
C型肝炎ウイルスワクチン
【課題】HCVに対するHCV特異的細胞性免疫(CMI)応答を生じさせるために広範囲の抗原を与えるアデノベクター、またはDNAプラスミドワクチンの提供。
【解決手段】Ad6ベクター及び、不活性NS5B−RNA依存性RNAポリメラーゼ領域を含むMet−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチドをコードする核酸。1つ以上のAD6領域及びAD6中に存在しない領域を含む組換え核酸であって、少なくとも1つのAD6領域がE1A、E1B、E2B、E2A、E4、L1、L2、L4及びL5からなる群から選択される、前記組換え核酸。
【解決手段】Ad6ベクター及び、不活性NS5B−RNA依存性RNAポリメラーゼ領域を含むMet−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチドをコードする核酸。1つ以上のAD6領域及びAD6中に存在しない領域を含む組換え核酸であって、少なくとも1つのAD6領域がE1A、E1B、E2B、E2A、E4、L1、L2、L4及びL5からなる群から選択される、前記組換え核酸。
【発明の詳細な説明】
【背景技術】
【0001】
本明細書中で引用した文献は本発明に対する従来技術とは認められない。
【0002】
世界の人口の約3%がC型肝炎ウイルス(HCV)に感染している(Wasleyら,Semin.Liver Dis.,20:1−16(2000))。HCVに曝されると、まれに顕性急性疾患にかかるが、多くの場合ウイルスは肝炎症を引き起こす慢性感染となり、ゆっくりと肝不全や肝硬変に進行する(Iwarson,FEMS Microbiol.Rev.,14:201−204(1994))。更に、疫学的調査から、HCVが肝細胞癌の発病において重要な役割をなしていることがわかった(Kew,FEMS Microbiol.Rev.,14:211−220(1994);Alter,Blood,85:1681−1695(1995))。
【0003】
1992年にHCVの血液スクリーニングがルーチンに実施されるまで、多くの患者は汚染されている血液、血液製剤または移植臓器にうっかり曝されたことにより感染していた。HCVの血液スクリーニングが実施されている地域では、HCVは主に感染血液に直接経皮的に暴露して、すなわち静脈薬の使用により感染している。まれな伝播方法には、周生期暴露、血液透析及びHCV感染者との性的接触が含まれる(Alterら,N.Engl.J.Med.,341(8):556−562(1999);Alter,J.Hepatol.,31(補遺):88−91(1999);Semin,Liver Dis.,201:1−16(2000))。
【0004】
HCVゲノムは、約3000アミノ酸の前駆体ポリタンパク質をコードする一本鎖RNA約9.5kbから構成されている(Chooら,Science,244:362−364(1989);Chooら,Science,244:359−362(1989);Takamizawaら,J.Virol.,65:1105−1113(1991))。HCVポリタンパク質は、C−E1−E2−p7−NS2−NS3−NS4A−NS4B−NS5A−NS5Bの順でウイルスタンパク質を含む。
【0005】
各ウイルスタンパク質は、HCVポリタンパク質のタンパク質分解により産生される。宿主細胞プロテアーゼは推定構造タンパク質C、E1、E2及びp7を放出し、アミノ酸810でNS2のN末端を形成する(Mizushimaら,J.Virol.,68:2731−2734(1994);Hijikataら,P.N.A.S.USA,90:10773−10777(1993))。
【0006】
非構造タンパク質NS3、NS4、NS4B、NS5A及びNS5Bは多分ウイルス複製機構を形成し、ポリタンパク質から放出される。NS2及びNS3のN末端に関連する亜鉛依存性プロテアーゼはNS2とNS3の間の開裂に関与する(Grakouiら,J.Virol.,67:1385−1395(1993);Hijikataら,P.N.A.S.USA,90:10773−10777(1993))。NS3のN末端ドメイン中に位置する別のセリンプロテアーゼはNS3/NS4A、NS4A/NS4B、NS4B/NS5A及びNS5A/NS5B接合部でのタンパク質分解開裂に関与している(Bartenschlagerら,J.Virol.,67:3835−3844(1993);Grakouiら,Proc.Natl.Acad.Sci.USA,90:10583−10587(1993);Tomeiら,J.Virol.,67:4017−4026(1993))。NS4AはNS3活性に対するコファクターを与える(Faillaら,J.Virol.,68:3753−3760(1994);DeFrancescoらの米国特許第5,739,002号明細書)。
【0007】
NS5Aはインターフェロン耐性を与える高リン酸化タンパク質である(DeFrancescoら,Semin.Liver Dis.,20(1):69−83(2000);Pawlotsky,Viral Hepat.,補遺1:47−48(1999))。
【0008】
NS5BはRNA依存性RNAポリメラーゼを与える(DeFrancescoらの国際特許出願公開第96/37619号パンフレット;Behrensら,EMBO,15:12−22(1996);Lohmannら,Virology,249:108−118(1998))。
【発明の概要】
【0009】
本発明は、Ad6ベクター、及び不活性NS5B RNA依存性RNAポリメラーゼ領域を含むMet−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチドをコードする核酸に関する。前記核酸は、HCVに対するHCV特異的細胞性免疫(CMI)応答を生じさせるために広範囲の抗原を与えるアデノベクターまたはDNAプラスミドワクチンの成分として特に有用である。
【0010】
HCV特異的CMI応答は、HCV抗原を認識する細胞毒性Tリンパ球及びTヘルパー細胞の産生を指す。CMI応答には非−HCV特異的免疫効果も含まれ得る。
【0011】
好ましい核酸は、配列番号1に実質的に類似し、配列番号1中に存在するNS5B領域に実質的に類似する少なくとも1つのポリペプチドを産生するように自己プロセシングするのに十分なプロテアーゼ活性を有するMet−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチドをコードする。NS5Bに対応する産生されたポリペプチドは酵素的に不活性である。より好ましくは、HCVポリペプチドは配列番号1中に存在するNS3、NS4A、NS4B、NS5A及びNS5B領域に実質的に類似するポリペプチドを産生するのに十分なプロテアーゼ活性を有する。
【0012】
「実質的に類似の配列」とは、参照配列に対する同一性が少なくとも約65%であることを示す。よって、例えば、配列番号1に実質的に類似のアミノ酸配列を有するポリペプチドは配列番号1に対して少なくとも約65%の全アミノ酸同一性を有する。
【0013】
NS3、NS4A、NS4B、NS5A及びNS5Bに対応するポリペプチドは、配列番号1中の対応領域に対して少なくとも約65%のアミノ酸配列同一性を有する。前記対応領域は、本明細書中NS3、NS4A、NS4B、NS5A及びNS5Bポリペプチドとも称される。
【0014】
従って、本発明の第1態様は、配列番号1に実質的に類似のMet−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチドをコードするヌクレオチド配列を含む核酸に関する。コードされるポリペプチドは、酵素的に不活性なNS5Bポリペプチドを産生するように自己プロセツシングするのに十分なプロテアーゼ活性を有する。
【0015】
好ましい実施態様において、前記核酸は、所望ヒト細胞においてMet−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチドを発現し得る発現ベクターである。ヒト細胞内での発現は、HCV感染を積極的に治療したり、HCV感染に対して予防的に治療する治療用途を有する。
【0016】
発現ベクターは、ポリペプチドをコードするヌクレオチド配列と共に適正な転写及びプロセシングのために調節要素を含む。任意に存在する調節要素には、前記ポリペプチドをコードするヌクレオチド配列に元々関連している調節要素及び前記ヌクレオチド配列に元々関連していない外因性調節要素が含まれる。外因性プロモーターのような外因性調節要素はヒト細胞のような特定宿主における発現のために有用であり得る。機能的発現のために有用な調節要素の例には、プロモーター、ターミネーター、リボソーム結合部位及びポリアデニル化シグナルが含まれる。
【0017】
本発明の別の態様は、ヒト細胞において配列番号1に実質的に類似のMet−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチドを発現し得る遺伝子発現カセットを含む核酸に関する。前記ポリペプチドは、酵素的に不活性なNS5Bタンパク質を産生するように自己プロセツシングし得る。前記遺伝子発現カセットは、少なくとも
a)ポリペプチドをコードするヌクレオチド配列に転写的にカップリングしているプロモーター、
b)前記ヌクレオチド配列に機能的にカップリングしている5’リボソーム結合部位、
c)前記ヌクレオチド配列の3’末端に連結しているターミネーター、及び
d)前記ヌクレオチド配列に機能的にカップリングしている3’ポリアデニル化シグナル
を含む。
【0018】
「転写的にカップリング」とは、ヌクレオチド配列がプロモーターで結合しているRNAポリメラーゼにより転写され得るようにプロモーターが位置づけられていることを指す。転写的カップリングでは、転写される配列がプロモーターに隣接している必要がない。
【0019】
「機能的カップリング」とは、ヌクレオチド配列への効果を媒介し得る能力を指す。機能的カップリングでは、カップリングされる配列が相互に隣接している必要はない。ヌクレオチド配列に機能的にカップリングしている3’ポリアデニル化シグナルにより、転写RNAの開裂及びポリアデニル化が促進される。ヌクレオチド配列に機能的にカップリングしている5’リボソーム結合部位により、リボソーム結合が促進される。
【0020】
好ましい実施態様では、核酸はHCVを治療する治療用途に適したまたは治療用ベクターの作成における中間体として適したDNAプラスミドベクターまたはアデノベクターである。HCVの治療には、HCV感染の積極的治療及びHCV感染に対する予防的治療が含まれる。
【0021】
本発明の別の態様は、(a)相同的組換え及び(b)アデノベクターレスキューを含む方法により産生される、配列番号1に実質的に類似のポリペプチドを発現し得るMet−NS3−NS4A−NS4B−NS5A−NS5B発現カセットを含むアデノベクターである。相同組換え段階により、アデノウイルスゲノムプラスミドが生ずる。アデノベクターレスキュー段階により、アデノゲノムプラスミドからアデノベクターが生ずる。
【0022】
本明細書に記載のアデノウイルスゲノムプラスミドは、E1領域及び場合によりE3領域が欠失されており、前記欠失領域の1つに遺伝子発現カセットが挿入されている組換えアデノウイルスゲノムを含む。組換えアデノウイルスゲノムは1つ以上のアデノウイルス血清タイプに実質的に類似の領域から構成されている。
【0023】
本発明の別の態様は、配列番号4の核酸配列またはその誘導体からなるアデノベクターに関し、前記誘導体では配列番号4中に存在するHCVポリタンパク質コード化配列が配列番号3、配列番号10または配列番号11のいずれかのHCVポリタンパク質コード化配列で置換されている。
【0024】
本発明の別の態様は、配列番号1に実質的に類似のMet−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチドをコードする配列を含む核酸を含む培養組換え細胞に関する。前記組換え細胞は、ベクター構築方法において前記ポリペプチドをコードする核酸を複製するために使用されるような各種用途を有する。
【0025】
本発明の別の態様は、配列番号1に実質的に類似のポリペプチドを発現し得るMet−NS3−NS4A−NS4B−NS5A−NS5B発現カセットを含むアデノベクターの作成方法に関する。前記方法は、(a)E1領域及びE3領域が欠失されており、前記欠失領域の1つに遺伝子発現カセットが挿入されている組換えアデノウイルスゲノムを含むアデノウイルスゲノムプラスミドを作成する段階、及び(b)前記アデノウイルスゲノムプラスミドからアデノベクターをレスキューする段階を含む。
【0026】
本発明の別の態様は、配列番号1に実質的に類似のMet−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチドを発現させるためのベクター及び医薬的に許容され得る担体を含む医薬組成物に関する。前記ベクターは患者における投与及びポリペプチド発現に適している。
【0027】
「患者」は、HCVに感染し得る哺乳動物を指す。患者はHCVに感染していてもいなくてもよい。患者の例はヒト及びチンパンジーである。
【0028】
本発明の別の態様は、配列番号1に実質的に類似のMet−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチドを発現するベクターを有効量患者に投与することを含む患者の治療方法に関する。前記ベクターは患者における投与及びポリペプチド発現に適している。
【0029】
治療を受ける患者はHCVに感染していてもいなくてもよい。HCV感染されている患者の場合、有効量はHCVの複製能力の低下、HCV負荷の低減、ウイルスクリアランスの上昇及び1つ以上のHCV特異的CMI応答の上昇という効果の1つ以上を達成するのに十分な量である。HCVに感染していない患者の場合、有効量はHCV感染に対するHCV特異的CMI応答の1つ以上の成分を産生する能力の上昇、HCV感染への感受性の低下及び感染ウイルスが慢性疾患に対する持続感染を樹立する能力の低下という効果の1つ以上を達成するのに十分な量である。
【0030】
本発明の別の態様は、Ad6領域及びAd6中に存在しない領域を含む組換え核酸に関する。「組換え」核酸とは、元々相互に関連しない2つ以上の核酸領域が存在することを意味する。Ad6組換え核酸はAd6領域及びAd6に非相同のポリペプチドをコードする遺伝子発現カセットを含むことが好ましい。
【0031】
本発明の他の特徴及び作用効果は、本明細書に記載されている複数の実施例を含めた追加記載から明らかである。ここに提示した実施例は本発明を実施するのに有用な成分及び方法を例示している。これらの実施例は本発明を限定しない。本明細書の記載に基づいて、当業者は本発明を実施するのに有用な他の成分及び方法を同定、使用することができる。
【図面の簡単な説明】
【0032】
【図1A】配列番号1を示す。
【図1B】配列番号1を示す。
【図2A】配列番号2を示す。配列番号2は、配列番号1をコードするヌクレオチド配列と共に最適化内部リボソームエントリー部位及びTAAA終止を規定する。ヌクレオチド1〜6は最適化内部リボソームエントリー部位を規定する。ヌクレオチド7〜5961はNos.5137〜5145にヌクレオチドを有するHCV Met−NS3−NS4A−NS4B−NS5A−NS5Bペリペプチドをコードし、アミノ酸No.1711〜1713にNS5Bを不活性とするAlaAlaGLy配列を規定する。ヌクレオチド5962〜5965はTAAA終止を規定する。
【図2B】配列番号2を示す。配列番号2は、配列番号1をコードするヌクレオチド配列と共に最適化内部リボソームエントリー部位及びTAAA終止を規定する。ヌクレオチド1〜6は最適化内部リボソームエントリー部位を規定する。ヌクレオチド7〜5961はNos.5137〜5145にヌクレオチドを有するHCV Met−NS3−NS4A−NS4B−NS5A−NS5Bペリペプチドをコードし、アミノ酸No.1711〜1713にNS5Bを不活性とするAlaAlaGLy配列を規定する。ヌクレオチド5962〜5965はTAAA終止を規定する。
【図2C】配列番号2を示す。配列番号2は、配列番号1をコードするヌクレオチド配列と共に最適化内部リボソームエントリー部位及びTAAA終止を規定する。ヌクレオチド1〜6は最適化内部リボソームエントリー部位を規定する。ヌクレオチド7〜5961はNos.5137〜5145にヌクレオチドを有するHCV Met−NS3−NS4A−NS4B−NS5A−NS5Bペリペプチドをコードし、アミノ酸No.1711〜1713にNS5Bを不活性とするAlaAlaGLy配列を規定する。ヌクレオチド5962〜5965はTAAA終止を規定する。
【図2D】配列番号2を示す。配列番号2は、配列番号1をコードするヌクレオチド配列と共に最適化内部リボソームエントリー部位及びTAAA終止を規定する。ヌクレオチド1〜6は最適化内部リボソームエントリー部位を規定する。ヌクレオチド7〜5961はNos.5137〜5145にヌクレオチドを有するHCV Met−NS3−NS4A−NS4B−NS5A−NS5Bペリペプチドをコードし、アミノ酸No.1711〜1713にNS5Bを不活性とするAlaAlaGLy配列を規定する。ヌクレオチド5962〜5965はTAAA終止を規定する。
【図3A】配列番号3を示す。配列番号3は配列番号2のコドン最適化バージョンである。ヌクレオチド7〜5961がHCV Met−NS3−NS4A−NS4B−NS5A−NS5Bペリペプチドをコードする。
【図3B】配列番号3を示す。配列番号3は配列番号2のコドン最適化バージョンである。ヌクレオチド7〜5961がHCV Met−NS3−NS4A−NS4B−NS5A−NS5Bペリペプチドをコードする。
【図3C】配列番号3を示す。配列番号3は配列番号2のコドン最適化バージョンである。ヌクレオチド7〜5961がHCV Met−NS3−NS4A−NS4B−NS5A−NS5Bペリペプチドをコードする。
【図3D】配列番号3を示す。配列番号3は配列番号2のコドン最適化バージョンである。ヌクレオチド7〜5961がHCV Met−NS3−NS4A−NS4B−NS5A−NS5Bペリペプチドをコードする。
【図4A】MRK Ad6−NSmut(配列番号4)を示す。配列番号4は配列番号1のポリペプチドが配列番号2によりコードされる発現カセットを含むアデノベクターである。塩基対1〜450はAd5 bp1−450に対応し、塩基対462〜1252はヒトCMVプロモーターに対応し、塩基対1258〜1267はコザック配列に対応し、塩基対1264〜7222はNS遺伝子に対応し、塩基対7231〜7451はBGHポリアデニル化シグナルに対応し、塩基対7469〜9506はAd5塩基対3511−5548に対応し、塩基対9507〜32121はAd6塩基対5542−28156に対応し、塩基対32122〜35117はAd6塩基対30789−33784に対応し、塩基対35118〜37089はAd5塩基対33967−35935に対応する。
【図4B】MRK Ad6−NSmut(配列番号4)を示す。配列番号4は配列番号1のポリペプチドが配列番号2によりコードされる発現カセットを含むアデノベクターである。塩基対1〜450はAd5 bp1−450に対応し、塩基対462〜1252はヒトCMVプロモーターに対応し、塩基対1258〜1267はコザック配列に対応し、塩基対1264〜7222はNS遺伝子に対応し、塩基対7231〜7451はBGHポリアデニル化シグナルに対応し、塩基対7469〜9506はAd5塩基対3511−5548に対応し、塩基対9507〜32121はAd6塩基対5542−28156に対応し、塩基対32122〜35117はAd6塩基対30789−33784に対応し、塩基対35118〜37089はAd5塩基対33967−35935に対応する。
【図4C】MRK Ad6−NSmut(配列番号4)を示す。配列番号4は配列番号1のポリペプチドが配列番号2によりコードされる発現カセットを含むアデノベクターである。塩基対1〜450はAd5 bp1−450に対応し、塩基対462〜1252はヒトCMVプロモーターに対応し、塩基対1258〜1267はコザック配列に対応し、塩基対1264〜7222はNS遺伝子に対応し、塩基対7231〜7451はBGHポリアデニル化シグナルに対応し、塩基対7469〜9506はAd5塩基対3511−5548に対応し、塩基対9507〜32121はAd6塩基対5542−28156に対応し、塩基対32122〜35117はAd6塩基対30789−33784に対応し、塩基対35118〜37089はAd5塩基対33967−35935に対応する。
【図4D】MRK Ad6−NSmut(配列番号4)を示す。配列番号4は配列番号1のポリペプチドが配列番号2によりコードされる発現カセットを含むアデノベクターである。塩基対1〜450はAd5 bp1−450に対応し、塩基対462〜1252はヒトCMVプロモーターに対応し、塩基対1258〜1267はコザック配列に対応し、塩基対1264〜7222はNS遺伝子に対応し、塩基対7231〜7451はBGHポリアデニル化シグナルに対応し、塩基対7469〜9506はAd5塩基対3511−5548に対応し、塩基対9507〜32121はAd6塩基対5542−28156に対応し、塩基対32122〜35117はAd6塩基対30789−33784に対応し、塩基対35118〜37089はAd5塩基対33967−35935に対応する。
【図4E】MRK Ad6−NSmut(配列番号4)を示す。配列番号4は配列番号1のポリペプチドが配列番号2によりコードされる発現カセットを含むアデノベクターである。塩基対1〜450はAd5 bp1−450に対応し、塩基対462〜1252はヒトCMVプロモーターに対応し、塩基対1258〜1267はコザック配列に対応し、塩基対1264〜7222はNS遺伝子に対応し、塩基対7231〜7451はBGHポリアデニル化シグナルに対応し、塩基対7469〜9506はAd5塩基対3511−5548に対応し、塩基対9507〜32121はAd6塩基対5542−28156に対応し、塩基対32122〜35117はAd6塩基対30789−33784に対応し、塩基対35118〜37089はAd5塩基対33967−35935に対応する。
【図4F】MRK Ad6−NSmut(配列番号4)を示す。配列番号4は配列番号1のポリペプチドが配列番号2によりコードされる発現カセットを含むアデノベクターである。塩基対1〜450はAd5 bp1−450に対応し、塩基対462〜1252はヒトCMVプロモーターに対応し、塩基対1258〜1267はコザック配列に対応し、塩基対1264〜7222はNS遺伝子に対応し、塩基対7231〜7451はBGHポリアデニル化シグナルに対応し、塩基対7469〜9506はAd5塩基対3511−5548に対応し、塩基対9507〜32121はAd6塩基対5542−28156に対応し、塩基対32122〜35117はAd6塩基対30789−33784に対応し、塩基対35118〜37089はAd5塩基対33967−35935に対応する。
【図4G】MRK Ad6−NSmut(配列番号4)を示す。配列番号4は配列番号1のポリペプチドが配列番号2によりコードされる発現カセットを含むアデノベクターである。塩基対1〜450はAd5 bp1−450に対応し、塩基対462〜1252はヒトCMVプロモーターに対応し、塩基対1258〜1267はコザック配列に対応し、塩基対1264〜7222はNS遺伝子に対応し、塩基対7231〜7451はBGHポリアデニル化シグナルに対応し、塩基対7469〜9506はAd5塩基対3511−5548に対応し、塩基対9507〜32121はAd6塩基対5542−28156に対応し、塩基対32122〜35117はAd6塩基対30789−33784に対応し、塩基対35118〜37089はAd5塩基対33967−35935に対応する。
【図4H】MRK Ad6−NSmut(配列番号4)を示す。配列番号4は配列番号1のポリペプチドが配列番号2によりコードされる発現カセットを含むアデノベクターである。塩基対1〜450はAd5 bp1−450に対応し、塩基対462〜1252はヒトCMVプロモーターに対応し、塩基対1258〜1267はコザック配列に対応し、塩基対1264〜7222はNS遺伝子に対応し、塩基対7231〜7451はBGHポリアデニル化シグナルに対応し、塩基対7469〜9506はAd5塩基対3511−5548に対応し、塩基対9507〜32121はAd6塩基対5542−28156に対応し、塩基対32122〜35117はAd6塩基対30789−33784に対応し、塩基対35118〜37089はAd5塩基対33967−35935に対応する。
【図4I】MRK Ad6−NSmut(配列番号4)を示す。配列番号4は配列番号1のポリペプチドが配列番号2によりコードされる発現カセットを含むアデノベクターである。塩基対1〜450はAd5 bp1−450に対応し、塩基対462〜1252はヒトCMVプロモーターに対応し、塩基対1258〜1267はコザック配列に対応し、塩基対1264〜7222はNS遺伝子に対応し、塩基対7231〜7451はBGHポリアデニル化シグナルに対応し、塩基対7469〜9506はAd5塩基対3511−5548に対応し、塩基対9507〜32121はAd6塩基対5542−28156に対応し、塩基対32122〜35117はAd6塩基対30789−33784に対応し、塩基対35118〜37089はAd5塩基対33967−35935に対応する。
【図4J】MRK Ad6−NSmut(配列番号4)を示す。配列番号4は配列番号1のポリペプチドが配列番号2によりコードされる発現カセットを含むアデノベクターである。塩基対1〜450はAd5 bp1−450に対応し、塩基対462〜1252はヒトCMVプロモーターに対応し、塩基対1258〜1267はコザック配列に対応し、塩基対1264〜7222はNS遺伝子に対応し、塩基対7231〜7451はBGHポリアデニル化シグナルに対応し、塩基対7469〜9506はAd5塩基対3511−5548に対応し、塩基対9507〜32121はAd6塩基対5542−28156に対応し、塩基対32122〜35117はAd6塩基対30789−33784に対応し、塩基対35118〜37089はAd5塩基対33967−35935に対応する。
【図4K】MRK Ad6−NSmut(配列番号4)を示す。配列番号4は配列番号1のポリペプチドが配列番号2によりコードされる発現カセットを含むアデノベクターである。塩基対1〜450はAd5 bp1−450に対応し、塩基対462〜1252はヒトCMVプロモーターに対応し、塩基対1258〜1267はコザック配列に対応し、塩基対1264〜7222はNS遺伝子に対応し、塩基対7231〜7451はBGHポリアデニル化シグナルに対応し、塩基対7469〜9506はAd5塩基対3511−5548に対応し、塩基対9507〜32121はAd6塩基対5542−28156に対応し、塩基対32122〜35117はAd6塩基対30789−33784に対応し、塩基対35118〜37089はAd5塩基対33967−35935に対応する。
【図4L】MRK Ad6−NSmut(配列番号4)を示す。配列番号4は配列番号1のポリペプチドが配列番号2によりコードされる発現カセットを含むアデノベクターである。塩基対1〜450はAd5 bp1−450に対応し、塩基対462〜1252はヒトCMVプロモーターに対応し、塩基対1258〜1267はコザック配列に対応し、塩基対1264〜7222はNS遺伝子に対応し、塩基対7231〜7451はBGHポリアデニル化シグナルに対応し、塩基対7469〜9506はAd5塩基対3511−5548に対応し、塩基対9507〜32121はAd6塩基対5542−28156に対応し、塩基対32122〜35117はAd6塩基対30789−33784に対応し、塩基対35118〜37089はAd5塩基対33967−35935に対応する。
【図4M】MRK Ad6−NSmut(配列番号4)を示す。配列番号4は配列番号1のポリペプチドが配列番号2によりコードされる発現カセットを含むアデノベクターである。塩基対1〜450はAd5 bp1−450に対応し、塩基対462〜1252はヒトCMVプロモーターに対応し、塩基対1258〜1267はコザック配列に対応し、塩基対1264〜7222はNS遺伝子に対応し、塩基対7231〜7451はBGHポリアデニル化シグナルに対応し、塩基対7469〜9506はAd5塩基対3511−5548に対応し、塩基対9507〜32121はAd6塩基対5542−28156に対応し、塩基対32122〜35117はAd6塩基対30789−33784に対応し、塩基対35118〜37089はAd5塩基対33967−35935に対応する。
【図5A】配列番号5及び6を示す。配列番号5は活性RNA依存性RNAポリメラーゼを有するHCV Met−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチドをコードする。配列番号6はポリペプチドのアミノ酸配列を規定する。
【図5B】配列番号5及び6を示す。配列番号5は活性RNA依存性RNAポリメラーゼを有するHCV Met−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチドをコードする。配列番号6はポリペプチドのアミノ酸配列を規定する。
【図5C】配列番号5及び6を示す。配列番号5は活性RNA依存性RNAポリメラーゼを有するHCV Met−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチドをコードする。配列番号6はポリペプチドのアミノ酸配列を規定する。
【図5D】配列番号5及び6を示す。配列番号5は活性RNA依存性RNAポリメラーゼを有するHCV Met−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチドをコードする。配列番号6はポリペプチドのアミノ酸配列を規定する。
【図5E】配列番号5及び6を示す。配列番号5は活性RNA依存性RNAポリメラーゼを有するHCV Met−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチドをコードする。配列番号6はポリペプチドのアミノ酸配列を規定する。
【図5F】配列番号5及び6を示す。配列番号5は活性RNA依存性RNAポリメラーゼを有するHCV Met−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチドをコードする。配列番号6はポリペプチドのアミノ酸配列を規定する。
【図5G】配列番号5及び6を示す。配列番号5は活性RNA依存性RNAポリメラーゼを有するHCV Met−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチドをコードする。配列番号6はポリペプチドのアミノ酸配列を規定する。
【図5H】配列番号5及び6を示す。配列番号5は活性RNA依存性RNAポリメラーゼを有するHCV Met−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチドをコードする。配列番号6はポリペプチドのアミノ酸配列を規定する。
【図5I】配列番号5及び6を示す。配列番号5は活性RNA依存性RNAポリメラーゼを有するHCV Met−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチドをコードする。配列番号6はポリペプチドのアミノ酸配列を規定する。
【図5J】配列番号5及び6を示す。配列番号5は活性RNA依存性RNAポリメラーゼを有するHCV Met−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチドをコードする。配列番号6はポリペプチドのアミノ酸配列を規定する。
【図5K】配列番号5及び6を示す。配列番号5は活性RNA依存性RNAポリメラーゼを有するHCV Met−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチドをコードする。配列番号6はポリペプチドのアミノ酸配列を規定する。
【図5L】配列番号5及び6を示す。配列番号5は活性RNA依存性RNAポリメラーゼを有するHCV Met−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチドをコードする。配列番号6はポリペプチドのアミノ酸配列を規定する。
【図5M】配列番号5及び6を示す。配列番号5は活性RNA依存性RNAポリメラーゼを有するHCV Met−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチドをコードする。配列番号6はポリペプチドのアミノ酸配列を規定する。
【図5N】配列番号5及び6を示す。配列番号5は活性RNA依存性RNAポリメラーゼを有するHCV Met−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチドをコードする。配列番号6はポリペプチドのアミノ酸配列を規定する。
【図5O】配列番号5及び6を示す。配列番号5は活性RNA依存性RNAポリメラーゼを有するHCV Met−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチドをコードする。配列番号6はポリペプチドのアミノ酸配列を規定する。
【図6A】pV1JnsAの核酸配列(配列番号7)を規定する。
【図6B】pV1JnsAの核酸配列(配列番号7)を規定する。
【図6C】pV1JnsAの核酸配列(配列番号7)を規定する。
【図7A】Ad6ゲノムの核酸配列(配列番号8)を規定する。
【図7B】Ad6ゲノムの核酸配列(配列番号8)を規定する。
【図7C】Ad6ゲノムの核酸配列(配列番号8)を規定する。
【図7D】Ad6ゲノムの核酸配列(配列番号8)を規定する。
【図7E】Ad6ゲノムの核酸配列(配列番号8)を規定する。
【図7F】Ad6ゲノムの核酸配列(配列番号8)を規定する。
【図7G】Ad6ゲノムの核酸配列(配列番号8)を規定する。
【図7H】Ad6ゲノムの核酸配列(配列番号8)を規定する。
【図7I】Ad6ゲノムの核酸配列(配列番号8)を規定する。
【図7J】Ad6ゲノムの核酸配列(配列番号8)を規定する。
【図7K】Ad6ゲノムの核酸配列(配列番号8)を規定する。
【図7L】Ad6ゲノムの核酸配列(配列番号8)を規定する。
【図7M】Ad6ゲノムの核酸配列(配列番号8)を規定する。
【図7N】Ad6ゲノムの核酸配列(配列番号8)を規定する。
【図7O】Ad6ゲノムの核酸配列(配列番号8)を規定する。
【図8A】Ad5ゲノムの核酸配列(配列番号9)を規定する。
【図8B】Ad5ゲノムの核酸配列(配列番号9)を規定する。
【図8C】Ad5ゲノムの核酸配列(配列番号9)を規定する。
【図8D】Ad5ゲノムの核酸配列(配列番号9)を規定する。
【図8E】Ad5ゲノムの核酸配列(配列番号9)を規定する。
【図8F】Ad5ゲノムの核酸配列(配列番号9)を規定する。
【図8G】Ad5ゲノムの核酸配列(配列番号9)を規定する。
【図8H】Ad5ゲノムの核酸配列(配列番号9)を規定する。
【図8I】Ad5ゲノムの核酸配列(配列番号9)を規定する。
【図8J】Ad5ゲノムの核酸配列(配列番号9)を規定する。
【図8K】Ad5ゲノムの核酸配列(配列番号9)を規定する。
【図9】Ad6ゲノムの各種領域を示す。線状(35759bp)ds DNAゲノムを2つの平行線で示し、100地図単位に分けられている。転写単位をゲノム中の位置及び方位に関して示す。初期遺伝子(E1A、E1B、E2A/B、E3及びE4)を灰色矢印で示す。黒色矢印で示す後期遺伝子(L1〜L5)は主要後期プロモーター(MLP)から生ずる転写物の選択的スプライシングにより産生され、いずれもその5’末端に三連リーダー(1、2、3)を含む。E1領域は約1.0〜11.5地図単位に位置し、E2領域は75.0〜11.5地図単位に位置し、E3領域は76.1〜86.7地図単位に位置し、E4領域は99.5〜91.2地図単位に位置している。主要後期転写単位は16.0〜91.2地図単位に位置している。
【図10】Ad6領域及びAd5領域を含むpAdE1−E3+を回収するための相同組換えを示す。
【図11】Ad6領域を含むpAdE1−E3+を回収するための相同組換えを示す。
【図12】各種HCV NSカセットを発現するプラスミドDNAをトランスフェクトした293細胞からの全細胞抽出物に対するウェスタンブロットを示す。成熟NS3及びNS5A産物を特異的抗体で検出した。“pV1Jns−NS”は、Met−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチドが配列番号5によりコードされ、配列番号5が配列番号7の塩基1881〜1912に挿入されているpV1JnsAプラスミドを指す。“pV1Jns−NSmut”は配列番号2が配列番号7の塩基1882〜1925に挿入されているpV1JnsAプラスミドを指す。“pV1Jns−NSOPTmut”は配列番号3が配列番号7の塩基1881〜1905に挿入されているpV1JnsAプラスミドを指す。
【図13A】C57black6マウス(A)及びBalbCマウス(B)において25μg及び50μgの各種HCV NSカセットをコードするプラスミドDNAの2回注射及び遺伝子エレクトロトランスファー(GET)により誘導したT細胞応答をIFNγELIspotにより示す。
【図13B】C57black6マウス(A)及びBalbCマウス(B)において25μg及び50μgの各種HCV NSカセットをコードするプラスミドDNAの2回注射及び遺伝子エレクトロトランスファー(GET)により誘導したT細胞応答をIFNγELIspotにより示す。
【図14】HeLa細胞の感染時の各種アデノベクターからのタンパク質発現を示す。MRKAd5−NSmutは、Ad5ゲノムが塩基対451〜3510のE1欠失、塩基対28134〜30817のE3欠失を有し、配列番号4の塩基対451〜7468のNS3−NS4A−NS4B−NS5A−NS5B発現カセットがNo.450〜3511に挿入されているAd5配列(配列番号9)をベースとするアデノベクターである。Ad5−NSは、塩基対342〜3523のE1欠失及び塩基対28134〜30817のE3欠失を有するAd5骨格をベースとし、配列番号5からのNS3−NS4A−NS4B−NS5A−NS5Bをコードする発現カセットを含むアデノベクターである。“MRKAd6−NSOPTmut”は、配列番号4の塩基1258〜7222が配列番号3で置換されている修飾配列番号4を有するアデノベクターを指す。
【図15】C57black6マウスにおいて109vpの各種HCV非構造遺伝子カセットを含むアデノベクターを2回注射することにより誘導したT細胞応答をINFγELIspotにより示す。
【図16A】アカゲザルにおいて1010vp(A)または1011vp(B)の各種HCV非構造遺伝子カセットを含むアデノベクターを1回または2回注射することにより誘導したT細胞応答をINFγELIspotにより示す。
【図16B】アカゲザルにおいて1010vp(A)または1011vp(B)の各種HCV非構造遺伝子カセットを含むアデノベクターを1回または2回注射することにより誘導したT細胞応答をINFγELIspotにより示す。
【図16C】アカゲザルにおいて1010vp(A)または1011vp(B)の各種HCV非構造遺伝子カセットを含むアデノベクターを1回または2回注射することにより誘導したT細胞応答をINFγELIspotにより示す。
【図16D】アカゲザルにおいて1010vp(A)または1011vp(B)の各種HCV非構造遺伝子カセットを含むアデノベクターを1回または2回注射することにより誘導したT細胞応答をINFγELIspotにより示す。
【図17A】アカゲザルにおいて1010vp(A)または1011vp(B)の各種HCV非構造遺伝子カセットを含むアデノベクターを2回注射することにより誘導したCD8+ T細胞応答をINFγICSにより示す。
【図17B】アカゲザルにおいて1010vp(A)または1011vp(B)の各種HCV非構造遺伝子カセットを含むアデノベクターを2回注射することにより誘導したCD8+ T細胞応答をINFγICSにより示す。
【図18A】アカゲザルにおいて1011vpのAd5−NS(A)、MRKAd5−NSmut(B)またはMRKAd6−NSmut(C)を2回注射することにより誘導したT細胞応答をバルクCTLアッセイにより示す。
【図18B】アカゲザルにおいて1011vpのAd5−NS(A)、MRKAd5−NSmut(B)またはMRKAd6−NSmut(C)を2回注射することにより誘導したT細胞応答をバルクCTLアッセイにより示す。
【図18C】アカゲザルにおいて1011vpのAd5−NS(A)、MRKAd5−NSmut(B)またはMRKAd6−NSmut(C)を2回注射することにより誘導したT細胞応答をバルクCTLアッセイにより示す。
【図18D】アカゲザルにおいて1011vpのAd5−NS(A)、MRKAd5−NSmut(B)またはMRKAd6−NSmut(C)を2回注射することにより誘導したT細胞応答をバルクCTLアッセイにより示す。
【図18E】アカゲザルにおいて1011vpのAd5−NS(A)、MRKAd5−NSmut(B)またはMRKAd6−NSmut(C)を2回注射することにより誘導したT細胞応答をバルクCTLアッセイにより示す。
【図18F】アカゲザルにおいて1011vpのAd5−NS(A)、MRKAd5−NSmut(B)またはMRKAd6−NSmut(C)を2回注射することにより誘導したT細胞応答をバルクCTLアッセイにより示す。
【図19】プラスミドpE2を示す。
【図20A】部分コドン最適化配列NSsuboptmut(配列番号10)を示す。Met−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチドのコード配列は塩基7〜5961である。
【図20B】部分コドン最適化配列NSsuboptmut(配列番号10)を示す。Met−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチドのコード配列は塩基7〜5961である。
【図20C】部分コドン最適化配列NSsuboptmut(配列番号10)を示す。Met−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチドのコード配列は塩基7〜5961である。
【図20D】部分コドン最適化配列NSsuboptmut(配列番号10)を示す。Met−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチドのコード配列は塩基7〜5961である。
【発明を実施するための形態】
【0033】
本発明は、Ad6ベクター、及び不活性NS5B領域を含むMet−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチドをコードする核酸に関する。不活性NS5B領域を設けると、活性ウイルス性RNAポリメラーゼによる副作用の可能性を低下させながらNS5B抗原が供給される。本発明の核酸の使用には、HCVに対するCMI応答を生じさせるべく広範囲の抗原を与えるHCVポリペプチドを細胞に導入するためのワクチン成分として及び前記ワクチン成分を作成するための中間体としての使用が含まれる。
【0034】
適応細胞免疫応答は、主要組織適合複合体(MHC)クラスI及びII発現が偏在分布しているために身体中のHCV感染細胞中でウイルス抗原を認識するように、免疫記憶を誘導するように、及び免疫記憶を維持するように機能し得る。これらの機能は抗原特異的CD4+ Tヘルパー(Th)及びCD8+ 細胞毒性T細胞(CTL)に起因する。
【0035】
特異的T細胞受容体を介して活性化すると、HCV特異的Th細胞は各種の免疫調節機能を実行し、その機能の多くはTh1及びTh2サイトカインにより媒介されている。HCV特異的Th細胞はB細胞の活性化及び分化、ウイルス特異的細胞毒性T細胞の誘導及び刺激を助ける。CTLと一緒になって、Th細胞は数種のウイルスの複製及び遺伝子発現を抑制するIFN−γ及びIFN−αも分泌し得る。更に、主要エフェクター細胞であるTh細胞及びCTLはウイルス感染細胞のアポトーシス及び溶解を誘導し得る。
【0036】
HCV特異的CTLは抗原提示細胞(pAPC)によりプロセシングされる抗原から生ずる。抗原はpAPC内で合成されたり、pAPCに導入されることもある。pAPC内での抗原合成は抗原をコードする発現カセットを細胞に導入することにより生起され得る。
【0037】
核酸ワクチンの好ましい投与ルートは筋肉内ルートである。筋肉内投与すると、核酸は体細胞及びpAPCへ導入、発現されるであろう。体細胞で産生されたHCV抗原はMHCクラスI分子の関係での提示のためにpAPCに移動し得る(Donnellyら,Annu.Rev.Immunol.,15:617−648(1997))。
【0038】
pAPCはより長い抗原をプロテアソーム複合体の形態のより小さいペプチド抗原にプロセシングする。前記抗原は、MHCクラスIタンパク質と結合するために小胞体/ゴルジ複合体分泌経路に転座する。CD8+ Tリンパ球は、T細胞受容体(TCR)及びCD8細胞表面タンパク質を介してクラス1 MHCに結合した抗原を認識する。
【0039】
Met−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチドをコードする核酸をワクチン成分として使用すると、単一ベクターからCMI応答を発生し得る広範囲の抗原を産生できる。前記ポリペプチドは少なくともNS5Bに対応する領域を生ずるように十分に自己プロセシングし得なければならない。好ましい核酸は、配列番号1中に存在するNS3、NS4A、NS4B、NS5A及びNS5B領域に実質的に類似している各HCVポリペプチドを産生するように自己プロセシングするのに十分なプロテアーゼ活性を有し、配列番号1に実質的に類似のアミノ酸配列をコードする。
【0040】
細胞中で自己プロセシングするのに十分なプロテアーゼ活性を有する配列番号1に実質的に類似のポリペプチドは、細胞に幾種のHCV株中に存在するT細胞エピトープを与える。プロテアーゼ活性はNS3及びNS3/NS4Aタンパク質により与えられ、適当な開裂部位でMet−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチドを消化すると、NS3、NS4A、NS4B、NS5A及びNS5Bに対応するポリペプチドを放出する。Met−NS3−NS4A−NS4B−NS5A−NS5Bの自己プロセシングにより、天然に存在するHCVポリペプチドに似ているポリペプチドが生ずる。
【0041】
本明細書に記載されているガイダンスに基づいて、患者において有利な効果を達成するのに十分に強い免疫応答を生じ得る。ここに記載されているガイダンスには、HCV配列選択、ベクター選択、ベクター作成、併用療法及び投与に関する情報が含まれている。
【0042】
I. HCV配列
多種多様の核酸配列が細胞にHCV Met−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチドを供給するためのワクチン成分として、またはワクチン成分を作成するための中間体として使用され得る。適当な核酸配列を得るための出発点としては、不活性NS5Bを産生するように修飾された天然NS3−NS4A−NS4B−NS5A−NS5Bポリペプチド配列が好ましい。
【0043】
CMI応答を生ずるようにHCV非構造抗原を与えるHCV核酸配列の使用は、Choら,Vaccine,17:1136−1144(1999);Paliardらの国際特許出願公開第01/30812号パンフレット(本発明に対する従来技術とは認められない)及びCoitらの国際特許出願公開第01/38360号パンフレット(本発明に対する従来技術とは認められない)に記載されている。前記文献は、例えば不活性NS5Bを産生するように自己プロセシングするポリペプチドもHCV配列とデリバリーベヒクルの特定組合せも全く記載していない。
【0044】
HCV Met−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチド配列への修飾は、コード化核酸を改変することによりなされ得る。欠失、挿入及び置換が生ずるように改変を実施し得る。
【0045】
小さな修飾はNS5B中で本質的に複製のためにモチーフを標的することにより不活性ポリメラーゼを生ずるようになされ得る。NS5B活性にとって重要なモチーフ及び不活性NS5Bを産生するようになされ得る修飾の例は、Lohmannら,Journal of Virology,74:8416−8426(1997)及びKolykhalovら,Journal of Virology,74:2046−2051(2000)に記載されている。
【0046】
HCV Met−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチド配列に修飾を加えるときに考慮すべき別の要因には、自己プロセシングする能力の維持及びT細胞抗原の維持が含まれる。HCVポリペプチドの自己プロセシング能力は、大体機能性NS3プロテアーゼにより決定される。NS3活性プロテアーゼ活性を維持する修飾は、NS3タンパク質、NS3に対するコファクターとして作用するNS4A、及びNS3−NS4A−NS4B−NS5A−NS5Bポリペプチド内に存在するNS3プロテアーゼ認識部位を考慮して得られ得る。
【0047】
広範囲のT細胞応答を誘発し得るポリペプチドを産生するために天然NS3−NS4A−NS4B−NS5A−NS5Bポリペプチド配列に対して各種修飾が加えられ得る。広範囲のT細胞応答を誘発するポリペプチドの能力に影響を及ぼす因子には、HCV特異的T細胞抗原領域の保存または導入、各種HCV単離物におけるT細胞抗原領域の優勢が含まれる。
【0048】
天然HCV単離物の多数の例が当業界で公知である。HCV単離物は1つ以上のサブタイプを含む以下の6つの主遺伝子型に分類され得る:HCV−1/(1a,1b,1c)、HCV−2/(2a,2b,2c)、HCV−3/(3a,3b,10c)、HCV−4/(4a)、HCV−5/(5a)及びHCV−6/(6a,6b,7b,8b,9b,11a)(Simmonds,J.Gen.Virol.,693−712(2001))。HCV−BK、HCV−J、HCV−N、HCV−Hのような特定HCV配列の例はGenBankに寄託されており、複数の文献に記載されている(例えば、Chamberlianら,J.Gen.Virol.,1341−1347(1997)参照)。
【0049】
HCV T細胞抗原は、例えば経験的実験により同定され得る。T細胞抗原を同定する1つの方法は、長ポリペプチドから一連の重複短ペプチドを作成し、その後ポジティブクローンについて感染患者からのT細胞集団をスクリーニングすることを含む。ポジティブクローンは特定ペプチドにより活性化/初期化される。ペプチド活性を測定するためにIFNγ−ELISPOT、IFNγ−細胞内染色及びバルクCTLアッセイのような方法を用いることができる。こうして同定されたペプチドは各病原のT細胞エピトープを表すと見做され得る。
【0050】
各種HCV単離物由来のHCV T細胞抗原領域は、例えば2つ以上の天然配列由来の領域を含むハイブリッドNS3−NS4A−NS4B−NS5A−NS5Bポリペプチドを作成することにより1つの配列に導入され得る。前記ハイブリッドは追加の修飾を含み得、この修飾がHCV CMI応答を生ずるポリペプチドの能力を低下させないことが好ましい。
【0051】
自己プロセシングし、CMI応答を生じさせる修飾Met−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチドの能力は、本明細書に記載されている方法または当業界で公知の方法により測定され得る。その方法には、HIV特異的CMI応答を測定するためのIFNγ−ELISPOT、IFNγ−細胞内染色及びバルクCTLアッセイの使用が含まれる。
【0052】
(A) Met−NS3−NS4A−NS4B−NS5A−NS5B配列
配列番号1は、好ましいMet−NS3−NS4A−NS4B−NS5A−NS5B配列を規定する。配列番号1は、数種のHCV単離物中に存在する多数のHCV特異的T細胞抗原を含む。配列番号1はHCV BK株ヌクレオチド配列(GenBank受託番号M58335)のNS3−NS4A−NS4B−NS5A−NS5B部分に類似している。
【0053】
配列番号1において、MHCクラスI分子による認識のために重要なアンカー位置は保存されているか、またはHCVポリタンパク質のNS3−NS4A−NS4B−NS5A−NS5B部分中の20個の公知T細胞エピトープのうちの18個の保存的置換を表す。残りの2個の公知T細胞エピトープのうち、1個のエピトープは異なるHLAスーパータイプにより認識され得る配列番号1中の非保存的アンカー置換を有し、1個のエピトープは保存されない1つのアンカー残基を有する。HCV T細胞エピトープはChisariら,Curr.Top.Microbiol.Immunol.,242:299−325(2000)及びLechnerら,J.Exp.Med.,9:1499−1512(2000)に記載されている。
【0054】
HCV BK NS3−NS4A−NS4B−NS5A−NS5Bヌクレオチド配列と配列番号1の違いとして、配列番号1中には5’末端にメチオニンが導入されており、修飾NS5B活性部位残基が存在していることが挙げられる。修飾により、GlyAspAspをAlaAlaGly(残基1711〜1713)で置換するとNS5Bが不活化される。
【0055】
コードされるHCV Met−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチドが配列番号1に実質的に類似のアミノ酸配列を有していることが好ましい。別の実施態様では、コードされるHCV Met−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチドは配列番号1に対して少なくとも65%、少なくとも75%、少なくとも85%、少なくとも95%、少なくとも99%または100%のアミノ酸同一性を有しているか、または配列番号1と1〜2、1〜3、1〜4、1〜5、1〜6、1〜7、1〜8、1〜9、1〜10、1〜11、1〜12、1〜13、1〜14、1〜15、1〜16、1〜17、1〜18、1〜19または1〜20アミノ酸だけ異なっている。
【0056】
Met−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチドと配列番号1のアミノ酸の違いは、2つの配列が異なるアミノ酸修飾の最低数を調べることにより計算される。アミノ酸修飾は欠失、付加、置換またはその組合せであり得る。
【0057】
アミノ酸配列同一性は、1つのポリペプチドのアミノ酸配列を第2ポリペプチドのアミノ酸配列と比較して、配列アラインメントを作成する当業界で公知の方法により測定される。アミノ酸同一性は、同一アミノ酸を有するアライン残基対の数をカウントすることにより前記アラインメントから算出される。
【0058】
配列同一性の測定方法には、G.D.Schuler,「生命情報科学:遺伝子及びタンパク質の分析に対する実践ガイド(Bioinformatics: A Pracitical Guide to the Analysis of Genes and Proteins)」,A.D.Baxevanis及びB.F.F.Ouelette編,John Wiley & Sons,Inc.(2001年)発行;Yonaら,「生命情報科学:配列、構造及びデータバンク(Bioinformatiacs: Sequence, structure and databanks)」,D.Higgins及びW.Taylor編,オックスフォード大学出版(2000年)発行;及び「生命情報科学:配列及びゲノム分析(Bioinformatics: Sequence and Genome analysis)」,D.W.Mount編,Cold Spring Harbor Laboratory Press(2001年)発行に記載されているものが含まれる。アミノ酸配列同一性を測定するための方法は、GAP(ウィスコンシン・パッケージ バージョン10.2;ウィスコンシン州マディソンに所在のGenetics Computer Group(GCG))、BLAST(Altschulら,J.Mol.Biol.,215(3):403−10(1990))及びFASTA(Pearson,Methods in Enzymology,R.F.Doolittle編,183:63−98(1990))のような公表されているコンピュータープログラムで体系化される。
【0059】
本発明の実施態様において、2つのポリペプチド間の配列同一性はGAPプログラム(ウィスコンシン・パッケージ バージョン10.2;ウィスコンシン州マディソンに所在のGenetics Computer Group(GCG))を用いて測定される。GAPはNeedleman及びWunschのアラインメント方法(Needlemanら,J.Mol.Biol.,48:443−453(1970))を利用している。GAPは2つの配列間の全ての可能性あるアラインメント及びギャップ位置を考察し、マッチド残基の数を最大限とする一方ギャップの数及び大きさを最小限とするグローバルアラインメントを作成する。シンボルマッチの値を割り当てるためにスコアリングマトリックスを用いる。加えて、アラインメントへのギャップの挿入を制限するためにはギャップ作成ペナルテイー及びギャップ延長ペナルティーが必要である。GAPを用いるポリペプチド比較のためのデフォルトプログラムパラメーターは、BLOSUM62(Henikoffら,Proc.Natl.Acad.Sci.USA,89:10915−10919(1992))アミノ酸スコアリングマトリックス(MATrix=blosum62.cmp)、ギャップ作成パラメーター(GAPweight=8)及びギャップ延長パラメーター(LENgthweight=2)である。
【0060】
全長に亘って配列番号1に実質的に類似していることに加えてより好ましいHCV Met−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチドは、配列番号1中に存在する対応領域に対して実質的に類似の各NS3、NS4A、NS4B、NS5A及びNS5B領域を生ずる。配列番号1中の対応領域は、Met−NS3アミノ酸1〜632、NS4Aアミノ酸633〜686、NS4Bアミノ酸687〜947、NS5Aアミノ酸948〜1394及びNS5Bアミノ酸1395〜1985である。
【0061】
別の実施態様では、NS3、NS4A、NS4B、NS5A及び/またはNS5B領域は配列番号1の対応領域に対して少なくとも65%、少なくとも75%、少なくとも85%、少なくとも95%、少なくとも99%または100%のアミノ酸同一性を有しているか、または1〜2、1〜3、1〜4、1〜5、1〜6、1〜7、1〜8、1〜9、1〜10、1〜11、1〜12、1〜13、1〜14、1〜15、1〜16、1〜17、1〜18、1〜19または1〜20アミノ酸だけ異なっている。
【0062】
好ましくは、配列番号1に対するアミノ酸修飾はT細胞抗原領域の全てまたは殆どを維持している。天然アミノ酸の違いは異なるアミノ酸側鎖(R基)による。R基はアミノ酸の諸特性、例えば物理的サイズ、電荷及び疎水性に影響を及ぼす。アミノ酸は以下のように複数のグループに分類され得る:中性で疎水性(アラニン、バリン、ロイシン、イソロイシン、プロリン、トリプトファン、フェニルアラニン及びメチオニン);中性で極性(グリシン、セリン、スレオニン、チロシン、システイン、アスパラギン及びグルタミン);塩基性(リシン、アルギニン及びヒスチジン);及び酸性(アスパラギン酸及びグルタミン酸)。
【0063】
通常、アミノ酸を置換するときには類似の特性を有するアミノ酸を交換することが好ましい。特定グループ内でアミノ酸を置換する、例えばロイシンをバリン、リシンをアルギニン、グルタミンをアスパラギンで置換することはポリペプチド3次構造を変化させない良好な候補である。
【0064】
特定のアミノ酸配列及び公知の遺伝子コードの縮重から出発して、多種多様のコード化核酸配列を得ることができる。遺伝子コードの縮重は、殆どすべてのアミノ酸がヌクレオチドトリプレット、すなわちコドンのいろいろな組合せによりコードされるために生ずる。特定コドンの特定アミノ酸への翻訳は当業界で公知である(例えば、Lewin,GENES IV,p.119,オックスフォード大学出版(1990年)発行参照)。アミノ酸は以下のようにコドンによりコードされる:
A=Ala=アラニン:コドン GCA,GCC,GCG,GCU;
C=Cys=システイン:コドン UGC,UGU;
D=Asp=アスパラギン酸:コドン GAC,GAU;
E=Glu=グルタミン酸:コドン GAA,GAG;
F=Phe=フェニルアラニン:コドン UUC,UUU;
G=Gly=グリシン:コドン GGA,GGC,GGG,GGU;
H=His=ヒスチジン:コドン CAC,CAU;
I=Ile=イソロイシン:コドン AUA,AUC,AUU;
K=Lys=リシン:コドン AAA,AAG;
L=Leu=ロイシン:コドン UUA,UUG,CUA,CUC,CUG,CUU;
M=Met=メチオニン:コドン AUG;
N=Asn=アスパラギン:コドン AAC,AAU;
P=Pro=プロリン:コドン CCA,CCC,CCG,CCU;
Q=Gln=グルタミン:コドン CAA,CAG;
R=Arg=アルギニン:コドン AGA,AGG,CGA,CGC,CGG,CGU;
S=Ser=セリン:コドン AGC,AGU,UCA,UCC,UCG,UCU;
T=Thr=スレオニン:コドン ACA,ACC,ACG,ACU;
V=Val=バリン:コドン GUA,GUC,GUG,GUU;
W=Trp=トリプトファン:コドン UCG;及び
Y=Tyr=チロシン:コドン UAC,UAU。
【0065】
核酸配列は、宿主における発現を強化しようとして最適化され得る。考慮すべき要因子には、G:C含量、好ましいコドン及び抑制性第2構造の回避が含まれる。これらの要因は特定宿主での高い発現を有する核酸配列を得るべくいろいろな方法で組合され得る(例えば、Donnellyらの国際特許出願公開第97/47358号パンフレット参照)。
【0066】
特定配列が特定宿主において高い発現を有する能力には幾つかの経験的実験を要する。前記実験には見込みのある核酸配列の発現を測定し、所要により前記配列を変化させることが含まれる。
【0067】
(B) コード化ヌクレオチド配列
