
ABCA1遺伝子の転写を調整する作用を有する物質のスクリーニング方法ならびにそのためのDNAおよびキット
【課題】 動脈硬化に伴う疾患の予防および/または治療を目的とした、肝ABCA1(ATP結合カセット膜トランスポーターA1)上昇作用を有する医薬をスクリーニングする新規な方法および該方法において用いられるDNAを提供する。
【解決手段】 ABCA1遺伝子のイントロン1中のSRE配列を含み、プロモーター活性を有するDNAを用いることを特徴とする、ABCA1遺伝子の転写を調整する作用を有する物質のスクリーニング方法が提供される。
【解決手段】 ABCA1遺伝子のイントロン1中のSRE配列を含み、プロモーター活性を有するDNAを用いることを特徴とする、ABCA1遺伝子の転写を調整する作用を有する物質のスクリーニング方法が提供される。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、肝臓においてABCA1遺伝子の発現を制御するプロモーター領域の発見に基づき、ABCA1遺伝子の転写を調整する作用を有する物質のスクリーニング方法ならびにそのためのDNAおよびキットに関するものであり、特に、プロモーター活性の増加を指標として、肝臓におけるABCA1発現の増加により動脈硬化性疾患を予防または治療する作用を有する物質をスクリーニングする方法に関するものである。
【背景技術】
【0002】
高密度リポ蛋白(HDL)は主に肝臓や小腸で合成、分泌され、リポ蛋白(アポA−IやアポA−IIなど)や脂質(リン脂質、コレステロール、トリグリセライド)から構成される複合分子である。HDLは、末梢細胞で過剰に蓄積したコレステロールを細胞膜表面から引き抜いて、胆汁酸への転換および排出が可能な肝臓へと輸送する、いわゆる「コレステロール逆輸送系」において重要な役割を担っている。血中HDL濃度が低いほど動脈硬化性疾患の発症率が高まる疫学的知見から、HDLは善玉コレステロールと呼ばれている。
【0003】
血中HDL濃度の低下は、動脈硬化症や高脂血症、肥満、糖尿病などの様々な疾患で認められる。家族性HDL欠損症であるタンジール病では、細胞内のコレステロールをアポA−Iを含むHDL粒子として放出する過程に障害を持つ。近年その原因遺伝子としてATP結合カセット膜トランスポーターA1(ABCA1)が同定され(非特許文献1、Nat.Genet.(1999)22, 347)、HDLの形成にABCA1タンパク質が必須の役割を担うことが明らかになった。
【0004】
培養細胞にABCA1を強制発現させるとHDL形成が増加する(非特許文献2、J.Clin.Invest.(1999)104, R25)。また、ABCA1過剰発現マウスでは血中HDL濃度が上昇し、動脈硬化に抵抗性を示すことが明らかにされている(非特許文献3、J.Biol.Chem.(2001)276, 33969)。したがって、ABCA1発現量の増加は、HDL濃度の上昇をもたらし、動脈硬化性疾患の予防および治療に有用であると考えられる。
【0005】
マクロファージや繊維芽細胞など末梢細胞におけるABCA1遺伝子の転写は、ABCA1遺伝子のエクソン1上流領域により制御されており、このABCA1遺伝子プロモーターの活性は複数の転写因子により調節されることが明らかにされている(非特許文献4、Biochem.Biophys.Res.Commun.(2000)274, 794:非特許文献5、J.Biol.Chem.(2002)277, 14443:非特許文献6、J.Lipd Res(2002)43, 297:非特許文献7、Mol.Cell.Biol.(2003)23,7756)。なかでも主要なものは、核内受容体LXRであり、RXRとのヘテロダイマーがLXR応答性配列に結合しABCA1の転写を活性化する。細胞内コレステロール濃度を感知するLXRは、ABCA1遺伝子の発現を促進し、HDL新生による細胞外へのコレステロール排出を促してコレステロール恒常性を保つ(非特許文献8、Proc.Natl.Acad.Sci.USA.(2000)97,12097)。
【0006】
【特許文献1】国際公開第00/078972号パンフレット
【非特許文献1】Nat.Genet.(1999)22, 347
【非特許文献2】J.Clin.Invest.(1999)104, R25
【非特許文献3】J.Biol.Chem.(2001)276, 33969
【非特許文献4】Biochem.Biophys.Res.Commun.(2000)274, 794
【非特許文献5】J.Biol.Chem.(2002)277, 14443
【非特許文献6】J.Lipd Res(2002)43, 297
【非特許文献7】Mol.Cell.Biol.(2003)23,7756
【非特許文献8】Proc.Natl.Acad.Sci.USA.(2000)97,12097
【非特許文献9】Biochem.Biophys.Res.Commun.(2004)324, 835
【非特許文献10】Arterioscler.Thromb.Vasc.Biol.(2004)24, 2365
【非特許文献11】J.Clin.Invest. (2005)115, 1333
【非特許文献12】J Biol Chem. 2005 Jun 10;280(23):22212-21
【発明の開示】
【発明が解決しようとする課題】
【0007】
スタチン系薬剤は、コレステロール合成の律速酵素であるHMG−CoA還元酵素の阻害剤であり、血中LDL濃度の低下薬として汎用されているが、血中HDL濃度の上昇効果も認められる。最近、スタチン系薬剤はヒト肝がん由来細胞HepG2においてABCA1遺伝子の発現促進作用を示すことが明らかにされた(非特許文献9、Biochem.Biophys.Res.Commun.(2004)324, 835)。一方、マクロファージ系の培養細胞THP−1やRAW264においては、ABCA1発現を著しく低下させる(非特許文献10、Arterioscler.Thromb.Vasc.Biol.(2004)24, 2365)。これは、コレステロール合成過程で生じるLXRの内因性リガンドがスタチン系薬剤の作用によって減少し、ABCA1遺伝子プロモーター活性が低下することによる。したがって、肝由来細胞のABCA1発現増加は、末梢細胞で機能する、上記のLXRが制御するプロモーター領域(エクソン1上流領域)とは別の発現調節系により引き起こされていると推定される。
【0008】
ABCA1は様々な組織に分布しているが、特に肝臓において強く発現する。マウスを用いた肝臓特異的なABCA1遺伝子のノックダウン解析では、血中HDLコレステロール量が20%以下まで低下し、肝臓のABCA1発現量は血中HDL量の決定に重要な役割を担うことが示されている(非特許文献11、J.Clin.Invest. (2005)115, 1333)。
【0009】
従って、HDL産生の主要臓器である肝臓においてABCA1遺伝子発現を増加させることは、動脈硬化性疾患を予防または治療する上で極めて重要となる。このような状況から、肝臓におけるABCA1遺伝子発現の制御機構の解明、さらにそれを利用した制御薬剤のスクリーニング方法が望まれていた。
【0010】
以上に鑑みて、本発明の課題は、肝臓のABCA1上昇作用を有し動脈硬化に伴う疾患の予防および/または治療に有用な医薬をスクリーニングする新規な方法および該方法において用いられるDNAを提供することにある。
【0011】
なお、本発明の方法において用いられるABCA1遺伝子は、Gene Bankデータベース上に「ATP結合カセット輸送体A1(ATP-binding cassette transporter A1)」として公開されている。また特許文献1(国際公開第00/078972号パンフレット)には「ABC1」をコードするものとして同遺伝子が、またそれと共に上記のエクソン1上流のプロモーターが開示されている。
【課題を解決するための手段】
【0012】
すなわち、本発明の一の側面によると、ABCA1遺伝子のイントロン1中のSRE配列を含み、プロモーター活性を有するDNAを用いることを特徴とする、ABCA1遺伝子の転写を調整する作用を有する物質のスクリーニング方法が提供される。
【0013】
また、本発明の他の側面によると、活性型SREBPの量を変化させる作用を有する物質(ピタバスタチン、シムバスタチンおよびアトルバスタチンを除く)を含有する、ABCA1遺伝子の転写調整剤が提供される。
【0014】
また、本発明の他の側面によると、ABCA1遺伝子のイントロン1中のSRE配列を含み、プロモーター活性を有する組換えDNAが提供される。
【0015】
また、本発明の他の側面によると、ABCA1遺伝子のイントロン1中のSRE配列を含み、プロモーター活性を有するDNAを有する組換えベクターが提供される。
【0016】
また、本発明の他の側面によると、ABCA1遺伝子のイントロン1中のSRE配列を含み、プロモーター活性を有する外因性のDNAに作動可能に連結した構造遺伝子を有する形質転換体が提供される。
【0017】
また、本発明の他の側面によると、上記組換えベクターおよび/または上記形質転換体を有する、ABCA1遺伝子の転写を調整する作用を有する物質をスクリーニングするためのキットが提供される。
【発明の効果】
【0018】
本発明は、ABCA1遺伝子の転写を調整する作用を有する物質、特に、動脈硬化性疾患の予防および/または治療剤を試験するための新規方法を確立する。また、本発明は、従来のABCA1遺伝子発現促進物質のスクリーニング方法に比べ、HDL上昇作用により効果的な分子を検出できる可能性がある。
【発明を実施するための最良の形態】
【0019】
以下に、本発明の実施の形態を説明する。もっとも、本発明は、以下に説明する実施の形態によって、限定されるものではない。
【0020】
本発明者らは、肝臓におけるABCA1遺伝子の発現調節に新たな制御機構が存在することを見出した。そこで、ABCA1遺伝子発現を調節する物質を選別または同定するための方法として、イントロン1に存在する新規肝臓型ABCA1プロモーター(本明細書中、肝臓型プロモーター、II型ABCA1プロモーター等とも記す。)を用いる方法を開発し、本発明を完成させた。
【0021】
本発明が開示する肝臓型プロモーターは、ABCA1遺伝子のエクソン1上流のプロモーター(特許文献1参照)とは全く異なる領域に存在し、DNA配列も異なる。この新規肝臓型プロモーターに関しては、ABCA1遺伝子転写制御、HDL形成との関連は報告されておらず、動脈硬化性疾患の治療または予防剤の試験法としての有用性も知られていない。上記したように、ピタバスタチン、シンバスタチン、アトルバスタチンがABCA1のmRNAを増加させることが示されている(非特許文献9、Biochem.Biophys.Res.Commun.(2004)324, 835)。しかし、その作用機序に関わるデータは無く、LXRが活性化される可能性が議論されている。また、アトルバスタチン、シンバスタチン、ピタバスタチンが(特定の条件下で)マクロファージのABCA1の発現を増加させること、その機序の一つとしてRhoAのゲラニル化が関与しているという報告がある(非特許文献12、J Biol Chem. 2005 Jun 10;280(23):22212-21)。しかし、これもPPAR gammaを介してLXRを活性化する(末梢型プロモーターを活性化する)メカニズムを想定している系であり、本発明にかかる新規肝臓型ABCA1プロモーターおよび当該プロモーターによる発現制御については記載も示唆もしていない。
【0022】
具体的には、本発明の一の側面によると、ABCA1遺伝子のイントロン1中のSRE配列を含み、プロモーター活性を有するDNAが提供される。当該DNAは、天然のものであってもよく、また人工的なものであってもよい。すなわち、本発明の他の側面によると、ABCA1遺伝子のイントロン1中のSRE配列を含み、プロモーター活性を有する組換えDNAが提供される。
【0023】
本発明にかかるDNAは、任意の生物に由来するDNAであってよく、また、人工的に設計および/または調製されたDNAであってもよい。具体的には、本発明にかかるDNAは、ヒト、サル、マウス、ラット、モルモット、ウサギ、ブタ、ウシ、ヒツジ、ニワトリなどの哺乳動物その他の恒温動物に由来するDNAであってよい。
【0024】
ABCA1遺伝子のイントロン1は、ABCA1遺伝子のエクソン1の下流かつエクソン2の上流に存在する領域である。具体的には、ヒトABCA1遺伝子のエクソン1、イントロン1およびエクソン2として、配列番号1に記載の配列の1〜219位、220〜24375位および24376〜24535位の配列をそれぞれ有するものが見出されている。マウスABCA1遺伝子のエクソン1、イントロン1およびエクソン2として、配列番号2に記載の配列の1〜215位、216〜15795位および15796〜15963位の配列をそれぞれ有するものが見出されている。ラットABCA1遺伝子のエクソン1、イントロン1およびエクソン2として、配列番号3に記載の配列の1〜220位、221〜16048位および16049〜16216位の配列をそれぞれ有するものが見出されている。また、ヒト、マウスおよびラットにおける上記イントロン1の相同体、変異体および誘導体の配列、ならびに、他の任意の生物における上記イントロン1の相同体ならびにその変異体および誘導体の配列は、当業者であれば、ヒト、マウスおよび/またはラットにおける上記イントロン1および/またはエクソン1、2の配列との相同性に基づき決定することができる。
【0025】
ABCA1遺伝子のイントロン1中のSRE配列を含み、プロモーター活性を有するDNAは、イントロン1中のSRE配列に加えて、エクソン2の翻訳開始点付近までを含むことが好ましい。なお、ヒト、マウスおよびラットのABCA1のエクソン2中の翻訳開始点は、それぞれ、配列番号1に記載の配列の24470位、配列番号2に記載の配列の15898位、および配列番号3に記載の配列の16151位である。プロモーター活性を有するのに必要な領域は、当業者であれば適宜決定することができる。プロモーター活性は、例えば、以下に詳細に説明するように、ルシフェラーゼアッセイ法、CAT法等により測定することができる。
【0026】
特に、ABCA1遺伝子のイントロン1中のSRE配列を含み、プロモーター活性を有するDNAは、(a)配列番号4〜6のいずれかに記載の配列を有するDNAの少なくとも一部とすることができる。配列番号4〜6に記載の配列を有するDNAは、それぞれ、ヒト、マウスおよびラットにおいて、配列番号1の22963〜24469位、配列番号2の14298〜15797位および配列番号3の15101〜16173位に見出される配列に対応する。さらに、ABCA1遺伝子のイントロン1中のSRE配列を含み、プロモーター活性を有するDNAは、以下に詳細に説明するように、(a)のDNAと実質的に同一の配列を有するDNAとすることができる。
