
Apo−2DcR
【課題】Apo-2リガンドに結合可能なApo-2DcRと命名された新規なポリペプチドの提供。
【解決手段】TNFRファミリーとの類似性を示す、新規ポリペプチド、Apo-2DcRをコード化したcDNAクローンを同定し、Apo-2リガンド(Apo-2L)に結合することを見出した。異種ポリペプチド又はアミノ酸配列と融合したApo-2DcRポリペプチドを含んでなるキメラ分子、Apo-2DcRをコードしている核酸、及びApo-2DcRに対する抗体を含む組成物。
【解決手段】TNFRファミリーとの類似性を示す、新規ポリペプチド、Apo-2DcRをコード化したcDNAクローンを同定し、Apo-2リガンド(Apo-2L)に結合することを見出した。異種ポリペプチド又はアミノ酸配列と融合したApo-2DcRポリペプチドを含んでなるキメラ分子、Apo-2DcRをコードしている核酸、及びApo-2DcRに対する抗体を含む組成物。
【発明の詳細な説明】
【発明の開示】
【0001】
発明の分野
本発明は、一般に、ここに「Apo-2DcR」と命名する新規ポリペプチドの同定、単離、及び組換え生産、及び抗Apo-2DcR抗体に関する。
【0002】
発明の背景
アポトーシス又は「プログラムされた細胞死」
哺乳動物における細胞数のコントロールは、細胞増殖と細胞死のバランスにより部分的に決定されると考えられている。しばしば壊死性細胞死と称される細胞死の一形態は、典型的には、ある種の外傷又は細胞傷害の結果生じる細胞死の病理的形態として特性付けられる。これに対して、通常は規則的又はコントロールされた状態で進行する細胞死の他の「生理的」形態がある。細胞死のこの規則的又はコントロールされた形態は、しばしば「アポトーシス」と称される[例えば、Barrら, Bio/Technology, 12:487-493(1994);Stellerら, Science, 267:1445-1449(1995)を参照]。アポトーシス性細胞死は、免疫系におけるクローン選択と胚の発達を含む多くの生理的プロセスにおいて自然に生じる[Itohら, Cell, 66:233-243(1991)]。アポトーシス性細胞死のレベルの減少は、癌、狼瘡、ヘルペスウイスル感染を含む種々の病理的条件に関連している[Thompson, Science, 267:1456-1462(1995)]。アポトーシス性細胞死のレベルの増加は、エイズ、アルツハイマー病、パーキンソン病、筋萎縮性側索硬化症、多発性硬化症、色素性網膜炎、小脳変性、無形成性貧血、心筋梗塞、脳卒中、再灌流傷害、及び毒素誘発性肝疾患を含む様々な他の病理状態に関連している[上掲のThompsonを参照]。
【0003】
典型的には、アポトーシス性細胞死には、細胞内における一又は複数の特徴的な形態学的及び生化学的変化、例えば細胞質の凝結、原形質膜の微絨毛の喪失、核の分節化、染色体DNAの分解又はミトコンドリア機能の喪失が伴う。様々な外因的及び内因的シグナルが、このような形態学的及び生化学的な細胞変化を惹起又は誘発すると考えられている[Raff, Nature, 356:397-400(1992);Steller, 上掲;Sachsら, Blood, 82:15(1993)]。例えば、ホルモンの刺激、例えば未成熟胸腺細胞に対する糖質コルチコイドホルモン、並びにある種の成長因子の退薬により惹起され得る[Watanabe-Fukunagaら, Nature, 356:314-317(1992)]。また、幾つかの同定された発癌遺伝子、例えばmyc、rel、及びE1A、及び腫瘍サプレッサー、例えばp53が、アポトーシスの誘発においてある役割を有していることも報告されている。ある種の化学療法薬及びある種の放射線も同様にアポトーシス誘発活性を有していることも見出されている[Thompson, 上掲]。
サイトカインのTNFファミリー
【0004】
様々な分子、例えば腫瘍壊死因子-α(「TNF-α」)、腫瘍壊死因子-β(「TNF-β」すなわち「リンホトキシン」)、CD30リガンド、CD27リガンド、CD40リガンド、OX-40リガンド、4-1BBリガンド、Apo-1リガンド(Fasリガンド又はCD95リガンドとも称される)、及びApo-2リガンド[トレイル(TRAIL)とも称される]が、サイトカインの腫瘍壊死因子(「TNF」)ファミリーのメンバーとして同定された[例えば、Gruss及びDower, Blood, 85:3378-3404(1995);Wileyら, Immunity, 3:673-682(1995);Pittiら, J. Biol. Chem., 271:12687-12690(1996)]。これらの分子のなかでも、TNF-α、TNF-β、CD30リガンド、4-1BBリガンド、Apo-1リガンド、及びApo-2リガンド(TRAIL)は、アポトーシス性細胞死に関与していることが報告されている。TNF-αとTNF-βの両方とも、感受性腫瘍細胞におけるアポトーシス性死を誘発することが報告されている[Schmidら, Proc. Natl. Acad. Sci., 83:1881(1986);Dealtryら, Eur. J. Immunol., 17:689(1987)]。ゼングらは、TNF-αがCD8ポジティブT細胞のポスト刺激性アポトーシスに関与していることを報告している[Zhengら, Nature, 377:348-351(1995)]。他の研究者は、CD30リガンドが胸腺における自己反応性T細胞の欠失に関与していることを報告している[Amakawaら, プログラム細胞死に関するコールドスプリングハーバー研究所のシンポジウム、要約集、第10巻、(1995)]。
【0005】
マウスのFas/Apo-1レセプター又はリガンド遺伝子(それぞれ1pr及びgldと呼称される)における変異が幾つかの自己免疫疾患に関連しており、Apo-1リガンドが末梢の自己反応性リンパ球のクローン除去の調節においてある役割を担っていることを示している[Krammerら, Curr. Op. Immunol., 6:279-289(1994);Nagataら, Science, 267:1449-1456(1995)]。また、Apo-1リガンドは、CD4ポジティブTリンパ球及びBリンパ球においてポスト刺激性アポトーシスを誘発することが報告されており、それらの機能がもはや必要でなくなった際の活性化リンパ球の除去に関与している[Krammerら, 上掲;Nagataら, 上掲]。Apo-1レセプターと特異的に結合するアゴニストのマウスモノクローナル抗体は、TNF-αに匹敵するか類似する細胞死滅活性を示すことが報告されている[Yoneharaら, J. Exp. Med., 169:1747-1756(1989)]。
レセプターのTNFファミリー
【0006】
このようなTNFファミリーのサイトカインが介在する種々の細胞反応誘導は、特異的な細胞レセプターに結合することにより開始されると考えられている。約55-kDa(TNFR1)と75-kDa(TNFR2)の2つの異なるTNFレセプターが同定されており[Hohmanら, J. Biol. Chem., 264:14927-14934(1989);Brockhausら, Proc. Natl. Acad. Sci., 87:3127-3131(1990);1991年3月20日に公開されたEP417563]、双方のレセプター型に対応するヒト及びマウスcDNAが単離され、特徴付けされている[Loetscherら, Cell, 61:351(1990);Schallら, Cell, 61:361(1990);Smithら, Science, 248:1019-1023(1990);Lewisら, Proc. Natl. Acad. Sci., 88:2830-2834(1991);Goodwinら, Mol. Cell. Biol., 11:3020-3026(1991)]。広範な多型性が、双方のTNFレセプター遺伝子に関連している[例えば、Takaoら, Immunogenetics, 37:199-203(1993)を参照]。双方のTNFRは細胞外、膜貫通及び細胞内領域を含む細胞表面レセプターの典型的な構造を共有する。双方のレセプターの細胞外部分はまた可溶性TNF結合タンパク質として天然に見出される[Nophar, Yら, EMBO J., 9:3269(1990);及びKohno, Tら, Proc. Natl. Acad. Sci. U.S.A., 87:8331(1990)]。更に最近になって、組換え体可溶性TNFレセプターのクローニングがヘイル(Hale)らにより報告されている[J. Cell. Biochem. 増補15F, 1991, p.113(P424)]。
【0007】
1型又は2型のTNFR(TNFR1及びTNFR2)の細胞外部分は、NH2末端から出発して、1〜4とされる4つのシステインに富んだドメイン(CRD)の反復アミノ酸配列パターンを含む。各CRDは約40のアミノ酸長のものであり、良好に保存された位置に4〜6のシステイン残基を含んでいる[Schallら, 上掲;Loetscherら, 上掲;Smithら, 上掲;Nopharら, 上掲;Kohnoら, 上掲]。TNFR1において、4つのCRDのおおよその境界は次の通りである:CRD1−14から約53までのアミノ酸;CRD2−約54から約97までのアミノ酸;CRD3−約98から約138までのアミノ酸;CRD4−約139から約167までのアミノ酸。TNFR2において、CRD1は、17〜約54までのアミノ酸を、CRD2は約55から約97までのアミノ酸を;CRD3は約98から約140までのアミノ酸を;CRD4は約141から約179までのアミノ酸を含む[Bannerら, Cell, 73:431-435(1993)]。また、リガンド結合におけるCRDの潜在的な役割は前掲のバナー(Banner)らにより記載されている。
【0008】
CRDの類似の反復パターンが、p75神経成長因子レセプター(NGFR)[Johnsonら, Cell, 47:545(1986);Radekeら, Nature, 325:593(1987)]、B細胞抗原CD40[Stamenkovicら, EMBO J., 8:1403(1989)]、T細胞抗原OX40[Malletら, EMBO J., 9:1063(1990)]及びFas抗原[Yoneharaら, 上掲、及びItohら, 上掲]を含む幾つかの他の細胞表面タンパク質に存在している。また、CRDはショープ(Shope)及び粘液腫ポックスウィルスの可溶型TNFR(sTNFR)様のT2タンパク質にも見出されている[Uptonら, Virology, 160:20-29(1987);Smithら, Biochem. Biophys. Res. Commun., 176:335(1991);Uptonら, Virology, 184:370(1991)]。これらの配列の最適なアラインメントは、システイン残基の位置が良好に保存されていることを示している。これらレセプターは、しばしば集合的に、TNF/NGFレセプタースーパーファミリーのメンバーと称される。p75NGFRに関する最近の研究では、CRD1の欠失[Welcher, A.A.ら, Proc. Natl. Acad. Sci. USA, 88:159-163(1991)]又はこのドメインにおける5-アミノ酸の挿入[Yan. H及びChao, M.V., J. Biol. Chem., 266:12099-12104(1991)]は、NGF結合にはほとんど又は全く影響を持たないことが示されている[Yan. H及びChao, M.V., 上掲]。p75NGFRはNGF結合に関与せず、そのCRD4と膜貫通領域の間に約60のアミノ酸のプロリンに富んだ伸展を含む[Peetre, Cら, Eur. J. Hematol.,41:414-419(1988);Seckinger, Pら, J. Biol. Chem., 264:11966-11973(1989);Yan. H及びChao, M.V., 上掲]。同様のプロリンに富んだ領域はTNFR2に見出されているが、TNFR1にはない。
【0009】
イトーらは、Apo-1レセプターが55-kDaのTNFR1によりシグナル化されるものと同様のアポトーシス性細胞死をシグナル化し得ることを開示している[Itohら, 上掲]。また、Apo-1抗原の発現は、細胞をTNF-α又は抗Apo-1のマウスモノクローナル抗体で処理した場合に、TNFR1のものと共にダウンレギュレーションされることが報告されている[Krammerら, 上掲;Nagataら, 上掲]。従って、Apo-1及びTNFR1レセプターの双方を同時発現する株化細胞が、共通のシグナル伝達経路を通して細胞死滅を媒介しているとの仮説を唱える研究者もいた[同]。
【0010】
リンホトキシン-αを除き、今日までに同定されているTNFファミリーのリガンドは、II型の膜貫通タンパク質であり、そのC末端は細胞外にある。これに対して、今日までに同定されているTNFレセプター(TNFR)ファミリーのレセプター類はI型の膜貫通タンパク質である。しかしながら、TNFリガンド及びレセプターファミリーの双方において、ファミリーメンバー間で同定された相同性は、主として細胞外ドメイン(「ECD」)において見出されている。TNF-α、Apo-1リガンド及びCD40リガンドを含むTNFファミリーサイトカインのいくつかは、細胞表面においてタンパク分解的に切断され;各場合に得られたタンパク質は、典型的には、可溶性サイトカインとして機能するホモ三量体分子を形成する。また、TNFレセプターファミリーのタンパク質は、通常、タンパク分解的に切断され、同族のサイトカインの阻害剤として機能し得る可溶性レセプターのECDを放出する。
【0011】
最近になって、TNFRファミリーの他のメンバーが同定されている。マースターズ(Marsters)ら、Curr. Biol., 6:750(1996)において、研究者は、細胞外のシステインに富んだ反復についてTNFRファミリーに対して類似性を示し、細胞質死亡ドメイン配列を含む点でTNFR1及びCD95に似ており、Apo-3と称される、ヒトのポリペプチド天然配列の全長を開示している[Marstersら,Curr. Biol., 6:1669(1996)もまた参照されたい]。他の研究者に依れば、Apo-3はDR3、wsl-1及びTRAMPとも称されている[Chinnaiyanら, Science, 274:990(1996);Kitsonら, Nature, 384:372(1996);Bodmerら, Immunity, 6:79(1997)]。
【0012】
パンらは、「DR4」と称される他のTNFレセプターファミリーのメンバーを開示している[Panら, Science, 276:111-113(1997)]。DR4は細胞自殺器を活動させる細胞質死亡ドメインを含むと報告されている。パンらは、DR4がApo-2リガンド又はTRAILとして知られているリガンドに対するレセプターであると考えられることを開示している。
【0013】
アポトーシス誘発性シグナル伝達複合体
現在理解されているように、細胞死プログラムは、少なくとも3つの重要な成分−活性化因子、阻害剤及びエフェクターを含む;線虫(C. elegans)において、これらの成分はそれぞれ3つの遺伝子、Ced-4、Ced-9及びCed-3によりコード化されている[Steller, Science, 267:1445(1995);Chinnaiyanら, Science, 275:1122-1126(1997)]。TNFRファミリーのメンバーの2つ、TNFR1及びFas/Apol(CD95)は、アポトーシス性細胞死を活性化し得る[Chinnaiyan及びDixit, Current Biology, 6:555-562(1996);Fraser及びEvan, Cell, 85:781-784(1996)]。また、TNFR1は転写因子、NF-κBの活性化を媒介することも知られている[Tartagliaら, Cell, 74:845-853(1993);Hsuら, Cell, 84:299-308(1996)]。ある程度のECD相同性に加えて、これら2つのレセプターは、死亡ドメインとして知られているオリゴマー形成界面の細胞内ドメイン(ICD)での相同性を共有する[Tartagliaら, 上掲;Nagata, Cell, 88:355(1997)]。また、死亡ドメインはアポトーシスを調節する幾つかの後生動物タンパク質、すなわちFADD/MORT1、TRADD及びRIPと称されるショウジョウバエタンパク質、リーパー(Reaper)及び哺乳動物タンパク質中においても見出されている[Cleaveland及びIhle, Cell, 81:479-482(1995)]。ラベンらは、酵母-二重ハイブリッド系を使用して、TNFR1死亡ドメインに結合するタンパク質wsl-1の同定を報告している[Ravenら, Programmed Cell Death Meeting, 9月20-24日,1995年,127頁の要約;Ravenら, European Cytokine Network, 7:210頁の要約82(1996年4-6月)]。wsl-1タンパク質はTNFR1に相同で(48%の同一性)、制限された組織分布を有すると記載されている。ラベン(Raven)らに依れば、wsl-1の組織分布は、TNFR1結合タンパク質、TRADDとは顕著に異なる。
【0014】
リガンド結合及びレセプターのクラスター形成の際に、TNFR1とCD95は死亡誘発性シグナル伝達複合体にFADDを補充するものと考えられている。言われるところによれば、CD95はFADDに直接結合する一方、TNFR1はTRADDを介して間接的にFADDに結合する[Chinnaiyanら, Cell, 81:505-512(1995);Boldinら, J. Biol. Chem., 270:387-391(1995);Hsuら, 上掲;Chinnaiyanら, J. Biol. Chem., 271:4961-4965(1996)]。FADDは、Ced-3-関連プロテアーゼ、MACHα/FLICE(カスパーゼ8)を、死亡シグナル伝達複合体に補充するアダプタータンパク質となることが報告されている[Boldinら, Cell, 85:803-815(1996);Muzioら, Cell, 85:817-827(1996)]。MACHα/FLICEは、細胞死プログラムの幾つかの重要な側面を実施し得る、インターロイキン-1β変換酵素(ICE)及びCPP32/Yamaを含むアポトーシス性プロテアーゼのカスケードを引き起こすトリガーであると思われる[Fraser及びEvan, 上掲]。
【0015】
プログラム細胞死が、線虫の細胞死遺伝子、ced-3、及び哺乳動物のIL-1-変換酵素、ICEに関連したシステインプロテアーゼファミリーのメンバーの活性に関与していることが最近開示された。ICE及びCPP32/Yamaプロテアーゼの活性は、牛痘ウイルス遺伝子、crmAの産物により阻害され得る[Rayら, Cell, 69:597-604(1992);Tewariら, Cell, 81:801-809(1995)]。最近の研究では、CrmAがTNFR1-及びCD95-誘発細胞死を阻害し得ることが示されている[Enariら, Nature, 375:78-81(1995);Tewariら, J. Biol. Chem., 270:3255-3260(1995)]。
【0016】
テワリ(Tewari)らにより最近レビューされているように、TNFR1、TNFR2及びCD40は、転写因子、NF-κBの活性化により、炎症誘発性及び同時刺激性サイトカイン、サイトカインレセプター、及び細胞接着分子の発現を変調する[Tewariら, Curr. Op. Genet. Develop., 6:39-44(1996)]。NF-κBは、そのサブユニットが保存Rel領域を含む二量体転写因子のファミリーの原型である[Vermaら, Genes Develop., 9:2723-2735(1996);Baldwin, Ann. Rev. Immunol., 14:649-681(1996)]。その潜伏形態において、NF-κBはIκB阻害剤ファミリーのメンバーと複合化しており;ある刺激に反応してのIκBの不活性化の際に、放出されたNF-κBが、特異的DNA配列と結合する核に転座して遺伝子転写を活性化する。
【0017】
サイトカインのTNFファミリー及びそれらのレセプターのレビューについては、上掲のグラス(Gruss)及びダゥアー(Dower)を参照されたい。
【0018】
発明の概要
本出願人は、本出願において「Apo-2DcR」と命名される新規ポリペプチドをコード化したcDNAクローンを同定した。Apo-2DcRはTNFRファミリーのメンバーであると信じられる;全長の天然配列ヒトApo-2DcRポリペプチドは、細胞外のシステインに富んだ反復部についてTNFRファミリーとの類似性を示す。本出願人は、Apo-2DcRがApo-2リガンド(Apo-2L)に結合することを見出した。
一実施態様において、本発明は、単離されたApo-2DcRポリペプチドを提供する。特に、本発明は、一実施態様においては図1Aの残基1〜259を含んでなるアミノ酸配列(配列番号:1)を含む、単離された天然配列Apo-2DcRポリペプチドを提供する。他の実施態様においては、単離されたApo-2DcRポリペプチドは、図1Aの残基1〜259(配列番号:1)を含んでなる天然配列Apo-2DcRポリペプチドと少なくとも約80%のアミノ酸配列同一性を有する。場合によっては、単離されたApo-2DcRポリペプチドは、−40〜259として図1Bにおいて同定された残基を含んでなるアミノ酸配列(配列番号:3)を含む。場合によっては、Apo-2DcRポリペプチドは、ATCC 209087として寄託されたベクターのcDNA挿入断片によりコード化されたポリペプチドを発現することにより得られた又は得られうるものである。
【0019】
他の実施態様において、本発明はApo-2DcRの単離された細胞外ドメイン(ECD)配列を提供する。場合によっては、単離された細胞外ドメイン配列は図1Aのアミノ酸残基1〜236(配列番号:1)又は図1Aの残基1〜161(配列番号:1)を含んでなる。場合によっては、単離された細胞外ドメイン配列は、ここにおいて同定されたApo-2DcR擬似反復の任意のものにおいて一又は複数のアミノ酸(図2参照)が欠失させられたアミノ酸配列を含んでなる。このような単離された細胞外ドメイン配列は、アミノ酸残基1〜X(ここで、Xは図1Aのアミノ酸残基161〜236(配列番号:1)の任意のものである)の配列を含んでなるポリペプチドを含んでもよい。
【0020】
他の実施態様において、本発明は、異種ポリペプチド又はアミノ酸配列と融合したApo-2DcRポリペプチドを含んでなるキメラ分子を提供する。このようなキメラ分子の例には、免疫グロブリン配列と融合したApo-2DcRが含まれる。他の例には、例えば免疫グロブリン配列のような、異種ポリペプチド又はアミノ酸配列と融合したApo-2DcRの細胞外ドメインが含まれる。
【0021】
他の実施態様において、本発明はApo-2DcRポリペプチドをコードしている単離核酸分子を提供する。一面において、該核酸分子は、Apo-2DcRポリペプチド又はApo-2DcRの特定のドメインをコード化するRNA又はDNAであるか、又はこのようなコード核酸配列に相補的であり、少なくとも中程度、場合によっては高いストリンジェント条件下でそれに安定して結合したままである。一実施態様において、核酸配列は:
(a)残基1〜残基259をコードする図1Aの核酸配列(配列番号:2)のコード領域(すなわち、ヌクレオチド193−195〜967−969);
(b)残基1〜残基236をコードする図1Aの核酸配列(配列番号:2)のコード領域(すなわち、ヌクレオチド193−195〜898−900);
(c)残基−40〜残基259をコードする図1Bの核酸配列(配列番号:4)のコード領域(すなわち、ヌクレオチド73−75〜967−969);
(d)遺伝コードの縮退の範囲内において(a)、(b)又は(c)配列に相当する配列;
から選択される。
【0022】
さらなる実施態様において、本発明はApo-2DcRポリペプチド又はApo-2DcRの特定のドメインをコード化した核酸分子を含んでなるベクターを提供する。また、ベクター又は核酸分子を含有する宿主細胞も提供する。Apo-2DcRの生産方法もさらに提供する。
【0023】
他の実施態様において、本発明はApo-2DcRに結合する抗体を提供する。抗体はアゴニスト、阻止又は中和抗体であってよい。
【0024】
他の実施態様において、本発明は非ヒト、トランスジェニック又はノックアウト動物を提供する。
【0025】
本発明のさらなる実施態様では、Apo-2DcR又はApo-2DcR抗体を含む製造品及びキットが提供される。
【0026】
好適な実施態様の詳細な説明
I.定義
ここで使用される際の「Apo-2DcRポリペプチド」及び「Apo-2DcR」という用語には、天然配列Apo-2DcR及びApo-2DcR変異体(ここでさらに定義される)が含まれる。これらの用語には、ヒトを含む種々の哺乳動物由来のApo-2DcRが含まれる。Apo-2DcRは種々の供給源、例えばヒト組織型又は他の供給源から単離されたもの、あるいは組換え又は合成法により調製されたものであってよい。
【0027】
「天然配列Apo-2DcR」には、天然由来のApo-2DcRと同一のアミノ酸配列を有するポリペプチドを含んでなる。しかして、天然配列Apo-2DcRは、任意の哺乳動物から自然に生じるApo-2DcRのアミノ酸配列を有し得る。このような天然配列Apo-2DcRは、自然から単離することもできるし、組換え又は合成手段により生産することもできる。「天然配列Apo-2DcR」という用語には、特に、Apo-2DcRの自然に生じる切断、分泌又は可溶型(例えば、細胞外ドメイン配列)、自然に生じる変異体型(例えば、選択的にスプライシングされた型)及びApo-2DcRの自然に生じる対立遺伝子変異体が含まれる。本発明の一実施態様において、天然配列Apo-2DcRは、図1A(配列番号:1)のアミノ酸1〜259又は図1B(配列番号:3)のアミノ酸−40〜259を含有する成熟又は全長天然配列Apo-2DcRである。場合によっては、Apo-2DcRポリペプチドは、ATCC 209087として寄託されているベクターのcDNA挿入物によりコードされたポリペプチドを発現させることにより得られた又は得られ得るものである。
【0028】
「Apo-2DcR細胞外ドメイン」すなわち「Apo-2DcRECD」は、膜貫通及び細胞質ドメインを本質的に有しないApo-2DcRの型を意味する。通常、Apo-2DcRECDは、膜貫通及び細胞質ドメインを1%未満、好ましくはそのようなドメインを0.5%未満だけ有する。場合によっては、Apo-2DcRECDは、図1A(配列番号:1)のアミノ酸残基1〜236又は図1A(配列番号:1)のアミノ酸残基1〜161を含むであろう。場合によっては、単離された細胞外ドメイン配列は、ここにおいて同定されたApo-2DcR擬似反復の任意のものにおいて同定される一又は複数のアミノ酸(図2参照)が欠失させられたアミノ酸配列を含んでなる。このような単離された細胞外ドメイン配列は、アミノ酸残基1〜X(ここで、Xは図1Aのアミノ酸残基161〜236(配列番号:1)の任意のものである)の配列を含んでなるポリペプチドを含んでもよい。
【0029】
「Apo-2DcR変異体」とは、全長天然配列ヒトApo-2DcRに対して図1Aに示されている推定アミノ酸配列(配列番号:1)又はここでApo-2DcRECDに対して同定された配列と少なくとも約80%のアミノ酸配列同一性を有する以下に定義する生物学的に活性なApo-2DcRを意味する。このようなApo-2DcR変異体には、例えば、図1Aの配列(配列番号:1)あるいはApo-2DcRECDに対してここで同定された配列のN又はC末端において一又は複数のアミノ酸残基が付加され、もしくは欠失されたApo-2DcRポリペプチドが含まれる。通常、Apo-2DcR変異体は、図1Aのアミノ酸配列(配列番号:1)と、少なくとも約80%のアミノ酸配列同一性、より好ましくは少なくとも約90%のアミノ酸配列同一性、さらにより好ましくは少なくとも約95%のアミノ酸配列同一性を有している。
【0030】
ここで同定されているApo-2DcR配列に対する「パーセント(%)アミノ酸配列同一性」は、配列を整列させ、最大のパーセント配列同一性を得るために必要ならば間隙を導入し、如何なる保存的置換も配列同一性の一部と考えないとした、Apo-2DcR配列のアミノ酸残基と同一である候補配列中のアミノ酸残基のパーセントとして定義される。パーセントアミノ酸配列同一性を決定する目的のためのアラインメントは、当業者の知る範囲にある種々の方法、例えばALIGN又はMegalign(DNASTAR)ソフトウエアのような公に入手可能なコンピュータソフトウエアを使用することにより達成可能である。当業者であれば、比較される配列の全長に対して最大のアラインメントを達成するために必要な任意のアルゴリズムを含む、アラインメントを測定するための適切なパラメータを決定することができる。
【0031】
「エピトープタグ」なる用語は、ここで用いられるときは、「タグポリペプチド」に融合したApo-2DcR、又はそれらのドメイン配列を含んでなるキメラポリペプチドを指す。タグポリペプチドは、その抗体が産生され得るエピトープを提供するに十分な数の残基を有しているが、その長さはApo-2DcRの活性を阻害しないよう充分に短い。また、タグポリペプチドは、好ましくは、抗体が他のエピトープと実質的に交差反応をしないようにかなり独特である。適切なタグポリペプチドは、一般に、少なくとも6のアミノ酸残基、通常は約8〜約50のアミノ酸残基(好ましくは約10〜約20の残基)を有する。
【0032】
「単離された」とは、ここで開示された種々のポリペプチドを記述するために使用するときは、その自然環境の成分から同定され分離され及び/又は回収されたポリペプチドを意味する。その自然環境の汚染成分とは、ポリペプチドの診断又は治療への使用を典型的には妨害する物質であり、酵素、ホルモン、及び他のタンパク質様又は非タンパク質様溶質が含まれる。好ましい実施態様において、ポリペプチドは、(1)スピニングカップシークエネーターを使用することにより、少なくとも15のN末端あるいは内部アミノ酸配列の残基を得るのに充分なほど、あるいは、(2)クーマシーブルーあるいは好ましくは銀染色を用いた非還元あるいは還元条件下でのSDS-PAGEによる均一性が得られるように充分なほど精製される。Apo-2DcRの自然環境の少なくとも1つの成分が存在しないため、単離されたポリペプチドには、組換え細胞内のインシトゥーのポリペプチドが含まれる。しかしながら、通常は、単離されたポリペプチドは少なくとも1つの精製工程により調製される。
【0033】
「単離された」Apo-2DcR核酸分子は、同定され、Apo-2DcR核酸の天然源に通常付随している少なくとも1つの汚染核酸分子から分離された核酸分子である。単離されたApo-2DcR核酸分子は、天然に見出される形態あるいは設定以外のものである。ゆえに、単離されたApo-2DcR核酸分子は、天然の細胞中に存在するApo-2DcR核酸細胞とは区別される。しかし、単離されたApo-2DcR核酸分子は、例えば、核酸分子が天然の細胞のものとは異なった染色体位置にあるApo-2DcRを通常発現する細胞に含まれるApo-2DcR核酸分子を含む。
【0034】
「対照配列」という表現は、特定の宿主生物において作用可能に関連付けられたコード配列を発現するために必要なDNA配列を指す。例えば原核生物に好適な対照配列は、プロモーター、場合によってはオペレータ配列、及びリボソーム結合部位を含む。真核生物の細胞は、プロモーター、ポリアデニル化シグナル及びエンハンサーを利用することが知られている。
【0035】
核酸は、他の核酸配列と機能的な関係にあるときに「作用可能に関連付けられ」ている。例えば、プレ配列あるいは分泌リーダーのDNAは、ポリペプチドの分泌に寄与するプレタンパク質として発現されているならそのポリペプチドのDNAに作用可能に関連付けられている;プロモーター又はエンハンサーは、配列の転写に影響を及ぼすならばコード配列に作用可能に関連付けられている;又はリボソーム結合部位は、もしそれが翻訳を容易にするような位置にあるならコード配列と作用可能に関連付けられている。一般的に、「作用可能に関連付けられる」とは、関連付けられたDNA配列が近接しており、分泌リーダーの場合には近接していて読みフェーズにある。しかし、エンハンサーは必ずしも近接しているわけではない。結合は簡便な制限部位でのライゲーションにより達成される。そのような部位が存在しない場合は、通常の手法にしたがって、合成されたオリゴヌクレオチドアダプターあるいはリンカーが使用される。
【0036】
「抗体」という用語は最も広い意味において使用され、特に抗Apo-2DcRモノクローナル抗体(アゴニスト、アンタゴニスト、及び中和抗体を含む)、及び多エピトープ特異性抗Apo-2DcR抗体組成物を包含している。
【0037】
ここで使用される「モノクローナル抗体」という用語は、実質的に均一な抗体の集団から得られる抗体を称する、すなわち、集団を構成する個々の抗体が、少量存在しうる自然に生じる可能な突然変異を除いて同一である。モノクローナル抗体は高度に特異的であり、一つの抗原部位に対応する。更に、異なる決定基(エピトープ)に対応する異なる抗体を典型的に含む通常の(ポリクローナル)抗体調製物とは異なり、各モノクローナル抗体は抗原の単一の決定基に対応する。
【0038】
ここで、モノクローナル抗体は、起源の種又は免疫グロブリンクラス又はサブクラスの命名に拘わらず、定常ドメインを有する抗Apo-2DcR抗体の可変(高頻度可変を含む)ドメイン、又は重鎖を有する軽鎖、又は他の種由来の鎖を有するある種由来の鎖、あるいは異種タンパク質との融合体をスプライシングすることによって得られるハイブリッド及び組換え抗体、並びにそれが所望の生物的活性を有する限り抗体断片(例えばFab、F(ab')2及びFv)を特に含む。例えば、米国特許第4816567号、及びMageら, Monoclonal Antibody Productuon Techniques and Applications, pp.79-97(Marcel Dekker, Inc.:ニューヨーク, 1987)を参照。
【0039】
従って、「モノクローナル」との形容は、実質的に均一な抗体集団から得られたという抗体の性質を示し、抗体を何か特定の方法で生産しなければならないことを意味するものではない。例えば、本発明に従って使用されるモノクローナル抗体は、最初にKohler及びMilsteinによって、Nature, 256:495 (1975)に記載されたハイブリドーマ法によって作ることができ、あるいは例えば米国特許第4816567号に記載された組換えDNA法によって作ることができる。「モノクローナル抗体」は、例えば、McCaffertyら, Nature, 348:552-554(1990)に記載された技術を用いてファージライブラリから単離することができる。
【0040】
非ヒト(例えばマウス)抗体の「ヒト化」形とは、キメラ免疫グロブリン、免疫グロブリン鎖、あるいはそれらの断片(例えばFv、Fab、Fab'、F(ab')2あるいは抗体の他の抗原結合サブ配列)であって、非ヒト免疫グロブリンに由来する最小配列を含むものである。大部分においてヒト化抗体はレシピエントの相補性決定領域(CDR)の残基が、マウス、ラット又はウサギのような所望の特異性、親和性及び能力を有する非ヒト(ドナー抗体)のCDRの残基によって置換されたヒト免疫グロブリン(レシピエント抗体)である。ある場合には、ヒト免疫グロブリンのFvフレームワーク領域(FR)残基は、対応する非ヒト残基によって置換されている。更に、ヒト化抗体は、レシピエント抗体にも、移入されたCDRもしくはフレームワーク配列にも見出されない残基を含んでもよい。これらの修飾は抗体の特性を更に洗練し、最適化するために行われる。一般に、ヒト化抗体は、全てあるいはほとんど全てのCDR領域が非ヒト免疫グロブリンのものに対応し、全てあるいはほとんど全てのFR領域がヒト免疫グロブリン配列のものである、少なくとも1つ、典型的には2つの可変ドメインの実質的に全てを含む。ヒト化抗体は、最適には免疫グロブリン定常領域又はドメイン(Fc)、典型的にはヒトの免疫グロブリンの定常領域又はドメインの少なくとも一部を含んでなる。
【0041】
ここで意図している「生物学的に活性な」及び「所望の生物学的活性」とは、(1)インビボ又はエキソビボで少なくとも一種類の哺乳動物細胞において(作用的又は刺激する形あるいは拮抗的すなわち阻止する形で)アポトーシスを変調する能力を有しているか;(2)Apo-2リガンドに結合する能力を有しているか;又は(3)Apo-2リガンドのシグナル伝達及びApo-2リガンド活性を変調する能力を有していることを意味する。
【0042】
「アポトーシス」及び「アポトーシス活性」という用語は広義に使用され、典型的には、細胞質の凝結、原形質膜の微絨毛の喪失、核の分節化、染色体DNAの分解又はミトコンドリア機能の喪失を含む一又は複数の特徴的な細胞変化を伴う、哺乳動物における細胞死の規則的又はコントロールされた形態を指す。この活性は、例えば細胞生死判別アッセイ、FACS分析又はDNA電気泳動法等、全て従来から知られている方法により決定し測定することができる。
【0043】
「癌」及び「癌性」という用語は、典型的には調節されない細胞成長を特徴とする、哺乳動物における生理学的状態を指すか記述する。癌の例には、これらに限定されるものではないが、癌腫、リンパ腫、芽細胞腫、肉腫、及び白血病が含まれる。このような癌のより特定の例には、扁平上皮細胞癌、小細胞肺癌、非小細胞肺癌、芽細胞腫、胃腸癌、腎臓癌、膵臓癌、神経膠芽細胞腫、神経芽腫、子宮頸管癌、卵巣癌、肝臓癌、胃癌、膀胱癌、肝細胞腫(hepatoma)、乳癌、大腸癌、結腸直腸癌、子宮体癌、唾液腺癌、腎臓癌、肝臓癌、前立腺癌、産卵口癌、甲状腺癌、肝癌(hepatic carcinoma)及び様々な種類の頭部及び頸部の癌が含まれる。
【0044】
ここで使用される「治療する」、「治療」及び「治療法」とは、治癒的療法、予防的療法及び防護的療法を称する。
【0045】
ここで使用される「哺乳動物」という用語は、ヒト、ウシ、ウマ、イヌ及びネコを含む哺乳動物として分類されるあらゆる動物を指す。本発明の好ましい実施態様においては、哺乳動物はヒトである。
【0046】
II. 本発明の組成物と方法
本発明は、新規に同定され、単離されたApo-2DcRポリペプチドを提供する。特に本出願人は、種々のヒトApo-2DcRポリペプチドを同定し、分離した。これらApo-2DcRポリペプチドの幾つかのものの性質と特徴は、以下の実施例においてさらに詳細に記載する。ここで開示されるApo-2DcRポリペプチドの性質と特徴に基づき、Apo-2DcRはTNFRファミリーのメンバーであるというのが本出願人の信ずるところである。
【0047】
Apo-2DcR、並びにApo-2DcRキメラ分子及び抗Apo-2DcR抗体を如何に調製するかにつき、以下に説明する。
【0048】
A. Apo-2DcRの調製
以下の説明は、主として、Apo-2DcR核酸を含むベクターで形質転換又はトランスフェクトされた細胞を培養してApo-2DcRを生産する方法に関する。もちろん、当該分野においてよく知られている他の方法を用いてApo-2DcRを調製することはできると考えられる。
【0049】
1. Apo-2DcRをコードするDNAの単離
Apo-2DcRをコードするDNAは、Apo-2DcRmRNAを保有していてそれを検出可能なレベルで発現すると考えられる組織から調製された任意のcDNAライブラリから得ることができる。従って、ヒトApo-2DcRDNAは、実施例1に記載されたヒトのcDNAのライブラリのように、ヒトの組織から調製されたcDNAライブラリから簡便に得ることができる。またApo-2DcRコード化遺伝子は、ゲノムライブラリから又はオリゴヌクレオチド合成により得ることもできる。
【0050】
ライブラリは、対象となる遺伝子あるいはその遺伝子によりコードされるタンパク質を同定するために設計された(Apo-2DcRに対する抗体又は少なくとも約20−80塩基のオリゴヌクレオチド等の)プローブによってスクリーニングできる。選択されたプローブによるcDNA又はゲノムライブラリのスクリーニングは、例えばSambrookら, Molecular Cloning: A Laboratory Manual(New York: Cold Spring Harbor Laboratory Press, 1989)に記載されている標準的な手順を使用して実施することができる。Apo-2DcRをコードする遺伝子を単離する他の方法はPCR法を使用するものである[Sambrookら,上掲;Dieffenbachら,PCR Primer:A Laboratory Manual(Cold Spring Harbor Laboratory Press, 1995)]。
【0051】
スクリーニングの好ましい方法は、種々のヒト組織からcDNAライブラリをスクリーニングするために選別したオリゴヌクレオチド配列を用いることである。以下の実施例1には、cDNAライブラリのスクリーニング技術を記載している。プローブとして選択されたオリゴヌクレオチド配列は、充分な長さで、疑陽性が最小化されるよう充分に明瞭でなければならない。オリゴヌクレオチドは、スクリーニングされるライブラリ内のDNAとのハイブリッド化時に検出可能であるように標識されていることが好ましい。標識化の方法は、当該分野において良く知られており、32P標識されたATPのような放射標識、ビオチン化あるいは酵素標識の使用が含まれる。中程度の厳密性及び高厳密性を含むハイブリッド化条件は、Sambrookら,上掲に提供されている。
【0052】
全てのタンパク質コード化配列を有する核酸は、初めてここで開示された推定アミノ酸配列を使用し、また必要ならば、cDNAに逆転写されなかったmRNAの生成中間体及び先駆物質を検出する上掲のSambrookらにより記述されている従来のプライマー伸展法を使用し、選択されたcDNA又はゲノムライブラリをスクリーニングすることにより得られる。
【0053】
Apo-2DcR変異体はApo-2DcRDNAに適切なヌクレオチド変化を導入するか、所望のApo-2DcRポリペプチドを合成することにより、調製することができる。アミノ酸の変化により、グリコシル化部位の数と位置の変化、膜係留特性の変更のように、Apo-2DcRの翻訳後過程を改変し得ることは当業者であれば理解されることである。
【0054】
ここで記載されている天然全長配列Apo-2DcR又はApo-2DcRの種々のドメインにおける変異は、例えば米国特許第5364934号に記載された保存的あるいは非保存的突然変異に関する技術とガイドラインの任意のものを使用して発生させることができる。変異は、Apo-2DcRをコードしている一又は複数のコドンの置換、欠失又は挿入であり、天然配列Apo-2DcRに対してApo-2DcRのアミノ酸配列に変化が生じている。場合によっては、変異は、Apo-2DcR分子の一又は複数のドメインにおいて、少なくとも1つのアミノ酸を任意の他のアミノ酸に置換することによりなされる。変異は、オリゴヌクレオチド媒介(部位特異的)突然変異誘発、アラニンスキャニング、及びPCR突然変異誘発のような、当該分野において既知の方法を用いて行うことができる。部位特異的突然変異誘発[Carterら, Nucl. Acids. Res., 13:4331(1986);Zollerら, Nucl. Acids. Res., 10:6487(1987)]、カセット突然変異[Wellsら, Gene, 34:315(1985)]、制限選択的突然変異誘発[Wellら, Philos. Trans. R. Soc. London. SerA, 317:415(1986)]又は他の既知の技術をクローンDNAに実施してApo-2DcR変異体のDNAを生産することができる。
【0055】
また、特定のリガンド又はレセプターとの相互作用に関与する、近接配列に沿った一又は複数のアミノ酸を同定するために、スキャニングアミノ酸分析法を使用することもできる。好ましいスキャニングアミノ酸は、比較的小さい中性アミノ酸である。このようなアミノ酸には、アラニン、グリシン、セリン及びシステインが含まれる。変異体の主鎖構造をあまり改変することなく、ベータ炭素を越えた側鎖が除去されるため、アラニンがこの群のなかで好ましいスキャニングアミノ酸である。またアラニンは最も一般的なアミノ酸であることも好ましい。さらに、アラニンは埋設及び露出位置の双方に頻繁に見出される[Creighton, The Proteins, (W.H. Freeman & Co., N.Y.);Chothia, J. Mol. Biol., 150:1(1976)]。アラニン置換により適切な量の変異体を生じない場合には、アイソテリックアミノ酸を使用することができる。
【0056】
一度選択されたApo-2DcR変異体が生産されると、それらを、例えばApo-2Lと接触させることができ、相互作用があれば測定することができる。Apo-2DcR変異体とApo-2Lとの相互作用は、例えば以下の実施例に記載したようなインビトロアッセイにより測定することができる。天然配列Apo-2DcRとApo-2DcR変異体の活性と性質を比較するため任意の数の分析的測定法を使用することができ、結合性に対して簡便なものは、天然配列Apo-2DcRの解離定数Kdと比較したApo-2DcR変異体とApo-2Lとの間に形成された複合体の解離定数Kdである。一般に置換残基当り≧3倍Kdが増加又は減少すると、置換残基がApo-2Lとの天然配列Apo-2DcRの相互作用において活性であることになる。
【0057】
場合によっては、突然変異(例えば一又は複数のアミノ酸の欠失)に好適なApo-2DcR配列中の代表的な部位には、細胞外ドメイン、特に一又は複数のシステインリッチドメイン、又は一又は複数の擬似反復内の部位が含まれる。このような変異は上述の方法を使用することにより達成される。
【0058】
2. 複製可能なベクターへの核酸の挿入
Apo-2DcRをコードした核酸(例えば、cDNA又はゲノムDNA)は、さらなるクローニング(DNAの増幅)又は発現のために、複製可能なベクター内に挿入される。様々なベクターが公的に入手可能である。ベクター成分としては、一般に、これらに制限されるものではないが、次のものの一又は複数が含まれる:シグナル配列、複製開始点、一又は複数のマーカー遺伝子、エンハンサーエレメント、プロモーター、及び転写終結配列であり、それぞれを以下に説明する。
【0059】
(i) シグナル配列成分
Apo-2DcRは直接的に組換え手法によって生産されるだけではなく、シグナル配列あるいは成熟タンパク質あるいはポリペプチドのN末端に特異的切断部位を有する他のポリペプチドである異種性ポリペプチドとの融合ペプチドとしても生産される。一般に、シグナル配列はベクターの成分であるか、ベクターに挿入されるApo-2DcRDNAの一部である。好ましく選択された異種シグナル配列は宿主細胞によって認識され加工される(すなわち、シグナルペプチダーゼによって切断される)ものである。シグナル配列は、例えばアルカリホスファターゼ、ペニシリナーゼ、lppあるいは熱安定なエンテロトキシンIIリーダーの群から選択される原核生物シグナル配列であってよい。酵母の分泌に関しては、シグナル配列は、酵母インベルターゼリーダー、アルファ因子リーダー(酵母菌属(Saccharomyces)及びクルイベロマイシス(Kluyveromyces)α因子リーダーを含み、後者は米国特許第5010182号に記載されている)、又は酸ホスフォターゼリーダー、白体(C.albicans)グルコアミラーゼリーダー(1990年4月4日発行のEP362179)、又は1990年11月15日に公開された国際特許出願第WO90/13646号に記載されているシグナルであり得る。哺乳動物細胞の発現においては、同一あるいは関連ある種の分泌ポリペプチド由来のシグナル配列、単純ヘルペスグリコタンパク質Dシグナルのようなウイルス分泌リーダーのような他の哺乳動物のシグナル配列をタンパク質の直接分泌に使用してもよいが、インビボにおけるヒト細胞の細胞膜へのApo-2DcRの挿入を通常指示する天然Apo-2DcRプレ配列が十分である。
