
B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体、それに関連する遺伝子およびペプチド、並びにB型肝炎ウイルスの検定方法、B型肝炎の診断方法、治療方法
【課題】全てのタイプのHBVに対して中和活性を有し得るHBs抗原特異的モノクローナル抗体を提供する。
【解決手段】HBs抗原特異的モノクローナル抗体。B型肝炎ウイルスのHBs抗原に対する単鎖抗体(scFv)、上記HBs抗原特異的モノクローナル抗体を構成する重鎖可変領域および軽鎖可変領域であるタンパク質、上記HBs抗原特異的モノクローナル抗体を構成する重鎖相補性決定領域1〜3および軽鎖相補性決定領域1〜3であるペプチド、上記HBs抗原特異的モノクローナル抗体を構成する重鎖可変領域および軽鎖可変領域をコードするDNA(遺伝子)、並びに上記HBs抗原特異的モノクローナル抗体を構成する重鎖相補性決定領域1〜3および軽鎖相補性決定領域1〜3をコードするDNA(遺伝子)。これらモノクローナル抗体等を利用するB型肝炎ウイルスの検定方法、B型肝炎の診断方法およびB型肝炎の治療方法。
【解決手段】HBs抗原特異的モノクローナル抗体。B型肝炎ウイルスのHBs抗原に対する単鎖抗体(scFv)、上記HBs抗原特異的モノクローナル抗体を構成する重鎖可変領域および軽鎖可変領域であるタンパク質、上記HBs抗原特異的モノクローナル抗体を構成する重鎖相補性決定領域1〜3および軽鎖相補性決定領域1〜3であるペプチド、上記HBs抗原特異的モノクローナル抗体を構成する重鎖可変領域および軽鎖可変領域をコードするDNA(遺伝子)、並びに上記HBs抗原特異的モノクローナル抗体を構成する重鎖相補性決定領域1〜3および軽鎖相補性決定領域1〜3をコードするDNA(遺伝子)。これらモノクローナル抗体等を利用するB型肝炎ウイルスの検定方法、B型肝炎の診断方法およびB型肝炎の治療方法。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の重鎖または軽鎖の可変領域または相補性決定領域をコードする遺伝子に関する。さらに本発明は、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の重鎖または軽鎖の可変領域または相補性決定領域であるペプチドに関する。さらに本発明は、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体、単鎖抗体(scFv)、その二量体および三量体、ならびにこのモノクローナル抗体、単鎖抗体(scFv)、その二量体および三量体、を用いるB型肝炎ウイルスの検定方法、B型肝炎の診断方法およびB型肝炎の治療方法に関する。
【背景技術】
【0002】
B型肝炎はB型肝炎ウイルスが肝細胞に感染することにより誘発され、肝不全、肝がんの誘発により死に至る病気である。B型肝炎ウイルスは病院等において、針刺し事故等により患者の血液が誤って体内に注入されることによりしばしば感染する。このような場合、現在ではHBワクチン接種等によりB型肝炎ウイルスに対する抗体を大量に持つようになった人の血液より抗血清を調製したものおよびHBワクチンを緊急に注射することにより、B型肝炎ウイルスの肝細胞への感染を抗体により中和(ブロック)することで感染を防ぐことが一般的に行われている。しかし、このように人から調製した抗血清は未知のウイルスの混入の可能性を除去することが難しく、人に用いる治療薬としては好ましいものではない。
【0003】
そこで、HBs抗原特異的抗体を作成し、それを中和(ブロック)に使用することが考えられている。
【0004】
[キメラ抗体、ヒト化抗体]
HBs抗原特異的抗体の作成方法(スクリーニング方法)としては、マウスをHBs抗原で免疫し、脾臓あるいはリンパ節のBリンパ球とミエローマ細胞を細胞融合法により融合し、HBs抗原特異的抗体を分泌しているハイブリドーマ細胞をELISA法等によりスクリーニングする。樹立したハイブリドーマよりマウス抗体遺伝子を回収し、マウスの定常部領域をヒトの定常部領域と置換(キメラ化)するか、マウスの抗体の抗原結合部位(相補性決定領域)(CDR1, 2, 3)をヒトの抗体のそれと置換(ヒト化)することにより、HBs抗原への結合性を保持しながら、ヒト抗体に近づける方法が知られている。[J Waters, M Pignatelli, S Galpin, K Ishihara, HC Thomas, Virus-neutralizing antibodies to hepatitis B virus: The nature of an immunogenic epitope on the S gene peptide. J gen Virol, 67:2767-2473, 1986(非特許文献1)、S Iwarson, E Tabor, HC Thomas, A Goodall, J Waters, P Snoy, JW-K Shih, RJ Gerety, Neutralization of hepatitis B virus infectivity by a murine monoclonal antibody: and experimental study in the Chimpanzee. J Med Virol, 16:89-96, 1985(非特許文献2)、CJ Ryu, P Gripon, HR Park, SS Park, YK Kim, C Guguen-Guillouzo, OJ Yoo, HJ Hong, In vitro neutralization of hepatitis B virus by monoclonal antibodies against the viral surface antigen. J Med Virol, 52:226-233, 1997(非特許文献5)、HJ Hong, CJ Ryu, H Hur, S Kim, HK Oh, MS Oh, SY Park. In vivo neutralization of hepatitis B virus infection by an anti-preS1 humanized antibody in chimpanzees. Virology, 318:134-141, 2004(非特許文献6)、CJ Ryu, EA Pallan, BR Jin, OJ Yoo, HJ Hong. A humanized antibody with specificity for hepatitis B surface antigen. Hum Antibo Hybridomas, 7:113-122, 1996(非特許文献7)]
【0005】
具体的なモノクローナル抗体としては、B型肝炎ウイルス表面抗原(HBs抗原)に結合できるヒトモノクローナル抗体が知られている。特表2000−513215号公報(特許文献1)には、B型肝炎ウイルス表面抗原(HBs抗原)に結合できるヒト抗体を産生するハイブリドーマ細胞系を得る方法、ならびにハイブリドーマ細胞系、およびその細胞系によって産生される抗体が開示されている。また、HBV感染の予防および処置における上記抗体の様々な使用が開示される。この発明では、抗HBs抗原抗体の高力価を有するヒトドナーから得られる末梢血リンパ球を、致死的に放射線を照射されSCID骨髄で保護された正常株マウスに移植する。このようなキメラマウスをHBs抗原で免疫したのち、そのマウス脾臓からヒト細胞を得て、異種骨髄腫細胞とインビトロ融合させ、HBs抗原に対して高い親和性および特異性を有するヒト抗体を分泌するハイブリドーマ細胞を発生させる。
【0006】
上記以外にもHBs抗原特異的抗体が報告され、以下の文献に記載されている。
(1)Maeda F et al. J Med Virol. 58:338-345, 1999(非特許文献8)
(2)Yano A et al. J Med Virol. 73:208-215, 2004(非特許文献9)
(3)日大医誌 41:1163-1171, 1982 (非特許文献10)
(4)Hybridoma 17 (6), 535-540 (1998) (非特許文献11)
(5)Zhonghua Gan Zang Bing Za Zhi 5 (3), 130-132 (2000) (非特許文献12)
(6)特開平9−20798号公報(特許文献2)
(7)Biochim. Biophys. Acta 1380 (2), 151-155 (1998) (非特許文献13)
(8)Proc. Natl. Acad. Sci. U.S.A. 89 (8), 3175-3179 (1992) (非特許文献14)
【0007】
また、HBs抗原特異的抗体の作成方法(スクリーニング方法)としては、以下の方法も知られている。
【0008】
[ヒトハイブリドーマ法]
末梢血Bリンパ球とミエローマ細胞を細胞融合法により融合し、HBs抗原特異的抗体を分泌しているハイブリドーマ細胞をELISA法等によりスクリーニングする。[H Sawada, S Iwasa, O Nishimura, K Kitano, Efficient production of anti-(hepatitis B virus) antibodies and their neutralizing activity in chimpanzees. Appl Microbiol Biotechnol, 43:445-451, 1995(非特許文献3)、Gary Witherell, XTL-001 XTL Pharmaceuticals. Current Opinion in Investigational Drugs, 3:684-692, 2002(非特許文献4)]
【0009】
[EBウイルス形質転換法]
末梢血ヒトBリンパ球にEBウイルスを感染させ不死化させB細胞株を確立し、HBs抗原特異的抗体を分泌しているB細胞株をELISA法等によりスクリーニングする。
【特許文献1】特表2000−513215号公報
【特許文献2】特開平9−20798号公報
【非特許文献1】J Waters, M Pignatelli, S Galpin, K Ishihara, HC Thomas, Virus-neutralizing antibodies to hepatitis B virus: The nature of an immunogenic epitope on the S gene peptide. J gen Virol, 67:2767-2473, 1986
【非特許文献2】S Iwarson, E Tabor, HC Thomas, A Goodall, J Waters, P Snoy, JW-K Shih, RJ Gerety, Neutralization of hepatitis B virus infectivity by a murine monoclonal antibody: and experimental study in the Chimpanzee. J Med Virol, 16:89-96, 1985
【非特許文献3】H Sawada, S Iwasa, O Nishimura, K Kitano, Efficient production of anti-(hepatitis B virus) antibodies and their neutralizing activity in chimpanzees. Appl Microbiol Biotechnol, 43:445-451, 1995
【非特許文献4】Gary Witherell, XTL-001 XTL Pharmaceuticals. Current Opinion in Investigational Drugs, 3:684-692, 2002
【非特許文献5】CJ Ryu, P Gripon, HR Park, SS Park, YK Kim, C Guguen-Guillouzo, OJ Yoo, HJ Hong, In vitro neutralization of hepatitis B virus by monoclonal antibodies against the viral surface antigen. J Med Virol, 52:226-233, 1997
【非特許文献6】HJ Hong, CJ Ryu, H Hur, S Kim, HK Oh, MS Oh, SY Park. In vivo neutralization of hepatitis B virus infection by an anti-preS1 humanized antibody in chimpanzees. Virology, 318:134-141, 2004
【非特許文献7】CJ Ryu, EA Pallan, BR Jin, OJ Yoo, HJ Hong. A humanized antibody with specificity for hepatitis B surface antigen. Hum Antibo Hybridomas, 7:113-122, 1996
【非特許文献8】Maeda F et al. J Med Virol. 58:338-345, 1999
【非特許文献9】Yano A et al. J Med Virol. 73:208-215, 2004
【非特許文献10】日大医誌 41:1163-1171, 1982
【非特許文献11】Hybridoma 17 (6), 535-540 (1998)
【非特許文献12】Zhonghua Gan Zang Bing Za Zhi 5 (3), 130-132 (2000)
【非特許文献13】Biochim. Biophys. Acta 1380 (2), 151-155 (1998)
【非特許文献14】Proc. Natl. Acad. Sci. U.S.A. 89 (8), 3175-3179 (1992)
【発明の開示】
【発明が解決しようとする課題】
【0010】
HBVとしては、アミノ酸の違いで10種類以上の異なるHBVが知られているが、特許文献1に記載の抗体はその一部しか認識できないという問題があった。さらに、特許文献1に記載の抗体は、中和実験の結果では、HBVを100%は中和できてはいないという問題もあった。
【0011】
また、非特許文献8〜10には、非特許文献8〜10に記載の抗体に中和があることは記載されているが、具体的なデータが示されていない。さらに、非特許文献11〜14および特許文献2には、そこに記載の抗体が中和活性を有するものであるかについての記載がない。
【0012】
そこで、本発明の目的は、全てのタイプのHBVに対して中和活性を有し得るHBs抗原特異的モノクローナル抗体を提供することにある。
【0013】
さらに本発明は、上記HBs抗原特異的モノクローナル抗体に加えて、
(1)B型肝炎ウイルスのHBs抗原に対する単鎖抗体(scFv)
(2)上記HBs抗原特異的モノクローナル抗体を構成する重鎖可変領域および軽鎖可変領域であるタンパク質、
(3)上記HBs抗原特異的モノクローナル抗体を構成する重鎖相補性決定領域1〜3および軽鎖相補性決定領域1〜3であるペプチド、
(4)上記HBs抗原特異的モノクローナル抗体を構成する重鎖可変領域および軽鎖可変領域をコードするDNA(遺伝子)、並びに
(5)上記HBs抗原特異的モノクローナル抗体を構成する重鎖相補性決定領域1〜3および軽鎖相補性決定領域1〜3をコードするDNA(遺伝子)
を提供することも目的とする。
【0014】
本発明者らは、B型肝炎ウイルスに対する抗体を細胞チップ技術および組換えDNA技術を用いて作製し、それを治療薬として用いることを考えた。すなわち、ワクチン接種した人の血液よりBリンパ球を調製し、それを本発明者の関係者らが開発したマイクロウェルアレイチップに播種し、B型肝炎ウイルス表面抗原(HBs抗原)で刺激し、スキャナーを用いてHBs抗原に反応するBリンパ球を検出した。検出したBリンパ球をチップより回収し、抗体遺伝子を回収し、発現ベクターに組込み動物細胞に導入することにより、組換え抗体を作製した。作製した組換え抗体は、HBs抗原に結合すること、さらには、ヒト肝細胞へのウイルスの感染をブロックすること(中和すること)が確認され、本発明を完成した。
【課題を解決するための手段】
【0015】
本発明は、以下の通りである。
[1]下記の何れかのDNAからなる遺伝子。
(1)配列番号1の塩基配列を有するDNA、
(2)配列番号1の塩基配列において1から数個の塩基の欠失、置換及び/又は付加を有する塩基配列を有し、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の重鎖可変領域をコードするDNA、および
(3)配列番号1の塩基配列またはこの塩基配列と相補的な塩基配列とストリンジェントな条件下でハイブリダイズする塩基配列を有し、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の重鎖可変領域をコードするDNA。
[2]下記の何れかのタンパク質をコードする遺伝子。
(1)配列番号2のアミノ酸配列を有するタンパク質、
(2)配列番号2のアミノ酸配列において1から数個のアミノ酸の欠失、置換及び/又は付加を有するアミノ酸配列を有し、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の重鎖可変領域であるタンパク質、および
(3)配列番号2のアミノ酸配列に対して90%以上の相同性を有するアミノ酸配列を有し、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の重鎖可変領域であるタンパク質。
[3]下記の何れかのDNAからなる遺伝子。
(1)配列番号3の配列を有するDNA、
(2)配列番号3の塩基配列において1から数個の塩基の欠失、置換及び/又は付加を有する塩基配列を有し、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の軽鎖可変領域をコードするDNA、および
(3)配列番号3の塩基配列またはこの塩基配列と相補的な塩基配列とストリンジェントな条件下でハイブリダイズする塩基配列を有し、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の軽鎖可変領域をコードするDNA。
[4]下記の何れかのタンパク質をコードする遺伝子。
(1)配列番号4のアミノ酸配列を有するタンパク質、
(2)配列番号4のアミノ酸配列において1から数個のアミノ酸の欠失、置換及び/又は付加を有するアミノ酸配列を有し、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の軽鎖可変領域であるタンパク質、および
(3)配列番号4のアミノ酸配列に対して90%以上の相同性を有するアミノ酸配列を有し、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の軽鎖可変領域であるタンパク質。
[5]下記の何れかのタンパク質。
(1)配列番号2のアミノ酸配列を有するタンパク質、
(2)配列番号2のアミノ酸配列において1から数個のアミノ酸の欠失、置換及び/又は付加を有するアミノ酸配列を有し、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の重鎖可変領域であるタンパク質、および
(3)配列番号2のアミノ酸配列に対して90%以上の相同性を有するアミノ酸配列を有し、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の重鎖可変領域であるタンパク質。
[6]下記の何れかのタンパク質。
(1)配列番号4のアミノ酸配列を有するタンパク質、
(2)配列番号4のアミノ酸配列において1から数個のアミノ酸の欠失、置換及び/又は付加を有するアミノ酸配列を有し、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の軽鎖可変領域であるタンパク質、および
(3)配列番号4のアミノ酸配列に対して90%以上の相同性を有するアミノ酸配列を有し、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の軽鎖可変領域であるタンパク質。
[7]下記の何れかの遺伝子。
(1)配列番号1の塩基配列における76〜105番目の塩基配列、または配列番号3のアミノ酸配列における26〜35番目のアミノ酸配列をコードする塩基配列を有する、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の重鎖相補性決定領域1(CDR−H1)をコードする遺伝子。
(2)配列番号1の塩基配列における148〜198の塩基配列、または配列番号3のアミノ酸配列における50〜66番目のアミノ酸配列をコードする塩基配列を有する、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の重鎖相補性決定領域2(CDR−H2)をコードする遺伝子。
(3)配列番号1の塩基配列における295〜333の塩基配列、または配列番号3のアミノ酸配列における99〜111番目のアミノ酸配列をコードする塩基配列を有する、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の重鎖相補性決定領域3(CDR−H3)をコードする遺伝子。
[8]下記の何れかの遺伝子。
(1)配列番号3の塩基配列における69〜102の塩基配列、または配列番号4のアミノ酸配列における24〜34のアミノ酸配列をコードする塩基配列を有する、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の軽鎖相補性決定領域1(CDR−L1)をコードする遺伝子。
(2)配列番号3の塩基配列における148〜168の塩基配列、または配列番号4のアミノ酸配列における50〜56のアミノ酸配列をコードする塩基配列を有する、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の軽鎖相補性決定領域2(CDR−L2)をコードする遺伝子。
(3)配列番号3の塩基配列における265〜291の塩基配列、または配列番号4のアミノ酸配列における89〜97のアミノ酸配列をコードする塩基配列を有する、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の軽鎖相補性決定領域3(CDR−L3)をコードする遺伝子。
[9]下記のいずれかのペプチド。
(1)配列番号2のアミノ酸配列における26〜35のアミノ酸配列を有する、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の重鎖相補性決定領域1(CDR−H1)であるペプチド、
(2)配列番号2のアミノ酸配列における50〜66のアミノ酸配列を有する、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の重鎖相補性決定領域2(CDR−H2)であるペプチド、
(3)配列番号2のアミノ酸配列における99〜111のアミノ酸配列を有する、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の重鎖相補性決定領域3(CDR−H3)であるペプチド。
[10]下記のいずれかのペプチド。
(1)配列番号4のアミノ酸配列における24〜34のアミノ酸配列を有する、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の軽鎖相補性決定領域1(CDR−L1)であるペプチド、
(2)配列番号4のアミノ酸配列における50〜56のアミノ酸配列を有する、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の軽鎖相補性決定領域2(CDR−L2)であるペプチド、
(3)配列番号4のアミノ酸配列における89〜97のアミノ酸配列を有する、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の軽鎖相補性決定領域3(CDR−L3)であるペプチド。
[11]配列番号2のアミノ酸配列を有するタンパク質を可変領域として含む重鎖と配列番号4のアミノ酸配列を有するタンパク質を可変領域として含む軽鎖とを含む、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体。
[12]配列番号2のアミノ酸配列における26〜35、50〜66および99〜111のアミノ酸配列を有するペプチドを相補性決定領域1〜3として含む重鎖と配列番号4のアミノ酸配列における24〜34、50〜56および89〜97のアミノ酸配列を有するペプチドを相補性決定領域1〜3として含む軽鎖とを含む、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体。
[13]配列番号2のアミノ酸配列を有するタンパク質と配列番号4のアミノ酸配列を有するタンパク質とをリンカーペプチドで結合した、B型肝炎ウイルスのHBs抗原に対する単鎖抗体。
[14]配列番号2のアミノ酸配列における26〜35、50〜66および99〜111のアミノ酸配列を有するペプチドを相補性決定領域1〜3として含むタンパク質と配列番号4のアミノ酸配列における24〜34、50〜56および89〜97のアミノ酸配列を有するペプチドを相補性決定領域1〜3として含むタンパク質とをリンカーペプチドで結合した、B型肝炎ウイルスのHBs抗原に対する単鎖抗体。
[15][13]または[14]に記載の単鎖抗体の二量体。
[16][13]または[14]に記載の単鎖抗体の三量体。
[17]配列番号2のアミノ酸配列を有するタンパク質と配列番号4のアミノ酸配列を有するタンパク質とをリンカーペプチドで結合し、かつ配列番号2のアミノ酸配列を有する前記タンパク質にCH3領域を結合した、B型肝炎ウイルスのHBs抗原に対するミニボディー。
[18]配列番号2のアミノ酸配列における26〜35、50〜66および99〜111のアミノ酸配列を有するペプチドを相補性決定領域1〜3として含むタンパク質と配列番号4のアミノ酸配列における24〜34、50〜56および89〜97のアミノ酸配列を有するペプチドを相補性決定領域1〜3として含むタンパク質とをリンカーペプチドで結合し、かつ配列番号2のアミノ酸配列における26〜35、50〜66および99〜111のアミノ酸配列を有するペプチドを相補性決定領域1〜3として含む前記タンパク質にCH3領域を結合した、B型肝炎ウイルスのHBs抗原に対するミニボディー(minibody)。
