
Abca12遺伝子機能欠損マウス
【課題】皮膚バリア機能障害を有する非ヒトモデル動物を提供すること、及び当該モデル動物を利用して皮膚バリア機能障害を治療若しくは改善、さらには予防することのできる薬物を探索する方法を提供すること。
【解決手段】Abca12遺伝子がノックアウトされた、皮膚バリア機能障害モデルマウス。本発明にかかる遺伝子欠損マウスは、重篤な皮膚バリア機能障害、特に魚鱗癬の症状を呈するモデル動物として利用可能であり、当該モデル動物を用いることで、皮膚バリア障害を治療又は改善することのできる物質を個体レベルでスクリーニングすることが可能となる。
【解決手段】Abca12遺伝子がノックアウトされた、皮膚バリア機能障害モデルマウス。本発明にかかる遺伝子欠損マウスは、重篤な皮膚バリア機能障害、特に魚鱗癬の症状を呈するモデル動物として利用可能であり、当該モデル動物を用いることで、皮膚バリア障害を治療又は改善することのできる物質を個体レベルでスクリーニングすることが可能となる。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、Abca12遺伝子の機能がノックアウトされた、皮膚バリア機能障害モデルマウス、及び乾癬などの皮膚疾患の治療、あるいは皮膚疾患によって失われた皮膚バリア機能の回復に有効な薬物、さらには乾癬などの皮膚疾患に対して有効な予防薬のスクリーニング方法または同定方法に関する。
【背景技術】
【0002】
近年の発生工学的技術の発展は、特定の外来遺伝子を導入したトランスジェニック動物や染色体上の特定の遺伝子の機能が破壊されたノックアウト動物の作製を可能にしてきた。この様な遺伝子操作動物は、導入あるいは破壊される遺伝子に依存した表現形質を示し得ることが知られている。そのため、遺伝子組み換え動物は、当該組み換えられる遺伝子と表現形質との関係を確認するために作製される他、疾患と密接な関係を有する表現形質すなわち病態を示す動物は、当該疾患モデル動物あるいは病態モデル動物としての価値を有する。
【0003】
一方、皮膚疾患に関しては、適切な疾患モデル動物或いは病態モデル動物が少なく、このことが皮膚疾患を治療する方法や、皮膚疾患の治療・予防薬の開発を困難なものとしている。中でも皮膚バリア障害は、患者に対して多くの場合致死的であり、したがってそのモデル動物を作製することは極めて困難であった。
【0004】
最も重篤な皮膚バリア機能障害を伴う疾患の一つに、道化師様魚鱗癬がある。この疾患は、常染色体劣性遺伝性の先天性魚鱗癬であり、患者は出生時から全身が板状の厚い鱗屑で覆われ、多くの場合、皮膚バリア機能障害に伴う呼吸不全、感染症、及び/又は水分多損失により、出生後1〜2週間以内に死亡する。そのため、皮膚バリア機能障害を改善することが、前記疾患患者の生存に不可欠である。
【0005】
近年、Akiyamaらによって、Abca12遺伝子の機能損失が道化師様魚鱗癬の原因であることが明らかにされた(非特許文献1、非特許文献2)。Abca12遺伝子は、ABC(ATP binding cassette transporter)をコードするスーパーファミリーに属する遺伝子の一種である。このスーパーファミリーは、細菌からヒトに至るまで進化中に非常によく保存されており、細胞外および細胞内の様々な物質の輸送、例えば塩素チャンネル、多剤耐性、胆汁酸塩輸送、グルタチオン抱合体輸送、HLAクラスI抗原輸送、スルホニル尿素受容体、オリゴA結合タンパク質、または脂質誘導(コレステロール、ホスファチジルセリン)輸送に関与していることが報告されている。
【0006】
ヒトのAbca12遺伝子は、特許文献1(特表2004−527237)にその塩基配列とそれにコードされるタンパク質のアミノ酸配列が記載され、またマウスのAbca12遺伝子は、Waterston R.H.ら(非特許文献3)に記載されている。
【0007】
前記Akiyamaらは、ヒトAbca12遺伝子がヒト表皮顆粒層細胞、特に層板顆粒において強く発現し、セラミド等の脂質輸送に関与していることを確認し、さらに、道化師様魚鱗癬疾患患者から分離して調製された患者培養表皮細胞に正常なヒトAbca12遺伝子を導入することで、前記患者培養表皮細胞のセラミド輸送障害を改善することを実験的に確認している。このことから、Abca12遺伝子又はその発現産物の機能欠失が、皮膚バリア機能に重要な角層細胞間脂質層の形成不全をもたらし、ヒトにおける道化師様魚鱗癬を発症する原因となっていると推察される。
【非特許文献1】Akiyamaら、J.Clin.Invest.、2005年、第115巻、第7号、第1777−1784頁
【非特許文献2】秋山真志ら、最新医学、2007年、第62巻、第11号、第2482−2488頁
【非特許文献3】Waterston R.H.ら、Nature、2002年、第420巻、第520−562頁
【特許文献1】特表2004−527237号
【発明の開示】
【発明が解決しようとする課題】
【0008】
本発明の目的は、皮膚バリア機能障害を有する非ヒトモデル動物を提供すること、及び当該モデル動物を利用して皮膚バリア機能障害を治療若しくは改善、さらには予防することのできる薬物を探索する方法を提供することにある。
【課題を解決するための手段】
【0009】
本発明者らは、Abca12遺伝子がノックアウトされたマウスを作出することに成功し、当該マウスが重篤な皮膚バリア機能障害を有することを見いだし、下記の各発明を完成した。
【0010】
(1)Abca12遺伝子の機能がノックアウトされた、皮膚バリア機能障害モデルマウス。
【0011】
(2)Abca12遺伝子のエクソン30を破壊することで当該遺伝子の機能がノックアウトされた、(1)に記載のモデルマウス。
【0012】
(3)ホモノックアウトマウスである、(1)又は(2)に記載のモデルマウス。
【0013】
(4)皮膚バリア機能障害が魚鱗癬による障害である、(1)〜(3)のいずれかに記載のモデルマウス。
【0014】
(5)(1)〜(4)の何れかに記載のモデルマウス又はその皮膚組織に物質を投与する工程、及び該モデルマウス又はその皮膚組織の皮膚バリア機能障害の改善効果を確認する工程を含む、皮膚バリア障害を治療又は改善することのできる物質をスクリーニングする方法。
【0015】
(6)(1)〜(4)の何れかに記載のモデルマウスを受胎した親マウスに物質を投与する工程、及び胎児マウスの出生率を測定する工程を含む、皮膚バリア障害を治療又は改善することのできる物質をスクリーニングする方法。
【発明の効果】
【0016】
本発明にかかる遺伝子欠損マウスは、これまでに報告のない重篤な皮膚バリア機能障害、特に道化師様魚鱗癬の症状を呈するモデル動物として利用可能であり、当該モデル動物を用いることで、皮膚バリア障害を治療又は改善することのできる物質を個体レベルでスクリーニングすることが可能となる。
【発明を実施するための最良の形態】
【0017】
