
Apo−2リガンド

【課題】哺乳動物細胞アポトーシスを誘導する「Apo-2リガンド」として表される新規なサイトカインの同定、単離、及び組換え生産、Apo-2リガンド抗体と該組成物を用いる方法を提供する。
【解決手段】哺乳動物細胞アポトーシスを誘導するApo-2リガンド(特定のアミノ酸配列を持つポリペプチド),Apo-2リガンドキメラ、Apo-2リガンドをコードする核酸、そしてApo-2リガンドに対する抗体を含む組成物。アポトーシスを誘導するための、そしてガンのような病理学上のコンディションを治療するためのApo-2リガンドの使用法。
【解決手段】哺乳動物細胞アポトーシスを誘導するApo-2リガンド(特定のアミノ酸配列を持つポリペプチド),Apo-2リガンドキメラ、Apo-2リガンドをコードする核酸、そしてApo-2リガンドに対する抗体を含む組成物。アポトーシスを誘導するための、そしてガンのような病理学上のコンディションを治療するためのApo-2リガンドの使用法。
【発明の詳細な説明】
【技術分野】
【0001】
関連出願
本出願は1996年1月9日に提出された米国特許第08/584,031号の一部継続出願であり、その内容は参考としてここで取り込まれる。
【0002】
本発明は一般的に哺乳動物細胞アポトーシスを誘導するここで「Apo-2リガンド」として表される新規なサイトカインの同定、単離、及び組換え生産に関し、およびApo-2リガンド抗体と該組成物を用いる方法に関する。
【背景技術】
【0003】
哺乳動物における細胞数のコントロールは、細胞増殖と細胞死の間のバランスによって決定されると考えられる。細胞死の一つの形態として、しばしば壊死性細胞死といわれるものは、ある外傷または細胞障害から由来する細胞死の病理学的な形態として典型的には特性指摘される。これに対して、通常規則的にまたはコントロールされた方法で進行する細胞死のもう一つの「生理学的な」形態が存在する。この規則的にまたはコントロールされた細胞死の形態は、しばしば「アポトーシス」といわれる[例えばBarr等,Bio/Technology,12:487-493(1994)参照]。アポトーシス性細胞死は多くの生理学的過程で天然で生じ、それには胚の発達及び免疫系でのクローン選択が含まれる[Itoh等,Cell,66:233-243(1991)]。しかしながら減少したレベルのアポトーシス性細胞死は、ガン、狼瘡、及びヘルペスウイルス感染を含む様々な病理学的コンディションと関連する[Thompson,Science,267:1456-1462(1995)]。
【0004】
アポトーシス性細胞死は細胞質の凝縮、細胞膜微絨毛の消失、核の断片化、染色体DNAの凝集、またはミトコンドリア機能の消失のような一つ以上の細胞内の形態学的及び生化学的変化を典型的に伴う。様々な外因性及び内因性のシグナルが、該形態学的及び生化学的細胞変化を引き起こす、または誘導すると考えられている[Raff,Nature,356:397-400(1992);Steller,Science,267:1445-1449(1995);Sachs等,Blood,82:15(1993)]。例えばそれらは非成熟胸腺細胞に対するグルココルチコイドホルモンのようなホルモン性の刺激によって引き起こされ、同様に特定の増殖因子の減衰によっても引き起こされる[Watanabe-Fukunaga等,Nature,356:314-317(1992)]。同様にmyc,rel及びE1Aのようないくつかのガン遺伝子、及びp53のような腫瘍サプレッサーが、アポトーシスを誘導するのに役割を担っていることが報告されている。特定の化学療法薬剤及びある形態の放射線も同様にアポトーシス誘発活性を持つことが観察されている[Thompson,上記参照]。
【0005】
腫瘍壊死因子-α(「TNF-α」)、腫瘍壊死因子-β(「TNF-β」または「リンホトキシン」)、CD30リガンド、CD27リガンド、CD40リガンド、OX-40リガンド、4-1BBリガンド、及びApo-1リガンド(FasリガンドまたはCD95リガンドとしてもいわれる)のような様々な分子が、サイトカインの腫瘍壊死因子(「TNF」)ファミリーのメンバーとして同定されている[例えばGruss及びDower,Blood,85:3378-3404(1995)参照]。これらの分子の中で、TNF-α、TNF-β,CD30リガンド,4-1BBリガンド及びApo-1リガンドは、アポトーシス性細胞死に関与することが報告されている。TNF-αとTNF-βは影響を受けやすい腫瘍細胞においてアポトーシス性の死を誘導することが報告されている[Schmid等,Proc.Natl.Acad.Sci.,83:1881(1986);Deaitry等,Eur.J.Immunol.,17:689(1987)]。Zheng等はTNF-αがCD8ポジティブT細胞の刺激後アポトーシスに関与することを報告している[Zheng等,Nature,377:348-351(1995)]。他の研究者はCD30リガンドが胸腺における活性化T細胞の除去に関与することを報告している[Amakawa等,Cold Spring Harbor Laboratory Symposium on Programmed Cell Death,Abstr.No.10,(1995)]。
【0006】
マウスFas/Apo-1受容体またはリガンド遺伝子(それぞれlpr及びgldと呼ばれる)におけるミューテーションはある自己免疫疾患と関連し、それはApo-1リガンドが末梢における活性化T細胞のクローン欠失を調節するのに役割を担っているであろうことを示す[Krammer等,Curr.Op.Immunol.,6:279-289(1994);Nagata等,Science,267:1449-1456(1995)]。Apo-1リガンドはまたCD4ポジティブTリンパ球及びBリンパ球における刺激後アポトーシスを誘導することが報告されており、そして活性化リンパ球の機能がもはや必要とされなくなった場合には該リンパ球の除去に関与するであろう[Krammer等,上記参照;Nagata等,上記参照]。Apo-1受容体に特異的に結合する作用剤マウスモノクローナル抗体は、TNF-αのものと比較可能なまたは同様な細胞殺傷活性を示すことが報告されている[Yonehara等,J.Exp.Med.,169:1747-1756(1989)]。
【0007】
該TNFファミリーサイトカインによって介在される様々な細胞応答の誘導は、特異的細胞受容体へのその結合によって開始されると考えられている。およそ55kDa(TNF-R1)及び75kDa(TNF-R2)の2の異なるTNF受容体が同定されており[Hohman等,J.Biol.Chem.,264:14927-14935(1989);Brockhaus等,Proc.Natl.Acad.Sci.,87:3127-3131(1990);1991年3月20日に印刷されたEP417,563]、両受容体タイプに相当するヒト及びマウスcDNAが単離され特性指摘されている[Loetscher等,Cell,61:351(1990);Schall等,Cell,61:361(1990);Smith等,Science,248:1019-1023(1990);Lewis等,Proc.Natl.Acad.Sci.,88:2830-2834(1991);Goodwin等,Mol.Cell.Biol.,11:3020-3026(1991)]。Itoh等はApo-1受容体が55kDaTNF-R1によって伝達されるものと同様なアポトーシス性細胞死を伝達し得ることを開示する[Itoh等,上記参照]。細胞がTNF-αまたは抗Apo-1マウスモノクローナル抗体のそれぞれで処理された場合、Apo-1抗原の発現がTNF-R1のものと共に下流調節されることが報告されている[Krammer等,上記参照;Nagata等,上記参照]。したがってある研究者はApo-1及びTNF-R1受容体の両者を共発現する細胞系が共通のシグナル経路を通して細胞殺傷を介在するであろうと仮説を立てている[Id.]。
【0008】
リンホトキシン-αの発現と共に今日同定されているTNFファミリーリガンドは、そのC末端が細胞外に存在するII型膜貫通タンパク質である。対照的に今日同定されているTNF受容体(TNFR)ファミリーにおける受容体は、I型膜貫通タンパク質である。しかしながらTNFリガンドと受容体ファミリーの両者において、ファミリーのメンバー間に同定される同一性は主に細胞外ドメイン(「ECD」)で見出されている。TNF-α、Apo-1リガンド及びCD40リガンドを含むTNFファミリーサイトカインのいくつかは、細胞表面でタンパク質分解的に切断される;それぞれの場合で結果として生じたタンパク質は可溶性サイトカインとして機能するホモ三量体分子を典型的に形成する。TNF受容体ファミリータンパク質はまた、同起源のサイトカインの阻害剤として機能し得る可溶性受容体ECDを放出するためにタンパク質分解的に通常切断される。サイトカインとその受容体のTNFファミリーのレビューとしては、Gruss及びDower,上記参照を参照。
【発明の開示】
【課題を解決するための手段】
【0009】
出願人は、「Apo-2リガンド」と呼ばれる新規なサイトカインをコードするcDNAクローンを同定した。Apo-2リガンドはTNFサイトカインファミリーのメンバーであると現在考えられている;Apo-2リガンドはApo-1リガンドを含むいくつかの周知のTNF関連タンパク質に対するアミノ酸配列において関連している。しかしながら出願人はApo-2リガンドがFas/Apo-1,TNF-R1,またはTNF-R2受容体のような周知の可溶性Apo-1またはTNF受容体によって明らかには阻害されないことを見出した。
【0010】
一つの実施態様として、本発明は単離された生物学的に活性なApo-2リガンドを提供する。特に本発明は図1Aの残基114-281を含むアミノ酸配列を含む単離された生物学的に活性なApo-2リガンドを提供する。もう一つの実施態様として、単離された生物学的に活性なApo-2リガンドは図1Aの残基1-281(SEQ ID NO:1)に示されたアミノ酸配列を含む。
【0011】
もう一つの実施態様として、本発明はもう一つの異種ポリペプチドに融合されたApo-2リガンドを含むキメラ分子を提供する。該キメラ分子の例としては、タグポリペプチド配列に融合されたApo-2リガンドが含まれる。
【0012】
もう一つの実施態様として、本発明はApo-2リガンドをコードする単離された核酸分子を提供する。一つの面として、核酸分子は生物学的に活性なApo-2リガンドをコードするRNAまたはDNAであり、あるいは該Apo-2リガンドをコードする核酸配列に相補的なものであり、そして少なくとも穏やかに厳格なコンディションの下でそれに安定に結合し続けるものである。一つの実施態様として該核酸配列は以下から選択される:
(a) 図1A(SEQ ID NO:2)に示された残基1から残基281(すなわち核酸91から281)のフルレングスタンパク質をコードする図1Aの核酸配列のコード領域、または残基41から281の細胞外タンパク質をコードする核酸211から933のコード領域、または残基114から281の細胞外タンパク質をコードする核酸430から933のコード領域;あるいは
(b) 遺伝学的コードの縮重の範囲内で(a)の配列に一致する配列。
【0013】
さらなる実施態様として、本発明はベクターでトランスフェクトまたはトランスフォームされたホスト細胞によって認識されるコントロール配列に実施可能にリンクしたApo-2リガンドをコードする核酸分子を含む複製可能なベクターを提供する。該ベクターまたは核酸分子を含むホスト細胞もまた提供される。該核酸分子を含むホスト細胞を培養すること、及び該ホスト細胞カルチャーからタンパク質を回収することを含むApo-2リガンドの生産法も、さらに提供される。
【0014】
もう一つの実施態様として、本発明はApo-2リガンドに結合する抗体を提供する。一つの面として、該抗体はApo-2リガンドに対する抗原特異性を持つモノクローナル抗体である。
【0015】
もう一つの実施態様として、本発明は生物学的に活性なApo-2リガンド及び製薬学的に許容できるキャリアーを含む組成物を提供する。該組成物はアポトーシスを誘導または刺激するのに有用な製薬学的組成物であろう。
【0016】
もう一つの実施態様として、本発明はアポトーシスを誘導するのに効果的なApo-2リガンドの量に、in vivoまたはex vivoで哺乳動物細胞をさらすことを含む哺乳動物細胞においてアポトーシスを誘導する方法を提供する。
【0017】
もう一つの実施態様として、本発明はガンを持つ哺乳動物の治療法を提供する。該方法においては、有効量のApo-2リガンドがガンを持つと診断された哺乳動物に投与される。Apo-2リガンドはまた化学療法、放射線療法、または抗腫瘍活性を発揮可能な他の試薬のような一つ以上の他の治療と共に哺乳動物に投与され得る。
本発明のさらなる実施態様として、Apo-2リガンドまたはApo-2リガンド抗体を含む製造品及びキットが提供される。製造品及びキットには容器、該容器についたラベル、容器内に含まれる組成物が含まれる。容器についたラベルは該組成物が特定の治療上のまたは非治療上の適用に用いられ得ることを示す。該組成物には、活性試薬、及びApo-2リガンドまたはApo-2リガンド抗体を含む活性試薬が含まれる。
【発明を実施するための最良の形態】
【0018】
I. 定義
「Apo-2リガンド」及び「Apo-2L」なる語は、図1Aに示されたアミノ酸配列のアミノ酸残基114-281、残基41-281、残基15-281、または残基1-281を含むポリペプチド配列をいうためにここで用いられ、同様に上記配列の生物学的に活性な欠失、挿入、または置換変異体をいうために用いられる。好ましい実施態様として、生物学的に活性な変異体は、上記配列のいずれか一つと少なくとも約80%の配列同一性、より好ましくは少なくとも約90%の配列同一性、そしてさらにより好ましくは少なくとも約95%の配列同一性を持つ。該定義はここで記述される(実施例8参照)ヒト組織タイプまたはもう一つのソースのようなApo-2リガンドソースから単離され、あるいは組換え法または合成法によって調製される。Apo-2リガンドの本定義は、Gen Bank HHEA47M,T90422,R31020,H43566,H44565,H44567,H54628,H44772,H54629,T82085,及びT10524のような周知のEST配列を除外する。
【0019】
ここで用いられている「タグを付けたエピトープ」なる語は、「タグポリペプチド」と融合したApo-2リガンドまたはそれらの一部を含むキメラポリペプチドを示す。タグポリペプチドは抗体を生産するときのエピトープを供給するために十分な残基をもち、しかしApo-2リガンドの活性を妨害しないよう十分に短い。タグポリペプチドはまた好ましくは抗体が他のエピトープと実質的に交差反応しないように全く独特でもある。適したタグポリペプチドは一般的には少なくとも6アミノ酸残基をもち、通常約8から約50の間のアミノ酸残基(好ましくは約10から約20の間の残基)をもつ。
【0020】
ここで開示されている様々なタンパク質を記述するのに用いられている「単離した」なる語は、自然界の構成要素から同定されて分離されおよび/または回収されたタンパク質を意味する。自然界の混合した構成要素は、タンパク質の診断または治療用途を典型的には妨害するであろう物質であり、酵素、ホルモンそして他のタンパク質性または非タンパク質性溶液を含むであろう。好ましい実施態様としては、タンパク質は(1)スピニングカップシークエネーターの使用によりN末端または内部のアミノ酸配列の少なくとも15残基を得るのに十分な程度に、(2)クマシーブルーまたは銀染色を用いる非還元または還元状態の下でのSDS-PAGEによって均質に精製されるであろう。Apo-2リガンド天然環境の少なくとも一つの構成要素が存在しないであろうから、単離したタンパク質は組換え細胞内でのIn situのタンパク質を含む。しかしながら普通は、単離したタンパク質は少なくとも一つの精製工程によって調製されるであろう。
【0021】
「単離した」Apo-2リガンド核酸分子は、通常はApo-2リガンド核酸の天然のソースとして関連している少なくとも一つの混合した核酸分子から同定され分離された核酸分子である。単離したApo-2リガンド核酸分子は天然で見出される形態と場所以外のものである。それゆえ単離したApo-2リガンド核酸分子は天然の細胞内に存在するようなApo-2リガンド核酸分子とは区別される。しかしながら単離したApo-2リガンド核酸分子は、例えば核酸分子が染色体の位置において天然の細胞のものとは異なるApo-2リガンドを通常は発現する細胞内に含まれるApo-2リガンド核酸分子を含む。
【0022】
「コントロール配列」なる語は、特定のホスト生物内で実施可能に結合したコード配列の発現のために必要なDNA配列を示す。原核生物に適したコントロール配列は、例えばポロモーター、必要であればオペレーター配列そしてリボソーム結合部位を含む。真核生物細胞はプロモーター、ポリアデニレーションシグナルそしてエンハンサーが知られている。
【0023】
核酸がさらなる核酸配列との機能的な関連内に置かれたとき、核酸は「実施可能に結合」している。例えばプレ配列または分泌リーダーのためのDNAは、もしそれが配列の転写に作用すれば、ポリペプチドのためのDNAと実施可能に結合している;またはリボソーム結合部位は、もしそれが翻訳を促進するために正しい場所に置かれていれば、コード配列と実施可能に結合している。一般的に「実施可能に結合」とは、結合されているDNA配列が隣接し、分泌リーダーの場合には隣接しそしてリーディングフレーム内にあることを意味する。しかしながらエンハンサーは隣接している必要はない。結合は簡便な制限部位でのライゲーションによって成し遂げられる。もし該部位が存在しなかったなら、合成オリゴヌクレオチドアダプターまたはリンカーをありきたりの実施にしたがって用いる。
【0024】
「抗体」なる語は、最も広い意味で用いられ、単一の請うApo-2リガンドモノクローナル抗体(作用剤および拮抗剤抗体を含む)およびポリエピトープ特異性を持つ請うApo-2リガンド抗体構成物をも特にカバーする。
【0025】
ここで用いられているような「モノクローナル抗体」なる語は、実質的に同質な抗体の集団から得られる抗体を示す、すなわち集団に含まれる個々の抗体が、少量存在するであろう自然に生じ得る突然変異を除いて同一なのである。モノクローナル抗体は高い特異性を持っており、単一の抗原部位に対して向けられている。さらに、異なる決定基(エピトープ)に対して向けられた異なる抗体を典型的に含むありきたりの(ポリクローナル)抗体調製物とは異なり、各モノクローナル抗体は抗原の単一の決定基に対して向けられている。
【0026】
ここではモノクローナル抗体は、由来する種や免疫グロブリンのクラスまたはサブクラスの名称に関わらず、抗Apo-2リガンド抗体の可変ドメイン(超可変ドメインを含む)を定常ドメインに接合したもの(例えば「ヒト化」抗体)、L鎖をH鎖に接合したもの、ある種の鎖をもう一つの種の鎖に接合したものまたは異種構造のタンパク質の融合によるものによって生産されるハイブリッド及び組換え抗体を含む。また望ましい活性を示す範囲で、抗体断片(例えばFab,F(ab')2およびFv)も含む。例えば、米国特許第4,816,567号及びMage等,Monoclonal Antibody Production Techniques and Applications内のpp.79-97(Marcel Dekker,Inc.,New York,1987)参照。
【0027】
それゆえ緩和した「モノクローナル」なる語は、実質的に同質の抗体の集団から得られたような抗体の性質を示し、いかなる特定の方法によって抗体を生産する必要があるとは解釈すべきではない。例えば本発明にしたがって用いられるモノクローナル抗体は、KohlerとMilstein,Nature 256:495(1975)によって最初に記述されたハイブリドーマ法によって得られ得るし、または米国特許第4,816,567号に記述されている組換えDNA法によっても得られ得る。「モノクローナル抗体」は例えばMcCafferty等,Nature,348:552-554(1990)に記述されている方法を用いて生産されるファージライブラリーからも単離し得る。
【0028】
非ヒト(例えばネズミ)抗体の「ヒト化」形態とは、非ヒト免疫グロブリンから由来する最小限の配列を含む、特異的なキメラ免疫グロブリン、免疫グロブリン鎖またはそれらの断片(Fv,Fab,Fab',F(ab')2または他の抗体の抗原結合部分配列)である。ほとんどの場合が、ヒト化抗体は受容者の相補性決定領域(CDR)由来の残基が、望ましい特異性、アフィニティーそして力量を持つマウス、ラットまたはウサギのような非ヒト種(提供者抗体)のCDR由来の残基に置き換えられているヒト免疫グロブリン(受容者抗体)である。ある例では、ヒト免疫グロブリンのFvフレームワーク領域(FR)残基が、相当する非ヒト残基で置き換えられている。さらに該ヒト化抗体は受容者抗体にもインポートCDRまたはフレームワーク配列にも見出されない残基をも含み得る。これらの変形は、抗体の能力をさらに洗練し最適化するためになされる。一般的にはヒト化抗体は、少なくとも一つ、そして典型的には二つの可変ドメインの実質的にすべてを含むであろうし、可変ドメイン内には非ヒト免疫グロブリンのものに相当するCDR領域の全てまたは実質的にすべておよびFR領域の全てまたは実質的に全てが、ヒト免疫グロブリンコンセンサス配列のものである。ヒト化抗体はまた最適には、免疫グロブリンコンセンサス領域(Fc)の少なくとも一部を含み、それは典型的にはヒト免疫グロブリンのものである。
【0029】
Apo-2リガンドを特徴付けるためにここで目的とされる「生物学的に活性」なる語は、in vivoまたはex vivoで哺乳動物細胞の少なくとも一タイプにおいてアポトーシスを誘導するまたは刺激する能力を持つことを意味する。
【0030】
「アポトーシス」及び「アポトーシス活性」なる語は広い意味で用いられ、細胞質の凝縮、細胞膜微絨毛の消失、核の断片化、染色体DNAの凝集、またはミトコンドリア機能の消失を含む一つ以上の特徴的な細胞変化を典型的に伴う、哺乳動物における規則的なまたはコントロールされた細胞死の形態をいう。この活性は例えば細胞生存力アッセイ、FACS分析またはDNA電気泳動によって決定され測定され得る。
【0031】
ここで用いられている「ガン」と「ガンの」なる語は、典型的に非制御細胞増殖によって特徴付けられる哺乳動物内の生理学的病気を示しまたは表す。ガンの例としては、ガン腫、リンパ腫、白血病、芽腫そして肉腫を含むがそれに限られない。該ガンのより好ましい例としては、扁平上皮細胞ガン腫、小細胞肺ガン、非小細胞肺ガン、神経芽腫、脾臓ガン、グリア芽腫多形、頚部ガン、胃ガン、膀胱ガン、肝腫瘍、乳ガン、大腸ガン腫そして頭と首のガンを含む。一つの実施態様として、該ガンには小胞リンパ腫、p53ミューテーションを伴うガン腫、または乳ガン、前立腺ガン、あるいは卵巣ガンのようなホルモン依存性のガンが含まれる。
【0032】
ここで用いられている「治療」なる語は、治癒力のある治療、予防治療そして予防的治療を示す。
【0033】
ここで用いられている「哺乳動物」なる語は、ヒト、ウシ、ウマ、イヌそしてネコを含む哺乳動物として分類されるいかなる動物をも示す。本発明の好ましい実施態様としては、哺乳動物はヒトである。
【0034】
II. 本発明の組成物及び方法
本発明はTNFリガンドファミリーに関連する新規なサイトカインを提供し、該サイトカインはここで「Apo-2リガンド」と同定される。ヒトApo-2リガンドの予想される成熟アミノ酸配列は281アミノ酸を含み、およそ32,5kDaの計算される分子量とおよそ7,63の等電点を持つ。ハイドロパシー分析では、残基15と40の間に疎水性領域の存在が示されるが、N末端に明らかなシグナル配列は存在しない。シグナル配列の不存在と内部疎水性領域の存在は、Apo-2リガンドがII型膜貫通タンパク質であることを示唆する。潜在的なNリンクグリコシル化部位は推定の細胞外領域の残基109に位置する。推定の細胞質領域は図1Aに示されるアミノ酸残基1-14を含み、膜貫通領域はアミノ酸残基14-40を含み、細胞外領域はアミノ酸残基41-281を含む。図1Aに示される細胞外領域のアミノ酸残基114-281を含むApo-2リガンドポリペプチドはまた以下の実施例に記述される。
【0035】
A. Apo-2リガンドの調製
以下の記述は主としてApo-2リガンド核酸を含むベクターを用いてトランスフォームまたはトランスフェクトされた細胞の培養と、細胞カルチャーからのポリぺプチドの回収によるApo-2リガンドの生産に関する。もちろん本分野でよく知られている代わりの方法が、Apo-2リガンドを調製するために用いられることは考えられる。
【0036】
1. Apo-2リガンドをコードするDNAの単離
Apo-2リガンドをコードするDNAは、Apo-2リガンドmRNAを持ち、検出可能なレベルにそれを発現すると考えられる組織から調製したcDNAライブラリーから得られるであろう。よって、ヒトApo-2リガンドDNAは実施例1に記述されているヒト胎盤cDNAのバクテリオファージライブラリーのようなヒト組織から調製されるcDNAライブラリーから便利に得ることができる。Apo-2リガンドをコードする遺伝子はまた、ゲノムライブラリーから、またはオリゴヌクレオチド合成によって得られるだろう。
【0037】
ライブラリーは(Apo-2リガンドまたは少なくとも約20-80塩基のオリゴヌクレオチドに対する抗体のような)プローブを用いてスクリーニングができ、そのプローブは興味ある遺伝子またはそれによってコードされるタンパク質を同定するためにデザインされている。オリゴヌクレオチドプローブの例は実施例1に提供される。選択されたプローブを用いてcDNAまたはゲノムのライブラリーをスクリーニングすることは、Sambrook等,Molecular Cloning:A Laboratory Manual(ニューヨーク:Cold Spring Harbor Laboratory Press,1989)に記述されているような標準的な方法を用いて実施されるであろう。Apo-2リガンドをコードする遺伝子の単離のための代わりの方法は、PCR方法体系を用いることである[Smabrook等,上記参照;Dieffenbach等,PCR Primer:A Laboratory Manual(Cold Spring Harbor Laboratory Press,1995)]。
【0038】
スクリーニングの一つの好ましい方法として、様々なヒト組織由来のcDNAライブラリーをスクリーニングするために、選択されたオリゴヌクレオチド配列を用いる。以下の実施例1は2の異なるオリゴヌクレオチドプローブを用いたcDNAライブラリーのスクリーニング法を記述する。プローブとして選択されたオリゴヌクレオチド配列は、偽のポジティブを最小化するのに十分な長さで十分に明白であるべきである。オリゴヌクレオチドはスクリーニングされるライブラリー内のDNAとのハイブリダイズに対して検出できるように好ましくはラベルされている。ラベルの方法は本技術ではよく知られており、32P-ラベルATP,ビオチン化または酵素ラベルのようなラジオラベルの使用が含まれる。全てのタンパク質をコードする配列を持つ核酸は、cDNAに逆転写されていないであろうmRNAの前駆体またはプロセッシングされた中間体を検出するために、ここで開示されるアミノ酸配列を用いて、およびもし必要ならば、Sambrook等,上記参照、に記述されているような一般的なプライマー伸長法を用いて、選択されたcDNAまたはゲノムライブラリーのスクリーニングによって得られるだろう。
【0039】
Apo-2リガンドのアミノ酸配列変異体は、Apo-2リガンドDNAに適した核酸の変化を導入することによって、または望ましいApo-2リガンドポリペプチドの合成によって調製することができる。該変異体は、細胞内領域、膜貫通領域、または細胞外領域、あるいは図1AのフルレングスApo-2リガンドに示されるアミノ酸配列の片方または両末端において、残基の挿入、置換及び/または欠失しているものをを表す。挿入、置換及び/または欠失のいかなる組み合わせも、最終構成物に到達するためになされ、ここで定義されるような望ましいアポトーシス活性を持つ最終構成物を提供することができる。好ましい実施態様としては、変異体はApo-2リガンドの細胞内、膜貫通、または細胞外領域、あるいはApo-2リガンドに対するフルレングス配列についてここで同定された配列と、少なくとも約80%の配列同一性、より好ましくは約90%の配列同一性、そしてさらにより好ましくは約95%の配列同一性をもつ。アミノ酸変化はまたグリコシル化部位の数または位置を変化する、または膜埋め込み性質を改変するようなApo-2リガンドの翻訳後プロセッシングを改変するであろう。
【0040】
上述のようなApo-2リガンドの配列のバリエーションは、米国特許第5,364,934号に示されている保存的および非保存的ミュータントに対する技術そしてガイドラインのいかなるものかを用いてなされうる。これらはオリゴヌクレオチド介在性(サイトディレクト)ミュータジェネシス、アラニンスキャニングそしてPCRミュータジェネシスを含む。
【0041】
2. 複製ベクターへの核酸の挿入
天然のまたは変異体Apo-2リガンドをコードする核酸(例えばcDNAまたはゲノムDNA)は、さらなるクローニング(DNAの増幅)のため、または発現のための複製ベクターに挿入することができる。様々なベクターが公に入手可能である。ベクター構成要素は一般的に、しかし限定することなく以下の一つまたはそれ以上を含む:シグナル配列、複製オリジン、一つかそれ以上のマーカー遺伝子、エンハンサーエレメント、プロモーターそして転写終結配列があり、それぞれが以下に記述されている。
【0042】
(i) シグナル配列構成要素
Apo-2リガンドは直接だけでなく、ヘテロ構造のポリペプチドを用いた融合ポリペプチドとしても組換え的に生産でき、そのヘテロ構造のポリペプチドは天然のタンパク質またはポリペプチドのN末端に特異的な切断部位を持つシグナル配列または他のポリペプチドであり得る。一般的には、シグナル配列はベクターの構成要素であり得、またはそれはベクターに挿入されるApo-2リガンドDNAの一部であり得る。選択されたヘテロ構造のシグナル配列は好ましくはホスト細胞によって認識されそしてプロセッシングされるものである(すなわち、シグナルぺプチダーゼによって切断される。シグナル配列は例えばアルカリホスファターゼ、ペニシリナーゼ、lppそして熱安定性エントロキシンIIリーダーから選択された原核生物のシグナル配列であり得る。酵母の分泌のためのシグナル配列は、例えば酵母インベルターゼリーダー、アルファーファクターリーダー(サッカロミセスとクルイベロミセスαファクターリーダーを含み、後者は米国特許第5,010,182号に記述されている)、または酸性ホスファターゼリーダー、C.albicansグルコアミラーゼリーダー(1990年4月4日に印刷されたEP 362,179)、または1990年11月15日に印刷されたWO 90/13646に記述されているシグナルであり得る。哺乳動物細胞発現では、哺乳動物のシグナル配列がタンパク質の分泌に向けられて用いることができ、それは同じまたは関連した種の分泌されたポリペプチド由来のシグナル配列のようなものから、例えば単純ヘルペス糖タンパク質Dシグナルのようなウイルス分泌リーダーでもよいが、in vivoでヒト細胞の細胞膜へのApo-2リガンドの挿入に直接的に向けられる天然のApo-2リガンドプレ配列でも十分である。
【0043】
該前駆体領域のためのDNAは好ましくはApo-2リガンドをコードするDNAに対するリーディングフレーム内にライゲーションされる。
【0044】
(ii) 複製オリジン構成要素
発現ベクターとクローニングベクターの両方が、一つかそれ以上の選択されたホスト細胞内でベクターの複製を可能にする核酸配列を含む。一般的にクローニングベクターではこの配列は、ベクターがホスト染色体DNAと非依存的に複製できるようにするものであり、複製のオリジンまたは自律複製配列を含む。該配列は様々な細菌、酵母そしてウイルスに対してよく知られている。プラスミドpBR322由来の複製オリジンは、ほとんどのグラムネガティブ細菌に対して適しており、2μプラスミドオリジンは酵母に適しており、そして様々なウイルスのオリジン(SV40,ポリオーマ,アデノウイルス,VSVまたはBPV)は哺乳動物細胞内でのクローニングベクターに対して有用である。一般的に複製構成物のオリジンは、哺乳動物発現ベクターに対しては必要でない(SV40オリジンは典型的にはそれが速いプロモーターを含むため用いられる)。
【0045】
ほとんどの発現ベクターは「シャトル」ベクターであり、すなわちそれらは少なくとも一つのクラスの生物内で複製できるが、発現のためもう一つの生物内にトランスフェクトすることができる。例えばベクターは大腸菌内でクローニングされ、そしてたとえそれがホスト細胞染色体と非依存的に複製できないとしても、それから同じベクターが酵母または哺乳動物細胞内にトランスフェクトされる。
【0046】
DNAはまたホストゲノム内に挿入されることによって増幅され得る。これはホストとしてバチルス種を用いれば容易に成し遂げられ、例えば、バチルスゲノムDNA内に見出される配列と相補的なDNA配列をベクター内に含ませることによって成し遂げられる。このベクターを用いたバチルスのトランスフェクションは、ゲノムとの相同的組換えおよびApo-2リガンドDNAの挿入の結果である。しかしながら、Apo-2リガンドをコードするゲノムDNAの回収は、Apo-2リガンドDNAを摘出するために制限酵素切断が必要とされるため、外来的に複製されるベクターの回収よりも複雑である。
【0047】
(iii) 選択遺伝子構成物
発現ベクターおよびクローニングベクターは典型的には、選択遺伝子、選択マーカーとも呼ばれるものを含む。この遺伝子は選択培地内で成長するトランスフォームされたホスト細胞の生存または成長のため必要なタンパク質をコードする。選択遺伝子を含まないベクターでトランスフォームされたホスト細胞は、培地内で生き残れないであろう。典型的な選択遺伝子は、(a)例えばアンピシリン、ネオマイシン、メトトレキセートまたはテトラサイクリンといった抗生物質または他の毒素に対して耐性を示すタンパク質、(b)栄養要求性欠乏を補足するタンパク質、または(c)例えばバチルスにとってのD-アラニンラセマーゼをコードする遺伝子といった複雑な培地からは入手できない重大な栄養素を供給するタンパク質をコードする。
【0048】
選択スキームの一つの例として、ホスト細胞の成長を抑える薬剤の使用がある。ヘテロ構造遺伝子を用いてうまくトランスフォームされた細胞は、薬剤耐性を示すタンパク質を生産し、それゆえ選択摂生を生き残れる。該ドミナント選択の例として、薬剤としてネオマイシン[Southern等,J.Molec.Appl.Genet.,1:327(1982)]、ミコフェノール酸[Mulligan等,Science,209:1422(1980)]またはハイグロマイシン[Sugden等,Mol.Cell.Biol.,5:410-413(1985)]を用いる。上述した該3例はそれぞれ、適切な薬剤G418またはネオマイシン(ジェネティシン)、xgpt(ミコフェノール酸)、またはハイグロマイシンに対する耐性を導くために、真核生物のコントロールの下で細菌の遺伝子を用いるものである。
【0049】
哺乳動物細胞に対して適した選択マーカーのもう一つの例は、DHFRまたはチミジンキナーゼのような、Apo-2リガンド核酸を取り込むのに適した細胞を同定できるものである。哺乳動物細胞トランスフォーマントは、トランスフォーマントだけがマーカーを取り込んだことによって唯一に生き残ることに適応する選択圧のもとに置かれる。選択圧は、培地中の選択試薬の濃度が継続的に変化するコンディションの下でトランスフォーマントを培養することによって押し付けられ、それによって選択遺伝子とApo-2リガンドをコードするDNAの両者の増幅が導かれる。増幅とは成長にとって重大なタンパク質の生産のため非常に大きな需要のある遺伝子が、組換え細胞の継続的な世代において染色体内でタンデムに繰り返されるプロセスである。Apo-2リガンドの増大した量は、増幅DNAから合成される。増幅可能な遺伝子の他の例として、メタロチオネイン-Iそして-II、アデノシンデアミナーゼそしてオルニチンデカルボキシラーゼが含まれる。
【0050】
DHFR選択遺伝子を用いてトランスフォームされた細胞は、DHFRの競合的な拮抗剤であるメトトレキセート(Mtx)を含む培地ですべてのトランスフォーマントを培養することによって最初に同定されるであろう。野生型DHFRを用いる場合に適したホスト細胞は、Urlaub等,Proc.Natl.Acad.Sci.USA,77:4216(1980)に記述されているように調製され普及している、DHFR活性に欠陥のあるチャイニーズハムスター卵巣(CHO)細胞系である。それからトランスフォームされた細胞を上昇した濃度のメトトレキセートにさらす。これによりDHFR遺伝子のマルチコピー、そしてそれに付随してApo-2リガンドをコードするDNAのような発現ベクターを含む他のDNAのマルチコピーの合成が導かる。もし例えばMtxに高い耐性であるミュータントDHFR遺伝子を用いるならば、外来性DHFRの存在にもかかわらず、この増幅法は例えばATCC番号CCL61CHO-K1などのいかなる他の適したホストを用いても使用できる(EP117,060)。
【0051】
代わりに、Apo-2リガンド、野生型DHFRタンパク質そしてアミノグリコシド3'-ホスホトランスフェラーゼ(APH)のようなもう一つの選択マーカーをコードするDNA配列を用いてトランスフォームまたはコトランスフォームされたホスト細胞(特に外来性DHFRを含む野生型ホスト)は、例えばカナマイシン、ネオマイシンまたはG418などのアミノグリコシド系抗生物質のような選択マーカーに対する選択試薬を含む培地中での細胞成長によって選択できる。米国特許第4,965,199号を参照。
【0052】
酵母を用いる場合に適した選択遺伝子は、酵母プラスミドYRp7内に存在するtrp1遺伝子である[Stinchcomb等,Nature,282:39(1979);Kingsman等,Gene,7:141(1979);Tschemper等,Gene,10:157(1980)]。trp1遺伝子は、例えばATCC番号44076またはPEP4-1のようなトリプトファン内で成長する能力を欠如した酵母のミュータント株のための選択マーカーを提供する[Jones,Genetics,85:12(1977)]。そこで酵母ホスト細胞ゲノム内のtrp1領域の存在は、トリプトファンの非存在下での成長によってトランスフォーメーションを検出するための効果的な環境を提供する。同様に、Leu2欠失酵母株(ATCC20,622または38,626)は、Leu2遺伝子をもつ既知のプラスミドによって相補される。
【0053】
加えて、1.6μm環状プラスミドpKD1由来のベクターは、クルイベロミセス酵母のトランスフォーメーションに対して用いられる[Bianchi等,Curr.Genet.,12:185(1987)]。さらに最近では、組換え仔ウシキモシンの大規模生産のための発現システムがK・ラクティスに対して報告されている[Van den Berg,Bio/Technology,8:135(1990)]。クルイベロミセスの工業的な株による成熟した組換えヒト血清アルブミンの分泌のための安定なマルチコピー発現ベクターもまた開示されている[Fleer等,Bio/Technology,9:968-975(1991)]。
【0054】
(iv) プロモーター構成要素
発現ベクターおよびクローニングベクターは、通常ホスト生物によって認識され、Apo-2リガンド核酸配列に実施可能に結合されたプロモーターを含む。プロモーターは構造遺伝子のスタートコドンの上流(5')に位置する(一般的には約100から1000bp内)非翻訳配列であり、それらが実施可能にリンクしているApo-2リガンド核酸配列のような特定の核酸配列の転写と翻訳をコントロールする。該プロモーターは典型的には、誘発性と構造性の2つに分類される。誘発性プロモーターは例えば栄養素の存在または非存在、あるいは温度変化といったいくつかの培地コンディションの変化に応じて、それのコントロールの下で増大したレベルのDNAからの転写を始めるプロモーターである。現在、様々な可能なホスト細胞によって認識される多くの数のプロモーターがよく知られている。これらのプロモーターは、制限酵素切断によって元となるDNAからプロモータを切り出し、ベクター内に単離したプロモーター配列を挿入することにより、Apo-2リガンドをコードするDNAに実施可能に結合される。天然のApo-2リガンドプロモーター配列と様々なヘテロ構造受容体プロモーターの両方が、Apo-2リガンドDNAの増幅及び/または発現に向けられて使用できるであろう。
【0055】
原核生物ホストを用いた使用にとって適したプロモーターは、β-ラクタマーゼとラクトースプロモーターシステム[Chang等,Nature,275:615(1978);Goeddel等,Nature,281:544(1979)]、アルカリホスファターゼ、トリプトファン(trp)プロモーターシステム[Goeddel,Nucleic Acids Res.,8:4057(1980);EP 36,776]そしてtacプロモーターのようなハイブリッドプロモーター[deBoer等,Proc.Natl.Acad.Sci.USA,80:21-25(1983)]を含む。しかしながら、他の既知の細菌プロモーターも適している。それらの核酸配列は印刷されており、それによって必要とされる制限部位を提供するためリンカーまたはアダプターを用いて、当業者はApo-2リガンドをコードするDNAにそれらを実施可能に結合できる[Siebenlist等,Cell,20:269(1980)]。細菌システムで用いられるプロモーターもまた、Apo-2リガンドをコードするDNAに実施可能に結合するシャインダルガノ(S.D.)配列を含むであろう。
【0056】
プロモーター配列は真核生物のためのものも知られている。事実上すべての真核生物の遺伝子が、転写開始部位からおよそ25から30塩基上流に位置するATリッチ領域を持っている。多くの遺伝子の転写の開始から70から80ベース上流に見出されるもう一つの配列は、CXCAAT領域であり、ここでXはいかなるヌクレオチドをもさす。ほとんどの真核生物遺伝子の3'末端には、AATAAA配列があり、それはコード配列の3'末端にポリAテールを付加するためのシグナルであり得る。これらの配列の全てが、真核生物発現ベクターに適するように挿入される。
【0057】
酵母ホストを用いる場合の適したプロモーター配列の例として、3-ホスホグリセリン酸キナーゼ[Hitzeman等,J.Biol.Chem.,255:2073(1980)]またはエノラーゼ、グリセルアルデヒド-3-リン酸デヒドロゲナーゼ、ヘクソキナーゼ、ピルビン酸デカルボキシラーゼ、ホスホフルクトキナーゼ、グルコース-6-リン酸イソメラーゼ、3-ホスホグリセリン酸ムターゼ、ピルビン酸キナーゼ、トリオースリン酸イソメラーゼ、ホスホグルコースイソメラーゼそしてグルコキナーゼのような他の解糖系の酵素[Hess等,J.Adv.Enzyme Reg.,7:149(1968);Holland,Biochemistry,17:4900(1978)]の各プロモーターが含まれる。
【0058】
成長コンディションによってコントロールされる転写という付加的な利点を有する誘発性プロモーターである他の酵母プロモーターは、アルコールデヒドロゲナーゼ2、イソシトクロムC、酸性ホスファターゼ、窒素代謝と関連する分解酵素、メタロチオネイン、グリセルアルデヒド-3-リン酸デヒドロゲナーゼそしてマルトースおよびガラクトース利用に関わる酵素に対する各プロモーター領域がある。酵母発現に用いる適したベクターおよびプロモーターは、さらにEP 73,657に記述されている。酵母エンハンサーもまた酵母プロモーターを用いて便利に使用される。
【0059】
哺乳動物ホスト細胞内のベクターからのApo-2リガンド転写は、例えばポリオーマイルス、鶏頭ウイルス(1989年7月5日に印刷されたUK 2,211,504)、アデノウイルス(アデノウイルス2のような)、ウシパピローマウイルス、トリ肉腫ウイルス、サイトメガロウイルス、B型肝炎ウイルスそして好ましくはシミアンウイルス40(SV40)のようなウイルスのゲノムから得られるプロモーター、以下のプロモーターがホスト細胞システムと両立できれば、例えばアクチンプロモーターまたは免疫グロブリンプロモーターなどのヘテロ構造哺乳動物プロモーター、またはヒートショックプロモーターから得られるプロモーター、あるいはApo-2リガンド配列と通常関連しているプロモーターから得られるプロモーターによってコントロールされる。
【0060】