配列番号2及び3は、Met−NS3−NS4A−NS4B−NS5A−NS5B配列をコードするヌクレオチド配列の2つの例を規定する。配列番号2のコード配列は、天然に存在するHCV−BK配列(GenBank受託番号M58335)のNS3−NS4A−NS4B−NS5A−NS5B領域に類似している(99.4%ヌクレオチド配列同一性)。配列番号3は配列番号2のコドン最適化バージョンである。配列番号2及び3は78.3%のヌクレオチド配列同一性を有する。
【0068】
HCV−BK NS3−NS4A−NS4B−NS5A−NS5Bヌクレオチド(GenBank受託番号M58335)と配列番号2の違いとして、配列番号2がリボソーム結合部位、ATGメチオニンコドン、修飾NS5B触媒ドメインをコードする領域、TAAAストップコドン及び更に30個のヌクレオチドの違いが挙げられる。修飾触媒ドメインはGlyAspAspではなくAlaAlaGly(残基1711〜1713)をコードして、NS5Bを不活化する。
【0069】
HCV Met−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチドをコードするヌクレオチド配列は、好ましくは配列番号2コード領域に実質的に類似している。別の実施態様において、HCV Met−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチドをコードするヌクレオチド配列は、配列番号2コード領域に対して少なくとも65%、少なくとも75%、少なくとも85%、少なくとも95%、少なくとも99%または100%のヌクレオチド配列同一性を有しているか、または配列番号2と1〜2、1〜3、1〜4、1〜5、1〜6、1〜7、1〜8、1〜9、1〜10、1〜11、1〜12、1〜13、1〜14、1〜15、1〜16、1〜17、1〜18、1〜19、1〜20、1〜25、1〜30、1〜35、1〜40、1〜45または1〜50ヌクレオチドだけ異なっている。
【0070】
Met−NS3−NS4A−NS4B−NS5A−NS5Bをコードする配列と配列番号2コード領域のヌクレオチドの違いは、2つの配列が異なるヌクレオチド修飾の最低数を調べることにより計算される。ヌクレオチド修飾は欠失、付加、置換またはその組合せであり得る。
【0071】
ヌクレオチド配列同一性は、1つの配列のヌクレオチド配列を第2配列のヌクレオチド配列と比較し、配列アラインメントを作成する当業界で公知の方法により測定される。配列同一性は、アラインさせた同一ヌクレオチドを有する位置の数をカウントすることにより前記アラインメントから算出される。
【0072】
2つのポリヌクレオチド間のヌクレオチド配列同一性の測定方法には、Schuler,「生命情報科学:遺伝子及びタンパク質の分析に対する実践ガイド(Bioinformatics: A Pracitical Guide to the Analysis of Genes and Proteins)」,A.D.Baxevanis及びB.F.F.Ouelette編,John Wiley & Sons,Inc.(2001年)発行;Yonaら,「生命情報科学:配列、構造及びデーターバンク(Bioinformatiacs: Sequence, structure and databanks)」,D.Higgins及びW.Taylor編,オックスフォード大学出版(2000年)発行;及び「生命情報科学:配列及びゲノム分析(Bioinformatics: Sequence and Genome analysis)」,D.W.Mount編,Cold Spring Harbor Laboratory Press(2001年)発行に記載されているものが含まれる。ヌクレオチド配列同一性を測定するための方法は、GAP(ウィスコンシン・パッケージ バージョン10.2;ウィスコンシン州マディソンに所在のGenetics Computer Group(GCG))、BLAST(Altschulら,J.Mol.Biol.,215(3):403−10(1990))及びFASTA(W.R.Person,Methods in Enzymology,R.F.Doolittle編,183:63−98(1990))のような公表されているコンピュータープログラムで体系化される。
【0073】
本発明の実施態様において、2つのポリヌクレオチド間の配列同一性はGAPプログラム(ウィスコンシン・パッケージ バージョン10.2;ウィスコンシン州マディソンに所在のGenetics Computer Group(GCG))を適用して測定される。GAPはNeedleman及びWunchのアラインメント方法(Needlemanら,J.Mol.Biol.,48:443−453(1970))を利用している。GAPは2つの配列間の全ての考えられるアラインメント及びギャップ位置を考察し、マッチド残基の数を最大限とする一方ギャップの数及び大きさを最小限とするグローバルアラインメントを形成する。シンボルマッチの値を割り当てるためにスコアリングマトリックスを用いる。加えて、アラインメントへのギャップの挿入を制限するためにはギャップ作成ペナルテイー及びギャップ延長ペナルティーが必要である。GAPを用いるポリペプチド比較のためのデフォルトプログラムパラメーターは、nwsgapda.cmpスコアリングマトリックス(MATrix=nwsgapda.cmp)、ギャップ作成パラメーター(GAPweight=50)及びギャップ延長パラメーター(LENgthweight=3)である。
【0074】
全長に亘って実質的に類似していることに加えてより好ましいHCV Met−NS3−NS4A−NS4B−NS5A−NS5Bヌクレオチド配列により、配列番号2中に存在する対応領域に対して実質的に類似の各NS3、NS4A、NS4B、NS5A及びNS5B領域が生ずる。配列番号2中の対応領域は、Met−NS3ヌクレオチド7〜1902、NS4Aヌクレオチド1903〜2064、NS4Bヌクレオチド2065〜2847、NS5Aヌクレオチド2848〜4188及びNS5Bヌクレオチド4189〜5661である。
【0075】
別の実施態様では、NS3、NS4A、NS4B、NS5A及び/またはNS5Bコード化領域は配列番号2中の対応領域に対して少なくとも65%、少なくとも75%、少なくとも85%、少なくとも95%、少なくとも99%または100%のヌクレオチド配列同一性を有しているか、または配列番号2と1〜2、1〜3、1〜4、1〜5、1〜6、1〜7、1〜8、1〜9、1〜10、1〜11、1〜12、1〜13、1〜14、1〜15、1〜16、1〜17、1〜18、1〜19、1〜20、1〜25、1〜30、1〜35、1〜40、1〜45または1〜50個のヌクレオチドだけ異なっている。
【0076】
(C) 遺伝子発現カセット
遺伝子発現カセットはポリペプチド発現に必要な要素を含んでいる。「ポリペプチド」はサイズを限定せず、タンパク質も含む。遺伝子発現カセット中に存在する調節要素は、通常(a)ポリペプチドをコードするヌクレオチド配列に転写的にカップリングされているプロモーター、(b)ヌクレオチド配列に機能的にカップリングされている5’リボソーム結合部位、(c)ヌクレオチド配列の3’末端に結合されているターミネーター、及び(d)ヌクレオチド配列に機能的にカップリングされている3’ポリアデニル化シグナルを含む。遺伝子発現またはポリペプチドプロセシングを強化または調節するのに有用な追加の調節要素を存在させてもよい。
【0077】
プロモーターは、RNAポリメラーゼにより認識され、下流領域の転写を媒介する遺伝子要素である。好ましいプロモーターは、転写レベルを向上させる強プロモーターである。強プロモーターの例は、前初期ヒトサイトメガロウイルスプロモーター(CMV)及びイントロンAを有するCMVである(Chapmanら,Nucl.Acids Res.,19:3979−3986(1991))。プロモーターの別の例には、EF1αプロモーター、マウスCMVプロモーター、ラウス肉腫ウイルスプロモーター、SV40初期/後期プロモーターやβ−アクチンプロモーターのような天然プロモーター;並びに合成筋肉特異的プロモーターやキメラ筋肉特異的/CMVプロモーターのような人工プロモーターが含まれる(Liら,Nat.Biotechnol.,17:241−245(1999);Hagstromら,Blood,95:2536−2542(2000))。
【0078】
リボソーム結合部位は開始コドンにまたはその近くに位置している。好ましいリボソーム結合部位の例には、CCACCAUGG、CCGCCAUGG及びACCAUGG(ここで、AUGは開始コドンである)が含まれる(Kozak,Cell,44:283−292(1986))。リボソーム結合部位の別の例はGCCACCAUGG(配列番号12)である。
【0079】
ポリアデニル化シグナルは、転写RNAの開裂及びポリ(A)テールのRNAへの付加に関与する。高級真核生物中のポリアデニル化シグナルは、ポリアデニル化付加部位から約11〜30ヌクレオチドにAAUAAA配列を含む。このAAUAAA配列はRNA開裂のシグナル化に関与する(Lewin,Genes IV,ニューヨークに所在のオックスフォード大学出版(1990年)出版)。ポリ(A)テールはmRNAプロセシングにとって重要である。
【0080】
遺伝子発現カセットの一部として使用され得るポリアデニル化シグナルは、最低の家兎β−グロビンポリアデニル化シグナル及びウシ成長ホルモンポリアデニル化(BGH)を含む(Xuら,Gene,272:149−156(2001);Postらの米国特許第5,122,458号明細書)を含む。更なる例には合成ポリアデニル化シグナル(SPA及びSV40ポリアデニル化シグナルが含まれる。SPA配列は、AAUAAAAGAUCUUUAUUUUCAUUAGAUCUGUGUGUUGGUUUUUUGUGUG(配列番号13)である。
【0081】
存在し得る遺伝子発現またはポリペプチドプロセシングを強化または調節するのに有用な追加の調節要素の例にはエンハンサー、リーダー配列及びオペレーターが含まれる。エンハンサー領域は転写を向上させる。エンハンサー領域の例にはCMVエンハンサー及びSV40エンハンサーが含まれる(Hittら,Methods in Molecular Genetics,7:13−30(1995);Xuら,Gene.272:149−156(2001))。エンハンサー領域はプロモーターに関連し得る。
【0082】
リーダー配列はポリペプチドをプロテオソームに指向させるポリペプチド上のアミノ酸領域である。リーダー配列をコードする核酸は構造遺伝子の5’であり、構造遺伝子に沿って転写される。リーダー配列の例はtPAである。
【0083】
遺伝子発現を調節するためにオペレーター配列を使用し得る。例えば、Tetオペレーター配列を遺伝子発現を抑制するために使用し得る。
【0084】
II. 治療用ベクター
Met−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチドをコードする核酸は、治療投与のために適したベクターを用いて患者に導入され得る。許容できない副作用を引き起こすことなく核酸を標的細胞にデリバーするベクターが適当であり得る。
【0085】
細胞発現は、Met−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチドをコードする遺伝子発現カセットを用いて達成される。遺伝子発現カセットは、有利な効果が得られるように標的細胞内で十分量の核酸を産生、プロセシングするために調節要素を含む。
【0086】
治療用途に使用され得るベクターの例には、第1世代及び第2世代アデノベクター、ヘルパー依存性アデノベクター、アデノ関連ウイルスベクター、レトロウイルスベクター、αウイルスベクター、ベネズエラウマ脳脊髄炎ウイルスベクター及びプラスミドベクターが含まれる(Hittら,Advances in Pharmacology,40:137−206(1997);Johnstonらの米国特許第6,156,588号明細書;Johnstonらの国際特許出願公開第95/32733号パンフレット)。Met−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチドを患者に導入するための好ましいベクターは第1世代アデノウイルスベクター及びプラスミドDNAベクターである。
【0087】
(A) 第1世代アデノウイルスベクター
遺伝子発現カセットを発現するための第1世代アデノベクターは、E1及び場合によりE3欠失組換えアデノウイルスゲノム中に発現カセットを含む。E1領域の欠失はアデノウイルスの複製に必要な要素を除去するのに十分に大きい。
【0088】
Met−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチドを発現するための第1世代アデノベクターはE1及びE3欠失組換えアデノウイルスゲノムを含む。E1領域の欠失はアデノウイルスの複製に必要な要素を除去するのに十分に大きい。E1及びE3領域の欠失の組合せは、Met−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチドをコードする遺伝子発現カセットを収容するのに十分な大きさである。
【0089】
アデノウイルスは両端に逆方向末端反復を有する二本鎖線状ゲノムを有する。ウイルス複製中、ゲノムはウイルスカプシド内部にパッケージされてビリオンを形成する。ウイルスはウイルス結合及びその後の内部移行によりその標的細胞に進入する(Hittら,Advances in Pharmacology,40:137−206(1997))。
【0090】
アデノベクターは、ヒトまたは動物中に見られるような各種アデノウイルス血清型をベースとしている。動物アデノウイルスの例にはウシ、ブタ、チンパンジー、マウス、イヌ及びトリが含まれる(CELO)。好ましいアデノベクターはヒト血清型、より好ましくはグループB、CまたはD血清型をベースとしている。ヒトアデノウイルスグループB、C、DまたはE血清型の例には、タイプ2(“Ad2”)、4(“Ad4”)、5(“Ad5”)、6(“Ad6”)、24(“Ad24”)、26(“Ad26”)、34(“Ad34”)及び35(“Ad35”)が含まれる。アデノベクターは1つのアデノウイルスに由来する領域または2つ以上のアデノウイルスに由来する領域を含み得る。
【0091】
別の実施態様において、アデノベクターはAd5、Ad6またはその組合せをベースとする。Ad5はChroboczekら,J.Virology,186:280−285(1992)に記載されている。Ad6は図7A〜7Nに記載されている。Ad5領域を含むAd6ベースのベクターは以下の実施例に記載されている。
【0092】
アデノベクターにおいてE1及びE3領域は完全に除去されていなくてもよい。むしろ、イントランスに供給されるE1タンパク質の非存在下でベクター複製を無能とするのに十分な量のE1領域が除去されている。E1欠失、またはE1欠失とE3欠失の組合せは遺伝子発現カセットを収容するのに十分な大きさである。
【0093】
E1欠失はAd5の塩基対約342〜塩基対約3523または他のアデノウイルスの対応領域から得られ得る。好ましくは、欠失領域はAd5の塩基対約450〜塩基対約3511の領域または他のアデノウイルスの対応領域の除去を含む。塩基対約341から始まる大きなE1領域が欠失されるとウイルスパッケージングを促進する要素が除去される。
【0094】
E3欠失はAd5の塩基対約27865〜塩基対約30995または他のアデノウイルスの対応領域から得られ得る。好ましくは、欠失領域はAd5の塩基対約28134〜塩基対約30817の領域または他のアデノウイルスの対応領域の除去を含む。
【0095】
E1領域及び場合によりE3領域の欠失の組合せは、遺伝子発現カセットを含む組換えゲノムの全サイズが野生型アデノウイルスゲノムの約105%を越えない程度に十分に大きくなければならない。例えば、組換えアデノウイルスAd5ゲノムが約105%を越える大きさになるとゲノムは不安定になる(Bettら,Journal of Virology,67:5911−5921(1993))。
【0096】
好ましくは、遺伝子発現カセットを含む組換えアデノウイルスゲノムの大きさは野生型アデノウイルスゲノムの大きさの約85%〜約105%である。別の実施態様では、発現カセットを含む組換えアデノウイルスゲノムの大きさは野生型アデノゲノムの大きさの約100%〜約105.2%、または100%である。
【0097】
E1及びE3欠失を有するアデノウイルスゲノムに約7500kbが挿入され得る。欠失なしの場合、Ad5ゲノムは35,935塩基対であり、Ad6ゲノムは35,759塩基対である。
【0098】
第1世代アデノベクターは、E1遺伝子産物をイントランスに供給することにより複製され得る。E1遺伝子産物はイントランスに、例えばアデノウイルスE1領域で形質転換させた細胞株を用いることにより供給され得る。アデノウイルスE1領域で形質転換させた細胞または細胞株の例はHEK 293細胞、911細胞、PERC.6(商標)細胞及びトランスフェクトした一次ヒトアミノサイト細胞である(Grahamら,Journal of Virology,36:59−72(1977);Schiednerら,Human Gene Therapy,11:2105−2116(2000);Fallauxら,Human Gene Therapy,9:1909−1917(1998);Boutらの米国特許第6,033,908号明細書)。
【0099】
Met−NS3−NS4A−NS4B−NS5A−NS5B発現カセットは、欠失E1領域または欠失E3領域に対応する領域で組換えアデノベクターゲノムに挿入されなければならない。発現カセットは平行または逆平行方位を有し得る。平行方位では、挿入遺伝子の転写方向は欠失E1またはE3領域と同じ方向である。逆平行方位転写では、逆鎖は鋳型として作用し、転写方向は逆方向である。
【0100】
本発明の実施態様では、アデノベクターはE1欠失領域に挿入された遺伝子発現カセットを有する。前記ベクターは、
a)Ad5またはAd6のいずれかに対応する塩基対約1〜塩基対約450の第1アデノウイルス領域、
b)前記第1領域に連結した、E1平行またはE1逆平行方位の遺伝子発現カセット、
c)前記発現カセットに連結した、Ad5に対応する塩基対約3511〜塩基対約5548またはAd6に対応する塩基対約3508〜塩基対約5541の第2アデノウイルス領域、
d)前記第2領域に連結した、Ad5に対応する塩基対約5549〜塩基対約28133またはAd6に対応する塩基対約5542〜塩基対約28156の第3アデノウイルス領域、
e)前記第3領域に連結した、Ad5に対応する塩基対約30818〜塩基対約33966またはAd6に対応する塩基対約30789〜塩基対約33784の第4アデノウイルス領域、及び
f)前記第4領域に連結した、Ad5に対応する塩基対約33967〜塩基対約35935またはAd6に対応する塩基対約33785〜塩基対約35759の第5アデノウイルス領域
を含む。
【0101】
本発明の別の実施態様では、アデノベクターはE3欠失領域に挿入された遺伝子発現カセットを有する。前記ベクターは、
a)Ad5またはAd6のいずれかに対応する塩基対約1〜塩基対約450の第1アデノウイルス領域、
b)前記第1領域に連結した、Ad5に対応する塩基対約3511〜塩基対約5548またはAd6に対応する塩基対約3508〜塩基対約5541の第2アデノウイルス領域、
c)前記第2領域に連結した、Ad5に対応する塩基対約5549〜塩基対約28133またはAd6に対応する塩基対約5542〜塩基対約28156の第3アデノウイルス領域、
d)前記第3領域に連結した、E3平行またはE3逆平行方位の遺伝子発現カセット、
e)前記遺伝子発現カセットに連結した、Ad5に対応する塩基対約30818〜塩基対約33966またはAd6に対応する塩基対約30789〜塩基対約33784の第4アデノウイルス領域、及び
f)前記第4領域に連結した、Ad5に対応する塩基対約33967〜塩基対約35935またはAd6に対応する塩基対約33785〜塩基対約35759の第5アデノウイルス領域
を含む。
【0102】
アデノウイルス領域に関する好ましい別の実施態様では、
(1)第1、第2、第3、第4及び第5領域はAd5に対応する;
(2)第1、第2、第3、第4及び第5領域はAd6に対応する;
(3)第1領域はAd5に対応し、第2領域はAd5に対応し、第3領域はAd6に対応し、第4領域はAd6に対応し、第5領域はAd5に対応する。
【0103】
(B) DNAプラスミドベクター
DNAワクチンプラスミドは遺伝子発現カセットと共に複製及び好ましくはベクター選択を容易とする要素を含む。好ましい要素は非哺乳動物細胞における複製及び選択マーカーを与える。前記ベクターは、ヒト細胞における複製またはヒト核酸への組込みを与える要素を含むべきではない。
【0104】
選択マーカーにより、マーカーを含む核酸の選択が容易となる。好ましい選択マーカーは抗生物質耐性を付与するものである。抗生物質選択遺伝子の例には、アンピシリン、ネオマイシン及びカナマイシンに対する耐性をコードする核酸が含まれる。
【0105】
好適なDNAワクチンベクターは細菌性複製起点及び選択マーカーを含むプラスミドから作成され得る。高い収率を与える細菌性複製起点にはColE1プラスミド誘導の細菌性複製起点が含まれる(Donnellyら,Annu.Rev.Immunol.,15:617−648(1997))。
【0106】
細菌性複製起点及び選択マーカーを存在させると、大腸菌のような細菌株においてDNAベクターが産生され得る。選択マーカーはDNAベクターを含まない細菌を除去するために使用される。
【0107】
III. Ad6組換え核酸
Ad6組換え核酸は、配列番号8中に存在するAd6領域に実質的に類似するAd6領域及びAd6核酸中に存在しない領域を含む。Ad6領域を含む組換え核酸は、別のAd6領域の産生において、Ad6ベースのベクターの作成における中間体として、または組換え遺伝子のデリバリー用ベクターとしてのように各種用途を有する。
【0108】
図9に示すように、Ad6のゲノム構成はAd5のゲノム構成に非常に類似している。Ad5とAd6のホモロジーは約98%である。
【0109】
別の実施態様において、Ad6組換え核酸はE1A、E1B、E2B、E2A、E3、E4、L1、L2、L3またはL4に実質的に類似しているヌクレオチド領域またはその組合せを含む。Ad6領域に実質的に類似の核酸領域は少なくとも65%、少なくとも75%、少なくとも85%、少なくとも95%、少なくとも99%または100%のヌクレオチド配列同一性を有するか、または1〜2、1〜3、1〜4、1〜5、1〜6、1〜7、1〜8、1〜9、1〜10、1〜11、1〜12、1〜13、1〜14、1〜15、1〜16、1〜17、1〜18、1〜19、1〜20、1〜25、1〜30、1〜35、1〜40、1〜45または1〜50個のヌクレオチドだけ異なっている。実質的に類似の核酸配列を調べるための方法及び実施態様は上記セクションI.B.に記載されている。
【0110】
好ましくは、組換えAd6核酸はAd6中に存在しないポリペプチドをコードする発現カセットを含む。発現カセットの例には、HCV領域をコードする発現カセット及び別のタイプのポリペプチドをコードする発現カセットが含まれる。
【0111】
別のタイプのアデノウイルスベクターは、異なる量の第1世代及び第2世代アデノベクターのようなAd6を導入して作成され得る。上記セクションII.A.に記載したように、第1世代アデノベクターはE1を欠き、E1をイントランスに供給したときに複製し得る。
【0112】
第2世代アデノベクターは第1世代ベクターよりも小さいアデノウイルスベクターを含み、アデノウイルスタンパク質を供給するヘルパーベクター及び/または補充細胞株と一緒に使用され得る。第2世代アデノベクターは複数の文献、例えばRusell,Journal of General Virology,81:2573−2604(2000);Hittら,「遺伝子導入のためのヒトAdベクター(Human Ad vectors for Gene
Transfer)」,Advances in Pharmacology,Vol.40,Academic Press発行に記載されている。
【0113】
本発明の実施態様では、Ad6組換え核酸はE1をイントランスに供給したときに複製し得るE1を欠くアデノウイルスベクターである。発現カセットは欠失E1領域及び/または欠失E3領域に挿入され得る。
【0114】
欠失E1領域に発現カセットを有するAd6ベースのアデノウイルスベクターの例は、
a)Ad5またはAd6のいずれかに対応する塩基対約1〜塩基対約450の第1アデノウイルス領域、
b)前記第1領域に連結した、E1平行またはE1逆平行方位の遺伝子発現カセット、
c)前記発現カセットに連結した、Ad5に対応する塩基対約3511〜塩基対約5548またはAd6に対応する塩基対約3508〜塩基対約5541の第2アデノウイルス領域、
d)前記第2領域に連結した、Ad5に対応する塩基対約5549〜塩基対約28133またはAd6に対応する塩基対約5542〜塩基対約28156の第3アデノウイルス領域、
e)場合によっては存在する、前記第3領域に連結した、Ad5に対応する塩基対約28134〜塩基対約30817またはAd6に対応する塩基対約28157〜塩基対約30788の第4アデノウイルス領域、
f)第4領域が存在するときにはその第4領域に連結し、第4領域が存在しないときには第3領域に連結した、Ad5に対応する塩基対約30818〜塩基対約33966またはAd6に対応する塩基対約30789〜塩基対約33784の第5アデノウイルス領域、及び
g)前記第5領域に連結した、Ad5に対応する塩基対約33967〜塩基対約35935またはAd6に対応する塩基対約33785〜塩基対約35759の第6アデノウイルス領域
を含むかまたはこれらから構成されるが、少なくとも1つのAd6領域は存在する。
【0115】
本発明の別の実施態様では、全ての領域がAd6に由来する;第1及び第2を除く全ての領域がAd6に由来する;第2、第3,第4及び第5領域から選択される1〜4個の領域がAd6に由来する。
【0116】
欠失E3領域に発現カセットを有するAd6ベースのアデノウイルスベクターの例は、
a)Ad5またはAd6のいずれかに対応する塩基対約1〜塩基対約450の第1アデノウイルス領域、
b)前記第1領域に連結した、Ad5に対応する塩基対約3511〜塩基対約5548またはAd6に対応する塩基対約3508〜塩基対約5541の第2アデノウイルス領域、
c)前記第2領域に連結した、Ad5に対応する塩基対約5549〜塩基対約28133またはAd6に対応する塩基対約5542〜塩基対約28156の第3アデノウイルス領域、
d)前記第3領域に連結した、E3平行またはE3逆平行方位の遺伝子発現カセット、
e)前記遺伝子発現カセットに連結した、Ad5に対応する塩基対約30818〜塩基対約33966またはAd6に対応する塩基対約30789〜塩基対約33784の第4アデノウイルス領域、及び
f)前記第4領域に連結した、Ad5に対応する塩基対約33967〜塩基対約35935またはAd6に対応する塩基対約33785〜塩基対約35759の第5アデノウイルス領域、
を含むかまたはこれらから構成されるが、少なくとも1つのAd6領域は存在する。
【0117】
本発明の別の実施態様では、全ての領域がAd6に由来する;第1及び第2を除く全ての領域がAd6に由来する;第2、第3,第4及び第5領域から選択される1〜4個の領域がAd6に由来する。
【0118】
IV. ベクター作成
ベクターは、制限酵素の使用、核酸ライゲーションや相同組換えを含むような組換え核酸方法を用いて作成され得る。組換え核酸方法は当業界で公知である(Ausubel,「分子生物学における現在のプロトコール(Current Protocols in Molecular Biology)」,John Wiley(1987−1998年)発行;Sambrookら,「分子クローニング,実験マニュアル(Molecular Cloning, A Laboratory Manual)」,第2版,Cold Spring Harbor Laboratory Press(1989年)発行)。
【0119】
中間体ベクターは治療ベクターを引き出すために、または発現カセットまたはその一部を1つのベクターから他のベクターに移動させるために使用される。中間体ベクターの例には、アデノウイルスゲノムプラスミド及びシャトルベクターが含まれる。
【0120】
中間体ベクター中の有用な要素には、複製起点、選択マーカー、相同組換え領域及び便利な制限部位が含まれる。便利な制限部位は核酸配列のクローニングまたは遊離を促進するために使用され得る。
【0121】
相同組換え領域は、別の核酸分子中の標的領域に相同な核酸領域である。相同領域は、標的領域に挿入される核酸配列に隣接している。別の実施態様では、相同領域は好ましくは約150〜600ヌクレオチド長、または約100〜500ヌクレオチド長である。
【0122】
本発明の実施態様は、Met−NS3−NS4A−NS4B−NS5A−NS5B発現カセット、選択マーカー、細菌性複製起点、第1アデノウイルス相同領域、及びE1領域に挿入またはE1領域を置換するために発現カセットを標的する第2アデノウイルス相同領域を含むシャトルベクターに関する。第1及び第2相同領域は発現カセットに隣接している。第1相同領域は、塩基対約4〜450の野生型アデノウイルス領域の少なくとも右端(3’端)に実質的に相同な少なくとも約100塩基対を含む。第2相同領域は、塩基対約3511〜5792のAd5の少なくとも左端(5’端)または他のアデノウイルス由来の対応領域に実質的に相同な少なくとも約100塩基対を含む。
【0123】
「実質的に相同な」は、標的領域と特異的に組み換えるのに十分に相同度を指す。別の実施態様では、実質的に相同とは少なくとも85%、少なくとも95%または100%の配列同一性を指す。配列同一性は上記セクションI.B.に記載されているように計算され得る。
【0124】
アデノベクターを産生するための1つの方法は、発現カセットを含むアデノウイルスゲノムプラスミドの作成を含む。プレアデノウイルスは、所望の相補(complimenting)細胞株での複製に必要な全てのアデノウイルス配列を含む。その後、プレアデノウイルスプラスミドを制限酵素で消化してウイルスITRを放出し、ウイルスレスキューのために相補細胞株にトランスフェクトする。ITRは複製が生ずるようにプラスミド配列から放出されなければならない。アデノベクターレスキューの結果、発現カセットを含むアデノベクターが生ずる。
【0125】
(A)アデノウイルスゲノムプラスミド
アデノウイルスゲノムプラスミドは、長いプラスミド(コスミドであってもよい)内にアデノベクター配列を含む。長いプラスミドは、プラスミドを産生及び維持するために使用される手順に応じて真核または細菌細胞における増殖及び選択を促進するような追加要素を含み得る。アデノウイルスゲノムプラスミドを産生するための方法には、シャトルベクター及び相同組換えの使用を含む技術及び遺伝子発現カセットのアデノウイルスコスミドへの挿入を含む技術が含まれる(Hittら,Methods in Molecular Genetics,7:13−30(1995);Danthinneら,Gene Therapy,7:1707−1714(2000))。
【0126】
アデノウイルスゲノムプラスミドがE1またはE3欠失領域に挿入された遺伝子発現カセットを有していることが好ましい。本発明の実施態様では、アデノウイルスゲノムプラスミドは、E1欠失領域に挿入された遺伝子発現カセット、複製起点、選択マーカー及び組換えアデノウイルス領域を含み、前記組換えアデノウイルス領域は
a)Ad5またはAd6のいずれかに対応する塩基対約1〜塩基対約450の第1アデノウイルス領域、
b)前記第1領域に連結した、E1平行またはE1逆平行方位の遺伝子発現カセット、
c)前記発現カセットに連結した、Ad5に対応する塩基対約3511〜塩基対約5548またはAd6に対応する塩基対約3508〜塩基対約5541の第2アデノウイルス領域、
d)前記第2領域に連結した、Ad5に対応する塩基対約5549〜塩基対約28133またはAd6に対応する塩基対約5542〜塩基対約28156の第3アデノウイルス領域、
e)前記第3領域に連結した、Ad5に対応する塩基対約30818〜塩基対約33966またはAd6に対応する塩基対約30789〜塩基対約33784の第4アデノウイルス領域、
f)前記第4領域に連結した、Ad5に対応する塩基対約33967〜塩基対約35935またはAd6に対応する塩基対約33785〜塩基対約35759の第5アデノウイルス領域、及び
g)場合によっては存在する、所望アデノベクターの全サイズを考慮してより小さなインサートのために存在し得る、Ad5またはAd6中に存在するE3領域の全部または一部に対応するE3領域
から構成される。
【0127】
本発明の別の実施態様では、組換えアデノウイルスゲノムプラスミドはE3欠失領域に挿入された遺伝子発現カセットを有する。前記ベクターは複製起点、選択マーカー、並びに
a)Ad5またはAd6のいずれかに対応する塩基対約1〜塩基対約450の第1アデノウイルス領域、
b)前記第1領域に連結した、Ad5に対応する塩基対約3511〜塩基対約5548またはAd6に対応する塩基対約3508〜塩基対約5541の第2アデノウイルス領域、
c)前記第2領域に連結した、Ad5に対応する塩基対約5549〜塩基対約28133またはAd6に対応する塩基対約5542〜塩基対約28156の第3アデノウイルス領域、
d)前記第3領域に連結した、E3平行またはE3逆平行方位の前記遺伝子発現カセット、
e)前記遺伝子発現カセットに連結した、Ad5に対応する塩基対約30818〜塩基対約33966またはAd6に対応する塩基対約30789〜塩基対約33784の第4アデノウイルス領域、及び
f)前記第4領域に連結した、Ad5に対応する塩基対約33967〜塩基対約35935またはAd6に対応する塩基対約33785〜塩基対約35759の第5アデノウイルス領域
を含む。
【0128】
存在するアデノウイルス領域に関する別の実施態様では
(1)第1、第2、第3、第4及び第5領域はAd5に対応する;
(2)第1、第2、第3、第4及び第5領域はAd6に対応する;
(3)第1領域はAd5に対応し、第2領域はAd5に対応し、第3領域はAd6に対応し、第4領域はAd6に対応し、第5領域はAd5に対応する。
【0129】
本発明の実施態様は、アデノウイルスゲノムプラスミドを産生するための相同組換え段階及びアデノウイルスレスキュー段階を含むアデノベクターの作成方法に関する。前記相同組換え段階は、アデノウイルス相同領域に隣接してMet−NS3−NS4A−NS4B−NS5A−NS5B発現カセットを含むシャトルベクターの使用を含む。前記アデノウイルス相同領域により、発現カセットはE1またはE3欠失領域に標的される。
【0130】
アデノウイルスゲノムプラスミドの産生に関する本発明の実施態様では、遺伝子発現カセットを、Ad5またはAd6のいずれかに対応する塩基対約1〜塩基対約450の第1アデノウイルス領域;前記第1領域に連結した、Ad5に対応する塩基対約3511〜塩基対約5548またはAd6に対応する塩基対約3508〜塩基対約5541の第2アデノウイルス領域;前記第2領域に連結した、Ad5に対応する塩基対約5549〜塩基対約28133またはAd6に対応する塩基対約5542〜塩基対約28156の第3アデノウイルス領域;前記第3領域に連結した、Ad5に対応する塩基対約30818〜塩基対約33966またはAd6に対応する塩基対約30789〜塩基対約33784の第4アデノウイルス領域;及び前記第4領域に連結した、Ad5に対応する塩基対約33967〜塩基対約35935またはAd6に対応する塩基対約33785〜塩基対約35759の第5アデノウイルス領域からなるベクターに挿入する。アデノウイルスゲノムプラスミドは複製起点及び選択マーカーを含んでいなければならず、Ad5またはAd6 E3領域の全部または一部を含んでいてもよい。
【0131】
存在するアデノウイルス領域に関する別の実施態様では、
(1)第1、第2、第3、第4及び第5領域はAd5に対応する;
(2)第1、第2、第3、第4及び第5領域はAd6に対応する;
(3)第1領域はAd5に対応し、第2領域はAd5に対応し、第3領域はAd6に対応し、第4領域はAd6に対応し、第5領域はAd5に対応する。
【0132】
(B) アデノベクターレスキュー
アデノベクターは、当業界で公知であるかまたは本明細書に記載されている方法を用いて組換えアデノウイルスゲノムプラスミドからレスキューされ得る。当業界で公知のアデノウイルスレスキューの方法の例は、Hittら,Methods in Molecular Genetics,7:13−30(1995)及びDanthinneら,Gene Therapy,7:1707−1714(2000)に記載されている。
【0133】
本明細書に記載されているアデノベクターをレスキューするための好ましい方法はアデノウイルス複製のブーストが含まれる。ブースティングアデノウイルス複製は、例えばE2タンパク質(ポリメラーゼ、プレ末端タンパク質及びDNA結合タンパク質)のようなアデノウイルス機能及びE4orf6を別のプラスミドに供給することにより実施され得る。以下の実施例10には、コドン最適化Met−NS3−NS4A−NS4B−NS5A−NS5B発現カセットを含むアデノベクターをレスキューするためのアデノウイルス複製のブーストを例示している。
【0134】
V. 部分最適化HCVコード配列
HCVポリタンパク質コード化核酸の部分最適化により、ヒトでの発現のために最適化されるコドンの量は完全最適化に比して少なくなる。全目的は、最適化コドンを有するHCVポリタンパク質コード化核酸を含むアデノベクターの作成を促進しながら、コドン最適化のために高い発現という効果を与えることである。
【0135】
HCVポリタンパク質コード化配列を完全最適化すると、各アミノ酸について最も頻繁に見られるヒトコドンが与えられる。完全最適化は、当業界で公知のコドン頻度表及びBLACKTRANSLATEプログラム(ウィスコンシンパッケージ バージョン10;ウィスコンシン州マディソンに所在のGenetics Computer Group(GCG))を用いて実施され得る。
【0136】
部分最適化は、存在する全HCVポリタンパク質コード化配列(例えば、NS3−NS5B)または存在する1つ以上の局部領域に対して実施され得る。別の実施態様では、存在する全HCVコード化ポリタンパク質のGC含量は少なくとも約65%以下であり、1つ以上の局部領域のGC含量は約70%以下である。
【0137】
局部領域はHCVコード化核酸中に存在する領域であり、サイズは異なり得る。例えば、局部領域は約60、約70、約80、約90または約100ヌクレオチド長であり得る。
【0138】
部分最適化は、最初天然に存在する配列に基づいて部分的に最適化されるHCVコード化ポリタンパク質配列を構築することにより達成され得る。或いは、最適化HCVコード化配列は部分最適化配列を作成するために比較に基づいて使用され得る。
【0139】
VI. HCV併用治療
HCV Met−NS3−NS4A−NS4B−NS5A−NS5Bワクチンは単独で患者を治療するために使用され得、他のHCV治療薬と一緒に使用され得、及び他のタイプの疾患を標的とする物質と一緒に使用され得る。追加の治療には、HCV及びHCV感染患者に広く蔓延している疾患を治療するための追加の治療薬が含まれる。他のタイプの疾患を標的とする物質にはHIV及びHBVに対するワクチンが含まれる。
【0140】
HCVを治療するための追加の治療薬にはワクチン及び非ワクチンが含まれる(Zein,Expert Opin.Investig.Drugs,10:1457−1469(2001))。追加のHCVワクタンの例には、HCVコア抗原及びHCV E1、E2またはp7領域に対する免疫応答を引き出すように設計されたワクチンが含まれる。ワクチン成分は天然に存在するHCVポリペプチド、HCVミモトープポリペプチドまたは前記ポリペプチドをコードする核酸であり得る。
【0141】
HCVミモトープポリペプチドはHCVエピトープを含むが、天然に存在するHCV抗原とは異なる配列を有する。HCVミモトープは天然に存在するHCV抗原に融合され得る。一般的なミモトープの作成方法及び各種HCVミモトープはFeliciらの米国特許第5,994,083号明細書及びNicosiaらの国際特許出願公開第99/60132号パンフレットに記載されている。
【0142】
VII. 医薬投与
HCVワクチンは、当業界で公知の方法を用い、本明細書に記載されているガイダンスを用いて処方、患者に投与される。医薬投与の一般的ガイドラインは、例えばいずれも援用により本明細書に含まれるとするModern Vaccinology,Kurstak編,Plenum Med.Co.(1994年)発行;Remington’s Pharmaceutical Science,第18版,Gennaro編,Mack Publishing(1990年)発行;及びModern Pharmaceutics,第2版,Banker及びRhodes編,Marcel Dekker,Inc.(1990年)発行に記載されている。
【0143】
HCVワクチンは、静脈内、腹腔内、皮下、筋肉内、皮内、皮膚を介する圧痕または鼻のような各種ルートにより投与され得る。好ましいルートは筋肉内である。
【0144】
筋肉内投与は各種方法を用いて、例えば1つ以上の電気パルスと共に又は電気パルスなしで注射により実施され得る。電気媒介導入は液性及び細胞免疫応答の両方を刺激することにより遺伝免疫化を助けることができる。
【0145】
ワクチン注射は各種方法を用いて、例えば針及び無針注射システムを用いることにより実施され得る。無針注射システムの例はジェット注射デバイスである(Donnellyらの国際特許出願公開第99/52463号パンフレット)。
【0146】
(A) 電気媒介導入
電気媒介導入、すなわち遺伝子エレクトロトランスファー(GET)は、核酸注射後適当な電気パルスをデリバリーすることにより実施され得る(Mathiesenの国際特許出願公開第98/43702号パンフレット参照)。プラスミド注射及びエレクトロポレーションはステンレス針を用いて実施され得る。針は2つ、3つまたはそれ以上の複雑なパターンで使用され得る。1つの構成では、針を、機械的支持体であり且つ適当なケーブルを用いて電場発生装置に針をつなげるプリント基板上にはんだ付けされている。
【0147】
電気刺激は電気パルスの形態で加えられる。パルスは各種形態(スクエア、シヌソイド、三角、指数関数的減衰)及び各種極性(正または負極の単極性、双極性)を有し得る。パルスは定電圧または定電流方式でデリバリーされ得る。
【0148】
HCVを含む核酸ワクチン及び他の核酸ワクチンを患者に導入するために多種多様な電気治療パターンが使用され得る。電気治療の可能なパターンには、
(治療1) パルス長0.2msec/フェーズ、周波数1000Hz、定電圧モード、45ボルト/フェーズ、浮動電流で1秒おきにデリバリーされる1000スクェア双極性パルス10列、
(治療2) パルス長2msec/フェーズ、周波数100Hz、定電流モード、100mA/フェーズ、浮動電圧で1秒おきにデリバリーされる1000スクェア双極性パルス2列、
(治療3) 約2msec/フェーズのパルス長で全部で3秒間の双極性パルス2列(組織に流れる実際の電流は約50mAに固定されている)
が含まれる。
【0149】
電気パルスは電場発生装置を介してデリバリーされる。好適な発生装置は、共通シャシに組み立てられ、駆動プログラムを実行するポータブルPCにより駆動される3つの独立したハードウェア部品から構成され得る。ソフトウェアは基本的及び補助持機能を管理する。前記デバイスの部品は、(1)マイクロプロセッサーにより駆動する信号発生器、(2)電力増幅器及び(3)デジタルオシロスコープである。
【0150】
信号発生器は、ソフトウェアのコントロール下で所定範囲の任意の周波数及び形状を有する信号を発する。同一ソフトウェアはデリバリーされる波形に対する対話式エディタを有する。前記発生器はデジタル制御の限流デバイス(最大電流出力を制御するための安全機構)である。電力増幅器は発生したシグナルを±150Vまで増幅させ得る。オシロスコープはデジタル式であり、増幅器によりデリバリーされる電流及び電圧の両方をサンプリングし得る。
【0151】
(B) 薬用担体
医薬的に許容され得る担体により、ワクチンの貯蔵及びワクチンの被験者への投与が容易となる。医薬的に許容され得る担体の例は本明細書に記載されている。別の医薬的に許容され得る担体は当業界で公知である。
【0152】
医薬的に許容され得る担体は各種成分、例えば緩衝液、通常生理食塩液、リン酸緩衝食塩液、スクロース、塩及びポリソルベートを含み得る。医薬的に許容され得る担体の例は次の通りである:2.5〜10mM トリス緩衝液、好ましくは約5mM トリス緩衝液;25〜100mM NaCl、好ましくは約75mM NaCl;2.5〜10% スクロース、好ましくは約5% スクロース;0.01〜2mM MgCl2;及び0.001〜0.01% ポリソルベート80(植物由来)。pHは好ましくは約7.0〜9.0、より好ましくは約8.0である。担体の特定例は、pH8.0で5mM トリス、75mM NaCl、5% スクロース、1mM MgCl2、0.005% ポリソルベート80を含む。
【0153】
(C) 投薬レジメ
好適な投与レジメは、特定ワクチンの効力及び諸要因(例えば、患者の年齢、体重、性別及び医学的状態;投与ルート;所望効果;及び投与回数)を考慮して決定され得る。特定ワクチンの効力は各種因子、例えば細胞において発現、プロセシングされ、MHCクラスI及びII複合体の関係で提示されるポリペプチドを産生する特定ワクチンの能力に依存する。
【0154】
患者に投与されるHCVコード化核酸は、アデノベクターのようなウイルスベクター及びDNAプラスミドワクチンを含めた各種タイプのベクターの一部であり得る。DNAプラスミドの投与に関する別の実施態様では、約0.1〜10mgのプラスミドが患者に投与され、約1〜5mgのプラスミドが患者に投与される。ウイルスベクター、好ましくはアデノウイルスベクターの投与に関する別の実施態様では、約105〜1011ウイルス粒子が患者に投与され、約107〜1010ウイルス粒子が患者に投与される。
【0155】
ウイルスベクターワクチン及びDNAプラスミドワクチンは単独で投与してもよく、または初期及び追加投与レジメの一部であってもよい。混合方式の初期及び追加接種としては、DNAワクチンを初期接種した後ウイルスベクターワクチンを追加接種したり、ウイルスベクターワクチンを初期接種した後DNAワクチンを追加接種することが挙げられる。
【0156】
初回接種を複数回、例えば約2〜4回またはそれ以上実施してもよい。初回接種と追加接種の間の期間は通常約4ヶ月〜1年の範囲で変更可能であるが、他の時間枠を使用してもよい。DNAワクチンを用いる初回接種レジメの使用は、ヒトが既に抗−アデノウイルス免疫応答を示している状況において好ましいことがある。
【0157】
本発明の実施態様では、1×107〜1〜1012粒子、好ましくは約1×1010〜1〜1011アデノベクター粒子を筋肉組織に直接投与する。初回ワクチン接種後、アデノベクターまたはDNAワクチンを用いて追加接種する。
【0158】
本発明の別の実施態様では、初回接種をDNAワクチンを用いて筋肉組織に直接実施する。初回接種後、アデノベクターまたはDNAワクチンを用いて追加接種する。
【0159】
免疫応答をブーストするためにインターロイキン−12、GM−CSF、B7−1、B7−2、IP10、Mig−1のような物質を同時投与することができる。前記物質はタンパク質として、または核酸ベクターを用いて同時投与され得る。
【0160】
(D) 非相同初回−追加接種
非相同初回−追加接種は、初回接種のために1タイプのウイルスベクター、追加接種のために別のタイプのウイルスベクターを使用することを含む混合方式である。非相同初回−追加接種では、異なるアデノウイルス血清型をベースとするベクターのような関連ベクター及びより遠縁のウイルス(例えば、アデノウイルス及びポックスウイルス)を使用し得る。マウスをマラリアから防御するためにポックスウイルス及びアデノウイルスベクターを使用することはGilbertら,Vaccine,20:1039−1045(2002)に記載されている。
【0161】
初回接種及び追加接種に関する別の実施態様では、Met−NS3−NS4A−NS4B−NS5A−NS5Bのような所望の抗原を発現する以下のタイプのベクターを使用する。すなわち、Ad5ベクター後Ad6ベクター;Ad6ベクター後Ad5ベクター;Ad5ベクター後ポックスウイルスベクター;ポックスウイルスベクター後Ad5ベクター;Ad6ベクター後ポックスウイルスベクター後;ポックスウイルスベクター後Ad6ベクター。
【0162】
初回接種と追加接種の間の期間は通常約4ヶ月〜1年の範囲で変更可能であるが、他の時間枠を使用してもよい。少なくとも時間枠は免疫学的に安静させるのに十分でなければならない。1つの実施態様では、この安静は少なくとも6ヶ月である。初回接種には1つのタイプのベクターを用いる複数回(例えば、2〜4回)の初回接種が含まれ得る。
【0163】
ポックスウイルスベクター中に存在する発現カセットは、当該ポックスウイルスまたは別のポックスウイルスメンバーに生まれつきであるかまたはそれから誘導されるプロモーターを含んでいなければならない。ワクシニアウイルス、修飾ワクシニアウイルス、アビポックスウイルス、アライグマポックスウイルス、修飾ワクシニアウイルスAnkara、カナリアポックスウイルス(例えば、ALVAC)、トリポックスウイルス、ウシポックスウイルス及びNYVACをベースとするベクターを含めた各種タイプのポックスウイルスベースのベクターを構築、使用するための諸戦略が当業界で公知である(Moss,Current Topics in Microbiology and Immunology,158:25−38(1982);Earlら,「分子生物学における現在のプロトコル(Current Protocols in Molecular Biology)」,Ausubelら編,ニューヨークのGreene Publishing Associates & Wiley Interscience(1991年)発行,16.16.1−16.16.7;Childら,Virology,174(2):625−9(1990);Tartagliaら,Virology,188:217−232(1992);米国特許第4,603,112号明細書、同第4,722,848号明細書、同第4,769,330号明細書、同第5,110,587号明細書、同第5,174,993号明細書、同第5,185,146号明細書、同第5,266,313号明細書、同第5,505,941号明細書、同第5,863,542号明細書及び同第5,942,235号明細書)。
【0164】
(E) アジュバント
HCVワクチンはアジュバントと一緒に処方され得る。アジュバントはDNAプラスミドワクチンのために特に有用である。アジュバントの例はミョウバン、AlPO4、アルヒドロゲル、リピド−A及びその誘導体または変異体、フロインド不完全アジュバント、中性レポソーム、ワクチン及びサイトカインを含むリポソーム、ノニオン性ブロックコポリマー及びケモカインである。
【0165】
ポリオキシエチレン(POE)及びポリオキシルプロピレン(POP)を含むノニオン性ブロックポリマー、例えばPOE−POP−POEブロックコポリマーがアジユバントとして使用され得る(Newmanら,Critical Reviews in Therapeutic Drug Carroer Systems,15:89−142(1998))。核酸の免疫応答はノニオン性ブロックコポリマーとアニオン性界面活性剤を併用して強化され得る。
【0166】
アジュバント処方物の特定例は、CRL−1005(CytRx Research Laboratoris)、DNA及びベンジルアルコニウムクロリド(BAK)を含むものである。前記処方物は、PBS中にプラスミドDNAを含む冷(<5℃)溶液に純粋なポリマーをポジティブ置換ビペットを用いて添加することにより調製され得る。次いで、前記溶液を撹拌して、ポリマーを可溶化する。ポリマーが完全溶解したら、ポリマーの曇点(から6−7℃)以下の温度で透明な溶液が得られる。次いで、PBS中に溶解したBAKの希溶液をゆっくり添加することにより約4mM BAKをPBS中DNA/CRL−1005溶液に添加する。ポリマー及びBAK添加前の初期DNA濃度は約6mg/mlであり、最終濃度は約5mg/mlである。BAKを添加後、処方物を十分に撹拌し、その間に温度は〜2℃から曇点以上に上昇する。次いで、処方物を氷上に置いて、温度を曇点以下に下げる。その後、処方物を撹拌すると、温度は〜2℃から曇点以上に上昇する。動的光散乱により測定して処方物の粒径が約200〜500nmに達するまで温度を〜2℃から曇点以上に上昇させながら冷却及び混合を数回繰り返す。次いで、溶液が透明になるまで処方物を氷上に保存した後、−70℃で保存する。使用前に、処方物を室温まで融解させる。
【0167】
(F) ワクチン貯蔵
アデノベクター及びDNAワクチンは各種タイプの緩衝液を用いて貯蔵され得る。例えば、以下の実施例9に記載の緩衝液A105をベクター貯蔵のために使用し得る。
【0168】
DNAの貯蔵は、微量金属イオンを除去またはキレート化することにより向上され得る。DNAワクチンの安定性を向上させるためにコハク酸やリンゴ酸のような試薬及びキレート剤を使用し得る。キレート剤の例には、複数のホスフェートリガンド及びEDTAが含まれる。DNAプラスミドのダメージを遊離ラジカル生成から保護するために非還元性フリーラジカルスカベンジャー(例えば、エタノールまたはグリセロール)の配合も有用であり得る。更に、緩衝液タイプ、pH、塩濃度、露光及びバイアルを作成するために使用される滅菌方法のタイプをDNAワクチンの安定性を最適化するために処方物中で調節してもよい。
【実施例】
【0169】
以下、本発明の各種要件を更に説明するために実施例を提示する。これらの実施例は本発明を実施するために有用な方法を例示する。これらの実施例は本発明を限定するものではない。
【0170】
(実施例1)
Met−NS3−NS4A−NS4B−NS5A−NS5B発現カセット
HCV NS3−NS4A−NS4B−NS5A−NS5Bをコードする各種遺伝子発現カセットを1bサブタイプHCV BK株をベースとして構築した。コードされる配列は、(1)活性NS5B配列(“NS”)、(2)不活性NS5B配列(“NSmut”)、または(3)不活性NS5B配列を有するコドン最適化配列(“NSOPTmut”)を有していた。前記発現カセットはCMVプロモーター/エンハンサー及びBGHポリアデニル化シグナルも含んでいた。
【0171】
NSヌクレオチド配列(配列番号5)はHCV BK株(GenBank受託番号M58335)と5952個のヌクレオチド中30個のみ異なっている。NSアミノ酸配列(配列番号6)は対応の1b遺伝子型HCV BK株と1984個のアミノ酸中7個のみ異なっている。翻訳を開始するために、ATGコドンがNS配列の5’末端に存在している。TGA終止配列がNS配列の3’末端に存在している。
【0172】
NSmutヌクレオチド配列(配列番号2,図2)はNS配列に類似している。NSmutとNSの違いとして、NSmutが改変NS5B触媒部位、5’末端に最適リボソーム結合部位及び3’末端にTAAA終止配列を有していることが挙げられる。NS5Bの改変はアミノ酸1711〜1713をコードする塩基5138〜5146を含む。この改変の結果、アミノ酸GlyAspAspがAlaAlaGlyに変化し、不活性形態NS5B RNA依存性RNAポリメラーゼNS5Bが生ずる。
【0173】
NSOPTmut配列(配列番号3,図3)をNSmutによりコードされるアミノ酸配列をベースとして設計した。NSmutアミノ酸配列をGCG(ウィスコンシンパッケージ バージョン10;ウィスコンシン州マディソンに所在のGenetics Computer Group(GCG))BLACKTRANSLATEプログラムを用いてヌクレオチド配列に逆翻訳した。各アミノ酸が対応する最も頻繁に見られるヒトコドンによりコードされるNSOPTmutヌクレオチド配列を作成するために、パラメーターとして最もありそうなヌクレオチド配列の作成を選択し、翻訳スキームとしてGCGパッケージ内で利用可能な高発現ヒト遺伝子のコドン出現頻度表(human_high.cod)を特定して前記プログラムを実行した。
【0174】
(実施例2)
NS、NSmutまたはNSOPTmut配列を有するpV1Jnsプラスミドの作成
NS配列、NSmut配列またはNSOPTmut配列のいずれかを含むpV1Jnsプラスミドを以下のように作成し、特徴づけた。
【0175】
NS配列を有するpV1Jnsプラスミド
HCV BKタイプ株由来のコード領域Met−NS3−NS4A−NS4B−NS5A及びコード領域Met−NS3−NS4A−NS4B−NS5A−NS5B(Tomeiら,J.Virol.,67:4017−4026(1993))をpcDNA3プラスミド(Invitrogen)にクローン化して、それぞれpcD3−5a及びpcD3−5bを作成した。PcD3−5AをHindIIIで消化し、クレノウ充填で平滑末端化し、その後XbaIで消化して、Met−NS3−NS4A−NS4B−NS5Aのコード領域に対応する断片を作成した。この断片をpV1Jns−polyにクローン化し、BglIIで消化し、クレノウ充填で平滑末端化し、その後XbaIで消化して、pV1JnsNS3−5Aを作成した。
【0176】
pV1Jns−polyは、XbaI、PmeI、PacIに対する認識部位を含むポリリンカーをユニークBglII及びNotI制限部位に挿入することにより修飾したpV1JnsAプラスミド(Montgomeryら,DNA and Cell Biol.,12:777−783(1993))の誘導体である。NS配列を有するpV1Jns(pV1JnsNS3−5B)は、XbaI及びNotI消化で直線化したpV1JN3−5Aと約200bpのNS5A、NS5Bコード領域及び約60bpのBGHポリアデニル化シグナルを含むPCR断片とを同時形質転換させる細菌株BJ5183への相同組換えにより得られた。生じたプラスミドをpV1Jns−NSと称する。
【0177】
pV1Jns−NSは以下のように要約され得る:
塩基 pV1JnsAの1〜1881、
追加 AGCTT、
次に Met−NS3−NS5B配列(配列番号5)、
次に wt TGAストップ、
追加 TCTAGAGCGTTTAAACCCTTAATTAAGG(配列番号14)、
塩基 pV1JnsAの1912〜4909。
【0178】
NSmut配列を有するpV1Jnsプラスミド
V1JnsNS3−5Aを、完全コザック配列の付加によりNS3コード配列の5’において修飾した。プラスミド(V1JNS3−5Aコザック)は、AflII消化により直線化したV1JNS3−5Aと、イントロンAの近位部分、制限部位BglII、完全コザック翻訳開始配列及びNS3コード配列の一部を含むPCR断片とを同時形質転換させる細菌株BJ5183への相同組換えにより得られた。
【0179】
生じたプラスミド(V1JNS3−5Aコザック)をXbaI消化により直線化し、約200bpのNS5A、NS5B変異配列、強翻訳停止TAAA及び約60bpのBGHポリアデニル化シグナルを含むPCR断片と共に細菌株BJ5183に同時形質転換した。増幅のために使用されるオリゴヌクレオチドにより変異を導入した22bp重複断片を2つ組合せることによりPCR断片を得た。生じたプラスミドをpV1Jns−NSmutと称する。
【0180】
pV1Jns−NSmutは以下のように要約され得る:
塩基 pV1JnsAの1〜1882、
次に コザックMet−NS3−NS5B(mut)TAAA配列(配列番号2)、
追加 TCTAGA、
塩基 pV1JnsAの1925〜4909。
【0181】
NSOPTmut配列を有するpV1Jnsプラスミド
酵素活性を阻害するための変異NS5B、完全コザック翻訳開始配列及び強翻訳停止を有するヒトコドン最適化合成遺伝子(NSOPTmut)を遺伝子の5’及び3’末端に存在するBamHI及びSalI制限部位で消化した。次いで、遺伝子をpV1JnsAプラスミドのポリリンカー中に存在するBglII及びSalI制限部位にクローン化して、pV1Jns−NSOPTmutを作成した。
【0182】
pV1Jns−NSOPTmutは以下のように要約され得る:
塩基 pV1JnsAの1〜1881、
追加 C、
次に コザックMet−NS3−NS5B(optmut)TAAA配列(配列番号3)、
追加 TTTAAATGTTTAAAC(配列番号15)、
塩基 pV1JnsAの1905〜4909。
【0183】
プラスミドの特徴づけ
L−グルタミン(最終4mM)を補充した10% FCS/DMEMにおいて増殖させたHEK293細胞のトランスフェクションによりHCV NSタンパク質の発現を試験した。トランスフェクションの24時間前に、トランスフェクション日に90〜95%集密度に達するまで細胞を直径35mmの6ウェルにおいて平板培養した。LIPOFECTAMINE 2000試薬を用いて、ラウス肉腫ウイルスプロモーターの制御下で40ngのプラスミドDNA(非飽和DNA量として前もって測定)を100ngのルシフェラーゼリポーター遺伝子を含むpRSV−Lucプラスミドと同時トランスフェクトした。細胞をCO2インキュベーター中37℃において48時間保持した。
【0184】
細胞抽出物を1% トリトン/TEN緩衝液中で調製した。この抽出物をルシフェラーゼ活性について正規化し、10% SDS−アクリルアミドゲル上で連続希釈で流した。タンパク質をニトロセルロースに移し、発現強度及び適正なタンパク質分解開裂を評価するためにNS3、NS5A及びNS5Bに対する抗体を用いてアッセイした。偽トランスフェクトした細胞をネガティブコントロールとして用いた。pV1JnsNS、pV1JnsNSmut及びpV1JnsNSOPTmutを試験した代表的な実験の結果を図12に示す。
【0185】
(実施例3)
プラスミドDNAベクターを用いるマウス免疫化
抗−HCV免疫応答を誘発する可能性を評価するために、各種マウス株にDNAプラスミドpV1Jns−NS、pV1Jns−NSmut及びpV1Jns−NSOPTmutを注射した。2種の株(Balb/C及びC57Black6,N=9−10)に対してDNA(25または50μg)を筋肉内注射した後電気パルスを加えた。各動物に対して3週間間隔で2回投与した。
【0186】
C57Black6マウスにおいてNS3タンパク質に対して誘発された体液性免疫応答を投与後の2つの血清中で細菌発現したNS3プロテアーゼドメインに対するELISAにより測定した。試験抗原に対して特異的な抗体が3つのすべてのベクターを用いて免疫化した動物で検出され、幾何平均(相乗平均)力価(GMT)は94000〜133000であった(表1〜3)。
【0187】
【表1】

