
C型肝炎ウイルス核酸ワクチン
本発明の主題は、HCV核酸ワクチンおよびワクチン成分として、または、HCVワクチンの製造に使用できる核酸構築物である。記載した構築物は、(1)第一のHCV菌株に基づくNS3−4A領域と第二の菌株に基づくNS3−NS4A−NS4B−NS5AまたはNS3−NS4A−NS4B−NS5A−NS5B領域とを含むキメラHCVポリペプチドをコードする構築物、および、(2)HCVポリペプチドをコードするチンパンジーに基づくアデノベクターを含む。

【発明の詳細な説明】
【背景技術】
【0001】
本願に引用した参考文献は特許請求の範囲に記載の発明に対する先行技術であると認められたものではない。
【0002】
全世界の人口の約3%がC型肝炎ウイルス(HCV)に感染している。(Wasleyら,Semin.Liver Dis.20,1−16,2000)。HCVに接触して明白な急性疾患が発症するパーセンテージは低いが、多くの場合にウイルスが慢性感染として定着し、肝臓の炎症を生じさせ、ゆっくりと肝不全および肝硬変に進行する。(Iwarson,FEMS Microbiol.Rev.14,201−204,1994)。さらに、疫学調査は、HCVが肝細胞癌の病理発生に重要な役割を演じることを示している。(Kew,FEMS Microbiol.Rev.14,211−220,1994,Alter,Blood 85,1681−1695,1995)。
【0003】
HCVに対する定例の血液スクリーニングが実施されるようになった1992年以前には、たいていの感染は、汚染血液、血液製品または移植臓器への迂闊な接触が原因となっていた。HCVの血液スクリーニングが実施されている地域では、HCVは主として静脈内薬物使用によって感染する。頻度は少ないが、周産期接触、血液透析およびHCV感染者との性的接触なども伝播方法に含まれる。(Alterら,N.Engl.J.Med.341(8),556−562,1999,Alter,J.Hepatol.31 Suppl.88−91,1999,Wasleyら,Semin.Liver.Dis.201,1−16,2000)。
【0004】
HCVゲノムは、約3000アミノ酸の前駆体ポリタンパク質をコードする約9.5kbの一本鎖RNAから成る。(Chooら,Science 244,362−364,1989,Chooら,Science 244,359−362,1989,Takamizawaら,J.Virol 65,1105−1113,1991)。HCVポリタンパク質はC−El−E2−p7−NS2−NS3−NS4A−NS4B−NS5A−NS5Bという序列のウイルスタンパク質を含有する。
【0005】
CMI応答を生じさせるために1つ以上のHCV非構造抗原を提供するHCV核酸配列を使用することは、例えば、Choら,Vaccine 77:1136−1144,1999;Paliardら,国際公開WO 01/30812;Coitら,国際公開WO 01/38360;およびEminiら,国際公開WO 03/031588によって言及されている。
【発明の開示】
【0006】
本発明の主題は、HCV核酸ワクチンおよびワクチン成分として、または、HCVワクチンの製造に使用できる核酸構築物である。記載した構築物は、(1)第一のHCV菌株に基づくNS3−4A領域と第二の菌株に基づくNS3−NS4A−NS4B−NS5AまたはNS3−NS4A−NS4B−NS5A−NS5B領域を含有するキメラHCVポリペプチドをコードする核酸、および、(2)チンパンジーに基づくHCVポリペプチドをコードするアデノベクターを含む。
【0007】
従って、本発明の第一の目的は、HCVキメラポリペプチドをコードするヌクレオチド配列を含む核酸配列を記載する。該ポリペプチドは、第一のHCV菌株に由来のHCV NS3−4aに実質的に相似のアミノ酸配列を含むHCV NS3−4a領域と、第二のHCV菌株に由来のHCV NS3−NS4A−NS4B−NS5Aに実質的に相似のアミノ酸配列を含むHCV NS3−NS4A−NS4B−NS5A領域とを含む。この場合、2つの領域に存在するNS3−4A配列は異なる配列を有している。第一領域は第二領域のアミノ側またはカルボキシル側に配置されている。
【0008】
アミノ酸配列に関する“実質的に相似の配列”という表現は、基準配列に対する少なくとも約70%の一致を意味する。基準配列に対するパーセント一致(一致パーセントと表すこともある)を決定するためには、最大数の一致アミノ酸が得られるようにポリペプチド領域を対応する基準領域に位置合せし、対応領域の一致アミノ酸の数を測定する。この数を基準領域のアミノ酸の総数で除算し、次いで100を乗算し、最も近い整数に丸める。例えば、HCV NS3−4A、NS3−NS4A−NS4B−NS5AおよびNS3−NS4A−NS4B−NS5A−NS5Bの領域が、異なるHCV菌株に存在する対応HCV領域でもよい。
【0009】
第一および第二の領域に異なるNS3−4A配列が存在することは、1つ以上のアミノ酸相違をもたらす。アミノ酸相違はおのおの独立に付加、置換または欠失である。
【0010】
好ましい実施態様では、核酸が、コードされたHCVポリペプチドをヒト細胞中で発現できる発現ベクターである。ヒト細胞中の発現は、HCV感染の能動的治療およびHCV感染に対する予防的処置という治療的用途を有している。
【0011】
発現ベクターは、ポリペプチドをコードするヌクレオチド配列を適正な転写およびプロセシング用の調節要素と共に含有する。存在し得る調節要素は、ポリペプチドをコードするヌクレオチド配列に天然に連携している調節要素およびヌクレオチド配列に天然に連携していない外来調節要素を含む。外来プロモーターのような外来調節要素はヒト細胞のような特定宿主中の発現に使用できる。
【0012】
好ましい発現ベクターは、1つ以上の欠失領域を有する組換えアデノウイルスゲノムである。組換えアデノウイルスゲノムは、1つ以上のアデノウイルス血清型に実質的に相似の種々の領域を含有し得る。欠失領域という表現は、指定した領域の全部または一部の欠失を表す。
【0013】
本発明の別の目的は、
a)HCV NS3−NS4A−NS4B−NS5Aを含むHCVポリペプチドをコードしている発現カセットと、
b)E1欠失、E3欠失および場合によりE4欠失を含む組換えアデノウイルスゲノムと、
を含み、該アデノウイルスゲノムが、(i)配列3または9に実質的に相似のアミノ酸配列をもつファイバー領域、(ii)配列5または11に実質的に相似のアミノ酸配列をもつヘキソン領域、および、(iii)配列7に実質的に相似のアミノ酸配列をもつペントン領域
の少なくとも1つをコードしている組換えアデノベクターを記載する。
【0014】
発現カセットはE1またはE3欠失に配置される。発現カセットの全部または一部が欠失領域に対応する位置に存在するならば発現カセットはE1またはE3欠失に配置されたと考えられる。
【0015】
HCVポリペプチドまたはアミノ酸配列という表現は、天然産生HCV配列、天然産生配列または指定配列に実質的に相似の天然産生配列の誘導体、および、キメラHCVポリペプチドを含む。キメラHCVポリペプチドは、上述の本発明の第一の目的に記載のHCVポリペプチドを含む。
【0016】
本発明の別の目的は組換えアデノウイルス粒子を記載する。該粒子は、本文中に記載の組換えアデノウイルスゲノムによってコードされ、該ゲノムのコピーをパッケージしている。
【0017】
本発明の別の目的は、(a)組換えアデノウイルスゲノムを発現させるためにE1相補細胞を使用して粒子を製造する段階と、(b)該粒子を実質的に精製する段階とを含む組換えアデノウイルス粒子の製造方法を記載する。粒子を実質的に精製するという表現は、粒子の製造に使用した細胞および細胞破片を完全にまたはほぼ完全に除去することを意味する。
【0018】
本発明の別の目的は、治療有効量のHCVポリペプチドをコードする核酸と医薬的に許容される担体とを含む医薬組成物を記載する。
【0019】
本発明の別の目的は、治療有効量のHCVポリペプチドをコードする核酸を患者に投与する段階を含む患者の治療方法を記載する。
【0020】
“治療する”という表現は、HCV感染患者の治療または活動性HCV感染の可能性もしくは重篤度を低下させるための患者の予防的処置を含む。“患者”という用語は、HCV感染の可能性のある哺乳動物を表す。患者はHCV感染している場合もあり非感染の場合もある。患者の例はヒトおよびチンパンジーである。
【0021】
HCV感染患者の場合、有効量は、以下の効果、すなわち、HCVの複製能力を低下させる効果、HCV荷重を減少させる効果、ウイルスクリアランスを増加させる効果、および、1つ以上のHCV特異的CMI応答を増加させる効果の1つ以上を発揮するために十分な量である:。
【0022】
HCV非感染患者の場合、有効量は、以下の効果、すなわち、HCV感染に対するHCV特異的CMI応答の1つ以上の成分の発生能力を増加させる効果、HCV感染に対する感受性を低下させる効果、および、慢性疾患に至る持続感染を定着させる感染ウイルスの能力を低下させる効果の1つ以上を達成するために十分な量である:。
【0023】
“含む”のような無制限的用語は、追加の要素または段階を許容する。追加の要素または段階の可能性を強調するために、ときには、“1つ以上”のような成句を無制限的用語と共にまたは単独で使用する。
【0024】
明白な記述がなければ、単数を表す不定冠詞は1に限定されない。例えば、“1つの細胞”は“複数の細胞”を排除しない。ときには、複数の存在が可能であることを強調するために、1つ以上という熟語を使用する。
【0025】
本発明のその他の特徴および利点は、様々な実施例を含む本文中の以後の記載より明らかであろう。与えられた実施例は本発明の実施に役立つ様々な構成要素および方法を例示する。実施例は特許請求の範囲に記載の本発明を限定しない。この開示に基づいて当業者は本発明の実施に役立つ他の構成要素および方法を鑑定し使用できるであろう。
【0026】
本発明は、種々のHCV菌株に基づくHCV非構造タンパク質を供給するHCVキメラポリペプチドをコードする核酸、および、HCVポリペプチドを発現するチンパンジーアデノベクターの使用を含む。記載した核酸の使用は、HCVに対するCMI応答を生じさせる広範囲の抗原を提供するHCVポリペプチドを細胞に導入するためのワクチン成分としての使用、および、このようなワクチン成分を製造する中間体としての使用を含む。
【0027】
適応型細胞性免疫応答は、主要組織適合複合体(MHC)クラスIおよびIIの遍在的分布によって全身のHCV感染細胞のウイルス抗原を認識し、免疫記憶を誘導し、免疫記憶を維持する機能を果たし得る。これらの機能を果たすのは、抗原特異的CD4+Tヘルパー細胞(Th)およびCD8+細胞障害性T細胞(CTL)である。
【0028】
特異的T細胞受容体を介して活性化されると、HCV特異的Th細胞は多様な免疫調節機能を発揮するが、たいていの機能にはTh1およびTh2サイトカインが介在する。HCV特異的Th細胞は、B細胞の活性化および分化ならびにウイルス特異的細胞障害性T細胞の誘導および刺激を助ける。Th細胞はまたCTLと共に、数種類のウイルスの複製および遺伝子発現を阻害するIFN−γおよびTNF−αを分泌し得る。さらに、Th細胞およびCTLは主要なエフェクター細胞であり、ウイルス感染細胞のアポトーシスおよび溶解を誘導できる。
【0029】
HCV特異的CTLは、専門的抗原提示細胞(pAPC)によってプロセシングされた抗原に応答して生じる。抗原はpAPC内部で合成されるかまたはpAPC導入される。抗原をコードする発現カセットを細胞に導入することによってpAPC内部で抗原を合成することができる。
【0030】
核酸ワクチンの好ましい投与経路は筋肉内経路である。筋肉内投与によって核酸が体細胞およびpAPCに導入され発現されると考えられる。体細胞で産生されたHCV抗原はpAPCに輸送され、MHCクラスI分子に関係して抗原提示される。(Donnellyら,Annu.Rev.Immunol.75:617−648,1997)。
【0031】
pAPCはプロテアソーム複合体において長い抗原をより短いペプチド抗原にプロセシングする。該抗原は小胞体/ゴルジ複合体分泌経路に転送されてMHCクラスIタンパク質に会合する。CD8+Tリンパ球は、MHCクラスIに会合した抗原をT細胞受容体(TCR)およびCD8細胞表面タンパク質を介して認識する。
【0032】
チンパンジーアデノウイルスに基づくベクターはHCV抗原導入ビヒクルとしてヒトアデノベクターに代替して使用できる。チンパンジーアデノウイルスに基づくベクターは多重免疫戦略で使用するのが特に有効である。この場合、治療される患者の体内では初回に使用したアデノベクターへの免疫応答が生じている。同種のアデノウイルスに基づくベクターとの接触が繰返されると、アデノウイルスタンパク質に対する免疫応答が原因でワクチンの有効性が低下するであろう。初回接触がアデノウイルス感染の結果という場合もあるが、このような感染は恐らく、HCV以外の疾患を治療するためにアデノベクターを使用したことによって生じたものであろう。
【0033】
ここに与える指針に基づいて、患者体内で有益な効果を発揮する十分に強力な免疫応答を発生できる。これらの指針は、HCV配列の選択、ベクター選択、ベクター産生、併用治療および投与に関する情報を含む。
【0034】
I.キメラ配列
種々のNS3−4A配列をもつ領域を提供するHCVキメラポリペプチドは、種々のHCV菌株を標的とする抗原を提供するワクチン成分として使用できる。HCVの主な特徴はそのゲノムの不均質性である。(Pawlotsky,Clin.Liver Dis,7:45−66,2003,Simmonds,J.General.Virol,85:3173−3188,2004)。さらに、RNAウイルスに共通の典型的な誤りがちのRNA−依存性RNAポリメラーゼおよび不十分なプルーフリーディング活性から生じた突然変異ならびに宿主から生じた突然変異は、HCVが偽種と呼ばれる複合ウイルス集団として宿主体内で循環する原因となる。(Blightら,Science 290:1972−1974,2000)。
【0035】
キメラ構築物の異なる領域に存在する異なるNS3−4A配列は、1つ以上のアミノ酸の違いとして表れる。アミノ酸の違いはおのおの独立に付加、置換または欠失である。種々の実施態様で、第一および第二の領域のNS3−4A配列は、少なくとも約5%、少なくとも10%、少なくとも15%だけ違っている。または、少なくとも1、5、10、15、20または25個のアミノ酸の変更が存在する。NS3−4A領域はNS3−4A配列に加えて、アミノ末端メチオニンのような付加アミノ酸および/または導入された開裂部位を含み得る。
【0036】
相違パーセントは配列一致を100から減算することによって決定できる。例えば、85%配列一致は15%相違を与える。
【0037】
キメラポリペプチドに存在する第一領域および第二ポリペプチド領域は、天然産生HCV単離物の多くの例に基づいて容易に作製できる。HCV単離物は1つ以上のサブタイプを含む以下の6つの主要遺伝子型に分類できる:HCV−1/(1a,1b,1c)、HCV−2/(2a,2b,2c)、HCV−3/(3a,3b,10a)、HCV−4/(4a)、HCV−5/(5a)およびHCV−6/(6a,6b,7b,8b,9a,11a)。(Simmonds,J.Gen.Virol,693−712,2001)。HCV−BK、HCV−J、HCV−NおよびHCV−Hのような特定のHCV配列はGenBankに寄託され種々の刊行物に記載されている。(例えば、Chamberlainら,J.Gen.Virol,1341−1347,1997)。
【0038】
好ましくは、第一領域および第二領域の双方がHCVプロテアーゼによってインビボでプロセシングされ、HCVキメラポリペプチドに存在する個々のタンパク質に対応する個々のタンパク質を提供する。個々のHCVタンパク質は細胞によってさらにプロセシングされ得る。
【0039】
第一領域に関する種々の実施態様では、該領域が、以下に存在するNS3−NS4A領域に実質的に相似のアミノ酸配列であるかまたは該アミノ酸配列を内包する:配列1のアミノ酸1−686;HCV 1a(Acc.No.M62321);HCV 2a(Acc.No.D00944;HCV 3a(Acc.No:D28917);HCV 4a(Acc.No:Y11604);HCV 5a(Acc.No:Y13184)またはHCV 6a(Acc.No:D84264)。
【0040】
第二領域に関する種々の実施態様では、該領域が、以下に存在する対応領域に実質的に相似のNS3−NS4A−NS4B−NS5AまたはNS3−NS4A−NS4B−NS5A−NS5B*配列であるかまたは該配列を内包する:配列1のアミノ酸686−2675、アミノ酸691−2675またはアミノ酸692−2675;HCV 1a(Acc.No.M62321);HCV 2a(Acc.No.D00944;HCV 3a(Acc.No:D28917);HCV 4a(Acc.No:Y11604);HCV 5a(Acc.No:Y13184)またはHCV 6a(Acc.No:D84264)。“NS5B*”という表記は不活性NS5Bを表す。
【0041】
好ましくは第二領域が、第一領域由来のプロテアーゼ活性に適合性のアミノ酸開裂部位を含む。開裂部位は既知の開裂配列に基づいて付加できる。
【0042】
アミノ酸配列に関する“実質的に相似の配列”という表現は、基準配列に対する少なくとも約70%の一致を意味する。基準配列に対するパーセント一致(一致パーセントと表すこともある)を決定するためには、最大数の一致アミノ酸が得られるようにポリペプチド領域を対応する基準領域に位置合せし、対応領域の一致アミノ酸の数を測定する。この数を基準領域のアミノ酸の総数で除算し、次いで100を乗算し、最も近い整数に丸める。
【0043】
種々の実施態様で、実質的に相似の配列は、少なくとも75%、少なくとも80%、少なくとも85%、少なくとも90%、少なくとも95%の一致を有しているか;または、0、1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19もしくは20個のアミノ酸の変更だけ違っている。変更はおのおの独立に挿入、置換または付加である。
【0044】
天然産生HCV配列に修飾を生じさせて実質的に相似の種々の配列を得ることもできる。天然産生アミノ酸の違いはアミノ酸側鎖(R基)の違いに起因する。R基は、物理的寸法、電荷および疎水性のようなアミノ酸の種々の特性を左右する。アミノ酸は以下のような種々のグループに分類できる:中性かつ疎水性(アラニン、バリン、ロイシン、イソロイシン、プロリン、トリプトファン、フェニルアラニン、メチオニン);中性かつ極性(グリシン、セリン、トレオニン、チロシン、システイン、アスパラギン、グルタミン);塩基性(リシン、アルギニン、ヒスチジン);酸性(アスパラギン酸、グルタミン酸)。
【0045】
活性を維持するために種々のアミノ酸を置換する場合、一般には同様の特性を有しているアミノ酸で置換するのが好ましい。バリンでロイシンを、アルギニンでリシンを、アスパラギンでグルタミンを置換するような特定グループ内部の異なるアミノ酸の置換が、ポリペプチド三次構造の変化を生じない良好な候補である。
【0046】
アミノ酸修飾はT細胞抗原領域を好ましく維持または追加する。広範囲のT細胞応答を誘発できるポリペプチドを産生するために天然産生HCVポリペプチドに様々な修飾を行うことが可能である。広いT細胞応答を誘発するポリペプチドの能力を左右する要因は、HCV特異的T細胞抗原の保存または導入、および、種々のHCV単離物中の種々のT細胞抗原領域の存在数である。
【0047】
HCV T細胞抗原は例えば実験作業によって同定できる。T細胞抗原を同定する1つの方法は、長いポリペプチドから一連の短いオーバーラップペプチドを作製し、次いで、感染患者に由来のT細胞集団を陽性クローンに基づいてスクリーニングする段階を含む。陽性クローンは特定のペプチドによって活性化/プライムされる。IFNγ−ELISPOT、IFNγ−細胞内染色およびバルクCTLアッセイのような技術がペプチド活性の測定に使用できる。このようにして同定したペプチドは、それぞれの病原体のT細胞エピトープを表すと考えることができる。
【0048】
自己をプロセシングしてCMI応答を生じるHCVポリペプチドの能力は、ここに記載の技術または当業界で公知の技術を使用して判定できる。(参照:例えばEminiら,国際公開WO 03/031588)。このような技術は、HCV特異的CMI応答を測定するためのIFNγ−ELISPOT、IFNγ−細胞内染色およびバルクCTLアッセイの使用を含む。
【0049】
複製に必須のモチーフを標的組換えすることによって不活性ポリメラーゼを産生するためにNS5Bに小さい修飾を行ってもよい。NS5B活性に必須のモチーフおよび不活性NS5Bを産生するために行うことができる修飾の例は、Lohmannら,Journal of Virology 77:8416−8426,1997、Kolykhalovら,Journal of Virology 74:2046−2051,2000およびEminiら,国際公開WO 03/031588に記載されている。
【0050】
キメラHCVポリペプチドを修飾するときに考慮すべき追加要因は、自己プロセシング能力の維持およびT細胞抗原の維持を含む。ポリペプチドの自己プロセシング能力は機能性プロテアーゼ活性によってほぼ判定できる。プロテアーゼ活性を維持する修飾は、NS3プロテアーゼ、NS3の補因子として役立つNS4A、および、HCVポリペプチド内部に存在するプロテアーゼ認識部位を考慮することによって得られる。
【0051】
II.キメラNS31−NS4A1−NS32−NS4A2−NS4B2−NS5A2−NS5B2*
好ましいキメラHCVポリペプチドは、NS31−NS4A1−NS32−NS4A2−NS4B2−NS5A2−NS5B2*である。下付き文字1および2は、種々のHCV菌株中の対応する領域に実質的に相似の配列をもつ領域を表し、“NS5B*”は酵素的に不活性のNS5Bを表す。NS31−NS4A1とNS32−NS4A2とは少なくとも1つのアミノ酸が違っている。種々の実施態様で、NS31−NS4A1とNS32−NS4A2とは少なくとも約5%、少なくとも約10%、少なくとも約15%違っている;または、少なくとも1、5、10、15、20または25個のアミノ酸の変更が存在する。
【0052】
好ましくは、NS31−NS4A1−NS32−NS4A2−NS4B2−NS5A2−NS5B2*ポリペプチドは、第一および第二領域のおのおのが1つ以上の個別HCVペプチドを産生するために十分なインビボのプロテアーゼ活性を提供する。好ましい実施態様で、ポリペプチドは、NS31、NS4A1、NS32、NS4A2、NS4B2、NS5A2、NS5B2*を個別ペプチドとして産生できる。
【0053】
種々のNS31−NS4A1およびNS32−NS4A2−NS4B2−NS5A2−NS5B2*領域は上述の項Iに記載のようにして供給される。種々の実施態様で、NS31−NS4A1領域は、配列1のアミノ酸1−686もしくはアミノ酸2−686に実質的に相似の配列であるかまたは該配列を内包する。NS32−NS4A2−NS4B2−NS5A2−NS5B2*領域は、配列1のアミノ酸687−2675、691−2675もしくは692−2675に実質的に相似の配列であるかまたは該配列を含む。または、NS31−NS4A1−NS32−NS4A2−NS4B2−NS5A2−NS5B2*が全体として配列1に実質的に相似である。種々の実施態様で、各領域は配列1の対応する領域に、少なくとも75%、少なくとも80%、少なくとも85%、少なくとも90%、少なくとも95%だけ実質的に相似であるか、または、0、1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19もしくは20個のアミノ酸の変更だけ違っている。
【0054】
III.遺伝子発現カセット
ポリペプチドをコードする遺伝子発現カセットは、ポリペプチド発現に必要な要素を含む。“ポリペプチド”という表現はキメラポリペプチドを含意する。
【0055】
III.A.コードされたポリペプチド配列
本文中に記載のHCVポリペプチドをコードした配列は種々のベクターに使用できる。特定例は、項IおよびIIに前述したようなキメラポリペプチド配列、および、配列16に実質的に相似の配列のようなHCVポリペプチド配列Met−NS3−NS4A−NS4B−NS5A−NS5B*を含む(参照:Eminiら,国際公開WO 03/031588)。配列16に実質的に相似の配列は、例えば項Vに後述するようなチンパンジーベクターに結合して使用できる。
【0056】
様々な実施態様で、配列16に実質的に相似の配列は、配列16に75%、少なくとも80%、少なくとも85%、少なくとも90%、少なくとも95%の配列一致を有しているか、または、0、1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19もしくは20個のアミノ酸の変更だけ違っている。
【0057】
III.B.コードされたポリペプチド配列
遺伝子発現カセット中に存在する調節要素は一般に、(a)ポリペプチドをコードするヌクレオチド配列に転写可能に連結されたプロモーターと、(b)ヌクレオチド配列に機能的に連結された5’リボソーム結合部位と、(c)ヌクレオチド配列の3’端に接合されたターミネーターと、(d)ヌクレオチド配列に機能的に連結された3’ポリアデニル化シグナルとを含む。遺伝子発現またはポリペプチドプロセシングに役立つ追加の調節要素が存在してもよい。
【0058】
プロモーターは、RNAポリメラーゼによって認識され、下流領域の転写を媒介する遺伝子要素である。好ましいプロモーターは、高い転写レベルを与える強プロモーターである。強プロモーターの例は、前初期ヒトサイトメガロウイルスプロモーター(CMV)およびイントロンAをもつCMVである。(Chapmanら,Nucl.Acids Res.19:3979−3986,1991)。プロモーターの追加例は、EF1アルファプロモーター、ネズミCMVプロモーター、ラウス肉腫ウイルスプロモーター、SV40初期/後期プロモーターおよびβ−アクチンプロモーターのような天然産生プロモーター、ならびに、合成の筋特異的プロモーター、キメラ筋特異的/CMVプロモーターのような人工プロモーターである。(Liら,Nat.Biotechnol.77:241−245,1999,Hagstromら,Blood 95:2536−2542,2000)。
【0059】
リボソーム結合部位は開始コドンまたはその近傍に局在する。リボソーム結合部位の例は、CCACCAUGG、CCGCCAUGGおよびACCAUGGを含み、ここにAUGは開始コドンである。(Kozak,Cell 44:283−292,1986)。リボソーム結合部位の別の例は、配列18によって示される。
【0060】
ポリアデニル化シグナルは、転写RNAを開裂しRNAにポリ(A)テールを付加する。高等真核生物のポリアデニル化シグナルは、ポリアデニル化付加部位から約11−30ヌクレオチドにAAUAAA配列を含んでいる。AAUAAA配列は、RNA開裂シグナルの伝達に関与する。(Lewin,Genes IV,Oxford University Press,NY,1990)。ポリ(A)テールはmRNAプロセシングに重要である。
【0061】
遺伝子発現カセットの部分として使用できるポリアデニル化シグナルは、ミニマルラビットβ−グロブリンポリアデニル化シグナルおよびウシ成長ホルモンポリアデニル化(BGH)シグナルを含む(Xuら,Gene 272:149−156,2001,Postら,米国特許No.5,122,458)。追加例は、合成ポリアデニル化シグナル(SPA)およびSV40ポリアデニル化シグナルを含む。SPA配列は配列19によって示されている。
【0062】
“転写可能に連結された”という表現は、プロモーターにRNAポリメラーゼが結合することによってヌクレオチド配列の転写が生じるようにプロモーターが配置されていることを表す。転写可能に連結されたとしても、転写される配列がプロモーターに隣接する必要はない。
【0063】
“機能的に連結された”という表現は、ヌクレオチド配列に間接的効果を与える能力を意味する。機能的に連結されたとしても、連結された配列が互いに隣接する必要はない。ヌクレオチド配列に機能的に連結された3’ポリアデニル化シグナルは、転写されたRNAの開裂およびポリアデニル化を促進する。ヌクレオチド配列に機能的に連結された5’リボソーム結合部位はリボソーム結合を促進する。
【0064】
遺伝子発現またはポリペプチドプロセシングを増進または調節するために有用な調節要素の追加例は、エンハンサー、リーダー配列およびオペレーターである。エンハンサー領域は転写を増進する。エンハンサー領域の例は、CMVエンハンサーおよびSV40エンハンサーである。(Hittら,Methods in Molecular Genetics 7:13−30,1995,Xuら,Gene 272:149−156, 2001)。エンハンサー領域はプロモーターに連携できる。
【0065】
リーダー配列は、ポリペプチドをプロテアソームに案内するポリペプチド上のアミノ酸領域である。リーダー配列をコードする核酸は構造遺伝子の5’であり、構造遺伝子と共に転写される。リーダー配列の一例はtPAである。
【0066】
オペレーター配列は遺伝子発現を調節するために使用できる。例えば、Tetオペレーター配列は遺伝子発現を抑制するために使用できる。
【0067】
IV.コーディング核酸配列
コーディング核酸配列は特定のアミノ酸配列をコードするコドンを提供する。特定のアミノ酸配列および遺伝子コードの既知の縮重性に基づいて異なる多数のコーディング核酸配列を得ることができる。遺伝子コードが縮重性をもつ理由は、ほぼすべてのアミノ酸がヌクレオチドトリプレットの様々な組合せすなわち“コドン”によってコードされているからである。
【0068】
特定のコドンが特定のアミノ酸に翻訳されることは当業界で公知である(参照:例えばLewin GENES IV,p.119,Oxford University Press,1990)。アミノ酸は以下のようなコドンによってコードされている:
A=Ala=アラニン:コドンGCA,GCC,GCG,GCU
C=Cys=システイン:コドンUGC,UGU
D=Asp=アスパラギン酸:コドンGAC,GAU
E=Glu=グルタミン酸:コドンGAA,GAG
F=Phe=フェニルアラニン:コドンUUC,UUU
G=Gly=グリシン:コドンGGA,GGC,GGG,GGU
H=His=ヒスチジン:コドンCAC,CAU
I=Ile=イソロイシン:コドンAUA,AUC,AUU
K=Lys=リシン:コドンAAA,AAG
L=Leu=ロイシン:コドンUUA,UUG,CUA,CUC,CUG,CUU
M=Met=メチオニン:コドンAUG
N=Asn=アスパラギン:コドンAAC,AAU
P=Pro=プロリン:コドンCCA,CCC,CCG,CCU
Q=Gln=グルタミン:コドンCAA,CAG
R=Arg=アルギニン:コドンAGA,AGG,CGA,CGC,CGG,CGU
S=Ser=セリン:コドンAGC,AGU,UCA,UCC,UCG,UCU
T=Thr=トレオニン:コドンACA,ACC,ACG,ACU
V=Val=バリン:コドンGUA,GUC,GUG,GUU
W=Trp=トリプトファン:コドンUGG
Y=Tyr=チロシン:コドンUAC,UAU。
【0069】
配列13のヌクレオチド1269−7223はNS3−NS4A−NS4B−NS5A−NS5B*配列の一例を示す。配列13のヌクレオチド1269−7223に実質的に相似の配列はワクチン成分の部分として使用できる。(Eminiら,国際公開WO 03/031588)。例えば、このような実質的に相似の配列は、ChA3またはChA63に基づくアデノベクターの部分として使用できる。
【0070】
ヌクレオチド配列に関する“実質的に相似の配列”という表現は、基準配列に対する少なくとも約70%の一致を表す。基準配列に対するパーセント一致(一致パーセントと表すこともある)を決定するためには、ヌクレオチド領域を最大一致が得られるように対応する基準領域に位置合せし、対応領域の一致ヌクレオチド数を測定する。この数を基準配列中のヌクレオチドの総数で除算し、100を乗算し、最も近い整数に丸める。
【0071】
核酸配列は宿主体内での発現を増進するように最適化できる。考察すべき要因は、C:G含量、好適なコドン、および、阻害性二次構造の回避を含む。特定宿主体内で発現増進を生じる核酸配列が得られるようにこれらの要因を様々に組合せる。(参照:例えばDonnellyら,国際公開WO 97/47358)。
【0072】
HCVコーディング核酸の最適化も、Eminiら,国際公開WO 03/031588らに記載されている。WO 03/031588は、NS3−NS4A−NS4B−NS5A−NS5B*をコードする種々の最適化配列の例を示している。
【0073】
NS31−NS4A1−NS32−NS4A2−NS4B2−NS5A2−NS5B2*のコドン最適化配列の一例は、配列2のヌクレオチド1891−9915によって示される。ヌクレオチド1891−1893は、NS31−NS4A1領域のメチオニンコドンを与える。ヌクレオチド3949−3960は、HCV 3aに基づく最初の4つのアミノ酸のあとにメチオニンが存在するNS32−NS4A2領域を与える。
【0074】
本発明の様々な実施態様で、NS31−NS4A1−NS32−NS4A2−NS4B2−NS5A2−NS5B2*コーディング領域は、配列2のヌクレオチド1891−9915または1894−9915に実質的に相似である。様々な実施態様でヌクレオチド配列は、配列2のヌクレオチド1891−9915または1894−9915に、少なくとも80%、少なくとも85%、少なくとも90%もしくは少なくとも95%の一致を有するか;または、0、1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20もしくは1−50ヌクレオチドだけ違っている。
【0075】
V.核酸ベクター
HCVポリペプチドをコードする核酸は、治療用ベクターとしてまたは治療用ベクターの製造に使用できる。治療用ベクターの製造に使用できるベクターはシャトルベクター、アデノウイルスゲノムプラスミドなどである。
【0076】