SV40の速いおよび遅いプロモーターはSV40ウイルス複製オリジンも含むSV40制限断片として便利に得られる[Fiers等,Nature,273:113(1978);MulliganとBerg,Science,209:1422-1427(1980);Pavlakis等,Proc.Natl.Acad.Sci.USA,78:7398-7402(1981)]。ヒトサイトメガロウイルスの極端に速いプロモーターはHindIIIE制限断片として便利に得られる[Greenaway等,Gene,18:355-360(1982)]。ベクターとしてウシパピローマウイルスを用いた哺乳動物ホスト内のDNA発現システムは、米国特許第4,419,446号に開示されている。このシステムの変形は米国特許第4,601,978号に記述されている[Gray等,Nature,295:503-508(1982)のサル細胞での免疫インターフェロンをコードするcDNAの発現;Reyes等,Nature,297:598-601(1982)の単純ヘルペス由来のチミジンキナーゼプロモーターのコントロールの下でのマウス細胞でのヒトβ-インターフェロンcDNAの発現;CanaaniとBerg,Proc.Natl.Acad.Sci.USA.79:5166-5170(1982)の培養マウスおよびウサギ細胞でのヒトインターフェロンβ1遺伝子の発現;そしてGorman等,Proc.Natl.Acad.Sci.USA,79:6777-6781(1982)のラウス肉腫ウイルスの長い末端反復配列をプロモーターとして用いた、CV-1サル腎細胞、ニワトリ胚繊維芽細胞、チャイニーズハムスター卵巣細胞、ヒーラ細胞そしてNIH-3T3細胞での細菌CAT配列の発現も参照]。
【0061】
(v) エンハンサーエレメント構成要素
高等哺乳動物による本発明のApo-2リガンドをコードするDNAの転写は、ベクターにエンハンサー配列を挿入することにより増大するであろう。エンハンサーはDNAのシス作用エレメントであり、通常約10から300bpであり、その転写を増大するためにプロモーターに基づいて作用する。エンハンサーは向きと位置に比較的非依存的であり、転写ユニットの5'[Laimins等,Proc.Natl.Acad.Sci.USA,78:993(1981)]そして3'[Lusky等,Mol.Cell Bio.,3:1108(1983)]、イントロン内[Banerji等,Cell,33:729(1983)]、同様にコード配列それ自体内[Osborne等,Mol.Cell Bio.,4:1293(1984)]に見出されている。多くのエンハンサー配列が哺乳動物遺伝子(グロビン、エラスターゼ、アルブミン、α-フェトプロテインそしてインスリン)から現在では知られている。しかしながら典型的には、当業者は真核生物細胞ウイルス由来のエンハンサーを用いる。例としては、複製オリジンの後ろ側(100-270bp)にSV40エンハンサー、サイトメガロウイルスの速いプロモーターエンハンサー、複製オリジンの後ろ側にポリオーマエンハンサーそしてアデノウイルスエンハンサーが含まれる。真核生物プロモーターの活性化のためのエンハンサーエレメントについてはYaniv,Nature,297:17-18(1982)も参照。エンハンサーはApo-2リガンド-コード配列に対して5'または3'の位置でベクターに挿入されるが、好ましくはプロモーターから5'部位に位置される。
【0062】
(vi) 転写終結構成要素
真核生物ホスト細胞(酵母、菌類、昆虫、植物、動物、ヒトまたは他の多核細胞生物由来の核のある細胞)内で用いられる発現ベクターは、また転写の終結とmRNAの安定化のために必要な配列を含むであろう。該配列は、真核生物またはウイルスのDNAまたはcDNAの非翻訳領域の5'そして時には3'から共通して入手可能である。これらの領域はApo-2リガンドをコードするmRNAの非翻訳領域内のポリアデニル化断片として転写される核酸部分を含む。
【0063】
(vii) ベクターの構築と解析
一つかそれ以上の上記リストの構成要素を含む適したベクターの構築は、標準的なライゲーション法を用いる。単離したプラスミドまたはDNA断片を切断し、調製し、そして必要とされるプラスミドを作製するために望ましい形に再ライゲーションする。
【0064】
構築したプラスミドが正しい配列か確かめるための解析として、ライゲーション混合物を大腸菌K12株294(ATCC31,446)をトランスフォームすることが用いられ、そして適当かどうかアンピシリンまたはテトラサイクリン耐性によって成功したトランスフォーマントを選択できる。トランスフォーマントからプラスミドを調製し、制限エンドヌクレアーゼ切断によって解析し、、および/またはMessing等,Nucleic Acids Res.,9:309(1981)の方法またはMaxam等,Methods in Enzymology,65:499(1980)の方法によってシークエンスする。
【0065】
(viii) 一過性発現ベクター
Apo-2リガンドをコードするDNAの哺乳動物細胞内での一過性発現のために提供される発現ベクターが用いられてもよい。一般的には一過性発現は、ホスト細胞が発現ベクターの多数のコピーを蓄積し、次には発現ベクターによってコードされている望ましいポリペプチドを高レベルで合成するように、ホスト細胞内で効率よく複製可能な発現ベクターの使用を含む[Sambrook等,上記参照]。適した発現ベクターとホスト細胞を含む一過性発現システムは、クローン化したDNAによってコードされるポリぺプチドの簡便な積極的な同定を可能にし、同様に望ましい生物学的または生理学的な性質のための該ポリペプチドの速いスクリーニングを可能にする。それゆえ一過性発現システムは生物学的に活性なApo-2リガンドであるApo-2リガンドの類似体及び変異体の同定の目的に対して、本発明では特に役に立つ。
【0066】
(ix) 適した具体例としての脊椎動物細胞ベクター
組換え脊椎動物細胞カルチャー内でのApo-2リガンドの合成に対する翻案のために適した他の方法、ベクターそしてホスト細胞は、Gething等,Nature,293:620-625(1981);Mantei等,Nature,281:40-46(1979);EP 117,060;そしてEP 117,058に記述されている。Apo-2リガンドの哺乳動物細胞カルチャー発現のための特に有用なプラスミドはpRK5[EP 307,247;実施例1にも記述されている]またはpSV16B[1991年6月13日印刷されたWO 91/08291]である。
【0067】
3. ホスト細胞の選択とトランスフォーメーション
ここでベクター内のDNAをクローン化または発現するための適したホスト細胞は、上述した原核生物、酵母、高等真核生物である。この目的のため適した原核生物は、グラムネガティブまたはグラムポジティブ生物のような真正細菌を限定なく含み、例えば大腸菌のようなエシェリキア、エンテロバクター、エルヴィニア、クレブシエラ、プロテウス、ネズミチフス菌のようなサルモネラ、霊菌のようなセラチアそしてシゲラといった腸内細菌、および枯草菌とバチルスリケニフォルミス(例えば1989年4月12日に印刷されたDD266,710に開示されているバチルスリケニフォルミス41P)のようなバチルス、緑膿菌のようなシュードモナスそしてストレプトミセスも同様に含む。好ましくはホスト細胞は、タンパク質分解酵素の最小限の量を分泌すべきである。
【0068】
原核生物に加えて、糸状菌または酵母のような真核生物細菌がApo-2リガンド-コードベクターのためのクローニングホストまたは発現ホストとして適している。サッカロミセスセレビシエまたは一般的なパン酵母は、低級真核生物ホスト微生物の間で最も広く用いられている。しかしながら、多くの他の属、種そして株が、広く入手でき、ここで有用である。
【0069】
グリコシル化Apo-2リガンドの発現にとって適したホスト細胞は、多細胞生物由来のものである。該ホスト細胞は複雑なプロセッシング及びグリコシル化活性が可能である。原則として、いかなる高等細胞カルチャーも、脊椎動物カルチャー由来から非脊椎動物カルチャー由来まで実施できる。非脊椎動物細胞の例として、植物細胞及び昆虫細胞が含まれる。非常に多くのバキュロウイルス株及び変異体そしてスポドプテラフルギペルダ(芋虫)、エデスエギプティ(カ)、エデスアルボピクタス(カ)、ドロソフィラメラノガスター(ショウジョウバエ)そしてカイコガのようなホスト由来の相当する複製を許容する昆虫ホスト細胞が、同定されている[Luckow等,Bio/Technology,6:47-55(1988);Setlow等編 Genetic Engineering、第8版(Plenum Pubulishing,1986),pp.277-279内のMiller等;そしてMaeda等,Nature,315:592-594(1985)]。トランスフェクションのための様々なウイルス株が広く入手可能であり、例えばオートグラファカリフォルニカNPVのL-1変異体およびカイコガNPVのBm-5株があり、特に実施例2に記述されているSpodoptera frugiperda(「Sf9」)細胞のトランスフェクションに対して、本発明にしたがったここでのウイルスのようなウイルスが用いられ得る。
【0070】
ワタ、トウモロコシ、ポテト、ソラマメ、ペチュニア、トマトそしてタバコの植物細胞が、ホストとして利用できる。典型的には植物細胞は、細菌アグロバクテリウムツメファシエンスの特定の株と共にインキュベーションによってトランスフェクトし、その細菌はApo-2リガンド-コードDNAを含むように前もって処理されている。A.ツメファシエンスと共に植物細胞をインキュベーションする間、Apo-2リガンドをコードするDNAは、トランスフェクトされるような植物細胞ホストにトランスファーされ、そして適したコンディションの下でApo-2リガンド-コードDNAを発現するであろう。加えて植物細胞で両立できる調節配列およびシグナル配列は、ノパリンシンテースプロモーターおよびポリアデニル化シグナル配列のように入手可能である[Depicker等,J.Mol.Appl.Gen.,1:561(1982)]。加えてT-DNA780遺伝子の上流領域から単離されたDNA領域により、組換えDNAを含む植物組織内で植物で発現できる遺伝子の活性化したまたは増大した転写レベルが可能である[1989年6月21日に印刷されたEP 321,196]。
【0071】
カルチャー内の脊椎動物細胞(組織細胞)の増殖もまた、本分野でよく知られている[例えばTissue Culture,Academic Press,KruseとPatterson編(1973)参照]。有用な哺乳動物ホスト細胞系の例として、SV40(COS-7,ATCC CRL 1651)によってトランスフォームされたサル腎CV1系;ヒト胚腎系(293細胞または懸濁液カルチャー内で成長のためサブクローン化された293細胞、Graham等,J.Gen Virol.,36:59(1977));仔ハムスター腎細胞(BHK,ATCC CCL 10);チャイニーズハムスター卵巣細胞/-DHFR(CHO,UrlaubとChasin,Proc.Natl.Acad.Sci.USA,77:4216(1980));マウスセルトリ細胞(TM4,Mather,Biol.Reprod.,23:243-251(1980));サル腎細胞(CV1 ATCC CCL 70);アフリカグリーンサル腎細胞(VERO-76,ATCCCRL-1587);ヒト子宮頚ガン細胞(HELA,ATCC CCL 2);イヌ腎細胞(MDCK,ATCC CCL 34);バッファローラット肝細胞(BRL 3A,ATCC CRL 1442);ヒト肺細胞(W138,ATCC CCL 75);ヒト肝細胞(Hep G2,HB 8065);マウス乳房腫瘍(MMT 060562,ATCC CCL 51);TRI細胞(Mather等,Annals N.Y.Acad.Sci.,383:44-68(1982));MRC5細胞;そしてFS4細胞がある。
【0072】
ホスト細胞はトランスフェクトされ、そして好ましくはApo-2リガンド生産のための上述の発現ベクターまたはクローニングベクターを用いてトランスフォームされ、そしてプロモーターの誘導、トランスフォーマントの選択または望ましい配列をコードする遺伝子の増幅のため適したように修飾されたありきたりの栄養素培地で培養される。
【0073】
トランスフェクションとは、いかなるコード配列が実際に発現されようとされまいと、ホスト細胞によって発現ベクターが取り込まれることを示す。非常に多くのトランスフェクションの方法が当業者に知られており、例えばCaPO4およびエレクトロポレーションがある。成功したトランスフェクションは一般的に、該ベクターの作用のいかなる徴候でもホスト細胞内で生じれば、認識される。
【0074】
トランスフォーメーションとは、染色体外エレメントとしてまたは染色体への組み込みによるかいずれでも、DNAを複製するために生物にDNAを導入することを示す。用いられるホスト細胞に依存して、トランスフォーメーションは該細胞に適した標準的な方法を用いてなされる。Sambrook等,上記参照内で記述されているような塩化カルシウムを用いたカルシウム処理またはエレクトロポレーションが、原核生物または実質的な細胞壁障害を持つ他の細胞のために使用される。アグロバクテリウムツメファシエンスを用いた感染は、Shaw等,Gene,23:315(1983)と1989年6月29日に印刷されたWO 89/05859に記述されているように、特定の植物細胞のトランスフォーメーションのために使用される。加えて植物は、1991年1月10日に印刷されたWO 91/00358に記述されているような超音波処理を用いてトランスフェクトしてもよい。
【0075】
該細胞壁を持たない哺乳動物細胞に対しては、Grahamとvan der Eb,Virology,52:456-457(1978)のリン酸カルシウム沈殿法が好ましい。哺乳動物細胞ホストシステムトランスフォーメーションの一般的な面としては、米国特許第4,399,216号に記述がある。酵母へのトランスフォーメーションは典型的には、Van Solingen等,J.Bact.,130:946(1977)とHsiao等,Proc.Natl.Acad.Sci.(USA),76:3829(1979)の方法にしたがって実施される。しかしながら、核マイクロインジェクション、エレクトロポレーション、完全な細胞への細菌プロトプラスト融合または例えばポリプレン、ポリオルニチンというポリカチオンのような、他の細胞へのDNA導入法もまた用いることができる。哺乳動物細胞をトランスフォームする様々な方法に対しては、Keown等,Methods in Enzymology,185:527-537(1990)とMansour等,Nature,336:348-352(1988)を参照。
【0076】
4. ホスト細胞の培養
Apo-2リガンドを生産するために用いられる原核生物細胞は、Sambrook等,上記参照で一般的に記述されている適した培地内で培養できる。
【0077】
Apo-2リガンドを生産するため用いられる哺乳動物ホスト細胞は、様々な培地で培養できる。商業的に入手可能な培地の例としては、Ham's F10(Sigma)、Minimal Essential Medium("MEM",Sigma)、PRMI-1640(Sigma)そしてダルベッコ変法イーグル培地("DMEM",Sigma)が含まれる。いかなる該培地も、ホルモンおよび/または増殖因子(インスリン、トランスフェリンまたは上皮増殖因子のような)、塩類(塩化ナトリウム、カルシウム、マグネシウムそしてリン酸塩のような)、バッファー(HEPESのような)、ヌクレオシド(アデノシン及びチミジンのような)、抗生物質(ゲンタマイシンTM薬剤のような)、微量元素(マイクロモーラー範囲で最終濃度に通常存在する無機化合物として定義される)及びグルコースまたは同等のエネルギー源が必要であるとして補われる。いかなる他のサプリメントであっても、当業者に既知であろう適した最終濃度で含んでよい。温度、pH、それに類するもののような培養条件は、発現のため選択されたホスト細胞に事前に用いたものであり、当業者には明白であろう。
【0078】
一般的に、哺乳動物細胞カルチャーの生産力を最大化するための原理、プロトコールそして実施上の技術は、Mammalian Cell Biotechnology:a Practical Approach,M.Butler編,(IRL Press,1991)に見出すことができる。
【0079】
この開示で示されるホスト細胞は、カルチャー内の細胞だけでなく、ホスト動物内にある細胞をも包含する。
【0080】
5. 遺伝子増幅/発現の検出
遺伝子増幅および/または発現は、ここで提供される配列に基づく適切なラベルプローブを用いて、例えばありきたりのサザンブロッティング、mRNAの転写量を測定するにはノーザンブロッティング[Thomas,Proc.Natl.Acad.Sci.USA,77:5201-5205(1980)]、ドットブロッティング(DNA解析)またはin situハイブリダイゼーションによって直接にサンプル内で測定することができる。様々なラベルが用いられうるが、最も一般的にはラジオアイソトープ、特に32Pがある。しかしながら、ポリヌクレオチドへの導入のためビオチン修飾ヌクレオチドを用いるように、他の方法もまた用いられる。すなわちビオチンは、アビジンまたは抗体に対する結合のための部位として用いられ、ラジオヌクレオチド、蛍光剤または酵素のような広く様々なラベルを用いてラベルすることが可能である。代わりにDNA二重鎖、RNA二重鎖そしてDNA-RNAハイブリッド二重鎖またはDNA-タンパク質二重鎖を含む、特異的な二重鎖を認識することができる抗体を用いてもよい。次に抗体はラベルされ、表面での二重鎖の形態により、二重鎖に結合した抗体の存在が検出できるために、二重鎖が表面のどこに結合しているかのアッセイを実施できる。
【0081】
代わりに遺伝子発現は、遺伝子産物の発現を直接測定するために、細胞または組織切片の免疫組織化学的染色および細胞カルチャーまたは体液のアッセイのような、免疫学的方法で測定することができる。免疫組織化学的染色法を用いると、細胞サンプルは典型的には、脱水および固定によって調製され、遺伝子産物と特異的に結合するラベル抗体を用いた反応が続くが、そのラベルは酵素的ラベル、蛍光ラベルまたは発光ラベル等のような通常視覚的に検出されるものである。
【0082】
免疫組織化学的染色および/またはサンプル体液のアッセイのために有効な抗体は、モノクローナルでもポリクローナルでもよく、いかなる動物内でも調製できる。簡便には該抗体は天然のApo-2リガンドポリペプチドに対して、またはここで提供されるDNA配列に基づく合成ペプチドに対して、あるいはApo-2リガンドDNAと融合され特異的抗体エピトープをコードする外因性配列に対して調製され得る。
【0083】
6. Apo-2リガンドポリペプチドの精製
Apo-2リガンドは分泌シグナル無しで直接生産されるとホスト細胞溶解産物から回収されるが、好ましくはHGF受容体拮抗剤は分泌ポリペプチドとして培地から回収される。もしApo-2リガンドが膜結合型であれば、適した洗浄剤溶液(例えばTriton-X100)を用いて膜からそれを引き離すことができ、あるいはその細胞外領域を酵素的な切断によって引き離すことができる。
【0084】
Apo-2リガンドがヒト由来の細胞以外の組換え細胞内で生産された場合には、Apo-2リガンドはヒト由来のタンパク質またはポリペプチドと関連がない。しかしながら、Apo-2リガンドとして実質的にホモ構造である調製を得るために、組換え細胞タンパク質またはポリペプチドからApo-2リガンドを精製することが望ましい。第一段階として、粒子状の細胞破片を除去するために、培地または溶解産物を遠心分離するとよい。その後Apo-2リガンドを水溶性タンパク質およびポリペプチドの混合物から精製するが、以下の方法は適した精製法の典型例である:イオン交換カラムを用いての分別;エタノール沈殿;逆相HPLC;シリカまたはDEAEのようなカチオン交換樹脂を用いてのクロマトグラフィー;等電点電気泳動;SDS-PAGE;硫安沈降;例えばSephadexG-75を用いたゲル濾過;そしてIgGのような不純物を除去するためのプロテインA Sepharoseカラム。
【0085】
好ましい実施態様として、Apo-2リガンドは実施例3に記述されるようなアフィニティークロマトグラフィーによって単離され得る。
【0086】
その残基が欠失、挿入、または置換されているApo-2リガンド変異体は、該変異値によって引き起こされる性質におけるいかなる実質的な変化をも考慮に入れて、天然のApo-2リガンドと同様な方式で回収される。
【0087】
フェニルメチルスルホニルフルオリド(PMSF)のようなプロテアーゼ阻害剤もまた、精製の間のタンパク質分解を阻害するのに有用であり、そして抗生物質も外来的な不純物の成長を妨げるために含まれよう。組換え細胞カルチャーでの発現に対してApo-2リガンドまたはその変異体の性質の変化を考慮して、天然のApo-2リガンドに適した精製方法が必要とされるであろうことは当業者に予測できよう。
【0088】
7. Apo-2リガンドポリペプチドの共有結合修飾
Apo-2リガンドの共有結合修飾は、本発明の範囲に含まれる。天然のApo-2リガンド及びApo-2リガンドのアミノ酸配列変異体の両者が共有結合で修飾され得る。Apo-2リガンドの共有結合の一つのタイプとして、Apo-2リガンドの選択された側鎖またはNあるいはC末端残基に反応可能な有機誘導体化試薬を用いて、Apo-2リガンドの目標とするアミノ酸に反応させて分子を導入することがある。
【0089】
二官能基を持つ誘導体化は、抗Apo-2リガンド抗体の精製の方法に用いる非水溶性支持体マトリックスまたは表面に対してApo-2リガンドを架橋するのに有用であり、逆もまた然りである。広く用いられている架橋試薬は、例えば1,1-ビス(ジアゾアセチル)-2-フェニルエタン、グルタルアルデヒド、例えば4-アジドサリチル酸のエステルといったN-ヒドロキシスクシンイミドエステル、3,3'-ジチオビス(スクシンイミジルプロピオネート)のようなジスクシンイミジルエステルを含む、ホモ二官能イミドエステル、そしてビス-N-マレイミド-1,8-オクタンのような二官能マレイミドを含む。メチル-3-[(p-アジドフェニル)ジチオ]プロピオイミデートのような誘導体化試薬は、光の存在の下で架橋を形成することが可能な光活性化中間体を生じる。代わりに、米国特許第3,969,287号;第3,691,016号;第4,195,128号;第4,247,642号;第4,229,537号そして第4,330,440号に記述されている臭化シアン-活性化炭化水素のような反応性非水溶性マトリックス及び反応基質が、タンパク質固定のために用いられる。
【0090】
他の修飾は、それぞれグルタミル残基およびアスパルチル残基に相当するグルタミニル残基およびアスパラギニル残基の脱アミド化、プロリンおよびリシンのヒドロキシル化、セリル残基またはトレオニル残基のヒドロキシル基のリン酸化、リシン、アルギニン及びヒスチジン側鎖のα-アミノ基のメチル化[T.E.Creighton,Proteins:Structure and Molecular Properties,W.H.Freeman & Co.,サンフランシスコ、pp.79-86(1983)]、N末端アミンのアセチル化およびC末端カルボキシル基の脱アミド化を含む。残基の修飾された形態は本発明の範囲に含まれる。
【0091】
本発明の範囲内に含まれるApo-2リガンドポリペプチドの共有結合の修飾のもう一つのタイプは、ポリペプチドの天然のグリコシル化パターンを変更することを含む。「天然のグリコシル化パターンを変更する」ということは、天然のApo-2リガンドに見出される一つかそれ以上の炭化水素部分を欠失すること、および/または天然のApo-2リガンドには存在しない一つかそれ以上のグリコシル化部位を加えることを意味するようここでの目的として向けられる。
【0092】
ポリペプチドのグリコシル化は、典型的にはN結合型とO結合型のいずれかである。N結合型とは、アスパラギン残基の側鎖に炭化水素部分が付着することを示す。アスパラギン-X-セリン及びアスパラギン-X-トレオニンというトリペプチドで、Xはプロリン以外のいかなるアミノ酸でもよいものが、アスパラギン側鎖に炭化水素部分を酵素的に付着するための認識配列である。それゆえポリペプチド内にこれらのトリペプチド配列のどちらかが存在すると、潜在的なグリコシル化部位を作り出し得る。O結合型グリコシル化は、ヒドロキシルアミノ酸、最も共通にはセリンまたはトレオニンに対して、N-アセチルガラクトサミン、ガラクトースまたはキシロースの糖類の一つが付着することを示すが、5-ヒドロキシプロリンまたはヒドロキシリシンもまた付着部位に使われ得る。
【0093】
Apo-2リガンドポリペプチドへのグリコシル化部位の付加は、(N結合型グリコシル化部位の場合)上述のトリペプチド配列の一つかそれ以上を含むようにアミノ酸配列を変更することによって成し遂げられる。変更は(O結合型グリコシル化部位の場合)、天然のApo-2リガンド配列に一つかそれ以上のセリンまたはトレオニン残基を付加または置換することによってもなし得る。Apo-2リガンドアミノ酸配列は、DNAレベルでの変化、特にコドンを望ましいアミノ酸に翻訳されるように選択するように、前もって選ばれた塩基にApo-2リガンドポリペプチドをコードするDNAを突然変異させることによって、任意に変化し得る。DNAミューテーション(類)は、上述した方法または米国特許第5,364,934号、上記参照内の方法を用いてなし得る。
【0094】
Apo-2リガンドポリペプチド内の炭化水素部分の数を増大するもう一つの方法は、ポリペプチドへのグリコシドの化学的または酵素的結合によるものである。用いられる結合方法に依存して、糖(類)は(a)アルギニンおよびヒスチジン、(b)フリーカルボキシル基、(c)システインのもののようなフリースルフヒドリル基、(d)セリン、トレオニンまたはヒドロキシプロリンのもののようなフリーヒドロキシル基、(e)フェニルアラニン、チロシンまたはトリプトファンのもののような芳香族残基、または(f)グルタミンのアミド基に付着し得る。これらの方法は1987年9月11日に印刷されたWO 87/05330およびAplinとWriston,CRC Crit.Rev.Biochem.,pp.259-306(1981)に記述されている。
【0095】
Apo-2リガンドポリペプチド内に存在する炭化水素部分の除去は、化学的にまたは酵素学的に、あるいはグリコシル化のターゲットとして機能するアミノ酸残基をコードするコドンの突然変異的置換によって成し遂げられうる。例えば化合物トリフルオロメタンスルホン酸または同等の化合物にポリペプチドをさらすことによる化学的な脱グリコシル化は、ポリペプチドは完全なままである一方、結合糖(N-アセチルグルコサミドまたはN-アセチルガラクトサミド)を除いて、ほとんどまたは全ての糖を切断することを引き起こす。化学的な脱グリコシル化については、Hakimuddin等,Arch.Biochem.Biophys.,259:52(1987)およびEdge等,Anal.Biochem.,118:131(1981)に記述がある。ポリペプチドの炭化水素部分の酵素的な切断は、Thotakura等,Meth.Enzymol.,138:350(1987)に記述されているように様々なエンドグリコシダーゼおよびエクソグリコシダーゼの使用により達成できる。
【0096】
潜在的グリコシル化部位のグリコシル化は、Duskin等,J.Biol.Chem.,257:3105(1982)に記述されているように化合物ツニカマイシンの使用により妨げられる。ツニカマイシンはタンパク質-N-グリコシド結合の形成をブロックする。
【0097】
Apo-2リガンドの共有結合修飾のもう一つのタイプは、米国特許第4,640,835号;第4,496,689号;第4,301,144号;第4,670,417号;第4,791,192号;または第4,179,337号に示された方法で、例えばポリエチレングリコール、ポリプロピレングリコールまたはポリオキシアルキレンといった様々な非タンパク質性ポリマーの一つにApo-2リガンドポリペプチドを結合することを含む。
【0098】
8. エピトープタグ化Apo-2リガンド
本発明はまた、もう一つのヘテロ構造ポリペプチドに融合されたApo-2リガンドを含むキメラポリペプチドをも提供する。一つの実施態様として、キメラポリペプチドはタグポリペプチドとのApo-2リガンドの融合を含み、該タグポリペプチドは抗タグ抗体が選択的に結合できるエピトープを提供する。エピトープタグは一般的にApo-2リガンドのアミノ末端またはカルボキシル末端に置かれる。Apo-2リガンドの該エピトープタグ型の存在は、タグポリペプチドに対する抗体を用いて検出できる。またエピトープタグの提供は、抗タグ抗体を用いるアフィニティー精製またはエピトープタグに結合するもう一つのタイプのアフィニティーマトリックスによって、Apo-2リガンドを容易に精製することを可能にする。
【0099】
様々なタグポリペプチドとそれら各自の抗体が本分野ではよく知られている。例として、flu HAタグポリペプチドとその抗体12CA5[Field等,Mol.Cell.Biol.,8:2159-2165(1988)];c-mycタグとそれらの抗体8F9,3C7,6E10,G4,B7そして9E10[Evan等,Molecular and Cellular Biology,5:3610-3616(1985)];そして単純ヘルペスウイルス糖タンパク質D(gD)タグとその抗体[Paborsky等,Protein Engineering,3(6):547-553(1990)]を含む。他のタグポリペプチドとしては、Flag-ペプチド[Hopp等,BioTechnology,6:1204-1210(1988)];KT3エピトープペプチド[Martin等,Science,255:192-194(1992)];αチューブリンエピトープペプチド[Skinner等,J.Biol.Chem.,266:15163-15166(1991)];そしてT7遺伝子10タンパク質ペプチドタグ[Lutz-Freyermuth等,Proc.Natl.Acad.Sci.USA,87:6393-6397(1990)]が含まれる。一度タグポリペプチドが選択されれば、それに対する抗体はここで開示される方法を用いて作製できる。
【0100】
一般的に、エピトープタグ化されたApo-2リガンドは、天然の及び変異体Apo-2リガンドに対して上述の方法にしたがって構築され生産され得る。Apo-2リガンド-タグポリぺプチ融合体は、好ましくはタグポリペプチドDNA配列にフレーム中でApo-2リガンド部分をコードするcDNA配列を融合し、そして適切なホスト細胞内で合成されたDNA融合合成物を発現することによって構築される。通常本発明のApo-2リガンド-タグポリペプチドキメラを調製する場合、タグポリペプチドのN末端をコードする核酸にApo-2リガンドの3'末端を融合するであろうが、しかしながら5'での融合もまた可能である。エピトープタグ化Apo-2リガンドの例は、以下の実施例2にさらに詳細に記述されている。
【0101】
エピトープタグ化Apo-2リガンドは、抗タグ抗体を用いたアフィニティークロマトグラフィーによって精製できる。アフィニティー抗体が付着しているマトリックスは、例えばアガロース、制御されたポアグラスまたはポリ(スチレンジビニル)ベンゼンを含むであろう。それからエピトープタグ化されたApo-2リガンドは、本分野で既知の方法を用いてアフィニティーカラムから溶出されることができる。
【0102】
B. Apo-2リガンドの治療上の使用
本明細書で開示されているApo-2リガンドは哺乳動物細胞においてアポトーシスを誘導するために治療上で用いられ得る。一般的に哺乳動物細胞においてアポトーシスを誘導するために方法には、細胞を有効量のApo-2リガンドにさらすことが含まれる。これは例えば以下に記述された方法及び実施例に記述された方法にしたがってin vivoまたはex vivoで成し遂げられ得る。アポトーシスを誘導する方法は、アポトーシスの減少したレベルによって特徴付けられる特定の生理学的コンディションに対する治療で用いられ得ることが予期される。該生理学的コンディションの例としては、狼瘡及び免疫介在性糸球体腎炎のような自己免疫疾患、及びガンが含まれる。ガンの治療に対するApo-2リガンドの治療上の応用は以下に詳細に記述されている。
【0103】
ガンの治療法において、Apo-2リガンドはガンを持つと診断された哺乳動物に投与される。Apo-2リガンドは他のアポトーシス誘導試薬、化学療法、放射線療法、及び手術を含むいかなる他の治療構成要素及び技術と組み合わせて用いることもできることはもちろん企図される。
【0104】
Apo-2リガンドは好ましくは、製薬学的に許容できるキャリー内で哺乳動物に投与される。適したキャリアーとその処方については、Oslo等によって編集されたRemington's Pharmaceutical Sciences,第16版、1980、Mark Publishing Co.に記述されている。典型的には適切な量の製薬学的に許容できる塩が、等浸透圧の処方にするための処方において用いられる。製薬学的に許容できるキャリアーの例として、塩水、リンガー溶液そしてブドウ糖溶液がある。溶液のpHは好ましくは約5から約8、そしてより好ましくは約7から約7.8である。特定のキャリアーが例えば投与の経路及び投与されるApo-2リガンドの濃度に依存してより好ましいということは、当業者には明らかであろう。
【0105】
Apo-2リガンドは注射(静脈、腹膜、皮下、筋肉の)、または効果的な形態で血流への輸送を確保する点滴のような他の方法によって、哺乳動物に投与される。Apo-2リガンドがin vivoまたはex vivoでの遺伝子治療によって投与され得ることもまた予期されよう。
【0106】
Apo-2リガンドを投与するための有効投与量とスケジュールは経験的に決まり、該決定をすることは当業者の手中にある。単独で用いられるApo-2リガンドの有効投与量または量は体重当たり1μg/kgから100mg/kgの範囲にあり、または一日当たりではそれ以上であろうと現在では考えられている。投与量の種間スケールは例えばMordenti等,Pharmaceut.Res.,8:1351(1991)に開示されているように本分野で周知の方法で実施され得る。投与されなければならないApo-2リガンドの投与量は、例えばApo-2リガンドを受けるであろう哺乳動物、投与経路、及び哺乳動物に投与される他の薬剤または治療に依存して変化するであろうことは当業者に理解されよう。
【0107】
哺乳動物に投与される一つ以上の他の治療は、化学療法及び/または放射線療法、免疫アジュバント、サイトカイン、及び抗体ベース治療を制限することなく含む。例として、インターロイキン(例えばIL-1,IL-2,IL-3,IL-6)、白血病阻害因子、インターフェロン、TGF-ベータ、エリトロポエチン、トロンボポエチン、抗VEGF抗体及びHER-2抗体が含まれる。哺乳動物細胞においてアポトーシスを誘導することが知られている他の試薬も用いられ得、該試薬にはTNF-α、TNF-β(リンホトキシン-α)、CD30リガンド、4-1BB、及びApo-1リガンドが含まれる。
【0108】
本発明によって企図される化学療法には、Doxorubicin,5-Fluorouracil("5-FU"),エトポシド,カンプトセシン(camptothecin),Leucovorin,Cytosine arabinoside("Ara-C"),Cyclophosphamide,Thiotepa,Busulfan,Cytoxin,Taxol,Methotrexate,Cisplatin,Melphalan,Vinblastine及びCarboplatinのような、本分野で周知であり、商業的に入手可能な化学的物質または薬剤が含まれる。該化学療法に対する調製と投与スケジュールは、製品の説明書にしたがって用いられるか、または当業者に経験的に決定されるであろう。該化学療法の調製と投与スケジュールはまたChemotherapy Service,M.C.Perry編,Williams & Wilkins,Baltimore,MD(1992)にも記述されている。
【0109】
化学療法はApo-2リガンドに対して上記したような製薬学的に許容できるキャリアーにおいて好ましくは投与される。化学療法の投与形態はApo-2リガンドに対して用いられるものと同様であり、または異なる形態でそれは哺乳動物に投与され得る。例えばApo-2リガンドは注射される一方で、化学療法は哺乳動物に経口で投与される。Apo-2リガンドと組み合わせた化学療法の投与形態は以下の実施例9-12にさらに詳細に記述されている。
【0110】
放射線治療は本分野で商業的に用いられ当業者に周知のプロトコールにしたがって哺乳動物に投与され得る。該治療にはセシウム、イリジウム、ヨウ素、またはコバルト放射線が含まれ得る。放射線治療は全身体的な照射によってなされ得、または体の内外の特異的な部位または組織に局所的に向けられ得る。典型的には放射線治療は約1から2週の期間にわたる間隔で投与される。しかしながら放射線治療はより長い期間にわたって投与され得る。任意に放射線治療は単一の投与または複数投与、連続的投与として投与され得る。
【0111】
Apo-2リガンドは及び一つ以上の他の治療は哺乳動物に同時的にまたは連続的に投与され得る。哺乳動物へのApo-2リガンド及び一つ以上の他の治療の投与に引き続いて、哺乳動物のガン及び生理学的なコンディションは当業者によく知られた様々な方法でモニターされ得る。例えば腫瘍の大きさは生検または標準的なx線イメージ法によって物理的に観察し得る。
【0112】
Apo-2リガンドはex vivoでガン細胞を治療するために用いられ得ることが企図される。該ex vivo治療は、骨髄移植および特に異種骨髄移植において有用であろう。例えばApo-2リガンドを用いて、及び場合により上記記述されているような一つ以上の他の治療薬をも用いて、ガン細胞を含む細胞または組織(類)の治療をすることは、受容者哺乳動物における移植前に、アポトーシスを誘導するため、そしてガン細胞を実質的に除去するために用いられ得る。
【0113】
ガン細胞を含む細胞または組織(類)は最初にドナー哺乳動物から得られる。細胞または組織(類)は外科的に得られ、好ましくは無菌的に得られる。移植のため骨髄を治療する方法では、骨髄は哺乳動物から針吸引によって得られる。それからガン細胞を含む細胞または組織(類)をApo-2リガンドを用いて治療し、任意に上述のような一つ以上の他の治療を伴う。骨髄は好ましくはApo-2リガンドを用いた治療の前に、単核細胞画分を得るために(ficoll-hypaque勾配による遠心分離によるように)好ましくは分画される。
【0114】
それから治療された細胞または組織(類)を受容者哺乳動物に注入または移植し得る。受容者哺乳動物はドナー哺乳動物と同じ個体であり得、またはもう一つの異種個体でもあり得る。自己骨髄移植に対しては、哺乳動物を移植の前に本分野で周知の、例えばAutologous Bone Marrow Transplantation: Proceedings of the Third International Symposium,Dicke等,編,University of Texas M.D. Anderson Hospital and Tumor Institute内に記述されている有効量の放射線または化学療法を用いて治療する。
【0115】
C. Apo-2リガンドの非治療上の使用
本発明のApo-2リガンドはまた非治療上の応用においても有用性を持つ。Apo-2リガンドをコードする核酸配列は組織特異的タイプ化のための診断薬として用い得る。例えばin situハイブリダイゼーション、ノーザン及びサザンブロッティング、及びPCR分析のような方法が、Apo-2リガンドをコードするDNA及び/またはRNAが試験される細胞タイプ(類)に存在するかどうかを調べるために用いられ得る。Apo-2リガンド核酸はまたここで記述される組換え法によってApo-2ポリペプチドの調製のためにも有用であろう。
【0116】
単離されたApo-2リガンドはApo-2リガンドの未知の量を含むサンプルが調整される際のコントロールとして定量的診断アッセイでも用いられ得る。Apo-2リガンド調製物はまた、Apo-2リガンドに対するアッセイ(例えばラジオイムノアッセイ、ラジオリセプターアッセイ、または酵素リンクイムノアッセイにおけるスタンダードとしての使用のためにApo-2リガンドをラベルすることによって)、例えばApo-2リガンドを結合する受容体を同定または単離する際のアフィニティー精製法、及び例えば放射性ヨウ素、酵素、またはフルオロフォアーでラベルした場合競合的タイプ受容体結合アッセイにおいて、スタンダードとして抗体を生産するのに有用でもある。
【0117】
Apo-2リガンドをコードする核酸はまたトランスジェニック動物または「ノックアウト」動物のそれぞれを生産するために用いられ得るが、該動物は各々治療上の有用な試薬の開発またはスクリーニングに有用である。トランスジェニック動物(例えばマウスまたはラット)は、トランス遺伝子を含む細胞を持つ動物であり、そのトランス遺伝子は動物内または例えば胚段階といった元となる動物の祖先に導入されたものである。トランス遺伝子はトランスジェニック動物が発達する細胞のゲノム内にインテグレートされたDNAである。一つの実施態様として、Apo-2リガンドをコードするcDNAまたはそれの適切な配列は、確立された方法にしたがってApo-2リガンドをコードするゲノムDNAをクローン化するために用いられ得、該ゲノム配列はApo-2リガンドをコードするDNAを発現する細胞を含むトランスジェニック動物を生産するために用いられる。トランスジェニック動物、特にマウスまたはラットのような動物を生産する方法は、本分野でありきたりのものになっており、例えば米国特許第4,736,866号及び第4,870,009号に記述されている。典型的には特定の細胞を組織特異的エンハンサーと共同してApo-2リガンドトランス遺伝子に対してターゲット化するであろう。胚段階で動物の生殖細胞系列内に導入されたApo-2リガンドをコードするトランス遺伝子の一コピーを含むトランスジェニック動物を、Apo-2リガンドをコードするDNAの増大した発現の影響を調べるため用いることができる。
【0118】
代わりに、Apo-2リガンドの非ヒト相同物を、Apo-2リガンドをコードする異種遺伝子と、動物の胚細胞内に導入されたApo-2リガンドをコードする改変されたゲノムDNAの間の相同的組換えの結果として、Apo-2リガンドをコードする不完全なまたは改変された遺伝子を持つApo-2リガンド「ノックアウト」動物を構築するために用いられ得る。例えばApo-2リガンドをコードするcDNAは、確立した方法にしたがってApo-2リガンドをコードするゲノムDNAをクローン化するために用いられ得る。Apo-2リガンドをコードするゲノムDNAの部分を、インテグレーションをモニターするために用いられ得る選択可能マーカーをコードする遺伝子のような、もう一つの遺伝子で欠失または置換し得る。典型的には非改変フランキングDNA(5'末端と3'末端の両方)の数キロベースを、ベクター内に取り込ませる[相同的組換えベクターの記述については、例えばThomas及びCapecchi,Cell,51:503(1987)参照]。該ベクターは胚幹細胞系(例えばエレクトロポレーションによって)内に導入され、導入されるDNAが内因性のDNAと相同的に組換える細胞が選択される[例えばLi等,Cell,69:915(1992)参照]。それから選択された細胞を集合キメラを形成するために動物(例えばマウスまたはラット)の胚胎盤内に注入する[例えばBradley,in Teratocarcinomas and Embryonic Stem Cells:A Practical Approach,E.J.Robertson,編(IRL,Oxford,1987),pp.113-152]。それからキメラの胚を適した偽妊娠メス里親動物内に移植し、該胚は「ノックアウト」動物を作製するための期間を運ぶ。その生殖細胞に相同的組換えDNAを運んでいる子孫を、標準的な方法で同定し、該動物の全ての細胞が相同的組換えDNAを含む動物を繁殖させるために用い得る。ノックアウト動物は例えば、特定の病理学上のコンディションに対して防御する能力によって、及びApo-2リガンドポリペプチドの不存在のため病理学的コンディションの発展によって特性指摘され得る。
【0119】
D. 抗Apo-2リガンド抗体調製
本発明はさらに抗Apo-2リガンド抗体を提供する。Apo-2リガンドに対する抗体は以下のように調製され得る。例示的な抗体には、ポリクローナル、モノクローナル、ヒト化、二重特異性、及びヘテロ接合の各抗体が含まれる。
【0120】
1. ポリクローナル抗体
Apo-2リガンド抗体にはポリクローナル抗体が含まれる。ポリクローナル抗体の調製法は当業者に周知である。ポリクローナル抗体は例えば免疫化試薬と必要であればアジュバントの一度以上の注射により哺乳動物において生産され得る。典型的には該免疫か試薬及び/またはアジュバントは、複数の皮下または腹膜内の注射により哺乳動物において注射されるであろう。免疫化試薬には、Apo-2リガンドポリペプチドまたはその融合タンパク質が含まれる。免疫化試薬を免疫化される哺乳動物において免疫原性であることが知られているタンパク質に接合することは、有用であろう。用いられるであろう該免疫原性タンパク質の例としては、キーホールリンペットヘモシアニン、血清アルブミン、ウシチログロブリン、及び大豆トリプシンインヒビターが制限されることなく含まれる。ミョウバンのような凝集試薬もまた、哺乳動物の免疫応答を増大するために用いられ得るであろう。用いられるであろうアジュバントの例としては、フロイント完全アジュバント及びMPL-TDMアジュバント(モノホスホリルリピドA,合成トレハロースジコリノミコレート)が含まれる。免疫化プロトコールは過度の実験なく当業者に選択され得る。それから該哺乳動物を採血し、抗体力価のため血清をアッセイし得る。必要であれば、哺乳動物を抗体力価が増大するまたは安定水準に達するまで追加免疫し得る。