【0027】
具体的には、ABCA1遺伝子のイントロン1中のSRE配列を含み、プロモーター活性を有するDNAは、(b)配列番号4〜6のいずれかに記載の配列において、1もしくは数個(好ましくは1〜80個、より好ましくは1〜40個、さらに好ましくは1〜20個、さらに好ましくは1〜10個)のヌクレオチドが置換、挿入または欠失された配列を有し、プロモーター活性を有するDNAの少なくとも一部とすることができる。当該技術分野における通常の知識を有する者であれば、野生型プロモーターのヌクレオチド配列の部分的な置換、挿入または欠失などの改変により、野生型プロモーターと同等またはそれ以上のプロモーター活性を有するDNAを調製することが可能である。
【0028】
また、ABCA1遺伝子のイントロン1中のSRE配列を含み、プロモーター活性を有するDNAは、(c)配列番号4〜6のいずれかに記載の配列と相同性が60%以上(好ましくは80%以上、さらに好ましくは90%以上)の配列を有し、プロモーター活性を有するDNAの少なくとも一部とすることができる。このようなDNAには、(a)のDNAの、自然界に存在する変異型遺伝子もしくは人為的に改変した変異型遺伝子または異種生物由来の相同遺伝子等が含まれる。
【0029】
相同性の比較は、通常、容易に入手できる配列比較プログラムを用いて行うことができる。商業的に入手可能なそのようなコンピュータープログラムは2以上の配列の間の%相同性を計算することができる。ほとんどの配列比較方法においては、全体としての相同性スコアに不当にペナルティを与えることなく可能な挿入および欠失を考慮に入れて、最適な整列を生じさせるよう設計される。これは、「ギャップ」を配列の整列に挿入して、局所的な相同性を最大化するよう努めることによって達成される。このような方法では、できる限り少数のギャップにおける配列の整列が、多くのギャップにおけるものより高いスコアを達成するように、各ギャップに「ギャップペナルティ」を割り当てて、相同性を計算する。ほとんどの整列プログラムはギャップペナルティを設定することができる。しかしながら、配列比較のためのそのようなソフトウェアを用いる場合、デフォルト値を用いるのが好ましい。このような配列比較を実行できるソフトウェアの例として、BLASTパッケージ(Ausubel et al.,1999 ibid-Chapter18参照)、FASTA(Atschul et al.,1990,J.Mol.Biol.,403-410)およびGENEWORKSスイートが挙げられる。
【0030】
また、ABCA1遺伝子のイントロン1中のSRE配列を含み、プロモーター活性を有するDNAは、(d)(a)のDNAとストリンジェントな条件でハイブリダイズし、プロモーター活性を有するDNAの少なくとも一部とすることができる。ここで、DNAは通常、互いに相補的な二本鎖DNA構造を有している。このため、(a)のDNAとハイブリダイズするDNAとは、(a)のDNAのいずれかの鎖とハイブリダイズする鎖を有するDNAをいうことに留意すべきである。
【0031】
なお、ストリンジェントな条件およびハイブリダイゼーションの方法は、例えば、J. Sambrook et al., Molecular Cloning, Cold Spring Harbour Laboratory Press, New York, USA, 第2版, 1989に従って設定することができる。具体的には、ストリンジェントな条件は、「1xSSC、0.1%SDS、37℃」とすることができ、よりストリンジェントな条件は、「0.5xSSC、0.1%SDS、42℃」とすることができ、さらにストリンジェントな条件は、「0.2xSSC、0.1%SDS、65℃」とすることができる(1xSSC:0.15MのNaCl、0.015Mのクエン酸ナトリウム、pH7.0)。ただし、これらの条件の組み合わせは例示であり、当業者であれば、ハイブリダイゼーションのストリンジェンシーを決定する上記または他の要素(例えば、プローブ濃度、プローブの長さ、ハイブリダイゼーション反応時間など)を適宜組み合わせることにより、上記と同様のストリンジェンシーを実現することが可能である。
【0032】
なお、(b)〜(d)のDNAのプロモーター活性は、例えば、以下に詳細に説明するように、ルシフェラーゼアッセイ法、CAT法等により測定することができる。また、(b)〜(d)のDNAは、(a)のDNAと「同様」のプロモーター活性を有することが好ましい。ここで、「同様」のプロモーター活性を有するとは、質的に類似のプロモーター活性を有することをいい、例えば、(b)〜(d)のDNAが、(a)のDNAと少なくとも1つの因子(活性型SREBP2等の転写因子等)について同一の制御を受ける場合が含まれる。一方で、「同様」のプロモーター活性を有するとは、量的に同程度のプロモーター活性を有することを意図するものではない。また、当業者には明らかなように、プロモーター活性を発揮するために、DNAは、配列番号4〜6のいずれかに記載の配列(またはこれと実質的に同一の配列)の全てを有する必要はなく、その一部であってもよい。上記したように、プロモーター活性を有するのに必要な領域は、当業者であれば、ルシフェラーゼアッセイ法、CAT法等により適宜決定することができる。
【0033】
SRE配列は、SREBPが特異的に結合する能力を有するいかなる配列でもよく、例えば、配列番号7〜10に記載のコンセンサス配列を有することが好ましい。また、SRE配列は、配列番号11、12に記載の、ラット肝臓型ABCA1プロモーター領域中のSREと相同なSREコンセンサス配列を有することが好ましい。換言すると、SRE配列は、配列番号7〜12に記載のコンセンサス配列を有し、SREBPが特異的に結合できる配列であることが好ましい。なお、配列番号11、12に記載のSREコンセンサス配列は、後述する配列番号14に記載のSRE配列に相補的な配列に相同な配列である。
【0034】
なお、配列番号7〜12に記載のSREコンセンサス配列を一塩基または多塩基置換することにより、SRE配列を不活化させることができる。(J Biol Chem. 1999 Feb 26;274(9):5285-91., PNAS. 2004 Aug 3;101(31):11245-50)変異の導入はクイックチェンジ(ストラタジェン)やオルタドサイトズ(プロメガ)やジーンエディター(プロメガ)などのキットを用いて行うことができる。また、制限酵素を用いてコンセンサス配列そのものを除去することで、SREを不活性化させるのと同様の効果を得ることもできる。
【0035】
SREBPが特異的に結合する能力の有無は、例えば、以下に詳細に説明するように、ゲルシフトアッセイ、DNase Iフットプリンティング法、クロマチン免疫沈降法(ChiPアッセイ)等により測定することができる。なお、「特異的に結合する」とは、好ましくは生理的条件下(好ましくは天然の細胞、組織または生物内と同等の条件下)で、当該タンパク質の目的のDNA配列への結合能が、当該タンパク質の他の無作為なDNA配列への結合能と比べて、有意に大きいことをいう。
【0036】
[ゲルシフトアッセイ]
ゲルシフトアッセイは、DNAとタンパク質との相互作用を検出する方法である。一般的には、32Pで標識したDNA断片と目的のタンパク質とを混合し低塩濃度の未変性ゲルで電気泳動しDNAをオートラジオグラフィーで検出する。タンパク質と結合したDNAは遊離DNAと比べゲル中で移動度が遅れるため検出が可能となる。
【0037】
[DNase Iフットプリンティング法]
DNase Iフットプリンティング法は、塩基配列特異的DNA分解酵素であるDNase IでDNAを部分的に消化する際、DNAに結合しているタンパク質は、結合部位のホスホジエステル結合をDNase Iによる攻撃から保護することを利用し、DNAとタンパク質との相互作用を検出する方法である。DNA断片の一端を放射性同位元素で標識しておき、DNase I反応後の生成物を高分解能ゲル電気泳動で分離した後、ゲルのオートラジオグラムを得ると、タンパク質結合部位のバンドが消失することになる。同じDNA断片をタンパク質非存在下においてDNase I処理したものとMaxam-Gilbertシークエンス反応(Maxam, A. & Gilbert, W., Methods Enzymol., 65, 499, 1980)したものを同時に電気泳動することにより、タンパク質の結合部位が同定できる。
【0038】
[クロマチン免疫沈降法(ChiPアッセイ)]
クロマチン免疫沈降法は、ホルムアルデヒド等を用いて細胞内でDNAとタンパク質を架橋し、目的のタンパク質に対する特異的な抗体を使った免疫沈降法と組み合わせることで、細胞内に実際に存在するクロマチンDNA上でのDNAとタンパク質の相互作用を検出する方法である。ホルムアルデヒド等によりDNAと目的のタンパク質の架橋を行い、目的のタンパク質に対する抗体で免疫沈降を行った後、可逆反応により架橋を外し、タンパク質を除き、DNA断片を回収する。回収されたDNAを鋳型にして、目的のDNA配列の特異的プライマーを用いてPCRまたはPCR/サザンブロットで解析を行う。
【0039】
具体的には、ヒトABCA1遺伝子のイントロン1におけるSRE配列として、TCTCAGCTGAG(配列番号13、配列番号1の24285〜24295位に対応)が見出されている。また、マウスおよびラットABCA1遺伝子のイントロン1におけるSRE配列として、GTCTGGTGAGA(配列番号14、配列番号2の15666〜15676位および配列番号3の15954〜15964位に対応)が見出されている。通常、SRE配列は、配列番号13または14に記載の野生型配列と50%以上の相同性を有し、好ましくは60%以上の相同性、さらに好ましくは80%以上の相同性を有する。
【0040】
また、上記したように、本発明の他の側面によると、ABCA1遺伝子のイントロン1中のSRE配列を含み、プロモーター活性を有する組換えDNAが提供される。本発明にかかる組換えDNAは、転写最小プロモーターをさらに有することが好ましい。最小プロモーターの例として、ヘルペスウイルスのチミジンキナーゼ(TK)遺伝子のプロモーター、CMV初期プロモーター、PGKプロモーター、SV40プロモーターなどが挙げられるが、これらに限定されるものではない。本発明にかかる組換えDNAは、協調因子の結合配列をさらに有することが好ましい。このような協調因子の例として、NF−YやSP1などが挙げられる。また、協調因子の結合配列とSRE配列との距離は、50塩基以内であることが好ましい。
【0041】
本発明にかかるDNAは、ゲノムライブラリーからの選別や、ゲノムDNAを鋳型としたポリメラーゼ連鎖反応(PCR)により取得できる。ゲノムライブラリーからの選別は、適当なベクターに組み込んだゲノムDNAと、目的のDNAの一部の配列を有する標識DNAとのハイブリダイゼーションによって行うことができる。ハイブリダイゼーションは、例えば、J. Sambrook et al., Molecular Cloning, Cold Spring Harbour Laboratory Press, New York, USA, 第2版, 1989に記載の方法などに従って行なうことができる。また、市販のライブラリーを使用する場合、添付の使用説明書に記載の方法に従って行なうことができる。また、ゲノムDNAを鋳型としたPCRは、目的のDNAの一部の配列を有するプライマーを用いて行うことができる。ヌクレオチド配列の改変は、例えば、制限酵素またはDNAエキソヌクレアーゼによる欠失導入、部位特異的変異誘発法による変異導入、変異プライマーを用いたPCR法による改変、合成変異DNAの直接導入などの方法により行うことができる。
【0042】
本発明の他の側面によると、ABCA1遺伝子のイントロン1中のSRE配列を含み、プロモーター活性を有するDNAを有する組換えベクターが提供される。組換えベクターは、当該プロモーター活性を有するDNAに作動可能に連結した構造遺伝子をさらに有してもよい。以下に詳細に説明するように、構造遺伝子は、例えば、組換えベクターをスクリーニングに用いる場合、その転写量および/または発現量を測定できるものであれば、任意の遺伝子でよい。
【0043】
本発明にあっては、ベクターは、任意のベクター、例えば、バクテリオファージ、プラスミド、コスミド、ファージミドなどであってよい。より具体的には、本発明に用いることができるベクターの例として、動物細胞用のベクターであるpCDベクター、cDM8ベクター、レトロウイルスベクター、アデノウイルスベクター、ヘルペスウイルスベクター、アデノアソシエイトウイルスベクターなどや、細菌用のベクターであるpUCなどを挙げることができる。構造遺伝子をベクター内に組み込むためには、構造遺伝子が正しく転写される方向に、さらにプロモーターが構造遺伝子に対して適正に機能する位置関係になる様に、プロモーター領域の下流に構造遺伝子を連結すればよい。プロモーターと構造遺伝子との連結は、制限酵素切断部位を利用して行うことができ、また、適当な制限酵素部位がなくてもリガーゼ反応を用いて行うことができる。本発明にかかる組換えベクターは、プロモーター活性を促進するエンハンサー領域等の転写調節領域、転写停止部位などその他の要素を有してもよい。
【0044】
なお、「作動可能に連結」とは、一の遺伝子要素が、他の遺伝子要素に対して予測されるように機能できるように配置されていることをいう。例えば、プロモーターが、構造遺伝子の転写の開始を助ける場合には、プロモーターは構造遺伝子に作動可能に結合している。この機能的な関係が維持される限り、プロモーターと構造遺伝子との間に残基が介在してもよい。
【0045】
本発明の他の側面によると、ABCA1遺伝子のイントロン1中のSRE配列を含み、プロモーター活性を有する外因性のDNAに作動可能に連結した構造遺伝子を有する形質転換体が提供される。本明細書において、形質転換体は、広く、外部からDNAが導入された結果、新しい形質を獲得した生物をいう。形質転換体は、任意の生物であってよく、また、任意の生物に由来する任意の細胞または組織であってよい。特に、本発明にかかる形質転換体は、上記組換えベクターで形質転換された形質転換体を含む。
【0046】
組換えベクターにより形質転換することができる宿主の例として、例えば、エシェリヒア属菌、バチルス属菌、酵母、昆虫細胞、昆虫、動物細胞などを挙げることができる。これらの宿主への形質転換の方法の例として、リン酸カルシウム法、リポフェクション法、エレクトロポレーション法、アデノウイルスベクターまたはレトロウイルスベクターによる遺伝子導入法、マイクロインジェクション法などを挙げることができる。