[19][5]若しくは[6]に記載のタンパク質、[9]若しくは[10]に記載のペプチド、[11]若しくは[12]に記載のモノクローナル抗体、[13]若しくは[14]に記載の単鎖抗体、[15]に記載の二量体、[16]に記載の三量体、[17]若しくは[18]に記載のミニボディーを用いるB型肝炎ウイルスの検定方法。
[20][5]若しくは[6]に記載のタンパク質、[9]若しくは[10]に記載のペプチド、[11]または[12]に記載のモノクローナル抗体、または[13]若しくは[14]に記載の単鎖抗体、[15]に記載の二量体、[16]に記載の三量体、[17]若しくは[18]に記載のミニボディーを用いるB型肝炎の診断方法。
[21][5]若しくは[6]に記載のタンパク質、[9]若しくは[10]に記載のペプチド、[11]または[12]に記載のモノクローナル抗体、または[13]若しくは[14]に記載の単鎖抗体、[15]に記載の二量体、[16]に記載の三量体、[17]若しくは[18]に記載のミニボディーを用いるB型肝炎の治療方法。
【発明の効果】
【0016】
本発明によれば、全てのタイプのHBVに対して中和活性を有し得るHBs抗原特異的モノクローナル抗体を提供することができる。
【発明を実施するための最良の形態】
【0017】
前述のように、本発明の組換え抗体は、ワクチン接種した人の血液より調製したBリンパ球をB型肝炎ウイルス表面抗原(HBs抗原)で刺激し、HBs抗原に反応する1個のBリンパ球を回収し、回収した1個のBリンパ球から抗体遺伝子を採取し、この抗体遺伝子を含む発現ベクターで形質転換した動物細胞を用いて作製した。この過程で、抗体遺伝子の塩基配列および抗体のアミノ酸配列を重鎖および軽鎖についてそれぞれ決定し、さらに、重鎖および軽鎖における相補性決定領域を決定した。
【0018】
本発明は、
(1)B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体、および単鎖抗体(scFv)
(2)このモノクローナル抗体を構成する重鎖可変領域および軽鎖可変領域であるタンパク質、
(3)このモノクローナル抗体を構成する重鎖相補性決定領域1〜3および軽鎖相補性決定領域1〜3であるペプチド、
(4)このモノクローナル抗体を構成する重鎖可変領域および軽鎖可変領域をコードするDNA(遺伝子)、並びに
(5)このモノクローナル抗体を構成する重鎖相補性決定領域1〜3および軽鎖相補性決定領域1〜3をコードするDNA(遺伝子)に関する。
【0019】
[可変領域をコードするDNA(遺伝子)]
本発明の重鎖可変領域をコードするDNA(遺伝子)は、下記の何れかのDNAからなる。
(1)配列番号1の塩基配列を有するDNA、
(2)配列番号1の塩基配列において1から数個の塩基の欠失、置換及び/又は付加を有する塩基配列を有し、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の重鎖可変領域をコードするDNA、および
(3)配列番号1の塩基配列またはこの塩基配列と相補的な塩基配列とストリンジェントな条件下でハイブリダイズする塩基配列を有し、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の重鎖可変領域をコードするDNA。
【0020】
さらに本発明の重鎖可変領域をコードするDNA(遺伝子)は、下記の何れかのタンパク質をコードする遺伝子である。
(1)配列番号2のアミノ酸配列を有するタンパク質、
(2)配列番号2のアミノ酸配列において1から数個のアミノ酸の欠失、置換及び/又は付加を有するアミノ酸配列を有し、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の重鎖可変領域であるタンパク質、および
(3)配列番号2のアミノ酸配列に対して90%以上の相同性を有するアミノ酸配列を有し、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の重鎖可変領域であるタンパク質。
【0021】
本発明の軽鎖可変領域をコードするDNA(遺伝子)は、下記の何れかのDNAからなる。
(1)配列番号3の配列を有するDNA、
(2)配列番号3の塩基配列において1から数個の塩基の欠失、置換及び/又は付加を有する塩基配列を有し、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の軽鎖可変領域をコードするDNA、および
(3)配列番号3の塩基配列またはこの塩基配列と相補的な塩基配列とストリンジェントな条件下でハイブリダイズする塩基配列を有し、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の軽鎖可変領域をコードするDNA。
【0022】
さらに、本発明の軽鎖可変領域をコードするDNA(遺伝子)は、下記の何れかのタンパク質をコードする遺伝子である。
(1)配列番号4のアミノ酸配列を有するタンパク質、
(2)配列番号4のアミノ酸配列において1から数個のアミノ酸の欠失、置換及び/又は付加を有するアミノ酸配列を有し、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の軽鎖可変領域であるタンパク質、および
(3)配列番号4のアミノ酸配列に対して90%以上の相同性を有するアミノ酸配列を有し、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の軽鎖可変領域であるタンパク質。
【0023】
配列番号1の塩基配列は、H鎖(重鎖)のV領域のcDNAの塩基配列であり、配列番号2のアミノ酸配列は、H鎖のV領域のアミノ酸である。さらに、配列番号3の塩基配列はL鎖(軽鎖)のV領域のcDNAの塩基配列であり、配列番号4のアミノ酸配列は、L鎖のV領域のアミノ酸配列である。これらの配列は、実施例で詳述されているように、HBs抗原に反応したBリンパ球を回収し、回収したBリンパ球からcDNAを合成し、合成したcDNAを用いて抗体のH鎖およびL鎖のV領域を増幅するようにPCR反応を行い、取得した細胞の抗体のH鎖およびL鎖のV領域のcDNAを作製し、作製したcDNAの塩基配列を決定し、さらに、このcDNAがタンパクに翻訳されることを確認して、決定した。その結果、本発明の抗体のH鎖およびL鎖のファミリー(サブクラス)は、H鎖は、VH3, DH, JH2であり、L鎖(κ鎖)はVκ3, Jκ4であった。
【0024】
本明細書で言う「1から数個の塩基の欠失、置換及び/又は付加を有する塩基配列」における「1から数個」の範囲は特には限定されないが、例えば、1から40個、好ましくは1から30個、より好ましくは1から20個、より好ましくは1から10個、さらに好ましくは1から5個、特に好ましくは1から3個程度を意味する。以下においても同様である。
【0025】
上記本発明のDNAが、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の重鎖または軽鎖の可変領域をコードするDNAであることは、このDNAを鋳型として作成されたタンパク質が、B型肝炎ウイルスのHBs抗原に対する反応性を有することで確認できる。B型肝炎ウイルスのHBs抗原に対する反応性は、例えば、酵素標識した二次抗体を用いるEnzyme linked immunosorbent assay (ELISA)、放射性同位元素で標識した二次抗体を用いるradio immuno assay (RIA)等により確認することができる。尚、DNAを鋳型として作成されるタンパク質が抗体の一部である場合、抗体と抗体の一部の抗原に対する反応性は同じようにELISA等で測定できる。ただし、例えばscFvの場合には抗体のFc部分が欠失しているので、二次抗体として酵素標識抗IgG抗体を用いることはできない。この場合はscFvにタグ(抗体のエピトープとなるペプチド)を付加して、そのタグに対する抗体等を二次抗体として用いることで、ELISA等で測定できる。
【0026】
ELISAは96穴プレートのウェルにHBs抗原をコートする。目的抗体をHBs抗原をコートしたウェルおよびしていないウェルに添加し、HBs抗原への目的抗体の結合を酵素標識した二次抗体の結合により酵素基質の発色を指標に確認することができる。この場合、抗原をコートしていないウェルには目的抗体は結合しないため発色せず、抗原をコートしたウェルには目的抗体が結合し発色する。RIAでは同様に96穴プレートにHBs抗原をコートする。目的抗体をHBs抗原をコートしたウェルおよびしていないウェルに添加し、HBs抗原への目的抗体の結合を放射性同位元素で標識した二次抗体の結合による放射能のカウントを指標に確認することができる。
【0027】
上記した「ストリンジェントな条件下でハイブリダイズする」とは、DNAをプローブとして使用し、コロニーハイブリダイゼーション法、プラークハイブリダイゼーション法、あるいはサザンブロットハイブリダイゼーション法等を用いることにより得られるDNAの塩基配列を意味し、例えば、コロニーあるいはプラーク由来のDNA又は該DNAの断片を固定化したフィルターを用いて、0.7〜1.0MのNaCl存在下、65℃でハイブリダイゼーションを行った後、0.1〜2×SSC溶液(1×SSC溶液は、150mM塩化ナトリウム、15mMクエン酸ナトリウム)を用い、65℃条件下でフィルターを洗浄することにより同定できるDNA等を挙げることができる。ハイブリダイゼーションは、モレキュラークローニング第2版等に記載されている方法に準じて行うことができる。
【0028】
ストリンジェントな条件下でハイブリダイズするDNAとしては、プローブとして使用するDNAの塩基配列と一定以上の相同性を有するDNAが挙げられ、例えば70%以上、好ましくは80%以上、より好ましくは90%以上、さらに好ましくは93%以上、特に好ましくは95%以上、最も好ましくは98%以上の相同性を有するDNAが挙げられる。
【0029】
本明細書で言う「1から数個のアミノ酸の欠失、置換及び/又は付加を有するアミノ酸配列」における「1から数個」の範囲は特には限定されないが、例えば、1から20個、好ましくは1から10個、より好ましくは1から7個、さらに好ましくは1から5個、特に好ましくは1から3個程度を意味する。
【0030】
本明細書で言う「配列表の配列番号2または4に記載のアミノ酸配列に対して90%以上の相同性を有するアミノ酸配列」における相同性は、90%以上であれば特に限定されないが、さらに好ましくは95%以上、さらに好ましくは97%以上、さらに好ましくは98%、特に好ましくは99%以上である。
【0031】
B型肝炎ウイルスのHBs抗原に対する反応性を有する抗体は既にいくつか報告されている。(特許文献1および2、非特許文献8〜14)それに対して、本発明の抗体のH鎖およびL鎖のファミリー(サブクラス)は、前述のとおりであり、本発明の抗体とH鎖およびL鎖のファミリー(サブクラス)が、H鎖についてVH3であり、かつL鎖(κ鎖)についてVκ3である抗体を、以下の表に示す。相同性は、本発明の抗体の該当する可変領域のアミノ酸配列との相同性である。
【0032】
【表1】

【0033】
本発明の遺伝子の取得方法は特に限定されない。本明細書中の配列表の配列番号1から4に記載したアミノ酸配列及び塩基配列の情報に基づいて適当なブローブやプライマーを調製し、それらを用いて抗体のcDNAライブラリーをスクリーニングすることにより本発明の遺伝子を単離することができる。単離した遺伝子が本発明の遺伝子であるかは、塩基配列の決定および遺伝子より発現させた抗体タンパクのHBs抗原への結合能をチェックすることにより調べることができる。cDNAライブラリーは、本発明の遺伝子を発現している細胞から常法により作製することができる。
【0034】
PCR法により本発明の遺伝子を取得することもできる。上記抗体の染色体DNAライブラリー又はcDNAライブラリーを鋳型として使用し、配列番号1又は3に記載した塩基配列を増幅できるように設計した1対のプライマーを用いてPCRを行う。PCRの反応条件は適宜設定することができ、例えば、94℃で30秒間(変性)、55℃で30秒〜1分間(アニーリング)、72℃で2分間(伸長)からなる反応工程を1サイクルとして、例えば30サイクル行った後、72℃で7分間反応させる条件などを挙げることができる。次いで、増幅されたDNA断片を、大腸菌等の宿主で増幅可能な適切なベクター中にクローニングすることができる。単離した遺伝子が本発明の遺伝子であるかは、塩基配列の決定および遺伝子より発現させた抗体タンパクのHBs抗原への結合能をチェックすることにより調べることができる。
【0035】
上記したプローブ又はプライマーの調製、cDNAライブラリーの構築、cDNAライブラリーのスクリーニング、並びに目的遺伝子のクローニング、塩基配列の決定、遺伝子からのタンパクの発現などの操作は当業者に既知であり、例えば、Molecular Cloning: A laboratory Mannual, 2nd Ed., Cold Spring Harbor Laboratory, Cold Spring Harbor, NY.,1989(以下、モレキュラークローニング第2版と略す)、Current Protocols in Molecular Biology, Supplement 1〜38, John Wiley & Sons (1987-1997)(以下、カレント・プロトコールズ・イン・モレキュラー・バイオロジーと略す)等に記載の方法に準じて行うことができる。
【0036】
上述した本発明の遺伝子の取得方法は、後述するCDRをコードする遺伝子の入手法にも適宜適用することができる。
【0037】
また、配列表の配列番号2または4に記載のアミノ酸配列において1から数個のアミノ酸の欠失、置換及び/又は付加を有するアミノ酸配列を有し、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の重鎖可変領域または軽鎖可変領域であるタンパク質をコードする遺伝子;並びに配列表の配列番号1または3に記載の塩基配列において1から数個の塩基の欠失、置換及び/又は付加を有する塩基配列を有し、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の重鎖可変領域または軽鎖可変領域をコードする遺伝子(以下、これらの遺伝子を変異遺伝子と称する)については、配列番号1〜4に記載のアミノ酸配列および塩基配列の情報に基づいて、化学合成、遺伝子工学的手法又は突然変異誘発などの当業者に既知の任意の方法で作製することができる。
【0038】
例えば、配列表の配列番号1または3に記載の塩基配列を有するDNAに対し、変異原となる薬剤と接触作用させる方法、紫外線を照射する方法、遺伝子工学的手法等を用いて行うことができる。遺伝子工学的手法の一つである部位特異的変異誘発法は特定の位置に特定の変異を導入できる手法であることから有用であり、モレキュラークローニング第2版、カレント・プロトコールズ・イン・モレキュラー・バイオロジー等に記載の方法に準じて行うことができる。
【0039】
[可変領域タンパク質]
本発明のB型肝炎ウイルスのHBs抗原に対するモノクローナル抗体を構成する重鎖可変領域であるタンパク質は、下記の何れかのタンパク質である。
(1)配列番号2のアミノ酸配列を有するタンパク質、
(2)配列番号2のアミノ酸配列において1から数個のアミノ酸の欠失、置換及び/又は付加を有するアミノ酸配列を有し、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の重鎖可変領域であるタンパク質、および
(3)配列番号2のアミノ酸配列に対して90%以上の相同性を有するアミノ酸配列を有し、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の重鎖可変領域であるタンパク質。
【0040】
本発明のB型肝炎ウイルスのHBs抗原に対するモノクローナル抗体を構成する軽鎖可変領域であるタンパク質は、下記の何れかのタンパク質である。
(1)配列番号4のアミノ酸配列を有するタンパク質、
(2)配列番号4のアミノ酸配列において1から数個のアミノ酸の欠失、置換及び/又は付加を有するアミノ酸配列を有し、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の軽鎖可変領域であるタンパク質、および
(3)配列番号4のアミノ酸配列に対して90%以上の相同性を有するアミノ酸配列を有し、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の軽鎖可変領域であるタンパク質。
【0041】
上述のように、配列番号2および4のアミノ酸配列は、HBs抗原に対するモノクローナル抗体の可変領域のcDNAがタンパクに翻訳されることを確認して決定したものである。
【0042】
本発明のタンパク質の取得方法は特に制限されず、化学合成により合成したタンパク質でもよいし、遺伝子組み換え技術により作製した組み換えタンパク質でもよい。組み換えタンパク質を作製する場合には、先ず、上記に記載した当該タンパク質をコードする遺伝子(DNA)を取得する。このDNAを適当な発現系に導入することにより、本発明のタンパク質を産生することができる。また、本発明のCDRペプチドも同様にして産生することができる。発現系でのタンパク質、ペプチドの発現については本明細書中後記する。
【0043】
[相補性決定領域DNA(遺伝子)]
本発明のモノクローナル抗体を構成する重鎖相補性決定領域1〜3をコードするDNA(遺伝子)は、下記の何れかの遺伝子である。
(1)配列番号1の塩基配列における76〜105番目の塩基配列(配列番号5)、または配列番号3のアミノ酸配列における26〜35番目のアミノ酸配列(配列番号11)をコードする塩基配列を有する、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の重鎖相補性決定領域1(CDR−H1)をコードする遺伝子。
(2)配列番号1の塩基配列における148〜198の塩基配列(配列番号6)、または配列番号3のアミノ酸配列における50〜66番目のアミノ酸配列(配列番号12)をコードする塩基配列を有する、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の重鎖相補性決定領域2(CDR−H2)をコードする遺伝子。
(3)配列番号1の塩基配列における295〜333の塩基配列(配列番号7)、または配列番号3のアミノ酸配列における99〜111番目のアミノ酸配列(配列番号13)をコードする塩基配列を有する、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の重鎖相補性決定領域3(CDR−H3)をコードする遺伝子。
【0044】
さらに本発明のモノクローナル抗体を構成する軽鎖相補性決定領域1〜3をコードするDNA(遺伝子)は、下記の何れかの遺伝子である。
(1)配列番号3の塩基配列における69〜102の塩基配列(配列番号8)、または配列番号4のアミノ酸配列における24〜34のアミノ酸配列(配列番号14)をコードする塩基配列を有する、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の軽鎖相補性決定領域1(CDR−L1)をコードする遺伝子。
(2)配列番号3の塩基配列における148〜168の塩基配列(配列番号9)、または配列番号4のアミノ酸配列における50〜56のアミノ酸配列(配列番号15)をコードする塩基配列を有する、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の軽鎖相補性決定領域2(CDR−L2)をコードする遺伝子。
(3)配列番号3の塩基配列における265〜291の塩基配列(配列番号10)、または配列番号4のアミノ酸配列における89〜97のアミノ酸配列(配列番号16)をコードする塩基配列を有する、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の軽鎖相補性決定領域3(CDR−L3)をコードする遺伝子。
【0045】
[相補性決定領域ペプチド]
本発明のモノクローナル抗体を構成する重鎖相補性決定領域1〜3であるペプチドは、下記のいずれかのペプチドである。
(1)配列番号2のアミノ酸配列における26〜35のアミノ酸配列を有する、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の重鎖相補性決定領域1(CDR−H1)であるペプチド、
(2)配列番号2のアミノ酸配列における50〜66のアミノ酸配列を有する、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の重鎖相補性決定領域2(CDR−H2)であるペプチド、
(3)配列番号2のアミノ酸配列における99〜111のアミノ酸配列を有する、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の重鎖相補性決定領域3(CDR−H3)であるペプチド。
【0046】
本発明のモノクローナル抗体を構成する軽鎖相補性決定領域1〜3であるペプチドは、下記のいずれかのペプチドである。
(1)配列番号4のアミノ酸配列における24〜34のアミノ酸配列を有する、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の軽鎖相補性決定領域1(CDR−L1)であるペプチド、
(2)配列番号4のアミノ酸配列における50〜56のアミノ酸配列を有する、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の軽鎖相補性決定領域2(CDR−L2)であるペプチド、
(3)配列番号4のアミノ酸配列における89〜97のアミノ酸配列を有する、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の軽鎖相補性決定領域3(CDR−L3)であるペプチド。
【0047】
本発明のモノクローナル抗体の重鎖相補性決定領域1〜3および軽鎖相補性決定領域1〜3は、Kabatの基準に照らして決定した。さらに、本発明のモノクローナル抗体の重鎖相補性決定領域1〜3および軽鎖相補性決定領域1〜3について、本発明のモノクローナル抗体と、H鎖およびL鎖のファミリー(サブクラス)が同一である、抗体(H鎖: VH、L鎖(κ鎖): Vκ3)のアミノ酸配列の対比およびアミノ酸配列の相同性を表2〜7に示す。
【0048】
【表2】
【0049】
【表3】
【0050】
【表4】
【0051】
【表5】
【0052】
【表6】
【0053】
【表7】
【0054】
[モノクローナル抗体]
本発明のB型肝炎ウイルスのHBs抗原に対するモノクローナル抗体は、配列番号2のアミノ酸配列を有するタンパク質を可変領域として含む重鎖と配列番号4のアミノ酸配列を有するタンパク質を可変領域として含む軽鎖とを含むものである。
【0055】
さらに本発明のB型肝炎ウイルスのHBs抗原に対するモノクローナル抗体は、配列番号2のアミノ酸配列における26〜35、50〜66および99〜111のアミノ酸配列を有するペプチドを相補性決定領域1〜3として含む重鎖と配列番号4のアミノ酸配列における24〜34、50〜56および89〜97のアミノ酸配列を有するペプチドを相補性決定領域1〜3として含む軽鎖とを含むものである。
【0056】
本発明のモノクローナル抗体は、以下の文献に記載の方法を利用して作成することができる [Kanda H, Mori K, Koga H, Taniguchi K, Kobayashi H, Sakahara H, Konishi J, Endo K, Watanabe T.Construction and expression of chimeric antibodies by a simple replacement of heavy and light chain V genes into a single cassette vector. Hybridoma. 13:359-66, 1994] 。具体的には、HBs抗原特異的Bリンパ球よりRT-PCRにより抗体遺伝子の重鎖可変領域および軽鎖可変領域の遺伝子を増幅する。増幅した遺伝子をpT7Blue-T vector (Novagen, A Brand of EMD Biosciences, Inc, An affiliate of Merck KGaA, Darmstadt, Germany)等にクローニングし、塩基配列を決定する。例えば、配列番号1および3に塩基配列を示す。その塩基配列を鋳型に制限酵素部位を含むプライマーを用いてPCRを行い、増幅させた遺伝子断片を制限酵素により切断し、その制限酵素部位を用いて抗体発現用ベクターに導入する。抗体発現用ベクターは抗体タンパク質の分泌に必要なシグナル配列に加え、抗体重鎖の定常部領域DNAあるいは抗体軽鎖の定常部領域DNAがクローニングされており、それぞれ抗体重鎖可変領域、抗体軽鎖可変領域遺伝子を導入することにより、完全長の抗体遺伝子を構築できる。
【0057】
構築した抗体重鎖遺伝子および抗体軽鎖遺伝子を同時にヒト胎児腎由来293T細胞などの動物細胞に導入し、2〜3日間培養することで、目的の抗体タンパクは培養上清に分泌される。
【0058】
CDR遺伝子を組み合わせて用い、抗体を調製する場合、以下のように行うことができる。抗体のV領域は、抗体の基本的な三次元構造を形作るフレームワーク部分(1〜4)と抗原に結合するCDR部分(1〜3)からなる。フレームワークの構造と任意のCDRを組み合わせることで任意の抗体のV領域を合成することが可能である。これをどのように組合せて1本のV領域遺伝子を合成するかは、図12に示すように「オーバーラップPCR」を用いる。たとえばFr1とCDR1の遺伝子配列がある時、ステプ1でFr1とCDR3遺伝子をPCRで合成する。この時、Fr1とCDR1の間のプライマー(P2、P3)はそれぞれCDR1あるいはFr1の塩基配列を加えて重なるようにしておく。ステップ1で出来上がったPCR産物は塩基配列が重なっており、これらのPCR産物を熱変性およびアニーリングを行うと、重なった部分でアニーリングする(ステップ2)。これをTaq DNAポリメラーゼで伸長させるとステップ3のようにFr1とCDR1がつながったものが出来上がる。これをプライマーP1およびP4を用いてPCRで増幅すればFr1とCDR1がつながったDNAが増幅される。ステップ2とステップ3は同時に行うことができる。実際にはFr2、CDR2、Fr3、CDR3、Fr4も同じように増幅してやり、ステップ3にてプライマーP1とP5を用いると抗体のV領域の遺伝子が増幅される。CDRの部分をランダムな塩基配列にしてやると、あらゆる抗原に結合する可能性のあるV領域のライブラリーを作成することができる。