【0188】
【表2】
【0189】
【表3】
【0190】
3週間間隔でプラスミドDNA(25μg)を2回筋肉内注射して免疫化したC57Black6マウスにおいてT細胞応答を調べた。定量ELIspotアッセイを実施して、NS3−MS5B配列を包含する10残基が重複している20merペプチドの5プールに応答してIFNγを分泌するT細胞の数を測定した。特異的CD8+応答を、C57Black6マウスの場合にはCD8+エピトープを包含する20merペプチド(pep1480)を用いて同一アッセイにより分析した。
【0191】
IFNγを抗原特異的に分泌する細胞を標準ELIspotアッセイを用いて検出した。3週間間隔でプラスミドDNA(50μg)を2回筋肉内注射して免疫化したC57Black6マウスにおけるT細胞応答を、NS3−NS5B配列を包含する10残基が重複している20merペプチドの5プールに応答してIFNγを分泌するT細胞の数を測定する同一のELIspotアッセイにより分析した。
【0192】
免疫化マウスから脾臓細胞を作成し、R10培地(10% FCS、2mM L−グルタミン、50U/ml〜50μg/ml ペニシリン/ストレプトマイシン、10mM Hepes、50μM 2−メルカプトエタノールを補充したRPMI 1640)に再懸濁した。マルチスクリーン96ウェル濾過プレート(Milliporeのカタログ番号MAIPS4510;マサチューセッツ州アッシュビー・ロード・ベッドフォードに所在のMillipore Corporation)を精製ラット抗−マウスIFN−γ抗体(PharMingenのカタログ番号18181D;カリフォルニア州サンジェゴに所在のPharmiMingen)で被覆した。一晩インキュベート後、プレートをPBS 1×/0.005% ツイーンで洗浄し、R10培地(250μl/ウェル)でブロックした。
【0193】
免疫化マウスから脾細胞を作成し、2.5×105/ウエルまたは5×105/ウエルの密度で10μM ペプチドの存在または非存在下で24時間インキュベートした。十分洗浄(PBS 1×/0.005% ツイーン)した後、ビオチニル化ラット抗−マウスIFNγ抗体(PharMingenのカタログ番号18112D;カリフォルニア州サンディェゴに所在のPharMingen)を添加し、4℃において一晩インキュベートした。展開のために、ストレプトアビジン−AKP(PharMingenのカタログ番号13043E;カリフォルニア州サンジェゴに所在のPharMingen)及び1−Step(商標)NBT−BCIP展開溶液(Pierceのカタログ番号34042;イリノイ州ロックフォードに所在のPierce)を添加した。
【0194】
HCV BK株NS3〜NS5Bの全配列を包含する20mer重複ペプチドのプールを用いて、HCV特異的IFNγ分泌T細胞を明らかとした。同様に、C57Black6マウスの場合には、CD8応答を検出するためにCD8+エピトープを包含する1つの20merペプチドを用いた。プラスミドベクターpV1Jns−NS、pV1Jns−NSmut及びpV1Jns−NSOPTmut(25〜50μg)を2回注射して免疫化したC57Black6及びBalb/Cマウス(N=9−10)のグループの代表的データーを図13A及び13Bに示す。
【0195】
(実施例4)
アカゲサルの免疫化
7.5mg/mlのCRL1005、0.6mM ベンザルコニウムクロリド中のプラスミドpV1Jns−NSOPTmut(5mg)を筋肉内注射することによりアカゲザル(N=3)を免疫化した。各動物の三角筋に0及び4週目に2回注射した。
【0196】
CMIをIFN−γ ELISPOTによりいろいろな時間に測定した。このアッセイはHCV抗原特異的CD8+及びCD4+ Tリンパ球応答を測定し、ヒト、アカゲザル、マウス及びラットのような各種哺乳動物に対して使用され得る。
【0197】
特定ペプチド及びペプチドプールを使用すると、CTL細胞毒性アッセイ、インターフェロン−γ ELISPOTアッセイ及びインターフェロン−γ細胞内染色アッセイにおける抗原提示を単純化し得る。各種HCVタンパク質(コア、E2、NS3、NS4A、NS4B、NS5A、NS5B)のアミノ酸配列をベースとするペプチドを、HCV DNA、アデノウイルスベクターワクチン接種したアカゲザル及びHCV感染ヒトにおける免疫応答を調べるために上記アッセイで使用するために作成した。各ペプチドは、10アミノ酸で埋め合わされる重複20merである。大プールのペプチドはHCVタンパク質に対する全応答を検出するために使用され得、小プールのペプチド及び各ペプチドは応答のエピトープ特異性を規定するために使用され得る。
【0198】
IFNγ−ELISPOT
IFNγ ELISPOTアッセイによりHCV特異的Tリンパ球応答が定量測定される。PBMCを連続希釈し、抗−アカゲザルIFN−γ抗体(MD−1 U−Cytech)を被覆したマイクロプレートウェルに置く。これらをHCVペプチドプールと20時間培養すると、前駆体細胞の再刺激及びIFN−γの分泌が生じる。細胞を洗浄すると、細胞が存在している濃厚域に抗体被覆ウェルに結合した分泌IFNが残る。捕捉IFNはビオチニル化抗−アカゲザルIFN抗体(デテクターAb U−Cytech)及びアルカリホスファターゼコンジュゲートストレプトアビジン(Pharmingen 13043E)を順次用いて検出する。不溶性アルカリホスファターゼ基質を添加すると、ウェル中の細胞が存在する場所に暗いスポットが生じ、IFN−γを分泌したT細胞毎に1つのスポットが残る。
【0199】
ウェルあたりのスポットの数は抗原特異的T細胞の前駆体頻度に直接関連している。γ−インターフェロンが最も一般的であり、活性化Tリンパ球により合成、分泌される最も豊富なサイトカインの1つであるので、これを(特定の抗−γインターフェロンモノクローナル抗体を用いる)本アッセイで可視化されるサイトカインとして選択した。このアッセイでは、100万個のPBMCあたりのスポット形成細胞(SFC)の数をペプチド抗原の存在及び非存在(培地コントロール)下でサンプル毎に測定する。2つの材料を投与後のPBMCについてのアカゲザルからのデーターを表4に示す。
【0200】
【表4】
【0201】
(実施例5)
Ad6プリアデノウイルスプラスミドの構築
Ad6プレアデノウイルスプラスミドを以下のように得た。
【0202】
pAd6 E1−E3+プレアデノウイルスプラスミドの構築
第1世代Ad6ベクターを作成するために使用され得るAd6ベースのプレアデノウイルスプラスミドを、Ad5とAd6間の広い配列同一性(約98%)を利用するかまたはAd6領域のみを含めて構築した。wtAd6配列を細菌プラスミドにクローン化するために相同組換えを使用した。
【0203】
Ad5領域及びAd6領域を含む細菌プラスミドとしてpAd6E1−E3+を回収するために使用される一般的方法を図10に示す。BJ5183細菌を精製wtAd6ウイルスDNA及びAd5 ITRカセットと称される第2DNA断片を用いて同時形質転換すると、相同組換えによりウイルスゲノムが回付(circularization)した。ITRカセットは、細菌性複製起点及びアンピシリン耐性遺伝子を含むプラスミド配列により分離されるAd5ゲノムの右端(bp33798〜35935)及び左端(bp1〜341及びbp3525〜5767)由来の配列を含む。ITRカセットはAd5 342〜3524のE1配列の欠失を含む。ITRカセット中のAd5配列により、組換えが起こり得る精製Ad6ウイルスDNAと相同な領域が生ずる。
【0204】
可能性あるクローンを制限分析によりスクリーニングし、1つのクローンをpAd6E1−E3+として選択した。次いで、このクローンを完全に配列分析した。pAd6E1−E3+は、bp1〜341及びbp3525〜5548のAd5配列、Ad6 bp5542〜33784、Ad5 bp33967〜35935を含む(bpの数はAd5及びAd6のwt配列を指す)。pAd6E1−E3+は、血清型特異性を構成する全てのAd6ビリオン構造タンパク質に対するコード配列を含んでいる。
【0205】
Ad6領域を含む細菌プラスミドとしてpAd6E1−E3+を回収するために使用される一般的方法を図11に示す。BJ5183細菌を精製wt Ad6ウイルスDNA及びAd6 ITRカセットと称される第2DNA断片を用いて同時形質転換すると、相同組換えによりウイルスゲノムが回付した。ITRカセットは、細菌性複製起点及びアンピシリン耐性遺伝子を含むプラスミド配列により分離されるAd6ゲノムの右端(bp35460〜35759)及び左端(bp1〜450及びbp3508〜3807)由来の配列を含んでいる。これら3つのセグメントをPCRにより作成し、pNEB193に順次クローン化して、pNEBAd6−3(ITRカセット)を作成した。このITRカセットはAd5 451〜3507のE1配列の欠失を含む。ITRカセット中のAd6配列により、組換えが起こり得る精製Ad6ウイルスDNAと相同な領域が生ずる。
【0206】
pAd6 E1−E3−プレアデノウイルスプラスミドの構築
A5領域を含み、E3領域が欠失しているAd6ベースのベクターを、Ad5領域を含むpAd6E1−E3+から始めて構築した。E3領域(Ad6 bp25871〜31192)を含むpAd6E1−E3+のA 5322bpサブ断片をpABS.3にサブクローン化して、pABSAd6E3を作成した。その後、このプラスミドにおいて3つのE3を欠失させて、3つの新しいプラスミドpABSAd6E3(1.8Kb)(Ad6 bp28602〜30440欠失)、pABSAd6E3(2.3Kb)(Ad6 bp28157〜30437欠失)及びpABSAd6E3(2.6Kb)(Ad6 bp28157〜30788欠失)を作成した。次いで、pAd6E1−E3+に3つのE3欠失部を逆置換するために細菌組換えを使用して、Ad6ゲノムプラスミドpAd6E1−E3−1.8Kb、pAd6E1−E3−2.3Kb及びpAd6E1−E3−2.6Kbを作成した。
【0207】
(実施例6)
NS配列を有するAd5ゲノムプラスミドの作成
コード領域NS3−NS4A−NS4B−NS5Aを含むpcDNA3プラスミド(Invitrogen)をXmuI及びNruI制限部位で消化し、CMVプロモーター、NS3−NS4A−NS4B−NS5Aコード領域及びウシ成長ホルモン(BGH)ポリアデニル化シグナルを含むDNA断片をシャトルベクターpDelE1 SpaのユニークEcorV制限部位にクローン化して、Sva3−5Aベクターを作成した。
【0208】
コード領域NS3−NS4A−NS4B−NS5A−NS5Bを含むpcDNA3プラスミドをXmnI及びEcorIで消化(部分消化)し、NS5Aの一部、NS5B遺伝子及びBGHポリアデニル化シグナルを含むDNA断片をSva3−5Aベクターにクローン化し、EcorI及びBglIIで消化し、クレノウで平滑末端化して、Sva3−5Bベクターを作成した。
【0209】
Sva3−5Bベクターは最後に消化したSspI及びBst1107I制限部位であり、アデノウイルス配列に隣接している発現カセット(CMVプロモーター,NS3−NS4A−NS4B−NS5A−NS5Bコード領域及びBGHポリアデニル化シグナル)を含むDNA断片をpAd5HVO(E1−,E3)ClaI直線化ゲノムプラスミドと共に細菌株BJ5183に同時形質転換して、pAd5HVONSを作成した。pAd5HVOはAd bp1〜341、bp3525〜28133及びbp30818〜35935を含んでいる。
【0210】
(実施例7)
NSmut配列を有するアデノウイルスゲノムプラスミドの作成
NS−mut配列を含むアデノウイルスゲノムプラスミドをAd5またはAd6バックグラウンドで作成した。Ad6バックグラウンドは塩基1〜450、3511〜5548及び33967〜35935でAd5領域を含んでいた。
【0211】
pV1JN3−5AコザックをBglII及びXbaI制限酵素で消化し、コザック配列及びNS3−NS4A−NS4B−NS5Aをコードする配列を含むDNA断片をBglII及びXbaI消化ポリpMRKpdelE1シャトルベクターにクローン化した。生じたベクターをshNS3−5Aコザックと称した。
【0212】
ポリpMRKpdelE1はBglII、PmeI、SwaI、XbaI、SalIに対する認識部位を含むポリリンカーをCMVプロモーターの下流に存在するユニークBglII制限部位に挿入することにより修飾したRKpdelE1(Pac/pIX/pack450)+CMVmin+BGHpA(str.)の誘導体である。MRKpdelE1(Pac/pIX/pack450)+CMVmin+BGHpA(str.)は、bp451〜3510のE1配列を欠失させたbp1〜5792のAd5配列を含む。ヒトCMVプロモーター及びBGHポリアデニル化シグナルをそれらを分離するユニークBglII部位とE1平行方位のE1欠失部に挿入した。
【0213】
酵素活性を阻害するように変異させた、3’末端に強翻訳停止を有するNS5B断片をアセンブリPCRにより得、相同組換えによりshNS3−5Aコザックベクターに挿入して、ポリpMRKpdelE1NSmutを作成した。ポリpMRKpdelE1NSmutでは、NS−mutコード配列はCMVプロモーターの制御下にあり、BGHポリアデニル化シグナルは下流に存在している。
【0214】
遺伝子発現カセット及び相同組換えし得るアデノウイルス配列を含む隣接領域をPacI及びBst1107I制限酵素を用いる消化により切り出し、pAd5HVO(E1−,E3−)またはpAd6E1−E3−2.6Kb ClaI直線化ゲノムプラスミドと共に細菌株BJ5183に同時形質転換して、それぞれpAd5HVONSmut及びpAd6E1−,E3−NSmutを作成した。
【0215】
pAd6E1−E3−2.6Kbは、Ad5 bp1〜341及びbp3525〜5548、Ad6 bp5542〜28157及びbp 30788〜33784、Ad5 bp33967〜35935を含んでいる(bpの数はAd5及びAd6の両方に対するwt配列を指す)。いずれのプラスミドでも、ウイルスITRは細菌性複製起点及びアンピシリン耐性遺伝子を含むプラスミド配列により連結されている。
【0216】
(実施例8)
NSOPTmutを有するアデノウイルスゲノムプラスミドの作成
pCRBluntベクター(Invitrogen)にクローン化した配列番号3のヒトコドン−最適化合成遺伝子(NSOPTmut)をBamH1及びSalI制限酵素で消化し、シャトルベクターポリpMRKpdelE1中に存在するBglIV及びSalI制限部位にクローン化した。生じたクローン(ポリpMRKpdelE1NSOPTmut)をPacI及びBst1107I制限酵素で消化し、pAd5HVO(E1−,E3−)またはpAd6E1−E3−2.6Kb ClaI直線化ゲノムプラスミドと共に細菌株BJ5183に同時形質転換して、それぞれpAd5HVONSOPTmut及びpAd6E1−,E3−NSOPTmutを作成した。
【0217】
(実施例9)
アデノウイルスベクターのレスキュー及び増幅
アデノベクターをPer.6細胞においてレスキューした。Per.C6を、L−グルタミン(最終4mM)、ペニシリン/ストレプトマイシン(最終100U/ml)及び10mM MgCl2を補充した10% FCS/DMEMにおいて増殖した。感染後、細胞を5% ウマ血清(HS)を補充した同一培地において維持した。ウイルスレスキューのために、2.5×106個のPer.C6を6cmΦペトリ皿において平板培養した。
【0218】
平板培養から24時間後、PacI直線化アデノウイルスDNA(10μg)を用いてリン酸カルシウム法により細胞をトランスフェクトした。DNA沈殿を細胞上に4時間放置した。培地を除去し、5% HS/DMEMを添加した。
【0219】
細胞変性効果が目に見えるまで(1週間)細胞をCO2インキュベーター中で維持した。細胞及び上清を回収し、凍結/融解サイクル(液体窒素/37℃の水浴)に3回かけた。ライセートを−4℃において3000rpmで20分間遠心し、回収した上清(細胞上に1回通したウイルスを含有する細胞ライセートに対応;P1)を10cmΦペトリ皿において80〜90%集密度のPer.C6を感染させるために1ml/皿の量で使用した。感染させた細胞を細胞変性効果が目に見えるまでインキュベートし、細胞及び上清を回収し、ライセートを上記したように作成した(P2)。
【0220】
2×15cmΦペトリ皿を感染させるためにP2ライセート(4ml)を使用した。この感染から回収したライセート(P3)を大きなウイルス調製物に対する出発点として使用されるウイルスストックとして複数のアリコートで−80℃において保存した。この場合、2×15cmΦペトリ皿を感染させるためには1mlのストックで十分であり、生じたライセート(P4)を大規模感染のためのペトリ皿の感染用に使用した。
【0221】
FCSを含まない培地で希釈し、10ml/皿の量で30×15cmΦペトリ皿(80〜90%集密度のPer.C6を含む)を感染させるために使用したP4ライセートから更なる増幅物を得た。細胞をCO2インキュベーター中で20分ごとに緩やかに混合しながら1時間インキュベートした。12ml/皿の5% HS/DMEMを添加し、細胞変性効果が目に見えるまで(約48時間)細胞をインキュベートした。
【0222】
細胞及び上清を集め、4℃において2000rpmで20分間遠心した。ペレットを0.1M トリス(pH8.0)(15ml)中に再懸濁した。細胞を凍結/融解サイクル(液体窒素/37℃の水浴)に3回かけて溶解した。2M MgCl2(150μl)及びDNAse(10mlの20mM トリス−HCl(pH7.4)、50mM NaCl、1mM ジチオトレイトール、0.1mg/ml ウシ血清アルブミン、50% グリセロール中10mg ウシ膵デオキシリボヌクレアーゼI)(75μl)を添加した。水浴において37℃において1時間インキュベート(15分毎に撹拌)した後、ライセートを4℃において4000rpmで15分間遠心した。回収した上清を直ちにCsCl勾配にかけた。
【0223】
CsCl勾配は、SW40無菌チューブにおいて次のように作成した:
1.5d CsCl(0.5ml)、
1.35d CsCl(3ml)、
1.25d CsCl(3ml)、
ウイルス上清(5ml/チューブ)を適用した。
【0224】
所要により、チューブの上部に0.1M トリス−Cl(pH8.0)を満たした。チューブを−10℃においてローターSW40を用いて35000rpmで1時間遠心した。(1.25/1.35インターフェースに位置する)ウイルスバンドを注射器により集めた。
【0225】
ウイルスを新しいSW40無菌チューブに移し、1.35d CsClをチューブの上部に満たした。10℃においてローターSW40において35000rpmで24時間遠心後、ウイルスをできるだけ少量で集め、緩衝液A 105(5mM トリス,5% スクロース,75mM NaCl,1mM MgCl2,0.005% ポリソルベート80(pH8.0))に対して徹底的に透析した。透析後、グリセロールを最終10%まで添加し、ウイルスを−80℃において複数のアリコートで保存した。
【0226】
(実施例10)
強化アデノベクターレスキュー
HCV NSOPTmut導入遺伝子を有する第1世代Ad5及びAd6ベクターはレスキューしがたいことが判明した。レスキュー過程の考えられる障害はアデノウイルスの複製機構のための最適以下鋳型であるプラスミドDNAの複製が効率的でないことに起因するであろう。ベクターのE1領域に挿入された導入遺伝子の非常に高いG−C含量に関連して、(通常ウイルスDNA中に存在する)DNAの5’末端に連結している末端タンパク質が不在であるために、プラスミド誘導アデノウイルスの複製速度が実質的に低減し得る。
【0227】
Adベクターをレスキューするためのより効率的で再現性の手順を確立するために、tet−誘導性プロモーターの制御下で全てのE2タンパク質(ポリメラーゼ、プレ末端タンパク質及びDNA結合タンパク質)及びE4 orf6を含む発現ベクター(pE2;図19)を用いた。PerC6及び293にpE2及び一般的なプレアデノプラスミドをトランスフェクトすることにより、Ad DNA複製が非常に向上し、完全な感染性アデノウイルス粒子がより効率的に生じる。
【0228】
プラスミド構築
pE2はクローニングベクターpBI(CLONTECH)をベースとし、細胞培養においてエピソームを複製し、選択するために2つの要素、すなわち(1)EBNA−1を発現させるときに細胞サイクルと同期してプラスミドを複製できるEBV−OriP(EBV[nt]7421−8042)領域及び(2)形質転換した細胞をポジティブ選択し得るハイグロマイシンBホスホトランスフェラーゼ(HPH)耐性遺伝子を加えた。アデノウイルス遺伝子E2a及びb及びE4−Orf6に対する2つの転写単位を構築し、下記するようにpE2において合わせた。
【0229】
Ad5−ポリメラーゼClaI/SphI断片及びAd5−pTP Acc65/EcoRV断片をpVac−Pol及びpVac−pTP(Stunnembergら,NAR,16:2431−2444(1988))から得た。両断片にクレノウを充填し、pBIのSalI(充填)及びEcoRV部位にクローン化して、それぞれpBI−Pol/pTPを得た。
【0230】
pCEP4(Invitrogen)由来のEBV−OriP要素をまずpJC13−1(Chungら,Cell,74(3):505−14(1993))のBamHI部位にクローン化することにより2つのトリβ−グロビン隔離体ダイマー内に挿入した。次いで、pJC13−OriP由来のHS4−OriP断片をpSA1mv(tk−Hygro−B耐性遺伝子発現カセット及びAd5複製起点を含むプラスミド)内部にクローン化した。ここで、ITRを頭−尾ジャンクションとして配置し、pFG140(Grahan,EMBO J.,3:2917−2922(1984))から5’−TCGAATCGATACGCGAACCTACGC−3’(配列番号16)及び5’−TCGACGTGTCGACTTCGAAGCGCACACCAAAAACGTC−3’(配列番号17)のプライマーを用いてPCRにより得た。こうして、pMVHS4Oripを作成した。次いで、隔離Orip、Ad5 ITRジャンクション及びtk−HygroBカセットを含むpMVHS4Orip由来のDNA断片をAseI/AatII制限pBI−Pol/pTPに挿入して、pBI−Pol/pTPHS4を作成した。
【0231】
Ad5−Orf6及びAd5−DBPを発現する第2転写単位を構築するために、PCRにより得たE4orf6(Ad5[nt]33193−34077)をまずpBIベクターに挿入して、pBI−Orf6を作成した。その後、DBPコーディングDNA配列(Ad5[nt]22443−24032)をpBI−Orf6に挿入して、第2の双方向的Tet調節発現ベクター(pBI−DBP/E4Orf6)を得た。pBI中に存在するオリジナルのポリAシグナルをBGH及びSV40ポリAで置換した。
【0232】
次いで、頭−尾ジャンクションで配置したAdeno5−ITR及びプラスミドpSA−1mvから得たハイグロマイシンB耐性遺伝子を含むDNA断片を挿入することによりpBI−DBP/E4orf6を修飾した。次いで、この新しいプラスミドpBI−DBP/E4orf6シャトルをドナープラスミドとして使用して、第2tet−調節転写単位を、pE2を保有する大腸菌株BJ5183を用いる相同組換えによりpBI−Pol/pTPHS4に挿入した。
【0233】
細胞株、トランスフェクション及びウイルス増幅
PerC6細胞を、10% ウシ胎仔血清(FBS)、10mM MgCl2、ペニシリン(100U/ml)、ストレプトマイシン(100μg/ml)及び2mM グルタミンを補充したダルベッコ改変イーグル培地(DMEM)において培養した。
【0234】
全ての一時的トランスフェクションをリポフェクタミン2000(Invitrogen)を製造業者が指示するように用いて実施した。6cmプレートにおいて平板培養した90%集密度のPERC.6(商標)をAd5/6NSOPTmutプレアデノプラスミド(3.5μg)でトランスフェクトし、PacIを単独でまたはpE2(5μg)+pUHD52.1(1μg)の組合せで用いて消化した。pUHD52.1は逆tetトランスアクティベーター2(rtTA2)(Urlinger,Proc.Natl.Acad.Sci.U.S.A.,97(14):7963−7968(2000))に対する発現ベクターである。トランスフェクション時に、pE2発現を活性化するために、細胞を1μg/mlのドキシサイクリンの存在下で培養した。トランスフェクションから7日後、細胞を回収し、細胞ライセートを凍結/融解サイクルを3回実施して得た。細胞ライセート(2ml)を使用して、第2のPerC6の6cm皿を感染させた。感染細胞を、完全CPEが観察されるまで培養した後、回収した。ウイルスを上記したように5回連続継代した後、CsCl勾配で精製した。精製ウイルスのDNA構造をエンドヌクレアーゼ消化及びアガロースゲル電気泳動分析によりコントロールし、元のプレアデノプラスミド制限パターンと比較した。
【0235】
(実施例11)
HCVポリタンパク質コード化核酸の部分最適化
ヒト宿主での発現のために最適化したコドンを含むアデノベクターの作成を促進するためにHCVポリタンパク質コード化核酸の部分最適化を実施した。総合的目的は、HCVポリタンパク質をコードするアデノベクターの作成を促進しながらコドン最適化により高い発現を与えることであった。
【0236】
ヒト宿主での発現のために最適化したコドンを含むHCVタンパク質をコードするアデノベクターを作成する際に幾つかの困難を経験した。最適化配列(配列番号3)を含むアデノベクターは、非最適化配列(配列番号2)を含むアデノベクターよりも合成及びレスキューが困難であることが判明した。
【0237】
配列番号3を含むアデノベクターを作成する際の困難は高いGC含量に起因していた。特に厄介な領域はNSOPTmut(配列番号3)の約No.3900の領域である。
【0238】
アデノベクター中での使用を容易とするために最適化HCVコード化核酸配列の別のバージョンを設計した。NSOPTmutと比較して、コード化ポリタンパク質及びそれぞれの開裂産物の発現を改善するように高レベルのコドン最適化を維持しながら低い全GC含量を有し、連続G’s及びC’sの潜在的に問題のモチーフの存在が低下/解消するように別のバージョンを設計した。
【0239】
コドンを次善に最適化した配列の作成の出発点はNSOPTmutヌクレオチド配列のコード領域(配列番号3の塩基7〜5961)である。コドン使用頻度の値(各アミノ酸につき1.0の全値に正規化した)をウィスコンシン・パッケージ バージョン10.3(Pharmacopeia,Inc.が100%出資の子会社のAccelrys Inc.)で販売しているファイルhuman_high.codから得た。
【0240】
局部及び全GC含量を低減するために、各アミノ酸毎に好ましいコドン置換を規定している表を手動で作成した。各アミノ酸について、1)最も一般的なコドンに比較して低いGC含量及び2)比較的に多く観察されるコドン使用頻度(human_high.codに定義されている)を有するコドンを置換コドンとして選択した。例えば、Argの場合、最高頻度のコドンはCGCである。Argをコードする他の5つの代替コドン(CGG、AGG、AGA、CGT、GGA)のうち、3つ(AGG、CGT、CGA)は1塩基により、1つ(AGA)は2塩基により、1つ(CGG)は0塩基によりGC含量を低減させる。AGAコドンはhuman_high.codに比較的低使用頻度(0.1)としてリストされているので、CGCを置換するコドンとして0.18の相対頻度を有するAGGが選択された。他のアミノ酸に対するコドン置換を確立するために同様の基準が適用され、表5に示すリストが作成された。以下の最適化手順で適用されるパラメーターは、生じる配列が十分に改善されたコドン使用(各アミノ酸について)を維持し、(全体及び保存G’s及び/またはC’sの局部ストレッチの形態の)GC含量が減少するように実験的に決定された。
【0241】
部分最適化HCVコード化配列の2つの例は配列番号10及び配列番号11である。配列番号10は全体的に部分最適化されているHCVコード化配列である。配列番号11は部分最適化されている領域を除いてコドン使用について完全に最適化されているHCVコード化配列である。
【0242】
コドン最適化は以下の手順を用いて実施した:
段階1)
インプット完全最適化NSOPTmut配列のコード領域を、各サイクル後1コドンだけスライディングウィンドをシフトする3コドン(9塩基)のスライディングウィンドを用いて分析した。5個以上の連続G’s及び/またはC’sを含むストレッチを前記ウィンドで検出したときにはいつも、以下の置換ルールを適用した。Let Nは既に実施したコドン置換の数を示す。Nが奇数のときにはウインド中の中間コドンを表5に特定したコドンで置換する。Nが偶数のときにはウィンドの第3末端コドンをhuman_high.codのようなコドン最適化表に特定されているコドンで置換する。LeuまたはValが第2または第3コドンに存在しているときには、非常に低い相対コドン使用頻度(例えば、human_high.cod参照)のLeuまたはValコドンが導入されないようにいずれの置換も適用しない。以下のサイクルで、シフトしたウインドの分析を以前のサイクルの置換を含む配列に適用した。
【0243】
3コドンウインド中の中間及び末端コドンの交互置換は、実験的に(手順後の最終配列から判断して)GC含量を低減させながら最適化コドン使用の全体維持をより満足させることが判明した。しかしながら、通常、正確な置換戦略は分析されるヌクレオチド配列によりコードされるアミノ酸配列に依存し、実験的に決定されるであろう。
【0244】
段階2)
段階1)で実施した全てのコドン置換を含む配列を21コドン(63塩基)長のスライディングウィンドを用いて更に分析した。調節可能なパラメーターに従ってウィンド中の全GC含量を測定した。ウィンド中のGC含量が70%以上ならば、以下のコドン置換戦略を適用した。このウィンドでは、アミノ酸Asn、Asp、Cys、Glu、His、Ile、Lys、Phe、Tyrのコドンを表5に示すコドンで置換する。この組のアミノ酸に対する置換の限定は、a)置換コドンがhuman_high.codにおける許容されるぐらい高い使用頻度を有している、及びb)置換コドンについてGUTG中の平均全ヒトコドン使用は最も高頻度くらい高いという事実により考えられた。以下のサイクルでは、シフトしたウインドの分析を以前のサイクルの置換を含む配列に適用される。
【0245】
各アミノ酸につきGC含量の全低減と高いコドン最適化の維持を妥協することにより閾値70%を実験的に決定した。段階1)と同様に、正確な置換戦略(アミノ酸の選択及びGC含量閾値)は分析されるヌクレオチド配列によりコードされるアミノ酸配列に依存し、実験的に決定されるであろう。
【0246】
段階3)
次いで、段階1)及び2)により作成した配列を手動で編集し、追加コドンを以下の基準に従って変化させた。21コドンのウィンドにおいてなお70%以上のGC含量を有している領域を手動で試験して、数個のコドンを表5に示すスキームに従って再び置換した。
【0247】
有用な制限部位を与えるため、相補鎖上の存在するオープンリーデイングフレームを除去するため、相同組換え領域を付加するために、コザックシグナルを付加するため及びターミネーターを付加するために以下の段階を実施した。これらの段階は段階4〜7である。
【0248】
段階4)
その後制限酵素のサブセットを用いてクローニングできるように、段階3で作成した配列を特定の制限部位(BglII、PmeI及びXbaI)の不在及び唯一のStuI部位の存在について試験した。2つの部位(BglIIに対する1部位及びStuIに対する1部位)を各認識部位の一部であるコドンを置換することにより配列から除去した。
【0249】
段階5)
次いで、段階1)〜4)により作成した配列をその後修飾NSOPTmut配列を(相同組換えにより)作成できるように修飾した。段階1)〜4)から得た配列では、塩基3556〜3755を含むセグメント及び塩基4456〜4656を含むセグメントをNSOPTmut由来の対応配列により置換した。相同組換えによりNSOPTmut中の問題の領域(No.3900あたり)を置換するために配列番号10の塩基3356〜4656を含むセグメントを使用して、配列番号11の配列を有するNSOPTmutの変異体を得ることができる。
【0250】
段階6)
段階1)〜5)で得た配列の分析から、相補鎖上の完全断片にほぼ広がっている可能性あるオープンリーディングフレームが示された。センス鎖からすべてのコドンCTA及びTTA(Leu)とTCA(Ser)を除去すると、相補鎖上の1つのリーディングフレームの1つ中の全てのストップコドンが有効的に除去された。この相補鎖オープンリーディングフレームの転写及びその後のタンパク質への翻訳の可能性は非常に少ないが、センス鎖上でコードされる配列の転写及びその後の翻訳との可能性ある干渉を避けるためにSerに対するTCAコドンをほぼ500塩基毎にセンス上に導入した。相同組換えを実施するために段階5)で導入したセグメントに変化を加えなかった。Serに対するTCAコドンはLeuに対するCTA及びTTAよりも好ましかった。なぜならば、human_high.codにおいてCTA(0.02)及びTTA(0.03)に比してTCA(0.05)は比較的により高い相対頻度であるからである。更に、CUTGからの平均ヒトコドン使用ではTCAが好ましかった(CTA及びTTAの0.07に対して0.14)。
【0251】
段階7)
最終段階では、最適化内部リボソームエントリー部位(コザックシグナル)を作成するためにGCCACCを配列の5’末端に付加し、3’にTAAAストップシグナルを付加した。NSsuboptmutの翻訳特性の開始を維持するために、コドン領域の最初の8コドンをNSOPTmut配列と同一に維持した。生じた配列を再びBglII、PmeI及びXbaI認識部位の不在及び唯一のStuI部位の存在についてチェックした。
【0252】
NSsuboptmut配列(配列番号10)は、NSOPTmut(70.3%)に比して全体的に低いGC含量(63.5%)を有しており、コドン使用最適化の十分に最適なレベルを維持している。NSsuboptmutのヌクレオチド同一性はNSmutに対して77.2%であった。
【0253】
【表5】
【0254】
(実施例12)
ウイルス特徴づけ
アデノベクターを、(a)物理的粒子/mlの測定、(b)TaqMan PCRアッセイの実行、及び(c)HeLa細胞の感染後のタンパク質発現のチェックにより特徴づけた。
【0255】
(a)物理的粒子の測定
CsCl精製ウイルスを0.1% SDS PBSで1/10及び1/100希釈した。コントロールとして、緩衝液A105を使用した。これらの希釈物を55℃において10分間インキュベートした。チューブを軽く回転させた後、260nmでODを測定した。ウイルス粒子の量を、1OD260nm=1.1×1012物理的粒子/mlに従って計算した。結果は典型的には5×1011〜1×1012物理的粒子/mlであった。
【0256】
(b)TaqMan PCRアッセイ
TaqMan PCRアッセイをアデノベクターゲノムの定量(Q−PCR粒子/ml)のために使用した。TaqMan PCRアッセイは、ABI Prism 7700−配列デテクターを用いて実施した。反応をアデノウイルス骨格に対して特異的なプローブ(最終200μMで)及びオリゴヌクレオチド(最終200nMで)の存在下で最終50μl容量で実施した。ウイルスを0.1% SDS PBSで1/10希釈し、55℃において10分間インキュベートした。チューブを軽く回転させた後、連続1/10希釈物(水中)を調製した。10−3、10−5及び10−7希釈物10μlをPCRアッセイの鋳型として使用した。
【0257】
各サンプル中に存在する粒子の量を、同一実験で作成した標準曲線に基づいて計算した。結果は典型的には1×1012〜3×1012Q−PCR粒子/mlであった。
【0258】
(c)HCV非構造タンパク質の発現
HCV NSタンパク質の発現をHeLa細胞を感染させることにより試験した。1.5×106細胞/皿(10cmΦペトリ皿)の感染前日に細胞を平板培養した。50、250及び1250pp/細胞のm.o.i.に対応する異なる量のCsCl精製ウイルスを最終容量5mlまで培地(FCS非含有)で希釈した。希釈ウイルスを細胞に添加し、CO2インキュベーター中37℃において1時間インキュベートした(20分毎にやさしく混合)。5% HS−DMEM(5ml)を添加し、細胞を37℃で48時間インキュベートした。
【0259】
1% トリトン/TEN緩衝液中で細胞抽出物を調製した。この抽出物を10% SDS−アクリルアミドゲル上に流し、ニトロセルロースにブロットし、NS3、NS5a及びNS5bに対する抗体を用いてアッセイして、適正なポリタンパク質開裂をチェックした。偽感染細胞をネガティブコントロールとして使用した。Ad5−NS、MRKAd5−NSmut、MRKAd6−NSmut及びMRKAd6−NSOPTmutを試験した代表的な実験の結果を図14に示す。
【0260】
(実施例13)
各種NSカセットをコードするアデノベクターを用いるマウス免疫化
アデノベクターAd5−NS、MRKAd5−NSmut、MRKAd6−NSmut及びMRKAd6−NSOPTmutをC57Black6マウス株に注射して、抗−HCV免疫応答を誘発する可能性を評価した。動物群(N=9−10)には109ppのCsCl精製ウイルスを筋肉内注射した。各動物に対して3週間間隔で2回投与した。
【0261】
NS3タンパク質に対する体液性免疫応答をC57Black6免疫化マウスからの投与後の2つの血清中で細菌発現させたNS3プロテアーゼドメインに対してELISAにより測定した。試験した抗原に対して特異的な抗体が100〜46000の幾何平均(相乗平均)力価(GMT)で検出された(表6、7、8及び9)。
【0262】
【表6】
【0263】
【表7】
【0264】
【表8】
【0265】
【表9】
【0266】
C57Black6マウスにおけるT細胞応答を、NS3−NS5B配列を包含する10残基が重複している20merペプチドの5プール(F〜L+Mの名称)に応じてIFNγを分泌するT細胞をの数を測定する定量ELISPOTアッセイにより分析した。C57Black6マウスにおいて誘導される特異的CD8−応答を、C57Black6マウスではCD8+エピトープを包含する20merペプチド(pep1480)を用いて同一アッセイにより分析した。抗原特異的にIFNγを分泌する細胞を標準ELIspotアッセイを用いて検出した。
【0267】
脾臓細胞、脾細胞及びペプチドを上記実施例3に記載されているように産生し、処理した。ベクターAd5−NS、MRKAd5−NSmut及びMRKAd6−NSmutの109ウイルス粒子を2回注射して免疫化したC57Black6マウスの群(N=9−10)からの代表的データーを図15に示す。
【0268】
(実施例14)
アデノベクターを用いるアカゲザルの免疫化
アカゲザル(N=3−4)を、CsCl精製Ad5−NS、MRKAd5−NSmut、MRKAd6−NSmutまたはMRKAd6−NSOPTmutウイルスを筋肉内注射することにより免疫化した。各動物の三角筋に1011または1010vpを0及び4週目に2回投与した。
【0269】
CMIを、a)IFN−γ ELISPOT(上記実施例3参照)、b)IFN−γ ICS及びc)バルクCTLアッセイにより異なる時間に測定した。これらのアッセイはHCV抗原特異的CD8+及びCD4+ Tリンパ球応答を測定し、ヒト、アカゲザル、マウス及びラットを含めた各種哺乳動物に対して使用され得る。
【0270】
特異的ペプチドまたはペプチドのプールの使用は、CTL細胞毒性アッセイ、インターフェロン−γ ELISPOTアッセイ及びインターフェロン−γ細胞内染色アッセイにおける抗原提示を単純化し得る。各種HCVタンパク質(コア、E2、NS3、NS4A、NS4B、NS5a、NS5b)のアミノ酸配列をベースとするペプチドを、HCV DNA、アデノウイルスベクターワクチン接種アカゲザル及びHCV感染ヒトにおける免疫応答を調べるための上記アッセイに使用するために作成した。各ペプチドは10アミノ酸で埋め合わされる重複20merである。大プールのペプチドをHCVタンパク質に対する全応答を検出するために使用することができ、小プールのペプチド及び各ペプチドは応答のエピトープ特異性を規定するために使用し得る。
【0271】
IFN−γ ICS
IFN−γ ICSのために、R10(10% FCSを補充したRPMI培地)(1ml)中の2×106PBMCをペプチドプール抗原で刺激した。各ペプチドの最終濃度は2μg/mlであった。細胞をCO2インキュベーター中37℃において1時間インキュベートした後、可溶性サイトカインの分泌を抑えるためにBrefeldin Aを10μg/mlの最終濃度まで添加した。細胞を37℃において更に14〜16時間インキュベートした。
【0272】
刺激を、共刺激抗体のCD28及びCD49d(抗−ヒトCD28 BD340975及び抗−ヒトCD49d BD340976)の存在下で実施した。インキュベート後、細胞を表面抗原に対する蛍光色素−コンジュゲート抗体の抗−CD3、抗−CD4、抗−CD8(CD3−APC Biosource APS0301、CD4−PE BD345769、CD8−PerCP BD345774)で染色した。
【0273】
細胞内サイトカインを検出するために、細胞をFACS透過化緩衝液2(BD340973)2×最終濃度で処理した。固定し、透過化した後、細胞をヒトIFN−γ、IFN−γFITCに対する抗体(Biosource AHC4338)とインキュベートした。
【0274】
細胞をPBS中の1% ホルムアルデヒド中に再懸濁し、24時間以内にFACSで分析した。4色FACS分析を2つのレーザーを備えたFACS Calibur装置(Becton Dickinson)を用いて実施した。CD3、CD8ポジティブ集団をカップリングしたForward対Side Scatterプロットにおいてリンパ球集団のゲーティングを得た。少なくとも3000事象のゲートが得られた。ポジティブ細胞を106リンパ球に対するIFN−γ発現細胞の数として表した。
【0275】
1010または1011vpの各種アデノベクターを1回または2回注射後の免疫化サルからのIFN−γ ELISPOT及びIFN−γ ICSデーターを図16A〜16D、17A及び17Bに示す。
【0276】
バルクCTLアッセイ
Tリンパ球の識別エフェクター機能は、この細胞集団のサブセットの適切なMHC−関連抗原ペプチドを示す細胞を直接溶解する能力である。この細胞毒性活性は、CD8+ Tリンパ球に最も頻繁に関係している。
【0277】
記憶T細胞の抗原再刺激及び増大を与えるように、PBMCサンプルにインビトロでHCV抗原を発現する組換えワクチンウイルスを約14日間感染させた。ペプチド抗原プールで処置した自己B細胞系に対する細胞毒性を試験した。
【0278】
培養物の溶解機能は、CTLエフェクター細胞との4時間インキュベーション中に標的細胞から遊離したクロムより生じた特異的溶解の%として測定する。特異的細胞毒性を測定し、無関係抗原または賦形剤処理B細胞系と比較した。このアッセイは半定量的であり、CTL応答がワクチンにより誘発されるかどうかを調べるための好ましい手段である。1011vp/投与のアデノベクターAd5−NS、MRKAd−NSmut及びMRKAd6−NSmutで免疫化したサルからの2回注射後のデーターを図18A〜18Fに示す。
【0279】
他の実施態様も請求の範囲内である。幾つかの実施態様を説明、例示してきたが、本発明の範囲及び趣旨を逸脱することなく各種修飾を加え得ることができる。