治療用ベクターは細胞にHCVポリペプチドを導入し発現させるために使用される。適正なベクターは、許容できない副作用を生じることなく標的細胞に核酸を送達できる。細胞発現は、HCVポリペプチドをコードする遺伝子発現カセットを使用して達成される。
【0077】
治療用途に使用できるベクターの例は、第一世代および第二世代のアデノベクター、ヘルパー依存性アデノベクター、アデノ関連ウイルスベクター、レトロウイルスベクター、アルファウイルスベクター、ベネズエラウマ脳炎ウイルスベクターおよびプラスミドベクターなどである。(Hittら,Advances in Pharmacology 40:131−206,1997,Johnstonら,米国特許6,156,588,Johnstonら,国際公開WO 95/32733,Eminiら,国際公開WO 03/031588)。
【0078】
V.A.アデノベクター
アデノベクターは、標的細胞中で所望のタンパク質またはポリペプチドを発現させるために組換えアデノウイルスゲノムを使用する。野生型アデノウイルスは、両端に逆方向末端反復配列をもつ二重鎖の直鎖状ゲノムを有している。ウイルス複製中にゲノムがウイルスキャプシド内部にパッケージされてビリオンを形成する。ウイルスはウイルス付着によって標的細胞に侵入し、次いでインターナリゼーションを達成する。(Hittら,Advances in Pharmacology 40:131−206,1997)。
【0079】
アデノウイルスゲノムは、アデノウイルスの複製およびプロセシングに必要な種々の要素を提供する。アデノウイルスゲノムの各末端は、逆方向末端反復配列(ITR)を含んでおり、これらはウイルス複製に必要である。該ウイルスはまた、感染性ビリオンの産生に必要なある種の構造タンパク質をプロセシングするために必要なプロテアーゼ活性をコードしている。
【0080】
アデノウイルスゲノムの構造は、宿主細胞に形質導入後のウイルス遺伝子の発現順序に基づいて記述できる。ウイルス遺伝子は、転写がDNA複製開始の以前に生じるかまたは以後に生じるかに従って初期(E)遺伝子または後期(L)遺伝子と呼ばれる。転写の初期段階で、E1、E2、E3およびE4遺伝子が発現されて宿主細胞のウイルス複製を準備する。ウイルスゲノムの必須初期領域1(E1)の欠失によってウイルスを複製欠陥にできる(Brodyら,Ann.N.Y.Acad.Sci. 7/6:90−101,1994)。
【0081】
後期段階で、ウイルス粒子の構造成分をコードする後期遺伝子L1−L5の発現がスイッチオンされる。すべての後期遺伝子が単一プロモーターのコントロール下にあり、ペントン(L2)、ヘキソン(L3)、100kDaのスカフォールドタンパク質およびファイバータンパク質(L5)を含むタンパク質をコードしており、これらは、アデノウイルスDNAをキャプシド封入した新しいウイルス粒子を形成する。最後に、野生型アデノウイルス複製プロセスが細胞溶解を惹起する。
【0082】
アデノベクターは、ヒトまたは動物に見出されるような種々のアデノウイルス血清型に基づく。動物アデノウイルスの例は、ウシ、ブタ、チンパンジー、ネズミ、イヌおよびトリ(CELO)を含む。ヒトアデノウイルスは、グループB、C、DまたはE血清型、例えば2型(“Ad2”)、4型(“Ad4”)、5型(“Ad5”)、6型(“Ad6”)、24型(“Ad24”)、26型(“Ad26”)、34型(“Ad34”)および35型(“Ad35”)を含む。アデノベクターは単一アデノウイルス由来または2種以上の異なるアデノウイルスに由来の領域を含有できる。
【0083】
様々な実施態様で、アデノベクターは、Ad5、Ad6、ChAd3、ChAd63またはそれらの組合せに基づく。Ad5は、Chroboczekら,J.Virology 756:280−285,1992に記載されている。Ad5およびAd6に基づくベクターは、Eminiら,国際公開WO 03/031588に記載されている。ChAd3の全長核酸配列を図6A−6Hに示す。ChAd63の全長核酸配列を図7A−7Hに示す。
【0084】
本発明の1つの実施態様では、アデノベクターが、表面に露出した1つ以上のチンパンジーChAd3またはChAd63構造タンパク質を含有する。表面露出タンパク質は、ファイバー、ヘキソンおよびペントンを含む。このようなタンパク質を含むアデノベクターは、ヒトアデノウイルスタンパク質と違って、患者が予めヒトアデノウイルスに接触したときの免疫応答の影響を受け難い。
【0085】
様々な実施態様で、組換えアデノベクターゲノムは、
a)配列3または9に実質的に相似のアミノ酸配列をもつファイバー領域、
b)配列5または11に実質的に相似のアミノ酸配列をもつヘキソン領域、および、
c)配列7に実質的に相似のアミノ酸配列をもつペントン領域
の少なくとも1つをコードしている。様々な追加実施態様では、配列相似が、少なくとも80%、少なくとも85%、少なくとも90%、少なくとも95%であるか、または、配列が、0、1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19もしくは20個のアミノ酸の変更だけ違っている。
【0086】
V.A.1.第一世代アデノベクター
第一世代アデノベクターは、E1欠失、場合によりE3欠失、場合によりE4欠失、および、発現カセットを有する組換えアデノウイルスゲノムを含有している。欠失の範囲および組合せは、ウイルスを複製不能にし、所望産物をコードする遺伝子発現カセットを収容するために十分な大きさである。ウイルスはE1欠失によって複製不能になる。
【0087】
E1欠失は、Ad5の塩基対約342を起点とし塩基対約3523に達する領域、または、他のアデノウイルスの対応する領域の欠失である。好ましくは、欠失領域が、Ad5の塩基対約450から塩基対約3511までの領域または他のアデノウイルスの対応する領域の除去を含む。塩基対約341を起点とするより大きいE1領域欠失は、ウイルスパッケージング促進要素を除去する。
【0088】
E3欠失は、Ad5の塩基対約27865を起点とし塩基対約30995に達する領域、または他のアデノベクターの対応する領域の欠失である。好ましくは、欠失領域が、Ad5の塩基対約28134から塩基対約30817までの領域または他のアデノベクターの対応する領域の除去を含む。
【0089】
E4欠失は、ChAd3の塩基対約34634を起点とし塩基対約37349に達する領域、または、他のアデノウイルスの対応する領域の欠失である。E4欠失は、先天性E4orf6を保持するかまたは別のアデノウイルス由来のE4orf6を挿入できる欠失でなければならない。Bertら,国際公開WO2004/018627は、このような異種E4orf6の使用を例示している。
【0090】
E1、E3およびE4の欠失の組合せは、遺伝子発現カセットを含有している組換えゲノムの全体寸法が野生型アデノウイルスゲノムの約105%を超過しない程度の十分な大きさでなければならない。例えば、組換えアデノウイルスAd5ゲノムの寸法が約105%を超過するとゲノムが不安定になる。(Bettら,Journal of Virology 67:5911−5921,1993)。
【0091】
好ましくは、遺伝子発現カセットを含有している組換えアデノウイルスゲノムの寸法は、野生型アデノウイルスゲノムの寸法の約85%から約105%までの範囲である。様々な実施態様で、発現カセットを含有している組換えアデノウイルスゲノムの寸法は、野生型ゲノムの寸法の約100%から約105.2%、または約100%である。
【0092】
E1およびE3欠失を有しているAd5またはAd6ゲノムには約7,500kbを挿入できる。全く欠失が全く存在しないとき、Ad5ゲノムは35,935塩基対であり、Ad6ゲノムは35,759塩基対である。
【0093】
ChAd3およびChAd63ベクターは、Ad5に比べて、ゲノムサイズがより大きく、また、より大きいE3領域が欠失可能領域として存在するので、核酸挿入容量が大きい。ChAd3ゲノムは、37,741塩基対であり、ChAd63ゲノムは36,643塩基対である。
【0094】
E1、E3が欠失しE4がAd5E4orf6で置換されたChAd3およびChAd63ベクターには約10,800bpを挿入できる。E4がAd5E4orf6によって置換されたとき、置換Ad5E4orf6が欠失領域よりも小さいので欠失および置換の双方である。これらのベクターには10,800bpを挿入すると野生型ゲノムの寸法の105%という限度に達する。
【0095】
第一世代のアデノベクターの複製は、E1遺伝子産物をトランスに供給することによって行うことができる。例えば、アデノウイルスE1領域で形質転換された細胞系を使用することによってE1遺伝子産物をトランスに供給できる。アデノウイルスE1領域で形質転換された細胞および細胞系の例は、HEK293細胞、911細胞、PERC.6TM細胞、および、トランスフェクトされた一次ヒトアミノサイト細胞である。(Grahamら,Journal of Virology 36:59−12,1977,Schiednerら,Human Gene Therapy 77:2105−2116,2000,Fallauxら, Human Gene Therapy 9:1909−1917,1998,Boutら,米国特許No.6,033,908)。
【0096】
チンパンジーアデノウイルスの先天性E4領域がAd5E4orf6によってシスに置換されると、Ad5相補細胞系に伝播した様々な血清型のチンパンジーアデノウイルスベクターの増殖が促進されおよび/または収量が増加する。293およびPER.C6細胞中のAd5E1配列は、チンパンジーアデノウイルスのようなグループCに所属するヒトアデノウイルス以外の血清型の複製を十分に相補しない。
【0097】
発現カセットは組換えアデノウイルスゲノムの欠失E1領域または欠失E3領域に対応する領域に挿入されなければならない。発現カセットは、平行配向でもよくまたは逆平行配向でもよい。平行配向では、挿入された遺伝子の転写方向が欠失したE1またはE3遺伝子と同方向である。逆平行配向転写では、対向鎖が鋳型として機能し、転写方向が対向方向である。
【0098】
本発明の1つの実施態様では、アデノベクターがE4欠失と、配列13のヌクレオチド34601−35482によって示されるAd5E4orf6配列に実質的に相似の配列の挿入とを含む。様々な実施態様で、配列一致は少なくとも75%、少なくとも80%、少なくとも85%、少なくとも90%、少なくとも95%であるか、または、配列5のヌクレオチド34601−35482から0、1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20もしくは1−50ヌクレオチドだけ違っている。
【0099】
本発明の1つの実施態様では、アデノウイルス主鎖が、配列13のヌクレオチド1−460および7468−35890または配列17のヌクレオチド1−454および7458−34658に少なくとも70%、少なくとも75%、少なくとも80%、少なくとも85%、少なくとも90%、少なくとも95%のヌクレオチド配列一致を有するか、または、配列13のヌクレオチド1−460および7468−35890または配列17のヌクレオチド1−454および7458−34658から0、1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20もしくは1−50ヌクレオチドだけ違っている。
【0100】
発現カセットを含有するアデノベクターの別の実施態様では、配列13または配列17に対するヌクレオチド配列一致が少なくとも70%、少なくとも75%、少なくとも80%、少なくとも85%、少なくとも90%、少なくとも95%であるか、または、配列13または配列17から0、1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20もしくは1−50ヌクレオチドだけ違っている。
【0101】
V.A.2.第二世代アデノベクター
第二世代のアデノベクターは、第一世代のベクターよりも小さいアデノウイルスゲノムを含有し、相補細胞系および/またはアデノウイルスタンパク質を供給するヘルパーベクターと共に使用できる。一般的な第二世代のアデノベクターは、Russell,Journal of General Virology 57:2573−2604,2000;Hittら,1997,Human Ad vectors for Gene Transfer,Advances in Pharmacology,Vol.40 Academic Press,Catalucciら.Journal of Virology 79:6400−6409,2005のような様々な参考文献に記載されている。第二世代のアデノベクターは、ヒトおよびチンパンジーアデノウイルスを含む様々な種類のアデノウイルスを基材にできる。
【0102】
V.B.DNAプラスミドベクター
DNAワクチンプラスミドベクターは、遺伝子発現カセットを複製促進要素および好ましくはベクー選択促進要素と共に含有する。好ましい要素は、非哺乳類細胞中の複製および選択可能マーカーを提供する。治療用ベクターは、ヒト細胞中の複製またはヒト核酸への組込みを提供する要素を含有してはならない。
【0103】
選択可能マーカーは、該マーカーを含有する核酸の選択を促進する。好ましい選択可能マーカーは、抗生物質抵抗性を与えるマーカーである。抗生物質選択遺伝子の例は、アンピシリン、ネオマイシンおよびカナマイシンに対する抵抗性をコードする核酸を含む。
【0104】
適当なDNAワクチンベクターは、細菌性複製起点と選択可能マーカーとを含有するプラスミドを出発材料として作製できる。より高い収率を与える細菌性複製起点の例は、ColE1プラスミド由来の細菌性複製起点を含む。(Donnellyら,Annu.Rev.Immunol.75:617−648,1997)。
【0105】
細菌性複製起点および選択可能マーカーの存在が大腸菌のような細菌株中のDNAベクターの産生を可能にする。選択可能マーカーは、DNAベクターを含有しない細菌を除去するために使用される。
【0106】
配列2は、HCVポリペプチドをコードする発現カセットを含有するプラスミドベクターの一例を示す。本発明の1つの実施態様で、プラスミドベクターは、配列2に少なくとも70%、少なくとも75%、少なくとも80%、少なくとも85%、少なくとも90%、少なくとも95%のヌクレオチド相似配列を有するか、または、0、1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20もしくは1−50ヌクレオチドだけ違っている。
【0107】
VI.ベクター作製
ベクターは、制限酵素、核酸結合および相同的組換えの使用を含む組換え核酸技術を使用して作製できる。組換え核酸技術は当業界で公知である。(Ausubel,Current Protocols in Molecular Biology,John Wiley,1987−1998、および、Sambrookら,Molecular Cloning,A Laboratory Manual,2nd Edition,Cold Spring Harbor Laboratory Press,1989)。
【0108】
治療用ベクターを得るためまたは発現カセットもしくはその一部分を1つのベクターから他のベクターに移すためには中間ベクターを使用する。中間ベクターの例は、アデノウイルスゲノムプラスミドおよびシャトルベクターを含む。
【0109】
中間ベクター中の有用な要素は、複製起点、選択可能マーカー、相同的組換え領域および適正な制限部位を含む。適正な制限部位は、核酸配列のクローニングまたは遊離を促進するために使用できる。
【0110】
相同的組換え領域は、別の核酸分子中の標的領域に相同的な核酸配列領域を供給する。相同的領域は標的領域に挿入される核酸配列に隣接する。様々な実施態様で、相同的領域は、約150から600ヌクレオチドの長さまたは約100から500ヌクレオチドの長さである。
【0111】
本発明の1つの実施態様は、HCVポリペプチドを発現する発現カセット、選択可能マーカー、細菌性複製起点、発現カセットを標的組換えによってE1領域に挿入または置換する第一のアデノウイルス相同性領域および第二のアデノウイルス相同性領域を含有するシャトルベクターを記載する。第一および第二の相同性領域は、発現カセットに隣接する。第一の相同性領域は、塩基対ほぼ4−450からの野生型アデノウイルスの領域の少なくとも右端(3’端)に実質的に相同の少なくとも約100塩基対を含有する。第二の相同性領域は、塩基対ほぼ3511−5792からのAd5の少なくとも左端(5’端)または別のアデノウイルスの対応する領域に実質的に相同の少なくとも約100塩基対を含有する。
【0112】
“実質的に相同の”という表現は、標的領域と特異的組換えを行うために十分な相同度を表す。様々な実施態様で、実質的相同は、少なくとも85%、少なくとも95%または100%の配列一致を表す。
【0113】
アデノベクターの1つの作製方法は、発現カセットを含有するプレアデノウイルスゲノムプラスミドの作製を経由する。該プレアデノウイルスプラスミドは、所望の相補細胞系中で複製されるために必要なすべてのアデノウイルス配列を含有する。次いでプレアデノウイルスプラスミドを制限酵素で消化してITRを遊離させ、相補細胞系にトランスフェクトしてウイルスを回収する。複製が生じ得るためにはITRがプラスミド配列から遊離されなければならない。アデノベクターの回収によって、発現カセットを含有するアデノベクターが得られる。(参照:例えばEminiら,国際公開WO 03/031588)。
【0114】
VI.A.アデノウイルスゲノムプラスミド
アデノウイルスゲノムプラスミドは、より長いプラスミド(コスミドでもよい)の内部にアデノベクター配列を含有している。より長いプラスミドは、プラスミドの製造および維持に使用される手順次第では真核細胞または原核細胞中の増殖および選択を促進する要素のような追加要素を含有している。アデノウイルスゲノムプラスミドは好ましくは、E1またはE3欠失領域に挿入された遺伝子発現カセットを有している。
【0115】
アデノウイルスゲノムプラスミド製造技術には、シャトルベクターおよび相同的組換えを使用する技術、アデノウイルスコスミドへの遺伝子発現カセットの挿入を含む技術などがある。(Hittら,Methods in Molecular Genetics 7:13−30,1995,Danthinneら,Gene Therapy 7:1707−1714,2000)。
【0116】
本発明の1つの実施態様は、アデノウイルスゲノムプラスミドを作製する相同的組換え段階と、アデノウイルス回収段階とを含むアデノベクターの製造方法を記載する。相同的組換え段階は、アデノウイルス相同領域に隣接されたHCVポリペプチド発現カセットを含有するシャトルベクターの使用を含む。アデノウイルス相同領域は、E1またはE3欠失領域に発現カセットを標的組換えする。
【0117】
VI.B.アデノベクター回収
アデノベクターは当業界で公知の方法またはここに記載の方法を使用してを組換えアデノウイルスゲノムプラスミドから回収できる。当業界で公知のアデノウイルス回収技術の例は、Hittら,Methods in Molecular Genetics 7:13−30,1995およびDanthinneら,Gene Therapy 7:1707−1714,2000に示されている。
【0118】
アデノベクター回収方法の一例は、アデノウイルス複製の増進を含む。増進は例えば、E2タンパク質(ポリメラーゼ、プレターミナルタンパク質およびDNA結合タンパク質)ならびにE4orf6のようなアデノウイルス機能を別のプラスミドから供給することによって実行できる(Eminiら,国際公開WO 03/031588)。
【0119】
VII.HCV併用治療
HCV核酸ワクチンは患者を治療するために単独で使用してもよく、他のHCV治療薬と併用してもよく、他の種類の疾患を対象とする薬剤と共に使用してもよい。追加治療薬は、HCVおよびHCV感染個体で有病率が高い疾患を治療する治療薬を含む。別種の疾患を対象とする薬剤は、HIVおよびHBCに特異的なワクチンを含む。
【0120】
HCVを治療するための追加治療薬はワクチンおよび非ワクチン薬を含む。(Zein,Expert Opin.Investig.Drugs 70:1457−1469,2001)。追加HCVワクチンの例は、HCVコア、E1、E2またはp7領域に対する免疫応答を誘発するように設計されたワクチンを含む。ワクチン成分の例は、天然産生ポリペプチド、HCVミモトープポリペプチド、または、このようなポリペプチドをコードする核酸を含む。
【0121】
一般的なミモトープ作製技術および種々のHCVミモトープを記載している参考文献は、Feliciら,米国特許No.5,994,083およびNicosiaら,国際出願WO 99/60132である。HCVミモトープは天然産生HCV抗原に融合できる。
【0122】
現在認可されている抗HCV薬はリバビリンと併用されるインターフェロンαである。組換えインターフェロンおよびPEG付加インターフェロンのような様々な形態のインターフェロンαをHCV感染の治療に使用できる。(De Francescoら,Antiviral Research 5S:1−16,2003,Walkerら,Antiviral Chemistry & Chemotherapy 14:1−21,2003)。
【0123】
異なる多様な抗HCV薬が様々な臨床開発段階にある。開発中の種々の抗HCV薬は、種々のHCV標的に特異的な薬剤を含む。種々のHCV標的の例は、HCVポリメラーゼおよびHCV NS3−NS4Aプロテアーゼを含む。(De Francescoら,Antiviral Research 58:1−16,2003,Walkerら,Antiviral Chemistry & Chemotherapy 14:1−21,2003)。
【0124】
VIII.医薬投与
HCVワクチンはここに提供した指針を当業界で公知の技術と共に使用して配合し患者に投与できる。一般的な医薬投与の指針は、Modern Vaccinology,Ed.Kurstak,Plenum Med.Co.1994;Remington’s Pharmaceutical Sciences 18th Edition,Ed.Gennaro,Mack Publishing,1990;およびModern Pharmaceutics 2nd Edition,Eds.Banker and Rhodes,Marcel Dekker,Inc.,1990に示されている。これらの文献はいずれも参照によって本発明に組込まれるものとする。
【0125】
HCVワクチンは、静脈内、腹腔内、皮下、筋肉内、経皮、皮膚圧入または鼻腔内のような種々の経路で投与できる。好ましい経路は筋肉内である。
【0126】
筋肉内投与は、1つ以上の電気パルスを伴うかまたは伴わない注射のような様々な技術を使用して行うことができる。電気介在移入は体液性および細胞性の双方の免疫応答を刺激することによって遺伝子免疫感作を補助する。
【0127】
ワクチン注射は、有針または無針の注射システムを使用する種々の技術を使用して行うことができる。無針注射システムの一例はジェット注射デバイスである。(Donnellyら,国際公開WO 99/52463)。
【0128】
電気介在移入すなわちジーン・エレクトロ・トランスファー(GET)は、核酸注入後に適正な電気パルスを送達することによって行うことができる。(参照:Mathiesen,国際公開WO 98/43702およびEminiら,国際公開WO 03/031588)
VIIIA.医薬担体
医薬的に許容される担体は、ワクチンの保存および対象へのワクチン投与を容易にする。医薬的に許容される担体の例をここに示す。医薬的に許容される追加の担体は当業界で公知である。
【0129】
医薬的に許容される担体は、バッファ、標準生理食塩水またはリン酸塩緩衝生理食塩水、ショ糖、塩およびポリソルベートのような様々な成分を含有し得る。医薬的に許容される担体の一例は、2.5−10mMのトリスバッファ、25−100mMのNaCl、2.5−10%ショ糖、0.01−2mMのMgCl2および0.001%−0.01%のポリソルベート80(植物由来)である。pHは約7.0−9.0でよい。担体の1つの具体例は、5mMのトリス、75mMのNaCl、5%ショ糖、1mMのMgCl2、0.005%のポリソルベート80をpH8.0で含有している。
【0130】
VIII.B.投薬計画
適正投薬計画は、特定ワクチンの効力、ならびに、患者の年齢、体重、性別および医学的状態、投与経路、所望の効果、投与回数などの要因を考慮して決定できる。特定ワクチンの効力は、細胞で発現およびプロセシングされMHCクラスIおよびII複合体に関係して提示されるポリペプチドを産生する特定ワクチンの能力のような様々な要因に依存する。
【0131】
患者に投与されたHCVポリペプチドをコードする核酸は、アデノベクターのようなウイルスベクターを含む様々なタイプのベクターおよびDNAプラスミドワクチンの部分を構成し得る。DNAプラスミドの投与に関する様々な実施態様では、約0.1から10mgのプラスミドを患者に投与し、また、約1から5mgのプラスミドを患者に投与する。ウイルスベクター好ましくはアデノウルイスベクターの投与に関する様々な実施態様では、約105から1011のウイルス粒子を患者に投与し、また107から1010のウイルス粒子を患者に投与する。
【0132】
ウイルスベクターワクチンおよびDNAプラスミドワクチンは、単独で投与してもよく、または、プライム(初回抗原刺激)およびブースト(追加抗原刺激)を含む投与計画の部分であってもよい。プライムおよびブーストの混合接種法は、DNAワクチンでプライムしウイルスベクターワクチンでブーストする方法、または、ウイルスベクターワクチンでプライムし、DNAワクチンでブーストする方法を含む。
【0133】
例えば約2から4回またはそれ以上の多数プライムを使用し得る。プライムとブーストとの間の時間の長さは、典型的には約4ヶ月から1年の範囲であるが、他の時間枠を使用してもよい。個体に抗アデノウイルス免疫応答が既に存在する場合にはDNAワクチンでプライムする投与計画の使用が好ましい。
【0134】
本発明の1つの実施態様では、初回ワクチン接種でDNAワクチンを筋肉組織に直接接種する。初回ワクチン接種後のブーストはアデノベクターまたはDNAワクチンで行う。
【0135】
免疫応答をブーストするために、インターロイキン−12、GM−CSF、B7−1、B7−2、IP10およびMig−1のような補助薬剤を併用投与できる。補助薬剤はタンパク質としてまたは核酸ベクターの使用を介して併用投与できる。
【0136】
VIII.C.異種プライム−ブースト
異種プライム−ブーストは、プライムに1つの種類のウイルスベクターを使用し、ブーストに別の種類のウイルスベクターを使用する混合方式である。異種プライム−ブーストは、種々のアデノウイルス血清型に基づくベクターのような近縁ベクター、ならびに、異なる動物のアデノウイルスおよびポックスウイルスのようなより遠縁のウイルスを含み得る。マウスをマラリアから守るためにポックスウイルスとアデノベクターとを使用することは、Gilbertら,Vaccine 20:1039−1045,2002によって示されている。HCVポリペプチドを発現するチンパンジーアデノベクターは、異種プライム−ブーストに使用できるベクターを提供する。
【0137】
プライムとブーストとの間の時間の長さは、典型的には約4ヶ月から1年の範囲であるが、他の時間枠を使用してもよい。最小時間枠は、免疫静止を考慮した十分な時間でなければならない。1つの実施態様では、この静止に少なくとも6ヶ月間を要する。プライムは1種類のベクターによる多回、例えば2から4回のプライムを含み得る。
【0138】
ポックスウイルスベクター中に存在する発現カセットは、当該ポックスウイルスまたは別のポックスウイルス構成員の先天的プロモーターまたはそれらに由来するプロモーターを含有しなければならない。ワクシニアウイルス、変性ワクシニアウイルス、アビポックスウイルス、アライグマポックスウイルス、変性ワクシニアウイルスAnkara、カナリアポックスウイルス(例えばALVAC)、トリポックスウイルス、ウシポックスウイルスおよびNYVACに基づくベクターのような様々な種類のポックスウイルスに基づくベクターを構築し使用する様々な戦略が当業界で公知である。(Moss,Current Topics in Microbiology and Immunology 755:25−38,1982;Earlら,In Current Protocols in Molecular Biology,Ausubelら,eds.,New York:Greene Publishing Associates & Wiley Interscience;1991:16.16.1−16.16.7;Childら,Virology 174(2):625−9,1990;Tartagliaら,Virology 188:217−232,1992;米国特許Nos.,4,603,112,4,722,848,4,769,330,5,110,587,5,174,993,5,185,146,5,266,313,5,505,941,5,863,542,および5,942,235)。
【0139】
VIII.D.アジュバント
HCVワクチンはアジュバントと共に配合できる。アジュバントは免疫原の免疫応答発生を補助する物質である。アジュバントは、生物学的または免疫学的半減期を延長する、免疫調節補助物質として作用する、または、免疫調節サイトカインの産生を誘導する、などの様々なメカニズムによって機能できる。種々のアジュバントを併用することもできる。
【0140】
HCVワクチンはアジュバントと共に配合できる。アジュバントの例は、ミョウバン、AlPO4、アルヒドロゲル、リピドAおよびそれらの誘導体または変異体、フロインド不完全アジュバント、中性リポソーム、ワクチンおよびサイトカインを内包するリポソーム、非イオン性ブロックコポリマー、ケモカイン、ならびに、免疫調節剤である。
【0141】
ポリオキシエチレン(POE)およびポリオキシプロピレン(POP)を含有する非イオン性ブロックコポリマー、例えば、POE−POP−POEブロックコポリマーはアジュバントとして使用し得る(Newmanら,Critical Reviews in Therapeutic Drug Carrier Systems 75:89−142,1998)。非イオン性ブロックコポリマーとアニオン性界面活性剤との併用によって核酸の免疫応答を増進できる。
【0142】
サイトカイン、ホルモン、脂質誘導体および低分子のような様々な種類の化合物を免疫調節剤として使用できる。免疫調節剤の例は、抗−CTLA−4、抗−CD137、抗−CD40、抗−CD28、抗−CD4、抗−CD25、抗PD1、抗−PD−Ll、抗−PD−L2、FOXP3−ブロッキング剤、Flt−3リガンド、イミキモッド、顆粒球−マクロファージコロニー−刺激因子(GM−CSF)、サルグラモスチン、トール様受容体(TLR)−7アゴニストおよびTLR−9アゴニストである。
【0143】
アジュバント配合物の具体例は、CRL−1005(CytRx Research Laboratories)、DNAおよびベンジルアルコニウムクロリド(BAK)を含有する配合物である。CRL−1005配合物は、例えばEminiら,国際公開WO 03/031588に記載されているように調製できる。
【0144】
VIII.E.ワクチン保存
ワクチンは様々な種類のバッファを使用して保存できる。例えば、Eminiら,国際公開WO 03/031588に記載されたバッファA105を使用できる。
【0145】
DNAの保存は微量金属イオンの除去またはキレート化によって強化できる。DNAワクチンの安定性を強化するためにコハク酸またはリンゴ酸のような試薬とキレート化剤とを使用できる。キレート化剤の例は、多くのリン酸塩リガンドおよびEDTAである。また、エタノールまたはグリセロールのような非還元性フリーラジカルスカベンジャーの含有はフリーラジカル発生によるDNAプラスミドの損傷を防止するために役立つ。さらに、DNAワクチンの安定性を最適化するために、配合物中のバッファの種類、pH、塩濃度、露光、バイアルの製造に使用した滅菌法などをコントロールするとよい。
【0146】
IX.実施例
本発明の様々な特徴をさらに詳細に説明するために実施例を以下に示す。実施例はまた本発明の有用な実施方法も示している。これらの実施例は特許請求の範囲に記載した本発明を限定しない。
【実施例1】
【0147】
ChAd63およびChAd3ゲノム配列の注釈
ChAd63およびChAd3を、“ヒトアデノウイルスC”群(HAdV−C;分類ID:129951)のすべてのタンパク質配列で作成されたローカルデータベースにブラストした。特定の分類IDをサーチすることによってタンパク質配列をNCBIサーバーからダウンロードした。ブラストサーチはblastxプログラムを使用して行った。列中に示す配列の数を1000に設定し、Filterをスイッチオフにした。次に、大規模配列相似解析用のBLAST強調ツールであるMSPcrunchによってブラスト結果を解析した。
【0148】
2つのゲノム配列について得られたCDS注釈のおのおのをATGおよび終結コドンの位置と必要ならばスプライシング部位の位置とを観察することによって手作業で確認した。このような産物の相同による予測の妥当性を検証するために、すべての産物を以前に作成されたアデノウイルスタンパク質データベースにblastpでサーチした。ChAd63およびChAd3の無加工のゲノム配列をMSPcrunch結果および手作業の校正に従ってVNTIで注釈した。遺伝子産物のChAd3およびChAd63を表1および2に示す。
【0149】
【表1】
【0150】
【表2】
【実施例2】
【0151】
ChAd3ベクター構築
ChAd3ΔE1,E3,E4,E4Ad5orf6ベクターの構築は以下の段階を含む。
【0152】
I.サブグループCシャトルベクターの構築
