
BTL−II核酸、タンパク質および抗体
本発明は、単離BTL−IIタンパク質、核酸、抗体、アンタゴニストおよびアゴニスト、そしてこれらを作製するおよび使用する方法を提供する。本発明の組成物を使用する、診断、スクリーニングおよび治療法が提供される。例えば、本発明の組成物は、炎症性腸疾患の診断および処置のため、そして抗原に対する粘膜免疫応答を促進するために使用することが可能である。
【発明の詳細な説明】
【技術分野】
【0001】
関連出願
本出願は、2002年12月23日に出願された米国仮出願第60/436,185号および2003年11月26日に出願された米国仮出願第60/525,298号の優先権を主張する。
【0002】
技術分野
本発明はブチロフィリン様タンパク質、特に、例えばB細胞および/またはT細胞のごとき免疫エフェクター細胞の機能を変調することが知られている、B7サブファミリーのブチロフィリン様タンパク質に関する。こうしたタンパク質をコードする核酸、こうしたタンパク質を産生するためのプロセス、こうしたタンパク質へ結合する抗体、こうしたタンパク質または抗体を含む医薬組成物、こうした核酸、タンパク質およびこうしたタンパク質に対する抗体を使用する方法もまた含まれている。
【背景技術】
【0003】
背景技術
免疫または炎症性応答の変調は、自己免疫疾患、異常な炎症および/または免疫応答により特徴付けられる疾患、並びに感染を含む、多様な種類の疾患の制御において、役に立つ手段であることが可能である。炎症性腸疾患および自己免疫または炎症性疾患のごとき、異常な炎症および/または免疫応答により特徴付けられる疾患の処置においては、免疫応答の下方変調が望まれる。他の事態、例えば、感染性疾患に対する免疫性を与えるために患者にワクチン接種する場合、免疫応答の刺激が望まれる。ワクチン設定において、同時投与される抗原に対する免疫応答を高めることが可能なアジュバントは、疾患に対する長期の保護を提供することにおいて役立つことが可能である。特に当該分野において欠けていることは、粘膜免疫応答を刺激することが可能なアジュバントである。全身性免疫応答に対する粘膜免疫応答は、まさしく共通の侵入地点、即ち、粘膜表面で感染を攻撃できるので、価値がある。本発明は、不適当なおよび/または異常な炎症および/または免疫応答により特徴付けられる疾患を診断そして処置するための治療剤、および免疫応答、特に粘膜免疫応答を刺激するためのアジュバントとして作用可能な治療剤を提供することにより、当該分野でのこれらの要求に取り組んでいる。
【発明の開示】
【発明が解決しようとする課題】
【0004】
概要
本発明は、単離BTL−IIタンパク質、核酸、抗体、BTL−II阻害剤およびアゴニスト、そしてこれらの組成物を使用するための方法を包含する。
【0005】
配列番号4のアミノ酸x−y、ここにおいてxは配列番号4の1から35位のいずれかのアミノ酸であり、そしてyは配列番号4の452−462位からのいずれかのアミノ酸である、から成るアミノ酸配列を含んでなる、単離BTL−IIタンパク質が提供される。こうしたBTL−IIタンパク質は、配列番号4のアミノ酸30から453、1から453、29から457、1から457、1から482および/または29から482を含むことができる。本発明はさらに、配列番号10、配列番号14または配列番号18のアミノ酸127から157、または配列番号16のアミノ酸126から156と少なくとも80%、所望により少なくとも85%、90%、92%、94%、96%、または98%同一であるアミノ酸配列から成るポリペプチドを含んでなる、単離BTL−IIタンパク質を提供し、ここにおいて、配列番号10、配列番号14または配列番号18のアミノ酸127から157、または配列番号16のアミノ酸126から156と並列された(aligned with)アミノ酸配列の同一性領域は少なくとも20、所望により少なくとも25または30アミノ酸長であり、そしてポリペプチドはB細胞またはT細胞上に発現された細胞表面レセプターへ結合可能であり、および/またはT細胞の増殖を阻害可能である。こうしたアミノ酸配列は少なくとも150アミノ酸長であり得、そして配列番号10、配列番号14、配列番号16または配列番号18のアミノ酸30から358と少なくとも80%、所望により少なくとも85%、90%、92%、94%、96%、98%、99%または99.5%同一であり得、ここにおいて、配列番号10、配列番号14、配列番号16または配列番号18のアミノ酸30から358と並列されたアミノ酸配列の同一性領域は少なくとも150、所望により少なくとも200、250または300アミノ酸長である。さらに、アミノ酸配列は、配列番号10、配列番号14、配列番号16または配列番号18のアミノ酸30から358と少なくとも90%、所望により少なくとも92%、94%、96%、98%、99%または99.5%同一であり得、および/または配列番号10、配列番号14、配列番号16または配列番号18のアミノ酸30から358を含んでなることが可能である。
【0006】
本発明の別の態様は、配列番号10、配列番号14、配列番号16または配列番号18のアミノ酸30から358と少なくとも80%、所望により少なくとも85%、90%、92%、94%、96%、98%、99%または99.5%同一である第一のアミノ酸配列から成る第一のポリペプチドを含んでなる、単離BTL−IIタンパク質を包含しており、ここにおいて、配列番号10、配列番号14、配列番号16または配列番号18のアミノ酸30から358と並列された第一のアミノ酸配列の同一性領域は少なくとも150アミノ酸であり、ここにおいて第一のポリペプチドは、配列番号10、配列番号14または配列番号18のアミノ酸127から157、または配列番号16のアミノ酸126から156と少なくとも80%、所望により少なくとも85%、90%、92%、94%、96%、98%、99%または99.5%同一である第二のアミノ酸配列から成る第二のポリペプチドを含んでなり、ここにおいて、配列番号10、配列番号14または配列番号18のアミノ酸127から157、または配列番号16のアミノ酸126から156と並列された、第二のアミノ酸配列の同一性領域は少なくとも20アミノ酸長であり、そしてここにおいて、第一のポリペプチドはT細胞の増殖を阻害可能である。第一のアミノ酸配列は、配列番号10、配列番号14または配列番号18のアミノ酸127から157、または配列番号16のアミノ酸126から156と同一であることが可能である。
【0007】
あるいは、本発明は、配列番号4のアミノ酸30から457と少なくとも80%、所望により少なくとも85%、90%、92%、94%、96%、98%、99%または99.5%同一であるアミノ酸配列から成るポリペプチドを含んでなる、単離されたBTL−IIタンパク質を提供し、ここにおいてポリペプチドはわずか2のまたはそれより少ないIg様ドメインを含んでなり、そしてここにおいて、ポリペプチドはT細胞の増殖を阻害可能である。アミノ酸配列は、配列番号8のアミノ酸30から247または配列番号12のアミノ酸30から243と、少なくとも80%、85%、90%、92%、94%、96%、98%、99%、99.5%または100%同一であり得る。アミノ酸配列は配列番号4のアミノ酸30から457と少なくとも90%、所望により少なくとも92%、94%、96%、98%、99%、99.5%または100%同一であり得る。
【0008】
あるいは、本発明のBTL−IIタンパク質は、配列番号4のアミノ酸247から452または配列番号6のアミノ酸248から447と、少なくとも80%、所望により少なくとも85%、90%、92%、94%、96%、98%、99%または99.5%同一である第一のアミノ酸配列から成る第一のポリペプチドを含んでなることが可能であり、ここにおいて第一のアミノ酸配列は、少なくとも25,所望により少なくとも50、75、100または150のアミノ酸の、配列番号4と並列された第一のアミノ酸配列との同一性領域を有する、配列番号4のアミノ酸32から232または配列番号6のアミノ酸27から232と、少なくとも80%、所望により少なくとも85%、90%、92%、94%、96%、98%、99%または99.5%同一であるアミノ酸配列を含んでおらず、そしてここにおいて、該タンパク質は、配列番号4のアミノ酸32から232または配列番号6のアミノ酸27から232と、少なくとも80%、所望により少なくとも85%、90%、92%、94%、96%、98%、99%または99.5%同一である第二のアミノ酸配列から成る第二のポリペプチドを含んでおらず、少なくとも25,所望により少なくとも50、75、100または150のアミノ酸の、配列番号4と並列された第二のアミノ酸配列との同一性領域を有し、そしてここにおいて、第一のポリペプチドはT細胞の増殖を阻害可能である。
【0009】
別の態様において、本発明の単離BTL−IIタンパク質は、配列番号8または配列番号12のアミノ酸32から242と、少なくとも80%、所望により少なくとも85%、90%、92%、94%、96%、98%、99%または99.5%同一である第一のアミノ酸配列から成る第一のポリペプチドを含んでなることが可能であり、ここにおいて、配列番号8または配列番号12と並列された第一のアミノ酸配列の同一性領域は少なくとも約50,所望により少なくとも約75、100、150または200アミノ酸長であり、ここにおいて、第一のポリペプチドは、配列番号8または配列番号12のアミノ酸10から40と、少なくとも80%、所望により少なくとも85%、90%、92%、94%、96%、98%、99%または99.5%同一である第二のアミノ酸配列から成る第二のポリペプチドを含んでなり、ここにおいて、配列番号8または配列番号12と並列された第二のアミノ酸配列の同一性領域は少なくとも約20,所望により少なくとも約25または30アミノ酸長であり、そしてここにおいて、第一のポリペプチドはT細胞の増殖を阻害可能である。
【0010】
さらに別の態様において、本発明は、配列番号10、配列番号14、配列番号16または配列番号18のアミノ酸30から358と、少なくとも80%、所望により少なくとも85%、90%、92%、94%、96%、98%、99%または99.5%同一であるアミノ酸配列から成るポリペプチドを含んでなる、単離BTL−IIタンパク質を包含しており、ここにおいて、配列番号10、配列番号14、配列番号16または配列番号18のアミノ酸30から358と並列されたアミノ酸配列の同一性領域は、少なくとも250、所望により少なくとも275または300アミノ酸であり、そしてここにおいて、ポリペプチドはT細胞の増殖を阻害可能である。
【0011】
本発明のBTL−IIタンパク質は、長さが長くて約480アミノ酸、約380アミノ酸、約270アミノ酸または約160アミノ酸であることができ、T細胞の増殖を阻害可能であるポリペプチドを含んでなることが可能である。
【0012】
本発明はさらに、配列番号10のアミノ酸32から358と、少なくとも80%、所望により少なくとも85%、90%、92%、94%、96%、98%、99%または99.5%同一である第一のアミノ酸配列から成る第一のポリペプチドを含んでなる、単離BTL−IIタンパク質を包含し、ここにおいて、配列番号10と並列された第一のアミノ酸配列の同一性領域は少なくとも約175、所望により約200、250、275または300アミノ酸長であり、ここにおいて、第一のアミノ酸配列は、長さが約380を超えず、所望により長くて約390、270または170アミノ酸であり、ここにおいて、第一のポリペプチドはT細胞の増殖を阻害可能であり、ここにおいて、第一のアミノ酸配列は、少なくとも約20、30、40、50、60または75アミノ酸の、配列番号4のアミノ酸148から232と並列された第一のアミノ酸配列の同一性領域を有する、配列番号4のアミノ酸148から232と少なくとも80%同一ではなく、そしてここにおいて、BTL−IIタンパク質は、少なくとも約20、30、40、50、60または75アミノ酸の、配列番号4のアミノ酸148から232と並列された第二のアミノ酸配列の同一性領域を有する、配列番号4のアミノ酸148から232と少なくとも80%同一である第二のアミノ酸配列を含んでいない。こうしたBTL−IIタンパク質は配列番号8または配列番号12のアミノ酸32から242を含んでなることができる。
【0013】
別の態様において、本発明は、BTL−IIタンパク質および、抗体のFc領域またはロイシンジッパーであることが可能な異種ポリペプチドを含んでなる、BTL−II組換え融合タンパク質を含んでなる。本発明はまた、少なくとも10アミノ酸長であり、そして配列番号4の360位に及ぶ断片へ特異的に結合する抗体を惹起可能である、配列番号4のアミノ酸29から457の免疫原性断片も包含する。少なくとも10アミノ酸長の、配列番号10、配列番号14または配列番号16、および配列番号18の免疫原性断片が提供され、ここにおいて、免疫原性断片は配列番号10、配列番号14、配列番号18または配列番号16の141から143位に及び、そして断片に特異的に結合する抗体を惹起可能である。あるいは、免疫原性は配列番号10、配列番号14または配列番号18の142位、または配列番号16の141位に及ぶことが可能である。
【0014】
代わりの態様において、マウスBTL−IIタンパク質を提供する。特に、本発明は、配列番号6のアミノ酸x−yから成るアミノ酸配列を含んでなる、単離BTL−IIタンパク質を提供し、ここにおいて、xは配列番号4の1から35位の任意のアミノ酸であり、そしてyは配列番号6の450−460位の任意のアミノ酸である。こうしたBTL−IIタンパク質は配列番号6のアミノ酸32−450、20から456および/または29から514を含んでなることが可能である。
【0015】
他の態様は、配列番号4のアミノ酸1−457、配列番号6のアミノ酸1−456、配列番号8のアミノ酸1−247、配列番号10のアミノ酸1−363、配列番号12のアミノ酸1−243、配列番号14のアミノ酸1−359、配列番号16のアミノ酸1−358または配列番号18のアミノ酸1−362から成るBTL−IIタンパク質に特異的に結合する単離された抗体を含む。こうした抗体は、モノクローナル抗体、ヒト化抗体またはヒト抗体であることが可能であり、そしてそのレセプターに対するBTL−IIの結合を阻害することができる。本発明は、こうした抗体をコードする核酸、そしてこうした抗体を産生することが可能な細胞を包含し、該細胞は、こうした抗体を産生するように遺伝子操作されたハイブリドーマ細胞または細胞であることができる。本発明はさらに、抗体を分泌することができるこうした細胞を培養することによる、抗体を産生する方法も包含する。
【0016】
他の態様はBTL−II核酸を含んでいる。本発明は、配列番号3のヌクレオチドxからyより成るポリヌクレオチドを含んでなる、または該ポリヌクレオチドの相補体を含んでなる単離されたBTL−II核酸を包含し、ここにおいて、xはヌクレオチド1から105であり、そしてyはヌクレオチド1345から1375である。こうした核酸は、配列番号3のヌクレオチド105から1345、または1から1371を含んでなることが可能である。さらに、いずれかの上記BTL−IIタンパク質をコードしているBTL−II核酸であるような、免疫原性断片をコードしている核酸が提供される。
【0017】
本発明はさらに、いずれかの上記BTL−II核酸または抗BTL−II抗体をコードしている核酸を含んでなるベクター、そしてこうしたベクターを含んでいる宿主細胞を提供する。あるいは、本発明は、BTL−IIタンパク質、BTL−IIの免疫原性断片、若しくはBTL−IIに対する抗体を発現するように遺伝子操作された宿主細胞を提供する。こうした宿主細胞は、CHO細胞を含む哺乳動物細胞であってよい。こうした宿主細胞を、BTL−IIタンパク質、免疫原性断片、若しくはBTL−IIに対する抗体の発現を可能にする条件下で培養することを含んでなる、BTL−IIタンパク質、免疫原性断片、若しくはBTL−IIに対する抗体を産生するための方法もまた本発明に包含される。この方法はさらに、宿主細胞または培地から、BTL−IIタンパク質、免疫原性断片、若しくは抗体を単離することを含んでなることができる。こうした方法により産生されたBTL−IIタンパク質、免疫原性断片、若しくは抗体もまた意図される。本発明はさらに、ハイブリドーマまたは骨髄腫細胞を含む、BTL−IIに対する抗体を産生する哺乳動物細胞、およびこれらの細胞を培養することによる、抗体を作製するための方法を包含する。
【0018】
本発明により包含される組成物を用いる、多様な治療法もまた意図される。本発明は、治療的に有効量のBTL−IIタンパク質、所望により可溶性BTL−IIタンパク質を投与することを含んでなる、炎症性腸疾患、所望によりクローン病かまたは潰瘍性大腸炎を患っている患者の腸管における炎症を軽減するための方法を提供する。治療的に有効量のBTL−IIアンタゴニストおよび抗原を投与することを含んでなる、全身性および/または粘膜免疫応答を含む、抗原に対する免疫応答を誘導するための方法もまた提供される。BTL−IIアンタゴニストは、抗体または小分子であることが可能であり、そして抗原は、粘膜表面に直接投与することが可能であり、若しくは全身的に投与することが可能である。さらに提供されるのは、BTL−II mRNAまたはタンパク質が過剰発現されているかどうかを決定するため、患者の腸管からの組織試料をアッセイすることを含んでなる、炎症性腸疾患を診断するまたは炎症性腸疾患の発症を予測するための方法である。組織は、抗BTL−II抗体を使用して、BTL−IIタンパク質発現をアッセイすることが可能である。本発明はさらに、抗原および可溶性BTL−IIタンパク質を同時投与することを含んでなる、抗原、特に自己免疫または炎症性疾患を患っている患者における自己抗原に対する免疫応答の勢いを弱めるための方法も提供する。抗原は、粘膜表面を経て投与することが可能である。
【0019】
さらなる態様において、本発明はT細胞の増殖およびサイトカイン産生を阻害するための方法を包含する。1つの態様において、本発明は、T細胞とBTL−IIポリペプチドを接触させることを含んでなる、T細胞増殖を阻害する阻害するために方法を含んでなる。T細胞はヒトT細胞であることが可能であり、そしてインビボでBTL−IIポリペプチドと接触させることが可能である。BTL−IIポリペプチドの代替物として、T細胞上に発現されたBTL−IIレセプターへ結合するアゴニスト性抗体(それがT細胞の増殖を阻害することが可能であれば)を使用することが可能である。別の態様において、本発明は、T細胞とBTL−IIポリペプチドを接触させることを含んでなる、T細胞によるサイトカイン産生を抑制するための方法を含んでいる。T細胞はヒトT細胞であることが可能であり、そしてインビボでBTL−IIポリペプチドと接触させることが可能である。サイトカインは、例えば、インターフェロンガンマ(IFNγ)、インターロイキン2(IL2)またはインターロイキン5(IL5)であることが可能である。
【課題を解決するための手段】
【0020】
配列表の簡単な説明
配列番号1は、寄託番号NM_019602でのアメリカ国立バイオテクノロジー情報センター(NCBI)エントリーからの、ヒトBTL−II cDNAのヌクレオチド配列である。
【0021】
配列番号2は、寄託番号NM_019602でのNCBIエントリーのcDNA配列から予測された、ヒトBTL−IIタンパク質のアミノ酸配列である。
配列番号3は、本発明の完全長ヒトBTL−II cDNAのヌクレオチド配列である。
【0022】
配列番号4は、配列番号3によりコードされた完全長ヒトBTL−IIタンパク質のアミノ酸配列である。
配列番号5は、本発明の完全長マウスBTL−II cDNAのヌクレオチド配列である。
【0023】
配列番号6は、配列番号5によりコードされた完全長マウスBTL−IIタンパク質のアミノ酸配列である。
配列番号7は、ヒトBTL−IIスプライス変異体の第一のカテゴリーの代表的メンバーからの、cDNAのヌクレオチド配列である(図3a)。
【0024】
配列番号8は、配列番号7によりコードされたアミノ酸配列である。
配列番号9は、ヒトBTL−IIスプライス変異体の第二のカテゴリーの代表的メンバーからの、cDNAのヌクレオチド配列である(図4a)。
【0025】
配列番号10は、配列番号9によりコードされたアミノ酸配列である。
配列番号11は、ヒトBTL−IIスプライス変異体の第三のカテゴリーの代表的メンバーからの、cDNAの部分ヌクレオチド配列である(図5a)。
【0026】
配列番号12は、配列番号11によりコードされたアミノ酸配列である。
配列番号13は、ヒトBTL−IIスプライス変異体の第四のカテゴリーの代表的メンバーからの、cDNAの部分ヌクレオチド配列である(図6a)。
【0027】
配列番号14は、配列番号13によりコードされたアミノ酸配列である。
配列番号15は、ヒトBTL−IIスプライス変異体の第五のカテゴリーの代表的メンバーからの、cDNAの部分ヌクレオチド配列である(図7a)。
【0028】
配列番号16は、配列番号15によりコードされたアミノ酸配列である。
配列番号17は、マウスBTL−IIスプライス変異体の第一のカテゴリーの代表的メンバーのヌクレオチド配列である(図9a)。
【0029】
配列番号18は、配列番号17によりコードされたアミノ酸配列である。
配列番号19は、実施例5に記載したBTL−II:Fc融合タンパク質をコードしているヌクレオチド配列である。
【0030】
配列番号20は、実施例5に記載したBTL−II:Fc融合タンパク質のアミノ酸配列である。
【0031】
発明の詳細な説明
本発明は、アゴニストまたはアンタゴニストであることが可能である、BTL−IIタンパク質、抗BTL−II抗体をコードしている組換えベクターを含む、BTL−IIタンパク質および核酸、ならびに、これらの分子を産生するそして使用するための方法、およびこれらを含んでいる医薬組成物を提供する。BTL−II発現は少数の組織型に限定されている。実施例4に例示されているように、BTL−IIは、マウス炎症性腸疾患モデルシステムにおいて、症状の発症に先立って、および症状段階の間に、腸管中で過剰発現される。それ故、BTL−II抗体は炎症性腸疾患を診断するため、若しくは発症の可能性を予測するために働くことが可能である。加えて、本発明はBTL−IIヌクレオチド配列の多数のアリル(allelic)変異体を提供する(図5aから7a)。これらは、炎症性腸疾患に対する感受性の予測における効用を見出すことが可能である。さらに、可溶性BTL−IIタンパク質はT細胞増殖およびサイトカイン産生を阻害することが可能であるので(実施例6−10)、BTL−IIタンパク質は自己免疫および炎症性疾患の処置に効用を見出すことが可能である。
【0032】
さらに、BTL−IIは、腸管における免疫試料採取に役割を演じていることが知られている特殊化した構造であるパイエル板で発現される。BTL−IIが好ましく発現されるのは、パイエル板に観察される他の細胞と比較して、他の樹状細胞を含んでも、パイエル板に観察されるCD11c+(低発現)CD8+B220+樹状細胞(形質細胞様樹状細胞とも称される)である。パイエル板樹状細胞は、T細胞分化に対するその影響力のため、粘膜表面での免疫寛容の誘導において役割を演じているとの仮説が立てられている。Weiner(2001),Nature Immunology 2(8):671−71;Weiner(2001),Immunol.Rev.182:207−14;IwasakiおよびKe1sall(1999),American Journal of Physiology−Gastrointestinal and Liver Physiology 276(5):G1074−78。それ故、抗BTL−II抗体は、パイエル板内のCD11c+(低発現)CD8+B220+樹状細胞を同定するために使用することが可能である。
【0033】
以下に説明するように、BTL−IIは、TおよびB細胞仲介応答の調節に役割を演じるブチロフィリン様タンパク質のB7サブファミリー内に存在する。BTL−IIは、特に腸管において、免疫システム仲介炎症の勢いを弱めるか、および/または促進することに役割を演じることができる。免疫システムの複雑さを仮定すれば、単一の細胞表面タンパク質は、ある場合には、エフェクター細胞上のいかなるレセプターが相互作用する分子に利用可能であるかに依存して、免疫エフェクター細胞による免疫応答を刺激する若しくは勢いを弱めるの両方を行うことができる。以下に議論されるB7−1およびB7−2タンパク質は、免疫エフェクター細胞、この場合T細胞に刺激性および勢いを弱める両方の効果を有する免疫調節細胞表面タンパク質の例である。この故に、BTL−IIはインビボで同様の二重の役割を果たすことができる。しかしながら、腸管は、食品抗原および共生微生物に対するその免疫寛容により証明されるように、全体的にみて外来抗原に対して高度に免疫寛容である。それ故、BTL−IIは、少なくともいくつかの状況において、インビボで免疫応答または炎症を弱めることに役割を演じることがありそうである。それ故、インビトロで選択されたBTL−IIアンタゴニスト(抗体を含むことが可能)結合タンパク質若しくは小分子は、抗原に対する粘膜免疫応答を刺激するように働くことができる。さらに、可溶性BTL−IIタンパク質、または機能的に均等な抗イディオタイプ抗体は、腸管における、または肺のごとき体内の他の粘膜表面における免疫応答を弱めることができる。
【0034】
本明細書において、“抗体”は、キメラ抗体であることが可能であり、単量体または一本鎖の、二量体の、三量体の、四量体のまたは多量体の抗体であることが可能であり、そして組換えタンパク質または非組換えタンパク質であることが可能である。“ドメイン”とは、一次配列モチーフおよび/または三次構造特性により区別することが可能な、タンパク質の一部または全てである。タンパク質ドメインを位置づけるために設計されたプログラムには、例えば、Pfam(Batemanら(1999),Nucleic Acids Res.27:260−62;Batemanら(1999),Nucleic Acids Res.28:263−66)、ProDom(Corpetら(1999),Nucleic Acids Res.27:263−67;Corpetら(1999),Nucleic Acids Res.28:267−69)、Domo(GracyおよびArgos(1998),Bioinformatics 14:164−87)およびSMART(Pontingら(1999),Nucleic Acids Res.27:229−32)が含まれる。三次構造は、例えば、X−線結晶学により経験的に決定することが可能であり、若しくはこうした使用のために設計されたコンピューターソフトウェアーを使用して予測することが可能である。例えば、構造データは、Molecular Modeling DatabaseからのNCBIのEntrezウェブサイトを通して(Wangら(2000),Nucleic Acids Res.28(1):243−45)、またはDALI(HolmおよびSander(1993),J.Mol.Biol.233:123−38)のごときソフトウェアーの使用により、アクセスすることが可能である。例えば、免疫グロブリン様ドメイン(Ig様)は、一次配列相同性よりもむしろ、その三次構造により主として区別される。例えば、Borkら(1994),J.Mol.Biol.242:309−20;HunkapillerおよびHood(1989),Adv.Immunol.44:1−63;WilliamsおよびBarclay(1988),Ann.Rev.Immunol.6:381−405、を参照されたい。しかしながら、IgVおよびIgCドメインは、それらの一次アミノ酸配列内の保存位置に出現するほんの少しの高度に保存されたアミノ酸を実際に含んでいる。例えば、Kabatら(1991)、免疫学的に興味が持たれるタンパク質の配列、米国保健福祉省、公衆衛生サービス、国立保健研究所、NIH出版番号91−3242、を参照されたい。適切な間隔で出現しているこうした高度に保存されたアミノ酸の存在は、IgC様またはIgV様ドメインの存在を示すことが可能である。
【0035】
タンパク質を“コードする”核酸とは、本明細書において、核酸またはその相補体がタンパク質をコードしているコドンを含んでなる場合を意味している。
タンパク質の発現を可能にする組換え核酸配列が、ウイルス感染、トランスフェクション、形質転換または電気穿孔法のごとき“遺伝子操作”の方法を使用して、細胞内へ導入された場合、細胞は特異的タンパク質を発現するように“遺伝子操作され”ている。例えば、Kaufmanら(1990),Meth.Enzymol.185:487−511、を参照されたい。このことは、例えば、細胞内へのタンパク質をコードしている核酸の導入、またはChappelの米国特許第5,272,071号に記載されているような、タンパク質をコードしている宿主遺伝子の発現を促進するための、調節配列の導入を含むことが可能である。“遺伝子操作”の方法は、限定するわけではないが、ポリメラーゼ連鎖反応を使用する核酸の増幅、大腸菌(E.coli)へそれらをクローン化することによる組換えDNA分子の組み立て、核酸の制限酵素消化、核酸のライゲーション、核酸のインビトロ合成、そして核酸の末端への塩基の移送、なかでも当該技術分野でよく知られている多数の方法、を含む多数の方法を包含している。例えば、Sambrookら,Molecular Cloning:A Laboratory Manual,第2版,1−3巻,Cold Spring Harbor Laboratory,1989、を参照されたい。
【0036】
“異種ポリペプチド”とは、本明細書で意味しているようなBTL−IIポリペプチドではなく、少なくとも3アミノ酸長である、任意のポリペプチドである。
ポリヌクレオチドまたはポリペプチドの配列同一性を決定するための比較に関連して、“同一性領域”により意味されることは、以下に述べたパラメーターを使用するコンピュータープログラムGAP(Devereuxら(1984),Nucleic Acids Res.12:387−95)により、別のポリヌクレオチドまたはポリペプチドと一致した(部分的にまたは正確に)ポリヌクレオチドまたはポリペプチドの一部である。例えば、20アミノ酸のポリペプチドが相当により長いタンパク質と並列された場合、最初の10アミノ酸がより長いタンパク質と正確に一致し、そして最後の10アミノ酸はより長いタンパク質と少しも一致しないとすれば、同一性領域は10アミノ酸である。一方、もし20アミノ酸ポリペプチドの最初および最後のアミノ酸がより長いタンパク質と一致し、そして八つの他の一致が間に散在すると、同一性領域は20アミノ酸である。しかしながら、少なくとも、例えば、20アミノ酸または60ヌクレオチドの同一のまたは保存的に置換されたアミノ酸または同一のヌクレオチドを有しない、並列された両鎖における長い伸展は、本明細書で意味するような同一性領域の終点を構成する。
【0037】
“Ig様”ドメインは免疫グロブリン様ドメインであり、そしてIgV様かまたはIgC様ドメインであってもよいし、または免疫グロブリン構造内へ折り畳むことが可能であるが、しかしIgV様かまたはIgC様としては明白に分類できないドメインであってもよい。
【0038】
“IgV様”ドメインは、免疫グロブリン可変領域ドメインに共通の特性で免疫グロブリン内へ折り畳むことが可能である、アミノ酸配列を有している。Borkら,上記文献;Millerら(1991),Proc.Natl.Acad.Sci.USA 88:4377−81;WilliamsおよびBarclay(1988),Ann.Rev.Immunol.6:381−405、を参照されたい。当業者は、保存された位置でのIgVドメイン中の高度に保存されたアミノ酸の存在から、ドメインをIgV様として同定することが可能であることを承知している。
【0039】
“IgC様”ドメインは、免疫グロブリン定常領域ドメインに共通の特性で免疫グロブリン内へ折り畳むことが可能である、アミノ酸配列を有している。Borkら,上記文献;WilliamsおよびBarclay,上記文献、を参照されたい。当業者は、保存された位置でのIgCドメイン中の高度に保存されたアミノ酸の存在から、ドメインをIgC様として同定することが可能であることを承知している。
【0040】
“炎症性腸疾患”は、クローン病、潰瘍性大腸炎、回腸炎、および胃腸管の慢性炎症により特徴付けられる他の疾患を含んでいる。
タンパク質は、二つのIg様ドメインを含み、そして別のIg様ドメインの、全てのまたはいくつかの認識可能な部分を含んでいない場合、“わずか2のまたはそれより少ないIg様ドメイン”を含んでなる。しかしながら、こうしたタンパク質は、Ig様ドメインではない他のアミノ酸配列、および“わずか2のまたはそれより少ないIg様ドメイン”をなお含むことが可能である。それ故、句“ほどのまたはそれより少ない”とは、Ig様ドメインのみを示し、タンパク質の一部であることができる他のアミノ酸配列を示してはいない。
【0041】
ポリペプチドが“T細胞の増殖を阻害する”ことが可能である、またはいくつかの他の生物学的機能を実行することが可能であると言われる場合、それはポリペプチドを含んでなるタンパク質が機能を実行できること、そして、生物学的機能を実行できない、少なくともいくつかのタンパク質へのポリペプチドの添加により、これらのタンパク質が機能を実行するのを可能にすることを意味している。いくつかの場合、生物学的機能を実行することが可能なポリペプチドは、追加の配列なしでそうすることが可能である。他の場合、生物学的機能を実行するには、他の配列(例えば、オリゴマー化配列)を必要とすることができる。一つのシナリオにおいて、一つのポリペプチドがFc領域またはロイシンジッパーまたはいくつかの二量体化ドメインに連結された場合、それは特定の生物学的機能を効率的に実行することが可能であるが、二量体化ドメインなしでは実行できない。本明細書中の意味として、そのようなポリペプチドは生物学的機能を実行することが可能である。
【0042】
“タンパク質”とは、少なくとも10アミノ酸、所望により少なくとも20、30、40、50、60、80、100、150、200、250および/または300アミノ酸を含んでなる任意のポリペプチドである。
【0043】
ポリペプチドまたはタンパク質に適用されるような“組換え”とは、タンパク質の産生が、核酸(タンパク質をコードしていてもしていなくてもよい)を、それらが天然には観察されない細胞内へ導入する、少なくとも一つの工程に依存していることを意味している。
【0044】
“組換え融合タンパク質”とは、天然には一緒に融合されることが観察されない、単一のポリペプチド鎖内へ融合された、少なくとも二つのタンパク質の一部または全てを含んでなる組換えタンパク質である。
【0045】
核酸配列における“サイレント(silent)突然変異”とは、核酸によりコードされたタンパク質の配列を変化させることなく、核酸の配列を変化させることである。
“可溶性”タンパク質とは、膜貫通ドメイン、または通常はタンパク質が膜へ埋め込まれるまたは膜に会合されることを引き起こす、GPIアンカー配列のごときいくつかの他のアミノ酸配列を欠いているタンパク質である。こうしたタンパク質は典型的には、膜貫通タンパク質の細胞外領域の全てまたは一部を含んでなることができる。
【0046】
本発明の目的のためには、二つのタンパク質または核酸は、もしそれらがアミノ酸または核酸配列において、少なくとも80%、所望により少なくとも85%、90%、95%、97%、98%、99%、99.5%または99.7%お互いに同一であり、そして非改変タンパク質の生物学的機能を維持しまたは望ましい様式に改変していれば、“実質的に類似”している。二つのアミノ酸または二つの核酸配列のパーセント同一性は、視覚的な点検および数学的計算により決定することが可能であり、またはより好ましくは、コンピュータープログラムを使用して配列情報を比較することにより比較を行う。コンピュータープログラムの例はGenetics Computer Group(GCG;Madison,WI)Wisconsinパッケージバージョン10.0プログラム,GAP(Devereuxら(1984),Nucleic Acids Res.12:387−95)である。GAPプログラムのための好ましいデフォルトパラメーターには以下のものが含まれる:(1)ヌクレオチドに関する単一(unary)比較マトリックス(同一に対し1および非同一に対し0の値を含む)、および「Atlas of Polypeptide Sequence and Structure」,SchwartzおよびDayhoff監修,National Biomedical Research Foundation,pp.353−358(1979)に記載されているような、GribskovおよびBurgess((1986)Nucleic Acids Res.14:6745)の加重アミノ酸比較マトリックス、または他の比較可能な比較マトリックスでのGCG実行;(2)各ギャップに対する8のペナルティおよびアミノ酸配列に対する各ギャップ中の各記号に対しさらに2のペナルティ、または各ギャップに対する50のペナルティおよびヌクレオチド配列に対する各ギャップ中の各記号に対しさらに3のペナルティ;(3)末端ギャップに対するペナルティなし、および(4)長いギャップに対し最大ペナルティなし。配列比較の分野で当業者により使用される他のプログラムもまた使用可能である。
【0047】
ブチロフィリン様タンパク質
現在まで報告されているブチロフィリン様タンパク質は、いくつかの構造特性を共有しており、そして多くは主要組織適合性座位(MHC)内にまたは隣接してコードされている。例えば、Henryら(1999),Immunology Today 20(6):285−88、を参照されたい。ブチロフィリンは、ウシミルクの脂肪小球に会合されている全タンパク質の40%を構成するタンパク質であり、そしてヒト相同体が存在する。Ruddyら(1997),Genome Research 7:441−56,JackおよびMather(1990),J.Biol.Chem.265:14481−86を引用している。ブチロフィリン様タンパク質間のアミノ酸配列レベルでの類似性は低くてもよいが、しかしドメイン構造は幾分保存されている。全てのメンバーは、通常IgV様ドメインであると報告されている、Ig様ドメインが続いたアミノ末端シグナルペプチドを含んでなる。多くの場合において、通常IgC様ドメインであると報告されている、別のIg様ドメイン、膜貫通ドメイン、そして細胞質ドメインが続いている。BTL−IIまたはB7−H3におけるように、二つのIg様ドメインはそれらの最初の出現に続いて直ちに反復されることが可能である。このドメイン構造は図1に例示されている。図1に図示されたタンパク質のほとんどは、ブチロフィリン様タンパク質のB7サブファミリーのメンバーである;三つ(ヒトBTN2A1、ヒトブチロフィリンおよびミエリンオリゴデンドロサイト糖タンパク質)のみがそうではない。
【0048】
Ig様ドメインは、その配列は非常に多岐にわたることが可能であるが、四つのβ鎖を含んでなる共通構造コアを全てが含む、多数の保存された折り畳みパターンの一つをそれでも保持している。免疫グロブリン定常および可変領域は、バレル(barrel)様形状を形成している、7から9の逆平行βストランドにより特徴付けられる三次構造を有している。Borkら(1994),J.Mol.Biol.242:309−20。IgVおよびIgC様免疫グロブリンドメインは各々、特有で、高度な保存残基をほんの少し含んでいる。HunkapillerおよびHood(1989),Adv.Immunol.44:1−63;Millerら(1991),Proc.Natl.Acad.Sci.USA 88:4377−81;WilliamsおよびBarclay(1988),Ann.Rev.Immunol.6:381−405。保存残基は多分構造または機能のために重要である。こうした保存残基は、BTL−IIのIg様ドメイン中に観察される。例えば、ヒトBTL−IIタンパク質の第一のIg様ドメイン(配列番号4)は、IgV様ドメインに特徴的な、こうした高度に保存された残基を、適切な位置、例えば、G43、C50、W65、L109、D118、G120、Y122およびC124位に含んでいる。表3を参照されたい。第三のIg様ドメインもまた、IgV様ドメインで保存されている残基に対応する残基を含んでいる。ヒトBTL−IIの第二および第四のIg様ドメインは、IgC様ドメインで高度に保存されている残基に対応する残基を含んでいる。表3。
【0049】
膜貫通および細胞質ドメインは、ブチロフィリン様タンパク質が第二のIg様ドメインを有しているかどうかに関わらず、ブチロフィリン様タンパク質中に出現する。膜貫通ドメインには、αへリックスコイルドコイル(coiled coil)モチーフに典型的な7アミノ酸繰り返しを思い出させる、一つまたはそれより多くの7アミノ酸単位が続いていてもよいし、続いていなくてもよい。7アミノ酸繰り返しはまた、ブチロフィリン様タンパク質中の別の位置に出現することもできる。7アミノ酸繰り返しの議論については、Millerら(1991),Proc.Natl.Acad.Sci.88:4377−81、を参照されたい。膜貫通ドメインを予測する方法は、当該技術分野でよく知られている。例えば、Ikedaら(2002),In Silico Biol.2(1):19−33;TusnadyおよびSimon(1998),J.Mol.Biol.283(2):489−506、を参照されたい。ブチロフィリン様タンパク質は細胞質ドメインを有することが可能である。こうした細胞質ドメインは、B30.2ドメインを含んでいても、または含んでいなくてもよい。多様なB30.2ドメインの配列が示されており、そしてB30.2ドメインの推定機能がHenryら((1998),Mol.Biol.Evol.15(12):1696−1705)により議論されている。B30.2ドメインはいろいろなタンパク質に観察され、そしてB30.2ドメインの機能は未知である。提案されたB30.2ドメインのリガンドはキサンチンオキシダーゼであり、それはブチロフィリンの細胞質ドメインと相互作用する。二つの異なったB30.2含有タンパク質のB30.2ドメインにおける突然変異は、二つの異なった疾患と関連付けられたが、突然変異および疾患表現型間の因果関係は確立されていない。Henryら、上記文献。
【0050】
ブチロフィリン様タンパク質のB7サブファミリー
BTL−IIは、例えば、T細胞、B細胞および骨髄細胞のごとき、免疫エフェクター細胞の調節に役割を演じる、B30.2ドメインを欠く多数のブチロフィリン様免疫調節タンパク質とドメイン構造を共有している。このサブファミリーは、ブチロフィリン様タンパク質の“B7サブファミリー”として本明細書において称される。本明細書で意味しているように、B7サブファミリーメンバーの特性には、限定ではなく、以下の項目が含まれる:(1)一つまたはそれより多くの細胞外Ig様ドメインを有し;(2)膜貫通および細胞質ドメインを有し;(3)B30.2ドメインを欠き;(4)抗原提示細胞上で発現されており;(5)活性化された免疫応答の間に発現の調節を起こし;そして(6)免疫応答を変調する。免疫応答を変調する、膜貫通ドメインを欠く、分泌、可溶性B7タンパク質は現在まで報告されていない。ほとんどのB7タンパク質は二つの細胞外Ig様ドメインを有しているが、B7−H3のいくつかのアイソフォームは四つ有している。図1を参照されたい。ヒトBTL−IIは、2から4の細胞外Ig様ドメインを有することが可能で、そして上にリストした、B7ファミリーメンバーの特性の全てを有している。それ故、我々はそれがブチロフィリン様タンパク質のB7サブファミリーのメンバーであると考えた。B7サブファミリーの全ての他の既知メンバーは、B細胞および/またはT細胞のごとき免疫エフェクター細胞上のレセプターと相互作用し、その相互作用は免疫応答を変調するためのシグナルとして働く。それ故、BTL−IIは、T細胞のごとき免疫エフェクター細胞上のレセプターと相互作用し、それ故免疫応答を変調すると予想される。
【0051】
B7ファミリーメンバーは免疫エフェクター細胞、特にT細胞の活性を変調することに役割を演じている。Henryら(1999),Immunology Today 20(6):285−88;SharpeおよびFreeman(2002),Nat.Rev.Immunol.2:116−26。最もよく特徴付けられた例は、CD80およびCD86(各々B7−1およびB7−2とも称される)であり、それらは抗原提示細胞上で発現され、T細胞の表面上に構成的に発現されるCD28と相互作用した場合、T細胞活性化を促進可能であり、またはT細胞の表面上にまた発現されるCTLA−4と相互作用した場合、T細胞活性化を阻害可能である。CTLA−4発現は構成的ではないが、しかしT細胞活性化に続いて迅速に上方調節される。例えば、Mastellerら(2000),J.Immunol.164:5319−27;Hehnerら(2000),J.Biol.Chem.275(24):18160−71;SharpeおよびFreeman(2002),Nat.Rev.Immunol.2:116−26、を参照されたい。さらに、CD80の両方のIg様ドメイン中の、多数の個々のアミノ酸残基がCTLA−4およびCD28への結合に重要であることが報告されている。Peachら(1995),J.Biol.Chem.270(36):21181−87。CD86は構成的に低レベルで発現され、そして活性化後に迅速に上方調節され、一方、CD80は、活性化後に誘導的に発現される。SharpeおよびFreeman(2002),Nature Reviews Immunology 2:116−26。
【0052】
本明細書においてB7RP−1と称される、別のT細胞調節分子は多血症(plethora)の名前を有している、KIAA0653(Ishikawaら(1998),DNA Res.5:169−76)、B7h(Swallowら(1999),Immunity 11:423−32)、GL50(Lingら(2000),J.Immunol.164:1653−57)、B7RP−1(Yoshinagaら(1999),Nature 402:827−32)、LICOS(Brodieら(2000),Curr.Biol.10:333−36)、B7−H2(Wangら(2000),Blood 96:2808−13)およびICOSL(SharpeおよびFreeman,上記文献)。B7RP−1は末梢リンパ組織、脾臓、リンパ節、肺、胸腺、脾細胞およびB細胞中で発現される。B7RP−1とT細胞の相互作用は、T細胞増殖およびサイトカイン産生を増加させることが可能である。B7RP−1は、活性化T細胞上に発現されるICOSレセプターを通してT細胞にシグナルを伝達する。Yoshinagaら(1999),Nature 402:827−32;Swallowら(1999),Immunity 11:423−32。B7RP−1は非刺激B細胞株で構成的に発現され、そしてその発現は、インターフェロンγにより単球中で誘導することが可能である。Aicherら(2000),J.Immunol.164:4689−96。調節T細胞の発生および調節機能は、B7RP−1およびT細胞上のそのレセプター間の相互作用に依存している。Akbariら(2002),Nature Medicine 8(9):1024−32。
【0053】
さらに、三つの他のT細胞調節分子、PD−L1(B7−H1とも称される)、PD−L2(B7−DCとも称される)およびB7−H4(B7S1およびB7xとも称される)は、T細胞増殖およびサイトカイン産生を阻害することが可能である。PD−L1およびPD−L2は、T細胞、B細胞および骨髄細胞上に発現される、それらの共通のレセプター、PD−1を通して作用する。Freemanら(2000),J.Exp.Med.192(7):1027−34;Dongら(1999),Nature Medicine 5(12):1365−69;Latchmanら(2001),Nature Immunology 2(3):261−68;Tamuraら(2001),Blood 97(6):1809−16;およびTsengら(2001),J.Exp.Med.193(7):839−45。PD−L1およびPD−L2の発現は、一般的な前炎症性サイトカインである、インターフェロンγにより誘導することが可能である。Latchmanら,上記文献。B7−H4はB細胞、マクロファージおよび樹状細胞上で発現され、BおよびT細胞上に発現される阻害的レセプターである、BTLAを通して作用するようである。Watanabeら(2003),Nature Immunology 4(7):670−79;Zangら(2003),Proc.Natl.Acad.Sci.100(18):10388−92;Sicaら(2003),Immunity 18:849−61;Prasadら(2003),Immunity 18:863−73;CarrenoおよびCollins(2003),Trends Immunol.24(10):524−27。
【0054】
T細胞を刺激することにおいて、同時刺激性役割を果たすさらに別のブチロフィリン様タンパク質はB7−H3である。Chapovalら(2001),Nature Immunol.2(3):269−74。他のB7ファミリーメンバーと同様に、B7−H3はシグナル配列、細胞外Ig様ドメイン、膜貫通ドメインおよび細胞質ドメインを有している。ヒトB7−H3遺伝子は、二つまたは四つの細胞外Ig様ドメインを有するアイソフォームをコードしている。Sunら(2002),J.Immunol.168:6294−97。B7−H3の発現は、炎症性サイトカインにより、樹状細胞上で誘導することが可能である。B7−H3は、T細胞の増殖および細胞毒性応答を刺激することが可能である。B7−H3は、CD28、CTLA−4、ICOSおよびPD−1とは異なる推定上のT細胞レセプターを通して作用する。Chapovalら、上記文献。
【0055】
BTL−IIタンパク質
ヒトおよびマウスBTL−IIタンパク質の存在は、ゲノム配列から予想されていた。Stammersら(2000),Immunogenetics 51:373−82。しかしながら、これらの著者は、ゲノム分析に基づいた、ヒトまたはマウスBTL−IIタンパク質中の膜貫通または細胞質ドメインの証拠、そしてマウスBTL−II mRNAを検出するために設計されたPCR実験において、エキソン1−4(各々、シグナルペプチド、二つのIg様ドメインおよび7アミノ酸繰り返し領域をコードする)とエキソン5および6(各々、別の二つのIg様ドメインをコードする)を連結している転写体の証拠を見出していない。Stammersら、上記文献。これらの知見に基づき、BTL−IIは、細胞表面、免疫変調タンパク質のB7サブファミリーには加えられなかった。これら同一の著者による、公共データベースへのその後の配列提出は、455アミノ酸をコードしているエキソン5および6を含んでおり、膜貫通ドメインおよび細胞質ドメインを欠いている、1368ヌクレオチドのヒトBTL−II mRNAを予測している(配列番号1(ヒトBTL−II cDNA配列)および配列番号2(ヒトBTL−IIタンパク質配列)を開示している、NCBI寄託番号第NM_019602号)。
【0056】
本発明のBTL−II核酸およびタンパク質配列は、多くの点でこれらの配列と異なっている。cDNA配列は、膜貫通または細胞質ドメインを含んでいない、以前に報告されているBTL−IIタンパク質配列とは違って、シグナル配列、細胞外ドメイン、膜貫通ドメイン、および細胞質ドメインをコードしている。BTL−IIの発現パターンと一緒に加えて、これらの特性のため、BTL−IIはブチロフィリン様タンパク質のB7サブファミリー内に加えられる。表1(下記)は、本発明のBTL−IIタンパク質および以前に報告されている配列間の相違を強調している。本発明のBTL−IIタンパク質は上段に示されており(配列番号4)、そしてNCBI寄託番号第NM_019602号に報告されているBTL−IIタンパク質は下段に示されている(配列番号2)。この比較から、二つの配列間には三つのミスマッチがあり(360、454および455位に)、そして配列番号4は、細胞外ドメインの追加の配列ならびに膜貫通および細胞質ドメインを構成する、さらに27のアミノ酸を有している。これらの配列は、上に列挙したパラメーターを使用するGAPプログラムに従うと、99.3%同一である。
【0057】
【表1】

【0058】
図1に例示したようなドメイン構造における総合的な類似性に加え、B7サブファミリータンパク質は、一次配列レベルでの類似性を有している。例えば、コンピュータープログラムGAPを使用して、対様式で並列した場合、ヒトBTL−IIタンパク質配列(配列番号4)は、下記表2に示されているように、他のB7サブファミリーメンバーに類似している。
【0059】
【表2】
【0060】
当業者は、任意のこれらの並列で保存されている残基は、保存されていない残基よりも、ヒトBTL−IIの構造または機能においておそらく本質的役割を演じることを了解するであろう。
【0061】
表3(下記)において、ヒトBTL−IIアミノ酸配列(配列番号4、上段)とマウスBTL−IIアミノ酸配列(配列番号6、下段)が並列されている。同一アミノ酸は垂直線で結ばれており、類似のアミノ酸は、それらの間に一つのまたは二つのドットを有している。GAP(上記)により決定された、これらの配列間のパーセント同一性は約62%であり、そしてパーセント類似性は約68%である。IgVまたはIgC様ドメイン、またはいわゆるIgV免疫グロブリンスーパーファミリーメンバーの“Iセット”中に観察される残基、またはこうした残基の保存的置換はボールド体で示されている。Peachら、上記文献;HarpazおよびChothia(1994),J.Mol.Biol.238:528−39。こうした残基は構造的に重要であるようであり、それ故、機能的効果を有することができる。適正な間隔をあけたこうしたアミノ酸のかなりの数の出現で、配列をIgV様またはIgC様として同定可能である。
【0062】
【表3】
【0063】