【0060】
このような前駆体領域のDNAは、好ましくは、Apo-2DcRをコードするDNAにリーディングフレームが結合される。
【0061】
(ii)複製開始点成分
発現及びクローニングベクターは共に一又は複数の選択された宿主細胞においてベクターの複製を可能にする核酸配列を含む。一般に、この配列はクローニングベクターにおいて、宿主染色体DNAとは独立にベクターが複製することを可能にするものであり、複製開始点又は自律的複製配列を含む。そのような配列は多くの細菌、酵母及びウイルスに対してよく知られている。プラスミドpBR322に由来する複製開始点は大部分のグラム陰性細菌に好適であり、2μプラスミド開始点は酵母に適しており、様々なウイルス開始点(SV40、ポリオーマ、アデノウイルス、VSV又はBPV)は哺乳動物細胞におけるクローニングベクターに有用である。一般には、哺乳動物の発現ベクターには複製開始点成分は不要である(SV40開始点が典型的には初期プロモーターを有しているため用いられる)。
【0062】
多くの発現ベクターは「シャトル」ベクターである、すなわち、それらは少なくとも一つのクラスの生物において複製可能であるが、発現のために他の生物に形質移入され得る。例えば、大腸菌においてベクターがクローン化され、そのベクターが酵母あるいは哺乳動物細胞に形質移入され、宿主細胞染色体と独立して複製することはできないとしても、発現する。
【0063】
DNAは宿主ゲノムに挿入することによって増幅され得る。これは、例えばベクターにバシラス(Bacillus)ゲノムDNAに見られる配列と相補的なDNA配列を含めることにより、宿主としてバシラス種を用いて容易に達成される。このベクターを用いたバシラスの形質移入は、ゲノムとの相同的組換え及びApo-2DcRDNAの挿入をもたらす。しかし、Apo-2DcRをコードするゲノムDNAの回収は、Apo-2DcRDNAを切除するのに制限酵素による消化を必要とするために、外来的に複製したベクターの場合よりも複雑である。
【0064】
(iii)選択遺伝子成分
発現及びクローニングベクターは、典型的には、選べるマーカーとも称される選択遺伝子を含む。この遺伝子は、選択的培地で成長させた形質転換宿主細胞の生存又は成長に必要なタンパク質をコードする。選択遺伝子を含むベクターと共に形質転換されていない宿主細胞は培地で生存しない。典型的な選択遺伝子は、(a)アンピシリン、ネオマイシン、メトトレキセートあるいはテトラサイクリンのような抗生物質あるいは他の毒素に耐性を与え、(b)栄養要求性欠陥を補い、(c)例えばバシリに対する遺伝子コードD-アラニンラセマーゼのような、複合培地から得られない重要な栄養素を供給する、タンパク質をコードする。
【0065】
選択技術の一例においては、宿主細胞の成長を抑止する薬物が用いられる。異種性遺伝子で首尾よく形質転換した細胞は、抗薬物性を付与し、選択工程を生存するタンパク質を生産する。このような優性選択の例としては、薬物ネオマイシン[Southernら, J. Molec. Appl. Genet., 1:327(1982)]、ミコフェノール酸[Mulliganら, Science, 209:1422(1980)]又はハイグロマイシン[Sugdenら, Mol. Cell. Biol., 5:410-413(1985)]が使用される。上述した3つの例は、真核生物のコントロール下で細菌遺伝子を用いて、適切な薬物G418又はネオマイシン(ジェネテシン)、xgpt(ミコフェノール酸)、又はハイグロマイシンにそれぞれ耐性を付与する。
【0066】
哺乳動物細胞に適切な選べるマーカーの他の例は、DHFRあるいはチミジンキナーゼのように、Apo-2DcR核酸を捕捉することのできる細胞成分を同定することのできるものである。哺乳動物細胞の形質転換細胞は、マーカーを捕捉することによって当該形質転換細胞のみが生存できるように独特に適応化された淘汰圧下に置かれる。淘汰圧は、培地中の選択剤の濃度が次第に変化する条件下で形質転換細胞を培養することにより課し、選択遺伝子とApo-2DcRをコードするDNAの双方を増幅させる。増幅は、成長に重要なタンパク質の生産に対する要求度が高い遺伝子が、組換え細胞の後の世代の染色体内に直列に反復されるプロセスである。増幅されたDNAから増大したApo-2DcR量が合成される。増幅可能な遺伝子のほかの例には、メタロチオネインI及びII、アデノシンデアミナーゼ、及びオルニチンデカルボキシラーゼが含まれる。
【0067】
DHFR選択遺伝子によって形質転換された細胞は、先ず、DHFRの競合的アンタゴニストであるメトトリキセート(Mtx)を含む培地において形質転換物の全てを培養することで同定される。野生型DHFRを用いた場合の好適な宿主細胞は、Urlaub ほかにより, Proc. Natl. Acad. Sci. USA, 77:7216 (1980)に記載されているようにして調製され増殖されたDHFR活性に欠陥のあるチャイニーズハムスター卵巣(CHO)株化細胞である。形質転換した細胞は次に濃度の高いメトトレキセートに接触させる。これによりDHFR遺伝子の複数コピーが合成され、同時に、Apo-2DcRをコードするDNAのような発現ベクターを含む他のDNAの複数コピーが作られる。この増幅方法は、もしMtxに高度に耐性である変異体DHFR遺伝子が使用されたような場合には内在性DHFRの存在にかかわらず、任意の他の適切な宿主、例えば、ATCC番号CCL61CHO−K1を使用することができる(EP117,060)。
【0068】
あるいは、Apo-2DcRをコードするDNA配列、野生型DHFRタンパク質、及びアミノグリコシド3'-ホスホトランスフェラーゼ(APH)のような他の選べるマーカーで形質転換あるいは同時形質転換した宿主細胞(特に、内在性DHFRを含む野生型宿主)は、カナマイシン、ネオマイシンあるいはG418のようなアミノグリコシド抗生物質のような選択可能マーカーの選択剤を含む培地における細胞増殖により選択することができる。米国特許第4965199号を参照。
【0069】
酵母中での使用に好適な選択遺伝子は酵母プラスミドYRp7に存在するtrp1遺伝子である[Stinchcombら, Nature, 282:39(1979);Kingmanら, Gene, 7:141(1979);Tschemperら, Gene, 10:157(1980)]。trp1遺伝子は、例えば、ATCC第44076号あるいはPEP4-1のようなトリプトファン内で成長する能力に欠ける酵母の突然変異株に対する選択マーカーを提供する[Jones, Genetics, 85:12 (1977)]。酵母宿主細胞ゲノムにtrp1破壊が存在することは、トリプトファンの不存在下における増殖による形質転換を検出するための有効な環境を提供する。同様に、Leu2欠陥酵母株(ATCC 20,622あるいは 38,626)は、Leu2遺伝子を有する既知のプラスミドによって補完される。
【0070】
更に、1.6μmの円形プラスミドpKD1由来のベクターは、クルイヴェロマイシス(Kluyveromyces)酵母の形質転換に用いることができる[Bianchi ほか., Curr. Genet., 12:185(1987)]。より最近では、組換え子ウシのキモシンの大量生産のための発現系がK.ラクティス(lactis)に対して報告されている[Van den Berg, Bio/Technology, 8:135 (1990)]。クルイヴェロマイシスの工業的な菌株からの、組換えによる成熟したヒト血清アルブミンを分泌する安定した複数コピー発現ベクターも開示されている[Fleer ほか, Bio/Technology,9:968-975 (1991)]。
【0071】
(iv)プロモーター成分
発現及びクローニングベクターは、通常、宿主生物によって認識され、Apo-2DcR核酸配列に作用可能に結びついているプロモーターを含む。プロモーターは、作用可能に結合しているApo-2DcR核酸配列のような特定の核酸配列の転写及び翻訳を制御する構造的な遺伝子(一般的に約100ないし1000塩基対)の開始コドンの上流側(5')に位置する未翻訳配列である。このようなプロモーターは典型的には、誘発的なクラス及び構成的なクラスの2つのクラスに属する。誘発的なプロモーターは、養分の存在あるいは不存在、温度変化等の培養条件のある変化に対応してその制御の下でDNAからの転写レベルを上昇させるプロモーターである。現時点において多種の可能な宿主細胞により認識される非常に多くのプロモーターがよく知られている。これらのプロモーターは、制限酵素の消化によって供給源DNAからプロモーターを排除し、ベクターに単離したプロモーター配列を挿入することで、Apo-2DcRをコードするDNAに作用的に結合している。天然のApo-2DcRプロモーター配列及び多くの異種性プロモーターはいずれもApo-2DcRDNAの直接増幅及び/又は発現に用いることができる。
【0072】
原核生物宿主での使用に好適なプロモーターはβラクタマーゼ及びラクトースプロモーター系[Cahng ほか, Nature, 275:615 (1978), Goeddel ほか, Nature, 281:544 (1979)]、アルカリホスファターゼ、トリプトファン(trp)プロモーター系[Goeddel, Nucleic Acids Res., 8:4057 (1980); EP 36,776]、及びハイブリッドプロモーター、例えばtacプロモーター[deBoer ほか, Proc. Natl. Acad. Sci. USA, 80:21-25 (1983)]を含む。しかし、他の既知の細菌プロモーターも好適である。これらのヌクレオチド配列は公表されており、よって当業者は、任意の所望の制限部位を提供するためにリンカーあるいはアダプターを使用することでApo-2DcRをコードするDNA[Siebenlist ほか, Cell, 20:269 (1980)]にそれらを作用可能に結合させることが可能である。細菌系で使用するプロモータもまたApo-2DcRをコードするDNAと作用可能に結合したシャイン・ダルガーノ(S.D.)配列を有する。
【0073】
真核生物に対してもプロモーター配列が知られている。実質的に全ての真核生物の遺伝子は、転写開始部位からおよそ25ないし30塩基上流に見出されるATリッチ領域を有する。多数の遺伝子の転写開始位置から70ないし80塩基上流に見出される他の配列は、Xが任意のヌクレオチドであるCXCAAT領域である。大部分の真核生物遺伝子の3'末端には、コード配列の3'末端へのポリA尾部の付加に対するシグナルであるAATAAA配列がある。これらの配列は全て真核生物の発現ベクターに適切に挿入される。
【0074】
酵母宿主と共に用いて好適なプロモーター配列の例としては、3-ホスホグリセラートキナーゼ[Hitzeman ほか, J. Biol. Chem., 255:2073 (1980)]又は他の糖分解酵素[Hess ほか, J. Adv. Enzyme Reg., 7:149 (1968);Holland, Biochemistry, 17:4900(1987)]、例えばエノラーゼ、グリセルアルデヒド-3-リン酸デヒドロゲナーゼ、ヘキソキナーゼ、ピルビン酸デカルボキシラーゼ、ホスホフルクトキナーゼ、グルコース-6-リン酸イソメラーゼ、3-ホスホグリセレートムターゼ、ピルビン酸キナーゼ、トリオセリン酸イソメラーゼ、ホスホグルコースイソメラーゼ、及びグルコキナーゼが含まれる。
【0075】
他の酵母プロモーターとしては、成長条件によって転写が制御される付加的効果を有する誘発的プロモーターであり、アルコールデヒドロゲナーゼ2、イソチトクロムC、酸ホスファターゼ、窒素代謝と関連する分解性酵素、メタロチオネイン、グリセルアルデヒド-3-リン酸デヒドロゲナーゼ、及びマルトース及びガラクトースの利用を支配する酵素のプロモーター領域がある。酵母の発現に好適に用いられるベクターとプロモータは欧州特許第73657号に更に記載されている。また酵母エンハンサーも酵母プロモーターと共に好適に用いられる。
【0076】
哺乳動物の宿主細胞におけるベクターからのApo-2DcR転写は、例えば、ポリオーマウィルス、伝染性上皮腫ウィルス(1989年7月5日公開のUK 2,211,504)、アデノウィルス(例えばアデノウィルス2)、ウシ乳頭腫ウィルス、トリ肉腫ウィルス、サイトメガロウィルス、レトロウィルス、B型肝炎ウィルス及び最も好ましくはサルウィルス40(SV40)のようなウィルスのゲノムから得られるプロモーター、異種性哺乳動物プロモーター、例えばアクチンプロモーター又は免疫グロブリンプロモーター、熱衝撃プロモーター、そしてApo-2DcR配列に通常付随するプロモーターによって、このようなプロモーターが宿主細胞系に適合し得る限り、調節される。
【0077】
SV40ウィルスの初期及び後期プロモーターは、SV40ウイルスの複製起点をさらに含むSV40制限断片として簡便に得られる[Fiersほか, Nature, 273:113 (1978);Mulligan及びBerg, Science, 209:1422-1427 (1980); Pavlakisほか, Proc. Natl. Acad. Sci. USA, 78:7398-7402 (1981)]。ヒトサイトメガロウィルスの最初期プロモーターは、HindIIIE制限断片として簡便に得られる[Greenawayほか, Gene, 18:355-360 (1982)]。ベクターとしてウシ乳頭腫ウィルスを用いて哺乳動物宿主でDNAを発現する系が、米国特許第4419446号に開示されている。この系の修飾は米国特許第4601978号に開示されている[また、サル細胞での免疫インターフェロンをコードしているcDNAの発現について、Grayら, Nature, 295:503-508(1982)を;単純ヘルペスウイルス由来のチミジンキナーゼプロモーターの調節下でのマウス細胞におけるヒトβインターフェロンcDNAの発現について、Reyesら, Nature, 297:598-601(1982)を;培養されたマウス及びウサギの細胞におけるヒトインターフェロンβ1遺伝子の発現について、Canaani及びBerg, Proc. Natl. Acad. Sci. USA, 79:5166-5170(1982)を;プロモーターとしてラウス肉腫ウィルスの長い末端反復を用いたCV-1サル腎臓細胞、ニワトリ胚線維芽細胞、チャイニーズハムスター卵巣細胞、HeLa細胞、及びマウスNIH-3T3細胞における細菌CAT配列の発現について、Gormanほか, Proc. Natl. Acas. Sci. USA, 79:6777-6781 (1982)を参照のこと]。
【0078】
(v)エンハンサーエレメント成分
より高等の真核生物による本発明のApo-2DcRをコードしているDNAの転写は、ベクター中にエンハンサー配列を挿入することによって増強され得る。エンハンサーは、通常は約10から300塩基対で、プロモーターに作用してその転写を増強するDNAのシス作動要素である。エンハンサーは、相対的に配向及び位置が独立しており、転写ユニットの5'[Laiminsほか, Proc. Natl. Acad. Sci. USA, 78:993 (1981)]及び 3'[Luskyほか, Mol. Cell Bio., 3:1108 (1983)]、イントロン内部[Banerjiほか, Cell, 33:729 (1983)]並びにコード配列自身の内部[Osbornほか, Mol. Cell Bio., 4:1293 (1984)]に見出されている。哺乳動物の遺伝子由来の多くのエンハンサー配列が現在知られている(グロビン、エラスターゼ、アルブミン、α-フェトプロテイン及びインスリン)。しかしながら、典型的には、真核細胞ウィルス由来のエンハンサーが用いられるであろう。例としては、複製起点の後期側のSV40エンハンサー(100-270塩基対)、サイトメガロウィルス初期プロモーターエンハンサー、複製起点の後期側のポリオーマエンハンサー及びアデノウィルスエンハンサーが含まれる。真核生物のプロモーターの活性化のための増強要素については、Yaniv, Nature, 297:17-18 (1982)もまた参照のこと。エンハンサーは、Apo-2DcRコード配列の5'又は3'位でベクター中にスプライシングされ得るが、好ましくはプロモーターから5'位に位置している。
【0079】
(vi)転写終結成分
真核生物宿主細胞(酵母、真菌、昆虫、植物、動物、ヒト、又は他の多細胞生物由来の有核細胞)に用いられる発現ベクターは、また転写の終結及びmRNAの安定化に必要な配列を含む。このような配列は、真核生物又はウィルスのDNA又はcDNAの5'、時には3'の非翻訳領域から一般に取得できる。これらの領域は、Apo-2DcRをコードしているmRNAの非翻訳部分にポリアデニル化断片として転写されるヌクレオチドセグメントを含む。
【0080】
(vii)ベクターの作成と分析
一又は複数の上に列挙した成分を含む適切なベクターの作成には標準的なライゲーション技術を用いる。分離されたプラスミド又はDNA断片を開裂させ、整え、そして必要とされるプラスミドの生成のために望ましい型に再ライゲーションする。
【0081】
作成されたプラスミドが正しい配列であることを確認する分析のために、ライゲーション混合物を用いて、大腸菌K12菌株294(ATCC 31446)を形質転換し、適当な場合にはアンピシリン又はテトラサイクリン耐性によって、形質転換細胞を好適に選択する。形質転換細胞からプラスミドを調製し、制限エンドヌクレアーゼ消化により分析し、及び/又はMessingほか, Nucleic Acids Res., 9:309 (1981)の方法又はMaximほか, Methods in Enzymology, 65:499 (1980)の方法によって配列決定する。
【0082】
(viii)一過性発現ベクター
Apo-2DcRをコードしているDNAの哺乳動物細胞における一過性発現をもたらす発現ベクターを使用することができる。一般に、一過性発現は、宿主細胞が発現ベクターの多くのコピーを蓄積し、次にその発現ベクターによってコードされている所望のポリペプチドを高レベルで合成するように、宿主細胞中で効果的に複製できる発現ベクターを使用することを含む[Sambrookほか,上掲]。一過性発現系は、適切な発現ベクターと宿主細胞を含むが、クローニングされたDNAによりコードされているポリペプチドの簡便で確実な同定並びに所望の生物学的又は生理学的性質についてのポリペプチドの迅速なスクリーニングを可能にする。したがって、一過性発現系は、本発明において、Apo-2DcR変異体を同定する目的のために特に有用である。
【0083】
(ix)適切な例示的脊椎動物細胞ベクター
組換え脊椎動物細胞培養でのApo-2DcRの合成に適応化するのに適切な他の方法、ベクター及び宿主細胞は、Gethingほか, Nature, 293:620-625 (1981); Manteiほか, Nature, 281:40-46 (1979); 欧州特許第117060号; 及び欧州特許第117058号に記載されている。
【0084】
3. 宿主細胞の選択及び形質転換
ここに記載のベクターにDNAをクローニングあるいは発現するために適切な宿主細胞は、上述の原核生物、酵母、又は高等真核生物細胞である。この目的にとって適切な原核生物は、限定するものではないが、真正細菌、例えばグラム陰性又はグラム陽性生物体、例えばエシェリチアのような腸内菌科、例えば大腸菌、エンテロバクター、エルウィニア(Erwinia)、クレブシエラ、プロテウス、サルモネラ、例えばネズミチフス菌、セラチア属、例えばセラチア・マルセスキャンス及び赤痢菌属、並びに桿菌、例えば枯草菌及びバシリ・リチェフォルミス(licheniformis)(例えば、1989年4月12日に公開された DD 266,710に開示されたバシリ・リチェニフォルミス41P)、シュードモナス属、例えば緑膿菌及びストレプトマイセス属を含む。好ましくは、宿主細胞は最小量のタンパク質分解酵素を分泌すべきである。
【0085】
原核生物に加えて、糸状菌又は酵母菌のような真核微生物は、Apo-2DcRをコードするベクターのための適切なクローニング又は発現宿主である。サッカロミセス・セレヴィシア、又は一般的なパン酵母は下等真核生物宿主微生物のなかで最も一般的に用いられる。しかしながら、多数の他の属、種及び菌株も、一般的に入手可能で有用である。
【0086】
グリコシル化Apo-2DcRの発現に適切な宿主細胞は、多細胞生物から誘導される。このような宿主細胞は、複雑なプロセシング及びグリコシル化活動が可能である。原則的には、脊椎動物であろうと無脊椎動物培養であろうと、任意のより高等の真核生物細胞培養が使用できる。無脊椎動物細胞の例としては植物及び昆虫細胞が含まれる。多数のバキュロウィルス株及び変異体及び対応する許容可能な昆虫宿主細胞、例えばスポドプテラ・フルギペルダ(毛虫)、アエデス・アエジプティ(蚊)、アエデス・アルボピクトゥス(蚊)、ドゥロソフィラ・メラノガスター(ショウジョウバエ)、及びボンビクス・モリが同定されている[例えば、Luckowほか, Bio/Technology, 6:47-55 (1988); Millerほか, Genetic Engineering, Setlowほか, eds., Vol. 8 (Plenum Publishing, 1986), pp.277-279; 及びMaedaほか, Nature, 315:592-594 (1985)を参照のこと]。トランスフェクションのための種々のウィルス株、例えば、オートグラファ・カリフォルニカNPVのL-1変異体とボンビクス・モリ NPVのBm-5株が公に利用できる。
【0087】
綿花、コーン、ジャガイモ、大豆、ペチュニア、トマト、及びタバコのような植物細胞培養を宿主として利用することができる。典型的には、植物細胞は、細菌アグロバクテリウム・トゥメファシエンスのある菌株と共にインキュベートすることによってトランスフェクトされる。A. トゥメファシエンスと共に植物細胞培養をインキュベートする間に、Apo-2DcRをコードしているDNAが、植物細胞宿主がトランスフェクトされるようにその植物細胞宿主に移され、そして適切な条件下でApo-2DcRをコードしているDNAを発現させる。加えて、例えば、ノパリンシンターゼプロモーター及びポリアデニル化シグナル配列のような、植物細胞と適合しうる調節及びシグナル配列が利用できる[Depickerほか, J. Mol. Appl. Gen., 1:561 (1982)]。また、T-DNA780遺伝子の上流領域から分離されるDNAセグメントは、組換えDNAを含む植物組織中の植物発現遺伝子の転写レベルを活性化又は増強しうる[1989年6月21日公開のEP 321,196]。
【0088】
培養(組織培養)中での脊椎動物細胞の増殖は、当該分野おいてよく知られている[例えば、Tissue Culture,Academic Press,編者Kruse及びPatterson(1973)を参照のこと]。有用な哺乳動物宿主株化細胞の例は、SV40によって形質転換されたサル腎臓CV1株 (COS-7, ATCC CRL 1651);ヒト胚腎臓株[293又は懸濁培養での増殖のためにサブクローン化された293細胞、Grahamほか, J. Gen Virol., 36:59 (1977)];ハムスター乳児腎細胞(BHK, ATCC CCL 10);チャイニーズハムスター卵巣細胞/-DHFR(CHO, Urlaub及びChasin, Proc. Natl. Acad. Sci. USA, 77:4216 (1980));マウスのセルトリ細胞[TM4, Mather, Biol. Reprod., 23:243-251 (1980)];サルの腎細胞 (CVI ATCC CCL 70); アフリカミドリザルの腎細胞(VERO-76, ATCC CRL-1587); ヒト子宮頸癌細胞 (HELA, ATCC CCL 2); イヌ腎細胞 (MDCK, ATCC CCL 34); バッファローラット肝細胞 (BRL 3A, ATCC CRL 1442); ヒト肺細胞 (W138, ATCC CCL 75); ヒト肝細胞 (Hep G2, HB 8065); マウス乳房腫瘍細胞 (MMT 060562, ATTC CCL51);TRI細胞[Motherほか, Annals N.Y. Acad. Sci., 383:44-68 (1982)];MRC5細胞;FS4細胞である。
【0089】
宿主細胞をトランスフェクトし、好ましくは上述のApo-2DcR生成のための発現又はクローニングベクターで形質転換し、プロモーターを誘導し、形質転換体を選択し、又は所望の配列をコードしている遺伝子を増幅するために適当に修飾された常套的栄養培地で培養する。
【0090】
トランスフェクションは、如何なるコード配列が実際に発現されるか否かにかかわらず、宿主細胞による発現ベクターの取り上げを意味する。多数のトランスフェクションの方法が当業者に知られている。例えば、CaPO4及びエレクトロポレーションである。このベクターの操作のあらゆる徴候が宿主細胞内で生じたときに成功したトランスフェクションが一般に認められる。
【0091】
形質転換は、染色体外のエレメントとしてであろうと染色体成分によってであろうと、DNAが複製可能であるように、生物体中にDNAを導入することを意味する。用いられる宿主細胞に応じて、そのような細胞に対して適した標準的な方法を用いて形質転換はなされる。前掲のSambrookほかにより記載された塩化カルシウムを用いるカルシウム処理又はエレクトロポレーションが、原核生物又は実質的な細胞壁障壁を含む他の細胞に対して用いられる。アグロバクテリウム・トゥメファシエンスによる感染が、Shawほか, Gene, 23:315 (1983)及び1989年6月29日公開の国際特許出願第WO89/05859号に記載されたように、ある種の植物細胞の形質転換に用いられる。加えて、1991年1月10日に公開された国際特許出願第WO91/00358号に記載されているように、超音波処理を用いて植物をトランスフェクトすることもできる。
【0092】
このような細胞壁のない哺乳動物の細胞に対しては、Graham及びvan der Eb, Virology, 52:456-457 (1978)のリン酸カルシウム沈殿法が好ましい。哺乳動物細胞の宿主系形質転換の一般的な側面は米国特許第4399216号に記載されている。酵母中の形質転換は、典型的には、Van solingenほか, J. Bact., 130:946 (1977)及びHsiaoほか, Proc. Natl. Acad. Sci. USA, 76:3829 (1979)の方法によって実施する。しかしながら、DNAを細胞中に導入する他の方法、例えば、核マイクロインジェクション、エレクトロポレーション、無傷の細胞、又はポリカチオン、例えばポリブレン、ポリオルニチン等を用いる細菌プロトプラスト融合もまた用いることもできる。哺乳動物細胞を形質転換するための種々の技術については、Keownほか, Methods in Enzymology, 185:527-537 (1990)及び Mansourほか, Nature, 336:348-352 (1988)を参照のこと。
【0093】
4.宿主細胞の培養
本発明のApo-2DcRポリペプチドを生成るために用いられる原核細胞は、前掲のSambrookほかにより記載されているような適切な培地で培養される。
【0094】
Apo-2DcRの生産に用いられる哺乳動物の宿主細胞は種々の培地において培養することができる。市販培地の例としては、ハム(Ham)のF10(シグマ)、最小必須培地(「MEM」,シグマ)、RPMI-1640(シグマ)及びダルベッコの改良イーグル培地(「DMEM」,シグマ)が含まれる。これらの培地はいずれも、ホルモン及び/又は他の成長因子(例えばインスリン、トランスフェリン、又は表皮成長因子)、塩類(例えば、塩化ナトリウム、カルシウム、マグネシウム及びリン酸塩)、バッファー(例えばHEPES)、ヌクレオシド(例えばアデノシン及びチミジン)、抗生物質(例えば、ゲンタマイシンTM薬)、微量元素(最終濃度がマイクロモル範囲で通常存在する無機化合物として定義される)及びグルコース又は同等のエネルギー源を必要に応じて補充することができる。任意の他の必要な補充物質もまた当業者に知られている適当な濃度で含むことができる。培養条件、例えば温度、pH等々は、発現のために選ばれた宿主細胞について以前から用いられているものであり、当業者には明らかであろう。
【0095】
一般に、哺乳動物の細胞培養の生産性を最大にするための原理、プロトコール、及び実用技術は、Mammalian Cell Biotechnology: a Practical Approach, M.Butler編 (IRL Press, 1991)に見出すことができる。
【0096】
この明細書において言及される宿主細胞は培養中の細胞並びに宿主動物内にある細胞を包含する。
【0097】
5. 遺伝子増幅/発現の検出
遺伝子の増幅及び/又は発現は、ここで提供された配列に基づき、適切に標識されたプローブを用い、例えば、従来よりのサザンブロット法、mRNAの転写を定量化するノーザンブロット法[Thomas,Proc. Natl. Acad. Sci. USA,77:5201-5205 (1980)]、ドットブロット法(DNA分析)、又はインシトゥハイブリッド形成法によって、直接的に試料中で測定することができる。種々の標識を用いることができ、最も一般的なものは放射性同位元素、特に32Pである。しかしながら、他の方法、例えばポリヌクレオチド中への導入のためのビオチン修飾されたヌクレオチドもまた使用することができる。ついで、このビオチンは、例えば放射性ヌクレオチド、蛍光剤又は酵素のような広範囲の標識で標識することができるアビジン又は抗体への結合部位として作用する。また、DNA二本鎖、RNA二本鎖及びDNA−RNAハイブリッド二本鎖又はDNA-タンパク二本鎖を含む、特異的二本鎖を認識することができる抗体を用いることもできる。ついで、抗体を標識し、アッセイを実施することができ、ここで二本鎖は表面に結合しており、その結果二本鎖の表面での形成の時点でその二本鎖に結合した抗体の存在を検出することができる。
【0098】
あるいは、遺伝子の発現は、遺伝子産物の発現を直接的に定量する免疫学的な方法、例えば細胞又は組織切片の免疫組織化学的染色及び細胞培養又は体液のアッセイによって、測定することもできる。免疫組織化学的染色技術では、細胞試料を、典型的には脱水と固定によって調製し、結合した遺伝子産物に対し特異的な標識化抗体と反応させるが、この標識は通常は視覚的に検出可能であり、例えば酵素的標識、蛍光標識、又はルミネサンス標識である。
【0099】
試料液の免疫組織化学的染色及び/又はアッセイに有用な抗体は、モノクローナルでもポリクローナルでもよく、任意の哺乳動物で調製することができる。簡便には、抗体は、天然配列Apo-2DcRポリペプチドに対して、又はここで提供されるDNA配列をベースとした合成ペプチドに対して、又はApo-2DcRDNAに融合し特異的抗体エピトープをコードする外因性配列に対して調製され得る。
【0100】
6. Apo-2DcRポリペプチドの精製
Apo-2DcRの形態は、培地又は宿主細胞の溶菌液から回収することができる。Apo-2DcRが膜結合性であるならば、適切な洗浄液(例えばトリトン-X100)を用いて膜から引き離すか、又はその細胞外ドメインを酵素的に切断する。Apo-2DcRはまた糖リン脂質膜アンカーの酵素的切断により細胞表面から引き離すことができる。
【0101】
Apo-2DcRがヒト起源のもの以外の組換え細胞でつくられるときは、Apo-2DcRはヒト起源のタンパク質又はポリペプチドを含んでいない。しかしながら、Apo-2DcRに関して実質的に相同である調製物を得るには、組換え細胞タンパク又はポリペプチドからApo-2DcRを精製することが望ましい。第一段階として、培地又は溶菌液を遠心分離して粒状の細胞屑を除去することができる。ついで、Apo-2DcRを、汚染した可溶性タンパク質及びポリペプチドから、適切な精製手順の例である次の手順により精製される:すなわち、イオン交換カラムでの分画;エタノール沈殿;逆相HPLC;シリカ又はカチオン交換樹脂、例えばDEAEによるクロマトグラフィー;クロマトフォーカシング;SDS-PAGE;硫酸アンモニウム沈殿;例えばセファデックスG-75を用いるゲル濾過;及びIgGのような汚染物を除くプロテインAセファロースカラムである。
【0102】
残基が欠失され、挿入され、又は置換されたApo-2DcR変異体は、その変異によってしばしば惹起された実質的な性質変化を考慮に入れて、天然配列Apo-2DcRと同じようにして回収することができる。例えば、他のタンパク質又はポリペプチド、例えば細菌性もしくはウイルス性抗原、免疫グロブリン配列、又はレセプター配列とApo-2DcRとの融合体の調製は精製を容易にする;配列に対する抗体を含む免疫アフィニティーカラムを、融合ポリペプチドを吸着するために使用することができる。他の種類のアフィニティーマトリックスもまた使用することもできる。
【0103】
例えばフェニルメチルスルホニルフロリド(PMSF)のようなプロテアーゼインヒビターもまた精製の間のタンパク分解を阻害するのに有用であり、偶発的な汚染物質の成長を防止するために抗生物質を含めることができる。天然配列Apo-2DcRに適切な精製方法は、組換え細胞培養の発現の際におけるApo-2DcR又はその変異体の特性の変化の起因となる改変が必要となることは、当業者であれば分かるであろう。
【0104】
7. Apo-2DcRポリペプチドの共有結合的修飾
Apo-2DcRの共有結合的修飾は本発明の範囲内に含まれる。Apo-2DcRの共有結合的修飾の一つの型は、Apo-2DcRの標的アミノ酸残基を、Apo-2DcRのN末端又はC末端残基、又は選択された側鎖と反応できる有機誘導体化剤と反応させることによって分子内に導入することができる。
【0105】
二官能性試薬による誘導体形成は、抗Apo-2DcR抗体を精製する方法に使用する水不溶性支持体マトリックス又は表面へのApo-2DcRの架橋に有用であり、またその逆も同様である。二官能性試薬による誘導体形成は、Apo-2DcR分子を架橋させてApo-2DcR二量体を産生するのにも有用である。このような二量体により結合アビディティーが増大させられ、インビボにおける分子の半減期が延びる。通常使用される架橋剤には、例えば、1,1-ビス(ジアゾアセチル)-2-フェニルエタン、グルタルアルデヒド、N-ヒドロキシスクシンイミドエステル、例えば、4-アジドサリチル酸とのエステル、3,3'-ジチオビス(スクシンイミジルプロピオナート)のようなジスクシンイミジルエステルを包含するホモ二官能性イミドエステル、及びビス-N-マレイミド-1,8-オクタンのような二官能性マレイミドが含まれる。メチル-3-[(p-アジドフェニル)ジチオ]プロピオイミダートのような誘導体化剤は、光の存在下で架橋を形成することができる光活性化中間体を生じる。また、臭化シアン活性化炭水化物のような反応性の水不溶性マトリックス及び米国特許第3969287号;3691016号;4195128号;4247642号;4229537号及び4330440号に記載されている反応性基質がタンパク固定に用いられる。
【0106】
その他の修飾は、それぞれ対応するグルタミル及びアスパルチル残基へのグルタミニル及びアスパラギニル残基の脱アミド化、プロリンとリジンのヒドロキシル化、セリル又はスレオニル残基のヒドロキシル基のリン酸化、リジン、アルギニン、及びヒスチジン側鎖のαアミノ基のメチル化[T.E. Creighton, Proteins: Structure and Molecular Properties, W.H. Freeman & Co., San Francisco, PP.79-86 (1983)]、N末端アミンのアセチル化、及び任意のC末端カルボキシル基のアミド化を含む。残基の修飾型も本発明の範囲に入る。
【0107】
本発明の範囲内に含まれるApo-2DcRポリペプチドの共有結合的修飾の他のタイプは、ポリペプチドの天然グリコシル化パターンを変更することを含む。「天然グリコシル化パターンの変更」とは、天然配列Apo-2DcRに見出される一又は複数の炭水化物部分の欠失、及び/又は天然配列Apo-2DcRに存在しない一又は複数のグリコシル化部位を付加することを意味することをここでは意図している。
【0108】
ポリペプチドのグリコシル化は、典型的には、N結合又はO結合の何れかである。N結合とは、アスパラギン残基の側鎖への炭水化物部分の結合を指す。アスパラギン-X-セリン及びアスパラギン-X-スレオニン(ここでXはプロリンを除く任意のアミノ酸)というトリペプチド配列は、アスパラギン側鎖への炭水化物部分の酵素的結合のための認識配列である。したがって、ポリペプチド中にこれらのトリペプチド配列の何れかが存在すると、可能性の有るグリコシル化部位が作り出される。O結合グリコシル化は、ヒドロキシルアミノ酸、最も一般的にはセリン又はスレオニン(5-ヒドロキシプロリン又は5-ヒドロキシリジンもまた用いられるが)に、糖類N-アセチルガラクトサミン、ガラクトース、又はキシロースの一つが結合することを意味する。
【0109】
Apo-2DcRポリペプチドへのグリコシル化部位の付加は、アミノ酸配列を、それが一又は複数の上述したトリペプチド配列(N結合グリコシル化部位のもの)を含むように変化させることによって達成される。この変化は、天然配列Apo-2DcRへの一又は複数のセリン又はスレオニン残基の付加、又はこれによる置換によってもなされる(O結合グリコシル化部位の場合)。場合によっては、Apo-2DcRアミノ酸はDNAレベルでの変化によって、特に、所望のアミノ酸に翻訳するコドンが産生されるように予め選んだ塩基でApo-2DcRポリペプチドをコードしているDNAを突然変異することによって変更される。このDNA突然変異は、上記に記載され前掲の米国特許第5364934号に記載された方法を用いてなされる。
【0110】
Apo-2DcRポリペプチド上の炭水化物部分の数を増加させる他の手段は、該ポリペプチドへのグリコシドの化学的又は酵素的結合による。用いられる結合形態に応じて、糖(類)は、(a)アルギニンとヒスチジンに、(b)遊離のカルボキシル基に、(c)遊離のスルフヒドリル基、例えばシステインのものに、(d)セリン、スレオニン又はヒドロキシプロリンのもののような遊離のヒドロキシル基に、(e)フェニルアラニン、チロシン又はトリプトファンのような芳香族残基、又は(f)グルタミンのアミノ基に結合される。これらの方法は1987年9月11日公開の国際特許出願第WO87/05330号及びAplin及びWriston, CRC Crit. Rev. Biochem., pp259-306 (1981)に記載されている。
【0111】
Apo-2DcRポリペプチド上に存在する炭水化物部分の除去は、化学的又は酵素的あるいはグリコシル化の標的となるアミノ酸残基をコードするコドンの突然変異的置換によりなされる。例えば、化学的脱グリコシル化は、化合物トリフルオロメタンスルホン酸、又は等価な化合物へ該ポリペプチドを曝露し、該ポリペプチドを無傷のまま残しながら、結合糖(N-アセチルグルコサミン又はN-アセチルガラクトサミン)を除く殆ど又は全ての糖を開裂させる。化学的脱グリコシル化は、Hakimuddinほか, Arch. Biochem Biophys., 259:52 (1987)及びEdgeほか, Anal. Biochem., 118:131 (1981)により記載されている。ポリペプチド上の炭水化物部分の酵素的開裂は、Thotakuraほか, Meth. Enzymol., 138:350 (1987)に記載されているように、種々のエンド及びエキソグリコシダーゼを使用して達成することができる。
【0112】
潜在的なグリコシル化部位でのグリコシル化は、Duskinほか, J. Biol. Chem., 257:3105 (1982)によって記載されているように、化合物ツニカマイシンを使用して防ぐことができる。ツニカマイシンはタンパク質-N-グルコシド結合の形成を阻害する。
【0113】
Apo-2DcRの共有結合的修飾の他のタイプは、米国特許番第4640835号;第4496689号;第4301144号;第4670417号;第4791192号又は第4179337号に記載されているように、Apo-2DcRポリペプチドを、種々の非タンパク性ポリマーの一つ、例えばポリエチレングリコール、ポリプロピレングリコール、又はポリオキシアルキレンに結合させることを含む。
【0114】
8. Apo-2DcRキメラ
また本発明は、他の異種性ポリペプチド又はアミノ酸配列と融合したApo-2DcRを含むキメラ分子を提供する。
【0115】
一実施態様では、キメラ分子は、抗タグ抗体が選択的に結合するエピトープを提供するタグポリペプチドとのApo-2DcRの融合体を含む。エピトープタグは一般にApo-2DcRのアミノ又はカルボキシル末端に位置させられる。Apo-2DcRのこのようなエピトープタグが付けられた形は、その存在をタグポリペプチドに対する抗体を用いて検出することができる。また、エピトープタグを供給すると、Apo-2DcRを抗タグ抗体又はエピトープタグに結合する他の種類のアフィニティーマトリックスを用いたアフィニティー精製によって直ぐに精製することができる。
【0116】
様々なタグポリペプチドとその各抗体は従来から良く知られている。例には、fluHAタグポリペプチドとその抗体12CA5[Fieldほか, Mol. Cell. Biol., 8:2159-2165 (1988)];c-mycタグとそれに対する8F9、3C7、6E10、G4、B7及び9E10抗体[Evanほか, Molecular and Cellular Biology, 5:3610-3616 (1985)];及び単純ヘルペスウィルス糖タンパクD(gD)タグとその抗体[Paborskyほか, Protein Engineering, 3(6):547-553 (1990)]が含まれる。他のタグポリペプチドには、フラッグ-ペプチド(Flag-peptide)[Hoppほか, BioTechnology, 6:1204-1210 (1988)];KT3エピトープペプチド[Martinほか, Science, 255:192-194 (1992)];αチューブリンエピトープペプチド[Skinnerほか, J. Biol. Chem., 266:15163-15166 (1991)];及びT7遺伝子10タンパクペプチドタグ[Lutz-Freyermuthほか, Proc. Natl. Acad. Sci. USA, 87:6393-6397 (1990)]が含まれる。ひとたびタグポリペプチドが選択されれば、それに対する抗体を、ここに開示した方法を用いて産生することができる。
【0117】
一般的に、エピトープタグApo-2DcRは、上述した方法に従い作成され生産される。Apo-2DcRタグポリペプチド融合体は、Apo-2DcRタンパク質をインフレームでコードしているcDNA配列をタグポリペプチドDNA配列に融合させ、得られたDNA融合作成物を適当な宿主細胞に発現させることによって好ましく作成される。通常は、本発明のApo-2DcRタグポリペプチドキメラを調製するときは、Apo-2DcRをコードしている核酸を、タグポリペプチドのN末端をコードしている核酸にその3'末端で融合させるが、5'融合もまた可能である。例えば、約5〜約10のヒスチジン残基を有するポリヒスチジン配列はN末端又はC末端で融合され、アフィニティークロマトグラフィーにおける精製ハンドルとして使用される。
【0118】
エピトープタグApo-2DcRは、抗タグ抗体を用いてアフィニティークロマトグラフィーによって精製することができる。アフィニティー抗体が付着されるマトリックスには、例えばアガロース、調整穴明きガラス又はポリ(スチレンジビニル)ベンゼンが含まれる。ついで、エピトープタグApo-2DcRは、当該分野において知られた技術を使用して、アフィニティーカラムから溶出することができる。
【0119】
他の実施態様において、キメラ分子は免疫グロブリン配列と融合したApo-2DcRポリペプチドを含む。キメラ分子は、また特定のApo-2DcRドメイン配列、例えば免疫グロブリン配列と融合したApo-2DcRの細胞外ドメイン配列も含む。これには、モノマー、又はホモ-又はヘテロマルチマー、そして特にはホモ-又はヘテロダイマー又は-テトラマー型のものが含まれ;場合によっては、キメラはダイマー型又はホモダイマー重鎖型であり得る。一般的に、これらの構築免疫グロブリンは以下の図に示されるような既知の単位構造を有している。

【0120】
基本的な4鎖の構造単位はIgG、IgD及びIgEが存在する型である。4鎖の単位がより大なる高分子量の免疫グロブリンにおいて繰り返される;IgMが一般にジスルフィド結合によって一緒になった基本的な4鎖単位のペンタマーとして存在する。IgAグロブリン、そして時折IgGグロブリンはまた血清中にマルチマー型で存在する。マルチマーの場合は、各4鎖単位は同一であるか異なっている。
【0121】
次の図は、幾つかの例示的なモノマー、ホモ-及びヘテロダイマー及びホモ-及びヘテロマルチマー構造を示している。これらの図は単に例証するためのものであって、マルチマーの鎖は、天然免疫グロブリンと同様にジスルフィド結合していると考えられる。
モノマー:
ホモダイマー:
ヘテロダイマー:
ホモテトラマー:
ヘテロテトラマー:
及び
【0122】
上図において、「A」はApo-2DcR配列又は異種性配列に融合したApo-2DcR配列を意味する;Xは、Aと同一でも異なっていてもよい付加的な薬剤、免疫グロブリンスーパーファミリーのメンバーの一部で、例えば天然又はキメラ免疫グロブリン可変領域を含む可変領域又は可変領域様ドメイン、例えばシュードモナス外毒素又はリシンのような毒素、又は他のサイトカイン(すなわち、IL-1、インターフェロン-γ)又は細胞表面分子(すなわち、NGFR、CD40、OX40、Fas抗原、ショープ及び粘液腫ポックスウィルスのT2タンパク質)のような他のタンパク質に機能的に結合している配列、又は定常ドメインと通常は結合していないポリペプチド治療剤である;Yはリンカー又は他のレセプター配列である;VL、VH、CL及びCHは免疫グロブリンの軽又は重鎖の可変又は定常ドメインを表す。「A」として少なくとも1つのApo-2DcR配列のCRDと、「X」としてCRDの反復性パターンを有する他の細胞表面タンパク質とを含む構造体が特に含まれる。
【0123】
上図は、単に本発明の可能性のあるキメラ構造を例証したものであって、全ての可能性を包括するものではない。例えば、これらの作成物のいずれにおいても、望ましくは幾つかの異なった「A」、「X」又は「Y」があるかもしれない。また、重又は軽鎖定常ドメインは、同一又は異なる免疫グロブリンから由来したものであってもよい。図示された類似の構造のあらゆる可能な組合わせは全て本発明の範囲に入る。
【0124】
一般的にキメラ分子は、ある種の抗体からの可変ドメインが他の種の可変ドメインで置換されたキメラ抗体と同様にして作成することができる。例えば、EP0125023;EP173494;Munro, Nature, 312:597(1984年12月13日);Neubergerら, Nature, 312:604-608(1984年12月13日);Sharonら, Nature, 309:364-367(1984年5月24日);Morrisonら, Proc. Nat'l. Acad. Sci. USA, 81:6851-6855(1984);Morrisonら, Science, 229:1202-1207(1985);Boulianneら, Nature, 312:643-646(1984年12月13日);Caponら, Nature, 337:525-531(1989);Trauneckerら, Nature, 339:68-70(1989)を参照のこと。
