
AAD−12イベント416、関連するトランスジェニックダイズ系統、およびそのイベント特異的な同定
本発明は、本明細書においてpDAB4468−0416と称される、ダイズ植物における除草剤耐性についての新規のaad−12形質転換イベントを包含する。本発明は、ダイズの細胞のゲノム内の特定の部位に挿入された異種ポリヌクレオチドを包含する。一部の実施形態では、前記イベント/ポリヌクレオチドに、例えば、他の除草剤耐性遺伝子(複数可)および/または昆虫抑制タンパク質を含めた他の形質を「積み重ねる」ことができる。さらに、本発明は、試料(例えばダイズ)中の本主題のイベントの存在を検出するためのアッセイを提供する。アッセイは、ダイズゲノムに挿入された組換え構築物のDNA配列、および挿入部位に隣接しているゲノム配列に基づくことができる。アッセイの実施において有用なキットおよび条件も提供される。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、AAD−12ダイズイベントおよびそれに由来する後代に関する。
【背景技術】
【0002】
aad−12遺伝子(元々はデルフチア・アシドボランス(Delftia acidovorans)由来)は、アリールオキシアルカノエートジオキシゲナーゼ(AAD−12)タンパク質をコードする。この形質により、例えば2,4−ジクロロフェノキシ酢酸に対する耐性、およびピリジルオキシ酢酸除草剤に対する耐性が付与される。aad−12遺伝子自体は、植物における除草剤耐性について、WO2007/053482に最初に開示された。
【0003】
植物における異種遺伝子または外来遺伝子の発現は、その外来遺伝子が染色体のどこに挿入されたかに影響される。これは、例えば、クロマチン構造(例えば、ヘテロクロマチン)のため、または組み込み部位に近い転写調節エレメント(例えば、エンハンサー)の近傍にあるためである可能性がある(Weisingら、Ann. Rev. Genet 22巻:421〜477頁、1988年)。同じ種類のトランスジェニック植物(または他の生物体)における同じ遺伝子が、異なるイベント間で発現レベルの広範な変動を示すことがある。発現の空間パターンまたは時間パターンにも差があることがある。例えば、種々の植物組織における導入遺伝子の相対的な発現量の差は、導入される遺伝子構築物中に存在する転写調節エレメントから予測されるパターンに対応しない可能性がある。
【0004】
したがって、多くの場合、所与の目的ために申し分のないレベルまで対象の導入された遺伝子を発現するイベントを特定するために、多数のイベントを創出し、スクリーニングする。商業目的のために、何百から何千もの異なるイベントを作出し、それらのイベントを、導入遺伝子の所望の発現レベルおよび発現パターンを有する単一のイベントについてスクリーニングすることが一般的である。導入遺伝子の発現の所望のレベルおよび/またはパターンを有するイベントは、従来の育種方法を使用して、有性異系交雑によって導入遺伝子を他の遺伝的背景に遺伝子移入するために有用である。そのような交雑の後代は、元の形質転換体の導入遺伝子の発現特性を維持する。この戦略を使用して、その土地の生長条件によく適合する多数の品種における信頼性の高い遺伝子発現を確実にする。
【0005】
米国特許出願公開第20090130071号は、ダイズイベントMON87701および検出方法に関する。米国特許出願公開第20090036308号および同第20080051288号は、ダイズイベント3560.4.3.5および検出方法に関する。米国特許出願第20080312082号は、ダイズイベントDP−305423−1および検出方法に関する。米国特許出願第20060282915号は、ダイズイベントMON89788および検出方法に関する。
【0006】
本明細書に開示されている特定のイベントを有するAAD−12ダイズは、これまでに開示されていない。
【発明の概要】
【0007】
本発明は、American Type Culture Collection(ATCC、アメリカの微生物系統保存機関)に受託番号PTA−10442で寄託されている種子を有する、DAS−68416−4と称されるAAD−12ダイズ(ダイズ(Glycine max))イベント、およびそれに由来する後代に関する。本発明の他の態様は、後代植物、ダイズ、種子、および/またはダイズイベントDAS−68416−4の植物および種子および後代の再生可能な部位、ならびにそのいずれかから製造された食品または飼料製品を含む。本発明は、花粉、胚珠、花、苗条、根、および葉、ならびに栄養細胞、花粉細胞、および卵細胞の核を含むが、これらに限定されない、ダイズイベントDAS−68416−4の植物の部位も包含する。本発明は、さらに、フェノキシオーキシン除草剤および/またはアリールオキシアルカノエート除草剤に対する耐性を有する(これらの除草剤を、ダイズ植物の上から施用するか、近接するまたは近くの土壌に施用するか、または近接するまたは近くの雑草に施用するかにかかわらず)ダイズ植物、ダイズイベントDAS−68416−4の新規の遺伝組成、およびダイズイベントDAS−68416−4を含むダイズ植物の農業生産力の側面に関する。
【0008】
本発明は、一部において、植物の育種および除草剤耐性植物に関する。本発明は、本明細書に記載の通り、ダイズの細胞のゲノム内の特定の部位に挿入されたポリヌクレオチド配列を含むダイズ植物における新規のaad−12形質転換イベントを包含する。
【0009】
一部の実施形態では、前記イベント/ポリヌクレオチド配列に、例えば、他の除草剤耐性遺伝子(複数可)および/または昆虫抑制タンパク質を含めた他の形質を「積み重ねる」ことができる。しかし、本発明は、本明細書に記載の通り、単一のイベントを有する植物を包含する。本発明の特定の実施形態では、種々の除草剤抵抗性の雑草(例えば、グリホサート抵抗性の雑草)を制御するために、1つまたは複数の除草剤耐性形質をダイズイベントDAS−68416−4に積み重ねる。
【0010】
さらに、本発明は、試料(例えば、ダイズの)中の本主題のイベントの存在を検出するためのアッセイを提供する。アッセイは、ダイズゲノムに挿入された組換え構築物のDNA配列、および挿入部位に隣接しているゲノム配列に基づくことができる。アッセイの実施において有用なキットおよび条件も提供される。
【0011】
したがって、本発明は、一部において、(トランスジェニックダイズ系統における)全aad−12挿入断片、およびその境界領域のDNA配列のクローニングおよび分析に関する。これらの配列は独特である。これらの挿入断片および境界の配列に基づいて、イベント特異的なプライマーを生成した。PCR分析により、これらのイベント特異的なプライマーセットを用いて生成したPCRアンプリコンを分析することによってこれらのイベントを同定することができることが実証された。したがって、これらおよび他の関連する手順を用いて、本発明のイベントを含むダイズ系統を一意的に同定することができる。
【図面の簡単な説明】
【0012】
【図1】pDAB4468のプラスミドマップを示す図である。
【図2】ダイズイベントのゲノムDNAを示す図であり、DAS−68416−4をEcoRVまたはPvuIIを用いて消化し、それを使用して対応するGENOMEWALKER(商標)ライブラリーを作製し、これを、標的DNA配列を増幅するために鋳型として使用した。
【図3】ダイズイベントDAS−68416−4の全長配列を5’端から3’端まで確認するためのプライマーの位置を示す図である。
【図4】ダイズイベントDAS−68416−4の全長配列を5’端から3’端まで確認するためのプライマーの位置を示す図である。
【図5】AAD−12ダイズイベントDAS−68416−4の挿入部位の配列を確認するためのプライマーの位置を示す図である。
【図6】植物の生活環を通した発現レベルを示す図である。
【0013】
[表の簡単な説明]
表1は、イベントDAS−68416−4についての挿入断片および隣接配列の配列番号1に関する残基の番号付けを示す。
表2は、サザン分析において使用したプローブの位置および長さを示す。
表3は、サザンブロット分析において予測されたハイブリダイズ断片および観察されたハイブリダイズ断片を示す。
表4は、隣接境界領域を増幅するためのダイズイベントDAS−68416−4のゲノムウォーキングの条件を示す。
表5は、ダイズイベントDAS−68416−4における境界領域およびイベント特異的な配列の標準のPCR増幅のための条件を示す。
表6は、T鎖挿入断片についてのアンプリコン1〜4に対するプライマーの説明を示す。
表7は、ダイズイベントDAS−68416−4における境界領域およびイベント特異的な配列の標準のPCR増幅のためのPCR混合物を示す。
表8は、2008年中に米国およびカナダにおいて生産されたダイズイベントDAS−68416−4から収集した組織中のAAD−12タンパク質レベルの概要を示す。
表9は、サザン分析において使用したプローブの位置および長さを示す。
表10は、サザンブロット分析において予測されたハイブリダイズ断片および観察されたハイブリダイズ断片を示す。
表11は、実験1において評価した農業形質パラメータを示す。
表12は、実験1からの農業特性の分析を示す。
表13は、2009年の農業試験および収量試験において収集されたデータを示す。
表14は、2009年の全ての地域にわたる農業特性の結果の概要を示す。
表15は、実験1からの病害発生率および虫害についての分析を示す。
表16は、DAS−68416−4および従来のダイズの試験において観察された病害ストレス要因および昆虫ストレス要因を示す。
表17は、温暖な条件下および寒冷な条件下でのダイズイベントDAS−68416−4種子の発芽を示す。
表18は、ダイズの茎葉の一般成分(proximate)分析、繊維分析およびミネラル分析の概要を示す。
表19は、ダイズの粒一般成分分析および繊維分析の概要を示す。
表20は、ダイズの粒のミネラル分析の概要を示す。
表21は、ダイズの粒のアミノ酸分析の概要を示す。
表22は、ダイズの粒の脂肪酸分析の概要を示す。
表23は、ダイズの粒のビタミン分析の概要を示す。
表24は、ダイズの粒のイソフラボン分析の概要を示す。
表25は、ダイズの粒の抗栄養因子分析の概要を示す。
表26は、2,4−D出芽前耐性試験についての現場および処理の情報を示す。
表27は、2,4−Dの出芽前施用に対するDAS−68416−4ダイズの耐性を例示している。
【0014】
[配列の簡単な説明]
配列番号1は、本主題のダイズイベントDAS−68416−4についての挿入断片配列および隣接配列を示す。
配列番号2〜28は、本明細書に記載のプライマーである。
配列番号29および30は、本明細書に記載の隣接SNPマーカーであるBARC−019093−03299およびBARC−044607−08736である。
【発明を実施するための形態】
【0015】
本発明は、一部において、植物の育種および除草剤耐性植物に関する。本発明は、本明細書に記載の通り、ダイズの細胞のゲノム内の特定の部位に挿入された、本主題のaad−12ポリヌクレオチド配列を含むダイズ植物(ダイズ)の新規の形質転換イベントを包含する。一部の実施形態では、前記ポリヌクレオチド配列に、他の形質を「積み重ねる」ことができる(例えば、他の除草剤耐性遺伝子(複数可)および/または昆虫抑制タンパク質をコードする遺伝子(複数可)など)。しかし、本発明は、本明細書に記載の単一のイベントを有する植物を包含する。
【0016】
さらに、本発明は、試料中の本主題のイベントの存在を検出するためのアッセイを提供する。本発明の態様は、本明細書において例証または提案される任意の診断用核酸分子、特に、完全にまたは部分的に本主題の隣接配列に基づく診断用核酸分子を設計および/または作出する方法を包含する。
【0017】
より詳細には、本発明は、一部において、トランスジェニックダイズイベントDAS−68416−4、これらのイベントを含む植物系統、ならびにこの挿入断片、および/またはその境界領域のDNA配列のクローニングおよび分析に関する。本発明の植物系統は、本明細書において開示され、提案されている配列を使用して検出することができる。
【0018】
一部の実施形態では、本発明は、除草剤耐性ダイズ系統、およびその同定に関する。本発明は、一部において、有性交雑の後代が対象のイベントを含有するかどうかを決定するために本主題のイベントの存在を検出することに関する。さらに、イベントを検出するための方法が包含され、それは、例えば、組換え作物に由来する食物の市販前承認および表示を必要とする規制に合致するために役立つ。任意の周知の核酸検出方法、例えばポリメラーゼ連鎖反応(PCR)または核酸プローブを使用したDNAのハイブリダイゼーションなどによって本主題のイベントの存在を検出することが可能である。イベント特異的なPCRアッセイは、例えば、Windelsら(Med. Fac. Landbouww、Univ. Gent 64/5b:459462、1999年)によって考察されている。これは、グリホサート耐性のダイズイベント40−3−2を、挿入断片と隣接DNAとの間の接合部にわたるプライマーセットを使用したPCRによって同定することに関する。より詳細には、1つのプライマーは挿入断片由来の配列を含み、第2のプライマーは隣接DNA由来の配列を含んだ。
【0019】
デルフチア・アシドボランス(Delftia acidovorans)に由来する、アリールオキシアルカノエートジオキシゲナーゼ(AAD−12)タンパク質をコードするaad−12遺伝子を挿入することによってダイズを改変した。この形質は、2,4−ジクロロフェノキシ酢酸除草剤およびピリジルオキシ酢酸除草剤に対する耐性を付与し、植物を形質転換する間、および育種苗床において選抜マーカーとして使用することができる。
【0020】
より詳細には、本明細書には、AAD12イベントpDAB4468−0416、ならびに、それを、代々、植物全体および分子レベルで安定性および発現について選抜し、特徴付けすることが記載されている。
【0021】
本発明に従って使用される対象の合成遺伝子(aad−12)は、デルフチア・アシドボランス(Delftia acidovorans)に由来し、フェノキシオーキシン(例えば、2,4−D、MCPA)、ならびにピリジルオキシオーキシン(例えば、フルロキシピル、トリクロピル)を含めたアリールオキシアルカノエート部分を持ついくつかの除草剤を不活性化することができる酵素をコードする。atUbi10プロモーターに駆動されるaad−12遺伝子を、ダイズ系統であるMaverickに、アグロバクテリウム・ツメファシエンス(Agrobacterium tumefaciens)法によって導入した。トランスジェニックT0植物を4〜6世代にわたって自家受粉させた。平行して、優良なダイズ品種にaad−12遺伝子の遺伝子移入も行った。全てのトランスジェニックイベントを、封じ込められ、規制された圃場苗床および研究室の環境において4〜5世代にわたって特徴付けた。
【0022】
イベントpDAB4468−0416挿入物の両末端について配列決定し、特徴付けた。イベント特異的なアッセイを開発した。これは、ダイズゲノム(ダイズの第4染色体)上にマッピングされた;隣接SNPマーカーは、本明細書に配列番号29および30として記載されている。イベントを別の優良な系統に遺伝子移入する。イベントにより、2,4−Dおよびグルホシネートに対する耐性がもたらされる。
【0023】
atUbi10プロモーターによって駆動し、アグロバクテリウム・ツメファシエンス(Agrobacterium tumefaciens)法によって100を超えるAAD12T1Maverickダイズイベントを生成した。イベントpDAB4468−0416を、5つの自家受粉させた世代およびいくらかの戻し交雑した世代の単一系列選抜によって選抜した。これは形態学的に正常であり、全ての自家受粉した世代を通じて、V3における2240g ae/haの2,4−D噴霧に対して耐性であった。イベントは、単一の優性遺伝子として遺伝性であり、その上、自家受粉した世代および戻し交雑した世代において正常なメンデル分離を有した。
【0024】
イベントpDAB4468−0416は、全長の植物転写単位(PTU)を伴う単一の組み込みを有することが見いだされた。ベクターの骨格からは抗生物質抵抗性遺伝子配列は見いだされず、また、aad−12遺伝子のホモ接合状態および半接合性状態において、数世代を通して遺伝子サイレンシングは検出されなかった。aad−12遺伝子は、2,4−D耐性のレベルをもたらす予測レベルで発現された。イベントは、aad−12遺伝子を安定に、複数の世代にわたって、および所与の世代において同胞系列の間で発現した。
【0025】
上の背景技術のセクションにおいて言及されている通り、導入遺伝子の植物ゲノムへの導入および組み込みは、いくつかのランダムなイベントを伴う(したがって発現される所与の挿入物について「イベント」と称する)。すなわち、アグロバクテリウム(Agrobacterium)形質転換、「遺伝子銃」、およびWHISKERなどの多くの形質転換技法を用いると、導入遺伝子がゲノム内のどこに挿入されるかは予測不可能である。したがって、挿入断片の両側の隣接植物ゲノムDNAを同定することは、所与の挿入イベントを有する植物を同定するために重要であり得る。例えば、挿入断片と宿主ゲノムとの接合領域にわたってPCRアンプリコンを生成するPCRプライマーを設計することができる。このPCRアンプリコンを使用して、独特のまたは別個の種類の挿入イベントを同定することができる。
【0026】
「イベント」は、元々ランダムなイベントであるので、本開示の一部として、イベントを含む少なくとも2500種子のダイズ系統がAmerican Type Culture Collection(ATCC)、10801 University Boulevard、Manassa、VA、20110に寄託され、制限なく一般に公開されている(しかし、特許権に支配される)。この寄託物は、ATCC寄託番号PTA−10442と称される。25個のダイズ(Glycine max)種子(AAD−12イベントpDAB4468−0416)のバイアルが、Dow AgroSciences LLC on October 22、2009を代表して寄託された。この寄託物は2009年11月2日に試験され、同日に種子は生育可能(viable)であった。この寄託物は、特許手続のための種子寄託物に関するブタペスト条約の条項に従い、かつその下で作製されたものであり、また、それに従い、かつその下で維持される。寄託物は、公共の受託所であるATCC受託所において、30年間、または最新の要求後5年間、または特許の有効期間のいずれかのより長い期間にわたって制限なく維持され、その期間中に生育不能になった場合は交換される。
【0027】
寄託された種子は本発明の一部である。明らかに、ダイズ植物をこれらの種子から生長させることができ、そのような植物は本発明の一部である。本発明は、これらのダイズ植物に含有される、これらの植物およびその後代を検出するために有用なDNA配列にも関する。本発明の検出方法およびキットは、試験の最終目的に応じて、これらのイベントの任意の1つ、2つ、またはさらには3つ全てを同定することを対象とすることができる。
【0028】
本発明の説明に役立つように、また当業者が本発明を実施する指針となるために、本明細書において定義および例が提供される。特に断りのない限り、用語は、当業者による従来の使用法に従って理解される。米国特許法施行規則第1.822条に明記されているDNA塩基の命名法を使用する。
【0029】
本明細書で使用される場合、用語「後代」は、AAD−12ダイズイベントDAS−68416−4を含む親植物の任意の世代の子孫を意味する。
【0030】
トランスジェニック「イベント」は、植物細胞を異種DNA、すなわち、対象の導入遺伝子を含む核酸構築物で形質転換すること、植物のゲノムに導入遺伝子を挿入することによって生じる植物の集団を再生させること、および特定のゲノム上の位置への挿入を特徴とする特定の植物を選択することによって作出される。用語「イベント」は、元の形質転換体および異種DNAを含むその形質転換体の後代を指す。用語「イベント」は、ゲノムDNA/導入遺伝子DNAを含む、形質転換体と別の品種との間の有性異系交雑によって作出される後代も指す。反復親との戻し交雑を繰り返した後であっても、形質転換された親由来の挿入された導入遺伝子DNAおよび隣接ゲノムDNA(ゲノムDNA/導入遺伝子DNA)は、交雑の後代において同じ染色体上の位置に存在する。用語「イベント」は、元の形質転換体由来のDNA、および挿入DNAを含む一方の親系統(例えば、元の形質転換体および自殖によって生じる後代)と挿入DNAを含有しない親系統との有性交雑の結果として、対象の導入遺伝子を含む挿入DNAを受け取る後代に伝達されることが予測され得る挿入DNAおよび挿入DNAのすぐ近接の隣接ゲノム配列を含むその後代も指す。
【0031】
「接合部配列」は、ゲノムに挿入されたDNAが、挿入点に隣接するダイズのネイティブなゲノム由来のDNAに連結している点にわたり、植物の遺伝物質中の一方または他方の接合部配列の特定または検出は、十分にイベントに特徴的である。本明細書に記載のダイズイベントにおける挿入物にわたるDNA配列および同様の長さの隣接DNAが包含される。そのような特徴的な配列の特定の例が本明細書において提供されるが、挿入物の接合部、または挿入物とゲノム配列との接合部とオーバーラップする他の配列も特徴的であり、本発明に従って使用することができる。
【0032】
本発明は、そのような隣接配列、接合部配列、および挿入断片配列を特定することに関する。関連するPCRプライマーおよびアンプリコンは、本発明に包含される。本発明に従って、挿入DNAおよびその端までわたるアンプリコンを使用するPCR分析方法を使用して、対象の登録商標を持つトランスジェニックダイズ系統に由来する商業化されたトランスジェニックダイズの品種または系統を検出または特定することができる。
【0033】
挿入断片を植物細胞のゲノムに導入するプロセスの間、挿入断片および/またはゲノム隣接配列のいくらかの欠失または他の変化が起こることは珍しくない。したがって、本明細書において提供される関連性のあるプラスミド配列のセグメントは、いくらかの軽微な変動を含む可能性がある。同じことが、本明細書において提供される隣接配列にも当てはまる。したがって、本主題の隣接配列および/または挿入断片配列といくらかの範囲の同一性を有するポリヌクレオチドを含む植物は、本発明の範囲内である。本発明の配列に対する同一性は、本明細書で例示または記載されている配列に対して少なくとも65%配列同一性、より好ましくは少なくとも70%配列同一性、より好ましくは少なくとも75%配列同一性、より好ましくは少なくとも80%同一性、およびより好ましくは少なくとも85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%配列同一性を有するポリヌクレオチド配列であってよい。本明細書において提供されるハイブリダイゼーションおよびハイブリダイゼーション条件は、そのような植物および本発明のポリヌクレオチド配列を定義するためにも使用することができる。隣接配列プラス挿入断片配列の配列は、寄託されている種子を照会して確認することができる。
【0034】
これらの挿入断片のそれぞれの全配列は、それぞれの隣接配列の部分と一緒に、本明細書において配列番号1として提供される。配列番号1(全部で10,212塩基対)に関してこのイベントについての挿入断片および隣接配列の位置が下の表1に示されている。これは、例えば、実施例3においてより詳細に考察されている。
【0035】
【表1】

【0036】
これらの配列(特に隣接配列)は独特である。これらの挿入断片および境界の配列に基づいて、イベント特異的なプライマーを生成した。PCR分析により、これらのイベント特異的なプライマーセットを用いて生成したPCRアンプリコンを分析することによって、異なるダイズ遺伝子型においてこれらのダイズ系統を同定することができることが実証された。したがって、これらおよび他の関連する手順を用いて、これらのダイズ系統を一意的に同定することができる。本明細書において同定される配列は独特である。
【0037】
本発明の検出技法は、植物育種と併せて、1つまたは複数の追加的な対象の形質を後代に与える取り組みにおいて、対象のイベントを含む親植物を別の植物系統と交雑した後に、どの後代植物が所与のイベントを含むかを決定するために特に有用である。これらのPCR分析方法は、ダイズの育種計画ならびに品質管理、特に商業化されたトランスジェニックダイズ種子に有益である。これらのトランスジェニックダイズ系統のPCR検出キットも、現在製造され、使用することができる。これは、製品登録および製品管理にも有益である。
【0038】
さらに、隣接ダイズ/ゲノム配列を使用して、それぞれの挿入断片の遺伝子位置を特異的に同定することができる。この情報を使用して、それぞれのイベントに特異的な分子マーカー系を作製することができる。これらは、育種戦略を加速させるために、および連鎖データを確立するために使用することができる。
【0039】
さらに、隣接配列の情報を使用して、導入遺伝子の組み込みプロセス、ゲノムの組み込み部位の特性、イベントの選別、導入遺伝子およびそれらの隣接配列の安定性、ならびに遺伝子発現(特に遺伝子サイレンシング、導入遺伝子のメチル化パターン、位置の影響、およびMARS[マトリックス付着領域]などの潜在的な発現関連エレメントなどに関連する)を試験し、特徴付けることができる。
【0040】
本開示全てに照らして、本発明は、ATCC寄託番号PTA−10442の下で入手可能な種子を包含することが明白でなければならない。本発明は、受託番号PTA−10442の下でATCCに寄託されている種子から生長させた除草剤耐性ダイズ植物も包含する。本発明は、さらに、前記植物の部位、例えば、葉、組織試料、前記植物によって生産される種子、花粉などを包含する。
【0041】
さらに、本発明は、寄託されている種子から生長させた植物の後裔植物および/または後代植物、好ましくは除草剤耐性ダイズ植物であって、本明細書に記載の検出可能な野生型ゲノムDNA/挿入DNA接合部配列を含むゲノムを有する植物を包含する。本明細書で使用される場合、用語「ダイズ」は、Glycine maxを意味し、ダイズ植物を用いて育種することができるその全品種を包含する。
【0042】
本発明は、本発明の植物を少なくとも一方の親として使用して交雑を行うプロセスをさらに包含する。例えば、本発明は、本明細書において例示されている植物のいずれかを一方の親または両親として有するF1ハイブリッド植物を包含する。また、本発明のそのようなF1ハイブリッドによって生産される種子も本発明の範囲内である。本発明は、例示した植物と、異なる(例えば、純系の親)植物を交雑し、得られたハイブリッド種子を収穫することによってF1ハイブリッド種子を作出するための方法を包含する。本発明は、雌親または雄親のいずれかである、例示した植物を包含する。得られる植物の特性は、親植物を慎重に考察することによって改良することができる。
【0043】
除草剤耐性ダイズ植物は、本明細書で言及されている系統の任意の1つの種子から生長させたダイズ植物からなる第1の親ダイズ植物と、第2の親ダイズ植物との有性交雑を最初に行い、それによって複数の第1の後代植物を作出すること;次いで、除草剤に対して抵抗性である(または、本発明のイベントの少なくとも1つを保有する)第1の後代植物を選択すること;第1の後代植物を自殖させ、それによって、複数の第2の後代植物を作出すること;次いで、第2の後代植物から、除草剤に対して抵抗性である(または、本発明のイベントの少なくとも1つを保有する)植物を選択することによって育種することができる。これらのステップは、第1の後代植物または第2の後代植物を第2の親ダイズ植物または第3の親ダイズ植物と戻し交雑することをさらに含んでよい。次いで本発明のダイズ種子を含むダイズ作物、またはその後代を植え付けることができる。
【0044】
2つの異なるトランスジェニック植物を交配して、それぞれ独立に分離して付加された2つの外因性遺伝子を含有する子孫を作出することもできることがまた理解される。適切な後代を自殖させることにより、付加された外因性遺伝子のどちらについてもホモ接合性である植物を作出することができる。栄養繁殖のように、親植物との戻し交雑および非トランスジェニック植物との異系交雑も意図されている。種々の形質および作物のために一般に使用される他の育種方法は当技術分野で公知である。反復親である望ましいホモ接合性の栽培品種または純系統に、単純に遺伝した、高度に遺伝性の形質の遺伝子を伝達するために戻し交雑育種が使用されてきた。伝達される形質の供給源は、供与親と称されている。生じた植物は、反復親(例えば、栽培品種)の属性および供与親から伝達された望ましい形質を有することが予測される。最初の交雑の後、供与親の表現型を保有する個体を選択し、反復親と繰り返し交雑(戻し交雑)する。生じた親は、反復親(例えば、栽培品種)の属性および供与親から伝達された望ましい形質を有することが予測される。
【0045】
本発明のDNA分子は、マーカー利用育種(MAB)法において分子マーカーとして使用することができる。本発明のDNA分子は、当技術分野で公知の通り、遺伝的に連鎖している作物学的に有用な形質を特定する方法(例えば、AFLPマーカー、RFLPマーカー、RAPDマーカー、SNPおよびSSRなど)において使用することができる。MAB法を使用して、本発明のダイズ植物(またはその後代および任意の他のダイズの栽培品種または品種)との交雑の後代において除草剤抵抗性形質を追跡することができる。DNA分子はこの形質についてのマーカーであり、当技術分野で周知のMAB法を使用して、本発明の少なくとも1つのダイズ系統、またはその後代が親または祖先であったダイズ植物における除草剤抵抗性形質(複数可)を追跡することができる。本発明の方法を使用して、対象イベントを有する任意のダイズ品種を特定することができる。
【0046】
本発明の方法は、本発明の植物を用いて育種するステップを含む、除草剤耐性ダイズ植物を作出する方法を包含する。より詳細には、前記方法は、本発明の2つの植物、または本発明の1つの植物と任意の他の植物とを交雑するステップを含んでよい。好ましい方法は、前記交雑の後代を、本発明に従って検出可能なイベントについて前記後代を分析することによって選択するステップをさらに含む。例えば、本発明を使用して、他の望ましい形質、例えば、作物学的形質など、例えば、本明細書において、種々の実施例において試験されるものなどを含む植物を用いた育種サイクルを通して対象イベントを追跡することができる。対象イベントおよび所望の形質を含む植物を検出し、特定し、選択し、例えば育種のさらなるラウンドに直ちに使用することができる。対象イベント/形質は、昆虫抵抗性形質(複数可)および/または別の除草剤耐性形質と育種を通じて組み合わせ、本発明に従って追跡することもできる。後者の好ましい一実施形態は、対象イベントを除草剤であるジカンバに対する抵抗性をコードする遺伝子と組み合わせて含む植物である。
【0047】
したがって、本発明は、例えば、グリホサート抵抗性(例えば、抵抗性植物または細菌のEPSPS、GOX、GAT)、グルホシネート抵抗性(例えば、Pat、bar)、アセト乳酸合成酵素(ALS)阻害性除草剤への抵抗性(例えば、イミダゾリノン[イマゼタピルなど]、スルホニル尿素系、トリアゾロピリミジンスルホンアニリド、ピリミジニルチオベンゾエート系、および他の化学物質[Csr1、SurAなど])、ブロモキシニル抵抗性(例えば、Bxn)、HPPD(4−ヒドロキシルフェニル−ピルビン酸−ジオキシゲナーゼ)酵素の阻害剤に対する抵抗性、フィトエンデサチュラーゼ(PDS)の阻害剤に対する抵抗性、光化学系II阻害性除草剤に対する抵抗性(例えば、psbA)、光化学系I阻害性除草剤に対する抵抗性、プロトポルフィリノーゲンオキシダーゼIX(PPO)阻害性除草剤に対する抵抗性(例えば、PPO−1)、フェニル尿素除草剤に対する抵抗性(例えば、CYP76B1)、ジカンバ分解酵素(例えば、US20030135879を参照されたい)をコードする形質と組み合わせることができ、その他のものを、単独で、または多数の組合せで積み重ねて、雑草のシフト(weed shift)および/または上述のクラスの任意の除草剤に対する抵抗性を有効に制御する、または妨げる能力をもたらすことができる。
【0048】
追加的な除草剤に関して、いくつかの追加的な好ましいALS(AHASとしても公知である)阻害剤としては、トリアゾロピリミジンスルホンアニリド系(例えば、クロランスラム−メチル、ジクロスラム、フロラスラム、フルメトスラム、メトスラム、およびペノキススラムなど)、ピリミジニルチオベンゾエート系(例えば、ビスピリバックおよびピリチオバックなど)、ならびにフルカルバゾンが挙げられる。いくつかの好ましいHPPD阻害剤としては、メソトリオン、イソキサフルトール、およびスルコトリオンが挙げられる。いくつかの好ましいPPO阻害剤としては、フルミクロラック、フルミオキサジン、フルフェンピル、ピラフルフェン、フルチアセット、ブタフェナシル、カルフェントラゾン、スルフェントラゾン、およびジフェニルエーテル系(例えば、アシフルオルフェン、ホメサフェン、ラクトフェン、およびオキシフルオルフェンなど)が挙げられる。
【0049】
さらに、AAD−12単独に、または1つもしくは複数の追加的なHTC形質を積み重ねたAAD−12に、1つまたは複数の追加的なインプット形質(例えば、昆虫抵抗性、真菌抵抗性、もしくはストレス耐性など)またはアウトプット形質(例えば、生産量の増加、油プロファイルの改良、繊維品質の改良など)を積み重ねることができる。したがって、本発明を使用して、任意の数の作物害虫を柔軟かつ費用効果的に制御する能力を伴う改良された作物の品質の完全な作物パッケージをもたらすことができる。
【0050】
対象のAAD−12酵素は、トランスジェニック発現を可能にして、ほぼ全ての広葉雑草およびイネ科雑草を制御し得る除草剤の組合せに対する耐性をもたらす。AAD−12は、例えば、他のHTC形質(例えば、グリホサート抵抗性、グルホシネート抵抗性、イミダゾリノン抵抗性、ブロモキシニル抵抗性など)、および昆虫抵抗性形質(Cry1F、Cry1Ab、Cry 34/45など)を積み重ねるための優れた除草剤耐性作物(HTC)形質としての機能を果たし得る。さらに、AAD−12は、第2の遺伝子または遺伝子群を用いて遺伝子操作した植物の一次形質転換体を選択するために役立つ選択マーカーとしての機能を果たし得る。
【0051】
本発明のAAD−12遺伝子は、フェノキシ酢酸オーキシン除草剤(例えば2,4−DB、MCPBなど)に変換される化合物に対する抵抗性ももたらす。2,4−DB除草剤に存在する酪酸部分は、β酸化によって植物毒性2,4−ジクロロフェノキシ酢酸に変換される。同様に、MCPBは、β酸化によって植物毒性MCPAに変換される。ブタン酸除草剤は、それ自体は非除草性である。ブタン酸除草剤は、感受性植物においてβ酸化によってそれらのそれぞれの酸に変換され、それは植物毒性である除草剤の酢酸の形態である。急速なβ酸化ができない植物は、ブタン酸除草剤による害を受けない。しかし、急速なβ酸化ができる植物およびブタン酸除草剤を酢酸形態に変換することができる植物は、その後にAAD−12によって保護される。
【0052】
除草剤を施用する方法は当技術分野で公知である。そのような施用は、2種以上の除草剤のタンク混合物を含んでよい。本発明に従って使用するためのいくつかの好ましい除草剤としては、2,4−D;2,4−DB;MCPA;MCPBなどのフェノキシオーキシン除草剤が挙げられる。これらに、1種または複数種の追加的な除草剤耐性遺伝子(複数可)および対応する除草剤(例えばグリホサートおよび/またはグルホシネート)を積み重ねることができる。1種、2種、3種、またはそれ以上の除草剤を、本開示の利益を有する当業者に明白になる有利な組合せで使用することができる。本主題の除草剤の1種または複数種を圃場/地域に、本発明の種子を植え付ける前に施用することができる。そのような施用は、例えば、植え付けの14日間以内であってよい。本主題の除草剤の1種または複数種は、植え付け時および/または植え付け後であるが出芽前に施用することもできる。本主題の除草剤の1種または複数種は、土に(雑草を制御するために)、または雑草および/または本発明のトランスジェニック植物の上から施用することもできる。本主題の除草剤は、例えば、ある除草剤には耐性であるが、別の除草剤には耐性でない可能性がある雑草を制御する、または防ぐために、交替させること、または組み合わせて使用することができる。本主題の3種類の除草剤について、種々の施用時間を当技術分野で公知の種々の方法で用いることができる。本発明は、AAD−12自生植物を制御するための方法ももたらす。発明の名称が「CONTROL OF AAD DICOT VOLUNTEERS IN MONOCOT CROPS」である同時に出願されたPCT出願を参照されたい。
【0053】
本発明のHTC形質は、他のHTC形質(これらに限定されないが、グリホサート耐性を含む)との新規の組合せにおいて使用することができる。これらの形質の組合せにより、除草剤(例えば、グリホサート)に対する新しく獲得した抵抗性または固有の耐性によって雑草(および同様のもの)種を制御する新規の方法が生じる。したがって、HTC形質に加えて、トランスジェニック作物において、前記酵素によって除草剤耐性を創出する、除草剤を使用して雑草を制御するための新規の方法は、本発明の範囲内である。
【0054】
さらに、世界中で栽培されているグリホサート耐性作物が一般的である。他のグリホサート耐性作物との輪作において多くの場合、グリホサート抵抗性自生植物を制御することは、輪作作物においては難しい。したがって、作物に個々に積み重ねた、または作物を個々に形質転換した対象のトランスジェニック形質の使用は、他のHTC自生作物を制御するためのツールを提供する。
【0055】
本発明の好ましい植物または種子は、そのゲノム内に、本明細書において同定される挿入断片配列と、本明細書において同定される、挿入断片の両側の少なくとも20〜500以上の連続した隣接ヌクレオチドを一緒に含む。別段の指定のない限り、隣接配列に言及する場合、それは配列番号1について特定されたものを指す(上の表1を参照されたい)。さらに、配列番号1は、元の形質転換体に挿入された異種DNAおよび挿入DNAのすぐ近接する例示的な隣接ゲノム配列を含む。これらの隣接配列の全部または一部は、イベントを含む親系統の有性交雑の結果として挿入DNAを受け取る後代に伝達されることが予測され得る。
【0056】
本発明は、本発明の植物の再生可能な細胞の組織培養物を包含する。また、そのような組織培養物から再生された植物も、特に前記植物が例示されている品種の形態学的性質および生理的性質の全てを発現することができる場合、包含される。本発明の好ましい植物は、寄託されている種子から生長させた植物の生理的特性および形態学的特性の全てを有する。本発明は、そのような種子および対象の品質形質を保有する種子の後代をさらに含む。
【0057】
植物または種子、またはその部位を操作(例えば、突然変異、さらなるトランスフェクション、およびさらに育種)することにより、「従属(本質的に由来する)」品種と称することができるものが創出され得る。植物新品種保護国際同盟(International Union for the Protection of New Varieties of Plants)(UPOV)は、ある品種が、保護された品種に由来しているかどうかを決定するための以下のガイドラインを提供した:
[A]以下の場合、品種は別の品種(「原品種」)に本質的に由来するとみなされるべきである
(i)原品種に主として由来する、または、それ自体が原品種に主として由来する品種に由来し、同時に原品種の遺伝子型または遺伝子型の組合せから生じる本質的な特性の発現を保持する;
(ii)原品種と明確に区別可能である;および
(iii)引き出す行為に由来する差異以外は、原品種の遺伝子型または遺伝子型の組合せから生じる本質的な特性の発現において原品種と一致する。
UPOVの国際機関の第6回会議、ジュネーブ、1992年10月30日;当該連合の事務局により作成された文書。
【0058】
本明細書で使用される場合、「系統」は、少なくとも1つの形質について、個体間で遺伝的変異をほとんど示さない、または示さない植物の群である。そのような系統は、何世代か自家受粉させ、選択すること、または組織または細胞の培養技法を使用して、単一の親から栄養繁殖させることによって創出することができる。
【0059】
本明細書で使用される場合、用語「栽培品種」および「品種」は同義であり、商業生産のために使用される系統を指す。
【0060】
「安定性」または「安定な」は、所与の構成要素に関しては、構成要素が代々、好ましくは、少なくとも3世代、実質的に同じレベルで、例えば、好ましくは±15%、より好ましくは±10%、最も好ましくは±5%で維持されることを意味する。安定性は、温度、位置、ストレスおよび植え付け時間に影響を受ける可能性がある。その後の世代を圃場条件下で比較することにより、構成要素が同様にもたらされるべきである。
【0061】
「商業的有用性」は、従来の農業設備を使用して農業者が作物を生産することができるように、および従来の圧搾および抽出用設備を使用して記載の構成要素を有する油を種子から抽出することができるように良好な植物生長力および高い稔性を有することと定義される。商業的に有用であるために、種子の重量、油の含有量、および1エーカー当たりの生産された油の総計によって測定した生産量は、同じ地域で栽培される高価値の形質を有さない、別の匹敵する市販のキャノーラ品種の平均生産量の15%以内である。
【0062】
「作物学的に優良」は、系統が対象イベント(複数可)に起因する昆虫抵抗性に加えて、例えば生産量、成熟度、病害抵抗性などの望ましい作物学的特性を有することを意味する。以下の実施例に記載の、本発明のイベントを含む植物に個々に、または任意の組合せで獲得される作物学的形質は、本発明の範囲内である。これらの作物学的特性およびデータポイントの全てを使用して、そのような植物を、そのような植物を定義するために使用する特性の範囲内の一点として、または一端もしくは両端で特定することができる。
【0063】
本開示に照らして当業者に理解されるように、検出キットの好ましい実施形態は、例えば、「接合部配列」または「移行部配列」(ダイズゲノムの隣接配列と挿入断片配列が接する場所)を対象とし、かつ/またはそれを含むプローブおよび/またはプライマーを含んでよい。これは、例えば、上記の表1に示されている一方のまたは両方の接合部配列(挿入断片と隣接配列が接する場所)を同定するために設計されたポリヌクレオチドプローブ、プライマー、および/またはアンプリコンを含む。1つの一般的な設計は、隣接領域内でハイブリダイズする1つのプライマー、および挿入断片内でハイブリダイズする1つのプライマーを有する。そのようなプライマーは、多くの場合、およそ、それぞれの長さが少なくとも約15残基である。この配置を用いて、プライマーを使用して、本発明のイベントの存在を示す検出可能なアンプリコンを生成/増幅することができる。これらのプライマーを使用して、上に示されている接合部配列にわたる(およびそれを含む)アンプリコンを生成することができる。
【0064】
隣接配列に「接している」プライマー(複数可)は、一般には、約200塩基を越えて、または接合部を越えてハイブリダイズするようには設計されない。したがって、典型的な隣接プライマーは、挿入断片の最初から隣接配列に入って200塩基の範囲内のどちらかの鎖の少なくとも15残基を含むように設計され得る。すなわち、配列番号1の残基約2530〜2730および/または約9122〜9322由来の(またはそれとハイブリダイズする)適切なサイズの配列を含むプライマーは、本発明の範囲内である。挿入プライマーは、挿入断片のどこにでも同様に設計することができるが、残基約2731〜2931および約8921〜9121を、例えば、そのようなプライマーの設計のために非排他的に使用することができる。
【0065】
当業者は、さまざまな標準のハイブリダイゼーションおよび/またはPCRの条件の下で、配列番号1(または相補物)のセグメント、およびその相補物とハイブリダイズするようにプライマーおよびプローブを設計することができ、ここでプライマーまたはプローブが例示された配列と完全に相補的ではないことも認識されよう。すなわち、ある程度のミスマッチが容認され得る。およそ20ヌクレオチドのプライマーについて、例えば、一般には、ミスマッチ塩基がアンプリコンと逆のプライマーの内部または末端にある場合、1または2ヌクレオチド程度は逆の鎖と結合しなくてよい。種々の適切なハイブリダイゼーション条件が以下に提供される。イノシンなどの合成ヌクレオチド類似体は、プローブにも使用することができる。ペプチド核酸(PNA)プローブ、ならびにDNAプローブおよびRNAプローブも使用することができる。重要なのは、そのようなプローブおよびプライマーが、本発明のイベントの存在に対して特徴的である(独自に特定し、区別することができる)ことである。
【0066】
PCR増幅において、例えば、軽微な配列決定のエラーを生じる可能性があるエラーが起こり得ることに留意するべきである。すなわち、別段の指定のない限り、本明細書において列挙されている配列は、ダイズのゲノムDNAから長いアンプリコンを生成し、次いでそのアンプリコンをクローニングし、配列決定することによって決定した。ゲノムDNAから配列決定するために十分なアンプリコンを生成するために必要な多数回の増幅を考慮すると、このように生成し、決定した配列においてわずかな差異および軽微な不一致が見いだされることは珍しいことではない。当業者は、これらの型の一般的な配列決定のエラーまたは不一致に起因して必要になる調整はいずれも本発明の範囲内であることを認識し、留意するべきである。
【0067】
例えば、イベントを創出する間に配列を挿入する場合、いくらかのゲノム配列が欠失することは珍しくないことにも留意すべきである。したがって、本主題の隣接配列と、例えばGENBANKに列挙されているゲノム配列との間にもいくらかの差異が出現する可能性がある。
【0068】
「挿入断片」の構成成分が図面に例示されており、以下の実施例においてより詳細に考察されている。これらの構成要素のDNAポリヌクレオチド配列、またはその断片を、本発明の方法においてDNAプライマーまたはプローブとして使用することができる。
【0069】
本発明の一部の実施形態では、ダイズ植物由来の植物および種子などにおける導入遺伝子/ゲノムの挿入領域の存在を検出するための組成物および方法が提供される。本明細書において提供される本主題の導入遺伝子/ゲノムの挿入領域の接合部配列(配列番号1の残基2730〜2731と残基9121〜9122の間)、そのセグメント、および例示された配列の相補物およびその任意のセグメントを含むDNA配列が提供される。挿入領域の接合部配列は、ゲノムに挿入された異種DNAと、挿入部位に隣接しているダイズの細胞由来のDNAの接合部にわたる。そのような配列は、所与のイベントに対して特徴的であり得る。
【0070】
これらの挿入断片および境界の配列に基づいて、イベント特異的なプライマーを生成することができる。PCR分析により、これらのイベント特異的なプライマーセットを用いて生成したPCRアンプリコンを分析することによって、異なるダイズ遺伝子型において本発明のダイズ系統を同定することができることが実証された。これらおよび他の関連する手順を用いて、これらのダイズ系統を一意的に同定することができる。したがって、そのようなプライマー対に由来するPCRアンプリコンは独特であり、これらのダイズ系統を同定するために使用することができる。
【0071】
一部の実施形態では、新規の導入遺伝子/ゲノムの挿入領域の連続した断片を含むDNA配列は本発明の態様である。導入遺伝子挿入断片配列の十分な長さのポリヌクレオチドおよび3つの上述のダイズ植物の1つまたは複数由来のダイズのゲノム配列の十分な長さのポリヌクレオチドを含むDNA配列および/またはこれらのダイズ植物の1つまたは複数に対して特徴的なアンプリコン産物を生成するためのプライマー配列として有用である配列が包含される。
【0072】
関連する実施形態は、本明細書において特定されるDNA配列(例えば、配列番号1およびそのセグメントなど)の導入遺伝子部分の少なくとも2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、またはそれ以上の連続したヌクレオチドを含むDNA配列、またはその相補物、およびこれらの配列由来の同様の長さのダイズの隣接DNA配列、またはその相補物に関する。そのような配列は、DNA増幅方法において、DNAプライマーとして有用である。これらのプライマーを使用して生成されるアンプリコンは、本明細書で言及されるダイズイベントのいずれかに対して特徴的である。したがって、本発明は、そのようなDNAプライマーおよび相同プライマーによって生成されるアンプリコンも包含する。
【0073】
本発明は、試料中の、本明細書で言及されるダイズイベントに対応するDNAの存在を検出する方法も包含する。そのような方法は、(a)DNAを含む試料を、これらのダイズイベントの少なくとも1つ由来のDNAを用いた核酸の増幅反応において使用したとき、前記イベント(複数可)に対して特徴的であるアンプリコンを生成するプライマーセットと接触させるステップと;(b)核酸の増幅反応を実施し、それによって、アンプリコンを生成するステップと;(c)アンプリコンを検出するステップとを含んでよい。
【0074】
本発明の別の検出方法は、試料中の、前記イベントの少なくとも1つに対応するDNAの存在を検出する方法であって、(a)DNAを含む試料を、ストリンジェントなハイブリダイゼーション条件下で前記ダイズイベントの少なくとも1つ由来のDNAとハイブリダイズさせ、ストリンジェントなハイブリダイゼーション条件下で対照のダイズ植物(対象のイベントがないDNA)とハイブリダイズしないプローブと接触させるステップと;(b)試料およびプローブをストリンジェントなハイブリダイゼーション条件に供するステップと;(c)プローブのDNAとのハイブリダイゼーションを検出するステップとを含む方法を包含する。
