】
なお各品種の育成については、以下の文献で報告されている。
【0079】
- 秣食豆公503[ダイズ遺伝資源由来]: Kitamura, K. et al. Mutant strains with low level of subunits of 7S globulin in soybean (Glycine max Mell.) seed. (1981) Japan. J. Breed. 31(4):353-359
- 毛振[ダイズ遺伝資源由来]: Kitamura, K. et al. Mutant strains with low level of subunits of 7S globulin in soybean (Glycine max Mell.) seed. (1981) Japan. J. Breed. 31(4):353-359
- タチユタカ: 橋本鋼二ほか.ダイズ新品種「タチユタカ」の育成.東北農試研報.77:27-44(1988)
- 刈系434号[(毛振//大館1号/秣食豆公503)/タチユタカ×2に由来]: Ogawa, T. et al. Genetic improvement of seed storage proteins using three variant alleles of 7S globulin subunits in soybean (Glycine max L.). (1989) Jpn. J. Breed. 39(2):137-147
- ゆめみのり[γ線照射した刈系434号に由来]: Takahashi, K. et al. Inheritance of α-subunit deficiency of β-conglycinin in soybean (Glycine max L. MERRILL) line induced by γ-ray irradiation. (1996) Breed.Sci. 46:251-255
【0080】
実施例1: ダイズ7Sグロブリンαサブユニット遺伝子の塩基配列解析
「刈系434号」、「毛振」及び「ゆめみのり」のそれぞれの幼葉約100mgから、PhytoPure DNA extraction kit(アマシャムバイオサイエンス)を用いて添付のプロトコールに従って、ゲノムDNAを抽出した。
【0081】
調製したゲノムDNAをテンプレートDNAとして、ポリメラーゼ連鎖反応(PCR)法によりDNA断片を増幅した。このPCRにおいては、国際塩基配列データベース(DDBJ/EMBL/GenBank)から登録番号AB051865(ゲノム配列;ダイズ品種「Forrest」由来)により入手できる7Sα遺伝子の既知塩基配列(配列番号5)に基づいて7Sα遺伝子全長を増幅するように設計された、以下のプライマーを用いた。
【0082】
・フォワードプライマー CG2TF: 5'-GGGTCATCGGGAACTAGTTCA-3'(AB051865の塩基配列上の第430位〜第452位の配列に相当;配列番号19)
・リバースプライマ− CG2TR: 5'-TTCATAGATGAAGCTCTAGTCATAGTTAC-3'(AB051865の塩基配列上の第2,919位〜第2,947位の配列に相当;配列番号20)
【0083】
PCR反応には、終濃度で1×バッファー、各0.2mM dNTP、1.5mM MgCl2、設定濃度のプライマー、1.25U DNAポリメラーゼ(ExTaq、TAKARA BIO INC.)、50〜100ngテンプレートDNAを含む25μlの反応液を用いた。PCR反応では、GeneAmp PCR system 9700(アプライドバイオシステムズ)を用いて、95℃で5分間の変性反応、続いて、95℃で30秒、62℃で30秒、72℃で1分を32回繰り返し、最後に72℃で7分間の伸長反応を行った。
【0084】
PCR反応後、反応物中のPCR増幅断片を、1%アガロースゲル(L03「TAKARA」、TAKARA BIO INC.)を用いて100Vで50分間の電気泳動によって分離し、エチジウムブロマイドによるゲル染色後、紫外線下で可視化した。サイズマーカーとしては2-Log DNA Ladder(New England BioLabs)を使用した(以下の実験でも同様)。
【0085】
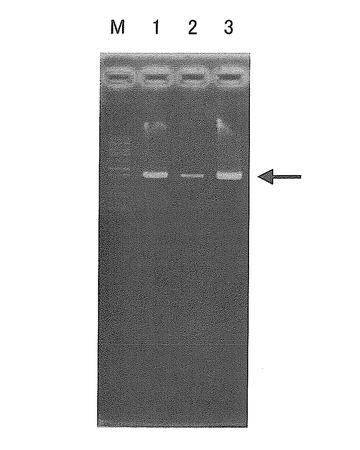
電気泳動の結果、いずれの品種・系統でも、AB051865の配列(ゲノム配列)から予測される通りの約2.5kbの増幅断片が確認された(図2)。図2中、M:サイズマーカー、1:「刈系434号」、2:「毛振」、3:「ゆめみのり」を示す。
【0086】
得られたDNA増幅断片を、核酸抽出キットQIAquick gel extraction kit(キアゲン)を用いてゲルから抽出し、それについてシークエンシング試薬BigDye terminatior v1.1 Cycle Sequencing kitおよびシークエンサーABI Prism 310 genetic analyzer(アプライドバイオシステムズ)を用いて、直接塩基配列を決定した。決定した配列については、オンラインソフトウェアCLUSTAL W(DNA Data Bank of Japan、http://www.ddbj.nig.ac.jp/)を用いて配列解析を行った。
【0087】
表1に示す通り、7Sグロブリンαサブユニット遺伝子として「刈系434号」はCG-2遺伝子のみを有し、「毛振」はCG-3遺伝子のみを有することから、本実施例で決定された塩基配列は「刈系434号」ではCG-2遺伝子、「毛振」ではCG-3遺伝子のものである。しかし配列解析の結果、「刈系434号」と「毛振」について決定された塩基配列は、全く同一であった。従って、本実施例で配列決定されたαサブユニット遺伝子の開始コドンから終止コドンまでを含む領域では、CG-2遺伝子とCG-3遺伝子は塩基配列上同一であった。
【0088】
さらに、「刈系434号」及び「毛振」について決定した配列は、AB051865(ダイズ品種「Forrest」)の第482位から第2,908位までの配列と、1塩基を除いて完全に一致しており、その違いは「刈系434号」及び「毛振」の配列においてAB051865の配列の2,564塩基目に当たるシトシンがアデニンに置換しているのみであった(図3)。
【0089】
一方、「ゆめみのり」の配列には、「刈系434号」の配列と比較して、4個のアデニン塩基の挿入が生じていた(図3)。その挿入位置は、AB051865配列の第704位〜第706位の3アデニン配列内に相当する。AB051865配列の第704位〜第706位は、AB051865配列にコードされる7Sα遺伝子の第1エキソン(野生型7Sα第1エキソン;配列番号1)の第215位〜第217位[第1エキソンの5'端の開始コドン「ATG」のAを第1位とした場合]に当たる。そして、このような4個のアデニン塩基の挿入により生じた塩基配列AAAAAAA、すなわち、「ゆめみのり」のCG-2遺伝子上の第1エキソンに相当する領域(挿入変異型7Sα第1エキソン;配列番号3)の第215位〜第221位の7アデニン配列は、「毛振」にもAB051865配列にも存在しなかった。
【0090】
次に、「ゆめみのり」について決定した配列から、それにコードされるアミノ酸配列を推定した。その結果、「ゆめみのり」の7Sαサブユニット遺伝子であるCG-2遺伝子においては、上記のような4個のアデニン塩基の挿入によりフレームシフトが生じ、第1エキソン中に終止コドンが生じていることが示された(図4、配列番号3及び4)。従って、この4塩基のアデニンの挿入が、「ゆめみのり」における7Sグロブリンαサブユニットの欠失の原因であると考えられた。
【0091】
実施例2: 7Sグロブリンαサブユニットの欠失性の検出
実施例1で確認された「ゆめみのり」のCG-2遺伝子(7Sグロブリンαサブユニット遺伝子)における4個のアデニン塩基の挿入の検出を、PCR増幅により行った。
【0092】
まず「タチユタカ」、「毛振」、「秣食豆公503」、「ゆめみのり」、「刈系434号」の幼葉から、実施例1と同様にしてゲノムDNAを調製し、それをテンプレートDNAとした。PCRには、以下のプライマーを設計して用いた。
【0093】
・欠失型CG-2特異的フォワードプライマー CG2MF:
5'-GAAGAATGTGAAGAAGGTGAAAAAAAT-3'[配列番号12]
(AB051865の塩基配列上の第685位〜第707位の配列+4アデニン塩基挿入に相当)
・CG-2/CG-3共通フォワードプライマー CG23F:
5'-AAGAAGAATGTGAAGAAGGTGAAAT-3'[配列番号13]
(AB051865の塩基配列上の第683位〜第707位の配列に相当)
・CG-2/CG-3共通リバースプライマー CG23R:
5'-GAATGCGACCATATGGTTTTTGA-3'[配列番号14]
(AB051865の塩基配列上の第1,109位〜第1,132位の配列に相当)
【0094】
ここで、欠失型CG-2特異的フォワードプライマー CG2MFは、4個のアデニン塩基の挿入配列を3'端に含む、欠失型CG-2遺伝子に特異的なプライマーである。
【0095】
PCR増幅は2つの反応系で行った。欠失型CG-2特異的フォワードプライマー CG2MFとCG-2/CG-3共通リバースプライマー CG23Rとを組み合わせたプライマーセット(欠失型CG-2特異的プライマーセット)を、1つ目のPCR増幅に用い、またCG-2/CG-3共通フォワードプライマー CG23FとCG-2/CG-3共通リバースプライマー CG23Rとを組み合わせたプライマーセット(野生型7Sα特異的プライマーセット)を、2つ目のPCR増幅に用いた。
【0096】
PCR反応には、終濃度で1×バッファー、各0.2mM dNTP、1.5mM MgCl2、各0.2μMのプライマー、1.25U DNAポリメラーゼ(ExTaq、TAKARA BIO INC.)、50〜100ngテンプレートDNAを含む25μlの反応液を用いた。PCR反応では、GeneAmp PCR system 9700(アプライドバイオシステムズ)を用いて、95℃で5分間の変性反応、続いて、95℃で30秒、62℃で30秒、72℃で1分を32回繰り返し、最後に72℃で7分間の伸長反応を行った。
【0097】
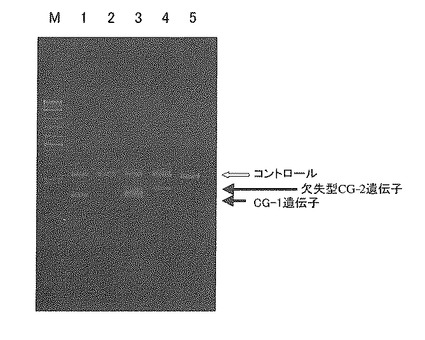
PCR反応後、4%アガロースゲルを用いて、その他は実施例1と同様にしてPCR増幅産物を電気泳動により分離し、可視化した。その結果を図5及び図6に示す。図5及び図6中、M:サイズマーカー、2:「タチユタカ」、3:「毛振」、4:「秣食豆公503」、5:「ゆめみのり」、6:「刈系434号」を示す。
【0098】
図5に示す通り、プライマーCG2MFとCG23Rを用いて行ったPCR増幅では、「ゆめみのり」のみ増幅断片が検出された。得られた増幅断片の長さは450bpであった。一方、プライマーCG23FとCG23Rを用いて行ったPCR増幅では、「ゆめみのり」以外の全品種・系統で、448bpの増幅断片が検出された(図6)。
【0099】
以上の通り、「ゆめみのり」のCG-2遺伝子内の4アデニン塩基挿入部位に設計した欠失型CG-2特異的プライマーセットおよび野生型7Sα特異的プライマーセットにより、「ゆめみのり」に認められるような7Sα欠失の有無を2回のPCRで判定できる共優性のマーカーを、開発することができた。
【0100】
実施例3: マルチプレックスPCRによる7Sα欠失性及び7Sα’欠失性の同時判定
ダイズ11Sグロブリンの含量を増加させるためには、7Sαと7Sα’を同時に欠失させることが有効である。そこで、7Sα’をコードするCG-1遺伝子に特異的なプライマーを設計して優性マーカーとし、実施例2で用いた7SαのマーカーとのマルチプレックスPCRにより7Sαと7Sα’の欠失の有無を同時に判定する方法を検討した。
