
AMV逆転写酵素遺伝子、該遺伝子によりコードされるAMV逆転写酵素及び該遺伝子を用いるAMV逆転写酵素の製造方法
【課題】工業的規模で発現することが可能な、新規なトリ骨髄芽球腫ウイルス(AMV)逆転写酵素遺伝子と、それを用いた該酵素の製造方法を提供する。
【解決手段】AMV逆転写酵素ベータ体をコードする遺伝子であって、特定の塩基配列からなるDNA、および該DNAのコードするAMV逆転写酵素ベータ。
【解決手段】AMV逆転写酵素ベータ体をコードする遺伝子であって、特定の塩基配列からなるDNA、および該DNAのコードするAMV逆転写酵素ベータ。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、新規なAMV逆転写酵素遺伝子及びそれを用いたAMV逆転写酵素の製造方法に関する。
【背景技術】
【0002】
逆転写酵素は、RNA依存性のDNAポリメラーゼであり、レトロウイルスの増殖に必須の因子として発見された。この酵素は一本鎖RNAを鋳型としてDNAを合成(逆転写)する活性を有しており、cDNA合成等に必須な遺伝子工学試薬あるいは遺伝子診断用酵素として利用されている。
【0003】
トリ骨髄芽球腫ウイルス(AMV)に由来する逆転写酵素は、約63kDaの分子量を有するアルファ鎖及び約95kDaの分子量を有するベータ鎖からなるヘテロダイマーであり、ASLVファミリーとして知られている。このアルファ鎖は、ベータ鎖のタンパク質分解性プロセシングにより形成される(非特許文献1)。この逆転写酵素は、所定のRTバリアントが大腸菌又は真核生物昆虫細胞において発現されるとの報告されており(特許文献1)、またシャペロンと共発現することで大腸菌において発現されると報告されているが(特許文献2)、工業的な利用の観点からはより大量の発現が必要とされている。
【0004】
【特許文献1】特表2002/534125号公報
【特許文献2】特開2002−315584号公報
【非特許文献1】Le Grice S.F.J.,1993 reverse Transcriptase, Cold Spring Harbor,New York:Cold Spring Harbor Laboratory Press,163
【発明の概要】
【発明が解決しようとする課題】
【0005】
そこで本発明の目的は、工業的規模で発現(工業的規模でAMV酵素を製造)することすることが可能な新規なAMV逆転写酵素遺伝子と、それを用いたAMV逆転写酵素の製造方法を提供することにある。
【課題を解決するための手段】
【0006】
本発明者らは、大腸菌を用いた工業的規模での発現(製造)を可能ならしめるべくAMV逆転写酵素の遺伝子配列を見直し、大腸菌が容易に翻訳するコドンに対応するようにDNA配列を変換することにより前記目的を達成し得ることを見出し、本発明を完成するに至った。すなわち本発明は、AMV逆転写酵素ベータ体をコードする遺伝子であって、配列番号3記載の塩基配列からなるDNA、それと実質的に相同なDNA又はこれらDNAとストリンジェントな条件下でハイブリダイズするDNAであり、AMV逆転写酵素アルファ体をコードする遺伝子であって、配列番号4記載の塩基配列からなるDNA、それと実質的に相同なDNA又はこれらDNAとストリンジェントな条件下でハイブリダイズするDNAであり、AMV逆転写酵素ベータ体のN末端アミノ酸をメチオニンとしたタンパク質をコードする遺伝子であって、配列番号5記載の塩基配列からなるDNA、それと実質的に相同なDNA又はこれらDNAとストリンジェントな条件下でハイブリダイズするDNAであり、AMV逆転写酵素アルファ体のN末端アミノ酸をメチオニンとしたタンパク質をコードする遺伝子であって、配列番号6記載の塩基配列からなるDNA、それと実質的に相同なDNA又はこれらDNAとストリンジェントな条件下でハイブリダイズするDNAである。
【0007】
また本発明は、配列番号3記載の塩基配列からなるDNA又はそれと実質的に相同なDNAによりコードされるAMV逆転写酵素ベータ体又はそれと実質的に相同なタンパク質であり、配列番号4記載の塩基配列からなるDNA又はそれと実質的に相同なDNAによりコードされるAMV逆転写酵素アルファ体又はそれと実質的に相同なタンパク質であり、配列番号5記載の塩基配列からなるDNA又はそれと実質的に相同なDNAによりコードされるAMV逆転写酵素ベータ体のN末端アミノ酸をメチオニンとしたタンパク質又はそれと実質的に相同なタンパク質であり、配列番号6記載の塩基配列からなるDNA又はそれと実質的に相同なDNAによりコードされるAMV逆転写酵素アルファ体のN末端アミノ酸をメチオニンとしたタンパク質又はそれと実質的に相同なタンパク質である。以下、本発明を詳細に説明する。
【0008】
本発明の「AMV逆転写酵素遺伝子」とは、AMV逆転写酵素をコードする遺伝子であり、その発現、即ちmRNAへの転写と転写されたmRNAの翻訳によってAMV逆転写酵素を製造し得るものである。なお天然のAMV逆転写酵素遺伝子は、全長が2.6kベースで、配列番号1記載の塩基配列を有するDNAである。この配列番号1記載の塩基配列を有するものはAMV逆転写酵素ベータ体をコードする遺伝子であるが、AMV逆転写酵素にはベータ体よりも低分子のアルファ体が存在し、アルファ体遺伝子は、前記配列番号1記載の塩基配列を有する遺伝子の5’末端側約1.7kベースからなるDNA(配列番号2)である。なお、これら遺伝子がコードするタンパク質の機能構造は、従来公的のデータベース(National Center for Biotechnology Information)に収録されている。
【0009】
本発明のAMV逆転写酵素遺伝子のうち、配列番号3記載の塩基配列からなるDNAは、AMV逆転写酵素ベータ体をコードする遺伝子であって、その塩基配列(アミノ酸残基をコードするコドン)が天然(ウイルス)型コドンから大腸菌型コドンに変換されたDNAである。配列番号4記載の塩基配列からなるDNAは、AMV逆転写酵素アルファ体をコードする遺伝子であって、その塩基配列(アミノ酸残基をコードするコドン)が天然(ウイルス)型コドンから大腸菌型コドンに変換されたDNAである。配列番号5記載の塩基配列からなるDNAは、AMV逆転写酵素ベータ体のN末端アミノ酸をメチオニンとしたタンパク質をコードする遺伝子であって、その塩基配列(アミノ酸残基をコードするコドン)が天然(ウイルス)型コドンから大腸菌型コドンに変換されたDNAである。そして配列番号6記載の塩基配列からなるDNAは、AMV逆転写酵素アルファ体のN末端アミノ酸をメチオニンとしたタンパク質をコードする遺伝子であって、その塩基配列(アミノ酸残基をコードするコドン)が天然(ウイルス)型コドンから大腸菌型コドンに変換されたDNAである。
【0010】
