
Bリンパ球上のヒトmIgEに結合可能な抗CεmX抗体
本発明は、ヒトBリンパ球の表面で発現する膜結合IgE(mIgE)のCεmXドメインに効率的に結合できる抗体の生産及び使用に関する。ヒト膜結合イプシロン鎖上のCH4ドメインとC末端膜アンカーペプチドとの間に位置する52アミノ酸のCεmXドメインは、mIgEを発現するB細胞の免疫標的化の抗原部位の一つであることが示唆されていた。CεmXのC末端でRADWPGPPペプチドと結合するa20を含む従来報告されていたモノクローナル抗体は、現在では、ヒトB細胞上のmIgEとの結合が弱いことが見出されている。出願人らは、CεmXのGLAGGSAQSQRAPDRVL及びHSGQQQGLPRAAGGSVPHPR等の所定の断片に特異的なモノクローナル抗体のみが、ヒトB細胞上のmIgEに有効に結合し得ること、ひいてはIgEが介在する疾患の処置のためにB細胞を標的化する用途に有用であることを見出した。
【発明の詳細な説明】
【技術分野】
【0001】
IgEは、アレルギー性喘息、アレルギー性鼻炎、アトピー性皮膚炎等を引き起こす原因である、I型過敏性反応の誘導に、中心的な役割を果たす。アレルギー反応は、イエダニ、樹木及び草花の花粉、特定の食物及び薬物、並びにヒアリの咬傷等の、低害性の環境物質に対する、免疫系の応答である。そのような応答において、アレルゲンと、好塩基球及びマスト細胞の表面上のIgEとの結合は、IgEの架橋、及びIgE.Fcの根本的な受容体である、I型IgE.Fc受容体、又はFcεRIの集合を引き起こす。この受容体の集合は、続いて、顆粒のエキソサイトーシス及び医薬的仲介物質、例えばヒスタミン、ロイコトリエン、トリプターゼ、サイトカイン及びケモカイン等の放出を引き起こす、シグナル経路を活性化する。このマスト細胞及び好塩基球からの仲介物質の放出は、様々なアレルギーの病理的兆候を引き起こす。
【背景技術】
【0002】
血中及び間質液中の遊離IgE、並びにB細胞中のmIgEと結合するが、好塩基球及びマスト細胞上のFcεRIに結合したIgEとは結合しない抗IgE抗体は、IgEが仲介するアレルギー性疾患の処置のために開発された。ヒト化抗IgE抗体のオマリズマブ(取引名Xolair)による処理は、様々なアレルギーの誘導におけるI型過敏性反応の低減において、複数の医薬的効果を示す。当該抗体は、FcεRIの結合部位と重複するFcのCH3ドメイン中の部位で、高い親和性でIgEと結合する。従って、この治療法は、当該抗体と、遊離IgE並びにBリンパ球及びメモリーB細胞上のmIgEとの結合に基づくもので、この結合は、血中及び間質液中の全体的な遊離IgEレベルの低下を引き起こす。
【0003】
抗IgEと遊離IgEとの結合は、更に、IgEが好塩基球及びマスト細胞の表面上のFcεRIと結合するのを妨げる。IgEが結合していないFcεRIは不安定であって、それらは内在化され(internalized)、分解されるので、遊離IgEと抗IgEとの結合の阻害は、好塩基球及びマスト細胞上のFcεRIを漸進的に下方制御する。当該抗体両方の他の効果の証拠として、サイトカイン活性の中和、全体的な炎症活性の緩和、及びIgE-抗IgE免疫複合体の蓄積によるアレルギーの掃引の可能性等が挙げられる。
【0004】
本発明の発明者の一人(T. W. Chang)は、オマリズマブが、IgEのCH3上の抗原部位に加えて、mIgEを発現するBリンパ球の標的化の為に、ヒトmIgE上に存在する、CεmXと称される他の抗原部位に結合することを見出した。は、ヒト膜結合ε鎖(mε)のCH4ドメインとC末端膜アンカー断片との間に位置する、52アミノ酸の断片である。研究対象とした殆どのヒトにおいて、CεmXを有しないmε(mεs)は僅かな割合を占め、一方CεmXを有するmε(mεL)が優勢に発現していることが示されている。遊離している分泌されたIgEのε鎖、並びにmIgEのmεs及びmεLのmRNAは、全て、εRNA転写産物の選択的スプライシングに由来する。CεmXのアミノ酸及びヌクレオチド配列は、全タンパク質及びDNAデータベース中で特異のものである。従って、CεmXは、mIgE及びmIgE発現B細胞を標的化するための特異的な抗原部位を提供する。
【0005】
Changの研究グループは、CεmX断片を含む組換えタンパク質、及びヒトmIgEを発現するヒトミエローマ由来細胞株のSK0−007細胞株の細胞、及びmεL(mεL(CH2-CM);CM:細胞質)の細胞質末端を通じてCH2ドメイン由来の断片に対応する遺伝子がトランスフェクションされたCHO細胞株の細胞と結合することができる、a20を含む幾つかのCεmX特異的マウスモノクローナル抗体の開発を既に報告している。抗体a20及びより初期に開発された全ての抗体は、8-a.a.のペプチド領域RADWPGPP、#45-52残基と、CεmXの52a.aのC末端で結合することが見出された。
【0006】
引用される関連特許文献
US5.091.313 2/1992 Chang
US5,254,671 10/1993 Chang
US5,260,416 11/1993 Chang
US5,274,075 12/1993 Chang
US5,292,867 3/1994 Chang
US5,342,924 8/1994 Chang
US2009/0010924A1 Wu
【0007】
引用される関連非特許文献
Davis FM, Gossett LA, Chang TW (1991) An epitope on membrane-bound but not secreted IgE: implications in isotype-specific regulation. Bio/Technology 9: 53-56.
Peng C, Davis FM, Sun LK, Liou RS, Kim YW, Chang TW (1992) A new isoform of human membrane-bound IgE. J Immunol 148: 129-136.
Chen, H.Y., Liu, F.T., Hou, C.M.H., Huang, J.S.W., Sharma, B.B., and Chang, TW. (2002) Monoclonal antibodies against CεmX domain in human membrane-bound IgE and their potential on targeting IgE-expressing B cells. Int. Archives Allergy & Immunol. 128, 315-324.