【0075】
さらに別の実施形態では、本発明は、本発明のaad−12イベントを含むダイズ植物を作出する方法であって、(a)第1の親ダイズ系統(前記系統の植物に前記除草剤抵抗性形質を付与する本発明の発現カセットを含む)と第2の親ダイズ系統(この除草剤耐性形質を欠く)を有性交雑し、それによって、複数の後代植物を作出するステップと;(b)分子マーカーを使用することによって後代植物を選択するステップとを含む方法を包含する。そのような方法は、場合によって、後代植物を第2の親ダイズ系統と戻し交雑して、前記昆虫耐性形質を含む真の育種(true-breeding)ダイズ植物を作出するさらなるステップを含んでよい。
【0076】
本発明の別の態様によると、前記3種のイベントの任意の1つ(または複数)を用いて交雑の後代の接合性を決定する方法が提供される。前記方法は、ダイズDNAを含む試料を本発明のプライマーセットと接触させるステップを含んでよい。前記プライマーは、前記ダイズイベントの少なくとも1つ由来のゲノムDNAを用いた核酸の増幅反応において使用したとき、前記ダイズイベントの少なくとも1つに対して特徴的である第1のアンプリコンを生成する。そのような方法は、核酸の増幅反応を実施し、それによって、第1のアンプリコンを生成するステップと;第1のアンプリコンを検出するステップと;ダイズDNAを含む試料を、ダイズ植物由来のゲノムDNAを用いた核酸の増幅反応において使用したとき、ダイズのゲノム領域と相同なネイティブなダイズのゲノムDNAを含む第2のアンプリコンを生成する前記プライマーセットと接触させるステップと;核酸の増幅反応を実施し、それによって、第2のアンプリコンを生成するステップとをさらに含む。この方法は、第2のアンプリコンを検出するステップと、試料中の第1のアンプリコンと第2のアンプリコンとを比較するステップであって、両方のアンプリコンが存在することにより、試料が、導入遺伝子挿入物についてヘテロ接合性であることが示されるステップとをさらに含む。
【0077】
DNA検出キットは、本明細書に開示されている組成物およびDNAの検出の技術分野で周知の方法を使用して開発することができる。このキットは、試料中の対象のダイズイベントDNAを特定するために有用であり、このDNAを含有するダイズ植物を育種するための方法に適用することができる。キットは、例えば、本明細書に開示されているアンプリコンと相同または相補的であるDNA配列、または対象イベントの導入遺伝子の遺伝エレメントに含有されるDNAと相同または相補的であるDNA配列を含有する。これらのDNA配列は、DNA増幅反応において、またはDNAのハイブリダイゼーション方法においてプローブとして、使用することができる。キットは、検出方法を実行するために必要な試薬および材料も含有することができる。
【0078】
「プローブ」は、従来の検出可能な標識またはレポーター分子(例えば、放射性同位元素、リガンド、化学発光剤、または酵素など)を付着させた単離された核酸分子である。そのようなプローブは、標的核酸の鎖、本発明の場合では、ダイズ植物由来であるかイベント由来のDNAを含む試料由来であるかにかかわらず、前記ダイズイベントの1つ由来のゲノムDNAの鎖と相補的である。本発明によるプローブは、デオキシリボ核酸またはリボ核酸だけでなく、標的DNA配列に特異的に結合し、その標的DNA配列の存在を検出するために使用することができるポリアミドおよび他のプローブ材料も含む。
【0079】
「プライマー」は、核酸ハイブリダイゼーションによって相補的な標的DNA鎖とアニーリングしてプライマーと標的DNA鎖との間のハイブリッドを形成し、次いでポリメラーゼ、例えば、DNAポリメラーゼによって標的DNA鎖に沿って伸長される、単離された/合成された核酸である。本発明のプライマー対は、例えば、ポリメラーゼ連鎖反応(PCR)または他の従来の核酸の増幅方法によって標的核酸配列を増幅するためのそれらの使用を指す。
【0080】
プローブおよびプライマーは、一般に、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35、36、37、38、39、40、41、42、43、44、45、46、47、48、49、50、51、52、53、54、55、56、57、58、59、60、61、62、63、64、65、66、67、68、69、70、71、72、73、74、75、76、77、78、79、80、81、82、83、84、85、86、87、88、89、90、91、92、93、94、95、96、97、98、99、100、101、102、103、104、105、106、107、108、109、110、111、112、113、114、115、116、117、118、119、120、121、122、123、124、125、126、127、128、129、130、131、132、133、134、135、136、137、138、139、140、141、142、143、144、145、146、147、148、149、150、151、152、153、154、155、156、157、158、159、160、161、162、163、164、165、166、167、168、169、170、171、172、173、174、175、176、177、178、179、180、181、182、183、184、185、186、187、188、189、190、191、192、193、194、195、196、197、198、199、200、201、202、203、204、205、206、207、208、209、210、211、212、213、214、215、216、217、218、219、220、221、222、223、224、225、226、227、228、229、230、231、232、233、234、235、236、237、238、239、240、241、242、243、244、245、246、247、248、249、250、251、252、253、254、255、256、257、258、259、260、261、262、263、264、265、266、267、268、269、270、271、272、273、274、275、276、277、278、279、280、281、282、283、284、285、286、287、288、289、290、291、292、293、294、295、296、297、298、299、300、301、302、303、304、305、306、307、308、309、310、311、312、313、314、315、316、317、318、319、320、321、322、323、324、325、326、327、328、329、330、331、332、333、334、335、336、337、338、339、340、341、342、343、344、345、346、347、348、349、350、351、352、353、354、355、356、357、358、359、360、361、362、363、364、365、366、367、368、369、370、371、372、373、374、375、376、377、378、379、380、381、382、383、384、385、386、387、388、389、390、391、392、393、394、395、396、397、398、399、400、401、402、403、404、405、406、407、408、409、410、411、412、413、414、415、416、417、418、419、420、421、422、423、424、425、426、427、428、429、430、431、432、433、434、435、436、437、438、439、440、441、442、443、444、445、446、447、448、449、450、451、452、453、454、455、456、457、458、459、460、461、462、463、464、465、466、467、468、469、470、471、472、473、474、475、476、477、478、479、480、481、482、483、484、485、486、487、488、489、490、491、492、493、494、495、496、497、498、499、または500ポリヌクレオチドまたはそれ以上の長さである。そのようなプローブおよびプライマーは、高ストリンジェンシーハイブリダイゼーション条件下で標的配列と特異的にハイブリダイズする。好ましくは、本発明によるプローブおよびプライマーは、標的配列と完全な配列類似性を有するが、標的配列とは異なり、標的配列とハイブリダイズする能力を保持するプローブを従来の方法によって設計することができる。
【0081】
プローブおよびプライマーを調製し、使用するための方法は、例えば、Molecular Cloning: A Laboratory Manual、第2版、1〜3巻Sambrookら編、Cold Spring Harbor Laboratory Press、Cold Spring Harbor、N.Y.、1989年に記載されている。PCRのプライマー対は、公知の配列から、例えば、その目的のためのコンピュータプログラムを使用することによって得ることができる。
【0082】
本明細書に開示されている隣接DNAおよび挿入断片の配列に基づくプライマーおよびプローブを使用して、従来の方法によって、例えば、そのような配列を再クローニングし、配列決定することによって、開示されている配列を確認すること(および、必要であれば、補正すること)ができる。
【0083】
本発明の核酸プローブおよびプライマーは、ストリンジェントな条件下で標的DNA配列とハイブリダイズする。任意の従来の核酸ハイブリダイゼーションまたは増幅の方法を使用して、試料中のトランスジェニックイベント由来のDNAの存在を特定することができる。核酸分子またはその断片は、ある特定の状況下で他の核酸分子と特異的にハイブリダイズすることができる。本明細書で使用される場合、2つの核酸分子は、2つの分子が、逆平行の、二本鎖核酸構造を形成することができる場合、互いと特異的にハイブリダイズすることができると言える。核酸分子は、別の核酸分子と完全な相補性を示す場合、その「相補物」であると言える。本明細書で使用される場合、分子は、分子の一方のあらゆるヌクレオチドが、他方のヌクレオチドと相補的である場合、「完全な相補性」を示すと言える。2つの分子は、少なくとも従来の「低ストリンジェンシー」条件下で、互いとアニーリングしたままであることを可能にするために十分な安定性で互いとハイブリダイズすることができる場合、「最小限相補的」であると言える。同様に、分子は、従来の「高ストリンジェンシー」条件下で、互いとアニーリングしたままであることを可能にするために十分な安定性で互いとハイブリダイズすることができる場合、「相補的」であると言える。従来のストリンジェンシー条件は、Sambrookら、1989年に記載されている。したがって、完全な相補性から逸脱することは、そのような逸脱により、二本鎖構造を形成する分子の能力が完全に妨げられない限りは許容できる。核酸分子がプライマーまたはプローブとしての機能を果たすためには、用いる特定の溶媒および塩濃度の下で安定な二本鎖構造を形成できるために十分に配列内で相補的であることのみが必要である。
【0084】
本明細書で使用される場合、実質的に相同な配列は、高ストリンジェンシー条件下で比較されている核酸配列の相補物と特異的にハイブリダイズし得る核酸配列である。用語「ストリンジェントな条件」は、Sambrookら、1989年、9.52〜9.55において考察されている特定のハイブリダイゼーション手順による、核酸プローブの標的核酸(すなわち、対象とする特定の核酸配列)とのハイブリダイゼーションに関して機能的に定義される。Sambrookら、1989年、9.47〜9.52および9.56〜9.58も参照されたい。したがって、本発明のヌクレオチド配列は、相補的なDNA断片のひと続きと2重鎖分子を選択的に形成するそれらの能力に関して使用することができる。
【0085】
構想される適用に応じて、プローブの標的配列に対する選択性のさまざまな程度を実現するためにハイブリダイゼーションのさまざまな条件を使用することができる。高い選択性を必要とする適用のために、一般には、ハイブリッドを形成するために比較的ストリンジェントな条件を用いることがある、例えば、約50℃〜約70℃の温度で約0.02M〜約0.15MのNaClによってもたらされるような比較的低塩かつ/または高温の条件を選択することがある。ストリンジェントな条件は、例えば、ハイブリダイゼーション濾過器を高ストリンジェンシー洗浄緩衝液(0.2×SSC、0.1%のSDS、65℃)で少なくとも2回洗浄することを伴ってよい。DNAのハイブリダイゼーションを促進する適切なストリンジェンシー条件、例えば、約45℃で6.0×塩化ナトリウム/クエン酸ナトリウム(SSC)、その後50℃で2.0×SSCで洗浄することは、当業者に公知である。例えば、洗浄ステップにおける塩濃度は、低ストリンジェンシーである50℃で約2.0×SSCから高ストリンジェンシーである50℃で約0.2×SSCまでから選択することができる。さらに、洗浄ステップにおける温度は、低ストリンジェンシー条件である室温、約22℃から高ストリンジェンシー条件である約65℃まで増加させることができる。温度と塩との両方が変動してよい、または、温度もしくは塩濃度のいずれかは、他方の変数が変化する一方で一定に保たれてよい。そのような選択的な条件は、もしあれば、プローブと鋳型または標的鎖との間のミスマッチを少し容認する。ハイブリダイゼーションによってDNA配列を検出することは当業者に周知であり、米国特許第4,965,188号および同第5,176,995号の教示は、ハイブリダイゼーション分析の方法の例である。
【0086】
特に好ましい実施形態では、本発明の核酸は、高ストリンジェンシー条件下で、本明細書において例証または提案される、その相補物および断片を含めたプライマー(またはアンプリコンもしくは他の配列)の1つまたは複数と特異的にハイブリダイズする。本発明の一態様では、本発明のマーカー核酸分子は、本明細書において例示された配列の1つに記載されている核酸配列、またはその相補物および/もしくは断片を有する。
【0087】
本発明の別の態様では、本発明のマーカー核酸分子は、そのような核酸配列と、80%から100%の間または90%から100%の間の配列同一性を共有する。本発明の別の態様では、本発明のマーカー核酸分子は、そのような配列と95%から100%の間の配列同一性を共有する。そのような配列は、遺伝的交雑の後代を特定するための植物育種方法において、マーカーとして使用することができる。プローブの標的DNA分子とのハイブリダイゼーションは、当業者に公知の任意の数の方法によって検出することができ、それらとしては、これらに限定されないが、蛍光タグ、放射性タグ、抗体ベースのタグおよび化学発光タグを挙げることができる。
【0088】
特定の増幅プライマー対を使用する標的核酸配列の増幅(例えば、PCRによる)に関して、「ストリンジェントな条件」は、プライマー対が、対応する野生型配列(またはその相補物)を有するプライマーが結合し得る標的核酸配列のみとハイブリダイズすること、および好ましくは特有の増幅産物、アンプリコンを生成することを可能にする条件である。
【0089】
用語「(標的配列)に特異的な」は、プローブまたはプライマーが、ストリンジェントなハイブリダイゼーション条件下で、標的配列を含む試料中の標的配列のみとハイブリダイズすることを示す。
【0090】
本明細書で使用される場合、「増幅されたDNA」または「アンプリコン」は、核酸鋳型の一部である標的核酸配列の核酸増幅産物を指す。例えば、有性交雑によって生じたダイズ植物が、本発明のダイズ植物由来のトランスジェニックイベントゲノムDNAを含有するかどうかを決定するために、ダイズ植物の組織試料から抽出したDNAを、挿入された異種DNAの挿入部位の近接する植物のゲノム内の隣接配列に由来するプライマー、および挿入された異種DNAに由来する第2のプライマーを含むプライマー対を使用して、イベントDNAの存在に対して特徴的であるアンプリコンを生成する核酸の増幅方法に供することができる。アンプリコンは、ある長さであり、同じくイベントに対して特徴的である配列を有する。アンプリコンの長さは、プライマー対の全て合わせた長さ足す1ヌクレオチド塩基対、および/またはプライマー対の全て合わせた長さ足す約2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35、36、37、38、39、40、41、42、43、44、45、46、47、48、49、50、51、52、53、54、55、56、57、58、59、60、61、62、63、64、65、66、67、68、69、70、71、72、73、74、75、76、77、78、79、80、81、82、83、84、85、86、87、88、89、90、91、92、93、94、95、96、97、98、99、100、101、102、103、104、105、106、107、108、109、110、111、112、113、114、115、116、117、118、119、120、121、122、123、124、125、126、127、128、129、130、131、132、133、134、135、136、137、138、139、140、141、142、143、144、145、146、147、148、149、150、151、152、153、154、155、156、157、158、159、160、161、162、163、164、165、166、167、168、169、170、171、172、173、174、175、176、177、178、179、180、181、182、183、184、185、186、187、188、189、190、191、192、193、194、195、196、197、198、199、200、201、202、203、204、205、206、207、208、209、210、211、212、213、214、215、216、217、218、219、220、221、222、223、224、225、226、227、228、229、230、231、232、233、234、235、236、237、238、239、240、241、242、243、244、245、246、247、248、249、250、251、252、253、254、255、256、257、258、259、260、261、262、263、264、265、266、267、268、269、270、271、272、273、274、275、276、277、278、279、280、281、282、283、284、285、286、287、288、289、290、291、292、293、294、295、296、297、298、299、300、301、302、303、304、305、306、307、308、309、310、311、312、313、314、315、316、317、318、319、320、321、322、323、324、325、326、327、328、329、330、331、332、333、334、335、336、337、338、339、340、341、342、343、344、345、346、347、348、349、350、351、352、353、354、355、356、357、358、359、360、361、362、363、364、365、366、367、368、369、370、371、372、373、374、375、376、377、378、379、380、381、382、383、384、385、386、387、388、389、390、391、392、393、394、395、396、397、398、399、400、401、402、403、404、405、406、407、408、409、410、411、412、413、414、415、416、417、418、419、420、421、422、423、424、425、426、427、428、429、430、431、432、433、434、435、436、437、438、439、440、441、442、443、444、445、446、447、448、449、450、451、452、453、454、455、456、457、458、459、460、461、462、463、464、465、466、467、468、469、470、471、472、473、474、475、476、477、478、479、480、481、482、483、484、485、486、487、488、489、490、491、492、493、494、495、496、497、498、499、または500、750、1000、1250、1500、1750、2000、またはそれ以上のヌクレオチド塩基対(上に列挙されている増大のいずれかを足すまたは引く)にわたってよい。あるいは、プライマー対は、挿入断片のヌクレオチド配列全体を含むアンプリコンが生成されるように、挿入DNAの両側の隣接配列に由来してもよい。植物のゲノム配列に由来するプライマー対のメンバーは、挿入DNA配列からある距離に位置してよい。この距離は、1ヌクレオチド塩基対から最大約2万ヌクレオチド塩基対までにわたることができる。用語「アンプリコン」の使用は、DNAの熱増幅反応において形成される可能性があるプライマー二量体を特に除外する。
【0091】
核酸の増幅は、ポリメラーゼ連鎖反応(PCR)を含めた当技術分野で公知の種々の核酸の増幅方法のいずれかによって実現することができる。種々の増幅方法が当技術分野で公知であり、とりわけ、米国特許第4,683,195号および米国特許第4,683,202号に記載されている。22kbに至るまでのゲノムDNAを増幅するためにPCR増幅方法が開発されてきた。これらの方法ならびにDNA増幅の技術分野で公知の他の方法を、本発明の実施において使用することができる。本明細書において提供される配列に由来するプライマーを使用してイベントからそのような配列を増幅し、その後PCRアンプリコンまたはクローニングされたDNAの標準のDNA配列決定を行うことにより、本主題のダイズイベント由来の異種導入遺伝子DNA挿入断片の配列または隣接ゲノム配列を検証すること(および必要であれば、補正すること)ができる。
【0092】
これらの方法によって生成されるアンプリコンは、複数の技法によって検出することができる。アガロースゲル電気泳動および臭化エチジウムを用いた染色は、DNAアンプリコンを検出する周知の一般的な方法である。別のそのような方法は、近接する隣接ゲノムDNA配列および挿入DNA配列の両方とオーバーラップするDNAオリゴヌクレオチドを設計する遺伝子ビット分析(Genetic Bit Analysis)である。オリゴヌクレオチドは、マイクロウェルプレートのウェル内に固定化されている。対象の領域のPCR後(挿入配列内の1つのプライマーおよび近接する隣接ゲノム配列内の1つのプライマーを使用する)、一本鎖のPCR産物は、固定化されたオリゴヌクレオチドとハイブリダイズし、DNAポリメラーゼおよび予測される次の塩基に特異的な標識したddNTPを使用する一塩基伸長反応の鋳型としての機能を果たし得る。読み取りは、蛍光またはELISAに基づいてよい。信号により、上首尾の増幅、ハイブリダイゼーション、および一塩基伸長に起因して、挿入断片/隣接配列が存在することが示される。
【0093】
別の方法は、Winge(Innov. Pharma. Tech. 00巻:18〜24頁、2000年)に記載されているようなパイロシークエンス技法である。この方法では近接するゲノムDNAと挿入DNAの接合部とオーバーラップするオリゴヌクレオチドを設計する。このオリゴヌクレオチドを、対象の領域(挿入配列内の1つのプライマーおよび隣接ゲノム配列内の1つのプライマー)由来の一本鎖のPCR産物とハイブリダイズさせ、DNAポリメラーゼ、ATP、スルフリラーゼ、ルシフェラーゼ、アピラーゼ、アデノシン5’−ホスホ硫酸およびルシフェリンの存在下でインキュベートする。dNTPを個々に加え、それが取り込まれた結果、測定される光信号がもたらされる。光信号により、増幅、ハイブリダイゼーション、および単一塩基または多塩基の伸長が上首尾であり、したがって導入遺伝子挿入断片/隣接配列が存在することが示される。
【0094】
蛍光偏光は、本発明のアンプリコンを検出するために使用することができる別の方法である。この方法に従って、オリゴヌクレオチドを、ゲノムの隣接DNAと挿入DNAの接合部とオーバーラップするように設計する。このオリゴヌクレオチドを、対象の領域(挿入DNA内の1つのプライマーおよび隣接ゲノムDNA配列内の1つのプライマー)由来の一本鎖のPCR産物とハイブリダイズさせ、DNAポリメラーゼおよび蛍光標識したddNTPの存在下でインキュベートする。一塩基伸長により、ddNTPが取り込まれる。取り込みは、蛍光光度計を使用して偏光の変化として測定することができる。偏光の変化は、増幅、ハイブリダイゼーション、および一塩基伸長の成功によって、導入遺伝子挿入断片/隣接配列が存在することを示す。
【0095】
TAQMAN(PE Applied Biosystem、Foster City、Calif.)は、DNA配列の存在を検出し、定量する方法である。簡単に述べると、ゲノムの隣接と挿入DNAの接合部とオーバーラップするFRETオリゴヌクレオチドプローブを設計する。FRETプローブおよびPCRプライマー(挿入DNA配列内の1つのプライマーおよび隣接ゲノム配列内の1つのプライマー)を、耐熱性ポリメラーゼおよびdNTPの存在下でサイクルにかける。特異的な増幅の間に、Taq DNAポリメラーゼにより、FRETプローブ上のクエンチング部分から蛍光部分が離れて除かれ、放出される。蛍光シグナルは、増幅およびハイブリダイゼーションの成功によって、隣接配列/導入遺伝子挿入断片配列が存在することを示す。
【0096】
配列の検出において使用するための分子ビーコンが記載されている。簡単に述べると、隣接ゲノムDNAと挿入DNAの接合部とオーバーラップするFRETオリゴヌクレオチドプローブを設計する。FRETプローブの独特の構造により、蛍光部分およびクエンチング部分が極めて近傍に保たれる二次構造が含有される。FRETプローブおよびPCRプライマー(挿入DNA配列内の1つのプライマーおよび隣接ゲノム配列内の1つのプライマー)を、耐熱性ポリメラーゼおよびdNTPの存在下でサイクルにかける。PCR増幅が上手くいった後、FRETプローブが標的配列にハイブリダイズすることにより、プローブの二次構造が除去され、蛍光部分およびクエンチング部分が空間的に分離される。蛍光シグナルが結果として生じる。蛍光シグナルは、増幅およびハイブリダイゼーションの成功によって、隣接ゲノム配列/導入遺伝子挿入断片配列が存在することを示す。
【0097】
ダイズゲノム内の挿入するために優れた位置を開示し、本発明は、この遺伝子位置の概ね近くに少なくとも1つの非aad12挿入断片を含むダイズ種子および/またはダイズ植物も含む。1つの選択肢は、本明細書において例示されているaad−12挿入断片を異なる挿入断片で置換することである。これらの一般的な点については、例えば、本発明に従って、標的化相同組換えを使用することができる。この種類の技術は、例えば、WO03/080809A2および対応する公開された米国特許出願(U.S.2003/0232410)の主題である。したがって、本発明は、本明細書において同定される隣接配列の全てまたは認識できる部分(例えば、配列番号1の残基1〜2730および残基9122〜10,212)が隣接する異種挿入断片(多コピーのaad−12の代わりにまたはそれと一緒に)を含む植物および植物細胞を包含する。aad−12遺伝子の1つの追加的なコピー(または複数の追加的なコピー)も、この/これらのように挿入の標的とすることができる。
【0098】
植物細胞の特定の染色体上の部位内に、相同組換えによってポリヌクレオチド配列を組み込むための方法は、当技術分野の範囲内で記載されている。例えば、米国特許出願公開第2009/0111188A1号に記載の部位特異的な組み込みでは、ドナーポリヌクレオチド配列の染色体上の標的への導入を媒介するためのリコンビナーゼまたはインテグラーゼの使用が記載されている。さらに、国際的な特許出願第WO2008/021207号には、1つまたは複数のドナーポリヌクレオチド配列をゲノムの特定の位置内に組み込むためのジンクフィンガー媒介相同組換えが記載されている。米国特許第6,720,475号に記載のFLP/FRTまたは米国特許第5,658,772号に記載のCRE/LOXなどのリコンビナーゼの使用を利用して、ポリヌクレオチド配列を特定の染色体上の部位に組み込むことができる。最終的に、ドナーポリヌクレオチドを特定の染色体上の位置に標的化するためのメガヌクレアーゼの使用が、Puchtaら、PNAS USA 93巻(1996年)5055〜5060頁に記載されている。
【0099】
植物細胞内に部位特異的に組み込むための他の種々の方法は、一般に、公知であり、適用可能である(Kumarら、Trands in Plant Sci. 6巻(4号)(2001年) 155〜159頁)。さらに、いくつかの原核生物および下等真核生物において同定された部位特異的な組換え系を植物における使用に適用することができる。そのような系の例としては、これらに限定されないが、酵母であるジゴサッカロマイセス・ロキシー(Zygosaccharomyces rouxii)のpSR1プラスミド由来のR/RSリコンビナーゼ系(Arakiら(1985年)J. Mol. Biol. 182巻:191〜203頁)、およびファージMuのGin/gix系(MaeserおよびKahlmann(1991年)Mol. Gen. Genet. 230巻:170〜176頁)が挙げられる。
【0100】
本発明の一部の実施形態では、既存のトランスジェニックイベントに近接して新しい導入遺伝子(複数可)を組み込む、または積み重ねることが望ましい場合がある。トランスジェニックイベントは、単一の挿入部位、正常なメンデル分離および安定発現、ならびに多数の環境区域における、およびそれ全てにわたる除草剤耐性および農業生産力を含めた効力の優れた組合せなどの独特の特性に基づいて選抜された好ましいゲノム遺伝子座と考えることができる。新しく組み込まれた導入遺伝子は、既存の形質転換体の導入遺伝子の発現特性を維持すべきである。さらに、新しく組み込まれたイベントのゲノム隣接配列および染色体上の位置はすでに同定されているので、新しく組み込まれたイベントを検出し、確認するためのアッセイの開発は克服されるであろう。最終的に、既存の導入遺伝子に連結された特定の染色体上の位置に新しい導入遺伝子を組み込むことにより、従来の育種方法を使用した有性異系交雑による導入遺伝子の他の遺伝的背景への遺伝子移入が促進される。
【0101】
本発明の一部の実施形態では、トランスジェニックイベントからポリヌクレオチド配列を削除することが望ましい場合がある。例えば、米国特許仮出願第61/297,628号に記載の導入遺伝子の削除では、染色体に組み込まれたトランスジェニックイベントから、遺伝子発現カセットからなるポリヌクレオチド配列を除去するためのジンクフィンガーヌクレアーゼの使用が記載されている。除去されるポリヌクレオチド配列は、選抜マーカーであってよい。ポリヌクレオチド配列を削除および除去したら、改変されたトランスジェニックイベントを、ポリヌクレオチド配列を挿入することによって再標的化することができる。ポリヌクレオチド配列を削除し、その後、改変されたトランスジェニックイベントを再標的化することにより、選抜マーカーの再使用、または特定の遺伝子が発現することによって生じる、植物のトランスクリプトームに対する意図されたものではない変化を克服する能力などの利点がもたらされる。
【0102】
本発明は、本明細書において、異種核酸を挿入するために優れた、ダイズゲノム内の第4染色体上の特定の部位を開示している。第4染色体上のターゲティング部位の位置を同定することにおいて有用な5’分子マーカー、3’分子マーカー、5’隣接配列、および3’隣接配列も開示されている。したがって、本発明は、対象の異種核酸をこの予め確立した標的部位に、またはこの標的部位の付近に導入するための方法を提供する。本発明は、開示されている標的部位またはそのような部位の概ね近くに挿入された任意の異種ヌクレオチド配列を含むダイズ種子および/またはダイズ植物も包含する。そのような標的化組み込みを実現するための1つの選択肢は、本明細書において例示されているpat発現カセットの代わりに異なる挿入断片を削除することおよび/または置換することである。この一般的な点について、例えば、またこれらに限定することなく、本発明に従って標的化相同組換えを使用することができる。
【0103】
本明細書で使用される場合、遺伝子、イベントまたは形質を「積み重ねること(stacking)」は、所望の形質を1つのトランスジェニック系統内に組み合わせることである。植物育種家は、それぞれが所望の形質を有する親同士の交雑を行い、次にこれらの所望の形質を両方とも有する子孫を同定することによってトランスジェニック形質を積み重ねる。遺伝子を積み重ねるための別の方法は、形質転換の間に、2種以上の遺伝子を同時に植物の細胞核に伝達することによる。遺伝子を積み重ねるための別の方法は、トランスジェニック植物を、対象とする別の遺伝子を用いて再形質転換することによる。例えば、遺伝子を積み重ねることを用いて、例えば、2種以上の異なる昆虫形質、昆虫抵抗性形質(複数可)と病害抵抗性形質(複数可)、2種以上の除草剤への抵抗性形質、および/または昆虫抵抗性形質(複数可)と除草剤抵抗性の形質(複数可)を含めた2種以上の異なる形質を組み合わせることができる。対象の遺伝子に加えて選抜マーカーを使用することも、遺伝子を積み重ねることと考えることができる。
【0104】
「相同組換え」とは、2種のヌクレオチド配列が相互作用して(組み換えられて)新しい組換えDNA配列を形成し得る類似したヌクレオチド配列を含有する対応する部位を有するヌクレオチド配列の任意の対の間の反応を指す。類似したヌクレオチド配列の部位は、本明細書ではそれぞれが「相同配列」と称される。一般に、相同組換えの頻度は、相同配列の長さが増加するにつれて増加する。したがって、相同組換えは、同一には満たない2つのヌクレオチド配列間で起こり得るが、組換え頻度(または効率)は、2つの配列間の相違が増加するにつれて減退する。組換えは、ドナー分子および標的分子のそれぞれの1つの相同配列を使用して実現することができ、それによって「単一乗換え」組換え産物を生成することができる。あるいは、標的ヌクレオチド配列およびドナーヌクレオチド配列のそれぞれに2つの相同配列を置くことができる。ドナー上の2つの相同配列と標的上の2つの相同配列との間の組換えにより、「2重乗換え」組換え産物が生成する。ドナー分子上の相同配列が、操作される配列(例えば、対象の配列)に隣接する場合、標的分子との2重乗換え組換えにより、対象の配列が元々標的分子上の相同配列の間にあったDNA配列と交換された組換え産物がもたらされる。2重乗換え組換えイベントによって標的とドナーとの間でDNA配列を交換することは、「配列交換」と称される。
【0105】
本明細書において言及または引用された全ての特許、特許出願、仮出願、および刊行物は、それらが本明細書の明確な教示と相反しない限りは、それらの全体が参照により本明細書に組み込まれる。
【0106】
以下の実施例は、本発明を実施するための手順を例示するため、および本発明のある特定の好ましい実施形態を実証するために含まれる。これらの実施例は、限定するものと解釈されるべきではない。当業者には、以下の実施例に開示されている技法は、それを実施するための好ましい方式を例示するために使用される特定の手法を表していることが理解されたい。しかし、当業者には、本開示に照らして、本発明の精神および範囲から逸脱することなく、なお同様または類似の結果を得ながら、これらの特定の実施形態において多くの変更を行うことができることが理解されたい。別段の指定のない限り、全ての百分率は重量により、特に断りのない限り、全ての溶媒混合物の割合は体積による。
【0107】
別段の指定のない限り、以下の略語が使用されている。
AAD−12 アリールオキシアルカノエートジオキシゲナーゼ−1
bp 塩基対
℃ 摂氏温度
DNA デオキシリボ核酸
DIG ジゴキシゲニン
EDTA エチレンジアミン四酢酸
kb キロベース
μg マイクログラム
μL マイクロリットル
mL ミリリットル
M モル質量
OLP オーバーラッププローブ
PCR ポリメラーゼ連鎖反応
PTU 植物転写単位
SDS ドデシル硫酸ナトリウム
SOP 標準操作手順
SSC 塩化ナトリウムとクエン酸ナトリウムとの混合物を含有する緩衝溶液、pH7.0
TBE トリス塩基、ホウ酸およびEDTAの混合物を含有する緩衝溶液、pH8.3
V ボルト
[実施例]
【実施例1】
【0108】
aad−12ダイズイベントDAS−68416−4の形質転換および選抜
トランスジェニックダイズ(Glycine max)イベントDAS−68416−4を、ダイズ子葉節の外植片をアグロバクテリウム媒介形質転換によって生成した。T鎖DNA領域内に選抜マーカー(pat)および対象の遺伝子(aad−12)と一緒にバイナリーベクターpDAB4468(図1)を保有する武装解除したアグロバクテリウム株EHA101(Hoodら、2006年)を使用して形質転換を開始した。
【0109】
Zengら(Zengら、2004年)の手順を改変して用いてアグロバクテリウム媒介形質転換を行った。簡単に述べると、ダイズ種子(Maverick栽培品種)を基本培地上で発芽させ、子葉節を単離し、アグロバクテリウムに感染させた。アグロバクテリウムを除去するために、苗条誘導(shoot initiation)培地、苗条伸長培地、および発根培地にセフォタキシム、チメンチンおよびバンコマイシンを補充した。グルホシネートによる選択を使用して、形質転換されていない苗条の生長を阻害した。選択された苗条を、根を発生させるために発根培地に移し、次に、小植物を順化させるために土壌混合物に移した。
【0110】
選択された小植物の頂小葉にグルホシネートを塗布して推定形質転換体についてスクリーニングした。スクリーニングされた小植物を温室に移して、気候順化させ、次に葉にグルホシネートを塗布して耐性を再確認し、推定形質転換体であるとみなした。スクリーニングされた植物の試料を採取し、選択マーカー遺伝子および/または対象の遺伝子を確認するための分子解析を行った。T0植物を温室内で自家受精させてT1種子を生じさせた。
【0111】
T1植物を戻し交雑し、優良な生殖質(Maverick)に遺伝子移入した。このイベント、ダイズイベントDAS−68416−4は、独立した形質転換分離株から生成した。イベントをその独特の特性、例えば、単一の挿入部位、正常なメンデル分離および安定発現、ならびに広範な遺伝子型背景および多数の環境全ての区域にわたる除草剤耐性および農業生産力を含めた効力の優れた組合せに基づいて選抜した。以下の実施例は、ダイズイベントDAS−68416−4を特徴付けるために使用したデータを含有する。
【実施例2】
【0112】
ダイズイベントDAS−68416−4のサザンブロットによる特徴付け
サザンブロット分析を使用して、ダイズイベントDAS−68418−4の組み込みパターンを確立した。これらの実験により、aad−12導入遺伝子のダイズゲノム内への組み込みおよびその完全性を実証するデータが生成した。ダイズイベントDAS−68418−4は、プラスミドpDAB4468由来のaad−12PTUの単一コピーを含有する全長の、単純な組み込みイベントであると特徴付けられた。
【0113】
サザンブロットのデータにより、pDAB4468T鎖断片がダイズイベントDAS−68418−4のゲノムに挿入されたことが示唆された。詳細なサザンブロット分析を、pDAB4468のT鎖組み込み領域に含有されるaad−12遺伝子に特異的なプローブ、およびプラスミド内に位置する切断部位を有し、プラスミドまたはプラスミドとダイズのゲノムDNAの接合部にわたる断片(境界断片)内にハイブリダイズ断片を生じる説明的な制限酵素を使用して行った。サザンハイブリダイゼーションによって示された制限酵素とプローブの組合せについての分子量は、このイベントに独特であり、その同定パターンを確立した。これらの分析により、aad−12PTUの再配置を伴わずにプラスミド断片がダイズのゲノムDNAに挿入されたことも示された。
【0114】
実施例2.1
ダイズの葉の試料の収集およびゲノムDNA(gDNA)の単離
ダイズイベントDAS−68416−4を含有する個々のダイズ植物から回収した葉組織からゲノムDNAを抽出した。さらに、aad−12遺伝子が存在しない物質系統の代表である遺伝的背景を含有する従来のダイズ植物であるMaverickからgDNAを単離した。
【0115】
標準のCTAB法の後、凍結乾燥した葉組織から個々のゲノムDNAを抽出した。抽出した後、DNAを、Pico Green試薬(Invitrogen、Carlsbad、CA)を使用して分光蛍光分析によって定量した。次いで、DNAをアガロースゲル上で可視化してPico Green分析からの値を確認し、DNAの質を決定した。
【0116】
実施例2.2
DNAの消化および分離
ダイズイベントDAS−68416−4をサザンブロットによって分子キャラクタリゼーションするために、10マイクログラム(10μg)のゲノムDNAを消化した。ダイズイベントDAS−68416−4および非トランスジェニックダイズ系統であるMaverick由来のゲノムDNAを、各DNA試料にDNA1μg当たりおよそ5ユニットの選択された制限酵素および対応する反応緩衝液を加えることによって消化した。各試料を、およそ37℃で一晩インキュベートした。消化のために、制限酵素SpeI、KpnI、およびPacIを個々に使用した(New EnglandBioLab、Ipswich、MA)。さらに、陽性ハイブリダイゼーション対照試料を、プラスミドDNAであるpDAB4468を非トランスジェニックダイズ品種であるMaverick由来のゲノムDNAと混ぜ合わせることによって調製した。プラスミドDNA/ゲノムDNA反応混液を、試験試料の場合と同じ手順および制限酵素を用いて消化した。消化物を一晩インキュベートした後、NaClを、最終濃度が0.1Mになるまで加え、消化されたDNA試料を、イソプロパノールを用いて沈殿させた。沈殿したDNAペレットを、1×ローディング緩衝液(0.1%ブロモフェノールブルー、100mMのEDTA、50%グリセロール、10mMのトリス、pH7.5)20μlに再懸濁させた。次いで、DNA試料および分子サイズマーカーを、0.4×TAE緩衝液(Fisher Scientific、Pittsburgh、PA)を用いた0.85%アガロースゲルにより、35ボルトでおよそ18〜22時間、電気泳動して断片の分離を実現した。ゲルを臭化エチジウム(Invitrogen、Carlsbad、CA)で染色し、DNAを紫外線(UV)の下で可視化した
【0117】
実施例2.3
サザン転写およびメンブレン処理
サザンブロット分析をSeversonら(1997年)に記載されている通り実施した。DNA断片を電気泳動によって分離し、UV光の下で可視化した後、ゲルを変性溶液(150mMのNaOH、3mMのEDTA)中におよそ20分間浸し、次いで、中和溶液(150mMのNaPO4、pH7.8)に移し少なくとも20分間置いた。ナイロンメンブレン(Roche Diagnostic、Indianapoli、IN)へのサザン転写を、転写緩衝液(25mMのピロリン酸ナトリウム、pH10)を伴うウィッキング系を用いて一晩実施した。転写後、メンブレンを65℃で約2時間、加熱乾燥することによってDNAをメンブレンに結合させた。このプロセスにより、ハイブリダイゼーションのためのサザンブロットメンブレンの準備ができた。
【0118】
実施例2.4
DNAプローブの標識化およびハイブリダイゼーション
標識したプローブを使用してナイロンメンブレンに結合したDNA断片を検出した。この実験のために使用したプローブは、プラスミドpDAB4468の特定のヌクレオチド領域に対するプライマーを使用したPCR増幅によって生成した。増幅されたPCR断片をアガロースゲルから単離し、精製し、ハイブリダイゼーションプローブのための鋳型として使用した。サザンブロットハイブリダイゼーションプローブを、GE Healthcare READY−TO−GO(商標)DNA Labeling Beads(GE Healthcare、Piscataway、NJ)を製造者の説明書に従って使用したランダムプライミングによってα32Pに特異的なヌクレオチドで標識し、PROBEQUANT(商標)G−50マイクロカラム(Amersham/Pharmacia、Piscataway、New Jersey、USA)によって精製した。この実験のために使用したプローブが記載された表は、表2に記載されている。プレハイブリダイゼーションを、ハイブリダイゼーション緩衝液(Sigma−Aldrich、St.Loui、MO)を使用して65℃で4時間行った。次いで、プレハイブリダイゼーション溶液をデカントし、水で5分間煮沸することによって予め変性させた所望の量の特異的なプローブを含有するハイブリダイゼーション溶液と交換した。ハイブリダイゼーション/プローブ混合物をナイロンメンブレンと一緒に65℃で一晩インキュベートした。
【0119】
ハイブリダイゼーション後、メンブレンを65oC、洗浄緩衝液(10mMのリン酸ナトリウム、2.5mMのピロリン酸ナトリウム、0.5mMのEDTA、0.1%SDS、リン酸を用いて調整pH7.8に調整した)で20分間、3回洗浄した。洗浄したフィルターをオートラジオグラフィーのためにPhosphorimagerスクリーンに曝露させ、画像をスキャンした。検出されたバンドの数およびサイズを、プローブについて実証した。さらに、サザンブロットにおいて分子量マーカーを使用してハイブリダイズ断片サイズを決定した。
【0120】
【表2】
【0121】
実施例2.5
サザンブロットの結果
aad−12PTUの公知の制限酵素部位に基づく、特定の消化物およびプローブに関して予測された断片サイズおよび観察された断片サイズが表3に示されている。予測された断片サイズは、pDAB4468のプラスミドマップ(図1)に基づき、観察された断片サイズは、これらの分析から概算したものであり、α32Pで標識されたMolecular Weight Marker II断片の指示サイズに基づく。
【0122】
これらの消化およびハイブリダイゼーションから2種類の断片を同定した:公知の酵素部位がプローブ領域に隣接し、aad−12PTUの挿入領域内に完全に含有されている内部の断片、および公知の酵素部位がプローブ領域の一端に位置し、第2の部位がダイズゲノム内にあると予測される境界断片。ほとんどの場合、DNA断片の組み込み部位はそれぞれのイベントに独特であるので、境界断片のサイズはイベントごとに変動する。境界断片により、組み込まれたDNAに対して制限酵素部位を位置づける手段およびDNA挿入物の数を評価する手段がもたらされる。イベントDAS−68416−4を含有するダイズの3世代に対して完了したサザンブロット分析により、プラスミドpDAB4468由来の低コピー、インタクトなaad−12PTUが、ダイズイベントDAS−68416−4のダイズゲノムに挿入されたことを示唆するデータがもたらされた。
【0123】
【表3】
【0124】
制限酵素SpeIおよびKpnIは、プラスミドpDAB4468に独特の制限部位を含有する。続いて、これらの酵素を、ダイズイベントDAS−68416−4におけるaad−12遺伝子挿入断片を特徴付けるために選択した。5,436bpより大きいまたは5,383bpより大きい境界断片は、それぞれSpeIおよびKpnIで消化した後、aad−12遺伝子プローブとハイブリダイズすることが予測された(表3)。それぞれSpeIおよびKpnIを使用したとき、約12,000bpおよび約16,000bpの単一のaad−12ハイブリダイゼーションバンドが観察された。プローブがこのサイズのバンドにハイブリダイズすることにより、ダイズイベントDAS−68416−4のダイズゲノム内にaad−12遺伝子に対する単一の挿入部位が存在することが示唆される。制限酵素PacIを、aad−12植物転写単位(PTU、プロモーター/遺伝子/ターミネーター)を含有する2,904bpの断片を放出させるために選択した(表3)。予測された2,904bpの断片は、PacI消化した後、aad−12遺伝子プローブを用いて観察された。全3種の酵素を用いてDAS−68416−4試料を消化し、その後aad−12遺伝子プローブとハイブリダイズさせることで得られた結果により、プラスミドpDAB4468由来のインタクトなaad−12PTUの単一コピーがダイズイベントDAS−68416−4のダイズゲノムに挿入されたことが示された。
【0125】
実施例2.6
骨格配列が存在しないこと
ダイズイベントDAS−68416−4にスペクチノマイシン抵抗性遺伝子が存在すること、または存在しないことをモニターするために、多重PCRアッセイを実施した。実験は、スペクチノマイシン抵抗性遺伝子コード配列の5つの異なる領域および内部標準として内在性のダイズレクチン遺伝子(GenBank ID No:K00821 M30884)配列内の407bp領域を検出するために設計した。さらに、以下の対照を含めた:(i)スペクチノマイシン抵抗性遺伝子を保有するプラスミドDNAを付加した形質転換されていないダイズのゲノムDNAを用いた陽性対照;(ii)形質転換されていないダイズ、Maverick由来のゲノムDNAを用いた陰性対照;および(iii)ゲノムDNAを有さないブランク。
【0126】
ダイズ由来のゲノムDNAを、CTAB法を使用して単離し、Pico Greenを使用して定量した。各試料のDNA濃度を100ng/μlに正規化した。PCR反応を、スペクチノマイシン抵抗性遺伝子コード配列およびレクチン遺伝子配列に特異的なプライマー配列を使用して実施した。この反応物を、試料当たりPCR産物20μlを2%Eゲルにローディングすることによって分析した。
【0127】