ここで、「天然型コドンから大腸菌型コドンに変換された」とは、いずれの遺伝子においても、その転写により、AMVが天然に有するAMV逆転写酵素遺伝子から転写されるmRNAにおいて見いだされる大腸菌レアコドン(rare codon)、すなわち大腸菌での使用頻度が低いコドンが、当該コドンがコードするアミノ酸を変更することなく大腸菌での使用頻度が高いコドンに置き換えられたmRNAを生成することを意味する。コドンの大腸菌における使用頻度の高低は、例えば公的データベース(http://www.kazusa.or.jp/codon/)を参考にゲノム遺伝子の解析を行うことによって推測することができるが、具体的に、大腸菌における使用頻度の高いコドンとして、アルギニン(Arg)コドンのAGA、AGG、CGG及びCGA、イソロイシン(Ile)コドンのAUA、ロイシン(Leu)コドンのCUA、グリシン(Gly)コドンのGGA、プロリン(Pro)コドンのCCCを例示することができる。このように大腸菌で使用頻度の高いコドンに置き換えられたmRNAを転写するよう塩基配列中のコドンを変更することで、本発明の遺伝子は大腸菌を用いることにより、工業的規模で発現することが可能である。
【0011】
コドンの変換は、当該コドンに対応する(当該コドンに転写される)DNAの塩基配列の部分(コドン)を変えることにより可能であるが、DNAの塩基配列の変換は、例えばSite−directed mutagenesis法等の従来公知の変異導入法を利用して行うことができる。中でも、合成オリゴヌクレオチドとPCRを組み合わせたDNAWorks法(Nucleic acid Res.,30,43,2002参照)がコドン変換のための特に好ましい方法として例示することができる。この方法は、アミノ酸配列から数十塩基からなるオリゴヌクレオチド群を合成し、PCRにより合成オリゴヌクレオチドをアッセンブリーさせることによって完全長の遺伝子を合成する方法である。
【0012】
本発明のAMV逆転写酵素遺伝子は、前記した4種類のDNAのほか、その一部が、その発現によってAMV逆転写酵素と実質的に同一な蛋白質であって、AMV逆転写酵素を形成するアミノ酸が欠失し、AMV逆転写酵素を形成するアミノ酸以外のアミノ酸が付加され、又は、AMV逆転写酵素を形成する所定のアミノ酸が他のアミノ酸へ置換されたAMV逆転写酵素様蛋白質をコードするDNA、また更には前記4種類のDNA又はAMV逆転写酵素様蛋白質をコードするDNAにストリンジェントな条件下でハイブリダイズするDNA(以下、これらDNAをまとめて前記4種類のDNAに「実質的に相同なDNA」と記載することがある)を含む。なお、「AMV逆転写酵素と実質的に同一な蛋白質」とは、具体的には、逆転写酵素活性、DNAポリメラーゼ活性又はRNaseH活性というAMV逆転写酵素が有する3つの主酵素活性のうち1つ以上がAMVと同等以上である蛋白質をいう。例えば、前記した各遺伝子との間で、公知の配列比較アルゴリズムを用いて解析した場合に、又は、手法自体が公知である手動整列及び目視試験により測定し、最大対応で整列化させた場合に、それぞれの塩基配列の75%以上、好ましくは85%、より好ましくは95%以上に同一性が見出される遺伝子が、その様な蛋白質をコードする遺伝子として例示できる。前記公知の解析方法として、例えば、解析局所相同性アルゴリズム(Smith等,Adv.Appl.Math.,2, 482,1981)、相同性整列化アルゴリズム(Needlman等,J.Mol. Biol.,48,443,1970)、類似性検索法(Person等,Proc. Natl.Acd.Sci.USA,85,2444,1988)等を例示できる。中でも、公知のPileup法やBLASTアルゴリズム(Altschul等,J.MOL.BIOL.,215,403,1990)が、本発明の実質的に相同なDNAを規定する上で好ましい方法として例示できる。そして「ストリンジェントな条件」とは、通常の状態と比較してDNA同士が二重鎖を形成し難い条件をいい、例えば、42℃で50%(v/v)ホルムアミド、0.1%ウシ血清アルブミン、0.1%フィコール、0.1%のポリビニルピロリドン、50mMリン酸ナトリウムバッファー(pH6.5)、150mMの塩化ナトリウム及び75mMクエン酸ナトリウムが共存する条件等が挙げられる。
【0013】
本発明のAMV逆転写酵素は、前記したAMV逆転写酵素遺伝子を発現させることにより得られる蛋白質(AMV逆転写酵素様蛋白質を含む)である。AMV逆転写酵素遺伝子の発現のためには従来公知の方法を使用することが可能であるが、かかる方法として例えば、本発明の遺伝子である前記DNAを適当な細胞に形質転換し、又は、プラスミドベクターに挿入して使用すれば良い。プラスミドベクターへの挿入は、そのプラスミドベクターの適当な位置に遺伝子工学的に挿入すればよい。ここでいう適当な位置とは、プラスミドベクターの複製機能、所望の抗生物質マーカー、あるいは伝達性に関わる領域を破壊しない範囲で自由に決定することが出来る。またプラスミドベクターに挿入する場合、例えばプロモータ機能を有するDNA配列を付加しても良い。大腸菌で機能するプロモータとして、例えばlacプロモータ、trcプロモータ又はT7プロモータ等を例示することができる。これらの中でも転写活性の調節が容易なlacプロモータは、好適なプロモータである。
【0014】
プラスミドベクターとしては形質転換する細胞内で安定に存在し、複製することができるものであれば特に制限はない。もっとも、本発明の遺伝子は、その塩基配列が大腸菌内で使用頻度の高いコドンに変換されているため、大腸菌内で安定に存在し複製できるプラスミドベクターとして知られているpUC系、pBR系、pET系、広宿主域(Broad−Host−Range)プラスミドベクター等の使用が好ましい。
【0015】
宿主としては前記の通り、K12系のE.coli JM109株に代表される大腸菌が好ましいが、変異処理した大腸菌変異株を使用することもできる。変異処理はニトロソグアニジン、メタンスルホン酸エチル、紫外線又は放射線等の、従来公知の変異手段を用いる方法を使用できる。
【0016】
以上に説明したプラスミドベクター等を公知の手法に従って大腸菌等に導入し、当該大腸菌を公知の手法に従って培養すればAMV逆転写酵素を製造することができる。プラスミドベクター等を大腸菌等に導入する方法としては、例えば、”Vectors for cloning genes”,Method in Enzymology,216,p .469−631,1992,Academic Press,や“Other Bacterial systems“,Method in Enzymology,204,p.305−636,1991,Academic Press)を参照することができる。
【0017】
大腸菌の培養方法としては、例えば必要な栄養源を補ったLB(Luria Broth)培地により培養する方法を例示できる。より具体的には、例えば、プラスミドベクターを含有した大腸菌のみを選択して培養するために、培地としてプラスミドベクターの構成を基にした選抜剤を含むものを使用することが例示できる。例えば、アンピシリン耐性遺伝子を有するプラスミドベクターを使用する場合には、培地にアンピシリンを添加することを例示できる。また培地には、炭素、窒素及び無機塩供給源の他に、適当な栄養源を添加しても良い。更にグルタチオン、システイン、シスタミン、チオグリコレート、ジチオスレイトールからなる群から選択される一種類以上の還元剤を添加しても良い。培養温度は、例えば約20℃から40℃、好ましくは25℃から35℃、より好ましくは約30℃とすることが例示できる。pHは約6.8から7.4、好ましくは7.0とすることが例示できる。
【0018】