【0101】
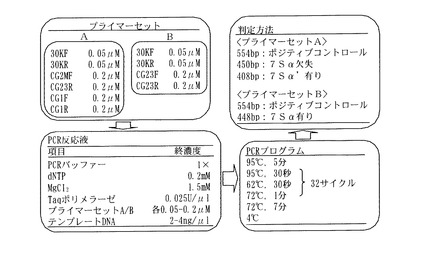
図7には、マルチプレックスPCRによる7Sα欠失性及び7Sα’欠失性の同時判定法の一例として行った以下の実験の具体的手順の概要を示す。
【0102】
まず、7Sグロブリンαサブユニット遺伝子のmRNA配列として報告されているGenBank登録番号X17698の塩基配列(配列番号7)と、7Sグロブリンα’サブユニット遺伝子(CG-1遺伝子)のmRNA配列として報告されているGenBank登録番号AB030838の塩基配列(配列番号11)とをアラインメントした(図8)。このアラインメントでは91%の高い相同性が示された。そこで、このアラインメントに基づき、CG-1遺伝子を特異的に増幅する以下のプライマーセット(CG-1特異的プライマーセット)を設計した。
【0103】
・CG-1特異的フォワードプライマー CG1F:
5'-GTCACAACAAGTGCCTCCGA-3'[配列番号15]
(AB030838の塩基配列上の第158位〜第177位の配列に相当)
・CG-1特異的リバースプライマー CG1R:
5'-GGTGCTTTTCCTGCTTGTG-3'[配列番号16]
(AB030838の塩基配列上の第547位〜第565位の配列に相当)
【0104】
マルチプレックスPCRには、このCG-1特異的プライマーセットに加えて、実施例2で用いた欠失型CG-2特異的プライマーセットを用いた。さらに、増幅断片が得られない場合にPCR反応自体が不成功に終わった可能性を除くため、ダイズタンパク質Gly-m Bd 30Kをコードする遺伝子の一部を増幅するプライマーセットも、ポジティブコントロールとして反応液に加えた。なおダイズタンパク質Gly-m Bd 30Kは、ダイズ貯蔵タンパク質に含まれる主要なアレルゲンであり、これまでの大規模なスクリーニングによっても欠失体は得られていない。このポジティブコントロール用プライマーセットは、以下のプライマーからなる。
【0105】
・ポジティブコントロール・フォワードプライマー30KF:
5'-AGCTCACTAATCACTATATATACGACATGCT-3'[配列番号17]
(GenBank登録番号AB013289の塩基配列上の第4,562位〜第4,592位の配列に相当)
・ポジティブコントロール・リバースプライマー30KR:
5'-CACAGCCCCCTTGGTACTTTAC-3'[配列番号18]
(GenBank登録番号AB013289の塩基配列上の第5,094位〜第5,115位の配列に相当)
【0106】
以上のCG-1特異的プライマーセット(CG1F及びCG1R)、欠失型CG-2特異的プライマーセット(CG2MF及びCG23R)、及びポジティブコントロール用プライマーセット(30KF及び30KR)は、まとめてプライマーセット(Aセット)として検出用に用いた。
【0107】
さらに、実施例2で用いた野生型7Sα特異的プライマーセット(CG23F及びCG23R;上記挿入変異を含まない野生型7Sα遺伝子を増幅する)と、上記ポジティブコントロール用プライマーセット(30KF及び30KR)を、まとめてプライマーセット(Bセット)としてコントロール用に用いた。
【0108】
表2には、本実施例におけるプライマーとその使用形態をまとめた。
【0109】
【表2】
【0110】
反応液は図7に記載された組成で全量25μlとして調製した。なおテンプレートDNAは実施例1と同様にして調製した。PCR反応は、GeneAmp PCR system 9700(アプライドバイオシステムズ)を用いて、以下の条件で行った:95℃で5分、続いて、95℃で30秒、62℃で30秒、72℃で1分を32サイクル、続いて72℃で7分の後、4℃で保持。
【0111】
PCR反応後、実施例2と同様にしてPCR増幅産物を電気泳動により分離し、可視化した。その結果を図9(Aセット使用)及び図10(Bセット使用)に示す。図9及び図10中、M:サイズマーカー、1:「タチユタカ」、2:「毛振」、3:「秣食豆公503」、4:「ゆめみのり」、5:「刈系434号」を示す。
【0112】
図9に示すように、プライマーセットAを用いて行ったPCR増幅では、450bpの増幅断片が「ゆめみのり」のみで得られ、この増幅断片長は欠失型CG-2特異的プライマーセットによる増幅サイズと一致していた。また「タチユタカ」及び「秣食豆公503」では、408bpの増幅断片が得られ、この増幅断片長はCG-1特異的プライマーセットによる増幅サイズと一致していた。さらに「毛振」及び「刈系434号」では、ポジティブコントロールプライマーによるもの以外の増幅断片は検出されなかった。なおポジティブコントロール用プライマーセットによる554bpの増幅断片は、全サンプルで検出された。
【0113】
一方、図10に示すように、プライマーセットBを用いて行ったコントロール用のPCR増幅では、「ゆめみのり」以外の品種で448bpの増幅断片が得られた。この増幅断片の長さは、野生型7Sグロブリンαサブユニット遺伝子を増幅する野生型7Sα特異的プライマーセットによる増幅サイズと一致していた。なおポジティブコントロール用プライマーセットによる554bpの増幅断片は、全サンプルで検出された。
【0114】
以上の結果は、上記表1に示した各品種・系統の特徴と完全に一致していた。このことから、これらのプライマーセットA、あるいはプライマーセットA及びBをPCR増幅に用いることにより、7Sαの欠失性及び7Sα’の欠失性を同時に判定できることが示された。
【0115】
また、これらのプライマーセットA及びBをPCR増幅に用いることにより、CG-1遺伝子の有無、野生型7Sα遺伝子の有無、欠失型CG-2遺伝子の有無から、ダイズ品種・系統が識別されることも示された。
【0116】
実施例4: 7Sα欠失性及び7Sα’欠失性の同時判定に基づくダイズの異品種・異系統混入検査
7Sグロブリンαサブユニット及びα’サブユニットの欠失性が異なるダイズ品種・系統の混入を、実施例3で述べた同時判定試験を利用して検査した。サンプルには、「ゆめみのり」(7Sα及び7Sα’の両方を欠失)のゲノムDNAに、「タチユタカ」(7Sα及び7Sα’の両方を有する)のゲノムDNAを混合した核酸試料を用いた。
【0117】
検出感度を確認するため、「ゆめみのり」のゲノムDNA量に対して0.1%、0.2%、0.5%、および1%となるように「タチユタカ」のゲノムDNAを加えたサンプルを用意し、それらをテンプレートDNAとした。
【0118】
各混入パーセンテージのテンプレートDNAを用い、実施例3と同様にして、Aセット及びBセットのプライマーセットを用いたPCR増幅を行った。PCR反応後、増幅産物を同様にして電気泳動し可視化した。その結果を図11(Aセット)及び図12(Bセット)に示す。
【0119】
図11に示される通り、プライマーセットAを用いて検出を行った場合、DNA重量換算で0.1%以上の混入量で、CG-1遺伝子を示す408bpの増幅断片が認められた。またプライマーセットBを用いて検出を行った場合、ゲノムDNA重量換算で0.2%以上の混入量で、野生型7Sα遺伝子を示す448bpの増幅断片が認められた(図12)。
【0120】
この同時判定法では、7Sα及び7Sα’の欠失性が異なるダイズ品種の混入を、混入率(DNA重量換算)0.1〜0.2%程度でも検出することができたことから、この方法は検出限界が非常に低く、高感度であることが示された。
【産業上の利用可能性】
【0121】
本発明の7Sグロブリンαサブユニット欠失ダイズの検出方法は、「ゆめみのり」又は「ゆめみのり」の兄弟系統、それらの子孫系統などの、7Sグロブリンαサブユニットを欠失したダイズ系統を簡便に検出するために使用することができる。この本発明の方法は、ダイズの混入検査法としても使用できる。
【図面の簡単な説明】
【0122】
【図1】図1は、ダイズアレルゲン7Sα欠失ダイズ「ゆめみのり」の系譜を示す図である。
【図2】図2は、7Sα遺伝子全長を含むDNA増幅断片の電気泳動写真である。M: マーカー、レーン1: 「刈系434号」、レーン2: 「毛振」、レーン3: 「ゆめみのり」。
【図3−1】図3は、7Sα遺伝子について各品種・系統で決定したゲノム配列のアラインメントを示す図である。基準配列として7Sα遺伝子の既知配列である登録番号AB051865の配列を一緒に示した。図中、「ゆめみのり(Yumeminori)」のみ、AB051865の配列の第704位〜第706位に相当する位置に、4個のアデニン塩基の挿入が見られる。
【図3−2】図3は、7Sα遺伝子について各品種・系統で決定したゲノム配列のアラインメントを示す図である。図3−2は、図3−1の続きである。AB051865の配列の第2564位に相当する位置において、「刈系434号」、「毛振」及び「ゆめみのり」ではシトシン(C)からアデニン(A)への置換が生じていた。
【図3−3】図3は、7Sα遺伝子について各品種・系統で決定したゲノム配列のアラインメントを示す図である。図3−3は、図3−1、図3−2の続きである。
【図4】図4は、「ゆめみのり」のCG-2遺伝子(7Sα遺伝子)においてフレームシフトが起こり、早期に終止コドンが生じたことを示す図である。
【図5】図5は、欠失型CG-2特異的プライマーセットを用いたPCR増幅の結果を示す電気泳動写真である。M: マーカー、レーン1: 「タチユタカ」、レーン2: 「毛振」、レーン3: 「秣食豆公503」、レーン4: 「ゆめみのり」、レーン5: 「刈系434号」。
【図6】図6は、野生型7Sα特異的プライマーセットを用いたPCR増幅の結果を示す電気泳動写真である。M: マーカー、レーン1: 「タチユタカ」、レーン2: 「毛振」、レーン3: 「秣食豆公503」、レーン4: 「ゆめみのり」、レーン5: 「刈系434号」。
【図7】図7は、7Sαと7Sα’の欠失性の同時判定方法の概要を示す図である。
【図8】図8は、登録番号X17698の配列とAB030838の配列とのアラインメントを示す図である。
【図9】図9は、プライマーセットAを用いた7Sα’および7Sα遺伝子検出用のPCR増幅の結果を示す電気泳動写真である。M: マーカー、レーン1: 「タチユタカ」、レーン2: 「毛振」、レーン3: 「秣食豆公503」、レーン4: 「ゆめみのり」、レーン5: 「刈系434号」。
【図10】図10は、プライマーセットBを用いたコントロール検出実験用のPCR増幅の結果を示す電気泳動写真である。M: マーカー、レーン1: 「タチユタカ」、レーン2: 「毛振」、レーン3: 「秣食豆公503」、レーン4: 「ゆめみのり」、レーン5: 「刈系434号」。
【図11】図11は、「ゆめみのり」への「タチユタカ」の混入を、プライマーセットAを用いて検出した結果を示す電気泳動写真である。
【図12】図12は、「ゆめみのり」への「タチユタカ」の混入を、プライマーセットBを用いて検出した結果を示す電気泳動写真である。
【配列表フリーテキスト】
【0123】
配列番号12〜20の配列はプライマーである。
【技術分野】
【0001】
本発明は、7Sグロブリンαサブユニット欠失ダイズを検出する方法、及びその方法に用いるプライマーに関する。
【背景技術】
【0002】
ダイズ貯蔵タンパク質のひとつである7Sグロブリン(別名:β-コングリシニン)は、α’、α及びβの3種類のサブユニットで構成されている。そのうち、7Sグロブリンのαサブユニット(以下、7Sαとも略記する)は、ダイズ陽性のアトピー性皮膚炎患者のうち25%の患者の血清によって検出された主要アレルゲンである。
【0003】