【0125】
あるいは、キメラ分子は以下のようにして作成することができる。所望の配列、例えばApo-2DcR及び/又はTNFR配列をコードする領域を含むDNAを、免疫グロブリン様ドメインをコードするドメインの3'末端又はその近傍で、及びApo-2DcR又はTNFRポリペプチドのN末端をコードするDNAもしくはその近傍の点(異なるリーダーの使用が想定される)で、又はTNFRのN末端コード領域もしくはその近傍(そこでは天然シグナルが使用される)で、制限酵素により開裂させる。ついで、このDNA断片を免疫グロブリン軽又は重鎖定常ドメインをコードするDNAの近傍に直ちに挿入し、必要ならば、得られた作成物を欠失変異誘導により整える。好ましくは、キメラ分子がヒトに対するインビボ治療法を意図したものである場合、Igはヒト免疫グロブリンである。免疫グロブリン軽又は重鎖定常領域をコードするDNAは既知であるか、cDNAライブラリから直ぐに入手できるか合成される。例えば、Adamsら, Biochemistry, 19:2711-2719(1980);Goughら, Biochemistry, 19:2702-2710(1980);Dolbyら, Proc. Natl. Acad. Sci.. USA, 77:6027-6031(1980);Riceら, Proc. Natl. Acad. Sci., 79:7862-7865(1982);Falknerら, Nature, 298:286-288(1982);及びMorrisonら, Ann. Rev. Immunol., 2:239-256(1984)を参照のこと。
【0126】
このような融合体の調製方法のさらなる詳細は、イムノアドヘシンの調製に関する刊行物に見出される。イムノアドヘシン一般、及び特にCD4-Ig融合分子は、1989年4月6日に公開された国際特許出願第WO89/02922号に開示されている。IgG重鎖定常領域に結合している、ヒト免疫不全ウイルス(HIV)のレセプター、CD4の細胞外部分を含む分子は当該分野において知られており、CD4の可溶性細胞外部分よりも著しく長い半減期とより低いクリアランスを有することが見出された[Caponら, 上掲;Byrnら, Nature, 344:667(1990)]。また、特異的キメラTNFR-IgG分子の作成が、Ashkenaziら, Proc. Natl. Acad. Sci., 88:10535-10539(1991),;Lesslauerら, [J. Cell. Biochem. Supplement 15F, 1991, p.115(P432)];及びPeppelとBeutler, J. Cell. Biochem. Supplement 15F, 1991, p.118(P439)]に記載されている。
【0127】
B. Apo-2DcRの治療的及び非治療的用途
本明細書に開示されているように、Apo-2DcRは、治療的に用いられて、哺乳動物細胞において、Apo-2LによるかApo-2DcRが結合する他のリガンドによるアポトーシス及び/又はNF-κB活性化を誘発する。この治療法は、例えばインビボ又はエキソビボの遺伝子療法を使用することによりなすことができ、ここで開示している死亡ドメイン配列の使用を含む。また、免疫グロブリン配列を含むApo-2DcRキメラ分子(Apo-2DcRの細胞外ドメイン配列を含むキメラ分子又は以下の実施例に記載されるApo-2DcRイムノアドヘシンを含む)も治療的に使用することができ、Apo-2Lの活性、例えばアポトーシス又はNF-κB誘導又はApo-2DcRが結合する他のリガンドの活性を阻害する。
【0128】
好適な担体とそれらの製剤は、Osloらにより編集されたRemington's Pharmaceutical Sciences, 16th ed., 1980, Mack Publishing Co.,に記載されている。典型的には、適量の製薬的に許容可能な塩が、製剤を等浸透圧にするために製剤において使用される。担体の例には、生理食塩水、リンガー液及びデキストロース液のようなバッファーが含まれる。溶液のpHは、好ましくは約5〜約8、さらに好ましくは約7.4〜約7.8である。例えば投与経路よっては、ある種の担体がより好ましくなることは、当業者には明らかである。
【0129】
哺乳動物への投与は、注射(例えば、静脈、腹腔内、皮下、筋内)、又は点滴のように、有効な形態で血流への送達を確実にする他の方法によりなすことができる。
【0130】
投与の有効な用量とスケジュールは経験的に決定することができ、このような決定は当業者の技量の範囲に含まれる。
【0131】
また、他の更なる治療法を哺乳動物に施してもよく、そのようなものには、限定されるものではないが、化学療法及び放射線療法、イムノアジュバント、サイトカイン、及び抗体ベース療法が含まれる。例としては、インターロイキン(例えば、IL-1、IL-2、IL-3、IL-6)、白血病阻害因子、インターフェロン、TGF-β、エリスロポイエチン、トロンボポイエチン、及びHER-2抗体が含まれる。哺乳動物細胞においてアポトーシスを誘発することが知られている他の薬剤を用いてもよく、そのような薬剤にはTNF-α、TNF-β(リンホトキシン-α)、CD30リガンド、4-1BBリガンド、及びApo-1リガンドが含まれる。
【0132】
本発明において考慮される化学療法には、当該分野において既知であって市販されている化学物質又は薬物、例えばドキソルビシン、5-フルオロウラシル、シトシンアラビノシド(「Ara-C」)、シクロホスファミド、チオテパ、ブスルファン、サイトキシン、タキソール、メトトレキセート、シスプラチン、メルファラン、ビンブラスチン及びカルボプラチンが含まれる。このような化学療法に対する調製法及び用量スケジュールは、製造者の指示に従って使用されるか、熟練した実務家により経験的に決定される。そのような化学療法に対する調製法及び用量スケジュールはまた化学療法サービス,M.C.Perry編, Williams & Wilkins, Baltimore, MD (1992)にも記載されている。化学療法剤は、上述されたもののような、製薬的に許容可能な担体で好ましく投与される。
【0133】
また、本発明のApo-2DcRは非治療的用途においても有用性を有している。Apo-2DcRをコードしている核酸配列を、組織特異性の分類のための診断に使用することもできる。例えば、インシトゥハイブリッド形成、ノーザン及びサザンブロット法、及びPCR分析のような手順を、Apo-2DcRをコードするDNA及び/又はRNAが評価されている細胞型中に存在しているか否かを決定するために使用することができる。Apo-2DcR核酸は、ここに記載されている組換え技術によるApo-2DcRの調製にもまた有用である。
【0134】
単離されたApo-2DcRは、未知量のApo-2DcRを含有するサンプルが調製された対照として、定量的診断アッセイに使用され得る。またApo-2DcR調製物は、抗体を産生する際、Apo-2DcRに対するアッセイにおける標準として(例えば、ラジオイムノアッセイ、ラジオレセプターアッセイ又は酵素結合免疫測定法における標準としての使用のためにApo-2DcRを標識することにより)、アフィニティー精製法において、及び例えば放射性ヨウ素、酵素又はフルオロフォアで標識された場合、競合型レセプター結合アッセイにおいて有用である。
【0135】
実施例に記載されたような、Apo-2DcRの単離された天然型を使用して、Apo-2DcRの交互型;例えばシグナル伝達経路に関与しうる細胞質ドメインを保有する型を同定してもよい。Apo-2DcRの修飾型、例えば前述のApo-2DcR-IgGキメラ分子(イムノアドヘシン)は、抗Apo-2DcR抗体の生産における免疫原として使用することができる。
【0136】
また、Apo-2DcR又はその修飾型をコードする核酸は、トランスジェニック動物か「ノックアウト」動物を産生するのに使用でき、これらは治療的に有用な試薬の開発やスクリーニングに有用である。トランスジェニック動物(例えばマウス又はラット)とは、出生前、例えば胚段階で、その動物又はその動物の祖先に導入された導入遺伝子を含む細胞を有する動物である。導入遺伝子とは、トランスジェニック動物が発生する細胞のゲノムに組み込まれたDNAである。一実施形態では、Apo-2DcRをコードするcDNA又はその適当な配列(例えば、Apo-2DcR-IgG)を、確立された技術によりApo-2DcRをコードするゲノムDNAをクローン化するために使用することができ、ゲノム配列を、Apo-2DcRをコードするDNAを発現する細胞を有するトランスジェニック動物を産生するために使用することができる。特にマウス又はラット等のトランスジェニック動物を産生する方法は当該分野において常套的になっており、例えば米国特許第4736866号や第4870009号に記述されている。典型的には、特定の細胞を組織特異的エンハンサーでのApo-2DcR導入遺伝子の導入の標的にする。胚段階で動物の生殖系列に導入されたApo-2DcRをコードする導入遺伝子のコピーを含むトランスジェニック動物はApo-2DcRをコードするDNAの増大した発現の影響を調べるために使用できる。このような動物は、例えば過度のアポトーシスを伴う病理的状態に対して保護をもたらすと思われる試薬のテスター動物として使用できる。発明のこの側面においては、動物を試薬で治療し、導入遺伝子を有する未治療の動物に比べ病状の発病率が低ければ、疾患に対する治療的処置の可能性が示される。他の実施態様において、可溶性型のApo-2DcR、例えばApo-2DcRECD又はこのような型の免疫グロブリンキメラを有するトランスジェニック動物を作成して、Apo-2DcRL、Apo-2DcRのリガンドの慢性的中和の効果をテストすることができる。
【0137】
あるいは、Apo-2DcRの非ヒト相同体は、動物の胚性細胞に導入されたApo-2DcRをコードする変更ゲノムDNAと、Apo-2DcRをコードする内在性遺伝子との間の相同的組換えによって、Apo-2DcRをコードする欠陥又は変更遺伝子を有する「ノックアウト」動物を作成するために使用できる。例えば、Apo-2DcRをコードするDNAは、確立された技術に従い、Apo-2DcRをコードするゲノムDNAのクローニングに使用できる。Apo-2DcRをコードするゲノムDNAの一部を欠失したり、組み込みを監視するために使用する選択可能なマーカーをコードする遺伝子等の他の遺伝子で置換することができる。典型的には、ベクターは無変化のフランキングDNA(5'と3'末端の両方)を数キロベース含む[例えば、相同的組換えベクターについてはThomas及びCapecchi, Cell, 51:503(1987)を参照のこと]。ベクターは胚性幹細胞に(例えば電気穿孔法等によって)導入し、導入されたDNAが内在性DNAと相同的に組換えられた細胞を選択する[例えば、Liほか, Cell, 69:915(1992)参照]。選択された細胞は次に動物(例えばマウス又はラット)の胚盤胞内に注入され、集合キメラを形成する[例えば、Bradley, Teratocarcinomas and Embryonic Stem Cells: A Practical Approach, E. J. Robertson, ed. (IRL, Oxford, 1987), pp. 113-152参照]。その後、キメラ性胚を適切な偽妊娠の雌性乳母に移植し、期間をおいて「ノックアウト」動物をつくり出す。胚細胞に相同的に組換えられたDNAを有する子孫は標準的な技術により同定され、それらを利用して動物の全細胞が相同的に組換えられたDNAを含む動物を繁殖させることができる。ノックアウト動物は、例えば腫瘍の発達を含む、Apo-2DcRポリペプチドが不在であることによるある種の病理的状態及びその病理的状態の発達に対する防御能力によって特徴付けられる。
【0138】
C. 抗Apo-2DcR抗体の調製
本発明は、さらに抗Apo-2DcR抗体を提供するものである。Apo-2DcRに対する抗体は以下のようにして調製することができる。抗体の例としては、ポリクローナル、モノクローナル、ヒト化、二重特異性及びヘテロ抱合体抗体が含まれる。
【0139】
1. ポリクローナル抗体
Apo-2DcR抗体はポリクローナル抗体を含む。ポリクローナル抗体の調製方法は当業者に知られている。哺乳動物においてポリクローナル抗体は、例えば免疫化剤、所望するのであればアジュバントを、一又は複数回注射することで発生させることができる。典型的には、免疫化剤及び/又はアジュバントを複数回皮下又は腹腔内注射により、哺乳動物に注射する。免疫化剤は、Apo-2DcRポリペプチド又はその融合タンパク質を含みうる。適切な免疫化剤の例は、Apo-2DcR-IgG融合タンパク質、又はキメラ分子(Apo-2DcREDC-IgG融合タンパク質を含む)である。また、その表面にApo-2DcRを発現する細胞を使用してもよい。免疫化されている哺乳動物において免疫原性であることが知られているタンパク質に免疫化剤を抱合させることが有用である。使用され得るそのような免疫原性タンパク質の例は、限定するものではないが、スカシガイヘモシアニン、血清アルブミン、ウシサイログロブリン及び大豆トリプシンインヒビターが含まれる。また、哺乳動物の免疫反応を増強するために、ミョウバンのような凝集剤を使用してもよい。使用され得るアジュバントの例には、フロイント完全アジュバント及びMPL-TDMアジュバント(モノホスホリル脂質A、合成トレハロースジコリノミコラート)が含まれる。免疫化プロトコールは、過度の実験なく当業者により選択されるであろう。哺乳動物から採血し、血清を検定して抗体価を求める。望まれるならば、抗体価が増加又はプラトーするまで哺乳動物に追加免疫を施す。
【0140】
2. モノクローナル抗体
あるいは、Apo-2DcR抗体はモノクローナル抗体であってもよい。モノクローナル抗体は、Kohler及びMilstein,上掲に記載されているようなハイブリドーマ法を使用することで調製することができる。ハイブリドーマ法では、マウス、ハムスター又は他の適切な宿主動物を典型的には免疫化剤により免疫化する(上述したようにして)ことで、免疫化剤に特異的に結合する抗体を生成するかあるいは生成可能なリンパ球を誘発する。また、リンパ球をインビトロで免疫化することもできる。
【0141】
免疫化剤は、典型的にはApo-2DcRポリペプチド又はその融合タンパク質を含む。適切な免疫化剤の例は、Apo-2DcR-IgG融合タンパク質又はキメラ分子である。Apo-2DcREDC-IgG免疫原の特定の例は、以下の実施例13に記載している。表面でApo-2DcRを発現する細胞もまた使用することができる。一般にヒト由来の細胞が望まれる場合には末梢血リンパ球(「PBL」)が使用され、あるいは非ヒト哺乳動物源が望まれている場合は、脾臓細胞又はリンパ節細胞が使用される。次いで、ポリエチレングリコール等の適当な融合剤を用いてリンパ球を不死化株化細胞と融合させ、ハイブリドーマ細胞を形成する[Goding, Monoclonal Antibodies: Principles and Practice, Academic Press, (1986) pp. 59-103]。不死化株化細胞は、通常は、形質転換した哺乳動物細胞、特に齧歯動物、ウシ、及びヒト由来の骨髄腫細胞である。通常、ラット及びマウスの骨髄腫細胞が使用される。ハイブリドーマ細胞は、好ましくは、未融合の不死化細胞の生存又は成長を阻害する一又は複数の物質を含有する適切な培地で培養される。例えば、親の形質転換細胞が、酵素のヒポキサンチングアニンホスホリボシルトランスフェラーゼ(HGPRT又はHPRT)を欠いていると、ハイブリドーマの培地は、典型的には、ヒポキサチン、アミノプチリン及びチミジン(「HAT培地」)を含み、この物質がHGPRT欠乏性細胞の増殖を阻止する。
【0142】
好ましい不死化株化細胞は、効率的に融合し、選択された抗体生成細胞による安定した高レベルの抗体発現を支援し、HAT培地のような培地に対して感受性であるものが望ましい。より好ましい不死化株化細胞はマウス骨髄腫株であり、これはカリフォルニア州サンディエゴのSalk Institute Cell Distribution Centerやバージニア州マナッサスのアメリカン・タイプ・カルチャー・コレクションより入手可能である。ヒトモノクローナル抗体を生成するためのヒト骨髄腫及びマウス-ヒト異種骨髄腫株化細胞も開示されている[Kozbor, J. Immunol., 133:3001 (1984)、Brodeurほか, Monoclonal Antibody Production Techniques and Applications, Marcel Dekker, Inc., New York, (1987) pp. 51-63]。
【0143】
ハイブリドーマ細胞を培養する培地を、Apo-2DcRに対するモノクローナル抗体の存在について検定する。好ましくは、ハイブリドーマ細胞によって生成されたモノクローナル抗体の結合特異性は免疫沈降又はラジオイムノアッセイ(RIA)や酵素結合免疫測定法(ELISA)等のインビトロ結合検定法によって測定する。このような技術及びアッセイは、当該分野において公知である。モノクローナル抗体の結合親和性は、例えばMunson及びPollard, Anal. Biochem., 107:220 (1980)によるスキャッチャード分析法によって測定することができる。
【0144】
所望のハイブリドーマ細胞が同定されたら、クローンを制限希釈工程を経てサブクローニングし、標準的な方法で増殖させることができる[Goding, 上掲]。この目的のための適当な培地には、例えば、ダルベッコの改変イーグル培地及びRPMI-1640倍地が含まれる。更に、ハイブリドーマ細胞は哺乳動物においてインビボで腹水として成長させることもできる。
【0145】
サブクローンによって分泌されるモノクローナル抗体は、例えばプロテインAセファロース法、ヒドロキシルアパタイトクロマトグラフィー法、ゲル電気泳動法、透析法又はアフィニティークロマトグラフィー等の従来の免疫グロブリン精製方法によって培地又は腹水液から分離又は精製される。
【0146】
また、モノクローナル抗体は、組換えDNA法、例えば米国特許第4816567号に記載された方法により作製することができる。本発明のモノクローナル抗体をコードするDNAは、常套的な方法によって(例えば、マウス抗体の重鎖及び軽鎖をコードする遺伝子に特異的に結合可能なオリゴヌクレオチドプローブを使用して)、容易に単離し配列決定することができる。本発明のハイブリドーマ細胞はそのようなDNAの好ましい供給源となる。ひとたび単離されたら、DNAは発現ベクター内に配することができ、これが宿主細胞、例えばサルCOS細胞、チャイニーズハムスター卵巣(CHO)細胞、あるいは免疫グロブリンタンパク質を生成等しない骨髄腫細胞内にトランスフェクトされ、組換え宿主細胞内でモノクローナル抗体の合成をすることができる。また、DNAは、例えば相同マウス配列に換えてヒト重鎖及び軽鎖定常ドメインのコード配列を置換することにより[US. Patent No.4816567;Morrisonほか, 上掲]、又は免疫グロブリンコード配列に非免疫グロブリンポリペプチドのコード配列の一部又は全部を共有結合することにより修飾することができる。このような非免疫グロブリンポリペプチドは、本発明の抗体の定常ドメインの代わりに置換するか、本発明の抗体の一つの抗原結合部位の可変ドメインの代わりに置換し、キメラ性二価抗体を産生することができる。
【0147】
以下の実施例に記載するようにして、抗Apo-2DcRモノクローナル抗体を調製した。これらの抗体のいくらかは、4G3.9.9、6D10.9.7、及び1C5.24.1としてATCCに寄託され、寄託受入番号 、 、及び がそれぞれ付与された。一実施態様において、本発明のモノクローナル抗体は、受入番号 、 、及び で寄託されたハイブリドーマ株化細胞により分泌される抗体の一又は複数と同じ生物学的特徴を有する。「生物学的特徴」という用語は、モノクローナル抗体のインビトロ及び/又はインビボの活性又は性質、例えばApo-2DcRに特異的に結合する能力又はApo-2DcR活性を実質的に阻止、誘導又は高める能力を指すために使用される。場合によっては、モノクローナル抗体はここで特に言及された3種の抗体の少なくとも1つと同じエピトープに結合する。そのようなエピトープ結合は、例えばここや実施例において記載したように、種々のアッセイを行うことにより測定することができる。
【0148】
本発明の抗体は一価抗体であってもよい。一価抗体の調製方法は当該分野においてよく知られてる。例えば、一つの方法は免疫グロブリン軽鎖と修飾重鎖の組換え発現を含む。重鎖は一般的に、重鎖の架橋を防止するようにFc領域の任意のポイントで切断される。あるいは、関連したシステイン残基を他のアミノ酸残基で置換するか欠失させて架橋を防止する。
【0149】
一価抗体の調製にはインビトロ法がまた適している。抗体の消化による、その断片、特にFab断片の調製は、当該分野において知られている慣用的技術を使用して達成できる。例えば、消化はパパインの使用により行うことができる。パパイン消化の例は、94/12/22に公開された国際特許出願第WO94/29348号、及び米国特許第4342566号に記載されている。抗体のパパイン消化は、典型的には、Fab断片と呼ばれ、各々が単一の抗原結合部位を有する2つの同一の抗原結合断片と、残りのFc断片を生成する。ペプシン処理により、2つの抗原結合部位を有し、抗原の架橋が尚も可能なF(ab')2断片が得られる。
【0150】
また、抗体の消化により生産されたFab断片は、軽鎖の定常ドメインと重鎖の第1定常ドメイン(CH1)を含む。Fab'断片は、抗体のヒンジ領域から一又は複数のシステインを含む重鎖CH1ドメインのカルボキシ末端に幾つかの残基が付加されているということで、Fab断片とは異なっている。Fab'-SHとは、定常ドメインのシステイン残基が遊離のチオール基を担持しているFab'に対するここでも命名である。F(ab’)2抗体断片は、本来は、それらの間にヒンジシステインを有するFab’断片の対として生産された。抗体断片の他の化学的結合もまた知られている。
【0151】
3. ヒト化抗体
本発明のApo-2DcR抗体は、さらにヒト化抗体又はヒト抗体を含む。非ヒト(例えばマウス)抗体のヒト化形とは、キメラ免疫グロブリン、免疫グロブリン鎖あるいはその断片(例えばFv、Fab、Fab'、F(ab')2あるいは抗体の他の抗原結合サブ配列)であって、非ヒト免疫グロブリンに由来する最小配列を含むものである。ヒト化抗体はレシピエントの相補性決定領域(CDR)の残基が、マウス、ラット又はウサギのような所望の特異性、親和性及び能力を有する非ヒト種(ドナー抗体)のCDRの残基によって置換されたヒト免疫グロブリン(レシピエント抗体)を含む。ある場合には、ヒト免疫グロブリンのFvフレームワーク残基は、対応する非ヒト残基によって置換されている。更に、ヒト化抗体は、レシピエント抗体にも、移入されたCDRもしくはフレームワーク配列にも見出されない残基を含んでいてもよい。一般に、ヒト化抗体は、全てあるいはほとんど全てのCDR領域が非ヒト免疫グロブリンのものに対応し、全てあるいはほとんど全てのFR領域がヒト免疫グロブリンコンセンサス配列のものである、少なくとも1つ、典型的には2つの可変ドメインの実質的に全てを含む。ヒト化抗体は、最適には免疫グロブリン定常領域(Fc)、典型的にはヒトの免疫グロブリンの定常領域の少なくとも一部を含んでなる[Jones ほか, Nature, 321:522-525 (1986); Riechmann ほか, Nature, 332:323-329 (1988); 及びPresta, Curr. Op Struct. Biol., 2:593-596 (1992)]。
【0152】
非ヒト抗体をヒト化する方法は従来からよく知られている。一般的に、ヒト化抗体には非ヒト由来の一又は複数のアミノ酸残基が導入される。これら非ヒトアミノ酸残基は、しばしば、典型的には「移入」可変ドメインから得られる「移入」残基と称される。ヒト化は基本的に齧歯動物のCDR又はCDR配列でヒト抗体の該当する配列を置換することによりウィンター及び共同研究者[Jonesほか, Nature, 321:522-525 (1986)、Riechmannほか, Nature, 332:323-327 (1988)、Verhoeyenほか, Science, 239:1534-1536 (1988)]の方法を使用して行える。よって、このような「ヒト化」抗体は、無傷のヒト可変ドメインより実質的に少ない分が非ヒト種由来の対応する配列で置換されたキメラ抗体(U.S. 4,816,567)である。実際には、ヒト化抗体は典型的にはある程度のCDR残基及び場合によってはいくらかのFR残基が齧歯類抗体の類似する部位からの残基によって置換されるヒト抗体である。
【0153】
抗原性を軽減するには、ヒト化抗体を生成するために使用するヒトの軽及び重可変ドメインの両方の選択が非常に重要である。「ベストフィット法」では、齧歯動物抗体の可変ドメインの配列を既知のヒト可変ドメイン配列ライブラリ全体に対してスクリーニングする。齧歯動物のものと最も近いヒトの配列を次にヒト化抗体のヒトフレームワーク(FR)として受け入れる[Simsほか, J. Immunol., 151:2296 (1993);Chothia及びLesk, J. Mol. Biol., 196:901 (1987)]。他の方法では、軽又は重鎖の特定のサブグループのヒト抗体全てのコンセンサス配列から誘導される特定のフレームワークを使用する。同じフレームワークを幾つかの異なるヒト化抗体に使用できる[Carterほか, Proc. Natl. Acad. Sci. USA, 89:4285 (1992);Prestaほか, J. Immunol., 151:2623 (1993)]。
【0154】
更に、抗体は、抗原に対する高親和性や他の好ましい生物学的性質を保持してヒト化することが重要である。この目標を達成するべく、好ましい方法では、親及びヒト化配列の三次元モデルを使用して、親配列及び様々な概念的ヒト化産物の分析工程を経てヒト化抗体を調製する。三次元免疫グロブリンモデルは一般的に入手可能であり、当業者にはよく知られている。選択された候補免疫グロブリン配列の推測三次元立体配座構造を図解し、表示するコンピュータプログラムは購入可能である。これら表示を見ることで、候補免疫グロブリン配列の機能における残基の役割の分析、すなわち候補免疫グログリンの抗原と結合する能力に影響を及ぼす残基の分析が可能となる。このようにして、例えば標的抗原に対する親和性を高めるといった、望ましい抗体特徴が得られるように、FR残基をコンセンサス及び移入配列から選択し、組み合わせることができる。一般的に、CDR残基は、直接かつ最も実質的に抗原結合性に影響を及ぼしている[1994年、3月3日に公開されたWO 94/04679を参照]。
【0155】
免疫化することで、内在性免疫グロブリンが生成されない状態でもヒト抗体の完全リパートリを生成することができるトランスジェニック動物(例えばマウス)を使用することが可能である。例えば、キメラ及び生殖系列変異マウスにおいて抗体重鎖結合領域(JH)遺伝子をホモ接合的に欠失させると内在性抗体生産の完全な阻害が生じることが記述されている。このような生殖系列変異マウスにヒト生殖系列免疫グロブリン遺伝子配列を移すと、抗原投与時にヒト抗体の生成が生じる[例えば、Jakobovitsほか, Proc. Natl. Acad. Sci. USA, 90:2551-255 (1993);Jakobovitsほか, Nature, 362:255-258 (1993);Bruggermanほか, Year in Immuno., 7:33 (1993)を参照]。また、ヒト抗体はファージ表示ライブラリ[Hoogenboom及びWinter, J. Mol. Biol., 227:381 (1992);Marksほか, J. Mol. Biol., 222:581 (1991)]において産生することもできる。また、Coleら及びBoernerらの方法も、ヒトモノクローナル抗体の調製に利用することができる[Coleら, Monoclonal Antibodies and Cancer Therapy, Alan R. Liss. p.77(1985)及びBoernerら, J. Immunol., 147(1):86-95(1991) ]。
【0156】
4. 二重特異性抗体
二重特異性抗体は、少なくとも2つの異なる抗原に対して結合特異性を有するモノクローナル抗体、好ましくはヒトもしくはヒト化抗体である。本発明の場合において、結合特異性の一方はApo-2DcRに対してであり、他方は任意の他の抗原、好ましくは細胞表面タンパク質又はレセプター又はレセプターサブユニットに対してである。
【0157】
二重特異性抗体を生成する方法は当該技術分野において周知である。伝統的には、二重特異性抗体の組換え生成方法は、二つの重鎖が異なる特異性を持つ二つの免疫グロブリン重鎖/軽鎖対の同時発現に基づく[Milstein及びCuello, Nature, 305:537-539 (1983)]。免疫グロブリンの重鎖と軽鎖を無作為に取り揃えたため、これらハイブリドーマ(クアドローマ)は10種の異なる抗体分子の潜在的混合物を生成でき、その内一種のみが正しい二重特異性構造を有する。正しい分子の精製は、アフィニティークロマトグラフィー工程によって通常達成される。同様の手順が1993年5月13日公開の国際特許出願第93/08829号、及びTrauneckerほか, EMBO J.,10:3655-3656 (1991)に開示されている。
【0158】
別のより好ましいアプローチによれば、所望の結合特異性(抗体-抗原組合せ部位)を有する抗体可変ドメインを免疫グロブリン定常ドメイン配列に融合する。融合は、好ましくは少なくともヒンジ部、CH2及びCH3領域の一部を含む免疫グロブリン重鎖定常ドメインとのものである。少なくとも一つの融合には軽鎖結合に必要な部位を含む第一の重鎖定常領域(CH1)が存在することが望ましい。免疫グロブリン重鎖融合をコードするDNA、及び望むのであれば免疫グロブリン軽鎖を、別々の発現ベクターに挿入し、適当な宿主生物に同時形質移入する。これは、作成に使用する三つのポリペプチド鎖が不等の比であるときに最高の収率が得られる実施態様において、三つのポリペプチド断片の相互比率の調節に大なるフレキシビリティをもたらす。しかし、少なくとも二つのポリペプチド鎖が同比率で発現すると高収率が得られる場合や比率が特に重要ではない場合には、一つの発現ベクターに二つ又は三つ全てのポリペプチド鎖のコード配列を挿入することができる。このアプローチの好適な形態では、二重特異性抗体は、一方のアームの第一結合特異性を有するハイブリッド免疫グロブリン重鎖と、他方のアームのハイブリッド免疫グロブリン重鎖/軽鎖対(第二の結合特異性をもたらす)からなる。このような非対称的構造は、二重特異性分子の半分にのみ免疫グロブリン軽鎖が存在すると容易な方法で分解できるので、所望の二重特異性化合物を不要な免疫グロブリン鎖の組合わせから分解し易くすることが見出された。このアプローチは1994年3月3日公開の国際特許出願第94/04690号によって開示されている。二重特異性抗体を生成するための更なる詳細については、例えばSureshほか,Methods in Enzymology, 121:210(1986)を参照されたい。
【0159】
5. ヘテロ抱合体抗体
ヘテロ抱合抗体もまた本発明の範囲に入る。ヘテロ抱合抗体は、2つの共有的に結合した抗体からなる。このような抗体は、例えば、免疫系細胞を不要な細胞に対してターゲティングさせるため(米国特許第4676980号)及びHIV感染の治療のために(WO91/00360、WO92/200373;EP03089)提案された。本抗体は、架橋剤に関連したものを含む合成タンパク化学における既知の方法を使用して、インビトロで調製することができると考えられる。例えば、ジスルフィド交換反応を使用するか又はチオエーテル結合を形成することにより、免疫毒素を作成することができる。この目的に対して好適な試薬の例には、イミノチオレート及びメチル-4-メルカプトブチリミデート、及び例えば米国特許第4676980号に開示されているものが含まれる。
【0160】
D. Apo-2DcR抗体の治療的及び非治療的用途
本発明のApo-2DcR抗体は治療的な有用性を有している。例えば、Apo-2リガンドに対する他のレセプターと交差反応するApo-2DcR抗体は過剰なアポトーシス(例えば神経変性疾患において)をブロックするか、潜在的な自己免疫/炎症効果をブロックするために使用することができる。場合によっては、Apo-2DcR阻止抗体は、Apo-2リガンド治療法と組合わせて使用することができる。Apo-2DcR抗体はApo-2DcRレセプターを阻止することができ、投与されたApo-2リガンドの生物利用能を増大させることができる。(Apo-2DcRに対しての上述のような)治療組成物と投与形態を用いることができる。
【0161】
さらに、Apo-2DcR抗体は、免疫組織化学染色検定法又はApo-2DcRの診断検定法、例えば特異的細胞、組織又は血清における発現を検出する検定法に使用することができる。当該分野において知られている様々な診断検定技術、例えば、競合的結合検定、直接的又は間接的サンドイッチ検定及び不均一又は均一相の何れにおいても実施される免疫沈降検定を使用することができる[Zola, Monoclonal Antibodies: A Manual of Techniques,CRC Press,Inc.,(1987) pp.147-158]。診断検定法に使用される抗体は、検出可能部分で標識することができる。検出可能成分は、直接的に又は間接的に検出可能なシグナルをつくりだすことができなければならない。例えば検出可能成分は、3H、14C、32P、35S又は125I等の放射性同位体、蛍光イソチオシアネート、ローダミン又はルシフェリン等の蛍光又は化学発光化合物、もしくはアルカリホスファターゼ、βガラクトシダーゼ又は西洋わさびペルオキシダーゼ等の酵素であってもよい。Hunterほか,Nature,144:945 (1962)、Davidほか, Biochemistry,13:1014(1974)、Painほか,J. Immunol. Meth.,40:219 (1981)及びNygrenほか,J. Histochem. and Cytochem.,30:407 (1982)等に記載されている方法を含み、検出可能部分に抗体を抱合させるための当該分野において知られている任意の方法を使用することができる。
【0162】
また、Apo-2DcR抗体は天然供給源又は組換え細胞培養からのApo-2DcRのアフィニティー精製にも有用である。この方法においては、Apo-2DcRに対する抗体を、当該分野でよく知られている方法を使用して、セファデックス樹脂や濾過紙のような適当な支持体に固定する。次に、固定された抗体を、精製するApo-2DcRを含む試料と接触させ、ついで、固定された抗体に結合したApo-2DcR以外の試料中の物質を実質的に全て除去する適当な溶媒で支持体を洗浄する。最後に、Apo-2DcRを抗体から離脱させる他の適当な溶媒で支持体を洗浄する。
【0163】
E. Apo-2DcR又はApo-2DcR抗体を含むキット
本発明の更なる実施態様においては、例えば上述の治療的又は非治療的用途に使用可能なApo-2DcR又はApo-2DcR抗体を含むキット及び製造品が提供される。製造品にはラベルが付された容器が含まれる。適切な容器には、例えばボトル、バイアル、及び試験管が含まれる。容器はガラス又はプラスチックのような種々の物質から形成できる。容器は、上述の治療的又は非治療的用途に有効な活性剤を含む組成物を収容する。組成物中の活性剤は、Apo-2DcR又はApo-2DcR抗体である。容器のラベルには、組成物が特定の治療的又は非治療的用途に使用されることが示され、また上述のもののような、インビボ又はインビトロのいずれかの使用の指示が示されている。
【0164】
本発明のキットは、典型的には、上述の容器と、商業上及び使用者の観点から望まれる、バッファー、希釈液、フィルター、針、注入器及び使用説明が記されたパッケージ挿入物を含む、材料を含む一又は複数の他の容器を含んでいる。
【0165】
以下の実施例は例示するためにのみ提供されるものであって、本発明の範囲を決して限定することを意図するものではない。
【0166】
本明細書で引用した全ての特許及び参考文献の全体を、出典明示によりここに取り込む。
【0167】
実施例
実施例において言及されている全ての制限酵素は、ニューイングランドバイオラボ社(New England Biolabs)から購入し、製造者の使用説明に従い使用した。実施例で言及されている全ての他の市販試薬は、特に示していない限りは、製造者の使用説明に従い使用した。ATCC受入番号により次の実施例及び明細書全体を通して特定している細胞の供給源はアメリカン・タイプ・カルチャー・コレクション(Mannasas,Virginia)である。
【0168】
実施例1
ヒトApo-2DcRをコードしているcDNAクローンの単離
【0169】
1.オリゴdTプライムcDNAライブラリ(「LIB111」)の調製
インビトロゲン、サンディエゴ、CAの試薬とプロトコールを使用してmRNAをヒト乳癌組織から単離した(高速トラック2)。ライフ・テクノロジーズ、ゲイザースバーグ、MDの試薬とプロトコール(スーパースクリプトプラスミド系)を使用し、ベクターpRK5D中にオリゴdTプライムcDNAライブラリ(「LIB111」)を産生するためにこのRNAを用いた。この手順において、二本鎖cDNAを1000塩基対よりも大きいサイズにし、Sa1I/NotIリンカーcDNAをXhoI/NotI切断ベクター内にクローニングした。pRK5Dはsp6転写開始部位にSfiI制限酵素部位が続き、その次にXhoI/NotIcDNAクローニング部位があるクローニングベクターである。
【0170】
2.ランダムプライムcDNAライブラリ(「LIB118」)の調製
一次cDNAクローンの5'末端を優先的に提示するために二次cDNAライブラリを産生した。sp6RNAを一次ライブラリ(上述のLIB111)から産生し、このRNAを用いて、ライフ・テクノロジーズの試薬とプロトコール(上述のスーパースクリプトプラスミド系)を使用してベクターpSST-AMY.0中にランダムプライムcDNAライブラリ(「LIB118」)を産生した。この手順において二本鎖cDNAを500−1000塩基対のサイズにし、NotIアダプターに平滑末端化してリンカーし、SfiIで切断し、SfiI/NotI切断ベクター中にクローニングした。pSST-AMY.0は、酵母アルコールデヒドロゲナーゼプロモータの後にcDNAクローニング部位とマウスアミラーゼ配列(分泌シグナルを持たない成熟配列)が続き、それに、酵母アルコールデヒドロゲナーゼターミネーターがクローニング部位後に続くクローニングベクターである。しかして、アミラーゼ配列とインフレームで融合されたこのベクター中にクローニングされたcDNAは、適切に形質移入された酵母コロニーからアミラーゼを分泌せしめる。
【0171】
3.形質転換と検出
LIB118からのDNAを氷上で冷却し、エレクトロコンピテントDH10B細菌 (Life Technologies,20ml)を添加した。ついで、細菌ベクター混合物を、製造者の推奨に従って電気穿孔法にかけた。続いてSOC培地(Life Technologies, 1ml)を添加し、混合物を37℃で30分間インキュベートした。ついで、形質転換体を、アンピシリンを含有する20の標準的な150mmLBプレートに蒔き、16時間(37℃)の間インキュベートした。陽性コロニーを擦り取り、CsCl勾配のような標準的なプロトコールを使用して、細菌ペレットからDNAを単離した。次に、精製したDNAについて以下の酵母プロトコールを進めた。
【0172】
本発明において使用される酵母方法は、3つの範疇に分けられた:(1)プラスミド/cDNA併用ベクターでの酵母の形質転換;(2)アミラーゼを分泌する酵母クローンの検出と単離;及び(3)酵母コロニーから直接の挿入断片のPCR増幅と配列決定及び更なる分析のためのDNAの精製である。
【0173】
安定な突然変異体ura3を含む任意の酵母株を本発明において使用することができるが、本発明の実施のために使用される好ましい酵母株はHD56-5A(ATCC-90785)であった。この株は次の遺伝子型:MATα、ura3-52、leu2-3、leu2-112、his3-11、his3-15、MAL+、SUC+、GAL+を有していた。
【0174】
形質転換を、Gietzら, Nucl. Acid. Res.,20:1425(1992) により概説されたプロトコールに基づいて実施した。この手順では、DNAマイクログラム当りおよそ1x105形質転換体の形質転換効率を得た。形質転換された細胞をついで寒天からYEPD複合培地ブロス(100ml)中に播種し、30℃で一晩増殖させた。YEPDブロスは、Kaiserら, Methods in Yeast Genetics, Cold Spring Harbor Press, Cold SpringHarbor, N.Y. USA, p207 (1994)に記載されているようにして調製した。ついで一晩の培養を約2x106細胞/ml(およそOD600=0.1)に希釈して新鮮なYEPDブロス(500ml)中に入れ、1x107細胞/ml(およそOD600=0.4−0.5)に再増殖させた。これには完了まで通常約3時間かかった。
【0175】
ついで、細胞を収集し、GS3ロータボトルに移しソーバル(Sorval)GS3ロータを5000rpmで5分間作動させて形質転換用に調製し、上清を捨て、ついで滅菌水に再懸濁させ、ベックマンGS-6KR遠心分離機で3500rpmにて50mlのファルコン管中で再び遠心分離した。上清を捨て、続いて細胞をLiAc/TE(10ml、10mMのトリス-HCl、1mMのEDTA、pH7.5、100mMのLi2OOCCH3)で洗浄し、LiAc/TE(2.5ml)中に再懸濁した。
【0176】
調製した細胞(100μl)を新鮮に変性した一本鎖サケ精巣DNA(Lofstrand Labs, Gaithersburg, MD, USA)と混合し微量遠心管中でDNA(1μg,容量<10μl)を形質転換することにより形質転換を行った。混合物をボルテックスすることにより簡単に混合し、ついで40%PEG/TE(600μl、40%ポリエチレングリコール-4000、10mMのトリス-HCl、1mMのEDTA、100mMのLi2OOCCH3、pH7.5)を添加した。この混合物を穏やかに混合し、30分間攪拌しながら30℃でインキュベートした。ついで細胞に42℃で15分間熱ショックをかけ、反応容器を微量遠心管中で5−10秒の間に12000rpmで遠心分離し、デカントし、TE(500μl、10mMのトリス-HCl、1mMのEDTA、pH7.5)に再懸濁させ、再び遠心分離した。ついで、細胞をTE(1ml)中に希釈し、一定分量(200μl)を、150mm成長板(VWR)に前もって調製していた選択培地上に広げた。
【0177】
あるいは、複数の小さな反応の代わりに、単一の大きな規模の反応で試薬量を多くしたものを使用して形質転換を行った。
【0178】
使用された選択培地は、Kaiserら,Methods in Yeast Genetics, Cold Spring Harbor Press, Cold Spring Harbor, N.Y. USA, p208-210 (1994)に記載されたようにして調製されたウラシルを欠く合成完全ブドウ糖寒天(SCD-Ura)であった。形質転換体は30℃で2−3日増殖させた。
【0179】
アミラーゼを分泌するコロニーの検出は選択増殖培地に赤色のデンプンを含めることにより実施した。Bielyら, Anal.Biochem.,172:176-179(1988) により記載された手順に従って、デンプンは赤色染料(リアクティブレッド-120、シグマ)に結合させた。結合されたデンプンは0.15%(w/v)の最終の濃度でSCD-Ura寒天プレート中に導入し、リン酸カルシウムで7.0のpHまで緩衝させた(50−100mMの最終濃度)。
【0180】
陽性のコロニーを取上げ、よく分離された同定可能なシングルコロニーを得るために新鮮選択培地(150mmプレート上)上に画線した。この工程はまた形質転換体の中でもプラスミドの維持を確実にした。アミラーゼ分泌に対して陽性のよく分離されたシングルコロニーを、緩衝化SCD-Ura寒天中への赤色デンプンの直接の導入により測定した。陽性のコロニーは直接可視化された陽性のコロニーの周りに明確なハローを生じるデンプンを破壊する能力により測定した。
【0181】
4.PCR増幅によるDNAの単離
陽性コロニーを単離したとき、その一部を爪楊枝により取上げ、滅菌水(30μl)中に希釈し、96ウェルプレート中に入れた。このとき、陽性コロニーは凍結して次の分析に対して保存するか、直ぐに増幅させた。一定分量の細胞(5μl)を、0.5μlのクレンタク(Klentaq)(クローンテック, Palo Alto, CA);4.0μlの10mMのdNTP's(パーキンエルマー-セタス);2.5μlのケンタク(Kentaq)バッファー(クローンテック);0.25μlのフォワードオリゴ1;0.25μlのリバースオリゴ2;12.5μlの蒸留水を含む25μl容量中でPCR反応に対する鋳型として使用した。フォワードオリゴヌクレオチド1の配列は:
TGTAAAACGACGGCCAGTTAAATAGACCTGCAATTATTAATCT (配列番号:5)
であった。リバースオリゴヌクレオチド2の配列は:
CAGGAAACAGCTATGACCACCTGCACACCTGCAAATCCATT(配列番号:6)
であった。
ついで、次のようにしてPCRを実施した:
a. 変性 92℃、5分
b. 3サイクルの変性 92℃、30秒
アニーリング 59℃、30秒
伸展 72℃、60秒
c. 3サイクルの変性 92℃、30秒
アニーリング 57℃、30秒
伸展 72℃、60秒
d. 25サイクルの変性 92℃、30秒
アニーリング 55℃、30秒
伸展 72℃、60秒
e. 保持 4℃
【0182】
オリゴヌクレオチドの下線領域はADHプロモータ領域とアミラーゼ領域にそれぞれアニーリングされ、挿入物が存在しないとき、ベクターpSST-AMY.0からの307塩基対領域を増幅した。典型的にはこれらのオリゴヌクレオチドの5'末端の最初の18ヌクレオチドはシーケンシングプライマーに対するアニーリング部位を含んでいた。従って、空のベクターからのPCR反応の総生成物は343塩基対であった。しかしながら、シグナル配列融合cDNAはかなり長い核酸配列を生じた。
【0183】
PCRに続いて、反応物の一定分量(5μl)を上掲のSambrookらにより記述されたようなトリス-ホウ酸塩-EDTA(TBE)緩衝系を使用する1%アガロース中でのアガロースゲル電気泳動により検査した。400塩基対よりも大きい単一の強いPCR生成物を生じるクローンを、96キアクイック(Qiaquick)PCR洗浄カラム(Qiagen Inc., Chatsworth, CA)での精製後にDNA配列決定により更に分析した。
【0184】
5.全長クローンの同定
上記スクリーニングにおいて分離されたcDNA配列(「DNA21705」)はヒトTNFR1とあるアミノ酸配列類似性又は相同性を有していることが分かった:
TNFR1 81 CRECESG-SFTASENHLRHCLSCSKCRKEMG
* * * .* . *. *. *. *. .
DNA21705 164 CNPCTEGVDYTNASNNEPSCFPCTVCKSD--
QVEISSCTVDRDTVCGCRK (配列番号:7)
* ****. ***** *..