【発明の概要】
【発明が解決しようとする課題】
【0008】
本発明は、ヒトmIgEのCεmXドメインに特異的で、ヒトBリンパ球上のmIgEに結合可能な抗体の開発及び同定に関する。また、アレルギー性疾患及びIgEにより仲介される他の疾患の処置におけるこれらの抗体の利用にも関する。
【課題を解決するための手段】
【0009】
Changの研究グループにより開発された抗CεmXモノクローナル抗体a20の研究において、a20は、Igα(CD79a)、Igβ(CD79b)、CD21、CD19、CD81及びB細胞受容体(BCR)に関連する他のタンパク質を発現しない、CHO細胞株又はNSO細胞株等の細胞株にmεL(CH2-CM)遺伝子をトランスフェクションしたものに、良好な結合を示すことを見出した。しかしながら、a20は、及び他のBCR関連タンパク質を発現する、例えばRamos細胞株等に、mεL(CH2-CM)遺伝子をトランスフェクションした細胞株に対しては、結合が弱いことが判明した。出願人らは、a20により認識されるCεmX上の抗原エピトープが、いずれかのBCR関連タンパク質によりブロックされていると推測した。故に、a20モノクローナル抗体及びそのキメラ及びヒト化版は、mIgEを発現するBリンパ球及びメモリー細胞をインビボで標的化する目的での使用には適しない。
【0010】
前記ペプチドエピトープRADWPGPPが抗体応答を誘導する唯一のエピトープであるとすると、ヒトCεmX含有タンパク質で免疫化したマウスを使用するハイブリドーマ法で生産したモノクローナル抗体は、全てこのペプチド領域に特異的なはずである。しかしながら、このエピトープがドミナントエピトープであるが、唯一の免疫原性エピトープではないとすると、CεmX上の他の抗原に特異的なモノクローナル抗体を開発する余地がある。BCR関連タンパク質により抗体の結合がブロックされないCεmXのエピトープが存在する可能性がある。もしこれが存在するのであれば、B細胞上のIgEに結合し、そのようなB細胞を標的化するのに使用できる、抗体を開発することができる。
【0011】
下記実施例において、出願人らは、RADWPGPPはドミナントエピトープであるが、CεmXの唯一の免疫原性及び抗原性エピトープではないことを示すのに成功した。更に、出願人らは、RADWPGPPの領域内に位置しない抗原性エピトープでCεmXに結合するモノクローナル抗体4B12及び26H2を見出した。これらのモノクローナル抗体は、CεmXとの結合において、a20抗体と競合しない。それらは、a20よりも一層強力にB細胞上のmIgEに結合し、a20よりも一層効果的にmIgE発現細胞の細胞溶解及びアポトーシスを引き起こす。
【0012】
この例は、4B12及び26H2等のモノクローナル抗体が、ヒトBリンパ球上のmIgEに結合可能で、IgE合成の下方制御のために、mIgEを発現するBリンパ球及びメモリーB細胞を標的化するための使用に適していることを示唆する。キメラ又はヒト化形態の前記抗体は、アレルギー性喘息、アレルギー性鼻炎及びアトピー性皮膚炎等の、IgE介在製のアレルギー性疾患に罹患した患者における使用に有用であり得る。抗IgEによるIgEの中和は、冷却誘導性のじんましん、慢性じんましん、コリン性じんましん、慢性鼻副鼻腔炎、全身性脂肪細胞症、皮膚脂肪細胞症、アレルギー性気管支肺アスペルギルス症、再発性特発性血管性浮腫、及び間質性膀胱炎、又は好酸球関連胃腸障害を効果的に処置することが示されていることから、4B12及び26H2等の抗体は、それらの様々な疾患を処置するのに利用される場合もある。
【0013】
更に、当該実施例は、CεmX、即ちmIgE発現B細胞に対する免疫応答の誘導における4B12及び26H2により認識されるペプチドの潜在的有用性を示唆する。類似の抗原的特徴、即ち、4B12及び26H2等の抗CεmX抗体に対する結合活性を有する前記ペプチド及びそれらの類似体が、個別に、又はT細胞のヘルプを誘導することが出来る部分を含有する分子構築物に組み合わせられて使用される場合もある。そのような構築物は、mIgE発現B細胞に対する強力な免疫化を誘導することができるため、全体のIgE合成の下方制御効果を実現する。
【図面の簡単な説明】
【0014】
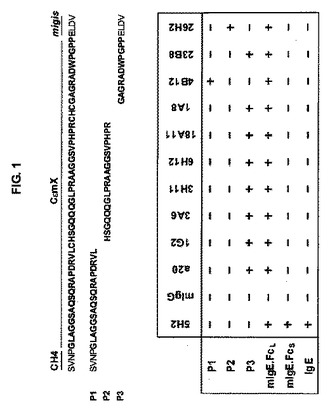
【図1】図1は、CεmXの連続的断片を有する3つの合成ペプチド、及びそれらのペプチドと様々な抗CεmX mAbsとの反応性を示す。CεmXドメインのアミノ酸残基は、太字で表記される。
【0015】
【図2】図2は、mIgE.FcL又はmIgE.Fcsを発現するCHO又はRamos細胞株に対する様々な抗CεmX mAbsの結合を示す。
【0016】
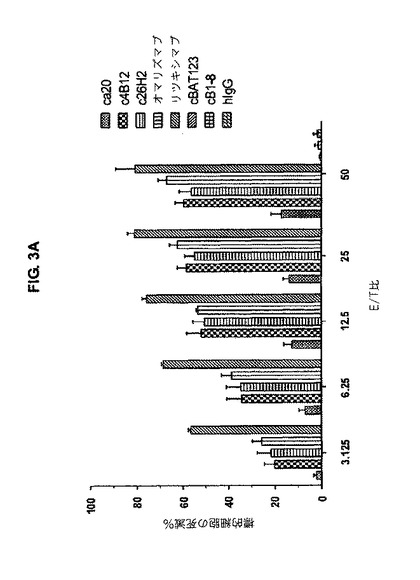
【図3A】図3Aは、キメラc4B12及びc26H2が、様々なE/T比で、mIgE.FcL発現Ramos細胞にADCCを誘導することを示す。
【0017】
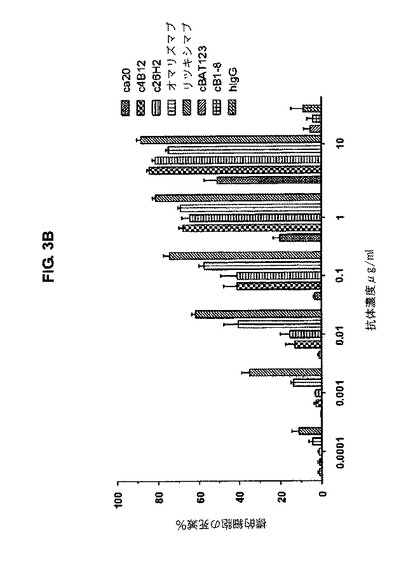
【図3B】図3Bは、mIgE.FcL発現Ramos細胞に対して、キメラc4B12及びc26H2が、用量依存的にADCCを誘導することを示す。
【0018】
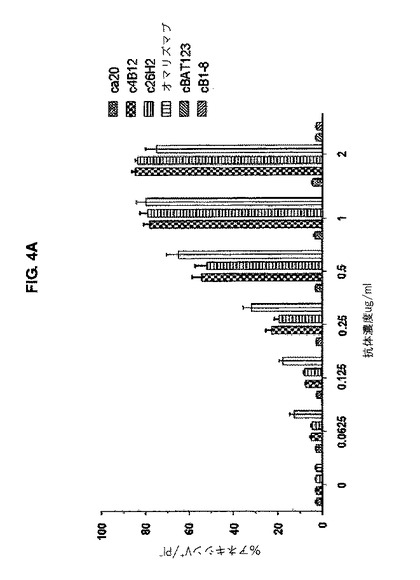
【図4A】図4Aは、mIgE.FcL発現Ramos細胞におけるキメラc4B12及びc26H2によるPSの露出が、用量依存的であることを示す。
【0019】
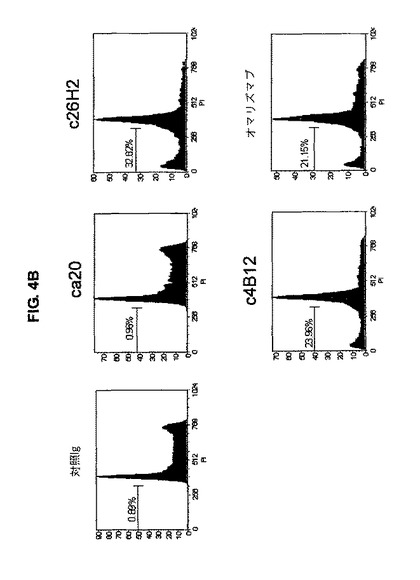
【図4B】図4Bは、アポトーシスを起こした核が、キメラc4B12及びc26H2で処理したmIgE.FcL発現Ramos細胞において観察されたことを示す。
【0020】
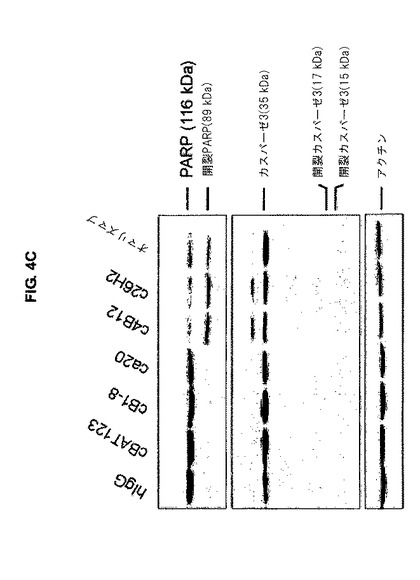
【図4C】図4Cは、カスパーゼ3位及びPARPの開裂が、キメラc4B12及びc26H2で処理したmIgE.FcL発現Ramos細胞において観察されたことを示す。
【0021】
【図5】図5は、親マウス4B12のVL及びVH、選択されたヒト生殖系列テンプレートKV2及びHV4のVL及びVH、並びにヒト化4B12(hu4B12)(アラインメント中「置換」と表記する)の、配列アラインメントを示す。このhu4B12は、キメラ4B12(c4B12)と同等の、CεmX組換えタンパク質、及びmIgE.FcL発現Ramos細胞に対する結合親和性を有する。
【実施例】
【0022】
実施例1:RADWPGPP以外の抗原性部位に結合する、新しい抗CεmXモノクローナル抗体
抗CεmX免疫応答を誘導するために、BALB/cマウスを、説明書に従って、TiterMax Gold adjuvant (Sigma-Aldrich)中で乳化した、50μgのn-ウンデシル−β−d−マルトピラノシド(UDM;Anatrace)可溶化mIgE.FcL組換えタンパク質を用いて、皮下投与により、2週間の間隔で、2回免疫化した。出願人らは、マウスが優勢なRADWPGPPエピトープに対してのみ抗体を産生することのないように、超免疫化(hyper-immunization)プロトコールの使用を避けた。0.1mgのUDM可溶化mIgE.FcL組換えタンパク質を、アジュバント無しで腹腔内投与して、最終的なブーストが与えられた。融合の1日前、NSO細胞を、10%ウシ胎児不活化血清(FBS; Invitrogen)及び1%ペニシリン−ストレプトマイシン(100x Pen-Strep solution; Invitrogen)混合物を添加した新鮮なDMEM培地(Invitrogen)中に、5x105 cells/mlの細胞密度で再播種した。最後のブーストの3日後、免疫化したマウス2頭の脾臓細胞を回収し、無血清DMEM培地で2回洗浄した。5x107個のNSO細胞を回収し、無血清DMEM培地で2回洗浄した。洗浄後、脾臓細胞及びNSO細胞に、予め温めておいた50%ポリエチレングリコール1500(PEG 1500, Roche Applied Science)を添加して融合させ、1分間ピペットチップを用いて静かに細胞を攪拌し、更に1分間攪拌し、2mlの予め温めておいた無血清DMEMを2分間かけて添加し、そして最後に8mlの予め温めておいた無血清DMEMを2分間かけて添加した。200xgで10分間遠心分離した後、融合した細胞を、600mlのHAT培地[2%ヒポキサンチン−アミノプテリン−チミジン混合物(50 x HAT solution; Invitrogen)、10% BM-Condimed H1(Roche Applied Science)、10%不活化FBS、及び1%ペニシリン−ストレプトマイシン混合物を添加したDMEM培地]に再懸濁し、200μl/ウェルずつ、96ウェル培養プレート30枚に分注した。3日目に、各ウェルに100μlのHAT培地を添加した。7日目及び10日目に、各ウェルの培地の半量を吸引し、同量のHAT培地を添加して、培地をリフレッシュした。14日目に、ハイブリドーマの上澄を用いて、酵素結合免疫吸着アッセイ(ELISA)により、UDM可溶化mIgE.FcL又はmIgE.Fcsタンパク質に結合する抗CεmX mAbsのスクリーニングを行った。