本発明は、Abca12遺伝子の機能がノックアウトされたマウスを、皮膚バリア機能障害モデル動物として使用することを特徴とする。ヒトのAbca12遺伝子にコードされるタンパク質は、2595アミノ酸残基からなるトランスポータータンパク質(以下、ヒトAbca12蛋白と表す)である。このヒトAbca12蛋白は、表皮細胞の層板顆粒へのセラミドなどの脂質輸送に関与しており、層板顆粒に輸送された脂質が角層細胞間脂質層を形成することで、皮膚バリア機能の要となることが、前記Akiyamaらによって明らかにされている。そして、このヒトAbca12遺伝子に何らかの変異が生じ、機能的なヒトAbca12蛋白が発現されないことによって、角層細胞間脂質層が十分に形成されず、皮膚バリア機能障害を来し、また魚鱗癬を発症することになる。
【0018】
一方、マウスにもヒトAbca12遺伝子に対応するマウスAbca12遺伝子が存在する。マウスAbca12遺伝子のcDNA及びゲノムDNAは既にクローニングされており、それらの塩基配列も、NCBIにアクセッション番号XM_001002308とNC_000067.5としてそれぞれ登録されている。アクセッション番号XM_001002308には、全2592アミノ酸残基からなるタンパク質(以下、マウスAbca12蛋白と表す)をコードする全7779bpのマウスAbca12遺伝子のcDNAの塩基配列情報が登録されている。このマウスAbca12蛋白のアミノ酸配列は、ヒトAbca12蛋白のアミノ酸配列に対して89%の同一性を有している。一方のアクセッション番号NC_000067.5には、52個のイントロンと53個のエクソンからなる構造を有する、約171kbpのゲノムDNAの塩基配列情報が登録されている。このマウスAbca12遺伝子のゲノムDNAは、マウスの第1染色体上に位置している。
【0019】
マウスにおけるAbca12遺伝子の機能は、ヒトのそれと比べて詳細には知られていないが、コードされる蛋白質間の同一性が89%と高いことから、マウスにおいてもヒトAbca12遺伝子と同様の役割を果たしているものと推察される。
【0020】
本発明における「Abca12遺伝子がノックアウトされたマウス」とは、前記のマウス染色体上のAbca12遺伝子に導入された変異により、変異が導入されていない正常な遺伝子と比較して、その機能的な遺伝子の転写発現が著しくあるいは完全に抑制されたマウス、あるいはマウスAbca12蛋白の活性の殆どが失われた変異Abca12蛋白を発現するマウスを意味する。特に好ましくは、機能的なAbca12遺伝子の転写発現が完全に抑制されるか、またはマウスAbca12蛋白の脂質輸送活性が完全に消失した変異Abca12蛋白を発現するマウスを意味する。
【0021】
かかる変異の導入は、ニトロソグアニジンのような化学的手段、放射線照射のような物理的手段、遺伝子組み換えのような生物学的手段など、如何なる手段によってもよいが、特に、マウス染色体上のAbca12遺伝子にターゲティングベクターを用いて外来遺伝子を挿入させる、いわゆる相同組み換え(homologous recombination)による導入が好ましい。相同組み換えによる外来遺伝子の挿入は、染色体上のマウスAbca12遺伝子の機能的な転写発現が阻害される限り、当該遺伝子の任意の位置に行ってもよく、好ましくは当該染色体上のマウスAbca12遺伝子のプロモーター領域又は当該遺伝子を構成する1又は2以上のエクソンの読み取り枠を破壊するように、外来遺伝子を導入することが好ましい。特にエクソン30を破壊する相同組み換えが好ましい。
【0022】
上記の相同組換えによる染色体上のAbca12遺伝子への変異の導入は、ターゲッティングベクターを用いてノックアウトマウスを作製する一般的な方法により、行うことができる。その典型例は、次の通りである。
【0023】
NCBIアクセッション番号XM_001002308及び/又はNC_000067.5として特定されるマウスAbca12遺伝子の塩基配列情報を基に、相同組換え用ターゲティングベクターを作製する。相同組換え用ターゲティングベクターは、染色体上のAbca12遺伝子の一部を欠いた塩基配列からなる核酸を有するように、もしくは染色体上のAbca12遺伝子の上流及び下流の塩基配列によってAbca12遺伝子とは異なる異種遺伝子の5’側及び3’側を囲んだ塩基配列からなる核酸を有するように、ベクター上の塩基配列を設計し、構築すればよい。
【0024】
ターゲティングベクターは、ベクターが取り込まれた細胞や、目的とする相同組換え体を選択するための適当なマーカー遺伝子を有することが好ましい。このようなマーカー遺伝子としては、例えばネオマイシン耐性遺伝子(neo)、ジフテリアトキシンAフラグメント遺伝子(DT−A)、ハイグロマイシン耐性遺伝子、アンピシリン耐性遺伝子、テトラサイクリン耐性遺伝子、ストレプトマイシン耐性遺伝子、ヘルペスウイルスチミジンキナーゼ遺伝子(HSV−tk)等、薬剤耐性選択に通常用いられる、それぞれ公知の薬剤耐性遺伝子を用いることができる。
【0025】
また、ターゲティングベクターは、相同的組み換えによって染色体上に組み込まれる位置にレポーター遺伝子を有していてもよい。この場合、例えばAbca12遺伝子の翻訳領域の配列を一部もしくは全部欠失させるかまたは非相同な別の塩基配列に置き換えた上、レポーター遺伝子の翻訳フレームがAbca12遺伝子の翻訳フレームと一致するように設計することが好ましい。レポーター遺伝子は、例えば大腸菌のβ−ガラクトシダーゼ遺伝子(LacZ)、β−グルコニダーゼ遺伝子(GUS)、ルシフェラーゼ遺伝子(Luc)、緑色蛍光タンパク質(GFP)、エクオリン遺伝子、タウマリン遺伝子等のような、一般に用いられるものであればいずれを用いてもよい。
【0026】
相同組換え用ターゲティングベクターの構築には、一般的な遺伝子工学的手法を用いて、上記の核酸、マーカー遺伝子、レポーター遺伝子等をPCRや合成リンカーDNA等を適宜用いて、通常のDNA組換え技術により容易に行うことができる。
【0027】
次に、相同組換え用ターゲティングベクターを適当な標的細胞へ導入する。この細胞へのターゲティングベクターの導入方法も、今や当業者に周知であり、その典型例はエレクトロポレーション法、マイクロインジェクション法などを挙げることができる。標的細胞は、胚性幹細胞(Embryonic Stem Cell、ES細胞)等、通常キメラ動物の作出に用いられるものが好ましい。マウスのES細胞としては、EK細胞(Evansら、Nature、1984年、第309巻、第255頁)、ES−D3細胞(Doetschmanら、J.Embryol.Exp.Molph.、1981年、第87巻、第27頁)、CCE細胞(Robertsonら、Nature、1986年、第323巻、第445頁)、129Sv/Ev由来マウスES細胞(文献出典・・・・・・)その他の既に樹立されたものを用いればよく、また上記の論文に記載された方法その他の既知の方法に準じて、新たにES細胞を樹立して用いてもよい。
【0028】