【背景技術】
【0001】
本明細書中で引用した文献は本発明に対する従来技術とは認められない。
【0002】
世界の人口の約3%がC型肝炎ウイルス(HCV)に感染している(Wasleyら,Semin.Liver Dis.,20:1−16(2000))。HCVに曝されると、まれに顕性急性疾患にかかるが、多くの場合ウイルスは肝炎症を引き起こす慢性感染となり、ゆっくりと肝不全や肝硬変に進行する(Iwarson,FEMS Microbiol.Rev.,14:201−204(1994))。更に、疫学的調査から、HCVが肝細胞癌の発病において重要な役割をなしていることがわかった(Kew,FEMS Microbiol.Rev.,14:211−220(1994);Alter,Blood,85:1681−1695(1995))。
【0003】
1992年にHCVの血液スクリーニングがルーチンに実施されるまで、多くの患者は汚染されている血液、血液製剤または移植臓器にうっかり曝されたことにより感染していた。HCVの血液スクリーニングが実施されている地域では、HCVは主に感染血液に直接経皮的に暴露して、すなわち静脈薬の使用により感染している。まれな伝播方法には、周生期暴露、血液透析及びHCV感染者との性的接触が含まれる(Alterら,N.Engl.J.Med.,341(8):556−562(1999);Alter,J.Hepatol.,31(補遺):88−91(1999);Semin,Liver Dis.,201:1−16(2000))。
【0004】
HCVゲノムは、約3000アミノ酸の前駆体ポリタンパク質をコードする一本鎖RNA約9.5kbから構成されている(Chooら,Science,244:362−364(1989);Chooら,Science,244:359−362(1989);Takamizawaら,J.Virol.,65:1105−1113(1991))。HCVポリタンパク質は、C−E1−E2−p7−NS2−NS3−NS4A−NS4B−NS5A−NS5Bの順でウイルスタンパク質を含む。
【0005】
各ウイルスタンパク質は、HCVポリタンパク質のタンパク質分解により産生される。宿主細胞プロテアーゼは推定構造タンパク質C、E1、E2及びp7を放出し、アミノ酸810でNS2のN末端を形成する(Mizushimaら,J.Virol.,68:2731−2734(1994);Hijikataら,P.N.A.S.USA,90:10773−10777(1993))。
【0006】
非構造タンパク質NS3、NS4、NS4B、NS5A及びNS5Bは多分ウイルス複製機構を形成し、ポリタンパク質から放出される。NS2及びNS3のN末端に関連する亜鉛依存性プロテアーゼはNS2とNS3の間の開裂に関与する(Grakouiら,J.Virol.,67:1385−1395(1993);Hijikataら,P.N.A.S.USA,90:10773−10777(1993))。NS3のN末端ドメイン中に位置する別のセリンプロテアーゼはNS3/NS4A、NS4A/NS4B、NS4B/NS5A及びNS5A/NS5B接合部でのタンパク質分解開裂に関与している(Bartenschlagerら,J.Virol.,67:3835−3844(1993);Grakouiら,Proc.Natl.Acad.Sci.USA,90:10583−10587(1993);Tomeiら,J.Virol.,67:4017−4026(1993))。NS4AはNS3活性に対するコファクターを与える(Faillaら,J.Virol.,68:3753−3760(1994);DeFrancescoらの米国特許第5,739,002号明細書)。
【0007】
NS5Aはインターフェロン耐性を与える高リン酸化タンパク質である(DeFrancescoら,Semin.Liver Dis.,20(1):69−83(2000);Pawlotsky,Viral Hepat.,補遺1:47−48(1999))。
【0008】
NS5BはRNA依存性RNAポリメラーゼを与える(DeFrancescoらの国際特許出願公開第96/37619号パンフレット;Behrensら,EMBO,15:12−22(1996);Lohmannら,Virology,249:108−118(1998))。
【発明の概要】
【0009】
本発明は、Ad6ベクター、及び不活性NS5B RNA依存性RNAポリメラーゼ領域を含むMet−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチドをコードする核酸に関する。前記核酸は、HCVに対するHCV特異的細胞性免疫(CMI)応答を生じさせるために広範囲の抗原を与えるアデノベクターまたはDNAプラスミドワクチンの成分として特に有用である。
【0010】
HCV特異的CMI応答は、HCV抗原を認識する細胞毒性Tリンパ球及びTヘルパー細胞の産生を指す。CMI応答には非−HCV特異的免疫効果も含まれ得る。
【0011】
好ましい核酸は、配列番号1に実質的に類似し、配列番号1中に存在するNS5B領域に実質的に類似する少なくとも1つのポリペプチドを産生するように自己プロセシングするのに十分なプロテアーゼ活性を有するMet−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチドをコードする。NS5Bに対応する産生されたポリペプチドは酵素的に不活性である。より好ましくは、HCVポリペプチドは配列番号1中に存在するNS3、NS4A、NS4B、NS5A及びNS5B領域に実質的に類似するポリペプチドを産生するのに十分なプロテアーゼ活性を有する。
【0012】
「実質的に類似の配列」とは、参照配列に対する同一性が少なくとも約65%であることを示す。よって、例えば、配列番号1に実質的に類似のアミノ酸配列を有するポリペプチドは配列番号1に対して少なくとも約65%の全アミノ酸同一性を有する。
【0013】
NS3、NS4A、NS4B、NS5A及びNS5Bに対応するポリペプチドは、配列番号1中の対応領域に対して少なくとも約65%のアミノ酸配列同一性を有する。前記対応領域は、本明細書中NS3、NS4A、NS4B、NS5A及びNS5Bポリペプチドとも称される。
【0014】
従って、本発明の第1態様は、配列番号1に実質的に類似のMet−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチドをコードするヌクレオチド配列を含む核酸に関する。コードされるポリペプチドは、酵素的に不活性なNS5Bポリペプチドを産生するように自己プロセツシングするのに十分なプロテアーゼ活性を有する。
【0015】
好ましい実施態様において、前記核酸は、所望ヒト細胞においてMet−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチドを発現し得る発現ベクターである。ヒト細胞内での発現は、HCV感染を積極的に治療したり、HCV感染に対して予防的に治療する治療用途を有する。
【0016】
発現ベクターは、ポリペプチドをコードするヌクレオチド配列と共に適正な転写及びプロセシングのために調節要素を含む。任意に存在する調節要素には、前記ポリペプチドをコードするヌクレオチド配列に元々関連している調節要素及び前記ヌクレオチド配列に元々関連していない外因性調節要素が含まれる。外因性プロモーターのような外因性調節要素はヒト細胞のような特定宿主における発現のために有用であり得る。機能的発現のために有用な調節要素の例には、プロモーター、ターミネーター、リボソーム結合部位及びポリアデニル化シグナルが含まれる。
【0017】
本発明の別の態様は、ヒト細胞において配列番号1に実質的に類似のMet−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチドを発現し得る遺伝子発現カセットを含む核酸に関する。前記ポリペプチドは、酵素的に不活性なNS5Bタンパク質を産生するように自己プロセツシングし得る。前記遺伝子発現カセットは、少なくとも
a)ポリペプチドをコードするヌクレオチド配列に転写的にカップリングしているプロモーター、
b)前記ヌクレオチド配列に機能的にカップリングしている5’リボソーム結合部位、
c)前記ヌクレオチド配列の3’末端に連結しているターミネーター、及び
d)前記ヌクレオチド配列に機能的にカップリングしている3’ポリアデニル化シグナル
を含む。
【0018】
「転写的にカップリング」とは、ヌクレオチド配列がプロモーターで結合しているRNAポリメラーゼにより転写され得るようにプロモーターが位置づけられていることを指す。転写的カップリングでは、転写される配列がプロモーターに隣接している必要がない。
【0019】
「機能的カップリング」とは、ヌクレオチド配列への効果を媒介し得る能力を指す。機能的カップリングでは、カップリングされる配列が相互に隣接している必要はない。ヌクレオチド配列に機能的にカップリングしている3’ポリアデニル化シグナルにより、転写RNAの開裂及びポリアデニル化が促進される。ヌクレオチド配列に機能的にカップリングしている5’リボソーム結合部位により、リボソーム結合が促進される。
【0020】
好ましい実施態様では、核酸はHCVを治療する治療用途に適したまたは治療用ベクターの作成における中間体として適したDNAプラスミドベクターまたはアデノベクターである。HCVの治療には、HCV感染の積極的治療及びHCV感染に対する予防的治療が含まれる。
【0021】
本発明の別の態様は、(a)相同的組換え及び(b)アデノベクターレスキューを含む方法により産生される、配列番号1に実質的に類似のポリペプチドを発現し得るMet−NS3−NS4A−NS4B−NS5A−NS5B発現カセットを含むアデノベクターである。相同組換え段階により、アデノウイルスゲノムプラスミドが生ずる。アデノベクターレスキュー段階により、アデノゲノムプラスミドからアデノベクターが生ずる。
【0022】
本明細書に記載のアデノウイルスゲノムプラスミドは、E1領域及び場合によりE3領域が欠失されており、前記欠失領域の1つに遺伝子発現カセットが挿入されている組換えアデノウイルスゲノムを含む。組換えアデノウイルスゲノムは1つ以上のアデノウイルス血清タイプに実質的に類似の領域から構成されている。
【0023】
本発明の別の態様は、配列番号4の核酸配列またはその誘導体からなるアデノベクターに関し、前記誘導体では配列番号4中に存在するHCVポリタンパク質コード化配列が配列番号3、配列番号10または配列番号11のいずれかのHCVポリタンパク質コード化配列で置換されている。
【0024】
本発明の別の態様は、配列番号1に実質的に類似のMet−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチドをコードする配列を含む核酸を含む培養組換え細胞に関する。前記組換え細胞は、ベクター構築方法において前記ポリペプチドをコードする核酸を複製するために使用されるような各種用途を有する。
【0025】
本発明の別の態様は、配列番号1に実質的に類似のポリペプチドを発現し得るMet−NS3−NS4A−NS4B−NS5A−NS5B発現カセットを含むアデノベクターの作成方法に関する。前記方法は、(a)E1領域及びE3領域が欠失されており、前記欠失領域の1つに遺伝子発現カセットが挿入されている組換えアデノウイルスゲノムを含むアデノウイルスゲノムプラスミドを作成する段階、及び(b)前記アデノウイルスゲノムプラスミドからアデノベクターをレスキューする段階を含む。
【0026】
本発明の別の態様は、配列番号1に実質的に類似のMet−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチドを発現させるためのベクター及び医薬的に許容され得る担体を含む医薬組成物に関する。前記ベクターは患者における投与及びポリペプチド発現に適している。
【0027】
「患者」は、HCVに感染し得る哺乳動物を指す。患者はHCVに感染していてもいなくてもよい。患者の例はヒト及びチンパンジーである。
【0028】
本発明の別の態様は、配列番号1に実質的に類似のMet−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチドを発現するベクターを有効量患者に投与することを含む患者の治療方法に関する。前記ベクターは患者における投与及びポリペプチド発現に適している。
【0029】
治療を受ける患者はHCVに感染していてもいなくてもよい。HCV感染されている患者の場合、有効量はHCVの複製能力の低下、HCV負荷の低減、ウイルスクリアランスの上昇及び1つ以上のHCV特異的CMI応答の上昇という効果の1つ以上を達成するのに十分な量である。HCVに感染していない患者の場合、有効量はHCV感染に対するHCV特異的CMI応答の1つ以上の成分を産生する能力の上昇、HCV感染への感受性の低下及び感染ウイルスが慢性疾患に対する持続感染を樹立する能力の低下という効果の1つ以上を達成するのに十分な量である。
【0030】
本発明の別の態様は、Ad6領域及びAd6中に存在しない領域を含む組換え核酸に関する。「組換え」核酸とは、元々相互に関連しない2つ以上の核酸領域が存在することを意味する。Ad6組換え核酸はAd6領域及びAd6に非相同のポリペプチドをコードする遺伝子発現カセットを含むことが好ましい。
【0031】
本発明の他の特徴及び作用効果は、本明細書に記載されている複数の実施例を含めた追加記載から明らかである。ここに提示した実施例は本発明を実施するのに有用な成分及び方法を例示している。これらの実施例は本発明を限定しない。本明細書の記載に基づいて、当業者は本発明を実施するのに有用な他の成分及び方法を同定、使用することができる。
【図面の簡単な説明】
【0032】
【図1A】配列番号1を示す。
【図1B】配列番号1を示す。
【図2A】配列番号2を示す。配列番号2は、配列番号1をコードするヌクレオチド配列と共に最適化内部リボソームエントリー部位及びTAAA終止を規定する。ヌクレオチド1〜6は最適化内部リボソームエントリー部位を規定する。ヌクレオチド7〜5961はNos.5137〜5145にヌクレオチドを有するHCV Met−NS3−NS4A−NS4B−NS5A−NS5Bペリペプチドをコードし、アミノ酸No.1711〜1713にNS5Bを不活性とするAlaAlaGLy配列を規定する。ヌクレオチド5962〜5965はTAAA終止を規定する。
【図2B】配列番号2を示す。配列番号2は、配列番号1をコードするヌクレオチド配列と共に最適化内部リボソームエントリー部位及びTAAA終止を規定する。ヌクレオチド1〜6は最適化内部リボソームエントリー部位を規定する。ヌクレオチド7〜5961はNos.5137〜5145にヌクレオチドを有するHCV Met−NS3−NS4A−NS4B−NS5A−NS5Bペリペプチドをコードし、アミノ酸No.1711〜1713にNS5Bを不活性とするAlaAlaGLy配列を規定する。ヌクレオチド5962〜5965はTAAA終止を規定する。
【図2C】配列番号2を示す。配列番号2は、配列番号1をコードするヌクレオチド配列と共に最適化内部リボソームエントリー部位及びTAAA終止を規定する。ヌクレオチド1〜6は最適化内部リボソームエントリー部位を規定する。ヌクレオチド7〜5961はNos.5137〜5145にヌクレオチドを有するHCV Met−NS3−NS4A−NS4B−NS5A−NS5Bペリペプチドをコードし、アミノ酸No.1711〜1713にNS5Bを不活性とするAlaAlaGLy配列を規定する。ヌクレオチド5962〜5965はTAAA終止を規定する。
【図2D】配列番号2を示す。配列番号2は、配列番号1をコードするヌクレオチド配列と共に最適化内部リボソームエントリー部位及びTAAA終止を規定する。ヌクレオチド1〜6は最適化内部リボソームエントリー部位を規定する。ヌクレオチド7〜5961はNos.5137〜5145にヌクレオチドを有するHCV Met−NS3−NS4A−NS4B−NS5A−NS5Bペリペプチドをコードし、アミノ酸No.1711〜1713にNS5Bを不活性とするAlaAlaGLy配列を規定する。ヌクレオチド5962〜5965はTAAA終止を規定する。
【図3A】配列番号3を示す。配列番号3は配列番号2のコドン最適化バージョンである。ヌクレオチド7〜5961がHCV Met−NS3−NS4A−NS4B−NS5A−NS5Bペリペプチドをコードする。
【図3B】配列番号3を示す。配列番号3は配列番号2のコドン最適化バージョンである。ヌクレオチド7〜5961がHCV Met−NS3−NS4A−NS4B−NS5A−NS5Bペリペプチドをコードする。
【図3C】配列番号3を示す。配列番号3は配列番号2のコドン最適化バージョンである。ヌクレオチド7〜5961がHCV Met−NS3−NS4A−NS4B−NS5A−NS5Bペリペプチドをコードする。
【図3D】配列番号3を示す。配列番号3は配列番号2のコドン最適化バージョンである。ヌクレオチド7〜5961がHCV Met−NS3−NS4A−NS4B−NS5A−NS5Bペリペプチドをコードする。
【図4A】MRK Ad6−NSmut(配列番号4)を示す。配列番号4は配列番号1のポリペプチドが配列番号2によりコードされる発現カセットを含むアデノベクターである。塩基対1〜450はAd5 bp1−450に対応し、塩基対462〜1252はヒトCMVプロモーターに対応し、塩基対1258〜1267はコザック配列に対応し、塩基対1264〜7222はNS遺伝子に対応し、塩基対7231〜7451はBGHポリアデニル化シグナルに対応し、塩基対7469〜9506はAd5塩基対3511−5548に対応し、塩基対9507〜32121はAd6塩基対5542−28156に対応し、塩基対32122〜35117はAd6塩基対30789−33784に対応し、塩基対35118〜37089はAd5塩基対33967−35935に対応する。
【図4B】MRK Ad6−NSmut(配列番号4)を示す。配列番号4は配列番号1のポリペプチドが配列番号2によりコードされる発現カセットを含むアデノベクターである。塩基対1〜450はAd5 bp1−450に対応し、塩基対462〜1252はヒトCMVプロモーターに対応し、塩基対1258〜1267はコザック配列に対応し、塩基対1264〜7222はNS遺伝子に対応し、塩基対7231〜7451はBGHポリアデニル化シグナルに対応し、塩基対7469〜9506はAd5塩基対3511−5548に対応し、塩基対9507〜32121はAd6塩基対5542−28156に対応し、塩基対32122〜35117はAd6塩基対30789−33784に対応し、塩基対35118〜37089はAd5塩基対33967−35935に対応する。
【図4C】MRK Ad6−NSmut(配列番号4)を示す。配列番号4は配列番号1のポリペプチドが配列番号2によりコードされる発現カセットを含むアデノベクターである。塩基対1〜450はAd5 bp1−450に対応し、塩基対462〜1252はヒトCMVプロモーターに対応し、塩基対1258〜1267はコザック配列に対応し、塩基対1264〜7222はNS遺伝子に対応し、塩基対7231〜7451はBGHポリアデニル化シグナルに対応し、塩基対7469〜9506はAd5塩基対3511−5548に対応し、塩基対9507〜32121はAd6塩基対5542−28156に対応し、塩基対32122〜35117はAd6塩基対30789−33784に対応し、塩基対35118〜37089はAd5塩基対33967−35935に対応する。
【図4D】MRK Ad6−NSmut(配列番号4)を示す。配列番号4は配列番号1のポリペプチドが配列番号2によりコードされる発現カセットを含むアデノベクターである。塩基対1〜450はAd5 bp1−450に対応し、塩基対462〜1252はヒトCMVプロモーターに対応し、塩基対1258〜1267はコザック配列に対応し、塩基対1264〜7222はNS遺伝子に対応し、塩基対7231〜7451はBGHポリアデニル化シグナルに対応し、塩基対7469〜9506はAd5塩基対3511−5548に対応し、塩基対9507〜32121はAd6塩基対5542−28156に対応し、塩基対32122〜35117はAd6塩基対30789−33784に対応し、塩基対35118〜37089はAd5塩基対33967−35935に対応する。
【図4E】MRK Ad6−NSmut(配列番号4)を示す。配列番号4は配列番号1のポリペプチドが配列番号2によりコードされる発現カセットを含むアデノベクターである。塩基対1〜450はAd5 bp1−450に対応し、塩基対462〜1252はヒトCMVプロモーターに対応し、塩基対1258〜1267はコザック配列に対応し、塩基対1264〜7222はNS遺伝子に対応し、塩基対7231〜7451はBGHポリアデニル化シグナルに対応し、塩基対7469〜9506はAd5塩基対3511−5548に対応し、塩基対9507〜32121はAd6塩基対5542−28156に対応し、塩基対32122〜35117はAd6塩基対30789−33784に対応し、塩基対35118〜37089はAd5塩基対33967−35935に対応する。
【図4F】MRK Ad6−NSmut(配列番号4)を示す。配列番号4は配列番号1のポリペプチドが配列番号2によりコードされる発現カセットを含むアデノベクターである。塩基対1〜450はAd5 bp1−450に対応し、塩基対462〜1252はヒトCMVプロモーターに対応し、塩基対1258〜1267はコザック配列に対応し、塩基対1264〜7222はNS遺伝子に対応し、塩基対7231〜7451はBGHポリアデニル化シグナルに対応し、塩基対7469〜9506はAd5塩基対3511−5548に対応し、塩基対9507〜32121はAd6塩基対5542−28156に対応し、塩基対32122〜35117はAd6塩基対30789−33784に対応し、塩基対35118〜37089はAd5塩基対33967−35935に対応する。
【図4G】MRK Ad6−NSmut(配列番号4)を示す。配列番号4は配列番号1のポリペプチドが配列番号2によりコードされる発現カセットを含むアデノベクターである。塩基対1〜450はAd5 bp1−450に対応し、塩基対462〜1252はヒトCMVプロモーターに対応し、塩基対1258〜1267はコザック配列に対応し、塩基対1264〜7222はNS遺伝子に対応し、塩基対7231〜7451はBGHポリアデニル化シグナルに対応し、塩基対7469〜9506はAd5塩基対3511−5548に対応し、塩基対9507〜32121はAd6塩基対5542−28156に対応し、塩基対32122〜35117はAd6塩基対30789−33784に対応し、塩基対35118〜37089はAd5塩基対33967−35935に対応する。
【図4H】MRK Ad6−NSmut(配列番号4)を示す。配列番号4は配列番号1のポリペプチドが配列番号2によりコードされる発現カセットを含むアデノベクターである。塩基対1〜450はAd5 bp1−450に対応し、塩基対462〜1252はヒトCMVプロモーターに対応し、塩基対1258〜1267はコザック配列に対応し、塩基対1264〜7222はNS遺伝子に対応し、塩基対7231〜7451はBGHポリアデニル化シグナルに対応し、塩基対7469〜9506はAd5塩基対3511−5548に対応し、塩基対9507〜32121はAd6塩基対5542−28156に対応し、塩基対32122〜35117はAd6塩基対30789−33784に対応し、塩基対35118〜37089はAd5塩基対33967−35935に対応する。
【図4I】MRK Ad6−NSmut(配列番号4)を示す。配列番号4は配列番号1のポリペプチドが配列番号2によりコードされる発現カセットを含むアデノベクターである。塩基対1〜450はAd5 bp1−450に対応し、塩基対462〜1252はヒトCMVプロモーターに対応し、塩基対1258〜1267はコザック配列に対応し、塩基対1264〜7222はNS遺伝子に対応し、塩基対7231〜7451はBGHポリアデニル化シグナルに対応し、塩基対7469〜9506はAd5塩基対3511−5548に対応し、塩基対9507〜32121はAd6塩基対5542−28156に対応し、塩基対32122〜35117はAd6塩基対30789−33784に対応し、塩基対35118〜37089はAd5塩基対33967−35935に対応する。
【図4J】MRK Ad6−NSmut(配列番号4)を示す。配列番号4は配列番号1のポリペプチドが配列番号2によりコードされる発現カセットを含むアデノベクターである。塩基対1〜450はAd5 bp1−450に対応し、塩基対462〜1252はヒトCMVプロモーターに対応し、塩基対1258〜1267はコザック配列に対応し、塩基対1264〜7222はNS遺伝子に対応し、塩基対7231〜7451はBGHポリアデニル化シグナルに対応し、塩基対7469〜9506はAd5塩基対3511−5548に対応し、塩基対9507〜32121はAd6塩基対5542−28156に対応し、塩基対32122〜35117はAd6塩基対30789−33784に対応し、塩基対35118〜37089はAd5塩基対33967−35935に対応する。
【図4K】MRK Ad6−NSmut(配列番号4)を示す。配列番号4は配列番号1のポリペプチドが配列番号2によりコードされる発現カセットを含むアデノベクターである。塩基対1〜450はAd5 bp1−450に対応し、塩基対462〜1252はヒトCMVプロモーターに対応し、塩基対1258〜1267はコザック配列に対応し、塩基対1264〜7222はNS遺伝子に対応し、塩基対7231〜7451はBGHポリアデニル化シグナルに対応し、塩基対7469〜9506はAd5塩基対3511−5548に対応し、塩基対9507〜32121はAd6塩基対5542−28156に対応し、塩基対32122〜35117はAd6塩基対30789−33784に対応し、塩基対35118〜37089はAd5塩基対33967−35935に対応する。
【図4L】MRK Ad6−NSmut(配列番号4)を示す。配列番号4は配列番号1のポリペプチドが配列番号2によりコードされる発現カセットを含むアデノベクターである。塩基対1〜450はAd5 bp1−450に対応し、塩基対462〜1252はヒトCMVプロモーターに対応し、塩基対1258〜1267はコザック配列に対応し、塩基対1264〜7222はNS遺伝子に対応し、塩基対7231〜7451はBGHポリアデニル化シグナルに対応し、塩基対7469〜9506はAd5塩基対3511−5548に対応し、塩基対9507〜32121はAd6塩基対5542−28156に対応し、塩基対32122〜35117はAd6塩基対30789−33784に対応し、塩基対35118〜37089はAd5塩基対33967−35935に対応する。
【図4M】MRK Ad6−NSmut(配列番号4)を示す。配列番号4は配列番号1のポリペプチドが配列番号2によりコードされる発現カセットを含むアデノベクターである。塩基対1〜450はAd5 bp1−450に対応し、塩基対462〜1252はヒトCMVプロモーターに対応し、塩基対1258〜1267はコザック配列に対応し、塩基対1264〜7222はNS遺伝子に対応し、塩基対7231〜7451はBGHポリアデニル化シグナルに対応し、塩基対7469〜9506はAd5塩基対3511−5548に対応し、塩基対9507〜32121はAd6塩基対5542−28156に対応し、塩基対32122〜35117はAd6塩基対30789−33784に対応し、塩基対35118〜37089はAd5塩基対33967−35935に対応する。
【図5A】配列番号5及び6を示す。配列番号5は活性RNA依存性RNAポリメラーゼを有するHCV Met−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチドをコードする。配列番号6はポリペプチドのアミノ酸配列を規定する。
【図5B】配列番号5及び6を示す。配列番号5は活性RNA依存性RNAポリメラーゼを有するHCV Met−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチドをコードする。配列番号6はポリペプチドのアミノ酸配列を規定する。
【図5C】配列番号5及び6を示す。配列番号5は活性RNA依存性RNAポリメラーゼを有するHCV Met−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチドをコードする。配列番号6はポリペプチドのアミノ酸配列を規定する。
【図5D】配列番号5及び6を示す。配列番号5は活性RNA依存性RNAポリメラーゼを有するHCV Met−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチドをコードする。配列番号6はポリペプチドのアミノ酸配列を規定する。
【図5E】配列番号5及び6を示す。配列番号5は活性RNA依存性RNAポリメラーゼを有するHCV Met−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチドをコードする。配列番号6はポリペプチドのアミノ酸配列を規定する。
【図5F】配列番号5及び6を示す。配列番号5は活性RNA依存性RNAポリメラーゼを有するHCV Met−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチドをコードする。配列番号6はポリペプチドのアミノ酸配列を規定する。
【図5G】配列番号5及び6を示す。配列番号5は活性RNA依存性RNAポリメラーゼを有するHCV Met−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチドをコードする。配列番号6はポリペプチドのアミノ酸配列を規定する。
【図5H】配列番号5及び6を示す。配列番号5は活性RNA依存性RNAポリメラーゼを有するHCV Met−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチドをコードする。配列番号6はポリペプチドのアミノ酸配列を規定する。
【図5I】配列番号5及び6を示す。配列番号5は活性RNA依存性RNAポリメラーゼを有するHCV Met−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチドをコードする。配列番号6はポリペプチドのアミノ酸配列を規定する。
【図5J】配列番号5及び6を示す。配列番号5は活性RNA依存性RNAポリメラーゼを有するHCV Met−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチドをコードする。配列番号6はポリペプチドのアミノ酸配列を規定する。
【図5K】配列番号5及び6を示す。配列番号5は活性RNA依存性RNAポリメラーゼを有するHCV Met−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチドをコードする。配列番号6はポリペプチドのアミノ酸配列を規定する。
【図5L】配列番号5及び6を示す。配列番号5は活性RNA依存性RNAポリメラーゼを有するHCV Met−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチドをコードする。配列番号6はポリペプチドのアミノ酸配列を規定する。
【図5M】配列番号5及び6を示す。配列番号5は活性RNA依存性RNAポリメラーゼを有するHCV Met−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチドをコードする。配列番号6はポリペプチドのアミノ酸配列を規定する。
【図5N】配列番号5及び6を示す。配列番号5は活性RNA依存性RNAポリメラーゼを有するHCV Met−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチドをコードする。配列番号6はポリペプチドのアミノ酸配列を規定する。
【図5O】配列番号5及び6を示す。配列番号5は活性RNA依存性RNAポリメラーゼを有するHCV Met−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチドをコードする。配列番号6はポリペプチドのアミノ酸配列を規定する。
【図6A】pV1JnsAの核酸配列(配列番号7)を規定する。
【図6B】pV1JnsAの核酸配列(配列番号7)を規定する。
【図6C】pV1JnsAの核酸配列(配列番号7)を規定する。
【図7A】Ad6ゲノムの核酸配列(配列番号8)を規定する。
【図7B】Ad6ゲノムの核酸配列(配列番号8)を規定する。
【図7C】Ad6ゲノムの核酸配列(配列番号8)を規定する。
【図7D】Ad6ゲノムの核酸配列(配列番号8)を規定する。
【図7E】Ad6ゲノムの核酸配列(配列番号8)を規定する。
【図7F】Ad6ゲノムの核酸配列(配列番号8)を規定する。
【図7G】Ad6ゲノムの核酸配列(配列番号8)を規定する。
【図7H】Ad6ゲノムの核酸配列(配列番号8)を規定する。
【図7I】Ad6ゲノムの核酸配列(配列番号8)を規定する。
【図7J】Ad6ゲノムの核酸配列(配列番号8)を規定する。
【図7K】Ad6ゲノムの核酸配列(配列番号8)を規定する。
【図7L】Ad6ゲノムの核酸配列(配列番号8)を規定する。
【図7M】Ad6ゲノムの核酸配列(配列番号8)を規定する。
【図7N】Ad6ゲノムの核酸配列(配列番号8)を規定する。
【図7O】Ad6ゲノムの核酸配列(配列番号8)を規定する。
【図8A】Ad5ゲノムの核酸配列(配列番号9)を規定する。
【図8B】Ad5ゲノムの核酸配列(配列番号9)を規定する。
【図8C】Ad5ゲノムの核酸配列(配列番号9)を規定する。
【図8D】Ad5ゲノムの核酸配列(配列番号9)を規定する。
【図8E】Ad5ゲノムの核酸配列(配列番号9)を規定する。
【図8F】Ad5ゲノムの核酸配列(配列番号9)を規定する。
【図8G】Ad5ゲノムの核酸配列(配列番号9)を規定する。
【図8H】Ad5ゲノムの核酸配列(配列番号9)を規定する。
【図8I】Ad5ゲノムの核酸配列(配列番号9)を規定する。
【図8J】Ad5ゲノムの核酸配列(配列番号9)を規定する。
【図8K】Ad5ゲノムの核酸配列(配列番号9)を規定する。
【図9】Ad6ゲノムの各種領域を示す。線状(35759bp)ds DNAゲノムを2つの平行線で示し、100地図単位に分けられている。転写単位をゲノム中の位置及び方位に関して示す。初期遺伝子(E1A、E1B、E2A/B、E3及びE4)を灰色矢印で示す。黒色矢印で示す後期遺伝子(L1〜L5)は主要後期プロモーター(MLP)から生ずる転写物の選択的スプライシングにより産生され、いずれもその5’末端に三連リーダー(1、2、3)を含む。E1領域は約1.0〜11.5地図単位に位置し、E2領域は75.0〜11.5地図単位に位置し、E3領域は76.1〜86.7地図単位に位置し、E4領域は99.5〜91.2地図単位に位置している。主要後期転写単位は16.0〜91.2地図単位に位置している。
【図10】Ad6領域及びAd5領域を含むpAdE1−E3+を回収するための相同組換えを示す。
【図11】Ad6領域を含むpAdE1−E3+を回収するための相同組換えを示す。
【図12】各種HCV NSカセットを発現するプラスミドDNAをトランスフェクトした293細胞からの全細胞抽出物に対するウェスタンブロットを示す。成熟NS3及びNS5A産物を特異的抗体で検出した。“pV1Jns−NS”は、Met−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチドが配列番号5によりコードされ、配列番号5が配列番号7の塩基1881〜1912に挿入されているpV1JnsAプラスミドを指す。“pV1Jns−NSmut”は配列番号2が配列番号7の塩基1882〜1925に挿入されているpV1JnsAプラスミドを指す。“pV1Jns−NSOPTmut”は配列番号3が配列番号7の塩基1881〜1905に挿入されているpV1JnsAプラスミドを指す。
【図13A】C57black6マウス(A)及びBalbCマウス(B)において25μg及び50μgの各種HCV NSカセットをコードするプラスミドDNAの2回注射及び遺伝子エレクトロトランスファー(GET)により誘導したT細胞応答をIFNγELIspotにより示す。
【図13B】C57black6マウス(A)及びBalbCマウス(B)において25μg及び50μgの各種HCV NSカセットをコードするプラスミドDNAの2回注射及び遺伝子エレクトロトランスファー(GET)により誘導したT細胞応答をIFNγELIspotにより示す。
【図14】HeLa細胞の感染時の各種アデノベクターからのタンパク質発現を示す。MRKAd5−NSmutは、Ad5ゲノムが塩基対451〜3510のE1欠失、塩基対28134〜30817のE3欠失を有し、配列番号4の塩基対451〜7468のNS3−NS4A−NS4B−NS5A−NS5B発現カセットがNo.450〜3511に挿入されているAd5配列(配列番号9)をベースとするアデノベクターである。Ad5−NSは、塩基対342〜3523のE1欠失及び塩基対28134〜30817のE3欠失を有するAd5骨格をベースとし、配列番号5からのNS3−NS4A−NS4B−NS5A−NS5Bをコードする発現カセットを含むアデノベクターである。“MRKAd6−NSOPTmut”は、配列番号4の塩基1258〜7222が配列番号3で置換されている修飾配列番号4を有するアデノベクターを指す。
【図15】C57black6マウスにおいて109vpの各種HCV非構造遺伝子カセットを含むアデノベクターを2回注射することにより誘導したT細胞応答をINFγELIspotにより示す。
【図16A】アカゲザルにおいて1010vp(A)または1011vp(B)の各種HCV非構造遺伝子カセットを含むアデノベクターを1回または2回注射することにより誘導したT細胞応答をINFγELIspotにより示す。
【図16B】アカゲザルにおいて1010vp(A)または1011vp(B)の各種HCV非構造遺伝子カセットを含むアデノベクターを1回または2回注射することにより誘導したT細胞応答をINFγELIspotにより示す。
【図16C】アカゲザルにおいて1010vp(A)または1011vp(B)の各種HCV非構造遺伝子カセットを含むアデノベクターを1回または2回注射することにより誘導したT細胞応答をINFγELIspotにより示す。
【図16D】アカゲザルにおいて1010vp(A)または1011vp(B)の各種HCV非構造遺伝子カセットを含むアデノベクターを1回または2回注射することにより誘導したT細胞応答をINFγELIspotにより示す。
【図17A】アカゲザルにおいて1010vp(A)または1011vp(B)の各種HCV非構造遺伝子カセットを含むアデノベクターを2回注射することにより誘導したCD8+ T細胞応答をINFγICSにより示す。
【図17B】アカゲザルにおいて1010vp(A)または1011vp(B)の各種HCV非構造遺伝子カセットを含むアデノベクターを2回注射することにより誘導したCD8+ T細胞応答をINFγICSにより示す。
【図18A】アカゲザルにおいて1011vpのAd5−NS(A)、MRKAd5−NSmut(B)またはMRKAd6−NSmut(C)を2回注射することにより誘導したT細胞応答をバルクCTLアッセイにより示す。
【図18B】アカゲザルにおいて1011vpのAd5−NS(A)、MRKAd5−NSmut(B)またはMRKAd6−NSmut(C)を2回注射することにより誘導したT細胞応答をバルクCTLアッセイにより示す。
【図18C】アカゲザルにおいて1011vpのAd5−NS(A)、MRKAd5−NSmut(B)またはMRKAd6−NSmut(C)を2回注射することにより誘導したT細胞応答をバルクCTLアッセイにより示す。
【図18D】アカゲザルにおいて1011vpのAd5−NS(A)、MRKAd5−NSmut(B)またはMRKAd6−NSmut(C)を2回注射することにより誘導したT細胞応答をバルクCTLアッセイにより示す。
【図18E】アカゲザルにおいて1011vpのAd5−NS(A)、MRKAd5−NSmut(B)またはMRKAd6−NSmut(C)を2回注射することにより誘導したT細胞応答をバルクCTLアッセイにより示す。
【図18F】アカゲザルにおいて1011vpのAd5−NS(A)、MRKAd5−NSmut(B)またはMRKAd6−NSmut(C)を2回注射することにより誘導したT細胞応答をバルクCTLアッセイにより示す。
【図19】プラスミドpE2を示す。
【図20A】部分コドン最適化配列NSsuboptmut(配列番号10)を示す。Met−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチドのコード配列は塩基7〜5961である。
【図20B】部分コドン最適化配列NSsuboptmut(配列番号10)を示す。Met−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチドのコード配列は塩基7〜5961である。
【図20C】部分コドン最適化配列NSsuboptmut(配列番号10)を示す。Met−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチドのコード配列は塩基7〜5961である。
【図20D】部分コドン最適化配列NSsuboptmut(配列番号10)を示す。Met−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチドのコード配列は塩基7〜5961である。
【発明を実施するための形態】
【0033】
本発明は、Ad6ベクター、及び不活性NS5B領域を含むMet−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチドをコードする核酸に関する。不活性NS5B領域を設けると、活性ウイルス性RNAポリメラーゼによる副作用の可能性を低下させながらNS5B抗原が供給される。本発明の核酸の使用には、HCVに対するCMI応答を生じさせるべく広範囲の抗原を与えるHCVポリペプチドを細胞に導入するためのワクチン成分として及び前記ワクチン成分を作成するための中間体としての使用が含まれる。
【0034】
適応細胞免疫応答は、主要組織適合複合体(MHC)クラスI及びII発現が偏在分布しているために身体中のHCV感染細胞中でウイルス抗原を認識するように、免疫記憶を誘導するように、及び免疫記憶を維持するように機能し得る。これらの機能は抗原特異的CD4+ Tヘルパー(Th)及びCD8+ 細胞毒性T細胞(CTL)に起因する。
【0035】
特異的T細胞受容体を介して活性化すると、HCV特異的Th細胞は各種の免疫調節機能を実行し、その機能の多くはTh1及びTh2サイトカインにより媒介されている。HCV特異的Th細胞はB細胞の活性化及び分化、ウイルス特異的細胞毒性T細胞の誘導及び刺激を助ける。CTLと一緒になって、Th細胞は数種のウイルスの複製及び遺伝子発現を抑制するIFN−γ及びIFN−αも分泌し得る。更に、主要エフェクター細胞であるTh細胞及びCTLはウイルス感染細胞のアポトーシス及び溶解を誘導し得る。
【0036】
HCV特異的CTLは抗原提示細胞(pAPC)によりプロセシングされる抗原から生ずる。抗原はpAPC内で合成されたり、pAPCに導入されることもある。pAPC内での抗原合成は抗原をコードする発現カセットを細胞に導入することにより生起され得る。
【0037】
核酸ワクチンの好ましい投与ルートは筋肉内ルートである。筋肉内投与すると、核酸は体細胞及びpAPCへ導入、発現されるであろう。体細胞で産生されたHCV抗原はMHCクラスI分子の関係での提示のためにpAPCに移動し得る(Donnellyら,Annu.Rev.Immunol.,15:617−648(1997))。