Eゲル上に多数のアンプリコンが存在すること(100bp、150bp、および407bpのバンド)は、ダイズイベントDAS−68416−4がスペクチノマイシン抵抗性遺伝子コード配列を含有することを示すことになる(スペクチノマイシン抵抗性遺伝子コード配列について、100bpおよび150bpの増幅されたバンドが予測される)。内部標準であるレクチン遺伝子配列に対応する407bpのアンプリコンのみが存在する場合、そのことは、ダイズイベントDAS−68416−4がスペクチノマイシン抵抗性コード配列を含有しないことを示す。
【0128】
ダイズイベントDAS−68416−4由来のDNA試料では、100bpまたは150bpの断片は増幅されなかった。407bp断片のみが増幅された。しかし、スペクチノマイシン抵抗性遺伝子を保有するプラスミドDNAをダイズのゲノムDNAに加えた陽性対照には100bp、150bp、および407bpの増幅断片が存在した。形質転換されていないダイズ由来のDNAを含有する陰性対照では、単一の407bp断片が増幅された。最終的に、いかなるゲノムDNAも含有しない反応物からは、いかなる増幅断片も産生されなかった。そのように、ダイズイベントDAS−68416−4においてスペクチノマイシン抵抗性コード配列は検出されなかった。
【実施例3】
【0129】
ダイズイベントDAS−68416−4の挿入断片および隣接境界領域におけるDNA配列のクローニングおよび特徴付け
ゲノムの挿入部位を特徴付け、説明するために、ダイズイベントDAS−68416−4のT鎖DNA挿入断片および隣接ゲノムDNA境界領域の配列を決定した。2,730bpの5’隣接境界配列、1,091bpの3’隣接境界配列、および6,391bpのT鎖挿入断片を含む、全部で10,212bpのダイズイベントDAS−68416−4ゲノム配列(配列番号1)を確認した。配列解析により、ダイズイベントDAS−68416−4が、予測されたT鎖挿入断片から配列が変動することなく、MARエレメント、aad−12発現カセット、およびpat発現カセットを含有するインタクトな導入遺伝子の単一コピーを含有することが検証された。
【0130】
ダイズイベントDAS−68416−4の挿入断片および境界の配列に基づくPCR増幅により、境界領域がダイズ起源であること、およびダイズイベントDAS−68416−4をイベント特異的に同定するために接合領域を使用することができることが検証された。隣接境界配列を含めた接合領域にわたる配列を分析することにより、T鎖の挿入によって生じるいかなる新規のオープンリーディングフレーム(ORF>=150 コドン)も同定されなかった。さらに、T鎖の挿入部位を、非トランスジェニックダイズのゲノムから同定された隣接境界配列の領域に対応するゲノムの断片をクローニングすることによって特徴付けた。ダイズイベントDAS−68416−4を野生型ゲノム配列と比較することにより、元の遺伝子座からの55bpの欠失およびイベントの3’組み込み接合部における9bpの挿入が明らかになった。全体的に、ダイズイベントDAS−68416−4の挿入断片および境界配列を特徴付けることにより、T鎖の単一の、インタクトなコピーがダイズゲノム内に存在することが示された。
【0131】
実施例3.1
ゲノムDNAの抽出および定量
凍結乾燥した葉組織または粉砕したての葉組織から、改変したCTAB法を使用してゲノムDNAを抽出した。ゲノムDNAを抽出した後、DNA試料を1×TE(10mMのトリス、pH8.0、1mMのEDTA)(Fluka、Sigma、St.Loui、MO)に溶解させ、Pico Green法を製造者(Molecular Probe、Eugene、OR)の説明書に従って使用して定量した。PCR分析のために、DNA試料を分子生物学グレードの水(5PRIME、Gaithersburg、MD)で希釈して、濃度を10〜100ng/μLにした。
【0132】
実施例3.2
PCRプライマー
表4には、ダイズイベントDAS−68416−4のDNA挿入断片および隣接境界領域をクローニングし、確認するために使用したプライマー配列が、図2で記した位置および説明とともに列挙されている。全てのプライマーは、Integrated DNA Technologie、Inc.(Coralville、IA)によって合成された。プライマーを、原液については水(5PRIME、Gaithersburg、MD)に溶解させて濃度を100μMにし、検量線用溶液については10μMの濃度まで水で希釈した。
【0133】
【表4】
【0134】
【表5】
【0135】
【表6】
【0136】
【表7】
【0137】
実施例3.3
ゲノムウォーキング
GENOMEWALKER(商標)Universal Kit(Clontech Laboratorie、Inc.、Mountain View、CA)を製造者の説明書に従って使用して、ダイズイベントDAS−68416−4についてpDAB4468T鎖挿入断片の5’隣接境界領域および3’隣接境界領域をクローニングした。ダイズイベントDAS−68416−4由来のゲノムDNAおよそ2μgをEcoRVおよびPvuIIを用いて一晩消化した(図2)。DNA消化物を、DNA Clean & Concentrator(商標)−25(Zymo Research、Orange、CA)を使用して精製し、その後GENOMEWALKER(商標)アダプターとライゲーションしてGENOMEWALKER(商標)ライブラリーを構築した。GENOMEWALKER(商標)ライブラリーのそれぞれを、アダプタープライマーAP1(このキットで提供される)および構築物特異的プライマーであるES_LEnd03またはES_PATEnd03(表4)を用いた一次PCR増幅のためのDNA鋳型として使用した。次に、一次PCR反応物1:25希釈物1マイクロリットル(1μL)を、このキットで提供されるネステッドアダプタープライマーAP2およびネステッド構築物特異的プライマーであるES_LEnd04またはES_PATEnd04(表4、7および図2)を用いた二次PCR増幅ための鋳型として使用した。
【0138】
実施例3.4
従来のPCR
標準のPCRを使用してダイズイベントDAS−68416−4の挿入断片および境界配列をクローニングし、確認した。従来のPCR増幅ために、TaKaRa LA TAQ(商標)(Takara Bio Inc、Shiga、Japan)、HOTSTARTAQ(商標)DNAポリメラーゼ(Qiagen、Valencia、CA)、HIGH FIDELITY(商標)PCRキット(Roche Diagnostic、Inc)、またはEASY−A(商標)高忠実度ポリメラーゼキット(Stratagene、LaJolla、CA)を製造者の推奨手順に従って使用した。特定のPCR条件およびアンプリコンの説明が表5、6、および7に列挙されている。
【0139】
実施例3.5
PCR産物の検出、精製、PCR産物のサブクローニング、および配列決定
PCR産物を1.2%または2%E−GEL(登録商標)(Invitrogen、Carlsbad、CA)を製品の説明書に従って使用して電気泳動によって検査した。DNAマーカーと比較することによって断片サイズを推定した。必要であれば、QIAquick Gel Extraction Kit(Qiagen、Valencia、CA)を使用して1%アガロースゲルから断片を切り取って1×TBE(89mMのトリス−ホウ酸、2mMのEDTA、pH8.3)に入れ、臭化エチジウムで染色することによってPCR断片を精製した。
【0140】
PCR断片を、配列決定するためにTOPO TA CLONING(登録商標)KIT(Invitrogen、Carlsbad、CA)を製品の説明書に従って使用してPCR(登録商標)4−TOPO(登録商標)ベクターにサブクローニングした。詳細には、製造者の説明書に従って、TOPO(登録商標)クローニング反応物2〜5マイクロリットルをOne Shot化学的コンピテントなTOP10細胞に導入し、形質転換した。クローニングされた断片を、プラスミドDNAをミニプレップした後に(QIAprep Spin Miniprep Kit、Qiagen、CA)、EcoRIで制限消化することによって、またはT3プライマーおよびT7プライマーを使用したダイレクトコロニーPCRによって検証した。次に、選択されたコロニーのプラスミドDNAまたはグリセロールストックを配列決定するために外部委託した。
【0141】
サブクローニングした後、予測されたDNA断片がクローニングされたことを確認するために、推定標的PCR産物について最初に配列決定した。予測されたDNA断片を含有するコロニーを選択して、プライマーウォーキングによる二本鎖の全長についての配列決定を完了した。配列決定は全て、Cogenics(Houston、TX)によって行われた。
【0142】
挿入断片および境界配列の最終的な組立てを、SEQUENCHER(登録商標)ソフトウェア(Gene Codes Corporation、Ann Arbor、MI)を使用して完了した。ダイズイベントDAS−68416−4の挿入断片およびその隣接境界配列のアノテーションを、ベクターNTI(バージョン10および11、Invitrogen、Carlsbad、CA)を使用して実施した。
【0143】
GenBankの非冗長ヌクレオチドデータベースに対してBLASTプログラムを使用して相同性検索を実施した。Vector NTI(バージョン11、Invitrogen)を使用したオープンリーディングフレーム(ORF)解析を実施して、ダイズイベントDAS−68416−4の完全な挿入断片および隣接境界配列、ならびに野生型Maverickダイズ系統の元の遺伝子座におけるORF(≧150コドン)を同定した。
【0144】
実施例3.6
5’末端境界配列
ダイズイベントDAS−68416−4のGENOMEWALKER(商標)ライブラリーのそれぞれから、導入遺伝子の5’末端に対して特異的なネステッドプライマーセットを使用してDNA断片を増幅した。EcoRV GENOMEWALKER(商標)ライブラリーから約1.8kbの断片、およびPvuII GENOMEWALKER(商標)ライブラリーから約3kbの断片が観察された。これらの断片を、PCR(登録商標)4−TOPO(登録商標)ベクターにクローニングした。ヌクレオチド配列のデータを生成するために、末端配列決定用にそれぞれのライブラリーについて5つのコロニーを無作為に選定した。プライマーウォーキングによって完全な配列を得るために、両方のPCRプライマーの配列を含有するコロニーを選択した。配列解析により、ダイズイベントDAS−68416−4のEcoRV GENOMEWALKER(商標)ライブラリーから増幅されたクローンが1,744bpのDNA断片を含有すること、およびダイズイベントDAS−68416−4のPvuII GENOMEWALKER(商標)ライブラリーから増幅されたクローンが3,047bpのDNA断片を含有することが明らかになった。配列解析により、EcoRV GENOMEWALKER(商標)ライブラリーから得られたDNA断片とPvuII GENOMEWALKER(商標)ライブラリークローンから得られたDNA断片が、プライマーES_LEnd04とEcoRV部位との間の領域においてオーバーラップしていることが明らかになった。これらのDNA断片は全て、導入遺伝子内にT鎖境界Bの5’末端接合部を含有し、これは、それらが、ダイズイベントDAS−68416−4の5’末端の導入遺伝子挿入断片およびその隣接境界の同じ領域から増幅されたことを示している。得られた2,730bpのダイズのゲノム配列は、GenBankにある配列との有意な相同性を有さないことが見いだされた。
【0145】
実施例3.7
3’末端境界配列
ダイズイベントDAS−68416−4のEcoRV GENOMEWALKER(商標)ライブラリーから、導入遺伝子の3’末端に対して特異的なネステッドプライマーセットを使用して、サイズが約1.3kbであるDNA断片を増幅した。次に、このDNA断片をPCR(登録商標)4−TOPO(登録商標)ベクターにクローニングした。末端配列決定のために5つのコロニーを無作為に選定した。5つのクローンは全て、プライマーAP2およびプライマーES_PATEnd04の両方の配列を含有した。これらのクローンについて完全に配列決定することにより、1,359bpのコンセンサスDNA断片が得られた。配列解析することにより、1,359bpの断片が、T鎖境界Aの3’末端領域由来の268bpの断片およびダイズのゲノムDNA由来の1,091bpの断片で構成されることが開示された。BLAST検索により、この1,091bpのダイズDNA配列とGenBankにある配列との間に有意な相同性は同定されなかった。
【0146】
実施例3.8
DNA挿入断片および接合部配列
以前に記載されているようなPCRに基づく方法を使用して、ダイズイベントDAS−68416−4からDNA挿入断片および隣接境界領域をクローニングした。5’隣接境界および3’隣接境界配列および予測された導入遺伝子配列を使用して、表6に列挙されているPCRプライマーを設計した。全部で、4つのオーバーラップDNA断片(978bpのアンプリコン1、2,414bpのアンプリコン2、1,834bpのアンプリコン3、および1,705bpのアンプリコン4)をクローニングし、配列決定した(図3)。挿入断片および隣接境界配列の全体を、4つの断片の間のオーバーラップ配列に基づいてアセンブリした。最終的にアセンブリした配列を解析することにより、pDAB4468の導入遺伝子に由来する6,391bpの断片が存在することが確認され、また、プラスミドpDAB4468から予測された配列と比較して、挿入DNA配列の塩基の変化は見られなかった。
【0147】
実施例3.9
ダイズのゲノム配列の確認
ダイズゲノム内のダイズイベントDAS−68416−4導入遺伝子の挿入部位を確認するために、異なるプライマー対を用いてPCRを行った(図4および表5)。ダイズイベントDAS−68416−4由来のゲノムDNAおよび他のトランスジェニックダイズ系統または非トランスジェニックダイズ系統由来のゲノムDNAを鋳型として使用した。したがって、得られた5’末端境界配列が正確であるかどうかを確認するために、aad−12遺伝子から5’末端境界配列にわたるDNAセグメントを増幅するためにaad−12特異的な2種のプライマー、例えば、AIILEnd05およびAIILEnd06、ならびに、16LEndG01および16LEndG02と称される5’末端境界配列に従って設計した2種のプライマーを使用した。同様に、クローニングされた3’末端境界配列を確認するために、pat特異的なプライマー、例えばPAT−End06、および16PATG01および16PATG02と称される、3’末端境界配列に従って設計した2種のプライマーを使用して、pat遺伝子から3’末端境界配列にわたるDNAセグメントを増幅した。予測されたサイズを有するDNA断片は、ダイズイベントDAS−68416−4の隣接境界に位置した1つのプライマー、導入遺伝子特異的な1つのプライマーの各プライマー対を用いたダイズイベントDAS−68416−4のゲノムDNAからのみ増幅されたが、他のトランスジェニックダイズ系統または非トランスジェニック対照由来のDNA試料からは増幅されなかった。結果は、クローニングされた5’境界配列および3’境界配列が、ダイズイベントDAS−68416−4におけるT鎖挿入断片の隣接境界配列であることを示している。
【0148】
ダイズゲノム内のDNA挿入物をさらに確認するために、2つのダイズ配列にわたるPCR増幅を完了した。5’末端境界配列に従って設計した2種のプライマー、16LEndG03および16LEndG04、ならびに3’末端境界配列に対する2種のプライマー、16PATG03および16PATG04を使用して、導入遺伝子、5’末端境界配列、および3’境界配列の全体を含有するDNAセグメントを増幅した。予測通り、16LEndG03および16PATG03のプライマー対を用いたPCR増幅により、ダイズイベントDAS−68416−4のゲノムDNAからおよそ9kbのDNA断片、および非トランスジェニックダイズ対照および他のダイズトランスジェニック系統から2.7kbのDNA断片が増幅された。同様に、16LEndG04および16PATG04のプライマー対を用いて完了したPCR反応により、ダイズイベントDAS−68416−4の試料からおよそ9kbのDNA断片、および他のダイズ対照系統全てから、同様に、2.8kbのDNA断片がもたらされた。両方のプライマー対をPCRに使用した場合、ダイズイベントDAS−68416−4以外の全てのダイズ試料において約6kbのサイズのかすかなバンドが目に見えたことが認められ、これは、このかすかなバンドが、このプライマー対でのダイズゲノムの非特異的な増幅に起因することを示唆している。
【0149】
実施例3.10
ダイズのゲノム配列の確認
16LEndG03と16PATG03のプライマー対と16LEndG04および16PATG04のプライマー対を使用して非トランスジェニックダイズ系統であるMaverickから増幅された2.7kbのDNA断片および2.8kbのDNA断片をクローニングし、配列決定した。それらの配列を互いに突き合せ、ダイズイベントDAS−68416−4からクローニングされた5’境界配列および3’境界配列とアラインメントした。これにより、クローニングされたDNA配列が、pDAB4468のT鎖がダイズイベントDAS−68416−4に組み込まれた遺伝子座を含有したことが実証された。アラインメント解析により、元の遺伝子座からの55bpの欠失および3’組み込み接合部における9bpの挿入物も明らかになった(図5)。クローニングされた元の遺伝子座のダイズのゲノム領域においてオープンリーディングフレーム(>/=450bp、150アミノ酸)は同定されなかった。
【実施例4】
【0150】
ダイズイベントDAS−68416−4の隣接SNPマーカーによるゲノムの特徴付け
ゲノムの挿入部位を特徴付け、説明するために、挿入断片の近傍に位置するマーカー配列を決定した。多型的な一塩基多型(SNP)マーカーのパネルを使用して導入遺伝子の位置を同定し、マッピングした。ダイズイベントDAS−68416−42は、第4染色体上の119.6cMに位置する。この位置は、2つの隣接SNPマーカー、BARC−044607−08736とBARC−019093−03299の間である。より詳細には、導入遺伝子の位置は、SNPマーカーBARC−019093−03299から1.3cM(480kb)離れてマッピングされた。
【0151】
実施例4.1
隣接境界領域の配列を用いたBLAST
ダイズイベントDAS−68416−4の隣接境界領域の配列(配列番号1)を、ダイズの全ゲノム配列のBLASTに使用した。BLASTの結果により、ダイズイベントDAS−68416−4の境界配列の両方が、連鎖群C1である第4染色体(Gm04)上に位置したことが示された。
【0152】
実施例4.2
SNPマッピングおよびBLASTの結果
境界配列を用いたBLASTおよびマッピングからの結果に基づいて、イベントは、第4染色体に割り当てられた。そのように、10種のSNPマーカーをダイズ遺伝連鎖マップから選択した。SNP配列を、Dr.Cregan、Beltsville Agricultural Research Center、およびUSDAによって開発されたSNPマーカーから選択した。これらのSNPマーカーは、第4染色体に対応する連鎖群C1に関連づけられる。ダイズイベントDAS−68416−4についてT鎖挿入断片の物理的な位置を決定するために、ダイズの全ゲノム配列のBLASTにSNP配列を使用した。
【0153】
実施例4.3
SNPマーカーの結果
ダイズイベントDAS−68416−4は、第4染色体上の119.6cMにマッピングされる。2つの隣接SNPマーカーは、BARC−044607−08736およびBARC−019093−03299である。導入遺伝子は、SNPマーカーBARC−019093−03299から1.3cM(480kb)離れており、SNPマーカーBARC−044607−08736とBARC−019093−03299の間のおよそ119.6cMである。
【実施例5】
【0154】
ダイズイベントDAS−68416−4におけるAAD−12タンパク質の特徴付け
組換えトランスジェニックダイズイベントDAS−68416−4に由来するAAD−12タンパク質の生化学的性質を特徴付けた。定量的酵素結合免疫吸着検定法(ELISA)、ドデシル硫酸ナトリウムポリアクリルアミドゲル電気泳動(SDS−PAGE、クーマシーブルー(Coomassie blue)を用いて染色し、糖タンパク質を検出する方法)、およびウェスタンブロット法を使用してタンパク質の生化学的性質を特徴付け、AAD−12タンパク質の発現を確認した。
【0155】
実施例5.1
植物組織におけるAAD−12タンパク質の発現
ダイズイベントDAS−68416−4におけるAAD−12タンパク質のレベルを決定した。可溶性の抽出可能なAAD−12タンパク質を、ダイズの葉における定量的酵素結合免疫吸着検定法(ELISA)方法を使用して測定した。
【0156】
ダイズ組織の試料を試験植物から単離し、発現を解析するために調製した。0.5%ウシ血清アルブミン(BSA)を含有する界面活性剤Tween−20(PBST)を含有するリン酸緩衝生理食塩水溶液を用いてダイズ植物の組織からAAD−12タンパク質を抽出した。植物組織を遠心分離し;水性上清を収集し、必要に応じて適切な緩衝液で希釈し、AAD−12ELISAキットをサンドイッチ形式で使用して解析した。キットは製造者の推奨プロトコールに従って使用した。
【0157】
ダイズイベントDAS−68416−4において、検出解析を実施して、発現の安定性および遺伝性を垂直方向(世代間)および水平方向(系列間)の両方で調査した。T5世代ダイズイベントDAS−68416−4において、発現は安定であり(分離性でない)、系列全てにわたって一貫した(図6)。
【0158】
種々の植物の段階における圃場発現レベル試験を、ダイズイベントDAS−68416−4において、V3前、V3後、R2前、およびR2後で実施した。噴霧処理全てについて、ならびに2,4−D除草剤を噴霧した小区画および噴霧していない小区画について、発現値は同様であった。2×噴霧率(2,240gm ae/haの2,4−D)を施用し、試験のどのポイントにおいても植物に対する害は確認されなかった。植物のv3前の段階における、系列全てにわたった平均の発現は300μg/cm2であった。2,4−Dを噴霧した後、発現は安定なままであり、系列にわたって平均400μg/cm2であった。ダイズがR2前に到達した時には、平均の発現は、平均200μg/cm2にわずかに降下した。2,4−Dを噴霧した後、R2後の発現は以前の平均400μg/cm2に戻った。図6を参照されたい。
【0159】
実施例5.2
植物組織におけるAAD−12タンパク質の発現
2008年中に米国およびカナダの6つの区域で追加的な圃場発現試験を行った。ダイズイベントDAS−68416−4の4つの処理(噴霧なし、2,4−Dを噴霧、グルホシネートを噴霧、または2,4−Dおよびグルホシネートの両方を噴霧)を試験した。試料採取した植物組織は、葉、粒、根、および茎葉を含んだ。発生のV5段階およびV10段階において葉組織を収集し、R3段階において根および茎葉を収集した。発生のR8段階において粒を収集した(Gaska、2006年)。可溶性の抽出可能なAAD−12タンパク質を、実施例5.1において前記されている通り、検証された酵素結合免疫吸着検定法(ELISA)方法を使用して測定した。全ての種類の組織について、AAD−12タンパク質レベルをng/乾燥重量(mg)に基づいて算出した。
【0160】
種々のダイズマトリックスにおけるAAD−12タンパク質の濃度(部位にわたって平均した)の概要が表8に示されている。平均の発現値は、R3段階の根における15.5ng/乾燥重量(mg)から、V5段階の葉組織における66.1ng/mgにわたった。発現値は、噴霧処理全てについて、ならびに2,4−D除草剤およびグルホシネート除草剤を噴霧した小区画および噴霧していない小区画について、同様であった。6つの区域全てにわたって、対照組織においてAAD−12タンパク質は検出されなかった。
【0161】
【表8】
【0162】
実施例5.3
AAD−12タンパク質のSDS PAGEおよびウェスタンブロット分析
安定剤を加えたPBST(0.05%Tween20、pH7.4を伴うリン酸緩衝生理食塩水)ベースの緩衝液中のダイズイベントDAS−68416−4の凍結乾燥した葉組織からAAD−12タンパク質を抽出し、可溶性タンパク質を遠心分離によって収集した。上清を濾過し、可溶性タンパク質をフェニルセファロース(PS)ビーズ(GE Healthcare、Piscataway、NJ)に結合させた。1時間インキュベートした後、PSビーズをPBSTで洗浄し、Milli−Q水を用いて結合したタンパク質を溶出した。伝導率を増加させるために、塩化ナトリウムを加え、PS精製されたタンパク質を、AAD−12特異的なポリクローナル抗体とコンジュゲートした抗AAD−12免疫親和性カラムにローディングした。結合していないタンパク質をカラムから収集し、カラムを予め冷却したPBS(リン酸緩衝生理食塩水、pH7.4)を用いて広範囲にわたって洗浄した。結合したタンパク質を、3.5MのNaSCN、50mMのトリス、pH8.0である緩衝液を用いてカラムから溶出させ、SDS−PAGEおよびウェスタンブロットによって検査した。同じプロトコールを使用して、対照ダイズ系統であるMaverickの葉組織からタンパク質を単離した。Maverickは、aad−12遺伝子を含有しないが、ダイズイベントDAS−68416−4植物を表す遺伝的背景を有する。
【0163】
ダイズイベントDAS−68416−4およびMaverick由来の凍結乾燥した葉組織を、約2.0%のプロテアーゼ阻害剤反応混液(Sigma、St.Loui、MO)を含有するPBST緩衝液と混合し、Geno−Grinderのボールベアリングを用いて粉砕することによってタンパク質を抽出した。試料を遠心分離し、上清を、Laemmli試料緩衝液と混合し、加熱し、短時間遠心分離した。試料をBio−Rad Criterion SDS−PAGEゲルに直接ローディングした。陽性参照標準物質、微生物由来のAAD−12タンパク質も、試料緩衝液と混合し、ゲルにローディングした。トリス/グリシン/SDS緩衝液(Bio−Rad、Hercule、CA)を用いて電気泳動を行った。電気泳動した後、ゲルを半分に切り、一方の半分はPierce GelCode Blueタンパク質染料で染料し、他方のゲルの半分は、ニトロセルロースメンブレンに電気ブロッティングした。次に、AAD−12特異的なポリクローナルウサギ抗体を用いてニトロセルロースメンブレンを探索した。化学発光基質を使用して、免疫反応性のバンドを可視化した。微生物由来のAAD−12では、クーマシー染色したSDS−PAGEゲルにおいて可視化された主要なタンパク質バンドは、およそ32kDaであった。予測通り、対応する植物由来のAAD−12タンパク質は、微生物由来のタンパク質とサイズが同一であった。予想通りに、植物の精製された画分は、AAD−12タンパク質に加えて、少量の非免疫反応性不純物を含有した。共精製されたタンパク質は、カラムマトリックスとの弱い相互作用によってカラム上に保持されたと思われる(Williamら、2006年、KennedyおよびBarne、1983年およびHolroydeら、1976年)。
【0164】
微生物由来のAAD−12およびDAS−68416−4植物組織抽出物は、抗AAD−12ポリクローナル抗体を使用したウェスタンブロットにおいて予測されるサイズの陽性シグナルを示した。AAD−12ウェスタンブロット分析では、対照であるMaverickの抽出物において免疫反応性タンパク質は観察されず、また、トランスジェニック植物由来の試料において代替のサイズのタンパク質(凝集体または分解生成物)は見られなかった。モノクローナル抗体により、微生物由来のタンパク質において少量のAAD−12二量体が検出された。これらの結果により、AAD−12タンパク質がダイズにおいて発現されているという証拠が追加される。
【実施例6】
【0165】
サザンブロットによるダイズイベントDAS−68416−4のメチル化検出解析
導入された導入遺伝子は、植物ゲノムに組み込まれた後にサイレンシングを受ける可能性がある。導入遺伝子の発現は、転写レベルおよび/または転写後レベルで阻害される可能性がある。転写レベルでの遺伝子サイレンシングは、導入遺伝子、そのプロモーターおよび他の関連性のある配列のメチル化に関連することが報告されている(Stemら、1997年)。特定の配列におけるメチル化を検出するために、ある方法では、メチル化感受性制限酵素を利用してDNAを消化し、その後DNA産物をサザンブロット分析する。特定の制限酵素部位がメチル化されていると、酵素はDNAを切断しない。制限部位がメチル化されていると、サザンブロットで検出可能な、分子量がより高いDNA断片がもたらされる。サザンブロットに基づくメチル化解析を実施して、ダイズイベントDAS−68416−4についてT鎖挿入断片のメチル化の状態を決定した。このアッセイは、aad−12遺伝子に特異的なプローブおよびそのプロモーターおよび2つのメチル化感受性制限酵素を使用して行った。aad−12発現カセットのメチル化は検出されなかった。
【0166】
6.1.ダイズの葉の試料収集およびゲノムDNA(gDNA)の単離
ゲノムDNAを、ダイズイベントDAS−68416−4および非トランスジェニックダイズ系統であるMaverickのそれぞれの植物の葉から調製した。伝統的なCTAB法を使用して、凍結乾燥した葉の試料からゲノムDNAを単離した。抽出後、DNAを、Pico Green試薬(Invitrogen、Carlsbad、CA)を使用して定量した。
【0167】
6.2.DNAの消化および分離
DNAの分子キャラクタリゼーションのために、10マイクログラム(10μg)のダイズイベントDAS−68416−4由来のゲノムDNAおよび非トランスジェニックダイズ系統であるMaverick由来のゲノムDNAを、DNA1μg当たりおよそ5ユニットの選択された制限酵素および対応する反応緩衝液を各DNA試料に加えることによって消化した。各試料を、およそ37℃で一晩インキュベートした。制限酵素AciIおよびHyp188IIIを使用して消化した(New EnglおよびBioLab、Ipswich、MA)。非トランスジェニックダイズであるMaverick由来のDNAを、試験試料と同じ手順および制限酵素を用いて消化して陰性対照として機能させた。消化されたDNA試料を、NaClを0.1Mの最終濃度まで加えた後、イソプロパノールを用いて沈殿させ、1×ローディング緩衝液(0.1%ブロモフェノールブルー、100mMのEDTA、50%グリセロール、10mMのトリス pH7.5)20μlに再懸濁させた。次いで、DNA試料および分子サイズマーカーを、0.4×TAE緩衝液(Fisher Scientific、Pittsburgh、PA)を35ボルトでおよそ18〜22時間用いて0.85%アガロースゲルによって電気泳動して断片の分離を実現した。ゲルを臭化エチジウム(Invitrogen、Carlsbad、CA)で染色し、DNAを紫外線(UV)の下で可視化した。
【0168】
6.3.サザン転写およびメンブレン処理
Seversonら、(1997年)に記載されている通りサザンブロット分析を実施した。簡単に述べると、DNA断片を電気泳動によって分離し、UV光の下で可視化した後、ゲルを変性溶液(150mMのNaOH、3mMのEDTA)におよそ20分間曝露させ、その後、中和溶液(150mMのNaPO4、pH7.8)に少なくとも20分間曝露させた。転写緩衝液(25mMのピロリン酸ナトリウム、pH10)を用いたウィッキング系を使用して、一晩、ナイロンメンブレン(Roche Diagnostic、Indianapoli、IN)にサザン転写を実施した。転写した後、メンブレンを65℃で約2時間加熱乾燥した。このプロセスにより、ハイブリダイゼーションのためのサザンブロットメンブレンの準備ができた。
【0169】
6.4.DNAプローブの標識化およびハイブリダイゼーション
ナイロンメンブレンに結合したDNA断片を、標識したプローブを使用して検出した。プローブは、プラスミドpDAB4468由来の特異的なプライマーを伴う増幅されたPCR断片として生成した。これらのPCR増幅された断片をアガロースゲルから切り取り、精製した。精製されたDNA断片を、ハイブリダイゼーションプローブを作製するための鋳型として使用した。ハイブリダイゼーションプローブを、GE Healthcare READY−TO−GO(商標)DNA Labeling Beads(GE Healthcare、Piscataway、NJ)を製造者の説明書に従って使用したランダムプライミングによってα32Pに特異的なヌクレオチドで標識し、PROBEQUANT(商標)G−50マイクロカラム(Amersham/Pharmacia、Piscataway、New Jersey、USA)によって精製した。この試験のために使用したプローブの一覧は、表9に記載されている。
【0170】
プレハイブリダイゼーションおよびハイブリダイゼーションを、それぞれ、65℃で4時間および一晩、ハイブリダイゼーション緩衝液(Sigma、St.Loui、MO)を使用して行った。ハイブリダイゼーション後、メンブレンを65℃、洗浄緩衝液(10mMのリン酸ナトリウム、2.5mMのピロリン酸ナトリウム、0.5mMのEDTA、0.1%SD、リン酸を用いてpH7.8に調整した)で20分間、3回洗浄した。洗浄したフィルターを、オートラジオグラフィーのためにPhosphorimagerスクリーンに曝露させ、画像をスキャンした。
【0171】
【表9】
【0172】
6.5 プローブの除去
サザンハイブリダイゼーションのデータを得た後、メンブレンブロットからDNAプローブを除去し、メンブレンブロットは、異なるDNAプローブを用いたハイブリダイゼーションに再利用することができた。簡単に述べると、曝露後、メンブレンブロットを室温で10分間、再生溶液1(30mMのNaOH、1mMのNa2EDTA)で洗浄し、65℃で30分間、再生溶液2(5mMのNaPO4、1mMのNa2EDTA、0.1%SDS)で洗浄した。次いで、メンブレンブロットを2×SSCで簡単に洗浄し、それで別のDNAプローブを用いたハイブリダイゼーションの準備ができた。メンブレンブロットをオートラジオグラフィーのためにPhosphorimagerスクリーンに曝露させて、次のハイブリダイゼーションに進む前にDNAプローブの全てを確実に除去した。
【0173】
6.6.サザンブロットの結果
この試験では、aad−12遺伝子およびそのプロモーターであるAtUbi10のメチル化の状態を決定するために、メチル化感受性制限酵素AciIおよびHyp188IIIを使用した。pDAB4468のT鎖DNAの公知の制限酵素部位に基づいて予測された特定の消化物およびプローブの断片サイズが表10で示されている。サザンブロットにおいて分子量がより高い断片が検出されることは、AciI制限酵素部位およびHyp188III制限酵素部位の認識配列内のシトシンがメチル化されていることを示すと思われる。そのように、メチル化により、制限酵素がゲノムDNAを消化することができなくなる。
【0174】
制限酵素AciIおよびHyp188IIIを使用してaad−12遺伝子のメチル化の状態を検査した。aad−12プローブを使用すると、予測されたサイズを有するハイブリダイゼーションバンドが観察された。このデータは、ダイズイベントDAS−68416−4において、AciIおよびHyp188IIIの認識部位におけるメチル化が起こっていないことを示唆している。同様に、Hyp188IIIで消化されたダイズイベントDAS−68416−4DNA試料において、AtUbi10プローブを使用すると、予測された分子量のバンドが検出された。このデータは、aad−12プロモーター配列における認識部位がメチル化されていなかったことを示している。
【0175】
【表10】
【実施例7】
【0176】
農業形質データ
2008年の米国およびカナダの6区域における組成研究の一部として、および2009年に米国およびカナダの8区域において行われた別々の試験においても、ダイズイベントDAS−68416−4を用いた農業試験を行った。これらの試験では、イベントDAS−68416−4ダイズ(2,4−D除草剤および/またはグルホシネート除草剤の施用を伴う、または伴わない)と、その非トランスジェニック近同質遺伝子対照(Maverick)を比較した。両方の試験の全てにわたる結果により、農業形質パラメータは、従来のダイズ系統について得られる範囲内であることが示された。
【0177】
実施例7.1.
2008年の農業形質データの生成
2008年に、アイオワ、イリノイ、インディアナ、ネブラスカおよびカナダのオンタリオ(2つの現場)に位置する6つの現場において、イベントDAS−68416−4ダイズおよび非トランスジェニック対照(Maverick品種)を用いた農業研究を行った。対照系統であるMaverickと比較して、ダイズイベントDAS−68416−4(除草剤処理を伴う、および除草剤処理を伴わない)の同等性を調査するために、群生/集団数、実生/植物生長力、植物の高さ、倒伏、病害発生率、虫害、および開花までの日数を含めた農業形質決定因子を評価した。この試験は、実験1と称される。
【0178】
試験ダイズ種子および対照ダイズ種子を、作条の間隔をおよそ30インチ(75cm)として、25ftの作条当たりおよそ112種子の播種密度で植えた。それぞれの現場において、それぞれの処理について3回反復の小区画を確立し、それぞれの小区画は2〜25ftの作条からなった。小区画を完全乱塊(randomized complete block)(RCB)計画に配置し、それぞれの現場において独特に無作為化した。それぞれのダイズの小区画には、同様の成熟度の非トランスジェニックダイズの2つの作条が隣接していた。試験現場全体を最低10ftの同様の相対的な成熟度の非トランスジェニックダイズで囲んだ。
【0179】
除草剤処理については、1エーカー当たりおよそ20ガロン(187L/ha)の噴霧体積を施用した。これらの施用は、商慣習の最大表示量を再現するように設計した。2,4−Dを、季節の合計3lb ae/Aで、上からの施用で3回散布して施用した。出芽前および生長段階のおよそV4およびR2において、それぞれ1.0lb ae A(1,120g/ha)の施用を行った。グルホシネートを、季節の合計0.74lb ai/A(828g ai/ha)、上からの施用で2回散布して施用した。生長段階のおよそV6およびR1において、それぞれ0.33lb ai/Aおよび0.41lb ai/A(374g ai/haおよび454g ai/ha)の施用を行った。
【0180】
圃場現場の全てにわたって、混合モデル(SAS バージョン8;SAS Institute 1999)を使用して農業形質データについて分散分析を行った。エントリーを固定効果とみなし、区域、区域内のブロック、区域×エントリー、およびエントリー×区域内のブロックを、変量効果として指定した。全体的な処理の効果の有意性を、F検定を用いて推定した。対照と、噴霧していないダイズイベントDAS−68416−4(噴霧なし)トランスジェニックエントリー、グルホシネートを噴霧したダイズイベントDAS−68416−4(ダイズイベントDAS−68416−4+グルホシネート)トランスジェニックエントリー、2,4−Dを噴霧したダイズイベントDAS−68416−4(ダイズイベントDAS−68416−4+2,4−D)トランスジェニックエントリーならびにグルホシネートおよび2,4−Dの両方を噴霧したダイズイベントDAS−68416−4(ダイズイベントDAS−68416−4+両方)トランスジェニックエントリーの間で、t検定を用いて対応のある対比を行った。多重性を制御するために、偽発見率(False Discovery Rate)(FDR)を使用して調整P値も算出した(BenjaminiおよびHochberg、1995年)。
【0181】
【表11】
【0182】
対照、噴霧していないダイズイベントDAS−68416−4、ダイズイベントDAS−68416−4+2,4−D、ダイズイベントDAS−68416−4+グルホシネート、およびダイズイベントDAS−68416−4+両方の除草剤から収集した農業形質データの分析を行った。群生数、初期集団、実生生長力、施用後の害、倒伏、最終的な群生数または開花までの日数について、統計的有意差は観察されなかった(表12)。高さについては、対照とダイズイベントDAS−68416−4+2,4−D噴霧との間に有意な対応のあるt検定が観察された。しかし、有意な全体的な処理の効果は観察されず、ダイズイベントDAS−68416−4処理と対照との間の差異は非常に小さく、また、差異は、異なるダイズイベントDAS−68416−4処理間で共有されなかった。これらの結果に基づいて、ダイズイベントDAS−68416−4は、近同質遺伝子非トランスジェニック対照と農業的に同等であった。
【0183】
【表12】
【0184】
実施例7.2.
2009年の農業形質データの生成
2009年に、アーカンソー、アイオワ、イリノイ、インディアナ、ミズーリ、およびネブラスカに位置する8か所の現場においてダイズイベントDAS−68416−4および非トランスジェニック対照(Maverick品種)を用いた農業研究行った。ダイズイベントDAS−68416−4ダイズ(除草剤処理を伴う、および除草剤処理を伴わない)の対照との同等性を調査するために、群生/集団数、実生/植物生長力、植物の高さ、病害発生率、虫害、および開花までの日数を含めた農業形質決定因子を評価した(表13)。
【0185】
【表13】
【0186】
試験には完全乱塊(randomized-complete-block)計画を用いた。エントリーは、ダイズイベントDAS−68416−4、Maverick対照系統、および市販されている非トランスジェニックダイズ系統であった。試験ダイズ種子、対照ダイズ種子および参照ダイズ種子を、作条の間隔をおよそ30インチ(75cm)として、作条当たりおよそ112種子の播種密度で植えた。それぞれの現場において、それぞれの処理について4回反復の小区画を確立し、各小区画は2〜25ftの作条からなった。それぞれのダイズの小区画には、非トランスジェニックダイズ(Maverick)の2つの作条が隣接していた。試験現場全体を、最低4つの作条(または10ft)の非トランスジェニックダイズ(Maverick)で囲んだ。適切な昆虫、雑草、および病害の制御慣習を施用して農業的に許容できる作物を生産した。
【0187】
商慣習の最大表示量(maximum label rate)を再現するように除草剤処理を適用した。処理は、噴霧しない対照および特定の生長段階において2,4−D、グルホシネート、2,4−D/グルホシネートを施用した除草剤施用からなった。2,4−D施用については、V4生長段階およびR2生長段階において除草剤を1.0lb ae/A(1,120g ae/ha)の比率で施用した。グルホシネート処理については、V4生長段階およびV6〜R2生長段階にある植物に施用を行った。両方の施用については、V4施用およびV6〜R2施用について、それぞれグルホシネートを0.33lb ai/A(374g ai/ha)および0.41lb ai/A(454 g ai/ha)の比率で施用した。両方の除草剤を施用するエントリーは、ダイズイベントDAS−68416−4および非トランスジェニックMaverickを含めた対照であった。Maverick小区画は、除草剤を施用した後に枯死することが予測された。
【0188】
圃場現場全てにわたって、混合モデル(SASバージョン8;SAS Institute 1999年)を用いて農業形質データについて分散分析を行った。エントリーを固定効果とみなし、区域、区域内のブロック、区域×エントリー、およびエントリー×区域内のブロックを、変量効果として指定した。個々の区域において、類似の様式で、エントリーを固定効果とし、ブロックおよびエントリー×ブロックを変量効果として分析を行った。データは統計分析には回さなかった。95%信頼水準において有意差が宣言され、F検定を用いて全体的な処理の効果の有意性を推定した。噴霧していないAAD−12(噴霧なし)トランスジェニックエントリー、グルホシネートを噴霧したAAD−12(AAD−12+グルホシネート)トランスジェニックエントリー、2,4−Dを噴霧したAAD−12(AAD−12+2,4−D)トランスジェニックエントリーならびにグルホシネートおよび2,4−Dの両方を噴霧したAAD−12(AAD−12+2,4−D+グルホシネート)トランスジェニックエントリーと対照エントリーとの間で、t検定を用いて対応のある対比を行った。
【0189】
この試験では多数の対比を行ったので、多重性が問題であった。予想外の効果を探すために単一の試験において多数の比較を行う場合、多重性が問題である。これらの条件下で、比較に関するp値に基づいて誤って差異が宣言される確率は非常に高い(1−0.95比較の数)。この試験では分析物当たり4つの比較があり(16種の農業形質についての分析所見)、農業形質について64の比較が生じる。したがって、調整していないp値に基づいて1つまたは複数の誤った差異が宣言される確率は、農業形質について99%であった(1−0.9564)。
【0190】
対照エントリー、AAD−12噴霧なしエントリー、AAD−12+グルホシネートエントリー、AAD−12+2,4−Dエントリー、およびAAD−12+2,4−D+グルホシネートエントリーから収集した農業形質データの分析を行った。現場全てにわたる分析について(表14)、実生生長力、最終集団、植物生長力/害(V4、R1)、倒伏、病害発生率、虫害、開花までの日数、成熟までの日数、莢の数、種子の数、生産量、および植物の高さについて統計的有意差は観察されなかった。群生数および初期集団については、対照とAAD−12+グルホシネートエントリーとの間に有意な対応のあるt検定が観察されが、有意な全体的な処理の効果またはFDR調整P値を伴わなかった。植物生長力/害(R2)については、対照とAAD−12+グルホシネートエントリーおよびAAD−12+2,4−D+グルホシネートエントリーのどちらとの間にも有意な対応のあるt検定および有意な全体的な処理の効果が観察されたが、有意なFDR調整P値を伴わなかった。これらの変数全てについて平均した結果も、この試験において試験された参照系統について見いだされた範囲内であった。
【0191】
【表14】
【0192】
実施例7.3.
生態学的評価
ダイズ栽培の実践に詳しい人(育種家、圃場の管理者、圃場の農学者、圃場の従業員)がダイズイベントDAS−68416−4の圃場試験をモニターし、観察した。圃場試験を行った人員は、ダイズイベントDAS−68416−4植物に対する植物病害および害虫の発生率を、同じ試験において従来のダイズ品種と比較して視覚的にモニターした。実施例7.1に記載の実験1の一部として、病害および虫害を、0%が病害発生率または昆虫抵抗性に起因する害を表す0〜100%の数値的な尺度で評価した。表15に、実験1に記載の6か所の現場全てにわたる結果が示されている。
【0193】
【表15】
【0194】
病害発生率については、統計的有意差は観察されなかった。虫害については、対照とダイズイベントDAS−68416−4+両方の除草剤との間に有意な対応のあるt検定が観察された。しかし有意な全体的な処理の効果は観察されず、ダイズイベントDAS−68416−4処理と対照との間の差異は小さく、また、差異は、異なるダイズイベントDAS−68416−4処理間で共有されなかった。
【0195】
2006年〜2008年に行われた全てのUSDA APHISに届け出た圃場試験からの生態学的な観察も行った。ダイズイベントDAS−68416−4植物の試験における病害の発生率および昆虫の存在を記録し、従来の対照と比較したダイズイベントDAS−68416−4植物の発生率または反応の差異を検査した。全ての場合において、ダイズイベントDAS−68416−4植物のいずれの試験でも従来の対照と比較して差異は見られなかった。ダイズイベントDAS−68416−4および従来のダイズの試験において観察された病害ストレス要因および昆虫ストレス要因は、表16に記載されている。これらの観察は、生態学的なストレス要因に対するダイズイベントDAS−68416−4の反応は、従来のダイズの反応と異ならないという結論を裏付ける。
【0196】
【表16】
【0197】
実施例7.4.
発芽(Germancy)および休眠の評価
ダイズイベントDAS−68416−4種子の発芽について、温暖な条件下および寒冷な条件下で近同質遺伝子型の比較器と比較して調べることによって種子の休眠特性の変化を評価した。
【0198】
温暖発芽試験については、ダイズイベントDAS−68416−4および対照ダイズの種子を、水を染み込ませた発芽用パッドを含有するペトリ皿に、プレート当たり25個置き、過剰な水を流し出した。プレートを25℃に置き、これらの条件下で5日間保持した。系統当たり16プレート(400種子)を調製した。5日後、発芽しなかった種子の数を記録した。
【0199】
寒冷発芽試験については、鉢植え用土を満たした平箱半分当たり種子100個の種子を植えた。平箱に多少水をまき、10℃で7日間保持し、その後25℃に5日間曝露し、その後、発芽しなかった種子の数を記録した。
【0200】
それぞれの試験からのデータを、反復当たり種子100個の4回の反復を伴う完全無作為計画を使用した分散分析(ANOVA)によって分析した。統計分析のために、発芽した種子の数を100で割った値の平方根のアークサインを使用してデータを変換した。発芽率(パーセント)が、表17に要約されている。
【0201】
【表17】
【0202】
温暖発芽実験または寒冷発芽実験のいずれにおいても、ダイズイベントDAS−68416−4の種子の発芽と対照ダイズの種子の発芽との間に有意差はなかった(それぞれPr>F=1.0および0.13)。これらの結果は、ダイズイベントDAS−68416−4において種子の休眠特性が変化してないことを示している。
【0203】
実施例7.5.