前記したlacプロモータ等の誘導性のプロモータを用い、誘導によってAMV逆転写酵素遺伝子を発現させる場合には、良好なAMV逆転写酵素の生産性が得られる程度にまで培養を行い、しかる後に誘導の操作を行うことが好ましい。その目安としては、例えば、培養液の濁度(660nmにおける吸光度)を測定し、約0.5から1.0を示す増殖期間に誘導の操作を行い、引き続き培養するが例示できる。lacプロモータの誘導剤としてはIPTGを例示することができるが、使用するプロモータに応じて適宜選択して使用すれば良い。lacプロモータを使用する場合のIPTGの添加濃度は終濃度で約0.1から1.0mMであれば良いが、好ましくは0.15mMである。
【0019】
培養液からAMV逆転写酵素を抽出する際には、発現の形態によって適宜抽出方法を選択すれば良い。例えば大腸菌のようにAMV逆転写酵素が宿主内に蓄積する場合は遠心分離操作等により菌体を集めた後、酵素処理剤や超音波破砕等により菌体を破砕して抽出することができる。この段階では種々の蛋白質が混在しているが、イオン交換クロマトグラフィー、疎水性相互作用クロマトグラフィー、ゲルろ過クロマトグラフィー又はアフィニティークロマトグラフィー等を単独又は組み合わせて適用することにより、AMV逆転写酵素のみを抽出することができる。
【発明の効果】
【0020】
AMV逆転写酵素遺伝子の天然(ウイルス)型コドンを大腸菌型コドンに変換したことにより、大腸菌を宿主として使用することによって工業的規模で発現(工業的規模でAMV逆転写酵素を製造)することが可能となる。
【図面の簡単な説明】
【0021】
【図1】図1から図6は、AMV逆転写酵素ベータ体の天然(ウィルス)型コドン及び大腸菌コドンの遺伝子配列を比較した図である。図中AMVbetaEcoliが大腸菌型コドンに変換したDNA配列の配列、AMVbetaが天然(ウィルス)型AMVベータ体の配列である。AMVアルファ体はその塩基番号1番から1716番目までである。
【図2】図1から図6は、AMV逆転写酵素ベータ体の天然(ウィルス)型コドン及び大腸菌コドンの遺伝子配列を比較した図である。
【図3】図1から図6は、AMV逆転写酵素ベータ体の天然(ウィルス)型コドン及び大腸菌コドンの遺伝子配列を比較した図である。
【図4】図1から図6は、AMV逆転写酵素ベータ体の天然(ウィルス)型コドン及び大腸菌コドンの遺伝子配列を比較した図である。
【図5】図1から図6は、AMV逆転写酵素ベータ体の天然(ウィルス)型コドン及び大腸菌コドンの遺伝子配列を比較した図である。
【図6】図1から図6は、AMV逆転写酵素ベータ体の天然(ウィルス)型コドン及び大腸菌コドンの遺伝子配列を比較した図である。
【図7】図7は、プラスミドベクターpTrcAMVBの構造を示す図である。
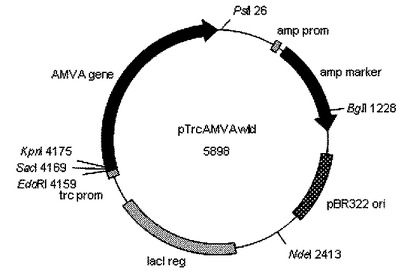
【図8】図8は、プラスミドベクターpTrcAMVAの構造を示す図である。
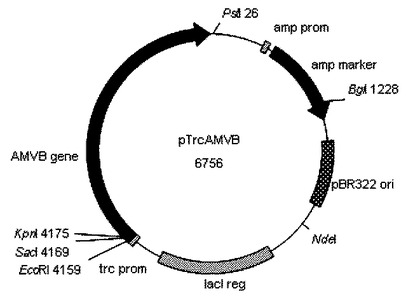
【図9】図9は、プラスミドベクターpTrcAMVBwildの構造を示す図である。
【図10】図10は、プラスミドベクターpTrcAMVAwildの構造を示す図である。
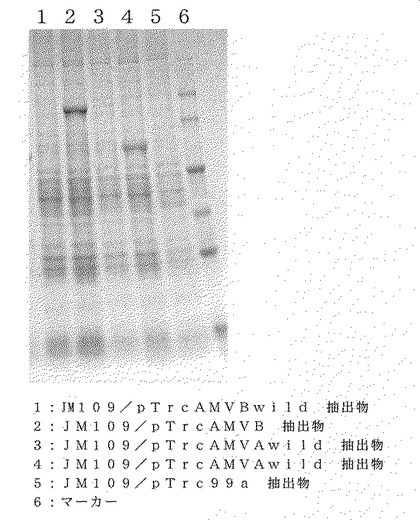
【図11】図11は、発現量の差を示す電気泳動像の結果を示す図である。
【発明を実施するための形態】
【0022】
以下、本発明の実施の形態について、実施例により詳細に説明する。
【実施例】
【0023】
以下の実施例は本発明の実施の一形態を説明するためのものであり、本発明を限定するものではない。
【0024】
実施例1 AMV逆転写酵素をコードするDNA配列の設計
配列番号1に記載したAMV逆転写酵素のアミノ酸配列に基づき、DNAWorks法により、コドンを大腸菌型に変換した。コドンを変換したDNA配列と天然(ウィルス)型コドンをアライメントした結果を図1から図6に示す。図1から図6の通り、アミノ酸配列はそのままにDNAの配列変換が行われ、そのDNA配列間の類似性は74%であった。
【0025】
実施例2 AMV逆転写酵素をコードするDNA配列の作製
天然(ウィルス)型コドンから大腸菌型コドンに変換したDNA配列を作製するために120種類のオリゴヌクレオチドを合成した。合成したオリゴヌクレオチドを配列番号7から126に示す。
【0026】
次いで、オリゴヌクレオチドから完全長のAMV逆転写酵素をコードするDNAを作製するために、二段階のPCR反応を行った。一段階目のPCR反応の反応液は表1の通りであり、94℃、5分の熱処理後、94℃、30秒間の第一ステップ、62℃、30秒間の第二ステップ、72℃、1分間の第三ステップを25サイクル行い、次いで、72℃、7分の第四ステップを行った。表1中のDNAミックスは、120種類の合成した50pmol/μLのオリゴヌクレオチドをそれぞれ一定量サンプリングし混合した溶液である。
【0027】
【表1】

第二段階目のPCR反応は、第一段階目のPCR反応の反応液を用いて、表2の反応液組成で行った。プライマーの配列は、配列番号7(5’―ATGACGGTGGCGTTACACCTG―3’)と配列番号126(5’―TTAGGCAAACAGCGGAGAGGCTTCGTCCTTCTTCGTCACTTCGTCT―3’)のオリゴヌクレオチドを用いた。94℃、5分の熱処理後、94℃、30秒間の第一ステップ、65℃、30秒間の第二ステップ、72℃、1分間の第三ステップを25サイクル行い、最後に、72℃、7分の第四ステップを行った。反応終了後、0.9%のアガロース電気泳動で確認したところ、設計通りのサイズのDNAバンドを確認することができた。
【0028】
【表2】
次いで、目的バンドから市販の試薬(Qiaquick Gelextraction kit:キアゲン社)を用いてDNAを抽出後、抽出したDNAの5’末端を市販の試薬(TAKARA BKL Kit:タカラバイオ社)を用いてリン酸化し、制限酵素SmaIで消化したpTrc99aプラスミドベクターに挿入しE.coli JM109株(タカラバイオ社)を形質転換した。調製したプラスミドベクターは制限解析及び配列決定により調査し、これをpTrcAMVBとした。
【0029】
さらに、pTrcAMVBを鋳型にAMV逆転写酵素アルファ体をコードする遺伝子をPCR法を用いて作製した。使用したPCRプライマーの配列は、配列番号7(5’―ATGACGGTGGCGTTACACCTG―3’)と配列番号127(5’―TTAATACGCTTGAAACGTGGCCTGGCTATCCGCC―3’)である。94℃、5分の熱処理後、94℃、30秒間の第一ステップ、65℃、30秒間の第二ステップ、72℃、1分間の第三ステップを25サイクル行い、最後に、72℃、7分の第四ステップを行った。反応終了後、0.9%のアガロース電気泳動で確認したところ、設計通りのサイズのDNAバンドを確認することができた。
【0030】
次いで、目的バンドから市販の試薬(Qiaquick Gelextraction kit:キアゲン社)を用いてDNAを抽出後、抽出したDNAの5’末端を市販の試薬(TAKARA BKL Kit:タカラバイオ社)を用いてリン酸化し、制限酵素SmaIで消化したpTrc99aプラスミドベクターに挿入しE.coli JM109株(タカラバイオ社)を形質転換した。調製したプラスミドベクターは制限解析及び配列決定により調査し、これをpTrcAMVAとした。図7にpTrcAMVB及び図8にpTrcAMVAの構造を示す。
