
B細胞の抗体遺伝子変異様式の転換方法
【課題】抗体をコードする遺伝子の点突然変異の頻度が上昇したB細胞由来の細胞の提供、及び抗体産生細胞であるB細胞由来の細胞において、抗体をコードする遺伝子の点突然変異の頻度を上昇させ、様々な特異性を有する抗体ライブラリーを形成させ、その中から所望の特異性を有する抗体を選択し製造する方法の提供。
【解決手段】XRCC3遺伝子の2つの対立遺伝子の片方のみが不活性化されたB細胞由来の細胞であって、抗体遺伝子への点突然変異の導入の頻度が両方のXRCC3遺伝子を有する細胞に比べ上昇した細胞。
【解決手段】XRCC3遺伝子の2つの対立遺伝子の片方のみが不活性化されたB細胞由来の細胞であって、抗体遺伝子への点突然変異の導入の頻度が両方のXRCC3遺伝子を有する細胞に比べ上昇した細胞。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、B細胞の抗体遺伝子変異様式を転換させ、点突然変異を有する抗体を製造する方法に関する。
【背景技術】
【0002】
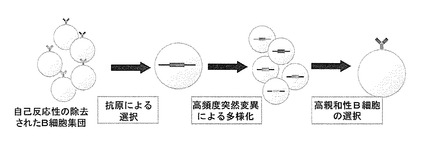
生体内では抗原による免疫後、産生される抗体の親和性が経時的に向上し、効率よく高親和性抗体が生み出される。この過程は親和性成熟と呼ばれている。親和性成熟は、抗原刺激されたB細胞の抗体遺伝子可変部に起こる高頻度突然変異により多様化したB細胞集団の中から、高親和性を獲得した変異B細胞が厳密に選択されて生存し、他の不要な細胞は死滅することにより進行する(図1)。この巧妙な仕組みは、一つの理想的な分子進化システムとみなすことができる。この原理により、生体への反復免疫により高特異性抗体を取得することができるが、この方法による抗体の取得には時間と労力を必要とする。
【0003】
図1に示すような生体内での抗体産生の過程を、in vitroの培養細胞系で再現できれば、迅速かつ効率的な抗体創製システムを構築することができる。この目的に使用できる高頻度変異機能を保持したB細胞株としては、ヒト、マウス、ニワトリ由来のB細胞株がいくつか知られているが、ニワトリB細胞株DT40は、以下のような優れた特性を有する。
1)培養中に自発的に抗体遺伝子に変異を導入し、多様な抗体ライブラリーを形成できる。
2)細胞表面にIgM抗体を膜結合型として発現し、また分泌している。
3)増殖速度が通常の動物細胞より2〜3倍速く培養が容易である。
4)動物細胞としては例外的に、外来遺伝子との相同組換え頻度が高いため、遺伝子ノックアウトなどの部位特異的遺伝子操作が容易に行える。
【0004】
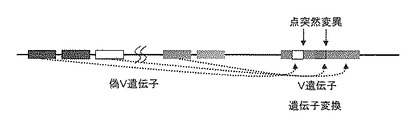
DT40細胞における抗体遺伝子の変異は主として遺伝子変換という機構で行われる。これは抗体可変部(V)遺伝子に、上流に配置された多くの偽V遺伝子の部分配列が遺伝子変換によって取り込まれることによって進行する(図2)。一塩基の置換による点突然変異も起こるが低頻度である。培養細胞の集団中には、これにより様々な特異性を持つ抗体ライブラリーが形成される。
【0005】
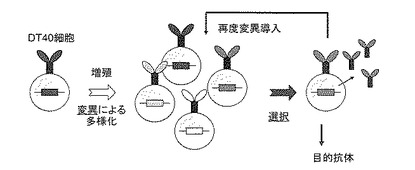
したがって培養したDT40細胞の集団から、目的抗原に結合する抗体を産生する細胞を単離することにより、必要な抗体を迅速かつ効率的に取得することができると考えられる(図3)。
【0006】
このような着想に基づき、本発明者は先に機能改変したDT40株DT40-SWを用いるin vitro抗体作成法を開発した(特許文献1を参照)。この技術のもう一つの重要な特徴は、目的抗体を産生する細胞を選択できた場合、図3に示すように再度その細胞クローンに変異を導入して多様化させ、選択を繰り返すことにより、更に高親和性の抗体へ進化させること(親和性成熟)が行える点にある。この親和性成熟を効率よく進行させるためには、次に述べるように更なる細胞の遺伝子レベルでの改変が必要である。
【0007】
一方、遺伝子変換型変異が優勢であるDT40細胞において、変異様式を遺伝子変換型から点突然変異型に転換させる方法については既に報告があり、DNAの損傷を修復するのに関与する因子XRCC2、XRCC3を欠損させることにより実現できることが知られていた(非特許文献1及び2を参照)。
【0008】
【特許文献1】特開2006-109711号公報
【非特許文献1】Sale, J. E. et al. Nature 412, 921-926(2001)
【非特許文献2】Arakawa, H. et al., PLOS Biol. 2, 967-974 (2004)
【発明の開示】
【発明が解決しようとする課題】
【0009】
本発明は、抗体をコードする遺伝子の点突然変異の頻度が上昇したB細胞由来の細胞の提供、及び抗体産生細胞であるB細胞由来の細胞において、抗体をコードする遺伝子の点突然変異の頻度を上昇させ、様々な特異性を有する抗体ライブラリーを形成させ、その中から所望の特異性を有する抗体を選択し製造する方法の提供を目的とする。
【課題を解決するための手段】
【0010】
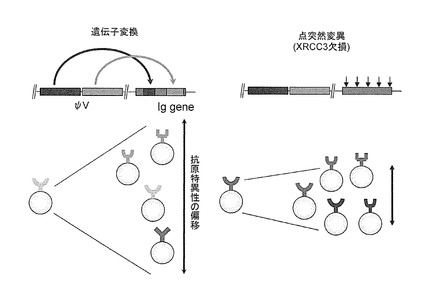
上記の親和性成熟を効率よく行うためには、変異による抗体ライブラリーの多様化が目的に適した形で進行することが重要である。図4(左)に示すように、DT40細胞においては、数十塩基程度の配列が一度に置き換わる遺伝子変換が変異様式として優位であり、これはニワトリ体内でB細胞の抗体多様性を生み出す機構と同じであるので、初期の抗体ライブラリーの拡大には有利であると言える。しかし、ニワトリ体内では、免疫後に高親和性抗体を生み出す親和性成熟の過程では、遺伝子変換と共に点突然変異の頻度が高まることが知られており、この変異様式が高親和性抗体を作るのに適していると考えられる。何故なら、一塩基置換である点突然変異では、元の特異性からより小さな偏移幅で抗原特異性が変化することが期待できるので、抗原特異性のより精密な改変が可能と考えられるからである(図4右)。
【0011】
そこで、抗体探索のためのライブラリー形成時には遺伝子変換型で変異が進行し、その中から目的抗体産生クローンが選択できた後には、点突然変異の頻度を上昇させるようにDT40を改変することが有効である。本発明者は、この問題を克服するための以下の検討を行い、有効な解決法を見いだした。
【0012】
本来、遺伝子変換型変異が優勢であるDT40細胞において、変異様式を遺伝子変換型から点突然変異型に転換させる方法については既に報告があり、DNAの損傷を修復するのに関与する因子XRCC2、XRCC3を欠損させることにより実現できることが知られている(図4右)(Sale, J. E. et al. Nature 412, 921-926(2001); Arakawa, H. et al., PLOS Biol. 2, 967-974 (2004))。
【0013】
しかし、これらの遺伝子を完全に欠損させると、1)細胞の増殖速度が低下する、2)点突然変異により効率に非機能的遺伝子が生じるため抗体を産生しない細胞が多く生じる、3)DT40の最大の長所である相同組換えが行えなくなり、遺伝子操作が困難となる、という問題が発生する。本発明者は、この問題を克服するために種々検討を重ねた結果、驚くべきことにXRCC3欠損細胞を作製する中間段階でできるXRCC3遺伝子の2つの対立遺伝子の片方のみを欠損させたヘテロノックアウト細胞、XRCC3(+/-)株が、上にあげた3つの問題点を克服するのに有用であることを見いだした。しかも、対立遺伝子の一方のみを破壊すればよいので、実験操作上極めて容易である。したがってこの方法はDT40を用いる抗体作製システムの性能向上に極めて有用である。
【0014】
すなわち、本発明の態様は以下のとおりである。
[1] XRCC3遺伝子の2つの対立遺伝子の片方のみが不活性化されたB細胞由来の細胞であって、抗体遺伝子への点突然変異の頻度が両方のXRCC3遺伝子を有する細胞に比べ上昇した細胞。
[2] XRCC3遺伝子の2つの対立遺伝子の片方の第6エキソンが不活性化された[1]の細胞。
[3] XRCC3遺伝子の2つの対立遺伝子の片方のみの不活性化が相同組換えにより行われる[1]又は[2]の細胞。
[4] B細胞由来の細胞がニワトリB細胞株DT40又はその変異株である、[1]〜[3]のいずれかの細胞。
[5] ニワトリB細胞株DT40の変異株が、
細胞外刺激により外因性Creリコンビナーゼ遺伝子の発現が誘導され、発現されたCreリコンビナーゼにより外因性AID(activation induced cytidine deaminase)遺伝子の向きを反転させることにより、AID発現を誘導すること及び停止させることが可能な変異株であって、以下の特徴:
1)内因性のAID遺伝子が機能的に破壊されており、内因性AID遺伝子発現によるAIDタンパク質は産生されないこと、