構造は配列よりも進化においてより保存されているので、非保存残基は保存残基よりも、BTL−IIタンパク質の全体の三次構造を決定することにおいて役割を演じそうもないことを当業者は理解するであろう。Borkら(1994),J.Mol.Biol.242:309−20。本明細書において、“非保存残基”とは、表3におけるように、ヒトおよびマウスBTL−IIタンパク質配列が比較された場合に保存されていない、BTL−IIタンパク質内のアミノ酸である。スプライス変異体によりコードされているBTL−IIタンパク質において、こうした残基は、配列内の異なった番号位置で出現するであろう。例えば、配列番号4の397位の非保存残基は、配列番号8の187位でみられる同一の非保存残基である。当業者は、タンパク質構造がタンパク質機能に影響可能であることを認識するであろう。さらに、非保存アミノ酸は、BTL−II機能において直接的な役割を演じることもありそうもない。例えば、残基4、6、25、26、35、36および多くの他の残基は、同一でもなくまたは類似してもいない。それ故、当業者は、こうした残基の改変がBTL−IIタンパク質機能に影響することは、保存されたまたは類似の残基の改変よりもありそうもないことを了解するであろう。さらに、保存的置換が、非保存的置換よりもタンパク質機能へ影響することは、よりありそうもないようである。生物学的活性へ影響することがありそうもない保存的置換であるアミノ酸置換の例には以下の置換が含まれる:Serに対するAla、Ileに対するVal、Gluに対するAsp、Serに対するThr、Glyに対するAla、Thrに対するAla、Asnに対するSer、Valに対するAla、Glyに対するSer、Pheに対するTyr、Proに対するAla、Argに対するLys、Asnに対するAsp、Ileに対するLeu、Valに対するLeu、Gluに対するAla、Glyに対するAsp、およびこれらの変化の逆。例えば、Neurathら,「The Proteins」,Academic Press,New York(1979)を参照されたい。さらに、グループ内の一つのアミノ酸の、同一のグループ内の別のアミノ酸への交換は保存的置換であり、ここでグループとは以下のようである:(1)アラニン、バリン、ロイシン、イソロイシン、メチオニン、ノルロイシンおよびフェニルアラニン;(2)ヒスチジン、アルギニン、リジン、グルタミンおよびアスパラギン;(3)アスパラギン酸およびグルタミン酸;(4)セリン、スレオニン、アラニン、チロシン、フェニルアラニン、トリプトファンおよびシステイン;(5)グリシン、プロリンおよびアラニン。
【0064】
ヒトBTL−IIタンパク質はいくつかの認識可能ドメインを含んでなる。第一は、配列番号4の残基1から、配列番号4の残基22から29付近までからの第二の位置に及ぶシグナル配列(エキソン1によりコードされている)である。次は、残基xから残基yに及ぶIg様ドメイン(エキソン2によりコードされている)であり、ここでxは配列番号4の残基22から32までからであり、そしてyは残基138から148までからである。これに続くのは、残基vから残基wに及ぶ別のIg様ドメイン(エキソン3によりコードされている)であり、ここでvは配列番号4の残基138から148までからであり、そしてwは残基232から242までからである。次は、残基tから残基uに及ぶ7アミノ酸反復領域(エキソン4によりコードされている)であり、ここでtは配列番号4の残基234から239までからであり、そしてuは残基240から247までからである。別のIg様領域が残基rから残基sに及んでおり、ここでrは配列番号4の残基240から247までからであり、そしてsは残基354から364までからである。第四のIg様領域が残基pから残基qに及んでおり、ここでpは配列番号4の残基355から365までからであり、そしてqは残基452から462までからである。ヒトBTL−IIのこれらの第一の6ドメインは、ヒトBTL−IIの細胞外領域を作り上げている。シグナル配列はタンパク質の分泌時にタンパク質の残り部分から切断されてもよいが、切断が必要とされるわけではない。膜貫通ドメインは残基nから残基oに及んでおり、ここでnは配列番号4の残基454から462までからであり、そしてoは残基473から481までからである。最後は、残基kから残基mに及ぶ細胞質ドメインであり、ここでkは配列番号4の残基474から481までからであり、そしてmは残基482付近である。
【0065】
マウスBTL−IIタンパク質は類似の組のドメインを含んでなる。第一は、配列番号6の残基1から始まり、残基20付近から残基27付近までからの位置で終わるシグナル配列である。第二は、残基xから残基yに及ぶIg様ドメインであり、ここでxは配列番号6の残基21から27までからであり、そしてyは残基138から148までからである。第三は、残基vから残基wの別のIg様ドメインであり、ここでvは配列番号6の残基139から148までからであり、そしてwは残基232から242までからである。7アミノ酸反復領域は残基tから残基uに及んでおり、ここでtは配列番号6の残基233から242までからであり、そしてuは残基238から248までからである。別のIg様領域が残基rからsに及んでおり、ここでrは配列番号6の残基239から248までからであり、そしてsは残基355から365までからである。最後のIg様領域が残基pからqに及んでおり、ここでpは配列番号6の残基356から365までからであり、そしてqは残基447から457までからである。これらの第一の6ドメインは、マウスBTL−IIの細胞外領域を作り上げている。膜貫通ドメインは残基nからoに及んでおり、ここでnは配列番号6の残基448から459までからであり、そしてoは残基470から478までからである。最後は、残基kからmに及ぶ細胞質ドメインであり、ここでは配列番号6の残基471から478までからであり、そしてmは残基514付近である。
【0066】
本発明はBTL−IIの分泌された、可溶性バージョン、ならびに、細胞表面上に発現することが可能である膜貫通ドメインを含んでなるバージョンを包含している。本発明はさらに、以下に記載するようなBTL−II核酸によりコードされたBTL−IIタンパク質も含んでいる。これらのタンパク質全ての組換え体バージョンは、本明細書で記載するように、抗体を産生するため、スクリーニングに、および/または治療剤として使用することが可能である。例えば、本発明は、配列番号4、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16および/または配列番号18の全て若しくは一部を含んでなるBTL−IIタンパク質を包含する。本発明のBTL−IIタンパク質は、一次アミノ酸配列の挿入、欠失、改変または置換により、配列番号4、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16または配列番号18とは異なったタンパク質を含んでいる。こうした変異体配列は、配列番号4、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16および/または配列番号18と少なくとも80%、85%、90%、95%、97%、98%、99%、99.5%または99.7%同一であり、そして上述の配列とGAPを使用して並列させた場合、10アミノ酸を超える内部ギャップを含んでいない。こうした配列の例には、図5b、6bおよび7bに示した、BTL−IIの天然に存在するヒトアリル(allelic)変異体が含まれる。もしこうした変異体配列が、配列番号4、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16および/または配列番号18と比較した場合にアミノ酸置換を含んでいれば、これらの置換は保存的アミノ酸置換であろう。さらに、変異体BTL−IIタンパク質は配列番号4、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16および/または配列番号18に関して、単一アミノ酸のわずか3、4、5、6、7、8、9、10、15、20または25の挿入、欠失または置換を含むことができる。
【0067】
本発明のBTL−IIタンパク質は、ヒトおよびマウスBTL−II mRNAの多様なスプライス変異体によりコードされているタンパク質を含む。こうした変異体ヒトBTL−IIタンパク質の配列は、配列番号8、配列番号10、配列番号12、配列番号14および配列番号16に示されている。変異体マウスBTL−IIタンパク質の配列は配列番号18に示されている。ヒトスプライス変異体は、エキソン3単独かまたはエキソン2および3の両方を欠いており、そして開示されているマウススプライス変異体は、エキソン3のみを欠いている。配列番号7、配列番号9、配列番号11、配列番号13、配列番号15および配列番号17;図3a、4a、5a、6a、7a、および9aを参照されたい。これらの配列および、10アミノ酸より長いギャップを含んでいないGAPを使用して、配列番号4、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16または配列番号18から成る群の少なくとも一つを有するタンパク質配列と並列した場合、実質的に類似の配列によりコードされているタンパク質は本発明に包含される。もしこれらのタンパク質が、配列番号4、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16および/または配列番号18と比較した場合にアミノ酸置換を含んでいれば、これらの置換は好ましくは保存的アミノ酸置換である。本発明のBTL−IIタンパク質は、T細胞の表面上のレセプターへ結合することが可能であり、そしてT細胞の増殖および/またはT細胞によるサイトカイン産生を阻害することが可能である。
【0068】
さらに、本発明はエキソン1および4(配列番号8および配列番号12)またはエキソン2および4(配列番号10、配列番号14、配列番号16および配列番号18)のスプライス接合部にまたがる核酸によりコードされているBTL−IIタンパク質、そしてT細胞の表面上に発現されたレセプターへ結合可能で、並びに/あるいは増殖および/またはT細胞によるサイトカイン産生を阻害可能な実質的に類似のタンパク質を提供する。これには特に、配列番号10、配列番号14または配列番号18のアミノ酸127から157、または配列番号16のアミノ酸126から156と少なくとも80%、85%、90%、95%、96%、97%、98%、99%、99.7%または100%同一であるアミノ酸配列から成るポリペプチドを含んでなるBTL−IIタンパク質が含まれる。配列番号10、配列番号14または配列番号18のアミノ酸127から157、または配列番号16のアミノ酸126から156と並列されたアミノ酸配列の同一性領域は、好ましくは少なくとも20、23、25、27、30、35または40アミノ酸長である。こうしたアミノ酸配列は少なくとも150アミノ酸長であることが可能であり、そして配列番号10、配列番号14、配列番号16または配列番号18のアミノ酸30から358と少なくとも80%、85%、90%、95%、96%、97%、98%、99%、99.7%または100%同一であることが可能である。配列番号10、配列番号14、配列番号16または配列番号18のアミノ酸30から358と並列されたアミノ酸配列の同一性領域は、少なくとも50、75、100、125、150、175、200または300アミノ酸であることが可能である。
【0069】
本発明はまた、わずか2のまたはそれより少ないIg様ドメインを含み、そしてB細胞またはT細胞上に発現された細胞表面レセプターへ結合可能である、配列番号4のアミノ酸30から457と少なくとも80%、85%、90%、95%、96%、97%、98%、99%、99.7%または100%同一であるアミノ酸配列から成るポリペプチドを含んでなるBTL−IIタンパク質も提供する。アミノ酸配列は、配列番号8のアミノ酸30から247または配列番号12のアミノ酸30から243と少なくとも80%、85%、90%、95%、96%、97%、98%、99%、99.7%または100%同一であることが可能である。
【0070】
さらなる態様において、本発明は配列番号11のヌクレオチド34から124から成るポリヌクレオチドと80%、85%、90%、95%、96%、97%、98%、99%、99.7%または100%同一であるポリヌクレオチドを含んでなる核酸によりコードされているタンパク質を提供し、ここにおいて同一性領域は少なくとも60、70、80、90または100ヌクレオチド長である。こうしたポリヌクレオチドによりコードされている成熟BTL−IIタンパク質は、このポリヌクレオチドにより少なくとも一部がコードされているシグナル配列(タンパク質の未成熟バージョンに存在する)を欠いていてもよいが、それは必須ではない。こうしたタンパク質はT細胞へ結合することが可能であり、および/またはT細胞の増殖および/またはT細胞によるサイトカイン産生を阻害することが可能である。
【0071】
BTL−IIタンパク質は異なる程度までグリコシル化されていてもよいし、またはグリコシル化されていなくてもよい。例として、本発明のBTL−IIタンパク質は、配列番号4、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16または配列番号18を含んでなるタンパク質中にすでに観察されるものに加え、一つまたはそれより多くのNまたはO連結グリコシル化部位を含んでなることができる。こうしたBTL−IIタンパク質は、それに結合されたより多いシアル酸部分を有することができるので、非改変タンパク質よりも長いインビボ半減期を有することが可能である。BTL−IIタンパク質はまた、ヒトまたはマウスBTL−IIのIg様ドメインまたは実質的に類似のドメインのどれか一つ、どれか二つ、どれか三つまたは四つ全てを含んでなるタンパク質も含んでいる。
【0072】
変異体
任意の型の改変(例えば、限定されるわけではないが、アミノ酸の挿入、欠失または置換;ポリペプチドのグリコシル化状態の変化;その三次元構造または自己会合状態を変化させるための再折り畳みまたは異性化;および他のポリペプチドまたは分子とのその会合の変化)により任意のBTL−IIタンパク質から誘導されたポリペプチドもまた、本明細書において意味するBTL−IIタンパク質である。本発明により提供されるBTL−IIタンパク質には、T細胞の表面に発現したレセプターに結合することが可能であり、および/またはT細胞の増殖および/またはT細胞によるサイトカイン産生を阻害することが可能である、BTL−IIタンパク質配列番号4、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16および/または配列番号18のアミノ酸配列と実質的に類似しているアミノ酸配列により特徴付けられるポリペプチドが含まれる。同一性の領域は、配列番号4、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16または配列番号18の30またはより高い位置から出発することが可能である。配列番号4、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16および/または配列番号18の少なくとも一つを有するこうした変異体タンパク質のGAP並列は、10アミノ酸より長い内部ギャップを有しないことができる。配列番号4、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16または配列番号18と実質的に類似しているBTL−IIタンパク質の部分は、少なくとも100、少なくとも125、少なくとも150、少なくとも175、少なくとも200または少なくとも250アミノ酸長であることが可能である。こうしたタンパク質における修飾は、天然に提供されるか、または故意に操作される。例えば、配列番号11(図6a)は、複数の位置を含んだほんの少しの位置での塩基変化を含んでいる、配列番号9(図4a)のアリルまたは多型性変異体である。
【0073】
本発明は、配列番号4、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16または配列番号18の配列に関して、単一アミノ酸の挿入、欠失または置換であることが可能である、単一または多アミノ酸改変を含む、BTL−IIタンパク質変異体を提供し、ここにおいて、BTL−II変異体タンパク質は、T細胞増殖および/またはサイトカイン産生を阻害することが可能である。改変は保存的アミノ酸置換であり得る。例えば、表3に示したヒトおよびマウスBTL−IIタンパク質の並列により、機能に影響することなくどのアミノ酸を変化させることが可能であるかについて、当業者は導かれる。ヒトおよびマウスBTL−IIにおいて同一または類似しているアミノ酸は、そうではないアミノ酸より機能のために重要であるようである。さらに、IgVまたはIgCドメインにおいて高度に保存されているアミノ酸(表3においてボールド体で示されている)もまた、機能的に重要であるようである。こうしたアリルまたは多型性変異体は、疾患の素因を決定するための診断剤として有用であることが可能である。配列番号3とは異なった配列を有するヒトアリル変異体は、図5a、5b、6a、6b、7aおよび7bに開示されている。例えば、炎症性腸疾患の症状がない正常の人々間で、配列番号3の配列を有するcDNAをコードしているBTL−II遺伝子を有している数と、図5a、6aおよび7aにおいて3−6でラベルされたアリル変異体突然変異をコードしているBTL−II遺伝子を有している数の間には特定の比が存在することが可能である。炎症性腸疾患の症状を有する患者の大部分が、図5a、6aおよび7aにおいて3−6でラベルされたアリル変異体突然変異を伴ったBTL−II遺伝子を有するように、これらのアリルの出現比率を患者間で改変することができる。患者の組織における多様なアリル変異体の存在は、例えば、多型性部位に広がっているDNAまたはRNAのセグメントのPCR増幅により決定することが可能である。上述のように、特にヒトおよびマウスBTL−IIタンパク質配列間で保存されていない部位での保存的アミノ酸置換は、保存的部位での非保存的置換よりも生物学的機能を保つようである。当業者はまた、例えば、DALI(HolmおよびSander(1993),J.Mol.Biol.233:123−38)のごときプログラムにより予測されるような、BTL−IIタンパク質の三次構造を実質的にひっくり返す置換もまた、機能を害しそうであることを理解するであろう。
【0074】
断片(fragment)
配列番号4、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16または配列番号18の断片、またはこれらの配列と少なくとも80%、85%、90%、95%、97%、98%、99%、99.5%または99.7%同一である断片を含んでなるタンパク質は、BTL−IIタンパク質に含まれている。好ましくは、こうした断片はT細胞の表面上に発現されたレセプターへ結合することが可能であり、そして少なくとも約50、60、70、80、90、100、150または200アミノ酸長である。好ましくは、こうした断片は水性溶液に可溶であり、そしてBTL−IIタンパク質の細胞外領域一部または全てを含んでなる。こうした断片を含んでなるタンパク質は分泌されることが可能である。例えば、上記ヒトまたはマウスBTL−IIタンパク質または実質的に類似したタンパク質のドメインの少なくとも一つを含んでなるBTL−IIタンパク質は本発明に包含され、ここにおいて該ドメインはBTL−IIタンパク質の少なくとも一つの生物学的特性を有している。例えば、こうしたタンパク質はT細胞の増殖および/またはサイトカイン産生を阻害可能である。
【0075】
さらに本発明に包含されるのは、ヒトおよびマウスBTL−IIタンパク質または実質的に類似のタンパク質の、以下の改変バージョンである:(1)エキソン3によりコードされている第二のIg様ドメイン(図1参照)を欠いているヒトおよびマウスBTL−IIタンパク質のバージョン;(2)エキソン2および3によりコードされている第一および第二のIg様ドメインを欠いているヒトおよびマウスBTL−IIタンパク質のバージョン;(3)エキソン5および6によりコードされている第三および第四のIg様ドメインを欠いているバージョン;(4)四つのIg様ドメインの任意の二つを欠いているバージョン;および(5)Ig様ドメインの任意の一つを欠いているバージョン。
【0076】
また本発明内に包含されるのは、配列番号4、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16または配列番号18の免疫原性断片であり、それは断片へ特異的に結合する抗体を惹起することが可能である。こうした断片は好ましくは少なくとも10アミノ酸長であり、そして好ましくは、上述の配列からの隣接アミノ酸残基を含んでなる。こうした断片で免疫化された動物により産生された抗体は、本出願の別のところで議論されているように、炎症性腸疾患の予測、診断および処置に有用であることが可能である。こうした断片はスプライス接合部によりコードされているこれらのタンパク質の領域に及ぶことが可能であり、スプライス変異体または完全長タンパク質によりコードされているタンパク質に特異的な抗体の発生に利点を有している。あるいは、エキソン3によりコードされているBTL−IIの一部に対する抗体は、完全長BTL−IIタンパク質によってのみ検出され、エキソン3を欠く全ての他のスプライス変異体によりコードされているタンパク質では検出されない。
【0077】
組換え融合タンパク質
本発明はさらに、上述の少なくとも一つのBTL−IIポリペプチド(BTL−IIタンパク質の一つである)、変異体または断片、そして少なくとも一つの他の部分を含んでなる融合タンパク質を包含する。他の部分は、異種ポリペプチド、即ち、BTL−IIポリペプチド以外のポリペプチドであることが可能である。他の部分はまた、例えば、ポリエチレングリコール(PEG)部分、または細胞毒性、細胞分裂抑制性、発光および/または放射活性部分のごとき、非タンパク質部分であることが可能である。PEGの結合は、少なくともいくつかのタンパク質のインビボ半減期を増加させることが知られている。さらに、細胞毒性、細胞分裂抑制性、発光および/または放射活性部分は、例えば、抗体が結合可能な細胞に位置させるための、細胞の増殖を阻害するための、または細胞を殺すための、診断または治療目的のために抗体へ融合されてきた。同様に、こうした部分へ融合されたBTL−IIポリペプチドは、BTL−IIが結合する細胞に位置させるために、細胞の増殖を阻害するために、または細胞を殺すために使用することが可能である。こうした細胞毒性、細胞分裂抑制性、発光および/または放射活性部分は、例えば、マイタンシン誘導体(DM1のごとき)、エンテロトキシン(ブドウ球菌エンテロトキシンのごとき)、ヨウ素同位元素(ヨード−125のごとき)、テクネチウム同位元素(Tc−99mのごとき)、シアニン蛍光色素(Cy5.5.18のごとき)、リボソーム不活性化タンパク質(ブーガニン、ゲロニンまたはサポリン−S6のごとき)、および商標MYLOTARGTM(Wyeth−Ayerst)の名で売買されている製品の一部である、細胞毒性物質のカリケアミシン、である。
【0078】
多様な異種ポリペプチドを、例えば、タンパク質のインビボ半減期を増加させるため、タンパク質の同定、単離および/または精製を容易にするため、タンパク質の活性を増加させるため、そしてタンパク質のオリゴマー化を促進するためのごとき、多様な目的のためにBTL−IIポリペプチドへ融合することが可能である。例えば、CD80のごときB7サブファミリーのいくつかのタンパク質は、主としてオリゴマー形でそれらのレセプターへ結合するので(CD80の場合は二量体、Collinsら(2002),Immunity 17:201−10、を参照されたい)、オリゴマー化は可溶性タンパク質の生物学的活性を保存するためには非常に重要であろう。
【0079】
多くの異種ポリペプチドは、それらがその一部である組換え融合タンパク質の同定および/または精製を容易にすることを可能にする。例には、ポリアルギニン、ポリヒスチジン、または、固定化された金属イオンに対して高い親和性を所有する非隣接ヒスチジンの天然に存在する配列であるHATTM(Clontech)が含まれる。これらの異種ポリペプチドを含んでなるタンパク質は、例えば、固定化されたニッケル、または固定化されたコバルトイオンを含んでなるTALONTM樹脂(Clontech)を使用するアフィニティークロマトグラフィーにより精製することが可能である。例えば、Knolら(1996),J.Biol.Chem.27(26):15358−15366、を参照されたい。ポリアルギニンを含んでなる異種ポリペプチドは、イオン交換クロマトグラフィーによる効率的な精製を可能にする。他の有用な異種ポリペプチドには、例えば、米国特許第5,011,912号およびHoppら(1988),Bio/Technology 6:1204、に記載されている抗原性同定ペプチドが含まれる。こうしたペプチドの一つはFLAG(登録商標)ペプチドであり、それは高度に抗原性であり、そして特異的モノクローナル抗体により可逆的に結合されるエピトープを提供し、発現された組換え融合タンパク質の、迅速なアッセイおよび容易な精製を可能にしている。4E11と称されるマウスハイブリドーマは、米国特許第5,011,912号に記載されているように、特定の二価金属カチオンの存在下でFLAG(登録商標)ペプチドを結合する、モノクローナル抗体を産生する。4E11ハイブリドーマ細胞株は、第HB9259号の寄託番号でAmerican Type Culture Collectionに寄託されている。FLAG(登録商標)ペプチドを結合するモノクローナル抗体は、FLAG(登録商標)ペプチドを含んでなるポリペプチド精製試薬を回収するための親和性試薬として使用することが可能である。他の適したタンパク質タグ(tag)および親和性試薬は以下のものである:1)固定化グルタチオンに対するグルタチオン−S−トランスフェラーゼ融合タンパク質の親和性を利用する、GST−BindTM)システム(Novagen)に記載されているもの;2)モノクローナル抗体に対するT7遺伝子10タンパク質のアミノ末端11アミノ酸の親和性を利用する、T7−Tag(登録商標)アフィニティー精製キット(Novagen)に記載されているもの;または3)タンパク質タグに対するストレプトアビジンの操作された形の親和性を利用する、Strep−tag(登録商標)システム(Novagen)に記載されているもの。上述のタンパク質タグのいくつか、並びにその他がSassenfeld(1990),TIBTECH 8:88−93,Brewerら,「Purification and Analysis of Recombinant Proteins」中,pp.239−266,SeetharamおよびSharma(監修),Marcel Dekker,Inc.(1991),およびBrewerおよびSassenfeld,「Protein Purification Applications」中,pp.91−111,HarrisおよびAngal(監修),Press,Inc.,Oxford England(1990)、に記載されている。さらに、二つまたはそれより多い上述のタグの融合、例えば、FLAGタグおよびポリヒスチジンタグの融合を本発明のBTL−IIタンパク質へ融合することが可能である。
【0080】
他の異種ポリペプチドを含んでなる組換え融合タンパク質は、例えば、二量体、三量体またはより高度な多量体を形成する性向、増加したインビボ半減期、および/または増加した生物学的活性のごとき、他の種類の独特な利点を有している。融合タンパク質を製造するための技術は既知であり、そして例えば、WO 99/31241およびCosmanら((2001),Immnunity 14:123−133)、に記載されている。例示として、IgG抗体のFc領域、または実質的に類似したタンパク質を含んでなる異種ポリペプチドを、BTL−IIポリペプチドまたは断片へ融合することが可能である。抗体のFc領域は、ヒトまたは動物起源の抗体からのCH2およびCH3ドメインまたはこれらに実質的に類似した免疫グロブリンドメインを含んでなるポリペプチドである。議論のためには、HasemannおよびCapra,免疫グロブリン:構造および機能,William E.Paul,監修,「Fundamental Immunology」中,第2版,212−213(1989)、を参照されたい。二量体化を促進するヒンジ領域を含んでなる、Fc領域の一部切断形もまた使用することが可能である。抗体および他の免疫グロブリンアイソタイプの他の部分を使用することが可能である。IgG抗体のFc領域を含んでなる組換え融合タンパク質は、二量体を形成するようである。抗体由来のタンパク質のいろいろな部分を含んでなる融合タンパク質は、Ashkenaziら((1991)Proc.Natl.Acad.Sci.USA 88:10535−39)、Byrnら((1990),Nature 344:677−70)、HollenbaughおよびAruffo(「免疫学における最新のプロトコール」中,補4,pp.10.19.1−10.19.11(1992))、Baumら((1994),EMBO J.13:3992−4001)および米国特許第5,457,035号およびWO 93/10151、に記載されている。いくつかの態様において、改変Fc領域は、野生型Fc領域と比較して、Fcレセプターに対するより低い親和性を有しているという利点を持つことが可能である。このことは、こうした組換え融合タンパク質が免疫エフェクター細胞により結合した細胞の溶解を少なくすることができるので都合がよい。実施例5は、ヒトFc領域へ融合されたマウスBTL−IIの細胞外領域を含んでいる融合タンパク質の製造を記載している。このタンパク質をコードしている核酸配列およびそのアミノ酸配列は、各々、配列番号19および配列番号20に開示されている。
【0081】
別の代替物として、本発明の組換え融合タンパク質は、ロイシンジッパーを含んでなる異種ポリペプチドを含んでなることが可能である。既知のロイシンジッパー配列の中で、二量体化を促進する配列および三量体化を促進する配列である。例えば、Landschulzら(1988),Science 240:1759−64、を参照されたい。ロイシンジッパーは、しばしば、他のアミノ酸と共に散らばる4または5のロイシン残基を持つ、7アミノ酸反復を含んでなる。ロイシンジッパーの使用および製造は、当該技術分野ではよく知られている。
【0082】
もしくは、組換え融合タンパク質の異種ポリペプチド形成部分は、二つまたはそれより多くのBTL−IIポリペプチドを連結している、一つまたはそれより多くのペプチドリンカーであり得る。一般に、ペプチドリンカーは、複数の同一の、類似のまたは異なったポリペプチドを連結するために働き、そしてタンパク質の連結された部分の望まれる機能に必要とされる可動性または強剛性を提供する、アミノ酸の伸長である。典型的には、ペプチドリンカーは、長さ約1および30アミノ酸の間である。ペプチドリンカーの例には、限定されるわけではないが、−−Gly−Gly−、GGGGS(配列番号1)、(GGGGS)n(配列番号2)、GKSSGSGSESKS(配列番号3)、GSTSGSGKSSEGKG(配列番号4)、GSTSGSGKSSEGSGSTKG(配列番号5)、GSTSGSGKSSEGKG(配列番号6)、GSTSGSGKPGSGEGSTKG(配列番号7)またはEGKSSGSGSESKEF(配列番号8)が含まれる。連結部分は、例えば、Huston,J.S.ら,Proc.Natl.Acad.Sci.85:5879−83(1988)、Whitlow,M.ら,Protein Engineering 6:989−95(1993)、およびNewton,D.L.ら,Biochemistry 35:545−53(1996)に記載されている。他の適切なペプチドリンカーは、米国特許第4,751,180号および第4,935,233号に記載されているものである。
【0083】
さらに、組換え融合タンパク質は、その正常なシグナル配列を欠き、そしてその代わりとして、それに置き換わる異種シグナル配列を有するBTL−IIタンパク質を含んでなることが可能である。シグナル配列の選択は、組換えタンパク質が産生されるべき宿主細胞の型に依存しており、そして天然のシグナル配列を異種シグナル配列に置き換えることが可能である。哺乳動物宿主細胞中で機能的であるシグナル配列には以下のものが含まれる:米国特許第4,965,195号に記載されているインターロイキン−7(IL−7)のためのシグナル配列;Cosmanら((1984),Nature 312:768)により記載されているインターロイキン−2レセプターのためのシグナル配列;EP特許第0 367 566号に記載されているインターロイキン−4レセプターシグナル配列;米国特許第4,968,607号に記載されているタイプI インターロイキン−1レセプターシグナル配列;EP特許第0 460 846号に記載されているタイプII インターロイキン−1レセプターシグナル配列。
【0084】
BTL−II核酸
本発明は、変異体、断片、組換え融合タンパク質、完全長タンパク質、可溶性タンパク質および分泌タンパク質を含む、上記のBTL−IIタンパク質、断片または免疫原性断片をコードする単離核酸を包含する。これらの核酸は、なかでも、組換えタンパク質を製造するため、および組織試料中のBTL−II核酸の存在を検出するために、例えば、診断使用のために有用である。こうした核酸は、ゲノムDNAまたはcDNAであり得る。核酸は、本発明のBTL−IIタンパク質をコードしている、中断されていないオープンリーディングフレーム(open reading frame)を含んでなることが可能である。本発明の核酸分子には、一本鎖および二本鎖形両方の形でのDNAおよびRNA、並びに、対応する相補的配列が含まれる。“単離核酸”とは、天然に存在する起源から単離された核酸の場合、該核酸が単離されたゲノムまたは生物体中に存在する、隣接する遺伝子配列から分離されている核酸である。オリゴヌクレオチドのごとく、化学的に、またはポリメラーゼ連鎖反応(PCR)生成物またはcDNAのごとく、鋳型から酵素的に合成された核酸の場合、こうしたプロセスから生じる核酸は単離核酸であることを理解されたい。単離核酸分子は、別々の断片の形での、またはより大きな核酸構築物の成分としての核酸分子も指している。
【0085】
さらに、本発明は、(1)例えば、サザンおよびノーザンブロッティング、ドットブロッティング、コロニーハイブリダイゼーションなど、当該技術分野ではよく知られた多数の方法によりBTL−II核酸を検出するためのプローブとして、(2)BTL−II核酸を増幅するためのポリメラーゼ連鎖反応(PCR)プライマーとして、または(3)例えば、アンチセンス核酸(ペプチド核酸を含んで)、リボザイム、三重らせん形成分子による発現の阻害を通して、または任意のこれらのRNAをコードするRNAまたはDNAを妨害することによる、BTL−II核酸の発現を調節するための手段として、働くことが可能であるBTL−IIをコードしている核酸の断片を包含する。PCRプライマーは、BTL−II核酸配列に加えて、増幅された核酸の使用を容易にする制限酵素切断部位のごとき他の配列を含んでなることが可能である。PCRは以下の文献に記載されている:Saikiら(1988),Science 239:487−91;「PCR Technology」,Erlich,監修,Stockton Press,(1989)。以下に説明するように、PCRはBTL−II mRNAの過剰発現を検出するために有用であることが可能であり、そしてPCRプライマーは、遺伝子の多様な部分から採ることが可能であり、そしてまた、異なったスプライス変異体間を区別するために選択することが可能である。アンチセンスRNA(およびそれらをコードしているDNA)、DNAまたは合成ヌクレオチド、そして発現を調節するためのそれらの使用は、当該技術分野ではよく知られており、例えば以下の文献に記載されている:IzantおよびWeintraub(1984),Cell 36(4):1007−15;IzantおよびWeintraub(1985),Science 229(4711):345−52;Hazel−Bellanら(1988),J.Exp.Med.168(6):2309−18;Sarinら(1988),Proc.Natl.Acad.Sci.USA 85(20):7448−51;Zon(1988),Pharm.Res.5(9):539−49;Harel−Bellanら(1988),J.Immunol.140(7):2431−35;Marcus−Sekuraら(1987),Nucleic Acids Res.15(14):5749−63;Gambari(2001),Curr.Pharm.Des.7(17):1839−62;およびLemaitreら(1987),Proc.Natl.Acad.Sci.USA 84(3):648−52。同様に、選択された遺伝子の発現を阻害するための干渉(interfering)RNA(およびそれらをコードしているDNA)そしてそれらの使用は、当該技術分野ではよく知られており、以下の文献に記載されている:Fjoseら(2001),Biotechnol.Ann.Rev.7:31−57;BosherおよびLabouesse(2000),Nature Cell Biol.2:E31−E36。さらに、リボザイムまたはDNAzymeは、特異的RNAを切断するように標的化することが可能であり、そして従って、LewinおよびHauswirth(2001)Trends Mol.Med.7(5):221−28;MenkeおよびHobom(1997),Mol.Biotechnol.8(1):17−33;Norrisら(2000),Adv.Exp.Med.Biol.465:293−301;Sioud(2001),Curr.Mol.Med.1(5):575−88;およびSantiagoおよびKhachigian(2001),J.Mol.Med.79(12):695−706、に記載されているように、遺伝子発現を阻害するために使用することが可能である。BTL−II発現を調節可能な核酸は、BTL−II機能のインビボまたはインビトロ研究における、または治療剤として、所望により遺伝子治療剤としての使用を見出すことが可能である。
【0086】
本発明はまた、中程度のストリンジェント条件下、より好ましくは高ストリンジェント条件下で、本明細書に記載したBTL−IIタンパク質をコードしている核酸へハイブリダイズする核酸も含んでいる。こうした核酸には、配列番号3、配列番号5、配列番号7、配列番号9、配列番号11、配列番号13、配列番号15または配列番号17が含まれる。好ましくは、こうした核酸は、T細胞の表面上のレセプターへ結合可能で、および/またはT細胞増殖および/またはサイトカイン産生を阻害可能なタンパク質をコードしている。ハイブリダイゼーション技術は当該技術分野ではよく知られており、そしてSambrook,J.,E.F.FritschおよびT.Maniatis(「Molecular Cloning:A Laboratory Manual」,Cold Spring Harbor Laboratory Press,コールドスプリングハーバー,NY,第9章および第11章,(1989))および「Current Protocols in Molecular Biology」(F.M.Ausubelら監修,John Wiley & Sons,Inc,2.10および6.3−6.4節(1995))により記載されている。中程度にストリンジェントな条件は、約50%ホルムアミド、6xSSC中、約42から55℃の温度でのハイブリダイゼーション、そして約60℃、0.5xSSC、0.1%SDS中での洗浄が含まれる。高ストリンジェントな条件は、上記のようなハイブリダイゼーション条件と定義されるが、およそ68℃、0.2xSSC、0.1% SDSでの洗浄を伴うと定義される。ハイブリダイゼーションおよび洗浄緩衝液において、SSPE(1xSSPEは、0.15M NaCl、10mM NaH2PO4、および1.26mM EDTA,pH7.4である)をSSC(1xSSCは、0.15M NaClおよび15mMクエン酸ナトリウムである)に対して置換することが可能である;洗浄は、好ましくは少なくとも2回、ハイブリダイゼーションが完了した後、15分間行う。
【0087】
当業者に知られるように、そして以下にさらに記載するように、ハイブリダイゼーション反応および二重鎖安定性を支配する基本原理を適用することによって、必要に応じて、望ましい度合いのストリンジェンシーを達成するよう、洗浄温度および洗浄塩濃度を調整可能であることを理解すべきである(例えば、Sambrookら、上記文献を参照されたい)。既知の配列の核酸をハイブリダイズさせる場合、ハイブリッド長は、核酸の配列を並列し、そして最適配列相補性を持つ、一つまたは複数の領域を同定することによって、決定可能である。長さが50塩基対未満であると予測されるハイブリッドのハイブリダイゼーション温度は、ハイブリッドの融解温度(Tm)より5から10℃低くなければならず、Tmは、以下の等式に従って決定する。長さ18塩基対未満のハイブリッドに関しては、Tm(度C)=2(A+T塩基数)+4(G+C塩基数)である。長さ18塩基対を超えるハイブリッドに関しては、Tm(度C)=81.5+16.6(log10[Na+])+0.41(%G+C)−(600/N)であり、式中、Nはハイブリッド中の塩基数であり、そして[Na+]はハイブリダイゼーション緩衝液中のナトリウムイオン濃度である(1xSSCの[Na+]=0.165M)。各々のこうしたハイブリダイズする核酸は、少なくとも15ヌクレオチド(または少なくとも18ヌクレオチド)、または少なくとも20、または少なくとも25、または少なくとも30、または少なくとも40、または少なくとも50、または少なくとも100である長さを有する。Sambrookら、上記文献。
【0088】
BTL−II核酸は、以下のポリヌクレオチドを含んでなる核酸を含んでいる:(1)配列番号3、配列番号5、配列番号7、配列番号9、配列番号11、配列番号13、配列番号15または配列番号17の全てまたは断片、ここにおいて、断片はT細胞の表面上に発現されたレセプターへ結合可能で、および/またはT細胞の増殖および/またはサイトカイン産生を阻害可能なBTL−IIタンパク質をコードしている;(2)少なくとも100、125、150、175、200、225、250、300、400、500、600、800、1000、1200、1400または1600ヌクレオチド長であり、そしてT細胞の表面上に発現されたレセプターへ結合可能で、および/またはT細胞の増殖および/またはサイトカイン産生を阻害可能なBTL−IIタンパク質をコードしている、配列番号3、配列番号5、配列番号7、配列番号9、配列番号11、配列番号13、配列番号15または配列番号17と少なくとも80%、85%、90%、95%、97%、98%、99%、99.5%または99.7%同一である配列;(3)本発明のBTL−IIタンパク質をコードしている核酸を検出するまたは増幅するために、またはBTL−II mRNAおよび/またはタンパク質の発現を調節するために有用である、配列番号3、配列番号5、配列番号7、配列番号9、配列番号11、配列番号13、配列番号15または配列番号17または実質的に類似した配列の断片;(4)配列番号7または配列番号11のヌクレオチド30から130から成るポリヌクレオチド、または配列番号9、配列番号13、配列番号15または配列番号17のヌクレオチド377−477と、少なくとも80%、85%、90%、95%、97%、98%、99%、99.5%または99.7%同一であるポリヌクレオチドを含んでなる核酸、ここにおいて、同一性の領域は少なくとも60,70,80,90または100ヌクレオチド長であり、そして核酸によりコードされているタンパク質は、T細胞の増殖および/またはサイトカイン産生を阻害可能である;および(5)配列番号3、配列番号5、配列番号7、配列番号9、配列番号11、配列番号13、配列番号15または配列番号17と比較して少なくとも1、2、3、4、6、8、10、15、20、25、30、35、40、50または75の改変を含んでなる核酸、ここにおいて、改変は、単一ヌクレオチドの挿入、欠失または置換であることが可能である。
【0089】
BTL−IIポリペプチドへ特異的に結合する抗体
変異体、断片および組換え融合タンパク質を含む、本発明のBTL−IIタンパク質へ特異的に結合する抗体は、本発明により包含される。本明細書において、別のタンパク質(抗体のごとき)によるBTL−II上のエピトープの特異的結合とは、同一のエピトープを含んでなる別のタンパク質により、特異的に結合されているタンパク質は、それが結合されているBTL−IIタンパク質の分子から置き換えられることが可能であるが、このエピトープを含んでいない別のタンパク質によっては置き換えられないことを意味している。多数の競合的結合アッセイが当該技術分野では知られている。エピトープは近接アミノ酸のみを含んでなることができるが、しかしまたBTL−IIタンパク質の三次の折り畳みにより近くにもたらされた非近接アミノ酸を含んでなることもできる。エピトープは当該技術分野では既知の方法により同定することが可能である。例えば、Leinonenら(2002),Clin.Chem.48(12):2208−16;Krogerら(2002),Biosens.Bioelectron.17(11−12):937−44;Zhuら(2001),Biochem.Biophys.Res.Commun.282(4):921−27、を参照されたい。本発明はまた、本明細書において免疫原性断片と称される、抗体を発生するために有用である、本明細書に記載したBTL−IIタンパク質のエピトープも包含する。免疫原性断片は、好ましくは少なくとも10アミノ酸長であり、そして好ましくは、配列番号4、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16または配列番号18からの近接アミノ酸を含んでなる。こうしたエピトープは、スプライス接合部によりコードされているBTL−IIタンパク質の領域に及んでおり、特異的スプライス変異体によりコードされているタンパク質への特異的結合の利点を有することができる。
【0090】
抗体はポリクローナルまたはモノクローナル抗体であることが可能であり、そして当該技術分野ではよく知られている方法により産生可能である。例えば、「Monoclonal Antibodies,Hybridomas:A New Dimension in Biological Analyses」,Kennetら(監修),Plenum Press,ニューヨーク(1980);および「Antibodies:A Laboratory Manual」,HarlowおよびLand(監修),Cold Spring Harbor Laboratory Press,コールドスプリングハーバー,NY,(1988);KohlerおよびMilstein(1980)Proc.Natl.Acad.Sci.USA,77:2197;Kozborら(1984),J.Immunol.133:3001−3005(ヒトB細胞ハイブリドーマ技術を記載している);Coleら,「Monoclonal Antibodies And Cancer Therapy」,Alan R.Liss.Inc.,pp.77−96(1985)(EBVハイブリドーマ技術を記載している);Kuby,「Immunology」,第2版,p.162−64,W.H.Freeman and Co.,ニューヨーク(1994)、を参照されたい。本発明のBTL−IIタンパク質に特異的なモノクローナル抗体を産生するハイブリドーマ細胞株もまた、本明細書に意図される。こうしたハイブリドーマは、慣用的技術によって産生しそして同定することが可能である。本発明のmAbを産生するハイブリドーマはインビトロまたはインビボで培養することが可能である。さらに、本発明の抗BTL−II抗体は、例えば、チャイニーズハムスター卵巣(CHO)、HeLa、VERO、BHK、Cos、MDCK、293、3T3、骨髄腫(例えば、NSO、NSI)またはWI38細胞、酵母細胞、昆虫細胞および大腸菌(Eschericha coli)を含む細菌細胞、を含む他の培養細胞中で産生することが可能である。こうした抗体は、抗体に加えてこれらの抗体の発現を可能にする核酸をコードしている核酸を所望の細胞内へ導入することにより産生することが可能である。抗体は、それから、これらの核酸が導入されている細胞を培養することにより産生することが可能である。モノクローナル抗体は、IgG、IgM、IgE、IgA、IgD、およびそのいかなるサブクラスも含む、いかなる免疫グロブリンクラスのものであってもよい。
【0091】
あるいは、抗体は重および軽鎖可変領域様ドメイン、そして所望により、また一つまたはそれより多くの定常領域様ドメインを含んでなる単鎖抗体(米国特許第4,946,778号;Birdら(1988),Science 242:423−26;Hustonら(1988),Proc.Natl.Acad.Sci.USA 85:5879−83)、二量体または多価抗体(例えば、Lanttoら(2002),J.Gen.Virol.83:2001−05;HudsonおよびSouriau(2001),Expert Opin.Biol.Ther.1(5):845−55)、四量体抗体(例えば、Janewayら,「Immunobiology:The Immune System in Health and Disease」,第5版,第II部,第3章,Garland Publishing(2001))、キメラ抗体(HudsonおよびSouriau,上記文献;Boulianneら(1984),Nature 312:643−46;Morrison(1984),Proc.Natl.Acad.Sci.USA 81:6851−55;Takedaら(1985),Nature 314:452−54;Neubergerら(1985),Nature 314:268−70)、異なったトランスジェニック動物で産生された完全ヒト抗体(例えば、米国特許第6,150,584に記載されている)またはインビトロ選択(米国特許出願第2002/0058033号)またはヒト化抗体(Morrisonら,上記文献;Takedaら,上記文献;Boulianneら,上記文献)であってもよい。さらに、抗体は、インビトロ選択スキームにより“成熟させる”ことが可能であり、例えば、それが結合するエピトープに対してより高い親和性のごとき、改変された特性を有する抗体を得る。例えば、Jacksonら(1995),J.Immunol.154(7):3310−19;PiniおよびBracci(2000),Curr.Protein Pept.Sci.1(2):155−69;E11markら(2002),Mol.Immunol.39(5−6):349;O’Connellら(2002),J.Mol.Biol.321(1):49−56;Hulsら(2001),Cancer Immunol.Immunother.50:163−71;HudsonおよびSouriau,上記文献;AdamsおよびSchier(1999),J.Immunol.Methods 231(1−2):249−60;Schmitzら(2000),Placenta 21 補A:S106−12、を参照されたい。あるいは、本発明のBTL−IIタンパク質へ特異的に結合可能な、例えば、Fab断片、F(ab’)2断片または単鎖Fv断片(scFv’s)のごとき抗体の断片も、本明細書において抗BTL−II抗体として意味されるものに包含される。FabおよびFv断片についての議論は、Kuby,上記文献,pp.109−112およびJanewayら,上記文献、を参照されたい。本発明はまた、BTL−IIタンパク質へ特異的に結合する抗体に特異的に結合する、そしてBTL−IIタンパク質の効果を模倣する、抗イディオタイプ抗体も包含する。こうした抗イディオタイプ抗体は、BTL−IIタンパク質としての同様の使用を発見した。抗イディオタイプ抗体を産生する方法は当該技術分野ではよく知られている。例えば、Kubyら,上記文献,371−72、を参照されたい。本発明のBTL−IIタンパク質および別のエピトープへ特異的に結合可能な、多様な種類の組換えおよび非組換え二重特異性抗体もまた意図される。多様な種類の二重特異性抗体およびそれらを作製する方法は、例えば、米国特許第4,474,893,6,060,285および6,106,833号に記載されている。
【0092】
抗BTL−II抗体は、BTL−IIのそのレセプターへの結合のごときBTL−IIの生物学的機能を遮断するアンタゴニスト抗体、若しくはBTL−IIの生物学的機能を促進するまたはBTL−IIの機能を模倣するアゴニスト抗体であることができる。アゴニスト抗体は、BTL−IIタンパク質の機能を模倣するアゴニスト性抗イディオタイプ抗体を含むことが可能である。BTL−II機能のためのアッセイは本明細書に記載されている。こうしたアッセイで決定された、BTL−IIの生物学的機能を遮断する抗BTL−II抗体は、本明細書において意味されるようにアンタゴニスト抗体である。アンタゴニスト抗体は、例えば、BTL−IIのそのレセプターへの結合を遮断することができる。こうしたアッセイへ加えた場合、BTL−IIの生物学的機能を促進するまたは増強する抗BTL−II抗体は、本明細書において意味されるようにアゴニスト抗体である。アンタゴニスト性抗BTL−II抗体は、例えば、粘膜免疫応答を増強するためのアジュバントとして使用可能である。さらに、BTL−IIレセプターに対するアゴニスト性抗は、例えば、クローン病または炎症性腸疾患のごとき、腸の不適切な炎症により特徴付けられる疾患を処置するため、粘膜免疫応答を抑制するために使用可能である。
【0093】
本発明の抗体はまた、インビトロまたはインビボの両方で、本発明のBTL−IIタンパク質の存在を検出するためのアッセイにも使用可能である。抗体はまた、免疫アフィニティークロマトグラフィーによる本発明のBTL−IIタンパク質の精製においても用いることが可能である。抗体はまた、免疫親和性クロマトグラフィーによる、本発明のBTL−IIタンパク質の精製に用いることが可能である。
【0094】
本発明は、本発明の抗体をコードしている核酸、およびこうした核酸を細胞に導入し、そして核酸を含んでいる細胞を培養することによる、抗体を産生するための方法を包含する。
【0095】
BTL−IIポリペプチドのアゴニストおよびアンタゴニスト
本発明はBTL−IIのアゴニストおよびアンタゴニスト、そしてアゴニストおよびアンタゴニストをスクリーニングするための方法、並びにアゴニストおよびアンタゴニストを使用するための方法を包含する。インビボまたはインビトロにおけるBTL−IIタンパク質の過剰または過少発現、またはインビボまたはインビトロにおけるBTL−II発現の完全非存在を含んでいる、例えば、細胞増殖アッセイ、サイトカイン分泌アッセイ、結合アッセイおよび遺伝子アッセイのごとき、BTL−II生物学的活性のためのアッセイが本明細書に記載されている。候補分子をこうしたアッセイに加えることが可能であり、BTL−IIタンパク質の生物学的活性に対するそれらの効果を決定する。BTL−IIアンタゴニストは、例えば、BTL−IIと、好ましくはB細胞またはT細胞上に発現されたそのレセプターとの相互作用を、遮断可能である。アンタゴニストはアンタゴニスト抗体を含み、そしてアゴニストはアゴニスト抗体を含む。加えて、アフィボディ(affibody)(Ronnmarkら(2002),J.Immunol.Methods 261(1−2):199−211)のような、本発明のBTL−IIタンパク質へ特異的に結合可能である他の抗体関連分子、および本発明のBTL−IIタンパク質に特異的に結合し、そしてBTL−IIタンパク質の生物学的活性を阻害する、WO 00/24782に記載されている生物学的に活性なペプチドが本発明に包含される。さらに、BTL−IIアンタゴニストには、例えば、干渉RNA(またはそれらをコードしているDNA)またはアンチセンスRNAまたはDNAのごとき、BTL−IIタンパク質および/またはmRNAの発現を変調するために有用である上述の核酸が含まれる。