【0121】
2. モノクローナル抗体
代わりにApo-2リガンド抗体はモノクローナル抗体であり得る。モノクローナル抗体はKohler及びMilstein,Nature,256:495(1975)に記述されているようなハイブリドーマ法を用いて調製され得る。ハイブリドーマ法においては、マウス、ハムスター、または他の適切なホスト動物が、免疫化試薬と特異的に結合するであろう抗体を生産するまたは生産し得るリンパ球を引き出すために免疫化試薬と(上述のように)典型的に免疫化される。代わりに、リンパ球はin vitroで免疫化され得る。
【0122】
免疫化試薬にはApo-2リガンドポリペプチドまたはその融合タンパク質が典型的には含まれる。その表面でApo-2リガンドを発現する細胞もまた用いられ得る。一般的にもしヒト起源の細胞が必要であれば、末梢血リンパ球(「PBL」)が用いられ、もし非ヒト哺乳動物ソースが必要であれば、脾臓細胞またはリンパ節細胞が用いられる。それからハイブリドーマ細胞を形成するためにポリエチレングリコールのような適当な融合試薬を用いて、該リンパ球を不朽化細胞系と融合する[Goding,Monoclonal Antibodies:Principles and Practice,Academic Press,(1986)pp.59-103]。不朽化細胞系は通常トランスフォームされた哺乳動物細胞であり、特にネズミ、ウシ、及びヒト起源のミエローマ細胞である。通常ラットまたはマウスミエローマ細胞系が用いられる。該ハイブリドーマ細胞は、非融合の不朽化細胞の成育と生存を阻害する一つかそれ以上の物質を好ましくは含む適当な培地において、培養され得る。例えばもし元となる細胞が、酵素ヒポキサンチングアニンホスホリボシルトランスフェラーゼ(HGPRTまたはHPRT)を欠失していたならば、ハイブリドーマに対する培地は典型的には、HGPRT欠失細胞の成長を妨げる物質であるヒポキサンチン、アミノプテリン及びチミジン(HAT培地)を含むであろう。
【0123】
好ましい不朽化細胞系は効率よく融合し、選択された抗体生産細胞による抗体の安定な高レベルの生産を維持し、そしてHAT培地のような培地に感受性なものである。より好ましい不朽化細胞系は、例えばSalk Institute Cell Distribution Center,San Diego,California及びAmerican Type Culture Collection,Rockville,Marylandから入手可能であるネズミミエローマ系である。ヒトミエローマ及びマウス-ヒトヘテロミエローマ細胞系もまた、ヒトモノクローナル抗体の生産のため記述されている[Kozbor,J.Immunol.,133:3001(1984);Brodeur等,Monoclonal Antibody Production Techniques and Applications,Marcel Dekker,Inc.,New York,(1987)pp.51-63]。
【0124】
それからハイブリドーマ細胞を培養する培地を、Apo-2リガンドに対して向けられたモノクローナル抗体の存在のためアッセイし得る。好ましくはハイブリドーマ細胞により生産されるモノクローナル抗体の結合特異性を、免疫沈降法によって、またはラジオイムノアッセイ(RIA)、蛍光活性化細胞ソーティング(FACS)、または固相酵素免疫検定法(ELISA)のようなin vitro結合アッセイによって測定する。モノクローナル抗体の結合アフィニティーは、例えばMunsonとPollard,Anal.Biochem.107:220(1980)のスキャッチャード分析によって測定される。
【0125】
望ましいハイブリドーマ細胞が同定された後、該クローンを限界希釈法によってサブクローン化し、標準的な方法[Goding,上記参照]によって成育させる。この目的のために適当な培地は、例えばダルベッコ変法イーグルス培地またはRPMI-1640培地である。代わりにハイブリドーマ細胞は、動物における腹水のようにin vivoで成育させることもできる。
【0126】
サブクローンにより分泌されるモノクローナル抗体は、例えばプロテインA-セファロース、ヒドロキシルアパタイトクロマトグラフィー、ゲル電気泳動、透析及びアフィニティークロマトグラフィーのようなありきたりの免疫グロブリン精製法によって、培地または腹水分泌液から単離され精製され得る。
【0127】
本発明の一つの実施態様として、モノクローナル抗体にはここと以下の実施例で記述される1D1,2G6,2E11または5C2抗体が含まれる。該モノクローナル抗体には、それぞれAmerican Type Culture Collection Accession Nos. ATCC HB-12556, HB-12557, HB-12558, またはHB-12559の下で寄託されているハイブリドーマ細胞系によって分泌される1D1,2G6,2E11または5C2モノクローナル抗体と同様の生物学的性質を持つ抗体も含まれる。「生物学的性質」なる語は、モノクローナル抗体のin vitro及び/またはin vivoでの活性、例えばApo-2リガンド誘発アポトーシスを実質的に減少する能力、またはApo-2リガンドのその受容体に対する結合を実質的に減少またはブロックする能力をいうために用いられる。該抗体は好ましくは、ここで開示される1D1,2G6,2E11または5C2抗体と同じエピトープ、または実質的に同じエピトープに結合する。これはここと実施例で記述されるアッセイを実施することによって測定され得る。
【0128】
該モノクローナル抗体は米国特許第4,816,567号に記述されるような組換えDNA法によっても作製され得る。本発明のモノクローナル抗体をコードするDNAは、ありきたりの方法を用いて容易に単離されシークエンスされ得る(例えばネズミ抗体のH鎖とL鎖をコードする遺伝子に特異的に結合できるオリゴヌクレオチドプローブを用いることによって)。本発明のハイブリドーマ細胞は上記DNAの好ましいソースとしても役に立つ。一度単離されると、DNAを発現ベクターにつなぎ、それからそれを組換えホスト細胞においてモノクローナル抗体の合成を得るために、サルCOS細胞、チャイニーズハムスター卵巣(CHO)細胞またはさもなければ免疫グロブリンタンパク質を生産しないミエローマ細胞のようなホスト細胞にトランスフェクトする。DNAはまた、例えば相同のネズミ配列の代わりにヒトH鎖およびL鎖定常ドメインのコード配列で置換することによって[米国特許第4,816,567号;Morrison等,上記参照]、あるいは非免疫グロブリンポリペプチドのコード配列を免疫グロブリンコード配列に共有結合でつなぐことによって修飾し得る。該非免疫グロブリンポリペプチドは本発明の抗体の定常ドメインの代わりに置換することができ、またはキメラ二価抗体を作製するために本発明の抗体の一つの抗原結合部位の可変ドメインの代わりに置換することができる。
【0129】
該抗体は一価抗体であり得る。一価抗体の調製法は本分野でよく知られている。例えば一つの方法には、免疫グロブリンL鎖と修飾H鎖の組換え発現が含まれる。H鎖は一般的にH鎖架橋を妨げるためにFc領域のいかなる部分でも切り詰められる。代わりに関連したシステイン残基をもう一つのアミノ酸残基で置換し、または架橋を妨げるために欠失する。
【0130】
in vitro法もまた一価抗体を調製するために適している。抗体の断片、特にFab断片を生産するための抗体の切断は、本分野で周知のありきたりの方法を用いて成し遂げられ得る。例えば切断はパパインを用いて実施され得る。パパイン切断の例は94年12月22日に印刷されたWO 94/29348及び米国特許第4,342,566号に記述されている。抗体のパパイン切断は典型的に、それぞれ単一の抗原結合部位を持つFab断片と呼ばれる2の同一の抗原結合断片と、残余のFc断片を生産する。パパイン処理は2の抗原結合部位をもち、依然として抗原を架橋できるF(ab')2断片を生ずる。
【0131】
抗体切断において生産されたFab断片はまた、L鎖の定常ドメインと、H鎖の第一の定常ドメイン(CH1)を含む。Fab'断片は、抗体ヒンジ領域由来の一つ以上のシステインを含むH鎖CH1ドメインのカルボキシ末端でのいくつかの残基の付加によって、Fab断片とは異なる。Fab'-SHは定常ドメインのシステイン残基(類)が遊離チオール基を持つFab'をここで表す。F(ab')2抗体断片はもともと、その間にヒンジシステインを持つFab'断片のペアとして生産された。他の化学的な抗体断片の結合もまた知られている。
【0132】
3. ヒト化抗体
本発明のApo-2リガンド抗体はさらに、ヒト化抗体またはヒト抗体を含む。非ヒト(例えばネズミ)抗体のヒト化形態は、キメラ免疫グロブリン、免疫グロブリン鎖または非ヒト免疫グロブリン由来の最小配列を含むその断片(Fv,Fab,Fab',F(ab')2または抗体の他の抗原結合サブ配列)である。ヒト化抗体には、受容者の相補性決定領域由来の残基が、望ましい特異性、アフィニティー及び力価を持つマウス、ラットまたはウサギのような非ヒト種(ドナー抗体)のCDR由来の残基で置換されているヒト免疫グロブリン(受容者抗体)が含まれる。例えばヒト免疫グロブリンのFvフレームワーク残基が相当する非ヒト残基で置換される。ヒト化抗体はまた、受容者抗体にもインポートCDRまたはフレームワーク配列にも見出されない残基をも含む。一般的に該ヒト化抗体は、少なくとも一つそして典型的には二つの可変ドメインの実質的にすべてを含み、該可変ドメインはCDR領域の全てまたは実質的に全てが非ヒト免疫グロブリンのものに相当し、FR領域の全てまたは実質的に全てがヒト免疫グロブリンコンセンサス配列のものである。該ヒト化抗体はまた適宜に、免疫グロブリン定常領域(Fc)の少なくとも一部を含み、典型的にはそれはヒト免疫グロブリンのものであろう[Jones等,Nature,321:522-525(1986);Reichmann等,Nature,332:323-329(1988);及びPresta,Curr.Op.Struct.Biol.2:593-596(1992)]。
【0133】
非ヒト抗体をヒト化する方法は本分野でよく知られている。一般的に、ヒト化抗体は非ヒトであるソースからそれに導入された一つかそれ以上のアミノ酸残基をもつ。これらの非ヒトアミノ酸残基は、しばしば「インポート残基」として示され、典型的には「インポート」可変ドメインから由来している。ヒト化は本質的に、Winterと共同研究者[Jones等,Nature 321:522-525(1986);Riechmann等,Nature 332:323-327(1988);Verhoeyen等,Science 239:1534-1536(1988)]の方法にしたがって実施され、ネズミCDRまたはCDR配列を、相当するヒト抗体の配列で置換することによるコンピューターモデルを用いたQueen等,Proc.Natl.Acad.Sci.86:10029-10033(1989)の方法によってなされる。したがって該「ヒト化」抗体はキメラ抗体であり(米国特許第4,816,567号)、そこでは実質的に完全なヒト可変ドメインより短い部分が、相当する非ヒト種由来の配列で置換されている。実際問題として、ヒト化抗体は典型的には、いくつかのCDR残基および可能ないくつかのFR残基がネズミ抗体の相似性の部位由来の残基によって置換されているヒト抗体である。
【0134】
ヒト化抗体を作製する場合に用いられるL鎖とH鎖の両方のヒト可変領域の選択は、抗原性を減少するために重要である。「ベスト-フィット」法にしたがって、ネズミ抗体の可変ドメインの配列が既知のヒト可変ドメイン配列の完全なライブラリーからスクリーニングされる。それからネズミのものに最も近いヒト配列が、ヒト化抗体のためにヒトフレームワーク(FR)として受け入れられる[Sims等,J.Immunol,151:2296(1993);ChothiaとLesk,J.Mol.Biol.196:901(1987)]。もう一つの方法は、L鎖とH鎖の特定のサブグループの全てのヒト抗体のコンセンサス配列由来の特定のフレームワークを用いる。同様のフレームワークはいくつかの異なるヒト化抗体のためにも用いられ得る[Carter等,Proc.Natl.Acad.Sci.USA.89:4285(1992);Presta等,J.Immunol.151:2623(1993)]。
【0135】
抗体は抗原に対する高アフィニティーおよび他の好ましい生物学的性質を保持したまま免疫化されることがさらに重要である。この目的を達成するため好ましい方法にしたがって、もととなるおよびヒト化配列の三次元モデルを用いてもととなる配列および様々な概念上のヒト化産物の分析の過程によって、ヒト化抗体を調製する。三次元免疫グロブリンモデルは、公に入手可能であり当業者には親しまれている。選択された免疫グロブリン配列の候補の可能な三次元構造を描写し表示するコンピュータープログラムも入手可能である。これらの表示を見ることによって、免疫グロブリン配列の候補の機能において残基の考え得る役割の分析、すなわち抗原に結合する場合に免疫グロブリン候補の能力に影響する残基の解析が可能となる。この方法では、FR残基が、標的抗原(類)に対する増大したアフィニティーのような望ましい抗体特性を達成するためのコンセンサス配列およびインポート配列から選択され結合され得る。一般的にCDR残基は直接にそして最も実質的に抗原結合を影響する部位に含まれる[1994年3月3日に印刷されたWO 94/04679参照]。
【0136】
免疫化により、内因性の免疫グロブリン生産の不存在下でヒト抗体のフルレパートリーを生産可能なトランスジェニック動物(例えばマウス)が用いられ得る。例えば、キメラで生殖系列ミュータントマウスにおける抗体H鎖結合領域(JH)遺伝子のホモ接合体欠失が、内因性抗体生産の完全な阻害を引き起こすことが記述されている。該生殖系列ミュータントマウスにおけるヒト生殖系列免疫グロブリン遺伝子配列のトランスファーは、抗原チャレンジに対するヒト抗体の生産を引き起こすであろう[例えばJakobovits等,Proc.Natl.Acad.Sci.USA,90:2551-255(1993);Jakobovits等,Nature,362:255-258(1993);Bruggermann等,Year in Immuno.,7:33(1993)]。ヒト抗体はまたファージディスプレーライブラリーにおいても生産し得る[Hoogenboom及びWinter,J.Mol.Biol.,227:381(1991);Marks等,J.Mol.Biol.,222:581(1991)]。Cote等及びBoerner等の方法もまた、ヒトモノクローナル抗体の調製のために有用である(Cote等,Monoclonal Antibodies and Cancer Therapy,Alan R.Liss,p.77(1985)及びBoerner等,J.Immunol.,147(1):86-95(1991)]。
【0137】
4. 二重特異性抗体
二重特異性抗体とは、少なくとも二つの異なる抗原に対する結合特異性を持つモノクローナルで、好ましくはヒトのまたはヒト化された抗体である。本ケースでは、結合特異性の一つはApo-2リガンドに対するものであり、もう一つはいかなる他の抗原に対するものでもよく、好ましくは細胞表面タンパク質または受容体あるいは受容体サブユニットに対するものである。
【0138】
二重特異性抗体を作成する方法は本分野で知られている。伝統的には、二重特異性抗体の組換え生産は、二つの免疫グロブリンH鎖/L鎖ペアの共同発現に基づいており、そこでは二つのH鎖は異なる特異性をもつ[MilsteinとCuello,Nature 305:537(1983)]。免疫グロブリンH鎖とL鎖のランダムな識別のため、これらのハイブリドーマ(クアッドローマ)は10種類の異なる抗体分子の混合物を潜在的に生産し、その中の一種だけが正しい二重特異性抗体なのである。正しい分子の精製は、通常アフィニティークロマトグラフィーステップによりなされる。同様の方法は1993年5月13日に印刷されたWO 93/08829およびTraunecker等,EMBO J.10:3655(1991)に記述されている。
【0139】
異なるそしてより好ましいアプローチにしたがうと、望ましい結合特異性を持つ抗体可変ドメイン(抗体抗原結合部位)を免疫グロブリン定常ドメイン配列に融合する。好ましくは融合は、少なくともヒンジ部、CH2、およびCH3を含む免疫グロブリンH鎖定常領域となされる。L鎖との結合に必要な部位を含む第一のH鎖定常領域(CH1)を融合物の少なくとも一つに存在させるのも好ましい。免疫グロブリンH鎖融合物をコードするDNAおよび必要であれば免疫グロブリンL鎖をコードするDNAを、別々の発現ベクターに組み込み、適したホスト生物内に共同してトランスフェクトする。構築に用いられる非等量割合の三つのポリペプチド鎖が最適条件収率を提供すると、これにより実施態様における三つのポリペプチド断片の相互の性質を調製する大きな柔軟性が提供される。しかしながら、等量割合で少なくとも二つのポリペプチド鎖の発現が高い収率を引き起こした場合、またはその割合が特定の重要性を持たない場合、一つの発現ベクターに二つまたは三つ全てのポリペプチド鎖をコードする配列を組み込むことが可能である。このアプローチの好ましい実施態様によると、二重特異性抗体は一つの腕の第一の結合特異性を持つハイブリッド免疫グロブリンH鎖と、もう一方の腕のハイブリッド免疫グロブリンH鎖/L鎖ペア(第二の結合特異性を提供する)より成る。この非対称の構造は、二重特異性分子の半分だけを占める免疫グロブリンL鎖の存在が選別の容易な方法を提供するように、望まない免疫グロブリン鎖の組み合わせから望ましい二重特異性化合物を選別するのを容易にすることが見出された。このアプローチは1994年3月3日に印刷されたWO 94/04690に開示されている。二重特異性抗体のさらに詳細を生産する方法については、例えばSuresh等,Methods in Enzymology 121:210(1986)を参照。
【0140】
5. ヘテロ接合抗体
ヘテロ接合抗体もまた本発明の範囲にある。ヘテロ接合抗体は二つの共有結合で結合された抗体より成る。該抗体は例えば、望まない細胞に対して免疫系細胞の標的にするため[米国特許第4,676,980号]、またはHIV感染の治療のため[WO 91/00360;WO 92/00373;そしてEP 03089])提案されている。該抗体は架橋試薬を含むもののような合成タンパク質化学において周知の方法を用いてin vitroで調製され得ることが企図されている。例えばイムノトキシンはジスルフィド交換反応を用いて、またはチオエーテル結合を形成することによって構築し得る。この目的に対する適した試薬の例としては、イミノチオレート及びメチル-4-メルカプトブチリミデートそして例えば米国特許第4,676,980号に開示されているものが含まれる。
【0141】
E. Apo-2リガンド抗体の使用
Apo-2リガンド抗体は例えば特異的細胞、組織、または血清における発現の検出といったApo-2リガンドに対する診断のアッセイに用いられ得る。本分野で周知の様々な診断のアッセイ法が用いられ得、その中には競合的結合アッセイ、直接または間接サンドイッチアッセイ及び異種または同種フェーズのそれぞれで実施される免疫沈降アッセイが含まれる[Zola,Monoclonal Antibodies:A Manual of Techniques,CRC Press,Inc.(1987)pp.147-158]。診断アッセイで用いられる抗体は検出可能な部分でラベルされ得る。検出可能な部分は直接または間接に検出可能なシグナルを生ずることが可能ないかなるものでもあり得る。例えば検出可能な部分は、3H,14C,32P,35Sまたは125Iのようなラジオアイソトープ、フルオレセイン、イソチオシアネート、ローダミンまたはルシフェリンのような蛍光化合物または化学ルミネセンス化合物、アルカリホスファターゼ、ベータ-ガラクトシダーゼまたはセイヨウワサビペルオキシダーゼのような酵素が挙げられる。検出可能な部分と抗体を別々に接合するための本分野で既知の方法も用いることができ、それにはHunter等,Nature 144:945(1962);David等,Biochemistry 13:1014(1974);Pain等,J.Immunol.Meth.40:219(1981);およびNygren,J.Histochem.and Cytochem.30:407(1982)に記述された方法が含まれる。
【0142】
Apo-2リガンド抗体はまた、組換え細胞カルチャーまたは天然のソースからApo-2リガンドのアフィニティー精製に対しても有用である。この工程においては、Apo-2リガンドに対する抗体が、本分野で周知の方法を用いて、Sephadex樹脂または濾紙のような適した支持体に固定化される。それから該固定化抗体を精製されるApo-2リガンドを含むサンプルと接触させ、その後該支持体を固定化抗体と結合したApo-2リガンドを除くサンプル中の実質的にすべての物質を除去するであろう適した溶媒を用いて洗浄する。最後に該支持体を抗体からApo-2リガンドを放出するであろうもう一つの適した溶媒で洗浄する。Apo-2リガンド抗体はまた可溶化Apo-2受容体のアフィニティー精製に対して、またはApo-2受容体の発現クローニングに対して有用である。
【0143】
ここで開示される抗体は治療薬としても用いられる。例えばApo-2リガンド活性(Apo-2リガンド誘発アポトーシスのような)をブロックする抗Apo-2リガンド抗体は、増大したアポトーシスと関連する病理学上のコンディションまたは疾患を治療するために用いられ得る[Thompson,上記参照参照]。
【0144】
F. Apo-2リガンドまたはApo-2リガンド抗体を含むキット
本発明の更なる実施態様において、例えば上述の治療上のまたは非治療上の応用に用いられ得るApo-2リガンドまたはApo-2リガンド抗体を含む製造物とキットが提供される。製造物としては、ラベルのついた容器が含まれる。適した容器は例えばボトル、ガラスビンそして試験管が含まれる。容器はガラスやプラスチックのような様々な物質から形成されるであろう。容器には、上述のような治療上または非治療上の応用に対して有効な活性試薬を含む構成物が入っている。構成物中の活性試薬はApo-2リガンドまたはApo-2リガンド抗体である。容器のラベルは構成物が、特異的な治療上または非治療上の応用に用いられ、上述したようなin vivoまたはin vitroでの各使用のための使用法も示されているであろう。
【0145】
本発明のキットは上述した容器を典型的には含み、商業的におよび使用者の見地から望ましい物質を含む一つ以上の他の容器を含み、その中には使用のための説明書と共に挿入された他のバッファー、希釈液、フィルター、針、シリンジそしてパッケージが含まれる。
【0146】
以下の実施例は説明のためのみを目的として提供され、いかなる方法においても本発明の範囲を制限することを企図しない。
【0147】
本明細書に引用される全ての参考文献は、ここで参考として全体として取り込まれる。
【実施例】
【0148】
本実施例で言及される全ての制限酵素は、New England Biolabsから購入され、製品の説明書にしたがって使用された。本実施例で言及される全ての他の商業的に入手可能な試薬は、もし他のいかなる指示もなければ、製品の説明書にしたがって用いられた。以下の実施例、及び明細書を通して、ATCC登録番号で示される細胞のソースは、American Type Culture Collection,Rockville,Marylandである。
【0149】
実施例1
ヒトApo-2リガンドをコードするcDNAクローンの単離
Apo-2リガンドに対するフルレングスcDNAを単離するために、ヒト胎盤cDNAのラムダgt11バクテリオファージライブラリー(約1×106クローン)(HL10756,Clontechから商業的に入手可能)を、ヒトFas/Apo-1リガンドとある程度の同一性を示すEST配列(GenBankローカスHMEA47M)に基づく合成オリゴヌクレオチドプローブを用いてハイブリダイゼーションによってスクリーニングした。HMEA47Mの該EST配列は390bpであり、その+3フレームで翻訳された場合、ヒトApo-1リガンドの34アミノ酸領域に対して16の同一性を示す。HMEA47Mの配列は以下のものである:
[配列1]

以下の配列を持つ60bpのオリゴヌクレオチドプローブをスクリーニングに用いた:
[配列2]
ハイブリダイゼーションは20%ホルムアミド、5×SSC、10%硫酸デキストラン、0.1%NaPiPO4、0.05MNaPO4、0.05mgサケ精子DNA、及び0.1%ドデシル硫酸ナトリウムを含むバッファーにおいて室温でオーバーナイトで実施し、引き続き5×SSCで42度での数回の洗浄をし、それから2×SSCで洗浄した。12のポジティブクローンをcDNAライブラリーで同定し、該ポジティブクローンを以下の配列を持つ第二の60bpのオリゴヌクレオチドプローブ(第一のプローブとオーバーラップしない)に対するハイブリダイゼーションによってスクリーニングした。
[配列3]
ハイブリダイゼーションを上記したように実施した。
【0150】
4の結果として生じたポジティブクローンを同定し、フランキング5'ベクター配列に基づき外来のClaI制限部位を加えたプライマーと、3'フランキングベクター配列に基づき外来のHindIII制限部位を加えたプライマーを用いたポリメラーゼ連鎖反応(PCR)により増幅した。PCR産物をゲル精製し、T-AライゲーションによりpGEM-T(Promegaから商業的に入手可能)内にサブクローン化した。それから異なるPCR由来の3の独立のクローンをディデオキシDNAシークエンシングに受けさせた。これらのクローンのDNA配列分析により、それらは本質的に同一であり、その5'領域でいくつかの長さのバリエーションを持つことが示された。
【0151】
Apo-2リガンドのコード領域の核酸配列は図1Aに示されている。該クローンの一つの下流3'末端配列のシークエンシングにより、特徴的なポリアデニル化部位が明らかになった(データは示していない)。該cDNAは核酸位91-93でATGコドンとして表される開始部位を持つ一つの長いオープンリーディングフレームを含んだ。この部位を取り囲む配列は開始部位に対して提案されているコンセンサス配列と理由のある一致を示す[Kozak,J.Cell.Biol.,115:887-903(1991)]。該オープンリーディングフレームは核酸位934-936の終始コドンTAAで終わる。
【0152】
ヒトApo-2リガンドの予想される成熟アミノ酸配列は281アミノ酸を含み、およそ32.5kDaの計算される分子量とおよそ7.63の等電点を持つ。ハイドロパシー分析(データは示していない)では残基15と40の間に疎水性領域の存在が示されるが、N末端の明らかなシグナル配列は存在しない。シグナル配列の不存在と内部の疎水性領域の存在は、Apo-2リガンドがII型膜貫通タンパク質であることを示唆する。推定の細胞内、膜貫通及び細胞外領域は、それぞれ14、26及び241アミノ酸長である。推定の膜貫通領域は図1Aにおいて下線が引かれている。潜在的なNリンクグリコシレーション部位は推定の細胞外ドメインの残基109に位置する。
【0153】
Apo-2リガンドのC末端領域のアミノ酸配列の他の周知のTNFサイトカインファミリーのメンバーとの比較により(AlignTMコンピュータープログラムを用いる)、C末端領域内でApo-2リガンドはApo-1リガンドと23.2%の同一性を示すことが示された(図1B)。比較分析は他のTNFファミリーメンバーとの比較的少ない程度の同一性を示した:CD40L(20.8%),LT-α(20.2%),LT-β(19.6%),TNF-α(19.0%),CD30L及びCD27L(15.5%),OX-40L(14.3%)及び4-1BBL(13.7%)。TNFサイトカインファミリーにおいて、TNF-α及びLT-αの結晶構造[Eck等,J.Bio.Chem.,264:17595-17605(1989);Eck等,J.Bio.Chem.,267:2119-2122(1992)]に基づいて、βストランドを形成すると予想される領域内の残基は、予想される結合ループにおける残基と比較して、他のTNFファミリーメンバーに関してより高く保存されている傾向にある。Apo-2リガンドは予想される結合ループにおける同一性と比較して、推定のβストランド領域で他のTNFファミリーメンバーとより大きい同一性を示すことが見出された。また、推定のβストランド、B及びB'を結合するループはApo-2リガンドにおいて著しくより長い。
【0154】
実施例2
ヒトApo-2リガンドの発現
A. フルレングスcDNA融合構築物
mycエピトープタグと融合したフルレングスApo-2リガンドcDNAを以下のように構築した。Apo-2リガンドcDNA挿入物をClaIおよびHindIII切断によって元となるpGEM-TApo-2リガンドプラスミド(実施例1に記述される)から摘出し、同じ制限酵素で札断されたpRK5哺乳動物発現プラスミド内に挿入した[Schall等,Cell,61:361-370(1990);Suva等,Science,237:893-896(1987)]。それから13アミノ酸mycエピトープタグをコードする配列
[配列4]
[Evan等,Mol.Cell.Biol.,5:3610-3616(1985)]を、プラスミドpRK5Apo-2リガンド-mycを作製するために、オリゴヌクレオチドディレクトミュータジェネシス[Zoller等,Nucleic Acids Res.,10:6487-6496(1982)]により、Apo-2リガンドコード配列の3'末端でコドン281と終始コドン(コドン282)の間に挿入した。
【0155】
pRK5Apo-2リガンド-mycプラスミドを、カルシウムリン酸沈降により、ネオマイシン耐性遺伝子を運ぶpRK5プラスミドと共にヒト293細胞(ATCCCRL1573)内にコトランスフェクトした。Apo-2リガンド-mycを発現する安定なクローンを抗生物質、G418(0.5mg/mL)(GIBCO)の存在下で50%HAM'sF12-50/50%DMEM(GIBCO)培地で成育する能力によって選択した。
【0156】
Apo-2リガンドのトポロジーを調べるために、G418耐性クローンを抗mycモノクローナル抗体(mAb)クローン9E10[Evan等,上記参照;Oncogene Scienceから商業的に入手可能]を用いて染色し、引き続きフィコエリスリン(PE)-接合ヤギ-マウス抗体(Jackson Immuno Researchから商業的に入手可能)を用いて染色した後、FACSにより分析した。FACS分析により偽のトランスフェクトされた細胞と比較して、特異的なポジティブ染色がApo-2リガンド-mycトランスフェクトクローンにおいてシフトすることが示され(図1C)、Apo-2リガンドはそのカルボキシ末端がさらされる形で細胞表面に発現されることが示された。したがってApo-2リガンドはII型膜貫通タンパク質であると考えられる。
【0157】
B. ECD融合構築物
2の可溶性Apo-2リガンド細胞外ドメイン(「ECD」)融合構築物を調製し、それにおいてもう一つの配列をApo-2リガンドのC末端領域の上流に融合した。
一つの構築において、ヘルペスウイルスグリコプロテインD(「gD」)シグナルペプチドの27アミノ酸[Lasky等,DNA,3:23-29(1984);Pennica等,Proc.Natl.Acad.Sci.,92:1142-1146(1995);Paborsky等,Protein Engineering,3:547-553(1990)に記述されている]及びエピトープタグ配列
[配列5]
をpRK5哺乳動物発現プラスミド内のApo-2リガンドのコドン114-281の上流に融合した。略記すると、gD配列を3'プライマーがApo-2リガンドのコドン114-121と同様にgD配列の3'領域に相補的であるPCRにおいて、元となるプラスミドpCHAD(Genentech,実質的にLasky等,Science,233:209-212(1986)に記述されているように調製した)から増幅した。該産物をpRK5Apo-2リガンドをテンプレートとして用いる連続的PCRにおけるApo-2リガンドコード領域の3'末端に相補的な3'プライマーと共に5'プライマーとして用いた。それからgD-Apo-2リガンドECD融合物をコードする該産物を、プラスミドpRK5gD-Apo-2リガンドECDを作製するためにpRK5プラスミド内にサブクローン化した。
【0158】
ヒト胎児腎293細胞(ATCC CRL 1573)をカルシウムリン酸沈降によってpRK5gD-Apo-2リガンドECDプラスミドまたはpRK5を用いて一過的にトランスフェクトした。可溶性gD-Apo-2リガンドタンパク質の発現を、35S-Cys及び35S-Metを用いたトランスフェクトされた細胞の代謝ラベルによって評価した。細胞上清を24時間後集め、遠心分離によってきれいにした。免疫沈降のため、5mlの上清を4℃でオーバーナイトで1μg/mlの濃度で5B6抗gDモノクローナル抗体(Genentech)と共にインキュベートした。それから25μlのPansorbin(Sigma)を4℃でさらに一時間加えた。熱したサンプルを再び回転させ、上清をSDS-PAGE及びオートラジオグラフィーにかけた。
【0159】
抗gD抗体を用いた免疫沈降により、gD-Apo-2リガンドプラスミドでトランスフェクトされた細胞の上清中に3の主要なバンドが現れた(図1E)。これらのバンドは23,48及び74kDaの相対的分子量(Mr)で移動した。成熟gD-Apo-2ポリペプチドの計算上の分子量は、およそ22.5kDaである;それゆえ観察されたバンドは、融合タンパク質の単量体(23kDa)、二量体(48kDa)、そして三量体(74kDa)の各形態を表し、Apo-2リガンドが哺乳動物細胞において分泌された可溶性gD融合タンパク質として発現され得ることを示すであろう。
【0160】
第二の構築において、12アミノ酸エンテロキナーゼ切断部位が引き続く、Met Gly His10配列(プラスミドpET19Bから由来する、Novagen)
[配列6]
をバキュロウイルス発現プラスミド(pVL1392,Pharmingen)内でApo-2リガンドのコドン114-281の上流に融合した。略記すると、Apo-2リガンドコドン114-281を、それぞれフランキングNdeIおよびBamHI制限部位を取り込んだ5'及び3'領域に相補的なプライマーを用いて、元となるpRK5Apo-2リガンドプラスミド(実施例1に記述されている)からPCRによって増幅した。該産物をT-AライゲーションによりpGEM-T(Rromega)内にサブクローン化し、DNA配列を確認した。それから挿入物をNdeIおよびBamHIを用いた切断により摘出し、アミノ末端Met Gly His10タグ及びエンテロキナーゼ切断部位を含む修飾バキュロウイルス発現ベクターpVL1392(Pharmingenから商業的に入手可能)内にサブクローン化した。
【0161】
組換えバキュロウイルスをリポフェクチン(GIBCO-BRLから商業的に入手可能)を用いて、Spodoptera frugiperda(「Sf9」)細胞(ATCC CRL 1711)内にHis10-Apo-2ECDプラスミドとBaculoGoldTMウイルスDNA(Pharmingen)をコトランスフェクトすることによって生産した。28℃でインキュベーションの4-5日後、放出されたウイルスを回収し、さらなる増幅に用いた。ウイルス感染とタンパク質発現を、O'Reilley等,Baculovirus expression vectors: A laboratory Manual,Oxford:Oxford University Press(1994)に記述されているように実施した。タンパク質を以下の実施例3に記述されているようにNi2+-キレートアフィニティークロマトグラフィーによって精製した。
【0162】
実施例3
組換えヒトApo-2リガンドの精製
Rupert等,Nature,362:175-179(1993)に記述されているように組換えウイルス感染及び偽の感染Sf9細胞(上記実施例2,セクションB参照)から抽出物を調製した。略記すると、Sf9細胞を洗浄し、ソニケーションバッファー(25mLHepes,pH7.9;12.5mMMgCl2;0.1mMEDTA;10%Glycerol;0.1%NP-40;0.4MKCl)に再懸濁し、そして氷上で20秒間2回ソニケートした。ソニケートされた物を遠心分離によりきれいにし、上清をローディングバッファー(50mMリン酸,300mMNaCl,10%Glycerol,pH7.8)で50倍に希釈し、0.45μmフィルターを通して濾過した。Ni2+-NTAアガロースカラム(Qiagenから商業的に入手可能)を5mLのベッド容量で調製し、25mLの水で洗浄し、25mLのローディングバッファーで平衡化した。濾過された細胞抽出物を分当たり0.5mLでカラムに乗せた。カラムをローディングバッファーを用いてベースラインA280に洗浄し、その時点で画分収集を開始した。次に、カラムを二次的洗浄バッファー(50mMリン酸;300mMNaCl,10%Glycerol,pH6.0)で洗浄し、それにより非特異的結合タンパク質が溶出した。再びA280ベースラインに到達した後、カラムを二次的洗浄バッファーにおける0から500mMのイミダゾール勾配にかけた。1mLの画分を回収し、SDS-PAHGE及び銀染色またはNi2+-NTA-接合アルカリホスファターゼ(Qiagen)を用いたウエスタンブロットによって分析した。溶出されたHis10-Apo-2リガンドタンパク質を含む画分をプールし、ローディングバッファーによって透析した。
【0163】
同一の方法をスタート物質として偽の感染Sf9細胞を用いて繰り返し、同じ画分をプールし、透析し、精製ヒトApo-2に対するコントロールとして用いた。
【0164】
精製タンパク質のSDS-PAGE分析により、分子量24kDaの主要なバンドが明らかになり、His10-Apo-2リガンド単量体に対する計算された分子量の22.4kDaに相当した(図1D,レーン3);タンパク質配列マイクロ分析(データは示していない)により、24kDaのバンドはHis10-Apo-2リガンドポリペプチドを表すことが確認された。少量の48kDa及び66kDaバンドもまた観察され、それらはおそらく可溶性Apo-2リガンドホモ二量体及びホモ三量体を表すであろう。スルホ-NHS(5mM)(Pierce Chemical)及び25mMと50mMでのEDC(Pierce Chemical)を用いたインキュベーションにより、精製His10-Apo-2リガンドの化学的架橋は(それぞれ図1D,レーン1と2)、タンパク質を主に66kDaバンドにシフトさせた。これらの結果により、溶液中のApo-2リガンドの主要な形態はホモ三量体であり、これらの三量体はSDSの存在下で二量体及び単量体に解離することが示唆される。
【0165】
実施例4
ヒトリンパ球細胞系に対するApo-2リガンドのアポトーシス活性
精製された可溶性Apo-2リガンド(実施例3に記述されている)のアポトーシス活性を、いくつかのヒトリンパ球細胞系を用いて調べた。第一の研究として、Epstein-Barrウイルス(EBV)トランスフォームヒト末梢血B細胞から由来する9D細胞(Genentech,Inc.)に対するApo-2リガンドの効果を調べた。9D細胞(10%胎児ウシ血清を加えたRPMI1640培地中に5×104細胞/ウェル)を、培地コントロール、Apo-2リガンド(3μg/ml、上述の実施例3に記述されているように調製された)、または抗Apo-1モノクローナル抗体、CH11(1μg/ml)のそれぞれで24時間インキュベートした[Yonehara等,J.Exp.Med.,169:1747-1756(1989)に記述されている;Medical and Biological Laboratories Co.から商業的に入手可能]。CH11抗Apo-1抗体は、fas/Apo-1リガンド活性をまねた作用剤抗体である。
【0166】
インキュベーション後、細胞をサイトスピンガラススライド上で回収し、倒立光学顕微鏡の下で写真を撮った。Apo-2リガンドと抗Apo-1モノクローナル抗体の両者は、同様のアポトーシス効果を誘導し、それは細胞質濃縮と細胞数の減少に特徴付けられた(図2A参照)。
【0167】
9D細胞に対するApo-2リガンドの効果と同様に、Raji細胞(ヒトバーキットリンパ腫B細胞系、ATCC CCL 86)及びJurkat細胞(ヒト急性T細胞白血病細胞系、ATCC TIB 152)に対する効果をさらにFACSによって分析した。該FACS分析は、アポトーシス化細胞死に対する確立された標準を用いて実施され、主に2のマーカー:(a) アポトーシス化細胞を染色するが生きた細胞はしないプロピジウムイオダイド(「PI」)色素、及び(b) アポトーシス化細胞の表面に見出されるさらされたホスファチジルセリンに結合するが、生きた細胞にはしないタンパク質の蛍光誘導体、アネキシンVを用いた細胞の蛍光染色に関するものであった[Darzynkiewicz等,Methods in Cell Biol.,41:15-38(1994);Fadok等,J.Immunol.,148:2207-2214(1992);Koopman等,Blood,84:1415-1420(1994)]。
【0168】
9D細胞(図2B)、Raji細胞(図2C)及びJurkat細胞(図2D)を、培地コントロール(左パネル)、Apo-2リガンド(3μg/ml、上述の実施例3に記述されているように調製された)(中央パネル)、または抗Apo-1モノクローナル抗体、CH11(1μg/ml)(右パネル)のそれぞれで24時間インキュベートした(1×106細胞/ウェル)。それから該細胞を洗浄し、PI及び蛍光チオシアネート(FITC)-接合アネキシンV(Brand Applicationsから購入)を用いて染色し、フローサイトメトリーで分析した。PIとアネキシンV染色(第3部分)の両者でネガティブな細胞は生きた細胞を表す;PI-ネガティブ、アネキシンV-ポジティブ染色細胞(第4部分)は早期アポトーシス化細胞を表す;PI-ポジティブ、アネキシンV-ネガティブ染色細胞(第2部分)は主にアポトーシスの後期段階の細胞を表す。
【0169】
Apo-2リガンドで処理された9D細胞は、上昇した細胞外アネキシンV結合を示し、同様に著しいPIの取り込みの増大を示し(図2B)、それはApo-2リガンドが細胞におけるアポトーシスを誘導したことを示す。抗Apo-1抗体、CH11(図2B)で比較可能な結果が得られた。Apo-2リガンドは抗Apo-1抗体がするように、Raji及びJurkat細胞において同様の応答を誘導した(図2C及び2D参照)。これらの細胞におけるコントロール及び抗Apo-1抗体と比較したApo-2リガンドによるアポトーシスの誘導(アポトーシス化細胞%ととして測定される)が、以下の表1にも示されている。
【0170】
Apo-2リガンドによる染色体内DNA断片化の活性化もまた分析した。Jurket細胞(左レーン)及び9D細胞(右レーン)を培地コントロールまたはApo-2リガンド(3μg/ml、実施例3に記述されているように調製した)と共に6時間インキュベートした(2×106細胞/ウェル)。それからDNAを細胞から抽出し、ターミナルトランスフェラーゼを用いて32P-ddATPを用いてラベルした。ラベル化DNAサンプルを2%アガロースゲルで電気泳動にかけ、その後オートラジオグラフィーにより分析した[Moore等,Cytotechnology,17:1-11(1995)]。Apo-2リガンドはJurkat細胞及び9D細胞の両者で染色体内DNA断片化を誘導した(図2E)。該DNA断片化はアポトーシスの特徴である[Cohen,Advances in Immunol.,50:55-85(1991)]。
【0171】
Apo-2リガンドアポトーシス活性のタイムコースを調べるために、9D細胞を0時間から50時間の範囲の期間、培地コントロールまたはApo-2リガンド(3μg/ml、実施例3に記述されているように調製される)を用いてミクロタイターディッシュ(5×104細胞/ウェル)でインキュベートした。インキュベーションに引き続き、死んだ細胞及び生きた細胞の数を血球計算板を用いて顕微鏡観察の下で測定した。
【0172】
図3Aに示されているように、細胞死の最大のレベルは24時間以内で9D細胞で誘導された。
【0173】
Apo-2リガンド誘導細胞死の量依存性を調べるために、9D細胞を培地コントロールまたはApo-2リガンド(実施例3に記述されているように調製された)の連続的な希釈物を用いて24時間でインキュベートした(5×104細胞/ウェル)。インキュベーションに引き続き死んだ細胞及び生きた細胞の数を上述のように測定した。その結果が図3Bに示されている。特異的アポトーシスはApo-2リガンド処理細胞におけるアポトーシス%からコントロールにおけるアポトーシス%を差し引くことによって計算された。アポトーシスの半最大活性化はおよそ0.1μg/ml(およそ1nM)で生じ、最大の誘導はおよそ1からおよそ3μg/ml(およそ10から30nM)で生じた。
【0174】
実施例5
ヒト非リンパ球腫瘍細胞系に対するApo-2リガンドのアポトーシス活性