【0031】
PCR法を用いて、プラスミド上にAMV逆転写酵素ベータ体をコードする遺伝子を保有する大腸菌クローン(ATCC 31990)からAMV逆転写酵素ベータ体をコードする遺伝子を取り出した。使用したPCRプライマーの配列は、配列番号128(5’― ATGACTGTTGCGCTACATCTGGCTATTCCGCTC―3’)と配列番号129(5’―TTATGCAAAAAGAGGGCTCGCCTCATC―3’)である。94℃、5分の熱処理後、94℃、30秒間の第一ステップ、65℃、30秒間の第二ステップ、72℃、1分間の第三ステップを25サイクル行い、最後に、72℃、7分の第四ステップを行った。反応終了後、0.9%のアガロース電気泳動で確認したところ、設計通りのサイズのDNAバンドを確認することができた。次いで、目的バンドから市販の試薬(Qiaquick Gelextraction kit:キアゲン社)を用いてDNAを抽出後、抽出したDNAの5’末端を市販の試薬(TAKARA BKL Kit:タカラバイオ社)を用いてリン酸化し、制限酵素SmaIで消化したpTrc99aプラスミドベクターに挿入し大腸菌E.coli JM109株(タカラバイオ社)を形質転換した。プラスミドベクターを複製させた後、複製したプラスミドベクターについて制限解析及び配列決定により調査し、これをpTrcAMVBwildとした。
【0032】
PCR法を用いて、プラスミド上にAMV逆転写酵素ベータ体をコードする遺伝子を保有する大腸菌クローン(ATCC 31990)からAMV逆転写酵素アルファ体をコードする遺伝子を取り出した。使用したPCRプライマーの配列は、配列番号128(5’―ATGACTGTTGCGCTACATCTGGCTATTCCGCTC―3’)と配列番号130(5’―TTAATACGCTTGAAAGGTGGCTTGGCTATCTGCC―3’)である。94℃、5分の熱処理後、94℃、30秒間の第一ステップ、65℃、30秒間の第二ステップ、72℃、1分間の第三ステップを25サイクル行い、最後に、72℃、7分の第四ステップを行った。反応終了後、0.9%のアガロース電気泳動で確認したところ、設計通りのサイズのDNAバンドを確認することができた。
【0033】
次いで、目的バンドから市販の試薬(Qiaquick Gelextraction kit:キアゲン社)を用いてDNAを抽出後、抽出したDNAの5’末端を市販の試薬(TAKARA BKL Kit:タカラバイオ社)を用いてリン酸化し、制限酵素SmaIで消化したpTrc99aプラスミドベクターに挿入しE.coli JM109株(タカラバイオ社)を形質転換した。形質転換した大腸菌を培養し、プラスミドベクターを複製させた後、複製したプラスミドベクターについて制限解析及び配列決定により調査し、これをpTrcAMVAwildとした。図9にpTrcAMVBwild及び図10にpTrcAMVAwildの構造を示す。
実施例3 配列の確認
実施例2において作製したプラスミドベクターpTrcAMVB及びpTrcAMVAに挿入したDNAの配列を、チェーンターミネータ法に基づく市販の試薬キット(Big Dye Terminator Cycle Sequencing FS read Reaction kit、PEアプライドバイオシステム社)を用いてサイクルシークエンス反応に供し、全自動DNAシークエンサーABI Prism 3700 DNA Analyzer(PEアプライドバイオシステム社)にて解析した。なお、シークエンス用プライマーには合成した120種類のオリゴヌクレオチドを適宜使用した。その結果、pTrcAMVB及びpTrcAMVAに挿入したDNAの塩基配列は設計通りであることを確認した。
【0034】
実施例4 AMV逆転写酵素の発現誘導
プラスミドベクターpTrcAMVBを用いて、50μg/mLの抗生物質アンピシリンを添加したLB寒天培地中でE.coli JM109(タカラバイオ社)を形質転換した。37℃で18時間培養後、出現した任意のコロニーをピックアップし50μg/mLの抗生物質アンピシリンを添加したLB液体培地に接種した。37℃で培養し、OD660nmの値が約0.5になったところで0.15mMのIPTG(タカラバイオ社製)を添加した。プラスミドベクターpTrcAMVBで形質転換した株をJM109/pTrcAMVBとした。プラスミドベクターpTrcAMVAについても同様に操作し、形質転換株をJM109/pTrcAMVAとした。
【0035】
同様の方法でプラスミドベクターpTrcAMVBwild及びプラスミドベクターpTrcAMVAwildも形質転換を行い、それぞれJM109/pTrcAMVBwild、JM109/pTrcAMVAwildとした。
【0036】
IPTG添加から適当な間隔で培養液をサンプリングし、それぞれの形質転換株から蛋白質溶液を調製した。調製は市販のキット(Bugbuster protein extraction Kit、ノバジェン社)により行い、調製した蛋白質溶液をSDS−PAGEに供した。電気泳動像の結果を図11に示す。図11の通り、発現するタンパク質のアミノ酸配列は同一であるものの、宿主である大腸菌に合わせたコドンでは天然(ウィルス)型コドンに比べて有意に発現量を増加していた。
【技術分野】
【0001】
本発明は、新規なAMV逆転写酵素遺伝子及びそれを用いたAMV逆転写酵素の製造方法に関する。
【背景技術】
【0002】
逆転写酵素は、RNA依存性のDNAポリメラーゼであり、レトロウイルスの増殖に必須の因子として発見された。この酵素は一本鎖RNAを鋳型としてDNAを合成(逆転写)する活性を有しており、cDNA合成等に必須な遺伝子工学試薬あるいは遺伝子診断用酵素として利用されている。
【0003】
トリ骨髄芽球腫ウイルス(AMV)に由来する逆転写酵素は、約63kDaの分子量を有するアルファ鎖及び約95kDaの分子量を有するベータ鎖からなるヘテロダイマーであり、ASLVファミリーとして知られている。このアルファ鎖は、ベータ鎖のタンパク質分解性プロセシングにより形成される(非特許文献1)。この逆転写酵素は、所定のRTバリアントが大腸菌又は真核生物昆虫細胞において発現されるとの報告されており(特許文献1)、またシャペロンと共発現することで大腸菌において発現されると報告されているが(特許文献2)、工業的な利用の観点からはより大量の発現が必要とされている。
【0004】
【特許文献1】特表2002/534125号公報
【特許文献2】特開2002−315584号公報
【非特許文献1】Le Grice S.F.J.,1993 reverse Transcriptase, Cold Spring Harbor,New York:Cold Spring Harbor Laboratory Press,163
【発明の概要】
【発明が解決しようとする課題】
【0005】
そこで本発明の目的は、工業的規模で発現(工業的規模でAMV酵素を製造)することすることが可能な新規なAMV逆転写酵素遺伝子と、それを用いたAMV逆転写酵素の製造方法を提供することにある。
【課題を解決するための手段】
【0006】