【0096】
アンタゴニストにはさらに、BTL−IIまたはそのレセプターへ結合するためにインビトロで選択されたアミノ酸配列を含んでなる、そして、所望により、BTL−IIまたはそのレセプターの相互作用を妨害可能な、タンパク質が含まれる。あるいは、こうしたタンパク質は、BTL−IIの生物学的機能を促進するまたは模倣するBTL−IIアゴニストであることが可能である。BTL−IIまたはそのレセプターへ結合するタンパク質は、BTL−IIとそのレセプターとの相互作用を妨害する能力でスクリーニングすることが可能であり、または、選択は、こうしたタンパク質を直接得るように設計することが可能である。
【0097】
タンパク質は、例えば、ファージディスプレイまたは細菌の表面のディスプレイのごとき、多くの方法により選択することができる。例えば、ParmleyおよびSmith(1989),Adv.Exp.Med.Biol.251:215−218;Luzzagoら(1995),Biotechnol.Annu.Rev.1:149−83;Luら(1995),Biotechnology(NY)13(4):366−372、を参照されたい。これらの方法において、結合ドメインのライブラリーの各々のメンバーは、個々のファージ粒子または細菌細胞上にディスプレイすることが可能であり、そして選ばれた条件下で目的のタンパク質へ結合する細菌またはファージを選択可能である。選択された結合ドメインをコードする核酸は、選択されたファージまたは細菌を成長させ、そしてそれらから核酸を単離することにより得ることが可能である。
【0098】
あるいは、タンパク質を全くインビトロで選択することが可能である。例えば、潜在的結合ドメインのライブラリー中の、個々のポリペプチドの各々を、それをコードする核酸へ付着可能であり、そして選ばれた条件下で目的のタンパク質へ結合するものを選択することが可能である。ポリペプチドはそれらをコードする核酸へ付着されているので、増幅、クローニング、または有効な結合ドメインをコードする核酸の配列決定のごとき、続いての操作が容易である。抗体−リボソーム−mRNA粒子、リボソームディスプレイ、共有結合的RNA−ペプチド融合、または共有結合的DNA−RNA−ペプチド融合を含む、こうした選択のための多様なスキームが当該技術分野では知られている。HeおよびTaussig(1997),Nucleic Acids.Res.25(24):5132−5134;HanesおよびPluckthun(1997),Proc.Natl.Acad.Sci.94:4937−4942;RobertsおよびSzostak(1997),Proc.Natl.Acad.Sci.94:12297−12302;LohseおよびWright(2001),Curr.Opin.Drug Discov.Devel.4(2):198−204;Kurzら(2000),Nucleic Acids Res.28(18):E83;Liuら(2000),Methods Enzymol.318:268−93;Nemotoら(1997),FEBS Lett.414(2):405−08;米国特許第6,261,804号;WO0032823;およびWO0034784。こうしたタンパク質はアンタゴニストまたはアゴニストであると選択することが可能である。
【0099】
BTL−IIタンパク質の生物学的活性のためのアッセイ
BTL−IIタンパク質の生物学的活性を検出するため、およびBTL−IIの結合パートナー(partner)を同定するために、多様なアッセイを使用することが可能である。BTL−IIタンパク質、BTL−IIのレセプター、および両方のアゴニストおよび/またはアンタゴニストを、こうしたアッセイに使用することが可能である。
【0100】
結合パートナーを同定するためのアッセイ
BTL−IIの結合パートナーは、最初にどの細胞の型に可溶性BTL−IIタンパク質が結合可能かを決定することにより同定可能である。ブチロフィリン様タンパク質のB7サブファミリーのメンバーのための既知レセプターはすべてT細胞上で発現されるので、構成的に(CD28)かまたは活性化後に(CTLA−4、ICOS、PD−1)、これらへBTL−IIが結合可能かどうかを決定するために、T細胞を試験することが可能である。BTL−IIの発現パターンのため、正常および炎症性腸から単離されたT細胞を、こうした試験に含むことが可能である。加えて、メモリーT細胞、ナイーブT細胞、αβT細胞、γδT細胞および多様な活性化状態のT細胞を含む、多様なサブセットのT細胞を試験することが可能である。
【0101】
こうした実験は、当該技術分野ではよく知られている方法を使用して実施可能である。例えば、マウスBTL−IIの細胞外ドメインに加えて抗体のFc領域を含んでなるBTL−II組換え融合タンパク質を、試験されている細胞へ結合させるために使用可能である。組換え融合タンパク質中のFc領域へ結合可能な蛍光標識抗体を、加えることが可能である。洗浄後、細胞は、BTL−IIが細胞へ結合可能かどうかを決定するため、蛍光表示式細胞分取(FACS)装置を使用して分析可能である。こうしたアッセイの一つはChapovalら((2001),Nature Immunol.2(3):269−74により記載されている。当該技術分野で既知の他の方法もまた、BTL−IIが特異的細胞集団へ結合するかどうかを決定するのに適切であり得る。
【0102】
さらに、B7サブファミリーメンバーの既知のレセプターは、BTL−IIがいかなるこれらのタンパク質にも結合するかどうかを決定するために試験されるであろう。B7サブファミリーメンバーの細胞外ドメイン(例えば、CTLA−4、B7−1およびB7−2のためのレセプター)および抗体のFc領域を含んでなる組換え融合タンパク質は、こうしたBTL−II融合タンパク質を作製するために使用した方法と類似の方法により作製可能である。細胞表面に発現される、膜貫通および細胞質ドメインを含むBTL−IIタンパク質をコードする、BTL−II核酸の完全長形で細胞をトランスフェクトすることが可能である。試験されるべきレセプター:Fc融合タンパク質を、Fc領域へ結合可能である蛍光標識抗体と一緒に、こうした細胞へ加えることが可能である。洗浄後、トランスフェクトされた細胞上に発現されたBTL−IIタンパク質へレセプター:Fc融合タンパク質が結合しているかどうかを決定するため、FACSにより細胞を分析することが可能である。逆実験もまた実行することが可能であり、ここでは、可溶性BTL−II:Fc融合タンパク質を使用し、そして完全長レセプタータンパク質を導入し、そしてトランスフェクションにより発現させる。こうした実験は、BTL−IIタンパク質および他のB7サブファミリーメンバーの既知のレセプター間の結合相互作用を明らかにすることが可能である。
【0103】
もし、BTL−IIが多様なT細胞の少なくとも一つとは結合するが、既知のレセプターへは結合しないとすれば、当該技術分野では既知の多様な発現クローニングまたはタンパク質精製法を、BTL−IIが結合するレセプターを同定するために使用可能である。例として、放射活性スライド(slide)結合cDNA発現クローニング法が使用可能である。簡単には、可溶性BTL−IIへ最も強い結合を有する細胞源を同定し、mRNAを単離し、そして哺乳動物発現ベクター中にcDNAライブラリーを構築する。スライド上で哺乳動物細胞をcDNAのプールでトランスフェクトし、そして発現を可能にするために適切にインキュベーションした後、可溶性BTL−II:Fc融合タンパク質を細胞へ結合させる。レセプターを運んでいる細胞への特異的結合を、以下の一連の工程により検出する:結合されたBTL−II:Fcタンパク質への放射活性抗Fc試薬の結合;スライドへのフィルム乳剤の適用;暴露させるためのインキュベーション;銀粒子を沈着させるためのフィルム現像;そして顕微鏡による粒子の検出。レセプター発現クローンは次ぎに、プールを副分画することによりプールから単離し、そして単一レセプタークローンを同定するためにスライド結合アッセイを反復する。こうした方法は、McMahonら(1991),EMBO J.10:2821−32、に記載されている。
【0104】
細胞への結合が達成された場合、多様な手段(動物免疫化またはファージディスプレイ技術を通して)を、BTL−IIを結合している抗体を単離するため、そして細胞へのその結合を破壊するために使用可能である。こうした抗体は、内在性BTL−IIの活性に拮抗するために、そして従って、免疫応答、および例えば、Cooperら((1993),Lab.Invest.69(2):238−49)およびTokoiら((1996),J.Gastroenterol.31(2):182−88)により記載されているような、炎症性腸疾患モデルのごとき疾患モデルにおいて、BTL−IIの有効量を低下させる効果を決定するためのアッセイに使用可能である。
【0105】
BTL−II機能を決定するためのアッセイ
B7サブファミリータンパク質は、“抗原”の用量を変化させ、T細胞レセプターを通してT細胞を活性化させることを含む、T細胞“同時刺激”アッセイにおいて活性を有することが示されている。B7サブファミリータンパク質の活性は、“最適以下”のT細胞レセプター刺激で、最も明らかである。例えば、Latchmanら(2001),Nature Immunol.2(3):261−68、を参照されたい。組織培養皿結合抗CD3ε抗体のごとき“代理(surrogate)抗原”または照射抗原提示細胞上のMHC分子により提示される抗原を使用可能である。可溶性BTL−IIタンパク質を、“抗原”についてのT細胞の応答に対するその効果を決定するために加えることが可能である。T細胞が刺激されているか、または抑制されているかを決定するため、多様なパラメーターが測定可能である。細胞増殖、細胞表面レセプター発現、そしてタンパク質および/またはmRNAレベルでの免疫変調分子(例えば、インターフェロンγおよび/またはIL−2のごとき)の発現レベル、を測定することが可能である。こうした方法は、例えば、Fitchら,「T Cell Subsets in Infectious and Autoimmune Diseases」,John Wiley and Sons,pp.68−85(1995);Freemanら(2000),J.Exp.Med.192(7):1027−34;Swallowら(1999),Immunity 11:423−32;Hutloffら(1999),Nature 397:263−66;Yoshinagaら(1999),Nature 402:827−32;Latchmanら,上記文献、に使用され、説明されている。BTL−IIの発現パターンを仮定すれば、こうしたアッセイは、粘膜組織またはこうした組織を排出するリンパ節に由来するT細胞を含むことが可能である。加えて、ナイーブおよびメモリーT細胞、CD4+およびCD8+T細胞、およびαβT細胞レセプター以外のT細胞レセプターを発現しているT細胞、を試験することが可能である。こうした実験から、BTL−IIが多数の異なった種類のT細胞の応答を改変できるかどうかを明らかにすることが可能である。こうした実験は以下に記載されており、そしてBTL−IIの可溶性バージョンが、細胞増殖を阻害する、およびインターフェロンガンマ(IFNγ)インターロイキン2(IL2)およびインターロイキン5(IL5)を含むサイトカインの産生を抑制可能であることを示している。実施例6−10;図12−17。
【0106】
この種類の実験の変法には、同時刺激性が可能である、他の可溶性B7サブファミリータンパク質または多様なTNFファミリーメンバーで観察される同時刺激応答について、BTL−IIタンパク質の可溶性形を含んだ効果を試験することが含まれる。このことは、BTL−IIが他の分子で見られる同時刺激を改変するかどうかを定義する助けとなるであろう。さらなる変法は、多様な種類の非照射抗原提示細胞の使用を含むことが可能である。このことは、抗原提示細胞の機能に対する、可溶性BTL−IIタンパク質の添加の効果を定義する助けとなるであろう。さらに別の変法においては、細胞へのBTL−II結合を遮断する抗体(上記参照)もまた、結合を防止する効果を決定するため、こうした同時刺激実験に導入可能である。
【0107】
別の種類の実験において、BTL−IIを発現する抗原提示細胞(例えば、パイエル板からの樹状細胞またはB細胞、または腸管上皮細胞(IEC)のごとき)をT細胞(上にリストした多様な種類のもののごとき)と組み合わせることが可能で、そして上にリストした一つまたはそれより多くのパラメーターを測定することが可能である。これらの細胞で得られた結果は、BTL−II発現を低くするために設計された、これらをコードしている干渉RNAまたはDNAが抗原提示細胞内へ導入された場合に得られた結果と比較可能である。
【0108】
T調節(T reg)細胞は、炎症性腸疾患モデルシステムにおいて、一部、T調節細胞の分化を誘導することにより、または調節機能を促進することにより、自己免疫応答を抑制するように、そしてそのようにすることを助けるように作用する。T reg細胞には、例えば、なかでも、Tr1細胞(Congら(2002),J.Immunol.169(11):6112−19;Grouxら(1997),Nature 389:737−42)、Th3細胞、CD4+Cd25+T reg細胞(例えば、MaloyおよびPowrie(2001),Nature Immunology 2(9):816−22)を参照されたい)、CD4+RbloCD25+T reg細胞、およびCD8+ T reg細胞(Allezら(2002),Gastroenterology 123:1516−26)が含まれる。従って、T調節細胞増殖および機能に対するBTL−IIの効果を評価することは、インビボにおけるBTL−IIの正確な役割を決定するために必要とされる。例えば、Tr1細胞存在下での、抗原特異的T細胞増殖および/またはT細胞によるサイトカイン産生を測定可能である。こうしたアッセイは、Congら、上記文献、に記載されている。BTL−IIの可溶性形、遮断抗体または干渉RNAを使用するBTL−II発現の阻害は、BTL−IIが、T調節細胞またはTr1細胞の増殖、維持、または他のT細胞の抗原活性を抑制する能力において役割を演じているかどうかを決定するために使用可能である。
【0109】
一つのT細胞サブセット、Th3細胞は、トランスフォーミング成長因子β(TGFβ)を産生し、そして粘膜抗原に対する経口免疫寛容と結びつけられている。例えば、Fukauraら(1996),J.Clin.Invest.98(1):70−77;Inobeら(1998),Eur.J.Immunol.28:2780−90、を参照されたい。こうした細胞は、TGFβ存在下、抗原による広範培養により、ナイーブT細胞から発生させることが可能である。可溶性BTL−IIタンパク質または遮断抗体は、上述の方法と同様の方法により、BTL−IIがこれらの細胞の増殖または機能を改変することが可能かどうかを問うために使用可能である。
【0110】
単純で明確な抗原(例えば、卵白アルブミンのごとき)または複雑な抗原(細菌またはウイルスを含んだ)を使用するモデルT細胞システムにおいて、可溶性BTL−IIタンパク質または拮抗性抗体を投与することの効果が確かめられるであろう。こうした抗原に対するT細胞応答(増殖および/またはインターフェロンγまたはIL−2のごとき分子の産生を含んで)を、BTL−IIタンパク質または抗体の存在下および非存在下で測定可能である。
【0111】
動物システムにおける同様の効果が試験可能である。初期の焦点は、特に粘膜免疫応答に関連するシステムであろう。これには、動物へ抗原を与えるモデルシステム(例えば、Yamashiroら(1994),Acta Paediatr.Jpn.36:550−56;Jainら(1996),Vaccine 14(13):1291−97;Chenら(2002),Immunology 105:171−80、を参照されたい)、デキストラン硫酸ナトリウム誘導炎症性腸疾患モデルにおけるごとき炎症性腸疾患システム(Cooperら(1993),Lab.Invest.69(2):238−49;Tokoiら(1996),J.Gastroentero1.31(2):182−88)、CD45RBhi/CD4+T細胞誘導萎縮病モデル(Morrisseyら(1993),J.Exp.Med.178:237−44)、および/またはマウス卵白アルブミン誘導喘息モデルのごとき、免疫応答を誘発するために気道内へ抗原を与えるまたは滴下するモデル喘息システム(Brusselleら(1994),Clin.Exp.Allergy 24(1):73−80)、が含まれる。特に、体液または細胞仲介応答の大きさが測定されるであろう、そして産生された免疫変調サイトカインの型が測定されるであろう。こうした実験は、増加したまたは減少したBTL−IIが、抗原(自己抗原を含んで)に対する応答を低下させることにおいて、または同種抗原に対する応答を増加させることにおいて役に立つことができるかどうかを明らかにすることが可能である。BTL−IIタンパク質または抗BTL−II抗体の効果はまた、炎症性疾患の他のモデルにおいても確かめることが可能である。
【0112】
T細胞または胸腺細胞増殖のためのアッセイには、以下のものが含まれるが、限定されるわけではない:「Current Protocols in Immunology」,Coliganら監修,Greene Publishing Associates and Wiley−Interscience(pp.3.1−3.19:マウスリンパ球機能のインビトロアッセイ;第7章:ヒトにおける免疫学的研究);Takaiら(1986),J.Immunol.137:3494−3500;Bertagnolliら(1990),J.Immunol.145:1706−1712;Bertagnolliら(1992),J.Immunol.149:3778−3783;Bowmanら(1994),J.Immunol.152:1756−1761。
【0113】
脾臓細胞、リンパ節細胞または胸腺細胞のサイトカイン産生および/または増殖のためのアッセイには、限定するわけではないが、以下のものが含まれる:KruisbeekおよびShevach(1994)ポリクローナルT細胞刺激,「Current Protocols in Immunology」中,Coliganら監修,第1巻,pp.3.12.1−3.12.14,John Wiley and Sons,トロント(1991);およびSchriber,(1994),マウスおよびヒトインターフェロンガンマの測定,「Current Protocols in Immunology」中,Coliganら監修,第1巻,pp.6.8.1−6.8.8,John Wiley and Sons,トロント(1991)。
【0114】
造血およびリンパ球新生細胞の増殖および分化のためのアッセイには、限定するわけではないが、以下のものが含まれる:Bottomlyら,1991,ヒトおよびマウスインターロイキン2およびインターロイキン4の測定,「Current Protocols in Immunology」中,Coliganら監修,第1巻,pp.6.3.1−6.3.12,John Wiley and Sons,トロント(1991);deVriesら(1991),J.Exp.Med.173:1205−1211;Moreauら(1988),Nature 336:690−692;Greenbergerら(1983),Proc Natl.Acad.Sci.USA 80:2931−2938;Nordan,マウスおよびヒトインターロイキン6の測定,「Current Protocols in Immunology」中,Coliganら監修,第1巻,pp.6.6.1−6.6.5,John Wiley and Sons,トロント(1991);Smithら(1986),Proc.Natl.Acad.Sci.USA 83:1857−1861;Bennettら,1991,ヒトインターロイキン11の測定,「Current Protocols in Immunology」中,Coliganら監修,第1巻,pp.6.15.1,John Wiley and Sons,トロント(1991);Ciarlettaら,マウスおよびヒトインターロイキン9の測定,「Current Protocols in Immunology」中,Coliganら監修,第1巻,pp.6.13,1,John Wiley and Sons,トロント(1991)。
【0115】
抗原に対するT細胞クローン応答のためのアッセイ(それは、なかでも、増殖およびサイトカイン産生を測定することにより、APC−T細胞相互作用に影響するポリペプチド、ならびに直接的T細胞効果を同定するであろう)には、限定するわけではないが、以下のものが含まれる:「Current Protocols in Immunology」,Coliganら監修,Greene Publishing Associates and Wiley−Interscience(第3章:マウスリンパ球機能のインビトロアッセイ;第6章:サイトカインおよびそれらの細胞レセプター;第7章:ヒトにおける免疫学的研究)(1991);Weinbergerら(1980),Proc.Natl.Acad.Sci.USA 77:6091−6095;Takaiら(1986),J.Immunol.137:3494−3500;Takaiら(1988),J.Immunol.140:508−512。
【0116】
胸腺細胞または脾細胞細胞毒性のためのアッセイには、限定するわけではないが、以下のものが含まれる:「Current Protocols in Immunology」,Coliganら監修,Greene Publishing Associates and Wiley−Interscience(第3章:マウスリンパ球機能のインビトロアッセイ,3.1−3.19;第7章:ヒトにおける免疫学的研究)(1991);HerrmannおよびMescher(1981),Proc.Natl.Acad.Sci.USA 78:2488−2492;Herrmannら(1982),J.Immunol.128:1968−1974;Handaら(1985),J.Immunol.135:1564−1572;Takaiら(1986),J.Immunol.137:3494−3500;Takaiら(1988),J.Immunol.140:508−512;Brownら(1994),J.Immunol.153:3079−3092。
【0117】
免疫グロブリン応答およびB細胞によるアイソタイプ転換のためのアッセイ(それは、なかでも、T細胞依存性抗体応答を変調し、そしてTh1/Th2プロフィールに影響するポリペプチドを同定するであろう)には、限定するわけではないが、以下のものが含まれる:Maliszewski(1990),J.Immunol.144:3028−3033;およびMondおよびBrunswick,B細胞機能のアッセイ:インビトロ抗体産生,「Current Protocols in Immunology」中,Coliganら監修,第1巻,pp.3.8.1−3.8.16,John Wiley and Sons,トロント(1991)。
【0118】
混合リンパ球反応(MLR)アッセイ(それは、なかでも、主としてTh1およびCTL応答を発生するポリペプチドを同定するであろう)には、限定するわけではないが、以下のものが含まれる:「Current Protocols in Immunology」,Coliganら監修,Greene Publishing Associates and Wiley−Interscience(第3章:マウスリンパ球機能のインビトロアッセイ,3.1−3.19;第7章:ヒトにおける免疫学的研究);Takaiら(1986),J.Immunol.137:3494−3500;Takaiら(1988),J.Immunol.140:508−512;Bertagnolliら(1992),J.Immunol.149:3778−3783。
【0119】
樹状細胞依存性アッセイ(それは、なかでも、ナイーブT細胞を活性化する樹状細胞により発現されるポリペプチドを同定するであろう)には、限定するわけではないが、以下のものが含まれる:GueryおよびAdorini(1995),J.Immunol.154:536−544;Inabaら(1991),J.Exp.Med.173:549−559;Macatoniaら(1995),J.Immunol.154:5071−5079;PorgadorおよびGilboa(1995),J.Exp.Med.182:255−260;Nairら(1993),J.Virology 67:4062−4069;Huangら(1994),Science 264:961−965;Macatoniaら(1989),J.Exp.Med.169:1255−1264;Bhardwajら(1994),J.Clin.Invest.94:797−807;およびInabaら(1990),J.Exp.Med.172:631−640。
【0120】
T細胞傾倒および発生の初期段階に影響するポリペプチドのためのアッセイには、限定するわけではないが、以下のものが含まれる:Anticaら(1994),Blood 84:111−117;Fineら(1994),Cell Immunol.155:111−122;Galyら(1995),Blood 85:2770−2778;Tokiら(1991),Proc.Natl.Acad.Sci.USA 88:7548−7551。
【0121】
レセプター−リガンド活性のためのアッセイには、限定するわけではないが、以下のものが含まれる:「Current Protocols in Immunology」,Coliganら監修,Greene Publishing Associates and Wiley−Interscience(第7.28章:静的条件下での細胞接着の測定 7.28.1−7.28.22);Takaiら(1987),Proc.Natl.Acad.Sci.USA 84:6864−6868;Biererら(1988),J.Exp.Med.168:1145−1156;Rosensteinら(1989),J.Exp.Med.169:149−160;Stoltenborgら(1994),J.Immunol.Methods 175:59−68;Stittら(1995),Cell 80:661−670。
【0122】
トランスジェニック動物を利用した機能のための遺伝子アッセイ
本明細書に開示したBTL−II核酸に対応する遺伝子(類)の複数のコピーを有し、好ましくは形質転換された細胞およびそれらの子孫内に安定的に維持されている遺伝子構築物を有する細胞の形質転換により産生されたトランスジェニック動物、好ましくはマウスを提供する。遺伝子発現レベルを増加させるまたは減少させる、または遺伝子発現の時間的または空間的パターンを変化させる、修飾された遺伝子制御領域を有するトランスジェニック動物もまた提供する(欧州特許第0 649 464 B1を参照されたい)。加えて、対応する遺伝子内への外来性配列の挿入を通して、または対応する遺伝子の全てまたは一部の欠失を通して、BTL−II遺伝子が部分的にまたは完全に不活性化されている生物体を提供する。部分的なまたは完全な遺伝子不活性化は、転位性要素の挿入(好ましくは続いての不正確な切除により)を通して(Plasterk(1992),Bioessays 14(9):629−633;Zwaalら(1993),Proc.Natl.Acad.Sci.USA 90(16):7431−7435;Clarkら(1994),Proc.Natl.Acad.Sci.USA 91(2):719−722)、または相同的組換え(好ましくは陽性/陰性遺伝子選択戦略により検出される)を通して(Mansourら(1988),Nature 336:348−352;米国特許第5,464,764;5,487,992;5,627,059;5,631,153;5,614,396;5,616,491;および5,679,523号)、達成可能である。代替法として、BTL−II遺伝子の発現は、トランスジェニック動物内へ導入されたDNAによりコードすることができる、干渉RNA、アンチセンスRNAまたはリボザイムの導入により、トランスジェニックまたは非トランスジェニック動物中で阻害可能である。こうした生物体の表現型は、BTL−II遺伝子のインビボ機能を解明することを可能にする。例えば、BTL−IIタンパク質を発現せずに、デキストラン硫酸ナトリウムの給餌に応答して炎症性腸疾患の症状を示す、トランスジェニック動物において増加した性向(野生型動物と比較して)から、BTL−IIは常態では腸管における炎症を低下させること、を示すことが可能である。あるいは、トランスジェニック動物はBTL−IIを過剰発現することが可能である。こうしたトランスジェニック動物の表現型もまた、BTL−IIタンパク質のインビボ機能に対する手掛かりを与えることが可能である。
【0123】
本発明のタンパク質、抗体、アンタゴニストおよびアゴニストの使用
特殊化した解剖学的構造、粘液膜の組内で作用する粘膜免疫システム、およびそれらが発生する免疫応答は、体内の他の解剖学的区画で発生する免疫応答からそれを区別する特性を有している。粘液膜はいくつかの独特な解剖学的区画のまさに一つであり、末梢リンパ節および脾臓、体腔(即ち、腹膜および胸膜)および免疫システムが活性である皮膚も含んでいる。粘膜表面は、肺、腸管、眼、鼻、口、のど、子宮および膣に観察される。粘膜表面は薄いシートまたは、体腔、仕切または隔壁、または二つの構造間の接合部の裏打ちのごとき、身体構造の被覆または外皮として働いている柔軟な組織の層である。粘膜表面は、大多数の感染性病原体の侵入経路であるので、粘膜免疫応答を増進可能であるアジュバントが特に望まれている。Janewayら,「Immunobiology:The Immune System in Health and Disease,第5版,第IV部,第10章,Garland Publishing,ニューヨークおよびロンドン(2001)。例えば、淋菌(Neisseria gonorrhoeae)のごとき、典型的には粘膜表面を通して侵入する感染性疾患は、しばしば1週間のみの粘膜免疫応答を発生させる。Russellら(1999),42(1):58−63。
【0124】
腸管はいくつかの様式で独特である。腸管を裏打ちしているのは、集合的に腸管付随リンパ組織(GALT)として知られている多数の特殊化した形のリンパ組織であり、以下のものが含まれる:扁桃腺およびアデノイド、それらは一緒になって口の奥の方でワルダイヤ輪を形成する;小腸中のパイエル板;虫垂;および大腸および直腸中の孤立リンパ濾胞。腸管中で外来抗原と遭遇することによるリンパ球の活性化は、粘液免疫システム中のいたるところでの抗原に対する適応性免疫応答の広がりを導くことが可能である。活性化リンパ球はリンパ系にそして、そこから血流へ入ることが可能である。血流は体中いたるところの粘膜部位へ活性化リンパ球を搬送可能であり、それは、粘膜組織で発現される粘膜アドレッシン(addressin)MAdCAM−1のごとき分子を使って、リンパ球により認識可能である。Janewayら、上記文献。腸管により産生される主抗体型はIgAであり、それは発現時に、腸管上皮の表面を覆う粘膜層中に主として観察される。加えて、多数の別々の種類のT細胞が腸管中に観察される。Janewayら、上記文献。
【0125】
外来抗原の腸管内への導入は、通常は免疫寛容を導くが、特異的免疫応答を導くこともできる。腸管は、普通は、外来抗原の莫大なアレイ、即ち、食物および腸管中に住む共生微生物、を受けおよび寛容する(免疫学的な意味で)。後でのタンパク質の注射が、アジュバント存在下でさえも、抗体応答を起こさないように、特別な外来タンパク質の給餌が、経口免疫寛容として知られている、そのタンパク質への特別な無応答性の状態を導くことが可能である。この現象は、脾臓およびリンパ節、ならびに粘膜免疫系が関与することができる。Gutgemannら(1998),Immunity 8:667−73、を参照されたい。しかしながら、例えば、サルモネラ−エルシニア(Salmonella Yersinia)またはエンタモエバ−ヒストリチカ(Entamoeba histolytica)のごとき腸病原体は、局所的(または全身的さえも)免疫応答を惹起することが可能である。腸管内への外来抗原の導入が免疫応答を惹起するかどうかを制御している因子は完全には理解されていない。Janewayら、上記文献、第5部、第14章。
【0126】
多くのワクチンは、抗原に対する免疫応答を促進するためにアジュバントを用いる。アジュバントは、抗原に対する免疫応答の特異性を強化するまたは広幅化する。免疫応答は、抗体力価の増加または抗原反応性T細胞数の増加を含むことができる。こうしたパラメーターを測定するための方法は当該技術分野に存在している。例えば、Zigtermanら(1988),J.Immunol.Methods 106(1):101−07。アジュバントが免疫応答を促進する機構は完全には理解されていないが、それらの使用は必須であろう。ごくわずかのワクチンが、選択された抗原に対する強い粘膜免疫応答を惹起可能である。
【0127】
本発明は、治療的有効量のBTL−IIのアンタゴニストおよび抗原を投与することを含んでなる、抗原に対する全身的または粘膜免疫応答を増進する方法を包含する。本発明を実施するために使用可能なBTL−IIのアンタゴニストには、BTL−IIの細胞外領域へ特異的に結合する拮抗的抗体またはインビトロ選択結合タンパク質、およびBTL−IIの生物学的活性を阻害可能な小分子が含まれる。所望により、BTL−IIタンパク質のアンタゴニストおよび抗原は、経口的に、経鼻的に、経膣的に、胃を経由して、または直腸を経由して、または吸入によるごとく、直接的に粘膜表面に投与可能である。例えば、少なくとも一つの場合においては、持続性免疫応答の誘導において膣投与よりも経鼻投与がより有効であることが報告されている。Russell(2002),Am.J.Reprod.Immunol.47(5):265−68。あるいは、BTL−IIアンタゴニストおよび/または抗原は、例えば、皮下、静脈内、筋肉内、動脈内または腹腔内に注射可能である。いくつかの態様において、抗体のごときBTL−IIアンタゴニストは注射可能であり、そして抗原は直接的に粘膜表面へ投与可能である。
【0128】
本発明の実施のために適した抗原には、任意の感染性病原体の全部もしくは一部、または感染性病原体に類似している病原体が含まれる。感染性病原体は、生きているまたは殺されたウイルス、細菌、およびアメーバ、鞭毛虫または蠕虫のごとき感染性真核動物を含むことが可能である。こうした感染性病原体に類似している病原体は、例えば、ワクチン接種されている哺乳動物へ感染可能なウイルスに類似しているが、しかし、それ自身ではワクチン接種されている哺乳動物へ感染できないウイルスであることができる。この例は、ワクチニアウイルス(ウシにおいては疾患を生じることが可能であるが、ヒトでは疾患を生じない)であり、それは天然痘(ヒトに疾患を生じる類似のウイルス)に対するワクチンを産生するためにジェンナーにより使用された。Janewayら、上記文献、第5部、第14章。表4は本発明を実施するために使用可能である、抗原の具体例を示している。
【0129】
【表4】
【0130】
【表5】
【0131】
あるいは、可溶性BTL−IIタンパク質を、自己免疫または炎症性疾患に関係づけられている抗原に対する寛容を促進するために使用可能である。例えば、実験的自己免疫脳脊髄炎(EAE)、多発性硬化症に多くの点で類似している状態を、例えば、ミエリン塩基性タンパク質またはミエリンオリゴデンドロサイト糖タンパク質(MOG)の多様なエピトープの注射により齧歯動物で誘導可能である。MOG誘導EAEは、いくつかの場合において、MOGまたはブチロフィリンの少量を前もって給餌することにより寛解可能である。Stefferlら(2000),J.Immunol.165:2859−65。可溶性BTL−IIタンパク質は、抗原に対する寛容性を促進し、そしてそれにより自己免疫疾患の症状を寛解するため、自己免疫疾患において標的とされていることが知られている抗原と同時投与することが可能である。所望により、抗原は、例えば、経鼻的に粘膜表面へ直接投与することが可能である。
【0132】
クローン病および潰瘍性大腸炎を含んだ炎症性腸疾患は、胃腸管の慢性炎症を含んでおり、多分、正常腸管フローラ(flora)の抗原に対する異常に促進された免疫応答のせいであろう。両方の疾患とも、発生は家族で集中発生する傾向があり、そしていくつかの遺伝子マーカーと関係づけることが可能であるので、少なくともいくらかの遺伝子的基盤を有しているようである。例えば、多薬剤耐性遺伝子(mdr1a)を発現しないマウスは自然発症的に大腸炎を発生する。Panwalaら(1998),J.Immunol.161:5733−44。クローン病および潰瘍性大腸炎両方の発生は、都市化した集団中で増加した発生が観察されるので、環境的因子にも影響されるようである。また、こうした疾患は、正常腸管フローラの非存在下では発生しない。
【0133】
クローン病には、口から肛門までの消化管のいずれかの部分の異常炎症が含まれるけれども、ほとんどの患者の異常炎症は回結腸、小腸および結腸−肛門直腸領域に限られている。典型的には、炎症は不連続である。共通の症状には、腹部痛、食欲不振、体重減少、発熱、下痢、腹部右下4分部位におけるふくらみおよび圧痛、便秘、嘔吐および肛門周囲不快および放出が含まれる。他の可能な症状には、なかでも、末梢関節炎、成長遅延、上強膜炎、アフタ性口内炎、結節性紅斑、壊疽性膿皮症、腎結石、尿希釈およびアルカリ化障害、吸収不良および胆石が含まれる。例えば、Stroberら,「Medical Immunology」,第10版,III節,第35章(2001);「Merck Manual of Diagnosis and Therapy」,第17版,3節,第31章,(1999)、を参照されたい。クローン病の患者から単離されたマクロファージは、増量されたIL−12、IFNγ、THFαおよび他の炎症性サイトカインを産生する。
【0134】
潰瘍性大腸炎は、しばしばクローン病と区別するのが難しいが、いくつかの点でクローン病とは性質が異なっている。第一に、それは一般に結腸に限定され、一方、クローン病は消化管を通して発生することができる。第二に、クローン病においては炎症が腸管または消化管の他の位置での壁を通してずっと入り込むことが可能であるのとは違って、潰瘍性大腸炎は主として腸管の浅層のみの炎症を含んでいる。最後に、潰瘍性大腸炎は、クローン病で典型的な炎症の非連続的部位よりもむしろ、炎症の連続的領域を典型的には含んでいる。クローン病同様に、潰瘍性大腸炎は主として都市領域で観察される。また、家族性集積の場合があるので、遺伝子的因子も潰瘍性大腸炎において役割を演じているらしい。自己抗体は、潰瘍性大腸炎患者において、クローン病患者よりもより頻繁に観察される。自己抗体はしばしば結腸上皮細胞成分に方向付けられる。最も普通であるのは、カタラーゼ、α−エノラーゼおよびラクトフェリンに特異性を有する、抗好中球細胞質抗体である。いくつかの場合、抗体は結腸微生物と交差反応する。
【0135】
潰瘍性大腸炎の症状は色々である。これらは、下痢、しぶり、腹部痙攣、便中の血液および粘液、発熱および直腸出血を含むことができる。結腸が6センチメーターを超えて拡張し、そしてその筋肉が緊張を失うおよび/または穿孔が起こってもよい、生命を脅かす可能性のある中毒性巨大結腸症もまた発生してもよい。潰瘍性大腸炎に伴ってもよい他の症状には、末梢関節炎、強直性脊髄炎、仙腸骨炎、前部ブドウ膜炎、結節性紅斑、壊疽性膿皮症、上強膜炎、自己免疫肝炎、原発性硬化性胆管炎、硬変症、および小児における成長および発育遅延を含むことができる。
【0136】
BTL−IIタンパク質へ特異的に結合する抗体またはインビトロ選択結合タンパク質は、炎症性腸疾患の発症を診断または予測するために使用可能である。下記実施例4に例示したように、炎症性腸疾患のモデルマウスにおいて、BTL−IIは、腸管において症状の発症に先立って、そして症状相の間に過剰発現される。従って、BTL−IIの過剰発現は、炎症性腸疾患の存在を示すことが可能であり、そしてその発症を予測可能である。抗BTL−II抗体は、ELISAアッセイまたは当該技術分野では既知の他の免疫に基づいたアッセイを使用し、患者の腸管からの組織試料をアッセイすることにより、BTL−IIの過剰発現を検出するために使用可能である。例えば、Reen(1994),酵素結合免疫吸着アッセイ(ELISA),「Basic Protein and Peptide Protocols,」中, Methods Mol.Biol.32:461−466、を参照されたい。過剰発現はまた、当該技術分野では既知のmRNA発現アッセイのなかでも、例えば、逆転写プラスPCR(RT−PCR)のごとき、BTL−II mRNA発現を測定するための、核酸に基づいた方法によっても検出可能である。例えば、Murphyら(1990),Biochemistry 29(45):10351−56、を参照されたい。
【0137】
別の態様において、本発明の可溶性BTL−IIタンパク質は、炎症性腸疾患を処置するために使用可能である。可溶性BTL−IIタンパク質は、B細胞またはT細胞上に発現される特異的レセプターへ結合可能であり、それにより免疫応答の下方調節を可能にしている。こうした下方調節は、例えば、CD4+T細胞によりマクロファージまたはB細胞の活性化を防止すること、または抗原によりT細胞の活性化を防止することが可能である。あるいは、こうした下方調節は、T細胞が、T細胞レセプターが特異的に結合可能な抗原に遭遇した場合、T細胞が反応不顕性(anergic)になることを起こすことが可能である。
【0138】
BTL−IIタンパク質へ特異的に結合する抗体またはインビトロ選択結合タンパク質はまた、炎症性腸疾患を患った、または炎症性腸疾患を発生する危険性のある患者を同定するための診断試薬としての使用も見いだせる。マウスモデルシステムにおいて、BTL−IIは炎症性腸疾患症状の発症に先立ってそしてその間に過剰発現されるので(実施例4)、BTL−II発現の異常に高いレベルは、腸管における炎症性疾患の存在、または腸管において炎症性疾患を発生する高い危険性を示すことが可能である。
【0139】
加えて、可溶性BTL−IIタンパク質は、移植(Manilayら,1998,Curr.Opin.Immunol.10:532−538)、移植片対宿主疾患、移植片拒絶、自己免疫または炎症性疾患、遺伝子治療(Hackettら,2000,Curr.Opin.Mol.Therap.2:376−382)などのごとき、免疫応答の下方調節が望まれる状態において有用であり得る。例えば、可溶性BTL−IIタンパク質は、哺乳動物への遺伝子治療ベクターの投与、移植、または所望の免疫抑制が適切な別の状態に先だって、ほとんど同時に(またはすぐ前かすぐ後に)、または同時に投与可能である。またこうした処置に適切であるのは、BTL−IIの機能を模倣した抗イディオタイプ抗体である。
【0140】
アゴニストBTL−II抗体、可溶性BTL−IIタンパク質または抗イディオタイプ抗体は、検出可能自己抗体の数を減少させるため、免疫エフェクター細胞の活性化を減少させるため、および/または疾患の症状を軽減または除去するため、自己免疫または炎症性疾患を患っている患者へ投与可能である。自己免疫および炎症性疾患には、患者自身の組織が患者の免疫系により引き起こされる有害な効果を受けやすい、すべての状態が含まれる。こうした効果は、可能性のなかでも、自己抗体により、および/または免疫エフェクター細胞の活性化により仲介可能である。自己免疫および炎症性疾患の原因は通常明らかではないが、多様な種類の感染と多様な自己免疫疾患の間の相関がいくつかの場合に確立されており、そして科学論文における議論の繰り返される主題である。例えば、Corapciogluら(2002),Thyroid 12:613−17;Sewellら(2002),Immunol.Lett.82:101−10;Rose(1998),Semin.Immunol.10(1):5−13;Matsiota−Bernard(1996),Cli.Exp.Immunol.104:228−35;およびMcMurrayおよびElboune(1997),Semin.Arthritis Rheum.26:690−701、を参照されたい。
【0141】
当業者は、自己免疫および炎症性疾患の症状が非常に広範で、そしてどの組織が患者の免疫系の標的にされているかに依存可能なことを認識するであろう。自己免疫および炎症性疾患は器官特異的または全身的であり得る。自己免疫および炎症性疾患には、多くの中でも、例えば、関節炎、アジソン病、インシュリン依存性糖尿病(タイプI糖尿病)、喘息、多発性内分泌障害症候群、全身性紅斑性エリテマトーデス、慢性活性肝炎、多様な形の甲状腺炎(ハシモト甲状腺炎、一過性甲状腺炎症候群およびグレーブス病を含んで)、リンパ球性腺下垂体炎、早期卵巣不全、特発性フィオパラサイロイディズム、悪性貧血、腎炎、自己免疫好中球減少症、グッドパスチャー症候群、多発性硬化症、白斑、重症筋無力症、リューマチ様関節炎、強皮症、原発性シェーグレン症候群、多発性筋炎、自己免疫溶血性貧血、炎症性腸疾患(クローン病および潰瘍性大腸炎を含んで)、乾癬、乾癬性関節炎、皮膚炎、自己免疫血小板減少性紫斑病、尋常性天疱瘡、急性リウマチ熱、混合本態性クリオグロブリン血症、および中程度自己免疫溶血性貧血が含まれる。
【0142】
ベクターおよび宿主細胞
本発明はまた、本発明の核酸を含んでいるベクター、ならびにこうしたベクターで形質転換した宿主細胞も提供する。本発明の核酸のいずれも、一般に選択可能マーカーおよび宿主中での増殖のための複製起点を含むベクターを含むことができる。ベクターはさらに、機能可能であるようにBTL−II核酸に連結された、哺乳動物、微生物、ウイルスまたは昆虫遺伝子からのもののごとき、適した転写または翻訳調節配列を含んでいる。こうした調節配列の例には、転写プロモーター、オペレーターまたはエンハンサー、mRNAリボソーム結合部位、および転写および翻訳を制御する適切な配列が含まれる。調節配列が、標的タンパク質をコードしているDNAへ機能的に関連している場合、ヌクレオチド配列は機能可能なように連結されている。それ故、もし、プロモーターヌクレオチド配列がBTL−II配列の転写を方向付けるならば、プロモーターヌクレオチド配列はBTL−II核酸配列へ機能可能なように連結されている。
【0143】
本発明の標的BTL−IIタンパク質をコードしているBTL−II核酸のクローニングのために適したベクターの選択は、ベクターが形質転換されるであろう宿主細胞、そして、当てはまる場合、標的ポリペプチドが発現されるべき宿主細胞に依存するであろう。BTL−IIタンパク質の発現に適した宿主細胞には、原核生物、酵母、昆虫そしてより高等な真核生物が含まれ、その各々が以下に議論されている。
【0144】
こうした宿主細胞中で発現されるべきBTL−IIタンパク質はまた、異種タンパク質からの領域を含む融合タンパク質であることができる。上で議論したように、こうした領域は、例えば、分泌、改良された安定性、容易にされた精製、標的化またはBTL−IIのオリゴマー化を可能にするために含ませることができる。例えば、適切なシグナル配列をコードしている核酸配列を発現ベクター内へ取り込むことが可能である。シグナル配列(分泌リーダー)をコードしている核酸配列は、BTL−IIがシグナル配列を含んでなる融合タンパク質として翻訳されるように、BTL−II配列へインフレーム(in−frame)で融合することができる。意図された宿主細胞内で機能的に可能であるシグナル配列は、BTL−IIタンパク質の細胞外分泌を促進することが可能である。異種シグナルペプチドは、天然のシグナル配列と置き換えることが可能である。哺乳動物宿主細胞中で機能的であるシグナルペプチドの例には、米国特許第4,965,195号に記載されているインターロイキン−7(IL−7)のためのシグナル配列、Cosmanら((1984),Nature 312:768)により記載されているインターロイキン−2レセプターのためのシグナル配列;欧州特許第0 367 566に記載されているインターロイキン−4レセプターシグナル配列;米国特許第4,968,607号に記載されているタイプIインターロイキン−1レセプターシグナル配列;欧州特許第0 460 846に記載されているタイプIIインターロイキン−1レセプターシグナル配列;ヒトIgKのシグナル配列(それはMETDTLLLWVLLLWVPGSTGである);およびヒト成長ホルモンのシグナル配列(それはMATGSRTSLLLAFGLLCLPWLQEGSAである)が含まれる。好ましくは、シグナル配列は、細胞からのBTL−IIタンパク質の分泌と同時にBTL−IIタンパク質から切断されるであろう。本発明の実施で使用可能である他のシグナル配列には、Sf9昆虫細胞中の、酵母α因子およびミツバチメラチンリーダーが含まれる。Brake(1989),Biotechnology 13:269−280;Homaら(1995),Protein Exp.Purif.6:141−148;Reavyら(2000),Protein Exp.Purif.6:221−228。
【0145】
本発明のタンパク質の発現に適した宿主細胞には、原核生物、酵母、そしてより高等な真核生物細胞が含まれる。これらのポリペプチドの発現に使用されるのに適した原核生物宿主には、エシェリキア(Escherichia)、バシラス(Bacillus)およびサルモネラ(Salmonella)属の細菌、ならびにシュードモナス(Pseudomonas)、ストレプトミセス(Streptomyces)およびスタフィロコッカス(Staphylococcus)属のメンバーが含まれる。原核生物細胞中、例えば、大腸菌中での発現のためには、BTL−IIタンパク質をコードしているポリヌクレオチド分子は、好ましくは、組換えポリペプチドの発現を容易にするために、N末端メチオニン残基を含んでいる。N末端Metは所望により発現されたポリペプチドから切断されてもよい。
【0146】
細胞性宿主中で使用するための発現ベクターは、一般に一つまたはそれより多くの表現型選択可能マーカー遺伝子を含んでなる。こうした遺伝子は、例えば、抗生物質耐性を与えるまたは栄養要求性要求を供給するタンパク質をコードしている。広範囲の種類のこうしたベクターは、商業源から容易に入手可能である。例には、pGEMベクター(Promega)、pSPORTベクターおよびpPROEXベクター(InVitrogen,Life Technologies,カールスバッド,CA),Bluescriptベクター(Stratagene)およびpQEベクター(Qiagen)が含まれる。
【0147】
BTL−IIはまた、サッカロミセス、ピキア(Pichia)およびクルベロミセス(Kluveromyces)を含む属からの酵母宿主細胞で発現可能である。好ましい酵母宿主は、S.セレビジエ(S.cerevisiae)およびP.パストリス(P.pastris)である。酵母ベクターはしばしば2μ酵母プラスミドからの複製配列起点、自己複製配列(ARS)、プロモーター領域、ポリアデニル化のための配列、転写終結のための配列、および選択可能マーカー遺伝子を含んでいるであろう。酵母および大腸菌の両方で複製可能なベクター(シャトルベクターと名付けられている)もまた使用することができる。酵母ベクターの上述の特性に加え、シャトルベクターはまた、大腸菌中での複製および選択のための配列を含んでいるであろう。酵母宿主中で発現された標的ポリペプチドの直接分泌は、BTL−IIコードヌクレオチド配列の5’末端に、酵母α因子リーダー配列をコードしているヌクレオチドの包含により達成することができる。Brake(1989),Biotechnology 13:269−280。
【0148】
昆虫宿主細胞培養系もまた、BTL−IIタンパク質の発現のために使用可能である。本発明のタンパク質は好ましくは、例えば、LuckowおよびSummers((1988),BioTechnology 6:47)による総説に記載されているような、バキュロウイルス発現系を使用して発現する。
【0149】
本発明のBTL−IIタンパク質は、哺乳動物宿主細胞においても発現可能である。適した哺乳動物宿主細胞の非制限例には、サル腎臓細胞のCOS−7株(Gluzmanら(1981),Cell 23:175−182)、チャイニーズハムスター卵巣(CHO)細胞(Puckら(1958),PNAS USA 60:1275−1281)CV−1(Fischerら(1970),Int.J.Cancer 5:21−27)およびヒト頚部癌腫細胞(HELA)(ATCC CCL2)が含まれる。
【0150】
本発明のBTL−IIタンパク質の発現に適した発現ベクターの選択は、使用されるべき特別の哺乳動物宿主細胞に依存するであろう。適した発現ベクターの例には、pcDNA3.1Hygro+(Invitrogen)、pDC409(McMahanら(1991),EMBO J.10:2821−2832)およびpSVL(Pharmacia Biotech)が含まれる。哺乳動物宿主細胞で使用するための発現ベクターは、ウイルスゲノムに由来する転写および翻訳制御配列を含むことが可能である。BTL−IIを発現するために使用可能な、普通に使用されるプロモーター配列およびエンハンサー配列には、限定されるわけではないが、ヒトサイトメガロウイルス(CMV)、アデノウイルス2、ポリオーマウイルスおよびサルウイルス40(SV40)に由来するものが含まれる。哺乳動物発現ベクターの構築法は、例えば、OkayamaおよびBerg((1982)Mol.Cell.Biol.2:161−170)、Cosmanら((1986)Mol.Immunol.23:935−941)、Cosmanら((1984)Nature 312:768−771)、EP−A−0367566およびWO 91/18982、に開示されている。
【0151】
特定のベクター内への挿入を容易にするためのBTL−II核酸の修飾(例えば、制限部位を修飾することにより)、特定の発現系または宿主における使用の容易さ(例えば、好ましい宿主コドンを使用して)などは既知であり、本発明での使用に意図されている。BTL−IIタンパク質の産生のための遺伝子操作法は、既知の方法に従った、無細胞発現系、細胞性宿主、組織および動物におけるポリヌクレオチド分子の発現を含んでいる。
【0152】
治療法
本明細書で言及したいかなる病気の“処置”も、疾患の少なくとも一つの症状の軽減、疾患の重度の軽減、または、いくつかの場合、疾患を伴うことができるより重度な症状への、または少なくとも一つの他の疾患への疾患進行の遅延または防止を包含している。処置は、疾患がすっかり治癒されることを意味する必要はない。有用な治療剤は、疾患の重度を軽減し、疾患またはその処置に付随する症状の重度を軽減し、または処置状態に続いていくらかの頻度で生じる可能性があるより重大な症状またはより重大な疾患の発症を遅延させることのみを必要とする。例えば、もし疾患が炎症性腸疾患であれば、治療剤は、腸管中の別々な部位の炎症の数を軽減し、影響を受けた腸管の全範囲を軽減し、痛みおよび/または膨潤を軽減し、下痢、便秘または嘔吐のごとき症状を軽減し、および/または腸管の穿孔を防止することができる。患者の状態は、バリウム浣腸または注腸に続いて実施されるX−線、内視鏡検査、大腸鏡検査および/または生検のごとき標準技術により評価可能である。適した方法は患者の状態および症状に従って変化する。