QKHKSSCTMTRDTVCQCKE (配列番号:8)
類似性に基づいて、DNA21705の配列からプローブを産生し、上述の第1節に記載したようにして調製したヒト胎児肺ライブラリ(「LIB25」)をスクリーニングするために使用した。クローニングベクターはpRK5B(pRK5BはSfiI部位を含まないpRK5Dの先駆体である)であり、切断されたcDNAサイズは2800塩基対未満であった。ヌクレオチド位置193−195に明らかな翻訳開始部位を持つ単一のオープンリーディングフレームを含み[Kozakら,上掲]、ヌクレオチド位置970−972に見出されるストップコドンで終端する(図1A;配列番号:2)全長クローンを同定した(DNA33085)(pRK-hApo-2DcR)(以下に示されるように、ATCC209087として寄託されたApo2-DcRとも呼ばれる)。予想されたポリペプチド先駆体は259アミノ酸長であり、およそ27.4kDaの計算分子量を有している。配列解析では、N末端シグナルペプチド、2つのシステインリッチドメイン、4のほぼ同一の15アミノ酸直列反復を含む配列、及び疎水性C末端領域が示された(図2及び図3)。C末端の疎水性配列には一対の小さいアミノ酸(Ala223及びAla224)が先行し;この構造と明白な細胞質ドメインがないことにより、Apo-2DcRがグリコシルホスファチジルイノシトール(GPI)アンカータンパク質でありうることが示唆される[Moran, J. Biol. Chem., 266: 1250- 1257(1991)を参照のこと]。Apo-2DcRは5つの潜在的なN結合グリコシル化部位を含んでいる(図2)。
【0185】
TNFレセプターファミリータンパク質は、典型的には、その細胞外領域に複数の(通常は4つの)システインに富んだドメインが存在していることを特徴としており、各システインリッチドメインは約45アミノ酸長であり、規則的に間隔を開けた約6つのシステイン残基を有している。1型TNFレセプターの結晶構造に基づき、各ドメインにおけるシステインは、典型的には3つのジスルフィド結合を形成し、通常システイン1と2、3と5、及び4と6が互いに対になっている。DR4及びApo-2(更に以下に記載)のように、Apo-2DcRは2つの細胞外のシステインに富んだ擬性反復部を含む(図2)が、これに対し他の同定されている哺乳動物TNFRファミリーのメンバーは3又はそれ以上のこのようなドメインを含む[Smithら, Cell, 76:959(1994)]。
【0186】
図1A(配列番号:1)に示す全長配列のアラインメント解析に基づいて、Apo-2DcRは、Apo-3、TNFR1、又はFas/Apo-1のような他のアポトーシス関連レセプターに対してよりも、DR4(60%)とApo-2(50%)に対してより高い配列同一性を示す。
【0187】
図1Bにおいて、出願人は、明らかな翻訳開始部位がヌクレオチド位置93−95(図1Bにアミノ酸残基−40として特定;配列番号:4)に交互にあてがわれうることを示した。図1Bに示すApo-2DcRはアミノ酸残基−40〜259を含む。
【0188】
実施例2
Apo-2Lに対するApo-2DcRの結合性とApo-2DcR活性に対するPI-PLCの影響
【0189】
Apo-2DcRがApo-2Lに結合するかどうかを試験するため、またApo-2DcRがGPI結合しているかどうかを評価するために、Apo-2DcRトランスフェクト293細胞への放射ヨウ素標識Apo-2Lの結合性を分析した。ホスファチジルイノシトール特異性ホスホリパーゼC(PI-PLC)での細胞の前処理の結合性に対する影響もまた分析した。
【0190】
ヒト293細胞(ATCC CRL 1573)を100mmプレートに蒔き(1x106細胞/プレート)、リン酸カルシウム沈殿法を使用して20μg/プレートのpRK5又は全長Apo-2DcR(実施例1に記載;ATCC寄託209087)をコードしているpRK5でトランスフェクトした。24時間後、細胞をPBS/10mMのEDTA中に収集し、リン酸緩衝食塩水(PBS)中で洗浄し、元のプレート当り2mlのPBS中に再懸濁し、トランスフェクション当り2組の1mlの一定分量に分けた。PI-PLC[Treanorら,Nature,382:80-83(1996)](1μg/ml)を、各トランスフェクションから取り出された2組の一定分量の1組に添加し、細胞を37℃で1時間インキュベートした。細胞を洗浄し、1%のBSA(シグマ)を含む1mlのPBS中に再懸濁させ、0.04mlの一定分量を3組のチューブに入れた。非特異的結合性の測定のために、0.005mlのおよそ20000cpm125I-Apo-2L(Apo-2Lは上掲のPittiらに記載され、通常のラクトペルオキシダーゼ法により放射ヨウ素標識した)を、0.005mlのPBS、又はPBSに入った0.005μlの非標識Apo-2L(最終濃度0.5μg/ml)と共にこれらのチューブに加えた。室温での1時間のインキュベーションの後、細胞を氷冷PBSで洗浄し、ペレット化し、放射能を計数した。
【0191】
Apo-2DcRによるトランスフェクションにより、特異的なApo-2Lの結合量が顕著に増大したが、これは、Apo-2DcRがApo-2Lに結合することを示している(図4)。PI-PLCでの処理により、Apo-2Lの結合が顕著に減少したが、これは、Apo-2DcRがGPIアンカーレセプターであることを示している(図4)。
【0192】
実施例3
全長Apo-2DcRによるApo-2L機能の阻害
【0193】
Apo-2DcRにおいて細胞質領域が無いことは、このレセプターが、Apo-2Lのシグナル伝達の伝達よりもむしろ変調に関与していることを示唆している。従って、Apo-2Lに対する細胞応答性に対するApo-2DcRトランスフェクションの影響を調べた。
【0194】
DR4とApo-2mRNAの両方を発現する(データは示さず)ヒト293細胞を、100mmプレートに蒔き(1x106細胞/プレート)、リン酸カルシウム沈殿法を使用して20μg/プレートのpRK5又はpRK5-hApo-2DcR(実施例2参照)と共にプレート当り3μgのpRKをコードしているグリーン蛍光タンパク質(GFP;クローンテックから購入)でトランスフェクトした。18時間後、細胞をPBS又はApo-2L(Pittiら,上掲,0.5μg/ml)で処理し、(プラスチック上の非固定細胞の明確な視覚化を可能にする)ホフマン光学系を備えた蛍光顕微鏡で6時間にわたって調べた。グリーン蛍光によりGFP陽性細胞を同定し、膜小疱形成(blebbing)及び細胞質凝結のような形態学基準によりアポトーシスを計数した。
【0195】
Apo-2DcRによるトランスフェクションにより、アポトーシス誘発により測定したApo-2Lに対する応答性が顕著に低減させられた(図5)。
【0196】
同様な実験において、pRK5又はpRK5-hApo-2DcR(20μg/プレート)により293細胞をトランスフェクトし、以下の実施例10におけるように、Apo-2L(0.5μg/ml)によるNF-κBの活性化を18時間後に分析した。その結果、Apo-2DcRは、アポトーシス誘発並びにNF-κB活性化により測定したApo-2L機能を阻害することが示された。
【0197】
実施例4
ノーザンブロット分析
【0198】
ヒト組織におけるApo-2DcRmRNAの発現を、ノーザンブロット分析によって検査した。ヒトRNAブロットを、全長Apo-2DcRcDNAをベースにした1.2キロベースの32P標識DNAプローブにハイブリダイズした;プローブはEcoRIでpRK5-Apo-2DcRプラスミドを消化させ、Apo-2DcRcDNA挿入断片を精製することにより産生した。ヒト胎児RNAブロットMTN(クローンテック)、ヒト成人RNAブロットMTN-II(クローンテック)及びヒト癌株化細胞RNAブロット(クローンテック)をDNAプローブと共にインキュベートした。ブロットをハイブリダイゼーション用バッファー(5X SSPE;2X デンハード溶液;100mg/mLの変性剪断されたサケ精子DNA;50%のホルムアミド;2%のSDS)に入ったプローブと共に、42℃で60時間インキュベートした。ブロットを2X SSC;0.05%のSDSを用い、室温で1時間、数回洗浄し、ついで0.1X SSC;0.1%のSDSを用い、50℃で30分間洗浄した。ブロットを一晩さらした後、リン光体イメージャー分析(Fuji)により展開させた。
【0199】
図7Aに示すように、幾つかのApo-2DcRmRNA転写物が検出された。相対的に高い発現が、成人の末梢血白血球(PBL)、脾臓、肺、肝臓及び胎盤において見られた。例えばPBL及び脾臓のようなApo-2DcRを発現する幾つかの成人組織は、Apo-2(以下の実施例11)及びDR4[Panら,上掲]を発現することが示された。
【0200】
図7Bに示されるように、Apo-2DcRメッセージは検査したヒト腫瘍株化細胞(すなわち、HL60前骨髄球性白血病、HeLaS3子宮頚癌、K562慢性骨髄性白血病、MOLT4急性リンパ芽球性白血病、SW480結腸直腸腺癌、A549肺癌、及びG361黒色腫)及び、特にApo-2DcRcDNAにサイズが対応するおよそ1.5kBの転写物の殆どにおいて存在しない。特定された腫瘍細胞タイプ以外の上述の正常なヒト組織におけるApo-2DcRの明らかな発現は、Apo-2DcRレセプターがApo-2リガンドにより癌細胞を(おそらくは癌細胞ではなく正常な細胞を保護することにより)優先的に死滅させることを示唆している。
【0201】
実施例5
ヒトApo-2をコードしているcDNAクローンの単離
【0202】
発現された配列タグ(EST)DNAデータベース(LIFESEQTM, Incyte Pharmaceuticals, Palo Alto, CA)を検索し、Apo-3レセプター[Marstersら, Curr. Biol., 6:750(1996)]の死亡ドメインに対して相同であったあるESTを同定した。ヒト膵臓(「LIB55」)及びヒト腎臓(「LIB28」)のcDNAライブラリ(pRK5ベクター中で上記の実施例1に記載されているようにして調製)を、ESTに基づく合成オリゴヌクレオチドプローブ:
GGGAGCCGCTCATGAGGAAGTTGGGCCTCATGGACAATGAGATAAAGGTGGCTAAAGCTG AGGCAGCGGG (配列番号:9)
でのハイブリダイゼーションによりスクリーニングした。
【0203】
3つのcDNAクローンの全体を配列決定した。cDNAの重なるコード領域はコドン410を除いて同一であった(図8の番号付け系を使用);おそらくは多型性のために、この位置が両方の膵臓cDNAにおけるロイシン残基(TTG)、そして腎臓cDNAにおけるメチオニン残基(ATG)をコード化した。
【0204】
Apo-2の全体核酸配列を図8に示す(配列番号:10)。(以下に示すようにATCC209021として寄託されたpRK5-Apo-2とも呼ばれる)クローン27868は、ヌクレオチド位置140-142に明らかな翻訳開始部位を有する単一のオープンリーディングフレームを含み[Kozakら,上掲]、ヌクレオチド部位1373-1375に見出される終止コドンで終了する(図8;配列番号10)。予想されるポリペプチド先駆体は411アミノ酸長で、I型膜貫通タンパク質であり、約45kDaの算定分子量を有する。ヒドロパシー分析(図示せず)により、シグナル配列(残基1-53)、続いて細胞外ドメイン(残基54-182)、膜貫通ドメイン(残基183-208)、及び細胞内ドメイン(残基209-411)の存在が示唆された(図9;配列番号:11)。293細胞に発現したApo-2-IgGのN末端アミノ酸配列分析により、成熟ポリペプチドがアミノ酸残基54で始まることが示され、これは、実際のシグナル配列が残基1-53を含むことが示している。
【0205】
DR4とApo-2DcRと同様に、Apo-2は2つの細胞外システインリッチ擬似反復を含む一方(図9)、他の同定された哺乳動物TNFRファミリーメンバーは3又はそれより多いそのようなドメインを含む[Smithら,Cell, 76:959(1994)]。
Apo-2の細胞質領域は、TNFR1(30%);CD95(19%);又はApo-3/DR3(29%)の死亡ドメインに対するよりもDR4(64%)の死亡ドメインに対して顕著により高いアミノ酸配列同一性を示す死亡ドメイン(図8に示すアミノ酸残基324−391;図2もまた参照のこと)を含む。TNFR1によるシグナル伝達に対して必要とされる6つの死亡ドメインアミノ酸のうち4つ[Tartagliaら,上掲]がApo-2に保存される一方、他の2つの残基は半保存される(図2参照)。
【0206】
全長配列の(ALIGNコンピュータプログラムを使用する)アラインメント解析に基づき、Apo-2は、例えばTNFR1(19%);CD95(17%);又はApo-3(DR3、WSL-1又はTRAMPとも呼ばれる)(29%)のような他のアポトーシス関連レセプターに対するよりもDR4(55%)に対してより高い配列同一性を示す。
【0207】
実施例6
A. Apo-2 ECDの発現
【0208】
可溶性の細胞外ドメイン(ECD)融合作成物を調製した。Apo-2ECD(図8に示すアミノ酸残基1-184)をPCR法により得て、C末端フラッグエピトープタグ(シグマ)に融合させた。(残基183及び184は膜貫通領域にあることが予想されていても、Apo-2ECD作成物は図8に示す残基183及び184を含んでいた)。ついで、フラッグエピトープタグ分子をpRK5に挿入し、ヒト293細胞(ATCC CRL 1573)に一過性トランスフェクションして発現させた。
【0209】
48時間インキュベートした後、細胞上清を収集し、共沈試験(実施例7を参照)に直接使用するか、製造者(シグマ)の説明に従い、抗フラッグアガロースビーズでのアフィニティークロマトグラフィーによるApo-2ECD-フラッグの精製にかけた。
【0210】
B. イムノアドヘシンとしてのApo-2ECDの発現
可溶性のApo-2ECDイムノアドヘシン作成物を調製した。Apo-2ECD(図8に示すアミノ酸1-184)を、以前に記載されているようにして[Ashkenaziら, Proc. Natl. Acad. Sci., 88:10535-10539(1991)]、pRK5のヒト免疫グロブリンG1重鎖のヒンジ及びFc領域に融合した。イムノアドヘシンは、ヒト293細胞に一過性トランスフェクションして発現させ、前掲のAshkenaziらにより記載されたプロテインAアフィニティークロマトグラフィーにより細胞上清から精製した。
【0211】
実施例7
Apo-2とApo-2リガンド間の結合相互作用を示す免疫沈降アッセイ
【0212】
Apo-2とApo-2Lが相互作用するか又は互いに結合するかを測定するために、擬似トランスフェクト293細胞又はApo-2ECD-フラッグ(上記の実施例6に記載されたもの)がトランスフェクトした293細胞からの上清(5ml)を、5μgのポリ-ヒスチジンタグ可溶性Apo-2L[Pittiら, 上掲]と室温で30分間インキュベートし、ついで共沈アッセイにより複合体形成について分析した。
【0213】
試料を25μlの抗フラッグ抱合アガロースビーズ(シグマ)又はニッケル抱合アガロースビーズ(Qiagen)を使用し、免疫沈降にかけた。4℃で1.5時間インキュベートした後、ビーズを遠心沈殿させ、リン酸緩衝食塩水(PBS)で4回洗浄した。抗フラッグアガロースを使用することにより、Apo-2LをフラッグタグApo-2ECDを通して沈殿させ;ニッケルアガロースを使用することにより、Apo-2ECDをHisタグApo-2Lを通して沈殿させた。沈殿したタンパク質を、SDS-PAGEバッファー中でビーズを5分間沸騰させることで放出させ、12%のポリアクリルアミドゲルでの電気泳動により分離し、ついでMarstersら, J. Biol. Chem.,272:14029-14032(1997)に記載されたようにして、抗Apo-2L又は抗フラッグ抗体(2μg/ml)での免疫ブロットにより検出した。
【0214】
図10に示した結果は、Apo-2ECDとApo-2Lが互いに結合可能であることを示している。
【0215】
さらに、結合相互作用を、抗フラッグビーズ(実施例6を参照)を用い、トランスフェクションした293細胞上清からApo-2ECDを精製し、ついでBIACORETM装置で試料を分析することにより、分析した。BIACORETM分析により、約1nMの解離定数(Kd)が示された。また、BIACORETM分析により、Apo-2ECDが他のアポトーシス誘発性TNFファミリーのメンバー、すなわちTNF-α(ジェネンテク・インク, Pennicaら, Nature, 312:712(1984))、リンホトキシン-α(ジェネンテク・インク), 又はFas/Apo-1リガンド(Alexis Biochemicals)と結合不可能であることが示された。よって、これらのデータは、Apo-2がApo-2Lに対する特異的なレセプターであることを示している。
【0216】
実施例8
Apo-2によるアポトーシスの誘発
【0217】
死亡ドメインはオリゴマー化界面として機能し得るため、死亡ドメインを含むレセプターの過剰発現により、リガンドの不在下におけるシグナル伝達が活性化する[Frazerら,上掲, Nagataら,上掲]。Apo-2が細胞死を誘発可能であるか否かを測定するために、ヒト293細胞又はHeLa細胞(ATCC CCL 2.2)をリン酸カルシウム沈殿法(293細胞)又は電気穿孔法(HeLa細胞)により、Apo-2及び/又はCrmAをコードしているpRK5ベクター又はpRK5系プラスミドで一過性トランスフェクションした。適用できる場合には、プラスミドDNAの全体量を、ベクターDNAを添加することにより調節した。アポトーシスを、形態学的(図11A)、DNA断片化(図11B)、又はMarstersら, Curr.Biol.,6:1669(1996)に記載されているようなホスファチジルセリン暴露のFACS分析(図11C)により、トランスフェクション24時間後に評価した。図11A及び11Bに示すように、Apo-2トランスフェクション293細胞には、顕著なアポトーシスが発生した。
【0218】
FACSにより分析される試料に対して、HeLa細胞にトランスフェクション用のマーカーとしてpRK5-CD4を同時形質移入させ、CD4発現細胞におけるアポトーシスを測定した;FADDをApo-2プラスミドで同時形質移入した;データは、Marstersら, Curr. Biol., 6:1669(1996)に記載されているように、少なくとも3回の実験の平均値±SEMである。カスパーゼ阻害剤、DEVD-fmk(Enzyme Systems)又はz-VAD-fmk(Research Biochemicals Intl.)を、トランスフェクション時に200μMで添加した。図11Cに示すように、カスパーゼ阻害剤CrmA、DEVD-fmk及びz-VAD-fmkが、Apo-2によるアポトーシスの誘発を阻害し、この反応におけるCed-3様プロテアーゼの関与が示された。
【0219】
FADDは、CD95、TNFR1及びApo-3/DR3によるアポトーシスの活性化を媒介するアダプタータンパク質である[Nagataら, 上掲]が、Apo-2L[Marstersら, 上掲]又はDR4[Panら, 上掲]によるアポトーシス誘導には必要であるとは思われない。CD95、TNFR1、又はApo-3/DR3によるアポトーシス誘発を阻害するFADDのドミナントネガティブ変異体型[Frazerら,上掲;Nagataら,上掲;Chinnayianら,上掲]は、HeLa細胞にApo-2を同時形質移入させた場合(図11C)に、Apo-2によるアポトーシス誘発を阻害しなかった。これらの結果より、Apo-2は、FADDとは独立して、アポトーシスのシグナル伝達をしていることが示唆される。この結論と一致して、Apo-2細胞質領域を含むグルタチオン-S-トランスフェラーゼ融合タンパク質は、インビトロで転写され翻訳されたFADDに結合しなかった(データは示さず)。
【0220】
実施例9
可溶性Apo-2ECDによるApo-2L活性の阻害
【0221】
可溶性Apo-2L(0.5μg/ml、Pittiら,上掲に記載されたようにして調製)を、抗フラッグ抗体(シグマ)(1μg/ml)と共に、PBSバッファー又はアフニティー精製されたApo-2ECD(5μg/ml)と室温で1時間、プレインキュベートし、HeLa細胞に添加した。インキュベート5時間後、細胞をFACS(上述)によりアポトーシスについて分析した(図11D)。
【0222】
Apo-2LはHeLa細胞において顕著なアポトーシスを誘発し、可溶性Apo-2ECDはApo-2Lの作用を阻止でき(図11D)、Apo-2LとApo-2との間に特異的な相互作用があることが確認された。同様の結果がApo-2ECDイムノアドヘシンでも得られた(図411E)。用量-反応分析では、約0.3nMのApo-2イムノアドヘシンで最大半減阻害が示されている(図11E)。
【0223】
実施例10
Apo-2によるNF-κBの活性化
【0224】
Apo-2がNF-κBを活性化させるか否かを測定するためのアッセイを行った。
HeLa細胞を、全長天然配列Apo-2、DR4又はApo-3をコードするpRK5発現プラスミドでトランスフェクションし、トランスフェクション24時間後に収集した。核抽出物を調製し、1μgの核タンパク質を、32P標識NF-κB特異性合成オリゴヌクレオチドプローブATCAGGGACTTTCCGCTGGGGACTTTCCG(配列番号:12)[MacKayら, J. Immunol., 153:5274-5284(1994)も参照のこと]と、単独又は50倍過剰の非標識プローブと共に、又は関連のない32P標識合成オリゴヌクレオチドAGGATGGGAAGTGTGTGATATATCCTTGAT(配列番号:13)と反応させた。幾つかの試料に、NF-κBのp65/Re1Aサブユニットに対する抗体(1μg/ml;Santa Cruz Biotechnology)を添加した。DNA結合性を、Hsuら,上掲;Marstersら,上掲、及びMacKayら,上掲に記載されたようにして、電気泳動易動度シフトアッセイにより分析した。
【0225】
結果を図12に示す。図12Aに示すように、電気泳動移動度シフトアッセイにより測定した場合、Apo-2とDR4の両方が、HeLa細胞へのトランスフェクションの際に、有意なNF-κβの活性化を誘発した;活性化レベルはApo-3/DR3で観察される活性化に匹敵するものであった。NF-κBのp65/Re1Aサブユニットに対する抗体は、NF-κBプローブの移動を阻害し、3つ全てのレセプターに対する反応においてp65を結び付けていた。
【0226】
また、Apo-2L自体がNF-κB活性を調節可能であるか否かを測定するアッセイを行った。HeLa細胞又はMCF7細胞(ヒト胸腺癌株化細胞、ATCC HTB 22)を、PBSバッファー、可溶性Apo-2L(Pittiら,上掲)又はTNF-α(ジェネンテク・インク, Pennicaら, Nature, 312:724(1984)を参照)(1μg/ml)で処理し、上述のようにしてNF-κB活性をアッセイした。結果を図12Bに示す。Apo-2Lは、処理されたHeLa細胞においては有意なNF-κB活性化を誘発したが、処理されたMCF7細胞においては誘発しなかった;TNF-αは両方の株化細胞においてより著しい活性化を誘発した。幾つかの研究において、TNFによるNF-κBの活性化により、TNF誘発性アポトーシスから細胞を保護できることが開示されている[Nagataら,上掲]。
【0227】
また、NF-κBインヒビターであるALLN(N-アセチル-ロイシン-ロイシン-ノルロイシン)及び転写インヒビターであるシクロヘキサミドの影響を試験した。HeLa細胞(6-ウェル皿に蒔いた)を、Apo-2L(1μg/ml)の添加前に、PBSバッファー、ALLN(Calbiochem)(40μg/ml)又はシクロヘキサミド(シグマ)(50μg/ml)と共に、1時間プレインキュベートした。インキュベート5時間後、FACSによりアポトーシスについて分析した(図12Cを参照)。
【0228】
結果を図12Cに示す。ALLN及びシクロヘキサミドの両方とも、HeLa細胞におけるApo-2L誘発性アポトーシスのレベルを増加させた。データは、Apo-2Lが保護性NF-κB依存性遺伝子を誘発可能であることを示している。また、データは、Apo-2Lがある種の株化細胞においてNF-κβを活性化することができ、Apo-2とDR4の両方がその機能を媒介し得ることを示している。
【0229】
実施例11
ノーザンブロット分析
【0230】
ヒト組織におけるApo-2mRNAの発現を、ノーザンブロット分析によって検査した。ヒトRNAブロットを、全長Apo-2cDNAをベースにした4.6キロベースの32P標識DNAプローブにハイブリダイズした;プローブはEcoRIでpRK5-Apo-2プラスミドを消化することにより産生した。ヒト胎児RNAブロットMTN(クローンテック)及びヒト成人RNAブロットMTN-II(クローンテック)をDNAプローブと共にインキュベートした。ブロットをハイブリダイゼーション用バッファー(5X SSPE;2Xデンハード溶液;100mg/mLの変性された剪断サケ精子DNA;50%のホルムアミド;2%のSDS)に入ったプローブと共に、42℃で60時間インキュベートした。ブロットを2X SSC;0.05%のSDSを用い、室温で1時間、数回洗浄し、ついで0.1X SSC;0.1%のSDSを用い、50℃で30分間洗浄した。ブロットを一晩さらした後、展開した。
【0231】
図13に示すように、約4.6キロベースの優性mRNAの転写物が複数の組織において検出された。発現は、胎児及び成人の肝臓及び肺、及び成人の卵巣及び末梢血白血球(PBL)において比較的高かったが、胎児及び成人の脳においてはmRNAの発現は検出されなかった。中レベルの発現は、成人の大腸、小腸、精巣、前立腺、胸腺、膵臓、腎臓、骨格筋、胎盤、及び心臓において見出された。Apo-2が発現した幾つかの成人組織、例えばPBL、卵巣及び脾臓は、DR4を発現することが以前に見出されているが[Panら,上掲]、各レセプターmRNAの発現の相対レベルは異なっているようである。
【0232】
実施例12
Apo-2、DR4及びApo-2DcR遺伝子の染色体局在化
【0233】
ヒトApo-2遺伝子の染色体局在化を、放射線ハイブリッド(RH)パネル分析により検査した。RHマッピングを、ヒト-マウス細胞の放射線ハイブリッドパネル(Research Genetics)とApo-2cDNAのコード領域に基づくプライマーを使用するPCRにより、実施した[Gelbら,Hum. Genet., 98:141(1996)]。スタンフォードヒトゲノムセンターデータベースを使用したPCRデータの解析では、Apo-2は11.05LODを有するマーカーD8S481に結合しており;D8S481はD8S2055と今度は結合しており、これがヒト染色体8p21に位置する。DR4の類似した分析においては、DR4が、ヒト染色体8p21にまた位置するマーカーD8S2127(13.00のLODを持つ)に結合していることが示された。Apo-2DcRの放射線ハイブリッドパネルを使用した解析では、Apo-2DcR遺伝子はマーカーWI-6536に結合し、これがD8S298についで結合しており、これがまたヒト染色体8p21に位置し、D8S2005とD8S2127の間に巣を形成している。従って、3つのApo-2Lレセプター、Apo-2、Apo-2DcR及びDR4に対するヒト遺伝子は、全て染色体8p21に位置している。
【0234】
本出願人の知る限りでは、今日まで、TNFR遺伝子ファミリーのメンバーで染色体8pに位置しているものはなかった。
【0235】
実施例13
Apo-2DcRに特異的なモノクローナル抗体の調製
【0236】
0.5μg/50μlのApo-2DcRイムノアドヘシンタンパク質(Ribi Immunochemical Research Inc., Hamilton, MTから購入したMPL-TDMアジュバントで希釈)を、各々の後足の裏の柔らかい部分に、3日の間隔で11回注射することで、Balb/cマウス(チャールズリヴァー研究所から得たもの)を免疫した。過去に開示されたように[Ashkenaziら、Proc. Natl. Acad. Sci., 88:10535-10539(1991)]、pRK5においてヒト免疫グロブリンG1重鎖のヒンジ及びFc領域に、Apo-2DcRのN末端領域を融合させることにより、Apo-2DcRイムノアドヘシンタンパク質を産生した。イムノアドヘシンタンパク質を、ヒト293細胞に一過性トランスフェクションして発現させ、前掲のAshkenaziらにより記載されたようにして、プロテインAアフィニティークロマトグラフィーにより細胞上清から精製した。
【0237】
最終の追加免疫から3日後に、膝窩リンパ節をマウスから取り除き、単細胞懸濁液を、1%のペニシリン-ストレプトマイシンが補填されたDMEM培地(Biowhitakker Corp.から得たもの)中で調製した。ついで、リンパ節細胞を35%のポリエチレングリコールを使用し、マウス骨髄腫細胞P3X63AgU.1(ATCC CRL 1597)と融合させ、96-ウェル培養プレートで培養した。融合の結果得られたハイブリドーマをHAT培地において選択した。融合から10日後に、ハイブリドーマ培養の上清をELISAでスクリーニングし、Apo-2DcRイムノアドヘシンタンパク質に結合するモノクローナル抗体の有無を試験した。
【0238】
ELISAにおいて、各々のウェルに、PBSに2μg/mlのヤギ抗ヒトIgG Fc(Cappel Laboratories社から購入)が入ったものを50μl添加して、96-ウェルマイクロタイタープレート(Maxisorb;Nunc, Kamstrup, Denmark)をコーティングし、4℃で一晩インキュベートした。ついでプレートを洗浄バッファー(0.05%のトゥイーン20を含有するPBS)で3回洗浄した。ついで、マイクロタイタープレートのウェルを、PBSに2.0%のウシ血清アルブミンが入ったもの200μlでブロックし、室温で1時間インキュベートした。ついでプレートを、洗浄バッファーで、再度3回洗浄した。
【0239】
洗浄工程後、アッセイ用バッファー(0.5%のBSAを含むPBS)に0.4μg/mlのApo-2DcRイムノアドヘシンタンパク質(上述のもの)が入ったもの50μlを各々のウェルに添加した。プレートを振盪装置上で、室温で1時間インキュベートし、続いて洗浄バッファーで3回洗浄した。
【0240】
洗浄工程に続き、100μlのハイブリドーマ上清又は精製した抗体(プロテインG-セファロースカラムを使用)(1μg/ml)を、アッセイ用バッファーの指定ウェルに添加した。100μlのP3X63AgU.1骨髄腫細胞の条件培地を、対照として他の指定ウェルに添加した。プレートを振盪装置上で、室温で1時間インキュベートし、続いて洗浄バッファーで3回洗浄した。
【0241】
次に、アッセイ用バッファーで1:1000に希釈されたHRP-抱合ヤギ抗マウスIgG Fc(Cappel Laboratories社から購入)を50μl、各々のウェルに添加し、プレートを振盪装置上で、室温で1時間インキュベートした。プレートを洗浄バッファーで3回洗浄し、ついで各々のウェルに50μlの基質(TMBマイクロウェル・ペルオキシダーゼ基質、Kirkegaard & Perry, Gaithersburg, MD)を添加し、室温で10分間インキュベートした。50μlのTMB1-成分終止溶液(ジエチレングリコール, Kirkegaard & Perry)を各々のウェルに添加して反応を終了させ、450nmでの吸光度を自動マイクロタイタープレート読取装置で読取った。
【0242】
ELISAによりスクリーニングされたハイブリドーマ上清の内、47の上清が陽性であると判断された(バックグラウンドのおよそ4倍高いと算出)。ELISAで陽性であると判断された上清を、さらにHUMEC細胞(Apo-2DcRを発現するヒト微小血管内皮株化細胞;Cell Syatems, Kirkland, WA)及びPE-抱合ヤギ抗マウスIgGを使用してFACS分析により分析した。この分析のため、細胞選別用バッファー(1%のFCSと0.02%のNaN3を含有するPBS)に懸濁させた25μlの細胞(4×106細胞/ml)をU字底のマイクロタイターウェルに添加し、100μlの培養上清又は精製した抗体(プロテインG-セファロースカラムで精製)(10μg/ml)が細胞選別用バッファーに入ったものと混合し、氷上で30分間インキュベートした。ついで、細胞を洗浄し、100μlのPE-抱合ヤギ抗マウスIgGと共に4℃で30分間インキュベートした。ついで細胞を2回洗浄し、200μlの細胞選別用バッファーに再懸濁させ、FACScan(Becton Dickinson, Mountain View, CA)により分析した。FACS分析により、12/35の上清が抗Apo-2抗体に対して陽性であることが示された。
【0243】
図14は、4G3.9.9;6D10.9.7;及び1C5.24.1と呼ばれるApo-2DcR抗体とインキュベートしたHUMEC細胞のFACS染色を示す。図14に示すように、各抗体は、HUMEC細胞において発現されるApo-2DcRレセプターを認識する。
【0244】
実施例14
他のApo-2リガンドレセプターへのApo-2DcR抗体の結合性をテストするためのELISAアッセイ
【0245】
実施例13に記載したモノクローナル抗体が、Apo-2DcR以外の他の既知のApo-2Lレセプターに結合することができたか否かを測定するためにELISAを実施した。特に、4G3.9.9;6D10.9.7;及び1C5.24.1抗体について、それぞれ、DR4[Panら,上掲]、Apo-2[上記の実施例に記載]、及びDcR2[Marstersら, Curr. Biol., 7:1003-1006(1997)]に対する結合性を調べた。ELISAは、本質的に上述の実施例13に記載したようにして実施した。
【0246】
結果を図15に示す。Apo-2DcR抗体4G3.9.9はApo-2DcRに結合した。また4G3.9.9抗体は、DR4とApo-2に対してはいくらかの交差反応性とDcR2に対してある限られた交差反応性を示した。6D10.9.7抗体はApo-2DcRに結合し、DR4、Apo-2及びDcR2に対してある限られた交差反応性を示した。最後に、1C5.24.1抗体はApo-2DcRに結合し、DR4に対して幾らかの交差反応性を示した。1C5.24.1抗体はApo-2とDcR2に対して些かより少ない交差反応性を示した。交差反応性のまとめをまた図16に示す。
【0247】
実施例15
抗体のアイソタイピング
【0248】
Apo-2DcR抗体(実施例13と14において上述したもの)のアイソタイプを、アイソタイプ特異的ヤギ抗マウスIg(Fisher Biotech, Pittsburgh, PA)で、マイクロタイタープレートを4℃で一晩かけてコーティングすることにより決定した。ついでプレートを洗浄バッファー(上述の実施例13に記載したもの)で洗浄した。ついで、マイクロタイタープレートのウェルを200μlの2%ウシ血清アルブミン(BSA)でブロックし、室温で1時間インキュベートした。プレートを、再度洗浄バッファーで3回洗浄した。次に、100μlのハイブリドーマ培養上清又は5μg/mlの精製抗体を指定ウェルに加えた。プレートを室温で30分インキュベートし、ついで50μlのHRP-抱合ヤギ抗マウスIgG(実施例13において上述したもの)を各々のウェルに添加した。プレートを室温で30分インキュベートした。プレートに結合したHRPのレベルを、上述のHRP基質を使用して検出した。
【0249】
アイソタイピング分析により、4G3.9.9及び1C5.24.1抗体はIgG1抗体であることが示された。またこの分析により、6D10.9.7抗体がIgG2b抗体であることが示された。これらの結果を図16にまた示す。
【0250】
材料の寄託
次の材料をアメリカン・タイプ・カルチャー・コレクション,10801 University Blvd., Manassas, Virginia, USA(ATCC)に寄託した:
材料 ATCC寄託番号 寄託日
pRK-Apo-2 209021 1997年5月8日
Apo2-DcR 209087 1997年5月30日
4G3.9.9 ___ _____________
6D10.9.7 ______ _____________
1C5.24.1 ______ _____________
【0251】
この寄託は、特許手続き上の微生物の寄託の国際的承認に関するブダペスト条約及びその規則(ブダペスト条約)の規定に従って行われた。これは、寄託の日付から30年間、寄託の生存可能な培養が維持されることを保証するものである。寄託物はブダペスト条約の条項に従い、またジェネンテク社とATCCとの間の合意に従い、ATCCから入手することができ、これは、どれが最初であろうとも、関連した米国特許の発行時又は任意の米国又は外国特許出願の公開時に、寄託培養物の後代を永久かつ非制限的に入手可能とすることを保証し、米国特許法第122条及びそれに従う特許庁長官規則(特に参照番号886OG638の37CFR第1.14条を含む)に従って権利を有すると米国特許庁長官が決定した者に後代を入手可能とすることを保証するものである。
【0252】
本出願の譲受人は、寄託した培養物が、適切な条件下で培養されていた場合に死亡もしくは損失又は破壊されたならば、材料は通知時に同一の他のもの即座に取り替えることに同意する。寄託物質の入手可能性は、特許法に従いあらゆる政府の権限下で認められた権利に違反して、本発明を実施するライセンスであるとみなされるものではない。
【0253】
上記の文書による明細書は、当業者に本発明を実施できるようにするために十分であると考えられる。寄託した態様は、本発明のある側面の一つの説明として意図されており、機能的に等価なあらゆる作成物がこの発明の範囲内にあるため、寄託された作成物により、本発明の範囲が限定されるものではない。ここでの物質の寄託は、文書による説明が、そのベストモードを含む、本発明の任意の側面の実施を可能にするために不十分であることを認めるものではないし、それが表す特定の例証に対して請求の範囲を制限するものと解釈されるものでもない。実際、ここに示し記載したものに加えて、本発明を様々に改変することは、前記の記載から当業者にとっては明らかなものであり、添付の請求の範囲内に入るものである。
【図面の簡単な説明】
【0254】
【図1A】図1Aは、天然配列ヒトApo-2DcRcDNAの核酸配列及びその誘導アミノ酸配列(開始部位は残基1にあてがわれる(ヌクレオチド193−195))を示す。
【図1B】図1Bは、天然配列ヒトApo-2DcRcDNAの核酸配列及びその誘導アミノ酸配列(開始部位は残基−40にあてがわれる(ヌクレオチド73−75))を示す。
【図2】図2は、Apo-2及びApo-2DcRの一次構造とmRNA発現を示す。この図は、全長DR4と整列化させたヒトApo-2及びApo-2DcRの推論アミノ酸配列を示す。Apo-2の死亡ドメインはDR4、Apo-3/DR3、TNFR1、及びCD95のものと整列化され;星印はTNFR1による死亡シグナル伝達に必須である残基を示す[Tartagliaら,上掲]。示されているのは、予想されたシグナルペプチド開裂部位(矢印)、2つのシステインリッチドメイン(CRD1、2)及びApo-2及びDR4の膜貫通ドメイン又はApo-2DcRの疎水性C末端(下線)である。また示されているのは、5つの潜在的なN結合グリコシル化部位(黒い箱)及びApo-2DcRの5つの配列擬似反復(括弧)である。
【図3】図3は、Apo-2とApo-2DcRのヒドロパシープロットを示す。上部の数はアミノ酸の位置を示している。
【図4】図4は、Apo-2DcRトランスフェクト細胞に対する放射ヨウ素標識Apo-2Lの結合と、PI-PLCでの細胞の前処理によるその阻害を示す。
【図5】図5はApo-2DcRによるアポトーシスのApo-2L誘発の阻害を示す。
【図6】図6はApo-2DcRによるNF-κBのApo-2L活性化の阻害を示す。
【図7】図7Aは、ヒト組織のポリA RNAブロットのノーザンハイブリダイゼーションにより分析されたヒト組織中でのApo-2DcRmRNAの発現を示す。図7Bは、ヒト癌株化細胞のポリA RNAブロットのノーザンハイブリダイゼーションにより分析されたヒト癌株化細胞中でのApo-2DcRmRNAの発現(の欠如)を示す。
【図8】図8は、天然配列ヒトApo-2cDNAの核酸配列とその誘導アミノ酸配列を示す。
【図9】図9は、天然配列ヒトApo-2の誘導アミノ酸配列を示し、推定シグナル配列には下線を引き、推定膜貫通ドメインは囲み、推定死亡ドメイン配列には破線の下線を付している。2つのシステインリッチドメインのシステインには個々に下線を引いている。
【図10】図10は、Apo-2LとのApo-2ECDの相互作用を示す。フラッグエピトープタグApo-2ECDでトランスフェクトされた293細胞又は擬似トランスフェクトされた293細胞の上清をポリ-His-タグApo-2Lと共にインキュベートし、抗フラッグ抱合又はニッケル抱合アガロースビーズで免疫沈降させた。沈殿したタンパク質をポリアクリルアミドゲルによる電気泳動法で分離し、抗Apo-2L又は抗フラッグ抗体を用いた免疫ブロットにより検出した。
【図11A】図11は、Apo-2によるアポトーシスの誘発と、可溶性Apo-2ECDによるApo-2L活性の阻害を示す。ヒト293細胞(A、B)又はHeLa細胞(C)を、Apo-2及び/又はCrmAをコードしたpRK5ベクター又はpRK5ベースプラスミドによりトランスフェクトした。アポトーシスを、形態学的(A)、DNA断片化(B)、又はFACS(C-E)により評価した。可溶性Apo-2Lを、バッファーあるいは抗フラッグ抗体又はApo-2DcRECDイムノアドヘシン又はDR4又はTNFR1イムノアドヘシンと共にアフィニティー精製されたApo-2ECDでプレインキュベートし、HeLa細胞に添加した。後で細胞のアポトーシスを分析した(D)。また、Apo-2ECDイムノアドヘシンを備えたApo-2Lを使用して用量反応分析を行った(E)。
【図11B】図11は、Apo-2によるアポトーシスの誘発と、可溶性Apo-2ECDによるApo-2L活性の阻害を示す。ヒト293細胞(A、B)又はHeLa細胞(C)を、Apo-2及び/又はCrmAをコードしたpRK5ベクター又はpRK5ベースプラスミドによりトランスフェクトした。アポトーシスを、形態学的(A)、DNA断片化(B)、又はFACS(C-E)により評価した。可溶性Apo-2Lを、バッファーあるいは抗フラッグ抗体又はApo-2DcRECDイムノアドヘシン又はDR4又はTNFR1イムノアドヘシンと共にアフィニティー精製されたApo-2ECDでプレインキュベートし、HeLa細胞に添加した。後で細胞のアポトーシスを分析した(D)。また、Apo-2ECDイムノアドヘシンを備えたApo-2Lを使用して用量反応分析を行った(E)。
【図11C】図11は、Apo-2によるアポトーシスの誘発と、可溶性Apo-2ECDによるApo-2L活性の阻害を示す。ヒト293細胞(A、B)又はHeLa細胞(C)を、Apo-2及び/又はCrmAをコードしたpRK5ベクター又はpRK5ベースプラスミドによりトランスフェクトした。アポトーシスを、形態学的(A)、DNA断片化(B)、又はFACS(C-E)により評価した。可溶性Apo-2Lを、バッファーあるいは抗フラッグ抗体又はApo-2DcRECDイムノアドヘシン又はDR4又はTNFR1イムノアドヘシンと共にアフィニティー精製されたApo-2ECDでプレインキュベートし、HeLa細胞に添加した。後で細胞のアポトーシスを分析した(D)。また、Apo-2ECDイムノアドヘシンを備えたApo-2Lを使用して用量反応分析を行った(E)。
【図11D】図11は、Apo-2によるアポトーシスの誘発と、可溶性Apo-2ECDによるApo-2L活性の阻害を示す。ヒト293細胞(A、B)又はHeLa細胞(C)を、Apo-2及び/又はCrmAをコードしたpRK5ベクター又はpRK5ベースプラスミドによりトランスフェクトした。アポトーシスを、形態学的(A)、DNA断片化(B)、又はFACS(C-E)により評価した。可溶性Apo-2Lを、バッファーあるいは抗フラッグ抗体又はApo-2DcRECDイムノアドヘシン又はDR4又はTNFR1イムノアドヘシンと共にアフィニティー精製されたApo-2ECDでプレインキュベートし、HeLa細胞に添加した。後で細胞のアポトーシスを分析した(D)。また、Apo-2ECDイムノアドヘシンを備えたApo-2Lを使用して用量反応分析を行った(E)。
【図11E】図11は、Apo-2によるアポトーシスの誘発と、可溶性Apo-2ECDによるApo-2L活性の阻害を示す。ヒト293細胞(A、B)又はHeLa細胞(C)を、Apo-2及び/又はCrmAをコードしたpRK5ベクター又はpRK5ベースプラスミドによりトランスフェクトした。アポトーシスを、形態学的(A)、DNA断片化(B)、又はFACS(C-E)により評価した。可溶性Apo-2Lを、バッファーあるいは抗フラッグ抗体又はApo-2DcRECDイムノアドヘシン又はDR4又はTNFR1イムノアドヘシンと共にアフィニティー精製されたApo-2ECDでプレインキュベートし、HeLa細胞に添加した。後で細胞のアポトーシスを分析した(D)。また、Apo-2ECDイムノアドヘシンを備えたApo-2Lを使用して用量反応分析を行った(E)。
【図12A】図12は、Apo-2、DR4及びApo-2LによるNF-κBの活性化を示す。(A)HeLa細胞を、示したタンパク質をコードする発現プラスミドでトランスフェクトした。核抽出物を調製し、電気泳動易動度シフトアッセイにより分析した。
【図12B】図12は、Apo-2、DR4及びApo-2LによるNF-κBの活性化を示す。(B)HeLa細胞又はMCF7細胞をバッファー、Apo-2L又はTNF-αで処理し、NF-κB活性をアッセイした。
【図12C】図12は、Apo-2、DR4及びApo-2LによるNF-κBの活性化を示す。(C)HeLa細胞をApo-2Lの添加前に、バッファー、ALLN又はシクロヘキサミドと共にプレインキュベートした。後にFACSによりアポトーシスを分析した。
【図13】図13は、ヒト組織のポリA RNAブロットのノーザンハイブリッド形成により分析した、ヒト組織におけるApo-2mRNAの発現を示す。
【図14A】図14は、IgG対照(点線)と比較したApo-2DcR抗体(太線で示す)のFASC分析を示す。抗体(1C5.24.1)は、HUMEC細胞において発現されたApo-2DcRレセプターを認識した。
【図14B】図14は、IgG対照(点線)と比較したApo-2DcR抗体(太線で示す)のFASC分析を示す。抗体(4G3.9.9)は、HUMEC細胞において発現されたApo-2DcRレセプターを認識した。
【図14C】図14は、IgG対照(点線)と比較したApo-2DcR抗体(太線で示す)のFASC分析を示す。抗体(6D10.9.7)は、HUMEC細胞において発現されたApo-2DcRレセプターを認識した。
【図15A】図15は、Apo-2DcR、及びDR4、Apo-2及びDcR2と称される他の既知のApo-2Lレセプターに対するApo-2DcR抗体1C5.24.の結合性をテストする酵素結合免疫吸着検定(ELISA)の結果を示すグラフを含む。
【図15B】図15は、Apo-2DcR、及びDR4、Apo-2及びDcR2と称される他の既知のApo-2Lレセプターに対するApo-2DcR抗体4G3.9.9の結合性をテストする酵素結合免疫吸着検定(ELISA)の結果を示すグラフを含む。
【図15C】図15は、Apo-2DcR、及びDR4、Apo-2及びDcR2と称される他の既知のApo-2Lレセプターに対するApo-2DcR抗体6D10.9.7の結合性をテストする酵素結合免疫吸着検定(ELISA)の結果を示すグラフを含む。
【図16】図16は、抗体1C5.24.1;4G3.9.9;及び6D10.9.7の交差反応性及びアイソタイプのまとめを提供する表である。
【発明の開示】
【0001】
発明の分野
本発明は、一般に、ここに「Apo-2DcR」と命名する新規ポリペプチドの同定、単離、及び組換え生産、及び抗Apo-2DcR抗体に関する。
【0002】
発明の背景
アポトーシス又は「プログラムされた細胞死」
哺乳動物における細胞数のコントロールは、細胞増殖と細胞死のバランスにより部分的に決定されると考えられている。しばしば壊死性細胞死と称される細胞死の一形態は、典型的には、ある種の外傷又は細胞傷害の結果生じる細胞死の病理的形態として特性付けられる。これに対して、通常は規則的又はコントロールされた状態で進行する細胞死の他の「生理的」形態がある。細胞死のこの規則的又はコントロールされた形態は、しばしば「アポトーシス」と称される[例えば、Barrら, Bio/Technology, 12:487-493(1994);Stellerら, Science, 267:1445-1449(1995)を参照]。アポトーシス性細胞死は、免疫系におけるクローン選択と胚の発達を含む多くの生理的プロセスにおいて自然に生じる[Itohら, Cell, 66:233-243(1991)]。アポトーシス性細胞死のレベルの減少は、癌、狼瘡、ヘルペスウイスル感染を含む種々の病理的条件に関連している[Thompson, Science, 267:1456-1462(1995)]。アポトーシス性細胞死のレベルの増加は、エイズ、アルツハイマー病、パーキンソン病、筋萎縮性側索硬化症、多発性硬化症、色素性網膜炎、小脳変性、無形成性貧血、心筋梗塞、脳卒中、再灌流傷害、及び毒素誘発性肝疾患を含む様々な他の病理状態に関連している[上掲のThompsonを参照]。
【0003】
典型的には、アポトーシス性細胞死には、細胞内における一又は複数の特徴的な形態学的及び生化学的変化、例えば細胞質の凝結、原形質膜の微絨毛の喪失、核の分節化、染色体DNAの分解又はミトコンドリア機能の喪失が伴う。様々な外因的及び内因的シグナルが、このような形態学的及び生化学的な細胞変化を惹起又は誘発すると考えられている[Raff, Nature, 356:397-400(1992);Steller, 上掲;Sachsら, Blood, 82:15(1993)]。例えば、ホルモンの刺激、例えば未成熟胸腺細胞に対する糖質コルチコイドホルモン、並びにある種の成長因子の退薬により惹起され得る[Watanabe-Fukunagaら, Nature, 356:314-317(1992)]。また、幾つかの同定された発癌遺伝子、例えばmyc、rel、及びE1A、及び腫瘍サプレッサー、例えばp53が、アポトーシスの誘発においてある役割を有していることも報告されている。ある種の化学療法薬及びある種の放射線も同様にアポトーシス誘発活性を有していることも見出されている[Thompson, 上掲]。
サイトカインのTNFファミリー
【0004】