本発明者らは、大腸菌を用いた工業的規模での発現(製造)を可能ならしめるべくAMV逆転写酵素の遺伝子配列を見直し、大腸菌が容易に翻訳するコドンに対応するようにDNA配列を変換することにより前記目的を達成し得ることを見出し、本発明を完成するに至った。すなわち本発明は、AMV逆転写酵素ベータ体をコードする遺伝子であって、配列番号3記載の塩基配列からなるDNA、それと実質的に相同なDNA又はこれらDNAとストリンジェントな条件下でハイブリダイズするDNAであり、AMV逆転写酵素アルファ体をコードする遺伝子であって、配列番号4記載の塩基配列からなるDNA、それと実質的に相同なDNA又はこれらDNAとストリンジェントな条件下でハイブリダイズするDNAであり、AMV逆転写酵素ベータ体のN末端アミノ酸をメチオニンとしたタンパク質をコードする遺伝子であって、配列番号5記載の塩基配列からなるDNA、それと実質的に相同なDNA又はこれらDNAとストリンジェントな条件下でハイブリダイズするDNAであり、AMV逆転写酵素アルファ体のN末端アミノ酸をメチオニンとしたタンパク質をコードする遺伝子であって、配列番号6記載の塩基配列からなるDNA、それと実質的に相同なDNA又はこれらDNAとストリンジェントな条件下でハイブリダイズするDNAである。
【0007】
また本発明は、配列番号3記載の塩基配列からなるDNA又はそれと実質的に相同なDNAによりコードされるAMV逆転写酵素ベータ体又はそれと実質的に相同なタンパク質であり、配列番号4記載の塩基配列からなるDNA又はそれと実質的に相同なDNAによりコードされるAMV逆転写酵素アルファ体又はそれと実質的に相同なタンパク質であり、配列番号5記載の塩基配列からなるDNA又はそれと実質的に相同なDNAによりコードされるAMV逆転写酵素ベータ体のN末端アミノ酸をメチオニンとしたタンパク質又はそれと実質的に相同なタンパク質であり、配列番号6記載の塩基配列からなるDNA又はそれと実質的に相同なDNAによりコードされるAMV逆転写酵素アルファ体のN末端アミノ酸をメチオニンとしたタンパク質又はそれと実質的に相同なタンパク質である。以下、本発明を詳細に説明する。
【0008】
本発明の「AMV逆転写酵素遺伝子」とは、AMV逆転写酵素をコードする遺伝子であり、その発現、即ちmRNAへの転写と転写されたmRNAの翻訳によってAMV逆転写酵素を製造し得るものである。なお天然のAMV逆転写酵素遺伝子は、全長が2.6kベースで、配列番号1記載の塩基配列を有するDNAである。この配列番号1記載の塩基配列を有するものはAMV逆転写酵素ベータ体をコードする遺伝子であるが、AMV逆転写酵素にはベータ体よりも低分子のアルファ体が存在し、アルファ体遺伝子は、前記配列番号1記載の塩基配列を有する遺伝子の5’末端側約1.7kベースからなるDNA(配列番号2)である。なお、これら遺伝子がコードするタンパク質の機能構造は、従来公的のデータベース(National Center for Biotechnology Information)に収録されている。
【0009】
本発明のAMV逆転写酵素遺伝子のうち、配列番号3記載の塩基配列からなるDNAは、AMV逆転写酵素ベータ体をコードする遺伝子であって、その塩基配列(アミノ酸残基をコードするコドン)が天然(ウイルス)型コドンから大腸菌型コドンに変換されたDNAである。配列番号4記載の塩基配列からなるDNAは、AMV逆転写酵素アルファ体をコードする遺伝子であって、その塩基配列(アミノ酸残基をコードするコドン)が天然(ウイルス)型コドンから大腸菌型コドンに変換されたDNAである。配列番号5記載の塩基配列からなるDNAは、AMV逆転写酵素ベータ体のN末端アミノ酸をメチオニンとしたタンパク質をコードする遺伝子であって、その塩基配列(アミノ酸残基をコードするコドン)が天然(ウイルス)型コドンから大腸菌型コドンに変換されたDNAである。そして配列番号6記載の塩基配列からなるDNAは、AMV逆転写酵素アルファ体のN末端アミノ酸をメチオニンとしたタンパク質をコードする遺伝子であって、その塩基配列(アミノ酸残基をコードするコドン)が天然(ウイルス)型コドンから大腸菌型コドンに変換されたDNAである。
【0010】
ここで、「天然型コドンから大腸菌型コドンに変換された」とは、いずれの遺伝子においても、その転写により、AMVが天然に有するAMV逆転写酵素遺伝子から転写されるmRNAにおいて見いだされる大腸菌レアコドン(rare codon)、すなわち大腸菌での使用頻度が低いコドンが、当該コドンがコードするアミノ酸を変更することなく大腸菌での使用頻度が高いコドンに置き換えられたmRNAを生成することを意味する。コドンの大腸菌における使用頻度の高低は、例えば公的データベース(http://www.kazusa.or.jp/codon/)を参考にゲノム遺伝子の解析を行うことによって推測することができるが、具体的に、大腸菌における使用頻度の高いコドンとして、アルギニン(Arg)コドンのAGA、AGG、CGG及びCGA、イソロイシン(Ile)コドンのAUA、ロイシン(Leu)コドンのCUA、グリシン(Gly)コドンのGGA、プロリン(Pro)コドンのCCCを例示することができる。このように大腸菌で使用頻度の高いコドンに置き換えられたmRNAを転写するよう塩基配列中のコドンを変更することで、本発明の遺伝子は大腸菌を用いることにより、工業的規模で発現することが可能である。
【0011】
コドンの変換は、当該コドンに対応する(当該コドンに転写される)DNAの塩基配列の部分(コドン)を変えることにより可能であるが、DNAの塩基配列の変換は、例えばSite−directed mutagenesis法等の従来公知の変異導入法を利用して行うことができる。中でも、合成オリゴヌクレオチドとPCRを組み合わせたDNAWorks法(Nucleic acid Res.,30,43,2002参照)がコドン変換のための特に好ましい方法として例示することができる。この方法は、アミノ酸配列から数十塩基からなるオリゴヌクレオチド群を合成し、PCRにより合成オリゴヌクレオチドをアッセンブリーさせることによって完全長の遺伝子を合成する方法である。
【0012】
本発明のAMV逆転写酵素遺伝子は、前記した4種類のDNAのほか、その一部が、その発現によってAMV逆転写酵素と実質的に同一な蛋白質であって、AMV逆転写酵素を形成するアミノ酸が欠失し、AMV逆転写酵素を形成するアミノ酸以外のアミノ酸が付加され、又は、AMV逆転写酵素を形成する所定のアミノ酸が他のアミノ酸へ置換されたAMV逆転写酵素様蛋白質をコードするDNA、また更には前記4種類のDNA又はAMV逆転写酵素様蛋白質をコードするDNAにストリンジェントな条件下でハイブリダイズするDNA(以下、これらDNAをまとめて前記4種類のDNAに「実質的に相同なDNA」と記載することがある)を含む。なお、「AMV逆転写酵素と実質的に同一な蛋白質」とは、具体的には、逆転写酵素活性、DNAポリメラーゼ活性又はRNaseH活性というAMV逆転写酵素が有する3つの主酵素活性のうち1つ以上がAMVと同等以上である蛋白質をいう。例えば、前記した各遺伝子との間で、公知の配列比較アルゴリズムを用いて解析した場合に、又は、手法自体が公知である手動整列及び目視試験により測定し、最大対応で整列化させた場合に、それぞれの塩基配列の75%以上、好ましくは85%、より好ましくは95%以上に同一性が見出される遺伝子が、その様な蛋白質をコードする遺伝子として例示できる。