ターゲティングベクターが取り込まれた細胞では、染色体の標的遺伝子、すなわちAbca12遺伝子が、ターゲティングベクター上の2つの相同な塩基配列部分との間で組換えを起こし、前記2つの相同な領域に囲まれたベクター上の核酸領域が染色体上に取り込まれ、変異が導入される。こうして変異が導入された細胞は、ベクター由来のマーカー遺伝子の発現に基づいて選択することができる。こうして選択された細胞のうち、所望の位置に変異が導入された細胞は、PCR法やサザンブロッティング法等により確認、選択することができる。また、核型分析により正常染色体数を確認し、キメラマウス作製に適した細胞を選択することができる。
【0029】
次に、上記方法で作製された変異が導入された細胞を用いて、注入法や凝集法その他の一般的な方法(例えば、細胞工学、第10巻、第403頁、1991年、又は「発生工学実験マニュアル」、野村達次監修、勝木元也編、講談社、1987年などを参照)に従ってキメラマウスを作出し、キメラ動物と適当な系統のマウスと交配させて産仔を得、得られる産仔の表現型、あるいはAbca12遺伝子座をPCR法やサザンブロッティング法等により解析して、その生殖細胞がES細胞に由来して、導入された変異を子孫に伝播することができる親キメラマウスを同定する。
【0030】
前記親キメラマウス同士を交配させることで、ヘテロ型のAbca12遺伝子ノックアウトマウス(以下、ヘテロ型Abcd12KOマウスと表す)を得ることができる。さらに得られたヘテロ型Abcd12KOマウス同士を交配させて、ホモ型のAbca12遺伝子ノックアウトマウス(以下、ホモ型Abca12KOマウスと表す)を得ることができる。Abca12遺伝子がヘテロ型もしくはホモ型にノックアウトされていることは、作出されたマウスの染色体DNAをPCR法やサザンブロッティング法等で解析することにより、確認することができる。本発明のマウスは、ヘテロ型Abcd12KOマウス及びホモ型Abca12KOマウスが挙げられるが、好ましくはホモ型Abca12KOマウスである。
【0031】
本発明のAbca12遺伝子がノックアウトされたマウス、特にホモ型Abca12KOマウスは、多くの場合、母体内で死亡し、出生率が極めて低い。また出生したマウスは、体重変化が確認できるほどの皮膚からの水分ロスが認められる他、トルイジンブルーを用いて染色するとわずか5分で染色されるなど、出生時から重篤な皮膚バリア機能障害を呈する。また、全身が紅色で、皮膚は鎧状で硬く柔軟性がなく、道化師様魚鱗癬と同様の外観を呈する。さらに、HE染色によりcompact hyperkeratosisが認められる一方、セラミド染色では角層の染色が観察されず、さらに皮膚組織の電子顕微鏡観察によって、角層内に多量の脂肪滴が観察される。
【0032】
したがって、本発明のAbca12遺伝子がノックアウトされたマウスに化合物を病態部位に対して外用的に投与するか、あるいは経口的に、または注射などにより投与し、ノックアウトマウスの皮膚バリア機能の改善効果を確認することにより、皮膚バリア障害を治療又は改善することのできる物質をスクリーニングすることができ、本発明は、かかるスクリーニング方法を提供するものである。また本発明のAbca12遺伝子がノックアウトされたマウスは、出生後の生存率を改善することのできる物質の探索、あるいは医学的なあるいは薬学的な方法の開発にも利用することができる。さらに、出生後のあるいは出生前の胎児の状態の本発明のAbca12遺伝子がノックアウトされたマウスから外科的に皮膚又は皮膚組織を取り出し、当該取り出した皮膚又は皮膚組織を用いて、またはそれらを正常マウスに移植して、物質のスクリーニングに使用してもよい。
【0033】
また、本発明のAbca12遺伝子がノックアウトされたマウスを受胎している親マウスに外用的に、経口的に、注射などにより、又は子宮内に物質を投与し、胎児マウスすなわちAbca12遺伝子がノックアウトされたマウスの出生率を測定することにより、多くの胎児マウスにとって致死的である皮膚バリア機能障害を治療又は改善することのできる物質をスクリーニングすることができ、本発明は、かかるスクリーニング方法も提供するものである。
【0034】
以下、実施例を挙げて本発明を具体的具体的に説明するが、本発明はこれらに限定されるものではない。なお、下記実施例において、特に明示がない限り、各操作はSambrook J.ら(Molecular Cloning、Cold Spring Harbor Laboratory Press、1989年発刊)その他の実験操作マニュアルに記載の方法、又は各種市販の試薬乃至キットに添付された指示書に従って行った。
【実施例】
【0035】
<実施例1>ノックアウトマウスの製造
BACPAC Resources Center(Children’s Hospital Oakland Research Institute、米国カリフォルニア州オークランド)から、129Sv/Evゲノムを含むBACクローンを購入し、10.6kbマウスゲノムDNAフラグメントをクローニングした。このフラグメントは、マウスAbca12のゲノムDNAのエクソン30を有する。
【0036】
エクソン30の47bp上流部位(イントロン29)から、エクソン30の203kb下流部位(イントロン30内部)までの領域を、ホスホグリセリン酸キナーゼプロモーター/ネオマイシン抵抗遺伝子カセット(PGK−neo)で置換した。置換された領域は、5’−CACTGATTGTGAACACAATAGCACC−3’(配列番号1)で開始しており、5’−TCACTGTGAGTTAAATTATCGA−3’(配列番号2)で終結している。
【0037】
このストラテジーによって、エクソン30のコード領域と、イントロン29および30の一部とが、PGK−neoで置換された改変フラグメントを得た。これにより、遺伝子転写がPGK−neoでトラップされ、エクソン30よりも下流のエクソン配列が発現されなかった。
【0038】
さらに改変フラグメントを、プラスミドベクターpSP72(プロメガ株式会社)の改変物に組み込んで、ターゲッティングベクターを得た。pSP72の改変物の塩基配列を、後述の配列表に示した(配列番号3)。ターゲッティングベクターの前記ターゲッティングベクターは、in Genious Targeting Laboratory, Inc.に委託して製造された。
【0039】
得られたターゲッティングベクターをNotIによって線状化して、129Sv/Ev胎生期幹細胞(WW1セルライン、米国National Institutes of Healthに登録されている)に、エレクトロポレーションによってトランスフェクトした。トランスフェクトされた細胞を、抗生物質G418を含む(500mg/ml)、ES細胞用の10%FCS−DMEM培地において37℃で培養し、組み換えクローンを選択した。
【0040】
選択された200の抵抗性コロニーを拡張させ、以下の条件でPCRを行い、組み換えクローンを同定した。まず、ターゲッティング構築物を得るために用いられる領域外のショートホモロジーアーム(short homology arm)の下流側(3’)へのプライマーA1(5’−TGAGACTTTCGTGCTGAATGGAG−3’)(配列番号4)と、Neoカセットの5’末端へのプライマーN1(5’−TGCGAGGCCAGAGGCCACTTGTGTAGC−3’)(配列番号5)を得た。プライマーA1とN1を用いて、0.35mMのMgCl2存在下、「95℃で30秒間、64℃で30秒間、さらに72℃で150秒間」35サイクルのPCR反応により、2.2kbフラグメントを増幅させた。