【0059】
[単鎖抗体]
本発明のB型肝炎ウイルスのHBs抗原に対する単鎖抗体は、配列番号2のアミノ酸配列を有するタンパク質と配列番号4のアミノ酸配列を有するタンパク質とをリンカーペプチドで結合したタンパク質である。さらに本発明のB型肝炎ウイルスのHBs抗原に対する単鎖抗体は、配列番号2のアミノ酸配列における26〜35、50〜66および99〜111のアミノ酸配列を有するペプチドを相補性決定領域1〜3として含むタンパク質と配列番号4のアミノ酸配列における24〜34、50〜56および89〜97のアミノ酸配列を有するペプチドを相補性決定領域1〜3として含むタンパク質とをリンカーペプチドで結合したタンパク質である。
【0060】
単鎖Fvタンパク(single chain Fv, scFv)は、抗体重鎖可変領域(VH)と抗体軽鎖可変領域(VL)を直列に結合したタンパク質であり、モノクローナル抗体と同様の方法で作製することができる。すなわち、上記モノクローナル抗体の作製方法で説明したpT7Blue-T vectorにクローニングした抗体重鎖可変領域遺伝子と抗体軽鎖可変領域遺伝子を鋳型として、この2つの領域をリンカー配列で結合させ、試験管内タンパク合成に適合するようT7プロモータ、リボソーム結合配列等を含む遺伝子をPCRを用いて作製する。作製した遺伝子を用いて、例えば無細胞系タンパク合成反応液PURESYSTEM S-S ((株)ポストゲノム研究所、東京)内でインキュベーションすることによりscFvタンパクを合成することができる。
【0061】
本発明は、上記単鎖抗体の二量体(diabody)および三量体(triabody)を包含する。さらに本発明は、上記単鎖抗体の配列番号2のアミノ酸配列を有するタンパク質または、配列番号2のアミノ酸配列における26〜35、50〜66および99〜111のアミノ酸配列を有するペプチドを相補性決定領域1〜3として含む前記タンパク質にCH3領域を結合した、B型肝炎ウイルスのHBs抗原に対するミニボディー(minibody)を包含する。
【0062】
単鎖Fvタンパクを構成するリンカーは、例えば、(GlyGlyGlyGlySer)を3つ連ねたものである。下記論文に記載のように、リンカーの長さを変えることによって、scFvが2つ組合わさった二量体(diabody)、3つ組合わさった三量体(triabody)が作製できる(Hudson PJ, Kortt AA, High avidity scFv multimers; diabodies and triabodies. J Immunol Methods, 231:177-189 (1999)参照、Edited by Frank Breitling, Sterfan Dubel "Recombinant Antibodies" (1999) A John Willey & Sons, Inc.(New York) and Spektrum Akademischer Verlag (Heidelberg) Copublication参照)。
【0063】
抗体を医薬として取り扱う場合、その分子量の大きさが問題となる。それを解決するために、scFv、二量体(diabody)、三量体(triabody)、ミニボディー(minibody)というような抗原結合能を保持したまま分子量を小さくした分子が作製され、抗体医薬としての評価が行われている。
【0064】
抗体遺伝子は図1のようにH鎖はVH遺伝子断片とCH1, (H), CH2, CH3、(CH4)遺伝子断片より、L鎖はVL遺伝子断片とCL遺伝子断片より構成されている。このうちのVHおよびVL遺伝子断片をリンカーで結合したものが単鎖Fv (scFv)である(図2)。scFvには5’-VH-VL-3’の構造のものと5’-VL-VH-3’のすなわち、scFvは抗体のVドメインより形成されC領域を有していない。これが二量体を形成したものがdiabody、三量体を形成したものがtriabodyである。これらはリンカーの長さを変えることで形成させることができるという報告がある(PJ Hudoson and AA Kortt, High avidity scFv multimers; diabodies and triabodies. J Immunol Methods 231:177-189, 1999)。scFvに比べるとdiabodyおよびtriabodyの方が総合的な結合力(avidity)は強くなる。また、scFvにIgGのCH3ドメインを結合したminibodyも抗原結合能を維持した抗体のミニチュア化の産物であり、CH3部分で二量体を形成する(LS Shahied, Y Tang, RK Alpaugh, R Somer, D Greenspon, LM Weiner.Bispecific minibodies targeting HER2/neu and CD16 exhibit improved tumor lysis when placed in a divalent tumor antigen binding format. J Biol Chem. 279:53907-53914)。ヒトIgGにはIgG1からIgG4の4つのバリエーションがあり、上記CH3部分は、IgG1からIgG4のいずれのCH3であることもできる。但し、IgG1のCH3部分であることが好ましい。
【0065】
[モノクローナル抗体の利用]
本発明は、上記本発明のモノクローナル抗体を用いるB型肝炎ウイルスの検定方法、B型肝炎の診断方法およびB型肝炎の治療方法を包含する。モノクローナル抗体の利用は、モノクローナル抗体そのもの以外に本発明のモノクローナル抗体の一部またはそれらのDNAを用いたもの、たとえばscFv単鎖抗体、単鎖抗体の二量体(diabody)、三量体(triabody)またはミニボディー(minibody)あるいはその遺伝子等を用いた利用も包含する。また、本発明のモノクローナル抗体の一部としては、例えば、このモノクローナル抗体を構成する重鎖可変領域および軽鎖可変領域であるタンパク質、並びに、このモノクローナル抗体を構成する重鎖相補性決定領域1〜3および軽鎖相補性決定領域1〜3であるペプチドを包含する。
【0066】
B型肝炎ウイルスの検定方法
本発明のモノクローナル抗体等を用いたB型肝炎ウイルスの検定方法は以下のように行うことができる。まず、96穴プレートのウェルに本発明のモノクローナル抗体を吸着させる。ブロッキングを行い非特異的吸着を抑えた後、B型肝炎ウイルスを含む検体をウェルに添加する。検体中にウイルスが含まれていれば、そのウイルスはウェルに吸着させた本発明のモノクローナル抗体に結合する。その後、ウェルを洗浄し、酵素標識した本発明モノクローナル抗体をウェルに添加する。ウェルを洗浄後、酵素基質をウェルに添加し、基質の発色で検体中のウイルスの存在を検出する。ウイルスが存在する検体を添加したウェルでは、ウェルに吸着させた抗体、それに結合したウイルス、さらにそのウイルスに結合した酵素標識抗体の複合体ができ、基質が発色する。
【0067】
また、本発明のモノクローナル抗体を用いたB型肝炎ウイルスの検定方法は以下のようにしても行うことができる。すなわち、本発明のモノクローナル抗体を96穴プレートあるいはビーズに固着させ、そこへB型肝炎ウイルスの存在をチェックする検体を添加する。B型肝炎ウイルスが存在すると、ウイルスが抗体に結合する。抗体に結合したウイルスよりウイルスDNAを抽出し、それを鋳型としてPCR等を行い、ウイルスDNAを増幅し、検出を行う。このようにしても検体中のウイルスの存在を検出することが可能である。
以上の方法によるウイルスの検出は本発明のモノクローナル抗体の一部、F(ab)’2, F(ab), scFv、単鎖抗体の二量体(diabody)、三量体(triabody)またはミニボディー(minibody)、このモノクローナル抗体を構成する重鎖可変領域および軽鎖可変領域であるタンパク質、並びに、このモノクローナル抗体を構成する重鎖相補性決定領域1〜3および軽鎖相補性決定領域1〜3であるペプチドなどを用いても行うことができる。
【0068】
B型肝炎の診断方法
本発明のモノクローナル抗体を用いたB型肝炎の診断は、上記検体として患者血清を調べることで、患者血清中のB型肝炎ウイルスを本発明のモノクローナル抗体を用いて検出することにより行うことができる。
また、患者の肝よりバイオプシーすることにより得た肝組織中のウイルスを本発明のモノクローナル抗体を用いて染色することにより診断することもできる。
以上の方法によるウイルスの診断は本発明のモノクローナル抗体の一部、F(ab)’2, F(ab), scFv、単鎖抗体の二量体(diabody)、三量体(triabody)またはミニボディー(minibody)、このモノクローナル抗体を構成する重鎖可変領域および軽鎖可変領域であるタンパク質、並びに、このモノクローナル抗体を構成する重鎖相補性決定領域1〜3および軽鎖相補性決定領域1〜3であるペプチドなどを用いても行うことができる。
【0069】
B型肝炎ウイルスの感染阻止およびB型肝炎の治療方法
本発明のモノクローナル抗体を用いたB型肝炎ウイルスの感染阻止は以下のように行うことができる。すなわち、B型肝炎ウイルスが混入している血液や体液、培養上清などが付着した注射針を誤って刺してしまったような、いわゆる針刺し事故においては、B型肝炎ウイルスワクチンをすぐに免疫すると同時にB型肝炎ウイルスに対する抗体価の高い抗血清を注入する治療法が一般的に行われている。すなわち、B型肝炎ウイルスに抗体が結合し細胞への感染を防ぐように作用する。しかし、人の血清製剤は献血由来の血清を材料とするため、未知のウイルスが混入する可能性もあり望ましいものではない。また、高力価で中和作用の優れた製剤を安定供給できるとは限らない。本発明のモノクローナル抗体を管理された環境で作製すれば、未知のウイルスの混入していないウイルス中和抗体を安定した品質および量で提供でき、上記ヒト由来抗血清の代わりに用いることができる。
【0070】
また、B型肝炎ウイルスが感染し、肝がん、肝硬変、肝不全に陥った患者で肝移植が治療法の選択肢となる場合がある。この場合、患者体内にB型肝炎ウイルスが残存していると移植した肝にB型肝炎ウイルスが再感染してしまい、治療効果が半減してしまうことになる。このような肝移植の場において、本発明のモノクローナル抗体を治療に用いることができる。すなわち、肝移植をするために患者の肝を除去した際に、本発明のモノクローナル抗体を単独あるいは他の抗体医薬や抗ウイルス剤と併用して患者に投与し、残存しているB型肝炎ウイルスを中和してやることにより、新たに移植した肝へのウイルスの感染を防ぐことができる。
【0071】
以上の方法による感染防止および治療は本発明のモノクローナル抗体の一部、F(ab)’2, F(ab), scFv、単鎖抗体の二量体(diabody)、三量体(triabody)またはミニボディー(minibody)、このモノクローナル抗体を構成する重鎖可変領域および軽鎖可変領域であるタンパク質、並びに、このモノクローナル抗体を構成する重鎖相補性決定領域1〜3および軽鎖相補性決定領域1〜3であるペプチドなどを用いても行うことができる。
【実施例】
【0072】
実施例1
[Bリンパ球の調製]
酵母菌で作製した組換えHBワクチンをHBs抗体陽性のボランティアに免疫し、免疫後9日目に末梢血よりリンパ球を調製した。すなわち、末梢血からヒトリンパ球をリンホセパールI液((株)免疫生物研究所、群馬県高崎市)を用いて遠心分離し、さらにAutoMACS (Miltenyi Biotec, Bergisch Gladbac, Germany)を用いてリンパ球画分から非Bリンパ球分画細胞を除去しBリンパ球画分を分離精製した。
【0073】
[Bリンパ球への蛍光色素の負荷]
調製した2 x 106個Bリンパ球を2μM Fluo-4/AM(カルシウム依存性蛍光色素、同仁、熊本)/1μM CellTracker Orange (Molecular Probe社)/0.04% pluronic F-127 (Molecular Probe社)/loading buffer (137 mM NaCl, 2.7 mM KCl,1.8 mM CaCl2, 1mM MgCl2, 1 mg/mLグルコース, 1mg/mL BSA, 20 mM Hepes (pH7.4))に懸濁し、ゆっくり震盪しながら室温で30分インキュベーションした。Loading bufferで細胞を洗浄し、細胞内に導入されなかった余分なFluo-4/AMを除いた。その後、細胞をRPMI1640/10% FCS溶液に懸濁した。なお、以下の全ての実験においてはPhenol Redを含まないRPMI1640を用いた。Fluo-4はカルシウムが結合すると蛍光強度が増加する蛍光色素であり、Bリンパ球の抗原による活性化の検出に用いる。CellTracker Orangeはカルシウムに関係なく蛍光を発する蛍光色素であり、マイクロアレイスキャナーを用いた細胞の認識あるいは蛍光顕微鏡下で細胞を回収する際の細胞の認識のために用いた。
【0074】
[マイクロウェルアレイチップ]
マイクロウェルアレイチップはシリコンを用いて作製されており(富山県工業技術センター製造)、直径10μm、深さ14μmのマイクロウェルが25μmのピッチ(ウェルの中心間の距離)で縦横に配列されている。マイクロウェルは30 x 30 (900ウェル)でひとつのクラスターを形成しており、それが縦に12クラスター、横に22クラスター並んでいるものを用いた。マイクロウェルアレイチップの両側には厚さ1mm、幅約1mm、長さ2cmのシールを貼った。
【0075】
[マイクロアレイスキャナー]
本装置は基本的に日立ソフトウェアエンジニアリング(株)(横浜市)のマイクロアレイスキャナー(CRBIO IIe-FITC)を用いており、473 nmおよび532 nmの波長の励起光を発することができるレーザーが搭載されている。また、2.5μmの解像度で解析が可能である。焦点深度は±50μmに設定されている。
【0076】
[マイクロウェルアレイチップへのリンパ球の播種]
マイクロウェルアレイチップのマイクロウェルへRPMI1640/10% FCS溶液を満たすために、マイクロウェルアレイチップをRPMI1640/10% FCS溶液に浸し、超音波をかけることでウェル中の空気を除くことにより、マイクロウェルへ溶液を満たした。マイクロウェルへのRPMI1640/10% FCS溶液の充填は以下の方法でも行った。マイクロウェルアレイチップへ100%アルコール液を添加する。アルコール液はマイクロウェルへ浸透していく。次に、アルコール液をPBSで置換し、さらにRPMI1640/10% FCS溶液で置換した。マイクロウェルへRPMI1640/10% FCS溶液を充填した後、チップ上の余分なバッファーを除き、そこへ上記細胞懸濁液を添加し、2〜5分間静置した。マイクロウェルに入らなかった細胞を回収し、チップ上へさらに細胞懸濁液を添加し、2〜5分間静置した。マイクロウェルに入らなかった細胞を回収し、チップ上へさらに細胞懸濁液を添加し、2〜5分間静置した。マイクロウェルに入らなかった細胞を回収し、さらにチップ上のマイクロウェルに入らなかった細胞をRPMI1640/10% FCS溶液を用いて洗い流した。リンパ球の直径は約8μmであり、使用するマイクロウェルの直径が10μmであるために一つのマイクロウェルにはリンパ球が1個入る。カバーグラスを上記シール上に置き、チップとカバーグラスの間にRPMI1640/10% FCS溶液を満たした。
【0077】
[マイクロウェルアレイチップを用いたHBs抗原特異的Bリンパ球の検出]
Bリンパ球を播種したマイクロウェルアレイチップをマイクロアレイスキャナー(CRBIO IIe-FITC)に挿入し、解像度2.5μmでスキャンし抗原刺激前のFluo-4の蛍光強度を測定しデータを保存した(抗原刺激前の蛍光データA)。次に、チップをスキャナーから取り出し、チップとカバーグラスの間のRPMI1640/10%FCS溶液を除き、そこへRPMI1640/10%FCS溶液に溶解させたHBs抗原(100μg/mL)を加えた。直ちにマイクロウェルアレイチップをマイクロアレイスキャナーに挿入し、抗原添加後約0.5〜2分の細胞内のFluo-4の蛍光強度を解像度2.5μmでスキャンし、データを保存した(抗原刺激後の蛍光データB)。スキャナーに付属の解析ソフトウェアを用いて刺激前後の蛍光強度の比(B/A)を計算し、比の大きいウェルを特定した。これらのウェルの中に抗原特異的Bリンパ球が存在する。
【0078】
[Bリンパ球のマイクロウェルからの回収]
検出したHBs抗原に反応し細胞内カルシウムが上昇したBリンパ球は蛍光顕微鏡下でマイクロマニピュレータを用いて回収した。すなわち、まず、カバーグラスとチップの間のRPMI1640/10%FCS溶液を取り除き、カバーグラスとチップの間に空気を入れることでカバーグラスを取り外した。チップが乾燥しないようにチップにRPMI1640/10%FCS溶液を加え、蛍光顕微鏡下で細胞のCellTracker Orangeの蛍光を観察しながら、マイクロマニピュレータで目的の細胞を回収した。
【0079】
[Bリンパ球からの抗体遺伝子の採取]
回収したBリンパ球をPCR用チューブに移す。細胞に15μLの細胞溶解液(25μLに希釈された時の最終濃度:1x 1ststrand buffer (GIBCO-BRL, SuperScript IIIに添付されている), 0.2 mM dNTP, 0.25% NP-40, 0.1 mg/mL BSA, 10 mM DTT, 0.05 μM Random Primer, 1U/μL RNasin (Promega, Madison, WI)を加え、そこへ逆転写酵素SuperScriptIII (GIBCO-BRL, Rockville, MD)を(5μL, 50U)添加し、37oC、1時間反応させmRNAよりcDNAを合成した。
【0080】
[抗体遺伝子の1回目の増幅]
合成したヒト抗体遺伝子のcDNAを鋳型に、抗体のH鎖およびL鎖のV領域を増幅するように設計された以下のプライマーを用いてPCR反応を行った。
【0081】
ヒト抗体(IgM およびIgG)のH鎖用のプライマー
PCR-1用
Primer1 5' mix
hVH17a.1 atggactgsayytggagvdtc(配列番号17)
hVH2a.1 tccacrctcctgctrctgac(配列番号18)
hVH3a.1 gggcygagstggvttttyct(配列番号19)
hVH4a.1 tcctcctsctggtggcagct(配列番号20)
hVH5.1 tcaaccgccatcctcgccct(配列番号21)
hVH6.1 ctccttcctcatcttcctgcc(配列番号22)
Primer1 3' mix
hIGHG1-4out agtccttgaccaggcagccca(配列番号23)
hIGHMout attctcacaggagacgagggg(配列番号24)
【0082】
ヒト抗体L鎖用のプライマー
PCR-1 用
Primer1 5’ mix
hKV12.1 atgaggstcccygctcagctc(配列番号25)
hKV3.1 ctcttcctcctgctactctggc(配列番号26)
hKV45.1 ctsttsctytggatctctg(配列番号27)
hKV6.1 tgggtttctgctgctctggg(配列番号28)
hKV7.1 atagggtccggggctcctttg(配列番号29)
hLV12.1 cykctsctcctcactctcctc(配列番号30)
hLV3.1 ttctcctcctcggcctcctct(配列番号31)
hLV4.2-2 ccagcytgtgctgactcaatc(配列番号32)
hLV789.2 tcycagmctgtgstgacycag(配列番号33)
hLV6.1 ttttatgctgactcagcccc(配列番号34)
hLV7.1 ggcctggactcctctctttctg(配列番号35)
hLV8.1 ggcctggatgatgcttctcctc(配列番号36)
hLV9.1 tcctctgctcctcaccctcct(配列番号37)
hLV10.1 cctgggtcatgctcctcctga(配列番号38)
hLV11.1 gcctgggctccactacttctc(配列番号39)
Primer1 3’ mix
hIGK1 ctgctcatcagatggcggga(配列番号40)
hIGL1 gacacacyagtgtggccttgt(配列番号41)
【0083】
PCR反応を行い、取得した細胞の抗体のH鎖およびL鎖のV領域のcDNAを増幅したH鎖およびL鎖は別個のチューブを用いて増幅した。PCR反応は、上記cDNA2μLを18μLのPCR mix1 (最終濃度:1x TAKARA ExTaq buffer, 0.25 mM dNTP, Primer1 5’ mix 0.5μM each, Primer1 3’ mix 0.5μM each, 0.05 U/μL ExTaq (Takara, 京都))に加え、94℃, 5 min; (94℃, 30 sec; 60℃, 1min; 72℃, 1 min 30 sec) x 40サイクル; 72℃, 5 min反応させる。このPCRの反応により抗体遺伝子のLeader配列から定常部領域までのDNAが増幅される。
【0084】
[抗体遺伝子の2回目の増幅]
1回のPCRではcDNAが十分増幅されないため、2回目のnested PCRを行った。2回目のPCRの反応は1回目に得られたPCRの反応溶液2μLを18μLのPCR mix2(最終濃度:1x TAKARA ExTaq buffer, 0.25 mM dNTP, Primer2 5’ mix 0.5μM each, Primer2 3’ mix 0.5μM each, 0.05 U/μL ExTaq)に加え、94 oC, 5 min; (94℃, 30 sec; 60℃, 1min; 72℃, 1 min 30 sec) x 40サイクル; 72℃, 5 min反応させる。2回目のPCRにより、1回目で増幅した抗体遺伝子の可変部領域から定常部領域までのcDNA配列が増幅される。以下に使用したプライマーの配列を示す。
【0085】
ヒト抗体(IgM およびIgG)のH鎖用のプライマー
PCR-2用
Primer2 5' mix
hVH17a.2 ggtgcagctkgtrcartctgg(配列番号42)
hVH2a.2 caccttgarggagtctggtcc(配列番号43)
hVH3a.2 aggtdcarctgktggagtcyg(配列番号44)
hVH4a.2 ggtcctgtctcaggtgcagct(配列番号45)
hVH5a.2 gtgcagctggtgcagtctgg(配列番号46)
hVH6.2 gcagcagtcaggtccaggact(配列番号47)
Primer2 3' mix
hIGHG1-4S aagacsgatgggcccttggtg(配列番号48)
hIGHMin aagggttgggcggatgcact(配列番号49)
【0086】
ヒト抗体L鎖用のプライマー
PCR-2 用
Primer1 5’ mix
hKV1.2 ccagatgacccagtctccatc(配列番号50)
hKV2.2 ccagtggggatattgtgatgac(配列番号51)
hKV3.2 cagtctccagccaccctgtct(配列番号52)
hKV4.2 gtgatgacccagtctccagac(配列番号53)
hKV 5.2 acactcacgcagtctccagca(配列番号54)
hKV67.2 ttgtgctgacycagtctccag(配列番号55)
hLV1.2 agtctgtgctgacgcagccgc(配列番号56)
hLV23.2 tgactcagccwcyctcmgtgtc(配列番号57)
hLV4.2-3 caatcatcctctgcmtctgc(配列番号58)
hLV5.2-2 gactcagccaacctccctctc(配列番号59)
hLV6.2 gactcagccccactctgtgtc(配列番号60)
hLV789.2 tcycagmctgtgstgacycag(配列番号61)
hLV1011.2 tgactcagccmcmctckgtgtc(配列番号62)
Primer1 3’ mix
hIGK2 gacagatggtgcagccacagt(配列番号63)
hIGL2 cttgragctcctcagaggaggg(配列番号64)
【0087】
[増幅した抗体遺伝子の塩基配列の決定]
PCR産物を、アガロースゲルを用いて解析、精製し、pT7Blue-T vector (Novagen, Madison, WI)にクローニングし、抗体遺伝子の塩基配列を決定し、タンパクに翻訳されることを確認した。H鎖のV領域のcDNAの塩基配列を配列番号1に、アミノ酸配列を配列番号2に示す。さらに、L鎖のV領域のcDNAの塩基配列を配列番号3に、アミノ酸配列を配列番号4に示す。
【0088】
実施例2
[抗体タンパクの産生、抗原結合能の解析]
RT-PCR法にて増幅した抗体のH鎖およびL鎖の可変領域遺伝子を抗体タンパク発現用ベクター(図3)に組込んだ。すなわち、増幅した抗体のH鎖可変領域の遺伝子断片は上記HEF-HBs-gγ1ベクターのVHの部分に、L鎖可変領域の遺伝子断片は上記HEFs-gκベクターの定常部領域(Human Cκ)の5’側に制限酵素部位を用いて挿入した。作製したH鎖およびL鎖の発現ベクターから抗体タンパクを発現させるために、両方の発現ベクターを同時にヒト胎児腎由来細胞293T細胞に遺伝子導入した。遺伝子導入は常法(細胞工学別冊8 新 細胞工学実験プロトコール、東京大学医科学研究所制癌研究部編、(株)秀潤社)に従い、リン酸カルシウム法を用いて行った。3日後に細胞上清を回収した。この培養上清中の抗体のHBs抗原への結合能を、HBs抗原をコートした96ウェルプレートを用いたELISA法により測定した。ELISAによる測定は以下のように行った。すなわち、96穴プレートにphosphate buffered saline (PBS)で希釈したHBs抗原(10μg/mL)を各ウェルに50μLずつ分注し、4℃で一晩反応させ吸着させた。PBSでウェルを洗浄し、非特異的結合を除去する為に3%BSA、0.05%Tween-20添加PBSを各ウェルに400μLずつ分注し、室温で2時間反応させブロッキングした。ネガティブコントロールのウェルとして、抗原を吸着させず、ブロッキングしたウェルも作成した。その後0.1% Tween-20添加PBS (PBS-T)でウェルを洗浄し、上記293T細胞の培養上清を各ウェルに50μLずつ分注し、室温で2時間反応させた。ウェルをPBS-Tで洗浄し、1000倍希釈したアルカリフォスファターゼ(ALP)標識抗ヒト免疫グロブリンを各ウェルに50μLずつ分注し、室温で2時間反応させた。ウェルをPBS-Tで洗浄し、ALP基質用緩衝液(100mM NaCl、5mM MgCl2 100mM Tris pH9.5)に溶解した1mg/mL p-nitrophenyl phosphate(ALP基質)を各ウェルに50μLずつ分注し、室温で20分反応後にマイクロプレートリーダーにて、405nmにおける吸光度を測定した。さらに、本発明のモノクローナル抗体の抗原への結合の特異性をチェックするために、抗体を含む細胞培養上清をウェルに添加する際に、可溶性のHBs抗原と培養上清とをプレインキュベーションさせた。その混合液をウェルに添加し、ウェルに吸着させたHBs抗原への抗体の結合が可溶性のHBs抗原により競合的に阻害されるかを検討した。結果を図4に示す。