前記公知の解析方法として、例えば、解析局所相同性アルゴリズム(Smith等,Adv.Appl.Math.,2, 482,1981)、相同性整列化アルゴリズム(Needlman等,J.Mol. Biol.,48,443,1970)、類似性検索法(Person等,Proc. Natl.Acd.Sci.USA,85,2444,1988)等を例示できる。中でも、公知のPileup法やBLASTアルゴリズム(Altschul等,J.MOL.BIOL.,215,403,1990)が、本発明の実質的に相同なDNAを規定する上で好ましい方法として例示できる。そして「ストリンジェントな条件」とは、通常の状態と比較してDNA同士が二重鎖を形成し難い条件をいい、例えば、42℃で50%(v/v)ホルムアミド、0.1%ウシ血清アルブミン、0.1%フィコール、0.1%のポリビニルピロリドン、50mMリン酸ナトリウムバッファー(pH6.5)、150mMの塩化ナトリウム及び75mMクエン酸ナトリウムが共存する条件等が挙げられる。
【0013】
本発明のAMV逆転写酵素は、前記したAMV逆転写酵素遺伝子を発現させることにより得られる蛋白質(AMV逆転写酵素様蛋白質を含む)である。AMV逆転写酵素遺伝子の発現のためには従来公知の方法を使用することが可能であるが、かかる方法として例えば、本発明の遺伝子である前記DNAを適当な細胞に形質転換し、又は、プラスミドベクターに挿入して使用すれば良い。プラスミドベクターへの挿入は、そのプラスミドベクターの適当な位置に遺伝子工学的に挿入すればよい。ここでいう適当な位置とは、プラスミドベクターの複製機能、所望の抗生物質マーカー、あるいは伝達性に関わる領域を破壊しない範囲で自由に決定することが出来る。またプラスミドベクターに挿入する場合、例えばプロモータ機能を有するDNA配列を付加しても良い。大腸菌で機能するプロモータとして、例えばlacプロモータ、trcプロモータ又はT7プロモータ等を例示することができる。これらの中でも転写活性の調節が容易なlacプロモータは、好適なプロモータである。
【0014】
プラスミドベクターとしては形質転換する細胞内で安定に存在し、複製することができるものであれば特に制限はない。もっとも、本発明の遺伝子は、その塩基配列が大腸菌内で使用頻度の高いコドンに変換されているため、大腸菌内で安定に存在し複製できるプラスミドベクターとして知られているpUC系、pBR系、pET系、広宿主域(Broad−Host−Range)プラスミドベクター等の使用が好ましい。
【0015】
宿主としては前記の通り、K12系のE.coli JM109株に代表される大腸菌が好ましいが、変異処理した大腸菌変異株を使用することもできる。変異処理はニトロソグアニジン、メタンスルホン酸エチル、紫外線又は放射線等の、従来公知の変異手段を用いる方法を使用できる。
【0016】
以上に説明したプラスミドベクター等を公知の手法に従って大腸菌等に導入し、当該大腸菌を公知の手法に従って培養すればAMV逆転写酵素を製造することができる。プラスミドベクター等を大腸菌等に導入する方法としては、例えば、”Vectors for cloning genes”,Method in Enzymology,216,p .469−631,1992,Academic Press,や“Other Bacterial systems“,Method in Enzymology,204,p.305−636,1991,Academic Press)を参照することができる。
【0017】
大腸菌の培養方法としては、例えば必要な栄養源を補ったLB(Luria Broth)培地により培養する方法を例示できる。より具体的には、例えば、プラスミドベクターを含有した大腸菌のみを選択して培養するために、培地としてプラスミドベクターの構成を基にした選抜剤を含むものを使用することが例示できる。例えば、アンピシリン耐性遺伝子を有するプラスミドベクターを使用する場合には、培地にアンピシリンを添加することを例示できる。また培地には、炭素、窒素及び無機塩供給源の他に、適当な栄養源を添加しても良い。更にグルタチオン、システイン、シスタミン、チオグリコレート、ジチオスレイトールからなる群から選択される一種類以上の還元剤を添加しても良い。培養温度は、例えば約20℃から40℃、好ましくは25℃から35℃、より好ましくは約30℃とすることが例示できる。pHは約6.8から7.4、好ましくは7.0とすることが例示できる。
【0018】
前記したlacプロモータ等の誘導性のプロモータを用い、誘導によってAMV逆転写酵素遺伝子を発現させる場合には、良好なAMV逆転写酵素の生産性が得られる程度にまで培養を行い、しかる後に誘導の操作を行うことが好ましい。その目安としては、例えば、培養液の濁度(660nmにおける吸光度)を測定し、約0.5から1.0を示す増殖期間に誘導の操作を行い、引き続き培養するが例示できる。lacプロモータの誘導剤としてはIPTGを例示することができるが、使用するプロモータに応じて適宜選択して使用すれば良い。lacプロモータを使用する場合のIPTGの添加濃度は終濃度で約0.1から1.0mMであれば良いが、好ましくは0.15mMである。
【0019】
培養液からAMV逆転写酵素を抽出する際には、発現の形態によって適宜抽出方法を選択すれば良い。例えば大腸菌のようにAMV逆転写酵素が宿主内に蓄積する場合は遠心分離操作等により菌体を集めた後、酵素処理剤や超音波破砕等により菌体を破砕して抽出することができる。この段階では種々の蛋白質が混在しているが、イオン交換クロマトグラフィー、疎水性相互作用クロマトグラフィー、ゲルろ過クロマトグラフィー又はアフィニティークロマトグラフィー等を単独又は組み合わせて適用することにより、AMV逆転写酵素のみを抽出することができる。
【発明の効果】
【0020】
AMV逆転写酵素遺伝子の天然(ウイルス)型コドンを大腸菌型コドンに変換したことにより、大腸菌を宿主として使用することによって工業的規模で発現(工業的規模でAMV逆転写酵素を製造)することが可能となる。
【図面の簡単な説明】
【0021】
【図1】図1から図6は、AMV逆転写酵素ベータ体の天然(ウィルス)型コドン及び大腸菌コドンの遺伝子配列を比較した図である。図中AMVbetaEcoliが大腸菌型コドンに変換したDNA配列の配列、AMVbetaが天然(ウィルス)型AMVベータ体の配列である。AMVアルファ体はその塩基番号1番から1716番目までである。
【図2】図1から図6は、AMV逆転写酵素ベータ体の天然(ウィルス)型コドン及び大腸菌コドンの遺伝子配列を比較した図である。
【図3】図1から図6は、AMV逆転写酵素ベータ体の天然(ウィルス)型コドン及び大腸菌コドンの遺伝子配列を比較した図である。
【図4】図1から図6は、AMV逆転写酵素ベータ体の天然(ウィルス)型コドン及び大腸菌コドンの遺伝子配列を比較した図である。
【図5】図1から図6は、AMV逆転写酵素ベータ体の天然(ウィルス)型コドン及び大腸菌コドンの遺伝子配列を比較した図である。
【図6】図1から図6は、AMV逆転写酵素ベータ体の天然(ウィルス)型コドン及び大腸菌コドンの遺伝子配列を比較した図である。