【0153】
本発明は、自己免疫疾患、移植片対宿主疾患および炎症性腸疾患を含んだ炎症性疾患を処置する方法を包含し、BTL−IIタンパク質または抗体の量を、特定の障害の重度または障害により引き起こされる症状の重度を反映する指標のベースラインを超えた持続性の改良を誘導するために、またはいくつかのまたは全ての場合において、処置された状態に続くより重大な疾患の発症を遅延させるまたは防止するために十分な時間使用する。本発明の処置は、問題とする障害のために通常使用する他の処置の前、後もしくは間に使用することができ、または他の処置なしでも使用することができる。例えば、クローン病および潰瘍性大腸炎は通常、スルファサラジン、5−アミノサリチル酸またはコルチコステロイドで処置する。これらの処置は、本発明の処置の前、間または後に使用することができる。
【0154】
上述の治療剤はいずれも組成物の形で、即ち、生理的に受容可能な坦体、賦形剤または希釈剤のごとき一つまたはそれより多くの追加の成分とともに投与可能である。例えば、組成物は本明細書に記載したような可溶性BTL−IIタンパク質に加えて、緩衝液、アスコルビン酸のごとき抗酸化剤、低分子量ポリペプチド(10より少ないアミノ酸を有しているもののごとき)、タンパク質、アミノ酸、グルコース、スクロースまたはデキストリンのごとき炭水化物、EDTAのごときキレート剤、グルタチオンおよび/または他の安定化剤、賦形剤および/または保存剤を含んでなる。組成物は液体または凍結乾燥物として処方することができる。医薬処方に用いることができる成分のさらなる例は、「Remington’s Pharmaceutical Sciences」,第16版,Mack Publishing Company,イーストン,Pa.(1980)、に示されている。
【0155】
上述の治療分子を含んでなる組成物は、限定するわけではないが、非経口、局所、経口、経鼻、経膣、直腸または肺(吸入により)投与を含んだ任意の適切な手段により投与可能である。もし注射するならば、組成物は関節内、静脈内、動脈内、筋肉内、腹腔内または皮下で、大量注射または連続的注入により投与可能である。経皮搬送および移植物からの徐放性放出、皮膚パッチまたは座剤であるような、局所投与(即ち、疾患の部位での)が意図される。吸入による搬送には、例えば、経鼻または経口吸入、噴霧器の使用、エアロゾル形での吸入などが含まれる。体腔内へ挿入された座剤を経る投与は、例えば、選ばれた体腔中に固形の組成物を挿入し、そしてそれを融解させることにより達成可能である。炎症性腸疾患を処置するための可溶性BTL−IIタンパク質またはアゴニストの場合、治療剤が適切に局在化するので、直腸座剤を経た投与が特に適切であることができる。他の代替物には、点眼、錠剤、ロゼンジ、シロップおよびチューインガムのごとき経口製剤、そしてローション、ゲル、スプレーおよび軟膏のごとき局所製剤が含まれる。ほとんどの場合、ポリペプチドである治療分子は、局所的または注射により、または吸入により投与可能である。
【0156】
上述の治療分子は、処置されている状態を処置するため、有効であり得るいかなる量、頻度および期間でも投与可能である。用量は、治療分子の分子的性質および処置されている障害の性質に依存する。処置は、所望の結果を達成するために必要な限り続けることができる。本発明の治療分子は、可能な投与計画の中でも、単一用量として、または1日数回、毎日、1日おき、週2回、週3回、毎週、1週おき、および毎月用量を含んだ、定期的に与える一連の用量として投与可能である。処置の定期性は、処置の期間を通して一定であってもよいし、一定でなくてもよい。例えば、処置を最初は週間隔で行い、そして後では1週おきに行うことができる。日、週、月または年の期間を有する処置が本発明に包含される。処置を中断し、その後再び始めることもできる。維持用量を最初の処置の後で投与することができる。
【0157】
用量は、体重キログラム当たりのミリグラムとして(mg/kg)、または皮膚表面の平方メーター当たりのミリグラムとして(mg/m2)、または身長または体重に関係なく固定用量として計ることができる。これらの全ては、当該技術分野における標準容量単位である。ヒト皮膚表面面積は、標準式を使用して、身長および体重から計算する。
本発明を特定の例を参照して説明してきた。これらの例はいかようにも本発明を限定することを意味していない。十分に本発明の範囲内である、多様な変更および修飾を本発明に行うことができることは、本開示の目的のためには言うまでもない。多くの他の変更を行うことができることを当業者は容易に思いつくであろうが、それは本明細書に開示されている、そして付随する特許請求の範囲に定義されているように、本発明の精神に包含される。
【0158】
本明細書は多くの特許、特許出願および出版物の引用を含んでいる。すべての目的のため、各々は本明細書において援用される。
【0159】
実施例1:ヒトBTL−II cDNAの単離
RNAはいくつかの起源、クローン病または潰瘍性大腸炎患者からのヒト結腸組織試料、ヒト結腸癌細胞株Caco−2(American Type Culture Collection(ATCC)番号HTB−37)およびT84と称される結腸上皮細胞株(ATCC番号CCL−248)、から単離した。RNAを逆転写し、NCBI寄託番号NM_019602に開示されている核酸配列に基づいて設計したプライマーを使用するPCRにより増幅した。これにより、完全長BTL−IIの配列の上流部分(配列番号3)および配列番号7、配列番号9、配列番号11、配列番号13および配列番号15に開示されているスプライス変異体配列を得た。BTL−II mRNAの3’末端を含んでいるcDNAの単離は、3’RACE(cDNA末端の迅速増幅)、即ち、本質的にFrohmanら((1988),Proc.Natl.Acad.Sci.USA 85(23):8998−9002)のプロトコールを使用して達成した。3’RACEによるBTL−II cDNAの多くの変異体の分析から、可溶性タンパク質をコードする変異体は膜貫通ドメインを欠いていないことを明らかにした。図5a、6aおよび7aに示したように、変異体の多くがBTL−II配列の多くの部位に配列多型を含んでいた(またはアリル変異体)。
【0160】
実施例2:マウスBTL−II cDNAの単離
マウスBTL−II cDNAは以下のように単離した。RNAは、RNeasyキット(Qiagen)を使用してマウス結腸および小腸から単離し、そして、製造元の推奨に従って、DNAse I(Ambion)で処理し、残余染色体DNAを除去した。精製RNAを、10mMトリス−HCl、pH8.3、50mM KCl、5mM MgCl2、各々1mMのdNTP、2.5μMランダムヘキサマープライマ一、lU/μl RNAse阻害剤、および2.5U/μl MuLV逆転写酵素(PE Biosystems)中に単離されたRNAを含んでいる反応混合物を使用して、cDNAに転写した。反応混合物を25℃で10分、続いて48℃で30分、続いて95℃で5分インキュベー卜した。BTL−II遺伝子のPCR増幅反応は、10mMトリス−HCl、pH8.3、50mM KCl、1.5mM MgCl2、各々200μMのdNTPおよび2.5U ampliTaq DNAポリメラーゼ(Perkin Elmer)、および両方25ピコモルの上流(5’−TTACTGAGAGAGGGAAACGGGCTGTTTTCTCC)および下流(5’−GGACTTCATTGGTGACTGATGCCATCCAC)プライマー、を含んでいる100μlの最終容量で実施した。増幅反応は、94℃で40秒、55℃で40秒および72℃で40秒を、35サイクル実施した。増幅生成物は2%アガロースゲルで分析し、エチジウムブロミドにより可視化し、そして配列決定した。視覚的判断によると、エキソン3を欠くスプライス変異体(図9a)はエキソン1から8を含んでいる変異体(図8)よりも豊富であった。
【0161】
実施例3:多様な起源の細胞および組織中でのBTL−IIの発現
BTL−II mRNAの発現は、本質的にHeidら((1996),Genome Res.6(10):986−94)のプロトコールに従ったリアルタイムPCRを使用し、そして後にPCRが続く標準逆転写(RT−PCR;例えば、Fuquaら(l990),Biotechniques 9(2):206−11、を参照されたい)を使用して測定した。試験された全ての細胞型は、マウスからであるバイエル板からのCD11c+CD8+B220+(または形質球様)樹状細胞を除いて、ヒト起源であった。BTL−II発現が検出された細胞は以下のものである:非刺激または殺したスタフィロコッカス アウレウス(Staphylococcus aureus)、CD40リガンドおよびインターロイキン4で刺激したヒトB細胞;インターフェロンγで刺激した正常ヒト気管支上皮細胞(NEBE細胞;Lecherら(1983),Cancer Res.43(12pt.1):5915−21、を参照されたい);Calu−3細胞、肺上皮細胞株(ATCC番号HTB−55)、非刺激;T84、結腸上皮細胞株、非刺激またはインターフェロンγで刺激;Caco−2細胞、ヒト結腸癌細胞株、非刺激;マウスパイエル板からのCD11c+(低発現)CD8+B220+細胞、それは主に樹状細胞である;およびマウス末梢血リンパ球。絶対スケール上、発現は、試験されたほとんどの細胞で低かった。BTL−II mRNAの発現は、樹状細胞型への分化を誘導するため、ヒト末梢血単球のインビトロ処理から生じた樹状細胞では検出されなかった。しかしながら、ヒト末梢血から精製された、ヒト血液から精製されたCD123+形質球様樹状細胞では、BTL−II発現を検出した。BTL−II mRNAは、パイエル板からのマウス樹状細胞のCD11c+(低発現)CD8+B220+サブセットにおいて高度に濃縮されていた。BTL−II mRNA発現を、脾臓、リンパ節、胃、腸管膜リンパ節、骨髄、小腸、盲腸、肺、大腸、パイエル板および胸腺を含んだ多くのマウス組織型で、同様の方法により検出した。最も高いレベルの発現は、小腸、パイエル板および盲腸組織で検出した。
【0162】
実施例4:炎症性腸疾患のためのマウスモデルにおけるBTL−IIの発現
mdr1a−/−マウスは、慢性炎症性腸疾患の研究のモデルシステムとなり得る。Panwalaら(1998),J.Immunol.161:5733−44。マウス多薬剤耐性遺伝子、mdr1aは、腸上皮細胞およびリンパ様細胞を含む多くの組織において発現される、170kDa膜貫通タンパク質をコードしている。mdr1aを欠失しているマウスは、調節不全の上皮細胞成長および大腸の粘膜固有層内への大量白血球湿潤により特徴付けられる、重度の自発的腸炎症を発生しやすい。経口抗生物質でmdr1a− /−マウスを処置すると、疾患の発生を防止し、そして活性な炎症を消散させる。活性な大腸炎を有するマウスから単離されたリンパ様細胞は、腸細菌抗原に対する促進された反応性を示した。mdr1aは上皮細胞および白血球の両方により発現されるけれども、大腸炎の発生は、上皮細胞上でのmdr1a発現の欠損と相関する。
【0163】
使用されたmdr1a−/−マウスは、FVB遺伝子バックグラウンドであった。典型的には、mdr1a−/−マウスの群の約20%が、18から20週齢で大腸炎を自発的に発生し、一方、mdr1a−/−マウスの残りのマウスは健康に残存し、そして大腸炎を発生しない。疾患を発生した動物のパーセンテージは、動物施設の清潔度に依存している。
【0164】
最初に、RNAを腸管組織から調製し、逆転写後に、得られたcDNA内へ蛍光標識を取り込ませ、使用して、Stammerら((2000),Immunogenetics 51:373−342)により発表されている配列に基づき、BTL−II mRNAを検出するために設計されたオリゴヌクレオチドを含んでいる、注文製作のAffymetrixチップ上のプローブのアレイヘハイブリダイズさせた。BTL−II mRNAの過剰発現を、炎症性腸疾患症状の発症前および症状の間の両方で、mdr1a−/−マウスにおいて検出した(FVB株の野生型個体と比較して)。こうしたデータの実例は、図10に示されている。これらのデータは、BTL−II mRNAは野生型FVB マウスよりもmdr1a−/−マウスにおいてより大きな程度で発現されていることを示している。さらに、より高いBTL−II mRNAの発現が、炎症性腸疾患の症状の発症に伴っている。
【0165】
これらの結果は、リアルタイムPCR技術(Heidら(1996),Genome Res.6(10):986−94)を使用する、同一RNAの分析により確認した。この分析は、2匹の健康な野生型FVBマウスと比較して、2匹の症状性mdr1a−/−マウスによるBTL−II mRNAの過剰発現を示した。このデータは図11に図示されている。2匹の親マウスは、図11の向かって左の対角の縞で印を付けられた2本の棒線で示されており、2匹の症状性mdr1a−/−は、図11の向かって右の市松模様(checkered pattern)で印を付けられた2本の棒線で示されている。これらのデータは、野生型および症状性mdr1a−/−マウス間の発現に約2から5倍の相違が存在することを示している。従って 、症状のない野生型マウスよりも、より高いレベルのBTL−II mRNAが炎症性腸疾患の症状を有するマウスで発現されている。
【0166】
実施例5:BTL−II:FC融合タンパク質の構築
ヒトFc領域へ融合されたマウスBTL−IIの細胞外領域から成る可溶性BTL−IIタンパク質(BTL−II:Fc)は以下の方法で製造した。BTL−II:Fcをコードしている融合cDNA構築物は、マウスBTL−IIの細胞外領域をコードしている核酸を、ヒトIgG1をコードしている核酸へ融合することにより(インフレーム)製造した。BTL−II:Fcタンパク質を製造するため、哺乳動物細胞を、LIPOFECTAMINETM2000トランスフェクション法(Invitrogen,カールスバッド,カリフォルニア,米国)を使用して、融合cDNAでトランスフェクトした。BTL−II:Fcタンパク質含有上清は、トランスフェクション6−7日後に収集し、そしてBTL−II:Fcタンパク質は、プロテインAカラムクロマトグラフィーにより精製した。BTL−II:Fcタンパク質をコードしている核酸配列およびBTL−II:Fcアミノ酸配列は、各々配列番号19および配列番号20にみられる。
【0167】
実施例6:BTL−II:FcによるヒトT細胞増殖の抑制
以下の実験は、BTL−IIの可溶性形が、他の同時刺激性分子とともに、またはなしで、モノクローナル抗CD3ε抗体に応答して、インビトロでのT細胞増殖を抑制可能かどうか試験している。
【0168】
BTL−II:Fcは実施例5に記載したように作製した。96ウェル、U底マイクロタイタープレートを、一つまたはそれより多くの他のタンパク質とともに、またはなしで、濃度を変化させた抗CD3ε抗体で被覆した。図12は、以下のような、何のタンパク質が各々の試料に使用されたかを示している:抗CD3ε抗体単独、
【0169】
【化1】
【0170】
;抗CD3ε抗体およびBTL−II:Fc、
【0171】
【化2】
【0172】
;抗CD3ε抗体およびB7RP−1:Fc、
【0173】
【化3】
【0174】
;抗CD3ε抗体、B7RP−1:FcおよびBTL−II:Fc、
【0175】
【化4】
【0176】
;抗CD3ε抗体およびB7−2:Fc、
【0177】
【化5】
【0178】
;抗CD3ε抗体、B7−2:FcおよびBTL−II:Fc、
【0179】
【化6】
【0180】
。
B7RP−1:Fcは、抗体のFc領域へ融合されたB7RP−1の細胞外領域(B7hとして知られている)から成り、そしてB7−2:Fcは、抗体のFc領域へ融合されたB7−2の細胞外領域から成っている。これらのタンパク質の両方とも、抗原に応答するT細胞を変調することが知られているB7ファミリーのタンパク質のメンバーである。これらの融合タンパク質は、例えば、R&D Systems(ミネアポリス,ミネソタ,米国)のごとき商業的製造供給元から購入することが可能であり、またはBTL−II:Fcタンパク質のために実施例5に記載したように単離可能である。B7−2はR&D Systemsから購入し、そしてB7RP−1は、B7−H2/Fcの名前でR&D Systemsから入手可能であるが、本質的に実施例5で記載したように作製した。マイクロタイタープレートウェルは、一つまたはそれより多くのタンパク質(それはBTL−II:Fc(10μg/ml)、B7RP1:Fc(10μg/ml)および/またはB7−2:Fc(2μg/ml)である)とともにまたはなしで、図12に示された抗CD3ε抗体の濃度を含んでいる、100μlのリン酸緩衝液(PBS)を加えることにより被覆した。プレートは4℃で一夜インキュベートし、その後PBSで2回洗浄した。ヒトT細胞は、磁気的標識により機能し、そしてCD4+T細胞以外の末梢血細胞を枯渇させ、比較的純粋なもとのままのCD4+T細胞の集団を生じる、Miltenyi Biotec(Bergisch Gladbach,ドイツ)からのCD4+T細胞単離キットを使用して、ヒト末梢血単核細胞から精製した。約lxl05精製T細胞を、200μlの容量の培養培地(l0%ウシ胎児血清を含むRPMI)中で各々のウェルへ加えた。細胞を総計で72時間インキュベートした。64時間目、1μCiの3H−チミジンを各々のウェルに加えた。72時間終了後、取り込まれなかったチミジンは、細胞をフィルター上に沈着し、そして培養培地を洗い流す、自動細胞収穫器(Tomtec,ハムデン、コネチカット,米国、から得た)を使用して除去した。その上に細胞を含むフィルターは次ぎに、どのくらいの放射活性を細胞が取り込んでいたかを決定するため、シンチレーションカウンター中でカウン卜した。結果は図l2aおよび12bに示されており、それらは、図12aが対数目盛りであり、図12bは直線目盛りであることのみが、異なっている。
【0181】
結果は、BTL−II:Fcが、抗CD3ε抗体により誘導されるT細胞増殖を抑制可能であることを示している。図12aおよび図12b。図12aにおいて、試験された最も高い抗体濃度では、BTL−IIプラス抗CD3ε抗体
【0182】
【化7】
【0183】
を含んでいる試料は、抗CD3ε抗体のみ
【0184】
【化8】
【0185】
を含んでいる試料よりも増殖が悪かった。さらに、B7−2
【0186】
【化9】
【0187】
およびB7RP−7
【0188】
【化10】
【0189】
の両方とも抗CD3ε抗体誘導T細胞増殖を刺激することは明らかである。B7−2:Fc添加によるT細胞増殖の効果を見るために必要とされる抗CD3ε抗体の濃度は、B7RP−1:Fcの添加による同様の効果を見るために必要とされる濃度よりも低い。さらに、BTL−IIはまた、B7−2:Fc
【0190】
【化11】
【0191】
およびB7RP−1:Fc
【0192】
【化12】
【0193】
の添加に応答した、この増強されたT細胞増殖も抑制することが可能である。B7−2:Fcプラス抗CD3ε抗体により誘導された増殖に対するBTL−II:Fcの効果は、試験された抗CD3ε抗体の最も低い濃度でのみ明らかであり、一方 、B7RP1:Fcにより誘導された増殖に対するBTL−II:Fcの効果は、試験された三つのより高い濃度でより明白である。従って、観察された効果は、抗CD3ε抗体の濃度に依存している。
【0194】
図13は、BTL−II:FcによるヒトT細胞増殖の抑制はまた、BTL−II:Fc濃度にも依存していることを示している。実験は、上述のように、ただし、抗CD3ε抗体濃度を0.5μg/mlで一定に保って実行した。BTL−II:Fc濃度は、図13に示されているように、0から10μg/mlへ変化させた。図13に示されている結果は、抗CD3ε誘導T細胞増殖の抑制が、BTL−II:Fcの濃度に依存していることを示している。
【0195】
実施例7:BTL−II:FcはマウスT細胞増殖を抑制する
この実験は、BTL−II:Fcがマウス、並びにヒトT細胞の増殖を抑制できるかどうかを決定するために行われた。T細胞増殖アッセイは、本質的に実施例6に記載したように行われ、ただし、Miltenyi Biotec GmbH(Bergisch Gladbach,ドイツ)から購入された磁気マイクロビーズを使用して精製されたマウスT細胞がヒトT細胞の代わりに使用された。トランスフェクトCHO細胞から作製された、ヒトFc領域から成るタンパク質が対照として働いた。ウェルは、図14に示した濃度の抗CD3ε抗体単独
【0196】
【化13】
【0197】
で、または、抗CD3ε抗体プラス10μg/mlのBTL−II:Fc
【0198】
【化14】
【0199】
かまたは10μg/mlのヒトFc領域から成るタンパク質
【0200】
【化15】
【0201】
で被覆した。結果は、BTL−II:Fcは抗CD3ε抗体に応答したマウスT細胞増殖を抑制できたが、ヒトFc領域から成るタンパク質単独では抑制できなかったことを示している。図14。
【0202】
実施例8:BTL−II:FcはマウスB細胞増殖を抑制しない
以下の実験は、ヒトBTL−II:Fcが、TALL−1およびIgM抗体により誘導されるマウスB細胞の増殖を阻害できるかどうかを決定するために設計された。実験は本質的に、Khareら((2000),Proc.Nad.Acad.Sci.97(7):3370−75)に記載されているように実行した。簡単には、マウスB細胞は、最初に密度勾配遠心分離によりリンパ球を精製し、次ぎにリンパ球は、単球/マクロファージおよびCD4+およびCD8+細胞を除去するB細胞カラム(Cedarlane,ウエストベリー,NY)を通過させることにより、脾臓から精製した。10%熱不活性化ウシ胎児血清を加えたMEM中の約1xl05精製B細胞を、ヤギF(ab’)2抗マウスIgM(2μg/ml)、ヒトTALL−1(10ng/ml)および/またはBTL−II:Fc(10μg/ml)とともに、またはなしで、96ウェルマイクロタイタープレート中、37℃にて4日インキュベー卜した。3H−チミジン(1μCi)は、インキュベーションの最後の8時間の間に加えた。4日の終了時に、実施例6に記載したように細胞を採取し、シンチレーションカウンターでカウントした。図15の印付けは、以下の細胞およびタンパク質の組み合わせを示している:B細胞単独、
【0203】
【化16】
【0204】
(これは図15に示したグラフ上、最も左の棒線であるが、検出可能なシグナルは存在しない);B細胞プラスTALL−1、
【0205】
【化17】
【0206】
;B細胞プラス抗IgM抗体、
【0207】
【化18】
【0208】
;B細胞プラスTALL−1および抗IgM抗体、
【0209】
【化19】
【0210】
;並びにB細胞プラスTALL−1、抗IgM抗体およびBTL−II:Fc、
【0211】
【化20】
【0212】
。結果は、BTL−II:FcはTALL−1および抗IgM抗体により誘導されたB細胞の増殖には効果がないことを示している。図15。それ故、BTL−IIはT細胞の増殖を阻害するが、B細胞の増殖は阻害しないと思われる。
【0213】
実施例9:T細胞によるサイトカイン産生に対するBTL−II:Fcの効果
以下の実験は、BTL−II:FcがT細胞によるサイトカイン産生に対して効果を有するかどうかを決定することを目的にしている。T細胞を精製し、マイクロタイタープレートを実施例6に記載したようにタンパク質で被覆した。約1x105の細胞を、200μl容量の培地(10%ウシ胎児血清を含むRPMI)を含む各々のウェルへ加えた。細胞は、37℃で64時間インキュベートした。次ぎに、サイトカイン濃度を決定するために150μlを除き、各々のウェル中に残っている50μlへ 3H−チミジンを加えた。マイクロタイタープレートは続いてさらに37℃で追加の8時間インキュベートし、続いて洗浄し、増殖の違いを確かめるため、実施例6に記載したようにカウントした(図16aに示されている)。何のタンパク質がウェルを被覆するために使用されたかを示す印付けは、図12aおよび図12bの通りである(実施例6に説明されている)。インターフェロンガンマ(IFNγ)、インターロイキン2(IL2)およびインターロイキン5(IL5)の濃度は、Meso Scale Discovery(IGEN International,Inc.の系列であるMSD,ガイサースブルグ,メリーランド ,米国)により販売されている多サイトカインの同時検出のための、電気化学に基づいたイムノアッセイシステムを使用して決定した。この種のサイトカイン検出の原理および操作は、例えば、Sennikovら(2003),J.Immunol.Methods 275:81−88、に説明されている。サイトカイン量の単位は、MSDの機械により発生された読み取り値から採られた。培地中のサイトカインの実際の濃度は、既知の濃度のタンパク質で発生させた検量線との比較なしでは、このデータから決定することはできず、それはこの特定の実験では付随していない。しかしながら、単一の実験内での読み取り値間の比較は、対照対実験試料に存在するサイトカインの相対量を提供することが可能である。
【0214】
結果は図16a−16dに示されている。図16aは、BTL−II:Fcは、抗CD3ε抗体または抗CD3ε抗体プラスB7RP−1:Fcに応答した増殖を阻害するが、使用された抗CD3ε抗体の濃度では、抗CD3ε抗体プラスB7−2:Fcに応答した増殖は阻害しないことを示している。しかしながら、実施例6で指摘したように(図12aおよび図12b)、より低い抗CD3ε抗体濃度、即ち、0.1μg/mlでは、抗CD3ε抗体プラスB7−2:Fcに応答した細胞増殖が、BTL−II:Fcにより阻害される。IFNγ、IL2およびIL5の産生は、抗CD3ε抗体単独で被覆されたウェルにおいては低く、そしてBTL−II:Fcの添加は、さらに有意にはそれを減少させなかった。図16a−16d。抗CD3ε抗体へのB7RP−l:FcかまたはB7−2:Fcの添加に応答したIFNγ産生の増加は、BTL−II:Fcにより阻害された。図16b。加えて、抗CD3ε抗体へのB7−2:Fcの添加に応答したIL2およびIL5産生の増加は、BTL−II:Fcにより阻害された。図16cおよび16d。試験された濃度では、抗CD3ε抗体へのB7RP−1:Fcの添加は、IL2またはIL5産生を評価できるほどは増加させなかった。図16cおよび16d。これらの結果は、BTL−II:Fcが、抗CD3ε抗体プラスB7−2:FcかまたはB7RP−1:Fcの組み合わせに応答した、少なくともいくつかのサイトカインの産生を阻害できることを示している。
【0215】
実施例10:BTL−II:FcによるT細胞の処置は、大量細胞死を生じさせない
以下の実験は、BTL−II:FcによるT細胞増殖の阻害が大量細胞死を伴うかどうかを決定するために行われた。T細胞増殖アッセイは本質的に実施例6に記載したように実施したが、ただし細胞は3H−チミジンでは標識しなかった。細胞は培養72時間後に、血球計数器でカウントし、そして細胞生存度は、トリパンブルー染色により決定した。マイクロタイタープレートウェルを被覆するために使用したタンパク質は、実施例6で説明し、そして図12aおよび図12bに示されているように図17でも示されている。抗CD3ε抗体は2μg/mlの濃度で使用した。結果は、各々のウェル中の死亡細胞の数は、ウェルを被覆するために使用されたタンパク質に関わらず、約0.5xl04から0.75xl04細胞の範囲であったことを示している。図17。それ故、BTL−II:Fc存在下で観察された細胞増殖の相違は、BTL−II:Fcの実質の毒性効果を反映していない。
【0216】
実施例11:マウスBTL−IIはマウスCTLA4、CD28、ICOSまたはPD−1とは結合しない
以下の組の実験は、BTL−IIが他のB7タンパク質の、いくつかの既知の結合パートナーの一つと結合するかどうかの疑問に取り組んでいる。例えば、B7RP−1はICOSと結合することが知られており(Yoshinagaら(1999),Nature402:827−32)、CD80(B7−1とも称されている)は、CD86(B7−2とも称されている)が行うように、CTLA4およびCD28へ結合することが知られている、例えば、SharpeおよびFreeman(2002),Nature Reviews Immunology 2:116−26)、そしてPD−L1およびPD−L2はPD−1へ結合することが知られている(Latchmanら(2001),Nature Immunology 2(3):261−68)。
【0217】
実験は以下のように行った。第一に、B7タンパク質へ結合することが知られているタンパク質の細胞外領域に加えてヒトIgG抗体のFc領域を含んでなる融合タンパク質を得た、または単離した。ヒト抗体のFc領域に加えてマウスCD28(mCD28−huFc)かまたはマウスPD−1(PD1−huFc)の細胞外領域を含んでなる融合タンパク質は、R&D Systems(ミネアポリス,ミネソタ,米国)から購入した。他の二つのB7結合タンパク質(CTLA4−huFcおよびICOS−huFc)もまたR&D Systemsから入手可能であるが、以下のように作製した。マウスCTLA4の細胞外領域をコードしているcDNAおよびマウスICOSの細胞外領域をコードしている別のcDNAを、哺乳動物細胞での発現に適したベクター中のヒトIgG抗体のFc領域をコードしているcDNAへ各々融合させた。これらの構築物の各々の細胞をトランスフェクトのために使用し、そして融合タンパク質は、プロテインAクロマトグラフィーにより、トランスフェクトされた細胞の培養培地から精製した。
【0218】
三つのマウスcDNAの各々の完全長バージョン(BTL−II、B7RP−1、CD80をコードしている)を発現に適したベクター内へ挿入した。これらの構築物の各々の約10μgを、空ベクターと共に、トランスフェクト293細胞に対して別々に使用した。トランスフェクション2日後、四つのトランスフェクションの各々から約100万細胞を、上述の融合タンパク質の各々で染色した。結合したタンパク質は、ヒトIgG Fc領域に対する蛍光標識抗体を使用して検出した。染色細胞はFACSにより分析した。結果は図18に示されている。
【0219】
予測されるように、空ベクターでトランスフェクトされた細胞(図18の上段)は、四つの融合タンパク質のいずれによっても染色されなかった。マウスBTL−IIでトランスフェクトされた細胞(図18の二段目)は同様に挙動し、融合タンパク質のどれもBTL−IIへ結合しないことを示している。予測されるように、マウスB7RP−1でトランスフェクトされた細胞はICOS−huFcで染色されたが、他の融合タンパク質のいずれでも染色されなかった。また予測されるように、マウスCD8Oでトランスフェク卜された細胞はCTLA4−huFcまたはmCD28−huFcで染色されたが、ICOS−huFcまたはPD−1−huFcでは染色されなかった。これらの結果は、BTL−IIは、四つのタンパク質、CTLA−4、PD−1、ICOSおよびCD28(その各々はB7ファミリー中の少なくとも一つのタンパク質へ結合することが知られている)へ結合できないことを示している。
【図面の簡単な説明】
【0220】
【図1】図1は、ブチロフィリン様タンパク質のファミリーの、選択されたメンバーのドメイン構造を示している。選択されたタンパク質のほとんどは、以下に議論するB7サブファミリーのメンバーである。各々のタンパク質の名前は、その構造が描かれている図の左に示されている。“Ig様”ドメインは以下に説明するように、免疫グロブリン様ドメインである。“7”とラベルされたドメインは以下に説明するように、7アミノ酸反復領域である。 “TM” とラベルされたドメインは膜貫通ドメインである。白四角は未同定の細胞質ドメインであり、ドメインの特別なファミリーの一部である。“B30.2” とラベルされたドメインは以下に説明するように、B30.2ドメインである。BTL−IIタンパク質は本明細書において、四つのIg様ドメインを有するように描かれているが、BTL−IIの形はまた、二つまたは三つのIg様ドメインを有するものも存在する。B7−H3は四つのIg様ドメインで描かれているが、それはまた二つのみのIg様ドメインを含んでいる形でも存在する。
【図2】図2は、ヒトBTL−II遺伝子およびmRNAの構造の図である。黒四角はエキソンを示している。四角の下に示されている水平線下の数字は、エキソンの配列番号3内の位置を示している。図の最下部の水平線は、細胞外ドメインおよび膜貫通および細胞質ドメイン(“TM/Cyto”)をコードし、そして3’非翻訳領域(“3UTR”)を形成している配列番号3の範囲を表示している。停止コドンは、日輪型(sunburst)または小さな爆発として記載することが可能な印により表示されている。
【図3a】図3aは、ヒトBTL−II mRNAのスプライス変異体の、第一のカテゴリーの構造の図である。記号は、図2で説明したものと同じである。エキソン2および3にかぶっている大きなXは、これらのエキソンが、スプライス変異体のこのカテゴリーでは失われていることを示している。
【図3b】図3bは、配列番号3の一部(上のライン)と並列された、スプライス変異体の第一のカテゴリーのメンバーの代表的配列(下のライン)である。失われたエキソン2および3により創造されたギャップ以外のミスマッチは存在しない。
【図4a】図4aは、ヒトBTL−II mRNAのスプライス変異体の、第二のカテゴリーの構造を示している。記号は、図2で説明したものと同じである。エキソン3にかぶっている大きなXは、このエキソンが、スプライス変異体のこのカテゴリーでは失われていることを示している。
【図4b】図4bは、配列番号3の一部(上のライン)と並列された、スプライス変異体の第二のカテゴリーのメンバーの代表的配列(下のライン)を示している。失われたエキソン3により創造されたギャップ以外のミスマッチは存在しない。
【図5a】図5aは、ヒトBTL−II mRNAのスプライス変異体の、第三のカテゴリーの構造を示している。記号は、図3で説明したものと同じである。エキソン2および3にかぶっている大きなXは、これらのエキソンが、スプライス変異体のこのカテゴリーでは失われていることを示している。黒四角に隣接する、数字を伴った星印は、スプライス変異体のこのカテゴリーに存在する配列多型の位置を示している。変異は、配列番号3内の以下の位置に存在する:変異2は1050位であり;変異3は1136および1140位であり;変異4は1178および1179位であり;変異5は1212位であり;および変異6は1242位である。
【図5b】図5bは、配列番号3の一部(上のライン)と並列された、スプライス変異体の第三のカテゴリーのメンバーの代表的配列(下のライン)を示している。ミスマッチ塩基はボールド体で示されている。
【図6a】図6aは、ヒトBTL−II mRNAのスプライス変異体の、第四のカテゴリーの構造を示している。エキソン3にかぶっている大きなXは、このエキソンが、スプライス変異体のこのカテゴリーでは失われていることを示している。配列多型は、図5aと同様に示されており、そして変異2から6は図5aと同様である。
【図6b】図6bは、配列番号3の一部(上のライン)と並列された、スプライス変異体の第四のカテゴリーのメンバーの代表的配列(下のライン)を示している。ミスマッチ塩基はボールド体で示されている。
【図7a】図7aは、ヒトBTL−II mRNAのスプライス変異体の、第五のカテゴリーの構造を示している。エキソン3にかぶっている大きなXは、このエキソンが、スプライス変異体のこのカテゴリーでは失われていることを示している。配列多型は、図5aと同様に示されており、そして変異2から6は図5aと同様である。変異1は、配列番号3のヌクレオチド78から80の欠失である。
【図7b】図7bは、配列番号3の一部(上のライン)と並列された、スプライス変異体の第五のカテゴリーのメンバーの代表的配列(下のライン)を示している。ミスマッチ塩基はボールド体で示されている。
【図8】図8はマウスBTL−II遺伝子およびmRNAの構造の図である。番号が配列番号5における位置を指していることを除いて、記号は図5で説明したものと同じである。
【図9a】図9aは、マウスBTL−II mRNAのスプライス変異体の、第一のカテゴリーの構造を示している。エキソン3にかぶっている大きなXは、このエキソンが、スプライス変異体のこのカテゴリーでは失われていることを示している。
【図9b】図9bは、配列番号5(上のライン)と並列された、マウススプライス変異体の第一のカテゴリーのメンバーの代表的配列(下のライン)を示している。失われたエキソン3により創造されたギャップ以外のミスマッチは存在しない。
【図10】図10は、非症状性、野生型の結腸組織におけるBTL−II mRNAの発現と比較した、炎症性腸疾患の症状を示していない場合(横縞)または炎症性腸疾患の症状を示している場合(市松模様)のmdr1a−/−マウス(FVBバックグラウンドにmdr1aヌル変異を運んでいる)の結腸組織におけるBTL−II mRNAの発現を表している。測定は、蛍光標識cDNAをBTL−II mRNAに相補的なオリゴヌクレオチドを含んでいるAffymetrixチップへハイブリダイズさせることにより行った。
【図11】図11は、FVB株の2匹の異なった野生型マウス(FVB#1および#2;対角の線)からの、および炎症性腸疾患の症状を示している2匹のmdr1a−/−マウス(Mdr1a−/−#1および#2;市松模様)からの結腸組織のリアルタイムPCRアッセイにより検出されたBTL−II mRNAの相対濃度を示しているグラフである。
【図12a】図12aは以下のタンパク質と培養された、精製ヒトT細胞の増殖(3H−チミジンの取り込みにより証明された)を示している棒グラフである:抗CD3ε抗体単独、
【0221】
【化21】
【0222】
;抗CD3ε抗体およびBTL−II:Fc、
【0223】
【化22】
【0224】
;抗CD3ε抗体およびB7RP−1:Fc、
【0225】
【化23】
【0226】
;抗CD3ε抗体、B7RP−1:FcおよびBTL−II:Fc、
【0227】
【化24】
【0228】
;抗CD3ε抗体およびB7−2:Fc、
【0229】
【化25】
【0230】
;および抗CD3ε抗体、B7−2:FcおよびBTL−II:Fc、
【0231】
【化26】
【0232】
。
【図12b】図12bは図12aと同一であるが、ただし対数目盛りではなく、直線目盛りが使用されている。
【図13】図13は、一定量の抗CD3ε抗体および示されているように量を変化させたBTL−II:Fcに応答した、精製ヒトT細胞の増殖を示している棒グラフである。
【図14】図14は、抗CD3ε抗体単独、
【0233】
【化27】
【0234】
、抗CD3ε抗体プラスBTL−II:Fc、
【0235】
【化28】
【0236】
、または抗CD3ε抗体プラスヒトFc領域から成る対照タンパク質、
【0237】
【化29】
【0238】
、に応答したマウスT細胞の増殖を示している棒グラフである。
【図15】図15は、タンパク質添加なし、
【0239】
【化30】
【0240】
、TALL−1単独、
【0241】
【化31】
【0242】
、抗IgM抗体単独、
【0243】
【化32】
【0244】
、TALL−1および抗IgM抗体、
【0245】
【化33】
【0246】
、およびTALL−1、抗IgM抗体およびBTL−II:Fc、
【0247】
【化34】
【0248】
、に応答したマウスB細胞の増殖を示している棒グラフである。
【図16a】図16aは、図12aに示したようなタンパク質の多様な組み合わせに応答した、精製ヒトT細胞の増殖を示している。
【図16b】図16bは、図12aに示したようなタンパク質の多様な組み合わせに応答した、相対インターフェロンガンマ(IFNγ)産生を示している。
【図16c】図16cは、図12aに示したようなタンパク質の多様な組み合わせに応答した、相対インターロイキン2(IL2)産生を示している。
【図16d】図16dは、図12aに示したようなタンパク質の多様な組み合わせに応答した、相対インターロイキン5(IL5)産生を示している。
【図17】図17は、図12aに示されているような、タンパク質の多様な組み合わせで培養された精製T細胞の培養において、死んだ細胞の総数を示している棒グラフである。
【図18】図18は、空ベクター(一番上の行)、かまたはマウスBTL−II(二番目の行)、マウスB7RP−1(三番目の行)またはマウスCD80(一番下の行)をコードしているcDNAを含んでいるベクターでトランスフェクされた細胞のFACSスキャンを示している。細胞は次のタンパク質で染色した:(1)IgG1抗体のヒトFc領域に融合されたマウスCTLA4の細胞外領域(一番目の列);(2)IgG1抗体のヒトFc領域に融合されたマウスCD28の細胞外領域(二番目の列);(3)IgG1抗体のヒトFc領域に融合されたマウスICOSの細胞外領域(三番目の列);および(4)IgG1抗体のヒトFc領域に融合されたPD1の細胞外領域(四番目の列)。各々のスキャンの縦軸(“カウント”とラベルされている)は細胞数を表し、そして横軸(“FL2−H”とラベルされている)は蛍光を表している。“M1”とラベルされた水平線は、“ゲート(gate)” がFACS機器に使用されたところを示している。このゲートに含まれる細胞が陽性と考えられる;全ての他の細胞は陰性と考えられる。各々のFACSスキャン上の文字は、個々の試料番号を示している。
【配列表】
【技術分野】
【0001】
関連出願
本出願は、2002年12月23日に出願された米国仮出願第60/436,185号および2003年11月26日に出願された米国仮出願第60/525,298号の優先権を主張する。
【0002】
技術分野
本発明はブチロフィリン様タンパク質、特に、例えばB細胞および/またはT細胞のごとき免疫エフェクター細胞の機能を変調することが知られている、B7サブファミリーのブチロフィリン様タンパク質に関する。こうしたタンパク質をコードする核酸、こうしたタンパク質を産生するためのプロセス、こうしたタンパク質へ結合する抗体、こうしたタンパク質または抗体を含む医薬組成物、こうした核酸、タンパク質およびこうしたタンパク質に対する抗体を使用する方法もまた含まれている。
【背景技術】
【0003】
背景技術
免疫または炎症性応答の変調は、自己免疫疾患、異常な炎症および/または免疫応答により特徴付けられる疾患、並びに感染を含む、多様な種類の疾患の制御において、役に立つ手段であることが可能である。炎症性腸疾患および自己免疫または炎症性疾患のごとき、異常な炎症および/または免疫応答により特徴付けられる疾患の処置においては、免疫応答の下方変調が望まれる。他の事態、例えば、感染性疾患に対する免疫性を与えるために患者にワクチン接種する場合、免疫応答の刺激が望まれる。ワクチン設定において、同時投与される抗原に対する免疫応答を高めることが可能なアジュバントは、疾患に対する長期の保護を提供することにおいて役立つことが可能である。特に当該分野において欠けていることは、粘膜免疫応答を刺激することが可能なアジュバントである。全身性免疫応答に対する粘膜免疫応答は、まさしく共通の侵入地点、即ち、粘膜表面で感染を攻撃できるので、価値がある。本発明は、不適当なおよび/または異常な炎症および/または免疫応答により特徴付けられる疾患を診断そして処置するための治療剤、および免疫応答、特に粘膜免疫応答を刺激するためのアジュバントとして作用可能な治療剤を提供することにより、当該分野でのこれらの要求に取り組んでいる。
【発明の開示】
【発明が解決しようとする課題】
【0004】
概要
本発明は、単離BTL−IIタンパク質、核酸、抗体、BTL−II阻害剤およびアゴニスト、そしてこれらの組成物を使用するための方法を包含する。
【0005】
配列番号4のアミノ酸x−y、ここにおいてxは配列番号4の1から35位のいずれかのアミノ酸であり、そしてyは配列番号4の452−462位からのいずれかのアミノ酸である、から成るアミノ酸配列を含んでなる、単離BTL−IIタンパク質が提供される。こうしたBTL−IIタンパク質は、配列番号4のアミノ酸30から453、1から453、29から457、1から457、1から482および/または29から482を含むことができる。本発明はさらに、配列番号10、配列番号14または配列番号18のアミノ酸127から157、または配列番号16のアミノ酸126から156と少なくとも80%、所望により少なくとも85%、90%、92%、94%、96%、または98%同一であるアミノ酸配列から成るポリペプチドを含んでなる、単離BTL−IIタンパク質を提供し、ここにおいて、配列番号10、配列番号14または配列番号18のアミノ酸127から157、または配列番号16のアミノ酸126から156と並列された(aligned with)アミノ酸配列の同一性領域は少なくとも20、所望により少なくとも25または30アミノ酸長であり、そしてポリペプチドはB細胞またはT細胞上に発現された細胞表面レセプターへ結合可能であり、および/またはT細胞の増殖を阻害可能である。こうしたアミノ酸配列は少なくとも150アミノ酸長であり得、そして配列番号10、配列番号14、配列番号16または配列番号18のアミノ酸30から358と少なくとも80%、所望により少なくとも85%、90%、92%、94%、96%、98%、99%または99.5%同一であり得、ここにおいて、配列番号10、配列番号14、配列番号16または配列番号18のアミノ酸30から358と並列されたアミノ酸配列の同一性領域は少なくとも150、所望により少なくとも200、250または300アミノ酸長である。さらに、アミノ酸配列は、配列番号10、配列番号14、配列番号16または配列番号18のアミノ酸30から358と少なくとも90%、所望により少なくとも92%、94%、96%、98%、99%または99.5%同一であり得、および/または配列番号10、配列番号14、配列番号16または配列番号18のアミノ酸30から358を含んでなることが可能である。
【0006】
本発明の別の態様は、配列番号10、配列番号14、配列番号16または配列番号18のアミノ酸30から358と少なくとも80%、所望により少なくとも85%、90%、92%、94%、96%、98%、99%または99.5%同一である第一のアミノ酸配列から成る第一のポリペプチドを含んでなる、単離BTL−IIタンパク質を包含しており、ここにおいて、配列番号10、配列番号14、配列番号16または配列番号18のアミノ酸30から358と並列された第一のアミノ酸配列の同一性領域は少なくとも150アミノ酸であり、ここにおいて第一のポリペプチドは、配列番号10、配列番号14または配列番号18のアミノ酸127から157、または配列番号16のアミノ酸126から156と少なくとも80%、所望により少なくとも85%、90%、92%、94%、96%、98%、99%または99.5%同一である第二のアミノ酸配列から成る第二のポリペプチドを含んでなり、ここにおいて、配列番号10、配列番号14または配列番号18のアミノ酸127から157、または配列番号16のアミノ酸126から156と並列された、第二のアミノ酸配列の同一性領域は少なくとも20アミノ酸長であり、そしてここにおいて、第一のポリペプチドはT細胞の増殖を阻害可能である。第一のアミノ酸配列は、配列番号10、配列番号14または配列番号18のアミノ酸127から157、または配列番号16のアミノ酸126から156と同一であることが可能である。
【0007】
あるいは、本発明は、配列番号4のアミノ酸30から457と少なくとも80%、所望により少なくとも85%、90%、92%、94%、96%、98%、99%または99.5%同一であるアミノ酸配列から成るポリペプチドを含んでなる、単離されたBTL−IIタンパク質を提供し、ここにおいてポリペプチドはわずか2のまたはそれより少ないIg様ドメインを含んでなり、そしてここにおいて、ポリペプチドはT細胞の増殖を阻害可能である。アミノ酸配列は、配列番号8のアミノ酸30から247または配列番号12のアミノ酸30から243と、少なくとも80%、85%、90%、92%、94%、96%、98%、99%、99.5%または100%同一であり得る。アミノ酸配列は配列番号4のアミノ酸30から457と少なくとも90%、所望により少なくとも92%、94%、96%、98%、99%、99.5%または100%同一であり得る。
【0008】
あるいは、本発明のBTL−IIタンパク質は、配列番号4のアミノ酸247から452または配列番号6のアミノ酸248から447と、少なくとも80%、所望により少なくとも85%、90%、92%、94%、96%、98%、99%または99.5%同一である第一のアミノ酸配列から成る第一のポリペプチドを含んでなることが可能であり、ここにおいて第一のアミノ酸配列は、少なくとも25,所望により少なくとも50、75、100または150のアミノ酸の、配列番号4と並列された第一のアミノ酸配列との同一性領域を有する、配列番号4のアミノ酸32から232または配列番号6のアミノ酸27から232と、少なくとも80%、所望により少なくとも85%、90%、92%、94%、96%、98%、99%または99.5%同一であるアミノ酸配列を含んでおらず、そしてここにおいて、該タンパク質は、配列番号4のアミノ酸32から232または配列番号6のアミノ酸27から232と、少なくとも80%、所望により少なくとも85%、90%、92%、94%、96%、98%、99%または99.5%同一である第二のアミノ酸配列から成る第二のポリペプチドを含んでおらず、少なくとも25,所望により少なくとも50、75、100または150のアミノ酸の、配列番号4と並列された第二のアミノ酸配列との同一性領域を有し、そしてここにおいて、第一のポリペプチドはT細胞の増殖を阻害可能である。
【0009】
別の態様において、本発明の単離BTL−IIタンパク質は、配列番号8または配列番号12のアミノ酸32から242と、少なくとも80%、所望により少なくとも85%、90%、92%、94%、96%、98%、99%または99.5%同一である第一のアミノ酸配列から成る第一のポリペプチドを含んでなることが可能であり、ここにおいて、配列番号8または配列番号12と並列された第一のアミノ酸配列の同一性領域は少なくとも約50,所望により少なくとも約75、100、150または200アミノ酸長であり、ここにおいて、第一のポリペプチドは、配列番号8または配列番号12のアミノ酸10から40と、少なくとも80%、所望により少なくとも85%、90%、92%、94%、96%、98%、99%または99.5%同一である第二のアミノ酸配列から成る第二のポリペプチドを含んでなり、ここにおいて、配列番号8または配列番号12と並列された第二のアミノ酸配列の同一性領域は少なくとも約20,所望により少なくとも約25または30アミノ酸長であり、そしてここにおいて、第一のポリペプチドはT細胞の増殖を阻害可能である。
【0010】
さらに別の態様において、本発明は、配列番号10、配列番号14、配列番号16または配列番号18のアミノ酸30から358と、少なくとも80%、所望により少なくとも85%、90%、92%、94%、96%、98%、99%または99.5%同一であるアミノ酸配列から成るポリペプチドを含んでなる、単離BTL−IIタンパク質を包含しており、ここにおいて、配列番号10、配列番号14、配列番号16または配列番号18のアミノ酸30から358と並列されたアミノ酸配列の同一性領域は、少なくとも250、所望により少なくとも275または300アミノ酸であり、そしてここにおいて、ポリペプチドはT細胞の増殖を阻害可能である。
【0011】
本発明のBTL−IIタンパク質は、長さが長くて約480アミノ酸、約380アミノ酸、約270アミノ酸または約160アミノ酸であることができ、T細胞の増殖を阻害可能であるポリペプチドを含んでなることが可能である。
【0012】
本発明はさらに、配列番号10のアミノ酸32から358と、少なくとも80%、所望により少なくとも85%、90%、92%、94%、96%、98%、99%または99.5%同一である第一のアミノ酸配列から成る第一のポリペプチドを含んでなる、単離BTL−IIタンパク質を包含し、ここにおいて、配列番号10と並列された第一のアミノ酸配列の同一性領域は少なくとも約175、所望により約200、250、275または300アミノ酸長であり、ここにおいて、第一のアミノ酸配列は、長さが約380を超えず、所望により長くて約390、270または170アミノ酸であり、ここにおいて、第一のポリペプチドはT細胞の増殖を阻害可能であり、ここにおいて、第一のアミノ酸配列は、少なくとも約20、30、40、50、60または75アミノ酸の、配列番号4のアミノ酸148から232と並列された第一のアミノ酸配列の同一性領域を有する、配列番号4のアミノ酸148から232と少なくとも80%同一ではなく、そしてここにおいて、BTL−IIタンパク質は、少なくとも約20、30、40、50、60または75アミノ酸の、配列番号4のアミノ酸148から232と並列された第二のアミノ酸配列の同一性領域を有する、配列番号4のアミノ酸148から232と少なくとも80%同一である第二のアミノ酸配列を含んでいない。こうしたBTL−IIタンパク質は配列番号8または配列番号12のアミノ酸32から242を含んでなることができる。
【0013】
別の態様において、本発明は、BTL−IIタンパク質および、抗体のFc領域またはロイシンジッパーであることが可能な異種ポリペプチドを含んでなる、BTL−II組換え融合タンパク質を含んでなる。本発明はまた、少なくとも10アミノ酸長であり、そして配列番号4の360位に及ぶ断片へ特異的に結合する抗体を惹起可能である、配列番号4のアミノ酸29から457の免疫原性断片も包含する。少なくとも10アミノ酸長の、配列番号10、配列番号14または配列番号16、および配列番号18の免疫原性断片が提供され、ここにおいて、免疫原性断片は配列番号10、配列番号14、配列番号18または配列番号16の141から143位に及び、そして断片に特異的に結合する抗体を惹起可能である。あるいは、免疫原性は配列番号10、配列番号14または配列番号18の142位、または配列番号16の141位に及ぶことが可能である。
【0014】
代わりの態様において、マウスBTL−IIタンパク質を提供する。特に、本発明は、配列番号6のアミノ酸x−yから成るアミノ酸配列を含んでなる、単離BTL−IIタンパク質を提供し、ここにおいて、xは配列番号4の1から35位の任意のアミノ酸であり、そしてyは配列番号6の450−460位の任意のアミノ酸である。こうしたBTL−IIタンパク質は配列番号6のアミノ酸32−450、20から456および/または29から514を含んでなることが可能である。
【0015】