【0047】
本発明の他の側面によると、ABCA1遺伝子の転写を調整する作用を有する物質をスクリーニングするためのキットが提供される。本発明にかかるキットは、動脈硬化性疾患の予防または治療に有用な物質をスクリーニングするために用いることができる。以下に詳細に説明するように、ABCA1遺伝子のイントロン1中のSRE配列を含み、プロモーター活性を有するDNAを用いることで、ABCA1遺伝子の転写を調整する作用を有する物質をスクリーニングすることができる。すなわち、本発明にかかるキットは、上記組換えベクターおよび/または上記形質転換体を有する。
【0048】
本発明にかかるキットには、スクリーニングの方法に応じて、酵素、酵素基質、緩衝液等の試薬、培地、道具等を組み合せることができる。酵素基質の例として、ルシフェリン、ATP、[14C]クロラムフェニコール、アセチルCoAを挙げることができる。緩衝液として、解析に用いる酵素の活性の維持に好適な緩衝液を適宜選択することができる。道具の例として、TLC用展開シート、メンブレン等を挙げることができる。
【0049】
本発明の他の側面によると、ABCA1遺伝子の転写を調整する作用を有する物質、特に、肝臓においてABCA1遺伝子の転写を調整する作用を有する物質のスクリーニング方法が提供される。特に、本発明にかかるスクリーニング方法によると、ABCA1遺伝子の転写を促進する物質をスクリーニングすることができる。本発明にかかるスクリーニング方法は、動脈硬化性疾患の予防または治療に有用な物質をスクリーニングするために用いることができる。本発明にかかるスクリーニング方法は、ABCA1遺伝子のイントロン1中のSRE配列を含み、プロモーター活性を有するDNAを用いることを特徴とする。当該DNAを用いることにより、本発明にかかるスクリーニング方法によると、例えば活性型SREBP量を増加させる作用を有する物質をスクリーニングすることができる。
【0050】
具体的には、本発明にかかるスクリーニング方法は、
被検物質を供するステップと、
ABCA1遺伝子のイントロン1中のSRE配列を含み、プロモーター活性を有するDNAに作動可能に連結した構造遺伝子を有するDNAを有する細胞または生物の複数の群を供するステップと、
前記被検物質を前記細胞または生物の少なくとも1つの群に接触させるステップと、
前記被検物質を接触させた群と接触させなかった群とにおいて、前記遺伝子の転写量および/または発現量を測定するステップと、
前記被検物質を接触させた群と接触させなかった群の前記転写量および/または発現量を比較するステップと
を含むことが好ましい。
【0051】
被検物質として、単一の化合物または組成物、天然または合成化合物、有機または無機化合物、タンパク質、ペプチド、オリゴヌクレオチド、ポリヌクレオチド、細胞抽出物、細胞培養上清、その他の任意の物質を供することができる。
【0052】
スクリーニングは、インビトロおよびインビボのいずれでも行うことができる。本発明にかかるスクリーニング方法において用いられる細胞または生物として、細菌等の原核生物(大腸菌等)、酵母(サッカロマイセス(Saccharomyces)属、クライヴェロマイセス(Kluyveromyces)属等)、昆虫、植物もしくは動物またはこれらに由来する細胞が挙げられる。動物の例として、恒温動物、特にヒト、サル、マウス、ラット、ウシ、ブタ、イヌ等の哺乳動物を挙げることができる。本発明で用いられる細胞は、実験室で培養することのできる培養細胞であることが好ましく、初代培養肝細胞であることがさらに好ましい。細胞は、恒温動物、特にヒト、サル、マウス、ラット、ウシ、ブタ、イヌ等の哺乳動物に由来することができ、ヒトに由来することが好ましい。また、細胞は、肝臓に由来することが好ましい。
【0053】
本発明に用いられる細胞または生物は、ABCA1遺伝子のイントロン1中のSRE配列を含み、プロモーター活性を有するDNAに作動可能に連結した構造遺伝子を有するDNAを有することが好ましい。当該DNAは、内因性であっても、または外因性であってもよい。外因性である場合、当該DNAは、任意の手段により細胞または生物に導入されてもよい。例えば、スクリーニングに培養細胞を用いる場合、培養細胞への当該DNAの導入方法として、本発明の属する技術分野において汎用される任意の方法を採用することができる。例えば、DNAの導入方法の例として、リン酸カルシウム−DNA共沈法、リポフェクション法、DEAE−デキストラン法、電気パルス穿孔法等が挙げられるが、これらに限定されない。いずれの方法においても、用いる細胞に応じて至適化されたトランスフェクション条件を用いることが好ましい。
【0054】
構造遺伝子は、その転写量および/または発現量を測定できるものであれば、任意の遺伝子でよい。当業者に明らかなように、上記構造遺伝子は、その転写量および/または発現量の測定方法に応じて、適宜選択することができる。上記構造遺伝子の例として、ABCA1、所定の活性を有するタンパク質(ルシフェラーゼ等)、所定の抗体が結合できるタンパク質、所定の物質が特異的に結合できるタンパク質(GST等)等をコードする遺伝子を挙げることができ、特に、ホタルルシフェラーゼ、クロラムフェニコールアセチルトランスフェラーゼ、β−ガラクトシダーゼ、分泌型アルカリホスファターゼおよび緑色蛍光蛋白質からなる群より選択されるタンパク質をコードする遺伝子(いわゆるレポーター遺伝子)を挙げることができる。
【0055】
被検物質を細胞または生物に接触させることは、当業者に明らかなように、任意の方法により行うことができる。例えば、培養細胞を用いる場合、被検物質を培地中に添加することで、接触させることができる。すなわち、培養細胞を用いる場合、細胞を、被検物質の存在下または非存在下で培養することが好ましい。また、動物を用いる場合は、任意の手段による投与により、例えば、任意の経路からの注射により、接触させることができる。被検物質を細胞または生物へ接触させる時間は、被験物質の効果、濃度、細胞または生物の種類および技術的考慮に応じて、適宜設定することができる。また、被験物質の濃度は、その毒性、透過性、細胞数、処理時間等に応じて、適宜設定することができ、通常1nM〜1mMであることが好ましい。培養細胞を用いる場合、一般的にこのような処理は、多くの試験を同時に実施できるマルチウェルプレートで実施することが好ましい。培養細胞を用いる場合、被験化合物の処理によるレポーター遺伝子の発現が測定可能な条件は、細胞が生存してレポーター遺伝子の発現産物が生産可能な条件であれば良い。特に好ましい条件にあっては、好ましくは使用される細胞株に適した培地を使用し、4〜6%の炭酸ガス存在下、36〜38℃で5〜72時間培養する。
【0056】
被検物質を接触させた群と接触させなかった群とにおいて、上記構造遺伝子の転写量および/または発現量を測定し、これらの転写量および/または発現量を比較することで、当該被検物質を、ABCA1遺伝子の転写を調整する作用を有する物質の候補物質とすることができるか否か判断することができる。具体的には、ある被検物質を接触させた群で、接触させなかった群と比べて、上記遺伝子の転写量および/または発現量が変化した場合、特に増加した場合、当該被検物質を候補物質とすることができる。
【0057】
被検物質を接触させた群と接触させなかった群とにおいて、上記構造遺伝子の転写量および/または発現量を測定するステップと、被検物質を接触させた群と接触させなかった群の転写量および/または発現量を比較するステップは、当業者に明らかなように、任意の方法により行うことができる。具体的には、ノーザンブロッティング法、DNAアレイ法等により、上記遺伝子の転写量を測定することができる。また、ルシフェラーゼアッセイ法やCAT法等により、上記遺伝子の発現量を測定することができる。さらに、上記所定の遺伝子に対する抗体が入手可能な場合は、ELISA法、免疫沈降法、ウエスタンブロッティング法により、当該遺伝子の発現量を測定することができる。また、上記所定の遺伝子として、GST等の所定の物質に特異的に結合するタンパク質の遺伝子を用いる場合は、当該タンパク質を沈降させ、沈降したタンパク質の量を測定することで、上記遺伝子の発現量を測定することができる。
【0058】
[ノーザンブロッティング法]
ノーザンブロッティング法は、変性したRNAを変性条件下のアガロース電気泳動で分離し、ニトロセルロースフィルターに移して、標識した特異的なプローブで検出するものである。RNAを変性させるのは、RNAは分子内に二次構造を持つため、そのままではサイズに従った正確な分離ができず、さらに、フィルターは一本鎖の核酸しか結合できないためである。
【0059】
[ルシフェラーゼアッセイ法、CAT法]
ルシフェラーゼアッセイ法およびCAT法は、目的とする遺伝子の転写調節領域の下流にレポーターとしてLUC(ルシフェラーゼ)またはCAT(クロラムフェニコールアセチルトランスフェラーゼ)遺伝子を組み込んだプラスミドを作成し、そのプラスミドを導入した細胞の酵素活性を測定するものである。具体的には、ルシフェラーゼアッセイ法は、マグネシウム存在下、ルシフェラーゼがルシフェリンとATPから酸化ルシフェリンとAMPを作る反応を触媒する際に発する波長560nmの光を、ルミノメーターを使って検出するものである。CAT法の場合、基質の[14C]クロラムフェニコールが、CATによりアセチル化されて、[14C]アセチルクロラムフェニコールが産生される。このサンプルを酢酸エチルで抽出し、薄層のシリカゲルプレート上に展開後、移動度の異なるスポットの放射活性を測定する。
【0060】
本発明の他の側面によると、上記スクリーニング方法によりスクリーニングされた、ABCA1遺伝子の転写を調整する作用を有する物質、特に、肝臓においてABCA1遺伝子の転写を調整する作用を有する物質が提供される。特に、上記したように、本発明にかかるスクリーニング方法によると、ABCA1遺伝子の転写を促進する物質をスクリーニングすることができる。このため、本発明の他の側面によると、上記スクリーニング方法によりスクリーニングされた、動脈硬化性疾患の予防または治療に有用な物質が提供される。
【0061】
本発明の他の側面によると、活性型SREBPの量を変化させる作用を有する物質を含有する、ABCA1遺伝子の転写調整剤が提供される。特に、活性型SREBPの量を変化させる作用を有する物質が、活性型SREBPの量を増加させる作用を有することが好ましい。ABCA1遺伝子の転写調整剤は、活性型SREBPの量を変化させる作用を有する物質を、ABCA1遺伝子の転写を調整するのに有効な量含有する。有効な量は、当業者であれば、転写調整剤を用いる目的、条件等に応じて、適宜設定することができる。なお、活性型SREBPはステロール応答性配列(SRE配列)を標的とする転写因子で、活性型SREBPを過剰発現させることでSRE配列下の転写が活性化されることが知られている。より具体的には、活性型SREBPは、以下のように制御されることが知られている。一般に、SREBPは小胞体膜に結合した形で存在している。細胞内コレステロール量が減少すると、SREBPはプロセシングを受けて膜から切り離され活性型SREBP(N末端480アミノ酸)として核内に移行し、標的遺伝子の転写を活性化あるいは抑制する。切断が行われるゴルジ体への輸送は、ステロールの低下により行われる。SREBPはSCAP(SREBP cleavage activating protein)と結合しINSIG−1によって小胞体に留められているが、コレステロール量が減少するとINSIG−1が解離し、SREBP−SCAP複合体がゴルジ体に移行する。
【0062】
また、活性型SREBPの量を変化させる作用を有する物質が、ピタバスタチン、シムバスタチンおよびアトルバスタチンを除く物質であることが好ましく、スタチン系薬剤を除く物質であることがさらに好ましい。活性型SREBPの量を変化させる作用を有する物質は、不活性型SREBPを活性型SREBPに(または活性型を不活性型に)変換する作用を有する物質であってもよく、また、例えば、活性型SREBPを発現させることで、活性型SREBPの量を直接変化させる作用を有する物質であってもよい。具体的には、活性型SREBPの量を変化させる作用を有する物質が、活性型SREBP発現ベクター、細胞内コレステロール量を減少させる作用を有する物質、SREBPの前駆体(不活性型SREBP)から核内移行型(活性型SREBP)へのプロセシングを促進する作用を有する物質、活性型SREBPの分解を抑制する作用を有する物質からなる群から選ばれる少なくとも1つであることが好ましい。より具体的には、細胞内コレステロール量を減少させる作用を有する物質の例として、コレステロール合成阻害剤、LDL受容体阻害剤、胆汁酸吸着樹脂が挙げられる。また、SREBPの前駆体から核内移行型へのプロセシングを促進する作用を有する物質の例として、インシュリンが挙げられる。その他、活性型SREBPの量を変化させる作用を有する物質の例として、ATPやインポーチンβ、ALLNなどのプロテアソーム阻害剤が挙げられる。
【実施例】
【0063】
本発明は以下の実施例においてより具体的に説明される。但し、本発明はこれらの実施例に限定されるものでない。
【0064】
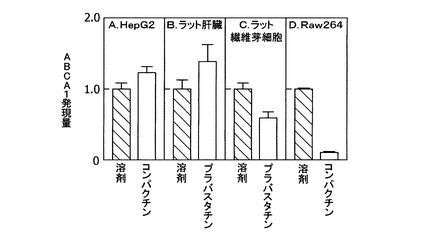
[実施例1:スタチン系薬剤のABCA1遺伝子発現に対する影響]
スタチン系薬剤がABCA1のmRNA発現に及ぼす影響を、以下の方法で調べた。ヒト肝ガン由来培養細胞HepG2およびラット繊維芽細胞Rat2を、10%FCSを含むDMEM培地で、マウス単球由来細胞Raw264を、10%FCSを含むDMEM/F−12培地でそれぞれ培養し、被験化合物を含む無血清培地(0.2%BSA)で24時間処理した。HepG2細胞およびRaw264細胞には、被験化合物として、コンパクチンを最終濃度50μMで添加した。Rat2細胞には、被験化合物として、プラバスタチンを、最終濃度50μMで添加した。それぞれ、溶剤のみを添加したものを対照とした。また、雄性ラットに被験化合物を経口投与し、その後該動物個体から肝臓を摘出した。被験化合物として、プラバスタチンを0.1%の濃度で飲料水に添加して1週間投与した。培養細胞およびラット組織からRNeasy(QIAGEN社製)を用いてRNAを調製した。7700 Sequence Detector(ABI社製)システムを用いたTaqManリアルタイム定量RT−PCRにより、調製したRNA中におけるABCA1のmRNA量を測定した。各試料中のmRNA量は、それぞれ18SRNAまたはGAPDHのmRNA量に対して正規化を行った。