様々な分子、例えば腫瘍壊死因子-α(「TNF-α」)、腫瘍壊死因子-β(「TNF-β」すなわち「リンホトキシン」)、CD30リガンド、CD27リガンド、CD40リガンド、OX-40リガンド、4-1BBリガンド、Apo-1リガンド(Fasリガンド又はCD95リガンドとも称される)、及びApo-2リガンド[トレイル(TRAIL)とも称される]が、サイトカインの腫瘍壊死因子(「TNF」)ファミリーのメンバーとして同定された[例えば、Gruss及びDower, Blood, 85:3378-3404(1995);Wileyら, Immunity, 3:673-682(1995);Pittiら, J. Biol. Chem., 271:12687-12690(1996)]。これらの分子のなかでも、TNF-α、TNF-β、CD30リガンド、4-1BBリガンド、Apo-1リガンド、及びApo-2リガンド(TRAIL)は、アポトーシス性細胞死に関与していることが報告されている。TNF-αとTNF-βの両方とも、感受性腫瘍細胞におけるアポトーシス性死を誘発することが報告されている[Schmidら, Proc. Natl. Acad. Sci., 83:1881(1986);Dealtryら, Eur. J. Immunol., 17:689(1987)]。ゼングらは、TNF-αがCD8ポジティブT細胞のポスト刺激性アポトーシスに関与していることを報告している[Zhengら, Nature, 377:348-351(1995)]。他の研究者は、CD30リガンドが胸腺における自己反応性T細胞の欠失に関与していることを報告している[Amakawaら, プログラム細胞死に関するコールドスプリングハーバー研究所のシンポジウム、要約集、第10巻、(1995)]。
【0005】
マウスのFas/Apo-1レセプター又はリガンド遺伝子(それぞれ1pr及びgldと呼称される)における変異が幾つかの自己免疫疾患に関連しており、Apo-1リガンドが末梢の自己反応性リンパ球のクローン除去の調節においてある役割を担っていることを示している[Krammerら, Curr. Op. Immunol., 6:279-289(1994);Nagataら, Science, 267:1449-1456(1995)]。また、Apo-1リガンドは、CD4ポジティブTリンパ球及びBリンパ球においてポスト刺激性アポトーシスを誘発することが報告されており、それらの機能がもはや必要でなくなった際の活性化リンパ球の除去に関与している[Krammerら, 上掲;Nagataら, 上掲]。Apo-1レセプターと特異的に結合するアゴニストのマウスモノクローナル抗体は、TNF-αに匹敵するか類似する細胞死滅活性を示すことが報告されている[Yoneharaら, J. Exp. Med., 169:1747-1756(1989)]。
レセプターのTNFファミリー
【0006】
このようなTNFファミリーのサイトカインが介在する種々の細胞反応誘導は、特異的な細胞レセプターに結合することにより開始されると考えられている。約55-kDa(TNFR1)と75-kDa(TNFR2)の2つの異なるTNFレセプターが同定されており[Hohmanら, J. Biol. Chem., 264:14927-14934(1989);Brockhausら, Proc. Natl. Acad. Sci., 87:3127-3131(1990);1991年3月20日に公開されたEP417563]、双方のレセプター型に対応するヒト及びマウスcDNAが単離され、特徴付けされている[Loetscherら, Cell, 61:351(1990);Schallら, Cell, 61:361(1990);Smithら, Science, 248:1019-1023(1990);Lewisら, Proc. Natl. Acad. Sci., 88:2830-2834(1991);Goodwinら, Mol. Cell. Biol., 11:3020-3026(1991)]。広範な多型性が、双方のTNFレセプター遺伝子に関連している[例えば、Takaoら, Immunogenetics, 37:199-203(1993)を参照]。双方のTNFRは細胞外、膜貫通及び細胞内領域を含む細胞表面レセプターの典型的な構造を共有する。双方のレセプターの細胞外部分はまた可溶性TNF結合タンパク質として天然に見出される[Nophar, Yら, EMBO J., 9:3269(1990);及びKohno, Tら, Proc. Natl. Acad. Sci. U.S.A., 87:8331(1990)]。更に最近になって、組換え体可溶性TNFレセプターのクローニングがヘイル(Hale)らにより報告されている[J. Cell. Biochem. 増補15F, 1991, p.113(P424)]。
【0007】
1型又は2型のTNFR(TNFR1及びTNFR2)の細胞外部分は、NH2末端から出発して、1〜4とされる4つのシステインに富んだドメイン(CRD)の反復アミノ酸配列パターンを含む。各CRDは約40のアミノ酸長のものであり、良好に保存された位置に4〜6のシステイン残基を含んでいる[Schallら, 上掲;Loetscherら, 上掲;Smithら, 上掲;Nopharら, 上掲;Kohnoら, 上掲]。TNFR1において、4つのCRDのおおよその境界は次の通りである:CRD1−14から約53までのアミノ酸;CRD2−約54から約97までのアミノ酸;CRD3−約98から約138までのアミノ酸;CRD4−約139から約167までのアミノ酸。TNFR2において、CRD1は、17〜約54までのアミノ酸を、CRD2は約55から約97までのアミノ酸を;CRD3は約98から約140までのアミノ酸を;CRD4は約141から約179までのアミノ酸を含む[Bannerら, Cell, 73:431-435(1993)]。また、リガンド結合におけるCRDの潜在的な役割は前掲のバナー(Banner)らにより記載されている。
【0008】
CRDの類似の反復パターンが、p75神経成長因子レセプター(NGFR)[Johnsonら, Cell, 47:545(1986);Radekeら, Nature, 325:593(1987)]、B細胞抗原CD40[Stamenkovicら, EMBO J., 8:1403(1989)]、T細胞抗原OX40[Malletら, EMBO J., 9:1063(1990)]及びFas抗原[Yoneharaら, 上掲、及びItohら, 上掲]を含む幾つかの他の細胞表面タンパク質に存在している。また、CRDはショープ(Shope)及び粘液腫ポックスウィルスの可溶型TNFR(sTNFR)様のT2タンパク質にも見出されている[Uptonら, Virology, 160:20-29(1987);Smithら, Biochem. Biophys. Res. Commun., 176:335(1991);Uptonら, Virology, 184:370(1991)]。これらの配列の最適なアラインメントは、システイン残基の位置が良好に保存されていることを示している。これらレセプターは、しばしば集合的に、TNF/NGFレセプタースーパーファミリーのメンバーと称される。p75NGFRに関する最近の研究では、CRD1の欠失[Welcher, A.A.ら, Proc. Natl. Acad. Sci. USA, 88:159-163(1991)]又はこのドメインにおける5-アミノ酸の挿入[Yan. H及びChao, M.V., J. Biol. Chem., 266:12099-12104(1991)]は、NGF結合にはほとんど又は全く影響を持たないことが示されている[Yan. H及びChao, M.V., 上掲]。p75NGFRはNGF結合に関与せず、そのCRD4と膜貫通領域の間に約60のアミノ酸のプロリンに富んだ伸展を含む[Peetre, Cら, Eur. J. Hematol.,41:414-419(1988);Seckinger, Pら, J. Biol. Chem., 264:11966-11973(1989);Yan. H及びChao, M.V., 上掲]。同様のプロリンに富んだ領域はTNFR2に見出されているが、TNFR1にはない。
【0009】
イトーらは、Apo-1レセプターが55-kDaのTNFR1によりシグナル化されるものと同様のアポトーシス性細胞死をシグナル化し得ることを開示している[Itohら, 上掲]。また、Apo-1抗原の発現は、細胞をTNF-α又は抗Apo-1のマウスモノクローナル抗体で処理した場合に、TNFR1のものと共にダウンレギュレーションされることが報告されている[Krammerら, 上掲;Nagataら, 上掲]。従って、Apo-1及びTNFR1レセプターの双方を同時発現する株化細胞が、共通のシグナル伝達経路を通して細胞死滅を媒介しているとの仮説を唱える研究者もいた[同]。
【0010】
リンホトキシン-αを除き、今日までに同定されているTNFファミリーのリガンドは、II型の膜貫通タンパク質であり、そのC末端は細胞外にある。これに対して、今日までに同定されているTNFレセプター(TNFR)ファミリーのレセプター類はI型の膜貫通タンパク質である。しかしながら、TNFリガンド及びレセプターファミリーの双方において、ファミリーメンバー間で同定された相同性は、主として細胞外ドメイン(「ECD」)において見出されている。TNF-α、Apo-1リガンド及びCD40リガンドを含むTNFファミリーサイトカインのいくつかは、細胞表面においてタンパク分解的に切断され;各場合に得られたタンパク質は、典型的には、可溶性サイトカインとして機能するホモ三量体分子を形成する。また、TNFレセプターファミリーのタンパク質は、通常、タンパク分解的に切断され、同族のサイトカインの阻害剤として機能し得る可溶性レセプターのECDを放出する。
【0011】
最近になって、TNFRファミリーの他のメンバーが同定されている。マースターズ(Marsters)ら、Curr. Biol., 6:750(1996)において、研究者は、細胞外のシステインに富んだ反復についてTNFRファミリーに対して類似性を示し、細胞質死亡ドメイン配列を含む点でTNFR1及びCD95に似ており、Apo-3と称される、ヒトのポリペプチド天然配列の全長を開示している[Marstersら,Curr. Biol., 6:1669(1996)もまた参照されたい]。他の研究者に依れば、Apo-3はDR3、wsl-1及びTRAMPとも称されている[Chinnaiyanら, Science, 274:990(1996);Kitsonら, Nature, 384:372(1996);Bodmerら, Immunity, 6:79(1997)]。
【0012】
パンらは、「DR4」と称される他のTNFレセプターファミリーのメンバーを開示している[Panら, Science, 276:111-113(1997)]。DR4は細胞自殺器を活動させる細胞質死亡ドメインを含むと報告されている。パンらは、DR4がApo-2リガンド又はTRAILとして知られているリガンドに対するレセプターであると考えられることを開示している。
【0013】
アポトーシス誘発性シグナル伝達複合体
現在理解されているように、細胞死プログラムは、少なくとも3つの重要な成分−活性化因子、阻害剤及びエフェクターを含む;線虫(C. elegans)において、これらの成分はそれぞれ3つの遺伝子、Ced-4、Ced-9及びCed-3によりコード化されている[Steller, Science, 267:1445(1995);Chinnaiyanら, Science, 275:1122-1126(1997)]。TNFRファミリーのメンバーの2つ、TNFR1及びFas/Apol(CD95)は、アポトーシス性細胞死を活性化し得る[Chinnaiyan及びDixit, Current Biology, 6:555-562(1996);Fraser及びEvan, Cell, 85:781-784(1996)]。また、TNFR1は転写因子、NF-κBの活性化を媒介することも知られている[Tartagliaら, Cell, 74:845-853(1993);Hsuら, Cell, 84:299-308(1996)]。ある程度のECD相同性に加えて、これら2つのレセプターは、死亡ドメインとして知られているオリゴマー形成界面の細胞内ドメイン(ICD)での相同性を共有する[Tartagliaら, 上掲;Nagata, Cell, 88:355(1997)]。また、死亡ドメインはアポトーシスを調節する幾つかの後生動物タンパク質、すなわちFADD/MORT1、TRADD及びRIPと称されるショウジョウバエタンパク質、リーパー(Reaper)及び哺乳動物タンパク質中においても見出されている[Cleaveland及びIhle, Cell, 81:479-482(1995)]。ラベンらは、酵母-二重ハイブリッド系を使用して、TNFR1死亡ドメインに結合するタンパク質wsl-1の同定を報告している[Ravenら, Programmed Cell Death Meeting, 9月20-24日,1995年,127頁の要約;Ravenら, European Cytokine Network, 7:210頁の要約82(1996年4-6月)]。wsl-1タンパク質はTNFR1に相同で(48%の同一性)、制限された組織分布を有すると記載されている。ラベン(Raven)らに依れば、wsl-1の組織分布は、TNFR1結合タンパク質、TRADDとは顕著に異なる。
【0014】
リガンド結合及びレセプターのクラスター形成の際に、TNFR1とCD95は死亡誘発性シグナル伝達複合体にFADDを補充するものと考えられている。言われるところによれば、CD95はFADDに直接結合する一方、TNFR1はTRADDを介して間接的にFADDに結合する[Chinnaiyanら, Cell, 81:505-512(1995);Boldinら, J. Biol. Chem., 270:387-391(1995);Hsuら, 上掲;Chinnaiyanら, J. Biol. Chem., 271:4961-4965(1996)]。FADDは、Ced-3-関連プロテアーゼ、MACHα/FLICE(カスパーゼ8)を、死亡シグナル伝達複合体に補充するアダプタータンパク質となることが報告されている[Boldinら, Cell, 85:803-815(1996);Muzioら, Cell, 85:817-827(1996)]。MACHα/FLICEは、細胞死プログラムの幾つかの重要な側面を実施し得る、インターロイキン-1β変換酵素(ICE)及びCPP32/Yamaを含むアポトーシス性プロテアーゼのカスケードを引き起こすトリガーであると思われる[Fraser及びEvan, 上掲]。
【0015】
プログラム細胞死が、線虫の細胞死遺伝子、ced-3、及び哺乳動物のIL-1-変換酵素、ICEに関連したシステインプロテアーゼファミリーのメンバーの活性に関与していることが最近開示された。ICE及びCPP32/Yamaプロテアーゼの活性は、牛痘ウイルス遺伝子、crmAの産物により阻害され得る[Rayら, Cell, 69:597-604(1992);Tewariら, Cell, 81:801-809(1995)]。最近の研究では、CrmAがTNFR1-及びCD95-誘発細胞死を阻害し得ることが示されている[Enariら, Nature, 375:78-81(1995);Tewariら, J. Biol. Chem., 270:3255-3260(1995)]。
【0016】
テワリ(Tewari)らにより最近レビューされているように、TNFR1、TNFR2及びCD40は、転写因子、NF-κBの活性化により、炎症誘発性及び同時刺激性サイトカイン、サイトカインレセプター、及び細胞接着分子の発現を変調する[Tewariら, Curr. Op. Genet. Develop., 6:39-44(1996)]。NF-κBは、そのサブユニットが保存Rel領域を含む二量体転写因子のファミリーの原型である[Vermaら, Genes Develop., 9:2723-2735(1996);Baldwin, Ann. Rev. Immunol., 14:649-681(1996)]。その潜伏形態において、NF-κBはIκB阻害剤ファミリーのメンバーと複合化しており;ある刺激に反応してのIκBの不活性化の際に、放出されたNF-κBが、特異的DNA配列と結合する核に転座して遺伝子転写を活性化する。
【0017】
サイトカインのTNFファミリー及びそれらのレセプターのレビューについては、上掲のグラス(Gruss)及びダゥアー(Dower)を参照されたい。
【0018】
発明の概要
本出願人は、本出願において「Apo-2DcR」と命名される新規ポリペプチドをコード化したcDNAクローンを同定した。Apo-2DcRはTNFRファミリーのメンバーであると信じられる;全長の天然配列ヒトApo-2DcRポリペプチドは、細胞外のシステインに富んだ反復部についてTNFRファミリーとの類似性を示す。本出願人は、Apo-2DcRがApo-2リガンド(Apo-2L)に結合することを見出した。
一実施態様において、本発明は、単離されたApo-2DcRポリペプチドを提供する。特に、本発明は、一実施態様においては図1Aの残基1〜259を含んでなるアミノ酸配列(配列番号:1)を含む、単離された天然配列Apo-2DcRポリペプチドを提供する。他の実施態様においては、単離されたApo-2DcRポリペプチドは、図1Aの残基1〜259(配列番号:1)を含んでなる天然配列Apo-2DcRポリペプチドと少なくとも約80%のアミノ酸配列同一性を有する。場合によっては、単離されたApo-2DcRポリペプチドは、−40〜259として図1Bにおいて同定された残基を含んでなるアミノ酸配列(配列番号:3)を含む。場合によっては、Apo-2DcRポリペプチドは、ATCC 209087として寄託されたベクターのcDNA挿入断片によりコード化されたポリペプチドを発現することにより得られた又は得られうるものである。
【0019】
他の実施態様において、本発明はApo-2DcRの単離された細胞外ドメイン(ECD)配列を提供する。場合によっては、単離された細胞外ドメイン配列は図1Aのアミノ酸残基1〜236(配列番号:1)又は図1Aの残基1〜161(配列番号:1)を含んでなる。場合によっては、単離された細胞外ドメイン配列は、ここにおいて同定されたApo-2DcR擬似反復の任意のものにおいて一又は複数のアミノ酸(図2参照)が欠失させられたアミノ酸配列を含んでなる。このような単離された細胞外ドメイン配列は、アミノ酸残基1〜X(ここで、Xは図1Aのアミノ酸残基161〜236(配列番号:1)の任意のものである)の配列を含んでなるポリペプチドを含んでもよい。
【0020】
他の実施態様において、本発明は、異種ポリペプチド又はアミノ酸配列と融合したApo-2DcRポリペプチドを含んでなるキメラ分子を提供する。このようなキメラ分子の例には、免疫グロブリン配列と融合したApo-2DcRが含まれる。他の例には、例えば免疫グロブリン配列のような、異種ポリペプチド又はアミノ酸配列と融合したApo-2DcRの細胞外ドメインが含まれる。
【0021】
他の実施態様において、本発明はApo-2DcRポリペプチドをコードしている単離核酸分子を提供する。一面において、該核酸分子は、Apo-2DcRポリペプチド又はApo-2DcRの特定のドメインをコード化するRNA又はDNAであるか、又はこのようなコード核酸配列に相補的であり、少なくとも中程度、場合によっては高いストリンジェント条件下でそれに安定して結合したままである。一実施態様において、核酸配列は:
(a)残基1〜残基259をコードする図1Aの核酸配列(配列番号:2)のコード領域(すなわち、ヌクレオチド193−195〜967−969);
(b)残基1〜残基236をコードする図1Aの核酸配列(配列番号:2)のコード領域(すなわち、ヌクレオチド193−195〜898−900);
(c)残基−40〜残基259をコードする図1Bの核酸配列(配列番号:4)のコード領域(すなわち、ヌクレオチド73−75〜967−969);
(d)遺伝コードの縮退の範囲内において(a)、(b)又は(c)配列に相当する配列;
から選択される。
【0022】
さらなる実施態様において、本発明はApo-2DcRポリペプチド又はApo-2DcRの特定のドメインをコード化した核酸分子を含んでなるベクターを提供する。また、ベクター又は核酸分子を含有する宿主細胞も提供する。Apo-2DcRの生産方法もさらに提供する。
【0023】
他の実施態様において、本発明はApo-2DcRに結合する抗体を提供する。抗体はアゴニスト、阻止又は中和抗体であってよい。
【0024】
他の実施態様において、本発明は非ヒト、トランスジェニック又はノックアウト動物を提供する。
【0025】
本発明のさらなる実施態様では、Apo-2DcR又はApo-2DcR抗体を含む製造品及びキットが提供される。
【0026】
好適な実施態様の詳細な説明
I.定義
ここで使用される際の「Apo-2DcRポリペプチド」及び「Apo-2DcR」という用語には、天然配列Apo-2DcR及びApo-2DcR変異体(ここでさらに定義される)が含まれる。これらの用語には、ヒトを含む種々の哺乳動物由来のApo-2DcRが含まれる。Apo-2DcRは種々の供給源、例えばヒト組織型又は他の供給源から単離されたもの、あるいは組換え又は合成法により調製されたものであってよい。
【0027】
「天然配列Apo-2DcR」には、天然由来のApo-2DcRと同一のアミノ酸配列を有するポリペプチドを含んでなる。しかして、天然配列Apo-2DcRは、任意の哺乳動物から自然に生じるApo-2DcRのアミノ酸配列を有し得る。このような天然配列Apo-2DcRは、自然から単離することもできるし、組換え又は合成手段により生産することもできる。「天然配列Apo-2DcR」という用語には、特に、Apo-2DcRの自然に生じる切断、分泌又は可溶型(例えば、細胞外ドメイン配列)、自然に生じる変異体型(例えば、選択的にスプライシングされた型)及びApo-2DcRの自然に生じる対立遺伝子変異体が含まれる。本発明の一実施態様において、天然配列Apo-2DcRは、図1A(配列番号:1)のアミノ酸1〜259又は図1B(配列番号:3)のアミノ酸−40〜259を含有する成熟又は全長天然配列Apo-2DcRである。場合によっては、Apo-2DcRポリペプチドは、ATCC 209087として寄託されているベクターのcDNA挿入物によりコードされたポリペプチドを発現させることにより得られた又は得られ得るものである。
【0028】
「Apo-2DcR細胞外ドメイン」すなわち「Apo-2DcRECD」は、膜貫通及び細胞質ドメインを本質的に有しないApo-2DcRの型を意味する。通常、Apo-2DcRECDは、膜貫通及び細胞質ドメインを1%未満、好ましくはそのようなドメインを0.5%未満だけ有する。場合によっては、Apo-2DcRECDは、図1A(配列番号:1)のアミノ酸残基1〜236又は図1A(配列番号:1)のアミノ酸残基1〜161を含むであろう。場合によっては、単離された細胞外ドメイン配列は、ここにおいて同定されたApo-2DcR擬似反復の任意のものにおいて同定される一又は複数のアミノ酸(図2参照)が欠失させられたアミノ酸配列を含んでなる。このような単離された細胞外ドメイン配列は、アミノ酸残基1〜X(ここで、Xは図1Aのアミノ酸残基161〜236(配列番号:1)の任意のものである)の配列を含んでなるポリペプチドを含んでもよい。
【0029】
「Apo-2DcR変異体」とは、全長天然配列ヒトApo-2DcRに対して図1Aに示されている推定アミノ酸配列(配列番号:1)又はここでApo-2DcRECDに対して同定された配列と少なくとも約80%のアミノ酸配列同一性を有する以下に定義する生物学的に活性なApo-2DcRを意味する。このようなApo-2DcR変異体には、例えば、図1Aの配列(配列番号:1)あるいはApo-2DcRECDに対してここで同定された配列のN又はC末端において一又は複数のアミノ酸残基が付加され、もしくは欠失されたApo-2DcRポリペプチドが含まれる。通常、Apo-2DcR変異体は、図1Aのアミノ酸配列(配列番号:1)と、少なくとも約80%のアミノ酸配列同一性、より好ましくは少なくとも約90%のアミノ酸配列同一性、さらにより好ましくは少なくとも約95%のアミノ酸配列同一性を有している。
【0030】
ここで同定されているApo-2DcR配列に対する「パーセント(%)アミノ酸配列同一性」は、配列を整列させ、最大のパーセント配列同一性を得るために必要ならば間隙を導入し、如何なる保存的置換も配列同一性の一部と考えないとした、Apo-2DcR配列のアミノ酸残基と同一である候補配列中のアミノ酸残基のパーセントとして定義される。パーセントアミノ酸配列同一性を決定する目的のためのアラインメントは、当業者の知る範囲にある種々の方法、例えばALIGN又はMegalign(DNASTAR)ソフトウエアのような公に入手可能なコンピュータソフトウエアを使用することにより達成可能である。当業者であれば、比較される配列の全長に対して最大のアラインメントを達成するために必要な任意のアルゴリズムを含む、アラインメントを測定するための適切なパラメータを決定することができる。
【0031】
「エピトープタグ」なる用語は、ここで用いられるときは、「タグポリペプチド」に融合したApo-2DcR、又はそれらのドメイン配列を含んでなるキメラポリペプチドを指す。タグポリペプチドは、その抗体が産生され得るエピトープを提供するに十分な数の残基を有しているが、その長さはApo-2DcRの活性を阻害しないよう充分に短い。また、タグポリペプチドは、好ましくは、抗体が他のエピトープと実質的に交差反応をしないようにかなり独特である。適切なタグポリペプチドは、一般に、少なくとも6のアミノ酸残基、通常は約8〜約50のアミノ酸残基(好ましくは約10〜約20の残基)を有する。
【0032】
「単離された」とは、ここで開示された種々のポリペプチドを記述するために使用するときは、その自然環境の成分から同定され分離され及び/又は回収されたポリペプチドを意味する。その自然環境の汚染成分とは、ポリペプチドの診断又は治療への使用を典型的には妨害する物質であり、酵素、ホルモン、及び他のタンパク質様又は非タンパク質様溶質が含まれる。好ましい実施態様において、ポリペプチドは、(1)スピニングカップシークエネーターを使用することにより、少なくとも15のN末端あるいは内部アミノ酸配列の残基を得るのに充分なほど、あるいは、(2)クーマシーブルーあるいは好ましくは銀染色を用いた非還元あるいは還元条件下でのSDS-PAGEによる均一性が得られるように充分なほど精製される。Apo-2DcRの自然環境の少なくとも1つの成分が存在しないため、単離されたポリペプチドには、組換え細胞内のインシトゥーのポリペプチドが含まれる。しかしながら、通常は、単離されたポリペプチドは少なくとも1つの精製工程により調製される。
【0033】
「単離された」Apo-2DcR核酸分子は、同定され、Apo-2DcR核酸の天然源に通常付随している少なくとも1つの汚染核酸分子から分離された核酸分子である。単離されたApo-2DcR核酸分子は、天然に見出される形態あるいは設定以外のものである。ゆえに、単離されたApo-2DcR核酸分子は、天然の細胞中に存在するApo-2DcR核酸細胞とは区別される。しかし、単離されたApo-2DcR核酸分子は、例えば、核酸分子が天然の細胞のものとは異なった染色体位置にあるApo-2DcRを通常発現する細胞に含まれるApo-2DcR核酸分子を含む。
【0034】
「対照配列」という表現は、特定の宿主生物において作用可能に関連付けられたコード配列を発現するために必要なDNA配列を指す。例えば原核生物に好適な対照配列は、プロモーター、場合によってはオペレータ配列、及びリボソーム結合部位を含む。真核生物の細胞は、プロモーター、ポリアデニル化シグナル及びエンハンサーを利用することが知られている。
【0035】
核酸は、他の核酸配列と機能的な関係にあるときに「作用可能に関連付けられ」ている。例えば、プレ配列あるいは分泌リーダーのDNAは、ポリペプチドの分泌に寄与するプレタンパク質として発現されているならそのポリペプチドのDNAに作用可能に関連付けられている;プロモーター又はエンハンサーは、配列の転写に影響を及ぼすならばコード配列に作用可能に関連付けられている;又はリボソーム結合部位は、もしそれが翻訳を容易にするような位置にあるならコード配列と作用可能に関連付けられている。一般的に、「作用可能に関連付けられる」とは、関連付けられたDNA配列が近接しており、分泌リーダーの場合には近接していて読みフェーズにある。しかし、エンハンサーは必ずしも近接しているわけではない。結合は簡便な制限部位でのライゲーションにより達成される。そのような部位が存在しない場合は、通常の手法にしたがって、合成されたオリゴヌクレオチドアダプターあるいはリンカーが使用される。
【0036】
「抗体」という用語は最も広い意味において使用され、特に抗Apo-2DcRモノクローナル抗体(アゴニスト、アンタゴニスト、及び中和抗体を含む)、及び多エピトープ特異性抗Apo-2DcR抗体組成物を包含している。
【0037】
ここで使用される「モノクローナル抗体」という用語は、実質的に均一な抗体の集団から得られる抗体を称する、すなわち、集団を構成する個々の抗体が、少量存在しうる自然に生じる可能な突然変異を除いて同一である。モノクローナル抗体は高度に特異的であり、一つの抗原部位に対応する。更に、異なる決定基(エピトープ)に対応する異なる抗体を典型的に含む通常の(ポリクローナル)抗体調製物とは異なり、各モノクローナル抗体は抗原の単一の決定基に対応する。
【0038】
ここで、モノクローナル抗体は、起源の種又は免疫グロブリンクラス又はサブクラスの命名に拘わらず、定常ドメインを有する抗Apo-2DcR抗体の可変(高頻度可変を含む)ドメイン、又は重鎖を有する軽鎖、又は他の種由来の鎖を有するある種由来の鎖、あるいは異種タンパク質との融合体をスプライシングすることによって得られるハイブリッド及び組換え抗体、並びにそれが所望の生物的活性を有する限り抗体断片(例えばFab、F(ab')2及びFv)を特に含む。例えば、米国特許第4816567号、及びMageら, Monoclonal Antibody Productuon Techniques and Applications, pp.79-97(Marcel Dekker, Inc.:ニューヨーク, 1987)を参照。
【0039】
従って、「モノクローナル」との形容は、実質的に均一な抗体集団から得られたという抗体の性質を示し、抗体を何か特定の方法で生産しなければならないことを意味するものではない。例えば、本発明に従って使用されるモノクローナル抗体は、最初にKohler及びMilsteinによって、Nature, 256:495 (1975)に記載されたハイブリドーマ法によって作ることができ、あるいは例えば米国特許第4816567号に記載された組換えDNA法によって作ることができる。「モノクローナル抗体」は、例えば、McCaffertyら, Nature, 348:552-554(1990)に記載された技術を用いてファージライブラリから単離することができる。
【0040】
非ヒト(例えばマウス)抗体の「ヒト化」形とは、キメラ免疫グロブリン、免疫グロブリン鎖、あるいはそれらの断片(例えばFv、Fab、Fab'、F(ab')2あるいは抗体の他の抗原結合サブ配列)であって、非ヒト免疫グロブリンに由来する最小配列を含むものである。大部分においてヒト化抗体はレシピエントの相補性決定領域(CDR)の残基が、マウス、ラット又はウサギのような所望の特異性、親和性及び能力を有する非ヒト(ドナー抗体)のCDRの残基によって置換されたヒト免疫グロブリン(レシピエント抗体)である。ある場合には、ヒト免疫グロブリンのFvフレームワーク領域(FR)残基は、対応する非ヒト残基によって置換されている。更に、ヒト化抗体は、レシピエント抗体にも、移入されたCDRもしくはフレームワーク配列にも見出されない残基を含んでもよい。これらの修飾は抗体の特性を更に洗練し、最適化するために行われる。一般に、ヒト化抗体は、全てあるいはほとんど全てのCDR領域が非ヒト免疫グロブリンのものに対応し、全てあるいはほとんど全てのFR領域がヒト免疫グロブリン配列のものである、少なくとも1つ、典型的には2つの可変ドメインの実質的に全てを含む。ヒト化抗体は、最適には免疫グロブリン定常領域又はドメイン(Fc)、典型的にはヒトの免疫グロブリンの定常領域又はドメインの少なくとも一部を含んでなる。
【0041】
ここで意図している「生物学的に活性な」及び「所望の生物学的活性」とは、(1)インビボ又はエキソビボで少なくとも一種類の哺乳動物細胞において(作用的又は刺激する形あるいは拮抗的すなわち阻止する形で)アポトーシスを変調する能力を有しているか;(2)Apo-2リガンドに結合する能力を有しているか;又は(3)Apo-2リガンドのシグナル伝達及びApo-2リガンド活性を変調する能力を有していることを意味する。
【0042】
「アポトーシス」及び「アポトーシス活性」という用語は広義に使用され、典型的には、細胞質の凝結、原形質膜の微絨毛の喪失、核の分節化、染色体DNAの分解又はミトコンドリア機能の喪失を含む一又は複数の特徴的な細胞変化を伴う、哺乳動物における細胞死の規則的又はコントロールされた形態を指す。この活性は、例えば細胞生死判別アッセイ、FACS分析又はDNA電気泳動法等、全て従来から知られている方法により決定し測定することができる。
【0043】
「癌」及び「癌性」という用語は、典型的には調節されない細胞成長を特徴とする、哺乳動物における生理学的状態を指すか記述する。癌の例には、これらに限定されるものではないが、癌腫、リンパ腫、芽細胞腫、肉腫、及び白血病が含まれる。このような癌のより特定の例には、扁平上皮細胞癌、小細胞肺癌、非小細胞肺癌、芽細胞腫、胃腸癌、腎臓癌、膵臓癌、神経膠芽細胞腫、神経芽腫、子宮頸管癌、卵巣癌、肝臓癌、胃癌、膀胱癌、肝細胞腫(hepatoma)、乳癌、大腸癌、結腸直腸癌、子宮体癌、唾液腺癌、腎臓癌、肝臓癌、前立腺癌、産卵口癌、甲状腺癌、肝癌(hepatic carcinoma)及び様々な種類の頭部及び頸部の癌が含まれる。
【0044】
ここで使用される「治療する」、「治療」及び「治療法」とは、治癒的療法、予防的療法及び防護的療法を称する。
【0045】
ここで使用される「哺乳動物」という用語は、ヒト、ウシ、ウマ、イヌ及びネコを含む哺乳動物として分類されるあらゆる動物を指す。本発明の好ましい実施態様においては、哺乳動物はヒトである。
【0046】
II. 本発明の組成物と方法
本発明は、新規に同定され、単離されたApo-2DcRポリペプチドを提供する。特に本出願人は、種々のヒトApo-2DcRポリペプチドを同定し、分離した。これらApo-2DcRポリペプチドの幾つかのものの性質と特徴は、以下の実施例においてさらに詳細に記載する。ここで開示されるApo-2DcRポリペプチドの性質と特徴に基づき、Apo-2DcRはTNFRファミリーのメンバーであるというのが本出願人の信ずるところである。
【0047】
Apo-2DcR、並びにApo-2DcRキメラ分子及び抗Apo-2DcR抗体を如何に調製するかにつき、以下に説明する。
【0048】
A. Apo-2DcRの調製
以下の説明は、主として、Apo-2DcR核酸を含むベクターで形質転換又はトランスフェクトされた細胞を培養してApo-2DcRを生産する方法に関する。もちろん、当該分野においてよく知られている他の方法を用いてApo-2DcRを調製することはできると考えられる。
【0049】
1. Apo-2DcRをコードするDNAの単離
Apo-2DcRをコードするDNAは、Apo-2DcRmRNAを保有していてそれを検出可能なレベルで発現すると考えられる組織から調製された任意のcDNAライブラリから得ることができる。従って、ヒトApo-2DcRDNAは、実施例1に記載されたヒトのcDNAのライブラリのように、ヒトの組織から調製されたcDNAライブラリから簡便に得ることができる。またApo-2DcRコード化遺伝子は、ゲノムライブラリから又はオリゴヌクレオチド合成により得ることもできる。
【0050】
ライブラリは、対象となる遺伝子あるいはその遺伝子によりコードされるタンパク質を同定するために設計された(Apo-2DcRに対する抗体又は少なくとも約20−80塩基のオリゴヌクレオチド等の)プローブによってスクリーニングできる。選択されたプローブによるcDNA又はゲノムライブラリのスクリーニングは、例えばSambrookら, Molecular Cloning: A Laboratory Manual(New York: Cold Spring Harbor Laboratory Press, 1989)に記載されている標準的な手順を使用して実施することができる。Apo-2DcRをコードする遺伝子を単離する他の方法はPCR法を使用するものである[Sambrookら,上掲;Dieffenbachら,PCR Primer:A Laboratory Manual(Cold Spring Harbor Laboratory Press, 1995)]。
【0051】
スクリーニングの好ましい方法は、種々のヒト組織からcDNAライブラリをスクリーニングするために選別したオリゴヌクレオチド配列を用いることである。以下の実施例1には、cDNAライブラリのスクリーニング技術を記載している。プローブとして選択されたオリゴヌクレオチド配列は、充分な長さで、疑陽性が最小化されるよう充分に明瞭でなければならない。オリゴヌクレオチドは、スクリーニングされるライブラリ内のDNAとのハイブリッド化時に検出可能であるように標識されていることが好ましい。標識化の方法は、当該分野において良く知られており、32P標識されたATPのような放射標識、ビオチン化あるいは酵素標識の使用が含まれる。中程度の厳密性及び高厳密性を含むハイブリッド化条件は、Sambrookら,上掲に提供されている。
【0052】
全てのタンパク質コード化配列を有する核酸は、初めてここで開示された推定アミノ酸配列を使用し、また必要ならば、cDNAに逆転写されなかったmRNAの生成中間体及び先駆物質を検出する上掲のSambrookらにより記述されている従来のプライマー伸展法を使用し、選択されたcDNA又はゲノムライブラリをスクリーニングすることにより得られる。
【0053】
Apo-2DcR変異体はApo-2DcRDNAに適切なヌクレオチド変化を導入するか、所望のApo-2DcRポリペプチドを合成することにより、調製することができる。アミノ酸の変化により、グリコシル化部位の数と位置の変化、膜係留特性の変更のように、Apo-2DcRの翻訳後過程を改変し得ることは当業者であれば理解されることである。
【0054】
ここで記載されている天然全長配列Apo-2DcR又はApo-2DcRの種々のドメインにおける変異は、例えば米国特許第5364934号に記載された保存的あるいは非保存的突然変異に関する技術とガイドラインの任意のものを使用して発生させることができる。変異は、Apo-2DcRをコードしている一又は複数のコドンの置換、欠失又は挿入であり、天然配列Apo-2DcRに対してApo-2DcRのアミノ酸配列に変化が生じている。場合によっては、変異は、Apo-2DcR分子の一又は複数のドメインにおいて、少なくとも1つのアミノ酸を任意の他のアミノ酸に置換することによりなされる。変異は、オリゴヌクレオチド媒介(部位特異的)突然変異誘発、アラニンスキャニング、及びPCR突然変異誘発のような、当該分野において既知の方法を用いて行うことができる。部位特異的突然変異誘発[Carterら, Nucl. Acids. Res., 13:4331(1986);Zollerら, Nucl. Acids. Res., 10:6487(1987)]、カセット突然変異[Wellsら, Gene, 34:315(1985)]、制限選択的突然変異誘発[Wellら, Philos. Trans. R. Soc. London. SerA, 317:415(1986)]又は他の既知の技術をクローンDNAに実施してApo-2DcR変異体のDNAを生産することができる。
【0055】
また、特定のリガンド又はレセプターとの相互作用に関与する、近接配列に沿った一又は複数のアミノ酸を同定するために、スキャニングアミノ酸分析法を使用することもできる。好ましいスキャニングアミノ酸は、比較的小さい中性アミノ酸である。このようなアミノ酸には、アラニン、グリシン、セリン及びシステインが含まれる。変異体の主鎖構造をあまり改変することなく、ベータ炭素を越えた側鎖が除去されるため、アラニンがこの群のなかで好ましいスキャニングアミノ酸である。またアラニンは最も一般的なアミノ酸であることも好ましい。さらに、アラニンは埋設及び露出位置の双方に頻繁に見出される[Creighton, The Proteins, (W.H. Freeman & Co., N.Y.);Chothia, J. Mol. Biol., 150:1(1976)]。アラニン置換により適切な量の変異体を生じない場合には、アイソテリックアミノ酸を使用することができる。
【0056】
一度選択されたApo-2DcR変異体が生産されると、それらを、例えばApo-2Lと接触させることができ、相互作用があれば測定することができる。Apo-2DcR変異体とApo-2Lとの相互作用は、例えば以下の実施例に記載したようなインビトロアッセイにより測定することができる。天然配列Apo-2DcRとApo-2DcR変異体の活性と性質を比較するため任意の数の分析的測定法を使用することができ、結合性に対して簡便なものは、天然配列Apo-2DcRの解離定数Kdと比較したApo-2DcR変異体とApo-2Lとの間に形成された複合体の解離定数Kdである。一般に置換残基当り≧3倍Kdが増加又は減少すると、置換残基がApo-2Lとの天然配列Apo-2DcRの相互作用において活性であることになる。
【0057】
場合によっては、突然変異(例えば一又は複数のアミノ酸の欠失)に好適なApo-2DcR配列中の代表的な部位には、細胞外ドメイン、特に一又は複数のシステインリッチドメイン、又は一又は複数の擬似反復内の部位が含まれる。このような変異は上述の方法を使用することにより達成される。
【0058】
2. 複製可能なベクターへの核酸の挿入
Apo-2DcRをコードした核酸(例えば、cDNA又はゲノムDNA)は、さらなるクローニング(DNAの増幅)又は発現のために、複製可能なベクター内に挿入される。様々なベクターが公的に入手可能である。ベクター成分としては、一般に、これらに制限されるものではないが、次のものの一又は複数が含まれる:シグナル配列、複製開始点、一又は複数のマーカー遺伝子、エンハンサーエレメント、プロモーター、及び転写終結配列であり、それぞれを以下に説明する。
【0059】
(i) シグナル配列成分
Apo-2DcRは直接的に組換え手法によって生産されるだけではなく、シグナル配列あるいは成熟タンパク質あるいはポリペプチドのN末端に特異的切断部位を有する他のポリペプチドである異種性ポリペプチドとの融合ペプチドとしても生産される。一般に、シグナル配列はベクターの成分であるか、ベクターに挿入されるApo-2DcRDNAの一部である。好ましく選択された異種シグナル配列は宿主細胞によって認識され加工される(すなわち、シグナルペプチダーゼによって切断される)ものである。シグナル配列は、例えばアルカリホスファターゼ、ペニシリナーゼ、lppあるいは熱安定なエンテロトキシンIIリーダーの群から選択される原核生物シグナル配列であってよい。酵母の分泌に関しては、シグナル配列は、酵母インベルターゼリーダー、アルファ因子リーダー(酵母菌属(Saccharomyces)及びクルイベロマイシス(Kluyveromyces)α因子リーダーを含み、後者は米国特許第5010182号に記載されている)、又は酸ホスフォターゼリーダー、白体(C.albicans)グルコアミラーゼリーダー(1990年4月4日発行のEP362179)、又は1990年11月15日に公開された国際特許出願第WO90/13646号に記載されているシグナルであり得る。哺乳動物細胞の発現においては、同一あるいは関連ある種の分泌ポリペプチド由来のシグナル配列、単純ヘルペスグリコタンパク質Dシグナルのようなウイルス分泌リーダーのような他の哺乳動物のシグナル配列をタンパク質の直接分泌に使用してもよいが、インビボにおけるヒト細胞の細胞膜へのApo-2DcRの挿入を通常指示する天然Apo-2DcRプレ配列が十分である。
【0060】
このような前駆体領域のDNAは、好ましくは、Apo-2DcRをコードするDNAにリーディングフレームが結合される。
【0061】
(ii)複製開始点成分
発現及びクローニングベクターは共に一又は複数の選択された宿主細胞においてベクターの複製を可能にする核酸配列を含む。一般に、この配列はクローニングベクターにおいて、宿主染色体DNAとは独立にベクターが複製することを可能にするものであり、複製開始点又は自律的複製配列を含む。そのような配列は多くの細菌、酵母及びウイルスに対してよく知られている。プラスミドpBR322に由来する複製開始点は大部分のグラム陰性細菌に好適であり、2μプラスミド開始点は酵母に適しており、様々なウイルス開始点(SV40、ポリオーマ、アデノウイルス、VSV又はBPV)は哺乳動物細胞におけるクローニングベクターに有用である。一般には、哺乳動物の発現ベクターには複製開始点成分は不要である(SV40開始点が典型的には初期プロモーターを有しているため用いられる)。
【0062】
多くの発現ベクターは「シャトル」ベクターである、すなわち、それらは少なくとも一つのクラスの生物において複製可能であるが、発現のために他の生物に形質移入され得る。例えば、大腸菌においてベクターがクローン化され、そのベクターが酵母あるいは哺乳動物細胞に形質移入され、宿主細胞染色体と独立して複製することはできないとしても、発現する。
【0063】
DNAは宿主ゲノムに挿入することによって増幅され得る。これは、例えばベクターにバシラス(Bacillus)ゲノムDNAに見られる配列と相補的なDNA配列を含めることにより、宿主としてバシラス種を用いて容易に達成される。このベクターを用いたバシラスの形質移入は、ゲノムとの相同的組換え及びApo-2DcRDNAの挿入をもたらす。しかし、Apo-2DcRをコードするゲノムDNAの回収は、Apo-2DcRDNAを切除するのに制限酵素による消化を必要とするために、外来的に複製したベクターの場合よりも複雑である。
【0064】
(iii)選択遺伝子成分
発現及びクローニングベクターは、典型的には、選べるマーカーとも称される選択遺伝子を含む。この遺伝子は、選択的培地で成長させた形質転換宿主細胞の生存又は成長に必要なタンパク質をコードする。選択遺伝子を含むベクターと共に形質転換されていない宿主細胞は培地で生存しない。典型的な選択遺伝子は、(a)アンピシリン、ネオマイシン、メトトレキセートあるいはテトラサイクリンのような抗生物質あるいは他の毒素に耐性を与え、(b)栄養要求性欠陥を補い、(c)例えばバシリに対する遺伝子コードD-アラニンラセマーゼのような、複合培地から得られない重要な栄養素を供給する、タンパク質をコードする。
【0065】
選択技術の一例においては、宿主細胞の成長を抑止する薬物が用いられる。異種性遺伝子で首尾よく形質転換した細胞は、抗薬物性を付与し、選択工程を生存するタンパク質を生産する。このような優性選択の例としては、薬物ネオマイシン[Southernら, J. Molec. Appl. Genet., 1:327(1982)]、ミコフェノール酸[Mulliganら, Science, 209:1422(1980)]又はハイグロマイシン[Sugdenら, Mol. Cell. Biol., 5:410-413(1985)]が使用される。上述した3つの例は、真核生物のコントロール下で細菌遺伝子を用いて、適切な薬物G418又はネオマイシン(ジェネテシン)、xgpt(ミコフェノール酸)、又はハイグロマイシンにそれぞれ耐性を付与する。
【0066】
哺乳動物細胞に適切な選べるマーカーの他の例は、DHFRあるいはチミジンキナーゼのように、Apo-2DcR核酸を捕捉することのできる細胞成分を同定することのできるものである。哺乳動物細胞の形質転換細胞は、マーカーを捕捉することによって当該形質転換細胞のみが生存できるように独特に適応化された淘汰圧下に置かれる。淘汰圧は、培地中の選択剤の濃度が次第に変化する条件下で形質転換細胞を培養することにより課し、選択遺伝子とApo-2DcRをコードするDNAの双方を増幅させる。増幅は、成長に重要なタンパク質の生産に対する要求度が高い遺伝子が、組換え細胞の後の世代の染色体内に直列に反復されるプロセスである。増幅されたDNAから増大したApo-2DcR量が合成される。増幅可能な遺伝子のほかの例には、メタロチオネインI及びII、アデノシンデアミナーゼ、及びオルニチンデカルボキシラーゼが含まれる。
【0067】
DHFR選択遺伝子によって形質転換された細胞は、先ず、DHFRの競合的アンタゴニストであるメトトリキセート(Mtx)を含む培地において形質転換物の全てを培養することで同定される。野生型DHFRを用いた場合の好適な宿主細胞は、Urlaub ほかにより, Proc. Natl. Acad. Sci. USA, 77:7216 (1980)に記載されているようにして調製され増殖されたDHFR活性に欠陥のあるチャイニーズハムスター卵巣(CHO)株化細胞である。形質転換した細胞は次に濃度の高いメトトレキセートに接触させる。これによりDHFR遺伝子の複数コピーが合成され、同時に、Apo-2DcRをコードするDNAのような発現ベクターを含む他のDNAの複数コピーが作られる。この増幅方法は、もしMtxに高度に耐性である変異体DHFR遺伝子が使用されたような場合には内在性DHFRの存在にかかわらず、任意の他の適切な宿主、例えば、ATCC番号CCL61CHO−K1を使用することができる(EP117,060)。
【0068】
あるいは、Apo-2DcRをコードするDNA配列、野生型DHFRタンパク質、及びアミノグリコシド3'-ホスホトランスフェラーゼ(APH)のような他の選べるマーカーで形質転換あるいは同時形質転換した宿主細胞(特に、内在性DHFRを含む野生型宿主)は、カナマイシン、ネオマイシンあるいはG418のようなアミノグリコシド抗生物質のような選択可能マーカーの選択剤を含む培地における細胞増殖により選択することができる。米国特許第4965199号を参照。
【0069】
酵母中での使用に好適な選択遺伝子は酵母プラスミドYRp7に存在するtrp1遺伝子である[Stinchcombら, Nature, 282:39(1979);Kingmanら, Gene, 7:141(1979);Tschemperら, Gene, 10:157(1980)]。trp1遺伝子は、例えば、ATCC第44076号あるいはPEP4-1のようなトリプトファン内で成長する能力に欠ける酵母の突然変異株に対する選択マーカーを提供する[Jones, Genetics, 85:12 (1977)]。酵母宿主細胞ゲノムにtrp1破壊が存在することは、トリプトファンの不存在下における増殖による形質転換を検出するための有効な環境を提供する。同様に、Leu2欠陥酵母株(ATCC 20,622あるいは 38,626)は、Leu2遺伝子を有する既知のプラスミドによって補完される。
【0070】
更に、1.6μmの円形プラスミドpKD1由来のベクターは、クルイヴェロマイシス(Kluyveromyces)酵母の形質転換に用いることができる[Bianchi ほか., Curr. Genet., 12:185(1987)]。より最近では、組換え子ウシのキモシンの大量生産のための発現系がK.ラクティス(lactis)に対して報告されている[Van den Berg, Bio/Technology, 8:135 (1990)]。クルイヴェロマイシスの工業的な菌株からの、組換えによる成熟したヒト血清アルブミンを分泌する安定した複数コピー発現ベクターも開示されている[Fleer ほか, Bio/Technology,9:968-975 (1991)]。