ダイズ貯蔵タンパク質には、7Sグロブリンの他、2Sグロブリン、11Sグロブリン、及び15Sグロブリンが知られている。7Sグロブリンは、アレルゲンであるだけでなく、11Sグロブリンに比べて栄養価や加工適性が劣るとされる。ダイズの7Sグロブリン含量は11Sグロブリンの含量との間で負の相関が認められることから、7Sグロブリンαサブユニット及びα’サブユニットを欠失させることによって相対的に11Sグロブリンの含量を高めることができる。そのため、7Sグロブリンαサブユニット及びα’サブユニットを欠失させて機能性を強化したダイズ品種の開発が進められている。
【0004】
7Sグロブリンのサブユニット変異体として、7Sグロブリンα’サブユニット(7Sα’)を欠失した「毛振(Keburi)」、7Sグロブリンαサブユニット及びβサブユニットの生成量が低下した「秣食豆公503(Moshidou Gong 503)」、秣食豆公503の7Sα及びβの低下性と毛振の7Sα’欠失が導入された「刈系434号(Kari-kei 434)」、刈系434号のγ線照射により作出された7Sα欠失品種「ゆめみのり(Yumeminori)」などを含む多様なダイズ品種・系統が知られている。これらのダイズ品種・系統の研究から、Region Cと呼ばれるダイズ染色体領域内のCG-1遺伝子が7Sグロブリンα’サブユニットをコードする遺伝子であること、及びRegion Aと呼ばれるダイズ染色体領域内のCG-2遺伝子及びCG-3遺伝子が7Sグロブリンαサブユニットをコードする遺伝子であることが明らかになった(非特許文献1及び2)。「毛振」ではCG-1遺伝子及びCG-2遺伝子が欠失していることが示されており(非特許文献2及び3)、「秣食豆公503」の7Sα低下性がCG-3遺伝子の欠失に起因することも報告されている(非特許文献3)。このようにダイズ7Sグロブリンサブユニットに関する研究は確実に進んできている。
【0005】
しかしながら、7Sグロブリンサブユニット変異の検出には、現在でも、手間を要するタンパク質レベルの手法が使用されている。そのような手法では、分析試料として、ダイズ種子粉末あるいは未分解のタンパク質を含むダイズ加工物の調製が必要である。また例えばタンパク質の検出の際に用いられるSDS-PAGEはゲルの作成に熟練を要するし、SDS-PAGEに続いて行うCBB法による検出は、感度が低いため異品種の僅かな混入などを検出できない場合がある。一方、SDS-PAGEに続いて行う銀染色法は、感度は高いが操作が煩雑であるため多数サンプルの分析が困難である。
【0006】
以上のような背景から、7Sグロブリンαサブユニット及びα’サブユニット欠失ダイズ品種の効率的な育成やそれらダイズ品種に関する異品種混入検査などに利用できる高感度な簡易判定技術の開発が求められている。
【0007】
【非特許文献1】Harada J.J. et al., Plant Cell (1989) 1: p.415-425
【非特許文献2】Ladin B.F. et al., J. Mol. Appl. Genet. (1984) 2: p.372-380
【非特許文献3】Yoshino et al., Breed Sci. (2002) 52: p.285-292
【発明の開示】
【発明が解決しようとする課題】
【0008】
本発明は、7Sグロブリンαサブユニット欠失ダイズの高感度な検出方法を提供することを課題とする。
【課題を解決するための手段】
【0009】
本発明者らは、上記課題を解決すべく鋭意研究を行った。その結果、7Sグロブリンαサブユニット(7Sα)を欠失したダイズ品種「ゆめみのり」において7Sα遺伝子であるCG-2遺伝子の第1エキソンに4個のアデニン塩基が挿入された変異が生じていることを見出した。「ゆめみのり」ではその4個のアデニン塩基の挿入によってCG-2遺伝子にフレームシフトが生じ、ごく早期に終止コドンが現れるようになっていることから、この挿入変異によって7Sグロブリンαサブユニットが生成されなくなったと考えられた。これらの知見に基づき、本発明者らは、「ゆめみのり」において見出されたCG-2遺伝子の第1エキソン中の4塩基のアデニンの挿入をダイズ核酸について検出することによって、7Sグロブリンαサブユニット欠失ダイズを検出する手法を開発し、本発明を完成するに至った。
【0010】
すなわち本発明は以下の通りである。
【0011】
[1] ダイズ由来の核酸試料について、CG−2遺伝子の第1エキソンの第215位〜第217位の配列AAA中への4個のアデニン塩基の挿入を検出する工程を特徴とする、7Sグロブリンαサブユニット欠失ダイズの検出方法。
この方法では、CG−2遺伝子の第1エキソンは、配列番号1で示す塩基配列からなるものであることが好ましい。
またこの方法の前記検出工程は、好ましくは、配列番号3で示す塩基配列上の第215〜第221位の配列AAAAAAAを含む少なくとも15塩基の連続した配列からなるDNAをプライマー又はプローブとして用いて検出する工程である。
【0012】
さらにその検出工程は、配列番号3で示す塩基配列上の第215〜第221位の配列AAAAAAAを含む15〜50塩基の連続した配列又はそれに相補的な配列からなるポリヌクレオチドプライマーを用いたPCR増幅により検出する工程であることが好ましい。
そのようなプライマーとしては、配列番号12で示す塩基配列からなるポリヌクレオチドプライマーが好適である。
さらに本発明では、配列番号12で示す塩基配列からなるポリヌクレオチドプライマーとともに、配列番号14で示す塩基配列からなるポリヌクレオチドプライマーをPCR増幅に用いることも好ましい。
この方法で検出される7Sグロブリンαサブユニット欠失ダイズは、好ましくは「ゆめみのり」、「ゆめみのり」の兄弟系統、又はそれらの子孫系統である。
【0013】
[2] 上記[1]に記載の方法において、前記プライマーとともに、CG−1遺伝子特異的プライマーセットをPCR増幅に用いることを特徴とする、ダイズの7Sグロブリンαサブユニット欠失性及び7Sグロブリンα’サブユニット欠失性の同時判定方法。
この方法で用いるCG−1遺伝子特異的プライマーセットとしては、配列番号15で示す塩基配列からなるポリヌクレオチドプライマーと、配列番号16で示す塩基配列からなるポリヌクレオチドプライマーとからなる組み合わせが好適である。
【0014】
[3] 上記[2]に記載の方法で得られた判定結果に基づいて、異系統のダイズの混在の有無を判定することを特徴とする、ダイズの混入検査法。
【0015】
[4] 配列番号3で示す塩基配列上の第215〜第221位の配列AAAAAAAを含む少なくとも15塩基の連続した配列からなるDNA。
【0016】
[5] 配列番号3で示す塩基配列上の第215〜第221位の配列AAAAAAAを含む15〜50塩基の連続した配列又はそれに相補的な配列からなるポリヌクレオチドプライマー。
【0017】
[6] 配列番号12で示す塩基配列からなるポリヌクレオチドプライマーと、配列番号14で示す塩基配列からなるポリヌクレオチドプライマーとからなる、プライマーセット。
【0018】
[7] 配列番号12〜14で示す塩基配列の各々からなる3種のポリヌクレオチドプライマーを含むキット。このキットは、好適には、「ゆめみのり」、「ゆめみのり」の兄弟系統、又はそれらの子孫系統である7Sグロブリンαサブユニット欠失ダイズを検出するためのキットである。
【0019】
[8] 配列番号12〜16で示す塩基配列の各々からなる5種のポリヌクレオチドプライマーを含むキット。このキットは、配列番号17及び18で示す塩基配列の各々からなる2種のポリヌクレオチドプライマーをさらに含むことが好ましい。このキットは、好適には、「ゆめみのり」、「ゆめみのり」の兄弟系統、又はそれらの子孫系統である7Sグロブリンαサブユニット欠失ダイズを検出するためのキットである。
【発明の効果】
【0020】
本発明の方法は、「ゆめみのり」、「ゆめみのり」の兄弟系統、又はそれらの子孫系統などの7Sグロブリンαサブユニットを欠失したダイズ系統を、高感度にかつ簡便に検出することができる。
【発明を実施するための最良の形態】
【0021】
1.7Sグロブリンαサブユニット欠失ダイズの検出
1)検出すべき変異
本発明の方法において検出すべき変異、すなわちダイズCG-2遺伝子の第1エキソン中への4個のアデニン塩基の挿入変異は、最初に、ダイズ品種「ゆめみのり」において見出されたものである。
【0022】
「ゆめみのり」は、ダイズの主要アレルゲンである7Sグロブリンαサブユニット(7Sα)が欠失しており、栄養価の高い11Sグロブリンの含有量が向上した、低アレルゲン性のダイズ品種である。「ゆめみのり」は、「毛振」の7Sα’欠失形質(すなわち、7Sα’が生成されない)と「秣食豆公503」の7Sα低下形質(すなわち、7Sαの生成量が低下する)とを導入した「刈系434号」に、γ線照射(200Gy)することによって得られた7Sα欠失変異体から確立した1系統に基づいて、品種登録された。図1に、ダイズ品種「ゆめみのり」の系譜を示す。「ゆめみのり」は、7Sα欠失形質(7Sαが生成されない)と7Sα’欠失形質(7Sα’が生成されない)の両方を示す。なおダイズ品種「ゆめみのり」は、独立行政法人 農業・生物系特定産業技術研究機構 東北農業研究センターから入手することができる。
【0023】
本発明者らは、「ゆめみのり」において7Sα遺伝子の1つであるCG-2遺伝子の第1エキソン中に4個のアデニン塩基の挿入が生じていること、その結果CG-2遺伝子にフレームシフトが起き、さらに終止コドンが早期に生じていることを見出した。「ゆめみのり」のCG-2遺伝子の第1エキソン配列を原品種「刈系434号」と比較すると、この4個のアデニン塩基の挿入箇所は、「刈系434号」のCG-2遺伝子第1エキソンの第215位〜第217位に位置する3アデニン配列(配列AAA)内に相当する。「ゆめみのり」においてはその3アデニン配列が、4個のアデニン塩基が挿入されたことにより、7アデニン配列(配列AAAAAAA)となっている(「ゆめみのり」のCG-2遺伝子の第1エキソンの第215位〜第221位)。「ゆめみのり」はもう1つの7Sα遺伝子であるCG-3遺伝子を完全欠失(遺伝子構造自体を完全に喪失)しているので、「ゆめみのり」の7Sα欠失形質が、CG-2遺伝子の第1エキソン中に4個のアデニン塩基が挿入された結果として引き起こされたことは明らかである。本発明では、第1エキソンの第215位〜第217位に位置する3アデニン配列(配列AAA)内に4個のアデニン塩基が挿入されたCG-2遺伝子を、便宜的に、欠失型CG-2遺伝子と呼ぶ。
【0024】
「ゆめみのり」のCG-2遺伝子の第1エキソン中への4個のアデニン塩基の挿入は、もともとは、「ゆめみのり」の育種過程で「刈系434号」にγ線照射(200Gy)することによって得られた7Sα欠失変異体に生じた変異である。この7Sα欠失変異体からは「ゆめみのり」と同様の形質をもった系統が他にも作成されており、それらの系統でもそのようなCG-2遺伝子の第1エキソン中への4個のアデニン塩基の挿入が認められる。本発明では、7Sα欠失変異体から作成したこれらの系統を便宜的に『「ゆめみのり」の兄弟系統』と呼ぶ。
【0025】
CG-2遺伝子の第1エキソン中への4個のアデニン塩基の挿入は、「ゆめみのり」又は「ゆめみのり」の兄弟系統の子孫系統にも受け継がれる。本発明において「ゆめみのり」又は「ゆめみのり」の兄弟系統の子孫系統とは、「ゆめみのり」又は「ゆめみのり」の兄弟系統を育種の母本として用いて、例えば突然変異誘発処理や他のダイズ系統・品種との交配によって作製された、後代のダイズを言う。「ゆめみのり」又は「ゆめみのり」の兄弟系統の子孫系統は、CG-2遺伝子の第1エキソン中への上記の4個のアデニン塩基の挿入をヘテロ接合又はホモ接合で有し得る。