ヒト非リンパ球腫瘍細胞系に対するApo-2リガンドの効果を、以下の細胞系を用いて調べた:HeLa(ヒト子宮頚ガン腫から由来する、ATCC CCL 22);ME-180(ヒト子宮頚ガン腫から由来する、ATCC HTB 33);MCF7(ヒト乳ガン腫から由来する、ATCC HTB 22);U-937(ヒト細網肉腫から由来する、ATCC CRL 1593);A549(ヒト肺ガン腫から由来する、ATCC CCL 185);及び293(アデノウイルストランスフォームヒト胎児腎細胞から由来する、ATCC CCL 1573)。該アッセイにおいては、それぞれの細胞系の1×106細胞を、培地コントロール、Apo-2リガンド(3μg/ml、実施例3に記述されているように調製した)またはApo-1モノクローナル抗体、CH11(1μg/ml)を用いて24時間でインキュベートした。該インキュベーションに引き続き、アポトーシスを実施例4に記述されているFACS分析によって測定した。その結果が以下の表1に示されている。
【0175】
【表1】
【0176】
HeLa細胞及びMCF7細胞は、CH11抗Apo-1抗体と比較してApo-2リガンドによってアポトーシスの誘導に等しく感受性であった。対照的にU-937細胞及びA549細胞は著しくApo-2リガンドによるアポトーシスの誘導により感受性であった。ME-180細胞はApo-2リガンドに対して大変感受性であるが、抗Apo-1抗体には比較的耐性であった。293細胞はApo-2リガンドに対しては耐性であり、抗Apo-1抗体に対しては弱く感受性であった。
【0177】
それゆえ、Apo-2リガンドはリンパ球起源の細胞と同様に、非リンパ球起源の細胞においてもアポトーシスを誘導することができる(実施例4参照)。また、十分に理解されておらず、いかなる特定の理論にも結び付けられることを期待しないが、出願人はApo-2リガンドがApo-1とは区別される受容体を介して機能すると現在考えている。この考えは、上述の細胞系がApo-2リガンドと抗Apo-1抗体に対する異なる感受性のパターンを示すということを表すここでのデータによって支持される。
【0178】
実施例6
ヒト末梢血単球に対するApo-2リガンドの効果
末梢血単核細胞(「PBMC」)を、Lymphocyte Separation Medium(LSM(登録商標)、Organon Teknika)を用いたFicoll密度勾配遠心分離によってヒトドナーの血液から単離した。単離されたT細胞の集団を、抗Igカラムに結合した表面Igを通してB細胞を除去し、Igカラムに結合しているFc受容体を通して単球を除去することによって、PBMCから調製した(R & D Systems)。単離されたB細胞の集団を、OKT3ミエローマ(ATCC CRL 8001)によって生産された抗cD3抗体と反応したT細胞の、及び4F2C13ハイブリドーマ(ATCC HB22)によって生産された単球特異的抗体と反応した単球の、それぞれ相補体介在性除去によりPBMCから調製した。さらなる単球の除去はプラスチックへの接着によって成し遂げられた。
【0179】
新鮮に単離された末梢血BまたはT細胞(1×106細胞/ウェル)を、培地コントロールまたはApo-2リガンド(3μg/ml、実施例3に記述されるように調製した)の存在下で3日間培養した。活性化のため、B細胞をリポ多糖(「LPS」、1μg/ml)で同時に処理し、T細胞をホルボールミリステートアセテート(「PMA」、10ng/ml)プラスイオノマイシン(1μg/ml)(Sigma)を用いて処理した。インターロイキン-2(「IL-2」)前処理のため、T細胞をApo-2リガンドにさらす前、IL-2(50U/ml)(Genzyme)の存在下で3-5日間培養した。アポトーシスを本質的に実施例4で上述されているようにFACS分析を用いて測定した。しかしながらB細胞は抗CD19/CD20抗体(Jackson Immunoresearch)によって捕らえられ、T細胞は抗CD4/CD8抗体(Jackson Immunoresearch)によって捕らえられた。その結果が以下の表2に示されており、それは独立の実験[Bリンパ球-9実験;Tリンパ球-8実験;Tリンパ球プラスIL-2-5実験]の平均±SEを表し、そこにおいては50,000細胞がデータポイント当たり分析された。統計的な分析はt検定を用いて実施した。表2において、それぞれのコントロールに対してa=p<0.05及びb=p<0.02である。
【0180】
【表2】
【0181】
Apo-2リガンドは非刺激B細胞、LPSにより活性化されたB細胞及びPMAとイオノマイシンを用いて活性化されたT細胞において有意にアポトーシスを誘導した。末梢T細胞はIL-2の存在下で細胞を培養することによってアポトーシスにかかりやすくなることができるということが以前に報告された[Lenardo等,Nature,353:858-861(1991)]。本研究はIL-2を用いた前処理がApo-2リガンド誘導死に対して末梢T細胞を感受性にすることを示した。
【0182】
実施例7
Fas/Apo-1及びTNF受容体を用いた阻害アッセイ
fas/Apo-1受容体、同様に1型及び2型TNF受容体(TNF-R1及びTNF-R2)の可溶性形態が精製された可溶性Apo-2リガンド(実施例3に記述されている)のアポトーシス活性を阻害できるかどうかを試験することによって、これらの受容体がApo-2リガンドのアポトーシス活性を介在することに関与しているかどうかを測定するためのアッセイを実施した。
【0183】
9D細胞をバッファーコントロール、CD4-IgGコントロール(25μg/ml)、可溶性Apo-1-IgG(25μg/ml)、可溶性TNFR1-IgG(25μg/ml)または可溶性TNFR2-IgG融合タンパク質(25μg/ml)の存在下で、培地コントロールまたはApo-2リガンド(0.3μg/ml、実施例3に記述されているように調製した)と共に24時間インキュベートした(5×104細胞/ウェル)。Fas/Apo-1、TNF-R1及びTNF-R2受容体の可溶性誘導体を、Ashkenazi等,Methods,8:104-115(1995)に記述されているようにIgG融合タンパク質として生産した。CD4-IgGを、Byrn等,Nature,344:667-670(1990)に記述されているようにIgG融合タンパク質として生産し、コントロールとして用いた。
【0184】
図3Cに示されているように、受容体-融合分子の全てが、9D細胞に対するApo-2リガンドアポトーシス活性を阻害しなかった。これらの結果から、Apo-2リガンドアポトーシス活性はfas/Apo-1及びTNF-R1とTNF-R2とは独立であることが示された。
【0185】
実施例8
哺乳動物組織におけるApo-2リガンドmRNAの発現
ヒト組織におけるApo-2リガンドmRNAの発現を、ノーザンブロット分析により調べた(図4)。ヒトRNAブロットはフルレングスApo-2リガンドcDNAに基づく32PラベルDNAプローブとハイブリダイズし、またはGenBank EST配列、HMEA47Mに基づく32PラベルRNAプローブとハイブリダイズした(実施例1参照)。ヒト胎児RNAブロットMTN(Clontech)及びヒト成人RNAブロットMTN-II(Clontech)を、DNAプローブとインキュベートし、一方でヒト成人RNAブロットMTN-I(Clontech)をRNAプローブとインキュベートした。ブロットをハイブリダイゼーションバッファー(5×SSPE;2×デンハルト溶液;100mg/mL変性切断サケ精子DNA;50%ホルムアミド;2%SDS)において42℃で16時間プローブとインキュベートした。該ブロットを、1×SSPEで数回;2%SDSで65℃で1時間及び50%新鮮な脱イオンホルムアミド;1×SSPE;0.2%SDSで65℃で30分洗浄した。該ブロットをホスホリムアガー(phosphorimager)(Fuji)を用いてオーバーナイトでさらした後解析した。
その結果が図4に示されている。胎児ヒト組織においては、Apo-2リガンドmRNA発現は肺、肝臓及び腎臓で検出され、脳組織ではされなかった。成人ヒト組織においては、Apo-2リガンドmRNA発現は脾臓、胸腺、前立腺、卵巣、小腸、末梢血リンパ球、心臓、胎盤、肺、腎臓で検出された。精巣、脳、骨格筋、及び膵臓ではほとんどまたは全く発現が検出されなかった。上述されているように、Apo-2リガンドに対して観察された発現プロフィールは、Apo-1リガンドのものとは同一ではなく、Apo-1リガンドは主にT細胞及び精巣で発現されている[Nagata等,上記参照]。
【0186】
実施例9
ヒト腫瘍細胞系に対するApo-2リガンドのアポトーシス活性
ヒト腫瘍細胞系に対するApo-2リガンド(実施例3に記述)のアポトーシス活性を、いくつかの化学療法試薬の一つの存在下または不存在下でさらに調べた。
以下のヒト腫瘍細胞系をアッセイした;A549(肺ガン腫、ATCC CCL 185);HCT116(大腸ガン腫、ATCC CCL 247);SW480(大腸腺ガン腫、ATCC CCL 228);MDA231(乳腺ガン腫、ATCC HTB 26);HeLa(子宮頚ガン腫、ATCC CCL 22);ME-180(子宮頚ガン腫,ATCC HTB 33);T24(膀胱ガン腫、ATCC HTB 4);SK-N-AS(神経芽腫、White等,Proc.Natl.Acad.Sci.,92:5520-5524(1995))。これらの細胞系のいくつかは、野生型p53を発現し、一方で他のものはミューテーションのため発現しておらず、それについては以下の表3を参照。該細胞を96穴プレートに2.5×105細胞/mlでまき、オーバーナイトでインキュベートした。該細胞をApo-2リガンドの2倍希釈物(100ng/mlから0.01ng/mlの範囲)の存在下で培養した。カルチャーのあるものには、化学療法試薬もまた24時間で加えた;シクロヘキサミド(「CHX」)(50μg/ml;Sigma Chemicals),ドキソルビシン(10-100μg/ml;Pharmacia)または5-FU(6mg/ml;Roche)。
【0187】
インキュベーションの24時間後、該細胞を20%メタノール中の0.5%クリスタルバイオレットで染色した。細胞生存力を0.1Mクエン酸ナトリウム(50%メタロール中に0.1Mクエン酸)を用いて染色細胞から色素を除去し、540nmの吸光度を測定することによって測定した。
その結果が以下の表に示されている。
【0188】
【表3】
+: 100ng/mlApo-2Lを用いた24時間後>35%が死
++: 100ng/mlApo-2Lを用いた24時間後>70%が死
+++: 10ng/mlApo-2Lを用いた24時間後>70%が死
これらの結果はApo-2リガンドが様々な腫瘍タイプから由来する腫瘍細胞系において細胞死を誘導し、Apo-2Lが腫瘍細胞のp53状態に独立して細胞死を誘導したことを示す。これらの結果はまたApo-2リガンド誘導細胞死がいくつかの異なる化学療法薬剤によって増加されることをも示す。
【0189】
実施例10
in vivoでのApo-2リガンドのアポトーシス活性
腫瘍を持つヌードマウスにおけるApo-2リガンドの効果を調べた。ヌードマウス(グループ当たり5-10マウス)(Harlan Sprague Dawleyから購入した)を、MDA231ヒト乳ガン腫細胞(ATCC HTB 23)(2×106細胞/マウス)で皮下に注射した(0日目)。それかが該腫瘍を14日間成長させた。14及び15日目に、2μg/0.05ml/マウスのApo-2リガンド(実施例3)及び/または10μg/0.05ml/マウスのドキソルビシン(Pharmacia)を、腫瘍部位に注射した。コントロール動物には0.05mlのPBSを同様に注射した。21日目に、該動物を殺し、腫瘍を調べ測量した(g)。
その結果が図5に示されている。該データにより、Apo-2リガンド処理はそれ自体で腫瘍成長を阻害し、そしてApo-2リガンドは腫瘍成長に対するドキスルビシンの阻害効果を増大することが示される。
【0190】
実施例11
in vivoでのApo-2リガンドのアポトーシス活性
Apo-2リガンドの抗腫瘍効果を、0日目にマウスをHCT116ヒト大腸ガン腫細胞(ATCC CCL 247)(2×106細胞/マウス)を用いて皮下に注射することを除いて、実施例10に記述されているように、腫瘍を持つヌードマウスで調べた。それから該腫瘍を14日間成長させた。14及び15日目に、2μg/0.05ml/マウスのApo-2リガンド及び/または10μg/0.05ml/マウスの5-FU(Roche)を、腫瘍部位に注射した。コントロール動物には0.05mlのPBSを同様に注射した。21日目に、該動物を殺し、腫瘍を調べ測量した(g)。
その結果が図6に示されている。これらの結果により、Apo-2リガンド処理はそれ自体で腫瘍成長を阻害し、そしてApo-2リガンドは腫瘍成長に対する5-FUの阻害効果を増大することが示される。
【0191】
実施例12
in vivoでのApo-2リガンドのアポトーシス活性
Apo-2リガンドの抗腫瘍効果を、1及び2日目に10μg/0.05ml/マウスのApo-2リガンド及び/または100μg/0.05ml/マウスの5-FU(Roche)を、腹膜内に注射することを除いて、実施例11に記述されているように、腫瘍を持つヌードマウスで調べた。コントロール動物には0.05mlのPBSを同様に注射した。それから腫瘍サイズ(mm2)を5,9,15日目に測定した。15日目に、該動物を殺し、腫瘍を調べ測量した(g)。
その結果が図7及び8に示されている。これらの結果により、Apo-2リガンドは腹膜内注射によって投与された場合でさえ、皮下腫瘍部位に到達し、抗腫瘍効果を発揮することができることが示される。また、これらの結果により、Apo-2リガンド処理はそれ自体で腫瘍成長を阻害し、そしてApo-2リガンドは腫瘍成長に対する5-FUの阻害効果を増大することが示される。
【0192】
実施例13
CrmAを用いた阻害アッセイ
ICE及びCPP32/YamaのようなプロテアーゼがApo-2リガンドによるアポトーシス誘導において役割を果たしているかどうかを調べるために、CrmAがApo-2リガンド誘導アポトーシスをブロックするかどうかを調べるためのアッセイを実施した[Marsters等,Current Biology,6:750-752(1996)]。CrmAは死プロテアーゼICE及びCPP/Yamaのポックスウイルス由来阻害剤であり、TNFR1及びFas/Apo-1による死シグナルをブロックする。加えて、Apo-1リガンド及びTNFによるアポトーシス誘導を介在するアダプタータンパク質、FADDを含む「死ドメイン」が、Apo-2リガンド誘導アポトーシスに関与するかどうかを調べるために[Chinnaiyan等,Cell,81:505-512(1995);Hsu等,Cell,84:299-308(1996)]、FADDのドミナントネガティブミュータント形態(FADD-DN)[Hsu等joukisannshou]が、Apo-2リガンド機能を阻害するかどうか[Marsters等,Current Biology,6:750-752(1996)]を調べるためのアッセイを実施した。
【0193】
HeLa-S3(ATCC CCL 22)細胞を、pRK5-CrmA発現プラスミド(CrmA配列はRay等,上記参照で報告された)またはpRK5-FADD-DN発現プラスミド(Hsu等,Cell,84:299-308(1996))を用いてトランスフェクトした。pRK5をコントロールとして用いた。該細胞をプラスミドDNAの取り込みに対するマーカーとしてpRK5-CD4(Smith等,Science,328:1704-1707(1988))と共にトランスフェクトした。トランスフェクトされた細胞をフィコエリスリン接合CD4抗体(Jackson Immunoresearch)を用いた染色によって同定し、アポトーシスを本質的に上記実施例4に記述されているようにFACSによって分析した。
【0194】
その結果が図9に示されている。CrmAは、抗Apo-1抗体により誘導されるアポトーシスと同様に、Apo-2リガンド誘導アポトーシスをブロックした。対照的に、FADD-DNは、Apo-2リガンド誘導アポトーシスに対してほとんど効果がないが、抗Apo-1抗体によるアポトーシス誘導を実質的にブロックした。したがって該アッセイの結果により、Apo-2リガンド、TNFR1及びFas/Apo-1は、アポトーシス性細胞死を活性化するための共通の末端シグナル経路を用いていることが示唆される。特に該結果は、ICE及びCPP/YamaのようなプロテアーゼがApo-2リガンド誘導アポトーシスに対して必要とされることを示唆する。対照的に、FADDはTNFR1及びFas/Apo-1による細胞死誘導に必要とされるが、Apo-2リガンドによるものには必要とされない。
【0195】
実施例14
A. 抗Apo-2リガンド抗体の調製
Balb/cマウス(Chales River Laboratoriesから得た)を、1μgApo-2リガンド(実施例3に記述されているように調製し、Ribi Immunochemical Research Inc.,ハミルトン、MTから購入したMPL-TDMアジュバントで希釈した)を1週間間隔で各後脚肉肢に10回注射することによって免疫化した。最終の追加免疫注射の3日後、マウスから膝窩リンパ節を取り出し、1%ペニシリン-ストレプトマイシンを補ったDMEM培地(Biowhitakker Corp.から得た)に単一細胞懸濁液を調製した。それから35%ポリエチレングリコール[Laskor等,Cell.Immunol.,55:251(1980)]を用いて、ネズミミエローマ細胞P3X63AgU.1(ATCC CRL 1597)とリンパ節細胞を融合し、96穴カルチャープレートで培養した。融合から由来するハイブリドーマをHAT培地で選択した。融合の10日後、c-Met-IgG融合タンパク質に結合するモノクローナル抗体の存在を試験するため、ハイブリドーマカルチャー上清をELISA[Kim等,J.Immunol.Meth.,156:9-17(1992)]でスクリーニングした。
【0196】
ELISAでは、PBS中の0.5μg/mlApo-2リガンド(実施例3参照)の50μlを各ウェルに加えることによって、96穴ミクロタイタープレートをコートし、4℃オーバーナイトでインキュベートした。それからプレートを洗浄バッファー(PBSプラス0.05%Tween20)で3回洗った。それから2%ウシ血清アルブミン(BSA)の200μlを用いて、ミクロタイター内のウェルをブロックし、1時間室温でインキュベートした。それからプレートを再び洗浄バッファーで3回洗った。
【0197】
洗浄ステップの後、2μg/mlのApo-2リガンド抗体の50μl、またはハイブリドーマカルチャー上清の100μlを各ウェルに加えた。培地に調和したP3X63AgU.1ミエローマ細胞の100μlをコントロールとして別の加工されたウェルに加えた。シェーカー装置で1時間室温でプレートをインキュベートし、それから洗浄バッファーで3回洗った。
【0198】
次に、アッセイバッファー(PBS中に0.5%ウシ血清アルブミン,0.05%Tween-20,0.01%Thimersol)で1:1000に希釈した50μlHRP結合ヤギ抗マウスIgG(Cappel Laboratoriesから購入した)を各ウェルに加え、シェーカー装置で室温で1時間プレートをインキュベートした。プレートを洗浄バッファーで3回洗い、引き続き基質(TMB,3,3',5,5'-テトラメチルベンジジンKirkegaard & Perry,Gaitherburg,MDから得た)の50μlを各ウェルに加え、10分間室温でインキュベートした。停止溶液(Kirkegaard & Perry)の50μlを各ウェルに加えることにより反応を止め、自動ミクロタイタープレートリーダーで450nmの吸光度を読み取った。
【0199】
ハイブリドーマ上清(選択された99)をApo-2リガンド誘導9D細胞殺傷をブロックする能力について試験した。活性は初めに、トリパンブルー色素溶出を用いて、処理された9D細胞の生存力%を調べることによって測定された。
【0200】
ブロッキング活性もまたFACS分析によって確認した。9D細胞(5×105細胞/0.5ml)を完全RPMI培地(RPMIプラス10%FCS、グルタミン、非必須アミノ酸、ペニシリン、ストレプトマイシン、ピルビン酸ナトリウム)に懸濁し、24穴マクロタイタープレート内にまいた。0.5mlのApo-2リガンド(1μg/ml)(実施例3に記述されているように調製した)を、完全RPMI培地内に懸濁し、10μgの精製モノクローナル抗体、または100μlの培養上清を用いてプレインキュベートし、それから9D細胞含有24マクロタイターウェル内に加えた。マクロタイタープレートを7%のCO2の存在下で37℃でオーバーナイトでインキュベートした。それからインキュベートされた細胞を回収し、PBSで一度洗浄した。細胞の生存力を製品の但し書き(Clontech)にしたがって、ホスファチジルセリンを結合したFITC-アネキシンVの染色により測定した。該細胞をPBSで洗浄し、200μlの結合バッファーに再懸濁した。10μlのアネキシンV-FITC(1μg/ml)及び10μlのプロピジウムイオダイドを該細胞に加えた。暗所で15分名インキュベーション後、該9d細胞をFACSにより分析した。
【0201】
8の潜在的なブロッキング抗体及び4の潜在的な非ブロッキング抗体を分泌する各ハイブリドーマを同定し、さらに限界希釈法によりクローン化(2回)した。
【0202】
モノクローナル抗体1D1,2G6,2E11及び5C2といわれる4の抗体のFACS分析が図10に示されている。(以下に示されるように、1D1,2G6,2E11及び5C2抗体はそれぞれハイブリドーマ1D1.12.4,2G6.3.4,2E11.5.5及び5C2.4.9によって生産され、それら全てがATCCに寄託されている)。Apo-2リガンドで処理された9D細胞(上部左図)は、非処理コントロール細胞(上部右図)より50%高いアポトーシス化細胞を示した。Apo-2リガンドプラス2E11,5C2,2G6または1D1抗体で処理された9D細胞は、非処理コントロールよりそれぞれ0%,6%,26&及び48%高いアポトーシス化細胞を示した。これらの結果は、5C2,2E11及び2G6抗体がブロッキング抗体である一方、1D1抗体は非ブロッキング抗体であることを示す。最も能力のあるブロッキング活性は、5C2抗体で観察された。
【0203】
4の抗体の抗原特異性もまたELISAで試験した。マイクロタイターウェルを2μg/mlリンホトキシン(Genentech,Inc.,EP 164,965、Gray等,Nature,312:721-724(1984)も参照)、TNF-アルファ(Genentech,Inc.,Pewnnica等,Nature,312:724-729(1984);Aggarwal等,J.Biol.Chem.,260:2345-2354(1985)も参照)、またはApo-2リガンド(実施例3参照)でコートした。モノクローナル抗体1D1,2G6,2E11及び5C2を10μg/mlの濃度で試験した。
【0204】
該アッセイの結果が図11に示されている。図11のデータは、モノクローナル抗体2G6,2E11及び5C2がApo-2リガンドに対して特異的である一方、モノクローナル抗体1D1はリンホトキシン及びTNF-アルファに対して弱い交差結合を示すことを表す。
【0205】
B. アイソタイピング
1D1,2G6,2E11及び5C2抗体(上記セクションAに記述されている)のアイソタイプを、アイソタイプ特異的ヤギ抗マウスIg(FisherBiotech,Pittsburgh,PA)でミクロタイタープレートを4℃でオーバーナイトでコートすることによって測定した。それから該プレートを上述の洗浄バッファーで洗浄した。それからミクロタイターのウェルを200μlの2%ウシ血清アルブミンでブロックし、室温で1時間インキュベートした。該プレートを洗浄バッファーで再び3回洗浄した。
【0206】
次に100μlの5μg/mlの精製Apo-2リガンド抗体、または100μlのハイブリドーマカルチャー上清を加工されたウェルに加えた。該プレートを室温で30分インキュベートし、それから50μlのHRP-接合ヤギ抗マウスIgG(上述のように)を各ウェルに加えた。該プレートを室温で30分インキュベートした。プレートに結合したHRPのレベルを上述のHRP基質を用いて検出した。
【0207】
アイソタイピング分析により、1D1及び2G6抗体はIgG2b抗体であり、そして2E11及び5C2抗体はIgG2a抗体であることが示された。
【0208】
C. エピトープマッピング
エピトープマッピングをビオチン化モノクローナル抗体を用いて、Kim等,上記参照に記述される競合的結合ELISAを用いて実施した。選択されたモノクローナル抗体を、Antibodies,A laboratory Manual,E.Harlow及びD.Lane,編、p.342に記述されているN-ヒドロキシルスクシンイミドを用いてビオチン化した。ミクロタイタープレートウェルを50μlのApo-2リガンド(実施例3参照、0.1μg/ml)でコートし、4℃でオーバーナイトで保存し、それから2%BSAで室温で1時間ブロックした。ミクロタイターウェルの洗浄後、所定最適化濃度のビオチン化抗体及び1000倍過剰の非ラベル抗体の混合物を、各ウェルに加えた。室温で1時間のインキュベーション後、該プレートを洗浄し、ビオチン化抗体の量をHRP-ストレプタビジンの添加によって検出し、そして該プレートをELISAプレートリーダーで490nmで読んだ。
【0209】
その結果が図12に示されている。該結果は、HRP接合抗体の結合がその抗体自身の過剰の量で効果的に阻害されるが、アッセイされた他の抗体では阻害されないことを示した。
【0210】
モノクローナル抗体により認識されるApo-2リガンドの領域を、Chuntharapai等,J.Immunol.,152:1783-1789(1994)に記述されているELISAで、合成ペプチド[図1Aに示されるApo-2リガンド配列のaa128-148(ペプチド「APO14」);aa144-159(ペプチド「APO15」);aa192-204(ペプチド「APO17」);aa230-238(ペプチド「APO18」);aa261-272(ペプチド「APO19」)]を用いて測定した。該結果が図13に示されている。1D1抗体はApo-2リガンドのアミノ酸残基192-204を含むAPO17ペプチドに結合することを示した。
【0211】
物質の寄託
以下の細胞系はAmerican Type Vulture Collection, 12301 Parklawn Drive,Rockville,MD,USA(ATCC)に寄託されている:
細胞系 ATCC寄託番号 寄託日
2935-pRK5-hApo-2L- CRL-12014 1996年1月3日
mycクローン2.1
1D1.12.4 HB-12557 1997年1月8日
2G6.3.4 HB-12559 1997年1月8日
2E11.5.5 HB-12556 1997年1月8日
5C2.4.9 HB-12558 1997年1月8日
【0212】
特許手続上の微生物の寄託の国際的承認に関するブタペスト条約の規定に基づいて寄託がなされた。これは寄託の日から30年間、寄託物の生存能力のあるカルチャーの管理を保証する。該寄託物はブタペスト条約の定める期間ATCCから入手可能であり、Genentech,Inc.とATCCの間の合意により、関係のある米国特許の発行か、米国または外国の特許出願の公開かの、いずれか早い時点で公衆に対して寄託物のカルチャーの子孫の永久に制限のない入手可能性が保証され、かつ米国特許法第122条ならびにそれに従う長官規則(特に886OG638に関係する37CFR1.14を含む)に従って米国特許商標長官によって権利を与えられることが決定された公衆に対する子孫の入手可能性が保障される。
【0213】
本出願の譲受人は、もし寄託した細胞系のカルチャーが適したコンディションの下で培養されながら死んだまたは失われたまたは破壊された場合、該細胞系を同じプラスミドのもう一つと共に告示して直ちに置き換えることを合意した。寄託された細胞系の利用可能性は、特許法にしたがっていかなる政府の権威に基づいて許可された権利に違反して、自由に本発明を実施できるというように解釈されない。
【0214】
先の文書の明細書が当業者をして本発明を実施せしめるのに十分であると考えられる。寄託された実施態様は本発明の特定の一面の単なる例証としてのつもりであり、機能的に均質であるいかなるカルチャーも本発明の範囲内にあるので、本発明は寄託されたカルチャーによって範囲を限定されることはない。ここで物質の寄託は、ここに含まれる記述されたものが、その最適のモードを含む本発明の任意の面の実施可能性に対して不十分であることの自認を構成するものではなく、存在する特別に説明された請求の範囲が表された特定の例に限定されるとみなされるものでもない。実際ここで示され記述されたものに加えて、本発明の様々な修飾が以前の記述から当業者に明らかになるであろうし、それらは付記される請求項の範囲内に存在する。
【図面の簡単な説明】
【0215】
【図1A】図1AはヒトApo-2リガンドcDNAの核酸配列及びそのコードするアミノ酸配列を示す。
【図1B】図1BはヒトTNFサイトカインファミリーの周知のメンバー、4-1BBL,OX40L,CD27L,CD30L,TNF-α,LT-β,LT-α,CD40L及びApo-1Lの相当する領域と共に、ヒトApo-2リガンドのC末端領域の配列を示す。
【図1C】図1Cは抗mycエピトープ抗体を用いるFACS分析によって測定された、ヒト293細胞で発現された組換えフルレングスC末端mycエピトープタグ化Apo-2リガンドの細胞トポロジーを示す。
【図1D】図1DはSDS-PAGE及び銀染色に引き続き、化学架橋の存在下(レーン2,3)または不存在下(レーン1)で測定された、組換えバキュロウイルス感染昆虫細胞で発現され、Ni2+キレートアフィニティークロマトグラフィーで精製された組換えHis10エピトープタグ化可溶性Apo-2のサイズ及びサブユニット構造を示す。
【図1E】図1EはSDS-PAGE及びオートラジオグラフィーに引き続き、抗gDエピトープ抗体を用いた免疫沈降によって測定された、代謝的にラベルされたヒト293細胞で発現された組換えgDエピトープタグ化可溶性Apo-2リガンドのサイズ及びサブユニット構造を示す。
【図2A】図2A-2EはApo-2リガンドによるB及びTリンパ球細胞系におけるアポトーシスの誘導を示す。アポトーシス化細胞は(A) 特徴的な形態変化によって;(B-D) フローサイトメトリーで測定された、プロピジウムイオダイド(PI)及びFITC接合アネキシンVを用いたポジティブ蛍光染色によって;及び(E) 染色体内DNA断片化の分析によって同定された。
【図2B】図2A-2EはApo-2リガンドによるB及びTリンパ球細胞系におけるアポトーシスの誘導を示す。アポトーシス化細胞は(A) 特徴的な形態変化によって;(B-D) フローサイトメトリーで測定された、プロピジウムイオダイド(PI)及びFITC接合アネキシンVを用いたポジティブ蛍光染色によって;及び(E) 染色体内DNA断片化の分析によって同定された。
【図2C】図2A-2EはApo-2リガンドによるB及びTリンパ球細胞系におけるアポトーシスの誘導を示す。アポトーシス化細胞は(A) 特徴的な形態変化によって;(B-D) フローサイトメトリーで測定された、プロピジウムイオダイド(PI)及びFITC接合アネキシンVを用いたポジティブ蛍光染色によって;及び(E) 染色体内DNA断片化の分析によって同定された。
【図2D】図2A-2EはApo-2リガンドによるB及びTリンパ球細胞系におけるアポトーシスの誘導を示す。アポトーシス化細胞は(A) 特徴的な形態変化によって;(B-D) フローサイトメトリーで測定された、プロピジウムイオダイド(PI)及びFITC接合アネキシンVを用いたポジティブ蛍光染色によって;及び(E) 染色体内DNA断片化の分析によって同定された。
【図2E】図2A-2EはApo-2リガンドによるB及びTリンパ球細胞系におけるアポトーシスの誘導を示す。アポトーシス化細胞は(A) 特徴的な形態変化によって;(B-D) フローサイトメトリーで測定された、プロピジウムイオダイド(PI)及びFITC接合アネキシンVを用いたポジティブ蛍光染色によって;及び(E) 染色体内DNA断片化の分析によって同定された。
【図3A】図3A-3CはApo-2リガンド誘発アポトーシスのタイムコースと量依存性、及びFas/Apo-1受容体、TNF-R1受容体またはTNF-R2受容体に基づく可溶性受容体IgG融合タンパク質による、Apo-2リガンド誘発アポトーシスの阻害の欠如を示す。
【図3B】図3A-3CはApo-2リガンド誘発アポトーシスのタイムコースと量依存性、及びFas/Apo-1受容体、TNF-R1受容体またはTNF-R2受容体に基づく可溶性受容体IgG融合タンパク質による、Apo-2リガンド誘発アポトーシスの阻害の欠如を示す。
【図3C】図3A-3CはApo-2リガンド誘発アポトーシスのタイムコースと量依存性、及びFas/Apo-1受容体、TNF-R1受容体またはTNF-R2受容体に基づく可溶性受容体IgG融合タンパク質による、Apo-2リガンド誘発アポトーシスの阻害の欠如を示す。
【図4】図4はノーザンブロット分析で測定された、ヒト胎児及びヒト成人組織におけるApo-2リガンドmRNAの発現を示す。
【図5】図5はヌードマウスにおいて成長したヒトMDA231乳ガンベース腫瘍の重量に対する、単独のまたはドキソルビシンと組み合わせた腫瘍内注射によって投与されたApo-2リガンドのin vivoでの効果を示す。
【図6】図6はヌードマウスにおいて成長したヒトHCT116大腸ガンベース腫瘍の重量に対する、単独のまたは5-FUと組み合わせた腫瘍内注射によって投与されたApo-2リガンドのin vivoでの効果を示す。
【図7】図7はヌードマウスにおいて成長したヒトHCT116大腸ガンベース腫瘍のサイズに対する、単独のまたは5-FUと組み合わせた腹膜内注射によって投与されたApo-2リガンドのin vivoでの効果を示す。
【図8】図8はヌードマウスにおいて成長したヒトHCT116大腸ガンベース腫瘍の重量に対する、単独のまたは5-FUと組み合わせた腹膜内注射によって投与されたApo-2リガンドのin vivoでの効果を示す。
【図9】図9はドミナントネガティブなFADDではないCrmAがHeLa-S3細胞においてApo-2リガンド誘発アポトーシスをブロックすることを示す棒グラフである。
【図10】図10はApo-2リガンドにより誘発されるアポトーシスのFACS分析及び4の抗Apo-2リガンド抗体:1D1,2G6,2E11及び5C2の効果を示す(FITC接合アネキシンVを用いて検出されたアポトーシス化9D細胞−太線;生きた非染色細胞-細線)。
【図11】図11はモノクローナル抗体1D1,2G6,2E11及び5C2の抗原特異性を示す棒グラフである。
【図12】図12はモノクローナル抗体1D1,2G6,2E11及び5C2のエピトープマッピングアッセイの結果を示す棒グラフである。
【図13】図13はApo-2リガンドの特異的アミノ酸領域より成るいくつかの異なる合成ペプチドに結合するモノクローナル抗体1D1の能力を試験するアッセイの結果を示す棒グラフである。
【技術分野】
【0001】
関連出願
本出願は1996年1月9日に提出された米国特許第08/584,031号の一部継続出願であり、その内容は参考としてここで取り込まれる。
【0002】
本発明は一般的に哺乳動物細胞アポトーシスを誘導するここで「Apo-2リガンド」として表される新規なサイトカインの同定、単離、及び組換え生産に関し、およびApo-2リガンド抗体と該組成物を用いる方法に関する。
【背景技術】
【0003】
哺乳動物における細胞数のコントロールは、細胞増殖と細胞死の間のバランスによって決定されると考えられる。細胞死の一つの形態として、しばしば壊死性細胞死といわれるものは、ある外傷または細胞障害から由来する細胞死の病理学的な形態として典型的には特性指摘される。これに対して、通常規則的にまたはコントロールされた方法で進行する細胞死のもう一つの「生理学的な」形態が存在する。この規則的にまたはコントロールされた細胞死の形態は、しばしば「アポトーシス」といわれる[例えばBarr等,Bio/Technology,12:487-493(1994)参照]。アポトーシス性細胞死は多くの生理学的過程で天然で生じ、それには胚の発達及び免疫系でのクローン選択が含まれる[Itoh等,Cell,66:233-243(1991)]。しかしながら減少したレベルのアポトーシス性細胞死は、ガン、狼瘡、及びヘルペスウイルス感染を含む様々な病理学的コンディションと関連する[Thompson,Science,267:1456-1462(1995)]。
【0004】
アポトーシス性細胞死は細胞質の凝縮、細胞膜微絨毛の消失、核の断片化、染色体DNAの凝集、またはミトコンドリア機能の消失のような一つ以上の細胞内の形態学的及び生化学的変化を典型的に伴う。様々な外因性及び内因性のシグナルが、該形態学的及び生化学的細胞変化を引き起こす、または誘導すると考えられている[Raff,Nature,356:397-400(1992);Steller,Science,267:1445-1449(1995);Sachs等,Blood,82:15(1993)]。例えばそれらは非成熟胸腺細胞に対するグルココルチコイドホルモンのようなホルモン性の刺激によって引き起こされ、同様に特定の増殖因子の減衰によっても引き起こされる[Watanabe-Fukunaga等,Nature,356:314-317(1992)]。同様にmyc,rel及びE1Aのようないくつかのガン遺伝子、及びp53のような腫瘍サプレッサーが、アポトーシスを誘導するのに役割を担っていることが報告されている。特定の化学療法薬剤及びある形態の放射線も同様にアポトーシス誘発活性を持つことが観察されている[Thompson,上記参照]。
【0005】
腫瘍壊死因子-α(「TNF-α」)、腫瘍壊死因子-β(「TNF-β」または「リンホトキシン」)、CD30リガンド、CD27リガンド、CD40リガンド、OX-40リガンド、4-1BBリガンド、及びApo-1リガンド(FasリガンドまたはCD95リガンドとしてもいわれる)のような様々な分子が、サイトカインの腫瘍壊死因子(「TNF」)ファミリーのメンバーとして同定されている[例えばGruss及びDower,Blood,85:3378-3404(1995)参照]。これらの分子の中で、TNF-α、TNF-β,CD30リガンド,4-1BBリガンド及びApo-1リガンドは、アポトーシス性細胞死に関与することが報告されている。TNF-αとTNF-βは影響を受けやすい腫瘍細胞においてアポトーシス性の死を誘導することが報告されている[Schmid等,Proc.Natl.Acad.Sci.,83:1881(1986);Deaitry等,Eur.J.Immunol.,17:689(1987)]。Zheng等はTNF-αがCD8ポジティブT細胞の刺激後アポトーシスに関与することを報告している[Zheng等,Nature,377:348-351(1995)]。他の研究者はCD30リガンドが胸腺における活性化T細胞の除去に関与することを報告している[Amakawa等,Cold Spring Harbor Laboratory Symposium on Programmed Cell Death,Abstr.No.10,(1995)]。
【0006】
マウスFas/Apo-1受容体またはリガンド遺伝子(それぞれlpr及びgldと呼ばれる)におけるミューテーションはある自己免疫疾患と関連し、それはApo-1リガンドが末梢における活性化T細胞のクローン欠失を調節するのに役割を担っているであろうことを示す[Krammer等,Curr.Op.Immunol.,6:279-289(1994);Nagata等,Science,267:1449-1456(1995)]。Apo-1リガンドはまたCD4ポジティブTリンパ球及びBリンパ球における刺激後アポトーシスを誘導することが報告されており、そして活性化リンパ球の機能がもはや必要とされなくなった場合には該リンパ球の除去に関与するであろう[Krammer等,上記参照;Nagata等,上記参照]。Apo-1受容体に特異的に結合する作用剤マウスモノクローナル抗体は、TNF-αのものと比較可能なまたは同様な細胞殺傷活性を示すことが報告されている[Yonehara等,J.Exp.Med.,169:1747-1756(1989)]。
【0007】
該TNFファミリーサイトカインによって介在される様々な細胞応答の誘導は、特異的細胞受容体へのその結合によって開始されると考えられている。およそ55kDa(TNF-R1)及び75kDa(TNF-R2)の2の異なるTNF受容体が同定されており[Hohman等,J.Biol.Chem.,264:14927-14935(1989);Brockhaus等,Proc.Natl.Acad.Sci.,87:3127-3131(1990);1991年3月20日に印刷されたEP417,563]、両受容体タイプに相当するヒト及びマウスcDNAが単離され特性指摘されている[Loetscher等,Cell,61:351(1990);Schall等,Cell,61:361(1990);Smith等,Science,248:1019-1023(1990);Lewis等,Proc.Natl.Acad.Sci.,88:2830-2834(1991);Goodwin等,Mol.Cell.Biol.,11:3020-3026(1991)]。Itoh等はApo-1受容体が55kDaTNF-R1によって伝達されるものと同様なアポトーシス性細胞死を伝達し得ることを開示する[Itoh等,上記参照]。細胞がTNF-αまたは抗Apo-1マウスモノクローナル抗体のそれぞれで処理された場合、Apo-1抗原の発現がTNF-R1のものと共に下流調節されることが報告されている[Krammer等,上記参照;Nagata等,上記参照]。したがってある研究者はApo-1及びTNF-R1受容体の両者を共発現する細胞系が共通のシグナル経路を通して細胞殺傷を介在するであろうと仮説を立てている[Id.]。
【0008】
リンホトキシン-αの発現と共に今日同定されているTNFファミリーリガンドは、そのC末端が細胞外に存在するII型膜貫通タンパク質である。対照的に今日同定されているTNF受容体(TNFR)ファミリーにおける受容体は、I型膜貫通タンパク質である。しかしながらTNFリガンドと受容体ファミリーの両者において、ファミリーのメンバー間に同定される同一性は主に細胞外ドメイン(「ECD」)で見出されている。TNF-α、Apo-1リガンド及びCD40リガンドを含むTNFファミリーサイトカインのいくつかは、細胞表面でタンパク質分解的に切断される;それぞれの場合で結果として生じたタンパク質は可溶性サイトカインとして機能するホモ三量体分子を典型的に形成する。TNF受容体ファミリータンパク質はまた、同起源のサイトカインの阻害剤として機能し得る可溶性受容体ECDを放出するためにタンパク質分解的に通常切断される。サイトカインとその受容体のTNFファミリーのレビューとしては、Gruss及びDower,上記参照を参照。
【発明の開示】
【課題を解決するための手段】
【0009】
出願人は、「Apo-2リガンド」と呼ばれる新規なサイトカインをコードするcDNAクローンを同定した。Apo-2リガンドはTNFサイトカインファミリーのメンバーであると現在考えられている;Apo-2リガンドはApo-1リガンドを含むいくつかの周知のTNF関連タンパク質に対するアミノ酸配列において関連している。しかしながら出願人はApo-2リガンドがFas/Apo-1,TNF-R1,またはTNF-R2受容体のような周知の可溶性Apo-1またはTNF受容体によって明らかには阻害されないことを見出した。
【0010】
一つの実施態様として、本発明は単離された生物学的に活性なApo-2リガンドを提供する。特に本発明は図1Aの残基114-281を含むアミノ酸配列を含む単離された生物学的に活性なApo-2リガンドを提供する。もう一つの実施態様として、単離された生物学的に活性なApo-2リガンドは図1Aの残基1-281(SEQ ID NO:1)に示されたアミノ酸配列を含む。
【0011】
もう一つの実施態様として、本発明はもう一つの異種ポリペプチドに融合されたApo-2リガンドを含むキメラ分子を提供する。該キメラ分子の例としては、タグポリペプチド配列に融合されたApo-2リガンドが含まれる。
【0012】
もう一つの実施態様として、本発明はApo-2リガンドをコードする単離された核酸分子を提供する。一つの面として、核酸分子は生物学的に活性なApo-2リガンドをコードするRNAまたはDNAであり、あるいは該Apo-2リガンドをコードする核酸配列に相補的なものであり、そして少なくとも穏やかに厳格なコンディションの下でそれに安定に結合し続けるものである。一つの実施態様として該核酸配列は以下から選択される:
(a) 図1A(SEQ ID NO:2)に示された残基1から残基281(すなわち核酸91から281)のフルレングスタンパク質をコードする図1Aの核酸配列のコード領域、または残基41から281の細胞外タンパク質をコードする核酸211から933のコード領域、または残基114から281の細胞外タンパク質をコードする核酸430から933のコード領域;あるいは
(b) 遺伝学的コードの縮重の範囲内で(a)の配列に一致する配列。
【0013】