2)互いに逆方向の2つのloxP配列で挟まれた外因性のAID遺伝子、及び該2つのloxP配列で挟まれた領域の上流側に存在する該動物細胞で機能し得るプロモーターを有し、上記AID遺伝子が該プロモーターに対して順方向に配置されている場合には、該プロモーターによるAID遺伝子の発現が可能であり、上記AID遺伝子が該プロモーターに対して逆方向に配置されている場合には、AID遺伝子の発現は停止すること、及び
3)Creリコンビナーゼ遺伝子が、細胞外刺激によりCreリコンビナーゼ活性化が可能な形で導入されており、Creリコンビナーゼ活性化により、上記外因性AID遺伝子を含む2つのloxP配列に挟まれた領域の方向が反転すること、
を有する変異株であり、細胞外刺激によりAID遺伝子の発現が停止し、変異機能が停止する変異株である、[4]の細胞。
[6] 外来性の抗体をコードする遺伝子が導入された[1]〜[5]のいずれかの細胞。
[7] 所望の特性を有する抗体を産生する抗体産生細胞を選択する方法であって、
(i) [1]〜[6]のいずれかの細胞を調製する工程、
(ii) 該細胞を一定期間培養する工程、
(iii) 該細胞が産生する抗体又は抗体をコードする遺伝子を単離して、その変異を検出する工程、並びに
(iv) 所望の変異を有する抗体を産生する細胞を選択する工程、を含む方法。
[8] 所望の特性を有する抗体を製造する方法であって、
(i) [1]〜[6]のいずれかの細胞を調製する工程、
(ii) 該細胞を一定期間培養する工程、
(iii) 該細胞が産生する抗体又は抗体をコードする遺伝子を単離して、その変異を検出する工程、
(iv) 所望の変異を有する抗体を産生する細胞を選択する工程、並びに
(v) 選択した所望の変異を有する細胞を培養して、抗体を生産する工程、を含む方法。
【発明の効果】
【0015】
本発明は、ニワトリB細胞由来のDT40細胞を用いる抗体作製システムであり、高親和性の抗体をより効率よく取得することを可能にする。また、本発明の抗体により、既存の抗体もXRCC3(+/-)変異株中で更に高性能な抗体に改良することが可能となる。特に、本発明の細胞を用いることにより、癌などの難治性疾患の治療に役立つ抗体の取得がより容易になる。
【発明を実施するための最良の形態】
【0016】
以下、本発明を詳細に説明する。
【0017】
本発明において用いるB細胞由来の細胞は、抗体を産生し得る限り、由来動物種、細胞株種は限定されない。ヒト、マウス、ヒツジ、ラット、ウサギ、ニワトリなどのB細胞若しくはその細胞株又はそれらの変異株を用いることができ、好ましくはニワトリB細胞由来のDT40細胞株を用いる。DT40細胞株は、Bリンパ腫由来の細胞であり、第2染色体がトリソミーという特徴をもつ(Baba, T.W., Giroir, B.P. and Humphries, E.H.:Virology 144 : 139-151,1985)。DT40株としては、野生型DT40株又は本発明者らが独自に樹立したDT40-SW株(変異機能を司る遺伝子AIDの発現を可逆的にスイッチすることにより、その抗体変異機能をON/OFF制御できる変異株(詳細はKanayama, N., Todo, K., Reth, M., Ohmori, H. Biochem. Biophys. Res. Commn. 327:70-75 (2005)及び特開2006-109711号公報に記載されている))を用いることができる。
【0018】
XRCC3(X-ray repair complementing defective repair in Chinese hamster cells 3)は、染色体の安定性維持とDNA損傷の修復に関与する遺伝子であり、細胞の相同組換えに関与する。ニワトリXRCC3遺伝子は、8個のエキソンを有する。ニワトリXRCC3遺伝子の塩基配列を配列番号1に示す。配列番号1中、8個のエキソンの位置は以下のとおりである。エキソン1: 1-215、エキソン2: 216-284、エキソン3: 285-422、エキソン4: 423-635、エキソン5: 636-790、エキソン6: 791-1003、エキソン7: 1004-1050、エキソン8: 1051-1935。
【0019】
本発明の細胞は、細胞の相同染色体のそれぞれに存在する2つのXRCC3対立遺伝子のうち、片方のみが不活性化されている。本発明の細胞は遺伝子型XRCC3(+/-)で表される。ここで、遺伝子の不活性化とは、遺伝子の塩基配列に部分的に欠失、置換、挿入、付加等が生じ遺伝子の発現が抑制されていることをいう。「遺伝子の発現を抑制」とは、遺伝子自体は発現するが、正常な機能を有する蛋白質をコードする遺伝子が発現されない場合も含まれる。「遺伝子が不活性化されている」ことを「遺伝子が破壊されている」、「遺伝子が欠損している」、「遺伝子がノックアウトされている」ということもある。また、遺伝子の不活性化は、遺伝子の発現が完全に抑制されている場合だけでなく、部分的に抑制されている場合も含まれる。本発明において、対立遺伝子の一方のみを不活性化するため、XRCC3遺伝子をヘテロでノックアウトするということがあり、得られた細胞をヘテロノックアウト細胞と呼ぶことがある。
【0020】
遺伝子の不活性化は、例えば、相同組換えにより行うことができる。相同組換えは、染色体上の遺伝子と外来DNAとの間で相同的遺伝子組換えを行い、目的の遺伝子を改変する方法をいう。相同組換えにおいては、タンパク質をコードする配列を分断する目的で、その遺伝子のエキソンに別のDNA配列を挿入する。相同組換えは、ジーンターゲティングベクターを用いて行う。相同組換え後、ジーンターゲティングベクターを持つ細胞を同定するために、目的の遺伝子を分断する配列には選択マーカー遺伝子を用いる。選択マーカー遺伝子としては、Blasticidin耐性遺伝子などの抗生物質耐性遺伝子、GFPなどの蛍光タンパク質遺伝子、色素タンパク質遺伝子等を用いることができる。
【0021】
本発明においては、ニワトリXRCC3遺伝子の8個のエキソンのうち、いずれのエキソンを分断してもよい。また、複数のエキソンを分断してもよい。好ましくは第6エキソンを分断する。
【0022】
ターゲッティングベクターは、ニワトリXRCC3遺伝子の配列情報に基づいて設計し、山村研一ら編 トランスジェニック動物 共立出版株式会社 1997年3月1日、相沢慎一 ジーンターゲティング ES細胞を用いた変異マウスの作製 バイオマニュアルシリーズ8 羊土社 1995、Hogan et al., Manipulating the Mouse Embryo, Cold Spring Hoarbor Laboratory Press (1944)、Joyner, A.L., Gene Targeting, A Practical Approach Series, IRL Press (1993)、松村正實ら編 実験医学別冊 新遺伝子工学ハンドブック(改定第3版) 羊土社 1999 等に記載の方法に従って作製することができる。ターゲティングベクターは挿入型及び置換型のどちらを用いてもよい。また、Cre-lox系を用いたターゲティングにより組換えを起こさせることもできる。Cre-lox系を用いたターゲティングは、例えば特表平11-503015号公報に記載の方法に従って行うことができる。相同組換えを起こした相同組換え体の選別方法としては、ポジティブ選択、プロモーター選択、ネガティブ選択、ポリA選択等の公知の選択方法を用いればよい。相同組換えを起こした細胞は、PCR法、サザンブロッティング法等の公知の方法で同定することができる。
【0023】
上記の方法で得られたB細胞由来の細胞のXRCC3(+/-)変異株は、抗体遺伝子の点突然変異が高頻度で起こる。また、本発明の細胞XRCC3(+/-)においては、野生型細胞XRCC3(+/+)において高頻度で生じる遺伝子変換も生じる。一方、2つのXRCC3対立遺伝子の両方を不活性化した細胞XRCC3(-/-)では、遺伝子変換の頻度は著しく低下する。従って、本発明の細胞は遺伝子変換と点突然変異の両方が生じやすく、多様な変異抗体を得ることができる。また、本発明の細胞XRCC3(+/-)の増殖速度は、野生型に対して若干低下するが、問題が生じるほどの低下は認められず、細胞密度は野生型と同様である。一方、2つのXRCC3対立遺伝子の両方を不活性化した細胞XRCC3(-/-)では、増殖速度及び細胞密度が野生型細胞XRCC(+/+)に比べ著しく低下する。さらに、XRCC3(-/-)では、抗体を産生できない株が高頻度で出現するのに対して、XRCC3(+/-)では抗体を産生できない株の出現率は低い。
【0024】