【0023】
ELISAによって抗CεmX mAbsを分泌するハイブリドーマをスクリーニングするために、0.1MのNaCO3 (pH 9.6)に溶解した精製UDM可溶化mIgE.FcL又はmIgE.Fcsタンパク質50 ng/ウェルで、96ウェルMaxiSorpプレート(Nunc)を、4℃で一昼夜コーティングした。コーティングされたウェルを、200μl/ウェルの1%BSAのPBS溶液により、室温で1時間ブロッキングした。0.05% Tween-20を加えたPBS 200μl/ウェルで、プレートを3回洗浄した。インキュベーションを、室温で2時間実行した。全てのウェルの培地を吸引し、0.05% Tween-20を加えたPBS 200μl/ウェルで、プレートを6回洗浄した。当該プレートに、1:10000で希釈したHRPコンジュゲートヤギ抗マウスIgG抗体(Chemicon)を加えて(100μl/ウェル)、1時間インキュベートした。続いて、全てのウェルの上澄を吸引し、0.05% Tween-20を加えたPBS 200μl/ウェルで、プレートを6回洗浄した。最後に、ウェルを、50μl/ウェルのテトラメチルベンジジン(TMB)基質溶液(SureBlue(商標), KPL)で現像し、50μl/ウェルの1N HClを加えて反応を停止させた。OD450の吸光度を、ELISAリーダー上で測定した。2つの細胞の融合体である4000超のハイブリドーマクローン中、17個のクローンは、ELISAにより、mIgE.Fcsよりも、UDM可溶化mIgE.FcLに特異的であることが示された。
【0024】
抗CεmX mAbsのCεmXに対する特異性を調査するために、前記複数のCεmX特異的クローンの、3つの合成ペプチドに対する反応性を試験した。当該3つの合成ペプチドは、#18残基に位置するC残基、及び#39-41残基に位置するCHC断片で分割した、CεmXの連続的な断片である。特に、P1ペプチドは、mεのCH4の最後の4アミノ酸残基、及びCεmXの最初の17アミノ酸残基(#1-17)、即ちGLAGGSAQSQRAPDRVLを含み;P2ペプチドは、CεmXの#19-38の20アミノ酸残基、即ちHSGQQQGLPRAAGGSVPHPRを含み;P3ペプチドは、CεmXの、末端の11アミノ酸残基(#42-52)、即ちGAGRADWPGPP、及び連続的ミギス(migis)領域の最初の4アミノ酸残基、即ちmε鎖の膜アンカーペプチドのN末端細胞外領域を含む。全てのペプチドは、Academia Sinica (Taipei, Taiwan)のGenomics Research Centerで合成された。これらのペプチドを、PBS中で、10mg/mlに再構築した。全てのペプチドを0.1MのNaCO3 (pH 9.6)に溶解し、500 ng/ウェルで、96ウェルMaxiSorpプレート(Nunc)を、4℃で一昼夜コーティングした。コーティングされたウェルを、200μl/ウェルの1%BSAのPBS溶液により、室温で1時間ブロッキングした。0.05% Tween-20を加えたPBS 200μl/ウェルで、プレートを3回洗浄し、続いてウェルに、1OOμlの、1μg/ml抗CεmX mAbs抗体を添加した。インキュベーションを、室温で2時間実行した。全てのウェルの培地を吸引し、0.05% Tween-20を加えたPBS 200μl/ウェルで、プレートを6回洗浄した。当該プレートに、1:10000で希釈したHRPコンジュゲートヤギ抗マウスIgG抗体を加えて、1時間インキュベートした。続いて、0.05% Tween-20を加えたPBS 200μl/ウェルで、プレートを6回洗浄した後、各ウェルに、50μl/ウェルのTMB基質溶液を添加した。当該反応を、50μl/ウェルの1N HClを加えて停止させた。OD450の吸光度を、ELISAリーダー上で測定した。出願人らの実験において調製した多数のCεmX特異的モノクローナル抗体の中で、4B12及び26H2のみ、RADWPGPPを含むP3ペプチドと反応しなかった。4B12はP1ペプチドと反応し、そして26H2はP2ペプチドと反応した。他のいずれのCεmX特異的モノクローナル抗体は、P3と反応した(図1)。従って、RADWPGPPは正しくドミナント免疫原エピトープである。しかしながら、唯一の免疫原エピトープではない。
【0025】
実施例2:mIgE発現B細胞上のmIgEへの4B12及び26H2の結合
出願人らは、更に、mεL(CH2-CM)又はmεs(CH2-CM)のいずれかをコードする組換えDNAをトランスフェクションしたCHO及びRamos細胞株に結合する多数のCεmX特異的モノクローナル抗体の性能を試験した。トランスフェクションされた2つのCHO細胞株はそれぞれmIgE.FcL又はmIgE.Fcsを生産したが、いずれもIgα及びIgβ等の副受容体と完全なB細胞受容体を形成しなかった。CHO細胞はそれらのタンパク質を発現しなかったためである。トランスフェクションされたRamos細胞株は、それぞれmIgE.FcL又はmIgE.Fcsを生産したが、いずれも内在(native)副受容体と複合体を形成した。抗CεmX mAbsの内在CεmXに対する結合を調査するために、mIgE.FcL又はmIgE.Fcsのいずれかを発現するCHO又はRamos細胞を、FACS緩衝剤[PBS、1% FBS、0.1%アジ化ナトリウム、及び2mM EDTA(pH8.0)]に、107 cells/mlの密度で再懸濁した。そして、106個の細胞を、100μlのハイブリドーマ上澄と、氷上で30分間インキュベーションし、続いてFACS緩衝剤で洗浄した。結合した抗体を、マウスIgGに対するFITC標識ウサギF(ab')2断片特異的抗体(AbD Secrotec)と氷上で30分間インキュベーションし、続いてFACS緩衝剤で2回洗浄したものを解析することにより検出した。FACSCanto IIフローサイトメーター(BD Bioscience)を使用してフローサイトメトリー試験を行い、そしてFCSExpressソフトウェア(De Novo Software)を使用して解析を行った。全てのCεmX特異的モノクローナル抗体において、mIgE.Fcsを発現するCHO及びRamos細胞に対する結合は確認されなかった。全てのCεmX特異的モノクローナル抗体は、mIgELを発現するCHO細胞に結合することが判明した。しかしながら、4B12及び26H2のみ、mIgE.FcLを発現するRamos細胞に結合したが、他の全てのCεmX特異的モノクローナル抗体は、を発現するRamos細胞に結合できなかった(図2)。
【0026】
実施例3:mIgE発現B細胞に対する4B12及び26H2による抗体依存的細胞毒性の誘導
キメラ抗CεmX mAbsのADCC活性を調査するために、出願人らは、末梢血単核細胞(PBMC)をエフェクター細胞として使用して、mIgE.FcLを発現するRamos細胞を標的化した。健康なドナーの軟膜(Taiwan Blood Service Foundation)から、Ficoll-Paque Plus (GE Healthcare)を用いた密度勾配遠心により、PBMCを精製し、これを90% FBS/10% DMSO (Hybri-Max(商標); Sigma- Aldrich)中で冷凍保存した。使用の前に、PBMCを融解し、2x106 cells/mlで、10%熱不活化FBS及び1%ペニシリン−ストレプトマイシン混合物を添加したMIDM培地(Invitrogen)中、一昼夜培養した。PBMCと共培養される標的細胞を特定するために、mIgE.FcL発現Ramos細胞を、0.1% BSA/PBS中の2.5μMの5−(及び−6)−カルボキシフルオレセインジアセテートスクシンイミジルエステル(CFDA, SE; Invitrogen)で、10分間、37℃で標識した。10%FBSを含有する冷却RPMI培地(Invitrogen)で3回洗浄した後、細胞を105 cells/mlに調整した。エフェクター−標的(E/T)比を滴定するために、200μlの完全RPMI培地中の20000個の標識した細胞を、1μg/mlの抗体に37℃で30分間曝露し、続いてこれを、同体積のPBMCと、50〜3.125の複数のE/T比で組み合わせた。抗体の滴定のために、完全RPMI培地200μl中の20000個のラベルした細胞を、様々な濃度の抗体(1000〜0.01 ng/ml)で、37℃で30分間オプソニン化して、続いて、E/T比25:1で、PBMCと組み合わせた。抗体非依存的な細胞死を測定するために、ラベルした標的細胞を、所定のE/T比で、抗体非存在下で、PBMCと組み合わせた。24時間のインキュベーションを終えて、氷上で15分間、死んだ細胞を2.5μg/mlの7−アミノアクチノマイシン(7-AAD; Invitrogen)で染色した。細胞を、Becton Dickinson FACSCanto IIフローサイトメーターで解析した。生きた標的細胞は、ドットプロット解析上で、CFSE-陽性/7-AAD-陰性のパーセンテージとして定義された。所定のE/T比で死んだ細胞のパーセンテージは、以下の式に従って計算された:
100x[(抗体非依存的対照中の生きた標的細胞の%−試料中の生きた標的細胞の%)/抗体非依存的対照中の生きた標的細胞の%]
c4B12、c26H2及びオマリズマブのADCC活性は、複数のE/T比で観察された。E/T比が50であるとき、c4B12、c26H2及びオマリズマブは60%の特異的溶解を引き起こし;一方、ca20の活性は低く、引き起こされた特異的溶解は僅か20%であった(図3A)。c4B12及びc26H2の濃度が0.01 μg/mlを超える場合、顕著なADCCが観察された。最大用量の10μg/mlにおいて、c4B12及びc26H2による標的細胞の特異的溶解は80〜90%で、一方ca20の特異的溶解は50%であった(図3B)。CD20を指向する陽性対照リツキシマブ及びオマリズマブは、複数のE/T比で、濃度依存的に、ADCCを効果的に誘導した。従って、出願人らは、c4B12及びc26H2は、ADCCの誘導において、ca20よりも強力な抗CεmX mAbsであって、インビボでmIgE発現B細胞を標的とするためのエフェクターを効率的に集合させることが出来る。
【0027】
実施例4:膜結合IgE.FcL発現Ramos細胞に対するキメラ抗CεmX mAbsによるアポトーシスの誘導
ホスファチジルセリン(PS)露出(exposure)を検出するために、mIgE.FcL-発現Ramos細胞(5x105 cell/ml)を、所定の濃度で、完全培養培地中、1時間、37℃で、キメラ抗CεmXmAbs、オマリズマブ又は対照抗体とインキュベーションした。続いて細胞を、濃度10μg/mlのヒトIgGのFc断片に特異的なヤギF(ab')2断片(Jackson ImmunoResearch Laboratories Inc.)で処理し、更に、37℃で24時間インキュベーションした。ホスファチジルセリン(PS)露出の検出は、1/200に希釈したフルオレセインイソチオシアネート(FITC)ラベルアネキシンV(Bio Vision)、及び2.5μg/mlプロビジウムヨーダイド(PI, Sigma- Aldrich)を含有する200μlのアネキシン緩衝剤中で、細胞を暗所、室温で15分間染色することにより評価された。細胞は、FACSCanto IIフローサイトメーター上で解析された。アポトーシス細胞は、ドットプロット解析上で、アネキシンV陰性/PI陽性細胞のパーセンテージとして検出された。mIgE.FcL発現Ramos細胞の約80%は、c4B12、c26H2、又はオマリズマブの濃度を増大させることによりアポトーシスで死滅したが、ca20の場合、最大で1μg/mlを与えてもそのようにならなかった(図4A)。
【0028】
アポトーシスを起こした核を検出するために、mIgE.FcL発現Ramos細胞(5x105 cell/ml)を、完全培養培地中1μg/mlのキメラ抗CεmX mAbs、オマリズマブ又は対照抗体と、37℃で1時間インキュベーションした。そして細胞を、最終濃度が10μg/mlとなるヒトIgGのFc断片に特異的なヤギF(ab')2断片で処理し、更に37℃で48時間インキュベーションした。5xlO5個の細胞を、0.5mlのプロビジウムヨーダイド(PI)/Triton溶液(PBS中0.1%酢酸ナトリウム、0.1%Triton X-100、15μg/ml PI、及び100μg/ml RNase A;全てSigma-Aldrich製)中で、氷上、暗所で1時間インキュベーションした。PIの蛍光は、FACSCanto IIフローサイトメーター上で決定した。無傷の核のDNAの内容は、リニアスケール上に記録された。低二倍(hypodiploid)DNAを含むアポトーシスを起こした核は、G0/G1ピークの下のチャンネルで蛍光を発し、全集団のパーセンテージとしてカウントされる。低二倍DNAを有する細胞集団の顕著な増大は、c4B12、c26H2又はオマリズマブで処理したmIgE.FcL発現Ramos細胞で観察された(図4B)。
【0029】
カスパーゼ3及びポリ(ADP-リボース)ポリメラーゼ(PARP)の開裂を検出するために、mIgE.FcL発現Ramos細胞を、37℃で1時間、完全培養培地中、1μg/mlの濃度で、キメラ抗CεmX mAbs、オマリズマブ又は対照抗体とインキュベーションした。そして細胞を、最終濃度が10μg/mlとなるヒトIgGのFc断片に特異的なヤギF(ab')2断片で処理し、更に37℃で24時間インキュベーションした。5xlO6個の細胞を氷冷PBSで洗浄し、100μlの氷冷調整RIPA緩衝剤[20 mM Tris (pH 7.4), 150 mM NaCl, 1% Triton-X 100, 0.5%デオキシコール酸塩, 0.1%ドデシル硫酸ナトリウム(SDS), 5 mM EDTA, 及びプロテアーゼインヒビター(Sigma-Aldrich)]中に再懸濁した。ライゼートを、20分間氷上でインキュベーションした。試料を、4℃で20分間16000xgで遠心分離した。上澄を新しい1.5mlチューブに移し、-80℃で保存した。清澄した各ライゼート中のタンパク質の量を、説明書に従い、Protein DCアッセイ(Bio-Rad Laboratories)を使用して定量した。各試料を全タンパク質量で平均化し、これをSDSポリアクリルアミドゲル電気泳動(SDS-PAGE)に供し、続いてPVDF膜(GE Healthcare)に転写した。カスパーゼ3及びPARPに対するウサギポリクローナル抗体をCell Signaling Techonologyから入手し、これを1:500に希釈した。HRPコンジュゲートヤギ抗ウサギIgG二次抗体(Sigma Aldrich)を、1:10000に希釈した。前記膜をECL(Immobilon(商標) Western; Millipore)試薬で現像した。等量のタンパク質がロードされたことは、β−アクチンに対する抗体(Sigma-Aldrich)を用いたウエスタンブロッティングにより評価された。c4B12、c26H2及びオマリズマブで処理したmIgE.FcL発現Ramos細胞は、ca20で処理したものと比べて、処理後24時間でのカスパーゼ3のMr 19及び17kDa断片への開裂がより顕著であった。一方、PARPの開裂は、Mx 116 kDa 完全PARP及びMx 89 kDa開裂産物を認識する抗体を使用して、c4B12、c26H2及びオマリズマブで処理したmIgE.FcL発現Ramos細胞において検出することが出来た(図4C)。
【技術分野】
【0001】
IgEは、アレルギー性喘息、アレルギー性鼻炎、アトピー性皮膚炎等を引き起こす原因である、I型過敏性反応の誘導に、中心的な役割を果たす。アレルギー反応は、イエダニ、樹木及び草花の花粉、特定の食物及び薬物、並びにヒアリの咬傷等の、低害性の環境物質に対する、免疫系の応答である。そのような応答において、アレルゲンと、好塩基球及びマスト細胞の表面上のIgEとの結合は、IgEの架橋、及びIgE.Fcの根本的な受容体である、I型IgE.Fc受容体、又はFcεRIの集合を引き起こす。この受容体の集合は、続いて、顆粒のエキソサイトーシス及び医薬的仲介物質、例えばヒスタミン、ロイコトリエン、トリプターゼ、サイトカイン及びケモカイン等の放出を引き起こす、シグナル経路を活性化する。このマスト細胞及び好塩基球からの仲介物質の放出は、様々なアレルギーの病理的兆候を引き起こす。
【背景技術】
【0002】
血中及び間質液中の遊離IgE、並びにB細胞中のmIgEと結合するが、好塩基球及びマスト細胞上のFcεRIに結合したIgEとは結合しない抗IgE抗体は、IgEが仲介するアレルギー性疾患の処置のために開発された。ヒト化抗IgE抗体のオマリズマブ(取引名Xolair)による処理は、様々なアレルギーの誘導におけるI型過敏性反応の低減において、複数の医薬的効果を示す。当該抗体は、FcεRIの結合部位と重複するFcのCH3ドメイン中の部位で、高い親和性でIgEと結合する。従って、この治療法は、当該抗体と、遊離IgE並びにBリンパ球及びメモリーB細胞上のmIgEとの結合に基づくもので、この結合は、血中及び間質液中の全体的な遊離IgEレベルの低下を引き起こす。