【0089】
図4に示されるように、作製した116抗体はHBs抗原に結合し、HBs抗原を結合させていないウェルには抗体は結合しなかった(データは示していない)。さらに、結合の特異性を検討するために116抗体と可溶性のHBs抗原を混合したものをHBs抗原が結合しているウェルに加え同様にELISAを行った。116抗体のウェルに結合したHBs抗原への結合が、可溶性HBs抗原により用量依存的、競合的に阻害されることが観察された。この結果より116抗体がHBs抗原に特異的に結合することが示された。
【0090】
116抗体が結合するエピトープの決定
ELISAの系を用いて、116抗体がadr型HBs抗原のa、d、rのどのエピトープに結合するかを、各エピトープに結合するモノクローナル抗体が116抗体のadr型HBs抗原への結合を競合的に阻害するかを調べることで検討した。まず、96穴プレートにリン酸緩衝生理食塩水(phosphate buffered saline (PBS))で希釈したadr型HBs抗原(10μg/mL)を各ウェルに50μLずつ分注し、4 oCで一晩反応させ吸着させた。PBSでウェルを洗浄し、非特異的結合を除去する為に3%BSA、0.05%Tween-20添加PBS(PBS-T/BSA)を各ウェルに400μLずつ分注し、室温で2時間反応させブロッキングした。ネガティブコントロールのウェルとして、抗原を吸着させず、ブロッキングしたウェルも作成した。その後0.1% Tween-20添加PBS (PBS-T)でウェルを洗浄し、PBS-T/BSAあるいは2μg/mLのエピトープaを認識するマウスモノクローナル抗体(抗a抗体)、エピトープdを認識するマウスモノクローナル抗体(抗d抗体)、あるいはエピトープrを認識するマウスモノクローナル抗体(抗r抗体)を各ウェルに50μLずつ分注し、室温で1時間反応させた。0.5μg/mLのビオチン化116抗体を各ウェルに5μLずつ分注し、さらに室温で2時間反応させた。ウェルをPBS-Tで洗浄し、1000倍希釈したアルカリフォスファターゼ(ALP)標識ストレプトアビジンを各ウェルに50μLずつ分注し、室温で2時間反応させた。ウェルをPBS-Tで洗浄し、ALP基質用緩衝液(100mM NaCl、5mM MgCl2、100mM Tris pH9.5)に溶解した1mg/mL p-ニトロフェニルフォスフェート(ALP基質)を各ウェルに50μLずつ分注し、室温で20分反応後にマイクロプレートリーダーにて、405nmにおける吸光度を測定した。結果を図5に示す。
【0091】
抗原を吸着させていないウェルでは発色せず(カラム1)、抗原を吸着させたウェルで発色した(カラム2)ことから、116抗体がadr型のHBs抗原に特異的に結合していることがわかる。抗a抗体は116抗体の抗原への結合を阻害したが(カラム3)、抗dおよび抗r抗体は116抗体の抗原への結合を阻害しなかった(カラム4、5)。これらの結果は、116抗体がエピトープdあるいはエピトープrを認識するのではなく、HBウイルスに共通のエピトープaを認識することを示している。
【0092】
本発明においては、116抗体を作製するにあたり、HBsワクチンはadrの型のものを用いており、スクリーニングもadrのHBs抗原を用いて行っている。したがって、116抗体はadr型のHBs抗原に結合するものである。エピトープa、エピトープd、エピトープrのどこを116抗体が認識するかについては、上記の様にadr型のHBs抗原への116抗体の結合をELISAで測定する時に、エピトープa、エピトープd、エピトープrに結合することがわかっている抗a抗体、抗d抗体、抗r抗体を同時に加えて競合させた時に、抗a抗体とは競合し結合が阻害されるが、抗d抗体や抗r抗体とは競合せず結合が阻害されないことから、ウイルスによって異なるエピトープdやエピトープrを認識するのではなく、全てのウイルスに共通のエピトープaを認識することがわかる。このように、本発明の抗体は、全てのタイプのHBVに対して中和活性を有し得るHBs抗原特異的抗体である。
【0093】
実施例3
[ヒト肝細胞キメラマウス]
ヒト肝細胞キメラマウスの作製法については以下の論文に詳述されている。[C Tateno, Y Yoshizane, N Saito, M Kataoka, R Utoh, C Yamasaki, A Tachibana, Y Soeno, K Asahina, H Hino, T Asahara, T Yokoki, T Furukawa, K Yoshizato. Near completely humanized liver in mice shows human-type metabolic responses to drugs. Am J Pathol, 165:901-912, 2004]簡単には、uPAマウス(B6SJL-TgN(Alb1Plau)144Bri, The Jackson Laboratory, Bar Harbor, ME)とscidマウス(Fox Chase SCID C.B-17/lcr-scid Jcl, Clea Japan inc., Tokyo, Japan)を掛け合わせuPA/SCIDマウスを作製した。ヒト肝細胞は、同意を得たドナーより採取した肝組織より小型肝細胞を含む肝非実質細胞を分離し移植に用いた。小型肝細胞移植1日前に、生後20〜30日のuPA/SCIDマウスに抗アシアロGM1抗体を腹腔内投与した。1日後、5〜7x105の生きた肝細胞を門脈を介して移植した。移植の成否はマウス血清中のヒトアルブミン値を測定することにより評価した。
【0094】
[抗HBs-抗体のウイルス中和能の解析]
HBVはヒトあるいはチンパンジーの肝細胞にしか感染しないため、抗HBs-抗体がHBVの感染をブロックできるかどうかは、これまでチンパンジーを用いて行われてきた。近年、フレッシュな肝臓組織をボランティアから採取し、その肝細胞を使ってin vitroにおいてHBVを感染させる実験系も報告されている。しかし、採取した肝細胞はin vitroで培養を続けることにより容易に感染感受性を失うため、常にフレッシュな肝細胞を用いる必要であり、人への負担から実際に行うことは困難であった。しかるに、上記ヒト肝細胞キメラマウスは移植した肝細胞のHBV感染感受性等の細胞の性質を失うことなく長期に肝細胞を保持できること、さらに、キメラマウス由来のヒト肝細胞を用いてキメラマウスを作製することも可能であることから、HBVの感染実験系を簡単に、安定に、低価格で供給することを可能にした。本実験では、ヒト肝細胞キメラマウスを用いてHBVの感染実験を行い、その実験系を用いて116抗体がHBVの感染をブロックすることができるかを検討した。実験の概略は以下の通りである。
【0095】
B型肝炎ウイルス感染患者の血清(1 x 108コピー/mLのB型肝炎ウイルス(HBV)のゲノムを含む)50μLと116抗体(20μg/匹)をプラスチックチューブ内で混合し室温で30分間反応させた後、その混合液をヒト肝細胞キメラマウスの鎖骨下静脈より注入した(2匹)。感染のポジティブコントロールとして患者血清のみをヒト肝細胞キメラマウス2匹に投与した。116抗体のネガティブコントロールとしてヒトIgG(20μg/匹)を患者血清とインキュベーションしたものをヒト肝細胞キメラマウス2匹に投与した。投与後、2週目、4週目、6週目にマウスの血清を採取し、B型肝炎ウイルスのゲノム遺伝子の存在を、定量PCR法を用いて定量した。
【0096】
結果を図6に示す。図6は、ヒトキメラマウス(1〜7)にHBVのみ(1〜3)、HBVとコントロールIgG(4、5)、HBVと116抗体(6、7)を注入し、2週目、4週目、6週目のマウス血清中のHBVゲノムDNAを定量PCR法にて測定した結果である。下欄にはヒト肝細胞キメラマウスの血清中のヒトアルブミンの量を示してあり、キメラマウスにおけるヒト肝細胞の存在を示す。
【0097】
図6に示すように患者血清(HBV)のみを投与した場合には投与2週後からウイルスゲノムDNAが検出され、4週目、6週目と経過するにつれ、DNA量は増加した。この結果は、HBVがマウス細胞には感染しないことから、移植されたヒト肝細胞にHBVが感染し、増殖していることを示す。ネガティブコントロール抗体としてヒトIgGを用いた場合も同様の結果を得た。一方、116抗体とHBVを混合したものをヒト肝細胞キメラマウスに投与した場合には、一匹のマウスにおいて2週目にウイルスゲノム遺伝子が微量検出されたが、4週目、6週目には2匹のマウスともにウイルスゲノム遺伝子は検出されなくなった。以上の結果は、116抗体がウイルスの細胞への感染を阻害する活性(中和活性)を保持していることを示す。
【0098】
実施例4
Single chain Fv (ScFv)
遺伝子の構築
116抗体遺伝子配列及びコントロール抗体遺伝子配列をそれぞれ鋳型とし、VH遺伝子の3’側及びVL遺伝子の5‘側をリンカー(GGGGS)3により結合した遺伝子をPCRにより構築した。さらに、このVH-(GGGGS)3-VL配列の5’側にT7プロモーター配列,翻訳効率を高めるためのε配列,リボソーム結合部位であるSD配列及び開始コドン、3’側にエピトープタグとしてHAタグ(アミノ酸配列:YPYDVPDYA)及び終止コドンをPCRにより付加した(図7および図8)。
【0099】
scFvタンパク合成
無細胞系タンパク合成反応液PURESYSTEM S-S ((株)ポストゲノム研究所、東京)50μLに上記PCR産物1pmolを鋳型として添加し37℃,60分間タンパク合成を行った.反応後チューブを氷上に移し,これをscFv116、及びscFvcont.として以下の実験に用いた。
【0100】
scFv116の抗原特異性の検討
scFvの抗原への結合をELISAにより検討した。96穴プレートにPBSで希釈したadr型HBs抗原(10μg/mL)を各ウェルに50μLずつ分注し、4 oCで一晩反応させ吸着させた。PBSでウェルを洗浄し、非特異的結合を除去する為に3%BSA、0.05%Tween-20添加PBS(PBS-T/BSA)を各ウェルに400μLずつ分注し、室温で2時間反応させてブロッキングした。0.1% Tween-20添加PBS (PBS-T)でウェルを洗浄後、各ウェルにPBS-T/BSAで希釈したscFv116あるいはscFvcontを50μLずつ加え、室温で1時間反応させた。その後、PBS-Tでウェルを洗浄後、1000倍希釈したマウス抗HAモノクローナル抗体を各ウェルに50μLずつ分注し、室温で1時間反応させた。PBS-Tで洗浄後、2000倍希釈したアルカリフォスファターゼ(ALP)標識ヤギ抗マウスIgG抗体を、各ウェルに50μLずつ分注し室温で2時間反応させた。ウェルをPBS-Tで洗浄し、ALP基質用緩衝液(100mM NaCl、5mM MgCl2、100mM Tris pH9.5)に溶解した1mg/mL p-nitrophenyl phosphate(ALP基質)を、各ウェルに50μLずつ分注し、室温で20分反応させた後、マイクロプレートリーダーにて405nmにおける吸光度を測定した。図9に示すように、コントロール遺伝子配列より構築したscFvタンパク(scFvcont)はHBs抗原とは全く結合せず、116遺伝子配列より構築したscFv(scFv116)は、濃度依存的にHBs抗原と結合することが示された。
【0101】
scFv116のHBs抗原への結合の特異性を検討するために、一定濃度のscFv116と種々の濃度 (0,10, 100,400,1,000 ng/mL) のHBs抗原と反応させた後に、上記ELISAを行った。図10に示すように、HBs抗原の濃度依存的にscFv116のHBs抗原への結合が阻害された。この結果は、scFv116とHBs抗原の結合が特異的であることを示す。
【0102】
さらに、scFv116と元のIgG116の認識するエピトープが同一であるか否かを検討するために、一定濃度のscFv116に対して種々の濃度 (0,40,200,1,000 ng/mL) のIgG116を加えて、上記ELISAを行った。その結果、添加したIgG116の濃度に依存的にscFv116の結合が阻害された(図11)。この結果は、HL116抗体のIgG型とscFv型は、同一のエピトープを認識することを示す。
以上、116抗体遺伝子を元に作製した
【産業上の利用可能性】
【0103】
このようなB型肝炎ウイルスに対する抗体(本発明の抗体)は、先に述べたような針刺し事故に対する治療に有効であることが期待されるが、肝移植時における効果も期待される。すなわち、B型肝炎ウイルスに感染している患者においては、肝移植を行ってもせっかく移植した新しい肝臓にB型肝炎ウイルスが再び感染することにより肝炎が発症し肝機能が低下する可能性が高く、肝移植の意義がなくなってしまうこともある。この場合、肝移植時にB型肝炎ウイルスに対する抗体で患者体内のB型肝炎ウイルスを中和・除去することで、移植した肝臓へのB型肝炎ウイルスの感染を防御できることが期待される。このように、B型肝炎ウイルスを中和する能力を持った抗体、特に、組換え抗体はB型肝炎ウイルスの感染阻止に非常に有用であると考えられる。
【図面の簡単な説明】
【0104】
【図1】抗体遺伝子の構造の模式図。
【図2】単鎖Fv (scFv)等の構造の模式図。
【図3】抗体γ鎖発現ベクターおよび抗体κ鎖発現ベクター。
【図4】116抗体のHBs抗原への特異的結合試験結果。
【図5】116抗体が認識するエピトープの試験結果。
【図6】116抗体のHBV中和能試験結果。
【図7】scFv遺伝子の構築およびscFvタンパクの合成スキーム。
【図8−1】116抗体scFv遺伝子の塩基配列。
【図8−2】116抗体scFv遺伝子の塩基配列。
【図9】scFvタンパク(scFvcont)のHBs抗原への結合試験結果。
【図10】scFv116のHBs抗原への特異性試験結果。
【図11】scFv116のIgG116への特異性試験結果。
【図12】CDR遺伝子を組み合わせて用いて抗体を調製する方法の説明図。
【技術分野】
【0001】
本発明は、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の重鎖または軽鎖の可変領域または相補性決定領域をコードする遺伝子に関する。さらに本発明は、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の重鎖または軽鎖の可変領域または相補性決定領域であるペプチドに関する。さらに本発明は、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体、単鎖抗体(scFv)、その二量体および三量体、ならびにこのモノクローナル抗体、単鎖抗体(scFv)、その二量体および三量体、を用いるB型肝炎ウイルスの検定方法、B型肝炎の診断方法およびB型肝炎の治療方法に関する。
【背景技術】
【0002】
B型肝炎はB型肝炎ウイルスが肝細胞に感染することにより誘発され、肝不全、肝がんの誘発により死に至る病気である。B型肝炎ウイルスは病院等において、針刺し事故等により患者の血液が誤って体内に注入されることによりしばしば感染する。このような場合、現在ではHBワクチン接種等によりB型肝炎ウイルスに対する抗体を大量に持つようになった人の血液より抗血清を調製したものおよびHBワクチンを緊急に注射することにより、B型肝炎ウイルスの肝細胞への感染を抗体により中和(ブロック)することで感染を防ぐことが一般的に行われている。しかし、このように人から調製した抗血清は未知のウイルスの混入の可能性を除去することが難しく、人に用いる治療薬としては好ましいものではない。
【0003】
そこで、HBs抗原特異的抗体を作成し、それを中和(ブロック)に使用することが考えられている。
【0004】
[キメラ抗体、ヒト化抗体]
HBs抗原特異的抗体の作成方法(スクリーニング方法)としては、マウスをHBs抗原で免疫し、脾臓あるいはリンパ節のBリンパ球とミエローマ細胞を細胞融合法により融合し、HBs抗原特異的抗体を分泌しているハイブリドーマ細胞をELISA法等によりスクリーニングする。樹立したハイブリドーマよりマウス抗体遺伝子を回収し、マウスの定常部領域をヒトの定常部領域と置換(キメラ化)するか、マウスの抗体の抗原結合部位(相補性決定領域)(CDR1, 2, 3)をヒトの抗体のそれと置換(ヒト化)することにより、HBs抗原への結合性を保持しながら、ヒト抗体に近づける方法が知られている。[J Waters, M Pignatelli, S Galpin, K Ishihara, HC Thomas, Virus-neutralizing antibodies to hepatitis B virus: The nature of an immunogenic epitope on the S gene peptide. J gen Virol, 67:2767-2473, 1986(非特許文献1)、S Iwarson, E Tabor, HC Thomas, A Goodall, J Waters, P Snoy, JW-K Shih, RJ Gerety, Neutralization of hepatitis B virus infectivity by a murine monoclonal antibody: and experimental study in the Chimpanzee. J Med Virol, 16:89-96, 1985(非特許文献2)、CJ Ryu, P Gripon, HR Park, SS Park, YK Kim, C Guguen-Guillouzo, OJ Yoo, HJ Hong, In vitro neutralization of hepatitis B virus by monoclonal antibodies against the viral surface antigen. J Med Virol, 52:226-233, 1997(非特許文献5)、HJ Hong, CJ Ryu, H Hur, S Kim, HK Oh, MS Oh, SY Park. In vivo neutralization of hepatitis B virus infection by an anti-preS1 humanized antibody in chimpanzees. Virology, 318:134-141, 2004(非特許文献6)、CJ Ryu, EA Pallan, BR Jin, OJ Yoo, HJ Hong. A humanized antibody with specificity for hepatitis B surface antigen. Hum Antibo Hybridomas, 7:113-122, 1996(非特許文献7)]
【0005】
具体的なモノクローナル抗体としては、B型肝炎ウイルス表面抗原(HBs抗原)に結合できるヒトモノクローナル抗体が知られている。特表2000−513215号公報(特許文献1)には、B型肝炎ウイルス表面抗原(HBs抗原)に結合できるヒト抗体を産生するハイブリドーマ細胞系を得る方法、ならびにハイブリドーマ細胞系、およびその細胞系によって産生される抗体が開示されている。また、HBV感染の予防および処置における上記抗体の様々な使用が開示される。この発明では、抗HBs抗原抗体の高力価を有するヒトドナーから得られる末梢血リンパ球を、致死的に放射線を照射されSCID骨髄で保護された正常株マウスに移植する。このようなキメラマウスをHBs抗原で免疫したのち、そのマウス脾臓からヒト細胞を得て、異種骨髄腫細胞とインビトロ融合させ、HBs抗原に対して高い親和性および特異性を有するヒト抗体を分泌するハイブリドーマ細胞を発生させる。
【0006】
上記以外にもHBs抗原特異的抗体が報告され、以下の文献に記載されている。
(1)Maeda F et al. J Med Virol. 58:338-345, 1999(非特許文献8)
(2)Yano A et al. J Med Virol. 73:208-215, 2004(非特許文献9)
(3)日大医誌 41:1163-1171, 1982 (非特許文献10)
(4)Hybridoma 17 (6), 535-540 (1998) (非特許文献11)
(5)Zhonghua Gan Zang Bing Za Zhi 5 (3), 130-132 (2000) (非特許文献12)
(6)特開平9−20798号公報(特許文献2)
(7)Biochim. Biophys. Acta 1380 (2), 151-155 (1998) (非特許文献13)
(8)Proc. Natl. Acad. Sci. U.S.A. 89 (8), 3175-3179 (1992) (非特許文献14)
【0007】
また、HBs抗原特異的抗体の作成方法(スクリーニング方法)としては、以下の方法も知られている。
【0008】
[ヒトハイブリドーマ法]
末梢血Bリンパ球とミエローマ細胞を細胞融合法により融合し、HBs抗原特異的抗体を分泌しているハイブリドーマ細胞をELISA法等によりスクリーニングする。[H Sawada, S Iwasa, O Nishimura, K Kitano, Efficient production of anti-(hepatitis B virus) antibodies and their neutralizing activity in chimpanzees. Appl Microbiol Biotechnol, 43:445-451, 1995(非特許文献3)、Gary Witherell, XTL-001 XTL Pharmaceuticals. Current Opinion in Investigational Drugs, 3:684-692, 2002(非特許文献4)]
【0009】
[EBウイルス形質転換法]
末梢血ヒトBリンパ球にEBウイルスを感染させ不死化させB細胞株を確立し、HBs抗原特異的抗体を分泌しているB細胞株をELISA法等によりスクリーニングする。
【特許文献1】特表2000−513215号公報
【特許文献2】特開平9−20798号公報
【非特許文献1】J Waters, M Pignatelli, S Galpin, K Ishihara, HC Thomas, Virus-neutralizing antibodies to hepatitis B virus: The nature of an immunogenic epitope on the S gene peptide. J gen Virol, 67:2767-2473, 1986
【非特許文献2】S Iwarson, E Tabor, HC Thomas, A Goodall, J Waters, P Snoy, JW-K Shih, RJ Gerety, Neutralization of hepatitis B virus infectivity by a murine monoclonal antibody: and experimental study in the Chimpanzee. J Med Virol, 16:89-96, 1985
【非特許文献3】H Sawada, S Iwasa, O Nishimura, K Kitano, Efficient production of anti-(hepatitis B virus) antibodies and their neutralizing activity in chimpanzees. Appl Microbiol Biotechnol, 43:445-451, 1995
【非特許文献4】Gary Witherell, XTL-001 XTL Pharmaceuticals. Current Opinion in Investigational Drugs, 3:684-692, 2002
【非特許文献5】CJ Ryu, P Gripon, HR Park, SS Park, YK Kim, C Guguen-Guillouzo, OJ Yoo, HJ Hong, In vitro neutralization of hepatitis B virus by monoclonal antibodies against the viral surface antigen. J Med Virol, 52:226-233, 1997
【非特許文献6】HJ Hong, CJ Ryu, H Hur, S Kim, HK Oh, MS Oh, SY Park. In vivo neutralization of hepatitis B virus infection by an anti-preS1 humanized antibody in chimpanzees. Virology, 318:134-141, 2004
【非特許文献7】CJ Ryu, EA Pallan, BR Jin, OJ Yoo, HJ Hong. A humanized antibody with specificity for hepatitis B surface antigen. Hum Antibo Hybridomas, 7:113-122, 1996
【非特許文献8】Maeda F et al. J Med Virol. 58:338-345, 1999
【非特許文献9】Yano A et al. J Med Virol. 73:208-215, 2004
【非特許文献10】日大医誌 41:1163-1171, 1982
【非特許文献11】Hybridoma 17 (6), 535-540 (1998)
【非特許文献12】Zhonghua Gan Zang Bing Za Zhi 5 (3), 130-132 (2000)
【非特許文献13】Biochim. Biophys. Acta 1380 (2), 151-155 (1998)