【0026】
本発明においては、限定するものではないが上述の「ゆめみのり」、「ゆめみのり」の兄弟系統、及びそれらの子孫系統のように、CG-2遺伝子の第1エキソンの第215位〜第217位に位置する3アデニン配列内への4個のアデニン塩基の挿入によってそのCG-2遺伝子の7Sグロブリンαサブユニット産生機能が失われたダイズを、「7Sグロブリンαサブユニット欠失ダイズ(7Sα欠失ダイズ)」と呼ぶ。7Sα欠失ダイズは、そのCG-2遺伝子中への4個のアデニン塩基の挿入変異をヘテロ接合又はホモ接合で有していてもよい。CG-2遺伝子とCG-3遺伝子は密接に連鎖していることから、この7Sグロブリンαサブユニット欠失ダイズは、通常は、「ゆめみのり」と同様にCG-3遺伝子を完全欠失している。
【0027】
多くのダイズ品種が、7Sグロブリンαサブユニット遺伝子としてCG-2およびCG-3の2つを保持している。しかし、「毛振」はCG-2遺伝子を完全欠失しており、7Sα遺伝子としてはCG-3遺伝子を保持する。一方、「秣食豆公503」及び「刈系434号」は、CG-3遺伝子を完全欠失しており、7Sα遺伝子としてはCG-2遺伝子を保持する。後述の実施例に示されるように、「毛振」のCG-3遺伝子と「刈系434号」のCG-2遺伝子とは、少なくとも開始コドンから終止コドンまでのゲノム配列領域内では同一の塩基配列を有しており、「ゆめみのり」のようなCG-2遺伝子における4個のアデニン塩基の挿入は含まない。本発明では、「ゆめみのり」で認められた上記の4個のアデニン塩基の挿入を含まない7Sα遺伝子、例えば「毛振」のCG-3遺伝子及び「刈系434号」のCG-2遺伝子などを、便宜的に、野生型7Sα遺伝子と呼ぶ。
【0028】
従って本発明では、ダイズ由来の核酸試料について、その核酸中に含まれるダイズ7Sα遺伝子であるCG-2遺伝子の第1エキソンの第215位〜第217位に位置する3アデニン塩基配列(配列AAA)中の4個のアデニン塩基の挿入を検出することにより、7Sグロブリンαサブユニット欠失ダイズを検出することができる。
【0029】
本発明におけるダイズのCG-2遺伝子は、限定するものではないが、例えばGenBank登録番号AB051865(ダイズ品種「Forrest」のゲノム配列;配列番号5)、同登録番号X17698(mRNA配列)、同登録番号AB030839(mRNA配列)に示された配列を有するものでありうる。本発明のダイズCG-2遺伝子は、便宜的には、登録番号AB051865に記載された塩基配列(配列番号5)の開始コドンから終止コドンまでの配列(第490位〜第2855位)を含むゲノムDNA領域、又は登録番号X17698(mRNA配列)の開始コドンから終止コドンまでの配列(第1位〜第1818位)を含むmRNA又はcDNA、あるいはそれらの塩基配列と非常に高い相同性(好ましくは98%以上、より好ましくは99%以上)を有する配列からなるゲノムDNA、mRNA又はcDNAとみなすこともできる。
【0030】
ダイズCG-2遺伝子は、通常はゲノム上で第1エキソン〜第6エキソンを含む。その第1エキソン(上記4アデニン塩基が挿入される)は、ダイズCG-2遺伝子中、登録番号AB051865に示す塩基配列(配列番号5)の第490位〜第1273位の領域に相当する。登録番号AB051865に示す塩基配列の第490位〜第1273位の領域は、「毛振」のCG-3遺伝子や「刈系434号」のCG-2遺伝子とも完全に一致していたことから、その領域を「野生型7Sα第1エキソン」と呼び、その野生型7Sα第1エキソンの塩基配列を配列番号1に示す。これに対し、「ゆめみのり」の、上記4アデニン塩基挿入部位を含む、CG-2遺伝子中の野生型7Sα第1エキソンに相当する領域(便宜的に、「挿入変異型7Sα第1エキソン」と呼ぶ)の塩基配列を、配列番号3に示す。
【0031】
2)挿入変異の検出
本発明では、7Sグロブリンαサブユニット欠失ダイズを検出するため、上記1)で述べたダイズCG-2遺伝子における挿入変異(CG-2遺伝子の第1エキソン中への4個のアデニン塩基の挿入)を、ダイズ由来の核酸を被験試料として検出する。
【0032】
この検出に供するダイズ(Glycine max Merrill)は、任意の品種又は系統のものであってよく、例えば、以下に限定するものではないが、「タチユタカ(Glycine max cv. Tachiyutaka)」、「毛振(Glycine max cv. Keburi)」、「秣食豆公503(Glycine max cv. Moshidou Gong 503」、「ゆめみのり(Glycine max cv. Yumeminori)」、「刈系434号(Glycine max, line Kari-kei434)」などが挙げられる。上記検出に供するダイズは、ダイズのゲノムを少なくとも1セット含む限り、例えばダイズと他の植物とのハイブリッド植物や遺伝子組換え植物などであってもよい。
【0033】
本発明において「ダイズ由来の核酸試料」とは、上記のような供試ダイズの植物体の一部(葉など)又は種子等から調製されたダイズ核酸(DNA又はRNA)を意味する。核酸試料を調製するためのダイズ試料の形態は特に限定されず、一般的に用いられている種子粉末等のタンパク質調製に適した試料だけでなく、ダイズから採取した葉(幼葉)、茎、花などの核酸抽出可能な任意の試料を用いることができる。本発明では、ダイズ由来の核酸試料として、ダイズから抽出されたゲノムDNA又はゲノムDNAをクローン化したゲノムライブラリーを用いることが好ましい。あるいは本発明におけるダイズ由来の核酸試料として、ダイズから抽出したmRNAから逆転写PCRによって合成されたcDNA、又はそのcDNAを組み込んだcDNAライブラリーを用いてもよい。本発明におけるダイズ由来の核酸試料は、PCR法などによって増幅した核酸増幅断片であってもよい。ダイズからの核酸の抽出、精製、調製などの実験手順は、分子生物学の分野で慣用されている手法に従えばよい。
【0034】
CG-2遺伝子中の挿入変異の検出は、SNP、塩基多型又は塩基変異を検出するために当業者が利用可能な公知の様々な方法を行うことができる(Orita et al., Proc. Natl. Acad. Sci., 86: 2766-2770等を参照されたい)。
【0035】
そのような変異検出法としては、より具体的には、例えば、dCAPS法〔Plant J 14, 381-385 (1998)〕、CAPS(Cleaved Amplified Polymorphic Sequence)法〔「モデル植物の実験プロトコール」細胞工学別冊 植物細胞工学シリーズ15、2001年4月2日発行、秀潤社、p.69-76〕、PCRダイレクトシークエンス法〔Biotechniques, 11, 246-249 (1991)〕、AP-PCR(Arbitrarily Primed-PCR)法〔Nucl. Acids Res., 18, 7213-7218 (1990)〕、PCR-SSCP(一本鎖DNA高次構造多型)法〔Biotechniques, 16,296-297 (1994), Biotechniques, 21, 510-514 (1996)〕、ASO(Allele Specific Oligonucleotide)ハイブリダイゼーション法〔Clin. Chim. Acta, 189, 153-157 (1990)〕、ARMS(Amplification Refracting Mutation System)法〔Nuc. Acids. Res., 19, 3561-3567 (1991), Nuc. Acids. Res., 20,4831-4837 (1992)〕、変性剤濃度勾配ゲル電気泳動(Denaturing Gradient Gel Electrophoresis; DGGE)法〔Biotechniqus, 27, 1016-1018 (1999)〕、RNaseA切断法〔DNA Cell. Biol., 14, 87-94 (1995)〕、化学切断法〔Biotechniques, 21, 216-218 (1996)〕、DOL(Dye-labeled Oligonucleotide Ligation)法〔Genome Res., 8, 549-556 (1998)〕、MALDI-TOF/MS(Matrix Assisted Laser Desorption-time of Flight/Mass Spectrometry)法〔Genome Res., 7, 378-388 (1997), Eur. J. Clin. Chem. Clin. Biochem., 35, 545-548 (1997)〕、TDI(Template-directed Dye-terminator Incorporation)法〔Proc. Natl. Acad. Sci. USA, 94, 10756-10761 (1997)〕、パドロック・プローブ(Padlock Probe)法〔Nat. Genet., 3, p225-232 (1998)、遺伝子医学, 4, p50-51 (2000)〕、モレキュラー・ビーコン(Molecular Beacons)法〔Nat. Biotechnol.,1, p49-53 (1998)、遺伝子医学、4, p46-48(2000)〕、TaqMan PCR法〔Genet. Anal., 14, 143-149 (1999), J. Clin. Microbiol., 34, 2933-2936 (1996)〕、インベーダー法〔Science, 5109, 778-783(1993), J. Biol. Chem., 30, 21387-21394 (1999), Nat. Biotechnol., 17, 292-296 (1999)〕、ダイナミック・アレル−スペシフィック・ハイブリダイゼーション(Dynamic Allele-Specific Hybridization (DASH))法〔Nat. Biotechnol.,1, p87-88, (1999)、遺伝子医学, 4, p47-48 (2000)〕、UCAN法〔タカラ酒造株式会社ホームページ(http://www.takara.co.jp)参照〕、及びDNAチップまたはDNAマイクロアレイを用いる方法〔Genomics 4, (1989), Drmanae, R., Labat, I., Brukner, I. and Crkvenjakov, R., p114-128、Bio Industry Vol.17 No.4, 「DNAチップ技術」 p5-11 (2000)〕等が挙げられるが、これらの方法に限定されるものではない。
【0036】
これらの方法において好適に用いられる本発明に係る挿入変異を検出するためのプライマー又はプローブは、上記挿入変異を含むCG-2遺伝子と、変異を含まない野生型7Sα遺伝子とを特異的に識別できるように、野生型7Sα第1エキソンの配列(配列番号1)及び「ゆめみのり」の挿入変異型7Sα第1エキソンの配列(配列番号3)に基づいて任意に設計することができる。プライマー又はプライマー対の設計は、当業者には公知の方法に従って行えばよい。プライマーの設計には、例えば、オンラインソフトウェアNetPrimer(PREMIER Biosoft International、http://www.premierbiosoft.com/)を利用することもできる。さらに本発明では、上記挿入変異を含む配列に特異的にハイブリダイズするプライマー又はプローブを設計して用いることがより好ましい。例えば、配列番号3で示す塩基配列上の第215〜第221位の配列AAAAAAAを含む少なくとも15塩基(15塩基〜783塩基)、例えば15塩基〜50塩基(プライマー用など)又は30塩基〜500塩基(プローブ用など)の連続した配列からなるDNAを、プライマー又はプローブとして好適に用いることができる。また、本発明の方法をより簡便な判定法とするためには、配列番号3で示す塩基配列上の第215〜第221位の配列AAAAAAAを含む15〜50塩基の連続した配列又はそれに相補的な配列からなるポリヌクレオチドプライマーを用いたPCR増幅を利用してその増幅断片の有無に基づいて検出することも好ましい。