【0071】
(iv)プロモーター成分
発現及びクローニングベクターは、通常、宿主生物によって認識され、Apo-2DcR核酸配列に作用可能に結びついているプロモーターを含む。プロモーターは、作用可能に結合しているApo-2DcR核酸配列のような特定の核酸配列の転写及び翻訳を制御する構造的な遺伝子(一般的に約100ないし1000塩基対)の開始コドンの上流側(5')に位置する未翻訳配列である。このようなプロモーターは典型的には、誘発的なクラス及び構成的なクラスの2つのクラスに属する。誘発的なプロモーターは、養分の存在あるいは不存在、温度変化等の培養条件のある変化に対応してその制御の下でDNAからの転写レベルを上昇させるプロモーターである。現時点において多種の可能な宿主細胞により認識される非常に多くのプロモーターがよく知られている。これらのプロモーターは、制限酵素の消化によって供給源DNAからプロモーターを排除し、ベクターに単離したプロモーター配列を挿入することで、Apo-2DcRをコードするDNAに作用的に結合している。天然のApo-2DcRプロモーター配列及び多くの異種性プロモーターはいずれもApo-2DcRDNAの直接増幅及び/又は発現に用いることができる。
【0072】
原核生物宿主での使用に好適なプロモーターはβラクタマーゼ及びラクトースプロモーター系[Cahng ほか, Nature, 275:615 (1978), Goeddel ほか, Nature, 281:544 (1979)]、アルカリホスファターゼ、トリプトファン(trp)プロモーター系[Goeddel, Nucleic Acids Res., 8:4057 (1980); EP 36,776]、及びハイブリッドプロモーター、例えばtacプロモーター[deBoer ほか, Proc. Natl. Acad. Sci. USA, 80:21-25 (1983)]を含む。しかし、他の既知の細菌プロモーターも好適である。これらのヌクレオチド配列は公表されており、よって当業者は、任意の所望の制限部位を提供するためにリンカーあるいはアダプターを使用することでApo-2DcRをコードするDNA[Siebenlist ほか, Cell, 20:269 (1980)]にそれらを作用可能に結合させることが可能である。細菌系で使用するプロモータもまたApo-2DcRをコードするDNAと作用可能に結合したシャイン・ダルガーノ(S.D.)配列を有する。
【0073】
真核生物に対してもプロモーター配列が知られている。実質的に全ての真核生物の遺伝子は、転写開始部位からおよそ25ないし30塩基上流に見出されるATリッチ領域を有する。多数の遺伝子の転写開始位置から70ないし80塩基上流に見出される他の配列は、Xが任意のヌクレオチドであるCXCAAT領域である。大部分の真核生物遺伝子の3'末端には、コード配列の3'末端へのポリA尾部の付加に対するシグナルであるAATAAA配列がある。これらの配列は全て真核生物の発現ベクターに適切に挿入される。
【0074】
酵母宿主と共に用いて好適なプロモーター配列の例としては、3-ホスホグリセラートキナーゼ[Hitzeman ほか, J. Biol. Chem., 255:2073 (1980)]又は他の糖分解酵素[Hess ほか, J. Adv. Enzyme Reg., 7:149 (1968);Holland, Biochemistry, 17:4900(1987)]、例えばエノラーゼ、グリセルアルデヒド-3-リン酸デヒドロゲナーゼ、ヘキソキナーゼ、ピルビン酸デカルボキシラーゼ、ホスホフルクトキナーゼ、グルコース-6-リン酸イソメラーゼ、3-ホスホグリセレートムターゼ、ピルビン酸キナーゼ、トリオセリン酸イソメラーゼ、ホスホグルコースイソメラーゼ、及びグルコキナーゼが含まれる。
【0075】
他の酵母プロモーターとしては、成長条件によって転写が制御される付加的効果を有する誘発的プロモーターであり、アルコールデヒドロゲナーゼ2、イソチトクロムC、酸ホスファターゼ、窒素代謝と関連する分解性酵素、メタロチオネイン、グリセルアルデヒド-3-リン酸デヒドロゲナーゼ、及びマルトース及びガラクトースの利用を支配する酵素のプロモーター領域がある。酵母の発現に好適に用いられるベクターとプロモータは欧州特許第73657号に更に記載されている。また酵母エンハンサーも酵母プロモーターと共に好適に用いられる。
【0076】
哺乳動物の宿主細胞におけるベクターからのApo-2DcR転写は、例えば、ポリオーマウィルス、伝染性上皮腫ウィルス(1989年7月5日公開のUK 2,211,504)、アデノウィルス(例えばアデノウィルス2)、ウシ乳頭腫ウィルス、トリ肉腫ウィルス、サイトメガロウィルス、レトロウィルス、B型肝炎ウィルス及び最も好ましくはサルウィルス40(SV40)のようなウィルスのゲノムから得られるプロモーター、異種性哺乳動物プロモーター、例えばアクチンプロモーター又は免疫グロブリンプロモーター、熱衝撃プロモーター、そしてApo-2DcR配列に通常付随するプロモーターによって、このようなプロモーターが宿主細胞系に適合し得る限り、調節される。
【0077】
SV40ウィルスの初期及び後期プロモーターは、SV40ウイルスの複製起点をさらに含むSV40制限断片として簡便に得られる[Fiersほか, Nature, 273:113 (1978);Mulligan及びBerg, Science, 209:1422-1427 (1980); Pavlakisほか, Proc. Natl. Acad. Sci. USA, 78:7398-7402 (1981)]。ヒトサイトメガロウィルスの最初期プロモーターは、HindIIIE制限断片として簡便に得られる[Greenawayほか, Gene, 18:355-360 (1982)]。ベクターとしてウシ乳頭腫ウィルスを用いて哺乳動物宿主でDNAを発現する系が、米国特許第4419446号に開示されている。この系の修飾は米国特許第4601978号に開示されている[また、サル細胞での免疫インターフェロンをコードしているcDNAの発現について、Grayら, Nature, 295:503-508(1982)を;単純ヘルペスウイルス由来のチミジンキナーゼプロモーターの調節下でのマウス細胞におけるヒトβインターフェロンcDNAの発現について、Reyesら, Nature, 297:598-601(1982)を;培養されたマウス及びウサギの細胞におけるヒトインターフェロンβ1遺伝子の発現について、Canaani及びBerg, Proc. Natl. Acad. Sci. USA, 79:5166-5170(1982)を;プロモーターとしてラウス肉腫ウィルスの長い末端反復を用いたCV-1サル腎臓細胞、ニワトリ胚線維芽細胞、チャイニーズハムスター卵巣細胞、HeLa細胞、及びマウスNIH-3T3細胞における細菌CAT配列の発現について、Gormanほか, Proc. Natl. Acas. Sci. USA, 79:6777-6781 (1982)を参照のこと]。
【0078】
(v)エンハンサーエレメント成分
より高等の真核生物による本発明のApo-2DcRをコードしているDNAの転写は、ベクター中にエンハンサー配列を挿入することによって増強され得る。エンハンサーは、通常は約10から300塩基対で、プロモーターに作用してその転写を増強するDNAのシス作動要素である。エンハンサーは、相対的に配向及び位置が独立しており、転写ユニットの5'[Laiminsほか, Proc. Natl. Acad. Sci. USA, 78:993 (1981)]及び 3'[Luskyほか, Mol. Cell Bio., 3:1108 (1983)]、イントロン内部[Banerjiほか, Cell, 33:729 (1983)]並びにコード配列自身の内部[Osbornほか, Mol. Cell Bio., 4:1293 (1984)]に見出されている。哺乳動物の遺伝子由来の多くのエンハンサー配列が現在知られている(グロビン、エラスターゼ、アルブミン、α-フェトプロテイン及びインスリン)。しかしながら、典型的には、真核細胞ウィルス由来のエンハンサーが用いられるであろう。例としては、複製起点の後期側のSV40エンハンサー(100-270塩基対)、サイトメガロウィルス初期プロモーターエンハンサー、複製起点の後期側のポリオーマエンハンサー及びアデノウィルスエンハンサーが含まれる。真核生物のプロモーターの活性化のための増強要素については、Yaniv, Nature, 297:17-18 (1982)もまた参照のこと。エンハンサーは、Apo-2DcRコード配列の5'又は3'位でベクター中にスプライシングされ得るが、好ましくはプロモーターから5'位に位置している。
【0079】
(vi)転写終結成分
真核生物宿主細胞(酵母、真菌、昆虫、植物、動物、ヒト、又は他の多細胞生物由来の有核細胞)に用いられる発現ベクターは、また転写の終結及びmRNAの安定化に必要な配列を含む。このような配列は、真核生物又はウィルスのDNA又はcDNAの5'、時には3'の非翻訳領域から一般に取得できる。これらの領域は、Apo-2DcRをコードしているmRNAの非翻訳部分にポリアデニル化断片として転写されるヌクレオチドセグメントを含む。
【0080】
(vii)ベクターの作成と分析
一又は複数の上に列挙した成分を含む適切なベクターの作成には標準的なライゲーション技術を用いる。分離されたプラスミド又はDNA断片を開裂させ、整え、そして必要とされるプラスミドの生成のために望ましい型に再ライゲーションする。
【0081】
作成されたプラスミドが正しい配列であることを確認する分析のために、ライゲーション混合物を用いて、大腸菌K12菌株294(ATCC 31446)を形質転換し、適当な場合にはアンピシリン又はテトラサイクリン耐性によって、形質転換細胞を好適に選択する。形質転換細胞からプラスミドを調製し、制限エンドヌクレアーゼ消化により分析し、及び/又はMessingほか, Nucleic Acids Res., 9:309 (1981)の方法又はMaximほか, Methods in Enzymology, 65:499 (1980)の方法によって配列決定する。
【0082】
(viii)一過性発現ベクター
Apo-2DcRをコードしているDNAの哺乳動物細胞における一過性発現をもたらす発現ベクターを使用することができる。一般に、一過性発現は、宿主細胞が発現ベクターの多くのコピーを蓄積し、次にその発現ベクターによってコードされている所望のポリペプチドを高レベルで合成するように、宿主細胞中で効果的に複製できる発現ベクターを使用することを含む[Sambrookほか,上掲]。一過性発現系は、適切な発現ベクターと宿主細胞を含むが、クローニングされたDNAによりコードされているポリペプチドの簡便で確実な同定並びに所望の生物学的又は生理学的性質についてのポリペプチドの迅速なスクリーニングを可能にする。したがって、一過性発現系は、本発明において、Apo-2DcR変異体を同定する目的のために特に有用である。
【0083】
(ix)適切な例示的脊椎動物細胞ベクター
組換え脊椎動物細胞培養でのApo-2DcRの合成に適応化するのに適切な他の方法、ベクター及び宿主細胞は、Gethingほか, Nature, 293:620-625 (1981); Manteiほか, Nature, 281:40-46 (1979); 欧州特許第117060号; 及び欧州特許第117058号に記載されている。
【0084】
3. 宿主細胞の選択及び形質転換
ここに記載のベクターにDNAをクローニングあるいは発現するために適切な宿主細胞は、上述の原核生物、酵母、又は高等真核生物細胞である。この目的にとって適切な原核生物は、限定するものではないが、真正細菌、例えばグラム陰性又はグラム陽性生物体、例えばエシェリチアのような腸内菌科、例えば大腸菌、エンテロバクター、エルウィニア(Erwinia)、クレブシエラ、プロテウス、サルモネラ、例えばネズミチフス菌、セラチア属、例えばセラチア・マルセスキャンス及び赤痢菌属、並びに桿菌、例えば枯草菌及びバシリ・リチェフォルミス(licheniformis)(例えば、1989年4月12日に公開された DD 266,710に開示されたバシリ・リチェニフォルミス41P)、シュードモナス属、例えば緑膿菌及びストレプトマイセス属を含む。好ましくは、宿主細胞は最小量のタンパク質分解酵素を分泌すべきである。
【0085】
原核生物に加えて、糸状菌又は酵母菌のような真核微生物は、Apo-2DcRをコードするベクターのための適切なクローニング又は発現宿主である。サッカロミセス・セレヴィシア、又は一般的なパン酵母は下等真核生物宿主微生物のなかで最も一般的に用いられる。しかしながら、多数の他の属、種及び菌株も、一般的に入手可能で有用である。
【0086】
グリコシル化Apo-2DcRの発現に適切な宿主細胞は、多細胞生物から誘導される。このような宿主細胞は、複雑なプロセシング及びグリコシル化活動が可能である。原則的には、脊椎動物であろうと無脊椎動物培養であろうと、任意のより高等の真核生物細胞培養が使用できる。無脊椎動物細胞の例としては植物及び昆虫細胞が含まれる。多数のバキュロウィルス株及び変異体及び対応する許容可能な昆虫宿主細胞、例えばスポドプテラ・フルギペルダ(毛虫)、アエデス・アエジプティ(蚊)、アエデス・アルボピクトゥス(蚊)、ドゥロソフィラ・メラノガスター(ショウジョウバエ)、及びボンビクス・モリが同定されている[例えば、Luckowほか, Bio/Technology, 6:47-55 (1988); Millerほか, Genetic Engineering, Setlowほか, eds., Vol. 8 (Plenum Publishing, 1986), pp.277-279; 及びMaedaほか, Nature, 315:592-594 (1985)を参照のこと]。トランスフェクションのための種々のウィルス株、例えば、オートグラファ・カリフォルニカNPVのL-1変異体とボンビクス・モリ NPVのBm-5株が公に利用できる。
【0087】
綿花、コーン、ジャガイモ、大豆、ペチュニア、トマト、及びタバコのような植物細胞培養を宿主として利用することができる。典型的には、植物細胞は、細菌アグロバクテリウム・トゥメファシエンスのある菌株と共にインキュベートすることによってトランスフェクトされる。A. トゥメファシエンスと共に植物細胞培養をインキュベートする間に、Apo-2DcRをコードしているDNAが、植物細胞宿主がトランスフェクトされるようにその植物細胞宿主に移され、そして適切な条件下でApo-2DcRをコードしているDNAを発現させる。加えて、例えば、ノパリンシンターゼプロモーター及びポリアデニル化シグナル配列のような、植物細胞と適合しうる調節及びシグナル配列が利用できる[Depickerほか, J. Mol. Appl. Gen., 1:561 (1982)]。また、T-DNA780遺伝子の上流領域から分離されるDNAセグメントは、組換えDNAを含む植物組織中の植物発現遺伝子の転写レベルを活性化又は増強しうる[1989年6月21日公開のEP 321,196]。
【0088】
培養(組織培養)中での脊椎動物細胞の増殖は、当該分野おいてよく知られている[例えば、Tissue Culture,Academic Press,編者Kruse及びPatterson(1973)を参照のこと]。有用な哺乳動物宿主株化細胞の例は、SV40によって形質転換されたサル腎臓CV1株 (COS-7, ATCC CRL 1651);ヒト胚腎臓株[293又は懸濁培養での増殖のためにサブクローン化された293細胞、Grahamほか, J. Gen Virol., 36:59 (1977)];ハムスター乳児腎細胞(BHK, ATCC CCL 10);チャイニーズハムスター卵巣細胞/-DHFR(CHO, Urlaub及びChasin, Proc. Natl. Acad. Sci. USA, 77:4216 (1980));マウスのセルトリ細胞[TM4, Mather, Biol. Reprod., 23:243-251 (1980)];サルの腎細胞 (CVI ATCC CCL 70); アフリカミドリザルの腎細胞(VERO-76, ATCC CRL-1587); ヒト子宮頸癌細胞 (HELA, ATCC CCL 2); イヌ腎細胞 (MDCK, ATCC CCL 34); バッファローラット肝細胞 (BRL 3A, ATCC CRL 1442); ヒト肺細胞 (W138, ATCC CCL 75); ヒト肝細胞 (Hep G2, HB 8065); マウス乳房腫瘍細胞 (MMT 060562, ATTC CCL51);TRI細胞[Motherほか, Annals N.Y. Acad. Sci., 383:44-68 (1982)];MRC5細胞;FS4細胞である。
【0089】
宿主細胞をトランスフェクトし、好ましくは上述のApo-2DcR生成のための発現又はクローニングベクターで形質転換し、プロモーターを誘導し、形質転換体を選択し、又は所望の配列をコードしている遺伝子を増幅するために適当に修飾された常套的栄養培地で培養する。
【0090】
トランスフェクションは、如何なるコード配列が実際に発現されるか否かにかかわらず、宿主細胞による発現ベクターの取り上げを意味する。多数のトランスフェクションの方法が当業者に知られている。例えば、CaPO4及びエレクトロポレーションである。このベクターの操作のあらゆる徴候が宿主細胞内で生じたときに成功したトランスフェクションが一般に認められる。
【0091】
形質転換は、染色体外のエレメントとしてであろうと染色体成分によってであろうと、DNAが複製可能であるように、生物体中にDNAを導入することを意味する。用いられる宿主細胞に応じて、そのような細胞に対して適した標準的な方法を用いて形質転換はなされる。前掲のSambrookほかにより記載された塩化カルシウムを用いるカルシウム処理又はエレクトロポレーションが、原核生物又は実質的な細胞壁障壁を含む他の細胞に対して用いられる。アグロバクテリウム・トゥメファシエンスによる感染が、Shawほか, Gene, 23:315 (1983)及び1989年6月29日公開の国際特許出願第WO89/05859号に記載されたように、ある種の植物細胞の形質転換に用いられる。加えて、1991年1月10日に公開された国際特許出願第WO91/00358号に記載されているように、超音波処理を用いて植物をトランスフェクトすることもできる。
【0092】
このような細胞壁のない哺乳動物の細胞に対しては、Graham及びvan der Eb, Virology, 52:456-457 (1978)のリン酸カルシウム沈殿法が好ましい。哺乳動物細胞の宿主系形質転換の一般的な側面は米国特許第4399216号に記載されている。酵母中の形質転換は、典型的には、Van solingenほか, J. Bact., 130:946 (1977)及びHsiaoほか, Proc. Natl. Acad. Sci. USA, 76:3829 (1979)の方法によって実施する。しかしながら、DNAを細胞中に導入する他の方法、例えば、核マイクロインジェクション、エレクトロポレーション、無傷の細胞、又はポリカチオン、例えばポリブレン、ポリオルニチン等を用いる細菌プロトプラスト融合もまた用いることもできる。哺乳動物細胞を形質転換するための種々の技術については、Keownほか, Methods in Enzymology, 185:527-537 (1990)及び Mansourほか, Nature, 336:348-352 (1988)を参照のこと。
【0093】
4.宿主細胞の培養
本発明のApo-2DcRポリペプチドを生成るために用いられる原核細胞は、前掲のSambrookほかにより記載されているような適切な培地で培養される。
【0094】
Apo-2DcRの生産に用いられる哺乳動物の宿主細胞は種々の培地において培養することができる。市販培地の例としては、ハム(Ham)のF10(シグマ)、最小必須培地(「MEM」,シグマ)、RPMI-1640(シグマ)及びダルベッコの改良イーグル培地(「DMEM」,シグマ)が含まれる。これらの培地はいずれも、ホルモン及び/又は他の成長因子(例えばインスリン、トランスフェリン、又は表皮成長因子)、塩類(例えば、塩化ナトリウム、カルシウム、マグネシウム及びリン酸塩)、バッファー(例えばHEPES)、ヌクレオシド(例えばアデノシン及びチミジン)、抗生物質(例えば、ゲンタマイシンTM薬)、微量元素(最終濃度がマイクロモル範囲で通常存在する無機化合物として定義される)及びグルコース又は同等のエネルギー源を必要に応じて補充することができる。任意の他の必要な補充物質もまた当業者に知られている適当な濃度で含むことができる。培養条件、例えば温度、pH等々は、発現のために選ばれた宿主細胞について以前から用いられているものであり、当業者には明らかであろう。
【0095】
一般に、哺乳動物の細胞培養の生産性を最大にするための原理、プロトコール、及び実用技術は、Mammalian Cell Biotechnology: a Practical Approach, M.Butler編 (IRL Press, 1991)に見出すことができる。
【0096】
この明細書において言及される宿主細胞は培養中の細胞並びに宿主動物内にある細胞を包含する。
【0097】
5. 遺伝子増幅/発現の検出
遺伝子の増幅及び/又は発現は、ここで提供された配列に基づき、適切に標識されたプローブを用い、例えば、従来よりのサザンブロット法、mRNAの転写を定量化するノーザンブロット法[Thomas,Proc. Natl. Acad. Sci. USA,77:5201-5205 (1980)]、ドットブロット法(DNA分析)、又はインシトゥハイブリッド形成法によって、直接的に試料中で測定することができる。種々の標識を用いることができ、最も一般的なものは放射性同位元素、特に32Pである。しかしながら、他の方法、例えばポリヌクレオチド中への導入のためのビオチン修飾されたヌクレオチドもまた使用することができる。ついで、このビオチンは、例えば放射性ヌクレオチド、蛍光剤又は酵素のような広範囲の標識で標識することができるアビジン又は抗体への結合部位として作用する。また、DNA二本鎖、RNA二本鎖及びDNA−RNAハイブリッド二本鎖又はDNA-タンパク二本鎖を含む、特異的二本鎖を認識することができる抗体を用いることもできる。ついで、抗体を標識し、アッセイを実施することができ、ここで二本鎖は表面に結合しており、その結果二本鎖の表面での形成の時点でその二本鎖に結合した抗体の存在を検出することができる。
【0098】
あるいは、遺伝子の発現は、遺伝子産物の発現を直接的に定量する免疫学的な方法、例えば細胞又は組織切片の免疫組織化学的染色及び細胞培養又は体液のアッセイによって、測定することもできる。免疫組織化学的染色技術では、細胞試料を、典型的には脱水と固定によって調製し、結合した遺伝子産物に対し特異的な標識化抗体と反応させるが、この標識は通常は視覚的に検出可能であり、例えば酵素的標識、蛍光標識、又はルミネサンス標識である。
【0099】
試料液の免疫組織化学的染色及び/又はアッセイに有用な抗体は、モノクローナルでもポリクローナルでもよく、任意の哺乳動物で調製することができる。簡便には、抗体は、天然配列Apo-2DcRポリペプチドに対して、又はここで提供されるDNA配列をベースとした合成ペプチドに対して、又はApo-2DcRDNAに融合し特異的抗体エピトープをコードする外因性配列に対して調製され得る。
【0100】
6. Apo-2DcRポリペプチドの精製
Apo-2DcRの形態は、培地又は宿主細胞の溶菌液から回収することができる。Apo-2DcRが膜結合性であるならば、適切な洗浄液(例えばトリトン-X100)を用いて膜から引き離すか、又はその細胞外ドメインを酵素的に切断する。Apo-2DcRはまた糖リン脂質膜アンカーの酵素的切断により細胞表面から引き離すことができる。
【0101】
Apo-2DcRがヒト起源のもの以外の組換え細胞でつくられるときは、Apo-2DcRはヒト起源のタンパク質又はポリペプチドを含んでいない。しかしながら、Apo-2DcRに関して実質的に相同である調製物を得るには、組換え細胞タンパク又はポリペプチドからApo-2DcRを精製することが望ましい。第一段階として、培地又は溶菌液を遠心分離して粒状の細胞屑を除去することができる。ついで、Apo-2DcRを、汚染した可溶性タンパク質及びポリペプチドから、適切な精製手順の例である次の手順により精製される:すなわち、イオン交換カラムでの分画;エタノール沈殿;逆相HPLC;シリカ又はカチオン交換樹脂、例えばDEAEによるクロマトグラフィー;クロマトフォーカシング;SDS-PAGE;硫酸アンモニウム沈殿;例えばセファデックスG-75を用いるゲル濾過;及びIgGのような汚染物を除くプロテインAセファロースカラムである。
【0102】
残基が欠失され、挿入され、又は置換されたApo-2DcR変異体は、その変異によってしばしば惹起された実質的な性質変化を考慮に入れて、天然配列Apo-2DcRと同じようにして回収することができる。例えば、他のタンパク質又はポリペプチド、例えば細菌性もしくはウイルス性抗原、免疫グロブリン配列、又はレセプター配列とApo-2DcRとの融合体の調製は精製を容易にする;配列に対する抗体を含む免疫アフィニティーカラムを、融合ポリペプチドを吸着するために使用することができる。他の種類のアフィニティーマトリックスもまた使用することもできる。
【0103】
例えばフェニルメチルスルホニルフロリド(PMSF)のようなプロテアーゼインヒビターもまた精製の間のタンパク分解を阻害するのに有用であり、偶発的な汚染物質の成長を防止するために抗生物質を含めることができる。天然配列Apo-2DcRに適切な精製方法は、組換え細胞培養の発現の際におけるApo-2DcR又はその変異体の特性の変化の起因となる改変が必要となることは、当業者であれば分かるであろう。
【0104】
7. Apo-2DcRポリペプチドの共有結合的修飾
Apo-2DcRの共有結合的修飾は本発明の範囲内に含まれる。Apo-2DcRの共有結合的修飾の一つの型は、Apo-2DcRの標的アミノ酸残基を、Apo-2DcRのN末端又はC末端残基、又は選択された側鎖と反応できる有機誘導体化剤と反応させることによって分子内に導入することができる。
【0105】
二官能性試薬による誘導体形成は、抗Apo-2DcR抗体を精製する方法に使用する水不溶性支持体マトリックス又は表面へのApo-2DcRの架橋に有用であり、またその逆も同様である。二官能性試薬による誘導体形成は、Apo-2DcR分子を架橋させてApo-2DcR二量体を産生するのにも有用である。このような二量体により結合アビディティーが増大させられ、インビボにおける分子の半減期が延びる。通常使用される架橋剤には、例えば、1,1-ビス(ジアゾアセチル)-2-フェニルエタン、グルタルアルデヒド、N-ヒドロキシスクシンイミドエステル、例えば、4-アジドサリチル酸とのエステル、3,3'-ジチオビス(スクシンイミジルプロピオナート)のようなジスクシンイミジルエステルを包含するホモ二官能性イミドエステル、及びビス-N-マレイミド-1,8-オクタンのような二官能性マレイミドが含まれる。メチル-3-[(p-アジドフェニル)ジチオ]プロピオイミダートのような誘導体化剤は、光の存在下で架橋を形成することができる光活性化中間体を生じる。また、臭化シアン活性化炭水化物のような反応性の水不溶性マトリックス及び米国特許第3969287号;3691016号;4195128号;4247642号;4229537号及び4330440号に記載されている反応性基質がタンパク固定に用いられる。
【0106】
その他の修飾は、それぞれ対応するグルタミル及びアスパルチル残基へのグルタミニル及びアスパラギニル残基の脱アミド化、プロリンとリジンのヒドロキシル化、セリル又はスレオニル残基のヒドロキシル基のリン酸化、リジン、アルギニン、及びヒスチジン側鎖のαアミノ基のメチル化[T.E. Creighton, Proteins: Structure and Molecular Properties, W.H. Freeman & Co., San Francisco, PP.79-86 (1983)]、N末端アミンのアセチル化、及び任意のC末端カルボキシル基のアミド化を含む。残基の修飾型も本発明の範囲に入る。
【0107】
本発明の範囲内に含まれるApo-2DcRポリペプチドの共有結合的修飾の他のタイプは、ポリペプチドの天然グリコシル化パターンを変更することを含む。「天然グリコシル化パターンの変更」とは、天然配列Apo-2DcRに見出される一又は複数の炭水化物部分の欠失、及び/又は天然配列Apo-2DcRに存在しない一又は複数のグリコシル化部位を付加することを意味することをここでは意図している。
【0108】
ポリペプチドのグリコシル化は、典型的には、N結合又はO結合の何れかである。N結合とは、アスパラギン残基の側鎖への炭水化物部分の結合を指す。アスパラギン-X-セリン及びアスパラギン-X-スレオニン(ここでXはプロリンを除く任意のアミノ酸)というトリペプチド配列は、アスパラギン側鎖への炭水化物部分の酵素的結合のための認識配列である。したがって、ポリペプチド中にこれらのトリペプチド配列の何れかが存在すると、可能性の有るグリコシル化部位が作り出される。O結合グリコシル化は、ヒドロキシルアミノ酸、最も一般的にはセリン又はスレオニン(5-ヒドロキシプロリン又は5-ヒドロキシリジンもまた用いられるが)に、糖類N-アセチルガラクトサミン、ガラクトース、又はキシロースの一つが結合することを意味する。
【0109】
Apo-2DcRポリペプチドへのグリコシル化部位の付加は、アミノ酸配列を、それが一又は複数の上述したトリペプチド配列(N結合グリコシル化部位のもの)を含むように変化させることによって達成される。この変化は、天然配列Apo-2DcRへの一又は複数のセリン又はスレオニン残基の付加、又はこれによる置換によってもなされる(O結合グリコシル化部位の場合)。場合によっては、Apo-2DcRアミノ酸はDNAレベルでの変化によって、特に、所望のアミノ酸に翻訳するコドンが産生されるように予め選んだ塩基でApo-2DcRポリペプチドをコードしているDNAを突然変異することによって変更される。このDNA突然変異は、上記に記載され前掲の米国特許第5364934号に記載された方法を用いてなされる。
【0110】
Apo-2DcRポリペプチド上の炭水化物部分の数を増加させる他の手段は、該ポリペプチドへのグリコシドの化学的又は酵素的結合による。用いられる結合形態に応じて、糖(類)は、(a)アルギニンとヒスチジンに、(b)遊離のカルボキシル基に、(c)遊離のスルフヒドリル基、例えばシステインのものに、(d)セリン、スレオニン又はヒドロキシプロリンのもののような遊離のヒドロキシル基に、(e)フェニルアラニン、チロシン又はトリプトファンのような芳香族残基、又は(f)グルタミンのアミノ基に結合される。これらの方法は1987年9月11日公開の国際特許出願第WO87/05330号及びAplin及びWriston, CRC Crit. Rev. Biochem., pp259-306 (1981)に記載されている。
【0111】
Apo-2DcRポリペプチド上に存在する炭水化物部分の除去は、化学的又は酵素的あるいはグリコシル化の標的となるアミノ酸残基をコードするコドンの突然変異的置換によりなされる。例えば、化学的脱グリコシル化は、化合物トリフルオロメタンスルホン酸、又は等価な化合物へ該ポリペプチドを曝露し、該ポリペプチドを無傷のまま残しながら、結合糖(N-アセチルグルコサミン又はN-アセチルガラクトサミン)を除く殆ど又は全ての糖を開裂させる。化学的脱グリコシル化は、Hakimuddinほか, Arch. Biochem Biophys., 259:52 (1987)及びEdgeほか, Anal. Biochem., 118:131 (1981)により記載されている。ポリペプチド上の炭水化物部分の酵素的開裂は、Thotakuraほか, Meth. Enzymol., 138:350 (1987)に記載されているように、種々のエンド及びエキソグリコシダーゼを使用して達成することができる。
【0112】
潜在的なグリコシル化部位でのグリコシル化は、Duskinほか, J. Biol. Chem., 257:3105 (1982)によって記載されているように、化合物ツニカマイシンを使用して防ぐことができる。ツニカマイシンはタンパク質-N-グルコシド結合の形成を阻害する。
【0113】
Apo-2DcRの共有結合的修飾の他のタイプは、米国特許番第4640835号;第4496689号;第4301144号;第4670417号;第4791192号又は第4179337号に記載されているように、Apo-2DcRポリペプチドを、種々の非タンパク性ポリマーの一つ、例えばポリエチレングリコール、ポリプロピレングリコール、又はポリオキシアルキレンに結合させることを含む。
【0114】
8. Apo-2DcRキメラ
また本発明は、他の異種性ポリペプチド又はアミノ酸配列と融合したApo-2DcRを含むキメラ分子を提供する。
【0115】
一実施態様では、キメラ分子は、抗タグ抗体が選択的に結合するエピトープを提供するタグポリペプチドとのApo-2DcRの融合体を含む。エピトープタグは一般にApo-2DcRのアミノ又はカルボキシル末端に位置させられる。Apo-2DcRのこのようなエピトープタグが付けられた形は、その存在をタグポリペプチドに対する抗体を用いて検出することができる。また、エピトープタグを供給すると、Apo-2DcRを抗タグ抗体又はエピトープタグに結合する他の種類のアフィニティーマトリックスを用いたアフィニティー精製によって直ぐに精製することができる。
【0116】
様々なタグポリペプチドとその各抗体は従来から良く知られている。例には、fluHAタグポリペプチドとその抗体12CA5[Fieldほか, Mol. Cell. Biol., 8:2159-2165 (1988)];c-mycタグとそれに対する8F9、3C7、6E10、G4、B7及び9E10抗体[Evanほか, Molecular and Cellular Biology, 5:3610-3616 (1985)];及び単純ヘルペスウィルス糖タンパクD(gD)タグとその抗体[Paborskyほか, Protein Engineering, 3(6):547-553 (1990)]が含まれる。他のタグポリペプチドには、フラッグ-ペプチド(Flag-peptide)[Hoppほか, BioTechnology, 6:1204-1210 (1988)];KT3エピトープペプチド[Martinほか, Science, 255:192-194 (1992)];αチューブリンエピトープペプチド[Skinnerほか, J. Biol. Chem., 266:15163-15166 (1991)];及びT7遺伝子10タンパクペプチドタグ[Lutz-Freyermuthほか, Proc. Natl. Acad. Sci. USA, 87:6393-6397 (1990)]が含まれる。ひとたびタグポリペプチドが選択されれば、それに対する抗体を、ここに開示した方法を用いて産生することができる。
【0117】
一般的に、エピトープタグApo-2DcRは、上述した方法に従い作成され生産される。Apo-2DcRタグポリペプチド融合体は、Apo-2DcRタンパク質をインフレームでコードしているcDNA配列をタグポリペプチドDNA配列に融合させ、得られたDNA融合作成物を適当な宿主細胞に発現させることによって好ましく作成される。通常は、本発明のApo-2DcRタグポリペプチドキメラを調製するときは、Apo-2DcRをコードしている核酸を、タグポリペプチドのN末端をコードしている核酸にその3'末端で融合させるが、5'融合もまた可能である。例えば、約5〜約10のヒスチジン残基を有するポリヒスチジン配列はN末端又はC末端で融合され、アフィニティークロマトグラフィーにおける精製ハンドルとして使用される。
【0118】
エピトープタグApo-2DcRは、抗タグ抗体を用いてアフィニティークロマトグラフィーによって精製することができる。アフィニティー抗体が付着されるマトリックスには、例えばアガロース、調整穴明きガラス又はポリ(スチレンジビニル)ベンゼンが含まれる。ついで、エピトープタグApo-2DcRは、当該分野において知られた技術を使用して、アフィニティーカラムから溶出することができる。
【0119】
他の実施態様において、キメラ分子は免疫グロブリン配列と融合したApo-2DcRポリペプチドを含む。キメラ分子は、また特定のApo-2DcRドメイン配列、例えば免疫グロブリン配列と融合したApo-2DcRの細胞外ドメイン配列も含む。これには、モノマー、又はホモ-又はヘテロマルチマー、そして特にはホモ-又はヘテロダイマー又は-テトラマー型のものが含まれ;場合によっては、キメラはダイマー型又はホモダイマー重鎖型であり得る。一般的に、これらの構築免疫グロブリンは以下の図に示されるような既知の単位構造を有している。
【0120】
基本的な4鎖の構造単位はIgG、IgD及びIgEが存在する型である。4鎖の単位がより大なる高分子量の免疫グロブリンにおいて繰り返される;IgMが一般にジスルフィド結合によって一緒になった基本的な4鎖単位のペンタマーとして存在する。IgAグロブリン、そして時折IgGグロブリンはまた血清中にマルチマー型で存在する。マルチマーの場合は、各4鎖単位は同一であるか異なっている。
【0121】
次の図は、幾つかの例示的なモノマー、ホモ-及びヘテロダイマー及びホモ-及びヘテロマルチマー構造を示している。これらの図は単に例証するためのものであって、マルチマーの鎖は、天然免疫グロブリンと同様にジスルフィド結合していると考えられる。
モノマー:
ホモダイマー:
ヘテロダイマー:
ホモテトラマー:
ヘテロテトラマー:
及び
【0122】
上図において、「A」はApo-2DcR配列又は異種性配列に融合したApo-2DcR配列を意味する;Xは、Aと同一でも異なっていてもよい付加的な薬剤、免疫グロブリンスーパーファミリーのメンバーの一部で、例えば天然又はキメラ免疫グロブリン可変領域を含む可変領域又は可変領域様ドメイン、例えばシュードモナス外毒素又はリシンのような毒素、又は他のサイトカイン(すなわち、IL-1、インターフェロン-γ)又は細胞表面分子(すなわち、NGFR、CD40、OX40、Fas抗原、ショープ及び粘液腫ポックスウィルスのT2タンパク質)のような他のタンパク質に機能的に結合している配列、又は定常ドメインと通常は結合していないポリペプチド治療剤である;Yはリンカー又は他のレセプター配列である;VL、VH、CL及びCHは免疫グロブリンの軽又は重鎖の可変又は定常ドメインを表す。「A」として少なくとも1つのApo-2DcR配列のCRDと、「X」としてCRDの反復性パターンを有する他の細胞表面タンパク質とを含む構造体が特に含まれる。
【0123】
上図は、単に本発明の可能性のあるキメラ構造を例証したものであって、全ての可能性を包括するものではない。例えば、これらの作成物のいずれにおいても、望ましくは幾つかの異なった「A」、「X」又は「Y」があるかもしれない。また、重又は軽鎖定常ドメインは、同一又は異なる免疫グロブリンから由来したものであってもよい。図示された類似の構造のあらゆる可能な組合わせは全て本発明の範囲に入る。
【0124】
一般的にキメラ分子は、ある種の抗体からの可変ドメインが他の種の可変ドメインで置換されたキメラ抗体と同様にして作成することができる。例えば、EP0125023;EP173494;Munro, Nature, 312:597(1984年12月13日);Neubergerら, Nature, 312:604-608(1984年12月13日);Sharonら, Nature, 309:364-367(1984年5月24日);Morrisonら, Proc. Nat'l. Acad. Sci. USA, 81:6851-6855(1984);Morrisonら, Science, 229:1202-1207(1985);Boulianneら, Nature, 312:643-646(1984年12月13日);Caponら, Nature, 337:525-531(1989);Trauneckerら, Nature, 339:68-70(1989)を参照のこと。
【0125】
あるいは、キメラ分子は以下のようにして作成することができる。所望の配列、例えばApo-2DcR及び/又はTNFR配列をコードする領域を含むDNAを、免疫グロブリン様ドメインをコードするドメインの3'末端又はその近傍で、及びApo-2DcR又はTNFRポリペプチドのN末端をコードするDNAもしくはその近傍の点(異なるリーダーの使用が想定される)で、又はTNFRのN末端コード領域もしくはその近傍(そこでは天然シグナルが使用される)で、制限酵素により開裂させる。ついで、このDNA断片を免疫グロブリン軽又は重鎖定常ドメインをコードするDNAの近傍に直ちに挿入し、必要ならば、得られた作成物を欠失変異誘導により整える。好ましくは、キメラ分子がヒトに対するインビボ治療法を意図したものである場合、Igはヒト免疫グロブリンである。免疫グロブリン軽又は重鎖定常領域をコードするDNAは既知であるか、cDNAライブラリから直ぐに入手できるか合成される。例えば、Adamsら, Biochemistry, 19:2711-2719(1980);Goughら, Biochemistry, 19:2702-2710(1980);Dolbyら, Proc. Natl. Acad. Sci.. USA, 77:6027-6031(1980);Riceら, Proc. Natl. Acad. Sci., 79:7862-7865(1982);Falknerら, Nature, 298:286-288(1982);及びMorrisonら, Ann. Rev. Immunol., 2:239-256(1984)を参照のこと。
【0126】
このような融合体の調製方法のさらなる詳細は、イムノアドヘシンの調製に関する刊行物に見出される。イムノアドヘシン一般、及び特にCD4-Ig融合分子は、1989年4月6日に公開された国際特許出願第WO89/02922号に開示されている。IgG重鎖定常領域に結合している、ヒト免疫不全ウイルス(HIV)のレセプター、CD4の細胞外部分を含む分子は当該分野において知られており、CD4の可溶性細胞外部分よりも著しく長い半減期とより低いクリアランスを有することが見出された[Caponら, 上掲;Byrnら, Nature, 344:667(1990)]。また、特異的キメラTNFR-IgG分子の作成が、Ashkenaziら, Proc. Natl. Acad. Sci., 88:10535-10539(1991),;Lesslauerら, [J. Cell. Biochem. Supplement 15F, 1991, p.115(P432)];及びPeppelとBeutler, J. Cell. Biochem. Supplement 15F, 1991, p.118(P439)]に記載されている。
【0127】
B. Apo-2DcRの治療的及び非治療的用途
本明細書に開示されているように、Apo-2DcRは、治療的に用いられて、哺乳動物細胞において、Apo-2LによるかApo-2DcRが結合する他のリガンドによるアポトーシス及び/又はNF-κB活性化を誘発する。この治療法は、例えばインビボ又はエキソビボの遺伝子療法を使用することによりなすことができ、ここで開示している死亡ドメイン配列の使用を含む。また、免疫グロブリン配列を含むApo-2DcRキメラ分子(Apo-2DcRの細胞外ドメイン配列を含むキメラ分子又は以下の実施例に記載されるApo-2DcRイムノアドヘシンを含む)も治療的に使用することができ、Apo-2Lの活性、例えばアポトーシス又はNF-κB誘導又はApo-2DcRが結合する他のリガンドの活性を阻害する。
【0128】
好適な担体とそれらの製剤は、Osloらにより編集されたRemington's Pharmaceutical Sciences, 16th ed., 1980, Mack Publishing Co.,に記載されている。典型的には、適量の製薬的に許容可能な塩が、製剤を等浸透圧にするために製剤において使用される。担体の例には、生理食塩水、リンガー液及びデキストロース液のようなバッファーが含まれる。溶液のpHは、好ましくは約5〜約8、さらに好ましくは約7.4〜約7.8である。例えば投与経路よっては、ある種の担体がより好ましくなることは、当業者には明らかである。
【0129】
哺乳動物への投与は、注射(例えば、静脈、腹腔内、皮下、筋内)、又は点滴のように、有効な形態で血流への送達を確実にする他の方法によりなすことができる。
【0130】
投与の有効な用量とスケジュールは経験的に決定することができ、このような決定は当業者の技量の範囲に含まれる。
【0131】
また、他の更なる治療法を哺乳動物に施してもよく、そのようなものには、限定されるものではないが、化学療法及び放射線療法、イムノアジュバント、サイトカイン、及び抗体ベース療法が含まれる。例としては、インターロイキン(例えば、IL-1、IL-2、IL-3、IL-6)、白血病阻害因子、インターフェロン、TGF-β、エリスロポイエチン、トロンボポイエチン、及びHER-2抗体が含まれる。哺乳動物細胞においてアポトーシスを誘発することが知られている他の薬剤を用いてもよく、そのような薬剤にはTNF-α、TNF-β(リンホトキシン-α)、CD30リガンド、4-1BBリガンド、及びApo-1リガンドが含まれる。
【0132】
本発明において考慮される化学療法には、当該分野において既知であって市販されている化学物質又は薬物、例えばドキソルビシン、5-フルオロウラシル、シトシンアラビノシド(「Ara-C」)、シクロホスファミド、チオテパ、ブスルファン、サイトキシン、タキソール、メトトレキセート、シスプラチン、メルファラン、ビンブラスチン及びカルボプラチンが含まれる。このような化学療法に対する調製法及び用量スケジュールは、製造者の指示に従って使用されるか、熟練した実務家により経験的に決定される。そのような化学療法に対する調製法及び用量スケジュールはまた化学療法サービス,M.C.Perry編, Williams & Wilkins, Baltimore, MD (1992)にも記載されている。化学療法剤は、上述されたもののような、製薬的に許容可能な担体で好ましく投与される。
【0133】
また、本発明のApo-2DcRは非治療的用途においても有用性を有している。Apo-2DcRをコードしている核酸配列を、組織特異性の分類のための診断に使用することもできる。例えば、インシトゥハイブリッド形成、ノーザン及びサザンブロット法、及びPCR分析のような手順を、Apo-2DcRをコードするDNA及び/又はRNAが評価されている細胞型中に存在しているか否かを決定するために使用することができる。Apo-2DcR核酸は、ここに記載されている組換え技術によるApo-2DcRの調製にもまた有用である。
【0134】
単離されたApo-2DcRは、未知量のApo-2DcRを含有するサンプルが調製された対照として、定量的診断アッセイに使用され得る。またApo-2DcR調製物は、抗体を産生する際、Apo-2DcRに対するアッセイにおける標準として(例えば、ラジオイムノアッセイ、ラジオレセプターアッセイ又は酵素結合免疫測定法における標準としての使用のためにApo-2DcRを標識することにより)、アフィニティー精製法において、及び例えば放射性ヨウ素、酵素又はフルオロフォアで標識された場合、競合型レセプター結合アッセイにおいて有用である。
【0135】
実施例に記載されたような、Apo-2DcRの単離された天然型を使用して、Apo-2DcRの交互型;例えばシグナル伝達経路に関与しうる細胞質ドメインを保有する型を同定してもよい。Apo-2DcRの修飾型、例えば前述のApo-2DcR-IgGキメラ分子(イムノアドヘシン)は、抗Apo-2DcR抗体の生産における免疫原として使用することができる。
【0136】
また、Apo-2DcR又はその修飾型をコードする核酸は、トランスジェニック動物か「ノックアウト」動物を産生するのに使用でき、これらは治療的に有用な試薬の開発やスクリーニングに有用である。トランスジェニック動物(例えばマウス又はラット)とは、出生前、例えば胚段階で、その動物又はその動物の祖先に導入された導入遺伝子を含む細胞を有する動物である。導入遺伝子とは、トランスジェニック動物が発生する細胞のゲノムに組み込まれたDNAである。一実施形態では、Apo-2DcRをコードするcDNA又はその適当な配列(例えば、Apo-2DcR-IgG)を、確立された技術によりApo-2DcRをコードするゲノムDNAをクローン化するために使用することができ、ゲノム配列を、Apo-2DcRをコードするDNAを発現する細胞を有するトランスジェニック動物を産生するために使用することができる。特にマウス又はラット等のトランスジェニック動物を産生する方法は当該分野において常套的になっており、例えば米国特許第4736866号や第4870009号に記述されている。典型的には、特定の細胞を組織特異的エンハンサーでのApo-2DcR導入遺伝子の導入の標的にする。胚段階で動物の生殖系列に導入されたApo-2DcRをコードする導入遺伝子のコピーを含むトランスジェニック動物はApo-2DcRをコードするDNAの増大した発現の影響を調べるために使用できる。このような動物は、例えば過度のアポトーシスを伴う病理的状態に対して保護をもたらすと思われる試薬のテスター動物として使用できる。発明のこの側面においては、動物を試薬で治療し、導入遺伝子を有する未治療の動物に比べ病状の発病率が低ければ、疾患に対する治療的処置の可能性が示される。他の実施態様において、可溶性型のApo-2DcR、例えばApo-2DcRECD又はこのような型の免疫グロブリンキメラを有するトランスジェニック動物を作成して、Apo-2DcRL、Apo-2DcRのリガンドの慢性的中和の効果をテストすることができる。
【0137】
あるいは、Apo-2DcRの非ヒト相同体は、動物の胚性細胞に導入されたApo-2DcRをコードする変更ゲノムDNAと、Apo-2DcRをコードする内在性遺伝子との間の相同的組換えによって、Apo-2DcRをコードする欠陥又は変更遺伝子を有する「ノックアウト」動物を作成するために使用できる。例えば、Apo-2DcRをコードするDNAは、確立された技術に従い、Apo-2DcRをコードするゲノムDNAのクローニングに使用できる。Apo-2DcRをコードするゲノムDNAの一部を欠失したり、組み込みを監視するために使用する選択可能なマーカーをコードする遺伝子等の他の遺伝子で置換することができる。典型的には、ベクターは無変化のフランキングDNA(5'と3'末端の両方)を数キロベース含む[例えば、相同的組換えベクターについてはThomas及びCapecchi, Cell, 51:503(1987)を参照のこと]。ベクターは胚性幹細胞に(例えば電気穿孔法等によって)導入し、導入されたDNAが内在性DNAと相同的に組換えられた細胞を選択する[例えば、Liほか, Cell, 69:915(1992)参照]。選択された細胞は次に動物(例えばマウス又はラット)の胚盤胞内に注入され、集合キメラを形成する[例えば、Bradley, Teratocarcinomas and Embryonic Stem Cells: A Practical Approach, E. J. Robertson, ed. (IRL, Oxford, 1987), pp. 113-152参照]。その後、キメラ性胚を適切な偽妊娠の雌性乳母に移植し、期間をおいて「ノックアウト」動物をつくり出す。胚細胞に相同的に組換えられたDNAを有する子孫は標準的な技術により同定され、それらを利用して動物の全細胞が相同的に組換えられたDNAを含む動物を繁殖させることができる。ノックアウト動物は、例えば腫瘍の発達を含む、Apo-2DcRポリペプチドが不在であることによるある種の病理的状態及びその病理的状態の発達に対する防御能力によって特徴付けられる。