【0065】
ヒト肝ガン由来培養細胞HepG2を50μMのコンパクチンで処理すると、ABCA1のmRNA量の増加が認められた(図1A)。また、雄性ラットにプラバスタチンを0.1%飲料水に添加して1週間投与した場合にも、肝臓のABCA1のmRNA量の増加が認められた(図1B)。しかし、ラット繊維芽細胞Rat2やマウス単球由来細胞RAW264のような末梢系細胞ではABCA1遺伝子の発現量は低下した(図1C,D)。したがって、肝臓のABCA1遺伝子は末梢組織とは異なる機構で発現制御されていると考えられる。
【0066】
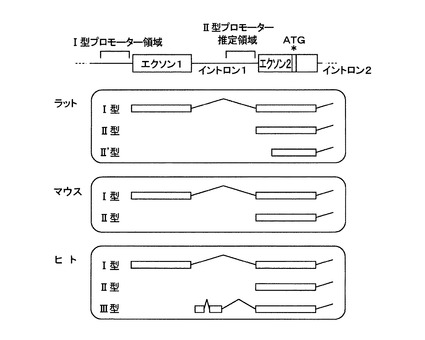
[実施例2:ヒト、マウスおよびラット肝臓で発現するABCA1のmRNA]
5’−RACE法により、ヒト、マウスおよびラット肝臓で発現しているABCA1のmRNAの5’末端領域を解析した。プライマーはABCA1のmRNAエクソン4および5の配列を基に設計した。配列番号15〜20に、ヒト、マウスおよびラット肝臓のそれぞれに対して用いた遺伝子特異的プライマーの配列を示す。これらのプライマーを用い、GeneRacerキット(Invitrogen社製)の添付プロトコールに従い、ヒト肝臓由来総RNA(Human Liver Total RNA;クロンテック社製)、マウス肝臓由来総RNA(Mouse Liver Total RNA;クロンテック社製)および雄性ウィスター系ラット肝臓より抽出した総RNAを鋳型としてcDNAを増幅した。
【0067】
ヒト、マウスおよびラット肝臓において、2または3種類の異なる転写開始点を持つABCA1のmRNAを検出した。配列番号21〜23にヒトI型〜III型のABCA1 mRNAを、配列番号24,25にマウスI型、II型のものを、配列番号26〜28にラットI型、II型、II’型のものの配列をそれぞれ示す。図2は、検出したABCA1のmRNAの5’末端の模式図である。ここでI型と仮称したmRNAは、これまで5’−RACE法を用いてヒト繊維芽細胞やヒト単球由来THP−1細胞から同定されたABCA1のmRNA[NM_005502]に相同する配列(ラット[NW_178095]およびマウス[NM_013454])であり、既報のプロモーター領域(エクソン1上流、Proc.Natl.Acad.Sci.(2000)97, 7987)により転写制御を受けると考えられる。他の2種類はI型とは異なる新規なABCA1 mRNAであった。特に、II型と命名したmRNAは、ヒト、マウス、ラットに共通に発現しており、I型が持つエクソン1領域を欠損しており、エクソン2から転写が開始されるmRNAである。ラット肝臓のII’型mRNAはII型の5’側が68塩基短縮されたmRNAに相当し、転写制御はII型と同一であると予測される。
【0068】
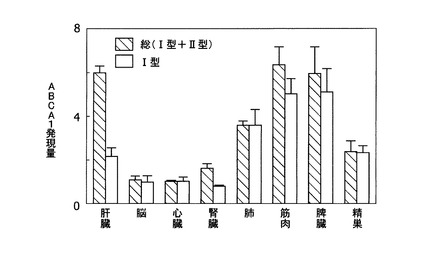
[実施例3:ラット組織におけるABCA1のmRNA発現分布]
ラットの各組織(肝臓、脳、心臓、腎臓、肺、筋肉、脾臓および精巣)から得られたmRNA(ラットMTCパネルI;クロンテック社製)を用い、各組織におけるI型およびII型ABCA1 mRNAの分布を調べた。ラットI型ABCA1 mRNA量はエクソン1領域(I型特有の非翻訳領域)を認識するプライマー・プローブ、また総ABCA1 mRNA量はタンパク質をコードする領域の配列を基に作成したプライマー・プローブを用いて、リアルタイムRT−PCRにより定量した。以下に、I型ABCA1検出用プローブおよび総ABCA1検出用プローブの配列を示す。
【0069】
・ラットI型ABCA1検出用標識プローブ(配列番号29)
FAM-ccctactttt ttctcccggt ttctgg-TAMRA
・ラットI型ABCA1検出用フォワードプライマー(配列番号30)
tctgctccct gtccccac
・ラットI型ABCA1検出用リバースプライマー(配列番号31)
ccaaccccta cacaaaccct
・ラット総ABCA1検出用標識プローブ(配列番号32)
FAM-tctttgctca gattgtcctg ccggc-TAMRA
・ラット総ABCA1検出用フォワードプライマー(配列番号33)
cccggcggag tagaaagg
・ラット総ABCA1検出用リバースプライマー(配列番号34)
agggcgatgc aaacaaagac
【0070】
上記の方法で、各組織についてI型と総ABCA1 mRNAの発現量を測定すると、I型とII型の発現は組織間で異なり、II型は主に肝臓および腎で発現していることが判明した(図3)。そこでII型を肝臓型、これに対しI型を末梢型と呼ぶことにした。
【0071】
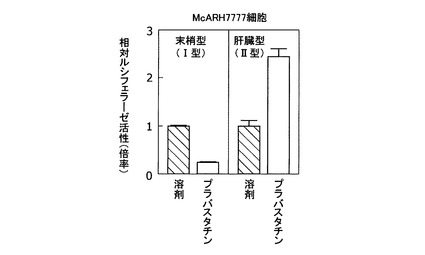
[実施例4:コンパクチンによる新規な肝臓型ABCA1遺伝子プロモーターの活性化]
肝臓型(II型)ABCA1 mRNAの発現調節領域の解析を行った。まず、ラット尾ゲノムDNAから末梢型(I型)および肝臓型(II型)ABCA1 mRNAの転写開始点上流領域約1000塩基をクローニングした。配列番号35に、ラット末梢型ABCA1転写開始点上流領域の配列を示す。配列番号6に、ラット肝臓型ABCA1転写開始点上流領域の配列を示す。得られた転写開始点上流領域を、ホタルルシフェラーゼベクターpGL3(プロメガ社製)に組み込み、レポータープラスミドを構築した。McARH7777細胞への一過性トランスフェクションにより、レポータープラスミドおよび遺伝子導入効率補正用のウミシイタケルシフェラーゼレポータープラスミドphRL−TK(プロメガ社製)を形質導入し、50μMプラバスタチンを含む10%脱脂血清培地で16時間処理した。その後、細胞を回収し、デュアルルシフェラーゼアッセイシステム(プロメガ社製)により細胞内ルシフェラーゼ活性を測定した。なお、転写活性は次のように定義して評価した。
(転写活性)=(ホタルルシフェラーゼ活性測定値)/(ウミシイタケルシフェラーゼ活性測定値)
【0072】
細胞をプラバスタチン処理することにより、末梢型転写開始点上流領域を組み込んだレポータープラスミドの転写活性は低下した(図4)。逆に、肝臓型転写開始点上流領域を組み込んだレポータープラスミドの転写活性は増大した。この結果は、スタチン系薬剤によるABCA1遺伝子発現の肝臓と末梢における応答性の違い(実施例1、図1)とよく一致した。従って、該肝臓型転写開始点上流領域が肝臓でのABCA1遺伝子発現を制御していることが示唆された。
【0073】
[実施例5:肝臓型ABCA1遺伝子プロモーター活性を調節するSRE配列]
スタチン系薬剤はステロール合成を阻害することにより、転写因子として機能する活性型(核内移行型)SREBPを二次的に増加させることが知られている。上記したように、活性型SREBP2はステロール応答性配列(SRE配列)を標的とする転写因子で、活性型SREBP2を過剰発現することでSRE配列下の転写が活性化されることが知られている。上記したように、SRE配列は、配列番号7〜12に記載のコンセンサス配列を有し、ラットの肝臓型ABCA1プロモーター領域に見出された(配列番号14)。
【0074】
そこで、肝臓型ABCA1プロモーター配列内に存在するSRE配列の変異解析を行った。実施例4で得られたラット肝臓型ABCA1プロモーターのSRE配列を、変異型SRE配列を含む合成オリゴヌクレオチドおよびクイックチェンジ(ストラタジェン社製)を用いて機能的に不活性化した。配列番号36に、変異型SRE配列の配列を示す。なお、この変異型SRE配列(GTGCGGCCGC)は、配列番号14に記載のSRE配列(GTCTGGTGAGA)にNotIの認識配列を導入したものであり、SREとして機能しないことが報告されている(J Biol Chem. 2003 Sep 19;278(38):36176-82.)。
【0075】
上述した方法(実施例4)に従い、野生型あるいは変異型SREを有する肝臓型ABCA1レポータープラスミドをMcARH7777細胞に形質導入した。実施例5にあっては、50μMプラスバスタチンの代わりに、50μMコンパクチンで処理し、転写活性を測定した。
【0076】
さらに、スタチン系薬剤で処理する代わりに、活性型SREBP2の過剰発現させた際の転写活性も測定した。活性型SREBP2の過剰発現実験においては、野生型あるいは変異型SREを有する肝臓型ABCA1レポータープラスミドに加えて、さらにpME−SREBP2(1−480)あるいはpME−18Sプラスミド(非特許文献13、J.Biol.Chem. (2003)36176)を共導入した。pME−SREBP2(1−480)プラスミドは、活性型SREBP2であるSREBP2のアミノ酸残基1〜480を発現することができるベクターであり、pME−18Sプラスミドは、その空ベクターである。
【0077】
コンパクチンによるラット肝臓型プロモーター転写活性促進作用は、配列中のSREの変異により消失した(図5)。活性型SREBP2を過剰発現した場合にも転写が活性化されるが、これもSRE変異により消失した(図5)。したがって、プロモーター内に存在するSRE配列が肝臓のABCA1遺伝子の転写調節に機能していることが判明した。
【0078】
[実施例6:ヒト肝臓型ABCA1遺伝子プロモーターのSREBP2による活性化]
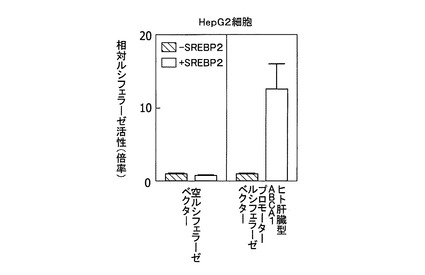
ヒトゲノムDNA(Human Genomic DNA;クロンテック社製)から、ヒト肝臓型ABCA1プロモーターをクローニングした。配列番号4に、ヒト肝臓型ABCA1転写開始点上流領域の配列を示す。得られた転写開始点上流領域を、ホタルルシフェラーゼレポーターベクターpGL3(プロメガ社製)に組み込んだ。ヒト肝臓型ABCA1レポータープラスミドおよびウミシイタケルシフェラーゼレポータープラスミドphRL−SV40(プロメガ社製)を、pME−18SまたはpME−SREBP2プラスミドと共に、HepG2細胞へ形質導入し、10%FCSを含むDMEM培地で16時間培養した。その後、細胞を回収し細胞内ルシフェラーゼ活性を測定した。
【0079】
ヒト肝臓型ABCA1プロモーターの活性は、活性型SREBP2の過剰発現により増大した(図6)。したがって、ヒト肝臓のABCA1遺伝子もラット肝臓と同様の転写制御を受けていると考えられる。
【図面の簡単な説明】
【0080】
【図1】図1Aは、コンパクチンで処理したヒト肝臓由来培養細胞HepG2におけるABCA1遺伝子の発現量変化を示すグラフである。図1Bは、プラバスタチンを投与したラットの肝臓におけるABCA1遺伝子の発現量変化を示すグラフである。図1Cは、プラバスタチンで処理したラット繊維芽細胞Rat2におけるABCA1遺伝子の発現量変化を示すグラフである。図1Dは、コンパクチンで処理したマウス単球由来細胞RAW264におけるABCA1遺伝子の発現量変化を示すグラフである。
【図2】図2は、5’−RACE法により解析したラット、マウスおよびヒト肝臓で発現するABCA1のmRNAの5’末端の模式図である。
【図3】図3は、ラット各組織(肝臓、脳、心臓、腎臓、肺、筋肉、脾臓および精巣)で発現する総ABCA1 mRNA量および末梢型(I型)ABCA1 mRNA量を示したグラフである。
【図4】図4Aは、プラバスタチンによるラット末梢型(I型)ABCA1遺伝子プロモーターの活性化を示したグラフである。図4Bは、プラバスタチンによるラット肝臓型(II型)ABCA1遺伝子プロモーターの活性化を示したグラフである。
【図5】図5は、ラット肝臓型(II型)ABCA1遺伝子プロモーターの活性がSRE配列の変異により消失したことを示すグラフである。
【図6】図6は、ヒト肝臓型(II型)ABCA1遺伝子プロモーターの活性がSREBP2の発現増加により活性化されることを示すグラフである。
【技術分野】
【0001】
本発明は、肝臓においてABCA1遺伝子の発現を制御するプロモーター領域の発見に基づき、ABCA1遺伝子の転写を調整する作用を有する物質のスクリーニング方法ならびにそのためのDNAおよびキットに関するものであり、特に、プロモーター活性の増加を指標として、肝臓におけるABCA1発現の増加により動脈硬化性疾患を予防または治療する作用を有する物質をスクリーニングする方法に関するものである。
【背景技術】
【0002】
高密度リポ蛋白(HDL)は主に肝臓や小腸で合成、分泌され、リポ蛋白(アポA−IやアポA−IIなど)や脂質(リン脂質、コレステロール、トリグリセライド)から構成される複合分子である。HDLは、末梢細胞で過剰に蓄積したコレステロールを細胞膜表面から引き抜いて、胆汁酸への転換および排出が可能な肝臓へと輸送する、いわゆる「コレステロール逆輸送系」において重要な役割を担っている。血中HDL濃度が低いほど動脈硬化性疾患の発症率が高まる疫学的知見から、HDLは善玉コレステロールと呼ばれている。
【0003】
血中HDL濃度の低下は、動脈硬化症や高脂血症、肥満、糖尿病などの様々な疾患で認められる。家族性HDL欠損症であるタンジール病では、細胞内のコレステロールをアポA−Iを含むHDL粒子として放出する過程に障害を持つ。近年その原因遺伝子としてATP結合カセット膜トランスポーターA1(ABCA1)が同定され(非特許文献1、Nat.Genet.(1999)22, 347)、HDLの形成にABCA1タンパク質が必須の役割を担うことが明らかになった。
【0004】
培養細胞にABCA1を強制発現させるとHDL形成が増加する(非特許文献2、J.Clin.Invest.(1999)104, R25)。また、ABCA1過剰発現マウスでは血中HDL濃度が上昇し、動脈硬化に抵抗性を示すことが明らかにされている(非特許文献3、J.Biol.Chem.(2001)276, 33969)。したがって、ABCA1発現量の増加は、HDL濃度の上昇をもたらし、動脈硬化性疾患の予防および治療に有用であると考えられる。
【0005】
マクロファージや繊維芽細胞など末梢細胞におけるABCA1遺伝子の転写は、ABCA1遺伝子のエクソン1上流領域により制御されており、このABCA1遺伝子プロモーターの活性は複数の転写因子により調節されることが明らかにされている(非特許文献4、Biochem.Biophys.Res.Commun.(2000)274, 794:非特許文献5、J.Biol.Chem.(2002)277, 14443:非特許文献6、J.Lipd Res(2002)43, 297:非特許文献7、Mol.Cell.Biol.(2003)23,7756)。なかでも主要なものは、核内受容体LXRであり、RXRとのヘテロダイマーがLXR応答性配列に結合しABCA1の転写を活性化する。細胞内コレステロール濃度を感知するLXRは、ABCA1遺伝子の発現を促進し、HDL新生による細胞外へのコレステロール排出を促してコレステロール恒常性を保つ(非特許文献8、Proc.Natl.Acad.Sci.USA.(2000)97,12097)。