【0041】
適切にターゲッティングされたマウス胚幹細胞を、C57BL/6Jマウス(Jackson Laboratories)から得られた胚盤胞にマイクロインジェクトして、キメラ胚を得た。得られたキメラ胚を、C57BL/6Jメスマウスに着床して、キメラマウスであるヘテロ型Abcd12KOマウス(C57BL/6Jメスと交雑されている)を発生させた。F1のヘテロ接合体を、6週齢を超えるC57BL/6Jと交雑させ、インタークロスさせてホモ型Abca12KOマウスであるABCA12−/−マウスを得た。
【0042】
成体マウス、および新生仔マウスについて、4つのPCRプライマー(A1、N1、WT1(5’−CACCTCACTGAAGACGTATC−3’、配列番号6)、WT2(5’−TCGAGTTATGACTGTGCCAT−3’、配列番号7))を用いて、その遺伝子型を同定した。A1およびN1のプライマーペアを用いたPCRでは2.2kbp変異バンドが増幅され、一方、WT1およびWT2のプライマーペアを用いたPCRでは、野生型ゲノムAbca12DNAの310bpフラグメントが増幅された。
【0043】
ヘテロ型Abcd12KOマウス、ホモ型Abca12KOマウス及び野生型マウスについて、Abca12遺伝子cDNAの部分配列をプローブとして用いて、Abca12遺伝子の発現をRT−PCR法により確認した。
【0044】
野生型マウスでは、PCR産物256bpのバンドが認められたが、ホモ型Abca12KOマウスでは全くシグナルは検出されず、タンパク質が翻訳され得ない状態にあることが確認された。また、ヘテロ型Abcd12KOマウスでは256bpのバンドを確認した。
【0045】
<実施例2>Abca12遺伝子がノックアウトされたマウスの表現型
C57BL/6Jマウス(野生型)の新生児マウスと比較したときの、実施例1で作成したヘテロ型Abcd12KOマウス及びホモ型Abca12KOマウスの特徴は、次のようにまとめられる。
【0046】
1)ヘテロ型Abcd12KOマウス)
・外見
野生型と比べて差異なし。
【0047】
・体重
野生型と比べて差異なし。
【0048】
・寿命
野生型と比べて差異なし。
【0049】
・光学顕微鏡病理所見
皮膚の構造について、及び他臓器にいずれも差を認められない。
【0050】
・電子顕微鏡所見
層板顆粒の形成、角質細胞間脂質の形成に差は認められない。
【0051】
2)ホモ型Abca12KOマウス
・外見
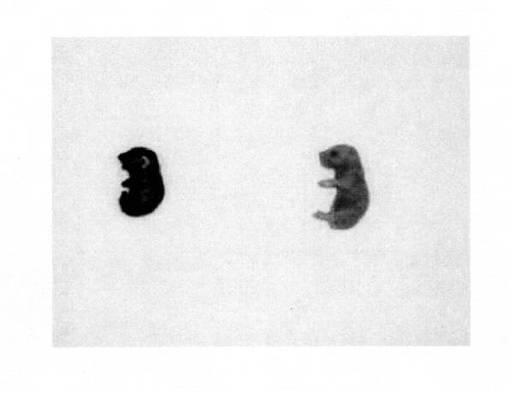
皮膚は硬く、鎧のような外見を呈しており、柔軟性がない(図1A)。また、四肢の短縮を認められる他、体に亀裂を生じたホモ型Abca12KOマウス(図1B)も認められた。これらはヒト道化師様魚鱗癬患者の特徴である。
【0052】
・体重
出生直後から3時間後の体重が、野生型は2%(95%信頼区間0−5%)の減少であるのに対して、ホモ型Abca12KOマウスは14%(95%信頼区間11−17%)減少と、著明な体重減少が認められた。
【0053】
・トルイジンブルー染色
出生直後の新生児を100%メタノールにて脱水後、0.1%トルイジンブルー液に全身を5分間浸けた結果、野生型マウスでは染色されないのに対して、ホモ型Abca12KOマウスは全身が青く染色され(図2)、KOマウスの皮膚透過性が亢進していることが確認された。
【0054】
・HE染色
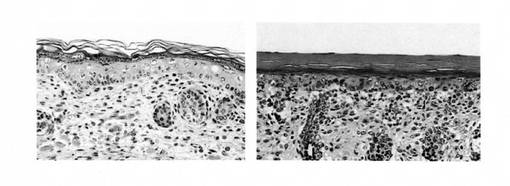
出生直後の新生児をホルマリン固定し、パラフィン包埋、ヘマトキシリン・エオジン染色を行った結果、野生型では正常角化(basket weave appearance)を認めたが、ホモ型Abca12KOマウスの皮膚では認められず、過角化を認め、道化師様魚鱗癬と同様の特徴を有することが確認された(図3)。一方、全身他臓器も観察を行ったが、野生型・ホモ型Abca12KOマウスの間に明らかな差を認めなかった。
【0055】
・セラミド染色
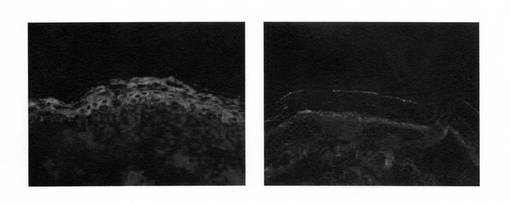
出生直後の新生児皮膚をOCT凍結包埋し、クリオスタットにて5μm厚切片を作成した。一次抗体ウサギ抗グルコシルセラミド抗を10倍稀釈、二次抗体Alexa488標識抗ウサギIgG抗体を50倍稀釈して、試料を染色した。その結果、野生型では角層での染色を認める一方、ホモ型Abca12KOマウスの皮膚では角層での染色が認められないという、道化師様魚鱗癬と同様の特徴が確認された。
【0056】
・電子顕微鏡観察
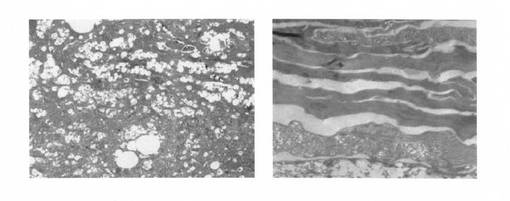
出生直後の新生児皮膚を5%グルタルアルデヒドにて固定、続いてルテニウム固定法を行い、電子顕微鏡用切片を作成し、電子顕微鏡で観察した。その結果、ホモ型Abca12KOマウスにおいてのみ、角層内に多量の脂肪滴が認められたが、角質細胞間脂肪層は形成されていないという、道化師様魚鱗癬と同様の特徴が確認された。
【0057】
・経表皮水分喪失量の測定
新生児マウスに対して、バポススキャンAS−VT100RS(ASAHIBIOMED)を使用して直接、経表皮水分喪失量を測定した。その結果、野生型マウスの水分損失は3.6(±1.5)g/h/m2であるのに対して、 ホモ型Abca12KOマウスのそれは46.2(±2.7)g/h/m2と、野生型に比べて著明に経表皮水分喪失量が上昇していることが確認された。
【図面の簡単な説明】
【0058】
【図1A】ホモ型Abca12KOマウス(左)と野生型マウス(右)の外見を示す写真である。
【図1B】体に亀裂を生じたホモ型Abca12KOマウスの外見を示す写真である。
【図2】ホモ型Abca12KOマウス(左)と野生型マウス(右)の新生児をトルイジンブルー染色した結果を示す。
【図3】ホモ型Abca12KOマウス(左)と野生型マウス(右)の新生児皮膚をHE染色した結果を示す。
【図4】ホモ型Abca12KOマウス(左)と野生型マウス(右)の新生児皮膚をセラミド染色した結果を示す。
【図5】ホモ型Abca12KOマウス(左)と野生型マウス(右)の新生児皮膚組織を電子顕微鏡観察した結果を示す。
【技術分野】
【0001】
本発明は、Abca12遺伝子の機能がノックアウトされた、皮膚バリア機能障害モデルマウス、及び乾癬などの皮膚疾患の治療、あるいは皮膚疾患によって失われた皮膚バリア機能の回復に有効な薬物、さらには乾癬などの皮膚疾患に対して有効な予防薬のスクリーニング方法または同定方法に関する。
【背景技術】
【0002】
近年の発生工学的技術の発展は、特定の外来遺伝子を導入したトランスジェニック動物や染色体上の特定の遺伝子の機能が破壊されたノックアウト動物の作製を可能にしてきた。この様な遺伝子操作動物は、導入あるいは破壊される遺伝子に依存した表現形質を示し得ることが知られている。そのため、遺伝子組み換え動物は、当該組み換えられる遺伝子と表現形質との関係を確認するために作製される他、疾患と密接な関係を有する表現形質すなわち病態を示す動物は、当該疾患モデル動物あるいは病態モデル動物としての価値を有する。