他の態様は、配列番号4のアミノ酸1−457、配列番号6のアミノ酸1−456、配列番号8のアミノ酸1−247、配列番号10のアミノ酸1−363、配列番号12のアミノ酸1−243、配列番号14のアミノ酸1−359、配列番号16のアミノ酸1−358または配列番号18のアミノ酸1−362から成るBTL−IIタンパク質に特異的に結合する単離された抗体を含む。こうした抗体は、モノクローナル抗体、ヒト化抗体またはヒト抗体であることが可能であり、そしてそのレセプターに対するBTL−IIの結合を阻害することができる。本発明は、こうした抗体をコードする核酸、そしてこうした抗体を産生することが可能な細胞を包含し、該細胞は、こうした抗体を産生するように遺伝子操作されたハイブリドーマ細胞または細胞であることができる。本発明はさらに、抗体を分泌することができるこうした細胞を培養することによる、抗体を産生する方法も包含する。
【0016】
他の態様はBTL−II核酸を含んでいる。本発明は、配列番号3のヌクレオチドxからyより成るポリヌクレオチドを含んでなる、または該ポリヌクレオチドの相補体を含んでなる単離されたBTL−II核酸を包含し、ここにおいて、xはヌクレオチド1から105であり、そしてyはヌクレオチド1345から1375である。こうした核酸は、配列番号3のヌクレオチド105から1345、または1から1371を含んでなることが可能である。さらに、いずれかの上記BTL−IIタンパク質をコードしているBTL−II核酸であるような、免疫原性断片をコードしている核酸が提供される。
【0017】
本発明はさらに、いずれかの上記BTL−II核酸または抗BTL−II抗体をコードしている核酸を含んでなるベクター、そしてこうしたベクターを含んでいる宿主細胞を提供する。あるいは、本発明は、BTL−IIタンパク質、BTL−IIの免疫原性断片、若しくはBTL−IIに対する抗体を発現するように遺伝子操作された宿主細胞を提供する。こうした宿主細胞は、CHO細胞を含む哺乳動物細胞であってよい。こうした宿主細胞を、BTL−IIタンパク質、免疫原性断片、若しくはBTL−IIに対する抗体の発現を可能にする条件下で培養することを含んでなる、BTL−IIタンパク質、免疫原性断片、若しくはBTL−IIに対する抗体を産生するための方法もまた本発明に包含される。この方法はさらに、宿主細胞または培地から、BTL−IIタンパク質、免疫原性断片、若しくは抗体を単離することを含んでなることができる。こうした方法により産生されたBTL−IIタンパク質、免疫原性断片、若しくは抗体もまた意図される。本発明はさらに、ハイブリドーマまたは骨髄腫細胞を含む、BTL−IIに対する抗体を産生する哺乳動物細胞、およびこれらの細胞を培養することによる、抗体を作製するための方法を包含する。
【0018】
本発明により包含される組成物を用いる、多様な治療法もまた意図される。本発明は、治療的に有効量のBTL−IIタンパク質、所望により可溶性BTL−IIタンパク質を投与することを含んでなる、炎症性腸疾患、所望によりクローン病かまたは潰瘍性大腸炎を患っている患者の腸管における炎症を軽減するための方法を提供する。治療的に有効量のBTL−IIアンタゴニストおよび抗原を投与することを含んでなる、全身性および/または粘膜免疫応答を含む、抗原に対する免疫応答を誘導するための方法もまた提供される。BTL−IIアンタゴニストは、抗体または小分子であることが可能であり、そして抗原は、粘膜表面に直接投与することが可能であり、若しくは全身的に投与することが可能である。さらに提供されるのは、BTL−II mRNAまたはタンパク質が過剰発現されているかどうかを決定するため、患者の腸管からの組織試料をアッセイすることを含んでなる、炎症性腸疾患を診断するまたは炎症性腸疾患の発症を予測するための方法である。組織は、抗BTL−II抗体を使用して、BTL−IIタンパク質発現をアッセイすることが可能である。本発明はさらに、抗原および可溶性BTL−IIタンパク質を同時投与することを含んでなる、抗原、特に自己免疫または炎症性疾患を患っている患者における自己抗原に対する免疫応答の勢いを弱めるための方法も提供する。抗原は、粘膜表面を経て投与することが可能である。
【0019】
さらなる態様において、本発明はT細胞の増殖およびサイトカイン産生を阻害するための方法を包含する。1つの態様において、本発明は、T細胞とBTL−IIポリペプチドを接触させることを含んでなる、T細胞増殖を阻害する阻害するために方法を含んでなる。T細胞はヒトT細胞であることが可能であり、そしてインビボでBTL−IIポリペプチドと接触させることが可能である。BTL−IIポリペプチドの代替物として、T細胞上に発現されたBTL−IIレセプターへ結合するアゴニスト性抗体(それがT細胞の増殖を阻害することが可能であれば)を使用することが可能である。別の態様において、本発明は、T細胞とBTL−IIポリペプチドを接触させることを含んでなる、T細胞によるサイトカイン産生を抑制するための方法を含んでいる。T細胞はヒトT細胞であることが可能であり、そしてインビボでBTL−IIポリペプチドと接触させることが可能である。サイトカインは、例えば、インターフェロンガンマ(IFNγ)、インターロイキン2(IL2)またはインターロイキン5(IL5)であることが可能である。
【課題を解決するための手段】
【0020】
配列表の簡単な説明
配列番号1は、寄託番号NM_019602でのアメリカ国立バイオテクノロジー情報センター(NCBI)エントリーからの、ヒトBTL−II cDNAのヌクレオチド配列である。
【0021】
配列番号2は、寄託番号NM_019602でのNCBIエントリーのcDNA配列から予測された、ヒトBTL−IIタンパク質のアミノ酸配列である。
配列番号3は、本発明の完全長ヒトBTL−II cDNAのヌクレオチド配列である。
【0022】
配列番号4は、配列番号3によりコードされた完全長ヒトBTL−IIタンパク質のアミノ酸配列である。
配列番号5は、本発明の完全長マウスBTL−II cDNAのヌクレオチド配列である。
【0023】
配列番号6は、配列番号5によりコードされた完全長マウスBTL−IIタンパク質のアミノ酸配列である。
配列番号7は、ヒトBTL−IIスプライス変異体の第一のカテゴリーの代表的メンバーからの、cDNAのヌクレオチド配列である(図3a)。
【0024】
配列番号8は、配列番号7によりコードされたアミノ酸配列である。
配列番号9は、ヒトBTL−IIスプライス変異体の第二のカテゴリーの代表的メンバーからの、cDNAのヌクレオチド配列である(図4a)。
【0025】
配列番号10は、配列番号9によりコードされたアミノ酸配列である。
配列番号11は、ヒトBTL−IIスプライス変異体の第三のカテゴリーの代表的メンバーからの、cDNAの部分ヌクレオチド配列である(図5a)。
【0026】
配列番号12は、配列番号11によりコードされたアミノ酸配列である。
配列番号13は、ヒトBTL−IIスプライス変異体の第四のカテゴリーの代表的メンバーからの、cDNAの部分ヌクレオチド配列である(図6a)。
【0027】
配列番号14は、配列番号13によりコードされたアミノ酸配列である。
配列番号15は、ヒトBTL−IIスプライス変異体の第五のカテゴリーの代表的メンバーからの、cDNAの部分ヌクレオチド配列である(図7a)。
【0028】
配列番号16は、配列番号15によりコードされたアミノ酸配列である。
配列番号17は、マウスBTL−IIスプライス変異体の第一のカテゴリーの代表的メンバーのヌクレオチド配列である(図9a)。
【0029】
配列番号18は、配列番号17によりコードされたアミノ酸配列である。
配列番号19は、実施例5に記載したBTL−II:Fc融合タンパク質をコードしているヌクレオチド配列である。
【0030】
配列番号20は、実施例5に記載したBTL−II:Fc融合タンパク質のアミノ酸配列である。
【0031】
発明の詳細な説明
本発明は、アゴニストまたはアンタゴニストであることが可能である、BTL−IIタンパク質、抗BTL−II抗体をコードしている組換えベクターを含む、BTL−IIタンパク質および核酸、ならびに、これらの分子を産生するそして使用するための方法、およびこれらを含んでいる医薬組成物を提供する。BTL−II発現は少数の組織型に限定されている。実施例4に例示されているように、BTL−IIは、マウス炎症性腸疾患モデルシステムにおいて、症状の発症に先立って、および症状段階の間に、腸管中で過剰発現される。それ故、BTL−II抗体は炎症性腸疾患を診断するため、若しくは発症の可能性を予測するために働くことが可能である。加えて、本発明はBTL−IIヌクレオチド配列の多数のアリル(allelic)変異体を提供する(図5aから7a)。これらは、炎症性腸疾患に対する感受性の予測における効用を見出すことが可能である。さらに、可溶性BTL−IIタンパク質はT細胞増殖およびサイトカイン産生を阻害することが可能であるので(実施例6−10)、BTL−IIタンパク質は自己免疫および炎症性疾患の処置に効用を見出すことが可能である。
【0032】
さらに、BTL−IIは、腸管における免疫試料採取に役割を演じていることが知られている特殊化した構造であるパイエル板で発現される。BTL−IIが好ましく発現されるのは、パイエル板に観察される他の細胞と比較して、他の樹状細胞を含んでも、パイエル板に観察されるCD11c+(低発現)CD8+B220+樹状細胞(形質細胞様樹状細胞とも称される)である。パイエル板樹状細胞は、T細胞分化に対するその影響力のため、粘膜表面での免疫寛容の誘導において役割を演じているとの仮説が立てられている。Weiner(2001),Nature Immunology 2(8):671−71;Weiner(2001),Immunol.Rev.182:207−14;IwasakiおよびKe1sall(1999),American Journal of Physiology−Gastrointestinal and Liver Physiology 276(5):G1074−78。それ故、抗BTL−II抗体は、パイエル板内のCD11c+(低発現)CD8+B220+樹状細胞を同定するために使用することが可能である。
【0033】
以下に説明するように、BTL−IIは、TおよびB細胞仲介応答の調節に役割を演じるブチロフィリン様タンパク質のB7サブファミリー内に存在する。BTL−IIは、特に腸管において、免疫システム仲介炎症の勢いを弱めるか、および/または促進することに役割を演じることができる。免疫システムの複雑さを仮定すれば、単一の細胞表面タンパク質は、ある場合には、エフェクター細胞上のいかなるレセプターが相互作用する分子に利用可能であるかに依存して、免疫エフェクター細胞による免疫応答を刺激する若しくは勢いを弱めるの両方を行うことができる。以下に議論されるB7−1およびB7−2タンパク質は、免疫エフェクター細胞、この場合T細胞に刺激性および勢いを弱める両方の効果を有する免疫調節細胞表面タンパク質の例である。この故に、BTL−IIはインビボで同様の二重の役割を果たすことができる。しかしながら、腸管は、食品抗原および共生微生物に対するその免疫寛容により証明されるように、全体的にみて外来抗原に対して高度に免疫寛容である。それ故、BTL−IIは、少なくともいくつかの状況において、インビボで免疫応答または炎症を弱めることに役割を演じることがありそうである。それ故、インビトロで選択されたBTL−IIアンタゴニスト(抗体を含むことが可能)結合タンパク質若しくは小分子は、抗原に対する粘膜免疫応答を刺激するように働くことができる。さらに、可溶性BTL−IIタンパク質、または機能的に均等な抗イディオタイプ抗体は、腸管における、または肺のごとき体内の他の粘膜表面における免疫応答を弱めることができる。
【0034】
本明細書において、“抗体”は、キメラ抗体であることが可能であり、単量体または一本鎖の、二量体の、三量体の、四量体のまたは多量体の抗体であることが可能であり、そして組換えタンパク質または非組換えタンパク質であることが可能である。“ドメイン”とは、一次配列モチーフおよび/または三次構造特性により区別することが可能な、タンパク質の一部または全てである。タンパク質ドメインを位置づけるために設計されたプログラムには、例えば、Pfam(Batemanら(1999),Nucleic Acids Res.27:260−62;Batemanら(1999),Nucleic Acids Res.28:263−66)、ProDom(Corpetら(1999),Nucleic Acids Res.27:263−67;Corpetら(1999),Nucleic Acids Res.28:267−69)、Domo(GracyおよびArgos(1998),Bioinformatics 14:164−87)およびSMART(Pontingら(1999),Nucleic Acids Res.27:229−32)が含まれる。三次構造は、例えば、X−線結晶学により経験的に決定することが可能であり、若しくはこうした使用のために設計されたコンピューターソフトウェアーを使用して予測することが可能である。例えば、構造データは、Molecular Modeling DatabaseからのNCBIのEntrezウェブサイトを通して(Wangら(2000),Nucleic Acids Res.28(1):243−45)、またはDALI(HolmおよびSander(1993),J.Mol.Biol.233:123−38)のごときソフトウェアーの使用により、アクセスすることが可能である。例えば、免疫グロブリン様ドメイン(Ig様)は、一次配列相同性よりもむしろ、その三次構造により主として区別される。例えば、Borkら(1994),J.Mol.Biol.242:309−20;HunkapillerおよびHood(1989),Adv.Immunol.44:1−63;WilliamsおよびBarclay(1988),Ann.Rev.Immunol.6:381−405、を参照されたい。しかしながら、IgVおよびIgCドメインは、それらの一次アミノ酸配列内の保存位置に出現するほんの少しの高度に保存されたアミノ酸を実際に含んでいる。例えば、Kabatら(1991)、免疫学的に興味が持たれるタンパク質の配列、米国保健福祉省、公衆衛生サービス、国立保健研究所、NIH出版番号91−3242、を参照されたい。適切な間隔で出現しているこうした高度に保存されたアミノ酸の存在は、IgC様またはIgV様ドメインの存在を示すことが可能である。
【0035】
タンパク質を“コードする”核酸とは、本明細書において、核酸またはその相補体がタンパク質をコードしているコドンを含んでなる場合を意味している。
タンパク質の発現を可能にする組換え核酸配列が、ウイルス感染、トランスフェクション、形質転換または電気穿孔法のごとき“遺伝子操作”の方法を使用して、細胞内へ導入された場合、細胞は特異的タンパク質を発現するように“遺伝子操作され”ている。例えば、Kaufmanら(1990),Meth.Enzymol.185:487−511、を参照されたい。このことは、例えば、細胞内へのタンパク質をコードしている核酸の導入、またはChappelの米国特許第5,272,071号に記載されているような、タンパク質をコードしている宿主遺伝子の発現を促進するための、調節配列の導入を含むことが可能である。“遺伝子操作”の方法は、限定するわけではないが、ポリメラーゼ連鎖反応を使用する核酸の増幅、大腸菌(E.coli)へそれらをクローン化することによる組換えDNA分子の組み立て、核酸の制限酵素消化、核酸のライゲーション、核酸のインビトロ合成、そして核酸の末端への塩基の移送、なかでも当該技術分野でよく知られている多数の方法、を含む多数の方法を包含している。例えば、Sambrookら,Molecular Cloning:A Laboratory Manual,第2版,1−3巻,Cold Spring Harbor Laboratory,1989、を参照されたい。
【0036】
“異種ポリペプチド”とは、本明細書で意味しているようなBTL−IIポリペプチドではなく、少なくとも3アミノ酸長である、任意のポリペプチドである。
ポリヌクレオチドまたはポリペプチドの配列同一性を決定するための比較に関連して、“同一性領域”により意味されることは、以下に述べたパラメーターを使用するコンピュータープログラムGAP(Devereuxら(1984),Nucleic Acids Res.12:387−95)により、別のポリヌクレオチドまたはポリペプチドと一致した(部分的にまたは正確に)ポリヌクレオチドまたはポリペプチドの一部である。例えば、20アミノ酸のポリペプチドが相当により長いタンパク質と並列された場合、最初の10アミノ酸がより長いタンパク質と正確に一致し、そして最後の10アミノ酸はより長いタンパク質と少しも一致しないとすれば、同一性領域は10アミノ酸である。一方、もし20アミノ酸ポリペプチドの最初および最後のアミノ酸がより長いタンパク質と一致し、そして八つの他の一致が間に散在すると、同一性領域は20アミノ酸である。しかしながら、少なくとも、例えば、20アミノ酸または60ヌクレオチドの同一のまたは保存的に置換されたアミノ酸または同一のヌクレオチドを有しない、並列された両鎖における長い伸展は、本明細書で意味するような同一性領域の終点を構成する。
【0037】
“Ig様”ドメインは免疫グロブリン様ドメインであり、そしてIgV様かまたはIgC様ドメインであってもよいし、または免疫グロブリン構造内へ折り畳むことが可能であるが、しかしIgV様かまたはIgC様としては明白に分類できないドメインであってもよい。
【0038】
“IgV様”ドメインは、免疫グロブリン可変領域ドメインに共通の特性で免疫グロブリン内へ折り畳むことが可能である、アミノ酸配列を有している。Borkら,上記文献;Millerら(1991),Proc.Natl.Acad.Sci.USA 88:4377−81;WilliamsおよびBarclay(1988),Ann.Rev.Immunol.6:381−405、を参照されたい。当業者は、保存された位置でのIgVドメイン中の高度に保存されたアミノ酸の存在から、ドメインをIgV様として同定することが可能であることを承知している。
【0039】
“IgC様”ドメインは、免疫グロブリン定常領域ドメインに共通の特性で免疫グロブリン内へ折り畳むことが可能である、アミノ酸配列を有している。Borkら,上記文献;WilliamsおよびBarclay,上記文献、を参照されたい。当業者は、保存された位置でのIgCドメイン中の高度に保存されたアミノ酸の存在から、ドメインをIgC様として同定することが可能であることを承知している。
【0040】
“炎症性腸疾患”は、クローン病、潰瘍性大腸炎、回腸炎、および胃腸管の慢性炎症により特徴付けられる他の疾患を含んでいる。
タンパク質は、二つのIg様ドメインを含み、そして別のIg様ドメインの、全てのまたはいくつかの認識可能な部分を含んでいない場合、“わずか2のまたはそれより少ないIg様ドメイン”を含んでなる。しかしながら、こうしたタンパク質は、Ig様ドメインではない他のアミノ酸配列、および“わずか2のまたはそれより少ないIg様ドメイン”をなお含むことが可能である。それ故、句“ほどのまたはそれより少ない”とは、Ig様ドメインのみを示し、タンパク質の一部であることができる他のアミノ酸配列を示してはいない。
【0041】
ポリペプチドが“T細胞の増殖を阻害する”ことが可能である、またはいくつかの他の生物学的機能を実行することが可能であると言われる場合、それはポリペプチドを含んでなるタンパク質が機能を実行できること、そして、生物学的機能を実行できない、少なくともいくつかのタンパク質へのポリペプチドの添加により、これらのタンパク質が機能を実行するのを可能にすることを意味している。いくつかの場合、生物学的機能を実行することが可能なポリペプチドは、追加の配列なしでそうすることが可能である。他の場合、生物学的機能を実行するには、他の配列(例えば、オリゴマー化配列)を必要とすることができる。一つのシナリオにおいて、一つのポリペプチドがFc領域またはロイシンジッパーまたはいくつかの二量体化ドメインに連結された場合、それは特定の生物学的機能を効率的に実行することが可能であるが、二量体化ドメインなしでは実行できない。本明細書中の意味として、そのようなポリペプチドは生物学的機能を実行することが可能である。
【0042】
“タンパク質”とは、少なくとも10アミノ酸、所望により少なくとも20、30、40、50、60、80、100、150、200、250および/または300アミノ酸を含んでなる任意のポリペプチドである。
【0043】
ポリペプチドまたはタンパク質に適用されるような“組換え”とは、タンパク質の産生が、核酸(タンパク質をコードしていてもしていなくてもよい)を、それらが天然には観察されない細胞内へ導入する、少なくとも一つの工程に依存していることを意味している。
【0044】
“組換え融合タンパク質”とは、天然には一緒に融合されることが観察されない、単一のポリペプチド鎖内へ融合された、少なくとも二つのタンパク質の一部または全てを含んでなる組換えタンパク質である。
【0045】
核酸配列における“サイレント(silent)突然変異”とは、核酸によりコードされたタンパク質の配列を変化させることなく、核酸の配列を変化させることである。
“可溶性”タンパク質とは、膜貫通ドメイン、または通常はタンパク質が膜へ埋め込まれるまたは膜に会合されることを引き起こす、GPIアンカー配列のごときいくつかの他のアミノ酸配列を欠いているタンパク質である。こうしたタンパク質は典型的には、膜貫通タンパク質の細胞外領域の全てまたは一部を含んでなることができる。
【0046】
本発明の目的のためには、二つのタンパク質または核酸は、もしそれらがアミノ酸または核酸配列において、少なくとも80%、所望により少なくとも85%、90%、95%、97%、98%、99%、99.5%または99.7%お互いに同一であり、そして非改変タンパク質の生物学的機能を維持しまたは望ましい様式に改変していれば、“実質的に類似”している。二つのアミノ酸または二つの核酸配列のパーセント同一性は、視覚的な点検および数学的計算により決定することが可能であり、またはより好ましくは、コンピュータープログラムを使用して配列情報を比較することにより比較を行う。コンピュータープログラムの例はGenetics Computer Group(GCG;Madison,WI)Wisconsinパッケージバージョン10.0プログラム,GAP(Devereuxら(1984),Nucleic Acids Res.12:387−95)である。GAPプログラムのための好ましいデフォルトパラメーターには以下のものが含まれる:(1)ヌクレオチドに関する単一(unary)比較マトリックス(同一に対し1および非同一に対し0の値を含む)、および「Atlas of Polypeptide Sequence and Structure」,SchwartzおよびDayhoff監修,National Biomedical Research Foundation,pp.353−358(1979)に記載されているような、GribskovおよびBurgess((1986)Nucleic Acids Res.14:6745)の加重アミノ酸比較マトリックス、または他の比較可能な比較マトリックスでのGCG実行;(2)各ギャップに対する8のペナルティおよびアミノ酸配列に対する各ギャップ中の各記号に対しさらに2のペナルティ、または各ギャップに対する50のペナルティおよびヌクレオチド配列に対する各ギャップ中の各記号に対しさらに3のペナルティ;(3)末端ギャップに対するペナルティなし、および(4)長いギャップに対し最大ペナルティなし。配列比較の分野で当業者により使用される他のプログラムもまた使用可能である。
【0047】
ブチロフィリン様タンパク質
現在まで報告されているブチロフィリン様タンパク質は、いくつかの構造特性を共有しており、そして多くは主要組織適合性座位(MHC)内にまたは隣接してコードされている。例えば、Henryら(1999),Immunology Today 20(6):285−88、を参照されたい。ブチロフィリンは、ウシミルクの脂肪小球に会合されている全タンパク質の40%を構成するタンパク質であり、そしてヒト相同体が存在する。Ruddyら(1997),Genome Research 7:441−56,JackおよびMather(1990),J.Biol.Chem.265:14481−86を引用している。ブチロフィリン様タンパク質間のアミノ酸配列レベルでの類似性は低くてもよいが、しかしドメイン構造は幾分保存されている。全てのメンバーは、通常IgV様ドメインであると報告されている、Ig様ドメインが続いたアミノ末端シグナルペプチドを含んでなる。多くの場合において、通常IgC様ドメインであると報告されている、別のIg様ドメイン、膜貫通ドメイン、そして細胞質ドメインが続いている。BTL−IIまたはB7−H3におけるように、二つのIg様ドメインはそれらの最初の出現に続いて直ちに反復されることが可能である。このドメイン構造は図1に例示されている。図1に図示されたタンパク質のほとんどは、ブチロフィリン様タンパク質のB7サブファミリーのメンバーである;三つ(ヒトBTN2A1、ヒトブチロフィリンおよびミエリンオリゴデンドロサイト糖タンパク質)のみがそうではない。
【0048】
Ig様ドメインは、その配列は非常に多岐にわたることが可能であるが、四つのβ鎖を含んでなる共通構造コアを全てが含む、多数の保存された折り畳みパターンの一つをそれでも保持している。免疫グロブリン定常および可変領域は、バレル(barrel)様形状を形成している、7から9の逆平行βストランドにより特徴付けられる三次構造を有している。Borkら(1994),J.Mol.Biol.242:309−20。IgVおよびIgC様免疫グロブリンドメインは各々、特有で、高度な保存残基をほんの少し含んでいる。HunkapillerおよびHood(1989),Adv.Immunol.44:1−63;Millerら(1991),Proc.Natl.Acad.Sci.USA 88:4377−81;WilliamsおよびBarclay(1988),Ann.Rev.Immunol.6:381−405。保存残基は多分構造または機能のために重要である。こうした保存残基は、BTL−IIのIg様ドメイン中に観察される。例えば、ヒトBTL−IIタンパク質の第一のIg様ドメイン(配列番号4)は、IgV様ドメインに特徴的な、こうした高度に保存された残基を、適切な位置、例えば、G43、C50、W65、L109、D118、G120、Y122およびC124位に含んでいる。表3を参照されたい。第三のIg様ドメインもまた、IgV様ドメインで保存されている残基に対応する残基を含んでいる。ヒトBTL−IIの第二および第四のIg様ドメインは、IgC様ドメインで高度に保存されている残基に対応する残基を含んでいる。表3。
【0049】
膜貫通および細胞質ドメインは、ブチロフィリン様タンパク質が第二のIg様ドメインを有しているかどうかに関わらず、ブチロフィリン様タンパク質中に出現する。膜貫通ドメインには、αへリックスコイルドコイル(coiled coil)モチーフに典型的な7アミノ酸繰り返しを思い出させる、一つまたはそれより多くの7アミノ酸単位が続いていてもよいし、続いていなくてもよい。7アミノ酸繰り返しはまた、ブチロフィリン様タンパク質中の別の位置に出現することもできる。7アミノ酸繰り返しの議論については、Millerら(1991),Proc.Natl.Acad.Sci.88:4377−81、を参照されたい。膜貫通ドメインを予測する方法は、当該技術分野でよく知られている。例えば、Ikedaら(2002),In Silico Biol.2(1):19−33;TusnadyおよびSimon(1998),J.Mol.Biol.283(2):489−506、を参照されたい。ブチロフィリン様タンパク質は細胞質ドメインを有することが可能である。こうした細胞質ドメインは、B30.2ドメインを含んでいても、または含んでいなくてもよい。多様なB30.2ドメインの配列が示されており、そしてB30.2ドメインの推定機能がHenryら((1998),Mol.Biol.Evol.15(12):1696−1705)により議論されている。B30.2ドメインはいろいろなタンパク質に観察され、そしてB30.2ドメインの機能は未知である。提案されたB30.2ドメインのリガンドはキサンチンオキシダーゼであり、それはブチロフィリンの細胞質ドメインと相互作用する。二つの異なったB30.2含有タンパク質のB30.2ドメインにおける突然変異は、二つの異なった疾患と関連付けられたが、突然変異および疾患表現型間の因果関係は確立されていない。Henryら、上記文献。
【0050】
ブチロフィリン様タンパク質のB7サブファミリー
BTL−IIは、例えば、T細胞、B細胞および骨髄細胞のごとき、免疫エフェクター細胞の調節に役割を演じる、B30.2ドメインを欠く多数のブチロフィリン様免疫調節タンパク質とドメイン構造を共有している。このサブファミリーは、ブチロフィリン様タンパク質の“B7サブファミリー”として本明細書において称される。本明細書で意味しているように、B7サブファミリーメンバーの特性には、限定ではなく、以下の項目が含まれる:(1)一つまたはそれより多くの細胞外Ig様ドメインを有し;(2)膜貫通および細胞質ドメインを有し;(3)B30.2ドメインを欠き;(4)抗原提示細胞上で発現されており;(5)活性化された免疫応答の間に発現の調節を起こし;そして(6)免疫応答を変調する。免疫応答を変調する、膜貫通ドメインを欠く、分泌、可溶性B7タンパク質は現在まで報告されていない。ほとんどのB7タンパク質は二つの細胞外Ig様ドメインを有しているが、B7−H3のいくつかのアイソフォームは四つ有している。図1を参照されたい。ヒトBTL−IIは、2から4の細胞外Ig様ドメインを有することが可能で、そして上にリストした、B7ファミリーメンバーの特性の全てを有している。それ故、我々はそれがブチロフィリン様タンパク質のB7サブファミリーのメンバーであると考えた。B7サブファミリーの全ての他の既知メンバーは、B細胞および/またはT細胞のごとき免疫エフェクター細胞上のレセプターと相互作用し、その相互作用は免疫応答を変調するためのシグナルとして働く。それ故、BTL−IIは、T細胞のごとき免疫エフェクター細胞上のレセプターと相互作用し、それ故免疫応答を変調すると予想される。
【0051】
B7ファミリーメンバーは免疫エフェクター細胞、特にT細胞の活性を変調することに役割を演じている。Henryら(1999),Immunology Today 20(6):285−88;SharpeおよびFreeman(2002),Nat.Rev.Immunol.2:116−26。最もよく特徴付けられた例は、CD80およびCD86(各々B7−1およびB7−2とも称される)であり、それらは抗原提示細胞上で発現され、T細胞の表面上に構成的に発現されるCD28と相互作用した場合、T細胞活性化を促進可能であり、またはT細胞の表面上にまた発現されるCTLA−4と相互作用した場合、T細胞活性化を阻害可能である。CTLA−4発現は構成的ではないが、しかしT細胞活性化に続いて迅速に上方調節される。例えば、Mastellerら(2000),J.Immunol.164:5319−27;Hehnerら(2000),J.Biol.Chem.275(24):18160−71;SharpeおよびFreeman(2002),Nat.Rev.Immunol.2:116−26、を参照されたい。さらに、CD80の両方のIg様ドメイン中の、多数の個々のアミノ酸残基がCTLA−4およびCD28への結合に重要であることが報告されている。Peachら(1995),J.Biol.Chem.270(36):21181−87。CD86は構成的に低レベルで発現され、そして活性化後に迅速に上方調節され、一方、CD80は、活性化後に誘導的に発現される。SharpeおよびFreeman(2002),Nature Reviews Immunology 2:116−26。
【0052】
本明細書においてB7RP−1と称される、別のT細胞調節分子は多血症(plethora)の名前を有している、KIAA0653(Ishikawaら(1998),DNA Res.5:169−76)、B7h(Swallowら(1999),Immunity 11:423−32)、GL50(Lingら(2000),J.Immunol.164:1653−57)、B7RP−1(Yoshinagaら(1999),Nature 402:827−32)、LICOS(Brodieら(2000),Curr.Biol.10:333−36)、B7−H2(Wangら(2000),Blood 96:2808−13)およびICOSL(SharpeおよびFreeman,上記文献)。B7RP−1は末梢リンパ組織、脾臓、リンパ節、肺、胸腺、脾細胞およびB細胞中で発現される。B7RP−1とT細胞の相互作用は、T細胞増殖およびサイトカイン産生を増加させることが可能である。B7RP−1は、活性化T細胞上に発現されるICOSレセプターを通してT細胞にシグナルを伝達する。Yoshinagaら(1999),Nature 402:827−32;Swallowら(1999),Immunity 11:423−32。B7RP−1は非刺激B細胞株で構成的に発現され、そしてその発現は、インターフェロンγにより単球中で誘導することが可能である。Aicherら(2000),J.Immunol.164:4689−96。調節T細胞の発生および調節機能は、B7RP−1およびT細胞上のそのレセプター間の相互作用に依存している。Akbariら(2002),Nature Medicine 8(9):1024−32。
【0053】
さらに、三つの他のT細胞調節分子、PD−L1(B7−H1とも称される)、PD−L2(B7−DCとも称される)およびB7−H4(B7S1およびB7xとも称される)は、T細胞増殖およびサイトカイン産生を阻害することが可能である。PD−L1およびPD−L2は、T細胞、B細胞および骨髄細胞上に発現される、それらの共通のレセプター、PD−1を通して作用する。Freemanら(2000),J.Exp.Med.192(7):1027−34;Dongら(1999),Nature Medicine 5(12):1365−69;Latchmanら(2001),Nature Immunology 2(3):261−68;Tamuraら(2001),Blood 97(6):1809−16;およびTsengら(2001),J.Exp.Med.193(7):839−45。PD−L1およびPD−L2の発現は、一般的な前炎症性サイトカインである、インターフェロンγにより誘導することが可能である。Latchmanら,上記文献。B7−H4はB細胞、マクロファージおよび樹状細胞上で発現され、BおよびT細胞上に発現される阻害的レセプターである、BTLAを通して作用するようである。Watanabeら(2003),Nature Immunology 4(7):670−79;Zangら(2003),Proc.Natl.Acad.Sci.100(18):10388−92;Sicaら(2003),Immunity 18:849−61;Prasadら(2003),Immunity 18:863−73;CarrenoおよびCollins(2003),Trends Immunol.24(10):524−27。
【0054】
T細胞を刺激することにおいて、同時刺激性役割を果たすさらに別のブチロフィリン様タンパク質はB7−H3である。Chapovalら(2001),Nature Immunol.2(3):269−74。他のB7ファミリーメンバーと同様に、B7−H3はシグナル配列、細胞外Ig様ドメイン、膜貫通ドメインおよび細胞質ドメインを有している。ヒトB7−H3遺伝子は、二つまたは四つの細胞外Ig様ドメインを有するアイソフォームをコードしている。Sunら(2002),J.Immunol.168:6294−97。B7−H3の発現は、炎症性サイトカインにより、樹状細胞上で誘導することが可能である。B7−H3は、T細胞の増殖および細胞毒性応答を刺激することが可能である。B7−H3は、CD28、CTLA−4、ICOSおよびPD−1とは異なる推定上のT細胞レセプターを通して作用する。Chapovalら、上記文献。
【0055】
BTL−IIタンパク質
ヒトおよびマウスBTL−IIタンパク質の存在は、ゲノム配列から予想されていた。Stammersら(2000),Immunogenetics 51:373−82。しかしながら、これらの著者は、ゲノム分析に基づいた、ヒトまたはマウスBTL−IIタンパク質中の膜貫通または細胞質ドメインの証拠、そしてマウスBTL−II mRNAを検出するために設計されたPCR実験において、エキソン1−4(各々、シグナルペプチド、二つのIg様ドメインおよび7アミノ酸繰り返し領域をコードする)とエキソン5および6(各々、別の二つのIg様ドメインをコードする)を連結している転写体の証拠を見出していない。Stammersら、上記文献。これらの知見に基づき、BTL−IIは、細胞表面、免疫変調タンパク質のB7サブファミリーには加えられなかった。これら同一の著者による、公共データベースへのその後の配列提出は、455アミノ酸をコードしているエキソン5および6を含んでおり、膜貫通ドメインおよび細胞質ドメインを欠いている、1368ヌクレオチドのヒトBTL−II mRNAを予測している(配列番号1(ヒトBTL−II cDNA配列)および配列番号2(ヒトBTL−IIタンパク質配列)を開示している、NCBI寄託番号第NM_019602号)。
【0056】
本発明のBTL−II核酸およびタンパク質配列は、多くの点でこれらの配列と異なっている。cDNA配列は、膜貫通または細胞質ドメインを含んでいない、以前に報告されているBTL−IIタンパク質配列とは違って、シグナル配列、細胞外ドメイン、膜貫通ドメイン、および細胞質ドメインをコードしている。BTL−IIの発現パターンと一緒に加えて、これらの特性のため、BTL−IIはブチロフィリン様タンパク質のB7サブファミリー内に加えられる。表1(下記)は、本発明のBTL−IIタンパク質および以前に報告されている配列間の相違を強調している。本発明のBTL−IIタンパク質は上段に示されており(配列番号4)、そしてNCBI寄託番号第NM_019602号に報告されているBTL−IIタンパク質は下段に示されている(配列番号2)。この比較から、二つの配列間には三つのミスマッチがあり(360、454および455位に)、そして配列番号4は、細胞外ドメインの追加の配列ならびに膜貫通および細胞質ドメインを構成する、さらに27のアミノ酸を有している。これらの配列は、上に列挙したパラメーターを使用するGAPプログラムに従うと、99.3%同一である。
【0057】
【表1】
【0058】
図1に例示したようなドメイン構造における総合的な類似性に加え、B7サブファミリータンパク質は、一次配列レベルでの類似性を有している。例えば、コンピュータープログラムGAPを使用して、対様式で並列した場合、ヒトBTL−IIタンパク質配列(配列番号4)は、下記表2に示されているように、他のB7サブファミリーメンバーに類似している。
【0059】
【表2】
【0060】
当業者は、任意のこれらの並列で保存されている残基は、保存されていない残基よりも、ヒトBTL−IIの構造または機能においておそらく本質的役割を演じることを了解するであろう。
【0061】
表3(下記)において、ヒトBTL−IIアミノ酸配列(配列番号4、上段)とマウスBTL−IIアミノ酸配列(配列番号6、下段)が並列されている。同一アミノ酸は垂直線で結ばれており、類似のアミノ酸は、それらの間に一つのまたは二つのドットを有している。GAP(上記)により決定された、これらの配列間のパーセント同一性は約62%であり、そしてパーセント類似性は約68%である。IgVまたはIgC様ドメイン、またはいわゆるIgV免疫グロブリンスーパーファミリーメンバーの“Iセット”中に観察される残基、またはこうした残基の保存的置換はボールド体で示されている。Peachら、上記文献;HarpazおよびChothia(1994),J.Mol.Biol.238:528−39。こうした残基は構造的に重要であるようであり、それ故、機能的効果を有することができる。適正な間隔をあけたこうしたアミノ酸のかなりの数の出現で、配列をIgV様またはIgC様として同定可能である。
【0062】
【表3】
【0063】
構造は配列よりも進化においてより保存されているので、非保存残基は保存残基よりも、BTL−IIタンパク質の全体の三次構造を決定することにおいて役割を演じそうもないことを当業者は理解するであろう。Borkら(1994),J.Mol.Biol.242:309−20。本明細書において、“非保存残基”とは、表3におけるように、ヒトおよびマウスBTL−IIタンパク質配列が比較された場合に保存されていない、BTL−IIタンパク質内のアミノ酸である。スプライス変異体によりコードされているBTL−IIタンパク質において、こうした残基は、配列内の異なった番号位置で出現するであろう。例えば、配列番号4の397位の非保存残基は、配列番号8の187位でみられる同一の非保存残基である。当業者は、タンパク質構造がタンパク質機能に影響可能であることを認識するであろう。さらに、非保存アミノ酸は、BTL−II機能において直接的な役割を演じることもありそうもない。例えば、残基4、6、25、26、35、36および多くの他の残基は、同一でもなくまたは類似してもいない。それ故、当業者は、こうした残基の改変がBTL−IIタンパク質機能に影響することは、保存されたまたは類似の残基の改変よりもありそうもないことを了解するであろう。さらに、保存的置換が、非保存的置換よりもタンパク質機能へ影響することは、よりありそうもないようである。生物学的活性へ影響することがありそうもない保存的置換であるアミノ酸置換の例には以下の置換が含まれる:Serに対するAla、Ileに対するVal、Gluに対するAsp、Serに対するThr、Glyに対するAla、Thrに対するAla、Asnに対するSer、Valに対するAla、Glyに対するSer、Pheに対するTyr、Proに対するAla、Argに対するLys、Asnに対するAsp、Ileに対するLeu、Valに対するLeu、Gluに対するAla、Glyに対するAsp、およびこれらの変化の逆。例えば、Neurathら,「The Proteins」,Academic Press,New York(1979)を参照されたい。さらに、グループ内の一つのアミノ酸の、同一のグループ内の別のアミノ酸への交換は保存的置換であり、ここでグループとは以下のようである:(1)アラニン、バリン、ロイシン、イソロイシン、メチオニン、ノルロイシンおよびフェニルアラニン;(2)ヒスチジン、アルギニン、リジン、グルタミンおよびアスパラギン;(3)アスパラギン酸およびグルタミン酸;(4)セリン、スレオニン、アラニン、チロシン、フェニルアラニン、トリプトファンおよびシステイン;(5)グリシン、プロリンおよびアラニン。
【0064】
ヒトBTL−IIタンパク質はいくつかの認識可能ドメインを含んでなる。第一は、配列番号4の残基1から、配列番号4の残基22から29付近までからの第二の位置に及ぶシグナル配列(エキソン1によりコードされている)である。次は、残基xから残基yに及ぶIg様ドメイン(エキソン2によりコードされている)であり、ここでxは配列番号4の残基22から32までからであり、そしてyは残基138から148までからである。これに続くのは、残基vから残基wに及ぶ別のIg様ドメイン(エキソン3によりコードされている)であり、ここでvは配列番号4の残基138から148までからであり、そしてwは残基232から242までからである。次は、残基tから残基uに及ぶ7アミノ酸反復領域(エキソン4によりコードされている)であり、ここでtは配列番号4の残基234から239までからであり、そしてuは残基240から247までからである。別のIg様領域が残基rから残基sに及んでおり、ここでrは配列番号4の残基240から247までからであり、そしてsは残基354から364までからである。第四のIg様領域が残基pから残基qに及んでおり、ここでpは配列番号4の残基355から365までからであり、そしてqは残基452から462までからである。ヒトBTL−IIのこれらの第一の6ドメインは、ヒトBTL−IIの細胞外領域を作り上げている。シグナル配列はタンパク質の分泌時にタンパク質の残り部分から切断されてもよいが、切断が必要とされるわけではない。膜貫通ドメインは残基nから残基oに及んでおり、ここでnは配列番号4の残基454から462までからであり、そしてoは残基473から481までからである。最後は、残基kから残基mに及ぶ細胞質ドメインであり、ここでkは配列番号4の残基474から481までからであり、そしてmは残基482付近である。
【0065】
マウスBTL−IIタンパク質は類似の組のドメインを含んでなる。第一は、配列番号6の残基1から始まり、残基20付近から残基27付近までからの位置で終わるシグナル配列である。第二は、残基xから残基yに及ぶIg様ドメインであり、ここでxは配列番号6の残基21から27までからであり、そしてyは残基138から148までからである。第三は、残基vから残基wの別のIg様ドメインであり、ここでvは配列番号6の残基139から148までからであり、そしてwは残基232から242までからである。7アミノ酸反復領域は残基tから残基uに及んでおり、ここでtは配列番号6の残基233から242までからであり、そしてuは残基238から248までからである。別のIg様領域が残基rからsに及んでおり、ここでrは配列番号6の残基239から248までからであり、そしてsは残基355から365までからである。最後のIg様領域が残基pからqに及んでおり、ここでpは配列番号6の残基356から365までからであり、そしてqは残基447から457までからである。これらの第一の6ドメインは、マウスBTL−IIの細胞外領域を作り上げている。膜貫通ドメインは残基nからoに及んでおり、ここでnは配列番号6の残基448から459までからであり、そしてoは残基470から478までからである。最後は、残基kからmに及ぶ細胞質ドメインであり、ここでは配列番号6の残基471から478までからであり、そしてmは残基514付近である。
【0066】
本発明はBTL−IIの分泌された、可溶性バージョン、ならびに、細胞表面上に発現することが可能である膜貫通ドメインを含んでなるバージョンを包含している。本発明はさらに、以下に記載するようなBTL−II核酸によりコードされたBTL−IIタンパク質も含んでいる。これらのタンパク質全ての組換え体バージョンは、本明細書で記載するように、抗体を産生するため、スクリーニングに、および/または治療剤として使用することが可能である。例えば、本発明は、配列番号4、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16および/または配列番号18の全て若しくは一部を含んでなるBTL−IIタンパク質を包含する。本発明のBTL−IIタンパク質は、一次アミノ酸配列の挿入、欠失、改変または置換により、配列番号4、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16または配列番号18とは異なったタンパク質を含んでいる。こうした変異体配列は、配列番号4、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16および/または配列番号18と少なくとも80%、85%、90%、95%、97%、98%、99%、99.5%または99.7%同一であり、そして上述の配列とGAPを使用して並列させた場合、10アミノ酸を超える内部ギャップを含んでいない。こうした配列の例には、図5b、6bおよび7bに示した、BTL−IIの天然に存在するヒトアリル(allelic)変異体が含まれる。もしこうした変異体配列が、配列番号4、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16および/または配列番号18と比較した場合にアミノ酸置換を含んでいれば、これらの置換は保存的アミノ酸置換であろう。さらに、変異体BTL−IIタンパク質は配列番号4、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16および/または配列番号18に関して、単一アミノ酸のわずか3、4、5、6、7、8、9、10、15、20または25の挿入、欠失または置換を含むことができる。
【0067】
本発明のBTL−IIタンパク質は、ヒトおよびマウスBTL−II mRNAの多様なスプライス変異体によりコードされているタンパク質を含む。こうした変異体ヒトBTL−IIタンパク質の配列は、配列番号8、配列番号10、配列番号12、配列番号14および配列番号16に示されている。変異体マウスBTL−IIタンパク質の配列は配列番号18に示されている。ヒトスプライス変異体は、エキソン3単独かまたはエキソン2および3の両方を欠いており、そして開示されているマウススプライス変異体は、エキソン3のみを欠いている。配列番号7、配列番号9、配列番号11、配列番号13、配列番号15および配列番号17;図3a、4a、5a、6a、7a、および9aを参照されたい。これらの配列および、10アミノ酸より長いギャップを含んでいないGAPを使用して、配列番号4、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16または配列番号18から成る群の少なくとも一つを有するタンパク質配列と並列した場合、実質的に類似の配列によりコードされているタンパク質は本発明に包含される。もしこれらのタンパク質が、配列番号4、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16および/または配列番号18と比較した場合にアミノ酸置換を含んでいれば、これらの置換は好ましくは保存的アミノ酸置換である。本発明のBTL−IIタンパク質は、T細胞の表面上のレセプターへ結合することが可能であり、そしてT細胞の増殖および/またはT細胞によるサイトカイン産生を阻害することが可能である。
【0068】
さらに、本発明はエキソン1および4(配列番号8および配列番号12)またはエキソン2および4(配列番号10、配列番号14、配列番号16および配列番号18)のスプライス接合部にまたがる核酸によりコードされているBTL−IIタンパク質、そしてT細胞の表面上に発現されたレセプターへ結合可能で、並びに/あるいは増殖および/またはT細胞によるサイトカイン産生を阻害可能な実質的に類似のタンパク質を提供する。これには特に、配列番号10、配列番号14または配列番号18のアミノ酸127から157、または配列番号16のアミノ酸126から156と少なくとも80%、85%、90%、95%、96%、97%、98%、99%、99.7%または100%同一であるアミノ酸配列から成るポリペプチドを含んでなるBTL−IIタンパク質が含まれる。配列番号10、配列番号14または配列番号18のアミノ酸127から157、または配列番号16のアミノ酸126から156と並列されたアミノ酸配列の同一性領域は、好ましくは少なくとも20、23、25、27、30、35または40アミノ酸長である。こうしたアミノ酸配列は少なくとも150アミノ酸長であることが可能であり、そして配列番号10、配列番号14、配列番号16または配列番号18のアミノ酸30から358と少なくとも80%、85%、90%、95%、96%、97%、98%、99%、99.7%または100%同一であることが可能である。配列番号10、配列番号14、配列番号16または配列番号18のアミノ酸30から358と並列されたアミノ酸配列の同一性領域は、少なくとも50、75、100、125、150、175、200または300アミノ酸であることが可能である。
【0069】
本発明はまた、わずか2のまたはそれより少ないIg様ドメインを含み、そしてB細胞またはT細胞上に発現された細胞表面レセプターへ結合可能である、配列番号4のアミノ酸30から457と少なくとも80%、85%、90%、95%、96%、97%、98%、99%、99.7%または100%同一であるアミノ酸配列から成るポリペプチドを含んでなるBTL−IIタンパク質も提供する。アミノ酸配列は、配列番号8のアミノ酸30から247または配列番号12のアミノ酸30から243と少なくとも80%、85%、90%、95%、96%、97%、98%、99%、99.7%または100%同一であることが可能である。
【0070】
さらなる態様において、本発明は配列番号11のヌクレオチド34から124から成るポリヌクレオチドと80%、85%、90%、95%、96%、97%、98%、99%、99.7%または100%同一であるポリヌクレオチドを含んでなる核酸によりコードされているタンパク質を提供し、ここにおいて同一性領域は少なくとも60、70、80、90または100ヌクレオチド長である。こうしたポリヌクレオチドによりコードされている成熟BTL−IIタンパク質は、このポリヌクレオチドにより少なくとも一部がコードされているシグナル配列(タンパク質の未成熟バージョンに存在する)を欠いていてもよいが、それは必須ではない。こうしたタンパク質はT細胞へ結合することが可能であり、および/またはT細胞の増殖および/またはT細胞によるサイトカイン産生を阻害することが可能である。
【0071】
BTL−IIタンパク質は異なる程度までグリコシル化されていてもよいし、またはグリコシル化されていなくてもよい。例として、本発明のBTL−IIタンパク質は、配列番号4、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16または配列番号18を含んでなるタンパク質中にすでに観察されるものに加え、一つまたはそれより多くのNまたはO連結グリコシル化部位を含んでなることができる。こうしたBTL−IIタンパク質は、それに結合されたより多いシアル酸部分を有することができるので、非改変タンパク質よりも長いインビボ半減期を有することが可能である。BTL−IIタンパク質はまた、ヒトまたはマウスBTL−IIのIg様ドメインまたは実質的に類似のドメインのどれか一つ、どれか二つ、どれか三つまたは四つ全てを含んでなるタンパク質も含んでいる。
【0072】
変異体
任意の型の改変(例えば、限定されるわけではないが、アミノ酸の挿入、欠失または置換;ポリペプチドのグリコシル化状態の変化;その三次元構造または自己会合状態を変化させるための再折り畳みまたは異性化;および他のポリペプチドまたは分子とのその会合の変化)により任意のBTL−IIタンパク質から誘導されたポリペプチドもまた、本明細書において意味するBTL−IIタンパク質である。本発明により提供されるBTL−IIタンパク質には、T細胞の表面に発現したレセプターに結合することが可能であり、および/またはT細胞の増殖および/またはT細胞によるサイトカイン産生を阻害することが可能である、BTL−IIタンパク質配列番号4、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16および/または配列番号18のアミノ酸配列と実質的に類似しているアミノ酸配列により特徴付けられるポリペプチドが含まれる。同一性の領域は、配列番号4、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16または配列番号18の30またはより高い位置から出発することが可能である。配列番号4、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16および/または配列番号18の少なくとも一つを有するこうした変異体タンパク質のGAP並列は、10アミノ酸より長い内部ギャップを有しないことができる。配列番号4、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16または配列番号18と実質的に類似しているBTL−IIタンパク質の部分は、少なくとも100、少なくとも125、少なくとも150、少なくとも175、少なくとも200または少なくとも250アミノ酸長であることが可能である。こうしたタンパク質における修飾は、天然に提供されるか、または故意に操作される。例えば、配列番号11(図6a)は、複数の位置を含んだほんの少しの位置での塩基変化を含んでいる、配列番号9(図4a)のアリルまたは多型性変異体である。