【0006】
【特許文献1】国際公開第00/078972号パンフレット
【非特許文献1】Nat.Genet.(1999)22, 347
【非特許文献2】J.Clin.Invest.(1999)104, R25
【非特許文献3】J.Biol.Chem.(2001)276, 33969
【非特許文献4】Biochem.Biophys.Res.Commun.(2000)274, 794
【非特許文献5】J.Biol.Chem.(2002)277, 14443
【非特許文献6】J.Lipd Res(2002)43, 297
【非特許文献7】Mol.Cell.Biol.(2003)23,7756
【非特許文献8】Proc.Natl.Acad.Sci.USA.(2000)97,12097
【非特許文献9】Biochem.Biophys.Res.Commun.(2004)324, 835
【非特許文献10】Arterioscler.Thromb.Vasc.Biol.(2004)24, 2365
【非特許文献11】J.Clin.Invest. (2005)115, 1333
【非特許文献12】J Biol Chem. 2005 Jun 10;280(23):22212-21
【発明の開示】
【発明が解決しようとする課題】
【0007】
スタチン系薬剤は、コレステロール合成の律速酵素であるHMG−CoA還元酵素の阻害剤であり、血中LDL濃度の低下薬として汎用されているが、血中HDL濃度の上昇効果も認められる。最近、スタチン系薬剤はヒト肝がん由来細胞HepG2においてABCA1遺伝子の発現促進作用を示すことが明らかにされた(非特許文献9、Biochem.Biophys.Res.Commun.(2004)324, 835)。一方、マクロファージ系の培養細胞THP−1やRAW264においては、ABCA1発現を著しく低下させる(非特許文献10、Arterioscler.Thromb.Vasc.Biol.(2004)24, 2365)。これは、コレステロール合成過程で生じるLXRの内因性リガンドがスタチン系薬剤の作用によって減少し、ABCA1遺伝子プロモーター活性が低下することによる。したがって、肝由来細胞のABCA1発現増加は、末梢細胞で機能する、上記のLXRが制御するプロモーター領域(エクソン1上流領域)とは別の発現調節系により引き起こされていると推定される。
【0008】
ABCA1は様々な組織に分布しているが、特に肝臓において強く発現する。マウスを用いた肝臓特異的なABCA1遺伝子のノックダウン解析では、血中HDLコレステロール量が20%以下まで低下し、肝臓のABCA1発現量は血中HDL量の決定に重要な役割を担うことが示されている(非特許文献11、J.Clin.Invest. (2005)115, 1333)。
【0009】
従って、HDL産生の主要臓器である肝臓においてABCA1遺伝子発現を増加させることは、動脈硬化性疾患を予防または治療する上で極めて重要となる。このような状況から、肝臓におけるABCA1遺伝子発現の制御機構の解明、さらにそれを利用した制御薬剤のスクリーニング方法が望まれていた。
【0010】
以上に鑑みて、本発明の課題は、肝臓のABCA1上昇作用を有し動脈硬化に伴う疾患の予防および/または治療に有用な医薬をスクリーニングする新規な方法および該方法において用いられるDNAを提供することにある。
【0011】
なお、本発明の方法において用いられるABCA1遺伝子は、Gene Bankデータベース上に「ATP結合カセット輸送体A1(ATP-binding cassette transporter A1)」として公開されている。また特許文献1(国際公開第00/078972号パンフレット)には「ABC1」をコードするものとして同遺伝子が、またそれと共に上記のエクソン1上流のプロモーターが開示されている。
【課題を解決するための手段】
【0012】
すなわち、本発明の一の側面によると、ABCA1遺伝子のイントロン1中のSRE配列を含み、プロモーター活性を有するDNAを用いることを特徴とする、ABCA1遺伝子の転写を調整する作用を有する物質のスクリーニング方法が提供される。
【0013】
また、本発明の他の側面によると、活性型SREBPの量を変化させる作用を有する物質(ピタバスタチン、シムバスタチンおよびアトルバスタチンを除く)を含有する、ABCA1遺伝子の転写調整剤が提供される。
【0014】
また、本発明の他の側面によると、ABCA1遺伝子のイントロン1中のSRE配列を含み、プロモーター活性を有する組換えDNAが提供される。
【0015】
また、本発明の他の側面によると、ABCA1遺伝子のイントロン1中のSRE配列を含み、プロモーター活性を有するDNAを有する組換えベクターが提供される。
【0016】
また、本発明の他の側面によると、ABCA1遺伝子のイントロン1中のSRE配列を含み、プロモーター活性を有する外因性のDNAに作動可能に連結した構造遺伝子を有する形質転換体が提供される。
【0017】
また、本発明の他の側面によると、上記組換えベクターおよび/または上記形質転換体を有する、ABCA1遺伝子の転写を調整する作用を有する物質をスクリーニングするためのキットが提供される。
【発明の効果】
【0018】
本発明は、ABCA1遺伝子の転写を調整する作用を有する物質、特に、動脈硬化性疾患の予防および/または治療剤を試験するための新規方法を確立する。また、本発明は、従来のABCA1遺伝子発現促進物質のスクリーニング方法に比べ、HDL上昇作用により効果的な分子を検出できる可能性がある。
【発明を実施するための最良の形態】
【0019】
以下に、本発明の実施の形態を説明する。もっとも、本発明は、以下に説明する実施の形態によって、限定されるものではない。
【0020】
本発明者らは、肝臓におけるABCA1遺伝子の発現調節に新たな制御機構が存在することを見出した。そこで、ABCA1遺伝子発現を調節する物質を選別または同定するための方法として、イントロン1に存在する新規肝臓型ABCA1プロモーター(本明細書中、肝臓型プロモーター、II型ABCA1プロモーター等とも記す。)を用いる方法を開発し、本発明を完成させた。
【0021】
本発明が開示する肝臓型プロモーターは、ABCA1遺伝子のエクソン1上流のプロモーター(特許文献1参照)とは全く異なる領域に存在し、DNA配列も異なる。この新規肝臓型プロモーターに関しては、ABCA1遺伝子転写制御、HDL形成との関連は報告されておらず、動脈硬化性疾患の治療または予防剤の試験法としての有用性も知られていない。上記したように、ピタバスタチン、シンバスタチン、アトルバスタチンがABCA1のmRNAを増加させることが示されている(非特許文献9、Biochem.Biophys.Res.Commun.(2004)324, 835)。しかし、その作用機序に関わるデータは無く、LXRが活性化される可能性が議論されている。また、アトルバスタチン、シンバスタチン、ピタバスタチンが(特定の条件下で)マクロファージのABCA1の発現を増加させること、その機序の一つとしてRhoAのゲラニル化が関与しているという報告がある(非特許文献12、J Biol Chem. 2005 Jun 10;280(23):22212-21)。しかし、これもPPAR gammaを介してLXRを活性化する(末梢型プロモーターを活性化する)メカニズムを想定している系であり、本発明にかかる新規肝臓型ABCA1プロモーターおよび当該プロモーターによる発現制御については記載も示唆もしていない。
【0022】
具体的には、本発明の一の側面によると、ABCA1遺伝子のイントロン1中のSRE配列を含み、プロモーター活性を有するDNAが提供される。当該DNAは、天然のものであってもよく、また人工的なものであってもよい。すなわち、本発明の他の側面によると、ABCA1遺伝子のイントロン1中のSRE配列を含み、プロモーター活性を有する組換えDNAが提供される。
【0023】
本発明にかかるDNAは、任意の生物に由来するDNAであってよく、また、人工的に設計および/または調製されたDNAであってもよい。具体的には、本発明にかかるDNAは、ヒト、サル、マウス、ラット、モルモット、ウサギ、ブタ、ウシ、ヒツジ、ニワトリなどの哺乳動物その他の恒温動物に由来するDNAであってよい。
【0024】
ABCA1遺伝子のイントロン1は、ABCA1遺伝子のエクソン1の下流かつエクソン2の上流に存在する領域である。具体的には、ヒトABCA1遺伝子のエクソン1、イントロン1およびエクソン2として、配列番号1に記載の配列の1〜219位、220〜24375位および24376〜24535位の配列をそれぞれ有するものが見出されている。マウスABCA1遺伝子のエクソン1、イントロン1およびエクソン2として、配列番号2に記載の配列の1〜215位、216〜15795位および15796〜15963位の配列をそれぞれ有するものが見出されている。ラットABCA1遺伝子のエクソン1、イントロン1およびエクソン2として、配列番号3に記載の配列の1〜220位、221〜16048位および16049〜16216位の配列をそれぞれ有するものが見出されている。また、ヒト、マウスおよびラットにおける上記イントロン1の相同体、変異体および誘導体の配列、ならびに、他の任意の生物における上記イントロン1の相同体ならびにその変異体および誘導体の配列は、当業者であれば、ヒト、マウスおよび/またはラットにおける上記イントロン1および/またはエクソン1、2の配列との相同性に基づき決定することができる。
【0025】
ABCA1遺伝子のイントロン1中のSRE配列を含み、プロモーター活性を有するDNAは、イントロン1中のSRE配列に加えて、エクソン2の翻訳開始点付近までを含むことが好ましい。なお、ヒト、マウスおよびラットのABCA1のエクソン2中の翻訳開始点は、それぞれ、配列番号1に記載の配列の24470位、配列番号2に記載の配列の15898位、および配列番号3に記載の配列の16151位である。プロモーター活性を有するのに必要な領域は、当業者であれば適宜決定することができる。プロモーター活性は、例えば、以下に詳細に説明するように、ルシフェラーゼアッセイ法、CAT法等により測定することができる。
【0026】
特に、ABCA1遺伝子のイントロン1中のSRE配列を含み、プロモーター活性を有するDNAは、(a)配列番号4〜6のいずれかに記載の配列を有するDNAの少なくとも一部とすることができる。配列番号4〜6に記載の配列を有するDNAは、それぞれ、ヒト、マウスおよびラットにおいて、配列番号1の22963〜24469位、配列番号2の14298〜15797位および配列番号3の15101〜16173位に見出される配列に対応する。さらに、ABCA1遺伝子のイントロン1中のSRE配列を含み、プロモーター活性を有するDNAは、以下に詳細に説明するように、(a)のDNAと実質的に同一の配列を有するDNAとすることができる。
【0027】
具体的には、ABCA1遺伝子のイントロン1中のSRE配列を含み、プロモーター活性を有するDNAは、(b)配列番号4〜6のいずれかに記載の配列において、1もしくは数個(好ましくは1〜80個、より好ましくは1〜40個、さらに好ましくは1〜20個、さらに好ましくは1〜10個)のヌクレオチドが置換、挿入または欠失された配列を有し、プロモーター活性を有するDNAの少なくとも一部とすることができる。当該技術分野における通常の知識を有する者であれば、野生型プロモーターのヌクレオチド配列の部分的な置換、挿入または欠失などの改変により、野生型プロモーターと同等またはそれ以上のプロモーター活性を有するDNAを調製することが可能である。
【0028】
また、ABCA1遺伝子のイントロン1中のSRE配列を含み、プロモーター活性を有するDNAは、(c)配列番号4〜6のいずれかに記載の配列と相同性が60%以上(好ましくは80%以上、さらに好ましくは90%以上)の配列を有し、プロモーター活性を有するDNAの少なくとも一部とすることができる。このようなDNAには、(a)のDNAの、自然界に存在する変異型遺伝子もしくは人為的に改変した変異型遺伝子または異種生物由来の相同遺伝子等が含まれる。
【0029】
相同性の比較は、通常、容易に入手できる配列比較プログラムを用いて行うことができる。商業的に入手可能なそのようなコンピュータープログラムは2以上の配列の間の%相同性を計算することができる。ほとんどの配列比較方法においては、全体としての相同性スコアに不当にペナルティを与えることなく可能な挿入および欠失を考慮に入れて、最適な整列を生じさせるよう設計される。これは、「ギャップ」を配列の整列に挿入して、局所的な相同性を最大化するよう努めることによって達成される。このような方法では、できる限り少数のギャップにおける配列の整列が、多くのギャップにおけるものより高いスコアを達成するように、各ギャップに「ギャップペナルティ」を割り当てて、相同性を計算する。ほとんどの整列プログラムはギャップペナルティを設定することができる。しかしながら、配列比較のためのそのようなソフトウェアを用いる場合、デフォルト値を用いるのが好ましい。このような配列比較を実行できるソフトウェアの例として、BLASTパッケージ(Ausubel et al.,1999 ibid-Chapter18参照)、FASTA(Atschul et al.,1990,J.Mol.Biol.,403-410)およびGENEWORKSスイートが挙げられる。
【0030】
また、ABCA1遺伝子のイントロン1中のSRE配列を含み、プロモーター活性を有するDNAは、(d)(a)のDNAとストリンジェントな条件でハイブリダイズし、プロモーター活性を有するDNAの少なくとも一部とすることができる。ここで、DNAは通常、互いに相補的な二本鎖DNA構造を有している。このため、(a)のDNAとハイブリダイズするDNAとは、(a)のDNAのいずれかの鎖とハイブリダイズする鎖を有するDNAをいうことに留意すべきである。
【0031】