【0003】
抗IgEと遊離IgEとの結合は、更に、IgEが好塩基球及びマスト細胞の表面上のFcεRIと結合するのを妨げる。IgEが結合していないFcεRIは不安定であって、それらは内在化され(internalized)、分解されるので、遊離IgEと抗IgEとの結合の阻害は、好塩基球及びマスト細胞上のFcεRIを漸進的に下方制御する。当該抗体両方の他の効果の証拠として、サイトカイン活性の中和、全体的な炎症活性の緩和、及びIgE-抗IgE免疫複合体の蓄積によるアレルギーの掃引の可能性等が挙げられる。
【0004】
本発明の発明者の一人(T. W. Chang)は、オマリズマブが、IgEのCH3上の抗原部位に加えて、mIgEを発現するBリンパ球の標的化の為に、ヒトmIgE上に存在する、CεmXと称される他の抗原部位に結合することを見出した。は、ヒト膜結合ε鎖(mε)のCH4ドメインとC末端膜アンカー断片との間に位置する、52アミノ酸の断片である。研究対象とした殆どのヒトにおいて、CεmXを有しないmε(mεs)は僅かな割合を占め、一方CεmXを有するmε(mεL)が優勢に発現していることが示されている。遊離している分泌されたIgEのε鎖、並びにmIgEのmεs及びmεLのmRNAは、全て、εRNA転写産物の選択的スプライシングに由来する。CεmXのアミノ酸及びヌクレオチド配列は、全タンパク質及びDNAデータベース中で特異のものである。従って、CεmXは、mIgE及びmIgE発現B細胞を標的化するための特異的な抗原部位を提供する。
【0005】
Changの研究グループは、CεmX断片を含む組換えタンパク質、及びヒトmIgEを発現するヒトミエローマ由来細胞株のSK0−007細胞株の細胞、及びmεL(mεL(CH2-CM);CM:細胞質)の細胞質末端を通じてCH2ドメイン由来の断片に対応する遺伝子がトランスフェクションされたCHO細胞株の細胞と結合することができる、a20を含む幾つかのCεmX特異的マウスモノクローナル抗体の開発を既に報告している。抗体a20及びより初期に開発された全ての抗体は、8-a.a.のペプチド領域RADWPGPP、#45-52残基と、CεmXの52a.aのC末端で結合することが見出された。
【0006】
引用される関連特許文献
US5.091.313 2/1992 Chang
US5,254,671 10/1993 Chang
US5,260,416 11/1993 Chang
US5,274,075 12/1993 Chang
US5,292,867 3/1994 Chang
US5,342,924 8/1994 Chang
US2009/0010924A1 Wu
【0007】
引用される関連非特許文献
Davis FM, Gossett LA, Chang TW (1991) An epitope on membrane-bound but not secreted IgE: implications in isotype-specific regulation. Bio/Technology 9: 53-56.
Peng C, Davis FM, Sun LK, Liou RS, Kim YW, Chang TW (1992) A new isoform of human membrane-bound IgE. J Immunol 148: 129-136.
Chen, H.Y., Liu, F.T., Hou, C.M.H., Huang, J.S.W., Sharma, B.B., and Chang, TW. (2002) Monoclonal antibodies against CεmX domain in human membrane-bound IgE and their potential on targeting IgE-expressing B cells. Int. Archives Allergy & Immunol. 128, 315-324.
【発明の概要】
【発明が解決しようとする課題】
【0008】
本発明は、ヒトmIgEのCεmXドメインに特異的で、ヒトBリンパ球上のmIgEに結合可能な抗体の開発及び同定に関する。また、アレルギー性疾患及びIgEにより仲介される他の疾患の処置におけるこれらの抗体の利用にも関する。
【課題を解決するための手段】
【0009】
Changの研究グループにより開発された抗CεmXモノクローナル抗体a20の研究において、a20は、Igα(CD79a)、Igβ(CD79b)、CD21、CD19、CD81及びB細胞受容体(BCR)に関連する他のタンパク質を発現しない、CHO細胞株又はNSO細胞株等の細胞株にmεL(CH2-CM)遺伝子をトランスフェクションしたものに、良好な結合を示すことを見出した。しかしながら、a20は、及び他のBCR関連タンパク質を発現する、例えばRamos細胞株等に、mεL(CH2-CM)遺伝子をトランスフェクションした細胞株に対しては、結合が弱いことが判明した。出願人らは、a20により認識されるCεmX上の抗原エピトープが、いずれかのBCR関連タンパク質によりブロックされていると推測した。故に、a20モノクローナル抗体及びそのキメラ及びヒト化版は、mIgEを発現するBリンパ球及びメモリー細胞をインビボで標的化する目的での使用には適しない。
【0010】
前記ペプチドエピトープRADWPGPPが抗体応答を誘導する唯一のエピトープであるとすると、ヒトCεmX含有タンパク質で免疫化したマウスを使用するハイブリドーマ法で生産したモノクローナル抗体は、全てこのペプチド領域に特異的なはずである。しかしながら、このエピトープがドミナントエピトープであるが、唯一の免疫原性エピトープではないとすると、CεmX上の他の抗原に特異的なモノクローナル抗体を開発する余地がある。BCR関連タンパク質により抗体の結合がブロックされないCεmXのエピトープが存在する可能性がある。もしこれが存在するのであれば、B細胞上のIgEに結合し、そのようなB細胞を標的化するのに使用できる、抗体を開発することができる。
【0011】
下記実施例において、出願人らは、RADWPGPPはドミナントエピトープであるが、CεmXの唯一の免疫原性及び抗原性エピトープではないことを示すのに成功した。更に、出願人らは、RADWPGPPの領域内に位置しない抗原性エピトープでCεmXに結合するモノクローナル抗体4B12及び26H2を見出した。これらのモノクローナル抗体は、CεmXとの結合において、a20抗体と競合しない。それらは、a20よりも一層強力にB細胞上のmIgEに結合し、a20よりも一層効果的にmIgE発現細胞の細胞溶解及びアポトーシスを引き起こす。
【0012】
この例は、4B12及び26H2等のモノクローナル抗体が、ヒトBリンパ球上のmIgEに結合可能で、IgE合成の下方制御のために、mIgEを発現するBリンパ球及びメモリーB細胞を標的化するための使用に適していることを示唆する。キメラ又はヒト化形態の前記抗体は、アレルギー性喘息、アレルギー性鼻炎及びアトピー性皮膚炎等の、IgE介在製のアレルギー性疾患に罹患した患者における使用に有用であり得る。抗IgEによるIgEの中和は、冷却誘導性のじんましん、慢性じんましん、コリン性じんましん、慢性鼻副鼻腔炎、全身性脂肪細胞症、皮膚脂肪細胞症、アレルギー性気管支肺アスペルギルス症、再発性特発性血管性浮腫、及び間質性膀胱炎、又は好酸球関連胃腸障害を効果的に処置することが示されていることから、4B12及び26H2等の抗体は、それらの様々な疾患を処置するのに利用される場合もある。
【0013】
更に、当該実施例は、CεmX、即ちmIgE発現B細胞に対する免疫応答の誘導における4B12及び26H2により認識されるペプチドの潜在的有用性を示唆する。類似の抗原的特徴、即ち、4B12及び26H2等の抗CεmX抗体に対する結合活性を有する前記ペプチド及びそれらの類似体が、個別に、又はT細胞のヘルプを誘導することが出来る部分を含有する分子構築物に組み合わせられて使用される場合もある。そのような構築物は、mIgE発現B細胞に対する強力な免疫化を誘導することができるため、全体のIgE合成の下方制御効果を実現する。
【図面の簡単な説明】
【0014】
【図1】図1は、CεmXの連続的断片を有する3つの合成ペプチド、及びそれらのペプチドと様々な抗CεmX mAbsとの反応性を示す。CεmXドメインのアミノ酸残基は、太字で表記される。