【0038】
pAPCはより長い抗原をプロテアソーム複合体の形態のより小さいペプチド抗原にプロセシングする。前記抗原は、MHCクラスIタンパク質と結合するために小胞体/ゴルジ複合体分泌経路に転座する。CD8+ Tリンパ球は、T細胞受容体(TCR)及びCD8細胞表面タンパク質を介してクラス1 MHCに結合した抗原を認識する。
【0039】
Met−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチドをコードする核酸をワクチン成分として使用すると、単一ベクターからCMI応答を発生し得る広範囲の抗原を産生できる。前記ポリペプチドは少なくともNS5Bに対応する領域を生ずるように十分に自己プロセシングし得なければならない。好ましい核酸は、配列番号1中に存在するNS3、NS4A、NS4B、NS5A及びNS5B領域に実質的に類似している各HCVポリペプチドを産生するように自己プロセシングするのに十分なプロテアーゼ活性を有し、配列番号1に実質的に類似のアミノ酸配列をコードする。
【0040】
細胞中で自己プロセシングするのに十分なプロテアーゼ活性を有する配列番号1に実質的に類似のポリペプチドは、細胞に幾種のHCV株中に存在するT細胞エピトープを与える。プロテアーゼ活性はNS3及びNS3/NS4Aタンパク質により与えられ、適当な開裂部位でMet−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチドを消化すると、NS3、NS4A、NS4B、NS5A及びNS5Bに対応するポリペプチドを放出する。Met−NS3−NS4A−NS4B−NS5A−NS5Bの自己プロセシングにより、天然に存在するHCVポリペプチドに似ているポリペプチドが生ずる。
【0041】
本明細書に記載されているガイダンスに基づいて、患者において有利な効果を達成するのに十分に強い免疫応答を生じ得る。ここに記載されているガイダンスには、HCV配列選択、ベクター選択、ベクター作成、併用療法及び投与に関する情報が含まれている。
【0042】
I. HCV配列
多種多様の核酸配列が細胞にHCV Met−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチドを供給するためのワクチン成分として、またはワクチン成分を作成するための中間体として使用され得る。適当な核酸配列を得るための出発点としては、不活性NS5Bを産生するように修飾された天然NS3−NS4A−NS4B−NS5A−NS5Bポリペプチド配列が好ましい。
【0043】
CMI応答を生ずるようにHCV非構造抗原を与えるHCV核酸配列の使用は、Choら,Vaccine,17:1136−1144(1999);Paliardらの国際特許出願公開第01/30812号パンフレット(本発明に対する従来技術とは認められない)及びCoitらの国際特許出願公開第01/38360号パンフレット(本発明に対する従来技術とは認められない)に記載されている。前記文献は、例えば不活性NS5Bを産生するように自己プロセシングするポリペプチドもHCV配列とデリバリーベヒクルの特定組合せも全く記載していない。
【0044】
HCV Met−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチド配列への修飾は、コード化核酸を改変することによりなされ得る。欠失、挿入及び置換が生ずるように改変を実施し得る。
【0045】
小さな修飾はNS5B中で本質的に複製のためにモチーフを標的することにより不活性ポリメラーゼを生ずるようになされ得る。NS5B活性にとって重要なモチーフ及び不活性NS5Bを産生するようになされ得る修飾の例は、Lohmannら,Journal of Virology,74:8416−8426(1997)及びKolykhalovら,Journal of Virology,74:2046−2051(2000)に記載されている。
【0046】
HCV Met−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチド配列に修飾を加えるときに考慮すべき別の要因には、自己プロセシングする能力の維持及びT細胞抗原の維持が含まれる。HCVポリペプチドの自己プロセシング能力は、大体機能性NS3プロテアーゼにより決定される。NS3活性プロテアーゼ活性を維持する修飾は、NS3タンパク質、NS3に対するコファクターとして作用するNS4A、及びNS3−NS4A−NS4B−NS5A−NS5Bポリペプチド内に存在するNS3プロテアーゼ認識部位を考慮して得られ得る。
【0047】
広範囲のT細胞応答を誘発し得るポリペプチドを産生するために天然NS3−NS4A−NS4B−NS5A−NS5Bポリペプチド配列に対して各種修飾が加えられ得る。広範囲のT細胞応答を誘発するポリペプチドの能力に影響を及ぼす因子には、HCV特異的T細胞抗原領域の保存または導入、各種HCV単離物におけるT細胞抗原領域の優勢が含まれる。
【0048】
天然HCV単離物の多数の例が当業界で公知である。HCV単離物は1つ以上のサブタイプを含む以下の6つの主遺伝子型に分類され得る:HCV−1/(1a,1b,1c)、HCV−2/(2a,2b,2c)、HCV−3/(3a,3b,10c)、HCV−4/(4a)、HCV−5/(5a)及びHCV−6/(6a,6b,7b,8b,9b,11a)(Simmonds,J.Gen.Virol.,693−712(2001))。HCV−BK、HCV−J、HCV−N、HCV−Hのような特定HCV配列の例はGenBankに寄託されており、複数の文献に記載されている(例えば、Chamberlianら,J.Gen.Virol.,1341−1347(1997)参照)。
【0049】
HCV T細胞抗原は、例えば経験的実験により同定され得る。T細胞抗原を同定する1つの方法は、長ポリペプチドから一連の重複短ペプチドを作成し、その後ポジティブクローンについて感染患者からのT細胞集団をスクリーニングすることを含む。ポジティブクローンは特定ペプチドにより活性化/初期化される。ペプチド活性を測定するためにIFNγ−ELISPOT、IFNγ−細胞内染色及びバルクCTLアッセイのような方法を用いることができる。こうして同定されたペプチドは各病原のT細胞エピトープを表すと見做され得る。
【0050】
各種HCV単離物由来のHCV T細胞抗原領域は、例えば2つ以上の天然配列由来の領域を含むハイブリッドNS3−NS4A−NS4B−NS5A−NS5Bポリペプチドを作成することにより1つの配列に導入され得る。前記ハイブリッドは追加の修飾を含み得、この修飾がHCV CMI応答を生ずるポリペプチドの能力を低下させないことが好ましい。
【0051】
自己プロセシングし、CMI応答を生じさせる修飾Met−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチドの能力は、本明細書に記載されている方法または当業界で公知の方法により測定され得る。その方法には、HIV特異的CMI応答を測定するためのIFNγ−ELISPOT、IFNγ−細胞内染色及びバルクCTLアッセイの使用が含まれる。
【0052】
(A) Met−NS3−NS4A−NS4B−NS5A−NS5B配列
配列番号1は、好ましいMet−NS3−NS4A−NS4B−NS5A−NS5B配列を規定する。配列番号1は、数種のHCV単離物中に存在する多数のHCV特異的T細胞抗原を含む。配列番号1はHCV BK株ヌクレオチド配列(GenBank受託番号M58335)のNS3−NS4A−NS4B−NS5A−NS5B部分に類似している。
【0053】
配列番号1において、MHCクラスI分子による認識のために重要なアンカー位置は保存されているか、またはHCVポリタンパク質のNS3−NS4A−NS4B−NS5A−NS5B部分中の20個の公知T細胞エピトープのうちの18個の保存的置換を表す。残りの2個の公知T細胞エピトープのうち、1個のエピトープは異なるHLAスーパータイプにより認識され得る配列番号1中の非保存的アンカー置換を有し、1個のエピトープは保存されない1つのアンカー残基を有する。HCV T細胞エピトープはChisariら,Curr.Top.Microbiol.Immunol.,242:299−325(2000)及びLechnerら,J.Exp.Med.,9:1499−1512(2000)に記載されている。
【0054】
HCV BK NS3−NS4A−NS4B−NS5A−NS5Bヌクレオチド配列と配列番号1の違いとして、配列番号1中には5’末端にメチオニンが導入されており、修飾NS5B活性部位残基が存在していることが挙げられる。修飾により、GlyAspAspをAlaAlaGly(残基1711〜1713)で置換するとNS5Bが不活化される。
【0055】
コードされるHCV Met−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチドが配列番号1に実質的に類似のアミノ酸配列を有していることが好ましい。別の実施態様では、コードされるHCV Met−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチドは配列番号1に対して少なくとも65%、少なくとも75%、少なくとも85%、少なくとも95%、少なくとも99%または100%のアミノ酸同一性を有しているか、または配列番号1と1〜2、1〜3、1〜4、1〜5、1〜6、1〜7、1〜8、1〜9、1〜10、1〜11、1〜12、1〜13、1〜14、1〜15、1〜16、1〜17、1〜18、1〜19または1〜20アミノ酸だけ異なっている。
【0056】
Met−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチドと配列番号1のアミノ酸の違いは、2つの配列が異なるアミノ酸修飾の最低数を調べることにより計算される。アミノ酸修飾は欠失、付加、置換またはその組合せであり得る。
【0057】
アミノ酸配列同一性は、1つのポリペプチドのアミノ酸配列を第2ポリペプチドのアミノ酸配列と比較して、配列アラインメントを作成する当業界で公知の方法により測定される。アミノ酸同一性は、同一アミノ酸を有するアライン残基対の数をカウントすることにより前記アラインメントから算出される。
【0058】
配列同一性の測定方法には、G.D.Schuler,「生命情報科学:遺伝子及びタンパク質の分析に対する実践ガイド(Bioinformatics: A Pracitical Guide to the Analysis of Genes and Proteins)」,A.D.Baxevanis及びB.F.F.Ouelette編,John Wiley & Sons,Inc.(2001年)発行;Yonaら,「生命情報科学:配列、構造及びデータバンク(Bioinformatiacs: Sequence, structure and databanks)」,D.Higgins及びW.Taylor編,オックスフォード大学出版(2000年)発行;及び「生命情報科学:配列及びゲノム分析(Bioinformatics: Sequence and Genome analysis)」,D.W.Mount編,Cold Spring Harbor Laboratory Press(2001年)発行に記載されているものが含まれる。アミノ酸配列同一性を測定するための方法は、GAP(ウィスコンシン・パッケージ バージョン10.2;ウィスコンシン州マディソンに所在のGenetics Computer Group(GCG))、BLAST(Altschulら,J.Mol.Biol.,215(3):403−10(1990))及びFASTA(Pearson,Methods in Enzymology,R.F.Doolittle編,183:63−98(1990))のような公表されているコンピュータープログラムで体系化される。
【0059】
本発明の実施態様において、2つのポリペプチド間の配列同一性はGAPプログラム(ウィスコンシン・パッケージ バージョン10.2;ウィスコンシン州マディソンに所在のGenetics Computer Group(GCG))を用いて測定される。GAPはNeedleman及びWunschのアラインメント方法(Needlemanら,J.Mol.Biol.,48:443−453(1970))を利用している。GAPは2つの配列間の全ての可能性あるアラインメント及びギャップ位置を考察し、マッチド残基の数を最大限とする一方ギャップの数及び大きさを最小限とするグローバルアラインメントを作成する。シンボルマッチの値を割り当てるためにスコアリングマトリックスを用いる。加えて、アラインメントへのギャップの挿入を制限するためにはギャップ作成ペナルテイー及びギャップ延長ペナルティーが必要である。GAPを用いるポリペプチド比較のためのデフォルトプログラムパラメーターは、BLOSUM62(Henikoffら,Proc.Natl.Acad.Sci.USA,89:10915−10919(1992))アミノ酸スコアリングマトリックス(MATrix=blosum62.cmp)、ギャップ作成パラメーター(GAPweight=8)及びギャップ延長パラメーター(LENgthweight=2)である。
【0060】
全長に亘って配列番号1に実質的に類似していることに加えてより好ましいHCV Met−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチドは、配列番号1中に存在する対応領域に対して実質的に類似の各NS3、NS4A、NS4B、NS5A及びNS5B領域を生ずる。配列番号1中の対応領域は、Met−NS3アミノ酸1〜632、NS4Aアミノ酸633〜686、NS4Bアミノ酸687〜947、NS5Aアミノ酸948〜1394及びNS5Bアミノ酸1395〜1985である。
【0061】
別の実施態様では、NS3、NS4A、NS4B、NS5A及び/またはNS5B領域は配列番号1の対応領域に対して少なくとも65%、少なくとも75%、少なくとも85%、少なくとも95%、少なくとも99%または100%のアミノ酸同一性を有しているか、または1〜2、1〜3、1〜4、1〜5、1〜6、1〜7、1〜8、1〜9、1〜10、1〜11、1〜12、1〜13、1〜14、1〜15、1〜16、1〜17、1〜18、1〜19または1〜20アミノ酸だけ異なっている。
【0062】
好ましくは、配列番号1に対するアミノ酸修飾はT細胞抗原領域の全てまたは殆どを維持している。天然アミノ酸の違いは異なるアミノ酸側鎖(R基)による。R基はアミノ酸の諸特性、例えば物理的サイズ、電荷及び疎水性に影響を及ぼす。アミノ酸は以下のように複数のグループに分類され得る:中性で疎水性(アラニン、バリン、ロイシン、イソロイシン、プロリン、トリプトファン、フェニルアラニン及びメチオニン);中性で極性(グリシン、セリン、スレオニン、チロシン、システイン、アスパラギン及びグルタミン);塩基性(リシン、アルギニン及びヒスチジン);及び酸性(アスパラギン酸及びグルタミン酸)。
【0063】
通常、アミノ酸を置換するときには類似の特性を有するアミノ酸を交換することが好ましい。特定グループ内でアミノ酸を置換する、例えばロイシンをバリン、リシンをアルギニン、グルタミンをアスパラギンで置換することはポリペプチド3次構造を変化させない良好な候補である。
【0064】
特定のアミノ酸配列及び公知の遺伝子コードの縮重から出発して、多種多様のコード化核酸配列を得ることができる。遺伝子コードの縮重は、殆どすべてのアミノ酸がヌクレオチドトリプレット、すなわちコドンのいろいろな組合せによりコードされるために生ずる。特定コドンの特定アミノ酸への翻訳は当業界で公知である(例えば、Lewin,GENES IV,p.119,オックスフォード大学出版(1990年)発行参照)。アミノ酸は以下のようにコドンによりコードされる:
A=Ala=アラニン:コドン GCA,GCC,GCG,GCU;
C=Cys=システイン:コドン UGC,UGU;
D=Asp=アスパラギン酸:コドン GAC,GAU;
E=Glu=グルタミン酸:コドン GAA,GAG;
F=Phe=フェニルアラニン:コドン UUC,UUU;
G=Gly=グリシン:コドン GGA,GGC,GGG,GGU;
H=His=ヒスチジン:コドン CAC,CAU;
I=Ile=イソロイシン:コドン AUA,AUC,AUU;
K=Lys=リシン:コドン AAA,AAG;
L=Leu=ロイシン:コドン UUA,UUG,CUA,CUC,CUG,CUU;
M=Met=メチオニン:コドン AUG;
N=Asn=アスパラギン:コドン AAC,AAU;
P=Pro=プロリン:コドン CCA,CCC,CCG,CCU;
Q=Gln=グルタミン:コドン CAA,CAG;
R=Arg=アルギニン:コドン AGA,AGG,CGA,CGC,CGG,CGU;
S=Ser=セリン:コドン AGC,AGU,UCA,UCC,UCG,UCU;
T=Thr=スレオニン:コドン ACA,ACC,ACG,ACU;
V=Val=バリン:コドン GUA,GUC,GUG,GUU;
W=Trp=トリプトファン:コドン UCG;及び
Y=Tyr=チロシン:コドン UAC,UAU。
【0065】
核酸配列は、宿主における発現を強化しようとして最適化され得る。考慮すべき要因子には、G:C含量、好ましいコドン及び抑制性第2構造の回避が含まれる。これらの要因は特定宿主での高い発現を有する核酸配列を得るべくいろいろな方法で組合され得る(例えば、Donnellyらの国際特許出願公開第97/47358号パンフレット参照)。
【0066】
特定配列が特定宿主において高い発現を有する能力には幾つかの経験的実験を要する。前記実験には見込みのある核酸配列の発現を測定し、所要により前記配列を変化させることが含まれる。
【0067】
(B) コード化ヌクレオチド配列
配列番号2及び3は、Met−NS3−NS4A−NS4B−NS5A−NS5B配列をコードするヌクレオチド配列の2つの例を規定する。配列番号2のコード配列は、天然に存在するHCV−BK配列(GenBank受託番号M58335)のNS3−NS4A−NS4B−NS5A−NS5B領域に類似している(99.4%ヌクレオチド配列同一性)。配列番号3は配列番号2のコドン最適化バージョンである。配列番号2及び3は78.3%のヌクレオチド配列同一性を有する。
【0068】
HCV−BK NS3−NS4A−NS4B−NS5A−NS5Bヌクレオチド(GenBank受託番号M58335)と配列番号2の違いとして、配列番号2がリボソーム結合部位、ATGメチオニンコドン、修飾NS5B触媒ドメインをコードする領域、TAAAストップコドン及び更に30個のヌクレオチドの違いが挙げられる。修飾触媒ドメインはGlyAspAspではなくAlaAlaGly(残基1711〜1713)をコードして、NS5Bを不活化する。
【0069】
HCV Met−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチドをコードするヌクレオチド配列は、好ましくは配列番号2コード領域に実質的に類似している。別の実施態様において、HCV Met−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチドをコードするヌクレオチド配列は、配列番号2コード領域に対して少なくとも65%、少なくとも75%、少なくとも85%、少なくとも95%、少なくとも99%または100%のヌクレオチド配列同一性を有しているか、または配列番号2と1〜2、1〜3、1〜4、1〜5、1〜6、1〜7、1〜8、1〜9、1〜10、1〜11、1〜12、1〜13、1〜14、1〜15、1〜16、1〜17、1〜18、1〜19、1〜20、1〜25、1〜30、1〜35、1〜40、1〜45または1〜50ヌクレオチドだけ異なっている。
【0070】
Met−NS3−NS4A−NS4B−NS5A−NS5Bをコードする配列と配列番号2コード領域のヌクレオチドの違いは、2つの配列が異なるヌクレオチド修飾の最低数を調べることにより計算される。ヌクレオチド修飾は欠失、付加、置換またはその組合せであり得る。
【0071】
ヌクレオチド配列同一性は、1つの配列のヌクレオチド配列を第2配列のヌクレオチド配列と比較し、配列アラインメントを作成する当業界で公知の方法により測定される。配列同一性は、アラインさせた同一ヌクレオチドを有する位置の数をカウントすることにより前記アラインメントから算出される。
【0072】
2つのポリヌクレオチド間のヌクレオチド配列同一性の測定方法には、Schuler,「生命情報科学:遺伝子及びタンパク質の分析に対する実践ガイド(Bioinformatics: A Pracitical Guide to the Analysis of Genes and Proteins)」,A.D.Baxevanis及びB.F.F.Ouelette編,John Wiley & Sons,Inc.(2001年)発行;Yonaら,「生命情報科学:配列、構造及びデーターバンク(Bioinformatiacs: Sequence, structure and databanks)」,D.Higgins及びW.Taylor編,オックスフォード大学出版(2000年)発行;及び「生命情報科学:配列及びゲノム分析(Bioinformatics: Sequence and Genome analysis)」,D.W.Mount編,Cold Spring Harbor Laboratory Press(2001年)発行に記載されているものが含まれる。ヌクレオチド配列同一性を測定するための方法は、GAP(ウィスコンシン・パッケージ バージョン10.2;ウィスコンシン州マディソンに所在のGenetics Computer Group(GCG))、BLAST(Altschulら,J.Mol.Biol.,215(3):403−10(1990))及びFASTA(W.R.Person,Methods in Enzymology,R.F.Doolittle編,183:63−98(1990))のような公表されているコンピュータープログラムで体系化される。
【0073】
本発明の実施態様において、2つのポリヌクレオチド間の配列同一性はGAPプログラム(ウィスコンシン・パッケージ バージョン10.2;ウィスコンシン州マディソンに所在のGenetics Computer Group(GCG))を適用して測定される。GAPはNeedleman及びWunchのアラインメント方法(Needlemanら,J.Mol.Biol.,48:443−453(1970))を利用している。GAPは2つの配列間の全ての考えられるアラインメント及びギャップ位置を考察し、マッチド残基の数を最大限とする一方ギャップの数及び大きさを最小限とするグローバルアラインメントを形成する。シンボルマッチの値を割り当てるためにスコアリングマトリックスを用いる。加えて、アラインメントへのギャップの挿入を制限するためにはギャップ作成ペナルテイー及びギャップ延長ペナルティーが必要である。GAPを用いるポリペプチド比較のためのデフォルトプログラムパラメーターは、nwsgapda.cmpスコアリングマトリックス(MATrix=nwsgapda.cmp)、ギャップ作成パラメーター(GAPweight=50)及びギャップ延長パラメーター(LENgthweight=3)である。
【0074】
全長に亘って実質的に類似していることに加えてより好ましいHCV Met−NS3−NS4A−NS4B−NS5A−NS5Bヌクレオチド配列により、配列番号2中に存在する対応領域に対して実質的に類似の各NS3、NS4A、NS4B、NS5A及びNS5B領域が生ずる。配列番号2中の対応領域は、Met−NS3ヌクレオチド7〜1902、NS4Aヌクレオチド1903〜2064、NS4Bヌクレオチド2065〜2847、NS5Aヌクレオチド2848〜4188及びNS5Bヌクレオチド4189〜5661である。
【0075】
別の実施態様では、NS3、NS4A、NS4B、NS5A及び/またはNS5Bコード化領域は配列番号2中の対応領域に対して少なくとも65%、少なくとも75%、少なくとも85%、少なくとも95%、少なくとも99%または100%のヌクレオチド配列同一性を有しているか、または配列番号2と1〜2、1〜3、1〜4、1〜5、1〜6、1〜7、1〜8、1〜9、1〜10、1〜11、1〜12、1〜13、1〜14、1〜15、1〜16、1〜17、1〜18、1〜19、1〜20、1〜25、1〜30、1〜35、1〜40、1〜45または1〜50個のヌクレオチドだけ異なっている。
【0076】
(C) 遺伝子発現カセット
遺伝子発現カセットはポリペプチド発現に必要な要素を含んでいる。「ポリペプチド」はサイズを限定せず、タンパク質も含む。遺伝子発現カセット中に存在する調節要素は、通常(a)ポリペプチドをコードするヌクレオチド配列に転写的にカップリングされているプロモーター、(b)ヌクレオチド配列に機能的にカップリングされている5’リボソーム結合部位、(c)ヌクレオチド配列の3’末端に結合されているターミネーター、及び(d)ヌクレオチド配列に機能的にカップリングされている3’ポリアデニル化シグナルを含む。遺伝子発現またはポリペプチドプロセシングを強化または調節するのに有用な追加の調節要素を存在させてもよい。
【0077】
プロモーターは、RNAポリメラーゼにより認識され、下流領域の転写を媒介する遺伝子要素である。好ましいプロモーターは、転写レベルを向上させる強プロモーターである。強プロモーターの例は、前初期ヒトサイトメガロウイルスプロモーター(CMV)及びイントロンAを有するCMVである(Chapmanら,Nucl.Acids Res.,19:3979−3986(1991))。プロモーターの別の例には、EF1αプロモーター、マウスCMVプロモーター、ラウス肉腫ウイルスプロモーター、SV40初期/後期プロモーターやβ−アクチンプロモーターのような天然プロモーター;並びに合成筋肉特異的プロモーターやキメラ筋肉特異的/CMVプロモーターのような人工プロモーターが含まれる(Liら,Nat.Biotechnol.,17:241−245(1999);Hagstromら,Blood,95:2536−2542(2000))。
【0078】
リボソーム結合部位は開始コドンにまたはその近くに位置している。好ましいリボソーム結合部位の例には、CCACCAUGG、CCGCCAUGG及びACCAUGG(ここで、AUGは開始コドンである)が含まれる(Kozak,Cell,44:283−292(1986))。リボソーム結合部位の別の例はGCCACCAUGG(配列番号12)である。
【0079】
ポリアデニル化シグナルは、転写RNAの開裂及びポリ(A)テールのRNAへの付加に関与する。高級真核生物中のポリアデニル化シグナルは、ポリアデニル化付加部位から約11〜30ヌクレオチドにAAUAAA配列を含む。このAAUAAA配列はRNA開裂のシグナル化に関与する(Lewin,Genes IV,ニューヨークに所在のオックスフォード大学出版(1990年)出版)。ポリ(A)テールはmRNAプロセシングにとって重要である。
【0080】
遺伝子発現カセットの一部として使用され得るポリアデニル化シグナルは、最低の家兎β−グロビンポリアデニル化シグナル及びウシ成長ホルモンポリアデニル化(BGH)を含む(Xuら,Gene,272:149−156(2001);Postらの米国特許第5,122,458号明細書)を含む。更なる例には合成ポリアデニル化シグナル(SPA及びSV40ポリアデニル化シグナルが含まれる。SPA配列は、AAUAAAAGAUCUUUAUUUUCAUUAGAUCUGUGUGUUGGUUUUUUGUGUG(配列番号13)である。
【0081】
存在し得る遺伝子発現またはポリペプチドプロセシングを強化または調節するのに有用な追加の調節要素の例にはエンハンサー、リーダー配列及びオペレーターが含まれる。エンハンサー領域は転写を向上させる。エンハンサー領域の例にはCMVエンハンサー及びSV40エンハンサーが含まれる(Hittら,Methods in Molecular Genetics,7:13−30(1995);Xuら,Gene.272:149−156(2001))。エンハンサー領域はプロモーターに関連し得る。
【0082】
リーダー配列はポリペプチドをプロテオソームに指向させるポリペプチド上のアミノ酸領域である。リーダー配列をコードする核酸は構造遺伝子の5’であり、構造遺伝子に沿って転写される。リーダー配列の例はtPAである。
【0083】
遺伝子発現を調節するためにオペレーター配列を使用し得る。例えば、Tetオペレーター配列を遺伝子発現を抑制するために使用し得る。
【0084】
II. 治療用ベクター
Met−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチドをコードする核酸は、治療投与のために適したベクターを用いて患者に導入され得る。許容できない副作用を引き起こすことなく核酸を標的細胞にデリバーするベクターが適当であり得る。
【0085】
細胞発現は、Met−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチドをコードする遺伝子発現カセットを用いて達成される。遺伝子発現カセットは、有利な効果が得られるように標的細胞内で十分量の核酸を産生、プロセシングするために調節要素を含む。
【0086】
治療用途に使用され得るベクターの例には、第1世代及び第2世代アデノベクター、ヘルパー依存性アデノベクター、アデノ関連ウイルスベクター、レトロウイルスベクター、αウイルスベクター、ベネズエラウマ脳脊髄炎ウイルスベクター及びプラスミドベクターが含まれる(Hittら,Advances in Pharmacology,40:137−206(1997);Johnstonらの米国特許第6,156,588号明細書;Johnstonらの国際特許出願公開第95/32733号パンフレット)。Met−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチドを患者に導入するための好ましいベクターは第1世代アデノウイルスベクター及びプラスミドDNAベクターである。
【0087】
(A) 第1世代アデノウイルスベクター
遺伝子発現カセットを発現するための第1世代アデノベクターは、E1及び場合によりE3欠失組換えアデノウイルスゲノム中に発現カセットを含む。E1領域の欠失はアデノウイルスの複製に必要な要素を除去するのに十分に大きい。
【0088】
Met−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチドを発現するための第1世代アデノベクターはE1及びE3欠失組換えアデノウイルスゲノムを含む。E1領域の欠失はアデノウイルスの複製に必要な要素を除去するのに十分に大きい。E1及びE3領域の欠失の組合せは、Met−NS3−NS4A−NS4B−NS5A−NS5Bポリペプチドをコードする遺伝子発現カセットを収容するのに十分な大きさである。
【0089】
アデノウイルスは両端に逆方向末端反復を有する二本鎖線状ゲノムを有する。ウイルス複製中、ゲノムはウイルスカプシド内部にパッケージされてビリオンを形成する。ウイルスはウイルス結合及びその後の内部移行によりその標的細胞に進入する(Hittら,Advances in Pharmacology,40:137−206(1997))。
【0090】
アデノベクターは、ヒトまたは動物中に見られるような各種アデノウイルス血清型をベースとしている。動物アデノウイルスの例にはウシ、ブタ、チンパンジー、マウス、イヌ及びトリが含まれる(CELO)。好ましいアデノベクターはヒト血清型、より好ましくはグループB、CまたはD血清型をベースとしている。ヒトアデノウイルスグループB、C、DまたはE血清型の例には、タイプ2(“Ad2”)、4(“Ad4”)、5(“Ad5”)、6(“Ad6”)、24(“Ad24”)、26(“Ad26”)、34(“Ad34”)及び35(“Ad35”)が含まれる。アデノベクターは1つのアデノウイルスに由来する領域または2つ以上のアデノウイルスに由来する領域を含み得る。
【0091】
別の実施態様において、アデノベクターはAd5、Ad6またはその組合せをベースとする。Ad5はChroboczekら,J.Virology,186:280−285(1992)に記載されている。Ad6は図7A〜7Nに記載されている。Ad5領域を含むAd6ベースのベクターは以下の実施例に記載されている。
【0092】
アデノベクターにおいてE1及びE3領域は完全に除去されていなくてもよい。むしろ、イントランスに供給されるE1タンパク質の非存在下でベクター複製を無能とするのに十分な量のE1領域が除去されている。E1欠失、またはE1欠失とE3欠失の組合せは遺伝子発現カセットを収容するのに十分な大きさである。
【0093】
E1欠失はAd5の塩基対約342〜塩基対約3523または他のアデノウイルスの対応領域から得られ得る。好ましくは、欠失領域はAd5の塩基対約450〜塩基対約3511の領域または他のアデノウイルスの対応領域の除去を含む。塩基対約341から始まる大きなE1領域が欠失されるとウイルスパッケージングを促進する要素が除去される。
【0094】
E3欠失はAd5の塩基対約27865〜塩基対約30995または他のアデノウイルスの対応領域から得られ得る。好ましくは、欠失領域はAd5の塩基対約28134〜塩基対約30817の領域または他のアデノウイルスの対応領域の除去を含む。
【0095】
E1領域及び場合によりE3領域の欠失の組合せは、遺伝子発現カセットを含む組換えゲノムの全サイズが野生型アデノウイルスゲノムの約105%を越えない程度に十分に大きくなければならない。例えば、組換えアデノウイルスAd5ゲノムが約105%を越える大きさになるとゲノムは不安定になる(Bettら,Journal of Virology,67:5911−5921(1993))。
【0096】
好ましくは、遺伝子発現カセットを含む組換えアデノウイルスゲノムの大きさは野生型アデノウイルスゲノムの大きさの約85%〜約105%である。別の実施態様では、発現カセットを含む組換えアデノウイルスゲノムの大きさは野生型アデノゲノムの大きさの約100%〜約105.2%、または100%である。
【0097】
E1及びE3欠失を有するアデノウイルスゲノムに約7500kbが挿入され得る。欠失なしの場合、Ad5ゲノムは35,935塩基対であり、Ad6ゲノムは35,759塩基対である。
【0098】
第1世代アデノベクターは、E1遺伝子産物をイントランスに供給することにより複製され得る。E1遺伝子産物はイントランスに、例えばアデノウイルスE1領域で形質転換させた細胞株を用いることにより供給され得る。アデノウイルスE1領域で形質転換させた細胞または細胞株の例はHEK 293細胞、911細胞、PERC.6(商標)細胞及びトランスフェクトした一次ヒトアミノサイト細胞である(Grahamら,Journal of Virology,36:59−72(1977);Schiednerら,Human Gene Therapy,11:2105−2116(2000);Fallauxら,Human Gene Therapy,9:1909−1917(1998);Boutらの米国特許第6,033,908号明細書)。
【0099】
Met−NS3−NS4A−NS4B−NS5A−NS5B発現カセットは、欠失E1領域または欠失E3領域に対応する領域で組換えアデノベクターゲノムに挿入されなければならない。発現カセットは平行または逆平行方位を有し得る。平行方位では、挿入遺伝子の転写方向は欠失E1またはE3領域と同じ方向である。逆平行方位転写では、逆鎖は鋳型として作用し、転写方向は逆方向である。
【0100】
本発明の実施態様では、アデノベクターはE1欠失領域に挿入された遺伝子発現カセットを有する。前記ベクターは、
a)Ad5またはAd6のいずれかに対応する塩基対約1〜塩基対約450の第1アデノウイルス領域、
b)前記第1領域に連結した、E1平行またはE1逆平行方位の遺伝子発現カセット、
c)前記発現カセットに連結した、Ad5に対応する塩基対約3511〜塩基対約5548またはAd6に対応する塩基対約3508〜塩基対約5541の第2アデノウイルス領域、
d)前記第2領域に連結した、Ad5に対応する塩基対約5549〜塩基対約28133またはAd6に対応する塩基対約5542〜塩基対約28156の第3アデノウイルス領域、
e)前記第3領域に連結した、Ad5に対応する塩基対約30818〜塩基対約33966またはAd6に対応する塩基対約30789〜塩基対約33784の第4アデノウイルス領域、及び
f)前記第4領域に連結した、Ad5に対応する塩基対約33967〜塩基対約35935またはAd6に対応する塩基対約33785〜塩基対約35759の第5アデノウイルス領域
を含む。
【0101】
本発明の別の実施態様では、アデノベクターはE3欠失領域に挿入された遺伝子発現カセットを有する。前記ベクターは、
a)Ad5またはAd6のいずれかに対応する塩基対約1〜塩基対約450の第1アデノウイルス領域、
b)前記第1領域に連結した、Ad5に対応する塩基対約3511〜塩基対約5548またはAd6に対応する塩基対約3508〜塩基対約5541の第2アデノウイルス領域、
c)前記第2領域に連結した、Ad5に対応する塩基対約5549〜塩基対約28133またはAd6に対応する塩基対約5542〜塩基対約28156の第3アデノウイルス領域、
d)前記第3領域に連結した、E3平行またはE3逆平行方位の遺伝子発現カセット、
e)前記遺伝子発現カセットに連結した、Ad5に対応する塩基対約30818〜塩基対約33966またはAd6に対応する塩基対約30789〜塩基対約33784の第4アデノウイルス領域、及び
f)前記第4領域に連結した、Ad5に対応する塩基対約33967〜塩基対約35935またはAd6に対応する塩基対約33785〜塩基対約35759の第5アデノウイルス領域
を含む。
【0102】
アデノウイルス領域に関する好ましい別の実施態様では、
(1)第1、第2、第3、第4及び第5領域はAd5に対応する;
(2)第1、第2、第3、第4及び第5領域はAd6に対応する;
(3)第1領域はAd5に対応し、第2領域はAd5に対応し、第3領域はAd6に対応し、第4領域はAd6に対応し、第5領域はAd5に対応する。
【0103】
(B) DNAプラスミドベクター
DNAワクチンプラスミドは遺伝子発現カセットと共に複製及び好ましくはベクター選択を容易とする要素を含む。好ましい要素は非哺乳動物細胞における複製及び選択マーカーを与える。前記ベクターは、ヒト細胞における複製またはヒト核酸への組込みを与える要素を含むべきではない。
【0104】
選択マーカーにより、マーカーを含む核酸の選択が容易となる。好ましい選択マーカーは抗生物質耐性を付与するものである。抗生物質選択遺伝子の例には、アンピシリン、ネオマイシン及びカナマイシンに対する耐性をコードする核酸が含まれる。
【0105】
好適なDNAワクチンベクターは細菌性複製起点及び選択マーカーを含むプラスミドから作成され得る。高い収率を与える細菌性複製起点にはColE1プラスミド誘導の細菌性複製起点が含まれる(Donnellyら,Annu.Rev.Immunol.,15:617−648(1997))。
【0106】
細菌性複製起点及び選択マーカーを存在させると、大腸菌のような細菌株においてDNAベクターが産生され得る。選択マーカーはDNAベクターを含まない細菌を除去するために使用される。
【0107】
III. Ad6組換え核酸
Ad6組換え核酸は、配列番号8中に存在するAd6領域に実質的に類似するAd6領域及びAd6核酸中に存在しない領域を含む。Ad6領域を含む組換え核酸は、別のAd6領域の産生において、Ad6ベースのベクターの作成における中間体として、または組換え遺伝子のデリバリー用ベクターとしてのように各種用途を有する。
【0108】
図9に示すように、Ad6のゲノム構成はAd5のゲノム構成に非常に類似している。Ad5とAd6のホモロジーは約98%である。
【0109】
別の実施態様において、Ad6組換え核酸はE1A、E1B、E2B、E2A、E3、E4、L1、L2、L3またはL4に実質的に類似しているヌクレオチド領域またはその組合せを含む。Ad6領域に実質的に類似の核酸領域は少なくとも65%、少なくとも75%、少なくとも85%、少なくとも95%、少なくとも99%または100%のヌクレオチド配列同一性を有するか、または1〜2、1〜3、1〜4、1〜5、1〜6、1〜7、1〜8、1〜9、1〜10、1〜11、1〜12、1〜13、1〜14、1〜15、1〜16、1〜17、1〜18、1〜19、1〜20、1〜25、1〜30、1〜35、1〜40、1〜45または1〜50個のヌクレオチドだけ異なっている。実質的に類似の核酸配列を調べるための方法及び実施態様は上記セクションI.B.に記載されている。
【0110】
好ましくは、組換えAd6核酸はAd6中に存在しないポリペプチドをコードする発現カセットを含む。発現カセットの例には、HCV領域をコードする発現カセット及び別のタイプのポリペプチドをコードする発現カセットが含まれる。
【0111】
別のタイプのアデノウイルスベクターは、異なる量の第1世代及び第2世代アデノベクターのようなAd6を導入して作成され得る。上記セクションII.A.に記載したように、第1世代アデノベクターはE1を欠き、E1をイントランスに供給したときに複製し得る。
【0112】
第2世代アデノベクターは第1世代ベクターよりも小さいアデノウイルスベクターを含み、アデノウイルスタンパク質を供給するヘルパーベクター及び/または補充細胞株と一緒に使用され得る。第2世代アデノベクターは複数の文献、例えばRusell,Journal of General Virology,81:2573−2604(2000);Hittら,「遺伝子導入のためのヒトAdベクター(Human Ad vectors for Gene
Transfer)」,Advances in Pharmacology,Vol.40,Academic Press発行に記載されている。
【0113】
本発明の実施態様では、Ad6組換え核酸はE1をイントランスに供給したときに複製し得るE1を欠くアデノウイルスベクターである。発現カセットは欠失E1領域及び/または欠失E3領域に挿入され得る。
【0114】
欠失E1領域に発現カセットを有するAd6ベースのアデノウイルスベクターの例は、
a)Ad5またはAd6のいずれかに対応する塩基対約1〜塩基対約450の第1アデノウイルス領域、
b)前記第1領域に連結した、E1平行またはE1逆平行方位の遺伝子発現カセット、
c)前記発現カセットに連結した、Ad5に対応する塩基対約3511〜塩基対約5548またはAd6に対応する塩基対約3508〜塩基対約5541の第2アデノウイルス領域、
d)前記第2領域に連結した、Ad5に対応する塩基対約5549〜塩基対約28133またはAd6に対応する塩基対約5542〜塩基対約28156の第3アデノウイルス領域、
e)場合によっては存在する、前記第3領域に連結した、Ad5に対応する塩基対約28134〜塩基対約30817またはAd6に対応する塩基対約28157〜塩基対約30788の第4アデノウイルス領域、
f)第4領域が存在するときにはその第4領域に連結し、第4領域が存在しないときには第3領域に連結した、Ad5に対応する塩基対約30818〜塩基対約33966またはAd6に対応する塩基対約30789〜塩基対約33784の第5アデノウイルス領域、及び
g)前記第5領域に連結した、Ad5に対応する塩基対約33967〜塩基対約35935またはAd6に対応する塩基対約33785〜塩基対約35759の第6アデノウイルス領域
を含むかまたはこれらから構成されるが、少なくとも1つのAd6領域は存在する。
【0115】