農業特性、病害特性、害虫特性、および発芽特性の概要
生育期全体を通して植物の生長特性を評価する農業形質データにより、ダイズイベントDAS−68416−4の従来の非トランスジェニックダイズとの同等性が実証されている。植物の生長および表現型の特性、病害および昆虫の圧力に対する感受性によって示される生態学的ストレス要因に対する反応、ならびに発芽および休眠の特性は、多様な環境にわたって、ダイズイベントDAS−68416−4植物と従来のダイズとの間で変化しなかった。したがって、これらのデータにより、ダイズイベントDAS−68416−4の農業特性、病害特性、および害虫特性は、従来のダイズの農業特性、病害特性、および害虫特性と有意に異ならず、また、ダイズイベントDAS−68416−4ダイズが植物害虫リスクの増加を示す徴候はないという結論が裏付けられる。Benjamini、Y.、Hochberg、Y.(1995年)Controling the false discovery rate:A practical and powerful approach to multiple testing. J. Royal Statistical Soc. B、57巻:289〜300頁。
【実施例8】
【0204】
粒および茎葉の組成
ダイズイベントDAS−68416−4(2,4−Dを噴霧、グルホシネートを噴霧、2,4−D+グルホシネートを噴霧、または2,4−Dまたはグルホシネートを噴霧していない)と従来のダイズとの間の同等性を調査するために、ダイズの茎葉および粒について組成分析を実施した。米国およびカナダの主要なダイズ生産地域内に位置する6か所の試験場において、ダイズイベントDAS−68416−4を伴う種子系統およびダイズイベントDAS−68416−4を伴わない種子系統を使用して試験を行った。試験場は、多様な農業習慣および環境条件の地域を代表し、タンパク質の発現解析および実施例7.1に記載の農業実験1のために使用したのと同じ現場であった。試験はアイオワ、イリノイ、インディアナ、ネブラスカ、およびカナダのオンタリオ(2現場)に位置した。
【0205】
ダイズの茎葉および粒の試料を、種々の試験を用いて栄養素量について分析した。茎葉について行った分析は、タンパク質、脂肪、灰分、水分、炭水化物、酸性デタージェント繊維(ADF)、中性デタージェント繊維(NDF)、カルシウムおよびリンを含んだ。粒について行った分析は、一般成分(灰分、全脂肪、水分、タンパク質、コレステロール、炭水化物)、繊維、ミネラル、アミノ酸、脂肪酸、ビタミン、抗栄養因子を含んだ。
【0206】
ダイズの茎葉および粒についての栄養分析の結果を文献において報告されている値と比較した。圃場現場の全てにわたって、混合モデルを使用して分散分析も行った。エントリーを固定効果とみなし、区域、区域内のブロック、および区域×エントリーを、変量効果として指定した。全体的な処理の効果の有意性を、F検定を用いて推定した。ダイズイベントDAS−68416−4トランスジェニックエントリー(噴霧していないAAD−12;2,4−Dまたはグルホシネートを噴霧していない)、グルホシネートを噴霧したダイズイベントDAS−68416−4トランスジェニックエントリー(ダイズイベントDAS−68416−4+グルホシネート)、2,4−Dを噴霧したダイズイベントDAS−68416−4トランスジェニックエントリー(ダイズイベントDAS−68416−4+2,4−D)、ならびにグルホシネートおよび2,4−Dの両方を噴霧したダイズイベントDAS−68416−4トランスジェニックエントリー(ダイズイベントDAS−68416−4+両方)と、対照エントリーとの間でt検定を用いて対応のある対比を行った。
【0207】
この試験では多数の対比を行ったので、多重性が問題であった。予想外の効果を探すために単一の試験において多数の比較を行う場合、多重性が問題である。これらの条件下で、比較に関するp値に基づいて誤って差異が宣言される確率は非常に高い(1−0.95比較の数)。この試験では分析物当たり4つの比較があり(75の定量化された分析物)、現場の組成分析全てにわたって300の比較が行われた。したがって、調整していないp値に基づいて1つまたは複数の誤った差異が宣言される確率は99.99%より高かった。
【0208】
多重性を説明するための1つの方法は、p値を調整して実験に関する誤差率(宣言された差異が全て有意である確率)を制御することであるが、試験において多くの比較を行う場合、特異的な効果を検出するための検定力は有意に低下する可能性がある。検定力がはるかに大きい代替法は、p値を調整してそれぞれの宣言された差異が有意である確率を制御することである。これは、偽発見率(FDR)手順(BenjaminiおよびHochberg、1995年)を用いて実現することができる。したがって、変量効果の処理間の真の差異の識別(偽陽性)を改善するために、FDRを用いてp値を調整した。
【0209】
実施例8.1.
ダイズの茎葉の組成分析
対照、噴霧していないダイズイベントDAS−68416−4、ダイズイベントDAS−68416−4+グルホシネート、ダイズイベントDAS−68416−4+2,4−DおよびダイズイベントDAS−68416−4+両方の除草剤由来のダイズの茎葉試料中のタンパク質、脂肪、灰分、水分、炭水化物、酸性デタージェント繊維(ADF)、中性デタージェント繊維(NDF)、カルシウムおよびリンの分析を実施した。全ての区域にわたる結果の概要が表18に示されている。
【0210】
タンパク質、脂肪、灰分、水分、炭水化物、ADF、NDF、カルシウムまたはリンについて、現場全てにわたる分析において、対照エントリーとトランスジェニックエントリーとの間に統計学的差異は観察されなかった。ダイズイベントDAS−68416−4+グルホシネートおよびダイズイベントDAS−68416−4+両方の除草剤についての現場全てにわたる平均灰分値は、ダイズイベントDAS−68416−4+グルホシネートおよびダイズイベントDAS−68416−4+2,4−DについてのNDF値と同様に、文献範囲の外側であった。非トランスジェニック対照を含めた全ての処理についてのADF値も、文献値の外側であった。茎葉におけるいかなる一般成分、繊維の種類、またはミネラルについても、平均値は、非トランスジェニック対照とトランスジェニックエントリーのいずれとの間でも有意に異ならなかった。これらの組成成分に基づいて、ダイズイベントDAS−68416−4ダイズ由来の茎葉は、近同質遺伝子非トランスジェニック対照の茎葉と実質的に同等であった。
【0211】
【表18】
【0212】
実施例8.2.
ダイズの粒の組成分析
実施例8.2.1
一般成分および繊維
対照、噴霧していないダイズイベントDAS−68416−4、ダイズイベントDAS−68416−4+グルホシネート、ダイズイベントDAS−68416−4+2,4−DおよびダイズイベントDAS−68416−4+両方の除草剤由来のダイズの粒試料中のタンパク質、脂肪、灰分、水分、コレステロール、炭水化物、ADF、NDFおよび総食物繊維の分析を実施した。全ての区域にわたる結果の概要が表19に示されている。
【0213】
脂肪、ADFまたは総食物繊維に関しては、現場全てにわたる分析において、対照エントリーとトランスジェニックエントリーとの間に統計学的差異は観察されなかった。しかし、ダイズイベントDAS−68416−4+2,4−Dエントリーについては、ADFが文献範囲よりもわずかに高かった。
【0214】
タンパク質レベルは、現場全てにわたる分析において、噴霧なし、ダイズイベントDAS−68416−4+2,4−D、およびダイズイベントDAS−68416−4+両方の除草剤について、調整していないP値に基づいて対照と比較して有意に異なった。しかし、FDRを調整した後、ダイズイベントDAS−68416−4+2,4−Dについてのp値のみが有意であり、また、全ての処理についての全体の平均のタンパク質の値は報告された文献値の範囲内であり、これは、差異が生物学的には意味がないことを示している。
【0215】
灰分の現場全てにわたる分析において、対照と2,4−Dを噴霧したダイズイベントDAS−68416−4処理との間に有意な調整していないP値が観察されたが、全体的な処理の効果または調整したP値は観察されなかった。灰分値も、報告された文献値の範囲内であり、これは、差異が生物学的には意味がないことを示している。
【0216】
水分レベルは、現場全てにわたる分析において、噴霧なし、ダイズイベントDAS−68416−4+2,4−D、およびダイズイベントDAS−68416−4+両方の除草剤について、調整していないP値に基づいて対照と比較して有意に異なった。しかし、水分についての全体的な処理の効果は有意でなく、ダイズイベントDAS−68416−4+2,4−D処理のみが有意なFDR調整したP値を有し、また、全ての処理について平均水分レベルは文献範囲内であった。このことにより、差異は生物学的には意味がないことが示された。
【0217】
コレステロール値は、全て定量限界未満(<LOQ)であり、また、報告された文献値はない。
【0218】
炭水化物レベルは、現場全てにわたる分析において、噴霧なし、ダイズイベントDAS−68416−4+グルホシネート、およびダイズイベントDAS−68416−4+2,4−Dについて、調整していないP値に基づいて対照と比較して有意に異なった。しかし、ダイズイベントDAS−68416−4+2,4−D処理のみが、FDR調整したP値に基づいて対照と有意に異なり、全ての処理の平均は報告された文献値の範囲内であり、これは、非トランスジェニックダイズとの同等性を示している。
【0219】
NDFレベルは、現場全てにわたる分析において、ダイズイベントDAS−68416−4+グルホシネートについて、調整していないP値に基づいて対照と比較して有意に異なっが、これは、有意な調整したP値または全体的な処理の効果を伴わなかった。現場全てにわたるNDFの値は、ダイズイベントDAS−68416−4+グルホシネートおよびダイズイベントDAS−68416−4+2,4−Dエントリーについて報告された文献値よりもわずかに高かったが、差異は、非トランスジェニック近同質遺伝子対照と比較して9%未満であった。
【0220】
これらの組成成分に基づいて、ダイズイベントDAS−68416−4由来の粒は、非トランスジェニックダイズの粒と実質的に同等であった。
【0221】
【表19】
【0222】
実施例8.2.2
ミネラル
対照、噴霧していないダイズイベントDAS−68416−4、ダイズイベントDAS−68416−4+グルホシネート、ダイズイベントDAS−68416−4+2,4−DおよびダイズイベントDAS−68416−4+両方の除草剤由来のダイズの粒試料中のカルシウム、クロム、銅、ヨウ素、鉄、マグネシウム、マンガン、モリブデン、リン、カリウム、セレン、ナトリウムおよび亜鉛の分析を実施した。全ての区域にわたる結果の概要が表20に示されている。
【0223】
クロム、銅、ヨウ素、鉄、マンガン、モリブデン、リン、セレンおよびナトリウム(検出されなかった)については、現場全てにわたる分析において、調整していないP値に基づいて、対照エントリーとトランスジェニックエントリーとの間に統計学的差異は観察されなかった。
【0224】
カルシウムは、現場全てにわたる分析におい て、ダイズイベントDAS−68416−4+2,4−Dについて、調整していないP値に基づいて有意差を有したが、これは、有意なFDR調整したP値または全体的な処理の効果を伴わず、全ての処理の平均は文献範囲内に入り、これは、差異が生物学的には意味がないことを示している。
【0225】
マグネシウムレベルは、現場全てにわたる分析において、ダイズイベントDAS−68416−4+両方の除草剤およびダイズイベントDAS−68416−4+グルホシネートについて、それぞれ、調整していないp値および調製したp値に基づいて対照と比較して有意に異なったが、全体的な処理の効果は有意でなかった。マグネシウムの現場全てにわたる平均値は、報告された文献値よりもわずかに低かったが、対照と比較して差異は小さく(<3%)、全てのダイズイベントDAS−68416−4エントリーが、対照と比較して、より文献値に近かった。
【0226】
全てのダイズイベントDAS−68416−4エントリーが、現場全てにわたる分析において、対照と比較して有意に高いカリウムの値を有した。しかし、対照と比較して差異は小さく(<5%)、全てのダイズイベントDAS−68416−4エントリーが、対照と比較して、より文献範囲に近かった。
【0227】
亜鉛レベルの差異は、現場全てにわたる分析において、ダイズイベントDAS−68416−4+両方の除草剤について、調整していないP値に基づいて有意であったが、これは、有意なFDR調整したP値または全体的な処理の効果を伴わず、また、差異は小さかった(<4%)。
【0228】
これらの組成成分に基づいて、ダイズイベントDAS−68416−4由来の粒は、非トランスジェニックダイズの粒と実質的に同等であった。
【0229】
【表20】
【0230】
実施例8.2.3
アミノ酸
対照、噴霧していないダイズイベントDAS−68416−4、ダイズイベントDAS−68416−4+グルホシネート、ダイズイベントDAS−68416−4+2,4−DおよびダイズイベントDAS−68416−4+両方の除草剤由来のダイズの粒試料中の以下のアミノ酸の分析を実施した:アラニン、アルギニン、アスパラギン酸、シスチン、グルタミン酸、グリシン、ヒスチジン、イソロイシン、ロイシン、リジン、メチオニン、フェニルアラニン、プロリン、セリン、トレオニン、トリプトファン、チロシン、およびバリン。全ての区域にわたる結果の概要が表21に示されている。
【0231】
システイン、メチオニン、プロリン、チロシンまたはトリプトファンについては、対照エントリーとトランスジェニックエントリーとの間に統計学的差異は観察されなかった。ダイズイベントDAS−68416−4+2,4−Dについてのイソロイシンレベルは、調整していないP値に基づいて対照と有意に異なったが、これは、有意なFDR調整したP値または有意な全体的な処理の効果を伴わなかった。残りの12種のアミノ酸のレベルは、ダイズイベントDAS−68416−4エントリーのうちの2つ以上について、対照と比較してわずかに低かったが(<7%)、全てが非トランスジェニックダイズについての文献範囲内に入った。全てのエントリーについて、全てのアミノ酸が文献範囲内であり、これは、差異が生物学的には意味がないことを示している。これらの組成成分に基づいて、ダイズイベントDAS−68416−4由来の粒は、非トランスジェニックダイズの粒と実質的に同等であった。
【0232】
【表21】
【0233】
実施例8.2.4
脂肪酸
対照、噴霧していないダイズイベントDAS−68416−4、ダイズイベントDAS−68416−4+グルホシネート、ダイズイベントDAS−68416−4+2,4−DおよびダイズイベントDAS−68416−4+両方の除草剤由来のダイズの粒試料中の22種の脂肪酸の分析を実施した。全ての区域にわたる結果の概要が表22に示されている。
【0234】
脂肪酸の10:0カプリン酸、15:0ペンタデカン酸、15:1ペンタデセン酸、20:3エイコサトリエン酸、20:4アラキドン酸、8:0カプリル酸、12:0ラウリン酸、14:0ミリスチン酸、14:1ミリストレイン酸、17:1ヘプタデセン酸、18:3ガンマリノレン酸、および20:2エイコサジエン酸を分析し、結果はLOQ未満であった。脂肪酸の16:0パルミチン酸、17:0ヘプタデカン酸、および20:1エイコセン酸は、対照とAAD−12エントリーとの間で有意に異ならなかったが、20:1エイコセン酸値は、AAD−12+グルホシネートおよびAAD−12+両方の除草剤について、報告された文献値よりも低かった。しかし、対照と比較して差異は小さかった(<5%)。
【0235】
16:1パルミトレイン酸のレベルは、対照と、噴霧していないダイズイベントDAS−68416−4、ダイズイベントDAS−68416−4+グルホシネート、ダイズイベントDAS−68416−4+2,4−D、およびダイズイベントDAS−68416−4+両方の除草剤との間で、調整していないp値に基づいて有意に異なった。しかし、噴霧していないダイズイベントDAS−68416−4エントリーのみが、16:1パルミトレイン酸について有意なFDR調整したP値を有した。この処理についての16:1パルミトレイン酸の現場全てにわたる値は報告された文献値と比較して低かったが、近同質遺伝子対照と比較して差異は小さかった(<13%)。
【0236】
18:0ステアリン酸のレベルは、対照と、噴霧なしおよびダイズイベントDAS−68416−4+グルホシネートとの間で、調整していないP値に基づいて有意に異なった。しかし、調整したp値または全体的な処理の効果に基づいて有意差は確認されず、また、全てのエントリーは報告された文献値の範囲内であり、これは、非トランスジェニックダイズとの同等性を示している。
【0237】
18:1オレイン酸のレベルは、対照と、噴霧していないダイズイベントDAS−68416−4、ダイズイベントDAS−68416−4+グルホシネート、ダイズイベントDAS−68416−4+2,4−D、およびダイズイベントDAS−68416−4+両方の除草剤との間で有意に異なった。しかし、18:1オレイン酸レベルは、全ての処理について報告された文献値の範囲内であり、これは、非トランスジェニックダイズとの同等性を示している。
【0238】
18:2リノール酸のレベルは、対照と、噴霧なしおよびダイズイベントDAS−68416−4+2,4−Dとの間で、調整していないP値に基づいて有意に異なった。しかし、調整したp値または全体的な処理の効果に有意差は確認されず、また、18:2リノール酸レベルは全ての処理について報告された文献値の範囲内であり、これは、非トランスジェニックダイズとの同等性を示している。
【0239】
18:3リノレン酸のレベルは、ダイズイベントDAS−68416−4エントリーのそれぞれと、対照との間で、調整していないp値に基づいて有意に異なり、また、調整したp値も、噴霧していないダイズイベントDAS−68416−4およびダイズイベントDAS−68416−4+両方の除草剤処理の間で対照と比較して有意であった。18:3リノレン酸について入手可能な文献値はないが、ダイズイベントDAS−68416−4と対照処理との間の差異は小さかった(<6%)。
【0240】
20:0アラキジン酸のレベルは、対照と、噴霧していないダイズイベントDAS−68416−4、ダイズイベントDAS−68416−4+グルホシネート、ダイズイベントDAS−68416−4+2,4−D、およびダイズイベントDAS−68416−4+両方の除草剤との間で、調整していないp値に基づいて有意に異なり、まら、20:0アラキジン酸は、現場全てにわたる分析において、噴霧なし、およびダイズイベントDAS−68416−4+グルホシネート処理について、調整したP値にも有意差を有した。しかし、20:0アラキジン酸レベルは全ての処理について報告された文献値の範囲内であり、これは、非トランスジェニックダイズとの同等性を示している。
【0241】
22:0ベヘン酸のレベルは、対照と、噴霧なし、ダイズイベントDAS−68416−4+グルホシネート、ダイズイベントDAS−68416−4+2,4−D、およびダイズイベントDAS−68416−4+両方の除草剤との間で、調整していないp値に基づいて有意に異なり、また、22:0ベヘン酸のレベルは、現場全てにわたる分析において、ダイズイベントDAS−68416−4+グルホシネートについて、調整したP値にも有意差を有した。しかし、有意な全体的な処理の効果はなく、また、22:0ベヘン酸レベルは全ての処理について報告された文献値の範囲内であり、これは、非トランスジェニックダイズとの同等性を示している。
【0242】
調査した22種の脂肪酸のうち、21種の脂肪酸について、4つのダイズイベントDAS−68416−4エントリーの全てが、対照と統計学的に区別できなかったか、または、文献値の範囲内であった。1つの場合(噴霧していないダイズイベントDAS−68416−4;16:1パルミトレイン酸)で、値は最小の文献値をわずかに下回り、対照と統計学的に異なった(<13%低かった)が、3種の噴霧処理の全ては、文献範囲内であった。これらの組成成分に基づいて、ダイズイベントDAS−68416由来の粒は、非トランスジェニックダイズの粒と実質的に同等であった。
【0243】
【表22】
【0244】
実施例8.2.5
ビタミン
対照、噴霧していないダイズイベントDAS−68416−4、ダイズイベントDAS−68416−4+グルホシネート、ダイズイベントDAS−68416−4+2,4−DおよびダイズイベントDAS−68416−4+両方の除草剤由来のダイズの粒試料中のビタミンの分析を実施した。全ての区域にわたる結果の概要が表23に示されている。
【0245】
ダイズの粒中のベータ−トコフェロール、デルタ−トコフェロール、ガンマ−トコフェロール、ビタミンA、ビタミンB5、ビタミンB6、ビタミンB12、ビタミンC、ビタミンDおよびナイアシンについての文献値は見つからなかった。ベータトコフェロール、ビタミンA、ビタミンB12およびビタミンDは全てLOQ未満であった。ビタミンB1、ビタミンB2、ビタミンB6、ビタミンC、ビタミンEまたはナイアシンについて、対照、噴霧していないAAD−12および処理したAAD−12の間に差異は観察されなかった。入手可能な文献範囲を有するこれらのビタミンの中で、近同質遺伝子対照を含めた全ての処理について値がその範囲を超えたビタミンB2を例外として、全ての処理がこれらの範囲内に入った。
【0246】
デルタ−トコフェロールレベルは、対照と、ダイズイベントDAS−68416−4+グルホシネートエントリーおよびダイズイベントDAS−68416−4+2,4−Dエントリーとの間で、調整していないp値に基づいて有意に異なった。しかし、これは、有意な調整したP値または全体的な処理の効果を伴わなかった。ガンマトコフェロールは、対照と、噴霧なしエントリーおよびダイズイベントDAS−68416−4+2,4−Dエントリーとの間で、調整していないp値および調整したp値に基づいて有意に異なった。しかし、ガンマトコフェロールは、ダイズイベントDAS−68416−4処理について、近同質遺伝子対照と比較して11%未満高かった。
【0247】
ビタミンB5レベルは、対照と、ダイズイベントDAS−68416−4+グルホシネートエントリーとの間で、調整したP値に基づいて有意に異なった。しかし、これは、有意な全体的な処理の効果を伴わなかった。
【0248】
葉酸は、対照と、噴霧なし、ダイズイベントDAS−68416−4+2,4−DおよびダイズイベントDAS−68416−4+両方の除草剤について、調整していないp値に基づいて有意に異なった。葉酸については、ダイズイベントDAS−68416−4エントリーの2種について、対照と比較して調整したp値にも有意差があった。しかし、葉酸レベルは全ての処理について報告された文献値の範囲内であり、ダイズイベントDAS−68416−4エントリーは、近同質遺伝子対照との差異が<9%であり、これは、非トランスジェニックダイズとの同等性を示している。
【0249】
これらの組成成分に基づいて、ダイズイベントDAS−68416−4由来の粒は、非トランスジェニックダイズの粒と実質的に同等であった。
【0250】
【表23】
【0251】
実施例8.2.5
イソフラボン
対照、噴霧していないダイズイベントDAS−68416−4、ダイズイベントDAS−68416−4+グルホシネート、ダイズイベントDAS−68416−4+2,4−DおよびダイズイベントDAS−68416−4+両方の除草剤由来のダイズの粒試料中のイソフラボンの分析を実施した。全ての区域にわたる結果の概要が表24に示されている。
【0252】
ゲニステインおよびグリシテインの結果は、処理した試料について、LOQを下回った。ダイジンレベルは、対照とダイズイベントDAS−68416−4+両方の除草剤エントリーとの間で、調整していないp値および調整したp値に基づいて有意に異なった。しかし、全体的な処理の効果は有意でなかった。報告された文献値はないが、ダイズイベントDAS−68416−4+両方の除草剤処理は、近同質遺伝子対照との差異が<9%であった。ゲニスチンレベルは、対照とダイズイベントDAS−68416−4+両方の除草剤エントリーとの間で、調整していないp値および調整したp値に基づいて有意に異なった。しかし、全体的な処理の効果は有意でなかった。ゲニスチン値は、全ての処理について、報告された文献値よりも高かったが、ダイズイベントDAS−68416−4処理は、近同質遺伝子対照と比較して差異が9%未満であった。グリシチンの値は対照とダイズイベントDAS−68416−4+両方の除草剤との間で、調整していないp値および調整したp値に基づいて有意に異なった。グリシチンについて報告された文献値はなかったが、全てのダイズイベントDAS−68416−4エントリーが、近同質遺伝子エントリーと比較して、差異が13%未満であった。
【0253】
これらの組成成分に基づいて、ダイズイベントDAS−68416−4ダイズ由来の粒は、非トランスジェニックダイズの粒と実質的に同等であった。
【0254】
【表24】
【0255】
実施例8.2.5
抗栄養因子
対照、噴霧していないダイズイベントDAS−68416−4、ダイズイベントDAS−68416−4+グルホシネート、ダイズイベントDAS−68416−4+2,4−DおよびダイズイベントDAS−68416−4+両方の除草剤由来のダイズの粒試料中の抗栄養因子の分析を実施した。全ての区域にわたる結果の概要が表25に示されている。
【0256】
レクチン、フィチン酸、またはトリプシン阻害物質について、対照エントリーとトランスジェニックエントリーとの間に統計学的差異は観察されなかった。これらの3種の抗栄養因子は全て文献範囲内でもあり、これは、非トランスジェニックダイズとの同等性を示している。
【0257】
ラフィノースは、ダイズイベントDAS−68416−4+グルホシネート処理について、対照と比較して調整していないp値に基づいて有意に低かった(<10%)。ラフィノースは、現場全てにわたる分析において、調整したP値または全体的な処理の効果に基づいて有意に異ならなかった。ラフィノースレベルは、全ての処理について報告された文献値の範囲内でもあり、これは、非トランスジェニックダイズとの同等性を示している。
【0258】
スタキオースは、対照およびダイズイベントDAS−68416−4+グルホシネートエントリーとの間で、調整していないP値に基づいて有意に異なった。スタキオースレベルは、現場全てにわたる分析において、調整したP値または全体的な処理の効果に基づいて有意に異ならなかった。スタキオースレベルは、全ての処理について報告された文献値の範囲内でもあり、これは、非トランスジェニックダイズとの同等性を示している。
【0259】
レクチン、フィチン酸、ラフィノース、スタキオースおよびトリプシン阻害物質についての抗栄養因子分析は、全て報告された文献値の範囲内であり、調整していないp値に基づく2つの有意差は、抗栄養因子のレベルがダイズイベントDAS−68416−4処理について対照と比較して低かった。
【0260】
これらの組成成分に基づいて、ダイズイベントDAS−68416−4ダイズ由来の粒は、非トランスジェニックダイズの粒と実質的に同等であった。
【0261】
【表25】
【0262】
実施例8.3
粒および茎葉の組成の概要
ダイズイベントDAS−68416−4の組成物は、近同質遺伝子系統と統計学的に区別できなかったか、または近同質遺伝子系統との差異が13%未満であったか、または非トランスジェニックダイズについての文献範囲内であった。したがって、ダイズイベントDAS−68416−4は、非トランスジェニックダイズと実質的に同等であることが見いだされた。組成の結果のプロットは、噴霧していないダイズイベントDAS−68416−4、ダイズイベントDAS−68416−4+グルホシネート、ダイズイベントDAS−68416−4+2,4−DおよびダイズイベントDAS−68416−4+両方ダイズと対照ダイズ系統との間に、生物学的に意味のある、処理に関連するいかなる組成上の差異も示していない。結論として、噴霧していないダイズイベントDAS−68416−4、ダイズイベントDAS−68416−4+グルホシネート、ダイズイベントDAS−68416−4+2,4−DおよびダイズイベントDAS−68416−4+両方の組成の結果により、ダイズイベントDAS−68416−4と従来のダイズの同等性が確認される。
【実施例9】
【0263】
植え付け前の、一掃するための施用および出芽前試験
この実施例では、本発明によって可能になる、ダイズ作物の植え付けおよび除草剤の使用の新しい方法を記載している。
【0264】
植え付け前の、一掃するための除草剤の施用は、所与の作物を植え付ける前に、冬または初春の間に現れた雑草を枯らすことを意図している。一般には、これらの施用は、植え付け前に雑草の物理的な除去が完了していない不耕起または低耕起の管理体制において適用する。したがって、不耕起または低耕起のダイズと併せて使用する除草剤計画によって、植え付け前に存在する非常に広範な広葉雑草およびイネ科雑草を制御しなければならない。
【0265】
グリホサート、グラモキソン、およびグルホシネートは、植え付け前の、一掃するための除草剤の施用のために広範に使用される非選択的な残留しない除草剤の例である。しかし、一部の雑草は、以下の因子の1つまたは複数に起因して、そのような除草剤で制御することが難しい:除草剤に対する雑草種または生物型の固有の非感受性、比較的大きなサイズの越冬一年生雑草、および除草剤の取り込みおよび活性を制限する寒冷な気候条件。非選択的な除草剤が不十分である場合に、これらの除草剤とタンク混合して雑草に対するスペクトルおよび活性を増加させるために、いくつかの除草剤の選択肢が利用可能である。1つの例は、ヒメムカシヨモギ(Conyza canadensis)(horseweed)を制御するために役立つ2,4−D+グリホサートのタンク混合の施用を用いることである。グリホサートは、存在する雑草の大部分を植え付け前に一掃制御するために420〜1680g ae/ha、より一般には、560〜840g ae/haで使用することができるが、多くの広葉雑草種(例えば、ヒメムカシヨモギ(horseweed))を制御することに役立つように、280〜1120g ae/haの2,4−Dをに施用することができる。
【0266】
2,4−Dは、非常に広範囲の広葉雑草(比較的低温においてさえも)に対して有効であり、かつ非常に安価であるので、一般的に好まれる植え付け前の除草剤である。しかし、一掃するための施用の後に植える作物が2,4−Dに対して感受性が高い場合(例えば、大部分の双子葉作物)、土壌に2,4−Dが残留すること(短期であっても)により、作物が害される恐れがある。ダイズに使用するために、多くの2,4−Dエステル除草剤の表示では、1lb ai/エーカーの一掃するための施用の少なくとも15日後までは新しい作物の植え付けが制限されている。2,4−Dの低揮発性エステル製剤の場合では、一掃するための最大0.5lb ai/エーカーまでの比率では、一般には、施用後7日を過ぎてから植え付けることが必要とされ、1.0lb ai/エーカーの比率では、一般には、施用後30日を過ぎてから植え付けることが必要とされる。水溶性である2,4−Dのアミン製品の場合のさらなる懸念は、除草剤が種子の帯域に浸出する恐れがあることである。しかし、aad−12遺伝子を含有する作物は、2,4−Dに対して耐性であり、土壌中に2,4−Dが残留することによって負の影響を受けない。この耐性に起因して、aad−12遺伝子を含有する植物は、aad−12遺伝子を含有しない植物よりも害を持続することなく、一掃後すぐに植えることができる。さらに、タンク混合(または市販のプレミックス)パートナーと用いる2,4−D除草剤の柔軟性が増し、費用が縮小することにより、雑草制御の選択肢が改善され、重要な不耕起および低耕の状況における一掃するための施用の頑強性が増す。
【0267】
2,4−Dアミンの出芽前施用を、植え付けの7日前、15日前または30日前に、1120g ae/ha、2240g ae/ha、4480g ae/haの比率で、aad−12遺伝子を含有するダイズ(イベントDAS−68416−4)および対照ダイズに適用する。出芽前施用を、当技術分野において認められている手順で、地理的に異なる地方に位置する圃場の小区画に適用する。出芽および早期生長の正確な評価をもたらすために、除草剤処理した小区画は、処理していない小区画と対になっている。植え付け後、および植え付けの7日前、15日前または30日前の2,4−Dの施用後;aad−12遺伝子を含有するダイズ(イベントDAS−68416−4)および対照ダイズの害を測定する。圃場試験の結果は、aad−12遺伝子を含有するダイズ(イベントDAS−68416−4)により、植え付けの7日前、15日前、または30日前の2,4−D除草剤の出芽前処理に頑強な耐性がもたらされることを示している。
【0268】
これらの実施例は、利用可能な多くの選択肢のうちのいくつかである。雑草制御の当業者は、例として、連邦の除草剤表示(CPR、2003年)に記載の製品およびAgriliance Crop Protection Guide(2003年)に記載の用法を利用することによって、これらに限定されないが、グラモキソン+2,4−Dまたはグルホシネート+2,4−Dを含めた種々の他の施用に注目されよう。上記の例を、安定に形質転換されていれば、AAD−12遺伝子によって保護される、2,4−D(または他のフェノキシオーキシン除草剤)に対する感受性が高い任意の作物に施用することができることも、当業者には認識されよう。
【0269】
DAS−68416−4ダイズは、2,4−Dに対して耐性である除草剤である。この実施例では、圃場条件下での、DAS−68416−4ダイズの除草剤耐性の特徴付けについて報告している。DAS−68416−4ダイズを、除草剤である2,4−Dに対する耐性について、米国で行った圃場試験において評価した。DAS−68416−4ダイズについて、提唱された最大の施用率は、2,4−Dについては1120g ae/haである。以下に考察するデータは、DAS−68416−4ダイズにより、これらの提唱された最大使用率の少なくとも4倍の比率において、許容できる耐性がもたらされることを示している。
【0270】
2,4−Dの解毒、およびこの化合物によって後に引き起こされる害からの保護に対するDAS−68416−4ダイズの効力を、圃場研究において特徴付けた。2,4−Dに対する出芽前の耐性を評価するために試験を設計した。出芽前試験を完全乱塊計画として行った。実験単位は、出芽前試験のための、長さおよそ3mの、作条が2つある小区画からなり、全ての試験は3回の反復を含有した。除草剤の施用は全て、噴霧体積およそ140〜190l/haを送達する、ガスで加圧された小さな小区画噴霧装置を用いて行った。使用した2,4−Dの製剤は、456g ae/リットルのジメチルアンモニウム塩であった。作物が出芽したおよそ7日後に、視覚的な害の評点を取得した。評点は、小区画内の植物全てにわたって観察された全ての害の症状の視覚的合成を反映する0〜100の尺度で取得し、0=処理していない小区画と比較して害なし、100=全ての植物が枯死とした。
【0271】
出芽前試験は、植え付け後だが、作物が出芽する前の、1120g ae/ha、2240g ae/ha、および4480g ae/haの施用からなった。出芽前試験に関する情報は、表26に提示されている。
【0272】
【表26】
【0273】
要約すると、2,4−D耐性の圃場評価を行った。ミシシッピ、インディアナ、およびミネソタの区域において、DAS−68416−4ダイズおよび形質転換されていないMaverickダイズの、出芽前の2,4−Dアミンの施用に対する反応を検査する試験を行った。この3区域全てにわたって平均すると、2,4−DアミンをDAS−68416−4に出芽前に施用することにより、施用率に関係なく、2%以下の害が生じた(表27)。形質転換されていないMaverickでは、同じ処理によって34〜60%の害が引き起こされた。2,4−Dを出芽前施用したものの害は、群生の減少および生長の低下からなった。
【0274】
【表27】
【実施例10】
【0275】
グリホサート耐性遺伝子+ダイズイベントDAS−68416−4の組合せを使用してグリホサート抵抗性の雑草を制御する方法
グリホサートは、非常に広範な広葉およびイネ科雑草種を制御するので、広範囲にわたって使用されている。しかし、GTCにおいて、および非作物施用においてグリホサートの使用を繰り返すことにより、雑草のシフトが自然により耐性の種またはグリホサート抵抗性の生物型に選択されてきており、またこれからもそれが続くであろう。大部分の除草剤抵抗性管理戦略では、抵抗性の雑草の出現を遅延させる方法として、同じ種の制御をもたらすが異なる作用様式を有する、効果的な比率で使用されるタンク混合の除草剤パートナーが処方される。416AAD−12イベントにグリホサート耐性形質(および/または他の除草剤耐性形質)を積み重ねることにより、グリホサート、フェノキシオーキシン(複数可)(例えば、2,4−D)およびピリジルオキシ酢酸オーキシン除草剤(例えば、トリクロピル)を同じ作物において選択的に使用することが可能になり、それによってGTCにおいてグリホサート抵抗性の双子葉雑草種を制御することが可能になる機構をもたらすことができた。これらの除草剤は、異なる作用様式の2つ以上の除草剤を含むタンク混合物中で同時に施用することができ;植え付け前、出芽前、または出芽後および分裂時のおよそ2時間からおよそ3カ月までにわたる施用のタイミングで、単一の除草剤の組成をそれぞれ逐次的に施用することができる;あるいは、それぞれの化学的クラスを代表する任意の数の除草剤の任意の組合せを、作物の植え付けの約7カ月以内、作物を収穫するまで(または、個々の除草剤について、いずれか最も短い収穫前の間隔)の任意のタイミングで施用することができる。
【0276】
広範なスペクトルのイネ科雑草および広葉雑草を制御することにおいて、施用のタイミング、個々の除草剤の比率、および厄介なまたは抵抗性の雑草を制御する能力の点で柔軟性を有することが重要である。グリホサート抵抗性遺伝子/AAD−12 416イベントの積み重ねを有する作物におけるグリホサートの施用は、約250〜2500g ae/haにわたってよく;(1種または複数種の)フェノキシオーキシン除草剤は、約25〜4000g ae/ha施用してよく;(1種または複数種の)ピリジルオキシ酢酸オーキシン除草剤は、25〜2000g ae/ha施用してよい。これらの施用(複数可)の最適な組合せ(複数可)およびタイミングは、特定の状況、種、および環境に左右され、雑草制御の当業者および本開示の利益を受ける当業者が決定することが最善である。
【0277】
北アメリカにおいて植えられている綿、キャノーラ、トウモロコシ、およびダイズの畑地の大部分は、グリホサート耐性(GT)形質を含有し、また、GTトウモロコシの採用は増えている。追加的なGT作物(例えば、コムギ、イネ、サトウダイコン、および芝生)が開発中であるが、これまで発売されていない。多くの他のグリホサート抵抗性種は、開発段階への実験中である(例えば、アルファルファ、サトウキビ、サンフラワー、ビート、エンドウ、ニンジン、キュウリ、レタス、タマネギ、イチゴ、トマト、およびタバコ;ポプラおよびモミジバフウのような森林種;ならびにマリーゴールド、ペチュニア、およびベゴニアのような園芸種;World Wide Web、isb.vt.edu/cfdocs/fieldtests1.cfm、2005年)。GTCは、このシステムによって提供される、雑草完全な広がりを制御するための価値があるツールであり、利便性および費用の有効性である。しかし、現在標準の基剤処理としてのグリホサートの有用性は、グリホサート抵抗性の雑草について選択されている。さらに、グリホサートのみの化学的プログラムが実施されている圃場において、グリホサートが本質的にあまり効果的でない雑草が優勢な種にシフトしている。AAD−12イベント416にGT形質を積み重ねることにより、従来の育種によってまたは新規の形質転換イベントと共同で、雑草を制御する効力、柔軟性、および雑草のシフトを管理する能力および除草剤抵抗性の発生を改良することができる。作物をAAD−12イベント416で形質転換することにより、単子葉作物は、フェノキシオーキシンまたはピリジルオキシオーキシンの安全性により高い余地を有することになり、双子葉作物にフェノキシオーキシンを選択的に施用することができる。AAD−12イベント416およびGT形質を任意の単子葉作物種または双子葉作物種に積み重ねた場合、改良された雑草制御の選択肢のためのいくつかのシナリオを構想することができる:
a)大部分のイネ科植物および広葉雑草種を制御するために、グリホサートを標準の出芽後の施用率(420〜2160g ae/ha、560〜840g ae/haが好ましい)で施用することができる。ヒメムカシヨモギ(Conyza canadensis)などのグリホサート抵抗性の広葉雑草またはグリホサートを用いて制御することが本質的に厄介な雑草(例えば、ツユクサ(Commelina)種、サツマイモ(Ipomoea)種など)を制御するために、280〜2240g ae/ha(560〜1120g ae/haが好ましい)の2,4−Dを、グリホサートと、逐次的に、タンク混合して、またはプレミックスとして施用して有効な制御をもたらすことができる。トリクロピルについては、施用率は一般には70〜1120g ae/ha、より一般には140〜420g ae/haの範囲になる。フルロキシピルについては、施用率は、一般には35〜560g ae/ha、より一般には70〜280 ae/haの範囲になる。
b)現在、GTCにおいて施用されるグリホサートの比率は、一般に、施用のタイミング当たり560〜2240g ae/haの範囲である。グリホサートは、広葉雑草種よりもイネ科植物種に対してはるかに効果的である。AAD−12イベント416+GTを積み重ねた形質により、イネ科植物に有効な比率のグリホサート(105−840g ae/ha、より好ましくは210−420g ae/ha)が可能になる。それから、2,4−D(280〜2240g ae/ha、より好ましくは560〜1120g ae/ha)を、イネ科植物に有効な比率のグリホサートと、逐次的に、タンク混合して、またはプレミックスとして施用して必要な広葉雑草の制御をもたらすことができる。上記の比率のトリクロピルおよびフルロキシピルは、処理レジメンにおいて許容できる構成成分になる。低率のグリホサートにより、広葉雑草の制御にもいくらかの利益がもたらされる;しかし、主要な制御は、2,4−D、トリクロピル、またはフルロキシピルによるものである。
【0278】
作物をAAD−12イベント416で形質転換することにより、1つまたは複数の商業的なアリールオキシオーキシン除草剤を単独で、または組み合わせて(逐次的にまたはそれぞれ独立に)使用することが可能になることが、雑草制御の当業者には認識されよう。これらの化学物質を代表する特定の比率の他の除草剤は、CPR(Crop Protection Reference)書籍または同様の編集物によって編集された除草剤の表示、オンライン(例えば、cdms.net/manuf/manuf.asp)で編集された表示、またはAgrilianceからのCrop Protection Guide(2005年)などの任意の商業的なまたは学術的な作物保護の手引きによって決定することができる。AAD−12イベント416によってHTCにおいて使用することが可能になったそれぞれの代替の除草剤は、単独で使用されようが、タンク混合して使用されようが、または逐次的に使用されようが、本発明の範囲内であるとみなされる。
(参考文献)
【技術分野】
【0001】
本発明は、AAD−12ダイズイベントおよびそれに由来する後代に関する。
【背景技術】
【0002】
aad−12遺伝子(元々はデルフチア・アシドボランス(Delftia acidovorans)由来)は、アリールオキシアルカノエートジオキシゲナーゼ(AAD−12)タンパク質をコードする。この形質により、例えば2,4−ジクロロフェノキシ酢酸に対する耐性、およびピリジルオキシ酢酸除草剤に対する耐性が付与される。aad−12遺伝子自体は、植物における除草剤耐性について、WO2007/053482に最初に開示された。
【0003】
植物における異種遺伝子または外来遺伝子の発現は、その外来遺伝子が染色体のどこに挿入されたかに影響される。これは、例えば、クロマチン構造(例えば、ヘテロクロマチン)のため、または組み込み部位に近い転写調節エレメント(例えば、エンハンサー)の近傍にあるためである可能性がある(Weisingら、Ann. Rev. Genet 22巻:421〜477頁、1988年)。同じ種類のトランスジェニック植物(または他の生物体)における同じ遺伝子が、異なるイベント間で発現レベルの広範な変動を示すことがある。発現の空間パターンまたは時間パターンにも差があることがある。例えば、種々の植物組織における導入遺伝子の相対的な発現量の差は、導入される遺伝子構築物中に存在する転写調節エレメントから予測されるパターンに対応しない可能性がある。
【0004】
したがって、多くの場合、所与の目的ために申し分のないレベルまで対象の導入された遺伝子を発現するイベントを特定するために、多数のイベントを創出し、スクリーニングする。商業目的のために、何百から何千もの異なるイベントを作出し、それらのイベントを、導入遺伝子の所望の発現レベルおよび発現パターンを有する単一のイベントについてスクリーニングすることが一般的である。導入遺伝子の発現の所望のレベルおよび/またはパターンを有するイベントは、従来の育種方法を使用して、有性異系交雑によって導入遺伝子を他の遺伝的背景に遺伝子移入するために有用である。そのような交雑の後代は、元の形質転換体の導入遺伝子の発現特性を維持する。この戦略を使用して、その土地の生長条件によく適合する多数の品種における信頼性の高い遺伝子発現を確実にする。
【0005】
米国特許出願公開第20090130071号は、ダイズイベントMON87701および検出方法に関する。米国特許出願公開第20090036308号および同第20080051288号は、ダイズイベント3560.4.3.5および検出方法に関する。米国特許出願第20080312082号は、ダイズイベントDP−305423−1および検出方法に関する。米国特許出願第20060282915号は、ダイズイベントMON89788および検出方法に関する。
【0006】
本明細書に開示されている特定のイベントを有するAAD−12ダイズは、これまでに開示されていない。
【発明の概要】
【0007】
本発明は、American Type Culture Collection(ATCC、アメリカの微生物系統保存機関)に受託番号PTA−10442で寄託されている種子を有する、DAS−68416−4と称されるAAD−12ダイズ(ダイズ(Glycine max))イベント、およびそれに由来する後代に関する。本発明の他の態様は、後代植物、ダイズ、種子、および/またはダイズイベントDAS−68416−4の植物および種子および後代の再生可能な部位、ならびにそのいずれかから製造された食品または飼料製品を含む。本発明は、花粉、胚珠、花、苗条、根、および葉、ならびに栄養細胞、花粉細胞、および卵細胞の核を含むが、これらに限定されない、ダイズイベントDAS−68416−4の植物の部位も包含する。本発明は、さらに、フェノキシオーキシン除草剤および/またはアリールオキシアルカノエート除草剤に対する耐性を有する(これらの除草剤を、ダイズ植物の上から施用するか、近接するまたは近くの土壌に施用するか、または近接するまたは近くの雑草に施用するかにかかわらず)ダイズ植物、ダイズイベントDAS−68416−4の新規の遺伝組成、およびダイズイベントDAS−68416−4を含むダイズ植物の農業生産力の側面に関する。
【0008】
本発明は、一部において、植物の育種および除草剤耐性植物に関する。本発明は、本明細書に記載の通り、ダイズの細胞のゲノム内の特定の部位に挿入されたポリヌクレオチド配列を含むダイズ植物における新規のaad−12形質転換イベントを包含する。
【0009】
一部の実施形態では、前記イベント/ポリヌクレオチド配列に、例えば、他の除草剤耐性遺伝子(複数可)および/または昆虫抑制タンパク質を含めた他の形質を「積み重ねる」ことができる。しかし、本発明は、本明細書に記載の通り、単一のイベントを有する植物を包含する。本発明の特定の実施形態では、種々の除草剤抵抗性の雑草(例えば、グリホサート抵抗性の雑草)を制御するために、1つまたは複数の除草剤耐性形質をダイズイベントDAS−68416−4に積み重ねる。
【0010】
さらに、本発明は、試料(例えば、ダイズの)中の本主題のイベントの存在を検出するためのアッセイを提供する。アッセイは、ダイズゲノムに挿入された組換え構築物のDNA配列、および挿入部位に隣接しているゲノム配列に基づくことができる。アッセイの実施において有用なキットおよび条件も提供される。
【0011】
したがって、本発明は、一部において、(トランスジェニックダイズ系統における)全aad−12挿入断片、およびその境界領域のDNA配列のクローニングおよび分析に関する。これらの配列は独特である。これらの挿入断片および境界の配列に基づいて、イベント特異的なプライマーを生成した。PCR分析により、これらのイベント特異的なプライマーセットを用いて生成したPCRアンプリコンを分析することによってこれらのイベントを同定することができることが実証された。したがって、これらおよび他の関連する手順を用いて、本発明のイベントを含むダイズ系統を一意的に同定することができる。
【図面の簡単な説明】
【0012】
【図1】pDAB4468のプラスミドマップを示す図である。