【0037】
本発明においてPCR増幅を用いた検出法を用いる場合、配列番号3で示す塩基配列上の第215〜第221位の配列AAAAAAAを3'末端付近に含む、15〜50塩基、より好ましくは18〜40塩基の連続した配列又はそれに相補的な配列からなるポリヌクレオチドプライマーを用いることが特に好ましい。このようなプライマーは、CG-2遺伝子において上記挿入変異を有するダイズ核酸のみを高感度に増幅することができる。
【0038】
配列AAAAAAAを3'末端付近に含むこのようなプライマーを用いた好適なプライマーセットとして、実施例に記載した欠失型CG-2特異的プライマーセット(プライマーCG2MF[配列番号12]及びCG23R[配列番号14])が挙げられる。この欠失型CG-2特異的プライマーセットを用いた場合、上記挿入変異を含むCG-2遺伝子のみが増幅され、上記挿入変異を含まないCG-2遺伝子やCG-3遺伝子(野生型7Sα遺伝子)は増幅されない。従って、この欠失型CG-2特異的プライマーセットを用いれば、CG-2遺伝子の上記挿入変異のみを特異的に検出することができる。さらにこのような検出において、挿入変異を含まない野生型7Sα遺伝子を特異的に検出できる、実施例に記載の野生型7Sα特異的プライマーセット(プライマーCG23F[配列番号13]及びCG23R[配列番号14])をコントロール試薬として用いることも好ましい。
【0039】
PCR増幅を用いた本発明の好適な検出法の具体例は、後述の実施例に記載している。
【0040】
本発明において用いるゲノムDNA及びmRNAの抽出及び精製、cDNAの作製(RT-PCR)、PCR、ベクター中へのライゲーション、細胞の形質転換及びライブラリーの作製、DNAの塩基配列決定、プライマーの合成、突然変異誘発、タンパク質の抽出などの分子生物学的・生化学的実験法は、基本的には、通常の実験書の記載に従って行うことができる。そのような実験書として、例えば、SambrookらのMolecular Cloning, A laboratory manual, 2001, Eds., Sambrook, J. & Russell, DW. Cold Spring Harbor Laboratory Pressを挙げることができる。
【0041】
3)7Sα欠失ダイズの検出及び識別
本発明の方法においては、以上のようにしてCG-2遺伝子の第1エキソン中への上記4アデニン塩基の挿入(挿入変異)を検出した結果から、ダイズの7Sα欠失性が示される。すなわち、CG-2遺伝子の第1エキソン中への上記4アデニン塩基の挿入が確認された場合、試験したダイズでは、少なくとも1つのCG-2遺伝子がフレームシフトを起こして7Sαタンパク質産生能を失っており、そのため7Sαタンパク質を(少なくとも部分的に)欠失していることが示される。
【0042】
従って、本発明の方法において、単一の品種又は系統のダイズ由来の核酸試料について、CG-2遺伝子の第1エキソン中への上記4アデニン塩基の挿入(挿入変異)が確認された場合には、そのダイズは7Sαタンパク質欠失ダイズであると判断することができる。
【0043】
また、本発明の方法において、複数の品種又は系統のダイズ由来の核酸を含む核酸試料について、CG-2遺伝子の第1エキソン中への上記4アデニン塩基の挿入(挿入変異)が確認された場合には、それらのダイズの一部が、7Sαタンパク質欠失ダイズであると判断することができる。
【0044】
本発明の方法においては、野生型7Sα遺伝子の特異的検出を並行して行うことにより、ダイズの7Sα欠失性をさらに詳細に判定することもできる。例えば、上記4アデニン塩基の挿入変異の存在が示され、かつ野生型7Sα遺伝子の存在が示されない場合は、供試ダイズは7Sαタンパク質を完全に欠失していると判定される。またこの場合、野生型7Sα遺伝子の存在が示されないことから、供試ダイズはCG-2遺伝子中の挿入変異についてホモ接合性であると判断できる。さらに、例えば「ゆめみのり」を交配親として作製した「ゆめみのり」の子孫系統について、上記の4アデニン塩基の挿入変異が検出され、かつ野生型7Sα遺伝子の存在が示されない場合には、その子孫系統は、7Sαを完全に欠失しており、「ゆめみのり」の7Sα欠失性の導入に成功したものと判断することができる。
【0045】
また、上記4アデニン塩基の挿入変異の存在が示される一方、野生型7Sα遺伝子の存在も示される場合には、7Sαタンパク質は部分的に欠失している。この場合、通常は、供試ダイズはCG-2遺伝子の挿入変異についてヘテロ接合性であると判定される。
【0046】
このように本発明では、ダイズの7Sα欠失性を判定した結果に基づいて、7Sグロブリンαサブユニット欠失ダイズを識別することもできる。本発明の方法を用いることにより、「ゆめみのり」、「ゆめみのり」の兄弟系統、又はそれらの子孫系統を用いて例えば7Sα欠失ダイズを育種しようとする際に、7Sα欠失形質を有するダイズを早期に選抜することもできる。そのため本発明は、7Sα欠失形質を有するダイズの効率的育種方法にも有用である。
【0047】
本発明において、7Sαの「欠失性」は、生成される7Sαタンパク質自体の欠失の有無だけでなく、7Sαを生成できない7Sα遺伝子、特に、「ゆめみのり」において認められる、第1エキソン中に4アデニン塩基の挿入変異を有し7Sαを産生できないCG-2遺伝子(「欠失型CG-2遺伝子」と呼ぶ)の有無として、理解してもよい。また、後述の7Sα’の「欠失性」も、同様に、生成される7Sα’タンパク質自体の欠失の有無だけでなく、7Sα’を生成できない7Sα’遺伝子、特に、その遺伝子構造全体が欠失しているために7Sα’を産生できないCG-1遺伝子(「欠失型7Sα’遺伝子」と呼ぶ)の有無として、理解することもできる。
【0048】
なお本発明において、ダイズの「系統」とは、品種登録されている純系のダイズ系統、及び品種登録されていない純系のダイズ系統を含むだけでなく、まだ純系として確立されていない育種過程にあるダイズ個体をも含む。
【0049】
2.7Sα欠失性及び7Sα’欠失性の同時判定、並びにダイズの混入検査法
本発明は、CG-2遺伝子の第1エキソン中への4アデニン塩基の挿入を検出する際に、7Sα’遺伝子の完全欠失(遺伝子構造自体の完全な喪失)の検出も同時に行う方法にも関する。7Sα’遺伝子の完全欠失の検出法としては、上記のCG-2遺伝子における挿入変異の検出と同様の検出手法を利用すればよいが、PCR増幅を利用して行う場合には、7Sα’遺伝子中の領域を特異的に増幅するプライマーセットを用いることによって行うことができる。例えば、7Sα’遺伝子の完全欠失は、7Sα’遺伝子を特異的に増幅可能なプライマー又はプローブを公知の7Sα’遺伝子(CG-1遺伝子)の塩基配列(例えばGenBank登録番号AB030838;配列番号11)に基づいて設計し、それを、CG-2遺伝子の第1エキソン中への4アデニン塩基の挿入を検出する工程において、その挿入変異を特異的に検出するためのプライマー又はプローブとともに(好ましくは混合して)使用して、検出することができる。
【0050】
7Sα’遺伝子を特異的に増幅可能なプライマーを含む特に好適なプライマーセットとしては、後述の実施例3に示したCG-1遺伝子特異的プライマーセット(CG1F[配列番号15]及びCG1R[配列番号16])が挙げられる。
【0051】
限定するものではないが、7Sα欠失性及び7Sα’欠失性の同時判定には、実施例3及び図7に記載したAセット及びBセットのプライマーセットが特に有用である。これらのプライマーセットを用いた同時判定法の手順は、実施例に詳述されている。
【0052】
7Sα欠失性及び7Sα’欠失性の同時判定法を用いれば、7Sα欠失性及び7Sα’欠失性の点で異なる性質を持つダイズ系統を、容易に識別することができる。このことは、7Sαの欠失による低アレルゲン性、及び7Sαと7Sα’の欠失による栄養価の高い11Sグロブリン含量の増加という有用な性質に関して、望ましい性質を持つダイズ系統を識別できることを意味する。
【0053】
さらに本発明は、7Sα及び7Sα’の欠失性が異なるダイズ系統の混入を、上記の同時判定試験を利用して検査する方法にも関する。ダイズ由来の核酸試料について、7Sα欠失性及び7Sα’欠失性の判定結果が、目的のダイズに関する結果と異なっていた場合には、その核酸試料の由来するダイズに、7Sα及び7Sα’の欠失性が異なる系統のダイズが混入していることが分かる。本発明のダイズの混入検査法は、検出限界が低く、高感度である。この混入検査法における検出限界は、核酸試料におけるDNA重量に換算した場合、混入率0.1%以上、好ましくは0.5%以上、好ましくは1%以上である。
【0054】
3.その他の実施形態
本発明は、CG-2遺伝子の第1エキソン中への4アデニン塩基の挿入変異を特異的に検出可能なDNA断片にも関する。これらのDNA断片は、プライマー又はプローブとして有用に用いることができる。
【0055】
具体的には、そのようなDNA断片は、「ゆめみのり」に見出されたCG-2遺伝子の挿入変異を含むDNAであって、配列番号3で示す塩基配列上の第215〜第221位の配列AAAAAAAを含む少なくとも15塩基(15塩基〜788塩基)、例えば15塩基〜50塩基(プライマー用など)又は30塩基〜500塩基(プローブ用など)の連続した配列からなるDNAであってよい。
【0056】
本発明は、「ゆめみのり」に見出されたCG-2遺伝子の挿入変異を含むプライマーであって、配列番号3で示す塩基配列上の第215〜第221位の配列AAAAAAAを含む15〜50塩基の連続した配列又はそれに相補的な配列からなるポリヌクレオチドプライマー、特に、配列番号3で示す塩基配列上の第215〜第221位の配列AAAAAAAを3'末端に含む、15〜50塩基、より好ましくは18〜40塩基の連続した配列又はそれに相補的な配列からなるポリヌクレオチドプライマーにも関する。この配列AAAAAAAを3'末端付近(3'末端又はその数塩基5'側に当たる3'末端近傍)に含むプライマーは、上記挿入変異を有するダイズ核酸のみを高感度に増幅することができるため、とりわけ有用である。このようなプライマーを用いた、欠失型CG-2特異的プライマーセット(プライマーCG2MF[配列番号12]及びCG23R[配列番号14])も本発明に含まれる。
【0057】
また本発明では、CG-2遺伝子中の挿入変異を特異的に検出できるプライマーに加えてコントロールプライマー等をさらに含む、検出に便利に使用できるプライマーセットにも関する。例えば、実施例3に記載した欠失型CG-2特異的プライマーセットと野生型7Sα特異的プライマーセットは、2回のPCRによって7Sαの有無を判定できる便利な共優性マーカーとして用いることができる。これらのマーカーを用いることによって、育種選抜において重要であるが従来のタンパク質での分析では検出不可能な、上記4アデニン塩基の挿入変異を有し7Sαを産生できないCG-2遺伝子(欠失型CG-2遺伝子)についてヘテロ接合であるダイズ系統を識別することができる。またこれらのマーカーを用いることによって、7Sα欠失ダイズ系統が野生型ダイズ系統に混入した場合の検出なども高感度に行うことができる。
【0058】