本発明の細胞XRCC(+/-)の継代培養を続けることにより、抗体遺伝子に種々の点突然変異を有する細胞集団を得ることができる。すなわち、細胞集団中に様々な特性を有する抗体ライブラリーが形成される。この細胞集団の中から所望の特性を有する抗体を選択すればよい。B細胞は表面に抗体が発現され、又は抗体を細胞外に分泌するため、細胞表面の抗体又は分泌された抗体の特性を調べて所望の抗体を選択することができる。細胞表面の抗体を選択し単離するためには、例えば抗原を固相化した磁気ビーズを用いればよい。すなわち、磁気ビーズに抗原を吸着により固相化させ、該抗原固相化磁気ビーズと細胞集団を混合し、所望の抗体を産生する細胞を磁気ビーズに結合させ、磁気を用いて磁気ビーズを回収することにより、所望の抗体を産生する細胞を回収・単離することができる。一方、培養液中に分泌された抗体は、ELISA(enzyme-linked immunosorbent assay)等により検出することができる。
【0025】
目的の抗原に結合し得る抗体を産生する細胞を単離し、さらに培養を行い変異を生じさせ、その中からより優れた特性を有する抗体を産生する細胞を選択するという操作を繰り返すことにより、さらに優れた特性を有する抗体を得ることができる。ここで、抗体の特性とは、目的の抗原に対する特異性、結合力の他、至適温度、至適pH、ADCC(antibody-dependent cellular cytotoxicity)、CDC(complement-dependent cytotoxicity)等の細胞傷害活性等を含む。特異性、結合力は、抗体の相補性決定領域(CDR)等の可変領域に変異が生じることにより改善される。また、特異性についても単に目的の抗原に対する特異性が向上した抗体を得るだけではなく、従来の動物に免疫する方法では作製が困難であった動物種間でアミノ酸配列が保存されたタンパク質に対しても特異性の高い抗体を得ることができる。
【0026】
本発明は、本発明の細胞を用いて所望の変異を有する抗体及び種々の変異を有する抗体ライブラリーを製造する方法を包含する。
【0027】
変異を有する抗体を製造する方法は、
B細胞の2つのXRCC3対立遺伝子の片方のみを不活性化されたB細胞を調製する工程、
該細胞を一定期間培養する工程、
該細胞が産生する抗体又は抗体をコードする遺伝子を単離して、その変異を検出する工程、
所望の変異を有する抗体を産生する細胞を選択する工程、並びに
選択した所望の変異を有する細胞を培養して、抗体を生産する工程、を含む。
【0028】
さらに、本発明の細胞を用いて既存の抗体に変異を起こさせ、より優れた特性を有する抗体を得ることができる。すなわち、本発明のXRCC3遺伝子の2つの対立遺伝子の片方のみが不活性化された細胞に既存の外来性の抗体をコードする遺伝子を発現可能な形態で導入し、該細胞のゲノムに組込み、該抗体遺伝子に変異を起こさせればよい。この場合、抗体は完全な抗体分子である必要はなく、抗原に結合するために必要な可変領域を含む抗体フラグメントを産生し得る細胞でもよく、またキメラ抗体またはヒト化抗体等、非天然型の抗原に結合し得る分子およびそのフラグメントを産生し得る細胞でもよい。
【0029】
本発明の方法に、本発明者らのKanayama, N., Todo, K.,Reth, M., Ohmori, H. Biochem. Biophys. Res. Commn. 327:70-75(2005)及び特開平2006-109711号公報に記載した、抗体遺伝子の変異機能を可逆的にON/OFF制御する方法を組み合わせるとDT40細胞を用いる培養系による革新的抗体作製技術が確立できる。本発明者らが開発した上記方法において用いられるDT40-SW細胞では、抗体遺伝子の変異を司るactivation-inducedcytidine deaminase(AID)遺伝子の発現を、外部から与えるエストロゲン誘導体等の刺激により可逆的にON/OFFすることができる。XRCC3遺伝子の2つの対立遺伝子の片方のみが不活性化されたXRCC3遺伝子へテロノックアウトDT40細胞から、目的クローンを単離できたとしても、そのクローンを増殖させる過程で更なる変異が導入される場合があり、この場合、有用な変異株を安定に保持することは困難になる。この問題を回避するためには、得られた目的とするクローンの変異機能を停止することが必要である。そこで、以下に述べるように、DT40-SW株において、XRCC3遺伝子の一方を不活性化させればよい。このXRCC3遺伝子へテロノックアウトDT40-SW細胞から選択されたクローンのAID発現を停止させることにより、目的抗体を産生する遺伝的に安定なクローンを得ることが可能となる。
【0030】
すなわち、本発明は、細胞外刺激により外因性Creリコンビナーゼが活性化され、活性化されたCreリコンビナーゼにより外因性AID(activation induced cytidine deaminase)遺伝子の向きを反転させることにより、AID発現を誘導すること及び停止させることが可能なB細胞由来の細胞であって、以下の特徴:
1)内因性のAID遺伝子が機能的に破壊されており、内因性AID遺伝子発現によるAIDタンパク質は産生されないこと、
2)互いに逆方向の2つのloxP配列で挟まれた外因性のAID遺伝子、及び該2つのloxP配列で挟まれた領域の上流側に存在する該細胞で機能し得るプロモーターを有し、上記AID遺伝子が該プロモーターに対して順方向に配置されている場合には、該プロモーターによるAID遺伝子の発現が可能であり、上記AID遺伝子が該プロモーターに対して逆方向に配置されている場合には、AID遺伝子の発現は停止すること、及び
3)Creリコンビナーゼ遺伝子が、細胞外刺激によりCreリコンビナーゼ活性化が可能な形で導入されており(具体的にはCreとエストロゲンレセプターの融合タンパク(Cre-ER)として発現させている。Cre-ERはエストロゲン誘導体が結合すると細胞質から核に移行し、基質であるloxPに挟まれたAID遺伝子に作用する(Zhang, Y. et al. Nucleic Acids Res. 24, 534-548 (1996)))、Creリコンビナーゼ活性化により、上記外因性AID遺伝子を含む2つのloxP配列に挟まれた領域の方向が反転すること、
を有する細胞において、2つのXRCC3対立遺伝子のうち、片方のみが不活性化された細胞を包含する。上記のプロモーターとしては、β-アクチンプロモーター、免疫グロブリンプロモーター、サイトメガロウイルスプロモーター、CAGプロモーター等が挙げられる。また、Creリコンビナーゼを活性化する細胞外刺激としては、例えば、Creリコンビナーゼが、エストロゲンレセプター、又はそのエストロゲン結合ドメインを含むタンパク質との融合タンパク質として発現し得るような形で、エストロゲンレセプター又はそのエストロゲン結合ドメインを含むタンパク質をコードする遺伝子とCreリコンビナーゼcDNAとをインフレームで連結したDNA構築物を細胞内に導入されている場合、エストロゲン誘導体刺激を細胞に与えると、Creリコンビナーゼの活性化が誘導される。
【0031】
さらに、本発明は細胞外刺激により外因性Creリコンビナーゼ遺伝子の発現が誘導され、発現されたCreリコンビナーゼにより外因性AID(activation induced cytidine deaminase)遺伝子の向きを反転させることにより、AID発現を誘導すること及び停止させることが可能なB細胞由来の細胞であって、以下の特徴:
1)内因性のAID遺伝子が機能的に破壊されており、内因性AID遺伝子発現によるAIDタンパク質は産生されないこと、
2)互いに逆方向の2つのloxP配列で挟まれた外因性のAID遺伝子、及び該2つのloxP配列で挟まれた領域の上流側に存在する該動物細胞で機能し得るプロモーターを有し、上記AID遺伝子が該プロモーターに対して順方向に配置されている場合には、該プロモーターによるAID遺伝子の発現が可能であり、上記AID遺伝子が該プロモーターに対して逆方向に配置されている場合には、AID遺伝子の発現は停止すること、及び
3)Creリコンビナーゼ遺伝子が、細胞外刺激によりCreリコンビナーゼ活性化が可能な形で導入されており、Creリコンビナーゼ活性化により、上記外因性AID遺伝子を含む2つのloxP配列に挟まれた領域の方向が反転すること、
を有する細胞において、2つのXRCC3対立遺伝子のうち、片方のみが不活性化された細胞を包含する。
【0032】
上記の細胞を用いて変異を有する抗体を得る方法は、
上記B細胞の2つのXRCC3対立遺伝子の片方のみを不活性化されたB細胞を調製する工程、
該細胞のAIDの発現が可能な状態にする工程、
該細胞を一定期間培養する工程、
該細胞にCreリコンビナーゼを活性化し得る細胞外刺激を与えて、AID遺伝子の向きを反転させて、AIDの発現を停止させる工程、
該細胞が産生する抗体又は抗体をコードする遺伝子を単離して、その変異を検出する工程、
所望の変異を有する抗体を産生する細胞を選択する工程、並びに
選択した所望の変異を有する細胞を培養して、抗体を生産する工程、を含む。
【実施例】
【0033】
本発明を以下の実施例によって具体的に説明するが、本発明はこれらの実施例によって限定されるものではない。
【0034】
1)XRCC3遺伝子の破壊によるXRCC3(+/-)、XRCC3(-/-)株の樹立