【0003】
一方、皮膚疾患に関しては、適切な疾患モデル動物或いは病態モデル動物が少なく、このことが皮膚疾患を治療する方法や、皮膚疾患の治療・予防薬の開発を困難なものとしている。中でも皮膚バリア障害は、患者に対して多くの場合致死的であり、したがってそのモデル動物を作製することは極めて困難であった。
【0004】
最も重篤な皮膚バリア機能障害を伴う疾患の一つに、道化師様魚鱗癬がある。この疾患は、常染色体劣性遺伝性の先天性魚鱗癬であり、患者は出生時から全身が板状の厚い鱗屑で覆われ、多くの場合、皮膚バリア機能障害に伴う呼吸不全、感染症、及び/又は水分多損失により、出生後1〜2週間以内に死亡する。そのため、皮膚バリア機能障害を改善することが、前記疾患患者の生存に不可欠である。
【0005】
近年、Akiyamaらによって、Abca12遺伝子の機能損失が道化師様魚鱗癬の原因であることが明らかにされた(非特許文献1、非特許文献2)。Abca12遺伝子は、ABC(ATP binding cassette transporter)をコードするスーパーファミリーに属する遺伝子の一種である。このスーパーファミリーは、細菌からヒトに至るまで進化中に非常によく保存されており、細胞外および細胞内の様々な物質の輸送、例えば塩素チャンネル、多剤耐性、胆汁酸塩輸送、グルタチオン抱合体輸送、HLAクラスI抗原輸送、スルホニル尿素受容体、オリゴA結合タンパク質、または脂質誘導(コレステロール、ホスファチジルセリン)輸送に関与していることが報告されている。
【0006】
ヒトのAbca12遺伝子は、特許文献1(特表2004−527237)にその塩基配列とそれにコードされるタンパク質のアミノ酸配列が記載され、またマウスのAbca12遺伝子は、Waterston R.H.ら(非特許文献3)に記載されている。
【0007】
前記Akiyamaらは、ヒトAbca12遺伝子がヒト表皮顆粒層細胞、特に層板顆粒において強く発現し、セラミド等の脂質輸送に関与していることを確認し、さらに、道化師様魚鱗癬疾患患者から分離して調製された患者培養表皮細胞に正常なヒトAbca12遺伝子を導入することで、前記患者培養表皮細胞のセラミド輸送障害を改善することを実験的に確認している。このことから、Abca12遺伝子又はその発現産物の機能欠失が、皮膚バリア機能に重要な角層細胞間脂質層の形成不全をもたらし、ヒトにおける道化師様魚鱗癬を発症する原因となっていると推察される。
【非特許文献1】Akiyamaら、J.Clin.Invest.、2005年、第115巻、第7号、第1777−1784頁
【非特許文献2】秋山真志ら、最新医学、2007年、第62巻、第11号、第2482−2488頁
【非特許文献3】Waterston R.H.ら、Nature、2002年、第420巻、第520−562頁
【特許文献1】特表2004−527237号
【発明の開示】
【発明が解決しようとする課題】
【0008】
本発明の目的は、皮膚バリア機能障害を有する非ヒトモデル動物を提供すること、及び当該モデル動物を利用して皮膚バリア機能障害を治療若しくは改善、さらには予防することのできる薬物を探索する方法を提供することにある。
【課題を解決するための手段】
【0009】
本発明者らは、Abca12遺伝子がノックアウトされたマウスを作出することに成功し、当該マウスが重篤な皮膚バリア機能障害を有することを見いだし、下記の各発明を完成した。
【0010】
(1)Abca12遺伝子の機能がノックアウトされた、皮膚バリア機能障害モデルマウス。
【0011】
(2)Abca12遺伝子のエクソン30を破壊することで当該遺伝子の機能がノックアウトされた、(1)に記載のモデルマウス。
【0012】
(3)ホモノックアウトマウスである、(1)又は(2)に記載のモデルマウス。
【0013】
(4)皮膚バリア機能障害が魚鱗癬による障害である、(1)〜(3)のいずれかに記載のモデルマウス。
【0014】
(5)(1)〜(4)の何れかに記載のモデルマウス又はその皮膚組織に物質を投与する工程、及び該モデルマウス又はその皮膚組織の皮膚バリア機能障害の改善効果を確認する工程を含む、皮膚バリア障害を治療又は改善することのできる物質をスクリーニングする方法。
【0015】
(6)(1)〜(4)の何れかに記載のモデルマウスを受胎した親マウスに物質を投与する工程、及び胎児マウスの出生率を測定する工程を含む、皮膚バリア障害を治療又は改善することのできる物質をスクリーニングする方法。
【発明の効果】
【0016】
本発明にかかる遺伝子欠損マウスは、これまでに報告のない重篤な皮膚バリア機能障害、特に道化師様魚鱗癬の症状を呈するモデル動物として利用可能であり、当該モデル動物を用いることで、皮膚バリア障害を治療又は改善することのできる物質を個体レベルでスクリーニングすることが可能となる。
【発明を実施するための最良の形態】
【0017】
本発明は、Abca12遺伝子の機能がノックアウトされたマウスを、皮膚バリア機能障害モデル動物として使用することを特徴とする。ヒトのAbca12遺伝子にコードされるタンパク質は、2595アミノ酸残基からなるトランスポータータンパク質(以下、ヒトAbca12蛋白と表す)である。このヒトAbca12蛋白は、表皮細胞の層板顆粒へのセラミドなどの脂質輸送に関与しており、層板顆粒に輸送された脂質が角層細胞間脂質層を形成することで、皮膚バリア機能の要となることが、前記Akiyamaらによって明らかにされている。そして、このヒトAbca12遺伝子に何らかの変異が生じ、機能的なヒトAbca12蛋白が発現されないことによって、角層細胞間脂質層が十分に形成されず、皮膚バリア機能障害を来し、また魚鱗癬を発症することになる。
【0018】
一方、マウスにもヒトAbca12遺伝子に対応するマウスAbca12遺伝子が存在する。マウスAbca12遺伝子のcDNA及びゲノムDNAは既にクローニングされており、それらの塩基配列も、NCBIにアクセッション番号XM_001002308とNC_000067.5としてそれぞれ登録されている。アクセッション番号XM_001002308には、全2592アミノ酸残基からなるタンパク質(以下、マウスAbca12蛋白と表す)をコードする全7779bpのマウスAbca12遺伝子のcDNAの塩基配列情報が登録されている。このマウスAbca12蛋白のアミノ酸配列は、ヒトAbca12蛋白のアミノ酸配列に対して89%の同一性を有している。一方のアクセッション番号NC_000067.5には、52個のイントロンと53個のエクソンからなる構造を有する、約171kbpのゲノムDNAの塩基配列情報が登録されている。このマウスAbca12遺伝子のゲノムDNAは、マウスの第1染色体上に位置している。
【0019】
マウスにおけるAbca12遺伝子の機能は、ヒトのそれと比べて詳細には知られていないが、コードされる蛋白質間の同一性が89%と高いことから、マウスにおいてもヒトAbca12遺伝子と同様の役割を果たしているものと推察される。