本発明は、CG-2遺伝子の上記挿入変異の有無と7Sα’遺伝子の有無をマルチプレックスPCRによって同時に検出することができる、実施例3に記載したCG-1特異的プライマーセットと欠失型CG-2特異的プライマーセットを含むプライマーセットにも関する。このプライマーセットには、実施例3に記載のようにさらにポジティブコントロール用プライマーセットを含んでもよい(実施例3のセットA)。本発明は、この検出のコントロール実験用に有用な、実施例3に記載の野生型7Sα特異的プライマーセットとポジティブコントロール用プライマーセットとを含むプライマーセット(実施例3のセットB)にも関する。これらのプライマーセットを用いて同時検出を行えば、欠失型CG-2遺伝子の有無と7Sα’遺伝子の有無が重要となる、7Sグロブリンαサブユニット変異体を利用した育種やダイズアレルゲン性の分析などにおいて、分析コストや労力を顕著に節減することができる。
【0059】
さらに、これらのプライマーセットは、ダイズの7Sα欠失性や7Sα’欠失性の検出用のキット、ダイズの識別用キット、ダイズの異系統混入検査キットなどに含めて提供することもありうる。このような各種キットも、本発明の範囲に含まれる。
【0060】
なお上記のようなDNA断片、並びにプライマー及びプローブなどは、当業者に公知の核酸調製法、例えば、限定するものではないが、化学合成法(液相法、固相法、カラム法及びバッチ法等)、クロマトグラフィー法による精製抽出法、PCR法、制限酵素消化等を利用して製造することができる。Applied Biosystems 3400 DNA合成機(Applied Biosystems)などの核酸合成装置も市販されている。
【0061】
4.配列の説明
・配列番号1: GenBank登録番号AB051865の配列中の、ダイズ7Sα遺伝子の第1エキソンに相当する塩基配列。野生型7Sα第1エキソンである。
【0062】
・配列番号2: 配列番号1の野生型7Sα第1エキソンにコードされるアミノ酸配列。
【0063】
・配列番号3: 「ゆめみのり」のダイズCG-2遺伝子(7Sα遺伝子)の、第1エキソンに相当する塩基配列。第215位〜第221位に挿入変異を含む配列AAAAAAAが存在し、その結果この第1エキソン領域内に終止コドンが生じている。
【0064】
・配列番号4: 配列番号3の配列にコードされるアミノ酸配列。
【0065】
・配列番号5: GenBank登録番号AB051865の塩基配列。ダイズ7Sα遺伝子を含むゲノム配列である。
【0066】
・配列番号6: 配列番号5の配列にコードされるアミノ酸配列。
【0067】
・配列番号7: GenBank登録番号X17698の塩基配列。野生型ダイズ7Sα遺伝子のmRNA配列である。
【0068】
・配列番号8: 刈系434号及び毛振の7Sα遺伝子を含むゲノム配列。
【0069】
・配列番号9: 刈系434号及び毛振の7Sα遺伝子にコードされるアミノ酸配列。
【0070】
・配列番号10:「ゆめみのり」のCG-2遺伝子(7Sα遺伝子)を含むゲノム配列。
【0071】
・配列番号11: GenBank登録番号AB051838の塩基配列。ダイズ7Sα’遺伝子(CG-1遺伝子)のmRNA配列である。
【0072】
・配列番号12: CG2MFプライマー
・配列番号13: CG23Fプライマー
・配列番号14: CG23Rプライマー
・配列番号15: CG1Fプライマー
・配列番号16: CG1Rプライマー
・配列番号17: 30KFプライマー
・配列番号18: 30KRプライマー
・配列番号19: CG2TFプライマー
・配列番号20: CG2TRプライマー
【実施例】
【0073】
以下、実施例及び図面を参照して本発明を説明するが、本発明はこれらの実施例に限定されるものではない。
【0074】
以下の実施例では、γ線照射によって7Sグロブリンα’サブユニット(7Sα’)および7Sグロブリンαサブユニット(7Sα)を欠失したダイズ品種「ゆめみのり」、「ゆめみのり」の原品種「刈系434号」、「ゆめみのり」の反復親「タチユタカ」、遺伝資源より見出された7Sサブユニットの変異体「毛振」および「秣食豆公503」を供試した。
【0075】
各品種における、7Sα’およびαをコードする遺伝子の有無は次の通りである。
【0076】
【表1】
【0077】
表1中、表現型について「欠失」とは、当該タンパク質の生成量がほぼゼロである完全欠失を意味し、「低下」とは当該タンパク質の生成量が野生型に比べて減少していることを意味し、「正常」とは当該タンパク質の生成量が野生型と同等であることを意味する。
【0078】
なお各品種の育成については、以下の文献で報告されている。
【0079】
- 秣食豆公503[ダイズ遺伝資源由来]: Kitamura, K. et al. Mutant strains with low level of subunits of 7S globulin in soybean (Glycine max Mell.) seed. (1981) Japan. J. Breed. 31(4):353-359
- 毛振[ダイズ遺伝資源由来]: Kitamura, K. et al. Mutant strains with low level of subunits of 7S globulin in soybean (Glycine max Mell.) seed. (1981) Japan. J. Breed. 31(4):353-359
- タチユタカ: 橋本鋼二ほか.ダイズ新品種「タチユタカ」の育成.東北農試研報.77:27-44(1988)
- 刈系434号[(毛振//大館1号/秣食豆公503)/タチユタカ×2に由来]: Ogawa, T. et al. Genetic improvement of seed storage proteins using three variant alleles of 7S globulin subunits in soybean (Glycine max L.). (1989) Jpn. J. Breed. 39(2):137-147
- ゆめみのり[γ線照射した刈系434号に由来]: Takahashi, K. et al. Inheritance of α-subunit deficiency of β-conglycinin in soybean (Glycine max L. MERRILL) line induced by γ-ray irradiation. (1996) Breed.Sci. 46:251-255
【0080】
実施例1: ダイズ7Sグロブリンαサブユニット遺伝子の塩基配列解析
「刈系434号」、「毛振」及び「ゆめみのり」のそれぞれの幼葉約100mgから、PhytoPure DNA extraction kit(アマシャムバイオサイエンス)を用いて添付のプロトコールに従って、ゲノムDNAを抽出した。
【0081】
調製したゲノムDNAをテンプレートDNAとして、ポリメラーゼ連鎖反応(PCR)法によりDNA断片を増幅した。このPCRにおいては、国際塩基配列データベース(DDBJ/EMBL/GenBank)から登録番号AB051865(ゲノム配列;ダイズ品種「Forrest」由来)により入手できる7Sα遺伝子の既知塩基配列(配列番号5)に基づいて7Sα遺伝子全長を増幅するように設計された、以下のプライマーを用いた。
【0082】
・フォワードプライマー CG2TF: 5'-GGGTCATCGGGAACTAGTTCA-3'(AB051865の塩基配列上の第430位〜第452位の配列に相当;配列番号19)
・リバースプライマ− CG2TR: 5'-TTCATAGATGAAGCTCTAGTCATAGTTAC-3'(AB051865の塩基配列上の第2,919位〜第2,947位の配列に相当;配列番号20)
【0083】
PCR反応には、終濃度で1×バッファー、各0.2mM dNTP、1.5mM MgCl2、設定濃度のプライマー、1.25U DNAポリメラーゼ(ExTaq、TAKARA BIO INC.)、50〜100ngテンプレートDNAを含む25μlの反応液を用いた。PCR反応では、GeneAmp PCR system 9700(アプライドバイオシステムズ)を用いて、95℃で5分間の変性反応、続いて、95℃で30秒、62℃で30秒、72℃で1分を32回繰り返し、最後に72℃で7分間の伸長反応を行った。
【0084】
PCR反応後、反応物中のPCR増幅断片を、1%アガロースゲル(L03「TAKARA」、TAKARA BIO INC.)を用いて100Vで50分間の電気泳動によって分離し、エチジウムブロマイドによるゲル染色後、紫外線下で可視化した。サイズマーカーとしては2-Log DNA Ladder(New England BioLabs)を使用した(以下の実験でも同様)。
【0085】
電気泳動の結果、いずれの品種・系統でも、AB051865の配列(ゲノム配列)から予測される通りの約2.5kbの増幅断片が確認された(図2)。図2中、M:サイズマーカー、1:「刈系434号」、2:「毛振」、3:「ゆめみのり」を示す。
【0086】
得られたDNA増幅断片を、核酸抽出キットQIAquick gel extraction kit(キアゲン)を用いてゲルから抽出し、それについてシークエンシング試薬BigDye terminatior v1.1 Cycle Sequencing kitおよびシークエンサーABI Prism 310 genetic analyzer(アプライドバイオシステムズ)を用いて、直接塩基配列を決定した。決定した配列については、オンラインソフトウェアCLUSTAL W(DNA Data Bank of Japan、http://www.ddbj.nig.ac.jp/)を用いて配列解析を行った。
【0087】
表1に示す通り、7Sグロブリンαサブユニット遺伝子として「刈系434号」はCG-2遺伝子のみを有し、「毛振」はCG-3遺伝子のみを有することから、本実施例で決定された塩基配列は「刈系434号」ではCG-2遺伝子、「毛振」ではCG-3遺伝子のものである。しかし配列解析の結果、「刈系434号」と「毛振」について決定された塩基配列は、全く同一であった。従って、本実施例で配列決定されたαサブユニット遺伝子の開始コドンから終止コドンまでを含む領域では、CG-2遺伝子とCG-3遺伝子は塩基配列上同一であった。
【0088】
さらに、「刈系434号」及び「毛振」について決定した配列は、AB051865(ダイズ品種「Forrest」)の第482位から第2,908位までの配列と、1塩基を除いて完全に一致しており、その違いは「刈系434号」及び「毛振」の配列においてAB051865の配列の2,564塩基目に当たるシトシンがアデニンに置換しているのみであった(図3)。
【0089】
一方、「ゆめみのり」の配列には、「刈系434号」の配列と比較して、4個のアデニン塩基の挿入が生じていた(図3)。その挿入位置は、AB051865配列の第704位〜第706位の3アデニン配列内に相当する。AB051865配列の第704位〜第706位は、AB051865配列にコードされる7Sα遺伝子の第1エキソン(野生型7Sα第1エキソン;配列番号1)の第215位〜第217位[第1エキソンの5'端の開始コドン「ATG」のAを第1位とした場合]に当たる。そして、このような4個のアデニン塩基の挿入により生じた塩基配列AAAAAAA、すなわち、「ゆめみのり」のCG-2遺伝子上の第1エキソンに相当する領域(挿入変異型7Sα第1エキソン;配列番号3)の第215位〜第221位の7アデニン配列は、「毛振」にもAB051865配列にも存在しなかった。
【0090】