【非特許文献14】Proc. Natl. Acad. Sci. U.S.A. 89 (8), 3175-3179 (1992)
【発明の開示】
【発明が解決しようとする課題】
【0010】
HBVとしては、アミノ酸の違いで10種類以上の異なるHBVが知られているが、特許文献1に記載の抗体はその一部しか認識できないという問題があった。さらに、特許文献1に記載の抗体は、中和実験の結果では、HBVを100%は中和できてはいないという問題もあった。
【0011】
また、非特許文献8〜10には、非特許文献8〜10に記載の抗体に中和があることは記載されているが、具体的なデータが示されていない。さらに、非特許文献11〜14および特許文献2には、そこに記載の抗体が中和活性を有するものであるかについての記載がない。
【0012】
そこで、本発明の目的は、全てのタイプのHBVに対して中和活性を有し得るHBs抗原特異的モノクローナル抗体を提供することにある。
【0013】
さらに本発明は、上記HBs抗原特異的モノクローナル抗体に加えて、
(1)B型肝炎ウイルスのHBs抗原に対する単鎖抗体(scFv)
(2)上記HBs抗原特異的モノクローナル抗体を構成する重鎖可変領域および軽鎖可変領域であるタンパク質、
(3)上記HBs抗原特異的モノクローナル抗体を構成する重鎖相補性決定領域1〜3および軽鎖相補性決定領域1〜3であるペプチド、
(4)上記HBs抗原特異的モノクローナル抗体を構成する重鎖可変領域および軽鎖可変領域をコードするDNA(遺伝子)、並びに
(5)上記HBs抗原特異的モノクローナル抗体を構成する重鎖相補性決定領域1〜3および軽鎖相補性決定領域1〜3をコードするDNA(遺伝子)
を提供することも目的とする。
【0014】
本発明者らは、B型肝炎ウイルスに対する抗体を細胞チップ技術および組換えDNA技術を用いて作製し、それを治療薬として用いることを考えた。すなわち、ワクチン接種した人の血液よりBリンパ球を調製し、それを本発明者の関係者らが開発したマイクロウェルアレイチップに播種し、B型肝炎ウイルス表面抗原(HBs抗原)で刺激し、スキャナーを用いてHBs抗原に反応するBリンパ球を検出した。検出したBリンパ球をチップより回収し、抗体遺伝子を回収し、発現ベクターに組込み動物細胞に導入することにより、組換え抗体を作製した。作製した組換え抗体は、HBs抗原に結合すること、さらには、ヒト肝細胞へのウイルスの感染をブロックすること(中和すること)が確認され、本発明を完成した。
【課題を解決するための手段】
【0015】
本発明は、以下の通りである。
[1]下記の何れかのDNAからなる遺伝子。
(1)配列番号1の塩基配列を有するDNA、
(2)配列番号1の塩基配列において1から数個の塩基の欠失、置換及び/又は付加を有する塩基配列を有し、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の重鎖可変領域をコードするDNA、および
(3)配列番号1の塩基配列またはこの塩基配列と相補的な塩基配列とストリンジェントな条件下でハイブリダイズする塩基配列を有し、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の重鎖可変領域をコードするDNA。
[2]下記の何れかのタンパク質をコードする遺伝子。
(1)配列番号2のアミノ酸配列を有するタンパク質、
(2)配列番号2のアミノ酸配列において1から数個のアミノ酸の欠失、置換及び/又は付加を有するアミノ酸配列を有し、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の重鎖可変領域であるタンパク質、および
(3)配列番号2のアミノ酸配列に対して90%以上の相同性を有するアミノ酸配列を有し、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の重鎖可変領域であるタンパク質。
[3]下記の何れかのDNAからなる遺伝子。
(1)配列番号3の配列を有するDNA、
(2)配列番号3の塩基配列において1から数個の塩基の欠失、置換及び/又は付加を有する塩基配列を有し、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の軽鎖可変領域をコードするDNA、および
(3)配列番号3の塩基配列またはこの塩基配列と相補的な塩基配列とストリンジェントな条件下でハイブリダイズする塩基配列を有し、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の軽鎖可変領域をコードするDNA。
[4]下記の何れかのタンパク質をコードする遺伝子。
(1)配列番号4のアミノ酸配列を有するタンパク質、
(2)配列番号4のアミノ酸配列において1から数個のアミノ酸の欠失、置換及び/又は付加を有するアミノ酸配列を有し、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の軽鎖可変領域であるタンパク質、および
(3)配列番号4のアミノ酸配列に対して90%以上の相同性を有するアミノ酸配列を有し、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の軽鎖可変領域であるタンパク質。
[5]下記の何れかのタンパク質。
(1)配列番号2のアミノ酸配列を有するタンパク質、
(2)配列番号2のアミノ酸配列において1から数個のアミノ酸の欠失、置換及び/又は付加を有するアミノ酸配列を有し、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の重鎖可変領域であるタンパク質、および
(3)配列番号2のアミノ酸配列に対して90%以上の相同性を有するアミノ酸配列を有し、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の重鎖可変領域であるタンパク質。
[6]下記の何れかのタンパク質。
(1)配列番号4のアミノ酸配列を有するタンパク質、
(2)配列番号4のアミノ酸配列において1から数個のアミノ酸の欠失、置換及び/又は付加を有するアミノ酸配列を有し、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の軽鎖可変領域であるタンパク質、および
(3)配列番号4のアミノ酸配列に対して90%以上の相同性を有するアミノ酸配列を有し、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の軽鎖可変領域であるタンパク質。
[7]下記の何れかの遺伝子。
(1)配列番号1の塩基配列における76〜105番目の塩基配列、または配列番号3のアミノ酸配列における26〜35番目のアミノ酸配列をコードする塩基配列を有する、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の重鎖相補性決定領域1(CDR−H1)をコードする遺伝子。
(2)配列番号1の塩基配列における148〜198の塩基配列、または配列番号3のアミノ酸配列における50〜66番目のアミノ酸配列をコードする塩基配列を有する、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の重鎖相補性決定領域2(CDR−H2)をコードする遺伝子。
(3)配列番号1の塩基配列における295〜333の塩基配列、または配列番号3のアミノ酸配列における99〜111番目のアミノ酸配列をコードする塩基配列を有する、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の重鎖相補性決定領域3(CDR−H3)をコードする遺伝子。
[8]下記の何れかの遺伝子。
(1)配列番号3の塩基配列における69〜102の塩基配列、または配列番号4のアミノ酸配列における24〜34のアミノ酸配列をコードする塩基配列を有する、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の軽鎖相補性決定領域1(CDR−L1)をコードする遺伝子。
(2)配列番号3の塩基配列における148〜168の塩基配列、または配列番号4のアミノ酸配列における50〜56のアミノ酸配列をコードする塩基配列を有する、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の軽鎖相補性決定領域2(CDR−L2)をコードする遺伝子。
(3)配列番号3の塩基配列における265〜291の塩基配列、または配列番号4のアミノ酸配列における89〜97のアミノ酸配列をコードする塩基配列を有する、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の軽鎖相補性決定領域3(CDR−L3)をコードする遺伝子。
[9]下記のいずれかのペプチド。
(1)配列番号2のアミノ酸配列における26〜35のアミノ酸配列を有する、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の重鎖相補性決定領域1(CDR−H1)であるペプチド、
(2)配列番号2のアミノ酸配列における50〜66のアミノ酸配列を有する、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の重鎖相補性決定領域2(CDR−H2)であるペプチド、
(3)配列番号2のアミノ酸配列における99〜111のアミノ酸配列を有する、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の重鎖相補性決定領域3(CDR−H3)であるペプチド。
[10]下記のいずれかのペプチド。
(1)配列番号4のアミノ酸配列における24〜34のアミノ酸配列を有する、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の軽鎖相補性決定領域1(CDR−L1)であるペプチド、
(2)配列番号4のアミノ酸配列における50〜56のアミノ酸配列を有する、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の軽鎖相補性決定領域2(CDR−L2)であるペプチド、
(3)配列番号4のアミノ酸配列における89〜97のアミノ酸配列を有する、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の軽鎖相補性決定領域3(CDR−L3)であるペプチド。
[11]配列番号2のアミノ酸配列を有するタンパク質を可変領域として含む重鎖と配列番号4のアミノ酸配列を有するタンパク質を可変領域として含む軽鎖とを含む、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体。
[12]配列番号2のアミノ酸配列における26〜35、50〜66および99〜111のアミノ酸配列を有するペプチドを相補性決定領域1〜3として含む重鎖と配列番号4のアミノ酸配列における24〜34、50〜56および89〜97のアミノ酸配列を有するペプチドを相補性決定領域1〜3として含む軽鎖とを含む、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体。
[13]配列番号2のアミノ酸配列を有するタンパク質と配列番号4のアミノ酸配列を有するタンパク質とをリンカーペプチドで結合した、B型肝炎ウイルスのHBs抗原に対する単鎖抗体。
[14]配列番号2のアミノ酸配列における26〜35、50〜66および99〜111のアミノ酸配列を有するペプチドを相補性決定領域1〜3として含むタンパク質と配列番号4のアミノ酸配列における24〜34、50〜56および89〜97のアミノ酸配列を有するペプチドを相補性決定領域1〜3として含むタンパク質とをリンカーペプチドで結合した、B型肝炎ウイルスのHBs抗原に対する単鎖抗体。
[15][13]または[14]に記載の単鎖抗体の二量体。
[16][13]または[14]に記載の単鎖抗体の三量体。
[17]配列番号2のアミノ酸配列を有するタンパク質と配列番号4のアミノ酸配列を有するタンパク質とをリンカーペプチドで結合し、かつ配列番号2のアミノ酸配列を有する前記タンパク質にCH3領域を結合した、B型肝炎ウイルスのHBs抗原に対するミニボディー。
[18]配列番号2のアミノ酸配列における26〜35、50〜66および99〜111のアミノ酸配列を有するペプチドを相補性決定領域1〜3として含むタンパク質と配列番号4のアミノ酸配列における24〜34、50〜56および89〜97のアミノ酸配列を有するペプチドを相補性決定領域1〜3として含むタンパク質とをリンカーペプチドで結合し、かつ配列番号2のアミノ酸配列における26〜35、50〜66および99〜111のアミノ酸配列を有するペプチドを相補性決定領域1〜3として含む前記タンパク質にCH3領域を結合した、B型肝炎ウイルスのHBs抗原に対するミニボディー(minibody)。
[19][5]若しくは[6]に記載のタンパク質、[9]若しくは[10]に記載のペプチド、[11]若しくは[12]に記載のモノクローナル抗体、[13]若しくは[14]に記載の単鎖抗体、[15]に記載の二量体、[16]に記載の三量体、[17]若しくは[18]に記載のミニボディーを用いるB型肝炎ウイルスの検定方法。
[20][5]若しくは[6]に記載のタンパク質、[9]若しくは[10]に記載のペプチド、[11]または[12]に記載のモノクローナル抗体、または[13]若しくは[14]に記載の単鎖抗体、[15]に記載の二量体、[16]に記載の三量体、[17]若しくは[18]に記載のミニボディーを用いるB型肝炎の診断方法。
[21][5]若しくは[6]に記載のタンパク質、[9]若しくは[10]に記載のペプチド、[11]または[12]に記載のモノクローナル抗体、または[13]若しくは[14]に記載の単鎖抗体、[15]に記載の二量体、[16]に記載の三量体、[17]若しくは[18]に記載のミニボディーを用いるB型肝炎の治療方法。
【発明の効果】
【0016】
本発明によれば、全てのタイプのHBVに対して中和活性を有し得るHBs抗原特異的モノクローナル抗体を提供することができる。
【発明を実施するための最良の形態】
【0017】
前述のように、本発明の組換え抗体は、ワクチン接種した人の血液より調製したBリンパ球をB型肝炎ウイルス表面抗原(HBs抗原)で刺激し、HBs抗原に反応する1個のBリンパ球を回収し、回収した1個のBリンパ球から抗体遺伝子を採取し、この抗体遺伝子を含む発現ベクターで形質転換した動物細胞を用いて作製した。この過程で、抗体遺伝子の塩基配列および抗体のアミノ酸配列を重鎖および軽鎖についてそれぞれ決定し、さらに、重鎖および軽鎖における相補性決定領域を決定した。
【0018】
本発明は、
(1)B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体、および単鎖抗体(scFv)
(2)このモノクローナル抗体を構成する重鎖可変領域および軽鎖可変領域であるタンパク質、
(3)このモノクローナル抗体を構成する重鎖相補性決定領域1〜3および軽鎖相補性決定領域1〜3であるペプチド、
(4)このモノクローナル抗体を構成する重鎖可変領域および軽鎖可変領域をコードするDNA(遺伝子)、並びに
(5)このモノクローナル抗体を構成する重鎖相補性決定領域1〜3および軽鎖相補性決定領域1〜3をコードするDNA(遺伝子)に関する。
【0019】
[可変領域をコードするDNA(遺伝子)]
本発明の重鎖可変領域をコードするDNA(遺伝子)は、下記の何れかのDNAからなる。
(1)配列番号1の塩基配列を有するDNA、
(2)配列番号1の塩基配列において1から数個の塩基の欠失、置換及び/又は付加を有する塩基配列を有し、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の重鎖可変領域をコードするDNA、および
(3)配列番号1の塩基配列またはこの塩基配列と相補的な塩基配列とストリンジェントな条件下でハイブリダイズする塩基配列を有し、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の重鎖可変領域をコードするDNA。
【0020】
さらに本発明の重鎖可変領域をコードするDNA(遺伝子)は、下記の何れかのタンパク質をコードする遺伝子である。
(1)配列番号2のアミノ酸配列を有するタンパク質、
(2)配列番号2のアミノ酸配列において1から数個のアミノ酸の欠失、置換及び/又は付加を有するアミノ酸配列を有し、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の重鎖可変領域であるタンパク質、および
(3)配列番号2のアミノ酸配列に対して90%以上の相同性を有するアミノ酸配列を有し、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の重鎖可変領域であるタンパク質。
【0021】
本発明の軽鎖可変領域をコードするDNA(遺伝子)は、下記の何れかのDNAからなる。
(1)配列番号3の配列を有するDNA、
(2)配列番号3の塩基配列において1から数個の塩基の欠失、置換及び/又は付加を有する塩基配列を有し、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の軽鎖可変領域をコードするDNA、および
(3)配列番号3の塩基配列またはこの塩基配列と相補的な塩基配列とストリンジェントな条件下でハイブリダイズする塩基配列を有し、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の軽鎖可変領域をコードするDNA。
【0022】
さらに、本発明の軽鎖可変領域をコードするDNA(遺伝子)は、下記の何れかのタンパク質をコードする遺伝子である。
(1)配列番号4のアミノ酸配列を有するタンパク質、
(2)配列番号4のアミノ酸配列において1から数個のアミノ酸の欠失、置換及び/又は付加を有するアミノ酸配列を有し、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の軽鎖可変領域であるタンパク質、および
(3)配列番号4のアミノ酸配列に対して90%以上の相同性を有するアミノ酸配列を有し、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の軽鎖可変領域であるタンパク質。
【0023】
配列番号1の塩基配列は、H鎖(重鎖)のV領域のcDNAの塩基配列であり、配列番号2のアミノ酸配列は、H鎖のV領域のアミノ酸である。さらに、配列番号3の塩基配列はL鎖(軽鎖)のV領域のcDNAの塩基配列であり、配列番号4のアミノ酸配列は、L鎖のV領域のアミノ酸配列である。これらの配列は、実施例で詳述されているように、HBs抗原に反応したBリンパ球を回収し、回収したBリンパ球からcDNAを合成し、合成したcDNAを用いて抗体のH鎖およびL鎖のV領域を増幅するようにPCR反応を行い、取得した細胞の抗体のH鎖およびL鎖のV領域のcDNAを作製し、作製したcDNAの塩基配列を決定し、さらに、このcDNAがタンパクに翻訳されることを確認して、決定した。その結果、本発明の抗体のH鎖およびL鎖のファミリー(サブクラス)は、H鎖は、VH3, DH, JH2であり、L鎖(κ鎖)はVκ3, Jκ4であった。
【0024】
本明細書で言う「1から数個の塩基の欠失、置換及び/又は付加を有する塩基配列」における「1から数個」の範囲は特には限定されないが、例えば、1から40個、好ましくは1から30個、より好ましくは1から20個、より好ましくは1から10個、さらに好ましくは1から5個、特に好ましくは1から3個程度を意味する。以下においても同様である。
【0025】
上記本発明のDNAが、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の重鎖または軽鎖の可変領域をコードするDNAであることは、このDNAを鋳型として作成されたタンパク質が、B型肝炎ウイルスのHBs抗原に対する反応性を有することで確認できる。B型肝炎ウイルスのHBs抗原に対する反応性は、例えば、酵素標識した二次抗体を用いるEnzyme linked immunosorbent assay (ELISA)、放射性同位元素で標識した二次抗体を用いるradio immuno assay (RIA)等により確認することができる。尚、DNAを鋳型として作成されるタンパク質が抗体の一部である場合、抗体と抗体の一部の抗原に対する反応性は同じようにELISA等で測定できる。ただし、例えばscFvの場合には抗体のFc部分が欠失しているので、二次抗体として酵素標識抗IgG抗体を用いることはできない。この場合はscFvにタグ(抗体のエピトープとなるペプチド)を付加して、そのタグに対する抗体等を二次抗体として用いることで、ELISA等で測定できる。
【0026】
ELISAは96穴プレートのウェルにHBs抗原をコートする。目的抗体をHBs抗原をコートしたウェルおよびしていないウェルに添加し、HBs抗原への目的抗体の結合を酵素標識した二次抗体の結合により酵素基質の発色を指標に確認することができる。この場合、抗原をコートしていないウェルには目的抗体は結合しないため発色せず、抗原をコートしたウェルには目的抗体が結合し発色する。RIAでは同様に96穴プレートにHBs抗原をコートする。目的抗体をHBs抗原をコートしたウェルおよびしていないウェルに添加し、HBs抗原への目的抗体の結合を放射性同位元素で標識した二次抗体の結合による放射能のカウントを指標に確認することができる。
【0027】
上記した「ストリンジェントな条件下でハイブリダイズする」とは、DNAをプローブとして使用し、コロニーハイブリダイゼーション法、プラークハイブリダイゼーション法、あるいはサザンブロットハイブリダイゼーション法等を用いることにより得られるDNAの塩基配列を意味し、例えば、コロニーあるいはプラーク由来のDNA又は該DNAの断片を固定化したフィルターを用いて、0.7〜1.0MのNaCl存在下、65℃でハイブリダイゼーションを行った後、0.1〜2×SSC溶液(1×SSC溶液は、150mM塩化ナトリウム、15mMクエン酸ナトリウム)を用い、65℃条件下でフィルターを洗浄することにより同定できるDNA等を挙げることができる。ハイブリダイゼーションは、モレキュラークローニング第2版等に記載されている方法に準じて行うことができる。
【0028】
ストリンジェントな条件下でハイブリダイズするDNAとしては、プローブとして使用するDNAの塩基配列と一定以上の相同性を有するDNAが挙げられ、例えば70%以上、好ましくは80%以上、より好ましくは90%以上、さらに好ましくは93%以上、特に好ましくは95%以上、最も好ましくは98%以上の相同性を有するDNAが挙げられる。
【0029】
本明細書で言う「1から数個のアミノ酸の欠失、置換及び/又は付加を有するアミノ酸配列」における「1から数個」の範囲は特には限定されないが、例えば、1から20個、好ましくは1から10個、より好ましくは1から7個、さらに好ましくは1から5個、特に好ましくは1から3個程度を意味する。
【0030】
本明細書で言う「配列表の配列番号2または4に記載のアミノ酸配列に対して90%以上の相同性を有するアミノ酸配列」における相同性は、90%以上であれば特に限定されないが、さらに好ましくは95%以上、さらに好ましくは97%以上、さらに好ましくは98%、特に好ましくは99%以上である。
【0031】
B型肝炎ウイルスのHBs抗原に対する反応性を有する抗体は既にいくつか報告されている。(特許文献1および2、非特許文献8〜14)それに対して、本発明の抗体のH鎖およびL鎖のファミリー(サブクラス)は、前述のとおりであり、本発明の抗体とH鎖およびL鎖のファミリー(サブクラス)が、H鎖についてVH3であり、かつL鎖(κ鎖)についてVκ3である抗体を、以下の表に示す。相同性は、本発明の抗体の該当する可変領域のアミノ酸配列との相同性である。
【0032】
【表1】
【0033】
本発明の遺伝子の取得方法は特に限定されない。本明細書中の配列表の配列番号1から4に記載したアミノ酸配列及び塩基配列の情報に基づいて適当なブローブやプライマーを調製し、それらを用いて抗体のcDNAライブラリーをスクリーニングすることにより本発明の遺伝子を単離することができる。単離した遺伝子が本発明の遺伝子であるかは、塩基配列の決定および遺伝子より発現させた抗体タンパクのHBs抗原への結合能をチェックすることにより調べることができる。cDNAライブラリーは、本発明の遺伝子を発現している細胞から常法により作製することができる。
【0034】
PCR法により本発明の遺伝子を取得することもできる。上記抗体の染色体DNAライブラリー又はcDNAライブラリーを鋳型として使用し、配列番号1又は3に記載した塩基配列を増幅できるように設計した1対のプライマーを用いてPCRを行う。PCRの反応条件は適宜設定することができ、例えば、94℃で30秒間(変性)、55℃で30秒〜1分間(アニーリング)、72℃で2分間(伸長)からなる反応工程を1サイクルとして、例えば30サイクル行った後、72℃で7分間反応させる条件などを挙げることができる。次いで、増幅されたDNA断片を、大腸菌等の宿主で増幅可能な適切なベクター中にクローニングすることができる。単離した遺伝子が本発明の遺伝子であるかは、塩基配列の決定および遺伝子より発現させた抗体タンパクのHBs抗原への結合能をチェックすることにより調べることができる。
【0035】
上記したプローブ又はプライマーの調製、cDNAライブラリーの構築、cDNAライブラリーのスクリーニング、並びに目的遺伝子のクローニング、塩基配列の決定、遺伝子からのタンパクの発現などの操作は当業者に既知であり、例えば、Molecular Cloning: A laboratory Mannual, 2nd Ed., Cold Spring Harbor Laboratory, Cold Spring Harbor, NY.,1989(以下、モレキュラークローニング第2版と略す)、Current Protocols in Molecular Biology, Supplement 1〜38, John Wiley & Sons (1987-1997)(以下、カレント・プロトコールズ・イン・モレキュラー・バイオロジーと略す)等に記載の方法に準じて行うことができる。
【0036】
上述した本発明の遺伝子の取得方法は、後述するCDRをコードする遺伝子の入手法にも適宜適用することができる。
【0037】