【図2】ダイズイベントのゲノムDNAを示す図であり、DAS−68416−4をEcoRVまたはPvuIIを用いて消化し、それを使用して対応するGENOMEWALKER(商標)ライブラリーを作製し、これを、標的DNA配列を増幅するために鋳型として使用した。
【図3】ダイズイベントDAS−68416−4の全長配列を5’端から3’端まで確認するためのプライマーの位置を示す図である。
【図4】ダイズイベントDAS−68416−4の全長配列を5’端から3’端まで確認するためのプライマーの位置を示す図である。
【図5】AAD−12ダイズイベントDAS−68416−4の挿入部位の配列を確認するためのプライマーの位置を示す図である。
【図6】植物の生活環を通した発現レベルを示す図である。
【0013】
[表の簡単な説明]
表1は、イベントDAS−68416−4についての挿入断片および隣接配列の配列番号1に関する残基の番号付けを示す。
表2は、サザン分析において使用したプローブの位置および長さを示す。
表3は、サザンブロット分析において予測されたハイブリダイズ断片および観察されたハイブリダイズ断片を示す。
表4は、隣接境界領域を増幅するためのダイズイベントDAS−68416−4のゲノムウォーキングの条件を示す。
表5は、ダイズイベントDAS−68416−4における境界領域およびイベント特異的な配列の標準のPCR増幅のための条件を示す。
表6は、T鎖挿入断片についてのアンプリコン1〜4に対するプライマーの説明を示す。
表7は、ダイズイベントDAS−68416−4における境界領域およびイベント特異的な配列の標準のPCR増幅のためのPCR混合物を示す。
表8は、2008年中に米国およびカナダにおいて生産されたダイズイベントDAS−68416−4から収集した組織中のAAD−12タンパク質レベルの概要を示す。
表9は、サザン分析において使用したプローブの位置および長さを示す。
表10は、サザンブロット分析において予測されたハイブリダイズ断片および観察されたハイブリダイズ断片を示す。
表11は、実験1において評価した農業形質パラメータを示す。
表12は、実験1からの農業特性の分析を示す。
表13は、2009年の農業試験および収量試験において収集されたデータを示す。
表14は、2009年の全ての地域にわたる農業特性の結果の概要を示す。
表15は、実験1からの病害発生率および虫害についての分析を示す。
表16は、DAS−68416−4および従来のダイズの試験において観察された病害ストレス要因および昆虫ストレス要因を示す。
表17は、温暖な条件下および寒冷な条件下でのダイズイベントDAS−68416−4種子の発芽を示す。
表18は、ダイズの茎葉の一般成分(proximate)分析、繊維分析およびミネラル分析の概要を示す。
表19は、ダイズの粒一般成分分析および繊維分析の概要を示す。
表20は、ダイズの粒のミネラル分析の概要を示す。
表21は、ダイズの粒のアミノ酸分析の概要を示す。
表22は、ダイズの粒の脂肪酸分析の概要を示す。
表23は、ダイズの粒のビタミン分析の概要を示す。
表24は、ダイズの粒のイソフラボン分析の概要を示す。
表25は、ダイズの粒の抗栄養因子分析の概要を示す。
表26は、2,4−D出芽前耐性試験についての現場および処理の情報を示す。
表27は、2,4−Dの出芽前施用に対するDAS−68416−4ダイズの耐性を例示している。
【0014】
[配列の簡単な説明]
配列番号1は、本主題のダイズイベントDAS−68416−4についての挿入断片配列および隣接配列を示す。
配列番号2〜28は、本明細書に記載のプライマーである。
配列番号29および30は、本明細書に記載の隣接SNPマーカーであるBARC−019093−03299およびBARC−044607−08736である。
【発明を実施するための形態】
【0015】
本発明は、一部において、植物の育種および除草剤耐性植物に関する。本発明は、本明細書に記載の通り、ダイズの細胞のゲノム内の特定の部位に挿入された、本主題のaad−12ポリヌクレオチド配列を含むダイズ植物(ダイズ)の新規の形質転換イベントを包含する。一部の実施形態では、前記ポリヌクレオチド配列に、他の形質を「積み重ねる」ことができる(例えば、他の除草剤耐性遺伝子(複数可)および/または昆虫抑制タンパク質をコードする遺伝子(複数可)など)。しかし、本発明は、本明細書に記載の単一のイベントを有する植物を包含する。
【0016】
さらに、本発明は、試料中の本主題のイベントの存在を検出するためのアッセイを提供する。本発明の態様は、本明細書において例証または提案される任意の診断用核酸分子、特に、完全にまたは部分的に本主題の隣接配列に基づく診断用核酸分子を設計および/または作出する方法を包含する。
【0017】
より詳細には、本発明は、一部において、トランスジェニックダイズイベントDAS−68416−4、これらのイベントを含む植物系統、ならびにこの挿入断片、および/またはその境界領域のDNA配列のクローニングおよび分析に関する。本発明の植物系統は、本明細書において開示され、提案されている配列を使用して検出することができる。
【0018】
一部の実施形態では、本発明は、除草剤耐性ダイズ系統、およびその同定に関する。本発明は、一部において、有性交雑の後代が対象のイベントを含有するかどうかを決定するために本主題のイベントの存在を検出することに関する。さらに、イベントを検出するための方法が包含され、それは、例えば、組換え作物に由来する食物の市販前承認および表示を必要とする規制に合致するために役立つ。任意の周知の核酸検出方法、例えばポリメラーゼ連鎖反応(PCR)または核酸プローブを使用したDNAのハイブリダイゼーションなどによって本主題のイベントの存在を検出することが可能である。イベント特異的なPCRアッセイは、例えば、Windelsら(Med. Fac. Landbouww、Univ. Gent 64/5b:459462、1999年)によって考察されている。これは、グリホサート耐性のダイズイベント40−3−2を、挿入断片と隣接DNAとの間の接合部にわたるプライマーセットを使用したPCRによって同定することに関する。より詳細には、1つのプライマーは挿入断片由来の配列を含み、第2のプライマーは隣接DNA由来の配列を含んだ。
【0019】
デルフチア・アシドボランス(Delftia acidovorans)に由来する、アリールオキシアルカノエートジオキシゲナーゼ(AAD−12)タンパク質をコードするaad−12遺伝子を挿入することによってダイズを改変した。この形質は、2,4−ジクロロフェノキシ酢酸除草剤およびピリジルオキシ酢酸除草剤に対する耐性を付与し、植物を形質転換する間、および育種苗床において選抜マーカーとして使用することができる。
【0020】
より詳細には、本明細書には、AAD12イベントpDAB4468−0416、ならびに、それを、代々、植物全体および分子レベルで安定性および発現について選抜し、特徴付けすることが記載されている。
【0021】
本発明に従って使用される対象の合成遺伝子(aad−12)は、デルフチア・アシドボランス(Delftia acidovorans)に由来し、フェノキシオーキシン(例えば、2,4−D、MCPA)、ならびにピリジルオキシオーキシン(例えば、フルロキシピル、トリクロピル)を含めたアリールオキシアルカノエート部分を持ついくつかの除草剤を不活性化することができる酵素をコードする。atUbi10プロモーターに駆動されるaad−12遺伝子を、ダイズ系統であるMaverickに、アグロバクテリウム・ツメファシエンス(Agrobacterium tumefaciens)法によって導入した。トランスジェニックT0植物を4〜6世代にわたって自家受粉させた。平行して、優良なダイズ品種にaad−12遺伝子の遺伝子移入も行った。全てのトランスジェニックイベントを、封じ込められ、規制された圃場苗床および研究室の環境において4〜5世代にわたって特徴付けた。
【0022】
イベントpDAB4468−0416挿入物の両末端について配列決定し、特徴付けた。イベント特異的なアッセイを開発した。これは、ダイズゲノム(ダイズの第4染色体)上にマッピングされた;隣接SNPマーカーは、本明細書に配列番号29および30として記載されている。イベントを別の優良な系統に遺伝子移入する。イベントにより、2,4−Dおよびグルホシネートに対する耐性がもたらされる。
【0023】
atUbi10プロモーターによって駆動し、アグロバクテリウム・ツメファシエンス(Agrobacterium tumefaciens)法によって100を超えるAAD12T1Maverickダイズイベントを生成した。イベントpDAB4468−0416を、5つの自家受粉させた世代およびいくらかの戻し交雑した世代の単一系列選抜によって選抜した。これは形態学的に正常であり、全ての自家受粉した世代を通じて、V3における2240g ae/haの2,4−D噴霧に対して耐性であった。イベントは、単一の優性遺伝子として遺伝性であり、その上、自家受粉した世代および戻し交雑した世代において正常なメンデル分離を有した。
【0024】
イベントpDAB4468−0416は、全長の植物転写単位(PTU)を伴う単一の組み込みを有することが見いだされた。ベクターの骨格からは抗生物質抵抗性遺伝子配列は見いだされず、また、aad−12遺伝子のホモ接合状態および半接合性状態において、数世代を通して遺伝子サイレンシングは検出されなかった。aad−12遺伝子は、2,4−D耐性のレベルをもたらす予測レベルで発現された。イベントは、aad−12遺伝子を安定に、複数の世代にわたって、および所与の世代において同胞系列の間で発現した。
【0025】
上の背景技術のセクションにおいて言及されている通り、導入遺伝子の植物ゲノムへの導入および組み込みは、いくつかのランダムなイベントを伴う(したがって発現される所与の挿入物について「イベント」と称する)。すなわち、アグロバクテリウム(Agrobacterium)形質転換、「遺伝子銃」、およびWHISKERなどの多くの形質転換技法を用いると、導入遺伝子がゲノム内のどこに挿入されるかは予測不可能である。したがって、挿入断片の両側の隣接植物ゲノムDNAを同定することは、所与の挿入イベントを有する植物を同定するために重要であり得る。例えば、挿入断片と宿主ゲノムとの接合領域にわたってPCRアンプリコンを生成するPCRプライマーを設計することができる。このPCRアンプリコンを使用して、独特のまたは別個の種類の挿入イベントを同定することができる。
【0026】
「イベント」は、元々ランダムなイベントであるので、本開示の一部として、イベントを含む少なくとも2500種子のダイズ系統がAmerican Type Culture Collection(ATCC)、10801 University Boulevard、Manassa、VA、20110に寄託され、制限なく一般に公開されている(しかし、特許権に支配される)。この寄託物は、ATCC寄託番号PTA−10442と称される。25個のダイズ(Glycine max)種子(AAD−12イベントpDAB4468−0416)のバイアルが、Dow AgroSciences LLC on October 22、2009を代表して寄託された。この寄託物は2009年11月2日に試験され、同日に種子は生育可能(viable)であった。この寄託物は、特許手続のための種子寄託物に関するブタペスト条約の条項に従い、かつその下で作製されたものであり、また、それに従い、かつその下で維持される。寄託物は、公共の受託所であるATCC受託所において、30年間、または最新の要求後5年間、または特許の有効期間のいずれかのより長い期間にわたって制限なく維持され、その期間中に生育不能になった場合は交換される。
【0027】
寄託された種子は本発明の一部である。明らかに、ダイズ植物をこれらの種子から生長させることができ、そのような植物は本発明の一部である。本発明は、これらのダイズ植物に含有される、これらの植物およびその後代を検出するために有用なDNA配列にも関する。本発明の検出方法およびキットは、試験の最終目的に応じて、これらのイベントの任意の1つ、2つ、またはさらには3つ全てを同定することを対象とすることができる。
【0028】
本発明の説明に役立つように、また当業者が本発明を実施する指針となるために、本明細書において定義および例が提供される。特に断りのない限り、用語は、当業者による従来の使用法に従って理解される。米国特許法施行規則第1.822条に明記されているDNA塩基の命名法を使用する。
【0029】
本明細書で使用される場合、用語「後代」は、AAD−12ダイズイベントDAS−68416−4を含む親植物の任意の世代の子孫を意味する。
【0030】
トランスジェニック「イベント」は、植物細胞を異種DNA、すなわち、対象の導入遺伝子を含む核酸構築物で形質転換すること、植物のゲノムに導入遺伝子を挿入することによって生じる植物の集団を再生させること、および特定のゲノム上の位置への挿入を特徴とする特定の植物を選択することによって作出される。用語「イベント」は、元の形質転換体および異種DNAを含むその形質転換体の後代を指す。用語「イベント」は、ゲノムDNA/導入遺伝子DNAを含む、形質転換体と別の品種との間の有性異系交雑によって作出される後代も指す。反復親との戻し交雑を繰り返した後であっても、形質転換された親由来の挿入された導入遺伝子DNAおよび隣接ゲノムDNA(ゲノムDNA/導入遺伝子DNA)は、交雑の後代において同じ染色体上の位置に存在する。用語「イベント」は、元の形質転換体由来のDNA、および挿入DNAを含む一方の親系統(例えば、元の形質転換体および自殖によって生じる後代)と挿入DNAを含有しない親系統との有性交雑の結果として、対象の導入遺伝子を含む挿入DNAを受け取る後代に伝達されることが予測され得る挿入DNAおよび挿入DNAのすぐ近接の隣接ゲノム配列を含むその後代も指す。
【0031】
「接合部配列」は、ゲノムに挿入されたDNAが、挿入点に隣接するダイズのネイティブなゲノム由来のDNAに連結している点にわたり、植物の遺伝物質中の一方または他方の接合部配列の特定または検出は、十分にイベントに特徴的である。本明細書に記載のダイズイベントにおける挿入物にわたるDNA配列および同様の長さの隣接DNAが包含される。そのような特徴的な配列の特定の例が本明細書において提供されるが、挿入物の接合部、または挿入物とゲノム配列との接合部とオーバーラップする他の配列も特徴的であり、本発明に従って使用することができる。
【0032】
本発明は、そのような隣接配列、接合部配列、および挿入断片配列を特定することに関する。関連するPCRプライマーおよびアンプリコンは、本発明に包含される。本発明に従って、挿入DNAおよびその端までわたるアンプリコンを使用するPCR分析方法を使用して、対象の登録商標を持つトランスジェニックダイズ系統に由来する商業化されたトランスジェニックダイズの品種または系統を検出または特定することができる。
【0033】
挿入断片を植物細胞のゲノムに導入するプロセスの間、挿入断片および/またはゲノム隣接配列のいくらかの欠失または他の変化が起こることは珍しくない。したがって、本明細書において提供される関連性のあるプラスミド配列のセグメントは、いくらかの軽微な変動を含む可能性がある。同じことが、本明細書において提供される隣接配列にも当てはまる。したがって、本主題の隣接配列および/または挿入断片配列といくらかの範囲の同一性を有するポリヌクレオチドを含む植物は、本発明の範囲内である。本発明の配列に対する同一性は、本明細書で例示または記載されている配列に対して少なくとも65%配列同一性、より好ましくは少なくとも70%配列同一性、より好ましくは少なくとも75%配列同一性、より好ましくは少なくとも80%同一性、およびより好ましくは少なくとも85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%配列同一性を有するポリヌクレオチド配列であってよい。本明細書において提供されるハイブリダイゼーションおよびハイブリダイゼーション条件は、そのような植物および本発明のポリヌクレオチド配列を定義するためにも使用することができる。隣接配列プラス挿入断片配列の配列は、寄託されている種子を照会して確認することができる。
【0034】
これらの挿入断片のそれぞれの全配列は、それぞれの隣接配列の部分と一緒に、本明細書において配列番号1として提供される。配列番号1(全部で10,212塩基対)に関してこのイベントについての挿入断片および隣接配列の位置が下の表1に示されている。これは、例えば、実施例3においてより詳細に考察されている。
【0035】
【表1】
【0036】
これらの配列(特に隣接配列)は独特である。これらの挿入断片および境界の配列に基づいて、イベント特異的なプライマーを生成した。PCR分析により、これらのイベント特異的なプライマーセットを用いて生成したPCRアンプリコンを分析することによって、異なるダイズ遺伝子型においてこれらのダイズ系統を同定することができることが実証された。したがって、これらおよび他の関連する手順を用いて、これらのダイズ系統を一意的に同定することができる。本明細書において同定される配列は独特である。
【0037】
本発明の検出技法は、植物育種と併せて、1つまたは複数の追加的な対象の形質を後代に与える取り組みにおいて、対象のイベントを含む親植物を別の植物系統と交雑した後に、どの後代植物が所与のイベントを含むかを決定するために特に有用である。これらのPCR分析方法は、ダイズの育種計画ならびに品質管理、特に商業化されたトランスジェニックダイズ種子に有益である。これらのトランスジェニックダイズ系統のPCR検出キットも、現在製造され、使用することができる。これは、製品登録および製品管理にも有益である。
【0038】
さらに、隣接ダイズ/ゲノム配列を使用して、それぞれの挿入断片の遺伝子位置を特異的に同定することができる。この情報を使用して、それぞれのイベントに特異的な分子マーカー系を作製することができる。これらは、育種戦略を加速させるために、および連鎖データを確立するために使用することができる。
【0039】
さらに、隣接配列の情報を使用して、導入遺伝子の組み込みプロセス、ゲノムの組み込み部位の特性、イベントの選別、導入遺伝子およびそれらの隣接配列の安定性、ならびに遺伝子発現(特に遺伝子サイレンシング、導入遺伝子のメチル化パターン、位置の影響、およびMARS[マトリックス付着領域]などの潜在的な発現関連エレメントなどに関連する)を試験し、特徴付けることができる。
【0040】
本開示全てに照らして、本発明は、ATCC寄託番号PTA−10442の下で入手可能な種子を包含することが明白でなければならない。本発明は、受託番号PTA−10442の下でATCCに寄託されている種子から生長させた除草剤耐性ダイズ植物も包含する。本発明は、さらに、前記植物の部位、例えば、葉、組織試料、前記植物によって生産される種子、花粉などを包含する。
【0041】
さらに、本発明は、寄託されている種子から生長させた植物の後裔植物および/または後代植物、好ましくは除草剤耐性ダイズ植物であって、本明細書に記載の検出可能な野生型ゲノムDNA/挿入DNA接合部配列を含むゲノムを有する植物を包含する。本明細書で使用される場合、用語「ダイズ」は、Glycine maxを意味し、ダイズ植物を用いて育種することができるその全品種を包含する。
【0042】
本発明は、本発明の植物を少なくとも一方の親として使用して交雑を行うプロセスをさらに包含する。例えば、本発明は、本明細書において例示されている植物のいずれかを一方の親または両親として有するF1ハイブリッド植物を包含する。また、本発明のそのようなF1ハイブリッドによって生産される種子も本発明の範囲内である。本発明は、例示した植物と、異なる(例えば、純系の親)植物を交雑し、得られたハイブリッド種子を収穫することによってF1ハイブリッド種子を作出するための方法を包含する。本発明は、雌親または雄親のいずれかである、例示した植物を包含する。得られる植物の特性は、親植物を慎重に考察することによって改良することができる。
【0043】
除草剤耐性ダイズ植物は、本明細書で言及されている系統の任意の1つの種子から生長させたダイズ植物からなる第1の親ダイズ植物と、第2の親ダイズ植物との有性交雑を最初に行い、それによって複数の第1の後代植物を作出すること;次いで、除草剤に対して抵抗性である(または、本発明のイベントの少なくとも1つを保有する)第1の後代植物を選択すること;第1の後代植物を自殖させ、それによって、複数の第2の後代植物を作出すること;次いで、第2の後代植物から、除草剤に対して抵抗性である(または、本発明のイベントの少なくとも1つを保有する)植物を選択することによって育種することができる。これらのステップは、第1の後代植物または第2の後代植物を第2の親ダイズ植物または第3の親ダイズ植物と戻し交雑することをさらに含んでよい。次いで本発明のダイズ種子を含むダイズ作物、またはその後代を植え付けることができる。
【0044】
2つの異なるトランスジェニック植物を交配して、それぞれ独立に分離して付加された2つの外因性遺伝子を含有する子孫を作出することもできることがまた理解される。適切な後代を自殖させることにより、付加された外因性遺伝子のどちらについてもホモ接合性である植物を作出することができる。栄養繁殖のように、親植物との戻し交雑および非トランスジェニック植物との異系交雑も意図されている。種々の形質および作物のために一般に使用される他の育種方法は当技術分野で公知である。反復親である望ましいホモ接合性の栽培品種または純系統に、単純に遺伝した、高度に遺伝性の形質の遺伝子を伝達するために戻し交雑育種が使用されてきた。伝達される形質の供給源は、供与親と称されている。生じた植物は、反復親(例えば、栽培品種)の属性および供与親から伝達された望ましい形質を有することが予測される。最初の交雑の後、供与親の表現型を保有する個体を選択し、反復親と繰り返し交雑(戻し交雑)する。生じた親は、反復親(例えば、栽培品種)の属性および供与親から伝達された望ましい形質を有することが予測される。
【0045】
本発明のDNA分子は、マーカー利用育種(MAB)法において分子マーカーとして使用することができる。本発明のDNA分子は、当技術分野で公知の通り、遺伝的に連鎖している作物学的に有用な形質を特定する方法(例えば、AFLPマーカー、RFLPマーカー、RAPDマーカー、SNPおよびSSRなど)において使用することができる。MAB法を使用して、本発明のダイズ植物(またはその後代および任意の他のダイズの栽培品種または品種)との交雑の後代において除草剤抵抗性形質を追跡することができる。DNA分子はこの形質についてのマーカーであり、当技術分野で周知のMAB法を使用して、本発明の少なくとも1つのダイズ系統、またはその後代が親または祖先であったダイズ植物における除草剤抵抗性形質(複数可)を追跡することができる。本発明の方法を使用して、対象イベントを有する任意のダイズ品種を特定することができる。
【0046】
本発明の方法は、本発明の植物を用いて育種するステップを含む、除草剤耐性ダイズ植物を作出する方法を包含する。より詳細には、前記方法は、本発明の2つの植物、または本発明の1つの植物と任意の他の植物とを交雑するステップを含んでよい。好ましい方法は、前記交雑の後代を、本発明に従って検出可能なイベントについて前記後代を分析することによって選択するステップをさらに含む。例えば、本発明を使用して、他の望ましい形質、例えば、作物学的形質など、例えば、本明細書において、種々の実施例において試験されるものなどを含む植物を用いた育種サイクルを通して対象イベントを追跡することができる。対象イベントおよび所望の形質を含む植物を検出し、特定し、選択し、例えば育種のさらなるラウンドに直ちに使用することができる。対象イベント/形質は、昆虫抵抗性形質(複数可)および/または別の除草剤耐性形質と育種を通じて組み合わせ、本発明に従って追跡することもできる。後者の好ましい一実施形態は、対象イベントを除草剤であるジカンバに対する抵抗性をコードする遺伝子と組み合わせて含む植物である。
【0047】
したがって、本発明は、例えば、グリホサート抵抗性(例えば、抵抗性植物または細菌のEPSPS、GOX、GAT)、グルホシネート抵抗性(例えば、Pat、bar)、アセト乳酸合成酵素(ALS)阻害性除草剤への抵抗性(例えば、イミダゾリノン[イマゼタピルなど]、スルホニル尿素系、トリアゾロピリミジンスルホンアニリド、ピリミジニルチオベンゾエート系、および他の化学物質[Csr1、SurAなど])、ブロモキシニル抵抗性(例えば、Bxn)、HPPD(4−ヒドロキシルフェニル−ピルビン酸−ジオキシゲナーゼ)酵素の阻害剤に対する抵抗性、フィトエンデサチュラーゼ(PDS)の阻害剤に対する抵抗性、光化学系II阻害性除草剤に対する抵抗性(例えば、psbA)、光化学系I阻害性除草剤に対する抵抗性、プロトポルフィリノーゲンオキシダーゼIX(PPO)阻害性除草剤に対する抵抗性(例えば、PPO−1)、フェニル尿素除草剤に対する抵抗性(例えば、CYP76B1)、ジカンバ分解酵素(例えば、US20030135879を参照されたい)をコードする形質と組み合わせることができ、その他のものを、単独で、または多数の組合せで積み重ねて、雑草のシフト(weed shift)および/または上述のクラスの任意の除草剤に対する抵抗性を有効に制御する、または妨げる能力をもたらすことができる。
【0048】
追加的な除草剤に関して、いくつかの追加的な好ましいALS(AHASとしても公知である)阻害剤としては、トリアゾロピリミジンスルホンアニリド系(例えば、クロランスラム−メチル、ジクロスラム、フロラスラム、フルメトスラム、メトスラム、およびペノキススラムなど)、ピリミジニルチオベンゾエート系(例えば、ビスピリバックおよびピリチオバックなど)、ならびにフルカルバゾンが挙げられる。いくつかの好ましいHPPD阻害剤としては、メソトリオン、イソキサフルトール、およびスルコトリオンが挙げられる。いくつかの好ましいPPO阻害剤としては、フルミクロラック、フルミオキサジン、フルフェンピル、ピラフルフェン、フルチアセット、ブタフェナシル、カルフェントラゾン、スルフェントラゾン、およびジフェニルエーテル系(例えば、アシフルオルフェン、ホメサフェン、ラクトフェン、およびオキシフルオルフェンなど)が挙げられる。
【0049】
さらに、AAD−12単独に、または1つもしくは複数の追加的なHTC形質を積み重ねたAAD−12に、1つまたは複数の追加的なインプット形質(例えば、昆虫抵抗性、真菌抵抗性、もしくはストレス耐性など)またはアウトプット形質(例えば、生産量の増加、油プロファイルの改良、繊維品質の改良など)を積み重ねることができる。したがって、本発明を使用して、任意の数の作物害虫を柔軟かつ費用効果的に制御する能力を伴う改良された作物の品質の完全な作物パッケージをもたらすことができる。
【0050】
対象のAAD−12酵素は、トランスジェニック発現を可能にして、ほぼ全ての広葉雑草およびイネ科雑草を制御し得る除草剤の組合せに対する耐性をもたらす。AAD−12は、例えば、他のHTC形質(例えば、グリホサート抵抗性、グルホシネート抵抗性、イミダゾリノン抵抗性、ブロモキシニル抵抗性など)、および昆虫抵抗性形質(Cry1F、Cry1Ab、Cry 34/45など)を積み重ねるための優れた除草剤耐性作物(HTC)形質としての機能を果たし得る。さらに、AAD−12は、第2の遺伝子または遺伝子群を用いて遺伝子操作した植物の一次形質転換体を選択するために役立つ選択マーカーとしての機能を果たし得る。
【0051】
本発明のAAD−12遺伝子は、フェノキシ酢酸オーキシン除草剤(例えば2,4−DB、MCPBなど)に変換される化合物に対する抵抗性ももたらす。2,4−DB除草剤に存在する酪酸部分は、β酸化によって植物毒性2,4−ジクロロフェノキシ酢酸に変換される。同様に、MCPBは、β酸化によって植物毒性MCPAに変換される。ブタン酸除草剤は、それ自体は非除草性である。ブタン酸除草剤は、感受性植物においてβ酸化によってそれらのそれぞれの酸に変換され、それは植物毒性である除草剤の酢酸の形態である。急速なβ酸化ができない植物は、ブタン酸除草剤による害を受けない。しかし、急速なβ酸化ができる植物およびブタン酸除草剤を酢酸形態に変換することができる植物は、その後にAAD−12によって保護される。
【0052】
除草剤を施用する方法は当技術分野で公知である。そのような施用は、2種以上の除草剤のタンク混合物を含んでよい。本発明に従って使用するためのいくつかの好ましい除草剤としては、2,4−D;2,4−DB;MCPA;MCPBなどのフェノキシオーキシン除草剤が挙げられる。これらに、1種または複数種の追加的な除草剤耐性遺伝子(複数可)および対応する除草剤(例えばグリホサートおよび/またはグルホシネート)を積み重ねることができる。1種、2種、3種、またはそれ以上の除草剤を、本開示の利益を有する当業者に明白になる有利な組合せで使用することができる。本主題の除草剤の1種または複数種を圃場/地域に、本発明の種子を植え付ける前に施用することができる。そのような施用は、例えば、植え付けの14日間以内であってよい。本主題の除草剤の1種または複数種は、植え付け時および/または植え付け後であるが出芽前に施用することもできる。本主題の除草剤の1種または複数種は、土に(雑草を制御するために)、または雑草および/または本発明のトランスジェニック植物の上から施用することもできる。本主題の除草剤は、例えば、ある除草剤には耐性であるが、別の除草剤には耐性でない可能性がある雑草を制御する、または防ぐために、交替させること、または組み合わせて使用することができる。本主題の3種類の除草剤について、種々の施用時間を当技術分野で公知の種々の方法で用いることができる。本発明は、AAD−12自生植物を制御するための方法ももたらす。発明の名称が「CONTROL OF AAD DICOT VOLUNTEERS IN MONOCOT CROPS」である同時に出願されたPCT出願を参照されたい。
【0053】
本発明のHTC形質は、他のHTC形質(これらに限定されないが、グリホサート耐性を含む)との新規の組合せにおいて使用することができる。これらの形質の組合せにより、除草剤(例えば、グリホサート)に対する新しく獲得した抵抗性または固有の耐性によって雑草(および同様のもの)種を制御する新規の方法が生じる。したがって、HTC形質に加えて、トランスジェニック作物において、前記酵素によって除草剤耐性を創出する、除草剤を使用して雑草を制御するための新規の方法は、本発明の範囲内である。
【0054】
さらに、世界中で栽培されているグリホサート耐性作物が一般的である。他のグリホサート耐性作物との輪作において多くの場合、グリホサート抵抗性自生植物を制御することは、輪作作物においては難しい。したがって、作物に個々に積み重ねた、または作物を個々に形質転換した対象のトランスジェニック形質の使用は、他のHTC自生作物を制御するためのツールを提供する。
【0055】
本発明の好ましい植物または種子は、そのゲノム内に、本明細書において同定される挿入断片配列と、本明細書において同定される、挿入断片の両側の少なくとも20〜500以上の連続した隣接ヌクレオチドを一緒に含む。別段の指定のない限り、隣接配列に言及する場合、それは配列番号1について特定されたものを指す(上の表1を参照されたい)。さらに、配列番号1は、元の形質転換体に挿入された異種DNAおよび挿入DNAのすぐ近接する例示的な隣接ゲノム配列を含む。これらの隣接配列の全部または一部は、イベントを含む親系統の有性交雑の結果として挿入DNAを受け取る後代に伝達されることが予測され得る。
【0056】
本発明は、本発明の植物の再生可能な細胞の組織培養物を包含する。また、そのような組織培養物から再生された植物も、特に前記植物が例示されている品種の形態学的性質および生理的性質の全てを発現することができる場合、包含される。本発明の好ましい植物は、寄託されている種子から生長させた植物の生理的特性および形態学的特性の全てを有する。本発明は、そのような種子および対象の品質形質を保有する種子の後代をさらに含む。
【0057】
植物または種子、またはその部位を操作(例えば、突然変異、さらなるトランスフェクション、およびさらに育種)することにより、「従属(本質的に由来する)」品種と称することができるものが創出され得る。植物新品種保護国際同盟(International Union for the Protection of New Varieties of Plants)(UPOV)は、ある品種が、保護された品種に由来しているかどうかを決定するための以下のガイドラインを提供した:
[A]以下の場合、品種は別の品種(「原品種」)に本質的に由来するとみなされるべきである
(i)原品種に主として由来する、または、それ自体が原品種に主として由来する品種に由来し、同時に原品種の遺伝子型または遺伝子型の組合せから生じる本質的な特性の発現を保持する;
(ii)原品種と明確に区別可能である;および
(iii)引き出す行為に由来する差異以外は、原品種の遺伝子型または遺伝子型の組合せから生じる本質的な特性の発現において原品種と一致する。
UPOVの国際機関の第6回会議、ジュネーブ、1992年10月30日;当該連合の事務局により作成された文書。
【0058】
本明細書で使用される場合、「系統」は、少なくとも1つの形質について、個体間で遺伝的変異をほとんど示さない、または示さない植物の群である。そのような系統は、何世代か自家受粉させ、選択すること、または組織または細胞の培養技法を使用して、単一の親から栄養繁殖させることによって創出することができる。
【0059】
本明細書で使用される場合、用語「栽培品種」および「品種」は同義であり、商業生産のために使用される系統を指す。
【0060】
「安定性」または「安定な」は、所与の構成要素に関しては、構成要素が代々、好ましくは、少なくとも3世代、実質的に同じレベルで、例えば、好ましくは±15%、より好ましくは±10%、最も好ましくは±5%で維持されることを意味する。安定性は、温度、位置、ストレスおよび植え付け時間に影響を受ける可能性がある。その後の世代を圃場条件下で比較することにより、構成要素が同様にもたらされるべきである。
【0061】
「商業的有用性」は、従来の農業設備を使用して農業者が作物を生産することができるように、および従来の圧搾および抽出用設備を使用して記載の構成要素を有する油を種子から抽出することができるように良好な植物生長力および高い稔性を有することと定義される。商業的に有用であるために、種子の重量、油の含有量、および1エーカー当たりの生産された油の総計によって測定した生産量は、同じ地域で栽培される高価値の形質を有さない、別の匹敵する市販のキャノーラ品種の平均生産量の15%以内である。
【0062】
「作物学的に優良」は、系統が対象イベント(複数可)に起因する昆虫抵抗性に加えて、例えば生産量、成熟度、病害抵抗性などの望ましい作物学的特性を有することを意味する。以下の実施例に記載の、本発明のイベントを含む植物に個々に、または任意の組合せで獲得される作物学的形質は、本発明の範囲内である。これらの作物学的特性およびデータポイントの全てを使用して、そのような植物を、そのような植物を定義するために使用する特性の範囲内の一点として、または一端もしくは両端で特定することができる。
【0063】
本開示に照らして当業者に理解されるように、検出キットの好ましい実施形態は、例えば、「接合部配列」または「移行部配列」(ダイズゲノムの隣接配列と挿入断片配列が接する場所)を対象とし、かつ/またはそれを含むプローブおよび/またはプライマーを含んでよい。これは、例えば、上記の表1に示されている一方のまたは両方の接合部配列(挿入断片と隣接配列が接する場所)を同定するために設計されたポリヌクレオチドプローブ、プライマー、および/またはアンプリコンを含む。1つの一般的な設計は、隣接領域内でハイブリダイズする1つのプライマー、および挿入断片内でハイブリダイズする1つのプライマーを有する。そのようなプライマーは、多くの場合、およそ、それぞれの長さが少なくとも約15残基である。この配置を用いて、プライマーを使用して、本発明のイベントの存在を示す検出可能なアンプリコンを生成/増幅することができる。これらのプライマーを使用して、上に示されている接合部配列にわたる(およびそれを含む)アンプリコンを生成することができる。
【0064】
隣接配列に「接している」プライマー(複数可)は、一般には、約200塩基を越えて、または接合部を越えてハイブリダイズするようには設計されない。したがって、典型的な隣接プライマーは、挿入断片の最初から隣接配列に入って200塩基の範囲内のどちらかの鎖の少なくとも15残基を含むように設計され得る。すなわち、配列番号1の残基約2530〜2730および/または約9122〜9322由来の(またはそれとハイブリダイズする)適切なサイズの配列を含むプライマーは、本発明の範囲内である。挿入プライマーは、挿入断片のどこにでも同様に設計することができるが、残基約2731〜2931および約8921〜9121を、例えば、そのようなプライマーの設計のために非排他的に使用することができる。
【0065】
当業者は、さまざまな標準のハイブリダイゼーションおよび/またはPCRの条件の下で、配列番号1(または相補物)のセグメント、およびその相補物とハイブリダイズするようにプライマーおよびプローブを設計することができ、ここでプライマーまたはプローブが例示された配列と完全に相補的ではないことも認識されよう。すなわち、ある程度のミスマッチが容認され得る。およそ20ヌクレオチドのプライマーについて、例えば、一般には、ミスマッチ塩基がアンプリコンと逆のプライマーの内部または末端にある場合、1または2ヌクレオチド程度は逆の鎖と結合しなくてよい。種々の適切なハイブリダイゼーション条件が以下に提供される。イノシンなどの合成ヌクレオチド類似体は、プローブにも使用することができる。ペプチド核酸(PNA)プローブ、ならびにDNAプローブおよびRNAプローブも使用することができる。重要なのは、そのようなプローブおよびプライマーが、本発明のイベントの存在に対して特徴的である(独自に特定し、区別することができる)ことである。
【0066】
PCR増幅において、例えば、軽微な配列決定のエラーを生じる可能性があるエラーが起こり得ることに留意するべきである。すなわち、別段の指定のない限り、本明細書において列挙されている配列は、ダイズのゲノムDNAから長いアンプリコンを生成し、次いでそのアンプリコンをクローニングし、配列決定することによって決定した。ゲノムDNAから配列決定するために十分なアンプリコンを生成するために必要な多数回の増幅を考慮すると、このように生成し、決定した配列においてわずかな差異および軽微な不一致が見いだされることは珍しいことではない。当業者は、これらの型の一般的な配列決定のエラーまたは不一致に起因して必要になる調整はいずれも本発明の範囲内であることを認識し、留意するべきである。
【0067】
例えば、イベントを創出する間に配列を挿入する場合、いくらかのゲノム配列が欠失することは珍しくないことにも留意すべきである。したがって、本主題の隣接配列と、例えばGENBANKに列挙されているゲノム配列との間にもいくらかの差異が出現する可能性がある。
【0068】
「挿入断片」の構成成分が図面に例示されており、以下の実施例においてより詳細に考察されている。これらの構成要素のDNAポリヌクレオチド配列、またはその断片を、本発明の方法においてDNAプライマーまたはプローブとして使用することができる。
【0069】
本発明の一部の実施形態では、ダイズ植物由来の植物および種子などにおける導入遺伝子/ゲノムの挿入領域の存在を検出するための組成物および方法が提供される。本明細書において提供される本主題の導入遺伝子/ゲノムの挿入領域の接合部配列(配列番号1の残基2730〜2731と残基9121〜9122の間)、そのセグメント、および例示された配列の相補物およびその任意のセグメントを含むDNA配列が提供される。挿入領域の接合部配列は、ゲノムに挿入された異種DNAと、挿入部位に隣接しているダイズの細胞由来のDNAの接合部にわたる。そのような配列は、所与のイベントに対して特徴的であり得る。
【0070】
これらの挿入断片および境界の配列に基づいて、イベント特異的なプライマーを生成することができる。PCR分析により、これらのイベント特異的なプライマーセットを用いて生成したPCRアンプリコンを分析することによって、異なるダイズ遺伝子型において本発明のダイズ系統を同定することができることが実証された。これらおよび他の関連する手順を用いて、これらのダイズ系統を一意的に同定することができる。したがって、そのようなプライマー対に由来するPCRアンプリコンは独特であり、これらのダイズ系統を同定するために使用することができる。
【0071】
一部の実施形態では、新規の導入遺伝子/ゲノムの挿入領域の連続した断片を含むDNA配列は本発明の態様である。導入遺伝子挿入断片配列の十分な長さのポリヌクレオチドおよび3つの上述のダイズ植物の1つまたは複数由来のダイズのゲノム配列の十分な長さのポリヌクレオチドを含むDNA配列および/またはこれらのダイズ植物の1つまたは複数に対して特徴的なアンプリコン産物を生成するためのプライマー配列として有用である配列が包含される。
【0072】
関連する実施形態は、本明細書において特定されるDNA配列(例えば、配列番号1およびそのセグメントなど)の導入遺伝子部分の少なくとも2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、またはそれ以上の連続したヌクレオチドを含むDNA配列、またはその相補物、およびこれらの配列由来の同様の長さのダイズの隣接DNA配列、またはその相補物に関する。そのような配列は、DNA増幅方法において、DNAプライマーとして有用である。これらのプライマーを使用して生成されるアンプリコンは、本明細書で言及されるダイズイベントのいずれかに対して特徴的である。したがって、本発明は、そのようなDNAプライマーおよび相同プライマーによって生成されるアンプリコンも包含する。
【0073】
本発明は、試料中の、本明細書で言及されるダイズイベントに対応するDNAの存在を検出する方法も包含する。そのような方法は、(a)DNAを含む試料を、これらのダイズイベントの少なくとも1つ由来のDNAを用いた核酸の増幅反応において使用したとき、前記イベント(複数可)に対して特徴的であるアンプリコンを生成するプライマーセットと接触させるステップと;(b)核酸の増幅反応を実施し、それによって、アンプリコンを生成するステップと;(c)アンプリコンを検出するステップとを含んでよい。
【0074】
本発明の別の検出方法は、試料中の、前記イベントの少なくとも1つに対応するDNAの存在を検出する方法であって、(a)DNAを含む試料を、ストリンジェントなハイブリダイゼーション条件下で前記ダイズイベントの少なくとも1つ由来のDNAとハイブリダイズさせ、ストリンジェントなハイブリダイゼーション条件下で対照のダイズ植物(対象のイベントがないDNA)とハイブリダイズしないプローブと接触させるステップと;(b)試料およびプローブをストリンジェントなハイブリダイゼーション条件に供するステップと;(c)プローブのDNAとのハイブリダイゼーションを検出するステップとを含む方法を包含する。
【0075】
さらに別の実施形態では、本発明は、本発明のaad−12イベントを含むダイズ植物を作出する方法であって、(a)第1の親ダイズ系統(前記系統の植物に前記除草剤抵抗性形質を付与する本発明の発現カセットを含む)と第2の親ダイズ系統(この除草剤耐性形質を欠く)を有性交雑し、それによって、複数の後代植物を作出するステップと;(b)分子マーカーを使用することによって後代植物を選択するステップとを含む方法を包含する。そのような方法は、場合によって、後代植物を第2の親ダイズ系統と戻し交雑して、前記昆虫耐性形質を含む真の育種(true-breeding)ダイズ植物を作出するさらなるステップを含んでよい。
【0076】
本発明の別の態様によると、前記3種のイベントの任意の1つ(または複数)を用いて交雑の後代の接合性を決定する方法が提供される。前記方法は、ダイズDNAを含む試料を本発明のプライマーセットと接触させるステップを含んでよい。前記プライマーは、前記ダイズイベントの少なくとも1つ由来のゲノムDNAを用いた核酸の増幅反応において使用したとき、前記ダイズイベントの少なくとも1つに対して特徴的である第1のアンプリコンを生成する。そのような方法は、核酸の増幅反応を実施し、それによって、第1のアンプリコンを生成するステップと;第1のアンプリコンを検出するステップと;ダイズDNAを含む試料を、ダイズ植物由来のゲノムDNAを用いた核酸の増幅反応において使用したとき、ダイズのゲノム領域と相同なネイティブなダイズのゲノムDNAを含む第2のアンプリコンを生成する前記プライマーセットと接触させるステップと;核酸の増幅反応を実施し、それによって、第2のアンプリコンを生成するステップとをさらに含む。この方法は、第2のアンプリコンを検出するステップと、試料中の第1のアンプリコンと第2のアンプリコンとを比較するステップであって、両方のアンプリコンが存在することにより、試料が、導入遺伝子挿入物についてヘテロ接合性であることが示されるステップとをさらに含む。
【0077】
DNA検出キットは、本明細書に開示されている組成物およびDNAの検出の技術分野で周知の方法を使用して開発することができる。このキットは、試料中の対象のダイズイベントDNAを特定するために有用であり、このDNAを含有するダイズ植物を育種するための方法に適用することができる。キットは、例えば、本明細書に開示されているアンプリコンと相同または相補的であるDNA配列、または対象イベントの導入遺伝子の遺伝エレメントに含有されるDNAと相同または相補的であるDNA配列を含有する。これらのDNA配列は、DNA増幅反応において、またはDNAのハイブリダイゼーション方法においてプローブとして、使用することができる。キットは、検出方法を実行するために必要な試薬および材料も含有することができる。
【0078】
「プローブ」は、従来の検出可能な標識またはレポーター分子(例えば、放射性同位元素、リガンド、化学発光剤、または酵素など)を付着させた単離された核酸分子である。そのようなプローブは、標的核酸の鎖、本発明の場合では、ダイズ植物由来であるかイベント由来のDNAを含む試料由来であるかにかかわらず、前記ダイズイベントの1つ由来のゲノムDNAの鎖と相補的である。本発明によるプローブは、デオキシリボ核酸またはリボ核酸だけでなく、標的DNA配列に特異的に結合し、その標的DNA配列の存在を検出するために使用することができるポリアミドおよび他のプローブ材料も含む。
【0079】
「プライマー」は、核酸ハイブリダイゼーションによって相補的な標的DNA鎖とアニーリングしてプライマーと標的DNA鎖との間のハイブリッドを形成し、次いでポリメラーゼ、例えば、DNAポリメラーゼによって標的DNA鎖に沿って伸長される、単離された/合成された核酸である。本発明のプライマー対は、例えば、ポリメラーゼ連鎖反応(PCR)または他の従来の核酸の増幅方法によって標的核酸配列を増幅するためのそれらの使用を指す。
【0080】
プローブおよびプライマーは、一般に、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35、36、37、38、39、40、41、42、43、44、45、46、47、48、49、50、51、52、53、54、55、56、57、58、59、60、61、62、63、64、65、66、67、68、69、70、71、72、73、74、75、76、77、78、79、80、81、82、83、84、85、86、87、88、89、90、91、92、93、94、95、96、97、98、99、100、101、102、103、104、105、106、107、108、109、110、111、112、113、114、115、116、117、118、119、120、121、122、123、124、125、126、127、128、129、130、131、132、133、134、135、136、137、138、139、140、141、142、143、144、145、146、147、148、149、150、151、152、153、154、155、156、157、158、159、160、161、162、163、164、165、166、167、168、169、170、171、172、173、174、175、176、177、178、179、180、181、182、183、184、185、186、187、188、189、190、191、192、193、194、195、196、197、198、199、200、201、202、203、204、205、206、207、208、209、210、211、212、213、214、215、216、217、218、219、220、221、222、223、224、225、226、227、228、229、230、231、232、233、234、235、236、237、238、239、240、241、242、243、244、245、246、247、248、249、250、251、252、253、254、255、256、257、258、259、260、261、262、263、264、265、266、267、268、269、270、271、272、273、274、275、276、277、278、279、280、281、282、283、284、285、286、287、288、289、290、291、292、293、294、295、296、297、298、299、300、301、302、303、304、305、306、307、308、309、310、311、312、313、314、315、316、317、318、319、320、321、322、323、324、325、326、327、328、329、330、331、332、333、334、335、336、337、338、339、340、341、342、343、344、345、346、347、348、349、350、351、352、353、354、355、356、357、358、359、360、361、362、363、364、365、366、367、368、369、370、371、372、373、374、375、376、377、378、379、380、381、382、383、384、385、386、387、388、389、390、391、392、393、394、395、396、397、398、399、400、401、402、403、404、405、406、407、408、409、410、411、412、413、414、415、416、417、418、419、420、421、422、423、424、425、426、427、428、429、430、431、432、433、434、435、436、437、438、439、440、441、442、443、444、445、446、447、448、449、450、451、452、453、454、455、456、457、458、459、460、461、462、463、464、465、466、467、468、469、470、471、472、473、474、475、476、477、478、479、480、481、482、483、484、485、486、487、488、489、490、491、492、493、494、495、496、497、498、499、または500ポリヌクレオチドまたはそれ以上の長さである。そのようなプローブおよびプライマーは、高ストリンジェンシーハイブリダイゼーション条件下で標的配列と特異的にハイブリダイズする。好ましくは、本発明によるプローブおよびプライマーは、標的配列と完全な配列類似性を有するが、標的配列とは異なり、標的配列とハイブリダイズする能力を保持するプローブを従来の方法によって設計することができる。