【0138】
C. 抗Apo-2DcR抗体の調製
本発明は、さらに抗Apo-2DcR抗体を提供するものである。Apo-2DcRに対する抗体は以下のようにして調製することができる。抗体の例としては、ポリクローナル、モノクローナル、ヒト化、二重特異性及びヘテロ抱合体抗体が含まれる。
【0139】
1. ポリクローナル抗体
Apo-2DcR抗体はポリクローナル抗体を含む。ポリクローナル抗体の調製方法は当業者に知られている。哺乳動物においてポリクローナル抗体は、例えば免疫化剤、所望するのであればアジュバントを、一又は複数回注射することで発生させることができる。典型的には、免疫化剤及び/又はアジュバントを複数回皮下又は腹腔内注射により、哺乳動物に注射する。免疫化剤は、Apo-2DcRポリペプチド又はその融合タンパク質を含みうる。適切な免疫化剤の例は、Apo-2DcR-IgG融合タンパク質、又はキメラ分子(Apo-2DcREDC-IgG融合タンパク質を含む)である。また、その表面にApo-2DcRを発現する細胞を使用してもよい。免疫化されている哺乳動物において免疫原性であることが知られているタンパク質に免疫化剤を抱合させることが有用である。使用され得るそのような免疫原性タンパク質の例は、限定するものではないが、スカシガイヘモシアニン、血清アルブミン、ウシサイログロブリン及び大豆トリプシンインヒビターが含まれる。また、哺乳動物の免疫反応を増強するために、ミョウバンのような凝集剤を使用してもよい。使用され得るアジュバントの例には、フロイント完全アジュバント及びMPL-TDMアジュバント(モノホスホリル脂質A、合成トレハロースジコリノミコラート)が含まれる。免疫化プロトコールは、過度の実験なく当業者により選択されるであろう。哺乳動物から採血し、血清を検定して抗体価を求める。望まれるならば、抗体価が増加又はプラトーするまで哺乳動物に追加免疫を施す。
【0140】
2. モノクローナル抗体
あるいは、Apo-2DcR抗体はモノクローナル抗体であってもよい。モノクローナル抗体は、Kohler及びMilstein,上掲に記載されているようなハイブリドーマ法を使用することで調製することができる。ハイブリドーマ法では、マウス、ハムスター又は他の適切な宿主動物を典型的には免疫化剤により免疫化する(上述したようにして)ことで、免疫化剤に特異的に結合する抗体を生成するかあるいは生成可能なリンパ球を誘発する。また、リンパ球をインビトロで免疫化することもできる。
【0141】
免疫化剤は、典型的にはApo-2DcRポリペプチド又はその融合タンパク質を含む。適切な免疫化剤の例は、Apo-2DcR-IgG融合タンパク質又はキメラ分子である。Apo-2DcREDC-IgG免疫原の特定の例は、以下の実施例13に記載している。表面でApo-2DcRを発現する細胞もまた使用することができる。一般にヒト由来の細胞が望まれる場合には末梢血リンパ球(「PBL」)が使用され、あるいは非ヒト哺乳動物源が望まれている場合は、脾臓細胞又はリンパ節細胞が使用される。次いで、ポリエチレングリコール等の適当な融合剤を用いてリンパ球を不死化株化細胞と融合させ、ハイブリドーマ細胞を形成する[Goding, Monoclonal Antibodies: Principles and Practice, Academic Press, (1986) pp. 59-103]。不死化株化細胞は、通常は、形質転換した哺乳動物細胞、特に齧歯動物、ウシ、及びヒト由来の骨髄腫細胞である。通常、ラット及びマウスの骨髄腫細胞が使用される。ハイブリドーマ細胞は、好ましくは、未融合の不死化細胞の生存又は成長を阻害する一又は複数の物質を含有する適切な培地で培養される。例えば、親の形質転換細胞が、酵素のヒポキサンチングアニンホスホリボシルトランスフェラーゼ(HGPRT又はHPRT)を欠いていると、ハイブリドーマの培地は、典型的には、ヒポキサチン、アミノプチリン及びチミジン(「HAT培地」)を含み、この物質がHGPRT欠乏性細胞の増殖を阻止する。
【0142】
好ましい不死化株化細胞は、効率的に融合し、選択された抗体生成細胞による安定した高レベルの抗体発現を支援し、HAT培地のような培地に対して感受性であるものが望ましい。より好ましい不死化株化細胞はマウス骨髄腫株であり、これはカリフォルニア州サンディエゴのSalk Institute Cell Distribution Centerやバージニア州マナッサスのアメリカン・タイプ・カルチャー・コレクションより入手可能である。ヒトモノクローナル抗体を生成するためのヒト骨髄腫及びマウス-ヒト異種骨髄腫株化細胞も開示されている[Kozbor, J. Immunol., 133:3001 (1984)、Brodeurほか, Monoclonal Antibody Production Techniques and Applications, Marcel Dekker, Inc., New York, (1987) pp. 51-63]。
【0143】
ハイブリドーマ細胞を培養する培地を、Apo-2DcRに対するモノクローナル抗体の存在について検定する。好ましくは、ハイブリドーマ細胞によって生成されたモノクローナル抗体の結合特異性は免疫沈降又はラジオイムノアッセイ(RIA)や酵素結合免疫測定法(ELISA)等のインビトロ結合検定法によって測定する。このような技術及びアッセイは、当該分野において公知である。モノクローナル抗体の結合親和性は、例えばMunson及びPollard, Anal. Biochem., 107:220 (1980)によるスキャッチャード分析法によって測定することができる。
【0144】
所望のハイブリドーマ細胞が同定されたら、クローンを制限希釈工程を経てサブクローニングし、標準的な方法で増殖させることができる[Goding, 上掲]。この目的のための適当な培地には、例えば、ダルベッコの改変イーグル培地及びRPMI-1640倍地が含まれる。更に、ハイブリドーマ細胞は哺乳動物においてインビボで腹水として成長させることもできる。
【0145】
サブクローンによって分泌されるモノクローナル抗体は、例えばプロテインAセファロース法、ヒドロキシルアパタイトクロマトグラフィー法、ゲル電気泳動法、透析法又はアフィニティークロマトグラフィー等の従来の免疫グロブリン精製方法によって培地又は腹水液から分離又は精製される。
【0146】
また、モノクローナル抗体は、組換えDNA法、例えば米国特許第4816567号に記載された方法により作製することができる。本発明のモノクローナル抗体をコードするDNAは、常套的な方法によって(例えば、マウス抗体の重鎖及び軽鎖をコードする遺伝子に特異的に結合可能なオリゴヌクレオチドプローブを使用して)、容易に単離し配列決定することができる。本発明のハイブリドーマ細胞はそのようなDNAの好ましい供給源となる。ひとたび単離されたら、DNAは発現ベクター内に配することができ、これが宿主細胞、例えばサルCOS細胞、チャイニーズハムスター卵巣(CHO)細胞、あるいは免疫グロブリンタンパク質を生成等しない骨髄腫細胞内にトランスフェクトされ、組換え宿主細胞内でモノクローナル抗体の合成をすることができる。また、DNAは、例えば相同マウス配列に換えてヒト重鎖及び軽鎖定常ドメインのコード配列を置換することにより[US. Patent No.4816567;Morrisonほか, 上掲]、又は免疫グロブリンコード配列に非免疫グロブリンポリペプチドのコード配列の一部又は全部を共有結合することにより修飾することができる。このような非免疫グロブリンポリペプチドは、本発明の抗体の定常ドメインの代わりに置換するか、本発明の抗体の一つの抗原結合部位の可変ドメインの代わりに置換し、キメラ性二価抗体を産生することができる。
【0147】
以下の実施例に記載するようにして、抗Apo-2DcRモノクローナル抗体を調製した。これらの抗体のいくらかは、4G3.9.9、6D10.9.7、及び1C5.24.1としてATCCに寄託され、寄託受入番号 、 、及び がそれぞれ付与された。一実施態様において、本発明のモノクローナル抗体は、受入番号 、 、及び で寄託されたハイブリドーマ株化細胞により分泌される抗体の一又は複数と同じ生物学的特徴を有する。「生物学的特徴」という用語は、モノクローナル抗体のインビトロ及び/又はインビボの活性又は性質、例えばApo-2DcRに特異的に結合する能力又はApo-2DcR活性を実質的に阻止、誘導又は高める能力を指すために使用される。場合によっては、モノクローナル抗体はここで特に言及された3種の抗体の少なくとも1つと同じエピトープに結合する。そのようなエピトープ結合は、例えばここや実施例において記載したように、種々のアッセイを行うことにより測定することができる。
【0148】
本発明の抗体は一価抗体であってもよい。一価抗体の調製方法は当該分野においてよく知られてる。例えば、一つの方法は免疫グロブリン軽鎖と修飾重鎖の組換え発現を含む。重鎖は一般的に、重鎖の架橋を防止するようにFc領域の任意のポイントで切断される。あるいは、関連したシステイン残基を他のアミノ酸残基で置換するか欠失させて架橋を防止する。
【0149】
一価抗体の調製にはインビトロ法がまた適している。抗体の消化による、その断片、特にFab断片の調製は、当該分野において知られている慣用的技術を使用して達成できる。例えば、消化はパパインの使用により行うことができる。パパイン消化の例は、94/12/22に公開された国際特許出願第WO94/29348号、及び米国特許第4342566号に記載されている。抗体のパパイン消化は、典型的には、Fab断片と呼ばれ、各々が単一の抗原結合部位を有する2つの同一の抗原結合断片と、残りのFc断片を生成する。ペプシン処理により、2つの抗原結合部位を有し、抗原の架橋が尚も可能なF(ab')2断片が得られる。
【0150】
また、抗体の消化により生産されたFab断片は、軽鎖の定常ドメインと重鎖の第1定常ドメイン(CH1)を含む。Fab'断片は、抗体のヒンジ領域から一又は複数のシステインを含む重鎖CH1ドメインのカルボキシ末端に幾つかの残基が付加されているということで、Fab断片とは異なっている。Fab'-SHとは、定常ドメインのシステイン残基が遊離のチオール基を担持しているFab'に対するここでも命名である。F(ab’)2抗体断片は、本来は、それらの間にヒンジシステインを有するFab’断片の対として生産された。抗体断片の他の化学的結合もまた知られている。
【0151】
3. ヒト化抗体
本発明のApo-2DcR抗体は、さらにヒト化抗体又はヒト抗体を含む。非ヒト(例えばマウス)抗体のヒト化形とは、キメラ免疫グロブリン、免疫グロブリン鎖あるいはその断片(例えばFv、Fab、Fab'、F(ab')2あるいは抗体の他の抗原結合サブ配列)であって、非ヒト免疫グロブリンに由来する最小配列を含むものである。ヒト化抗体はレシピエントの相補性決定領域(CDR)の残基が、マウス、ラット又はウサギのような所望の特異性、親和性及び能力を有する非ヒト種(ドナー抗体)のCDRの残基によって置換されたヒト免疫グロブリン(レシピエント抗体)を含む。ある場合には、ヒト免疫グロブリンのFvフレームワーク残基は、対応する非ヒト残基によって置換されている。更に、ヒト化抗体は、レシピエント抗体にも、移入されたCDRもしくはフレームワーク配列にも見出されない残基を含んでいてもよい。一般に、ヒト化抗体は、全てあるいはほとんど全てのCDR領域が非ヒト免疫グロブリンのものに対応し、全てあるいはほとんど全てのFR領域がヒト免疫グロブリンコンセンサス配列のものである、少なくとも1つ、典型的には2つの可変ドメインの実質的に全てを含む。ヒト化抗体は、最適には免疫グロブリン定常領域(Fc)、典型的にはヒトの免疫グロブリンの定常領域の少なくとも一部を含んでなる[Jones ほか, Nature, 321:522-525 (1986); Riechmann ほか, Nature, 332:323-329 (1988); 及びPresta, Curr. Op Struct. Biol., 2:593-596 (1992)]。
【0152】
非ヒト抗体をヒト化する方法は従来からよく知られている。一般的に、ヒト化抗体には非ヒト由来の一又は複数のアミノ酸残基が導入される。これら非ヒトアミノ酸残基は、しばしば、典型的には「移入」可変ドメインから得られる「移入」残基と称される。ヒト化は基本的に齧歯動物のCDR又はCDR配列でヒト抗体の該当する配列を置換することによりウィンター及び共同研究者[Jonesほか, Nature, 321:522-525 (1986)、Riechmannほか, Nature, 332:323-327 (1988)、Verhoeyenほか, Science, 239:1534-1536 (1988)]の方法を使用して行える。よって、このような「ヒト化」抗体は、無傷のヒト可変ドメインより実質的に少ない分が非ヒト種由来の対応する配列で置換されたキメラ抗体(U.S. 4,816,567)である。実際には、ヒト化抗体は典型的にはある程度のCDR残基及び場合によってはいくらかのFR残基が齧歯類抗体の類似する部位からの残基によって置換されるヒト抗体である。
【0153】
抗原性を軽減するには、ヒト化抗体を生成するために使用するヒトの軽及び重可変ドメインの両方の選択が非常に重要である。「ベストフィット法」では、齧歯動物抗体の可変ドメインの配列を既知のヒト可変ドメイン配列ライブラリ全体に対してスクリーニングする。齧歯動物のものと最も近いヒトの配列を次にヒト化抗体のヒトフレームワーク(FR)として受け入れる[Simsほか, J. Immunol., 151:2296 (1993);Chothia及びLesk, J. Mol. Biol., 196:901 (1987)]。他の方法では、軽又は重鎖の特定のサブグループのヒト抗体全てのコンセンサス配列から誘導される特定のフレームワークを使用する。同じフレームワークを幾つかの異なるヒト化抗体に使用できる[Carterほか, Proc. Natl. Acad. Sci. USA, 89:4285 (1992);Prestaほか, J. Immunol., 151:2623 (1993)]。
【0154】
更に、抗体は、抗原に対する高親和性や他の好ましい生物学的性質を保持してヒト化することが重要である。この目標を達成するべく、好ましい方法では、親及びヒト化配列の三次元モデルを使用して、親配列及び様々な概念的ヒト化産物の分析工程を経てヒト化抗体を調製する。三次元免疫グロブリンモデルは一般的に入手可能であり、当業者にはよく知られている。選択された候補免疫グロブリン配列の推測三次元立体配座構造を図解し、表示するコンピュータプログラムは購入可能である。これら表示を見ることで、候補免疫グロブリン配列の機能における残基の役割の分析、すなわち候補免疫グログリンの抗原と結合する能力に影響を及ぼす残基の分析が可能となる。このようにして、例えば標的抗原に対する親和性を高めるといった、望ましい抗体特徴が得られるように、FR残基をコンセンサス及び移入配列から選択し、組み合わせることができる。一般的に、CDR残基は、直接かつ最も実質的に抗原結合性に影響を及ぼしている[1994年、3月3日に公開されたWO 94/04679を参照]。
【0155】
免疫化することで、内在性免疫グロブリンが生成されない状態でもヒト抗体の完全リパートリを生成することができるトランスジェニック動物(例えばマウス)を使用することが可能である。例えば、キメラ及び生殖系列変異マウスにおいて抗体重鎖結合領域(JH)遺伝子をホモ接合的に欠失させると内在性抗体生産の完全な阻害が生じることが記述されている。このような生殖系列変異マウスにヒト生殖系列免疫グロブリン遺伝子配列を移すと、抗原投与時にヒト抗体の生成が生じる[例えば、Jakobovitsほか, Proc. Natl. Acad. Sci. USA, 90:2551-255 (1993);Jakobovitsほか, Nature, 362:255-258 (1993);Bruggermanほか, Year in Immuno., 7:33 (1993)を参照]。また、ヒト抗体はファージ表示ライブラリ[Hoogenboom及びWinter, J. Mol. Biol., 227:381 (1992);Marksほか, J. Mol. Biol., 222:581 (1991)]において産生することもできる。また、Coleら及びBoernerらの方法も、ヒトモノクローナル抗体の調製に利用することができる[Coleら, Monoclonal Antibodies and Cancer Therapy, Alan R. Liss. p.77(1985)及びBoernerら, J. Immunol., 147(1):86-95(1991) ]。
【0156】
4. 二重特異性抗体
二重特異性抗体は、少なくとも2つの異なる抗原に対して結合特異性を有するモノクローナル抗体、好ましくはヒトもしくはヒト化抗体である。本発明の場合において、結合特異性の一方はApo-2DcRに対してであり、他方は任意の他の抗原、好ましくは細胞表面タンパク質又はレセプター又はレセプターサブユニットに対してである。
【0157】
二重特異性抗体を生成する方法は当該技術分野において周知である。伝統的には、二重特異性抗体の組換え生成方法は、二つの重鎖が異なる特異性を持つ二つの免疫グロブリン重鎖/軽鎖対の同時発現に基づく[Milstein及びCuello, Nature, 305:537-539 (1983)]。免疫グロブリンの重鎖と軽鎖を無作為に取り揃えたため、これらハイブリドーマ(クアドローマ)は10種の異なる抗体分子の潜在的混合物を生成でき、その内一種のみが正しい二重特異性構造を有する。正しい分子の精製は、アフィニティークロマトグラフィー工程によって通常達成される。同様の手順が1993年5月13日公開の国際特許出願第93/08829号、及びTrauneckerほか, EMBO J.,10:3655-3656 (1991)に開示されている。
【0158】
別のより好ましいアプローチによれば、所望の結合特異性(抗体-抗原組合せ部位)を有する抗体可変ドメインを免疫グロブリン定常ドメイン配列に融合する。融合は、好ましくは少なくともヒンジ部、CH2及びCH3領域の一部を含む免疫グロブリン重鎖定常ドメインとのものである。少なくとも一つの融合には軽鎖結合に必要な部位を含む第一の重鎖定常領域(CH1)が存在することが望ましい。免疫グロブリン重鎖融合をコードするDNA、及び望むのであれば免疫グロブリン軽鎖を、別々の発現ベクターに挿入し、適当な宿主生物に同時形質移入する。これは、作成に使用する三つのポリペプチド鎖が不等の比であるときに最高の収率が得られる実施態様において、三つのポリペプチド断片の相互比率の調節に大なるフレキシビリティをもたらす。しかし、少なくとも二つのポリペプチド鎖が同比率で発現すると高収率が得られる場合や比率が特に重要ではない場合には、一つの発現ベクターに二つ又は三つ全てのポリペプチド鎖のコード配列を挿入することができる。このアプローチの好適な形態では、二重特異性抗体は、一方のアームの第一結合特異性を有するハイブリッド免疫グロブリン重鎖と、他方のアームのハイブリッド免疫グロブリン重鎖/軽鎖対(第二の結合特異性をもたらす)からなる。このような非対称的構造は、二重特異性分子の半分にのみ免疫グロブリン軽鎖が存在すると容易な方法で分解できるので、所望の二重特異性化合物を不要な免疫グロブリン鎖の組合わせから分解し易くすることが見出された。このアプローチは1994年3月3日公開の国際特許出願第94/04690号によって開示されている。二重特異性抗体を生成するための更なる詳細については、例えばSureshほか,Methods in Enzymology, 121:210(1986)を参照されたい。
【0159】
5. ヘテロ抱合体抗体
ヘテロ抱合抗体もまた本発明の範囲に入る。ヘテロ抱合抗体は、2つの共有的に結合した抗体からなる。このような抗体は、例えば、免疫系細胞を不要な細胞に対してターゲティングさせるため(米国特許第4676980号)及びHIV感染の治療のために(WO91/00360、WO92/200373;EP03089)提案された。本抗体は、架橋剤に関連したものを含む合成タンパク化学における既知の方法を使用して、インビトロで調製することができると考えられる。例えば、ジスルフィド交換反応を使用するか又はチオエーテル結合を形成することにより、免疫毒素を作成することができる。この目的に対して好適な試薬の例には、イミノチオレート及びメチル-4-メルカプトブチリミデート、及び例えば米国特許第4676980号に開示されているものが含まれる。
【0160】
D. Apo-2DcR抗体の治療的及び非治療的用途
本発明のApo-2DcR抗体は治療的な有用性を有している。例えば、Apo-2リガンドに対する他のレセプターと交差反応するApo-2DcR抗体は過剰なアポトーシス(例えば神経変性疾患において)をブロックするか、潜在的な自己免疫/炎症効果をブロックするために使用することができる。場合によっては、Apo-2DcR阻止抗体は、Apo-2リガンド治療法と組合わせて使用することができる。Apo-2DcR抗体はApo-2DcRレセプターを阻止することができ、投与されたApo-2リガンドの生物利用能を増大させることができる。(Apo-2DcRに対しての上述のような)治療組成物と投与形態を用いることができる。
【0161】
さらに、Apo-2DcR抗体は、免疫組織化学染色検定法又はApo-2DcRの診断検定法、例えば特異的細胞、組織又は血清における発現を検出する検定法に使用することができる。当該分野において知られている様々な診断検定技術、例えば、競合的結合検定、直接的又は間接的サンドイッチ検定及び不均一又は均一相の何れにおいても実施される免疫沈降検定を使用することができる[Zola, Monoclonal Antibodies: A Manual of Techniques,CRC Press,Inc.,(1987) pp.147-158]。診断検定法に使用される抗体は、検出可能部分で標識することができる。検出可能成分は、直接的に又は間接的に検出可能なシグナルをつくりだすことができなければならない。例えば検出可能成分は、3H、14C、32P、35S又は125I等の放射性同位体、蛍光イソチオシアネート、ローダミン又はルシフェリン等の蛍光又は化学発光化合物、もしくはアルカリホスファターゼ、βガラクトシダーゼ又は西洋わさびペルオキシダーゼ等の酵素であってもよい。Hunterほか,Nature,144:945 (1962)、Davidほか, Biochemistry,13:1014(1974)、Painほか,J. Immunol. Meth.,40:219 (1981)及びNygrenほか,J. Histochem. and Cytochem.,30:407 (1982)等に記載されている方法を含み、検出可能部分に抗体を抱合させるための当該分野において知られている任意の方法を使用することができる。
【0162】
また、Apo-2DcR抗体は天然供給源又は組換え細胞培養からのApo-2DcRのアフィニティー精製にも有用である。この方法においては、Apo-2DcRに対する抗体を、当該分野でよく知られている方法を使用して、セファデックス樹脂や濾過紙のような適当な支持体に固定する。次に、固定された抗体を、精製するApo-2DcRを含む試料と接触させ、ついで、固定された抗体に結合したApo-2DcR以外の試料中の物質を実質的に全て除去する適当な溶媒で支持体を洗浄する。最後に、Apo-2DcRを抗体から離脱させる他の適当な溶媒で支持体を洗浄する。
【0163】
E. Apo-2DcR又はApo-2DcR抗体を含むキット
本発明の更なる実施態様においては、例えば上述の治療的又は非治療的用途に使用可能なApo-2DcR又はApo-2DcR抗体を含むキット及び製造品が提供される。製造品にはラベルが付された容器が含まれる。適切な容器には、例えばボトル、バイアル、及び試験管が含まれる。容器はガラス又はプラスチックのような種々の物質から形成できる。容器は、上述の治療的又は非治療的用途に有効な活性剤を含む組成物を収容する。組成物中の活性剤は、Apo-2DcR又はApo-2DcR抗体である。容器のラベルには、組成物が特定の治療的又は非治療的用途に使用されることが示され、また上述のもののような、インビボ又はインビトロのいずれかの使用の指示が示されている。
【0164】
本発明のキットは、典型的には、上述の容器と、商業上及び使用者の観点から望まれる、バッファー、希釈液、フィルター、針、注入器及び使用説明が記されたパッケージ挿入物を含む、材料を含む一又は複数の他の容器を含んでいる。
【0165】
以下の実施例は例示するためにのみ提供されるものであって、本発明の範囲を決して限定することを意図するものではない。
【0166】
本明細書で引用した全ての特許及び参考文献の全体を、出典明示によりここに取り込む。
【0167】
実施例
実施例において言及されている全ての制限酵素は、ニューイングランドバイオラボ社(New England Biolabs)から購入し、製造者の使用説明に従い使用した。実施例で言及されている全ての他の市販試薬は、特に示していない限りは、製造者の使用説明に従い使用した。ATCC受入番号により次の実施例及び明細書全体を通して特定している細胞の供給源はアメリカン・タイプ・カルチャー・コレクション(Mannasas,Virginia)である。
【0168】
実施例1
ヒトApo-2DcRをコードしているcDNAクローンの単離
【0169】
1.オリゴdTプライムcDNAライブラリ(「LIB111」)の調製
インビトロゲン、サンディエゴ、CAの試薬とプロトコールを使用してmRNAをヒト乳癌組織から単離した(高速トラック2)。ライフ・テクノロジーズ、ゲイザースバーグ、MDの試薬とプロトコール(スーパースクリプトプラスミド系)を使用し、ベクターpRK5D中にオリゴdTプライムcDNAライブラリ(「LIB111」)を産生するためにこのRNAを用いた。この手順において、二本鎖cDNAを1000塩基対よりも大きいサイズにし、Sa1I/NotIリンカーcDNAをXhoI/NotI切断ベクター内にクローニングした。pRK5Dはsp6転写開始部位にSfiI制限酵素部位が続き、その次にXhoI/NotIcDNAクローニング部位があるクローニングベクターである。
【0170】
2.ランダムプライムcDNAライブラリ(「LIB118」)の調製
一次cDNAクローンの5'末端を優先的に提示するために二次cDNAライブラリを産生した。sp6RNAを一次ライブラリ(上述のLIB111)から産生し、このRNAを用いて、ライフ・テクノロジーズの試薬とプロトコール(上述のスーパースクリプトプラスミド系)を使用してベクターpSST-AMY.0中にランダムプライムcDNAライブラリ(「LIB118」)を産生した。この手順において二本鎖cDNAを500−1000塩基対のサイズにし、NotIアダプターに平滑末端化してリンカーし、SfiIで切断し、SfiI/NotI切断ベクター中にクローニングした。pSST-AMY.0は、酵母アルコールデヒドロゲナーゼプロモータの後にcDNAクローニング部位とマウスアミラーゼ配列(分泌シグナルを持たない成熟配列)が続き、それに、酵母アルコールデヒドロゲナーゼターミネーターがクローニング部位後に続くクローニングベクターである。しかして、アミラーゼ配列とインフレームで融合されたこのベクター中にクローニングされたcDNAは、適切に形質移入された酵母コロニーからアミラーゼを分泌せしめる。
【0171】
3.形質転換と検出
LIB118からのDNAを氷上で冷却し、エレクトロコンピテントDH10B細菌 (Life Technologies,20ml)を添加した。ついで、細菌ベクター混合物を、製造者の推奨に従って電気穿孔法にかけた。続いてSOC培地(Life Technologies, 1ml)を添加し、混合物を37℃で30分間インキュベートした。ついで、形質転換体を、アンピシリンを含有する20の標準的な150mmLBプレートに蒔き、16時間(37℃)の間インキュベートした。陽性コロニーを擦り取り、CsCl勾配のような標準的なプロトコールを使用して、細菌ペレットからDNAを単離した。次に、精製したDNAについて以下の酵母プロトコールを進めた。
【0172】
本発明において使用される酵母方法は、3つの範疇に分けられた:(1)プラスミド/cDNA併用ベクターでの酵母の形質転換;(2)アミラーゼを分泌する酵母クローンの検出と単離;及び(3)酵母コロニーから直接の挿入断片のPCR増幅と配列決定及び更なる分析のためのDNAの精製である。
【0173】
安定な突然変異体ura3を含む任意の酵母株を本発明において使用することができるが、本発明の実施のために使用される好ましい酵母株はHD56-5A(ATCC-90785)であった。この株は次の遺伝子型:MATα、ura3-52、leu2-3、leu2-112、his3-11、his3-15、MAL+、SUC+、GAL+を有していた。
【0174】
形質転換を、Gietzら, Nucl. Acid. Res.,20:1425(1992) により概説されたプロトコールに基づいて実施した。この手順では、DNAマイクログラム当りおよそ1x105形質転換体の形質転換効率を得た。形質転換された細胞をついで寒天からYEPD複合培地ブロス(100ml)中に播種し、30℃で一晩増殖させた。YEPDブロスは、Kaiserら, Methods in Yeast Genetics, Cold Spring Harbor Press, Cold SpringHarbor, N.Y. USA, p207 (1994)に記載されているようにして調製した。ついで一晩の培養を約2x106細胞/ml(およそOD600=0.1)に希釈して新鮮なYEPDブロス(500ml)中に入れ、1x107細胞/ml(およそOD600=0.4−0.5)に再増殖させた。これには完了まで通常約3時間かかった。
【0175】
ついで、細胞を収集し、GS3ロータボトルに移しソーバル(Sorval)GS3ロータを5000rpmで5分間作動させて形質転換用に調製し、上清を捨て、ついで滅菌水に再懸濁させ、ベックマンGS-6KR遠心分離機で3500rpmにて50mlのファルコン管中で再び遠心分離した。上清を捨て、続いて細胞をLiAc/TE(10ml、10mMのトリス-HCl、1mMのEDTA、pH7.5、100mMのLi2OOCCH3)で洗浄し、LiAc/TE(2.5ml)中に再懸濁した。
【0176】
調製した細胞(100μl)を新鮮に変性した一本鎖サケ精巣DNA(Lofstrand Labs, Gaithersburg, MD, USA)と混合し微量遠心管中でDNA(1μg,容量<10μl)を形質転換することにより形質転換を行った。混合物をボルテックスすることにより簡単に混合し、ついで40%PEG/TE(600μl、40%ポリエチレングリコール-4000、10mMのトリス-HCl、1mMのEDTA、100mMのLi2OOCCH3、pH7.5)を添加した。この混合物を穏やかに混合し、30分間攪拌しながら30℃でインキュベートした。ついで細胞に42℃で15分間熱ショックをかけ、反応容器を微量遠心管中で5−10秒の間に12000rpmで遠心分離し、デカントし、TE(500μl、10mMのトリス-HCl、1mMのEDTA、pH7.5)に再懸濁させ、再び遠心分離した。ついで、細胞をTE(1ml)中に希釈し、一定分量(200μl)を、150mm成長板(VWR)に前もって調製していた選択培地上に広げた。
【0177】
あるいは、複数の小さな反応の代わりに、単一の大きな規模の反応で試薬量を多くしたものを使用して形質転換を行った。
【0178】
使用された選択培地は、Kaiserら,Methods in Yeast Genetics, Cold Spring Harbor Press, Cold Spring Harbor, N.Y. USA, p208-210 (1994)に記載されたようにして調製されたウラシルを欠く合成完全ブドウ糖寒天(SCD-Ura)であった。形質転換体は30℃で2−3日増殖させた。
【0179】
アミラーゼを分泌するコロニーの検出は選択増殖培地に赤色のデンプンを含めることにより実施した。Bielyら, Anal.Biochem.,172:176-179(1988) により記載された手順に従って、デンプンは赤色染料(リアクティブレッド-120、シグマ)に結合させた。結合されたデンプンは0.15%(w/v)の最終の濃度でSCD-Ura寒天プレート中に導入し、リン酸カルシウムで7.0のpHまで緩衝させた(50−100mMの最終濃度)。
【0180】
陽性のコロニーを取上げ、よく分離された同定可能なシングルコロニーを得るために新鮮選択培地(150mmプレート上)上に画線した。この工程はまた形質転換体の中でもプラスミドの維持を確実にした。アミラーゼ分泌に対して陽性のよく分離されたシングルコロニーを、緩衝化SCD-Ura寒天中への赤色デンプンの直接の導入により測定した。陽性のコロニーは直接可視化された陽性のコロニーの周りに明確なハローを生じるデンプンを破壊する能力により測定した。
【0181】
4.PCR増幅によるDNAの単離
陽性コロニーを単離したとき、その一部を爪楊枝により取上げ、滅菌水(30μl)中に希釈し、96ウェルプレート中に入れた。このとき、陽性コロニーは凍結して次の分析に対して保存するか、直ぐに増幅させた。一定分量の細胞(5μl)を、0.5μlのクレンタク(Klentaq)(クローンテック, Palo Alto, CA);4.0μlの10mMのdNTP's(パーキンエルマー-セタス);2.5μlのケンタク(Kentaq)バッファー(クローンテック);0.25μlのフォワードオリゴ1;0.25μlのリバースオリゴ2;12.5μlの蒸留水を含む25μl容量中でPCR反応に対する鋳型として使用した。フォワードオリゴヌクレオチド1の配列は:
TGTAAAACGACGGCCAGTTAAATAGACCTGCAATTATTAATCT (配列番号:5)
であった。リバースオリゴヌクレオチド2の配列は:
CAGGAAACAGCTATGACCACCTGCACACCTGCAAATCCATT(配列番号:6)
であった。
ついで、次のようにしてPCRを実施した:
a. 変性 92℃、5分
b. 3サイクルの変性 92℃、30秒
アニーリング 59℃、30秒
伸展 72℃、60秒
c. 3サイクルの変性 92℃、30秒
アニーリング 57℃、30秒
伸展 72℃、60秒
d. 25サイクルの変性 92℃、30秒
アニーリング 55℃、30秒
伸展 72℃、60秒
e. 保持 4℃
【0182】
オリゴヌクレオチドの下線領域はADHプロモータ領域とアミラーゼ領域にそれぞれアニーリングされ、挿入物が存在しないとき、ベクターpSST-AMY.0からの307塩基対領域を増幅した。典型的にはこれらのオリゴヌクレオチドの5'末端の最初の18ヌクレオチドはシーケンシングプライマーに対するアニーリング部位を含んでいた。従って、空のベクターからのPCR反応の総生成物は343塩基対であった。しかしながら、シグナル配列融合cDNAはかなり長い核酸配列を生じた。
【0183】
PCRに続いて、反応物の一定分量(5μl)を上掲のSambrookらにより記述されたようなトリス-ホウ酸塩-EDTA(TBE)緩衝系を使用する1%アガロース中でのアガロースゲル電気泳動により検査した。400塩基対よりも大きい単一の強いPCR生成物を生じるクローンを、96キアクイック(Qiaquick)PCR洗浄カラム(Qiagen Inc., Chatsworth, CA)での精製後にDNA配列決定により更に分析した。
【0184】
5.全長クローンの同定
上記スクリーニングにおいて分離されたcDNA配列(「DNA21705」)はヒトTNFR1とあるアミノ酸配列類似性又は相同性を有していることが分かった:
TNFR1 81 CRECESG-SFTASENHLRHCLSCSKCRKEMG
* * * .* . *. *. *. *. .
DNA21705 164 CNPCTEGVDYTNASNNEPSCFPCTVCKSD--
QVEISSCTVDRDTVCGCRK (配列番号:7)
* ****. ***** *..
QKHKSSCTMTRDTVCQCKE (配列番号:8)
類似性に基づいて、DNA21705の配列からプローブを産生し、上述の第1節に記載したようにして調製したヒト胎児肺ライブラリ(「LIB25」)をスクリーニングするために使用した。クローニングベクターはpRK5B(pRK5BはSfiI部位を含まないpRK5Dの先駆体である)であり、切断されたcDNAサイズは2800塩基対未満であった。ヌクレオチド位置193−195に明らかな翻訳開始部位を持つ単一のオープンリーディングフレームを含み[Kozakら,上掲]、ヌクレオチド位置970−972に見出されるストップコドンで終端する(図1A;配列番号:2)全長クローンを同定した(DNA33085)(pRK-hApo-2DcR)(以下に示されるように、ATCC209087として寄託されたApo2-DcRとも呼ばれる)。予想されたポリペプチド先駆体は259アミノ酸長であり、およそ27.4kDaの計算分子量を有している。配列解析では、N末端シグナルペプチド、2つのシステインリッチドメイン、4のほぼ同一の15アミノ酸直列反復を含む配列、及び疎水性C末端領域が示された(図2及び図3)。C末端の疎水性配列には一対の小さいアミノ酸(Ala223及びAla224)が先行し;この構造と明白な細胞質ドメインがないことにより、Apo-2DcRがグリコシルホスファチジルイノシトール(GPI)アンカータンパク質でありうることが示唆される[Moran, J. Biol. Chem., 266: 1250- 1257(1991)を参照のこと]。Apo-2DcRは5つの潜在的なN結合グリコシル化部位を含んでいる(図2)。
【0185】
TNFレセプターファミリータンパク質は、典型的には、その細胞外領域に複数の(通常は4つの)システインに富んだドメインが存在していることを特徴としており、各システインリッチドメインは約45アミノ酸長であり、規則的に間隔を開けた約6つのシステイン残基を有している。1型TNFレセプターの結晶構造に基づき、各ドメインにおけるシステインは、典型的には3つのジスルフィド結合を形成し、通常システイン1と2、3と5、及び4と6が互いに対になっている。DR4及びApo-2(更に以下に記載)のように、Apo-2DcRは2つの細胞外のシステインに富んだ擬性反復部を含む(図2)が、これに対し他の同定されている哺乳動物TNFRファミリーのメンバーは3又はそれ以上のこのようなドメインを含む[Smithら, Cell, 76:959(1994)]。
【0186】
図1A(配列番号:1)に示す全長配列のアラインメント解析に基づいて、Apo-2DcRは、Apo-3、TNFR1、又はFas/Apo-1のような他のアポトーシス関連レセプターに対してよりも、DR4(60%)とApo-2(50%)に対してより高い配列同一性を示す。
【0187】
図1Bにおいて、出願人は、明らかな翻訳開始部位がヌクレオチド位置93−95(図1Bにアミノ酸残基−40として特定;配列番号:4)に交互にあてがわれうることを示した。図1Bに示すApo-2DcRはアミノ酸残基−40〜259を含む。
【0188】
実施例2
Apo-2Lに対するApo-2DcRの結合性とApo-2DcR活性に対するPI-PLCの影響
【0189】
Apo-2DcRがApo-2Lに結合するかどうかを試験するため、またApo-2DcRがGPI結合しているかどうかを評価するために、Apo-2DcRトランスフェクト293細胞への放射ヨウ素標識Apo-2Lの結合性を分析した。ホスファチジルイノシトール特異性ホスホリパーゼC(PI-PLC)での細胞の前処理の結合性に対する影響もまた分析した。
【0190】
ヒト293細胞(ATCC CRL 1573)を100mmプレートに蒔き(1x106細胞/プレート)、リン酸カルシウム沈殿法を使用して20μg/プレートのpRK5又は全長Apo-2DcR(実施例1に記載;ATCC寄託209087)をコードしているpRK5でトランスフェクトした。24時間後、細胞をPBS/10mMのEDTA中に収集し、リン酸緩衝食塩水(PBS)中で洗浄し、元のプレート当り2mlのPBS中に再懸濁し、トランスフェクション当り2組の1mlの一定分量に分けた。PI-PLC[Treanorら,Nature,382:80-83(1996)](1μg/ml)を、各トランスフェクションから取り出された2組の一定分量の1組に添加し、細胞を37℃で1時間インキュベートした。細胞を洗浄し、1%のBSA(シグマ)を含む1mlのPBS中に再懸濁させ、0.04mlの一定分量を3組のチューブに入れた。非特異的結合性の測定のために、0.005mlのおよそ20000cpm125I-Apo-2L(Apo-2Lは上掲のPittiらに記載され、通常のラクトペルオキシダーゼ法により放射ヨウ素標識した)を、0.005mlのPBS、又はPBSに入った0.005μlの非標識Apo-2L(最終濃度0.5μg/ml)と共にこれらのチューブに加えた。室温での1時間のインキュベーションの後、細胞を氷冷PBSで洗浄し、ペレット化し、放射能を計数した。
【0191】
Apo-2DcRによるトランスフェクションにより、特異的なApo-2Lの結合量が顕著に増大したが、これは、Apo-2DcRがApo-2Lに結合することを示している(図4)。PI-PLCでの処理により、Apo-2Lの結合が顕著に減少したが、これは、Apo-2DcRがGPIアンカーレセプターであることを示している(図4)。
【0192】
実施例3
全長Apo-2DcRによるApo-2L機能の阻害
【0193】
Apo-2DcRにおいて細胞質領域が無いことは、このレセプターが、Apo-2Lのシグナル伝達の伝達よりもむしろ変調に関与していることを示唆している。従って、Apo-2Lに対する細胞応答性に対するApo-2DcRトランスフェクションの影響を調べた。
【0194】
DR4とApo-2mRNAの両方を発現する(データは示さず)ヒト293細胞を、100mmプレートに蒔き(1x106細胞/プレート)、リン酸カルシウム沈殿法を使用して20μg/プレートのpRK5又はpRK5-hApo-2DcR(実施例2参照)と共にプレート当り3μgのpRKをコードしているグリーン蛍光タンパク質(GFP;クローンテックから購入)でトランスフェクトした。18時間後、細胞をPBS又はApo-2L(Pittiら,上掲,0.5μg/ml)で処理し、(プラスチック上の非固定細胞の明確な視覚化を可能にする)ホフマン光学系を備えた蛍光顕微鏡で6時間にわたって調べた。グリーン蛍光によりGFP陽性細胞を同定し、膜小疱形成(blebbing)及び細胞質凝結のような形態学基準によりアポトーシスを計数した。
【0195】
Apo-2DcRによるトランスフェクションにより、アポトーシス誘発により測定したApo-2Lに対する応答性が顕著に低減させられた(図5)。
【0196】
同様な実験において、pRK5又はpRK5-hApo-2DcR(20μg/プレート)により293細胞をトランスフェクトし、以下の実施例10におけるように、Apo-2L(0.5μg/ml)によるNF-κBの活性化を18時間後に分析した。その結果、Apo-2DcRは、アポトーシス誘発並びにNF-κB活性化により測定したApo-2L機能を阻害することが示された。
【0197】
実施例4
ノーザンブロット分析
【0198】
ヒト組織におけるApo-2DcRmRNAの発現を、ノーザンブロット分析によって検査した。ヒトRNAブロットを、全長Apo-2DcRcDNAをベースにした1.2キロベースの32P標識DNAプローブにハイブリダイズした;プローブはEcoRIでpRK5-Apo-2DcRプラスミドを消化させ、Apo-2DcRcDNA挿入断片を精製することにより産生した。ヒト胎児RNAブロットMTN(クローンテック)、ヒト成人RNAブロットMTN-II(クローンテック)及びヒト癌株化細胞RNAブロット(クローンテック)をDNAプローブと共にインキュベートした。ブロットをハイブリダイゼーション用バッファー(5X SSPE;2X デンハード溶液;100mg/mLの変性剪断されたサケ精子DNA;50%のホルムアミド;2%のSDS)に入ったプローブと共に、42℃で60時間インキュベートした。ブロットを2X SSC;0.05%のSDSを用い、室温で1時間、数回洗浄し、ついで0.1X SSC;0.1%のSDSを用い、50℃で30分間洗浄した。ブロットを一晩さらした後、リン光体イメージャー分析(Fuji)により展開させた。
【0199】
図7Aに示すように、幾つかのApo-2DcRmRNA転写物が検出された。相対的に高い発現が、成人の末梢血白血球(PBL)、脾臓、肺、肝臓及び胎盤において見られた。例えばPBL及び脾臓のようなApo-2DcRを発現する幾つかの成人組織は、Apo-2(以下の実施例11)及びDR4[Panら,上掲]を発現することが示された。
【0200】
図7Bに示されるように、Apo-2DcRメッセージは検査したヒト腫瘍株化細胞(すなわち、HL60前骨髄球性白血病、HeLaS3子宮頚癌、K562慢性骨髄性白血病、MOLT4急性リンパ芽球性白血病、SW480結腸直腸腺癌、A549肺癌、及びG361黒色腫)及び、特にApo-2DcRcDNAにサイズが対応するおよそ1.5kBの転写物の殆どにおいて存在しない。特定された腫瘍細胞タイプ以外の上述の正常なヒト組織におけるApo-2DcRの明らかな発現は、Apo-2DcRレセプターがApo-2リガンドにより癌細胞を(おそらくは癌細胞ではなく正常な細胞を保護することにより)優先的に死滅させることを示唆している。
【0201】
実施例5
ヒトApo-2をコードしているcDNAクローンの単離
【0202】
発現された配列タグ(EST)DNAデータベース(LIFESEQTM, Incyte Pharmaceuticals, Palo Alto, CA)を検索し、Apo-3レセプター[Marstersら, Curr. Biol., 6:750(1996)]の死亡ドメインに対して相同であったあるESTを同定した。ヒト膵臓(「LIB55」)及びヒト腎臓(「LIB28」)のcDNAライブラリ(pRK5ベクター中で上記の実施例1に記載されているようにして調製)を、ESTに基づく合成オリゴヌクレオチドプローブ:
GGGAGCCGCTCATGAGGAAGTTGGGCCTCATGGACAATGAGATAAAGGTGGCTAAAGCTG AGGCAGCGGG (配列番号:9)
でのハイブリダイゼーションによりスクリーニングした。
【0203】
3つのcDNAクローンの全体を配列決定した。cDNAの重なるコード領域はコドン410を除いて同一であった(図8の番号付け系を使用);おそらくは多型性のために、この位置が両方の膵臓cDNAにおけるロイシン残基(TTG)、そして腎臓cDNAにおけるメチオニン残基(ATG)をコード化した。
【0204】
Apo-2の全体核酸配列を図8に示す(配列番号:10)。(以下に示すようにATCC209021として寄託されたpRK5-Apo-2とも呼ばれる)クローン27868は、ヌクレオチド位置140-142に明らかな翻訳開始部位を有する単一のオープンリーディングフレームを含み[Kozakら,上掲]、ヌクレオチド部位1373-1375に見出される終止コドンで終了する(図8;配列番号10)。予想されるポリペプチド先駆体は411アミノ酸長で、I型膜貫通タンパク質であり、約45kDaの算定分子量を有する。ヒドロパシー分析(図示せず)により、シグナル配列(残基1-53)、続いて細胞外ドメイン(残基54-182)、膜貫通ドメイン(残基183-208)、及び細胞内ドメイン(残基209-411)の存在が示唆された(図9;配列番号:11)。293細胞に発現したApo-2-IgGのN末端アミノ酸配列分析により、成熟ポリペプチドがアミノ酸残基54で始まることが示され、これは、実際のシグナル配列が残基1-53を含むことが示している。
【0205】
DR4とApo-2DcRと同様に、Apo-2は2つの細胞外システインリッチ擬似反復を含む一方(図9)、他の同定された哺乳動物TNFRファミリーメンバーは3又はそれより多いそのようなドメインを含む[Smithら,Cell, 76:959(1994)]。
Apo-2の細胞質領域は、TNFR1(30%);CD95(19%);又はApo-3/DR3(29%)の死亡ドメインに対するよりもDR4(64%)の死亡ドメインに対して顕著により高いアミノ酸配列同一性を示す死亡ドメイン(図8に示すアミノ酸残基324−391;図2もまた参照のこと)を含む。TNFR1によるシグナル伝達に対して必要とされる6つの死亡ドメインアミノ酸のうち4つ[Tartagliaら,上掲]がApo-2に保存される一方、他の2つの残基は半保存される(図2参照)。
【0206】
全長配列の(ALIGNコンピュータプログラムを使用する)アラインメント解析に基づき、Apo-2は、例えばTNFR1(19%);CD95(17%);又はApo-3(DR3、WSL-1又はTRAMPとも呼ばれる)(29%)のような他のアポトーシス関連レセプターに対するよりもDR4(55%)に対してより高い配列同一性を示す。
【0207】
実施例6
A. Apo-2 ECDの発現
【0208】
可溶性の細胞外ドメイン(ECD)融合作成物を調製した。Apo-2ECD(図8に示すアミノ酸残基1-184)をPCR法により得て、C末端フラッグエピトープタグ(シグマ)に融合させた。(残基183及び184は膜貫通領域にあることが予想されていても、Apo-2ECD作成物は図8に示す残基183及び184を含んでいた)。ついで、フラッグエピトープタグ分子をpRK5に挿入し、ヒト293細胞(ATCC CRL 1573)に一過性トランスフェクションして発現させた。
【0209】
48時間インキュベートした後、細胞上清を収集し、共沈試験(実施例7を参照)に直接使用するか、製造者(シグマ)の説明に従い、抗フラッグアガロースビーズでのアフィニティークロマトグラフィーによるApo-2ECD-フラッグの精製にかけた。
【0210】
B. イムノアドヘシンとしてのApo-2ECDの発現
可溶性のApo-2ECDイムノアドヘシン作成物を調製した。Apo-2ECD(図8に示すアミノ酸1-184)を、以前に記載されているようにして[Ashkenaziら, Proc. Natl. Acad. Sci., 88:10535-10539(1991)]、pRK5のヒト免疫グロブリンG1重鎖のヒンジ及びFc領域に融合した。イムノアドヘシンは、ヒト293細胞に一過性トランスフェクションして発現させ、前掲のAshkenaziらにより記載されたプロテインAアフィニティークロマトグラフィーにより細胞上清から精製した。
【0211】
実施例7
Apo-2とApo-2リガンド間の結合相互作用を示す免疫沈降アッセイ
【0212】
Apo-2とApo-2Lが相互作用するか又は互いに結合するかを測定するために、擬似トランスフェクト293細胞又はApo-2ECD-フラッグ(上記の実施例6に記載されたもの)がトランスフェクトした293細胞からの上清(5ml)を、5μgのポリ-ヒスチジンタグ可溶性Apo-2L[Pittiら, 上掲]と室温で30分間インキュベートし、ついで共沈アッセイにより複合体形成について分析した。