【図7】図7は、プラスミドベクターpTrcAMVBの構造を示す図である。
【図8】図8は、プラスミドベクターpTrcAMVAの構造を示す図である。
【図9】図9は、プラスミドベクターpTrcAMVBwildの構造を示す図である。
【図10】図10は、プラスミドベクターpTrcAMVAwildの構造を示す図である。
【図11】図11は、発現量の差を示す電気泳動像の結果を示す図である。
【発明を実施するための形態】
【0022】
以下、本発明の実施の形態について、実施例により詳細に説明する。
【実施例】
【0023】
以下の実施例は本発明の実施の一形態を説明するためのものであり、本発明を限定するものではない。
【0024】
実施例1 AMV逆転写酵素をコードするDNA配列の設計
配列番号1に記載したAMV逆転写酵素のアミノ酸配列に基づき、DNAWorks法により、コドンを大腸菌型に変換した。コドンを変換したDNA配列と天然(ウィルス)型コドンをアライメントした結果を図1から図6に示す。図1から図6の通り、アミノ酸配列はそのままにDNAの配列変換が行われ、そのDNA配列間の類似性は74%であった。
【0025】
実施例2 AMV逆転写酵素をコードするDNA配列の作製
天然(ウィルス)型コドンから大腸菌型コドンに変換したDNA配列を作製するために120種類のオリゴヌクレオチドを合成した。合成したオリゴヌクレオチドを配列番号7から126に示す。
【0026】
次いで、オリゴヌクレオチドから完全長のAMV逆転写酵素をコードするDNAを作製するために、二段階のPCR反応を行った。一段階目のPCR反応の反応液は表1の通りであり、94℃、5分の熱処理後、94℃、30秒間の第一ステップ、62℃、30秒間の第二ステップ、72℃、1分間の第三ステップを25サイクル行い、次いで、72℃、7分の第四ステップを行った。表1中のDNAミックスは、120種類の合成した50pmol/μLのオリゴヌクレオチドをそれぞれ一定量サンプリングし混合した溶液である。
【0027】
【表1】
第二段階目のPCR反応は、第一段階目のPCR反応の反応液を用いて、表2の反応液組成で行った。プライマーの配列は、配列番号7(5’―ATGACGGTGGCGTTACACCTG―3’)と配列番号126(5’―TTAGGCAAACAGCGGAGAGGCTTCGTCCTTCTTCGTCACTTCGTCT―3’)のオリゴヌクレオチドを用いた。94℃、5分の熱処理後、94℃、30秒間の第一ステップ、65℃、30秒間の第二ステップ、72℃、1分間の第三ステップを25サイクル行い、最後に、72℃、7分の第四ステップを行った。反応終了後、0.9%のアガロース電気泳動で確認したところ、設計通りのサイズのDNAバンドを確認することができた。
【0028】
【表2】
次いで、目的バンドから市販の試薬(Qiaquick Gelextraction kit:キアゲン社)を用いてDNAを抽出後、抽出したDNAの5’末端を市販の試薬(TAKARA BKL Kit:タカラバイオ社)を用いてリン酸化し、制限酵素SmaIで消化したpTrc99aプラスミドベクターに挿入しE.coli JM109株(タカラバイオ社)を形質転換した。調製したプラスミドベクターは制限解析及び配列決定により調査し、これをpTrcAMVBとした。
【0029】
さらに、pTrcAMVBを鋳型にAMV逆転写酵素アルファ体をコードする遺伝子をPCR法を用いて作製した。使用したPCRプライマーの配列は、配列番号7(5’―ATGACGGTGGCGTTACACCTG―3’)と配列番号127(5’―TTAATACGCTTGAAACGTGGCCTGGCTATCCGCC―3’)である。94℃、5分の熱処理後、94℃、30秒間の第一ステップ、65℃、30秒間の第二ステップ、72℃、1分間の第三ステップを25サイクル行い、最後に、72℃、7分の第四ステップを行った。反応終了後、0.9%のアガロース電気泳動で確認したところ、設計通りのサイズのDNAバンドを確認することができた。
【0030】
次いで、目的バンドから市販の試薬(Qiaquick Gelextraction kit:キアゲン社)を用いてDNAを抽出後、抽出したDNAの5’末端を市販の試薬(TAKARA BKL Kit:タカラバイオ社)を用いてリン酸化し、制限酵素SmaIで消化したpTrc99aプラスミドベクターに挿入しE.coli JM109株(タカラバイオ社)を形質転換した。調製したプラスミドベクターは制限解析及び配列決定により調査し、これをpTrcAMVAとした。図7にpTrcAMVB及び図8にpTrcAMVAの構造を示す。
【0031】
PCR法を用いて、プラスミド上にAMV逆転写酵素ベータ体をコードする遺伝子を保有する大腸菌クローン(ATCC 31990)からAMV逆転写酵素ベータ体をコードする遺伝子を取り出した。使用したPCRプライマーの配列は、配列番号128(5’― ATGACTGTTGCGCTACATCTGGCTATTCCGCTC―3’)と配列番号129(5’―TTATGCAAAAAGAGGGCTCGCCTCATC―3’)である。94℃、5分の熱処理後、94℃、30秒間の第一ステップ、65℃、30秒間の第二ステップ、72℃、1分間の第三ステップを25サイクル行い、最後に、72℃、7分の第四ステップを行った。反応終了後、0.9%のアガロース電気泳動で確認したところ、設計通りのサイズのDNAバンドを確認することができた。次いで、目的バンドから市販の試薬(Qiaquick Gelextraction kit:キアゲン社)を用いてDNAを抽出後、抽出したDNAの5’末端を市販の試薬(TAKARA BKL Kit:タカラバイオ社)を用いてリン酸化し、制限酵素SmaIで消化したpTrc99aプラスミドベクターに挿入し大腸菌E.coli JM109株(タカラバイオ社)を形質転換した。プラスミドベクターを複製させた後、複製したプラスミドベクターについて制限解析及び配列決定により調査し、これをpTrcAMVBwildとした。
【0032】
PCR法を用いて、プラスミド上にAMV逆転写酵素ベータ体をコードする遺伝子を保有する大腸菌クローン(ATCC 31990)からAMV逆転写酵素アルファ体をコードする遺伝子を取り出した。使用したPCRプライマーの配列は、配列番号128(5’―ATGACTGTTGCGCTACATCTGGCTATTCCGCTC―3’)と配列番号130(5’―TTAATACGCTTGAAAGGTGGCTTGGCTATCTGCC―3’)である。94℃、5分の熱処理後、94℃、30秒間の第一ステップ、65℃、30秒間の第二ステップ、72℃、1分間の第三ステップを25サイクル行い、最後に、72℃、7分の第四ステップを行った。反応終了後、0.9%のアガロース電気泳動で確認したところ、設計通りのサイズのDNAバンドを確認することができた。
【0033】
次いで、目的バンドから市販の試薬(Qiaquick Gelextraction kit:キアゲン社)を用いてDNAを抽出後、抽出したDNAの5’末端を市販の試薬(TAKARA BKL Kit:タカラバイオ社)を用いてリン酸化し、制限酵素SmaIで消化したpTrc99aプラスミドベクターに挿入しE.coli JM109株(タカラバイオ社)を形質転換した。形質転換した大腸菌を培養し、プラスミドベクターを複製させた後、複製したプラスミドベクターについて制限解析及び配列決定により調査し、これをpTrcAMVAwildとした。図9にpTrcAMVBwild及び図10にpTrcAMVAwildの構造を示す。