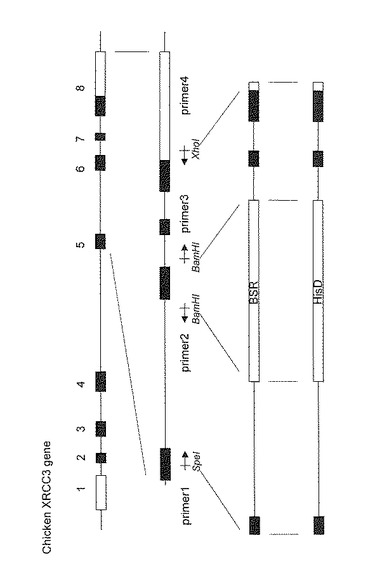
DT40細胞のXRCC3遺伝子の破壊のために、図5に示すようにXRCC3遺伝子の第6エキソン内部にブラスチサイジン耐性遺伝子又はヒスチジノールデヒドロゲナーゼ遺伝子を挿入した2種類のターゲッティングベクター(それぞれBsr、HisDと呼ぶ)を次の手順で作製した。SpeIサイトを含むprimer1とBamHIサイトを含むprimer2を用いて、PCRにより5’側の遺伝子断片を得た。また、BamHIサイトを含むprimer3とXhoIサイトを含むprimer4を用いて、PCRにより3’側の遺伝子断片を得た。Primer1〜4の配列は、それぞれ配列表の配列番号2〜5に示す。そこに、薬剤耐性マーカーとしてHisDとBSR遺伝子を挿入し、ターゲッティングベクターを作製した。
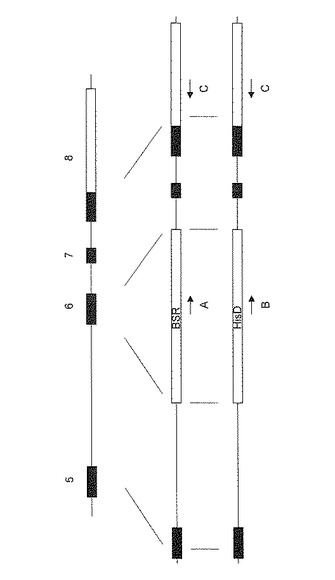
【0035】
最初に、Bsrを導入して、標的相同組換えにより一方のXRCC3を破壊し、XRCC3(+/-)株を得た。続いて、XRCC3(+/-)株にHisDを導入し、残りのXRCC3を破壊し、XRCC3(-/-)株を樹立した。ターゲッティングベクターを導入したDT40細胞クローンよりDNAを抽出し、PCRにより標的相同組換えが正しく行なわれたことを確認した。PCRには図6に示すプライマーA、B、Cを用いた(それぞれの配列は配列表の配列番号6〜8に示す)。Bsrの場合はプライマーAとC、HisDの場合はプライマーBとCを用いて相同組換えを確認した。図7に示すように、XRCC3の標的相同組換えによる遺伝子破壊の結果、XRCC3(+/-)、XRCC3(-/-)株が正しく作製されたことが確認された。第二段階の遺伝子導入も成功して、XRCC3(-/-)が取得できたことは、XRCC3(+/-)株において、相同組換え機能が保持されていることを示しており、XRCC3(+/-)株の有用性を更に裏付けるものである。
【0036】
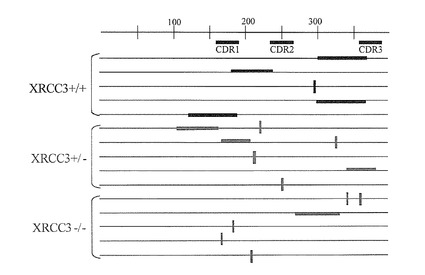
2)XRCC3(+/+)、(+/-)、(-/-)における変異パターンの比較
元株であるXRCC3(+/+)とXRCC3を部分的あるいは完全に破壊したXRCC3(+/-)、XRCC3(-/-)の各細胞を4週間継続的に培養し、抗体L鎖遺伝子の配列解析により、変異導入効率と変異様式を評価した。図8及び表1に示すように、元株XRCC3(+/+)では遺伝子変換が優位に起こっているが、XRCC3(-/-)株では、点突然変異型に大きく転換していた。一方、驚くべきことに、XRCC3(+/-)株においては、遺伝子変換がやや低下するだけで、点突然変異は、XRCC3(-/-)と同程度まで増加していた。
【0037】
更に、遺伝子変換と点突然変異を合わせた、全体の変異頻度ではXRCC3(+/-)が最も高かった。これは、XRCC3(+/-)株が遺伝子変換と点突然変異の長所を併せ持つ優良な変異株であることを示している。
【0038】
【表1】

【0039】
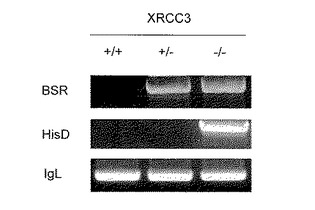
3)表面IgM消失率及び増殖速度の比較
XRCC3(-/-)株では、点突然変異は増加するがそれに伴って、ストップコドンやフレームシフトなどが生じるために、抗体タンパクを作れない変異細胞(表面IgM陰性細胞)が増加することが報告されている(Sale, J. E. et al. Nature 412, 921-926(2001))。また、XRCC3の欠損に伴い、細胞の増殖速度が低下することも知られている(同文献)これは抗体探索システムとしては重大な欠点となる。
【0040】
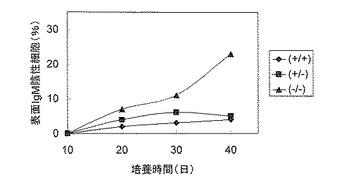
そこで、XRCC3(+/+)、(+/-)、(-/-)の3種類の細胞株を図9に示すように、長期間培養し、表面IgM抗体の発現をフローサイトメトリーにより測定した。報告されているように、XRCC3(-/-)株では表面IgM陰性の細胞が時間とともに増加したが、XRCC3(+/+)では増加しなかった。驚くべきことに、点突然変異効率が上昇しているXRCC3(+/-)においても表面IgM陰性細胞の出現率は低かった。このことは、点突然変異による抗体遺伝子の機能喪失が遺伝子変換によって修復されていることを示唆し、XRCC3(+/-)株の有用性をさらに支持するものである。
【0041】
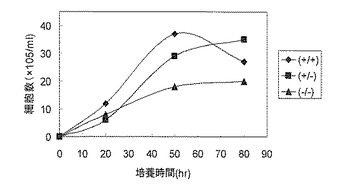
XRCC3(+/+)、(+/-)、(-/-)の3種類の細胞株の増殖速度を比較したところ、XRCC3(+/+)株の増殖が最も速く、XRCC3(+/-)の増殖はやや遅かったが問題となる程度ではなく、最大の細胞密度もXRCC3(+/+)株とほぼ同程度であった。一方、XRCC3(-/-)株では、増殖速度、最大細胞密度ともに低下していた(図10)。この結果もXRCC3(+/-)株の有用性を示している。
【図面の簡単な説明】
【0042】
【図1】免疫系における高特異性抗体の産生(親和性成熟)の原理を示す図である。
【図2】ニワトリ抗体遺伝子の多様化機構を示す図である。
【図3】ニワトリB細胞株DT40を用いる抗体作製システムを示す図である。
【図4】XRCC3遺伝子の欠損による遺伝子変換から点突然変異への変異様式の転換を示す図である。
【図5】XRCC3ターゲッティングベクターの構築図である。
【図6】標的相同組換えによるXRCC3遺伝子の不活性化の方法及び相同組換え確認のためのプライマーの位置を示す図である。
【図7】XRCC3の標的相同組換えによるXRCC3遺伝子の不活性化の確認の結果を示す図である。
【図8】XRCC3遺伝子の不活性化による点突然変異の増加を示す図である。各変異細胞の代表的な5クローンの結果を示し、太い横線は遺伝子変換で挿入された配列、縦の太線は点突然変異の場所を示す。
【図9】XRCC3(+/+)、XRCC3(+/-)及びXRCC3(-/-)における表面IgMの消失率を示す図である。
【図10】XRCC3(+/+)、XRCC3(+/-)及びXRCC3(-/-)における細胞増殖速度を示す図である。
【配列表フリーテキスト】
【0043】
配列番号2〜8 プライマー
【技術分野】
【0001】
本発明は、B細胞の抗体遺伝子変異様式を転換させ、点突然変異を有する抗体を製造する方法に関する。
【背景技術】
【0002】
生体内では抗原による免疫後、産生される抗体の親和性が経時的に向上し、効率よく高親和性抗体が生み出される。この過程は親和性成熟と呼ばれている。親和性成熟は、抗原刺激されたB細胞の抗体遺伝子可変部に起こる高頻度突然変異により多様化したB細胞集団の中から、高親和性を獲得した変異B細胞が厳密に選択されて生存し、他の不要な細胞は死滅することにより進行する(図1)。この巧妙な仕組みは、一つの理想的な分子進化システムとみなすことができる。この原理により、生体への反復免疫により高特異性抗体を取得することができるが、この方法による抗体の取得には時間と労力を必要とする。
【0003】
図1に示すような生体内での抗体産生の過程を、in vitroの培養細胞系で再現できれば、迅速かつ効率的な抗体創製システムを構築することができる。この目的に使用できる高頻度変異機能を保持したB細胞株としては、ヒト、マウス、ニワトリ由来のB細胞株がいくつか知られているが、ニワトリB細胞株DT40は、以下のような優れた特性を有する。
1)培養中に自発的に抗体遺伝子に変異を導入し、多様な抗体ライブラリーを形成できる。
2)細胞表面にIgM抗体を膜結合型として発現し、また分泌している。
3)増殖速度が通常の動物細胞より2〜3倍速く培養が容易である。
4)動物細胞としては例外的に、外来遺伝子との相同組換え頻度が高いため、遺伝子ノックアウトなどの部位特異的遺伝子操作が容易に行える。
【0004】
DT40細胞における抗体遺伝子の変異は主として遺伝子変換という機構で行われる。これは抗体可変部(V)遺伝子に、上流に配置された多くの偽V遺伝子の部分配列が遺伝子変換によって取り込まれることによって進行する(図2)。一塩基の置換による点突然変異も起こるが低頻度である。培養細胞の集団中には、これにより様々な特異性を持つ抗体ライブラリーが形成される。
【0005】
したがって培養したDT40細胞の集団から、目的抗原に結合する抗体を産生する細胞を単離することにより、必要な抗体を迅速かつ効率的に取得することができると考えられる(図3)。
【0006】
このような着想に基づき、本発明者は先に機能改変したDT40株DT40-SWを用いるin vitro抗体作成法を開発した(特許文献1を参照)。この技術のもう一つの重要な特徴は、目的抗体を産生する細胞を選択できた場合、図3に示すように再度その細胞クローンに変異を導入して多様化させ、選択を繰り返すことにより、更に高親和性の抗体へ進化させること(親和性成熟)が行える点にある。この親和性成熟を効率よく進行させるためには、次に述べるように更なる細胞の遺伝子レベルでの改変が必要である。