【0213】
試料を25μlの抗フラッグ抱合アガロースビーズ(シグマ)又はニッケル抱合アガロースビーズ(Qiagen)を使用し、免疫沈降にかけた。4℃で1.5時間インキュベートした後、ビーズを遠心沈殿させ、リン酸緩衝食塩水(PBS)で4回洗浄した。抗フラッグアガロースを使用することにより、Apo-2LをフラッグタグApo-2ECDを通して沈殿させ;ニッケルアガロースを使用することにより、Apo-2ECDをHisタグApo-2Lを通して沈殿させた。沈殿したタンパク質を、SDS-PAGEバッファー中でビーズを5分間沸騰させることで放出させ、12%のポリアクリルアミドゲルでの電気泳動により分離し、ついでMarstersら, J. Biol. Chem.,272:14029-14032(1997)に記載されたようにして、抗Apo-2L又は抗フラッグ抗体(2μg/ml)での免疫ブロットにより検出した。
【0214】
図10に示した結果は、Apo-2ECDとApo-2Lが互いに結合可能であることを示している。
【0215】
さらに、結合相互作用を、抗フラッグビーズ(実施例6を参照)を用い、トランスフェクションした293細胞上清からApo-2ECDを精製し、ついでBIACORETM装置で試料を分析することにより、分析した。BIACORETM分析により、約1nMの解離定数(Kd)が示された。また、BIACORETM分析により、Apo-2ECDが他のアポトーシス誘発性TNFファミリーのメンバー、すなわちTNF-α(ジェネンテク・インク, Pennicaら, Nature, 312:712(1984))、リンホトキシン-α(ジェネンテク・インク), 又はFas/Apo-1リガンド(Alexis Biochemicals)と結合不可能であることが示された。よって、これらのデータは、Apo-2がApo-2Lに対する特異的なレセプターであることを示している。
【0216】
実施例8
Apo-2によるアポトーシスの誘発
【0217】
死亡ドメインはオリゴマー化界面として機能し得るため、死亡ドメインを含むレセプターの過剰発現により、リガンドの不在下におけるシグナル伝達が活性化する[Frazerら,上掲, Nagataら,上掲]。Apo-2が細胞死を誘発可能であるか否かを測定するために、ヒト293細胞又はHeLa細胞(ATCC CCL 2.2)をリン酸カルシウム沈殿法(293細胞)又は電気穿孔法(HeLa細胞)により、Apo-2及び/又はCrmAをコードしているpRK5ベクター又はpRK5系プラスミドで一過性トランスフェクションした。適用できる場合には、プラスミドDNAの全体量を、ベクターDNAを添加することにより調節した。アポトーシスを、形態学的(図11A)、DNA断片化(図11B)、又はMarstersら, Curr.Biol.,6:1669(1996)に記載されているようなホスファチジルセリン暴露のFACS分析(図11C)により、トランスフェクション24時間後に評価した。図11A及び11Bに示すように、Apo-2トランスフェクション293細胞には、顕著なアポトーシスが発生した。
【0218】
FACSにより分析される試料に対して、HeLa細胞にトランスフェクション用のマーカーとしてpRK5-CD4を同時形質移入させ、CD4発現細胞におけるアポトーシスを測定した;FADDをApo-2プラスミドで同時形質移入した;データは、Marstersら, Curr. Biol., 6:1669(1996)に記載されているように、少なくとも3回の実験の平均値±SEMである。カスパーゼ阻害剤、DEVD-fmk(Enzyme Systems)又はz-VAD-fmk(Research Biochemicals Intl.)を、トランスフェクション時に200μMで添加した。図11Cに示すように、カスパーゼ阻害剤CrmA、DEVD-fmk及びz-VAD-fmkが、Apo-2によるアポトーシスの誘発を阻害し、この反応におけるCed-3様プロテアーゼの関与が示された。
【0219】
FADDは、CD95、TNFR1及びApo-3/DR3によるアポトーシスの活性化を媒介するアダプタータンパク質である[Nagataら, 上掲]が、Apo-2L[Marstersら, 上掲]又はDR4[Panら, 上掲]によるアポトーシス誘導には必要であるとは思われない。CD95、TNFR1、又はApo-3/DR3によるアポトーシス誘発を阻害するFADDのドミナントネガティブ変異体型[Frazerら,上掲;Nagataら,上掲;Chinnayianら,上掲]は、HeLa細胞にApo-2を同時形質移入させた場合(図11C)に、Apo-2によるアポトーシス誘発を阻害しなかった。これらの結果より、Apo-2は、FADDとは独立して、アポトーシスのシグナル伝達をしていることが示唆される。この結論と一致して、Apo-2細胞質領域を含むグルタチオン-S-トランスフェラーゼ融合タンパク質は、インビトロで転写され翻訳されたFADDに結合しなかった(データは示さず)。
【0220】
実施例9
可溶性Apo-2ECDによるApo-2L活性の阻害
【0221】
可溶性Apo-2L(0.5μg/ml、Pittiら,上掲に記載されたようにして調製)を、抗フラッグ抗体(シグマ)(1μg/ml)と共に、PBSバッファー又はアフニティー精製されたApo-2ECD(5μg/ml)と室温で1時間、プレインキュベートし、HeLa細胞に添加した。インキュベート5時間後、細胞をFACS(上述)によりアポトーシスについて分析した(図11D)。
【0222】
Apo-2LはHeLa細胞において顕著なアポトーシスを誘発し、可溶性Apo-2ECDはApo-2Lの作用を阻止でき(図11D)、Apo-2LとApo-2との間に特異的な相互作用があることが確認された。同様の結果がApo-2ECDイムノアドヘシンでも得られた(図411E)。用量-反応分析では、約0.3nMのApo-2イムノアドヘシンで最大半減阻害が示されている(図11E)。
【0223】
実施例10
Apo-2によるNF-κBの活性化
【0224】
Apo-2がNF-κBを活性化させるか否かを測定するためのアッセイを行った。
HeLa細胞を、全長天然配列Apo-2、DR4又はApo-3をコードするpRK5発現プラスミドでトランスフェクションし、トランスフェクション24時間後に収集した。核抽出物を調製し、1μgの核タンパク質を、32P標識NF-κB特異性合成オリゴヌクレオチドプローブATCAGGGACTTTCCGCTGGGGACTTTCCG(配列番号:12)[MacKayら, J. Immunol., 153:5274-5284(1994)も参照のこと]と、単独又は50倍過剰の非標識プローブと共に、又は関連のない32P標識合成オリゴヌクレオチドAGGATGGGAAGTGTGTGATATATCCTTGAT(配列番号:13)と反応させた。幾つかの試料に、NF-κBのp65/Re1Aサブユニットに対する抗体(1μg/ml;Santa Cruz Biotechnology)を添加した。DNA結合性を、Hsuら,上掲;Marstersら,上掲、及びMacKayら,上掲に記載されたようにして、電気泳動易動度シフトアッセイにより分析した。
【0225】
結果を図12に示す。図12Aに示すように、電気泳動移動度シフトアッセイにより測定した場合、Apo-2とDR4の両方が、HeLa細胞へのトランスフェクションの際に、有意なNF-κβの活性化を誘発した;活性化レベルはApo-3/DR3で観察される活性化に匹敵するものであった。NF-κBのp65/Re1Aサブユニットに対する抗体は、NF-κBプローブの移動を阻害し、3つ全てのレセプターに対する反応においてp65を結び付けていた。
【0226】
また、Apo-2L自体がNF-κB活性を調節可能であるか否かを測定するアッセイを行った。HeLa細胞又はMCF7細胞(ヒト胸腺癌株化細胞、ATCC HTB 22)を、PBSバッファー、可溶性Apo-2L(Pittiら,上掲)又はTNF-α(ジェネンテク・インク, Pennicaら, Nature, 312:724(1984)を参照)(1μg/ml)で処理し、上述のようにしてNF-κB活性をアッセイした。結果を図12Bに示す。Apo-2Lは、処理されたHeLa細胞においては有意なNF-κB活性化を誘発したが、処理されたMCF7細胞においては誘発しなかった;TNF-αは両方の株化細胞においてより著しい活性化を誘発した。幾つかの研究において、TNFによるNF-κBの活性化により、TNF誘発性アポトーシスから細胞を保護できることが開示されている[Nagataら,上掲]。
【0227】
また、NF-κBインヒビターであるALLN(N-アセチル-ロイシン-ロイシン-ノルロイシン)及び転写インヒビターであるシクロヘキサミドの影響を試験した。HeLa細胞(6-ウェル皿に蒔いた)を、Apo-2L(1μg/ml)の添加前に、PBSバッファー、ALLN(Calbiochem)(40μg/ml)又はシクロヘキサミド(シグマ)(50μg/ml)と共に、1時間プレインキュベートした。インキュベート5時間後、FACSによりアポトーシスについて分析した(図12Cを参照)。
【0228】
結果を図12Cに示す。ALLN及びシクロヘキサミドの両方とも、HeLa細胞におけるApo-2L誘発性アポトーシスのレベルを増加させた。データは、Apo-2Lが保護性NF-κB依存性遺伝子を誘発可能であることを示している。また、データは、Apo-2Lがある種の株化細胞においてNF-κβを活性化することができ、Apo-2とDR4の両方がその機能を媒介し得ることを示している。
【0229】
実施例11
ノーザンブロット分析
【0230】
ヒト組織におけるApo-2mRNAの発現を、ノーザンブロット分析によって検査した。ヒトRNAブロットを、全長Apo-2cDNAをベースにした4.6キロベースの32P標識DNAプローブにハイブリダイズした;プローブはEcoRIでpRK5-Apo-2プラスミドを消化することにより産生した。ヒト胎児RNAブロットMTN(クローンテック)及びヒト成人RNAブロットMTN-II(クローンテック)をDNAプローブと共にインキュベートした。ブロットをハイブリダイゼーション用バッファー(5X SSPE;2Xデンハード溶液;100mg/mLの変性された剪断サケ精子DNA;50%のホルムアミド;2%のSDS)に入ったプローブと共に、42℃で60時間インキュベートした。ブロットを2X SSC;0.05%のSDSを用い、室温で1時間、数回洗浄し、ついで0.1X SSC;0.1%のSDSを用い、50℃で30分間洗浄した。ブロットを一晩さらした後、展開した。
【0231】
図13に示すように、約4.6キロベースの優性mRNAの転写物が複数の組織において検出された。発現は、胎児及び成人の肝臓及び肺、及び成人の卵巣及び末梢血白血球(PBL)において比較的高かったが、胎児及び成人の脳においてはmRNAの発現は検出されなかった。中レベルの発現は、成人の大腸、小腸、精巣、前立腺、胸腺、膵臓、腎臓、骨格筋、胎盤、及び心臓において見出された。Apo-2が発現した幾つかの成人組織、例えばPBL、卵巣及び脾臓は、DR4を発現することが以前に見出されているが[Panら,上掲]、各レセプターmRNAの発現の相対レベルは異なっているようである。
【0232】
実施例12
Apo-2、DR4及びApo-2DcR遺伝子の染色体局在化
【0233】
ヒトApo-2遺伝子の染色体局在化を、放射線ハイブリッド(RH)パネル分析により検査した。RHマッピングを、ヒト-マウス細胞の放射線ハイブリッドパネル(Research Genetics)とApo-2cDNAのコード領域に基づくプライマーを使用するPCRにより、実施した[Gelbら,Hum. Genet., 98:141(1996)]。スタンフォードヒトゲノムセンターデータベースを使用したPCRデータの解析では、Apo-2は11.05LODを有するマーカーD8S481に結合しており;D8S481はD8S2055と今度は結合しており、これがヒト染色体8p21に位置する。DR4の類似した分析においては、DR4が、ヒト染色体8p21にまた位置するマーカーD8S2127(13.00のLODを持つ)に結合していることが示された。Apo-2DcRの放射線ハイブリッドパネルを使用した解析では、Apo-2DcR遺伝子はマーカーWI-6536に結合し、これがD8S298についで結合しており、これがまたヒト染色体8p21に位置し、D8S2005とD8S2127の間に巣を形成している。従って、3つのApo-2Lレセプター、Apo-2、Apo-2DcR及びDR4に対するヒト遺伝子は、全て染色体8p21に位置している。
【0234】
本出願人の知る限りでは、今日まで、TNFR遺伝子ファミリーのメンバーで染色体8pに位置しているものはなかった。
【0235】
実施例13
Apo-2DcRに特異的なモノクローナル抗体の調製
【0236】
0.5μg/50μlのApo-2DcRイムノアドヘシンタンパク質(Ribi Immunochemical Research Inc., Hamilton, MTから購入したMPL-TDMアジュバントで希釈)を、各々の後足の裏の柔らかい部分に、3日の間隔で11回注射することで、Balb/cマウス(チャールズリヴァー研究所から得たもの)を免疫した。過去に開示されたように[Ashkenaziら、Proc. Natl. Acad. Sci., 88:10535-10539(1991)]、pRK5においてヒト免疫グロブリンG1重鎖のヒンジ及びFc領域に、Apo-2DcRのN末端領域を融合させることにより、Apo-2DcRイムノアドヘシンタンパク質を産生した。イムノアドヘシンタンパク質を、ヒト293細胞に一過性トランスフェクションして発現させ、前掲のAshkenaziらにより記載されたようにして、プロテインAアフィニティークロマトグラフィーにより細胞上清から精製した。
【0237】
最終の追加免疫から3日後に、膝窩リンパ節をマウスから取り除き、単細胞懸濁液を、1%のペニシリン-ストレプトマイシンが補填されたDMEM培地(Biowhitakker Corp.から得たもの)中で調製した。ついで、リンパ節細胞を35%のポリエチレングリコールを使用し、マウス骨髄腫細胞P3X63AgU.1(ATCC CRL 1597)と融合させ、96-ウェル培養プレートで培養した。融合の結果得られたハイブリドーマをHAT培地において選択した。融合から10日後に、ハイブリドーマ培養の上清をELISAでスクリーニングし、Apo-2DcRイムノアドヘシンタンパク質に結合するモノクローナル抗体の有無を試験した。
【0238】
ELISAにおいて、各々のウェルに、PBSに2μg/mlのヤギ抗ヒトIgG Fc(Cappel Laboratories社から購入)が入ったものを50μl添加して、96-ウェルマイクロタイタープレート(Maxisorb;Nunc, Kamstrup, Denmark)をコーティングし、4℃で一晩インキュベートした。ついでプレートを洗浄バッファー(0.05%のトゥイーン20を含有するPBS)で3回洗浄した。ついで、マイクロタイタープレートのウェルを、PBSに2.0%のウシ血清アルブミンが入ったもの200μlでブロックし、室温で1時間インキュベートした。ついでプレートを、洗浄バッファーで、再度3回洗浄した。
【0239】
洗浄工程後、アッセイ用バッファー(0.5%のBSAを含むPBS)に0.4μg/mlのApo-2DcRイムノアドヘシンタンパク質(上述のもの)が入ったもの50μlを各々のウェルに添加した。プレートを振盪装置上で、室温で1時間インキュベートし、続いて洗浄バッファーで3回洗浄した。
【0240】
洗浄工程に続き、100μlのハイブリドーマ上清又は精製した抗体(プロテインG-セファロースカラムを使用)(1μg/ml)を、アッセイ用バッファーの指定ウェルに添加した。100μlのP3X63AgU.1骨髄腫細胞の条件培地を、対照として他の指定ウェルに添加した。プレートを振盪装置上で、室温で1時間インキュベートし、続いて洗浄バッファーで3回洗浄した。
【0241】
次に、アッセイ用バッファーで1:1000に希釈されたHRP-抱合ヤギ抗マウスIgG Fc(Cappel Laboratories社から購入)を50μl、各々のウェルに添加し、プレートを振盪装置上で、室温で1時間インキュベートした。プレートを洗浄バッファーで3回洗浄し、ついで各々のウェルに50μlの基質(TMBマイクロウェル・ペルオキシダーゼ基質、Kirkegaard & Perry, Gaithersburg, MD)を添加し、室温で10分間インキュベートした。50μlのTMB1-成分終止溶液(ジエチレングリコール, Kirkegaard & Perry)を各々のウェルに添加して反応を終了させ、450nmでの吸光度を自動マイクロタイタープレート読取装置で読取った。
【0242】
ELISAによりスクリーニングされたハイブリドーマ上清の内、47の上清が陽性であると判断された(バックグラウンドのおよそ4倍高いと算出)。ELISAで陽性であると判断された上清を、さらにHUMEC細胞(Apo-2DcRを発現するヒト微小血管内皮株化細胞;Cell Syatems, Kirkland, WA)及びPE-抱合ヤギ抗マウスIgGを使用してFACS分析により分析した。この分析のため、細胞選別用バッファー(1%のFCSと0.02%のNaN3を含有するPBS)に懸濁させた25μlの細胞(4×106細胞/ml)をU字底のマイクロタイターウェルに添加し、100μlの培養上清又は精製した抗体(プロテインG-セファロースカラムで精製)(10μg/ml)が細胞選別用バッファーに入ったものと混合し、氷上で30分間インキュベートした。ついで、細胞を洗浄し、100μlのPE-抱合ヤギ抗マウスIgGと共に4℃で30分間インキュベートした。ついで細胞を2回洗浄し、200μlの細胞選別用バッファーに再懸濁させ、FACScan(Becton Dickinson, Mountain View, CA)により分析した。FACS分析により、12/35の上清が抗Apo-2抗体に対して陽性であることが示された。
【0243】
図14は、4G3.9.9;6D10.9.7;及び1C5.24.1と呼ばれるApo-2DcR抗体とインキュベートしたHUMEC細胞のFACS染色を示す。図14に示すように、各抗体は、HUMEC細胞において発現されるApo-2DcRレセプターを認識する。
【0244】
実施例14
他のApo-2リガンドレセプターへのApo-2DcR抗体の結合性をテストするためのELISAアッセイ
【0245】
実施例13に記載したモノクローナル抗体が、Apo-2DcR以外の他の既知のApo-2Lレセプターに結合することができたか否かを測定するためにELISAを実施した。特に、4G3.9.9;6D10.9.7;及び1C5.24.1抗体について、それぞれ、DR4[Panら,上掲]、Apo-2[上記の実施例に記載]、及びDcR2[Marstersら, Curr. Biol., 7:1003-1006(1997)]に対する結合性を調べた。ELISAは、本質的に上述の実施例13に記載したようにして実施した。
【0246】
結果を図15に示す。Apo-2DcR抗体4G3.9.9はApo-2DcRに結合した。また4G3.9.9抗体は、DR4とApo-2に対してはいくらかの交差反応性とDcR2に対してある限られた交差反応性を示した。6D10.9.7抗体はApo-2DcRに結合し、DR4、Apo-2及びDcR2に対してある限られた交差反応性を示した。最後に、1C5.24.1抗体はApo-2DcRに結合し、DR4に対して幾らかの交差反応性を示した。1C5.24.1抗体はApo-2とDcR2に対して些かより少ない交差反応性を示した。交差反応性のまとめをまた図16に示す。
【0247】
実施例15
抗体のアイソタイピング
【0248】
Apo-2DcR抗体(実施例13と14において上述したもの)のアイソタイプを、アイソタイプ特異的ヤギ抗マウスIg(Fisher Biotech, Pittsburgh, PA)で、マイクロタイタープレートを4℃で一晩かけてコーティングすることにより決定した。ついでプレートを洗浄バッファー(上述の実施例13に記載したもの)で洗浄した。ついで、マイクロタイタープレートのウェルを200μlの2%ウシ血清アルブミン(BSA)でブロックし、室温で1時間インキュベートした。プレートを、再度洗浄バッファーで3回洗浄した。次に、100μlのハイブリドーマ培養上清又は5μg/mlの精製抗体を指定ウェルに加えた。プレートを室温で30分インキュベートし、ついで50μlのHRP-抱合ヤギ抗マウスIgG(実施例13において上述したもの)を各々のウェルに添加した。プレートを室温で30分インキュベートした。プレートに結合したHRPのレベルを、上述のHRP基質を使用して検出した。
【0249】
アイソタイピング分析により、4G3.9.9及び1C5.24.1抗体はIgG1抗体であることが示された。またこの分析により、6D10.9.7抗体がIgG2b抗体であることが示された。これらの結果を図16にまた示す。
【0250】
材料の寄託
次の材料をアメリカン・タイプ・カルチャー・コレクション,10801 University Blvd., Manassas, Virginia, USA(ATCC)に寄託した:
材料 ATCC寄託番号 寄託日
pRK-Apo-2 209021 1997年5月8日
Apo2-DcR 209087 1997年5月30日
4G3.9.9 ___ _____________
6D10.9.7 ______ _____________
1C5.24.1 ______ _____________
【0251】
この寄託は、特許手続き上の微生物の寄託の国際的承認に関するブダペスト条約及びその規則(ブダペスト条約)の規定に従って行われた。これは、寄託の日付から30年間、寄託の生存可能な培養が維持されることを保証するものである。寄託物はブダペスト条約の条項に従い、またジェネンテク社とATCCとの間の合意に従い、ATCCから入手することができ、これは、どれが最初であろうとも、関連した米国特許の発行時又は任意の米国又は外国特許出願の公開時に、寄託培養物の後代を永久かつ非制限的に入手可能とすることを保証し、米国特許法第122条及びそれに従う特許庁長官規則(特に参照番号886OG638の37CFR第1.14条を含む)に従って権利を有すると米国特許庁長官が決定した者に後代を入手可能とすることを保証するものである。
【0252】
本出願の譲受人は、寄託した培養物が、適切な条件下で培養されていた場合に死亡もしくは損失又は破壊されたならば、材料は通知時に同一の他のもの即座に取り替えることに同意する。寄託物質の入手可能性は、特許法に従いあらゆる政府の権限下で認められた権利に違反して、本発明を実施するライセンスであるとみなされるものではない。
【0253】
上記の文書による明細書は、当業者に本発明を実施できるようにするために十分であると考えられる。寄託した態様は、本発明のある側面の一つの説明として意図されており、機能的に等価なあらゆる作成物がこの発明の範囲内にあるため、寄託された作成物により、本発明の範囲が限定されるものではない。ここでの物質の寄託は、文書による説明が、そのベストモードを含む、本発明の任意の側面の実施を可能にするために不十分であることを認めるものではないし、それが表す特定の例証に対して請求の範囲を制限するものと解釈されるものでもない。実際、ここに示し記載したものに加えて、本発明を様々に改変することは、前記の記載から当業者にとっては明らかなものであり、添付の請求の範囲内に入るものである。
【図面の簡単な説明】
【0254】
【図1A】図1Aは、天然配列ヒトApo-2DcRcDNAの核酸配列及びその誘導アミノ酸配列(開始部位は残基1にあてがわれる(ヌクレオチド193−195))を示す。
【図1B】図1Bは、天然配列ヒトApo-2DcRcDNAの核酸配列及びその誘導アミノ酸配列(開始部位は残基−40にあてがわれる(ヌクレオチド73−75))を示す。
【図2】図2は、Apo-2及びApo-2DcRの一次構造とmRNA発現を示す。この図は、全長DR4と整列化させたヒトApo-2及びApo-2DcRの推論アミノ酸配列を示す。Apo-2の死亡ドメインはDR4、Apo-3/DR3、TNFR1、及びCD95のものと整列化され;星印はTNFR1による死亡シグナル伝達に必須である残基を示す[Tartagliaら,上掲]。示されているのは、予想されたシグナルペプチド開裂部位(矢印)、2つのシステインリッチドメイン(CRD1、2)及びApo-2及びDR4の膜貫通ドメイン又はApo-2DcRの疎水性C末端(下線)である。また示されているのは、5つの潜在的なN結合グリコシル化部位(黒い箱)及びApo-2DcRの5つの配列擬似反復(括弧)である。
【図3】図3は、Apo-2とApo-2DcRのヒドロパシープロットを示す。上部の数はアミノ酸の位置を示している。
【図4】図4は、Apo-2DcRトランスフェクト細胞に対する放射ヨウ素標識Apo-2Lの結合と、PI-PLCでの細胞の前処理によるその阻害を示す。
【図5】図5はApo-2DcRによるアポトーシスのApo-2L誘発の阻害を示す。
【図6】図6はApo-2DcRによるNF-κBのApo-2L活性化の阻害を示す。
【図7】図7Aは、ヒト組織のポリA RNAブロットのノーザンハイブリダイゼーションにより分析されたヒト組織中でのApo-2DcRmRNAの発現を示す。図7Bは、ヒト癌株化細胞のポリA RNAブロットのノーザンハイブリダイゼーションにより分析されたヒト癌株化細胞中でのApo-2DcRmRNAの発現(の欠如)を示す。
【図8】図8は、天然配列ヒトApo-2cDNAの核酸配列とその誘導アミノ酸配列を示す。
【図9】図9は、天然配列ヒトApo-2の誘導アミノ酸配列を示し、推定シグナル配列には下線を引き、推定膜貫通ドメインは囲み、推定死亡ドメイン配列には破線の下線を付している。2つのシステインリッチドメインのシステインには個々に下線を引いている。
【図10】図10は、Apo-2LとのApo-2ECDの相互作用を示す。フラッグエピトープタグApo-2ECDでトランスフェクトされた293細胞又は擬似トランスフェクトされた293細胞の上清をポリ-His-タグApo-2Lと共にインキュベートし、抗フラッグ抱合又はニッケル抱合アガロースビーズで免疫沈降させた。沈殿したタンパク質をポリアクリルアミドゲルによる電気泳動法で分離し、抗Apo-2L又は抗フラッグ抗体を用いた免疫ブロットにより検出した。
【図11A】図11は、Apo-2によるアポトーシスの誘発と、可溶性Apo-2ECDによるApo-2L活性の阻害を示す。ヒト293細胞(A、B)又はHeLa細胞(C)を、Apo-2及び/又はCrmAをコードしたpRK5ベクター又はpRK5ベースプラスミドによりトランスフェクトした。アポトーシスを、形態学的(A)、DNA断片化(B)、又はFACS(C-E)により評価した。可溶性Apo-2Lを、バッファーあるいは抗フラッグ抗体又はApo-2DcRECDイムノアドヘシン又はDR4又はTNFR1イムノアドヘシンと共にアフィニティー精製されたApo-2ECDでプレインキュベートし、HeLa細胞に添加した。後で細胞のアポトーシスを分析した(D)。また、Apo-2ECDイムノアドヘシンを備えたApo-2Lを使用して用量反応分析を行った(E)。
【図11B】図11は、Apo-2によるアポトーシスの誘発と、可溶性Apo-2ECDによるApo-2L活性の阻害を示す。ヒト293細胞(A、B)又はHeLa細胞(C)を、Apo-2及び/又はCrmAをコードしたpRK5ベクター又はpRK5ベースプラスミドによりトランスフェクトした。アポトーシスを、形態学的(A)、DNA断片化(B)、又はFACS(C-E)により評価した。可溶性Apo-2Lを、バッファーあるいは抗フラッグ抗体又はApo-2DcRECDイムノアドヘシン又はDR4又はTNFR1イムノアドヘシンと共にアフィニティー精製されたApo-2ECDでプレインキュベートし、HeLa細胞に添加した。後で細胞のアポトーシスを分析した(D)。また、Apo-2ECDイムノアドヘシンを備えたApo-2Lを使用して用量反応分析を行った(E)。
【図11C】図11は、Apo-2によるアポトーシスの誘発と、可溶性Apo-2ECDによるApo-2L活性の阻害を示す。ヒト293細胞(A、B)又はHeLa細胞(C)を、Apo-2及び/又はCrmAをコードしたpRK5ベクター又はpRK5ベースプラスミドによりトランスフェクトした。アポトーシスを、形態学的(A)、DNA断片化(B)、又はFACS(C-E)により評価した。可溶性Apo-2Lを、バッファーあるいは抗フラッグ抗体又はApo-2DcRECDイムノアドヘシン又はDR4又はTNFR1イムノアドヘシンと共にアフィニティー精製されたApo-2ECDでプレインキュベートし、HeLa細胞に添加した。後で細胞のアポトーシスを分析した(D)。また、Apo-2ECDイムノアドヘシンを備えたApo-2Lを使用して用量反応分析を行った(E)。
【図11D】図11は、Apo-2によるアポトーシスの誘発と、可溶性Apo-2ECDによるApo-2L活性の阻害を示す。ヒト293細胞(A、B)又はHeLa細胞(C)を、Apo-2及び/又はCrmAをコードしたpRK5ベクター又はpRK5ベースプラスミドによりトランスフェクトした。アポトーシスを、形態学的(A)、DNA断片化(B)、又はFACS(C-E)により評価した。可溶性Apo-2Lを、バッファーあるいは抗フラッグ抗体又はApo-2DcRECDイムノアドヘシン又はDR4又はTNFR1イムノアドヘシンと共にアフィニティー精製されたApo-2ECDでプレインキュベートし、HeLa細胞に添加した。後で細胞のアポトーシスを分析した(D)。また、Apo-2ECDイムノアドヘシンを備えたApo-2Lを使用して用量反応分析を行った(E)。
【図11E】図11は、Apo-2によるアポトーシスの誘発と、可溶性Apo-2ECDによるApo-2L活性の阻害を示す。ヒト293細胞(A、B)又はHeLa細胞(C)を、Apo-2及び/又はCrmAをコードしたpRK5ベクター又はpRK5ベースプラスミドによりトランスフェクトした。アポトーシスを、形態学的(A)、DNA断片化(B)、又はFACS(C-E)により評価した。可溶性Apo-2Lを、バッファーあるいは抗フラッグ抗体又はApo-2DcRECDイムノアドヘシン又はDR4又はTNFR1イムノアドヘシンと共にアフィニティー精製されたApo-2ECDでプレインキュベートし、HeLa細胞に添加した。後で細胞のアポトーシスを分析した(D)。また、Apo-2ECDイムノアドヘシンを備えたApo-2Lを使用して用量反応分析を行った(E)。
【図12A】図12は、Apo-2、DR4及びApo-2LによるNF-κBの活性化を示す。(A)HeLa細胞を、示したタンパク質をコードする発現プラスミドでトランスフェクトした。核抽出物を調製し、電気泳動易動度シフトアッセイにより分析した。
【図12B】図12は、Apo-2、DR4及びApo-2LによるNF-κBの活性化を示す。(B)HeLa細胞又はMCF7細胞をバッファー、Apo-2L又はTNF-αで処理し、NF-κB活性をアッセイした。
【図12C】図12は、Apo-2、DR4及びApo-2LによるNF-κBの活性化を示す。(C)HeLa細胞をApo-2Lの添加前に、バッファー、ALLN又はシクロヘキサミドと共にプレインキュベートした。後にFACSによりアポトーシスを分析した。
【図13】図13は、ヒト組織のポリA RNAブロットのノーザンハイブリッド形成により分析した、ヒト組織におけるApo-2mRNAの発現を示す。
【図14A】図14は、IgG対照(点線)と比較したApo-2DcR抗体(太線で示す)のFASC分析を示す。抗体(1C5.24.1)は、HUMEC細胞において発現されたApo-2DcRレセプターを認識した。
【図14B】図14は、IgG対照(点線)と比較したApo-2DcR抗体(太線で示す)のFASC分析を示す。抗体(4G3.9.9)は、HUMEC細胞において発現されたApo-2DcRレセプターを認識した。
【図14C】図14は、IgG対照(点線)と比較したApo-2DcR抗体(太線で示す)のFASC分析を示す。抗体(6D10.9.7)は、HUMEC細胞において発現されたApo-2DcRレセプターを認識した。
【図15A】図15は、Apo-2DcR、及びDR4、Apo-2及びDcR2と称される他の既知のApo-2Lレセプターに対するApo-2DcR抗体1C5.24.の結合性をテストする酵素結合免疫吸着検定(ELISA)の結果を示すグラフを含む。
【図15B】図15は、Apo-2DcR、及びDR4、Apo-2及びDcR2と称される他の既知のApo-2Lレセプターに対するApo-2DcR抗体4G3.9.9の結合性をテストする酵素結合免疫吸着検定(ELISA)の結果を示すグラフを含む。
【図15C】図15は、Apo-2DcR、及びDR4、Apo-2及びDcR2と称される他の既知のApo-2Lレセプターに対するApo-2DcR抗体6D10.9.7の結合性をテストする酵素結合免疫吸着検定(ELISA)の結果を示すグラフを含む。
【図16】図16は、抗体1C5.24.1;4G3.9.9;及び6D10.9.7の交差反応性及びアイソタイプのまとめを提供する表である。
【特許請求の範囲】
【請求項1】
図1A(配列番号:1)のアミノ酸残基1〜259を含んでなる天然配列Apo-2DcRポリペプチドと、少なくとも約80%のアミノ酸配列同一性を有する単離Apo-2DcRポリペプチド。
【請求項2】
前記Apo-2DcRポリペプチドが少なくとも約90%のアミノ酸配列同一性を有する請求項1に記載のApo-2DcRポリペプチド。
【請求項3】
前記Apo-2DcRポリペプチドが少なくとも約95%のアミノ酸配列同一性を有する請求項2に記載のApo-2DcRポリペプチド。
【請求項4】
図1A(配列番号:1)のアミノ酸残基1〜259を含んでなる単離された天然配列Apo-2DcRポリペプチド。
【請求項5】
図1A(配列番号:1)のアミノ酸残基1〜161を含んでなるApo-2DcRポリペプチドの単離された細胞外ドメイン配列。
【請求項6】
図1A(配列番号:1)のアミノ酸残基1〜165を含んでなる請求項5に記載の細胞外ドメイン配列。
【請求項7】
図1A(配列番号:1)のアミノ酸残基1〜236を含んでなる請求項5に記載の細胞外ドメイン配列。
【請求項8】
Xが図1A(配列番号:1)のアミノ酸残基161〜236の何れか一つであるアミノ酸残基1〜Xを含んでなるApo-2DcRポリペプチドの単離された細胞外ドメイン配列。
【請求項9】
図1B(配列番号:3)のアミノ酸残基−40〜259を含んでなる単離された天然配列Apo-2DcRポリペプチド。
【請求項10】
異種性アミノ酸配列に融合した請求項5に記載の細胞外ドメイン配列又は請求項1に記載のApo-2DcRポリペプチドを含んでなるキメラ分子。
【請求項11】
前記異種性アミノ酸配列がエピトープタグ配列である請求項10に記載のキメラ分子。
【請求項12】
前記異種性アミノ酸配列が免疫グロブリン配列である請求項10に記載のキメラ分子。
【請求項13】
前記免疫グロブリン配列がIgGである請求項12に記載のキメラ分子。
【請求項14】
前記細胞外ドメイン配列が図1A(配列番号:1)のアミノ酸残基1〜165を含んでなる請求項12に記載のキメラ分子。
【請求項15】
請求項1に記載のApo-2DcRポリペプチド又は請求項5に記載の細胞外ドメイン配列に結合する抗体。
【請求項16】
前記抗体がモノクローナル抗体である請求項15に記載の抗体。
【請求項17】
阻止抗体を含んでなる請求項15に記載の抗体。
【請求項18】
Apo-2DcRポリペプチドへの結合に加えて、他のApo-2リガンドレセプターに結合する抗体を含んでなる請求項15に記載の抗体。
【請求項19】
キメラ抗体を含んでなる請求項15に記載の抗体。
【請求項20】
ヒト抗体を含んでなる請求項15に記載の抗体。
【請求項21】
IgG抗体を含んでなる請求項15に記載の抗体。
【請求項22】
ATCC寄託番号___として寄託されたハイブリドーマ株化細胞により産生された4G3.9.9モノクローナル抗体の生物学的性質を有する請求項16に記載の抗体。
【請求項23】
ATCC寄託番号___として寄託されたハイブリドーマ株化細胞により産生された6D10.9.7モノクローナル抗体の生物学的性質を有する請求項16に記載の抗体。
【請求項24】
ATCC寄託番号___として寄託されたハイブリドーマ株化細胞により産生された1C5.24.1モノクローナル抗体の生物学的性質を有する請求項16に記載の抗体。
【請求項25】
ATCC寄託番号___として寄託されたハイブリドーマ株化細胞により産生された4G3.9.9モノクローナル抗体が結合するエピトープと同じエピトープに抗体が結合する請求項16に記載の抗体。
【請求項26】
ATCC寄託番号___として寄託されたハイブリドーマ株化細胞により産生された6D10.9.7モノクローナル抗体が結合するエピトープと同じエピトープに抗体が結合する請求項16に記載の抗体。
【請求項27】
ATCC寄託番号___として寄託されたハイブリドーマ株化細胞により産生された1C5.24.1モノクローナル抗体が結合するエピトープと同じエピトープに抗体が結合する請求項16に記載の抗体。
【請求項28】
請求項16に記載の抗体を産生するハイブリドーマ株化細胞。
【請求項29】
ATCC寄託番号___として寄託されたハイブリドーマ株化細胞。
【請求項30】
ATCC寄託番号___として寄託されたハイブリドーマ株化細胞。
【請求項31】
ATCC寄託番号___として寄託されたハイブリドーマ株化細胞。
【請求項32】
ATCC寄託番号___として寄託されたハイブリドーマ株化細胞により産生された4G3.9.9モノクローナル抗体。
【請求項33】
ATCC寄託番号___として寄託されたハイブリドーマ株化細胞により産生された6D10.9.7モノクローナル抗体。
【請求項34】
ATCC寄託番号___として寄託されたハイブリドーマ株化細胞により産生された1C5.24.1モノクローナル抗体。
【請求項35】
請求項5に記載の細胞外ドメイン配列又は請求項1に記載のApo-2DcRポリペプチドをコード化している核酸配列を含んでなる単離された核酸。
【請求項36】
前記核酸配列が図1A(配列番号:1)のアミノ酸残基を含んでなる天然配列Apo-2DcRポリペプチドをコード化する請求項35に記載の核酸。
【請求項37】
前記核酸配列が図1A(配列番号:2)のヌクレオチド193〜969を含んでなる請求項36に記載の核酸。
【請求項38】
請求項35に記載の核酸を含んでなるベクター。
【請求項39】
ベクターで形質転換した宿主細胞により認識される対照配列に作用可能に結合した請求項38に記載のベクター。
【請求項40】
請求項38に記載のベクターを含んでなる宿主細胞。
【請求項41】
CHO細胞を含んでなる請求項40に記載の宿主細胞。
【請求項42】
酵母細胞を含んでなる請求項40に記載の宿主細胞。
【請求項43】
大腸菌を含んでなる請求項40に記載の宿主細胞。
【請求項44】
Apo-2DcRポリペプチドをコードしている核酸分子を使用してApo-2DcRポリペプチドの生産を行う方法において、請求項40に記載の宿主細胞を培養することを含んでなる方法。
【請求項45】
Apo-2DcRポリペプチドをコードする核酸を発現する細胞を含む非ヒト、トランスジェニック動物。
【請求項46】
マウス又はラットである請求項45に記載の動物。
【請求項47】
Apo-2DcRポリペプチドをコードする変更遺伝子を有する細胞を含む、非ヒト、ノックアウト動物。
【請求項48】
マウス又はラットである請求項47に記載の動物。
【請求項49】
請求項1又は5に記載のApo-2DcRと担体を含んでなる組成物。
【請求項50】
請求項15に記載のApo-2DcR抗体と担体を含んでなる組成物。
【請求項51】
容器と該容器に入れられる組成物を含んでなり、組成物がApo-2DcRポリペプチド又はApo-2DcR抗体を含む製造品。
【請求項52】
インビボ又はエキソビボにおいてApo-2DcRポリペプチド又はApo-2DcR抗体を使用するための使用説明書をさらに含んでなる請求項51に記載の製造品。
【請求項53】
哺乳動物細胞におけるアポトーシスを変調する方法において、Apo-2DcRポリペプチドに前記細胞を暴露させることを含んでなる方法。
【請求項54】
前記細胞が更にApo-2リガンドに暴露される請求項53に記載の方法。
【請求項1】
図1A(配列番号:1)のアミノ酸残基1〜259を含んでなる天然配列Apo-2DcRポリペプチドと、少なくとも約80%のアミノ酸配列同一性を有する単離Apo-2DcRポリペプチド。
【請求項2】
前記Apo-2DcRポリペプチドが少なくとも約90%のアミノ酸配列同一性を有する請求項1に記載のApo-2DcRポリペプチド。
【請求項3】
前記Apo-2DcRポリペプチドが少なくとも約95%のアミノ酸配列同一性を有する請求項2に記載のApo-2DcRポリペプチド。
【請求項4】
図1A(配列番号:1)のアミノ酸残基1〜259を含んでなる単離された天然配列Apo-2DcRポリペプチド。
【請求項5】
図1A(配列番号:1)のアミノ酸残基1〜161を含んでなるApo-2DcRポリペプチドの単離された細胞外ドメイン配列。
【請求項6】
図1A(配列番号:1)のアミノ酸残基1〜165を含んでなる請求項5に記載の細胞外ドメイン配列。
【請求項7】
図1A(配列番号:1)のアミノ酸残基1〜236を含んでなる請求項5に記載の細胞外ドメイン配列。
【請求項8】
Xが図1A(配列番号:1)のアミノ酸残基161〜236の何れか一つであるアミノ酸残基1〜Xを含んでなるApo-2DcRポリペプチドの単離された細胞外ドメイン配列。
【請求項9】
図1B(配列番号:3)のアミノ酸残基−40〜259を含んでなる単離された天然配列Apo-2DcRポリペプチド。
【請求項10】
異種性アミノ酸配列に融合した請求項5に記載の細胞外ドメイン配列又は請求項1に記載のApo-2DcRポリペプチドを含んでなるキメラ分子。
【請求項11】
前記異種性アミノ酸配列がエピトープタグ配列である請求項10に記載のキメラ分子。
【請求項12】
前記異種性アミノ酸配列が免疫グロブリン配列である請求項10に記載のキメラ分子。
【請求項13】
前記免疫グロブリン配列がIgGである請求項12に記載のキメラ分子。
【請求項14】
前記細胞外ドメイン配列が図1A(配列番号:1)のアミノ酸残基1〜165を含んでなる請求項12に記載のキメラ分子。
【請求項15】
請求項1に記載のApo-2DcRポリペプチド又は請求項5に記載の細胞外ドメイン配列に結合する抗体。
【請求項16】
前記抗体がモノクローナル抗体である請求項15に記載の抗体。
【請求項17】
阻止抗体を含んでなる請求項15に記載の抗体。
【請求項18】
Apo-2DcRポリペプチドへの結合に加えて、他のApo-2リガンドレセプターに結合する抗体を含んでなる請求項15に記載の抗体。
【請求項19】
キメラ抗体を含んでなる請求項15に記載の抗体。
【請求項20】
ヒト抗体を含んでなる請求項15に記載の抗体。
【請求項21】
IgG抗体を含んでなる請求項15に記載の抗体。
【請求項22】
ATCC寄託番号___として寄託されたハイブリドーマ株化細胞により産生された4G3.9.9モノクローナル抗体の生物学的性質を有する請求項16に記載の抗体。
【請求項23】
ATCC寄託番号___として寄託されたハイブリドーマ株化細胞により産生された6D10.9.7モノクローナル抗体の生物学的性質を有する請求項16に記載の抗体。
【請求項24】
ATCC寄託番号___として寄託されたハイブリドーマ株化細胞により産生された1C5.24.1モノクローナル抗体の生物学的性質を有する請求項16に記載の抗体。
【請求項25】
ATCC寄託番号___として寄託されたハイブリドーマ株化細胞により産生された4G3.9.9モノクローナル抗体が結合するエピトープと同じエピトープに抗体が結合する請求項16に記載の抗体。
【請求項26】
ATCC寄託番号___として寄託されたハイブリドーマ株化細胞により産生された6D10.9.7モノクローナル抗体が結合するエピトープと同じエピトープに抗体が結合する請求項16に記載の抗体。
【請求項27】
ATCC寄託番号___として寄託されたハイブリドーマ株化細胞により産生された1C5.24.1モノクローナル抗体が結合するエピトープと同じエピトープに抗体が結合する請求項16に記載の抗体。
【請求項28】
請求項16に記載の抗体を産生するハイブリドーマ株化細胞。
【請求項29】
ATCC寄託番号___として寄託されたハイブリドーマ株化細胞。
【請求項30】
ATCC寄託番号___として寄託されたハイブリドーマ株化細胞。
【請求項31】
ATCC寄託番号___として寄託されたハイブリドーマ株化細胞。
【請求項32】
ATCC寄託番号___として寄託されたハイブリドーマ株化細胞により産生された4G3.9.9モノクローナル抗体。
【請求項33】
ATCC寄託番号___として寄託されたハイブリドーマ株化細胞により産生された6D10.9.7モノクローナル抗体。
【請求項34】
ATCC寄託番号___として寄託されたハイブリドーマ株化細胞により産生された1C5.24.1モノクローナル抗体。
【請求項35】
請求項5に記載の細胞外ドメイン配列又は請求項1に記載のApo-2DcRポリペプチドをコード化している核酸配列を含んでなる単離された核酸。
【請求項36】
前記核酸配列が図1A(配列番号:1)のアミノ酸残基を含んでなる天然配列Apo-2DcRポリペプチドをコード化する請求項35に記載の核酸。
【請求項37】
前記核酸配列が図1A(配列番号:2)のヌクレオチド193〜969を含んでなる請求項36に記載の核酸。
【請求項38】
請求項35に記載の核酸を含んでなるベクター。
【請求項39】
ベクターで形質転換した宿主細胞により認識される対照配列に作用可能に結合した請求項38に記載のベクター。
【請求項40】
請求項38に記載のベクターを含んでなる宿主細胞。
【請求項41】
CHO細胞を含んでなる請求項40に記載の宿主細胞。
【請求項42】
酵母細胞を含んでなる請求項40に記載の宿主細胞。
【請求項43】
大腸菌を含んでなる請求項40に記載の宿主細胞。
【請求項44】
Apo-2DcRポリペプチドをコードしている核酸分子を使用してApo-2DcRポリペプチドの生産を行う方法において、請求項40に記載の宿主細胞を培養することを含んでなる方法。
【請求項45】
Apo-2DcRポリペプチドをコードする核酸を発現する細胞を含む非ヒト、トランスジェニック動物。
【請求項46】
マウス又はラットである請求項45に記載の動物。
【請求項47】
Apo-2DcRポリペプチドをコードする変更遺伝子を有する細胞を含む、非ヒト、ノックアウト動物。
【請求項48】
マウス又はラットである請求項47に記載の動物。
【請求項49】
請求項1又は5に記載のApo-2DcRと担体を含んでなる組成物。
【請求項50】
請求項15に記載のApo-2DcR抗体と担体を含んでなる組成物。
【請求項51】
容器と該容器に入れられる組成物を含んでなり、組成物がApo-2DcRポリペプチド又はApo-2DcR抗体を含む製造品。
【請求項52】
インビボ又はエキソビボにおいてApo-2DcRポリペプチド又はApo-2DcR抗体を使用するための使用説明書をさらに含んでなる請求項51に記載の製造品。
【請求項53】
哺乳動物細胞におけるアポトーシスを変調する方法において、Apo-2DcRポリペプチドに前記細胞を暴露させることを含んでなる方法。
【請求項54】
前記細胞が更にApo-2リガンドに暴露される請求項53に記載の方法。
【図1A】


【図1B】

【図2】

【図3】
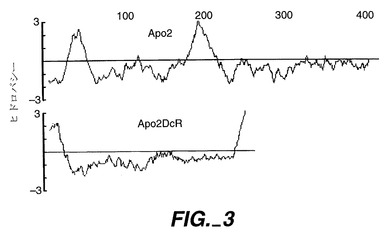
【図4】
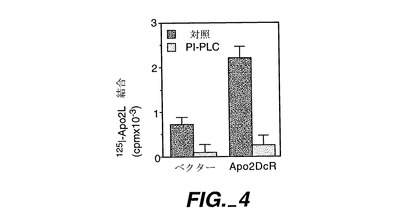
【図5】
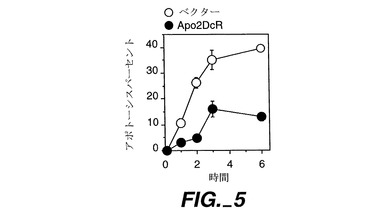
【図6】
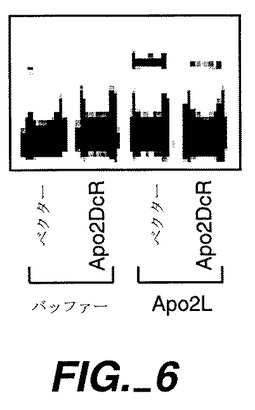
【図7】
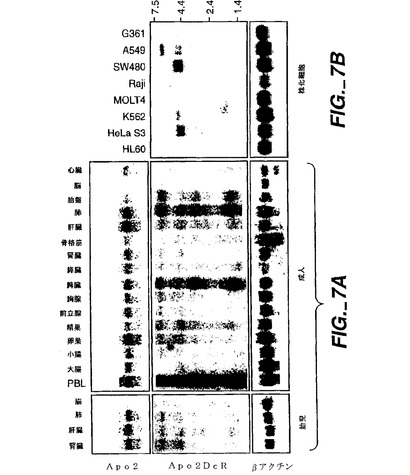
【図8】

【図9】

【図10】
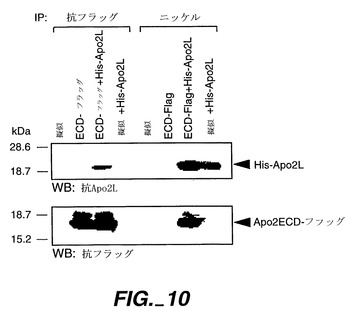
【図11A】
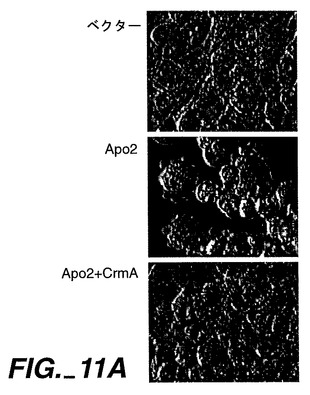
【図11B】
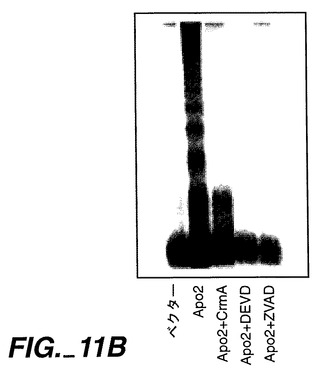
【図11C】
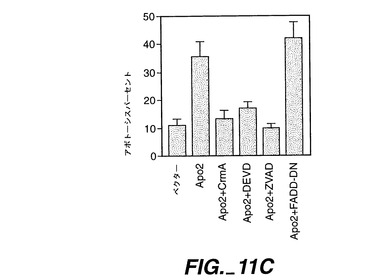
【図11D】
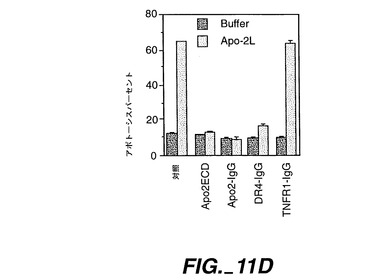
【図11E】
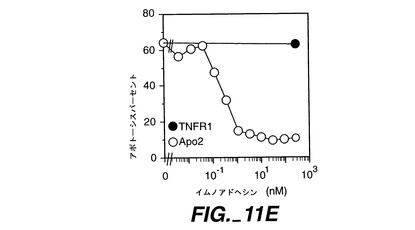
【図12A】
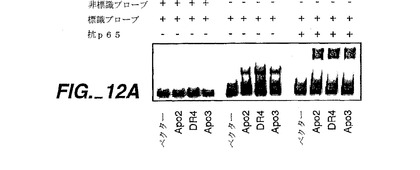
【図12B】
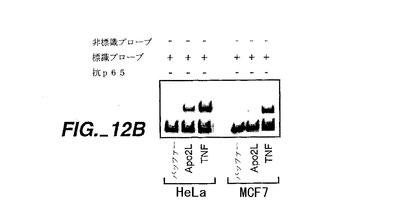
【図12C】
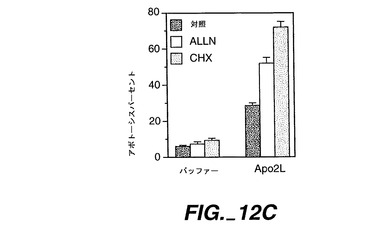
【図13】
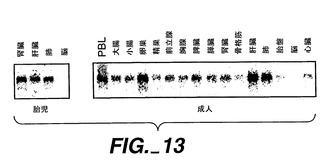
【図14A】
【図14B】
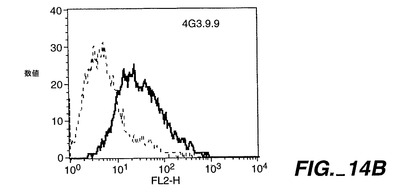
【図14C】
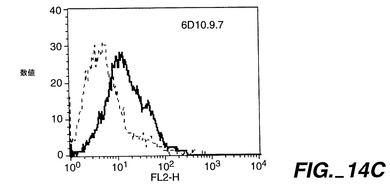
【図15A】
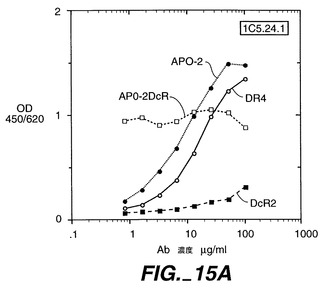
【図15B】
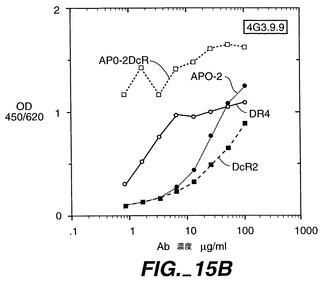
【図15C】
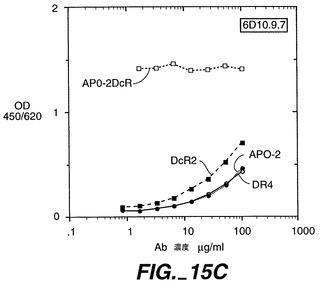
【図16】
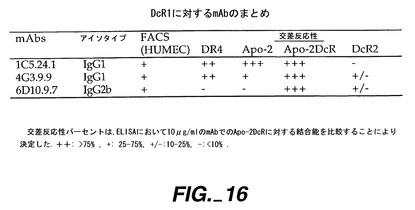
【図1B】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11A】
【図11B】
【図11C】
【図11D】
【図11E】
【図12A】
【図12B】
【図12C】
【図13】
【図14A】
【図14B】
【図14C】
【図15A】
【図15B】
【図15C】
【図16】
【公開番号】特開2010−13451(P2010−13451A)
【公開日】平成22年1月21日(2010.1.21)
【国際特許分類】
【外国語出願】
【出願番号】特願2009−171161(P2009−171161)
【出願日】平成21年7月22日(2009.7.22)
【分割の表示】特願2008−95736(P2008−95736)の分割
【原出願日】平成10年6月12日(1998.6.12)
【出願人】(596168317)ジェネンテック・インコーポレーテッド (372)
【氏名又は名称原語表記】GENENTECH,INC.
【Fターム(参考)】
【公開日】平成22年1月21日(2010.1.21)
【国際特許分類】
【出願番号】特願2009−171161(P2009−171161)
【出願日】平成21年7月22日(2009.7.22)
【分割の表示】特願2008−95736(P2008−95736)の分割
【原出願日】平成10年6月12日(1998.6.12)
【出願人】(596168317)ジェネンテック・インコーポレーテッド (372)
【氏名又は名称原語表記】GENENTECH,INC.
【Fターム(参考)】
[ Back to top ]