【0015】
【図2】図2は、mIgE.FcL又はmIgE.Fcsを発現するCHO又はRamos細胞株に対する様々な抗CεmX mAbsの結合を示す。
【0016】
【図3A】図3Aは、キメラc4B12及びc26H2が、様々なE/T比で、mIgE.FcL発現Ramos細胞にADCCを誘導することを示す。
【0017】
【図3B】図3Bは、mIgE.FcL発現Ramos細胞に対して、キメラc4B12及びc26H2が、用量依存的にADCCを誘導することを示す。
【0018】
【図4A】図4Aは、mIgE.FcL発現Ramos細胞におけるキメラc4B12及びc26H2によるPSの露出が、用量依存的であることを示す。
【0019】
【図4B】図4Bは、アポトーシスを起こした核が、キメラc4B12及びc26H2で処理したmIgE.FcL発現Ramos細胞において観察されたことを示す。
【0020】
【図4C】図4Cは、カスパーゼ3位及びPARPの開裂が、キメラc4B12及びc26H2で処理したmIgE.FcL発現Ramos細胞において観察されたことを示す。
【0021】
【図5】図5は、親マウス4B12のVL及びVH、選択されたヒト生殖系列テンプレートKV2及びHV4のVL及びVH、並びにヒト化4B12(hu4B12)(アラインメント中「置換」と表記する)の、配列アラインメントを示す。このhu4B12は、キメラ4B12(c4B12)と同等の、CεmX組換えタンパク質、及びmIgE.FcL発現Ramos細胞に対する結合親和性を有する。
【実施例】
【0022】
実施例1:RADWPGPP以外の抗原性部位に結合する、新しい抗CεmXモノクローナル抗体
抗CεmX免疫応答を誘導するために、BALB/cマウスを、説明書に従って、TiterMax Gold adjuvant (Sigma-Aldrich)中で乳化した、50μgのn-ウンデシル−β−d−マルトピラノシド(UDM;Anatrace)可溶化mIgE.FcL組換えタンパク質を用いて、皮下投与により、2週間の間隔で、2回免疫化した。出願人らは、マウスが優勢なRADWPGPPエピトープに対してのみ抗体を産生することのないように、超免疫化(hyper-immunization)プロトコールの使用を避けた。0.1mgのUDM可溶化mIgE.FcL組換えタンパク質を、アジュバント無しで腹腔内投与して、最終的なブーストが与えられた。融合の1日前、NSO細胞を、10%ウシ胎児不活化血清(FBS; Invitrogen)及び1%ペニシリン−ストレプトマイシン(100x Pen-Strep solution; Invitrogen)混合物を添加した新鮮なDMEM培地(Invitrogen)中に、5x105 cells/mlの細胞密度で再播種した。最後のブーストの3日後、免疫化したマウス2頭の脾臓細胞を回収し、無血清DMEM培地で2回洗浄した。5x107個のNSO細胞を回収し、無血清DMEM培地で2回洗浄した。洗浄後、脾臓細胞及びNSO細胞に、予め温めておいた50%ポリエチレングリコール1500(PEG 1500, Roche Applied Science)を添加して融合させ、1分間ピペットチップを用いて静かに細胞を攪拌し、更に1分間攪拌し、2mlの予め温めておいた無血清DMEMを2分間かけて添加し、そして最後に8mlの予め温めておいた無血清DMEMを2分間かけて添加した。200xgで10分間遠心分離した後、融合した細胞を、600mlのHAT培地[2%ヒポキサンチン−アミノプテリン−チミジン混合物(50 x HAT solution; Invitrogen)、10% BM-Condimed H1(Roche Applied Science)、10%不活化FBS、及び1%ペニシリン−ストレプトマイシン混合物を添加したDMEM培地]に再懸濁し、200μl/ウェルずつ、96ウェル培養プレート30枚に分注した。3日目に、各ウェルに100μlのHAT培地を添加した。7日目及び10日目に、各ウェルの培地の半量を吸引し、同量のHAT培地を添加して、培地をリフレッシュした。14日目に、ハイブリドーマの上澄を用いて、酵素結合免疫吸着アッセイ(ELISA)により、UDM可溶化mIgE.FcL又はmIgE.Fcsタンパク質に結合する抗CεmX mAbsのスクリーニングを行った。
【0023】
ELISAによって抗CεmX mAbsを分泌するハイブリドーマをスクリーニングするために、0.1MのNaCO3 (pH 9.6)に溶解した精製UDM可溶化mIgE.FcL又はmIgE.Fcsタンパク質50 ng/ウェルで、96ウェルMaxiSorpプレート(Nunc)を、4℃で一昼夜コーティングした。コーティングされたウェルを、200μl/ウェルの1%BSAのPBS溶液により、室温で1時間ブロッキングした。0.05% Tween-20を加えたPBS 200μl/ウェルで、プレートを3回洗浄した。インキュベーションを、室温で2時間実行した。全てのウェルの培地を吸引し、0.05% Tween-20を加えたPBS 200μl/ウェルで、プレートを6回洗浄した。当該プレートに、1:10000で希釈したHRPコンジュゲートヤギ抗マウスIgG抗体(Chemicon)を加えて(100μl/ウェル)、1時間インキュベートした。続いて、全てのウェルの上澄を吸引し、0.05% Tween-20を加えたPBS 200μl/ウェルで、プレートを6回洗浄した。最後に、ウェルを、50μl/ウェルのテトラメチルベンジジン(TMB)基質溶液(SureBlue(商標), KPL)で現像し、50μl/ウェルの1N HClを加えて反応を停止させた。OD450の吸光度を、ELISAリーダー上で測定した。2つの細胞の融合体である4000超のハイブリドーマクローン中、17個のクローンは、ELISAにより、mIgE.Fcsよりも、UDM可溶化mIgE.FcLに特異的であることが示された。
【0024】
抗CεmX mAbsのCεmXに対する特異性を調査するために、前記複数のCεmX特異的クローンの、3つの合成ペプチドに対する反応性を試験した。当該3つの合成ペプチドは、#18残基に位置するC残基、及び#39-41残基に位置するCHC断片で分割した、CεmXの連続的な断片である。特に、P1ペプチドは、mεのCH4の最後の4アミノ酸残基、及びCεmXの最初の17アミノ酸残基(#1-17)、即ちGLAGGSAQSQRAPDRVLを含み;P2ペプチドは、CεmXの#19-38の20アミノ酸残基、即ちHSGQQQGLPRAAGGSVPHPRを含み;P3ペプチドは、CεmXの、末端の11アミノ酸残基(#42-52)、即ちGAGRADWPGPP、及び連続的ミギス(migis)領域の最初の4アミノ酸残基、即ちmε鎖の膜アンカーペプチドのN末端細胞外領域を含む。全てのペプチドは、Academia Sinica (Taipei, Taiwan)のGenomics Research Centerで合成された。これらのペプチドを、PBS中で、10mg/mlに再構築した。全てのペプチドを0.1MのNaCO3 (pH 9.6)に溶解し、500 ng/ウェルで、96ウェルMaxiSorpプレート(Nunc)を、4℃で一昼夜コーティングした。コーティングされたウェルを、200μl/ウェルの1%BSAのPBS溶液により、室温で1時間ブロッキングした。0.05% Tween-20を加えたPBS 200μl/ウェルで、プレートを3回洗浄し、続いてウェルに、1OOμlの、1μg/ml抗CεmX mAbs抗体を添加した。インキュベーションを、室温で2時間実行した。全てのウェルの培地を吸引し、0.05% Tween-20を加えたPBS 200μl/ウェルで、プレートを6回洗浄した。当該プレートに、1:10000で希釈したHRPコンジュゲートヤギ抗マウスIgG抗体を加えて、1時間インキュベートした。続いて、0.05% Tween-20を加えたPBS 200μl/ウェルで、プレートを6回洗浄した後、各ウェルに、50μl/ウェルのTMB基質溶液を添加した。当該反応を、50μl/ウェルの1N HClを加えて停止させた。OD450の吸光度を、ELISAリーダー上で測定した。出願人らの実験において調製した多数のCεmX特異的モノクローナル抗体の中で、4B12及び26H2のみ、RADWPGPPを含むP3ペプチドと反応しなかった。4B12はP1ペプチドと反応し、そして26H2はP2ペプチドと反応した。他のいずれのCεmX特異的モノクローナル抗体は、P3と反応した(図1)。従って、RADWPGPPは正しくドミナント免疫原エピトープである。しかしながら、唯一の免疫原エピトープではない。
【0025】
実施例2:mIgE発現B細胞上のmIgEへの4B12及び26H2の結合