【0020】
本発明における「Abca12遺伝子がノックアウトされたマウス」とは、前記のマウス染色体上のAbca12遺伝子に導入された変異により、変異が導入されていない正常な遺伝子と比較して、その機能的な遺伝子の転写発現が著しくあるいは完全に抑制されたマウス、あるいはマウスAbca12蛋白の活性の殆どが失われた変異Abca12蛋白を発現するマウスを意味する。特に好ましくは、機能的なAbca12遺伝子の転写発現が完全に抑制されるか、またはマウスAbca12蛋白の脂質輸送活性が完全に消失した変異Abca12蛋白を発現するマウスを意味する。
【0021】
かかる変異の導入は、ニトロソグアニジンのような化学的手段、放射線照射のような物理的手段、遺伝子組み換えのような生物学的手段など、如何なる手段によってもよいが、特に、マウス染色体上のAbca12遺伝子にターゲティングベクターを用いて外来遺伝子を挿入させる、いわゆる相同組み換え(homologous recombination)による導入が好ましい。相同組み換えによる外来遺伝子の挿入は、染色体上のマウスAbca12遺伝子の機能的な転写発現が阻害される限り、当該遺伝子の任意の位置に行ってもよく、好ましくは当該染色体上のマウスAbca12遺伝子のプロモーター領域又は当該遺伝子を構成する1又は2以上のエクソンの読み取り枠を破壊するように、外来遺伝子を導入することが好ましい。特にエクソン30を破壊する相同組み換えが好ましい。
【0022】
上記の相同組換えによる染色体上のAbca12遺伝子への変異の導入は、ターゲッティングベクターを用いてノックアウトマウスを作製する一般的な方法により、行うことができる。その典型例は、次の通りである。
【0023】
NCBIアクセッション番号XM_001002308及び/又はNC_000067.5として特定されるマウスAbca12遺伝子の塩基配列情報を基に、相同組換え用ターゲティングベクターを作製する。相同組換え用ターゲティングベクターは、染色体上のAbca12遺伝子の一部を欠いた塩基配列からなる核酸を有するように、もしくは染色体上のAbca12遺伝子の上流及び下流の塩基配列によってAbca12遺伝子とは異なる異種遺伝子の5’側及び3’側を囲んだ塩基配列からなる核酸を有するように、ベクター上の塩基配列を設計し、構築すればよい。
【0024】
ターゲティングベクターは、ベクターが取り込まれた細胞や、目的とする相同組換え体を選択するための適当なマーカー遺伝子を有することが好ましい。このようなマーカー遺伝子としては、例えばネオマイシン耐性遺伝子(neo)、ジフテリアトキシンAフラグメント遺伝子(DT−A)、ハイグロマイシン耐性遺伝子、アンピシリン耐性遺伝子、テトラサイクリン耐性遺伝子、ストレプトマイシン耐性遺伝子、ヘルペスウイルスチミジンキナーゼ遺伝子(HSV−tk)等、薬剤耐性選択に通常用いられる、それぞれ公知の薬剤耐性遺伝子を用いることができる。
【0025】
また、ターゲティングベクターは、相同的組み換えによって染色体上に組み込まれる位置にレポーター遺伝子を有していてもよい。この場合、例えばAbca12遺伝子の翻訳領域の配列を一部もしくは全部欠失させるかまたは非相同な別の塩基配列に置き換えた上、レポーター遺伝子の翻訳フレームがAbca12遺伝子の翻訳フレームと一致するように設計することが好ましい。レポーター遺伝子は、例えば大腸菌のβ−ガラクトシダーゼ遺伝子(LacZ)、β−グルコニダーゼ遺伝子(GUS)、ルシフェラーゼ遺伝子(Luc)、緑色蛍光タンパク質(GFP)、エクオリン遺伝子、タウマリン遺伝子等のような、一般に用いられるものであればいずれを用いてもよい。
【0026】
相同組換え用ターゲティングベクターの構築には、一般的な遺伝子工学的手法を用いて、上記の核酸、マーカー遺伝子、レポーター遺伝子等をPCRや合成リンカーDNA等を適宜用いて、通常のDNA組換え技術により容易に行うことができる。
【0027】
次に、相同組換え用ターゲティングベクターを適当な標的細胞へ導入する。この細胞へのターゲティングベクターの導入方法も、今や当業者に周知であり、その典型例はエレクトロポレーション法、マイクロインジェクション法などを挙げることができる。標的細胞は、胚性幹細胞(Embryonic Stem Cell、ES細胞)等、通常キメラ動物の作出に用いられるものが好ましい。マウスのES細胞としては、EK細胞(Evansら、Nature、1984年、第309巻、第255頁)、ES−D3細胞(Doetschmanら、J.Embryol.Exp.Molph.、1981年、第87巻、第27頁)、CCE細胞(Robertsonら、Nature、1986年、第323巻、第445頁)、129Sv/Ev由来マウスES細胞(文献出典・・・・・・)その他の既に樹立されたものを用いればよく、また上記の論文に記載された方法その他の既知の方法に準じて、新たにES細胞を樹立して用いてもよい。
【0028】
ターゲティングベクターが取り込まれた細胞では、染色体の標的遺伝子、すなわちAbca12遺伝子が、ターゲティングベクター上の2つの相同な塩基配列部分との間で組換えを起こし、前記2つの相同な領域に囲まれたベクター上の核酸領域が染色体上に取り込まれ、変異が導入される。こうして変異が導入された細胞は、ベクター由来のマーカー遺伝子の発現に基づいて選択することができる。こうして選択された細胞のうち、所望の位置に変異が導入された細胞は、PCR法やサザンブロッティング法等により確認、選択することができる。また、核型分析により正常染色体数を確認し、キメラマウス作製に適した細胞を選択することができる。
【0029】
次に、上記方法で作製された変異が導入された細胞を用いて、注入法や凝集法その他の一般的な方法(例えば、細胞工学、第10巻、第403頁、1991年、又は「発生工学実験マニュアル」、野村達次監修、勝木元也編、講談社、1987年などを参照)に従ってキメラマウスを作出し、キメラ動物と適当な系統のマウスと交配させて産仔を得、得られる産仔の表現型、あるいはAbca12遺伝子座をPCR法やサザンブロッティング法等により解析して、その生殖細胞がES細胞に由来して、導入された変異を子孫に伝播することができる親キメラマウスを同定する。
【0030】
前記親キメラマウス同士を交配させることで、ヘテロ型のAbca12遺伝子ノックアウトマウス(以下、ヘテロ型Abcd12KOマウスと表す)を得ることができる。さらに得られたヘテロ型Abcd12KOマウス同士を交配させて、ホモ型のAbca12遺伝子ノックアウトマウス(以下、ホモ型Abca12KOマウスと表す)を得ることができる。Abca12遺伝子がヘテロ型もしくはホモ型にノックアウトされていることは、作出されたマウスの染色体DNAをPCR法やサザンブロッティング法等で解析することにより、確認することができる。本発明のマウスは、ヘテロ型Abcd12KOマウス及びホモ型Abca12KOマウスが挙げられるが、好ましくはホモ型Abca12KOマウスである。
【0031】