さらなる実施態様として、本発明はベクターでトランスフェクトまたはトランスフォームされたホスト細胞によって認識されるコントロール配列に実施可能にリンクしたApo-2リガンドをコードする核酸分子を含む複製可能なベクターを提供する。該ベクターまたは核酸分子を含むホスト細胞もまた提供される。該核酸分子を含むホスト細胞を培養すること、及び該ホスト細胞カルチャーからタンパク質を回収することを含むApo-2リガンドの生産法も、さらに提供される。
【0014】
もう一つの実施態様として、本発明はApo-2リガンドに結合する抗体を提供する。一つの面として、該抗体はApo-2リガンドに対する抗原特異性を持つモノクローナル抗体である。
【0015】
もう一つの実施態様として、本発明は生物学的に活性なApo-2リガンド及び製薬学的に許容できるキャリアーを含む組成物を提供する。該組成物はアポトーシスを誘導または刺激するのに有用な製薬学的組成物であろう。
【0016】
もう一つの実施態様として、本発明はアポトーシスを誘導するのに効果的なApo-2リガンドの量に、in vivoまたはex vivoで哺乳動物細胞をさらすことを含む哺乳動物細胞においてアポトーシスを誘導する方法を提供する。
【0017】
もう一つの実施態様として、本発明はガンを持つ哺乳動物の治療法を提供する。該方法においては、有効量のApo-2リガンドがガンを持つと診断された哺乳動物に投与される。Apo-2リガンドはまた化学療法、放射線療法、または抗腫瘍活性を発揮可能な他の試薬のような一つ以上の他の治療と共に哺乳動物に投与され得る。
本発明のさらなる実施態様として、Apo-2リガンドまたはApo-2リガンド抗体を含む製造品及びキットが提供される。製造品及びキットには容器、該容器についたラベル、容器内に含まれる組成物が含まれる。容器についたラベルは該組成物が特定の治療上のまたは非治療上の適用に用いられ得ることを示す。該組成物には、活性試薬、及びApo-2リガンドまたはApo-2リガンド抗体を含む活性試薬が含まれる。
【発明を実施するための最良の形態】
【0018】
I. 定義
「Apo-2リガンド」及び「Apo-2L」なる語は、図1Aに示されたアミノ酸配列のアミノ酸残基114-281、残基41-281、残基15-281、または残基1-281を含むポリペプチド配列をいうためにここで用いられ、同様に上記配列の生物学的に活性な欠失、挿入、または置換変異体をいうために用いられる。好ましい実施態様として、生物学的に活性な変異体は、上記配列のいずれか一つと少なくとも約80%の配列同一性、より好ましくは少なくとも約90%の配列同一性、そしてさらにより好ましくは少なくとも約95%の配列同一性を持つ。該定義はここで記述される(実施例8参照)ヒト組織タイプまたはもう一つのソースのようなApo-2リガンドソースから単離され、あるいは組換え法または合成法によって調製される。Apo-2リガンドの本定義は、Gen Bank HHEA47M,T90422,R31020,H43566,H44565,H44567,H54628,H44772,H54629,T82085,及びT10524のような周知のEST配列を除外する。
【0019】
ここで用いられている「タグを付けたエピトープ」なる語は、「タグポリペプチド」と融合したApo-2リガンドまたはそれらの一部を含むキメラポリペプチドを示す。タグポリペプチドは抗体を生産するときのエピトープを供給するために十分な残基をもち、しかしApo-2リガンドの活性を妨害しないよう十分に短い。タグポリペプチドはまた好ましくは抗体が他のエピトープと実質的に交差反応しないように全く独特でもある。適したタグポリペプチドは一般的には少なくとも6アミノ酸残基をもち、通常約8から約50の間のアミノ酸残基(好ましくは約10から約20の間の残基)をもつ。
【0020】
ここで開示されている様々なタンパク質を記述するのに用いられている「単離した」なる語は、自然界の構成要素から同定されて分離されおよび/または回収されたタンパク質を意味する。自然界の混合した構成要素は、タンパク質の診断または治療用途を典型的には妨害するであろう物質であり、酵素、ホルモンそして他のタンパク質性または非タンパク質性溶液を含むであろう。好ましい実施態様としては、タンパク質は(1)スピニングカップシークエネーターの使用によりN末端または内部のアミノ酸配列の少なくとも15残基を得るのに十分な程度に、(2)クマシーブルーまたは銀染色を用いる非還元または還元状態の下でのSDS-PAGEによって均質に精製されるであろう。Apo-2リガンド天然環境の少なくとも一つの構成要素が存在しないであろうから、単離したタンパク質は組換え細胞内でのIn situのタンパク質を含む。しかしながら普通は、単離したタンパク質は少なくとも一つの精製工程によって調製されるであろう。
【0021】
「単離した」Apo-2リガンド核酸分子は、通常はApo-2リガンド核酸の天然のソースとして関連している少なくとも一つの混合した核酸分子から同定され分離された核酸分子である。単離したApo-2リガンド核酸分子は天然で見出される形態と場所以外のものである。それゆえ単離したApo-2リガンド核酸分子は天然の細胞内に存在するようなApo-2リガンド核酸分子とは区別される。しかしながら単離したApo-2リガンド核酸分子は、例えば核酸分子が染色体の位置において天然の細胞のものとは異なるApo-2リガンドを通常は発現する細胞内に含まれるApo-2リガンド核酸分子を含む。
【0022】
「コントロール配列」なる語は、特定のホスト生物内で実施可能に結合したコード配列の発現のために必要なDNA配列を示す。原核生物に適したコントロール配列は、例えばポロモーター、必要であればオペレーター配列そしてリボソーム結合部位を含む。真核生物細胞はプロモーター、ポリアデニレーションシグナルそしてエンハンサーが知られている。
【0023】
核酸がさらなる核酸配列との機能的な関連内に置かれたとき、核酸は「実施可能に結合」している。例えばプレ配列または分泌リーダーのためのDNAは、もしそれが配列の転写に作用すれば、ポリペプチドのためのDNAと実施可能に結合している;またはリボソーム結合部位は、もしそれが翻訳を促進するために正しい場所に置かれていれば、コード配列と実施可能に結合している。一般的に「実施可能に結合」とは、結合されているDNA配列が隣接し、分泌リーダーの場合には隣接しそしてリーディングフレーム内にあることを意味する。しかしながらエンハンサーは隣接している必要はない。結合は簡便な制限部位でのライゲーションによって成し遂げられる。もし該部位が存在しなかったなら、合成オリゴヌクレオチドアダプターまたはリンカーをありきたりの実施にしたがって用いる。
【0024】
「抗体」なる語は、最も広い意味で用いられ、単一の請うApo-2リガンドモノクローナル抗体(作用剤および拮抗剤抗体を含む)およびポリエピトープ特異性を持つ請うApo-2リガンド抗体構成物をも特にカバーする。
【0025】
ここで用いられているような「モノクローナル抗体」なる語は、実質的に同質な抗体の集団から得られる抗体を示す、すなわち集団に含まれる個々の抗体が、少量存在するであろう自然に生じ得る突然変異を除いて同一なのである。モノクローナル抗体は高い特異性を持っており、単一の抗原部位に対して向けられている。さらに、異なる決定基(エピトープ)に対して向けられた異なる抗体を典型的に含むありきたりの(ポリクローナル)抗体調製物とは異なり、各モノクローナル抗体は抗原の単一の決定基に対して向けられている。
【0026】
ここではモノクローナル抗体は、由来する種や免疫グロブリンのクラスまたはサブクラスの名称に関わらず、抗Apo-2リガンド抗体の可変ドメイン(超可変ドメインを含む)を定常ドメインに接合したもの(例えば「ヒト化」抗体)、L鎖をH鎖に接合したもの、ある種の鎖をもう一つの種の鎖に接合したものまたは異種構造のタンパク質の融合によるものによって生産されるハイブリッド及び組換え抗体を含む。また望ましい活性を示す範囲で、抗体断片(例えばFab,F(ab')2およびFv)も含む。例えば、米国特許第4,816,567号及びMage等,Monoclonal Antibody Production Techniques and Applications内のpp.79-97(Marcel Dekker,Inc.,New York,1987)参照。
【0027】
それゆえ緩和した「モノクローナル」なる語は、実質的に同質の抗体の集団から得られたような抗体の性質を示し、いかなる特定の方法によって抗体を生産する必要があるとは解釈すべきではない。例えば本発明にしたがって用いられるモノクローナル抗体は、KohlerとMilstein,Nature 256:495(1975)によって最初に記述されたハイブリドーマ法によって得られ得るし、または米国特許第4,816,567号に記述されている組換えDNA法によっても得られ得る。「モノクローナル抗体」は例えばMcCafferty等,Nature,348:552-554(1990)に記述されている方法を用いて生産されるファージライブラリーからも単離し得る。
【0028】
非ヒト(例えばネズミ)抗体の「ヒト化」形態とは、非ヒト免疫グロブリンから由来する最小限の配列を含む、特異的なキメラ免疫グロブリン、免疫グロブリン鎖またはそれらの断片(Fv,Fab,Fab',F(ab')2または他の抗体の抗原結合部分配列)である。ほとんどの場合が、ヒト化抗体は受容者の相補性決定領域(CDR)由来の残基が、望ましい特異性、アフィニティーそして力量を持つマウス、ラットまたはウサギのような非ヒト種(提供者抗体)のCDR由来の残基に置き換えられているヒト免疫グロブリン(受容者抗体)である。ある例では、ヒト免疫グロブリンのFvフレームワーク領域(FR)残基が、相当する非ヒト残基で置き換えられている。さらに該ヒト化抗体は受容者抗体にもインポートCDRまたはフレームワーク配列にも見出されない残基をも含み得る。これらの変形は、抗体の能力をさらに洗練し最適化するためになされる。一般的にはヒト化抗体は、少なくとも一つ、そして典型的には二つの可変ドメインの実質的にすべてを含むであろうし、可変ドメイン内には非ヒト免疫グロブリンのものに相当するCDR領域の全てまたは実質的にすべておよびFR領域の全てまたは実質的に全てが、ヒト免疫グロブリンコンセンサス配列のものである。ヒト化抗体はまた最適には、免疫グロブリンコンセンサス領域(Fc)の少なくとも一部を含み、それは典型的にはヒト免疫グロブリンのものである。
【0029】
Apo-2リガンドを特徴付けるためにここで目的とされる「生物学的に活性」なる語は、in vivoまたはex vivoで哺乳動物細胞の少なくとも一タイプにおいてアポトーシスを誘導するまたは刺激する能力を持つことを意味する。
【0030】
「アポトーシス」及び「アポトーシス活性」なる語は広い意味で用いられ、細胞質の凝縮、細胞膜微絨毛の消失、核の断片化、染色体DNAの凝集、またはミトコンドリア機能の消失を含む一つ以上の特徴的な細胞変化を典型的に伴う、哺乳動物における規則的なまたはコントロールされた細胞死の形態をいう。この活性は例えば細胞生存力アッセイ、FACS分析またはDNA電気泳動によって決定され測定され得る。
【0031】
ここで用いられている「ガン」と「ガンの」なる語は、典型的に非制御細胞増殖によって特徴付けられる哺乳動物内の生理学的病気を示しまたは表す。ガンの例としては、ガン腫、リンパ腫、白血病、芽腫そして肉腫を含むがそれに限られない。該ガンのより好ましい例としては、扁平上皮細胞ガン腫、小細胞肺ガン、非小細胞肺ガン、神経芽腫、脾臓ガン、グリア芽腫多形、頚部ガン、胃ガン、膀胱ガン、肝腫瘍、乳ガン、大腸ガン腫そして頭と首のガンを含む。一つの実施態様として、該ガンには小胞リンパ腫、p53ミューテーションを伴うガン腫、または乳ガン、前立腺ガン、あるいは卵巣ガンのようなホルモン依存性のガンが含まれる。
【0032】
ここで用いられている「治療」なる語は、治癒力のある治療、予防治療そして予防的治療を示す。
【0033】
ここで用いられている「哺乳動物」なる語は、ヒト、ウシ、ウマ、イヌそしてネコを含む哺乳動物として分類されるいかなる動物をも示す。本発明の好ましい実施態様としては、哺乳動物はヒトである。
【0034】
II. 本発明の組成物及び方法
本発明はTNFリガンドファミリーに関連する新規なサイトカインを提供し、該サイトカインはここで「Apo-2リガンド」と同定される。ヒトApo-2リガンドの予想される成熟アミノ酸配列は281アミノ酸を含み、およそ32,5kDaの計算される分子量とおよそ7,63の等電点を持つ。ハイドロパシー分析では、残基15と40の間に疎水性領域の存在が示されるが、N末端に明らかなシグナル配列は存在しない。シグナル配列の不存在と内部疎水性領域の存在は、Apo-2リガンドがII型膜貫通タンパク質であることを示唆する。潜在的なNリンクグリコシル化部位は推定の細胞外領域の残基109に位置する。推定の細胞質領域は図1Aに示されるアミノ酸残基1-14を含み、膜貫通領域はアミノ酸残基14-40を含み、細胞外領域はアミノ酸残基41-281を含む。図1Aに示される細胞外領域のアミノ酸残基114-281を含むApo-2リガンドポリペプチドはまた以下の実施例に記述される。
【0035】
A. Apo-2リガンドの調製
以下の記述は主としてApo-2リガンド核酸を含むベクターを用いてトランスフォームまたはトランスフェクトされた細胞の培養と、細胞カルチャーからのポリぺプチドの回収によるApo-2リガンドの生産に関する。もちろん本分野でよく知られている代わりの方法が、Apo-2リガンドを調製するために用いられることは考えられる。
【0036】
1. Apo-2リガンドをコードするDNAの単離
Apo-2リガンドをコードするDNAは、Apo-2リガンドmRNAを持ち、検出可能なレベルにそれを発現すると考えられる組織から調製したcDNAライブラリーから得られるであろう。よって、ヒトApo-2リガンドDNAは実施例1に記述されているヒト胎盤cDNAのバクテリオファージライブラリーのようなヒト組織から調製されるcDNAライブラリーから便利に得ることができる。Apo-2リガンドをコードする遺伝子はまた、ゲノムライブラリーから、またはオリゴヌクレオチド合成によって得られるだろう。
【0037】
ライブラリーは(Apo-2リガンドまたは少なくとも約20-80塩基のオリゴヌクレオチドに対する抗体のような)プローブを用いてスクリーニングができ、そのプローブは興味ある遺伝子またはそれによってコードされるタンパク質を同定するためにデザインされている。オリゴヌクレオチドプローブの例は実施例1に提供される。選択されたプローブを用いてcDNAまたはゲノムのライブラリーをスクリーニングすることは、Sambrook等,Molecular Cloning:A Laboratory Manual(ニューヨーク:Cold Spring Harbor Laboratory Press,1989)に記述されているような標準的な方法を用いて実施されるであろう。Apo-2リガンドをコードする遺伝子の単離のための代わりの方法は、PCR方法体系を用いることである[Smabrook等,上記参照;Dieffenbach等,PCR Primer:A Laboratory Manual(Cold Spring Harbor Laboratory Press,1995)]。
【0038】
スクリーニングの一つの好ましい方法として、様々なヒト組織由来のcDNAライブラリーをスクリーニングするために、選択されたオリゴヌクレオチド配列を用いる。以下の実施例1は2の異なるオリゴヌクレオチドプローブを用いたcDNAライブラリーのスクリーニング法を記述する。プローブとして選択されたオリゴヌクレオチド配列は、偽のポジティブを最小化するのに十分な長さで十分に明白であるべきである。オリゴヌクレオチドはスクリーニングされるライブラリー内のDNAとのハイブリダイズに対して検出できるように好ましくはラベルされている。ラベルの方法は本技術ではよく知られており、32P-ラベルATP,ビオチン化または酵素ラベルのようなラジオラベルの使用が含まれる。全てのタンパク質をコードする配列を持つ核酸は、cDNAに逆転写されていないであろうmRNAの前駆体またはプロセッシングされた中間体を検出するために、ここで開示されるアミノ酸配列を用いて、およびもし必要ならば、Sambrook等,上記参照、に記述されているような一般的なプライマー伸長法を用いて、選択されたcDNAまたはゲノムライブラリーのスクリーニングによって得られるだろう。
【0039】
Apo-2リガンドのアミノ酸配列変異体は、Apo-2リガンドDNAに適した核酸の変化を導入することによって、または望ましいApo-2リガンドポリペプチドの合成によって調製することができる。該変異体は、細胞内領域、膜貫通領域、または細胞外領域、あるいは図1AのフルレングスApo-2リガンドに示されるアミノ酸配列の片方または両末端において、残基の挿入、置換及び/または欠失しているものをを表す。挿入、置換及び/または欠失のいかなる組み合わせも、最終構成物に到達するためになされ、ここで定義されるような望ましいアポトーシス活性を持つ最終構成物を提供することができる。好ましい実施態様としては、変異体はApo-2リガンドの細胞内、膜貫通、または細胞外領域、あるいはApo-2リガンドに対するフルレングス配列についてここで同定された配列と、少なくとも約80%の配列同一性、より好ましくは約90%の配列同一性、そしてさらにより好ましくは約95%の配列同一性をもつ。アミノ酸変化はまたグリコシル化部位の数または位置を変化する、または膜埋め込み性質を改変するようなApo-2リガンドの翻訳後プロセッシングを改変するであろう。
【0040】
上述のようなApo-2リガンドの配列のバリエーションは、米国特許第5,364,934号に示されている保存的および非保存的ミュータントに対する技術そしてガイドラインのいかなるものかを用いてなされうる。これらはオリゴヌクレオチド介在性(サイトディレクト)ミュータジェネシス、アラニンスキャニングそしてPCRミュータジェネシスを含む。
【0041】
2. 複製ベクターへの核酸の挿入
天然のまたは変異体Apo-2リガンドをコードする核酸(例えばcDNAまたはゲノムDNA)は、さらなるクローニング(DNAの増幅)のため、または発現のための複製ベクターに挿入することができる。様々なベクターが公に入手可能である。ベクター構成要素は一般的に、しかし限定することなく以下の一つまたはそれ以上を含む:シグナル配列、複製オリジン、一つかそれ以上のマーカー遺伝子、エンハンサーエレメント、プロモーターそして転写終結配列があり、それぞれが以下に記述されている。
【0042】
(i) シグナル配列構成要素
Apo-2リガンドは直接だけでなく、ヘテロ構造のポリペプチドを用いた融合ポリペプチドとしても組換え的に生産でき、そのヘテロ構造のポリペプチドは天然のタンパク質またはポリペプチドのN末端に特異的な切断部位を持つシグナル配列または他のポリペプチドであり得る。一般的には、シグナル配列はベクターの構成要素であり得、またはそれはベクターに挿入されるApo-2リガンドDNAの一部であり得る。選択されたヘテロ構造のシグナル配列は好ましくはホスト細胞によって認識されそしてプロセッシングされるものである(すなわち、シグナルぺプチダーゼによって切断される。シグナル配列は例えばアルカリホスファターゼ、ペニシリナーゼ、lppそして熱安定性エントロキシンIIリーダーから選択された原核生物のシグナル配列であり得る。酵母の分泌のためのシグナル配列は、例えば酵母インベルターゼリーダー、アルファーファクターリーダー(サッカロミセスとクルイベロミセスαファクターリーダーを含み、後者は米国特許第5,010,182号に記述されている)、または酸性ホスファターゼリーダー、C.albicansグルコアミラーゼリーダー(1990年4月4日に印刷されたEP 362,179)、または1990年11月15日に印刷されたWO 90/13646に記述されているシグナルであり得る。哺乳動物細胞発現では、哺乳動物のシグナル配列がタンパク質の分泌に向けられて用いることができ、それは同じまたは関連した種の分泌されたポリペプチド由来のシグナル配列のようなものから、例えば単純ヘルペス糖タンパク質Dシグナルのようなウイルス分泌リーダーでもよいが、in vivoでヒト細胞の細胞膜へのApo-2リガンドの挿入に直接的に向けられる天然のApo-2リガンドプレ配列でも十分である。
【0043】
該前駆体領域のためのDNAは好ましくはApo-2リガンドをコードするDNAに対するリーディングフレーム内にライゲーションされる。
【0044】
(ii) 複製オリジン構成要素
発現ベクターとクローニングベクターの両方が、一つかそれ以上の選択されたホスト細胞内でベクターの複製を可能にする核酸配列を含む。一般的にクローニングベクターではこの配列は、ベクターがホスト染色体DNAと非依存的に複製できるようにするものであり、複製のオリジンまたは自律複製配列を含む。該配列は様々な細菌、酵母そしてウイルスに対してよく知られている。プラスミドpBR322由来の複製オリジンは、ほとんどのグラムネガティブ細菌に対して適しており、2μプラスミドオリジンは酵母に適しており、そして様々なウイルスのオリジン(SV40,ポリオーマ,アデノウイルス,VSVまたはBPV)は哺乳動物細胞内でのクローニングベクターに対して有用である。一般的に複製構成物のオリジンは、哺乳動物発現ベクターに対しては必要でない(SV40オリジンは典型的にはそれが速いプロモーターを含むため用いられる)。
【0045】
ほとんどの発現ベクターは「シャトル」ベクターであり、すなわちそれらは少なくとも一つのクラスの生物内で複製できるが、発現のためもう一つの生物内にトランスフェクトすることができる。例えばベクターは大腸菌内でクローニングされ、そしてたとえそれがホスト細胞染色体と非依存的に複製できないとしても、それから同じベクターが酵母または哺乳動物細胞内にトランスフェクトされる。
【0046】
DNAはまたホストゲノム内に挿入されることによって増幅され得る。これはホストとしてバチルス種を用いれば容易に成し遂げられ、例えば、バチルスゲノムDNA内に見出される配列と相補的なDNA配列をベクター内に含ませることによって成し遂げられる。このベクターを用いたバチルスのトランスフェクションは、ゲノムとの相同的組換えおよびApo-2リガンドDNAの挿入の結果である。しかしながら、Apo-2リガンドをコードするゲノムDNAの回収は、Apo-2リガンドDNAを摘出するために制限酵素切断が必要とされるため、外来的に複製されるベクターの回収よりも複雑である。
【0047】
(iii) 選択遺伝子構成物
発現ベクターおよびクローニングベクターは典型的には、選択遺伝子、選択マーカーとも呼ばれるものを含む。この遺伝子は選択培地内で成長するトランスフォームされたホスト細胞の生存または成長のため必要なタンパク質をコードする。選択遺伝子を含まないベクターでトランスフォームされたホスト細胞は、培地内で生き残れないであろう。典型的な選択遺伝子は、(a)例えばアンピシリン、ネオマイシン、メトトレキセートまたはテトラサイクリンといった抗生物質または他の毒素に対して耐性を示すタンパク質、(b)栄養要求性欠乏を補足するタンパク質、または(c)例えばバチルスにとってのD-アラニンラセマーゼをコードする遺伝子といった複雑な培地からは入手できない重大な栄養素を供給するタンパク質をコードする。
【0048】
選択スキームの一つの例として、ホスト細胞の成長を抑える薬剤の使用がある。ヘテロ構造遺伝子を用いてうまくトランスフォームされた細胞は、薬剤耐性を示すタンパク質を生産し、それゆえ選択摂生を生き残れる。該ドミナント選択の例として、薬剤としてネオマイシン[Southern等,J.Molec.Appl.Genet.,1:327(1982)]、ミコフェノール酸[Mulligan等,Science,209:1422(1980)]またはハイグロマイシン[Sugden等,Mol.Cell.Biol.,5:410-413(1985)]を用いる。上述した該3例はそれぞれ、適切な薬剤G418またはネオマイシン(ジェネティシン)、xgpt(ミコフェノール酸)、またはハイグロマイシンに対する耐性を導くために、真核生物のコントロールの下で細菌の遺伝子を用いるものである。
【0049】
哺乳動物細胞に対して適した選択マーカーのもう一つの例は、DHFRまたはチミジンキナーゼのような、Apo-2リガンド核酸を取り込むのに適した細胞を同定できるものである。哺乳動物細胞トランスフォーマントは、トランスフォーマントだけがマーカーを取り込んだことによって唯一に生き残ることに適応する選択圧のもとに置かれる。選択圧は、培地中の選択試薬の濃度が継続的に変化するコンディションの下でトランスフォーマントを培養することによって押し付けられ、それによって選択遺伝子とApo-2リガンドをコードするDNAの両者の増幅が導かれる。増幅とは成長にとって重大なタンパク質の生産のため非常に大きな需要のある遺伝子が、組換え細胞の継続的な世代において染色体内でタンデムに繰り返されるプロセスである。Apo-2リガンドの増大した量は、増幅DNAから合成される。増幅可能な遺伝子の他の例として、メタロチオネイン-Iそして-II、アデノシンデアミナーゼそしてオルニチンデカルボキシラーゼが含まれる。
【0050】
DHFR選択遺伝子を用いてトランスフォームされた細胞は、DHFRの競合的な拮抗剤であるメトトレキセート(Mtx)を含む培地ですべてのトランスフォーマントを培養することによって最初に同定されるであろう。野生型DHFRを用いる場合に適したホスト細胞は、Urlaub等,Proc.Natl.Acad.Sci.USA,77:4216(1980)に記述されているように調製され普及している、DHFR活性に欠陥のあるチャイニーズハムスター卵巣(CHO)細胞系である。それからトランスフォームされた細胞を上昇した濃度のメトトレキセートにさらす。これによりDHFR遺伝子のマルチコピー、そしてそれに付随してApo-2リガンドをコードするDNAのような発現ベクターを含む他のDNAのマルチコピーの合成が導かる。もし例えばMtxに高い耐性であるミュータントDHFR遺伝子を用いるならば、外来性DHFRの存在にもかかわらず、この増幅法は例えばATCC番号CCL61CHO-K1などのいかなる他の適したホストを用いても使用できる(EP117,060)。
【0051】
代わりに、Apo-2リガンド、野生型DHFRタンパク質そしてアミノグリコシド3'-ホスホトランスフェラーゼ(APH)のようなもう一つの選択マーカーをコードするDNA配列を用いてトランスフォームまたはコトランスフォームされたホスト細胞(特に外来性DHFRを含む野生型ホスト)は、例えばカナマイシン、ネオマイシンまたはG418などのアミノグリコシド系抗生物質のような選択マーカーに対する選択試薬を含む培地中での細胞成長によって選択できる。米国特許第4,965,199号を参照。
【0052】
酵母を用いる場合に適した選択遺伝子は、酵母プラスミドYRp7内に存在するtrp1遺伝子である[Stinchcomb等,Nature,282:39(1979);Kingsman等,Gene,7:141(1979);Tschemper等,Gene,10:157(1980)]。trp1遺伝子は、例えばATCC番号44076またはPEP4-1のようなトリプトファン内で成長する能力を欠如した酵母のミュータント株のための選択マーカーを提供する[Jones,Genetics,85:12(1977)]。そこで酵母ホスト細胞ゲノム内のtrp1領域の存在は、トリプトファンの非存在下での成長によってトランスフォーメーションを検出するための効果的な環境を提供する。同様に、Leu2欠失酵母株(ATCC20,622または38,626)は、Leu2遺伝子をもつ既知のプラスミドによって相補される。
【0053】
加えて、1.6μm環状プラスミドpKD1由来のベクターは、クルイベロミセス酵母のトランスフォーメーションに対して用いられる[Bianchi等,Curr.Genet.,12:185(1987)]。さらに最近では、組換え仔ウシキモシンの大規模生産のための発現システムがK・ラクティスに対して報告されている[Van den Berg,Bio/Technology,8:135(1990)]。クルイベロミセスの工業的な株による成熟した組換えヒト血清アルブミンの分泌のための安定なマルチコピー発現ベクターもまた開示されている[Fleer等,Bio/Technology,9:968-975(1991)]。
【0054】
(iv) プロモーター構成要素
発現ベクターおよびクローニングベクターは、通常ホスト生物によって認識され、Apo-2リガンド核酸配列に実施可能に結合されたプロモーターを含む。プロモーターは構造遺伝子のスタートコドンの上流(5')に位置する(一般的には約100から1000bp内)非翻訳配列であり、それらが実施可能にリンクしているApo-2リガンド核酸配列のような特定の核酸配列の転写と翻訳をコントロールする。該プロモーターは典型的には、誘発性と構造性の2つに分類される。誘発性プロモーターは例えば栄養素の存在または非存在、あるいは温度変化といったいくつかの培地コンディションの変化に応じて、それのコントロールの下で増大したレベルのDNAからの転写を始めるプロモーターである。現在、様々な可能なホスト細胞によって認識される多くの数のプロモーターがよく知られている。これらのプロモーターは、制限酵素切断によって元となるDNAからプロモータを切り出し、ベクター内に単離したプロモーター配列を挿入することにより、Apo-2リガンドをコードするDNAに実施可能に結合される。天然のApo-2リガンドプロモーター配列と様々なヘテロ構造受容体プロモーターの両方が、Apo-2リガンドDNAの増幅及び/または発現に向けられて使用できるであろう。
【0055】
原核生物ホストを用いた使用にとって適したプロモーターは、β-ラクタマーゼとラクトースプロモーターシステム[Chang等,Nature,275:615(1978);Goeddel等,Nature,281:544(1979)]、アルカリホスファターゼ、トリプトファン(trp)プロモーターシステム[Goeddel,Nucleic Acids Res.,8:4057(1980);EP 36,776]そしてtacプロモーターのようなハイブリッドプロモーター[deBoer等,Proc.Natl.Acad.Sci.USA,80:21-25(1983)]を含む。しかしながら、他の既知の細菌プロモーターも適している。それらの核酸配列は印刷されており、それによって必要とされる制限部位を提供するためリンカーまたはアダプターを用いて、当業者はApo-2リガンドをコードするDNAにそれらを実施可能に結合できる[Siebenlist等,Cell,20:269(1980)]。細菌システムで用いられるプロモーターもまた、Apo-2リガンドをコードするDNAに実施可能に結合するシャインダルガノ(S.D.)配列を含むであろう。
【0056】
プロモーター配列は真核生物のためのものも知られている。事実上すべての真核生物の遺伝子が、転写開始部位からおよそ25から30塩基上流に位置するATリッチ領域を持っている。多くの遺伝子の転写の開始から70から80ベース上流に見出されるもう一つの配列は、CXCAAT領域であり、ここでXはいかなるヌクレオチドをもさす。ほとんどの真核生物遺伝子の3'末端には、AATAAA配列があり、それはコード配列の3'末端にポリAテールを付加するためのシグナルであり得る。これらの配列の全てが、真核生物発現ベクターに適するように挿入される。
【0057】
酵母ホストを用いる場合の適したプロモーター配列の例として、3-ホスホグリセリン酸キナーゼ[Hitzeman等,J.Biol.Chem.,255:2073(1980)]またはエノラーゼ、グリセルアルデヒド-3-リン酸デヒドロゲナーゼ、ヘクソキナーゼ、ピルビン酸デカルボキシラーゼ、ホスホフルクトキナーゼ、グルコース-6-リン酸イソメラーゼ、3-ホスホグリセリン酸ムターゼ、ピルビン酸キナーゼ、トリオースリン酸イソメラーゼ、ホスホグルコースイソメラーゼそしてグルコキナーゼのような他の解糖系の酵素[Hess等,J.Adv.Enzyme Reg.,7:149(1968);Holland,Biochemistry,17:4900(1978)]の各プロモーターが含まれる。
【0058】
成長コンディションによってコントロールされる転写という付加的な利点を有する誘発性プロモーターである他の酵母プロモーターは、アルコールデヒドロゲナーゼ2、イソシトクロムC、酸性ホスファターゼ、窒素代謝と関連する分解酵素、メタロチオネイン、グリセルアルデヒド-3-リン酸デヒドロゲナーゼそしてマルトースおよびガラクトース利用に関わる酵素に対する各プロモーター領域がある。酵母発現に用いる適したベクターおよびプロモーターは、さらにEP 73,657に記述されている。酵母エンハンサーもまた酵母プロモーターを用いて便利に使用される。
【0059】
哺乳動物ホスト細胞内のベクターからのApo-2リガンド転写は、例えばポリオーマイルス、鶏頭ウイルス(1989年7月5日に印刷されたUK 2,211,504)、アデノウイルス(アデノウイルス2のような)、ウシパピローマウイルス、トリ肉腫ウイルス、サイトメガロウイルス、B型肝炎ウイルスそして好ましくはシミアンウイルス40(SV40)のようなウイルスのゲノムから得られるプロモーター、以下のプロモーターがホスト細胞システムと両立できれば、例えばアクチンプロモーターまたは免疫グロブリンプロモーターなどのヘテロ構造哺乳動物プロモーター、またはヒートショックプロモーターから得られるプロモーター、あるいはApo-2リガンド配列と通常関連しているプロモーターから得られるプロモーターによってコントロールされる。
【0060】
SV40の速いおよび遅いプロモーターはSV40ウイルス複製オリジンも含むSV40制限断片として便利に得られる[Fiers等,Nature,273:113(1978);MulliganとBerg,Science,209:1422-1427(1980);Pavlakis等,Proc.Natl.Acad.Sci.USA,78:7398-7402(1981)]。ヒトサイトメガロウイルスの極端に速いプロモーターはHindIIIE制限断片として便利に得られる[Greenaway等,Gene,18:355-360(1982)]。ベクターとしてウシパピローマウイルスを用いた哺乳動物ホスト内のDNA発現システムは、米国特許第4,419,446号に開示されている。このシステムの変形は米国特許第4,601,978号に記述されている[Gray等,Nature,295:503-508(1982)のサル細胞での免疫インターフェロンをコードするcDNAの発現;Reyes等,Nature,297:598-601(1982)の単純ヘルペス由来のチミジンキナーゼプロモーターのコントロールの下でのマウス細胞でのヒトβ-インターフェロンcDNAの発現;CanaaniとBerg,Proc.Natl.Acad.Sci.USA.79:5166-5170(1982)の培養マウスおよびウサギ細胞でのヒトインターフェロンβ1遺伝子の発現;そしてGorman等,Proc.Natl.Acad.Sci.USA,79:6777-6781(1982)のラウス肉腫ウイルスの長い末端反復配列をプロモーターとして用いた、CV-1サル腎細胞、ニワトリ胚繊維芽細胞、チャイニーズハムスター卵巣細胞、ヒーラ細胞そしてNIH-3T3細胞での細菌CAT配列の発現も参照]。
【0061】
(v) エンハンサーエレメント構成要素
高等哺乳動物による本発明のApo-2リガンドをコードするDNAの転写は、ベクターにエンハンサー配列を挿入することにより増大するであろう。エンハンサーはDNAのシス作用エレメントであり、通常約10から300bpであり、その転写を増大するためにプロモーターに基づいて作用する。エンハンサーは向きと位置に比較的非依存的であり、転写ユニットの5'[Laimins等,Proc.Natl.Acad.Sci.USA,78:993(1981)]そして3'[Lusky等,Mol.Cell Bio.,3:1108(1983)]、イントロン内[Banerji等,Cell,33:729(1983)]、同様にコード配列それ自体内[Osborne等,Mol.Cell Bio.,4:1293(1984)]に見出されている。多くのエンハンサー配列が哺乳動物遺伝子(グロビン、エラスターゼ、アルブミン、α-フェトプロテインそしてインスリン)から現在では知られている。しかしながら典型的には、当業者は真核生物細胞ウイルス由来のエンハンサーを用いる。例としては、複製オリジンの後ろ側(100-270bp)にSV40エンハンサー、サイトメガロウイルスの速いプロモーターエンハンサー、複製オリジンの後ろ側にポリオーマエンハンサーそしてアデノウイルスエンハンサーが含まれる。真核生物プロモーターの活性化のためのエンハンサーエレメントについてはYaniv,Nature,297:17-18(1982)も参照。エンハンサーはApo-2リガンド-コード配列に対して5'または3'の位置でベクターに挿入されるが、好ましくはプロモーターから5'部位に位置される。
【0062】
(vi) 転写終結構成要素
真核生物ホスト細胞(酵母、菌類、昆虫、植物、動物、ヒトまたは他の多核細胞生物由来の核のある細胞)内で用いられる発現ベクターは、また転写の終結とmRNAの安定化のために必要な配列を含むであろう。該配列は、真核生物またはウイルスのDNAまたはcDNAの非翻訳領域の5'そして時には3'から共通して入手可能である。これらの領域はApo-2リガンドをコードするmRNAの非翻訳領域内のポリアデニル化断片として転写される核酸部分を含む。
【0063】
(vii) ベクターの構築と解析
一つかそれ以上の上記リストの構成要素を含む適したベクターの構築は、標準的なライゲーション法を用いる。単離したプラスミドまたはDNA断片を切断し、調製し、そして必要とされるプラスミドを作製するために望ましい形に再ライゲーションする。
【0064】
構築したプラスミドが正しい配列か確かめるための解析として、ライゲーション混合物を大腸菌K12株294(ATCC31,446)をトランスフォームすることが用いられ、そして適当かどうかアンピシリンまたはテトラサイクリン耐性によって成功したトランスフォーマントを選択できる。トランスフォーマントからプラスミドを調製し、制限エンドヌクレアーゼ切断によって解析し、、および/またはMessing等,Nucleic Acids Res.,9:309(1981)の方法またはMaxam等,Methods in Enzymology,65:499(1980)の方法によってシークエンスする。
【0065】
(viii) 一過性発現ベクター
Apo-2リガンドをコードするDNAの哺乳動物細胞内での一過性発現のために提供される発現ベクターが用いられてもよい。一般的には一過性発現は、ホスト細胞が発現ベクターの多数のコピーを蓄積し、次には発現ベクターによってコードされている望ましいポリペプチドを高レベルで合成するように、ホスト細胞内で効率よく複製可能な発現ベクターの使用を含む[Sambrook等,上記参照]。適した発現ベクターとホスト細胞を含む一過性発現システムは、クローン化したDNAによってコードされるポリぺプチドの簡便な積極的な同定を可能にし、同様に望ましい生物学的または生理学的な性質のための該ポリペプチドの速いスクリーニングを可能にする。それゆえ一過性発現システムは生物学的に活性なApo-2リガンドであるApo-2リガンドの類似体及び変異体の同定の目的に対して、本発明では特に役に立つ。
【0066】
(ix) 適した具体例としての脊椎動物細胞ベクター
組換え脊椎動物細胞カルチャー内でのApo-2リガンドの合成に対する翻案のために適した他の方法、ベクターそしてホスト細胞は、Gething等,Nature,293:620-625(1981);Mantei等,Nature,281:40-46(1979);EP 117,060;そしてEP 117,058に記述されている。Apo-2リガンドの哺乳動物細胞カルチャー発現のための特に有用なプラスミドはpRK5[EP 307,247;実施例1にも記述されている]またはpSV16B[1991年6月13日印刷されたWO 91/08291]である。
【0067】
3. ホスト細胞の選択とトランスフォーメーション
ここでベクター内のDNAをクローン化または発現するための適したホスト細胞は、上述した原核生物、酵母、高等真核生物である。この目的のため適した原核生物は、グラムネガティブまたはグラムポジティブ生物のような真正細菌を限定なく含み、例えば大腸菌のようなエシェリキア、エンテロバクター、エルヴィニア、クレブシエラ、プロテウス、ネズミチフス菌のようなサルモネラ、霊菌のようなセラチアそしてシゲラといった腸内細菌、および枯草菌とバチルスリケニフォルミス(例えば1989年4月12日に印刷されたDD266,710に開示されているバチルスリケニフォルミス41P)のようなバチルス、緑膿菌のようなシュードモナスそしてストレプトミセスも同様に含む。好ましくはホスト細胞は、タンパク質分解酵素の最小限の量を分泌すべきである。
【0068】
原核生物に加えて、糸状菌または酵母のような真核生物細菌がApo-2リガンド-コードベクターのためのクローニングホストまたは発現ホストとして適している。サッカロミセスセレビシエまたは一般的なパン酵母は、低級真核生物ホスト微生物の間で最も広く用いられている。しかしながら、多くの他の属、種そして株が、広く入手でき、ここで有用である。
【0069】
グリコシル化Apo-2リガンドの発現にとって適したホスト細胞は、多細胞生物由来のものである。該ホスト細胞は複雑なプロセッシング及びグリコシル化活性が可能である。原則として、いかなる高等細胞カルチャーも、脊椎動物カルチャー由来から非脊椎動物カルチャー由来まで実施できる。非脊椎動物細胞の例として、植物細胞及び昆虫細胞が含まれる。非常に多くのバキュロウイルス株及び変異体そしてスポドプテラフルギペルダ(芋虫)、エデスエギプティ(カ)、エデスアルボピクタス(カ)、ドロソフィラメラノガスター(ショウジョウバエ)そしてカイコガのようなホスト由来の相当する複製を許容する昆虫ホスト細胞が、同定されている[Luckow等,Bio/Technology,6:47-55(1988);Setlow等編 Genetic Engineering、第8版(Plenum Pubulishing,1986),pp.277-279内のMiller等;そしてMaeda等,Nature,315:592-594(1985)]。トランスフェクションのための様々なウイルス株が広く入手可能であり、例えばオートグラファカリフォルニカNPVのL-1変異体およびカイコガNPVのBm-5株があり、特に実施例2に記述されているSpodoptera frugiperda(「Sf9」)細胞のトランスフェクションに対して、本発明にしたがったここでのウイルスのようなウイルスが用いられ得る。
【0070】
ワタ、トウモロコシ、ポテト、ソラマメ、ペチュニア、トマトそしてタバコの植物細胞が、ホストとして利用できる。典型的には植物細胞は、細菌アグロバクテリウムツメファシエンスの特定の株と共にインキュベーションによってトランスフェクトし、その細菌はApo-2リガンド-コードDNAを含むように前もって処理されている。A.ツメファシエンスと共に植物細胞をインキュベーションする間、Apo-2リガンドをコードするDNAは、トランスフェクトされるような植物細胞ホストにトランスファーされ、そして適したコンディションの下でApo-2リガンド-コードDNAを発現するであろう。加えて植物細胞で両立できる調節配列およびシグナル配列は、ノパリンシンテースプロモーターおよびポリアデニル化シグナル配列のように入手可能である[Depicker等,J.Mol.Appl.Gen.,1:561(1982)]。加えてT-DNA780遺伝子の上流領域から単離されたDNA領域により、組換えDNAを含む植物組織内で植物で発現できる遺伝子の活性化したまたは増大した転写レベルが可能である[1989年6月21日に印刷されたEP 321,196]。
【0071】
カルチャー内の脊椎動物細胞(組織細胞)の増殖もまた、本分野でよく知られている[例えばTissue Culture,Academic Press,KruseとPatterson編(1973)参照]。有用な哺乳動物ホスト細胞系の例として、SV40(COS-7,ATCC CRL 1651)によってトランスフォームされたサル腎CV1系;ヒト胚腎系(293細胞または懸濁液カルチャー内で成長のためサブクローン化された293細胞、Graham等,J.Gen Virol.,36:59(1977));仔ハムスター腎細胞(BHK,ATCC CCL 10);チャイニーズハムスター卵巣細胞/-DHFR(CHO,UrlaubとChasin,Proc.Natl.Acad.Sci.USA,77:4216(1980));マウスセルトリ細胞(TM4,Mather,Biol.Reprod.,23:243-251(1980));サル腎細胞(CV1 ATCC CCL 70);アフリカグリーンサル腎細胞(VERO-76,ATCCCRL-1587);ヒト子宮頚ガン細胞(HELA,ATCC CCL 2);イヌ腎細胞(MDCK,ATCC CCL 34);バッファローラット肝細胞(BRL 3A,ATCC CRL 1442);ヒト肺細胞(W138,ATCC CCL 75);ヒト肝細胞(Hep G2,HB 8065);マウス乳房腫瘍(MMT 060562,ATCC CCL 51);TRI細胞(Mather等,Annals N.Y.Acad.Sci.,383:44-68(1982));MRC5細胞;そしてFS4細胞がある。
【0072】