出願人らは、更に、mεL(CH2-CM)又はmεs(CH2-CM)のいずれかをコードする組換えDNAをトランスフェクションしたCHO及びRamos細胞株に結合する多数のCεmX特異的モノクローナル抗体の性能を試験した。トランスフェクションされた2つのCHO細胞株はそれぞれmIgE.FcL又はmIgE.Fcsを生産したが、いずれもIgα及びIgβ等の副受容体と完全なB細胞受容体を形成しなかった。CHO細胞はそれらのタンパク質を発現しなかったためである。トランスフェクションされたRamos細胞株は、それぞれmIgE.FcL又はmIgE.Fcsを生産したが、いずれも内在(native)副受容体と複合体を形成した。抗CεmX mAbsの内在CεmXに対する結合を調査するために、mIgE.FcL又はmIgE.Fcsのいずれかを発現するCHO又はRamos細胞を、FACS緩衝剤[PBS、1% FBS、0.1%アジ化ナトリウム、及び2mM EDTA(pH8.0)]に、107 cells/mlの密度で再懸濁した。そして、106個の細胞を、100μlのハイブリドーマ上澄と、氷上で30分間インキュベーションし、続いてFACS緩衝剤で洗浄した。結合した抗体を、マウスIgGに対するFITC標識ウサギF(ab')2断片特異的抗体(AbD Secrotec)と氷上で30分間インキュベーションし、続いてFACS緩衝剤で2回洗浄したものを解析することにより検出した。FACSCanto IIフローサイトメーター(BD Bioscience)を使用してフローサイトメトリー試験を行い、そしてFCSExpressソフトウェア(De Novo Software)を使用して解析を行った。全てのCεmX特異的モノクローナル抗体において、mIgE.Fcsを発現するCHO及びRamos細胞に対する結合は確認されなかった。全てのCεmX特異的モノクローナル抗体は、mIgELを発現するCHO細胞に結合することが判明した。しかしながら、4B12及び26H2のみ、mIgE.FcLを発現するRamos細胞に結合したが、他の全てのCεmX特異的モノクローナル抗体は、を発現するRamos細胞に結合できなかった(図2)。
【0026】
実施例3:mIgE発現B細胞に対する4B12及び26H2による抗体依存的細胞毒性の誘導
キメラ抗CεmX mAbsのADCC活性を調査するために、出願人らは、末梢血単核細胞(PBMC)をエフェクター細胞として使用して、mIgE.FcLを発現するRamos細胞を標的化した。健康なドナーの軟膜(Taiwan Blood Service Foundation)から、Ficoll-Paque Plus (GE Healthcare)を用いた密度勾配遠心により、PBMCを精製し、これを90% FBS/10% DMSO (Hybri-Max(商標); Sigma- Aldrich)中で冷凍保存した。使用の前に、PBMCを融解し、2x106 cells/mlで、10%熱不活化FBS及び1%ペニシリン−ストレプトマイシン混合物を添加したMIDM培地(Invitrogen)中、一昼夜培養した。PBMCと共培養される標的細胞を特定するために、mIgE.FcL発現Ramos細胞を、0.1% BSA/PBS中の2.5μMの5−(及び−6)−カルボキシフルオレセインジアセテートスクシンイミジルエステル(CFDA, SE; Invitrogen)で、10分間、37℃で標識した。10%FBSを含有する冷却RPMI培地(Invitrogen)で3回洗浄した後、細胞を105 cells/mlに調整した。エフェクター−標的(E/T)比を滴定するために、200μlの完全RPMI培地中の20000個の標識した細胞を、1μg/mlの抗体に37℃で30分間曝露し、続いてこれを、同体積のPBMCと、50〜3.125の複数のE/T比で組み合わせた。抗体の滴定のために、完全RPMI培地200μl中の20000個のラベルした細胞を、様々な濃度の抗体(1000〜0.01 ng/ml)で、37℃で30分間オプソニン化して、続いて、E/T比25:1で、PBMCと組み合わせた。抗体非依存的な細胞死を測定するために、ラベルした標的細胞を、所定のE/T比で、抗体非存在下で、PBMCと組み合わせた。24時間のインキュベーションを終えて、氷上で15分間、死んだ細胞を2.5μg/mlの7−アミノアクチノマイシン(7-AAD; Invitrogen)で染色した。細胞を、Becton Dickinson FACSCanto IIフローサイトメーターで解析した。生きた標的細胞は、ドットプロット解析上で、CFSE-陽性/7-AAD-陰性のパーセンテージとして定義された。所定のE/T比で死んだ細胞のパーセンテージは、以下の式に従って計算された:
100x[(抗体非依存的対照中の生きた標的細胞の%−試料中の生きた標的細胞の%)/抗体非依存的対照中の生きた標的細胞の%]
c4B12、c26H2及びオマリズマブのADCC活性は、複数のE/T比で観察された。E/T比が50であるとき、c4B12、c26H2及びオマリズマブは60%の特異的溶解を引き起こし;一方、ca20の活性は低く、引き起こされた特異的溶解は僅か20%であった(図3A)。c4B12及びc26H2の濃度が0.01 μg/mlを超える場合、顕著なADCCが観察された。最大用量の10μg/mlにおいて、c4B12及びc26H2による標的細胞の特異的溶解は80〜90%で、一方ca20の特異的溶解は50%であった(図3B)。CD20を指向する陽性対照リツキシマブ及びオマリズマブは、複数のE/T比で、濃度依存的に、ADCCを効果的に誘導した。従って、出願人らは、c4B12及びc26H2は、ADCCの誘導において、ca20よりも強力な抗CεmX mAbsであって、インビボでmIgE発現B細胞を標的とするためのエフェクターを効率的に集合させることが出来る。
【0027】
実施例4:膜結合IgE.FcL発現Ramos細胞に対するキメラ抗CεmX mAbsによるアポトーシスの誘導
ホスファチジルセリン(PS)露出(exposure)を検出するために、mIgE.FcL-発現Ramos細胞(5x105 cell/ml)を、所定の濃度で、完全培養培地中、1時間、37℃で、キメラ抗CεmXmAbs、オマリズマブ又は対照抗体とインキュベーションした。続いて細胞を、濃度10μg/mlのヒトIgGのFc断片に特異的なヤギF(ab')2断片(Jackson ImmunoResearch Laboratories Inc.)で処理し、更に、37℃で24時間インキュベーションした。ホスファチジルセリン(PS)露出の検出は、1/200に希釈したフルオレセインイソチオシアネート(FITC)ラベルアネキシンV(Bio Vision)、及び2.5μg/mlプロビジウムヨーダイド(PI, Sigma- Aldrich)を含有する200μlのアネキシン緩衝剤中で、細胞を暗所、室温で15分間染色することにより評価された。細胞は、FACSCanto IIフローサイトメーター上で解析された。アポトーシス細胞は、ドットプロット解析上で、アネキシンV陰性/PI陽性細胞のパーセンテージとして検出された。mIgE.FcL発現Ramos細胞の約80%は、c4B12、c26H2、又はオマリズマブの濃度を増大させることによりアポトーシスで死滅したが、ca20の場合、最大で1μg/mlを与えてもそのようにならなかった(図4A)。
【0028】
アポトーシスを起こした核を検出するために、mIgE.FcL発現Ramos細胞(5x105 cell/ml)を、完全培養培地中1μg/mlのキメラ抗CεmX mAbs、オマリズマブ又は対照抗体と、37℃で1時間インキュベーションした。そして細胞を、最終濃度が10μg/mlとなるヒトIgGのFc断片に特異的なヤギF(ab')2断片で処理し、更に37℃で48時間インキュベーションした。5xlO5個の細胞を、0.5mlのプロビジウムヨーダイド(PI)/Triton溶液(PBS中0.1%酢酸ナトリウム、0.1%Triton X-100、15μg/ml PI、及び100μg/ml RNase A;全てSigma-Aldrich製)中で、氷上、暗所で1時間インキュベーションした。PIの蛍光は、FACSCanto IIフローサイトメーター上で決定した。無傷の核のDNAの内容は、リニアスケール上に記録された。低二倍(hypodiploid)DNAを含むアポトーシスを起こした核は、G0/G1ピークの下のチャンネルで蛍光を発し、全集団のパーセンテージとしてカウントされる。低二倍DNAを有する細胞集団の顕著な増大は、c4B12、c26H2又はオマリズマブで処理したmIgE.FcL発現Ramos細胞で観察された(図4B)。
【0029】