本発明の別の実施態様では、全ての領域がAd6に由来する;第1及び第2を除く全ての領域がAd6に由来する;第2、第3,第4及び第5領域から選択される1〜4個の領域がAd6に由来する。
【0116】
欠失E3領域に発現カセットを有するAd6ベースのアデノウイルスベクターの例は、
a)Ad5またはAd6のいずれかに対応する塩基対約1〜塩基対約450の第1アデノウイルス領域、
b)前記第1領域に連結した、Ad5に対応する塩基対約3511〜塩基対約5548またはAd6に対応する塩基対約3508〜塩基対約5541の第2アデノウイルス領域、
c)前記第2領域に連結した、Ad5に対応する塩基対約5549〜塩基対約28133またはAd6に対応する塩基対約5542〜塩基対約28156の第3アデノウイルス領域、
d)前記第3領域に連結した、E3平行またはE3逆平行方位の遺伝子発現カセット、
e)前記遺伝子発現カセットに連結した、Ad5に対応する塩基対約30818〜塩基対約33966またはAd6に対応する塩基対約30789〜塩基対約33784の第4アデノウイルス領域、及び
f)前記第4領域に連結した、Ad5に対応する塩基対約33967〜塩基対約35935またはAd6に対応する塩基対約33785〜塩基対約35759の第5アデノウイルス領域、
を含むかまたはこれらから構成されるが、少なくとも1つのAd6領域は存在する。
【0117】
本発明の別の実施態様では、全ての領域がAd6に由来する;第1及び第2を除く全ての領域がAd6に由来する;第2、第3,第4及び第5領域から選択される1〜4個の領域がAd6に由来する。
【0118】
IV. ベクター作成
ベクターは、制限酵素の使用、核酸ライゲーションや相同組換えを含むような組換え核酸方法を用いて作成され得る。組換え核酸方法は当業界で公知である(Ausubel,「分子生物学における現在のプロトコール(Current Protocols in Molecular Biology)」,John Wiley(1987−1998年)発行;Sambrookら,「分子クローニング,実験マニュアル(Molecular Cloning, A Laboratory Manual)」,第2版,Cold Spring Harbor Laboratory Press(1989年)発行)。
【0119】
中間体ベクターは治療ベクターを引き出すために、または発現カセットまたはその一部を1つのベクターから他のベクターに移動させるために使用される。中間体ベクターの例には、アデノウイルスゲノムプラスミド及びシャトルベクターが含まれる。
【0120】
中間体ベクター中の有用な要素には、複製起点、選択マーカー、相同組換え領域及び便利な制限部位が含まれる。便利な制限部位は核酸配列のクローニングまたは遊離を促進するために使用され得る。
【0121】
相同組換え領域は、別の核酸分子中の標的領域に相同な核酸領域である。相同領域は、標的領域に挿入される核酸配列に隣接している。別の実施態様では、相同領域は好ましくは約150〜600ヌクレオチド長、または約100〜500ヌクレオチド長である。
【0122】
本発明の実施態様は、Met−NS3−NS4A−NS4B−NS5A−NS5B発現カセット、選択マーカー、細菌性複製起点、第1アデノウイルス相同領域、及びE1領域に挿入またはE1領域を置換するために発現カセットを標的する第2アデノウイルス相同領域を含むシャトルベクターに関する。第1及び第2相同領域は発現カセットに隣接している。第1相同領域は、塩基対約4〜450の野生型アデノウイルス領域の少なくとも右端(3’端)に実質的に相同な少なくとも約100塩基対を含む。第2相同領域は、塩基対約3511〜5792のAd5の少なくとも左端(5’端)または他のアデノウイルス由来の対応領域に実質的に相同な少なくとも約100塩基対を含む。
【0123】
「実質的に相同な」は、標的領域と特異的に組み換えるのに十分に相同度を指す。別の実施態様では、実質的に相同とは少なくとも85%、少なくとも95%または100%の配列同一性を指す。配列同一性は上記セクションI.B.に記載されているように計算され得る。
【0124】
アデノベクターを産生するための1つの方法は、発現カセットを含むアデノウイルスゲノムプラスミドの作成を含む。プレアデノウイルスは、所望の相補(complimenting)細胞株での複製に必要な全てのアデノウイルス配列を含む。その後、プレアデノウイルスプラスミドを制限酵素で消化してウイルスITRを放出し、ウイルスレスキューのために相補細胞株にトランスフェクトする。ITRは複製が生ずるようにプラスミド配列から放出されなければならない。アデノベクターレスキューの結果、発現カセットを含むアデノベクターが生ずる。
【0125】
(A)アデノウイルスゲノムプラスミド
アデノウイルスゲノムプラスミドは、長いプラスミド(コスミドであってもよい)内にアデノベクター配列を含む。長いプラスミドは、プラスミドを産生及び維持するために使用される手順に応じて真核または細菌細胞における増殖及び選択を促進するような追加要素を含み得る。アデノウイルスゲノムプラスミドを産生するための方法には、シャトルベクター及び相同組換えの使用を含む技術及び遺伝子発現カセットのアデノウイルスコスミドへの挿入を含む技術が含まれる(Hittら,Methods in Molecular Genetics,7:13−30(1995);Danthinneら,Gene Therapy,7:1707−1714(2000))。
【0126】
アデノウイルスゲノムプラスミドがE1またはE3欠失領域に挿入された遺伝子発現カセットを有していることが好ましい。本発明の実施態様では、アデノウイルスゲノムプラスミドは、E1欠失領域に挿入された遺伝子発現カセット、複製起点、選択マーカー及び組換えアデノウイルス領域を含み、前記組換えアデノウイルス領域は
a)Ad5またはAd6のいずれかに対応する塩基対約1〜塩基対約450の第1アデノウイルス領域、
b)前記第1領域に連結した、E1平行またはE1逆平行方位の遺伝子発現カセット、
c)前記発現カセットに連結した、Ad5に対応する塩基対約3511〜塩基対約5548またはAd6に対応する塩基対約3508〜塩基対約5541の第2アデノウイルス領域、
d)前記第2領域に連結した、Ad5に対応する塩基対約5549〜塩基対約28133またはAd6に対応する塩基対約5542〜塩基対約28156の第3アデノウイルス領域、
e)前記第3領域に連結した、Ad5に対応する塩基対約30818〜塩基対約33966またはAd6に対応する塩基対約30789〜塩基対約33784の第4アデノウイルス領域、
f)前記第4領域に連結した、Ad5に対応する塩基対約33967〜塩基対約35935またはAd6に対応する塩基対約33785〜塩基対約35759の第5アデノウイルス領域、及び
g)場合によっては存在する、所望アデノベクターの全サイズを考慮してより小さなインサートのために存在し得る、Ad5またはAd6中に存在するE3領域の全部または一部に対応するE3領域
から構成される。
【0127】
本発明の別の実施態様では、組換えアデノウイルスゲノムプラスミドはE3欠失領域に挿入された遺伝子発現カセットを有する。前記ベクターは複製起点、選択マーカー、並びに
a)Ad5またはAd6のいずれかに対応する塩基対約1〜塩基対約450の第1アデノウイルス領域、
b)前記第1領域に連結した、Ad5に対応する塩基対約3511〜塩基対約5548またはAd6に対応する塩基対約3508〜塩基対約5541の第2アデノウイルス領域、
c)前記第2領域に連結した、Ad5に対応する塩基対約5549〜塩基対約28133またはAd6に対応する塩基対約5542〜塩基対約28156の第3アデノウイルス領域、
d)前記第3領域に連結した、E3平行またはE3逆平行方位の前記遺伝子発現カセット、
e)前記遺伝子発現カセットに連結した、Ad5に対応する塩基対約30818〜塩基対約33966またはAd6に対応する塩基対約30789〜塩基対約33784の第4アデノウイルス領域、及び
f)前記第4領域に連結した、Ad5に対応する塩基対約33967〜塩基対約35935またはAd6に対応する塩基対約33785〜塩基対約35759の第5アデノウイルス領域
を含む。
【0128】
存在するアデノウイルス領域に関する別の実施態様では
(1)第1、第2、第3、第4及び第5領域はAd5に対応する;
(2)第1、第2、第3、第4及び第5領域はAd6に対応する;
(3)第1領域はAd5に対応し、第2領域はAd5に対応し、第3領域はAd6に対応し、第4領域はAd6に対応し、第5領域はAd5に対応する。
【0129】
本発明の実施態様は、アデノウイルスゲノムプラスミドを産生するための相同組換え段階及びアデノウイルスレスキュー段階を含むアデノベクターの作成方法に関する。前記相同組換え段階は、アデノウイルス相同領域に隣接してMet−NS3−NS4A−NS4B−NS5A−NS5B発現カセットを含むシャトルベクターの使用を含む。前記アデノウイルス相同領域により、発現カセットはE1またはE3欠失領域に標的される。
【0130】
アデノウイルスゲノムプラスミドの産生に関する本発明の実施態様では、遺伝子発現カセットを、Ad5またはAd6のいずれかに対応する塩基対約1〜塩基対約450の第1アデノウイルス領域;前記第1領域に連結した、Ad5に対応する塩基対約3511〜塩基対約5548またはAd6に対応する塩基対約3508〜塩基対約5541の第2アデノウイルス領域;前記第2領域に連結した、Ad5に対応する塩基対約5549〜塩基対約28133またはAd6に対応する塩基対約5542〜塩基対約28156の第3アデノウイルス領域;前記第3領域に連結した、Ad5に対応する塩基対約30818〜塩基対約33966またはAd6に対応する塩基対約30789〜塩基対約33784の第4アデノウイルス領域;及び前記第4領域に連結した、Ad5に対応する塩基対約33967〜塩基対約35935またはAd6に対応する塩基対約33785〜塩基対約35759の第5アデノウイルス領域からなるベクターに挿入する。アデノウイルスゲノムプラスミドは複製起点及び選択マーカーを含んでいなければならず、Ad5またはAd6 E3領域の全部または一部を含んでいてもよい。
【0131】
存在するアデノウイルス領域に関する別の実施態様では、
(1)第1、第2、第3、第4及び第5領域はAd5に対応する;
(2)第1、第2、第3、第4及び第5領域はAd6に対応する;
(3)第1領域はAd5に対応し、第2領域はAd5に対応し、第3領域はAd6に対応し、第4領域はAd6に対応し、第5領域はAd5に対応する。
【0132】
(B) アデノベクターレスキュー
アデノベクターは、当業界で公知であるかまたは本明細書に記載されている方法を用いて組換えアデノウイルスゲノムプラスミドからレスキューされ得る。当業界で公知のアデノウイルスレスキューの方法の例は、Hittら,Methods in Molecular Genetics,7:13−30(1995)及びDanthinneら,Gene Therapy,7:1707−1714(2000)に記載されている。
【0133】
本明細書に記載されているアデノベクターをレスキューするための好ましい方法はアデノウイルス複製のブーストが含まれる。ブースティングアデノウイルス複製は、例えばE2タンパク質(ポリメラーゼ、プレ末端タンパク質及びDNA結合タンパク質)のようなアデノウイルス機能及びE4orf6を別のプラスミドに供給することにより実施され得る。以下の実施例10には、コドン最適化Met−NS3−NS4A−NS4B−NS5A−NS5B発現カセットを含むアデノベクターをレスキューするためのアデノウイルス複製のブーストを例示している。
【0134】
V. 部分最適化HCVコード配列
HCVポリタンパク質コード化核酸の部分最適化により、ヒトでの発現のために最適化されるコドンの量は完全最適化に比して少なくなる。全目的は、最適化コドンを有するHCVポリタンパク質コード化核酸を含むアデノベクターの作成を促進しながら、コドン最適化のために高い発現という効果を与えることである。
【0135】
HCVポリタンパク質コード化配列を完全最適化すると、各アミノ酸について最も頻繁に見られるヒトコドンが与えられる。完全最適化は、当業界で公知のコドン頻度表及びBLACKTRANSLATEプログラム(ウィスコンシンパッケージ バージョン10;ウィスコンシン州マディソンに所在のGenetics Computer Group(GCG))を用いて実施され得る。
【0136】
部分最適化は、存在する全HCVポリタンパク質コード化配列(例えば、NS3−NS5B)または存在する1つ以上の局部領域に対して実施され得る。別の実施態様では、存在する全HCVコード化ポリタンパク質のGC含量は少なくとも約65%以下であり、1つ以上の局部領域のGC含量は約70%以下である。
【0137】
局部領域はHCVコード化核酸中に存在する領域であり、サイズは異なり得る。例えば、局部領域は約60、約70、約80、約90または約100ヌクレオチド長であり得る。
【0138】
部分最適化は、最初天然に存在する配列に基づいて部分的に最適化されるHCVコード化ポリタンパク質配列を構築することにより達成され得る。或いは、最適化HCVコード化配列は部分最適化配列を作成するために比較に基づいて使用され得る。
【0139】
VI. HCV併用治療
HCV Met−NS3−NS4A−NS4B−NS5A−NS5Bワクチンは単独で患者を治療するために使用され得、他のHCV治療薬と一緒に使用され得、及び他のタイプの疾患を標的とする物質と一緒に使用され得る。追加の治療には、HCV及びHCV感染患者に広く蔓延している疾患を治療するための追加の治療薬が含まれる。他のタイプの疾患を標的とする物質にはHIV及びHBVに対するワクチンが含まれる。
【0140】
HCVを治療するための追加の治療薬にはワクチン及び非ワクチンが含まれる(Zein,Expert Opin.Investig.Drugs,10:1457−1469(2001))。追加のHCVワクタンの例には、HCVコア抗原及びHCV E1、E2またはp7領域に対する免疫応答を引き出すように設計されたワクチンが含まれる。ワクチン成分は天然に存在するHCVポリペプチド、HCVミモトープポリペプチドまたは前記ポリペプチドをコードする核酸であり得る。
【0141】
HCVミモトープポリペプチドはHCVエピトープを含むが、天然に存在するHCV抗原とは異なる配列を有する。HCVミモトープは天然に存在するHCV抗原に融合され得る。一般的なミモトープの作成方法及び各種HCVミモトープはFeliciらの米国特許第5,994,083号明細書及びNicosiaらの国際特許出願公開第99/60132号パンフレットに記載されている。
【0142】
VII. 医薬投与
HCVワクチンは、当業界で公知の方法を用い、本明細書に記載されているガイダンスを用いて処方、患者に投与される。医薬投与の一般的ガイドラインは、例えばいずれも援用により本明細書に含まれるとするModern Vaccinology,Kurstak編,Plenum Med.Co.(1994年)発行;Remington’s Pharmaceutical Science,第18版,Gennaro編,Mack Publishing(1990年)発行;及びModern Pharmaceutics,第2版,Banker及びRhodes編,Marcel Dekker,Inc.(1990年)発行に記載されている。
【0143】
HCVワクチンは、静脈内、腹腔内、皮下、筋肉内、皮内、皮膚を介する圧痕または鼻のような各種ルートにより投与され得る。好ましいルートは筋肉内である。
【0144】
筋肉内投与は各種方法を用いて、例えば1つ以上の電気パルスと共に又は電気パルスなしで注射により実施され得る。電気媒介導入は液性及び細胞免疫応答の両方を刺激することにより遺伝免疫化を助けることができる。
【0145】
ワクチン注射は各種方法を用いて、例えば針及び無針注射システムを用いることにより実施され得る。無針注射システムの例はジェット注射デバイスである(Donnellyらの国際特許出願公開第99/52463号パンフレット)。
【0146】
(A) 電気媒介導入
電気媒介導入、すなわち遺伝子エレクトロトランスファー(GET)は、核酸注射後適当な電気パルスをデリバリーすることにより実施され得る(Mathiesenの国際特許出願公開第98/43702号パンフレット参照)。プラスミド注射及びエレクトロポレーションはステンレス針を用いて実施され得る。針は2つ、3つまたはそれ以上の複雑なパターンで使用され得る。1つの構成では、針を、機械的支持体であり且つ適当なケーブルを用いて電場発生装置に針をつなげるプリント基板上にはんだ付けされている。
【0147】
電気刺激は電気パルスの形態で加えられる。パルスは各種形態(スクエア、シヌソイド、三角、指数関数的減衰)及び各種極性(正または負極の単極性、双極性)を有し得る。パルスは定電圧または定電流方式でデリバリーされ得る。
【0148】
HCVを含む核酸ワクチン及び他の核酸ワクチンを患者に導入するために多種多様な電気治療パターンが使用され得る。電気治療の可能なパターンには、
(治療1) パルス長0.2msec/フェーズ、周波数1000Hz、定電圧モード、45ボルト/フェーズ、浮動電流で1秒おきにデリバリーされる1000スクェア双極性パルス10列、
(治療2) パルス長2msec/フェーズ、周波数100Hz、定電流モード、100mA/フェーズ、浮動電圧で1秒おきにデリバリーされる1000スクェア双極性パルス2列、
(治療3) 約2msec/フェーズのパルス長で全部で3秒間の双極性パルス2列(組織に流れる実際の電流は約50mAに固定されている)
が含まれる。
【0149】
電気パルスは電場発生装置を介してデリバリーされる。好適な発生装置は、共通シャシに組み立てられ、駆動プログラムを実行するポータブルPCにより駆動される3つの独立したハードウェア部品から構成され得る。ソフトウェアは基本的及び補助持機能を管理する。前記デバイスの部品は、(1)マイクロプロセッサーにより駆動する信号発生器、(2)電力増幅器及び(3)デジタルオシロスコープである。
【0150】
信号発生器は、ソフトウェアのコントロール下で所定範囲の任意の周波数及び形状を有する信号を発する。同一ソフトウェアはデリバリーされる波形に対する対話式エディタを有する。前記発生器はデジタル制御の限流デバイス(最大電流出力を制御するための安全機構)である。電力増幅器は発生したシグナルを±150Vまで増幅させ得る。オシロスコープはデジタル式であり、増幅器によりデリバリーされる電流及び電圧の両方をサンプリングし得る。
【0151】
(B) 薬用担体
医薬的に許容され得る担体により、ワクチンの貯蔵及びワクチンの被験者への投与が容易となる。医薬的に許容され得る担体の例は本明細書に記載されている。別の医薬的に許容され得る担体は当業界で公知である。
【0152】
医薬的に許容され得る担体は各種成分、例えば緩衝液、通常生理食塩液、リン酸緩衝食塩液、スクロース、塩及びポリソルベートを含み得る。医薬的に許容され得る担体の例は次の通りである:2.5〜10mM トリス緩衝液、好ましくは約5mM トリス緩衝液;25〜100mM NaCl、好ましくは約75mM NaCl;2.5〜10% スクロース、好ましくは約5% スクロース;0.01〜2mM MgCl2;及び0.001〜0.01% ポリソルベート80(植物由来)。pHは好ましくは約7.0〜9.0、より好ましくは約8.0である。担体の特定例は、pH8.0で5mM トリス、75mM NaCl、5% スクロース、1mM MgCl2、0.005% ポリソルベート80を含む。
【0153】
(C) 投薬レジメ
好適な投与レジメは、特定ワクチンの効力及び諸要因(例えば、患者の年齢、体重、性別及び医学的状態;投与ルート;所望効果;及び投与回数)を考慮して決定され得る。特定ワクチンの効力は各種因子、例えば細胞において発現、プロセシングされ、MHCクラスI及びII複合体の関係で提示されるポリペプチドを産生する特定ワクチンの能力に依存する。
【0154】
患者に投与されるHCVコード化核酸は、アデノベクターのようなウイルスベクター及びDNAプラスミドワクチンを含めた各種タイプのベクターの一部であり得る。DNAプラスミドの投与に関する別の実施態様では、約0.1〜10mgのプラスミドが患者に投与され、約1〜5mgのプラスミドが患者に投与される。ウイルスベクター、好ましくはアデノウイルスベクターの投与に関する別の実施態様では、約105〜1011ウイルス粒子が患者に投与され、約107〜1010ウイルス粒子が患者に投与される。
【0155】
ウイルスベクターワクチン及びDNAプラスミドワクチンは単独で投与してもよく、または初期及び追加投与レジメの一部であってもよい。混合方式の初期及び追加接種としては、DNAワクチンを初期接種した後ウイルスベクターワクチンを追加接種したり、ウイルスベクターワクチンを初期接種した後DNAワクチンを追加接種することが挙げられる。
【0156】
初回接種を複数回、例えば約2〜4回またはそれ以上実施してもよい。初回接種と追加接種の間の期間は通常約4ヶ月〜1年の範囲で変更可能であるが、他の時間枠を使用してもよい。DNAワクチンを用いる初回接種レジメの使用は、ヒトが既に抗−アデノウイルス免疫応答を示している状況において好ましいことがある。
【0157】
本発明の実施態様では、1×107〜1〜1012粒子、好ましくは約1×1010〜1〜1011アデノベクター粒子を筋肉組織に直接投与する。初回ワクチン接種後、アデノベクターまたはDNAワクチンを用いて追加接種する。
【0158】
本発明の別の実施態様では、初回接種をDNAワクチンを用いて筋肉組織に直接実施する。初回接種後、アデノベクターまたはDNAワクチンを用いて追加接種する。
【0159】
免疫応答をブーストするためにインターロイキン−12、GM−CSF、B7−1、B7−2、IP10、Mig−1のような物質を同時投与することができる。前記物質はタンパク質として、または核酸ベクターを用いて同時投与され得る。
【0160】
(D) 非相同初回−追加接種
非相同初回−追加接種は、初回接種のために1タイプのウイルスベクター、追加接種のために別のタイプのウイルスベクターを使用することを含む混合方式である。非相同初回−追加接種では、異なるアデノウイルス血清型をベースとするベクターのような関連ベクター及びより遠縁のウイルス(例えば、アデノウイルス及びポックスウイルス)を使用し得る。マウスをマラリアから防御するためにポックスウイルス及びアデノウイルスベクターを使用することはGilbertら,Vaccine,20:1039−1045(2002)に記載されている。
【0161】
初回接種及び追加接種に関する別の実施態様では、Met−NS3−NS4A−NS4B−NS5A−NS5Bのような所望の抗原を発現する以下のタイプのベクターを使用する。すなわち、Ad5ベクター後Ad6ベクター;Ad6ベクター後Ad5ベクター;Ad5ベクター後ポックスウイルスベクター;ポックスウイルスベクター後Ad5ベクター;Ad6ベクター後ポックスウイルスベクター後;ポックスウイルスベクター後Ad6ベクター。
【0162】
初回接種と追加接種の間の期間は通常約4ヶ月〜1年の範囲で変更可能であるが、他の時間枠を使用してもよい。少なくとも時間枠は免疫学的に安静させるのに十分でなければならない。1つの実施態様では、この安静は少なくとも6ヶ月である。初回接種には1つのタイプのベクターを用いる複数回(例えば、2〜4回)の初回接種が含まれ得る。
【0163】
ポックスウイルスベクター中に存在する発現カセットは、当該ポックスウイルスまたは別のポックスウイルスメンバーに生まれつきであるかまたはそれから誘導されるプロモーターを含んでいなければならない。ワクシニアウイルス、修飾ワクシニアウイルス、アビポックスウイルス、アライグマポックスウイルス、修飾ワクシニアウイルスAnkara、カナリアポックスウイルス(例えば、ALVAC)、トリポックスウイルス、ウシポックスウイルス及びNYVACをベースとするベクターを含めた各種タイプのポックスウイルスベースのベクターを構築、使用するための諸戦略が当業界で公知である(Moss,Current Topics in Microbiology and Immunology,158:25−38(1982);Earlら,「分子生物学における現在のプロトコル(Current Protocols in Molecular Biology)」,Ausubelら編,ニューヨークのGreene Publishing Associates & Wiley Interscience(1991年)発行,16.16.1−16.16.7;Childら,Virology,174(2):625−9(1990);Tartagliaら,Virology,188:217−232(1992);米国特許第4,603,112号明細書、同第4,722,848号明細書、同第4,769,330号明細書、同第5,110,587号明細書、同第5,174,993号明細書、同第5,185,146号明細書、同第5,266,313号明細書、同第5,505,941号明細書、同第5,863,542号明細書及び同第5,942,235号明細書)。
【0164】
(E) アジュバント
HCVワクチンはアジュバントと一緒に処方され得る。アジュバントはDNAプラスミドワクチンのために特に有用である。アジュバントの例はミョウバン、AlPO4、アルヒドロゲル、リピド−A及びその誘導体または変異体、フロインド不完全アジュバント、中性レポソーム、ワクチン及びサイトカインを含むリポソーム、ノニオン性ブロックコポリマー及びケモカインである。
【0165】
ポリオキシエチレン(POE)及びポリオキシルプロピレン(POP)を含むノニオン性ブロックポリマー、例えばPOE−POP−POEブロックコポリマーがアジユバントとして使用され得る(Newmanら,Critical Reviews in Therapeutic Drug Carroer Systems,15:89−142(1998))。核酸の免疫応答はノニオン性ブロックコポリマーとアニオン性界面活性剤を併用して強化され得る。
【0166】
アジュバント処方物の特定例は、CRL−1005(CytRx Research Laboratoris)、DNA及びベンジルアルコニウムクロリド(BAK)を含むものである。前記処方物は、PBS中にプラスミドDNAを含む冷(<5℃)溶液に純粋なポリマーをポジティブ置換ビペットを用いて添加することにより調製され得る。次いで、前記溶液を撹拌して、ポリマーを可溶化する。ポリマーが完全溶解したら、ポリマーの曇点(から6−7℃)以下の温度で透明な溶液が得られる。次いで、PBS中に溶解したBAKの希溶液をゆっくり添加することにより約4mM BAKをPBS中DNA/CRL−1005溶液に添加する。ポリマー及びBAK添加前の初期DNA濃度は約6mg/mlであり、最終濃度は約5mg/mlである。BAKを添加後、処方物を十分に撹拌し、その間に温度は〜2℃から曇点以上に上昇する。次いで、処方物を氷上に置いて、温度を曇点以下に下げる。その後、処方物を撹拌すると、温度は〜2℃から曇点以上に上昇する。動的光散乱により測定して処方物の粒径が約200〜500nmに達するまで温度を〜2℃から曇点以上に上昇させながら冷却及び混合を数回繰り返す。次いで、溶液が透明になるまで処方物を氷上に保存した後、−70℃で保存する。使用前に、処方物を室温まで融解させる。
【0167】
(F) ワクチン貯蔵
アデノベクター及びDNAワクチンは各種タイプの緩衝液を用いて貯蔵され得る。例えば、以下の実施例9に記載の緩衝液A105をベクター貯蔵のために使用し得る。
【0168】
DNAの貯蔵は、微量金属イオンを除去またはキレート化することにより向上され得る。DNAワクチンの安定性を向上させるためにコハク酸やリンゴ酸のような試薬及びキレート剤を使用し得る。キレート剤の例には、複数のホスフェートリガンド及びEDTAが含まれる。DNAプラスミドのダメージを遊離ラジカル生成から保護するために非還元性フリーラジカルスカベンジャー(例えば、エタノールまたはグリセロール)の配合も有用であり得る。更に、緩衝液タイプ、pH、塩濃度、露光及びバイアルを作成するために使用される滅菌方法のタイプをDNAワクチンの安定性を最適化するために処方物中で調節してもよい。
【実施例】
【0169】
以下、本発明の各種要件を更に説明するために実施例を提示する。これらの実施例は本発明を実施するために有用な方法を例示する。これらの実施例は本発明を限定するものではない。
【0170】
(実施例1)
Met−NS3−NS4A−NS4B−NS5A−NS5B発現カセット
HCV NS3−NS4A−NS4B−NS5A−NS5Bをコードする各種遺伝子発現カセットを1bサブタイプHCV BK株をベースとして構築した。コードされる配列は、(1)活性NS5B配列(“NS”)、(2)不活性NS5B配列(“NSmut”)、または(3)不活性NS5B配列を有するコドン最適化配列(“NSOPTmut”)を有していた。前記発現カセットはCMVプロモーター/エンハンサー及びBGHポリアデニル化シグナルも含んでいた。
【0171】
NSヌクレオチド配列(配列番号5)はHCV BK株(GenBank受託番号M58335)と5952個のヌクレオチド中30個のみ異なっている。NSアミノ酸配列(配列番号6)は対応の1b遺伝子型HCV BK株と1984個のアミノ酸中7個のみ異なっている。翻訳を開始するために、ATGコドンがNS配列の5’末端に存在している。TGA終止配列がNS配列の3’末端に存在している。
【0172】
NSmutヌクレオチド配列(配列番号2,図2)はNS配列に類似している。NSmutとNSの違いとして、NSmutが改変NS5B触媒部位、5’末端に最適リボソーム結合部位及び3’末端にTAAA終止配列を有していることが挙げられる。NS5Bの改変はアミノ酸1711〜1713をコードする塩基5138〜5146を含む。この改変の結果、アミノ酸GlyAspAspがAlaAlaGlyに変化し、不活性形態NS5B RNA依存性RNAポリメラーゼNS5Bが生ずる。
【0173】
NSOPTmut配列(配列番号3,図3)をNSmutによりコードされるアミノ酸配列をベースとして設計した。NSmutアミノ酸配列をGCG(ウィスコンシンパッケージ バージョン10;ウィスコンシン州マディソンに所在のGenetics Computer Group(GCG))BLACKTRANSLATEプログラムを用いてヌクレオチド配列に逆翻訳した。各アミノ酸が対応する最も頻繁に見られるヒトコドンによりコードされるNSOPTmutヌクレオチド配列を作成するために、パラメーターとして最もありそうなヌクレオチド配列の作成を選択し、翻訳スキームとしてGCGパッケージ内で利用可能な高発現ヒト遺伝子のコドン出現頻度表(human_high.cod)を特定して前記プログラムを実行した。
【0174】
(実施例2)
NS、NSmutまたはNSOPTmut配列を有するpV1Jnsプラスミドの作成
NS配列、NSmut配列またはNSOPTmut配列のいずれかを含むpV1Jnsプラスミドを以下のように作成し、特徴づけた。
【0175】
NS配列を有するpV1Jnsプラスミド
HCV BKタイプ株由来のコード領域Met−NS3−NS4A−NS4B−NS5A及びコード領域Met−NS3−NS4A−NS4B−NS5A−NS5B(Tomeiら,J.Virol.,67:4017−4026(1993))をpcDNA3プラスミド(Invitrogen)にクローン化して、それぞれpcD3−5a及びpcD3−5bを作成した。PcD3−5AをHindIIIで消化し、クレノウ充填で平滑末端化し、その後XbaIで消化して、Met−NS3−NS4A−NS4B−NS5Aのコード領域に対応する断片を作成した。この断片をpV1Jns−polyにクローン化し、BglIIで消化し、クレノウ充填で平滑末端化し、その後XbaIで消化して、pV1JnsNS3−5Aを作成した。
【0176】
pV1Jns−polyは、XbaI、PmeI、PacIに対する認識部位を含むポリリンカーをユニークBglII及びNotI制限部位に挿入することにより修飾したpV1JnsAプラスミド(Montgomeryら,DNA and Cell Biol.,12:777−783(1993))の誘導体である。NS配列を有するpV1Jns(pV1JnsNS3−5B)は、XbaI及びNotI消化で直線化したpV1JN3−5Aと約200bpのNS5A、NS5Bコード領域及び約60bpのBGHポリアデニル化シグナルを含むPCR断片とを同時形質転換させる細菌株BJ5183への相同組換えにより得られた。生じたプラスミドをpV1Jns−NSと称する。
【0177】
pV1Jns−NSは以下のように要約され得る:
塩基 pV1JnsAの1〜1881、
追加 AGCTT、
次に Met−NS3−NS5B配列(配列番号5)、
次に wt TGAストップ、
追加 TCTAGAGCGTTTAAACCCTTAATTAAGG(配列番号14)、
塩基 pV1JnsAの1912〜4909。
【0178】
NSmut配列を有するpV1Jnsプラスミド
V1JnsNS3−5Aを、完全コザック配列の付加によりNS3コード配列の5’において修飾した。プラスミド(V1JNS3−5Aコザック)は、AflII消化により直線化したV1JNS3−5Aと、イントロンAの近位部分、制限部位BglII、完全コザック翻訳開始配列及びNS3コード配列の一部を含むPCR断片とを同時形質転換させる細菌株BJ5183への相同組換えにより得られた。
【0179】
生じたプラスミド(V1JNS3−5Aコザック)をXbaI消化により直線化し、約200bpのNS5A、NS5B変異配列、強翻訳停止TAAA及び約60bpのBGHポリアデニル化シグナルを含むPCR断片と共に細菌株BJ5183に同時形質転換した。増幅のために使用されるオリゴヌクレオチドにより変異を導入した22bp重複断片を2つ組合せることによりPCR断片を得た。生じたプラスミドをpV1Jns−NSmutと称する。
【0180】
pV1Jns−NSmutは以下のように要約され得る:
塩基 pV1JnsAの1〜1882、
次に コザックMet−NS3−NS5B(mut)TAAA配列(配列番号2)、
追加 TCTAGA、
塩基 pV1JnsAの1925〜4909。
【0181】
NSOPTmut配列を有するpV1Jnsプラスミド
酵素活性を阻害するための変異NS5B、完全コザック翻訳開始配列及び強翻訳停止を有するヒトコドン最適化合成遺伝子(NSOPTmut)を遺伝子の5’及び3’末端に存在するBamHI及びSalI制限部位で消化した。次いで、遺伝子をpV1JnsAプラスミドのポリリンカー中に存在するBglII及びSalI制限部位にクローン化して、pV1Jns−NSOPTmutを作成した。
【0182】
pV1Jns−NSOPTmutは以下のように要約され得る:
塩基 pV1JnsAの1〜1881、
追加 C、
次に コザックMet−NS3−NS5B(optmut)TAAA配列(配列番号3)、
追加 TTTAAATGTTTAAAC(配列番号15)、
塩基 pV1JnsAの1905〜4909。
【0183】
プラスミドの特徴づけ
L−グルタミン(最終4mM)を補充した10% FCS/DMEMにおいて増殖させたHEK293細胞のトランスフェクションによりHCV NSタンパク質の発現を試験した。トランスフェクションの24時間前に、トランスフェクション日に90〜95%集密度に達するまで細胞を直径35mmの6ウェルにおいて平板培養した。LIPOFECTAMINE 2000試薬を用いて、ラウス肉腫ウイルスプロモーターの制御下で40ngのプラスミドDNA(非飽和DNA量として前もって測定)を100ngのルシフェラーゼリポーター遺伝子を含むpRSV−Lucプラスミドと同時トランスフェクトした。細胞をCO2インキュベーター中37℃において48時間保持した。
【0184】
細胞抽出物を1% トリトン/TEN緩衝液中で調製した。この抽出物をルシフェラーゼ活性について正規化し、10% SDS−アクリルアミドゲル上で連続希釈で流した。タンパク質をニトロセルロースに移し、発現強度及び適正なタンパク質分解開裂を評価するためにNS3、NS5A及びNS5Bに対する抗体を用いてアッセイした。偽トランスフェクトした細胞をネガティブコントロールとして用いた。pV1JnsNS、pV1JnsNSmut及びpV1JnsNSOPTmutを試験した代表的な実験の結果を図12に示す。
【0185】
(実施例3)
プラスミドDNAベクターを用いるマウス免疫化
抗−HCV免疫応答を誘発する可能性を評価するために、各種マウス株にDNAプラスミドpV1Jns−NS、pV1Jns−NSmut及びpV1Jns−NSOPTmutを注射した。2種の株(Balb/C及びC57Black6,N=9−10)に対してDNA(25または50μg)を筋肉内注射した後電気パルスを加えた。各動物に対して3週間間隔で2回投与した。
【0186】
C57Black6マウスにおいてNS3タンパク質に対して誘発された体液性免疫応答を投与後の2つの血清中で細菌発現したNS3プロテアーゼドメインに対するELISAにより測定した。試験抗原に対して特異的な抗体が3つのすべてのベクターを用いて免疫化した動物で検出され、幾何平均(相乗平均)力価(GMT)は94000〜133000であった(表1〜3)。
【0187】
【表1】
【0188】
【表2】
【0189】
【表3】
【0190】
3週間間隔でプラスミドDNA(25μg)を2回筋肉内注射して免疫化したC57Black6マウスにおいてT細胞応答を調べた。定量ELIspotアッセイを実施して、NS3−MS5B配列を包含する10残基が重複している20merペプチドの5プールに応答してIFNγを分泌するT細胞の数を測定した。特異的CD8+応答を、C57Black6マウスの場合にはCD8+エピトープを包含する20merペプチド(pep1480)を用いて同一アッセイにより分析した。
【0191】
IFNγを抗原特異的に分泌する細胞を標準ELIspotアッセイを用いて検出した。3週間間隔でプラスミドDNA(50μg)を2回筋肉内注射して免疫化したC57Black6マウスにおけるT細胞応答を、NS3−NS5B配列を包含する10残基が重複している20merペプチドの5プールに応答してIFNγを分泌するT細胞の数を測定する同一のELIspotアッセイにより分析した。
【0192】
免疫化マウスから脾臓細胞を作成し、R10培地(10% FCS、2mM L−グルタミン、50U/ml〜50μg/ml ペニシリン/ストレプトマイシン、10mM Hepes、50μM 2−メルカプトエタノールを補充したRPMI 1640)に再懸濁した。マルチスクリーン96ウェル濾過プレート(Milliporeのカタログ番号MAIPS4510;マサチューセッツ州アッシュビー・ロード・ベッドフォードに所在のMillipore Corporation)を精製ラット抗−マウスIFN−γ抗体(PharMingenのカタログ番号18181D;カリフォルニア州サンジェゴに所在のPharmiMingen)で被覆した。一晩インキュベート後、プレートをPBS 1×/0.005% ツイーンで洗浄し、R10培地(250μl/ウェル)でブロックした。
【0193】
免疫化マウスから脾細胞を作成し、2.5×105/ウエルまたは5×105/ウエルの密度で10μM ペプチドの存在または非存在下で24時間インキュベートした。十分洗浄(PBS 1×/0.005% ツイーン)した後、ビオチニル化ラット抗−マウスIFNγ抗体(PharMingenのカタログ番号18112D;カリフォルニア州サンディェゴに所在のPharMingen)を添加し、4℃において一晩インキュベートした。展開のために、ストレプトアビジン−AKP(PharMingenのカタログ番号13043E;カリフォルニア州サンジェゴに所在のPharMingen)及び1−Step(商標)NBT−BCIP展開溶液(Pierceのカタログ番号34042;イリノイ州ロックフォードに所在のPierce)を添加した。
【0194】
HCV BK株NS3〜NS5Bの全配列を包含する20mer重複ペプチドのプールを用いて、HCV特異的IFNγ分泌T細胞を明らかとした。同様に、C57Black6マウスの場合には、CD8応答を検出するためにCD8+エピトープを包含する1つの20merペプチドを用いた。プラスミドベクターpV1Jns−NS、pV1Jns−NSmut及びpV1Jns−NSOPTmut(25〜50μg)を2回注射して免疫化したC57Black6及びBalb/Cマウス(N=9−10)のグループの代表的データーを図13A及び13Bに示す。
【0195】
(実施例4)
アカゲサルの免疫化
7.5mg/mlのCRL1005、0.6mM ベンザルコニウムクロリド中のプラスミドpV1Jns−NSOPTmut(5mg)を筋肉内注射することによりアカゲザル(N=3)を免疫化した。各動物の三角筋に0及び4週目に2回注射した。
【0196】
CMIをIFN−γ ELISPOTによりいろいろな時間に測定した。このアッセイはHCV抗原特異的CD8+及びCD4+ Tリンパ球応答を測定し、ヒト、アカゲザル、マウス及びラットのような各種哺乳動物に対して使用され得る。
【0197】
特定ペプチド及びペプチドプールを使用すると、CTL細胞毒性アッセイ、インターフェロン−γ ELISPOTアッセイ及びインターフェロン−γ細胞内染色アッセイにおける抗原提示を単純化し得る。各種HCVタンパク質(コア、E2、NS3、NS4A、NS4B、NS5A、NS5B)のアミノ酸配列をベースとするペプチドを、HCV DNA、アデノウイルスベクターワクチン接種したアカゲザル及びHCV感染ヒトにおける免疫応答を調べるために上記アッセイで使用するために作成した。各ペプチドは、10アミノ酸で埋め合わされる重複20merである。大プールのペプチドはHCVタンパク質に対する全応答を検出するために使用され得、小プールのペプチド及び各ペプチドは応答のエピトープ特異性を規定するために使用され得る。
【0198】
IFNγ−ELISPOT
IFNγ ELISPOTアッセイによりHCV特異的Tリンパ球応答が定量測定される。PBMCを連続希釈し、抗−アカゲザルIFN−γ抗体(MD−1 U−Cytech)を被覆したマイクロプレートウェルに置く。これらをHCVペプチドプールと20時間培養すると、前駆体細胞の再刺激及びIFN−γの分泌が生じる。細胞を洗浄すると、細胞が存在している濃厚域に抗体被覆ウェルに結合した分泌IFNが残る。捕捉IFNはビオチニル化抗−アカゲザルIFN抗体(デテクターAb U−Cytech)及びアルカリホスファターゼコンジュゲートストレプトアビジン(Pharmingen 13043E)を順次用いて検出する。不溶性アルカリホスファターゼ基質を添加すると、ウェル中の細胞が存在する場所に暗いスポットが生じ、IFN−γを分泌したT細胞毎に1つのスポットが残る。
【0199】
ウェルあたりのスポットの数は抗原特異的T細胞の前駆体頻度に直接関連している。γ−インターフェロンが最も一般的であり、活性化Tリンパ球により合成、分泌される最も豊富なサイトカインの1つであるので、これを(特定の抗−γインターフェロンモノクローナル抗体を用いる)本アッセイで可視化されるサイトカインとして選択した。このアッセイでは、100万個のPBMCあたりのスポット形成細胞(SFC)の数をペプチド抗原の存在及び非存在(培地コントロール)下でサンプル毎に測定する。2つの材料を投与後のPBMCについてのアカゲザルからのデーターを表4に示す。
【0200】
【表4】
【0201】
(実施例5)
Ad6プリアデノウイルスプラスミドの構築
Ad6プレアデノウイルスプラスミドを以下のように得た。
【0202】
pAd6 E1−E3+プレアデノウイルスプラスミドの構築
第1世代Ad6ベクターを作成するために使用され得るAd6ベースのプレアデノウイルスプラスミドを、Ad5とAd6間の広い配列同一性(約98%)を利用するかまたはAd6領域のみを含めて構築した。wtAd6配列を細菌プラスミドにクローン化するために相同組換えを使用した。