ホスト細胞はトランスフェクトされ、そして好ましくはApo-2リガンド生産のための上述の発現ベクターまたはクローニングベクターを用いてトランスフォームされ、そしてプロモーターの誘導、トランスフォーマントの選択または望ましい配列をコードする遺伝子の増幅のため適したように修飾されたありきたりの栄養素培地で培養される。
【0073】
トランスフェクションとは、いかなるコード配列が実際に発現されようとされまいと、ホスト細胞によって発現ベクターが取り込まれることを示す。非常に多くのトランスフェクションの方法が当業者に知られており、例えばCaPO4およびエレクトロポレーションがある。成功したトランスフェクションは一般的に、該ベクターの作用のいかなる徴候でもホスト細胞内で生じれば、認識される。
【0074】
トランスフォーメーションとは、染色体外エレメントとしてまたは染色体への組み込みによるかいずれでも、DNAを複製するために生物にDNAを導入することを示す。用いられるホスト細胞に依存して、トランスフォーメーションは該細胞に適した標準的な方法を用いてなされる。Sambrook等,上記参照内で記述されているような塩化カルシウムを用いたカルシウム処理またはエレクトロポレーションが、原核生物または実質的な細胞壁障害を持つ他の細胞のために使用される。アグロバクテリウムツメファシエンスを用いた感染は、Shaw等,Gene,23:315(1983)と1989年6月29日に印刷されたWO 89/05859に記述されているように、特定の植物細胞のトランスフォーメーションのために使用される。加えて植物は、1991年1月10日に印刷されたWO 91/00358に記述されているような超音波処理を用いてトランスフェクトしてもよい。
【0075】
該細胞壁を持たない哺乳動物細胞に対しては、Grahamとvan der Eb,Virology,52:456-457(1978)のリン酸カルシウム沈殿法が好ましい。哺乳動物細胞ホストシステムトランスフォーメーションの一般的な面としては、米国特許第4,399,216号に記述がある。酵母へのトランスフォーメーションは典型的には、Van Solingen等,J.Bact.,130:946(1977)とHsiao等,Proc.Natl.Acad.Sci.(USA),76:3829(1979)の方法にしたがって実施される。しかしながら、核マイクロインジェクション、エレクトロポレーション、完全な細胞への細菌プロトプラスト融合または例えばポリプレン、ポリオルニチンというポリカチオンのような、他の細胞へのDNA導入法もまた用いることができる。哺乳動物細胞をトランスフォームする様々な方法に対しては、Keown等,Methods in Enzymology,185:527-537(1990)とMansour等,Nature,336:348-352(1988)を参照。
【0076】
4. ホスト細胞の培養
Apo-2リガンドを生産するために用いられる原核生物細胞は、Sambrook等,上記参照で一般的に記述されている適した培地内で培養できる。
【0077】
Apo-2リガンドを生産するため用いられる哺乳動物ホスト細胞は、様々な培地で培養できる。商業的に入手可能な培地の例としては、Ham's F10(Sigma)、Minimal Essential Medium("MEM",Sigma)、PRMI-1640(Sigma)そしてダルベッコ変法イーグル培地("DMEM",Sigma)が含まれる。いかなる該培地も、ホルモンおよび/または増殖因子(インスリン、トランスフェリンまたは上皮増殖因子のような)、塩類(塩化ナトリウム、カルシウム、マグネシウムそしてリン酸塩のような)、バッファー(HEPESのような)、ヌクレオシド(アデノシン及びチミジンのような)、抗生物質(ゲンタマイシンTM薬剤のような)、微量元素(マイクロモーラー範囲で最終濃度に通常存在する無機化合物として定義される)及びグルコースまたは同等のエネルギー源が必要であるとして補われる。いかなる他のサプリメントであっても、当業者に既知であろう適した最終濃度で含んでよい。温度、pH、それに類するもののような培養条件は、発現のため選択されたホスト細胞に事前に用いたものであり、当業者には明白であろう。
【0078】
一般的に、哺乳動物細胞カルチャーの生産力を最大化するための原理、プロトコールそして実施上の技術は、Mammalian Cell Biotechnology:a Practical Approach,M.Butler編,(IRL Press,1991)に見出すことができる。
【0079】
この開示で示されるホスト細胞は、カルチャー内の細胞だけでなく、ホスト動物内にある細胞をも包含する。
【0080】
5. 遺伝子増幅/発現の検出
遺伝子増幅および/または発現は、ここで提供される配列に基づく適切なラベルプローブを用いて、例えばありきたりのサザンブロッティング、mRNAの転写量を測定するにはノーザンブロッティング[Thomas,Proc.Natl.Acad.Sci.USA,77:5201-5205(1980)]、ドットブロッティング(DNA解析)またはin situハイブリダイゼーションによって直接にサンプル内で測定することができる。様々なラベルが用いられうるが、最も一般的にはラジオアイソトープ、特に32Pがある。しかしながら、ポリヌクレオチドへの導入のためビオチン修飾ヌクレオチドを用いるように、他の方法もまた用いられる。すなわちビオチンは、アビジンまたは抗体に対する結合のための部位として用いられ、ラジオヌクレオチド、蛍光剤または酵素のような広く様々なラベルを用いてラベルすることが可能である。代わりにDNA二重鎖、RNA二重鎖そしてDNA-RNAハイブリッド二重鎖またはDNA-タンパク質二重鎖を含む、特異的な二重鎖を認識することができる抗体を用いてもよい。次に抗体はラベルされ、表面での二重鎖の形態により、二重鎖に結合した抗体の存在が検出できるために、二重鎖が表面のどこに結合しているかのアッセイを実施できる。
【0081】
代わりに遺伝子発現は、遺伝子産物の発現を直接測定するために、細胞または組織切片の免疫組織化学的染色および細胞カルチャーまたは体液のアッセイのような、免疫学的方法で測定することができる。免疫組織化学的染色法を用いると、細胞サンプルは典型的には、脱水および固定によって調製され、遺伝子産物と特異的に結合するラベル抗体を用いた反応が続くが、そのラベルは酵素的ラベル、蛍光ラベルまたは発光ラベル等のような通常視覚的に検出されるものである。
【0082】
免疫組織化学的染色および/またはサンプル体液のアッセイのために有効な抗体は、モノクローナルでもポリクローナルでもよく、いかなる動物内でも調製できる。簡便には該抗体は天然のApo-2リガンドポリペプチドに対して、またはここで提供されるDNA配列に基づく合成ペプチドに対して、あるいはApo-2リガンドDNAと融合され特異的抗体エピトープをコードする外因性配列に対して調製され得る。
【0083】
6. Apo-2リガンドポリペプチドの精製
Apo-2リガンドは分泌シグナル無しで直接生産されるとホスト細胞溶解産物から回収されるが、好ましくはHGF受容体拮抗剤は分泌ポリペプチドとして培地から回収される。もしApo-2リガンドが膜結合型であれば、適した洗浄剤溶液(例えばTriton-X100)を用いて膜からそれを引き離すことができ、あるいはその細胞外領域を酵素的な切断によって引き離すことができる。
【0084】
Apo-2リガンドがヒト由来の細胞以外の組換え細胞内で生産された場合には、Apo-2リガンドはヒト由来のタンパク質またはポリペプチドと関連がない。しかしながら、Apo-2リガンドとして実質的にホモ構造である調製を得るために、組換え細胞タンパク質またはポリペプチドからApo-2リガンドを精製することが望ましい。第一段階として、粒子状の細胞破片を除去するために、培地または溶解産物を遠心分離するとよい。その後Apo-2リガンドを水溶性タンパク質およびポリペプチドの混合物から精製するが、以下の方法は適した精製法の典型例である:イオン交換カラムを用いての分別;エタノール沈殿;逆相HPLC;シリカまたはDEAEのようなカチオン交換樹脂を用いてのクロマトグラフィー;等電点電気泳動;SDS-PAGE;硫安沈降;例えばSephadexG-75を用いたゲル濾過;そしてIgGのような不純物を除去するためのプロテインA Sepharoseカラム。
【0085】
好ましい実施態様として、Apo-2リガンドは実施例3に記述されるようなアフィニティークロマトグラフィーによって単離され得る。
【0086】
その残基が欠失、挿入、または置換されているApo-2リガンド変異体は、該変異値によって引き起こされる性質におけるいかなる実質的な変化をも考慮に入れて、天然のApo-2リガンドと同様な方式で回収される。
【0087】
フェニルメチルスルホニルフルオリド(PMSF)のようなプロテアーゼ阻害剤もまた、精製の間のタンパク質分解を阻害するのに有用であり、そして抗生物質も外来的な不純物の成長を妨げるために含まれよう。組換え細胞カルチャーでの発現に対してApo-2リガンドまたはその変異体の性質の変化を考慮して、天然のApo-2リガンドに適した精製方法が必要とされるであろうことは当業者に予測できよう。
【0088】
7. Apo-2リガンドポリペプチドの共有結合修飾
Apo-2リガンドの共有結合修飾は、本発明の範囲に含まれる。天然のApo-2リガンド及びApo-2リガンドのアミノ酸配列変異体の両者が共有結合で修飾され得る。Apo-2リガンドの共有結合の一つのタイプとして、Apo-2リガンドの選択された側鎖またはNあるいはC末端残基に反応可能な有機誘導体化試薬を用いて、Apo-2リガンドの目標とするアミノ酸に反応させて分子を導入することがある。
【0089】
二官能基を持つ誘導体化は、抗Apo-2リガンド抗体の精製の方法に用いる非水溶性支持体マトリックスまたは表面に対してApo-2リガンドを架橋するのに有用であり、逆もまた然りである。広く用いられている架橋試薬は、例えば1,1-ビス(ジアゾアセチル)-2-フェニルエタン、グルタルアルデヒド、例えば4-アジドサリチル酸のエステルといったN-ヒドロキシスクシンイミドエステル、3,3'-ジチオビス(スクシンイミジルプロピオネート)のようなジスクシンイミジルエステルを含む、ホモ二官能イミドエステル、そしてビス-N-マレイミド-1,8-オクタンのような二官能マレイミドを含む。メチル-3-[(p-アジドフェニル)ジチオ]プロピオイミデートのような誘導体化試薬は、光の存在の下で架橋を形成することが可能な光活性化中間体を生じる。代わりに、米国特許第3,969,287号;第3,691,016号;第4,195,128号;第4,247,642号;第4,229,537号そして第4,330,440号に記述されている臭化シアン-活性化炭化水素のような反応性非水溶性マトリックス及び反応基質が、タンパク質固定のために用いられる。
【0090】
他の修飾は、それぞれグルタミル残基およびアスパルチル残基に相当するグルタミニル残基およびアスパラギニル残基の脱アミド化、プロリンおよびリシンのヒドロキシル化、セリル残基またはトレオニル残基のヒドロキシル基のリン酸化、リシン、アルギニン及びヒスチジン側鎖のα-アミノ基のメチル化[T.E.Creighton,Proteins:Structure and Molecular Properties,W.H.Freeman & Co.,サンフランシスコ、pp.79-86(1983)]、N末端アミンのアセチル化およびC末端カルボキシル基の脱アミド化を含む。残基の修飾された形態は本発明の範囲に含まれる。
【0091】
本発明の範囲内に含まれるApo-2リガンドポリペプチドの共有結合の修飾のもう一つのタイプは、ポリペプチドの天然のグリコシル化パターンを変更することを含む。「天然のグリコシル化パターンを変更する」ということは、天然のApo-2リガンドに見出される一つかそれ以上の炭化水素部分を欠失すること、および/または天然のApo-2リガンドには存在しない一つかそれ以上のグリコシル化部位を加えることを意味するようここでの目的として向けられる。
【0092】
ポリペプチドのグリコシル化は、典型的にはN結合型とO結合型のいずれかである。N結合型とは、アスパラギン残基の側鎖に炭化水素部分が付着することを示す。アスパラギン-X-セリン及びアスパラギン-X-トレオニンというトリペプチドで、Xはプロリン以外のいかなるアミノ酸でもよいものが、アスパラギン側鎖に炭化水素部分を酵素的に付着するための認識配列である。それゆえポリペプチド内にこれらのトリペプチド配列のどちらかが存在すると、潜在的なグリコシル化部位を作り出し得る。O結合型グリコシル化は、ヒドロキシルアミノ酸、最も共通にはセリンまたはトレオニンに対して、N-アセチルガラクトサミン、ガラクトースまたはキシロースの糖類の一つが付着することを示すが、5-ヒドロキシプロリンまたはヒドロキシリシンもまた付着部位に使われ得る。
【0093】
Apo-2リガンドポリペプチドへのグリコシル化部位の付加は、(N結合型グリコシル化部位の場合)上述のトリペプチド配列の一つかそれ以上を含むようにアミノ酸配列を変更することによって成し遂げられる。変更は(O結合型グリコシル化部位の場合)、天然のApo-2リガンド配列に一つかそれ以上のセリンまたはトレオニン残基を付加または置換することによってもなし得る。Apo-2リガンドアミノ酸配列は、DNAレベルでの変化、特にコドンを望ましいアミノ酸に翻訳されるように選択するように、前もって選ばれた塩基にApo-2リガンドポリペプチドをコードするDNAを突然変異させることによって、任意に変化し得る。DNAミューテーション(類)は、上述した方法または米国特許第5,364,934号、上記参照内の方法を用いてなし得る。
【0094】
Apo-2リガンドポリペプチド内の炭化水素部分の数を増大するもう一つの方法は、ポリペプチドへのグリコシドの化学的または酵素的結合によるものである。用いられる結合方法に依存して、糖(類)は(a)アルギニンおよびヒスチジン、(b)フリーカルボキシル基、(c)システインのもののようなフリースルフヒドリル基、(d)セリン、トレオニンまたはヒドロキシプロリンのもののようなフリーヒドロキシル基、(e)フェニルアラニン、チロシンまたはトリプトファンのもののような芳香族残基、または(f)グルタミンのアミド基に付着し得る。これらの方法は1987年9月11日に印刷されたWO 87/05330およびAplinとWriston,CRC Crit.Rev.Biochem.,pp.259-306(1981)に記述されている。
【0095】
Apo-2リガンドポリペプチド内に存在する炭化水素部分の除去は、化学的にまたは酵素学的に、あるいはグリコシル化のターゲットとして機能するアミノ酸残基をコードするコドンの突然変異的置換によって成し遂げられうる。例えば化合物トリフルオロメタンスルホン酸または同等の化合物にポリペプチドをさらすことによる化学的な脱グリコシル化は、ポリペプチドは完全なままである一方、結合糖(N-アセチルグルコサミドまたはN-アセチルガラクトサミド)を除いて、ほとんどまたは全ての糖を切断することを引き起こす。化学的な脱グリコシル化については、Hakimuddin等,Arch.Biochem.Biophys.,259:52(1987)およびEdge等,Anal.Biochem.,118:131(1981)に記述がある。ポリペプチドの炭化水素部分の酵素的な切断は、Thotakura等,Meth.Enzymol.,138:350(1987)に記述されているように様々なエンドグリコシダーゼおよびエクソグリコシダーゼの使用により達成できる。
【0096】
潜在的グリコシル化部位のグリコシル化は、Duskin等,J.Biol.Chem.,257:3105(1982)に記述されているように化合物ツニカマイシンの使用により妨げられる。ツニカマイシンはタンパク質-N-グリコシド結合の形成をブロックする。
【0097】
Apo-2リガンドの共有結合修飾のもう一つのタイプは、米国特許第4,640,835号;第4,496,689号;第4,301,144号;第4,670,417号;第4,791,192号;または第4,179,337号に示された方法で、例えばポリエチレングリコール、ポリプロピレングリコールまたはポリオキシアルキレンといった様々な非タンパク質性ポリマーの一つにApo-2リガンドポリペプチドを結合することを含む。
【0098】
8. エピトープタグ化Apo-2リガンド
本発明はまた、もう一つのヘテロ構造ポリペプチドに融合されたApo-2リガンドを含むキメラポリペプチドをも提供する。一つの実施態様として、キメラポリペプチドはタグポリペプチドとのApo-2リガンドの融合を含み、該タグポリペプチドは抗タグ抗体が選択的に結合できるエピトープを提供する。エピトープタグは一般的にApo-2リガンドのアミノ末端またはカルボキシル末端に置かれる。Apo-2リガンドの該エピトープタグ型の存在は、タグポリペプチドに対する抗体を用いて検出できる。またエピトープタグの提供は、抗タグ抗体を用いるアフィニティー精製またはエピトープタグに結合するもう一つのタイプのアフィニティーマトリックスによって、Apo-2リガンドを容易に精製することを可能にする。
【0099】
様々なタグポリペプチドとそれら各自の抗体が本分野ではよく知られている。例として、flu HAタグポリペプチドとその抗体12CA5[Field等,Mol.Cell.Biol.,8:2159-2165(1988)];c-mycタグとそれらの抗体8F9,3C7,6E10,G4,B7そして9E10[Evan等,Molecular and Cellular Biology,5:3610-3616(1985)];そして単純ヘルペスウイルス糖タンパク質D(gD)タグとその抗体[Paborsky等,Protein Engineering,3(6):547-553(1990)]を含む。他のタグポリペプチドとしては、Flag-ペプチド[Hopp等,BioTechnology,6:1204-1210(1988)];KT3エピトープペプチド[Martin等,Science,255:192-194(1992)];αチューブリンエピトープペプチド[Skinner等,J.Biol.Chem.,266:15163-15166(1991)];そしてT7遺伝子10タンパク質ペプチドタグ[Lutz-Freyermuth等,Proc.Natl.Acad.Sci.USA,87:6393-6397(1990)]が含まれる。一度タグポリペプチドが選択されれば、それに対する抗体はここで開示される方法を用いて作製できる。
【0100】
一般的に、エピトープタグ化されたApo-2リガンドは、天然の及び変異体Apo-2リガンドに対して上述の方法にしたがって構築され生産され得る。Apo-2リガンド-タグポリぺプチ融合体は、好ましくはタグポリペプチドDNA配列にフレーム中でApo-2リガンド部分をコードするcDNA配列を融合し、そして適切なホスト細胞内で合成されたDNA融合合成物を発現することによって構築される。通常本発明のApo-2リガンド-タグポリペプチドキメラを調製する場合、タグポリペプチドのN末端をコードする核酸にApo-2リガンドの3'末端を融合するであろうが、しかしながら5'での融合もまた可能である。エピトープタグ化Apo-2リガンドの例は、以下の実施例2にさらに詳細に記述されている。
【0101】
エピトープタグ化Apo-2リガンドは、抗タグ抗体を用いたアフィニティークロマトグラフィーによって精製できる。アフィニティー抗体が付着しているマトリックスは、例えばアガロース、制御されたポアグラスまたはポリ(スチレンジビニル)ベンゼンを含むであろう。それからエピトープタグ化されたApo-2リガンドは、本分野で既知の方法を用いてアフィニティーカラムから溶出されることができる。
【0102】
B. Apo-2リガンドの治療上の使用
本明細書で開示されているApo-2リガンドは哺乳動物細胞においてアポトーシスを誘導するために治療上で用いられ得る。一般的に哺乳動物細胞においてアポトーシスを誘導するために方法には、細胞を有効量のApo-2リガンドにさらすことが含まれる。これは例えば以下に記述された方法及び実施例に記述された方法にしたがってin vivoまたはex vivoで成し遂げられ得る。アポトーシスを誘導する方法は、アポトーシスの減少したレベルによって特徴付けられる特定の生理学的コンディションに対する治療で用いられ得ることが予期される。該生理学的コンディションの例としては、狼瘡及び免疫介在性糸球体腎炎のような自己免疫疾患、及びガンが含まれる。ガンの治療に対するApo-2リガンドの治療上の応用は以下に詳細に記述されている。
【0103】
ガンの治療法において、Apo-2リガンドはガンを持つと診断された哺乳動物に投与される。Apo-2リガンドは他のアポトーシス誘導試薬、化学療法、放射線療法、及び手術を含むいかなる他の治療構成要素及び技術と組み合わせて用いることもできることはもちろん企図される。
【0104】
Apo-2リガンドは好ましくは、製薬学的に許容できるキャリー内で哺乳動物に投与される。適したキャリアーとその処方については、Oslo等によって編集されたRemington's Pharmaceutical Sciences,第16版、1980、Mark Publishing Co.に記述されている。典型的には適切な量の製薬学的に許容できる塩が、等浸透圧の処方にするための処方において用いられる。製薬学的に許容できるキャリアーの例として、塩水、リンガー溶液そしてブドウ糖溶液がある。溶液のpHは好ましくは約5から約8、そしてより好ましくは約7から約7.8である。特定のキャリアーが例えば投与の経路及び投与されるApo-2リガンドの濃度に依存してより好ましいということは、当業者には明らかであろう。
【0105】
Apo-2リガンドは注射(静脈、腹膜、皮下、筋肉の)、または効果的な形態で血流への輸送を確保する点滴のような他の方法によって、哺乳動物に投与される。Apo-2リガンドがin vivoまたはex vivoでの遺伝子治療によって投与され得ることもまた予期されよう。
【0106】
Apo-2リガンドを投与するための有効投与量とスケジュールは経験的に決まり、該決定をすることは当業者の手中にある。単独で用いられるApo-2リガンドの有効投与量または量は体重当たり1μg/kgから100mg/kgの範囲にあり、または一日当たりではそれ以上であろうと現在では考えられている。投与量の種間スケールは例えばMordenti等,Pharmaceut.Res.,8:1351(1991)に開示されているように本分野で周知の方法で実施され得る。投与されなければならないApo-2リガンドの投与量は、例えばApo-2リガンドを受けるであろう哺乳動物、投与経路、及び哺乳動物に投与される他の薬剤または治療に依存して変化するであろうことは当業者に理解されよう。
【0107】
哺乳動物に投与される一つ以上の他の治療は、化学療法及び/または放射線療法、免疫アジュバント、サイトカイン、及び抗体ベース治療を制限することなく含む。例として、インターロイキン(例えばIL-1,IL-2,IL-3,IL-6)、白血病阻害因子、インターフェロン、TGF-ベータ、エリトロポエチン、トロンボポエチン、抗VEGF抗体及びHER-2抗体が含まれる。哺乳動物細胞においてアポトーシスを誘導することが知られている他の試薬も用いられ得、該試薬にはTNF-α、TNF-β(リンホトキシン-α)、CD30リガンド、4-1BB、及びApo-1リガンドが含まれる。
【0108】
本発明によって企図される化学療法には、Doxorubicin,5-Fluorouracil("5-FU"),エトポシド,カンプトセシン(camptothecin),Leucovorin,Cytosine arabinoside("Ara-C"),Cyclophosphamide,Thiotepa,Busulfan,Cytoxin,Taxol,Methotrexate,Cisplatin,Melphalan,Vinblastine及びCarboplatinのような、本分野で周知であり、商業的に入手可能な化学的物質または薬剤が含まれる。該化学療法に対する調製と投与スケジュールは、製品の説明書にしたがって用いられるか、または当業者に経験的に決定されるであろう。該化学療法の調製と投与スケジュールはまたChemotherapy Service,M.C.Perry編,Williams & Wilkins,Baltimore,MD(1992)にも記述されている。
【0109】
化学療法はApo-2リガンドに対して上記したような製薬学的に許容できるキャリアーにおいて好ましくは投与される。化学療法の投与形態はApo-2リガンドに対して用いられるものと同様であり、または異なる形態でそれは哺乳動物に投与され得る。例えばApo-2リガンドは注射される一方で、化学療法は哺乳動物に経口で投与される。Apo-2リガンドと組み合わせた化学療法の投与形態は以下の実施例9-12にさらに詳細に記述されている。
【0110】
放射線治療は本分野で商業的に用いられ当業者に周知のプロトコールにしたがって哺乳動物に投与され得る。該治療にはセシウム、イリジウム、ヨウ素、またはコバルト放射線が含まれ得る。放射線治療は全身体的な照射によってなされ得、または体の内外の特異的な部位または組織に局所的に向けられ得る。典型的には放射線治療は約1から2週の期間にわたる間隔で投与される。しかしながら放射線治療はより長い期間にわたって投与され得る。任意に放射線治療は単一の投与または複数投与、連続的投与として投与され得る。
【0111】
Apo-2リガンドは及び一つ以上の他の治療は哺乳動物に同時的にまたは連続的に投与され得る。哺乳動物へのApo-2リガンド及び一つ以上の他の治療の投与に引き続いて、哺乳動物のガン及び生理学的なコンディションは当業者によく知られた様々な方法でモニターされ得る。例えば腫瘍の大きさは生検または標準的なx線イメージ法によって物理的に観察し得る。
【0112】
Apo-2リガンドはex vivoでガン細胞を治療するために用いられ得ることが企図される。該ex vivo治療は、骨髄移植および特に異種骨髄移植において有用であろう。例えばApo-2リガンドを用いて、及び場合により上記記述されているような一つ以上の他の治療薬をも用いて、ガン細胞を含む細胞または組織(類)の治療をすることは、受容者哺乳動物における移植前に、アポトーシスを誘導するため、そしてガン細胞を実質的に除去するために用いられ得る。
【0113】
ガン細胞を含む細胞または組織(類)は最初にドナー哺乳動物から得られる。細胞または組織(類)は外科的に得られ、好ましくは無菌的に得られる。移植のため骨髄を治療する方法では、骨髄は哺乳動物から針吸引によって得られる。それからガン細胞を含む細胞または組織(類)をApo-2リガンドを用いて治療し、任意に上述のような一つ以上の他の治療を伴う。骨髄は好ましくはApo-2リガンドを用いた治療の前に、単核細胞画分を得るために(ficoll-hypaque勾配による遠心分離によるように)好ましくは分画される。
【0114】
それから治療された細胞または組織(類)を受容者哺乳動物に注入または移植し得る。受容者哺乳動物はドナー哺乳動物と同じ個体であり得、またはもう一つの異種個体でもあり得る。自己骨髄移植に対しては、哺乳動物を移植の前に本分野で周知の、例えばAutologous Bone Marrow Transplantation: Proceedings of the Third International Symposium,Dicke等,編,University of Texas M.D. Anderson Hospital and Tumor Institute内に記述されている有効量の放射線または化学療法を用いて治療する。
【0115】
C. Apo-2リガンドの非治療上の使用
本発明のApo-2リガンドはまた非治療上の応用においても有用性を持つ。Apo-2リガンドをコードする核酸配列は組織特異的タイプ化のための診断薬として用い得る。例えばin situハイブリダイゼーション、ノーザン及びサザンブロッティング、及びPCR分析のような方法が、Apo-2リガンドをコードするDNA及び/またはRNAが試験される細胞タイプ(類)に存在するかどうかを調べるために用いられ得る。Apo-2リガンド核酸はまたここで記述される組換え法によってApo-2ポリペプチドの調製のためにも有用であろう。
【0116】
単離されたApo-2リガンドはApo-2リガンドの未知の量を含むサンプルが調整される際のコントロールとして定量的診断アッセイでも用いられ得る。Apo-2リガンド調製物はまた、Apo-2リガンドに対するアッセイ(例えばラジオイムノアッセイ、ラジオリセプターアッセイ、または酵素リンクイムノアッセイにおけるスタンダードとしての使用のためにApo-2リガンドをラベルすることによって)、例えばApo-2リガンドを結合する受容体を同定または単離する際のアフィニティー精製法、及び例えば放射性ヨウ素、酵素、またはフルオロフォアーでラベルした場合競合的タイプ受容体結合アッセイにおいて、スタンダードとして抗体を生産するのに有用でもある。
【0117】
Apo-2リガンドをコードする核酸はまたトランスジェニック動物または「ノックアウト」動物のそれぞれを生産するために用いられ得るが、該動物は各々治療上の有用な試薬の開発またはスクリーニングに有用である。トランスジェニック動物(例えばマウスまたはラット)は、トランス遺伝子を含む細胞を持つ動物であり、そのトランス遺伝子は動物内または例えば胚段階といった元となる動物の祖先に導入されたものである。トランス遺伝子はトランスジェニック動物が発達する細胞のゲノム内にインテグレートされたDNAである。一つの実施態様として、Apo-2リガンドをコードするcDNAまたはそれの適切な配列は、確立された方法にしたがってApo-2リガンドをコードするゲノムDNAをクローン化するために用いられ得、該ゲノム配列はApo-2リガンドをコードするDNAを発現する細胞を含むトランスジェニック動物を生産するために用いられる。トランスジェニック動物、特にマウスまたはラットのような動物を生産する方法は、本分野でありきたりのものになっており、例えば米国特許第4,736,866号及び第4,870,009号に記述されている。典型的には特定の細胞を組織特異的エンハンサーと共同してApo-2リガンドトランス遺伝子に対してターゲット化するであろう。胚段階で動物の生殖細胞系列内に導入されたApo-2リガンドをコードするトランス遺伝子の一コピーを含むトランスジェニック動物を、Apo-2リガンドをコードするDNAの増大した発現の影響を調べるため用いることができる。
【0118】
代わりに、Apo-2リガンドの非ヒト相同物を、Apo-2リガンドをコードする異種遺伝子と、動物の胚細胞内に導入されたApo-2リガンドをコードする改変されたゲノムDNAの間の相同的組換えの結果として、Apo-2リガンドをコードする不完全なまたは改変された遺伝子を持つApo-2リガンド「ノックアウト」動物を構築するために用いられ得る。例えばApo-2リガンドをコードするcDNAは、確立した方法にしたがってApo-2リガンドをコードするゲノムDNAをクローン化するために用いられ得る。Apo-2リガンドをコードするゲノムDNAの部分を、インテグレーションをモニターするために用いられ得る選択可能マーカーをコードする遺伝子のような、もう一つの遺伝子で欠失または置換し得る。典型的には非改変フランキングDNA(5'末端と3'末端の両方)の数キロベースを、ベクター内に取り込ませる[相同的組換えベクターの記述については、例えばThomas及びCapecchi,Cell,51:503(1987)参照]。該ベクターは胚幹細胞系(例えばエレクトロポレーションによって)内に導入され、導入されるDNAが内因性のDNAと相同的に組換える細胞が選択される[例えばLi等,Cell,69:915(1992)参照]。それから選択された細胞を集合キメラを形成するために動物(例えばマウスまたはラット)の胚胎盤内に注入する[例えばBradley,in Teratocarcinomas and Embryonic Stem Cells:A Practical Approach,E.J.Robertson,編(IRL,Oxford,1987),pp.113-152]。それからキメラの胚を適した偽妊娠メス里親動物内に移植し、該胚は「ノックアウト」動物を作製するための期間を運ぶ。その生殖細胞に相同的組換えDNAを運んでいる子孫を、標準的な方法で同定し、該動物の全ての細胞が相同的組換えDNAを含む動物を繁殖させるために用い得る。ノックアウト動物は例えば、特定の病理学上のコンディションに対して防御する能力によって、及びApo-2リガンドポリペプチドの不存在のため病理学的コンディションの発展によって特性指摘され得る。
【0119】
D. 抗Apo-2リガンド抗体調製
本発明はさらに抗Apo-2リガンド抗体を提供する。Apo-2リガンドに対する抗体は以下のように調製され得る。例示的な抗体には、ポリクローナル、モノクローナル、ヒト化、二重特異性、及びヘテロ接合の各抗体が含まれる。
【0120】
1. ポリクローナル抗体
Apo-2リガンド抗体にはポリクローナル抗体が含まれる。ポリクローナル抗体の調製法は当業者に周知である。ポリクローナル抗体は例えば免疫化試薬と必要であればアジュバントの一度以上の注射により哺乳動物において生産され得る。典型的には該免疫か試薬及び/またはアジュバントは、複数の皮下または腹膜内の注射により哺乳動物において注射されるであろう。免疫化試薬には、Apo-2リガンドポリペプチドまたはその融合タンパク質が含まれる。免疫化試薬を免疫化される哺乳動物において免疫原性であることが知られているタンパク質に接合することは、有用であろう。用いられるであろう該免疫原性タンパク質の例としては、キーホールリンペットヘモシアニン、血清アルブミン、ウシチログロブリン、及び大豆トリプシンインヒビターが制限されることなく含まれる。ミョウバンのような凝集試薬もまた、哺乳動物の免疫応答を増大するために用いられ得るであろう。用いられるであろうアジュバントの例としては、フロイント完全アジュバント及びMPL-TDMアジュバント(モノホスホリルリピドA,合成トレハロースジコリノミコレート)が含まれる。免疫化プロトコールは過度の実験なく当業者に選択され得る。それから該哺乳動物を採血し、抗体力価のため血清をアッセイし得る。必要であれば、哺乳動物を抗体力価が増大するまたは安定水準に達するまで追加免疫し得る。
【0121】
2. モノクローナル抗体
代わりにApo-2リガンド抗体はモノクローナル抗体であり得る。モノクローナル抗体はKohler及びMilstein,Nature,256:495(1975)に記述されているようなハイブリドーマ法を用いて調製され得る。ハイブリドーマ法においては、マウス、ハムスター、または他の適切なホスト動物が、免疫化試薬と特異的に結合するであろう抗体を生産するまたは生産し得るリンパ球を引き出すために免疫化試薬と(上述のように)典型的に免疫化される。代わりに、リンパ球はin vitroで免疫化され得る。
【0122】
免疫化試薬にはApo-2リガンドポリペプチドまたはその融合タンパク質が典型的には含まれる。その表面でApo-2リガンドを発現する細胞もまた用いられ得る。一般的にもしヒト起源の細胞が必要であれば、末梢血リンパ球(「PBL」)が用いられ、もし非ヒト哺乳動物ソースが必要であれば、脾臓細胞またはリンパ節細胞が用いられる。それからハイブリドーマ細胞を形成するためにポリエチレングリコールのような適当な融合試薬を用いて、該リンパ球を不朽化細胞系と融合する[Goding,Monoclonal Antibodies:Principles and Practice,Academic Press,(1986)pp.59-103]。不朽化細胞系は通常トランスフォームされた哺乳動物細胞であり、特にネズミ、ウシ、及びヒト起源のミエローマ細胞である。通常ラットまたはマウスミエローマ細胞系が用いられる。該ハイブリドーマ細胞は、非融合の不朽化細胞の成育と生存を阻害する一つかそれ以上の物質を好ましくは含む適当な培地において、培養され得る。例えばもし元となる細胞が、酵素ヒポキサンチングアニンホスホリボシルトランスフェラーゼ(HGPRTまたはHPRT)を欠失していたならば、ハイブリドーマに対する培地は典型的には、HGPRT欠失細胞の成長を妨げる物質であるヒポキサンチン、アミノプテリン及びチミジン(HAT培地)を含むであろう。
【0123】
好ましい不朽化細胞系は効率よく融合し、選択された抗体生産細胞による抗体の安定な高レベルの生産を維持し、そしてHAT培地のような培地に感受性なものである。より好ましい不朽化細胞系は、例えばSalk Institute Cell Distribution Center,San Diego,California及びAmerican Type Culture Collection,Rockville,Marylandから入手可能であるネズミミエローマ系である。ヒトミエローマ及びマウス-ヒトヘテロミエローマ細胞系もまた、ヒトモノクローナル抗体の生産のため記述されている[Kozbor,J.Immunol.,133:3001(1984);Brodeur等,Monoclonal Antibody Production Techniques and Applications,Marcel Dekker,Inc.,New York,(1987)pp.51-63]。
【0124】
それからハイブリドーマ細胞を培養する培地を、Apo-2リガンドに対して向けられたモノクローナル抗体の存在のためアッセイし得る。好ましくはハイブリドーマ細胞により生産されるモノクローナル抗体の結合特異性を、免疫沈降法によって、またはラジオイムノアッセイ(RIA)、蛍光活性化細胞ソーティング(FACS)、または固相酵素免疫検定法(ELISA)のようなin vitro結合アッセイによって測定する。モノクローナル抗体の結合アフィニティーは、例えばMunsonとPollard,Anal.Biochem.107:220(1980)のスキャッチャード分析によって測定される。
【0125】
望ましいハイブリドーマ細胞が同定された後、該クローンを限界希釈法によってサブクローン化し、標準的な方法[Goding,上記参照]によって成育させる。この目的のために適当な培地は、例えばダルベッコ変法イーグルス培地またはRPMI-1640培地である。代わりにハイブリドーマ細胞は、動物における腹水のようにin vivoで成育させることもできる。
【0126】
サブクローンにより分泌されるモノクローナル抗体は、例えばプロテインA-セファロース、ヒドロキシルアパタイトクロマトグラフィー、ゲル電気泳動、透析及びアフィニティークロマトグラフィーのようなありきたりの免疫グロブリン精製法によって、培地または腹水分泌液から単離され精製され得る。
【0127】
本発明の一つの実施態様として、モノクローナル抗体にはここと以下の実施例で記述される1D1,2G6,2E11または5C2抗体が含まれる。該モノクローナル抗体には、それぞれAmerican Type Culture Collection Accession Nos. ATCC HB-12556, HB-12557, HB-12558, またはHB-12559の下で寄託されているハイブリドーマ細胞系によって分泌される1D1,2G6,2E11または5C2モノクローナル抗体と同様の生物学的性質を持つ抗体も含まれる。「生物学的性質」なる語は、モノクローナル抗体のin vitro及び/またはin vivoでの活性、例えばApo-2リガンド誘発アポトーシスを実質的に減少する能力、またはApo-2リガンドのその受容体に対する結合を実質的に減少またはブロックする能力をいうために用いられる。該抗体は好ましくは、ここで開示される1D1,2G6,2E11または5C2抗体と同じエピトープ、または実質的に同じエピトープに結合する。これはここと実施例で記述されるアッセイを実施することによって測定され得る。
【0128】
該モノクローナル抗体は米国特許第4,816,567号に記述されるような組換えDNA法によっても作製され得る。本発明のモノクローナル抗体をコードするDNAは、ありきたりの方法を用いて容易に単離されシークエンスされ得る(例えばネズミ抗体のH鎖とL鎖をコードする遺伝子に特異的に結合できるオリゴヌクレオチドプローブを用いることによって)。本発明のハイブリドーマ細胞は上記DNAの好ましいソースとしても役に立つ。一度単離されると、DNAを発現ベクターにつなぎ、それからそれを組換えホスト細胞においてモノクローナル抗体の合成を得るために、サルCOS細胞、チャイニーズハムスター卵巣(CHO)細胞またはさもなければ免疫グロブリンタンパク質を生産しないミエローマ細胞のようなホスト細胞にトランスフェクトする。DNAはまた、例えば相同のネズミ配列の代わりにヒトH鎖およびL鎖定常ドメインのコード配列で置換することによって[米国特許第4,816,567号;Morrison等,上記参照]、あるいは非免疫グロブリンポリペプチドのコード配列を免疫グロブリンコード配列に共有結合でつなぐことによって修飾し得る。該非免疫グロブリンポリペプチドは本発明の抗体の定常ドメインの代わりに置換することができ、またはキメラ二価抗体を作製するために本発明の抗体の一つの抗原結合部位の可変ドメインの代わりに置換することができる。
【0129】
該抗体は一価抗体であり得る。一価抗体の調製法は本分野でよく知られている。例えば一つの方法には、免疫グロブリンL鎖と修飾H鎖の組換え発現が含まれる。H鎖は一般的にH鎖架橋を妨げるためにFc領域のいかなる部分でも切り詰められる。代わりに関連したシステイン残基をもう一つのアミノ酸残基で置換し、または架橋を妨げるために欠失する。
【0130】
in vitro法もまた一価抗体を調製するために適している。抗体の断片、特にFab断片を生産するための抗体の切断は、本分野で周知のありきたりの方法を用いて成し遂げられ得る。例えば切断はパパインを用いて実施され得る。パパイン切断の例は94年12月22日に印刷されたWO 94/29348及び米国特許第4,342,566号に記述されている。抗体のパパイン切断は典型的に、それぞれ単一の抗原結合部位を持つFab断片と呼ばれる2の同一の抗原結合断片と、残余のFc断片を生産する。パパイン処理は2の抗原結合部位をもち、依然として抗原を架橋できるF(ab')2断片を生ずる。
【0131】
抗体切断において生産されたFab断片はまた、L鎖の定常ドメインと、H鎖の第一の定常ドメイン(CH1)を含む。Fab'断片は、抗体ヒンジ領域由来の一つ以上のシステインを含むH鎖CH1ドメインのカルボキシ末端でのいくつかの残基の付加によって、Fab断片とは異なる。Fab'-SHは定常ドメインのシステイン残基(類)が遊離チオール基を持つFab'をここで表す。F(ab')2抗体断片はもともと、その間にヒンジシステインを持つFab'断片のペアとして生産された。他の化学的な抗体断片の結合もまた知られている。
【0132】
3. ヒト化抗体
本発明のApo-2リガンド抗体はさらに、ヒト化抗体またはヒト抗体を含む。非ヒト(例えばネズミ)抗体のヒト化形態は、キメラ免疫グロブリン、免疫グロブリン鎖または非ヒト免疫グロブリン由来の最小配列を含むその断片(Fv,Fab,Fab',F(ab')2または抗体の他の抗原結合サブ配列)である。ヒト化抗体には、受容者の相補性決定領域由来の残基が、望ましい特異性、アフィニティー及び力価を持つマウス、ラットまたはウサギのような非ヒト種(ドナー抗体)のCDR由来の残基で置換されているヒト免疫グロブリン(受容者抗体)が含まれる。例えばヒト免疫グロブリンのFvフレームワーク残基が相当する非ヒト残基で置換される。ヒト化抗体はまた、受容者抗体にもインポートCDRまたはフレームワーク配列にも見出されない残基をも含む。一般的に該ヒト化抗体は、少なくとも一つそして典型的には二つの可変ドメインの実質的にすべてを含み、該可変ドメインはCDR領域の全てまたは実質的に全てが非ヒト免疫グロブリンのものに相当し、FR領域の全てまたは実質的に全てがヒト免疫グロブリンコンセンサス配列のものである。該ヒト化抗体はまた適宜に、免疫グロブリン定常領域(Fc)の少なくとも一部を含み、典型的にはそれはヒト免疫グロブリンのものであろう[Jones等,Nature,321:522-525(1986);Reichmann等,Nature,332:323-329(1988);及びPresta,Curr.Op.Struct.Biol.2:593-596(1992)]。
【0133】
非ヒト抗体をヒト化する方法は本分野でよく知られている。一般的に、ヒト化抗体は非ヒトであるソースからそれに導入された一つかそれ以上のアミノ酸残基をもつ。これらの非ヒトアミノ酸残基は、しばしば「インポート残基」として示され、典型的には「インポート」可変ドメインから由来している。ヒト化は本質的に、Winterと共同研究者[Jones等,Nature 321:522-525(1986);Riechmann等,Nature 332:323-327(1988);Verhoeyen等,Science 239:1534-1536(1988)]の方法にしたがって実施され、ネズミCDRまたはCDR配列を、相当するヒト抗体の配列で置換することによるコンピューターモデルを用いたQueen等,Proc.Natl.Acad.Sci.86:10029-10033(1989)の方法によってなされる。したがって該「ヒト化」抗体はキメラ抗体であり(米国特許第4,816,567号)、そこでは実質的に完全なヒト可変ドメインより短い部分が、相当する非ヒト種由来の配列で置換されている。実際問題として、ヒト化抗体は典型的には、いくつかのCDR残基および可能ないくつかのFR残基がネズミ抗体の相似性の部位由来の残基によって置換されているヒト抗体である。
【0134】