ChAd3ウイルスゲノムを完全に配列決定し(配列14)、その情報を使用して、相同的組換えによって完全ゲノムを容易にクローニングできるシャトルベクターを構築した。要約すると、ここでpChAd3EGFPと呼ぶサブグループCチンパンジーアデノウイルス3をクローニングするために使用したシャトルベクターを以下の手順で構築した。pIXコーディング領域を含有しているChAd3 DNAフラグメント(nt3542−4105)を配列20および21のプライマーと共にPCR増幅し、Sgf1−AscIで消化し、次いで、SgfI−AscIで消化したpARSCV32−3にクローニングして、pARS−ChAd3Dを作製した。ChAd3右端(nt37320−7441)を配列22および23のプライマーと共にPCR増幅し、XbaIおよびBamHIで消化し、次いで、XbaIおよびBamHIで制限したpARS−ChAd3Dに結合して、pARS−ChAd3RDを作製した。ChAd3ウイルスDNA左端(nt1−460)を配列24および25のプライマーと共にPCR増幅し、EcoRIおよびSgfIで消化し、次いで、EcoRIおよびSgfIで消化したpARS−ChAd3RDにクローニングし、このようにしてpARS−ChAd3RLDを作製した。このウイルスDNAカセットはまた、消化によってプラスミドDNAからウイルスDNAが遊離されるように双方のITRの末端に局在する制限酵素部位(PmeI)も含むように設計した。
【0153】
II.ΔE1 ChAd3ベクターの構築
ChAd3ベクターはBJ5183大腸菌株中の相同的組換えによって構築した。BJ5183細胞を、ChAd3精製ウイルスDNAとBstEIIおよびBst11071で消化したpChAd3EGFPシャトルベクターとによって同時形質転換した。pIX遺伝子、直鎖化pChAd3EGFPの末端に存在する右ITR DNA配列とウイルスゲノムDNAとの間の相同的組換えによってウイルスゲノムDNAがプラスミドベクターに挿入され、同時に、E1領域が欠失してEGFP発現カセットで置換された。ヒトサイトメガロウイルス(HCMV)プロモーターおよびウシ成長ホルモンポリアデニル化シグナル(Bgh ポリA)に基づくHCV NS領域発現カセットを、Eminiら,国際公開WO 03/031588に記載された手順で構築し、HCMVおよびBgh−ポリAのDNA配列間の相同性を利用した相同的組換えによってBJ5183大腸菌株中のChAd3ΔE1EGFPベクターに挿入した。
【0154】
III.E3領域欠失
ChAd3ベクター主鎖に完全なE3領域の欠失を導入するために、E3遺伝子に隣接する2つのDNA領域をPCR増幅して2つのDNAフラグメントを作製した。486bpのフラグメントはnt28159からnt28644(pVIII遺伝子の3’、E3領域の上流)の範囲であり、474bpのDNAフラグメントは、ファイバー遺伝子の3’端(bp32633からbp33106まで、E3領域の下流)を含んでいる。第一のDNAフラグメントの3’端および第二のフラグメントの5’端にEcoRI制限部位を導入した。2つのPCRフラグメントをEcoRIで消化し、インビトロ結合によって接合した。得られたDNAフラグメント(988bp)を次に、pVIII順方向オリゴおよびファイバー逆方向オリゴを使用してさらに増幅した。
【0155】
互いに接合した3’および5’のDNAフランキング領域を含有している988bpのE3領域のDNAフラグメントを、BJ5183細胞の同時形質転換によって、HpaI(野生型ChAd3の32384bpでE3領域切断)で直鎖化したpChAd3ΔE1/EGFPに組換えて、E3欠失を導入した。最終組換え産物はpChAd3ΔE1,E3/EGFPプレアデノプラスミドであった。
【0156】
IV.E4領域の欠失およびAd5E4orf6の挿入
ChAd3E4領域をAd5E4orf6で置換するために、Ad5E4orf6を、ChAd3ゲノムの右端に由来の最後の393bp(bp37349からbp37741まで)を含有するシャトルプラスミドに導入した。引き続いて、ファイバー3’端に由来しE4ポリAを含有している144bpのDNAフラグメント(ChAd3マップのbp34491からbp34634まで)をAd5E4orf6の下流に導入して、プラスミドpARSChAd3Ad5E4orf6−2を作製した。
【0157】
最後にBJ5183大腸菌株の同時形質転換によって、境界にファイバー3’端/E4ポリAおよびChAd3右端を含むpARSChAd3Ad5E4orf6−2に由来のDNAフラグメントを、PacI制限酵素(PacI部位、野生型ChAd3のnt36924)で直鎖化したpChAd3ΔE1,E3/EGFPに相同的組換えによって導入し、pChAd3ΔE1,3,4Ad5orf6EGFPを作製した。
【0158】
この戦略によって、ChAd3E4コーディング領域が完全に欠失し、ChAd3E4プロモーターのコントロール下に推定E4TATAシグナルの62bp下流にクローニングされたAd5E4orf6遺伝子で置換された。
【実施例3】
【0159】
ChAd63ベクター構築
ChAd3ΔE1,E3,E4,E4Ad5orf6ベクターに類似のChAd63ベクターは以下のように構築した。
【0160】
I.サブグループEシャトルベクターの構築
ChAd63ウイルスゲノムを完全に配列決定し、その情報を使用して、相同的組換えによって完全ゲノムを容易にクローニングできるシャトルベクターを構築した。要約すると、ここでpARSChAd63_EGFPと呼ぶサブグループEチンパンジーアデノウイルス63をクローニングするために使用したベクターを以下に記載の手順で構築した。
【0161】
ChAd63右端(nt36216−36643)を配列26および27のプライマーと共にPCR増幅し、XbaIおよびBamHIで消化し、次いで、XbaIおよびBamHIで制限したpARSChAd3−RLDに結合して、pARS−ChAd63Rを作製した。pIXコーディング領域を含有しているChAd63 DNAフラグメント(nt3422−3814)を配列28および29のプライマーと共にPCR増幅し、SgfI−AscIで消化し、次いで、SgfI−AscIで消化したpARS−ChAd63Rにクローニングして、pARS−ChAd63RDを作製した。ChAd63ウイルスDNA左端(nt1−455)をプライマー30および31と共にPCR増幅し、EcoRIおよびEcoRVで消化し、次いで、EcoRIおよびEcoRVで消化したpARS−ChAd63RDにクローニングし、このようにしてpARS−ChAd63RLDを作製した。HCMV−EGFP−bghポリAカセットを、配列32および33のプライマーと共にPCR増幅し、EcoRVで消化し、次いで、EcoRVで消化したpARS−ChAd63RLDにクローニングして、pARS−ChAd63RLD−EGFPを作製した。このウイルスDNAカセットはまた、消化によってプラスミドDNAからウイルスDNAが遊離されるように双方のITRの末端に局在する制限酵素部位(PmeI)も含むように設計した。
【0162】
II.ΔE1 ChAd63ベクターの構築
ChAd63ベクターはBJ5183大腸菌株中の相同的組換えによって構築した。BJ5183細胞を、ChAd63精製ウイルスDNAとAscIで消化したpARS−ChAd63RLD−EGFPとによって同時形質転換した。pIX遺伝子、直鎖化pARS−ChAd63RLD−EGFPの末端に存在する右ITR DNA配列とウイルスゲノムDNAとの間の相同的組換えによってウイルスゲノムDNAがプラスミドベクターに挿入され、同時に、E1領域が欠失してEGFP発現カセットで置換された。
【0163】
III.E3領域欠失およびChAd63NSmutベクター構築
ChAd63ベクター主鎖に完全なE3領域欠失を導入するために、E3遺伝子に隣接する2つのDNA領域をPCR増幅して2つのDNAフラグメントを作製した。567bpのフラグメントはnt26665からnt27207まで(pVIII遺伝子の3’端,E3領域の上流)の範囲であり、563bpのDNAフラグメントはファイバー遺伝子の3’端(bp31788からbp32326まで,E3領域の下流)を含んでいる。第一のDNAフラグメントの3’端および第二フラグメントの5’にPacI制限部位を導入した。2つのPCRフラグメントをPacIで消化し、インビトロ結合によって接合した。得られたDNAフラグメント(1112bp)を次に、pVIII順方向オリゴヌクレオチドおよびファイバー逆方向オリゴヌクレオチドを使用してさらに増幅した。
【0164】
互いに接合した3’および5’のDNAフランキング領域を含有している1112bpのE3領域のDNAフラグメントを、BJ5183細胞中の同時形質転換によって、HpaI(野生型ChAd63の30168bpでE3領域切断)で直鎖化したpChAd63ΔE1/EGFPと組換えて、E3欠失を導入した。最終組換え産物はpChAd63ΔE1,E3/EGFPプレアデノプラスミドであった。
【0165】
ヒトサイトメガロウイルスプロモーター(HCMV)およびウシ成長ホルモンポリアデニル化シグナル(Bgh ポリA)に基づくHCV NS領域発現カセットをEminiら,国際公開WO 03/031588に記載された手順で構築し、HCMVおよびBghポリAのDNA配列間の相同性を利用した相同的組換えによってBJ5183大腸菌株中のpChAd63ΔE1,E3/EGFPベクターに挿入し、ChAd63NSmutを作製した。
【0166】
IV.E4領域の欠失およびAd5E4orf6の挿入
ChAd63E4領域をAd5E4orf6で置換するために、Ad5E4orf6を、ChAd63ゲノムの右端に由来の428bp(bp36216からbp36643まで)下流でpARS−ChAd63RLD−EGFPに導入した。引き続いて、ファイバー3’端に由来しE4ポリAを含有している200bpのDNAフラグメント(ChAd63マップのbp33624からbp33823まで)をAd5E4orf6の下流に導入して、プラスミドpARSChAd63Ad5E4orf6−2を作製した。最後に、BJ5183大腸菌株の同時形質転換によって、境界にファイバー3’端/E4ポリAおよびChAd63左端を含むpARSChAd63Ad5E4orf6−2に由来のDNAフラグメントを、PmeI制限酵素(プラスミドDNAからウイルスDNAを遊離する)で消化したpChAd63ΔE1,E3/EGFPに相同的組換えによって導入し、このようにしてpChAd63ΔE1,3,4Ad5orf6EGFPを作製した。
【0167】
この戦略によって、ChAd63E4コーディング領域が完全に欠失し、ChAd63E4プロモーターのコントロール下に推定E4TATAシグナルの131bp下流にクローニングされたAd5E4orf6遺伝子で置換された。
【実施例4】
【0168】
ChAd3NSmut(配列13)の発現
Catalucciら,Journal of Virology 79:6400−6409,2005に記載の手順に準拠した技術を使用してChAd3NSmutのHCVタンパク質発現を試験した。HeLa細胞にChAd3NSmutおよびMRKAd6NSmutを感染させた。MRKAd6NSmutはEminiら,国際公開WO 03/031588に記載されている。細胞抽出物を抗NS5Aモノクローナル抗体によるイムノブロットで分析した。図8に示すように、ChAd3NSmutは、ヒトAd6に基づくベクター(MRKAd6NSmut)と同様にHCVタンパク質を発現する。
【実施例5】
【0169】
ChAd3NSmut(配列13)の安定性
Catalucciら,Journal of Virology 79:6400−6409,2005に記載の手順に準拠した技術を使用してChAd3NSmutの遺伝的安定性を検証した。5つの独立クローン(継代10)から抽出したウイルスDNAの制限分析を行った。プレChAd3NSmutプラスミドを陽性対照として含ませた。ChAd3NSmutはPerC.6細胞までの継代期間は遺伝的に安定であった。
【実施例6】
【0170】
ChAd3NSmut(配列13)およびChAd63NSmut(配列17)はマウス体内でCMIを誘導した。
【0171】
Eminiら,国際公開WO 03/031588に記載の手順に準拠した技術を使用してChAd3NSmutおよびChAd63NSmutの細胞性免疫誘導能力をC57/B6マウスで試験した。図9は、C57/B6マウス体内でのChAd3NSmut(配列13)、ChAd63NSmut(配列17)およびMRKAd6NSmutの細胞性免疫誘導能力の比較を示す。図9は、注入3週後に行ったIFNγ ELIspot実験(H2Kb制限ペプチド使用、NS3プロテアーゼ中でマッピング)を示す(データはN=5の平均として示す)。108および109の用量のChAd3NSmutおよびChAd63NSmutによってマウス体内でMRKAd6NSmutと同等のCMIが誘発される。
【実施例7】
【0172】
ChAd3NSmutおよびChAd63NSmutはアカゲザル体内でCMIを誘導した。
【0173】
ChAd3NSmut(配列13)およびChAd63NSmut(配列17)のCMI誘導能力をヒト以外の霊長類で確認するために、Eminiら,国際公開WO 03/031588およびCirilloら,国際公開WO 2005/071093に記載の手順に準拠した技術を使用してアカゲザルを免疫感作した。異なる3つの非交差反応性ベクターの連続注射に基づく異種プライム/ブースト法で3匹1グループのサルを免疫感作してベクターを評価した。3匹の動物は、0および4週目に1匹あたり1010vpの用量のChAd3NSmutを2回注射してプライムし、次いで22週目にMRKAd6NSmut、42週目にChAd63NSmutを注射した。IFN−γ ELISPOTによって測定した免疫応答の時間経過を図11に報告するが、これは、所与のいずれかの時点で種々のHCV NSペプチドプールに観察された応答の和を表している。この結果は、ChAd3NS注射によって全部の動物に効率的なプライム効果が得られたこと、および、MRKAd6NSmutおよびChAd63NSmutの双方の投与によってCMIが強力にブーストされることを示す。
【実施例8】
【0174】
キメラHCVポリペプチドをコードするプラスミドDNA(配列1)の構築
HCV 3aに基づくNS3−4A領域とHCV 1bに基づくNS3−NS4A−NS4B−NS5A−NS5B領域とを含み、ここでpV1JnsNSOPTmut3a−1bと呼ぶキメラHCVポリペプチドをコードするプラスミド(図12および13)はBJ5183大腸菌株中の相同的組換えによって得られた。
【0175】
最適翻訳開始(コザック)配列と、成熟NS3配列の第一アミノ酸に融合したメチオニン開始コドンとをもつHCV 3a由来の完全コドン最適化NS3−4aをコードするプラスミドを合成的に作製した。NS3−4aコーディング配列には、pV1JnsNSOPTmutb受容体プラスミドに挿入するためにイントロンA配列とNS3(HCV)コーディング配列の起点とに相同の2つの組換え領域が隣接している(Eminiら,国際公開WO 03/031588)。親プラスミドからインサートを切除するHindIII制限部位を新しいNS3−4a配列の両端に導入した。
【0176】
このpV1JnsOPTmutプラスミドをHpaI非反復部位消化によって直鎖化した。直鎖化したpV1JnsOPTmutプラスミドおよびHindIII消化したNS3−4a(3a)インサートでBJ5183菌株を同時形質転換して、pV1JnsNSOPTmut3a−1bを作製した。図13に示す得られたpV1JnsNSOPTmut3a−lbの遺伝子構造を制限酵素およびDNA配列解析によって検証した。
【実施例9】
【0177】
キメラHCVポリペプチドをコードするプラスミドDNAはマウス体内でDMIを誘導した。
【0178】
キメラHCVポリペプチドをコードするプラスミドDNAの種々のHCV遺伝子型に対する細胞性免疫誘導能力をマウスで試験した。キメラポリペプチド(配列1)は、HCV 3aに基づくNS3−4A領域およびHCV 1bに基づくNS3−NS4A−NS4B−NS5A−NS5B領域を含んでいた(pV1Jns−NSOPTmut3a−1b)。
【0179】
異なる3つのマウス株(2つの同系交配:Balb/c,C57B1/6および1つの異系交配:CD1)に、50μgのDNAを筋肉内注射し次いで電気パルスを与えた。各動物に、キメラプラスミド(pV1Jns−NSOPTmut3a−1b)、または、HCV 1bに基づくNS3−NS4A−NS4B−NS5A−NS5B領域をコードするpV1Jns−NSOPTmutプラスミド(Eminiら,国際公開WO 03/031588)を2回与えた。HCV 1bおよび3aに由来のウイルスタンパク質に特異的なCMIを、Eminiら,国際公開WO 03/031588に記載された技術を使用して測定した。図14は、CD1マウス(異系交配株)の体内でHCV 1bおよび3aに由来のNS3タンパク質に応答してIFN−γを分泌するT細胞の数を示す(1,000,000脾細胞あたりのスポット形成細胞として表す)。1b NS3タンパク質に対する特異的応答は双方のプラスミドで同等であるが、3a NS3タンパク質に対してはキメラ構築物のほうが高い応答を誘導する(スチューデントT検定でp=0.04)。2つの同系交配株のマウス(Balb/cおよびC57B1/6)の体内でHCV 1bおよび3aに由来のNS3タンパク質に対して誘導されたCMIは双方の構築物で同等であった。
【0180】
特許請求の範囲は他の実施態様も包含する。いくつかの実施態様を提示して説明したが、本発明の要旨および範囲を逸脱することなく様々な変更を行うことが可能であろう。
【図面の簡単な説明】
【0181】
【図1】配列1のアミノ酸配列を示す。アミノ酸1−686は、HCV 3aに基づくNS3−NS4A領域を与える。アミノ酸687−690は、HCV 3a NS4B領域の最初の4つのアミノ酸を構成し、また、HCV 3a NS3−NS4A領域の開裂接合を与える。アミノ酸691−2675は、HCV 1bに基づくNS3−NS4A−NS4B−NS5A−NS5B領域を与える。NS3−NS4A領域およびNS3−NS4A−NS4B−NS5A−NS5B領域はまた開始メチオニンの付加を含む。
【図2A】図2A−2Cは配列2の核酸配列を示す。ヌクレオチド318−10182は以下の要素を含む発現カセットを構成する:HCMVプロモーター:nt318−905Int A:nt1040−1865コザック配列:nt1885−1890HCV Met−NS3−NS4A(3aに基づく、最適化):nt1891−3948HCV NS4Bの最初の4アミノ酸(3aに基づく):nt3949−3960HCV Met−NS3−5B(1bに基づく、Bk菌株、最適化):nt3961−9915TAAAターミネーター:nt9916−9919BGH:nt9956−10179。
【図2B】図2A−2Cは配列2の核酸配列を示す。ヌクレオチド318−10182は以下の要素を含む発現カセットを構成する:HCMVプロモーター:nt318−905Int A:nt1040−1865コザック配列:nt1885−1890HCV Met−NS3−NS4A(3aに基づく、最適化):nt1891−3948HCV NS4Bの最初の4アミノ酸(3aに基づく):nt3949−3960HCV Met−NS3−5B(1bに基づく、Bk菌株、最適化):nt3961−9915TAAAターミネーター:nt9916−9919BGH:nt9956−10179。
【図2C】図2A−2Cは配列2の核酸配列を示す。ヌクレオチド318−10182は以下の要素を含む発現カセットを構成する:HCMVプロモーター:nt318−905Int A:nt1040−1865コザック配列:nt1885−1890HCV Met−NS3−NS4A(3aに基づく、最適化):nt1891−3948HCV NS4Bの最初の4アミノ酸(3aに基づく):nt3949−3960HCV Met−NS3−5B(1bに基づく、Bk菌株、最適化):nt3961−9915TAAAターミネーター:nt9916−9919BGH:nt9956−10179。
【図3】met−NS3−5B配列(配列16)を示す。
【図4A】図4A−4Jは、ChAd3ファイバー(図4A、配列3)、ChAd3ヘキソン(図4B、配列5)、ChAd3ペントン(図4C、配列7)、ChAd63ファイバー(図4D、配列9)、ChAd63ヘキソン(図4E、配列11)のアミノ酸配列、ならびに、ChAd3ファイバー(図4F、配列4)、ChAd3ヘキソン(図4G、配列6)、ChAd3ペントン(図4H、配列8)、ChAd63ファイバー(図4I、配列10)およびChAd63ヘキソン(図4J、配列12)のコーディング核酸を表す。
【図4B】図4A−4Jは、ChAd3ファイバー(図4A、配列3)、ChAd3ヘキソン(図4B、配列5)、ChAd3ペントン(図4C、配列7)、ChAd63ファイバー(図4D、配列9)、ChAd63ヘキソン(図4E、配列11)のアミノ酸配列、ならびに、ChAd3ファイバー(図4F、配列4)、ChAd3ヘキソン(図4G、配列6)、ChAd3ペントン(図4H、配列8)、ChAd63ファイバー(図4I、配列10)およびChAd63ヘキソン(図4J、配列12)のコーディング核酸を表す。
【図4C】図4A−4Jは、ChAd3ファイバー(図4A、配列3)、ChAd3ヘキソン(図4B、配列5)、ChAd3ペントン(図4C、配列7)、ChAd63ファイバー(図4D、配列9)、ChAd63ヘキソン(図4E、配列11)のアミノ酸配列、ならびに、ChAd3ファイバー(図4F、配列4)、ChAd3ヘキソン(図4G、配列6)、ChAd3ペントン(図4H、配列8)、ChAd63ファイバー(図4I、配列10)およびChAd63ヘキソン(図4J、配列12)のコーディング核酸を表す。
【図4D】図4A−4Jは、ChAd3ファイバー(図4A、配列3)、ChAd3ヘキソン(図4B、配列5)、ChAd3ペントン(図4C、配列7)、ChAd63ファイバー(図4D、配列9)、ChAd63ヘキソン(図4E、配列11)のアミノ酸配列、ならびに、ChAd3ファイバー(図4F、配列4)、ChAd3ヘキソン(図4G、配列6)、ChAd3ペントン(図4H、配列8)、ChAd63ファイバー(図4I、配列10)およびChAd63ヘキソン(図4J、配列12)のコーディング核酸を表す。
【図4E】図4A−4Jは、ChAd3ファイバー(図4A、配列3)、ChAd3ヘキソン(図4B、配列5)、ChAd3ペントン(図4C、配列7)、ChAd63ファイバー(図4D、配列9)、ChAd63ヘキソン(図4E、配列11)のアミノ酸配列、ならびに、ChAd3ファイバー(図4F、配列4)、ChAd3ヘキソン(図4G、配列6)、ChAd3ペントン(図4H、配列8)、ChAd63ファイバー(図4I、配列10)およびChAd63ヘキソン(図4J、配列12)のコーディング核酸を表す。
【図4F】図4A−4Jは、ChAd3ファイバー(図4A、配列3)、ChAd3ヘキソン(図4B、配列5)、ChAd3ペントン(図4C、配列7)、ChAd63ファイバー(図4D、配列9)、ChAd63ヘキソン(図4E、配列11)のアミノ酸配列、ならびに、ChAd3ファイバー(図4F、配列4)、ChAd3ヘキソン(図4G、配列6)、ChAd3ペントン(図4H、配列8)、ChAd63ファイバー(図4I、配列10)およびChAd63ヘキソン(図4J、配列12)のコーディング核酸を表す。
【図4G】図4A−4Jは、ChAd3ファイバー(図4A、配列3)、ChAd3ヘキソン(図4B、配列5)、ChAd3ペントン(図4C、配列7)、ChAd63ファイバー(図4D、配列9)、ChAd63ヘキソン(図4E、配列11)のアミノ酸配列、ならびに、ChAd3ファイバー(図4F、配列4)、ChAd3ヘキソン(図4G、配列6)、ChAd3ペントン(図4H、配列8)、ChAd63ファイバー(図4I、配列10)およびChAd63ヘキソン(図4J、配列12)のコーディング核酸を表す。
【図4H】図4A−4Jは、ChAd3ファイバー(図4A、配列3)、ChAd3ヘキソン(図4B、配列5)、ChAd3ペントン(図4C、配列7)、ChAd63ファイバー(図4D、配列9)、ChAd63ヘキソン(図4E、配列11)のアミノ酸配列、ならびに、ChAd3ファイバー(図4F、配列4)、ChAd3ヘキソン(図4G、配列6)、ChAd3ペントン(図4H、配列8)、ChAd63ファイバー(図4I、配列10)およびChAd63ヘキソン(図4J、配列12)のコーディング核酸を表す。
【図4I】図4A−4Jは、ChAd3ファイバー(図4A、配列3)、ChAd3ヘキソン(図4B、配列5)、ChAd3ペントン(図4C、配列7)、ChAd63ファイバー(図4D、配列9)、ChAd63ヘキソン(図4E、配列11)のアミノ酸配列、ならびに、ChAd3ファイバー(図4F、配列4)、ChAd3ヘキソン(図4G、配列6)、ChAd3ペントン(図4H、配列8)、ChAd63ファイバー(図4I、配列10)およびChAd63ヘキソン(図4J、配列12)のコーディング核酸を表す。
【図4J】図4A−4Jは、ChAd3ファイバー(図4A、配列3)、ChAd3ヘキソン(図4B、配列5)、ChAd3ペントン(図4C、配列7)、ChAd63ファイバー(図4D、配列9)、ChAd63ヘキソン(図4E、配列11)のアミノ酸配列、ならびに、ChAd3ファイバー(図4F、配列4)、ChAd3ヘキソン(図4G、配列6)、ChAd3ペントン(図4H、配列8)、ChAd63ファイバー(図4I、配列10)およびChAd63ヘキソン(図4J、配列12)のコーディング核酸を表す。
【図5A】図5A−5Hは、ChAd3ΔE1,3,4,Ad5E4orf6NSmut−35,890bp(ChAd3NSmut、配列13)の核酸配列を表す。野生型ゲノムの欠失の組合せは、nt461からnt3541まで(3080bp)のEl欠失、nt28644からnt32633まで(3989bp)のE3欠失、nt34634からnt37349まで(2715bp)のE4欠失である。様々な領域は以下の場所に存在する:ChAd3左側ITR+パッケージングシグナル:nt1−460HCMVプロモーター:nt467−1257コザックコンセンサス配列:nt1263−1268HCV NS3−5B(BK菌株):nt1269−7223TAAターミネーター:nt7224−7226BGHポリA:nt7234−7452ChAd3主鎖:nt7468−35890Ad5E4orf6:nt34601−35482。
【図5B】図5A−5Hは、ChAd3ΔE1,3,4,Ad5E4orf6NSmut−35,890bp(ChAd3NSmut、配列13)の核酸配列を表す。野生型ゲノムの欠失の組合せは、nt461からnt3541まで(3080bp)のEl欠失、nt28644からnt32633まで(3989bp)のE3欠失、nt34634からnt37349まで(2715bp)のE4欠失である。様々な領域は以下の場所に存在する:ChAd3左側ITR+パッケージングシグナル:nt1−460HCMVプロモーター:nt467−1257コザックコンセンサス配列:nt1263−1268HCV NS3−5B(BK菌株):nt1269−7223TAAターミネーター:nt7224−7226BGHポリA:nt7234−7452ChAd3主鎖:nt7468−35890Ad5E4orf6:nt34601−35482。
【図5C】図5A−5Hは、ChAd3ΔE1,3,4,Ad5E4orf6NSmut−35,890bp(ChAd3NSmut、配列13)の核酸配列を表す。野生型ゲノムの欠失の組合せは、nt461からnt3541まで(3080bp)のEl欠失、nt28644からnt32633まで(3989bp)のE3欠失、nt34634からnt37349まで(2715bp)のE4欠失である。様々な領域は以下の場所に存在する:ChAd3左側ITR+パッケージングシグナル:nt1−460HCMVプロモーター:nt467−1257コザックコンセンサス配列:nt1263−1268HCV NS3−5B(BK菌株):nt1269−7223TAAターミネーター:nt7224−7226BGHポリA:nt7234−7452ChAd3主鎖:nt7468−35890Ad5E4orf6:nt34601−35482。
【図5D】図5A−5Hは、ChAd3ΔE1,3,4,Ad5E4orf6NSmut−35,890bp(ChAd3NSmut、配列13)の核酸配列を表す。野生型ゲノムの欠失の組合せは、nt461からnt3541まで(3080bp)のEl欠失、nt28644からnt32633まで(3989bp)のE3欠失、nt34634からnt37349まで(2715bp)のE4欠失である。様々な領域は以下の場所に存在する:ChAd3左側ITR+パッケージングシグナル:nt1−460HCMVプロモーター:nt467−1257コザックコンセンサス配列:nt1263−1268HCV NS3−5B(BK菌株):nt1269−7223TAAターミネーター:nt7224−7226BGHポリA:nt7234−7452ChAd3主鎖:nt7468−35890Ad5E4orf6:nt34601−35482。
【図5E】図5A−5Hは、ChAd3ΔE1,3,4,Ad5E4orf6NSmut−35,890bp(ChAd3NSmut、配列13)の核酸配列を表す。野生型ゲノムの欠失の組合せは、nt461からnt3541まで(3080bp)のEl欠失、nt28644からnt32633まで(3989bp)のE3欠失、nt34634からnt37349まで(2715bp)のE4欠失である。様々な領域は以下の場所に存在する:ChAd3左側ITR+パッケージングシグナル:nt1−460HCMVプロモーター:nt467−1257コザックコンセンサス配列:nt1263−1268HCV NS3−5B(BK菌株):nt1269−7223TAAターミネーター:nt7224−7226BGHポリA:nt7234−7452ChAd3主鎖:nt7468−35890Ad5E4orf6:nt34601−35482。
【図5F】図5A−5Hは、ChAd3ΔE1,3,4,Ad5E4orf6NSmut−35,890bp(ChAd3NSmut、配列13)の核酸配列を表す。野生型ゲノムの欠失の組合せは、nt461からnt3541まで(3080bp)のEl欠失、nt28644からnt32633まで(3989bp)のE3欠失、nt34634からnt37349まで(2715bp)のE4欠失である。様々な領域は以下の場所に存在する:ChAd3左側ITR+パッケージングシグナル:nt1−460HCMVプロモーター:nt467−1257コザックコンセンサス配列:nt1263−1268HCV NS3−5B(BK菌株):nt1269−7223TAAターミネーター:nt7224−7226BGHポリA:nt7234−7452ChAd3主鎖:nt7468−35890Ad5E4orf6:nt34601−35482。
【図5G】図5A−5Hは、ChAd3ΔE1,3,4,Ad5E4orf6NSmut−35,890bp(ChAd3NSmut、配列13)の核酸配列を表す。野生型ゲノムの欠失の組合せは、nt461からnt3541まで(3080bp)のEl欠失、nt28644からnt32633まで(3989bp)のE3欠失、nt34634からnt37349まで(2715bp)のE4欠失である。様々な領域は以下の場所に存在する:ChAd3左側ITR+パッケージングシグナル:nt1−460HCMVプロモーター:nt467−1257コザックコンセンサス配列:nt1263−1268HCV NS3−5B(BK菌株):nt1269−7223TAAターミネーター:nt7224−7226BGHポリA:nt7234−7452ChAd3主鎖:nt7468−35890Ad5E4orf6:nt34601−35482。
【図5H】図5A−5Hは、ChAd3ΔE1,3,4,Ad5E4orf6NSmut−35,890bp(ChAd3NSmut、配列13)の核酸配列を表す。野生型ゲノムの欠失の組合せは、nt461からnt3541まで(3080bp)のEl欠失、nt28644からnt32633まで(3989bp)のE3欠失、nt34634からnt37349まで(2715bp)のE4欠失である。様々な領域は以下の場所に存在する:ChAd3左側ITR+パッケージングシグナル:nt1−460HCMVプロモーター:nt467−1257コザックコンセンサス配列:nt1263−1268HCV NS3−5B(BK菌株):nt1269−7223TAAターミネーター:nt7224−7226BGHポリA:nt7234−7452ChAd3主鎖:nt7468−35890Ad5E4orf6:nt34601−35482。
【図6A】図6A−6Hは野生型ChAd3の核酸配列(配列14)を示す。
【図6B】図6A−6Hは野生型ChAd3の核酸配列(配列14)を示す。
【図6C】図6A−6Hは野生型ChAd3の核酸配列(配列14)を示す。
【図6D】図6A−6Hは野生型ChAd3の核酸配列(配列14)を示す。
【図6E】図6A−6Hは野生型ChAd3の核酸配列(配列14)を示す。
【図6F】図6A−6Hは野生型ChAd3の核酸配列(配列14)を示す。
【図6G】図6A−6Hは野生型ChAd3の核酸配列(配列14)を示す。
【図6H】図6A−6Hは野生型ChAd3の核酸配列(配列14)を示す。
【図7A】図7A−7Hは野生型ChAd63の核酸配列(配列15)を示す。
【図7B】図7A−7Hは野生型ChAd63の核酸配列(配列15)を示す。
【図7C】図7A−7Hは野生型ChAd63の核酸配列(配列15)を示す。
【図7D】図7A−7Hは野生型ChAd63の核酸配列(配列15)を示す。
【図7E】図7A−7Hは野生型ChAd63の核酸配列(配列15)を示す。
【図7F】図7A−7Hは野生型ChAd63の核酸配列(配列15)を示す。
【図7G】図7A−7Hは野生型ChAd63の核酸配列(配列15)を示す。
【図7H】図7A−7Hは野生型ChAd63の核酸配列(配列15)を示す。
【図8】ChAd3NSmut(配列13)およびMRKAd6NSmutに感染したHeLa細胞中のHCV NSタンパク質の発現の比較を示す。
【図9】C57/B6マウス体内で細胞性免疫を誘導するChAd3NSmut(配列13)およびMRKAd6NSmutの能力の比較を示す。
【図10A】図10A−10HはChAd63ΔE1,3,4,Ad5E4orf6,NSmut(配列17)の核酸配列を示す。野生型ゲノムの欠失の組合せは、nt455からnt3421まで(2967bp)のEl欠失、nt27207からnt31778まで(4582bp)のE3欠失、nt33825からnt36215まで(2390bp)のE4欠失である。様々な領域は以下の場所に存在する:ChAd63左側ITR+パッケージングシグナル:nt1−454HCMVプロモーター:nt458−1248コザックコンセンサス配列:nt1254−1259HCV NS3−5B(BK菌株):nt1260−7214TAAターミネーター:nt7215−7217BGHポリA:nt7227−7447ChAd63主鎖:nt7458−34658Ad5E4orf6:nt33316−34197。