実施例3 配列の確認
実施例2において作製したプラスミドベクターpTrcAMVB及びpTrcAMVAに挿入したDNAの配列を、チェーンターミネータ法に基づく市販の試薬キット(Big Dye Terminator Cycle Sequencing FS read Reaction kit、PEアプライドバイオシステム社)を用いてサイクルシークエンス反応に供し、全自動DNAシークエンサーABI Prism 3700 DNA Analyzer(PEアプライドバイオシステム社)にて解析した。なお、シークエンス用プライマーには合成した120種類のオリゴヌクレオチドを適宜使用した。その結果、pTrcAMVB及びpTrcAMVAに挿入したDNAの塩基配列は設計通りであることを確認した。
【0034】
実施例4 AMV逆転写酵素の発現誘導
プラスミドベクターpTrcAMVBを用いて、50μg/mLの抗生物質アンピシリンを添加したLB寒天培地中でE.coli JM109(タカラバイオ社)を形質転換した。37℃で18時間培養後、出現した任意のコロニーをピックアップし50μg/mLの抗生物質アンピシリンを添加したLB液体培地に接種した。37℃で培養し、OD660nmの値が約0.5になったところで0.15mMのIPTG(タカラバイオ社製)を添加した。プラスミドベクターpTrcAMVBで形質転換した株をJM109/pTrcAMVBとした。プラスミドベクターpTrcAMVAについても同様に操作し、形質転換株をJM109/pTrcAMVAとした。
【0035】
同様の方法でプラスミドベクターpTrcAMVBwild及びプラスミドベクターpTrcAMVAwildも形質転換を行い、それぞれJM109/pTrcAMVBwild、JM109/pTrcAMVAwildとした。
【0036】
IPTG添加から適当な間隔で培養液をサンプリングし、それぞれの形質転換株から蛋白質溶液を調製した。調製は市販のキット(Bugbuster protein extraction Kit、ノバジェン社)により行い、調製した蛋白質溶液をSDS−PAGEに供した。電気泳動像の結果を図11に示す。図11の通り、発現するタンパク質のアミノ酸配列は同一であるものの、宿主である大腸菌に合わせたコドンでは天然(ウィルス)型コドンに比べて有意に発現量を増加していた。
【特許請求の範囲】
【請求項1】
AMV逆転写酵素ベータ体をコードする遺伝子であって、配列番号3記載の塩基配列からなるDNA、それと実質的に相同なDNA又はこれらDNAとストリンジェントな条件下でハイブリダイズするDNA。
【請求項2】
AMV逆転写酵素アルファ体をコードする遺伝子であって、配列番号4記載の塩基配列からなるDNA、それと実質的に相同なDNA又はこれらDNAとストリンジェントな条件下でハイブリダイズするDNA。
【請求項3】
AMV逆転写酵素ベータ体のN末端アミノ酸をメチオニンとしたタンパク質をコードする遺伝子であって、配列番号5記載の塩基配列からなるDNA、それと実質的に相同なDNA又はこれらDNAとストリンジェントな条件下でハイブリダイズするDNA。
【請求項4】
AMV逆転写酵素アルファ体のN末端アミノ酸をメチオニンとしたタンパク質をコードする遺伝子であって、配列番号6記載の塩基配列からなるDNA、それと実質的に相同なDNA又はこれらDNAとストリンジェントな条件下でハイブリダイズするDNA。
【請求項5】
配列番号3記載の塩基配列からなるDNA又はそれと実質的に相同なDNAによりコードされるAMV逆転写酵素ベータ体又はそれと実質的に相同なタンパク質。
【請求項6】
配列番号4記載の塩基配列からなるDNA又はそれと実質的に相同なDNAによりコードされるAMV逆転写酵素アルファ体又はそれと実質的に相同なタンパク質。
【請求項7】
配列番号5記載の塩基配列からなるDNA又はそれと実質的に相同なDNAによりコードされるAMV逆転写酵素ベータ体のN末端アミノ酸をメチオニンとしたタンパク質又はそれと実質的に相同なタンパク質。
【請求項8】
配列番号6記載の塩基配列からなるDNA又はそれと実質的に相同なDNAによりコードされるAMV逆転写酵素アルファ体のN末端アミノ酸をメチオニンとしたタンパク質又はそれと実質的に相同なタンパク質。
【請求項1】
AMV逆転写酵素ベータ体をコードする遺伝子であって、配列番号3記載の塩基配列からなるDNA、それと実質的に相同なDNA又はこれらDNAとストリンジェントな条件下でハイブリダイズするDNA。
【請求項2】
AMV逆転写酵素アルファ体をコードする遺伝子であって、配列番号4記載の塩基配列からなるDNA、それと実質的に相同なDNA又はこれらDNAとストリンジェントな条件下でハイブリダイズするDNA。
【請求項3】
AMV逆転写酵素ベータ体のN末端アミノ酸をメチオニンとしたタンパク質をコードする遺伝子であって、配列番号5記載の塩基配列からなるDNA、それと実質的に相同なDNA又はこれらDNAとストリンジェントな条件下でハイブリダイズするDNA。
【請求項4】
AMV逆転写酵素アルファ体のN末端アミノ酸をメチオニンとしたタンパク質をコードする遺伝子であって、配列番号6記載の塩基配列からなるDNA、それと実質的に相同なDNA又はこれらDNAとストリンジェントな条件下でハイブリダイズするDNA。
【請求項5】
配列番号3記載の塩基配列からなるDNA又はそれと実質的に相同なDNAによりコードされるAMV逆転写酵素ベータ体又はそれと実質的に相同なタンパク質。
【請求項6】
配列番号4記載の塩基配列からなるDNA又はそれと実質的に相同なDNAによりコードされるAMV逆転写酵素アルファ体又はそれと実質的に相同なタンパク質。
【請求項7】
配列番号5記載の塩基配列からなるDNA又はそれと実質的に相同なDNAによりコードされるAMV逆転写酵素ベータ体のN末端アミノ酸をメチオニンとしたタンパク質又はそれと実質的に相同なタンパク質。
【請求項8】
配列番号6記載の塩基配列からなるDNA又はそれと実質的に相同なDNAによりコードされるAMV逆転写酵素アルファ体のN末端アミノ酸をメチオニンとしたタンパク質又はそれと実質的に相同なタンパク質。
【図1】


【図2】

【図3】

【図4】

【図5】

【図6】

【図7】
【図8】

【図9】

【図10】
【図11】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【公開番号】特開2012−120506(P2012−120506A)
【公開日】平成24年6月28日(2012.6.28)
【国際特許分類】
【出願番号】特願2010−275557(P2010−275557)
【出願日】平成22年12月10日(2010.12.10)
【出願人】(000003300)東ソー株式会社 (1,901)
【Fターム(参考)】
【公開日】平成24年6月28日(2012.6.28)
【国際特許分類】
【出願日】平成22年12月10日(2010.12.10)
【出願人】(000003300)東ソー株式会社 (1,901)
【Fターム(参考)】
[ Back to top ]