【0007】
一方、遺伝子変換型変異が優勢であるDT40細胞において、変異様式を遺伝子変換型から点突然変異型に転換させる方法については既に報告があり、DNAの損傷を修復するのに関与する因子XRCC2、XRCC3を欠損させることにより実現できることが知られていた(非特許文献1及び2を参照)。
【0008】
【特許文献1】特開2006-109711号公報
【非特許文献1】Sale, J. E. et al. Nature 412, 921-926(2001)
【非特許文献2】Arakawa, H. et al., PLOS Biol. 2, 967-974 (2004)
【発明の開示】
【発明が解決しようとする課題】
【0009】
本発明は、抗体をコードする遺伝子の点突然変異の頻度が上昇したB細胞由来の細胞の提供、及び抗体産生細胞であるB細胞由来の細胞において、抗体をコードする遺伝子の点突然変異の頻度を上昇させ、様々な特異性を有する抗体ライブラリーを形成させ、その中から所望の特異性を有する抗体を選択し製造する方法の提供を目的とする。
【課題を解決するための手段】
【0010】
上記の親和性成熟を効率よく行うためには、変異による抗体ライブラリーの多様化が目的に適した形で進行することが重要である。図4(左)に示すように、DT40細胞においては、数十塩基程度の配列が一度に置き換わる遺伝子変換が変異様式として優位であり、これはニワトリ体内でB細胞の抗体多様性を生み出す機構と同じであるので、初期の抗体ライブラリーの拡大には有利であると言える。しかし、ニワトリ体内では、免疫後に高親和性抗体を生み出す親和性成熟の過程では、遺伝子変換と共に点突然変異の頻度が高まることが知られており、この変異様式が高親和性抗体を作るのに適していると考えられる。何故なら、一塩基置換である点突然変異では、元の特異性からより小さな偏移幅で抗原特異性が変化することが期待できるので、抗原特異性のより精密な改変が可能と考えられるからである(図4右)。
【0011】
そこで、抗体探索のためのライブラリー形成時には遺伝子変換型で変異が進行し、その中から目的抗体産生クローンが選択できた後には、点突然変異の頻度を上昇させるようにDT40を改変することが有効である。本発明者は、この問題を克服するための以下の検討を行い、有効な解決法を見いだした。
【0012】
本来、遺伝子変換型変異が優勢であるDT40細胞において、変異様式を遺伝子変換型から点突然変異型に転換させる方法については既に報告があり、DNAの損傷を修復するのに関与する因子XRCC2、XRCC3を欠損させることにより実現できることが知られている(図4右)(Sale, J. E. et al. Nature 412, 921-926(2001); Arakawa, H. et al., PLOS Biol. 2, 967-974 (2004))。
【0013】
しかし、これらの遺伝子を完全に欠損させると、1)細胞の増殖速度が低下する、2)点突然変異により効率に非機能的遺伝子が生じるため抗体を産生しない細胞が多く生じる、3)DT40の最大の長所である相同組換えが行えなくなり、遺伝子操作が困難となる、という問題が発生する。本発明者は、この問題を克服するために種々検討を重ねた結果、驚くべきことにXRCC3欠損細胞を作製する中間段階でできるXRCC3遺伝子の2つの対立遺伝子の片方のみを欠損させたヘテロノックアウト細胞、XRCC3(+/-)株が、上にあげた3つの問題点を克服するのに有用であることを見いだした。しかも、対立遺伝子の一方のみを破壊すればよいので、実験操作上極めて容易である。したがってこの方法はDT40を用いる抗体作製システムの性能向上に極めて有用である。
【0014】
すなわち、本発明の態様は以下のとおりである。
[1] XRCC3遺伝子の2つの対立遺伝子の片方のみが不活性化されたB細胞由来の細胞であって、抗体遺伝子への点突然変異の頻度が両方のXRCC3遺伝子を有する細胞に比べ上昇した細胞。
[2] XRCC3遺伝子の2つの対立遺伝子の片方の第6エキソンが不活性化された[1]の細胞。
[3] XRCC3遺伝子の2つの対立遺伝子の片方のみの不活性化が相同組換えにより行われる[1]又は[2]の細胞。
[4] B細胞由来の細胞がニワトリB細胞株DT40又はその変異株である、[1]〜[3]のいずれかの細胞。
[5] ニワトリB細胞株DT40の変異株が、
細胞外刺激により外因性Creリコンビナーゼ遺伝子の発現が誘導され、発現されたCreリコンビナーゼにより外因性AID(activation induced cytidine deaminase)遺伝子の向きを反転させることにより、AID発現を誘導すること及び停止させることが可能な変異株であって、以下の特徴:
1)内因性のAID遺伝子が機能的に破壊されており、内因性AID遺伝子発現によるAIDタンパク質は産生されないこと、
2)互いに逆方向の2つのloxP配列で挟まれた外因性のAID遺伝子、及び該2つのloxP配列で挟まれた領域の上流側に存在する該動物細胞で機能し得るプロモーターを有し、上記AID遺伝子が該プロモーターに対して順方向に配置されている場合には、該プロモーターによるAID遺伝子の発現が可能であり、上記AID遺伝子が該プロモーターに対して逆方向に配置されている場合には、AID遺伝子の発現は停止すること、及び
3)Creリコンビナーゼ遺伝子が、細胞外刺激によりCreリコンビナーゼ活性化が可能な形で導入されており、Creリコンビナーゼ活性化により、上記外因性AID遺伝子を含む2つのloxP配列に挟まれた領域の方向が反転すること、
を有する変異株であり、細胞外刺激によりAID遺伝子の発現が停止し、変異機能が停止する変異株である、[4]の細胞。
[6] 外来性の抗体をコードする遺伝子が導入された[1]〜[5]のいずれかの細胞。
[7] 所望の特性を有する抗体を産生する抗体産生細胞を選択する方法であって、
(i) [1]〜[6]のいずれかの細胞を調製する工程、
(ii) 該細胞を一定期間培養する工程、
(iii) 該細胞が産生する抗体又は抗体をコードする遺伝子を単離して、その変異を検出する工程、並びに
(iv) 所望の変異を有する抗体を産生する細胞を選択する工程、を含む方法。
[8] 所望の特性を有する抗体を製造する方法であって、
(i) [1]〜[6]のいずれかの細胞を調製する工程、
(ii) 該細胞を一定期間培養する工程、
(iii) 該細胞が産生する抗体又は抗体をコードする遺伝子を単離して、その変異を検出する工程、
(iv) 所望の変異を有する抗体を産生する細胞を選択する工程、並びに
(v) 選択した所望の変異を有する細胞を培養して、抗体を生産する工程、を含む方法。
【発明の効果】
【0015】
本発明は、ニワトリB細胞由来のDT40細胞を用いる抗体作製システムであり、高親和性の抗体をより効率よく取得することを可能にする。また、本発明の抗体により、既存の抗体もXRCC3(+/-)変異株中で更に高性能な抗体に改良することが可能となる。特に、本発明の細胞を用いることにより、癌などの難治性疾患の治療に役立つ抗体の取得がより容易になる。
【発明を実施するための最良の形態】
【0016】
以下、本発明を詳細に説明する。
【0017】
本発明において用いるB細胞由来の細胞は、抗体を産生し得る限り、由来動物種、細胞株種は限定されない。ヒト、マウス、ヒツジ、ラット、ウサギ、ニワトリなどのB細胞若しくはその細胞株又はそれらの変異株を用いることができ、好ましくはニワトリB細胞由来のDT40細胞株を用いる。DT40細胞株は、Bリンパ腫由来の細胞であり、第2染色体がトリソミーという特徴をもつ(Baba, T.W., Giroir, B.P. and Humphries, E.H.:Virology 144 : 139-151,1985)。DT40株としては、野生型DT40株又は本発明者らが独自に樹立したDT40-SW株(変異機能を司る遺伝子AIDの発現を可逆的にスイッチすることにより、その抗体変異機能をON/OFF制御できる変異株(詳細はKanayama, N., Todo, K., Reth, M., Ohmori, H. Biochem. Biophys. Res. Commn. 327:70-75 (2005)及び特開2006-109711号公報に記載されている))を用いることができる。
【0018】
XRCC3(X-ray repair complementing defective repair in Chinese hamster cells 3)は、染色体の安定性維持とDNA損傷の修復に関与する遺伝子であり、細胞の相同組換えに関与する。ニワトリXRCC3遺伝子は、8個のエキソンを有する。ニワトリXRCC3遺伝子の塩基配列を配列番号1に示す。配列番号1中、8個のエキソンの位置は以下のとおりである。エキソン1: 1-215、エキソン2: 216-284、エキソン3: 285-422、エキソン4: 423-635、エキソン5: 636-790、エキソン6: 791-1003、エキソン7: 1004-1050、エキソン8: 1051-1935。