【0203】
Ad5領域及びAd6領域を含む細菌プラスミドとしてpAd6E1−E3+を回収するために使用される一般的方法を図10に示す。BJ5183細菌を精製wtAd6ウイルスDNA及びAd5 ITRカセットと称される第2DNA断片を用いて同時形質転換すると、相同組換えによりウイルスゲノムが回付(circularization)した。ITRカセットは、細菌性複製起点及びアンピシリン耐性遺伝子を含むプラスミド配列により分離されるAd5ゲノムの右端(bp33798〜35935)及び左端(bp1〜341及びbp3525〜5767)由来の配列を含む。ITRカセットはAd5 342〜3524のE1配列の欠失を含む。ITRカセット中のAd5配列により、組換えが起こり得る精製Ad6ウイルスDNAと相同な領域が生ずる。
【0204】
可能性あるクローンを制限分析によりスクリーニングし、1つのクローンをpAd6E1−E3+として選択した。次いで、このクローンを完全に配列分析した。pAd6E1−E3+は、bp1〜341及びbp3525〜5548のAd5配列、Ad6 bp5542〜33784、Ad5 bp33967〜35935を含む(bpの数はAd5及びAd6のwt配列を指す)。pAd6E1−E3+は、血清型特異性を構成する全てのAd6ビリオン構造タンパク質に対するコード配列を含んでいる。
【0205】
Ad6領域を含む細菌プラスミドとしてpAd6E1−E3+を回収するために使用される一般的方法を図11に示す。BJ5183細菌を精製wt Ad6ウイルスDNA及びAd6 ITRカセットと称される第2DNA断片を用いて同時形質転換すると、相同組換えによりウイルスゲノムが回付した。ITRカセットは、細菌性複製起点及びアンピシリン耐性遺伝子を含むプラスミド配列により分離されるAd6ゲノムの右端(bp35460〜35759)及び左端(bp1〜450及びbp3508〜3807)由来の配列を含んでいる。これら3つのセグメントをPCRにより作成し、pNEB193に順次クローン化して、pNEBAd6−3(ITRカセット)を作成した。このITRカセットはAd5 451〜3507のE1配列の欠失を含む。ITRカセット中のAd6配列により、組換えが起こり得る精製Ad6ウイルスDNAと相同な領域が生ずる。
【0206】
pAd6 E1−E3−プレアデノウイルスプラスミドの構築
A5領域を含み、E3領域が欠失しているAd6ベースのベクターを、Ad5領域を含むpAd6E1−E3+から始めて構築した。E3領域(Ad6 bp25871〜31192)を含むpAd6E1−E3+のA 5322bpサブ断片をpABS.3にサブクローン化して、pABSAd6E3を作成した。その後、このプラスミドにおいて3つのE3を欠失させて、3つの新しいプラスミドpABSAd6E3(1.8Kb)(Ad6 bp28602〜30440欠失)、pABSAd6E3(2.3Kb)(Ad6 bp28157〜30437欠失)及びpABSAd6E3(2.6Kb)(Ad6 bp28157〜30788欠失)を作成した。次いで、pAd6E1−E3+に3つのE3欠失部を逆置換するために細菌組換えを使用して、Ad6ゲノムプラスミドpAd6E1−E3−1.8Kb、pAd6E1−E3−2.3Kb及びpAd6E1−E3−2.6Kbを作成した。
【0207】
(実施例6)
NS配列を有するAd5ゲノムプラスミドの作成
コード領域NS3−NS4A−NS4B−NS5Aを含むpcDNA3プラスミド(Invitrogen)をXmuI及びNruI制限部位で消化し、CMVプロモーター、NS3−NS4A−NS4B−NS5Aコード領域及びウシ成長ホルモン(BGH)ポリアデニル化シグナルを含むDNA断片をシャトルベクターpDelE1 SpaのユニークEcorV制限部位にクローン化して、Sva3−5Aベクターを作成した。
【0208】
コード領域NS3−NS4A−NS4B−NS5A−NS5Bを含むpcDNA3プラスミドをXmnI及びEcorIで消化(部分消化)し、NS5Aの一部、NS5B遺伝子及びBGHポリアデニル化シグナルを含むDNA断片をSva3−5Aベクターにクローン化し、EcorI及びBglIIで消化し、クレノウで平滑末端化して、Sva3−5Bベクターを作成した。
【0209】
Sva3−5Bベクターは最後に消化したSspI及びBst1107I制限部位であり、アデノウイルス配列に隣接している発現カセット(CMVプロモーター,NS3−NS4A−NS4B−NS5A−NS5Bコード領域及びBGHポリアデニル化シグナル)を含むDNA断片をpAd5HVO(E1−,E3)ClaI直線化ゲノムプラスミドと共に細菌株BJ5183に同時形質転換して、pAd5HVONSを作成した。pAd5HVOはAd bp1〜341、bp3525〜28133及びbp30818〜35935を含んでいる。
【0210】
(実施例7)
NSmut配列を有するアデノウイルスゲノムプラスミドの作成
NS−mut配列を含むアデノウイルスゲノムプラスミドをAd5またはAd6バックグラウンドで作成した。Ad6バックグラウンドは塩基1〜450、3511〜5548及び33967〜35935でAd5領域を含んでいた。
【0211】
pV1JN3−5AコザックをBglII及びXbaI制限酵素で消化し、コザック配列及びNS3−NS4A−NS4B−NS5Aをコードする配列を含むDNA断片をBglII及びXbaI消化ポリpMRKpdelE1シャトルベクターにクローン化した。生じたベクターをshNS3−5Aコザックと称した。
【0212】
ポリpMRKpdelE1はBglII、PmeI、SwaI、XbaI、SalIに対する認識部位を含むポリリンカーをCMVプロモーターの下流に存在するユニークBglII制限部位に挿入することにより修飾したRKpdelE1(Pac/pIX/pack450)+CMVmin+BGHpA(str.)の誘導体である。MRKpdelE1(Pac/pIX/pack450)+CMVmin+BGHpA(str.)は、bp451〜3510のE1配列を欠失させたbp1〜5792のAd5配列を含む。ヒトCMVプロモーター及びBGHポリアデニル化シグナルをそれらを分離するユニークBglII部位とE1平行方位のE1欠失部に挿入した。
【0213】
酵素活性を阻害するように変異させた、3’末端に強翻訳停止を有するNS5B断片をアセンブリPCRにより得、相同組換えによりshNS3−5Aコザックベクターに挿入して、ポリpMRKpdelE1NSmutを作成した。ポリpMRKpdelE1NSmutでは、NS−mutコード配列はCMVプロモーターの制御下にあり、BGHポリアデニル化シグナルは下流に存在している。
【0214】
遺伝子発現カセット及び相同組換えし得るアデノウイルス配列を含む隣接領域をPacI及びBst1107I制限酵素を用いる消化により切り出し、pAd5HVO(E1−,E3−)またはpAd6E1−E3−2.6Kb ClaI直線化ゲノムプラスミドと共に細菌株BJ5183に同時形質転換して、それぞれpAd5HVONSmut及びpAd6E1−,E3−NSmutを作成した。
【0215】
pAd6E1−E3−2.6Kbは、Ad5 bp1〜341及びbp3525〜5548、Ad6 bp5542〜28157及びbp 30788〜33784、Ad5 bp33967〜35935を含んでいる(bpの数はAd5及びAd6の両方に対するwt配列を指す)。いずれのプラスミドでも、ウイルスITRは細菌性複製起点及びアンピシリン耐性遺伝子を含むプラスミド配列により連結されている。
【0216】
(実施例8)
NSOPTmutを有するアデノウイルスゲノムプラスミドの作成
pCRBluntベクター(Invitrogen)にクローン化した配列番号3のヒトコドン−最適化合成遺伝子(NSOPTmut)をBamH1及びSalI制限酵素で消化し、シャトルベクターポリpMRKpdelE1中に存在するBglIV及びSalI制限部位にクローン化した。生じたクローン(ポリpMRKpdelE1NSOPTmut)をPacI及びBst1107I制限酵素で消化し、pAd5HVO(E1−,E3−)またはpAd6E1−E3−2.6Kb ClaI直線化ゲノムプラスミドと共に細菌株BJ5183に同時形質転換して、それぞれpAd5HVONSOPTmut及びpAd6E1−,E3−NSOPTmutを作成した。
【0217】
(実施例9)
アデノウイルスベクターのレスキュー及び増幅
アデノベクターをPer.6細胞においてレスキューした。Per.C6を、L−グルタミン(最終4mM)、ペニシリン/ストレプトマイシン(最終100U/ml)及び10mM MgCl2を補充した10% FCS/DMEMにおいて増殖した。感染後、細胞を5% ウマ血清(HS)を補充した同一培地において維持した。ウイルスレスキューのために、2.5×106個のPer.C6を6cmΦペトリ皿において平板培養した。
【0218】
平板培養から24時間後、PacI直線化アデノウイルスDNA(10μg)を用いてリン酸カルシウム法により細胞をトランスフェクトした。DNA沈殿を細胞上に4時間放置した。培地を除去し、5% HS/DMEMを添加した。
【0219】
細胞変性効果が目に見えるまで(1週間)細胞をCO2インキュベーター中で維持した。細胞及び上清を回収し、凍結/融解サイクル(液体窒素/37℃の水浴)に3回かけた。ライセートを−4℃において3000rpmで20分間遠心し、回収した上清(細胞上に1回通したウイルスを含有する細胞ライセートに対応;P1)を10cmΦペトリ皿において80〜90%集密度のPer.C6を感染させるために1ml/皿の量で使用した。感染させた細胞を細胞変性効果が目に見えるまでインキュベートし、細胞及び上清を回収し、ライセートを上記したように作成した(P2)。
【0220】
2×15cmΦペトリ皿を感染させるためにP2ライセート(4ml)を使用した。この感染から回収したライセート(P3)を大きなウイルス調製物に対する出発点として使用されるウイルスストックとして複数のアリコートで−80℃において保存した。この場合、2×15cmΦペトリ皿を感染させるためには1mlのストックで十分であり、生じたライセート(P4)を大規模感染のためのペトリ皿の感染用に使用した。
【0221】
FCSを含まない培地で希釈し、10ml/皿の量で30×15cmΦペトリ皿(80〜90%集密度のPer.C6を含む)を感染させるために使用したP4ライセートから更なる増幅物を得た。細胞をCO2インキュベーター中で20分ごとに緩やかに混合しながら1時間インキュベートした。12ml/皿の5% HS/DMEMを添加し、細胞変性効果が目に見えるまで(約48時間)細胞をインキュベートした。
【0222】
細胞及び上清を集め、4℃において2000rpmで20分間遠心した。ペレットを0.1M トリス(pH8.0)(15ml)中に再懸濁した。細胞を凍結/融解サイクル(液体窒素/37℃の水浴)に3回かけて溶解した。2M MgCl2(150μl)及びDNAse(10mlの20mM トリス−HCl(pH7.4)、50mM NaCl、1mM ジチオトレイトール、0.1mg/ml ウシ血清アルブミン、50% グリセロール中10mg ウシ膵デオキシリボヌクレアーゼI)(75μl)を添加した。水浴において37℃において1時間インキュベート(15分毎に撹拌)した後、ライセートを4℃において4000rpmで15分間遠心した。回収した上清を直ちにCsCl勾配にかけた。
【0223】
CsCl勾配は、SW40無菌チューブにおいて次のように作成した:
1.5d CsCl(0.5ml)、
1.35d CsCl(3ml)、
1.25d CsCl(3ml)、
ウイルス上清(5ml/チューブ)を適用した。
【0224】
所要により、チューブの上部に0.1M トリス−Cl(pH8.0)を満たした。チューブを−10℃においてローターSW40を用いて35000rpmで1時間遠心した。(1.25/1.35インターフェースに位置する)ウイルスバンドを注射器により集めた。
【0225】
ウイルスを新しいSW40無菌チューブに移し、1.35d CsClをチューブの上部に満たした。10℃においてローターSW40において35000rpmで24時間遠心後、ウイルスをできるだけ少量で集め、緩衝液A 105(5mM トリス,5% スクロース,75mM NaCl,1mM MgCl2,0.005% ポリソルベート80(pH8.0))に対して徹底的に透析した。透析後、グリセロールを最終10%まで添加し、ウイルスを−80℃において複数のアリコートで保存した。
【0226】
(実施例10)
強化アデノベクターレスキュー
HCV NSOPTmut導入遺伝子を有する第1世代Ad5及びAd6ベクターはレスキューしがたいことが判明した。レスキュー過程の考えられる障害はアデノウイルスの複製機構のための最適以下鋳型であるプラスミドDNAの複製が効率的でないことに起因するであろう。ベクターのE1領域に挿入された導入遺伝子の非常に高いG−C含量に関連して、(通常ウイルスDNA中に存在する)DNAの5’末端に連結している末端タンパク質が不在であるために、プラスミド誘導アデノウイルスの複製速度が実質的に低減し得る。
【0227】
Adベクターをレスキューするためのより効率的で再現性の手順を確立するために、tet−誘導性プロモーターの制御下で全てのE2タンパク質(ポリメラーゼ、プレ末端タンパク質及びDNA結合タンパク質)及びE4 orf6を含む発現ベクター(pE2;図19)を用いた。PerC6及び293にpE2及び一般的なプレアデノプラスミドをトランスフェクトすることにより、Ad DNA複製が非常に向上し、完全な感染性アデノウイルス粒子がより効率的に生じる。
【0228】
プラスミド構築
pE2はクローニングベクターpBI(CLONTECH)をベースとし、細胞培養においてエピソームを複製し、選択するために2つの要素、すなわち(1)EBNA−1を発現させるときに細胞サイクルと同期してプラスミドを複製できるEBV−OriP(EBV[nt]7421−8042)領域及び(2)形質転換した細胞をポジティブ選択し得るハイグロマイシンBホスホトランスフェラーゼ(HPH)耐性遺伝子を加えた。アデノウイルス遺伝子E2a及びb及びE4−Orf6に対する2つの転写単位を構築し、下記するようにpE2において合わせた。
【0229】
Ad5−ポリメラーゼClaI/SphI断片及びAd5−pTP Acc65/EcoRV断片をpVac−Pol及びpVac−pTP(Stunnembergら,NAR,16:2431−2444(1988))から得た。両断片にクレノウを充填し、pBIのSalI(充填)及びEcoRV部位にクローン化して、それぞれpBI−Pol/pTPを得た。
【0230】
pCEP4(Invitrogen)由来のEBV−OriP要素をまずpJC13−1(Chungら,Cell,74(3):505−14(1993))のBamHI部位にクローン化することにより2つのトリβ−グロビン隔離体ダイマー内に挿入した。次いで、pJC13−OriP由来のHS4−OriP断片をpSA1mv(tk−Hygro−B耐性遺伝子発現カセット及びAd5複製起点を含むプラスミド)内部にクローン化した。ここで、ITRを頭−尾ジャンクションとして配置し、pFG140(Grahan,EMBO J.,3:2917−2922(1984))から5’−TCGAATCGATACGCGAACCTACGC−3’(配列番号16)及び5’−TCGACGTGTCGACTTCGAAGCGCACACCAAAAACGTC−3’(配列番号17)のプライマーを用いてPCRにより得た。こうして、pMVHS4Oripを作成した。次いで、隔離Orip、Ad5 ITRジャンクション及びtk−HygroBカセットを含むpMVHS4Orip由来のDNA断片をAseI/AatII制限pBI−Pol/pTPに挿入して、pBI−Pol/pTPHS4を作成した。
【0231】
Ad5−Orf6及びAd5−DBPを発現する第2転写単位を構築するために、PCRにより得たE4orf6(Ad5[nt]33193−34077)をまずpBIベクターに挿入して、pBI−Orf6を作成した。その後、DBPコーディングDNA配列(Ad5[nt]22443−24032)をpBI−Orf6に挿入して、第2の双方向的Tet調節発現ベクター(pBI−DBP/E4Orf6)を得た。pBI中に存在するオリジナルのポリAシグナルをBGH及びSV40ポリAで置換した。
【0232】
次いで、頭−尾ジャンクションで配置したAdeno5−ITR及びプラスミドpSA−1mvから得たハイグロマイシンB耐性遺伝子を含むDNA断片を挿入することによりpBI−DBP/E4orf6を修飾した。次いで、この新しいプラスミドpBI−DBP/E4orf6シャトルをドナープラスミドとして使用して、第2tet−調節転写単位を、pE2を保有する大腸菌株BJ5183を用いる相同組換えによりpBI−Pol/pTPHS4に挿入した。
【0233】
細胞株、トランスフェクション及びウイルス増幅
PerC6細胞を、10% ウシ胎仔血清(FBS)、10mM MgCl2、ペニシリン(100U/ml)、ストレプトマイシン(100μg/ml)及び2mM グルタミンを補充したダルベッコ改変イーグル培地(DMEM)において培養した。
【0234】
全ての一時的トランスフェクションをリポフェクタミン2000(Invitrogen)を製造業者が指示するように用いて実施した。6cmプレートにおいて平板培養した90%集密度のPERC.6(商標)をAd5/6NSOPTmutプレアデノプラスミド(3.5μg)でトランスフェクトし、PacIを単独でまたはpE2(5μg)+pUHD52.1(1μg)の組合せで用いて消化した。pUHD52.1は逆tetトランスアクティベーター2(rtTA2)(Urlinger,Proc.Natl.Acad.Sci.U.S.A.,97(14):7963−7968(2000))に対する発現ベクターである。トランスフェクション時に、pE2発現を活性化するために、細胞を1μg/mlのドキシサイクリンの存在下で培養した。トランスフェクションから7日後、細胞を回収し、細胞ライセートを凍結/融解サイクルを3回実施して得た。細胞ライセート(2ml)を使用して、第2のPerC6の6cm皿を感染させた。感染細胞を、完全CPEが観察されるまで培養した後、回収した。ウイルスを上記したように5回連続継代した後、CsCl勾配で精製した。精製ウイルスのDNA構造をエンドヌクレアーゼ消化及びアガロースゲル電気泳動分析によりコントロールし、元のプレアデノプラスミド制限パターンと比較した。
【0235】
(実施例11)
HCVポリタンパク質コード化核酸の部分最適化
ヒト宿主での発現のために最適化したコドンを含むアデノベクターの作成を促進するためにHCVポリタンパク質コード化核酸の部分最適化を実施した。総合的目的は、HCVポリタンパク質をコードするアデノベクターの作成を促進しながらコドン最適化により高い発現を与えることであった。
【0236】
ヒト宿主での発現のために最適化したコドンを含むHCVタンパク質をコードするアデノベクターを作成する際に幾つかの困難を経験した。最適化配列(配列番号3)を含むアデノベクターは、非最適化配列(配列番号2)を含むアデノベクターよりも合成及びレスキューが困難であることが判明した。
【0237】
配列番号3を含むアデノベクターを作成する際の困難は高いGC含量に起因していた。特に厄介な領域はNSOPTmut(配列番号3)の約No.3900の領域である。
【0238】
アデノベクター中での使用を容易とするために最適化HCVコード化核酸配列の別のバージョンを設計した。NSOPTmutと比較して、コード化ポリタンパク質及びそれぞれの開裂産物の発現を改善するように高レベルのコドン最適化を維持しながら低い全GC含量を有し、連続G’s及びC’sの潜在的に問題のモチーフの存在が低下/解消するように別のバージョンを設計した。
【0239】
コドンを次善に最適化した配列の作成の出発点はNSOPTmutヌクレオチド配列のコード領域(配列番号3の塩基7〜5961)である。コドン使用頻度の値(各アミノ酸につき1.0の全値に正規化した)をウィスコンシン・パッケージ バージョン10.3(Pharmacopeia,Inc.が100%出資の子会社のAccelrys Inc.)で販売しているファイルhuman_high.codから得た。
【0240】
局部及び全GC含量を低減するために、各アミノ酸毎に好ましいコドン置換を規定している表を手動で作成した。各アミノ酸について、1)最も一般的なコドンに比較して低いGC含量及び2)比較的に多く観察されるコドン使用頻度(human_high.codに定義されている)を有するコドンを置換コドンとして選択した。例えば、Argの場合、最高頻度のコドンはCGCである。Argをコードする他の5つの代替コドン(CGG、AGG、AGA、CGT、GGA)のうち、3つ(AGG、CGT、CGA)は1塩基により、1つ(AGA)は2塩基により、1つ(CGG)は0塩基によりGC含量を低減させる。AGAコドンはhuman_high.codに比較的低使用頻度(0.1)としてリストされているので、CGCを置換するコドンとして0.18の相対頻度を有するAGGが選択された。他のアミノ酸に対するコドン置換を確立するために同様の基準が適用され、表5に示すリストが作成された。以下の最適化手順で適用されるパラメーターは、生じる配列が十分に改善されたコドン使用(各アミノ酸について)を維持し、(全体及び保存G’s及び/またはC’sの局部ストレッチの形態の)GC含量が減少するように実験的に決定された。
【0241】
部分最適化HCVコード化配列の2つの例は配列番号10及び配列番号11である。配列番号10は全体的に部分最適化されているHCVコード化配列である。配列番号11は部分最適化されている領域を除いてコドン使用について完全に最適化されているHCVコード化配列である。
【0242】
コドン最適化は以下の手順を用いて実施した:
段階1)
インプット完全最適化NSOPTmut配列のコード領域を、各サイクル後1コドンだけスライディングウィンドをシフトする3コドン(9塩基)のスライディングウィンドを用いて分析した。5個以上の連続G’s及び/またはC’sを含むストレッチを前記ウィンドで検出したときにはいつも、以下の置換ルールを適用した。Let Nは既に実施したコドン置換の数を示す。Nが奇数のときにはウインド中の中間コドンを表5に特定したコドンで置換する。Nが偶数のときにはウィンドの第3末端コドンをhuman_high.codのようなコドン最適化表に特定されているコドンで置換する。LeuまたはValが第2または第3コドンに存在しているときには、非常に低い相対コドン使用頻度(例えば、human_high.cod参照)のLeuまたはValコドンが導入されないようにいずれの置換も適用しない。以下のサイクルで、シフトしたウインドの分析を以前のサイクルの置換を含む配列に適用した。
【0243】
3コドンウインド中の中間及び末端コドンの交互置換は、実験的に(手順後の最終配列から判断して)GC含量を低減させながら最適化コドン使用の全体維持をより満足させることが判明した。しかしながら、通常、正確な置換戦略は分析されるヌクレオチド配列によりコードされるアミノ酸配列に依存し、実験的に決定されるであろう。
【0244】
段階2)
段階1)で実施した全てのコドン置換を含む配列を21コドン(63塩基)長のスライディングウィンドを用いて更に分析した。調節可能なパラメーターに従ってウィンド中の全GC含量を測定した。ウィンド中のGC含量が70%以上ならば、以下のコドン置換戦略を適用した。このウィンドでは、アミノ酸Asn、Asp、Cys、Glu、His、Ile、Lys、Phe、Tyrのコドンを表5に示すコドンで置換する。この組のアミノ酸に対する置換の限定は、a)置換コドンがhuman_high.codにおける許容されるぐらい高い使用頻度を有している、及びb)置換コドンについてGUTG中の平均全ヒトコドン使用は最も高頻度くらい高いという事実により考えられた。以下のサイクルでは、シフトしたウインドの分析を以前のサイクルの置換を含む配列に適用される。
【0245】
各アミノ酸につきGC含量の全低減と高いコドン最適化の維持を妥協することにより閾値70%を実験的に決定した。段階1)と同様に、正確な置換戦略(アミノ酸の選択及びGC含量閾値)は分析されるヌクレオチド配列によりコードされるアミノ酸配列に依存し、実験的に決定されるであろう。
【0246】
段階3)
次いで、段階1)及び2)により作成した配列を手動で編集し、追加コドンを以下の基準に従って変化させた。21コドンのウィンドにおいてなお70%以上のGC含量を有している領域を手動で試験して、数個のコドンを表5に示すスキームに従って再び置換した。
【0247】
有用な制限部位を与えるため、相補鎖上の存在するオープンリーデイングフレームを除去するため、相同組換え領域を付加するために、コザックシグナルを付加するため及びターミネーターを付加するために以下の段階を実施した。これらの段階は段階4〜7である。
【0248】
段階4)
その後制限酵素のサブセットを用いてクローニングできるように、段階3で作成した配列を特定の制限部位(BglII、PmeI及びXbaI)の不在及び唯一のStuI部位の存在について試験した。2つの部位(BglIIに対する1部位及びStuIに対する1部位)を各認識部位の一部であるコドンを置換することにより配列から除去した。
【0249】
段階5)
次いで、段階1)〜4)により作成した配列をその後修飾NSOPTmut配列を(相同組換えにより)作成できるように修飾した。段階1)〜4)から得た配列では、塩基3556〜3755を含むセグメント及び塩基4456〜4656を含むセグメントをNSOPTmut由来の対応配列により置換した。相同組換えによりNSOPTmut中の問題の領域(No.3900あたり)を置換するために配列番号10の塩基3356〜4656を含むセグメントを使用して、配列番号11の配列を有するNSOPTmutの変異体を得ることができる。
【0250】
段階6)
段階1)〜5)で得た配列の分析から、相補鎖上の完全断片にほぼ広がっている可能性あるオープンリーディングフレームが示された。センス鎖からすべてのコドンCTA及びTTA(Leu)とTCA(Ser)を除去すると、相補鎖上の1つのリーディングフレームの1つ中の全てのストップコドンが有効的に除去された。この相補鎖オープンリーディングフレームの転写及びその後のタンパク質への翻訳の可能性は非常に少ないが、センス鎖上でコードされる配列の転写及びその後の翻訳との可能性ある干渉を避けるためにSerに対するTCAコドンをほぼ500塩基毎にセンス上に導入した。相同組換えを実施するために段階5)で導入したセグメントに変化を加えなかった。Serに対するTCAコドンはLeuに対するCTA及びTTAよりも好ましかった。なぜならば、human_high.codにおいてCTA(0.02)及びTTA(0.03)に比してTCA(0.05)は比較的により高い相対頻度であるからである。更に、CUTGからの平均ヒトコドン使用ではTCAが好ましかった(CTA及びTTAの0.07に対して0.14)。
【0251】
段階7)
最終段階では、最適化内部リボソームエントリー部位(コザックシグナル)を作成するためにGCCACCを配列の5’末端に付加し、3’にTAAAストップシグナルを付加した。NSsuboptmutの翻訳特性の開始を維持するために、コドン領域の最初の8コドンをNSOPTmut配列と同一に維持した。生じた配列を再びBglII、PmeI及びXbaI認識部位の不在及び唯一のStuI部位の存在についてチェックした。
【0252】
NSsuboptmut配列(配列番号10)は、NSOPTmut(70.3%)に比して全体的に低いGC含量(63.5%)を有しており、コドン使用最適化の十分に最適なレベルを維持している。NSsuboptmutのヌクレオチド同一性はNSmutに対して77.2%であった。
【0253】
【表5】
【0254】
(実施例12)
ウイルス特徴づけ
アデノベクターを、(a)物理的粒子/mlの測定、(b)TaqMan PCRアッセイの実行、及び(c)HeLa細胞の感染後のタンパク質発現のチェックにより特徴づけた。
【0255】
(a)物理的粒子の測定
CsCl精製ウイルスを0.1% SDS PBSで1/10及び1/100希釈した。コントロールとして、緩衝液A105を使用した。これらの希釈物を55℃において10分間インキュベートした。チューブを軽く回転させた後、260nmでODを測定した。ウイルス粒子の量を、1OD260nm=1.1×1012物理的粒子/mlに従って計算した。結果は典型的には5×1011〜1×1012物理的粒子/mlであった。
【0256】
(b)TaqMan PCRアッセイ
TaqMan PCRアッセイをアデノベクターゲノムの定量(Q−PCR粒子/ml)のために使用した。TaqMan PCRアッセイは、ABI Prism 7700−配列デテクターを用いて実施した。反応をアデノウイルス骨格に対して特異的なプローブ(最終200μMで)及びオリゴヌクレオチド(最終200nMで)の存在下で最終50μl容量で実施した。ウイルスを0.1% SDS PBSで1/10希釈し、55℃において10分間インキュベートした。チューブを軽く回転させた後、連続1/10希釈物(水中)を調製した。10−3、10−5及び10−7希釈物10μlをPCRアッセイの鋳型として使用した。
【0257】
各サンプル中に存在する粒子の量を、同一実験で作成した標準曲線に基づいて計算した。結果は典型的には1×1012〜3×1012Q−PCR粒子/mlであった。
【0258】
(c)HCV非構造タンパク質の発現
HCV NSタンパク質の発現をHeLa細胞を感染させることにより試験した。1.5×106細胞/皿(10cmΦペトリ皿)の感染前日に細胞を平板培養した。50、250及び1250pp/細胞のm.o.i.に対応する異なる量のCsCl精製ウイルスを最終容量5mlまで培地(FCS非含有)で希釈した。希釈ウイルスを細胞に添加し、CO2インキュベーター中37℃において1時間インキュベートした(20分毎にやさしく混合)。5% HS−DMEM(5ml)を添加し、細胞を37℃で48時間インキュベートした。
【0259】
1% トリトン/TEN緩衝液中で細胞抽出物を調製した。この抽出物を10% SDS−アクリルアミドゲル上に流し、ニトロセルロースにブロットし、NS3、NS5a及びNS5bに対する抗体を用いてアッセイして、適正なポリタンパク質開裂をチェックした。偽感染細胞をネガティブコントロールとして使用した。Ad5−NS、MRKAd5−NSmut、MRKAd6−NSmut及びMRKAd6−NSOPTmutを試験した代表的な実験の結果を図14に示す。
【0260】
(実施例13)
各種NSカセットをコードするアデノベクターを用いるマウス免疫化
アデノベクターAd5−NS、MRKAd5−NSmut、MRKAd6−NSmut及びMRKAd6−NSOPTmutをC57Black6マウス株に注射して、抗−HCV免疫応答を誘発する可能性を評価した。動物群(N=9−10)には109ppのCsCl精製ウイルスを筋肉内注射した。各動物に対して3週間間隔で2回投与した。
【0261】
NS3タンパク質に対する体液性免疫応答をC57Black6免疫化マウスからの投与後の2つの血清中で細菌発現させたNS3プロテアーゼドメインに対してELISAにより測定した。試験した抗原に対して特異的な抗体が100〜46000の幾何平均(相乗平均)力価(GMT)で検出された(表6、7、8及び9)。
【0262】
【表6】
【0263】
【表7】
【0264】
【表8】
【0265】
【表9】
【0266】
C57Black6マウスにおけるT細胞応答を、NS3−NS5B配列を包含する10残基が重複している20merペプチドの5プール(F〜L+Mの名称)に応じてIFNγを分泌するT細胞をの数を測定する定量ELISPOTアッセイにより分析した。C57Black6マウスにおいて誘導される特異的CD8−応答を、C57Black6マウスではCD8+エピトープを包含する20merペプチド(pep1480)を用いて同一アッセイにより分析した。抗原特異的にIFNγを分泌する細胞を標準ELIspotアッセイを用いて検出した。
【0267】
脾臓細胞、脾細胞及びペプチドを上記実施例3に記載されているように産生し、処理した。ベクターAd5−NS、MRKAd5−NSmut及びMRKAd6−NSmutの109ウイルス粒子を2回注射して免疫化したC57Black6マウスの群(N=9−10)からの代表的データーを図15に示す。
【0268】
(実施例14)
アデノベクターを用いるアカゲザルの免疫化
アカゲザル(N=3−4)を、CsCl精製Ad5−NS、MRKAd5−NSmut、MRKAd6−NSmutまたはMRKAd6−NSOPTmutウイルスを筋肉内注射することにより免疫化した。各動物の三角筋に1011または1010vpを0及び4週目に2回投与した。
【0269】
CMIを、a)IFN−γ ELISPOT(上記実施例3参照)、b)IFN−γ ICS及びc)バルクCTLアッセイにより異なる時間に測定した。これらのアッセイはHCV抗原特異的CD8+及びCD4+ Tリンパ球応答を測定し、ヒト、アカゲザル、マウス及びラットを含めた各種哺乳動物に対して使用され得る。
【0270】
特異的ペプチドまたはペプチドのプールの使用は、CTL細胞毒性アッセイ、インターフェロン−γ ELISPOTアッセイ及びインターフェロン−γ細胞内染色アッセイにおける抗原提示を単純化し得る。各種HCVタンパク質(コア、E2、NS3、NS4A、NS4B、NS5a、NS5b)のアミノ酸配列をベースとするペプチドを、HCV DNA、アデノウイルスベクターワクチン接種アカゲザル及びHCV感染ヒトにおける免疫応答を調べるための上記アッセイに使用するために作成した。各ペプチドは10アミノ酸で埋め合わされる重複20merである。大プールのペプチドをHCVタンパク質に対する全応答を検出するために使用することができ、小プールのペプチド及び各ペプチドは応答のエピトープ特異性を規定するために使用し得る。
【0271】
IFN−γ ICS
IFN−γ ICSのために、R10(10% FCSを補充したRPMI培地)(1ml)中の2×106PBMCをペプチドプール抗原で刺激した。各ペプチドの最終濃度は2μg/mlであった。細胞をCO2インキュベーター中37℃において1時間インキュベートした後、可溶性サイトカインの分泌を抑えるためにBrefeldin Aを10μg/mlの最終濃度まで添加した。細胞を37℃において更に14〜16時間インキュベートした。
【0272】
刺激を、共刺激抗体のCD28及びCD49d(抗−ヒトCD28 BD340975及び抗−ヒトCD49d BD340976)の存在下で実施した。インキュベート後、細胞を表面抗原に対する蛍光色素−コンジュゲート抗体の抗−CD3、抗−CD4、抗−CD8(CD3−APC Biosource APS0301、CD4−PE BD345769、CD8−PerCP BD345774)で染色した。
【0273】
細胞内サイトカインを検出するために、細胞をFACS透過化緩衝液2(BD340973)2×最終濃度で処理した。固定し、透過化した後、細胞をヒトIFN−γ、IFN−γFITCに対する抗体(Biosource AHC4338)とインキュベートした。
【0274】
細胞をPBS中の1% ホルムアルデヒド中に再懸濁し、24時間以内にFACSで分析した。4色FACS分析を2つのレーザーを備えたFACS Calibur装置(Becton Dickinson)を用いて実施した。CD3、CD8ポジティブ集団をカップリングしたForward対Side Scatterプロットにおいてリンパ球集団のゲーティングを得た。少なくとも3000事象のゲートが得られた。ポジティブ細胞を106リンパ球に対するIFN−γ発現細胞の数として表した。
【0275】
1010または1011vpの各種アデノベクターを1回または2回注射後の免疫化サルからのIFN−γ ELISPOT及びIFN−γ ICSデーターを図16A〜16D、17A及び17Bに示す。
【0276】
バルクCTLアッセイ
Tリンパ球の識別エフェクター機能は、この細胞集団のサブセットの適切なMHC−関連抗原ペプチドを示す細胞を直接溶解する能力である。この細胞毒性活性は、CD8+ Tリンパ球に最も頻繁に関係している。
【0277】
記憶T細胞の抗原再刺激及び増大を与えるように、PBMCサンプルにインビトロでHCV抗原を発現する組換えワクチンウイルスを約14日間感染させた。ペプチド抗原プールで処置した自己B細胞系に対する細胞毒性を試験した。
【0278】
培養物の溶解機能は、CTLエフェクター細胞との4時間インキュベーション中に標的細胞から遊離したクロムより生じた特異的溶解の%として測定する。特異的細胞毒性を測定し、無関係抗原または賦形剤処理B細胞系と比較した。このアッセイは半定量的であり、CTL応答がワクチンにより誘発されるかどうかを調べるための好ましい手段である。1011vp/投与のアデノベクターAd5−NS、MRKAd−NSmut及びMRKAd6−NSmutで免疫化したサルからの2回注射後のデーターを図18A〜18Fに示す。
【0279】
他の実施態様も請求の範囲内である。幾つかの実施態様を説明、例示してきたが、本発明の範囲及び趣旨を逸脱することなく各種修飾を加え得ることができる。
【特許請求の範囲】
【請求項1】
1つ以上のAD6領域及びAD6中に存在しない領域を含む組換え核酸であって、少なくとも1つのAD6領域がE1A、E1B、E2B、E2A、E4、L1、L2、L4及びL5からなる群から選択されることを特徴とする前記組換え核酸。
【請求項2】
AD6中に存在しない前記領域がAD6中に存在しないポリペプチドをコードする発現カセットであることを特徴とする請求の範囲第1項に記載の組換え核酸。
【請求項3】
前記組換え核酸が、E1をイントランスで供給したときに複製できるE1を少なくとも欠くアデノウイルスベクターであることを特徴とする請求の範囲第2項に記載の組換え核酸。
【請求項4】
前記ベクターが
a)Ad5またはAd6のいずれかに対応する塩基対約1〜塩基対約450の第1アデノウイルス領域、
b)前記第1領域に連結した、E1平行またはE1逆平行方位の前記遺伝子発現カセット、
c)前記遺伝子発現カセットに連結した、Ad5に対応する塩基対約3511〜塩基対約5548またはAd6に対応する塩基対約3508〜塩基対約5541の第2アデノウイルス領域、
d)前記第2領域に連結した、Ad5に対応する塩基対約5549〜塩基対約28133またはAd6に対応する塩基対約5542〜塩基対約28156の第3アデノウイルス領域、
e)場合によっては存在する、前記第3領域に連結した、Ad5に対応する塩基対約28134〜塩基対約30817またはAd6に対応する塩基対約28157〜塩基対約30789の第4アデノウイルス領域、
f)前記第4領域が存在するときにはその第4領域に連結し、前記第4領域が存在しないときには前記第3領域に連結した、Ad5に対応する塩基対約30818〜塩基対約33966またはAd6に対応する塩基対約30789〜塩基対約33784の第5アデノウイルス領域、及び
g)前記第4領域に連結した、Ad5に対応する塩基対約33967〜塩基対約35935またはAd6に対応する塩基対約33785〜塩基対約35759の第6アデノウイルス領域、
から構成され、ただし前記した第2、第3及び第5領域の少なくとも1つがAd6由来であることを特徴とする請求の範囲第3項に記載の組換え核酸。
【請求項5】
前記ベクターが
a)Ad5またはAd6のいずれかに対応する塩基対約1〜塩基対約450の第1アデノウイルス領域、
b)前記第1領域に連結した、Ad5に対応する塩基対約3511〜塩基対約5548またはAd6に対応する塩基対約3508〜塩基対約5541の第2アデノウイルス領域、
c)前記第2領域に連結した、Ad5に対応する塩基対約5549〜塩基対約28133またはAd6に対応する塩基対約5542〜塩基対約28156の第3アデノウイルス領域、
d)前記第3領域に連結した、E3平行またはE3逆平行方位の前記遺伝子発現カセット、
e)前記遺伝子発現カセットに連結した、Ad5に対応する塩基対約30818〜塩基対約33966またはAd6に対応する塩基対約30789〜塩基対約33784の第4アデノウイルス領域、及び
f)前記第4領域に連結した、Ad5に対応する塩基対約33967〜塩基対約35935またはAd6に対応する塩基対約33785〜塩基対約35759の第5アデノウイルス領域
から構成され、ただし前記した第2、第3及び第4領域の少なくとも1つがAd6由来であることを特徴とする請求の範囲第3項に記載の組換え核酸。
【請求項1】
1つ以上のAD6領域及びAD6中に存在しない領域を含む組換え核酸であって、少なくとも1つのAD6領域がE1A、E1B、E2B、E2A、E4、L1、L2、L4及びL5からなる群から選択されることを特徴とする前記組換え核酸。
【請求項2】
AD6中に存在しない前記領域がAD6中に存在しないポリペプチドをコードする発現カセットであることを特徴とする請求の範囲第1項に記載の組換え核酸。
【請求項3】
前記組換え核酸が、E1をイントランスで供給したときに複製できるE1を少なくとも欠くアデノウイルスベクターであることを特徴とする請求の範囲第2項に記載の組換え核酸。
【請求項4】
前記ベクターが
a)Ad5またはAd6のいずれかに対応する塩基対約1〜塩基対約450の第1アデノウイルス領域、
b)前記第1領域に連結した、E1平行またはE1逆平行方位の前記遺伝子発現カセット、
c)前記遺伝子発現カセットに連結した、Ad5に対応する塩基対約3511〜塩基対約5548またはAd6に対応する塩基対約3508〜塩基対約5541の第2アデノウイルス領域、
d)前記第2領域に連結した、Ad5に対応する塩基対約5549〜塩基対約28133またはAd6に対応する塩基対約5542〜塩基対約28156の第3アデノウイルス領域、
e)場合によっては存在する、前記第3領域に連結した、Ad5に対応する塩基対約28134〜塩基対約30817またはAd6に対応する塩基対約28157〜塩基対約30789の第4アデノウイルス領域、
f)前記第4領域が存在するときにはその第4領域に連結し、前記第4領域が存在しないときには前記第3領域に連結した、Ad5に対応する塩基対約30818〜塩基対約33966またはAd6に対応する塩基対約30789〜塩基対約33784の第5アデノウイルス領域、及び
g)前記第4領域に連結した、Ad5に対応する塩基対約33967〜塩基対約35935またはAd6に対応する塩基対約33785〜塩基対約35759の第6アデノウイルス領域、
から構成され、ただし前記した第2、第3及び第5領域の少なくとも1つがAd6由来であることを特徴とする請求の範囲第3項に記載の組換え核酸。
【請求項5】
前記ベクターが
a)Ad5またはAd6のいずれかに対応する塩基対約1〜塩基対約450の第1アデノウイルス領域、
b)前記第1領域に連結した、Ad5に対応する塩基対約3511〜塩基対約5548またはAd6に対応する塩基対約3508〜塩基対約5541の第2アデノウイルス領域、
c)前記第2領域に連結した、Ad5に対応する塩基対約5549〜塩基対約28133またはAd6に対応する塩基対約5542〜塩基対約28156の第3アデノウイルス領域、
d)前記第3領域に連結した、E3平行またはE3逆平行方位の前記遺伝子発現カセット、
e)前記遺伝子発現カセットに連結した、Ad5に対応する塩基対約30818〜塩基対約33966またはAd6に対応する塩基対約30789〜塩基対約33784の第4アデノウイルス領域、及び
f)前記第4領域に連結した、Ad5に対応する塩基対約33967〜塩基対約35935またはAd6に対応する塩基対約33785〜塩基対約35759の第5アデノウイルス領域
から構成され、ただし前記した第2、第3及び第4領域の少なくとも1つがAd6由来であることを特徴とする請求の範囲第3項に記載の組換え核酸。
【図1A】