次に、「ゆめみのり」について決定した配列から、それにコードされるアミノ酸配列を推定した。その結果、「ゆめみのり」の7Sαサブユニット遺伝子であるCG-2遺伝子においては、上記のような4個のアデニン塩基の挿入によりフレームシフトが生じ、第1エキソン中に終止コドンが生じていることが示された(図4、配列番号3及び4)。従って、この4塩基のアデニンの挿入が、「ゆめみのり」における7Sグロブリンαサブユニットの欠失の原因であると考えられた。
【0091】
実施例2: 7Sグロブリンαサブユニットの欠失性の検出
実施例1で確認された「ゆめみのり」のCG-2遺伝子(7Sグロブリンαサブユニット遺伝子)における4個のアデニン塩基の挿入の検出を、PCR増幅により行った。
【0092】
まず「タチユタカ」、「毛振」、「秣食豆公503」、「ゆめみのり」、「刈系434号」の幼葉から、実施例1と同様にしてゲノムDNAを調製し、それをテンプレートDNAとした。PCRには、以下のプライマーを設計して用いた。
【0093】
・欠失型CG-2特異的フォワードプライマー CG2MF:
5'-GAAGAATGTGAAGAAGGTGAAAAAAAT-3'[配列番号12]
(AB051865の塩基配列上の第685位〜第707位の配列+4アデニン塩基挿入に相当)
・CG-2/CG-3共通フォワードプライマー CG23F:
5'-AAGAAGAATGTGAAGAAGGTGAAAT-3'[配列番号13]
(AB051865の塩基配列上の第683位〜第707位の配列に相当)
・CG-2/CG-3共通リバースプライマー CG23R:
5'-GAATGCGACCATATGGTTTTTGA-3'[配列番号14]
(AB051865の塩基配列上の第1,109位〜第1,132位の配列に相当)
【0094】
ここで、欠失型CG-2特異的フォワードプライマー CG2MFは、4個のアデニン塩基の挿入配列を3'端に含む、欠失型CG-2遺伝子に特異的なプライマーである。
【0095】
PCR増幅は2つの反応系で行った。欠失型CG-2特異的フォワードプライマー CG2MFとCG-2/CG-3共通リバースプライマー CG23Rとを組み合わせたプライマーセット(欠失型CG-2特異的プライマーセット)を、1つ目のPCR増幅に用い、またCG-2/CG-3共通フォワードプライマー CG23FとCG-2/CG-3共通リバースプライマー CG23Rとを組み合わせたプライマーセット(野生型7Sα特異的プライマーセット)を、2つ目のPCR増幅に用いた。
【0096】
PCR反応には、終濃度で1×バッファー、各0.2mM dNTP、1.5mM MgCl2、各0.2μMのプライマー、1.25U DNAポリメラーゼ(ExTaq、TAKARA BIO INC.)、50〜100ngテンプレートDNAを含む25μlの反応液を用いた。PCR反応では、GeneAmp PCR system 9700(アプライドバイオシステムズ)を用いて、95℃で5分間の変性反応、続いて、95℃で30秒、62℃で30秒、72℃で1分を32回繰り返し、最後に72℃で7分間の伸長反応を行った。
【0097】
PCR反応後、4%アガロースゲルを用いて、その他は実施例1と同様にしてPCR増幅産物を電気泳動により分離し、可視化した。その結果を図5及び図6に示す。図5及び図6中、M:サイズマーカー、2:「タチユタカ」、3:「毛振」、4:「秣食豆公503」、5:「ゆめみのり」、6:「刈系434号」を示す。
【0098】
図5に示す通り、プライマーCG2MFとCG23Rを用いて行ったPCR増幅では、「ゆめみのり」のみ増幅断片が検出された。得られた増幅断片の長さは450bpであった。一方、プライマーCG23FとCG23Rを用いて行ったPCR増幅では、「ゆめみのり」以外の全品種・系統で、448bpの増幅断片が検出された(図6)。
【0099】
以上の通り、「ゆめみのり」のCG-2遺伝子内の4アデニン塩基挿入部位に設計した欠失型CG-2特異的プライマーセットおよび野生型7Sα特異的プライマーセットにより、「ゆめみのり」に認められるような7Sα欠失の有無を2回のPCRで判定できる共優性のマーカーを、開発することができた。
【0100】
実施例3: マルチプレックスPCRによる7Sα欠失性及び7Sα’欠失性の同時判定
ダイズ11Sグロブリンの含量を増加させるためには、7Sαと7Sα’を同時に欠失させることが有効である。そこで、7Sα’をコードするCG-1遺伝子に特異的なプライマーを設計して優性マーカーとし、実施例2で用いた7SαのマーカーとのマルチプレックスPCRにより7Sαと7Sα’の欠失の有無を同時に判定する方法を検討した。
【0101】
図7には、マルチプレックスPCRによる7Sα欠失性及び7Sα’欠失性の同時判定法の一例として行った以下の実験の具体的手順の概要を示す。
【0102】
まず、7Sグロブリンαサブユニット遺伝子のmRNA配列として報告されているGenBank登録番号X17698の塩基配列(配列番号7)と、7Sグロブリンα’サブユニット遺伝子(CG-1遺伝子)のmRNA配列として報告されているGenBank登録番号AB030838の塩基配列(配列番号11)とをアラインメントした(図8)。このアラインメントでは91%の高い相同性が示された。そこで、このアラインメントに基づき、CG-1遺伝子を特異的に増幅する以下のプライマーセット(CG-1特異的プライマーセット)を設計した。
【0103】
・CG-1特異的フォワードプライマー CG1F:
5'-GTCACAACAAGTGCCTCCGA-3'[配列番号15]
(AB030838の塩基配列上の第158位〜第177位の配列に相当)
・CG-1特異的リバースプライマー CG1R:
5'-GGTGCTTTTCCTGCTTGTG-3'[配列番号16]
(AB030838の塩基配列上の第547位〜第565位の配列に相当)
【0104】
マルチプレックスPCRには、このCG-1特異的プライマーセットに加えて、実施例2で用いた欠失型CG-2特異的プライマーセットを用いた。さらに、増幅断片が得られない場合にPCR反応自体が不成功に終わった可能性を除くため、ダイズタンパク質Gly-m Bd 30Kをコードする遺伝子の一部を増幅するプライマーセットも、ポジティブコントロールとして反応液に加えた。なおダイズタンパク質Gly-m Bd 30Kは、ダイズ貯蔵タンパク質に含まれる主要なアレルゲンであり、これまでの大規模なスクリーニングによっても欠失体は得られていない。このポジティブコントロール用プライマーセットは、以下のプライマーからなる。
【0105】
・ポジティブコントロール・フォワードプライマー30KF:
5'-AGCTCACTAATCACTATATATACGACATGCT-3'[配列番号17]
(GenBank登録番号AB013289の塩基配列上の第4,562位〜第4,592位の配列に相当)
・ポジティブコントロール・リバースプライマー30KR:
5'-CACAGCCCCCTTGGTACTTTAC-3'[配列番号18]
(GenBank登録番号AB013289の塩基配列上の第5,094位〜第5,115位の配列に相当)
【0106】
以上のCG-1特異的プライマーセット(CG1F及びCG1R)、欠失型CG-2特異的プライマーセット(CG2MF及びCG23R)、及びポジティブコントロール用プライマーセット(30KF及び30KR)は、まとめてプライマーセット(Aセット)として検出用に用いた。
【0107】
さらに、実施例2で用いた野生型7Sα特異的プライマーセット(CG23F及びCG23R;上記挿入変異を含まない野生型7Sα遺伝子を増幅する)と、上記ポジティブコントロール用プライマーセット(30KF及び30KR)を、まとめてプライマーセット(Bセット)としてコントロール用に用いた。
【0108】
表2には、本実施例におけるプライマーとその使用形態をまとめた。
【0109】
【表2】
【0110】
反応液は図7に記載された組成で全量25μlとして調製した。なおテンプレートDNAは実施例1と同様にして調製した。PCR反応は、GeneAmp PCR system 9700(アプライドバイオシステムズ)を用いて、以下の条件で行った:95℃で5分、続いて、95℃で30秒、62℃で30秒、72℃で1分を32サイクル、続いて72℃で7分の後、4℃で保持。
【0111】
PCR反応後、実施例2と同様にしてPCR増幅産物を電気泳動により分離し、可視化した。その結果を図9(Aセット使用)及び図10(Bセット使用)に示す。図9及び図10中、M:サイズマーカー、1:「タチユタカ」、2:「毛振」、3:「秣食豆公503」、4:「ゆめみのり」、5:「刈系434号」を示す。
【0112】
図9に示すように、プライマーセットAを用いて行ったPCR増幅では、450bpの増幅断片が「ゆめみのり」のみで得られ、この増幅断片長は欠失型CG-2特異的プライマーセットによる増幅サイズと一致していた。また「タチユタカ」及び「秣食豆公503」では、408bpの増幅断片が得られ、この増幅断片長はCG-1特異的プライマーセットによる増幅サイズと一致していた。さらに「毛振」及び「刈系434号」では、ポジティブコントロールプライマーによるもの以外の増幅断片は検出されなかった。なおポジティブコントロール用プライマーセットによる554bpの増幅断片は、全サンプルで検出された。
【0113】
一方、図10に示すように、プライマーセットBを用いて行ったコントロール用のPCR増幅では、「ゆめみのり」以外の品種で448bpの増幅断片が得られた。この増幅断片の長さは、野生型7Sグロブリンαサブユニット遺伝子を増幅する野生型7Sα特異的プライマーセットによる増幅サイズと一致していた。なおポジティブコントロール用プライマーセットによる554bpの増幅断片は、全サンプルで検出された。
【0114】
以上の結果は、上記表1に示した各品種・系統の特徴と完全に一致していた。このことから、これらのプライマーセットA、あるいはプライマーセットA及びBをPCR増幅に用いることにより、7Sαの欠失性及び7Sα’の欠失性を同時に判定できることが示された。
【0115】
また、これらのプライマーセットA及びBをPCR増幅に用いることにより、CG-1遺伝子の有無、野生型7Sα遺伝子の有無、欠失型CG-2遺伝子の有無から、ダイズ品種・系統が識別されることも示された。
【0116】
実施例4: 7Sα欠失性及び7Sα’欠失性の同時判定に基づくダイズの異品種・異系統混入検査
7Sグロブリンαサブユニット及びα’サブユニットの欠失性が異なるダイズ品種・系統の混入を、実施例3で述べた同時判定試験を利用して検査した。サンプルには、「ゆめみのり」(7Sα及び7Sα’の両方を欠失)のゲノムDNAに、「タチユタカ」(7Sα及び7Sα’の両方を有する)のゲノムDNAを混合した核酸試料を用いた。
【0117】
検出感度を確認するため、「ゆめみのり」のゲノムDNA量に対して0.1%、0.2%、0.5%、および1%となるように「タチユタカ」のゲノムDNAを加えたサンプルを用意し、それらをテンプレートDNAとした。
【0118】
各混入パーセンテージのテンプレートDNAを用い、実施例3と同様にして、Aセット及びBセットのプライマーセットを用いたPCR増幅を行った。PCR反応後、増幅産物を同様にして電気泳動し可視化した。その結果を図11(Aセット)及び図12(Bセット)に示す。
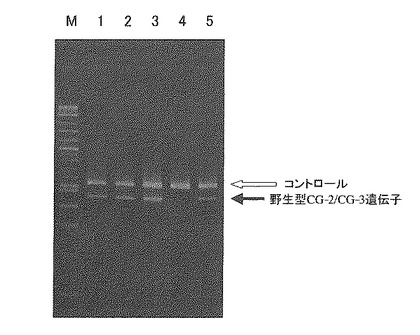
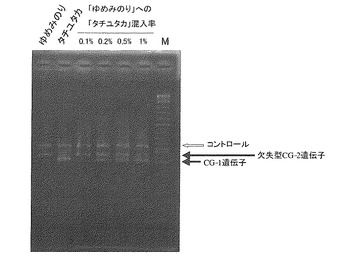
【0119】
図11に示される通り、プライマーセットAを用いて検出を行った場合、DNA重量換算で0.1%以上の混入量で、CG-1遺伝子を示す408bpの増幅断片が認められた。またプライマーセットBを用いて検出を行った場合、ゲノムDNA重量換算で0.2%以上の混入量で、野生型7Sα遺伝子を示す448bpの増幅断片が認められた(図12)。