また、配列表の配列番号2または4に記載のアミノ酸配列において1から数個のアミノ酸の欠失、置換及び/又は付加を有するアミノ酸配列を有し、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の重鎖可変領域または軽鎖可変領域であるタンパク質をコードする遺伝子;並びに配列表の配列番号1または3に記載の塩基配列において1から数個の塩基の欠失、置換及び/又は付加を有する塩基配列を有し、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の重鎖可変領域または軽鎖可変領域をコードする遺伝子(以下、これらの遺伝子を変異遺伝子と称する)については、配列番号1〜4に記載のアミノ酸配列および塩基配列の情報に基づいて、化学合成、遺伝子工学的手法又は突然変異誘発などの当業者に既知の任意の方法で作製することができる。
【0038】
例えば、配列表の配列番号1または3に記載の塩基配列を有するDNAに対し、変異原となる薬剤と接触作用させる方法、紫外線を照射する方法、遺伝子工学的手法等を用いて行うことができる。遺伝子工学的手法の一つである部位特異的変異誘発法は特定の位置に特定の変異を導入できる手法であることから有用であり、モレキュラークローニング第2版、カレント・プロトコールズ・イン・モレキュラー・バイオロジー等に記載の方法に準じて行うことができる。
【0039】
[可変領域タンパク質]
本発明のB型肝炎ウイルスのHBs抗原に対するモノクローナル抗体を構成する重鎖可変領域であるタンパク質は、下記の何れかのタンパク質である。
(1)配列番号2のアミノ酸配列を有するタンパク質、
(2)配列番号2のアミノ酸配列において1から数個のアミノ酸の欠失、置換及び/又は付加を有するアミノ酸配列を有し、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の重鎖可変領域であるタンパク質、および
(3)配列番号2のアミノ酸配列に対して90%以上の相同性を有するアミノ酸配列を有し、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の重鎖可変領域であるタンパク質。
【0040】
本発明のB型肝炎ウイルスのHBs抗原に対するモノクローナル抗体を構成する軽鎖可変領域であるタンパク質は、下記の何れかのタンパク質である。
(1)配列番号4のアミノ酸配列を有するタンパク質、
(2)配列番号4のアミノ酸配列において1から数個のアミノ酸の欠失、置換及び/又は付加を有するアミノ酸配列を有し、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の軽鎖可変領域であるタンパク質、および
(3)配列番号4のアミノ酸配列に対して90%以上の相同性を有するアミノ酸配列を有し、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の軽鎖可変領域であるタンパク質。
【0041】
上述のように、配列番号2および4のアミノ酸配列は、HBs抗原に対するモノクローナル抗体の可変領域のcDNAがタンパクに翻訳されることを確認して決定したものである。
【0042】
本発明のタンパク質の取得方法は特に制限されず、化学合成により合成したタンパク質でもよいし、遺伝子組み換え技術により作製した組み換えタンパク質でもよい。組み換えタンパク質を作製する場合には、先ず、上記に記載した当該タンパク質をコードする遺伝子(DNA)を取得する。このDNAを適当な発現系に導入することにより、本発明のタンパク質を産生することができる。また、本発明のCDRペプチドも同様にして産生することができる。発現系でのタンパク質、ペプチドの発現については本明細書中後記する。
【0043】
[相補性決定領域DNA(遺伝子)]
本発明のモノクローナル抗体を構成する重鎖相補性決定領域1〜3をコードするDNA(遺伝子)は、下記の何れかの遺伝子である。
(1)配列番号1の塩基配列における76〜105番目の塩基配列(配列番号5)、または配列番号3のアミノ酸配列における26〜35番目のアミノ酸配列(配列番号11)をコードする塩基配列を有する、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の重鎖相補性決定領域1(CDR−H1)をコードする遺伝子。
(2)配列番号1の塩基配列における148〜198の塩基配列(配列番号6)、または配列番号3のアミノ酸配列における50〜66番目のアミノ酸配列(配列番号12)をコードする塩基配列を有する、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の重鎖相補性決定領域2(CDR−H2)をコードする遺伝子。
(3)配列番号1の塩基配列における295〜333の塩基配列(配列番号7)、または配列番号3のアミノ酸配列における99〜111番目のアミノ酸配列(配列番号13)をコードする塩基配列を有する、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の重鎖相補性決定領域3(CDR−H3)をコードする遺伝子。
【0044】
さらに本発明のモノクローナル抗体を構成する軽鎖相補性決定領域1〜3をコードするDNA(遺伝子)は、下記の何れかの遺伝子である。
(1)配列番号3の塩基配列における69〜102の塩基配列(配列番号8)、または配列番号4のアミノ酸配列における24〜34のアミノ酸配列(配列番号14)をコードする塩基配列を有する、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の軽鎖相補性決定領域1(CDR−L1)をコードする遺伝子。
(2)配列番号3の塩基配列における148〜168の塩基配列(配列番号9)、または配列番号4のアミノ酸配列における50〜56のアミノ酸配列(配列番号15)をコードする塩基配列を有する、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の軽鎖相補性決定領域2(CDR−L2)をコードする遺伝子。
(3)配列番号3の塩基配列における265〜291の塩基配列(配列番号10)、または配列番号4のアミノ酸配列における89〜97のアミノ酸配列(配列番号16)をコードする塩基配列を有する、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の軽鎖相補性決定領域3(CDR−L3)をコードする遺伝子。
【0045】
[相補性決定領域ペプチド]
本発明のモノクローナル抗体を構成する重鎖相補性決定領域1〜3であるペプチドは、下記のいずれかのペプチドである。
(1)配列番号2のアミノ酸配列における26〜35のアミノ酸配列を有する、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の重鎖相補性決定領域1(CDR−H1)であるペプチド、
(2)配列番号2のアミノ酸配列における50〜66のアミノ酸配列を有する、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の重鎖相補性決定領域2(CDR−H2)であるペプチド、
(3)配列番号2のアミノ酸配列における99〜111のアミノ酸配列を有する、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の重鎖相補性決定領域3(CDR−H3)であるペプチド。
【0046】
本発明のモノクローナル抗体を構成する軽鎖相補性決定領域1〜3であるペプチドは、下記のいずれかのペプチドである。
(1)配列番号4のアミノ酸配列における24〜34のアミノ酸配列を有する、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の軽鎖相補性決定領域1(CDR−L1)であるペプチド、
(2)配列番号4のアミノ酸配列における50〜56のアミノ酸配列を有する、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の軽鎖相補性決定領域2(CDR−L2)であるペプチド、
(3)配列番号4のアミノ酸配列における89〜97のアミノ酸配列を有する、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の軽鎖相補性決定領域3(CDR−L3)であるペプチド。
【0047】
本発明のモノクローナル抗体の重鎖相補性決定領域1〜3および軽鎖相補性決定領域1〜3は、Kabatの基準に照らして決定した。さらに、本発明のモノクローナル抗体の重鎖相補性決定領域1〜3および軽鎖相補性決定領域1〜3について、本発明のモノクローナル抗体と、H鎖およびL鎖のファミリー(サブクラス)が同一である、抗体(H鎖: VH、L鎖(κ鎖): Vκ3)のアミノ酸配列の対比およびアミノ酸配列の相同性を表2〜7に示す。
【0048】
【表2】
【0049】
【表3】
【0050】
【表4】
【0051】
【表5】
【0052】
【表6】
【0053】
【表7】
【0054】
[モノクローナル抗体]
本発明のB型肝炎ウイルスのHBs抗原に対するモノクローナル抗体は、配列番号2のアミノ酸配列を有するタンパク質を可変領域として含む重鎖と配列番号4のアミノ酸配列を有するタンパク質を可変領域として含む軽鎖とを含むものである。
【0055】
さらに本発明のB型肝炎ウイルスのHBs抗原に対するモノクローナル抗体は、配列番号2のアミノ酸配列における26〜35、50〜66および99〜111のアミノ酸配列を有するペプチドを相補性決定領域1〜3として含む重鎖と配列番号4のアミノ酸配列における24〜34、50〜56および89〜97のアミノ酸配列を有するペプチドを相補性決定領域1〜3として含む軽鎖とを含むものである。
【0056】
本発明のモノクローナル抗体は、以下の文献に記載の方法を利用して作成することができる [Kanda H, Mori K, Koga H, Taniguchi K, Kobayashi H, Sakahara H, Konishi J, Endo K, Watanabe T.Construction and expression of chimeric antibodies by a simple replacement of heavy and light chain V genes into a single cassette vector. Hybridoma. 13:359-66, 1994] 。具体的には、HBs抗原特異的Bリンパ球よりRT-PCRにより抗体遺伝子の重鎖可変領域および軽鎖可変領域の遺伝子を増幅する。増幅した遺伝子をpT7Blue-T vector (Novagen, A Brand of EMD Biosciences, Inc, An affiliate of Merck KGaA, Darmstadt, Germany)等にクローニングし、塩基配列を決定する。例えば、配列番号1および3に塩基配列を示す。その塩基配列を鋳型に制限酵素部位を含むプライマーを用いてPCRを行い、増幅させた遺伝子断片を制限酵素により切断し、その制限酵素部位を用いて抗体発現用ベクターに導入する。抗体発現用ベクターは抗体タンパク質の分泌に必要なシグナル配列に加え、抗体重鎖の定常部領域DNAあるいは抗体軽鎖の定常部領域DNAがクローニングされており、それぞれ抗体重鎖可変領域、抗体軽鎖可変領域遺伝子を導入することにより、完全長の抗体遺伝子を構築できる。
【0057】
構築した抗体重鎖遺伝子および抗体軽鎖遺伝子を同時にヒト胎児腎由来293T細胞などの動物細胞に導入し、2〜3日間培養することで、目的の抗体タンパクは培養上清に分泌される。
【0058】
CDR遺伝子を組み合わせて用い、抗体を調製する場合、以下のように行うことができる。抗体のV領域は、抗体の基本的な三次元構造を形作るフレームワーク部分(1〜4)と抗原に結合するCDR部分(1〜3)からなる。フレームワークの構造と任意のCDRを組み合わせることで任意の抗体のV領域を合成することが可能である。これをどのように組合せて1本のV領域遺伝子を合成するかは、図12に示すように「オーバーラップPCR」を用いる。たとえばFr1とCDR1の遺伝子配列がある時、ステプ1でFr1とCDR3遺伝子をPCRで合成する。この時、Fr1とCDR1の間のプライマー(P2、P3)はそれぞれCDR1あるいはFr1の塩基配列を加えて重なるようにしておく。ステップ1で出来上がったPCR産物は塩基配列が重なっており、これらのPCR産物を熱変性およびアニーリングを行うと、重なった部分でアニーリングする(ステップ2)。これをTaq DNAポリメラーゼで伸長させるとステップ3のようにFr1とCDR1がつながったものが出来上がる。これをプライマーP1およびP4を用いてPCRで増幅すればFr1とCDR1がつながったDNAが増幅される。ステップ2とステップ3は同時に行うことができる。実際にはFr2、CDR2、Fr3、CDR3、Fr4も同じように増幅してやり、ステップ3にてプライマーP1とP5を用いると抗体のV領域の遺伝子が増幅される。CDRの部分をランダムな塩基配列にしてやると、あらゆる抗原に結合する可能性のあるV領域のライブラリーを作成することができる。
【0059】
[単鎖抗体]
本発明のB型肝炎ウイルスのHBs抗原に対する単鎖抗体は、配列番号2のアミノ酸配列を有するタンパク質と配列番号4のアミノ酸配列を有するタンパク質とをリンカーペプチドで結合したタンパク質である。さらに本発明のB型肝炎ウイルスのHBs抗原に対する単鎖抗体は、配列番号2のアミノ酸配列における26〜35、50〜66および99〜111のアミノ酸配列を有するペプチドを相補性決定領域1〜3として含むタンパク質と配列番号4のアミノ酸配列における24〜34、50〜56および89〜97のアミノ酸配列を有するペプチドを相補性決定領域1〜3として含むタンパク質とをリンカーペプチドで結合したタンパク質である。
【0060】
単鎖Fvタンパク(single chain Fv, scFv)は、抗体重鎖可変領域(VH)と抗体軽鎖可変領域(VL)を直列に結合したタンパク質であり、モノクローナル抗体と同様の方法で作製することができる。すなわち、上記モノクローナル抗体の作製方法で説明したpT7Blue-T vectorにクローニングした抗体重鎖可変領域遺伝子と抗体軽鎖可変領域遺伝子を鋳型として、この2つの領域をリンカー配列で結合させ、試験管内タンパク合成に適合するようT7プロモータ、リボソーム結合配列等を含む遺伝子をPCRを用いて作製する。作製した遺伝子を用いて、例えば無細胞系タンパク合成反応液PURESYSTEM S-S ((株)ポストゲノム研究所、東京)内でインキュベーションすることによりscFvタンパクを合成することができる。
【0061】
本発明は、上記単鎖抗体の二量体(diabody)および三量体(triabody)を包含する。さらに本発明は、上記単鎖抗体の配列番号2のアミノ酸配列を有するタンパク質または、配列番号2のアミノ酸配列における26〜35、50〜66および99〜111のアミノ酸配列を有するペプチドを相補性決定領域1〜3として含む前記タンパク質にCH3領域を結合した、B型肝炎ウイルスのHBs抗原に対するミニボディー(minibody)を包含する。
【0062】
単鎖Fvタンパクを構成するリンカーは、例えば、(GlyGlyGlyGlySer)を3つ連ねたものである。下記論文に記載のように、リンカーの長さを変えることによって、scFvが2つ組合わさった二量体(diabody)、3つ組合わさった三量体(triabody)が作製できる(Hudson PJ, Kortt AA, High avidity scFv multimers; diabodies and triabodies. J Immunol Methods, 231:177-189 (1999)参照、Edited by Frank Breitling, Sterfan Dubel "Recombinant Antibodies" (1999) A John Willey & Sons, Inc.(New York) and Spektrum Akademischer Verlag (Heidelberg) Copublication参照)。
【0063】
抗体を医薬として取り扱う場合、その分子量の大きさが問題となる。それを解決するために、scFv、二量体(diabody)、三量体(triabody)、ミニボディー(minibody)というような抗原結合能を保持したまま分子量を小さくした分子が作製され、抗体医薬としての評価が行われている。
【0064】
抗体遺伝子は図1のようにH鎖はVH遺伝子断片とCH1, (H), CH2, CH3、(CH4)遺伝子断片より、L鎖はVL遺伝子断片とCL遺伝子断片より構成されている。このうちのVHおよびVL遺伝子断片をリンカーで結合したものが単鎖Fv (scFv)である(図2)。scFvには5’-VH-VL-3’の構造のものと5’-VL-VH-3’のすなわち、scFvは抗体のVドメインより形成されC領域を有していない。これが二量体を形成したものがdiabody、三量体を形成したものがtriabodyである。これらはリンカーの長さを変えることで形成させることができるという報告がある(PJ Hudoson and AA Kortt, High avidity scFv multimers; diabodies and triabodies. J Immunol Methods 231:177-189, 1999)。scFvに比べるとdiabodyおよびtriabodyの方が総合的な結合力(avidity)は強くなる。また、scFvにIgGのCH3ドメインを結合したminibodyも抗原結合能を維持した抗体のミニチュア化の産物であり、CH3部分で二量体を形成する(LS Shahied, Y Tang, RK Alpaugh, R Somer, D Greenspon, LM Weiner.Bispecific minibodies targeting HER2/neu and CD16 exhibit improved tumor lysis when placed in a divalent tumor antigen binding format. J Biol Chem. 279:53907-53914)。ヒトIgGにはIgG1からIgG4の4つのバリエーションがあり、上記CH3部分は、IgG1からIgG4のいずれのCH3であることもできる。但し、IgG1のCH3部分であることが好ましい。
【0065】
[モノクローナル抗体の利用]
本発明は、上記本発明のモノクローナル抗体を用いるB型肝炎ウイルスの検定方法、B型肝炎の診断方法およびB型肝炎の治療方法を包含する。モノクローナル抗体の利用は、モノクローナル抗体そのもの以外に本発明のモノクローナル抗体の一部またはそれらのDNAを用いたもの、たとえばscFv単鎖抗体、単鎖抗体の二量体(diabody)、三量体(triabody)またはミニボディー(minibody)あるいはその遺伝子等を用いた利用も包含する。また、本発明のモノクローナル抗体の一部としては、例えば、このモノクローナル抗体を構成する重鎖可変領域および軽鎖可変領域であるタンパク質、並びに、このモノクローナル抗体を構成する重鎖相補性決定領域1〜3および軽鎖相補性決定領域1〜3であるペプチドを包含する。
【0066】
B型肝炎ウイルスの検定方法
本発明のモノクローナル抗体等を用いたB型肝炎ウイルスの検定方法は以下のように行うことができる。まず、96穴プレートのウェルに本発明のモノクローナル抗体を吸着させる。ブロッキングを行い非特異的吸着を抑えた後、B型肝炎ウイルスを含む検体をウェルに添加する。検体中にウイルスが含まれていれば、そのウイルスはウェルに吸着させた本発明のモノクローナル抗体に結合する。その後、ウェルを洗浄し、酵素標識した本発明モノクローナル抗体をウェルに添加する。ウェルを洗浄後、酵素基質をウェルに添加し、基質の発色で検体中のウイルスの存在を検出する。ウイルスが存在する検体を添加したウェルでは、ウェルに吸着させた抗体、それに結合したウイルス、さらにそのウイルスに結合した酵素標識抗体の複合体ができ、基質が発色する。
【0067】
また、本発明のモノクローナル抗体を用いたB型肝炎ウイルスの検定方法は以下のようにしても行うことができる。すなわち、本発明のモノクローナル抗体を96穴プレートあるいはビーズに固着させ、そこへB型肝炎ウイルスの存在をチェックする検体を添加する。B型肝炎ウイルスが存在すると、ウイルスが抗体に結合する。抗体に結合したウイルスよりウイルスDNAを抽出し、それを鋳型としてPCR等を行い、ウイルスDNAを増幅し、検出を行う。このようにしても検体中のウイルスの存在を検出することが可能である。
以上の方法によるウイルスの検出は本発明のモノクローナル抗体の一部、F(ab)’2, F(ab), scFv、単鎖抗体の二量体(diabody)、三量体(triabody)またはミニボディー(minibody)、このモノクローナル抗体を構成する重鎖可変領域および軽鎖可変領域であるタンパク質、並びに、このモノクローナル抗体を構成する重鎖相補性決定領域1〜3および軽鎖相補性決定領域1〜3であるペプチドなどを用いても行うことができる。
【0068】
B型肝炎の診断方法
本発明のモノクローナル抗体を用いたB型肝炎の診断は、上記検体として患者血清を調べることで、患者血清中のB型肝炎ウイルスを本発明のモノクローナル抗体を用いて検出することにより行うことができる。
また、患者の肝よりバイオプシーすることにより得た肝組織中のウイルスを本発明のモノクローナル抗体を用いて染色することにより診断することもできる。
以上の方法によるウイルスの診断は本発明のモノクローナル抗体の一部、F(ab)’2, F(ab), scFv、単鎖抗体の二量体(diabody)、三量体(triabody)またはミニボディー(minibody)、このモノクローナル抗体を構成する重鎖可変領域および軽鎖可変領域であるタンパク質、並びに、このモノクローナル抗体を構成する重鎖相補性決定領域1〜3および軽鎖相補性決定領域1〜3であるペプチドなどを用いても行うことができる。
【0069】
B型肝炎ウイルスの感染阻止およびB型肝炎の治療方法
本発明のモノクローナル抗体を用いたB型肝炎ウイルスの感染阻止は以下のように行うことができる。すなわち、B型肝炎ウイルスが混入している血液や体液、培養上清などが付着した注射針を誤って刺してしまったような、いわゆる針刺し事故においては、B型肝炎ウイルスワクチンをすぐに免疫すると同時にB型肝炎ウイルスに対する抗体価の高い抗血清を注入する治療法が一般的に行われている。すなわち、B型肝炎ウイルスに抗体が結合し細胞への感染を防ぐように作用する。しかし、人の血清製剤は献血由来の血清を材料とするため、未知のウイルスが混入する可能性もあり望ましいものではない。また、高力価で中和作用の優れた製剤を安定供給できるとは限らない。本発明のモノクローナル抗体を管理された環境で作製すれば、未知のウイルスの混入していないウイルス中和抗体を安定した品質および量で提供でき、上記ヒト由来抗血清の代わりに用いることができる。
【0070】
また、B型肝炎ウイルスが感染し、肝がん、肝硬変、肝不全に陥った患者で肝移植が治療法の選択肢となる場合がある。この場合、患者体内にB型肝炎ウイルスが残存していると移植した肝にB型肝炎ウイルスが再感染してしまい、治療効果が半減してしまうことになる。このような肝移植の場において、本発明のモノクローナル抗体を治療に用いることができる。すなわち、肝移植をするために患者の肝を除去した際に、本発明のモノクローナル抗体を単独あるいは他の抗体医薬や抗ウイルス剤と併用して患者に投与し、残存しているB型肝炎ウイルスを中和してやることにより、新たに移植した肝へのウイルスの感染を防ぐことができる。
【0071】
以上の方法による感染防止および治療は本発明のモノクローナル抗体の一部、F(ab)’2, F(ab), scFv、単鎖抗体の二量体(diabody)、三量体(triabody)またはミニボディー(minibody)、このモノクローナル抗体を構成する重鎖可変領域および軽鎖可変領域であるタンパク質、並びに、このモノクローナル抗体を構成する重鎖相補性決定領域1〜3および軽鎖相補性決定領域1〜3であるペプチドなどを用いても行うことができる。
【実施例】
【0072】
実施例1
[Bリンパ球の調製]
酵母菌で作製した組換えHBワクチンをHBs抗体陽性のボランティアに免疫し、免疫後9日目に末梢血よりリンパ球を調製した。すなわち、末梢血からヒトリンパ球をリンホセパールI液((株)免疫生物研究所、群馬県高崎市)を用いて遠心分離し、さらにAutoMACS (Miltenyi Biotec, Bergisch Gladbac, Germany)を用いてリンパ球画分から非Bリンパ球分画細胞を除去しBリンパ球画分を分離精製した。
【0073】
[Bリンパ球への蛍光色素の負荷]
調製した2 x 106個Bリンパ球を2μM Fluo-4/AM(カルシウム依存性蛍光色素、同仁、熊本)/1μM CellTracker Orange (Molecular Probe社)/0.04% pluronic F-127 (Molecular Probe社)/loading buffer (137 mM NaCl, 2.7 mM KCl,1.8 mM CaCl2, 1mM MgCl2, 1 mg/mLグルコース, 1mg/mL BSA, 20 mM Hepes (pH7.4))に懸濁し、ゆっくり震盪しながら室温で30分インキュベーションした。Loading bufferで細胞を洗浄し、細胞内に導入されなかった余分なFluo-4/AMを除いた。その後、細胞をRPMI1640/10% FCS溶液に懸濁した。なお、以下の全ての実験においてはPhenol Redを含まないRPMI1640を用いた。Fluo-4はカルシウムが結合すると蛍光強度が増加する蛍光色素であり、Bリンパ球の抗原による活性化の検出に用いる。CellTracker Orangeはカルシウムに関係なく蛍光を発する蛍光色素であり、マイクロアレイスキャナーを用いた細胞の認識あるいは蛍光顕微鏡下で細胞を回収する際の細胞の認識のために用いた。
【0074】
[マイクロウェルアレイチップ]
マイクロウェルアレイチップはシリコンを用いて作製されており(富山県工業技術センター製造)、直径10μm、深さ14μmのマイクロウェルが25μmのピッチ(ウェルの中心間の距離)で縦横に配列されている。マイクロウェルは30 x 30 (900ウェル)でひとつのクラスターを形成しており、それが縦に12クラスター、横に22クラスター並んでいるものを用いた。マイクロウェルアレイチップの両側には厚さ1mm、幅約1mm、長さ2cmのシールを貼った。
【0075】
[マイクロアレイスキャナー]