本発明のAbca12遺伝子がノックアウトされたマウス、特にホモ型Abca12KOマウスは、多くの場合、母体内で死亡し、出生率が極めて低い。また出生したマウスは、体重変化が確認できるほどの皮膚からの水分ロスが認められる他、トルイジンブルーを用いて染色するとわずか5分で染色されるなど、出生時から重篤な皮膚バリア機能障害を呈する。また、全身が紅色で、皮膚は鎧状で硬く柔軟性がなく、道化師様魚鱗癬と同様の外観を呈する。さらに、HE染色によりcompact hyperkeratosisが認められる一方、セラミド染色では角層の染色が観察されず、さらに皮膚組織の電子顕微鏡観察によって、角層内に多量の脂肪滴が観察される。
【0032】
したがって、本発明のAbca12遺伝子がノックアウトされたマウスに化合物を病態部位に対して外用的に投与するか、あるいは経口的に、または注射などにより投与し、ノックアウトマウスの皮膚バリア機能の改善効果を確認することにより、皮膚バリア障害を治療又は改善することのできる物質をスクリーニングすることができ、本発明は、かかるスクリーニング方法を提供するものである。また本発明のAbca12遺伝子がノックアウトされたマウスは、出生後の生存率を改善することのできる物質の探索、あるいは医学的なあるいは薬学的な方法の開発にも利用することができる。さらに、出生後のあるいは出生前の胎児の状態の本発明のAbca12遺伝子がノックアウトされたマウスから外科的に皮膚又は皮膚組織を取り出し、当該取り出した皮膚又は皮膚組織を用いて、またはそれらを正常マウスに移植して、物質のスクリーニングに使用してもよい。
【0033】
また、本発明のAbca12遺伝子がノックアウトされたマウスを受胎している親マウスに外用的に、経口的に、注射などにより、又は子宮内に物質を投与し、胎児マウスすなわちAbca12遺伝子がノックアウトされたマウスの出生率を測定することにより、多くの胎児マウスにとって致死的である皮膚バリア機能障害を治療又は改善することのできる物質をスクリーニングすることができ、本発明は、かかるスクリーニング方法も提供するものである。
【0034】
以下、実施例を挙げて本発明を具体的具体的に説明するが、本発明はこれらに限定されるものではない。なお、下記実施例において、特に明示がない限り、各操作はSambrook J.ら(Molecular Cloning、Cold Spring Harbor Laboratory Press、1989年発刊)その他の実験操作マニュアルに記載の方法、又は各種市販の試薬乃至キットに添付された指示書に従って行った。
【実施例】
【0035】
<実施例1>ノックアウトマウスの製造
BACPAC Resources Center(Children’s Hospital Oakland Research Institute、米国カリフォルニア州オークランド)から、129Sv/Evゲノムを含むBACクローンを購入し、10.6kbマウスゲノムDNAフラグメントをクローニングした。このフラグメントは、マウスAbca12のゲノムDNAのエクソン30を有する。
【0036】
エクソン30の47bp上流部位(イントロン29)から、エクソン30の203kb下流部位(イントロン30内部)までの領域を、ホスホグリセリン酸キナーゼプロモーター/ネオマイシン抵抗遺伝子カセット(PGK−neo)で置換した。置換された領域は、5’−CACTGATTGTGAACACAATAGCACC−3’(配列番号1)で開始しており、5’−TCACTGTGAGTTAAATTATCGA−3’(配列番号2)で終結している。
【0037】
このストラテジーによって、エクソン30のコード領域と、イントロン29および30の一部とが、PGK−neoで置換された改変フラグメントを得た。これにより、遺伝子転写がPGK−neoでトラップされ、エクソン30よりも下流のエクソン配列が発現されなかった。
【0038】
さらに改変フラグメントを、プラスミドベクターpSP72(プロメガ株式会社)の改変物に組み込んで、ターゲッティングベクターを得た。pSP72の改変物の塩基配列を、後述の配列表に示した(配列番号3)。ターゲッティングベクターの前記ターゲッティングベクターは、in Genious Targeting Laboratory, Inc.に委託して製造された。
【0039】
得られたターゲッティングベクターをNotIによって線状化して、129Sv/Ev胎生期幹細胞(WW1セルライン、米国National Institutes of Healthに登録されている)に、エレクトロポレーションによってトランスフェクトした。トランスフェクトされた細胞を、抗生物質G418を含む(500mg/ml)、ES細胞用の10%FCS−DMEM培地において37℃で培養し、組み換えクローンを選択した。
【0040】
選択された200の抵抗性コロニーを拡張させ、以下の条件でPCRを行い、組み換えクローンを同定した。まず、ターゲッティング構築物を得るために用いられる領域外のショートホモロジーアーム(short homology arm)の下流側(3’)へのプライマーA1(5’−TGAGACTTTCGTGCTGAATGGAG−3’)(配列番号4)と、Neoカセットの5’末端へのプライマーN1(5’−TGCGAGGCCAGAGGCCACTTGTGTAGC−3’)(配列番号5)を得た。プライマーA1とN1を用いて、0.35mMのMgCl2存在下、「95℃で30秒間、64℃で30秒間、さらに72℃で150秒間」35サイクルのPCR反応により、2.2kbフラグメントを増幅させた。
【0041】
適切にターゲッティングされたマウス胚幹細胞を、C57BL/6Jマウス(Jackson Laboratories)から得られた胚盤胞にマイクロインジェクトして、キメラ胚を得た。得られたキメラ胚を、C57BL/6Jメスマウスに着床して、キメラマウスであるヘテロ型Abcd12KOマウス(C57BL/6Jメスと交雑されている)を発生させた。F1のヘテロ接合体を、6週齢を超えるC57BL/6Jと交雑させ、インタークロスさせてホモ型Abca12KOマウスであるABCA12−/−マウスを得た。
【0042】
成体マウス、および新生仔マウスについて、4つのPCRプライマー(A1、N1、WT1(5’−CACCTCACTGAAGACGTATC−3’、配列番号6)、WT2(5’−TCGAGTTATGACTGTGCCAT−3’、配列番号7))を用いて、その遺伝子型を同定した。A1およびN1のプライマーペアを用いたPCRでは2.2kbp変異バンドが増幅され、一方、WT1およびWT2のプライマーペアを用いたPCRでは、野生型ゲノムAbca12DNAの310bpフラグメントが増幅された。
【0043】
ヘテロ型Abcd12KOマウス、ホモ型Abca12KOマウス及び野生型マウスについて、Abca12遺伝子cDNAの部分配列をプローブとして用いて、Abca12遺伝子の発現をRT−PCR法により確認した。
【0044】
野生型マウスでは、PCR産物256bpのバンドが認められたが、ホモ型Abca12KOマウスでは全くシグナルは検出されず、タンパク質が翻訳され得ない状態にあることが確認された。また、ヘテロ型Abcd12KOマウスでは256bpのバンドを確認した。