なお、ストリンジェントな条件およびハイブリダイゼーションの方法は、例えば、J. Sambrook et al., Molecular Cloning, Cold Spring Harbour Laboratory Press, New York, USA, 第2版, 1989に従って設定することができる。具体的には、ストリンジェントな条件は、「1xSSC、0.1%SDS、37℃」とすることができ、よりストリンジェントな条件は、「0.5xSSC、0.1%SDS、42℃」とすることができ、さらにストリンジェントな条件は、「0.2xSSC、0.1%SDS、65℃」とすることができる(1xSSC:0.15MのNaCl、0.015Mのクエン酸ナトリウム、pH7.0)。ただし、これらの条件の組み合わせは例示であり、当業者であれば、ハイブリダイゼーションのストリンジェンシーを決定する上記または他の要素(例えば、プローブ濃度、プローブの長さ、ハイブリダイゼーション反応時間など)を適宜組み合わせることにより、上記と同様のストリンジェンシーを実現することが可能である。
【0032】
なお、(b)〜(d)のDNAのプロモーター活性は、例えば、以下に詳細に説明するように、ルシフェラーゼアッセイ法、CAT法等により測定することができる。また、(b)〜(d)のDNAは、(a)のDNAと「同様」のプロモーター活性を有することが好ましい。ここで、「同様」のプロモーター活性を有するとは、質的に類似のプロモーター活性を有することをいい、例えば、(b)〜(d)のDNAが、(a)のDNAと少なくとも1つの因子(活性型SREBP2等の転写因子等)について同一の制御を受ける場合が含まれる。一方で、「同様」のプロモーター活性を有するとは、量的に同程度のプロモーター活性を有することを意図するものではない。また、当業者には明らかなように、プロモーター活性を発揮するために、DNAは、配列番号4〜6のいずれかに記載の配列(またはこれと実質的に同一の配列)の全てを有する必要はなく、その一部であってもよい。上記したように、プロモーター活性を有するのに必要な領域は、当業者であれば、ルシフェラーゼアッセイ法、CAT法等により適宜決定することができる。
【0033】
SRE配列は、SREBPが特異的に結合する能力を有するいかなる配列でもよく、例えば、配列番号7〜10に記載のコンセンサス配列を有することが好ましい。また、SRE配列は、配列番号11、12に記載の、ラット肝臓型ABCA1プロモーター領域中のSREと相同なSREコンセンサス配列を有することが好ましい。換言すると、SRE配列は、配列番号7〜12に記載のコンセンサス配列を有し、SREBPが特異的に結合できる配列であることが好ましい。なお、配列番号11、12に記載のSREコンセンサス配列は、後述する配列番号14に記載のSRE配列に相補的な配列に相同な配列である。
【0034】
なお、配列番号7〜12に記載のSREコンセンサス配列を一塩基または多塩基置換することにより、SRE配列を不活化させることができる。(J Biol Chem. 1999 Feb 26;274(9):5285-91., PNAS. 2004 Aug 3;101(31):11245-50)変異の導入はクイックチェンジ(ストラタジェン)やオルタドサイトズ(プロメガ)やジーンエディター(プロメガ)などのキットを用いて行うことができる。また、制限酵素を用いてコンセンサス配列そのものを除去することで、SREを不活性化させるのと同様の効果を得ることもできる。
【0035】
SREBPが特異的に結合する能力の有無は、例えば、以下に詳細に説明するように、ゲルシフトアッセイ、DNase Iフットプリンティング法、クロマチン免疫沈降法(ChiPアッセイ)等により測定することができる。なお、「特異的に結合する」とは、好ましくは生理的条件下(好ましくは天然の細胞、組織または生物内と同等の条件下)で、当該タンパク質の目的のDNA配列への結合能が、当該タンパク質の他の無作為なDNA配列への結合能と比べて、有意に大きいことをいう。
【0036】
[ゲルシフトアッセイ]
ゲルシフトアッセイは、DNAとタンパク質との相互作用を検出する方法である。一般的には、32Pで標識したDNA断片と目的のタンパク質とを混合し低塩濃度の未変性ゲルで電気泳動しDNAをオートラジオグラフィーで検出する。タンパク質と結合したDNAは遊離DNAと比べゲル中で移動度が遅れるため検出が可能となる。
【0037】
[DNase Iフットプリンティング法]
DNase Iフットプリンティング法は、塩基配列特異的DNA分解酵素であるDNase IでDNAを部分的に消化する際、DNAに結合しているタンパク質は、結合部位のホスホジエステル結合をDNase Iによる攻撃から保護することを利用し、DNAとタンパク質との相互作用を検出する方法である。DNA断片の一端を放射性同位元素で標識しておき、DNase I反応後の生成物を高分解能ゲル電気泳動で分離した後、ゲルのオートラジオグラムを得ると、タンパク質結合部位のバンドが消失することになる。同じDNA断片をタンパク質非存在下においてDNase I処理したものとMaxam-Gilbertシークエンス反応(Maxam, A. & Gilbert, W., Methods Enzymol., 65, 499, 1980)したものを同時に電気泳動することにより、タンパク質の結合部位が同定できる。
【0038】
[クロマチン免疫沈降法(ChiPアッセイ)]
クロマチン免疫沈降法は、ホルムアルデヒド等を用いて細胞内でDNAとタンパク質を架橋し、目的のタンパク質に対する特異的な抗体を使った免疫沈降法と組み合わせることで、細胞内に実際に存在するクロマチンDNA上でのDNAとタンパク質の相互作用を検出する方法である。ホルムアルデヒド等によりDNAと目的のタンパク質の架橋を行い、目的のタンパク質に対する抗体で免疫沈降を行った後、可逆反応により架橋を外し、タンパク質を除き、DNA断片を回収する。回収されたDNAを鋳型にして、目的のDNA配列の特異的プライマーを用いてPCRまたはPCR/サザンブロットで解析を行う。
【0039】
具体的には、ヒトABCA1遺伝子のイントロン1におけるSRE配列として、TCTCAGCTGAG(配列番号13、配列番号1の24285〜24295位に対応)が見出されている。また、マウスおよびラットABCA1遺伝子のイントロン1におけるSRE配列として、GTCTGGTGAGA(配列番号14、配列番号2の15666〜15676位および配列番号3の15954〜15964位に対応)が見出されている。通常、SRE配列は、配列番号13または14に記載の野生型配列と50%以上の相同性を有し、好ましくは60%以上の相同性、さらに好ましくは80%以上の相同性を有する。
【0040】
また、上記したように、本発明の他の側面によると、ABCA1遺伝子のイントロン1中のSRE配列を含み、プロモーター活性を有する組換えDNAが提供される。本発明にかかる組換えDNAは、転写最小プロモーターをさらに有することが好ましい。最小プロモーターの例として、ヘルペスウイルスのチミジンキナーゼ(TK)遺伝子のプロモーター、CMV初期プロモーター、PGKプロモーター、SV40プロモーターなどが挙げられるが、これらに限定されるものではない。本発明にかかる組換えDNAは、協調因子の結合配列をさらに有することが好ましい。このような協調因子の例として、NF−YやSP1などが挙げられる。また、協調因子の結合配列とSRE配列との距離は、50塩基以内であることが好ましい。
【0041】
本発明にかかるDNAは、ゲノムライブラリーからの選別や、ゲノムDNAを鋳型としたポリメラーゼ連鎖反応(PCR)により取得できる。ゲノムライブラリーからの選別は、適当なベクターに組み込んだゲノムDNAと、目的のDNAの一部の配列を有する標識DNAとのハイブリダイゼーションによって行うことができる。ハイブリダイゼーションは、例えば、J. Sambrook et al., Molecular Cloning, Cold Spring Harbour Laboratory Press, New York, USA, 第2版, 1989に記載の方法などに従って行なうことができる。また、市販のライブラリーを使用する場合、添付の使用説明書に記載の方法に従って行なうことができる。また、ゲノムDNAを鋳型としたPCRは、目的のDNAの一部の配列を有するプライマーを用いて行うことができる。ヌクレオチド配列の改変は、例えば、制限酵素またはDNAエキソヌクレアーゼによる欠失導入、部位特異的変異誘発法による変異導入、変異プライマーを用いたPCR法による改変、合成変異DNAの直接導入などの方法により行うことができる。
【0042】
本発明の他の側面によると、ABCA1遺伝子のイントロン1中のSRE配列を含み、プロモーター活性を有するDNAを有する組換えベクターが提供される。組換えベクターは、当該プロモーター活性を有するDNAに作動可能に連結した構造遺伝子をさらに有してもよい。以下に詳細に説明するように、構造遺伝子は、例えば、組換えベクターをスクリーニングに用いる場合、その転写量および/または発現量を測定できるものであれば、任意の遺伝子でよい。
【0043】
本発明にあっては、ベクターは、任意のベクター、例えば、バクテリオファージ、プラスミド、コスミド、ファージミドなどであってよい。より具体的には、本発明に用いることができるベクターの例として、動物細胞用のベクターであるpCDベクター、cDM8ベクター、レトロウイルスベクター、アデノウイルスベクター、ヘルペスウイルスベクター、アデノアソシエイトウイルスベクターなどや、細菌用のベクターであるpUCなどを挙げることができる。構造遺伝子をベクター内に組み込むためには、構造遺伝子が正しく転写される方向に、さらにプロモーターが構造遺伝子に対して適正に機能する位置関係になる様に、プロモーター領域の下流に構造遺伝子を連結すればよい。プロモーターと構造遺伝子との連結は、制限酵素切断部位を利用して行うことができ、また、適当な制限酵素部位がなくてもリガーゼ反応を用いて行うことができる。本発明にかかる組換えベクターは、プロモーター活性を促進するエンハンサー領域等の転写調節領域、転写停止部位などその他の要素を有してもよい。
【0044】
なお、「作動可能に連結」とは、一の遺伝子要素が、他の遺伝子要素に対して予測されるように機能できるように配置されていることをいう。例えば、プロモーターが、構造遺伝子の転写の開始を助ける場合には、プロモーターは構造遺伝子に作動可能に結合している。この機能的な関係が維持される限り、プロモーターと構造遺伝子との間に残基が介在してもよい。
【0045】
本発明の他の側面によると、ABCA1遺伝子のイントロン1中のSRE配列を含み、プロモーター活性を有する外因性のDNAに作動可能に連結した構造遺伝子を有する形質転換体が提供される。本明細書において、形質転換体は、広く、外部からDNAが導入された結果、新しい形質を獲得した生物をいう。形質転換体は、任意の生物であってよく、また、任意の生物に由来する任意の細胞または組織であってよい。特に、本発明にかかる形質転換体は、上記組換えベクターで形質転換された形質転換体を含む。
【0046】
組換えベクターにより形質転換することができる宿主の例として、例えば、エシェリヒア属菌、バチルス属菌、酵母、昆虫細胞、昆虫、動物細胞などを挙げることができる。これらの宿主への形質転換の方法の例として、リン酸カルシウム法、リポフェクション法、エレクトロポレーション法、アデノウイルスベクターまたはレトロウイルスベクターによる遺伝子導入法、マイクロインジェクション法などを挙げることができる。
【0047】
本発明の他の側面によると、ABCA1遺伝子の転写を調整する作用を有する物質をスクリーニングするためのキットが提供される。本発明にかかるキットは、動脈硬化性疾患の予防または治療に有用な物質をスクリーニングするために用いることができる。以下に詳細に説明するように、ABCA1遺伝子のイントロン1中のSRE配列を含み、プロモーター活性を有するDNAを用いることで、ABCA1遺伝子の転写を調整する作用を有する物質をスクリーニングすることができる。すなわち、本発明にかかるキットは、上記組換えベクターおよび/または上記形質転換体を有する。
【0048】
本発明にかかるキットには、スクリーニングの方法に応じて、酵素、酵素基質、緩衝液等の試薬、培地、道具等を組み合せることができる。酵素基質の例として、ルシフェリン、ATP、[14C]クロラムフェニコール、アセチルCoAを挙げることができる。緩衝液として、解析に用いる酵素の活性の維持に好適な緩衝液を適宜選択することができる。道具の例として、TLC用展開シート、メンブレン等を挙げることができる。
【0049】
本発明の他の側面によると、ABCA1遺伝子の転写を調整する作用を有する物質、特に、肝臓においてABCA1遺伝子の転写を調整する作用を有する物質のスクリーニング方法が提供される。特に、本発明にかかるスクリーニング方法によると、ABCA1遺伝子の転写を促進する物質をスクリーニングすることができる。本発明にかかるスクリーニング方法は、動脈硬化性疾患の予防または治療に有用な物質をスクリーニングするために用いることができる。本発明にかかるスクリーニング方法は、ABCA1遺伝子のイントロン1中のSRE配列を含み、プロモーター活性を有するDNAを用いることを特徴とする。当該DNAを用いることにより、本発明にかかるスクリーニング方法によると、例えば活性型SREBP量を増加させる作用を有する物質をスクリーニングすることができる。
【0050】
具体的には、本発明にかかるスクリーニング方法は、
被検物質を供するステップと、
ABCA1遺伝子のイントロン1中のSRE配列を含み、プロモーター活性を有するDNAに作動可能に連結した構造遺伝子を有するDNAを有する細胞または生物の複数の群を供するステップと、
前記被検物質を前記細胞または生物の少なくとも1つの群に接触させるステップと、
前記被検物質を接触させた群と接触させなかった群とにおいて、前記遺伝子の転写量および/または発現量を測定するステップと、
前記被検物質を接触させた群と接触させなかった群の前記転写量および/または発現量を比較するステップと
を含むことが好ましい。
【0051】
被検物質として、単一の化合物または組成物、天然または合成化合物、有機または無機化合物、タンパク質、ペプチド、オリゴヌクレオチド、ポリヌクレオチド、細胞抽出物、細胞培養上清、その他の任意の物質を供することができる。
【0052】
スクリーニングは、インビトロおよびインビボのいずれでも行うことができる。本発明にかかるスクリーニング方法において用いられる細胞または生物として、細菌等の原核生物(大腸菌等)、酵母(サッカロマイセス(Saccharomyces)属、クライヴェロマイセス(Kluyveromyces)属等)、昆虫、植物もしくは動物またはこれらに由来する細胞が挙げられる。動物の例として、恒温動物、特にヒト、サル、マウス、ラット、ウシ、ブタ、イヌ等の哺乳動物を挙げることができる。本発明で用いられる細胞は、実験室で培養することのできる培養細胞であることが好ましく、初代培養肝細胞であることがさらに好ましい。細胞は、恒温動物、特にヒト、サル、マウス、ラット、ウシ、ブタ、イヌ等の哺乳動物に由来することができ、ヒトに由来することが好ましい。また、細胞は、肝臓に由来することが好ましい。