本装置は基本的に日立ソフトウェアエンジニアリング(株)(横浜市)のマイクロアレイスキャナー(CRBIO IIe-FITC)を用いており、473 nmおよび532 nmの波長の励起光を発することができるレーザーが搭載されている。また、2.5μmの解像度で解析が可能である。焦点深度は±50μmに設定されている。
【0076】
[マイクロウェルアレイチップへのリンパ球の播種]
マイクロウェルアレイチップのマイクロウェルへRPMI1640/10% FCS溶液を満たすために、マイクロウェルアレイチップをRPMI1640/10% FCS溶液に浸し、超音波をかけることでウェル中の空気を除くことにより、マイクロウェルへ溶液を満たした。マイクロウェルへのRPMI1640/10% FCS溶液の充填は以下の方法でも行った。マイクロウェルアレイチップへ100%アルコール液を添加する。アルコール液はマイクロウェルへ浸透していく。次に、アルコール液をPBSで置換し、さらにRPMI1640/10% FCS溶液で置換した。マイクロウェルへRPMI1640/10% FCS溶液を充填した後、チップ上の余分なバッファーを除き、そこへ上記細胞懸濁液を添加し、2〜5分間静置した。マイクロウェルに入らなかった細胞を回収し、チップ上へさらに細胞懸濁液を添加し、2〜5分間静置した。マイクロウェルに入らなかった細胞を回収し、チップ上へさらに細胞懸濁液を添加し、2〜5分間静置した。マイクロウェルに入らなかった細胞を回収し、さらにチップ上のマイクロウェルに入らなかった細胞をRPMI1640/10% FCS溶液を用いて洗い流した。リンパ球の直径は約8μmであり、使用するマイクロウェルの直径が10μmであるために一つのマイクロウェルにはリンパ球が1個入る。カバーグラスを上記シール上に置き、チップとカバーグラスの間にRPMI1640/10% FCS溶液を満たした。
【0077】
[マイクロウェルアレイチップを用いたHBs抗原特異的Bリンパ球の検出]
Bリンパ球を播種したマイクロウェルアレイチップをマイクロアレイスキャナー(CRBIO IIe-FITC)に挿入し、解像度2.5μmでスキャンし抗原刺激前のFluo-4の蛍光強度を測定しデータを保存した(抗原刺激前の蛍光データA)。次に、チップをスキャナーから取り出し、チップとカバーグラスの間のRPMI1640/10%FCS溶液を除き、そこへRPMI1640/10%FCS溶液に溶解させたHBs抗原(100μg/mL)を加えた。直ちにマイクロウェルアレイチップをマイクロアレイスキャナーに挿入し、抗原添加後約0.5〜2分の細胞内のFluo-4の蛍光強度を解像度2.5μmでスキャンし、データを保存した(抗原刺激後の蛍光データB)。スキャナーに付属の解析ソフトウェアを用いて刺激前後の蛍光強度の比(B/A)を計算し、比の大きいウェルを特定した。これらのウェルの中に抗原特異的Bリンパ球が存在する。
【0078】
[Bリンパ球のマイクロウェルからの回収]
検出したHBs抗原に反応し細胞内カルシウムが上昇したBリンパ球は蛍光顕微鏡下でマイクロマニピュレータを用いて回収した。すなわち、まず、カバーグラスとチップの間のRPMI1640/10%FCS溶液を取り除き、カバーグラスとチップの間に空気を入れることでカバーグラスを取り外した。チップが乾燥しないようにチップにRPMI1640/10%FCS溶液を加え、蛍光顕微鏡下で細胞のCellTracker Orangeの蛍光を観察しながら、マイクロマニピュレータで目的の細胞を回収した。
【0079】
[Bリンパ球からの抗体遺伝子の採取]
回収したBリンパ球をPCR用チューブに移す。細胞に15μLの細胞溶解液(25μLに希釈された時の最終濃度:1x 1ststrand buffer (GIBCO-BRL, SuperScript IIIに添付されている), 0.2 mM dNTP, 0.25% NP-40, 0.1 mg/mL BSA, 10 mM DTT, 0.05 μM Random Primer, 1U/μL RNasin (Promega, Madison, WI)を加え、そこへ逆転写酵素SuperScriptIII (GIBCO-BRL, Rockville, MD)を(5μL, 50U)添加し、37oC、1時間反応させmRNAよりcDNAを合成した。
【0080】
[抗体遺伝子の1回目の増幅]
合成したヒト抗体遺伝子のcDNAを鋳型に、抗体のH鎖およびL鎖のV領域を増幅するように設計された以下のプライマーを用いてPCR反応を行った。
【0081】
ヒト抗体(IgM およびIgG)のH鎖用のプライマー
PCR-1用
Primer1 5' mix
hVH17a.1 atggactgsayytggagvdtc(配列番号17)
hVH2a.1 tccacrctcctgctrctgac(配列番号18)
hVH3a.1 gggcygagstggvttttyct(配列番号19)
hVH4a.1 tcctcctsctggtggcagct(配列番号20)
hVH5.1 tcaaccgccatcctcgccct(配列番号21)
hVH6.1 ctccttcctcatcttcctgcc(配列番号22)
Primer1 3' mix
hIGHG1-4out agtccttgaccaggcagccca(配列番号23)
hIGHMout attctcacaggagacgagggg(配列番号24)
【0082】
ヒト抗体L鎖用のプライマー
PCR-1 用
Primer1 5’ mix
hKV12.1 atgaggstcccygctcagctc(配列番号25)
hKV3.1 ctcttcctcctgctactctggc(配列番号26)
hKV45.1 ctsttsctytggatctctg(配列番号27)
hKV6.1 tgggtttctgctgctctggg(配列番号28)
hKV7.1 atagggtccggggctcctttg(配列番号29)
hLV12.1 cykctsctcctcactctcctc(配列番号30)
hLV3.1 ttctcctcctcggcctcctct(配列番号31)
hLV4.2-2 ccagcytgtgctgactcaatc(配列番号32)
hLV789.2 tcycagmctgtgstgacycag(配列番号33)
hLV6.1 ttttatgctgactcagcccc(配列番号34)
hLV7.1 ggcctggactcctctctttctg(配列番号35)
hLV8.1 ggcctggatgatgcttctcctc(配列番号36)
hLV9.1 tcctctgctcctcaccctcct(配列番号37)
hLV10.1 cctgggtcatgctcctcctga(配列番号38)
hLV11.1 gcctgggctccactacttctc(配列番号39)
Primer1 3’ mix
hIGK1 ctgctcatcagatggcggga(配列番号40)
hIGL1 gacacacyagtgtggccttgt(配列番号41)
【0083】
PCR反応を行い、取得した細胞の抗体のH鎖およびL鎖のV領域のcDNAを増幅したH鎖およびL鎖は別個のチューブを用いて増幅した。PCR反応は、上記cDNA2μLを18μLのPCR mix1 (最終濃度:1x TAKARA ExTaq buffer, 0.25 mM dNTP, Primer1 5’ mix 0.5μM each, Primer1 3’ mix 0.5μM each, 0.05 U/μL ExTaq (Takara, 京都))に加え、94℃, 5 min; (94℃, 30 sec; 60℃, 1min; 72℃, 1 min 30 sec) x 40サイクル; 72℃, 5 min反応させる。このPCRの反応により抗体遺伝子のLeader配列から定常部領域までのDNAが増幅される。
【0084】
[抗体遺伝子の2回目の増幅]
1回のPCRではcDNAが十分増幅されないため、2回目のnested PCRを行った。2回目のPCRの反応は1回目に得られたPCRの反応溶液2μLを18μLのPCR mix2(最終濃度:1x TAKARA ExTaq buffer, 0.25 mM dNTP, Primer2 5’ mix 0.5μM each, Primer2 3’ mix 0.5μM each, 0.05 U/μL ExTaq)に加え、94 oC, 5 min; (94℃, 30 sec; 60℃, 1min; 72℃, 1 min 30 sec) x 40サイクル; 72℃, 5 min反応させる。2回目のPCRにより、1回目で増幅した抗体遺伝子の可変部領域から定常部領域までのcDNA配列が増幅される。以下に使用したプライマーの配列を示す。
【0085】
ヒト抗体(IgM およびIgG)のH鎖用のプライマー
PCR-2用
Primer2 5' mix
hVH17a.2 ggtgcagctkgtrcartctgg(配列番号42)
hVH2a.2 caccttgarggagtctggtcc(配列番号43)
hVH3a.2 aggtdcarctgktggagtcyg(配列番号44)
hVH4a.2 ggtcctgtctcaggtgcagct(配列番号45)
hVH5a.2 gtgcagctggtgcagtctgg(配列番号46)
hVH6.2 gcagcagtcaggtccaggact(配列番号47)
Primer2 3' mix
hIGHG1-4S aagacsgatgggcccttggtg(配列番号48)
hIGHMin aagggttgggcggatgcact(配列番号49)
【0086】
ヒト抗体L鎖用のプライマー
PCR-2 用
Primer1 5’ mix
hKV1.2 ccagatgacccagtctccatc(配列番号50)
hKV2.2 ccagtggggatattgtgatgac(配列番号51)
hKV3.2 cagtctccagccaccctgtct(配列番号52)
hKV4.2 gtgatgacccagtctccagac(配列番号53)
hKV 5.2 acactcacgcagtctccagca(配列番号54)
hKV67.2 ttgtgctgacycagtctccag(配列番号55)
hLV1.2 agtctgtgctgacgcagccgc(配列番号56)
hLV23.2 tgactcagccwcyctcmgtgtc(配列番号57)
hLV4.2-3 caatcatcctctgcmtctgc(配列番号58)
hLV5.2-2 gactcagccaacctccctctc(配列番号59)
hLV6.2 gactcagccccactctgtgtc(配列番号60)
hLV789.2 tcycagmctgtgstgacycag(配列番号61)
hLV1011.2 tgactcagccmcmctckgtgtc(配列番号62)
Primer1 3’ mix
hIGK2 gacagatggtgcagccacagt(配列番号63)
hIGL2 cttgragctcctcagaggaggg(配列番号64)
【0087】
[増幅した抗体遺伝子の塩基配列の決定]
PCR産物を、アガロースゲルを用いて解析、精製し、pT7Blue-T vector (Novagen, Madison, WI)にクローニングし、抗体遺伝子の塩基配列を決定し、タンパクに翻訳されることを確認した。H鎖のV領域のcDNAの塩基配列を配列番号1に、アミノ酸配列を配列番号2に示す。さらに、L鎖のV領域のcDNAの塩基配列を配列番号3に、アミノ酸配列を配列番号4に示す。
【0088】
実施例2
[抗体タンパクの産生、抗原結合能の解析]
RT-PCR法にて増幅した抗体のH鎖およびL鎖の可変領域遺伝子を抗体タンパク発現用ベクター(図3)に組込んだ。すなわち、増幅した抗体のH鎖可変領域の遺伝子断片は上記HEF-HBs-gγ1ベクターのVHの部分に、L鎖可変領域の遺伝子断片は上記HEFs-gκベクターの定常部領域(Human Cκ)の5’側に制限酵素部位を用いて挿入した。作製したH鎖およびL鎖の発現ベクターから抗体タンパクを発現させるために、両方の発現ベクターを同時にヒト胎児腎由来細胞293T細胞に遺伝子導入した。遺伝子導入は常法(細胞工学別冊8 新 細胞工学実験プロトコール、東京大学医科学研究所制癌研究部編、(株)秀潤社)に従い、リン酸カルシウム法を用いて行った。3日後に細胞上清を回収した。この培養上清中の抗体のHBs抗原への結合能を、HBs抗原をコートした96ウェルプレートを用いたELISA法により測定した。ELISAによる測定は以下のように行った。すなわち、96穴プレートにphosphate buffered saline (PBS)で希釈したHBs抗原(10μg/mL)を各ウェルに50μLずつ分注し、4℃で一晩反応させ吸着させた。PBSでウェルを洗浄し、非特異的結合を除去する為に3%BSA、0.05%Tween-20添加PBSを各ウェルに400μLずつ分注し、室温で2時間反応させブロッキングした。ネガティブコントロールのウェルとして、抗原を吸着させず、ブロッキングしたウェルも作成した。その後0.1% Tween-20添加PBS (PBS-T)でウェルを洗浄し、上記293T細胞の培養上清を各ウェルに50μLずつ分注し、室温で2時間反応させた。ウェルをPBS-Tで洗浄し、1000倍希釈したアルカリフォスファターゼ(ALP)標識抗ヒト免疫グロブリンを各ウェルに50μLずつ分注し、室温で2時間反応させた。ウェルをPBS-Tで洗浄し、ALP基質用緩衝液(100mM NaCl、5mM MgCl2 100mM Tris pH9.5)に溶解した1mg/mL p-nitrophenyl phosphate(ALP基質)を各ウェルに50μLずつ分注し、室温で20分反応後にマイクロプレートリーダーにて、405nmにおける吸光度を測定した。さらに、本発明のモノクローナル抗体の抗原への結合の特異性をチェックするために、抗体を含む細胞培養上清をウェルに添加する際に、可溶性のHBs抗原と培養上清とをプレインキュベーションさせた。その混合液をウェルに添加し、ウェルに吸着させたHBs抗原への抗体の結合が可溶性のHBs抗原により競合的に阻害されるかを検討した。結果を図4に示す。
【0089】
図4に示されるように、作製した116抗体はHBs抗原に結合し、HBs抗原を結合させていないウェルには抗体は結合しなかった(データは示していない)。さらに、結合の特異性を検討するために116抗体と可溶性のHBs抗原を混合したものをHBs抗原が結合しているウェルに加え同様にELISAを行った。116抗体のウェルに結合したHBs抗原への結合が、可溶性HBs抗原により用量依存的、競合的に阻害されることが観察された。この結果より116抗体がHBs抗原に特異的に結合することが示された。
【0090】
116抗体が結合するエピトープの決定
ELISAの系を用いて、116抗体がadr型HBs抗原のa、d、rのどのエピトープに結合するかを、各エピトープに結合するモノクローナル抗体が116抗体のadr型HBs抗原への結合を競合的に阻害するかを調べることで検討した。まず、96穴プレートにリン酸緩衝生理食塩水(phosphate buffered saline (PBS))で希釈したadr型HBs抗原(10μg/mL)を各ウェルに50μLずつ分注し、4 oCで一晩反応させ吸着させた。PBSでウェルを洗浄し、非特異的結合を除去する為に3%BSA、0.05%Tween-20添加PBS(PBS-T/BSA)を各ウェルに400μLずつ分注し、室温で2時間反応させブロッキングした。ネガティブコントロールのウェルとして、抗原を吸着させず、ブロッキングしたウェルも作成した。その後0.1% Tween-20添加PBS (PBS-T)でウェルを洗浄し、PBS-T/BSAあるいは2μg/mLのエピトープaを認識するマウスモノクローナル抗体(抗a抗体)、エピトープdを認識するマウスモノクローナル抗体(抗d抗体)、あるいはエピトープrを認識するマウスモノクローナル抗体(抗r抗体)を各ウェルに50μLずつ分注し、室温で1時間反応させた。0.5μg/mLのビオチン化116抗体を各ウェルに5μLずつ分注し、さらに室温で2時間反応させた。ウェルをPBS-Tで洗浄し、1000倍希釈したアルカリフォスファターゼ(ALP)標識ストレプトアビジンを各ウェルに50μLずつ分注し、室温で2時間反応させた。ウェルをPBS-Tで洗浄し、ALP基質用緩衝液(100mM NaCl、5mM MgCl2、100mM Tris pH9.5)に溶解した1mg/mL p-ニトロフェニルフォスフェート(ALP基質)を各ウェルに50μLずつ分注し、室温で20分反応後にマイクロプレートリーダーにて、405nmにおける吸光度を測定した。結果を図5に示す。
【0091】
抗原を吸着させていないウェルでは発色せず(カラム1)、抗原を吸着させたウェルで発色した(カラム2)ことから、116抗体がadr型のHBs抗原に特異的に結合していることがわかる。抗a抗体は116抗体の抗原への結合を阻害したが(カラム3)、抗dおよび抗r抗体は116抗体の抗原への結合を阻害しなかった(カラム4、5)。これらの結果は、116抗体がエピトープdあるいはエピトープrを認識するのではなく、HBウイルスに共通のエピトープaを認識することを示している。
【0092】
本発明においては、116抗体を作製するにあたり、HBsワクチンはadrの型のものを用いており、スクリーニングもadrのHBs抗原を用いて行っている。したがって、116抗体はadr型のHBs抗原に結合するものである。エピトープa、エピトープd、エピトープrのどこを116抗体が認識するかについては、上記の様にadr型のHBs抗原への116抗体の結合をELISAで測定する時に、エピトープa、エピトープd、エピトープrに結合することがわかっている抗a抗体、抗d抗体、抗r抗体を同時に加えて競合させた時に、抗a抗体とは競合し結合が阻害されるが、抗d抗体や抗r抗体とは競合せず結合が阻害されないことから、ウイルスによって異なるエピトープdやエピトープrを認識するのではなく、全てのウイルスに共通のエピトープaを認識することがわかる。このように、本発明の抗体は、全てのタイプのHBVに対して中和活性を有し得るHBs抗原特異的抗体である。
【0093】
実施例3
[ヒト肝細胞キメラマウス]
ヒト肝細胞キメラマウスの作製法については以下の論文に詳述されている。[C Tateno, Y Yoshizane, N Saito, M Kataoka, R Utoh, C Yamasaki, A Tachibana, Y Soeno, K Asahina, H Hino, T Asahara, T Yokoki, T Furukawa, K Yoshizato. Near completely humanized liver in mice shows human-type metabolic responses to drugs. Am J Pathol, 165:901-912, 2004]簡単には、uPAマウス(B6SJL-TgN(Alb1Plau)144Bri, The Jackson Laboratory, Bar Harbor, ME)とscidマウス(Fox Chase SCID C.B-17/lcr-scid Jcl, Clea Japan inc., Tokyo, Japan)を掛け合わせuPA/SCIDマウスを作製した。ヒト肝細胞は、同意を得たドナーより採取した肝組織より小型肝細胞を含む肝非実質細胞を分離し移植に用いた。小型肝細胞移植1日前に、生後20〜30日のuPA/SCIDマウスに抗アシアロGM1抗体を腹腔内投与した。1日後、5〜7x105の生きた肝細胞を門脈を介して移植した。移植の成否はマウス血清中のヒトアルブミン値を測定することにより評価した。
【0094】
[抗HBs-抗体のウイルス中和能の解析]
HBVはヒトあるいはチンパンジーの肝細胞にしか感染しないため、抗HBs-抗体がHBVの感染をブロックできるかどうかは、これまでチンパンジーを用いて行われてきた。近年、フレッシュな肝臓組織をボランティアから採取し、その肝細胞を使ってin vitroにおいてHBVを感染させる実験系も報告されている。しかし、採取した肝細胞はin vitroで培養を続けることにより容易に感染感受性を失うため、常にフレッシュな肝細胞を用いる必要であり、人への負担から実際に行うことは困難であった。しかるに、上記ヒト肝細胞キメラマウスは移植した肝細胞のHBV感染感受性等の細胞の性質を失うことなく長期に肝細胞を保持できること、さらに、キメラマウス由来のヒト肝細胞を用いてキメラマウスを作製することも可能であることから、HBVの感染実験系を簡単に、安定に、低価格で供給することを可能にした。本実験では、ヒト肝細胞キメラマウスを用いてHBVの感染実験を行い、その実験系を用いて116抗体がHBVの感染をブロックすることができるかを検討した。実験の概略は以下の通りである。
【0095】
B型肝炎ウイルス感染患者の血清(1 x 108コピー/mLのB型肝炎ウイルス(HBV)のゲノムを含む)50μLと116抗体(20μg/匹)をプラスチックチューブ内で混合し室温で30分間反応させた後、その混合液をヒト肝細胞キメラマウスの鎖骨下静脈より注入した(2匹)。感染のポジティブコントロールとして患者血清のみをヒト肝細胞キメラマウス2匹に投与した。116抗体のネガティブコントロールとしてヒトIgG(20μg/匹)を患者血清とインキュベーションしたものをヒト肝細胞キメラマウス2匹に投与した。投与後、2週目、4週目、6週目にマウスの血清を採取し、B型肝炎ウイルスのゲノム遺伝子の存在を、定量PCR法を用いて定量した。
【0096】
結果を図6に示す。図6は、ヒトキメラマウス(1〜7)にHBVのみ(1〜3)、HBVとコントロールIgG(4、5)、HBVと116抗体(6、7)を注入し、2週目、4週目、6週目のマウス血清中のHBVゲノムDNAを定量PCR法にて測定した結果である。下欄にはヒト肝細胞キメラマウスの血清中のヒトアルブミンの量を示してあり、キメラマウスにおけるヒト肝細胞の存在を示す。
【0097】
図6に示すように患者血清(HBV)のみを投与した場合には投与2週後からウイルスゲノムDNAが検出され、4週目、6週目と経過するにつれ、DNA量は増加した。この結果は、HBVがマウス細胞には感染しないことから、移植されたヒト肝細胞にHBVが感染し、増殖していることを示す。ネガティブコントロール抗体としてヒトIgGを用いた場合も同様の結果を得た。一方、116抗体とHBVを混合したものをヒト肝細胞キメラマウスに投与した場合には、一匹のマウスにおいて2週目にウイルスゲノム遺伝子が微量検出されたが、4週目、6週目には2匹のマウスともにウイルスゲノム遺伝子は検出されなくなった。以上の結果は、116抗体がウイルスの細胞への感染を阻害する活性(中和活性)を保持していることを示す。
【0098】
実施例4
Single chain Fv (ScFv)
遺伝子の構築
116抗体遺伝子配列及びコントロール抗体遺伝子配列をそれぞれ鋳型とし、VH遺伝子の3’側及びVL遺伝子の5‘側をリンカー(GGGGS)3により結合した遺伝子をPCRにより構築した。さらに、このVH-(GGGGS)3-VL配列の5’側にT7プロモーター配列,翻訳効率を高めるためのε配列,リボソーム結合部位であるSD配列及び開始コドン、3’側にエピトープタグとしてHAタグ(アミノ酸配列:YPYDVPDYA)及び終止コドンをPCRにより付加した(図7および図8)。
【0099】
scFvタンパク合成
無細胞系タンパク合成反応液PURESYSTEM S-S ((株)ポストゲノム研究所、東京)50μLに上記PCR産物1pmolを鋳型として添加し37℃,60分間タンパク合成を行った.反応後チューブを氷上に移し,これをscFv116、及びscFvcont.として以下の実験に用いた。
【0100】
scFv116の抗原特異性の検討
scFvの抗原への結合をELISAにより検討した。96穴プレートにPBSで希釈したadr型HBs抗原(10μg/mL)を各ウェルに50μLずつ分注し、4 oCで一晩反応させ吸着させた。PBSでウェルを洗浄し、非特異的結合を除去する為に3%BSA、0.05%Tween-20添加PBS(PBS-T/BSA)を各ウェルに400μLずつ分注し、室温で2時間反応させてブロッキングした。0.1% Tween-20添加PBS (PBS-T)でウェルを洗浄後、各ウェルにPBS-T/BSAで希釈したscFv116あるいはscFvcontを50μLずつ加え、室温で1時間反応させた。その後、PBS-Tでウェルを洗浄後、1000倍希釈したマウス抗HAモノクローナル抗体を各ウェルに50μLずつ分注し、室温で1時間反応させた。PBS-Tで洗浄後、2000倍希釈したアルカリフォスファターゼ(ALP)標識ヤギ抗マウスIgG抗体を、各ウェルに50μLずつ分注し室温で2時間反応させた。ウェルをPBS-Tで洗浄し、ALP基質用緩衝液(100mM NaCl、5mM MgCl2、100mM Tris pH9.5)に溶解した1mg/mL p-nitrophenyl phosphate(ALP基質)を、各ウェルに50μLずつ分注し、室温で20分反応させた後、マイクロプレートリーダーにて405nmにおける吸光度を測定した。図9に示すように、コントロール遺伝子配列より構築したscFvタンパク(scFvcont)はHBs抗原とは全く結合せず、116遺伝子配列より構築したscFv(scFv116)は、濃度依存的にHBs抗原と結合することが示された。
【0101】
scFv116のHBs抗原への結合の特異性を検討するために、一定濃度のscFv116と種々の濃度 (0,10, 100,400,1,000 ng/mL) のHBs抗原と反応させた後に、上記ELISAを行った。図10に示すように、HBs抗原の濃度依存的にscFv116のHBs抗原への結合が阻害された。この結果は、scFv116とHBs抗原の結合が特異的であることを示す。
【0102】
さらに、scFv116と元のIgG116の認識するエピトープが同一であるか否かを検討するために、一定濃度のscFv116に対して種々の濃度 (0,40,200,1,000 ng/mL) のIgG116を加えて、上記ELISAを行った。その結果、添加したIgG116の濃度に依存的にscFv116の結合が阻害された(図11)。この結果は、HL116抗体のIgG型とscFv型は、同一のエピトープを認識することを示す。
以上、116抗体遺伝子を元に作製した
【産業上の利用可能性】
【0103】
このようなB型肝炎ウイルスに対する抗体(本発明の抗体)は、先に述べたような針刺し事故に対する治療に有効であることが期待されるが、肝移植時における効果も期待される。すなわち、B型肝炎ウイルスに感染している患者においては、肝移植を行ってもせっかく移植した新しい肝臓にB型肝炎ウイルスが再び感染することにより肝炎が発症し肝機能が低下する可能性が高く、肝移植の意義がなくなってしまうこともある。この場合、肝移植時にB型肝炎ウイルスに対する抗体で患者体内のB型肝炎ウイルスを中和・除去することで、移植した肝臓へのB型肝炎ウイルスの感染を防御できることが期待される。このように、B型肝炎ウイルスを中和する能力を持った抗体、特に、組換え抗体はB型肝炎ウイルスの感染阻止に非常に有用であると考えられる。