【図10B】図10A−10HはChAd63ΔE1,3,4,Ad5E4orf6,NSmut(配列17)の核酸配列を示す。野生型ゲノムの欠失の組合せは、nt455からnt3421まで(2967bp)のEl欠失、nt27207からnt31778まで(4582bp)のE3欠失、nt33825からnt36215まで(2390bp)のE4欠失である。様々な領域は以下の場所に存在する:ChAd63左側ITR+パッケージングシグナル:nt1−454HCMVプロモーター:nt458−1248コザックコンセンサス配列:nt1254−1259HCV NS3−5B(BK菌株):nt1260−7214TAAターミネーター:nt7215−7217BGHポリA:nt7227−7447ChAd63主鎖:nt7458−34658Ad5E4orf6:nt33316−34197。
【図10C】図10A−10HはChAd63ΔE1,3,4,Ad5E4orf6,NSmut(配列17)の核酸配列を示す。野生型ゲノムの欠失の組合せは、nt455からnt3421まで(2967bp)のEl欠失、nt27207からnt31778まで(4582bp)のE3欠失、nt33825からnt36215まで(2390bp)のE4欠失である。様々な領域は以下の場所に存在する:ChAd63左側ITR+パッケージングシグナル:nt1−454HCMVプロモーター:nt458−1248コザックコンセンサス配列:nt1254−1259HCV NS3−5B(BK菌株):nt1260−7214TAAターミネーター:nt7215−7217BGHポリA:nt7227−7447ChAd63主鎖:nt7458−34658Ad5E4orf6:nt33316−34197。
【図10D】図10A−10HはChAd63ΔE1,3,4,Ad5E4orf6,NSmut(配列17)の核酸配列を示す。野生型ゲノムの欠失の組合せは、nt455からnt3421まで(2967bp)のEl欠失、nt27207からnt31778まで(4582bp)のE3欠失、nt33825からnt36215まで(2390bp)のE4欠失である。様々な領域は以下の場所に存在する:ChAd63左側ITR+パッケージングシグナル:nt1−454HCMVプロモーター:nt458−1248コザックコンセンサス配列:nt1254−1259HCV NS3−5B(BK菌株):nt1260−7214TAAターミネーター:nt7215−7217BGHポリA:nt7227−7447ChAd63主鎖:nt7458−34658Ad5E4orf6:nt33316−34197。
【図10E】図10A−10HはChAd63ΔE1,3,4,Ad5E4orf6,NSmut(配列17)の核酸配列を示す。野生型ゲノムの欠失の組合せは、nt455からnt3421まで(2967bp)のEl欠失、nt27207からnt31778まで(4582bp)のE3欠失、nt33825からnt36215まで(2390bp)のE4欠失である。様々な領域は以下の場所に存在する:ChAd63左側ITR+パッケージングシグナル:nt1−454HCMVプロモーター:nt458−1248コザックコンセンサス配列:nt1254−1259HCV NS3−5B(BK菌株):nt1260−7214TAAターミネーター:nt7215−7217BGHポリA:nt7227−7447ChAd63主鎖:nt7458−34658Ad5E4orf6:nt33316−34197。
【図10F】図10A−10HはChAd63ΔE1,3,4,Ad5E4orf6,NSmut(配列17)の核酸配列を示す。野生型ゲノムの欠失の組合せは、nt455からnt3421まで(2967bp)のEl欠失、nt27207からnt31778まで(4582bp)のE3欠失、nt33825からnt36215まで(2390bp)のE4欠失である。様々な領域は以下の場所に存在する:ChAd63左側ITR+パッケージングシグナル:nt1−454HCMVプロモーター:nt458−1248コザックコンセンサス配列:nt1254−1259HCV NS3−5B(BK菌株):nt1260−7214TAAターミネーター:nt7215−7217BGHポリA:nt7227−7447ChAd63主鎖:nt7458−34658Ad5E4orf6:nt33316−34197。
【図10G】図10A−10HはChAd63ΔE1,3,4,Ad5E4orf6,NSmut(配列17)の核酸配列を示す。野生型ゲノムの欠失の組合せは、nt455からnt3421まで(2967bp)のEl欠失、nt27207からnt31778まで(4582bp)のE3欠失、nt33825からnt36215まで(2390bp)のE4欠失である。様々な領域は以下の場所に存在する:ChAd63左側ITR+パッケージングシグナル:nt1−454HCMVプロモーター:nt458−1248コザックコンセンサス配列:nt1254−1259HCV NS3−5B(BK菌株):nt1260−7214TAAターミネーター:nt7215−7217BGHポリA:nt7227−7447ChAd63主鎖:nt7458−34658Ad5E4orf6:nt33316−34197。
【図10H】図10A−10HはChAd63ΔE1,3,4,Ad5E4orf6,NSmut(配列17)の核酸配列を示す。野生型ゲノムの欠失の組合せは、nt455からnt3421まで(2967bp)のEl欠失、nt27207からnt31778まで(4582bp)のE3欠失、nt33825からnt36215まで(2390bp)のE4欠失である。様々な領域は以下の場所に存在する:ChAd63左側ITR+パッケージングシグナル:nt1−454HCMVプロモーター:nt458−1248コザックコンセンサス配列:nt1254−1259HCV NS3−5B(BK菌株):nt1260−7214TAAターミネーター:nt7215−7217BGHポリA:nt7227−7447ChAd63主鎖:nt7458−34658Ad5E4orf6:nt33316−34197。
【図11】IFN−γ ELISPOTによって測定した免疫応答の時間経過を、いずれか所与の時点に種々のHCV NSペプチドプールで観察した応答の和として表す。
【図12】pV1JnsNSOPTmut 3a−1bの種々の構成要素を示す。
【図13】キメラHCVポリペプチドをコードするpV1JnsNSOPTmut 3a−lb発現カセットの遺伝子構造を示す。様々な領域は以下の場所に存在する:−ヒトCMVプロモーター:318−905;−イントロンA:1040−1865;−コザック配列:1885−1894;−HCV MetNS3−NS4A(遺伝子型3a):1891−3960;−HCV MetNS3−NS5BOPTmut(遺伝子型1b):3961−9915;−TAAAターミネーター:9916−9919;−BGHポリA:9965−10182。
【図14】キメラプラスミド(pV1Jns−NSOPTmut 3a−1b)またはpV1Jns−NSOPTmutで免疫感作した動物(CD1マウス)体内のHCV 1bおよび3a由来のNS3タンパク質に応答してIFN−γを分泌するT細胞の数を示す(脾細胞1,000,000個あたりのスポット形成細胞として表す)。
【背景技術】
【0001】
本願に引用した参考文献は特許請求の範囲に記載の発明に対する先行技術であると認められたものではない。
【0002】
全世界の人口の約3%がC型肝炎ウイルス(HCV)に感染している。(Wasleyら,Semin.Liver Dis.20,1−16,2000)。HCVに接触して明白な急性疾患が発症するパーセンテージは低いが、多くの場合にウイルスが慢性感染として定着し、肝臓の炎症を生じさせ、ゆっくりと肝不全および肝硬変に進行する。(Iwarson,FEMS Microbiol.Rev.14,201−204,1994)。さらに、疫学調査は、HCVが肝細胞癌の病理発生に重要な役割を演じることを示している。(Kew,FEMS Microbiol.Rev.14,211−220,1994,Alter,Blood 85,1681−1695,1995)。
【0003】
HCVに対する定例の血液スクリーニングが実施されるようになった1992年以前には、たいていの感染は、汚染血液、血液製品または移植臓器への迂闊な接触が原因となっていた。HCVの血液スクリーニングが実施されている地域では、HCVは主として静脈内薬物使用によって感染する。頻度は少ないが、周産期接触、血液透析およびHCV感染者との性的接触なども伝播方法に含まれる。(Alterら,N.Engl.J.Med.341(8),556−562,1999,Alter,J.Hepatol.31 Suppl.88−91,1999,Wasleyら,Semin.Liver.Dis.201,1−16,2000)。
【0004】
HCVゲノムは、約3000アミノ酸の前駆体ポリタンパク質をコードする約9.5kbの一本鎖RNAから成る。(Chooら,Science 244,362−364,1989,Chooら,Science 244,359−362,1989,Takamizawaら,J.Virol 65,1105−1113,1991)。HCVポリタンパク質はC−El−E2−p7−NS2−NS3−NS4A−NS4B−NS5A−NS5Bという序列のウイルスタンパク質を含有する。
【0005】
CMI応答を生じさせるために1つ以上のHCV非構造抗原を提供するHCV核酸配列を使用することは、例えば、Choら,Vaccine 77:1136−1144,1999;Paliardら,国際公開WO 01/30812;Coitら,国際公開WO 01/38360;およびEminiら,国際公開WO 03/031588によって言及されている。
【発明の開示】
【0006】
本発明の主題は、HCV核酸ワクチンおよびワクチン成分として、または、HCVワクチンの製造に使用できる核酸構築物である。記載した構築物は、(1)第一のHCV菌株に基づくNS3−4A領域と第二の菌株に基づくNS3−NS4A−NS4B−NS5AまたはNS3−NS4A−NS4B−NS5A−NS5B領域を含有するキメラHCVポリペプチドをコードする核酸、および、(2)チンパンジーに基づくHCVポリペプチドをコードするアデノベクターを含む。
【0007】
従って、本発明の第一の目的は、HCVキメラポリペプチドをコードするヌクレオチド配列を含む核酸配列を記載する。該ポリペプチドは、第一のHCV菌株に由来のHCV NS3−4aに実質的に相似のアミノ酸配列を含むHCV NS3−4a領域と、第二のHCV菌株に由来のHCV NS3−NS4A−NS4B−NS5Aに実質的に相似のアミノ酸配列を含むHCV NS3−NS4A−NS4B−NS5A領域とを含む。この場合、2つの領域に存在するNS3−4A配列は異なる配列を有している。第一領域は第二領域のアミノ側またはカルボキシル側に配置されている。
【0008】
アミノ酸配列に関する“実質的に相似の配列”という表現は、基準配列に対する少なくとも約70%の一致を意味する。基準配列に対するパーセント一致(一致パーセントと表すこともある)を決定するためには、最大数の一致アミノ酸が得られるようにポリペプチド領域を対応する基準領域に位置合せし、対応領域の一致アミノ酸の数を測定する。この数を基準領域のアミノ酸の総数で除算し、次いで100を乗算し、最も近い整数に丸める。例えば、HCV NS3−4A、NS3−NS4A−NS4B−NS5AおよびNS3−NS4A−NS4B−NS5A−NS5Bの領域が、異なるHCV菌株に存在する対応HCV領域でもよい。
【0009】
第一および第二の領域に異なるNS3−4A配列が存在することは、1つ以上のアミノ酸相違をもたらす。アミノ酸相違はおのおの独立に付加、置換または欠失である。
【0010】
好ましい実施態様では、核酸が、コードされたHCVポリペプチドをヒト細胞中で発現できる発現ベクターである。ヒト細胞中の発現は、HCV感染の能動的治療およびHCV感染に対する予防的処置という治療的用途を有している。
【0011】
発現ベクターは、ポリペプチドをコードするヌクレオチド配列を適正な転写およびプロセシング用の調節要素と共に含有する。存在し得る調節要素は、ポリペプチドをコードするヌクレオチド配列に天然に連携している調節要素およびヌクレオチド配列に天然に連携していない外来調節要素を含む。外来プロモーターのような外来調節要素はヒト細胞のような特定宿主中の発現に使用できる。
【0012】
好ましい発現ベクターは、1つ以上の欠失領域を有する組換えアデノウイルスゲノムである。組換えアデノウイルスゲノムは、1つ以上のアデノウイルス血清型に実質的に相似の種々の領域を含有し得る。欠失領域という表現は、指定した領域の全部または一部の欠失を表す。
【0013】
本発明の別の目的は、
a)HCV NS3−NS4A−NS4B−NS5Aを含むHCVポリペプチドをコードしている発現カセットと、
b)E1欠失、E3欠失および場合によりE4欠失を含む組換えアデノウイルスゲノムと、
を含み、該アデノウイルスゲノムが、(i)配列3または9に実質的に相似のアミノ酸配列をもつファイバー領域、(ii)配列5または11に実質的に相似のアミノ酸配列をもつヘキソン領域、および、(iii)配列7に実質的に相似のアミノ酸配列をもつペントン領域
の少なくとも1つをコードしている組換えアデノベクターを記載する。
【0014】
発現カセットはE1またはE3欠失に配置される。発現カセットの全部または一部が欠失領域に対応する位置に存在するならば発現カセットはE1またはE3欠失に配置されたと考えられる。
【0015】
HCVポリペプチドまたはアミノ酸配列という表現は、天然産生HCV配列、天然産生配列または指定配列に実質的に相似の天然産生配列の誘導体、および、キメラHCVポリペプチドを含む。キメラHCVポリペプチドは、上述の本発明の第一の目的に記載のHCVポリペプチドを含む。
【0016】
本発明の別の目的は組換えアデノウイルス粒子を記載する。該粒子は、本文中に記載の組換えアデノウイルスゲノムによってコードされ、該ゲノムのコピーをパッケージしている。
【0017】
本発明の別の目的は、(a)組換えアデノウイルスゲノムを発現させるためにE1相補細胞を使用して粒子を製造する段階と、(b)該粒子を実質的に精製する段階とを含む組換えアデノウイルス粒子の製造方法を記載する。粒子を実質的に精製するという表現は、粒子の製造に使用した細胞および細胞破片を完全にまたはほぼ完全に除去することを意味する。
【0018】
本発明の別の目的は、治療有効量のHCVポリペプチドをコードする核酸と医薬的に許容される担体とを含む医薬組成物を記載する。
【0019】
本発明の別の目的は、治療有効量のHCVポリペプチドをコードする核酸を患者に投与する段階を含む患者の治療方法を記載する。
【0020】
“治療する”という表現は、HCV感染患者の治療または活動性HCV感染の可能性もしくは重篤度を低下させるための患者の予防的処置を含む。“患者”という用語は、HCV感染の可能性のある哺乳動物を表す。患者はHCV感染している場合もあり非感染の場合もある。患者の例はヒトおよびチンパンジーである。
【0021】
HCV感染患者の場合、有効量は、以下の効果、すなわち、HCVの複製能力を低下させる効果、HCV荷重を減少させる効果、ウイルスクリアランスを増加させる効果、および、1つ以上のHCV特異的CMI応答を増加させる効果の1つ以上を発揮するために十分な量である:。
【0022】
HCV非感染患者の場合、有効量は、以下の効果、すなわち、HCV感染に対するHCV特異的CMI応答の1つ以上の成分の発生能力を増加させる効果、HCV感染に対する感受性を低下させる効果、および、慢性疾患に至る持続感染を定着させる感染ウイルスの能力を低下させる効果の1つ以上を達成するために十分な量である:。
【0023】
“含む”のような無制限的用語は、追加の要素または段階を許容する。追加の要素または段階の可能性を強調するために、ときには、“1つ以上”のような成句を無制限的用語と共にまたは単独で使用する。
【0024】
明白な記述がなければ、単数を表す不定冠詞は1に限定されない。例えば、“1つの細胞”は“複数の細胞”を排除しない。ときには、複数の存在が可能であることを強調するために、1つ以上という熟語を使用する。
【0025】
本発明のその他の特徴および利点は、様々な実施例を含む本文中の以後の記載より明らかであろう。与えられた実施例は本発明の実施に役立つ様々な構成要素および方法を例示する。実施例は特許請求の範囲に記載の本発明を限定しない。この開示に基づいて当業者は本発明の実施に役立つ他の構成要素および方法を鑑定し使用できるであろう。
【0026】
本発明は、種々のHCV菌株に基づくHCV非構造タンパク質を供給するHCVキメラポリペプチドをコードする核酸、および、HCVポリペプチドを発現するチンパンジーアデノベクターの使用を含む。記載した核酸の使用は、HCVに対するCMI応答を生じさせる広範囲の抗原を提供するHCVポリペプチドを細胞に導入するためのワクチン成分としての使用、および、このようなワクチン成分を製造する中間体としての使用を含む。
【0027】
適応型細胞性免疫応答は、主要組織適合複合体(MHC)クラスIおよびIIの遍在的分布によって全身のHCV感染細胞のウイルス抗原を認識し、免疫記憶を誘導し、免疫記憶を維持する機能を果たし得る。これらの機能を果たすのは、抗原特異的CD4+Tヘルパー細胞(Th)およびCD8+細胞障害性T細胞(CTL)である。
【0028】
特異的T細胞受容体を介して活性化されると、HCV特異的Th細胞は多様な免疫調節機能を発揮するが、たいていの機能にはTh1およびTh2サイトカインが介在する。HCV特異的Th細胞は、B細胞の活性化および分化ならびにウイルス特異的細胞障害性T細胞の誘導および刺激を助ける。Th細胞はまたCTLと共に、数種類のウイルスの複製および遺伝子発現を阻害するIFN−γおよびTNF−αを分泌し得る。さらに、Th細胞およびCTLは主要なエフェクター細胞であり、ウイルス感染細胞のアポトーシスおよび溶解を誘導できる。
【0029】
HCV特異的CTLは、専門的抗原提示細胞(pAPC)によってプロセシングされた抗原に応答して生じる。抗原はpAPC内部で合成されるかまたはpAPC導入される。抗原をコードする発現カセットを細胞に導入することによってpAPC内部で抗原を合成することができる。
【0030】
核酸ワクチンの好ましい投与経路は筋肉内経路である。筋肉内投与によって核酸が体細胞およびpAPCに導入され発現されると考えられる。体細胞で産生されたHCV抗原はpAPCに輸送され、MHCクラスI分子に関係して抗原提示される。(Donnellyら,Annu.Rev.Immunol.75:617−648,1997)。
【0031】
pAPCはプロテアソーム複合体において長い抗原をより短いペプチド抗原にプロセシングする。該抗原は小胞体/ゴルジ複合体分泌経路に転送されてMHCクラスIタンパク質に会合する。CD8+Tリンパ球は、MHCクラスIに会合した抗原をT細胞受容体(TCR)およびCD8細胞表面タンパク質を介して認識する。
【0032】
チンパンジーアデノウイルスに基づくベクターはHCV抗原導入ビヒクルとしてヒトアデノベクターに代替して使用できる。チンパンジーアデノウイルスに基づくベクターは多重免疫戦略で使用するのが特に有効である。この場合、治療される患者の体内では初回に使用したアデノベクターへの免疫応答が生じている。同種のアデノウイルスに基づくベクターとの接触が繰返されると、アデノウイルスタンパク質に対する免疫応答が原因でワクチンの有効性が低下するであろう。初回接触がアデノウイルス感染の結果という場合もあるが、このような感染は恐らく、HCV以外の疾患を治療するためにアデノベクターを使用したことによって生じたものであろう。
【0033】
ここに与える指針に基づいて、患者体内で有益な効果を発揮する十分に強力な免疫応答を発生できる。これらの指針は、HCV配列の選択、ベクター選択、ベクター産生、併用治療および投与に関する情報を含む。
【0034】
I.キメラ配列
種々のNS3−4A配列をもつ領域を提供するHCVキメラポリペプチドは、種々のHCV菌株を標的とする抗原を提供するワクチン成分として使用できる。HCVの主な特徴はそのゲノムの不均質性である。(Pawlotsky,Clin.Liver Dis,7:45−66,2003,Simmonds,J.General.Virol,85:3173−3188,2004)。さらに、RNAウイルスに共通の典型的な誤りがちのRNA−依存性RNAポリメラーゼおよび不十分なプルーフリーディング活性から生じた突然変異ならびに宿主から生じた突然変異は、HCVが偽種と呼ばれる複合ウイルス集団として宿主体内で循環する原因となる。(Blightら,Science 290:1972−1974,2000)。
【0035】
キメラ構築物の異なる領域に存在する異なるNS3−4A配列は、1つ以上のアミノ酸の違いとして表れる。アミノ酸の違いはおのおの独立に付加、置換または欠失である。種々の実施態様で、第一および第二の領域のNS3−4A配列は、少なくとも約5%、少なくとも10%、少なくとも15%だけ違っている。または、少なくとも1、5、10、15、20または25個のアミノ酸の変更が存在する。NS3−4A領域はNS3−4A配列に加えて、アミノ末端メチオニンのような付加アミノ酸および/または導入された開裂部位を含み得る。
【0036】
相違パーセントは配列一致を100から減算することによって決定できる。例えば、85%配列一致は15%相違を与える。
【0037】
キメラポリペプチドに存在する第一領域および第二ポリペプチド領域は、天然産生HCV単離物の多くの例に基づいて容易に作製できる。HCV単離物は1つ以上のサブタイプを含む以下の6つの主要遺伝子型に分類できる:HCV−1/(1a,1b,1c)、HCV−2/(2a,2b,2c)、HCV−3/(3a,3b,10a)、HCV−4/(4a)、HCV−5/(5a)およびHCV−6/(6a,6b,7b,8b,9a,11a)。(Simmonds,J.Gen.Virol,693−712,2001)。HCV−BK、HCV−J、HCV−NおよびHCV−Hのような特定のHCV配列はGenBankに寄託され種々の刊行物に記載されている。(例えば、Chamberlainら,J.Gen.Virol,1341−1347,1997)。
【0038】
好ましくは、第一領域および第二領域の双方がHCVプロテアーゼによってインビボでプロセシングされ、HCVキメラポリペプチドに存在する個々のタンパク質に対応する個々のタンパク質を提供する。個々のHCVタンパク質は細胞によってさらにプロセシングされ得る。
【0039】
第一領域に関する種々の実施態様では、該領域が、以下に存在するNS3−NS4A領域に実質的に相似のアミノ酸配列であるかまたは該アミノ酸配列を内包する:配列1のアミノ酸1−686;HCV 1a(Acc.No.M62321);HCV 2a(Acc.No.D00944;HCV 3a(Acc.No:D28917);HCV 4a(Acc.No:Y11604);HCV 5a(Acc.No:Y13184)またはHCV 6a(Acc.No:D84264)。
【0040】
第二領域に関する種々の実施態様では、該領域が、以下に存在する対応領域に実質的に相似のNS3−NS4A−NS4B−NS5AまたはNS3−NS4A−NS4B−NS5A−NS5B*配列であるかまたは該配列を内包する:配列1のアミノ酸686−2675、アミノ酸691−2675またはアミノ酸692−2675;HCV 1a(Acc.No.M62321);HCV 2a(Acc.No.D00944;HCV 3a(Acc.No:D28917);HCV 4a(Acc.No:Y11604);HCV 5a(Acc.No:Y13184)またはHCV 6a(Acc.No:D84264)。“NS5B*”という表記は不活性NS5Bを表す。
【0041】
好ましくは第二領域が、第一領域由来のプロテアーゼ活性に適合性のアミノ酸開裂部位を含む。開裂部位は既知の開裂配列に基づいて付加できる。
【0042】
アミノ酸配列に関する“実質的に相似の配列”という表現は、基準配列に対する少なくとも約70%の一致を意味する。基準配列に対するパーセント一致(一致パーセントと表すこともある)を決定するためには、最大数の一致アミノ酸が得られるようにポリペプチド領域を対応する基準領域に位置合せし、対応領域の一致アミノ酸の数を測定する。この数を基準領域のアミノ酸の総数で除算し、次いで100を乗算し、最も近い整数に丸める。
【0043】
種々の実施態様で、実質的に相似の配列は、少なくとも75%、少なくとも80%、少なくとも85%、少なくとも90%、少なくとも95%の一致を有しているか;または、0、1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19もしくは20個のアミノ酸の変更だけ違っている。変更はおのおの独立に挿入、置換または付加である。
【0044】
天然産生HCV配列に修飾を生じさせて実質的に相似の種々の配列を得ることもできる。天然産生アミノ酸の違いはアミノ酸側鎖(R基)の違いに起因する。R基は、物理的寸法、電荷および疎水性のようなアミノ酸の種々の特性を左右する。アミノ酸は以下のような種々のグループに分類できる:中性かつ疎水性(アラニン、バリン、ロイシン、イソロイシン、プロリン、トリプトファン、フェニルアラニン、メチオニン);中性かつ極性(グリシン、セリン、トレオニン、チロシン、システイン、アスパラギン、グルタミン);塩基性(リシン、アルギニン、ヒスチジン);酸性(アスパラギン酸、グルタミン酸)。
【0045】
活性を維持するために種々のアミノ酸を置換する場合、一般には同様の特性を有しているアミノ酸で置換するのが好ましい。バリンでロイシンを、アルギニンでリシンを、アスパラギンでグルタミンを置換するような特定グループ内部の異なるアミノ酸の置換が、ポリペプチド三次構造の変化を生じない良好な候補である。
【0046】
アミノ酸修飾はT細胞抗原領域を好ましく維持または追加する。広範囲のT細胞応答を誘発できるポリペプチドを産生するために天然産生HCVポリペプチドに様々な修飾を行うことが可能である。広いT細胞応答を誘発するポリペプチドの能力を左右する要因は、HCV特異的T細胞抗原の保存または導入、および、種々のHCV単離物中の種々のT細胞抗原領域の存在数である。
【0047】
HCV T細胞抗原は例えば実験作業によって同定できる。T細胞抗原を同定する1つの方法は、長いポリペプチドから一連の短いオーバーラップペプチドを作製し、次いで、感染患者に由来のT細胞集団を陽性クローンに基づいてスクリーニングする段階を含む。陽性クローンは特定のペプチドによって活性化/プライムされる。IFNγ−ELISPOT、IFNγ−細胞内染色およびバルクCTLアッセイのような技術がペプチド活性の測定に使用できる。このようにして同定したペプチドは、それぞれの病原体のT細胞エピトープを表すと考えることができる。
【0048】
自己をプロセシングしてCMI応答を生じるHCVポリペプチドの能力は、ここに記載の技術または当業界で公知の技術を使用して判定できる。(参照:例えばEminiら,国際公開WO 03/031588)。このような技術は、HCV特異的CMI応答を測定するためのIFNγ−ELISPOT、IFNγ−細胞内染色およびバルクCTLアッセイの使用を含む。
【0049】
複製に必須のモチーフを標的組換えすることによって不活性ポリメラーゼを産生するためにNS5Bに小さい修飾を行ってもよい。NS5B活性に必須のモチーフおよび不活性NS5Bを産生するために行うことができる修飾の例は、Lohmannら,Journal of Virology 77:8416−8426,1997、Kolykhalovら,Journal of Virology 74:2046−2051,2000およびEminiら,国際公開WO 03/031588に記載されている。
【0050】
キメラHCVポリペプチドを修飾するときに考慮すべき追加要因は、自己プロセシング能力の維持およびT細胞抗原の維持を含む。ポリペプチドの自己プロセシング能力は機能性プロテアーゼ活性によってほぼ判定できる。プロテアーゼ活性を維持する修飾は、NS3プロテアーゼ、NS3の補因子として役立つNS4A、および、HCVポリペプチド内部に存在するプロテアーゼ認識部位を考慮することによって得られる。
【0051】
II.キメラNS31−NS4A1−NS32−NS4A2−NS4B2−NS5A2−NS5B2*
好ましいキメラHCVポリペプチドは、NS31−NS4A1−NS32−NS4A2−NS4B2−NS5A2−NS5B2*である。下付き文字1および2は、種々のHCV菌株中の対応する領域に実質的に相似の配列をもつ領域を表し、“NS5B*”は酵素的に不活性のNS5Bを表す。NS31−NS4A1とNS32−NS4A2とは少なくとも1つのアミノ酸が違っている。種々の実施態様で、NS31−NS4A1とNS32−NS4A2とは少なくとも約5%、少なくとも約10%、少なくとも約15%違っている;または、少なくとも1、5、10、15、20または25個のアミノ酸の変更が存在する。
【0052】
好ましくは、NS31−NS4A1−NS32−NS4A2−NS4B2−NS5A2−NS5B2*ポリペプチドは、第一および第二領域のおのおのが1つ以上の個別HCVペプチドを産生するために十分なインビボのプロテアーゼ活性を提供する。好ましい実施態様で、ポリペプチドは、NS31、NS4A1、NS32、NS4A2、NS4B2、NS5A2、NS5B2*を個別ペプチドとして産生できる。
【0053】
種々のNS31−NS4A1およびNS32−NS4A2−NS4B2−NS5A2−NS5B2*領域は上述の項Iに記載のようにして供給される。種々の実施態様で、NS31−NS4A1領域は、配列1のアミノ酸1−686もしくはアミノ酸2−686に実質的に相似の配列であるかまたは該配列を内包する。NS32−NS4A2−NS4B2−NS5A2−NS5B2*領域は、配列1のアミノ酸687−2675、691−2675もしくは692−2675に実質的に相似の配列であるかまたは該配列を含む。または、NS31−NS4A1−NS32−NS4A2−NS4B2−NS5A2−NS5B2*が全体として配列1に実質的に相似である。種々の実施態様で、各領域は配列1の対応する領域に、少なくとも75%、少なくとも80%、少なくとも85%、少なくとも90%、少なくとも95%だけ実質的に相似であるか、または、0、1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19もしくは20個のアミノ酸の変更だけ違っている。
【0054】
III.遺伝子発現カセット
ポリペプチドをコードする遺伝子発現カセットは、ポリペプチド発現に必要な要素を含む。“ポリペプチド”という表現はキメラポリペプチドを含意する。
【0055】
III.A.コードされたポリペプチド配列
本文中に記載のHCVポリペプチドをコードした配列は種々のベクターに使用できる。特定例は、項IおよびIIに前述したようなキメラポリペプチド配列、および、配列16に実質的に相似の配列のようなHCVポリペプチド配列Met−NS3−NS4A−NS4B−NS5A−NS5B*を含む(参照:Eminiら,国際公開WO 03/031588)。配列16に実質的に相似の配列は、例えば項Vに後述するようなチンパンジーベクターに結合して使用できる。
【0056】
様々な実施態様で、配列16に実質的に相似の配列は、配列16に75%、少なくとも80%、少なくとも85%、少なくとも90%、少なくとも95%の配列一致を有しているか、または、0、1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19もしくは20個のアミノ酸の変更だけ違っている。
【0057】
III.B.コードされたポリペプチド配列
遺伝子発現カセット中に存在する調節要素は一般に、(a)ポリペプチドをコードするヌクレオチド配列に転写可能に連結されたプロモーターと、(b)ヌクレオチド配列に機能的に連結された5’リボソーム結合部位と、(c)ヌクレオチド配列の3’端に接合されたターミネーターと、(d)ヌクレオチド配列に機能的に連結された3’ポリアデニル化シグナルとを含む。遺伝子発現またはポリペプチドプロセシングに役立つ追加の調節要素が存在してもよい。
【0058】
プロモーターは、RNAポリメラーゼによって認識され、下流領域の転写を媒介する遺伝子要素である。好ましいプロモーターは、高い転写レベルを与える強プロモーターである。強プロモーターの例は、前初期ヒトサイトメガロウイルスプロモーター(CMV)およびイントロンAをもつCMVである。(Chapmanら,Nucl.Acids Res.19:3979−3986,1991)。プロモーターの追加例は、EF1アルファプロモーター、ネズミCMVプロモーター、ラウス肉腫ウイルスプロモーター、SV40初期/後期プロモーターおよびβ−アクチンプロモーターのような天然産生プロモーター、ならびに、合成の筋特異的プロモーター、キメラ筋特異的/CMVプロモーターのような人工プロモーターである。(Liら,Nat.Biotechnol.77:241−245,1999,Hagstromら,Blood 95:2536−2542,2000)。
【0059】
リボソーム結合部位は開始コドンまたはその近傍に局在する。リボソーム結合部位の例は、CCACCAUGG、CCGCCAUGGおよびACCAUGGを含み、ここにAUGは開始コドンである。(Kozak,Cell 44:283−292,1986)。リボソーム結合部位の別の例は、配列18によって示される。
【0060】
ポリアデニル化シグナルは、転写RNAを開裂しRNAにポリ(A)テールを付加する。高等真核生物のポリアデニル化シグナルは、ポリアデニル化付加部位から約11−30ヌクレオチドにAAUAAA配列を含んでいる。AAUAAA配列は、RNA開裂シグナルの伝達に関与する。(Lewin,Genes IV,Oxford University Press,NY,1990)。ポリ(A)テールはmRNAプロセシングに重要である。
【0061】