【0045】
<実施例2>Abca12遺伝子がノックアウトされたマウスの表現型
C57BL/6Jマウス(野生型)の新生児マウスと比較したときの、実施例1で作成したヘテロ型Abcd12KOマウス及びホモ型Abca12KOマウスの特徴は、次のようにまとめられる。
【0046】
1)ヘテロ型Abcd12KOマウス)
・外見
野生型と比べて差異なし。
【0047】
・体重
野生型と比べて差異なし。
【0048】
・寿命
野生型と比べて差異なし。
【0049】
・光学顕微鏡病理所見
皮膚の構造について、及び他臓器にいずれも差を認められない。
【0050】
・電子顕微鏡所見
層板顆粒の形成、角質細胞間脂質の形成に差は認められない。
【0051】
2)ホモ型Abca12KOマウス
・外見
皮膚は硬く、鎧のような外見を呈しており、柔軟性がない(図1A)。また、四肢の短縮を認められる他、体に亀裂を生じたホモ型Abca12KOマウス(図1B)も認められた。これらはヒト道化師様魚鱗癬患者の特徴である。
【0052】
・体重
出生直後から3時間後の体重が、野生型は2%(95%信頼区間0−5%)の減少であるのに対して、ホモ型Abca12KOマウスは14%(95%信頼区間11−17%)減少と、著明な体重減少が認められた。
【0053】
・トルイジンブルー染色
出生直後の新生児を100%メタノールにて脱水後、0.1%トルイジンブルー液に全身を5分間浸けた結果、野生型マウスでは染色されないのに対して、ホモ型Abca12KOマウスは全身が青く染色され(図2)、KOマウスの皮膚透過性が亢進していることが確認された。
【0054】
・HE染色
出生直後の新生児をホルマリン固定し、パラフィン包埋、ヘマトキシリン・エオジン染色を行った結果、野生型では正常角化(basket weave appearance)を認めたが、ホモ型Abca12KOマウスの皮膚では認められず、過角化を認め、道化師様魚鱗癬と同様の特徴を有することが確認された(図3)。一方、全身他臓器も観察を行ったが、野生型・ホモ型Abca12KOマウスの間に明らかな差を認めなかった。
【0055】
・セラミド染色
出生直後の新生児皮膚をOCT凍結包埋し、クリオスタットにて5μm厚切片を作成した。一次抗体ウサギ抗グルコシルセラミド抗を10倍稀釈、二次抗体Alexa488標識抗ウサギIgG抗体を50倍稀釈して、試料を染色した。その結果、野生型では角層での染色を認める一方、ホモ型Abca12KOマウスの皮膚では角層での染色が認められないという、道化師様魚鱗癬と同様の特徴が確認された。
【0056】
・電子顕微鏡観察
出生直後の新生児皮膚を5%グルタルアルデヒドにて固定、続いてルテニウム固定法を行い、電子顕微鏡用切片を作成し、電子顕微鏡で観察した。その結果、ホモ型Abca12KOマウスにおいてのみ、角層内に多量の脂肪滴が認められたが、角質細胞間脂肪層は形成されていないという、道化師様魚鱗癬と同様の特徴が確認された。
【0057】
・経表皮水分喪失量の測定
新生児マウスに対して、バポススキャンAS−VT100RS(ASAHIBIOMED)を使用して直接、経表皮水分喪失量を測定した。その結果、野生型マウスの水分損失は3.6(±1.5)g/h/m2であるのに対して、 ホモ型Abca12KOマウスのそれは46.2(±2.7)g/h/m2と、野生型に比べて著明に経表皮水分喪失量が上昇していることが確認された。
【図面の簡単な説明】
【0058】
【図1A】ホモ型Abca12KOマウス(左)と野生型マウス(右)の外見を示す写真である。
【図1B】体に亀裂を生じたホモ型Abca12KOマウスの外見を示す写真である。
【図2】ホモ型Abca12KOマウス(左)と野生型マウス(右)の新生児をトルイジンブルー染色した結果を示す。
【図3】ホモ型Abca12KOマウス(左)と野生型マウス(右)の新生児皮膚をHE染色した結果を示す。
【図4】ホモ型Abca12KOマウス(左)と野生型マウス(右)の新生児皮膚をセラミド染色した結果を示す。
【図5】ホモ型Abca12KOマウス(左)と野生型マウス(右)の新生児皮膚組織を電子顕微鏡観察した結果を示す。
【特許請求の範囲】
【請求項1】
Abca12遺伝子の機能がノックアウトされた、皮膚バリア機能障害モデルマウス。
【請求項2】
Abca12遺伝子のエクソン30を破壊することで当該遺伝子の機能がノックアウトされた、請求項1に記載のモデルマウス。
【請求項3】
ホモノックアウトマウスである、請求項1又は2に記載のモデルマウス。
【請求項4】
皮膚バリア機能障害が魚鱗癬による障害である、請求項1〜3のいずれかに記載のモデルマウス。
【請求項5】
請求項1〜4の何れかに記載のモデルマウス又はその皮膚組織に物質を投与する工程、及び該モデルマウス又はその皮膚組織の皮膚バリア機能障害の改善効果を確認する工程を含む、皮膚バリア障害を治療又は改善することのできる物質をスクリーニングする方法。
【請求項6】
請求項1〜4の何れかに記載のモデルマウスを受胎した親マウスに物質を投与する工程、及び胎児マウスの出生率を測定する工程を含む、皮膚バリア障害を治療又は改善することのできる物質をスクリーニングする方法。
【請求項1】
Abca12遺伝子の機能がノックアウトされた、皮膚バリア機能障害モデルマウス。
【請求項2】
Abca12遺伝子のエクソン30を破壊することで当該遺伝子の機能がノックアウトされた、請求項1に記載のモデルマウス。
【請求項3】
ホモノックアウトマウスである、請求項1又は2に記載のモデルマウス。
【請求項4】
皮膚バリア機能障害が魚鱗癬による障害である、請求項1〜3のいずれかに記載のモデルマウス。
【請求項5】
請求項1〜4の何れかに記載のモデルマウス又はその皮膚組織に物質を投与する工程、及び該モデルマウス又はその皮膚組織の皮膚バリア機能障害の改善効果を確認する工程を含む、皮膚バリア障害を治療又は改善することのできる物質をスクリーニングする方法。
【請求項6】
請求項1〜4の何れかに記載のモデルマウスを受胎した親マウスに物質を投与する工程、及び胎児マウスの出生率を測定する工程を含む、皮膚バリア障害を治療又は改善することのできる物質をスクリーニングする方法。
【図1A】



【図1B】

【図2】
【図3】
【図4】
【図5】
【図1B】
【図2】
【図3】
【図4】
【図5】
【公開番号】特開2009−225697(P2009−225697A)
【公開日】平成21年10月8日(2009.10.8)
【国際特許分類】
【出願番号】特願2008−72866(P2008−72866)
【出願日】平成20年3月21日(2008.3.21)
【出願人】(504173471)国立大学法人 北海道大学 (971)
【Fターム(参考)】
【公開日】平成21年10月8日(2009.10.8)
【国際特許分類】
【出願日】平成20年3月21日(2008.3.21)
【出願人】(504173471)国立大学法人 北海道大学 (971)
【Fターム(参考)】
[ Back to top ]