【0073】
本発明は、配列番号4、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16または配列番号18の配列に関して、単一アミノ酸の挿入、欠失または置換であることが可能である、単一または多アミノ酸改変を含む、BTL−IIタンパク質変異体を提供し、ここにおいて、BTL−II変異体タンパク質は、T細胞増殖および/またはサイトカイン産生を阻害することが可能である。改変は保存的アミノ酸置換であり得る。例えば、表3に示したヒトおよびマウスBTL−IIタンパク質の並列により、機能に影響することなくどのアミノ酸を変化させることが可能であるかについて、当業者は導かれる。ヒトおよびマウスBTL−IIにおいて同一または類似しているアミノ酸は、そうではないアミノ酸より機能のために重要であるようである。さらに、IgVまたはIgCドメインにおいて高度に保存されているアミノ酸(表3においてボールド体で示されている)もまた、機能的に重要であるようである。こうしたアリルまたは多型性変異体は、疾患の素因を決定するための診断剤として有用であることが可能である。配列番号3とは異なった配列を有するヒトアリル変異体は、図5a、5b、6a、6b、7aおよび7bに開示されている。例えば、炎症性腸疾患の症状がない正常の人々間で、配列番号3の配列を有するcDNAをコードしているBTL−II遺伝子を有している数と、図5a、6aおよび7aにおいて3−6でラベルされたアリル変異体突然変異をコードしているBTL−II遺伝子を有している数の間には特定の比が存在することが可能である。炎症性腸疾患の症状を有する患者の大部分が、図5a、6aおよび7aにおいて3−6でラベルされたアリル変異体突然変異を伴ったBTL−II遺伝子を有するように、これらのアリルの出現比率を患者間で改変することができる。患者の組織における多様なアリル変異体の存在は、例えば、多型性部位に広がっているDNAまたはRNAのセグメントのPCR増幅により決定することが可能である。上述のように、特にヒトおよびマウスBTL−IIタンパク質配列間で保存されていない部位での保存的アミノ酸置換は、保存的部位での非保存的置換よりも生物学的機能を保つようである。当業者はまた、例えば、DALI(HolmおよびSander(1993),J.Mol.Biol.233:123−38)のごときプログラムにより予測されるような、BTL−IIタンパク質の三次構造を実質的にひっくり返す置換もまた、機能を害しそうであることを理解するであろう。
【0074】
断片(fragment)
配列番号4、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16または配列番号18の断片、またはこれらの配列と少なくとも80%、85%、90%、95%、97%、98%、99%、99.5%または99.7%同一である断片を含んでなるタンパク質は、BTL−IIタンパク質に含まれている。好ましくは、こうした断片はT細胞の表面上に発現されたレセプターへ結合することが可能であり、そして少なくとも約50、60、70、80、90、100、150または200アミノ酸長である。好ましくは、こうした断片は水性溶液に可溶であり、そしてBTL−IIタンパク質の細胞外領域一部または全てを含んでなる。こうした断片を含んでなるタンパク質は分泌されることが可能である。例えば、上記ヒトまたはマウスBTL−IIタンパク質または実質的に類似したタンパク質のドメインの少なくとも一つを含んでなるBTL−IIタンパク質は本発明に包含され、ここにおいて該ドメインはBTL−IIタンパク質の少なくとも一つの生物学的特性を有している。例えば、こうしたタンパク質はT細胞の増殖および/またはサイトカイン産生を阻害可能である。
【0075】
さらに本発明に包含されるのは、ヒトおよびマウスBTL−IIタンパク質または実質的に類似のタンパク質の、以下の改変バージョンである:(1)エキソン3によりコードされている第二のIg様ドメイン(図1参照)を欠いているヒトおよびマウスBTL−IIタンパク質のバージョン;(2)エキソン2および3によりコードされている第一および第二のIg様ドメインを欠いているヒトおよびマウスBTL−IIタンパク質のバージョン;(3)エキソン5および6によりコードされている第三および第四のIg様ドメインを欠いているバージョン;(4)四つのIg様ドメインの任意の二つを欠いているバージョン;および(5)Ig様ドメインの任意の一つを欠いているバージョン。
【0076】
また本発明内に包含されるのは、配列番号4、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16または配列番号18の免疫原性断片であり、それは断片へ特異的に結合する抗体を惹起することが可能である。こうした断片は好ましくは少なくとも10アミノ酸長であり、そして好ましくは、上述の配列からの隣接アミノ酸残基を含んでなる。こうした断片で免疫化された動物により産生された抗体は、本出願の別のところで議論されているように、炎症性腸疾患の予測、診断および処置に有用であることが可能である。こうした断片はスプライス接合部によりコードされているこれらのタンパク質の領域に及ぶことが可能であり、スプライス変異体または完全長タンパク質によりコードされているタンパク質に特異的な抗体の発生に利点を有している。あるいは、エキソン3によりコードされているBTL−IIの一部に対する抗体は、完全長BTL−IIタンパク質によってのみ検出され、エキソン3を欠く全ての他のスプライス変異体によりコードされているタンパク質では検出されない。
【0077】
組換え融合タンパク質
本発明はさらに、上述の少なくとも一つのBTL−IIポリペプチド(BTL−IIタンパク質の一つである)、変異体または断片、そして少なくとも一つの他の部分を含んでなる融合タンパク質を包含する。他の部分は、異種ポリペプチド、即ち、BTL−IIポリペプチド以外のポリペプチドであることが可能である。他の部分はまた、例えば、ポリエチレングリコール(PEG)部分、または細胞毒性、細胞分裂抑制性、発光および/または放射活性部分のごとき、非タンパク質部分であることが可能である。PEGの結合は、少なくともいくつかのタンパク質のインビボ半減期を増加させることが知られている。さらに、細胞毒性、細胞分裂抑制性、発光および/または放射活性部分は、例えば、抗体が結合可能な細胞に位置させるための、細胞の増殖を阻害するための、または細胞を殺すための、診断または治療目的のために抗体へ融合されてきた。同様に、こうした部分へ融合されたBTL−IIポリペプチドは、BTL−IIが結合する細胞に位置させるために、細胞の増殖を阻害するために、または細胞を殺すために使用することが可能である。こうした細胞毒性、細胞分裂抑制性、発光および/または放射活性部分は、例えば、マイタンシン誘導体(DM1のごとき)、エンテロトキシン(ブドウ球菌エンテロトキシンのごとき)、ヨウ素同位元素(ヨード−125のごとき)、テクネチウム同位元素(Tc−99mのごとき)、シアニン蛍光色素(Cy5.5.18のごとき)、リボソーム不活性化タンパク質(ブーガニン、ゲロニンまたはサポリン−S6のごとき)、および商標MYLOTARGTM(Wyeth−Ayerst)の名で売買されている製品の一部である、細胞毒性物質のカリケアミシン、である。
【0078】
多様な異種ポリペプチドを、例えば、タンパク質のインビボ半減期を増加させるため、タンパク質の同定、単離および/または精製を容易にするため、タンパク質の活性を増加させるため、そしてタンパク質のオリゴマー化を促進するためのごとき、多様な目的のためにBTL−IIポリペプチドへ融合することが可能である。例えば、CD80のごときB7サブファミリーのいくつかのタンパク質は、主としてオリゴマー形でそれらのレセプターへ結合するので(CD80の場合は二量体、Collinsら(2002),Immunity 17:201−10、を参照されたい)、オリゴマー化は可溶性タンパク質の生物学的活性を保存するためには非常に重要であろう。
【0079】
多くの異種ポリペプチドは、それらがその一部である組換え融合タンパク質の同定および/または精製を容易にすることを可能にする。例には、ポリアルギニン、ポリヒスチジン、または、固定化された金属イオンに対して高い親和性を所有する非隣接ヒスチジンの天然に存在する配列であるHATTM(Clontech)が含まれる。これらの異種ポリペプチドを含んでなるタンパク質は、例えば、固定化されたニッケル、または固定化されたコバルトイオンを含んでなるTALONTM樹脂(Clontech)を使用するアフィニティークロマトグラフィーにより精製することが可能である。例えば、Knolら(1996),J.Biol.Chem.27(26):15358−15366、を参照されたい。ポリアルギニンを含んでなる異種ポリペプチドは、イオン交換クロマトグラフィーによる効率的な精製を可能にする。他の有用な異種ポリペプチドには、例えば、米国特許第5,011,912号およびHoppら(1988),Bio/Technology 6:1204、に記載されている抗原性同定ペプチドが含まれる。こうしたペプチドの一つはFLAG(登録商標)ペプチドであり、それは高度に抗原性であり、そして特異的モノクローナル抗体により可逆的に結合されるエピトープを提供し、発現された組換え融合タンパク質の、迅速なアッセイおよび容易な精製を可能にしている。4E11と称されるマウスハイブリドーマは、米国特許第5,011,912号に記載されているように、特定の二価金属カチオンの存在下でFLAG(登録商標)ペプチドを結合する、モノクローナル抗体を産生する。4E11ハイブリドーマ細胞株は、第HB9259号の寄託番号でAmerican Type Culture Collectionに寄託されている。FLAG(登録商標)ペプチドを結合するモノクローナル抗体は、FLAG(登録商標)ペプチドを含んでなるポリペプチド精製試薬を回収するための親和性試薬として使用することが可能である。他の適したタンパク質タグ(tag)および親和性試薬は以下のものである:1)固定化グルタチオンに対するグルタチオン−S−トランスフェラーゼ融合タンパク質の親和性を利用する、GST−BindTM)システム(Novagen)に記載されているもの;2)モノクローナル抗体に対するT7遺伝子10タンパク質のアミノ末端11アミノ酸の親和性を利用する、T7−Tag(登録商標)アフィニティー精製キット(Novagen)に記載されているもの;または3)タンパク質タグに対するストレプトアビジンの操作された形の親和性を利用する、Strep−tag(登録商標)システム(Novagen)に記載されているもの。上述のタンパク質タグのいくつか、並びにその他がSassenfeld(1990),TIBTECH 8:88−93,Brewerら,「Purification and Analysis of Recombinant Proteins」中,pp.239−266,SeetharamおよびSharma(監修),Marcel Dekker,Inc.(1991),およびBrewerおよびSassenfeld,「Protein Purification Applications」中,pp.91−111,HarrisおよびAngal(監修),Press,Inc.,Oxford England(1990)、に記載されている。さらに、二つまたはそれより多い上述のタグの融合、例えば、FLAGタグおよびポリヒスチジンタグの融合を本発明のBTL−IIタンパク質へ融合することが可能である。
【0080】
他の異種ポリペプチドを含んでなる組換え融合タンパク質は、例えば、二量体、三量体またはより高度な多量体を形成する性向、増加したインビボ半減期、および/または増加した生物学的活性のごとき、他の種類の独特な利点を有している。融合タンパク質を製造するための技術は既知であり、そして例えば、WO 99/31241およびCosmanら((2001),Immnunity 14:123−133)、に記載されている。例示として、IgG抗体のFc領域、または実質的に類似したタンパク質を含んでなる異種ポリペプチドを、BTL−IIポリペプチドまたは断片へ融合することが可能である。抗体のFc領域は、ヒトまたは動物起源の抗体からのCH2およびCH3ドメインまたはこれらに実質的に類似した免疫グロブリンドメインを含んでなるポリペプチドである。議論のためには、HasemannおよびCapra,免疫グロブリン:構造および機能,William E.Paul,監修,「Fundamental Immunology」中,第2版,212−213(1989)、を参照されたい。二量体化を促進するヒンジ領域を含んでなる、Fc領域の一部切断形もまた使用することが可能である。抗体および他の免疫グロブリンアイソタイプの他の部分を使用することが可能である。IgG抗体のFc領域を含んでなる組換え融合タンパク質は、二量体を形成するようである。抗体由来のタンパク質のいろいろな部分を含んでなる融合タンパク質は、Ashkenaziら((1991)Proc.Natl.Acad.Sci.USA 88:10535−39)、Byrnら((1990),Nature 344:677−70)、HollenbaughおよびAruffo(「免疫学における最新のプロトコール」中,補4,pp.10.19.1−10.19.11(1992))、Baumら((1994),EMBO J.13:3992−4001)および米国特許第5,457,035号およびWO 93/10151、に記載されている。いくつかの態様において、改変Fc領域は、野生型Fc領域と比較して、Fcレセプターに対するより低い親和性を有しているという利点を持つことが可能である。このことは、こうした組換え融合タンパク質が免疫エフェクター細胞により結合した細胞の溶解を少なくすることができるので都合がよい。実施例5は、ヒトFc領域へ融合されたマウスBTL−IIの細胞外領域を含んでいる融合タンパク質の製造を記載している。このタンパク質をコードしている核酸配列およびそのアミノ酸配列は、各々、配列番号19および配列番号20に開示されている。
【0081】
別の代替物として、本発明の組換え融合タンパク質は、ロイシンジッパーを含んでなる異種ポリペプチドを含んでなることが可能である。既知のロイシンジッパー配列の中で、二量体化を促進する配列および三量体化を促進する配列である。例えば、Landschulzら(1988),Science 240:1759−64、を参照されたい。ロイシンジッパーは、しばしば、他のアミノ酸と共に散らばる4または5のロイシン残基を持つ、7アミノ酸反復を含んでなる。ロイシンジッパーの使用および製造は、当該技術分野ではよく知られている。
【0082】
もしくは、組換え融合タンパク質の異種ポリペプチド形成部分は、二つまたはそれより多くのBTL−IIポリペプチドを連結している、一つまたはそれより多くのペプチドリンカーであり得る。一般に、ペプチドリンカーは、複数の同一の、類似のまたは異なったポリペプチドを連結するために働き、そしてタンパク質の連結された部分の望まれる機能に必要とされる可動性または強剛性を提供する、アミノ酸の伸長である。典型的には、ペプチドリンカーは、長さ約1および30アミノ酸の間である。ペプチドリンカーの例には、限定されるわけではないが、−−Gly−Gly−、GGGGS(配列番号1)、(GGGGS)n(配列番号2)、GKSSGSGSESKS(配列番号3)、GSTSGSGKSSEGKG(配列番号4)、GSTSGSGKSSEGSGSTKG(配列番号5)、GSTSGSGKSSEGKG(配列番号6)、GSTSGSGKPGSGEGSTKG(配列番号7)またはEGKSSGSGSESKEF(配列番号8)が含まれる。連結部分は、例えば、Huston,J.S.ら,Proc.Natl.Acad.Sci.85:5879−83(1988)、Whitlow,M.ら,Protein Engineering 6:989−95(1993)、およびNewton,D.L.ら,Biochemistry 35:545−53(1996)に記載されている。他の適切なペプチドリンカーは、米国特許第4,751,180号および第4,935,233号に記載されているものである。
【0083】
さらに、組換え融合タンパク質は、その正常なシグナル配列を欠き、そしてその代わりとして、それに置き換わる異種シグナル配列を有するBTL−IIタンパク質を含んでなることが可能である。シグナル配列の選択は、組換えタンパク質が産生されるべき宿主細胞の型に依存しており、そして天然のシグナル配列を異種シグナル配列に置き換えることが可能である。哺乳動物宿主細胞中で機能的であるシグナル配列には以下のものが含まれる:米国特許第4,965,195号に記載されているインターロイキン−7(IL−7)のためのシグナル配列;Cosmanら((1984),Nature 312:768)により記載されているインターロイキン−2レセプターのためのシグナル配列;EP特許第0 367 566号に記載されているインターロイキン−4レセプターシグナル配列;米国特許第4,968,607号に記載されているタイプI インターロイキン−1レセプターシグナル配列;EP特許第0 460 846号に記載されているタイプII インターロイキン−1レセプターシグナル配列。
【0084】
BTL−II核酸
本発明は、変異体、断片、組換え融合タンパク質、完全長タンパク質、可溶性タンパク質および分泌タンパク質を含む、上記のBTL−IIタンパク質、断片または免疫原性断片をコードする単離核酸を包含する。これらの核酸は、なかでも、組換えタンパク質を製造するため、および組織試料中のBTL−II核酸の存在を検出するために、例えば、診断使用のために有用である。こうした核酸は、ゲノムDNAまたはcDNAであり得る。核酸は、本発明のBTL−IIタンパク質をコードしている、中断されていないオープンリーディングフレーム(open reading frame)を含んでなることが可能である。本発明の核酸分子には、一本鎖および二本鎖形両方の形でのDNAおよびRNA、並びに、対応する相補的配列が含まれる。“単離核酸”とは、天然に存在する起源から単離された核酸の場合、該核酸が単離されたゲノムまたは生物体中に存在する、隣接する遺伝子配列から分離されている核酸である。オリゴヌクレオチドのごとく、化学的に、またはポリメラーゼ連鎖反応(PCR)生成物またはcDNAのごとく、鋳型から酵素的に合成された核酸の場合、こうしたプロセスから生じる核酸は単離核酸であることを理解されたい。単離核酸分子は、別々の断片の形での、またはより大きな核酸構築物の成分としての核酸分子も指している。
【0085】
さらに、本発明は、(1)例えば、サザンおよびノーザンブロッティング、ドットブロッティング、コロニーハイブリダイゼーションなど、当該技術分野ではよく知られた多数の方法によりBTL−II核酸を検出するためのプローブとして、(2)BTL−II核酸を増幅するためのポリメラーゼ連鎖反応(PCR)プライマーとして、または(3)例えば、アンチセンス核酸(ペプチド核酸を含んで)、リボザイム、三重らせん形成分子による発現の阻害を通して、または任意のこれらのRNAをコードするRNAまたはDNAを妨害することによる、BTL−II核酸の発現を調節するための手段として、働くことが可能であるBTL−IIをコードしている核酸の断片を包含する。PCRプライマーは、BTL−II核酸配列に加えて、増幅された核酸の使用を容易にする制限酵素切断部位のごとき他の配列を含んでなることが可能である。PCRは以下の文献に記載されている:Saikiら(1988),Science 239:487−91;「PCR Technology」,Erlich,監修,Stockton Press,(1989)。以下に説明するように、PCRはBTL−II mRNAの過剰発現を検出するために有用であることが可能であり、そしてPCRプライマーは、遺伝子の多様な部分から採ることが可能であり、そしてまた、異なったスプライス変異体間を区別するために選択することが可能である。アンチセンスRNA(およびそれらをコードしているDNA)、DNAまたは合成ヌクレオチド、そして発現を調節するためのそれらの使用は、当該技術分野ではよく知られており、例えば以下の文献に記載されている:IzantおよびWeintraub(1984),Cell 36(4):1007−15;IzantおよびWeintraub(1985),Science 229(4711):345−52;Hazel−Bellanら(1988),J.Exp.Med.168(6):2309−18;Sarinら(1988),Proc.Natl.Acad.Sci.USA 85(20):7448−51;Zon(1988),Pharm.Res.5(9):539−49;Harel−Bellanら(1988),J.Immunol.140(7):2431−35;Marcus−Sekuraら(1987),Nucleic Acids Res.15(14):5749−63;Gambari(2001),Curr.Pharm.Des.7(17):1839−62;およびLemaitreら(1987),Proc.Natl.Acad.Sci.USA 84(3):648−52。同様に、選択された遺伝子の発現を阻害するための干渉(interfering)RNA(およびそれらをコードしているDNA)そしてそれらの使用は、当該技術分野ではよく知られており、以下の文献に記載されている:Fjoseら(2001),Biotechnol.Ann.Rev.7:31−57;BosherおよびLabouesse(2000),Nature Cell Biol.2:E31−E36。さらに、リボザイムまたはDNAzymeは、特異的RNAを切断するように標的化することが可能であり、そして従って、LewinおよびHauswirth(2001)Trends Mol.Med.7(5):221−28;MenkeおよびHobom(1997),Mol.Biotechnol.8(1):17−33;Norrisら(2000),Adv.Exp.Med.Biol.465:293−301;Sioud(2001),Curr.Mol.Med.1(5):575−88;およびSantiagoおよびKhachigian(2001),J.Mol.Med.79(12):695−706、に記載されているように、遺伝子発現を阻害するために使用することが可能である。BTL−II発現を調節可能な核酸は、BTL−II機能のインビボまたはインビトロ研究における、または治療剤として、所望により遺伝子治療剤としての使用を見出すことが可能である。
【0086】
本発明はまた、中程度のストリンジェント条件下、より好ましくは高ストリンジェント条件下で、本明細書に記載したBTL−IIタンパク質をコードしている核酸へハイブリダイズする核酸も含んでいる。こうした核酸には、配列番号3、配列番号5、配列番号7、配列番号9、配列番号11、配列番号13、配列番号15または配列番号17が含まれる。好ましくは、こうした核酸は、T細胞の表面上のレセプターへ結合可能で、および/またはT細胞増殖および/またはサイトカイン産生を阻害可能なタンパク質をコードしている。ハイブリダイゼーション技術は当該技術分野ではよく知られており、そしてSambrook,J.,E.F.FritschおよびT.Maniatis(「Molecular Cloning:A Laboratory Manual」,Cold Spring Harbor Laboratory Press,コールドスプリングハーバー,NY,第9章および第11章,(1989))および「Current Protocols in Molecular Biology」(F.M.Ausubelら監修,John Wiley & Sons,Inc,2.10および6.3−6.4節(1995))により記載されている。中程度にストリンジェントな条件は、約50%ホルムアミド、6xSSC中、約42から55℃の温度でのハイブリダイゼーション、そして約60℃、0.5xSSC、0.1%SDS中での洗浄が含まれる。高ストリンジェントな条件は、上記のようなハイブリダイゼーション条件と定義されるが、およそ68℃、0.2xSSC、0.1% SDSでの洗浄を伴うと定義される。ハイブリダイゼーションおよび洗浄緩衝液において、SSPE(1xSSPEは、0.15M NaCl、10mM NaH2PO4、および1.26mM EDTA,pH7.4である)をSSC(1xSSCは、0.15M NaClおよび15mMクエン酸ナトリウムである)に対して置換することが可能である;洗浄は、好ましくは少なくとも2回、ハイブリダイゼーションが完了した後、15分間行う。
【0087】
当業者に知られるように、そして以下にさらに記載するように、ハイブリダイゼーション反応および二重鎖安定性を支配する基本原理を適用することによって、必要に応じて、望ましい度合いのストリンジェンシーを達成するよう、洗浄温度および洗浄塩濃度を調整可能であることを理解すべきである(例えば、Sambrookら、上記文献を参照されたい)。既知の配列の核酸をハイブリダイズさせる場合、ハイブリッド長は、核酸の配列を並列し、そして最適配列相補性を持つ、一つまたは複数の領域を同定することによって、決定可能である。長さが50塩基対未満であると予測されるハイブリッドのハイブリダイゼーション温度は、ハイブリッドの融解温度(Tm)より5から10℃低くなければならず、Tmは、以下の等式に従って決定する。長さ18塩基対未満のハイブリッドに関しては、Tm(度C)=2(A+T塩基数)+4(G+C塩基数)である。長さ18塩基対を超えるハイブリッドに関しては、Tm(度C)=81.5+16.6(log10[Na+])+0.41(%G+C)−(600/N)であり、式中、Nはハイブリッド中の塩基数であり、そして[Na+]はハイブリダイゼーション緩衝液中のナトリウムイオン濃度である(1xSSCの[Na+]=0.165M)。各々のこうしたハイブリダイズする核酸は、少なくとも15ヌクレオチド(または少なくとも18ヌクレオチド)、または少なくとも20、または少なくとも25、または少なくとも30、または少なくとも40、または少なくとも50、または少なくとも100である長さを有する。Sambrookら、上記文献。
【0088】
BTL−II核酸は、以下のポリヌクレオチドを含んでなる核酸を含んでいる:(1)配列番号3、配列番号5、配列番号7、配列番号9、配列番号11、配列番号13、配列番号15または配列番号17の全てまたは断片、ここにおいて、断片はT細胞の表面上に発現されたレセプターへ結合可能で、および/またはT細胞の増殖および/またはサイトカイン産生を阻害可能なBTL−IIタンパク質をコードしている;(2)少なくとも100、125、150、175、200、225、250、300、400、500、600、800、1000、1200、1400または1600ヌクレオチド長であり、そしてT細胞の表面上に発現されたレセプターへ結合可能で、および/またはT細胞の増殖および/またはサイトカイン産生を阻害可能なBTL−IIタンパク質をコードしている、配列番号3、配列番号5、配列番号7、配列番号9、配列番号11、配列番号13、配列番号15または配列番号17と少なくとも80%、85%、90%、95%、97%、98%、99%、99.5%または99.7%同一である配列;(3)本発明のBTL−IIタンパク質をコードしている核酸を検出するまたは増幅するために、またはBTL−II mRNAおよび/またはタンパク質の発現を調節するために有用である、配列番号3、配列番号5、配列番号7、配列番号9、配列番号11、配列番号13、配列番号15または配列番号17または実質的に類似した配列の断片;(4)配列番号7または配列番号11のヌクレオチド30から130から成るポリヌクレオチド、または配列番号9、配列番号13、配列番号15または配列番号17のヌクレオチド377−477と、少なくとも80%、85%、90%、95%、97%、98%、99%、99.5%または99.7%同一であるポリヌクレオチドを含んでなる核酸、ここにおいて、同一性の領域は少なくとも60,70,80,90または100ヌクレオチド長であり、そして核酸によりコードされているタンパク質は、T細胞の増殖および/またはサイトカイン産生を阻害可能である;および(5)配列番号3、配列番号5、配列番号7、配列番号9、配列番号11、配列番号13、配列番号15または配列番号17と比較して少なくとも1、2、3、4、6、8、10、15、20、25、30、35、40、50または75の改変を含んでなる核酸、ここにおいて、改変は、単一ヌクレオチドの挿入、欠失または置換であることが可能である。
【0089】
BTL−IIポリペプチドへ特異的に結合する抗体
変異体、断片および組換え融合タンパク質を含む、本発明のBTL−IIタンパク質へ特異的に結合する抗体は、本発明により包含される。本明細書において、別のタンパク質(抗体のごとき)によるBTL−II上のエピトープの特異的結合とは、同一のエピトープを含んでなる別のタンパク質により、特異的に結合されているタンパク質は、それが結合されているBTL−IIタンパク質の分子から置き換えられることが可能であるが、このエピトープを含んでいない別のタンパク質によっては置き換えられないことを意味している。多数の競合的結合アッセイが当該技術分野では知られている。エピトープは近接アミノ酸のみを含んでなることができるが、しかしまたBTL−IIタンパク質の三次の折り畳みにより近くにもたらされた非近接アミノ酸を含んでなることもできる。エピトープは当該技術分野では既知の方法により同定することが可能である。例えば、Leinonenら(2002),Clin.Chem.48(12):2208−16;Krogerら(2002),Biosens.Bioelectron.17(11−12):937−44;Zhuら(2001),Biochem.Biophys.Res.Commun.282(4):921−27、を参照されたい。本発明はまた、本明細書において免疫原性断片と称される、抗体を発生するために有用である、本明細書に記載したBTL−IIタンパク質のエピトープも包含する。免疫原性断片は、好ましくは少なくとも10アミノ酸長であり、そして好ましくは、配列番号4、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16または配列番号18からの近接アミノ酸を含んでなる。こうしたエピトープは、スプライス接合部によりコードされているBTL−IIタンパク質の領域に及んでおり、特異的スプライス変異体によりコードされているタンパク質への特異的結合の利点を有することができる。
【0090】
抗体はポリクローナルまたはモノクローナル抗体であることが可能であり、そして当該技術分野ではよく知られている方法により産生可能である。例えば、「Monoclonal Antibodies,Hybridomas:A New Dimension in Biological Analyses」,Kennetら(監修),Plenum Press,ニューヨーク(1980);および「Antibodies:A Laboratory Manual」,HarlowおよびLand(監修),Cold Spring Harbor Laboratory Press,コールドスプリングハーバー,NY,(1988);KohlerおよびMilstein(1980)Proc.Natl.Acad.Sci.USA,77:2197;Kozborら(1984),J.Immunol.133:3001−3005(ヒトB細胞ハイブリドーマ技術を記載している);Coleら,「Monoclonal Antibodies And Cancer Therapy」,Alan R.Liss.Inc.,pp.77−96(1985)(EBVハイブリドーマ技術を記載している);Kuby,「Immunology」,第2版,p.162−64,W.H.Freeman and Co.,ニューヨーク(1994)、を参照されたい。本発明のBTL−IIタンパク質に特異的なモノクローナル抗体を産生するハイブリドーマ細胞株もまた、本明細書に意図される。こうしたハイブリドーマは、慣用的技術によって産生しそして同定することが可能である。本発明のmAbを産生するハイブリドーマはインビトロまたはインビボで培養することが可能である。さらに、本発明の抗BTL−II抗体は、例えば、チャイニーズハムスター卵巣(CHO)、HeLa、VERO、BHK、Cos、MDCK、293、3T3、骨髄腫(例えば、NSO、NSI)またはWI38細胞、酵母細胞、昆虫細胞および大腸菌(Eschericha coli)を含む細菌細胞、を含む他の培養細胞中で産生することが可能である。こうした抗体は、抗体に加えてこれらの抗体の発現を可能にする核酸をコードしている核酸を所望の細胞内へ導入することにより産生することが可能である。抗体は、それから、これらの核酸が導入されている細胞を培養することにより産生することが可能である。モノクローナル抗体は、IgG、IgM、IgE、IgA、IgD、およびそのいかなるサブクラスも含む、いかなる免疫グロブリンクラスのものであってもよい。
【0091】
あるいは、抗体は重および軽鎖可変領域様ドメイン、そして所望により、また一つまたはそれより多くの定常領域様ドメインを含んでなる単鎖抗体(米国特許第4,946,778号;Birdら(1988),Science 242:423−26;Hustonら(1988),Proc.Natl.Acad.Sci.USA 85:5879−83)、二量体または多価抗体(例えば、Lanttoら(2002),J.Gen.Virol.83:2001−05;HudsonおよびSouriau(2001),Expert Opin.Biol.Ther.1(5):845−55)、四量体抗体(例えば、Janewayら,「Immunobiology:The Immune System in Health and Disease」,第5版,第II部,第3章,Garland Publishing(2001))、キメラ抗体(HudsonおよびSouriau,上記文献;Boulianneら(1984),Nature 312:643−46;Morrison(1984),Proc.Natl.Acad.Sci.USA 81:6851−55;Takedaら(1985),Nature 314:452−54;Neubergerら(1985),Nature 314:268−70)、異なったトランスジェニック動物で産生された完全ヒト抗体(例えば、米国特許第6,150,584に記載されている)またはインビトロ選択(米国特許出願第2002/0058033号)またはヒト化抗体(Morrisonら,上記文献;Takedaら,上記文献;Boulianneら,上記文献)であってもよい。さらに、抗体は、インビトロ選択スキームにより“成熟させる”ことが可能であり、例えば、それが結合するエピトープに対してより高い親和性のごとき、改変された特性を有する抗体を得る。例えば、Jacksonら(1995),J.Immunol.154(7):3310−19;PiniおよびBracci(2000),Curr.Protein Pept.Sci.1(2):155−69;E11markら(2002),Mol.Immunol.39(5−6):349;O’Connellら(2002),J.Mol.Biol.321(1):49−56;Hulsら(2001),Cancer Immunol.Immunother.50:163−71;HudsonおよびSouriau,上記文献;AdamsおよびSchier(1999),J.Immunol.Methods 231(1−2):249−60;Schmitzら(2000),Placenta 21 補A:S106−12、を参照されたい。あるいは、本発明のBTL−IIタンパク質へ特異的に結合可能な、例えば、Fab断片、F(ab’)2断片または単鎖Fv断片(scFv’s)のごとき抗体の断片も、本明細書において抗BTL−II抗体として意味されるものに包含される。FabおよびFv断片についての議論は、Kuby,上記文献,pp.109−112およびJanewayら,上記文献、を参照されたい。本発明はまた、BTL−IIタンパク質へ特異的に結合する抗体に特異的に結合する、そしてBTL−IIタンパク質の効果を模倣する、抗イディオタイプ抗体も包含する。こうした抗イディオタイプ抗体は、BTL−IIタンパク質としての同様の使用を発見した。抗イディオタイプ抗体を産生する方法は当該技術分野ではよく知られている。例えば、Kubyら,上記文献,371−72、を参照されたい。本発明のBTL−IIタンパク質および別のエピトープへ特異的に結合可能な、多様な種類の組換えおよび非組換え二重特異性抗体もまた意図される。多様な種類の二重特異性抗体およびそれらを作製する方法は、例えば、米国特許第4,474,893,6,060,285および6,106,833号に記載されている。
【0092】
抗BTL−II抗体は、BTL−IIのそのレセプターへの結合のごときBTL−IIの生物学的機能を遮断するアンタゴニスト抗体、若しくはBTL−IIの生物学的機能を促進するまたはBTL−IIの機能を模倣するアゴニスト抗体であることができる。アゴニスト抗体は、BTL−IIタンパク質の機能を模倣するアゴニスト性抗イディオタイプ抗体を含むことが可能である。BTL−II機能のためのアッセイは本明細書に記載されている。こうしたアッセイで決定された、BTL−IIの生物学的機能を遮断する抗BTL−II抗体は、本明細書において意味されるようにアンタゴニスト抗体である。アンタゴニスト抗体は、例えば、BTL−IIのそのレセプターへの結合を遮断することができる。こうしたアッセイへ加えた場合、BTL−IIの生物学的機能を促進するまたは増強する抗BTL−II抗体は、本明細書において意味されるようにアゴニスト抗体である。アンタゴニスト性抗BTL−II抗体は、例えば、粘膜免疫応答を増強するためのアジュバントとして使用可能である。さらに、BTL−IIレセプターに対するアゴニスト性抗は、例えば、クローン病または炎症性腸疾患のごとき、腸の不適切な炎症により特徴付けられる疾患を処置するため、粘膜免疫応答を抑制するために使用可能である。
【0093】
本発明の抗体はまた、インビトロまたはインビボの両方で、本発明のBTL−IIタンパク質の存在を検出するためのアッセイにも使用可能である。抗体はまた、免疫アフィニティークロマトグラフィーによる本発明のBTL−IIタンパク質の精製においても用いることが可能である。抗体はまた、免疫親和性クロマトグラフィーによる、本発明のBTL−IIタンパク質の精製に用いることが可能である。
【0094】
本発明は、本発明の抗体をコードしている核酸、およびこうした核酸を細胞に導入し、そして核酸を含んでいる細胞を培養することによる、抗体を産生するための方法を包含する。
【0095】
BTL−IIポリペプチドのアゴニストおよびアンタゴニスト
本発明はBTL−IIのアゴニストおよびアンタゴニスト、そしてアゴニストおよびアンタゴニストをスクリーニングするための方法、並びにアゴニストおよびアンタゴニストを使用するための方法を包含する。インビボまたはインビトロにおけるBTL−IIタンパク質の過剰または過少発現、またはインビボまたはインビトロにおけるBTL−II発現の完全非存在を含んでいる、例えば、細胞増殖アッセイ、サイトカイン分泌アッセイ、結合アッセイおよび遺伝子アッセイのごとき、BTL−II生物学的活性のためのアッセイが本明細書に記載されている。候補分子をこうしたアッセイに加えることが可能であり、BTL−IIタンパク質の生物学的活性に対するそれらの効果を決定する。BTL−IIアンタゴニストは、例えば、BTL−IIと、好ましくはB細胞またはT細胞上に発現されたそのレセプターとの相互作用を、遮断可能である。アンタゴニストはアンタゴニスト抗体を含み、そしてアゴニストはアゴニスト抗体を含む。加えて、アフィボディ(affibody)(Ronnmarkら(2002),J.Immunol.Methods 261(1−2):199−211)のような、本発明のBTL−IIタンパク質へ特異的に結合可能である他の抗体関連分子、および本発明のBTL−IIタンパク質に特異的に結合し、そしてBTL−IIタンパク質の生物学的活性を阻害する、WO 00/24782に記載されている生物学的に活性なペプチドが本発明に包含される。さらに、BTL−IIアンタゴニストには、例えば、干渉RNA(またはそれらをコードしているDNA)またはアンチセンスRNAまたはDNAのごとき、BTL−IIタンパク質および/またはmRNAの発現を変調するために有用である上述の核酸が含まれる。
【0096】
アンタゴニストにはさらに、BTL−IIまたはそのレセプターへ結合するためにインビトロで選択されたアミノ酸配列を含んでなる、そして、所望により、BTL−IIまたはそのレセプターの相互作用を妨害可能な、タンパク質が含まれる。あるいは、こうしたタンパク質は、BTL−IIの生物学的機能を促進するまたは模倣するBTL−IIアゴニストであることが可能である。BTL−IIまたはそのレセプターへ結合するタンパク質は、BTL−IIとそのレセプターとの相互作用を妨害する能力でスクリーニングすることが可能であり、または、選択は、こうしたタンパク質を直接得るように設計することが可能である。
【0097】
タンパク質は、例えば、ファージディスプレイまたは細菌の表面のディスプレイのごとき、多くの方法により選択することができる。例えば、ParmleyおよびSmith(1989),Adv.Exp.Med.Biol.251:215−218;Luzzagoら(1995),Biotechnol.Annu.Rev.1:149−83;Luら(1995),Biotechnology(NY)13(4):366−372、を参照されたい。これらの方法において、結合ドメインのライブラリーの各々のメンバーは、個々のファージ粒子または細菌細胞上にディスプレイすることが可能であり、そして選ばれた条件下で目的のタンパク質へ結合する細菌またはファージを選択可能である。選択された結合ドメインをコードする核酸は、選択されたファージまたは細菌を成長させ、そしてそれらから核酸を単離することにより得ることが可能である。
【0098】
あるいは、タンパク質を全くインビトロで選択することが可能である。例えば、潜在的結合ドメインのライブラリー中の、個々のポリペプチドの各々を、それをコードする核酸へ付着可能であり、そして選ばれた条件下で目的のタンパク質へ結合するものを選択することが可能である。ポリペプチドはそれらをコードする核酸へ付着されているので、増幅、クローニング、または有効な結合ドメインをコードする核酸の配列決定のごとき、続いての操作が容易である。抗体−リボソーム−mRNA粒子、リボソームディスプレイ、共有結合的RNA−ペプチド融合、または共有結合的DNA−RNA−ペプチド融合を含む、こうした選択のための多様なスキームが当該技術分野では知られている。HeおよびTaussig(1997),Nucleic Acids.Res.25(24):5132−5134;HanesおよびPluckthun(1997),Proc.Natl.Acad.Sci.94:4937−4942;RobertsおよびSzostak(1997),Proc.Natl.Acad.Sci.94:12297−12302;LohseおよびWright(2001),Curr.Opin.Drug Discov.Devel.4(2):198−204;Kurzら(2000),Nucleic Acids Res.28(18):E83;Liuら(2000),Methods Enzymol.318:268−93;Nemotoら(1997),FEBS Lett.414(2):405−08;米国特許第6,261,804号;WO0032823;およびWO0034784。こうしたタンパク質はアンタゴニストまたはアゴニストであると選択することが可能である。
【0099】
BTL−IIタンパク質の生物学的活性のためのアッセイ
BTL−IIタンパク質の生物学的活性を検出するため、およびBTL−IIの結合パートナー(partner)を同定するために、多様なアッセイを使用することが可能である。BTL−IIタンパク質、BTL−IIのレセプター、および両方のアゴニストおよび/またはアンタゴニストを、こうしたアッセイに使用することが可能である。
【0100】
結合パートナーを同定するためのアッセイ
BTL−IIの結合パートナーは、最初にどの細胞の型に可溶性BTL−IIタンパク質が結合可能かを決定することにより同定可能である。ブチロフィリン様タンパク質のB7サブファミリーのメンバーのための既知レセプターはすべてT細胞上で発現されるので、構成的に(CD28)かまたは活性化後に(CTLA−4、ICOS、PD−1)、これらへBTL−IIが結合可能かどうかを決定するために、T細胞を試験することが可能である。BTL−IIの発現パターンのため、正常および炎症性腸から単離されたT細胞を、こうした試験に含むことが可能である。加えて、メモリーT細胞、ナイーブT細胞、αβT細胞、γδT細胞および多様な活性化状態のT細胞を含む、多様なサブセットのT細胞を試験することが可能である。
【0101】
こうした実験は、当該技術分野ではよく知られている方法を使用して実施可能である。例えば、マウスBTL−IIの細胞外ドメインに加えて抗体のFc領域を含んでなるBTL−II組換え融合タンパク質を、試験されている細胞へ結合させるために使用可能である。組換え融合タンパク質中のFc領域へ結合可能な蛍光標識抗体を、加えることが可能である。洗浄後、細胞は、BTL−IIが細胞へ結合可能かどうかを決定するため、蛍光表示式細胞分取(FACS)装置を使用して分析可能である。こうしたアッセイの一つはChapovalら((2001),Nature Immunol.2(3):269−74により記載されている。当該技術分野で既知の他の方法もまた、BTL−IIが特異的細胞集団へ結合するかどうかを決定するのに適切であり得る。
【0102】
さらに、B7サブファミリーメンバーの既知のレセプターは、BTL−IIがいかなるこれらのタンパク質にも結合するかどうかを決定するために試験されるであろう。B7サブファミリーメンバーの細胞外ドメイン(例えば、CTLA−4、B7−1およびB7−2のためのレセプター)および抗体のFc領域を含んでなる組換え融合タンパク質は、こうしたBTL−II融合タンパク質を作製するために使用した方法と類似の方法により作製可能である。細胞表面に発現される、膜貫通および細胞質ドメインを含むBTL−IIタンパク質をコードする、BTL−II核酸の完全長形で細胞をトランスフェクトすることが可能である。試験されるべきレセプター:Fc融合タンパク質を、Fc領域へ結合可能である蛍光標識抗体と一緒に、こうした細胞へ加えることが可能である。洗浄後、トランスフェクトされた細胞上に発現されたBTL−IIタンパク質へレセプター:Fc融合タンパク質が結合しているかどうかを決定するため、FACSにより細胞を分析することが可能である。逆実験もまた実行することが可能であり、ここでは、可溶性BTL−II:Fc融合タンパク質を使用し、そして完全長レセプタータンパク質を導入し、そしてトランスフェクションにより発現させる。こうした実験は、BTL−IIタンパク質および他のB7サブファミリーメンバーの既知のレセプター間の結合相互作用を明らかにすることが可能である。
【0103】
もし、BTL−IIが多様なT細胞の少なくとも一つとは結合するが、既知のレセプターへは結合しないとすれば、当該技術分野では既知の多様な発現クローニングまたはタンパク質精製法を、BTL−IIが結合するレセプターを同定するために使用可能である。例として、放射活性スライド(slide)結合cDNA発現クローニング法が使用可能である。簡単には、可溶性BTL−IIへ最も強い結合を有する細胞源を同定し、mRNAを単離し、そして哺乳動物発現ベクター中にcDNAライブラリーを構築する。スライド上で哺乳動物細胞をcDNAのプールでトランスフェクトし、そして発現を可能にするために適切にインキュベーションした後、可溶性BTL−II:Fc融合タンパク質を細胞へ結合させる。レセプターを運んでいる細胞への特異的結合を、以下の一連の工程により検出する:結合されたBTL−II:Fcタンパク質への放射活性抗Fc試薬の結合;スライドへのフィルム乳剤の適用;暴露させるためのインキュベーション;銀粒子を沈着させるためのフィルム現像;そして顕微鏡による粒子の検出。レセプター発現クローンは次ぎに、プールを副分画することによりプールから単離し、そして単一レセプタークローンを同定するためにスライド結合アッセイを反復する。こうした方法は、McMahonら(1991),EMBO J.10:2821−32、に記載されている。
【0104】
細胞への結合が達成された場合、多様な手段(動物免疫化またはファージディスプレイ技術を通して)を、BTL−IIを結合している抗体を単離するため、そして細胞へのその結合を破壊するために使用可能である。こうした抗体は、内在性BTL−IIの活性に拮抗するために、そして従って、免疫応答、および例えば、Cooperら((1993),Lab.Invest.69(2):238−49)およびTokoiら((1996),J.Gastroenterol.31(2):182−88)により記載されているような、炎症性腸疾患モデルのごとき疾患モデルにおいて、BTL−IIの有効量を低下させる効果を決定するためのアッセイに使用可能である。
【0105】
BTL−II機能を決定するためのアッセイ
B7サブファミリータンパク質は、“抗原”の用量を変化させ、T細胞レセプターを通してT細胞を活性化させることを含む、T細胞“同時刺激”アッセイにおいて活性を有することが示されている。B7サブファミリータンパク質の活性は、“最適以下”のT細胞レセプター刺激で、最も明らかである。例えば、Latchmanら(2001),Nature Immunol.2(3):261−68、を参照されたい。組織培養皿結合抗CD3ε抗体のごとき“代理(surrogate)抗原”または照射抗原提示細胞上のMHC分子により提示される抗原を使用可能である。可溶性BTL−IIタンパク質を、“抗原”についてのT細胞の応答に対するその効果を決定するために加えることが可能である。T細胞が刺激されているか、または抑制されているかを決定するため、多様なパラメーターが測定可能である。細胞増殖、細胞表面レセプター発現、そしてタンパク質および/またはmRNAレベルでの免疫変調分子(例えば、インターフェロンγおよび/またはIL−2のごとき)の発現レベル、を測定することが可能である。こうした方法は、例えば、Fitchら,「T Cell Subsets in Infectious and Autoimmune Diseases」,John Wiley and Sons,pp.68−85(1995);Freemanら(2000),J.Exp.Med.192(7):1027−34;Swallowら(1999),Immunity 11:423−32;Hutloffら(1999),Nature 397:263−66;Yoshinagaら(1999),Nature 402:827−32;Latchmanら,上記文献、に使用され、説明されている。BTL−IIの発現パターンを仮定すれば、こうしたアッセイは、粘膜組織またはこうした組織を排出するリンパ節に由来するT細胞を含むことが可能である。加えて、ナイーブおよびメモリーT細胞、CD4+およびCD8+T細胞、およびαβT細胞レセプター以外のT細胞レセプターを発現しているT細胞、を試験することが可能である。こうした実験から、BTL−IIが多数の異なった種類のT細胞の応答を改変できるかどうかを明らかにすることが可能である。こうした実験は以下に記載されており、そしてBTL−IIの可溶性バージョンが、細胞増殖を阻害する、およびインターフェロンガンマ(IFNγ)インターロイキン2(IL2)およびインターロイキン5(IL5)を含むサイトカインの産生を抑制可能であることを示している。実施例6−10;図12−17。
【0106】
この種類の実験の変法には、同時刺激性が可能である、他の可溶性B7サブファミリータンパク質または多様なTNFファミリーメンバーで観察される同時刺激応答について、BTL−IIタンパク質の可溶性形を含んだ効果を試験することが含まれる。このことは、BTL−IIが他の分子で見られる同時刺激を改変するかどうかを定義する助けとなるであろう。さらなる変法は、多様な種類の非照射抗原提示細胞の使用を含むことが可能である。このことは、抗原提示細胞の機能に対する、可溶性BTL−IIタンパク質の添加の効果を定義する助けとなるであろう。さらに別の変法においては、細胞へのBTL−II結合を遮断する抗体(上記参照)もまた、結合を防止する効果を決定するため、こうした同時刺激実験に導入可能である。
【0107】
別の種類の実験において、BTL−IIを発現する抗原提示細胞(例えば、パイエル板からの樹状細胞またはB細胞、または腸管上皮細胞(IEC)のごとき)をT細胞(上にリストした多様な種類のもののごとき)と組み合わせることが可能で、そして上にリストした一つまたはそれより多くのパラメーターを測定することが可能である。これらの細胞で得られた結果は、BTL−II発現を低くするために設計された、これらをコードしている干渉RNAまたはDNAが抗原提示細胞内へ導入された場合に得られた結果と比較可能である。
【0108】
T調節(T reg)細胞は、炎症性腸疾患モデルシステムにおいて、一部、T調節細胞の分化を誘導することにより、または調節機能を促進することにより、自己免疫応答を抑制するように、そしてそのようにすることを助けるように作用する。T reg細胞には、例えば、なかでも、Tr1細胞(Congら(2002),J.Immunol.169(11):6112−19;Grouxら(1997),Nature 389:737−42)、Th3細胞、CD4+Cd25+T reg細胞(例えば、MaloyおよびPowrie(2001),Nature Immunology 2(9):816−22)を参照されたい)、CD4+RbloCD25+T reg細胞、およびCD8+ T reg細胞(Allezら(2002),Gastroenterology 123:1516−26)が含まれる。従って、T調節細胞増殖および機能に対するBTL−IIの効果を評価することは、インビボにおけるBTL−IIの正確な役割を決定するために必要とされる。例えば、Tr1細胞存在下での、抗原特異的T細胞増殖および/またはT細胞によるサイトカイン産生を測定可能である。こうしたアッセイは、Congら、上記文献、に記載されている。BTL−IIの可溶性形、遮断抗体または干渉RNAを使用するBTL−II発現の阻害は、BTL−IIが、T調節細胞またはTr1細胞の増殖、維持、または他のT細胞の抗原活性を抑制する能力において役割を演じているかどうかを決定するために使用可能である。