ヒト化抗体を作製する場合に用いられるL鎖とH鎖の両方のヒト可変領域の選択は、抗原性を減少するために重要である。「ベスト-フィット」法にしたがって、ネズミ抗体の可変ドメインの配列が既知のヒト可変ドメイン配列の完全なライブラリーからスクリーニングされる。それからネズミのものに最も近いヒト配列が、ヒト化抗体のためにヒトフレームワーク(FR)として受け入れられる[Sims等,J.Immunol,151:2296(1993);ChothiaとLesk,J.Mol.Biol.196:901(1987)]。もう一つの方法は、L鎖とH鎖の特定のサブグループの全てのヒト抗体のコンセンサス配列由来の特定のフレームワークを用いる。同様のフレームワークはいくつかの異なるヒト化抗体のためにも用いられ得る[Carter等,Proc.Natl.Acad.Sci.USA.89:4285(1992);Presta等,J.Immunol.151:2623(1993)]。
【0135】
抗体は抗原に対する高アフィニティーおよび他の好ましい生物学的性質を保持したまま免疫化されることがさらに重要である。この目的を達成するため好ましい方法にしたがって、もととなるおよびヒト化配列の三次元モデルを用いてもととなる配列および様々な概念上のヒト化産物の分析の過程によって、ヒト化抗体を調製する。三次元免疫グロブリンモデルは、公に入手可能であり当業者には親しまれている。選択された免疫グロブリン配列の候補の可能な三次元構造を描写し表示するコンピュータープログラムも入手可能である。これらの表示を見ることによって、免疫グロブリン配列の候補の機能において残基の考え得る役割の分析、すなわち抗原に結合する場合に免疫グロブリン候補の能力に影響する残基の解析が可能となる。この方法では、FR残基が、標的抗原(類)に対する増大したアフィニティーのような望ましい抗体特性を達成するためのコンセンサス配列およびインポート配列から選択され結合され得る。一般的にCDR残基は直接にそして最も実質的に抗原結合を影響する部位に含まれる[1994年3月3日に印刷されたWO 94/04679参照]。
【0136】
免疫化により、内因性の免疫グロブリン生産の不存在下でヒト抗体のフルレパートリーを生産可能なトランスジェニック動物(例えばマウス)が用いられ得る。例えば、キメラで生殖系列ミュータントマウスにおける抗体H鎖結合領域(JH)遺伝子のホモ接合体欠失が、内因性抗体生産の完全な阻害を引き起こすことが記述されている。該生殖系列ミュータントマウスにおけるヒト生殖系列免疫グロブリン遺伝子配列のトランスファーは、抗原チャレンジに対するヒト抗体の生産を引き起こすであろう[例えばJakobovits等,Proc.Natl.Acad.Sci.USA,90:2551-255(1993);Jakobovits等,Nature,362:255-258(1993);Bruggermann等,Year in Immuno.,7:33(1993)]。ヒト抗体はまたファージディスプレーライブラリーにおいても生産し得る[Hoogenboom及びWinter,J.Mol.Biol.,227:381(1991);Marks等,J.Mol.Biol.,222:581(1991)]。Cote等及びBoerner等の方法もまた、ヒトモノクローナル抗体の調製のために有用である(Cote等,Monoclonal Antibodies and Cancer Therapy,Alan R.Liss,p.77(1985)及びBoerner等,J.Immunol.,147(1):86-95(1991)]。
【0137】
4. 二重特異性抗体
二重特異性抗体とは、少なくとも二つの異なる抗原に対する結合特異性を持つモノクローナルで、好ましくはヒトのまたはヒト化された抗体である。本ケースでは、結合特異性の一つはApo-2リガンドに対するものであり、もう一つはいかなる他の抗原に対するものでもよく、好ましくは細胞表面タンパク質または受容体あるいは受容体サブユニットに対するものである。
【0138】
二重特異性抗体を作成する方法は本分野で知られている。伝統的には、二重特異性抗体の組換え生産は、二つの免疫グロブリンH鎖/L鎖ペアの共同発現に基づいており、そこでは二つのH鎖は異なる特異性をもつ[MilsteinとCuello,Nature 305:537(1983)]。免疫グロブリンH鎖とL鎖のランダムな識別のため、これらのハイブリドーマ(クアッドローマ)は10種類の異なる抗体分子の混合物を潜在的に生産し、その中の一種だけが正しい二重特異性抗体なのである。正しい分子の精製は、通常アフィニティークロマトグラフィーステップによりなされる。同様の方法は1993年5月13日に印刷されたWO 93/08829およびTraunecker等,EMBO J.10:3655(1991)に記述されている。
【0139】
異なるそしてより好ましいアプローチにしたがうと、望ましい結合特異性を持つ抗体可変ドメイン(抗体抗原結合部位)を免疫グロブリン定常ドメイン配列に融合する。好ましくは融合は、少なくともヒンジ部、CH2、およびCH3を含む免疫グロブリンH鎖定常領域となされる。L鎖との結合に必要な部位を含む第一のH鎖定常領域(CH1)を融合物の少なくとも一つに存在させるのも好ましい。免疫グロブリンH鎖融合物をコードするDNAおよび必要であれば免疫グロブリンL鎖をコードするDNAを、別々の発現ベクターに組み込み、適したホスト生物内に共同してトランスフェクトする。構築に用いられる非等量割合の三つのポリペプチド鎖が最適条件収率を提供すると、これにより実施態様における三つのポリペプチド断片の相互の性質を調製する大きな柔軟性が提供される。しかしながら、等量割合で少なくとも二つのポリペプチド鎖の発現が高い収率を引き起こした場合、またはその割合が特定の重要性を持たない場合、一つの発現ベクターに二つまたは三つ全てのポリペプチド鎖をコードする配列を組み込むことが可能である。このアプローチの好ましい実施態様によると、二重特異性抗体は一つの腕の第一の結合特異性を持つハイブリッド免疫グロブリンH鎖と、もう一方の腕のハイブリッド免疫グロブリンH鎖/L鎖ペア(第二の結合特異性を提供する)より成る。この非対称の構造は、二重特異性分子の半分だけを占める免疫グロブリンL鎖の存在が選別の容易な方法を提供するように、望まない免疫グロブリン鎖の組み合わせから望ましい二重特異性化合物を選別するのを容易にすることが見出された。このアプローチは1994年3月3日に印刷されたWO 94/04690に開示されている。二重特異性抗体のさらに詳細を生産する方法については、例えばSuresh等,Methods in Enzymology 121:210(1986)を参照。
【0140】
5. ヘテロ接合抗体
ヘテロ接合抗体もまた本発明の範囲にある。ヘテロ接合抗体は二つの共有結合で結合された抗体より成る。該抗体は例えば、望まない細胞に対して免疫系細胞の標的にするため[米国特許第4,676,980号]、またはHIV感染の治療のため[WO 91/00360;WO 92/00373;そしてEP 03089])提案されている。該抗体は架橋試薬を含むもののような合成タンパク質化学において周知の方法を用いてin vitroで調製され得ることが企図されている。例えばイムノトキシンはジスルフィド交換反応を用いて、またはチオエーテル結合を形成することによって構築し得る。この目的に対する適した試薬の例としては、イミノチオレート及びメチル-4-メルカプトブチリミデートそして例えば米国特許第4,676,980号に開示されているものが含まれる。
【0141】
E. Apo-2リガンド抗体の使用
Apo-2リガンド抗体は例えば特異的細胞、組織、または血清における発現の検出といったApo-2リガンドに対する診断のアッセイに用いられ得る。本分野で周知の様々な診断のアッセイ法が用いられ得、その中には競合的結合アッセイ、直接または間接サンドイッチアッセイ及び異種または同種フェーズのそれぞれで実施される免疫沈降アッセイが含まれる[Zola,Monoclonal Antibodies:A Manual of Techniques,CRC Press,Inc.(1987)pp.147-158]。診断アッセイで用いられる抗体は検出可能な部分でラベルされ得る。検出可能な部分は直接または間接に検出可能なシグナルを生ずることが可能ないかなるものでもあり得る。例えば検出可能な部分は、3H,14C,32P,35Sまたは125Iのようなラジオアイソトープ、フルオレセイン、イソチオシアネート、ローダミンまたはルシフェリンのような蛍光化合物または化学ルミネセンス化合物、アルカリホスファターゼ、ベータ-ガラクトシダーゼまたはセイヨウワサビペルオキシダーゼのような酵素が挙げられる。検出可能な部分と抗体を別々に接合するための本分野で既知の方法も用いることができ、それにはHunter等,Nature 144:945(1962);David等,Biochemistry 13:1014(1974);Pain等,J.Immunol.Meth.40:219(1981);およびNygren,J.Histochem.and Cytochem.30:407(1982)に記述された方法が含まれる。
【0142】
Apo-2リガンド抗体はまた、組換え細胞カルチャーまたは天然のソースからApo-2リガンドのアフィニティー精製に対しても有用である。この工程においては、Apo-2リガンドに対する抗体が、本分野で周知の方法を用いて、Sephadex樹脂または濾紙のような適した支持体に固定化される。それから該固定化抗体を精製されるApo-2リガンドを含むサンプルと接触させ、その後該支持体を固定化抗体と結合したApo-2リガンドを除くサンプル中の実質的にすべての物質を除去するであろう適した溶媒を用いて洗浄する。最後に該支持体を抗体からApo-2リガンドを放出するであろうもう一つの適した溶媒で洗浄する。Apo-2リガンド抗体はまた可溶化Apo-2受容体のアフィニティー精製に対して、またはApo-2受容体の発現クローニングに対して有用である。
【0143】
ここで開示される抗体は治療薬としても用いられる。例えばApo-2リガンド活性(Apo-2リガンド誘発アポトーシスのような)をブロックする抗Apo-2リガンド抗体は、増大したアポトーシスと関連する病理学上のコンディションまたは疾患を治療するために用いられ得る[Thompson,上記参照参照]。
【0144】
F. Apo-2リガンドまたはApo-2リガンド抗体を含むキット
本発明の更なる実施態様において、例えば上述の治療上のまたは非治療上の応用に用いられ得るApo-2リガンドまたはApo-2リガンド抗体を含む製造物とキットが提供される。製造物としては、ラベルのついた容器が含まれる。適した容器は例えばボトル、ガラスビンそして試験管が含まれる。容器はガラスやプラスチックのような様々な物質から形成されるであろう。容器には、上述のような治療上または非治療上の応用に対して有効な活性試薬を含む構成物が入っている。構成物中の活性試薬はApo-2リガンドまたはApo-2リガンド抗体である。容器のラベルは構成物が、特異的な治療上または非治療上の応用に用いられ、上述したようなin vivoまたはin vitroでの各使用のための使用法も示されているであろう。
【0145】
本発明のキットは上述した容器を典型的には含み、商業的におよび使用者の見地から望ましい物質を含む一つ以上の他の容器を含み、その中には使用のための説明書と共に挿入された他のバッファー、希釈液、フィルター、針、シリンジそしてパッケージが含まれる。
【0146】
以下の実施例は説明のためのみを目的として提供され、いかなる方法においても本発明の範囲を制限することを企図しない。
【0147】
本明細書に引用される全ての参考文献は、ここで参考として全体として取り込まれる。
【実施例】
【0148】
本実施例で言及される全ての制限酵素は、New England Biolabsから購入され、製品の説明書にしたがって使用された。本実施例で言及される全ての他の商業的に入手可能な試薬は、もし他のいかなる指示もなければ、製品の説明書にしたがって用いられた。以下の実施例、及び明細書を通して、ATCC登録番号で示される細胞のソースは、American Type Culture Collection,Rockville,Marylandである。
【0149】
実施例1
ヒトApo-2リガンドをコードするcDNAクローンの単離
Apo-2リガンドに対するフルレングスcDNAを単離するために、ヒト胎盤cDNAのラムダgt11バクテリオファージライブラリー(約1×106クローン)(HL10756,Clontechから商業的に入手可能)を、ヒトFas/Apo-1リガンドとある程度の同一性を示すEST配列(GenBankローカスHMEA47M)に基づく合成オリゴヌクレオチドプローブを用いてハイブリダイゼーションによってスクリーニングした。HMEA47Mの該EST配列は390bpであり、その+3フレームで翻訳された場合、ヒトApo-1リガンドの34アミノ酸領域に対して16の同一性を示す。HMEA47Mの配列は以下のものである:
[配列1]
以下の配列を持つ60bpのオリゴヌクレオチドプローブをスクリーニングに用いた:
[配列2]
ハイブリダイゼーションは20%ホルムアミド、5×SSC、10%硫酸デキストラン、0.1%NaPiPO4、0.05MNaPO4、0.05mgサケ精子DNA、及び0.1%ドデシル硫酸ナトリウムを含むバッファーにおいて室温でオーバーナイトで実施し、引き続き5×SSCで42度での数回の洗浄をし、それから2×SSCで洗浄した。12のポジティブクローンをcDNAライブラリーで同定し、該ポジティブクローンを以下の配列を持つ第二の60bpのオリゴヌクレオチドプローブ(第一のプローブとオーバーラップしない)に対するハイブリダイゼーションによってスクリーニングした。
[配列3]
ハイブリダイゼーションを上記したように実施した。
【0150】
4の結果として生じたポジティブクローンを同定し、フランキング5'ベクター配列に基づき外来のClaI制限部位を加えたプライマーと、3'フランキングベクター配列に基づき外来のHindIII制限部位を加えたプライマーを用いたポリメラーゼ連鎖反応(PCR)により増幅した。PCR産物をゲル精製し、T-AライゲーションによりpGEM-T(Promegaから商業的に入手可能)内にサブクローン化した。それから異なるPCR由来の3の独立のクローンをディデオキシDNAシークエンシングに受けさせた。これらのクローンのDNA配列分析により、それらは本質的に同一であり、その5'領域でいくつかの長さのバリエーションを持つことが示された。
【0151】
Apo-2リガンドのコード領域の核酸配列は図1Aに示されている。該クローンの一つの下流3'末端配列のシークエンシングにより、特徴的なポリアデニル化部位が明らかになった(データは示していない)。該cDNAは核酸位91-93でATGコドンとして表される開始部位を持つ一つの長いオープンリーディングフレームを含んだ。この部位を取り囲む配列は開始部位に対して提案されているコンセンサス配列と理由のある一致を示す[Kozak,J.Cell.Biol.,115:887-903(1991)]。該オープンリーディングフレームは核酸位934-936の終始コドンTAAで終わる。
【0152】
ヒトApo-2リガンドの予想される成熟アミノ酸配列は281アミノ酸を含み、およそ32.5kDaの計算される分子量とおよそ7.63の等電点を持つ。ハイドロパシー分析(データは示していない)では残基15と40の間に疎水性領域の存在が示されるが、N末端の明らかなシグナル配列は存在しない。シグナル配列の不存在と内部の疎水性領域の存在は、Apo-2リガンドがII型膜貫通タンパク質であることを示唆する。推定の細胞内、膜貫通及び細胞外領域は、それぞれ14、26及び241アミノ酸長である。推定の膜貫通領域は図1Aにおいて下線が引かれている。潜在的なNリンクグリコシレーション部位は推定の細胞外ドメインの残基109に位置する。
【0153】
Apo-2リガンドのC末端領域のアミノ酸配列の他の周知のTNFサイトカインファミリーのメンバーとの比較により(AlignTMコンピュータープログラムを用いる)、C末端領域内でApo-2リガンドはApo-1リガンドと23.2%の同一性を示すことが示された(図1B)。比較分析は他のTNFファミリーメンバーとの比較的少ない程度の同一性を示した:CD40L(20.8%),LT-α(20.2%),LT-β(19.6%),TNF-α(19.0%),CD30L及びCD27L(15.5%),OX-40L(14.3%)及び4-1BBL(13.7%)。TNFサイトカインファミリーにおいて、TNF-α及びLT-αの結晶構造[Eck等,J.Bio.Chem.,264:17595-17605(1989);Eck等,J.Bio.Chem.,267:2119-2122(1992)]に基づいて、βストランドを形成すると予想される領域内の残基は、予想される結合ループにおける残基と比較して、他のTNFファミリーメンバーに関してより高く保存されている傾向にある。Apo-2リガンドは予想される結合ループにおける同一性と比較して、推定のβストランド領域で他のTNFファミリーメンバーとより大きい同一性を示すことが見出された。また、推定のβストランド、B及びB'を結合するループはApo-2リガンドにおいて著しくより長い。
【0154】
実施例2
ヒトApo-2リガンドの発現
A. フルレングスcDNA融合構築物
mycエピトープタグと融合したフルレングスApo-2リガンドcDNAを以下のように構築した。Apo-2リガンドcDNA挿入物をClaIおよびHindIII切断によって元となるpGEM-TApo-2リガンドプラスミド(実施例1に記述される)から摘出し、同じ制限酵素で札断されたpRK5哺乳動物発現プラスミド内に挿入した[Schall等,Cell,61:361-370(1990);Suva等,Science,237:893-896(1987)]。それから13アミノ酸mycエピトープタグをコードする配列
[配列4]
[Evan等,Mol.Cell.Biol.,5:3610-3616(1985)]を、プラスミドpRK5Apo-2リガンド-mycを作製するために、オリゴヌクレオチドディレクトミュータジェネシス[Zoller等,Nucleic Acids Res.,10:6487-6496(1982)]により、Apo-2リガンドコード配列の3'末端でコドン281と終始コドン(コドン282)の間に挿入した。
【0155】
pRK5Apo-2リガンド-mycプラスミドを、カルシウムリン酸沈降により、ネオマイシン耐性遺伝子を運ぶpRK5プラスミドと共にヒト293細胞(ATCCCRL1573)内にコトランスフェクトした。Apo-2リガンド-mycを発現する安定なクローンを抗生物質、G418(0.5mg/mL)(GIBCO)の存在下で50%HAM'sF12-50/50%DMEM(GIBCO)培地で成育する能力によって選択した。
【0156】
Apo-2リガンドのトポロジーを調べるために、G418耐性クローンを抗mycモノクローナル抗体(mAb)クローン9E10[Evan等,上記参照;Oncogene Scienceから商業的に入手可能]を用いて染色し、引き続きフィコエリスリン(PE)-接合ヤギ-マウス抗体(Jackson Immuno Researchから商業的に入手可能)を用いて染色した後、FACSにより分析した。FACS分析により偽のトランスフェクトされた細胞と比較して、特異的なポジティブ染色がApo-2リガンド-mycトランスフェクトクローンにおいてシフトすることが示され(図1C)、Apo-2リガンドはそのカルボキシ末端がさらされる形で細胞表面に発現されることが示された。したがってApo-2リガンドはII型膜貫通タンパク質であると考えられる。
【0157】
B. ECD融合構築物
2の可溶性Apo-2リガンド細胞外ドメイン(「ECD」)融合構築物を調製し、それにおいてもう一つの配列をApo-2リガンドのC末端領域の上流に融合した。
一つの構築において、ヘルペスウイルスグリコプロテインD(「gD」)シグナルペプチドの27アミノ酸[Lasky等,DNA,3:23-29(1984);Pennica等,Proc.Natl.Acad.Sci.,92:1142-1146(1995);Paborsky等,Protein Engineering,3:547-553(1990)に記述されている]及びエピトープタグ配列
[配列5]
をpRK5哺乳動物発現プラスミド内のApo-2リガンドのコドン114-281の上流に融合した。略記すると、gD配列を3'プライマーがApo-2リガンドのコドン114-121と同様にgD配列の3'領域に相補的であるPCRにおいて、元となるプラスミドpCHAD(Genentech,実質的にLasky等,Science,233:209-212(1986)に記述されているように調製した)から増幅した。該産物をpRK5Apo-2リガンドをテンプレートとして用いる連続的PCRにおけるApo-2リガンドコード領域の3'末端に相補的な3'プライマーと共に5'プライマーとして用いた。それからgD-Apo-2リガンドECD融合物をコードする該産物を、プラスミドpRK5gD-Apo-2リガンドECDを作製するためにpRK5プラスミド内にサブクローン化した。
【0158】
ヒト胎児腎293細胞(ATCC CRL 1573)をカルシウムリン酸沈降によってpRK5gD-Apo-2リガンドECDプラスミドまたはpRK5を用いて一過的にトランスフェクトした。可溶性gD-Apo-2リガンドタンパク質の発現を、35S-Cys及び35S-Metを用いたトランスフェクトされた細胞の代謝ラベルによって評価した。細胞上清を24時間後集め、遠心分離によってきれいにした。免疫沈降のため、5mlの上清を4℃でオーバーナイトで1μg/mlの濃度で5B6抗gDモノクローナル抗体(Genentech)と共にインキュベートした。それから25μlのPansorbin(Sigma)を4℃でさらに一時間加えた。熱したサンプルを再び回転させ、上清をSDS-PAGE及びオートラジオグラフィーにかけた。
【0159】
抗gD抗体を用いた免疫沈降により、gD-Apo-2リガンドプラスミドでトランスフェクトされた細胞の上清中に3の主要なバンドが現れた(図1E)。これらのバンドは23,48及び74kDaの相対的分子量(Mr)で移動した。成熟gD-Apo-2ポリペプチドの計算上の分子量は、およそ22.5kDaである;それゆえ観察されたバンドは、融合タンパク質の単量体(23kDa)、二量体(48kDa)、そして三量体(74kDa)の各形態を表し、Apo-2リガンドが哺乳動物細胞において分泌された可溶性gD融合タンパク質として発現され得ることを示すであろう。
【0160】
第二の構築において、12アミノ酸エンテロキナーゼ切断部位が引き続く、Met Gly His10配列(プラスミドpET19Bから由来する、Novagen)
[配列6]
をバキュロウイルス発現プラスミド(pVL1392,Pharmingen)内でApo-2リガンドのコドン114-281の上流に融合した。略記すると、Apo-2リガンドコドン114-281を、それぞれフランキングNdeIおよびBamHI制限部位を取り込んだ5'及び3'領域に相補的なプライマーを用いて、元となるpRK5Apo-2リガンドプラスミド(実施例1に記述されている)からPCRによって増幅した。該産物をT-AライゲーションによりpGEM-T(Rromega)内にサブクローン化し、DNA配列を確認した。それから挿入物をNdeIおよびBamHIを用いた切断により摘出し、アミノ末端Met Gly His10タグ及びエンテロキナーゼ切断部位を含む修飾バキュロウイルス発現ベクターpVL1392(Pharmingenから商業的に入手可能)内にサブクローン化した。
【0161】
組換えバキュロウイルスをリポフェクチン(GIBCO-BRLから商業的に入手可能)を用いて、Spodoptera frugiperda(「Sf9」)細胞(ATCC CRL 1711)内にHis10-Apo-2ECDプラスミドとBaculoGoldTMウイルスDNA(Pharmingen)をコトランスフェクトすることによって生産した。28℃でインキュベーションの4-5日後、放出されたウイルスを回収し、さらなる増幅に用いた。ウイルス感染とタンパク質発現を、O'Reilley等,Baculovirus expression vectors: A laboratory Manual,Oxford:Oxford University Press(1994)に記述されているように実施した。タンパク質を以下の実施例3に記述されているようにNi2+-キレートアフィニティークロマトグラフィーによって精製した。
【0162】
実施例3
組換えヒトApo-2リガンドの精製
Rupert等,Nature,362:175-179(1993)に記述されているように組換えウイルス感染及び偽の感染Sf9細胞(上記実施例2,セクションB参照)から抽出物を調製した。略記すると、Sf9細胞を洗浄し、ソニケーションバッファー(25mLHepes,pH7.9;12.5mMMgCl2;0.1mMEDTA;10%Glycerol;0.1%NP-40;0.4MKCl)に再懸濁し、そして氷上で20秒間2回ソニケートした。ソニケートされた物を遠心分離によりきれいにし、上清をローディングバッファー(50mMリン酸,300mMNaCl,10%Glycerol,pH7.8)で50倍に希釈し、0.45μmフィルターを通して濾過した。Ni2+-NTAアガロースカラム(Qiagenから商業的に入手可能)を5mLのベッド容量で調製し、25mLの水で洗浄し、25mLのローディングバッファーで平衡化した。濾過された細胞抽出物を分当たり0.5mLでカラムに乗せた。カラムをローディングバッファーを用いてベースラインA280に洗浄し、その時点で画分収集を開始した。次に、カラムを二次的洗浄バッファー(50mMリン酸;300mMNaCl,10%Glycerol,pH6.0)で洗浄し、それにより非特異的結合タンパク質が溶出した。再びA280ベースラインに到達した後、カラムを二次的洗浄バッファーにおける0から500mMのイミダゾール勾配にかけた。1mLの画分を回収し、SDS-PAHGE及び銀染色またはNi2+-NTA-接合アルカリホスファターゼ(Qiagen)を用いたウエスタンブロットによって分析した。溶出されたHis10-Apo-2リガンドタンパク質を含む画分をプールし、ローディングバッファーによって透析した。
【0163】
同一の方法をスタート物質として偽の感染Sf9細胞を用いて繰り返し、同じ画分をプールし、透析し、精製ヒトApo-2に対するコントロールとして用いた。
【0164】
精製タンパク質のSDS-PAGE分析により、分子量24kDaの主要なバンドが明らかになり、His10-Apo-2リガンド単量体に対する計算された分子量の22.4kDaに相当した(図1D,レーン3);タンパク質配列マイクロ分析(データは示していない)により、24kDaのバンドはHis10-Apo-2リガンドポリペプチドを表すことが確認された。少量の48kDa及び66kDaバンドもまた観察され、それらはおそらく可溶性Apo-2リガンドホモ二量体及びホモ三量体を表すであろう。スルホ-NHS(5mM)(Pierce Chemical)及び25mMと50mMでのEDC(Pierce Chemical)を用いたインキュベーションにより、精製His10-Apo-2リガンドの化学的架橋は(それぞれ図1D,レーン1と2)、タンパク質を主に66kDaバンドにシフトさせた。これらの結果により、溶液中のApo-2リガンドの主要な形態はホモ三量体であり、これらの三量体はSDSの存在下で二量体及び単量体に解離することが示唆される。
【0165】
実施例4
ヒトリンパ球細胞系に対するApo-2リガンドのアポトーシス活性
精製された可溶性Apo-2リガンド(実施例3に記述されている)のアポトーシス活性を、いくつかのヒトリンパ球細胞系を用いて調べた。第一の研究として、Epstein-Barrウイルス(EBV)トランスフォームヒト末梢血B細胞から由来する9D細胞(Genentech,Inc.)に対するApo-2リガンドの効果を調べた。9D細胞(10%胎児ウシ血清を加えたRPMI1640培地中に5×104細胞/ウェル)を、培地コントロール、Apo-2リガンド(3μg/ml、上述の実施例3に記述されているように調製された)、または抗Apo-1モノクローナル抗体、CH11(1μg/ml)のそれぞれで24時間インキュベートした[Yonehara等,J.Exp.Med.,169:1747-1756(1989)に記述されている;Medical and Biological Laboratories Co.から商業的に入手可能]。CH11抗Apo-1抗体は、fas/Apo-1リガンド活性をまねた作用剤抗体である。
【0166】
インキュベーション後、細胞をサイトスピンガラススライド上で回収し、倒立光学顕微鏡の下で写真を撮った。Apo-2リガンドと抗Apo-1モノクローナル抗体の両者は、同様のアポトーシス効果を誘導し、それは細胞質濃縮と細胞数の減少に特徴付けられた(図2A参照)。
【0167】
9D細胞に対するApo-2リガンドの効果と同様に、Raji細胞(ヒトバーキットリンパ腫B細胞系、ATCC CCL 86)及びJurkat細胞(ヒト急性T細胞白血病細胞系、ATCC TIB 152)に対する効果をさらにFACSによって分析した。該FACS分析は、アポトーシス化細胞死に対する確立された標準を用いて実施され、主に2のマーカー:(a) アポトーシス化細胞を染色するが生きた細胞はしないプロピジウムイオダイド(「PI」)色素、及び(b) アポトーシス化細胞の表面に見出されるさらされたホスファチジルセリンに結合するが、生きた細胞にはしないタンパク質の蛍光誘導体、アネキシンVを用いた細胞の蛍光染色に関するものであった[Darzynkiewicz等,Methods in Cell Biol.,41:15-38(1994);Fadok等,J.Immunol.,148:2207-2214(1992);Koopman等,Blood,84:1415-1420(1994)]。
【0168】
9D細胞(図2B)、Raji細胞(図2C)及びJurkat細胞(図2D)を、培地コントロール(左パネル)、Apo-2リガンド(3μg/ml、上述の実施例3に記述されているように調製された)(中央パネル)、または抗Apo-1モノクローナル抗体、CH11(1μg/ml)(右パネル)のそれぞれで24時間インキュベートした(1×106細胞/ウェル)。それから該細胞を洗浄し、PI及び蛍光チオシアネート(FITC)-接合アネキシンV(Brand Applicationsから購入)を用いて染色し、フローサイトメトリーで分析した。PIとアネキシンV染色(第3部分)の両者でネガティブな細胞は生きた細胞を表す;PI-ネガティブ、アネキシンV-ポジティブ染色細胞(第4部分)は早期アポトーシス化細胞を表す;PI-ポジティブ、アネキシンV-ネガティブ染色細胞(第2部分)は主にアポトーシスの後期段階の細胞を表す。
【0169】
Apo-2リガンドで処理された9D細胞は、上昇した細胞外アネキシンV結合を示し、同様に著しいPIの取り込みの増大を示し(図2B)、それはApo-2リガンドが細胞におけるアポトーシスを誘導したことを示す。抗Apo-1抗体、CH11(図2B)で比較可能な結果が得られた。Apo-2リガンドは抗Apo-1抗体がするように、Raji及びJurkat細胞において同様の応答を誘導した(図2C及び2D参照)。これらの細胞におけるコントロール及び抗Apo-1抗体と比較したApo-2リガンドによるアポトーシスの誘導(アポトーシス化細胞%ととして測定される)が、以下の表1にも示されている。
【0170】
Apo-2リガンドによる染色体内DNA断片化の活性化もまた分析した。Jurket細胞(左レーン)及び9D細胞(右レーン)を培地コントロールまたはApo-2リガンド(3μg/ml、実施例3に記述されているように調製した)と共に6時間インキュベートした(2×106細胞/ウェル)。それからDNAを細胞から抽出し、ターミナルトランスフェラーゼを用いて32P-ddATPを用いてラベルした。ラベル化DNAサンプルを2%アガロースゲルで電気泳動にかけ、その後オートラジオグラフィーにより分析した[Moore等,Cytotechnology,17:1-11(1995)]。Apo-2リガンドはJurkat細胞及び9D細胞の両者で染色体内DNA断片化を誘導した(図2E)。該DNA断片化はアポトーシスの特徴である[Cohen,Advances in Immunol.,50:55-85(1991)]。
【0171】
Apo-2リガンドアポトーシス活性のタイムコースを調べるために、9D細胞を0時間から50時間の範囲の期間、培地コントロールまたはApo-2リガンド(3μg/ml、実施例3に記述されているように調製される)を用いてミクロタイターディッシュ(5×104細胞/ウェル)でインキュベートした。インキュベーションに引き続き、死んだ細胞及び生きた細胞の数を血球計算板を用いて顕微鏡観察の下で測定した。
【0172】
図3Aに示されているように、細胞死の最大のレベルは24時間以内で9D細胞で誘導された。
【0173】
Apo-2リガンド誘導細胞死の量依存性を調べるために、9D細胞を培地コントロールまたはApo-2リガンド(実施例3に記述されているように調製された)の連続的な希釈物を用いて24時間でインキュベートした(5×104細胞/ウェル)。インキュベーションに引き続き死んだ細胞及び生きた細胞の数を上述のように測定した。その結果が図3Bに示されている。特異的アポトーシスはApo-2リガンド処理細胞におけるアポトーシス%からコントロールにおけるアポトーシス%を差し引くことによって計算された。アポトーシスの半最大活性化はおよそ0.1μg/ml(およそ1nM)で生じ、最大の誘導はおよそ1からおよそ3μg/ml(およそ10から30nM)で生じた。
【0174】
実施例5
ヒト非リンパ球腫瘍細胞系に対するApo-2リガンドのアポトーシス活性
ヒト非リンパ球腫瘍細胞系に対するApo-2リガンドの効果を、以下の細胞系を用いて調べた:HeLa(ヒト子宮頚ガン腫から由来する、ATCC CCL 22);ME-180(ヒト子宮頚ガン腫から由来する、ATCC HTB 33);MCF7(ヒト乳ガン腫から由来する、ATCC HTB 22);U-937(ヒト細網肉腫から由来する、ATCC CRL 1593);A549(ヒト肺ガン腫から由来する、ATCC CCL 185);及び293(アデノウイルストランスフォームヒト胎児腎細胞から由来する、ATCC CCL 1573)。該アッセイにおいては、それぞれの細胞系の1×106細胞を、培地コントロール、Apo-2リガンド(3μg/ml、実施例3に記述されているように調製した)またはApo-1モノクローナル抗体、CH11(1μg/ml)を用いて24時間でインキュベートした。該インキュベーションに引き続き、アポトーシスを実施例4に記述されているFACS分析によって測定した。その結果が以下の表1に示されている。
【0175】
【表1】
【0176】
HeLa細胞及びMCF7細胞は、CH11抗Apo-1抗体と比較してApo-2リガンドによってアポトーシスの誘導に等しく感受性であった。対照的にU-937細胞及びA549細胞は著しくApo-2リガンドによるアポトーシスの誘導により感受性であった。ME-180細胞はApo-2リガンドに対して大変感受性であるが、抗Apo-1抗体には比較的耐性であった。293細胞はApo-2リガンドに対しては耐性であり、抗Apo-1抗体に対しては弱く感受性であった。
【0177】
それゆえ、Apo-2リガンドはリンパ球起源の細胞と同様に、非リンパ球起源の細胞においてもアポトーシスを誘導することができる(実施例4参照)。また、十分に理解されておらず、いかなる特定の理論にも結び付けられることを期待しないが、出願人はApo-2リガンドがApo-1とは区別される受容体を介して機能すると現在考えている。この考えは、上述の細胞系がApo-2リガンドと抗Apo-1抗体に対する異なる感受性のパターンを示すということを表すここでのデータによって支持される。
【0178】
実施例6
ヒト末梢血単球に対するApo-2リガンドの効果
末梢血単核細胞(「PBMC」)を、Lymphocyte Separation Medium(LSM(登録商標)、Organon Teknika)を用いたFicoll密度勾配遠心分離によってヒトドナーの血液から単離した。単離されたT細胞の集団を、抗Igカラムに結合した表面Igを通してB細胞を除去し、Igカラムに結合しているFc受容体を通して単球を除去することによって、PBMCから調製した(R & D Systems)。単離されたB細胞の集団を、OKT3ミエローマ(ATCC CRL 8001)によって生産された抗cD3抗体と反応したT細胞の、及び4F2C13ハイブリドーマ(ATCC HB22)によって生産された単球特異的抗体と反応した単球の、それぞれ相補体介在性除去によりPBMCから調製した。さらなる単球の除去はプラスチックへの接着によって成し遂げられた。
【0179】
新鮮に単離された末梢血BまたはT細胞(1×106細胞/ウェル)を、培地コントロールまたはApo-2リガンド(3μg/ml、実施例3に記述されるように調製した)の存在下で3日間培養した。活性化のため、B細胞をリポ多糖(「LPS」、1μg/ml)で同時に処理し、T細胞をホルボールミリステートアセテート(「PMA」、10ng/ml)プラスイオノマイシン(1μg/ml)(Sigma)を用いて処理した。インターロイキン-2(「IL-2」)前処理のため、T細胞をApo-2リガンドにさらす前、IL-2(50U/ml)(Genzyme)の存在下で3-5日間培養した。アポトーシスを本質的に実施例4で上述されているようにFACS分析を用いて測定した。しかしながらB細胞は抗CD19/CD20抗体(Jackson Immunoresearch)によって捕らえられ、T細胞は抗CD4/CD8抗体(Jackson Immunoresearch)によって捕らえられた。その結果が以下の表2に示されており、それは独立の実験[Bリンパ球-9実験;Tリンパ球-8実験;Tリンパ球プラスIL-2-5実験]の平均±SEを表し、そこにおいては50,000細胞がデータポイント当たり分析された。統計的な分析はt検定を用いて実施した。表2において、それぞれのコントロールに対してa=p<0.05及びb=p<0.02である。
【0180】
【表2】
【0181】
Apo-2リガンドは非刺激B細胞、LPSにより活性化されたB細胞及びPMAとイオノマイシンを用いて活性化されたT細胞において有意にアポトーシスを誘導した。末梢T細胞はIL-2の存在下で細胞を培養することによってアポトーシスにかかりやすくなることができるということが以前に報告された[Lenardo等,Nature,353:858-861(1991)]。本研究はIL-2を用いた前処理がApo-2リガンド誘導死に対して末梢T細胞を感受性にすることを示した。
【0182】
実施例7
Fas/Apo-1及びTNF受容体を用いた阻害アッセイ
fas/Apo-1受容体、同様に1型及び2型TNF受容体(TNF-R1及びTNF-R2)の可溶性形態が精製された可溶性Apo-2リガンド(実施例3に記述されている)のアポトーシス活性を阻害できるかどうかを試験することによって、これらの受容体がApo-2リガンドのアポトーシス活性を介在することに関与しているかどうかを測定するためのアッセイを実施した。
【0183】
9D細胞をバッファーコントロール、CD4-IgGコントロール(25μg/ml)、可溶性Apo-1-IgG(25μg/ml)、可溶性TNFR1-IgG(25μg/ml)または可溶性TNFR2-IgG融合タンパク質(25μg/ml)の存在下で、培地コントロールまたはApo-2リガンド(0.3μg/ml、実施例3に記述されているように調製した)と共に24時間インキュベートした(5×104細胞/ウェル)。Fas/Apo-1、TNF-R1及びTNF-R2受容体の可溶性誘導体を、Ashkenazi等,Methods,8:104-115(1995)に記述されているようにIgG融合タンパク質として生産した。CD4-IgGを、Byrn等,Nature,344:667-670(1990)に記述されているようにIgG融合タンパク質として生産し、コントロールとして用いた。
【0184】
図3Cに示されているように、受容体-融合分子の全てが、9D細胞に対するApo-2リガンドアポトーシス活性を阻害しなかった。これらの結果から、Apo-2リガンドアポトーシス活性はfas/Apo-1及びTNF-R1とTNF-R2とは独立であることが示された。
【0185】
実施例8
哺乳動物組織におけるApo-2リガンドmRNAの発現
ヒト組織におけるApo-2リガンドmRNAの発現を、ノーザンブロット分析により調べた(図4)。ヒトRNAブロットはフルレングスApo-2リガンドcDNAに基づく32PラベルDNAプローブとハイブリダイズし、またはGenBank EST配列、HMEA47Mに基づく32PラベルRNAプローブとハイブリダイズした(実施例1参照)。ヒト胎児RNAブロットMTN(Clontech)及びヒト成人RNAブロットMTN-II(Clontech)を、DNAプローブとインキュベートし、一方でヒト成人RNAブロットMTN-I(Clontech)をRNAプローブとインキュベートした。ブロットをハイブリダイゼーションバッファー(5×SSPE;2×デンハルト溶液;100mg/mL変性切断サケ精子DNA;50%ホルムアミド;2%SDS)において42℃で16時間プローブとインキュベートした。該ブロットを、1×SSPEで数回;2%SDSで65℃で1時間及び50%新鮮な脱イオンホルムアミド;1×SSPE;0.2%SDSで65℃で30分洗浄した。該ブロットをホスホリムアガー(phosphorimager)(Fuji)を用いてオーバーナイトでさらした後解析した。
その結果が図4に示されている。胎児ヒト組織においては、Apo-2リガンドmRNA発現は肺、肝臓及び腎臓で検出され、脳組織ではされなかった。成人ヒト組織においては、Apo-2リガンドmRNA発現は脾臓、胸腺、前立腺、卵巣、小腸、末梢血リンパ球、心臓、胎盤、肺、腎臓で検出された。精巣、脳、骨格筋、及び膵臓ではほとんどまたは全く発現が検出されなかった。上述されているように、Apo-2リガンドに対して観察された発現プロフィールは、Apo-1リガンドのものとは同一ではなく、Apo-1リガンドは主にT細胞及び精巣で発現されている[Nagata等,上記参照]。
【0186】
実施例9
ヒト腫瘍細胞系に対するApo-2リガンドのアポトーシス活性
ヒト腫瘍細胞系に対するApo-2リガンド(実施例3に記述)のアポトーシス活性を、いくつかの化学療法試薬の一つの存在下または不存在下でさらに調べた。
以下のヒト腫瘍細胞系をアッセイした;A549(肺ガン腫、ATCC CCL 185);HCT116(大腸ガン腫、ATCC CCL 247);SW480(大腸腺ガン腫、ATCC CCL 228);MDA231(乳腺ガン腫、ATCC HTB 26);HeLa(子宮頚ガン腫、ATCC CCL 22);ME-180(子宮頚ガン腫,ATCC HTB 33);T24(膀胱ガン腫、ATCC HTB 4);SK-N-AS(神経芽腫、White等,Proc.Natl.Acad.Sci.,92:5520-5524(1995))。これらの細胞系のいくつかは、野生型p53を発現し、一方で他のものはミューテーションのため発現しておらず、それについては以下の表3を参照。該細胞を96穴プレートに2.5×105細胞/mlでまき、オーバーナイトでインキュベートした。該細胞をApo-2リガンドの2倍希釈物(100ng/mlから0.01ng/mlの範囲)の存在下で培養した。カルチャーのあるものには、化学療法試薬もまた24時間で加えた;シクロヘキサミド(「CHX」)(50μg/ml;Sigma Chemicals),ドキソルビシン(10-100μg/ml;Pharmacia)または5-FU(6mg/ml;Roche)。
【0187】
インキュベーションの24時間後、該細胞を20%メタノール中の0.5%クリスタルバイオレットで染色した。細胞生存力を0.1Mクエン酸ナトリウム(50%メタロール中に0.1Mクエン酸)を用いて染色細胞から色素を除去し、540nmの吸光度を測定することによって測定した。
その結果が以下の表に示されている。
【0188】
【表3】
+: 100ng/mlApo-2Lを用いた24時間後>35%が死
++: 100ng/mlApo-2Lを用いた24時間後>70%が死
+++: 10ng/mlApo-2Lを用いた24時間後>70%が死
これらの結果はApo-2リガンドが様々な腫瘍タイプから由来する腫瘍細胞系において細胞死を誘導し、Apo-2Lが腫瘍細胞のp53状態に独立して細胞死を誘導したことを示す。これらの結果はまたApo-2リガンド誘導細胞死がいくつかの異なる化学療法薬剤によって増加されることをも示す。
【0189】
実施例10
in vivoでのApo-2リガンドのアポトーシス活性
腫瘍を持つヌードマウスにおけるApo-2リガンドの効果を調べた。ヌードマウス(グループ当たり5-10マウス)(Harlan Sprague Dawleyから購入した)を、MDA231ヒト乳ガン腫細胞(ATCC HTB 23)(2×106細胞/マウス)で皮下に注射した(0日目)。それかが該腫瘍を14日間成長させた。14及び15日目に、2μg/0.05ml/マウスのApo-2リガンド(実施例3)及び/または10μg/0.05ml/マウスのドキソルビシン(Pharmacia)を、腫瘍部位に注射した。コントロール動物には0.05mlのPBSを同様に注射した。21日目に、該動物を殺し、腫瘍を調べ測量した(g)。