遺伝子発現カセットの部分として使用できるポリアデニル化シグナルは、ミニマルラビットβ−グロブリンポリアデニル化シグナルおよびウシ成長ホルモンポリアデニル化(BGH)シグナルを含む(Xuら,Gene 272:149−156,2001,Postら,米国特許No.5,122,458)。追加例は、合成ポリアデニル化シグナル(SPA)およびSV40ポリアデニル化シグナルを含む。SPA配列は配列19によって示されている。
【0062】
“転写可能に連結された”という表現は、プロモーターにRNAポリメラーゼが結合することによってヌクレオチド配列の転写が生じるようにプロモーターが配置されていることを表す。転写可能に連結されたとしても、転写される配列がプロモーターに隣接する必要はない。
【0063】
“機能的に連結された”という表現は、ヌクレオチド配列に間接的効果を与える能力を意味する。機能的に連結されたとしても、連結された配列が互いに隣接する必要はない。ヌクレオチド配列に機能的に連結された3’ポリアデニル化シグナルは、転写されたRNAの開裂およびポリアデニル化を促進する。ヌクレオチド配列に機能的に連結された5’リボソーム結合部位はリボソーム結合を促進する。
【0064】
遺伝子発現またはポリペプチドプロセシングを増進または調節するために有用な調節要素の追加例は、エンハンサー、リーダー配列およびオペレーターである。エンハンサー領域は転写を増進する。エンハンサー領域の例は、CMVエンハンサーおよびSV40エンハンサーである。(Hittら,Methods in Molecular Genetics 7:13−30,1995,Xuら,Gene 272:149−156, 2001)。エンハンサー領域はプロモーターに連携できる。
【0065】
リーダー配列は、ポリペプチドをプロテアソームに案内するポリペプチド上のアミノ酸領域である。リーダー配列をコードする核酸は構造遺伝子の5’であり、構造遺伝子と共に転写される。リーダー配列の一例はtPAである。
【0066】
オペレーター配列は遺伝子発現を調節するために使用できる。例えば、Tetオペレーター配列は遺伝子発現を抑制するために使用できる。
【0067】
IV.コーディング核酸配列
コーディング核酸配列は特定のアミノ酸配列をコードするコドンを提供する。特定のアミノ酸配列および遺伝子コードの既知の縮重性に基づいて異なる多数のコーディング核酸配列を得ることができる。遺伝子コードが縮重性をもつ理由は、ほぼすべてのアミノ酸がヌクレオチドトリプレットの様々な組合せすなわち“コドン”によってコードされているからである。
【0068】
特定のコドンが特定のアミノ酸に翻訳されることは当業界で公知である(参照:例えばLewin GENES IV,p.119,Oxford University Press,1990)。アミノ酸は以下のようなコドンによってコードされている:
A=Ala=アラニン:コドンGCA,GCC,GCG,GCU
C=Cys=システイン:コドンUGC,UGU
D=Asp=アスパラギン酸:コドンGAC,GAU
E=Glu=グルタミン酸:コドンGAA,GAG
F=Phe=フェニルアラニン:コドンUUC,UUU
G=Gly=グリシン:コドンGGA,GGC,GGG,GGU
H=His=ヒスチジン:コドンCAC,CAU
I=Ile=イソロイシン:コドンAUA,AUC,AUU
K=Lys=リシン:コドンAAA,AAG
L=Leu=ロイシン:コドンUUA,UUG,CUA,CUC,CUG,CUU
M=Met=メチオニン:コドンAUG
N=Asn=アスパラギン:コドンAAC,AAU
P=Pro=プロリン:コドンCCA,CCC,CCG,CCU
Q=Gln=グルタミン:コドンCAA,CAG
R=Arg=アルギニン:コドンAGA,AGG,CGA,CGC,CGG,CGU
S=Ser=セリン:コドンAGC,AGU,UCA,UCC,UCG,UCU
T=Thr=トレオニン:コドンACA,ACC,ACG,ACU
V=Val=バリン:コドンGUA,GUC,GUG,GUU
W=Trp=トリプトファン:コドンUGG
Y=Tyr=チロシン:コドンUAC,UAU。
【0069】
配列13のヌクレオチド1269−7223はNS3−NS4A−NS4B−NS5A−NS5B*配列の一例を示す。配列13のヌクレオチド1269−7223に実質的に相似の配列はワクチン成分の部分として使用できる。(Eminiら,国際公開WO 03/031588)。例えば、このような実質的に相似の配列は、ChA3またはChA63に基づくアデノベクターの部分として使用できる。
【0070】
ヌクレオチド配列に関する“実質的に相似の配列”という表現は、基準配列に対する少なくとも約70%の一致を表す。基準配列に対するパーセント一致(一致パーセントと表すこともある)を決定するためには、ヌクレオチド領域を最大一致が得られるように対応する基準領域に位置合せし、対応領域の一致ヌクレオチド数を測定する。この数を基準配列中のヌクレオチドの総数で除算し、100を乗算し、最も近い整数に丸める。
【0071】
核酸配列は宿主体内での発現を増進するように最適化できる。考察すべき要因は、C:G含量、好適なコドン、および、阻害性二次構造の回避を含む。特定宿主体内で発現増進を生じる核酸配列が得られるようにこれらの要因を様々に組合せる。(参照:例えばDonnellyら,国際公開WO 97/47358)。
【0072】
HCVコーディング核酸の最適化も、Eminiら,国際公開WO 03/031588らに記載されている。WO 03/031588は、NS3−NS4A−NS4B−NS5A−NS5B*をコードする種々の最適化配列の例を示している。
【0073】
NS31−NS4A1−NS32−NS4A2−NS4B2−NS5A2−NS5B2*のコドン最適化配列の一例は、配列2のヌクレオチド1891−9915によって示される。ヌクレオチド1891−1893は、NS31−NS4A1領域のメチオニンコドンを与える。ヌクレオチド3949−3960は、HCV 3aに基づく最初の4つのアミノ酸のあとにメチオニンが存在するNS32−NS4A2領域を与える。
【0074】
本発明の様々な実施態様で、NS31−NS4A1−NS32−NS4A2−NS4B2−NS5A2−NS5B2*コーディング領域は、配列2のヌクレオチド1891−9915または1894−9915に実質的に相似である。様々な実施態様でヌクレオチド配列は、配列2のヌクレオチド1891−9915または1894−9915に、少なくとも80%、少なくとも85%、少なくとも90%もしくは少なくとも95%の一致を有するか;または、0、1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20もしくは1−50ヌクレオチドだけ違っている。
【0075】
V.核酸ベクター
HCVポリペプチドをコードする核酸は、治療用ベクターとしてまたは治療用ベクターの製造に使用できる。治療用ベクターの製造に使用できるベクターはシャトルベクター、アデノウイルスゲノムプラスミドなどである。
【0076】
治療用ベクターは細胞にHCVポリペプチドを導入し発現させるために使用される。適正なベクターは、許容できない副作用を生じることなく標的細胞に核酸を送達できる。細胞発現は、HCVポリペプチドをコードする遺伝子発現カセットを使用して達成される。
【0077】
治療用途に使用できるベクターの例は、第一世代および第二世代のアデノベクター、ヘルパー依存性アデノベクター、アデノ関連ウイルスベクター、レトロウイルスベクター、アルファウイルスベクター、ベネズエラウマ脳炎ウイルスベクターおよびプラスミドベクターなどである。(Hittら,Advances in Pharmacology 40:131−206,1997,Johnstonら,米国特許6,156,588,Johnstonら,国際公開WO 95/32733,Eminiら,国際公開WO 03/031588)。
【0078】
V.A.アデノベクター
アデノベクターは、標的細胞中で所望のタンパク質またはポリペプチドを発現させるために組換えアデノウイルスゲノムを使用する。野生型アデノウイルスは、両端に逆方向末端反復配列をもつ二重鎖の直鎖状ゲノムを有している。ウイルス複製中にゲノムがウイルスキャプシド内部にパッケージされてビリオンを形成する。ウイルスはウイルス付着によって標的細胞に侵入し、次いでインターナリゼーションを達成する。(Hittら,Advances in Pharmacology 40:131−206,1997)。
【0079】
アデノウイルスゲノムは、アデノウイルスの複製およびプロセシングに必要な種々の要素を提供する。アデノウイルスゲノムの各末端は、逆方向末端反復配列(ITR)を含んでおり、これらはウイルス複製に必要である。該ウイルスはまた、感染性ビリオンの産生に必要なある種の構造タンパク質をプロセシングするために必要なプロテアーゼ活性をコードしている。
【0080】
アデノウイルスゲノムの構造は、宿主細胞に形質導入後のウイルス遺伝子の発現順序に基づいて記述できる。ウイルス遺伝子は、転写がDNA複製開始の以前に生じるかまたは以後に生じるかに従って初期(E)遺伝子または後期(L)遺伝子と呼ばれる。転写の初期段階で、E1、E2、E3およびE4遺伝子が発現されて宿主細胞のウイルス複製を準備する。ウイルスゲノムの必須初期領域1(E1)の欠失によってウイルスを複製欠陥にできる(Brodyら,Ann.N.Y.Acad.Sci. 7/6:90−101,1994)。
【0081】
後期段階で、ウイルス粒子の構造成分をコードする後期遺伝子L1−L5の発現がスイッチオンされる。すべての後期遺伝子が単一プロモーターのコントロール下にあり、ペントン(L2)、ヘキソン(L3)、100kDaのスカフォールドタンパク質およびファイバータンパク質(L5)を含むタンパク質をコードしており、これらは、アデノウイルスDNAをキャプシド封入した新しいウイルス粒子を形成する。最後に、野生型アデノウイルス複製プロセスが細胞溶解を惹起する。
【0082】
アデノベクターは、ヒトまたは動物に見出されるような種々のアデノウイルス血清型に基づく。動物アデノウイルスの例は、ウシ、ブタ、チンパンジー、ネズミ、イヌおよびトリ(CELO)を含む。ヒトアデノウイルスは、グループB、C、DまたはE血清型、例えば2型(“Ad2”)、4型(“Ad4”)、5型(“Ad5”)、6型(“Ad6”)、24型(“Ad24”)、26型(“Ad26”)、34型(“Ad34”)および35型(“Ad35”)を含む。アデノベクターは単一アデノウイルス由来または2種以上の異なるアデノウイルスに由来の領域を含有できる。
【0083】
様々な実施態様で、アデノベクターは、Ad5、Ad6、ChAd3、ChAd63またはそれらの組合せに基づく。Ad5は、Chroboczekら,J.Virology 756:280−285,1992に記載されている。Ad5およびAd6に基づくベクターは、Eminiら,国際公開WO 03/031588に記載されている。ChAd3の全長核酸配列を図6A−6Hに示す。ChAd63の全長核酸配列を図7A−7Hに示す。
【0084】
本発明の1つの実施態様では、アデノベクターが、表面に露出した1つ以上のチンパンジーChAd3またはChAd63構造タンパク質を含有する。表面露出タンパク質は、ファイバー、ヘキソンおよびペントンを含む。このようなタンパク質を含むアデノベクターは、ヒトアデノウイルスタンパク質と違って、患者が予めヒトアデノウイルスに接触したときの免疫応答の影響を受け難い。
【0085】
様々な実施態様で、組換えアデノベクターゲノムは、
a)配列3または9に実質的に相似のアミノ酸配列をもつファイバー領域、
b)配列5または11に実質的に相似のアミノ酸配列をもつヘキソン領域、および、
c)配列7に実質的に相似のアミノ酸配列をもつペントン領域
の少なくとも1つをコードしている。様々な追加実施態様では、配列相似が、少なくとも80%、少なくとも85%、少なくとも90%、少なくとも95%であるか、または、配列が、0、1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19もしくは20個のアミノ酸の変更だけ違っている。
【0086】
V.A.1.第一世代アデノベクター
第一世代アデノベクターは、E1欠失、場合によりE3欠失、場合によりE4欠失、および、発現カセットを有する組換えアデノウイルスゲノムを含有している。欠失の範囲および組合せは、ウイルスを複製不能にし、所望産物をコードする遺伝子発現カセットを収容するために十分な大きさである。ウイルスはE1欠失によって複製不能になる。
【0087】
E1欠失は、Ad5の塩基対約342を起点とし塩基対約3523に達する領域、または、他のアデノウイルスの対応する領域の欠失である。好ましくは、欠失領域が、Ad5の塩基対約450から塩基対約3511までの領域または他のアデノウイルスの対応する領域の除去を含む。塩基対約341を起点とするより大きいE1領域欠失は、ウイルスパッケージング促進要素を除去する。
【0088】
E3欠失は、Ad5の塩基対約27865を起点とし塩基対約30995に達する領域、または他のアデノベクターの対応する領域の欠失である。好ましくは、欠失領域が、Ad5の塩基対約28134から塩基対約30817までの領域または他のアデノベクターの対応する領域の除去を含む。
【0089】
E4欠失は、ChAd3の塩基対約34634を起点とし塩基対約37349に達する領域、または、他のアデノウイルスの対応する領域の欠失である。E4欠失は、先天性E4orf6を保持するかまたは別のアデノウイルス由来のE4orf6を挿入できる欠失でなければならない。Bertら,国際公開WO2004/018627は、このような異種E4orf6の使用を例示している。
【0090】
E1、E3およびE4の欠失の組合せは、遺伝子発現カセットを含有している組換えゲノムの全体寸法が野生型アデノウイルスゲノムの約105%を超過しない程度の十分な大きさでなければならない。例えば、組換えアデノウイルスAd5ゲノムの寸法が約105%を超過するとゲノムが不安定になる。(Bettら,Journal of Virology 67:5911−5921,1993)。
【0091】
好ましくは、遺伝子発現カセットを含有している組換えアデノウイルスゲノムの寸法は、野生型アデノウイルスゲノムの寸法の約85%から約105%までの範囲である。様々な実施態様で、発現カセットを含有している組換えアデノウイルスゲノムの寸法は、野生型ゲノムの寸法の約100%から約105.2%、または約100%である。
【0092】
E1およびE3欠失を有しているAd5またはAd6ゲノムには約7,500kbを挿入できる。全く欠失が全く存在しないとき、Ad5ゲノムは35,935塩基対であり、Ad6ゲノムは35,759塩基対である。
【0093】
ChAd3およびChAd63ベクターは、Ad5に比べて、ゲノムサイズがより大きく、また、より大きいE3領域が欠失可能領域として存在するので、核酸挿入容量が大きい。ChAd3ゲノムは、37,741塩基対であり、ChAd63ゲノムは36,643塩基対である。
【0094】
E1、E3が欠失しE4がAd5E4orf6で置換されたChAd3およびChAd63ベクターには約10,800bpを挿入できる。E4がAd5E4orf6によって置換されたとき、置換Ad5E4orf6が欠失領域よりも小さいので欠失および置換の双方である。これらのベクターには10,800bpを挿入すると野生型ゲノムの寸法の105%という限度に達する。
【0095】
第一世代のアデノベクターの複製は、E1遺伝子産物をトランスに供給することによって行うことができる。例えば、アデノウイルスE1領域で形質転換された細胞系を使用することによってE1遺伝子産物をトランスに供給できる。アデノウイルスE1領域で形質転換された細胞および細胞系の例は、HEK293細胞、911細胞、PERC.6TM細胞、および、トランスフェクトされた一次ヒトアミノサイト細胞である。(Grahamら,Journal of Virology 36:59−12,1977,Schiednerら,Human Gene Therapy 77:2105−2116,2000,Fallauxら, Human Gene Therapy 9:1909−1917,1998,Boutら,米国特許No.6,033,908)。
【0096】
チンパンジーアデノウイルスの先天性E4領域がAd5E4orf6によってシスに置換されると、Ad5相補細胞系に伝播した様々な血清型のチンパンジーアデノウイルスベクターの増殖が促進されおよび/または収量が増加する。293およびPER.C6細胞中のAd5E1配列は、チンパンジーアデノウイルスのようなグループCに所属するヒトアデノウイルス以外の血清型の複製を十分に相補しない。
【0097】
発現カセットは組換えアデノウイルスゲノムの欠失E1領域または欠失E3領域に対応する領域に挿入されなければならない。発現カセットは、平行配向でもよくまたは逆平行配向でもよい。平行配向では、挿入された遺伝子の転写方向が欠失したE1またはE3遺伝子と同方向である。逆平行配向転写では、対向鎖が鋳型として機能し、転写方向が対向方向である。
【0098】
本発明の1つの実施態様では、アデノベクターがE4欠失と、配列13のヌクレオチド34601−35482によって示されるAd5E4orf6配列に実質的に相似の配列の挿入とを含む。様々な実施態様で、配列一致は少なくとも75%、少なくとも80%、少なくとも85%、少なくとも90%、少なくとも95%であるか、または、配列5のヌクレオチド34601−35482から0、1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20もしくは1−50ヌクレオチドだけ違っている。
【0099】
本発明の1つの実施態様では、アデノウイルス主鎖が、配列13のヌクレオチド1−460および7468−35890または配列17のヌクレオチド1−454および7458−34658に少なくとも70%、少なくとも75%、少なくとも80%、少なくとも85%、少なくとも90%、少なくとも95%のヌクレオチド配列一致を有するか、または、配列13のヌクレオチド1−460および7468−35890または配列17のヌクレオチド1−454および7458−34658から0、1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20もしくは1−50ヌクレオチドだけ違っている。
【0100】
発現カセットを含有するアデノベクターの別の実施態様では、配列13または配列17に対するヌクレオチド配列一致が少なくとも70%、少なくとも75%、少なくとも80%、少なくとも85%、少なくとも90%、少なくとも95%であるか、または、配列13または配列17から0、1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20もしくは1−50ヌクレオチドだけ違っている。
【0101】
V.A.2.第二世代アデノベクター
第二世代のアデノベクターは、第一世代のベクターよりも小さいアデノウイルスゲノムを含有し、相補細胞系および/またはアデノウイルスタンパク質を供給するヘルパーベクターと共に使用できる。一般的な第二世代のアデノベクターは、Russell,Journal of General Virology 57:2573−2604,2000;Hittら,1997,Human Ad vectors for Gene Transfer,Advances in Pharmacology,Vol.40 Academic Press,Catalucciら.Journal of Virology 79:6400−6409,2005のような様々な参考文献に記載されている。第二世代のアデノベクターは、ヒトおよびチンパンジーアデノウイルスを含む様々な種類のアデノウイルスを基材にできる。
【0102】
V.B.DNAプラスミドベクター
DNAワクチンプラスミドベクターは、遺伝子発現カセットを複製促進要素および好ましくはベクー選択促進要素と共に含有する。好ましい要素は、非哺乳類細胞中の複製および選択可能マーカーを提供する。治療用ベクターは、ヒト細胞中の複製またはヒト核酸への組込みを提供する要素を含有してはならない。
【0103】
選択可能マーカーは、該マーカーを含有する核酸の選択を促進する。好ましい選択可能マーカーは、抗生物質抵抗性を与えるマーカーである。抗生物質選択遺伝子の例は、アンピシリン、ネオマイシンおよびカナマイシンに対する抵抗性をコードする核酸を含む。
【0104】
適当なDNAワクチンベクターは、細菌性複製起点と選択可能マーカーとを含有するプラスミドを出発材料として作製できる。より高い収率を与える細菌性複製起点の例は、ColE1プラスミド由来の細菌性複製起点を含む。(Donnellyら,Annu.Rev.Immunol.75:617−648,1997)。
【0105】
細菌性複製起点および選択可能マーカーの存在が大腸菌のような細菌株中のDNAベクターの産生を可能にする。選択可能マーカーは、DNAベクターを含有しない細菌を除去するために使用される。
【0106】
配列2は、HCVポリペプチドをコードする発現カセットを含有するプラスミドベクターの一例を示す。本発明の1つの実施態様で、プラスミドベクターは、配列2に少なくとも70%、少なくとも75%、少なくとも80%、少なくとも85%、少なくとも90%、少なくとも95%のヌクレオチド相似配列を有するか、または、0、1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20もしくは1−50ヌクレオチドだけ違っている。
【0107】
VI.ベクター作製
ベクターは、制限酵素、核酸結合および相同的組換えの使用を含む組換え核酸技術を使用して作製できる。組換え核酸技術は当業界で公知である。(Ausubel,Current Protocols in Molecular Biology,John Wiley,1987−1998、および、Sambrookら,Molecular Cloning,A Laboratory Manual,2nd Edition,Cold Spring Harbor Laboratory Press,1989)。
【0108】
治療用ベクターを得るためまたは発現カセットもしくはその一部分を1つのベクターから他のベクターに移すためには中間ベクターを使用する。中間ベクターの例は、アデノウイルスゲノムプラスミドおよびシャトルベクターを含む。
【0109】
中間ベクター中の有用な要素は、複製起点、選択可能マーカー、相同的組換え領域および適正な制限部位を含む。適正な制限部位は、核酸配列のクローニングまたは遊離を促進するために使用できる。
【0110】
相同的組換え領域は、別の核酸分子中の標的領域に相同的な核酸配列領域を供給する。相同的領域は標的領域に挿入される核酸配列に隣接する。様々な実施態様で、相同的領域は、約150から600ヌクレオチドの長さまたは約100から500ヌクレオチドの長さである。
【0111】
本発明の1つの実施態様は、HCVポリペプチドを発現する発現カセット、選択可能マーカー、細菌性複製起点、発現カセットを標的組換えによってE1領域に挿入または置換する第一のアデノウイルス相同性領域および第二のアデノウイルス相同性領域を含有するシャトルベクターを記載する。第一および第二の相同性領域は、発現カセットに隣接する。第一の相同性領域は、塩基対ほぼ4−450からの野生型アデノウイルスの領域の少なくとも右端(3’端)に実質的に相同の少なくとも約100塩基対を含有する。第二の相同性領域は、塩基対ほぼ3511−5792からのAd5の少なくとも左端(5’端)または別のアデノウイルスの対応する領域に実質的に相同の少なくとも約100塩基対を含有する。
【0112】
“実質的に相同の”という表現は、標的領域と特異的組換えを行うために十分な相同度を表す。様々な実施態様で、実質的相同は、少なくとも85%、少なくとも95%または100%の配列一致を表す。
【0113】
アデノベクターの1つの作製方法は、発現カセットを含有するプレアデノウイルスゲノムプラスミドの作製を経由する。該プレアデノウイルスプラスミドは、所望の相補細胞系中で複製されるために必要なすべてのアデノウイルス配列を含有する。次いでプレアデノウイルスプラスミドを制限酵素で消化してITRを遊離させ、相補細胞系にトランスフェクトしてウイルスを回収する。複製が生じ得るためにはITRがプラスミド配列から遊離されなければならない。アデノベクターの回収によって、発現カセットを含有するアデノベクターが得られる。(参照:例えばEminiら,国際公開WO 03/031588)。
【0114】
VI.A.アデノウイルスゲノムプラスミド
アデノウイルスゲノムプラスミドは、より長いプラスミド(コスミドでもよい)の内部にアデノベクター配列を含有している。より長いプラスミドは、プラスミドの製造および維持に使用される手順次第では真核細胞または原核細胞中の増殖および選択を促進する要素のような追加要素を含有している。アデノウイルスゲノムプラスミドは好ましくは、E1またはE3欠失領域に挿入された遺伝子発現カセットを有している。
【0115】
アデノウイルスゲノムプラスミド製造技術には、シャトルベクターおよび相同的組換えを使用する技術、アデノウイルスコスミドへの遺伝子発現カセットの挿入を含む技術などがある。(Hittら,Methods in Molecular Genetics 7:13−30,1995,Danthinneら,Gene Therapy 7:1707−1714,2000)。
【0116】
本発明の1つの実施態様は、アデノウイルスゲノムプラスミドを作製する相同的組換え段階と、アデノウイルス回収段階とを含むアデノベクターの製造方法を記載する。相同的組換え段階は、アデノウイルス相同領域に隣接されたHCVポリペプチド発現カセットを含有するシャトルベクターの使用を含む。アデノウイルス相同領域は、E1またはE3欠失領域に発現カセットを標的組換えする。
【0117】
VI.B.アデノベクター回収
アデノベクターは当業界で公知の方法またはここに記載の方法を使用してを組換えアデノウイルスゲノムプラスミドから回収できる。当業界で公知のアデノウイルス回収技術の例は、Hittら,Methods in Molecular Genetics 7:13−30,1995およびDanthinneら,Gene Therapy 7:1707−1714,2000に示されている。
【0118】
アデノベクター回収方法の一例は、アデノウイルス複製の増進を含む。増進は例えば、E2タンパク質(ポリメラーゼ、プレターミナルタンパク質およびDNA結合タンパク質)ならびにE4orf6のようなアデノウイルス機能を別のプラスミドから供給することによって実行できる(Eminiら,国際公開WO 03/031588)。
【0119】
VII.HCV併用治療
HCV核酸ワクチンは患者を治療するために単独で使用してもよく、他のHCV治療薬と併用してもよく、他の種類の疾患を対象とする薬剤と共に使用してもよい。追加治療薬は、HCVおよびHCV感染個体で有病率が高い疾患を治療する治療薬を含む。別種の疾患を対象とする薬剤は、HIVおよびHBCに特異的なワクチンを含む。
【0120】
HCVを治療するための追加治療薬はワクチンおよび非ワクチン薬を含む。(Zein,Expert Opin.Investig.Drugs 70:1457−1469,2001)。追加HCVワクチンの例は、HCVコア、E1、E2またはp7領域に対する免疫応答を誘発するように設計されたワクチンを含む。ワクチン成分の例は、天然産生ポリペプチド、HCVミモトープポリペプチド、または、このようなポリペプチドをコードする核酸を含む。
【0121】
一般的なミモトープ作製技術および種々のHCVミモトープを記載している参考文献は、Feliciら,米国特許No.5,994,083およびNicosiaら,国際出願WO 99/60132である。HCVミモトープは天然産生HCV抗原に融合できる。
【0122】
現在認可されている抗HCV薬はリバビリンと併用されるインターフェロンαである。組換えインターフェロンおよびPEG付加インターフェロンのような様々な形態のインターフェロンαをHCV感染の治療に使用できる。(De Francescoら,Antiviral Research 5S:1−16,2003,Walkerら,Antiviral Chemistry & Chemotherapy 14:1−21,2003)。
【0123】
異なる多様な抗HCV薬が様々な臨床開発段階にある。開発中の種々の抗HCV薬は、種々のHCV標的に特異的な薬剤を含む。種々のHCV標的の例は、HCVポリメラーゼおよびHCV NS3−NS4Aプロテアーゼを含む。(De Francescoら,Antiviral Research 58:1−16,2003,Walkerら,Antiviral Chemistry & Chemotherapy 14:1−21,2003)。
【0124】
VIII.医薬投与
HCVワクチンはここに提供した指針を当業界で公知の技術と共に使用して配合し患者に投与できる。一般的な医薬投与の指針は、Modern Vaccinology,Ed.Kurstak,Plenum Med.Co.1994;Remington’s Pharmaceutical Sciences 18th Edition,Ed.Gennaro,Mack Publishing,1990;およびModern Pharmaceutics 2nd Edition,Eds.Banker and Rhodes,Marcel Dekker,Inc.,1990に示されている。これらの文献はいずれも参照によって本発明に組込まれるものとする。
【0125】
HCVワクチンは、静脈内、腹腔内、皮下、筋肉内、経皮、皮膚圧入または鼻腔内のような種々の経路で投与できる。好ましい経路は筋肉内である。
【0126】
筋肉内投与は、1つ以上の電気パルスを伴うかまたは伴わない注射のような様々な技術を使用して行うことができる。電気介在移入は体液性および細胞性の双方の免疫応答を刺激することによって遺伝子免疫感作を補助する。
【0127】
ワクチン注射は、有針または無針の注射システムを使用する種々の技術を使用して行うことができる。無針注射システムの一例はジェット注射デバイスである。(Donnellyら,国際公開WO 99/52463)。
【0128】
電気介在移入すなわちジーン・エレクトロ・トランスファー(GET)は、核酸注入後に適正な電気パルスを送達することによって行うことができる。(参照:Mathiesen,国際公開WO 98/43702およびEminiら,国際公開WO 03/031588)
VIIIA.医薬担体
医薬的に許容される担体は、ワクチンの保存および対象へのワクチン投与を容易にする。医薬的に許容される担体の例をここに示す。医薬的に許容される追加の担体は当業界で公知である。
【0129】
医薬的に許容される担体は、バッファ、標準生理食塩水またはリン酸塩緩衝生理食塩水、ショ糖、塩およびポリソルベートのような様々な成分を含有し得る。医薬的に許容される担体の一例は、2.5−10mMのトリスバッファ、25−100mMのNaCl、2.5−10%ショ糖、0.01−2mMのMgCl2および0.001%−0.01%のポリソルベート80(植物由来)である。pHは約7.0−9.0でよい。担体の1つの具体例は、5mMのトリス、75mMのNaCl、5%ショ糖、1mMのMgCl2、0.005%のポリソルベート80をpH8.0で含有している。
【0130】
VIII.B.投薬計画
適正投薬計画は、特定ワクチンの効力、ならびに、患者の年齢、体重、性別および医学的状態、投与経路、所望の効果、投与回数などの要因を考慮して決定できる。特定ワクチンの効力は、細胞で発現およびプロセシングされMHCクラスIおよびII複合体に関係して提示されるポリペプチドを産生する特定ワクチンの能力のような様々な要因に依存する。
【0131】
患者に投与されたHCVポリペプチドをコードする核酸は、アデノベクターのようなウイルスベクターを含む様々なタイプのベクターおよびDNAプラスミドワクチンの部分を構成し得る。DNAプラスミドの投与に関する様々な実施態様では、約0.1から10mgのプラスミドを患者に投与し、また、約1から5mgのプラスミドを患者に投与する。ウイルスベクター好ましくはアデノウルイスベクターの投与に関する様々な実施態様では、約105から1011のウイルス粒子を患者に投与し、また107から1010のウイルス粒子を患者に投与する。
【0132】
ウイルスベクターワクチンおよびDNAプラスミドワクチンは、単独で投与してもよく、または、プライム(初回抗原刺激)およびブースト(追加抗原刺激)を含む投与計画の部分であってもよい。プライムおよびブーストの混合接種法は、DNAワクチンでプライムしウイルスベクターワクチンでブーストする方法、または、ウイルスベクターワクチンでプライムし、DNAワクチンでブーストする方法を含む。
【0133】
例えば約2から4回またはそれ以上の多数プライムを使用し得る。プライムとブーストとの間の時間の長さは、典型的には約4ヶ月から1年の範囲であるが、他の時間枠を使用してもよい。個体に抗アデノウイルス免疫応答が既に存在する場合にはDNAワクチンでプライムする投与計画の使用が好ましい。
【0134】
本発明の1つの実施態様では、初回ワクチン接種でDNAワクチンを筋肉組織に直接接種する。初回ワクチン接種後のブーストはアデノベクターまたはDNAワクチンで行う。
【0135】
免疫応答をブーストするために、インターロイキン−12、GM−CSF、B7−1、B7−2、IP10およびMig−1のような補助薬剤を併用投与できる。補助薬剤はタンパク質としてまたは核酸ベクターの使用を介して併用投与できる。
【0136】
VIII.C.異種プライム−ブースト
異種プライム−ブーストは、プライムに1つの種類のウイルスベクターを使用し、ブーストに別の種類のウイルスベクターを使用する混合方式である。異種プライム−ブーストは、種々のアデノウイルス血清型に基づくベクターのような近縁ベクター、ならびに、異なる動物のアデノウイルスおよびポックスウイルスのようなより遠縁のウイルスを含み得る。マウスをマラリアから守るためにポックスウイルスとアデノベクターとを使用することは、Gilbertら,Vaccine 20:1039−1045,2002によって示されている。HCVポリペプチドを発現するチンパンジーアデノベクターは、異種プライム−ブーストに使用できるベクターを提供する。
【0137】