【0109】
一つのT細胞サブセット、Th3細胞は、トランスフォーミング成長因子β(TGFβ)を産生し、そして粘膜抗原に対する経口免疫寛容と結びつけられている。例えば、Fukauraら(1996),J.Clin.Invest.98(1):70−77;Inobeら(1998),Eur.J.Immunol.28:2780−90、を参照されたい。こうした細胞は、TGFβ存在下、抗原による広範培養により、ナイーブT細胞から発生させることが可能である。可溶性BTL−IIタンパク質または遮断抗体は、上述の方法と同様の方法により、BTL−IIがこれらの細胞の増殖または機能を改変することが可能かどうかを問うために使用可能である。
【0110】
単純で明確な抗原(例えば、卵白アルブミンのごとき)または複雑な抗原(細菌またはウイルスを含んだ)を使用するモデルT細胞システムにおいて、可溶性BTL−IIタンパク質または拮抗性抗体を投与することの効果が確かめられるであろう。こうした抗原に対するT細胞応答(増殖および/またはインターフェロンγまたはIL−2のごとき分子の産生を含んで)を、BTL−IIタンパク質または抗体の存在下および非存在下で測定可能である。
【0111】
動物システムにおける同様の効果が試験可能である。初期の焦点は、特に粘膜免疫応答に関連するシステムであろう。これには、動物へ抗原を与えるモデルシステム(例えば、Yamashiroら(1994),Acta Paediatr.Jpn.36:550−56;Jainら(1996),Vaccine 14(13):1291−97;Chenら(2002),Immunology 105:171−80、を参照されたい)、デキストラン硫酸ナトリウム誘導炎症性腸疾患モデルにおけるごとき炎症性腸疾患システム(Cooperら(1993),Lab.Invest.69(2):238−49;Tokoiら(1996),J.Gastroentero1.31(2):182−88)、CD45RBhi/CD4+T細胞誘導萎縮病モデル(Morrisseyら(1993),J.Exp.Med.178:237−44)、および/またはマウス卵白アルブミン誘導喘息モデルのごとき、免疫応答を誘発するために気道内へ抗原を与えるまたは滴下するモデル喘息システム(Brusselleら(1994),Clin.Exp.Allergy 24(1):73−80)、が含まれる。特に、体液または細胞仲介応答の大きさが測定されるであろう、そして産生された免疫変調サイトカインの型が測定されるであろう。こうした実験は、増加したまたは減少したBTL−IIが、抗原(自己抗原を含んで)に対する応答を低下させることにおいて、または同種抗原に対する応答を増加させることにおいて役に立つことができるかどうかを明らかにすることが可能である。BTL−IIタンパク質または抗BTL−II抗体の効果はまた、炎症性疾患の他のモデルにおいても確かめることが可能である。
【0112】
T細胞または胸腺細胞増殖のためのアッセイには、以下のものが含まれるが、限定されるわけではない:「Current Protocols in Immunology」,Coliganら監修,Greene Publishing Associates and Wiley−Interscience(pp.3.1−3.19:マウスリンパ球機能のインビトロアッセイ;第7章:ヒトにおける免疫学的研究);Takaiら(1986),J.Immunol.137:3494−3500;Bertagnolliら(1990),J.Immunol.145:1706−1712;Bertagnolliら(1992),J.Immunol.149:3778−3783;Bowmanら(1994),J.Immunol.152:1756−1761。
【0113】
脾臓細胞、リンパ節細胞または胸腺細胞のサイトカイン産生および/または増殖のためのアッセイには、限定するわけではないが、以下のものが含まれる:KruisbeekおよびShevach(1994)ポリクローナルT細胞刺激,「Current Protocols in Immunology」中,Coliganら監修,第1巻,pp.3.12.1−3.12.14,John Wiley and Sons,トロント(1991);およびSchriber,(1994),マウスおよびヒトインターフェロンガンマの測定,「Current Protocols in Immunology」中,Coliganら監修,第1巻,pp.6.8.1−6.8.8,John Wiley and Sons,トロント(1991)。
【0114】
造血およびリンパ球新生細胞の増殖および分化のためのアッセイには、限定するわけではないが、以下のものが含まれる:Bottomlyら,1991,ヒトおよびマウスインターロイキン2およびインターロイキン4の測定,「Current Protocols in Immunology」中,Coliganら監修,第1巻,pp.6.3.1−6.3.12,John Wiley and Sons,トロント(1991);deVriesら(1991),J.Exp.Med.173:1205−1211;Moreauら(1988),Nature 336:690−692;Greenbergerら(1983),Proc Natl.Acad.Sci.USA 80:2931−2938;Nordan,マウスおよびヒトインターロイキン6の測定,「Current Protocols in Immunology」中,Coliganら監修,第1巻,pp.6.6.1−6.6.5,John Wiley and Sons,トロント(1991);Smithら(1986),Proc.Natl.Acad.Sci.USA 83:1857−1861;Bennettら,1991,ヒトインターロイキン11の測定,「Current Protocols in Immunology」中,Coliganら監修,第1巻,pp.6.15.1,John Wiley and Sons,トロント(1991);Ciarlettaら,マウスおよびヒトインターロイキン9の測定,「Current Protocols in Immunology」中,Coliganら監修,第1巻,pp.6.13,1,John Wiley and Sons,トロント(1991)。
【0115】
抗原に対するT細胞クローン応答のためのアッセイ(それは、なかでも、増殖およびサイトカイン産生を測定することにより、APC−T細胞相互作用に影響するポリペプチド、ならびに直接的T細胞効果を同定するであろう)には、限定するわけではないが、以下のものが含まれる:「Current Protocols in Immunology」,Coliganら監修,Greene Publishing Associates and Wiley−Interscience(第3章:マウスリンパ球機能のインビトロアッセイ;第6章:サイトカインおよびそれらの細胞レセプター;第7章:ヒトにおける免疫学的研究)(1991);Weinbergerら(1980),Proc.Natl.Acad.Sci.USA 77:6091−6095;Takaiら(1986),J.Immunol.137:3494−3500;Takaiら(1988),J.Immunol.140:508−512。
【0116】
胸腺細胞または脾細胞細胞毒性のためのアッセイには、限定するわけではないが、以下のものが含まれる:「Current Protocols in Immunology」,Coliganら監修,Greene Publishing Associates and Wiley−Interscience(第3章:マウスリンパ球機能のインビトロアッセイ,3.1−3.19;第7章:ヒトにおける免疫学的研究)(1991);HerrmannおよびMescher(1981),Proc.Natl.Acad.Sci.USA 78:2488−2492;Herrmannら(1982),J.Immunol.128:1968−1974;Handaら(1985),J.Immunol.135:1564−1572;Takaiら(1986),J.Immunol.137:3494−3500;Takaiら(1988),J.Immunol.140:508−512;Brownら(1994),J.Immunol.153:3079−3092。
【0117】
免疫グロブリン応答およびB細胞によるアイソタイプ転換のためのアッセイ(それは、なかでも、T細胞依存性抗体応答を変調し、そしてTh1/Th2プロフィールに影響するポリペプチドを同定するであろう)には、限定するわけではないが、以下のものが含まれる:Maliszewski(1990),J.Immunol.144:3028−3033;およびMondおよびBrunswick,B細胞機能のアッセイ:インビトロ抗体産生,「Current Protocols in Immunology」中,Coliganら監修,第1巻,pp.3.8.1−3.8.16,John Wiley and Sons,トロント(1991)。
【0118】
混合リンパ球反応(MLR)アッセイ(それは、なかでも、主としてTh1およびCTL応答を発生するポリペプチドを同定するであろう)には、限定するわけではないが、以下のものが含まれる:「Current Protocols in Immunology」,Coliganら監修,Greene Publishing Associates and Wiley−Interscience(第3章:マウスリンパ球機能のインビトロアッセイ,3.1−3.19;第7章:ヒトにおける免疫学的研究);Takaiら(1986),J.Immunol.137:3494−3500;Takaiら(1988),J.Immunol.140:508−512;Bertagnolliら(1992),J.Immunol.149:3778−3783。
【0119】
樹状細胞依存性アッセイ(それは、なかでも、ナイーブT細胞を活性化する樹状細胞により発現されるポリペプチドを同定するであろう)には、限定するわけではないが、以下のものが含まれる:GueryおよびAdorini(1995),J.Immunol.154:536−544;Inabaら(1991),J.Exp.Med.173:549−559;Macatoniaら(1995),J.Immunol.154:5071−5079;PorgadorおよびGilboa(1995),J.Exp.Med.182:255−260;Nairら(1993),J.Virology 67:4062−4069;Huangら(1994),Science 264:961−965;Macatoniaら(1989),J.Exp.Med.169:1255−1264;Bhardwajら(1994),J.Clin.Invest.94:797−807;およびInabaら(1990),J.Exp.Med.172:631−640。
【0120】
T細胞傾倒および発生の初期段階に影響するポリペプチドのためのアッセイには、限定するわけではないが、以下のものが含まれる:Anticaら(1994),Blood 84:111−117;Fineら(1994),Cell Immunol.155:111−122;Galyら(1995),Blood 85:2770−2778;Tokiら(1991),Proc.Natl.Acad.Sci.USA 88:7548−7551。
【0121】
レセプター−リガンド活性のためのアッセイには、限定するわけではないが、以下のものが含まれる:「Current Protocols in Immunology」,Coliganら監修,Greene Publishing Associates and Wiley−Interscience(第7.28章:静的条件下での細胞接着の測定 7.28.1−7.28.22);Takaiら(1987),Proc.Natl.Acad.Sci.USA 84:6864−6868;Biererら(1988),J.Exp.Med.168:1145−1156;Rosensteinら(1989),J.Exp.Med.169:149−160;Stoltenborgら(1994),J.Immunol.Methods 175:59−68;Stittら(1995),Cell 80:661−670。
【0122】
トランスジェニック動物を利用した機能のための遺伝子アッセイ
本明細書に開示したBTL−II核酸に対応する遺伝子(類)の複数のコピーを有し、好ましくは形質転換された細胞およびそれらの子孫内に安定的に維持されている遺伝子構築物を有する細胞の形質転換により産生されたトランスジェニック動物、好ましくはマウスを提供する。遺伝子発現レベルを増加させるまたは減少させる、または遺伝子発現の時間的または空間的パターンを変化させる、修飾された遺伝子制御領域を有するトランスジェニック動物もまた提供する(欧州特許第0 649 464 B1を参照されたい)。加えて、対応する遺伝子内への外来性配列の挿入を通して、または対応する遺伝子の全てまたは一部の欠失を通して、BTL−II遺伝子が部分的にまたは完全に不活性化されている生物体を提供する。部分的なまたは完全な遺伝子不活性化は、転位性要素の挿入(好ましくは続いての不正確な切除により)を通して(Plasterk(1992),Bioessays 14(9):629−633;Zwaalら(1993),Proc.Natl.Acad.Sci.USA 90(16):7431−7435;Clarkら(1994),Proc.Natl.Acad.Sci.USA 91(2):719−722)、または相同的組換え(好ましくは陽性/陰性遺伝子選択戦略により検出される)を通して(Mansourら(1988),Nature 336:348−352;米国特許第5,464,764;5,487,992;5,627,059;5,631,153;5,614,396;5,616,491;および5,679,523号)、達成可能である。代替法として、BTL−II遺伝子の発現は、トランスジェニック動物内へ導入されたDNAによりコードすることができる、干渉RNA、アンチセンスRNAまたはリボザイムの導入により、トランスジェニックまたは非トランスジェニック動物中で阻害可能である。こうした生物体の表現型は、BTL−II遺伝子のインビボ機能を解明することを可能にする。例えば、BTL−IIタンパク質を発現せずに、デキストラン硫酸ナトリウムの給餌に応答して炎症性腸疾患の症状を示す、トランスジェニック動物において増加した性向(野生型動物と比較して)から、BTL−IIは常態では腸管における炎症を低下させること、を示すことが可能である。あるいは、トランスジェニック動物はBTL−IIを過剰発現することが可能である。こうしたトランスジェニック動物の表現型もまた、BTL−IIタンパク質のインビボ機能に対する手掛かりを与えることが可能である。
【0123】
本発明のタンパク質、抗体、アンタゴニストおよびアゴニストの使用
特殊化した解剖学的構造、粘液膜の組内で作用する粘膜免疫システム、およびそれらが発生する免疫応答は、体内の他の解剖学的区画で発生する免疫応答からそれを区別する特性を有している。粘液膜はいくつかの独特な解剖学的区画のまさに一つであり、末梢リンパ節および脾臓、体腔(即ち、腹膜および胸膜)および免疫システムが活性である皮膚も含んでいる。粘膜表面は、肺、腸管、眼、鼻、口、のど、子宮および膣に観察される。粘膜表面は薄いシートまたは、体腔、仕切または隔壁、または二つの構造間の接合部の裏打ちのごとき、身体構造の被覆または外皮として働いている柔軟な組織の層である。粘膜表面は、大多数の感染性病原体の侵入経路であるので、粘膜免疫応答を増進可能であるアジュバントが特に望まれている。Janewayら,「Immunobiology:The Immune System in Health and Disease,第5版,第IV部,第10章,Garland Publishing,ニューヨークおよびロンドン(2001)。例えば、淋菌(Neisseria gonorrhoeae)のごとき、典型的には粘膜表面を通して侵入する感染性疾患は、しばしば1週間のみの粘膜免疫応答を発生させる。Russellら(1999),42(1):58−63。
【0124】
腸管はいくつかの様式で独特である。腸管を裏打ちしているのは、集合的に腸管付随リンパ組織(GALT)として知られている多数の特殊化した形のリンパ組織であり、以下のものが含まれる:扁桃腺およびアデノイド、それらは一緒になって口の奥の方でワルダイヤ輪を形成する;小腸中のパイエル板;虫垂;および大腸および直腸中の孤立リンパ濾胞。腸管中で外来抗原と遭遇することによるリンパ球の活性化は、粘液免疫システム中のいたるところでの抗原に対する適応性免疫応答の広がりを導くことが可能である。活性化リンパ球はリンパ系にそして、そこから血流へ入ることが可能である。血流は体中いたるところの粘膜部位へ活性化リンパ球を搬送可能であり、それは、粘膜組織で発現される粘膜アドレッシン(addressin)MAdCAM−1のごとき分子を使って、リンパ球により認識可能である。Janewayら、上記文献。腸管により産生される主抗体型はIgAであり、それは発現時に、腸管上皮の表面を覆う粘膜層中に主として観察される。加えて、多数の別々の種類のT細胞が腸管中に観察される。Janewayら、上記文献。
【0125】
外来抗原の腸管内への導入は、通常は免疫寛容を導くが、特異的免疫応答を導くこともできる。腸管は、普通は、外来抗原の莫大なアレイ、即ち、食物および腸管中に住む共生微生物、を受けおよび寛容する(免疫学的な意味で)。後でのタンパク質の注射が、アジュバント存在下でさえも、抗体応答を起こさないように、特別な外来タンパク質の給餌が、経口免疫寛容として知られている、そのタンパク質への特別な無応答性の状態を導くことが可能である。この現象は、脾臓およびリンパ節、ならびに粘膜免疫系が関与することができる。Gutgemannら(1998),Immunity 8:667−73、を参照されたい。しかしながら、例えば、サルモネラ−エルシニア(Salmonella Yersinia)またはエンタモエバ−ヒストリチカ(Entamoeba histolytica)のごとき腸病原体は、局所的(または全身的さえも)免疫応答を惹起することが可能である。腸管内への外来抗原の導入が免疫応答を惹起するかどうかを制御している因子は完全には理解されていない。Janewayら、上記文献、第5部、第14章。
【0126】
多くのワクチンは、抗原に対する免疫応答を促進するためにアジュバントを用いる。アジュバントは、抗原に対する免疫応答の特異性を強化するまたは広幅化する。免疫応答は、抗体力価の増加または抗原反応性T細胞数の増加を含むことができる。こうしたパラメーターを測定するための方法は当該技術分野に存在している。例えば、Zigtermanら(1988),J.Immunol.Methods 106(1):101−07。アジュバントが免疫応答を促進する機構は完全には理解されていないが、それらの使用は必須であろう。ごくわずかのワクチンが、選択された抗原に対する強い粘膜免疫応答を惹起可能である。
【0127】
本発明は、治療的有効量のBTL−IIのアンタゴニストおよび抗原を投与することを含んでなる、抗原に対する全身的または粘膜免疫応答を増進する方法を包含する。本発明を実施するために使用可能なBTL−IIのアンタゴニストには、BTL−IIの細胞外領域へ特異的に結合する拮抗的抗体またはインビトロ選択結合タンパク質、およびBTL−IIの生物学的活性を阻害可能な小分子が含まれる。所望により、BTL−IIタンパク質のアンタゴニストおよび抗原は、経口的に、経鼻的に、経膣的に、胃を経由して、または直腸を経由して、または吸入によるごとく、直接的に粘膜表面に投与可能である。例えば、少なくとも一つの場合においては、持続性免疫応答の誘導において膣投与よりも経鼻投与がより有効であることが報告されている。Russell(2002),Am.J.Reprod.Immunol.47(5):265−68。あるいは、BTL−IIアンタゴニストおよび/または抗原は、例えば、皮下、静脈内、筋肉内、動脈内または腹腔内に注射可能である。いくつかの態様において、抗体のごときBTL−IIアンタゴニストは注射可能であり、そして抗原は直接的に粘膜表面へ投与可能である。
【0128】
本発明の実施のために適した抗原には、任意の感染性病原体の全部もしくは一部、または感染性病原体に類似している病原体が含まれる。感染性病原体は、生きているまたは殺されたウイルス、細菌、およびアメーバ、鞭毛虫または蠕虫のごとき感染性真核動物を含むことが可能である。こうした感染性病原体に類似している病原体は、例えば、ワクチン接種されている哺乳動物へ感染可能なウイルスに類似しているが、しかし、それ自身ではワクチン接種されている哺乳動物へ感染できないウイルスであることができる。この例は、ワクチニアウイルス(ウシにおいては疾患を生じることが可能であるが、ヒトでは疾患を生じない)であり、それは天然痘(ヒトに疾患を生じる類似のウイルス)に対するワクチンを産生するためにジェンナーにより使用された。Janewayら、上記文献、第5部、第14章。表4は本発明を実施するために使用可能である、抗原の具体例を示している。
【0129】
【表4】
【0130】
【表5】
【0131】
あるいは、可溶性BTL−IIタンパク質を、自己免疫または炎症性疾患に関係づけられている抗原に対する寛容を促進するために使用可能である。例えば、実験的自己免疫脳脊髄炎(EAE)、多発性硬化症に多くの点で類似している状態を、例えば、ミエリン塩基性タンパク質またはミエリンオリゴデンドロサイト糖タンパク質(MOG)の多様なエピトープの注射により齧歯動物で誘導可能である。MOG誘導EAEは、いくつかの場合において、MOGまたはブチロフィリンの少量を前もって給餌することにより寛解可能である。Stefferlら(2000),J.Immunol.165:2859−65。可溶性BTL−IIタンパク質は、抗原に対する寛容性を促進し、そしてそれにより自己免疫疾患の症状を寛解するため、自己免疫疾患において標的とされていることが知られている抗原と同時投与することが可能である。所望により、抗原は、例えば、経鼻的に粘膜表面へ直接投与することが可能である。
【0132】
クローン病および潰瘍性大腸炎を含んだ炎症性腸疾患は、胃腸管の慢性炎症を含んでおり、多分、正常腸管フローラ(flora)の抗原に対する異常に促進された免疫応答のせいであろう。両方の疾患とも、発生は家族で集中発生する傾向があり、そしていくつかの遺伝子マーカーと関係づけることが可能であるので、少なくともいくらかの遺伝子的基盤を有しているようである。例えば、多薬剤耐性遺伝子(mdr1a)を発現しないマウスは自然発症的に大腸炎を発生する。Panwalaら(1998),J.Immunol.161:5733−44。クローン病および潰瘍性大腸炎両方の発生は、都市化した集団中で増加した発生が観察されるので、環境的因子にも影響されるようである。また、こうした疾患は、正常腸管フローラの非存在下では発生しない。
【0133】
クローン病には、口から肛門までの消化管のいずれかの部分の異常炎症が含まれるけれども、ほとんどの患者の異常炎症は回結腸、小腸および結腸−肛門直腸領域に限られている。典型的には、炎症は不連続である。共通の症状には、腹部痛、食欲不振、体重減少、発熱、下痢、腹部右下4分部位におけるふくらみおよび圧痛、便秘、嘔吐および肛門周囲不快および放出が含まれる。他の可能な症状には、なかでも、末梢関節炎、成長遅延、上強膜炎、アフタ性口内炎、結節性紅斑、壊疽性膿皮症、腎結石、尿希釈およびアルカリ化障害、吸収不良および胆石が含まれる。例えば、Stroberら,「Medical Immunology」,第10版,III節,第35章(2001);「Merck Manual of Diagnosis and Therapy」,第17版,3節,第31章,(1999)、を参照されたい。クローン病の患者から単離されたマクロファージは、増量されたIL−12、IFNγ、THFαおよび他の炎症性サイトカインを産生する。
【0134】
潰瘍性大腸炎は、しばしばクローン病と区別するのが難しいが、いくつかの点でクローン病とは性質が異なっている。第一に、それは一般に結腸に限定され、一方、クローン病は消化管を通して発生することができる。第二に、クローン病においては炎症が腸管または消化管の他の位置での壁を通してずっと入り込むことが可能であるのとは違って、潰瘍性大腸炎は主として腸管の浅層のみの炎症を含んでいる。最後に、潰瘍性大腸炎は、クローン病で典型的な炎症の非連続的部位よりもむしろ、炎症の連続的領域を典型的には含んでいる。クローン病同様に、潰瘍性大腸炎は主として都市領域で観察される。また、家族性集積の場合があるので、遺伝子的因子も潰瘍性大腸炎において役割を演じているらしい。自己抗体は、潰瘍性大腸炎患者において、クローン病患者よりもより頻繁に観察される。自己抗体はしばしば結腸上皮細胞成分に方向付けられる。最も普通であるのは、カタラーゼ、α−エノラーゼおよびラクトフェリンに特異性を有する、抗好中球細胞質抗体である。いくつかの場合、抗体は結腸微生物と交差反応する。
【0135】
潰瘍性大腸炎の症状は色々である。これらは、下痢、しぶり、腹部痙攣、便中の血液および粘液、発熱および直腸出血を含むことができる。結腸が6センチメーターを超えて拡張し、そしてその筋肉が緊張を失うおよび/または穿孔が起こってもよい、生命を脅かす可能性のある中毒性巨大結腸症もまた発生してもよい。潰瘍性大腸炎に伴ってもよい他の症状には、末梢関節炎、強直性脊髄炎、仙腸骨炎、前部ブドウ膜炎、結節性紅斑、壊疽性膿皮症、上強膜炎、自己免疫肝炎、原発性硬化性胆管炎、硬変症、および小児における成長および発育遅延を含むことができる。
【0136】
BTL−IIタンパク質へ特異的に結合する抗体またはインビトロ選択結合タンパク質は、炎症性腸疾患の発症を診断または予測するために使用可能である。下記実施例4に例示したように、炎症性腸疾患のモデルマウスにおいて、BTL−IIは、腸管において症状の発症に先立って、そして症状相の間に過剰発現される。従って、BTL−IIの過剰発現は、炎症性腸疾患の存在を示すことが可能であり、そしてその発症を予測可能である。抗BTL−II抗体は、ELISAアッセイまたは当該技術分野では既知の他の免疫に基づいたアッセイを使用し、患者の腸管からの組織試料をアッセイすることにより、BTL−IIの過剰発現を検出するために使用可能である。例えば、Reen(1994),酵素結合免疫吸着アッセイ(ELISA),「Basic Protein and Peptide Protocols,」中, Methods Mol.Biol.32:461−466、を参照されたい。過剰発現はまた、当該技術分野では既知のmRNA発現アッセイのなかでも、例えば、逆転写プラスPCR(RT−PCR)のごとき、BTL−II mRNA発現を測定するための、核酸に基づいた方法によっても検出可能である。例えば、Murphyら(1990),Biochemistry 29(45):10351−56、を参照されたい。
【0137】
別の態様において、本発明の可溶性BTL−IIタンパク質は、炎症性腸疾患を処置するために使用可能である。可溶性BTL−IIタンパク質は、B細胞またはT細胞上に発現される特異的レセプターへ結合可能であり、それにより免疫応答の下方調節を可能にしている。こうした下方調節は、例えば、CD4+T細胞によりマクロファージまたはB細胞の活性化を防止すること、または抗原によりT細胞の活性化を防止することが可能である。あるいは、こうした下方調節は、T細胞が、T細胞レセプターが特異的に結合可能な抗原に遭遇した場合、T細胞が反応不顕性(anergic)になることを起こすことが可能である。
【0138】
BTL−IIタンパク質へ特異的に結合する抗体またはインビトロ選択結合タンパク質はまた、炎症性腸疾患を患った、または炎症性腸疾患を発生する危険性のある患者を同定するための診断試薬としての使用も見いだせる。マウスモデルシステムにおいて、BTL−IIは炎症性腸疾患症状の発症に先立ってそしてその間に過剰発現されるので(実施例4)、BTL−II発現の異常に高いレベルは、腸管における炎症性疾患の存在、または腸管において炎症性疾患を発生する高い危険性を示すことが可能である。
【0139】
加えて、可溶性BTL−IIタンパク質は、移植(Manilayら,1998,Curr.Opin.Immunol.10:532−538)、移植片対宿主疾患、移植片拒絶、自己免疫または炎症性疾患、遺伝子治療(Hackettら,2000,Curr.Opin.Mol.Therap.2:376−382)などのごとき、免疫応答の下方調節が望まれる状態において有用であり得る。例えば、可溶性BTL−IIタンパク質は、哺乳動物への遺伝子治療ベクターの投与、移植、または所望の免疫抑制が適切な別の状態に先だって、ほとんど同時に(またはすぐ前かすぐ後に)、または同時に投与可能である。またこうした処置に適切であるのは、BTL−IIの機能を模倣した抗イディオタイプ抗体である。
【0140】
アゴニストBTL−II抗体、可溶性BTL−IIタンパク質または抗イディオタイプ抗体は、検出可能自己抗体の数を減少させるため、免疫エフェクター細胞の活性化を減少させるため、および/または疾患の症状を軽減または除去するため、自己免疫または炎症性疾患を患っている患者へ投与可能である。自己免疫および炎症性疾患には、患者自身の組織が患者の免疫系により引き起こされる有害な効果を受けやすい、すべての状態が含まれる。こうした効果は、可能性のなかでも、自己抗体により、および/または免疫エフェクター細胞の活性化により仲介可能である。自己免疫および炎症性疾患の原因は通常明らかではないが、多様な種類の感染と多様な自己免疫疾患の間の相関がいくつかの場合に確立されており、そして科学論文における議論の繰り返される主題である。例えば、Corapciogluら(2002),Thyroid 12:613−17;Sewellら(2002),Immunol.Lett.82:101−10;Rose(1998),Semin.Immunol.10(1):5−13;Matsiota−Bernard(1996),Cli.Exp.Immunol.104:228−35;およびMcMurrayおよびElboune(1997),Semin.Arthritis Rheum.26:690−701、を参照されたい。
【0141】
当業者は、自己免疫および炎症性疾患の症状が非常に広範で、そしてどの組織が患者の免疫系の標的にされているかに依存可能なことを認識するであろう。自己免疫および炎症性疾患は器官特異的または全身的であり得る。自己免疫および炎症性疾患には、多くの中でも、例えば、関節炎、アジソン病、インシュリン依存性糖尿病(タイプI糖尿病)、喘息、多発性内分泌障害症候群、全身性紅斑性エリテマトーデス、慢性活性肝炎、多様な形の甲状腺炎(ハシモト甲状腺炎、一過性甲状腺炎症候群およびグレーブス病を含んで)、リンパ球性腺下垂体炎、早期卵巣不全、特発性フィオパラサイロイディズム、悪性貧血、腎炎、自己免疫好中球減少症、グッドパスチャー症候群、多発性硬化症、白斑、重症筋無力症、リューマチ様関節炎、強皮症、原発性シェーグレン症候群、多発性筋炎、自己免疫溶血性貧血、炎症性腸疾患(クローン病および潰瘍性大腸炎を含んで)、乾癬、乾癬性関節炎、皮膚炎、自己免疫血小板減少性紫斑病、尋常性天疱瘡、急性リウマチ熱、混合本態性クリオグロブリン血症、および中程度自己免疫溶血性貧血が含まれる。
【0142】
ベクターおよび宿主細胞
本発明はまた、本発明の核酸を含んでいるベクター、ならびにこうしたベクターで形質転換した宿主細胞も提供する。本発明の核酸のいずれも、一般に選択可能マーカーおよび宿主中での増殖のための複製起点を含むベクターを含むことができる。ベクターはさらに、機能可能であるようにBTL−II核酸に連結された、哺乳動物、微生物、ウイルスまたは昆虫遺伝子からのもののごとき、適した転写または翻訳調節配列を含んでいる。こうした調節配列の例には、転写プロモーター、オペレーターまたはエンハンサー、mRNAリボソーム結合部位、および転写および翻訳を制御する適切な配列が含まれる。調節配列が、標的タンパク質をコードしているDNAへ機能的に関連している場合、ヌクレオチド配列は機能可能なように連結されている。それ故、もし、プロモーターヌクレオチド配列がBTL−II配列の転写を方向付けるならば、プロモーターヌクレオチド配列はBTL−II核酸配列へ機能可能なように連結されている。
【0143】
本発明の標的BTL−IIタンパク質をコードしているBTL−II核酸のクローニングのために適したベクターの選択は、ベクターが形質転換されるであろう宿主細胞、そして、当てはまる場合、標的ポリペプチドが発現されるべき宿主細胞に依存するであろう。BTL−IIタンパク質の発現に適した宿主細胞には、原核生物、酵母、昆虫そしてより高等な真核生物が含まれ、その各々が以下に議論されている。
【0144】
こうした宿主細胞中で発現されるべきBTL−IIタンパク質はまた、異種タンパク質からの領域を含む融合タンパク質であることができる。上で議論したように、こうした領域は、例えば、分泌、改良された安定性、容易にされた精製、標的化またはBTL−IIのオリゴマー化を可能にするために含ませることができる。例えば、適切なシグナル配列をコードしている核酸配列を発現ベクター内へ取り込むことが可能である。シグナル配列(分泌リーダー)をコードしている核酸配列は、BTL−IIがシグナル配列を含んでなる融合タンパク質として翻訳されるように、BTL−II配列へインフレーム(in−frame)で融合することができる。意図された宿主細胞内で機能的に可能であるシグナル配列は、BTL−IIタンパク質の細胞外分泌を促進することが可能である。異種シグナルペプチドは、天然のシグナル配列と置き換えることが可能である。哺乳動物宿主細胞中で機能的であるシグナルペプチドの例には、米国特許第4,965,195号に記載されているインターロイキン−7(IL−7)のためのシグナル配列、Cosmanら((1984),Nature 312:768)により記載されているインターロイキン−2レセプターのためのシグナル配列;欧州特許第0 367 566に記載されているインターロイキン−4レセプターシグナル配列;米国特許第4,968,607号に記載されているタイプIインターロイキン−1レセプターシグナル配列;欧州特許第0 460 846に記載されているタイプIIインターロイキン−1レセプターシグナル配列;ヒトIgKのシグナル配列(それはMETDTLLLWVLLLWVPGSTGである);およびヒト成長ホルモンのシグナル配列(それはMATGSRTSLLLAFGLLCLPWLQEGSAである)が含まれる。好ましくは、シグナル配列は、細胞からのBTL−IIタンパク質の分泌と同時にBTL−IIタンパク質から切断されるであろう。本発明の実施で使用可能である他のシグナル配列には、Sf9昆虫細胞中の、酵母α因子およびミツバチメラチンリーダーが含まれる。Brake(1989),Biotechnology 13:269−280;Homaら(1995),Protein Exp.Purif.6:141−148;Reavyら(2000),Protein Exp.Purif.6:221−228。
【0145】
本発明のタンパク質の発現に適した宿主細胞には、原核生物、酵母、そしてより高等な真核生物細胞が含まれる。これらのポリペプチドの発現に使用されるのに適した原核生物宿主には、エシェリキア(Escherichia)、バシラス(Bacillus)およびサルモネラ(Salmonella)属の細菌、ならびにシュードモナス(Pseudomonas)、ストレプトミセス(Streptomyces)およびスタフィロコッカス(Staphylococcus)属のメンバーが含まれる。原核生物細胞中、例えば、大腸菌中での発現のためには、BTL−IIタンパク質をコードしているポリヌクレオチド分子は、好ましくは、組換えポリペプチドの発現を容易にするために、N末端メチオニン残基を含んでいる。N末端Metは所望により発現されたポリペプチドから切断されてもよい。
【0146】
細胞性宿主中で使用するための発現ベクターは、一般に一つまたはそれより多くの表現型選択可能マーカー遺伝子を含んでなる。こうした遺伝子は、例えば、抗生物質耐性を与えるまたは栄養要求性要求を供給するタンパク質をコードしている。広範囲の種類のこうしたベクターは、商業源から容易に入手可能である。例には、pGEMベクター(Promega)、pSPORTベクターおよびpPROEXベクター(InVitrogen,Life Technologies,カールスバッド,CA),Bluescriptベクター(Stratagene)およびpQEベクター(Qiagen)が含まれる。
【0147】
BTL−IIはまた、サッカロミセス、ピキア(Pichia)およびクルベロミセス(Kluveromyces)を含む属からの酵母宿主細胞で発現可能である。好ましい酵母宿主は、S.セレビジエ(S.cerevisiae)およびP.パストリス(P.pastris)である。酵母ベクターはしばしば2μ酵母プラスミドからの複製配列起点、自己複製配列(ARS)、プロモーター領域、ポリアデニル化のための配列、転写終結のための配列、および選択可能マーカー遺伝子を含んでいるであろう。酵母および大腸菌の両方で複製可能なベクター(シャトルベクターと名付けられている)もまた使用することができる。酵母ベクターの上述の特性に加え、シャトルベクターはまた、大腸菌中での複製および選択のための配列を含んでいるであろう。酵母宿主中で発現された標的ポリペプチドの直接分泌は、BTL−IIコードヌクレオチド配列の5’末端に、酵母α因子リーダー配列をコードしているヌクレオチドの包含により達成することができる。Brake(1989),Biotechnology 13:269−280。
【0148】
昆虫宿主細胞培養系もまた、BTL−IIタンパク質の発現のために使用可能である。本発明のタンパク質は好ましくは、例えば、LuckowおよびSummers((1988),BioTechnology 6:47)による総説に記載されているような、バキュロウイルス発現系を使用して発現する。
【0149】
本発明のBTL−IIタンパク質は、哺乳動物宿主細胞においても発現可能である。適した哺乳動物宿主細胞の非制限例には、サル腎臓細胞のCOS−7株(Gluzmanら(1981),Cell 23:175−182)、チャイニーズハムスター卵巣(CHO)細胞(Puckら(1958),PNAS USA 60:1275−1281)CV−1(Fischerら(1970),Int.J.Cancer 5:21−27)およびヒト頚部癌腫細胞(HELA)(ATCC CCL2)が含まれる。
【0150】
本発明のBTL−IIタンパク質の発現に適した発現ベクターの選択は、使用されるべき特別の哺乳動物宿主細胞に依存するであろう。適した発現ベクターの例には、pcDNA3.1Hygro+(Invitrogen)、pDC409(McMahanら(1991),EMBO J.10:2821−2832)およびpSVL(Pharmacia Biotech)が含まれる。哺乳動物宿主細胞で使用するための発現ベクターは、ウイルスゲノムに由来する転写および翻訳制御配列を含むことが可能である。BTL−IIを発現するために使用可能な、普通に使用されるプロモーター配列およびエンハンサー配列には、限定されるわけではないが、ヒトサイトメガロウイルス(CMV)、アデノウイルス2、ポリオーマウイルスおよびサルウイルス40(SV40)に由来するものが含まれる。哺乳動物発現ベクターの構築法は、例えば、OkayamaおよびBerg((1982)Mol.Cell.Biol.2:161−170)、Cosmanら((1986)Mol.Immunol.23:935−941)、Cosmanら((1984)Nature 312:768−771)、EP−A−0367566およびWO 91/18982、に開示されている。
【0151】
特定のベクター内への挿入を容易にするためのBTL−II核酸の修飾(例えば、制限部位を修飾することにより)、特定の発現系または宿主における使用の容易さ(例えば、好ましい宿主コドンを使用して)などは既知であり、本発明での使用に意図されている。BTL−IIタンパク質の産生のための遺伝子操作法は、既知の方法に従った、無細胞発現系、細胞性宿主、組織および動物におけるポリヌクレオチド分子の発現を含んでいる。
【0152】
治療法
本明細書で言及したいかなる病気の“処置”も、疾患の少なくとも一つの症状の軽減、疾患の重度の軽減、または、いくつかの場合、疾患を伴うことができるより重度な症状への、または少なくとも一つの他の疾患への疾患進行の遅延または防止を包含している。処置は、疾患がすっかり治癒されることを意味する必要はない。有用な治療剤は、疾患の重度を軽減し、疾患またはその処置に付随する症状の重度を軽減し、または処置状態に続いていくらかの頻度で生じる可能性があるより重大な症状またはより重大な疾患の発症を遅延させることのみを必要とする。例えば、もし疾患が炎症性腸疾患であれば、治療剤は、腸管中の別々な部位の炎症の数を軽減し、影響を受けた腸管の全範囲を軽減し、痛みおよび/または膨潤を軽減し、下痢、便秘または嘔吐のごとき症状を軽減し、および/または腸管の穿孔を防止することができる。患者の状態は、バリウム浣腸または注腸に続いて実施されるX−線、内視鏡検査、大腸鏡検査および/または生検のごとき標準技術により評価可能である。適した方法は患者の状態および症状に従って変化する。
【0153】
本発明は、自己免疫疾患、移植片対宿主疾患および炎症性腸疾患を含んだ炎症性疾患を処置する方法を包含し、BTL−IIタンパク質または抗体の量を、特定の障害の重度または障害により引き起こされる症状の重度を反映する指標のベースラインを超えた持続性の改良を誘導するために、またはいくつかのまたは全ての場合において、処置された状態に続くより重大な疾患の発症を遅延させるまたは防止するために十分な時間使用する。本発明の処置は、問題とする障害のために通常使用する他の処置の前、後もしくは間に使用することができ、または他の処置なしでも使用することができる。例えば、クローン病および潰瘍性大腸炎は通常、スルファサラジン、5−アミノサリチル酸またはコルチコステロイドで処置する。これらの処置は、本発明の処置の前、間または後に使用することができる。
【0154】
上述の治療剤はいずれも組成物の形で、即ち、生理的に受容可能な坦体、賦形剤または希釈剤のごとき一つまたはそれより多くの追加の成分とともに投与可能である。例えば、組成物は本明細書に記載したような可溶性BTL−IIタンパク質に加えて、緩衝液、アスコルビン酸のごとき抗酸化剤、低分子量ポリペプチド(10より少ないアミノ酸を有しているもののごとき)、タンパク質、アミノ酸、グルコース、スクロースまたはデキストリンのごとき炭水化物、EDTAのごときキレート剤、グルタチオンおよび/または他の安定化剤、賦形剤および/または保存剤を含んでなる。組成物は液体または凍結乾燥物として処方することができる。医薬処方に用いることができる成分のさらなる例は、「Remington’s Pharmaceutical Sciences」,第16版,Mack Publishing Company,イーストン,Pa.(1980)、に示されている。
【0155】
上述の治療分子を含んでなる組成物は、限定するわけではないが、非経口、局所、経口、経鼻、経膣、直腸または肺(吸入により)投与を含んだ任意の適切な手段により投与可能である。もし注射するならば、組成物は関節内、静脈内、動脈内、筋肉内、腹腔内または皮下で、大量注射または連続的注入により投与可能である。経皮搬送および移植物からの徐放性放出、皮膚パッチまたは座剤であるような、局所投与(即ち、疾患の部位での)が意図される。吸入による搬送には、例えば、経鼻または経口吸入、噴霧器の使用、エアロゾル形での吸入などが含まれる。体腔内へ挿入された座剤を経る投与は、例えば、選ばれた体腔中に固形の組成物を挿入し、そしてそれを融解させることにより達成可能である。炎症性腸疾患を処置するための可溶性BTL−IIタンパク質またはアゴニストの場合、治療剤が適切に局在化するので、直腸座剤を経た投与が特に適切であることができる。他の代替物には、点眼、錠剤、ロゼンジ、シロップおよびチューインガムのごとき経口製剤、そしてローション、ゲル、スプレーおよび軟膏のごとき局所製剤が含まれる。ほとんどの場合、ポリペプチドである治療分子は、局所的または注射により、または吸入により投与可能である。
【0156】
上述の治療分子は、処置されている状態を処置するため、有効であり得るいかなる量、頻度および期間でも投与可能である。用量は、治療分子の分子的性質および処置されている障害の性質に依存する。処置は、所望の結果を達成するために必要な限り続けることができる。本発明の治療分子は、可能な投与計画の中でも、単一用量として、または1日数回、毎日、1日おき、週2回、週3回、毎週、1週おき、および毎月用量を含んだ、定期的に与える一連の用量として投与可能である。処置の定期性は、処置の期間を通して一定であってもよいし、一定でなくてもよい。例えば、処置を最初は週間隔で行い、そして後では1週おきに行うことができる。日、週、月または年の期間を有する処置が本発明に包含される。処置を中断し、その後再び始めることもできる。維持用量を最初の処置の後で投与することができる。
【0157】
用量は、体重キログラム当たりのミリグラムとして(mg/kg)、または皮膚表面の平方メーター当たりのミリグラムとして(mg/m2)、または身長または体重に関係なく固定用量として計ることができる。これらの全ては、当該技術分野における標準容量単位である。ヒト皮膚表面面積は、標準式を使用して、身長および体重から計算する。
本発明を特定の例を参照して説明してきた。これらの例はいかようにも本発明を限定することを意味していない。十分に本発明の範囲内である、多様な変更および修飾を本発明に行うことができることは、本開示の目的のためには言うまでもない。多くの他の変更を行うことができることを当業者は容易に思いつくであろうが、それは本明細書に開示されている、そして付随する特許請求の範囲に定義されているように、本発明の精神に包含される。
【0158】
本明細書は多くの特許、特許出願および出版物の引用を含んでいる。すべての目的のため、各々は本明細書において援用される。
【0159】
実施例1:ヒトBTL−II cDNAの単離
RNAはいくつかの起源、クローン病または潰瘍性大腸炎患者からのヒト結腸組織試料、ヒト結腸癌細胞株Caco−2(American Type Culture Collection(ATCC)番号HTB−37)およびT84と称される結腸上皮細胞株(ATCC番号CCL−248)、から単離した。RNAを逆転写し、NCBI寄託番号NM_019602に開示されている核酸配列に基づいて設計したプライマーを使用するPCRにより増幅した。これにより、完全長BTL−IIの配列の上流部分(配列番号3)および配列番号7、配列番号9、配列番号11、配列番号13および配列番号15に開示されているスプライス変異体配列を得た。BTL−II mRNAの3’末端を含んでいるcDNAの単離は、3’RACE(cDNA末端の迅速増幅)、即ち、本質的にFrohmanら((1988),Proc.Natl.Acad.Sci.USA 85(23):8998−9002)のプロトコールを使用して達成した。3’RACEによるBTL−II cDNAの多くの変異体の分析から、可溶性タンパク質をコードする変異体は膜貫通ドメインを欠いていないことを明らかにした。図5a、6aおよび7aに示したように、変異体の多くがBTL−II配列の多くの部位に配列多型を含んでいた(またはアリル変異体)。
【0160】
実施例2:マウスBTL−II cDNAの単離
マウスBTL−II cDNAは以下のように単離した。RNAは、RNeasyキット(Qiagen)を使用してマウス結腸および小腸から単離し、そして、製造元の推奨に従って、DNAse I(Ambion)で処理し、残余染色体DNAを除去した。精製RNAを、10mMトリス−HCl、pH8.3、50mM KCl、5mM MgCl2、各々1mMのdNTP、2.5μMランダムヘキサマープライマ一、lU/μl RNAse阻害剤、および2.5U/μl MuLV逆転写酵素(PE Biosystems)中に単離されたRNAを含んでいる反応混合物を使用して、cDNAに転写した。反応混合物を25℃で10分、続いて48℃で30分、続いて95℃で5分インキュベー卜した。BTL−II遺伝子のPCR増幅反応は、10mMトリス−HCl、pH8.3、50mM KCl、1.5mM MgCl2、各々200μMのdNTPおよび2.5U ampliTaq DNAポリメラーゼ(Perkin Elmer)、および両方25ピコモルの上流(5’−TTACTGAGAGAGGGAAACGGGCTGTTTTCTCC)および下流(5’−GGACTTCATTGGTGACTGATGCCATCCAC)プライマー、を含んでいる100μlの最終容量で実施した。増幅反応は、94℃で40秒、55℃で40秒および72℃で40秒を、35サイクル実施した。増幅生成物は2%アガロースゲルで分析し、エチジウムブロミドにより可視化し、そして配列決定した。視覚的判断によると、エキソン3を欠くスプライス変異体(図9a)はエキソン1から8を含んでいる変異体(図8)よりも豊富であった。
【0161】
実施例3:多様な起源の細胞および組織中でのBTL−IIの発現
BTL−II mRNAの発現は、本質的にHeidら((1996),Genome Res.6(10):986−94)のプロトコールに従ったリアルタイムPCRを使用し、そして後にPCRが続く標準逆転写(RT−PCR;例えば、Fuquaら(l990),Biotechniques 9(2):206−11、を参照されたい)を使用して測定した。試験された全ての細胞型は、マウスからであるバイエル板からのCD11c+CD8+B220+(または形質球様)樹状細胞を除いて、ヒト起源であった。BTL−II発現が検出された細胞は以下のものである:非刺激または殺したスタフィロコッカス アウレウス(Staphylococcus aureus)、CD40リガンドおよびインターロイキン4で刺激したヒトB細胞;インターフェロンγで刺激した正常ヒト気管支上皮細胞(NEBE細胞;Lecherら(1983),Cancer Res.43(12pt.1):5915−21、を参照されたい);Calu−3細胞、肺上皮細胞株(ATCC番号HTB−55)、非刺激;T84、結腸上皮細胞株、非刺激またはインターフェロンγで刺激;Caco−2細胞、ヒト結腸癌細胞株、非刺激;マウスパイエル板からのCD11c+(低発現)CD8+B220+細胞、それは主に樹状細胞である;およびマウス末梢血リンパ球。絶対スケール上、発現は、試験されたほとんどの細胞で低かった。BTL−II mRNAの発現は、樹状細胞型への分化を誘導するため、ヒト末梢血単球のインビトロ処理から生じた樹状細胞では検出されなかった。しかしながら、ヒト末梢血から精製された、ヒト血液から精製されたCD123+形質球様樹状細胞では、BTL−II発現を検出した。BTL−II mRNAは、パイエル板からのマウス樹状細胞のCD11c+(低発現)CD8+B220+サブセットにおいて高度に濃縮されていた。BTL−II mRNA発現を、脾臓、リンパ節、胃、腸管膜リンパ節、骨髄、小腸、盲腸、肺、大腸、パイエル板および胸腺を含んだ多くのマウス組織型で、同様の方法により検出した。最も高いレベルの発現は、小腸、パイエル板および盲腸組織で検出した。
【0162】
実施例4:炎症性腸疾患のためのマウスモデルにおけるBTL−IIの発現
mdr1a−/−マウスは、慢性炎症性腸疾患の研究のモデルシステムとなり得る。Panwalaら(1998),J.Immunol.161:5733−44。マウス多薬剤耐性遺伝子、mdr1aは、腸上皮細胞およびリンパ様細胞を含む多くの組織において発現される、170kDa膜貫通タンパク質をコードしている。mdr1aを欠失しているマウスは、調節不全の上皮細胞成長および大腸の粘膜固有層内への大量白血球湿潤により特徴付けられる、重度の自発的腸炎症を発生しやすい。経口抗生物質でmdr1a− /−マウスを処置すると、疾患の発生を防止し、そして活性な炎症を消散させる。活性な大腸炎を有するマウスから単離されたリンパ様細胞は、腸細菌抗原に対する促進された反応性を示した。mdr1aは上皮細胞および白血球の両方により発現されるけれども、大腸炎の発生は、上皮細胞上でのmdr1a発現の欠損と相関する。
【0163】
使用されたmdr1a−/−マウスは、FVB遺伝子バックグラウンドであった。典型的には、mdr1a−/−マウスの群の約20%が、18から20週齢で大腸炎を自発的に発生し、一方、mdr1a−/−マウスの残りのマウスは健康に残存し、そして大腸炎を発生しない。疾患を発生した動物のパーセンテージは、動物施設の清潔度に依存している。
【0164】
最初に、RNAを腸管組織から調製し、逆転写後に、得られたcDNA内へ蛍光標識を取り込ませ、使用して、Stammerら((2000),Immunogenetics 51:373−342)により発表されている配列に基づき、BTL−II mRNAを検出するために設計されたオリゴヌクレオチドを含んでいる、注文製作のAffymetrixチップ上のプローブのアレイヘハイブリダイズさせた。BTL−II mRNAの過剰発現を、炎症性腸疾患症状の発症前および症状の間の両方で、mdr1a−/−マウスにおいて検出した(FVB株の野生型個体と比較して)。こうしたデータの実例は、図10に示されている。これらのデータは、BTL−II mRNAは野生型FVB マウスよりもmdr1a−/−マウスにおいてより大きな程度で発現されていることを示している。さらに、より高いBTL−II mRNAの発現が、炎症性腸疾患の症状の発症に伴っている。
【0165】
これらの結果は、リアルタイムPCR技術(Heidら(1996),Genome Res.6(10):986−94)を使用する、同一RNAの分析により確認した。この分析は、2匹の健康な野生型FVBマウスと比較して、2匹の症状性mdr1a−/−マウスによるBTL−II mRNAの過剰発現を示した。このデータは図11に図示されている。2匹の親マウスは、図11の向かって左の対角の縞で印を付けられた2本の棒線で示されており、2匹の症状性mdr1a−/−は、図11の向かって右の市松模様(checkered pattern)で印を付けられた2本の棒線で示されている。これらのデータは、野生型および症状性mdr1a−/−マウス間の発現に約2から5倍の相違が存在することを示している。従って 、症状のない野生型マウスよりも、より高いレベルのBTL−II mRNAが炎症性腸疾患の症状を有するマウスで発現されている。
【0166】
実施例5:BTL−II:FC融合タンパク質の構築