【0081】
プローブおよびプライマーを調製し、使用するための方法は、例えば、Molecular Cloning: A Laboratory Manual、第2版、1〜3巻Sambrookら編、Cold Spring Harbor Laboratory Press、Cold Spring Harbor、N.Y.、1989年に記載されている。PCRのプライマー対は、公知の配列から、例えば、その目的のためのコンピュータプログラムを使用することによって得ることができる。
【0082】
本明細書に開示されている隣接DNAおよび挿入断片の配列に基づくプライマーおよびプローブを使用して、従来の方法によって、例えば、そのような配列を再クローニングし、配列決定することによって、開示されている配列を確認すること(および、必要であれば、補正すること)ができる。
【0083】
本発明の核酸プローブおよびプライマーは、ストリンジェントな条件下で標的DNA配列とハイブリダイズする。任意の従来の核酸ハイブリダイゼーションまたは増幅の方法を使用して、試料中のトランスジェニックイベント由来のDNAの存在を特定することができる。核酸分子またはその断片は、ある特定の状況下で他の核酸分子と特異的にハイブリダイズすることができる。本明細書で使用される場合、2つの核酸分子は、2つの分子が、逆平行の、二本鎖核酸構造を形成することができる場合、互いと特異的にハイブリダイズすることができると言える。核酸分子は、別の核酸分子と完全な相補性を示す場合、その「相補物」であると言える。本明細書で使用される場合、分子は、分子の一方のあらゆるヌクレオチドが、他方のヌクレオチドと相補的である場合、「完全な相補性」を示すと言える。2つの分子は、少なくとも従来の「低ストリンジェンシー」条件下で、互いとアニーリングしたままであることを可能にするために十分な安定性で互いとハイブリダイズすることができる場合、「最小限相補的」であると言える。同様に、分子は、従来の「高ストリンジェンシー」条件下で、互いとアニーリングしたままであることを可能にするために十分な安定性で互いとハイブリダイズすることができる場合、「相補的」であると言える。従来のストリンジェンシー条件は、Sambrookら、1989年に記載されている。したがって、完全な相補性から逸脱することは、そのような逸脱により、二本鎖構造を形成する分子の能力が完全に妨げられない限りは許容できる。核酸分子がプライマーまたはプローブとしての機能を果たすためには、用いる特定の溶媒および塩濃度の下で安定な二本鎖構造を形成できるために十分に配列内で相補的であることのみが必要である。
【0084】
本明細書で使用される場合、実質的に相同な配列は、高ストリンジェンシー条件下で比較されている核酸配列の相補物と特異的にハイブリダイズし得る核酸配列である。用語「ストリンジェントな条件」は、Sambrookら、1989年、9.52〜9.55において考察されている特定のハイブリダイゼーション手順による、核酸プローブの標的核酸(すなわち、対象とする特定の核酸配列)とのハイブリダイゼーションに関して機能的に定義される。Sambrookら、1989年、9.47〜9.52および9.56〜9.58も参照されたい。したがって、本発明のヌクレオチド配列は、相補的なDNA断片のひと続きと2重鎖分子を選択的に形成するそれらの能力に関して使用することができる。
【0085】
構想される適用に応じて、プローブの標的配列に対する選択性のさまざまな程度を実現するためにハイブリダイゼーションのさまざまな条件を使用することができる。高い選択性を必要とする適用のために、一般には、ハイブリッドを形成するために比較的ストリンジェントな条件を用いることがある、例えば、約50℃〜約70℃の温度で約0.02M〜約0.15MのNaClによってもたらされるような比較的低塩かつ/または高温の条件を選択することがある。ストリンジェントな条件は、例えば、ハイブリダイゼーション濾過器を高ストリンジェンシー洗浄緩衝液(0.2×SSC、0.1%のSDS、65℃)で少なくとも2回洗浄することを伴ってよい。DNAのハイブリダイゼーションを促進する適切なストリンジェンシー条件、例えば、約45℃で6.0×塩化ナトリウム/クエン酸ナトリウム(SSC)、その後50℃で2.0×SSCで洗浄することは、当業者に公知である。例えば、洗浄ステップにおける塩濃度は、低ストリンジェンシーである50℃で約2.0×SSCから高ストリンジェンシーである50℃で約0.2×SSCまでから選択することができる。さらに、洗浄ステップにおける温度は、低ストリンジェンシー条件である室温、約22℃から高ストリンジェンシー条件である約65℃まで増加させることができる。温度と塩との両方が変動してよい、または、温度もしくは塩濃度のいずれかは、他方の変数が変化する一方で一定に保たれてよい。そのような選択的な条件は、もしあれば、プローブと鋳型または標的鎖との間のミスマッチを少し容認する。ハイブリダイゼーションによってDNA配列を検出することは当業者に周知であり、米国特許第4,965,188号および同第5,176,995号の教示は、ハイブリダイゼーション分析の方法の例である。
【0086】
特に好ましい実施形態では、本発明の核酸は、高ストリンジェンシー条件下で、本明細書において例証または提案される、その相補物および断片を含めたプライマー(またはアンプリコンもしくは他の配列)の1つまたは複数と特異的にハイブリダイズする。本発明の一態様では、本発明のマーカー核酸分子は、本明細書において例示された配列の1つに記載されている核酸配列、またはその相補物および/もしくは断片を有する。
【0087】
本発明の別の態様では、本発明のマーカー核酸分子は、そのような核酸配列と、80%から100%の間または90%から100%の間の配列同一性を共有する。本発明の別の態様では、本発明のマーカー核酸分子は、そのような配列と95%から100%の間の配列同一性を共有する。そのような配列は、遺伝的交雑の後代を特定するための植物育種方法において、マーカーとして使用することができる。プローブの標的DNA分子とのハイブリダイゼーションは、当業者に公知の任意の数の方法によって検出することができ、それらとしては、これらに限定されないが、蛍光タグ、放射性タグ、抗体ベースのタグおよび化学発光タグを挙げることができる。
【0088】
特定の増幅プライマー対を使用する標的核酸配列の増幅(例えば、PCRによる)に関して、「ストリンジェントな条件」は、プライマー対が、対応する野生型配列(またはその相補物)を有するプライマーが結合し得る標的核酸配列のみとハイブリダイズすること、および好ましくは特有の増幅産物、アンプリコンを生成することを可能にする条件である。
【0089】
用語「(標的配列)に特異的な」は、プローブまたはプライマーが、ストリンジェントなハイブリダイゼーション条件下で、標的配列を含む試料中の標的配列のみとハイブリダイズすることを示す。
【0090】
本明細書で使用される場合、「増幅されたDNA」または「アンプリコン」は、核酸鋳型の一部である標的核酸配列の核酸増幅産物を指す。例えば、有性交雑によって生じたダイズ植物が、本発明のダイズ植物由来のトランスジェニックイベントゲノムDNAを含有するかどうかを決定するために、ダイズ植物の組織試料から抽出したDNAを、挿入された異種DNAの挿入部位の近接する植物のゲノム内の隣接配列に由来するプライマー、および挿入された異種DNAに由来する第2のプライマーを含むプライマー対を使用して、イベントDNAの存在に対して特徴的であるアンプリコンを生成する核酸の増幅方法に供することができる。アンプリコンは、ある長さであり、同じくイベントに対して特徴的である配列を有する。アンプリコンの長さは、プライマー対の全て合わせた長さ足す1ヌクレオチド塩基対、および/またはプライマー対の全て合わせた長さ足す約2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35、36、37、38、39、40、41、42、43、44、45、46、47、48、49、50、51、52、53、54、55、56、57、58、59、60、61、62、63、64、65、66、67、68、69、70、71、72、73、74、75、76、77、78、79、80、81、82、83、84、85、86、87、88、89、90、91、92、93、94、95、96、97、98、99、100、101、102、103、104、105、106、107、108、109、110、111、112、113、114、115、116、117、118、119、120、121、122、123、124、125、126、127、128、129、130、131、132、133、134、135、136、137、138、139、140、141、142、143、144、145、146、147、148、149、150、151、152、153、154、155、156、157、158、159、160、161、162、163、164、165、166、167、168、169、170、171、172、173、174、175、176、177、178、179、180、181、182、183、184、185、186、187、188、189、190、191、192、193、194、195、196、197、198、199、200、201、202、203、204、205、206、207、208、209、210、211、212、213、214、215、216、217、218、219、220、221、222、223、224、225、226、227、228、229、230、231、232、233、234、235、236、237、238、239、240、241、242、243、244、245、246、247、248、249、250、251、252、253、254、255、256、257、258、259、260、261、262、263、264、265、266、267、268、269、270、271、272、273、274、275、276、277、278、279、280、281、282、283、284、285、286、287、288、289、290、291、292、293、294、295、296、297、298、299、300、301、302、303、304、305、306、307、308、309、310、311、312、313、314、315、316、317、318、319、320、321、322、323、324、325、326、327、328、329、330、331、332、333、334、335、336、337、338、339、340、341、342、343、344、345、346、347、348、349、350、351、352、353、354、355、356、357、358、359、360、361、362、363、364、365、366、367、368、369、370、371、372、373、374、375、376、377、378、379、380、381、382、383、384、385、386、387、388、389、390、391、392、393、394、395、396、397、398、399、400、401、402、403、404、405、406、407、408、409、410、411、412、413、414、415、416、417、418、419、420、421、422、423、424、425、426、427、428、429、430、431、432、433、434、435、436、437、438、439、440、441、442、443、444、445、446、447、448、449、450、451、452、453、454、455、456、457、458、459、460、461、462、463、464、465、466、467、468、469、470、471、472、473、474、475、476、477、478、479、480、481、482、483、484、485、486、487、488、489、490、491、492、493、494、495、496、497、498、499、または500、750、1000、1250、1500、1750、2000、またはそれ以上のヌクレオチド塩基対(上に列挙されている増大のいずれかを足すまたは引く)にわたってよい。あるいは、プライマー対は、挿入断片のヌクレオチド配列全体を含むアンプリコンが生成されるように、挿入DNAの両側の隣接配列に由来してもよい。植物のゲノム配列に由来するプライマー対のメンバーは、挿入DNA配列からある距離に位置してよい。この距離は、1ヌクレオチド塩基対から最大約2万ヌクレオチド塩基対までにわたることができる。用語「アンプリコン」の使用は、DNAの熱増幅反応において形成される可能性があるプライマー二量体を特に除外する。
【0091】
核酸の増幅は、ポリメラーゼ連鎖反応(PCR)を含めた当技術分野で公知の種々の核酸の増幅方法のいずれかによって実現することができる。種々の増幅方法が当技術分野で公知であり、とりわけ、米国特許第4,683,195号および米国特許第4,683,202号に記載されている。22kbに至るまでのゲノムDNAを増幅するためにPCR増幅方法が開発されてきた。これらの方法ならびにDNA増幅の技術分野で公知の他の方法を、本発明の実施において使用することができる。本明細書において提供される配列に由来するプライマーを使用してイベントからそのような配列を増幅し、その後PCRアンプリコンまたはクローニングされたDNAの標準のDNA配列決定を行うことにより、本主題のダイズイベント由来の異種導入遺伝子DNA挿入断片の配列または隣接ゲノム配列を検証すること(および必要であれば、補正すること)ができる。
【0092】
これらの方法によって生成されるアンプリコンは、複数の技法によって検出することができる。アガロースゲル電気泳動および臭化エチジウムを用いた染色は、DNAアンプリコンを検出する周知の一般的な方法である。別のそのような方法は、近接する隣接ゲノムDNA配列および挿入DNA配列の両方とオーバーラップするDNAオリゴヌクレオチドを設計する遺伝子ビット分析(Genetic Bit Analysis)である。オリゴヌクレオチドは、マイクロウェルプレートのウェル内に固定化されている。対象の領域のPCR後(挿入配列内の1つのプライマーおよび近接する隣接ゲノム配列内の1つのプライマーを使用する)、一本鎖のPCR産物は、固定化されたオリゴヌクレオチドとハイブリダイズし、DNAポリメラーゼおよび予測される次の塩基に特異的な標識したddNTPを使用する一塩基伸長反応の鋳型としての機能を果たし得る。読み取りは、蛍光またはELISAに基づいてよい。信号により、上首尾の増幅、ハイブリダイゼーション、および一塩基伸長に起因して、挿入断片/隣接配列が存在することが示される。
【0093】
別の方法は、Winge(Innov. Pharma. Tech. 00巻:18〜24頁、2000年)に記載されているようなパイロシークエンス技法である。この方法では近接するゲノムDNAと挿入DNAの接合部とオーバーラップするオリゴヌクレオチドを設計する。このオリゴヌクレオチドを、対象の領域(挿入配列内の1つのプライマーおよび隣接ゲノム配列内の1つのプライマー)由来の一本鎖のPCR産物とハイブリダイズさせ、DNAポリメラーゼ、ATP、スルフリラーゼ、ルシフェラーゼ、アピラーゼ、アデノシン5’−ホスホ硫酸およびルシフェリンの存在下でインキュベートする。dNTPを個々に加え、それが取り込まれた結果、測定される光信号がもたらされる。光信号により、増幅、ハイブリダイゼーション、および単一塩基または多塩基の伸長が上首尾であり、したがって導入遺伝子挿入断片/隣接配列が存在することが示される。
【0094】
蛍光偏光は、本発明のアンプリコンを検出するために使用することができる別の方法である。この方法に従って、オリゴヌクレオチドを、ゲノムの隣接DNAと挿入DNAの接合部とオーバーラップするように設計する。このオリゴヌクレオチドを、対象の領域(挿入DNA内の1つのプライマーおよび隣接ゲノムDNA配列内の1つのプライマー)由来の一本鎖のPCR産物とハイブリダイズさせ、DNAポリメラーゼおよび蛍光標識したddNTPの存在下でインキュベートする。一塩基伸長により、ddNTPが取り込まれる。取り込みは、蛍光光度計を使用して偏光の変化として測定することができる。偏光の変化は、増幅、ハイブリダイゼーション、および一塩基伸長の成功によって、導入遺伝子挿入断片/隣接配列が存在することを示す。
【0095】
TAQMAN(PE Applied Biosystem、Foster City、Calif.)は、DNA配列の存在を検出し、定量する方法である。簡単に述べると、ゲノムの隣接と挿入DNAの接合部とオーバーラップするFRETオリゴヌクレオチドプローブを設計する。FRETプローブおよびPCRプライマー(挿入DNA配列内の1つのプライマーおよび隣接ゲノム配列内の1つのプライマー)を、耐熱性ポリメラーゼおよびdNTPの存在下でサイクルにかける。特異的な増幅の間に、Taq DNAポリメラーゼにより、FRETプローブ上のクエンチング部分から蛍光部分が離れて除かれ、放出される。蛍光シグナルは、増幅およびハイブリダイゼーションの成功によって、隣接配列/導入遺伝子挿入断片配列が存在することを示す。
【0096】
配列の検出において使用するための分子ビーコンが記載されている。簡単に述べると、隣接ゲノムDNAと挿入DNAの接合部とオーバーラップするFRETオリゴヌクレオチドプローブを設計する。FRETプローブの独特の構造により、蛍光部分およびクエンチング部分が極めて近傍に保たれる二次構造が含有される。FRETプローブおよびPCRプライマー(挿入DNA配列内の1つのプライマーおよび隣接ゲノム配列内の1つのプライマー)を、耐熱性ポリメラーゼおよびdNTPの存在下でサイクルにかける。PCR増幅が上手くいった後、FRETプローブが標的配列にハイブリダイズすることにより、プローブの二次構造が除去され、蛍光部分およびクエンチング部分が空間的に分離される。蛍光シグナルが結果として生じる。蛍光シグナルは、増幅およびハイブリダイゼーションの成功によって、隣接ゲノム配列/導入遺伝子挿入断片配列が存在することを示す。
【0097】
ダイズゲノム内の挿入するために優れた位置を開示し、本発明は、この遺伝子位置の概ね近くに少なくとも1つの非aad12挿入断片を含むダイズ種子および/またはダイズ植物も含む。1つの選択肢は、本明細書において例示されているaad−12挿入断片を異なる挿入断片で置換することである。これらの一般的な点については、例えば、本発明に従って、標的化相同組換えを使用することができる。この種類の技術は、例えば、WO03/080809A2および対応する公開された米国特許出願(U.S.2003/0232410)の主題である。したがって、本発明は、本明細書において同定される隣接配列の全てまたは認識できる部分(例えば、配列番号1の残基1〜2730および残基9122〜10,212)が隣接する異種挿入断片(多コピーのaad−12の代わりにまたはそれと一緒に)を含む植物および植物細胞を包含する。aad−12遺伝子の1つの追加的なコピー(または複数の追加的なコピー)も、この/これらのように挿入の標的とすることができる。
【0098】
植物細胞の特定の染色体上の部位内に、相同組換えによってポリヌクレオチド配列を組み込むための方法は、当技術分野の範囲内で記載されている。例えば、米国特許出願公開第2009/0111188A1号に記載の部位特異的な組み込みでは、ドナーポリヌクレオチド配列の染色体上の標的への導入を媒介するためのリコンビナーゼまたはインテグラーゼの使用が記載されている。さらに、国際的な特許出願第WO2008/021207号には、1つまたは複数のドナーポリヌクレオチド配列をゲノムの特定の位置内に組み込むためのジンクフィンガー媒介相同組換えが記載されている。米国特許第6,720,475号に記載のFLP/FRTまたは米国特許第5,658,772号に記載のCRE/LOXなどのリコンビナーゼの使用を利用して、ポリヌクレオチド配列を特定の染色体上の部位に組み込むことができる。最終的に、ドナーポリヌクレオチドを特定の染色体上の位置に標的化するためのメガヌクレアーゼの使用が、Puchtaら、PNAS USA 93巻(1996年)5055〜5060頁に記載されている。
【0099】
植物細胞内に部位特異的に組み込むための他の種々の方法は、一般に、公知であり、適用可能である(Kumarら、Trands in Plant Sci. 6巻(4号)(2001年) 155〜159頁)。さらに、いくつかの原核生物および下等真核生物において同定された部位特異的な組換え系を植物における使用に適用することができる。そのような系の例としては、これらに限定されないが、酵母であるジゴサッカロマイセス・ロキシー(Zygosaccharomyces rouxii)のpSR1プラスミド由来のR/RSリコンビナーゼ系(Arakiら(1985年)J. Mol. Biol. 182巻:191〜203頁)、およびファージMuのGin/gix系(MaeserおよびKahlmann(1991年)Mol. Gen. Genet. 230巻:170〜176頁)が挙げられる。
【0100】
本発明の一部の実施形態では、既存のトランスジェニックイベントに近接して新しい導入遺伝子(複数可)を組み込む、または積み重ねることが望ましい場合がある。トランスジェニックイベントは、単一の挿入部位、正常なメンデル分離および安定発現、ならびに多数の環境区域における、およびそれ全てにわたる除草剤耐性および農業生産力を含めた効力の優れた組合せなどの独特の特性に基づいて選抜された好ましいゲノム遺伝子座と考えることができる。新しく組み込まれた導入遺伝子は、既存の形質転換体の導入遺伝子の発現特性を維持すべきである。さらに、新しく組み込まれたイベントのゲノム隣接配列および染色体上の位置はすでに同定されているので、新しく組み込まれたイベントを検出し、確認するためのアッセイの開発は克服されるであろう。最終的に、既存の導入遺伝子に連結された特定の染色体上の位置に新しい導入遺伝子を組み込むことにより、従来の育種方法を使用した有性異系交雑による導入遺伝子の他の遺伝的背景への遺伝子移入が促進される。
【0101】
本発明の一部の実施形態では、トランスジェニックイベントからポリヌクレオチド配列を削除することが望ましい場合がある。例えば、米国特許仮出願第61/297,628号に記載の導入遺伝子の削除では、染色体に組み込まれたトランスジェニックイベントから、遺伝子発現カセットからなるポリヌクレオチド配列を除去するためのジンクフィンガーヌクレアーゼの使用が記載されている。除去されるポリヌクレオチド配列は、選抜マーカーであってよい。ポリヌクレオチド配列を削除および除去したら、改変されたトランスジェニックイベントを、ポリヌクレオチド配列を挿入することによって再標的化することができる。ポリヌクレオチド配列を削除し、その後、改変されたトランスジェニックイベントを再標的化することにより、選抜マーカーの再使用、または特定の遺伝子が発現することによって生じる、植物のトランスクリプトームに対する意図されたものではない変化を克服する能力などの利点がもたらされる。
【0102】
本発明は、本明細書において、異種核酸を挿入するために優れた、ダイズゲノム内の第4染色体上の特定の部位を開示している。第4染色体上のターゲティング部位の位置を同定することにおいて有用な5’分子マーカー、3’分子マーカー、5’隣接配列、および3’隣接配列も開示されている。したがって、本発明は、対象の異種核酸をこの予め確立した標的部位に、またはこの標的部位の付近に導入するための方法を提供する。本発明は、開示されている標的部位またはそのような部位の概ね近くに挿入された任意の異種ヌクレオチド配列を含むダイズ種子および/またはダイズ植物も包含する。そのような標的化組み込みを実現するための1つの選択肢は、本明細書において例示されているpat発現カセットの代わりに異なる挿入断片を削除することおよび/または置換することである。この一般的な点について、例えば、またこれらに限定することなく、本発明に従って標的化相同組換えを使用することができる。
【0103】
本明細書で使用される場合、遺伝子、イベントまたは形質を「積み重ねること(stacking)」は、所望の形質を1つのトランスジェニック系統内に組み合わせることである。植物育種家は、それぞれが所望の形質を有する親同士の交雑を行い、次にこれらの所望の形質を両方とも有する子孫を同定することによってトランスジェニック形質を積み重ねる。遺伝子を積み重ねるための別の方法は、形質転換の間に、2種以上の遺伝子を同時に植物の細胞核に伝達することによる。遺伝子を積み重ねるための別の方法は、トランスジェニック植物を、対象とする別の遺伝子を用いて再形質転換することによる。例えば、遺伝子を積み重ねることを用いて、例えば、2種以上の異なる昆虫形質、昆虫抵抗性形質(複数可)と病害抵抗性形質(複数可)、2種以上の除草剤への抵抗性形質、および/または昆虫抵抗性形質(複数可)と除草剤抵抗性の形質(複数可)を含めた2種以上の異なる形質を組み合わせることができる。対象の遺伝子に加えて選抜マーカーを使用することも、遺伝子を積み重ねることと考えることができる。
【0104】
「相同組換え」とは、2種のヌクレオチド配列が相互作用して(組み換えられて)新しい組換えDNA配列を形成し得る類似したヌクレオチド配列を含有する対応する部位を有するヌクレオチド配列の任意の対の間の反応を指す。類似したヌクレオチド配列の部位は、本明細書ではそれぞれが「相同配列」と称される。一般に、相同組換えの頻度は、相同配列の長さが増加するにつれて増加する。したがって、相同組換えは、同一には満たない2つのヌクレオチド配列間で起こり得るが、組換え頻度(または効率)は、2つの配列間の相違が増加するにつれて減退する。組換えは、ドナー分子および標的分子のそれぞれの1つの相同配列を使用して実現することができ、それによって「単一乗換え」組換え産物を生成することができる。あるいは、標的ヌクレオチド配列およびドナーヌクレオチド配列のそれぞれに2つの相同配列を置くことができる。ドナー上の2つの相同配列と標的上の2つの相同配列との間の組換えにより、「2重乗換え」組換え産物が生成する。ドナー分子上の相同配列が、操作される配列(例えば、対象の配列)に隣接する場合、標的分子との2重乗換え組換えにより、対象の配列が元々標的分子上の相同配列の間にあったDNA配列と交換された組換え産物がもたらされる。2重乗換え組換えイベントによって標的とドナーとの間でDNA配列を交換することは、「配列交換」と称される。
【0105】
本明細書において言及または引用された全ての特許、特許出願、仮出願、および刊行物は、それらが本明細書の明確な教示と相反しない限りは、それらの全体が参照により本明細書に組み込まれる。
【0106】
以下の実施例は、本発明を実施するための手順を例示するため、および本発明のある特定の好ましい実施形態を実証するために含まれる。これらの実施例は、限定するものと解釈されるべきではない。当業者には、以下の実施例に開示されている技法は、それを実施するための好ましい方式を例示するために使用される特定の手法を表していることが理解されたい。しかし、当業者には、本開示に照らして、本発明の精神および範囲から逸脱することなく、なお同様または類似の結果を得ながら、これらの特定の実施形態において多くの変更を行うことができることが理解されたい。別段の指定のない限り、全ての百分率は重量により、特に断りのない限り、全ての溶媒混合物の割合は体積による。
【0107】
別段の指定のない限り、以下の略語が使用されている。
AAD−12 アリールオキシアルカノエートジオキシゲナーゼ−1
bp 塩基対
℃ 摂氏温度
DNA デオキシリボ核酸
DIG ジゴキシゲニン
EDTA エチレンジアミン四酢酸
kb キロベース
μg マイクログラム
μL マイクロリットル
mL ミリリットル
M モル質量
OLP オーバーラッププローブ
PCR ポリメラーゼ連鎖反応
PTU 植物転写単位
SDS ドデシル硫酸ナトリウム
SOP 標準操作手順
SSC 塩化ナトリウムとクエン酸ナトリウムとの混合物を含有する緩衝溶液、pH7.0
TBE トリス塩基、ホウ酸およびEDTAの混合物を含有する緩衝溶液、pH8.3
V ボルト
[実施例]
【実施例1】
【0108】
aad−12ダイズイベントDAS−68416−4の形質転換および選抜
トランスジェニックダイズ(Glycine max)イベントDAS−68416−4を、ダイズ子葉節の外植片をアグロバクテリウム媒介形質転換によって生成した。T鎖DNA領域内に選抜マーカー(pat)および対象の遺伝子(aad−12)と一緒にバイナリーベクターpDAB4468(図1)を保有する武装解除したアグロバクテリウム株EHA101(Hoodら、2006年)を使用して形質転換を開始した。
【0109】
Zengら(Zengら、2004年)の手順を改変して用いてアグロバクテリウム媒介形質転換を行った。簡単に述べると、ダイズ種子(Maverick栽培品種)を基本培地上で発芽させ、子葉節を単離し、アグロバクテリウムに感染させた。アグロバクテリウムを除去するために、苗条誘導(shoot initiation)培地、苗条伸長培地、および発根培地にセフォタキシム、チメンチンおよびバンコマイシンを補充した。グルホシネートによる選択を使用して、形質転換されていない苗条の生長を阻害した。選択された苗条を、根を発生させるために発根培地に移し、次に、小植物を順化させるために土壌混合物に移した。
【0110】
選択された小植物の頂小葉にグルホシネートを塗布して推定形質転換体についてスクリーニングした。スクリーニングされた小植物を温室に移して、気候順化させ、次に葉にグルホシネートを塗布して耐性を再確認し、推定形質転換体であるとみなした。スクリーニングされた植物の試料を採取し、選択マーカー遺伝子および/または対象の遺伝子を確認するための分子解析を行った。T0植物を温室内で自家受精させてT1種子を生じさせた。
【0111】
T1植物を戻し交雑し、優良な生殖質(Maverick)に遺伝子移入した。このイベント、ダイズイベントDAS−68416−4は、独立した形質転換分離株から生成した。イベントをその独特の特性、例えば、単一の挿入部位、正常なメンデル分離および安定発現、ならびに広範な遺伝子型背景および多数の環境全ての区域にわたる除草剤耐性および農業生産力を含めた効力の優れた組合せに基づいて選抜した。以下の実施例は、ダイズイベントDAS−68416−4を特徴付けるために使用したデータを含有する。
【実施例2】
【0112】
ダイズイベントDAS−68416−4のサザンブロットによる特徴付け
サザンブロット分析を使用して、ダイズイベントDAS−68418−4の組み込みパターンを確立した。これらの実験により、aad−12導入遺伝子のダイズゲノム内への組み込みおよびその完全性を実証するデータが生成した。ダイズイベントDAS−68418−4は、プラスミドpDAB4468由来のaad−12PTUの単一コピーを含有する全長の、単純な組み込みイベントであると特徴付けられた。
【0113】
サザンブロットのデータにより、pDAB4468T鎖断片がダイズイベントDAS−68418−4のゲノムに挿入されたことが示唆された。詳細なサザンブロット分析を、pDAB4468のT鎖組み込み領域に含有されるaad−12遺伝子に特異的なプローブ、およびプラスミド内に位置する切断部位を有し、プラスミドまたはプラスミドとダイズのゲノムDNAの接合部にわたる断片(境界断片)内にハイブリダイズ断片を生じる説明的な制限酵素を使用して行った。サザンハイブリダイゼーションによって示された制限酵素とプローブの組合せについての分子量は、このイベントに独特であり、その同定パターンを確立した。これらの分析により、aad−12PTUの再配置を伴わずにプラスミド断片がダイズのゲノムDNAに挿入されたことも示された。
【0114】
実施例2.1
ダイズの葉の試料の収集およびゲノムDNA(gDNA)の単離
ダイズイベントDAS−68416−4を含有する個々のダイズ植物から回収した葉組織からゲノムDNAを抽出した。さらに、aad−12遺伝子が存在しない物質系統の代表である遺伝的背景を含有する従来のダイズ植物であるMaverickからgDNAを単離した。
【0115】
標準のCTAB法の後、凍結乾燥した葉組織から個々のゲノムDNAを抽出した。抽出した後、DNAを、Pico Green試薬(Invitrogen、Carlsbad、CA)を使用して分光蛍光分析によって定量した。次いで、DNAをアガロースゲル上で可視化してPico Green分析からの値を確認し、DNAの質を決定した。
【0116】
実施例2.2
DNAの消化および分離
ダイズイベントDAS−68416−4をサザンブロットによって分子キャラクタリゼーションするために、10マイクログラム(10μg)のゲノムDNAを消化した。ダイズイベントDAS−68416−4および非トランスジェニックダイズ系統であるMaverick由来のゲノムDNAを、各DNA試料にDNA1μg当たりおよそ5ユニットの選択された制限酵素および対応する反応緩衝液を加えることによって消化した。各試料を、およそ37℃で一晩インキュベートした。消化のために、制限酵素SpeI、KpnI、およびPacIを個々に使用した(New EnglandBioLab、Ipswich、MA)。さらに、陽性ハイブリダイゼーション対照試料を、プラスミドDNAであるpDAB4468を非トランスジェニックダイズ品種であるMaverick由来のゲノムDNAと混ぜ合わせることによって調製した。プラスミドDNA/ゲノムDNA反応混液を、試験試料の場合と同じ手順および制限酵素を用いて消化した。消化物を一晩インキュベートした後、NaClを、最終濃度が0.1Mになるまで加え、消化されたDNA試料を、イソプロパノールを用いて沈殿させた。沈殿したDNAペレットを、1×ローディング緩衝液(0.1%ブロモフェノールブルー、100mMのEDTA、50%グリセロール、10mMのトリス、pH7.5)20μlに再懸濁させた。次いで、DNA試料および分子サイズマーカーを、0.4×TAE緩衝液(Fisher Scientific、Pittsburgh、PA)を用いた0.85%アガロースゲルにより、35ボルトでおよそ18〜22時間、電気泳動して断片の分離を実現した。ゲルを臭化エチジウム(Invitrogen、Carlsbad、CA)で染色し、DNAを紫外線(UV)の下で可視化した
【0117】
実施例2.3
サザン転写およびメンブレン処理
サザンブロット分析をSeversonら(1997年)に記載されている通り実施した。DNA断片を電気泳動によって分離し、UV光の下で可視化した後、ゲルを変性溶液(150mMのNaOH、3mMのEDTA)中におよそ20分間浸し、次いで、中和溶液(150mMのNaPO4、pH7.8)に移し少なくとも20分間置いた。ナイロンメンブレン(Roche Diagnostic、Indianapoli、IN)へのサザン転写を、転写緩衝液(25mMのピロリン酸ナトリウム、pH10)を伴うウィッキング系を用いて一晩実施した。転写後、メンブレンを65℃で約2時間、加熱乾燥することによってDNAをメンブレンに結合させた。このプロセスにより、ハイブリダイゼーションのためのサザンブロットメンブレンの準備ができた。
【0118】
実施例2.4
DNAプローブの標識化およびハイブリダイゼーション
標識したプローブを使用してナイロンメンブレンに結合したDNA断片を検出した。この実験のために使用したプローブは、プラスミドpDAB4468の特定のヌクレオチド領域に対するプライマーを使用したPCR増幅によって生成した。増幅されたPCR断片をアガロースゲルから単離し、精製し、ハイブリダイゼーションプローブのための鋳型として使用した。サザンブロットハイブリダイゼーションプローブを、GE Healthcare READY−TO−GO(商標)DNA Labeling Beads(GE Healthcare、Piscataway、NJ)を製造者の説明書に従って使用したランダムプライミングによってα32Pに特異的なヌクレオチドで標識し、PROBEQUANT(商標)G−50マイクロカラム(Amersham/Pharmacia、Piscataway、New Jersey、USA)によって精製した。この実験のために使用したプローブが記載された表は、表2に記載されている。プレハイブリダイゼーションを、ハイブリダイゼーション緩衝液(Sigma−Aldrich、St.Loui、MO)を使用して65℃で4時間行った。次いで、プレハイブリダイゼーション溶液をデカントし、水で5分間煮沸することによって予め変性させた所望の量の特異的なプローブを含有するハイブリダイゼーション溶液と交換した。ハイブリダイゼーション/プローブ混合物をナイロンメンブレンと一緒に65℃で一晩インキュベートした。
【0119】
ハイブリダイゼーション後、メンブレンを65oC、洗浄緩衝液(10mMのリン酸ナトリウム、2.5mMのピロリン酸ナトリウム、0.5mMのEDTA、0.1%SDS、リン酸を用いて調整pH7.8に調整した)で20分間、3回洗浄した。洗浄したフィルターをオートラジオグラフィーのためにPhosphorimagerスクリーンに曝露させ、画像をスキャンした。検出されたバンドの数およびサイズを、プローブについて実証した。さらに、サザンブロットにおいて分子量マーカーを使用してハイブリダイズ断片サイズを決定した。
【0120】
【表2】
【0121】
実施例2.5
サザンブロットの結果
aad−12PTUの公知の制限酵素部位に基づく、特定の消化物およびプローブに関して予測された断片サイズおよび観察された断片サイズが表3に示されている。予測された断片サイズは、pDAB4468のプラスミドマップ(図1)に基づき、観察された断片サイズは、これらの分析から概算したものであり、α32Pで標識されたMolecular Weight Marker II断片の指示サイズに基づく。
【0122】
これらの消化およびハイブリダイゼーションから2種類の断片を同定した:公知の酵素部位がプローブ領域に隣接し、aad−12PTUの挿入領域内に完全に含有されている内部の断片、および公知の酵素部位がプローブ領域の一端に位置し、第2の部位がダイズゲノム内にあると予測される境界断片。ほとんどの場合、DNA断片の組み込み部位はそれぞれのイベントに独特であるので、境界断片のサイズはイベントごとに変動する。境界断片により、組み込まれたDNAに対して制限酵素部位を位置づける手段およびDNA挿入物の数を評価する手段がもたらされる。イベントDAS−68416−4を含有するダイズの3世代に対して完了したサザンブロット分析により、プラスミドpDAB4468由来の低コピー、インタクトなaad−12PTUが、ダイズイベントDAS−68416−4のダイズゲノムに挿入されたことを示唆するデータがもたらされた。
【0123】
【表3】
【0124】
制限酵素SpeIおよびKpnIは、プラスミドpDAB4468に独特の制限部位を含有する。続いて、これらの酵素を、ダイズイベントDAS−68416−4におけるaad−12遺伝子挿入断片を特徴付けるために選択した。5,436bpより大きいまたは5,383bpより大きい境界断片は、それぞれSpeIおよびKpnIで消化した後、aad−12遺伝子プローブとハイブリダイズすることが予測された(表3)。それぞれSpeIおよびKpnIを使用したとき、約12,000bpおよび約16,000bpの単一のaad−12ハイブリダイゼーションバンドが観察された。プローブがこのサイズのバンドにハイブリダイズすることにより、ダイズイベントDAS−68416−4のダイズゲノム内にaad−12遺伝子に対する単一の挿入部位が存在することが示唆される。制限酵素PacIを、aad−12植物転写単位(PTU、プロモーター/遺伝子/ターミネーター)を含有する2,904bpの断片を放出させるために選択した(表3)。予測された2,904bpの断片は、PacI消化した後、aad−12遺伝子プローブを用いて観察された。全3種の酵素を用いてDAS−68416−4試料を消化し、その後aad−12遺伝子プローブとハイブリダイズさせることで得られた結果により、プラスミドpDAB4468由来のインタクトなaad−12PTUの単一コピーがダイズイベントDAS−68416−4のダイズゲノムに挿入されたことが示された。
【0125】
実施例2.6
骨格配列が存在しないこと
ダイズイベントDAS−68416−4にスペクチノマイシン抵抗性遺伝子が存在すること、または存在しないことをモニターするために、多重PCRアッセイを実施した。実験は、スペクチノマイシン抵抗性遺伝子コード配列の5つの異なる領域および内部標準として内在性のダイズレクチン遺伝子(GenBank ID No:K00821 M30884)配列内の407bp領域を検出するために設計した。さらに、以下の対照を含めた:(i)スペクチノマイシン抵抗性遺伝子を保有するプラスミドDNAを付加した形質転換されていないダイズのゲノムDNAを用いた陽性対照;(ii)形質転換されていないダイズ、Maverick由来のゲノムDNAを用いた陰性対照;および(iii)ゲノムDNAを有さないブランク。
【0126】
ダイズ由来のゲノムDNAを、CTAB法を使用して単離し、Pico Greenを使用して定量した。各試料のDNA濃度を100ng/μlに正規化した。PCR反応を、スペクチノマイシン抵抗性遺伝子コード配列およびレクチン遺伝子配列に特異的なプライマー配列を使用して実施した。この反応物を、試料当たりPCR産物20μlを2%Eゲルにローディングすることによって分析した。
【0127】
Eゲル上に多数のアンプリコンが存在すること(100bp、150bp、および407bpのバンド)は、ダイズイベントDAS−68416−4がスペクチノマイシン抵抗性遺伝子コード配列を含有することを示すことになる(スペクチノマイシン抵抗性遺伝子コード配列について、100bpおよび150bpの増幅されたバンドが予測される)。内部標準であるレクチン遺伝子配列に対応する407bpのアンプリコンのみが存在する場合、そのことは、ダイズイベントDAS−68416−4がスペクチノマイシン抵抗性コード配列を含有しないことを示す。
【0128】
ダイズイベントDAS−68416−4由来のDNA試料では、100bpまたは150bpの断片は増幅されなかった。407bp断片のみが増幅された。しかし、スペクチノマイシン抵抗性遺伝子を保有するプラスミドDNAをダイズのゲノムDNAに加えた陽性対照には100bp、150bp、および407bpの増幅断片が存在した。形質転換されていないダイズ由来のDNAを含有する陰性対照では、単一の407bp断片が増幅された。最終的に、いかなるゲノムDNAも含有しない反応物からは、いかなる増幅断片も産生されなかった。そのように、ダイズイベントDAS−68416−4においてスペクチノマイシン抵抗性コード配列は検出されなかった。
【実施例3】
【0129】
ダイズイベントDAS−68416−4の挿入断片および隣接境界領域におけるDNA配列のクローニングおよび特徴付け
ゲノムの挿入部位を特徴付け、説明するために、ダイズイベントDAS−68416−4のT鎖DNA挿入断片および隣接ゲノムDNA境界領域の配列を決定した。2,730bpの5’隣接境界配列、1,091bpの3’隣接境界配列、および6,391bpのT鎖挿入断片を含む、全部で10,212bpのダイズイベントDAS−68416−4ゲノム配列(配列番号1)を確認した。配列解析により、ダイズイベントDAS−68416−4が、予測されたT鎖挿入断片から配列が変動することなく、MARエレメント、aad−12発現カセット、およびpat発現カセットを含有するインタクトな導入遺伝子の単一コピーを含有することが検証された。
【0130】
ダイズイベントDAS−68416−4の挿入断片および境界の配列に基づくPCR増幅により、境界領域がダイズ起源であること、およびダイズイベントDAS−68416−4をイベント特異的に同定するために接合領域を使用することができることが検証された。隣接境界配列を含めた接合領域にわたる配列を分析することにより、T鎖の挿入によって生じるいかなる新規のオープンリーディングフレーム(ORF>=150 コドン)も同定されなかった。さらに、T鎖の挿入部位を、非トランスジェニックダイズのゲノムから同定された隣接境界配列の領域に対応するゲノムの断片をクローニングすることによって特徴付けた。ダイズイベントDAS−68416−4を野生型ゲノム配列と比較することにより、元の遺伝子座からの55bpの欠失およびイベントの3’組み込み接合部における9bpの挿入が明らかになった。全体的に、ダイズイベントDAS−68416−4の挿入断片および境界配列を特徴付けることにより、T鎖の単一の、インタクトなコピーがダイズゲノム内に存在することが示された。
【0131】
実施例3.1
ゲノムDNAの抽出および定量
凍結乾燥した葉組織または粉砕したての葉組織から、改変したCTAB法を使用してゲノムDNAを抽出した。ゲノムDNAを抽出した後、DNA試料を1×TE(10mMのトリス、pH8.0、1mMのEDTA)(Fluka、Sigma、St.Loui、MO)に溶解させ、Pico Green法を製造者(Molecular Probe、Eugene、OR)の説明書に従って使用して定量した。PCR分析のために、DNA試料を分子生物学グレードの水(5PRIME、Gaithersburg、MD)で希釈して、濃度を10〜100ng/μLにした。
【0132】
実施例3.2
PCRプライマー
表4には、ダイズイベントDAS−68416−4のDNA挿入断片および隣接境界領域をクローニングし、確認するために使用したプライマー配列が、図2で記した位置および説明とともに列挙されている。全てのプライマーは、Integrated DNA Technologie、Inc.(Coralville、IA)によって合成された。プライマーを、原液については水(5PRIME、Gaithersburg、MD)に溶解させて濃度を100μMにし、検量線用溶液については10μMの濃度まで水で希釈した。
【0133】
【表4】
【0134】
【表5】
【0135】
【表6】
【0136】
【表7】
【0137】
実施例3.3
ゲノムウォーキング
GENOMEWALKER(商標)Universal Kit(Clontech Laboratorie、Inc.、Mountain View、CA)を製造者の説明書に従って使用して、ダイズイベントDAS−68416−4についてpDAB4468T鎖挿入断片の5’隣接境界領域および3’隣接境界領域をクローニングした。ダイズイベントDAS−68416−4由来のゲノムDNAおよそ2μgをEcoRVおよびPvuIIを用いて一晩消化した(図2)。DNA消化物を、DNA Clean & Concentrator(商標)−25(Zymo Research、Orange、CA)を使用して精製し、その後GENOMEWALKER(商標)アダプターとライゲーションしてGENOMEWALKER(商標)ライブラリーを構築した。GENOMEWALKER(商標)ライブラリーのそれぞれを、アダプタープライマーAP1(このキットで提供される)および構築物特異的プライマーであるES_LEnd03またはES_PATEnd03(表4)を用いた一次PCR増幅のためのDNA鋳型として使用した。次に、一次PCR反応物1:25希釈物1マイクロリットル(1μL)を、このキットで提供されるネステッドアダプタープライマーAP2およびネステッド構築物特異的プライマーであるES_LEnd04またはES_PATEnd04(表4、7および図2)を用いた二次PCR増幅ための鋳型として使用した。
【0138】
実施例3.4
従来のPCR
標準のPCRを使用してダイズイベントDAS−68416−4の挿入断片および境界配列をクローニングし、確認した。従来のPCR増幅ために、TaKaRa LA TAQ(商標)(Takara Bio Inc、Shiga、Japan)、HOTSTARTAQ(商標)DNAポリメラーゼ(Qiagen、Valencia、CA)、HIGH FIDELITY(商標)PCRキット(Roche Diagnostic、Inc)、またはEASY−A(商標)高忠実度ポリメラーゼキット(Stratagene、LaJolla、CA)を製造者の推奨手順に従って使用した。特定のPCR条件およびアンプリコンの説明が表5、6、および7に列挙されている。
【0139】
実施例3.5