プライムとブーストとの間の時間の長さは、典型的には約4ヶ月から1年の範囲であるが、他の時間枠を使用してもよい。最小時間枠は、免疫静止を考慮した十分な時間でなければならない。1つの実施態様では、この静止に少なくとも6ヶ月間を要する。プライムは1種類のベクターによる多回、例えば2から4回のプライムを含み得る。
【0138】
ポックスウイルスベクター中に存在する発現カセットは、当該ポックスウイルスまたは別のポックスウイルス構成員の先天的プロモーターまたはそれらに由来するプロモーターを含有しなければならない。ワクシニアウイルス、変性ワクシニアウイルス、アビポックスウイルス、アライグマポックスウイルス、変性ワクシニアウイルスAnkara、カナリアポックスウイルス(例えばALVAC)、トリポックスウイルス、ウシポックスウイルスおよびNYVACに基づくベクターのような様々な種類のポックスウイルスに基づくベクターを構築し使用する様々な戦略が当業界で公知である。(Moss,Current Topics in Microbiology and Immunology 755:25−38,1982;Earlら,In Current Protocols in Molecular Biology,Ausubelら,eds.,New York:Greene Publishing Associates & Wiley Interscience;1991:16.16.1−16.16.7;Childら,Virology 174(2):625−9,1990;Tartagliaら,Virology 188:217−232,1992;米国特許Nos.,4,603,112,4,722,848,4,769,330,5,110,587,5,174,993,5,185,146,5,266,313,5,505,941,5,863,542,および5,942,235)。
【0139】
VIII.D.アジュバント
HCVワクチンはアジュバントと共に配合できる。アジュバントは免疫原の免疫応答発生を補助する物質である。アジュバントは、生物学的または免疫学的半減期を延長する、免疫調節補助物質として作用する、または、免疫調節サイトカインの産生を誘導する、などの様々なメカニズムによって機能できる。種々のアジュバントを併用することもできる。
【0140】
HCVワクチンはアジュバントと共に配合できる。アジュバントの例は、ミョウバン、AlPO4、アルヒドロゲル、リピドAおよびそれらの誘導体または変異体、フロインド不完全アジュバント、中性リポソーム、ワクチンおよびサイトカインを内包するリポソーム、非イオン性ブロックコポリマー、ケモカイン、ならびに、免疫調節剤である。
【0141】
ポリオキシエチレン(POE)およびポリオキシプロピレン(POP)を含有する非イオン性ブロックコポリマー、例えば、POE−POP−POEブロックコポリマーはアジュバントとして使用し得る(Newmanら,Critical Reviews in Therapeutic Drug Carrier Systems 75:89−142,1998)。非イオン性ブロックコポリマーとアニオン性界面活性剤との併用によって核酸の免疫応答を増進できる。
【0142】
サイトカイン、ホルモン、脂質誘導体および低分子のような様々な種類の化合物を免疫調節剤として使用できる。免疫調節剤の例は、抗−CTLA−4、抗−CD137、抗−CD40、抗−CD28、抗−CD4、抗−CD25、抗PD1、抗−PD−Ll、抗−PD−L2、FOXP3−ブロッキング剤、Flt−3リガンド、イミキモッド、顆粒球−マクロファージコロニー−刺激因子(GM−CSF)、サルグラモスチン、トール様受容体(TLR)−7アゴニストおよびTLR−9アゴニストである。
【0143】
アジュバント配合物の具体例は、CRL−1005(CytRx Research Laboratories)、DNAおよびベンジルアルコニウムクロリド(BAK)を含有する配合物である。CRL−1005配合物は、例えばEminiら,国際公開WO 03/031588に記載されているように調製できる。
【0144】
VIII.E.ワクチン保存
ワクチンは様々な種類のバッファを使用して保存できる。例えば、Eminiら,国際公開WO 03/031588に記載されたバッファA105を使用できる。
【0145】
DNAの保存は微量金属イオンの除去またはキレート化によって強化できる。DNAワクチンの安定性を強化するためにコハク酸またはリンゴ酸のような試薬とキレート化剤とを使用できる。キレート化剤の例は、多くのリン酸塩リガンドおよびEDTAである。また、エタノールまたはグリセロールのような非還元性フリーラジカルスカベンジャーの含有はフリーラジカル発生によるDNAプラスミドの損傷を防止するために役立つ。さらに、DNAワクチンの安定性を最適化するために、配合物中のバッファの種類、pH、塩濃度、露光、バイアルの製造に使用した滅菌法などをコントロールするとよい。
【0146】
IX.実施例
本発明の様々な特徴をさらに詳細に説明するために実施例を以下に示す。実施例はまた本発明の有用な実施方法も示している。これらの実施例は特許請求の範囲に記載した本発明を限定しない。
【実施例1】
【0147】
ChAd63およびChAd3ゲノム配列の注釈
ChAd63およびChAd3を、“ヒトアデノウイルスC”群(HAdV−C;分類ID:129951)のすべてのタンパク質配列で作成されたローカルデータベースにブラストした。特定の分類IDをサーチすることによってタンパク質配列をNCBIサーバーからダウンロードした。ブラストサーチはblastxプログラムを使用して行った。列中に示す配列の数を1000に設定し、Filterをスイッチオフにした。次に、大規模配列相似解析用のBLAST強調ツールであるMSPcrunchによってブラスト結果を解析した。
【0148】
2つのゲノム配列について得られたCDS注釈のおのおのをATGおよび終結コドンの位置と必要ならばスプライシング部位の位置とを観察することによって手作業で確認した。このような産物の相同による予測の妥当性を検証するために、すべての産物を以前に作成されたアデノウイルスタンパク質データベースにblastpでサーチした。ChAd63およびChAd3の無加工のゲノム配列をMSPcrunch結果および手作業の校正に従ってVNTIで注釈した。遺伝子産物のChAd3およびChAd63を表1および2に示す。
【0149】
【表1】
【0150】
【表2】
【実施例2】
【0151】
ChAd3ベクター構築
ChAd3ΔE1,E3,E4,E4Ad5orf6ベクターの構築は以下の段階を含む。
【0152】
I.サブグループCシャトルベクターの構築
ChAd3ウイルスゲノムを完全に配列決定し(配列14)、その情報を使用して、相同的組換えによって完全ゲノムを容易にクローニングできるシャトルベクターを構築した。要約すると、ここでpChAd3EGFPと呼ぶサブグループCチンパンジーアデノウイルス3をクローニングするために使用したシャトルベクターを以下の手順で構築した。pIXコーディング領域を含有しているChAd3 DNAフラグメント(nt3542−4105)を配列20および21のプライマーと共にPCR増幅し、Sgf1−AscIで消化し、次いで、SgfI−AscIで消化したpARSCV32−3にクローニングして、pARS−ChAd3Dを作製した。ChAd3右端(nt37320−7441)を配列22および23のプライマーと共にPCR増幅し、XbaIおよびBamHIで消化し、次いで、XbaIおよびBamHIで制限したpARS−ChAd3Dに結合して、pARS−ChAd3RDを作製した。ChAd3ウイルスDNA左端(nt1−460)を配列24および25のプライマーと共にPCR増幅し、EcoRIおよびSgfIで消化し、次いで、EcoRIおよびSgfIで消化したpARS−ChAd3RDにクローニングし、このようにしてpARS−ChAd3RLDを作製した。このウイルスDNAカセットはまた、消化によってプラスミドDNAからウイルスDNAが遊離されるように双方のITRの末端に局在する制限酵素部位(PmeI)も含むように設計した。
【0153】
II.ΔE1 ChAd3ベクターの構築
ChAd3ベクターはBJ5183大腸菌株中の相同的組換えによって構築した。BJ5183細胞を、ChAd3精製ウイルスDNAとBstEIIおよびBst11071で消化したpChAd3EGFPシャトルベクターとによって同時形質転換した。pIX遺伝子、直鎖化pChAd3EGFPの末端に存在する右ITR DNA配列とウイルスゲノムDNAとの間の相同的組換えによってウイルスゲノムDNAがプラスミドベクターに挿入され、同時に、E1領域が欠失してEGFP発現カセットで置換された。ヒトサイトメガロウイルス(HCMV)プロモーターおよびウシ成長ホルモンポリアデニル化シグナル(Bgh ポリA)に基づくHCV NS領域発現カセットを、Eminiら,国際公開WO 03/031588に記載された手順で構築し、HCMVおよびBgh−ポリAのDNA配列間の相同性を利用した相同的組換えによってBJ5183大腸菌株中のChAd3ΔE1EGFPベクターに挿入した。
【0154】
III.E3領域欠失
ChAd3ベクター主鎖に完全なE3領域の欠失を導入するために、E3遺伝子に隣接する2つのDNA領域をPCR増幅して2つのDNAフラグメントを作製した。486bpのフラグメントはnt28159からnt28644(pVIII遺伝子の3’、E3領域の上流)の範囲であり、474bpのDNAフラグメントは、ファイバー遺伝子の3’端(bp32633からbp33106まで、E3領域の下流)を含んでいる。第一のDNAフラグメントの3’端および第二のフラグメントの5’端にEcoRI制限部位を導入した。2つのPCRフラグメントをEcoRIで消化し、インビトロ結合によって接合した。得られたDNAフラグメント(988bp)を次に、pVIII順方向オリゴおよびファイバー逆方向オリゴを使用してさらに増幅した。
【0155】
互いに接合した3’および5’のDNAフランキング領域を含有している988bpのE3領域のDNAフラグメントを、BJ5183細胞の同時形質転換によって、HpaI(野生型ChAd3の32384bpでE3領域切断)で直鎖化したpChAd3ΔE1/EGFPに組換えて、E3欠失を導入した。最終組換え産物はpChAd3ΔE1,E3/EGFPプレアデノプラスミドであった。
【0156】
IV.E4領域の欠失およびAd5E4orf6の挿入
ChAd3E4領域をAd5E4orf6で置換するために、Ad5E4orf6を、ChAd3ゲノムの右端に由来の最後の393bp(bp37349からbp37741まで)を含有するシャトルプラスミドに導入した。引き続いて、ファイバー3’端に由来しE4ポリAを含有している144bpのDNAフラグメント(ChAd3マップのbp34491からbp34634まで)をAd5E4orf6の下流に導入して、プラスミドpARSChAd3Ad5E4orf6−2を作製した。
【0157】
最後にBJ5183大腸菌株の同時形質転換によって、境界にファイバー3’端/E4ポリAおよびChAd3右端を含むpARSChAd3Ad5E4orf6−2に由来のDNAフラグメントを、PacI制限酵素(PacI部位、野生型ChAd3のnt36924)で直鎖化したpChAd3ΔE1,E3/EGFPに相同的組換えによって導入し、pChAd3ΔE1,3,4Ad5orf6EGFPを作製した。
【0158】
この戦略によって、ChAd3E4コーディング領域が完全に欠失し、ChAd3E4プロモーターのコントロール下に推定E4TATAシグナルの62bp下流にクローニングされたAd5E4orf6遺伝子で置換された。
【実施例3】
【0159】
ChAd63ベクター構築
ChAd3ΔE1,E3,E4,E4Ad5orf6ベクターに類似のChAd63ベクターは以下のように構築した。
【0160】
I.サブグループEシャトルベクターの構築
ChAd63ウイルスゲノムを完全に配列決定し、その情報を使用して、相同的組換えによって完全ゲノムを容易にクローニングできるシャトルベクターを構築した。要約すると、ここでpARSChAd63_EGFPと呼ぶサブグループEチンパンジーアデノウイルス63をクローニングするために使用したベクターを以下に記載の手順で構築した。
【0161】
ChAd63右端(nt36216−36643)を配列26および27のプライマーと共にPCR増幅し、XbaIおよびBamHIで消化し、次いで、XbaIおよびBamHIで制限したpARSChAd3−RLDに結合して、pARS−ChAd63Rを作製した。pIXコーディング領域を含有しているChAd63 DNAフラグメント(nt3422−3814)を配列28および29のプライマーと共にPCR増幅し、SgfI−AscIで消化し、次いで、SgfI−AscIで消化したpARS−ChAd63Rにクローニングして、pARS−ChAd63RDを作製した。ChAd63ウイルスDNA左端(nt1−455)をプライマー30および31と共にPCR増幅し、EcoRIおよびEcoRVで消化し、次いで、EcoRIおよびEcoRVで消化したpARS−ChAd63RDにクローニングし、このようにしてpARS−ChAd63RLDを作製した。HCMV−EGFP−bghポリAカセットを、配列32および33のプライマーと共にPCR増幅し、EcoRVで消化し、次いで、EcoRVで消化したpARS−ChAd63RLDにクローニングして、pARS−ChAd63RLD−EGFPを作製した。このウイルスDNAカセットはまた、消化によってプラスミドDNAからウイルスDNAが遊離されるように双方のITRの末端に局在する制限酵素部位(PmeI)も含むように設計した。
【0162】
II.ΔE1 ChAd63ベクターの構築
ChAd63ベクターはBJ5183大腸菌株中の相同的組換えによって構築した。BJ5183細胞を、ChAd63精製ウイルスDNAとAscIで消化したpARS−ChAd63RLD−EGFPとによって同時形質転換した。pIX遺伝子、直鎖化pARS−ChAd63RLD−EGFPの末端に存在する右ITR DNA配列とウイルスゲノムDNAとの間の相同的組換えによってウイルスゲノムDNAがプラスミドベクターに挿入され、同時に、E1領域が欠失してEGFP発現カセットで置換された。
【0163】
III.E3領域欠失およびChAd63NSmutベクター構築
ChAd63ベクター主鎖に完全なE3領域欠失を導入するために、E3遺伝子に隣接する2つのDNA領域をPCR増幅して2つのDNAフラグメントを作製した。567bpのフラグメントはnt26665からnt27207まで(pVIII遺伝子の3’端,E3領域の上流)の範囲であり、563bpのDNAフラグメントはファイバー遺伝子の3’端(bp31788からbp32326まで,E3領域の下流)を含んでいる。第一のDNAフラグメントの3’端および第二フラグメントの5’にPacI制限部位を導入した。2つのPCRフラグメントをPacIで消化し、インビトロ結合によって接合した。得られたDNAフラグメント(1112bp)を次に、pVIII順方向オリゴヌクレオチドおよびファイバー逆方向オリゴヌクレオチドを使用してさらに増幅した。
【0164】
互いに接合した3’および5’のDNAフランキング領域を含有している1112bpのE3領域のDNAフラグメントを、BJ5183細胞中の同時形質転換によって、HpaI(野生型ChAd63の30168bpでE3領域切断)で直鎖化したpChAd63ΔE1/EGFPと組換えて、E3欠失を導入した。最終組換え産物はpChAd63ΔE1,E3/EGFPプレアデノプラスミドであった。
【0165】
ヒトサイトメガロウイルスプロモーター(HCMV)およびウシ成長ホルモンポリアデニル化シグナル(Bgh ポリA)に基づくHCV NS領域発現カセットをEminiら,国際公開WO 03/031588に記載された手順で構築し、HCMVおよびBghポリAのDNA配列間の相同性を利用した相同的組換えによってBJ5183大腸菌株中のpChAd63ΔE1,E3/EGFPベクターに挿入し、ChAd63NSmutを作製した。
【0166】
IV.E4領域の欠失およびAd5E4orf6の挿入
ChAd63E4領域をAd5E4orf6で置換するために、Ad5E4orf6を、ChAd63ゲノムの右端に由来の428bp(bp36216からbp36643まで)下流でpARS−ChAd63RLD−EGFPに導入した。引き続いて、ファイバー3’端に由来しE4ポリAを含有している200bpのDNAフラグメント(ChAd63マップのbp33624からbp33823まで)をAd5E4orf6の下流に導入して、プラスミドpARSChAd63Ad5E4orf6−2を作製した。最後に、BJ5183大腸菌株の同時形質転換によって、境界にファイバー3’端/E4ポリAおよびChAd63左端を含むpARSChAd63Ad5E4orf6−2に由来のDNAフラグメントを、PmeI制限酵素(プラスミドDNAからウイルスDNAを遊離する)で消化したpChAd63ΔE1,E3/EGFPに相同的組換えによって導入し、このようにしてpChAd63ΔE1,3,4Ad5orf6EGFPを作製した。
【0167】
この戦略によって、ChAd63E4コーディング領域が完全に欠失し、ChAd63E4プロモーターのコントロール下に推定E4TATAシグナルの131bp下流にクローニングされたAd5E4orf6遺伝子で置換された。
【実施例4】
【0168】
ChAd3NSmut(配列13)の発現
Catalucciら,Journal of Virology 79:6400−6409,2005に記載の手順に準拠した技術を使用してChAd3NSmutのHCVタンパク質発現を試験した。HeLa細胞にChAd3NSmutおよびMRKAd6NSmutを感染させた。MRKAd6NSmutはEminiら,国際公開WO 03/031588に記載されている。細胞抽出物を抗NS5Aモノクローナル抗体によるイムノブロットで分析した。図8に示すように、ChAd3NSmutは、ヒトAd6に基づくベクター(MRKAd6NSmut)と同様にHCVタンパク質を発現する。
【実施例5】
【0169】
ChAd3NSmut(配列13)の安定性
Catalucciら,Journal of Virology 79:6400−6409,2005に記載の手順に準拠した技術を使用してChAd3NSmutの遺伝的安定性を検証した。5つの独立クローン(継代10)から抽出したウイルスDNAの制限分析を行った。プレChAd3NSmutプラスミドを陽性対照として含ませた。ChAd3NSmutはPerC.6細胞までの継代期間は遺伝的に安定であった。
【実施例6】
【0170】
ChAd3NSmut(配列13)およびChAd63NSmut(配列17)はマウス体内でCMIを誘導した。
【0171】
Eminiら,国際公開WO 03/031588に記載の手順に準拠した技術を使用してChAd3NSmutおよびChAd63NSmutの細胞性免疫誘導能力をC57/B6マウスで試験した。図9は、C57/B6マウス体内でのChAd3NSmut(配列13)、ChAd63NSmut(配列17)およびMRKAd6NSmutの細胞性免疫誘導能力の比較を示す。図9は、注入3週後に行ったIFNγ ELIspot実験(H2Kb制限ペプチド使用、NS3プロテアーゼ中でマッピング)を示す(データはN=5の平均として示す)。108および109の用量のChAd3NSmutおよびChAd63NSmutによってマウス体内でMRKAd6NSmutと同等のCMIが誘発される。
【実施例7】
【0172】
ChAd3NSmutおよびChAd63NSmutはアカゲザル体内でCMIを誘導した。
【0173】
ChAd3NSmut(配列13)およびChAd63NSmut(配列17)のCMI誘導能力をヒト以外の霊長類で確認するために、Eminiら,国際公開WO 03/031588およびCirilloら,国際公開WO 2005/071093に記載の手順に準拠した技術を使用してアカゲザルを免疫感作した。異なる3つの非交差反応性ベクターの連続注射に基づく異種プライム/ブースト法で3匹1グループのサルを免疫感作してベクターを評価した。3匹の動物は、0および4週目に1匹あたり1010vpの用量のChAd3NSmutを2回注射してプライムし、次いで22週目にMRKAd6NSmut、42週目にChAd63NSmutを注射した。IFN−γ ELISPOTによって測定した免疫応答の時間経過を図11に報告するが、これは、所与のいずれかの時点で種々のHCV NSペプチドプールに観察された応答の和を表している。この結果は、ChAd3NS注射によって全部の動物に効率的なプライム効果が得られたこと、および、MRKAd6NSmutおよびChAd63NSmutの双方の投与によってCMIが強力にブーストされることを示す。
【実施例8】
【0174】
キメラHCVポリペプチドをコードするプラスミドDNA(配列1)の構築
HCV 3aに基づくNS3−4A領域とHCV 1bに基づくNS3−NS4A−NS4B−NS5A−NS5B領域とを含み、ここでpV1JnsNSOPTmut3a−1bと呼ぶキメラHCVポリペプチドをコードするプラスミド(図12および13)はBJ5183大腸菌株中の相同的組換えによって得られた。
【0175】
最適翻訳開始(コザック)配列と、成熟NS3配列の第一アミノ酸に融合したメチオニン開始コドンとをもつHCV 3a由来の完全コドン最適化NS3−4aをコードするプラスミドを合成的に作製した。NS3−4aコーディング配列には、pV1JnsNSOPTmutb受容体プラスミドに挿入するためにイントロンA配列とNS3(HCV)コーディング配列の起点とに相同の2つの組換え領域が隣接している(Eminiら,国際公開WO 03/031588)。親プラスミドからインサートを切除するHindIII制限部位を新しいNS3−4a配列の両端に導入した。
【0176】
このpV1JnsOPTmutプラスミドをHpaI非反復部位消化によって直鎖化した。直鎖化したpV1JnsOPTmutプラスミドおよびHindIII消化したNS3−4a(3a)インサートでBJ5183菌株を同時形質転換して、pV1JnsNSOPTmut3a−1bを作製した。図13に示す得られたpV1JnsNSOPTmut3a−lbの遺伝子構造を制限酵素およびDNA配列解析によって検証した。
【実施例9】
【0177】
キメラHCVポリペプチドをコードするプラスミドDNAはマウス体内でDMIを誘導した。
【0178】
キメラHCVポリペプチドをコードするプラスミドDNAの種々のHCV遺伝子型に対する細胞性免疫誘導能力をマウスで試験した。キメラポリペプチド(配列1)は、HCV 3aに基づくNS3−4A領域およびHCV 1bに基づくNS3−NS4A−NS4B−NS5A−NS5B領域を含んでいた(pV1Jns−NSOPTmut3a−1b)。
【0179】
異なる3つのマウス株(2つの同系交配:Balb/c,C57B1/6および1つの異系交配:CD1)に、50μgのDNAを筋肉内注射し次いで電気パルスを与えた。各動物に、キメラプラスミド(pV1Jns−NSOPTmut3a−1b)、または、HCV 1bに基づくNS3−NS4A−NS4B−NS5A−NS5B領域をコードするpV1Jns−NSOPTmutプラスミド(Eminiら,国際公開WO 03/031588)を2回与えた。HCV 1bおよび3aに由来のウイルスタンパク質に特異的なCMIを、Eminiら,国際公開WO 03/031588に記載された技術を使用して測定した。図14は、CD1マウス(異系交配株)の体内でHCV 1bおよび3aに由来のNS3タンパク質に応答してIFN−γを分泌するT細胞の数を示す(1,000,000脾細胞あたりのスポット形成細胞として表す)。1b NS3タンパク質に対する特異的応答は双方のプラスミドで同等であるが、3a NS3タンパク質に対してはキメラ構築物のほうが高い応答を誘導する(スチューデントT検定でp=0.04)。2つの同系交配株のマウス(Balb/cおよびC57B1/6)の体内でHCV 1bおよび3aに由来のNS3タンパク質に対して誘導されたCMIは双方の構築物で同等であった。
【0180】
特許請求の範囲は他の実施態様も包含する。いくつかの実施態様を提示して説明したが、本発明の要旨および範囲を逸脱することなく様々な変更を行うことが可能であろう。
【図面の簡単な説明】
【0181】
【図1】配列1のアミノ酸配列を示す。アミノ酸1−686は、HCV 3aに基づくNS3−NS4A領域を与える。アミノ酸687−690は、HCV 3a NS4B領域の最初の4つのアミノ酸を構成し、また、HCV 3a NS3−NS4A領域の開裂接合を与える。アミノ酸691−2675は、HCV 1bに基づくNS3−NS4A−NS4B−NS5A−NS5B領域を与える。NS3−NS4A領域およびNS3−NS4A−NS4B−NS5A−NS5B領域はまた開始メチオニンの付加を含む。
【図2A】図2A−2Cは配列2の核酸配列を示す。ヌクレオチド318−10182は以下の要素を含む発現カセットを構成する:HCMVプロモーター:nt318−905Int A:nt1040−1865コザック配列:nt1885−1890HCV Met−NS3−NS4A(3aに基づく、最適化):nt1891−3948HCV NS4Bの最初の4アミノ酸(3aに基づく):nt3949−3960HCV Met−NS3−5B(1bに基づく、Bk菌株、最適化):nt3961−9915TAAAターミネーター:nt9916−9919BGH:nt9956−10179。
【図2B】図2A−2Cは配列2の核酸配列を示す。ヌクレオチド318−10182は以下の要素を含む発現カセットを構成する:HCMVプロモーター:nt318−905Int A:nt1040−1865コザック配列:nt1885−1890HCV Met−NS3−NS4A(3aに基づく、最適化):nt1891−3948HCV NS4Bの最初の4アミノ酸(3aに基づく):nt3949−3960HCV Met−NS3−5B(1bに基づく、Bk菌株、最適化):nt3961−9915TAAAターミネーター:nt9916−9919BGH:nt9956−10179。
【図2C】図2A−2Cは配列2の核酸配列を示す。ヌクレオチド318−10182は以下の要素を含む発現カセットを構成する:HCMVプロモーター:nt318−905Int A:nt1040−1865コザック配列:nt1885−1890HCV Met−NS3−NS4A(3aに基づく、最適化):nt1891−3948HCV NS4Bの最初の4アミノ酸(3aに基づく):nt3949−3960HCV Met−NS3−5B(1bに基づく、Bk菌株、最適化):nt3961−9915TAAAターミネーター:nt9916−9919BGH:nt9956−10179。
【図3】met−NS3−5B配列(配列16)を示す。
【図4A】図4A−4Jは、ChAd3ファイバー(図4A、配列3)、ChAd3ヘキソン(図4B、配列5)、ChAd3ペントン(図4C、配列7)、ChAd63ファイバー(図4D、配列9)、ChAd63ヘキソン(図4E、配列11)のアミノ酸配列、ならびに、ChAd3ファイバー(図4F、配列4)、ChAd3ヘキソン(図4G、配列6)、ChAd3ペントン(図4H、配列8)、ChAd63ファイバー(図4I、配列10)およびChAd63ヘキソン(図4J、配列12)のコーディング核酸を表す。
【図4B】図4A−4Jは、ChAd3ファイバー(図4A、配列3)、ChAd3ヘキソン(図4B、配列5)、ChAd3ペントン(図4C、配列7)、ChAd63ファイバー(図4D、配列9)、ChAd63ヘキソン(図4E、配列11)のアミノ酸配列、ならびに、ChAd3ファイバー(図4F、配列4)、ChAd3ヘキソン(図4G、配列6)、ChAd3ペントン(図4H、配列8)、ChAd63ファイバー(図4I、配列10)およびChAd63ヘキソン(図4J、配列12)のコーディング核酸を表す。
【図4C】図4A−4Jは、ChAd3ファイバー(図4A、配列3)、ChAd3ヘキソン(図4B、配列5)、ChAd3ペントン(図4C、配列7)、ChAd63ファイバー(図4D、配列9)、ChAd63ヘキソン(図4E、配列11)のアミノ酸配列、ならびに、ChAd3ファイバー(図4F、配列4)、ChAd3ヘキソン(図4G、配列6)、ChAd3ペントン(図4H、配列8)、ChAd63ファイバー(図4I、配列10)およびChAd63ヘキソン(図4J、配列12)のコーディング核酸を表す。
【図4D】図4A−4Jは、ChAd3ファイバー(図4A、配列3)、ChAd3ヘキソン(図4B、配列5)、ChAd3ペントン(図4C、配列7)、ChAd63ファイバー(図4D、配列9)、ChAd63ヘキソン(図4E、配列11)のアミノ酸配列、ならびに、ChAd3ファイバー(図4F、配列4)、ChAd3ヘキソン(図4G、配列6)、ChAd3ペントン(図4H、配列8)、ChAd63ファイバー(図4I、配列10)およびChAd63ヘキソン(図4J、配列12)のコーディング核酸を表す。
【図4E】図4A−4Jは、ChAd3ファイバー(図4A、配列3)、ChAd3ヘキソン(図4B、配列5)、ChAd3ペントン(図4C、配列7)、ChAd63ファイバー(図4D、配列9)、ChAd63ヘキソン(図4E、配列11)のアミノ酸配列、ならびに、ChAd3ファイバー(図4F、配列4)、ChAd3ヘキソン(図4G、配列6)、ChAd3ペントン(図4H、配列8)、ChAd63ファイバー(図4I、配列10)およびChAd63ヘキソン(図4J、配列12)のコーディング核酸を表す。
【図4F】図4A−4Jは、ChAd3ファイバー(図4A、配列3)、ChAd3ヘキソン(図4B、配列5)、ChAd3ペントン(図4C、配列7)、ChAd63ファイバー(図4D、配列9)、ChAd63ヘキソン(図4E、配列11)のアミノ酸配列、ならびに、ChAd3ファイバー(図4F、配列4)、ChAd3ヘキソン(図4G、配列6)、ChAd3ペントン(図4H、配列8)、ChAd63ファイバー(図4I、配列10)およびChAd63ヘキソン(図4J、配列12)のコーディング核酸を表す。
【図4G】図4A−4Jは、ChAd3ファイバー(図4A、配列3)、ChAd3ヘキソン(図4B、配列5)、ChAd3ペントン(図4C、配列7)、ChAd63ファイバー(図4D、配列9)、ChAd63ヘキソン(図4E、配列11)のアミノ酸配列、ならびに、ChAd3ファイバー(図4F、配列4)、ChAd3ヘキソン(図4G、配列6)、ChAd3ペントン(図4H、配列8)、ChAd63ファイバー(図4I、配列10)およびChAd63ヘキソン(図4J、配列12)のコーディング核酸を表す。
【図4H】図4A−4Jは、ChAd3ファイバー(図4A、配列3)、ChAd3ヘキソン(図4B、配列5)、ChAd3ペントン(図4C、配列7)、ChAd63ファイバー(図4D、配列9)、ChAd63ヘキソン(図4E、配列11)のアミノ酸配列、ならびに、ChAd3ファイバー(図4F、配列4)、ChAd3ヘキソン(図4G、配列6)、ChAd3ペントン(図4H、配列8)、ChAd63ファイバー(図4I、配列10)およびChAd63ヘキソン(図4J、配列12)のコーディング核酸を表す。
【図4I】図4A−4Jは、ChAd3ファイバー(図4A、配列3)、ChAd3ヘキソン(図4B、配列5)、ChAd3ペントン(図4C、配列7)、ChAd63ファイバー(図4D、配列9)、ChAd63ヘキソン(図4E、配列11)のアミノ酸配列、ならびに、ChAd3ファイバー(図4F、配列4)、ChAd3ヘキソン(図4G、配列6)、ChAd3ペントン(図4H、配列8)、ChAd63ファイバー(図4I、配列10)およびChAd63ヘキソン(図4J、配列12)のコーディング核酸を表す。
【図4J】図4A−4Jは、ChAd3ファイバー(図4A、配列3)、ChAd3ヘキソン(図4B、配列5)、ChAd3ペントン(図4C、配列7)、ChAd63ファイバー(図4D、配列9)、ChAd63ヘキソン(図4E、配列11)のアミノ酸配列、ならびに、ChAd3ファイバー(図4F、配列4)、ChAd3ヘキソン(図4G、配列6)、ChAd3ペントン(図4H、配列8)、ChAd63ファイバー(図4I、配列10)およびChAd63ヘキソン(図4J、配列12)のコーディング核酸を表す。
【図5A】図5A−5Hは、ChAd3ΔE1,3,4,Ad5E4orf6NSmut−35,890bp(ChAd3NSmut、配列13)の核酸配列を表す。野生型ゲノムの欠失の組合せは、nt461からnt3541まで(3080bp)のEl欠失、nt28644からnt32633まで(3989bp)のE3欠失、nt34634からnt37349まで(2715bp)のE4欠失である。様々な領域は以下の場所に存在する:ChAd3左側ITR+パッケージングシグナル:nt1−460HCMVプロモーター:nt467−1257コザックコンセンサス配列:nt1263−1268HCV NS3−5B(BK菌株):nt1269−7223TAAターミネーター:nt7224−7226BGHポリA:nt7234−7452ChAd3主鎖:nt7468−35890Ad5E4orf6:nt34601−35482。
【図5B】図5A−5Hは、ChAd3ΔE1,3,4,Ad5E4orf6NSmut−35,890bp(ChAd3NSmut、配列13)の核酸配列を表す。野生型ゲノムの欠失の組合せは、nt461からnt3541まで(3080bp)のEl欠失、nt28644からnt32633まで(3989bp)のE3欠失、nt34634からnt37349まで(2715bp)のE4欠失である。様々な領域は以下の場所に存在する:ChAd3左側ITR+パッケージングシグナル:nt1−460HCMVプロモーター:nt467−1257コザックコンセンサス配列:nt1263−1268HCV NS3−5B(BK菌株):nt1269−7223TAAターミネーター:nt7224−7226BGHポリA:nt7234−7452ChAd3主鎖:nt7468−35890Ad5E4orf6:nt34601−35482。
【図5C】図5A−5Hは、ChAd3ΔE1,3,4,Ad5E4orf6NSmut−35,890bp(ChAd3NSmut、配列13)の核酸配列を表す。野生型ゲノムの欠失の組合せは、nt461からnt3541まで(3080bp)のEl欠失、nt28644からnt32633まで(3989bp)のE3欠失、nt34634からnt37349まで(2715bp)のE4欠失である。様々な領域は以下の場所に存在する:ChAd3左側ITR+パッケージングシグナル:nt1−460HCMVプロモーター:nt467−1257コザックコンセンサス配列:nt1263−1268HCV NS3−5B(BK菌株):nt1269−7223TAAターミネーター:nt7224−7226BGHポリA:nt7234−7452ChAd3主鎖:nt7468−35890Ad5E4orf6:nt34601−35482。