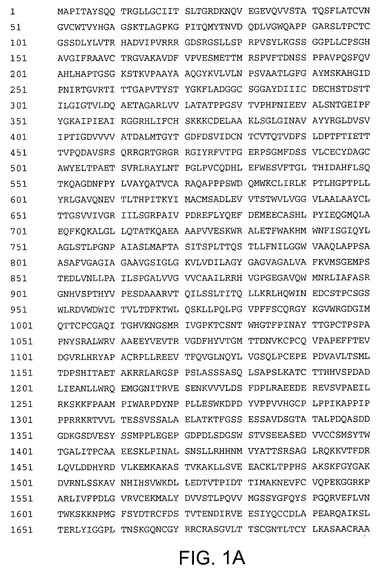
【図1B】

【図2A】
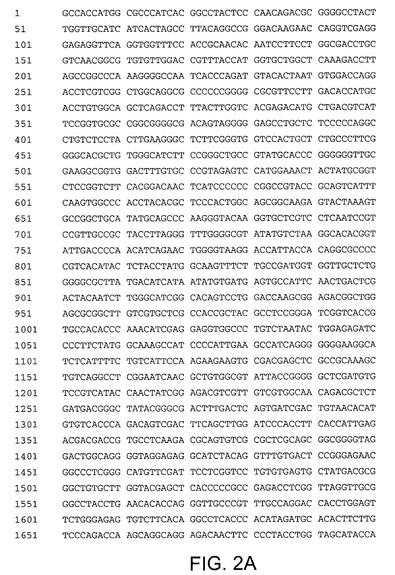
【図2B】
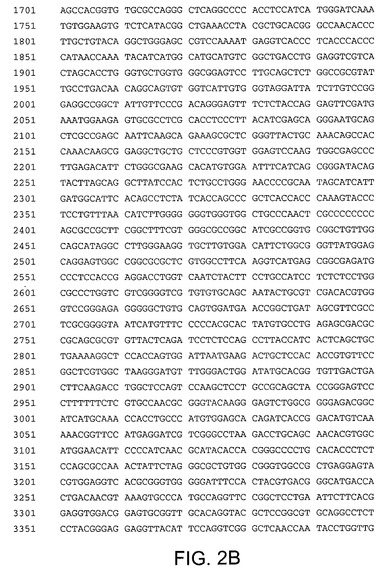
【図2C】

【図2D】

【図3A】
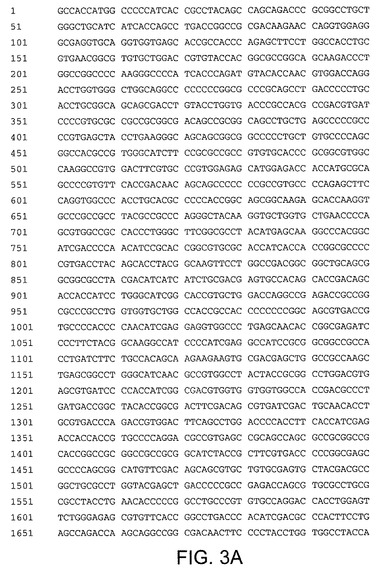
【図3B】
【図3C】
【図3D】

【図4A】
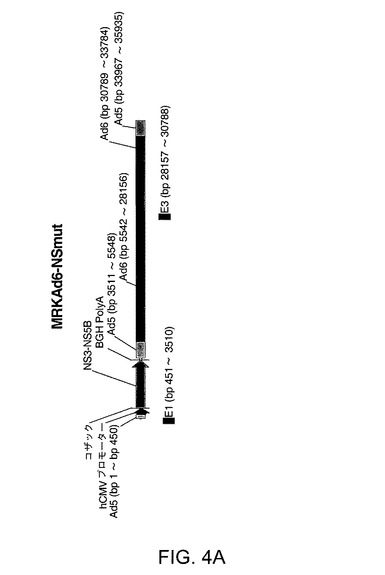
【図4B】

【図4C】

【図4D】

【図4E】

【図4F】

【図4G】

【図4H】

【図4I】

【図4J】

【図4K】

【図4L】

【図4M】

【図5A】
【図5B】
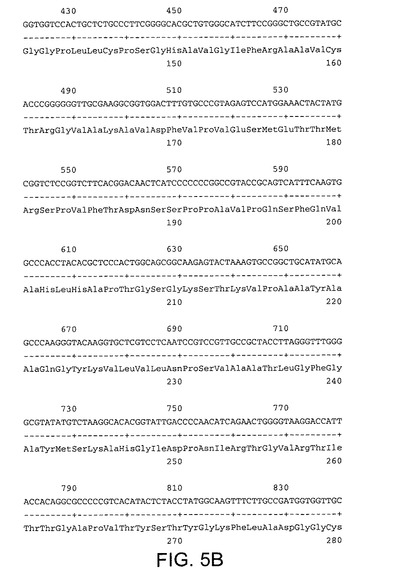
【図5C】

【図5D】

【図5E】

【図5F】

【図5G】

【図5H】

【図5I】

【図5J】

【図5K】
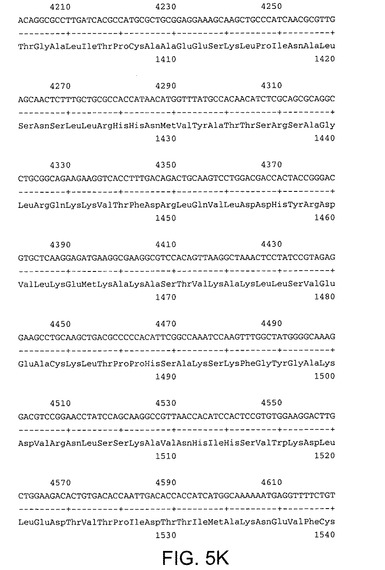
【図5L】

【図5M】

【図5N】

【図5O】
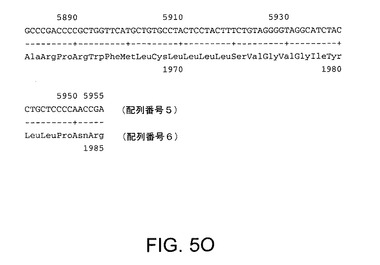
【図6A】

【図6B】
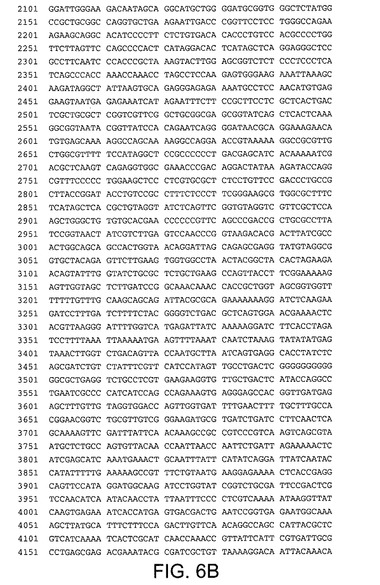
【図6C】

【図7A】

【図7B】

【図7C】

【図7D】
【図7E】

【図7F】

【図7G】

【図7H】
【図7I】

【図7J】

【図7K】

【図7L】

【図7M】

【図7N】

【図7O】

【図8A】
【図8B】
【図8C】
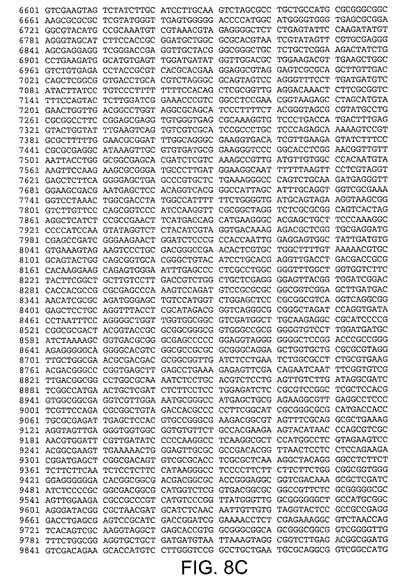
【図8D】
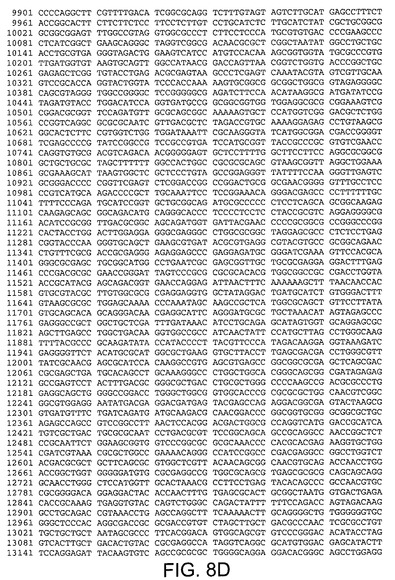
【図8E】
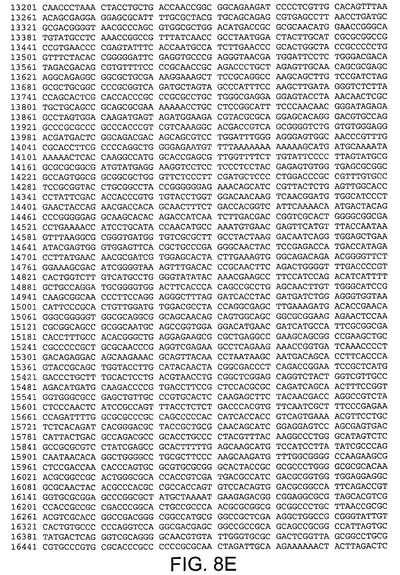
【図8F】
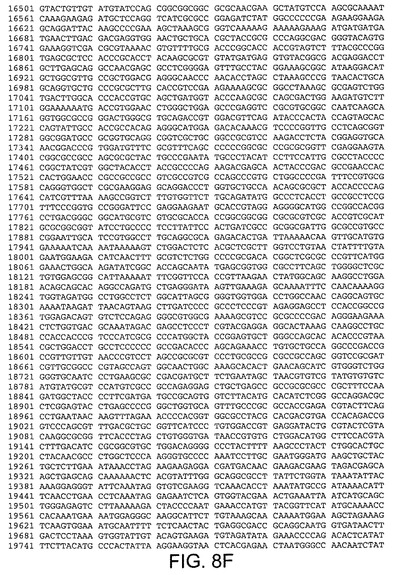
【図8G】
【図8H】
【図8I】
【図8J】

【図8K】
【図9】
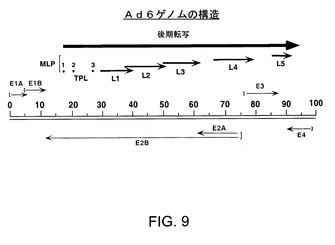
【図10】
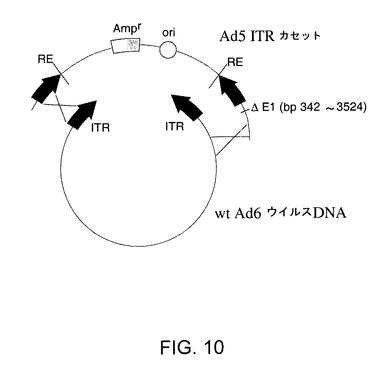
【図11】
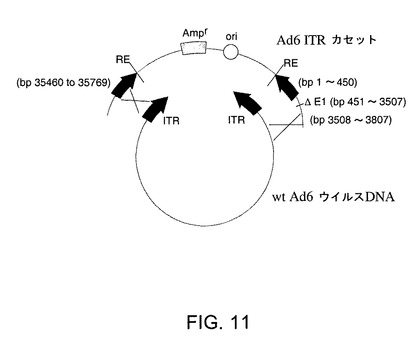
【図12】
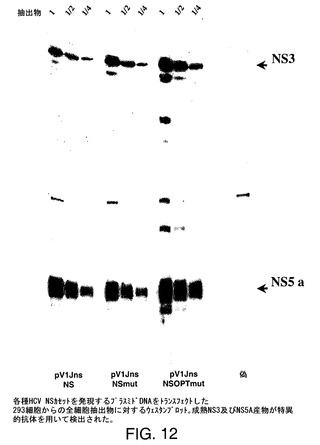
【図13A】
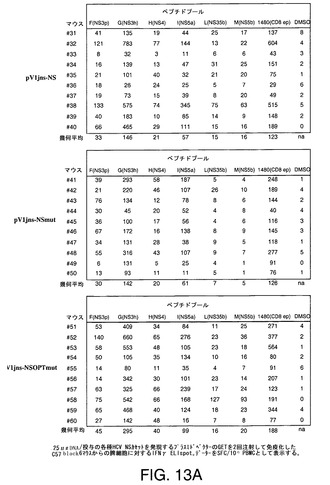
【図13B】
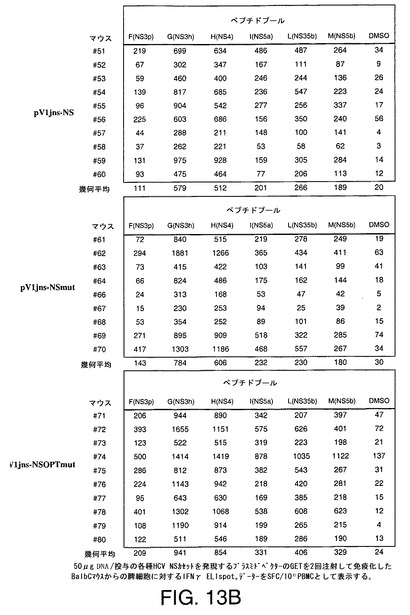
【図14】
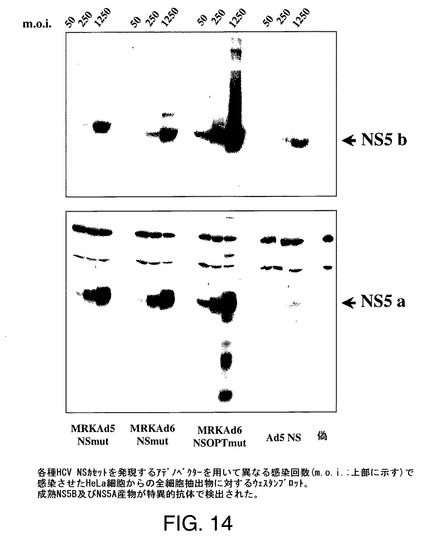
【図15】
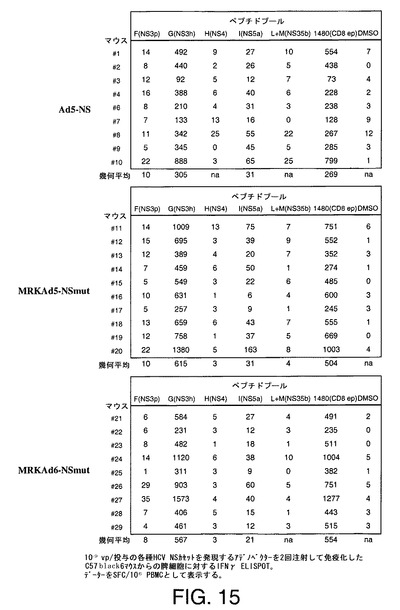
【図16A】
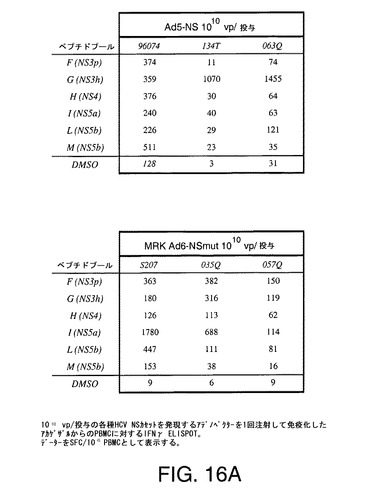
【図16B】

【図16C】
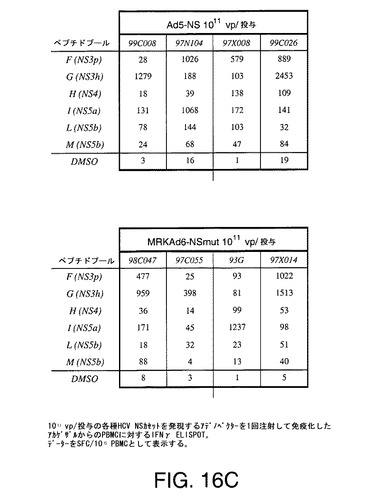
【図16D】

【図17A】
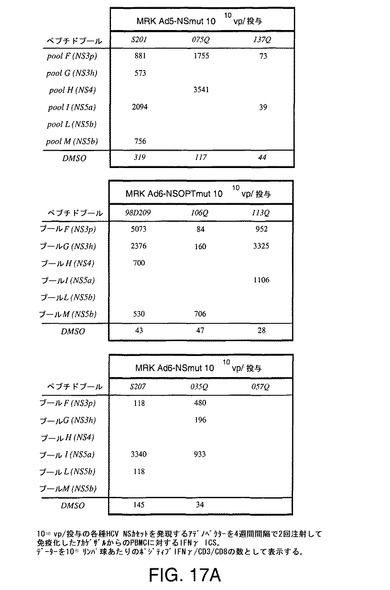
【図17B】

【図18A】
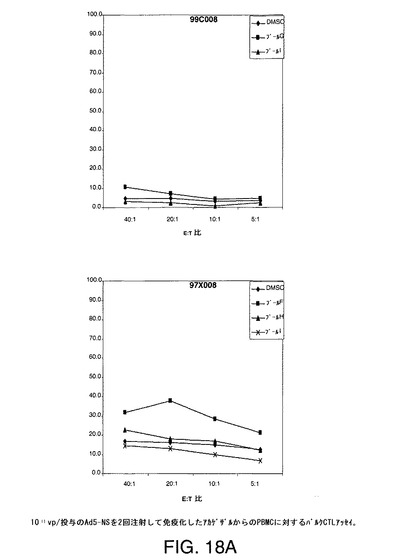
【図18B】
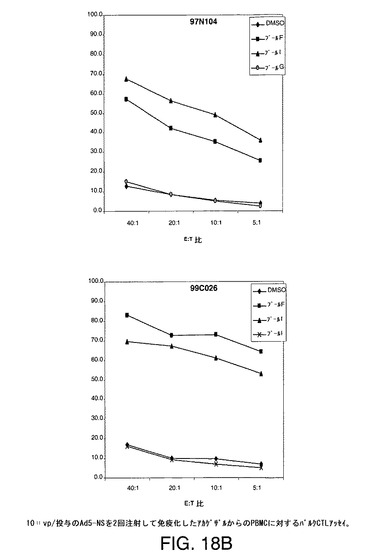
【図18C】
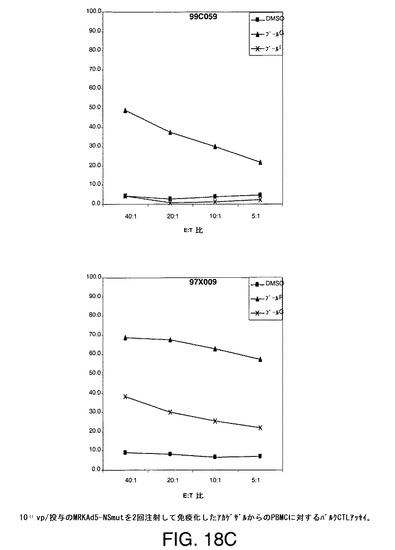
【図18D】
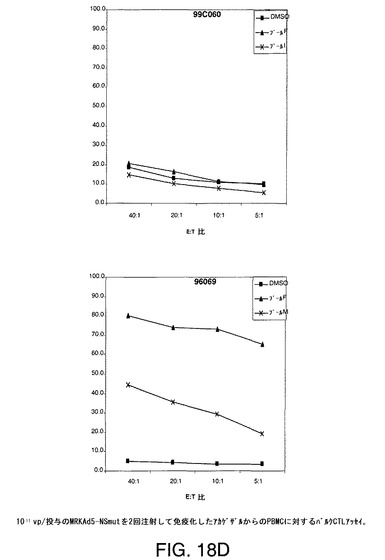
【図18E】
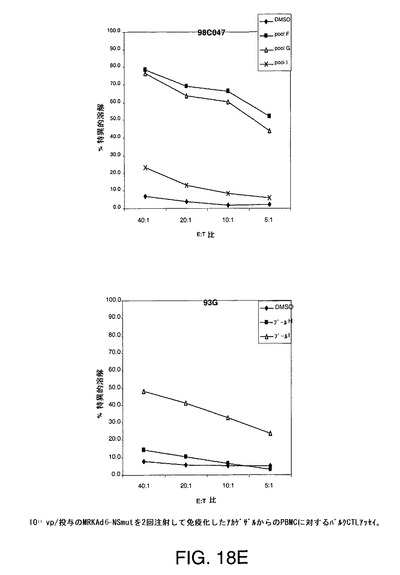
【図18F】
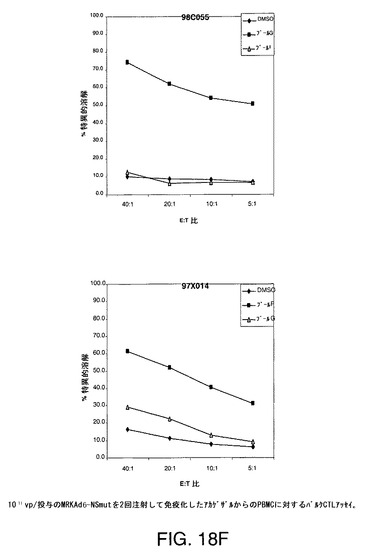
【図19】
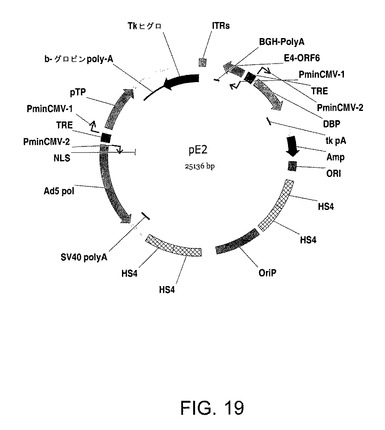
【図20A】

【図20B】

【図20C】

【図20D】

【図1B】
【図2A】
【図2B】
【図2C】
【図2D】
【図3A】
【図3B】
【図3C】
【図3D】
【図4A】
【図4B】
【図4C】
【図4D】
【図4E】
【図4F】
【図4G】
【図4H】
【図4I】
【図4J】
【図4K】
【図4L】
【図4M】
【図5A】
【図5B】
【図5C】
【図5D】
【図5E】
【図5F】
【図5G】
【図5H】
【図5I】
【図5J】
【図5K】
【図5L】
【図5M】
【図5N】
【図5O】
【図6A】
【図6B】
【図6C】
【図7A】
【図7B】
【図7C】
【図7D】
【図7E】
【図7F】
【図7G】
【図7H】
【図7I】
【図7J】
【図7K】
【図7L】
【図7M】
【図7N】
【図7O】
【図8A】
【図8B】
【図8C】
【図8D】
【図8E】
【図8F】
【図8G】
【図8H】
【図8I】
【図8J】
【図8K】
【図9】
【図10】
【図11】
【図12】
【図13A】
【図13B】
【図14】
【図15】
【図16A】
【図16B】
【図16C】
【図16D】
【図17A】
【図17B】
【図18A】
【図18B】
【図18C】
【図18D】
【図18E】
【図18F】
【図19】
【図20A】
【図20B】
【図20C】
【図20D】
【公開番号】特開2009−183295(P2009−183295A)
【公開日】平成21年8月20日(2009.8.20)
【国際特許分類】
【出願番号】特願2009−80838(P2009−80838)
【出願日】平成21年3月30日(2009.3.30)
【分割の表示】特願2003−534559(P2003−534559)の分割
【原出願日】平成14年10月10日(2002.10.10)
【出願人】(390023526)メルク エンド カムパニー インコーポレーテッド (924)
【氏名又は名称原語表記】MERCK & COMPANY INCOPORATED
【出願人】(501209427)イステイチユート・デイ・リチエルケ・デイ・ビオロジア・モレコラーレ・ピ・アンジエレツテイ・エツセ・ピー・アー (90)
【Fターム(参考)】
【公開日】平成21年8月20日(2009.8.20)
【国際特許分類】
【出願日】平成21年3月30日(2009.3.30)
【分割の表示】特願2003−534559(P2003−534559)の分割
【原出願日】平成14年10月10日(2002.10.10)
【出願人】(390023526)メルク エンド カムパニー インコーポレーテッド (924)
【氏名又は名称原語表記】MERCK & COMPANY INCOPORATED
【出願人】(501209427)イステイチユート・デイ・リチエルケ・デイ・ビオロジア・モレコラーレ・ピ・アンジエレツテイ・エツセ・ピー・アー (90)
【Fターム(参考)】
[ Back to top ]