【0053】
本発明に用いられる細胞または生物は、ABCA1遺伝子のイントロン1中のSRE配列を含み、プロモーター活性を有するDNAに作動可能に連結した構造遺伝子を有するDNAを有することが好ましい。当該DNAは、内因性であっても、または外因性であってもよい。外因性である場合、当該DNAは、任意の手段により細胞または生物に導入されてもよい。例えば、スクリーニングに培養細胞を用いる場合、培養細胞への当該DNAの導入方法として、本発明の属する技術分野において汎用される任意の方法を採用することができる。例えば、DNAの導入方法の例として、リン酸カルシウム−DNA共沈法、リポフェクション法、DEAE−デキストラン法、電気パルス穿孔法等が挙げられるが、これらに限定されない。いずれの方法においても、用いる細胞に応じて至適化されたトランスフェクション条件を用いることが好ましい。
【0054】
構造遺伝子は、その転写量および/または発現量を測定できるものであれば、任意の遺伝子でよい。当業者に明らかなように、上記構造遺伝子は、その転写量および/または発現量の測定方法に応じて、適宜選択することができる。上記構造遺伝子の例として、ABCA1、所定の活性を有するタンパク質(ルシフェラーゼ等)、所定の抗体が結合できるタンパク質、所定の物質が特異的に結合できるタンパク質(GST等)等をコードする遺伝子を挙げることができ、特に、ホタルルシフェラーゼ、クロラムフェニコールアセチルトランスフェラーゼ、β−ガラクトシダーゼ、分泌型アルカリホスファターゼおよび緑色蛍光蛋白質からなる群より選択されるタンパク質をコードする遺伝子(いわゆるレポーター遺伝子)を挙げることができる。
【0055】
被検物質を細胞または生物に接触させることは、当業者に明らかなように、任意の方法により行うことができる。例えば、培養細胞を用いる場合、被検物質を培地中に添加することで、接触させることができる。すなわち、培養細胞を用いる場合、細胞を、被検物質の存在下または非存在下で培養することが好ましい。また、動物を用いる場合は、任意の手段による投与により、例えば、任意の経路からの注射により、接触させることができる。被検物質を細胞または生物へ接触させる時間は、被験物質の効果、濃度、細胞または生物の種類および技術的考慮に応じて、適宜設定することができる。また、被験物質の濃度は、その毒性、透過性、細胞数、処理時間等に応じて、適宜設定することができ、通常1nM〜1mMであることが好ましい。培養細胞を用いる場合、一般的にこのような処理は、多くの試験を同時に実施できるマルチウェルプレートで実施することが好ましい。培養細胞を用いる場合、被験化合物の処理によるレポーター遺伝子の発現が測定可能な条件は、細胞が生存してレポーター遺伝子の発現産物が生産可能な条件であれば良い。特に好ましい条件にあっては、好ましくは使用される細胞株に適した培地を使用し、4〜6%の炭酸ガス存在下、36〜38℃で5〜72時間培養する。
【0056】
被検物質を接触させた群と接触させなかった群とにおいて、上記構造遺伝子の転写量および/または発現量を測定し、これらの転写量および/または発現量を比較することで、当該被検物質を、ABCA1遺伝子の転写を調整する作用を有する物質の候補物質とすることができるか否か判断することができる。具体的には、ある被検物質を接触させた群で、接触させなかった群と比べて、上記遺伝子の転写量および/または発現量が変化した場合、特に増加した場合、当該被検物質を候補物質とすることができる。
【0057】
被検物質を接触させた群と接触させなかった群とにおいて、上記構造遺伝子の転写量および/または発現量を測定するステップと、被検物質を接触させた群と接触させなかった群の転写量および/または発現量を比較するステップは、当業者に明らかなように、任意の方法により行うことができる。具体的には、ノーザンブロッティング法、DNAアレイ法等により、上記遺伝子の転写量を測定することができる。また、ルシフェラーゼアッセイ法やCAT法等により、上記遺伝子の発現量を測定することができる。さらに、上記所定の遺伝子に対する抗体が入手可能な場合は、ELISA法、免疫沈降法、ウエスタンブロッティング法により、当該遺伝子の発現量を測定することができる。また、上記所定の遺伝子として、GST等の所定の物質に特異的に結合するタンパク質の遺伝子を用いる場合は、当該タンパク質を沈降させ、沈降したタンパク質の量を測定することで、上記遺伝子の発現量を測定することができる。
【0058】
[ノーザンブロッティング法]
ノーザンブロッティング法は、変性したRNAを変性条件下のアガロース電気泳動で分離し、ニトロセルロースフィルターに移して、標識した特異的なプローブで検出するものである。RNAを変性させるのは、RNAは分子内に二次構造を持つため、そのままではサイズに従った正確な分離ができず、さらに、フィルターは一本鎖の核酸しか結合できないためである。
【0059】
[ルシフェラーゼアッセイ法、CAT法]
ルシフェラーゼアッセイ法およびCAT法は、目的とする遺伝子の転写調節領域の下流にレポーターとしてLUC(ルシフェラーゼ)またはCAT(クロラムフェニコールアセチルトランスフェラーゼ)遺伝子を組み込んだプラスミドを作成し、そのプラスミドを導入した細胞の酵素活性を測定するものである。具体的には、ルシフェラーゼアッセイ法は、マグネシウム存在下、ルシフェラーゼがルシフェリンとATPから酸化ルシフェリンとAMPを作る反応を触媒する際に発する波長560nmの光を、ルミノメーターを使って検出するものである。CAT法の場合、基質の[14C]クロラムフェニコールが、CATによりアセチル化されて、[14C]アセチルクロラムフェニコールが産生される。このサンプルを酢酸エチルで抽出し、薄層のシリカゲルプレート上に展開後、移動度の異なるスポットの放射活性を測定する。
【0060】
本発明の他の側面によると、上記スクリーニング方法によりスクリーニングされた、ABCA1遺伝子の転写を調整する作用を有する物質、特に、肝臓においてABCA1遺伝子の転写を調整する作用を有する物質が提供される。特に、上記したように、本発明にかかるスクリーニング方法によると、ABCA1遺伝子の転写を促進する物質をスクリーニングすることができる。このため、本発明の他の側面によると、上記スクリーニング方法によりスクリーニングされた、動脈硬化性疾患の予防または治療に有用な物質が提供される。
【0061】
本発明の他の側面によると、活性型SREBPの量を変化させる作用を有する物質を含有する、ABCA1遺伝子の転写調整剤が提供される。特に、活性型SREBPの量を変化させる作用を有する物質が、活性型SREBPの量を増加させる作用を有することが好ましい。ABCA1遺伝子の転写調整剤は、活性型SREBPの量を変化させる作用を有する物質を、ABCA1遺伝子の転写を調整するのに有効な量含有する。有効な量は、当業者であれば、転写調整剤を用いる目的、条件等に応じて、適宜設定することができる。なお、活性型SREBPはステロール応答性配列(SRE配列)を標的とする転写因子で、活性型SREBPを過剰発現させることでSRE配列下の転写が活性化されることが知られている。より具体的には、活性型SREBPは、以下のように制御されることが知られている。一般に、SREBPは小胞体膜に結合した形で存在している。細胞内コレステロール量が減少すると、SREBPはプロセシングを受けて膜から切り離され活性型SREBP(N末端480アミノ酸)として核内に移行し、標的遺伝子の転写を活性化あるいは抑制する。切断が行われるゴルジ体への輸送は、ステロールの低下により行われる。SREBPはSCAP(SREBP cleavage activating protein)と結合しINSIG−1によって小胞体に留められているが、コレステロール量が減少するとINSIG−1が解離し、SREBP−SCAP複合体がゴルジ体に移行する。
【0062】
また、活性型SREBPの量を変化させる作用を有する物質が、ピタバスタチン、シムバスタチンおよびアトルバスタチンを除く物質であることが好ましく、スタチン系薬剤を除く物質であることがさらに好ましい。活性型SREBPの量を変化させる作用を有する物質は、不活性型SREBPを活性型SREBPに(または活性型を不活性型に)変換する作用を有する物質であってもよく、また、例えば、活性型SREBPを発現させることで、活性型SREBPの量を直接変化させる作用を有する物質であってもよい。具体的には、活性型SREBPの量を変化させる作用を有する物質が、活性型SREBP発現ベクター、細胞内コレステロール量を減少させる作用を有する物質、SREBPの前駆体(不活性型SREBP)から核内移行型(活性型SREBP)へのプロセシングを促進する作用を有する物質、活性型SREBPの分解を抑制する作用を有する物質からなる群から選ばれる少なくとも1つであることが好ましい。より具体的には、細胞内コレステロール量を減少させる作用を有する物質の例として、コレステロール合成阻害剤、LDL受容体阻害剤、胆汁酸吸着樹脂が挙げられる。また、SREBPの前駆体から核内移行型へのプロセシングを促進する作用を有する物質の例として、インシュリンが挙げられる。その他、活性型SREBPの量を変化させる作用を有する物質の例として、ATPやインポーチンβ、ALLNなどのプロテアソーム阻害剤が挙げられる。
【実施例】
【0063】
本発明は以下の実施例においてより具体的に説明される。但し、本発明はこれらの実施例に限定されるものでない。
【0064】
[実施例1:スタチン系薬剤のABCA1遺伝子発現に対する影響]
スタチン系薬剤がABCA1のmRNA発現に及ぼす影響を、以下の方法で調べた。ヒト肝ガン由来培養細胞HepG2およびラット繊維芽細胞Rat2を、10%FCSを含むDMEM培地で、マウス単球由来細胞Raw264を、10%FCSを含むDMEM/F−12培地でそれぞれ培養し、被験化合物を含む無血清培地(0.2%BSA)で24時間処理した。HepG2細胞およびRaw264細胞には、被験化合物として、コンパクチンを最終濃度50μMで添加した。Rat2細胞には、被験化合物として、プラバスタチンを、最終濃度50μMで添加した。それぞれ、溶剤のみを添加したものを対照とした。また、雄性ラットに被験化合物を経口投与し、その後該動物個体から肝臓を摘出した。被験化合物として、プラバスタチンを0.1%の濃度で飲料水に添加して1週間投与した。培養細胞およびラット組織からRNeasy(QIAGEN社製)を用いてRNAを調製した。7700 Sequence Detector(ABI社製)システムを用いたTaqManリアルタイム定量RT−PCRにより、調製したRNA中におけるABCA1のmRNA量を測定した。各試料中のmRNA量は、それぞれ18SRNAまたはGAPDHのmRNA量に対して正規化を行った。
【0065】
ヒト肝ガン由来培養細胞HepG2を50μMのコンパクチンで処理すると、ABCA1のmRNA量の増加が認められた(図1A)。また、雄性ラットにプラバスタチンを0.1%飲料水に添加して1週間投与した場合にも、肝臓のABCA1のmRNA量の増加が認められた(図1B)。しかし、ラット繊維芽細胞Rat2やマウス単球由来細胞RAW264のような末梢系細胞ではABCA1遺伝子の発現量は低下した(図1C,D)。したがって、肝臓のABCA1遺伝子は末梢組織とは異なる機構で発現制御されていると考えられる。
【0066】
[実施例2:ヒト、マウスおよびラット肝臓で発現するABCA1のmRNA]
5’−RACE法により、ヒト、マウスおよびラット肝臓で発現しているABCA1のmRNAの5’末端領域を解析した。プライマーはABCA1のmRNAエクソン4および5の配列を基に設計した。配列番号15〜20に、ヒト、マウスおよびラット肝臓のそれぞれに対して用いた遺伝子特異的プライマーの配列を示す。これらのプライマーを用い、GeneRacerキット(Invitrogen社製)の添付プロトコールに従い、ヒト肝臓由来総RNA(Human Liver Total RNA;クロンテック社製)、マウス肝臓由来総RNA(Mouse Liver Total RNA;クロンテック社製)および雄性ウィスター系ラット肝臓より抽出した総RNAを鋳型としてcDNAを増幅した。
【0067】
ヒト、マウスおよびラット肝臓において、2または3種類の異なる転写開始点を持つABCA1のmRNAを検出した。配列番号21〜23にヒトI型〜III型のABCA1 mRNAを、配列番号24,25にマウスI型、II型のものを、配列番号26〜28にラットI型、II型、II’型のものの配列をそれぞれ示す。図2は、検出したABCA1のmRNAの5’末端の模式図である。ここでI型と仮称したmRNAは、これまで5’−RACE法を用いてヒト繊維芽細胞やヒト単球由来THP−1細胞から同定されたABCA1のmRNA[NM_005502]に相同する配列(ラット[NW_178095]およびマウス[NM_013454])であり、既報のプロモーター領域(エクソン1上流、Proc.Natl.Acad.Sci.(2000)97, 7987)により転写制御を受けると考えられる。他の2種類はI型とは異なる新規なABCA1 mRNAであった。特に、II型と命名したmRNAは、ヒト、マウス、ラットに共通に発現しており、I型が持つエクソン1領域を欠損しており、エクソン2から転写が開始されるmRNAである。ラット肝臓のII’型mRNAはII型の5’側が68塩基短縮されたmRNAに相当し、転写制御はII型と同一であると予測される。
【0068】
[実施例3:ラット組織におけるABCA1のmRNA発現分布]
ラットの各組織(肝臓、脳、心臓、腎臓、肺、筋肉、脾臓および精巣)から得られたmRNA(ラットMTCパネルI;クロンテック社製)を用い、各組織におけるI型およびII型ABCA1 mRNAの分布を調べた。ラットI型ABCA1 mRNA量はエクソン1領域(I型特有の非翻訳領域)を認識するプライマー・プローブ、また総ABCA1 mRNA量はタンパク質をコードする領域の配列を基に作成したプライマー・プローブを用いて、リアルタイムRT−PCRにより定量した。以下に、I型ABCA1検出用プローブおよび総ABCA1検出用プローブの配列を示す。
【0069】
・ラットI型ABCA1検出用標識プローブ(配列番号29)
FAM-ccctactttt ttctcccggt ttctgg-TAMRA
・ラットI型ABCA1検出用フォワードプライマー(配列番号30)
tctgctccct gtccccac
・ラットI型ABCA1検出用リバースプライマー(配列番号31)