PCR産物の検出、精製、PCR産物のサブクローニング、および配列決定
PCR産物を1.2%または2%E−GEL(登録商標)(Invitrogen、Carlsbad、CA)を製品の説明書に従って使用して電気泳動によって検査した。DNAマーカーと比較することによって断片サイズを推定した。必要であれば、QIAquick Gel Extraction Kit(Qiagen、Valencia、CA)を使用して1%アガロースゲルから断片を切り取って1×TBE(89mMのトリス−ホウ酸、2mMのEDTA、pH8.3)に入れ、臭化エチジウムで染色することによってPCR断片を精製した。
【0140】
PCR断片を、配列決定するためにTOPO TA CLONING(登録商標)KIT(Invitrogen、Carlsbad、CA)を製品の説明書に従って使用してPCR(登録商標)4−TOPO(登録商標)ベクターにサブクローニングした。詳細には、製造者の説明書に従って、TOPO(登録商標)クローニング反応物2〜5マイクロリットルをOne Shot化学的コンピテントなTOP10細胞に導入し、形質転換した。クローニングされた断片を、プラスミドDNAをミニプレップした後に(QIAprep Spin Miniprep Kit、Qiagen、CA)、EcoRIで制限消化することによって、またはT3プライマーおよびT7プライマーを使用したダイレクトコロニーPCRによって検証した。次に、選択されたコロニーのプラスミドDNAまたはグリセロールストックを配列決定するために外部委託した。
【0141】
サブクローニングした後、予測されたDNA断片がクローニングされたことを確認するために、推定標的PCR産物について最初に配列決定した。予測されたDNA断片を含有するコロニーを選択して、プライマーウォーキングによる二本鎖の全長についての配列決定を完了した。配列決定は全て、Cogenics(Houston、TX)によって行われた。
【0142】
挿入断片および境界配列の最終的な組立てを、SEQUENCHER(登録商標)ソフトウェア(Gene Codes Corporation、Ann Arbor、MI)を使用して完了した。ダイズイベントDAS−68416−4の挿入断片およびその隣接境界配列のアノテーションを、ベクターNTI(バージョン10および11、Invitrogen、Carlsbad、CA)を使用して実施した。
【0143】
GenBankの非冗長ヌクレオチドデータベースに対してBLASTプログラムを使用して相同性検索を実施した。Vector NTI(バージョン11、Invitrogen)を使用したオープンリーディングフレーム(ORF)解析を実施して、ダイズイベントDAS−68416−4の完全な挿入断片および隣接境界配列、ならびに野生型Maverickダイズ系統の元の遺伝子座におけるORF(≧150コドン)を同定した。
【0144】
実施例3.6
5’末端境界配列
ダイズイベントDAS−68416−4のGENOMEWALKER(商標)ライブラリーのそれぞれから、導入遺伝子の5’末端に対して特異的なネステッドプライマーセットを使用してDNA断片を増幅した。EcoRV GENOMEWALKER(商標)ライブラリーから約1.8kbの断片、およびPvuII GENOMEWALKER(商標)ライブラリーから約3kbの断片が観察された。これらの断片を、PCR(登録商標)4−TOPO(登録商標)ベクターにクローニングした。ヌクレオチド配列のデータを生成するために、末端配列決定用にそれぞれのライブラリーについて5つのコロニーを無作為に選定した。プライマーウォーキングによって完全な配列を得るために、両方のPCRプライマーの配列を含有するコロニーを選択した。配列解析により、ダイズイベントDAS−68416−4のEcoRV GENOMEWALKER(商標)ライブラリーから増幅されたクローンが1,744bpのDNA断片を含有すること、およびダイズイベントDAS−68416−4のPvuII GENOMEWALKER(商標)ライブラリーから増幅されたクローンが3,047bpのDNA断片を含有することが明らかになった。配列解析により、EcoRV GENOMEWALKER(商標)ライブラリーから得られたDNA断片とPvuII GENOMEWALKER(商標)ライブラリークローンから得られたDNA断片が、プライマーES_LEnd04とEcoRV部位との間の領域においてオーバーラップしていることが明らかになった。これらのDNA断片は全て、導入遺伝子内にT鎖境界Bの5’末端接合部を含有し、これは、それらが、ダイズイベントDAS−68416−4の5’末端の導入遺伝子挿入断片およびその隣接境界の同じ領域から増幅されたことを示している。得られた2,730bpのダイズのゲノム配列は、GenBankにある配列との有意な相同性を有さないことが見いだされた。
【0145】
実施例3.7
3’末端境界配列
ダイズイベントDAS−68416−4のEcoRV GENOMEWALKER(商標)ライブラリーから、導入遺伝子の3’末端に対して特異的なネステッドプライマーセットを使用して、サイズが約1.3kbであるDNA断片を増幅した。次に、このDNA断片をPCR(登録商標)4−TOPO(登録商標)ベクターにクローニングした。末端配列決定のために5つのコロニーを無作為に選定した。5つのクローンは全て、プライマーAP2およびプライマーES_PATEnd04の両方の配列を含有した。これらのクローンについて完全に配列決定することにより、1,359bpのコンセンサスDNA断片が得られた。配列解析することにより、1,359bpの断片が、T鎖境界Aの3’末端領域由来の268bpの断片およびダイズのゲノムDNA由来の1,091bpの断片で構成されることが開示された。BLAST検索により、この1,091bpのダイズDNA配列とGenBankにある配列との間に有意な相同性は同定されなかった。
【0146】
実施例3.8
DNA挿入断片および接合部配列
以前に記載されているようなPCRに基づく方法を使用して、ダイズイベントDAS−68416−4からDNA挿入断片および隣接境界領域をクローニングした。5’隣接境界および3’隣接境界配列および予測された導入遺伝子配列を使用して、表6に列挙されているPCRプライマーを設計した。全部で、4つのオーバーラップDNA断片(978bpのアンプリコン1、2,414bpのアンプリコン2、1,834bpのアンプリコン3、および1,705bpのアンプリコン4)をクローニングし、配列決定した(図3)。挿入断片および隣接境界配列の全体を、4つの断片の間のオーバーラップ配列に基づいてアセンブリした。最終的にアセンブリした配列を解析することにより、pDAB4468の導入遺伝子に由来する6,391bpの断片が存在することが確認され、また、プラスミドpDAB4468から予測された配列と比較して、挿入DNA配列の塩基の変化は見られなかった。
【0147】
実施例3.9
ダイズのゲノム配列の確認
ダイズゲノム内のダイズイベントDAS−68416−4導入遺伝子の挿入部位を確認するために、異なるプライマー対を用いてPCRを行った(図4および表5)。ダイズイベントDAS−68416−4由来のゲノムDNAおよび他のトランスジェニックダイズ系統または非トランスジェニックダイズ系統由来のゲノムDNAを鋳型として使用した。したがって、得られた5’末端境界配列が正確であるかどうかを確認するために、aad−12遺伝子から5’末端境界配列にわたるDNAセグメントを増幅するためにaad−12特異的な2種のプライマー、例えば、AIILEnd05およびAIILEnd06、ならびに、16LEndG01および16LEndG02と称される5’末端境界配列に従って設計した2種のプライマーを使用した。同様に、クローニングされた3’末端境界配列を確認するために、pat特異的なプライマー、例えばPAT−End06、および16PATG01および16PATG02と称される、3’末端境界配列に従って設計した2種のプライマーを使用して、pat遺伝子から3’末端境界配列にわたるDNAセグメントを増幅した。予測されたサイズを有するDNA断片は、ダイズイベントDAS−68416−4の隣接境界に位置した1つのプライマー、導入遺伝子特異的な1つのプライマーの各プライマー対を用いたダイズイベントDAS−68416−4のゲノムDNAからのみ増幅されたが、他のトランスジェニックダイズ系統または非トランスジェニック対照由来のDNA試料からは増幅されなかった。結果は、クローニングされた5’境界配列および3’境界配列が、ダイズイベントDAS−68416−4におけるT鎖挿入断片の隣接境界配列であることを示している。
【0148】
ダイズゲノム内のDNA挿入物をさらに確認するために、2つのダイズ配列にわたるPCR増幅を完了した。5’末端境界配列に従って設計した2種のプライマー、16LEndG03および16LEndG04、ならびに3’末端境界配列に対する2種のプライマー、16PATG03および16PATG04を使用して、導入遺伝子、5’末端境界配列、および3’境界配列の全体を含有するDNAセグメントを増幅した。予測通り、16LEndG03および16PATG03のプライマー対を用いたPCR増幅により、ダイズイベントDAS−68416−4のゲノムDNAからおよそ9kbのDNA断片、および非トランスジェニックダイズ対照および他のダイズトランスジェニック系統から2.7kbのDNA断片が増幅された。同様に、16LEndG04および16PATG04のプライマー対を用いて完了したPCR反応により、ダイズイベントDAS−68416−4の試料からおよそ9kbのDNA断片、および他のダイズ対照系統全てから、同様に、2.8kbのDNA断片がもたらされた。両方のプライマー対をPCRに使用した場合、ダイズイベントDAS−68416−4以外の全てのダイズ試料において約6kbのサイズのかすかなバンドが目に見えたことが認められ、これは、このかすかなバンドが、このプライマー対でのダイズゲノムの非特異的な増幅に起因することを示唆している。
【0149】
実施例3.10
ダイズのゲノム配列の確認
16LEndG03と16PATG03のプライマー対と16LEndG04および16PATG04のプライマー対を使用して非トランスジェニックダイズ系統であるMaverickから増幅された2.7kbのDNA断片および2.8kbのDNA断片をクローニングし、配列決定した。それらの配列を互いに突き合せ、ダイズイベントDAS−68416−4からクローニングされた5’境界配列および3’境界配列とアラインメントした。これにより、クローニングされたDNA配列が、pDAB4468のT鎖がダイズイベントDAS−68416−4に組み込まれた遺伝子座を含有したことが実証された。アラインメント解析により、元の遺伝子座からの55bpの欠失および3’組み込み接合部における9bpの挿入物も明らかになった(図5)。クローニングされた元の遺伝子座のダイズのゲノム領域においてオープンリーディングフレーム(>/=450bp、150アミノ酸)は同定されなかった。
【実施例4】
【0150】
ダイズイベントDAS−68416−4の隣接SNPマーカーによるゲノムの特徴付け
ゲノムの挿入部位を特徴付け、説明するために、挿入断片の近傍に位置するマーカー配列を決定した。多型的な一塩基多型(SNP)マーカーのパネルを使用して導入遺伝子の位置を同定し、マッピングした。ダイズイベントDAS−68416−42は、第4染色体上の119.6cMに位置する。この位置は、2つの隣接SNPマーカー、BARC−044607−08736とBARC−019093−03299の間である。より詳細には、導入遺伝子の位置は、SNPマーカーBARC−019093−03299から1.3cM(480kb)離れてマッピングされた。
【0151】
実施例4.1
隣接境界領域の配列を用いたBLAST
ダイズイベントDAS−68416−4の隣接境界領域の配列(配列番号1)を、ダイズの全ゲノム配列のBLASTに使用した。BLASTの結果により、ダイズイベントDAS−68416−4の境界配列の両方が、連鎖群C1である第4染色体(Gm04)上に位置したことが示された。
【0152】
実施例4.2
SNPマッピングおよびBLASTの結果
境界配列を用いたBLASTおよびマッピングからの結果に基づいて、イベントは、第4染色体に割り当てられた。そのように、10種のSNPマーカーをダイズ遺伝連鎖マップから選択した。SNP配列を、Dr.Cregan、Beltsville Agricultural Research Center、およびUSDAによって開発されたSNPマーカーから選択した。これらのSNPマーカーは、第4染色体に対応する連鎖群C1に関連づけられる。ダイズイベントDAS−68416−4についてT鎖挿入断片の物理的な位置を決定するために、ダイズの全ゲノム配列のBLASTにSNP配列を使用した。
【0153】
実施例4.3
SNPマーカーの結果
ダイズイベントDAS−68416−4は、第4染色体上の119.6cMにマッピングされる。2つの隣接SNPマーカーは、BARC−044607−08736およびBARC−019093−03299である。導入遺伝子は、SNPマーカーBARC−019093−03299から1.3cM(480kb)離れており、SNPマーカーBARC−044607−08736とBARC−019093−03299の間のおよそ119.6cMである。
【実施例5】
【0154】
ダイズイベントDAS−68416−4におけるAAD−12タンパク質の特徴付け
組換えトランスジェニックダイズイベントDAS−68416−4に由来するAAD−12タンパク質の生化学的性質を特徴付けた。定量的酵素結合免疫吸着検定法(ELISA)、ドデシル硫酸ナトリウムポリアクリルアミドゲル電気泳動(SDS−PAGE、クーマシーブルー(Coomassie blue)を用いて染色し、糖タンパク質を検出する方法)、およびウェスタンブロット法を使用してタンパク質の生化学的性質を特徴付け、AAD−12タンパク質の発現を確認した。
【0155】
実施例5.1
植物組織におけるAAD−12タンパク質の発現
ダイズイベントDAS−68416−4におけるAAD−12タンパク質のレベルを決定した。可溶性の抽出可能なAAD−12タンパク質を、ダイズの葉における定量的酵素結合免疫吸着検定法(ELISA)方法を使用して測定した。
【0156】
ダイズ組織の試料を試験植物から単離し、発現を解析するために調製した。0.5%ウシ血清アルブミン(BSA)を含有する界面活性剤Tween−20(PBST)を含有するリン酸緩衝生理食塩水溶液を用いてダイズ植物の組織からAAD−12タンパク質を抽出した。植物組織を遠心分離し;水性上清を収集し、必要に応じて適切な緩衝液で希釈し、AAD−12ELISAキットをサンドイッチ形式で使用して解析した。キットは製造者の推奨プロトコールに従って使用した。
【0157】
ダイズイベントDAS−68416−4において、検出解析を実施して、発現の安定性および遺伝性を垂直方向(世代間)および水平方向(系列間)の両方で調査した。T5世代ダイズイベントDAS−68416−4において、発現は安定であり(分離性でない)、系列全てにわたって一貫した(図6)。
【0158】
種々の植物の段階における圃場発現レベル試験を、ダイズイベントDAS−68416−4において、V3前、V3後、R2前、およびR2後で実施した。噴霧処理全てについて、ならびに2,4−D除草剤を噴霧した小区画および噴霧していない小区画について、発現値は同様であった。2×噴霧率(2,240gm ae/haの2,4−D)を施用し、試験のどのポイントにおいても植物に対する害は確認されなかった。植物のv3前の段階における、系列全てにわたった平均の発現は300μg/cm2であった。2,4−Dを噴霧した後、発現は安定なままであり、系列にわたって平均400μg/cm2であった。ダイズがR2前に到達した時には、平均の発現は、平均200μg/cm2にわずかに降下した。2,4−Dを噴霧した後、R2後の発現は以前の平均400μg/cm2に戻った。図6を参照されたい。
【0159】
実施例5.2
植物組織におけるAAD−12タンパク質の発現
2008年中に米国およびカナダの6つの区域で追加的な圃場発現試験を行った。ダイズイベントDAS−68416−4の4つの処理(噴霧なし、2,4−Dを噴霧、グルホシネートを噴霧、または2,4−Dおよびグルホシネートの両方を噴霧)を試験した。試料採取した植物組織は、葉、粒、根、および茎葉を含んだ。発生のV5段階およびV10段階において葉組織を収集し、R3段階において根および茎葉を収集した。発生のR8段階において粒を収集した(Gaska、2006年)。可溶性の抽出可能なAAD−12タンパク質を、実施例5.1において前記されている通り、検証された酵素結合免疫吸着検定法(ELISA)方法を使用して測定した。全ての種類の組織について、AAD−12タンパク質レベルをng/乾燥重量(mg)に基づいて算出した。
【0160】
種々のダイズマトリックスにおけるAAD−12タンパク質の濃度(部位にわたって平均した)の概要が表8に示されている。平均の発現値は、R3段階の根における15.5ng/乾燥重量(mg)から、V5段階の葉組織における66.1ng/mgにわたった。発現値は、噴霧処理全てについて、ならびに2,4−D除草剤およびグルホシネート除草剤を噴霧した小区画および噴霧していない小区画について、同様であった。6つの区域全てにわたって、対照組織においてAAD−12タンパク質は検出されなかった。
【0161】
【表8】
【0162】
実施例5.3
AAD−12タンパク質のSDS PAGEおよびウェスタンブロット分析
安定剤を加えたPBST(0.05%Tween20、pH7.4を伴うリン酸緩衝生理食塩水)ベースの緩衝液中のダイズイベントDAS−68416−4の凍結乾燥した葉組織からAAD−12タンパク質を抽出し、可溶性タンパク質を遠心分離によって収集した。上清を濾過し、可溶性タンパク質をフェニルセファロース(PS)ビーズ(GE Healthcare、Piscataway、NJ)に結合させた。1時間インキュベートした後、PSビーズをPBSTで洗浄し、Milli−Q水を用いて結合したタンパク質を溶出した。伝導率を増加させるために、塩化ナトリウムを加え、PS精製されたタンパク質を、AAD−12特異的なポリクローナル抗体とコンジュゲートした抗AAD−12免疫親和性カラムにローディングした。結合していないタンパク質をカラムから収集し、カラムを予め冷却したPBS(リン酸緩衝生理食塩水、pH7.4)を用いて広範囲にわたって洗浄した。結合したタンパク質を、3.5MのNaSCN、50mMのトリス、pH8.0である緩衝液を用いてカラムから溶出させ、SDS−PAGEおよびウェスタンブロットによって検査した。同じプロトコールを使用して、対照ダイズ系統であるMaverickの葉組織からタンパク質を単離した。Maverickは、aad−12遺伝子を含有しないが、ダイズイベントDAS−68416−4植物を表す遺伝的背景を有する。
【0163】
ダイズイベントDAS−68416−4およびMaverick由来の凍結乾燥した葉組織を、約2.0%のプロテアーゼ阻害剤反応混液(Sigma、St.Loui、MO)を含有するPBST緩衝液と混合し、Geno−Grinderのボールベアリングを用いて粉砕することによってタンパク質を抽出した。試料を遠心分離し、上清を、Laemmli試料緩衝液と混合し、加熱し、短時間遠心分離した。試料をBio−Rad Criterion SDS−PAGEゲルに直接ローディングした。陽性参照標準物質、微生物由来のAAD−12タンパク質も、試料緩衝液と混合し、ゲルにローディングした。トリス/グリシン/SDS緩衝液(Bio−Rad、Hercule、CA)を用いて電気泳動を行った。電気泳動した後、ゲルを半分に切り、一方の半分はPierce GelCode Blueタンパク質染料で染料し、他方のゲルの半分は、ニトロセルロースメンブレンに電気ブロッティングした。次に、AAD−12特異的なポリクローナルウサギ抗体を用いてニトロセルロースメンブレンを探索した。化学発光基質を使用して、免疫反応性のバンドを可視化した。微生物由来のAAD−12では、クーマシー染色したSDS−PAGEゲルにおいて可視化された主要なタンパク質バンドは、およそ32kDaであった。予測通り、対応する植物由来のAAD−12タンパク質は、微生物由来のタンパク質とサイズが同一であった。予想通りに、植物の精製された画分は、AAD−12タンパク質に加えて、少量の非免疫反応性不純物を含有した。共精製されたタンパク質は、カラムマトリックスとの弱い相互作用によってカラム上に保持されたと思われる(Williamら、2006年、KennedyおよびBarne、1983年およびHolroydeら、1976年)。
【0164】
微生物由来のAAD−12およびDAS−68416−4植物組織抽出物は、抗AAD−12ポリクローナル抗体を使用したウェスタンブロットにおいて予測されるサイズの陽性シグナルを示した。AAD−12ウェスタンブロット分析では、対照であるMaverickの抽出物において免疫反応性タンパク質は観察されず、また、トランスジェニック植物由来の試料において代替のサイズのタンパク質(凝集体または分解生成物)は見られなかった。モノクローナル抗体により、微生物由来のタンパク質において少量のAAD−12二量体が検出された。これらの結果により、AAD−12タンパク質がダイズにおいて発現されているという証拠が追加される。
【実施例6】
【0165】
サザンブロットによるダイズイベントDAS−68416−4のメチル化検出解析
導入された導入遺伝子は、植物ゲノムに組み込まれた後にサイレンシングを受ける可能性がある。導入遺伝子の発現は、転写レベルおよび/または転写後レベルで阻害される可能性がある。転写レベルでの遺伝子サイレンシングは、導入遺伝子、そのプロモーターおよび他の関連性のある配列のメチル化に関連することが報告されている(Stemら、1997年)。特定の配列におけるメチル化を検出するために、ある方法では、メチル化感受性制限酵素を利用してDNAを消化し、その後DNA産物をサザンブロット分析する。特定の制限酵素部位がメチル化されていると、酵素はDNAを切断しない。制限部位がメチル化されていると、サザンブロットで検出可能な、分子量がより高いDNA断片がもたらされる。サザンブロットに基づくメチル化解析を実施して、ダイズイベントDAS−68416−4についてT鎖挿入断片のメチル化の状態を決定した。このアッセイは、aad−12遺伝子に特異的なプローブおよびそのプロモーターおよび2つのメチル化感受性制限酵素を使用して行った。aad−12発現カセットのメチル化は検出されなかった。
【0166】
6.1.ダイズの葉の試料収集およびゲノムDNA(gDNA)の単離
ゲノムDNAを、ダイズイベントDAS−68416−4および非トランスジェニックダイズ系統であるMaverickのそれぞれの植物の葉から調製した。伝統的なCTAB法を使用して、凍結乾燥した葉の試料からゲノムDNAを単離した。抽出後、DNAを、Pico Green試薬(Invitrogen、Carlsbad、CA)を使用して定量した。
【0167】
6.2.DNAの消化および分離
DNAの分子キャラクタリゼーションのために、10マイクログラム(10μg)のダイズイベントDAS−68416−4由来のゲノムDNAおよび非トランスジェニックダイズ系統であるMaverick由来のゲノムDNAを、DNA1μg当たりおよそ5ユニットの選択された制限酵素および対応する反応緩衝液を各DNA試料に加えることによって消化した。各試料を、およそ37℃で一晩インキュベートした。制限酵素AciIおよびHyp188IIIを使用して消化した(New EnglおよびBioLab、Ipswich、MA)。非トランスジェニックダイズであるMaverick由来のDNAを、試験試料と同じ手順および制限酵素を用いて消化して陰性対照として機能させた。消化されたDNA試料を、NaClを0.1Mの最終濃度まで加えた後、イソプロパノールを用いて沈殿させ、1×ローディング緩衝液(0.1%ブロモフェノールブルー、100mMのEDTA、50%グリセロール、10mMのトリス pH7.5)20μlに再懸濁させた。次いで、DNA試料および分子サイズマーカーを、0.4×TAE緩衝液(Fisher Scientific、Pittsburgh、PA)を35ボルトでおよそ18〜22時間用いて0.85%アガロースゲルによって電気泳動して断片の分離を実現した。ゲルを臭化エチジウム(Invitrogen、Carlsbad、CA)で染色し、DNAを紫外線(UV)の下で可視化した。
【0168】
6.3.サザン転写およびメンブレン処理
Seversonら、(1997年)に記載されている通りサザンブロット分析を実施した。簡単に述べると、DNA断片を電気泳動によって分離し、UV光の下で可視化した後、ゲルを変性溶液(150mMのNaOH、3mMのEDTA)におよそ20分間曝露させ、その後、中和溶液(150mMのNaPO4、pH7.8)に少なくとも20分間曝露させた。転写緩衝液(25mMのピロリン酸ナトリウム、pH10)を用いたウィッキング系を使用して、一晩、ナイロンメンブレン(Roche Diagnostic、Indianapoli、IN)にサザン転写を実施した。転写した後、メンブレンを65℃で約2時間加熱乾燥した。このプロセスにより、ハイブリダイゼーションのためのサザンブロットメンブレンの準備ができた。
【0169】
6.4.DNAプローブの標識化およびハイブリダイゼーション
ナイロンメンブレンに結合したDNA断片を、標識したプローブを使用して検出した。プローブは、プラスミドpDAB4468由来の特異的なプライマーを伴う増幅されたPCR断片として生成した。これらのPCR増幅された断片をアガロースゲルから切り取り、精製した。精製されたDNA断片を、ハイブリダイゼーションプローブを作製するための鋳型として使用した。ハイブリダイゼーションプローブを、GE Healthcare READY−TO−GO(商標)DNA Labeling Beads(GE Healthcare、Piscataway、NJ)を製造者の説明書に従って使用したランダムプライミングによってα32Pに特異的なヌクレオチドで標識し、PROBEQUANT(商標)G−50マイクロカラム(Amersham/Pharmacia、Piscataway、New Jersey、USA)によって精製した。この試験のために使用したプローブの一覧は、表9に記載されている。
【0170】
プレハイブリダイゼーションおよびハイブリダイゼーションを、それぞれ、65℃で4時間および一晩、ハイブリダイゼーション緩衝液(Sigma、St.Loui、MO)を使用して行った。ハイブリダイゼーション後、メンブレンを65℃、洗浄緩衝液(10mMのリン酸ナトリウム、2.5mMのピロリン酸ナトリウム、0.5mMのEDTA、0.1%SD、リン酸を用いてpH7.8に調整した)で20分間、3回洗浄した。洗浄したフィルターを、オートラジオグラフィーのためにPhosphorimagerスクリーンに曝露させ、画像をスキャンした。
【0171】
【表9】
【0172】
6.5 プローブの除去
サザンハイブリダイゼーションのデータを得た後、メンブレンブロットからDNAプローブを除去し、メンブレンブロットは、異なるDNAプローブを用いたハイブリダイゼーションに再利用することができた。簡単に述べると、曝露後、メンブレンブロットを室温で10分間、再生溶液1(30mMのNaOH、1mMのNa2EDTA)で洗浄し、65℃で30分間、再生溶液2(5mMのNaPO4、1mMのNa2EDTA、0.1%SDS)で洗浄した。次いで、メンブレンブロットを2×SSCで簡単に洗浄し、それで別のDNAプローブを用いたハイブリダイゼーションの準備ができた。メンブレンブロットをオートラジオグラフィーのためにPhosphorimagerスクリーンに曝露させて、次のハイブリダイゼーションに進む前にDNAプローブの全てを確実に除去した。
【0173】
6.6.サザンブロットの結果
この試験では、aad−12遺伝子およびそのプロモーターであるAtUbi10のメチル化の状態を決定するために、メチル化感受性制限酵素AciIおよびHyp188IIIを使用した。pDAB4468のT鎖DNAの公知の制限酵素部位に基づいて予測された特定の消化物およびプローブの断片サイズが表10で示されている。サザンブロットにおいて分子量がより高い断片が検出されることは、AciI制限酵素部位およびHyp188III制限酵素部位の認識配列内のシトシンがメチル化されていることを示すと思われる。そのように、メチル化により、制限酵素がゲノムDNAを消化することができなくなる。
【0174】
制限酵素AciIおよびHyp188IIIを使用してaad−12遺伝子のメチル化の状態を検査した。aad−12プローブを使用すると、予測されたサイズを有するハイブリダイゼーションバンドが観察された。このデータは、ダイズイベントDAS−68416−4において、AciIおよびHyp188IIIの認識部位におけるメチル化が起こっていないことを示唆している。同様に、Hyp188IIIで消化されたダイズイベントDAS−68416−4DNA試料において、AtUbi10プローブを使用すると、予測された分子量のバンドが検出された。このデータは、aad−12プロモーター配列における認識部位がメチル化されていなかったことを示している。
【0175】
【表10】
【実施例7】
【0176】
農業形質データ
2008年の米国およびカナダの6区域における組成研究の一部として、および2009年に米国およびカナダの8区域において行われた別々の試験においても、ダイズイベントDAS−68416−4を用いた農業試験を行った。これらの試験では、イベントDAS−68416−4ダイズ(2,4−D除草剤および/またはグルホシネート除草剤の施用を伴う、または伴わない)と、その非トランスジェニック近同質遺伝子対照(Maverick)を比較した。両方の試験の全てにわたる結果により、農業形質パラメータは、従来のダイズ系統について得られる範囲内であることが示された。
【0177】
実施例7.1.
2008年の農業形質データの生成
2008年に、アイオワ、イリノイ、インディアナ、ネブラスカおよびカナダのオンタリオ(2つの現場)に位置する6つの現場において、イベントDAS−68416−4ダイズおよび非トランスジェニック対照(Maverick品種)を用いた農業研究を行った。対照系統であるMaverickと比較して、ダイズイベントDAS−68416−4(除草剤処理を伴う、および除草剤処理を伴わない)の同等性を調査するために、群生/集団数、実生/植物生長力、植物の高さ、倒伏、病害発生率、虫害、および開花までの日数を含めた農業形質決定因子を評価した。この試験は、実験1と称される。
【0178】
試験ダイズ種子および対照ダイズ種子を、作条の間隔をおよそ30インチ(75cm)として、25ftの作条当たりおよそ112種子の播種密度で植えた。それぞれの現場において、それぞれの処理について3回反復の小区画を確立し、それぞれの小区画は2〜25ftの作条からなった。小区画を完全乱塊(randomized complete block)(RCB)計画に配置し、それぞれの現場において独特に無作為化した。それぞれのダイズの小区画には、同様の成熟度の非トランスジェニックダイズの2つの作条が隣接していた。試験現場全体を最低10ftの同様の相対的な成熟度の非トランスジェニックダイズで囲んだ。
【0179】
除草剤処理については、1エーカー当たりおよそ20ガロン(187L/ha)の噴霧体積を施用した。これらの施用は、商慣習の最大表示量を再現するように設計した。2,4−Dを、季節の合計3lb ae/Aで、上からの施用で3回散布して施用した。出芽前および生長段階のおよそV4およびR2において、それぞれ1.0lb ae A(1,120g/ha)の施用を行った。グルホシネートを、季節の合計0.74lb ai/A(828g ai/ha)、上からの施用で2回散布して施用した。生長段階のおよそV6およびR1において、それぞれ0.33lb ai/Aおよび0.41lb ai/A(374g ai/haおよび454g ai/ha)の施用を行った。
【0180】
圃場現場の全てにわたって、混合モデル(SAS バージョン8;SAS Institute 1999)を使用して農業形質データについて分散分析を行った。エントリーを固定効果とみなし、区域、区域内のブロック、区域×エントリー、およびエントリー×区域内のブロックを、変量効果として指定した。全体的な処理の効果の有意性を、F検定を用いて推定した。対照と、噴霧していないダイズイベントDAS−68416−4(噴霧なし)トランスジェニックエントリー、グルホシネートを噴霧したダイズイベントDAS−68416−4(ダイズイベントDAS−68416−4+グルホシネート)トランスジェニックエントリー、2,4−Dを噴霧したダイズイベントDAS−68416−4(ダイズイベントDAS−68416−4+2,4−D)トランスジェニックエントリーならびにグルホシネートおよび2,4−Dの両方を噴霧したダイズイベントDAS−68416−4(ダイズイベントDAS−68416−4+両方)トランスジェニックエントリーの間で、t検定を用いて対応のある対比を行った。多重性を制御するために、偽発見率(False Discovery Rate)(FDR)を使用して調整P値も算出した(BenjaminiおよびHochberg、1995年)。
【0181】
【表11】
【0182】
対照、噴霧していないダイズイベントDAS−68416−4、ダイズイベントDAS−68416−4+2,4−D、ダイズイベントDAS−68416−4+グルホシネート、およびダイズイベントDAS−68416−4+両方の除草剤から収集した農業形質データの分析を行った。群生数、初期集団、実生生長力、施用後の害、倒伏、最終的な群生数または開花までの日数について、統計的有意差は観察されなかった(表12)。高さについては、対照とダイズイベントDAS−68416−4+2,4−D噴霧との間に有意な対応のあるt検定が観察された。しかし、有意な全体的な処理の効果は観察されず、ダイズイベントDAS−68416−4処理と対照との間の差異は非常に小さく、また、差異は、異なるダイズイベントDAS−68416−4処理間で共有されなかった。これらの結果に基づいて、ダイズイベントDAS−68416−4は、近同質遺伝子非トランスジェニック対照と農業的に同等であった。
【0183】
【表12】
【0184】
実施例7.2.
2009年の農業形質データの生成
2009年に、アーカンソー、アイオワ、イリノイ、インディアナ、ミズーリ、およびネブラスカに位置する8か所の現場においてダイズイベントDAS−68416−4および非トランスジェニック対照(Maverick品種)を用いた農業研究行った。ダイズイベントDAS−68416−4ダイズ(除草剤処理を伴う、および除草剤処理を伴わない)の対照との同等性を調査するために、群生/集団数、実生/植物生長力、植物の高さ、病害発生率、虫害、および開花までの日数を含めた農業形質決定因子を評価した(表13)。
【0185】
【表13】
【0186】
試験には完全乱塊(randomized-complete-block)計画を用いた。エントリーは、ダイズイベントDAS−68416−4、Maverick対照系統、および市販されている非トランスジェニックダイズ系統であった。試験ダイズ種子、対照ダイズ種子および参照ダイズ種子を、作条の間隔をおよそ30インチ(75cm)として、作条当たりおよそ112種子の播種密度で植えた。それぞれの現場において、それぞれの処理について4回反復の小区画を確立し、各小区画は2〜25ftの作条からなった。それぞれのダイズの小区画には、非トランスジェニックダイズ(Maverick)の2つの作条が隣接していた。試験現場全体を、最低4つの作条(または10ft)の非トランスジェニックダイズ(Maverick)で囲んだ。適切な昆虫、雑草、および病害の制御慣習を施用して農業的に許容できる作物を生産した。
【0187】
商慣習の最大表示量(maximum label rate)を再現するように除草剤処理を適用した。処理は、噴霧しない対照および特定の生長段階において2,4−D、グルホシネート、2,4−D/グルホシネートを施用した除草剤施用からなった。2,4−D施用については、V4生長段階およびR2生長段階において除草剤を1.0lb ae/A(1,120g ae/ha)の比率で施用した。グルホシネート処理については、V4生長段階およびV6〜R2生長段階にある植物に施用を行った。両方の施用については、V4施用およびV6〜R2施用について、それぞれグルホシネートを0.33lb ai/A(374g ai/ha)および0.41lb ai/A(454 g ai/ha)の比率で施用した。両方の除草剤を施用するエントリーは、ダイズイベントDAS−68416−4および非トランスジェニックMaverickを含めた対照であった。Maverick小区画は、除草剤を施用した後に枯死することが予測された。
【0188】
圃場現場全てにわたって、混合モデル(SASバージョン8;SAS Institute 1999年)を用いて農業形質データについて分散分析を行った。エントリーを固定効果とみなし、区域、区域内のブロック、区域×エントリー、およびエントリー×区域内のブロックを、変量効果として指定した。個々の区域において、類似の様式で、エントリーを固定効果とし、ブロックおよびエントリー×ブロックを変量効果として分析を行った。データは統計分析には回さなかった。95%信頼水準において有意差が宣言され、F検定を用いて全体的な処理の効果の有意性を推定した。噴霧していないAAD−12(噴霧なし)トランスジェニックエントリー、グルホシネートを噴霧したAAD−12(AAD−12+グルホシネート)トランスジェニックエントリー、2,4−Dを噴霧したAAD−12(AAD−12+2,4−D)トランスジェニックエントリーならびにグルホシネートおよび2,4−Dの両方を噴霧したAAD−12(AAD−12+2,4−D+グルホシネート)トランスジェニックエントリーと対照エントリーとの間で、t検定を用いて対応のある対比を行った。
【0189】
この試験では多数の対比を行ったので、多重性が問題であった。予想外の効果を探すために単一の試験において多数の比較を行う場合、多重性が問題である。これらの条件下で、比較に関するp値に基づいて誤って差異が宣言される確率は非常に高い(1−0.95比較の数)。この試験では分析物当たり4つの比較があり(16種の農業形質についての分析所見)、農業形質について64の比較が生じる。したがって、調整していないp値に基づいて1つまたは複数の誤った差異が宣言される確率は、農業形質について99%であった(1−0.9564)。
【0190】
対照エントリー、AAD−12噴霧なしエントリー、AAD−12+グルホシネートエントリー、AAD−12+2,4−Dエントリー、およびAAD−12+2,4−D+グルホシネートエントリーから収集した農業形質データの分析を行った。現場全てにわたる分析について(表14)、実生生長力、最終集団、植物生長力/害(V4、R1)、倒伏、病害発生率、虫害、開花までの日数、成熟までの日数、莢の数、種子の数、生産量、および植物の高さについて統計的有意差は観察されなかった。群生数および初期集団については、対照とAAD−12+グルホシネートエントリーとの間に有意な対応のあるt検定が観察されが、有意な全体的な処理の効果またはFDR調整P値を伴わなかった。植物生長力/害(R2)については、対照とAAD−12+グルホシネートエントリーおよびAAD−12+2,4−D+グルホシネートエントリーのどちらとの間にも有意な対応のあるt検定および有意な全体的な処理の効果が観察されたが、有意なFDR調整P値を伴わなかった。これらの変数全てについて平均した結果も、この試験において試験された参照系統について見いだされた範囲内であった。
【0191】
【表14】
【0192】
実施例7.3.
生態学的評価
ダイズ栽培の実践に詳しい人(育種家、圃場の管理者、圃場の農学者、圃場の従業員)がダイズイベントDAS−68416−4の圃場試験をモニターし、観察した。圃場試験を行った人員は、ダイズイベントDAS−68416−4植物に対する植物病害および害虫の発生率を、同じ試験において従来のダイズ品種と比較して視覚的にモニターした。実施例7.1に記載の実験1の一部として、病害および虫害を、0%が病害発生率または昆虫抵抗性に起因する害を表す0〜100%の数値的な尺度で評価した。表15に、実験1に記載の6か所の現場全てにわたる結果が示されている。
【0193】
【表15】
【0194】
病害発生率については、統計的有意差は観察されなかった。虫害については、対照とダイズイベントDAS−68416−4+両方の除草剤との間に有意な対応のあるt検定が観察された。しかし有意な全体的な処理の効果は観察されず、ダイズイベントDAS−68416−4処理と対照との間の差異は小さく、また、差異は、異なるダイズイベントDAS−68416−4処理間で共有されなかった。
【0195】
2006年〜2008年に行われた全てのUSDA APHISに届け出た圃場試験からの生態学的な観察も行った。ダイズイベントDAS−68416−4植物の試験における病害の発生率および昆虫の存在を記録し、従来の対照と比較したダイズイベントDAS−68416−4植物の発生率または反応の差異を検査した。全ての場合において、ダイズイベントDAS−68416−4植物のいずれの試験でも従来の対照と比較して差異は見られなかった。ダイズイベントDAS−68416−4および従来のダイズの試験において観察された病害ストレス要因および昆虫ストレス要因は、表16に記載されている。これらの観察は、生態学的なストレス要因に対するダイズイベントDAS−68416−4の反応は、従来のダイズの反応と異ならないという結論を裏付ける。
【0196】
【表16】
【0197】
実施例7.4.
発芽(Germancy)および休眠の評価
ダイズイベントDAS−68416−4種子の発芽について、温暖な条件下および寒冷な条件下で近同質遺伝子型の比較器と比較して調べることによって種子の休眠特性の変化を評価した。
【0198】
温暖発芽試験については、ダイズイベントDAS−68416−4および対照ダイズの種子を、水を染み込ませた発芽用パッドを含有するペトリ皿に、プレート当たり25個置き、過剰な水を流し出した。プレートを25℃に置き、これらの条件下で5日間保持した。系統当たり16プレート(400種子)を調製した。5日後、発芽しなかった種子の数を記録した。
【0199】
寒冷発芽試験については、鉢植え用土を満たした平箱半分当たり種子100個の種子を植えた。平箱に多少水をまき、10℃で7日間保持し、その後25℃に5日間曝露し、その後、発芽しなかった種子の数を記録した。
【0200】
それぞれの試験からのデータを、反復当たり種子100個の4回の反復を伴う完全無作為計画を使用した分散分析(ANOVA)によって分析した。統計分析のために、発芽した種子の数を100で割った値の平方根のアークサインを使用してデータを変換した。発芽率(パーセント)が、表17に要約されている。
【0201】
【表17】
【0202】
温暖発芽実験または寒冷発芽実験のいずれにおいても、ダイズイベントDAS−68416−4の種子の発芽と対照ダイズの種子の発芽との間に有意差はなかった(それぞれPr>F=1.0および0.13)。これらの結果は、ダイズイベントDAS−68416−4において種子の休眠特性が変化してないことを示している。
【0203】
実施例7.5.
農業特性、病害特性、害虫特性、および発芽特性の概要
生育期全体を通して植物の生長特性を評価する農業形質データにより、ダイズイベントDAS−68416−4の従来の非トランスジェニックダイズとの同等性が実証されている。植物の生長および表現型の特性、病害および昆虫の圧力に対する感受性によって示される生態学的ストレス要因に対する反応、ならびに発芽および休眠の特性は、多様な環境にわたって、ダイズイベントDAS−68416−4植物と従来のダイズとの間で変化しなかった。したがって、これらのデータにより、ダイズイベントDAS−68416−4の農業特性、病害特性、および害虫特性は、従来のダイズの農業特性、病害特性、および害虫特性と有意に異ならず、また、ダイズイベントDAS−68416−4ダイズが植物害虫リスクの増加を示す徴候はないという結論が裏付けられる。Benjamini、Y.、Hochberg、Y.(1995年)Controling the false discovery rate:A practical and powerful approach to multiple testing. J. Royal Statistical Soc. B、57巻:289〜300頁。
【実施例8】
【0204】
粒および茎葉の組成
ダイズイベントDAS−68416−4(2,4−Dを噴霧、グルホシネートを噴霧、2,4−D+グルホシネートを噴霧、または2,4−Dまたはグルホシネートを噴霧していない)と従来のダイズとの間の同等性を調査するために、ダイズの茎葉および粒について組成分析を実施した。米国およびカナダの主要なダイズ生産地域内に位置する6か所の試験場において、ダイズイベントDAS−68416−4を伴う種子系統およびダイズイベントDAS−68416−4を伴わない種子系統を使用して試験を行った。試験場は、多様な農業習慣および環境条件の地域を代表し、タンパク質の発現解析および実施例7.1に記載の農業実験1のために使用したのと同じ現場であった。試験はアイオワ、イリノイ、インディアナ、ネブラスカ、およびカナダのオンタリオ(2現場)に位置した。
【0205】
ダイズの茎葉および粒の試料を、種々の試験を用いて栄養素量について分析した。茎葉について行った分析は、タンパク質、脂肪、灰分、水分、炭水化物、酸性デタージェント繊維(ADF)、中性デタージェント繊維(NDF)、カルシウムおよびリンを含んだ。粒について行った分析は、一般成分(灰分、全脂肪、水分、タンパク質、コレステロール、炭水化物)、繊維、ミネラル、アミノ酸、脂肪酸、ビタミン、抗栄養因子を含んだ。
【0206】
ダイズの茎葉および粒についての栄養分析の結果を文献において報告されている値と比較した。圃場現場の全てにわたって、混合モデルを使用して分散分析も行った。エントリーを固定効果とみなし、区域、区域内のブロック、および区域×エントリーを、変量効果として指定した。全体的な処理の効果の有意性を、F検定を用いて推定した。ダイズイベントDAS−68416−4トランスジェニックエントリー(噴霧していないAAD−12;2,4−Dまたはグルホシネートを噴霧していない)、グルホシネートを噴霧したダイズイベントDAS−68416−4トランスジェニックエントリー(ダイズイベントDAS−68416−4+グルホシネート)、2,4−Dを噴霧したダイズイベントDAS−68416−4トランスジェニックエントリー(ダイズイベントDAS−68416−4+2,4−D)、ならびにグルホシネートおよび2,4−Dの両方を噴霧したダイズイベントDAS−68416−4トランスジェニックエントリー(ダイズイベントDAS−68416−4+両方)と、対照エントリーとの間でt検定を用いて対応のある対比を行った。
【0207】
この試験では多数の対比を行ったので、多重性が問題であった。予想外の効果を探すために単一の試験において多数の比較を行う場合、多重性が問題である。これらの条件下で、比較に関するp値に基づいて誤って差異が宣言される確率は非常に高い(1−0.95比較の数)。この試験では分析物当たり4つの比較があり(75の定量化された分析物)、現場の組成分析全てにわたって300の比較が行われた。したがって、調整していないp値に基づいて1つまたは複数の誤った差異が宣言される確率は99.99%より高かった。
【0208】
多重性を説明するための1つの方法は、p値を調整して実験に関する誤差率(宣言された差異が全て有意である確率)を制御することであるが、試験において多くの比較を行う場合、特異的な効果を検出するための検定力は有意に低下する可能性がある。検定力がはるかに大きい代替法は、p値を調整してそれぞれの宣言された差異が有意である確率を制御することである。これは、偽発見率(FDR)手順(BenjaminiおよびHochberg、1995年)を用いて実現することができる。したがって、変量効果の処理間の真の差異の識別(偽陽性)を改善するために、FDRを用いてp値を調整した。
【0209】
実施例8.1.
ダイズの茎葉の組成分析
対照、噴霧していないダイズイベントDAS−68416−4、ダイズイベントDAS−68416−4+グルホシネート、ダイズイベントDAS−68416−4+2,4−DおよびダイズイベントDAS−68416−4+両方の除草剤由来のダイズの茎葉試料中のタンパク質、脂肪、灰分、水分、炭水化物、酸性デタージェント繊維(ADF)、中性デタージェント繊維(NDF)、カルシウムおよびリンの分析を実施した。全ての区域にわたる結果の概要が表18に示されている。
【0210】
タンパク質、脂肪、灰分、水分、炭水化物、ADF、NDF、カルシウムまたはリンについて、現場全てにわたる分析において、対照エントリーとトランスジェニックエントリーとの間に統計学的差異は観察されなかった。ダイズイベントDAS−68416−4+グルホシネートおよびダイズイベントDAS−68416−4+両方の除草剤についての現場全てにわたる平均灰分値は、ダイズイベントDAS−68416−4+グルホシネートおよびダイズイベントDAS−68416−4+2,4−DについてのNDF値と同様に、文献範囲の外側であった。非トランスジェニック対照を含めた全ての処理についてのADF値も、文献値の外側であった。茎葉におけるいかなる一般成分、繊維の種類、またはミネラルについても、平均値は、非トランスジェニック対照とトランスジェニックエントリーのいずれとの間でも有意に異ならなかった。これらの組成成分に基づいて、ダイズイベントDAS−68416−4ダイズ由来の茎葉は、近同質遺伝子非トランスジェニック対照の茎葉と実質的に同等であった。
【0211】
【表18】
【0212】
実施例8.2.