【0019】
本発明の細胞は、細胞の相同染色体のそれぞれに存在する2つのXRCC3対立遺伝子のうち、片方のみが不活性化されている。本発明の細胞は遺伝子型XRCC3(+/-)で表される。ここで、遺伝子の不活性化とは、遺伝子の塩基配列に部分的に欠失、置換、挿入、付加等が生じ遺伝子の発現が抑制されていることをいう。「遺伝子の発現を抑制」とは、遺伝子自体は発現するが、正常な機能を有する蛋白質をコードする遺伝子が発現されない場合も含まれる。「遺伝子が不活性化されている」ことを「遺伝子が破壊されている」、「遺伝子が欠損している」、「遺伝子がノックアウトされている」ということもある。また、遺伝子の不活性化は、遺伝子の発現が完全に抑制されている場合だけでなく、部分的に抑制されている場合も含まれる。本発明において、対立遺伝子の一方のみを不活性化するため、XRCC3遺伝子をヘテロでノックアウトするということがあり、得られた細胞をヘテロノックアウト細胞と呼ぶことがある。
【0020】
遺伝子の不活性化は、例えば、相同組換えにより行うことができる。相同組換えは、染色体上の遺伝子と外来DNAとの間で相同的遺伝子組換えを行い、目的の遺伝子を改変する方法をいう。相同組換えにおいては、タンパク質をコードする配列を分断する目的で、その遺伝子のエキソンに別のDNA配列を挿入する。相同組換えは、ジーンターゲティングベクターを用いて行う。相同組換え後、ジーンターゲティングベクターを持つ細胞を同定するために、目的の遺伝子を分断する配列には選択マーカー遺伝子を用いる。選択マーカー遺伝子としては、Blasticidin耐性遺伝子などの抗生物質耐性遺伝子、GFPなどの蛍光タンパク質遺伝子、色素タンパク質遺伝子等を用いることができる。
【0021】
本発明においては、ニワトリXRCC3遺伝子の8個のエキソンのうち、いずれのエキソンを分断してもよい。また、複数のエキソンを分断してもよい。好ましくは第6エキソンを分断する。
【0022】
ターゲッティングベクターは、ニワトリXRCC3遺伝子の配列情報に基づいて設計し、山村研一ら編 トランスジェニック動物 共立出版株式会社 1997年3月1日、相沢慎一 ジーンターゲティング ES細胞を用いた変異マウスの作製 バイオマニュアルシリーズ8 羊土社 1995、Hogan et al., Manipulating the Mouse Embryo, Cold Spring Hoarbor Laboratory Press (1944)、Joyner, A.L., Gene Targeting, A Practical Approach Series, IRL Press (1993)、松村正實ら編 実験医学別冊 新遺伝子工学ハンドブック(改定第3版) 羊土社 1999 等に記載の方法に従って作製することができる。ターゲティングベクターは挿入型及び置換型のどちらを用いてもよい。また、Cre-lox系を用いたターゲティングにより組換えを起こさせることもできる。Cre-lox系を用いたターゲティングは、例えば特表平11-503015号公報に記載の方法に従って行うことができる。相同組換えを起こした相同組換え体の選別方法としては、ポジティブ選択、プロモーター選択、ネガティブ選択、ポリA選択等の公知の選択方法を用いればよい。相同組換えを起こした細胞は、PCR法、サザンブロッティング法等の公知の方法で同定することができる。
【0023】
上記の方法で得られたB細胞由来の細胞のXRCC3(+/-)変異株は、抗体遺伝子の点突然変異が高頻度で起こる。また、本発明の細胞XRCC3(+/-)においては、野生型細胞XRCC3(+/+)において高頻度で生じる遺伝子変換も生じる。一方、2つのXRCC3対立遺伝子の両方を不活性化した細胞XRCC3(-/-)では、遺伝子変換の頻度は著しく低下する。従って、本発明の細胞は遺伝子変換と点突然変異の両方が生じやすく、多様な変異抗体を得ることができる。また、本発明の細胞XRCC3(+/-)の増殖速度は、野生型に対して若干低下するが、問題が生じるほどの低下は認められず、細胞密度は野生型と同様である。一方、2つのXRCC3対立遺伝子の両方を不活性化した細胞XRCC3(-/-)では、増殖速度及び細胞密度が野生型細胞XRCC(+/+)に比べ著しく低下する。さらに、XRCC3(-/-)では、抗体を産生できない株が高頻度で出現するのに対して、XRCC3(+/-)では抗体を産生できない株の出現率は低い。
【0024】
本発明の細胞XRCC(+/-)の継代培養を続けることにより、抗体遺伝子に種々の点突然変異を有する細胞集団を得ることができる。すなわち、細胞集団中に様々な特性を有する抗体ライブラリーが形成される。この細胞集団の中から所望の特性を有する抗体を選択すればよい。B細胞は表面に抗体が発現され、又は抗体を細胞外に分泌するため、細胞表面の抗体又は分泌された抗体の特性を調べて所望の抗体を選択することができる。細胞表面の抗体を選択し単離するためには、例えば抗原を固相化した磁気ビーズを用いればよい。すなわち、磁気ビーズに抗原を吸着により固相化させ、該抗原固相化磁気ビーズと細胞集団を混合し、所望の抗体を産生する細胞を磁気ビーズに結合させ、磁気を用いて磁気ビーズを回収することにより、所望の抗体を産生する細胞を回収・単離することができる。一方、培養液中に分泌された抗体は、ELISA(enzyme-linked immunosorbent assay)等により検出することができる。
【0025】
目的の抗原に結合し得る抗体を産生する細胞を単離し、さらに培養を行い変異を生じさせ、その中からより優れた特性を有する抗体を産生する細胞を選択するという操作を繰り返すことにより、さらに優れた特性を有する抗体を得ることができる。ここで、抗体の特性とは、目的の抗原に対する特異性、結合力の他、至適温度、至適pH、ADCC(antibody-dependent cellular cytotoxicity)、CDC(complement-dependent cytotoxicity)等の細胞傷害活性等を含む。特異性、結合力は、抗体の相補性決定領域(CDR)等の可変領域に変異が生じることにより改善される。また、特異性についても単に目的の抗原に対する特異性が向上した抗体を得るだけではなく、従来の動物に免疫する方法では作製が困難であった動物種間でアミノ酸配列が保存されたタンパク質に対しても特異性の高い抗体を得ることができる。
【0026】
本発明は、本発明の細胞を用いて所望の変異を有する抗体及び種々の変異を有する抗体ライブラリーを製造する方法を包含する。
【0027】
変異を有する抗体を製造する方法は、
B細胞の2つのXRCC3対立遺伝子の片方のみを不活性化されたB細胞を調製する工程、
該細胞を一定期間培養する工程、
該細胞が産生する抗体又は抗体をコードする遺伝子を単離して、その変異を検出する工程、
所望の変異を有する抗体を産生する細胞を選択する工程、並びに
選択した所望の変異を有する細胞を培養して、抗体を生産する工程、を含む。
【0028】
さらに、本発明の細胞を用いて既存の抗体に変異を起こさせ、より優れた特性を有する抗体を得ることができる。すなわち、本発明のXRCC3遺伝子の2つの対立遺伝子の片方のみが不活性化された細胞に既存の外来性の抗体をコードする遺伝子を発現可能な形態で導入し、該細胞のゲノムに組込み、該抗体遺伝子に変異を起こさせればよい。この場合、抗体は完全な抗体分子である必要はなく、抗原に結合するために必要な可変領域を含む抗体フラグメントを産生し得る細胞でもよく、またキメラ抗体またはヒト化抗体等、非天然型の抗原に結合し得る分子およびそのフラグメントを産生し得る細胞でもよい。
【0029】
本発明の方法に、本発明者らのKanayama, N., Todo, K.,Reth, M., Ohmori, H. Biochem. Biophys. Res. Commn. 327:70-75(2005)及び特開平2006-109711号公報に記載した、抗体遺伝子の変異機能を可逆的にON/OFF制御する方法を組み合わせるとDT40細胞を用いる培養系による革新的抗体作製技術が確立できる。本発明者らが開発した上記方法において用いられるDT40-SW細胞では、抗体遺伝子の変異を司るactivation-inducedcytidine deaminase(AID)遺伝子の発現を、外部から与えるエストロゲン誘導体等の刺激により可逆的にON/OFFすることができる。XRCC3遺伝子の2つの対立遺伝子の片方のみが不活性化されたXRCC3遺伝子へテロノックアウトDT40細胞から、目的クローンを単離できたとしても、そのクローンを増殖させる過程で更なる変異が導入される場合があり、この場合、有用な変異株を安定に保持することは困難になる。この問題を回避するためには、得られた目的とするクローンの変異機能を停止することが必要である。そこで、以下に述べるように、DT40-SW株において、XRCC3遺伝子の一方を不活性化させればよい。このXRCC3遺伝子へテロノックアウトDT40-SW細胞から選択されたクローンのAID発現を停止させることにより、目的抗体を産生する遺伝的に安定なクローンを得ることが可能となる。