カスパーゼ3及びポリ(ADP-リボース)ポリメラーゼ(PARP)の開裂を検出するために、mIgE.FcL発現Ramos細胞を、37℃で1時間、完全培養培地中、1μg/mlの濃度で、キメラ抗CεmX mAbs、オマリズマブ又は対照抗体とインキュベーションした。そして細胞を、最終濃度が10μg/mlとなるヒトIgGのFc断片に特異的なヤギF(ab')2断片で処理し、更に37℃で24時間インキュベーションした。5xlO6個の細胞を氷冷PBSで洗浄し、100μlの氷冷調整RIPA緩衝剤[20 mM Tris (pH 7.4), 150 mM NaCl, 1% Triton-X 100, 0.5%デオキシコール酸塩, 0.1%ドデシル硫酸ナトリウム(SDS), 5 mM EDTA, 及びプロテアーゼインヒビター(Sigma-Aldrich)]中に再懸濁した。ライゼートを、20分間氷上でインキュベーションした。試料を、4℃で20分間16000xgで遠心分離した。上澄を新しい1.5mlチューブに移し、-80℃で保存した。清澄した各ライゼート中のタンパク質の量を、説明書に従い、Protein DCアッセイ(Bio-Rad Laboratories)を使用して定量した。各試料を全タンパク質量で平均化し、これをSDSポリアクリルアミドゲル電気泳動(SDS-PAGE)に供し、続いてPVDF膜(GE Healthcare)に転写した。カスパーゼ3及びPARPに対するウサギポリクローナル抗体をCell Signaling Techonologyから入手し、これを1:500に希釈した。HRPコンジュゲートヤギ抗ウサギIgG二次抗体(Sigma Aldrich)を、1:10000に希釈した。前記膜をECL(Immobilon(商標) Western; Millipore)試薬で現像した。等量のタンパク質がロードされたことは、β−アクチンに対する抗体(Sigma-Aldrich)を用いたウエスタンブロッティングにより評価された。c4B12、c26H2及びオマリズマブで処理したmIgE.FcL発現Ramos細胞は、ca20で処理したものと比べて、処理後24時間でのカスパーゼ3のMr 19及び17kDa断片への開裂がより顕著であった。一方、PARPの開裂は、Mx 116 kDa 完全PARP及びMx 89 kDa開裂産物を認識する抗体を使用して、c4B12、c26H2及びオマリズマブで処理したmIgE.FcL発現Ramos細胞において検出することが出来た(図4C)。
【特許請求の範囲】
【請求項1】
ヒトBリンパ球上の膜結合IgEと結合可能で、かつRADWPGPPペプチドと結合不可能な、CεmX-特異的抗体。
【請求項2】
マウスモノクローナル抗体である、請求項1に記載の抗体。
【請求項3】
マウスモノクローナル抗体の可変領域及びヒト抗体の定常領域を含むキメラ抗体である、請求項1に記載の抗体。
【請求項4】
本質的にマウスモノクローナル抗体の超可変領域、並びにヒト抗体のフレームワーク領域及び定常領域を含む、請求項1に記載の抗体。
【請求項5】
ヒト抗体である、請求項1に記載の抗体。
【請求項6】
請求項1に記載のCεmX-特異的抗体の断片であって、ヒトBリンパ球上の膜結合IgEと結合可能で、かつRADWPGPPペプチドと結合不可能な断片。
【請求項7】
Fab、F(ab)'2、又は単鎖Fvである、請求項6に記載の抗体の断片。
【請求項8】
請求項1に記載の抗体を用いてIgE介在性疾患を処置する治療方法。
【請求項9】
前記IgE介在性疾患がアレルギー性喘息、アレルギー性鼻炎、又はアトピー性皮膚炎である、請求項8に記載の治療方法。
【請求項10】
前記IgE介在性疾患が、冷却誘導性のじんましん、慢性じんましん、コリン性じんましん、慢性鼻副鼻腔炎、全身性脂肪細胞症、皮膚脂肪細胞症、アレルギー性気管支肺アスペルギルス症、再発性特発性血管性浮腫、及び間質性膀胱炎、又は好酸球関連胃腸障害である、請求項8に記載の治療方法。
【請求項11】
GLAGGSAQSQRAPDRVL、又は同様の抗原的特徴を有する類似体と結合する、請求項1に記載の抗体。
【請求項12】
HSGQQQGLPRAAGGSVPHPR、又は同様の抗原的特徴を有する類似体と結合する、請求項1に記載の抗体。
【請求項13】
GLAGGSAQSQRAPDRVLを含む免疫原又は同様の抗原的特徴を有する類似体を用いて、インビボで患者の免疫応答を誘導する治療方法。
【請求項14】
HSGQQQGLPRAAGGSVPHPRを含む免疫原又は同様の抗原的特徴を有する類似体を用いて、インビボで患者の免疫応答を誘導する治療方法。
【請求項15】
GLAGGSAQSQRAPDRVLを含む免疫原又は同様の抗原的特徴を有する類似体、及びHSGQQQGLPRAAGGSVPHPRを含む免疫原又は同様の抗原的特徴を有する類似体を用いて、インビボで患者の免疫応答を誘導する治療方法。
【請求項1】
ヒトBリンパ球上の膜結合IgEと結合可能で、かつRADWPGPPペプチドと結合不可能な、CεmX-特異的抗体。
【請求項2】
マウスモノクローナル抗体である、請求項1に記載の抗体。
【請求項3】
マウスモノクローナル抗体の可変領域及びヒト抗体の定常領域を含むキメラ抗体である、請求項1に記載の抗体。
【請求項4】
本質的にマウスモノクローナル抗体の超可変領域、並びにヒト抗体のフレームワーク領域及び定常領域を含む、請求項1に記載の抗体。
【請求項5】
ヒト抗体である、請求項1に記載の抗体。
【請求項6】
請求項1に記載のCεmX-特異的抗体の断片であって、ヒトBリンパ球上の膜結合IgEと結合可能で、かつRADWPGPPペプチドと結合不可能な断片。
【請求項7】
Fab、F(ab)'2、又は単鎖Fvである、請求項6に記載の抗体の断片。
【請求項8】
請求項1に記載の抗体を用いてIgE介在性疾患を処置する治療方法。
【請求項9】
前記IgE介在性疾患がアレルギー性喘息、アレルギー性鼻炎、又はアトピー性皮膚炎である、請求項8に記載の治療方法。
【請求項10】
前記IgE介在性疾患が、冷却誘導性のじんましん、慢性じんましん、コリン性じんましん、慢性鼻副鼻腔炎、全身性脂肪細胞症、皮膚脂肪細胞症、アレルギー性気管支肺アスペルギルス症、再発性特発性血管性浮腫、及び間質性膀胱炎、又は好酸球関連胃腸障害である、請求項8に記載の治療方法。
【請求項11】
GLAGGSAQSQRAPDRVL、又は同様の抗原的特徴を有する類似体と結合する、請求項1に記載の抗体。
【請求項12】
HSGQQQGLPRAAGGSVPHPR、又は同様の抗原的特徴を有する類似体と結合する、請求項1に記載の抗体。
【請求項13】
GLAGGSAQSQRAPDRVLを含む免疫原又は同様の抗原的特徴を有する類似体を用いて、インビボで患者の免疫応答を誘導する治療方法。
【請求項14】
HSGQQQGLPRAAGGSVPHPRを含む免疫原又は同様の抗原的特徴を有する類似体を用いて、インビボで患者の免疫応答を誘導する治療方法。
【請求項15】
GLAGGSAQSQRAPDRVLを含む免疫原又は同様の抗原的特徴を有する類似体、及びHSGQQQGLPRAAGGSVPHPRを含む免疫原又は同様の抗原的特徴を有する類似体を用いて、インビボで患者の免疫応答を誘導する治療方法。
【図1】


【図2】

【図3A】
【図3B】
【図4A】
【図4B】
【図4C】
【図5】

【図2】
【図3A】
【図3B】
【図4A】
【図4B】
【図4C】
【図5】
【公表番号】特表2013−520396(P2013−520396A)
【公表日】平成25年6月6日(2013.6.6)
【国際特許分類】
【出願番号】特願2011−551394(P2011−551394)
【出願日】平成22年2月25日(2010.2.25)
【国際出願番号】PCT/CN2010/000232
【国際公開番号】WO2010/097012
【国際公開日】平成22年9月2日(2010.9.2)
【出願人】(596118493)アカデミア シニカ (33)
【氏名又は名称原語表記】ACADEMIA SINICA
【住所又は居所原語表記】128 Sec 2,Academia Road,Nankang,Taipei 11529 TW
【Fターム(参考)】
【公表日】平成25年6月6日(2013.6.6)
【国際特許分類】
【出願日】平成22年2月25日(2010.2.25)
【国際出願番号】PCT/CN2010/000232
【国際公開番号】WO2010/097012
【国際公開日】平成22年9月2日(2010.9.2)
【出願人】(596118493)アカデミア シニカ (33)
【氏名又は名称原語表記】ACADEMIA SINICA
【住所又は居所原語表記】128 Sec 2,Academia Road,Nankang,Taipei 11529 TW
【Fターム(参考)】
[ Back to top ]