ダイズの粒の組成分析
実施例8.2.1
一般成分および繊維
対照、噴霧していないダイズイベントDAS−68416−4、ダイズイベントDAS−68416−4+グルホシネート、ダイズイベントDAS−68416−4+2,4−DおよびダイズイベントDAS−68416−4+両方の除草剤由来のダイズの粒試料中のタンパク質、脂肪、灰分、水分、コレステロール、炭水化物、ADF、NDFおよび総食物繊維の分析を実施した。全ての区域にわたる結果の概要が表19に示されている。
【0213】
脂肪、ADFまたは総食物繊維に関しては、現場全てにわたる分析において、対照エントリーとトランスジェニックエントリーとの間に統計学的差異は観察されなかった。しかし、ダイズイベントDAS−68416−4+2,4−Dエントリーについては、ADFが文献範囲よりもわずかに高かった。
【0214】
タンパク質レベルは、現場全てにわたる分析において、噴霧なし、ダイズイベントDAS−68416−4+2,4−D、およびダイズイベントDAS−68416−4+両方の除草剤について、調整していないP値に基づいて対照と比較して有意に異なった。しかし、FDRを調整した後、ダイズイベントDAS−68416−4+2,4−Dについてのp値のみが有意であり、また、全ての処理についての全体の平均のタンパク質の値は報告された文献値の範囲内であり、これは、差異が生物学的には意味がないことを示している。
【0215】
灰分の現場全てにわたる分析において、対照と2,4−Dを噴霧したダイズイベントDAS−68416−4処理との間に有意な調整していないP値が観察されたが、全体的な処理の効果または調整したP値は観察されなかった。灰分値も、報告された文献値の範囲内であり、これは、差異が生物学的には意味がないことを示している。
【0216】
水分レベルは、現場全てにわたる分析において、噴霧なし、ダイズイベントDAS−68416−4+2,4−D、およびダイズイベントDAS−68416−4+両方の除草剤について、調整していないP値に基づいて対照と比較して有意に異なった。しかし、水分についての全体的な処理の効果は有意でなく、ダイズイベントDAS−68416−4+2,4−D処理のみが有意なFDR調整したP値を有し、また、全ての処理について平均水分レベルは文献範囲内であった。このことにより、差異は生物学的には意味がないことが示された。
【0217】
コレステロール値は、全て定量限界未満(<LOQ)であり、また、報告された文献値はない。
【0218】
炭水化物レベルは、現場全てにわたる分析において、噴霧なし、ダイズイベントDAS−68416−4+グルホシネート、およびダイズイベントDAS−68416−4+2,4−Dについて、調整していないP値に基づいて対照と比較して有意に異なった。しかし、ダイズイベントDAS−68416−4+2,4−D処理のみが、FDR調整したP値に基づいて対照と有意に異なり、全ての処理の平均は報告された文献値の範囲内であり、これは、非トランスジェニックダイズとの同等性を示している。
【0219】
NDFレベルは、現場全てにわたる分析において、ダイズイベントDAS−68416−4+グルホシネートについて、調整していないP値に基づいて対照と比較して有意に異なっが、これは、有意な調整したP値または全体的な処理の効果を伴わなかった。現場全てにわたるNDFの値は、ダイズイベントDAS−68416−4+グルホシネートおよびダイズイベントDAS−68416−4+2,4−Dエントリーについて報告された文献値よりもわずかに高かったが、差異は、非トランスジェニック近同質遺伝子対照と比較して9%未満であった。
【0220】
これらの組成成分に基づいて、ダイズイベントDAS−68416−4由来の粒は、非トランスジェニックダイズの粒と実質的に同等であった。
【0221】
【表19】
【0222】
実施例8.2.2
ミネラル
対照、噴霧していないダイズイベントDAS−68416−4、ダイズイベントDAS−68416−4+グルホシネート、ダイズイベントDAS−68416−4+2,4−DおよびダイズイベントDAS−68416−4+両方の除草剤由来のダイズの粒試料中のカルシウム、クロム、銅、ヨウ素、鉄、マグネシウム、マンガン、モリブデン、リン、カリウム、セレン、ナトリウムおよび亜鉛の分析を実施した。全ての区域にわたる結果の概要が表20に示されている。
【0223】
クロム、銅、ヨウ素、鉄、マンガン、モリブデン、リン、セレンおよびナトリウム(検出されなかった)については、現場全てにわたる分析において、調整していないP値に基づいて、対照エントリーとトランスジェニックエントリーとの間に統計学的差異は観察されなかった。
【0224】
カルシウムは、現場全てにわたる分析におい て、ダイズイベントDAS−68416−4+2,4−Dについて、調整していないP値に基づいて有意差を有したが、これは、有意なFDR調整したP値または全体的な処理の効果を伴わず、全ての処理の平均は文献範囲内に入り、これは、差異が生物学的には意味がないことを示している。
【0225】
マグネシウムレベルは、現場全てにわたる分析において、ダイズイベントDAS−68416−4+両方の除草剤およびダイズイベントDAS−68416−4+グルホシネートについて、それぞれ、調整していないp値および調製したp値に基づいて対照と比較して有意に異なったが、全体的な処理の効果は有意でなかった。マグネシウムの現場全てにわたる平均値は、報告された文献値よりもわずかに低かったが、対照と比較して差異は小さく(<3%)、全てのダイズイベントDAS−68416−4エントリーが、対照と比較して、より文献値に近かった。
【0226】
全てのダイズイベントDAS−68416−4エントリーが、現場全てにわたる分析において、対照と比較して有意に高いカリウムの値を有した。しかし、対照と比較して差異は小さく(<5%)、全てのダイズイベントDAS−68416−4エントリーが、対照と比較して、より文献範囲に近かった。
【0227】
亜鉛レベルの差異は、現場全てにわたる分析において、ダイズイベントDAS−68416−4+両方の除草剤について、調整していないP値に基づいて有意であったが、これは、有意なFDR調整したP値または全体的な処理の効果を伴わず、また、差異は小さかった(<4%)。
【0228】
これらの組成成分に基づいて、ダイズイベントDAS−68416−4由来の粒は、非トランスジェニックダイズの粒と実質的に同等であった。
【0229】
【表20】
【0230】
実施例8.2.3
アミノ酸
対照、噴霧していないダイズイベントDAS−68416−4、ダイズイベントDAS−68416−4+グルホシネート、ダイズイベントDAS−68416−4+2,4−DおよびダイズイベントDAS−68416−4+両方の除草剤由来のダイズの粒試料中の以下のアミノ酸の分析を実施した:アラニン、アルギニン、アスパラギン酸、シスチン、グルタミン酸、グリシン、ヒスチジン、イソロイシン、ロイシン、リジン、メチオニン、フェニルアラニン、プロリン、セリン、トレオニン、トリプトファン、チロシン、およびバリン。全ての区域にわたる結果の概要が表21に示されている。
【0231】
システイン、メチオニン、プロリン、チロシンまたはトリプトファンについては、対照エントリーとトランスジェニックエントリーとの間に統計学的差異は観察されなかった。ダイズイベントDAS−68416−4+2,4−Dについてのイソロイシンレベルは、調整していないP値に基づいて対照と有意に異なったが、これは、有意なFDR調整したP値または有意な全体的な処理の効果を伴わなかった。残りの12種のアミノ酸のレベルは、ダイズイベントDAS−68416−4エントリーのうちの2つ以上について、対照と比較してわずかに低かったが(<7%)、全てが非トランスジェニックダイズについての文献範囲内に入った。全てのエントリーについて、全てのアミノ酸が文献範囲内であり、これは、差異が生物学的には意味がないことを示している。これらの組成成分に基づいて、ダイズイベントDAS−68416−4由来の粒は、非トランスジェニックダイズの粒と実質的に同等であった。
【0232】
【表21】
【0233】
実施例8.2.4
脂肪酸
対照、噴霧していないダイズイベントDAS−68416−4、ダイズイベントDAS−68416−4+グルホシネート、ダイズイベントDAS−68416−4+2,4−DおよびダイズイベントDAS−68416−4+両方の除草剤由来のダイズの粒試料中の22種の脂肪酸の分析を実施した。全ての区域にわたる結果の概要が表22に示されている。
【0234】
脂肪酸の10:0カプリン酸、15:0ペンタデカン酸、15:1ペンタデセン酸、20:3エイコサトリエン酸、20:4アラキドン酸、8:0カプリル酸、12:0ラウリン酸、14:0ミリスチン酸、14:1ミリストレイン酸、17:1ヘプタデセン酸、18:3ガンマリノレン酸、および20:2エイコサジエン酸を分析し、結果はLOQ未満であった。脂肪酸の16:0パルミチン酸、17:0ヘプタデカン酸、および20:1エイコセン酸は、対照とAAD−12エントリーとの間で有意に異ならなかったが、20:1エイコセン酸値は、AAD−12+グルホシネートおよびAAD−12+両方の除草剤について、報告された文献値よりも低かった。しかし、対照と比較して差異は小さかった(<5%)。
【0235】
16:1パルミトレイン酸のレベルは、対照と、噴霧していないダイズイベントDAS−68416−4、ダイズイベントDAS−68416−4+グルホシネート、ダイズイベントDAS−68416−4+2,4−D、およびダイズイベントDAS−68416−4+両方の除草剤との間で、調整していないp値に基づいて有意に異なった。しかし、噴霧していないダイズイベントDAS−68416−4エントリーのみが、16:1パルミトレイン酸について有意なFDR調整したP値を有した。この処理についての16:1パルミトレイン酸の現場全てにわたる値は報告された文献値と比較して低かったが、近同質遺伝子対照と比較して差異は小さかった(<13%)。
【0236】
18:0ステアリン酸のレベルは、対照と、噴霧なしおよびダイズイベントDAS−68416−4+グルホシネートとの間で、調整していないP値に基づいて有意に異なった。しかし、調整したp値または全体的な処理の効果に基づいて有意差は確認されず、また、全てのエントリーは報告された文献値の範囲内であり、これは、非トランスジェニックダイズとの同等性を示している。
【0237】
18:1オレイン酸のレベルは、対照と、噴霧していないダイズイベントDAS−68416−4、ダイズイベントDAS−68416−4+グルホシネート、ダイズイベントDAS−68416−4+2,4−D、およびダイズイベントDAS−68416−4+両方の除草剤との間で有意に異なった。しかし、18:1オレイン酸レベルは、全ての処理について報告された文献値の範囲内であり、これは、非トランスジェニックダイズとの同等性を示している。
【0238】
18:2リノール酸のレベルは、対照と、噴霧なしおよびダイズイベントDAS−68416−4+2,4−Dとの間で、調整していないP値に基づいて有意に異なった。しかし、調整したp値または全体的な処理の効果に有意差は確認されず、また、18:2リノール酸レベルは全ての処理について報告された文献値の範囲内であり、これは、非トランスジェニックダイズとの同等性を示している。
【0239】
18:3リノレン酸のレベルは、ダイズイベントDAS−68416−4エントリーのそれぞれと、対照との間で、調整していないp値に基づいて有意に異なり、また、調整したp値も、噴霧していないダイズイベントDAS−68416−4およびダイズイベントDAS−68416−4+両方の除草剤処理の間で対照と比較して有意であった。18:3リノレン酸について入手可能な文献値はないが、ダイズイベントDAS−68416−4と対照処理との間の差異は小さかった(<6%)。
【0240】
20:0アラキジン酸のレベルは、対照と、噴霧していないダイズイベントDAS−68416−4、ダイズイベントDAS−68416−4+グルホシネート、ダイズイベントDAS−68416−4+2,4−D、およびダイズイベントDAS−68416−4+両方の除草剤との間で、調整していないp値に基づいて有意に異なり、まら、20:0アラキジン酸は、現場全てにわたる分析において、噴霧なし、およびダイズイベントDAS−68416−4+グルホシネート処理について、調整したP値にも有意差を有した。しかし、20:0アラキジン酸レベルは全ての処理について報告された文献値の範囲内であり、これは、非トランスジェニックダイズとの同等性を示している。
【0241】
22:0ベヘン酸のレベルは、対照と、噴霧なし、ダイズイベントDAS−68416−4+グルホシネート、ダイズイベントDAS−68416−4+2,4−D、およびダイズイベントDAS−68416−4+両方の除草剤との間で、調整していないp値に基づいて有意に異なり、また、22:0ベヘン酸のレベルは、現場全てにわたる分析において、ダイズイベントDAS−68416−4+グルホシネートについて、調整したP値にも有意差を有した。しかし、有意な全体的な処理の効果はなく、また、22:0ベヘン酸レベルは全ての処理について報告された文献値の範囲内であり、これは、非トランスジェニックダイズとの同等性を示している。
【0242】
調査した22種の脂肪酸のうち、21種の脂肪酸について、4つのダイズイベントDAS−68416−4エントリーの全てが、対照と統計学的に区別できなかったか、または、文献値の範囲内であった。1つの場合(噴霧していないダイズイベントDAS−68416−4;16:1パルミトレイン酸)で、値は最小の文献値をわずかに下回り、対照と統計学的に異なった(<13%低かった)が、3種の噴霧処理の全ては、文献範囲内であった。これらの組成成分に基づいて、ダイズイベントDAS−68416由来の粒は、非トランスジェニックダイズの粒と実質的に同等であった。
【0243】
【表22】
【0244】
実施例8.2.5
ビタミン
対照、噴霧していないダイズイベントDAS−68416−4、ダイズイベントDAS−68416−4+グルホシネート、ダイズイベントDAS−68416−4+2,4−DおよびダイズイベントDAS−68416−4+両方の除草剤由来のダイズの粒試料中のビタミンの分析を実施した。全ての区域にわたる結果の概要が表23に示されている。
【0245】
ダイズの粒中のベータ−トコフェロール、デルタ−トコフェロール、ガンマ−トコフェロール、ビタミンA、ビタミンB5、ビタミンB6、ビタミンB12、ビタミンC、ビタミンDおよびナイアシンについての文献値は見つからなかった。ベータトコフェロール、ビタミンA、ビタミンB12およびビタミンDは全てLOQ未満であった。ビタミンB1、ビタミンB2、ビタミンB6、ビタミンC、ビタミンEまたはナイアシンについて、対照、噴霧していないAAD−12および処理したAAD−12の間に差異は観察されなかった。入手可能な文献範囲を有するこれらのビタミンの中で、近同質遺伝子対照を含めた全ての処理について値がその範囲を超えたビタミンB2を例外として、全ての処理がこれらの範囲内に入った。
【0246】
デルタ−トコフェロールレベルは、対照と、ダイズイベントDAS−68416−4+グルホシネートエントリーおよびダイズイベントDAS−68416−4+2,4−Dエントリーとの間で、調整していないp値に基づいて有意に異なった。しかし、これは、有意な調整したP値または全体的な処理の効果を伴わなかった。ガンマトコフェロールは、対照と、噴霧なしエントリーおよびダイズイベントDAS−68416−4+2,4−Dエントリーとの間で、調整していないp値および調整したp値に基づいて有意に異なった。しかし、ガンマトコフェロールは、ダイズイベントDAS−68416−4処理について、近同質遺伝子対照と比較して11%未満高かった。
【0247】
ビタミンB5レベルは、対照と、ダイズイベントDAS−68416−4+グルホシネートエントリーとの間で、調整したP値に基づいて有意に異なった。しかし、これは、有意な全体的な処理の効果を伴わなかった。
【0248】
葉酸は、対照と、噴霧なし、ダイズイベントDAS−68416−4+2,4−DおよびダイズイベントDAS−68416−4+両方の除草剤について、調整していないp値に基づいて有意に異なった。葉酸については、ダイズイベントDAS−68416−4エントリーの2種について、対照と比較して調整したp値にも有意差があった。しかし、葉酸レベルは全ての処理について報告された文献値の範囲内であり、ダイズイベントDAS−68416−4エントリーは、近同質遺伝子対照との差異が<9%であり、これは、非トランスジェニックダイズとの同等性を示している。
【0249】
これらの組成成分に基づいて、ダイズイベントDAS−68416−4由来の粒は、非トランスジェニックダイズの粒と実質的に同等であった。
【0250】
【表23】
【0251】
実施例8.2.5
イソフラボン
対照、噴霧していないダイズイベントDAS−68416−4、ダイズイベントDAS−68416−4+グルホシネート、ダイズイベントDAS−68416−4+2,4−DおよびダイズイベントDAS−68416−4+両方の除草剤由来のダイズの粒試料中のイソフラボンの分析を実施した。全ての区域にわたる結果の概要が表24に示されている。
【0252】
ゲニステインおよびグリシテインの結果は、処理した試料について、LOQを下回った。ダイジンレベルは、対照とダイズイベントDAS−68416−4+両方の除草剤エントリーとの間で、調整していないp値および調整したp値に基づいて有意に異なった。しかし、全体的な処理の効果は有意でなかった。報告された文献値はないが、ダイズイベントDAS−68416−4+両方の除草剤処理は、近同質遺伝子対照との差異が<9%であった。ゲニスチンレベルは、対照とダイズイベントDAS−68416−4+両方の除草剤エントリーとの間で、調整していないp値および調整したp値に基づいて有意に異なった。しかし、全体的な処理の効果は有意でなかった。ゲニスチン値は、全ての処理について、報告された文献値よりも高かったが、ダイズイベントDAS−68416−4処理は、近同質遺伝子対照と比較して差異が9%未満であった。グリシチンの値は対照とダイズイベントDAS−68416−4+両方の除草剤との間で、調整していないp値および調整したp値に基づいて有意に異なった。グリシチンについて報告された文献値はなかったが、全てのダイズイベントDAS−68416−4エントリーが、近同質遺伝子エントリーと比較して、差異が13%未満であった。
【0253】
これらの組成成分に基づいて、ダイズイベントDAS−68416−4ダイズ由来の粒は、非トランスジェニックダイズの粒と実質的に同等であった。
【0254】
【表24】
【0255】
実施例8.2.5
抗栄養因子
対照、噴霧していないダイズイベントDAS−68416−4、ダイズイベントDAS−68416−4+グルホシネート、ダイズイベントDAS−68416−4+2,4−DおよびダイズイベントDAS−68416−4+両方の除草剤由来のダイズの粒試料中の抗栄養因子の分析を実施した。全ての区域にわたる結果の概要が表25に示されている。
【0256】
レクチン、フィチン酸、またはトリプシン阻害物質について、対照エントリーとトランスジェニックエントリーとの間に統計学的差異は観察されなかった。これらの3種の抗栄養因子は全て文献範囲内でもあり、これは、非トランスジェニックダイズとの同等性を示している。
【0257】
ラフィノースは、ダイズイベントDAS−68416−4+グルホシネート処理について、対照と比較して調整していないp値に基づいて有意に低かった(<10%)。ラフィノースは、現場全てにわたる分析において、調整したP値または全体的な処理の効果に基づいて有意に異ならなかった。ラフィノースレベルは、全ての処理について報告された文献値の範囲内でもあり、これは、非トランスジェニックダイズとの同等性を示している。
【0258】
スタキオースは、対照およびダイズイベントDAS−68416−4+グルホシネートエントリーとの間で、調整していないP値に基づいて有意に異なった。スタキオースレベルは、現場全てにわたる分析において、調整したP値または全体的な処理の効果に基づいて有意に異ならなかった。スタキオースレベルは、全ての処理について報告された文献値の範囲内でもあり、これは、非トランスジェニックダイズとの同等性を示している。
【0259】
レクチン、フィチン酸、ラフィノース、スタキオースおよびトリプシン阻害物質についての抗栄養因子分析は、全て報告された文献値の範囲内であり、調整していないp値に基づく2つの有意差は、抗栄養因子のレベルがダイズイベントDAS−68416−4処理について対照と比較して低かった。
【0260】
これらの組成成分に基づいて、ダイズイベントDAS−68416−4ダイズ由来の粒は、非トランスジェニックダイズの粒と実質的に同等であった。
【0261】
【表25】
【0262】
実施例8.3
粒および茎葉の組成の概要
ダイズイベントDAS−68416−4の組成物は、近同質遺伝子系統と統計学的に区別できなかったか、または近同質遺伝子系統との差異が13%未満であったか、または非トランスジェニックダイズについての文献範囲内であった。したがって、ダイズイベントDAS−68416−4は、非トランスジェニックダイズと実質的に同等であることが見いだされた。組成の結果のプロットは、噴霧していないダイズイベントDAS−68416−4、ダイズイベントDAS−68416−4+グルホシネート、ダイズイベントDAS−68416−4+2,4−DおよびダイズイベントDAS−68416−4+両方ダイズと対照ダイズ系統との間に、生物学的に意味のある、処理に関連するいかなる組成上の差異も示していない。結論として、噴霧していないダイズイベントDAS−68416−4、ダイズイベントDAS−68416−4+グルホシネート、ダイズイベントDAS−68416−4+2,4−DおよびダイズイベントDAS−68416−4+両方の組成の結果により、ダイズイベントDAS−68416−4と従来のダイズの同等性が確認される。
【実施例9】
【0263】
植え付け前の、一掃するための施用および出芽前試験
この実施例では、本発明によって可能になる、ダイズ作物の植え付けおよび除草剤の使用の新しい方法を記載している。
【0264】
植え付け前の、一掃するための除草剤の施用は、所与の作物を植え付ける前に、冬または初春の間に現れた雑草を枯らすことを意図している。一般には、これらの施用は、植え付け前に雑草の物理的な除去が完了していない不耕起または低耕起の管理体制において適用する。したがって、不耕起または低耕起のダイズと併せて使用する除草剤計画によって、植え付け前に存在する非常に広範な広葉雑草およびイネ科雑草を制御しなければならない。
【0265】
グリホサート、グラモキソン、およびグルホシネートは、植え付け前の、一掃するための除草剤の施用のために広範に使用される非選択的な残留しない除草剤の例である。しかし、一部の雑草は、以下の因子の1つまたは複数に起因して、そのような除草剤で制御することが難しい:除草剤に対する雑草種または生物型の固有の非感受性、比較的大きなサイズの越冬一年生雑草、および除草剤の取り込みおよび活性を制限する寒冷な気候条件。非選択的な除草剤が不十分である場合に、これらの除草剤とタンク混合して雑草に対するスペクトルおよび活性を増加させるために、いくつかの除草剤の選択肢が利用可能である。1つの例は、ヒメムカシヨモギ(Conyza canadensis)(horseweed)を制御するために役立つ2,4−D+グリホサートのタンク混合の施用を用いることである。グリホサートは、存在する雑草の大部分を植え付け前に一掃制御するために420〜1680g ae/ha、より一般には、560〜840g ae/haで使用することができるが、多くの広葉雑草種(例えば、ヒメムカシヨモギ(horseweed))を制御することに役立つように、280〜1120g ae/haの2,4−Dをに施用することができる。
【0266】
2,4−Dは、非常に広範囲の広葉雑草(比較的低温においてさえも)に対して有効であり、かつ非常に安価であるので、一般的に好まれる植え付け前の除草剤である。しかし、一掃するための施用の後に植える作物が2,4−Dに対して感受性が高い場合(例えば、大部分の双子葉作物)、土壌に2,4−Dが残留すること(短期であっても)により、作物が害される恐れがある。ダイズに使用するために、多くの2,4−Dエステル除草剤の表示では、1lb ai/エーカーの一掃するための施用の少なくとも15日後までは新しい作物の植え付けが制限されている。2,4−Dの低揮発性エステル製剤の場合では、一掃するための最大0.5lb ai/エーカーまでの比率では、一般には、施用後7日を過ぎてから植え付けることが必要とされ、1.0lb ai/エーカーの比率では、一般には、施用後30日を過ぎてから植え付けることが必要とされる。水溶性である2,4−Dのアミン製品の場合のさらなる懸念は、除草剤が種子の帯域に浸出する恐れがあることである。しかし、aad−12遺伝子を含有する作物は、2,4−Dに対して耐性であり、土壌中に2,4−Dが残留することによって負の影響を受けない。この耐性に起因して、aad−12遺伝子を含有する植物は、aad−12遺伝子を含有しない植物よりも害を持続することなく、一掃後すぐに植えることができる。さらに、タンク混合(または市販のプレミックス)パートナーと用いる2,4−D除草剤の柔軟性が増し、費用が縮小することにより、雑草制御の選択肢が改善され、重要な不耕起および低耕の状況における一掃するための施用の頑強性が増す。
【0267】
2,4−Dアミンの出芽前施用を、植え付けの7日前、15日前または30日前に、1120g ae/ha、2240g ae/ha、4480g ae/haの比率で、aad−12遺伝子を含有するダイズ(イベントDAS−68416−4)および対照ダイズに適用する。出芽前施用を、当技術分野において認められている手順で、地理的に異なる地方に位置する圃場の小区画に適用する。出芽および早期生長の正確な評価をもたらすために、除草剤処理した小区画は、処理していない小区画と対になっている。植え付け後、および植え付けの7日前、15日前または30日前の2,4−Dの施用後;aad−12遺伝子を含有するダイズ(イベントDAS−68416−4)および対照ダイズの害を測定する。圃場試験の結果は、aad−12遺伝子を含有するダイズ(イベントDAS−68416−4)により、植え付けの7日前、15日前、または30日前の2,4−D除草剤の出芽前処理に頑強な耐性がもたらされることを示している。
【0268】
これらの実施例は、利用可能な多くの選択肢のうちのいくつかである。雑草制御の当業者は、例として、連邦の除草剤表示(CPR、2003年)に記載の製品およびAgriliance Crop Protection Guide(2003年)に記載の用法を利用することによって、これらに限定されないが、グラモキソン+2,4−Dまたはグルホシネート+2,4−Dを含めた種々の他の施用に注目されよう。上記の例を、安定に形質転換されていれば、AAD−12遺伝子によって保護される、2,4−D(または他のフェノキシオーキシン除草剤)に対する感受性が高い任意の作物に施用することができることも、当業者には認識されよう。
【0269】
DAS−68416−4ダイズは、2,4−Dに対して耐性である除草剤である。この実施例では、圃場条件下での、DAS−68416−4ダイズの除草剤耐性の特徴付けについて報告している。DAS−68416−4ダイズを、除草剤である2,4−Dに対する耐性について、米国で行った圃場試験において評価した。DAS−68416−4ダイズについて、提唱された最大の施用率は、2,4−Dについては1120g ae/haである。以下に考察するデータは、DAS−68416−4ダイズにより、これらの提唱された最大使用率の少なくとも4倍の比率において、許容できる耐性がもたらされることを示している。
【0270】
2,4−Dの解毒、およびこの化合物によって後に引き起こされる害からの保護に対するDAS−68416−4ダイズの効力を、圃場研究において特徴付けた。2,4−Dに対する出芽前の耐性を評価するために試験を設計した。出芽前試験を完全乱塊計画として行った。実験単位は、出芽前試験のための、長さおよそ3mの、作条が2つある小区画からなり、全ての試験は3回の反復を含有した。除草剤の施用は全て、噴霧体積およそ140〜190l/haを送達する、ガスで加圧された小さな小区画噴霧装置を用いて行った。使用した2,4−Dの製剤は、456g ae/リットルのジメチルアンモニウム塩であった。作物が出芽したおよそ7日後に、視覚的な害の評点を取得した。評点は、小区画内の植物全てにわたって観察された全ての害の症状の視覚的合成を反映する0〜100の尺度で取得し、0=処理していない小区画と比較して害なし、100=全ての植物が枯死とした。
【0271】
出芽前試験は、植え付け後だが、作物が出芽する前の、1120g ae/ha、2240g ae/ha、および4480g ae/haの施用からなった。出芽前試験に関する情報は、表26に提示されている。
【0272】
【表26】
【0273】
要約すると、2,4−D耐性の圃場評価を行った。ミシシッピ、インディアナ、およびミネソタの区域において、DAS−68416−4ダイズおよび形質転換されていないMaverickダイズの、出芽前の2,4−Dアミンの施用に対する反応を検査する試験を行った。この3区域全てにわたって平均すると、2,4−DアミンをDAS−68416−4に出芽前に施用することにより、施用率に関係なく、2%以下の害が生じた(表27)。形質転換されていないMaverickでは、同じ処理によって34〜60%の害が引き起こされた。2,4−Dを出芽前施用したものの害は、群生の減少および生長の低下からなった。
【0274】
【表27】
【実施例10】
【0275】
グリホサート耐性遺伝子+ダイズイベントDAS−68416−4の組合せを使用してグリホサート抵抗性の雑草を制御する方法
グリホサートは、非常に広範な広葉およびイネ科雑草種を制御するので、広範囲にわたって使用されている。しかし、GTCにおいて、および非作物施用においてグリホサートの使用を繰り返すことにより、雑草のシフトが自然により耐性の種またはグリホサート抵抗性の生物型に選択されてきており、またこれからもそれが続くであろう。大部分の除草剤抵抗性管理戦略では、抵抗性の雑草の出現を遅延させる方法として、同じ種の制御をもたらすが異なる作用様式を有する、効果的な比率で使用されるタンク混合の除草剤パートナーが処方される。416AAD−12イベントにグリホサート耐性形質(および/または他の除草剤耐性形質)を積み重ねることにより、グリホサート、フェノキシオーキシン(複数可)(例えば、2,4−D)およびピリジルオキシ酢酸オーキシン除草剤(例えば、トリクロピル)を同じ作物において選択的に使用することが可能になり、それによってGTCにおいてグリホサート抵抗性の双子葉雑草種を制御することが可能になる機構をもたらすことができた。これらの除草剤は、異なる作用様式の2つ以上の除草剤を含むタンク混合物中で同時に施用することができ;植え付け前、出芽前、または出芽後および分裂時のおよそ2時間からおよそ3カ月までにわたる施用のタイミングで、単一の除草剤の組成をそれぞれ逐次的に施用することができる;あるいは、それぞれの化学的クラスを代表する任意の数の除草剤の任意の組合せを、作物の植え付けの約7カ月以内、作物を収穫するまで(または、個々の除草剤について、いずれか最も短い収穫前の間隔)の任意のタイミングで施用することができる。
【0276】
広範なスペクトルのイネ科雑草および広葉雑草を制御することにおいて、施用のタイミング、個々の除草剤の比率、および厄介なまたは抵抗性の雑草を制御する能力の点で柔軟性を有することが重要である。グリホサート抵抗性遺伝子/AAD−12 416イベントの積み重ねを有する作物におけるグリホサートの施用は、約250〜2500g ae/haにわたってよく;(1種または複数種の)フェノキシオーキシン除草剤は、約25〜4000g ae/ha施用してよく;(1種または複数種の)ピリジルオキシ酢酸オーキシン除草剤は、25〜2000g ae/ha施用してよい。これらの施用(複数可)の最適な組合せ(複数可)およびタイミングは、特定の状況、種、および環境に左右され、雑草制御の当業者および本開示の利益を受ける当業者が決定することが最善である。
【0277】
北アメリカにおいて植えられている綿、キャノーラ、トウモロコシ、およびダイズの畑地の大部分は、グリホサート耐性(GT)形質を含有し、また、GTトウモロコシの採用は増えている。追加的なGT作物(例えば、コムギ、イネ、サトウダイコン、および芝生)が開発中であるが、これまで発売されていない。多くの他のグリホサート抵抗性種は、開発段階への実験中である(例えば、アルファルファ、サトウキビ、サンフラワー、ビート、エンドウ、ニンジン、キュウリ、レタス、タマネギ、イチゴ、トマト、およびタバコ;ポプラおよびモミジバフウのような森林種;ならびにマリーゴールド、ペチュニア、およびベゴニアのような園芸種;World Wide Web、isb.vt.edu/cfdocs/fieldtests1.cfm、2005年)。GTCは、このシステムによって提供される、雑草完全な広がりを制御するための価値があるツールであり、利便性および費用の有効性である。しかし、現在標準の基剤処理としてのグリホサートの有用性は、グリホサート抵抗性の雑草について選択されている。さらに、グリホサートのみの化学的プログラムが実施されている圃場において、グリホサートが本質的にあまり効果的でない雑草が優勢な種にシフトしている。AAD−12イベント416にGT形質を積み重ねることにより、従来の育種によってまたは新規の形質転換イベントと共同で、雑草を制御する効力、柔軟性、および雑草のシフトを管理する能力および除草剤抵抗性の発生を改良することができる。作物をAAD−12イベント416で形質転換することにより、単子葉作物は、フェノキシオーキシンまたはピリジルオキシオーキシンの安全性により高い余地を有することになり、双子葉作物にフェノキシオーキシンを選択的に施用することができる。AAD−12イベント416およびGT形質を任意の単子葉作物種または双子葉作物種に積み重ねた場合、改良された雑草制御の選択肢のためのいくつかのシナリオを構想することができる:
a)大部分のイネ科植物および広葉雑草種を制御するために、グリホサートを標準の出芽後の施用率(420〜2160g ae/ha、560〜840g ae/haが好ましい)で施用することができる。ヒメムカシヨモギ(Conyza canadensis)などのグリホサート抵抗性の広葉雑草またはグリホサートを用いて制御することが本質的に厄介な雑草(例えば、ツユクサ(Commelina)種、サツマイモ(Ipomoea)種など)を制御するために、280〜2240g ae/ha(560〜1120g ae/haが好ましい)の2,4−Dを、グリホサートと、逐次的に、タンク混合して、またはプレミックスとして施用して有効な制御をもたらすことができる。トリクロピルについては、施用率は一般には70〜1120g ae/ha、より一般には140〜420g ae/haの範囲になる。フルロキシピルについては、施用率は、一般には35〜560g ae/ha、より一般には70〜280 ae/haの範囲になる。
b)現在、GTCにおいて施用されるグリホサートの比率は、一般に、施用のタイミング当たり560〜2240g ae/haの範囲である。グリホサートは、広葉雑草種よりもイネ科植物種に対してはるかに効果的である。AAD−12イベント416+GTを積み重ねた形質により、イネ科植物に有効な比率のグリホサート(105−840g ae/ha、より好ましくは210−420g ae/ha)が可能になる。それから、2,4−D(280〜2240g ae/ha、より好ましくは560〜1120g ae/ha)を、イネ科植物に有効な比率のグリホサートと、逐次的に、タンク混合して、またはプレミックスとして施用して必要な広葉雑草の制御をもたらすことができる。上記の比率のトリクロピルおよびフルロキシピルは、処理レジメンにおいて許容できる構成成分になる。低率のグリホサートにより、広葉雑草の制御にもいくらかの利益がもたらされる;しかし、主要な制御は、2,4−D、トリクロピル、またはフルロキシピルによるものである。
【0278】
作物をAAD−12イベント416で形質転換することにより、1つまたは複数の商業的なアリールオキシオーキシン除草剤を単独で、または組み合わせて(逐次的にまたはそれぞれ独立に)使用することが可能になることが、雑草制御の当業者には認識されよう。これらの化学物質を代表する特定の比率の他の除草剤は、CPR(Crop Protection Reference)書籍または同様の編集物によって編集された除草剤の表示、オンライン(例えば、cdms.net/manuf/manuf.asp)で編集された表示、またはAgrilianceからのCrop Protection Guide(2005年)などの任意の商業的なまたは学術的な作物保護の手引きによって決定することができる。AAD−12イベント416によってHTCにおいて使用することが可能になったそれぞれの代替の除草剤は、単独で使用されようが、タンク混合して使用されようが、または逐次的に使用されようが、本発明の範囲内であるとみなされる。
(参考文献)
【特許請求の範囲】
【請求項1】
配列番号1を含むゲノムを含むトランスジェニックダイズ植物。
【請求項2】
American Type Culture Collection(ATCC)に受託番号PTA−10442の下で寄託されている代表的な種子に存在するAAD−12イベントpDAB4468−0416を含むゲノムを含むダイズ種子。
【請求項3】
配列番号1を含むゲノムを含むダイズ種子。
【請求項4】
配列番号1を含む、請求項2に記載の種子を生長させることによって作出されるダイズ植物。
【請求項5】
AAD−12イベントpDAB4468−0416を含む、請求項4に記載のダイズ植物の後代植物。
【請求項6】
配列番号1を含む、請求項1に記載のダイズ植物の除草剤耐性後代植物。
【請求項7】
配列番号29および配列番号30のSNPマーカー間の第4染色体に位置するダイズ染色体標的部位に導入遺伝子挿入断片を含むトランスジェニックダイズ植物であって、前記標的部位が異種核酸を含む、トランスジェニックダイズ植物。
【請求項8】
配列番号29および配列番号30のSNPマーカー間の第4染色体上の位置において異種核酸を挿入するステップを含むトランスジェニックダイズ植物を作製する方法であって、前記標的部位が異種核酸を含む方法。
【請求項9】
花粉、胚珠、花、莢、リント、苗条、根、および葉からなる群から選択され、配列番号1を含む、請求項4に記載の植物の一部。
【請求項10】
配列番号1の残基1〜2730および配列番号1の残基9122〜10,212からなる群から選択されるゲノム配列中に、またはそれに隣接して導入遺伝子挿入断片を含むトランスジェニックダイズ植物。
【請求項11】
少なくとも15ヌクレオチドを含み、ストリンジェントな洗浄条件下で配列番号1の残基1〜2730および配列番号1の残基9122〜10,212からなる群から選択される核酸配列とのハイブリダイゼーションを維持する、単離されたポリヌクレオチド分子。
【請求項12】
配列番号2〜28からなる群から選択されるヌクレオチド配列を含む、単離されたポリヌクレオチド。
【請求項13】
ストリンジェントな洗浄条件下で、配列番号1の残基2720〜2740、配列番号1の残基9112〜9132、およびその相補物からなる群から選択されるヌクレオチド配列とハイブリダイズする、単離されたポリヌクレオチド。
【請求項14】
ポリメラーゼ連鎖反応によって生成するアンプリコンである、請求項13に記載のポリヌクレオチド。
【請求項15】
配列番号1を含む、ポリヌクレオチド。
【請求項16】
請求項15に記載のポリヌクレオチドを作出する方法。
【請求項17】
導入遺伝子をダイズゲノムのDNAセグメントに挿入する方法であって、前記DNAセグメントが、配列番号1の残基1〜2730を含む5’末端および配列番号1の残基9122〜10,212を含む3’末端を含む方法。
【請求項18】
ダイズ植物を育種する方法であって、配列番号1を含む第1のダイズ植物を第2のダイズ植物と交雑してゲノムを含む第3のダイズ植物を作出するステップと、前記第3のダイズ植物を、前記ゲノム内に配列番号1が存在することについてアッセイするステップとを含む方法。
【請求項19】
除草剤耐性形質をダイズ植物に遺伝子移入する方法であって、配列番号1を含む第1のダイズ植物を第2のダイズ植物と交雑してゲノムを含む第3のダイズ植物を作出するステップと、前記第3のダイズ植物を、前記ゲノム内に配列番号1が存在することについてアッセイするステップとを含む方法。
【請求項20】
雑草を制御する方法であって、請求項1の植物を含む圃場にアリールオキシアルカノエート除草剤を施用するステップを含む方法。
【請求項21】
前記除草剤が、2,4−D;2,4−DB;MCPA;およびMCPBからなる群から選択される、請求項20に記載の方法。
【請求項22】
前記圃場に第2の除草剤を施用するステップを含む、請求項20に記載の方法。
【請求項23】
前記第2の除草剤が、グリホサートおよびジカンバからなる群から選択される、請求項22に記載の方法。
【請求項24】
雑草を制御する方法であって、圃場にアリールオキシアルカノエート除草剤を施用するステップと、前記除草剤を施用してから14日間以内に前記圃場に請求項3の種子を植え付けるステップとを含む方法。
【請求項25】
ダイズDNAを含む試料中のダイズイベントを検出する方法であって、前記試料を、American Type Culture Collection(ATCC)に受託番号PTA−10442の下で寄託されている代表的な種子に存在するAAD−12イベントpDAB4468−0416に特徴的な少なくとも1つのポリヌクレオチドと接触させるステップを含む方法。
【請求項26】
前記試料を、
a.配列番号1の残基1〜2730、配列番号1の残基9122〜10,212、およびその相補物からなる群から選択される隣接配列に結合する第1のプライマー、および
b.配列番号1の残基2731〜9121またはその相補物を含む挿入断片配列に結合する第2のプライマー
と接触させるステップと、
前記試料をポリメラーゼ連鎖反応に供するステップと、前記プライマーの間で生成されたアンプリコンについてアッセイするステップとを含む、請求項25に記載の方法。
【請求項27】
前記プライマーが、配列番号2〜7からなる群から選択される、請求項26に記載の方法。
【請求項28】
前記ポリヌクレオチドが、少なくとも30ヌクレオチドを含み、かつストリンジェントな条件下で配列番号1の残基2720〜2740、配列番号1の残基9112〜9132、およびその相補物からなる群から選択される配列とハイブリダイズし、前記方法が、前記試料および前記ポリヌクレオチドをストリンジェントなハイブリダイゼーション条件に供するステップと、前記試料を、前記ポリヌクレオチドの前記DNAへのハイブリダイゼーションについてアッセイするステップとをさらに含む、請求項25に記載の方法。
【請求項29】
請求項27に記載の第1のプライマーおよび第2のプライマーを含むDNA検出キット。
【請求項30】
請求項28に記載の方法を実施するためのDNA検出キット。
【請求項31】
少なくとも30ヌクレオチドを含み、かつストリンジェントな条件下で配列番号1の残基2720〜2740、配列番号1の残基9112〜9132、およびその相補物からなる群から選択される配列とハイブリダイズするポリヌクレオチドを含むDNA検出キット。
【請求項32】
除草剤抵抗性の雑草を処理する、または防ぐために使用される、請求項20に記載の方法。
【請求項33】
前記除草剤が、前記植物を植え付ける前に施用される、請求項20に記載の方法。
【請求項34】
グリホサート耐性形質をさらに含む少なくとも1つの請求項1に記載の植物を含む地域においてグリホサート抵抗性の雑草を制御する方法であって、アリールオキシアルカノエート除草剤を前記地域の少なくとも一部に施用するステップを含む方法。
【請求項35】
前記除草剤がフェノキシオーキシンである、請求項34に記載の方法。
【請求項36】
前記除草剤がグリホサートとのタンク混合物から施用される、請求項34に記載の方法。
【請求項37】
前記雑草の少なくとも1種が、前記植物と異なる種のグリホサート抵抗性の自生植物である、請求項20に記載の方法。
【請求項1】
配列番号1を含むゲノムを含むトランスジェニックダイズ植物。
【請求項2】
American Type Culture Collection(ATCC)に受託番号PTA−10442の下で寄託されている代表的な種子に存在するAAD−12イベントpDAB4468−0416を含むゲノムを含むダイズ種子。
【請求項3】
配列番号1を含むゲノムを含むダイズ種子。
【請求項4】
配列番号1を含む、請求項2に記載の種子を生長させることによって作出されるダイズ植物。
【請求項5】
AAD−12イベントpDAB4468−0416を含む、請求項4に記載のダイズ植物の後代植物。
【請求項6】
配列番号1を含む、請求項1に記載のダイズ植物の除草剤耐性後代植物。
【請求項7】
配列番号29および配列番号30のSNPマーカー間の第4染色体に位置するダイズ染色体標的部位に導入遺伝子挿入断片を含むトランスジェニックダイズ植物であって、前記標的部位が異種核酸を含む、トランスジェニックダイズ植物。
【請求項8】
配列番号29および配列番号30のSNPマーカー間の第4染色体上の位置において異種核酸を挿入するステップを含むトランスジェニックダイズ植物を作製する方法であって、前記標的部位が異種核酸を含む方法。
【請求項9】
花粉、胚珠、花、莢、リント、苗条、根、および葉からなる群から選択され、配列番号1を含む、請求項4に記載の植物の一部。
【請求項10】
配列番号1の残基1〜2730および配列番号1の残基9122〜10,212からなる群から選択されるゲノム配列中に、またはそれに隣接して導入遺伝子挿入断片を含むトランスジェニックダイズ植物。
【請求項11】
少なくとも15ヌクレオチドを含み、ストリンジェントな洗浄条件下で配列番号1の残基1〜2730および配列番号1の残基9122〜10,212からなる群から選択される核酸配列とのハイブリダイゼーションを維持する、単離されたポリヌクレオチド分子。
【請求項12】
配列番号2〜28からなる群から選択されるヌクレオチド配列を含む、単離されたポリヌクレオチド。
【請求項13】
ストリンジェントな洗浄条件下で、配列番号1の残基2720〜2740、配列番号1の残基9112〜9132、およびその相補物からなる群から選択されるヌクレオチド配列とハイブリダイズする、単離されたポリヌクレオチド。
【請求項14】
ポリメラーゼ連鎖反応によって生成するアンプリコンである、請求項13に記載のポリヌクレオチド。
【請求項15】
配列番号1を含む、ポリヌクレオチド。
【請求項16】
請求項15に記載のポリヌクレオチドを作出する方法。
【請求項17】
導入遺伝子をダイズゲノムのDNAセグメントに挿入する方法であって、前記DNAセグメントが、配列番号1の残基1〜2730を含む5’末端および配列番号1の残基9122〜10,212を含む3’末端を含む方法。
【請求項18】
ダイズ植物を育種する方法であって、配列番号1を含む第1のダイズ植物を第2のダイズ植物と交雑してゲノムを含む第3のダイズ植物を作出するステップと、前記第3のダイズ植物を、前記ゲノム内に配列番号1が存在することについてアッセイするステップとを含む方法。
【請求項19】
除草剤耐性形質をダイズ植物に遺伝子移入する方法であって、配列番号1を含む第1のダイズ植物を第2のダイズ植物と交雑してゲノムを含む第3のダイズ植物を作出するステップと、前記第3のダイズ植物を、前記ゲノム内に配列番号1が存在することについてアッセイするステップとを含む方法。
【請求項20】
雑草を制御する方法であって、請求項1の植物を含む圃場にアリールオキシアルカノエート除草剤を施用するステップを含む方法。
【請求項21】
前記除草剤が、2,4−D;2,4−DB;MCPA;およびMCPBからなる群から選択される、請求項20に記載の方法。
【請求項22】
前記圃場に第2の除草剤を施用するステップを含む、請求項20に記載の方法。
【請求項23】
前記第2の除草剤が、グリホサートおよびジカンバからなる群から選択される、請求項22に記載の方法。
【請求項24】
雑草を制御する方法であって、圃場にアリールオキシアルカノエート除草剤を施用するステップと、前記除草剤を施用してから14日間以内に前記圃場に請求項3の種子を植え付けるステップとを含む方法。
【請求項25】
ダイズDNAを含む試料中のダイズイベントを検出する方法であって、前記試料を、American Type Culture Collection(ATCC)に受託番号PTA−10442の下で寄託されている代表的な種子に存在するAAD−12イベントpDAB4468−0416に特徴的な少なくとも1つのポリヌクレオチドと接触させるステップを含む方法。
【請求項26】
前記試料を、
a.配列番号1の残基1〜2730、配列番号1の残基9122〜10,212、およびその相補物からなる群から選択される隣接配列に結合する第1のプライマー、および
b.配列番号1の残基2731〜9121またはその相補物を含む挿入断片配列に結合する第2のプライマー
と接触させるステップと、
前記試料をポリメラーゼ連鎖反応に供するステップと、前記プライマーの間で生成されたアンプリコンについてアッセイするステップとを含む、請求項25に記載の方法。
【請求項27】
前記プライマーが、配列番号2〜7からなる群から選択される、請求項26に記載の方法。
【請求項28】
前記ポリヌクレオチドが、少なくとも30ヌクレオチドを含み、かつストリンジェントな条件下で配列番号1の残基2720〜2740、配列番号1の残基9112〜9132、およびその相補物からなる群から選択される配列とハイブリダイズし、前記方法が、前記試料および前記ポリヌクレオチドをストリンジェントなハイブリダイゼーション条件に供するステップと、前記試料を、前記ポリヌクレオチドの前記DNAへのハイブリダイゼーションについてアッセイするステップとをさらに含む、請求項25に記載の方法。
【請求項29】
請求項27に記載の第1のプライマーおよび第2のプライマーを含むDNA検出キット。
【請求項30】
請求項28に記載の方法を実施するためのDNA検出キット。
【請求項31】
少なくとも30ヌクレオチドを含み、かつストリンジェントな条件下で配列番号1の残基2720〜2740、配列番号1の残基9112〜9132、およびその相補物からなる群から選択される配列とハイブリダイズするポリヌクレオチドを含むDNA検出キット。
【請求項32】
除草剤抵抗性の雑草を処理する、または防ぐために使用される、請求項20に記載の方法。
【請求項33】
前記除草剤が、前記植物を植え付ける前に施用される、請求項20に記載の方法。
【請求項34】
グリホサート耐性形質をさらに含む少なくとも1つの請求項1に記載の植物を含む地域においてグリホサート抵抗性の雑草を制御する方法であって、アリールオキシアルカノエート除草剤を前記地域の少なくとも一部に施用するステップを含む方法。
【請求項35】
前記除草剤がフェノキシオーキシンである、請求項34に記載の方法。
【請求項36】
前記除草剤がグリホサートとのタンク混合物から施用される、請求項34に記載の方法。
【請求項37】
前記雑草の少なくとも1種が、前記植物と異なる種のグリホサート抵抗性の自生植物である、請求項20に記載の方法。
【図1】

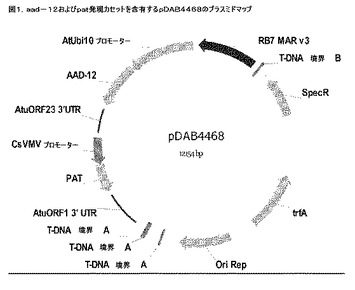
【図2】
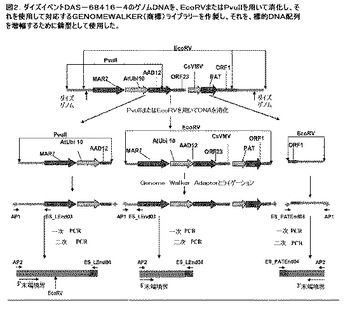
【図3】
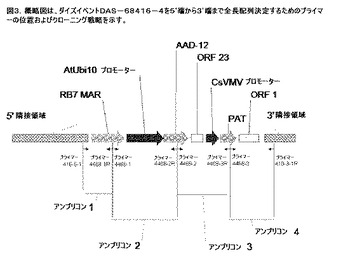
【図4】
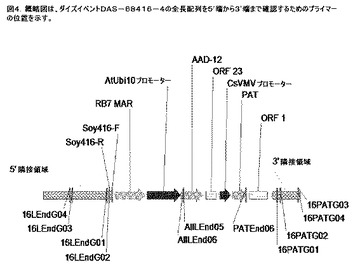
【図5】

【図6】
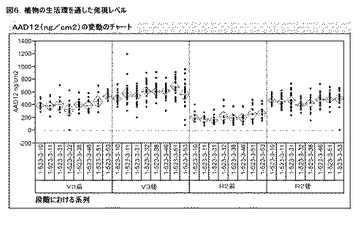
【図2】
【図3】
【図4】
【図5】
【図6】
【公表番号】特表2013−511985(P2013−511985A)
【公表日】平成25年4月11日(2013.4.11)
【国際特許分類】
【出願番号】特願2012−541192(P2012−541192)
【出願日】平成22年11月24日(2010.11.24)
【国際出願番号】PCT/US2010/058001
【国際公開番号】WO2011/066384
【国際公開日】平成23年6月3日(2011.6.3)
【出願人】(501035309)ダウ アグロサイエンシィズ エルエルシー (197)
【Fターム(参考)】
【公表日】平成25年4月11日(2013.4.11)
【国際特許分類】
【出願日】平成22年11月24日(2010.11.24)
【国際出願番号】PCT/US2010/058001
【国際公開番号】WO2011/066384
【国際公開日】平成23年6月3日(2011.6.3)
【出願人】(501035309)ダウ アグロサイエンシィズ エルエルシー (197)
【Fターム(参考)】
[ Back to top ]