ヒトFc領域へ融合されたマウスBTL−IIの細胞外領域から成る可溶性BTL−IIタンパク質(BTL−II:Fc)は以下の方法で製造した。BTL−II:Fcをコードしている融合cDNA構築物は、マウスBTL−IIの細胞外領域をコードしている核酸を、ヒトIgG1をコードしている核酸へ融合することにより(インフレーム)製造した。BTL−II:Fcタンパク質を製造するため、哺乳動物細胞を、LIPOFECTAMINETM2000トランスフェクション法(Invitrogen,カールスバッド,カリフォルニア,米国)を使用して、融合cDNAでトランスフェクトした。BTL−II:Fcタンパク質含有上清は、トランスフェクション6−7日後に収集し、そしてBTL−II:Fcタンパク質は、プロテインAカラムクロマトグラフィーにより精製した。BTL−II:Fcタンパク質をコードしている核酸配列およびBTL−II:Fcアミノ酸配列は、各々配列番号19および配列番号20にみられる。
【0167】
実施例6:BTL−II:FcによるヒトT細胞増殖の抑制
以下の実験は、BTL−IIの可溶性形が、他の同時刺激性分子とともに、またはなしで、モノクローナル抗CD3ε抗体に応答して、インビトロでのT細胞増殖を抑制可能かどうか試験している。
【0168】
BTL−II:Fcは実施例5に記載したように作製した。96ウェル、U底マイクロタイタープレートを、一つまたはそれより多くの他のタンパク質とともに、またはなしで、濃度を変化させた抗CD3ε抗体で被覆した。図12は、以下のような、何のタンパク質が各々の試料に使用されたかを示している:抗CD3ε抗体単独、
【0169】
【化1】
【0170】
;抗CD3ε抗体およびBTL−II:Fc、
【0171】
【化2】
【0172】
;抗CD3ε抗体およびB7RP−1:Fc、
【0173】
【化3】
【0174】
;抗CD3ε抗体、B7RP−1:FcおよびBTL−II:Fc、
【0175】
【化4】
【0176】
;抗CD3ε抗体およびB7−2:Fc、
【0177】
【化5】
【0178】
;抗CD3ε抗体、B7−2:FcおよびBTL−II:Fc、
【0179】
【化6】
【0180】
。
B7RP−1:Fcは、抗体のFc領域へ融合されたB7RP−1の細胞外領域(B7hとして知られている)から成り、そしてB7−2:Fcは、抗体のFc領域へ融合されたB7−2の細胞外領域から成っている。これらのタンパク質の両方とも、抗原に応答するT細胞を変調することが知られているB7ファミリーのタンパク質のメンバーである。これらの融合タンパク質は、例えば、R&D Systems(ミネアポリス,ミネソタ,米国)のごとき商業的製造供給元から購入することが可能であり、またはBTL−II:Fcタンパク質のために実施例5に記載したように単離可能である。B7−2はR&D Systemsから購入し、そしてB7RP−1は、B7−H2/Fcの名前でR&D Systemsから入手可能であるが、本質的に実施例5で記載したように作製した。マイクロタイタープレートウェルは、一つまたはそれより多くのタンパク質(それはBTL−II:Fc(10μg/ml)、B7RP1:Fc(10μg/ml)および/またはB7−2:Fc(2μg/ml)である)とともにまたはなしで、図12に示された抗CD3ε抗体の濃度を含んでいる、100μlのリン酸緩衝液(PBS)を加えることにより被覆した。プレートは4℃で一夜インキュベートし、その後PBSで2回洗浄した。ヒトT細胞は、磁気的標識により機能し、そしてCD4+T細胞以外の末梢血細胞を枯渇させ、比較的純粋なもとのままのCD4+T細胞の集団を生じる、Miltenyi Biotec(Bergisch Gladbach,ドイツ)からのCD4+T細胞単離キットを使用して、ヒト末梢血単核細胞から精製した。約lxl05精製T細胞を、200μlの容量の培養培地(l0%ウシ胎児血清を含むRPMI)中で各々のウェルへ加えた。細胞を総計で72時間インキュベートした。64時間目、1μCiの3H−チミジンを各々のウェルに加えた。72時間終了後、取り込まれなかったチミジンは、細胞をフィルター上に沈着し、そして培養培地を洗い流す、自動細胞収穫器(Tomtec,ハムデン、コネチカット,米国、から得た)を使用して除去した。その上に細胞を含むフィルターは次ぎに、どのくらいの放射活性を細胞が取り込んでいたかを決定するため、シンチレーションカウンター中でカウン卜した。結果は図l2aおよび12bに示されており、それらは、図12aが対数目盛りであり、図12bは直線目盛りであることのみが、異なっている。
【0181】
結果は、BTL−II:Fcが、抗CD3ε抗体により誘導されるT細胞増殖を抑制可能であることを示している。図12aおよび図12b。図12aにおいて、試験された最も高い抗体濃度では、BTL−IIプラス抗CD3ε抗体
【0182】
【化7】
【0183】
を含んでいる試料は、抗CD3ε抗体のみ
【0184】
【化8】
【0185】
を含んでいる試料よりも増殖が悪かった。さらに、B7−2
【0186】
【化9】
【0187】
およびB7RP−7
【0188】
【化10】
【0189】
の両方とも抗CD3ε抗体誘導T細胞増殖を刺激することは明らかである。B7−2:Fc添加によるT細胞増殖の効果を見るために必要とされる抗CD3ε抗体の濃度は、B7RP−1:Fcの添加による同様の効果を見るために必要とされる濃度よりも低い。さらに、BTL−IIはまた、B7−2:Fc
【0190】
【化11】
【0191】
およびB7RP−1:Fc
【0192】
【化12】
【0193】
の添加に応答した、この増強されたT細胞増殖も抑制することが可能である。B7−2:Fcプラス抗CD3ε抗体により誘導された増殖に対するBTL−II:Fcの効果は、試験された抗CD3ε抗体の最も低い濃度でのみ明らかであり、一方 、B7RP1:Fcにより誘導された増殖に対するBTL−II:Fcの効果は、試験された三つのより高い濃度でより明白である。従って、観察された効果は、抗CD3ε抗体の濃度に依存している。
【0194】
図13は、BTL−II:FcによるヒトT細胞増殖の抑制はまた、BTL−II:Fc濃度にも依存していることを示している。実験は、上述のように、ただし、抗CD3ε抗体濃度を0.5μg/mlで一定に保って実行した。BTL−II:Fc濃度は、図13に示されているように、0から10μg/mlへ変化させた。図13に示されている結果は、抗CD3ε誘導T細胞増殖の抑制が、BTL−II:Fcの濃度に依存していることを示している。
【0195】
実施例7:BTL−II:FcはマウスT細胞増殖を抑制する
この実験は、BTL−II:Fcがマウス、並びにヒトT細胞の増殖を抑制できるかどうかを決定するために行われた。T細胞増殖アッセイは、本質的に実施例6に記載したように行われ、ただし、Miltenyi Biotec GmbH(Bergisch Gladbach,ドイツ)から購入された磁気マイクロビーズを使用して精製されたマウスT細胞がヒトT細胞の代わりに使用された。トランスフェクトCHO細胞から作製された、ヒトFc領域から成るタンパク質が対照として働いた。ウェルは、図14に示した濃度の抗CD3ε抗体単独
【0196】
【化13】
【0197】
で、または、抗CD3ε抗体プラス10μg/mlのBTL−II:Fc
【0198】
【化14】
【0199】
かまたは10μg/mlのヒトFc領域から成るタンパク質
【0200】
【化15】
【0201】
で被覆した。結果は、BTL−II:Fcは抗CD3ε抗体に応答したマウスT細胞増殖を抑制できたが、ヒトFc領域から成るタンパク質単独では抑制できなかったことを示している。図14。
【0202】
実施例8:BTL−II:FcはマウスB細胞増殖を抑制しない
以下の実験は、ヒトBTL−II:Fcが、TALL−1およびIgM抗体により誘導されるマウスB細胞の増殖を阻害できるかどうかを決定するために設計された。実験は本質的に、Khareら((2000),Proc.Nad.Acad.Sci.97(7):3370−75)に記載されているように実行した。簡単には、マウスB細胞は、最初に密度勾配遠心分離によりリンパ球を精製し、次ぎにリンパ球は、単球/マクロファージおよびCD4+およびCD8+細胞を除去するB細胞カラム(Cedarlane,ウエストベリー,NY)を通過させることにより、脾臓から精製した。10%熱不活性化ウシ胎児血清を加えたMEM中の約1xl05精製B細胞を、ヤギF(ab’)2抗マウスIgM(2μg/ml)、ヒトTALL−1(10ng/ml)および/またはBTL−II:Fc(10μg/ml)とともに、またはなしで、96ウェルマイクロタイタープレート中、37℃にて4日インキュベー卜した。3H−チミジン(1μCi)は、インキュベーションの最後の8時間の間に加えた。4日の終了時に、実施例6に記載したように細胞を採取し、シンチレーションカウンターでカウントした。図15の印付けは、以下の細胞およびタンパク質の組み合わせを示している:B細胞単独、
【0203】
【化16】
【0204】
(これは図15に示したグラフ上、最も左の棒線であるが、検出可能なシグナルは存在しない);B細胞プラスTALL−1、
【0205】
【化17】
【0206】
;B細胞プラス抗IgM抗体、
【0207】
【化18】
【0208】
;B細胞プラスTALL−1および抗IgM抗体、
【0209】
【化19】
【0210】
;並びにB細胞プラスTALL−1、抗IgM抗体およびBTL−II:Fc、
【0211】
【化20】
【0212】
。結果は、BTL−II:FcはTALL−1および抗IgM抗体により誘導されたB細胞の増殖には効果がないことを示している。図15。それ故、BTL−IIはT細胞の増殖を阻害するが、B細胞の増殖は阻害しないと思われる。
【0213】
実施例9:T細胞によるサイトカイン産生に対するBTL−II:Fcの効果
以下の実験は、BTL−II:FcがT細胞によるサイトカイン産生に対して効果を有するかどうかを決定することを目的にしている。T細胞を精製し、マイクロタイタープレートを実施例6に記載したようにタンパク質で被覆した。約1x105の細胞を、200μl容量の培地(10%ウシ胎児血清を含むRPMI)を含む各々のウェルへ加えた。細胞は、37℃で64時間インキュベートした。次ぎに、サイトカイン濃度を決定するために150μlを除き、各々のウェル中に残っている50μlへ 3H−チミジンを加えた。マイクロタイタープレートは続いてさらに37℃で追加の8時間インキュベートし、続いて洗浄し、増殖の違いを確かめるため、実施例6に記載したようにカウントした(図16aに示されている)。何のタンパク質がウェルを被覆するために使用されたかを示す印付けは、図12aおよび図12bの通りである(実施例6に説明されている)。インターフェロンガンマ(IFNγ)、インターロイキン2(IL2)およびインターロイキン5(IL5)の濃度は、Meso Scale Discovery(IGEN International,Inc.の系列であるMSD,ガイサースブルグ,メリーランド ,米国)により販売されている多サイトカインの同時検出のための、電気化学に基づいたイムノアッセイシステムを使用して決定した。この種のサイトカイン検出の原理および操作は、例えば、Sennikovら(2003),J.Immunol.Methods 275:81−88、に説明されている。サイトカイン量の単位は、MSDの機械により発生された読み取り値から採られた。培地中のサイトカインの実際の濃度は、既知の濃度のタンパク質で発生させた検量線との比較なしでは、このデータから決定することはできず、それはこの特定の実験では付随していない。しかしながら、単一の実験内での読み取り値間の比較は、対照対実験試料に存在するサイトカインの相対量を提供することが可能である。
【0214】
結果は図16a−16dに示されている。図16aは、BTL−II:Fcは、抗CD3ε抗体または抗CD3ε抗体プラスB7RP−1:Fcに応答した増殖を阻害するが、使用された抗CD3ε抗体の濃度では、抗CD3ε抗体プラスB7−2:Fcに応答した増殖は阻害しないことを示している。しかしながら、実施例6で指摘したように(図12aおよび図12b)、より低い抗CD3ε抗体濃度、即ち、0.1μg/mlでは、抗CD3ε抗体プラスB7−2:Fcに応答した細胞増殖が、BTL−II:Fcにより阻害される。IFNγ、IL2およびIL5の産生は、抗CD3ε抗体単独で被覆されたウェルにおいては低く、そしてBTL−II:Fcの添加は、さらに有意にはそれを減少させなかった。図16a−16d。抗CD3ε抗体へのB7RP−l:FcかまたはB7−2:Fcの添加に応答したIFNγ産生の増加は、BTL−II:Fcにより阻害された。図16b。加えて、抗CD3ε抗体へのB7−2:Fcの添加に応答したIL2およびIL5産生の増加は、BTL−II:Fcにより阻害された。図16cおよび16d。試験された濃度では、抗CD3ε抗体へのB7RP−1:Fcの添加は、IL2またはIL5産生を評価できるほどは増加させなかった。図16cおよび16d。これらの結果は、BTL−II:Fcが、抗CD3ε抗体プラスB7−2:FcかまたはB7RP−1:Fcの組み合わせに応答した、少なくともいくつかのサイトカインの産生を阻害できることを示している。
【0215】
実施例10:BTL−II:FcによるT細胞の処置は、大量細胞死を生じさせない
以下の実験は、BTL−II:FcによるT細胞増殖の阻害が大量細胞死を伴うかどうかを決定するために行われた。T細胞増殖アッセイは本質的に実施例6に記載したように実施したが、ただし細胞は3H−チミジンでは標識しなかった。細胞は培養72時間後に、血球計数器でカウントし、そして細胞生存度は、トリパンブルー染色により決定した。マイクロタイタープレートウェルを被覆するために使用したタンパク質は、実施例6で説明し、そして図12aおよび図12bに示されているように図17でも示されている。抗CD3ε抗体は2μg/mlの濃度で使用した。結果は、各々のウェル中の死亡細胞の数は、ウェルを被覆するために使用されたタンパク質に関わらず、約0.5xl04から0.75xl04細胞の範囲であったことを示している。図17。それ故、BTL−II:Fc存在下で観察された細胞増殖の相違は、BTL−II:Fcの実質の毒性効果を反映していない。
【0216】
実施例11:マウスBTL−IIはマウスCTLA4、CD28、ICOSまたはPD−1とは結合しない
以下の組の実験は、BTL−IIが他のB7タンパク質の、いくつかの既知の結合パートナーの一つと結合するかどうかの疑問に取り組んでいる。例えば、B7RP−1はICOSと結合することが知られており(Yoshinagaら(1999),Nature402:827−32)、CD80(B7−1とも称されている)は、CD86(B7−2とも称されている)が行うように、CTLA4およびCD28へ結合することが知られている、例えば、SharpeおよびFreeman(2002),Nature Reviews Immunology 2:116−26)、そしてPD−L1およびPD−L2はPD−1へ結合することが知られている(Latchmanら(2001),Nature Immunology 2(3):261−68)。
【0217】
実験は以下のように行った。第一に、B7タンパク質へ結合することが知られているタンパク質の細胞外領域に加えてヒトIgG抗体のFc領域を含んでなる融合タンパク質を得た、または単離した。ヒト抗体のFc領域に加えてマウスCD28(mCD28−huFc)かまたはマウスPD−1(PD1−huFc)の細胞外領域を含んでなる融合タンパク質は、R&D Systems(ミネアポリス,ミネソタ,米国)から購入した。他の二つのB7結合タンパク質(CTLA4−huFcおよびICOS−huFc)もまたR&D Systemsから入手可能であるが、以下のように作製した。マウスCTLA4の細胞外領域をコードしているcDNAおよびマウスICOSの細胞外領域をコードしている別のcDNAを、哺乳動物細胞での発現に適したベクター中のヒトIgG抗体のFc領域をコードしているcDNAへ各々融合させた。これらの構築物の各々の細胞をトランスフェクトのために使用し、そして融合タンパク質は、プロテインAクロマトグラフィーにより、トランスフェクトされた細胞の培養培地から精製した。
【0218】
三つのマウスcDNAの各々の完全長バージョン(BTL−II、B7RP−1、CD80をコードしている)を発現に適したベクター内へ挿入した。これらの構築物の各々の約10μgを、空ベクターと共に、トランスフェクト293細胞に対して別々に使用した。トランスフェクション2日後、四つのトランスフェクションの各々から約100万細胞を、上述の融合タンパク質の各々で染色した。結合したタンパク質は、ヒトIgG Fc領域に対する蛍光標識抗体を使用して検出した。染色細胞はFACSにより分析した。結果は図18に示されている。
【0219】
予測されるように、空ベクターでトランスフェクトされた細胞(図18の上段)は、四つの融合タンパク質のいずれによっても染色されなかった。マウスBTL−IIでトランスフェクトされた細胞(図18の二段目)は同様に挙動し、融合タンパク質のどれもBTL−IIへ結合しないことを示している。予測されるように、マウスB7RP−1でトランスフェクトされた細胞はICOS−huFcで染色されたが、他の融合タンパク質のいずれでも染色されなかった。また予測されるように、マウスCD8Oでトランスフェク卜された細胞はCTLA4−huFcまたはmCD28−huFcで染色されたが、ICOS−huFcまたはPD−1−huFcでは染色されなかった。これらの結果は、BTL−IIは、四つのタンパク質、CTLA−4、PD−1、ICOSおよびCD28(その各々はB7ファミリー中の少なくとも一つのタンパク質へ結合することが知られている)へ結合できないことを示している。
【図面の簡単な説明】
【0220】
【図1】図1は、ブチロフィリン様タンパク質のファミリーの、選択されたメンバーのドメイン構造を示している。選択されたタンパク質のほとんどは、以下に議論するB7サブファミリーのメンバーである。各々のタンパク質の名前は、その構造が描かれている図の左に示されている。“Ig様”ドメインは以下に説明するように、免疫グロブリン様ドメインである。“7”とラベルされたドメインは以下に説明するように、7アミノ酸反復領域である。 “TM” とラベルされたドメインは膜貫通ドメインである。白四角は未同定の細胞質ドメインであり、ドメインの特別なファミリーの一部である。“B30.2” とラベルされたドメインは以下に説明するように、B30.2ドメインである。BTL−IIタンパク質は本明細書において、四つのIg様ドメインを有するように描かれているが、BTL−IIの形はまた、二つまたは三つのIg様ドメインを有するものも存在する。B7−H3は四つのIg様ドメインで描かれているが、それはまた二つのみのIg様ドメインを含んでいる形でも存在する。
【図2】図2は、ヒトBTL−II遺伝子およびmRNAの構造の図である。黒四角はエキソンを示している。四角の下に示されている水平線下の数字は、エキソンの配列番号3内の位置を示している。図の最下部の水平線は、細胞外ドメインおよび膜貫通および細胞質ドメイン(“TM/Cyto”)をコードし、そして3’非翻訳領域(“3UTR”)を形成している配列番号3の範囲を表示している。停止コドンは、日輪型(sunburst)または小さな爆発として記載することが可能な印により表示されている。
【図3a】図3aは、ヒトBTL−II mRNAのスプライス変異体の、第一のカテゴリーの構造の図である。記号は、図2で説明したものと同じである。エキソン2および3にかぶっている大きなXは、これらのエキソンが、スプライス変異体のこのカテゴリーでは失われていることを示している。
【図3b】図3bは、配列番号3の一部(上のライン)と並列された、スプライス変異体の第一のカテゴリーのメンバーの代表的配列(下のライン)である。失われたエキソン2および3により創造されたギャップ以外のミスマッチは存在しない。
【図4a】図4aは、ヒトBTL−II mRNAのスプライス変異体の、第二のカテゴリーの構造を示している。記号は、図2で説明したものと同じである。エキソン3にかぶっている大きなXは、このエキソンが、スプライス変異体のこのカテゴリーでは失われていることを示している。
【図4b】図4bは、配列番号3の一部(上のライン)と並列された、スプライス変異体の第二のカテゴリーのメンバーの代表的配列(下のライン)を示している。失われたエキソン3により創造されたギャップ以外のミスマッチは存在しない。
【図5a】図5aは、ヒトBTL−II mRNAのスプライス変異体の、第三のカテゴリーの構造を示している。記号は、図3で説明したものと同じである。エキソン2および3にかぶっている大きなXは、これらのエキソンが、スプライス変異体のこのカテゴリーでは失われていることを示している。黒四角に隣接する、数字を伴った星印は、スプライス変異体のこのカテゴリーに存在する配列多型の位置を示している。変異は、配列番号3内の以下の位置に存在する:変異2は1050位であり;変異3は1136および1140位であり;変異4は1178および1179位であり;変異5は1212位であり;および変異6は1242位である。
【図5b】図5bは、配列番号3の一部(上のライン)と並列された、スプライス変異体の第三のカテゴリーのメンバーの代表的配列(下のライン)を示している。ミスマッチ塩基はボールド体で示されている。
【図6a】図6aは、ヒトBTL−II mRNAのスプライス変異体の、第四のカテゴリーの構造を示している。エキソン3にかぶっている大きなXは、このエキソンが、スプライス変異体のこのカテゴリーでは失われていることを示している。配列多型は、図5aと同様に示されており、そして変異2から6は図5aと同様である。
【図6b】図6bは、配列番号3の一部(上のライン)と並列された、スプライス変異体の第四のカテゴリーのメンバーの代表的配列(下のライン)を示している。ミスマッチ塩基はボールド体で示されている。
【図7a】図7aは、ヒトBTL−II mRNAのスプライス変異体の、第五のカテゴリーの構造を示している。エキソン3にかぶっている大きなXは、このエキソンが、スプライス変異体のこのカテゴリーでは失われていることを示している。配列多型は、図5aと同様に示されており、そして変異2から6は図5aと同様である。変異1は、配列番号3のヌクレオチド78から80の欠失である。
【図7b】図7bは、配列番号3の一部(上のライン)と並列された、スプライス変異体の第五のカテゴリーのメンバーの代表的配列(下のライン)を示している。ミスマッチ塩基はボールド体で示されている。
【図8】図8はマウスBTL−II遺伝子およびmRNAの構造の図である。番号が配列番号5における位置を指していることを除いて、記号は図5で説明したものと同じである。
【図9a】図9aは、マウスBTL−II mRNAのスプライス変異体の、第一のカテゴリーの構造を示している。エキソン3にかぶっている大きなXは、このエキソンが、スプライス変異体のこのカテゴリーでは失われていることを示している。
【図9b】図9bは、配列番号5(上のライン)と並列された、マウススプライス変異体の第一のカテゴリーのメンバーの代表的配列(下のライン)を示している。失われたエキソン3により創造されたギャップ以外のミスマッチは存在しない。
【図10】図10は、非症状性、野生型の結腸組織におけるBTL−II mRNAの発現と比較した、炎症性腸疾患の症状を示していない場合(横縞)または炎症性腸疾患の症状を示している場合(市松模様)のmdr1a−/−マウス(FVBバックグラウンドにmdr1aヌル変異を運んでいる)の結腸組織におけるBTL−II mRNAの発現を表している。測定は、蛍光標識cDNAをBTL−II mRNAに相補的なオリゴヌクレオチドを含んでいるAffymetrixチップへハイブリダイズさせることにより行った。
【図11】図11は、FVB株の2匹の異なった野生型マウス(FVB#1および#2;対角の線)からの、および炎症性腸疾患の症状を示している2匹のmdr1a−/−マウス(Mdr1a−/−#1および#2;市松模様)からの結腸組織のリアルタイムPCRアッセイにより検出されたBTL−II mRNAの相対濃度を示しているグラフである。
【図12a】図12aは以下のタンパク質と培養された、精製ヒトT細胞の増殖(3H−チミジンの取り込みにより証明された)を示している棒グラフである:抗CD3ε抗体単独、
【0221】
【化21】
【0222】
;抗CD3ε抗体およびBTL−II:Fc、
【0223】
【化22】
【0224】
;抗CD3ε抗体およびB7RP−1:Fc、
【0225】
【化23】
【0226】
;抗CD3ε抗体、B7RP−1:FcおよびBTL−II:Fc、
【0227】
【化24】
【0228】
;抗CD3ε抗体およびB7−2:Fc、
【0229】
【化25】
【0230】
;および抗CD3ε抗体、B7−2:FcおよびBTL−II:Fc、
【0231】
【化26】
【0232】
。
【図12b】図12bは図12aと同一であるが、ただし対数目盛りではなく、直線目盛りが使用されている。
【図13】図13は、一定量の抗CD3ε抗体および示されているように量を変化させたBTL−II:Fcに応答した、精製ヒトT細胞の増殖を示している棒グラフである。
【図14】図14は、抗CD3ε抗体単独、
【0233】
【化27】
【0234】
、抗CD3ε抗体プラスBTL−II:Fc、
【0235】
【化28】
【0236】
、または抗CD3ε抗体プラスヒトFc領域から成る対照タンパク質、
【0237】
【化29】
【0238】
、に応答したマウスT細胞の増殖を示している棒グラフである。
【図15】図15は、タンパク質添加なし、
【0239】
【化30】
【0240】
、TALL−1単独、
【0241】
【化31】
【0242】
、抗IgM抗体単独、
【0243】
【化32】
【0244】
、TALL−1および抗IgM抗体、
【0245】
【化33】
【0246】
、およびTALL−1、抗IgM抗体およびBTL−II:Fc、
【0247】
【化34】
【0248】
、に応答したマウスB細胞の増殖を示している棒グラフである。
【図16a】図16aは、図12aに示したようなタンパク質の多様な組み合わせに応答した、精製ヒトT細胞の増殖を示している。
【図16b】図16bは、図12aに示したようなタンパク質の多様な組み合わせに応答した、相対インターフェロンガンマ(IFNγ)産生を示している。
【図16c】図16cは、図12aに示したようなタンパク質の多様な組み合わせに応答した、相対インターロイキン2(IL2)産生を示している。
【図16d】図16dは、図12aに示したようなタンパク質の多様な組み合わせに応答した、相対インターロイキン5(IL5)産生を示している。
【図17】図17は、図12aに示されているような、タンパク質の多様な組み合わせで培養された精製T細胞の培養において、死んだ細胞の総数を示している棒グラフである。
【図18】図18は、空ベクター(一番上の行)、かまたはマウスBTL−II(二番目の行)、マウスB7RP−1(三番目の行)またはマウスCD80(一番下の行)をコードしているcDNAを含んでいるベクターでトランスフェクされた細胞のFACSスキャンを示している。細胞は次のタンパク質で染色した:(1)IgG1抗体のヒトFc領域に融合されたマウスCTLA4の細胞外領域(一番目の列);(2)IgG1抗体のヒトFc領域に融合されたマウスCD28の細胞外領域(二番目の列);(3)IgG1抗体のヒトFc領域に融合されたマウスICOSの細胞外領域(三番目の列);および(4)IgG1抗体のヒトFc領域に融合されたPD1の細胞外領域(四番目の列)。各々のスキャンの縦軸(“カウント”とラベルされている)は細胞数を表し、そして横軸(“FL2−H”とラベルされている)は蛍光を表している。“M1”とラベルされた水平線は、“ゲート(gate)” がFACS機器に使用されたところを示している。このゲートに含まれる細胞が陽性と考えられる;全ての他の細胞は陰性と考えられる。各々のFACSスキャン上の文字は、個々の試料番号を示している。
【配列表】
【特許請求の範囲】
【請求項1】
単離BTL−IIタンパク質であって、配列番号4のアミノ酸xからy、から成るアミノ酸配列、ここにおいて、xは配列番号4の1から35位までからのいずれかのアミノ酸であり、そしてyは配列番号4の452から462位までからのいずれかのアミノ酸である、を含んでなる、前記BTL−IIタンパク質。
【請求項2】
BTL−IIタンパク質が配列番号4のアミノ酸30から453を含んでなる、請求項1に記載のBTL−IIタンパク質。
【請求項3】
BTL−IIタンパク質が配列番号4のアミノ酸29から457を含んでなる、請求項2に記載のBTL−IIタンパク質。
【請求項4】
配列番号4のアミノ酸29から482を含んでなる、請求項1に記載のBTL−IIタンパク質。
【請求項5】
単離BTL−IIタンパク質であって、配列番号10、配列番号14、配列番号16または配列番号18のアミノ酸30から358と少なくとも80%同一であるアミノ酸配列から成るポリペプチド、ここにおいて配列番号10、配列番号14、配列番号16または配列番号18のアミノ酸30から358と並列されたアミノ酸配列の同一性領域は少なくとも150アミノ酸であり、
ここにおいてアミノ酸配列は、配列番号10、配列番号14または配列番号18のアミノ酸127から157と、または配列番号16のアミノ酸126から156と少なくとも80%同一であり、ここにおいて配列番号10、配列番号14または配列番号18のアミノ酸127から157と、または配列番号16のアミノ酸126から156と並列されたアミノ酸配列の同一性領域は少なくとも20アミノ酸長であり、そして
ここにおいてポリペプチドはT細胞の増殖を阻害可能である、
を含んでなる、前記BTL−IIタンパク質。
【請求項6】
アミノ酸配列が、配列番号10、配列番号14、配列番号16または配列番号18のアミノ酸30から358と少なくとも90%同一である、請求項5に記載のBTL−IIタンパク質。
【請求項7】
アミノ酸配列が、配列番号10、配列番号14または配列番号18のアミノ酸127から157,または配列番号16のアミノ酸126から156と少なくとも90%同一である、請求項5または6に記載のBTL−IIタンパク質。
【請求項8】
配列番号10、配列番号14、配列番号16または配列番号18のアミノ酸30から358を含んでなる、請求項5から7のいずれか一項に記載のBTL−IIタンパク質。
【請求項9】
単離BTL−IIタンパク質であって、配列番号10、配列番号14、配列番号16または配列番号18のアミノ酸32から358と少なくとも80%同一であるアミノ酸配列から成るポリペプチド、
ここにおいて配列番号10、配列番号14、配列番号16または配列番号18のアミノ酸32から358と並列されたアミノ酸配列の同一性領域は少なくとも275アミノ酸であり、そして
ここにおいてポリペプチドはT細胞の増殖を阻害可能である、
を含んでなる、前記BTL−IIタンパク質。
【請求項10】
アミノ酸配列が、配列番号10、配列番号14、配列番号16または配列番号18のアミノ酸32から358と少なくとも90%同一である、請求項9に記載のBTL−IIタンパク質。
【請求項11】
アミノ酸配列が、配列番号10、配列番号14、配列番号16または配列番号18のアミノ酸32から358を含んでなる、請求項10に記載のBTL−IIタンパク質。
【請求項12】
単離BTL−IIタンパク質であって、配列番号10のアミノ酸32から358と少なくとも80%同一である第一のアミノ酸配列から成る第一のポリペプチド、ここにおいて配列番号10に並列された第一のアミノ酸配列の同一性領域は少なくとも約175アミノ酸長であり、
ここにおいて第一のアミノ酸配列は長さが約390アミノ酸を超えず、
ここにおいて第一のポリペプチドはT細胞の増殖を阻害可能であり、
ここにおいて第一のアミノ酸配列は、配列番号4のアミノ酸148から232と少なくとも80%同一ではなく、少なくとも約40アミノ酸の、配列番号4のアミノ酸148から232と並列された第一のアミノ酸配列の同一性領域を有し、そして
ここにおいてBTL−IIタンパク質は、配列番号4のアミノ酸148から232と少なくとも80%同一で、少なくとも約40アミノ酸の、配列番号4のアミノ酸148から232と並列された第二のアミノ酸配列の同一性領域を有する、第二のアミノ酸配列を含んでいない、
を含んでなる、前記BTL−IIタンパク質。
【請求項13】
第一のアミノ酸配列は長さが約270アミノ酸を超えない、請求項12に記載のBTL−IIタンパク質。
【請求項14】
第一のアミノ酸配列が、配列番号10と少なくとも90%同一である、請求項12または13に記載のBTL−IIタンパク質。
【請求項15】
配列番号8または配列番号12のアミノ酸32から242を含んでなる、請求項12から14のいずれか一項に記載のBTL−IIタンパク質。
【請求項16】
異種ポリペプチドをさらに含んでなる、請求項1、5、9または12に記載のBTL−IIタンパク質。
【請求項17】
異種ポリペプチドが、抗体のFc領域である、請求項16に記載のBTL−IIタンパク質。
【請求項18】
配列番号4の免疫原性断片であって、
ここにおいて免疫原性断片は、免疫原性断片へ特異的に結合する抗体を惹起することが可能であり、
ここにおいて免疫原性断片は少なくとも10アミノ酸長であり、そして
ここにおいて免疫原性断片は配列番号4の360位に及ぶ、
前記断片。
【請求項19】
配列番号10、配列番号14、配列番号16または配列番号18から成る群より選択される配列の免疫原性断片であって、
ここにおいて免疫原性断片は配列番号10、配列番号14、配列番号16または配列番号18の141から143位に及び、
ここにおいて免疫原性断片は、免疫原性断片へ特異的に結合する抗体を惹起することが可能であり、
ここにおいて免疫原性断片は少なくとも10アミノ酸長である、
前記断片。
【請求項20】
単離BTL−IIタンパク質であって、配列番号6のアミノ酸xからyまでから成るアミノ酸配列、ここにおいてxは配列番号6の1から35位までからのいずれかのアミノ酸であり、そしてyは配列番号6の450から460位までからのいずれかのアミノ酸である、を含んでなる、前記BTL−IIタンパク質。
【請求項21】
配列番号6のアミノ酸32から450を含んでなる、請求項20に記載のBTL−IIタンパク質。
【請求項22】
配列番号6のアミノ酸29から456を含んでなる、請求項21に記載のBTL−IIタンパク質。
【請求項23】
配列番号6のアミノ酸29から514を含んでなる、請求項22に記載のBTL−IIタンパク質。
【請求項24】
配列番号4のアミノ酸1から457、から成る、BTL−IIタンパク質へ特異的に結合する単離抗体。
【請求項25】
配列番号6のアミノ酸1から456、から成る、BTL−IIタンパク質へ特異的に結合する単離抗体。
【請求項26】
配列番号8のアミノ酸1から247;
配列番号10のアミノ酸1から363;
配列番号12のアミノ酸1から243;
配列番号14のアミノ酸1から359;
配列番号16のアミノ酸1から358;および
配列番号18のアミノ酸1から362
から成る群より選択されるアミノ酸配列から成るBTL−IIタンパク質へ特異的に結合する単離抗体。
【請求項27】
抗体がモノクローナル抗体である、請求項24から26のいずれか一項に記載の抗体。
【請求項28】
抗体がヒト抗体である、請求項24から27のいずれか一項に記載の抗体。
【請求項29】
抗体がBTL−IIのアンタゴニストである、請求項24から28のいずれか一項に記載の抗体。
【請求項30】
単離BTL−II核酸であって、配列番号3のヌクレオチドxからy、ここにおいてxはヌクレオチド1から105よりであり、そしてyはヌクレオチド1345から1375よりである、より成るポリヌクレオチドまたはその相補体を含んでなる、前記BTL−II核酸。
【請求項31】
配列番号3のヌクレオチド105から1345を含んでなる、請求項30に記載のBTL−II核酸。
【請求項32】
配列番号3のヌクレオチド1から1371を含んでなる、請求項31に記載のBTL−II核酸。
【請求項33】
BTL−II核酸が、請求項1から23のいずれか一項のBTL−IIタンパク質または免疫原性断片をコードする、単離BTL−II核酸またはその相補体。
【請求項34】
請求項30から33のいずれか一項に記載のBTL−II核酸を含んでなるベクター。
【請求項35】
請求項34に記載のベクターを含んでいる宿主細胞。
【請求項36】
BTL−IIタンパク質または免疫原性断片を産生するための方法であって、BTL−IIタンパク質または免疫原性断片の発現を可能にする条件下、請求項35に記載の宿主細胞を培養することを含んでなる、前記方法。
【請求項37】
請求項1から23のいずれかの一項に記載のBTL−IIタンパク質または免疫原性断片を発現するように遺伝子操作された宿主細胞。
【請求項38】
宿主細胞が哺乳動物細胞である、請求項37に記載の宿主細胞。
【請求項39】
宿主細胞がCHO細胞である、請求項38に記載の宿主細胞。
【請求項40】
BTL−IIタンパク質または免疫原性断片を産生するための方法であって、BTL−IIタンパク質または免疫原性断片の発現を可能にする条件下、請求項37の宿主細胞を培養することを含んでなる、前記方法。
【請求項41】
宿主細胞または培地から、BTL−IIタンパク質を単離することをさらに含んでなる、請求項36または40に記載の方法。
【請求項42】
炎症性腸疾患を患っている患者に、治療的に有効量の、請求項1から17のいずれか一項に記載のBTL−IIタンパク質を投与することを含んでなる方法。
【請求項43】
患者がクローン病を患っている、請求項42に記載の方法。
【請求項44】
患者が潰瘍性大腸炎を患っている、請求項42に記載の方法。
【請求項45】
抗原に対する免疫応答を誘導するための方法であって、治療的に有効量のBTL−IIアンタゴニストおよび抗原を哺乳動物に投与することを含んでなる、前記方法。
【請求項46】
免疫応答が、抗原に対する粘膜免疫応答を含んでなる、請求項45に記載の方法。
【請求項47】
アンタゴニストが請求項29の抗体である、請求項45に記載の方法。
【請求項48】
抗原が粘膜表面を経て投与される、請求項45から47のいずれか一項に記載の方法。
【請求項49】
抗原に対する免疫応答を弱めるための方法であって、抗原および請求項1から17および20から23のいずれか一項に記載のBTL−IIタンパク質を哺乳動物に投与することを含んでなる、前記方法。
【請求項50】
抗原が粘膜表面を経て投与される、請求項49に記載の方法。
【請求項51】
T細胞増殖を阻害するための方法であって、T細胞と請求項1から17および20から23のいずれか一項に記載のBTL−IIタンパク質を接触させることを含んでなる、前記方法。
【請求項52】
T細胞をインビボでBTL−IIタンパク質と接触させる、請求項51に記載の方法。
【請求項53】
T細胞がヒトT細胞である、請求項51または52に記載の方法。
【請求項54】
T細胞によるサイトカイン産生を阻害するための方法であって、T細胞と請求項1から17および20から23のいずれか一項に記載のポリペプチドを接触させることを含んでなる、前記方法。
【請求項55】
T細胞をインビボでポリペプチドと接触させる、請求項54に記載の方法。
【請求項56】
T細胞がヒトT細胞である、請求項54または55に記載の方法。
【請求項57】
インターフェロンガンマ産生が阻害される、請求項54から56のいずれか一項に記載の方法。
【請求項58】
IL2産生が阻害される、請求項54から57のいずれか一項に記載の方法。
【請求項59】
IL5産生が阻害される、請求項54から58のいずれか一項に記載の方法。
【請求項1】
単離BTL−IIタンパク質であって、配列番号4のアミノ酸xからy、から成るアミノ酸配列、ここにおいて、xは配列番号4の1から35位までからのいずれかのアミノ酸であり、そしてyは配列番号4の452から462位までからのいずれかのアミノ酸である、を含んでなる、前記BTL−IIタンパク質。
【請求項2】
BTL−IIタンパク質が配列番号4のアミノ酸30から453を含んでなる、請求項1に記載のBTL−IIタンパク質。
【請求項3】
BTL−IIタンパク質が配列番号4のアミノ酸29から457を含んでなる、請求項2に記載のBTL−IIタンパク質。
【請求項4】
配列番号4のアミノ酸29から482を含んでなる、請求項1に記載のBTL−IIタンパク質。
【請求項5】
単離BTL−IIタンパク質であって、配列番号10、配列番号14、配列番号16または配列番号18のアミノ酸30から358と少なくとも80%同一であるアミノ酸配列から成るポリペプチド、ここにおいて配列番号10、配列番号14、配列番号16または配列番号18のアミノ酸30から358と並列されたアミノ酸配列の同一性領域は少なくとも150アミノ酸であり、
ここにおいてアミノ酸配列は、配列番号10、配列番号14または配列番号18のアミノ酸127から157と、または配列番号16のアミノ酸126から156と少なくとも80%同一であり、ここにおいて配列番号10、配列番号14または配列番号18のアミノ酸127から157と、または配列番号16のアミノ酸126から156と並列されたアミノ酸配列の同一性領域は少なくとも20アミノ酸長であり、そして
ここにおいてポリペプチドはT細胞の増殖を阻害可能である、
を含んでなる、前記BTL−IIタンパク質。
【請求項6】
アミノ酸配列が、配列番号10、配列番号14、配列番号16または配列番号18のアミノ酸30から358と少なくとも90%同一である、請求項5に記載のBTL−IIタンパク質。
【請求項7】
アミノ酸配列が、配列番号10、配列番号14または配列番号18のアミノ酸127から157,または配列番号16のアミノ酸126から156と少なくとも90%同一である、請求項5または6に記載のBTL−IIタンパク質。
【請求項8】
配列番号10、配列番号14、配列番号16または配列番号18のアミノ酸30から358を含んでなる、請求項5から7のいずれか一項に記載のBTL−IIタンパク質。
【請求項9】
単離BTL−IIタンパク質であって、配列番号10、配列番号14、配列番号16または配列番号18のアミノ酸32から358と少なくとも80%同一であるアミノ酸配列から成るポリペプチド、
ここにおいて配列番号10、配列番号14、配列番号16または配列番号18のアミノ酸32から358と並列されたアミノ酸配列の同一性領域は少なくとも275アミノ酸であり、そして
ここにおいてポリペプチドはT細胞の増殖を阻害可能である、
を含んでなる、前記BTL−IIタンパク質。
【請求項10】
アミノ酸配列が、配列番号10、配列番号14、配列番号16または配列番号18のアミノ酸32から358と少なくとも90%同一である、請求項9に記載のBTL−IIタンパク質。
【請求項11】
アミノ酸配列が、配列番号10、配列番号14、配列番号16または配列番号18のアミノ酸32から358を含んでなる、請求項10に記載のBTL−IIタンパク質。
【請求項12】
単離BTL−IIタンパク質であって、配列番号10のアミノ酸32から358と少なくとも80%同一である第一のアミノ酸配列から成る第一のポリペプチド、ここにおいて配列番号10に並列された第一のアミノ酸配列の同一性領域は少なくとも約175アミノ酸長であり、
ここにおいて第一のアミノ酸配列は長さが約390アミノ酸を超えず、
ここにおいて第一のポリペプチドはT細胞の増殖を阻害可能であり、
ここにおいて第一のアミノ酸配列は、配列番号4のアミノ酸148から232と少なくとも80%同一ではなく、少なくとも約40アミノ酸の、配列番号4のアミノ酸148から232と並列された第一のアミノ酸配列の同一性領域を有し、そして
ここにおいてBTL−IIタンパク質は、配列番号4のアミノ酸148から232と少なくとも80%同一で、少なくとも約40アミノ酸の、配列番号4のアミノ酸148から232と並列された第二のアミノ酸配列の同一性領域を有する、第二のアミノ酸配列を含んでいない、
を含んでなる、前記BTL−IIタンパク質。
【請求項13】
第一のアミノ酸配列は長さが約270アミノ酸を超えない、請求項12に記載のBTL−IIタンパク質。
【請求項14】
第一のアミノ酸配列が、配列番号10と少なくとも90%同一である、請求項12または13に記載のBTL−IIタンパク質。
【請求項15】
配列番号8または配列番号12のアミノ酸32から242を含んでなる、請求項12から14のいずれか一項に記載のBTL−IIタンパク質。
【請求項16】
異種ポリペプチドをさらに含んでなる、請求項1、5、9または12に記載のBTL−IIタンパク質。
【請求項17】
異種ポリペプチドが、抗体のFc領域である、請求項16に記載のBTL−IIタンパク質。
【請求項18】
配列番号4の免疫原性断片であって、
ここにおいて免疫原性断片は、免疫原性断片へ特異的に結合する抗体を惹起することが可能であり、
ここにおいて免疫原性断片は少なくとも10アミノ酸長であり、そして
ここにおいて免疫原性断片は配列番号4の360位に及ぶ、
前記断片。
【請求項19】
配列番号10、配列番号14、配列番号16または配列番号18から成る群より選択される配列の免疫原性断片であって、
ここにおいて免疫原性断片は配列番号10、配列番号14、配列番号16または配列番号18の141から143位に及び、
ここにおいて免疫原性断片は、免疫原性断片へ特異的に結合する抗体を惹起することが可能であり、
ここにおいて免疫原性断片は少なくとも10アミノ酸長である、
前記断片。
【請求項20】
単離BTL−IIタンパク質であって、配列番号6のアミノ酸xからyまでから成るアミノ酸配列、ここにおいてxは配列番号6の1から35位までからのいずれかのアミノ酸であり、そしてyは配列番号6の450から460位までからのいずれかのアミノ酸である、を含んでなる、前記BTL−IIタンパク質。
【請求項21】
配列番号6のアミノ酸32から450を含んでなる、請求項20に記載のBTL−IIタンパク質。
【請求項22】
配列番号6のアミノ酸29から456を含んでなる、請求項21に記載のBTL−IIタンパク質。
【請求項23】
配列番号6のアミノ酸29から514を含んでなる、請求項22に記載のBTL−IIタンパク質。
【請求項24】
配列番号4のアミノ酸1から457、から成る、BTL−IIタンパク質へ特異的に結合する単離抗体。
【請求項25】
配列番号6のアミノ酸1から456、から成る、BTL−IIタンパク質へ特異的に結合する単離抗体。
【請求項26】
配列番号8のアミノ酸1から247;
配列番号10のアミノ酸1から363;
配列番号12のアミノ酸1から243;
配列番号14のアミノ酸1から359;
配列番号16のアミノ酸1から358;および
配列番号18のアミノ酸1から362
から成る群より選択されるアミノ酸配列から成るBTL−IIタンパク質へ特異的に結合する単離抗体。
【請求項27】
抗体がモノクローナル抗体である、請求項24から26のいずれか一項に記載の抗体。
【請求項28】
抗体がヒト抗体である、請求項24から27のいずれか一項に記載の抗体。
【請求項29】
抗体がBTL−IIのアンタゴニストである、請求項24から28のいずれか一項に記載の抗体。
【請求項30】
単離BTL−II核酸であって、配列番号3のヌクレオチドxからy、ここにおいてxはヌクレオチド1から105よりであり、そしてyはヌクレオチド1345から1375よりである、より成るポリヌクレオチドまたはその相補体を含んでなる、前記BTL−II核酸。
【請求項31】
配列番号3のヌクレオチド105から1345を含んでなる、請求項30に記載のBTL−II核酸。
【請求項32】
配列番号3のヌクレオチド1から1371を含んでなる、請求項31に記載のBTL−II核酸。
【請求項33】
BTL−II核酸が、請求項1から23のいずれか一項のBTL−IIタンパク質または免疫原性断片をコードする、単離BTL−II核酸またはその相補体。
【請求項34】
請求項30から33のいずれか一項に記載のBTL−II核酸を含んでなるベクター。
【請求項35】
請求項34に記載のベクターを含んでいる宿主細胞。
【請求項36】
BTL−IIタンパク質または免疫原性断片を産生するための方法であって、BTL−IIタンパク質または免疫原性断片の発現を可能にする条件下、請求項35に記載の宿主細胞を培養することを含んでなる、前記方法。
【請求項37】
請求項1から23のいずれかの一項に記載のBTL−IIタンパク質または免疫原性断片を発現するように遺伝子操作された宿主細胞。
【請求項38】
宿主細胞が哺乳動物細胞である、請求項37に記載の宿主細胞。
【請求項39】
宿主細胞がCHO細胞である、請求項38に記載の宿主細胞。
【請求項40】
BTL−IIタンパク質または免疫原性断片を産生するための方法であって、BTL−IIタンパク質または免疫原性断片の発現を可能にする条件下、請求項37の宿主細胞を培養することを含んでなる、前記方法。
【請求項41】
宿主細胞または培地から、BTL−IIタンパク質を単離することをさらに含んでなる、請求項36または40に記載の方法。
【請求項42】
炎症性腸疾患を患っている患者に、治療的に有効量の、請求項1から17のいずれか一項に記載のBTL−IIタンパク質を投与することを含んでなる方法。
【請求項43】
患者がクローン病を患っている、請求項42に記載の方法。
【請求項44】
患者が潰瘍性大腸炎を患っている、請求項42に記載の方法。
【請求項45】
抗原に対する免疫応答を誘導するための方法であって、治療的に有効量のBTL−IIアンタゴニストおよび抗原を哺乳動物に投与することを含んでなる、前記方法。
【請求項46】
免疫応答が、抗原に対する粘膜免疫応答を含んでなる、請求項45に記載の方法。
【請求項47】
アンタゴニストが請求項29の抗体である、請求項45に記載の方法。
【請求項48】
抗原が粘膜表面を経て投与される、請求項45から47のいずれか一項に記載の方法。
【請求項49】
抗原に対する免疫応答を弱めるための方法であって、抗原および請求項1から17および20から23のいずれか一項に記載のBTL−IIタンパク質を哺乳動物に投与することを含んでなる、前記方法。
【請求項50】
抗原が粘膜表面を経て投与される、請求項49に記載の方法。
【請求項51】
T細胞増殖を阻害するための方法であって、T細胞と請求項1から17および20から23のいずれか一項に記載のBTL−IIタンパク質を接触させることを含んでなる、前記方法。
【請求項52】
T細胞をインビボでBTL−IIタンパク質と接触させる、請求項51に記載の方法。
【請求項53】
T細胞がヒトT細胞である、請求項51または52に記載の方法。
【請求項54】
T細胞によるサイトカイン産生を阻害するための方法であって、T細胞と請求項1から17および20から23のいずれか一項に記載のポリペプチドを接触させることを含んでなる、前記方法。
【請求項55】
T細胞をインビボでポリペプチドと接触させる、請求項54に記載の方法。
【請求項56】
T細胞がヒトT細胞である、請求項54または55に記載の方法。
【請求項57】
インターフェロンガンマ産生が阻害される、請求項54から56のいずれか一項に記載の方法。
【請求項58】
IL2産生が阻害される、請求項54から57のいずれか一項に記載の方法。
【請求項59】
IL5産生が阻害される、請求項54から58のいずれか一項に記載の方法。
【図1】

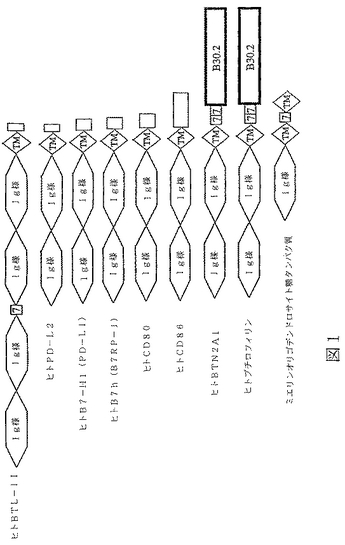
【図2】
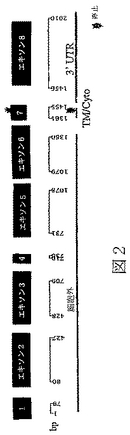
【図3a】
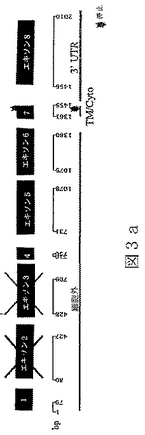
【図3b】

【図4a】
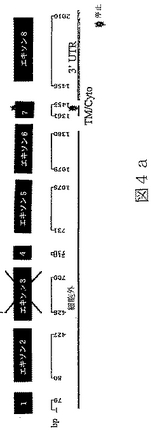
【図4b】

【図5a】
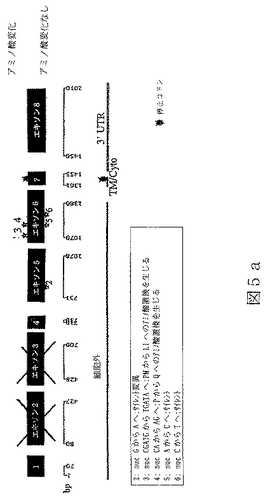
【図5b】
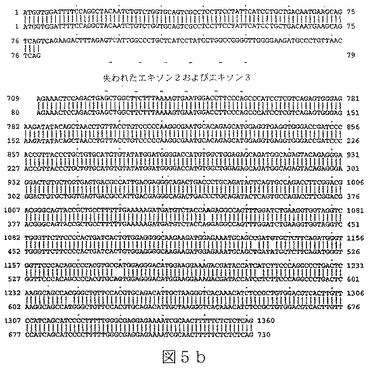
【図6a】
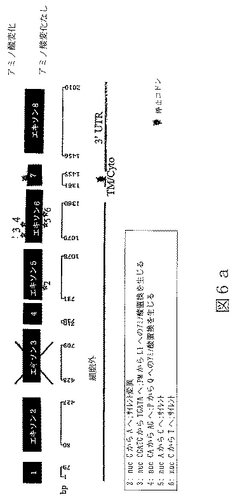
【図6b】
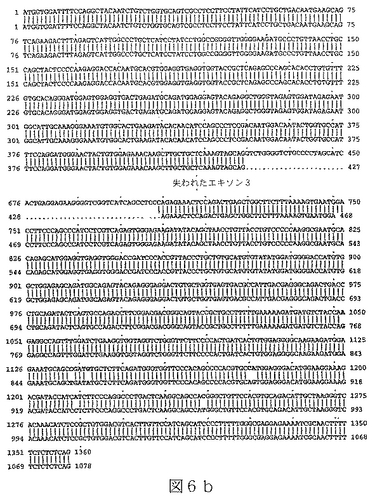
【図7a】
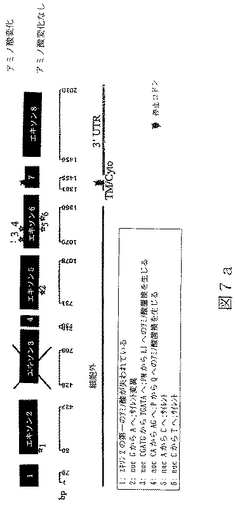
【図7b】
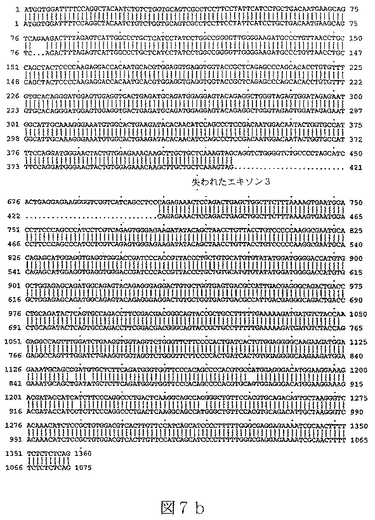
【図8】
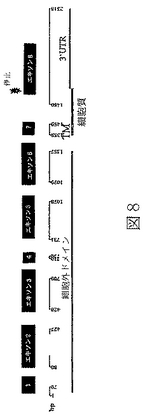
【図9a】
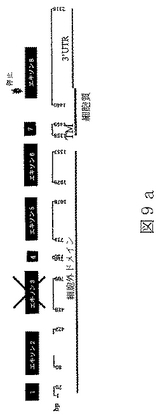
【図9b】
【図10】
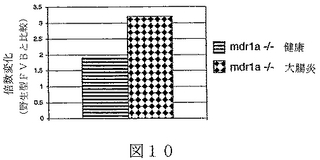
【図11】
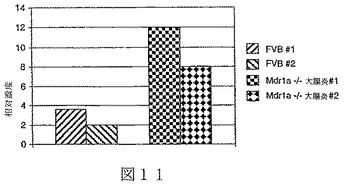
【図12a】
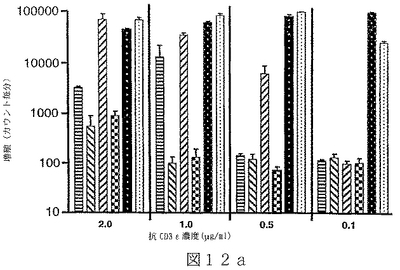
【図12b】
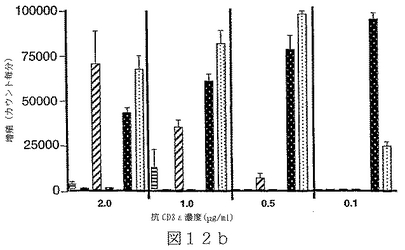
【図13】
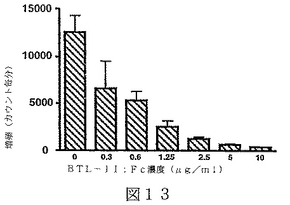
【図14】
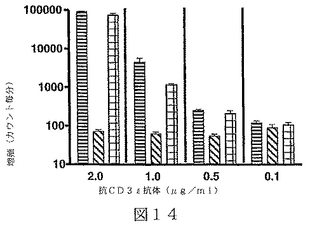
【図15】
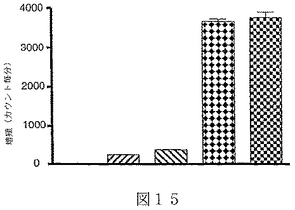
【図16a】
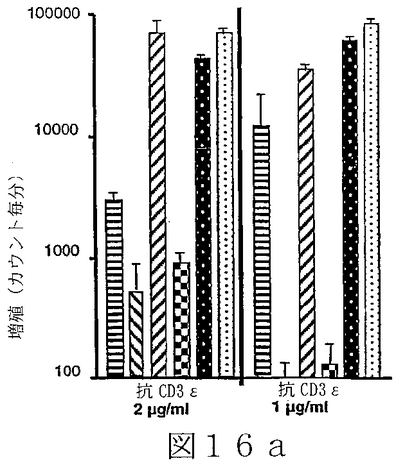
【図16b】
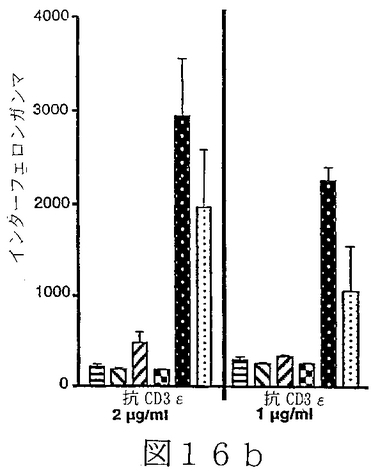
【図16c】
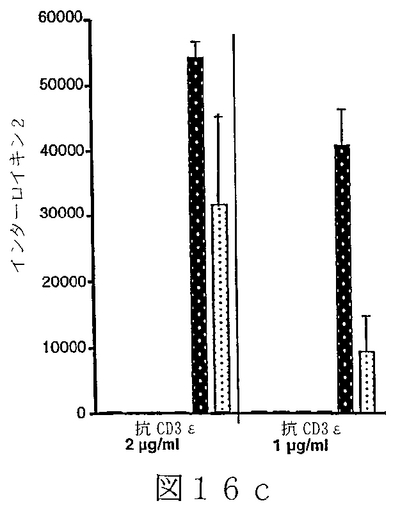
【図16d】
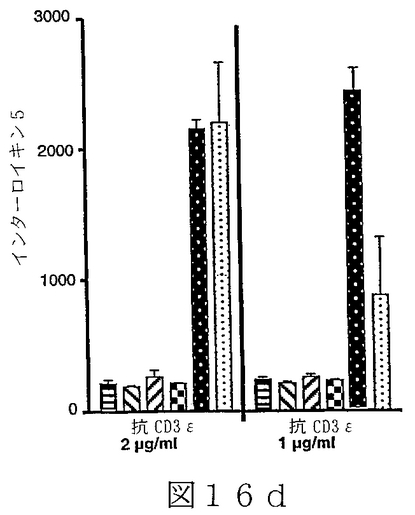
【図17】
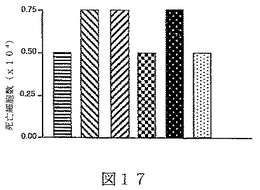
【図18】
【図2】
【図3a】
【図3b】
【図4a】
【図4b】
【図5a】
【図5b】
【図6a】
【図6b】
【図7a】
【図7b】
【図8】
【図9a】
【図9b】
【図10】
【図11】
【図12a】
【図12b】
【図13】
【図14】
【図15】
【図16a】
【図16b】
【図16c】
【図16d】
【図17】
【図18】
【公表番号】特表2006−525786(P2006−525786A)
【公表日】平成18年11月16日(2006.11.16)
【国際特許分類】
【出願番号】特願2004−563747(P2004−563747)
【出願日】平成15年12月19日(2003.12.19)
【国際出願番号】PCT/US2003/040338
【国際公開番号】WO2004/058986
【国際公開日】平成16年7月15日(2004.7.15)
【出願人】(591123609)イミュネックス・コーポレーション (24)
【氏名又は名称原語表記】IMMUNEX CORPORATION
【Fターム(参考)】
【公表日】平成18年11月16日(2006.11.16)
【国際特許分類】
【出願日】平成15年12月19日(2003.12.19)
【国際出願番号】PCT/US2003/040338
【国際公開番号】WO2004/058986
【国際公開日】平成16年7月15日(2004.7.15)
【出願人】(591123609)イミュネックス・コーポレーション (24)
【氏名又は名称原語表記】IMMUNEX CORPORATION
【Fターム(参考)】
[ Back to top ]