【図面の簡単な説明】
【0104】
【図1】抗体遺伝子の構造の模式図。
【図2】単鎖Fv (scFv)等の構造の模式図。
【図3】抗体γ鎖発現ベクターおよび抗体κ鎖発現ベクター。
【図4】116抗体のHBs抗原への特異的結合試験結果。
【図5】116抗体が認識するエピトープの試験結果。
【図6】116抗体のHBV中和能試験結果。
【図7】scFv遺伝子の構築およびscFvタンパクの合成スキーム。
【図8−1】116抗体scFv遺伝子の塩基配列。
【図8−2】116抗体scFv遺伝子の塩基配列。
【図9】scFvタンパク(scFvcont)のHBs抗原への結合試験結果。
【図10】scFv116のHBs抗原への特異性試験結果。
【図11】scFv116のIgG116への特異性試験結果。
【図12】CDR遺伝子を組み合わせて用いて抗体を調製する方法の説明図。
【特許請求の範囲】
【請求項1】
下記の何れかのDNAからなる遺伝子。
(1)配列番号1の塩基配列を有するDNA、
(2)配列番号1の塩基配列において1から数個の塩基の欠失、置換及び/又は付加を有する塩基配列を有し、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の重鎖可変領域をコードするDNA、および
(3)配列番号1の塩基配列またはこの塩基配列と相補的な塩基配列とストリンジェントな条件下でハイブリダイズする塩基配列を有し、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の重鎖可変領域をコードするDNA。
【請求項2】
下記の何れかのタンパク質をコードする遺伝子。
(1)配列番号2のアミノ酸配列を有するタンパク質、
(2)配列番号2のアミノ酸配列において1から数個のアミノ酸の欠失、置換及び/又は付加を有するアミノ酸配列を有し、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の重鎖可変領域であるタンパク質、および
(3)配列番号2のアミノ酸配列に対して90%以上の相同性を有するアミノ酸配列を有し、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の重鎖可変領域であるタンパク質。
【請求項3】
下記の何れかのDNAからなる遺伝子。
(1)配列番号3の配列を有するDNA、
(2)配列番号3の塩基配列において1から数個の塩基の欠失、置換及び/又は付加を有する塩基配列を有し、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の軽鎖可変領域をコードするDNA、および
(3)配列番号3の塩基配列またはこの塩基配列と相補的な塩基配列とストリンジェントな条件下でハイブリダイズする塩基配列を有し、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の軽鎖可変領域をコードするDNA。
【請求項4】
下記の何れかのタンパク質をコードする遺伝子。
(1)配列番号4のアミノ酸配列を有するタンパク質、
(2)配列番号4のアミノ酸配列において1から数個のアミノ酸の欠失、置換及び/又は付加を有するアミノ酸配列を有し、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の軽鎖可変領域であるタンパク質、および
(3)配列番号4のアミノ酸配列に対して90%以上の相同性を有するアミノ酸配列を有し、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の軽鎖可変領域であるタンパク質。
【請求項5】
下記の何れかのタンパク質。
(1)配列番号2のアミノ酸配列を有するタンパク質、
(2)配列番号2のアミノ酸配列において1から数個のアミノ酸の欠失、置換及び/又は付加を有するアミノ酸配列を有し、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の重鎖可変領域であるタンパク質、および
(3)配列番号2のアミノ酸配列に対して90%以上の相同性を有するアミノ酸配列を有し、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の重鎖可変領域であるタンパク質。
【請求項6】
下記の何れかのタンパク質。
(1)配列番号4のアミノ酸配列を有するタンパク質、
(2)配列番号4のアミノ酸配列において1から数個のアミノ酸の欠失、置換及び/又は付加を有するアミノ酸配列を有し、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の軽鎖可変領域であるタンパク質、および
(3)配列番号4のアミノ酸配列に対して90%以上の相同性を有するアミノ酸配列を有し、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の軽鎖可変領域であるタンパク質。
【請求項7】
下記の何れかの遺伝子。
(1)配列番号1の塩基配列における76〜105番目の塩基配列、または配列番号3のアミノ酸配列における26〜35番目のアミノ酸配列をコードする塩基配列を有する、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の重鎖相補性決定領域1(CDR−H1)をコードする遺伝子。
(2)配列番号1の塩基配列における148〜198の塩基配列、または配列番号3のアミノ酸配列における50〜66番目のアミノ酸配列をコードする塩基配列を有する、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の重鎖相補性決定領域2(CDR−H2)をコードする遺伝子。
(3)配列番号1の塩基配列における295〜333の塩基配列、または配列番号3のアミノ酸配列における99〜111番目のアミノ酸配列をコードする塩基配列を有する、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の重鎖相補性決定領域3(CDR−H3)をコードする遺伝子。
【請求項8】
下記の何れかの遺伝子。
(1)配列番号3の塩基配列における69〜102の塩基配列、または配列番号4のアミノ酸配列における24〜34のアミノ酸配列をコードする塩基配列を有する、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の軽鎖相補性決定領域1(CDR−L1)をコードする遺伝子。
(2)配列番号3の塩基配列における148〜168の塩基配列、または配列番号4のアミノ酸配列における50〜56のアミノ酸配列をコードする塩基配列を有する、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の軽鎖相補性決定領域2(CDR−L2)をコードする遺伝子。
(3)配列番号3の塩基配列における265〜291の塩基配列、または配列番号4のアミノ酸配列における89〜97のアミノ酸配列をコードする塩基配列を有する、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の軽鎖相補性決定領域3(CDR−L3)をコードする遺伝子。
【請求項9】
下記のいずれかのペプチド。
(1)配列番号2のアミノ酸配列における26〜35のアミノ酸配列を有する、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の重鎖相補性決定領域1(CDR−H1)であるペプチド、
(2)配列番号2のアミノ酸配列における50〜66のアミノ酸配列を有する、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の重鎖相補性決定領域2(CDR−H2)であるペプチド、
(3)配列番号2のアミノ酸配列における99〜111のアミノ酸配列を有する、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の重鎖相補性決定領域3(CDR−H3)であるペプチド。
【請求項10】
下記のいずれかのペプチド。
(1)配列番号4のアミノ酸配列における24〜34のアミノ酸配列を有する、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の軽鎖相補性決定領域1(CDR−L1)であるペプチド、
(2)配列番号4のアミノ酸配列における50〜56のアミノ酸配列を有する、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の軽鎖相補性決定領域2(CDR−L2)であるペプチド、
(3)配列番号4のアミノ酸配列における89〜97のアミノ酸配列を有する、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の軽鎖相補性決定領域3(CDR−L3)であるペプチド。
【請求項11】
配列番号2のアミノ酸配列を有するタンパク質を可変領域として含む重鎖と配列番号4のアミノ酸配列を有するタンパク質を可変領域として含む軽鎖とを含む、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体。
【請求項12】
配列番号2のアミノ酸配列における26〜35、50〜66および99〜111のアミノ酸配列を有するペプチドを相補性決定領域1〜3として含む重鎖と配列番号4のアミノ酸配列における24〜34、50〜56および89〜97のアミノ酸配列を有するペプチドを相補性決定領域1〜3として含む軽鎖とを含む、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体。
【請求項13】
配列番号2のアミノ酸配列を有するタンパク質と配列番号4のアミノ酸配列を有するタンパク質とをリンカーペプチドで結合した、B型肝炎ウイルスのHBs抗原に対する単鎖抗体。
【請求項14】
配列番号2のアミノ酸配列における26〜35、50〜66および99〜111のアミノ酸配列を有するペプチドを相補性決定領域1〜3として含むタンパク質と配列番号4のアミノ酸配列における24〜34、50〜56および89〜97のアミノ酸配列を有するペプチドを相補性決定領域1〜3として含むタンパク質とをリンカーペプチドで結合した、B型肝炎ウイルスのHBs抗原に対する単鎖抗体。
【請求項15】
請求項13または14に記載の単鎖抗体の二量体。
【請求項16】
請求項13または14に記載の単鎖抗体の三量体。
【請求項17】
配列番号2のアミノ酸配列を有するタンパク質と配列番号4のアミノ酸配列を有するタンパク質とをリンカーペプチドで結合し、かつ配列番号2のアミノ酸配列を有する前記タンパク質にCH3領域を結合した、B型肝炎ウイルスのHBs抗原に対するミニボディー。
【請求項18】
配列番号2のアミノ酸配列における26〜35、50〜66および99〜111のアミノ酸配列を有するペプチドを相補性決定領域1〜3として含むタンパク質と配列番号4のアミノ酸配列における24〜34、50〜56および89〜97のアミノ酸配列を有するペプチドを相補性決定領域1〜3として含むタンパク質とをリンカーペプチドで結合し、かつ配列番号2のアミノ酸配列における26〜35、50〜66および99〜111のアミノ酸配列を有するペプチドを相補性決定領域1〜3として含む前記タンパク質にCH3領域を結合した、B型肝炎ウイルスのHBs抗原に対するミニボディー(minibody)。
【請求項19】
請求項5若しくは6に記載のタンパク質、請求項9若しくは10に記載のペプチド、請求項11若しくは12に記載のモノクローナル抗体、請求項13若しくは14に記載の単鎖抗体、請求項15に記載の二量体、請求項16に記載の三量体、請求項17若しくは18に記載のミニボディーを用いるB型肝炎ウイルスの検定方法。
【請求項20】
請求項5若しくは6に記載のタンパク質、請求項9若しくは10に記載のペプチド、請求項11または12に記載のモノクローナル抗体、または請求項13若しくは14に記載の単鎖抗体、請求項15に記載の二量体、請求項16に記載の三量体、請求項17若しくは18に記載のミニボディーを用いるB型肝炎の診断方法。
【請求項21】
請求項5若しくは6に記載のタンパク質、請求項9若しくは10に記載のペプチド、請求項11または12に記載のモノクローナル抗体、または請求項13若しくは14に記載の単鎖抗体、請求項15に記載の二量体、請求項16に記載の三量体、請求項17若しくは18に記載のミニボディーを用いるB型肝炎の治療方法。
【請求項1】
下記の何れかのDNAからなる遺伝子。
(1)配列番号1の塩基配列を有するDNA、
(2)配列番号1の塩基配列において1から数個の塩基の欠失、置換及び/又は付加を有する塩基配列を有し、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の重鎖可変領域をコードするDNA、および
(3)配列番号1の塩基配列またはこの塩基配列と相補的な塩基配列とストリンジェントな条件下でハイブリダイズする塩基配列を有し、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の重鎖可変領域をコードするDNA。
【請求項2】
下記の何れかのタンパク質をコードする遺伝子。
(1)配列番号2のアミノ酸配列を有するタンパク質、
(2)配列番号2のアミノ酸配列において1から数個のアミノ酸の欠失、置換及び/又は付加を有するアミノ酸配列を有し、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の重鎖可変領域であるタンパク質、および
(3)配列番号2のアミノ酸配列に対して90%以上の相同性を有するアミノ酸配列を有し、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の重鎖可変領域であるタンパク質。
【請求項3】
下記の何れかのDNAからなる遺伝子。
(1)配列番号3の配列を有するDNA、
(2)配列番号3の塩基配列において1から数個の塩基の欠失、置換及び/又は付加を有する塩基配列を有し、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の軽鎖可変領域をコードするDNA、および
(3)配列番号3の塩基配列またはこの塩基配列と相補的な塩基配列とストリンジェントな条件下でハイブリダイズする塩基配列を有し、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の軽鎖可変領域をコードするDNA。
【請求項4】
下記の何れかのタンパク質をコードする遺伝子。
(1)配列番号4のアミノ酸配列を有するタンパク質、
(2)配列番号4のアミノ酸配列において1から数個のアミノ酸の欠失、置換及び/又は付加を有するアミノ酸配列を有し、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の軽鎖可変領域であるタンパク質、および
(3)配列番号4のアミノ酸配列に対して90%以上の相同性を有するアミノ酸配列を有し、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の軽鎖可変領域であるタンパク質。
【請求項5】
下記の何れかのタンパク質。
(1)配列番号2のアミノ酸配列を有するタンパク質、
(2)配列番号2のアミノ酸配列において1から数個のアミノ酸の欠失、置換及び/又は付加を有するアミノ酸配列を有し、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の重鎖可変領域であるタンパク質、および
(3)配列番号2のアミノ酸配列に対して90%以上の相同性を有するアミノ酸配列を有し、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の重鎖可変領域であるタンパク質。
【請求項6】
下記の何れかのタンパク質。
(1)配列番号4のアミノ酸配列を有するタンパク質、
(2)配列番号4のアミノ酸配列において1から数個のアミノ酸の欠失、置換及び/又は付加を有するアミノ酸配列を有し、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の軽鎖可変領域であるタンパク質、および
(3)配列番号4のアミノ酸配列に対して90%以上の相同性を有するアミノ酸配列を有し、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の軽鎖可変領域であるタンパク質。
【請求項7】
下記の何れかの遺伝子。
(1)配列番号1の塩基配列における76〜105番目の塩基配列、または配列番号3のアミノ酸配列における26〜35番目のアミノ酸配列をコードする塩基配列を有する、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の重鎖相補性決定領域1(CDR−H1)をコードする遺伝子。
(2)配列番号1の塩基配列における148〜198の塩基配列、または配列番号3のアミノ酸配列における50〜66番目のアミノ酸配列をコードする塩基配列を有する、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の重鎖相補性決定領域2(CDR−H2)をコードする遺伝子。
(3)配列番号1の塩基配列における295〜333の塩基配列、または配列番号3のアミノ酸配列における99〜111番目のアミノ酸配列をコードする塩基配列を有する、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の重鎖相補性決定領域3(CDR−H3)をコードする遺伝子。
【請求項8】
下記の何れかの遺伝子。
(1)配列番号3の塩基配列における69〜102の塩基配列、または配列番号4のアミノ酸配列における24〜34のアミノ酸配列をコードする塩基配列を有する、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の軽鎖相補性決定領域1(CDR−L1)をコードする遺伝子。
(2)配列番号3の塩基配列における148〜168の塩基配列、または配列番号4のアミノ酸配列における50〜56のアミノ酸配列をコードする塩基配列を有する、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の軽鎖相補性決定領域2(CDR−L2)をコードする遺伝子。
(3)配列番号3の塩基配列における265〜291の塩基配列、または配列番号4のアミノ酸配列における89〜97のアミノ酸配列をコードする塩基配列を有する、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の軽鎖相補性決定領域3(CDR−L3)をコードする遺伝子。
【請求項9】
下記のいずれかのペプチド。
(1)配列番号2のアミノ酸配列における26〜35のアミノ酸配列を有する、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の重鎖相補性決定領域1(CDR−H1)であるペプチド、
(2)配列番号2のアミノ酸配列における50〜66のアミノ酸配列を有する、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の重鎖相補性決定領域2(CDR−H2)であるペプチド、
(3)配列番号2のアミノ酸配列における99〜111のアミノ酸配列を有する、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の重鎖相補性決定領域3(CDR−H3)であるペプチド。
【請求項10】
下記のいずれかのペプチド。
(1)配列番号4のアミノ酸配列における24〜34のアミノ酸配列を有する、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の軽鎖相補性決定領域1(CDR−L1)であるペプチド、
(2)配列番号4のアミノ酸配列における50〜56のアミノ酸配列を有する、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の軽鎖相補性決定領域2(CDR−L2)であるペプチド、
(3)配列番号4のアミノ酸配列における89〜97のアミノ酸配列を有する、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体の軽鎖相補性決定領域3(CDR−L3)であるペプチド。
【請求項11】
配列番号2のアミノ酸配列を有するタンパク質を可変領域として含む重鎖と配列番号4のアミノ酸配列を有するタンパク質を可変領域として含む軽鎖とを含む、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体。
【請求項12】
配列番号2のアミノ酸配列における26〜35、50〜66および99〜111のアミノ酸配列を有するペプチドを相補性決定領域1〜3として含む重鎖と配列番号4のアミノ酸配列における24〜34、50〜56および89〜97のアミノ酸配列を有するペプチドを相補性決定領域1〜3として含む軽鎖とを含む、B型肝炎ウイルスのHBs抗原に対するモノクローナル抗体。
【請求項13】
配列番号2のアミノ酸配列を有するタンパク質と配列番号4のアミノ酸配列を有するタンパク質とをリンカーペプチドで結合した、B型肝炎ウイルスのHBs抗原に対する単鎖抗体。
【請求項14】
配列番号2のアミノ酸配列における26〜35、50〜66および99〜111のアミノ酸配列を有するペプチドを相補性決定領域1〜3として含むタンパク質と配列番号4のアミノ酸配列における24〜34、50〜56および89〜97のアミノ酸配列を有するペプチドを相補性決定領域1〜3として含むタンパク質とをリンカーペプチドで結合した、B型肝炎ウイルスのHBs抗原に対する単鎖抗体。
【請求項15】
請求項13または14に記載の単鎖抗体の二量体。
【請求項16】
請求項13または14に記載の単鎖抗体の三量体。
【請求項17】
配列番号2のアミノ酸配列を有するタンパク質と配列番号4のアミノ酸配列を有するタンパク質とをリンカーペプチドで結合し、かつ配列番号2のアミノ酸配列を有する前記タンパク質にCH3領域を結合した、B型肝炎ウイルスのHBs抗原に対するミニボディー。
【請求項18】
配列番号2のアミノ酸配列における26〜35、50〜66および99〜111のアミノ酸配列を有するペプチドを相補性決定領域1〜3として含むタンパク質と配列番号4のアミノ酸配列における24〜34、50〜56および89〜97のアミノ酸配列を有するペプチドを相補性決定領域1〜3として含むタンパク質とをリンカーペプチドで結合し、かつ配列番号2のアミノ酸配列における26〜35、50〜66および99〜111のアミノ酸配列を有するペプチドを相補性決定領域1〜3として含む前記タンパク質にCH3領域を結合した、B型肝炎ウイルスのHBs抗原に対するミニボディー(minibody)。
【請求項19】
請求項5若しくは6に記載のタンパク質、請求項9若しくは10に記載のペプチド、請求項11若しくは12に記載のモノクローナル抗体、請求項13若しくは14に記載の単鎖抗体、請求項15に記載の二量体、請求項16に記載の三量体、請求項17若しくは18に記載のミニボディーを用いるB型肝炎ウイルスの検定方法。
【請求項20】
請求項5若しくは6に記載のタンパク質、請求項9若しくは10に記載のペプチド、請求項11または12に記載のモノクローナル抗体、または請求項13若しくは14に記載の単鎖抗体、請求項15に記載の二量体、請求項16に記載の三量体、請求項17若しくは18に記載のミニボディーを用いるB型肝炎の診断方法。
【請求項21】
請求項5若しくは6に記載のタンパク質、請求項9若しくは10に記載のペプチド、請求項11または12に記載のモノクローナル抗体、または請求項13若しくは14に記載の単鎖抗体、請求項15に記載の二量体、請求項16に記載の三量体、請求項17若しくは18に記載のミニボディーを用いるB型肝炎の治療方法。
【図7】

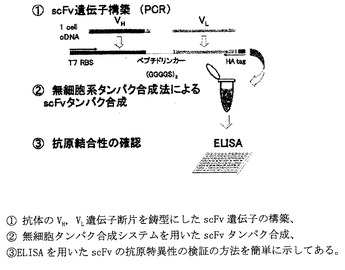
【図8−1】
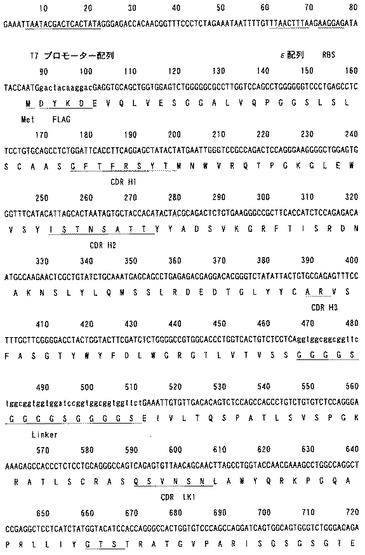
【図8−2】
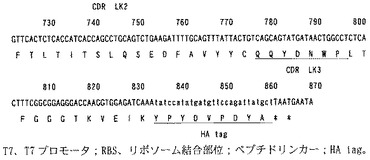
【図11】

【図1】
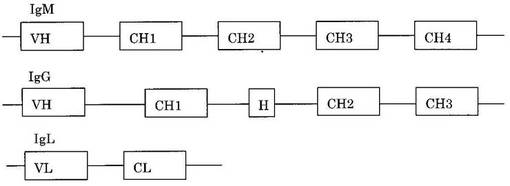
【図2】
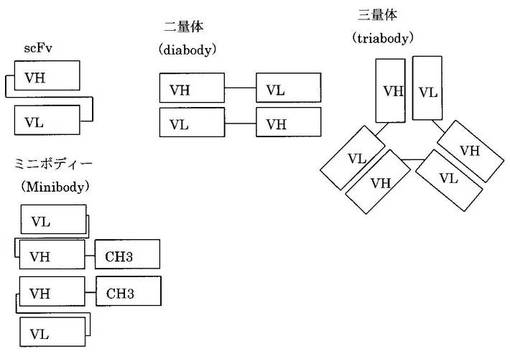
【図3】
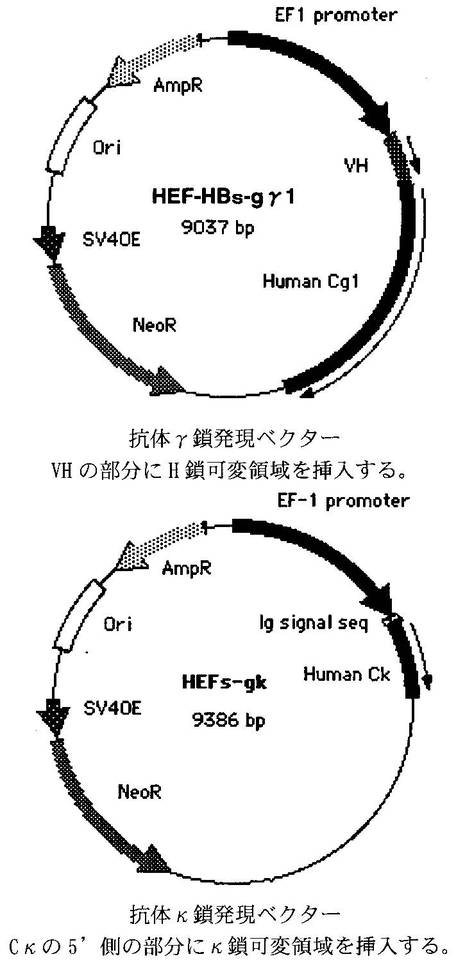
【図4】
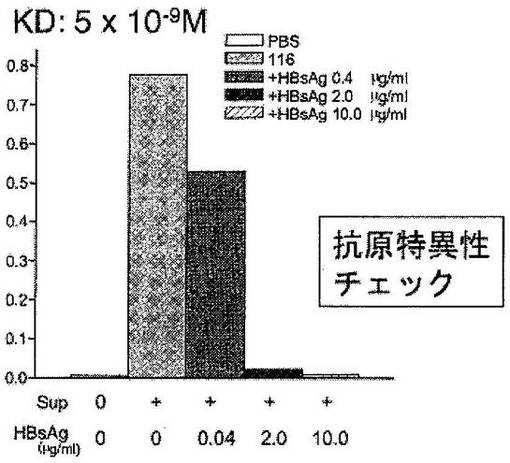
【図5】
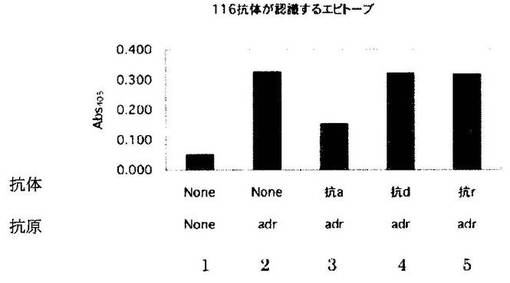
【図6】
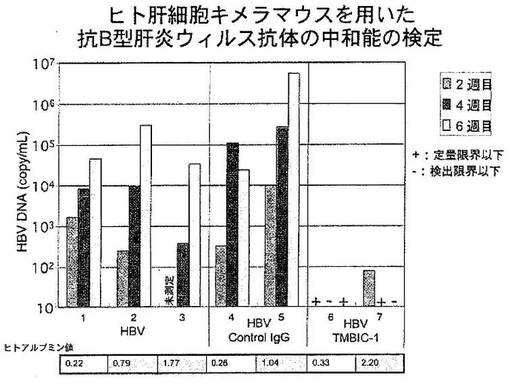
【図9】
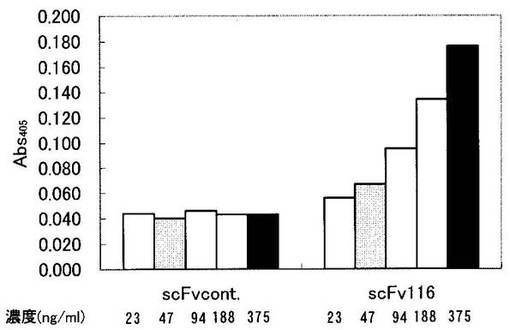
【図10】
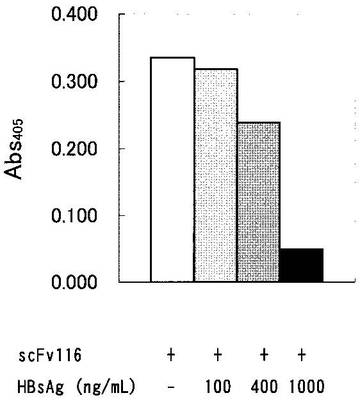
【図12】
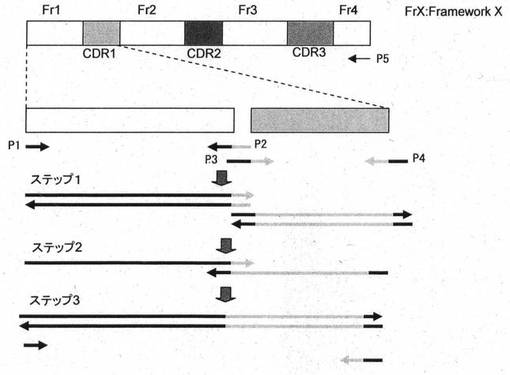
【図8−1】
【図8−2】
【図11】
【図1】
【図2】
【図3】
【図4】
【図5】
【図6】
【図9】
【図10】
【図12】
【公開番号】特開2007−14267(P2007−14267A)
【公開日】平成19年1月25日(2007.1.25)
【国際特許分類】
【出願番号】特願2005−199147(P2005−199147)
【出願日】平成17年7月7日(2005.7.7)
【出願人】(305060567)国立大学法人富山大学 (194)
【出願人】(803000104)財団法人ひろしま産業振興機構 (70)
【出願人】(000003311)中外製薬株式会社 (228)
【上記2名の代理人】
【識別番号】100089705
【弁理士】
【氏名又は名称】社本 一夫
【Fターム(参考)】
【公開日】平成19年1月25日(2007.1.25)
【国際特許分類】
【出願日】平成17年7月7日(2005.7.7)
【出願人】(305060567)国立大学法人富山大学 (194)
【出願人】(803000104)財団法人ひろしま産業振興機構 (70)
【出願人】(000003311)中外製薬株式会社 (228)
【上記2名の代理人】
【識別番号】100089705
【弁理士】
【氏名又は名称】社本 一夫
【Fターム(参考)】
[ Back to top ]