【図5D】図5A−5Hは、ChAd3ΔE1,3,4,Ad5E4orf6NSmut−35,890bp(ChAd3NSmut、配列13)の核酸配列を表す。野生型ゲノムの欠失の組合せは、nt461からnt3541まで(3080bp)のEl欠失、nt28644からnt32633まで(3989bp)のE3欠失、nt34634からnt37349まで(2715bp)のE4欠失である。様々な領域は以下の場所に存在する:ChAd3左側ITR+パッケージングシグナル:nt1−460HCMVプロモーター:nt467−1257コザックコンセンサス配列:nt1263−1268HCV NS3−5B(BK菌株):nt1269−7223TAAターミネーター:nt7224−7226BGHポリA:nt7234−7452ChAd3主鎖:nt7468−35890Ad5E4orf6:nt34601−35482。
【図5E】図5A−5Hは、ChAd3ΔE1,3,4,Ad5E4orf6NSmut−35,890bp(ChAd3NSmut、配列13)の核酸配列を表す。野生型ゲノムの欠失の組合せは、nt461からnt3541まで(3080bp)のEl欠失、nt28644からnt32633まで(3989bp)のE3欠失、nt34634からnt37349まで(2715bp)のE4欠失である。様々な領域は以下の場所に存在する:ChAd3左側ITR+パッケージングシグナル:nt1−460HCMVプロモーター:nt467−1257コザックコンセンサス配列:nt1263−1268HCV NS3−5B(BK菌株):nt1269−7223TAAターミネーター:nt7224−7226BGHポリA:nt7234−7452ChAd3主鎖:nt7468−35890Ad5E4orf6:nt34601−35482。
【図5F】図5A−5Hは、ChAd3ΔE1,3,4,Ad5E4orf6NSmut−35,890bp(ChAd3NSmut、配列13)の核酸配列を表す。野生型ゲノムの欠失の組合せは、nt461からnt3541まで(3080bp)のEl欠失、nt28644からnt32633まで(3989bp)のE3欠失、nt34634からnt37349まで(2715bp)のE4欠失である。様々な領域は以下の場所に存在する:ChAd3左側ITR+パッケージングシグナル:nt1−460HCMVプロモーター:nt467−1257コザックコンセンサス配列:nt1263−1268HCV NS3−5B(BK菌株):nt1269−7223TAAターミネーター:nt7224−7226BGHポリA:nt7234−7452ChAd3主鎖:nt7468−35890Ad5E4orf6:nt34601−35482。
【図5G】図5A−5Hは、ChAd3ΔE1,3,4,Ad5E4orf6NSmut−35,890bp(ChAd3NSmut、配列13)の核酸配列を表す。野生型ゲノムの欠失の組合せは、nt461からnt3541まで(3080bp)のEl欠失、nt28644からnt32633まで(3989bp)のE3欠失、nt34634からnt37349まで(2715bp)のE4欠失である。様々な領域は以下の場所に存在する:ChAd3左側ITR+パッケージングシグナル:nt1−460HCMVプロモーター:nt467−1257コザックコンセンサス配列:nt1263−1268HCV NS3−5B(BK菌株):nt1269−7223TAAターミネーター:nt7224−7226BGHポリA:nt7234−7452ChAd3主鎖:nt7468−35890Ad5E4orf6:nt34601−35482。
【図5H】図5A−5Hは、ChAd3ΔE1,3,4,Ad5E4orf6NSmut−35,890bp(ChAd3NSmut、配列13)の核酸配列を表す。野生型ゲノムの欠失の組合せは、nt461からnt3541まで(3080bp)のEl欠失、nt28644からnt32633まで(3989bp)のE3欠失、nt34634からnt37349まで(2715bp)のE4欠失である。様々な領域は以下の場所に存在する:ChAd3左側ITR+パッケージングシグナル:nt1−460HCMVプロモーター:nt467−1257コザックコンセンサス配列:nt1263−1268HCV NS3−5B(BK菌株):nt1269−7223TAAターミネーター:nt7224−7226BGHポリA:nt7234−7452ChAd3主鎖:nt7468−35890Ad5E4orf6:nt34601−35482。
【図6A】図6A−6Hは野生型ChAd3の核酸配列(配列14)を示す。
【図6B】図6A−6Hは野生型ChAd3の核酸配列(配列14)を示す。
【図6C】図6A−6Hは野生型ChAd3の核酸配列(配列14)を示す。
【図6D】図6A−6Hは野生型ChAd3の核酸配列(配列14)を示す。
【図6E】図6A−6Hは野生型ChAd3の核酸配列(配列14)を示す。
【図6F】図6A−6Hは野生型ChAd3の核酸配列(配列14)を示す。
【図6G】図6A−6Hは野生型ChAd3の核酸配列(配列14)を示す。
【図6H】図6A−6Hは野生型ChAd3の核酸配列(配列14)を示す。
【図7A】図7A−7Hは野生型ChAd63の核酸配列(配列15)を示す。
【図7B】図7A−7Hは野生型ChAd63の核酸配列(配列15)を示す。
【図7C】図7A−7Hは野生型ChAd63の核酸配列(配列15)を示す。
【図7D】図7A−7Hは野生型ChAd63の核酸配列(配列15)を示す。
【図7E】図7A−7Hは野生型ChAd63の核酸配列(配列15)を示す。
【図7F】図7A−7Hは野生型ChAd63の核酸配列(配列15)を示す。
【図7G】図7A−7Hは野生型ChAd63の核酸配列(配列15)を示す。
【図7H】図7A−7Hは野生型ChAd63の核酸配列(配列15)を示す。
【図8】ChAd3NSmut(配列13)およびMRKAd6NSmutに感染したHeLa細胞中のHCV NSタンパク質の発現の比較を示す。
【図9】C57/B6マウス体内で細胞性免疫を誘導するChAd3NSmut(配列13)およびMRKAd6NSmutの能力の比較を示す。
【図10A】図10A−10HはChAd63ΔE1,3,4,Ad5E4orf6,NSmut(配列17)の核酸配列を示す。野生型ゲノムの欠失の組合せは、nt455からnt3421まで(2967bp)のEl欠失、nt27207からnt31778まで(4582bp)のE3欠失、nt33825からnt36215まで(2390bp)のE4欠失である。様々な領域は以下の場所に存在する:ChAd63左側ITR+パッケージングシグナル:nt1−454HCMVプロモーター:nt458−1248コザックコンセンサス配列:nt1254−1259HCV NS3−5B(BK菌株):nt1260−7214TAAターミネーター:nt7215−7217BGHポリA:nt7227−7447ChAd63主鎖:nt7458−34658Ad5E4orf6:nt33316−34197。
【図10B】図10A−10HはChAd63ΔE1,3,4,Ad5E4orf6,NSmut(配列17)の核酸配列を示す。野生型ゲノムの欠失の組合せは、nt455からnt3421まで(2967bp)のEl欠失、nt27207からnt31778まで(4582bp)のE3欠失、nt33825からnt36215まで(2390bp)のE4欠失である。様々な領域は以下の場所に存在する:ChAd63左側ITR+パッケージングシグナル:nt1−454HCMVプロモーター:nt458−1248コザックコンセンサス配列:nt1254−1259HCV NS3−5B(BK菌株):nt1260−7214TAAターミネーター:nt7215−7217BGHポリA:nt7227−7447ChAd63主鎖:nt7458−34658Ad5E4orf6:nt33316−34197。
【図10C】図10A−10HはChAd63ΔE1,3,4,Ad5E4orf6,NSmut(配列17)の核酸配列を示す。野生型ゲノムの欠失の組合せは、nt455からnt3421まで(2967bp)のEl欠失、nt27207からnt31778まで(4582bp)のE3欠失、nt33825からnt36215まで(2390bp)のE4欠失である。様々な領域は以下の場所に存在する:ChAd63左側ITR+パッケージングシグナル:nt1−454HCMVプロモーター:nt458−1248コザックコンセンサス配列:nt1254−1259HCV NS3−5B(BK菌株):nt1260−7214TAAターミネーター:nt7215−7217BGHポリA:nt7227−7447ChAd63主鎖:nt7458−34658Ad5E4orf6:nt33316−34197。
【図10D】図10A−10HはChAd63ΔE1,3,4,Ad5E4orf6,NSmut(配列17)の核酸配列を示す。野生型ゲノムの欠失の組合せは、nt455からnt3421まで(2967bp)のEl欠失、nt27207からnt31778まで(4582bp)のE3欠失、nt33825からnt36215まで(2390bp)のE4欠失である。様々な領域は以下の場所に存在する:ChAd63左側ITR+パッケージングシグナル:nt1−454HCMVプロモーター:nt458−1248コザックコンセンサス配列:nt1254−1259HCV NS3−5B(BK菌株):nt1260−7214TAAターミネーター:nt7215−7217BGHポリA:nt7227−7447ChAd63主鎖:nt7458−34658Ad5E4orf6:nt33316−34197。
【図10E】図10A−10HはChAd63ΔE1,3,4,Ad5E4orf6,NSmut(配列17)の核酸配列を示す。野生型ゲノムの欠失の組合せは、nt455からnt3421まで(2967bp)のEl欠失、nt27207からnt31778まで(4582bp)のE3欠失、nt33825からnt36215まで(2390bp)のE4欠失である。様々な領域は以下の場所に存在する:ChAd63左側ITR+パッケージングシグナル:nt1−454HCMVプロモーター:nt458−1248コザックコンセンサス配列:nt1254−1259HCV NS3−5B(BK菌株):nt1260−7214TAAターミネーター:nt7215−7217BGHポリA:nt7227−7447ChAd63主鎖:nt7458−34658Ad5E4orf6:nt33316−34197。
【図10F】図10A−10HはChAd63ΔE1,3,4,Ad5E4orf6,NSmut(配列17)の核酸配列を示す。野生型ゲノムの欠失の組合せは、nt455からnt3421まで(2967bp)のEl欠失、nt27207からnt31778まで(4582bp)のE3欠失、nt33825からnt36215まで(2390bp)のE4欠失である。様々な領域は以下の場所に存在する:ChAd63左側ITR+パッケージングシグナル:nt1−454HCMVプロモーター:nt458−1248コザックコンセンサス配列:nt1254−1259HCV NS3−5B(BK菌株):nt1260−7214TAAターミネーター:nt7215−7217BGHポリA:nt7227−7447ChAd63主鎖:nt7458−34658Ad5E4orf6:nt33316−34197。
【図10G】図10A−10HはChAd63ΔE1,3,4,Ad5E4orf6,NSmut(配列17)の核酸配列を示す。野生型ゲノムの欠失の組合せは、nt455からnt3421まで(2967bp)のEl欠失、nt27207からnt31778まで(4582bp)のE3欠失、nt33825からnt36215まで(2390bp)のE4欠失である。様々な領域は以下の場所に存在する:ChAd63左側ITR+パッケージングシグナル:nt1−454HCMVプロモーター:nt458−1248コザックコンセンサス配列:nt1254−1259HCV NS3−5B(BK菌株):nt1260−7214TAAターミネーター:nt7215−7217BGHポリA:nt7227−7447ChAd63主鎖:nt7458−34658Ad5E4orf6:nt33316−34197。
【図10H】図10A−10HはChAd63ΔE1,3,4,Ad5E4orf6,NSmut(配列17)の核酸配列を示す。野生型ゲノムの欠失の組合せは、nt455からnt3421まで(2967bp)のEl欠失、nt27207からnt31778まで(4582bp)のE3欠失、nt33825からnt36215まで(2390bp)のE4欠失である。様々な領域は以下の場所に存在する:ChAd63左側ITR+パッケージングシグナル:nt1−454HCMVプロモーター:nt458−1248コザックコンセンサス配列:nt1254−1259HCV NS3−5B(BK菌株):nt1260−7214TAAターミネーター:nt7215−7217BGHポリA:nt7227−7447ChAd63主鎖:nt7458−34658Ad5E4orf6:nt33316−34197。
【図11】IFN−γ ELISPOTによって測定した免疫応答の時間経過を、いずれか所与の時点に種々のHCV NSペプチドプールで観察した応答の和として表す。
【図12】pV1JnsNSOPTmut 3a−1bの種々の構成要素を示す。
【図13】キメラHCVポリペプチドをコードするpV1JnsNSOPTmut 3a−lb発現カセットの遺伝子構造を示す。様々な領域は以下の場所に存在する:−ヒトCMVプロモーター:318−905;−イントロンA:1040−1865;−コザック配列:1885−1894;−HCV MetNS3−NS4A(遺伝子型3a):1891−3960;−HCV MetNS3−NS5BOPTmut(遺伝子型1b):3961−9915;−TAAAターミネーター:9916−9919;−BGHポリA:9965−10182。
【図14】キメラプラスミド(pV1Jns−NSOPTmut 3a−1b)またはpV1Jns−NSOPTmutで免疫感作した動物(CD1マウス)体内のHCV 1bおよび3a由来のNS3タンパク質に応答してIFN−γを分泌するT細胞の数を示す(脾細胞1,000,000個あたりのスポット形成細胞として表す)。
【特許請求の範囲】
【請求項1】
第一HCV領域と第二HCV領域とを含み、
前記第一HCV領域が、第一のHCV菌株由来のHCV NS3−4Aに実質的に相似のHCV NS3−4Aアミノ酸配列を含み、
前記第二HCV領域が、第二のHCV菌株に由来のHCV NS3−NS4A−NS4B−NS5Aに実質的に相似のHCV NS3−NS4A−NS4B−NS5Aアミノ酸配列を含み、
前記第一領域のHCV NS3−4Aアミノ酸配列が前記第二領域のHCV NS3−4Aアミノ酸配列とは違っており、
前記第一領域が前記第二領域のアミノまたはカルボキシル側に配置されたHCVポリペプチドをコードするヌクレオチド配列を含む核酸。
【請求項2】
前記第一領域のHCV NS3−4Aアミノ酸配列が前記第一HCV菌株由来のHCV NS3−4Aに少なくとも85%の配列一致を有しており、前記第二領域のHCV NS3−NS4A−NS4B−NS5Aアミノ酸が前記第二菌株由来のHCV NS3−NS4A−NS4B−NS5Aに少なくとも85%の配列一致を有しており、さらに、前記第一領域のHCV NS3−4Aアミノ酸配列と前記第二領域のHCV NS3−4Aアミノ酸配列が少なくとも5%だけ違っている請求項1に記載の核酸。
【請求項3】
前記第二HCV領域はさらに、前記第二領域がNS3−NS4A−NS4B−NS5A−NS5Bを供給するようにHCV NS5Bを含んでおり、前記第二領域がさらに、インビボで自己をプロセシングして前記NS5Bを産生するために十分なプロテアーゼ活性を有しており、前記NS5Bタンパク質が酵素的に不活性である請求項2に記載の核酸。
【請求項4】
前記第一領域が前記第二領域のアミノ側に存在し、前記第一領域が自己をプロセシングしてHCV NS3を産生するために十分なプロテアーゼ活性を有している請求項3に記載の核酸。
【請求項5】
前記HCVポリペプチドが配列1に実質的に相似である請求項4に記載の核酸。
【請求項6】
前記HCVポリペプチドが配列1から成る請求項5に記載の核酸。
【請求項7】
前記ヌクレオチド配列が配列2の塩基1891−9915に実質的に相似である請求項5に記載の核酸。
【請求項8】
前記核酸が前記ヌクレオチド配列をコードする発現カセットを含む請求項1から7のいずれか一項に記載の核酸。
【請求項9】
前記核酸が、選択可能マーカーと、複製起点と、前記発現カセットに隣接する第一のアデノウイルス相同領域および第二のアデノウイルス相同領域とをさらに含むシャトルベクターであり、前記第一相同領域が野生型アデノウイルス領域の少なくとも右端に実質的に相同の少なくとも約100の塩基対を有しており、前記第二相同領域が野生型アデノウイルス領域の少なくとも左端に実質的に相同の少なくとも約100の塩基対を有している請求項8に記載の核酸。
【請求項10】
前記核酸が、選択可能マーカーと、複製起点と、E1欠失、E3欠失および場合によりE4欠失を含む組換えアデノベクターゲノムとを含むアデノウイルスゲノムプラスミドであり、前記発現カセットがE1またはE3欠失に配置されている請求項8に記載の核酸。
【請求項11】
前記組換えアデノベクターゲノムが、
a)配列3または9に実質的に相似のアミノ酸配列をもつファイバー領域、
b)配列5または11に実質的に相似のアミノ酸配列をもつヘキソン領域、および、
c)配列7に実質的に相似のアミノ酸配列をもつペントン領域
の少なくとも1つをコードしている請求項10に記載の核酸。
【請求項12】
前記核酸が発現ベクターである請求項8に記載の核酸。
【請求項13】
前記発現ベクターがプラスミドである請求項12に記載の核酸。
【請求項14】
前記プラスミドがさらに原核性複製起点と選択可能マーカーとを含む請求項13に記載の核酸。
【請求項15】
前記核酸が配列2の核酸に実質的に相似である請求項14に記載の核酸。
【請求項16】
前記核酸が配列2から成る請求項15に記載の核酸。
【請求項17】
前記発現ベクターが組換えアデノベクターであり、前記アデノベクターが組換えアデノウイルスゲノムを含む請求項12に記載の核酸。
【請求項18】
前記組換えアデノウイルスゲノムがE1欠失、E3欠失および場合によりE4欠失を含み、前記発現カセットがE1またはE3欠失に配置されている請求項17に記載の核酸。
【請求項19】
前記組換えアデノベクターゲノムが、
a)配列3または9に実質的に相似のアミノ酸配列をもつファイバー領域、
b)配列5または11に実質的に相似のアミノ酸配列をもつヘキソン領域、および、
c)配列7に実質的に相似のアミノ酸配列をもつペントン領域
の少なくとも1つをコードしている請求項18に記載の核酸。
【請求項20】
前記アデノウイルスゲノムが配列14または15の対応領域に実質的に相似である請求項19に記載の核酸。
【請求項21】
a)HCV NS3−NS4A−NS5Aを含むHCVポリペプチドをコードしている発現カセットと、
b)E1欠失、E3欠失および場合によりE4欠失を含むアデノウイルスゲノムと、
を含み、前記ゲノムが、
i)配列3または9に実質的に相似のアミノ酸配列をもつファイバー領域、
ii)配列5または11に実質的に相似のアミノ酸配列をもつヘキソン領域、および、
iii)配列7に実質的に相似のアミノ酸配列をもつペントン領域
の少なくとも1つをコードしており、
前記発現カセットがE1またはE3欠失に配置されている組換えアデノベクター。
【請求項22】
前記HCVポリペプチドがHCV NS3−NS4A−NS5A−NS5Bを含み、インビボで自己をプロセシングしてNS5Bタンパク質を産生するために十分な活性を有しており、前記NS5Bタンパク質が酵素的に不活性であり、前記アデノウイルスゲノムが、E4欠失と、配列13のヌクレオチド34601−35482に実質的に相似のAd5E4orf6配列の挿入とを含む請求項21に記載のベクター。
【請求項23】
前記ベクターが実質的に配列13または17である請求項21に記載のベクター。
【請求項24】
前記ベクターが配列13または17の核酸配列から成る請求項23に記載のベクター。
【請求項25】
請求項17に記載の組換えアデノウイルスゲノムを含み、前記組換えアデノウイルスゲノムによってコードされている組換えアデノウイルス粒子。
【請求項26】
請求項18に記載の組換えアデノウイルスゲノムを含み、前記組換えアデノウイルスゲノムによってコードされている組換えアデノウイルス粒子。
【請求項27】
請求項19に記載の組換えアデノウイルスゲノムを含み、前記組換えアデノウイルスゲノムによってコードされている組換えアデノウイルス粒子。
【請求項28】
請求項22に記載の組換えアデノウイルスゲノムを含み、前記組換えアデノウイルスゲノムによってコードされている組換えアデノウイルス粒子。
【請求項29】
請求項23に記載の組換えアデノウイルスゲノムを含み、前記組換えアデノウイルスゲノムによってコードされている組換えアデノウイルス粒子。
【請求項30】
請求項24に記載の組換えアデノウイルスゲノムを含み、前記組換えアデノウイルスゲノムによってコードされている組換えアデノウイルス粒子。
【請求項31】
a)請求項17に記載の組換えアデノウイルスゲノムを発現させるためにE1相補細胞を使用して粒子を製造する段階と、
b)前記粒子を実質的に精製する段階と、
を含む組換えアデノウイルス粒子の製造方法。
【請求項32】
a)請求項18に記載の組換えアデノウイルスゲノムを発現させるためにE1相補細胞を使用して粒子を製造する段階と、
b)前記粒子を実質的に精製する段階と、
を含む組換えアデノウイルス粒子の製造方法。
【請求項33】
a)請求項19に記載の組換えアデノウイルスゲノムを発現させるためにE1相補細胞を使用して粒子を製造する段階と、
b)前記粒子を実質的に精製する段階と、
を含む組換えアデノウイルス粒子の製造方法。
【請求項34】
a)請求項22に記載の組換えアデノウイルスゲノムを発現させるためにE1相補細胞を使用して粒子を製造する段階と、
b)前記粒子を実質的に精製する段階と、
を含む組換えアデノウイルス粒子の製造方法。
【請求項35】
a)請求項23に記載の組換えアデノウイルスゲノムを発現させるためにE1相補細胞を使用して粒子を製造する段階と、
b)前記粒子を実質的に精製する段階と、
を含む組換えアデノウイルス粒子の製造方法。
【請求項36】
a)請求項24に記載の組換えアデノウイルスゲノムを発現させるためにE1相補細胞を使用して粒子を製造する段階と、
b)前記粒子を実質的に精製する段階と、
を含む組換えアデノウイルス粒子の製造方法。
【請求項37】
治療有効量の請求項12に記載の核酸と医薬的に許容される担体とを含む医薬配合物。
【請求項38】
治療有効量の請求項14に記載の核酸と医薬的に許容される担体とを含む医薬配合物。
【請求項39】
治療有効量の請求項15に記載の核酸と医薬的に許容される担体とを含む医薬配合物。
【請求項40】
治療有効量の請求項17に記載の核酸と医薬的に許容される担体とを含む医薬配合物。
【請求項41】
治療有効量の請求項18に記載の核酸と医薬的に許容される担体とを含む医薬配合物。
【請求項42】
治療有効量の請求項19に記載の核酸と医薬的に許容される担体とを含む医薬配合物。
【請求項43】
治療有効量の請求項21から24のいずれか一項に記載の核酸と医薬的に許容される担体とを含む医薬配合物。
【請求項44】
治療有効量の請求項12に記載の核酸を患者に投与する段階を含む患者の治療方法。
【請求項45】
治療有効量の請求項14に記載の核酸を患者に投与する段階を含む患者の治療方法。
【請求項46】
治療有効量の請求項15に記載の核酸を患者に投与する段階を含む患者の治療方法。
【請求項47】
治療有効量の請求項17に記載の核酸を患者に投与する段階を含む患者の治療方法。
【請求項48】
治療有効量の請求項18に記載の核酸を患者に投与する段階を含む患者の治療方法。
【請求項49】
治療有効量の請求項19に記載の核酸を患者に投与する段階を含む患者の治療方法。
【請求項50】
治療有効量の請求項21から24のいずれか一項に記載の核酸を患者に投与する段階を含む患者の治療方法。
【請求項1】
第一HCV領域と第二HCV領域とを含み、
前記第一HCV領域が、第一のHCV菌株由来のHCV NS3−4Aに実質的に相似のHCV NS3−4Aアミノ酸配列を含み、
前記第二HCV領域が、第二のHCV菌株に由来のHCV NS3−NS4A−NS4B−NS5Aに実質的に相似のHCV NS3−NS4A−NS4B−NS5Aアミノ酸配列を含み、
前記第一領域のHCV NS3−4Aアミノ酸配列が前記第二領域のHCV NS3−4Aアミノ酸配列とは違っており、
前記第一領域が前記第二領域のアミノまたはカルボキシル側に配置されたHCVポリペプチドをコードするヌクレオチド配列を含む核酸。
【請求項2】
前記第一領域のHCV NS3−4Aアミノ酸配列が前記第一HCV菌株由来のHCV NS3−4Aに少なくとも85%の配列一致を有しており、前記第二領域のHCV NS3−NS4A−NS4B−NS5Aアミノ酸が前記第二菌株由来のHCV NS3−NS4A−NS4B−NS5Aに少なくとも85%の配列一致を有しており、さらに、前記第一領域のHCV NS3−4Aアミノ酸配列と前記第二領域のHCV NS3−4Aアミノ酸配列が少なくとも5%だけ違っている請求項1に記載の核酸。
【請求項3】
前記第二HCV領域はさらに、前記第二領域がNS3−NS4A−NS4B−NS5A−NS5Bを供給するようにHCV NS5Bを含んでおり、前記第二領域がさらに、インビボで自己をプロセシングして前記NS5Bを産生するために十分なプロテアーゼ活性を有しており、前記NS5Bタンパク質が酵素的に不活性である請求項2に記載の核酸。
【請求項4】
前記第一領域が前記第二領域のアミノ側に存在し、前記第一領域が自己をプロセシングしてHCV NS3を産生するために十分なプロテアーゼ活性を有している請求項3に記載の核酸。
【請求項5】
前記HCVポリペプチドが配列1に実質的に相似である請求項4に記載の核酸。
【請求項6】
前記HCVポリペプチドが配列1から成る請求項5に記載の核酸。
【請求項7】
前記ヌクレオチド配列が配列2の塩基1891−9915に実質的に相似である請求項5に記載の核酸。
【請求項8】
前記核酸が前記ヌクレオチド配列をコードする発現カセットを含む請求項1から7のいずれか一項に記載の核酸。
【請求項9】
前記核酸が、選択可能マーカーと、複製起点と、前記発現カセットに隣接する第一のアデノウイルス相同領域および第二のアデノウイルス相同領域とをさらに含むシャトルベクターであり、前記第一相同領域が野生型アデノウイルス領域の少なくとも右端に実質的に相同の少なくとも約100の塩基対を有しており、前記第二相同領域が野生型アデノウイルス領域の少なくとも左端に実質的に相同の少なくとも約100の塩基対を有している請求項8に記載の核酸。
【請求項10】
前記核酸が、選択可能マーカーと、複製起点と、E1欠失、E3欠失および場合によりE4欠失を含む組換えアデノベクターゲノムとを含むアデノウイルスゲノムプラスミドであり、前記発現カセットがE1またはE3欠失に配置されている請求項8に記載の核酸。
【請求項11】
前記組換えアデノベクターゲノムが、
a)配列3または9に実質的に相似のアミノ酸配列をもつファイバー領域、
b)配列5または11に実質的に相似のアミノ酸配列をもつヘキソン領域、および、
c)配列7に実質的に相似のアミノ酸配列をもつペントン領域
の少なくとも1つをコードしている請求項10に記載の核酸。
【請求項12】
前記核酸が発現ベクターである請求項8に記載の核酸。
【請求項13】
前記発現ベクターがプラスミドである請求項12に記載の核酸。
【請求項14】
前記プラスミドがさらに原核性複製起点と選択可能マーカーとを含む請求項13に記載の核酸。
【請求項15】
前記核酸が配列2の核酸に実質的に相似である請求項14に記載の核酸。
【請求項16】
前記核酸が配列2から成る請求項15に記載の核酸。
【請求項17】
前記発現ベクターが組換えアデノベクターであり、前記アデノベクターが組換えアデノウイルスゲノムを含む請求項12に記載の核酸。
【請求項18】
前記組換えアデノウイルスゲノムがE1欠失、E3欠失および場合によりE4欠失を含み、前記発現カセットがE1またはE3欠失に配置されている請求項17に記載の核酸。
【請求項19】
前記組換えアデノベクターゲノムが、
a)配列3または9に実質的に相似のアミノ酸配列をもつファイバー領域、
b)配列5または11に実質的に相似のアミノ酸配列をもつヘキソン領域、および、
c)配列7に実質的に相似のアミノ酸配列をもつペントン領域
の少なくとも1つをコードしている請求項18に記載の核酸。
【請求項20】
前記アデノウイルスゲノムが配列14または15の対応領域に実質的に相似である請求項19に記載の核酸。
【請求項21】
a)HCV NS3−NS4A−NS5Aを含むHCVポリペプチドをコードしている発現カセットと、
b)E1欠失、E3欠失および場合によりE4欠失を含むアデノウイルスゲノムと、
を含み、前記ゲノムが、
i)配列3または9に実質的に相似のアミノ酸配列をもつファイバー領域、
ii)配列5または11に実質的に相似のアミノ酸配列をもつヘキソン領域、および、
iii)配列7に実質的に相似のアミノ酸配列をもつペントン領域
の少なくとも1つをコードしており、
前記発現カセットがE1またはE3欠失に配置されている組換えアデノベクター。
【請求項22】
前記HCVポリペプチドがHCV NS3−NS4A−NS5A−NS5Bを含み、インビボで自己をプロセシングしてNS5Bタンパク質を産生するために十分な活性を有しており、前記NS5Bタンパク質が酵素的に不活性であり、前記アデノウイルスゲノムが、E4欠失と、配列13のヌクレオチド34601−35482に実質的に相似のAd5E4orf6配列の挿入とを含む請求項21に記載のベクター。
【請求項23】
前記ベクターが実質的に配列13または17である請求項21に記載のベクター。
【請求項24】
前記ベクターが配列13または17の核酸配列から成る請求項23に記載のベクター。
【請求項25】
請求項17に記載の組換えアデノウイルスゲノムを含み、前記組換えアデノウイルスゲノムによってコードされている組換えアデノウイルス粒子。
【請求項26】
請求項18に記載の組換えアデノウイルスゲノムを含み、前記組換えアデノウイルスゲノムによってコードされている組換えアデノウイルス粒子。
【請求項27】
請求項19に記載の組換えアデノウイルスゲノムを含み、前記組換えアデノウイルスゲノムによってコードされている組換えアデノウイルス粒子。
【請求項28】
請求項22に記載の組換えアデノウイルスゲノムを含み、前記組換えアデノウイルスゲノムによってコードされている組換えアデノウイルス粒子。
【請求項29】
請求項23に記載の組換えアデノウイルスゲノムを含み、前記組換えアデノウイルスゲノムによってコードされている組換えアデノウイルス粒子。
【請求項30】
請求項24に記載の組換えアデノウイルスゲノムを含み、前記組換えアデノウイルスゲノムによってコードされている組換えアデノウイルス粒子。
【請求項31】
a)請求項17に記載の組換えアデノウイルスゲノムを発現させるためにE1相補細胞を使用して粒子を製造する段階と、
b)前記粒子を実質的に精製する段階と、
を含む組換えアデノウイルス粒子の製造方法。
【請求項32】
a)請求項18に記載の組換えアデノウイルスゲノムを発現させるためにE1相補細胞を使用して粒子を製造する段階と、
b)前記粒子を実質的に精製する段階と、
を含む組換えアデノウイルス粒子の製造方法。
【請求項33】
a)請求項19に記載の組換えアデノウイルスゲノムを発現させるためにE1相補細胞を使用して粒子を製造する段階と、
b)前記粒子を実質的に精製する段階と、
を含む組換えアデノウイルス粒子の製造方法。
【請求項34】
a)請求項22に記載の組換えアデノウイルスゲノムを発現させるためにE1相補細胞を使用して粒子を製造する段階と、
b)前記粒子を実質的に精製する段階と、
を含む組換えアデノウイルス粒子の製造方法。
【請求項35】
a)請求項23に記載の組換えアデノウイルスゲノムを発現させるためにE1相補細胞を使用して粒子を製造する段階と、
b)前記粒子を実質的に精製する段階と、
を含む組換えアデノウイルス粒子の製造方法。
【請求項36】
a)請求項24に記載の組換えアデノウイルスゲノムを発現させるためにE1相補細胞を使用して粒子を製造する段階と、
b)前記粒子を実質的に精製する段階と、
を含む組換えアデノウイルス粒子の製造方法。
【請求項37】
治療有効量の請求項12に記載の核酸と医薬的に許容される担体とを含む医薬配合物。
【請求項38】
治療有効量の請求項14に記載の核酸と医薬的に許容される担体とを含む医薬配合物。
【請求項39】
治療有効量の請求項15に記載の核酸と医薬的に許容される担体とを含む医薬配合物。
【請求項40】
治療有効量の請求項17に記載の核酸と医薬的に許容される担体とを含む医薬配合物。
【請求項41】
治療有効量の請求項18に記載の核酸と医薬的に許容される担体とを含む医薬配合物。
【請求項42】
治療有効量の請求項19に記載の核酸と医薬的に許容される担体とを含む医薬配合物。
【請求項43】
治療有効量の請求項21から24のいずれか一項に記載の核酸と医薬的に許容される担体とを含む医薬配合物。
【請求項44】
治療有効量の請求項12に記載の核酸を患者に投与する段階を含む患者の治療方法。
【請求項45】
治療有効量の請求項14に記載の核酸を患者に投与する段階を含む患者の治療方法。
【請求項46】
治療有効量の請求項15に記載の核酸を患者に投与する段階を含む患者の治療方法。
【請求項47】
治療有効量の請求項17に記載の核酸を患者に投与する段階を含む患者の治療方法。
【請求項48】
治療有効量の請求項18に記載の核酸を患者に投与する段階を含む患者の治療方法。
【請求項49】
治療有効量の請求項19に記載の核酸を患者に投与する段階を含む患者の治療方法。
【請求項50】
治療有効量の請求項21から24のいずれか一項に記載の核酸を患者に投与する段階を含む患者の治療方法。
【図1】