その結果が図5に示されている。該データにより、Apo-2リガンド処理はそれ自体で腫瘍成長を阻害し、そしてApo-2リガンドは腫瘍成長に対するドキスルビシンの阻害効果を増大することが示される。
【0190】
実施例11
in vivoでのApo-2リガンドのアポトーシス活性
Apo-2リガンドの抗腫瘍効果を、0日目にマウスをHCT116ヒト大腸ガン腫細胞(ATCC CCL 247)(2×106細胞/マウス)を用いて皮下に注射することを除いて、実施例10に記述されているように、腫瘍を持つヌードマウスで調べた。それから該腫瘍を14日間成長させた。14及び15日目に、2μg/0.05ml/マウスのApo-2リガンド及び/または10μg/0.05ml/マウスの5-FU(Roche)を、腫瘍部位に注射した。コントロール動物には0.05mlのPBSを同様に注射した。21日目に、該動物を殺し、腫瘍を調べ測量した(g)。
その結果が図6に示されている。これらの結果により、Apo-2リガンド処理はそれ自体で腫瘍成長を阻害し、そしてApo-2リガンドは腫瘍成長に対する5-FUの阻害効果を増大することが示される。
【0191】
実施例12
in vivoでのApo-2リガンドのアポトーシス活性
Apo-2リガンドの抗腫瘍効果を、1及び2日目に10μg/0.05ml/マウスのApo-2リガンド及び/または100μg/0.05ml/マウスの5-FU(Roche)を、腹膜内に注射することを除いて、実施例11に記述されているように、腫瘍を持つヌードマウスで調べた。コントロール動物には0.05mlのPBSを同様に注射した。それから腫瘍サイズ(mm2)を5,9,15日目に測定した。15日目に、該動物を殺し、腫瘍を調べ測量した(g)。
その結果が図7及び8に示されている。これらの結果により、Apo-2リガンドは腹膜内注射によって投与された場合でさえ、皮下腫瘍部位に到達し、抗腫瘍効果を発揮することができることが示される。また、これらの結果により、Apo-2リガンド処理はそれ自体で腫瘍成長を阻害し、そしてApo-2リガンドは腫瘍成長に対する5-FUの阻害効果を増大することが示される。
【0192】
実施例13
CrmAを用いた阻害アッセイ
ICE及びCPP32/YamaのようなプロテアーゼがApo-2リガンドによるアポトーシス誘導において役割を果たしているかどうかを調べるために、CrmAがApo-2リガンド誘導アポトーシスをブロックするかどうかを調べるためのアッセイを実施した[Marsters等,Current Biology,6:750-752(1996)]。CrmAは死プロテアーゼICE及びCPP/Yamaのポックスウイルス由来阻害剤であり、TNFR1及びFas/Apo-1による死シグナルをブロックする。加えて、Apo-1リガンド及びTNFによるアポトーシス誘導を介在するアダプタータンパク質、FADDを含む「死ドメイン」が、Apo-2リガンド誘導アポトーシスに関与するかどうかを調べるために[Chinnaiyan等,Cell,81:505-512(1995);Hsu等,Cell,84:299-308(1996)]、FADDのドミナントネガティブミュータント形態(FADD-DN)[Hsu等joukisannshou]が、Apo-2リガンド機能を阻害するかどうか[Marsters等,Current Biology,6:750-752(1996)]を調べるためのアッセイを実施した。
【0193】
HeLa-S3(ATCC CCL 22)細胞を、pRK5-CrmA発現プラスミド(CrmA配列はRay等,上記参照で報告された)またはpRK5-FADD-DN発現プラスミド(Hsu等,Cell,84:299-308(1996))を用いてトランスフェクトした。pRK5をコントロールとして用いた。該細胞をプラスミドDNAの取り込みに対するマーカーとしてpRK5-CD4(Smith等,Science,328:1704-1707(1988))と共にトランスフェクトした。トランスフェクトされた細胞をフィコエリスリン接合CD4抗体(Jackson Immunoresearch)を用いた染色によって同定し、アポトーシスを本質的に上記実施例4に記述されているようにFACSによって分析した。
【0194】
その結果が図9に示されている。CrmAは、抗Apo-1抗体により誘導されるアポトーシスと同様に、Apo-2リガンド誘導アポトーシスをブロックした。対照的に、FADD-DNは、Apo-2リガンド誘導アポトーシスに対してほとんど効果がないが、抗Apo-1抗体によるアポトーシス誘導を実質的にブロックした。したがって該アッセイの結果により、Apo-2リガンド、TNFR1及びFas/Apo-1は、アポトーシス性細胞死を活性化するための共通の末端シグナル経路を用いていることが示唆される。特に該結果は、ICE及びCPP/YamaのようなプロテアーゼがApo-2リガンド誘導アポトーシスに対して必要とされることを示唆する。対照的に、FADDはTNFR1及びFas/Apo-1による細胞死誘導に必要とされるが、Apo-2リガンドによるものには必要とされない。
【0195】
実施例14
A. 抗Apo-2リガンド抗体の調製
Balb/cマウス(Chales River Laboratoriesから得た)を、1μgApo-2リガンド(実施例3に記述されているように調製し、Ribi Immunochemical Research Inc.,ハミルトン、MTから購入したMPL-TDMアジュバントで希釈した)を1週間間隔で各後脚肉肢に10回注射することによって免疫化した。最終の追加免疫注射の3日後、マウスから膝窩リンパ節を取り出し、1%ペニシリン-ストレプトマイシンを補ったDMEM培地(Biowhitakker Corp.から得た)に単一細胞懸濁液を調製した。それから35%ポリエチレングリコール[Laskor等,Cell.Immunol.,55:251(1980)]を用いて、ネズミミエローマ細胞P3X63AgU.1(ATCC CRL 1597)とリンパ節細胞を融合し、96穴カルチャープレートで培養した。融合から由来するハイブリドーマをHAT培地で選択した。融合の10日後、c-Met-IgG融合タンパク質に結合するモノクローナル抗体の存在を試験するため、ハイブリドーマカルチャー上清をELISA[Kim等,J.Immunol.Meth.,156:9-17(1992)]でスクリーニングした。
【0196】
ELISAでは、PBS中の0.5μg/mlApo-2リガンド(実施例3参照)の50μlを各ウェルに加えることによって、96穴ミクロタイタープレートをコートし、4℃オーバーナイトでインキュベートした。それからプレートを洗浄バッファー(PBSプラス0.05%Tween20)で3回洗った。それから2%ウシ血清アルブミン(BSA)の200μlを用いて、ミクロタイター内のウェルをブロックし、1時間室温でインキュベートした。それからプレートを再び洗浄バッファーで3回洗った。
【0197】
洗浄ステップの後、2μg/mlのApo-2リガンド抗体の50μl、またはハイブリドーマカルチャー上清の100μlを各ウェルに加えた。培地に調和したP3X63AgU.1ミエローマ細胞の100μlをコントロールとして別の加工されたウェルに加えた。シェーカー装置で1時間室温でプレートをインキュベートし、それから洗浄バッファーで3回洗った。
【0198】
次に、アッセイバッファー(PBS中に0.5%ウシ血清アルブミン,0.05%Tween-20,0.01%Thimersol)で1:1000に希釈した50μlHRP結合ヤギ抗マウスIgG(Cappel Laboratoriesから購入した)を各ウェルに加え、シェーカー装置で室温で1時間プレートをインキュベートした。プレートを洗浄バッファーで3回洗い、引き続き基質(TMB,3,3',5,5'-テトラメチルベンジジンKirkegaard & Perry,Gaitherburg,MDから得た)の50μlを各ウェルに加え、10分間室温でインキュベートした。停止溶液(Kirkegaard & Perry)の50μlを各ウェルに加えることにより反応を止め、自動ミクロタイタープレートリーダーで450nmの吸光度を読み取った。
【0199】
ハイブリドーマ上清(選択された99)をApo-2リガンド誘導9D細胞殺傷をブロックする能力について試験した。活性は初めに、トリパンブルー色素溶出を用いて、処理された9D細胞の生存力%を調べることによって測定された。
【0200】
ブロッキング活性もまたFACS分析によって確認した。9D細胞(5×105細胞/0.5ml)を完全RPMI培地(RPMIプラス10%FCS、グルタミン、非必須アミノ酸、ペニシリン、ストレプトマイシン、ピルビン酸ナトリウム)に懸濁し、24穴マクロタイタープレート内にまいた。0.5mlのApo-2リガンド(1μg/ml)(実施例3に記述されているように調製した)を、完全RPMI培地内に懸濁し、10μgの精製モノクローナル抗体、または100μlの培養上清を用いてプレインキュベートし、それから9D細胞含有24マクロタイターウェル内に加えた。マクロタイタープレートを7%のCO2の存在下で37℃でオーバーナイトでインキュベートした。それからインキュベートされた細胞を回収し、PBSで一度洗浄した。細胞の生存力を製品の但し書き(Clontech)にしたがって、ホスファチジルセリンを結合したFITC-アネキシンVの染色により測定した。該細胞をPBSで洗浄し、200μlの結合バッファーに再懸濁した。10μlのアネキシンV-FITC(1μg/ml)及び10μlのプロピジウムイオダイドを該細胞に加えた。暗所で15分名インキュベーション後、該9d細胞をFACSにより分析した。
【0201】
8の潜在的なブロッキング抗体及び4の潜在的な非ブロッキング抗体を分泌する各ハイブリドーマを同定し、さらに限界希釈法によりクローン化(2回)した。
【0202】
モノクローナル抗体1D1,2G6,2E11及び5C2といわれる4の抗体のFACS分析が図10に示されている。(以下に示されるように、1D1,2G6,2E11及び5C2抗体はそれぞれハイブリドーマ1D1.12.4,2G6.3.4,2E11.5.5及び5C2.4.9によって生産され、それら全てがATCCに寄託されている)。Apo-2リガンドで処理された9D細胞(上部左図)は、非処理コントロール細胞(上部右図)より50%高いアポトーシス化細胞を示した。Apo-2リガンドプラス2E11,5C2,2G6または1D1抗体で処理された9D細胞は、非処理コントロールよりそれぞれ0%,6%,26&及び48%高いアポトーシス化細胞を示した。これらの結果は、5C2,2E11及び2G6抗体がブロッキング抗体である一方、1D1抗体は非ブロッキング抗体であることを示す。最も能力のあるブロッキング活性は、5C2抗体で観察された。
【0203】
4の抗体の抗原特異性もまたELISAで試験した。マイクロタイターウェルを2μg/mlリンホトキシン(Genentech,Inc.,EP 164,965、Gray等,Nature,312:721-724(1984)も参照)、TNF-アルファ(Genentech,Inc.,Pewnnica等,Nature,312:724-729(1984);Aggarwal等,J.Biol.Chem.,260:2345-2354(1985)も参照)、またはApo-2リガンド(実施例3参照)でコートした。モノクローナル抗体1D1,2G6,2E11及び5C2を10μg/mlの濃度で試験した。
【0204】
該アッセイの結果が図11に示されている。図11のデータは、モノクローナル抗体2G6,2E11及び5C2がApo-2リガンドに対して特異的である一方、モノクローナル抗体1D1はリンホトキシン及びTNF-アルファに対して弱い交差結合を示すことを表す。
【0205】
B. アイソタイピング
1D1,2G6,2E11及び5C2抗体(上記セクションAに記述されている)のアイソタイプを、アイソタイプ特異的ヤギ抗マウスIg(FisherBiotech,Pittsburgh,PA)でミクロタイタープレートを4℃でオーバーナイトでコートすることによって測定した。それから該プレートを上述の洗浄バッファーで洗浄した。それからミクロタイターのウェルを200μlの2%ウシ血清アルブミンでブロックし、室温で1時間インキュベートした。該プレートを洗浄バッファーで再び3回洗浄した。
【0206】
次に100μlの5μg/mlの精製Apo-2リガンド抗体、または100μlのハイブリドーマカルチャー上清を加工されたウェルに加えた。該プレートを室温で30分インキュベートし、それから50μlのHRP-接合ヤギ抗マウスIgG(上述のように)を各ウェルに加えた。該プレートを室温で30分インキュベートした。プレートに結合したHRPのレベルを上述のHRP基質を用いて検出した。
【0207】
アイソタイピング分析により、1D1及び2G6抗体はIgG2b抗体であり、そして2E11及び5C2抗体はIgG2a抗体であることが示された。
【0208】
C. エピトープマッピング
エピトープマッピングをビオチン化モノクローナル抗体を用いて、Kim等,上記参照に記述される競合的結合ELISAを用いて実施した。選択されたモノクローナル抗体を、Antibodies,A laboratory Manual,E.Harlow及びD.Lane,編、p.342に記述されているN-ヒドロキシルスクシンイミドを用いてビオチン化した。ミクロタイタープレートウェルを50μlのApo-2リガンド(実施例3参照、0.1μg/ml)でコートし、4℃でオーバーナイトで保存し、それから2%BSAで室温で1時間ブロックした。ミクロタイターウェルの洗浄後、所定最適化濃度のビオチン化抗体及び1000倍過剰の非ラベル抗体の混合物を、各ウェルに加えた。室温で1時間のインキュベーション後、該プレートを洗浄し、ビオチン化抗体の量をHRP-ストレプタビジンの添加によって検出し、そして該プレートをELISAプレートリーダーで490nmで読んだ。
【0209】
その結果が図12に示されている。該結果は、HRP接合抗体の結合がその抗体自身の過剰の量で効果的に阻害されるが、アッセイされた他の抗体では阻害されないことを示した。
【0210】
モノクローナル抗体により認識されるApo-2リガンドの領域を、Chuntharapai等,J.Immunol.,152:1783-1789(1994)に記述されているELISAで、合成ペプチド[図1Aに示されるApo-2リガンド配列のaa128-148(ペプチド「APO14」);aa144-159(ペプチド「APO15」);aa192-204(ペプチド「APO17」);aa230-238(ペプチド「APO18」);aa261-272(ペプチド「APO19」)]を用いて測定した。該結果が図13に示されている。1D1抗体はApo-2リガンドのアミノ酸残基192-204を含むAPO17ペプチドに結合することを示した。
【0211】
物質の寄託
以下の細胞系はAmerican Type Vulture Collection, 12301 Parklawn Drive,Rockville,MD,USA(ATCC)に寄託されている:
細胞系 ATCC寄託番号 寄託日
2935-pRK5-hApo-2L- CRL-12014 1996年1月3日
mycクローン2.1
1D1.12.4 HB-12557 1997年1月8日
2G6.3.4 HB-12559 1997年1月8日
2E11.5.5 HB-12556 1997年1月8日
5C2.4.9 HB-12558 1997年1月8日
【0212】
特許手続上の微生物の寄託の国際的承認に関するブタペスト条約の規定に基づいて寄託がなされた。これは寄託の日から30年間、寄託物の生存能力のあるカルチャーの管理を保証する。該寄託物はブタペスト条約の定める期間ATCCから入手可能であり、Genentech,Inc.とATCCの間の合意により、関係のある米国特許の発行か、米国または外国の特許出願の公開かの、いずれか早い時点で公衆に対して寄託物のカルチャーの子孫の永久に制限のない入手可能性が保証され、かつ米国特許法第122条ならびにそれに従う長官規則(特に886OG638に関係する37CFR1.14を含む)に従って米国特許商標長官によって権利を与えられることが決定された公衆に対する子孫の入手可能性が保障される。
【0213】
本出願の譲受人は、もし寄託した細胞系のカルチャーが適したコンディションの下で培養されながら死んだまたは失われたまたは破壊された場合、該細胞系を同じプラスミドのもう一つと共に告示して直ちに置き換えることを合意した。寄託された細胞系の利用可能性は、特許法にしたがっていかなる政府の権威に基づいて許可された権利に違反して、自由に本発明を実施できるというように解釈されない。
【0214】
先の文書の明細書が当業者をして本発明を実施せしめるのに十分であると考えられる。寄託された実施態様は本発明の特定の一面の単なる例証としてのつもりであり、機能的に均質であるいかなるカルチャーも本発明の範囲内にあるので、本発明は寄託されたカルチャーによって範囲を限定されることはない。ここで物質の寄託は、ここに含まれる記述されたものが、その最適のモードを含む本発明の任意の面の実施可能性に対して不十分であることの自認を構成するものではなく、存在する特別に説明された請求の範囲が表された特定の例に限定されるとみなされるものでもない。実際ここで示され記述されたものに加えて、本発明の様々な修飾が以前の記述から当業者に明らかになるであろうし、それらは付記される請求項の範囲内に存在する。
【図面の簡単な説明】
【0215】
【図1A】図1AはヒトApo-2リガンドcDNAの核酸配列及びそのコードするアミノ酸配列を示す。
【図1B】図1BはヒトTNFサイトカインファミリーの周知のメンバー、4-1BBL,OX40L,CD27L,CD30L,TNF-α,LT-β,LT-α,CD40L及びApo-1Lの相当する領域と共に、ヒトApo-2リガンドのC末端領域の配列を示す。
【図1C】図1Cは抗mycエピトープ抗体を用いるFACS分析によって測定された、ヒト293細胞で発現された組換えフルレングスC末端mycエピトープタグ化Apo-2リガンドの細胞トポロジーを示す。
【図1D】図1DはSDS-PAGE及び銀染色に引き続き、化学架橋の存在下(レーン2,3)または不存在下(レーン1)で測定された、組換えバキュロウイルス感染昆虫細胞で発現され、Ni2+キレートアフィニティークロマトグラフィーで精製された組換えHis10エピトープタグ化可溶性Apo-2のサイズ及びサブユニット構造を示す。
【図1E】図1EはSDS-PAGE及びオートラジオグラフィーに引き続き、抗gDエピトープ抗体を用いた免疫沈降によって測定された、代謝的にラベルされたヒト293細胞で発現された組換えgDエピトープタグ化可溶性Apo-2リガンドのサイズ及びサブユニット構造を示す。
【図2A】図2A-2EはApo-2リガンドによるB及びTリンパ球細胞系におけるアポトーシスの誘導を示す。アポトーシス化細胞は(A) 特徴的な形態変化によって;(B-D) フローサイトメトリーで測定された、プロピジウムイオダイド(PI)及びFITC接合アネキシンVを用いたポジティブ蛍光染色によって;及び(E) 染色体内DNA断片化の分析によって同定された。
【図2B】図2A-2EはApo-2リガンドによるB及びTリンパ球細胞系におけるアポトーシスの誘導を示す。アポトーシス化細胞は(A) 特徴的な形態変化によって;(B-D) フローサイトメトリーで測定された、プロピジウムイオダイド(PI)及びFITC接合アネキシンVを用いたポジティブ蛍光染色によって;及び(E) 染色体内DNA断片化の分析によって同定された。
【図2C】図2A-2EはApo-2リガンドによるB及びTリンパ球細胞系におけるアポトーシスの誘導を示す。アポトーシス化細胞は(A) 特徴的な形態変化によって;(B-D) フローサイトメトリーで測定された、プロピジウムイオダイド(PI)及びFITC接合アネキシンVを用いたポジティブ蛍光染色によって;及び(E) 染色体内DNA断片化の分析によって同定された。
【図2D】図2A-2EはApo-2リガンドによるB及びTリンパ球細胞系におけるアポトーシスの誘導を示す。アポトーシス化細胞は(A) 特徴的な形態変化によって;(B-D) フローサイトメトリーで測定された、プロピジウムイオダイド(PI)及びFITC接合アネキシンVを用いたポジティブ蛍光染色によって;及び(E) 染色体内DNA断片化の分析によって同定された。
【図2E】図2A-2EはApo-2リガンドによるB及びTリンパ球細胞系におけるアポトーシスの誘導を示す。アポトーシス化細胞は(A) 特徴的な形態変化によって;(B-D) フローサイトメトリーで測定された、プロピジウムイオダイド(PI)及びFITC接合アネキシンVを用いたポジティブ蛍光染色によって;及び(E) 染色体内DNA断片化の分析によって同定された。
【図3A】図3A-3CはApo-2リガンド誘発アポトーシスのタイムコースと量依存性、及びFas/Apo-1受容体、TNF-R1受容体またはTNF-R2受容体に基づく可溶性受容体IgG融合タンパク質による、Apo-2リガンド誘発アポトーシスの阻害の欠如を示す。
【図3B】図3A-3CはApo-2リガンド誘発アポトーシスのタイムコースと量依存性、及びFas/Apo-1受容体、TNF-R1受容体またはTNF-R2受容体に基づく可溶性受容体IgG融合タンパク質による、Apo-2リガンド誘発アポトーシスの阻害の欠如を示す。
【図3C】図3A-3CはApo-2リガンド誘発アポトーシスのタイムコースと量依存性、及びFas/Apo-1受容体、TNF-R1受容体またはTNF-R2受容体に基づく可溶性受容体IgG融合タンパク質による、Apo-2リガンド誘発アポトーシスの阻害の欠如を示す。
【図4】図4はノーザンブロット分析で測定された、ヒト胎児及びヒト成人組織におけるApo-2リガンドmRNAの発現を示す。
【図5】図5はヌードマウスにおいて成長したヒトMDA231乳ガンベース腫瘍の重量に対する、単独のまたはドキソルビシンと組み合わせた腫瘍内注射によって投与されたApo-2リガンドのin vivoでの効果を示す。
【図6】図6はヌードマウスにおいて成長したヒトHCT116大腸ガンベース腫瘍の重量に対する、単独のまたは5-FUと組み合わせた腫瘍内注射によって投与されたApo-2リガンドのin vivoでの効果を示す。
【図7】図7はヌードマウスにおいて成長したヒトHCT116大腸ガンベース腫瘍のサイズに対する、単独のまたは5-FUと組み合わせた腹膜内注射によって投与されたApo-2リガンドのin vivoでの効果を示す。
【図8】図8はヌードマウスにおいて成長したヒトHCT116大腸ガンベース腫瘍の重量に対する、単独のまたは5-FUと組み合わせた腹膜内注射によって投与されたApo-2リガンドのin vivoでの効果を示す。
【図9】図9はドミナントネガティブなFADDではないCrmAがHeLa-S3細胞においてApo-2リガンド誘発アポトーシスをブロックすることを示す棒グラフである。
【図10】図10はApo-2リガンドにより誘発されるアポトーシスのFACS分析及び4の抗Apo-2リガンド抗体:1D1,2G6,2E11及び5C2の効果を示す(FITC接合アネキシンVを用いて検出されたアポトーシス化9D細胞−太線;生きた非染色細胞-細線)。
【図11】図11はモノクローナル抗体1D1,2G6,2E11及び5C2の抗原特異性を示す棒グラフである。
【図12】図12はモノクローナル抗体1D1,2G6,2E11及び5C2のエピトープマッピングアッセイの結果を示す棒グラフである。
【図13】図13はApo-2リガンドの特異的アミノ酸領域より成るいくつかの異なる合成ペプチドに結合するモノクローナル抗体1D1の能力を試験するアッセイの結果を示す棒グラフである。
【特許請求の範囲】
【請求項1】
哺乳動物ガン細胞にアポトーシスを誘導するのに有効量のApo-2リガンドに対して前記細胞をさらす工程を含む、哺乳動物ガン細胞におけるin vitroでのアポトーシスの誘導方法であって、前記Apo-2リガンドが、
(a)図1A(配列番号1)のアミノ酸残基41-281を含むポリペプチド;
(b)図1A(配列番号1)のアミノ酸残基114-281を含むポリペプチド;
(c)図1A(配列番号1)のアミノ酸残基114-281からなるポリペプチド;
(d)図1A(配列番号1)のアミノ酸残基1-281からなるポリペプチド;及び
(e)(a)、(b)、(c)、または(d)の断片であるポリペプチド;
からなる群から選択されるポリペプチドであり、
前記哺乳動物ガン細胞が、乳ガン細胞、肺ガン細胞、大腸ガン細胞、前立腺ガン細胞、卵巣ガン細胞、及びグリア芽腫ガン細胞からなる群から選択される方法。
【請求項2】
前記Apo-2リガンドポリペプチドが図1A(配列番号1)のアミノ酸残基114-281からなる、請求項1に記載の方法。
【請求項3】
前記哺乳動物ガン細胞が放射線療法または化学療法にもさらされる、請求項1または2に記載の方法。
【請求項4】
前記化学療法がドキソルビシン(Doxorubicin)、5-フルオロウラシル(5-Fluorouracil)、シトシンアラビノシド(Cytosine arabinoside)、シクロホスファミド(Cyclophosphamide)、チオテパ(Thiotepa)、ブスルファン(Busulfan)、サイトキシン(Cytoxin)、タキソール(Taxol)、メトトレキサート(Methotrexate)、シスプラチン(Cisplatin)、メルファラン(Melphalan)、ビンブラスチン(Vinblastine)、及びカルボプラチン(Carboplatin)からなる群から選択される、請求項3に記載の方法。
【請求項5】
前記Apo-2リガンドポリペプチドがポリエチレングリコール、ポリプロピレングリコール、及びポリオキシアルキレンからなる群から選択される一つ以上の非タンパク質性ポリマーに結合されている、請求項1または2に記載の方法。
【請求項6】
前記Apo-2リガンドポリペプチドがグリコシル化されていない、請求項1または2に記載の方法。
【請求項7】
前記Apo-2リガンドポリペプチドが大腸菌で生産される、請求項6に記載の方法。
【請求項8】
哺乳動物のガン細胞にアポトーシスを誘導するのに有効量のApo-2リガンドをを含む、哺乳動物におけるガンの治療剤であって、前記Apo-2リガンドが、
(a)図1A(配列番号1)のアミノ酸残基41-281を含むポリペプチド;
(b)図1A(配列番号1)のアミノ酸残基114-281を含むポリペプチド;
(c)図1A(配列番号1)のアミノ酸残基114-281からなるポリペプチド;
(d)図1A(配列番号1)のアミノ酸残基1-281からなるポリペプチド;及び
(e)(a)、(b)、(c)、または(d)の断片であるポリペプチド;
からなる群から選択されるポリペプチドであり、
前記哺乳動物のガン細胞が、乳ガン細胞、肺ガン細胞、大腸ガン細胞、前立腺ガン細胞、卵巣ガン細胞、及びグリア芽腫ガン細胞からなる群から選択される治療剤。
【請求項9】
前記Apo-2リガンドポリペプチドが図1A(配列番号1)のアミノ酸残基114-281からなる、請求項8に記載の治療剤。
【請求項10】
更に放射線療法または化学療法の前記哺乳動物への投与と組み合わされて使用される、請求項8または9に記載の治療剤。
【請求項11】
前記化学療法がドキソルビシン(Doxorubicin)、5-フルオロウラシル(5-Fluorouracil)、シトシンアラビノシド(Cytosine arabinoside)、シクロホスファミド(Cyclophosphamide)、チオテパ(Thiotepa)、ブスルファン(Busulfan)、サイトキシン(Cytoxin)、タキソール(Taxol)、メトトレキサート(Methotrexate)、シスプラチン(Cisplatin)、メルファラン(Melphalan)、ビンブラスチン(Vinblastine)、及びカルボプラチン(Carboplatin)からなる群から選択される、請求項10に記載の治療剤。
【請求項12】
前記Apo-2リガンドポリペプチドがポリエチレングリコール、ポリプロピレングリコール、及びポリオキシアルキレンからなる群から選択される一つ以上の非タンパク質性ポリマーに結合されている、請求項8または9に記載の治療剤。
【請求項13】
前記Apo-2リガンドポリペプチドがグリコシル化されていない、請求項8または9に記載の治療剤。
【請求項14】
前記Apo-2リガンドポリペプチドが大腸菌で生産される、請求項13に記載の治療剤。
【請求項15】
American Type Culture Collection Accession Number ATCC HB-12258の下で寄託されたハイブリドーマ細胞系。
【請求項16】
American Type Culture Collection Accession Number ATCC HB-12257の下で寄託されたハイブリドーマ細胞系。
【請求項17】
American Type Culture Collection Accession Number ATCC HB-12256の下で寄託されたハイブリドーマ細胞系。
【請求項18】
American Type Culture Collection Accession Number ATCC HB-12259の下で寄託されたハイブリドーマ細胞系。
【請求項19】
American Type Culture Collection Accession Number ATCC HB-12257の下で寄託されたハイブリドーマ細胞系によって生産される、Apo-2リガンドモノクローナル抗体。
【請求項20】
American Type Culture Collection Accession Number ATCC HB-12259の下で寄託されたハイブリドーマ細胞系によって生産される、Apo-2リガンドモノクローナル抗体。
【請求項21】
American Type Culture Collection Accession Number ATCC HB-12256の下で寄託されたハイブリドーマ細胞系によって生産される、Apo-2リガンドモノクローナル抗体。
【請求項22】
American Type Culture Collection Accession Number ATCC HB-12258の下で寄託されたハイブリドーマ細胞系によって生産される、Apo-2リガンドモノクローナル抗体。
【請求項23】
(a)図1A(配列番号1)のアミノ酸114から281を含むApo-2リガンドポリペプチドに結合し;及び
(b)American Type Culture Collection Accession Number ATCC HB-12257の下で寄託されたハイブリドーマ細胞系によって生産される1D1モノクローナル抗体が結合するのと同じApo-2リガンドエピトープに結合する;
単離されたApo-2リガンドモノクローナル抗体。
【請求項24】
図1A(配列番号1)のアミノ酸192から204を含むApo-2リガンドポリペプチドのエピトープに結合する、請求項23に記載の抗体。
【請求項25】
(a)図1A(配列番号1)のアミノ酸114から281を含むApo-2リガンドポリペプチドに結合し;及び
(b)American Type Culture Collection Accession Number ATCC HB-12259の下で寄託されたハイブリドーマ細胞系によって生産される2G6モノクローナル抗体が結合するのと同じApo-2リガンドエピトープに結合する;
単離されたApo-2リガンドモノクローナル抗体。
【請求項26】
Apo-2リガンドレセプターに対するApo-2リガンドの結合をブロックする、または哺乳動物細胞においてApo-2リガンドが誘導したアポトーシスをブロックする、請求項25に記載の抗体。
【請求項27】
(a)図1A(配列番号1)のアミノ酸114から281を含むApo-2リガンドポリペプチドに結合し;及び
(b)American Type Culture Collection Accession Number ATCC HB-12256の下で寄託されたハイブリドーマ細胞系によって生産される2E11モノクローナル抗体が結合するのと同じApo-2リガンドエピトープに結合する;
単離されたApo-2リガンドモノクローナル抗体。
【請求項28】
Apo-2リガンドレセプターに対するApo-2リガンドの結合をブロックする、または哺乳動物細胞においてApo-2リガンドが誘導したアポトーシスをブロックする、請求項27に記載の抗体。
【請求項29】
(a)図1A(配列番号1)のアミノ酸114から281を含むApo-2リガンドポリペプチドに結合し;及び
(b)American Type Culture Collection Accession Number ATCC HB-12258の下で寄託されたハイブリドーマ細胞系によって生産される5C2モノクローナル抗体が結合するのと同じApo-2リガンドエピトープに結合する;
単離されたApo-2リガンドモノクローナル抗体。
【請求項30】
Apo-2リガンドレセプターに対するApo-2リガンドの結合をブロックする、または哺乳動物細胞においてApo-2リガンドが誘導したアポトーシスをブロックする、請求項29に記載の抗体。
【請求項1】
哺乳動物ガン細胞にアポトーシスを誘導するのに有効量のApo-2リガンドに対して前記細胞をさらす工程を含む、哺乳動物ガン細胞におけるin vitroでのアポトーシスの誘導方法であって、前記Apo-2リガンドが、
(a)図1A(配列番号1)のアミノ酸残基41-281を含むポリペプチド;
(b)図1A(配列番号1)のアミノ酸残基114-281を含むポリペプチド;
(c)図1A(配列番号1)のアミノ酸残基114-281からなるポリペプチド;
(d)図1A(配列番号1)のアミノ酸残基1-281からなるポリペプチド;及び
(e)(a)、(b)、(c)、または(d)の断片であるポリペプチド;
からなる群から選択されるポリペプチドであり、
前記哺乳動物ガン細胞が、乳ガン細胞、肺ガン細胞、大腸ガン細胞、前立腺ガン細胞、卵巣ガン細胞、及びグリア芽腫ガン細胞からなる群から選択される方法。
【請求項2】
前記Apo-2リガンドポリペプチドが図1A(配列番号1)のアミノ酸残基114-281からなる、請求項1に記載の方法。
【請求項3】
前記哺乳動物ガン細胞が放射線療法または化学療法にもさらされる、請求項1または2に記載の方法。
【請求項4】
前記化学療法がドキソルビシン(Doxorubicin)、5-フルオロウラシル(5-Fluorouracil)、シトシンアラビノシド(Cytosine arabinoside)、シクロホスファミド(Cyclophosphamide)、チオテパ(Thiotepa)、ブスルファン(Busulfan)、サイトキシン(Cytoxin)、タキソール(Taxol)、メトトレキサート(Methotrexate)、シスプラチン(Cisplatin)、メルファラン(Melphalan)、ビンブラスチン(Vinblastine)、及びカルボプラチン(Carboplatin)からなる群から選択される、請求項3に記載の方法。
【請求項5】
前記Apo-2リガンドポリペプチドがポリエチレングリコール、ポリプロピレングリコール、及びポリオキシアルキレンからなる群から選択される一つ以上の非タンパク質性ポリマーに結合されている、請求項1または2に記載の方法。
【請求項6】
前記Apo-2リガンドポリペプチドがグリコシル化されていない、請求項1または2に記載の方法。
【請求項7】
前記Apo-2リガンドポリペプチドが大腸菌で生産される、請求項6に記載の方法。
【請求項8】
哺乳動物のガン細胞にアポトーシスを誘導するのに有効量のApo-2リガンドをを含む、哺乳動物におけるガンの治療剤であって、前記Apo-2リガンドが、
(a)図1A(配列番号1)のアミノ酸残基41-281を含むポリペプチド;
(b)図1A(配列番号1)のアミノ酸残基114-281を含むポリペプチド;
(c)図1A(配列番号1)のアミノ酸残基114-281からなるポリペプチド;
(d)図1A(配列番号1)のアミノ酸残基1-281からなるポリペプチド;及び
(e)(a)、(b)、(c)、または(d)の断片であるポリペプチド;
からなる群から選択されるポリペプチドであり、
前記哺乳動物のガン細胞が、乳ガン細胞、肺ガン細胞、大腸ガン細胞、前立腺ガン細胞、卵巣ガン細胞、及びグリア芽腫ガン細胞からなる群から選択される治療剤。
【請求項9】
前記Apo-2リガンドポリペプチドが図1A(配列番号1)のアミノ酸残基114-281からなる、請求項8に記載の治療剤。
【請求項10】
更に放射線療法または化学療法の前記哺乳動物への投与と組み合わされて使用される、請求項8または9に記載の治療剤。
【請求項11】
前記化学療法がドキソルビシン(Doxorubicin)、5-フルオロウラシル(5-Fluorouracil)、シトシンアラビノシド(Cytosine arabinoside)、シクロホスファミド(Cyclophosphamide)、チオテパ(Thiotepa)、ブスルファン(Busulfan)、サイトキシン(Cytoxin)、タキソール(Taxol)、メトトレキサート(Methotrexate)、シスプラチン(Cisplatin)、メルファラン(Melphalan)、ビンブラスチン(Vinblastine)、及びカルボプラチン(Carboplatin)からなる群から選択される、請求項10に記載の治療剤。
【請求項12】
前記Apo-2リガンドポリペプチドがポリエチレングリコール、ポリプロピレングリコール、及びポリオキシアルキレンからなる群から選択される一つ以上の非タンパク質性ポリマーに結合されている、請求項8または9に記載の治療剤。
【請求項13】
前記Apo-2リガンドポリペプチドがグリコシル化されていない、請求項8または9に記載の治療剤。
【請求項14】
前記Apo-2リガンドポリペプチドが大腸菌で生産される、請求項13に記載の治療剤。
【請求項15】
American Type Culture Collection Accession Number ATCC HB-12258の下で寄託されたハイブリドーマ細胞系。
【請求項16】
American Type Culture Collection Accession Number ATCC HB-12257の下で寄託されたハイブリドーマ細胞系。
【請求項17】
American Type Culture Collection Accession Number ATCC HB-12256の下で寄託されたハイブリドーマ細胞系。
【請求項18】
American Type Culture Collection Accession Number ATCC HB-12259の下で寄託されたハイブリドーマ細胞系。
【請求項19】
American Type Culture Collection Accession Number ATCC HB-12257の下で寄託されたハイブリドーマ細胞系によって生産される、Apo-2リガンドモノクローナル抗体。
【請求項20】
American Type Culture Collection Accession Number ATCC HB-12259の下で寄託されたハイブリドーマ細胞系によって生産される、Apo-2リガンドモノクローナル抗体。
【請求項21】
American Type Culture Collection Accession Number ATCC HB-12256の下で寄託されたハイブリドーマ細胞系によって生産される、Apo-2リガンドモノクローナル抗体。
【請求項22】
American Type Culture Collection Accession Number ATCC HB-12258の下で寄託されたハイブリドーマ細胞系によって生産される、Apo-2リガンドモノクローナル抗体。
【請求項23】
(a)図1A(配列番号1)のアミノ酸114から281を含むApo-2リガンドポリペプチドに結合し;及び
(b)American Type Culture Collection Accession Number ATCC HB-12257の下で寄託されたハイブリドーマ細胞系によって生産される1D1モノクローナル抗体が結合するのと同じApo-2リガンドエピトープに結合する;
単離されたApo-2リガンドモノクローナル抗体。
【請求項24】
図1A(配列番号1)のアミノ酸192から204を含むApo-2リガンドポリペプチドのエピトープに結合する、請求項23に記載の抗体。
【請求項25】
(a)図1A(配列番号1)のアミノ酸114から281を含むApo-2リガンドポリペプチドに結合し;及び
(b)American Type Culture Collection Accession Number ATCC HB-12259の下で寄託されたハイブリドーマ細胞系によって生産される2G6モノクローナル抗体が結合するのと同じApo-2リガンドエピトープに結合する;
単離されたApo-2リガンドモノクローナル抗体。
【請求項26】
Apo-2リガンドレセプターに対するApo-2リガンドの結合をブロックする、または哺乳動物細胞においてApo-2リガンドが誘導したアポトーシスをブロックする、請求項25に記載の抗体。
【請求項27】
(a)図1A(配列番号1)のアミノ酸114から281を含むApo-2リガンドポリペプチドに結合し;及び
(b)American Type Culture Collection Accession Number ATCC HB-12256の下で寄託されたハイブリドーマ細胞系によって生産される2E11モノクローナル抗体が結合するのと同じApo-2リガンドエピトープに結合する;
単離されたApo-2リガンドモノクローナル抗体。
【請求項28】
Apo-2リガンドレセプターに対するApo-2リガンドの結合をブロックする、または哺乳動物細胞においてApo-2リガンドが誘導したアポトーシスをブロックする、請求項27に記載の抗体。
【請求項29】
(a)図1A(配列番号1)のアミノ酸114から281を含むApo-2リガンドポリペプチドに結合し;及び
(b)American Type Culture Collection Accession Number ATCC HB-12258の下で寄託されたハイブリドーマ細胞系によって生産される5C2モノクローナル抗体が結合するのと同じApo-2リガンドエピトープに結合する;
単離されたApo-2リガンドモノクローナル抗体。
【請求項30】
Apo-2リガンドレセプターに対するApo-2リガンドの結合をブロックする、または哺乳動物細胞においてApo-2リガンドが誘導したアポトーシスをブロックする、請求項29に記載の抗体。
【図1A】


【図1B】

【図1C】

【図1D】

【図1E】
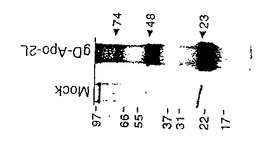
【図2A】

【図2B】

【図2C】

【図2D】
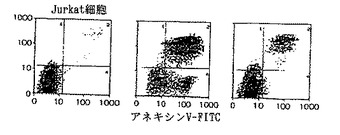
【図2E】

【図3A】
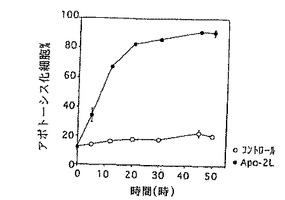
【図3B】
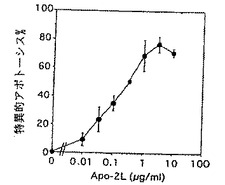
【図3C】
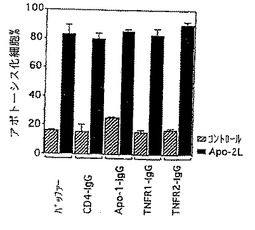
【図4】
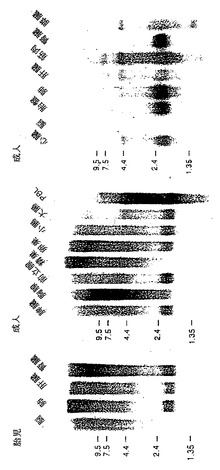
【図5】
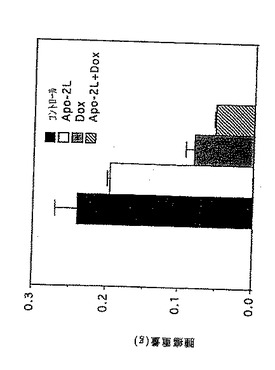
【図6】
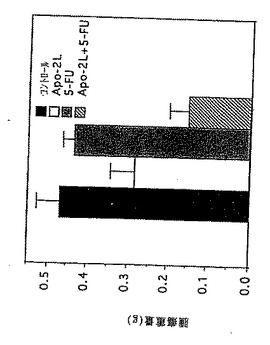
【図7】

【図8】
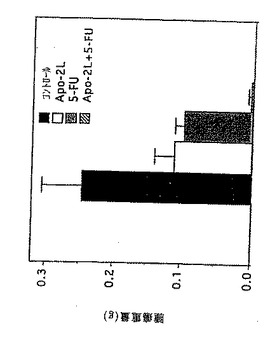
【図9】
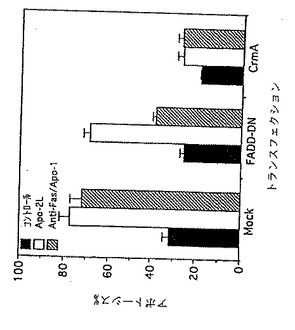
【図10】
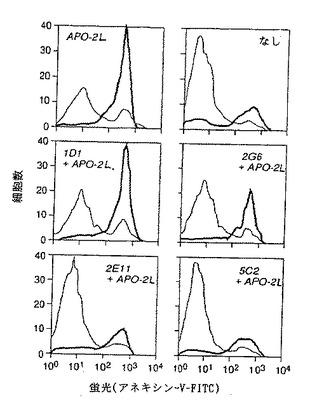
【図11】
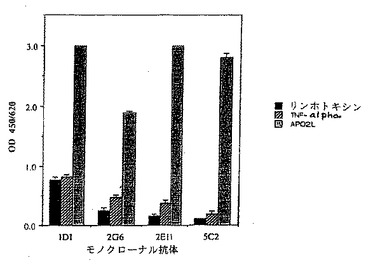
【図12】
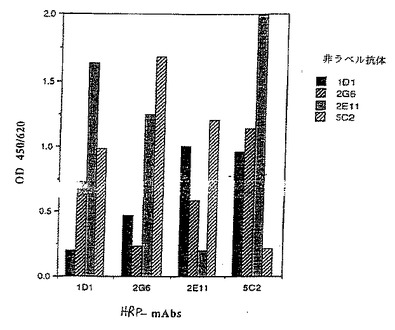
【図13】

【図1B】
【図1C】
【図1D】
【図1E】
【図2A】
【図2B】
【図2C】
【図2D】
【図2E】
【図3A】
【図3B】
【図3C】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【公開番号】特開2007−169280(P2007−169280A)
【公開日】平成19年7月5日(2007.7.5)
【国際特許分類】
【出願番号】特願2006−343397(P2006−343397)
【出願日】平成18年12月20日(2006.12.20)
【分割の表示】特願2004−328142(P2004−328142)の分割
【原出願日】平成9年1月8日(1997.1.8)
【出願人】(596168317)ジェネンテック・インコーポレーテッド (372)
【氏名又は名称原語表記】GENENTECH,INC.
【Fターム(参考)】
【公開日】平成19年7月5日(2007.7.5)
【国際特許分類】
【出願日】平成18年12月20日(2006.12.20)
【分割の表示】特願2004−328142(P2004−328142)の分割
【原出願日】平成9年1月8日(1997.1.8)
【出願人】(596168317)ジェネンテック・インコーポレーテッド (372)
【氏名又は名称原語表記】GENENTECH,INC.
【Fターム(参考)】
[ Back to top ]