【0120】
この同時判定法では、7Sα及び7Sα’の欠失性が異なるダイズ品種の混入を、混入率(DNA重量換算)0.1〜0.2%程度でも検出することができたことから、この方法は検出限界が非常に低く、高感度であることが示された。
【産業上の利用可能性】
【0121】
本発明の7Sグロブリンαサブユニット欠失ダイズの検出方法は、「ゆめみのり」又は「ゆめみのり」の兄弟系統、それらの子孫系統などの、7Sグロブリンαサブユニットを欠失したダイズ系統を簡便に検出するために使用することができる。この本発明の方法は、ダイズの混入検査法としても使用できる。
【図面の簡単な説明】
【0122】
【図1】図1は、ダイズアレルゲン7Sα欠失ダイズ「ゆめみのり」の系譜を示す図である。
【図2】図2は、7Sα遺伝子全長を含むDNA増幅断片の電気泳動写真である。M: マーカー、レーン1: 「刈系434号」、レーン2: 「毛振」、レーン3: 「ゆめみのり」。
【図3−1】図3は、7Sα遺伝子について各品種・系統で決定したゲノム配列のアラインメントを示す図である。基準配列として7Sα遺伝子の既知配列である登録番号AB051865の配列を一緒に示した。図中、「ゆめみのり(Yumeminori)」のみ、AB051865の配列の第704位〜第706位に相当する位置に、4個のアデニン塩基の挿入が見られる。
【図3−2】図3は、7Sα遺伝子について各品種・系統で決定したゲノム配列のアラインメントを示す図である。図3−2は、図3−1の続きである。AB051865の配列の第2564位に相当する位置において、「刈系434号」、「毛振」及び「ゆめみのり」ではシトシン(C)からアデニン(A)への置換が生じていた。
【図3−3】図3は、7Sα遺伝子について各品種・系統で決定したゲノム配列のアラインメントを示す図である。図3−3は、図3−1、図3−2の続きである。
【図4】図4は、「ゆめみのり」のCG-2遺伝子(7Sα遺伝子)においてフレームシフトが起こり、早期に終止コドンが生じたことを示す図である。
【図5】図5は、欠失型CG-2特異的プライマーセットを用いたPCR増幅の結果を示す電気泳動写真である。M: マーカー、レーン1: 「タチユタカ」、レーン2: 「毛振」、レーン3: 「秣食豆公503」、レーン4: 「ゆめみのり」、レーン5: 「刈系434号」。
【図6】図6は、野生型7Sα特異的プライマーセットを用いたPCR増幅の結果を示す電気泳動写真である。M: マーカー、レーン1: 「タチユタカ」、レーン2: 「毛振」、レーン3: 「秣食豆公503」、レーン4: 「ゆめみのり」、レーン5: 「刈系434号」。
【図7】図7は、7Sαと7Sα’の欠失性の同時判定方法の概要を示す図である。
【図8】図8は、登録番号X17698の配列とAB030838の配列とのアラインメントを示す図である。
【図9】図9は、プライマーセットAを用いた7Sα’および7Sα遺伝子検出用のPCR増幅の結果を示す電気泳動写真である。M: マーカー、レーン1: 「タチユタカ」、レーン2: 「毛振」、レーン3: 「秣食豆公503」、レーン4: 「ゆめみのり」、レーン5: 「刈系434号」。
【図10】図10は、プライマーセットBを用いたコントロール検出実験用のPCR増幅の結果を示す電気泳動写真である。M: マーカー、レーン1: 「タチユタカ」、レーン2: 「毛振」、レーン3: 「秣食豆公503」、レーン4: 「ゆめみのり」、レーン5: 「刈系434号」。
【図11】図11は、「ゆめみのり」への「タチユタカ」の混入を、プライマーセットAを用いて検出した結果を示す電気泳動写真である。
【図12】図12は、「ゆめみのり」への「タチユタカ」の混入を、プライマーセットBを用いて検出した結果を示す電気泳動写真である。
【配列表フリーテキスト】
【0123】
配列番号12〜20の配列はプライマーである。
【特許請求の範囲】
【請求項1】
ダイズ由来の核酸試料について、CG−2遺伝子の第1エキソンの第215位〜第217位の配列AAA中への4個のアデニン塩基の挿入を検出する工程を特徴とする、7Sグロブリンαサブユニット欠失ダイズの検出方法。
【請求項2】
CG−2遺伝子の第1エキソンが、配列番号1で示す塩基配列からなる、請求項1に記載の方法。
【請求項3】
前記検出工程が、配列番号3で示す塩基配列上の第215〜第221位の配列AAAAAAAを含む少なくとも15塩基の連続した配列からなるDNAをプライマー又はプローブとして用いて検出する工程である、請求項1又は2に記載の方法。
【請求項4】
前記検出工程が、配列番号3で示す塩基配列上の第215〜第221位の配列AAAAAAAを含む15〜50塩基の連続した配列又はそれに相補的な配列からなるポリヌクレオチドプライマーを用いたPCR増幅により検出する工程である、請求項1又は2に記載の方法。
【請求項5】
前記プライマーが、配列番号12で示す塩基配列からなるポリヌクレオチドプライマーである、請求項4に記載の方法。
【請求項6】
配列番号12で示す塩基配列からなるポリヌクレオチドプライマーとともに、配列番号14で示す塩基配列からなるポリヌクレオチドプライマーをPCR増幅に用いる、請求項5に記載の方法。
【請求項7】
7Sグロブリンαサブユニット欠失ダイズが、「ゆめみのり」、「ゆめみのり」の兄弟系統、又はそれらの子孫系統である、請求項1〜6のいずれか1項に記載の方法。
【請求項8】
請求項4〜7のいずれか1項に記載の方法において、前記プライマーとともに、CG−1遺伝子特異的プライマーセットをPCR増幅に用いることを特徴とする、ダイズの7Sグロブリンαサブユニット欠失性及び7Sグロブリンα’サブユニット欠失性の同時判定方法。
【請求項9】
CG−1遺伝子特異的プライマーセットが、配列番号15で示す塩基配列からなるポリヌクレオチドプライマーと、配列番号16で示す塩基配列からなるポリヌクレオチドプライマーとからなる、請求項8に記載の方法。
【請求項10】
請求項8又は9に記載の方法で得られた判定結果に基づいて、異系統のダイズの混在の有無を判定することを特徴とする、ダイズの混入検査法。
【請求項11】
配列番号3で示す塩基配列上の第215〜第221位の配列AAAAAAAを含む少なくとも15塩基の連続した配列からなるDNA。
【請求項12】
配列番号3で示す塩基配列上の第215〜第221位の配列AAAAAAAを含む15〜50塩基の連続した配列又はそれに相補的な配列からなるポリヌクレオチドプライマー。
【請求項13】
配列番号12で示す塩基配列からなるポリヌクレオチドプライマーと、配列番号14で示す塩基配列からなるポリヌクレオチドプライマーとからなる、プライマーセット。
【請求項14】
配列番号12〜14で示す塩基配列の各々からなる3種のポリヌクレオチドプライマーを含むキット。
【請求項15】
配列番号12〜16で示す塩基配列の各々からなる5種のポリヌクレオチドプライマーを含むキット。
【請求項16】
配列番号17及び18で示す塩基配列の各々からなる2種のポリヌクレオチドプライマーをさらに含む、請求項15に記載のキット。
【請求項17】
「ゆめみのり」、「ゆめみのり」の兄弟系統、又はそれらの子孫系統である7Sグロブリンαサブユニット欠失ダイズを検出するための、請求項14〜16のいずれか1項に記載のキット。
【請求項1】
ダイズ由来の核酸試料について、CG−2遺伝子の第1エキソンの第215位〜第217位の配列AAA中への4個のアデニン塩基の挿入を検出する工程を特徴とする、7Sグロブリンαサブユニット欠失ダイズの検出方法。
【請求項2】
CG−2遺伝子の第1エキソンが、配列番号1で示す塩基配列からなる、請求項1に記載の方法。
【請求項3】
前記検出工程が、配列番号3で示す塩基配列上の第215〜第221位の配列AAAAAAAを含む少なくとも15塩基の連続した配列からなるDNAをプライマー又はプローブとして用いて検出する工程である、請求項1又は2に記載の方法。
【請求項4】
前記検出工程が、配列番号3で示す塩基配列上の第215〜第221位の配列AAAAAAAを含む15〜50塩基の連続した配列又はそれに相補的な配列からなるポリヌクレオチドプライマーを用いたPCR増幅により検出する工程である、請求項1又は2に記載の方法。
【請求項5】
前記プライマーが、配列番号12で示す塩基配列からなるポリヌクレオチドプライマーである、請求項4に記載の方法。
【請求項6】
配列番号12で示す塩基配列からなるポリヌクレオチドプライマーとともに、配列番号14で示す塩基配列からなるポリヌクレオチドプライマーをPCR増幅に用いる、請求項5に記載の方法。
【請求項7】
7Sグロブリンαサブユニット欠失ダイズが、「ゆめみのり」、「ゆめみのり」の兄弟系統、又はそれらの子孫系統である、請求項1〜6のいずれか1項に記載の方法。
【請求項8】
請求項4〜7のいずれか1項に記載の方法において、前記プライマーとともに、CG−1遺伝子特異的プライマーセットをPCR増幅に用いることを特徴とする、ダイズの7Sグロブリンαサブユニット欠失性及び7Sグロブリンα’サブユニット欠失性の同時判定方法。
【請求項9】
CG−1遺伝子特異的プライマーセットが、配列番号15で示す塩基配列からなるポリヌクレオチドプライマーと、配列番号16で示す塩基配列からなるポリヌクレオチドプライマーとからなる、請求項8に記載の方法。
【請求項10】
請求項8又は9に記載の方法で得られた判定結果に基づいて、異系統のダイズの混在の有無を判定することを特徴とする、ダイズの混入検査法。
【請求項11】
配列番号3で示す塩基配列上の第215〜第221位の配列AAAAAAAを含む少なくとも15塩基の連続した配列からなるDNA。
【請求項12】
配列番号3で示す塩基配列上の第215〜第221位の配列AAAAAAAを含む15〜50塩基の連続した配列又はそれに相補的な配列からなるポリヌクレオチドプライマー。
【請求項13】
配列番号12で示す塩基配列からなるポリヌクレオチドプライマーと、配列番号14で示す塩基配列からなるポリヌクレオチドプライマーとからなる、プライマーセット。
【請求項14】
配列番号12〜14で示す塩基配列の各々からなる3種のポリヌクレオチドプライマーを含むキット。
【請求項15】
配列番号12〜16で示す塩基配列の各々からなる5種のポリヌクレオチドプライマーを含むキット。
【請求項16】
配列番号17及び18で示す塩基配列の各々からなる2種のポリヌクレオチドプライマーをさらに含む、請求項15に記載のキット。
【請求項17】
「ゆめみのり」、「ゆめみのり」の兄弟系統、又はそれらの子孫系統である7Sグロブリンαサブユニット欠失ダイズを検出するための、請求項14〜16のいずれか1項に記載のキット。
【図1】

【図2】
【図3−1】

【図3−2】

【図3−3】

【図4】

【図5】

【図6】

【図7】
【図8】

【図9】
【図10】
【図11】
【図12】
【図2】
【図3−1】
【図3−2】
【図3−3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【公開番号】特開2007−53(P2007−53A)
【公開日】平成19年1月11日(2007.1.11)
【国際特許分類】
【出願番号】特願2005−182469(P2005−182469)
【出願日】平成17年6月22日(2005.6.22)
【出願人】(501203344)独立行政法人農業・食品産業技術総合研究機構 (827)
【Fターム(参考)】
【公開日】平成19年1月11日(2007.1.11)
【国際特許分類】
【出願日】平成17年6月22日(2005.6.22)
【出願人】(501203344)独立行政法人農業・食品産業技術総合研究機構 (827)
【Fターム(参考)】
[ Back to top ]