【図2A】

【図2B】

【図2C】

【図3】

【図4A】

【図4B】

【図4C】

【図4D】

【図4E】

【図4F】

【図4G】

【図4H】
【図4I】

【図4J】

【図5A】

【図5B】

【図5C】

【図5D】

【図5E】

【図5F】

【図5G】

【図5H】

【図6A】

【図6B】

【図6C】

【図6D】

【図6E】

【図6F】

【図6G】

【図6H】

【図7A】

【図7B】

【図7C】

【図7D】

【図7E】

【図7F】

【図7G】

【図7H】

【図8】
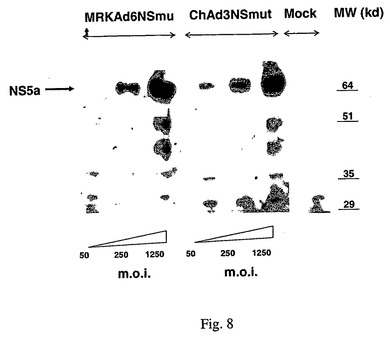
【図9】
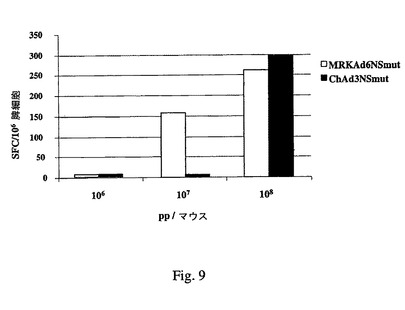
【図10A】

【図10B】

【図10C】

【図10D】

【図10E】

【図10F】

【図10G】

【図10H】

【図11】
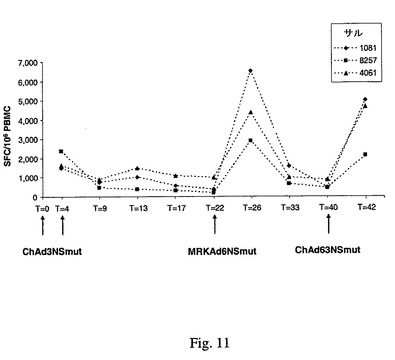
【図12】
【図13】
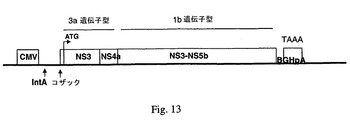
【図14】
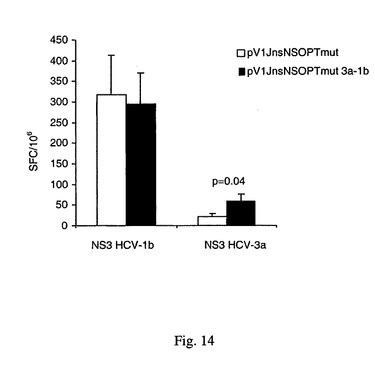
【図2A】
【図2B】
【図2C】
【図3】
【図4A】
【図4B】
【図4C】
【図4D】
【図4E】
【図4F】
【図4G】
【図4H】
【図4I】
【図4J】
【図5A】
【図5B】
【図5C】
【図5D】
【図5E】
【図5F】
【図5G】
【図5H】
【図6A】
【図6B】
【図6C】
【図6D】
【図6E】
【図6F】
【図6G】
【図6H】
【図7A】
【図7B】
【図7C】
【図7D】
【図7E】
【図7F】
【図7G】
【図7H】
【図8】
【図9】
【図10A】
【図10B】
【図10C】
【図10D】
【図10E】
【図10F】
【図10G】
【図10H】
【図11】
【図12】
【図13】
【図14】
【公表番号】特表2008−543295(P2008−543295A)
【公表日】平成20年12月4日(2008.12.4)
【国際特許分類】
【出願番号】特願2008−516223(P2008−516223)
【出願日】平成18年6月13日(2006.6.13)
【国際出願番号】PCT/EP2006/005697
【国際公開番号】WO2006/133911
【国際公開日】平成18年12月21日(2006.12.21)
【出願人】(501209427)イステイチユート・デイ・リチエルケ・デイ・ビオロジア・モレコラーレ・ピ・アンジエレツテイ・エツセ・ピー・アー (90)
【Fターム(参考)】
【公表日】平成20年12月4日(2008.12.4)
【国際特許分類】
【出願日】平成18年6月13日(2006.6.13)
【国際出願番号】PCT/EP2006/005697
【国際公開番号】WO2006/133911
【国際公開日】平成18年12月21日(2006.12.21)
【出願人】(501209427)イステイチユート・デイ・リチエルケ・デイ・ビオロジア・モレコラーレ・ピ・アンジエレツテイ・エツセ・ピー・アー (90)
【Fターム(参考)】
[ Back to top ]