【0030】
すなわち、本発明は、細胞外刺激により外因性Creリコンビナーゼが活性化され、活性化されたCreリコンビナーゼにより外因性AID(activation induced cytidine deaminase)遺伝子の向きを反転させることにより、AID発現を誘導すること及び停止させることが可能なB細胞由来の細胞であって、以下の特徴:
1)内因性のAID遺伝子が機能的に破壊されており、内因性AID遺伝子発現によるAIDタンパク質は産生されないこと、
2)互いに逆方向の2つのloxP配列で挟まれた外因性のAID遺伝子、及び該2つのloxP配列で挟まれた領域の上流側に存在する該細胞で機能し得るプロモーターを有し、上記AID遺伝子が該プロモーターに対して順方向に配置されている場合には、該プロモーターによるAID遺伝子の発現が可能であり、上記AID遺伝子が該プロモーターに対して逆方向に配置されている場合には、AID遺伝子の発現は停止すること、及び
3)Creリコンビナーゼ遺伝子が、細胞外刺激によりCreリコンビナーゼ活性化が可能な形で導入されており(具体的にはCreとエストロゲンレセプターの融合タンパク(Cre-ER)として発現させている。Cre-ERはエストロゲン誘導体が結合すると細胞質から核に移行し、基質であるloxPに挟まれたAID遺伝子に作用する(Zhang, Y. et al. Nucleic Acids Res. 24, 534-548 (1996)))、Creリコンビナーゼ活性化により、上記外因性AID遺伝子を含む2つのloxP配列に挟まれた領域の方向が反転すること、
を有する細胞において、2つのXRCC3対立遺伝子のうち、片方のみが不活性化された細胞を包含する。上記のプロモーターとしては、β-アクチンプロモーター、免疫グロブリンプロモーター、サイトメガロウイルスプロモーター、CAGプロモーター等が挙げられる。また、Creリコンビナーゼを活性化する細胞外刺激としては、例えば、Creリコンビナーゼが、エストロゲンレセプター、又はそのエストロゲン結合ドメインを含むタンパク質との融合タンパク質として発現し得るような形で、エストロゲンレセプター又はそのエストロゲン結合ドメインを含むタンパク質をコードする遺伝子とCreリコンビナーゼcDNAとをインフレームで連結したDNA構築物を細胞内に導入されている場合、エストロゲン誘導体刺激を細胞に与えると、Creリコンビナーゼの活性化が誘導される。
【0031】
さらに、本発明は細胞外刺激により外因性Creリコンビナーゼ遺伝子の発現が誘導され、発現されたCreリコンビナーゼにより外因性AID(activation induced cytidine deaminase)遺伝子の向きを反転させることにより、AID発現を誘導すること及び停止させることが可能なB細胞由来の細胞であって、以下の特徴:
1)内因性のAID遺伝子が機能的に破壊されており、内因性AID遺伝子発現によるAIDタンパク質は産生されないこと、
2)互いに逆方向の2つのloxP配列で挟まれた外因性のAID遺伝子、及び該2つのloxP配列で挟まれた領域の上流側に存在する該動物細胞で機能し得るプロモーターを有し、上記AID遺伝子が該プロモーターに対して順方向に配置されている場合には、該プロモーターによるAID遺伝子の発現が可能であり、上記AID遺伝子が該プロモーターに対して逆方向に配置されている場合には、AID遺伝子の発現は停止すること、及び
3)Creリコンビナーゼ遺伝子が、細胞外刺激によりCreリコンビナーゼ活性化が可能な形で導入されており、Creリコンビナーゼ活性化により、上記外因性AID遺伝子を含む2つのloxP配列に挟まれた領域の方向が反転すること、
を有する細胞において、2つのXRCC3対立遺伝子のうち、片方のみが不活性化された細胞を包含する。
【0032】
上記の細胞を用いて変異を有する抗体を得る方法は、
上記B細胞の2つのXRCC3対立遺伝子の片方のみを不活性化されたB細胞を調製する工程、
該細胞のAIDの発現が可能な状態にする工程、
該細胞を一定期間培養する工程、
該細胞にCreリコンビナーゼを活性化し得る細胞外刺激を与えて、AID遺伝子の向きを反転させて、AIDの発現を停止させる工程、
該細胞が産生する抗体又は抗体をコードする遺伝子を単離して、その変異を検出する工程、
所望の変異を有する抗体を産生する細胞を選択する工程、並びに
選択した所望の変異を有する細胞を培養して、抗体を生産する工程、を含む。
【実施例】
【0033】
本発明を以下の実施例によって具体的に説明するが、本発明はこれらの実施例によって限定されるものではない。
【0034】
1)XRCC3遺伝子の破壊によるXRCC3(+/-)、XRCC3(-/-)株の樹立
DT40細胞のXRCC3遺伝子の破壊のために、図5に示すようにXRCC3遺伝子の第6エキソン内部にブラスチサイジン耐性遺伝子又はヒスチジノールデヒドロゲナーゼ遺伝子を挿入した2種類のターゲッティングベクター(それぞれBsr、HisDと呼ぶ)を次の手順で作製した。SpeIサイトを含むprimer1とBamHIサイトを含むprimer2を用いて、PCRにより5’側の遺伝子断片を得た。また、BamHIサイトを含むprimer3とXhoIサイトを含むprimer4を用いて、PCRにより3’側の遺伝子断片を得た。Primer1〜4の配列は、それぞれ配列表の配列番号2〜5に示す。そこに、薬剤耐性マーカーとしてHisDとBSR遺伝子を挿入し、ターゲッティングベクターを作製した。
【0035】
最初に、Bsrを導入して、標的相同組換えにより一方のXRCC3を破壊し、XRCC3(+/-)株を得た。続いて、XRCC3(+/-)株にHisDを導入し、残りのXRCC3を破壊し、XRCC3(-/-)株を樹立した。ターゲッティングベクターを導入したDT40細胞クローンよりDNAを抽出し、PCRにより標的相同組換えが正しく行なわれたことを確認した。PCRには図6に示すプライマーA、B、Cを用いた(それぞれの配列は配列表の配列番号6〜8に示す)。Bsrの場合はプライマーAとC、HisDの場合はプライマーBとCを用いて相同組換えを確認した。図7に示すように、XRCC3の標的相同組換えによる遺伝子破壊の結果、XRCC3(+/-)、XRCC3(-/-)株が正しく作製されたことが確認された。第二段階の遺伝子導入も成功して、XRCC3(-/-)が取得できたことは、XRCC3(+/-)株において、相同組換え機能が保持されていることを示しており、XRCC3(+/-)株の有用性を更に裏付けるものである。
【0036】
2)XRCC3(+/+)、(+/-)、(-/-)における変異パターンの比較
元株であるXRCC3(+/+)とXRCC3を部分的あるいは完全に破壊したXRCC3(+/-)、XRCC3(-/-)の各細胞を4週間継続的に培養し、抗体L鎖遺伝子の配列解析により、変異導入効率と変異様式を評価した。図8及び表1に示すように、元株XRCC3(+/+)では遺伝子変換が優位に起こっているが、XRCC3(-/-)株では、点突然変異型に大きく転換していた。一方、驚くべきことに、XRCC3(+/-)株においては、遺伝子変換がやや低下するだけで、点突然変異は、XRCC3(-/-)と同程度まで増加していた。
【0037】
更に、遺伝子変換と点突然変異を合わせた、全体の変異頻度ではXRCC3(+/-)が最も高かった。これは、XRCC3(+/-)株が遺伝子変換と点突然変異の長所を併せ持つ優良な変異株であることを示している。
【0038】
【表1】
【0039】
3)表面IgM消失率及び増殖速度の比較
XRCC3(-/-)株では、点突然変異は増加するがそれに伴って、ストップコドンやフレームシフトなどが生じるために、抗体タンパクを作れない変異細胞(表面IgM陰性細胞)が増加することが報告されている(Sale, J. E. et al. Nature 412, 921-926(2001))。また、XRCC3の欠損に伴い、細胞の増殖速度が低下することも知られている(同文献)これは抗体探索システムとしては重大な欠点となる。
【0040】
そこで、XRCC3(+/+)、(+/-)、(-/-)の3種類の細胞株を図9に示すように、長期間培養し、表面IgM抗体の発現をフローサイトメトリーにより測定した。報告されているように、XRCC3(-/-)株では表面IgM陰性の細胞が時間とともに増加したが、XRCC3(+/+)では増加しなかった。驚くべきことに、点突然変異効率が上昇しているXRCC3(+/-)においても表面IgM陰性細胞の出現率は低かった。このことは、点突然変異による抗体遺伝子の機能喪失が遺伝子変換によって修復されていることを示唆し、XRCC3(+/-)株の有用性をさらに支持するものである。
【0041】