ccaaccccta cacaaaccct
・ラット総ABCA1検出用標識プローブ(配列番号32)
FAM-tctttgctca gattgtcctg ccggc-TAMRA
・ラット総ABCA1検出用フォワードプライマー(配列番号33)
cccggcggag tagaaagg
・ラット総ABCA1検出用リバースプライマー(配列番号34)
agggcgatgc aaacaaagac
【0070】
上記の方法で、各組織についてI型と総ABCA1 mRNAの発現量を測定すると、I型とII型の発現は組織間で異なり、II型は主に肝臓および腎で発現していることが判明した(図3)。そこでII型を肝臓型、これに対しI型を末梢型と呼ぶことにした。
【0071】
[実施例4:コンパクチンによる新規な肝臓型ABCA1遺伝子プロモーターの活性化]
肝臓型(II型)ABCA1 mRNAの発現調節領域の解析を行った。まず、ラット尾ゲノムDNAから末梢型(I型)および肝臓型(II型)ABCA1 mRNAの転写開始点上流領域約1000塩基をクローニングした。配列番号35に、ラット末梢型ABCA1転写開始点上流領域の配列を示す。配列番号6に、ラット肝臓型ABCA1転写開始点上流領域の配列を示す。得られた転写開始点上流領域を、ホタルルシフェラーゼベクターpGL3(プロメガ社製)に組み込み、レポータープラスミドを構築した。McARH7777細胞への一過性トランスフェクションにより、レポータープラスミドおよび遺伝子導入効率補正用のウミシイタケルシフェラーゼレポータープラスミドphRL−TK(プロメガ社製)を形質導入し、50μMプラバスタチンを含む10%脱脂血清培地で16時間処理した。その後、細胞を回収し、デュアルルシフェラーゼアッセイシステム(プロメガ社製)により細胞内ルシフェラーゼ活性を測定した。なお、転写活性は次のように定義して評価した。
(転写活性)=(ホタルルシフェラーゼ活性測定値)/(ウミシイタケルシフェラーゼ活性測定値)
【0072】
細胞をプラバスタチン処理することにより、末梢型転写開始点上流領域を組み込んだレポータープラスミドの転写活性は低下した(図4)。逆に、肝臓型転写開始点上流領域を組み込んだレポータープラスミドの転写活性は増大した。この結果は、スタチン系薬剤によるABCA1遺伝子発現の肝臓と末梢における応答性の違い(実施例1、図1)とよく一致した。従って、該肝臓型転写開始点上流領域が肝臓でのABCA1遺伝子発現を制御していることが示唆された。
【0073】
[実施例5:肝臓型ABCA1遺伝子プロモーター活性を調節するSRE配列]
スタチン系薬剤はステロール合成を阻害することにより、転写因子として機能する活性型(核内移行型)SREBPを二次的に増加させることが知られている。上記したように、活性型SREBP2はステロール応答性配列(SRE配列)を標的とする転写因子で、活性型SREBP2を過剰発現することでSRE配列下の転写が活性化されることが知られている。上記したように、SRE配列は、配列番号7〜12に記載のコンセンサス配列を有し、ラットの肝臓型ABCA1プロモーター領域に見出された(配列番号14)。
【0074】
そこで、肝臓型ABCA1プロモーター配列内に存在するSRE配列の変異解析を行った。実施例4で得られたラット肝臓型ABCA1プロモーターのSRE配列を、変異型SRE配列を含む合成オリゴヌクレオチドおよびクイックチェンジ(ストラタジェン社製)を用いて機能的に不活性化した。配列番号36に、変異型SRE配列の配列を示す。なお、この変異型SRE配列(GTGCGGCCGC)は、配列番号14に記載のSRE配列(GTCTGGTGAGA)にNotIの認識配列を導入したものであり、SREとして機能しないことが報告されている(J Biol Chem. 2003 Sep 19;278(38):36176-82.)。
【0075】
上述した方法(実施例4)に従い、野生型あるいは変異型SREを有する肝臓型ABCA1レポータープラスミドをMcARH7777細胞に形質導入した。実施例5にあっては、50μMプラスバスタチンの代わりに、50μMコンパクチンで処理し、転写活性を測定した。
【0076】
さらに、スタチン系薬剤で処理する代わりに、活性型SREBP2の過剰発現させた際の転写活性も測定した。活性型SREBP2の過剰発現実験においては、野生型あるいは変異型SREを有する肝臓型ABCA1レポータープラスミドに加えて、さらにpME−SREBP2(1−480)あるいはpME−18Sプラスミド(非特許文献13、J.Biol.Chem. (2003)36176)を共導入した。pME−SREBP2(1−480)プラスミドは、活性型SREBP2であるSREBP2のアミノ酸残基1〜480を発現することができるベクターであり、pME−18Sプラスミドは、その空ベクターである。
【0077】
コンパクチンによるラット肝臓型プロモーター転写活性促進作用は、配列中のSREの変異により消失した(図5)。活性型SREBP2を過剰発現した場合にも転写が活性化されるが、これもSRE変異により消失した(図5)。したがって、プロモーター内に存在するSRE配列が肝臓のABCA1遺伝子の転写調節に機能していることが判明した。
【0078】
[実施例6:ヒト肝臓型ABCA1遺伝子プロモーターのSREBP2による活性化]
ヒトゲノムDNA(Human Genomic DNA;クロンテック社製)から、ヒト肝臓型ABCA1プロモーターをクローニングした。配列番号4に、ヒト肝臓型ABCA1転写開始点上流領域の配列を示す。得られた転写開始点上流領域を、ホタルルシフェラーゼレポーターベクターpGL3(プロメガ社製)に組み込んだ。ヒト肝臓型ABCA1レポータープラスミドおよびウミシイタケルシフェラーゼレポータープラスミドphRL−SV40(プロメガ社製)を、pME−18SまたはpME−SREBP2プラスミドと共に、HepG2細胞へ形質導入し、10%FCSを含むDMEM培地で16時間培養した。その後、細胞を回収し細胞内ルシフェラーゼ活性を測定した。
【0079】
ヒト肝臓型ABCA1プロモーターの活性は、活性型SREBP2の過剰発現により増大した(図6)。したがって、ヒト肝臓のABCA1遺伝子もラット肝臓と同様の転写制御を受けていると考えられる。
【図面の簡単な説明】
【0080】
【図1】図1Aは、コンパクチンで処理したヒト肝臓由来培養細胞HepG2におけるABCA1遺伝子の発現量変化を示すグラフである。図1Bは、プラバスタチンを投与したラットの肝臓におけるABCA1遺伝子の発現量変化を示すグラフである。図1Cは、プラバスタチンで処理したラット繊維芽細胞Rat2におけるABCA1遺伝子の発現量変化を示すグラフである。図1Dは、コンパクチンで処理したマウス単球由来細胞RAW264におけるABCA1遺伝子の発現量変化を示すグラフである。
【図2】図2は、5’−RACE法により解析したラット、マウスおよびヒト肝臓で発現するABCA1のmRNAの5’末端の模式図である。
【図3】図3は、ラット各組織(肝臓、脳、心臓、腎臓、肺、筋肉、脾臓および精巣)で発現する総ABCA1 mRNA量および末梢型(I型)ABCA1 mRNA量を示したグラフである。
【図4】図4Aは、プラバスタチンによるラット末梢型(I型)ABCA1遺伝子プロモーターの活性化を示したグラフである。図4Bは、プラバスタチンによるラット肝臓型(II型)ABCA1遺伝子プロモーターの活性化を示したグラフである。
【図5】図5は、ラット肝臓型(II型)ABCA1遺伝子プロモーターの活性がSRE配列の変異により消失したことを示すグラフである。
【図6】図6は、ヒト肝臓型(II型)ABCA1遺伝子プロモーターの活性がSREBP2の発現増加により活性化されることを示すグラフである。
【特許請求の範囲】
【請求項1】
ABCA1遺伝子のイントロン1中のSRE配列を含み、プロモーター活性を有するDNAを用いることを特徴とする、ABCA1遺伝子の転写を調整する作用を有する物質のスクリーニング方法。
【請求項2】
前記ABCA1遺伝子の転写を調整する作用を有する物質が、動脈硬化性疾患の予防または治療に有用な物質である、請求項1に記載の方法。
【請求項3】
前記プロモーター活性を有するDNAが、
(a)配列番号4〜6のいずれかに記載の配列を有するDNAと、
(b)配列番号4〜6のいずれかに記載の配列において、1もしくは数個のヌクレオチドが置換、挿入もしくは欠失された配列を有し、プロモーター活性を有するDNAと、
(c)配列番号4〜6のいずれかに記載の配列と相同性が60%以上の配列を有し、プロモーター活性を有するDNAと、
(d)(a)のDNAとストリンジェントな条件でハイブリダイズし、プロモーター活性を有するDNAと
からなる群から選ばれるDNAの少なくとも一部である、請求項1または2に記載の方法。
【請求項4】
前記SRE配列が、配列番号7〜12のいずれかに記載の配列である請求項1〜3のいずれかに記載の方法。
【請求項5】
被検物質を供するステップと、
前記プロモーター活性を有するDNAに作動可能に連結した構造遺伝子を有するDNAを有する細胞または生物の複数の群を供するステップと、
前記被検物質を前記細胞または生物の少なくとも1つの群に接触させるステップと、
前記被検物質を接触させた群と接触させなかった群とにおいて、前記遺伝子の転写量および/または発現量を測定するステップと、
前記被検物質を接触させた群と接触させなかった群の前記転写量および/または発現量を比較するステップと
を含む請求項1〜4のいずれかに記載の方法。
【請求項6】
活性型SREBPの量を変化させる作用を有する物質(ピタバスタチン、シムバスタチンおよびアトルバスタチンを除く)を含有する、ABCA1遺伝子の転写調整剤。
【請求項7】
前記活性型SREBPの量を変化させる作用を有する物質が、スタチン系薬剤を除く物質である、請求項6に記載のABCA1遺伝子の転写調整剤。
【請求項8】
ABCA1遺伝子のイントロン1中のSRE配列を含み、プロモーター活性を有する組換えDNA。
【請求項9】
ABCA1遺伝子のイントロン1中のSRE配列を含み、プロモーター活性を有するDNAを有する組換えベクター。
【請求項10】
前記プロモーター活性を有するDNAに作動可能に連結した構造遺伝子をさらに有する請求項9に記載の組換えベクター。
【請求項11】
ABCA1遺伝子のイントロン1中のSRE配列を含み、プロモーター活性を有する外因性のDNAに作動可能に連結した構造遺伝子を有する形質転換体。
【請求項12】
請求項10に記載の組換えベクターおよび/または請求項11に記載の形質転換体を有する、ABCA1遺伝子の転写を調整する作用を有する物質をスクリーニングするためのキット。
【請求項13】
前記ABCA1遺伝子の転写を調整する作用を有する物質が、動脈硬化性疾患の予防または治療に有用な物質である、請求項12に記載のキット。
【請求項1】
ABCA1遺伝子のイントロン1中のSRE配列を含み、プロモーター活性を有するDNAを用いることを特徴とする、ABCA1遺伝子の転写を調整する作用を有する物質のスクリーニング方法。
【請求項2】
前記ABCA1遺伝子の転写を調整する作用を有する物質が、動脈硬化性疾患の予防または治療に有用な物質である、請求項1に記載の方法。
【請求項3】
前記プロモーター活性を有するDNAが、
(a)配列番号4〜6のいずれかに記載の配列を有するDNAと、
(b)配列番号4〜6のいずれかに記載の配列において、1もしくは数個のヌクレオチドが置換、挿入もしくは欠失された配列を有し、プロモーター活性を有するDNAと、
(c)配列番号4〜6のいずれかに記載の配列と相同性が60%以上の配列を有し、プロモーター活性を有するDNAと、
(d)(a)のDNAとストリンジェントな条件でハイブリダイズし、プロモーター活性を有するDNAと
からなる群から選ばれるDNAの少なくとも一部である、請求項1または2に記載の方法。
【請求項4】
前記SRE配列が、配列番号7〜12のいずれかに記載の配列である請求項1〜3のいずれかに記載の方法。
【請求項5】
被検物質を供するステップと、
前記プロモーター活性を有するDNAに作動可能に連結した構造遺伝子を有するDNAを有する細胞または生物の複数の群を供するステップと、
前記被検物質を前記細胞または生物の少なくとも1つの群に接触させるステップと、
前記被検物質を接触させた群と接触させなかった群とにおいて、前記遺伝子の転写量および/または発現量を測定するステップと、
前記被検物質を接触させた群と接触させなかった群の前記転写量および/または発現量を比較するステップと
を含む請求項1〜4のいずれかに記載の方法。
【請求項6】
活性型SREBPの量を変化させる作用を有する物質(ピタバスタチン、シムバスタチンおよびアトルバスタチンを除く)を含有する、ABCA1遺伝子の転写調整剤。
【請求項7】
前記活性型SREBPの量を変化させる作用を有する物質が、スタチン系薬剤を除く物質である、請求項6に記載のABCA1遺伝子の転写調整剤。
【請求項8】
ABCA1遺伝子のイントロン1中のSRE配列を含み、プロモーター活性を有する組換えDNA。
【請求項9】
ABCA1遺伝子のイントロン1中のSRE配列を含み、プロモーター活性を有するDNAを有する組換えベクター。
【請求項10】
前記プロモーター活性を有するDNAに作動可能に連結した構造遺伝子をさらに有する請求項9に記載の組換えベクター。
【請求項11】
ABCA1遺伝子のイントロン1中のSRE配列を含み、プロモーター活性を有する外因性のDNAに作動可能に連結した構造遺伝子を有する形質転換体。
【請求項12】
請求項10に記載の組換えベクターおよび/または請求項11に記載の形質転換体を有する、ABCA1遺伝子の転写を調整する作用を有する物質をスクリーニングするためのキット。
【請求項13】
前記ABCA1遺伝子の転写を調整する作用を有する物質が、動脈硬化性疾患の予防または治療に有用な物質である、請求項12に記載のキット。
【図1】


【図2】
【図3】
【図4】
【図5】

【図6】
【図2】
【図3】
【図4】
【図5】
【図6】
【公開番号】特開2007−82445(P2007−82445A)
【公開日】平成19年4月5日(2007.4.5)
【国際特許分類】
【出願番号】特願2005−273693(P2005−273693)
【出願日】平成17年9月21日(2005.9.21)
【出願人】(803000056)財団法人ヒューマンサイエンス振興財団 (341)
【Fターム(参考)】
【公開日】平成19年4月5日(2007.4.5)
【国際特許分類】
【出願日】平成17年9月21日(2005.9.21)
【出願人】(803000056)財団法人ヒューマンサイエンス振興財団 (341)
【Fターム(参考)】
[ Back to top ]