XRCC3(+/+)、(+/-)、(-/-)の3種類の細胞株の増殖速度を比較したところ、XRCC3(+/+)株の増殖が最も速く、XRCC3(+/-)の増殖はやや遅かったが問題となる程度ではなく、最大の細胞密度もXRCC3(+/+)株とほぼ同程度であった。一方、XRCC3(-/-)株では、増殖速度、最大細胞密度ともに低下していた(図10)。この結果もXRCC3(+/-)株の有用性を示している。
【図面の簡単な説明】
【0042】
【図1】免疫系における高特異性抗体の産生(親和性成熟)の原理を示す図である。
【図2】ニワトリ抗体遺伝子の多様化機構を示す図である。
【図3】ニワトリB細胞株DT40を用いる抗体作製システムを示す図である。
【図4】XRCC3遺伝子の欠損による遺伝子変換から点突然変異への変異様式の転換を示す図である。
【図5】XRCC3ターゲッティングベクターの構築図である。
【図6】標的相同組換えによるXRCC3遺伝子の不活性化の方法及び相同組換え確認のためのプライマーの位置を示す図である。
【図7】XRCC3の標的相同組換えによるXRCC3遺伝子の不活性化の確認の結果を示す図である。
【図8】XRCC3遺伝子の不活性化による点突然変異の増加を示す図である。各変異細胞の代表的な5クローンの結果を示し、太い横線は遺伝子変換で挿入された配列、縦の太線は点突然変異の場所を示す。
【図9】XRCC3(+/+)、XRCC3(+/-)及びXRCC3(-/-)における表面IgMの消失率を示す図である。
【図10】XRCC3(+/+)、XRCC3(+/-)及びXRCC3(-/-)における細胞増殖速度を示す図である。
【配列表フリーテキスト】
【0043】
配列番号2〜8 プライマー
【特許請求の範囲】
【請求項1】
XRCC3遺伝子の2つの対立遺伝子の片方のみが不活性化されたB細胞由来の細胞であって、抗体遺伝子への点突然変異導入の頻度が両方のXRCC3遺伝子を有する細胞に比べ上昇した細胞。
【請求項2】
XRCC3遺伝子の2つの対立遺伝子の片方の第6エキソンが不活性化された請求項1に記載の細胞。
【請求項3】
XRCC3遺伝子の2つの対立遺伝子の片方のみの不活性化が相同組換えにより行われる請求項1又は2に記載の細胞。
【請求項4】
B細胞由来の細胞がニワトリB細胞株DT40又はその変異株である、請求項1〜3のいずれか1項に記載の細胞。
【請求項5】
ニワトリB細胞株DT40の変異株が、細胞外刺激により外因性Creリコンビナーゼ遺伝子の発現が誘導され、発現されたCreリコンビナーゼにより外因性AID(activation induced cytidine deaminase)遺伝子の向きを反転させることにより、AID発現を誘導すること及び停止させることが可能な変異株であって、以下の特徴:
1)内因性のAID遺伝子が機能的に破壊されており、内因性AID遺伝子発現によるAIDタンパク質は産生されないこと、
2)互いに逆方向の2つのloxP配列で挟まれた外因性のAID遺伝子、及び該2つのloxP配列で挟まれた領域の上流側に存在する該動物細胞で機能し得るプロモーターを有し、上記AID遺伝子が該プロモーターに対して順方向に配置されている場合には、該プロモーターによるAID遺伝子の発現が可能であり、上記AID遺伝子が該プロモーターに対して逆方向に配置されている場合には、AID遺伝子の発現は停止すること、及び
3)Creリコンビナーゼ遺伝子が、細胞外刺激によりCreリコンビナーゼ活性化が可能な形で導入されており、Creリコンビナーゼ活性化により、上記外因性AID遺伝子を含む2つのloxP配列に挟まれた領域の方向が反転すること、
を有する変異株であり、細胞外刺激によりAID遺伝子の発現が停止し、変異機能が停止する変異株である、請求項4記載の細胞。
【請求項6】
外来性の抗体をコードする遺伝子が導入された請求項1〜5のいずれか1項に記載の細胞。
【請求項7】
所望の特性を有する抗体を産生する抗体産生細胞を選択する方法であって、
(i) 請求項1〜6のいずれか1項に記載の細胞を調製する工程、
(ii) 該細胞を一定期間培養する工程、
(iii) 該細胞が産生する抗体又は抗体をコードする遺伝子を単離して、その変異を検出する工程、並びに
(iv) 所望の変異を有する抗体を産生する細胞を選択する工程、を含む方法。
【請求項8】
所望の特性を有する抗体を製造する方法であって、
(i) 請求項1〜6のいずれか1項に記載の細胞を調製する工程、
(ii) 該細胞を一定期間培養する工程、
(iii) 該細胞が産生する抗体又は抗体をコードする遺伝子を単離して、その変異を検出する工程、
(iv) 所望の変異を有する抗体を産生する細胞を選択する工程、並びに
(v) 選択した所望の変異を有する細胞を培養して、抗体を生産する工程、を含む方法。
【請求項1】
XRCC3遺伝子の2つの対立遺伝子の片方のみが不活性化されたB細胞由来の細胞であって、抗体遺伝子への点突然変異導入の頻度が両方のXRCC3遺伝子を有する細胞に比べ上昇した細胞。
【請求項2】
XRCC3遺伝子の2つの対立遺伝子の片方の第6エキソンが不活性化された請求項1に記載の細胞。
【請求項3】
XRCC3遺伝子の2つの対立遺伝子の片方のみの不活性化が相同組換えにより行われる請求項1又は2に記載の細胞。
【請求項4】
B細胞由来の細胞がニワトリB細胞株DT40又はその変異株である、請求項1〜3のいずれか1項に記載の細胞。
【請求項5】
ニワトリB細胞株DT40の変異株が、細胞外刺激により外因性Creリコンビナーゼ遺伝子の発現が誘導され、発現されたCreリコンビナーゼにより外因性AID(activation induced cytidine deaminase)遺伝子の向きを反転させることにより、AID発現を誘導すること及び停止させることが可能な変異株であって、以下の特徴:
1)内因性のAID遺伝子が機能的に破壊されており、内因性AID遺伝子発現によるAIDタンパク質は産生されないこと、
2)互いに逆方向の2つのloxP配列で挟まれた外因性のAID遺伝子、及び該2つのloxP配列で挟まれた領域の上流側に存在する該動物細胞で機能し得るプロモーターを有し、上記AID遺伝子が該プロモーターに対して順方向に配置されている場合には、該プロモーターによるAID遺伝子の発現が可能であり、上記AID遺伝子が該プロモーターに対して逆方向に配置されている場合には、AID遺伝子の発現は停止すること、及び
3)Creリコンビナーゼ遺伝子が、細胞外刺激によりCreリコンビナーゼ活性化が可能な形で導入されており、Creリコンビナーゼ活性化により、上記外因性AID遺伝子を含む2つのloxP配列に挟まれた領域の方向が反転すること、
を有する変異株であり、細胞外刺激によりAID遺伝子の発現が停止し、変異機能が停止する変異株である、請求項4記載の細胞。
【請求項6】
外来性の抗体をコードする遺伝子が導入された請求項1〜5のいずれか1項に記載の細胞。
【請求項7】
所望の特性を有する抗体を産生する抗体産生細胞を選択する方法であって、
(i) 請求項1〜6のいずれか1項に記載の細胞を調製する工程、
(ii) 該細胞を一定期間培養する工程、
(iii) 該細胞が産生する抗体又は抗体をコードする遺伝子を単離して、その変異を検出する工程、並びに
(iv) 所望の変異を有する抗体を産生する細胞を選択する工程、を含む方法。
【請求項8】
所望の特性を有する抗体を製造する方法であって、
(i) 請求項1〜6のいずれか1項に記載の細胞を調製する工程、
(ii) 該細胞を一定期間培養する工程、
(iii) 該細胞が産生する抗体又は抗体をコードする遺伝子を単離して、その変異を検出する工程、
(iv) 所望の変異を有する抗体を産生する細胞を選択する工程、並びに
(v) 選択した所望の変異を有する細胞を培養して、抗体を生産する工程、を含む方法。
【図1】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【公開番号】特開2009−60850(P2009−60850A)
【公開日】平成21年3月26日(2009.3.26)
【国際特許分類】
【出願番号】特願2007−231741(P2007−231741)
【出願日】平成19年9月6日(2007.9.6)
【国等の委託研究の成果に係る記載事項】(出願人による申告)平成18年度、独立行政法人新エネルギー・産業技術総合開発機構「新機能抗体創製技術開発」に係る委託研究、産業技術力強化法第19条の適用を受ける特許出願
【出願人】(504147243)国立大学法人 岡山大学 (444)
【Fターム(参考)】
【公開日】平成21年3月26日(2009.3.26)
【国際特許分類】
【出願日】平成19年9月6日(2007.9.6)
【国等の委託研究の成果に係る記載事項】(出願人による申告)平成18年度、独立行政法人新エネルギー・産業技術総合開発機構「新機能抗体創製技術開発」に係る委託研究、産業技術力強化法第19条の適用を受ける特許出願
【出願人】(504147243)国立大学法人 岡山大学 (444)
【Fターム(参考)】
[ Back to top ]